/



























































































































































































































































































































































































































































































































































































Автор: Зитте П. Вайлер Э.В. Кадерайт Й.В. Брезински А.Б. Кёрнер К.
Теги: ботаника
ISBN: 978-5-7695-2741-8
Год: 2007




Текст
П.Зитте, Э. В. Вайлер,
W
И.В.Кадерайт, А. Брезински, К. Кёрнер
Ботаника
Эволюция и систематика
Lehrbuch der
Botanik
fur Hochschulen
Begrundet von
E. Strasburger • F. Noll
H. Schenck A. F. W. Schimper
35. Auflage
neubearbeitet von
Peter Sitte
Elmar W.Weiler
Joachim W. Kacereit
Andreas Bresinsky
Christian Komer
Spektrum Akaaemischer Verlag Heidelberg • Berlin
Ботаника
Учебник для вузов
На основе учебника
Э. Страсбургера, Ф.Нолля, Г. Шенка, А. Ф. В.Шимпера
35-е издание
Издание переработано
П.Зитте, Э. В. Вайлером, Й. В. Кадерайтом,
А. Брезински, К. Кёрнером
ACADEMIA
Москва
Издательский центр «Академия»
2007
П.ЗИТТЕ, Э. В. ВАЙЛЕР. Й. В. КАДЕРАЙТ, А.БРЕЗИНСКИ, К. КЁРНЕР
Ботаника
Учебник для вузов
На основе учебника
Э. Страсбургера, Ф. Нолля, Г. Шенка, А. Ф. В. Шимпера
В четырех томах
Перевод с немецкого
Под редакцией А. Г. Е~еневского, В. Н. Павлова, А. К. Тимонина, И И Сидоровой, В В.Чуба
academ’a
Москва
Издательский центр «Академия»
2007
УДК 58(075.8)
ББК 28.5я73
Б86
Рецензенты:
проф. Г. Н. Огуреева (Московский государственный университет им. М. В. Ломоносова); чл.-кор. РАН, проф. Р. В. Камелин
Ботаника. Учебник для вузов : в 4 т.: / П. Зитте, Э. В. Вайлер, Й. В. Каде-Б86 райт, А.Брезински, К. Кёрнер ; на основе учебника Э. Страсбургера [и др.] ; пер. с нем. Е. Б. Поспеловой, К. Л.Тарасова, Н. В.Хмелевской. — М.: Издательский центр «Академия», 2007. — 576 с.
Т. 3. Эволюция и систематика / под ред. А. К.Тимонина, И. И. Сидоровой
ISBN 978-5-7695-2741-8 (рус.)
ISBN 978-5-7695-2746-3 (Т. 3) (рус.)
ISBN 3-8274-1010-Х (Elsevier GmbH)
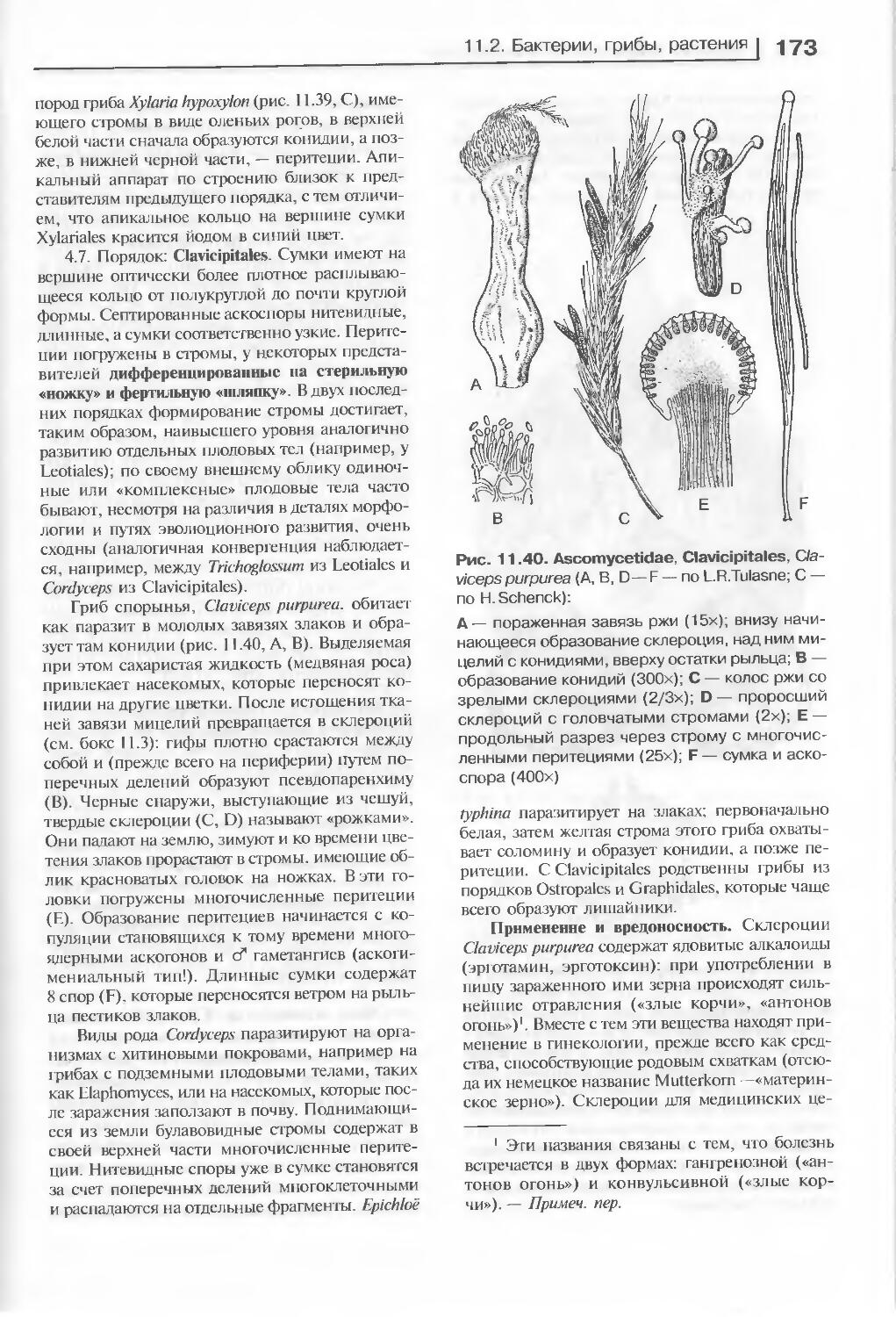
Многократно переиздававшийся в Германии, переведенный на многие языки учебник «Ботаника» Э. Страсбургера уникален своей исторической преемственностью, широтой охвата материала и ультрасовременностью приводимых данных: каждое издание, в том числе и последнее, полностью перерабатывается и обновляется по сравнению с предыдущим.
На русском языке учебник выходит в четырех томах. Третий том посвящен эволюции, систематике и филогении растений.
Для студентов высших учебных заведений. Может быть полезен преподавателям и научным работникам.
УДК 58(075.8)
ББК 28.5я73
Данное произведение является переводом Strasburger Lehrbuch der Botanik. 35-е издание.
Elsevier GmbH и ИЦ «Академия» не несут ответственности за ущерб, который может быть причинен в результате использования материалов, содержащихся в книге, за их достоверность, а также за возможные нарушения авторских прав третьих лиц и права на неприкосновенность частной жизни.
Авторские права П. Зитте, Э. В. Вайлера, Й. В. Кадерайта, А. Брезински, К. Кёрнера защищены в этом и во всех последующих изданиях.
Оригинал-макет данного издания является собственностью Издательского центра «Академия», и его воспроизведение любым способом без согласия правообладателя запрещается
ISBN 978-5-7695-2741-8 (рус.) ©2002 Elsevier GmbH, Spektrum Akademischer Verlag, Heidelberg ISBN 978-5-7695-2746-3 (T. 3) (рус.) © Издание на русском языке, перевод на русский язык, ISBN 3-8274-1010-Х (Elsevier GmbH) оформление. Издательский центр «Академия», 2007
Предисловие к 35-му немецкому изданию
Учебник «Ботаника» для высшей школы (35-е издание) Эдуарда Страсбургера вступает в новую фазу своей более чем 100-летней истории. Два соавтора, Хуберт Циглер и Фридрих Эрендорфер, активно участвовавшие соответственно более двадцати и тридцати лет в создании данного труда, выбыли из страсбургерской команды по возрасту. Мы, а также большая армия читателей курируемых Циглером и Эрендорфером разделов благодарны им за их выдающиеся достижения. Теперь авторами этих разделов стали более молодые эксперты. Часть материала по физиологии переработана и дополнена главой по алле-лофизиологии Эльмаром В. Вайлером (Бохум), который, кроме того, обновил главу по молекулярным основам в структурной части. Материал в области эволюции и методы систематики переработал Йоахим В. Ка-дерайт (Майнц), взявший на себя также обзор семенных растений и историю растений. Наконец, Кристианом Кёрнером (Базель) в значительной мере заново составлены главы по экологии (экология растений, растения в биосфере, популяционная и вегетативная экология).
Новая команда старалась соответствовать задачам академического преподавания ботаники — представить на хорошем уровне все существенные основы этого предмета, а также проинформировать читателей о достаточно достоверных новых результатах исследований и их применении (например, трансгенные растения или биомасса и продуктивность). Неизменной осталась и цель — достойно отразить все разделы науки о растениях и смежные области. В соответствии с этим в учебнике уделяется большое внимание не только фотоавтотрофным, «зеленым», организмам, но и всем тем гетеротрофным группам, ко
торые происходят от автотрофных или важны для понимания филогении, физиологии и экологии автотрофов (прокариоты, грибы). Кроме того, мы стремились улучшить и так богатый иллюстративный материал (например, с помощью четырехцветных иллюстраций, унификации всех химических формул), а также структуру учебника (например, используя численное членение, обзоры содержания в начале каждой главы, ссылки, вставки по важным специальным темам и, наконец, перечни вставок и таблиц в оглавлении). Ссылки на новую литературу делают возможным углубление в предмет там, где более подробное обсуждение выходит за рамки учебника. Многочисленные ссылки экономят время студента на обращение к указателю, но в основном обозначают связи между дисциплинами и создают систему материала, вынужденно предложенного линейно. Наконец, набор некоторого материала мелким шрифтом позволяет «перепрыгнуть» те места в книге, содержание которых менее важно, прежде всего для начинающего. Поэтому учебник представлен как бы в двух вариантах: один для младших курсов, другой для старших.
Сердечно благодарим тех, кто помогал нам конструктивной критикой и предоставлением иллюстраций. (Их имена частично раскрываются в благодарностях, помещенных в начале текстовой части, и в подписях к рисункам. Иллюстрации, авторы которых не названы, делали мы сами.) В работе над книгой нам основательно помогал доктор Андреас Буль (Халле). Решающей помощью было для нас необычное приглашение к работе лектора Инги Айкен (Штутгарт) и госпожи Эльке Литтманн из производственного отдела издательства. Издательство Spektrum Verlag, возглавля
8 | ПРЕДИСЛОВИЕ К 35-МУ НЕМЕЦКОМУ ИЗДАНИЮ
емое доктором Ульрихом Г. Мольтманном, несмотря на давление постоянно обостряющейся конкуренции, не только полностью поддержало новое оформление традиционного труда, но и со своей стороны энергично продвигало его вперед. Обновление в этой области касается пакетных предложений, благодаря которым в Германии книга может быть получена вместе с уже зарекомендовавшей себя брошюрой «Помощь в обучении. Ботаника», а также со «Словарем по ботанике» Г. Вагенитца (Гёттинген)
на CD-ROM и/или постером «Систематика: Ботаника» А.Брезински и Й. В.Каде-райта.
Мы желаем всем читателям успешно освоить с помощью учебника «Ботаника» эту науку, важность которой растет вместе с ее комплексностью.
Фрайбург, Бохум, Майнц, Регенсбург и Базель, февраль 2002
Авторы
Предисловие к 1 -му немецкому изданию
Авторы учебника «Ботаника» уже много лет работают доцентами ботаники в университете г. Бонна. Они постоянно обмениваются научными мыслями и методическими предложениями. Теперь авторы пытаются совместно изложить в книге свой научный опыт, накопленный в течение жизни. Материал учебника был распределен так: Эдуард Страсбургер взял на себя обязанность по написанию введения и морфологии, Фритц Нолль — физиологии, Генрих Шенк— материала о споровых растениях, А. Ф. В. Шимпер — явнобрачных растениях.
Даже если каждый автор несет научную ответственность только за написанную им часть, единообразие всех частей обеспечивалось постоянным согласованием. Поэтому книгу, несмотря на то что у нее есть несколько авторов, можно считать единым произведением.
Учебник «Ботаника» предназначен для студентов вузов и должен прежде всего
пробудить у них научный интерес, стимулировать научные знания и выводы. Но одновременно авторы обращают внимание на практические требования обучения и удовлетворяют потребности медиков и фармацевтов. Так, медик сможет из цветных иллюстраций получить сведения о ядовитых растениях, важных для него, фармацевт — найти в книге необходимые указания на лекарственные растения и наркотики.
Многочисленные иллюстрации были подготовлены в основном авторами учебника, ряд иллюстраций имеют других авторов.
Нельзя недооценить любезность господина издателя, который не жалел расходов на цветные иллюстрации в тексте и прилагал все усилия, чтобы придать книге законченный вид.
Бонн, июль 1894 Авторы
Эдуард Страсбургер
*01.02.1844, Варшава — 119.05.1912, Бонн основоположник учебника для высшей школы «Ботаника»
После изучения естественных наук в Париже, Бонне и Йене, а также написания докторской диссертации в Йене Эдуард Страсбургер защитил докторскую диссертацию в 1867 году в Варшаве и в 1869 году в возрасте 25 лет был приглашен в качестве профессора ботаники в университет Йены, а в 1881 году — в университет Бонна. Под его руководством Ботанический институт в Поппельсдорфском замке вошел в список международных центров ботаники. Здесь Э. Страсбургер со своими сотрудниками Ф. Ноллем, Г. Шенком и А. Ф.В. Шимпером написал в 1894 году учебник для высшей школы «Ботаника» (раньше коротко называемый «Боннский учебник»). Этот учебник вместе с много
кратно издававшимся «Малым ботаническим практикумом» и более обширным «Ботаническим практикумом» отразили бо-таническо-микроскопическую практику того времени в вузах. Исследовательская работа Страсбургера в первую очередь была важна и для истории развития цитологии. Ученый установил, что процессы деления ядра (образование, расщепление и движение хромосом) у растений протекают так же, как у животных, т.е. одинаково у всех организмов (1875). Он впервые наблюдал у цветковых растений процессы оплодотворения и слияния мужского ядра с ядром яйцеклетки и сделал вывод, что клеточное ядро — важнейший носитель наследственных структур (1884).
Авторы учебника «Ботаника»
Учебник «Ботаника» был написан в 1894 году учеными-ботаниками, работавшими в Бонне: Эдуардом Страсбургером, Фритцем Ноллем, Генрихом Шенком, А. Ф. Вильгельмом Шимпером, и в последующее время ими, а также ниже названными авторами переиздание учебника было продолжено.
Введение и морфология, или структура:
1 — 11-е издания (1894—1911) — Эдуард Сграс-бургер
12 — 26-е издания (1913—1954)— Ганс Фиттинг
27 —32-е издания (1958—1983) — Дитрих фон Денффер
33 — 35-е издания (1991—2002) — Петер Зитге
Физиология:
1 -9-е издания (1894—1908) — Фритц Нолль 10— 16-е издания (1909—1923) — Людвиг Йост 17—21-е издания (1928—1939)— Германн Зирп 22 —30-е издания (1944—1971) — Вальтер Шумахер
31 -34-е издания (1978—1998) — Хуберт Циглер 35-е издание (2002) — Эльмар
В.Вайлер
Эволюция и систематика, общие основы:
30—34-е издания (1971 —1998) — Фридрих
Эрендорфер
35-е издание (2002) — Йоахим
В. Кадерайт
Низшие растения:
1 — 16-е издания (1894—1923)— Генрих Шенк 17—28-е издания (1928—1962)— Рихард Хардер 29 — 31-е издания (1967—1978)— Карл Мэгде-фрау
32 —35-е издания (1983 — 2002) — Андреас Бре-зински
Семенные растения:
1—5-е издания (1894—1901)— А.Ф.В.Шимпер 6 — 19-е издания (1904—1936)— Георг Карстен 20—29-е изда- (1939—1967)— Франц Фирбас ния
30—34-е изда- (1971 — 1998) — Фридрих
ния Эрендорфер
35-е издание (2002) — Йоахим
Кадерайт
География растений, геоботаника или экология:
20—29-е издания (1939—1967) — Франц Фирбас
30—34-е издания (1971 —1998) — Фридрих
Эрендорфер
35-е издание (2002) — Кристиан
Кёрнер
Издания на иностранных языках
Английский:
Лондон: 1896,1902,1907,1911,1920,1930,1965, 1971,1975
Итальянский:
Милан: 1896,1913,1921,1928,1954,1965,1982,
2002
Польский:
Варшава: 1960, ND 1962,1967,1971, ND 1973
Испанский:
Барселона: 1923, 1935,1943,1953,1960,1974, 1986,1994
Сербско-хорватский:
Загреб: 1980, 1982,1988, ND 1991
Турецкий:
Стамбул: 1998
Хронология
ок. 300 до н.э.
1151-
1158
после 1530
1533 1583
1590
1665
1675
1682
1683
1694
1735
1779
1790
1793
1804
1805
1809
1822
1831
«Естественная история растений»: Теофрастос Эрезиос (371 -286 до н.э.)
Описание 300 лекарственных и сельскохозяйственных растений, пряностей и наркотиков «De plan-tis», «De arboribus»: Хильдегард фон Бинген
Старейшая «Книга трав»: Отто Брунфельс, Иеронимус Бок, Леонхарт Фукс
Первая кафедра ботаники в Падуе Первый общий учебник ботаники «De Plantis»: Андреа Чезальпино Изобретение микроскопа: Йоханнес и Захариас Янссен Открытие клеточного строения тканей «Micrographia»: Роберт Гук «Anatome plantarum»: Марселло Мальпиги
«Анатомия растений»: Неемия Грю Первое изображение бактерий: Антониус ван Левенгук
Наличие полов у растений: Рудольф Якоб Камерариус Бинарная номенклатура. «Systema naturae»; «Species plantarum» (1753): Карл фон Линней (Каро-лус Линнеус, 1707 — 1778): Открытие фотосинтеза: Ян Инген-хаусп
«Метаморфоз растений»: Иоганн Вольфганг фон Гёте
Основание экологии цветков: Кристиан Конрад Шпренгель Открытие растительного газообмена: Николя Теодор де Соссюр Основание географии растений: Александр фон Гумбольдт «Philosophic zoologique», учение о происхождении видов: Жан Батист де Ламарк
Открытие осмоса: Анри Дютроше Открытие клеточного ядра: Роберт Браун
1835 Деление клетки у растении: Гуго
фон Моль
1838 Основание клеточной теории:
Маттиас Якоб Шляйден совместно с анатомом и физиологом Теодором Шванном
1839 Минеральное питание растений, опровержение гумусовой теории: Юстус фон Либих
1846 Термин «протоплазма»: Гуго фон Моль
1851 Сходства в смене поколений у растений: Вильгельм Хофмайстер
1855 «Omnis cellula е cellula»: Рудольф
Вирхов
1858 Мицеллярная теория: Карл
Нэгели
1859 «Происхождение видов...»:
Чарлз Дарвин
1860 Водная культура: Юлиус Сакс
1860 Опровсрже! i не теории абиогене-
за: Херманн Хоффманн, Луи Пастер
1862 Крахмал как продукт фото-
синтеза: Юлиус Сакс
1866 «Опыты с растительными гибри-
дами», правила наследования: Грегор Мендель (1822—1884)
1866 Концепция экологии: Эрнст Геккель
1867—1869 Двойственная природа лишайников: Симон Швенденер
1869 Открытие ДНК, фосфорсодержащий «нуклеин»: Фридрих Мишер
1875 Открытие деления ядра у растений: Эдуард Страсбургер
1877 «Осмотические исследования»:
Вильгельм П феффер
1883 Пластиды как самореплицирую-
щиеся органеллы, возможные потомки внутриклеточных симбионтов: Андреас Ф. В. Шимпер, Ф. Шмитц
ХРОНОЛОГИЯ | -|3
1884
1884
1884
1887
1888
1894
1897
1900
1901
1902
1904
1909
1910
1913
1913
1916
1917
1920
после 1920 1922
1925
1926
«Физиологическая анатомия растений»: Готглиб Хаберландт «Сравнительная морфология и биология грибов, миксомицетов и бактерий»: Антон де Бари Открытие слияния ядер при оплодотворении цветковых растений: Эдуард Страсбургер Мейоз: Теодор Бовери Функция корневых клубеньков бобовых: X. Хелльригель и X. Вил fa-фарт, М.В. Байеринк, А. Празмов-ски
Первое издание учебника Эдуарда Страсбургера «Ботаника» Сбраживание с помощью бесклеточного дрожжевого экстракта: Эдуард Бухнер
Переоткрытие менделевских правил наследования: Эрих Чермак фон Сейсенегт, Карл Корре нс и Гуго де Фриз
Мутационная теория: Гуго де Фриз Симбиогенез. пластиды как потомки цианобактерий: Константин Мережковский
Концепция экосистем: Т.А.Тэнсли Пластиды как носители наследственных структур: Карл Корре нс и Эрвин Баур
Полиплоидия: Эдуард Страсбургер Объяснение структуры хлорофилла: Рихард Вилльштеттер «Микрохимия растений»: Ганс Молиш
Экспериментальное производство полиплоидного томата: Ганс Винклер
Математика формообразования, аллометрия: «О росте и форме»: Д'Арсп В. Томпсон
Первые систематические исследования фотопериодизма: В. Гарнер и X. А. Аллард Макромолекулярная химия: X. Штаудингер
Генотипическая концепция растительной адаптации: Г.Турессон Двухслойная модель биомембран: Э. Гортер, Ф. Грендель Доказательство образования фактора роста (гиббереллин) Gibberella fujikuroi: Э. Куросава
1928 Открытие пенициллина: А. Фле-
минг
1928 Трансформация пневмококков.
Ф. Гриффит
1928 Эу- и гетерохроматин: Э.Хайтц
1930 Теория флоэмного транспорта:
Э. Мюнх
1930 Экспериментальный ресинтез
аллотетраплоидного гибридного вида Galeopsis tetrahit: А. Мюнтцинг
1930—1934 Физический анализ транспирации, транспирационные сопротивления: А. Зейбольд
1930—1950 Синтез генетики и эволюционной теории: Р. А. Фишер, Дж. С. Холдейн, Ф. Г. Добжан-ски, Э. Майр, Д. С. Хаксли, Дж. Г. Симпсон, Дж.Л. Стеббинс
1931 Фотосинтетический О2 происхо-
дит из воды: К. ван Нил
1931 Первый электронный микро-
скоп: Э.Руска; с 1939 г. коммерческое изготовление «сверхмикроскопов» по Э.Руске и Б.фон Боррису на заводе Siemens, по X. Малю и др. на заводе AEG
1933 Теория клеточного дыхания: X.Виланд
1934 Концепция ниш сосуществова-
ния организмов: Г. Ф. Гаузе
1935 Физиологические основы про-
изводства лесов: П.Бойсен-Йенсен
1935 Кристаллизация вируса табачной
мозаики: У. М. Стэнли
1935 Первое использование изотопов
для исследований обмена веществ: Р. Шёнхаймер и Д. Рит-тенберг
1937 Цикл лимонной кислоты:
X. А. Кребс
1937 Фотолиз воды с помощью изоли-
рованных хлоропластов: Р. Хилл
1937— 1943 «Сравнительная морфология высших растений»: В. Тролль
1938 «Субмикроскопическая морфология протоплазмы и ее производных»: А. Фрай-Висслинг
1938— 1947 Цитогенетическая биосистематика и эволюционные исследования у сосудистых растений: Э. Б. Бэбкок, Дж.Л. Стеббинс
14 | хронология
1939—1941 Центральная роль АТФ в энергетическом балансе клетки: Ф.Липманн
1939—1953 Изменение 13С у растений:
А. Нир и Э. Гульбрансон, X. К.Урей, М. Кальвин, И. В. Вайгель, П. Берчи
1941 Данные по живым экземплярам Metasequoia, которая до этого была известна только как ископаемая: Т. Кан, В. Вонг, К. By. Описание М. glyptostroboides в 1948 г. X. X. Ху и В. К. Ченг
1943 Доказательство генетического действия ДНК; О. Т. Эвери, К. М. МакЛеод и М. МакКарти 1947—1949 САМ-метаболизм:
В. и И. Боннер, М. Томас
1950 Прыгающие гены у кукурузы:
Барбара МакКлинток
1950 Кладистические методы био-
систематики: В. Хенних
1952 9 + 2-строение жгутиков:
Ирэне Мантон
1952 Доказательство трансдукции
наследственных структур у бактерий: Й.Ледерберг
1952 —1953 Методы фиксирования и тонких срезов для электронной микроскопии: К. Р. Портер, Ф. С. Сёстранд, Г. Э. Паладе
1952—1954 Фитохромная система:
X. А. Бортовик, С. Б. Хендрикс
1953 Производство аминокислот в условиях первичной земли: С. Миллер
1953 Двуспиральная модель ДНК:
Дж. Д. Уотсон, Ф.Х. К. Крик
1953 Закономерности использова-
ния света в растительных травостоях: М. Монси, Т. Сэки
1954 Фотофосфорилирование:
Д.Арнон
1954 Инфракрасный газоанализатор
для непрерывного измерения фотосинтеза: К. Эгле и А. Эрнст
1954 Выделение веществ с цитокининовым действием: Ф.Скруг, К. О. Миллер
1954 — 1966 Открытие С4-фотосинтеза: X. П. Корчак, Й. С. Карпилов, М. Д. Хач и К. Р. Слэйк
1955 Первое доказательство «self-asscmbly «/самосборки (у ВТМ): X. Френкель-Конрат и Р. Вилльямс
1957 Цикл фотосинтеза: М. Кальвин
1958 Экспериментальное доказа-
тельство полуконсервативной репликации ДНК: М. Мезель-сон и Ф. В. Шталь
1960 Выделение протопластов:
Э. К. Кокинг
1960—1961 Две световые реакции в эукариотических фототрофных организмах: Р. Хилл, Л.Н.М.Дуйсенс, Х.Т. Витт, Б. Кок
1961 Хемиосмагическая теория
образования АТФ: П.Д. Митчелл
1961 Генетический код объяснен: М. В. Ниренберг, Й. X. Матгэи и др.; универсальность кода: Ф.Х.К.Крик, Л.Барнетт, С. Бреннер и Р. Й. Уоттс-Тобин
1961 Модель регуляции активности
генов: Ф. Жакоб и Й. Моно
1961 «Жизнь, ее природа, проис-
хождение и развитие»: А. И. Опарин
1961 ДНК-гибридизация: С. Шпи-
гельман
1962 Фотодыхание: Н.Е. Толберт
1962 Хемотаксономия растений;
Р. Хегнауэр
1963 — 1964 Открытие абсцизовой кислоты: П. Ф. Уореинг и Ф. Т. Эд-дикотт
1964 Закономерности компартмен-тации у настоящих клеточных: Э. Шнепф
1964—1966 Гаплонтные культуры; С. Гупта и С. К. Магешвари
1965 Первый коммерческий растровый электронный микроскоп: К.Оатс, Cambridge Instr.
1968 Повторяющиеся последовательности в составе генов эукариот: Р. Й. Бриттен и Д. Э. Коне
1970 Про- и эукариоты как отдельные империи организмов: Р. Й. Станир
1970 Современная формулировка эндосимбиотической теории: Линн Маргулис
хронология | is
1970 Родословные древа последо- 1986
вательностей: Маргарет О.Дейхофф
1971 Получение высших растений из
протопластов листа: И.Такебе и Г. Мельхерс 1991
1971-1972 Сигнальные последовательности при транспорте протеинов через мембраны: Г.Блобел и Б.Доб- 1993
берштайн, К.Милыптайн
1972 Жидкостно-мозаичная модель биомембраны: С. Й. Сингер и Г. Л. Николсон
1974 Рестрикционные эндонуклеазы как инструменты анализа ДНК: В.Эрбер 1995
1976 Пэтч-кламп-техника для изучения ионных каналов в мембранах: Э. Неер, Б. Сакманн 1996
1977 Секвенирование ДНК: В. Гилберт, Ф. Сэнгер
1977 Особое положение архебакте-рий: К. Р. Вёзе, О. Кандлер
1977 Мозаичные гены, интрон/экзон-ная структура генов: С. Хогнесс,
Й.Л. Мандель, П.Чамбон 1999
1979 Arabidopsis thaliana как модельное растение для молекулярной биологии («растительная дрозофила»): К. Р. Сомервиль,
Э. М. Мейеровитн и др. 2000
1979 Agrobacterium tumefaciens как переносчик генов: Й.Шелл, М.ван Монтагю и др.
1980 Реконструкция гаметофита псилофитов: В. Реми
1982 Объяснение структуры бакте-
риального фотосинтетического реакционного центра: Й. Дай-зенхофер, X. Михель, Р. Хубер 2001
1982 «Рибозимы», РНК как энзимы: Т.Р.Чех, С. Альтман
1985 Полимеразная цепная реакция: К. Муллис
Первые полные секвенирования хлоропластной ДНК (/Vz-cotiana’. М.Сугиура с сотр.; Marchantia. К. Охайама с сотр.) Генетическое программирование образования цветка гомеотическими генами, «ABC-модель»: Э.М.Мейе-ровитц, Э. С. Коэн, X. Седлер Молекулярная кладограмма покрытосеменных на основе ДНК-последовательностей хлоропластного гена rbcL: М. Чейз с сотр.
Первые полные ДНК-последовательности геномов бактерий {Haemophilus influenza и Mycoplasma genitaliunr. И. К. Вентер с сотр.)
Первые полные ДНК-последовательности геномов архебакте-рий {Methanococcus jannaschii'. Й.К.Вентер) и эукариот (дрожжи, Saccharomyces cerevisiae: участвовали свыше ИХ) лабораторий)
Идентификация Amborellaceae как базальной группы покрытосеменных: С. Мэтьюс и М. Донохью; П.С. Солтис с сотр.; Й.-Л. Кью с сотр.
Первая полная ДНК-последовательность высшего растения резушки Таля Arabidopsis thali-апа\ The Arabidopsis Genome Initiative, участвовали 27 лабораторий в СШа, Европе и Японии
«Золотой рис»: первое введение пути биосинтеза (провитамина А) в особенно важную для человеческого питания растительную ткань, эндосперм риса, путем трансформации: И. Пот-рикус и П. Бейер
Предисловие к русскому изданию
Предлагаемая вниманию читателей книга Э. Страсбургера «Ботаника» впервые вышла в свет более 100 лет назад и быстро получила признание мировой научной общественности за фундаментальность и широту охвата материала, оригинальность и наглядность его представления, доступность изложения, став классическим пособием для многих поколений исследователей. Жизнь растений во всем многообразии ее проявлений, сложность структурно-функциональных взаимосвязей, обеспечивающих роль и место растений в биосфере, становились более понятными и зримыми после прочтения книги. Переиздание книги после кончины Э. Страсбургера поддерживается международными коллективами авторитетных ученых, которые, сохраняя принципы построения книги, наполняют ее новым фактическим материалом, поддерживают теоретический уровень изложения согласно времени переиздания.
Постоянное обновление издания обеспечило ему необычно долгую жизнь. И настоящее издание очень отличается от предыдущих: в переработке четырех основных разделов из шести приняли участие новые авторы. Перевод 35-го издания этой оригинальной книги, без сомнения, классической сводки по ботанике, должен стать полезным и необходимым пособием для русскоязычных читателей, изучающих ботанику как интегральную науку о жизни растений. Его нельзя рассматривать в качестве стандартного учебника. Это пособие для всех тех, кто выбрал для себя растительный мир как интересный объект исследования и хочет получить о нем новейшие научные представления, что важно и для начинающего ботаника, и для опытного натуралиста-исследователя, и особенно для студентов, аспирантов, преподавателей вузов.
Книга начинается с описания структуры и свойств воды и биохимии полимеров растительной клетки (нуклеиновых кислот, белков, полисахаридов, липидов), что является достаточно обоснованным и позволяет при изучении последующих разделов — цитологического, морфологических и анатомических — лучше понять принципы функциональной организации клетки и растительного организма. В разделе о строении клетки дается описание методологии ее изучения, приводятся современные представления о биохимии, физико-химии и работе мембран, о компартментации и клеточных компартментах, их эволюции.
Значительная часть книги — это описание физиологических функций растительного организма, их метаболических систем, составляющих основу жизнедеятельности и обеспечивающих существование организма в разнообразных условиях среды. Такие метаболические системы находятся в сложных взаимоотношениях друг с другом, и координация их функционирования во времени и пространстве представлена на современном уровне.
Обменные реакции в организме требуют значительных затрат энергии, источником которой служат универсальные для клеток бактерий, хлоропластов и митохондрий растений и животных процессы трансформации энергии. Во всех этих системах световая энергия, или энергия окисляемых субстратов, используется для организации электронного транспорта в мембране, сопряженного с созданием протонного градиента, который применяется для синтеза молекул АТФ. Поэтому нужно понять общие принципы организации элек-трон-транспортных цепей в сопрягающих мембранах и дать характеристику основным компонентам, участвующим в переносе
Предисловие | 17
электрона. Это возможно при внимательном изучении соответствующих глав. Важную роль в жизни растительного организма играют взаимодействия митохондрий и хлоропластов, во время которых между ними устанавливаются системы прямых и обратных связей, имеющих существенное значение для поддержания клетки как целостной функциональной единицы, гибко приспосабливающей интенсивность дыхания к действию различных эндогенных и экзогенных факторов. Важнейшей задачей для понимания сущности процессов жизнедеятельности растения является изучение современных представлений в области фотосинтеза как физиологической функции, составляющей основу биоэнергетики. Фундаментальные процессы фотосинтеза — структурно-функциональная организация фотосинтетического аппарата, молекулярная структура и физико-химические свойства пигментных систем, механизм первичных процессов преобразования энергии и представления о структуре и функционировании реакционных центров — подверглись в последние годы пе-реисследованию, что нашло отражение в соответствующих главах.
Растения — это компоненты биосферы, которые выполняют огромную геохимическую работу, обусловленную прежде всего их способом минерального питания и водообмена. За последние годы существенно изменились наши представления о поступлении ионов и воды в растение, значительно расширились знания о функциях элементов минерального питания. Большой объем материала о механизмах поступления ионов и воды в апопласт. системах их транспорта через мембраны, транспорте веществ на ближние и дальние расстояния и их круговороте по растению, изложенный в книге, позволяет понять роль этих процессов в формировании функциональноцелостной системы.
Рост и развитие растений представляют собой интегральные физиологические функции, в которых раскрываются генетические возможности организма, обеспечивающие адаптивный и репродуктивный потенциал вида. Изучение принципов гормональной, световой, термо- и других видов
регуляции роста и развития растений составляет важнейший раздел ботаники. Жизнь растительного организма протекает в постоянном взаимодействии как с абиотическими (физическими, химическими), так и с биотическими (другими растениями, животными, микроорганизмами) факторами внешней среды. И эти взаимодействия могут существенно модифицировать развитие растения и его потенции к воспроизведению. Очевидны возможные изменения в структуре ценозов, обусловленные гипервлиянием абиотических и биотических факторов. Эволюция выработала множество механизмов, позволяющих растению преодолеть неблагоприятные воздействия и сохранить потенциал размножения и распространения. Действие каждого стресс-фактора рассматривается как с позиций его повреждающего эффекта, так и с точки зрения ответных реакций растительного организма, направленных на формирование механизмов адаптации, которые позволяют преодолевать или избегать неблагоприятные воздействия.
Помимо текста несомненным достоинством книги является широкое использование многочисленных схем, таблиц и разнообразных иллюстраций: рисунков, микрофотографий высокого разрешения, карт и фотографий ландшафтной растительности. Можно отметить существенное увеличение объема раздела «Экология», включившего в этом издании все основные сведения современной науки о растительном покрове Земли, а также систематику растений в рамках системы, основанной на учете новейших молекулярно-генетических исследований родственных связей между различными таксонами. Разработанные ботаниками в последние годы подходы позволили значительно продвинуться в понимании принципов функционирования растительного организма или растительных сообществ как целостных биологических систем, что отражено в данной книге и несомненно будет интересно широкому кругу читателей.
Профессор Московского государственного университета им. М. В. Ломоносова —----------------------И. П. Ермаков
БИБЛаК)' А БгУТ
Предисловие научного редактора
Ботаническая систематика в естественных учебных курсах традиционно понимается как описание признаков таксонов крупных рангов (не ниже рода), т.е. фактически как описание биоразнообразия. Следует заметить, что учебная литература по зоологии не несет в заглавиях слова «систематика», а именуется соответственно «Зоология позвоночных», «Зоология беспозвоночных», «Энтомология» ит.д. Дело, конечно, не в заголовках, а в том, что задачи систематики не исчерпываются классификацией. Систематика помимо описания таксонов, а это ее важнейшая задача, должна включать разделы, которые могут быть объединены рубрикой «теоретическая систематика». Данный пробел в отечественной ботанической литературе недавно был до известной степени заполнен книгой Р. В. Камелина «Лекции по систематике растений» (2004), представляющей собой обработку лекционного курса. Хотя в книгах некоторые проблемы Р. В. Камелина и Э. Страсбургера неизбежно оказались общими (например, систематика и филогения), подходы к их решению и базовый материал весьма различны. Впрочем, вряд ли стоит еще раз подчеркивать, что «Теоретическая систематика» столь многогранна, что можно радоваться каждой новой публикации.
В предлагаемом вашему вниманию учебнике «Ботаника» удачно совмещаются оба глобальных раздела систематики: описание биоразнообразия и теоретические аспекты. Среди последних можно выделить описание методов нумерической систематики (фенетики) и принципов кладистики, бурно развивающейся у нас в стране особенно в последние три десятилетия. В учебнике большое внимание уделяется подходам к определению монофилии так
сонов. Здесь же встает вопрос о признании (или непризнании) парафилетических групп, что активно дискутируется в современной науке.
В отдельную главу выделена эволюция. В ней прежде всего изложены генетические аспекты систематики, механизмы изменчивости, различные варианты мутаций, рекомбинаций, даны представления о потоке генов, генетическом дрейфе. Все это подводит к проблемам видообразования, которое также рассматривается в учебнике. Однако некоторые понятия, такие как перепатрическое и парапатрическое видообразование, малоупотребительны среди отечественных специалистов-биологов. Особое внимание справедливо уделяется гибридизации и в конечном счете аллопо-липлоидии. Следует подчеркнуть важность данного раздела для лиц, стремящихся к глубокому изучению систематики растений.
Разделы главы «Эволюция» и теоретические разделы главы «Систематика» столь тесно связаны между собой, что их следовало бы объединить рубрикой «общая часть», предпослав ее «специальной части» — описанию таксонов.
Несколько замечаний по поводу «специальной части» учебника. Порядок расположения таксонов традиционен: бактерии—грибы—водоросли—мохообразные— папоротникообразные—голосеменные— покрытосеменные. Таксоны рассматриваются, как правило, не ниже ранга порядков (у покрытосеменных — семейств). Характеристика, по необходимости, максимально лаконична, однако содержит число родов и видов в семействе, а также географию, что в целом придает характеристике таксонов определенность и информативность. Систематика покрытосемен
Предисловие I -| g
ных строится в соответствии с известным филогенетическим древом по генно-молекулярным данным (Чейз и др., 1993). Это отражает наиболее распространенную в наше время точку зрения на Amborella как анцестральное цветковое растение и на Acorus как однодольное, сестринское по отношению ко всем однодольным. Следует заметить, что авторы 35-го издания учебника «Ботаника» прекрасно знакомы с современной научной литературой. Это ощущение возникает при ознакомлении с самыми разными группами. Конечно, при таком массиве таксонов и признаков невозможно избежать неточностей и даже ошибок. Кроме того, в случаях, когда возможна
различная трактовка природы органов (например, цветок и зародыш злаков), автор вправе придерживаться своей концепции, хотя она может разделяться лишь немногими.
В заключение следует отметить, что классический учебник Э. Страсбургера всегда пользовался высоким авторитетом у нашей ботанической общественности, с интересом встречавшей каждое его издание. Нет сомнения, что перевод книги на русский язык существенно расширит читательскую аудиторию.
Доктор биологических наук, профессор А. Г. Еленевский
Г>
(se ijiopilosa)
oriidiunp
ict
is.
tipi)
inatae)
I MecowOjpsi» pcutinulptt^. ^Meconopsis dtlavay^ =|| I Meconopsis horrik^hf^/ \: Meconopsis aculeatja-^ !
^;(sect
?Cj^4econel la) feonella)
Месой|Ж
urn)
"• J '>' tsrs
ipsis s
tf-
л
•Л-?
| Meconnpsin in \Meeonvpsis ptin^pea •
—I Mteconopsis disc
Meeonopsis
iMectfniy 1|
Arpemone mexieana lisehst'noljtia caltfnrnica
- Pteridophyllum. ract'm-osum
•lia
Введение
Поразительное разнообразие организмов, живших в прошлом и живущих в настоящее время, с их самыми различными деталями строения и многообразными функциями возникло в течение истории Земли в ходе эволюции.
Глава 10 «Эволюция» и глава 11 «Систематика» посвящены этому эволюционному процессу и примерам организменного разнообразия, появившегося в результате данного процесса.
В главе 10 показано, как взаимодействие генетических изменений и естественного отбора ведет к изменению генетического состава следующих одно за другим поколений, которое в конечном счете приводит к возникновению новых видов. При этом естественный отбор является результатом взаимоотношений индивидуума с его абиотической и биотической средой обитания. Отношения организмов и их среды подробно представлены также в экологическом разделе учебника.
Хотя процесс эволюции непрерывен и потому происходит и в настоящее время, все же многие его составляющие не поддаются непосредственному наблюдению, и их приходится объяснять, исходя из свойств ныне живущих организмов. Согласно эволюционной теории, считается, что процессы, наблюдаемые в настоящее время или
выявленные на основании изучения современных организмов, действовали и в прошлом подобным образом и что иные процессы в то время не оказывали принципиального влияния.
Глава 11 показывает, как могут быть систематизированы современные (а частично также и жившие ранее) организмы. Важнейшей целью систематики является установление порядка, т. е. составление системы, которая отражает родственные связи между организмами и, соответственно, их происхождение (филогению). При этом систематика использует большое количество признаков, из которых особенно важна информация, содержащаяся непосредственно в наследственном материале. Одновременно систематике требуются также сложные методы, чтобы конструировать филогенетические деревья (эволюционные деревья), прежде всего на основе данных, полученных при изучении ныне живущих организмов.
Поскольку процесс формирования филогенетического разнообразия происходил в далеком прошлом и потому — как уже сказано выше при упоминании некоторых аспектов эволюционного процесса — недоступен для непосредственного наблюдения, реконструированные филогенетические деревья всегда гипотетичны.
<1
Разнообразие организмов, живших в прошлом и живущих в настоящее время, возникло в процессе эволюции. Поэтому систематика старается расположить возникшие таким образом организмы согласно их предполагаемому родству и представить в виде филогенетического дерева (дендрограммы). Хотя систематик использует при этом все признаки, наблюдающиеся у организмов, особая важность в настоящее время придается анализу содержащейся в ДНК информации. Это иллюстрируется примером. Определение последовательности определенных генов в ДНК различных маков (Papaver) и близких к ним родов (Meconopsis, Roemeria, Stylomecori) позволяет составить филогенетическое дерево, представленное на рисунке.
Представленная здесь система не только охватывает многообразие растений (Glaueobionta, Rhodobionta — красные водоросли, Chlorobionta — зеленые растения),
но и включает также прокариотические организмы (Bacteria, Archaea), слизевики (Acrasiobionta, Heterokontobionta) и хитиновые грибы (Mycobionta).
Благодарности
Раздел систематики бактерий, грибов и низших растений подготовлен в представленной форме не без помощи многочисленных коллег. Со времени 32-го издания учебника было унаследовано от Карла Мегдефрау (Тюбинген, Мюнхен, 1907 — 1999) его направление в изучении низших растений. Автор считает своим долгом подчеркнуть, что большие части текста, относящиеся к морфологии и развитию отдельных рассматриваемых организмов, были взяты в исходном изложении у К. Мегдефрау. Так последователь смог продолжить строение на достижениях своего предшественника, на его чрезвычайно основательных разработках. За достижения, которые позволили объединить равным образом непрерывность преемственности и согласование с успехами науки, К. Мегдефрау выражается глубокая благодарность.
В окончательном редактировании предлагаемого текста особое участие принимали в свое время Йозеф Поэлт (Грац, 1924 — 1995), Фридрих Эрендорфер (Вена), Дитрих фон Денффср (Гиссен) и Хуберт Циглер (Мюнхен), за что я и сегодня, оглядываясь назад, весьма им признателен. За ценные указания благодарю Гельмута Весла (Регенсбург), Бургхарда Бюделя (Карлсруэ), Карла Эссера (Бохум), Вольфганга Фрея (Берлин), Герхарда Юритца (Карлсруэ), Отто Людвига Ланге (Вюрцбург), Михаэля Мелконяна (Дюссельдорф), Франца Обервин-клера (Тюбинген), Аннемари Шмид (Зальцбург), Герхарда Вагенитца (Гёттинген) и многих рецензентов. Спасибо всем соавторам за коллегиальную работу в дружеском
сообществе над рядом изданий учебника на протяжении нескольких лет.
Ханс Пенглер (Регенсбург) исполнил заново значительную часть рисунков в следующих разделах: «Грибы», «Водоросли», «Мхи» и «Папоротникообразные», а также частично предложил новые. Благодарю и его за эту деятельность, которая была столь полезна при работе над учебником начиная с 33-го издания.
Регенсбург, февраль 2002 А. Брезински
Даже если структура и содержание составленной мной главы во многих местах заметно отклоняются от текста предшествующего издания «Страсбургера», все же эта глава основывается на работе моего предшественника в коллективе составителей учебника Фридриха Эрендорфера (Вена). За это ему особая признательность.
Сердечной благодарности заслуживают также Фолькер Биттрих (Кампинас), Гудрун Клаузинг (Майнц), Ганс-Петер Комес (Майнц), Петер Эндресс (Цюрих), Бернхард фон Хаген (Галле), Петер Ляйнс (Гейдельберг), Йенс Ровер (Гамбург), Герхард Ваге-нитц (Гёттинген) за критическое прочтение рукописи, Барбара Ал1>брехт (Майнц), Ганс-Петер Комес (Майнц), Штефан Гляй-сберг (Майнц) и Натали Шмальц (Майнц) за помощь в литературном поиске, а также Вольфганг Лихт (Майнц) за помощь в корректуре и работе над указателем.
Майнц, февраль 2002 Й. В. Кадерайт
Эволюция
10.1. Изменчивость...............25
10.1.1. Онтогенетическая изменчивость
и фенотипическая пластичность.....27
10.1.2. Генетическая изменчивость.28
10.1.2.1. Генные мутации..........29
10.1.2.2. Хромосомные мутации.....33
10.1.2.3. Геномные мутации........37
10.1.2.4. Рекомбинация............40
10.1.2.5. Внеядерное наследование......................45
10.1.3. Рекомбинативная система ..46
10.1.3.1. Система оплодотворения..47
10.1.3.2. Опыление................54
10.1.3.3. Система размножения.....55
10.1.3.4. Поток генов и жизненная форма 58
10.2. Типы и причины естественной изменчивости......................60
10.2.1. Естественный отбор........60
10.2.2. Дрейф генов ..............67
10.3. Видообразование............69
10.3.1. Определения вида.........69
10.3.2. Дивергентное видообразование..................71
10.3.2.1. Аллопатрическое видообразование..................71
10.3.2.2. Репродуктивная изоляция .... 73
10.3.2.3. Перипатрическое, парапатрическое, симпатрическое видообразование и эффект
основателя.......................74
10.3.2.4. Генетика видовых различий.........................77
10.3.3. Гибридизация и гибридное видообразование..................80
10.3.3.1. Гибридизация ..........80
10.3.3.2. Гомоплоидное гибридное видообразование..................82
10.3.3.3. Интрогрессивная гибридизация ................... 83
10.3.3.4. Аллополиплоидия .......84
10.4. Макроэволюция..............91
Наша Земля населена огромным числом организмов самой разнообразной формы и образа жизни. Только в царствах растений и грибов в настоящее время известно около 360000 видов и, вероятно, куда большее их число еще не описано. Все это многообразие появилось в результате эволюции после, скорее всего, однократного возникновения жизни более 3,5 млрд лет назад, т.е. благодаря тому что виды изменялись и одни виды возникали от других. «Ничто в биологии не может быть понято вне эволюционного контекста» — это часто цитируемое высказывание Ф.Доб-жанского метко отмечает ключевое значение эволюционного процесса для биоло
гии. В этой главе представлены важнейшие аспекты эволюционного процесса.
Очень упрощенно эволюционный процесс представляют следующим образом. Благодаря производству обычно многочисленного потомства каждый вид потенциально способен увеличить свою численность в геометрической прогрессии. Так, например, одно дерево секвойи вечнозеленой {Sequoia sempervirens) в течение своей продолжительной, несколько сотен лет, жизни приносит 109—1О10 семян. И даже живущий всего несколько недель однолетник, например мятлик однолетний {Роа annua), к концу своей жизни производит около 100 семян. Этот потенциальный гео
24 I ГЛАВА 10. эволюция
метрический прирост численности, однако, ограничен биотическими и абиотическими факторами окружающей среды, так что число индивидуумов определенного вида в течение сравнительно короткого геологического промежутка времени остается более или менее постоянным. Из этого следует, что в чреде последовательных поколений статистически один родительский индивидуум заменяется одним потомком. Какие индивидуумы при этом выживут и смогут размножаться, чаще всего зависит не от слепого случая, а от особенностей конкурирующих индивидуумов. Ведь потомство оказывается неидентичным, потому что благодаря как мутациям, так и связанным с парасексуальным и половым процессами рекомбинациям возникает генетическая и, следовательно, наследственная изменчивость. Различные индивидуумы соответственно своим генетическим свойствам более или менее успешны в условиях данной окружающей среды. Больший «успех» (приспособленность) заключается в сравнительно высокой вероятности выживания и репродукции. Результатом этой дифференциальной выживаемости и репродуктивного успеха различных индивидуумов и является естественный отбор. Следствием совместного действия мутаций, рекомбинаций и естественного отбора может быть изменение генетического состава последующих поколений, а именно эволюция. Кроме названных процессов для эволюции важны и случайные изменения частоты генов.
В настоящее время никто из ученых не оспаривает существования эволюции, она доказана множеством наблюдений. Это и прямые наблюдения изменений видов в природе со временем (самый известный, наверное, пример — изменение относительных частот темной и светлой форм бабочки березовой пяденицы в процессе индустриализации в Великобритании, которая вызвала почернение стволов берез из-за копоти), и обнаружение широкого распространения внутривидовых вариаций и почти непрерывных переходов между многими видами, и наблюдение гомологического сходства между самыми различными организмами по морфологическим и молекулярным признакам, и иерархическая структура этих сходств (т.е. уменьшения сходства в ряду:
виды одного рода -> роды одного семейства -> -> семейства одного отряда и т.д.). и постепенное возникновение ныне существующих организмов, документированное ископаемыми остатками, а также многократно использованные человеком возможности экспериментально изменять виды в результате селекции.
Еще до опубликования в 1859 г. Чарлзом Дарвином своего эпохального труда «Происхождение видов путем естественного отбора...» не раз предполагали, что виды не постоянны, а изменчивы и что все жившие в прошлом и ныне существующие виды произошли от одного общего предка. Заслуга Дарвина состоит прежде всего в том, что ему удалось предложить механизм эволюционного изменения, в целом принимаемый и сегодня. Несмотря на то что феномен эволюции как процесса изменения видов был воспринят быстро, процесс естественного отбора долго оспаривали, вплоть до работ А. Ф.Л. Вейсмана в конце 19-го столетия и даже еще позже. Многие принимали, что важную роль в изменении видов играют описанное Ж. Б. Ламарком еще в 1809 г. «внутреннее стремление к совершенству», а также наследование благоприобретенных свойств. Даже Ч.Дарвин признавал значение этих механизмов. Самым большим недостатком эволюционной теории Дарвина было отсутствие убедительной теории наследственности. Основополагающие представления Г. Менделя о механизмах наследственности, описанные в его «Опытах над растительными гибридами» (1866), остались неизвестными Дарвину. Лишь после так называемого переоткрытия менделевских законов наследования Г. М. де Фризом, С. Е. Коррен-сом и Эдлером фон Чермаком (в случае с де Фризом и Э. Чермаком существует, правда, вполне серьезное сомнение в том, что они открыли заново законы Менделя, не зная его работу) на рубеже столетий началось объединение эволюционной теории и генетики. Сначала, впрочем, ранние генетики, работавшие в основном с сильно различающимися признаками, не признавали предложенную Дарвином модель постепенных превращений и предполагали, что эволюция происходит скорее скачкообразно, путем так называемых макромутаций. В 1918 г. Р. А. Фишер впервые показал, что и непрерывная изменчивость может быть результатом менделевского наследования, а Р. А. Фишер (1930), Дж. Б. С. Холдейн (1932) и С.Райт (1931) показали, что естественный отбор может привести к генетическим изменениям в соответствии с принципами менделевской генетики. Предложенный этими авторами «современный синтез» был продолжен прежде всего в работах Ф. Добжанского (1937), Дж.С.Хаксли (1942), Э.Майра (1942),
10.1. Изменчивость J 25
Дж.Дж.Симпсона (1944) и Дж.Л.Стебинса в области ботаники «Изменчивость и эволюция у растений» (1950), что сделало идеи синтетической эволюции широко доступными научной общественности. С тех пор современная эволюционная теория получает все новые подтверждения, уточнения и дополнения.
10.1. Изменчивость
Почти все признаки растений, такие, например, как форма и величина листьев (рис. 10.1), даже при сравнении их у одного растения или всего у двух индивидуумов в большинстве случаев оказываются вовсе не одинаковыми, а различными, и этим демонстрируют изменчивость. Масса семян фасоли с одного поля (рис. 10.2) варьирует непрерывно, т.е, в определенных интервалах могут быть найдены фактически любые значения массы. Прерывисто (или ме-ристически) варьирует число семян в бобе. Здесь всегда речь идет о целых числах, а не о промежуточных значениях.
Особая форма прерывистой изменчивости — слабые, чаше всего качественные прояв-
Рис. 10.1. Внутри- и межпопуляционная изменчивость прикорневых листьев у 6 популяций калифорнийского сложноцветного Layia gaillardio-ides (по J. Clausen).
Слева — популяции с влажных внешних, справа — сухих внутренних прибрежных гор. Все растения культивировали в одинаковых условиях; каждый лист принадлежит отдельному индивидууму
Рис. 10.2. Непрерывная вариационная кривая массы 712 семян фасоли от нескольких генетических одинаковых индивидуумов (по W. Johannsen):
А — масса 0,1 г; В — число семян в каждом из весовых классов по 0,05 г; 1 — фактические вариации; 2 — теоретическая кривая случайного распределения. Средние значения встречаются значительно чаще, чем крайние
ления признака у одного вида (например, семена с гладкой или бородавчатой поверхностью у торицы полевой (Spergula arvensis), особей с белыми или красными цветками у хохлатки полой (Corydalis cava)). Если такая форма изменчивости удовлетворяет определенным генетическим критериям, то говорят о полиморфизме.
Сравнение признаков и наблюдение таким образом за изменчивостью возможны на весьма различных уровнях организации. Можно сравнивать признаки одного индивидуума (внутри-организменная изменчивость) или разных индивидуумов (межорганизменная изменчивость), относящихся к одной популяции — группе инди
26 I ГЛАВА 10, ЭВОЛЮЦИЯ
видуумов, растущих в одном месте и свободно скрещивающихся между собой (внутрипопуля-ционная изменчивость), или к различным популяциям одного вида (межпопуляционная изменчивость), а также к разным видам (межвидовая изменчивость) ит.д.
Причиной изменчивости можно, по всей вероятности, считать три фактора. Первый фактор — онтогенетическая изменчивость. В этом случае гомологические структуры в ходе развития индивидуума образуются по-разному. Пример — изменение формы листьев вдоль побега
(см. рис. 4.66, 4.68). Второй фактор кроется во взаимодействии генетической конституции индивидуума (генотипа) с окружающей средой. Вследствие этого в зависимости от условий окружающей среды генотип может проявляться в виде различных фенотипов. Если разделить один экземпляр тысячелистника (Achillea millefolium) на две половины и выращивать их, к примеру, на равнине и в горах, то довольно скоро эти генетически идентичные растения станут выглядеть по-разному (рис. 10.3). Такую форму изменчивое-
см см
Рис. 10.3. Экспериментально полученные модификации у калифорнийского тысячелистника (Achillea millefolium agg.: A.lanulosa, тетраплоид) (no J.Clausen, D. D. Keck и W. M. Hiesey).
Вегетативно размноженные части (клоны) 7 индивидуумов из одной горной популяции в Сьерра-Неваде (Mather), выращенные в трех экспериментальных садах: Stanford (30 м над уровнем моря), Mather (1 400 м над уровнем моря) и Timberline (3 050 м над уровнем моря). Наследственные различия между индивидуумами, в том числе различная норма реакции каждого индивидуума, на разной высоте i :ад уровнем моря
10.1. Изменчивость J 27
ти называют модификациями, а растения демонстрируют фенотипическую пластичность (см. 12.2). Наконец, наблюдаемые вариации могут определяться и тем, что различные индивидуумы обычно генетически различаются (генетическая изменчивость). В то время как онтогенетические вариации имеют место только в пределах организма, а генетические различия проявляются лишь при сравнении разных индивидуумов, часть модификаций представляет собой внутриорганизменную изменчивость, а часть — межорганизменную.
10.1.1. Онтогенетическая изменчивость
и фенотипическая пластичность
Онтогенетическая изменчивость — результат различного развития гомологичных структур у одного организма в зависимости от внутренних и/или внешних условий. Впечатляющие примеры онтогенетической изменчивости демонстрируют изменение формы плесневых грибов или образование листьев различной формы в индивидуальном развитии семенных растений (см. рис. 4.68).
Плющ (Hedera helix) образует в период вегетативного развития характерные лопастные листья, а с начала репродуктивного периода — цельные. Если использовать для вегетативного размножения участок побега с цельными листьями, то возврата к лопастным листьям не происходит.
Изменение формы в ходе чередования поколений (гаметофита и спорофита) можно также рассматривать как онтогенетическую изменчивость, хотя здесь имеется не один индивидуум, а много генетически очень сходных, сменяющих друг друга индивидуумов. Изменение формы в ходе чередования поколений привело, например, к тому, что гаметофит и спорофит некоторых бурых водорослей были описаны как самостоятельные роды.
Фенотипическая пластичность означает, что генотип (т.е. индивидуум определен
ной генетической конституции1) в зависимости от условий среды может образовать различные фенотипы. Поскольку модификации возникают в ходе развития организма, то очевидно, что фенотипическую пластичность нелегко бывает отделить от онтогенетической изменчивости. Последняя имеет место только тогда, когда онтогенетические вариации являются результатом осуществления программы развития, регулируемой исключительно эндогенно. Но чаще и онтогенетическая изменчивость зависит от восприятия сигналов из окружающей среды. Комплексное совместное действие генотипа и влияния среды на образование конкретного фенотипа показывает, что генотип определяет в конечном счете не признаки, а скорее различные возможности реализации фенотипа в некоторых границах, т.е. норму реакции.
Важное различие между генотипом и фенотипом было установлено датским ботаником В. Л. Иогансеном. Ученый работал с семенами от полученных инбридингом гомозиготных индивидуумов фасоли обыкновенной (Phaseolus vulgaris). Из различных по массе семян одного индивидуума он отбирал для следующей генерации самые легкие и самые тяжелые. Так как средняя масса семян от полученных подобным образом растений второго поколения была идентичной, Иогансен сделал вывод, что различие в массе семян, образованных одним индивидуумом, должно быть результатом влияния условий среды. Обнаруженные различия в средней массе семян от индивидуумов, происходящих от разных родителей, он отнес на счет генетических различий этих индивидуумов. Таким образом, изменчивость массы семян одного индивидуума чисто модификационная, тогда как вариабельность массы семян двух генетически различных индивидуумов является и генетической.
Фенотипическую пластичность легко обнаружить экспериментально, если, например, полученные вегетативным размножением и, соответственно, генетически идентичные индивидуумы выращивать в раз
1 Генотип — это наследственная информация, содержащаяся в ДНК (редко — в РНК). Его никак нельзя отождествлять с индивидуумом. — Примеч. ред.
28 I ГЛАВА 10. эволюция
личных условиях (см. рис. 10.3). Если вегетативное размножение невозможно (как, например, у многих однолетних видов), приходится работать с генетически близкими индивидуумами, например с сестринскими экземплярами. В такой ситуации, конечно, учитывают, что наблюдаемые различия могут иметь (хотя и малый) генетический компонент.
Многочисленные эксперименты привели к следующим общим выводам о фенотипической пластичности.
• Модификации не наследственны. Если, к примеру, потомство очень маленького, выросшего в плохих условиях растения выращивают в хороших условиях, то потомки станут не маленькими, а крупными в пределах нормы реакции.
• Разные признаки одного растения проявляют разную степень фенотипической пластичности и не коррелируют друг с другом. В целом вегетативные структуры (например, высота растения, форма и размер листьев) пластичнее, чем репродуктивные (к примеру, размер цветков, масса семян). Правда, отмечены существенная пластичность и репродуктивных структур. Таково, например, образование клейстогамных цветков у фиалки душистой (Viola odorata) в конце вегетационного периода или образование коричневых или черных семян в зависимости от длины дня у видов сведы (Suaeda).
Фенотипическая пластичность не ограничивается морфолого-анатомическими признаками, она может проявляться также и в физиологических и биохимических признаках.
• Модифицирующее влияние окружающей среды может вызывать различия между одинаковыми органами одного индивидуума. У бука лесного (Fagus sylvatica) слабая освещенность приводит к образованию теневых листьев (см. рис. 7.73). Если условия освещенности изменятся, то на том же побеге за теневыми листьями развиваются световые.
• Разные модификации вызваны своими специфическими влияниями среды. У хлопчатника, например, число междоузлий коррелирует с обеспеченностью азотом и не зависит от водообеспеченности,
однако длина междоузлий зависит от обеспеченности водой.
• Индивидуумы одного вида могут различаться по степени фенотипической пластичности анализируемого признака. Экспериментальное культивирование восьми клонов 192 генотипов костра мягкого (Вго-mus hordeaceus) в различных условиях состава почвы, удобрений и фотопериода показало, что, к примеру, степень пластичности признаков длительности развития и величины соцветия соответствует нормальному распределению, т. е. относительно немногие из 192 генотипов проявляют малую или высокую пластичность наблюдаемых признаков, а большинство генотипов показывают среднюю пластичность. Точно так же можно наблюдать различия в пластичности гомологичных признаков между разными популяциями одного вида (и еще более выраженные — между разными видами).
• Степень пластичности признака наследственна и поддается отбору.
Два последних результата подтверждают, что способность к фенотипической пластичности имеет генетическую основу.
Фенотипическая пластичность вносит свой вклад как во внутриорганизменную, так и в межорганизменную изменчивость. Так, одно и то же растение лютика водного {Ranunculus aquatilis) образует в зависимости от длины дня и расположения листового зачатка под или над поверхностью воды листья разной формы: под водой сильно рассеченные, а плавающие по поверхности воды и надводные листья обычно только лопастные (см. рис. 4.68). Если сравнивать листья двух индивидуумов лютика водного, занимающих одинаковое положение на побеге, то может оказаться, что у одного растения такой лист тонко рассеченный (если он начал развиваться под водой при соответствующей длине дня), а у другого — только лопастной.
10.1.2. Генетическая изменчивость
Хотя онтогенетическая изменчивость и вариабельность в результате фенотипической пластичности важны для приспособ
10.1. Изменчивость J 29
ления генотипа1 к окружающей среде и потому имеют определенное эволюционное значение, наибольшую значимость для эволюционных преобразований имеет генетическая изменчивость. Генетические вариации фенотипов индивидуумов, к примеру, одной популяции или вида, можно обнаружить, если выращивать их в идентичных условиях и сравнивать онтогенетически гомологичные структуры или иные свойства. Только таким путем можно узнать, каков вклад генетических различий в наблюдаемые различия. Главный источник генетической изменчивости — мутации и рекомбинации.
Мугация как спонтанное (или индуцированное экспериментально) изменение наследственного материала может произойти на самых различных уровнях и во всех геномах растительной клетки. Так, могут измениться последовательность нуклеотидов ДНК одного гена (генная мутация), структура хромосомы (хромосомная мутация) и, наконец, весь геном (геномная мутация). Следует четко знать, что все мутации случайны, т. е. нет никакой возможности предсказать их вид и место, и что они ненаправленны и не связаны с условиями отбора, которому подвергался индивидуум.
Один из примеров ненаправленного характера мутаций — эволюция устойчивости растений к гербицидам. Многие гербициды применяют в различных частях Земли (например, в Европе и Северной Америке) примерно с одного и того же времени и приблизительно в одинаковых концентрациях. Несмотря на эти сходные условия отбора (по крайней мере в отношении этого фактора), можно видеть, что один и тот же вид в одной области выработал устойчивые к гербицидам генотипы, а в другой — нет. Например, повсеместно распространенная пастушья сумка обыкновенная (Capsella bursa-pastoris) приобрела устойчивость к гербицидам в Польше с 1984 г., но больше нигде. В Северной Америке ежовник обыкновенный (Echinochloa crus-galli) стал устойчивым к гербицидам благодаря мутации пластидного генома, а в Европе — благодаря, очевидно, мутации ядерного генома. Это доказывает, что сходное давление отбора совсем не обязательно приводит к сходным мутациям.
1 К среде приспособлены организмы, т.е. реализованные в фенотипах генотипы, а не непосредственно генотипы. — Примеч. ред.
10.1.2.1. Генные мутации
Генные мутации бывают точковыми, нарушениями рамки считывания, либо возникают вследствие активности подвижных генетических элементов, так называемых транспозонов (рис. 10.4). В случае точковой мутации один нуклеотид заменяется другим. Если при этом пуриновый нуклеотид заменяется на пуриновый или пиримидиновый на пиримидиновый, то говорят о транзиции, если пуриновый нуклеотид заменяется пиримидиновым или наоборот, такой обмен называют трансверсией. При нарушении рамки считывания в существующую последовательность вставляется один или несколько нуклеотидов (инсер-ция) или нуклеотиды утрачиваются (деле-
А гп
AUG АЦА| GGA CUА । GUU CGA GGC AUAAUU.
I I I I I I I I I
Met lie Gly Leu Vai Arg Gly lie lie
Субституция отмеченного основания
AUGAUG GGA CUA GUU CGA GGC AUA AUU...
I I I I I I I I I
Met Met Gly Leu Val Arg Gly lie He
AUG Alj^GGA CUA GUU CGA GGC AUA AUU..
1 1 1 i i T ; i
Met Met Gly Leu Val Arg Gly lie lie
Делеция
отмеченного основания
AUG AUGGAC UAG UUC GAGGCA UAA UU...
I'll h”
Met Met Asp Stop
Рис. 10.4. Точковая мутация (А) и нарушение рамки считывания (В) (по K.-F. Fischbach; из W. Seyffert)-
А — замена отмеченного в цепочке нуклеотида (G вместо А), приводящая к встраиванию другой аминокислоты (метионина (Met) вместо изолейцина (Не); В — потеря (делеция) отмеченного нуклеотида, изменяющая порядок считывания последующих триплетов и, соответственно, последовательность аминокислот. Появление стоп-кодона в новом порядке прочтения вызывает обрыв белкового синтеза
30 I ГЛАВА 10, ЭВОЛЮЦИЯ
ция). Следствием инсерции или делеции оказывается сдвиг триплетов в последующем участке ДНК и совершенно иное ее прочтение. Причиной такого рода мутаций бывают случайные ошибки в репликации ДНК при отсутствии ее репарации.
Транзиции могут происходить потому, что вместо обычных аминных форм аденина или цитозина в последовательность встраиваются весьма редкие таутомерные иминные формы, а вместо кетонных форм гуанина и тимина — еще более редкие енольные формы (рис. I0.5). Так, например, имипоформа аденина может связываться с цитозином вместо тимина, а енольная форма гуанина — с тимином вместо цитозина. Трансверсии могут встречаться в тех случаях, если из-за потери нуклеотидов (депуринизация, де-пиримидинизация) возникают пропуски в нуклеотидной последовательности. Когда пропуск возникает из-за утраты гуанина, в это место обычно встраивается аденин. Если пропуск в цепочке, потерявшей гуанин, заполняется тимином, комплементарным аденину, то результатом будет трансверсия от GC к АТ (см. рис. 10.4, рис. 10.5). Мутации ошибки считывания накапливаются в та
ких участках нуклеотидной цепочки, где несколько одинаковых нуклеотидов следуют друг за другом. «Ошибки» ДНК-полимеразы могут во время синтеза ДНК привести к пропуску одного нуклеотида в кодирующей цепочке ДНК. В результате в следующем цикле репликации соответствующий нуклеотид выпадает, т.е. происходит деления. Может также случиться, что во вновь синтезируемую цепочку ДНК встраивается дополнительный нуклеотид, благодаря чему возникает инсерция.
Транспозоны — генетические элементы, автономно размножающиеся и способные менять местоположение в геноме. Эта способность достигается благодаря тому, что транспозоны содержат генетическую информацию для фермента (транспозазы), который может узнавать как целевую последовательность в геноме, так и конец транспозона и вырезать его. К тому же транспозоны в состоянии использовать ферменты клетки, необходимые для репликации. Когда транспозон встраивается в ген, он может нарушить его функцию и таким образом привести к возникновению мутации.
Благодаря мутациям генов возникают аллели, которые можно определить как воз
N —С N — H = N СН
/ \ / \ / dR N = C C-N
\ // \ N-H“O dR
Аденин (аминоформа)
Тимин (кетоформа)
Гуанин Цитозин
(кетоформа) (аминоформа)
Н Н
\ / N=H-N ""С С-С С-СН
\ И \ // W
N — С N-Н= N СН
/ \ / \ /
dR N = C C-N
\ // \
Н О dR
N-C N = H-N СН
/ \ / \ /
dR N = C C-N
\ // \
N — Н = О dR
Аденин (иминоформа)
Цитозин (аминоформа)
Гуанин Тимин
(енольная форма) (кетоформа)
Рис. 10.5. Амино- и иминоформы аденина (А) и кетоформа и енольная форма гуанина (В) (по K.-F. Fischbach; из W. Seyffert).
Редкая иминоформа аденина спаривается с цитозином вместо тимина, редкая енольная форма гуанина соединяется с тимином вместо цитозина
10.1. Изменчивость I 31
никшие одна из другой различающиеся формы гена. Если диплоидный индивидуум (имеющий по паре гомологичных хромосом) содержит два одинаковых аллеля, то по рассматриваемому гену он гомозиготен. Если два аллеля неодинаковы, он гетерозиготен. В то время как в диплоидном индивидууме может быть только два аллеля одного гена, популяция может содержать несколько аллелей (множественные аллели). Аллели одного гена бывают полностью доминантными (такой аллель определяет фенотип) или рецессивными (такой аллель фенотипически не проявляется1), а также возможно их неполное доминирование (проявление в фенотипе в различном соотношении обоих аллелей), причем промежуточные свойства фенотипа по наблюдаемому признаку являются особым случаем неполного доминирования. Кодоминант-ность (полное проявление обоих аллелей в фенотипе) чаще всего обнаруживают на уровне белков.
В старой литературе гены и их аллели часто обозначали буквами. При этом прописными буквами отмечали доминантность аллеля, а строчными — рецессивность. Так, аллель доминантного красного цвета лепестков львиного зева (Antirrhinum majus) обозначали буквой R, а рецессивного белого — г. В новой литературе чаще используют трехбуквенное сокращение для обозначения гена и его аллелей. Независимо от доминантности и рецессивности прописными буквами обозначают аллели дикого типа, а строчными — мутантные аллели. В любом случае для обозначения генов и аллелей используют курсив.
Считается, что у эукариот средняя частота мутирования гена составляет I мутацию на ген, приходящуюся на 105—106 гамет. Однако при точном изучении отдельного признака можно обнаружить изменчивость частоты его мутирования. У кукурузы (Zea mays) ферменты биосинтеза антоциана мутируют с частотой в 4,92 х ИГ4 гамет, но при образовании морщинистых семянок вместо гладких частота мутаций 1,2 х КГ6. При этом оказывается, что фено
1 Рецессивный аллель не проявляется фенотипически только в гетерозиготном состоянии. — Примеч. ред.
типически наблюдаемые мутации генетически совершенно не обязательно гомологичны, следовательно, необязательно изменен один и тот же участок ДНК одинаковым образом. Эти данные о частоте мутирования, величина которой сильно зависит также от примененных методов наблюдения, относятся к так называемым спонтанным мутациям, для которых невозможно установить какие-либо внешние причины. Более высокие частоты мутирования можно вызвать, к примеру, ионизирующим излучением, ультрафиолетовым светом и различными мутагенными веществами.
Частоту фенотипически проявляющихся мутаций можно определить путем скрещивания индивидуума, гомозиготного по доминантному гену, с индивидуумом, гомозиготным по тому же рецессивному гену (ААхаа). В отсутствие мутаций следует ожидать, что все гибридные индивидуумы будут иметь генетическую конституцию Аа, а по фенотипу соответствовать гомозиготному доминантному родителю. Если все же обнаружатся индивидуумы с фенотипом гомозиготного рецессивного родителя, то их появление следует объяснить слиянием мутантной гаметы (А —> а) гомозиготного доминантного родителя с гаметой гомозиготного рецессивного родителя. Частота таких рецессивных фенотипов позволяет рассчитать частоту мутирования.
Исходя из средней частоты мутирования 1 х 10 5 и принимая во внимание, что высшие растения согласно лучшим для резушки Таля (Arabidopsis thaliand) оценкам имеют около 25 500 генов (см. бокс 7.1, см. рис. 7.4), можно сделать вывод, что около 20 % гамет являются носителями мутаций. Это значение, как бы сильно оно ни менялось для различных генов, отчетливо показывает, что мутационная генетическая изменчивость— самое обычное явление. Вероятность мутирования не одинакова для любого участка ДНК; имеются зоны, более и менее часто мутирующие. По-видимому, это обусловлено тем, что вероятность мутирования нуклеотида зависит от его окружения.
Поскольку толковые мутации и нарушения рамки считывания в основном зависят от точности репликации ДНК и эффективности механизмов ее репарации, сама частота мутаций находится под генетическим контролем. Мутации в фермен
32 I ГЛАВА 10, эволюция
тах-репликаторах ДНК и ферменгах-репа-раторах могут влиять на частоту мутирования. Соответственно гены этих ферментов и названы генами-мутаторами.
Частота мутаций в пластидных и митохондриальных геномах меньше, чем в ядерном геноме. Замена нуклеотида, измеряемая как субституция в одной позиции в год, в ядерном геноме происходит с частотой в среднем (5 — 30)10-9, в пластидном — (I —3)10 9 и в митохондриальном геноме — (0,2— 1)10 9.
Эффект мутаций на кодируемый геном продукт бывает весьма различным. Скрытые мутации вследствие вырожденности генетического кода не вызывают замены аминокислоты и, соответственно, не производят никакого эффекта. То же присуще нейтральным мутациям, когда замена аминокислоты хотя и происходит, но не изменяет функцию затронутого белка. Если же замена аминокислоты влияет на функцию продукта данного гена, то говорят о смысловой мутации. Сильный эффект на продукт гена оказывают, к примеру, такие мутации, когда кодирующий аминокислоту
Рис. 10.6. Генные мутанты львиного зева большого (Antirrhinum majus) (по H.Stubbe).
Общее развитие растения: А— нормальное; В — карликовое; С — рано цветущее. Форма цветков: D — нормальная зигоморфная; Е — радиальная; F— со шпорцем
триплет мутирует в стоп-кодон (бессмысленная мутация) или возникает нарушение порядка считывания, при котором в результате мутации получается совершенно другой продукт гена. В обоих случаях образуется нефункционирующий генный продукт.
Проявятся ли мутации через воздействие на продукт гена также и в фенотипе растения (рис. 10.6), зависит от того, находится ли мутантный ген в гаплоидном или диплоидном состоянии. В диплоидной спо-рофитной генерации многие мутации фенотипически не проявляются, потому что они рецессивны. Это можно объяснить тем, что в диплоидном организме каждый ген представлен дважды. После мутации одного гена из такой пары немутированный аллель может образовать неизмененный генный продукт.
Эффект мутаций зависит и от того, какие функции имеет ген в иерархической организации, к примеру, обмена веществ или процесса развития. Если ген осуществляет регуляторную функцию на высоком уровне, то эффект мутации может быть драматическим.
Хорошо исследованный в последние годы преимущественно на резушке Таля (Arabidopsis thaliana) и львином зеве большом (Antirrhinum majus) пример такого эффекта показал ген, определяющий идентичность частей цветка. Мутации гена, кодирующего факторы транскрипции и этим вмешивающегося в функции подчиненных генов, могут привести к тому, что вместо нормальной последовательности органов цветка (чашечка, венчик, тычинки, пестик) получаются цветки с нарушенной последовательностью частей, например пестики, тычинки, тычинки, пестики или чашечка, чашечка, пестики, пестики (см. 7.4.3).
Как иерархическая организация генома и связанное с этим существование цепей генного действия и цепей биосинтеза, так и распределение продукта гена по различным структурам растения приводит к тому, что один ген может влиять на несколько признаков фенотипа (плейотропия). Примером этого служит действие гена окраски цветков левкоя седого (Matthiola incana) на характер опушения растения (гомози
10.1. Изменчивость J 33
готность рецессивного аллеля, приводящего к выпадению биосинтеза красящего вещества (пигмента) приводит и к отсутствию волосков на растении), а также действие гена антопиана гороха на окраску цветков, бобов, семян и прилистников. И наоборот, часто на признаки влияют несколько генов (полигеиия). Наконец, есть взаимодействия различного рода между негомологичными генами, которые объединены под понятием «эпистаз».
Очень большое число мутаций имеет негативный эффект, т.е. снижает приспособленность мутантов. Это понятно, ведь структура гена является результатом долгой адаптивной эволюции, так что вероятность его улучшения благодаря случайным и ненаправленным мутациям мала.
Хорошо известная мутация пластидного генома с большим фенотипическим эффектом определяет механизм устойчивости к гербицидам. Триазиновые гербициды воздействуют на растения, связывая белок (QB) фотосистемы II и тем самым прерывая фотосинтетический транспорт электронов. Резистентность к гербицидам возникла, к примеру, у мари белой (Chenopodium album) и мятлика однолетнего (Роа annua) благодаря точковой мутации в пластидном гене psb-A, где произошла замена аминокислоты (глицин вместо серина) в позиции 264 белка QB, что резко снизило связывание гербицида с этим белком.
Мутации митохондриального генома могут быть причиной часто встречающейся у растений спонтанной стерильности пыльцы. При этих мутациях, правда, имеют место не изменения генов, а перестройки всего митохондриального генома.
Транспозонная мутация ответственна за наблюдавшиеся Г. Менделем и генетически проанализированные им различия между гладкими и морщинистыми семенами у гороха посевного (Pisum sativum). Здесь из-за внедрения транспозона был поврежден ген, кодирующий разветвленность молекул крахмала и тем самым содержание воды в семенах. Более высокое содержание воды в семенах мутантных индивидуумов приводило к более сильному усыханию и сморщиванию поверхности созревающих семян. Точно так же результатом транспозонных мутаций являются светлые цветки с красными секторами у львиного зева. Красные сектора возникают в тех участках лепестков, где восстанавливается биосинтез вследствие потери транспозона ТатЗ, нарушающего этот биосинтез.
Последний пример ясно показывает, что транспозонные мутации могут приводить к генетической изменчивости тканей одного индивидуума. Таким образом, даже неизменность генотипа внутри одного индивидуума оказывается относительной.
10.1.2.2. Хромосомные мутации
Причиной хромосомных мутаций являются разрывы хромосом, возникающие спонтанно и благодаря активности транспозонов. Как и в случае генных мутаций, частоту хромосомных мутаций можно повысить экспериментально. В зависимости от числа встречающихся разрывов и поведения возникших таким путем хромосомных фрагментов различают следующие типы хромосомных мутапий (рис. 10.7).
• Делецией называют потерю концевого участка хромосомы. Следствием потери теломеры оказывается слияние измененными концами сестринских хроматид, образовавшихся после репликации мутантной хромосомы. В итоге возникает хромосома с двумя центромерами, которая при следующем клеточном делении разрывается. Продолжение этого процесса слияния и разрыва известно как цикл «разрыв — слияние—мост». В результате к стабильному изменению структуры хромосом деле-ции чаще всего не ведут.
• Если в хромосоме происходят два разрыва и теряется участок между ними, то говорят о нехватке (дефиции). У гетерозиготного по этой мутации индивидуума (имеющего одну мутантную и одну неизмененную хромосому) можно распознать нехватку определенной величины в бивалентах мейотических хромосом вследствие образования петлеобразного выпячивания одной из них. Эта петля содержит участок неизмененной хромосомы, утраченный в мутантной хромосоме, которому теперь не с чем конъюгировать.
• Если хромосомный фрагмент, возникший после двух разрывов, не теряется, как при дефиции, а встраивается в другую хромосому в месте ее одинарного разрыва, то получается дупликация (удвоение). При этом фрагмент может встроиться не только в гомологичную хромосому, но и в не-
34 I ГЛАВА 10. эволюция
гомологичную. Если дуплицированные области в гомологичной хромосоме следуют непосредственно друг за другом, то они могут иметь либо одинаковую (тандемная дупликация), либо противоположную ориентацию (инвертированная дупликация). Дупликации также возможно распознать
у гетерозиготного индивидуума (при встраивании в гомологичную хромосому) при мейоти ческой конъюгации хромосом по образованию петлевидного выпячивания.
• Инверсией называют хромосомную мутацию, при которой хромосомный
Делеция abc defg|hi к
a b с d е f g
Нехватка ab cdlefg|hi к
а b с d h । к
a b f е deg h
a b с d h I к
Транслокация
abc de|f v w|x у z
abc dexyz v wf
Дупликация
a L с , d е f ( g h i К
ab cghidefghik
...
I
Рис. 10.7. Хромосомные мутации (no O.Hess; из W.Seyffert):
А — делеция: потеря концевого участка хромосомы (от h до к); В — нехватка: потеря промежуточного участка хромосомы (от е до д). У гетерозиготного индивидуума по этому признаку нехватка проявляется в петлеобразном выпячивании соответствующего участка немутантной хромосомы; С — дупликация: удвоение промежуточного участка хромосомы (от g до i). У гетерозиготного организма по этому признаку дупликация различима благодаря петлеобразному выпячиванию участка мутантной хромосомы; D — инверсия: встраивание промежуточного участка хромосомы (от с до f) в противоположной ориентации. У гетерозиготного индивидуума по инверсии это приводит к образованию инверсионной петли; Е — транслокация: реципрокный перенос концевого участка хромосомы (f и соответственно х—z) в негомологичные хромосомы. У гетерозиготного по реципрокной транслокации индивидуума при мейозе возникает крестообразная фигура конъюгирующих хромосом
e f g h i к
h
10.1. Изменчивость J 35
фрагмент, возникший вследствие двух разрывов, вновь встраивается в то же место, но ориентирован противоположно. В случае перицентрической инверсии центромера он оказывается частью инвертированной области, при парацентрической инверсии это не случается. Инверсия у гетерозиготного индивидуума проявляется в мейозе как характерная инверсионная петля.
• При транслокациях хромосомный фрагмент переносится в другую, негомологичную, хромосому. Если транслокация ре-ципрокна, т.е. между двумя негомологичными хромосомами произошел обмен фрагментами, у гетерозиготного индивидуума в мейозе образуются крестообразные фигуры при конъюгации четырех хромосом. Особый случай транслокации — слияние двух акроцентрических хромосом (робертсоновская транслокация).
Непосредственный фенотипический эффект хромосомных мутаций может быть весьма различным. Делеции и дефиции в зависимости от функции затронутого ими гена приводят к образованию либо нежизнеспособных гамет, либо нежизнеспособного диплоидного организма, гомозиготного по мутантной хромосоме. При гетерозиготности по мутантной хромосоме не только делеция и нехватка [дефиция], но и дупликация генетической информации могут привести к нарушению генного баланса.
Уровни экспрессии генов одного генома тонко сбалансированы между собой. Так как количество генного продукта пропорционально числу копий гена и аллелей, то следствием уменьшения или увеличения их числа бывает нарушение этого баланса.
Наконец, бывает и так, что экспрессия гена зависит от его положения в геноме. Изменение положения в результате хромосомной мутации может повлиять на фенотип как эффект положения, если экспрессия гена нарушена, например из-за нового соседства с гетерохроматиновыми участками хромосомы.
Кроме непосредственного фенотипического проявления в мутантном организме хромосомные мутации имеют и далеко
идущие эволюционные последствия. Дупликация генов может привести к возникновению семейств генов. Семейства генов, с одной стороны, могут облегчить синтез большого количества генного продукта, а с другой стороны, делают возможной диверсификацию функций кодируемых белков.
Пример большой потребности в продукте — запасные белки в семенах. Почти половина их у кукурузы (Zea mays) — это зеин. Зеин состоит, помимо прочего, из полипептидов с молекулярной массой 19 000 или 22000. Вероятно, что не менее 54 генных копий кодируют меньший полипептид и 24 генных копий — больший. Эти гены распределены по меньшей мере по трем хромосомам. У многочисленных генов, составляющих семейство белков, связывающихся с хлорофиллами а или Ь. обнаружена сильная диверсификация функции связывания пигментов. Дивергенция последовательностей нуклеотидов в кодирующих участках этих генов может составлять 55 %.
Изменение пространственного расположения генов путем хромосомных мутаций влияет различным образом на дальнейшую рекомбинацию генов. Так, функционально связанные гены могут благодаря хромосомным мутациям попасть в непосредственное соседство, что снижает вероятность их рекомбинации. Пример этого — локус самонесовместимости у гетероморфных примул, который фактически содержит три гена (см. 10.1.3.1). Рекомбинационные возможности ограничены, например, когда индивидуум гетерозиготен по перицентрической инверсии. Если в этом случае в области инверсионной петли происходит кроссинговер, возникают одна ацентрическая и одна дицентрическая хромосомы, которые затем обе утрачиваются в дальнейшем ходе мейоза. Поскольку таким путем возникают хромосомно несбалансированные и, вероятно, нежизнеспособные гаметы, инвертированный участок хромосомы в результате оказывается защищенным от рекомбинации (рис. 10.8).
Ограничение рекомбинации благодаря хромосомным мутациям, а именно транслокациям, находят и у некоторых ослинников (Oenothera)'. вследствие многочисленных транслокаций, затрагивающих все хромосомы, возникают такие хромосомы, которые при гетерозиготности в
36 I ГЛАВА 10. ЭВОЛЮЦИЯ
В
Рис. 10.8. Барьерный эффект хромосомной мутации: инверсия (по G.L. Stebbins):
А — схема измененной пары хромосом (исходная форма — черная, мутантная — серая) с некоторыми нанесенными генами-маркерами (a, b, с, d), местами разрывов и разворотом поврежденного участка хромосомы; В — мейоз в F,: конъюгация различных по структуре хромосом и кроссинговер в инвертированном участке; С — возникновение в анафазе I моста с двумя центромерами и фрагмента без центромеры. Обе структуры элиминируются, жизнеспособны только гаметы с неизмененными хромосомами исходной формы или мутанты
Рис. 10.9. Перманентная комплексная гетерозиготность.
У некоторых ослинников (Oenothera) благодаря многочисленным охватывающим все хромосомы транслокациям возникают хромосомы, которые при гетерозиготности в мейозе не образуют бивалентов, а располагаются цепочками вследствие гомологии концевых участков различных хромосом. Так возникают гаметы либо только с материнским, либо только с отцовским набором хромосом
Рис. 10.10. Изменение числа хромосом путем хромосомной мутации (А— по D.W. Kyhos; В, С — по F. Ehrendorfer):
А— гаплоидные кариограммы двух близкородственных видов Chaenactis (Asteraceae) с 2n = 12 и 2n = = 10; В — схема дифференцирующей реципрокной транслокации и выпадения фрагмента; С — мейо-тическая конъюгация хромосом в F,
10.1. Изменчивость J 37
мейозе не образуют бивалентов, а напротив, из-за гомологии концевых участков различных хромосом располагаются цепочками (рис. 10.9). Таким образом возникают гаметы либо только с материнским, либо только с отцовским набором хромосом. Эта перманентная комплексная гетерозиготность поддерживается благодаря тому, что встреча гамет с одинаковой хромосомной конфигурацией приводит к нежизнеспособным зиготам в результате действия летальных факторов.
Наконец, хромосомные мутации (особенно транслокации) могут стать причиной изменения числа хромосом (рис. 10.10). Этот механизм изменения числа хромосом называют также дисплоидией.
Частота хромосомных мутаций, по-ви-димому, весьма различается в разных таксонах. Сравнение по молекулярным признакам установленных генетических карт сцепления показывает, что хромосомы пшеницы, ячменя и ржи в высшей степени ко-линеарны, т.е. линейное расположение генов в них мало изменено и почти одинаково. В противоположность этому два близкородственных вида подсолнечника различаются уже десятью хромосомными мутациями.
Хромосомные мутации известны не только в ядерном геноме, но и в геномах органелл. В то время как в геномах пластид они сравнительно редки и по этой причине обычно хорошо маркируют таксоны, перестройки в митохондриальном геноме чрезвычайно часты. Причина этого в том, что митохондриальный геном в отличие от пластидного содержит очень много последовательностей в нескольких копиях. Это обеспечивает конъюгацию и тем самым рекомбинацию внутри митохондриального генома.
10.1.2.3. Геномные мутации
Изменение числа хромосом благодаря механизмам, отличным от вышеописанной дисплоидии, называют геномными мутациями. Чаще всего они возникают при митотическом или мейотическом делении ядра, когда нарушается распределение хроматид или хромосом по дочерним клеткам. Если при этом затронут не весь геном, а только какая-либо одна или немногие хромосомы, говорят об анеуплоидии. Например, если в мейозе II не происходит разделения хроматид одной хромосомы («non-disjunction»),
то в результате одна гаплоидная клетка содержит лишнюю хромосому, отсутствующую во второй гаплоидной клетке (рис. 1-0.10).
Этот механизм может привести к изменению числа хромосом у вида. Чтобы различить дисплоидию и анеуплоидию, вызвавшие изменение числа хромосом, необходимо изучить конъюгацию хромосом в мейозе подходящего гибридного индивидуума.
Если все хромосомы не расходятся при клеточном делении, то происходит эупло-идное изменение их числа. Самая обычная форма эуплоидных геномных мутаций — полиплоидия.
Гаплоидное число хромосом организма принято обозначать как х. Диплоидные индивидуумы соответственно имеют 2х хромосом, а полиплоидные — 4х (тетраплоиды), 6х (гексаплоиды), 8х (октоплоиды) и т.д. Кратное четное увеличение числа хромосом называют ортоплоидией. Конечно, возможно возникновение анортошюидного генома с числом хромосом Зх (триплоиды), 5х (пентаплоиды) и т.д. Геном тетраплоидного организма с основным гаплоидным числом хромосом х = 7 обычно обозначают 2п = 4х = 28. Таким образом показывают, что мейоз у этого организма протекает нормально и в нем можно наблюдать только биваленты.
Соматическая полиплоидия имеет место в том случае, если при митозе клетки хромосомы реплицируются, но деления ядра и клетки не происходит. Это приводит к образованию реституционных ядер с удвоенным числом хромосом.
Данный феномен может быть вызван и экспериментально путем применения, например, колхицина — алкалоида безвременника осеннего. Колхицин подавляет образование веретена деления, но не удвоение хромосом.
Соматическая полиплоидия может привести и к образованию эидополиплоцдной ткани у индивидуума. Пример — тапетум пыльников, который обычно эндополиплоиден и клетки которого могут содержать многочисленные наборы хромосом. Если хроматиды хромосомы не расходятся, возникают так называемые политенные (гигантские) хромосомы. Такие структуры обычные, например у двукрылых (Diptera), бывают иногда и в клетках зародышевого мешка растений.
Эволюционное значение соматическая полиплоидия приобретает тогда, когда по-
38 I ГЛАВА 10. эволюция
липлоидизированные ткани участвуют в развитии репродуктивных органов, что может привести к образованию гамет с удвоенным числом хромосом.
Это было обнаружено у Primula х kewensis, стерильного гибрида между Р. verticillata и P.flo-ribunda. Причиной спонтанного образования фертильного соцветия на остальном стерильном индивидууме этого гибрида была как раз соматическая полиплоидизация.
В случае генеративной полиплоидии происходит слияние нередуцированных гамет. Такие гаметы из-за ошибок в мейозе оказываются диплоидными и встречаются у всех растений с малой частотой (в среднем у исследованных видов 0,57 %; частота диплоидных пыльцевых зерен и яйцеклеток, по всей видимости, не различается). У некоторых сортов кукурузы (Zea mays) нередуцированные пыльцевые зерна наблюдали с частотой по крайней мере 3,5 %. При этом частота образования диплоидных гамет контролируется как генетически, так и условиями внешней среды. Например, высокие или низкие температуры, а также недостаток питательных веществ приводят к возрастанию этой частоты. При слиянии двух нередуцированных гамет сразу возникает тетраплоидный организм. Чаще всего из-за малой частоты появления нередуцированных гамет генеративная полиплоидизация проходит в два этапа. На первом этапе возникает триплоидный (Зх) индивидуум благодаря слиянию нормальной редуцированной (гаплоидной х) гаметы с нередуцированной (2х). При этом нередуцированной чаще оказывается яйцеклетка. Если триплоидная гамета такого организма сольется с нормальной редуцированной гаметой, то возникнет тетраплоидный (4х) индивидуум.
Частота образования нередуцированных триплоидных гамет у триплоидных растений значительно выше (около 5 %), чем частота образования нередуцированных гамет у диплоидных индивидуумов. Однако триплоидные растения образуют также и гаплоидные, и диплоидные гаметы, так что большое число их гамет содержит больше или меньше половины трипло-идного набора хромосом.
Различают несколько форм полиплоидии в зависимости от уровня гомологии
комбинирующихся геномов полиплоидного индивидуума. Если скомбинированные геномы гомологичны, говорят об аутополиплоидии, если же геномы различны, речь идет об аллополиплоидии (см. 10.3.3.4).
Аутополиплоидия и аллополиплоидия — крайние формы непрерывного ряда сходства генома, а не объективные категории. В то время как возникновение полиплоидного потомства благодаря соматической полиплоидии, самооплодотворению или скрещиванию двух индивидуумов одной популяции однозначно попадает в категорию аутополиплоидии, некоторая дивергенция геномов наблюдается уже при сравнении индивидуумов разных популяций одного вида, еще более выражена она у двух индивидуумов различных подвидов и т.д. Учитывая невозможность объективно разграничить ауто- и аллополиплоидию, целесообразно установить различия между ними на видовых границах. Тогда полиплоидия внутри одного вида будет аутополиплоидией, а полиплоидия вследствие скрещивания индивидуумов разных видов — аллополи-плоидией. Однако в действительности проблема остается нерешенной, так как и сами виды объективно не определимы и биологически не эквивалентны (см. 10.3.1). Иногда используют понятие «сегментарной аллополиплоидии» как промежуточную категорию между ауто- и аллополи плоидией.
Относительная частота образования полиплоидов путем прямого слияния двух нередуцированных гамет или через промежуточную триплоидную фазу может быть различной при ауто- и аллополиплоидии. В то время как путь к аутополиплоидии, пожалуй, чаще всего идет через промежуточную триплоидную фазу, путь к аллополиплоидии чаще всего, вероятно, должен быть прямым. Это объясняют обычно тем, что частота образования нередуцированных гамет у гибридов значительно выше (в среднем около 27 %), чем частота образования нередуцированных гамет у негибридных индивидуумов (0,57 %).
В принципе возможно не только увеличение, но и гаплоидизация генома. Это бывает благодаря тому, что яйцеклетка может развиваться партеногенетически, т. е. без оплодотворения (см. 10.1.3.3). Случается, что у диплоидного растения появляется гаплоидное потомство. Если этот процесс происходит у полиплоидного растения, то получается полигаплоцдное потомство. И хотя у нормально размножающихся половым путем видов часто наблюдают появление гаплоидного или полиплоидного потомства,
10.1. Изменчивость J 39
неясно, каково возможное значение таких геномных мутаций для эволюции растений.
Обычно наблюдаемый непосредственный эффект полиплоидизации — увеличение всех частей растения. Этот так называемый эффект гигантизма вызван тем, что увеличение числа хромосом приводит к увеличению ядра клетки, а величина ядра, в свою очередь, коррелирует с величиной клетки (рис. 10.11). Так, полиплоидные растения обычно имеют более толстые листья и лепестки, чем их диплоидные предки, однако часто они менее ветвисты и отличаются удлиненным сроком развития. Большое эволюционное значение имеет тот факт, что у аутополиплоидных растений велика доля различных нарушений мейоза. Они возникают благодаря тому, что гомологичные хромосомы имеются теперь не в двойном, а в четверном числе, например у тетраплоидов. Это приводит к тому, что в мейозе образуются не только биваленты, но и мультиваленты из нескольких гомологичных хромосом, а отдельные хромосомы остаются без пары — униваленты (рис. 10.12). И тогда в результате расхождения хромосом в мейозе образуются недостаточно жизнеспособные или даже вообще стерильные гаметы с лишними или недостающими хромосомами. Таким образом, фертильность аутоплоидных растений чаще всего понижена.
Вследствие наличия у ауто полиплоидно го растения в наборах по четыре гомологичные хромосомы процесс наследования проходит иначе, чем у диплоидного индивидуума. Обнаружение тетрасомической вместо дисомической наследственности часто используют в качестве критерия для интерпретации вида как ауто- или аллополи плоидного.
Мейотический эффект при аллополи-плоидии другой. Здесь диплоидные гибриды обычно имеют пониженную фертильность из-за недостатка гомологичных хромосом в геноме, полученном от родительских видов. Благодаря полиплоидии, однако, возникают гомологичные хромосомы, и фертильность восстанавливается (см. 10.3.3.4).
Частоту полиплоидии оценивают в зависимости от того, какое число хромосом признают полиплоидным. Если в качестве полиплоидов рассматривать только те виды,
Рис. 10.11. Эффект гигантизма (по В. Bamford и F. В.Winkler; из G. L. Stebbins).
Полиплоидизация может привести к увеличению клеточного ядра, клеток и всего растения. Устьица, пыльцевые зерна и хромосомы диплоидного и тетраплоидного Antirrhinum majus
у которых обнаружено число хромосом, кратное самому малому из известных для представителей рода, то среди цветковых растений 30 — 35% видов полиплоидны. Если же принять, что и самое малое число хромосом, встречающееся в каком-либо роде, уже может быть полиплоидным, и исходить из того, что все гаплоидные числа х> 9 полиплоидны, то доля полиплоидных
Рис. 10.12. Образование тетравалентов у ауто-тетраплоидной Nasturtium officinale (по I. Manton; из D. Briggs и S. М.Walters)
40 | ГЛАВА 10. эволюция
цветковых растений возрастет до 70—80 %. В любом случае очевидно, что полиплоидия у цветковых растений — широко распространенное явление и потому полипло-идизация — важный эволюционный процесс. У папоротников и родственных им растений доля полиплоидов оценивается даже в 95 %, в то время как среди нецветковых семенных растений встречается всего около 5% полиплоидов.
Хотя аутополиплоидные индивидуумы возникают, очевидно, чаще, чем аллополи-плоидные, выявить их значительно труднее вследствие нерегулярности мейоза. Кроме того, благодаря гибридному происхождению аллополиплоиды характеризуются большей генетической изменчивостью (см. 10.3.3.4), поэтому обычно принято считать, что преобладающая часть полиплоидных растений возникает путем аллополиплоидии и соответственно восходит к аллополиплоид-ным предкам. С другой стороны, аутополиплоидия хорошо документирована, например для подорожника среднего (Plantago media), ежи сборной (Dactylisglomerata) или видов рода Heuchera.
10.1.2.4. Рекомбинация
Генетическая изменчивость возникает вследствие не только мутаций, но и перемешивания наследственного материала различных индивидуумов. Этот процесс, названный рекомбинацией, связан у эукариотических организмов с половым процессом. Рекомбинация родительского наследственного материала происходит, с одной стороны, из-за случайности слияния половых клеток (сингамии), а с другой стороны, благодаря процессу мейотического деления при образовании гамет следующего поколения.
Несмотря на отсутствие полового процесса у бактерий и архей, у этих форм существует возможность генетического обмена и таким путем рекомбинации наследственного материала. Обмен ДНК может осуществляться при прямом контакте клеток (конъюгация), путем переноса ДНК бактериофагами (трансдукция) или переноса свободной ДНК (трансформация). Эти процессы объединяют под общим названием «парасексуальность».
Процессы рекомбинации можно выявить в закономерностях наследования.
В 1866 г. Грегор Мендель в «Опытах над растительными гибридами» впервые количественно сформулировал закономерности наследственности. Достижения Менделя, чрезвычайно важные не только для генетики, но и для эволюционной биологии, в то время остались незамеченными. Лишь после так называемого пе-реоткрытия установленных им правил наследования Г. М. де Фризом, С. Е. Корренсом и А. Эллером фон Чермаком в 1900 г. началось стремительное развитие генетики. Основным объектом исследования Менделя был горох посевной (Pisum sativum), наличие многочисленных сортов которого, четко различающихся по многим признакам и к тому же гомозиготных благодаря постоянному самоопылению, позволило ученому получить наглядные и количест венно интерпретируемые результаты.
Следующие эксперименты привели Г. Менделя к постулированию известных закономерностей, которые сейчас называют обычно законами Менделя. При скрещивании двух индивидуумов (Р), различающихся между собой только по одному признаку (моногибридное скрещивание), первое дочернее поколение (F,) оказывается единообразным. В представленном на рис. 10.13 примере с ночной красавицей (Mirabilis jalapa) родительские индивидуумы имеют красные или белые цветки, а все потомки F, проявляют промежуточное состояние признака — у них цветки розовые. Окажется ли F, промежуточным по исследуемому признаку или похожим на одного из родителей, как при скрещивании двух индивидуумов крапивы шариконосной (Urtica pilulifera) с зубчатыми и цельнокрайними листьями (рис. 10.14), зависит от экспрессии аллелей этого признака. В случае доми-нантности/рецессивности аллелей F, окажется подобным родителю с доминантным аллелем, а в случае неполного доминирования F, может оказаться промежуточным. Сходство всех потомков первого поколения (F,) описано в первом законе Менделя — законе единообразия F(. Правда, его можно наблюдать только тогда, когда родительские индивидуумы гомозиготны по рассматриваемому гену. Единообразие F, при выполнении этой предпосылки проявляется независимо от направления скрещива-
10.1. Изменчивость J 41
Рис. 10.13. Дисомическое наследование окраски цветков у ночной красавицы (Mirabilis Jalapa) (по С. E.Correns).
Моногибридное скрещивание родительских растений (Р) с белыми и красными цветками; три поколения их потомства (F,, F2, F3), гетерозиготные индивидуумы с промежуточной (розовой) окраской цветков. Приведена аллельная конституция (г— белые, R— красные) диплоидных растений и гаплоидных гамет
Рис. 10.14. Наследование зубчатости края листа у Urtica pilulifera. Моногибридное скрещивание родительских растений (Р) с острозубчатым (pilulifera) или почти ровным (dodartii) краем листа; их потомки трех поколений (F,, F2, F3) (по С. Е. Correns).
Показана аллельная конституция (Z — острозубчатые листья; z — почти цельнокрайные листья) диплоидных растений
42 | ГЛАВА 10. эволюция____________
ния: неважно, какой из генотипов используется как материнский, а какой — как отцовский1. Если у ночной красавицы затем скрещивают два индивидуума из Fb то получают второе дочернее поколение (F2), в котором индивидуумы с белыми, розовыми и красными цветками встречаются в соотношении 1:2:1. В результате скрещивания между собой индивидуумов F, у Urtica piluli-fera получается F2, в котором индивидуумы с зубчатыми и целънокрайными листьями встречаются в соотношении 3:1. В обоих случаях наблюдается расщепление признака в F2, из-за чего это явление известно как второй закон Менделя — закон расщепления в F2. Для объяснения соответствующих результатов при скрещивании различных сортов гороха, к примеру с гладкими и морщинистыми или желтыми и зелеными семенами, Г. Мендель высказал предположение, что каждый исследованный признак определяется двумя наследственными факторами, которые сегодня называют аллелями одного гена. В случае с ночной красавицей результат скрещивания RR (красные цветки; каждый диплоидный индивидуум содержит два аллеля одного гена, гаплоидные гаметы получают один аллель Rj'xrr (белые цветки; гаплоидные гаметы содержат аллель г) проявляется в единообразии F| при аллельном составе Rr. Каждый индивидуум F] образует одинаковое число гамет с Л и г. Случайный процесс оплодотворения приводит к тому, что в F2 встречаются три различных генотипа (RR, Rr, rr) в соотношении 1:2:1. У Urtica при скрещивании ZZxzz в F, содержат генотипы Zz, а в F2 — генотипы ZZ, Zz и zz в соотношении 1:2:1, но из-за полного доминирования Z соответствующие фенотипы встречаются в соотношении 3 (листья с зубчатым краем) : 1 (целыю-крайные листья). Встречаемость родительских гено- и фенотипов в F2 ясно показывает, что наследственные факторы дискретны, т.е. в F, они хотя и комбинируются, но не смешиваются.
Выявление дискретности наследственных факторов стало значительным шагом вперед по
1 Скрещивать возможно не генотипы, а организмы с определенными генотипами. — Примеч. ред.
сравнению с представлениями Ч. Дарвина, предполагавшего, что наследственные факторы смешиваются («blending inheritance»). Это было самым слабым местом в теории эволюции Дарвина, что понимали уже его современники, поскольку полезные мутации вряд ли тогда могли бы сохраняться из-за постоянного «разбавления» в скрещиваниях с немутантными индивидами.
В названных примерах наблюдался процесс наследования у диплоидного споро-фитного поколения исследуемого обьекта. Такое наследование называют диплогено-типическим. Применительно к организмам вроде зеленой водоросли Chlamydomonas, у которой митозы, вегетативное размножение и дифференцировка признаков происходят в гаплоидной фазе, а диплоидна только зигота, говорят о гаплогенотипическом наследовании.
Генотипические различия идентичных по фенотипу индивидуумов F2 с зубчатым краем листа Urtica (ZZ\ Zz. в соотношении 1:2) можно выявить, получив от каждого из них следующее поколение F3 путем самоопыления или скрещивания каждого индивидуума F2 с гомозиготным рецессивным родительским организмом (<ф, т.е. провести реципрокное (анализирующее) скрещивание. В случае скрещивания Z^x^ индивиды реципрокного поколения (R) с зубчатыми (Zz) и цельнокрайними (zz) листьями оказываются в соотношении 1:1, в то время как при скрещивании ZZxzz. все индивиды имеют зубчатые (Zz) листья.
Индивидуумы могут различаться не по одному, а по двум или большему числу признаков (дигибридные или полигибридные скрещивания). Тогда можно наблюдать и другую закономерность. Скрещивание сортов львиного зева большого Antirrhinum ntajus с красными и актиноморфными цветками (RRzz) и с белыми и зигоморфными цветками (rrZZ) (рис. 10.15) приводит и здесь к проявлению первого правила Менделя о единообразии F, — все растения оказываются с красными и зигоморфными цветками. Если скрестить два индивидуума из F), то в F2 встречаются фенотипы: цветки красные и зигоморфные, красные и актиноморфные, белые и зигоморфные, а также белые и актиноморфные в соотношении 9:3:3: 1. Эти данные можно объяс
10.1. Изменчивость |
43
нить тем, что красный цвет (R) и зигоморфная форма (Z) доминантны и что единообразное первое поколение с генотипом RrZz образует четыре различных типа гамет. Случайные комбинации этих четырех типов гамет RZ, Rz, rZ и rz образуют 16 возможных сочетаний (число возможных комбинаций можно рассчитать как комбинационное число 4П, где п — число исследуемых генов, среди которых имеется 9 генотипов (1 х RRZZ, 2 х RRZz, 2 х RrZZ, 4 х RrZz, 1 х RRzz, 2 х Rrzz, 1 х rrZZ, 2 х rrZz, 1 хгггг). Согласно правилу доминирования, в исследованном примере эти девять генотипов попадают в четыре фенотипических класса {RRZZ, RRZz, RrZZ, RrZz — красные зигоморфные; RRzz, Rrzz — красные актиноморфные; rrZZ. rrZz — белые зигоморфные; rrzz — белые актиноморфные). Примечательно, что в этом случае в поколении F2 встречаются комбинации признаков, которых не было ни у родительского поколения, пи в F|. На уровне фенотипов — это красные зигоморфные и белые актиноморфные цветки, а на уровне генотипов— все комбинации, отличающиеся от RRzz, rrZ/.и RrZz. Таким образом, очевидно, что генетическая рекомбинация способствует возникновению генетической изменчивости. Наследственные задатки двух анализируемых признаков не остаются в сочетаниях, свойственных родителям, а образуют комбинации между собой независимо. Эти данные соответствуют третьему закону Мевделя — закону независимой комбинации разных наследственных задатков.
Только что описанная независимая комбинация наследственных задатков во многих случаях, однако, не проявляется. При скрещивании растений гороха с прямыми зелеными бобами и с изогнутыми восковожелтыми бобами во втором поколении не обнаруживается ожидаемое расщепление в соотношении 9:3:3;1, а комбинации признаков родительских форм наблюдаются значительно чаще, чем новые комбинации. Таким образом, два рассматриваемых гена, определяющих форму плода и его окраску, не независимы друг от друга, а сцеплены.
Все до сих пор описанные процессы наследования, включая и последнее отклонение от третьего закона Менделя, можно
Рис. 10.15. Схема дигибридного скрещивания у Antirrhinum majus. Родительские растения с красными актиноморфными и белыми зигоморфными цветками; их потомки в F, и F2; б" и <[> гаметы.
Гены (каждый с двумя аллелями, доминанты обозначены заглавными буквами, рецессивные— строчными), определяющие окраску цветков (Я — красные, г — белые) и форму цветков (Z — зигоморфные, z — актиноморфные) находятся в разных хромосомах (и соответственно не сцеплены). 9 генотипов F2 (RRZZ, RRZz, RrZZ, RrZz, RRzz, Rrzz, rrZZ, rrZz, rrzz) попадают в 4 фенотипических класса (RRZZ, RRZz, RrZZ, RrZz — красные зигоморфные; RRzz, Rrzz — красные актиноморфные; rrZZ, rrZz — белые зигоморфные; rrzz — белые актиноморфные
объяснить процессами слияния гамет и мейозом, а также организацией генов в клеточном ядре. Единообразие первого поколения гибридов F| возникает благодаря тому, что в диплоидном организме каждая хромосома имеется в двойном числе и соответственно каждый ген представлен двумя аллелями. Потомство (F,) гомозиготных родителей оказывается однотипным — гетерозиготным. Так как в мейозе число хромосом уменьшается вдвое, образующиеся гаметы несут только один аллель. Их случайное объединение при слиянии половых клеток при моногибридном скрещивании приводит во втором поколении (F2) к образованию трех различных генотипов в соотношении 1:2:1, т. е. наблюдается расщепление в F2. Описываемое третьим законом
44 1ПЛАВА 10. эволюция
Менделя независимое комбинирование наследственных задатков обнаруживается в случаях, если анализируемые гены находятся в разных хромосомах. Их независимость определяется тем, что в первом делении мейоза располагающиеся попарно бивалентами гомологичные хромосомы родителей, как правило, расходятся случайным образом, так что не все хромосомы одного родителя попадают в одну дочернюю клетку, а другого — в другую. Таким образом, благодаря случайной ориентации бивалентов возникает перемешивание родительских хромосом. Этот процесс называют также интерхромосомной рекомбинацией, потому что перемешивание родительских хромосом хотя и происходит, но сами хромосомы остаются интактными. Отклонения от независимого комбинирования наследственных задатков встречаются, когда анализируемые гены находятся в одной и той же хромосоме и физически связаны друг с другом. Наблюдение того, что такие гены не всегда остаются вместе, а с различной частотой оказываются отделены друг от друга, объясняется тем, что родительские хромосомы в мейозе могут обмениваться участками благодаря кроссин-говеру. Так возникают хромосомы, состоящие из отцовской и материнской частей. В результате гены одной хромосомы могут происходить соответственно от отца и от матери. Этот второй процесс рекомбинации называют интрахромосомной рекомбинацией.
Масштаб интер- и интрахромосомных рекомбинаций сильно зависит от свойственного виду числа хромосом и их размера. Более высокое хромосомное число увеличивает возможности комбинирования материнских и отцовских хромосом, а частота кроссинговера выше в более крупных хромосомах, чем в мелких, из-за чисто пространственных соотношений.
Частота, с которой расположенные в одной хромосоме гены отделяются друг от друга, зависит от расстояния между ними. Большое расстояние чаще приводит к разделению, так как вероятность кроссинговера между генами в этих случаях велика. Если же два гена расположены по соседству, то они разделяются редко, потому что кроссинговер на очень
коротком участке хромосомы маловероятен.
Частоту рекомбинаций можно использовать для составления генетических карт сцепления (рис. 10.16), на которых представлено линейное расположение генов в хромосоме. Правда, частота рекомбинаций далеко расположенных генов всегда ниже значения, которое получается суммированием рекомбинационных частот генов, расположенных между ними. Это можно объяснить тем, что при очень больших расстояниях частота рекомбинации вновь падает вследствие двойного или многократного кроссинговера. Место гена на генетической карте сцепления называется локусом.
До сих пор были использованы примеры. в которых один признак кодируется одним геном с двумя аллелями. Примеры, когда очень многие и в особенности непрерывно варьирующие признаки, такие, как высота растения или длина листовой пластинки, не расщепляются в F2 на дискретные классы признаков, а скорее характеризуются непрерывным вариационным рядом, приводят нас к заключению, что эти признаки, вероятно, по-лигенны, т.е. кодируются многими генами. Распознавание дискретных классов признака в расщепляющемся поколении может быть дополнительно затруднено и тем, что на экспрессию гена влияет окружающая среда, которая затушевывает имеющиеся дискретные классы признака. Процесс наследования количественных признаков невозможно анализировать методами менделевской генетики, его исследуют методами количественной генетики (см. рис. 10.16).
В последнее время доступность молекулярных методов позволила применить новые подходы в генетическом анализе количественных признаков. При этом на первом этапе составляют генетические карты сцепления молекулярных признаков. На втором этапе пытаются найти сходное расщепление молекулярных и фенотипических признаков. Если, к примеру, какой-то фенотипический признак индивидуумов расщепляющегося второго поколения (F2) статистически достоверно коррелирует с определенным молекулярным признаком (это называют косегрегацией), то делают вывод, что определяющий фенотип ген расположен по соседству с геном молекулярного признака. Таким путем можно оценить число генов, вли-
10.1. Изменчивость J 45
Бобы: совместно с N прямые I изогнутые (ср)
Бобы: средней ширины / более узкие (ten) —
Окраска бобов: зеленая / восково-желтая (др)----
Листочки: овальные / заостренные морщинистые (ел)
Окраска цветков: совместно с А Ат Аг В се-------'
пурпурная / буровато-пурпурная (ст)
Бобы: средней ширины / явно узкие (te) ---------
Форма семян: шаровидные / сплющенные (сот) 1
Семенная кожура (теста): совместно с А z тр Dem Lob Ve тип частичной окраски fusca — calvitium (cal)
Форма семян; шаровидные / бороздчатые.
углубление от зародышевого корешка (sul)
Окраска цветков: совместно с А Ат Аг В Сг пурпурная / розовая (се)
Семенная кожура (теста): совместно с АВ темная,_।
сине-фиолетовая, в крапинку / без крапинок.
совместно с АЬ обычно незаметные крапинки (fs)
Семенная кожура (теста): совместно с А черно-фиолетовая (U), сине-фиолетовая '
полосатая (Ust) / неокрашенная (и)
Рис. 10.16. Положение некоторых генов (ср, ten, др и т.д.) в хромосоме V гороха посевного (Pisum sativum) (по Н. Lamprecht; из Е. Gunther).
Слева — фенотип при нормальном или мутантном состоянии гена; частичное проявление только при совместном действии с другими генами (например, с А — основным геном для образования антоциана). Справа — частоты рекомбинаций
яющих на какой-то признак, их положение в геноме и их относительный эффект. Эти методы известны как QTL-картирование (Quantitative Trait Loci).
Генетическая рекомбинация потенциально ведет к возникновению крайне большого числа новых генотипов. Число генотипов (g) в поколении F2 рассчитывают как g = Зп, где п — число независимо сегрегирующих генов, каждый из которых имеет два аллеля. В группе индивидуумов, в которой имеется не два, а несколько аллелей, число новых генотипов для F2 рассчитывают по формуле
g = (r(r+l)/2)* (10.1)
где г — число аллелей гена; п — число независимо сегрегирующих генов. Только для пяти генов с четырьмя аллелями каждый возможно 100000 комбинаций. Правда, при таких расчетах следует учитывать, что не все гены могут свободно комбинироваться, поскольку они иногда расположены в одной хромосоме. Тем не менее такая коли
чественная оценка показывает, сколь велико возникновение новых генотипов при рекомбинации.
10.1.2.5. Внеядерное наследование
Пластиды и митохондрии (органеллы растительных клеток) благодаря своему симбиотическому происхождению имеют собственный геном, соответственно пластидный и митохондриальный. Наследование признаков, кодируемых этими геномами, не подчиняется законам Менделя. Особенность такого внеядерного (экстрахромо-сомного) наследования объясняют тем, что зигота получает свои органеллы чаще всего не от обоих родителей, а только от материнского организма (материнское наследование) и что слияние органелл как первая предпосылка рекомбинации наследственного материала встречается редко. Слияние пластид наблюдали у Chlamydomonas, а митохондрий — у дрожжей.
46 I ГЛАВА 10. ЭВОЛЮЦИЯ
Рис. 10.17. Внехромосомное наследование зелено-белой пятнистости. Материнское наследование у ночной красавицы Mirabilis jalapa (по С. Е.Сог-rens)
Типичный для пластид ход наследования можно наблюдать у ночной красавицы (Mirabilis jalapa) (рис. IO. 17). Здесь имеются, как и у многих других видов, индивидуумы с нормальными зелеными листьями и с пятнистыми бело-зелеными. Белые участки в ткани листа возникают благодаря тому, что клетки в них имеют только бесцветные пластиды из-за дефекта в образовании хлорофилла. Скрещивание нормального зеленого индивидуума ночной красавицы с пятнистым приводит в зависимости от направления скрещивания к различным результатам, существенно отличающимся от менделевских правил. Если зеленое растение используют как материнское, то все потомки оказываются зелеными. Если же в качестве материнского берут пятнистое растение, большинство потомков имеют пятнистые листья. Эти результаты объясняются тем, что у ночной красавицы пластиды наследуются только через яйцеклетку, поэтому потомки имеют пластидные признаки только женского родителя. Случаи, когда при использовании пятнистого растения в качестве женского родителя, а также при самоопылении пятнистого растения встречаются и нормальные зеленые потомки, обусловлены тем, что либо какая-то яйцеклетка случайно содержит только нормальные пластиды, либо во время развития эмбриона из зиготы с нормальными и дефектными пластидами имеет место такое случайное распределение пластид, что в клетки листьев попадают лишь нормальные пластиды.
При материнском наследовании пластиды исключаются из мужских половых клеток во время развития пыльцы, созревания спермиев или только при оплодотворении, или же пластиды, полученные от мужских половых клеток, дегенерируют. Хотя у цветковых растений законо
мерно наследование пластид по материнской линии, у них известно и бипарентальное наследование пластид, например у Pelargonium и Hypericum. Наследование пластид по отцовской линии описано для некоторых хвойных, например у сосны (Pinus) и лиственницы (Larix), но также и у киви (Actinidia). Митохондрии наследуются чаше всего по материнской линии, но и для этих органелл известны случаи бипарентального и отцовского наследования.
10.1.3. Рекомбинативная система
Из формулы (Ю.1) расчета числа возможных рекомбинаций в зависимости от числа исследуемых генов и аллелей (см. 10.1.2.4) следует, что при отсутствии аллельных вариаций (г = 1; сомозиготность по каждому гену) новые рекомбинации не возникают. Хотя клеточные механизмы рекомбинаций имеют место и здесь, скрещивание генетически идентичных индивидуумов не приводит к генетическим новообразованиям в потомстве. Мера генетических рекомбинаций, способствующих возникновению генетических вариаций, зависит от того, насколько велико генетическое сходство скрещиваемых индивидуумов. Генетическое сходство индивидуумов репродуктивного сообщества определяется системой оплодотворения (самооплодотворение/пе-рекрестное оплодотворение и опыление), системой размножения (половое/бесполое),
10.1. Изменчивость J 47
жизненной формой, дальностью разноса пыльцы и соответственно семян или плодов (поток генов). Совокупность этих факторов можно назвать рекомбинативной системой вида.
10.1.3.1. Система оплодотворения
Двудомность и другие половые типы
Большинство цветковых растений имеет обоеполые (гермафродитные) цветки, в строении которых заложена возможность самоопыления и самооплодотворения. Постоянное скрещивание индивидуума самого с собой и последующее самооплодотворение увеличивают гомозиготность потомства (рис. 10.18), соответственно уменьшая генетическую изменчивость. Повышение уровня гомозиготности приводит к тому, что рецессивные аллели, став гомозиготными, проявляют свои часто негативные качества. Связанные с этим фенотипические особенности, ухудшающие физическое состояние потомства, называют депрессией инбридинга (англ, inbreeding depression). Самая простая возможность воспрепятствовать самооплодотворению — двудомность, т. е. развитие однополых цветков на разных индивидуумах. В то время как эта форма разделения полов встречается у множества животных, для цветковых растений она большая редкость и встречается только у 5% видов. Двудомные виды чаще встречаются, например, среди тропических древесных растений и на океанических островах.
Так, на Гавайских островах двудомны около 15 % видов. Определение пола у двудомных цветковых растений первично диплогенотипическое, т.е. генетическая конституция спорофитов является решающим фактором для образования или только мужских, или только женских гаметофитов в цветках конкретного индивидуума. Например, у смолевки обыкновенной {Silene latifolia) половые хромосомы есть (рис. 10.19).
Половые хромосомы противопоставляются как гетеросомы остальным хромосомам генома — аутосомам. В данном случае мужские растения имеют хромосомную Конституцию XY и характеризуются как
гетерогаметные. поскольку их гаметы получают X- или Y-хромосому, а женские растения имеют хромосомную конституцию XX и как гомогаметные образуют гаметы только одного типа. У большинства растений, как и у многих животных, мужские организмы гетерогаметны, а женские — гомогаметны. При скрещивании женских и мужских индивидуумов это приводит к образованию мужского и женского потомства в соотношении 1: 1 (см. рис. 10.19). Однако у большинства видов растений обнаружены отклонения от этого численного соотношения. Это означает, что определение пола может быть обусловлено не только генотипически, но и модифи-капионно (фенотипически). Такие факторы, как температура, длина светового дня или доступность воды, оказывают экспериментально доказанное влияние на определение пола у двудомных растений. У двудомной конопли {Cannabis sativa) определение пола может быть изменено использованием абсцизовой кислоты, ауксина, гиббереллина, цитокинина, высокой концентрации бора, моноокиси углерода (СО) или выращиванием при длинном световом дне и повреждением растения.
Гетерогаметный женский пол известен, к примеру, у Fragaria, Potentilla и Cotula. Половые
Рис. 10.18. Инбридинг и гомозиготизация (по D. Lewis).
Самоопыление и сестринское скрещивание приводят за несколько поколений к полной гомозиготности
48 I ГЛАВА 10. эволюция
Бокс 1и.1. Уне? и анализ фенигипй-«доий и генАги-^хий иллчкнчмвости
Генетическую изменчивость вида можно наблюдать на разных уровнях различными методами. Изменчивость фенотипа имеет как генетическую, так и модификационную компоненту (см. 10.1.1). Так как модификационную компоненту полностью исключить нельзя, для определения доли генетической изменчивости используют результаты экспериментов, в которых все анализируемые генотипы исследуют в одинаковых условиях среды. Для этого растительный материал различного происхождения выращивают из семян в достаточном для статистического анализа количестве при единообразии условий среды. В таком сравнительном эксперименте в культуре (англ, common garden trial) можно распознать, в какой мере представители одного вида отличаются друг от друга генетически, поскольку сохраняющиеся в культуре при одинаковых условиях различия можно объяснить только генетическими отличиями. При этом можно наблюдать все признаки фенотипа — морфологические, анатомические, физиологические или экологические особенности. Следует иметь в виду, что из-за фенотипической пластичности признаков генетические различия вовсе не всегда проявляются при специфических условиях культивирования и что случайно могут встретиться и такие свойства, которые не характерны для вида в естественных условиях произрастания. Сравнительное культивирование при различных условиях и сравнение вариаций в эксперименте с вариациями, наблюдаемыми в природе, позволяют распознать такие случаи. Изменчивость фенотипических особенностей обычно соответствует нормальному распределению. При этом малые и высокие значения признака редки, значительно преобладают средние значения (см. рис. 10.2). Для описания изменчивости используют важные статистические величины, такие, как среднее значение, дисперсия и стандартное отклонение. При этом среднее значение х рассчитывают как частное от деления суммы ^х всех измеряемых показате-„ (_ У х4) леи на число п этих показателей х = — .
У и ) Дисперсия и стандартное отклонение — это меры для описания разброса данных, который не отражает среднее значение. Данные различных распределений могут иметь одинаковые средние значения (рис. А). Дисперсию s2 рассчитывают как частное от деления суммы квадратов отклонений каждого измеряемого
показателя от среднего значения ]Г(х - х) на число измеряемых показателей минус 1(п - 1). Таким образом, дисперсия
g2 = X(x-x)2 п - 1
И наконец, стандартное отклонение s — это корень квадратный из дисперсии s =-JsT В эволюционно-биологических задачах обычно важно сравнить, например, изменчивость какого-либо признака в двух популяциях, чтобы выяснить отсутствие или наличие статистически достоверных различий. Для этого можно воспользоваться анализом дисперсий. Проверяют, достоверно ли больше дисперсия между популяциями, чем дисперсии внутри популяций. Если хотят исследовать не один, а несколько признаков, то используют методы многомерной статистики.
В биологическом материале обычны отклонения от нормального распределения — положительно или отрицательно смешенные распределения, в которых высокие или низкие значения измеряемых величин встречаются чаще, чем средние.
Генетическую изменчивость можно обнаружить и на уровне белков и ДНК. Для характеристики внутривидовой изменчивости из белков обычно исследуют алло- или изоэнзимы. При этом чаще имеют дело с самыми обычными энзимами первичного метаболизма растений. Энзимы, кодируемые одним генетическим локусом с различными аллелями, — это аллоэнзимы. Если для энзима имеется несколько локусов, говорят об изоэнзимах. Аллели одного локуса или разных локусов одной ферментной системы можно обнаружить благодаря тому, что они разделяются при электрофорезе из-за различий в их электрических зарядах и впоследствии могут быть окрашены специальными методами1.
Большое преимущество алло- и изоэнзимов, объединяемых под понятием «изозимы», состоит в том, что оба аллеля одного локуса чаще всего экспрессируются кодоминантно, т.е. оба образуют белок. Это позволяет распознать гетерозиготные индивидуумы без проведения скрещиваний.
1 Разные заряды имеют белковые продукты разных аллелей; их, а не сами аллели, разделяют и выявляют при электрофорезе. — Примеч. ред.
10.1. Изменчивость J 49
Рис. А. Две популяции (А и В) с различным распределением мерных признаков могут иметь одинаковые средние значения в популяциях (по A. M.Srb и R.D.Owen; из D. Briggs и S.M. Walters)
Часто используемые для внутривидовых анализов методы анализа ДНК можно объединить в так называемый метод отпечатков. Примерами могут служить методы RAPD (Random Amplified Polymorphic DNAs), AFLP (Amplified Fragment Length Polymorphism), ISSR (Inter Simple Sequence Repeats) и анализы мини- и микросаттелитной ДНК (VNTRs: Variable Number Tandem Repeats). Эти техники основаны на полимеразной цепной реакции (PCR: Polymerase Chain Reaction) и/или рестрикционном анализе (RFLP: Restriction Fragment Length Polymorphism).
В описании генетической изменчивости по таким признакам, которые позволяют распознать генные локусы и их аллели, указывают обычно различные количественные величины, разрешающие сравнивать виды с различными свойствами, например процентное соотношение полиморфных локусов и число ал
лелей на локус. Локус считают полиморфным в том случае, если в исследуемом материале обнаружено более одного аллеля или если для самого обычного аллеля показана частота менее 0,99. Другая важная величина — средний уровень гетерозиготности. Для отдельного локуса среднюю гетерозиготность h рассчитывают по формуле
h = !-£>?, (10.3)
где х, — частота i-ro аллеля; m — число аллелей. Средняя гетерозиготность Н всех локусов — среднее значение всех h. В дальнейшем анализе структуры генетической изменчивости внутри вида общую вариабельность И, можно расчленить на две составляющие. Из них Н, описывает изменчивость внутри популяций, a Gs, (по М. Нею) или FSI (по С. Райту) — изменчивость между популяциями.
Половые клетки
Рис. 10.19. Половые хромосомы и диплогенетическое определение пола (А, В — 1 800х; из К. Belar; С — из W. Schumacher).
Мейоз (метафаза 1) в материнской клетке пыльцевого зерна (А) и материнской клетке зародышевого мешка ( I) у двудомного растения Silene latifolia; С — схема диплогенетического определения пола; X, Y — половые хромосомы
50 I ГЛАВА 10. эволюция
фомосомы содержат регуляторные гены, которые влияют на образование мужских и женских органов цветка. Однако гены, отвечающие за структуру этих органов, локализованы и в аутосомах. Причиной возникновения половых хромосом скорее всего было то, что обычно полная стерильность, сопровождающая рекомбинацию морфологически различных хромосом, случайно оказалась подавленной. Половые хромосомы наблюдали далеко не у всех двудомных видов.
Если роль двудомности как препятствия самооплодотворению очевидна, то ее возникновение в результате отбора на перекрестное опыление часто вызывает сомнения. Альтернативная гипотеза состоит в том, что при отсутствии мужских органов в цветке можно вкладывать больше ресурсов в семена и плоды. Это могло бы объяснить, почему двудомность нередко коррелирует с образованием мясистых и потому требующих «больших затрат» плодов.
Двудомность, вероятно, чаще всего возникает через гинодиецию, когда на одном растении есть только женские цветки, а на другом — обоеполые. Двудомность и гинодиеция — не единственные варианты распределения половых функций между разными цветками и индивидуумами. Кроме того, имеются однодомность, гиномо-ноеция, андромоноеция, возможно, андродиеция и полигамия (табл. 10.1).
Среди других групп семенных растений двудомны Ginkgo, все Cycadopsida и большинство Gnetopsida. Половые хромосомы можно найти и у некоторых их представителей. Среди Conifero-psida двудомность весьма редка. В европейской флоре, например, двудомны тис ягодный (Тахиз baccata) и можжевельник обыкновенный (Juni-perus communis). Среди споровых растений раздельнополость присуща лишь гаметофитам, но не спорофитам. У папоротников и родственных им растений разнополые гаметофиты встреча-
Таблица 10.1. Приблизительные частоты различных форм распределения полов у покрытосеменных растений
Распределение полов Пол цветка Пол индивидуума Прибл. частота, %
Гермафродитизм <?И ! с? и ¥ 71
Однодомность сГ ИЛИ ¥ с? И ¥ 5
Гиномоноеция <? И ¥/¥ сГ И ¥ 2,8
Андромоноеция <? и ¥ 1,7
Двудомность сГ ИЛИ ¥ сГ ИЛИ ¥ 5
Гинодиеция с? И ¥/¥ <? и ¥/¥ 7
Андродиеция с? И ¥ /о” с? и ¥/о" 0
Полигамия d И ¥/<?/¥ о' И ¥/<?/¥ 3,6
ются почти у одних только разноспоровых таксонов. Однако гаметофиты разного пола всегда образованы одним и тем же спорофитом. У равноспоровых папоротников однополые гаметофиты могут возникать благодаря модификационному определению пола. Гаметофиты либо мужского, либо женского пола известны у мхов и водорослей, а печеночник Sphaerocarpos имеет также и половые хромосомы.
Система несовместимости
Самонесовместимость позволяет избежать самооплодотворения в обоеполых цветках (рис. 10.19): оплодотворение спер-миями собственной пыльцы невозможно. Независимо от того как функционируют конкретные системы несовместимости, генетический принцип, лежащий в их основе, всегда один и тот же. Если в пыльцевом зерне или мужском гаметофите и на рыльце экспрессируется один и тот же аллель самонесовместимости (5), то оплодотворения не происходит. В зависимости от детерминации поведения пыльцы генотипом самого мужского гаметофита или генотипом образовавшего его спорофита, а также в зависимости от морфологии цветка, связанной с системой самонесовместимости, можно выделить три системы самонесовместимости: гаметофитную (ГС) гомоморфную, спорофитную (СС) гомоморфную и гетероморфную.
При ГС реакция пыльцы определяется генотипом пыльцевого зерна, т. е. образованные гетерозиготным индивидуумом пыльцевые зерна попадают в два реакционных класса в соответствии с содержащимся в них аллелем 5, а экспрессия двух аллелей 5 в столбике кодоминантна. При самоопылении индивидуума с аллельной конституцией S|.S2 (и при скрещивании двух индивидуумов с обоими этими аллелями) ни пыльца ни пыльца S2 не приводят к оплодотворению (рис. 10.20), так как оба аллеля экспрессированы и на рыльце. При скрещивании А, S3 (материнское растение) xS|S2 (отцовское растение) пыльца S2 осуществляет оплодотворение, так как аллель Л2 отсутствует в пестике. При скрещивании S3S4xS|S2 оба типа пыльцевых зерен могут осуществить оплодотворение.
10.1. Изменчивость 51
ГС с одним геном 5 известна во многих семействах растений, например у Papaveraceae, Rosaceae, Solanaceae и Scrophulariaceae. Число различных аллелей гена 5 колеблется от 20 до 70. ГС с двумя генами (5 и Z) отмечена у злаков (Роасеае). Здесь реакция несовместимости проявляется в том случае, если аллели обоих генов идентичны как в пыльце, так и в пестике. Три гена самонесовместимости обнаружены у лютика едкого (Ranunculus acris), а четыре — у сахарной свеклы (Beta vulgaris).
При СС реакция пыльцевого зерна определяется не его собственным аллелем, а генотипом образовавшего его индивидуума. В этом случае все пыльцевые зерна одного индивидуума показывают одинаковую реакцию, хотя и содержат различные аллели.
При СС следует ожидать большой изменчивости в экспрессии аллелей в пыльце и пестике. Чаще всего (см. рис. 10.20) один из аллелей доминантен в реакции пыльцы, а два аллеля кодоминантны в пестике.
При скрещивании 5'|52 (материнское растение) хЛ'|5'з (отцовское растение) в
случае СС при доминантности S, проявляется полная реакция самонесовместимости в отличие от отношений при ГС (при ГС пыльца 53 осуществляет оплодотворение), так как 5, определяет реакцию и пыльцы, и пестика при кодоминантной экспрессии (см. рис. 10.20). В случае доминантности в пыльце 5'з реакция самонесовместимости вообще не возникнет, и пыльца с аллелем Sj может обеспечить оплодотворение, поскольку реакция пыльцы всецело определена аллелем 53.
СС лучше всего изучена у Asteraceae и Brassicaceae, а вообще известна в восьми семействах. Число аллелей гена 5 почти такое же, как и при ГС.
Разумеется, пыльца как мужской гаметофит семенных растений имеет признаки, управляемые собственным гаплоидным геномом. Случаи, когда некоторые признаки пыльцы зависят от генотипа образовавшего ее родительского индивидуума, можно объяснить тем, что тапетум пыльника участвует в образовании оболочки пыльцевого зерна Эффектный и рано исследованный пример — это форма пыльцевых зерен
Гаметофитная самонесовместимость
Спорофитная самонесовместимость
Рис. 10.20. Несовместимость при опылении и оплодотворении у покрытосеменных. S-аллели (S,, S2, S3, S4) пыльцевых зерен на рыльце (сверху) и в материнской ткани столбика и завязи (снизу), пыльцевые зерна и пыльцевые трубки (обозначены светлыми линиями) (по F. Ehrendorfer).
При гаметофитной самонесовместимости реакция зависит от генотипа гаплоидных пыльцевых зерен. При спорофитной самонесовместимости решающим оказывается генотип индивидуума, образующего пыльцу (в скобках). (Генотипы S|S3 и S3S4 проявляются в пестике, а не в пыльце. — Примеч. пер.)
52 | ГЛАВА 10. эволюция
у гороха (Pi im sativum). Когда скрещивают его индивидуум с удлиненными пыльцевыми зернами (LL) с индивидуумами с округлыми пыльцевыми зернами (//), то гетерозиготное (LI) F, образует только удлиненные пыльцевые зерна, хотя половина пыльцы содержит аллель круглых пыльцевых зерен I. Напрогив, наличие или отсутствие крахмала в пыльце кукурузы (Zea mays) является чисто гаметофитным признаком. Гетерозиготные индивидуумы, полученные от скрещивания «крахмалоносных» и «бескрахмаль-ных» растений, образуют в соотношении 1 : 1 пыльцевые зерна с крахмалом и без него.
Гомоморфные ГС и СС в большинстве случаев коррелируют с определенными другими признаками. При ГС пыльца к моменту опыления чаше всего двухклеточная, кутикула рыльца разорванная и рыльце влажное, а процессу самооплодотворения препятствует остановка роста пыльцевой трубки в столбике. В противоположность этому при СС пыльца в процессе опыления чаще всего трехклеточная, кутикула рыльца сплошная и рыльце сухое, так что пыльцевая трубка просто не может внедриться в рыльце. Исключение представляет ГС злаков, морфологически соответствующая СС.
При гетероморфоной самонесовмести-мости генетически детерминированные типы скрещивания проявляются и в морфологии. Примером может служить примула (Primula). У большинства видов этого рода находят два типа цветков (рис. 10.21). Так, есть индивидуум с длинным столбиком и низко сидящими в трубке венчика
пыльниками и индивидуум с короткими столбиками и высоко сидяшими пыльниками. Обе эти формы различаются также величиной сосочков рыльца и пыльцевыми зернами. Этот феномен назван дисти-лией или, более общо, гетеростилией.
Гетероморфии сходного типа известны примерно в 25 семействах цветковых растений и 155 родах. Водном только Rubiaceae имеется 91 гетероморфный род. К морфологической дифференцировке типов цветков можно отнести дополнительно или альтернативно к названным признакам также опушение столбика (например, у OxaUs) и его окраску (например, у Eichhornia), размер пыльников (к примеру, у Lithospermum, Pulmonaria) или скульптуру экзины (например, у Armeria, Limonium. Linum). Не только две. но и три формы цветка (тристилия) находят, например, у дербенника иволистного (Lythrum salicaria), в роде Eichhornia (Poniederiaceae) и у Narcissus (см. рис. 10.21).
Гетеростилия связана с генетической самонесовместимостью. У Primula индивидуумы с короткими столбиками гетерозиготны по 5х, а формы с длинными столбиками гомозиготны по хх. При доминировании S и спорофитной реакции самонесов-местимости успешное оплодотворение возможно только между двумя этими морфами цветка. Оплодотворения внутри цветка или между цветками одного морфотипа не происходит.
Если бы .$ был не доминантным, а кодоминантным с х, то пыльца индивидуумов с длинными столбиками не могла бы прорастать на рыльце цветка с коротким столбиком (5s), так
Рис. 10.21. Гетеростилия у Primula sinensis. Рыльца (G) и пыльники (S) располагаются на разной высоте (А, В — небольшое увеличение; Р, N, р, п — 80х; по F.Noll):
А — цветок с длинным столбиком и крупными сосочками на рыльце (N) и мелкими пыльцевыми зернами (р); в- цветок с коротким столбиком, маленькими сосочками рыльца (п) и крупными зернами пыльцы (Р)
10.1. Изменчивость J 53
как оба растения содержали бы один и тот же аллель. Если бы реакция самонесовместимости была гаметофитной, то при доминантности Л' было бы возможно самооплодотворение индивидуумов с короткими столбиками, так как половина их пыльцы содержит аллель s, который не проявляется в рыльце.
В противоположность ГС и гомоморфной СС ген S при гетероморфной системе имеет только два аллеля. Соответственно в популяции статистически успешно только каждое второе скрещивание, так как в идеальном случае обе формы цветка встречаются в соотношении 1:1. При очень большом числе аллелей в случаях ГС и СС процент успешных скрещиваний в популяции значительно выше.
Функция гетероморфности, дополнительная к осуществлению генетической самонесовместимости, состоит, вероятно, в том, чтобы уменьшить частоту иллегитимного опыления (внутри одного цветка или между двумя цветками одинакового строения). У Primula пыльца прилипает к брюшку насекомого-опылителя при посещении им цветка с коротким столбиком. Эта пыльца с большей вероятностью попадет на рыльце цветка с длинным столбиком, нежели с коротким. При иллегитимном опылении рыльце может оказаться заблокированным для легитимной пыльцы или иллегитимная пыльца может замедлить прорастание легатимной пыльцы. Вопрос об эволюционной последовательности возникновения гетероморфности и генетической системы несовместимости остается спорным.
Ген 5 у Primula — это в действительности не единичный ген, а группа из трех очень тесно сцепленных генов. Первый ген ((?/§) кодирует длину столбика, величину сосочков рыльца и реакцию несовместимости столбика; второй (Р/р) кодирует величину пыльцевого зерна и его реакцию несовместимости; третий (A/а) кодирует положение пыльников. Редкая рекомбинация внутри этого комплекса генов в гетерозиготных индивидуумах с короткими столбиками в определенных скрещиваниях может привести к потомству с низко расположенными пыльниками и коротким столбиком или высоко расположенными пыльниками и длинным столбиком. Оба этих гомоморфных рекомбинантов самосов-местимы и вследствие взаимного расположения пыльников и рыльца могут легко самоопыляться. Этот генный комплекс у Primula— пример того, как возникшее, вероятно, благодаря хромосомным мутациям тесное сцепление трех генов может привести к ослаблению генетической рекомбинации. В этом примере таким путем отчет
ливо снижена частота разрывов внутри функционального генного комплекса.
В дополнение к уже описанным системам самонесовместимости иногда признают еще как особую систему отсроченной самонесовместимости. При всех различиях в деталях общее для этих явлений то, что реакция несовместимости проявляется лишь после прорастания пыльцевой трубки через столбик и чаще всего выражается в отмирании семязачатков.
Лежащие в основе реакций самонесовместимости биохимические процессы должны быть, по-видимому, очень разными в различных системах. В то время как при ГС, возможно, гликопротеины с рибонуклеазной активностью узнают аллелеспецифичную пыльцевую РНК и разрушают ее, для СС у Brassicaceae известно, что в реакции самонесовместимости участвует взаимодействие выделенных пыльцой гликопротеинов и рецепторных протеин-киназ.
Из всех исследованных цветковых растений около 50 % самонесовместимы. Распределение различных систем самонесовместимости по системе, а также сравнение их функций на биохимическом уровне позволяют думать, что три описанные системы возникли независимо друг от друга. Далее можно принять, что ГС и гетероморфная спорофитная самонесов-местимость возникали многократно параллельно, чаще всего, возможно, от самосовместимых предков. Это подразумевает, что спорофитная реакция пыльцы в гетероморфных таксонах не гомологична спорофитной реакции пыльцы при ГС. Вопрос о том, возникала ли широко распространенная ГС многократно или нет, остается открытым. Сравнение нуклеотидных последовательностей гликопротеинов с рибонуклеазной активностью, оперирующих в этой системе, пока не дало окончательного ответа на этот вопрос. Вопреки ранним предположениям в настоящее время преобладает мнение, что первые покрытосеменные растения были самосовместимыми. Нередок, правда, и вторичный переход от самонесовместимости к само-совместимости (см. 10.1.3.2).
В селекции растений система совместимости вида имеет большое практическое значение. Так, для скрещивания самосовместимых и сильно самоопыляющихся видов требуется применение особых методов, а у самонесовместимых видов создание «чистых» инцухтных линий обходится дорого.
54 I ГЛАВА 10. эволюция
10.1.3.2. Опыление
Самосовместимые гермафродитные цветки ие обязательно должны быть само-оплодотворяющимися. Воспрепятствовать самооплодотворению (автогамии) и стимулировать оплодотворение чужеродными гаметами (аллогамию) возможно, затруднив или полностью исключив самоопыление. Этого можно достичь либо разделением по времени созревания тычинок и плодолистиков, либо их пространственным разделением в цветке. При временном разделении (дихогамия; рис. 10.22) либо андроцей созревает раньше гинецея (протерандрия, или протандрия, например у Asteraceae), либо гинецей — раньше андроцея (см. рис. 10.22) (протерогиния, или протогииия, например у многих Ranunculaceae).
В то время как более редкая протогиния обычно бывает связана с самосовместимостью, протандрию часто находят в самонесовместимых цветках. Здесь функция протандрии состоит, возможно, в том, чтобы препятствовать блокированию рыльца собственной пыльцой или чтобы уменьшить невыгодное для чужой пыльцы взаимодействие с собственной пыльцой.
При пространственном разделении (гер-когамия) тычинки и плодолистики распо
ложены в цветке так, что самоопыления не происходит.
Примером могут служить цветки ириса (Iris', рис. 10.22), в которых каждое рыльце, расположенное над тычинкой, прикрыто выростом лепестковидной лопасти столбика. Когда нагруженное пыльцой насекомое покидает цветок, эта лопасть прижимается к поверхности столбика и прикрывает рыльце. При посещении насекомым следующего цветка такая же лопасть счищает с него пыльцу, которая теперь может попасть на рыльце пестика.
Хотя дихогамия и геркогамия могут эффективно препятствовать самоопылению в отдельном цветке, они ие могут исключить опыления между цветками одного соцветия. Опыление между цветками одного индивидуума (гейтоногамия) генетически точно так же является самоопылением.
Если последовательность расцветания цветков в соцветии (например, снизу вверх) и перемещение по нему опылителя (например, снизу вверх) жестко детерминированы, то опылитель передвигается главным образом от функционально женских цветков к функционально мужским. В результате дихогамия по типу протерандрии может препятствовать гейтоногамии.
Поскольку большинство видов цветковых растений имеют гермафродитные цвет
Рис. 10.22. Дихогамия (А, В) и геркогамия (С—Е). (А, В— по F.E. Clements и F. L. Long; С—Е— по W. Troll).
Протерандрия у Epilobium angustifolium. Цветок в мужской (А) и женской (В) стадиях развития (в натуральную величину). В цветках Iris pseudacorus пыльники и рыльце прост >анственно отделены друг от друга и рыльце (п) прикрыто листовидной лопастью рыльца (С — общий вид; D — продольный срез цветка; Е — схематический поперечный срез в области пыльников)
10.1. Изменчивость J 55
ки, а генетическая система самоиесовмес-тимости свойственна немногим из них и механизмы биологии цветения не всегда могут воспрепятствовать самоопылению или опылению между цветками одного индивидуума, приходится признать, что самоопыление и самооплодотворение у растений — обычное явление. Подсчитано, что в мире около 40 % цветковых растений способны к самоопылению и самооплодотворению. Указывают даже, что во флорах умеренного климата (например, Британских островов) около 2/3 видов имеют эту возможность. Так как эффект продолжительного самооплодотворения негативен (потеря генетического разнообразия, или депрессия инбридинга), требуется найти объяснение для столь немалой частоты самооп-лодотворяющихся видов. Утрата генетической изменчивости вследствие возрастания гомозиготности уменьшается благодаря тому, что популяция самооплодотворяюще-гося вида обычно не только состоит из потомства одного индивидуума, но и содержит многочисленные, отличающиеся друг от друга, самооплодотворяющиеся генотипы1. Кроме того, даже низкая частота перекрестного оплодотворения эффективно препятствует полной гомозиготизации. Если, к примеру, только каждое десятое оплодотворение перекрестное, то после нескольких самооплодотворяющихся поколений аллель А, исходно встречавшийся в популяции с частотой 0,5 (50 % всех имевшихся в популяции аллелей одного гена принадлежали к А), сохраняется тем не менее в равновесной частоте =0,1 (10 % А). Депрессия инбридинга может быть преодолена благодаря тому, что в течение немногих поколений генотипы с гомозиготными рецессивными аллелями, обладающими неблагоприятным эффектом, элиминируются отбором. Возможное преимущество самооплодотворения заключается в повышенной эффективности размножения и ускорении развития.
Например, в роде Leavenworthia (Brassicaceae) в самонесовместимых популяциях развиваются в семена 52—62 % семязачатков, а в самосовме-
1 Самооплодотворяться могут растения, а не генотипы. — Примеч. ред.
стимых популяциях — 73—94%. При рассмотрении множества видов эти показатели составляют около 22 % для самонесовместимых и 75 % для самосовместимых популяций.
Эффективность репродукции может быть нарушена просто из-за отсутствия опылителя или партнера для скрещивания. Нехватка опылителей обычно имеет место в экстремальных условиях, например в постоянно холодных и сырых местообитаниях. Для видов, однократно цветущих в своей жизни, существует опасность, что в этот обычно короткий промежуток времени опылителей окажется мало или не будет вовсе. С проблемой отсутствия партнера для скрещивания особенно часто сталкиваются виды-колонизаторы. Если, например, заселяется оставленное под паром поле, то развитие на нем популяций вселяющихся видов обычно начинается с появления только одного индивидуума. Успех колонизации гарантирован, если этот индивидуум может оплодотворить себя сам. Соответственно многие сорняки, колонизирующие нарушенные человеком местообитания, в массе своей — самооплодотворяющиеся растения. Так как обычное самооплодотворение и отсутствие необходимости привлекать опылителей приводят у многих сорняков к уменьшению размера цветков, сокращению производства нектара и снижению количества пыльцы, развитие этих растений ускоряется путем укорочения репродуктивной фазы жизни. Это повышает их успех в колонизации, в частности, благодаря тому, что за один вегетационный период они могут образовать несколько поколений. Повышенная скорость развития, однако, может потребоваться и в условиях очень короткого вегетационного периода.
10.1.3.3. Система размножения
Растения, как и многие другие организмы, способны не только к половому воспроизведению, ио и к бесполому размножению. О размножении говорят тогда, когда число индивидуумов в последовательных поколениях возрастает. Размножение и воспроизведение почти всегда связаны между собой. Так как при бесполом раз
56 I ГЛАВА 10. ЭВОЛЮЦИЯ
множении возникает потомство, генетически идентичное своим родителям, генетическая изменчивость вида или популяции в значительной мере зависит от относительной частоты бесполого размножения. Бесполое размножение, которое также называют апомиксисом, может быть или вегетативным, или семенным (благодаря агамоспермии — образованию семян без полового процесса).
Вегетативное размножение заключается в образовании потомства из соматических тканей при полном отсутствии каких-либо элементов полового процесса. Потомство возникает исключительно благодаря митотическим делениям клеток и потому без изменения йдериой фазы. Это обычный феномен у грибов, водорослей, мхов, папоротников и покрытосеменных растений. У голосеменных растений вегетативное размножение встречается редко. Для флоры цветковых растений Британских островов показано, что около 46 % видов обладают способностью к вегетативному размножению. При этом речь идет чаще всего о многолетних травах, ио иногда и о кустарниках. Одно- и двулетние растения не могут размножаться вегетативно, а у деревьев способность к вегетативному размножению встречается редко. Примеры такой способности — тополя {Populus), вязы (Ulmus), а также растения рода Prunus. Особенно выражена способность к вегетативному размножению, например, у злаков и водных растений. Произошедшие в результате вегетативного размножения и больше не связанные физически с материнским растением индивидуумы называют также раметами. Совокупность рамет, возникших от одного генетически индивидуума (гене-ты), составляет клон.
У сосудистых растений органами вегетативного размножения чаще всего являются побеги или модифицированные побеги. Это может быть простая фрагментация обычного надземного (например, отделение после укоренения верхушки побега у Rubus) или водного побега (например, у Elodea), уса (например, у Fragaria) и корневища (у многих злаков). Но бывает, что образуются особые многоклеточные выводковые тела. Это, к примеру, выводковые почки (бульбочки у наземных растений, например у Allium, Polygonum viviparum, турионы — зимующие почки водных
растений, к примеру у Elodea, Flydrocharis), корневые клубни (например, у Ranunculus ficaria), клубни на подземных столонах (например, у картофеля), луковички (например, у Galanthus) или выводковые почки на месте колосков (например, у Роа alpina и Р. bulbosa). Выводковые тела могут образовываться также и у водорослей, лишайников и мхов. Образование побегов на корнях с последующей фрагментацией корней — самый обычный механизм вегетативного размножения у деревьев (корневая поросль). В некоторых случаях и листья могут образовать «детки» для вегетативного размножения (например, у Asplenium, Kalanchoe', см. рис. 4.32). У водорослей, лишайников и мхов важнейшими механизмами вегетативного размножения являются деление и почкование клеток (например, у дрожжей), распадение на отдельные клетки или фрагментация таллома, а также образование особых выводковых телец.
Вклад вегетативного размножения в поддержание популяции может быть значительным. Так, например, подсчитано, что более 99 % индивидуумов в популяции лютика ползучего (Ranunculus repens) произошли благодаря вегетативному размножению. В ходе вегетативного размножения генетические индивидуумы — генеты — могут достигать значительной величины и иногда неожиданно большого возраста.
Использование методов молекулярной генетики для определения генетической идентичности индивидуумов при исследовании, к примеру, североамериканского вида тополя (Populus tremuloides) показало, что один клон занимает 43 га и в целом объединяет около 47 006 рамет. Но и у травянистых растений вроде овсяницы красной (Festuca rubra) было обнаружено, что индивидуумы одного клона могут расти на расстоянии до 220 м друг от друга. При наблюдении современного темпа роста этого вида можно рассчитать, что один такой клон достигает возраста от нескольких сотен до примерно 1 000 лет. Эти результаты показывают, что в популяции число генетических индивидуумов может быть намного меньше, чем число физически независимых индивидуумов. Очевидно, что это сильно затрагивает масштаб генетической изменчивости в популяции.
Агамоспермией называют образование семян без участия полового процесса. Этот феномен известен для представителей около 34 семейств и особенно часто встречается у Asteraceae, Роасеае и Rosaceae. Так как мнения о разграничении агамосперм-
10.1. Изменчивость 57
ных видов очень различны, невозможно сделать надежные выводы об их числе. Механизмы образования агамоспермных семян весьма различны. У представителей родов Citrus, Opuntia или Nigrttella агамо-спермные зародыши возникают в ткани семязачатка без образования зародышевого мешка. При этой так называемой адвентивной эмбрионии (= спорофитная агамоспермия) образованию зародыша не препятствует половой процесс. Так, и половые, и агамоспермные зародыши бывают на одном растении, иногда даже в одном семени. В этом случае агамоспермия факультативна. Однако несмотря на отсутствие полового процесса для образования семян, как правило, необходимы опыление и оплодотворение (псевдогамия), запускающие процесс образования эндосперма. При гаметофитной агамоспермии в семязачатке образуется зародышевый мешок со спорофит-ным, т. е. нередуцированным, числом хромосом. Если этот зародышевый мешок возникает независимо от наличия обычного гаплоидного зародышевого мешка, то мы имеем дело с апоспорией. Если же такой зародышевый мешок замещает обычный зародышевый мешок, то говорят о дипло-спории. Оба эти механизма агамоспермии чаще всего сочетаются с полиплоидией. Зародыш в обоих случаях может развиться или из неоплодотворенной яйцеклетки с нередуцированным числом хромосом (партеногенез), или из какой-либо другой клетки нередуцированного зародышевого мешка (апогамия)'. Аиоспорические виды вроде мятлика лугового (Роа pratensis), лапчатки Неймана (Potentilia neumanniana) или лютика золотистого (Ranunculus auricomus), как и виды с адвентивной эмбрионией, факультативно агамоспермны, так как у них существует возможность образования типичного зародышевого мешка. Здесь также чаще всего необходимы опыление и оплодотворение для образования эндосперма. В противоположность этому диплоспори-ческие виды, например ястребинки (Hieracium), одуванчики (Taraxacum), вейники
' В России обычно используют название «апогаметия», этимологически более правильное. — Примеч. ред.
(Calamagrostis) или белоус (Nardus), чаще всего облигатно агамоспермны, так как у них при образовании зародышей бесполым путем есть ткань, подобная той, из которой у видов с половым процессом возникает зародышевый мешок. При диплоспории для развития эндосперма псевдогамия чаще всего не требуется.
Разные механизмы агамоспермии возможно четко различить по цитологическим процессам. Но так как различные механизмы могут встречаться у близкородственных форм (например, Hieracium подрод Hieracium. диплоспория, подрод Pilosella-. апоспория), можно предположить, что все они тесно связаны по генезису.
Методически трудно выявить генетическую основу агамоспермии. У агамоспермных видов, с одной стороны, иногда имеет место половой процесс, с другой — существование близкородственных половых видов с той же плоидностью, с которыми они могут скрещиваться. Адвентивная эмбриония. по-видимому, регулируется только одним-единственным генетическим локусом. В случае апоспории и диплоспории достоверно участвуют по крайней мере два гена. У Taraxacum, например, один ген обусловливает подавление мейоза при развитии зародышевого мешка, а другой ген индуцирует партеногенетическое развитие яйцеклетки. У различных злаков (например, родов Panicum, Brachiaha, Tripsacum) гаме-тофитная агамоспермия регулируется только одним геном. Правда, картирование генома показало, что за этим «геном», возможно, скрывается более крупный участок хромосомы, который отличается сильно сниженной рекомбинационной способностью или ее совершенным отсутствием. Еще не ясно, как много генов фактически содержатся в этом участке и принимают участие в регуляции агамоспермии.
Ожидаемого полного отсутствия генетической изменчивости у агамоспермных видов нет. Наблюдаемые у них генетические вариации могут иметь разные причины.
• Агамоспермия редко строго облигат-на и по меньшей мере случайный половой процесс может вызывать изменчивость.
• Образование нередуцированиого зародышевого мешка у диплоспорических видов может начинаться с мейоза, но не проходит до конца, и в результате образуется нередуцированное так называемое реституционное ядро.
• Соматические мутации могут накапливаться в течение нескольких поколений
58 I ГЛАВА 10. эволюция_______________
• У агамоспермных видов, вероятно, часто встречаются соматические рекомбинации в виде обусловленных транспозонами хромосомных перестроек в соматических клетках.
Несмотря на эти механизмы возникновения генетической изменчивости, уровень ее у агамоспермных видов сильно снижен по сравнению с их близкими родственниками, характеризующимися наличием полового процесса.
Агамоспермия с обычно сниженной генетической изменчивостью ведет к тому, что и самые мелкие различия признаков остаются более или менее постоянными в череде поколений. Так как постоянство различий признаков является основой для разграничения видов при использовании морфологической концепции вида (см. 10.3.1), у агамоспермных форм можно признать большое число агамных видов (микрс-виды). Например, в центральноевропейской флоре, по мнению некоторых авторов, роды Taraxacum и Hieracium включают около 250 и 190 таких микровидов.
10.1.3.4. Поток генов и жизненная форма
Генетическое сходство скрещивающихся индивидуумов зависит от расстояния, на которое разносятся пыльца и диаспоры (споры, семена, плоды). Если бы пыльца и диаспоры распространялись, как правило, лишь на очень короткие расстояния, то очень велика вероятность скрещивания между родителями и потомками или потомками одного индивидуума, т.е. между генетически очень сходными индивидуумами. Зато эффект генетических рекомбинаций тогда был бы мал. С увеличением расстояния транспортировки пыльцы и диаспор возрастает вероятность скрещивания генетически различающихся индивидуумов. Так как с пыльцой и диаспорами перемешается и наследственный материал, то в тех случаях, когда опыление приводит к оплодотворению, а диаспоры прорастают и возникшие растения скрещиваются с другими индивидуумами, оба эти явления можно объединить под общим понятием «поток генов». Учитывая многообразие механизмов опыления и
распространения диаспор (см. 11.2), трудно прийти к реалистичным опенкам дистанций потока генов.
Для опыления считается твердо установленным факт, что частота опыления пыльцой какого-либо индивидуума с удалением от него экспоненциально уменьшается (рис. 10.23) и что опыление чаще всего происходит между индивидуумами, находящимися на расстоянии от нескольких дециметров до нескольких десятков и значительно реже — нескольких сотен метров. Однако наблюдали опылителей, преодолевших и значительно большие дистанции. Некоторые южноамериканские пчелы совершают перелеты до 23 км, во время которых осуществляют опыление. Галловые осы, опыляющие тропические фикусы, регулярно совершают перелеты от 6 до 14 км, а позвоночные животные (например, птицы и летучие мыши) могут в качестве опылителей преодолевать в среднем большие расстояния, чем насекомые.
Перенос пыльцы можно учитывать непосредственно, например, при зоофилии — наблюдая за передвижениями животных-опылителей, а при опылении ветром (анемофилии) — подсчитывая количество оседающей пыльцы на разном удалении от ее источника. Прямое наблюдение не дает, однако, прямых данных о том, завершилось ли отмеченное опыление оплодотворением. Лучше всего это можно установить путем генетического сравнения родителей с их потомками при знании генетической конституции других потенциальных родителей. Экспериментально созданные популяции губасти-ка (Mimulus} (см. рис. 10.23), в которых была известна генетическая конституция каждого индивидуума, позволили определить отцовство каждого образованного в популяции семени и таким образом максимально подробно проанализировать поток генов с переносимой пыльцой. Исследование этого вида показало, что при опылении животными расстояние генного потока обычно больше, чем дистанция перелета опылителей между двумя индивидуумами. Данные наблюдения можно объяснить тем, что обычно не вся собранная на одном цветке пыльца остается на следующем цветке, часть ее может быть занесена и значительно дальше. Это явление называют «сапу over». Наоборот, при опылении ветром дистанция генного потока обычно меньше, чем расстояние разноса пыльцы. И здесь объяснение кроется в методике наблюдений, так как на большом расстоянии от
10.1. Изменчивость
59
источника пыльцы ее плотность в воздухе очень мала, а следовательно, очень мала и вероятность опыления и оплодотворения.
Детальные знания о дистанциях переноса пыльцы важны при оценке потенциальной опасности от измененных с помощью генной инженерии культурных растений, выращиваемых неподалеку от близкородственных и скрещивающихся с ними дикорастущих видов.
Генетически несходные индивидуумы как партнеры для скрещивания могут оказаться поблизости друг от друга благодаря тому, что диаспоры могут быть перенесены на весьма большие расстояния. Все сказанное о переносе пыльцы в принципе относится и к транспорту диаспор. С удалением от индивидуума плотность производимых им диаспор экспоненциально снижается (рис. 10.24), и расстояния, на которые распространяются семена и плоды цветковых растений, составляют от нескольких дециметров до нескольких сотен метров. При этом есть исключения, имеющие эволюционно-биологическое значение.
Так, например, известно, что океанические острова вулканического происхождения (например, Кракатау, Сортсей) сравнительно быстро были заселены растениями. Многочисленные анализы родства в комбинации с датировками, основанными на генетических дистанциях, позволяют предположить, что дальний транспорт диаспор даже между континентами и островами Южного полушария в геологическом масштабе времени — нередкое явление. Факт дальнего распространения, к примеру, между Авст-ралией/Новой Зеландией и Южной Америкой можно считать доказанным для Taraxacum и Gentianella.
В дискуссии о самооплодотворении и перекрестном оплодотворении было показано, что и случайное перекрестное оплодотворение может в значительной мере способствовать поддержанию генетической изменчивости. Аналогично это касается и возможного участия в поддержании или увеличении генетической изменчивости случайного потока генов на большие расстояния в ходе переноса как пыльцы, так и диаспор. Благодаря этому труднонаблюдаемые исключительные случаи переноса пыльцы и диаспор имеют большое значение.
Другим аспектом взаимосвязи между потоком генов и генетической изменчивостью в популяциях является жизненная
Рис. 10.23. Распределение числа шмелей (Bombus sp.), посещающих Primula veris в Нортумберленде (Англия), в зависимости от расстояния (по A. J. Richards)
Рис. 10.24. Распределение обилия семян Ver-bascum thapsus в зависимости от дистанции распространения (по E.J. Salisbury; из J.L. Harper)
форма исследуемого вида. Продолжительность жизни как один из аспектов жизненной формы важна, поскольку обмен индивидуумами между популяциями многолетних видов имеет большую вероятность,
60 | ГЛАВА 10. ЭВОЛЮЦИЯ_________
чем между популяциями однолетников. Это приводит к существенному возрастанию генетической изменчивости у многолетних видов.
По продолжительности жизни различают однолетники, двулетники и многолетники. Среди однолетников в умеренном климате можно выделить зимующие однолетники и летники (яровые, или не зимующие, однолетники). В то время как цикл развития зимующих однолетников чаще всего начинается осенью и заканчивается с приходом весны или в начале лета (т.е. цикл развития захватывает два календарных года, продолжаясь менее одного года), цикл развития летников укладывается в один вегетационный период. Продолжительность жизни многолетников варьирует в широких пределах — от немногих лет до нескольких тысяч лет Кроме продолжительности жизни в различие жизненных форм могут внести свой вклад и особенности размножения. Растения, которые независимо от продолжительности жизни размножаются всего один раз. называют монокарпическими, растения с многократной репродукцией — поликарпическими.
10.2. Типы и причины естественной изменчивости
10.2.1. Естественный отбор
Наблюдаемые в природе примеры внутривидовой генетической изменчивости чаще всего не случайны, а коррелируют с особенностями окружающей среды и свойствами исследуемых видов растений. Уже с начала XVIII в. была известна внутривидовая изменчивость, например, сроков цветения лесных деревьев северного и южного происхождения. Однако до экспериментов шведского ботаника Г.Турессона в 20-е годы XX столетия не было единого мнения, имеет ли такая внутривидовая изменчивость генетическую основу или она в большей мере результат условий местообитания. Культивируя при одинаковых условиях растения различного происхождения, относящиеся к широко распространенным и встречающимся в разных экологических условиях видам, Турессои смог показать, что наблюдаемые в естественных местообита
ниях различия в большинстве случаев хотя бы частично сохраняются в культуре. Из этого он сделал вывод, что внутри вида существует генетическая изменчивость изучаемых признаков. Так как материал из различных мест со сходными экологическими условиями (рис. 10.25) (например, дюнные популяции ястребинки зонтичной Hieracium umbellatum] всегда демонстрировал сходные признаки и отличался от материала из мест с другими условиями (например, из популяций на прибрежных скалах), Турессон установил, что внутривидовая генетическая изменчивое! ь коррелирует с условиями местообитания. Данные и выводы ученого удалось подтвердить и уточнить в большом числе сравнительных исследований. Соответственно можно уверенно считать, что коррелирующая с условиями среды внутривидовая генетическая изменчивость оказывается очень распространенным явлением.
Вдоль транссекты с запада на восток через Калифорнию от побережья до высокогорий калифорнийских гор Сьерра-Невада и пограничного с ней Большого Бассейна улалось зарегистрировать генетическую изменчивость многих видов, коррелирующую с климатическими факторами. Например, растения тысячелистника Achillea lanulosa, происходящие с различных высотных уровней хр. Сьерра-Невада, отличаются высотой от растений того же вида, произрастающих в Большом Бассейне (см. рис. 10.25). Исследование злака Andropogon scoparius, широко распространенного в Северной Америке, показало, что северным формам для стимуляции цветения требуется более длительное освещение (15 ч в день), чем растениям южного происхождения (14 ч в день). Для понимания этого наблюдения необходимо знать, что длина дня летом возрастает с широтой, а запуск цветения у многих растений зависит от длины дня. Корреляцию генетической изменчивости с различными свойствами почвы смогли показать у Achillea borealis. В эксперименте формы, взятые с серпентиновых почв, хорошо растут на серпентиновых почвах (серпентин — горная порода, на которой возникают богатые магнием и бедные известью почвы). Но формы того же вида, которые в природе не встречаются на серпентиновых почвах, в экспериментальной культуре на серпентиновых почвах растут плохо. Удалось показать, что растения Anthoxanthum odoratum из бедных известью местообитаний нуждаются для роста в значительно меньшем количестве извести, чем расте-
10.2. Типы и причины естественной изменчивости | 61
Рис. 10.25. Экологические расы калифорнийского тысячелистника (Achillea lanulosa, тетраплоид) с различной высоты над уровнем моря (1 400—3 350—2 100 м) вдоль транссекты длиной около 60 км через Сьерра-Неваду и примыкающий район Большого Бассейна (примерно 38" с. ш.) (по J. Clausen, D. D. Keck и W. М. Hiesey).
Около 60 индивидов из каждой популяции были выращены из семян в Стэнфорде (30 м). Диаграммы показывают наследственную изменчивость высоты побега, среднее значение (стрелками) и рисунок типичного индивида из каждой популяции
Рис. 10.26. Клинальная изменчивость у клевера ползучего (Tnfolium repens) (из H.Daday).
Частота аллеля А, ответственного за образование цианогенного гликозида (альтернативный аллель а) в популяциях (кружки с черными (= А) и белыми (= а) секторами), коррелирует с январскими изотермами от Средиземноморья до Северной Европы (карта слева), в Альпах — с высотой над уровнем моря (круговые диаграммы справа)
62 I ГЛАВА 10. ЭВОЛЮЦИЯ
ния того же вида родом из местообитаний, богатых известью. Многочисленные виды (например, Agrostis capillaris, Anthoxanthum odoratum, Festuca ovina, Mimulus guttatus, Plantago lanceolata, Rumex acetosa) имеют генотипы, которые позволяют им расти на почвах, загрязненных тяжелыми металлами, например кадмием, медью, цинком, в то время как другие их генотипы не в состоянии это обеспечить. Внутривидовая дифференциация может коррелировать не только с абиотическими факторами, но и с биотическими. Как и многие другие виды, клевер ползучий (Tn’folium repens) содержит цианогенный гликозид, ферментативное расщепление которого, например, при повреждении тканей приводит к высвобождению синильной кислоты. Данный вид полиморфен по этому признаку. Можно показать, что относительная частота цианогенных (с цианогенным гликозидом) и ацианогенных (без этого гликозида) генотипов коррелирует не только с температурой местообитания (рис. 10.26) при мелкомасштабном рассмотрении, но и с частотой улиток, поедающих этот клевер (при крупномасштабном рассмотрении). Plantago lanceolata с сильно вытаптываемых местообитаний в отличие от подорожника из мест, где ходят мало, имеет не прямостоячую форму роста, а стелющуюся или приподнимающуюся. Внутривидовая дифференциация, коррелирующая с факторами окружающей среды кроме вышеназванных примеров показа
на также и в отношении доступного для фотосинтеза света, засухоустойчивости, морозоустойчивости, устойчивости к нападению паразитов и травоядных животных, привлечения опылителей, конкуренции с окружающими растениями и т. п. (рис. 10.27).
Детальное пространственное размещение генетических вариаций в природе может быть разных типов. Если коррелирующий с каким-либо признаком растения фактор окружающей среды распределен в природе прерывисто (к примеру, щелочные и кислые почвы над известковыми и силикатными породами в Альпах), растения могут показывать более или менее дискретную изменчивость. Если же, напротив, фактор среды изменяется непрерывно (например, снижение средних температур с увеличением высоты над уровнем моря), то и в исследуемом растительном материале надо ожидать непрерывной генетической изменчивости. Оба эти типа внутривидовой изменчивости давно уже различали как экотипическую (экотип) и экоклинальную (экоклина) изменчивость. Разграничение этих типов бывает затруднено тем, что анализируемый признак
0-4 5-9 10-14 15-19 20-24 25-29 30-34 35-39 40-44
Температура, °C
Рис. 10.27. Экологическая дифференциация у семенных растений (А — по Н.А. Mooney nW. D. Billings В — по О. Langlet):
А— экотипы кисличника двухстолбчатого (Oxyria digyna, Polygonaceae) и их различные физиологические нормы реакции: средние показатели фотосинтеза (кружки) и дыхания (треугольники), измеренные в мг СО2 на квадратный дециметр поверхности листа, в зависимости от температуры у юж ной альпийской (1) и северной арктической (2) рас; В— клинальная изменчивость сосны обыкновенной (Pinus sylvestris): при одинаковых условиях выращивания 52 растения европейского происхождения показывают тесную корреляцию массы сухого вещества хвои (как меры устойчивости к холоду) со временем суток в первый весенний день (средняя температура +6 °C) в их естественном местообитании (как мера географической широты, континентальности и длины вегетационного периода)
13 14 15 16 17 18 19 20 21 22 23 24
Время суток
10.2. Типы и причины естественной изменчивости | 63
может коррелировать не с одним, а с двумя или несколькими факторами окружающей среды, изменяющимися в пространстве разнонаправленно.
Обнаруженные типы внутривидовой генетической изменчивости зависят и от многих экспериментальных параметров. Так, сравнение пространственно далеких популяций чаще всего показывает более четкие различия, чем сравнение соседних популяций, а сравнение выращенного из семян потомства представителей двух популяций будет демонстрировать менее выраженные различия, нежели сравнение взрослых растений, взятых из естественных местообитаний.
Естественный отбор общепризнан как важнейшая причина внутривидовой изменчивости, коррелирующей с факторами окружающей среды. Причиной естественного отбора является то, что разные индивидуумы (генотипы) популяции генетически различаются по успешности размножения в данной среде. Различия в успешности размножения проявляются в разной вероятности выживания до репродуктивного возраста, а после достижения репродуктивной фазы — в разном репродуктивном успехе. Разные шансы выжить и различный репродуктивный успех можно в совокупности назвать приспособленностью генотипа. Таким образом, объектом естественного отбора оказывается индивидуальный генотип, точнее, реализованный последним фенотип, т. е. индивидуум. Если разные генотипы популяции имеют разную приспособленность, то возникает естественный отбор. В результате естественного отбора может измениться относительная частота аллелей как на разных стадиях развития одного поколения, так и у последовательных поколений. Изменение частот аллелей в последовательности поколений можно назвать эволюцией. Конкретным (но мысленным) примером эволюции путем естественного отбора может служить один из видов тысячелистника — Achillea lanulosa, произрастающий вдоль транссекты через Калифорнию. Если две идентичные по генетическому составу популяции с генетически изменчивой высотой растений (с чем связаны число цветков и скорость развития до первого
цветения) вырастить на побережье и в высокогорье, то их развитие в течение нескольких поколений, вероятно, пошло бы разными путями. В прибрежной популяции высокорослые растения с большим числом цветков дали бы в среднем больше потомков, чем низкорослые растения с немногими цветками. При более или менее постоянной численности популяции это приведет к увеличению относительной частоты высокорослых растений. В горах, где высокие растения из-за короткого вегетационного периода, возможно, даже не успеют зацвести или лишь самые первые их цветки смогут образовать семена, увеличилась бы относительная частота низкорослых растений. Таким образом, естественный отбор объясняет как эволюцию, так и генетическое приспособление (адаптацию), ибо, по-видимому, рационально считать вариабельность высоты растений адаптацией к условиям произрастания. Итак, изменчивость, коррелирующую с окружающей средой, в большинстве случаев следует понимать как адаптивную изменчивость.
Признак можно интерпретировать как генетическую адаптацию в тех случаях, если он наследуется и способствует повышению вероятности выживания или успеха в размножении, т. е. повышению приспособленности индивидуума. Термином «адаптация» обозначают как процесс приспособления, так и состояние приспособленности. Процесс генетической адаптации начинается с того, что какое-то свойство возникает благодаря мутации, т.е. совершенно случайно. Если новое свойство индивидуума повышает его приспособленность, то вторым шагом возникновения генетической адаптации становится распространение этого свойства в популяции благодаря естественному отбору.
Для доказательства роли естественного отбора как причины естественной генетической изменчивости недостаточно наблюдать корреляцию генетической изменчивости с определенной вариабельностью среды. Е1еобходимо многократно показать, что разные генотипы в разных условиях среды имеют различную приспособленность или что генотип в своей естественной среде обитания имеет более высокую приспособленность, чем в чуждой ему среде. Это можно осуществить,
64 | ГЛАВА 10, ЭВОЛЮЦИЯ
либо подвергнув разные генотипы1 действию определенного решающего фактора среды и измерив их приспособленность, либо произведя в природе реципрокную пересадку материала различного происхождения. Наконец, можно определить различия приспособленности индивидуумов одной популяции в естественных условиях.
Первый вариант исследования был реализован на подорожнике большом (Plantago major). Отмечено, что растения в местах, где регулярно ходят, имеют более или менее распростертые цветоносы, а растения из мест, где вытаптывания нет, итйеют прямостоячие цветоносы. Это различие сохраняется и в культуре при одинаковых условиях выращивания. Экспериментальное вытаптывание растений различного происхождения (для этого на них многократно опускали металлические гири подходящего размера так, чтобы производимое ими давление на единицу поверхности примерно соответствовало давлению, оказываемому взрослым человеком) показало, что стелющиеся цветоносы по приспособленности (измеренной как сухой вес репродуктивных органов) статистически достоверно слабее реагируют на искусственное вытаптывание, чем прямостоячие. В контроле (без вытаптывания) приспособленность распростертых и прямостоячих форм оказалась почти одинаковой. В эксперименте с реципрокными пересадками поменяли местами популяции одного вида из контрастных местообитаний. Так, например, прибрежные популяции калифорнийского Achillea lanulosa были пересажены в горы, а высокогорные — на побережье. В обоих случаях было обнаружено, что приспособленность популяций в чуждом местообитании ниже, чем в естественном. Оба эксперимента позволяют сделать вывод, что наблюдаемые в природе различия между популяциями, обитающими в разных экологических условиях, являются результатом естественного отбора и могут интерпретироваться как адаптивные различия. Реципрокное пересаживание имеет то преимущество перед экспериментальным анализом только одного фактора окружающей среды, которое позволяет измерить приспособленность ко всей среде в целом, не устанавливая часто трудно идентифицируемого фактора среды, предположительно наиболее важного для изменчивости.
1 Все эксперименты возможны не непосредственно с генотипами, а с организмами, т.е. с фенотипами, обладающими разными генотипами. — Примеч. ред.
Совершенно не ясно, можно ли каждый фенотипический признак организма интерпретировать как адаптацию. Существует альтернативная вероятность, что возникающие благодаря мутациям признаки селективно нейтральны и сохраняются благодаря особым генетическим процессам (например, при плейотропии, генетическом сцеплении). Можно ожидать, что мутации, приводящие к селективно нейтральному признаку (поскольку они не влияют на приспособленность), происходят сравнительно часто и медленно отсеиваются естественным отбором. Известное постоянство признаков указывало бы, таким образом, на то, что ограниченная изменчивость признака — результат естественного отбора. Соответственно нельзя с уверенностью принять, что каждое свойство организма является совершенной адаптацией. Следует думать, что структура или свойство чаще всего имеет многообразные связи с окружающей средой и, соответственно, может быть понята как результат действовавшего в различных направлениях естественного отбора, т.е. как некий компромисс. Одно какое-либо свойство, кодируемое плей-отропно действующим геном или комплексом тесно сцепленных генов, может поддерживаться естественным отбором, тогда как другое свойство, кодируемое тем же геном или тем же комплексом, — нет. Наличие последнего тогда не адаптивно. Наконец, может быть и так, что какое-то свойство является приспособлением к уже не существующим условиям среды, а времени для его эволюционного изменения прошло слишком мало. Так, например, очень крупные и жесткие плоды многочисленных южно-американских видов деревьев (например, Crescentia alata, Bignoniaceae; Schee-lea rostrata, Arecaceae) интерпретируют как приспособление к крупным травоядным животным, вымершим примерно 10 000 лет назад.
В зависимости от эффекта эволюционного изменения признака различают три формы естественного отбора: движущий, дизруптивный и стабилизирующий (рис. 10.28). В случае движущего отбора в популяции с нормальным распределением генетической изменчивости наиболее
10.2. Типы и причины естественной изменчивости | 65
приспособленными оказываются одни из крайних вариантов. Это приводит в последовательных поколениях к направленному смещению изменчивости вариационной кривой в сторону этой краевой области. При дизруптивном отборе в популяции генотипы обеих краевых зон вариационной кривой имеют более высокую приспособленность, чем генотипы индивидов средней части спектра изменчивости. В результате начинается разделение популяции на две. В случае стабилизирующего отбора, наоборот, генотипы в области средних популяционных показателей имеют наибольшую приспособленность, а генотипы в обеих краевых зонах— низкую. Такое распределение уровня приспособленности индивидуумов не изменяет генетическую изменчивость в ряду последовательных поколений. На примере стабилизирующего отбора очевидно, что эволюционным результатом естественного отбора не всегда должно быть изменение частот аллелей.
Внутривидовая генетическая изменчивость коррелирует не только с факторами окружающей среды. Анализ генетической изменчивости, в особенности на молекулярном уровне (например, изоферменты), показал, что данный тип генетической изменчивости частично коррелируют и обусловлен теми свойствами, которые в совокупности составляют рекомбинационную систему вида. Здесь прежде всего интересно сравнение внутрипопуляционной и межпопуляционной изменчивости. Система опыления в целом действует так, что самоопыляющимся видам в основном присуща сравнительно малая внутрипопуляци-онная, сравнительно большая межпопуляционная изменчивость. У перекрестноопы-ляющихся видов соотношения обратные — внутри популяций изменчивость выше, чем между популяциями. Объясняются эти примеры, с одной стороны, генетическим эффектом постоянного самооплодотворения, с другой стороны, различной интенсивностью генетического обмена (потока генов) между популяциями у само- и перекрест ноопыляющихся видов. Самооплодотворение ведет к утрате генетической изменчивости внутри популяции и одновременно
Рис. 10.28. Стабилизирующий, движущий и дизруптивный отбор (по К. Mather).
Размах вариации (абсцисса) исходных популяций (внизу) обусловлен частотой индивидуумов с различной наследственностью: он либо сохраняется (А), либо сдвигается (В), либо разделяется на две части (С) благодаря разным формам отбора (стрелки)
способствует изоляции соседних популяций. Влияние потока генов на структуру внутривидовой генетической изменчивости ясно также при сравнении разных механизмов переноса пыльцы и диаспор. Ветроопыляемым, распространяемым ветром видам свойственны сравнительно большая внутрипопуляционная и сравнительно меньшая межпопуляционная изменчивость, чем видам, опыляемым и распространяемым животными, и видам без особых приспособлений для распространения. Создается впечатление, что дистанции потока генов и соответственно генетический обмен между популяциями у ветроопыляемых и распространяемых ветром видов в среднем больше, чем у видов, опыляемых и распространяемых животными. На проявление изменчивости влияет и жизненная форма: однолетние виды демонстрируют меньшую изменчивость внутри популяций и большую между популяциями, нежели короткоживущие травянистые многолетники, а последние, в свою очередь, — меньшую внутрипопуляционную и большую межпопуляционную изменчивость, чем долгоживущие древесные многолетние виды. Возможное объяснение этих примеров лежит, с одной стороны, в известной корреляции самоопыления и малого потока генов с короткой продолжительностью жизни, а перекрестного опыления и более сильного потока генов с большой продолжительно-
66 | ГЛАВА 10. ЭВОЛЮЦИЯ
---- “ ’ ''------°
Бркг 1 rj_^_ Популяционная генетика
Изменение частот аллей в последовательных поколениях количественно исследуют в популяционной генетике. Популяционная генетика пытается таким образом ответить на вопрос: какими станут частоты генотипов или соответственно аллелей в каком-либо поколении, если известны их частоты в предыдущем поколении? Частоту аллелей можно выяснить из рассчитываемой частоты генотипов. В случае единственного генетического локуса с аллелями Лио имеется три генотипа — АА, Аа и аа с частотами Р, Q и R. Частоту р аллеля Л рассчитывают делением суммы удвоенной частоты гомозиготных генотипов АА и частоты гетерозиготных генотипов Аа на удвоенное число индивидуумов в популяции:
2P + Q
2(Р + Q + R)'
(10.4)
Следует использовать удвоенную частоту генотипа АА и удвоенное число индивидуумов, потому что каждый диплоидный индивидуум содержит два аллеля, так что число аллелей в популяции вдвое больше числа индивидуумов. Так как частоты Р, Q и R трех генотипов в сумме дают частоту всех индивидуумов в популяции и равны 1, данную выше формулу можно упростить: р = Р + Q/2. Частоту q аллеля а рассчитывают аналогичным образом по формуле q = R + Q/2. Поскольку частоты обоих аллелей в сумме равны l(p + q= l),q можно вычислять так: q = = 1 - р. Напротив, частоту генотипов по частотам аллелей можно вычислить только при определенных условиях. Рассчитать частоту генотипов в следующем поколении по частотам аллелей в предыдущем поколении возможно, если:
• приспособленность трех генотипов АА, Аа и аа (как в примере) одинакова,
• генотипы скрещиваются случайно,
• не появляются новые аллели (из-за мутаций или потока генов),
• популяция достаточно многочисленна, чтобы исключить случайные флуктуации частот аллелей.
В этом случае частота АА равна р2, частота Аа равна 2pq и частота аа равна q2. Эта основополагающая для популяционной генетики связь между частотой аллеля и частотой генотипа известна как закон Харди—Вайнберга. Он означает, что при постоянных условиях в череде последовательных поколений не происходит сдвига частот аллелей и генотипов и,
соответственно, нет эволюции. Однако можно проверить, находится ли популяция в равновесии или нет. Из частоты имеющихся в популяции генотипов АА, Аа и аа можно рассчитать частоты р аллеля А и q аллеля а. Если популяция находится в равновесии, то частоты генотипов АА, Аа и аа должны оказаться р2, 2pq и q2. Если наблюдаемые частоты генотипов отклоняются от ожидаемых согласно закону Харди—Вайнберга, то популяция не находится в равновесии. Причиной этого могла бы быть неодинаковая приспособленность трех генотипов. Можно рассчитать, как неодинаковая приспособленность влияет на частоту аллелей в последующих поколениях. Для простоты можно принять, что приспособленности А А и Аа одинаковы и равны 1, но приспособленность аа меньше: 1 - s. Буквой s обозначают коэффициент отбора, который равен единице минус приспособленность, причем в простейшем случае приспособленность можно рассчитать как отношение уровня выживания данного генотипа к уровню выживания наиболее приспособленного генотипа. В нашем примере генотипы АА и Аа имеют приспособленность 1, а уровень выживания генотипа аа мог быть 90 % от таковой АА и Аа. Тогда его приспособленность была бы 0,9, а коэффициент отбора s = 0, Е В исходной популяции частоты гентипов составляют АА = р2, Аа = 2pq и аа = q2(l - s). Общая величина популяции: р2 + 2pq + q2(l - s). Если в этой формуле заменить р на 1 - q (р + q = 1, откуда р = 1 - q), то выражение для общей величины популяции можно упростить: 1 - sq2. Тогда частоты генотипов в популяции будут:
.. Р2 Л 2РЧ
/1/1 = —-—=-, Аа = —i-Ar-, 1 - sq2 1 - sq2
аа =
?2(l-s) 1 — sq2
Частота рн изменившаяся под действием о. бора, будет:
Pi =
Р2 +Р9
1 - sq2
(10.5)
(Частота АЛ + половина частоты Аа как доли общей популяции.) Изменение частоты в ходе отбора:
Ap=pi_p=pl±M-p=p2+pq-p+spq2
1 - sq2
p(p + q-l + sq2) 1-sq2
1 - sq2
(Ю.6)
1 - sq2
10.2, Типы и причины естественной изменчивости | 67
Если первоначально р = q = 0,5 и s = 0,1, то д 0,1 0,5 0,52 ,
Др = —-------Ц- = 0,0128. Таким образом,
1-0,10,52 частота р, равна 0,5128, а частота q, равна 0,4872. Эта формула дает возможность рассчитать и предсказать изменение частоты аллеля в последовательных поколениях при случайном скрещивании и разной приспособленности генотипов. Если принять, что коэффициент отбора гомозиготного рецессивного генотипа аа = 1, т. е. индивидуумы этого генотипа никогда не достигают репродуктивного состояния, то по исходной частоте qc аллеля а можно рассчитать частоту a q„ через п поколений:
q =-5^-
1 + nq0
(Ю.7)
При исходной частоте q0 = 0,5 частота qn аллеля а после, к примеру, 10 поколений (п = = 10) постоянной элиминации станет q1() = = 0.083. Отсюда ясно, что даже очень низкая при
способленность гомозиготного фенотипа не может привести к полной элиминации обусловливающего его аллеля, так как данный аллель содержится в гетерозиготных генотипах. Это подтверждает большое эволюционное значение диплоидности и связанной с ней гетерозиготности большого числа организмов. Сложнее сделать подобные расчеты в тех случаях, когда генотипы различаются не уровнем выживания, а репродуктивным успехом, когда скрещивания в популяции неслучайны, когда имеет место поток генов между популяциями или когда приспособленность генотипов изменяется в ряду поколений, например, в зависимости от их частоты. Еще более усложняется количественный анализ, когда наблюдают отбор не по одному генетическому локусу, а по двум или более локусам, взаимно влияющим друг на друга. В то же время это более соответствует реальной ситуации, так как, пожалуй, лишь немногие фенотипические особенности кодируются только одним генетическим локусом.
стью жизни, а с другой стороны, в том, что со временем популяции долгоживущих видов с большей вероятностью обогащаются генетически благодаря притоку генов из других популяций, чем популяции короткоживущих видов. Наконец, у видов с бесполым или частично бесполым размножением генетическая изменчивость существенно меньше, чем у видов только с половым размножением.
10.2.2. Дрейф генов
Другой важный для генетической структуры видов фактор — случайность. Случайные сдвиги частот аллелей в следующих друг за другом поколениях называют дрейфом генов. К таким событиям может, в частности, привести резкое сокращение размера популяции, причем с уменьшением популяции возрастает вероятность случайного сдвига частот аллелей. В результате дрейфа генов может произойти утрата одного из аллелей, и благодаря этому по другому аллелю одного генетического локуса все индивидуумы оказываются гомозиготными (фиксация). Правда, генетический эффект уменьшения численности популяции ста
новится заметным только тогда, когда ее величина становится меньше 10 индивидуумов и в течение нескольких поколений остается низкой, вследствие чего в основном происходит инбридинг, а один из аллелей локуса был редким в исходной популяции. Величина популяции может быть малой, например, из-за того, что немногие индивидуумы или даже всего один индивидуум (гермафродитный и самооплодот-воряющийся) оказались на острове в результате дальнего заноса или случайного заноса человеком и основали новую популяцию (эффект основателя, англ, founder effect). Резко сократить численность популяции также могут и драматические изменения среды (эффект бутылочного горлышка, англ, bottleneck).
Пример эффекта основателя — занесенный человеком из Северной Америки в Австралию злак Echinochloa microstachya. В то время как в Северной Америке каждая популяция этого вида отличается от других характерной для нее комбинацией аллелей, 18 из 20 изученных в Австралии популяций оказались генетически идентичными. Однако подобное единообразие встречается не всегда. Так, занесенная из Европы в Северную Америку метлица (Apera spica-venti) демонстрирует на обоих континентах сходный размах генетической изменчивости.
68 I ГЛАВА 10. ЭВОЛЮЦИЯ
Рис. 10.29. Географическое распространение 11 различных типов пластид Alnus glutinosa (по R. A. King и С. Ferris).
В ледниковых рефугиумах в Южной Европе находят большее генетическое разнообразие, чем во вновь заселенных оттуда областях. Кружки с черными и белыми сегментами указывают на гетерогенность популяций по типам пластид
Постоянное изменение высоты ледяного покрова в четвертичном периоде и связанные с этим климатические изменения сильно влияли на величину популяций всех живших тогда организмов. Этим можно объяснить, что в областях-убежищах (рефугиумах) многочисленные виды деревьев характеризуются сравнительно высокой генетической изменчивостью, а в колонизированных ими в конце ледникового периода областях на Севере популяции генетически обеднены (рис. 10.29). Такой тип генетической изменчивости мог возникнуть благодаря тому, что повторное заселение подвергшейся оледенению территории не шло путем плавного расширения ареала прежде отступивших популяций. В уже заново заселенных областях всегда происходили колебания численности, обусловленные быстрыми изменениями климата, либо единичные или немногие индивидуумы могли натурализоваться далеко впереди от наступающей популяции.
Наблюдения, позволяющие объяснить характер генетической изменчивости определенными особенностями растений (на
пример, системой оплодотворения) или случайными событиями, поднимают вопрос о роли естественного отбора. Более общий вопрос — в какой мере генетическая изменчивость нейтральна, т.е. в какой мере различные генотипы имеют одинаковую приспособленность. Относительно фенотипических признаков можно в целом принять, что различные генотипы нередко имеют и разную приспособленность. Поэтому характер генетической изменчивости по фенотипическим признакам часто является результатом естественного отбора. Относительно молекулярных признаков1 было постулировано, прежде всего М. Кимурой, что большинство имеющихся в популяции генотипов обладают одинаковой приспособленностью и потому селективно нейтральны. В то же время нет никакого сомне
1 Исключение молекулярных признаков из числа фенотипических — ошибка. — Примеч. ред.
10.3. Видообразование | 69
ния в том, что эффект большинства новых мутаций (см. рис. 10.29) отрицательный, так как эти мутации элиминируются отбором. Кимура предположил: генетическая изменчивость на молекулярном уровне — скорее результат дрейфа генов, нежели естественного отбора.
Эта теория нейтральной эволюции основывается на слишком высоких для результатов естественного отбора темпах эволюции протеинов, слишком большом многообразии протеинов внутри видов, на известном постоянстве темпов эволюции протеинов в разных эволюционных линиях и на более высоких темпах эволюции нефункциональных частей ферментов в сравнении с функциональными. Аргументация, первоначально базировавшаяся на анализе протеиновых последовательностей, позднее была изменена благодаря несколько иным результатам анализа нуклеотидных последовательностей ДНК.
Ответ на вопрос, что влияет сильнее на молекулярную эволюцию — дрейф генов или естественный отбор,зависит в значительной мере от представлений о размере популяций, темпах мутационного процесса и коэффициентах отбора. Поскольку эти величины обычно не удается определить количественно и им приходится давать только приблизительную оценку, то в зависимости от принятых оценок определенные варианты генетической изменчивости можно объяснить в соответствии с нейтралистской и селекционист-ской моделями эволюции. Соответственно нельзя окончательно ответить на вопрос об относительной роли дрейфа и отбора. Достоверно лишь то, что, к примеру, скрытые мутации или некоторые нетранскрибируемые последовательности ДНК эволюируют нейтрально и что на молекулярную эволюцию, следовательно, влияют как дрейф, так и отбор. Но и в фенотипической эволюции может проявляться дрейф генов.
10.3. Видообразование
10.3.1. Определения вида
Обсуждение генетической изменчивости внутри вида и механизмов видообразования с очевидностью требует определения вида. Дать непротиворечивое определение вида чрезвычайно трудно. Несомненно, в практической систематике большинство видов описано на основе наблюдаемой морфологической изменчивости и, та
ким образом, использована морфологическая (таксономическая или фенотипическая) концепция вида. При этом систематик ищет скоррелированную прерывистость в изменении разных фенотипических, главным образом морфологических, признаков (рис. 10.30). Эту прерывистость рассматривают как границу между видами. Изменчивость внутри видов непрерывна, а изменчивость между видами прерывиста. Однако следует иметь в виду, что в особых случаях и внутривидовая изменчивость бывает прерывистой в форме полиморфизма (например, гетеростилия у Primula, двудомность; см. 10.1.3). В подобных ситуациях для разграничения видов требуются дополнительные критерии. Именно по этой причине в понимании морфологического вида сильна субъективная компонента, ибо невозможно установить объективные критерии величины фенотипической преры вистости, необходимой для разграничения видов.
Видовые границы могут быть по-разному выражены в разных пространственных масштабах. Разграничение видов в узкоограниченной области чаще всего вызывает немного проблем, но они существенно возрастают при рассмотрении всей области распространения вида (см. рис. 10.30).
.- Длина листа (см)
Длина плода (мм)
Рис. 10.30. Схематическое изображение дискретной изменчивости между видами по количественным и качественным признакам.
Виды А и В различаются по окраске цветков, опушению цветоносов, а также по длине листьев и плодов
70 I ГЛАВА 10, эволюция
В то время как морфологическое определение вида в принципе не связано с представлениями об эволюционном процессе, в других определениях вида пытаются учесть процесс его эволюционного образования. Сформулированная Э. Майром биологическая концепция вида определяет виды как группы свободно скрещивающихся природных популяций, которые репродуктивно изолированы от других групп популяций (т.е. от других видов). Репродуктивная изоляция имеет генетическую основу. Это определение подразумевает, что виды распознаваемы потому, что они не гибридизируют с другими видами. Гибридизация как критерий распознавания вида уже давно и неоднократно обсуждалась и использовалась Э. Майром.
В биологической концепции вида подчеркивается именно репродуктивная изоляция одного вида от другого. Очень похожая концепция взаи-моузнавания конспецифичных партнеров (англ, recognition species concept), напротив, подчеркивает взаимное узнавание партнеров по скрещиванию внутри вида. Экологическая концепция вида определяет вид как группу популяций, которые занимают одинаковые экологические ниши. Принимается, что интеграция видов возникает на основе сходной экологии принадлежащих к ним индивидуумов и популяций и соответственно сходного направления отбора. Генетическая концепция вцда пытается объяснить существование видов тем, что они могут изменяться только в определенных границах, определяемых их собственной генетической структурой.
В то время как вышеназванные концепции вида применимы к ныне существующим организмам, другие концепции пытаются определить вид в его историческом существовании. Эволюционная концепция вида Дж. Симпсона определяет вид как линию развития (т. е. последовательность возникающих друг от друга популяций), которая эволюирует независимо от других подобных линий развития и имеет собственную эволюционную роль и тенденцию развития.
И наконец, в различных вариантах филогенетической (кладистической) концепции вида в один вид объединяют всех произошедших от одного предка членов одной (монофилетической) эволюционной линии развития — от ее возникновения (образования вида) до конца (т. е. образования следующих видов). Таким образом, здесь в основу для распознавания видов положено общее происхождение от одного предка и неявно предполагается, что виды возможно различать только благодаря их собственным признакам (аутапоморфиям).
Очевидно, что различные концепции вида были сформулированы на основе разных, но отнюдь не взаимоисключающих теоретических
положений. Независимо от их теоретической ценности они в очень разной степени годятся для практического использования. Выделенные согласно разным концепциям виды наверняка нередко широко перекрываются. Можно заключить, что морфологические виды существуют потому, что принадлежащие к ним индивидуумы скрещиваются между собой, но репродуктивно изолированы от других видов, что они образовались при одинаковом давлении отбора, являются результатом независимой эволюции и возникли от одного общего предка. Поскольку нет общепринятого критерия видового ранга, морфологическую концепцию вида иногда рассматривают как ненаучную.
Прежде чем давать определение вида, необходимо ответить на вопрос о правомерности объединения индивидуумов и популяций в виды как реально существующие в природе основные единицы биологической изменчивости. В зависимости от той или иной точки зрения на этот вопрос можно от ветить по-разному. Так как основной единицей эволюционного процесса является популяция как группа живущих на определенной территории и скрещивающихся между' собой индивидуумов — ибо именно в популяциях имеет место сдвиг частот аллелей в череде поколений, — можно сделать вывод, что сам ход эволюционного процесса не позволяет признать виды за естественные целостности, и виды, так же как и таксономические единицы более высокого ранга (роды, семейства и т.п.), — категории искусственные. К этому выводу пришел уже Ч. Дарвин. С другой стороны, правда и то, что существование скрещивающихся между собой популяций и их репродуктивная изоляция от других популяций — биологическая реальность, что несколько популяций подвержены сходному давлению отбора благодаря сходной экологии и что несколько популяций, обладающих общими, только им свойственными признаками, произошли от общего предка. При таком подходе объединение индивидуумов и популяций в виды отражает биологическую реальность.
Подводя итог, можно сказать, что систематическая категория «вид» независимо от трудности или даже невозможности ее удовлетворительного определения является до сих пор незаменимым основным понятием для оперирования с научными наблюдениями. Так как на практике большинство видов можно опознать по морфологическим признакам и очень многие «морфологические» виды соответствуют также и критериям других концепций вида, здесь следует понимать виды именно как морфологиче
10.3. Видообразование | 71
ские, которые обычно изолированы друг от друга генетическими механизмами. При этом нельзя отрицать, что морфологическая концепция вида в известной мере субъективна и теоретически недостаточна. Особая проблема распознавания видов возникает у агамоспермных форм (см. 10.1.3.3).
Таким образом, дискуссия о процессе видообразования должна объяснить, как в эволюции возникает, с одной стороны, морфологическое разнообразие, а с другой — репродуктивная изоляция.
10.3.2. Дивергентное видообразование
10.3.2.1. Аллопатрическое видообразование
Внутривидовая изменчивость возникает преимущественно благодаря мутациям, естественному отбору и дрейфу генов. Если
генетический обмен между популяциями или популяционными группами одного вида прерывается, например из-за их пространственного разделения или вымирания промежуточных популяций, то процесс внутривидовой дифференциации в генетически изолированных частях вида может продолжаться благодаря различным мутациям, приспособлению к разным условиям среды в ходе естественного отбора и/или случайной фиксации разных аллелей и привести к возникновению новых видов. Эволюционный процесс разделения одного вида на два (или несколько) новых называется видообразованием (англ, speciation). Просгейшая возможность прекращения потока генов между популяциями одного вида — пространственное разделение. Соответственно этот механизм видообразования называют также географическим, или аллопатрическим, видообразованием. В данном случае усиливающаяся дивергенция изолированных популя-
Рис. 10.31. Географическая дифференциация круга горных средиземноморских форм сосны черной (Pinus nigra) (по W. B.Critchfield и E.L. Little; H.Meusel, Е. Jager и Е.Weinert; Н. Niklfeld).
Подвиды поименованы, соподчиненные локальные расы обозначены цифрами
72 I ГЛАВА 10, эволюция
Рис. 10.32. Видообразование и географическое распространение в одной из групп видов рода Gilia (Polemoniaceae) на юго-западе Северной Америки (по V.Grant).
Существуют географически граничащие друг с другом (А — G. latiflora) или изолированные (В — G. leptantha) подвиды внутри видов, близкородственные и частично перекрывающиеся виды (С— G. lenti-flora, G. latiflora, G. leptantha), менее близкородственные либо аллопатрические (D — G. ochroleuca, G. mexicana) либо симпатрические виды (E — G. ochroleuca, G. tenuiflora, G. leptantha, G. latiflora). Этот пример географического распространения и различной степени родства интерпретируют как видообразование на различных стадиях
ций происходит постепенно, и принято считать, что дивергенция обусловлена преимущественно естественным отбором.
Пространственное разделение и связанная с ним генетическая изоляция популяций одного вида могут, к примеру, возникнуть вследствие разделения массивов суши при движении континентов, возникновения горных систем, фрагментации ареалов (при четвертичных оледенениях) или разделения массивов суши водными преградами из-за потепления климата в после-ледниковье (например, Британские острова/Ев-ропейский континент). Геологические и климатические изменения в истории Земли бессчетное число раз вызывали расчленение прежде
непрерывных ареалов и создавали тем самым предпосылки для видообразования.
Ввиду почти полной невозможности непосредственно наблюдать процесс видообразования в природе важнейшим указанием на большое значение аллопатрического видообразования оказывается географическая специфика внутривидовой изменчивости, соответственно — географическое распространение близкородственных видов. Так, например, у обитающей в Средиземноморье сосны черной (Pinus nigra) обнаружены многочисленные географически изолированные подвиды (рис. 10.31), а внутри подвидов от
10.3. Видообразование | 73
мечена географическая дифференциация. Дифференциацию внутри подвидов и сами подвиды можно рассматривать как разные стадии непрерывного процесса аллопатрического видообразования.
Части вида называют подвидами в том случае, если большинство индивидов можно отнести к тому или иному подвиду (см. рис. 10.31), но существуют и редкие промежуточные формы. Таким образом, изменчивость между подвидами не полностью прерывиста. Кроме того, важно, что подвиды либо обитают в разных областях, либо различаются экологией.
В группе видов Gilia (Polemoniaceae), распространенных на юго-западе Северной Америки, находят географически обособленные (С. leptanlha) или граничащие друг с другом (С. latiflora) близкородственные виды с частично перекрывающимися ареалами (С. tenuiflora, G. latiflora, G. leptanlha), а также менее близкородственные аллопатрические (G. ochroleuca, G. mexicana) либо симпатрические виды (G. ochroleuca, G. tenuiflora, G. leptantha, G. latiflora) (рис. 10.32).
Такой характер географического распространения и разной степени родства интерпретирую! как разные стадии видообразования. Подобные типы распространения и родственных связей можно найти почти повсеместно. Gilia ochroleuca не способна скрещиваться с встречающимися в той же области G. tenuiflora, G. leptantha, G. latiflora (см. рис. 10.32). Но последние три вида с частично перекрывающимися ареалами время от времени скрещиваются между собой. Эти наблюдения показывают, что алло-патрически возникающие виды могут унаследовать механизмы репродуктивной изоляции, которые позволяют совместно обитать близкородственным видам.
Правда, следует задаться вопросом: действительно ли такое распространение родственных видов отражает процесс аллопатрического видообразования и им обусловлено, разве не могло оно возникнуть вследствие совсем других причин уже после образования видов?
Постепенность аллопатрического ви- . дообразования проявляется, например, в более или менее непрерывном увеличении генетического расстояния, выявляемого при сравнении изоферментов, между популяциями одного подвида, подвидами одного вида или близкородственными видами, более далекими видами и т.д.
10.3.2.2. Репродуктивная изоляция
Возможные механизмы репродуктивной изоляции, обусловленные своеобразием растений, можно сгруппировать в зависимости от момента их действия. При этом самые важные моменты — это опыление и образование зиготы. Уже попадание на рыльце индивида большого количества пыльцы другого вида (рис. 10.32) может препятствовать его репродуктивному успеху. Образование зиготы при оплодотворении яйцеклетки спермием чужого вида ведет к бесплодию и тем самым снижает приспособленность индивидуума. Действенные механизмы репродуктивной изоляции, включающиеся перед опылением (англ, pre-mating) и оплодотворением (пре-зиготические), являются важной предпосылкой аллопатрического видообразования наряду с уже обсуждавшейся пространственной изоляцией.
• Экологическая изоляция. Почти полностью перекрывающиеся ареалами виды гравилат городской (Geum urbanum) и гравилат речной (G. rivale) встречаются соответственно во влажных смешанных лесах, кустарниковых зарослях, по опушкам и в сырых местообитаниях различного рода. Если их местообитания оказываются рядом, то виды часто гибридизируют.
• Временная изоляция. Время цветения близкородственных и встречающихся вместе видов может быть разным. Пример суточного различия времени цветения — цветущий весной Lactuca graminifolia и цветущий летом L. canadensis на юго-востоке США. Пример суточного различия цветения — Silene latifolia (цветки раскрываются ночью) и A. dioica (цветки раскрываются днем); они также изолированы благодаря разным опылителям.
• Изоляция экологией цветения. Гибридизации близкородственных видов могут препятствовать специфические опылители. Яркий пример — орхидеи рода Ophrys, строение и аромат цветков разных видов которых имитируют самок разных перепончатокрылых и их феромоны и потому посещаются самцами разных видов. Близкородственные губастик красный (Mimulus саг-
74 I ГЛАВА 10. эволюция
dinalis) и губастик Льюиса (M.lewisii) (Scro-phulariaceae) в Калифорнии опыляют соответственно колибри и шмели. С различием опылителей обычно связаны также и морфологические различия цветков, которые более или менее исключают опыление неспецифичным опылителем. Данное явление называют также механической изоляцией.
Другой случай изоляции экологией цветения — связанная с видообразованием (или внутривидовой дифференциацией) замена системы самонесовместимости и перекрестного опыления на самосовместимость и самооплодотворение. Это особенно четко выявлено на примере эволюции от связанной с тристилией самонссовместимости к почти гомостилии и самосовместимости в роде Eichhornia (Pontedcriaceae). Здесь репродуктивную изоляцию между видами обеспечивают самоопыление и самооплодотворение.
Успешному оплодотворению и образованию зиготы после опыления (англ, postmating) может препятствовать так называемая гибридная несовместимость.
• Гибридная несовместимость. Ее обеспечивают механизмы, препятствующие прорастанию пыльцы, росту пыльцевой трубки через столбик, выходу ядра спермия из пыльцевой трубки или оплодотворению вторичного ядра зародышевого мешка, необходимого для образования эндосперма у цветковых растений.
Другой аспект роста пыльцевых трубок состоит в том, что при попадании на рыльце собственной и чужой пыльцы пыльцевые трубки собственной пыльцы обычно растут быстрее, чем пыльцевые трубки чужеродной пыльцы. В зависимости от количественных соотношений между собственной и чужой пыльцой в той или мере снижается успех оплодотворения пыльцой чужого вида.
После образования зиготы включаются постзиготические механизмы.
• Сниженная жизнеспособность гибридов Fj. Пониженная жизнеспособность гибридных индивидуумов может проявлят ься на разных стадиях— от первого деления зиготы до цветущего и плодоносящего растения.
• Стерильность гибридов. Фертильность гибридов может быть снижена благодаря генным, хромосомным или цитоплазматиче
ским различиям между родительскими видами. Различия по одному или многим генам могут привести при их комбинировании у гибридов к снижению фертильности.
Пример хромосомной мутации (инверсия) как причины сниженной фертильности гибридов между индивидуумами с мутацией и без нее представлен на рис. 10.8. Удалось показать, что сниженная фертильность гибридов между двумя видами подсолнечника (Helianthus annuus и Н. argophytlus) обусловлена различием родительских видов по двум реципрокным транслокациям. Пониженную фертильность гибридов могут обусловить также различия в числе хромосом, возникшие в результате анеуплоидии, дисплои-дии или полиплоидии. Понятно, что возникновение хромосомных различий внутри вида представляет исходный момент для видообразования. При сравнении близкородственных видов во многих случаях четко выявлены хромосомные мутации. Проявляющиеся в стерильности гибридов генные и хромосомные различия чаще всего вызывают нарушения в образовании бивалентов в мейозе у гибридных индивидуумов.
• Гибридный распад. Снижение жизнеспособности или фертильности проявляется не в первом гибридном поколении, а в последующих.
Концепция аллопатрического видообразования предполагает, что названные механизмы изоляции являются побочным продуктом возрастающей генетической дивергенции образующихся видов.
Разную степень репродуктивной изоляции использовали в экспериментальной систематике в трудоемких опытах по скрещиванию для определения степени родства между видами. При этом, правда, стало очевидно, что эволюционная дивергенция по морфологическим признакам и репродуктивная изоляция вовсе не всегда тесно сцеплены. Известны примеры, когда морфологически очень разные виды могут легко скрещиваться и давать фертильные гибриды, а также примеры, где едва различимые формы репродуктивно изолированы друг от друга.
10.3.2.3. Перипатрическое, парапатрическое, симпатрическое видообразование и эффект основателя
Серьезная критика модели аллопатрического видообразования основана на представлении о том, что весьма трудно идеи-
10.3. Видообразование | 75
тифицировать факторы, ответственные за общую эволюцию множества популяций, населяющих часть области распространения вида. Эта общность эволюции была бы возможной при условии потока генов между популяциями и одинакового направления отбора во всей данной области. На это следует возразить, поскольку дистанции потока генов в природе чаще всего малы (см. 10.1.3.4), а условия окружающей среды обычно варьируют на небольших расстояниях, популяции приспособлены к локальным условиям. На основе данного соображения предложены модели видообразования, в которых за исходную точку берут малые, но пространственно изолированные популяции вида. Один из примеров — пе-рипатрическое видообразование (по-английски иногда его называют quantum speciation), при котором обитающие на краю ареала популяции из-за отклоняющихся условий среды оказываются изолированными и мотут дать начало новому виду. Важнейшие аргументы в пользу этой модели видообразования — опять же специфика географического распространения внутривидовых вариаций или близкородственных видов. В принципе похожая модель видообразования, названная эффектом основателя (англ, founder effect speciation), не содержит указаний на то, как малая популяция стала изолированной. Здесь, в частности, возможно, что диаспоры вида попали в географически изолированную область, например на остров. Для обеих моделей общим является внимание к эволюционным процессам при возникновении и после возникновения малых популяций: к уменьшению генетической изменчивости при образовании малой популяции (дрейф генов; см. 10.2.2), а также к быстрой эволюции и возникновению новых особенностей благодаря инбридингу после образования популяции. Это подразумевает, что при видообразовании в малых популяциях случайные процессы имеют большее значение, чем естественный отбор.
Примеры возможного перипатрического видообразования — пары видов Lasthenia minor/ L. maritima (Asteraceae) и Salix alaxensis / S. sili-cicola (Salicaceae). Lasthenia minor населяет материковую часть Калифорнии, тогда как L. maritima
чаше всего растет на скалистых островках с птичьими базарами у калифорнийского побережья. Предположили, что L. maritima возникла из краевых популяций L. minor, изолированных от материка в конце четвертичного периода в результате подъема уровня моря. В то время как арктический вид Salix alaxensis широко распространен в западной Канаде и на Аляске, 5. silicicola имеет узкий ареал в дюнах, образовавшихся после ледникового периода по южному краю озера Атабаска в Канаде. Ареалы обоих видов в настоящее время разделяет примерно 300 км. Возможно, 5. silicicola возникла после отступления ледника в конце четвертичного периода из сохранившихся у озера Атабаска популяций мигрировавшей на северо-запад 5. alaxensis. Как L. maritima. так и 5. silicicola отличаются генетически от их предполагаемых предков L. minor и 5. alaxensis прежде всего утратой генетической изменчивости.
Примеры эффекта основателя — виды Cir sium canescens /С. pitcheri (Asteraceae) и Sanicula (Apiaceae) на Гавайских островах. Подобно Lasthenia и Salix распространенный по берегам Великих озер Северной Америки С. pitcheri проявляет лишь малую часть генетической изменчивости, характерной для широкораспространенного в Небраске С. canescens. Предположили, что предок С. pitcheri мигрировал во время последнего оледенения вдоль края ледника к Великим озерам и таким образом оказался изолированным от С. canescens. Sanicula на Гавайях — один из многих примеров видообразования после однократного заселения островов немногими диаспорами. Четыре вида Sanicula, встречающиеся на трех островах Гавайского архипелага, близкородственны калифорнийским видам этого рода. Обнаружение сниженной по сравнению с материковыми видами генетической изменчивости как у всей группы четырех гавайских видов, так и у каждого из них. а также интерпретация кла-дистических расстояний последовательностей ДПК как молекулярных часов позволяют предположить, что около 8,9 млн лет назад архипелага достигли всего один раз немногие диаспоры (или даже единственная) и что дальнейшее видообразование происходило на архипелаге по модели эффекта основателя примерно 0,9 млн лет назад. Видообразование на архипелаге — пример адаптивной радиации, при которой за колонизацией нового жизненного пространства чаще всего быстро следует эволюционная диверсификация.
Если бы видообразование происходило только в малых популяциях, то географическое распространение, например подвидов Pinus nigra (см. рис. 10.31), можно было
76 I ГЛАВА 10. ЭВОЛЮЦИЯ
бы объяснить и тем, что новые таксоны, возникнув в малых популяциях путем пе-рипатрического образования подвидов или видов или благодаря эффекту основателя, увеличили свои ареалы.
Бесспорно, географически, а тем самым и генетически изолированные части вида могут дать начало новым видам. Однако спорным остается вопрос о том, мотут ли стать новыми видами непосредственно граничащие друг с другом (парапатрические)
или распространенные на одной и той же территории (симпатрические) популяции одного вида. Поскольку прекращение потока генов — важная предпосылка для дивергентной эволюции, в таких случаях был бы необходим механизм репродуктивной изоляции. Этот механизм может возникнуть, если гибриды между соседними, но адаптированными к разным условиям среды популяциями обладают резко пониженной присп особленностью.
Исходный материал
Время цветения
Время цветения
После отбора против гибридизации
67,0
Дни прорастания до цветения (со средними значениями)
Рис. 10.33. Симпатрическое видообразование (no Е. Paterniani).
В эксперименте с кукурузой (Zea mays) интенсивный отбор против гибридизации между двумя сортами (White flint. Yellow sweet) с одинаковыми сроками цветения после 6 поколений приводит к отчетливому расхождению по срокам цветения
10.3. Видообразование | 77
Возникновение репродуктивной изоляции в симпатрических популяциях благодаря сдвигу во времени цветения было показано в селекционных экспериментах с двумя сортами кукурузы (Zea mays) (рис. 10.33). Исходным материалом были сорта кукурузы с желтыми (Yellow sweet) и белыми (White flint) зерновками. Зерновки межсортовых гибридов можно было различить по их окраске. Опыление растений сорта White flint пыльцой растений сорта Yellow sweet приводит к образованию желтых плодов. В эксперименте выращивали смесь из индивидуумов обоих сортов. В начале эксперимента в среднем 35,8 % зерновок, образованных сортом White flint, были желтыми, а 46,7 % зерновок, образованных Yellow sweet, — белыми. Для скрещиваний в последующих поколениях высевали зерновки тех индивидуумов, которые имели самый низкий процент гибридных плодов. Таким образом проводили отбор против । ибридизапии. Через шесть поколений процент гибридных зерновок снизился соответственно до 4,9% (White flint) и 3,4 % (Yellow sweet). Уменьшение гибридизации стало возможным, потому что снизилось перекрывание сроков цветения обоих сортов. Признаки этого явления наблюдали и в природе. У Agrostis capillaris известны генотипы, толерантные к тяжелым металлам. Популяции с этими генотипами часто непосредственно соседствуют с таковыми без устойчивости к тяжелым металлам, например по краю рудничных отвалов. На медном руднике в Уэльсе удалось показать, что толерантные к тяжелым металлам растения на нормальной почве имеют пониженную приспособленность, а нетолерантные растения почти не способны расти на почвах с медью. Гибридное потомство между толерантными и нетолерантными индивидуумами имеет бцтее низкую приспособленность, чем потомство от индивидуумов с одинаковыми генотипами. Это обусловливает (см. рис. 10.33) появление естественного отбора против гибридов. Наблюдали, что толерантные растения Agrostis capillaris начинают цвести примерно на неделю раньше нетолерантных индивидуумов. Эти различия в сроках цветения сохраняются и при культивировании в одинаковых условиях. Так как в целом сроки цветения значительно (на несколько недель) перекрываются, здесь можно предположить по крайнем мере начало эволюционного становления репродуктивной изоляции путем сдвига сроков цветения.
При очень сильном отборе против гибридов, т. е. при сильно сниженной приспособленности гибридных индивидуумов, возможны пара- или симпатрическое возникновение механизмов изоляции и тем са
мым пара- или симпатрическое видообразование. Правда, не ясно, существует ли в природе достаточно сильный отбор этого типа и быстрее ли он ведет к репродуктивной изоляции, чем генетический обмен к гомогенизации генетически дифференцированных популяций. По крайней мере, теоретическая возможность эволюции репродуктивной изоляции при пара- или симпатрии показывает, что репродуктивная изоляция не только бывает побочным продуктом эволюционной дивергенции, но и может возникать вследствие естественного отбора на изоляцию. Это важно и в тех случаях, когда начинают вторично контактировать неполностью репродуктивно изолированные близкородственные виды. В такой ситуации существующие несовершенные барьеры, препятствующие потоку генов, могут быть усилены естественным отбором (англ, reinforcement).
Обычно области гибридизации между видами известны как гибридные зоны. Такие гибридные зоны могут возникать либо вторично благодаря встрече ранее аллопатрически дифференцированных видов, либо, возможно, первично — в ходе пара- или симпатрического видообразования. В обоих случаях они функционируют как барьеры потоку генов, потому что пространственно изолируют друг от друга виды, например отсеивают чужеродную пыльцу.
10.3.2.4. Генетика видовых различий
Аллопатрическое видообразование чаще всего рассматривают как постепенный процесс на основе наблюдений уменьшающегося фенотипического и генетического сходства и постепенно возрастающей репродуктивной изоляции между все менее близкородственными подвидами, видами и т.д. Эта гипотеза подразумевает, что и морфологические изменения — процесс постепенный. Генетические исследования видовых различий многократно показали (рис. 10.34), что изменчивость анализируемых признаков в любом выбранном поколении непрерывна; четкие классы признаков чаще всего выделить невозможно. Поэтому пришли к выводу, что на морфологические призна-
78 I ГЛАВА 10. эволюция
10.3. Видообразование | 79
ки влияют многие гены. Мутации некоторых из многочисленных генов должны были бы тогда приводить к мелким изменениям морфологического признака. Методы молекулярной генетики принесли новые результаты. Стало возможно идентифицировать и анализировать морфогенетические гены. Далее путем анализа распределения морфологических и молекулярных признаков, локализованных на генетической карте сцеплений, можно выявить, сколько генов и с каким относительным эффектом участвует в формировании фенотипа (см. 10.1.2.4).
Особенно хорошо исследована в этом отношении кукуруза. Культивируемая кукуруза (Zea mays subsp. mays) происходит от теосинте (Z mays subsp. parviglumis) и, вероятно, впервые появилась около 10 тыс. лет назад. Важное различие между двумя этими подвидами — строение побеговой системы и расположение женских и мужских соцветий (см. рис. 10.34). У кукурузы главный побег заканчивается мужским соцветием, а боковые ветви первого порядка имеют очень короткие междоузлия и несут верхушечные початки. У теосинте главный побег также заканчивается мужским соцветием. Однако междоузлия боковых побегов первого порядка не укорочены, и побеги заканчиваются также мужскими соцветиями. Женские соцветия следует искать на верхушках боковых побегов второго порядка. Генетический и молекулярный анализ этих различий показал, что они обусловлены всего одним геном. Этот ген, известный у одного мутанта кукурузы как teosinte branched (tbl), влияет на длину междоузлий боковых побегов и пол соцветий. Ген tbl проявляется у теосинте только в пазушных почках боковых побегов первого порядка, из которых образуются женские соцветия. У мутантов культурной кукурузы, напротив, tbl проявляется уже в пазушных почках главного побега. Это при
водит к образованию укороченных боковых побегов с верхушечными женскими соцветиями. Экспрессия этого гена у кукурузы отчетливо сильнее, чем у теосинте. Эволюция здесь, похоже, проявляется главным образом в изменении генной регуляции. tbl — регуляторный ген, влияющий, очевидно, на силу апикального доминирования.
Кукуруза и теосинте различаются и по структуре женских соцветий. У теосинте колоски расположены в два ряда. В каждом углублении на оси соцветия по одному колоску с очень жесткими колосковыми чешуями, охватывающими весь колосок. При созревании женские колоски разрываются. Напротив, у культурной кукурузы колоски расположены в многочисленных рядах. Углубления на оси початка мелкие, каждое содержит по два колоска. Колосковые чешуи маленькие, сравнительно мягкие и не охватываю! колоски. При созревании колоски остаются неповрежденными. Удалось показать, что один ген (teosinte glume architecture (tgal)) влияет на жесткость колосковых чешуй, регулируя отложение в них кремния и лигнина, величину колосковых чешуй и глубину углублений на оси соцветия.
Другой пример большого эффекта генов дают виды губастика (Mimulus). Опыляемый шмелями Mimulus lewisii образует небольшое количество (около 0,5 мкл) очень концентрированного нектара. Опыляемый колибри М. саг-dinalis, напротив, имеет много (около 40 мкл) сравнительно жидкого нектара. Это различие, по-видимому, регулируется преимущественно одним геном.
Такие различия показывают, что, несмотря на влияние на структуру многочисленных генов, морфологическая эволюция может идти путем глубоких преобразований благодаря изменениям регуляторного гена, занимающего более высокое положение в функциональной иерархии генов.
<1
Рис. 10.34. Теосинте (А—С, Z. mays subsp. parviglumis) и дикая форма культурной кукурузы (D — F, Z mays subsp. mays). Различаются по архитектуре побеговой системы и расположению мужских и женских соцветий (по J.Doebley, A.Stec и Ch.Gustus):
А— теосинте: боковые побеги с верхушечными мужскими соцветиями; D— кукуруза: боковые побеги с верхушечными женскими соцветиями (различна также структура женских соцветий); В, С — теосинте: колоски в два ряда, 1 колосок в обертке; Е, F— кукуруза: многочисленные ряды колосков, 2 колоска в обертке. Эти различия определяются в основном двумя генами (teosinte branched 1, tb1 и teosinte glume architecture, tga1)
80
| ГЛАВА 10. ЭВОЛЮЦИЯ
10.3.3. Гибридизация и гибридное видообразование
10.3.3.1. Гибридизация
При всей значимости репродуктивной изоляции для видообразования многие виды растений репродуктивно не изолированы друг от друга и могут гибридизировать. Понятие «гибридизация» здесь следует применять к скрещиванию между видами, но в расширенном смысле его можно использовать для обозначения скрещивания между генетически дифференцированными популяциями, подвидами и другими группировками внутри вида.
Частоту скрещивания между видами в природе можно наблюдать на различных уровнях. С одной стороны, анализ пяти хорошо известных флор умеренного климата показал, что гибридны от 5,8 % («межгорная» флора Кордильер, США) до 22% (Британские острова) от общего числа видов высших растений этих флор. С другой стороны, анализ смешанных популяций гибридизирующих видов показал, что гибриды составляют от <1 % (например, Senecio vernalise S. vulgaris) до 31% (например, у Quercus) индивидуумов этих популяций.
Гибридизация часто бывает асимметричной, когда она успешна только в тех случаях, если два вида выступают в качестве материнского и отцовского растений в определенной комбина
ции. Например, скрещивание между двумя видами примул — Primula vulgaris и Р. veris — бывает успешным, если только в качестве материнского растения выступает Р. veris. Очень часто скрещивание между самосовместимым и самонесов-местимым видами приводит к гибридному потомству лишь в тех случаях, когда материнским растением был самосовместимый вид.
Поскольку частота гибридизации между видами зависит от мощности пре- и постзиготических изоляционных барьеров, она связана и с относительной частотой гибридизирующих видов в их смешанной популяции. Частота гибридизации обычно увеличивается, если один из двух видов встречается в значительно меньшем числе. Это можно объяснить среди прочих причин тем, что вероятность опыления чужеродной пыльцой вида, представленного лишь немногими индивидуумами, в такой ситуации выше, чем при равной численности обоих видов.
Наконец, наличие гибридизации в значительной мере коррелирует с нарушениями окружающей среды либо вследствие естественных причин (например, климатические изменения в четвертичном периоде), либо под воздействием человека. Эффект нарушения состоит в том, что он допускает взаимные встречи видов, географически или экологически разделенных в ненарушенной среде, чем повышается вероятность появления гибридных индивидуумов; если в биотопе, заселенном родительскими видами, гибриды чаще всего оказываются угнетенными, то в так называемом гибридном местообитании или во вновь заселяемых биотопах они могут иметь преимущество перед родительскими видами.
2
2 1.25
1 00
0,75-
| 0.50
X
X 0,25 S
S 3
10
20 30 40 50 60
Длина рахиса, см
Листья: О короткие длинные Облиственные боковые оси соцветия из корзинок:
О 0 О' 1-2 (У 3-6
Срединные листья:
ос Y) без широкого основания листа
Низовые листья:
6 с о без черешковидной зоны
Язычковые цветки:
О белые О светло- • розовые розовые
Опушение:
о густое д рыхлое О скудное
Рис. 10.35. Диаграмма гибридологического анализа. Экспериментальное скрещивание диплоидных видов Achillea setacea (Ps) и A. aspleniifolia (Ра) (по F. Ehrendorfer).
Разные индивидуумы р! от реципрокных скрещиваний (s х а, а х s) обведены пунктирной линией, родительские индивидуумы — штриховой линией. Все остальные точки представляют собой субвитальное F2. Спонтанно возникший аллотетраплоидный индивидуум показан серым цветом
10.3. Видообразование | 81
Рис. 10.36. Возникновение и анализ полиплоидного комплекса у папоротников (северо-американ-ские виды Asplenium) (по W Н.Wagner, D.M.Smith и D. A. Levin).
При подсчетах хромосомных чисел и пар конъюгирующих хромосом у гибридов установлены формулы геномов: диплоидные исходные виды A. platyneuron (РР), A. rhizophyllum (HR), A. montanum (ММ), ди-, три- или тетраплоидные гибриды: RP, RMM, RMP и RMPM, а также аллотетраплоидные дочерние виды A. ebenoides (RRPP), A. pinnatifidum (RRMM) и A. bradleyi (РРММ); история их возникновения подтверждена морфологией, сравнительной фитохимией (фенольные соединения: ксантоны, по результатам двумерной бумажной хроматографии), а также анализом образцов изо- и аллозимов
Гибриды по морфологическим признакам промежуточны между родительскими видами, i ю иногда имеют и новые признаки, т.е. (рис. 10.35) не встречающиеся у родительских видов. Новые признаки у гибридов могут получиться как благодаря новым комбинациям генов, не встречающимся у родительских видов, так и благодаря гетерозиготности отдельных генных локусов.
Распределение морфологических признаков предположительно гибридной популяции (или гибридного вида) в сравнении с таковым родительских видов представлено на диаграмме (см. рис. 10.35). Оно может быть важным инструментом для подтверждения предполагаемого гибридного происхождения. Гибридные индивидуумы часто содержат вторичные вещества, характерные для обоих родительских форм (рис. 10.36),
но нередко и в этих случаях встречаются новые вещества. На уровне генетических признаков свойства гибрида зависят и от хода наследования рассматриваемого признака. В то время как у гибридов, по крайней мере первого поколения, при бипарентальном наследовании ДНК находят признаки ДНК обоих родительских видов, гибрид при наследовании ДНК по материнской или отповской линии может обладать ДНК только одного из родителей.
В ходе дальнейшей эволюции гибридов может наблюдаться гомогенизация последовательности ДНК в направлении одного из родительских видов, как, например, обычно используемого в систематике внутреннего транскрибируемого участка рибосомальной ДНК (ITS). В результате через несколько поколений у гибридов возможно обнаружить ITS-последовательность только одного из родителей. Этот феномен — । появление так называемой согласованной эволюции, которая особенно характерна для повто
82 I ГЛАВА 10. эволюция
ряющихся последовательностей и включает также гомогенизацию многочисленных повторов.
Часто гибриды, по крайней мере между близкородственными видами, отличаются от родителей более мощным ростом. Это явление, хорошо известное как гетерозис, отражает гетерозиготность многочисленных локусов у гибридных индивидуумов.
Данную особенность используют в сельском хозяйстве. Урожайность многих культурных растений (например, кукурузы, сахарной свеклы) можно повысить, используя посевной материал, полученный от скрещивания «чистых» (инбред-ных) линий и дающий сильный эффект гетерозиса. Получение такого посевного материала облегчает использование одной из родительских линий со стерильной пыльцой.
Фертильность гибридов совершенно не связана с гетерозисом. В зависимости от мощности постзиготических изолирующих барьеров гибридные индивидуумы F1 нередко демонстрируют пониженную фертильность. Но так как фертильность разных гибридных индивидуумов, получившихся в результате одного и того же скрещивания, вариабельна, существует возможность того, что отдельные гибридные индивидуумы мало и совсем не отличаются по фертильности от родительских индивидуумов. В то время как приспособленность гибридов в местообитании родительских видов чаще всего ниже, чем у родительских растений, приспособленность гибридов в «гибридном» местообитании может оказаться даже выше, чем у их родителей.
Обычно существование гибридов ограничено во времени и пространстве, но существует возможность того, что они становятся исходным пунктом дальнейшей эволюции и возникновения новых видов. Предпосылкой этого является неполная стерильность гибридных индивидуумов. Для успешного появления гибридного потомства важно быстрое восстановление неограниченной способности к размножению. Оно может происходить или без изменения числа хромосом (гомоплоидное гибридное видообразование и интрогрессивная гибридизация), или в ходе полиплои-дизации (аллополиплоидное гибридное видообразование). Наконец, полностью сте
рильные гомоплоидные или полиплоидные гибриды могут размножаться апомикти-чески.
10.3.3.2. Гомоплоидное гибридное видообразование
Условием восстановления полной фертильности гибридов без изменения числа хромосом (гомоплоидное гибридное видообразование) является рекомбинация в гибридном потомстве. Для этого необходимо, чтобы гибридные индивидуумы могли размножаться либо путем самооплодотворения, либо благодаря скрещиванию друг с другом или возвратному скрещиванию с родительскими видами. При отборе на повышение фертильности результат рекомбинации должен состоять в том, чтобы гибридное потомство становилось гомозиготным по таким факторам, по которым различаются родительские виды и которые как элементы постзиготических механизмов изоляции вызывают пониженную фертильность гибридов. Этими факторами являются генные или чаще хромосомные мутации, по которым различаются родительские виды (см. 10.1.2.2). Поэтому гомоплоидное видообразование часто называют также рекомбинационным видообразованием. Соответственно частоте хромосомных мутаций у гибридизирующих видов гомоплоидное видообразование обычно связано с быстрым изменением структуры хромосом благодаря последующим хромосомным мутациям. Эта хромосомная эволюция у гибридного потомства одновременно приводит к установлению репродуктивной изоляции гибридов от родительских видов. Далее для существования фертильного гибридного потомства важно, чтобы для него нашлось подходящее местообитание.
Процесс гомоплоидного видообразования в связи с хромосомными мутациями и заселением нового местообитания особенно хорошо описан у подсолнечников (Helianthus) в Северной Америке. От гибридизации Helianthus annuus, чаще всего обитающего на тяжелых глинистых почвах, и Н. petiolaris с песчаных почв возникли три гибридных вида: Н. anomalus, Н. deserticola и Н. paradoxus. В противоположность родительским видам Н. anomalus и Н. deserticola заселяют край
10.3. Видообразование | 83
не сухие почвы, а Н. paradoxus— влажные солонцы. Сопоставление последовательности генов в хромосомах Helianthus annuus и Н. petiolaris как родительских видов и Н. anomalus как гибридного потомка показало, что родительские виды с гаплоидным числом хромосом 17 различаются по меньшей мере десятью хромосомными мутациями (три инверсии, семь транслокаций). Гибридный потомок Н. anomalus имеет в шести хромосомах такое же расположение генов, как у обоих родительских видов, в четырех других хромосомах гены расположены либо как у одного, либо как у другого родительского вида, а семь из 17 хромосом отличаются от хромосом обоих родителей (рис. 10.37). Эти преобразования, по-видимому, могли произойти очень быстро. Экспериментальное получение гибридов между Н. annuus и Н. petiolaris с последующим отбором на фертильность привело к ее росту от <10 % у гибридов Fj к почти полной фертильности после пяти поколений. Структура хромосом этих искусственных гибридов была удивительно похожа на хромосомы встречающегося в природе Н. anomalus.
Быстрое восстановление высокой фертильности показано и в селекционных экспериментах с гибридами, например Elymus и Gilia. Наряду с Helianthus гомоплоидное видообраование хорошо документировано также в родах Iris, Paeonia, Pinus и Stephanomeria. Удалось показать, что в возникновении Iris nelsonii принимали участие даже три вида (I.fulva, I. hexa уста, I. brevicaulis).
10.3.3.3. Интрогрессивная гибридизация
В зависимости от относительной частоты самооплодотворения (насколько она вообще возможна) у гибридов, их скрещивания друг с другом и возвратного скрещивания с родительскими видами гибридное потомство в дополнение к случайно появившимся новым признакам будет иметь более или менее промежуточное строение, т.е. проявит большее или меньшее сходство с родительскими видами. Регулярное или по крайней мере сравнительно частое возвратное скрещивание гибридного потомства с одним из родительских видов может привести к тому, что только относительно немногие признаки одного вида постоянно переходят к другому виду. Этот процесс, названный интрогрессивной гибридизацией, или интрогрессией, в принципе соответствует традиционной селек-
Helianthus annuus anomalus petiolaris annuus anomalus petiolaris
Рис. 10.37. Рекомбинация у гомоплоидного гибридного вида (по L. Н. Rieseberg, С. van Fossen и A. Desrochers).
При гаплоидном числе х = 17 подсолнечники Helianthus annuus и Н. petiolaris различаются по меньшей мере 10 хромосомными мутациями (3 инверсии, 7 транслокаций). Гомоплоидный гибридный потомок Н. anomalus в 6 хромосомах (А—F) имеет такое же расположение генов, как у обоих родительских видов, 4 хромосомы (L/M, N Т, U) демонстрируют расположение генов как у одного из родителей, а остальные 7 хромосом отличаются от обоих родительских видов
ции растений, когда путем скрещиваний пытаются передать культурным растениям желаемые особенности дикорастущих видов (например, устойчивость к грибным заболеваниям).
Интрогрессия выявлена, например, у крестовника обыкновенного (Senecio vulgaris) в Великобритании (рис. 10.38). В 1875 г. у этого вида, в норме не имеющего язычковых цветков в корзинках (var. vulgaris), была описана разновидность (var. hibernicus) с короткими язычковыми цветками. Путем экспериментального ресинтеза и
84 | ГЛАВА 10. эволюция
Рис. 10.38. Интрогрессивная гибридизация (по S. Ross-Craig).
Короткие язычковые цветки в корзинках Senecio vulgaris var. hibernicus (С) получились благодаря интрогрессивной гибридизации между S. vulgaris var. vulgaris (В) без язычковых цветков и S. squalidus (А) с длинными язычковыми цветками
использования молекулярных признаков удалось показать, что данный признак возник благодаря интрогрессивной гибридизации между занесенным в Великобританию в XVIII столетии 5. squa-lidus с крупными язычковыми цветками и 5. vulgaris. S. squalidus и сам является гомоплоидным гибридным видом, возникшим из сицилианских 5. aethnensis и 5. chrysanthemifolius.
Как показывает пример широко распространенной в Великобритании var. hibernicus. интрогрессия не ограничена узкими пространственными пределами.
Сравнение пластидных и ядерных ДНК привело к выводу, что интрогрессия пластидных ДНК встречается значительно чаще, нежели интрогрессия ядерных ДНК. Так, нередко обнаруживают, что индивидуумы одного вида обладают пластидным геномом другого вида, а в их ядерной ДНК нет признаков гибридизации. Этот феномен также известен как хлоропластный захват («chloroplast capture»).
Из 141 исследованного в Южной Калифорнии индивидуума Helianthus petiolaris 137 обладали пластидным геномом Н. annuus. Только два индивидуума Н. petiolaris имели признаки I/. annuus и в ядерном геноме. Так как Н. petiolaris был занесен в Калифорнию всего около 50 лет назад, этот пример иллюстрирует и потенциально высокую скорость интрогрессивной гибридизации.
Успешное появление гомоплоидных гибридных видов или возникающих путем интрогрессии новых форм одного вида ос
новано преимущественно на том, что гибриды имеют повышенную генетическую изменчивость благодаря комбинации родительских геномов. Рекомбинация этих вариаций в последующих гибридных поколениях при гомоплоидном гибридном видообразовании может привести к возникновению новых признаков.
Однако гибридизация и интрогрессия ведут также и к снижению репродуктивной изоляции. Это позволяет 5. vulgaris var. hibernicus, возникшей путем интрогрессии между Senecio vulgaris и .S'. squalidus, легче скрещиваться с .S'. squalidus, чем с незатронутой гибридизацией var. vulgaris, что возможно объяснить большим генетическим сходством var. hibernicus с .S', squalidus, генетический материал которого она инкорпорировала, чем сходством var. vulgaris с этим видом. Уменьшение репродуктивной изоляции может привести в конце концов к слиянию двух видов путем гибридизации. Показано, что редкому Канарскому локальному эндемику Argyranthemum coronopifo-Нит как виду угрожает исчезновение вследствие гибридизации с быстро распространяющимся на Канарских островах под влиянием человека A. frutescens. Таким образом возникает важный аспект охраны редких видов — снижение гибридизации между редкими и обычными видами.
10.3.3.4. Аллополиплоидия
Фертильность гибридов, чаще всего пониженная благодаря постзиготическим механизмам изоляции, может восстановить
10.3. Видообразование | 85
ся путем полиплоидизации. Для пояснения обозначим разные гаплоидные геномы гибридизирующих видов как А и В (или А и X при менее выраженных различиях): диплоидные гибридные индивидуумы имеют тогда геномный состав АВ (или АА’)-Из-за недостаточной гомологии хромосом вследствие генных либо хромосомных раз-.ичий образование бивалентов в мейозе более или менее нарушено, что и снижает фертильность (см. 2.2.3.7). Если в ходе гибридизации благодаря слиянию нередуцированных гамет непосредственно или через анортоплоидную промежуточную стадию (см. 10.1.2.3) происходит удвоение генома, то получаются тетраплоидные гибриды с геномным составом ААВВ или ААА’А’, у которых в мейозе каждая хромосома находит себе гомологичного партнера для конъюгации. Это приводит к правильному образованию бивалентов и чаще всего имеет следствием полное восстановление фертильности. Данная модель особенно убедительна, если гибридизирующие виды различаются хромосомными мутациями.
Воздействие полиплоидизации на фертильность более или менее стерильных диплоидных гибридов доказано многократно. В качестве примера можно привести еще раз гибрид между Primula floribunda и Р. verticillata (Р. х kewensis), у которого соматическая полиплоидизации обычного стерильного индивидуума привела к образованию единственного фертильного соцветия. Регулярное образование бивалентов у полиплоидных гибридных растений является, однако, функцией не только гомологии между хромосомами от одного родителя, но и отсутствия гомологии между хромосомами разных родителей. Путем последовательной элиминации 21 пары различающихся хромосом аллогексаплоидной (2п = 6х = 42) мягкой пшеницы (Triticum sativum) далось показать, что при отсутствии 5-й хромосомы В-генома регулярное образование бива-1ентов нарушается и имеет место мультивалентность. Это позволяет сделать вывод, что образование бивалентов также находится под генетическим контролем, причем в данном случае важные для этого гены локализованы в названной хромосоме.
Полиплоидные гибриды в известной мере репродуктивно изолированы от родительских видов вследствие измененного числа хромосом. Возвратное скрещивание, например, тетраплоидного (4х) гибрида с
одним из диплоидных родителей (2х) ведет к образованию триплоидных индивидуумов (Зх), мейоз у которых существенно нарушен из-за невозможности конъюгации одного из трех гаплоидных геномов и которые в соответствии с этим имеют пониженную фертильность.
Однако поток генов между индивидуумами с разной плоидностью не исключен полностью, так как, например, триплоидные растения все же образуют небольшое число диплоидных гамет (см. 10.1.2.3), которые при слиянии с диплоидными гаметами тетраплоидных гибридов могут привести к фертильному потомству. В качестве одного из многих примеров потока генов между растениями различной плоидности можно сослаться на интрогрессивную гибридизацию между Senecio vulgaris и 5. squalidus, поскольку 5. squalidus — диплоид (2n = 20), а 5. vulgaris — тетра-плоид (2п = 40).
Считается, что шансы возникших полиплоидных гибридов могут сохраниться, как и у гомоплоидных гибридов они увеличиваются, если в их распоряжении оказываются нарушенное «гибридное» местообитание либо новый биотоп для заселения. Часто это можно заметить в географическом распространении диплоидных и полиплоидных видов одного рода.
Диплоидные виды и подвиды Biscutella ser. Laevigatae произрастают далеко за пределами области, подвергшейся оледенению в четвертичном периоде, в то время как тетраплоидные таксоны можно встретить главным образом внутри этой области (рис. 10.39). Эффект естественного нарушения местообитаний из-за постоянно меняющегося в четвертичном периоде положения ледника состоял, с одной стороны, в том, что климатические изменения допускали тогда контакты гибридизирующих таксонов, а с другой стороны, в том, что отступление ледника освобождало огромные пространства, которые могли заселять новообразующиеся полиплоидные таксоны. Аналогичный пример дают североамериканские ирисы Iris virginica (2n = 72), /. setosa subsp. interior (2n = 72) и их аллополиплоидный гибрид /. versicolor (2n = 108). В то время как /. virginica и I. setosa встречаются в далеких областях, не подвергавшихся оледенению (юго-восток Северной Америки и Аляска), I. versicolor растет почти исключительно в прежде покрытых ледником областях северо-востока Северной Америки.
Уже давно известный факт, что частота полиплоидных растений возрастает с уве-
86 I ГЛАВА 10. ЭВОЛЮЦИЯ
Рис. 10.39. Распространение диплоидных и тетраплоидных форм Biscutella ser. Laevigatae в Центральной Европе (по I. Manton и др., схематизировано и дополнено C.Konig).
Диплоиды предпочли неоледеневавшие области в эпоху вюрмско! о оледенения, тетраплоиды можно найти также в подвергшихся сильному оледенению Альпах
Рис. 10.40. Геномная гибридизация in-situ («genome painting») соматических хромосом у аллотетра-плоидного табака (Nicotiana tabacum, 2n = 48) (по Е. Moscone):
А — фото; В — поясняющий рисунок. Специфическое связывание ДНК и флуорохромная окраска позволяют различить хромосомные наборы обоих диплоидных родительских видов, белым показан N. otophora (2п = 24), серым — N. sylvestris (2п = 24). Пары хромосом 1 —3 имеют смешанную окраску: здесь произошла транслокация между хромосомами родительских видов
10.3. Видообразование | 87
личением географической широты (доля полиплоидов в скандинавской флоре составляет 56—72 %, а в североафриканской, средиземноморской и Канарской флорах — 23 — 34%), лучше всего можно объяснить изменениями климата в палеоген-неогено-вый и преимущественно в четвертичный периоды. Климатически обусловленные сдвиги ареалов и возникновение незаселенных территорий привели, как и в случае с американскими Biscutella, к появлению полиплоидных гибридных видов.
На возникновение полиплоидного гибридного вида может указывать комбинация родительских признаков, выявляемая морфологическим, фитохимиче-ским, кариологическим и молекулярным методами.
Особенно показательны для кариологиче-ских признаков методы видоспецифичной диф-ференцальной окраски хромосом. К примеру, хромосомные препараты у полиплоидного гибридного вида можно обработать видоспецифическими зондами ДНК, связанными с флуоресцентными красителями, и сделать таким образом узнаваемыми хромосомы его родителей (рис. 10.40). Эта техника известна и как метод хромосомных отпечатков («chromosome painting») или FISH (Fluorescence In Situ Hybridisation). Гибридизацию геномных ДНК на клеточных препаратах в целом называют GISH-техникой (Genomic In Situ Hybridisation).
Кроме этих приемов, можно скрещивать полиплоидные гибридные виды и с их предполагаемыми диплоидными родительскими видами. В мейозе у такого триплоидного индивидуума в самом благоприятном случае удается наблюдать, что два из трех имеющихся гаплоидных хромосомных наборов образуют биваленты, а третий хромосомный набор в конъюгации участия не принимает. Это может означать, что использованный диплоидный вид действительно был родителем гибридного вида. Если образования бивалентов не происходит, то использованный в скрещивании диплоидный вид можно исключить из числа предполагаемых родителей. У триплоидного гибрида между аллотетраплоидным папоротником Asplenium pinnatifldum с генной формулой RRMM и одним из диплоидных родительских видов (A. montanum: ММ) можно наблюдать конъюгацию хромосом только М-ге-нома. Однако возможности этой техники анализа полиплоидных гибридов ограничены, так как конъюгация хромосом не только зависит от их гомологии, но и находится под генетическим контролем, а для отсутствия бивалентов чаще всего
требуется сильная дивергенция хромосом обоих родительских видов.
Наконец, важным средством доказательства является экспериментальный ресинтез полиплоидного гибридного вида из предполагаемых родительских видов. Первый такой ресинтез был осуществлен в 1930 г. А. Мюнтцингом. Мюнтцинг предположил, что пикульник обыкновенный (Galeopsis tetrahif) мог возникнуть как полиплоидный гибридный вид (2п = 4х = 32) из пикуль-ника пушистого (G. pubescens; 2п = 2х =16) и пикульника красивого (6'. speciosa; 2п = 2х = 16). Для доказательства этой гипотезы он скрестил оба последних вида. Самоопыление почти полностью стерильных гибридов дало потомство, в котором был обнаружен единственный трипло-идный индивидуум. Скрещивание этого растения с G. pubescens привело к появлению тетра-плоидных растений, морфологически очень похожих на G. tetrahit и успешно скрещивавшихся с данным видом.
В других случаях был выбран прямой путь экспериментального ресинтеза. Так, табак деревенский, или махорка (Nicotiana rustica; 2n = 4х = = 28), был получен в результате полиплоидиза-ции с помощью колхицина гибрида между N. ра-niculata и N. undulata (оба имели 2п = 2х = 24), а рапс (Brassica napus; 2n = 2х = 38) — в результате полиплоидизации гибрида между капустой (В. oleracea; 2п = 2 х = 18) и репой (В. гара; 2п = = 2х = 20).
Из многочисленных хорошо исследованных примеров возникновения полиплоидных гибридных видов следует еще кратко описать папоротник многоножку (Polypodium), пшеницу мягкую (Triticum aestivum) и козлобородник (Tragopogori). Использовав очень много признаков, удалось показать, что семь диплоидных видов Polypodium принимали участие в возникновении шести тетраплоидных и одного гексаплоидно-го вида (см. рис. 10.40, рис. 10.41). При этом тетраплоидная многоножка обыкновенная (Р. vulgare) возникла вследствие гибридизации диплоидных Р. glycyrrhiza и Р. sibiricum и в свою очередь вместе с диплоидной Р. australe участвовала в возникновении гексаплоидной Р. intetjectum. Благодаря анализу у наследуемой по материнской линии пластидной ДНК удалось надежно показать, что некоторые виды Polypodium (Р. са-lirhiza, Р. hesperium, Р. virginianum) содержат пластидную ДНК обоих родителей и потому, очевидно, возникали по меньшей мере дважды.
88 I ГЛАВА 10. эволюция
P glycyrrhiza 2n
P calirhiza
4n
P califomicum 2n
Р amorphum 2n
P hesperium P saximontanum
4n 4n
P vulgare 4n
P sibiricum 2n
P "okiense”
4n
Р faurici 2n
P interjectum 6n
P virginianum 4n
P australe
2n
P appalachianum 2n
Рис. 10.41. Полиплоидный комплекс в роде Polypodium (по C.H.Haufler, M.D.Windham и E.W. Rabe) 7 диплоидных видов рода Polypodium участвовали в образовании 6 тетраплоидных и 1 гексаплоид-ного вида. Тетраплоидная многоножка обыкновенная (Р. vulgare) возникла в результате гибридизации диплоидных Р. glycyrrhiza и Р. sibiricum. Сравнительный анализ признаков, унаследованных от материнской формы и от обоих родителей, показал, что некоторые виды возникали по крайней мере дважды (Р. calirhiza, Р, hesperium, Р virginianum)
Гексаплоидная (2п = 6х = 42) мягкая пшеница возникла из трех разных видов, из которых, правда, сегодня можно назвать только два (рис. 10.42). Вначале Triticum urartu (геномная формула АА) гибридизировала с неизвестным видом (ВВ). Возникшая в результате Triticum turgidum (ААВВ)
бх
4х Т. turgidum
AABB
Рис. 10.42. Возникновение гексаплоидной пшеницы мягкой (2п = 6х = 42) путем аллополип-лоидии (по F. Ehrendorfer).
Гибридизация Triticum urartu (формула генома АА) с неизвестным видом (ВВ) привела к Triticum turgidum (ААВВ), которая в качестве материнского вида после скрещивания с Aegilops squarrosa (DD) и дала современную мягкую пшеницу с геномным составом AABBDD
в конце третьего тысячелетия до нашей эры вновь гибридизировала в качестве материнского организма с Aegilops squarrosa (DD), дав современную мягкую пшеницу с геномным составом AABBDD. Археологические находки четко показывают, что люди культивировали уже и диплоидные и тетраплоид-ные виды — соответственно, однозернянку и эммер (рис. 10.42).
Как ь мягкая пшеница, многие другие культурные растения также имеют полиплоидное и гибридное происхождение. К ним относятся, например, уже названные рапс (Brassica), табак (Nicotiana), овес (Диепа), виноград (Vitis) и т.д. Полиплоидия у культурных растений, впрочем, не более обычна, чем у родственных им дикорастущих видов. Это можно объяснить, вероятно, тем, что нарушение естественной растительности и культивирование растений человеком аналогично эффекту климатических изменений в четвертичном периоде привели к тому, что географически и экологически изолированные в природе виды встретились и могли гибридизировать, причем уже на ранней земледельческой стадии возделывание более или менее освобожденных от конкуренции культурных растений допускало выживание полиплоидных гибридных видов.
В роде козлобородник (Tragopogori) три европейских вида — Т. dubius, Т. porrifolius
10.3. Видообразование | 89
и Т. pratensis, которые были занесены в Северную Америку, от скрещивания между соответственно Т. dubius и Т. porrifolius и Т. dubius и Т. pratensis возникли тетраплоидные гибридные виды Т. minis и Т. mis-cellus. Путем анализа как ядерной, так и пластидной ДНК было показано, что оба тетраплоидных вида подобно видам Polypodium возникали повторно. Так, Т. miscellus в одном случае имел в качестве материнского растения Т. dubius, а во всех других — Т. pratensis. Повторное возникновение ал-лополиплоидных видов удавалось доказать неоднократно.
Тот факт, что аллотетраплоидные Т. minis и Т. miscellus возникли только в Америке, но не на своей европейской родине, еще раз показывает, что для выживания гибридных видов требуется измененная окружающая среда. Вероятно, отсутствие гибридных видов в Европе связано не с тем, что они там не возникали, а с тем, что они не смогли там закрепиться.
Как и диплоидные гибридные растения, полиплоидные гибриды проявляют большую генетическую изменчивость. Она выражается в повышенной гетерозиготности (в зависимости от аллельной конституции родительских видов) вследствие большого числа аллелей в локусе и большого числа гетерозиготных локусов, а также в образовании новых комбинаций генов.
В то время как у диплоидных гибридов гетерозиготность преходящая вследствие сегрегации аллелей в чреде поколений, у полиплоидов гетерозиготность может фиксироваться. Это происходит, когда гомологичные локусы содержат разные аллели в хромосомах, полученных от обоих различающихся родителей, когда соответствующие родительские хромосомы не образуют биваленты и, соответст венно, отсутствует сегрегация геномов. Удвоение всех общих обоим родителям генов при полиплоидизации позволяет, так же как и удвоение генов при хромосомных мутациях (см. 10.1.2.2), изменить функцию одного из удвоенных генов в последующей эволюции полиплоидов. С другой стороны, известно, что многократно повторяющиеся гены могут утратить свою функцию. Этот феномен известен как «молчащие гены» (gene silencing). Хотя у полиплоидных гибридных растений чаще всего, по-видимому, конъюгируют гомологичные хромосомы одного родительского генома, не исключена и конъюгация гомологичных хромосом, унаследованных от разных родителей, и
таким путем — рекомбинация между разными родительскими геномами (межгеномная рекомбинация). Показано, что геномы поколений F2 и Fs, полученных от самооплодотворения полиплоидных гибридов F,, искусственно созданных в ходе скрещиваний в разных комбинациях Brassica гара, В. nigra и В. oleracea, различаются по 38—96 признакам. Сделан вывод, что эти различия, установленные молекулярными методами, обусловлены перестройками хромосом. Частота геномных изменений в этом примере увеличивалась с возрастанием различий между родительскими геномами. Методом хромосомных отпечатков удалось показать межгеномные транслокации и у табака (Nicotiana) (см. рис. 10.40). Как и у гомоплоидных гибридов, у полиплоидных гибридов рекомбинация высвобождает дополнительную генетическую изменчивость.
Этим увеличением генетической изменчивости, вероятно, и обусловлен очевидный успех полиплоидных гибридных видов в эволюции почти всех групп растений.
На примере Polypodium и Tragopogon было показано, что один диплоидный вид может участвовать в образовании нескольких полиплоидных гибридных видов. Благодаря этому разные полиплоидные гибридные виды имеют общий геном, полученный от одного и того же диплоидного вида, поэтому репродуктивные барьеры между аллополиплоидами обычно слабее, чем между исходными диплоидными видами. Вследствие гибридизации аллополи-плоидов друг с другом или с диплоидами образуются полиплоцдные ряды.
Примеры таких рядов — упомянутые многоножка {Polypodiunr. 2х, 4х, 6х) и пшеница (Triticum-. 2х, 4х, 6х). Еще более длинные полиплоидные ряды обнаружены, например, у подмаренника (Galium anisophyllum: от 2х до 10х) или в роде щавель (Rurnex: от 2х до 20х)
Возраст полиплоидов оценивают в зависимости от относительной частоты диплоидных и полиплоидных видов в полиплоидных рядах. Если в обширном круге родственных форм лишь немногие виды полиплоидные, то говорят о неополиплоидии. Если же, напротив, имеется много полиплоидов порой с очень большим числом хромосом при совершенном отсутствии диплоидных форм, то такой случай называют палеополиплоцдией.
90 | ГЛАВА 10. эволюция
Крайний случай палеополиплоидии — род ужовник (Ophioglossum), в котором у О. reticulatum известен хромосомный набор 2п = 1440 (96-пло-ид). Магнолиевые (Magnoliaceae) с гаплоидным набором хромосом х = 19 также считают палеополиплоидами. Нсо- и палеополиплоидии связаны в непрерывный ряд промежуточными состояниями плоидности и значениями хромосомных чисел.
Как гомоплоидное, так и полиплоидное гибридное видообразование начинается с гибридизации видов, совместно встречающихся в одной области. И если впоследствии гибридные виды благополучно закрепляются, заселяют местообитание, отличающееся от родительских местообитаний, и наконец иную область, то само гибридное видообразование происходит симпат-рически. Этим гибридное видообразование отличается от наиболее обычного аллопатрического видообразования путем эволюционной дивергенции в географически разделенных популяциях. Симпатрия родительских видов и гибридного потомства возможна благодаря тому, что между ними возникает сильная (хотя и не всегда полная) репродуктивная изоляция вследствие рекомбинации в случае гомоплоидного гибридного видообразования и вследствие изменения числа хромосом в случае полиплоидного гибридного видообразования. Полиплоидное гибридное видообразование особенно примечательно тем, что оно более или менее прерывисто и потенциально может произойти за одно поколение. Как и гомоплоидному видообразованию, ему приписывают высокую скорость. Это еще одно отличие от постепенной эволюционной дивергенции географически разделенных популяций.
Очевидно, что гибридизация с се различными возможностями стабилизации гибридного потомства — весьма обычный процесс эволюционного изменения и видообразования в растительном мире. Возможность широкого участия гибридизационных процессов в эволюции растений (такая эволюция называется также сетчатой) всегда следует принимать во внимание при реконструкции филогенетических отношений.
Гибриды с пониженной фертильностью могут сохраниться благодаря агамоспермии
или вегетативному размножению. Полиплоидные и обычно анортоплоидные (например, Зх, 5х) хромосомные числа имеют преимущественно формы с гаметофитной агамоспермией (см. 10.1.3.3). Уже не раз предполагали, что агамоспермные виды возникли как более или менее стерильные гибриды.
В роде рябина (Sorbus) есть три широко распространенных амфимиктических вида — ария (Л. aria), рябина обыкновенная (S. aucuparia) и глоговина (S. to/minalis) с диплоидным хромосомным числом 2п = 2х =34. В результате гибридизации этих трех видов в различных комбинациях и частично при участии нередуцированных гамет возникли многочисленные агамоспермные группы видов. Так, эндемичная для западной Англии S. bristolensis с триплоидным хромосомным числом 2п = Зх = 51 происходит, по-ви-димому, от скрещивания 5. torminalis х 5. aria. По-видимому, из этого же скрещивания следует выводить и агамоспермный, но диплоидный сборный вид .S', latifolia, состоящий из множества узко распространенных мелких видов.
Признают также гибридогенное возникновение форм мятлика альпийского (Роа alpina), представленных амфимикгическимм диплоида-ми и тетраплоидами и апомиктическими (живородящими или агамоспермными) полиплоидами и анеуплоидами (2п = 31 —61), агамосперм-ной лапчатки Нойманна (сборный вид Potentilla neumanniana; 4х— 12х) и многих агамоспермных видов из родов малина (Rub us), манжетка (Alche-milla), ястребинка (Hieradum) и одуванчик (Taraxacum). Так как агамоспермия сравнительно редко полностью облигатна (см. 10.1.3.3), половой процесс, происходящий время от времени у преимущественно агамоспермных гибридогенных видов, может привести благодаря дальнейшей гибридизации к чрезвычайно сложным и динамично изменяющимся комплексам форм. Особенно яркий пример этого — малина и ежевика (Rubus).
Стерильные гибриды могут сохраняться и благодаря вегетативному размножению. Пример — гибрид (Circaea х intermedia) между двулепестниками альпийским (С. alpina) и парижским (С. lutetiana), который несмотря на почти полную стерильность широко распространился благодаря вегетативному размножению.
Гибрид между европейской Spartina maritime (2n = 60) и завезенной из Северной Америки 5. alternifolia (2п = 62) закрепился, с одной стороны, благодаря вегетативному размножению без изменения числа хромосом (5. х toumendii), а с
10.4. Макроэволюция | 91
другой стороны, благодаря полиплоидии как фертильный вид (.S', anglicar, 2n =120, 122). У чистяка (Ranunculus ficaria) диплоидные формы размножаются половым путем, но триплоидные и тет-раплоидные формы — посредством образующихся в пазухах листьев выводковых клубеньков.
10.4. Макроэволюция
Описанных совместных действий случайных мутаций, генетической рекомбинации, дрейфа генов, репродуктивной изоляции и естественного отбора как компонентов неодарвинистических представлений об эволюционном процессе достаточно для понимания внутривидовой дифференциации и различных процессов видообразования. Этот уровень эволюции называют микроэволюцией. Но наверняка существуют и редкие единичные события с необычно существенными эволюционными последствиями. К их числу относятся, например, возникновение автотрофной растительной клетки путем эндосимбиоза гетеротрофного эвкариотного организма с автотрофными бактериями или массовые вымирания типа того, что случилось на границе мела и палеогена, вероятно, в результате падения астероида. Эндосимбиоз можно уподобить очень крупной мутации, а массовое вымирание — это ведь не что иное, как внезапное и резкое изменение условий отбора. Таким образом, оба события подходят под общепринятое понимание эволюционного процесса. Тем не менее возникает вопрос: можно ли объяснить механизмами эволюционных изменений, происходящих на уровне вида, примеры эволюционных изменений, наблюдаемых преимущественно методами палеоботаники и сравнительной морфологии и происходящих в течение длительных геологических промежутков времени (макроэволюция)?
Неодарвинизм на данный вопрос чаще всего отвечает положительно: принято считать, что длительность действия отмеченных выше эволюционных механизмов способна создать наблюдаемые крупные эволюционные изменения. Лежащее в основе этого утверждения представление, что ныне наблюдаемые механизмы эволюции не отличаются от механизмов, действо
вавших в далеком прошлом, восходит к английскому геологу Чарлзу Лайелю, который в своих «Принципах геологии» (Ch. Lyell «Principles of Geology», 1830—1833) объяснял геологическое и геоморфологическое строение Земли именно принципом актуализма, отвергая, таким образом, популярную в его время теорию катастроф Ж. Кювье. Чтение этого сочинения Лайеля во время кругосветного плавания сильно повлияло на Ч. Дарвина.
В качестве примеров макроэволюции следует привести глубокие различия между группами организмов высшего систематического ранга, так называемые эволюционные тренды (главные направления развития): возникновение, длительное существование и вымирание групп организмов.
Заселение суши в ордовике, силуре и девоне приспособленными к жизни в воде растениями — пример огромного эволюционного изменения, потребовавшего множества морфологических, анатомических и физиологических изменений. Растениям потребовалась непроницаемая для воды кутикула, чтобы предотвратить неконтролируемую потерю воды. Так как с появлением кутикулы стало невозможным поглощение воды всей поверхностью растения, возникли ризоиды и корни как особые органы поглощения воды. Необходимость транспорта воды от этих, чаще всего погруженных в почву, органов в другие части растения даже у мелких растений было эффективно только благодаря возникновению особых проводящих элементов (гидроидов, трахеид, сосудов). Возвышающиеся над землей части растений нуждались, кроме того, в укрепляющих структурах. Газообмен, также затрудненный из-за образования кутикулы, был обеспечен благодаря возникновению устьиц, а транспорт газов внутри растения потребовал появления системы внутренних полостей (межклетников). Соответственно этим приспособлениям к жизни на суше очень велики морфолого-анатомические различия между первично-водными и первично-наземными растениями. Палеоботаника показала, что различные структуры наземных растений в истории Земли появлялись последовательно (например, первые фрагменты кутикулы известны из ордовика, растения
92 | ГЛАВА 10. эволюция__________
с устьицами — из силура, а прямостоячие оси с трахеидами — из девона). Далее все вышеназванные структуры можно убедительно интерпретировать как приспособления. Эти два факта указывают на то, что возникновение наземных растений несмотря на их современные большие отличия от первично-водных растений, было постепенным процессом приспособления к измененным условиям среды, который не ускоряли никакие другие механизмы эволюции, кроме уже известных.
Неоднократно мож! ю наблюдать эволюционные тренды — эволюцию признака или комплекса признаков, проходящую в течение долгого времени только в одном направлении. Особенно яркий пример направленной эволюции на высоком систематическом уровне — это усиливающаяся редукция гаметофитного поколения в ходе эволюции наземных растений от мхов с их доминированием гаметофита через папоротники и голосеменные к покрытосеменным, у которых как мужские, так и женские гаметофиты состоят лишь из немногих клеток. Столь же заметно при сравнении разных групп наземных растений усиливающееся укрытие женского гаметофита (см. 11.2). Возможное объяснение эволюционного тренда состоит в допущении направленного изменения среды и тем самым направленного отбора в течение длительного времени. Однако большой вопрос: насколько корректно такое допущение, учитывая, что история климата Земли была скорее турбулентной с сильными колебаниями климата, например, в четвертичном и более ранних периодах? Другое допущение заключается в принятии направленности эволюции в сторону прогресса (прогрессивная эволюция). Применительно к эволюционной редукции гаметофитного поколения и связанного с ней возрастающего доминирования спорофитного поколения можно было бы утверждать, что при изначальном сходстве обоих поколений одно из них более жизнеспособно. Это, как полагают, было бы именно спорофитное поколение, поскольку у него как у диплоидного поколения в противоположность гаплоидному гаметофитному поколению неблагоприятные мутации проявляются
только тогда, когда они гомозиготны. То, что тренд возможно было бы рассматривать как прогрессивную эволюцию, вовсе не означает, что представители ранних фаз этой эволюции были неприспособленными к окружающей среде.
Вместо того чтобы объяснять эволюционные тренды естественным отбором, был предложен процесс, альтернативный признаваемым неодарвинизмом процессам, — межвидовой отбор (англ, species selection). При этом предполагают, что признак, например разная величина, связан с различными темпами видообразования или вымирания. Так, виды, состоящие из более крупных индивидуумов, могли бы иметь более высокий темп видообразования или более низкую скорость вымирания. С течением времени это привело бы к увеличению числа видов из более крупных индивидуумов относительно числа видов из более мелких индивидуумов, а также к возрастанию средних размеров индивидуумов. При этом важно, что размер непосредственно не отбирается естественным отбором. Существование межвидового отбора как процесса, не зависимого от естественного отбора, спорно.
Палеонтологическая «летопись» ясно показывает, что группы организмов возникают, существуют различное время в большем или меньшем видовом разнообразии и затем вымирают или остаются в виде очень немногих видов. Примерами мотуг служить Lycopodiopsida или Equisetopsida, которые в карбоне были представлены несколькими линиями древесных форм и господствовали в тогдашней растительности, тогда как сегодня сохранилось сравнительно немного их видов как примесь к доминирующим в растительности цветковым растениям. Объяснение этого феномена могло бы состоять в том, что одна группа, лучше приспособленная, вытесняет другую. При этом возможны две ситуации: с одной стороны, в неизменной среде возникают прогрессивные признаки, которые ведут к выживанию новообразовавшейся группы организмов, с другой стороны, в измененной среде группы, приспособившиеся к новым условиям, заменяют группы, приспособленные к старой среде.
Большие различия между группами организмов, эволюционных трендов, возникновения и вымирания групп организмов можно без больших проблем понимать
10.4. Макроэволюция | 93
как возникшие благодаря естественному отбору более или менее постепенные адаптивные изменения. И хотя далеко не каждому известному примеру макроэволюци-онных изменений легко найти удовлетворительное объяснение в рамках известных эволюционных процессов, до сих пор не известно более совершенных или общепринятых альтернативных объяснений.
Для описания характера эволюционных изменений в геологическом масштабе времени введено много понятий. Если со временем какая-либо линия развития эволюционно изменяется, то говорят об анагенезе; если в течение длительного времени она не меняется, говорят о стазисе. Эволюционные изменения в одном направлении (эволюционный тренд) называют ортогенезом. Дифференциация одной линии развития и разделение ее на несколько новых известны как кладогенез. Особым случаем кладогенеза можно считать адаптивную радиацию, которая определяется как экологическая дифференциация одной линии развития. Самый известный пример адаптивной радиации — диверсификация, следующая за первичным вселением на архипелаг островов. Хотя адаптивная радиация обычно протекает сравнительно быстро, ее скорость не входит в определение адаптивной радиации. В палентологической «летописи» обычно наблюдают сравнительно долгие периоды стазиса, сменяющиеся короткими фазами интенсивного кладогенеза. Такой ход эволюции называют прерывистым равновесием (англ, punctuated equilibrium).
В эволюции признаков можно отличать гомологии от конвергенций и параллелизмов1. Гомологию следует здесь определить как соответствие признака у разных организмов вследствие его унаследования от общего предка. В противоположность этому признаки называют конвергентными, если они возникли в разных неблизкородственных линиях развития при приспособлении к сходным условиям среды. Обычно конвергентно возникшие сходные признаки можно легко установить на основе их детальной структуры или функции. Часто называемые примеры — это суккулен-тные стебли кактусов Нового Света и молочаев Старого Света (см. рис. 4.35) или редукция околоцветника, возникновение тычинок с подвижными нитями и большими пыльниками с сухой пыльцой, рылец с большой поверхностью и т.п. у ветроопыляемых растений самого разного родства (см. 11.2, опыление). В отличие от вышесказанного при параллелизме речь идет о независимом возникновении сходного признака в близкородственных линиях развития. Параллелизм обычно можно распознать не по особенностям признака, а только благодаря анализу родства. Такой анализ должен показать, что признак общий двум организмам, не выводится непосредственно от их общего предка. Пример такого рода — крестовники обыкновенный (Senecio vulgaris} и лесной (.S'. sylvaticus). Полная или почти полная редукция язычковых цветков в соцветиях этих видов произошла независимо и, соответственно, параллельно в ходе их возникновения от разных предков с нормально развитыми язычковыми цветками. В основе параллельной эволюции лежит как приспособление к сходным условиям среды, так и вероятная канализация эволюционного изменения, обусловленная особенностями общего предка.
1 Гомологию возможно отличать от аналогии, тогда как конвергенцию и параллелизм можно противопоставить дивергенции. — Примеч. ред.
11
Систематика и филогения
11.1. Методы систематики..........95
11.1.1. Распознавание видов.......95
11.1.2. Монографии, флоры и определительные ключи...........95
11.1.3. Исследование родства......96
11.1.3.1 . Признаки...............97
11.1.3.2 . «Конфликты» признаков .99
11.1.3.3 . Нумерическая систематика..................... 100
11.1.3.4 . Филогенетическая систематика..................... 101
11.1.3.5 . Метод максимального подобия..........................103
11.1.3.6 . Статистический анализ гипотез
о родстве........................104
11.1.4 . Филогения и классификация ..104
11.1.5 . Номенклатура ............ 107
11.2. Бактерии, грибы, растения ... 108
11.3. Филогенез растений и история растительности ....................513
11.3.1. Обзор филогенеза организмов.........................513
11.3.2. История растительности.....516
11.3.2.1. Методы...................516
11.3.2.2. Докембрий и палеозой (4 000 — 245 млн лет)..............520
11.3.2.3. Мезозой (245 —
65 млн лет)........................524
11.3.2.4. Кайнозой (65 млн лет — по настоящее время)................526
Цель систематического исследования — упорядочить бесконечное многообразие организмов самых разных форм и образа жизни. Для этого необходимо умение распознавать виды и объединять их в систематические группы более высокого ранга (роды, семейства и т.д.). В задачу систематики входит также описание отдельных видов и более высоких систематических групп, присвоение им особых названий и создание определительных ключей для их идентификации. Опираясь на учение о биологической эволюции, созданное Чарлзом Дарвином (см. гл. 10), систематика добивается такого объединения видов в роды, родов в семейства и т.д., которое отражает естественные родственные связи между организмами и группами организмов. Единственное об ъективное основание для группирования, представляемое самой природой, — история происхождения организмов. В последние годы цель выявления родства
(родственных связей) групп стала значительно ближе, так как систематический анализ все в большей мере опирается на информацию, содержащуюся в ДНК организмов.. Эту информацию в сочетании с данными о «традиционных» признаках, например морфологических, в наше время успешно интерпретируют систематически, используя сильно математизированные и потому результативные методы, а также статистически проверяя качество предлагаемых систематических гипотез. Благодаря этому систематика стала методически вполне современной наукой.
Систематика— первый и важнейший шаг в исследовании биологического разнообразия. Она предоставляет справочную основу для всех остальных биологических дисциплин и всех тех частей (отраслей) нашего хозяйства, которые имеют дело с организмами или их продуктами, обеспечивая точную идентификацию и найме-
11.1. Методы систематики | gg
нование каждого организма и тем самым — ясные связи между организмами. Систематика является также незаменимой основой для понимания эволюционных взаимосвязей биологических явлений. Если, к примеру, цитолог исследует эволюционное происхождение ядерной мембраны, генетик задается вопросом о возникновении интрона, морфолог пытается реконструировать структуру самых первых цветков, физиолог занимается эволюцией различных путей фотосинтеза, а эколог анализирует происхождение разнообразных потребностей растений в составе почвы, то реконструированный систематиком филогенез, в котором определены исходные и производные организмы, представляет собой важнейшее обобщенное знание объекта.
В этой главе сначала представлены методы систематического исследования, а затем дан обзор строения и систематики бактерий, грибов и растений.
11.1. Методы систематики
11.1.1. Распознавание видов
Вид (species) — основная единица систематики. Это не означает, что вид должен быть и основной единицей эволюционных изменений (см. 10.3). Существует множество различных концепций вида (см. 10.3.1), однако большинство видов на практике распознают на основе прерывистых фенотипических вариаций. Таким образом, если можно отчетливо разделить какую-то группу индивидуумов, например по длине листа, характеру опушения цветоножки, окраске цветков и по размеру плодов (см. рис. 10.30), на две подгруппы, причем вариации рассматриваемых признаков внутри этих подгрупп оказываются непрерывными, то данные подгруппы можно рассматривать как различные виды. Этот метод настолько объективен, насколько надежно может быть вычислена и повторно подтверждена наблюдаемая прерывистость мерных и счетных признаков. Во всех этих случаях необходимо прежде всего пока
зать, что мы работаем с однородным или сопоставимым репрезентативным материалом. Впрочем, требуемую для распознавания видов меру фенотипической прерывистости устанавливают субъективно. Фенотипическая прерывистость — это документируемый факт, тогда как отнесение обнаруженных подгрупп исследуемой группы к вариациям, подвидам или видам оказывается лишь формальной проблемой номенклатуры. Для видов общепринято, что относящиеся к ним индивидуумы происходят от одного общего предка и потому не полифилетичны.
Используемые для распознавания видов фенотипические признаки в принципе те же самые (см. 11.1.3.1), что применяются для объединения видов в систематические группы более высокого ранга. На практике большинство признаков анатомо-морфологические, так как первоначальное определение видов чаще всего ведется при работе с музейным материалом — гербарием, и четкая выраженность таких признаков как в природе, так и в коллекциях музеев упрощает отнесение исследуемого индивидуума к тому или иному виду. Огромное значение ботанических музеев (гербариев) состоит прежде всего в том, что только в этих, порой сотни лет собираемых коллекциях исследователь имеет в распоряжении достаточно репрезентативный материал исследуемой группы растений. Самые крупные гербарии мира содержат примерно 6 млн экземпляров растений, и благодаря принятому межгербарному обмену материалом исследователь может получить на время необходимый материал из различных коллекций. Вместе с тем крайне важно исследовать и живые растения, так как определенные признаки, например число хромосом, можно изучить только на таком материале.
11.1.2. Монографии, флоры и определительные ключи
Важным продуктом систематического исследования на видовом уровне являются монографии как систематические обработки определенного круга родственных форм (например, монография рода Primula) и флоры как систематические обработки растений, населяющих какую-либо географическую область (к примеру, флора Германии). Монографии и флоры, с
96 | ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
одной стороны, содержат описания изученных растений, а с другой стороны, делают доступной идентификацию растения благодаря определительным ключам. В монографиях дополнительно содержатся все формальные аспекты систематической обработки (ботаник, первым описавший таксон, — автор вида, рода и т.п., время описания и название публикации, синонимы ит. п.). Описания могут быть весьма различной детализации, но должны ясно показать как изменчивость внутри вида или группы, так и границы с другими видами и группами.
Определительный ключ может быть построен по дихотомическому или политомическому принципу. Возьмем, к примеру, три признака с разными состояниями (см. 11.1.3.2):
1-0 Лепестки красные
1-1 Лепестки желтые
2-0 Плод ягода
2-1 Плод коробочка
3-0 Растение однолетнее
3-1 Растение многолетнее
и распределение признаков по видам:
Вид Окраска лепестков Плод Жизненная форма
А Красная Ягода Многолетник
В Красная Коробочка Однолетник
С Желтая Ягода Однолетник
D Желтая Коробочка Многолетник
В этом случае дихотомический ключ — наиболее часто используемая форма определительных ключей — выглядит следующим образом:
1. Лепестки красные 2.
Лепестки желтые 3.
2. Плод ягода вид А
Плод коробочка вид В
3. Растение однолетнее вид С
Растение многолетнее вид D
В политомическом ключе все признаки для одного вида собраны вместе:
Вид Комбинация признаков
А 1-0; 2-0; 3-1
В 1-0; 2-1; 3-0
С 1-1; 2-0; 3-0
D 1-1; 2-1; 3-1
Преимущество политомического ключа в том, что он позволяет определить вид и тогда, когда, к примеру, окраска цветков неизвестна, так как уже одна комбинация типа плода и жизненной
формы дает возможность различить все четыре вида. В дихотомическом ключе, напротив, без знания окраски цветков нельзя различить виды А и С или В и D. Правда, данный недостаток можно уменьшить, если включить в тезы и антитезы ключа вопрос не об одном, а о двух или большем числе признаков. Это всегда и делается в хороших ключах, но и здесь может встретиться случай, когда невозможно наблюдать запрашиваемый признак. Политомические ключи все чаще предлагают в электронной форме. Например, предлагаемые в компьютерной программе ‘Intkey’ (M.J.Dallwitz) так называемые интерактивные ключи имеют многочисленные преимущества по сравнению с дихотомическими ключами. Прежде всего они состоят в том, что работа по определению может быть начата с любого признака или с любой комбинации признаков. Содержащаяся в монографиях пли флорах информация также частично уже доступна в форме компьютерной базы данных.
11.1.3. Исследование родства
Каждую систематическую группу независимо от ее ранга обычно называют таксоном. Степень родства между таксонами определяют по относительному возрасту последнего общего предка. Так, таксон В имеет более тесное родство с таксоном С, чем с таксоном А, если последний общий предок В и С моложе общего предка А и В (рис. 11.1).
Знание родственных взаимоотношений между таксонами важно с трех точек зрения. Во-первых, родство (о котором недавно можно было сформулировать лишь более или менее хорошо обоснованную гипотезу) — это единственная объективная основа классификации и потому имеет для систематика решающее значение. Во-вторых, классификация, базирующаяся на родстве, имеет известную прогностическую силу. Это означает, что определенное свойство вида может быть с большей вероятностью обнаружено у близких, нежели у дальних родственников этого вида. Так, отдельные алкалоиды группы морфинов мака снотворного (Papaver somniferum) известны только для очень близкородственного мака изящного (Papaver gracile) и для чуть менее близкого мака прицветникового (Papaver bracteatum), но не характерны для более далеких видов мака. Однако родство не всегда имеет прогностическую силу, так как признаки могут быть утрачены. В-третьих, знание родства — единственная возможность реконструировать эволюцию
11.1. Методы систематики! gy
признаков любого вида. Если, например, физио-югию растений интересует вопрос, возникли ли среди амарантовых (Amaranthaceae) С4-растения из С3-растений только один раз или несколько раз, то ответить на него возможно, рассматривая не сам признак, а родственные отношения таксонов с таким свойством.
11.1.3.1. Признаки
Группы признаков, используемые для определения видов и, более того, для исследования родства, т.е. для объединения видов в таксон более высокого ранга, крайне разнообразны.
Историческое развитие систематики показывает, что расширение области наблюдаемых признаков тесно связано с развитием новых методов наблюдения и возрастанием легкости их использования, а также с общим состоянием теоретических знаний. С античных времен важнейшую базу сравнений составляли признаки габитуса растения, затем с XVI—XVII вв. до К.Линнея и далее вплоть до XIX в. — макроскопические признаки цветков и плодов. С широкого использования микроскопа в XIX в. началось исследование слоевищных растений и их oprai юв размножения (Е. М. Fries, Н. A. de Baty, A. Pascher, W. Hofmeister, J.-В. Payer и др.) и, кроме того, анатомических признаков побеговых растений (к примеру, В. Н. Solereder, С. R. Metcalfe). После утверждения эволюционной теории все более важным источником исследования родства становится палеоботаника (H.Solms-Laubach, R. Kid-ston, W. Zimmermann и др.). Еще во второй половине XIX в. выяснили и значение особенностей ареалов для исследования родства (A. Kerner V. Marilaun, R.v. Wettstein и др.). В 20-е годы XX столетия в систематике начали использовать сведения о числе хромосом и их структуре (например, Е. М. East, Е. В. Babcock), а в начале второй половины XX в. большее значение приобрети данные фитохимии (R.E. Alston, R.Hegnauer и др.) и электронной микроскопии (I. Manton, к примеру, в отношении водорослей). В два последние десятилетия прошлого века все более важными становились данные о строении белков (например, L. D. Gottlieb — изоферменты) и нуклеиновых кислот (например, C.R.Woese, J. D. Palmer, М. W. Chase, D. E. & P. S. Soltis), доминирующие в современной систематике.
Большое значение для систематики имеет морфология (см. гл. 4), которая описывает внешнее строение растений. Примеры разнообразия морфологических при
знаков: клеточная организация зеленых водорослей (монадная, коккоидная, нитчатая, тканевая, талломная и т.п.; см. гл. 5; 11.2, Chlorobionta), супротивное или очередное расположение листьев, число и расположение тычинок в цветке (см. 11.2, расположение частей цветка). Для оценки структурных гомологий морфологических признаков часто необходимо изучить их развитие (онтогенез). Так, например, многочленные андроцеи могут возникать совершенно разным способом (см. рис. 11.173). Признаки внутреннего строения исследует анатомия. При этом гистология (см. гл. 3) занимается структурой тканей, как, например, коллатеральным или биколлатеральным расположением ксилемы и флоэмы в проводящих пучках покрытосеменных растений или наличием настоящих тканей или плектенхимы у зеленых и красных водорослей (см. 11.2, Chlorobionta, Rhodophyta). Цитология (см. гл. 2), еще одна часть анатомии, имеет дело с тонким строением клеток. Так, большое значение для систематики имеют структура базального тельца жгутиков (см. рис. 11.88) и детальный ход клеточного деления у зеленых водорослей или электронно-микроскопическая структура пластид ситовидных трубок у покрытосеменных (см. рис. 11.167). Кариология как часть цитологии исследует хромосомы клеточного ядра. При этом выявленная с помощью различных методов окрашивания структура хро-
Рис. 11.1. Родство.
Степень родства определяется относительным возрастом последнего общего предка. Родство таксона В с таксоном С ближе, чем с таксоном А, поскольку последний общий предок (2) В и С моложе, чем последний общий предок (1) А и В
98 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
мосом в интерфазе и преимущественно в метафазе, а также весь хромосомный набор клетки оказываются вариабельны при сравнении различных таксонов. Разделами анатомии можно также считать палинологию и эмбриологию. Палинология занимается структурой спор и пыльцевых зерен (см. рис. 11.174,11.175,11.176). Здесь выявлены различия в характере слоев стенки спор, видимой при электронной микроскопии скульптуры поверхности спор, в числе, форме и расположении апертур. Развитие спорангиев, гаметофитов, гаметангиев, эндосперма и зародыша — область эмбриологии. Различия в возникновении выстилающего слоя (тапетума) спорангиев лежат, к примеру, в основе подразделения папоротников на группы, а развитие зародышевого мешка у покрытосеменных варьирует по числу участвующих в его образовании мегаспор и конечному числу слагающих клеток (см. рис. 11.187). Структуру содержащихся в растениях веществ изучает фитохимия. При этом речь может идти о пигментах фотосинтеза (различные хлорофиллы и дополнительные пигменты; см. 6.4.2), запасных углеводах (крахмал, инулин) или о таких общих для большого круга покрытосеменных растений классов веществ, как алкалоиды, терпеноиды или флавоноиды (см. рис. 11.168). Систематик может получить, кроме того, важные признаки также из области физиологии (например, С3-, С4-фотосинтез; см. 6.5), экологии (например, требования к почве), географического распространения таксона, выявляемого географией растений, или из фитопатологии (например, в отношении специфичности патогенных грибов или травоядных животных; см. 9.3,9.4); особенно важно при изучении родства принимать во внимание исследованные палеоботаниками ископаемые формы. В настоящее время утрачивает свое значение экспериментальная систематика, занимающаяся гибридизацией и интерпретирующая успех скрещивания как критерий родства. Напротив, сегодня для исследования родства особенное значение приобрел анализ протеинов и нуклеиновых кислот. В то время как серологический анализ протеинов (сравнительное осаждение протеинов специфичес
кими антителами) и анализ аминокислотных последовательностей в них сейчас также теряют свое значение, анализ изоферментов (выявление числа генетических локусов и числа и качества аллелей ферментов путем электрофореза; см. бокс 10.1), как и прежде, остается обычным приемом для характеристики генетической конституции популяций или близкородственных видов. Анализ РНК и главным образом ДНК начался с практически утратившего сейчас значение метода гибрцдизации ДНК, когда заключение о степени родства основывали на фотометрически регистрируемой скорости образования двойных цепочек ДНК из отдельных участков ДНК, выделенных из различных видов. Анализ ДНК методом обработки ферментами-рестриктазами, разрезающими ее на специфические участки (например, из 6 или 4 пар оснований) (RFLP), также отходит на задний план. При RFLP-анализе ДНК разных видов нарезают с помощью одинаковых рестриктаз. Различия в получаемых фрагментах обусловлены мутациями в области нарезанных участков. Эти мутации используют как признаки. При преобладающем теперь секвенировании РНК/ДНК определяют последовательность нуклеотидов в нуклеиновой кислоте и каждую позицию нуклеотида рассматривают как признак. Сегодня в систематике растений наиболее часто используют, пожалуй, нуклеотидные последовательности генов или участков генов пластидной rbcL (большая субъединица рибу-лозо-1,5 - би фосфаткарбоксилазы; другие все чаще используемые гены пластид — это, к примеру, matK и ndKF или же определенные пластидные нитроны и спейсеры) и рибосомальной ДНК клеточного ядра. К рибосомальным ДНК клеточного ядра принадлежат как ITS -последовательности (внутренний транскрибируемый спейсер), так и 16/18S последовательности РНК, которые кодируют малую субъединицу рибосомы. Начали также использовать и митохондриальные гены. Предпочтительное применение пластидной ДНК в сравнении с ядерной обусловлено тем, что большинство ядерных генов представлено целыми семействами, вследствие чего при их гомологизации возникают проблемы, в то вре
11.1. Методы систематики | gg
мя как пластидные гены чаще всего имеются в единственном числе. Правда, пластидная ДНК чаще всего наследуется только по материнской линии (см. 10.1.2.5). Особенно информативными могут быть выяв-[яемые при секвенировании или RFLP-анализе макромутации ДНК типа инсер-пий или инверсий, поскольку они редко возникают параллельно у разных видов. Для анализа ДНК на низком таксономическом уровне сегодня еще используют множество тетодов так называемых отпечатков (см. бокс 10.1), с помощью которых, чаще всего с применением полимеразной цепной реакции (PCR), иногда удается даже различить отдельные индивидуумы.
Строение ДНК во многих отношениях имеет преимущество перед другими признаками растений. Это определяется тем. что данные о ДНК возможно однозначно кодировать. В последовательности ДНК можно точно определить положение нуклеотидов. Четко отнести морфологический признак к одному из альтернативных состояний, напротив, часто очень непросто или даже невозможно из-за существования переходных форм. Это особенно проявляется в работе с близкородственными видами, где различия часто только количественные. Другое преимущество данных о ДНК в том, что они позволяют сравнивать друг с другом организмы, имеющие мало общего в фенотипических признаках (например, одноклеточные водоросли и семенные растения), но с одинаковыми генами (например. rbcL). Из строения ДНК можно получить, наконец, очень много признаков, но при этом следует учитывать, что при секвенировании одного гена ана-шзируют множество нуклеотидных последовательностей, хотя рассматривают только один ген. В противоположность этому анализ фенотипов включает обширную область различных признаков и тем самым — также очень большое и совершенно неизвестное число генов.
оценки признаков следует из того, что чаще всего не все признаки подтверждают одинаковое группирование объектов. При рассмотрении нескольких признаков в сущности всегда встречается «конфликт» признаков.
При рассмотрении, например, трех таксонов — А, В и С и двух признаков, имеющих состояния 0 и 1, может оказаться, что признак 1 противопоставляет группу А + В таксону С, а признак 2 — таксон А группе В + С (рис. 11.2).
«Конфликты» признаков имеют место, во-первых, из-за того, что при сравнении двух (и более) таксонов идентичные проявления признака могут быть результатом конвергенции или параллелизма (см. 10.4), т.е. возникали в процессе эволюции неоднократно и независимо друг от друга, что вовсе не очевидно из структуры самого признака. В то время как для обоснования соответствия у двух таксонов какого-либо структурного признака, например морфологического, дополнительно к его особому качеству можно привлечь его относительное положение и онтогенез, для оценки идентичного нуклеотида, находящегося в одинаковой позиции в ДНК двух таксонов, таких дополнительных критериев нет. «Конфликты» признаков возникают, во-вторых, потому что значимость признаков для выявления родства различается в зависимости от времени их возникновения относительно времени возникновения исследуемой группы (см. 11.1.3.4).
Для разрешения «конфликтов» признаков используют разные пути. Первая возможность состоит в том, чтобы приписать различным признакам разный «вес» и таким образом решить проблему. Насколько несомненно то, что разные признаки име-
11.1.3.2. «Конфликты» признаков
В зависимости от исследуемого таксона и его систематического ранга используют разные группы признаков. Для дальнейшего обсуждения ценности признаков имеет смысл разграничить признак и его состояние (англ, character, character state). Так, например, окраска цветка — признак, а красная, белая, синяя и т.д. — состояния этого признака. Необходимость развития методов
Признаки таксоны Группирование
А В С
1 0 0 1 А, В/С
2 0 1 1 А/В, С
Рис. 11.2. «Конфликт» признаков.
Признаки 1 и 2 со своими состояниями 1-0, 1 -1, 2-0 и 2-1 по-разному указывают на родство между таксонами А, В и С: признак 1 показывает группирование А с В, признак 2 — В с С
100 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
ют разные «веса» вследствие своей разной сложности, настолько же трудно объективно оценить их различные «веса». В результате возникают варианты классификаций, имеющие сильный субъективный компонент. Соответственно они могут сильно различаться, несмотря на одинаковый набор используемых признаков. Объективизацией представлений о классификации организмов занимаются, с одной стороны, ну-мерическая систематика, а с другой стороны — филогенетическая систематика.
11.1.3.3. Нумерическая систематика
Нумерическая систематика (фенетика) занимается парными сравнениями всех таксонов для установления сходства их друг с другом и расчета структуры сходств в исследуемой группе. В нумерической систематике анализируемые таксоны обычно называют операциональными таксономическими единицами, «operational taxonomic unit» (OTU). Конкретный способ действий состоит в том, чтобы с самого начала (как и во всех других методах обработки данных) выяснить выраженность признаков у всех таксонов исследуемой группы. Затем можно закодировать их в так называемой бинарной матрице данных, в которой таксоны помещают напротив признаков (рис. 11.3, А). Так, например, для признака «окраска цветков» проявления признака красный цвет или белый цвет кодируют произвольно как 0 и 1. На следующем этапе попарного сравнения всех таксонов рассчитывают сходство (или дистанцию, принимая 1 за сходство). I [ростой способ его вычисления состоит в том, что число признаков, общих двум таксонам, делят на общее число наблюдаемых признаков (рис. 11.3, В). Рассчитанный таким образом коэффициент сходства известен и как коэффициент простого совпадения («simple matching coefficient»). Наконец, на третьем этапе, используя вес коэффициенты сходства, таксоны объединяют в группы со все уменьшающимся сходством. Представленная на рис. 11.3, С фенограмма является результатом так называемого кластерного
анализа, в котором таксоны располагаются в иерархическом порядке. Различные группы, или «ветви», расходящиеся от места разветвления, называют также кластерами.
Только что описанный метод можно видоизменять во многих местах или использовать различные опции. Так, некий признак (например, окраска цветков) может иметь не два, а большее
А В с Г п Ш М Q L J F
А 8/10 5/10 6/10 3/10 2/10
В 0,8 7/10 6/10 1/10 0/10
С 0.5 0,7 9/10 2/10 3/10
D 0,6 0,6 0,9 3/10 4/10
Е 0,3 0,1 0,2 0,3 9/10
F 0,2 0 0,3 0,4 0,9
Рис. 11.3. Нумерическая систематика (по О Spring и Н. Buschmann):
А — матрица данных для таксонов от А до F и 10 признаков, имеющих состояния 0 и 1 каждый; В — парные сходства между таксонами (отношение числа общих признаков к общему числу признаков); С — рассчитанная по коэффициентам фенограмма
11.1. Методы систематики
J 101
число состояний (например, красный, белый, желтый (англ, multistate character). Этот вариант можно рассчитать, как и признаки, только с двумя состояниями. При кодировке количественных признаков, таких, как длина листа, следует создать классы значений, которые обозначают, например, все листья короче 10 см как 0, а все листья длиннее 10 см— как 1. Такие классы обычно задают произвольно. Но в принципе существует возможность отказаться от кодирования таких количественных признаков и рассматривать простую разность между выраженностью признака у двух таксонов как меру сходства (дистанции) по этому признаку. Кроме очень простого коэффициента простого совпадения можно использовать большое число других коэффициентов сходства или дистанции. С их помощью можно, например, учитывать, обусловлена ли общность между двумя таксонами отсутствием или наличием проявления признака. Коэффициент Жаккара не учитывает сходство по отсутствующим признакам. При рассмотрении, например, признаков последовательности ДНК не учитывают увеличение вероятности многократных и конвергентных изменений одного нуклеотида с увеличением дистанции между двумя таксонами (например, дистанция Кантора, дистанция Кимуры). Наконец, есть много разных приемов кластерного анализа и еще так называемый ординационный анализ. Например, метод главных компонентов («principal component analysis», РСА) в отличие от кластерного анализа дает неиерархическое представление отношений сходства. Обычно применяют — в особенности для молекулярных данных — метод UPGMA (метод невзвешенного парно-группового среднего, «unweighted pair group method using arithmetic averages») и соседнего связывания («neighbour joining», NJ). В то время как UPGMA поставляет «правильные» результаты только когда темп эволюции во всех эволюционных линиях одинаков, NJ может справиться и с различными темпами эволюции благодаря постоянному пересчету исходной матрицы дистанций.
Претензия фенетики на объективность во многих отношениях ограничена. Как и все другие методы, она начинает с отбора признаков и их оценки, для чего нет никаких объекгивных критериев. Специфическая проблема фонетических методов состоит в том, что как для вычисления коэффициентов сходства, так и для дальнейших расчетов парного сходства есть множество различных способов, которые могут привести к разным результатам, но нет объективного критерия выбора этих способов. Исходя из этого, представляется принципиальным вопрос, можно ли отождествлять сходство и родство, так как именно родство является единственным объективным кри-
А В С
• У .
Сходство
Рис. 11.4. Сходство и родство (по М. Ridley). При различной скорости эволюции сходство не отражает родство. Хотя А и В более сходны, чем В и С, более близкородственными оказываются В и С
терием группирования. Сходство и родство тождественны только в том случае, когда темп эволюции во всех эволюционных линиях одинаков и нет параллелизмов и конвергенций. Если ускоренная эволюция в одной линии развития, например при завоевании совершенно нового жизненного пространства, ведет к сильной фенотипической дивергенции, то сходство будет неверно отражать родственные отношения (рис. 11.4). Первоначально, впрочем, фенетика и не претендовала на реконструкцию родства.
11.1.3.4 . Филогенетическая систематика
Метод филогенетической систематики (кладистики) разработан энтомологом В. Хеннигом (W. Hennig). На первом этапе отбирают все признаки. Второй этап анализа заключается в оценке признаков по относительному времени их появления. Те признаки, которые встречаются только внутри исследуемой группы (англ.: ingroup) (рис. 11.5), называют относительно продвинутыми, или апоморфными (апоморфия), а те, которые имелись уже вне исследуемой группы, считают исходными, или плезио-морфными (плезиоморфия). В зависимости от того, имеется ли апоморфия только у одного или у нескольких таксонов, говорят об аут- или синапоморфии, а при плезио-морфии используют соответственно понятие симплезиоморфия. Оценка состояния
102 | ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Рис. 11.5. Сравнение с сестринской группой. Если в исследуемой группе таксонов А, В, С проявления признака для Абудут 1-0, для В — 1 -0 и для С — 1 -1, а проявление признака 1 -0 у сестринской группы S считать плезиоморфным состоянием, то придется постулировать только одну трансформацию признака: 1-0 —> 1-1 в линии, ведущей к С. Если же принять за исходное состояние (1-1), то потребуются две трансформации признака: или трансформация признака (1-1 —> 1-0) имела место как при возникновении А, так и при возникновении В, или эта трансформация ((1-1 -> 1-0)) произошла у общего предка А, В, С уже после возникновения S, а при возникновении С случилась обратная трансформация: ((1-0 -> 1-1)). Сравнение с сестринской группой служит основой принципа экономии при распознавании апоморфных признаков
признака как апоморфного или плезиомор-фного относительна, потому что она меняется с объемом исследуемой группы. Так, обладание обоеполыми цветками для покрытосеменных в целом — плезиоморфный признак (в сравнении с однополыми цветками), так как обоеполые цветки — это исходное состояние у этих растений (см. 11.2. Цветок). Для всех семенных растений, однако, обоеполые цветки — это апоморфия, так как это состояние признака встречается только у покрытосеменных и может быть использовано для обоснования монофилии данной группы. Важнейшее достижение Хеннига состояло в обосновании того, что для установления родства годятся только апоморфные, но не плезиоморфные признаки. Так, если бы, например, при сравнении автотрофной астры (Asteraceae/Astera-les) с автотрофным коровяком (Scrophula-riaceae/Lamiales) и паразитической зара
зихой (Orobanchaceae/Lamiales) пришли к выводу о более близком родстве астры и коровяка на основании наличия у обоих хлорофилла, то этот вывод был бы ошибочным. Причина в том, что наличие хлорофилла — исходное свойство зеленых растений и как плезиоморфия не может служить для установления родства в одной маленькой их подгруппе. У астры и коровяка хлорофилл сохранился, а у заразихи был утрачен.
Для решения вопроса, является ли состояние признака апоморфным или плезиоморфным, важнейшим методом служит сравнение с сестринской группой (англ, sister group comparison). При этом принято считать, что состояние признака, встречающееся у ближайшего родственника исследуемой группы (сестринской группы), плезио-морфно. Причина такого допущения в том, что оно экономнее (англ, more parsimonious), чем альтернативное допущение о возможности апоморфного проявления признака в несестринской группе1. Если в исследуемой группе из трех таксонов А, В и С состояния признака следующие: А: 1-0, В: 1-0 и С 1-1, а состояние признака 1-0 у сестринской группы S считать плезиоморфным, то требуется только одна трансформация признака (англ, character transformation) от 1-0 к 1-1 в линии, ведущей к С (рис. 11.5). Если же принять, что исходным состоянием было 1-1, потребовались бы две трансформации признака: либо трансформация от 1-1 к 1-0 имела место дважды при возникновении А и В, или же эта трансформация имела место у общего предка А, В и С после возникновения S, а при возникновении С произошла обратная тра! кгформа-ции от 1-0 к 1-1 (рис. 11.5). Так как лишь апоморфный признак дает возможность судить о родстве, одинаковое состояние признака 1-0 у А и Вне свидетельствует о тесном родстве этих таксонов. Скорее у А и В сохранилось состояние признака общего предка.
1 Авторы нечетко представляют кладистичес-кий принцип парсимонии. Речь должна идти о независимом появлении одинаковых состояний признака в исследуемой и сестринской ей группах. — Примеч. ред.
11.1. Методы систематики | Q3
Под экономностью (англ, parsimony) как причиной выбора информативного признака не следует подразумевать, что и сама эволюция должна была бы быть экономной и всегда идти кратчайшим путем. Экономность — скорее общий научный принцип, который требует минимального числа гипотез, необходимых для объяснения.
Даже если известен самый близкий родственник исследуемой группы, необходимо кроме сестринской группы дополнительно провести сравнение ее с другими родст венными группами. Сестринская группа и эти другие родичи вместе взятые образуют внешнюю труппу (англ, outgroup). Понятие «внешняя группа» более общее, чем понятие «сестринская группа». Проблематичность сравнения с сестринской и внешней группами в том, что принимается неизменность раз возникших признаков в этих группах. Так как сест ринская группа геологически того же возраста, что и исследуемая группа, а остальные таксоны внешней группы геологически старше, они имели столько же (сестринская группа) или даже больше времени (другие таксоны внешней группы) для эволюционного преобразования, чем сама исследуемая группа. Поэтому просто невероятно, чтобы с момента возникновения исследуемой группы ее сестринская или внешняя группа не изменились.
После того как на втором этапе анализа признак оценен как апоморфный или плезиоморфный, на следующем этапе для установления родства оставляют только апоморфные признаки. Если при первичном отборе признаков правильно распознаны все случаи параллельного развития — параллелизмы (англ, parallelism), а также случаи обратного развития признаков — реверсии (англ, reversal), то апоморфные состояния признаков должны быть непротиворечивыми. Однако так почти никогда не бывает. Противоречивость признаков, вызванную параллелизмами или реверсиями, называют гомоплазией (англ, homoplasy), состояние признака называют соответственно гомопластичным, если он в исследуемой группе возникал более одного раза или после возникновения вернулся к исходному состоянию. Существующие «конфликты» признаков разрешают таким образом, чтобы, используя подходящие методы вычисления и следуя принципу экономии, минимизировать число необходимых в целом трансформаций признаков.
Этот последний шаг — важнейший принцип используемого сегодня чаще всего максимально экономного кладизма, который практически заменил филогенетическую систематику в ее первоначальной только что описанной форме и который использован в компьютерных программах, например PAUP (‘phylogenetic analysis using parsimony’). Принцип максимальной экономии состоит в отказе от предварительной оценки состояния признака как апо-морфного или плезиоморфного и тем самым от сравнения с сестринской внешней группой; все признаки рассчитывают так, чтобы число необходимых трансформаций признака в полученных кладограммах было минимальным. Несмотря на это, сестринс-кую/внешнюю группу включают в анализ. Предпочтение отдают самой экономной (англ, most parsimonious) кладограмме, которая первоначально не имеет исходной точки («неукорененная» кладограмма — англ, unrooted). «Корень» как исходную точку такой кладограммы располагают соответственно результатам сравнения исследуемой и внешней групп. Оценка признаков как апо- или плезиоморфных при использовании метода максимальной экономии получается лишь после того, как кладограмма рассчитана и «укоренена». Различные группы, расходящиеся от точки разветвления, или «ветви» кладограммы, называют также «кладами» (‘clades’).
Так называемая эволюционная систематика пытается, хотя и без формализованных методик, отразить в классификации как сходство, так и родство. Еще и сегодня часто используемые системы покрытосеменных растений, например системы А. Сгоп-quist, R.M.T. Dahlgren, А.Л.Тахтаджана и R. F. Thome, представляют как раз эту эволюционную систематику.
11.1.3.5 . Метод максимального подобия
В интерпретации признаков последовательности нуклеотидов ДНК все чаще применяют метод, названный методом максимального подобия (maximum likelihood). Исходным пунктом этого метода является формулировка модели эволюции последо
104 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
вательности нуклеотидов, специфичной для исследуемой группы. С одной стороны, такая модель учитывает вариации последовательности в исследуемой группе, к примеру, оценивая частоту субституций на основе наблюдаемой изменчивости в каждой позиции или из вариаций последовательности, делая вывод об относительной частоте транзиций и трансверсий. С другой стороны, в модели эколюции последовательности дополнительно принимают общие допущения, например, о независимости друг от друга позиций последовательности. Затем рассчитывают родословное древо, которое объясняет с максимальной вероятностью (maximum likelihood) наблюдаемые в исследуемой группе вариации последовательности на основе принятой модели эволюции последовательности, специфической для исследуемой группы.
11.1.3.6 . Статистический анализ гипотез о родстве
Гипотезы о родстве, рассчитанные с помощью фенетики, кладистики или метода максимальной экономии, в известной мере доступны статистической проверке их стабильности. Один из таких методов — бут-стреп-анализ. Путем случайного выбора из оригинальных данных и многократных повторений составляют новую матрицу данных, а затем анализируют ее. Эта новая матрица имеет такой же размер, как и оригинальная матрица, но может отличаться от нее тем, что один признак, например, использован трижды, зато два другие вообще не включены в матрицу. Величина бутст-реп-индекса для одной клады 90 означает, что в 90 % повторных анализов различных матриц данных положение таксонов на этой кладе идентично с положением в анализе матрицы оригинальных данных. В кладистике обычно используют Decay-индекс (а также Bremer-Index). Здесь определяют число дополнительных шагов (трансформаций признаков), необходимых для того, чтобы разрешенная клада (т.е. показывающая родственные отношения между А, В и С; см. рис. 11.1) преобразовалась в политомию (когда невозможно различить родственные связи А, В и С друг с другом).
Значения Decay-индекса 1, к примеру, означало бы, что одна из разрешенных ветвей (клад) в максимально экономном родословном древе становится неразрешенной в древе, которое только на один шаг длиннее максимально экономного. Увеличение Decay-индекса интерпретируют как увеличение обоснованности гипотезы о родстве.
11.1.4. Филогения и классификация
Выше описанные возможности систематического анализа ведут к созданию гипотез о родственных отношениях между таксонами, т.е. филогении таксонов. Особая проблема, равно присущая фенетическим и кладистическим методам, состоит в том, что они с трудом разрешают сетчатую эволюцию (т.е. возникновение нового таксона путем гибридизации дивергировавших линий развития), столь обычную у растений (см. 10.3.3). Как фенограмма, так и кладограмма не являются собственно родословным древом, потому что не дают сведений о предках или потомках. Филогению таксона можно использовать для того, чтобы выявить эволюцию признака. Простое нанесение признаков на фенограмму или кладограмму показывает, как изменялся признак в процессе филогенетического развития таксона. Это представлено на примере форм, родственных чистотелу большому (Chelidonium majus'. Papaveraceae-Chelidonioideae) (рис. 11.6). Фенограмма или кладограмма должны быть основаны на таких признаках, которые независимы от тех, чью филогению собираются реконструировать.
При переводе фенограмм и кладограмм в формальную классификацию следуют многократно принятому принципу филогенетической систематики, согласно которому надо признавать только монофилетические группы. Монофилия (рис. 11.7), по определению, — это тот случай, когда таксон содержит всех прямых потомков одного общего предка. Парафилия — это тот случай, когда все включенные в таксон группы происходят от одного общего пред-
11.1. Методы систематики | "|Q5
Рис. 11.6. Эволюция признаков (по F. R.BIattner и J.W. Kadereit).
Нанесение признаков на кладограмму показывает, как мог бы меняться признак в филогенезе таксона. В круге родства чистотела (Chelidonium majus) многопоровые пыльцевые зерна возникли у общего предка Sanguinaria, Еотесоп, Macleaya и Bocconia, а имеющиеся у общего предка Hylomecon, Stylophorum и Chelidonium трехбороздные пыльцевые зерна стали многобороздными у предка Stylophorum и Chelidonium. При возникновении Chelidonium вновь появились трехбороздные пыльцевые зерна
Рис. 11.7. Моно-, пара- и полифилия (по М. Ridley).
Монофилетический таксон (3-4, 2-4, 1 -4) содержит всех потомков одного непосредственного общего предка. Парафилия наблюдается тогда, когда все включенные в таксон группы (1-2, 1-3) имеют одного общего предка, но не все потомки (3-4, 4) этого предка включены в данный таксон. Полифилетический таксон содержит подгруппы (1 и 3), которые произошли от разных предков
1 06 | ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
ка, но не все потомки этого предка включены в данный таксон. При полифилии в один таксон включают прямых потомков разных предков.
Один из многих известных примеров парафилии — это два североамериканских рода: Clarkia и Heterogaura (Onagraceae). В то время как Clarkia обладает двумя кругами
Clarkia (А—С) и Heterogaura (D — F) различаются числом фертильных тычинок, формой рыльца и типом плода. Анализ родства показывает, что выделение Heterogaura heterandra в самостоятельный род ведет к признанию парафилии рода Clarkia
11.1. Методы систематики
фертильных тычинок, раздельным рыльцем и плодом коробочкой, Heterogaura имеет только один круг фертильных тычинок, головчатое рыльце и плод орех, благодаря чему этот таксон и был описан как особый род. Молекулярные анализы ясно показали, что Heterogaura близкородственна некоторым видам Clarkia и, таким образом, возникла из рода Clarkia (рис. 11.8). Если сохранить Heterogaura как род, тогда Clarkia по отношению к Heterogaura парафилетична, так как, хотя все представители Clarkia и сводятся к одному прямому общему предку, но вследствие исключения Heterogaura не все потомки этого предка включены в один таксон. Отказ от парафилетических таксонов прежде всего предполагает, что критерием для классификации должно быть не сходство, а родство. Поэтому Heterogaura, явно не похожую на виды рода Clarkia, следует классифицировать как представителя рода Clarkia, потому что ее самые близкие родичи относятся к Clarkia'. Молекулярные анализы показали также, что двудольные растения пара-филетичны относительно однодольных (см. рис. 11.213). Однако многие оспаривают требование филогенетической систематики не учитывать в классификации плезиоморф-ные признаки.
11.1.5. Номенклатура
Для наименования и классификации растений существует много формальных правил, которые содержатся в регулярно перерабатываемом «Международном кодексе ботанической номенклатуры». Формальные аспекты систематического исследования называют также таксономией. Из-за крайне неоднозначного определения этого понятия в литературе мы здесь от него полностью отказались. Правда, понятия «систематика» и «таксономия» обычно рассматривают как синонимы.
В системе растений используют ряд обязательных таксономических рангов, или категорий. При этом речь идет об абстрактных понятиях упо-
1 Этот таксон целесообразно включать в род Clarkia потому, что некоторые виды последнего более близкородственны Heterogaum, чем другим видам Clarkia. — Примеч. ред.
рядочивания, которые в иерархии занимают совершенно определенные положения. Название вида представляет так называемый бином (двойное название), состоящий из названия рода и видового эпитета (например, Achillea millefolium). Полное видовое название включает также фамилию автора, который описал этот вид. У Achillea millefolium L. буква L означает Линней. Названия надвидовых таксонов состоят из одного слова (уни-номинальны) (например, Achillea). В табл. 11.1 представлены важнейшие таксономические категории, их общепринятые окончания, а также соответствующие названия конкретных таксонов на примере тысячелистника обыкновенного (Achillea millefolium). Новый таксон в ботанике тогда считают действительно обнародованным (англ, validly published), когда его описание удовлетворяет определенным критериям. Это выбор названия в соответст вии с твердо установленными правилами, наличие диагноза, публикация в одном из общедоступных литературных источников, а также указание так называемого типа (см. ниже), который должен быть сохранен и общедоступен. Условия доступности для всех исследователей литературного источника и типа определены в «Кодексе». Все научные названия растений используют в латинизированной форме. Для родовых названий, а также названий таксонов более высокого ранга используют имена существительные, а для видовых эпитетов (и других названий внутривидовых категорий) чаще всего употребляют прилагательные. Пер-воописание в ботанике должно быть составлено на латинском языке. Благодаря типифика-ции описанное название типа однозначно связано с конкретным растительным индивидуумом. Тип названия вида или внутривидового таксона чаще всего представлен растением, сохраняемым в гербарии. Только в исключительных случаях соответственно исторической традиции типом можно считать изображение растения или его описание. При этом тип вовсе не всегда типичен для таксона. Род типизирован видом, а семейство — родом. Если для таксона существует несколько законно опубликованных названий, то вступает в силу правило приоритета. Используют то название, которое было законно опубликовано первым. Это правило не относится к таксонам выше ранга семейства, а иногда могут быть признаны законными («законсервированы») и не приоритетные названия. Позднее опубликованные названия получают статус синонимов законного названия.
В недавно широко обсуждавшейся «филогенетической номенклатуре», представленной коротко в «Филокодексе» (‘PhyloCode’), при наименовании организмов ориентируются исключительно на филогенетическое родство.
1 08 | ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Таблица 11.1. Иерархический перечень основных таксономических категорий, их нормативные окончания, а также таксономическое положение в системе тысячелистника обыкновенного {Achillea millefolium)
Таксономический ранг (русское и латинское название) Окончания Таксономическое положение (примеры, синонимы)
Царство (regnum) Подцарство (subregnum) Отдел, или тип (divisio/ phylum) Подтип (subphylum) Класс (classis) Подкласс (subclassis) Надпорядок (superordo) Порядок (ordo) Семейство (familia) Подсемейство (subfamilia) Триба (tribus) Род (genus) Секция (sectio) Серия (series) Группа видов (aggregatum) Вид (species) Подвид (subspecies) Вариация (varietas) Форма (forma) -bionta -phyta, -mycota -phytina, mycotina -phyceae, -mycetes или -opsida (или -atae) -idae -anae -ales -aceae -oideae -eae Eucarya Chlorobionta Streptophyta Spermatophytina Magnoliopsida Rosidae Asterales Asteraceae (= Compositae) Asteroideae Anthemideae Achillea Achillea sect. Achillea Achillea millefolium agg. Achillea millefolium A. m. subsp. sudetica A. m. subsp. s. f. rosea
11.2. Бактерии, грибы, растения
I. Царство (домен): Bacteria.......114
Первый отдел: грамположительные бактерии (Posibacteriota)........121
Второй отдел: грамотрицательные бактерии (Negibacteriota)........122
Третий отдел: синезеленые водоросли, или цианобактерии (Cyanobacteriota, Cyanoprokaryota, Cyanophyta)......................127
Четвертый отдел: прохлорофиты (Prochlorobacteriota, Prochlorophyta)..................132
II. Царство (домен): археи (Archaea)..........................132
Первый отдел: Crenarchaeota......133
Второй отдел: Euryarchaeota......133
III. Царство (домен): эукариоты (Eucarya, Eukaryoten)..............134
Размножение, смена ядерных фаз и поколений..................... 135
Первое подцарство: Acrasiobionta. 137
Единственный отдел: Acrasiomycota.................. 137
Второе подцарство: Myxobionta .... 137
Первый отдел: Myxomycota....... 138
Второй отдел: плазмодиофоромицеты (Plasmodiophoromycota)......... 140
Третье подцарство: Heterckontobionta............... 142
Первый отдел: лабиринтуловые (Labyrinthulomycota)........... 142
Второй отдел: Оомицеты, или целлюлозные грибы (Oomycota)... 142
11.2. Бактерии, грибы, растения | -|Qg
Третий отдел: Heterokontophyta ... 150 Четвертое подцарство: хитиновые грибы (Mycobionta).............. 150
Единственный отдел: Eumycota... 150 1. Класс: хитридиомицеты (Chytridiomycetes).............151
2. Класс: зигомицеты (Zygomycetes).................156
3. Класс: сумчатые грибы, или аскомицеты (Ascomycetes)... 160
1. Подкласс: тафриномицетиды (Taphrinomycetidae)......... 161
2. Подкласс: эндомицетиды (Endomycetidae)............. 161
3. Подкласс: лабульбениомицетиды (Laboulbeniomycetidae)....... 163
4. Подкласс: аскомицетиды (Ascomycetidae)............. 164
4. Класс: базидиомицеты (Basidiomycetes) ............ 176
1. Подкласс: устомицетиды (Ustomycetidae)..............177
2. Подкласс: уредомицетиды (Uredomycetidae) ............180
3. Подкласс: Hymenomycetidae .............185
1. Приложение к Mycobionta: несовершенные грибы (Deuteromycetes) .............203
2. Приложение к Mycobionta: лишайники (Lichenes)..........203
Пятое подцарство: Glaucobionta.... 209 Шестое подцарство:
Rhodobionta .....................210
Единственный отдел: красные водоросли (Rhodophyta)...........210
1. Подкласс: Bangiophycidae..217
2. Подкласс:
Florideophycidae.............217
1. Приложение к Rhodobionta: отдел криптофиты (Cryptophyta) ........218
2. Приложение к Rhodobionta: отдел динофиты (Dinophyta, Pyrrhophyta, Dinoflagellate)..................219
3. Приложение к Rhodobionta:
отдел Haptophyta.................222
4. Приложение к Rhodobionta: отдел Heterokontophyta
(= Chrysophyta, Chromophyta).....223
1. Класс:
Chloromonadophyceae..........224
2. Класс: желтозеленые водоросли (Xanthophyceae)....225
3. Класс: золотистые водоросли (Chrysophyceae)..............229
4. Класс: диатомовые, или кремнеземные водоросли
(Bacillariophyceae = Diatomeae).................231
5. Класс: бурые водоросли (Phaeophyceae)...............235
Седьмое подцарство: хлоробионты (Chlorobionta, «Viridiplantae»)..245
Первый отдел: зеленые водоросли I (Chlorophyta)........246
1. Класс: Prasinophyceae.....248
2. Класс: Ulvophyceae........249
3. Класс: Trebouxiophyceae...250
4. Класс: Cladophorophyceae............251
5. Класс: Bryopsidophyceae
(= сифоновые)................252
6. Класс: Dasycladophyceae...253
7. Класс: Trentepohliophyceae..........254
8. Класс: Chlorophyceae .....255
1. Приложение к Chlorophyta: Chlorarachniophyta...............265
2. Приложение к Chlorophyta: отдел эвгленовые (Euglenophyta)........265
Второй отдел: Streptophyta ......267
Первый подотдел: Streptophytina (зеленые водоросли II)........267
1. Класс:
Mesostigmatophyceae..........267
2. Класс: конъюгаты, или сцеплянки (Zygnematophyceae, Conjugatae)..................267
3. Класс: Klebsormidiophyceae
(Coleochaetophyceae).........270
4. Класс: харовые, или лучицы
(Charophyceae)...............271
Второй подотдел: мхи (Bryophytina) ................277
1. Класс: Marchantiopsida (= Hepaticae р.р., талломные печеночники).................281
2. Класс: Jungermanniopsida
(= Hepaticae р.р., преимущественно листостебельные
печеночники).................286
3. Класс: Bryopsida (= Musci, листостебельные мхи).........288
110 | ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
1. Подкласс: торфяные мхи (Sphagnidae)...............293
2. Подкласс: Andreaeidae ..294
3. Подкласс: Bryidae.......294
4. Класс: антоцеротовые (Anthocerotopsida) ...........300
Третий подотдел: папоротникообразные, птеридофиты (Pteridophytina) .... 305
1. Класс: псилофиты (Psilophytopsida) .............310
2. Класс: плауны (Lycopodiopsida) ..............312
3. Класс: хвощевидные (Equisetopsida
= Sphenopsida).................322
4. Класс: псилотовые (Psilotopsida).................328
5. Класс: папоротники (Pteridopsida) (= Filicopsida).329
Ступень развития: первопапоротники (Primofilices = = Protopteridiidae)........330
Ступень развития: эуспорангиатные папоротники (Eusporangiatae =
= Ophioglossidae)..........333
Ступень развития: лептоспорангиатные папоротники (Leptosporangiatae = = Pterididae)..............335
Ступень развития: водные папоротники (Hydropterides = = Salviniidae) ............342
Четвертый подотдел: семенные растения (Spermatophytina)....350
Чередование поколений.........350
Вегетативные органы...........353
Вторичные метаболиты..........357
Цветки........................358
Околоцветник..................359
Микроспорофиллы...............360
Пыльца.....................364
Мужской гаметофит..........367
Мегаспорофиллы ...............369
Семязачатки................373
Женский гаметофит..........375
Нектарники.................377
Расположение элементов цветка.....................377
Соцветия.....................379
Опыление.....................379
Оплодотворение...............386
Семя.........................387
Плоды .......................393
Вскрывающиеся плоды........394
Невскрывающиеся плоды .....395
Соплодия...................395
Распространение плодов и семян......................396
Прорастание семян............398
Система Spermatophytina......399
1. Класс: саговники (Cycadopsida) (Palmfarne)..399
2. Класс: Ginkgopsida (Ginkgo)...................402
3. Класс: хвойные (Coniferopsida)............403
4. Класс: Gnetopsida......411
5. Класс: цветковые растения, покрытосеменные (Magnoliopsida)
(Angiospermae).............415
Эволюция и филогения покрытосеменных (обзор)......416
Система покрытосеменных......420
1. Подкласс: Magnoliidae..420
2. Подкласс: Liliidae (Monocotyledonae)..........426
«Lilianae»...............432
«Коммелиноидные»
однодольные..............438
3. Подкласс: «настоящие двудольные»
(Rosidae)..................446
Основные «настоящие двудольные» ...............450
«Розиды».................458
«Эурозиды I».............459
«Эурозиды II»............478
«Астериды»...............485
«Эвастериды I»...........488
«Эвастериды II»..........499
Происхождение и родственные связи семенных растений............508
11.2. Бактерии, грибы, растения | -| -| -|
Понятия «животные» и «растения» исходно приравнивали к двум основным систематическим группам (таксонам) живых организмов («regnum vegetabile» и «regnum animate» соответственно). В настоящее время известно, что мы имеем здесь дело с типами организации, различающимися по физиологии питания, а не с естественными группами, связанными общим родством. При этом «растительное царство» даже не представляет собой определенной эволюционной линии и соответственно не является каким бы то ни было таксоном. Растения можно определить как фотоавтотрофные организмы. Соответственно ботаника — это биология фотоавтотрофов. В рамках ботаники рассматриваются все фотоавтотрофные организмы, однако также и те гетеротрофные группы, которые происходят от автотрофов или важны для понимания филогении автотрофов.
В самом широком смысле, следовательно, и грибы (лишайниковый симбиоз!), и прокариоты (гипотеза эндосимбионтов!) являются предметом ботаники и соответственно этой книги. Грибы имеют с типичными растениями некоторые конвер-гентно возникшие общие признаки (см. ниже).
Ультраструктурное исследование строения клетки выявило у живых организмов два принципиально различных, в настоящее время не связанных между собой переходными формами плана строения: протоциты и эуциты. Эти два плана строения клетки сравнительно недавно были положены в основу подразделения всего живого на две основные группы (царства): Procaryota (прокариоты) и Eucaryota (эукариоты). Недавние исследования с привлечением методов молекулярной биологии свидетельствуют в пользу факта очень раннего расхождения на три основные группы: археи (Archaea), бактерии (Bacteria) и эукарии (Eucarya), причем Archaea на филогенетическом древе (рис. I1.9) оказываются ближе к Eucarya, чем к Bacteria. Однако на основе строения клетки Archaea и Bacteria как прокариоты противопоставляются Eucarya (эукариотам). Все три основные группы (Bacteria, Archaea, Eucarya)
рассматриваются здесь в ранге царств. Чтобы избежать путаницы с применявшимся ранее понятием «царство» («растительное царство», противопоставляемое «животному царству»), вместо понятия «царство» при филогенетической классификации живых организмов все больше используется категория «домен» (домен Bacteria ит.д.).
В приводимой ниже системе рассмотрим Archaea и Bacteria, а из Eucarya — все автотрофные и произошедшие от них вторично гетеротрофные растения (в узком смысле слова), а также гетеротрофные грибы (в широком смысле слова). Как правило, это организмы, ведущие прикрепленный образ жизни, с жесткими клеточными стенками; они могут получать питательные вещества только в растворенном (или же газообразном) виде. Таким образом, сюда не включаются одно- и многоклеточные животные (Archaeozoa, Protozoa и Metazoa = Zoobionta). При этом необходимо учитывать, что разграничение растений, грибов и животных у низкоорганизованных эукариот (= Protobionta, например слизевики и эвглены, рис. 11.9) связано с немалыми трудностями.
В пределах рассматриваемых царств (regna, домены) обширные, однако однозначно произошедшие от одной предковой группы (монофилетические) таксоны трактуются в ранге подцарств (subregna, субдомены) и отделов (phyla). Их названия в случае подцарств оканчиваются на -bionta, в случае отделов автотрофных эукариот на -phyta, а в случае грибов — на -mycota.
Взаимоотношения и возможное родство большинства описываемых в этой книге групп суммируются в кратком обзоре, приводимом после соответствующего раздела. Общий обзор принятой авторами книги системы можно видеть в оглавлении и на рис. 11.9.
Более ранние названия крупных групп (например, водоросли, грибы, семенные растения) в свете современных исследований родственных связей в значительной мере не годятся для обозначения монофилетических эволюционных линий. В крайнем случае такие названия соответствуют
112 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
a LU
4 Gymnospermae
filicopsida
Magnowdae
Equisetopsida
Lycopodiopsida
Streptophyte
Labyrsnthulomycolai
МИТ0Х0НДРИ1
Negibactenote
Proch oiwactenote
turyarchaeota
7
Bryophytma
Anthocerotopsida
Bryopsidj
Archaea
Crenarchaeota
Cyanobactenote
Bacteria
Ptendophytma
Psilotopsida
Первичные пластиды
Posibactenota
(Angiospermae)
Jungermanniopsida
Marchantiopsida
Streptophytma
Klebsormidicphyceae
Charophyceae Zygnematophyceae Mesostfgmatophyceae
Chlorophyta-Chlofophytina
Chlorophyceae
Ulvophyceae
7 „^‘ТгеЬоихюрЬусеае
у Cladophorophyceae
>1 Bryopsidophi'cea^’’**?
Glaucobionta Glaticophyte
•Dasydadophyceae
Trentepohljophyceae
Rhodobionta Rhodophyto
Mycobionta
BasidiomyceSes
Zygomycetes
Animaha (Metazoa)
Lichenes
Ascomycetes
Chytridiomycetes
Heterokontobionta
Heterokontophyta ptaeophycea.
Baallariophyceae
Chrysophyccae
Xanthophyceae «—
Chloromonadoptiyceae
Oomycota
| 'K’f Chloraracfmiophyta
Dinophyta
Cryptophyte
Haptophyte
Progenota
• Acrasiobionta
Рис. 11.9. Филогенетическое древо растений и грибов (по A. Bresinsky и J.W.Kadereit):
Стрелками указаны случаи возникновения эндосимбиозов и как результат этого —возникновение митохондрий и пластид. Стрелки, идущие к лишайникам (Lichenes), показывают установление эктосимбиоза с цианобактериями и Chlorophyta:
1 — возникновение; 2 — приобретение митохондрий в ходе эндосимбиоза; 3 — цианобактерии как первичные эндосимбионты, приобретение «первичных пластид»; 4 — красные водоросли как вторичные симбионты, приобретение «вторичных пластид»; 5 — зеленые водоросли как вторичные симби-оны, приобретение «вторичных пластид»; 6 — фотоавтотрофность вследствие эктосимбиоза
11.2. Бактерии, грибы, растения | -| -| 3
типам организации. Однако для соответствующего большим группам плана строения (типа организации) или одного уровня развития (уровня организации) эти понятия все еще сохраняют определенное значение при обозначении конкретных общностей и для легкого охвата многообразия форм. Поэтому более ранние обозначения типов организации объяснены в этой книге в специальных вставках— боксах (J 1.1 —11.3; 11.5; 11.8—11.10), хотя они и не составляют ныне основу для систематического подразделения.
К одному типу организации (любого объема) относят группы организмов, которые по особенностям своей внешней (т.е. морфологической) или также и внутренней (т. е. анатомической и цитоло!ической) организации в значительной мере совпадают. Типы организации часто соответствуют уровням развития и как таковые являются выражением неоднократно возникавших приспособлений к определенным условиям жизни или же общего прогрессивного развития организации.
Таким образом, типы организации охватывают и явно гетерогенные группы. Однако, с другой стороны, они нередко разделяют родственные группы, которые с точки зрения эволюции и систематики должны быть помещены близко друг от друга.
История системы «растительного царства» связана со сменой лежащих в ее основе фундаментальных принципов. Наиболее известная из искусственных систем та, которую предложил К.Линней (1735) на основе половых структур. Он противопоставил 23 классам цветковых растений 24-й класс — Ciyptogamia («тайнобрачные»), к которым отнес не только тогда еще малоизученные папоротники, мхи, водоросли и грибы, но и некоторые высшие растения с трудно распознаваемыми цветками (Ficus, Lemna) и даже кораллы и губки. Подразделения (таксоны) цветковых растений (Phanerogamia, «явнобрачных») Линней различал прежде всего по распределению пола в цветках и по числу, срастанию, расположению и соотношениям длин тычинок. Тайнобрачные ныне можно называть споровыми растениями, так как у них развитие новых особей происходит чаще всего из одноклеточных диаспор (например, спор), а явнобрачные —
цветковыми, или, лучше, семенными растениями.
Уже Линней попытался создать естественную систему растений, однако только А.Л. де Жюссье (1789), О. П.Декандоль (1819), Ст.Эндлихер (1836) идр. могут считаться создателями важнейших формальных систем. Даже после появления эволюционного учения в системах А. Брауна (1864), Дж. Бентама и Дж. Д. Гукера (1862 — 1883), А. В. Эйхлера (1883) и особенно в широко используемой и сейчас системе А. Энглера для выделения таксонов сохраняется использование уровней организации и уровней развития.
Первая попытка создания филогенетической системы была сделана Р. фон Вет-тштейном (1901 — 1908). Современные общепринятые системы представляют собой различные этапы на пути от формальной к филогенетической и синтетической группировкам.
Даже при самом общем рассмотрении всевозможных ныне конкурирующих между собой систем можно увидеть многие, часто глубокие различия. Это показывает, в какой степени систематика все еще находится в состоянии становления. Правда, в последние годы вследствие сравнения и кладистической обработки данных по последовательностям различных гомологичных участков ДНК из хлоропластного, митохондриального и ядерного генома сделан существенный скачок в выяснении родственных связей всех групп организмов.
Таким образом, можно надеяться достичь в обозримом будущем общепризнанной приемлемой естественной группировки.
Точно так же и приводимая здесь система представляет собой попытку в какой-то мере представить в общих чертах родственные связи между большими группами. Однако учитывая учебный характер данной книги, мы сделали в описании определенные упрощения.
К настоящему времени известно около 500000 ныне живущих растений. Из них более двух третей относятся к семенным растениям (около 700 голосеменных и 240000 покрытосеменных), около 10000 —
114 | ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
к папоротникообразным и 24 000 — ко мхам. Среди Protobionta число описанных водорослей оценивается приблизительно в 23 000, грибов — около 100000, лишайников — примерно 20 000. Наконец, следует учитывать еще 3 000 видов бактерий и 2 000 — синезеленых водорослей. Каждый год описываются все новые виды (особенно грибов и покрытосеменных!), поэтому вполне можно предположить, что еще далеко не законченная инвентаризация растительного царства приведет к тому, что число его видов значительно превысит полмиллиона!
I. ЦАРСТВО (ДОМЕН): BACTERIA
Царство Bacteria (бактерии, цианобактерии. прохлоробактерии) включает прокариотные организмы (бокс 11.1) с особой химической структурой клеточной стенки (муреиновый мешок).
Два рассматриваемых далее отдела — Posibacteriota и Negibacteriota — образую! группу бактерий в узком смысле слова, они называются также эубактериями (Eubacte-ria; в противоположность цианобактериям и т.д.).
Эубактерии (см. рис. 2.93) — это прокариоты, подавляющее большинство видов которых гетеротрофны (см. 9.1); кроме того, они очень мелкие (см. рис. 1.1) и морфологически малодифференцированны. Различные формы клеток бактерий могут быть выведены из исходной формы шара, а также прямого или изогнутого цилиндра (см. рис. 11.10).
Различают: шаровидные кокки, которые могут объединяться в простые группы типа колоний; палочки, спорообразующие формы которых называют бациллами; формы от изогнутых (вибрионы) до винтообразно извитых палочек (спириллы).
У некоторых бактерий клетки после деления остаются соединенными между собой, образуя скопления из клеток, пакеты (сарцины, рис. 11.10, F), нити (рис. 11.10, Н) или сети. Уже у бактерий можно, таким образом, констатировать тенденцию к развитию более сложных структур. Однако у таких многоклеточных форм принцип разделения функций осуществляется редко (ср. Chlorochromatium).
Состоящие из суммы клеток нити могут быть простыми или разветвленными; некоторые из них погружены в слизистые влагалища или имеют жгутики. Аналогичную конвергенцию с мицелием
Бокс 11.1. Типы организации прокариот
Прокариотные клетки (протоциты) не имеют настоящего, т.е. ограниченного оболочкой, ядра (отсюда возникло их прежнее название — Anucleobionta. безъядерные), однако у них есть от одного до нескольких эквивалентов ядра — нуклеоид(ы). ДНК свободно располагается как генофор в так называемой нуклеоплазме. Митоз и мейоз отсутствуют. Подразделение клетки на реакционные пространства (компарт-менты) выражено меньше, чем у эукариот: отсутствуют хлоропласты и митохондрии. У некоторых прокариот имеются двигательные органеллы, но их структура существенно отличается от структуры аналогичных органелл у эукариот. Стенка прокариотной клетки состоит из гетерополимерных веществ, которые
пока не удалось выявить ни у каких эукари-отных организмов. Клеточная стенка представляет собой гигантскую мешковидную полисахаридную молекулу различного химического состава, за счет основных валентностей связанную в сетевидное образование (см. 2.3.3).
Если эукариоты очень зависимы в своей жизни от кислорода, отношение к этому веществу у разных прокариот разное. У них наблюдается переход от полной непереносимости кислорода до обязательной потребности в нем. Только у прокариот имеется широко встречающаяся здесь способность связывать атмосферный азот.
К прокариотам относятся бактерии, цианобактерии, прохлоробактерии и археи.
11.2. Бактерии, грибы, растения | -|-(5
Рис. 11.10. Bacteria. Формы бактерий (А — по Umeda; В — по O.Kandler; С — по Stolp; D — по Krieg; Е— по Houwink; F— по Beveridge; G—J— по Brock и Honinger; К— по Н. G.Schlegel; L—N — по Grillone):
A — Staphylococcus; В — Lactobacillus; С — Bdellovibrio; t> — Spirillum; Е — Caulobacter (4 000х); F — Sarcina; G—J — Sphaerotilus: G — подвижная стадия (700x), H — форма Sphaerotilus (330x), J — начало разделения клеток (800x); К — Streptomyces; L—N — Chondromyces: L — палочки (200x), M, N — плодовые тела (ЗОх)
эукариотных грибов представляют разветвленные, многоклеточные нити актино-мицетов (рис. 11.10, К). Миксобактерии — это гибкие, ползающие по поверхности субстрата прокариоты. Аналогично некоторым эукариотным миксомицетам, отдельные клетки некоторых из таких форм сползаются в плодовые тела размером до 1 мм (рис. 11.10, L—N).
Морфологическая дифференцировка свидетельствует об эволюции в сторону более комплексных структур. Хотя здесь ни в коей мере не достигается комплексность, свойственная эукариотам, все же встречаются уже соответствующие эукариотам формы, выработавшиеся в связи с приспособлением к определенным условиям жизни (колонии, нити, разветвленные нити, мицелий, плодовые тела, споры, жгутики). Од
нако встречаются и явления редукции, вплоть до размеров вирусов.
Нуклеоид и плазмиды. ДНК бактерий распределена в цитоплазме не диффузно, а локализована в определенной части клетки — нуклеоплазме. Нуклеоид представляет собой клубок из тонких нитей, граничащий непосредственно с цитоплазмой; ядер-ная оболочка отсутствует. Из-за деления ге-нофора, опережающего клеточное деление, в бактериальной клетке часто встречаются 2—4 нуклеоида. У Escherichia coli генофор состоит из одной-единственной кольцевидно замкнутой нити ДНК длиной 1,4 мм. Делению генофора у бактерий, очевидно, предшествует прикрепление его к клеточной мембране (никаких митоза и мейоза!). Кроме генофора в бактериальных клетках встречаются кольца ДНК меньшего разме
116 | ГЛАВА 11, СИСТЕМАТИКА И ФИЛОГЕНИЯ
ра, способные к самостоятельной репликации — так называемые плазмиды.
Ввиду многих особенностей у бактерий, в отличие от эукариот, ни в коем случае не следует говорить о «клеточных ядрах» и «хромосомах», что, к сожалению, сейчас еще часто делается. Напротив, ДНК бактерий по строению и функциям в значительной мере соответствует ДНК всех других организмов (см. 1.2). При быстрой репликации скорость новообразования ДНК достигает 33 мкм (длина цепочки) в минуту. Бактерии — это те организмы, на которых были получены наиболее существенные дан
ные молекулярной генетики. Для Е. coli, Salmonella typhimurium и других бактерий уже созданы генетические карты.
Цитоплазма отграничена от клеточной стенки цитоплазматической мембраной, которая, как и у всех других организмов, многослойная (= плазмалемма). В цитоплазме находятся нуклеоид (иногда несколько), различные мембранные системы и клеточные включения. Рибосомы бактерий, размер которых составляет 16x18 нм, состоят примерно на 60% из РНК и на 40% из
Рис. 11.11. Bacteria. Расположение жгутиков у бактерий (А — по van Iterson; В — по Labaw и Mosley; С — по Ziegler; D — по Scanga; Е — по Houwink и van Iterson):
А — Монотрих (Vibrio metchnikovii, 1000х); В — часть жгутика (Bordetella bronchiseptica, 60 000х); С — базальное зерно у места прикрепления жгутика (Rhizobium radicicola, 20 000х); D — лофотрих (Spirillum undula, 8 000х); Е — перитих (Proteus vulgaris, содержимое клетки частично автолизировано, 10 ОООх)
Рис. 11.12. Bacteria. Фотоавтотрофная бактерия Rhodopseudomonas с тилакоидами (по Drews и Giesbrecht):
СМ — цитоплазматическая мембрана; Ро — полифосфатное тельце; W — клеточная стенка
11.2. Бактерии, грибы, растения | -| 17
белка. Они содержатся в клетке в числе примерно 5 000—50 000. Рибосомы осаждаются в ультрацентрифуге при 70S (S — единица Сведберга, коэффициент для определения молекулярной массы). Напротив, эукариоты имеют в цитоплазме 808-рибо-сомы, а в митохондриях и хлоропластах — 708-рибосомы. Мембраны внутри плазмы бактерий образуют (насколько это было исследовано) сеть.
Мезосомы возникают из впячиваний цитоплазматической мембраны. Они были описаны, в частности, как мезосомы и тубулярные тельца. У фототрофных бактерий отмечены также фотосинтетически активные пузырьки, аналогичные соответствующим структурам в хлоропластах зеленых растений — тилакоидам (рис. 11.12); у некоторых видов они даже сходным образом сгруппированы в стопки. Мембраны тилакоидов — носители поглощающих свет пигментов (бактериохлорофиллов и каротиноидов), а также компонентов систем транспорта электронов и фосфорилирования при фотосинтезе. Правда, тилакоиды у бактерий никогда не бывают окружены общей оболочкой; таким образом, здесь мы имеем дело не с настоящими пластидами. У некоторых бактерий встречаются также газовые пузырьки (например, Chromatiaceae).
Компоненты клеточной стенки. Откладывающиеся в клетке вещества отчасти являются запасными. Многие бактерии запасают полисахариды типа гликогена. Липофильные граны состоят из пол и-lira дрокси масля ной кислоты. Mycobacterium и Actinomyces накапливают преимущественно нейтральные жиры и воски. Фосфорная кислота накапливается в форме полифосфатных гранул («волютина») (см. рис. 2.93).
Клеточная стенка бактерий (см. рис. 2.98) имеет толщину примерно 20 нм. У нее нет фибриллярной структуры, как у целлюлозной стенки клеток высших растений. Ее механическая прочность достигается, как правило, покровом — мешком. Последний состоит преимущественно из полимера муреина, образованного из субъединиц N-ane-тилмураминовой кислоты и N-ацетилглю-козамина; они в чередующейся |3-1,4-гли-козидной последовательности соединены
в полигликановые тяжи. За счет соединения тяжей с короткими пептидами (тетра- или пентапептидами, содержащими D-или L-аминокислоты) возникает сетевидная макромолекула, «муреиновый мешок». Пептидоглюкан у всех бактерий (в том числе и цианобактерий) в более чем 100 вариантах (типы пептидоглюкана) является компонентом клеточной оболочки. У некоторых бактерий на поверхности образуется мощный слой слизи (зооглеи) или «капсулы» различного состава (большей частью это полисахариды или полипептиды). Клетки Acetobacter xylinum соединяются целлюлозой в пленку; у Sarcina ventriculi клетки также связаны между собой целлюлозой.
Движение бактерий осуществляется за счет очень нежных плазматических жгутиков (см. 2.3.2; рис. 2.96), которые встречаются на определенных стадиях развития многих бактерий и обеспечивают активное движение клетки в том или ином направлении. При электронной микроскопии эти бактериальные жгутики показывают винтообразную поверхностную структуру (рис. 11.11, В); они составлены из нескольких крайне тонких продольных фибрилл, скрученных между собой. Структуры «2 + 9», как у настоящих жгутиков эукариот, у них нет (см. рис. 2.16). Способность к движению обеспечивает сократительный белок (фла-геллин), сходный с миозином мышечных клеток. Диаметр жгутиков составляет чаще всего 10 — 20 нм, длина — до 20 мкм. Они расположены в клетке по-разному: в единственном числе на конце клетки (моно-трихи, рис. 11.11, А) или пучками (лофо-трихи, как Spirillum, рис. 11.11, D); они могут быть равномерно распределены по всей поверхности клетки (перитрихи, рис. 11.11, Е). Прикрепление к клетке может быть полярным (рис. 11.11, С), биполярным, латеральным (сбоку) или немного ниже конца клетки (субполярным). Каждый жгутик отходит (насколько известно) от базального тельца (рис. 11.11, С), погруженного в оболочку клетки (см. рис. 2.97). Число жгутиков может зависеть от внешних условий: так, Proteus vulgaris при скудном питании имеет 2 субполярных
118 | ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
жгутика вместо многих распределенных по всей поверхности клетки жгутиков в обычных условиях. В пучках насчитывается 2—50 жгутиков (политрихи).
Движение. Скорость движения с помощью жгутиков составляет, например, у Bacillus megatherium до 200 мкм/с — это примерно 50-кратная длина самой клетки. Spirillum за одну секунду может 13 раз обернуться вокруг своей оси. При этом жгутики совершают 40 оборотов, что соответствует числу оборотов электромотора. Движение происходит, как правило, за счет толчка, как у корабельного винта, однако может заменяться подтягивающим движением по типу самолетного пропеллера. Оно осуществляется чаще всего внутри жидких сред, реже по влажным поверхностям (так движется по агару имеющий перитрихиальное жгутование Proteus vulgaris). В зависимости от действующего фактора движение представляет собой хемотаксис (см. 8.2.1.1), аэротаксис, фототаксис (см. 8.2.1.2) и магнитотаксис (см. 8.2.1.3). Движения в ответ на то или иное раздражение позволяют подвижным формам скапливаться в местах с оптимальным наличием полезных для них веществ. Подвижность имеющей жгутики прокариотной клетки можно в плане строения и эволюции рассматривать как пример конвергенции с аналогичными клетками у эукариот. К скользящему движению способны лишенные жтугиков, похожие на цианобактерии, но гетеротрофные бактерии. Ползающее движение очень медленное (примерно 250 мкм/мин) и связано с выделением слизистых чехлов. О способности к движению миксобактерий по типу голых протопластов упоминалось выше.
Помимо жгутиков у некоторых бактерий встречаются многочисленные более тонкие нити («фимбрии», или «pili»), функция которых в значительной степени пока неизвестна. У Escherichia coli так называемые F-, или половые, фимбрии обеспечивают парасексуальную конъюгацию (рис. 11.13, А, В).
Физиология. Питание бактерий может различаться в зависимости от источника энергии, донора электронов и источника углерода. Получение энергии происходит
либо путем разложения веществ в субстрате (хемотрофия, см. 6.9), либо за счет использования световой энергии (фототрофия, см. 6.4). В качестве донора электронов служат органические (органотрофия) или неорганические вещества, например NH3, H2S или Fe4-4 (автотрофия). Автотрофные бактерии в зависимости от донора электронов и источника энергии бывают хемолитотрофа-ми или фототрофами. Строго анаэробные виды в присутствии кислорода не могут ни расти, ни размножаться. Факультативные анаэробы могут существовать и без кислорода; микроаэрофильные формы выдерживают лишь незначительные концентрации кислорода. Для облигатно аэробных бактерий кислород совершенно необходим. Распространена — и встречается только у прокариот— способность к фиксации атмосферного азота (см. 9.2.1).
Размножение происходит, как правило, путем деления клеток на две, у вытянутых форм всегда перпендикулярно продольной оси. При этом сначала образуется от края клетки к середине (центрипе-тально) поперечная перегородка, по которой клетки отделяются друг от друга (отсюда старое название «дробянки» — Schizophyta). У почти всех исследованных до настоящего времени бактерий пепти-доглюкановая капсула (насколько она имеется) с самого начала принимает участие в образовании перегородки. Клетки могут после деления оставаться соединенными в рыхлые цепочки (например. Streptococcus).
Для переживания неблагоприятных условий некоторые формы образуют покоящиеся клетки, или споры. У некоторых палочковидных бактерий споры закладываются внутри клеток как эндоспоры; они отличаются от вегетативных клеток меньшей способностью к окрашиванию и сильным преломлением света. Значение эндоспор состоит прежде всего в их устойчивости к нагреванию, благодаря чему они могут сохраняться неповрежденными, например при длительном кипячении. Вегетативные клетки спорообразующих форм, наоборот, погибают уже при пастеризации (нагревании в течение 10 мин до 80 °C). Спорообразование внутри бактериальной
11.2. Бактерии, грибы, растения
119
Рис. 11.13. Escherichia coli. Перенос ДНК от клетки-донора к клетке-реципиенту (А, В). Бактериофаги (С—Е) (А — по Brinton и Carnahan; В — по W. Nultsch; С, D — по F. Ehrendorfer; Е — по Kellenberger и Arber):
А— электронная микрофотография. Клетка-донор и клетка-реципиент (последняя слева) посредством полового волоска (копуляционной фимбрии) соединены между собой (3 500х); В— схема переноса ДНК. Вверху — половой волосок, внизу — реципиент. Донорная ДНК (показана черным цветом) разделена на две цепи, из которых одна проникает в клетку-реципиент. Репликация (светлая линия) одноцепочечной ДНК (темная линия) происходит в половом волоске и реципиенте. Некоторые локусы генов отмечены кружками и буквами (Cal, Lac, Т'ь Az', L); С, D — размножение бактериофагов в клетке хозяина с ее разрушением (лизогенный цикл). При смене назад к литическому циклу может происходить трансдукция, т. е. перенос бактериальной ДНК на освободившиеся фаги; Е — отдельные фаги Т2 (40 ОООх)
120 | ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
клетки начинается с различных химических превращений в материнской клетке, причем разлагается 75 % ее белков. Затем следует деление материнской клетки на две дочерние клетки неравной величины. Спорообразование завершается формированием вокруг меньшей клетки, становящейся спорой, толстой стенки, которая может составлять до 50 % ее объема и сухой массы. У терморезистентных спор накапливается специфичная для них дипиколиновая кислота. При этом усиливаются терморезистентность и светопреломление.
Спорообразование определяется внешними условиями и осуществляется, например, при недостатке питательных веществ. Готовность к прорастанию спор повышается при хранении и нагревании. Из датированных частиц почвы, которые содержали бактериальные споры (например, почва на гербарных растениях) можно было еще прорастить споры после 200 — 320 лет хранения в сухих условиях. Правда, при хранении в сухом состоянии почвенного образца 90 % спор теряли жизнеспособность в течение 50 лет.
У бактерий возможен частичный перенос генетического материала («парасексуальность»): фрагменты ДНК могут передаваться от клетки-донора к клетке-реципиенту непосредственно путем конъюгации, с помощью бактериофагов или в экстрагированной форме путем трансформации (рис. 11.13, В; относительно функции половых волосков, или пилей: рис. 11.13, А).
В противоположность чаще всего более крупным бактериям (см. 1.2.5), вирусы, имеющие значительно меньшие размеры и проходящие через бактериальные фильтры, не являются самостоятельными организмами. Они развиваются из генетического материала клеток. Вирусы представляют собой как бы ставшие самостоятельными гены, которые взяли на себя функцию управлять обменом веществ клетки-хозяина для собственного синтеза. Возможно, хотя бы некоторые вирусы даже могли возникнуть путем крайней редукции из патогенных бактерий. Если бактерии содержат ДНК и РНК в соотноше
нии примерно 1: 3,5 %, то вирусы всегда содержат только один тип нуклеиновых кислот — либо ДНК, либо РНК. Вирусы могут воспроизводиться только в живых клетках; они не показывают ни роста, ни деления и нечувствительны к пенициллину и сульфонамидам. На электронной микрофотографии отсутствуют (несмотря на иногда весьма высокую морфологическую организацию) все структуры, характерные для бактерий. Известные примеры болезней, вызываемых вирусами: мозаика табака (ср. рис. 3, т. 1), ящур, бешенство, желтая лихорадка, гепатит, грипп, оспа, герпес. Ретровирусы — РНК-вирусы, у которых одноцепочечная РНК транскрибируется на двойной спирали ДНК (т.е. противоположно обычной транскрипции с ДНК на РНК). К этой группе относятся, в частности, возбудители иммунодефицита (СПИД, ВИЧ) и вызывающие опухоли онковирусы.
Бактериофаги представляют собой особенно высокоорганизованные, относительно крупные вирусы (длина 150—110 мкм). Они состоят из «головки», содержащей ДНК, оболочки и белкового «хвоста» (рис. 11.13, Е). Кончик хвоста прикрепляется к поверхности бактериальной клетки, и только после этого содержащаяся в головке ДНК внедряется в тело бактерии через полый хвост. Спустя несколько минут становятся заметны первые признаки образуемых заново частей фага, а еще примерно через такое же время несколько сотен новых фагов высвобождаются из бактериальной клетки в результате ее разрушения (лизиса). Они возникают не путем деления, а путем образования заново из плазмы бактерий. Это явление основано на том, что ДНК фага включается в обмен веществ хозяина, воздействует на его генетический аппарат, и вместо нормальных компонентов бактерии синтезируются специфические компоненты фага. Фаги могут внутри клетки-хозяина изменять за счет мутаций свои биохимические свойства; их можно скрещивать и рекомбинировать. Прежде думали, что фаги, возможно, являются начальным этапом жизни. Однако у них нет собственного обмена веществ и энергии (например, отсутствует
11.2. Бактерии, грибы, растения
дыхание), и поэтому в настоящее время их считают обособившимися частями бактериальной ДНК, которые приобрели способность к самовоспроизведению и даже к сохранению вне клетки в полностью неактивном (латентном) состоянии, пока опять не включатся в обмен веществ хозяина. Существенным подтверждением такого предположения было, в частности, открытие того, что не все бактериофаги смертельны для бактерий, а ДНК так называемых «умеренных» фагов без вреда для хозяина может длительное время реплицироваться бактериальной ДНК (рис. 11.13, С, D). «Генетическое вещество» фагов и бактерий частично очень сходно с таковым бактерии.
Систематическое подразделение эубак-терий. При преобладающей бедности морфологических признаков для подразделения бактерий согласно предполагаемым родственным связям наряду с доступными морфологическими данными особенно большое значение имеют также биохимические и физиологические критерии. Важную роль играет сравнение последовательностей нуклеиновых кислот (см. 1.2).
Бактерии подразделяются на грамотри-цательные и грамположительные — это группы, которые могут рассматриваться в ранге отдела. Для их идентификации используется так называемая окраска по Граму (предложена X. К. И. Грамом, 1853 — 1938). Грамположительными называются такие бактерии, у которых анилиновый краситель сохраняется в клетке, а у грамот-рицательных бактерий он может вымываться. Для лишенных клеточной стенки бактерий, которые могут возникать как спонтанно, так и в эксперименте (ср. микоплазмы), окраска по Граму не применяется.
Первый отдел: грамположительные бактерии (Posibacteriota)
Из многослойного муреинового покрова гоамположительных эубактерий при окраске по Граму краситель не вымывается. Их клеточная оболочка (см. рис. 2.98, А)
отличается следующими особенностями. Многослойная муреиновая сеть составляет до 30—70 % сухой массы клеточной стенки. Особенностью аминокислотного состава является то, что диаминопимелиновая кислота часто заменяется лизином. Полисахариды отсутствуют или ковалентно связаны. Содержание белка меньше. Часто встречаются тейхоевые кислоты — полимеры рибитфосфорной кислоты или гли-церофосфорной кислоты, которые через фосфодиэфирные связи соединены с му-рамовой кислотой. Среди грамположитель-ных бактерий широко представлены всевозможные морфологические и физиологические типы, известные у бактерий. Наиболее сложной морфологической организации достигли актиномицеты, вегетативное тело которых представляет собой ветвящийся мицелий. Однако образование плодовых тел, как у миксобактерий, а также способность к фотосинтезу отсутствуют. Образование эндоспор встречается только у некоторых палочковидных грам-положительных бактерий. Представление о многообразии относящихся к этому отделу форм дает подразделение их на следующие искусственные группы.
1. Кокки. Грамположительные кокки (аналогично рис.-11.10, А, С) могут представлять собой либо одиночные клетки, либо сгруппированные попарно, тетрадами, а также образуют скопления в виде пакетов (у Sarcina ventriculi до 64 клеток, скрепленных вместе целлюлозой). Это облигатные или факультативные анаэробы, иногда аэробы. Пример: молочно-кислые бактерии (такие, как Streptococcus lactis) из семейства Streptococ-сасеае. Сюда же относится Deinococcus (см. 7.7.2.3).
2. Неспорообразующие палочки. Сюда относятся палочковидные (см. рис. 11.10, В) анаэробные или факультативно аэробные молочнокислые бактерии из семейства Lactobacillaceae (Lactobacillus).
3. Спорообразующие палочки могут образовывать эндоспоры. Они относятся к одному-единственному семейству — Clostridiaceae (Bacillus, Clostridium). Это либо неподвижные, либо (за счет наличия латерально или же перитрихиаль-но расположенных жгутиков) подвижные, аэробные, живущие в почве, или факультативно анаэробные бактерии. Многие из них образуют также цепочки из клеток или нити.
4. Коринеформные бгктерии (включая Actino-mycetales). Коринеформными называют грам-
122 I ГЛАВА 11, СИСТЕМАТИКА И ФИЛОГЕНИЯ
положительные бактерии с сильно меняющимся обликом, т.е. палочки могут становиться булавовидными, короткими, кокками или слабо разветвленными формами. Образование эндоспор полностью отсутствует. К ним относятся пропионовокислые бактерии (Propionibacteriaceae), которые в качестве анаэробных организмов встречаются в рубце и кишечнике жвачных животных. У Mycobacteriaceae, относящихся уже к Actinomycetales, тенденция к ветвлению более выражена по сравнению с ранее рассмотренными формами. Если настоящее ветвление у Mycobacterium встречается только в молодых культурах, то у так называемых «лучистых грибков», или актиномицетов (Actinomycetaceae, Streptomycetaceae, Nocardiaceae), оно является правилом. Многочисленные виды почвенных актиномицетов развивают в искусственной культуре чаще всего колонии диаметром до нескольких сантиметров. Они образуются «мицелием», который часто состоит из одной-един-ственной клетки, лишенной поперечных перегородок, крайне нежной и, как правило, часто обильно разветвленной (диаметр нити 0,5 — 1 мкм; см. рис. 11.10, К). Ни хитин, ни целлюлоза в клеточной стенке не встречаются. Иногда нити становятся многоклеточными и могут легко распадаться на палочки, которые чрезвычайно похожи на некоторые палочковидные бактерии. Кроме того, они образуют (особенно на воздухе) различного рода экзоспоры, располагающиеся в цепочках.
Streptomyces scabies вызывает паршу картофеля и других овощных культур, внешне проявляющуюся в образовании коростинок на пораженной части растений. В корневых клубеньках ольхи и видов некоторых других родов обитает относящийся к актиномицетам симбионт (Fran-kia alni), который ассимилирует здесь свободный атмосферный азот (см. 9.2.1f). Виды Nocardia, как и представители рода Mycobacterium, могут окислять этан (ср. Flavobacteriurri). Виды Thermohiono-spora и Thermoactinomyces растут при высоких температурах. Продукты выделения некоторых актиномицетов находят применение в медицине как антибиотики для борьбы с инфекциями, вызываемыми патогенными бактериями (актиномицин, стрептомицин и т. д.). В естественных условиях они, вероятно, важны доя противостояния микроорганизмам-конкурентам. К группе коринеформных бактерий могут быть отнесены также Clavibacter (см. 9.3.2) и Rhodococcus (см. 7.6.2.1).
5. Микоплазмы (например, Mycoplasma), раньше обозначавшиеся также PPLO (от англ, pleuropneumonia-like organisms — организмы, сходные с возбудителями плевропневмонии), не имеют клеточной стенки и соответственно по
стоянной формы, поэтому их нельзя отнести к грамположительным бактериям на основании окрашивания по Граму. Однако данные по последовательностям 16S-pPHK свидетельствуют о том, что они произошли от грамположительных бактерий. Сюда относится, очевидно, также род Metallogenium, виды которого участвуют в окислении марганца в водоемах (марганцевые бактерии, см. 6.9.1).
Второй отдел: грамотрицательные бактерии (Negibacteriota)
У грамотрицательных бактерий муреиновая сеть тонкая, однослойная и составляет меньше 10% сухой массы клеточной стенки; вещества, поступающие при окраске по Граму, могут легко вымываться. Внешняя мембрана состоит из откладывающихся, однако в любом случае не связанных ковалентно липопротеинов, липополисахаридов и других липидов, которые составляют до 80 % сухой массы клеточной стенки. Ионы Са2+ повышают стабильность липопротеидного слоя. Тей-хоевые кислоты (см. рис. 2.90, В) не обнаружены.
К этому отделу относятся кокки, палочки, вибрионы, спириллы, спирохеты и скользящие формы. По способу получения энергии это фототрофы или хемотрофы. В отличие от грамположительных бактерий некоторые представители отдела способны к фотосинтезу, причем здесь он происходит без выделения кислорода (в отличие от цианобактерий и эукариот). Родственные связи фототрофных и нефототрофных грамотрицательных бактерий чаще всего выражены значительно больше, чем связи с цианобактериями, которые при фотосинтезе используют воду в качестве донора электронов. Среди хемотрофных групп различают хемолитотрофные и хемооргано-трофные.
Классификация отдела находится в периоде становления и лишь частично использует возможные родственные связи. Наряду с порядками, в значительной мере являющимися естественными, имеются также различные семейства, объединенные в искусственные группы (1 — 12) по сходству
11.2. Бактерии, грибы, растения . -|23
облика и обмена веществ. Мы начнем с кокков и/или палочек и рассмотрим их по группам анаэробов (1), факультативных анаэробов (2) и аэробов (3). Далее будут следовать вытянутые и одновременно спирально закрученные формы, а именно жесткие спириллы (4) и гибкие спирохеты (5), а также бактерии, несущие придатки (6). Исходя из особенностей морфологии, в специальные группы выделяются хламидобак-терии (7) и способные к скользящему движению виды из Cytophagales (8) и Мухо-bacterales (9); последние образуют структуры типа плодовых тел. По обмену веществ можно различать хемолитоавтотрофные (11) и фотоавтотрофные (12), а по образу жизни — облигатно паразитические бактерии (10).
1. Анаэробные кокки и палочки объединяются в семейства Veillonellaceae и Bacteroidaceae. К последнему семейству примыкают организмы пока еще неясного систематического положения из рода Desulfovibrio. Это полярные моно-трихи или политрихи, представляющие собой по облику вибрионы (палочки, изогнутые в виде запятой) или спириллы (см. ниже, 4). Они относятся к небольшой группе восстановителей сульфатов до сульфитов, способных к «дыханию за счет сульфата» (см. 6.7), и живут как хемолитогетеротрофы. Виды Desulfovibrio— обитатели сапропеля («гнилого ила»).
2. К факультативно анаэробным палочкам относятся, в частности, организмы семейства Enterobacteriaceae (Enterobacter, Erwinia, Klebsiella и часто живущая в кишечнике теплокровных организмов и широко используемая в экспериментальных исследованиях кишечная палочка — Escherichia coli). Виды родов Photobacterium и Вепеккеа из семейства Vibrionaceae приспособились к жизни в морской воде и известны как светящиеся бактерии («биолюминесценция»). Рыбы и головоногие моллюски используют эти виды в своих органах свечения в качестве симбионтов для ориентации и отпугивания. Сюда примыкают роды Haemophilus, Yersinia (бокс 9.2) и Zymomonas. Elavobacterium, также примыкающий к этой группе, отличается способностью к окислению этана.
3. Аэробные кокки и палочки представлены среди грамотрицательных бактерий в большом разнообразии. К ним относятся, в частности, семейства Azotobacteraceae (Azotobacter) и Rhizo-biaceae, виды которых часто способны к связыванию свободного атмосферного азота. Первые живут свободно и могут связывать до 20 мг азо
та на 1 г использованного сахара. Представители Rhizobiaceae (Rhizohium. Azo-. Brady-, Meso-. Sinorhizobium) заражают корни бобовых растений, которые реагирую! на это образованием корневых клубеньков (см. 9.2.1). Phyllobacterium rubiacearum — вид, относящийся к тому же семейству, — является симбионтом в листьях различных Rubiaceae (Psychotria, Pavetta) и Ardisia из Myrsinaceae.
Род Agrobacterium не способен к связыванию молекулярного азота. Agrobacterium tumefaciens образует галлы на цветковых растениях (бокс 9.2, рис. А). Он использовался для получения трансгенных растений (см. хронологическую таблицу). Сюда же относятся семейство Pseudomona-daceae (с родами Pseudomonas и Xanthomonas), уксуснокислые бактерии (например, Acetobacter aceti), а также роды Bordetella, Citrobacterw Methy-lomonas.
4. Спириллы (Spirillales с родом Spirillum) — спирально извитые, жесткие палочки с разным количеством витков — от нескольких до множества. По типу жгутикового аппарата это биполярные политрихи. Чаще всего они являются аэробами, реже — факультативными анаэробами.
5. Спирохеты (Spirochaetales) — чрезвычайно длинные (до 500 мкм!) и тонкие (диаметр 0,1—0,6 мм) анаэробные или аэробные бактерии, спирально извитые, как и спириллы. В противоположность последним они гибкие: тонкие клеточные стенки позволяют им при отсутствии жгутиков активно двигаться за счет сокращения осевой нити, находящейся внутри клеток.
6. Бактерии с придатками — формы, имеющие различное эволюционное родство, которые при делении дают дочерние клетки неравной величины, а также образуют придатки в форме палочек и выростов. Палочки состоят из слизи, нитевидные выросты образуются в результате вырастания клетки. Gallionella femigi-пеа известна как железобактерия, которая весной в водоемах, содержащих железо, образует ржаво-коричневые массы. Pedomicrobium тап-ganicum окисляет марганец (марганцевые бактерии, см. 6.9.1).
7. Хламидобактерии обладают трубковидными чехлами (влагалищами), которые удерживают клетки вместе в цепочках. Широко известен так называемый «гриб сточных вод» Sphaerotilus natans (см. рис. 11.10, G—J). Эта бактерия (!) растет в сильно загрязненных водах, например в отстойниках сахарных заводов. Образуя нити, хлопья и даже наросты, напоминающие густой мех. она может забивать трубы и сточные канавы. Сюда относится также Leptothrix.
124 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
8. Порядок Cytophagales. Представители порядка производят скользящие движения. Оба относящиеся сюда рода — Cytophaga и Sporocyto-phaga — вместе с рассматривающимися дальше миксобактериями являются аэробными почвенными бактериями, разлагающими целлюлозу. В порядке Cytophagales в отличие от мик-собактерий не образуется никаких плодовых тел. В качестве скользящих нитчатых форм сюда могут быть присоединены Thiothrix (см. 6.9.1) и Beggiatoaceae. О Chloroflexus см. ниже (группа 12).
9. Порядок миксобактерии (Myxobacterales) составляет основную часть группы скользящих бактерий. Однако в эволюционном плане они продвинулись дальше предыдущей группы, поскольку обладают комплексной организацией, при которой одноклеточные формы могут образовывать плодовые тела. Красные или другого цвета скопления клеток миксобактерий, живущие на почве или в помете животных, представляют собой колонии («псевдоплазмодий») из мелких, лишенных клеточной стенки, активно изгибающихся палочек без жгутиков, которые за счет активных сокращений клеток могут успешно совершать скользящие движения. У некоторых видов палочки при совместном сползании собираются в определенных местах и образуют характерные, различающиеся в зависимости от рода по облику и окраске, иногда связанные слизью общие скопления — так называемые плодовые тельца, или цистофоры. Из их внутренней части снова могут образовываться подвижные клетки (например, у распространенных видов Myxococcus и у Chondrotnyces, рис. 11.10, L—N). В культуре некоторые миксобактерии могут питаться живыми микроорганизмами (например, бактериями). В жизненном цикле здесь наблюдается, таким образом, примечательная конвергенция с эукариотными Acrasiobionta.
10. Облигатно паразитические бактерии объединяются в порядок риккетсии (Rickettsiales). Будучи облигатными паразитами, риккетсии не могут культивироваться вне живых клеток. Они очень мелкие, от вирусов отличаются соотношением ДНК/РНК (1:3,5); кроме того, клеточная стенка здесь чувствительна к лизозиму и содержит мураминовую кислоту.
С риккетсиями, возможно, могут быть сближены долго считавшиеся вирусами возбудители пситтакозов (болезней попугаев). Это приспособившиеся к паразитическому образу жизни бактерии, имеющие ДНК и РНК, а также содержащие специфические вещества (например, мурамовую кислоту).
II. У хемолитотрофпых бактерий — в противоположность гетеротрофным бактериям (см.,
например, группу 1) — имеется обязательная взаимосвязь хемолитотрофии с автотрофной фиксацией СО2 (см. 6.9). Аэробные Nitrobacte-гасеае окисляют аммиак до нитрита (Nitroso-monas) или нитрит до нитрата (Nitrobacter). Морфологически это могут быть кокки, палочки или спириллы; если у них имеются жгутики, то они прикрепляются субполярно или же клетки являются перитрихами. С этим семейством могут быть сближены бактерии, окисляющие восстановленные соединения серы (отчасти Thiobacillus) или Fe2+ до Fe1' (отчасти Thiobacillus и Siderocapsaceae). Наконец, стоит упомянуть водородные бактерии, или бактерии гремучего газа (например, Alcaligenes eutrophus), которые только факультативно автотрофны. Они могут, с одной стороны, лучше развиваться на органических питательных средах, однако, с другой стороны, могут также окислять молекулярный водород с помощью гидрогеназ. Таким путем эти бактерии получают энергию, осуществляют процессы восстановительного синтеза и синтезируют специфические углеводы, фиксируя СО2.
12. Фотоавтотрофные Rhodospirillales — в значительной мере анаэробы. Они отличаются наличием различных фотосинтетических пигментов (бактериохлорофиллы а — е) и каротинов, которые придают им характерную пурпурно-фиолетовую, красноватую, коричневую, оливковую или зеленую окраску. Кислород ингибирует синтез и функционирование бактериохлорофиллов, которые этим отличаются также от хлорофилла а цианобактерий (см. рис. 6.45) и эукариот. В качестве доноров электронов используются, в частности, органические соединения (Rhodospirillaceae). Эти приспособившиеся к использованию света как источника энергии фотоавтотрофные бактерии, филогенетически крайне не родственные, встречаются в виде кокков, палочек или спирилл. Если имеются жгутики, то они располагаются полярно или биполярно.
Семейство Rhodospirillaceae — лишенные серы пурпурные бактерии. Как и последующее семейство, отличается наличием преимущественно бактериохлорофилла а или b на цитоплазматической мембранной системе. Элементарная сера ими, как правило, не окисляется. Наиболее известные представители относятся к родам Rhodospirillum, Rhodobacter и Rhodomicro-bium.
У представителей обоих последующих семейств в качестве донора электронов используется элементарная сера или сероводород. Chromatiaceae накапливают в клетках или на их поверхности серу. Чаще всего они имеют пурпурную окраску, поэтому их называют
11.2. Бактерии, грибы, растения
серосодержащими пурпурными бактериями. Такими являются виды Chromatium и Thiospirillum, клетки которых достигают значительных размеров (20 — 40 х 3,5 — 4 мкм), а также Thiocapsa. Зеленые серные бактерии, относящиеся к семейству Chlorobiaceae (род Chlorobium и некоторые другие), не могут ни накапливать, ни откладывать серу. Они содержат бактериохлорофиллы (преимущественно с или rf; у некоторых также есть бактериохлорофилл а в незначительных количествах) в пузырьках, которые расположены вблизи цитоплазматической мембраны или прикрепляются к ней. Эта особенность отличает их от обоих предыдущих семейств. Особые формы известны под родовым названием Chlorochromatium. Они выглядят как скопления из нескольких неподвижных зеленых серобактерий и расположенной в центре бесцветной бактерии с полярными жгутиками; такое образование движется в пространстве как единое целое. Chloroflexaceae (лишенные серы зеленые бактерии, объединяемые в род Chloroflexus), производят скользящие движения и поэтому представляют собой нечто среднее между группами 8 и 12.
Фотоавтотрофные бактерии обитают в анаэробных зонах в пресноводных лужах, прудах и озерах, в медленно текущих водотоках, а также в морских бухтах. Серосодержащие пурпурные бактерии образуют, например, розоватые или винно-красные налеты на разлагающихся частях растений у дна водоемов. Иногда наблюдается их массовое развитие («цветение воды») в более глубоких анаэробных зонах озер. Это происходит при определенных температурных условиях — при достаточно высоких концентрациях сероводорода, диоксида углерода и органических соединений. Благодаря высокому содержанию каротиноидов пурпурные бактерии могут улавливать проникающий в глубину коротковолновый свет и используют его для фотосинтеза. Соответственно на больших глубинах водоемов преобладают пурпурные бактерии, а из серобактерий — имеющие бурую окраску из-за большего содержания каротиноидов.
Встречаемость и образ жизни бактерий
Бактерии, представленные многими видами (около 3 000) и неизмеримым числом особей, встречаются по всему земному шару: в воде, почве, а с пылью — повсюду в атмосфере и на всех предметах. Их широ
кому распространению способствуют главным образом следующие факторы: во-первых, малый размер и связанная с ним очень большая поверхность по сравнению с массой тела, за счет чего становятся возможными очень высокая физиологическая активность и интенсивность обмена веществ (например, способность к очень быстрому размножению); во-вторых, устойчивость их вегетативных клеток и особенно спор к неблагоприятным внешним воздействиям, а также многообразие способов питания. При оптимальных условиях некоторые виды (например, Vibrio cholerae) могут делиться несколько раз в час, так что от одной бактериальной клетки за 24 часа могут возникнуть несколько миллиардов новых клеток.
Споры бактерий очень устойчивы к высыханию и экстремальным температурам; некоторые выдерживают пребывание в течение нескольких часов в кипящей воде (максимально 30 ч), а также при глубоком холоде. Также и вегетативные клетки многих видов особенно устойчивы к высыханию. Некоторые могут жить при высокой температуре (90—НО °C), например в горячих источниках, а некоторые сами активно выделяют значительное количество теплоты («самонагревание» более чем до 60 °C сена, навоза, табака, хлопка, например такими видами, как Bacillus stearothermophilus или видами родов Thermomonospora и Thermoactinomyces).
Термофилия. Термофильные бактерии, например виды родов Bacillus, Clostridium, различные микобактерии, а также рассматриваемые далее археи, не только выносят высокие температуры, но и нуждаются в них для оптимального роста. Настоящая термофилия в этом смысле встречается только у прокариот.
К умеренным термофилам относятся бактерии, которые высвобождают тепловую энергию при обмене веществ.
Термофильные бактерии используют термостабильные белки и ферменты, которые отличаются высокими температурными оптимумами. Стабильность белков повышается, в частности, ионами металлов или соединением с клеточными мембранами, а также особым аминокислотным составом;
126 | ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
так, термостабильные белки содержат больше аргининовых остатков, чем термолабильные.
Обмен веществ. У прокариот, и именно уже только среди эубактерий, встречается большее число типов обмена веществ, чем среди эукариот. Большинство бактерий живет гетеротрофно — как сапротрофы или паразиты. Однако облигатный паразитизм (например, у риккетсий) редок; большинство патогенных видов может размножаться и вне растительного или животного организма. Поэтому их культивирование на соответствующих питательных растворах (например, мясном отваре с пептоном) не представляет никакой сложности. На твердых питательных средах (агаре, желатине) бактерии часто образуют слизистые скопления разного облика — «колонии» (см. 5.1), которые чаще всего бесцветны, но могут быть и окрашены за счет выделения пигментов. Пигменты в клетках (в тилакоидах, см. рис. 11.12, или же в цитоплазматической мембране) имеют только фотосинтезирующие зеленые бактерии или пурпурные бактерии — аналогично галобактери-ям из архей.
За счет выделяемых ферментов бактерии вызывают интенсивное разложение субстрата либо в анаэробных, либо в аэробных условиях. Как особенности физиологии обмена веществ разных бактерий можно, в частности, назвать: автотрофность — за счет либо фотосинтеза (красные и зеленые серобактерии), либо хемосинтеза (см. 6.9); гетеротрофность у сапротрофов, у паразитов или в симбиозе; кислородный или бескислородный энергетический обмен веществ; денитрификацию или десульфурификаиию (см. 6.6; 6.7); связывание молекулярного азота (см. 9.2.2). Бактерии осуществляют многие процессы брожения: молочно-кислое и масляно-кислое брожение, сбраживание целлюлозы, пектина и белка, аэробное уксуснокислое брожение (см. 6.10.2.2). Бактерии могут разлагать почти все природные вещества, даже нефть, парафины, асфальт. Углеводороды разлагаются тем хуже, чем короче их цепи; этан и метан используются только особыми специализированными видами (см. 6.9.1). К бактериаль
ному разложению в значительной мере устойчивы лишь некоторые искусственные смолы и пластиковые материалы, а также особенно устойчивый спорополленин (см. 2.2.7.6).
Симбионты. Среди симбиотических бактерий особенно важны для многих сосудистых растений (Fabaceae, Alnus, Hip-pophad, Ardisia, Pavetta, Psychotria', см. 9.2.1) виды, фиксирующие атмосферный азот, в том числе из семейств Rhizobiaceae (Rhi-zobium, Phyllobacterium) и Actinomycetaceae (Frankia). Полагают, что митохондрии эука-риотных клеток возникли путем эндосимбиоза (исходно эндосимбионтами были организмы типа риккетсий).
Возбудители болезней. Многочисленные виды бактерий вызывают болезни животных и человека. Профилактика таких болезней возможна путем иммунизации (вакцинации): при активной иммунизации человеку (или животному) вводится ослабленный возбудитель или его яды, чтобы стимулировать образование антител; при пассивной иммунизации вводят антитела из иммунизированных животных.
Примерами болезней человека, вызываемых грамположительными бактериями, могут быть: гнойные инфекции (Staphylococcus), сибирская язва (Bacillus anthracis), столбняк (Clostridium tetani). дифтерия (Corynebacterium diphtheriae). туберкулез (Mycobacterium tuberculosis), угри (Pro-pionibacterium acni), актиномикозы (Actinomyces bovi). Близка к грамположительным бактериям Mycoplasma pneumoniae, вызывающая заболевания легких. Грамотрицательные бактерии вызывают воспаление легких и инфекции дыхательных путей (Klebsiella pneumoniae; Bordetella bronchiseptica; Haemophilus influenzae), тиф (Salmonella typhi), паратиф (Salmonella paratyphi), пищевые отравления (Salmonella typhimurium), чуму (Yersinia pestis), холеру (Vibrio cholerae), венерические заболевания (гонорея — Neisseria gono-rhoeae; сифилис — Treponema pallidum), воспаление головного мозга (Neisseria meningitides). пятнистая лихорадка (Rickettsia sp.).
Фитопатогенные виды внедряются в растение либо через устьица, гидатоды и т. п. (особенно виды Pseudomonas и Xantho-monas), либо инфицируют раны (морозо-бойные трещины, повреждения насекомыми и т.п., например Erwinia carotovora). Па
11.2. Бактерии, грибы, растения
тогенные бактерии отравляют организм хозяина токсинами. Наличие или отсутствие жгутиков не играет для патогенности никакой роли; странным образом для растений патогенны только палочковидные, неспорообразующие формы. Патогенные бактерии обитают чаще всего в межклетниках и оттуда растворяют срединные пластинки (см. 2.2.7.1), так что отделенные друг от друга клетки отмирают; иногда этот процесс ускоряется еще токсинами. Ткань хозяина превращается при этом в кашеобразную гнилую массу (мокрые гнили). В живые клетки внедряются относительно немногие бактерии (в том числе Pseudomonas tabaci). Изредка бактерии закупоривают сосуды и таким путем вызывают увядание и отмирание растения, чему чаше всего еще способствуют токсины увядания (например, Corynebacterium michiga-nense). Известно свыше 200 бактериозов растений.
Биотехнология. Бактерии и другие микроорганизмы могут применяться в технических процессах и промышленном производстве. Это, например, такие области, как генная инженерия (трансформация), получение антибиотиков (также для отщепления боковых цепей синтетически полученных предшественников), ферментов и других белков; разложение отходов (например, метанное сбраживание ила сточных вод); обогащение металлов путем микробного выщелачивания (перевод трудно растворимых соединений меди и урана в растворимые в воде сульфаты видами Thiobacillus). Бактерии, растущие на содержащих нефть субстратах, могут служить индикаторами при поиске новых месторождений.
Третий отдел: синезеленые водоросли, или цианобактерии (Cyanobacteriota, Cyanoprokaryota, Cyanophyta)
Представители этого отдела напоминают водоросли (отсюда название — синезеленые водоросли; прокариотные водоросли) — это морфологически просто устро
енные орагнизмы, осуществляющие фотосинтез. При фотосинтезе высвобождается кислород (оксигенный фотосинтез). Однако в отличие от эукариотных водорослей строение клетки в основных чертах соответствует ранее рассмотренным бактериям, т.е. это прокариоты, и поэтому вместо старого названия «синезеленые водоросли» лучше употреблять название «цианобактерии».
Отличия от эубактерий. Среди прокариот цианобактерии представляют собой относительно гомогенную группу. Определение нуклеотидных последовательностей 16S рРНК показывает более высокий уровень гомогенности между родами цианобактерий, чем между ними и остальными бактериями. От фототрофных родов эубактерий цианобактерии отличаются другими фотосинтетическими пигментами (хлорофилл а вместо бактериохлорофилла) и высвобождением кислорода при фотосинтезе; однако наряду с оксигенным при определенных условиях может осуществляться и аноксигенный фотосинтез. Клетка цианобактерий в среднем в 5—10 раз крупнее бактериальной.
Отличия от эукариотных водорослей. Фототрофные цианобактерии (синезеленые водоросли), часто образующие простые или разветвленные нити, будучи прокариотами, отличаются от эукариотных водорослей (см. бокс 11.5) по следующим признакам. В клетках отсутствует оформленное клеточное ядро, митохондрии, лизосомы, эндоплазматическая сеть, ограниченные мембранами хлоропласты и окруженные тонопластом вакуоли с клеточным соком; правда, некоторые цианобактерии имеют, как и некоторые эубактерий, наполненные газом пузырьки — газовые вакуоли. Однако в противоположность всем эукариотам некоторые цианобактерии способны, как и некоторые эубактерий, связывать свободный атмосферный азот (N2). Эта способность обусловлена прежде всего наличием гетероцист (см. 9.2.1), которые отличаются от остальных клеток утратой пигментации, наличием целлюлозы, а также светопреломляющих полярных телец (рис. 11.15, F). Очевидно, образующиеся в гетероцистах соединения азота че
1 28 | ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
рез тонкие каналы в полярных тельцах переходят в соседние клетки.
Строение клетки. В центральной, бесцветной части клеток (нуклео-, или цент-роплазме) располагаются элементы в виде гран, палочек, сети или нитей, которые содержат ДНК. Их совокупность, так называемый хроматиновый аппарат, представляет собой эквивалент ядра. При делении клетки весь этот комплекс делится надвое (рис. 11.15, N). Центроплазма окружена нечетко отграниченной периферической окрашенной хроматоплазмой, в зависимости от формы клетки имеющей вид полого шара или цилиндра. Хроматоплазма очень вязкая и в отличие от протоплазмы эукариотных клеток не находится в движении. В диффузно рассеянных рибосомах она содержит рибонуклеиновую кислоту, а в тилакоидах находится ассимиляционный пигмент — хлорофиллы а и Ь, а в отдельных случаях также хлорофиллы с и d. В качестве дополнительных пигментов встречаются наряду с каротиноидами (особенно Р-каротином, но также и зеаксантином, эхиненоном и миксоксантофиллом, но не лютеином) два водорастворимых хромопротеида (фикоби-липротеида), простетические группы которых (преобладающий здесь фикоцианин, а также фикоэритрин) носят название фикобилины. Фикобилины родственны желчным пигментам и в незначительно отличающейся форме встречаются также у отделов эукариотных водорослей — Crypto-
phyta и Rhodophyta. Фикобилипротеиды у цианобактерий, как и у красных водорослей, локализованы в тельцах — фикобили-сомах (см. рис. 2.89), которые расположены на тилакоидах (рис. 11.14, А). Последние находятся на примерно равных расстояниях друг от друга и не сгруппированы по два, по три или в стопки.
Запасные вещества. Цианобактериальный (цианофициевый) крахмал запасается в виде не различимых при световой микроскопии частиц между тилакоидами. Это глюкан, близкий к гликогену и родственный багрянковому крахмалу красных водорослей. Кроме того, встречаются цианофициновые зернышки, видимые под микроскопом — немного угловатые мелкие тельца, которые состоят из полимеров аминокислот аргинина и аспарагина; очевидно, это азотсодержащие запасные вещества. Фосфор в виде высокополимерных фосфатов накапливается в волютиновых тельцах, построенных из нуклеопротеидов. Возможно, эти тельца также служат как накопители энергии (АТФ).
Жесткая клеточная стенка (= защитный слой) состоит из муреина: целлюлоза практически отсутствует (однако см. выше о гетероцистах). Снаружи у цианобактерий часто встречаются слизистые чехлы (влагалища), которые, как показывают электронно-микроскопические исследования, имеют волокнистую структуру и наряду с аминокислотами и жирными кислотами содер
Рис. 11.14. Cyanobacteriota (А — по Hall и Claus; В — по H.Drawert):
А — концентрические тилакоиды (25 000х); В — Cylindrospermum, поровый пояс на поперечной перегородке (26 000х); L— липидное тельце; Р — фосфатное тельце
11.2. Бактерии, грибы, растения | -|29
жат полисахариды. Клеточная стенка состоит из четырех слоев и разрушается лизоцимом. По ультраструктуре и химизму клеточные стенки цианобактерий занимают промежуточное положение между стенками грамотрицательных и грамположитель-ных эубактерий.
Морфология. Некоторые цианобактерии одноклеточные (например, Dermocarpa). Остальные формы представляют собой колонии, содержащие от нескольких до многих клеток (Chroococcus, Meristnopedia), неразвет-вленные нити без (Oscillatoria) или с гетероцистами (Nostoc, Anabaena) и нити с ге-терополярной дифференциацией (Rivularia), нити с ложным ветвлением (Tolypothrix, Scytonema) или настоящим ветвлением (Hapalosiphon). Ложное ветвление возникав ет в результате того, что материнская нить разрывается, а конец дочерней нити, прорывая слизистый чехол, продолжает расти вбок, образуя ветвь (рис. 11.15, К). В противоположность этому настоящее ветвление происходит за счет изменения плоскости деления. Ветвление начинается с клеток, которые за счет деления в другом направлении образуются параллельно продольной оси нити и далее сохраняют такую ориентацию деления. Некоторые нитчатые цианобактерии имеют на продольном и поперечном срезах от нескольких до многих клеток и при этом обладают настоящим ветвлением (Stigonema, некоторые виды с верхушечным ростом; Fischerella). Здесь клетки делятся в основном более чем в одном направлении. У всех многоклеточных видов цианобактерий фактически мы имеем дело с колониями, в которых отдельные клетки рыхло расположены внутри выделяемой ими совместно слизи или в исходной клеточной стенке (см. 5.1).
Движение. Талломы многих видов, чаще всего нитчатых, совершают скользящие движения (2—11 мкм/с). Движение возможно только на твердом и к тому же влажном субстрате. Оно основано не только на простом выделении слизи (ср. движение десмидиевых водорослей, Zygnematophyceae), но и, по-видимому, на работе микрофибрилл. Эти структуры обвивают нить или клетку поверх муреинового слоя и за счет трения о субстрат сообщают вращательное
движение. Для упора им служит собственная слизь, выделяемая через тончайшие поры диаметром в 10 нм (рис. 11.14, В) в клеточной стенке. К вращательному движению с перемещением вперед способны только нитчатые Oscillatoriaceae, тогда как представители других групп двигаются вперед без одновременного вращения.
Размножение цианобактерий осуществляется путем клеточного деления. Подвижные стадии со жгутиками отсутствуют. Нитчатые синезеленые водоросли растут ин-теркалярно путем деления любых клеток в нити. Поперечные перегородки образуются при этом нентрипетально наподобие ирисовой диафрагмы (рис. 11.15, N). Они состоят только из материала опорного слоя. Размножение происходит за счет неспецифической фрагментации нитей или за счет обособления гормогониев из небольшого числа клеток (рис. 11.15, L). Гормогонии — фрагменты нитей, состоящие из молодых, неспециализированных клеток, — отделяются от материнской нити, уползают и разрастаются в новые нити. У некоторых одноклеточных форм содержимое увеличивающейся в размере материнской клетки последовательно делится на большое число округлых эндоспор, каждая из которых после выхода из материнской клетки вырастает в новую особь. У некоторых видов с вытянутыми в длину клетками базальная часть остается стерильной, тогда как апикальная часть постоянно регенерирует для образования спор (рис. 11.15, D). Эндоспоры цианобактерий отличаются от эндоспор эубактерий своей структурой и развитием. Встречаются также и экзоспоры; они от-шнуровываются от материнской клетки. Все эти виды спор не имеют жгутиков. Для переживания неблагоприятных периодов образуются (особенно у гормогониевых) покоящиеся клетки — акинеты. В них превращаются отдельные клетки, в которых откладываются запасные вещества, при этом размер их увеличивается, а стенки сильно утолщаются (рис. 11.15, J). Акинеты прорастают в гормогонии. Однако может быть и так, что боковые ответвления нитей целиком окружаются прочной стенкой и превращаются в покоящийся орган — гармонисту. Таким образом, имеются различные
130 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
f 00 00 j GOOD t ©ООО i 00 ~ ‘ t СС И;С
00
00 GC
00 00 - ис
00 ОС
СЭ со
СС 00 00
С
Рис. 11.15. Cyanobacteriota (A, D, Е, J, М — по L.Geitler; В — по К. Magdefrau; С— по Smith; F, G — по G.G.Thuret; Н — по Brehm; К, L— по O.Kirchner; N — по M.M.Gomont):
А — Chroococcus turgidus (400х); В — Aphanocapsa pulchra (500х); С — Merismopedia punctata (бООх); D— Dermocarpa clavata, образование эндоспор; Е, F — Nostoc commune: Е— колония (1х), F — то же, нить с 4 гетероцистами (400х); G — Rivularia polyotis, часть колонии (200х); Н — Rivularia haematites, часть колонии на поперечном срезе, с отложениями извести и годичными кольцами (15х); J— Cylindrospermum stagnate, с удлиненной покоящейся клеткой и шаровидной гетероцистой у конца нити (500х); К— Plectonema wollei с ложным ветвлением (200х); L— Lyngbya aestuarii, образование гормогониев (500х); М — Stigonema mamillosum, конец нити (250х); N — Oscillatoria princeps, конец нити (ЗООх)
возможности не только для вегетативного размножения, но и для образования покоящихся органов, которые могут встречаться в жизненном цикле при определенных (т.е. неблагоприятных) условиях.
Половое размножение неизвестно. Неясно, основан ли наблюдаемый иногда обмен генетическим материалом (в частности, могла происходить рекомбинация факторов устойчивости к разным антибиотикам двух штаммов в одном-единственном штамме) на парасексуальных процессах.
Систематика. Основанное на морфологических признаках подразделение совпадает с молекулярно-филогенетическими данными лишь частично. Cyanoprokaryota (т.е. цианобактерии) составляют единственный класс (Cyanophyceae) и в зависимости от ступени организации подразделяются на несколько порядков.
1. Порядок: Chroococcales. К нему относятся одноклеточные формы или простые колонии (шары; таблички; короткие неразветвленные нити). Неразветвленные короткие нити уже могут объединяться в своего рода «псевдопаренхиму». Размножение происходит путем клеточного деления, у некоторых образуются также эндоспоры (например, Dermocarpa', рис. 11.15, D) и экзоспоры. Если при этом дочерние клетки не вырастают до нормального размера материнских клеток, говорят о размножении посредством наноцитов. Synechococcus и Synechocystis — одноклеточные формы. У Chroococcus и Gloeo-capsa (см. рис. 5.2) клетки после деления остаются объединенными в колонии по 2, 4 или 8 внутри общих слизистых обверток, которые могут быть слоистыми (см. 5.1). У Chroococcus молодые дочерние клетки полушаровидные (рис. 11.15, А), тогда как у Gloeocapsa они яйцевидно закругленные и располагаются в очень толстой слизистой обвертке. Виды обоих родов встречаются чаще всего в слизистых налетах на
11.2. Бактерии, грибы, растения | -|3-|
влажных скалах и стенах. У Aphanocapsa (рис. 11.15. В), Aphanotheca, Microcystis и Merismopedia (рис. 11.15, С) число клеток, входящих в колонию, больше. Колонии в виде табличек у видов Merismopedia (рис. 11.15, С) возникают в результате клеточных делений строго в одной плоскости; эти виды обитают в пресных волах, некоторые — в морях.
2. Порядок: Oscillatoriales. У этих нитчатых форм еще отсутствуют гетероцисты и акинеты, только конечные клетки отличаются по облику от остальных клеток нити. Деление клеток всегда происходит в одном направлении, поэтому ветвление отсутствует. Нити часто встречающихся в воде и иле видов Oscillatoria состоят, из однородных, часто дисковидных клеток (рис. 11.15, М). Рост осуществляется ин-теркалярно, а размножение гормогониями. К этому же порядку относятся роды Phormidium, Schizothrix, Spirulina, Trichndesmium, Plectonema и Lyngbya.
3. Порядок: Nostocales. Здесь клеточное деление происходит, как и у представителей пре-дьщущего порядка, но перпендикулярно продольной оси нитей (см. рис. 5.1). Размножение происходит, как и у Oscillatoriales, гормогониями. В качестве особых, хорошо заметных форм клеток регулярно встречаются гетероцисты и иногда акинеты. Виды рода Nostoc, которые образуют в воде или на влажной почве округлые или аморфные лопастные колонии, представлены четковидными нитями, погруженными в общую полисахаридную слизь (рис. 11.15, F). Виды родов Cylindrospermum, Aphanizomenon и Anabaena, часть которых — планктонные формы, образуют покоящиеся клетки (рис. 11.15, J). У видов Rivularia (рис. 11.15, G, Н) в колониях, прикрепляющихся к водным растениям и камням, имеется четкое различие межу основанием и концом нити: на нижнем конце находится гетероциста, а кверху нить постепенно вытягивается в бесцветный волосок; таким образом, здесь имеется дифференциация, обусловленная планом строения. Виды Tolypothrix и Scytonema отличаются наличием ложного ветвления. К этому же порядку относятся роды Anabaenopsis, Calothrix и Aulosira (см. 9.2.1).
4. Порядок: Stigonematales. К порядку относятся наиболее дифференцированные формы. В результате поперечных и продольных клеточных делений для них возможны и характерны настоящее ветвление и наличие многорядных нитей. Размножение осуществляется гормогониями. Могут встречаться гетероцисты или акинеты. У Stigonema (рис. 11.15, М) таллом подразделяется на основание и вершину; верхушечная клетка отчленяет вниз сегменты, которые дальше делятся продольными и поперечными перегородками; многорядные нити могут образовы
вать также боковые ветви. К этому же порядку относятся роды Fischerella, Hapalosiphon и Ма-stigodadus.
Встречаемость и образ жизни цианобактерий (синезеленых водорослей)
Цианобактерии, насчитывающие порядка 2 000 видов, распространены по всему земному шару. Часто их можно видеть невооруженным глазом как слизистую массу, тонко-нитчатые налеты, «цветение воды» и т. д. Они обитают прежде всего в пресной воде (даже в горячих источниках с температурой порядка 75 °C), но встречаются также на влажных и сухих почвах, на коре деревьев и скалах — до Арктики и Антарктики. Таким образом, некоторые виды приспособились к жизни вне воды.
Разрушение и образование горных пород. Синезеленые водоросли, обитающие на известковых породах, подвержены воздействию больших колебаний температуры и влажности. Они живут здесь отчасти на поверхности (эпилитно), отчасти в тончайших трещинках (эндолитно) и нередко образуют черные полосы (чернильные штрихи). Некоторые эндолитные виды могут растворять известковые породы, у других же (например, Rivularia, Schizothrix) известь откладывается в слизистых чехлах (рис. 11.15, Н), что в пресной воде приводит к образованию мела и известкового туфа, а в приливно-отливной зоне теплых морей — к отложению слоистых известковых корок (строматолитов). Ископаемые строматолиты обнаружены в докембрийских отложениях, и полагают, что образовавшие их синезеленыс водоросли уже в ту геологическую эпоху покрывали большие площади и были широко распространены.
Виды, встречающиеся в больших массах у поверхности пресных и соленых вод, могут вызывать так называемое «цветение воды». Oscillatoria rubescens вызывает в эвтрофных водоемах красное «цветение» и известна как водоросль «бургундского цветения». Другие виды, такие, как Microcystis aeruginosa и Aphanizomenon flos-aquae, образуют ядовитые пептиды, что может вызывать гибель рыбы в пресных водах. Цвете
132 | ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
ние воды, вызываемое Spirulina platensis в соленых озерах Южной Африки, — основной источник питания малого фламинго. При биологическом анализе воды обилие цианобактерий означает критическую степень эвтрофирования (бокс 11.7).
Азотфиксация. Во многих родах (Nostoc, Anabaena и др.) есть вилы, которые связывают свободный атмосферный азот (см. 9.2.1). На заливаемых водой рисовых полях цианобактерии ежегодно связывают до 50 кг азота на гектар. В противоположность некоторым эубактериям (Rhizobiaceae) цианобактерии, фиксирующие атмосферный азот, тем самым полностью приспособлены к свободному образу жизни. Вклад синезеленых водорослей в функционировании экосистем при этом, по-видимому, больше, чем азотфиксируюших эубактерий. Среди цианобактерий также больше число азотфиксирующих родов и видов.
Представители нескольких родов вступают в симбиотические отношения с другими организмами. Водоросли некоторых лишайников (см. Mycobionta, приложение 2) — цианобактерии. Некоторые формы живут как эндофиты в полостях тканей других растений, например Anabaena — в листьях Azolla (см. рис. 11.159, D), Nostoc — в талломе некоторых печеночников (Blasio, Anthocems. см. рис. 11.127, D), в корнях Cycas и корневище Gunnera (покрытосеменного растения). В этих симбиотических ассоциациях цианобактерии снабжают своих партнеров азотом.
Четвертый отдел: прохлорофиты (Prochlorobacteriota, Prochlorophyta)
Прохлорофиты — одноклеточные, живущие в симбиозе с морскими асцидиями водоросли, у которых отсутствуют фикобилины и к тому же наряду с хлорофиллом а имеется хлорофилл Ь. По строению клетки это типичные прокариоты: ограниченное мембранами ядро отсутствует, клеточные стенки состоят из муреина. В культуре они размножаются самостоятельно. К этому отделу относятся также свободноживущие
нитчатые пресноводные формы, по остальным признакам соответствующие диагнозу Prochlorophyta1.
II. ЦАРСТВО (ДОМЕН): АРХЕИ (ARCHAEA)
Археи внешне напоминают бактерии, несмотря на существеннейшие отличия от них (почему их раньше называли архебак-терии). Однако у архей (насчитывающих около 80 видов) имеется большое разнообразие в строении клеточных стенок и мембран, причем мурамовая кислота — типичный компонент клеточной стенки эубактерий — всегда отсутствует. В зависимости от рода образуются белковые и полисахаридные покровы, белковые чехлы (влагалища), а также клеточные стенки, иногда с псевдомуреином, компонентом которого является не мурамовая кислота, а L-талозами-нуроновая. Удалось выявить только L-ами-нокислоты. Благодаря другой клеточной структуре археи устойчивы к пенициллину, D-циклосерину и другим антибиотикам, нарушающим биосинтез муреина. Другие специфичные вещества — разветвленные, содержащие фитан липиды с эфирными связями, РНК-полимеразы комплексного строения и большая доля модифицированных нуклеотидов в рибосомных нуклеиновых кислотах. Результаты анализа нуклеотидных последовательностей рРНК свидетельствуют о большой обособленности архей от эубактерий.
Сходные и даже совпадаюшие облик и особенности жизнедеятельности у архей и эубактерий возникли, очевидно, в ходе независимого эволюционного развития. В обеих группах встречаются кокки, палочки. сарпины. спириллы и нитчатые формы.
1 Прохлорофиты были открыты сравнительно недавно (в 1970-е гг.). и это открытие было тогда настоящей сенсацией. Однако данные молекулярной биологии говорят о близком родстве их к некоторым цианобактериям, так что во многих современных системах их как таксон вообще не рассматривают, а интегрируют в систему цианобактерий: одноклеточные формы помещают в Chroococcales, а нитчатые — в Osci-llatoriales (см. выше). — Примеч. пер.
11.2. Бактерии, грибы, растения | 133
У архей встречаются еще клетки в форме пластинок и часто имеются переходы от одной формы к другой. Некоторые представители способны к активному движению, чаше всего за счет наличия одного жгутика (например, Methanobacterium mobile). Как и у бактерий, среди архей есть аэробные и анаэробные, гетеротрофные, серозависимые и фототрофные формы. Некоторые представители являются при этом экстремально термофильными, ацидофильными или галофильными. Таксоны архей в соответствии с традициями и установленными правилами номенклатуры часто имеют вводящие в заблуждение концовки названий типа -bacterium или -bacteriales.
На основании различий в составе 16S рРНК современные исследования отдельных представителей дают основание предполагать сильную дивергенцию в ходе эволюции архей, бактерий и эукариот (не считая их хлоропластов и митохондрий) (см. рис. 11.9). Однако по ряду биохимических признаков археи показывают большее родство с эукариотами, чем с бактериями, что еще больше подчеркивает их обособленное положение среди прокариот.
Эволюционное обособление архей произошло, по-видимому, примерно 4 млрд лет назад, так как древнейшие известные ископаемые находки цианобактерий имеют возраст около 3 млрд лет, а археи, исходя из данных по 16S рРНК, должны были возникнуть до разделения на эу- и цианобактерии. На этом раннем временном отрезке дифференцировки жизни (более 3 млрд лет назад) в атмосфере Земли, носившей в значительной степени восстановительный характер, могли существовать метановые бактерии (Н2 — из атмосферы; СО2 — из примитивных процессов брожения в древних морях).
Археи включают древние эколого-физиологические приспособительные типы, которые (вероятно, в ходе прогрессивного эволюционного развития) сохранились до настоящего времени в соответствующих биотопах (например, метанобактерии в сапропеле и рубце жвачных животных). В пользу таксономического разделения на рано и сильно разошедшиеся эволюционные линии говорит реликтовая природа доживших до наших дней представителей этой группы.
Первый отдел: Crenarchaeota
Чаше всего термофильные (термоацидофильные) и зависимые от серы представители этого отдела, по-видимому, наиболее близки к предкам архей. С позиций молекулярной филогенетики они характеризуются своеобразной структурой РНК в рибосомах, отличаясь от второго отдела (Euryarchaeota).
Верхняя температурная граница для этих организмов, приспособившихся к архаичным условиям жизни, определяется доступностью воды, а также стабильностью компонентов клетки. Клетки Pyrodictium occidtum (Sulfolobales) диаметром 0,3 — 2,5 мкм имеют форму от дисковидной до чашевидной; они заключены в сеть из полых фибрилл; оптимальная для их роста температура 100 °C, верхняя температурная граница роста ПО "С. Вода при подобных высоких температурах остается жидкой только под давлением, например у морского дна или в глубине сольфатаровых полей. Pyrodictium был открыт в глубине морских вод с вулканическим нагревом. Это крайне анаэробный организм, восстанавливающий серу в сероводород. Sulfolobus acidocal-darius— аэробный и крайне ацидо-термофиль-ный вид; он факультативно автотрофный, отчасти использующий для дыхания органический субстрат и отчасти окисляющий серу до серной кислоты с использованием кислорода и воды. Температурный минимум для его роста 60 °C (у Pyrodiction occultum 82 °C). Acidothennus infernus в зависимости от наличия кислорода может образовывать сероводород или серную кислоту. Thermoplasma acidophdum (Thermoplasmales) — организм, который был изолирован из дымящихся угольных отвалов и горячих источников. Эти археи, лишенные клеточной стенки, в нейтральной среде погибают; оптимальные для их роста значения pH I —2, а температуры 59 °C. Их клетки, размножающиеся почкованием, очень вариабельны: от сильно вытянутых до неправильно-коккоидных. Были также открыты формы с пучком жгутиков. Дисковидные клетки видов Ther-moproteus и Thermofilum (Thermoproteales) могут даже вырастать в нити длиной свыше 100 мкм.
Второй отдел: Euryarchaeota
Представители этого отдела обитают в широком спектре экстремальных биотопов. Они отличаются с позиций молекулярной филогенетики (рРНК) от видов предыдущего отдела.
134 I ГЛАВА 11, СИСТЕМАТИКА И ФИЛОГЕНИЯ
1. Метаногенные археи — анаэробы, продуцирующие метан; при попадании на воздух они погибают быстрее, чем анаэробные бактерии. Спорообразование не установлено. Это морфологически разнообразная, но физиологически весьма однородная группа, способная к автотрофному образу жизни с использованием СО2 и Н2 в качестве единственных источников углерода и энергии. Как альтернативные источники углерода могут использоваться также простые карбоновые кислоты и спирт. Кроме того, они отличаются наличием двух специфических, больше нигде не встречающихся кофакторов, из которых один (СоМ, 2-меркаптоэтансульфоновая кислота) участвует как переносчик метана при его образовании, а другой (F420) действует как переносчик водорода. Формы Methano-bacterium (Methanobacteriaies) бывают от кок-коидных до узко-палочковидных; кроме того, все виды грамположительные и имеют псевдомуреиновый чехол. Последующие роды грамот-рицательны. Род Methanospirillum (Methano-microbiales) объединяет организмы в виде длинных закрученных палочек; их белковое влагалище не участвует в образовании перегородок (ср. образование перегородок у эубактерий). Methanosarcina имеет необычно крупные, сгруппированные в пакеты клетки с гетерополиса-харидным чехлом. Methanococcus (не путать с Methylococcus) — типовой род Methanococcales — образует кокки; вместо жесткой клеточной стенки имеется поверхностный слой из белковых компонентов. Таким образом, большинство родов «метанобактерий» по-разному решило проблему построения клеточного покрова. К тому же эти формы практически не имеют сходства по общему химическому составу, не отмечено их сходство и с эубактериями.
2. Галофильные археи (Halobacteriales) выживают даже в высушенной соли и встречаются в сильно соленых водоемах. Halobacterium halobium может расти даже при 12%-й концентрации NaCl. При значении pH 5,5 галобактерии жить уже не могут; их температурный оптимум находится в пределах 40—45 °C. При определенных условиях они могут осуществлять фосфорилирование (см. 6.4.9) и вследствие содержания каротиноидов окрашивать соленую воду в красный цвет.
III. ЦАРСТВО (ДОМЕН): ЭУКАРИОТЫ (EUCARYA, EUKARYOTEN)
Эукариоты по числу видов и биомассе составляют большую часть ныне живущих организмов. Эукариотная клетка (эуцит)
отличается наличием истинного клеточного ядра, которое отграничено от цитоплазмы оболочкой из двух мембран, имеющей поры (см. рис. 2.26). Кроме того, важная особенность эукариот— наличие разнообразных клеточных органелл, которые, как и ядро, четко отграничены от основной плазмы. К ним относятся эндоплазматическая сеть, диктиосомы (аппарат Гольджи), митохондрии и микротельца (см. 2.2.8, 2.2.6.6). Для фотоавтотрофных эукариотных растений характерны хлоропласты, окруженные двумя или большим числом мембран (см. 2.2.9.1). Жгутики эукариотных клеток построены по одному плану: они имеют две центральные простые микротрубочки и 9 периферических двойных (структура 2 + + 9!). Если имеется клеточная стенка, то она образована сплетением макромолекул, связанных только соседними валентностями (целлюлоза, хитин и т.д.). В целом эукариоты четко отличаются от прокариот большей комплексностью клеток. Этим различием обусловлена глубокая пропасть между всеми ископаемыми и ныне живущими прокариотами — с одной стороны, и эукариотами — с другой (ср. с динофитами, см. приложение 2 к Rhodobionta). Аминокислотные последовательности ферментных белков со сходными функциями у про- и эукариот совпадают в значительно меньшей степени, чем у представителей любой из этих обеих групп. Так, элементы цепочки цитохрома с бактерий и эукариот имеют до 60 % различных аминокислот, тогда как соответствующее значение при сравнении человека и растений пшеницы дает 45 %, млекопитающих и птиц — 12 %, а человека и шимпанзе — 0 %.
Возникновение эукариот и образование при этом ограниченных мембранами органелл в эуците в настоящее время чаще всего рассматриваются в свете эндосимбио-тической теории (см. 2.4; рис. 11.9). В основе ее лежит признание многократно и независимо осуществлявшихся поглощений симбионтов примитивными эуци-тами.
Ископаемые находки позволяют приблизительно определить время эволюционного обособления эукариот. Древнейшие одноклеточные организмы из архея (свыше
11.2. Бактерии, грибы, растения 1 -|35
3,4 млрд лет назад) имеют средний размер 5 мкм и, таким образом, соответствуют современным прокариотам. В дальнейшем, около 1,4 млрд лет назад, начинают преобладать клетки более крупных размеров — в среднем 13 мкм, что характерно для эукариот. Поэтому есть основание считать, что разделение про- и эукариот могло произойти примерно 2 млрд лет назад, тогда как разделение растений и животных — «только» 1,1 млрд лет назад.
Размножение, смена ядерных фаз и поколений
Клеточное ядро в типе делится путем митоза. В ходе бесполого (вегетативного) размножения происходит митотическое деление ядер. При половом размножении сливаются цитоплазма и ядра двух часто специализированных в качестве гамет клеток (плазмо- и кариогамия = сингамия); закономерно следующий мейоз ведет от диплофазы к гаплофазе и, таким образом, обеспечивает характерную для эукариот смену ядерных фаз.
Сингамия. Связанная с половым размножением рекомбинация получаемого от родителей наследственного материала (генетическая рекомбинация) решительным образом способствовала эволюции эукариот. Соответственно у них половое размножение — за исключением лишь немногих групп, где оно, по-видимому, полностью отсутствует (например, у Euglenophyta), — прочно закрепилось и выработалось уже на ранних этапах эволюции. При этом надежность сингамии все больше подкреплялась образованием большого числа половых клеток или половых ядер, чем обеспечивалось большое число диплоидных ядер, возникавших в результате сингамии. Благодаря этому значительно интенсивнее стала происходить генетическая рекомбинация. В целом сингамия осуществляется путем слияния двух половых клеток (гамет; га-метогамия). Они могут быть одинакового вида, в некоторых случаях со жгутиками (изогамия), или же разного вида: меньшие по размеру и имеющие жгутики — мужские, более крупные и также имеющие жгу
тики — женские (анизогамия1), или же жгутики имеют только мужские клетки, а женские лишены их (оогамия) (см., например, Chlamydornonadaceae). Если гаметангии (клетки, отличающиеся от остальных и содержащие сначала половые ядра, а потом иногда и гаметы) сливаются друг с другом непосредственно, не высвобождая гамет, то имеет место гаметангиогамия (см., например, Oomycota). Если вегетативные клетки сливаются друг с другом без особой дифференцировки. этот процесс называют со-матогамия (см., например, Hymenomyceti-dae). При этом группируются попарно и сливаются друг с другом половые ядра, а не гаметы, как это имеет место при гаме-тогамии.
Смена ядерных фаз и поколений. Исходные эукариоты были (если они уже могли размножаться половым путем) чистые гаплонты с жизненным циклом, полностью проходящим в гаплофазе: после слияния половых клеток (гамет) или половых ядер в одну диплоидную клетку (зиготу) последняя непосредственно претерпевает мейоз. Возникающие в результате гаплоидные клетки образуют при митотических ядерных и клеточных делениях гаплоидную конечную стадию в онтогенезе. Диплоидна только зигота. Смена ядерных фаз (переход от дипло- к гаплофазе и наоборот) обозначается здесь как зиготическая, так как она связана с зиготой (например, Ulothrix, см. рис. 11.101, А1 2). Возникающее за счет сингамии ядро зиготы у гаплодиплонтов (см. рис. 11.101, В) на протяжении собственного диплоидного отрезка жизненного цикла размножается митотически (спорофит), пока возникшие таким образом многочисленные диплоидные ядра не примут участие в мейозе и, таким образом, не дадут
1 В отечественной литературе чаще используется термин гетерогамия. — Примеч. пер.
2 Пример неудачен, так как у Ulothrix зигота не непосредственно претерпевает мейоз, а предварительно прорастает в маленькое растеньице грушевидной формы; в него переходит диплоидное ядро и уже здесь происходит мейоз. Это растеньипе часто трактуют как диплоидную стадию водоросли (СЬЛо/ши-стадия), и тогда уже нельзя говорить о зиготической смене ядерных фаз. — Примеч. пер.
136 I ГЛАВА 11, СИСТЕМАТИКА И ФИЛОГЕНИЯ
начало многочисленным рекомбинантным гаплоидным репродуктивным структурам (мейоспорам). С них начинается гаплоидный отрезок жизненного цикла: вырастает гаплоидный организм (гаметофит), производящий гаметы. Смена ядерных фаз здесь занимает промежуточное положение между образованием зиготы и образованием гамет.
Гаплодиплонты отличаются сменой поколений, т.е. регулярной последовательностью поколений в онтогенезе; они размножаются по-разному и соответственно в ходе развития ограничены разными репродуктивными клетками. У растений гаплоидное поколение называют гаметофит. Последний возникает из мейоспор и заканчивает раз витие образованием гаплоидных гамет. Диплоидный спорофит возникает из зиготы (до этого возникшей из слившихся гамет) и заканчивает развитие образованием мейоспор, которые из-за непосредственно предшествующего мейоза диплоидных ядер гаплоидны. Обозначать диплоидное спорофитное поколение как отрезок жизненного цикла с бесполым размножением, строго говоря, некорректно, так как мейоз и связанное с ним образование мейоспор всегда неразделимы с половым размножением. Разве только спорофитное поколение может дополнительно размножаться бесполым 1гутем с образованием диплоидных митоспор. Если гаметофит и спорофит имеют в значительной мере одинаковый облик, говорят об изоморфной (например, Cladophora, см. рис. 11.101, В), если различаются по облику — об анизоморфной (или гетероморфной) смене поколений (Cutleria, Laminaria, см. рис. 11.87). В целом смена поколений является двучленной, а гаметофит и спорофит представляют собой самостоятельно живущие организмы.
При более редкой трехчленной смене поколений за гаметофитом следуют два по-разному размножающихся спорофитных поколения (Rhodophyta. см. рис. 11.65). В этом особом случае со сменой ядерных фаз связаны только переходы от гаметофита к первому спорофитному поколению и от второго спорофитного поколения к гаметофиту. Переход от первого ко второму спорофитному поколению происходит,
однако, в одной и той же ядерной фазе. Это показывает, что смена поколений не всегда должна сопровождаться сменой ядерных фаз. Два поколения могут быть связаны между собой и таким образом, что образуют одну особь. Если при этом одно поколение питается за счет другого, говорят о гонотрофии (Bryophytina, см. рис. 11.108, D). Гаплодикарионты (например, Hymenomyce-tidae, см. рис. 11.51, В; Derbesia', рис. 11.101, С) в значительной мере соответствуют гап-лодиплонтам, однако вместо диплофазы у них имеется дикариофаза, в которой половые ядра хотя и попадают в одну клетку (зиготу), но на длительном отрезке жизненного цикла делятся порознь, в результате чего возникает и поддерживается двухъядерная фаза (дикарион). Только непосредственно перед мейозом половые ядра сливаются в диплоидные. У диплонтов (например, Oomycota, см. рис. 11.20, D) весь жизненный цикл проходит в диплофазе. Возникающие в результате мейоза гаметы гаплоидны; смена ядерных фаз является, таким образом, гаметической. Среди растений диплонты возникли путем все большей редукции гаметофита и интеграции немногих клеток, оставшихся от гаметофита, в спорофит (Fucus, см. рис. 11.87). Таким образом, диплонты имеют часто лишь скрытую смену поколений (семенные растения; см. бокс 11.10).
Эволюция эукариот характеризуется прогрессирующими комплексностью, дифференцировкой и функциональной специализацией органов, а также приспособлением к различным стратегиям питания и местообитаниям. Так возникли уровни (типы) организации, которые чаще всего должны рассматриваться не как эволюционно родственные группы, а именно (за исключением не рассматриваемых здесь животных) слизевики (бокс 11.2), грибы (см. бокс 11.3), лишайники (Mycobionta, приложение 2), водоросли (см. бокс 11.5), Embryo-phyta (см. бокс 11.8) и сосудистые растения (см. бокс 11.9).
Особенно обращает внимание многократное возникновение одного и того же типа организации у лишайников путем независимого друг от друга образования симбиоза различных водорослей и грибов. Так
11.2. Бактерии, грибы, растения | -(37
же и грибы I щедставляют собой скорее пат раллельные линии, эволюционировавшие по пути приспособления к гетеротрофному образу жизни, чем филогенетически единую ipynny. То же относится и к рано разделившимся эволюционным линиям эукариотных водорослей, по меньшей мере в плане учета фотоавтотрофных эндосимбионтов.
Первое подцарство: Acrasiobionta
Это подцарство включает слизевиков, у которых на всех стадиях жизненного цикла отсутствуют клетки со жгутиками1. Их агрегирующие плазмодии образуются путем агрегации отдельных клеток без слияния.
Единственный отдел: Acrasiomycota
У Acrasiobionta, к которым относится единственный отдел и единственный класс акразиомицеты (Acrasiomycetes)1 2, миксаме-бы сползаются в агрегационный плазмодий (также называемый псевдоплазмодий; см. рис. 5.3), не сливаясь между собой. Мик-софлагелляты, имеющие жгутики, отсутствуют. Клеточные стенки состоят из целлюлозы.
Жизненный цикл. В фазе бесполого размножения амебы делятся до тех пор, пока не исчерпаны источники питания. Питание у них, как и у миксомииетов, фаготрофное; пищей служат преимущественно бактерии. Если пищи больше не хватает, то начинается сползание определенной
1 У некоторых представителей собственно ак-разиевых (Acrasiales в этой книге) жгутиковые стадии имеются. — Примеч. пер.
2 В настоящее время практически общепризнанно, что Acrasiomycetes («клеточные слизевики») в таком понимании, как здесь, — исключительно гетерогенная группа, которая не может считаться каким-либо естественным таксоном: собственно акразиевые (здесь — Acrasiales) — очень обособленная группа, показывающая отдаленное родство с эвгленами, тогда как диктиостелиевые (здесь — Dictyosteliales) — сородичи миксомине-тов (Myxomycota). — Примеч. пер.
группы амеб к некоторому центру (см. рис. 5.3). Амебы, сползающиеся в агрегационный плазмодий, не сливаясь при этом между собой, привлекаются друг к другу веществом акразином (см. 8.2.1.1; рис. 8.5).
Агрегационный плазмодий может переползать по субстрату в том или ином направлении, пока не наступит фаза кульминации, когда он станет подниматься вверх в виде колонки. И теперь сохраняется индивидуальность одноклеточных амеб, хотя и становятся явными определенные процессы дифференцировки при происходящем теперь формировании плодовых тел. В центральной части зачатка плодового тела (см. рис. 5.3) амебы вскоре останавливаются, одеваются жесткой клеточной стенкой и образуют ножку из отдельных клеток; отсюда происходит название «клеточные слизевики», даваемое акразиомицетам. При добавлении других амеб снизу ножка удлиняется. На нее наползают новые амебы, образуя наверху собственно спорокарп («спорангий»). В головке периферические клетки становятся корой, внутренние клетки принимают округлую форму и превращаются в гаплоидные споры (цисты); образуется также колонка как продолжение ножки. После споруляции ножка и коровые клетки отмирают.
Вопрос о половом процессе путем слияния амеб в диплоидные мегацисты и следующем после этого мейозе долгое время оставался спорным. В настоящее время считается достоверным половое размножение, по крайней мере, у агрегационного плазмодия Polysphondylium (Dictyosteliales). Широко известный объект лабораторных исследований — Dictyostelium. У Acrasis (Acrasiales), в противоположность представителям предыдущего рода, амебы не образуют в пределах агрегационного плазмодия потоков в направлении центра образования спо-роношения.
Второе подцарство: Myxobionta
Отличающие этот таксон плазмодии возникают путем слияния миксофлагел-лят или миксамеб или же развиваются из отдельных клеток без предшествующего полового процесса. В жизненном цикле встречаются подвижные стадии со жгутиками. Клеточные стенки, насколько они различимы на определенных стадиях развития, построены из галактозамина и целлюлозы.
138 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Первый отдел: Myxomycota
I. Класс: миксомицсты (Myxomycetcs). Вегетативная (= соматическая) фаза представляет собой диплоидный, многоядерный, не подразделенный на отдельные клетки настоящий плазмодий (см. рис. 2.9; 11.16), питающийся фаготрофно. Из плазмодиев развиваются плодовые тела, причем часть плазмы отвердевает и создает характерные структуры, тогда как другая часть, содержащая ядра, превращается в мейоспоры. Последние имеют клеточную стенку, состоящую не менее чем из двух слоев и, согласно новейшим исследованиям, не содержащую в своем составе ни целлюлозы, ни хитина, алишь преимущественно полимерный галактозамин. В качестве запасного питательного вещества образуется гликоген. Плазмодии и особенно плодовые тела часто окрашены в яркие цвета. Химическая структура пигментов изучена лишь отчасти; она иная, чем у грибов.
Жизненный цикл. Споры прорастают в воде или на влажном субстрате. Всхожесть их часто сохраняется очень долго. Так, удалось прорастить споры из гербарного образца, хранившегося свыше 70 лет. При этом споры высвобождают или одноядерные, голые амебоидные подвижные миксамебы, или миксофлагелляты, имеющие жгутики (рис. 11.17, А). Миксофлагелляты чаще всего двужгутиковые, причем один жгутик часто сильно редуцирован или полностью отсутствует. В последнем случае, по крайней мере, имеется второй, утративший свою
Рис. 11.16. Myxomycota. Край плазмодия у
Badhamia utricularis (2х) (по E.Jahn)
функцию, блефаропласт. Подвижные клетки могут за счет утраты жгутиков превращаться в миксамебы. Последние размножаются, как и подвижные клетки, путем деления. Миксамебы или миксофлагелляты сливаются попарно друг с другом в аме-бозиготу (или имеющую жгутики плано-зиготу), в которой затем сливаются ядра (плазмогамия, затем кариогамия). Получающаяся диплоидная структура претерпевает многочисленные митотические ядерные деления и развивается в крупный многоядерный плазмодий (см. рис. П.16). Такие плазмодии могут в свою очередь сливаться между собой. Митозы закрытые и происходят у всех ядер одного плазмодия синхронно. В плазмодиях плазма находится в интенсивном движении. Плазмодии развиваются при большой влажности воздуха в лесной почве, в соломе, среди травы, мхов или в гнилой древесине. Позже они изменяют форму и медленно выползают на поверхность субстрата. Клеточные стенки отсутствуют как на поверхности, так и внутри плазмодия. Его передняя часть (см. рис. 11.16) состоит из более плотной плазмы; ближе к задней части он часто выглядит как разделенный на сеть из отдельных тяжей. Диаметр плазмодия у некоторых видов может превышать 20 см (например, Fuligo, Brefeldia).
Образование плодовых тел начинается при определенных, еще недостаточно изученных экзогенных условиях (исчерпание субстрата, свет, температура, pH); возможно, решающую роль играют также эндогенные факторы. Перед этим меняется поведение плазмодия: он выползает из влажного субстрата к свету и превращается при сильной потере воды в многочисленные спорокарпы (спорангии, см. рис. 11.17, D). Эти плодовые тела имеют внешнюю, часто содержащую известь стенку — перидий, а нередко также и ножку, которая может продолжаться внутрь спорокарпа как колонка, а кроме того, нередко и систему нитей— капиллиций. Структуры формируются из затвердевшей безъядерной плазмы, которая не идет на образование спор. Возникновение капиллиция, очевидно, начинается с отложения строительного материала в особых пузырьках. Содержащая
11.2. Бактерии, грибы, растения | 13g
Рис. 11.17. Myxomycota (A — no Gilbert; В — по Н.А. von Stosch, R. von Wettstein; C — no C.G. Elliot; D, E, F — по H. Schenck; G, H — no A. L. Lister):
A—С — миксофлагелляты: А (и В) — короткий жгутик не изображен, В — копуляция (1 500х), С — показаны длинный и короткий жгутик; D — Leocarpus fragilis, многочисленные плодовые тела на мхе (нат. величина); Е— Comatricha typhoides, часть капиллиция (1В0х); F— Stemonitis fusca, плодовое тело (5х); G — Cribraria rufa, плодовое тело (ЗОх); Н — Trichia varia; нить капиллиция и споры (ЗООх)
ядра плазма путем разделения образует одноядерные, сначала диплоидные споры. В результате последующего мейоза в каждой споре возникает 4 гаплоидных ядра, из которых три ядра исчезают и остается только одно. При созревании плодового тела перидий разрывается, и споры выбрасываются из сплетения капиллиция. У некоторых видов капиллиций, подобно
элатерам печеночников, за счет гигроскопических изменений формы способствует освобождению спорокарпа. В жизненном цикле гаплоидны миксофлагелляты и не копулировавшие миксамебы, диплоидными являются плазмодии, плодовые тела и молодые споры, а зрелые споры опять гаплоидны. Таким образом, диплоидная фаза в ходе развития преобладает.
Бокс 11.2. Типы организации слизевиков
Организмы обоих подцарств — Acrasiobionta и Myxobionta — называются слизевиками. Слизевики отличаются вегетативным телом, представляющим собой лишенную клеточной стенки, многоядерную амебоидную подвижную плазменную массу — плазмодий. Он возникает;
• как агрегационный плазмодий; миксамебы сползаются в плазменные скопления, не теряя своей индивидуальности;
• как настоящий плазмодий: миксамебы или миксофлагеллаты должны слиться между собой, образуя диплоидный многоядерный плазмодий.
Еще одна возможность— возникновение плазмодия бесполым путем из одной-един-ственной клетки с делением ядер (но без клеточного деления).
Имеющиеся в жизненном цикле плазмодии различны, таким образом, по своей природе и аналогичны прокариотическим плазмодиям миксобактерий (Myxobacteriales). Размножение осуществляется спорами, которые возникают в особых плодовых телах (см. рис. 11.17, G), если
это не паразитические формы, обитающие внутри хозяина. У подвижных стадий имеется два гладких жгутика чаще всего неравной длины, реже один жгутик редуцирован. Между разными отделами слизевиков близкого эволюционного родства нет. Очень своеобразные по своей форме и образу жизни слизевики имеют много общих признаков с представителями выделяемой зоологами группы простейших, с которыми они близкородственны, но отличаются образованием плодовых тел и спор. Сходство состоит, в частности, в следующем;
• гетеротрофность — большинство форм питается, как и животные, фаготрофно, поглощая целые твердые частицы;
• наличие амебоидных стадий, входящих в жизненный цикл;
• отсутствие клеточных стенок, по крайней мере, в вегетативных фазах развития.
Это сходство с простейшими побудило многих исследователей к тому, чтобы относить слизевиков под названием Mycetozoa к царству животных.
140 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Питание плазмодиев или предшествующих им одноклеточных стадий осуществляется в природе, вероятно, всегда за счет поглощения различных микроорганизмов, таких, как бактерии, простейшие, споры, дрожжевые клетки, гифы грибов и т.д. Частины пищи заключаются в пищеварительные вакуоли и ферментативно перевариваются; непереваренные остатки через некоторое время выбрасываются. В культуре большинство видов может поддерживаться только тогда, когда им скармливают живые микроорганизмы (например, бактерии). Некоторые виды могут также поддерживаться в чисто сапротрофных условиях на питательных средах определенного состава.
В условиях культуры плазмодии некоторых видов могут захватывать и накапливать внутри себя одноклеточные зеленые водоросли, не переваривая их. Становясь таким образом зелеными, плазмодии приобретают фотосинтетическую активность. Хотя зеленые водоросли и попадают в плодовые тела, в естественных условиях такой симбиоз не может поддерживаться из поколения в поколение.
Систематика. Подразделение миксомицетов основывается прежде всего на различиях во внешнем виде плодовых тел. В более примитивных группах отсутствует капиллиций. Эволюция должна была идти также от сидячих плодовых тел к плодовым телам на ножках и от одиночных плодовых тел к комплексным. Примерно 500 известных видов подразделяются на следующие порядки.
1. Порядок: Ceratiomyxales. Спорообразование осуществляется экзогенно. На поверхности имеющего вид колонки плодового тела отчленяются на ножках споры (вероятно, односпоровые спорокарпы). Каждая спора дает при прорастании плазматический вырост с 4 гаплоидными ядрами, из которых после митоза образуется 8 гаплоидных подвижных клеток. В порядок входит только один род Ceratiomyxa с одним видом, представленным многочисленными формами и встречающимся по всему земному шару на гнилой древесине.
Во всех последующих порядках (2 — 6) споры образуются эндогенно внутри плодовых тел.
Плодовые тела у следующих 4 порядков (2 — 5) формируются из полукруглых выпуклостей плазмодия, которые сначала соединены с ним и друг с друтом плазматическими тяжами, однако позже становятся изолированными. Прилегающий к субстрату, составляющий основу слой
плазмодия («гипоталлус») или вообще не сохраняется при созревании плодовых тел, или от него остается только слизистая кожица.
2. Порядок: Liceales. В противоположность дальнейшим порядкам (3 — 6) здесь отсутствуют капиллиций и колонка (например, Lycogala, Cribraria, рис. 11.17, G).
3. Порядок: Echinosteliales. Имеется колонка.
4. Порядок: Trichiales. Колонка отсутствует. Капиллиций состоит из нитей со свободными концами, как у Trichia' (рис. 11.17, Н).
У всех трех предыдущих порядков (2 — 4) споровая масса светлых тонов.
5. Порядок: Physarales. Споровая масса черная или разных тонов — от темно-фиолетовых до ржавых. На перидии и часто также на капиллиции видны белые отложения извести (например, Didymium). Сюда относятся также Leocarpus (рис. 11.17, D); Badhamia (рис. 11.16); Fuligo septica с комплексным плодовым телом (эталием).
6. Порядок: Stemonitales. Плодовые тела, имеющие размер от 0,5 до 1 — 2 см, развиваются на «гипоталлусе». Внутри они дифференцированы на колонку, капиллиций и споры. У Stemonitis капиллиций образует по периферии замкнутую сеть, у Comatricha нити имеют свободные концы (рис. 11.7, Е). Lamproderma отличается перидием с металлическим блеском, у Brefeldia плоское, крупное плодовое тело составлено из многих споро-карпов (эталий).
Практическое значение миксомицетов невелико. Большую популярность как тест-объект при физиологических и биохимических исследованиях получил Physarum (Physarales).
В следующем классе плазмодии возникают без предшествующего полового процесса, непосредственно из одиночных клеток (также и без агрегации плазмодиев).
II. Класс: нротостелиомицеты (Protoste-liomycetes). Многоядерные сетчатые плазмодии образуются из клеток со жгутиками или без них. На тонких ножках экзогенно отчленяются от одной до четырех спор.
Второй отдел: плазмодиофоромицеты (Plasmodiophoromycota)
Этот отдел отличается от всех ранее рассмотренных слизевиков наличием хитиновых клеточных стенок, а также одной осо-
1 У видов рода Trichia нити капиллиция не-разветвленные; у видов других родов они могут быть разветвленными. — Примеч. пер.
11.2. Бактерии, грибы, растения | -| 41
бенностью ядерного деления: в метафазе хроматиновые массы располагаются перпендикулярно к обеим сторонам крупного, несколько вытянутого ядрышка, так что возникает крестообразная фигура внутри ядер-ной мембраны. Остается подвопросом, правомерно ли рассматривать Plasmodiopho-romycota как производные миксомицетов, перешедшие к эндопаразитизму. Правда, у этих двух групп имеются сходные зооспоры с двумя жгутиками неравной длины. В цикле развития участвуют гаплоидные и диплоидные плазмодии; у миксомицетов они всегда диплоидные, у протостелиоми-цетов — гаплоидные.
Известный представитель единственного класса Plasmodiophoromycetes — Plasmodiophora brassicae — возбудитель килы капусты1 (рис. 11.18).
Жизненный цикл. Перезимовавшие покоящиеся споры (гипноспоры) паразита прорастают весной в почве двужгутиковыми гаплоидными зооспорами, которые после утраты жгутиков внедряются, как амебоиды, в корневые волоски молодых растений капус ты. Здесь каждая внедрившаяся амеба образует многоядерный гаплоидный плазмодий (рис. 11.18, В). Он может распадаться на многоядерные частицы, которые в свою очередь (после разрушения стенок в тканях хозяина) перемещаются дальше от клетки к клетке и таким образом быстро расширяют очаг инфекции. Позже, после распада плазмодия на сначала одноядерные, потом многоядерные фрагменты, возникают многоядерные гаметангии. Последние распадаются на двужгутиковые гаметы, число которых соответствует числу ядер. После разрушения ткани хозяина они высвобождаются и копулируют между собой в почве.
Диплоидные планозиготы после отбрасывания жгутиков снова внедряются в корни уже развившегося к тому времени растения капусты (теперь уже не исключительно через корневые волоски), где они разрастаются в многоядерные протопласты без клеточных стенок — диплоидные плазмодии. Растение-хозяин отвечает на это образованием опухолей (рис. 11.18. А). Затем после мейоза в клетках растения-хозяина возникают покоящиеся толстостенные гаплоидные мейоспоры (покоящиеся споры, гипноспоры), которые зимуют вместе с пораженным растением и весной, после сгнивания ткани опухоли, снова попадают в почву.
1 Более корректно называть эту болезнь «кила крестоцветных», поскольку поражается не только капуста. — Примеч. пер.
Рис. 11.18. Plasmodiophoromycota, Plastrodio-phora brassicae (A — no Ross; В — no C. Chupp; C, D — no M.Woronin):
A— кила на корнях растения кольраби (1/Зх); В — плазмодии в корневом волоске (ЗООх); С — клетки коры корня со спорами (520х); D — прорастание спор (1 240х)
Зооспоры и гаметы несут на переднем конце два очень различающихся по длине жгутика без мастигонем. Смена гаплоидного и диплоидного плазмодиев соответствует смене генераций, однако места кариогамии и мейоза в жизненном цикле еще окончательно не выяснены.
Виды некоторых родственных родов (например, Polymyxa) — паразиты разных наземных и водных растений, которые образуют похожие разрастания органов (60 облигатно эндопаразитических видов, поражающих сосудистые растения, водоросли и грибы).
Обзор слизевиков. Слизевики (Acrasio-bionta и Myxobionta; около 600 видов) находятся в основании эволюционного развития гетеротрофных эукариот. Протосте -лиомицеты и миксомицеты возникли, скорее всего, от бесцветных жгутиковых, а ак-разиомицеты — от каких-то амеб. Эти группы из-за особенностей морфологии и цик ла развития занимают очень обособленное положение («Mycetozoa»). Что касается плазмодиофоромипетов, то остается неясным, возник ли у них плазмодий исходно или только вторично в связи с эндосимби-отическим образом жизни. Во втором случае следовало бы присоединить этот класс к рассматриваемым далее грибам и там
1 42 | ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
сближать с группами, характеризующимися сходными признаками. Правда, там нигде не встречаются подвижные стадии с двумя гладкими жгутиками неравной длины, как у миксомицетов, протостелиомицетов и плазмодиофоромицетов.
Третье подцарство: Heterokontobionta
Организмы, относящиеся к этой большой группе, отличаются гетероконтными подвижными стадиями, т.е. они снабжены одним направленным вперед перистым жгутиком и одним направленным назад гладким жгутиком, который служит рулем (см. рис. 11.20, А, С; 11.72, F). Перистый жгутик имеет два ряда жестких волосков. Последние состоят из основания, трубчатого ствола и терминальных волосков (одного или нескольких). Волоски, встречающиеся иногда на жгутиках среди представителей других эволюционных групп (например, Glaucobionta, Chlorobionta), никогда не бывают ни жесткими, ни трубчатыми.
В этом подцарстве от одних предков возникли как гетеротрофные грибоподобные организмы (Labyrinthulomycota; Oomy-cota), так и автотрофные водоросли (I letero-kontophyta). Сначала рассмотрим гетеротрофные группы, которые по типу организации соответствуют грибам (бокс 11.3).
Первый отдел: лабиринтуловые (Labyrinthulomycota)
Сетчатые слизевики, объединяемые в класс Labyrinthulomycetes, имеют еще организацию слизевиков (см. бокс 11.2). Однако из-за гетероконтного типа организации жгутикового аппарата они сближаются с рассматриваемыми далее Oomycota. Сюда относятся виды, которые поражают как эндопаразиты морские растения (например, Zostera, Laminaria)1. Для них характерны
1 Среди лабиринтуловых известны также паразиты некоторых пресноводных водорослей (например, видов Cladophora и Vaucheria). Известны виды и из наземных биотопов, например паразиты корней пшеницы. — Примеч. пер.
многоклеточные сетчатые плазмодии; последние возникают путем деления двужгутиковых подвижных клеток внутри увеличивающейся в размере слизистой оболочки. Сетчатые слизевики представляют собой исходную группу среди Heterokontobionta. Сходные с ними организмы были предками рассматриваемых далее гетеротрофных Oomycota, а также ставших в результате эндоцитобиоза автотрофными Heterokontophyta2.
Второй отдел: Оомицеты, или целлюлозные грибы (Oomycota)
Этот отдел, включающий около 500 видов, хотя имеет определенное сходство с грибами (бокс 11.3), по ряду признаков отличается от всех типичных грибов. Таллом,
•’ К этому же отделу обычно относят и еще одну очень важную группу — класс траустохит-рневые (Thraustochytrioniycetes). Внешне их талломы конвергентно сходны с некоторыми хит-ридиомицетами (например, типа Rhizophydium, см. ниже), что и отразилось в их названии. Клетки у них, как и у лабиринтуловых, окружены слизистым чехлом. Эта эктоплазматическая сеть развита слабее, чем у лабиринтуловых, и напоминает ризомицелий у хитридиевых грибов. Она служит только для обмена веществ между клетками и окружающей средой, а также для прикрепления к субстрату. Размножение — посредством зооспор (изредка апланоспор и амебоидных клеток). Зооспоры образуются в большом количестве (иногда свыше 100) в зооспорангиях. Цикл развития от зооспоры до нового зооспорангия может занимать всего 2 — 3 ч. Поэтому траусто-хитриевые стали одними из самых обильных морских организмов, в частности в планктоне и нейстоне (например, в гипонейстоне их численность может составлять 2—3 млн на I мл воды), и играют важную роль в питании морских жи-вотных-фильтраторов. Есть указания на наличие у траустохитриевых полового процесса, но доказательства отсутствуют. Траустохитриевые — облигатно морские и галофильные сапротрофы, встречающиеся по всему земному шару. После бактерий это наиболее массовые гетеротрофные организмы в морях и соленых внутренних водоемах. Некоторые известны как паразиты ряда морских беспозвоночных и рыб. Их насчитывается свыше 40 видов. Ряд видов хорошо развиваются на пыльце сосны, помещенной в морскую воду, так что их можно «выманить» для дальнейших исследований. — Примеч. пер.
11.2. Бактерии, грибы, растения | ( 43
реже одноклеточный (Lagenidiales) и чаще всего сифональный, имеет почти всегда стенки, содержащие целлюлозу. Размножение осуществляется путем слияния сГ гаметангиев с оогониями (гаметангиогамия); первые образуют оплодотворяющие выросты, внедряющиеся в оогонии. После оплодотворения в оогониях развиваются зиготы в числе от одной до нескольких (так называемые «ооспоры»). Возникающие при бесполом размножении зооспоры гетеро-контные: они несут направленный вперед перистый жгутик и направленный назад гладкий жгутик, чаще всего несколько более длинный (см. рис. 11.20, В). Кроме того, оомицеты (единственный класс отдела), согласно всем имеющимся данным, диплонты, так что мейоз у них происходит перед образованием гамет (гаметическая смена ядерных фаз). Сплетения гиф и плодовые тела никогда не образуются.
Что касается обмена веществ, то оомицеты помимо химизма клеточной стенки имеют следующие особенности. Белок клеточной стенки содержит гидроксипролин. Биосинтез лизина осуществляется после связывания пирувата и аспартата в дигидродипиколиновую кислоту по пути диаминопимелиновой кислоты (у других грибов через аминадипиновую кислоту). Никотиновая кислота синтезируется не из триптофана (как у животных и других грибов), а из СЗ-соединений. Ферменты, участвующие в синтезе триптофана, особого, нигде больше не встречающегося типа. Молекулярная масса рибосомной РНК (258-фракция) иная, чем у других грибов (за исключением слизевиков). Оомицеты практически все бесцветны; из них не удалось выделить никаких пигментов.
Все названные морфологические и химические признаки, а также диплоидный жизненный цикл говорят об обособленности Oomycota от хитиновых грибов (Eumy-cota).
Образ жизни. Виды, обладающие признаками, считающимися исходными, обитают в воде чаше всего как сапротрофы. Эволюционно более подвинутые обитатели наземной среды — паразиты высших растений. Это подразделение на группы в зависимости от образа жизни соответствует двум важнейшим (1,3) порядкам оомицетов.
1. Порядок: Saprolegniales. Лишенный поперечных перегородок многоядерный
мицелий (см. рис. 11.21, С) обитает в воде, чаще всего в пресной, у некоторых видов также в солоноватой, чаше всего сапро-трофно на погруженных туда гниющих частях растений и трупах насекомых; реже он паразитирует на ослабленных живых рыбах. При бесполом размножении концы гиф расширяются в слабо выраженные булавовидные зооспороцисты и отграничиваются перегородкой от несущей гифы. Путем деления плазмы в них возникают одноядерные грушевидные митозооспоры с двумя апикальными жгутиками неравной длины, из которых один несет два ряда волосков (см. рис. 11.21, А; 11.20, А). После плавания в течение некоторого времени жгутики впячиваются. Споры, становящиеся округлыми и одетыми стенкой, образуют на подходящем субстрате проростковую трубку и развиваются в новый организм.
У некоторых оомицетов сначала образуются еще другие зооспоры. Эти вторые подвижные стадии (рис. 11.21. В) отличаются от образуемых первоначально внешним видом и местом прикрепления жгутиков (рис. 11.21, А): они почковидные, и жгутики прикрепляются сбоку (рис. 11.21, В). Это явление дипланетизма отличает род Sapro-legnia от других родов1. У других родов первичные зооспоры встречаются только внутри или же в непосредственной близости от спороцисты (Achlya) или вообще не встречаются (Thrausto-theca, Dictyuchus)', в последнем случае диплане -тизм отсутствует. У Aplanes зооспор нет; споры, инцистирующиеся внутри спороцисты, дают про-ростковые трубки через ее стенку. Аналогично ведет себя Geolegnia, но с той разницей, что споры высвобождаются из спороцисты до образования проростковой трубки. Для Saprolegnia характерно то, что гифа, несущая спороцисту, после выхода зооспор прорастает новой спороцистой внутрь освободившейся.
Гаметангии отделяются поперечной перегородкой от несущих их гиф. Шаровидные оогонии содержат сначала много ядер, большинство из которых, однако, дегенерирует, после чего вокруг каждого из оставшихся ядер группируется плазма (оосфе-
1 Более корректно называть дипланетизмом способность зооспор инцистироваться, а потом снова существовать в виде зооспоры; явление изменения морфологии зооспор правильнее называть диморфизмом. — Примеч. пер.
144 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Бокс 11.3. Типы организации грибов
Грибы в более узком смысле слова, как и слизевики, не имеют пластин и хлорофилла; они живут как сапротрофы или паразиты в пресной воде или на суше, реже в морях. Часто их можно культивировать на подходящих питательных средах — в основном это сапротроф-ные, но также и некоторые паразитные формы; они гетеротрофны не только по отношению к углероду, но и в отношении азота и определенных активных веществ. Эти эукариот-ные организмы, образующие таллом (слоевище), хотя и рассматриваются традиционно вместе с растениями, занимают особое положение в системе живых организмов. В противоположность слизевикам у них не бывает плазмодия, а таллом чаше всего не является амебоидным, одет клеточной стенкой из хитина, глюканов и т.д. Вегетативное тело редко бывает в виде пузыря или капли, чаще всего оно нитевидное. Отдельные нити гриба называются гифами, а совокупность гиф вне плодовых тел — мицелием. В плодовом теле гифы сплетаются в ложную ткань — плектенхиму.
У грибов можно различить следующие уровни организации:
• голые паразитирующие протопласты (например, Olpidium)', во всех последующих случаях вегетативная фаза отличается наличием клеточной стенки;
• ризомицелий: содержащий ядро пузырь выпускает в субстрат нитчатые выросты, лишенные ядер (например, Rhizophydium, см. рис. 11.24, А);
• почкующийся мицелий: таллом состоит из каплевидных или несколько вытянутых клеток, которые путем почкования образуют дочерние клетки; в результате их неполного отчлене-ния могут возникать короткие цепочки из клеток (например, у дрожжей; см. рис. 2.34); от этой формы можно вывести
• псевдомицелий: здесь возникшие в результате почкования клетки остаются соединенными в разветвленные цепочки (аналогично см. рис. 11.30, G);
• мицелий из гиф и плектенхима плодовых тел: таллом образуется из нитевидных клеток; последние чаще всего разветвленные, у некоторых грибов не разделенные перегородками (си-фональные формы; см. рис. 11.28, D), у других они закономерно разделены септами (трихаль-ные формы; см. рис. 11.51, А); гифы часто плотно собраны вместе или сгруппированы в плодовое тело (см. рис. 11.51, В).
Грибы с талломами в форме пузыревидных одиночных клеток или несептированных гиф
раньше трактовались как фикомицеты («грибы-водоросли») и противопоставлялись грибам с септированным мицелием. Поперечные стенки последних имеют центральную, простую или комплексную, пору. Пора чаще всего сквозная и может обеспечивать миграцию плазмы и ядер. Плазма находится внутри гиф в интенсивном движении.
Название «фикомицеты» («грибы-водоросли») следовало бы оставить за грибами, для которых установлено родство с водорослями (т.е. за Oomycota; см. рис. 11.9).
В качестве запасных веществ широко встречаются гликоген и жир, реже маннит и другие вещества; крахмал как запасное вещество у грибов не встречается.
Размножение осуществляется разного рода специальными клетками, которые при эндогенном возникновении называются спорами. Конидии образуются всегда экзогенно и служат для бесполого размножения, как исключение — для переноса сГ ядер при половом размножении. У грибов, обитающих в воде, споры часто голые, со жгутиками (зооспоры, планоспоры). у обитающих на суше они одеты клеточной стенкой и лишены жгутиков (апланоспоры). Споры могут при наличии полового процесса возникать после мейоза (мейоспоры) или образовываться после митотических ядерных делений (митоспоры). Некоторые грибы могут также размножаться путем распада мицелия на отдельные клетки (оидии). Часто возникают покоящиеся структуры в форме жестких клубневидных сплетений гиф (склероциев). Примечательна группировка мицелия в шнуровидные тяжи длиной в несколько метров (ризоморфы), которые служат для распространения (например, у Amillaria melted).
При половом размножении копулируют гаметы (изо-, анизо- или оогамия), целые гаметангии (гаметангиогамия), гаметы или же конидии с гаметангиями (гамето- или же кони-дио-гаметангиогамия) или две клетки таллома, не дифференцированные как специальные половые клетки (соматогамия). Гаметангии (если они имеются) никогда не бывают окружены покровом из многих клеток, поэтому их по аналогии с водорослями называют не анте-ридиями и оогониями (ср. Bryophytina), а в зависимости от дифференцировки, способа образования и дальнейшего развития — просто как сГили <j> гаметангии, как спермогонии (6"), сперматангии (сГ), оогонии (<j>) и аскогоны (О); имеются и другие обозначения.
11.2. Бактерии, грибы, растения
145
Двудомность Однодомность
Гетероталличная Гомоталличная Гетероталличная
Совместимость Гомогенная несовместимость
Рис. 11.19. Взаимодействие полов у грибов (по К. Esser).
Прямоугольники означают мицелии с б" (отдающими ядра) и <j> (воспринимающими ядра) органами. Двудомность: б" и <j> органы на разных мицелиях. Однодомность: o' и <j> органы на одном и том же мицелии. Гетероталлизм: отдельный изолированный мицелий не может образовывать зиготы. У однодомных грибов гетероталлизм может быть обусловлен гомогенной несовместимостью, т. е. только такие б" ядра совместимы с ядрами, которые различаются с ними по фактору скрещивания: - б" х +<j> и +б" х - <j>, но, например, не -б" х - <j>
Органы, образующие споры, у грибов (как и у водорослей) также никогда не бывают окружены покровами из стерильных клеток. Поэтому в настоящее время их называют не спорангиями, а спороцистами, в отличие от органов с теми же функциями у высших наземных растений (например, мхов, папоротников).
Специальное название для лишенных клеточных покровов структур, которые образуют гаметы или ядра гамет (т.е. название «гамето-цисты» вместо названия «гаметангии»), излишне. Скорее следовало бы сохранить установившееся понятие «гаметангий» и производные аг него термины, такие, как «гаметангиогамия» и т.д. Следует также в дальнейшем придерживаться принципа: не вводить без крайней необходимости новых понятий. Гаметангии, защищенные стерильными клетками, напротив, должны называться антеридиями и архегониями.
Часто преобладает бесполое размножение, в некоторых случаях половое размножение неизвестно или же в ходе эволюционного развития было утрачено. Те части таллома, которые образуют репродуктивные структуры без смены адер-ных фаз (митоспоры, конидии и т.д.), у грибов называются «побочными спороношениями» (анаморфы). В противоположность им основное спороношение (телеоморфа) состоит из частей таллома, в которых происходит слияние ядер (кариогамия) и смена ядерных фаз (мейоз).
Разделение полов и дифференцировку на б" и <j> органы или же репродуктивные клетки часто бывает трудно выявить. Тем не менее
отдаваемое ядро можно трактовать как б", воспринимающее — как . Исходя из такой предпосылки, можно различать однодомное и двудомное разделение полов (рис. 11.19). Двудомность имеет место тогда, когда один мицелий является либо донором ядер, либо же их реципиентом (рис. 11.19, слева). При однодомности каждый мицелий может быть как донором, так и реципиентом ядер.
Следующая пара понятий, часто используемая при описании полового размножения грибов, не совсем соответствует рассмотренным выше обозначениям и основана на других генетических особенностях. Гомоталлич-ные грибы в культурах образуют из единственных спор зиготы и соответственно плодовые тела, тогда как у гетероталличных видов для этого необходимы два мицелия различных типов скрещивания (например, «+» и «-»).
У однодомно-гетероталличных грибов слияние ядер одного-единственного мицелия невозможно (как в случае двудомности). Генетически эта несовместимость основывается на наличии по крайней мере двух аллелей одного фактора скрещиваемости, которые обозначают «+» и «-»(или другими символами). Ядра с идентичными знаками (например, «+» и «+») несовместимы и друг с другом не сливаются — в таком случае говорят о гомогенной несовместимости (рис. 11.19, справа). Гетерогенная несовместимость установлена при скрещивании географических рас одного вида; она основывается на несовместимости разных знаков.
146 | ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Подвижные клетки (зооспоры, гаметы) грибов могут иметь разные типы жгутикового аппарата: опистоконтный — с единственным гладким задним жгутиком; акро-
контный — с единственным перистым передним жгутиком; гетероконтный — с двумя жгутиками, из которых один гладкий, а другой перистый.
ра), которая сокращается, превращаясь в шаровидную голую яйцеклетку. Эти яйцеклетки, от одной до нескольких, свободно располагаются в оогонии, т.е. не окружены периплазмой. Многоядерные о" гаметангии закладываются как целая струк~ура, не образующая половых клеток. Она хемотропически (за счет антеридиола, оогониола) привлекается к оогонию и дает простые или разветвленные оплодотворяющие выросты внутрь оогония, которые достигают яйцеклеток (гаметангиогамия: рис. 11.21, Е, F) аналогично оплодотворению пыльцевой трубкой у семенных растений. В каждую яйцеклетку поступает только одно ядро, которое и сливается с ядром яйцеклетки. После этого яйцеклетка превращается в
зиготу с жесткой оболочкой, устойчивой к микробной инфекции.
Зиготы после периода покоя прорастают без редукционного деления многоядерной проростковой трубкой, которая чаще всего вскоре образует проростковую спороцисту (см. рис. 11.21,1). Имеются однодомные (о" гаметангии и оогонии на одном и том же талломе) и двудомные виды.
2. Порядок: Leptomitales. Это живущие в воде сапротрофы с регулярно перешнурованными, однако не септированными гифами и пузыревидными спороцистами. В оогонии развивается, как и у представителей следующего порядка, ооспора, окруженная периплазмой. Leptomitus — обитатель сильно загрязненных (сточных) вод.
Рис. 11.20. Oomycota (А— по Kole, Horstra; В, С — по Kole, Gielink; J.N. Couch):
А — Oomycetes, перистый жгутик (слева) и гладкий жгутик (справа) зооспоры Phytophthora infestans (8 000х); В — Achlya (Oomycetes, гетероконтная зооспора с перистым передним и гладким задним жгутиком); С — Rhizidiomyces (Hyphochytridiomycetes, зооспора с единственным передним перистым жгутиком); D— Saprolegniales, жизненный цикл. Светлые линии — гаплофаза, темные — диплофаза; R! — редукционное деление. Диплогенотипическое определение пола (о = <j> ядро; • = б" ядро)
11.2. Бактерии, грибы, растения | -|47
3. Порядок: Peronosporales. Сюда относятся паразиты, которые поражают преимущественно наземные высшие растения, вызывая у них заболевание, известное как ложная мучнистая роса (особенно Регопо-sporaceae). Растущие между клетками в ткани хозяина гифы гриба посылают в живые клетки короткие выросты — гаустории (рис. 11.22, D). Чаще всего мицелий выходит из устьиц хозяина и образует здесь разветвленные спороцистоносцы (рис. 11.22, А), в совокупности видимые невооруженным глазом как плесневые налеты. Спороцисты отличаются от таковых у Saprolegniales тем, что отрываются от несущих их гиф обычно в виде округлых или эллипсоидных образований. Чаше всего (например, у Plasmo-pard) целые (') спороцисты переносятся ветром на листья других растений, где они при наличии водяных капель (дождь, роса)
высвобождают свое разделившееся тем временем содержимое в виде нескольких почковидных зооспор (соответствующих вторичным зооспорам Saprolegnia, рис. 11.22, С; здесь, таким образом, дипланетизма нет).
В связи с прогрессирующей адаптацией к наземному образу жизни (ср. Trentepohliophyceae среди водорослей; см. рис. 11.93) зооспороцисты Peronosporales все больше превращаются в конидии.
У Pythium зооспороцисты, фиксированные на несущих их гифах, всегда высвобождают зооспоры. Спороцисты очень сходны с вегетативными гифами. У Phytophthora, Plasmopara и Pseudcpero-nospora спороцисты отрываются и переносятся ветром; они прорастают, как правило, зооспорами, однако при особых внешних условиях (малая влажность!) образуют также проростковые трубки. У Phytophthora спороцистоносцы уже четко отличаются от остальных гиф; после отделения спороцист они могут расти дальше как вегета
Рис. 11.21. Oomycota, Saprolegniales (A, D— по G. Klebs; В— noW. Hohnk; С — по Schrader; Е— G — по Moreau; Н, J — по A.W.Ziegler):
А — спорангий, который дает зооспоры s с двумя жгутиками на переднем конце (200х); В — второй тип спор с боковым расположением жгутиков (примерно 350х); С — участок несептированной гифы с многочисленными ядрами (500х); D — гифа с половыми органами: а — о" гаметангий, который дал оплодотворяющие отростки внутрь оогония, о— оплодотворенные яйцеклетки (бООх); Е— оплодотворяющий отросток с О" ядрами; F — 6" ядро внедряется в яйцеклетку; G — зигота со слившимися ядрами (Е—G — бООх); Н — проростковая трубка; J — проростковый спорангий с еще неподвижными зооспорами (Н, J — 1 400х). А, В, D — Saprolegnia mixta; С — Thraustotheca; Е—G — Achlya flagellata; Н — Isoachlya intermedia; J — Thraustothreca primoachlya
148 | ГJАВА 11 • СИСТЕМАТИКА И ФИЛОГЕНИЯ
тивные ги<|и1. У Plasmopara, Pseudoperonospora и Peronospora спороцистоносцы (конидиеносцы) чорфологически дифференцированы специфическим для каждого рода образом; нарастания их после споруляции не происходит. Спороцисты Peronospora прорастают только инфекционными гифами; гомологичные спороцистам органы стали, таким образом, полностью конидиями, которые за счет изменения формы несущей их структуры при уменьшении влажности могут активно отбрасываться.
Половое размножение. Половые органы возникают внутри растения-хозяина: оогонии — как округлые вздутия концов гиф, О* гаметангии — выросты в виде трубок (рис. 11.22, Е). И те, и другие органы отграничиваются поперечной перегородкой и содержат много ядер. Резкое разграниче
ние d" гамет отсутствует; яйцеклетка каждого оогония (как правило, единственная) окружена периплазмой. Оплодотворение и формирование зиготы в оогонии представлены на рис. 11.22, Е—G, на примере родов Peronospora и Albugo.
Число способных к оплодотворению ядер в d" гаметангии и в оогонии варьирует от многих до одного в зависимости от рода. В оплодотворенных оогониях возникает по одной ооспоре, которая у некоторых видов может содержать много диплоидных ядер (це-нозигота). У некоторых Peronosporaceae (например, Basidiophora entospora) способные к функционированию сГ гаметангии полностью отсутствуют; происходит попарное слияние ядер в одном оогонии (автогамия).
Рис. 11.22. Oomycota, Peronosporales. Plasmopara, Р. viticola (A—D) (А, В — по Millardet; С — по Arens; Е—G — по Н.Wager; Н — по С. Drechsler);
А — спорангиеносцы, выходящие из устьиц; В — оогонии (с о" гаметангием) и зиготами (100х); С — образование и выход зооспор (бООх); D — прорастание зооспор (z) через устьица в межклетники (250х); Е — Peronospora, Р. parasitica, молодой многоядерный оогоний (о) и б" гаметангий (g); F, G — Albugo, A. Candida: F — оогоний с оплодотворяющим отростком (Ь) о" гаметангия, который вводит о" ядро, G — зигота в оогонии, окруженная молодой стенкой и периплазмой (р) (Е—G — бООх); Н — Pythium, Р. ultimum, зигота, прорастающая зооспорами (800х); oz — центральная одноядерная часть оогония; 1,2,3 — стадии развития
11.2. Бактерии, грибы, растения | -|4д
Некоторые виды размножаются на значительной части своего ареала только бесполым путем. Гетероталличный возбудитель фитофтороза картофеля (Phytophthora infe-stans) у себя на родине в Южной и Центральной Америке представлен обоими типами скрещивания, так что там установлено и половое размножение. Однако в Европу, Северную Америку и др., очевидно, проник только один из двух типов скрещивания; здесь размножение осуществляется исключительно бесполым путем1.
Прорастание зигот осуществляется либо непосредственно путем образования зооспор, либо, чаще, путем образования проро-стковой трубки, которая на конпе образует спороцисту с зооспорами (рис. 11.22, Н). У более эволюционно продвинутых форм про-ростковая трубка, не отчленяя спороцисты, непосредственно внедряется в ткань хозяина (аналогично первращению зооспороцист в конидии).
В пределах родов (особенно Peronospora) виды часто приурочены к одному или немногим хозяевам. Видообразование связано с различным выбором хозяев и первоначально более или менее непрерывным, а по мере дальнейшей эволюции все более дискретным изменением морфологических признаков (например, размера конидий). В то же время эти признаки обусловлены вариабельностью внешней среды (например, влияние на размер конидий помимо возраста также температуры, влажности, субстрата), поэтому генетически обусловленный процесс видообразования и связанное с ним разделение видов могут маскироваться модификациями.
Образ жизни и вредоносность Peronospo-rales. Лишь немногие представители порядка обитают в пресной воле или почве (некоторые представители Pythiaceae). Будучи в большинстве паразитами наземных растений (например, Peronosporaceae, Pythiaceae), они могут вызывать многочисленные заболевания культурных растений. Эти организмы могут быть распространены по всему земному шару, однако для их развития требуется высокая влажность.
Опасный паразит картофеля — Phytophthora infestans (Pythiaceae); гриб сначала
1 В настоящее время в Евразии представлены оба типа скрешивания, так что стал возможен половой процесс. — Примеч. пер.
вызывает гниль ботвы и переходит также на клубни, так как во время дождя спороцисты смываются в почву и заражают клубни через чечевички. Зооспоры хемотактически привлекаются корнями; у видов грибов, поражающих определенные виды растений-хозяев, это вызывается только соответствующим видом растения-хозяина. В дождливые годы может быть уничтожено более 20 % урожая картофеля. В XIX столетии эпифитотии фитофтороза оставляли без пищи население целых больших регионов. Так, в 1845—1846гг. фитофтороз картофеля привел к большому голоду в Ирландии, сильно уменьшившему ее население, после чего последовала волна эмиграции в США. В Ирландии и сейчас не достигнута прежняя численность населения — 8 млн человек.
В хозяйственной деятельности человека значение имеет также вызываемая Plas-mopara viticola (Peronosporaceae) ложная мучнистая роса (милдью) виноградной лозы (рис. 11.22, А), при сырой погоде поражение приобретает характер эпифитотии. Ягоды превращаются в кожистые гнилые образования. Из урожая винограда ежегодно уничтожается примерно 20 % этой и другими, менее важными, грибными болезнями (остальные 20 % — вредителями). Поражения ложной мучнистой росой встречаются также на сахарной свекле, луке, хмеле и других культурных растениях. В 1959 г. в Европе впервые появился (ранее встречалась в Америке и Австралии) возбудитель перонос-пороза табака — Peronospora tabacina (эту болезнь еще называют голубой плесенью из-за беловато-голубоватой окраски массы конидий). В дождливое лето 1960 г. гриб уничтожил значительную часть плантаций табака в Средней Европе. Pythium debaryanum — гриб, широко распространенный в почве, вызывает полегание («черную ножку») у сеянцев различных растений, отчего они гибнут. Для борьбы с грибами ложной мучнистой росы (настоящая мучнистая роса — см. Erysipha-les) используют опрыскивание листьев содержащими медь фунгицидами (исходно медно-известковую смесь), что подавляет прорастание спороцист.
Обзор оомицетов. Объединяемые в этом классе порядки показывают переход от вод
150 | ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
ного к наземному образу жизни, постепенную замену зооспор конидиями и переход от гидрохорного к анемохорному распространению. С этими эволюционными явлениями, которые достигают вершины у Peronosporaceae. связаны повышение биологических потребностей и все большее проявление паразитических свойств. Это выразилось в переходе от сапротрофизма к паразитизму, специализации относительно определенных хозяев и их органов, а также, наконец, в том. что хозяин начинает поражаться лишь частично. В азотном питании наблюдается прогрессирующее (не связанное с вышеназванными тенденциями) ограничение органическими соединениями азота. Если некоторые Peronosporales могут использовать наряду с аммонийным также и нитратный азот, то Saprolegniales и Leptomitales не способны к этому, а последние, кроме того, не усваивают и аммонийный азот и используют только его органические соединения. В размножении участвуют лишь определенные части таллома, а остальные продолжают рост («эукарпия»); только у самых примитивных форм (например, Lagenisma из Lagenidiales; Thrausto-chytridiales1) в кчестве гаметангия функционирует таллом в целом («холокарпия»).
Филогения. Для оомицетов, родственных автотрофным водорослям из типа Heterokontophyta (см. приложение 4 к Rhodobionta; рис. 11.9). общими признаками являются гетероконтные жгутиковые стадии, сифональное строение таллома и целлюлоза в клеточных стенках. Этим они отличаются от всех прочих грибов. Правда, тип размножения оомицетов — гаметангиогамия и связанная с ней утрата имеющих жгутики гамет сильно видоизменены. В качестве подвижных стадий выступают только служащие для бесполого размножения зооспоры. Последние, за исключением весьма продвинутых в эволюционном отношении форм (например, Peronospora), составляют значительную долю отдела.
Небольшой класс гифохитридиомицеты (Ну-phochytridioinycctes) (15 видов) отличается признаками, частично сходными с оомицетами. Хотя их подвижные стадии обладают только одним, направленным вперед по ходу движения жгутиком, он. как и у оомицетов. перистый (рис. 11.20, С). В кле
1 Траустохитриевые в настоящее время относят не к оомицетам, а к отделу либиринтуло-вые. — Примеч. пер.
точных стенках наряду с хитином содержится целлюлоза. Однако этих признаков недостаточно для однозначного определения родственных связей гифохитридиомицетов. Они обитают в пресной и морской воде как паразиты водорослей и грибов или сапротрофы на остатках растений и насекомых. Виды этого класса, например Anisolpidium ectocarpi с простыми бутылевидными клетками внутри клеток хозяина (бурой водоросли Ectocarpus), чаще всего «холокарпные».
Третий отдел: Heterokontophyta
Относимые сюда группы из Heterokon-tobionta представлены фотоавтотрофами. Фотоавтотрофность возникла на ранних этапах эволюции у исходно гетеротрофных клеток в результате захвата ими в качестве эндосимбионтов фотосинтезирующих организмов, в этом случае уже эукариотных. Возникшая таким образом новая фототрофная форма организации имеет пока примитивное и чаще всего мало расчлененное строение водорослевого типа организации (см. бокс 11.5). Этот отдел рассмотрен ниже, вместе с прочими фотоавтотрофными эукариотами (см. приложение 4 к Rhodobionta).
Четвертое подцарство: хитиновые грибы (Mycobionta)
Гетеротрофные организмы, эволюционно близкие определенным группам животных (исходным Metazoa из типов губок или стрекающих), в клеточных стенках которых вместо целлюлозы содержится чаще всего хитин, образуют большую группу грибов в более узком смысле. Они не относятся к растениям и даже не родственны им. однако традиционно остаются объектом ботаники.
Единственный отдел: Eumycota
В связи с прогрессирующей адаптацией к жизни вне воды более подвинутые классы «настоящих грибов» (Eumycota) полностью утрачивают снабженные жгутиками зооспоры и гаметы. Они встречаются у наиболее примитивных представителей, но их органелла движения представляет собой единственный гладкий жгутик, на
11.2. Бактерии, грибы, растения | -j 54
правленный назад по ходу движения (опи-стоконтные подвижные стадии). Что касается строения таллома, то в этом, рассматриваемом как монофилетичный, отделе (см. рис. 11.9) достигнуты все известные для грибов уровни организации. В богатых видами более высокоорганизованных классах на определенных фазах развития широко встречаются специальные плектен-химные структуры (плодовые тела).
Клеточная стенка содержит в качестве главного компонента почти всегда хитин (часто совместно с глюканами), целлюлоза практически полностью отсутствует. Представители некоторых групп содержат в стенке маннан-р-глюканы (Saccharomyces) или галактозамингалактаны (Trichomycetes). Некоторые немногие формы, приспособившиеся к паразитическому образу жизни, полностью утратили клеточную стенку, так что в цикле развития встречаются вторично голые протопласты (например, Olpidium, см. рис. 11.23, С).
Оплодотворение может происходить в виде изогамии, анизогамии, оогамии (редко), гаметангиогамии и соматогамии. При га-метангиогамии почти никогда не участвуют оогонии с яйцеклетками. Покоящиеся структуры никогда не возникают внутри оогониев (ср. Oomycota). Большинство представителей — гаплонты, гаплодиплонты или гаплодикарионты; диплонты составляют исключение. В цикле развития все большую и большую часть занимает дикариофаза.
Многие виды образуют каротины (известные у водорослей как дополнительные пигменты); они выполняют функцию фоторецепторов при фо-тотропных изгибах (Pilobolus). Наряду с ними встречаются многие другие пигменты, которые относятся к самым различным структурным типам. Часто присутствуют фенольные пигменты (см. рис. 11.60), также известны азотсодержащие гетероциклы, однако антоцианы и флавоны, широко распространенные у последующих отделов, практически полностью отсутствуют.
1. Класс: хитридиомицеты (Chytridiomycetes)
Хитридиомицеты существуют в виде отдельных клеток или образуют многоядерный таллом без поперечных перегородок (сифональный). Подвижные клетки (гаме
ты и зооспоры) опистоконтные (см. рис. 11.25, F). Вместо ядрышка чаще всего встречается богатый РНК «ядерный колпачок» (см. рис. 11.25, Е).
Особенности размножения можно рассмотреть на примере Allomyces (Blastocla-diales). Этот «эукарпный» почвенный гриб уже имеет обильно разветвленные, многоядерные гифы. Жизненный цикл проходит как изоморфная смена генераций (у группы «Еи-AUomyces», см. рис. 11.25, Q).
Гаметофит образует на концах гиф отделенные септами гаметангии; гаметангий чаще всего сидит непосредственно на 0я гаметангии (однодомность). Оба через специальные поры высвобождают гаметы; 6я гаметы мельче и окрашены у-кароти-ном в оранжево-красный цвет; гаметы бесцветные; гаметы выделяют сиренин (см. 8.2.1.1) и путем хемотаксиса привлекают 6я гаметы. После анизогамии возникает диплоидная зигота, только в самом начале имеющая жгутики. Из нее прорастает спорофит, который по размеру и общему виду сходен с гаметофитом. На отдельных участках гиф спорофита развиваются спороцисты двух разных типов. Расположенные по бокам гиф, чаще всего попарно друг над другом, тонкостенные ми-тоспороцисты, открывающиеся сосочком, высвобождают после исключительно митотических ядерных делений диплоидные митозооспоры; последние прорастают снова в диплоидный спорофит, так что смена генераций нарушается за счет интенсивного бесполого размножения спорофита (побочное спороношение с митоспороци-стами). На концах гиф, часто поодиночке, обособляются толстостенные, имеющие орнаментацию в виде пчелиных сот, тем-ноокрашенные мейоспороцисты, которые отпадают целиком. После периода покоя (как гипноспороцисты) они претерпевают мейоз и высвобождают гаплоидные мей-озооспоры, прорастающие в гаметофиты.
Группа «Ей-Allomyces» представлена видами A. arbuscula (с? гаметангии расположены на <j>) и A. macrogynus (<j> на О"). Гибрид этих двух видов, встречающихся в разных вариантах пло-идности (A. arbuscukr. п = 8,16, 24, 32; A. macrogynus: п = 14, 28, 56), — А. х javanicus — дает мейоспоры, имеющие меньший процент прорастания (0,1 —
152 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
3,2 %) и образующие гаметофиты с различным расположением гаметангиев. От полной смены генераций имеются отклонения с редукцией гаметофита (например, Brachy-Allomyces).
Три порядка хитридиомицетов отличаются строением таллома, характером бесполого размножения и ультраструктурой зооспор. Известно около 500 видов. Большинство видов живет в воде, некоторые также в почве или как паразиты в клетках высших растений1.
1. Порядок: Chytridiales. Таллом грибов, относящихся к этому порядку, развит слабо. Чаще всего он одноклеточный, в виде шара или пузыря: мицелий из гиф не образуется, однако часто имеются тонкие придатки одной-единственной клетки, лишенные собственных ядер (ризоми-целий) Половое размножение происходит как изогамия, анизогамия или гаметангиогамия. Определение пола генотипическое (Olpidiaceae: Rozella) или модификационное (Synchytrium). Обычно в образовании спор или гамет участвует весь таллом («холокарпия»); более подвинутые формы на ризомипелии развивают специальные собирательные клетки, которые могут давать начало репродуктивным клеткам («эукар-пия»). Зооспоры содержат одну хорошо заметную каплю масла.
Цикл развития низших Chytridiales можно рассмотреть на примере Olpidiaceae. Голый протопласт ведет паразитический образ жизни в клетке растения-хозяина, увеличивается в размере и окружается хитиновой стенкой (рис. 11.23, С, D), претерпевает ядерные деления и деление цитоплазмы и образует в результате опистоконт-ные подвижные стадии; на их образование идет
1 К сожалению, здесь совершенно не рассматривается крайне интересный порядок Neocalli-masticales, ныне также относимый к хитридио-мицетам. Виды этого порядка — облигатные анаэробы (этим данный порядок уникален среди всех грибов в целом), обитающие в рубце и слепой кишке жвачных животных. Некоторые виды имеют типичные для хитридиомицетов одножгутиковые зооспоры, у некоторых жгутиков более 10. Митохондрий у этих грибов нет (они считаются утраченными в ходе эволюции) — это естественная ситуация для анаэробных организмов. Для некоторых видов показана способность образовывать покоящиеся споры, которые могут сохраняться в высушенном помете в аэробных условиях. Неокаллимастиксовые — обязательный компонент рубца жвачных. Считается, что они играют важную роль в переваривании пищи. — Примеч. пер.
весь протопласт. Эти подвижные стадии либо инфицируют новые клетки хозяина, как зооспоры, или копулируют, как гаметы, попарно друг с другом (факультативное определение функции; изогамия) с образованием голых двужгутиковых планозигот (рис. 11.23, F). Последние внедряются в клетки хозяина и позже превращаются там в гипнозиготы с жесткой стенкой; два половых ядра сливаются только следующей весной (прообраз дикариофазы; рис. 11.23, G, Н), а затем образуют (очевидно, после редукционного деления) многочисленные подвижные стадии, которые высвобождаются через специальный сосочек.
Olpidium brassicae — возбудитель «черной ножки» проростков капусты. Родственные Syn-chytriaceae живут как эндопаразиты в цветковых растениях, где могут вызывать разрастания типа галл; Synchytrium endobioticum вызывает рак картофеля. Все эти грибы развиваются внутри тканей хозяина — в полностью пораженных клетках; кроме того, они «холокарпные».
У последующих семейств не весь таллом идет на образование репродуктивных клеток («эукар-пия»).
Rhizidiaceae — часто встречающиеся паразиты планктонных водорослей и пыльцы (Rhizo-phydiunr, рис. 11.24, А). Таллом «функционально» разделяется на репродуктивный пузырек, расположенный вне субстрата, и питающий ризоид, который внедряется в клетку хозяина. Центр мо-нопентрического таллома у Polyphagus euglenae образует один-единственный «центральный пузырь», от которого отходит несколько ризоидов. Этот гриб питается за счет того, что он своими ризоидальными выростами внедряется в водоросли из рода Euglena и высасывает их содержимое. Один экземпляр Polyphagus euglenae может поразить свыше 50 эвглен (рис. 11.62, А). Половое размножение — анизогаметангиогамия: более мелкие сГ особи посылают «поисковый ризоид»; встретившись с центральным пузырем <j> особи, он сливается с ней, и в место слияния переходят соответственно О" и <j> ядра копулирующих грибов (см. рис. 11.42, Е, F). Возникающая зигота имеет толстую шиповатую стенку и претерпевает состояние покоя как дикариоти-ческая гипнозигота («покоящаяся спора», G). Слияние ядер (кариогамия) и, вероятно, мейоз происходят при прорастании гипнозиготы (Н); в проростковой трубке в результате ядерных делений и одновременного распадения цитоплазмы образуются многочисленные зооспоры (вероятно, мейозооспоры). После освобождения они садятся на эвглены, инцистируются, а затем циста прорастает и возникает «центральный пузырь» с ризоидами, которые дальше поражают клетки эвглен.
11.2. Бактерии, грибы, растения
153
.© Q Э о/
Le®c 6
XX? ф
Рис. 11.23. Eumycota, Chytridiomycetes, Chytridiales, Olpidium viciae (no S. Kusano):
A — зооспоры; В — внедрение в клетку хозяина; С — голый протопласт гриба в клетке хозяина; D — зооспорангий, могущий служить как гаметангий; Е — зооспорангий опорожненный; F — копуляция двух опистоконтных гамет (А—F — 500х); G — молодая, еще двухъядерная зигота (бООх); Н — инци-стированная зигота; I — инцистированная зигота, прорастающая (Н, J — 120х)
В обоих последующих семействах для выхода зогхзпор у спороцисты образуется крышечка (оперкулятный тип — в противоположность ино-перкулятному у предыдущих семейств).
У Chytridiaceae формируется моноцентриче-ский, у Megachytriaceae чаще всего полицентрический таллом с несколькими пузыревидными зооспороцистами, связанными ризоидальными тяжами. Половое размножение в немногих исследованных случаях представляет собой гаме-тангиогамию. У Zygochytrium (Megachytriaceae) две копуляционные ветви, в данном случае равнозначные, растут навстречу друг другу, и в месте слияния их концы (рис. 11.24, К) образуют толстостенную гипнозиготу; о поведении ядер до образования зиготы ничего не известно.
Имеются случаи, когда копулирующие с образованием или без образования трубки гаметангии многоядерны. Такие процессы очень на, изминают картину, с которой мы позже столкнемся у зиго-мицетов. Если у Chytridiales гетерофазная смена генераций еще не встречается (только у Physoderma' есть указания на нее), в следующем порядке она достаточно выражена.
2. Порядок: Blastocladiales. Представители порядка образуют чаще всего таллом, состоящий из гиг)), которые обособляют несколько вмести
1 В настоящее время род Physoderma обычно относят к следующему порядку — Blastocladiales. — Примеч. пер.
лищ репродукт ивных клеток путем образования поперечных перегородок на своих концах. К субстрату таллом прикрепляется ризоидоподобными выростами. Наиболее примитивные представители внешне близки к Chytridiales, например обитающая в почве «холокарпная» Blastocladiella (рис. 11.25, А—D). Зооспоры содержат несколько не особенно крупных капелек масла и прорастают двумя проростковыми трубками. Большинство представителей обитает саттротрофно в почве, воде, часто на остатках растений и животных. Два вида Blastocladiella поражают синезеленые водоросли. Развитие Allotnyces было рассмотрено выше (введение к хитридиомицетам). Blastocladiella (рис. 11.25, А) образует талломы с вместилищами репродуктивных клеток только одного типа: каждый гаметофит развивает один гаметангий; сливающиеся изогаметы различаются только окраской (каротин). Спорофит несет либо одну митоспороцисту, либо одну мейоспо-роцисту.
3. Порядок Monoblepharidalcs. Порядок отличается от предыдущих наличием четко отграниченной базальной части таллома, несущей ризоиды, а также прежде всего оогамным размножением. гаметы представляют собой лишенные жгутиков яйцеклетки, которые после оплодотворения имеющими жгутик сГ гаметами (оогамия) развиваются дальше в ооспоры. Несмотря на более прогрессивную, чем у Blastocla-diales, форму размножения, они остались гаплон-
154 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Рис. 11.24. Chytridiomycetes Chytridiales (А — по Е. R. Uebelmesser; В — по в.,е11о; С—J — по Н. Wagner; К—Р— по N.Sorokin):
А — Rhizophydium halophilum, зооспорангий с сосочками, через которые выходят опистоконтные зооспоры, на пылинке Pinus с гаусториями внутри; В — Polychitrium aggregatum, небольшой многоядерный неклеточный мицелий с развитыми по-разному спорангиями и двумя опистоконтными зооспорами (А, В — 400х); С—J — Polyphagus euglenae (примерно 450х): С — зооспора, D — таллом, образующий ризоиды, Е — копуляция между мелкой сГ и более крупной особями, F — d" ядро в будущей зиготе, G — зигота с еще не слившимися сГ и <j> ядрами, Н, J — развитие и освобождение от зооспор (s) зооспороцисты (z); К — Р — Zygochytrium aurantiacum (350х): К — таллом с двумя расположенными на концах гиф, уже опустевшими зооспорангиями и двумя копулирующими гаметангиями (g), L— зооспорангий в процессе освобождения от зооспор, М — Р— образование зиготы (z) из копулировавших гаметангиев Р — зрелая гипнозигота (z) (зигоспора)
тами: мейоз у них происходит при прорастании зиготы (зиготическая смена ядерных фаз); смена генераций отсутствует.
Для полового размножения служат вздутые одноядерные оогонии (рис. 11.25, К), расположенные чаще всего на концах гиф; их содержимое редуцировано до одной-единственной одноядерной яйцеклетки1. Расположенные под оогонием сперматогонии высвобождают некоторое коли
1 У некоторых представителей Momoblepha-ridales яйцеклеток может быть несколько — Примеч. пер.
чество одноядерных и одножгугиковых сперматозоидов (К). Последние проникают через специальное отверстие в оогоний и оплодотворяют яйцеклетку (L, М), которая к этому времени либо остается внутри оогония, либо (у большинства видов) выталкивается через специальное отверстие из оогония (N) и здесь становится толстостенной шиповатой гипнозиготой (О); или зигота даже уплывает оттуда с помощью сохраняющегося д" жгутика. Зиготы прорастают не зооспорами, а проростковой трубкой (Р), что считается более прогрессивным признаком. Бесполое размножение осуществляется зооспорами (J).
11.2. Бактерии, грибы, растения
Представители порядка обитают как сапротрофы на растительных остатках в воде.
При обзоре хитрцдиомицетов можно отметить некоторые важные прогрессивные тенденции. Вместо исходной «холокарпии» все больше развивается «эукарпия». Наблюдается переход от изогамии с факультативным определением функции репродуктивных клеток через изогамию с генотипическим определением пола гамет к анизогамии, оогамии (Monoblepharidales) и гаме-тангиогамии. В отдельных случаях кариога-
мия происходит не сразу после копуляции гамет и связанной с ней плазмогамией; следствием является включение в цикл развития дикариофазы (Olpidium, Polyphagus). Наряду с однодомностью встречается и двудомность; в отдельных группах наблюдаются смена генераций и ее дальнейшее развитие в диплонтный жизненный цикл (Allomyces). Голые талломы приспособились к существованию в качестве эндопаразитов. Истинные наземные формы с воздушным мицелием в этой группе еще не возникли.
Рис. 11.25. Chytridiomycetes (А—D— по R. Harder и G.Soergel; Е—Н— по H.Kniep; J —О — по M.Woronin; Р — по Laibach):
А— Н — Blastocladiales, А— D — Blastocladiella variabilis: А — спорофит, В — зооспорангий в процессе выхода зооспор, С — с покоящимся спорангием (А—С ЗЗх), D — зооспора (450х); Е—Н — Allomyces javanicus (1 ОООх), Е— <j>, F — сГ опистоконтная гамета; G — копуляция гамет; Н — планозигота; J—Р— Monoblepharidales, Monoblepharis (ЗООх); J — М. macrandra, спорангий с выходящими из него зооспорами, К—Р — М. sphaerica: К — конец гифы с оогонием и расположенным под ним спермогонием, из которого выходит сперматозоид, L— сперматозиод внедряется через апикальное отверстие оогония к яйцеклетке и сливается с ней, М — слияние совершилось, N — оплодотворенное яйцо выползает из оогония, О — гипнозигота с толстой шиповатой стенкой, Р — прорастание зиготы; Q — Blastocladiales, Allomyces: схема смены поколений, светлые линии — гаплофаза, темные — диплофаза, G — гаметофит, R! — редукционное деление, S — спорофит
|56 | ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Во всех следующих далее классах (2—4) имеющие жгутики подвижные стадии (гаметы, зооспоры) полностью отсутствуют. Широко развивалось приспособление к наземному образу жизни.
2. Класс: зигомицеты (Zygomycetes)
Зигомицеты имеют чаще всего развитый мицелий из гиф, обычно не септированных и многоядерных (ценоцитный мицелий, который соответствует сифонально-му типу организации водорослей); у некоторых форм имеются поперечные перегородки. При половом размножении никогда не образуется гамет: всегда копулируют
друг с другом два гаметангия, чаще всего многоядерные, нередко имеющие одинаковый облик. Они обособляются от концов растущих навстречу друг другу гиф (гаме-тангиогамия) и дают покоящуюся зиготу. Эта так называемая «зигоспора» — результат полового процесса. Она прорастает, претерпевая мейоз, в зародышевую спороцисту, в которой за счет разделения многоядерного плазматического содержимого эндогенно в большом числе возникают мейоспоры.
Бесполое размножение приспособлено к наземному образу жизни, однако несколько иначе, чем у оомицетов. Если у оомице-тов спороциста отрывалась, чтобы образо
Рис. 11.26. Zygomycetes, Mucorales (А — по О. Brefeld; В — по R. A. Harper; С — по Moreau; D — по J. Webster):
А — спорангий на продольном оптическом разрезе Mucor mucedo (225х); В — разрез через зрелый спорангий с многоядерными митоспорами Sporodinia grandis; С — Cunninghamella echinulata: 1 — образование конидий (370х), 2 — конидия (1 ОООх); D — Thamnidium elegans, разветвления, несущие спо-рангиоли (200х); Е— жизненный цикл: светлые детали рисунка— гаплофаза, темный круг вверху — диплофаза; с — колонка; s — спорангиоспоры; w — стенка; R! — редукционное деление
11.2. Бактерии, грибы, растения
вать зооспоры на месте своего прорастания, то у зигомицетов окруженные клеточными стенками споры возникают (за счет деления содержимого) эндогенно, внутри спороцист. Высвобождаясь из спороцисты, они распространяются по воздуху. Аналогичным образом у зигомицетов спороцисты превращаются в конидии, дающие про-ростковую трубку.
Половое размножение происходит при встрече мицелиев противоположных типов скрещивания (+ и -). Тогда оба мицелия под воздействием выделяемых гамонов образуют булавовидные гаметангии, изгибающиеся навстречу друг другу и, наконец, смыкающиеся концами. Они многоядерные и отделяются от несущих гиф (суспензоров) поперечной перегородкой. Разделительная двойная стенка между гаметангиями исчезает (см. рис. 11.27, В), и оба гаметангия участвуют в формировании возникающей зиготы. Последняя представляет собой покоящуюся гипнозиготу с толстой, многослойной, снаружи бородавчатой стенкой («зигоспора» D), в которой попарно сближаются многочисленные половые ядра (+, -). В конце периода покоя немногие ядра, иногда всего одна-единственная пара, претерпевают слияние (кариогамия), тогда как другие пары ядер погибают. Таким образом, в оплодотворении участвуют исключительно гаметные ядра, свободных гамет не бывает. «Зигоспора» прорастает, претерпевая мейоз, проростковой трубкой, причем дальнейшее митотическое деление происходит лишь в одном-единственном из получившихся гаплоидных ядер (остальные продукты мейоза дегенерируют): все ядра, следовательно, генотипически одинаковы. На конце проростковой трубки образуется проростковая спороциста (J), которая содержит многочисленные мейоспоры, принадлежащие какому-либо одному (+ или -) типу скрещивания. Внешне она сходна со спороцистой бесполого размножения, однако возникающие здесь споры представляют собой одноядерные мейоспоры одного полового знака (+ или -). В обоих случаях спороцисты отграничиваются от несущей ее гифы поперечной перегородкой, которая в виде так называемой колонки вдается внутрь спороцисты
(рис. 11.26, А). Многоядерная плазма спороцист распадается либо на многоядерные гаплоидные митоспоры, либо на одноядерные гаплоидные мейоспоры.
К этому классу относится примерно 500 видов, преимущественно сапротрофных. Они распределяются по нескольким порядкам.
1. Порядок: Mucorales. Споры образуются в спороцистах, причем последние чаще всего вскрываются и высвобождают споры; реже спороцисты содержат немного спор или всего одну спору и отваливаются целиком; их называют спо-ронистиоли. «Зигоспоры» возникают как часть сливающихся гаметангиев. К этому порядку относятся наземные плесневые грибы, которые ведут преимущественно сапротрофный образ жизни, реже паразитируют на растениях и животных.
Один из наиболее широко распространенных видов — головчатая плесень (Mucor mucedo). Сильно разветвленный, лишенный поперечных перегородок мицелий этого гриба образует белые плесневые налеты на помете животных, хлебе и т.д. Выходящие из субстрата гифы, служащие для поглощения питательных веществ, поднимают вверх вертикальные выросты мицелия, каждый из которых несет на конце округлую спороцисту (рис. 11.26, А). Внутри нее возникают многочисленные приспособленные к наземной жизни споры, имеющие клеточную стенку и способные переносить высыхание. Они округлые, многоядерные; будучи митоспорами, относятся к побочным спороношениям и долгое время сохраняют всхожесть.
У различных видов существует много отклонений в строении спороцист, гаметангиев и зигоспор, а также в особенностях размножения. В частности, имеются такие типы спороцист, которые, каки зооспорангии некоторых хитридио-мицетов, открываются крышечкой (Saksenaea), тогда как обычно споры освобождаются за счет разрыва стенки спороцисты. Среди побочных спороношений четко прослеживается тенденция к сильному уменьшению числа спор в спороцисте, а также к тому, чтобы сильно уменьшившаяся в размере спороциста распространялась целиком (при утрате колонки, например у Thamnidium). В конце этого ряда мы встречаем такие спороцисты, которые соответствуют экзогенно отделяющимся конидиям, в которых способность к образованию спор сильно подавлена (например, Haplosporangium, Blakeslea, Choanephora) или полностью пропадает (Си-nninghamella). У Choanephora в зависимости от внешних условий (питание, температура) образуются «конидии» или же спороцисты со спо-
| 58 | ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
роцистоспорами. Некоторые виды имеют только конидии. Нередко встречающийся на конском помете пилоболус (Pilobolus) за счет тур-горного давления отбрасывает целиком спороцисту черного цвета, расположенную на конце несущей клетки, способной к положительным фототропным изгибам (см. рис. 11.28, А). Дальность полета составляет до 1,8 м в высоту и до 2,4 м в сторону при начальной скорости примерно 10 м/с. При гаметангиогамии у двудомных (рис. 11.27, А) или однодомных видов (Е) участвующие в этом процессе гаметангии обычно имеют одинаковый облик (изогаметан-гиогамия; A, G) или же отличаются по размеру и поведению (анизогаметангиогамия; Е). Гипнозиготы нередко защищены гифами ^бверт-ки, которые вырастают из суспензоров. Здесь можно видеть прообраз формирования плодовых тел, столь характерных для аскомицетов и базидиомицетов. Под плодовыми телами мы понимаем видимые невооруженным глазом структуры из гиф основного спороношения.
У Absidia (анизогаметангиогамия) только суспензор, соответствующий более крупному гаметангию, образует неразветвленные, длинные, коричневые выросты, закрывающие собой зигоспору. У Phycomyces (изогаметангиогамия) зиготу обрастают идущие от обоих суспензоров почти черные, разветвленные придатки (G). У Mortierella развивается вокруг каждой зиготы, а у Endogone (следующий порядок) — вокруг нескольких зигот защитная структура из плотно переплетенных гиф.
2. Порядок: Endogonales (с родом Endogone и еще несколькими родами). Виды рода Endogone, имеющие клубневидные плодовые тела размером с лесной орех, развиваются под землей и часто образуют микоризу.
Гаметангии после копуляции формируют структуры в виде моста (рис. 11.28, J), из вершины которых выпячиваются шаровидные зиготы (К) Мицелий сначала многоядерный и несептированный, позже появляются поперечные перегородки.
Рис. 11.27. Zygomycetes, Mucorales (А— D — по М. L. Keene; Е, F — по Green; G — по Н. Gwynne Vaughan; Н — по Moreau; J, К — по O.Brefeld):
А—D — оплодотворяющий орган и образованье гипнозиготы у Sporodinia grandis (50х); Е, F — то же, у Zygorrhynchus moelleri (75х); G — Phycomyces blakesleeanus, зигота с обрастающими ее придатками, отходящими от суспензоров (ЗОх); Н — Mucor hiemalis, зигота с гаплоидными ядрами, слияние ядер и диплоидные ядра (550х); J — Mucor mucedo, проростковый спорангий (60х); К — Chaetocladium jonesii, прорастание зиготы конидиеносцем (75х); g— гаметангий; к— копуляционная ветвь; s — суспензор; z — зигота
11.2. Бактерии, грибы, растения I -| gg
Рис. 11.28. Zygomycetes (А — по J. Webster; В, С — по A. H.R. Buller; D—G — по L.S. Olive; Н — по F. Dangeard; J, К— по R.Thaxter; L, М — по Buchholz):
А—С — Mucorales, Pilobolus crystallinus: А — несущая клетка (спорангиеносец) со спороцистой (sp); первая покрыта выделяющимися каплями жидкости (t) (20х), В, С —отброшенная спороциста соответственно незадолго до и после столкновения с препятствием; D — Н — Entomophthorales: D — F — Entomophthora muscae (450х): D — конец гифы из мухи, Е— возникающий из нее, прорывающийся наружу конидиеносец, F— образование конидии, G — молодая гифа Entomophthora sciarea (180х); Н — Ancylistes closterii, оплодотворение между соседними клетками (500х); J—М — Endogonales, Endogone, оплодотворение: J — копуляция, К — готовая зигота £ pisiformis, L — выпячивание зиготы после перехода ядра из сГ (gm) в <j> (gw) гаметангий, М — готовая зигота (z) Е. lactiflua (ЗООх); р — слизистое кольцо; s — суспензор
3. Порядок: Entomophthorales. Виды порядка размножаются бесполым путем почти исключительно конидиями, которые, как и у Mucorales, произошли из спороцист. Копуляция гаметангиев осуществляется между двумя гифами или может быть также боковой между соседними клетками одной и той же гифы (рис. 11.28, Н). Гипнозигота, как и у Endogone, образуется в виде выроста слившихся гаметангиев. В гифах встречаются поперечные перегородки; образующиеся таким образом обособленные участки бывают от многоядерных с самым разным числом ядер до одноядерных (рис. 11.28, G), у Basi-diobolus ranarum они в основном только одноядерные. У наиболее известного представителя — гриба Entomophthora muscae, вызывающего эпизоотии — заболевания комнатной мухи, многоядерная конидия (рис. 11.28, Е, F) отбрасывается от несущей ее клетки. Попав на муху, конидия образует проростковую трубку, которая внедряется внутрь тела животного и там разви
вается мицелий, убивающий муху. Из трупа в массе вырастают конидиеносцы. От них активно отбрасываются конидии, которые (например, на оконном стекле) окружают мертвую муху светлым венцом. В высохших мухах возникают внутри гиф толстостенные цисты, которые, возможно, следует рассматривать как партеногенетические зиготы.
Близкие к Entomophthorales представители порядка Zoopagales (4-й порядок) паразитируют на амебах и нематодах, образуя ветви гиф, превращающиеся в гаустории (см. рис. 11.62, G, бокс 11.4)1.
Спорным является сближение с зигомице-тами видов (около 60), объединяемых в класс три-хомицеты (Trichomycetes). Они обитают как па
1 К этому же порядку относят и семейство Piptocephalidaceae, представители которого паразитируют на других зигомицетах (Mucorales). — Примеч. пер.
160 | ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
разиты на поверхности или внутри тела насекомых (особенно водных) и имеют сильно редуцированный таллом, клеточные стенки которого состоят из полигалактозамина и галактана. Возникающие в результате гаметангиогамии зигоспоры и отсутствие гамет говорят об их родстве с зигомицетами, которые, однако, имеют в клеточной стенке хитин. Иногда встречающиеся амебоидные стадии свидетельствуют об обособленности этой группы. Примерами родов могут служить Amoebidiwn, развивающиеся на водных насекомых, и Harpella — в кишечнике мошек (Simuliidae)1.
3. Класс: сумчатые грибы, или аскомицеты (Ascomycetes)
Аскомицеты живут, как и базидиомице-ты (см. ниже), преимущественно в наземных местобитаниях. Некоторые виды ветре-
1 К большому сожалению, здесь не рассматриваются грибы порядка гломусовых (Glomerales, или — неграмотно — Glomales), относимого к зи-гомицетам или выделяемого в отдельный класс гломеромицеты (Glomeromycetes). Это грибы, образующие наиболее распространенный тип микориз — везикулярно-арбускулярные микоризы (ВАМ). Из числа видов растений, имеющих микоризы, свыше 80 % имеют именно такие микоризы. Представители этого класса, в частности, образуют микоризу с наиболее важными сельскохозяйственными растениями — такими, как злаки. ВАМ относятся к эндомикоризам, т.е. в отличие от эктомикориз, имеющих хорошо выраженный наружный чехол и сеть гиф в межклетниках коры (сеть Гартига), грибы ВАМ сильно не изменяют внешнюю морфологию корней. Только иногда на поверхности корня встречается сетка гиф, соединяющая внутритканевой мицелий с грибом, находящимся в почве. Гриб проникает внутрь клеток корня растения, образуя там древовидно-ветвящиеся образования — ар-бускулы. Помимо них внутри клеток или за их пределами обычно встречаются округлые вздутия — везикулы. Питание растений осуществляется за счет переваривания арбускулов, а иногда к тому же везикулов и гиф. Известны случаи, когда грибная инфекция сосредоточена в специальных отдельных клубеньках. Для воспроизведения и перенесения неблагоприятных условий у этих грибов образуются покоящиеся споры, представляющие собой азигоспоры со сложной 6-слойной стенкой, содержащей хитин и целлюлозу. Они образуются одиночно или рыхлыми группами вне корней растений или, реже, внутри них. — Примеч. пер.
чаются в пресных водах или в морях. По большей же части это паразиты растений или сапротрофы на отмерших растительных тканях и в соках растений. Вегетативное тело — как правило, обильно разветвленный мицелий из септированных гиф; в их поперечных перегородках имеются простые поры. Формы, приспособившиеся к определенному характеру питания, имеют вегетативное тело в виде почкующихся клеток по типу дрожжей. Клеточные стенки состоят из хитина и глюканов (у дрожжей, Endomycetidae, доля хитина в стенках незначительна или он отсутствует совсем). При сильном увеличении (электронная микроскопия) видно, что стенки двухслойные: внутренний слой светлый, толстый и бесструктурный, внешний слой темный и тонкий.
Половое размножение приводит к образованию сумки, или аска, — характерной мейоспороцисты, часто имеющей мешковидную форму. В сумке происходит: слияние половых ядер (например, «+» и «-»-ядер; кариогамия), редукционное деление (мейоз), а также эндогенное образование мейоспор (= аскош юр) в виде свободных клеток. Кроме того, сумки часто приспособлены к активному выбрасыванию аскоспор. Имеющие жгутики репродуктивные клетки отсутствуют, как у зигомицетов и у полностью приспособившихся к наземной жизни базидио-мицетов. В жизненном цикле аскомицетов появляется двухъядерная фаза (дикарион); она характерна для всех аскомицетов (кроме Endomycetidae) и базидиомицетов.
Аскомицеты, или сумчатые грибы, насчитывая примерно 30 000 известных видов, составляют порядка 30% всех описанных к настоящему времени грибов. Если сюда причислить еше по большей части происшедшие от аскомицетов несовершенные грибы (Deuteromycetes, приложение 1 к Mycobionta), то доля аскомицетов в общем числе грибов повысится до 60%. Систематическое подразделение основывается на различном возникновении сумок в жизненном цикле, на строении и типе вскрытия сумок, а также на форме и характере развития плодовых тел. Системы, предлагаемые разными авторами, различаются в зависимости от оценки ими тех или иных признаков. В дальнейшем мы будем исходить из того, что эво
11.2. Бактерии, грибы, растения | -j g-j
люция грибов шла от хитридиомицетов через зигомицеты к аскомицетам.
В первых трех подклассах плодовые тела не образуются.
1. ПОДКЛАСС: ТАФРИНОМИЦЕТИДЫ (TAPHRINOMYCETIDAE)
Представители этой группы аскомице-тов паразитируют на растениях. Плодовые тела не образуются.
Виды Taphrina могут вызывать у растений различные деформации. Некоторые виды провоцируют образование «ведьминых мётел» на косточковых, березах и грабах; Т. deformans обусловливает курчавость листьев персика; Т. pruni превращает завязи сливы в полые внутри, лишенные косточки галлы, так называемые кармашки. Сумки возникают из коротких, сначала двухъядерных, затем одноядерных диплоидных клеток гиф между кутикулой и эпидермисом хозяина, после редукционного деления и нескольких митозов. Они прорываются между клетками эпидермиса растения-хозяина, образуют палисадный слой (рис. 11.29) и открываются на вершине простой трещиной, через которую освобождаются аскоспоры. При прорастании аскоспоры почкуются, напоминая этим клетки дрожжей. Затем на поверхности растения-хозяина возникает сначала гаплоидный, сапротрофно живущий почкующийся мицелий. При слиянии вегетативных клеток или автогамной попарной группировки ядер формируется стадия дикариотичного мицелия, который теперь внедряется как паразит в межклетники хозяина. Его дальнейшее развитие осуществляется независимо от гаплоидного мицелия. В этом плане Taphrinomycetidae сходны скорее с базидиоми-цетами, имеющими самостоятельную дикариофазу, чем с аскомицетами (рассматриваются ниже).
К Taphrinomycetidae близки следующие порядки.
Protomycetales. Сюда относятся некоторые паразиты цветковых растений, которые вызывают характерные изменения окраски или пузырчатые вздутия на своих хозяевах.
Ascosphaerales. Ascosphaera apis вызывает болезнь пчел.
2. ПОДКЛАСС: ЭНДОМИЦЕТИДЫ (ENDOMYCETIDAE)
В этом подклассе объединены дрожжевидные аскомицеты. Дрожжи — это грибы, которые размножаются почкованием, как пекарские дрожжи (см. рис. 2.34); другой от-
Рис. 11.29. Ascomycetes, Taphrinomycetidae, Taphrina deformans: кариогамия и зрелые сумки (800х) (по Е. М. Martin)
личительной особенностью их может быть размножение фрагментами гиф (артроспорами). Сумки возникают непосредственно из зигот или других отдельных клеток, не после Дикариофазы и не в плодовых телах. Стенка сумки распадается или ослизняется при созревании спор, таким образом аскоспоры не отбрасываются. Таллом у некоторых представителей распадается на отдельные клетки. Обычно он представляет собой почкующийся мицелий, реже — септированный нитевидный мицелий. Грибы, относящиеся к этому подклассу, часто живут в богатых сахарами субстратах (например, в истечениях сока деревьев и кустарников, в нектаре).
К роду Saccharomyces и близко родственным родам (Hansenula, Schizosaccharomyces') относятся широко известные, часто применяемые на практике и в научных исследованиях дрожжи. Их округлые или овальные одноядерные клетки размножаются чаще всего почкованием (см. рис. 2.34) и отчасти остаются в разветвленных цепочках разной
1 В современных системах делящиеся дрожжи типа Schizosaccharomyces обычно считаются далеко отстоящими в эволюционном плане от почкующихся дрожжей типа Saccharomyces и сближаются с тафриновыми. — Примеч. пер.
1 62 | ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Рис. 11.30. Ascomycetes, Endomycetidae, Saccharoinycetales (А, V — по J. Lodder, N.J.W. Kreger; В — F, H — U— no A.Guillermond; G — no G. Lindau; W—Y — no H.O.Juel):
A—F — Schizosaccharomyces, S. octosporus (350x): A — группа соединенных клеток, В —F — копуляция и образование сумки; G —L— Saccharomyces, S. cerevisiae: G— цепочки почкующихся клеток (200х), Н — L— образование сумки (550х); М —S — Saccharomycodes, S. ludwigii (375x): M — P— копуляция прорастающих мейоспор в сумке, Q — прорастание диплоидной клетки, R, S — образование аскоспор; Т, U — Endomyces, Е. magnusii, копуляция и образование сумки (375х); V — Candida, С. reukaufii (375х); W—Y — Dipodascus, D. albidus, копуляция и образование сумки (275х)
длины (рис. 11.30, G); у Schizosaccharomyces клетки размножаются за счет образования noi юречных перегородок (рис. 11.30, А). Большинство дрожжей (например, пекарские, винные и пивные1) почкуются с образованием рубца. Дочерние клетки возникают здесь таким образом, что у материнской клетки в одном месте образуется почковидное вздутие. Разрастающаяся дочерняя клетка, куда заходит ядро, отрывается после образования разделительной перегородки от материнской клетки. И у материнской, и у дочерней клетки в месте разделения остается рубен. Каждая дрожжевая клетка
1 По современным представлениям, пекарские, винные и пивные дрожжи, ранее рассматривавшиеся как самостоятельные вилы, представляют собой один вид — Saccharomyces cerevisiae. — Примеч. пер.
имеет один рубец, возникший при ее образовании, и много (до 32) рубцов там, где возникали дочерние клетки. В качестве запасного вещества клетки откладывают гликоген и содержат многочисленные витамины, особенно группы В.
Пекарские дрожжи (Saccharomyces cerevisiae) были первым эукариотным организмом, у которого была полностью секвениро-вана последовательность нуклеотидов ДНК. При половом размножении (рис. 11.30) две клетки копулируют друг с другом (иногда через короткий копуляционный мостик). Если смешиваются суспензии дрожжей различных типов скрещивания, происходит выпадение осадка в виде облака. Эта агглютинация обусловлена специфическими белками клеточной стенки одного типа скрещивания и полисахаридами клеточной стенки другого типа скрещивания. Соеди
11.2. Бактерии, грибы, растения | -J63
нение в белково-полисахаридный комплекс является причиной агглютинации. Зигота непосредственно или после промежуточной почкующейся стадии становится сумкой, образуя после мейоза 4 или 8 аскоспор, которые после разрыва стенки получившейся сумки освобождаются и прорастают в новые вегетативные клетки.
Можно выделить три типа процесса развития у дрожжей. При гаплонтном типе (Schizosaccharornyces, рис. 11.30, А— F) ядро зиготы делится мейотически непосредственно после ее образования и зигота превращается в сумку; вегетативное размножение осуществляется в гаплофазе. При гаплодиплонтном типе (Saccharomyces cerevisiae, рис. 11.30, G— L) зигота дает начало почкующимся клеткам, в некоторых из них после мейоза возникают аскоспоры. Таким образом, здесь имеет место промежуточная смена ядерных фаз. Из гаплоидных аскоспор снова развиваются почкующиеся клетки, теперь они гаплоидные. Наконец, при диплонтном типе (Saccha-romycodes ludwigii, рис. 11.30, М —S) аскоспоры сливаются попарно друг с другом уже в сумке; вегетативное почкование осуществляется исключительно в диплофазе.
Дрожжи (Saccharomyces) находят разнообразное применение как возбудители спиртового брожения, причем как конечный продукт в одних случаях используется спирт (вино, пиво и т.д.), в других — СО2 (для разрыхления хлебного теста). Если винные дрожжи (Saccharomyces edipsoideus = S. vim) встречаются и в диком виде на ягодах, то пивные дрожжи (A cerevisiae и 5. carlsbergensis с многочисленными расами) известны только в культуре. При брожении более крупные клетки дрожжей оседают. а клетки меньшего размера остаются парящими в жидкости. В сортах пива «поверхностного брожения» (например, белое пиво) большая часть дрожжевых клеток отводится с пеной (в противоположность нормальным сортам 1 шва «низового брожения»). Используемое в хлебопечении кислое тесто содержит кроме дрожжей также молочно-кислые бактерии. Так называемый чайный гриб представляет собой смесь из дрожжей и бактерий; он превращает черный чай в кислый на вкус напиток.
Единственный порядок: Endomycetales. Представители порядка имеют типичное для подкласса строение. Помимо Saccharomyces, это строение можно рассмотреть на примере родов Dipodascus и Endomyces. Все эти роды можно рассматривать как самостоятельные семейства. Di-podascaceae: клетки гиф Dipodascus образуют длинные цепочки. Эти клетки могут быть одно-(D. uninucleatus) и многоядерными (например, D. alhidus). У D. albidus, который встречается в слизистых истечениях деревьев, образуются клювовидные гаметангии (рис. 11.30, W). которые сливаются концами и затем отграничиваются в основаниях поперечными перегородками (рис. 11.3. X). Ядра одного гаметангия (б") переходят в несколько более крупный другой (£). Слияние, однако, происходит только у одной пары ядер. Женский гаметангий вытягивается в длинную сумку. Не слившиеся ядра отмирают, а из диплоидного ядра слияния после мейоза возникают многочисленные гаплоидные ядра, из которых каждое становится ядром аскоспоры. D. uninucleatus обитает на мертвых насекомых. При его размножении сливаются две соседние клетки, становящиеся таким образом гаметангиями; ядра у них крупнее, чем у остальных клеток. Представители семейств Endomycetaceae и Saccharomycetaceae содержат в сумках не более восьми аскоспор.
Endomyces образует нитевидный мицелий. У Е. magnusii, который обитает в слизистых истечениях (возникающих из экссудата флоэмы) дуба, размеры мужской и женской копуляпи-онных ветвей значительно различаются; последняя после слияния ядер и следующего за ним мейоза становится четырехспоровой сумкой (рис. 11.30, Т, С).
Дрожжевидные талломы встречаются не только у Endomycetidae; они имеются (по крайней мере, на определенных стадиях развития) также у других аскомицетов (Taphrinomycetidae), у базидиомицетов (например, Sporobolomyceta-ceae, Exobasidiaceae, Ustilaginaceae) и у несовершенных грибов (например, Cryptococcaceae).
3. ПОДКЛАСС: ЛАБУЛЬБЕНИОМИЦЕТИ-ДЫ (LABOULBENIOMYCETIDAE)
Мы лишь вкратце коснемся этой группы грибов — паразитов насекомых, в ходе эволюции рано отделившейся от остальных аскомицетов. Для них характерны редуцированные талломы, но строго фиксированное строение мелких органов спороношения (рис. 11.31). Эти грибы чаще всего проникают лишь короткой, темноокрашен-ной «ножкой» в хитиновый покров своих хозяев. Многие виды (из общего числа примерно I 500) очень специфичны в отношении хозяев. Талломы развивают в специальном вместилище («пе-
164 | ГЛАВА 11 СИСТЕМАТИКА И ФИЛОГЕНИЯ
Рис. 11.31. Ascomycetes, Laboulbeniomyce-tidae. Stigmatomyces baerii (no R.Thaxter).
Верхний ряд: развитие до образования сперма-тангиев (ап) и материнской клетки аскогона (гл). Нижний ряд: стадия оплодотворения (ад — аскогон, t — трихогина), наполовину зрелый перитеций с молодыми сумками (а), сумка с 4 двухклеточными спорами (400х)
ритеции») — женский половой орган (аскогон) с трихогиной. Он оплодотворяется спермация-ми (не гаметангиогамия!), которые высвобождаются из мелких бутылевидных сперматангиев. После оплодотворения возникают сумки с нежными стенками, которые содержат одно- или двухклеточные аскоспоры. Время от заражения хозяина до созревания спор составляет лишь 10— 20 дней. Единственный порядок Laboulbeniales этого подкласса показывает определенные родственные связи с Ascomycetidae; согласно другой точке зрения он занимает очень изолированное положение в системе.
4. ПОДКЛАСС: АСКОМИЦЕТИДЫ (ASCOMYCETIDAE)
Представители этого подкласса отличаются нитевидным гаплоидным мицелием в вегетативной фазе, гифами с парами ядер (аскогенными гифами) в генеративной стадии, а также плодовыми телами, в которых перемешаны гаплоидные и дикариотичные
гифы. Дикариотичные гифы связаны с гаплоидным мицелием пространственно и в отношении физиологии питания. Группировка ядер в пары происходит после различных процессов оплодотворения.
Размножение. При гаметангиогамии, например у Ругопета confluens', в молодых зачатках плодовых тел на некоторых концах гиф возникают $ органы; они состоят из клетки-ножки, расширенного многоядерного $ гаметангия — аскогона (рис. 11.32, В: ag; А) и расположенного на вершине аскогона изогнутого многоядерного придатка — трихогины (1). В непосредственной близости от аскогона возникает (также из гаплоидных гиф) булавовидный многоядерный д' гаметангий (g). Разные половые органы встречаются иногда группами и растут навстречу друг другу, так что сГ гаметангий сливается с трихогиной (гаметангиогамия). Трихогина открывается в месте их соприкосновения (ее ядра при этом дегенерируют); d" ядра мигрируют из сГ гаметангия в трихогину и оттуда через временно открывающуюся пору в аскогон (плазмогамия). Там ядра о" и $ группируются попарно (рис. 11.32, С). После этого из аскогона образуются многочисленные выросты, в которые переходят пары ядер: аскогенные гифы, которые растут, претерпевая клеточные деления, и ветвятся. При всех клеточных делениях в каждой клетке остаются попарно сгруппированные ядра, так как деления ядер происходят конъю-гированно (т.е. одновременно). Так возникают клетки дикариофазы с двумя ядрами разных типов скрещивания (здесь d" и $). У других аскомицетов перенос сГ ядер в аскогон может происходить не из сГ гаметангиев, а в результате образования многоядерных или одноядерных конидий (гаме-то-гаметангиогамия), а также через гаплоидные гифы. При соматогамии гаметангиев нет, а сливаются обычные, особо не дифференцированные гифы. В результате слияния (после плазмогамии) и здесь возникает дикариотичный мицелий. Часто половая функция бывает подавлена. Говорят: о
1 Правильное современное научное название этого гриба — Ругопета omphalodes. — Примеч. пер.
11.2. Бактерии, грибы, растения 1 -| je.
Рис. 11.32. Ascomycetes, Ascomycetidae (А — по R. Harder; В—D — по Р. Claussen; Е — по A. de Вагу;
F—J — по R. A. Harper; К — no Р. Claussen):
А — плодовое тело однодомного дискомицета (схема), гаплофаза, дикариофаза, диплофаза (крючки не изображены); В —J — Pyronema confluens: В — зачаток плодового тела (450х), С — попарная группировка d” и <j> ядер в аскогоне, D — переход дикарионов в вырастающие из аскогона аскогенные гифы (С, D ЮООх), Е — аскогон с аскогенными гифами (150х), F—J — развитие сумки; К — Boudiera, молодая сумка (с) с аскоспорами (F — К — ЮООх); а — будущая сумка; ад — аскогон с трихогиной t; д — сГ гаметангии; h — крючок; s — клетка-ножка
партеногамии, когда внутри аскогона происходит попарная координация ядер без предшествующего оплодотворения б" ядрами; об автогамии — когда образование дикариона происходит где-либо без участия аскогонов; об апомиксисе — когда половая функция утрачена и развитие проходит в гаплофазе. Дикариотичные (аскогенные) гифы у аскомицетов часто отличаются наличием у поперечных перегородок своеобразных крючков, которые возникают следующим образом (рис. 11.32, F—J). Растущая конечная клетка образует сбоку, несколько ниже конца гифы (субтерминально), отходящий вниз и в сторону, противоположную направлению роста, крючковидный вырост. Одновременно ядра дикариона делятся, причем одно из образовавшихся дочерних ядер мигрирует в крючковидный вырост. Затем верхняя пара ядер отделяется поперечными перегородками (G), тогда как крючок на конце сливается с
основной гифой и находящееся в нем ядро возвращается туда (J). Образование крючка повторяется при каждом новом делении конечной клетки, пока в ней не произойдет кариогамия и не начнется образование сумки (Н). Из конечных клеток аскогенных гиф, после того, как там произойдут кариогамия и мейоз, возникают сумки. Молодые зачатки сумок первоначально двуядерные (рис. 11.32, F, G). После слияния ядер (Н) конечная клетка превращается в булавовидную, первоначально еще одноядерную диплоидную спороцисту. Из ядра слияния после трехкратного деления (имеет место мейоз) возникает 8 ядер, которые в результате свободного деления клеток (см. Ascomycetes) отграничиваются стенками. Таким образом, сумка — это мейоспороциста, в которой к двум делениям мейоза добавляется митоз.
Плазма, не используемая при спорообразовании, — периплазма — часто находит
166 | ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
применение для отложения еще одного, разнообразно скулы ггурированного слоя на стенках спор. Сумки развиваются, как правило, внутри плодовых тел, реже — свободно, на незащищенных гифах зачатка плодового тела. Аскогимениальным называют такое развитие, при котором только аскогенные дикариотичные гифы окружаются оболочкой (перидием); таким образом, начало образования плодового тела определяется оплодотворением. При асколокуляр-ном типе развития зачатки плодовых тел или сплетения гиф, которые впоследствии должны превратиться во вместилища сумок, закладываются уже до оплодотворения, а аскогенные гифы врастают в образующиеся после оплодотворения полости (ло-кулы). Систематическое подразделение основывается здесь наряду с прочими признаками на разном онтогенезе и разном строении плодовых тел и сумок. Различают несколько видов плодовых тел: округлое закрытое — клейстотеций; блюдце видное открытое — апотеций; бутылевидное, с выводным отверстием — перитеций, при ас-когимениальном, и псевдотеций, при аско-локулярном развитии. Псевдотеции могут широко раскрываться или, как клейстотеции, разрываться пассивно.
Рис. 11.33. Ascomycetidae, Eurotiales (А — по L. Кпу; В, D — по О. Brefeld; С, Е — по A. De Вагу): А — Aspergillus glaucus, леечник, конидиеносец (ЗООх); В — Penicillium glaucum, «кистевик», конидиеносец (ЗООх): р — фиалида, m — метула; С— Eurotium, спирально закрученный аскогон, охватывающий сГ гаметангий (450х); D — Talaromyces, обвивающие друг друга гаметангии (500х); Е — Eurotium, клейстотеций в поперечном разреза (250х)
I. Надпорядок: Eurotianae. Стенки (про-тотуникатных) сумок недифференцированные, тонкие. Часто они ослизняются до созревания спор, так что аскоспоры освобождаются пассивно.
I.I. Порядок: Eurotiales. Сюда относятся грибы, у которых часто подавлено или отсутствует половое спороношение. Однако характеристика и положение порядка в системе аскомицетов основаны на признаках половой стадии (рис. 11.33, С—Е). Она образуется, например, у Eurotiaceae после слияния булавовидных гаметангиев (аскогона, cf гаметангия). У поперечных перегородок возникающих после этого дикарио-тичных аскогенных гиф крючки отсутствуют. Булавовидные сумки закладываются внутри закрытых шаровидных плодовых тел. Сумки содержат каждая по 4 или 8 аскоспор, часто дисковидных, и беспорядочно располагаются в большом количестве в плодовом теле, плектенхиматическая стенка которого должна разрушаться, чтобы сумки и аскоспоры могли распространяться. Соответственно плодовые тела здесь — клейстотеции без специально образующегося выводного отверстия. Характерны и бесполые спороношения (рис. 11.33, А, В). Некоторые имеют названия Aspergillus и Penicillium, они относятся к наиболее частым плесневым грибам («плесень» — не систематическое понятие, а общее обозначение для поверхностно растущих мицелиев грибов). Размножение осуществляется бесполым путем — конидиями (см. бокс 11.3), которые образуются на расположенных близко друг к другу конидиеносцах и часто бывают окрашены в сине-зеленый цвет.
У леечника, или аспергилла {Aspergillus), конидии расположены на конидиогенных клетках (фиалидах), расходящихся во все стороны от шаровидного вздутия на вершине конидиеносца. Фиалиды последовательно отшнуровывают конидии, которые связаны друг с другом в виде цепочек. У ки-стевика, или пеницилла {Penicillium), конидии, также располагающиеся цепочками, возникают на разветвленных конидиеносцах, причем ветви, образующие конидии, называются фиалиды, а расположенные под ними — метулы. Систематические единицы в пределах Eurotiales называют по
11.2. Бактерии, грибы, растения | -|g7
половой стадии, а когда она отсутствует — по конидиальной стадии. Например, кони-диальная стадия у Eurotium, Sartorya — Aspergillus; у Talaromyces, Carpenteles— Pe-nicillium.
К отдельному семейству относятся обитающие под землей (гипогейно) оленьи трюфели — Elaphomyces (Elaphomycetaceae). Их клубневидные плодовые тела размером 1 — 4 см для человека несъедобны, однако выкапываются и съедаются дикими животными; таким путем распространяются споры.
Применение и наносимый ущерб. Из Penicillium notatum, Р. chrysogenum' и других видов получают антибиотик пенициллин, подавляющий синтез клеточной стенки бактерий. Антибиотик выделяется [рибом в питательную среду ergillus Р. roquefortii и Р. camembertii необходимы для изготовления определенных сортов сыра. Asp. wentii проду! щрует амилазы и протеазы и поэтому используется в биотехнологии; Asp. fla-vus образует афлатоксины, которые вызывают рак и повреждения печени. Asp. fumi-gatus вызывает заболевания легких и бронхов у человека. Другие опасные возбудители грибных заболеваний человека и животных — микозы — также относятся к этому порядку или должны быть отнесены сюда же на основании единственно известных анаморф, сходных с конидиальными стадиями Eurotiales.
К прототуникатным аскомицетам относятся и другие, меньшие порядки, перечисленные ниже.
1.2. Порядок: Microascales. К порядку относятся возбудители широко известного отмирания вязов из рода Ophiostoma. Гриб обитает в личиночных ходах жуков-заболонников, поражающих вязы. Длинные шейки их перитециев далеко выступают в личиночные ходы. Анаморфы развивают конидиеносцы. Аскоспоры и конидии собираются в каплевидных выделениях, которые захватываются личинками жуков и распространяются. Отмирание вязов представляет собой болезнь увядания, вызываемую закупоркой сосудов молодых ветвей и выделением токсина, убивающего листья (цератоульмина). Только после заноса вирулентного штамма из Север-
1 По современным представлениям, Р. notatum и Р. chrysogenum — один вид (для которого должно быть оставлено второе название). — Примеч. пер.
ной Америки (Ophiostoma novo-ulmi) болезнь распространилась в виде эпифиготии и по Европе. Другие порядки — Onygenales (Onygena equina на копытах лошадей), чаще всего образующие лишайники Caliciales, а также Coronophorales и Ме-liolales.
Стенка сумки у всех последующих порядков эутуникатная, т.е. четко различима как более толстый слой, прочная и снабженная приспособлениями для выброса аскоспор. Сумки первоначально однослойные (у порядков 2—4— унитуникатные).
2. Надпорядок: Erysiphales. Эти паразитические грибы, как и возбудители настоящей мучнистой росы, живут на растениях. Пораженные растения выглядят как бы обсыпанные мукой. Такое впечатление создается из-за белого поверхностного мицелия, который летом образует в больших количествах конидии (рис. 11.34, А). Через гаустории, которые проникают в клетки эпидермиса хозяина (A: h), гриб получает питательные вещества. Половое спороноше-ние представляет собой мелкие клейстотеции от коричневого до черного цвета, видимые невооруженным глазом как точки поверх белого налета мицелия и кониди-ального спороношения.
Половое размножение. Мужская копуляци-онная ветвь, которая делится на клетку-ножку и одноядерный сГ гаметангий, прикладывается к также одноядерному аскогону (рис. 11.34, D). Половое ядро о" переходит в аскогон (Е). После этого из каждого оплодотворенного аскогона возникает сумка без промежуточного образования аскогенных гиф или же аскогон (рис. 11.35) дает начало аскогенным гифам, конечные клетки которых образуют сумки. В первом случае пара ядер после синхронного деления (G) сливается в диплоидное ядро зиготы, которое после мейоза дает от 4 до 8 ядер аскоспор. Во втором случае развитие сумок соответствует описанному выше нормальному варианту, но поперечные стенки аскогенных гиф не имеют крючков.
Одновременно с образованием и оплодотворением аскогона он оплетается гифами оболочки, которые в конечном счете образуют светлый основной слой и темный перидий клейстотеция. Последний при созревании разрывается трещиной под давлением набухающих сумок. Чаше всего у основания клейстотеция возникает венец из гиф, часто на конце дихотомически ветвящихся или крючковидно загнутых. Они. по-видимому, способствуют распространению гриба (В, С). Сумки в клейстотеции расположены
168 | ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Рис. 11.34. Ascomycetidae, Erysiphales (А, В — по Р. Sorauer; С — по S. Blumer; D—G — по Bergman):
А — Uncinula necator, образование конидий (100х); В — то же, клейстотеций с придатками (ЗОх); С — Microsphaera alphitoides, клейстотеций с придатками (ЗОх); D—G— оплодотворение у Sphaerotheca fuliginosa (250х); а — аскогон; g — d" гаметангий; h — гаусторий; hh — покровные гифы; к — конидия; s — клетка-ножка
пучком (если сумка не одна) и открываются (иногда небольшой крышечкой), отбрасывая аскоспоры в воздух на расстояние до 2 см.
Uncinula necator (А, В) поражает листья и ягоды винограда (конидиальная стадия: Oidium tuckeri). Sphaerotheca mors-uvae (с одной сумкой в клейстотеции) поражает крыжовник; Sphaerotheca pannosa — розы; Microsphaera alphitoides (С) обитает на листьях дуба. Blumeria graminis (Erysi-phe graminis) — паразит дикорастущих и хлебных злаков. Для борьбы с «настоящей мучнистой росой» применяются препараты серы.
Рис. 11.35. Ascomycetidae, сумки до и после выбрасывания спор (A, D — по Е Oberwinkler; В, С — по A. Beckett):
А—С — иноперкулятные сумки, вершины сумок с апикальными аппаратами до (В) и после (С) выхода спор; > — оперкулятная сумка, отверстие с крышечкой
3. Надпорядок: Pczizales. Этот очень разнообразный по особенностям развития и строения порядок насчитывает примерно 1 000 видов, в основном сапротрофов. Типичная форма плодового тела у представителей Pezizaceae и близких семейств — чашевидный до дисковидного апотеций (например, Pevza), на верхней стороне которого в виде палисадного слоя расположен гимений, состоящий из сумок и гаплоидных стерильных парафиз. Сумки открываются на вершине крышечкой; таким образом, они упитуникатнысоперкулятные. как в предыдущем порядке (рис. 11.35). Споры часто отбрасываются на большие расстояния (Dasyobolus, см. 8.3.2.7).
Оплодотворение и образование сумок было впервые открыто и детально изучено у Ругопета (см. рис. 11.32). Р. confluens образует относительно мелкие дисковидные плодовые тела. Они часто плотно прижаты друг к другу, образуя почти корковидные налеты на местах, подвергшихся воздействию огня, или на почве. Уже перед копуляцией половые органы оплетаются рыхлым слоем гаплоидных покровных гиф. После оплодотворения (плазмогамии) возникают аскогенные гифы; монокариотич-ные гаплоидные гифы и дикариотичные аскогенные гифы, чаще всего образующие крючки, переплетаются и совместно образуют плодовое тело.
Образование плодового тела связано с половым процессом, который происходит
11.2. Бактерии, грибы, растения i -jgg
одновременно или в заранее возникающих зачатках плодовых тел. Гимений у Ругопета развивается с самого начала открыто на поверхности плодового тела (гимнокарпный тип). У других родов (например, Ascophanus) гимений возникает сначала внутри шаровидного зачатка плодового тела, поверхностный слой которого позже наверху разрывается, так что гимений становится открытым (гемиангиокарпный тип). Размер плодового тела в зависимости от вида — от нескольких миллиметров до более одного дециметра (Sarcosphaera).
Некоторые представители обладают более крупными апотециями на ножке. Ножка может иметь продольные бороздки (например, Helvetia), а поверхность самого апотеция, края которого отогнуты вниз, может быть подразделена на отдельные камеры (например, Morchella). Увеличение площади гимения и подъем его на ножке обеспечивают более действенное распространение спор. У некоторых родов (например, Helvetia, Gyromitra) не образуется ни аскогонов, ни о" гаметангиев. Сливаются друг с другом вегетативные гифы совместимых типов скрещивания (соматогамия). У Morchella часто
встречается слияние гиф только одного и того же вида (автогамия) (рис. 11.36).
Чаще всего развивающиеся в лесных почвах плодовые тела настоящих трюфелей (Tuberaceae) могут быть выведены из открытых блюдцевидных форм и отнесены к Pezizales в связи с наличием переходов. Однако их плодовые тела остаются в почве и замкнуты (гипогейный образ жизни). Освобождение аскоспор происходит в значительной мере за счет питающихся этими грибами животных или при разрушение плодового тела. Плодовые тела, чаще всего имеющие клубневидную форму, пронизаны ходами, которые, по крайней мере на ранней стадии развития, выходят наружу и покрыты своего рода гимением (см. рис. 11.37). Эти ходы представляют собой впячи-вания гимения внутрь. В широких булавовидных сумках, которые возникают после со-матогамии (автогамии) из конечных клеток аскогенных гиф, несущих пряжки (см. Ну-menomycetidae), располагается 1—5скульп-турированных аскоспор коричневого цвета. Исходно оперкулятное строение сумок почти не различимо, так как их нежная стенка не дифференцирована.
Рис. 11.36. Ascomycetidae, Pezizales (А — по Е. Michael; В — по J. Sachs; D — по G. Bresadola; Е — по Н. Schenck):
A — Otidea (Peziza) leporina (2/Зх); В — Pulvmula convexula, разрез через апотеций; на верхней стороне расположен гиме11ий (20х); С — Morchella esculenta, плодовое тело (3/4х); D — Helvetia pezizoides (3/4х);
Е — Morchella esculenta (3/4х); а — сумка; р — парафиза; sh — субгимениальное сплетение гиф
170 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Рис. 11.37. Pezizales, Tuberaceae, Tuber return (no L. R.Tulasne):
А — плодовое тело, вертикальный разрез (Зх); В — разрез части гимения (ЗООх); а — темные вены из плотного сплетения гиф: g — рыхлая плектенхима; h — гимений; г — кора
Более крупные представители Pezizales (например, Morchella, рис. 11.36, С) находят применение как съедобные грибы. Известны также и ядовитые виды. К ним относится Sarcosphaera crassa. Строчок (Gyromitra esculenta), яд которого разрушается при нагревании, иногда используется в пищу после отваривания (отвар сливают), однако из-за опасности отравления использовать его в пищу не рекомендуется. Из трюфелей, живущих в симбиозе с лесными породами деревьев, несколько видов (особенно Tuber magnatum — пьемонтский трюфель и Tuber melanosporum — перигорский трюфель) издревле высоко ценились как съедобные грибы благодаря летучим ароматическим веществам, придающим блюдам из них особую привлекательность. Подземные плодовые тела находят с помощью специально натасканных для этого свиней или собак. Трюфели имеют чрезвычайно высокую цену на рынке («черные бриллианты»)1.
1 1 кг перигорского трюфеля стоит в Европе порядка I 500 евро. — Примеч. пер.
4. Надпорядок: Leotianae. На вершине унитуникатной и иноперкулятной сумки (см. рис. 11.35) образуется отверстие в виде поры, окруженное или простым расплывающимся валиком, или дополнительно апикальным кольцом, или верхушечным валиком. Последние при использовании реактива, содержащего йод, часто окрашиваются в синий цвет и тогда получают название амилоидных. Механизм выбрасывания спор, вызываемого светом или изменением влажности, полностью не выяснен. По-видимому, разбухание валика или же апикального кольца играет при этом решающую роль, как и тургор внутри сумки. Плодовые тела— преимущественно апотеции (4.1-4.2).
4.1. Порядок: Leotiales. Эти грибы имеют плодовые тела от бокаловидной до блюдцевидной формы, возникающие аскогимениальным путем, однако их размеры, а также размеры сумок и спор часто меньше, чем у Pezizales. Наряду с типичными апотециями здесь встречаются, как и у Pezizales, другие производные формы плодовых тел. например булавовидные — у Trichoglo-ssum, блюдцевидные на ножках — у Sclerotinia или дифференцированные на шляпку и ножку — у Cudonia (результат конвергентной эволюции плодовых тел с Pezizales и т.д.). Большинство видов многочисленных родов живет сапротроф-но, однако некоторые — паразиты: например, Trichoscyphella willkommii — возбудитель рака лиственницы, Pseudopeziza trifolii — возбудитель болезней клевера. Sclerotinia fructigena поражает яблоки и груши; сначала развиваются кониди-альные пустулы, часто расположенные в виде концентрических окружностей (что обусловлено ежедневными сменами светового и темнового периодов) со спороношениями типа «Monilia», а весной на мумифицированных плодах возникают апотеции на длинных ножках (рис. 11.38). Конидиальная стадия гриба Sclerotinia fuckeliana, называемая Botrytis cinerea, в дождливые годы вызывает опадание ягод винограда, а при сухой погоде — «благородную гниль» ягод, особенно при большом содержании в них сахара. Обитающий сапротрофно на гнилой древесине Chlorosplenium aeruginosum окрашивает субстрат в интенсивно сине-зеленый цвет.
4.2. Порядок; Phacidiales (включая Rhytis-matales). Эта группа, ранее объединявшаяся с предыдущим порядком, отличается от него ас-колокулярным развитием плодового тела. Плоские плодовые тела открываются трещиной или продольными разрывами. Представи-
11.2. Бактерии, грибы, растения | -jy-j
Рис. 11.38. Ascomycetidae, Leotia-les, Sclerotinia fructigena (A — no Honey; В — no W. Kotte):
A — плодовое тело на мумифицированном персике (3/4х); В — конидиаль-ная стадия Monilia на груше. Мицелий образует конидии концентрическими кругами (1/2х)
тели порядка Phacidiales ведут преимущественно паразитический образ жизни. Сюда (Rhytis-matales) относятся возбудитель черной пятнистости клена, Rhytisma acerinum, который вызывает осенью черные пятна на листьях (апотеции образуются весной), и Lophodermium seditiosum на сосновых иглах, вызывающий болезнь «шютте».
4.3. Порядок: Lecanorales. Представители порядка составляют основную часть грибов, входящих в состав лишайников наших широт, и поэтому будут рассмотрены в соответствующем разделе (см. приложение 2 к Mycobionta). Грибы, живущие в лишайниковом симбиозе, образуют
апотеции, которые между парафизами (см. рис. 11.36, С) содержат особого строения сумки с утолщенными вершинами. Эти сумки булавовидные, толстостенные, иногда их стенка состоит из нескольких слоев (причем слои, в отличие от тех. что наблюдаются у битуникатных сумок, имеют одинаковую эластичность) и имеют вокруг поры на вершине верхушечный валик, окрашивающийся йодом в синий цвет.
Последующие порядки (4.4 —4.7) характеризуются бутылевидными плодовыми телами (перитециями) с направленным вверх выводным отверстием (ostiolum) на вершине; тип развития у них аскогимениальный. Сумки совместно
Рис. 11.39. Ascomycetidae (А — по С. Т. Ingold; В — по Brauns и Riehm; С — по К. Magdefrau):
А, В — Sphaeriales: А — Podospora fimiseda, перитеций (90х); В — нектриозный рак на ветке плодового дерева (возбудитель — Nectria galligena) (в натур, величину); С — Xylariales, Xylaria hypoxylon (натуральная величина); а — сумки; kb — область конидиального спороношения; m — гифы мицелия; р — перифизы; pb — область перитециев
172 | ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
с многочисленными гаплоидными гифами (парафизами) образуют палисадный плодущий слой (гимений), который выстилает основание и стенки заранее образующейся полости плодового тела (рис. 11.39, А). При созревании одна сумка за другой по очереди выдвигается до тех пор, пока ее вершина не войдет в выводное отверстие перитеция, после чего сумка сразу выбрасывает все 8 спор. Высота, на которую они выбрасываются, может составлять 20 см и более. После этого сумка спадается, так что отверстие перитеция освобождается для следующей сумки.
4.4. Порядок: Sphaeriales1. Тупые наверху сумки представителей этого порядка имеют в верхней части сумки образующийся вокруг апикальной поры валик; апикальный аппарат выглядит чаще всего как замок поры в форме пластинки. Как представитель этого порядка может быть упомянута прежде всего Neurospora. N. sitophila и N. crassa вызывают красную хлебную плесень и выносят высокие температуры (до 75 °C). Виды Neurospora образуют на каждом отдельном мицелии как аскогоны, так и клетки, которые предназначены для переноса сГ ядер. В качестве переносчиков d" ядер на трихогину аскогона служат многоядерные конидии (мегаконидии), одноядерные спермации или микроконидии, а также соматические гифы. Спермации представляют собой специализированные клетки, предназначенные для этой цели; мега- и микроконидии могут также образовывать проростковую трубку и соответственно давать начало новому мицелию. Перекрестное оплодотворение достигается за счет того, что аскогоны могут оплодотворяться только ядрами противоположного типа скрещивания (гомогенная несовместимость; см. рис. 11.19). После оплодотворения (плазмогамии) из аскогона вырастают обычным путем дикари-отичные аскогенные гифы, образующие крючки.
Большое значение для генетических исследований приобрели обитающие в природе на навозе Podospora anserina, Sordaria fimicola и 5. macrospora. Сумки этих видов Sordaria содержат по 8 аскоспор, тогда как у Р. anserina они 4-споровые. У Р. anserina мегаконидии отсутствуют; О* ядра переносятся спермациями. У обоих упомянутых видов Sordaria мега- и микроконидии отсутствуют, аскогоны не образуют трихо-гины, а группировка ядер в дикарион осуществляется партеногенетически; таким образом, перитеции развиваются в результате самооплодотворения. Правда, у мутантов может происходить также соматогамия между разными мицелиями.
1 В настоящее время в литературе этот порядок чаще разделяется на несколько порядков: Sordariales, Xylariales, Hypocreales и др. — Примеч. пер.
В области конфронтации возникают в этом случае перитеции как результат скрещивания, в их сумках может наблюдаться предредукция или же постредукция.
Перитеции Sphaeriales, часто едва достигающие размера 0,5 мм, располагаются обычно поодиночке, но иногда и группами, которые могут находиться на более или менее четко выраженном сплетении (строме). Строма — это, как правило, твердое образование типа склероция, в которое погружены отдельные перитеции или их группы (тогда это образование представляет собой как бы комплексное плодовое тело). Перитеции Nectria cinnaharina сидят на подушковидных киноварно-красных стромах; последние развивают сначала конидии, позже перитеции; в обеих формах они видны как красные подушечки на отмерших ветвях. Nectria galligena (с бесцветными конидиями) обитает как паразит в коре и вызывает рак плодовых деревьев. Отмершие под влиянием гриба части коры обрастаю! раневым каллусом, который, в свою очередь, сам отмирает, что приводит к беспорядочным волнистым разрастаниям (рис. 11.39, В) и может, наконец, погубить все дерево. Из культуральных фильтратов паразитирующей на растениях риса Gibberella было выделено ростовое вещество гиббереллин (см. 7.6.3). Из этого же порядка можно еще упомянуть роды Cochliobolus (см. 9.3.3), Daldinia, Gaeumannomyces и Hypoxylon.
4.5. Порядок: Diaporthales. Пора сумки в дополнение к апикальному валику окружена оптически плотным кольцом, которое хорошо красится анилиновым синим. Сумки часто растворяются и под конец вместе со спорами выдавливаются из выводного отверстия плодового тела. В остальном этот порядок, представленный родами Diaporthe, Diaporthella и др., очень сходен с предыдущим. Перитеции возникают внутри стро-матического сплетения гиф. Виды, многие из которых — паразиты, внедряются своими стромами в ткань хозяина, а из нее обычно несколько выступают наружу только длинные шейки перитециев. Endothia parasitica' была занесена в Северную Америку и практически полностью уничтожила на всем материке большие массивы настоящего каштана (Castanea dentata, см. 13.8). На территории Европы (например, в Тичино) ущерб от этого гриба также очень велик.
4.6. Порядок: Xylariales. Стромы представляют собой более крупные образования в форме подушек, шаров, булав или оленьих рогов, в которые погружены многочисленные перитеции. У часто встречающегося на пнях лиственных
1 В настоящее время в литературе этот вид чаще встречается под названием Cryphonectria parasitica. — Примеч. пер.
11.2. Бактерии, грибы, растения
173
пород гриба Xylaria hypoxylon (рис. 11.39, С), имеющего стромы в виде оленьих рогов, в верхней белой части сначала образуются конидии, а позже, в нижней черной части, — перитеции. Апикальный аппарат по строению близок к представителям предыдущего порядка, с тем отличием, что апикальное кольцо на вершине сумки Xylariales красится йодом в синий цвет.
4.7. Порядок: Clavicipitales. Сумки имеют на вершине оптически более плотное расплывающееся кольцо от полукруглой до почти круглой формы. Септированные аскоспоры нитевидные, длинные, а сумки соответственно узкие. Перитеции погружены в стромы, у некоторых представителей дифференцированные на стерильную «ножку» и ферт ильную «шляпку». В двух последних порядках формирование стромы достигает, таким образом, наивысшего уровня аналогично развитию отдельных плодовых тел (например, у Leotiales); по своему внешнему облику одиночные или «комплексные» плодовые тела часто бывают, несмотря на различия в деталях морфологии и путях эволюционного развития, очень сходны (аналогичная конвергенция наблюдается, например, между Trichoglossum из Leotiales и Cordyceps из Clavicipitales).
Гриб спорынья, Claviceps purpurea, обитает как паразит в молодых завязях злаков и образует там конидии (рис. 11.40, А, В). Выделяемая при этом сахаристая жидкость (медвяная роса) привлекает насекомых, которые переносят конидии на другие цветки. После истощения тканей завязи мицелий превращается в склероций (см. бокс 11.3): гифы плотно срастаются между собой и (прежде всего на периферии) пугем поперечных делений образуют псевдопаренхиму (В). Черные снаружи, выступающие из чешуй, твердые склероции (С, D) называю! «рожками». Они падают на землю, зимуют и ко времени цветения злаков прорастают в стромы, имеющие облик красноватых головок на ножках. В эти головки погружены многочисленные перитеции (Е). Образование перитециев начинается с копуляции становящихся к тому времени многоядерными аскогонов и 0я гаметангиев (аскоги-мениальный тип!). Длинные сумки содержат 8 спор (F). которые переносятся ветром на рыльца пестиков злаков.
Виды рода Cordyceps паразитируют на организмах с хитиновыми покровами, например на грибах с подземными плодовыми телами, таких как Elaphomyces, или на насекомых, которые после заражения заползают в почву. Поднимающиеся из земли булавовидные стромы содержат в своей верхней части многочисленные перитеции. Нитевидные споры уже в сумке становятся за счет поперечных делений многоклеточными и распадаются на отдельные фрагменты. Epichloe
Рис. 11.40. Ascomycetidae, Clavicipitales, Claviceps purpurea (A, В, D—F — no L.R.Tulasne; C — по H. Schenck):
A— пораженная завязь ржи (15x); внизу начинающееся образование склероция, над ним мицелий с конидиями, вверху остатки рыльца; F — образование конидий (ЗООх); С — колос ржи со зрелыми склероциями (2/Зх); О — проросший склероций с головчатыми стромами (2х); Е — продольный разрез через строму с многочисленными перитециями (25х); F — сумка и аскоспора (400х)
typhina паразитирует на злаках; первоначально белая, затем желтая строма этого гриба охватывает соломину и образует конидии, а позже перитеции. С Clavicipitales родственны грибы из порядков Ostropales и Graphidales, которые чаще всего образуют лишайники.
Применение и вредоносность. Склероции Claviceps purpurea содержат ядовитые алкалоиды (эрготамин, эрготоксин): при употреблении в пищу зараженного ими зерна происходят сильнейшие отравления («злые корчи», «антонов огонь»)'. Вместе с тем эти вещества находят применение в гинекологии, прежде всего как средства, способствующие родовым схваткам (отсюда их немецкое название Mutterkorn —«материнское зерно»). Склероции для медицинских це
1 Эти названия связаны с тем, что болезнь встречается в двух формах: гангренозной («антонов огонь») и конвульсивной («злые корчи»), — Примеч. пер.
174 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
лей выращиваются в широком масштабе, например, путем специального заражения ржи; в настоящее время налажено получение алкалоидов из мицелия в ферментерах. Из конидиальной стадии вида Cordyceps (называемой Tolypodadiuni inflatum) был выделен циклоспорин, который, в
Рис. 11.41. Ascomycetidae, Dothideales (А—
Е — по N Pringsheim; F—Н— по Higgins):
А—Е— Pyrenophora, Р. scirpi, способ вскрытия битуникатной сумки (400>): А— зрелая сумка с 8 четырехклеточными спорами, В — то же, внешняя стенка сумки разорвалась, внутренняя растягивается, С — последняя спора непосредственно перед выбросом, D — освободившаяся сумка, Е — прорастающая спора; F—Н — Мусо-sphaerella, М. tulipifera (175х), развитие псевдо-теция: F — ранняя стадия с разветвленным аскогоном, G — с сумками разного возраста, Н — зрелый псевдотеций
частности, применяется при пересадке органов для предупреждения их отторжения.
5. Надпорядок; Dothideales. Стенка сумки у представителей порядка состоит из двух слоев, по-разному растяжимых; таким образом, в отличие от ранее рассмотренных порядков, она битуникатная. Внешний тонкий слой относительно нерастяжим и разрывается на вершине при увеличении тургорного давления внутри сумки. Толстая внутренняя, растяжимая оболочка сумки после этого сильно вытягивается, причем из-за нарастающего давления аскоспоры одна за другой выбрасываются наружу, сначала замыкая собой пору на вершине (рис. 11.41, С). Повышение осмотического давления внутри сумки обусловлено превращением осмотически неактивных веществ в активные (возможно, гликогена в сахар). Бутылевидные плодовые тела с образующимся в них выводным отверстием чаще всего возникают по аско-локулярному типу (см. Ascomycetidae, введение). Будучи внешне сходными с перитециями, из-за различия в характере развития эти плодовые тела получили название псевдотеции (рис. 11.41).
К этому порядку относятся возбудители многих болезней растений. Виды рода Venturia (кони-диальная стадия: Fusidadium) вызывают паршу яблок и груш, образуя темные пятна на опавших или находящихся на дереве плодах (рис. 11.42). Виды рода Capnodium вызывают буро-черный налет — («чернь») на листьях; будучи сапротро-фами, эти грибы используют для питания выделения листьев или тлей. Виды рода Herpotrichia в альпийской зоне обволакивают покрытые снегом ветви хвойных деревьев буро-черным сплетением гиф и вызывают отмирание хвои. Сородичами Dothideales являются и виды, образующие лишайники, в том числе относящиеся к порядку Vernicariales.
Обзор аскомицетов. Среди образующих плодовые тела аскомицетов (Ascomycetidae) в параллельных эволюционных рядах (параллелизм, см. 4.1.1), часто с использованием разных принципов строения, выработались сходные формы, обеспечивающие активное распространение спор, например булавовидные, бутылевидные плодовые тела или плодовые тела, состоящие из ножки и шляпки. Разные эволюционные линии чет
11.2. Бактерии, грибы, растения | -,75
ко представляют аскогимениальный (перитеций) и асколокулярный (псевдотеций) типы развития. Если не брать во внимание уклоняющиеся Laboulbeniomycetidae и Endomycetidae, аскомицеты показывают принципиальную общность циклов развития. Половой акт часто осуществляется в аскогонах путем принятия д" ядер. Половые ядра партнеров по скрещиванию сразу не сливаются, а сгруппированные парами переходят в аскогенные гифы, размножаются здесь путем сопряженного деления и сливаются в диплоидное ядро только в зачатке сумки (= конечных клетках аскогенных гиф). Плазмогамия и кариогамия разделены далеко в пространстве и во времени двухъядерной стадией (дикариофазой). Дикариотичные клетки функционально уже диплоидны, только их ядра еще индивидуализированы. Типичные аскомицеты — гаплонты с вклинивающимся в жизненный цикл дикари-онтом, питание которого, однако, остается зависимым от предшествующего гаплоидного поколения (исключение: тафриномице-тиды). Гаплоидный мицелий, чаще всего образующий 6я гаметангии и аскогоны, может рассматриваться как гаметофит, дикариотичные аскогенные гифы — как спорофит. Фаза дикариотичного спорофита заканчивается образованием сумок (мейоспоро-цист); в них после слияния ядер и редукционного деления формируются гаплоидные мейоспоры (= аскоспоры). Обычно в каждой сумке возникает 8 аскороспор, однако некоторые виды обладают 1-, 2-, 4-или многоспоровыми сумками. Плектенхима плодового тела состоит из сильно развитых гаплоидных гиф гаметофита, в которые вплетены дикариотичные гифы. Аскогоны и д’ гаметангии или дающие д' ядра клетки (т. е. мегаконидии, микроконидии, соматические гифы) образуются на одном и том же мицелии (однодомность; см. рис. 11.32, В). Самооплодотворению часто препятствует биполярная гомогенная несовместимость (см. рис. 11.19). Среди аскомицетов можно проследить редукцию гаметангиев. Редукция или полная утрата половой функции может наблюдаться у партеногенетических или автогамных видов, которые могут быть выведены от форм, размножающихся половым путем.
Рис. 11.42. Ascomycetidae, Dothideales: парша на груше, вызываемая Fusicladium (по Kirchner и Boltshauser)
Филогения и родственные связи. Вопрос о происхождении аскомицетов остается спорным. Происхождение их от красных водорослей (Rhodophyta) в настоящее время практически уже не обсуждается (его рассматривали ранее на основе сходства формы аскогона и карпого-на и деталей развития после оплодотворения).
Отличия от красных водорослей проявляются в отсутствии ассимиляционных пигментов и, что еше более важно, в наличии дикариофазы, в химизме клеточной стенки, а также в аппарате Гольджи. Против признания такого происхождения аскомицетов может говорить и тот факт, что Rhodophyta как очень специализированные водоросли встречаются только с пермского периода. тогда как аскомицеты в формах, соответствующих современным таксонам, известны уже с карбона.
176 | ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
За выведение аскомицетов из грибов со свойствами, характерными для современных зи-гомицетов и хитридиомицетов, говорят уже имеющаяся у них выраженная или частично сходная гаметангиогамия и в отдельных случаях намечающаяся дикариофаза, состав клеточной стенки из хитина и соответствующих глюканов (с р-1,3-, Р-1,4-, р-1,6-связями), синтез лизина через аминоадипиновую кислоту, остающаяся интактной ядерная оболочка при делении ядра. Утрата активно передвигающихся репродуктивных клеток у аско- и зигомицетов в отличие от хитридиомицетов может быть связана с наземным образом жизни. В таком же плане могут быть истолкованы последовательное усовершенствование плодовых тел и септированность гиф. По морфологическим признакам к аско-мицетам ближе зигомицеты. по биохимическим — хитридиомицеты.
Выведение аскомицетов от низших грибов делает рациональным такую их классификацию, при которой тафриномицетиды и эндомицети-ды стоят в начале системы. Правда, остается еще в значительной мере сомнительным, можно ли выводить слияние копулирующих клеток, которое можно наблюдать у Saccharomycetales, из зигогамии зигомицетов. Признаки Endomyceti-dae, прежде всего отсутствие дикариофазы, свидетельствует в пользу признания их обособленности, которой иногда придают столь большое значение, что их выделяют из аскомицетов в самостоятельный класс. У некоторых Endomy-cetidae зигота прорастает в небольшой диплоидный мицелий, и только на нем образуются сумки. Остальные группы аскомицетов, у которых появилась задержка слияния ядер и соответственно более или менее выраженная дикариофаза, отделились в ходе эволюции достаточно рано.
Это обособление могло произойти от предков с признаками Taphrinomycetidae, которые, однако, по сравнению с Ascomycetidae, имеют более простое строение (отсутствие плодовых тел), но уже обладают дикариофазой (дикарио-тичный мицелий самостоятелен в плане питания), как все другие аско- и базидиомицеты. Наличие у них дрожжевидных стадий в ходе развития свидетельствует еще о родственных связях с Endomycctidae. Отсюда выводятся эволюционные линии, с одной стороны, к остальным аскомицетам, с другой стороны — к LJstomycetidae из базидиомицетов. Если у последних дикарио-тичный мицелий самостоятелен, то у обеих первых названных групп грибов он находится в физиологической зависимости от гаплоидного первичного мицелия. В основание системы аскомицетов должны быть поставлены (после грибов с особенностями Taphrinomycetidae) формы с
признаками современных Eurotiales и низших Pezizales (Ascomycetidae). Дальнейшая эволюция шла здесь прежде всего по пути усовершенствования сумок и плодовых тел.
4. Класс: базидиомицеты (Basidiomycetes)
Дня базидиомицетов (к ним относится около 30 000 видов т.е. 30% всех грибов) характерна мейоспороциста — базцдия, которая в типичном случае отшнуровывает от себя 4 раздельно расположенных мейоспоры. В базидии, как и в сумке, непосредственно друг за другом следуют кариогамия и мейоз (см. рис. 11.52). Возникшие в результате редукционного деления гаплоидные ядра (обычно их 4) мигрируют в кончики палочковидных выростов базидии (стеригмы, см. рис. 11.52,9) и только здесь происходит «экзогенное» образование базидиоспор. Жизненный цикл, отчасти различающийся в разных группах базидиомицетов, протекает по следующей, приблизительно соответствующей шляпочным грибам, схеме: базидиоспоры прорастают (см. рис. 11.45) в мицелий с одноядерными клетками, способный практически к неограниченному росту; гаметангии, как и у эволюционно продвинутых аскомицетов, не образуются. Если встречаются мицелии противоположных типов спаривания (например,«+» и «-»), то две соприкоснувшиеся вегетативные клетки сливаются между собой (соматогамия, см. рис. 11.52, 3), причем их ядра попарно лишь сближаются, но не сливаются между собой. Возникающая таким образом дикариотичная клетка дает начало дикариотичному мицелию, способному к самостоятельному питанию и не зависящему от гаплоидных и мо-нокариотичных гиф.
Кроме того, базидиомицеты отличаются от аскомицетов и строением поры в поперечной перегородке гифы. Если у аскомицетов это всегда простое отверстие, то у базидиомицетов, у которых также встречаются простые поры, по большей части возле поры имеется расширение в виде бочонка («долипора», лат. dolium — сосуд для вина), покрытое с каждой стороны парентосомой (греч. parenthesis — вдвинутый; soma — тело), которая образуется эндоплазматической се
11.2. Бактерии, грибы, растения
177
тью (рис. 11.43). Клеточная стенка базидио-мицетов показывает слоистую ультраструктуру (ср. Ascomycetes, см. введение).
Базидия может быть септированной (фрагмобазидия, рис. 11.44, А, В) или булавовидной и одноклеточной (холобазидия, рис. 11.44, F). Соответственно этому различию в строении раньше разделяли фраг-мобазидиомицеты и холобазидиомицеты.
Представители двух первых подклассов, которые рассмотрены ниже, раньше объединялись в подкласс Heterobasidiomy-cetidae. Однако, согласно молекулярным и ультраструктурным признакам, предлагается другая группировка, отчасти с другим объединением представителей (например, Auriculariales, Tremellales, Microbotiyales). Поперечные стенки гиф часто имеют простые поры (см. рис. 11.43, А); пряжки (см. Hymeno-mycetidae) у значительного большинства отсутствуют.
1. ПОДКЛАСС: УСТОМИЦЕТИДЫ (USTOMYCETIDAE)
В этом подклассе объединяются облигатные паразиты растений. У тех представителей, у которых базидия разделена поперечными перегородками (фрагмобазидия), предшествующая дикариофаза не размножается дикариотичными спорами. Виды этого подкласса — возбудители головни
различных растений («головневые грибы»). Такие названия и заболевания, и грибы получили потому, что пораженные части растений выглядят как бы обугленными.
I. Порядок: Tilletiales. У базидий отсутствуют поперечные перегородки. Базидии
Рис. 11.44. Basidiomycetes. Формы базидий (750х) (по F. Oberwinkler):
А— Platygloea (Auriculariales); В— Bourdotia (Tremellales); С — Tulasnella (Tulasnellales); D — Dacrymyces (Dacrymycetales); E— Sistotrema (Poriales); F — Hyphoderma (Poriales); G — Exobasidium (Exobasidiales); H — Xenasma (Proto-hymeniales); J — Repetobasidium (Poriales); К — Scleroderma (Boletales)
Рис. 11.43. Basidiomycetes. Поперечные перегородки гифы базидиомицетов (В — по R. Т Moore и J.H.McAlear):
А— с простой порой; В — с долипорой: d — долипора. р — парентосома; w — клеточная стенка
Рис. 11.45. Basidiomycetes. Прорастающие споры (1 OOOx) (A— no F. Oberwinkler; В — no O.Brefeld):
A — Exidiopsis effusa co вторичной спорой; В — Auricularia auricula-judae: 1 —3 — базидиоспоры (2 — базидиоспора с поперечными перегородками, 3— базидиоспора с конидиями); С — Pleurotus ostreatus с проростковой гифой
178 | ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
образуют на вершине чаще всего 4 или 8 удлиненных мейоспор. Только для разграничения с «головневыми спорами» образуются одна или несколько септ (рис. 11.46, J).
Между мейоспорами разных типов скрещивания возникают — часто уже тогда, когда они сидят на спороцистах, — копуляционные мостики (К), через которые плазма и ядра одной споры мигрируют в другую.
От начинающего после этого расти дикарио-тичного мицелия отчленяются дикариотичные конидии, которые активно отбрасываются (баллис-токонидии). Как дикариотичная, так и гаплоидная стадия могут размножаться конидиями. У пшеницы Tilletia caries вызывает твердую, или вонючую, головню, — Urocystis tritici — головню стеблей или полосатую головню листьев. Виды Entyloma поражают преимущественно сложноцветные растения. С Tilletia caries можно успешно бороться путем кратковременного «протравливания» посевного материала высокой температурой или ядовитыми веществами либо путем опыления веществами, которые убивают приставшие к семенам головневые споры. На пораженном растении злака образуется несколько миллионов головневых спор, которые при молотьбе напыляются на посевной материал и после посева заражают молодые про
ростки (проростковая инфекция), поэтому болезнь может легко распространиться на многие растения; вследствие этого раньше погибало до 20 % (иногда даже 60 %) урожая зерна.
От рассматриваемого дальше порядка Ustilaginales порядок Tilletiales отличается, кроме строения базидии, также тем, что в головневых спорах происходит не только кариогамия, но и чаще всего уже мейоз.
2. Порядок: Ustilaginales. Не имеющие плодовых тел устиляговые обитают как паразиты обычно в межклетниках высших растений и развивают в определенных органах своих хозяев (например, корнях, осях побегов, завязях плодов, тычинках) толстостенные споры. Грибы этого порядка, как и предыдущего, привлекают особое внимание как патогенные для растений организмы. Ustilago maydis образует на соцветиях (и других частях) кукурузы желваки и пузыри размером с кулак, которые заполнены головневыми спорами. Другие виды рода Ustilago заполняют завязи и частично прилегающие структуры соцветий ячменя и
Рис. 11.46. Ustomycetidae (А, В— по R. A. Harper; С, К— по F. Rawitscher; D— по W.Stempell; Е— G — по R.Seyfert; Н, J — по O.Brefeld):
А—Н — Ustilaginales: А, В — Ustilago scabiosae, проросшие головневые споры и образование мейоспор (s) на четырехклеточной базидии (110х); С — Ustilago carbo, копулирующие базидиоспоры (1 200х); D — Entyloma calendulae, мицелий с пряжками и дикарионами; Е—G — Ustilago vuijckii, образование головневых спор, дикариотичные и диплоидные головневые споры; Н — Ustilago sp., прорастающие в питательном растворе головневые спорвы (350х); J, К — Tilletiales, Tilletia caries'. J — проросшая из головневой споры базидия с четырьмя парами расположенных на ее вершине мейоспор (ЗООх); К — две копулировавшие базидиоспоры, прорастающие в дикариотичный мицелий, с конидией (к) (650х)
11.2. Бактерии, грибы, растения 1 -(79
пшеницы пылящим порошком головневых спор (пыльная головня — например, U. avenae вызывает пыльную головню овса). В случае пыльной головни ячменя (U. hor~ del) и пшеницы (U. tritici) головневые споры образуются еще до раскрытия цветков в молодых завязях и распыляются, когда растения находятся в полном цвету. Споры переносятся ветром и прорастают в том же году между чешуями здоровых цветков (цветковая инфекция). Развивающийся из базидиоспор мицелий врастает затем в образующееся семя и зимует в его зародыше.
Жизненный цикл. Из базидиоспор, имеющих половой знак «+» или «-» (биполярный гетероталлизм), при благоприятных условиях вырастает дрожжевидный псевдомицелий; он гаплоиден и может жить только сапротрофно. Его можно также культивировать на искусственных питательных средах. Если генотипически различные клетки «+» и «-» псевдомицелиев встречаются между собой, то через копуляционный мостик происходит слияние плазматического содержимого (плазмогамия) и группировка ядер в дикарион. Содержимое одной клетки переходит в другую, поэтому воспринимающая клетка становится двухъядерной. Она вырастает в дикариотичную гифу, которая отныне способна заражать растение-хозяина. Способность к паразитизму ограничена, таким образом, ди-кариофазой. Дикариотичный мицелий распространяется в хозяине и образует там в определенных органах головневые споры, в которых осуществляется кариогамия. В деталях проходят следующие стадии: дикариотичный мицелий, который у ряда видов имеет пряжки (рис. 11.46, D, Е), проникает в проростке растения-хозяина по межклетникам до апикальной меристемы и растет сначала вместе с растением, не вызывая внешних симптомов болезни, пока не достигнет определенных мест, например тычинок или завязей. Ткани хозяина полностью разрушаются, а в плотном сплетении мицелия образуются расположенные цепочками наподобие бус вздутия, которые окружаются толстой стенкой коричнево-черного цвета (рис. 11.46, Е) и отделяются от образовавшей их гифы. Они распыляются из места образования, как зола, поэтому их называют «головневые споры». В этой форме головневые споры гомологичны молодым базидиям, поскольку здесь также происходит кариогамия. Однако сами головневые споры еще не представляют собой спороцисты типа базидии, поэтому их называют пробазидии. Головневые споры прорастают (чаше всего только после перезимовки) в гифенный вырост, впос-
Рис. 11.47. Ustomycetidae, Exobasidiales, Exobasidium vaccinii (A — по K. Magdefrau; В — no M.Woronin; C — no F. Oberwinkler):
A — Vaccirium vitis-idaea с тремя листьями, пораженными Exobasidium (2/Зх); В — поперечный разрез через пораженный лист; слева нормальное образование листовой ткани, справа развитие ткани, гипертрофированной из-за поражения грибом (60х); С — прорывающийся между клетками эпидермиса мицелий с базидиями и прорастание базидиоспор (ЗЗОх)
ледствии поперечно септированный (рис. 11.46, А, В). При этом происходит мейоз, так что теперь в каждой из 4 отгороженных поперечной перегородкой клеток находится одно гаплоидное ядро. На этой стадии вырост— пробазидия — соответствует септированной фрагмоба-зидии. Эта пробазидия отшнуровывает вбок гаплоидные споры (так называемые споридии, В), причем возникающие в результате мейоза дочерние ядра мигрируют обратно в базидию. Они имеют генетически детерминированные половые знаки «+» или «-» в соотношении 1:1. При хорошем питании от базидии могут многократно отшнуровываться новые базидиоспоры. Цикл развития гаплодикариотичный: он аналогичен гаплодиплоидному циклу дрожжей.
Как и у дрожжей, некоторые формы (например, у Ustilago maydis — возбудителя пузырчатой головни кукурузы) чисто гаплоидны. Встречаются также другие отклонения. Некоторые формы уже тетраполярны (см. рис. 11.55).
1 80 |ГЛАВА 11 • СИСТЕМАТИКА И ФИЛОГЕНИЯ
3. Порядок: Exobasidiales. Представители порядка паразитирует, как и Taphrinales из аскомицетов, на цветковых растениях (в Европе прежде всего на вересковых) и не образуют плодовых тел. Часто они вызывают галлоподобные деформации на пораженных частях растения-хозяина (рис. 11.47, А), которые возникают из-за гипертрофии мезофилла (рис. 11.47, В). Мицелий пронизывает ткань растения, проходя по межклетникам и через клетки. Гифы паразита выходят на поверхность хозяина через устьица или между клетками эпидермиса и образуют там несептированные базидии (рис. 11.47, С) На тупых, сильно растопыренных стеригмах располагаются рядом друг с другом изогнутые споры, которые пассивно отпадают, образуют поперечные перегородки и прорастают конидями (рис. 11.47, С). В чистой культуре из спор вырастают скопления клеток, почкующихся наподобие дрожжей. На основании особенностей ультраструктуры и биохимии Exobasidiales относят, как делаем здесь и мы, к Ustomycetidae
2. ПОДКЛАСС: УРЕДОМИЦЕТИДЫ (UREDOMYCETIDAE)
К этому подклассу относятся паразиты, обычно еще не образующие плодовых тел. Характерная особенность — хорошо выраженная дикариофаза, воспроизводимая ди-кариоспорами (уредоспорами). Она предшествует образованию четырехклеточных фрагмобазидий с поперечными перегородками (см. рис. 11.49, D, F). Последние возникают всегда из округлых пробазидий; такие структуры часто отсутствуют в других подклассах базидиомицетов (например, среди Ustomycetidae у Tilletiales и Exobasidiales; среди Hymenomycetidae — у подавляющего большинства). Этот подкласс, насчитывающий несколько тысяч видов, включает грибы-возбудители широко распространенного заболевания растений — ржавчины.
1. Порядок: Uredinales (ржавчинные). Ржавчинные грибы живут как паразиты, прежде всего в межклетниках, не убивая пораженные ткани. В клетки хозяина внедряются гаустории (рис. 11.48, А). Изредка мицелий обильно разрастается по всему рас
тению, но чаще всего он распространяется лишь вблизи места заражения. На дикарио-тичных гифах пряжки отсутствуют. С аско-мицетами ржавчинные сходны по следующим признакам: биполярный гетероталлизм, наличие спермациев и воспринимающих гиф в качестве половых органов, септы с простыми порами, хорошо представленные анаморфы. За редкими исключениями, ржавчинные грибы не образуют каких-либо заметных плодовых тел. Это можно считать приспособлением к паразитированию на чаще всего недолговечных вегетативных органах высших растений. Ржавчинные грибы характеризуются большим разнообразием типов спор (полный цикл развития охватывает 5 разных типов спор; см. рис. 11.50), которые закономерно сменяют друг друга и связаны со сменой ядерных фаз, а часто и со сменой хозяев.
Дикариотичные гифы возникают из базальных клеток зачатка эцидия, когда они получат ядро одним из двух возможных путей. При переносе ядер спермациями важную роль играют вышеназванные пикниды. Их кувшиновидные плектенхиматические мицелиальные тела при созревании прорывают верхний эпидермис пораженных листьев барбариса и становятся видны как желтоватые пустулы (рис. 11.48, А). Они содержат, кроме стерильных гиф возле выводного отверстия (перифиз), короткие, плотно прижатые друг к другу гифы в центре, которые отчленяют мелкие одноядерные эллиптические спермации (так называемые пикноспоры, рис. 11.48, D). Последние, однако, прорастают на питательном растворе в короткую проростковую трубку, а при переносе на здоровый лист не могут заражать его. Их функция состоит в том, чтобы перенести свое ядро на так называемые воспринимающие гифы (рис. 11.48, A: eh). Это ответвления гаплоидного мицелия, которые выходят между клетками эпидермиса и перифизами и выдаются над поверхностью листа. Поперечных перегородок у них нет. Спермации сливаются только с воспринимающими гифами противоположных типов скрещивания (+ х -), что при смешанной (+, -) инфекции не составляет сложности. Кроме того, пикниды выделяют пахучую жидкость («нектар»), которую собирают на-
11.2. Бактерии, грибы, растения
181
Рис. 11.48. Uredomycetidae, Uredinales (А — по А. Н. R. Buller; В — по Н. Schenck; С — по V.H.Blackman; D — по R.H.Colley; Е— по А. Н. Christmann; F— по V. Н. Blackman):
А, В — Puccinia graminis: А— пикнида на барбарисе на продольном разрезе (140х), В — эцидий на барбарисе (140х); С— Gymnosporangium clavariaeforme, пикнида на листе боярышника, прорывающая эпидермис на верхней стороне (450х); D— Peridermium strobi, отшнуровывание одноядерных спермациев (1 200х); Е— Phragmidium speciosum, базальные клетки (Ь) с копуляционным мостиком (1 200х); F— Phragmidium violaceum, зрелая эцидиоспора (800х); а,, а2— двухъядерные эцидиоспоры; ак — цепочки дикариотичных эцидиоспор; е — эпидермис; eh — воспринимающие гифы; h — гаусторий; m— гаплоидный межклеточный мицелий; р— перифизы; рр— псевдоперидий; pz— палисадные клетки с гаусторием (h); s— спермации; z,, z2— промежуточные клетки
секомые, так что спермации могут быть перенесены ими и на другие листья, первоначально инфицированные только одним типом скрещивания.
Внедрившееся в воспринимающие гифы ядро мигрирует (в примыкающие к ним гифы) от клетки к клетке через перфорации в клеточной стенке до зачатка эцидия, где в базальных клетках возникает стадия дикариона. Второй путь состоит в том, что перенос ядра происходит при соматогамии: сливаются простые «+»- и «-»-гифы в ткани хозяина, если имела место смешанная
инфекция. Базальные клетки зачатка эцидия, ставшие теперь дикариотичными, разрастаются в бокаловидные эцидии ярко-оранжевой окраски, которые прорываются на нижней стороне листа. В них возникают многочисленные цепочки, состоящие чаще всего из правильно чередующихся между собой дикариотичных эцидиоспор и мелких, позже расплывающихся и исчезающих промежуточных клеток (рис. 11.48, Е: У ви-
дов некоторых родов (например, Puccinia) самые конечные споры каждой цепочки, а также все споры периферических цепочек
182 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
утрачивают репродуктивную функцию и склеиваются друг с другом в плотный покров (псевдоперидий, рис. 11.48, В: рр). За счет давления постоянно образующихся у основания цепочек новых спор (у Puccinia graminis свыше К) ООО в одном эцидии) псевдоперидий и эпидермис разрываются, так что споры (сначала угловатые из-за давления со всех сторон, потом округлые) могут разноситься ветром.
Со сменой ядерной фазы (с гаплоидной на дикариотичную) изменяется также круг растений-хозяев. Эцидиоспоры
прорастают только на хлебных и дикорастущих злаках. Их проростковая трубка внедряется через устьице в ткань этого второго хозяина и развивается в межклеточный (интерпеллюлярный), локально ограниченный мицелий. Он содержит дикарионы, но не имеет пряжек. Вскоре мицетий переходит к интенсивному образованию дикари-отнчных конидий, которые здесь называются уредоспоры, или «летние споры» (рис. 11.49, Е). Последние возникают поодиночке из вздувающихся конечных клеток несущих их гиф. Внешне это спороношение выгля
Рис. 11.49. Uredomycetidae, Uredinales (А, В — по A. De Вагу; С — по F.Tavel: D, F — по L. R.Tulasne;
Е — по Р. Sappin-Trouffy; G — по V. Н. Blackman):
А—D — Puccinia graminis: А— группа уредоспор (и) с одной толстостенной двухклеточной телейтоспорой (t) между ними, В — прорастающая уредсспора (А, В — ЗООх), С — стебель хлебного злака с телейтоспороношением (150х); D — двухклеточная телейтоспора, прорастающая двумя базидиями (ЗООх); Е— Phragmidium rubi, край уредоспороношения после разрыва отброшенного влево эпидермиса (е) растения-хозяина; споры разной степени зрелости (565х); F, G — телейтоспоры (500х): F — Uromyces appendicuiatus, одноклеточная телейтоспора, проросшая базидией (ядра не показаны), G — Phragmidium violaceum; внизу с парными ядрами, которые в обеих верхних клетках уже слились; к — проростковые поры; р — парафиза
11.2. Бактерии, грибы, растения I -| gg
дит как мелкие черточки ржавой окраски (отсюда название — ржавчинные грибы!), прорывающие эпидермис. Они обеспечивают распространение гриба летом (перенос на другие экземпляры того же хозяина). Каждая отдельная уредопустула образует очень много (на восприимчивом хозяине — миллионы) уредоспор. Они сразу же инфицируют другие растения злаков, на которых уже через 3 недели после заражения развиваются новые уредопустулы. Таким путем болезнь очень быстро распространяется, причем и на большие расстояния.
К осени дикариотичный мицелий в уре-допустулах или в других местах дает еще одну форму спороношения — двуклеточные телейтоспоры (рис. 11.49, A: t; С). В клетках этих спор пары ядер сливаются между собой (кариогамия). Телейтоспоры имеют толстые стенки, устойчивы к высыханию и холоду и служат для перезимовки в состоянии покоя. Следующей весной каждая из двух диплоидных клеток (= пробазидия) телейтоспоры претерпевает мейоз и прорастает через образуемую заранее пророс -тковую пору (рис. 11.49, D: к) в вытянутую базидию (D, F). Между четырьмя гаплоидными ядрами образуются поперечные перегородки, и из каждой из этих четырех клеток вырастает базидиоспора (мейоспора), в которую переходит ядро (D). Базидиоспоры отбрасываются и переносятся ветром на первого хозяина — барбарис. Этим замыкается цикл развития, в ходе которого имеются монокариотичные гаплоидные базидиоспоры (1) и гаплоидные спермации (2), дикариотичные эцидио- (3) и уредо-споры (4), а также сначала дикариотичные, а потом диплоидные телейтоспоры (5) — всего пять различных типов спор.
При так называемом Eu-типе никл развития полный и имеется смена поколений. Стадия гаплоидного гаметофита подавлена, а дикариотич-ная спорофитная фаза преобладает; эта тенденция еще усиливается за счет повторного образования уредоспор (рис. 11.50), реже — эцидиев (виды Cronartium). Наряду с видами, имеющими облигатную смену хозяев (разнохозяинность) и регулярную последовательность гаплоидной и дикариотичной фаз, есть такие виды, жизненный цикл которых проходит на одном и том же хозяине (однохозяинность). За счет подавления той или иной формы спороношения жизненный
Смена хозяина
Рис. 11.50. Uredomycetidae, Uredinales. Схема развития Puccinia graminis.
Светлые линии: гаплоидная фаза; двойные темные линии: дикариотическая фаза; жирные темные линии: диплоидная фаза. Мелкие клетки — барбарис, крупные — злак в качестве растений-хозяев (ср. рис. 11.48, 11.49). Аес— эцидий; В — базальная клетка; Рус — пикнида; R! — редукционное деление; So — соматогамия; Sp — оплодотворение спермациями; Т — телейтоспо-роношение (число спор, как у U, уменьшено); U — уредоспороношение
184 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
цикл также упрощается; могут выпадать: эцидиоспоры (Brachy-тип, например, Uromyces fa-bae); эцидио- и уредоспоры при наличии периода покоя у телейтоспор (Micro-тип, например, Tranzschelia fusca) или без него (Lepto-тип, например, Puccinia malvacearumY, уредоспоры (Opsis-тип, например Gymnosporangium juniperinum)', уре-до- и телейтоспоры (Endo-тип, например, Endo-phyllum sempervivi); эцидио-, телейто- и базидиоспоры (несовершенные ржавчинные грибы). При этом всегда сохраняется выраженная дикарио-фаза. В местностях с коротким вегетационным периодом такие укороченные типы развития, ограниченные одним хозяином, имеют преимущество.
Так же, как и возбудитель стеблевой ржавчины злаков, возбудитель широко распространенной ржавчины гороха (Uromyces pisi) относится к Eu-типу. Пикниды и эцидии встречаются на видах молочая (Euphorbia cyparissias и др.). Пораженные растения остаются неветвящими-ся, имеют желтоватые, короткие, толстые листья и чаще всего не цветут. Уредо- и (здесь одноклеточные) телейтоспоры развиваются на листьях гороха (Pisum sativum) и видах Lathyrus. На одном хозяине проходит все развитие возбудителя ржавчины фасоли (Uromyces phaseoli). Пикниды и эцидии встречаются летом на несколько гипертрофированных, горбатых листочках фасоли, на которых осенью образуются уредоспоры и, наконец, телейтоспоры. Так же ведут себя паразитирующие на листьях Rosaceae виды рода Phragmidium (см. рис. 1L49, Е). Эцидии здесь не окружены псевдоперидием, как у возбудителя стеблевой ржавчины злаков, а цепочки эцидиоспор прорывают, часто на обширной площади, эпидермис таким же образом, как и уредоспоры (тип Саеоша). Некое подобие плодовых тел представляют собой слизистые скопления телейтоспор, приподнимающиеся вверх, как палочка, у видов Gymnosporangium, и образования из цепочек телейтоспор у видов Cronartium.
Филогения. Ржавчинные грибы представляют собой эволюционно очень древнюю группу и встречаются уже в карбоне как паразиты на папоротниках. Образующие базидии Uredinales и образующие сумки Taphrinomycetidae, возможно, произошли от одной предковой группы. Их общий предок должен был иметь самостоятельно питающуюся дикариофазу. И у тафриномицетид паразитирующие на папоротниках представители свидетельствуют о древности этой группы. В мезозое Uredinales перешли на голосеменные, особенно хвойные, и, наконец, с верхнего мела — на покрытосеменные. С проникновением в области более холодного климата у них, возможно, и возникли покоящиеся пробазидии, т.е. телейтоспоры, с толстыми стенками.
Ржавчинные грибы — опасные возбудители болезней растений. Особенно значительны вызываемые ими потери урожая зерновых (в отдельных случаях до 25 %, однако в целом они заметно не превышают примерно 5%). По всему земному шару распространен вид Puccinia graminis, вызывающий стеблевую ржавчину, которую из-за темной окраски скопления телейтоспор называют также черной ржавчиной. Он поражает все наши виды хлебных злаков и многочисленные дикорастущие злаки.
В Средней Европе наносимый им ущерб не столь велик, как в более теплых странах, так как там развитие этого относительно теплолюбивого гриба протекает быстрее. В Германии особенно опасна желтая ржавчина (возбудитель — Р. striiformis со скоплениями уредоспор от светло-желтого до оранжевого цвета), поражение которой может носить характер эпифитотий, особенно пшеницы, а также ржи, ячменя и различных дикорастущих злаков. Промежуточный хозяин гриба неизвестен.
Р. coronata — возбудитель корончатой ржавчины овса и некоторых дикорастущих злаков — имеет промежуточным хозяином Rhamnus cathartica и еще некоторые виды, а Р. hordei, возбудитель карликовой ржавчины ячменя, образует эцидии на видах Ornithogalum.
Другие виды Puccinia встречаются на спарже, моркови, луке, крыжовнике и других культурных растениях; виды Uromyces — на горохе, фасоли и сахарной свекле; Gymnosporangium — на листьях груши («сетчатая ржавчина»). К другим семействам ржавчинных грибов относятся Melampsora Uni — однохозяинный возбудитель ржавчины льна, который разрушает его лубяные волокна; имеющие значение для лесного хозяйства грибы Melampsorella caryophylla-сеагит («ведьмины метлы» и рак пихты белой, уредо- и телейтоспоры на гвоздичных); Cronartium ribicola (его эцидиальная стадия поражает веймутову сосну и часто вызывает ее гибель; эцидии прорываются как крупные пузыревидные скопления из коры дерева; другой хозяин в жизненном цикле — смородина).
Надежда на то, что с разнохозяинными ржавчинными грибами можно бороться
11.2. Бактерии, грибы, растения I -| 85
путем устранения промежуточных хозяев, может оправдываться лишь очень ограниченно, поскольку у подавляющего большинства видов могут зимовать и уредо-споры или уже осенью заражаются молодые всходы озимых злаков, а также различные кормовые и декоративные злаки (в случае желтой ржавчины — также пырей); кроме того, уредоспоры могут заноситься из чрезвычайно отдаленных регионов (в Германию — даже через Альпы). Поскольку применение химических средств борьбы небезопасно, пытаются выводить устойчивые к ржавчине сорта культурных растений. Это имеет свои трудности, так как у каждого вида ржавчинных грибов насчитывается большое число физиологических рас, морфологически чаще всего не различимых, которые специализируются относительно различных сортов культурных растений, а за счет мутаций и рекомбинаций постоянно возникают все новые расы. Подобное расообразование у грибов-возбудителей болезней растений играет в фитопатологии очень большую роль, поэтому работа селекционеров в области выведения устойчивых сортов никогда не закончится.
Сюда примыкает порядок Septobasidiales, представители которого живут совместно со щитовками (симбиоз?).
На основании анализа ДНК и особенностей ультраструктуры скорее сюда, а не к головневым грибам, следует отнести грибы-возбудители головни пыльников гвоздичных. Виды рода Mi-crobotrium (Microbotryales) заполняют пыльники хозяина черно-фиолетовыми головневыми спорами вместо пыльцы. У женских растений раздельнополых смолевок (Silene alba и 5. dioica) при инфекции таким грибом стимулируется образование пыльников, в которых затем развиваются споры гриба.
3. ПОДКЛАСС: HYMENOMYCETIDAE
У представителей этого подкласса мицелий чаще всего образует плодовые тела — макроскопические структуры из гиф, на которых базидии располагаются палисадным слоем (гимением). Ход развития совпадает с тем, который имеет место у пластинчатых грибов; на гифах возле септ часто образуются пряжки (рис. 11.51).
В жизненном цикле, который сходен у всех образующих плодовые тела базидиомицетов (рис. 11.51), базидиоспоры прорастают в гаплоидные гифы, состоящие из одноядерных клеток. После слияния гиф (со-матогамии) тотчас же образуется дикари-отичный мицелий, часто несущий пряжки (см. рис. 4.6). Слияние гиф, партнеров скрещивания, контролируется генетически.
Биполярная и теграполярная несовместимость. У многих базидиомицетов пол oi 1ределяется не одним геном (пара аллелей «+» и «-»), а двумя не-сцепленными генами. Аллели одного фактора обозначаются Д и А2, другого — В\ и В2. Диплоидное ядро зиготы содержит, таким образом, At/A2 и (!) BJB2. После редукционного деления одно плодовое тело содержит базидиоспоры 4 типов: Ах В,, А2В2, AiB2 или Л2В|. Только скрещивание с различными А- и В-факторами может продолжить жизненный цикл, дав начало дикариотичному мицелию (часто с пряжками у септ гиф); так, сочетание А, В, х х А2В2 совместимо, но AtB} х или AtBt х Л,В2, напротив, несовместимы (рис. 11.55).
Здесь говорят о тетраполярной гомогенной несовместимости. С ней связан тот факт, что при встрече гаплоидных монокариотичных гиф, которые прорастают из спор одного плодового тела, фактически только в 25 % случаев возникает мицелий с пряжками. Генетической рекомбинации мицелиев разного географического происхождения способствует явление множественных аллелей, так как в ней могут участвовать и другие аллели, например А3В3 и А3Ва. Поэтому скрещивания с факторами А\/А2 или BJВ, х Д/Д или В3/Д) (например, ДВ, х А3В3) на 100% совместимы. Механизм биполярной или тетраполярной несовместимости, который преимущественно связан с наличием множественных аллелей, способствует таким путем перекрестному оплодотворению (аутбридингу), причем этот эффект при тетраполярной несовместимости сильнее (25%: 100%), чем при биполярной (50%: 100%). Если у аскомицетов встречается только биполярная несовместимость, то у базидиомицетов отмечается все больше случаев тет-раполярного механизма.
Пряжки гомологичны крючку аскомицетов. Они принимают в себя временно одно из двух дочерних ядер, возникших при делении клетки, с тем, чтобы оно потом опять перешло в ту из дочерних клеток, которая расположена дальше от конца гифы (рис. 11.51, 5 —7).
Процесс образования пряжки повторяется при каждом клеточном делении,
186 | ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Рис. 11.51. Hymenomycetidae (по R. Harder):
А — развитие пряжкового мицелия: 1 — генотипически различные (+, -) споры, 2 — прорастание спор в мицелий без пряжек, 3—4 — копуляция, 5—7 — образование первой пряжки, 8 — образование следующей пряжки, 9 — пряжковый мицелий с дикариотичной материнской клеткой базидии (а), молодой базидией с диплоидным ядром (Ь) и зрелой базидией (с) со спорами разного типа скрещивания (с); В - схема развития шляпочного гриба. Светлыми линиями показана гаплоидная фаза; тонкими темными — дикариотичная фаза; жирными темными — диплоидная фаза. Пряжки не изображены, базидии изображены непропорционально крупными по отношению к шляпке
Рис. 11.52. Hymenomycetidae, Agaricales. Образование пряжек и развитие базидии (1 — 7 — по Н. Kniep; 8, 9 — по W. Ruhland):
1—7 — Oudemansiella mucida (620х): 1 — начало образования пряжки в двухъядерной конечной клетке, 2 — клеточное ядро отходит в пряжку, 3 — одновременное ядерное деление, 4 — образование стенки внутри пряжки и рядом с ней, материнская клетка базидии отделяется от несущей ее клетки (клетки-ножки), 5 — слияние пряжки с клеткой-ножкой, 6 — два гаплоидных ядра материнской клетки базидии сливаются в диплоидное ядро, 7 — молодая базидия с четырьмя ядрами будущих базидиоспор, образовавшимися в результате мейоза (вверху четыре зачатка стеригм); 8, 9— Psathyrella (1 500х): 8 — базидия с 4 ядрами перед их переходом в молодые базидиоспоры, расположенные на вершине базидии, 9 — переход клеточного ядра через стеригму в базидиоспору
11.2. Бактерии, грибы, растения
так что возникает обильно разветвленный «пряжковый мицелий», содержащий в каждой клетке дикарионы и имеющий пряжки у каждой поперечной перегородки (рис. 11.51, 8, 9). Устанавливающаяся таким образом дикариотичная фаза (в отличие от аскомицетов) способна к самостоятельному существованию. Такой мицелий может годами разрастаться в почве, древесине и других субстратах и претерпевать бесчисленные дальнейшие клеточные деления с конъюгированным делением ядер дикариона, пока под влиянием еще не известных нам условий не разовьются плодовые тела. В противоположность аскоми-цетам (см. рис. 11.32) плодовое тело бази-диомицетов состоит исключительно из дикариотичных гиф (рис. 11.51). Его возникновение, таким образом, не связано, как у аскомицетов, каждый раз с новым актом плазмогамии при половом процессе. Последний у базидиомицетов происходит только один раз для возникновения многолетнего дикариотичного мицелия, способного,
в отличие от аскомицетов, из года в год давать плодовые тела. На поверхности или внутри плодовых тел (чаще всего на их нижней стороне) булавовидно вздутые конечные клетки дикариотичных гиф располагаются в виде палисадного слоя — гимения (рис. 11.51, В). Только в этих конечных клетках — молодых базидиях — два ядра дикариона сливаются между собой (кариогамия, рис. 11.52,6), после чего сразу же происходит мейоз (с определением типа скрещивания) и образуются четыре гаплоидные мейоспоры — базидиоспоры. Развивающиеся из базидиоспор гаплоидные мицелии соответствуют гаметофитам; возникающий из них в результате соматогам-ной копуляции дикариотичный мицелий может рассматриваться как дикариотичный спорофит.
При развитии базидиоспор концы стеригм вздуваются, образуя споровые мешочки (рис. 11.52, 8). Из 4 гаплоидных ядер каждое протискивается через свою стеригму (рис. 11.52, 9), и теперь в каждом споро-
Рис. 11.53. Hymenomycetidae (по F. Oberwinkler):
А, В — Auricularia auricula-judae: А— плодовое тело (натуральная величина), В — продольный разрез через гимений (400х); С, D— Tremella mesenterica: С— плодовое тело (натуральная величина), D — поперечный разрез через гимений (400х)
188 I ГЛАВА 11 СИСТЕМАТИКА И ФИЛОГЕНИЯ
А
А
Рис. 11.54. Hymenomycetidae (А—К— по А. Н. R. Buller; L — по К. Wells):
A—L— Dacrymycetales: Calocera cornea (900х), отбрасывание базидиоспоры: А, В — вытягивание стеригмы, С—F — отшнуровывание базидиоспоры (продолжительность примерно 40 мин), G, Н — образование капельки жидкости (t) у места прикрепления споры (длительность примерно 10с), I — отбрасывание споры вместе с капелькой (t), J, К — опадание стеригмы; L — Schi-zophyllales, Schizophyllum commune (1 500x), место прикрепления базидиоспоры к стеригме; bw— стенка базидии; sw— стенка споры
вом мешочке образуется одна спора. Однако почти всегда стенка споры рано сливается со стенкой мешочка, так что двойная природа споровой оболочки становится незаметной; стенка мешочка образует при этом периспорий. Таким образом,
мейоспоры только внешне кажутся образующимися экзогенно. Они преимущественно эллипсоидные и уплощены с одной стороны. Споры отбрасываются лишь на короткое расстояние за счет того, что находящаяся в состоянии сильного тургора базидия выдавливает из вершины стеригмы каплю, которая увлекает с собой спору (рис. 11.53, 11.54).
К рассматриваемой группе относится большинство организмов, известных в обиходе как грибы (см. рис. 11.57; 11.59; 11.61). Их мицелий часто можно культивировать, однако в культуре плодовые тела образуются редко. Многие виды часто имеют быстрорастущие плодовые тела, особенно в конце лета и осенью. Можно различать гимениальные и гастроидные плодовые тела.
У гимениальных плодовых тел гимено-мицетов (рис. 11.55— 11.57) в более узком смысле слова гимений в ходе развития становится открытым, а базидиоспоры активно отбрасываются от базидий. Гимений содержит базидии и в отдельных случаях цистилы, расположенные в виде палисадного слоя (рис. 11.56, 11.59, С). Гименофор представляет собой макроскопическую структуру, увеличивающую спороносную поверхность. Морфология гимениальных плодовых тел весьма разнообразна — от корко
is 1w
Рис. 11.55. Hymenomycetidae, Pleurotus ostreatus.
Обеспечение перекрестного оплодотворения путем гомогенной несовместимости (тетраполярный механизм, множественные аллели). Плодовые тела с несколько различающимися по окраске шляпками — штаммы одного и того же вида разного географического происхождения. А. В — факторы скрещивания используемых для скрещивания монокариотичных мицелиев; — скрещивание дает ди-кариотичный пряжковый мицелий
11.2. Бактерии, грибы, растения | -|g
видных или булавовидных до сильно разветвленных, консолевидных или дифференцированных на шляпку и ножку («шляпочные грибы»). Столь же разнообразно и строение гименофора: он может быть гладким или складчатым, иметь вид пчелиных сот, пор, трубочек, шипов или пластинок. Существуют почти все возможные комбинации форм плодовых тел и типов гименофора, причем в ходе эволюции достигалось максимально возможное число спор при наиболее благоприятном пути их распространения. Плодовое тело гимениального типа развивается различным образом. У гимнокарпного плодового тела гимений (гименофор) с самого начала закладывается на открытых внешних поверхностях. В случае гемиангиокарпного развития (рис. 11.59, А, В) гимений образуется сначала внутри еще молодых плодовых тел. Однако при вытягивании ножки и раскрытии шляпки исходная обвертка разрывается. Ее остатки часто сохраняются и на зрелом плодовом теле как части общего и/или частного покрывала. Частное покрывало (velum partiale — vp) образует в таком случае кольцо (т) или еще недолгое время соединяет шляпку и ножку в виде паутинистой вуали (cortina) либо же полностью исчезает. Общее покрывало (velum universale — vu) оставляет обвертки в виде влагалища у основания ножки (volva — v; на-
Рис. 11.56. Hymenomycetidae. Формы цистид (Poriales, 750х) (по F. Oberwinkler):
A — Tubuliciunr, В — Vara ria; С — Peniophora; D — Tubulicrinis', E — Stereum; F — Hyphoderma
пример, у бледной поганки) и/или белые лоскутья и клочья на поверхности шляпки (например, у Amanita muscaria — мухомора).
Псевдоангиокарпное развитие первоначально сходно с гимнокарпным, однако край шляпки сильно завертывается внутрь,
Рис. 11.57. Hymenomycetidae. Различные гимениальные плодовые тела (А— по Е. Schild; В — по Е Oberwinkler; С— по G.Bresadola; D— по Н. Schenck; Е— по R. Harder):
А— Flamaria botrytis (1/2х); В — Stereum hirsutum (12х); С — Dendropolyporus umbellatus (1/Зх); D — Hydnum repandum (1 /2х); Е — Phellinus igniarius, многолетнее плодовое тело с годичными зонами (1 /2х)
190 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Рис. 11.58. Hymenomycetidae, Poriales, Tra-metes versicolor (по E. J. H. Corner).
Разрез через край растущего плодового тела с двумя зачатками трубочек гименофора. Толстостенные: скелетные гифы; тонкостенные: гифы, образующие базидии; полностью черные: связующие гифы (150х)
так что его гифы сплетаются с гифами коры ножки.
Отбрасывание спор. Споры отбрасываются от базидий на небольшое расстояние. Оно составляет у грибов с трубчатым гименофором примерно половину диаметра трубочки (у грибов с пластинчатым гименофором — половину расстояния между пластинками). Полет быстро превращается в падение по вертикали за пределы трубочки или пространства между пластин
ками, где споры подхватываются движением воздуха. Собственно распространение спор осуществляется воздушными потоками. Если положить зрелую шляпку пластинчатого гриба пластинками вниз на лист бумаги, то уже через несколько часов выпадающие споры образуют на нем четкий отпечаток пластинок. Зрелое плодовое тело шампиньона полевого (Agaricus campestris) диаметром 10 см имеет поверхность гимения 1 200 см2, которая в совокупности производит около 1,2 млрд спор; за один час отбрасывается приблизительно 40 млн спор.
В гимении наряду со зрелыми находятся и молодые базидии, а также стерильные гифы с дегенерировавшими дикарионами и более крупные, также стерильные конечные гифы разной формы — цистиды (см. рис. 11.56). Последние выполняют функцию защиты и выделения (например, у Poriales) или, возможно, препятствуют слипанию пластинок (например, у Coprinus). Они важны для систематического подразделения и определения видов. Плектенхима плодового тела — трама (рис. 11.58) — также дифференцируется: например, толстостенные «скелетные гифы» служат для прочности; толстостенные разветвленные «связующие гифы» схватывают другие гифы; тонкостенные «генеративные гифы» образуют базидии.
Рис. 11.59. Hymenomycetidae, Agaricales. Схематические продольные разрезы через плодовые тела с покрывалами (А, В — по Е. Fischer):
А — частное покрывало (velum partiale, vp); В — общее покрывало (velum universale, vu) и частное покрывало, слева — у молодого, справа — у зрелого плодового тела; С— разрез через гимений Ну-pholoma (1 000х); Ь — базидия; Ьо — молодая базидия; с — цистида; f — остатки общего покрывала на шляпке; m — кольцо, остаток частного покрывала; s — базидиоспора; st — стеригма; sh — субгимений; t — трама; v — обвертка (volva), остаток общего покрывала у основания ножки
11.2. Бактерии, грибы, растения | ig-|
Гастроидные плодовые тела «гастероми-цетов» (рис. 11.61) образуют базидии внутри себя; первоначально закладывающийся на ранних стадиях развития гимений разрушается ко времени созревания спор или вообще не образуется. Развитие плодовых тел ангиокарпное или гемиангиокарпное. Плодовые тела могут быть закрытыми, подразделенными или неподразделенными внутри на отдельные камеры, или же споровая масса выносится вверх на способном к растяжению элементе (рецептаку-ле) из оболочки (перидия) плодового тела (рис. 11.61, А—С). Споры не отбрасываются от базидий. Распространение спор осуществляется ветром, а в отдельных случаях также насекомыми и млекопитающими.
Между гимениальными и гастроидны-ми плодовыми телами существуют переходы, так что большинство «гастеромице-тов» имеет тесную родственную связь с типичными гименомицетами, поэтому сейчас
Оксидаза, О?
Синее окрашивание
Рис. 11.60. Hymenomycetidae, Boletales (по
W. Steglich).
Вариегатовая кислота (производное пульвино-вой кислоты) из Suillus variegates: вверху — нео-кисленная (желтая) форма, внизу — окисленный (синий) анион; R = ОН
Рис. 11.61. Hymenomycetidae, «Gasteromycetes» (А— по J.E. Lange; В, D, G— по J. Poelt, H.Jahn и С. Caspari; С — по V. Fayod; Е — no Е. Gramberg; F — Е. Michael и В. Hennig):
А—С — Phallales: А — Phallus impudicus, зрелое плодовое тело с каплями оплывающей глебы на шляпке и молодое плодовое тело на продольном срезе (1/2х), В — Anthurus archer! (1/2х), С— Clathrus ruber (1/2х); D — Boletales (ранее относили к Sclerodermatales), Scleroderma citrinum, на срезе видна глеба с белыми бесплодными участками в виде прожилок (1/2х); Е, F — Nidulariales: Е — Cyathus striatus (натуральная величина), F — Sphaerobolus stellatus, справа — отбрасывание эндоперидия (Зх); G — Geastrales, Geastrum quadnfidum (1/2х)
192 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
его помешают в общий подкласс — Hymenomycetidae.
Сначала выстраиваются преимущественно порядки, представители которых имеют базидии с поперечными или продольными перегородками (фрагмобазидии). Базидиоспоры прорастают конидиями, вторичными спорами или дрожжевидными клетками (см. рис. 11.45). Вторичные споры — однократно отшнуровывающиеся структуры, в которые переходит единственное ядро базидиоспоры.
Морфологическая взаимосвязь между Ure-domycetidae (Uredinales, Septobasidiales) и следующих за ними порядков (Tremellales, Auricula-riales) становится очевидной благодаря наличию переходных форм. Так, представители некоторых семейств Uredinales (например, Chrysomyxac эпи-диями на Picea и уредо- и телейтоспорами на Ericales) вместо толстостенных телейтоспор развивают базидии типа Auricularia. Uredinella coccidiophaga обитает аналогично Septobasidiales на щитовках, однако имеет пробазидии, сходные с телейтоспорами. Patouillardina (Tremellales) с беспорядочно септированными базидиями является промежуточным звеном между Auricula-riales и Tremellales. У Bourdotia (Tremellales) продольные перегородки в базидиях не доходят до их основания; такие формы представляют собой переход от фрагмобазидии к холобазидии. Если у Uredinales базидии лишь в отдельных случаях образуются на своего рода примитивных плодовых телах, то Auriculariales и Tremellales, за исключением немногих форм без плодовых тел, показывают усложняющийся ряд разнообразнейших типов плодовых тел: плоские налеты, булавовидные, состоящие из ножки и головки, консолевидные, состоящие из ножки и шляпки. Сходные формы плодовых тел, возникавшие в ходе параллельного эволюционного развития, будут нам встречаться неоднократно и среди других групп базидиомицетов (см. рис. 11.57). Поверхность гимения может увеличиваться за счет образования складок, структур типа пчелиных сот или зубчиков. В пределах одного вида Auricularia auricula-judae можно наблюдать переходы от гладкой спороносной поверхности через складчатую к поверхности, имеющей облик сот или почти трубочек (= тенденция к выработке гименофора). У Auriculariales (частично) и Tremellales (закономерно) впервые в пределах упомянутых эволюционных рядов встречается пряжковый мицелий; он возникает после соматогамии и отличает дикариофазу.
1. Порядок: Tremellales, дрожалковые. Фрагмобазидии этих грибов разделены продольными
перегородками, расположенными крест-накрест (см. рис. 11.53, D; 11.44, В). Они обитают преимущественно на отмершей древесине, изредка на других субстратах или же паразитируют на других грибах. У наиболее примитивных представителей плодовые тела отсутствуют. Виды рода Tremella образуют студенистые плодовые тела от мозговидной до листовидной формы, имеющие желтую, буроватую или черную окраску (см. рис. 11.53, С). Студенистая «ткань» плодовых тел служит для накапливания влаги. Шляпковидное плодовое тело Pseudohydnum, имеющее боковую ножку, несет на нижней стороне шипы, которые покрыты гимением. Exidiopsis — ср. с рис. 11.45.
2. Порядок: Dacrymycetales. Эти грибы отличаются 2-споровой холобазидией в форме камертона (Dacrymyces, см. рис. 11.44, D), чаще всего септированными базидиоспорами, прорастающими конидиями, и тем, что они обитают как сапротро-фы на древесине. Плодовые тела имеют вид простых корочек или пустул либо представляют собой образования от состоящих из ножки и головки, бокаловидных до разветвленно-булавовидных. Таким образом, здесь мы имеем набор вариантов, конвергентно сходных с имеющимися в других эволюционных линиях. Плодовые тела, окрашенные каротиноидами, различны — чаще всего от мягких до вязко-студенистых. Порядок показывает определенные общие черты с уже обсуждавшимися паразитическими Exobasidiales. В частности, это касается прорастания базидиоспор, имеющих поперечные перегородки, крючковидно изогнутыми конидиями — эта особенность не встречается у прочих базидиомицетов, имеющих холобазидии. С другой стороны, здесь имеется также родство с Tulasnellales (С).
3. Порядок: Auriculariales (уховидные грибы). С представителями Uredomycetidae виды этого порядка сближает наличие фрагмобазидии с поперечными перегородками. Существование родов, виды которых являются паразитами без плодовых тел (например, Herpobasidium, Helicobasidi-ит), указывает на возможную родственную связь с Uredinales. Иудино ухо (Auricularia auricula-judae, см. рис. 11.53, А) — вид со студенистыми темно-коричневыми плодовыми телами в форме ушной раковины, вырастающими из стволов бузины, может рассматриваться как эволюционно продвинутый. На своей вогнутой поверхности, которая может варьировать от гладкой до складчатой, они несут гимений. Базидии, как у всех Auriculariales. разделены поперечными перегородками на расположенные друг над другом 4 клетки, от каждой из которых вырастает вбок стеригма с мейоспорой (см. рис. 11.53, В; 11.44, А). У некоторых Auriculariales базидия имеет в основании шаровидное вздутие. В этой «пробазидии», которая некоторое время представляет собой
11.2. Бактерии, грибы, растения | 193
конец дикариотичной гифы, происходит слияние ядер. После мейоза из нее вырастает собственно базидия. Базидиоспоры имеют ко времени прорастания (например, у Auricularia) несколько поперечных перегородок и образуют от каждой возникшей таким образом клетки множество конидий. К этому порядку примыкают Tulasnellales (см. рис. 11.44, С).
Последующие порядки базидиомицетов, имеющие исключительно холобазидии, раньше объединялись в группу Holobasi-diomycetidae. Споры всегда прорастают гифами. Парентосомы у долипоровых септ пронизаны порами и образуют, таким образом, ситовидную структуру. Форма и размер холобазидии разнообразны (см. рис. 11.44). Наряду с широко распространенной булавовидной формой (F) встречаются, например, сильновыпуклые (Е), возникающие сбоку на несущей гифе плевробазидии (Н), многократно прорастающие друг из друга репетобазидии (J). Имеется большое разнообразие форм плодовых тел и характера поверхности слоя, несущего гимений (гименофора). Как и у рассмотренных ранее порядков (в том числе характеризующихся наличием фрагмобазидии), здесь также в разных эволюционных линиях происходит выработка сходных типов плодовых тел, так что у этих грибов, как и у водорослей, можно видеть разные типы дифференциации. Внешняя форма плодовых тел здесь часто служит критерием для различения родов и семейств того или иного порядка.
По внешней форме плодовых тел раньше различали искусственные (т.е. не отражающие родственных связей) группы афиллофоровых (Poriales = Aphyllophorales в широком смысле), агариковых, или пластинчатых (Agaricales в широком смысле), и гастеромицетов (Gasteromyce-tales). В настоящее время при систематизации этих организмов пытаются отражать родственные связи, а именно исходить из сходства как можно большего числа признаков разного рода: например, таких микроскопических структур, как строение гимения и плектенхимы плодового тела (= трамы), состава метаболитов, ДНК и т.д. При группировке таксонов более низкого ранга в таксоны более высокого ранга (например, порядки) важную роль играют переходные формы, которые, с одной стороны, имеют признаки разных таксонов, а с другой — показывают переходы признаков. Из-за наличия плавных переходов между вышеназванными формами пло
довых тел афиллофоровых, агариковых и гастеромицетов системы, построенные на особенностях плодовых тел, оказались несостоятельными.
4. Порядок: Cantharellales. Наиболее известный представитель порядка — лисичка настоящая (Cantharellus cibarius, Cantharellaceae), ценный съедобный гриб. Его состоящее из ножки и шляпки плодовое тело имеет гименофор из толстых, часто анастомозирующих между собой складок, и гимений с удлиненными стихически-ми базидиями, т.е. в них оси веретен ядерного деления располагаются по продольной оси базидии. В качестве пигментов имеются и каротиноиды, которые вообще редки у высших холо-базидиомииетов. Некоторые виды имеют почти гладкий гименофор, как, в частности, виды рода Clavulina. имеющие плодовые тела в форме оленьих рогов с булавовидными расширениями на концах. Ежовик желтый (Hydnum repandum. см. рис. 11.57, D) отличается состоящими из шляпки и ножки плодовыми телами с шиповидным гименофором.
5. Группа порядков: Gomphanae. Выделение этой группы порядков основано на данных анализа ДНК. Морфологически они весьма гетеро-генны: с одной стороны, сюда относятся виды с плодовыми телами от булавовидных до имеющих форму оленьих рогов (Gomphales с родами Gomphus, Ramana, см. рис. 11.57, А), а с другой стороны — также виды с гастроидными плодовыми телами (Geastrales, Pliallales, см. рис. 11.61, А— С, G).
5.1. Geastrales. Плодовые тела земляных звездочек (Geastrum, см. рис. 11.61, G) имеют характерный облик: экзоперидий разрывается на лопасти, которые отгибаются вниз и освобождают бумагообразный эндоперидий с заключенной в нем глебой (G). Гифы имеют возле септ пряжки (в отличие от Lycoperdales — см. ниже). Базидии выпуклые; на них на коротких стеригмах образуются споры (часто их больше 4).
5.2. Phallales. Плодовые тела на ранних стадиях развития окружены слизистой оболочкой, которая позже разрывается и становится сходной с вольвой (velum universale) некоторых пластинчатых шляпочных грибов. Первоначально заключенная внутри оболочки глеба (см. порядок 10.4) подразделяется на камеры и при созревании образует слизистую массу с сильным неприятным запахом, содержащую базидиоспоры. Эта масса у многих представителей порядка выносится вверх способным к растяжению осевым элементом (рецептакулом). Распространение спор осуществляется насекомыми, которые привлекаются запахом глебы, а у некоторых видов еще и яркой окраской рецептакула. Броские формы (так называемые «грибы-цветы») распространены особенно в тропиках. Обитающая
194 | ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
в умеренной зоне веселка (Phallus impudicus, см. рис. 11.61, А) внешне несколько напоминает сморчок, относящийся к аскомицетам, но имеет совершенно другой тип развития и структуру (конвергентное сходство). Молодое плодовое тело, окруженное мягкой белой оболочкой, в Западной Европе называют «ведьмино яйцо». Вольва состоит из внешнего и кожистого внутреннего перидия и студенистого промежуточного слоя. Развитие плодового тела можно наблюдать на извлеченном из земли снаружи мягком, а внутри твердом «ведьмином яйце»: рецептакул — уже заложившийся внутри молодого плодового тела— вытягивается в течение нескольких часов до длины примерно в 15 см, разрывая при этом оболочку, которая остается в виде бокала, вынося наверх «шляпку». Последняя закладывается уже в «ведьмином яйце» в виде окружающего ножку слоя, имеющего форму колокола; снаружи она примыкает к вольве. «Шляпка» состоит из кожистого несущего слоя, разделенного на камеры, и расположенной на нем чернозеленоватой слизистой споровой массы, имеющей неприятный запах. Эта «шляпка» согласно одной точке зрения в целом соответствует глебе, согласно другой — она отчасти представляет собой вырост рецептакула (несущий слой), а отчасти соответствует глебе (споровая масса). Споровая масса расплывается и капает с ячеистой поверхности «шляпки». Мухи (синие и зеленые падальные) распространяют споры на поверхности своего тела. У видов тропического рода Dictyophora от вершины ножки отходит ажурная сеточка, которая сначала находится между «шляпкой» и ножкой, а потом конусообразно спускается вниз (почему немцы называют этот гриб «дама с вуалью»).
Рещеточник (Clathrus, см. рис. 11.61, С) и цве-тохвостник (Anthurus. В) имеют сходный тип развития плодовых тел с тем только различием, что у первого рода окрашенный в красный цвет рецептакул имеет форму сетки, а у второго на вершине расходится несколькими лопастями.
На основании строения и развития плодовых тел и отсутствия гимения оба рассмотренных выше порядка могут рассматриваться как весьма изолированные среди Hymenomycetidae.
6. Порядок: Hymenochaetales. Гифы окрашены коричневыми пигментами (стирилпиронами), локализованными в мембранах. Пряжки у септ гиф отсутствуют, а в гимении обращают на себя внимание чаще всего заостренные, толстостенные, окрашенные в коричневый цвет цистиды (= щетинки). Споры гладкостенные.
Подразделение на роды отражает различия в формах плодовых тел. Гладкие корочки, которые частично в виде ребрышек отстают от субстрата, представлены в роде Hymenochaete. Мно
голетние консолевидные плодовые тела со слоистым трубчатым гименофором отличают род Phellinus. По чередующимся слоям трубочек и трамы на поперечном срезе можно судить о возрасте плодового тела (см. рис. 11.57, Е). Трубочки с узкими выходными отверстиями в виде пор показывают у таких плодовых тел (как и в других порядках) положительное геотропическое направление роста с высочайшей точностью, что обеспечивает в подобном случае выход спор. Coltricia имеет жесткое, хорошо сохраняющееся плодовое тело с трубчатым гименофором, состоящее из шляпки и ножки.
7. Порядок: Russulalcs. Грибы, известные как сыроежки и млечники, однозначно отличаются часто амилоидной (см. объяснение при описании Leotianae) орнаментацией спор, а также чаще всего группами округлых клеток между вы-тянутыми гифами и органами выделения, проводящими терпеноиды (млечные гифы, синеющие в ванилин-Н2БО4 цистиды). Сыроежки (Russula) с их хрупкими, легко разламывающимися пластинками имеют в качестве пигментов азотсодержащие, растворимые в воде руссугпе-ридины. Плодовые тела млечников (Lactarius) выделяют при повреждении млечный сок, иногда несколько окрашенный. В пределах порядка наблюдаются плавные переходы от форм с плодовыми телами, дифференцированными на шляпку и ножку, с развитым гимением к формам с подземными гастроидными плодовыми телами. Сюда же примыкают растущие на гнилой древесине виды Stereum (Stereaceae). У них многослойные распростертые плодовые тела частично отстают от субстрата, а гименофор гладкий (см. рис. 11.57, В). Сходное изменение трактовки родственных связей необходимо принять также для Heterobasidion (см. Poriales) и Albatrellus (овечьего трутовика).
8. Группа порядков: Aphyllophoranae. Представители группы образуют трубчатый гименофор.
8.1. Polyporales. Жесткомясистые, состоящие из шляпки и ножки плодовые тела видов рода Polyporus, давшего название порядку, могут обрастать своим исключительно трубчатым гименофором посторонние предметы (например, ветки, соломины злаков). В культурах были получены плодовые тела также из монокариотичного мицелия, в данном случае гаплоидного. У Dendro-polyporus (см. рис. 11.57, В) возникает несколько шляпок на общей сильноразветвлснной ножке.
8.2. Poriales. В настоящее время эта группа пока еще выделяется в порядок. Большинство видов способно тем или иным образом обрастать своими плодовыми телами посторонние предметы. Гимений располагается свободно на плодовом теле, он образуется рано, по гимнокарпному типу и по мере увеличения плодового
11.2. Бактерии, грибы, растения i -j gjj
тела дает все новый и новый прирост. Виды типового рода Porta (Poriaceae) образуют плоские, прижатые к субстрату корки, которые несут на верхней стороне трубчатый гименофор. У Corti-cium (Corticiaceae) плодовые тела также прижатые, с гладким гименофором без особого пространственного членения. Часто встречающиеся на пнях деревьев виды рода Trametes (например, Т. versicolor) имеют отходящие от субстрата кожисто-корковые плодовые тела с трубчатым гименофором. Такой же гименофор образуют консолевидные многолетние плодовые тела настоящего трутовика (Femes fomentarius). который в Германии особенно часто встречается как паразит на буке. У березового трутовика (березовой губки, Piptoporus betulinus) внутренняя часть однолетнего плодового тела относительно мягкая. У других трутовиков гименофор переходного вида — от лабиринтовидного (Daedalea quercina, дубовая губка) до сходного с пластинчатым (Daedaleopsis tricolor). Различные плодовые тела и типы гименофора образуются также в пределах рода Gloephyllum; плодовые тела заборного гриба (G. septarium) растут в виде отстающих от субстрата корок и имеют гименофор типа пластинчатого, тогда как пахучий трутовик (G.odoratum) с запахом, напоминающим запах фенхеля, с нижней стороны консолевидного плодового тела имеет трубчатый гименофор. Оранжево-бурая окраска плодовых тел обоих видов обусловлена трамети-ном. Вид Heterobasidion annosus — корневая губка — вызывает гниль (особая форма белой гнили, см. бокс 11.4, с. 124) и по признакам микроструктуры и характеристикам ДНК должен быть отнесен скорее к Russulales.
9. Порядок: Thelcphorales. Представители порядка отличаются от Poriales и Hymenochaetales пигментированными спорами с бугорчатой поверхностью, имеющими чаще всего попарно расположенные бородавки или шипы. Эти грибы накапливают телефоровую кислоту в форме пигментированных отложений на гифах трамы. В пределах порядка существуют переходы от плоских, прилегающих к субстрату плодовых тел с гладким или шиповатым гименофором (Tomentella) к булавовидно-разветвленным или шляпковидным плодовым телам (Thelephora). Плодовые тела, состоящие из ножки и шляпки с шиповатым гименофором, отличают, в частности, род Sarcodon (например, саркодон черепитчатый, или ежовик пестрый, 5. imbricatus), а трубчатый гименофор — род Boletopsis. Ежовик желтый (Hydnum repandum, Cantharellalcs), имея внешне сходное плодовое тело с шиповатым гименофором, однако отличающийся совершенно другими микроскопическими и химическими признаками, не может быть отнесен к этому порядку (конвергенция!).
10. Группа порядков: Agaricanae (включая Lycoperdanae). Представители этой группы — прежде всего грибы, имеющие недолговечные плодовые тела с четким разделением на ножку и шляпку, не способные обрастать посторонние предметы и образующие с нижней стороны пластинчатый гименофор. Четкие родственные связи таких типичных пластинчатых грибов с родами, отличающимися другой формой плодовых тел, в частности афиллофороидного или гасте-роидного типа, привели к новой концепции порядков в данном случае.
10.1. Порядок: Schizophyllales. Щелелистник (Schizophyllum) — объект, часто используемый в экспериментальных исследованиях (см. рис. 11.54, L); гименофор здесь в форме жестких, продольно расщепленных пластинок, деформирующихся при изменении влажности. Плодовые тела могут рассматриваться как комплексные, составленные из простых бокаловидных плодовых тел (какие сохранились еще у Cyphellopsis).
10.2. Порядок: Agaricales. Как и Poriales, этот порядок выделен временно. Его окончательный состав (в Средней Европе около 2000 видов) определяется соответствием набора признаков типовому роду Agaricus (шампиньон). У многих представителей этой группы гимений образуется внутри плодовых тел в схизогенных полостях, но при их раскрытии он становится свободным. Однако наряду с гемиангиокарпным развитием встречаются также псевдоангиокарпное и гимнокарпное развитие. Плодовые тела недолговечные. У представителей некоторых родов (например, навозник, или чернильный гриб, Coprinus) они разрушаются уже через несколько часов после возникновения. Заложение гименофора осуществляется не постепенно и не от центра к периферии (как у Aphyllophoranae), а единовременно. Гименофор чаще всего состоит из радиально расходящихся вертикальных пластин, у зрелых плодовых тел выстилающих нижнюю сторону шляпки, расположенной на ножке. Основное сплетение гиф пластинок — трама — несет на внешней стороне слой, состоящий из округлых клеток — субгимений, который покрывается гимением. Гимений состоит главным образом из базидий разной степени зрелости и часто также из цистид (см. рис. 11.59, С). Веретено деления ядра располагается перпендикулярно продольной оси базидий (хиастобазидии).
К этому порядку относятся как съедобные грибы, подлежащие охране, так и опасные ядовитые грибы. В широком масштабе культивируется шампиньон двуспоровый (Agaricus bisporus). Две (не 4!) споры на его базидиях содержат по два совместимых («+» и «-») ядра; таким образом здесь в отличие от большинства других Agaricales не требуется половой процесс для на
196 | ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
чала нового жизненного никла, и при культивировании этот гриб можно размножать посредством дикариотичного посевного материала. Опасные и коварные ядовитые грибы (симптомы отравления появляются только по прошествии от 6 до 24 часов после употребления в пищу!) — различные виды рода Amanita, особенно бледная и весенняя поганки (соответственно A. phalloides и A. virosa). Они содержат ядовитые циклические пептиды (ама- и фаллотоксины). Их плодовые тела имеют свободно закругленные, т.е. не приросшие к ножке (в отличие от шампиньона) пластинки, остающиеся белыми или беловатыми, а также мешковидную обвертку у основания ножки — остаток общего покрывала. Пластинчатые грибы, имеющие определенное сходство с Polyporales (см. 8.1), такие, как пи-лолистник (Lentinus) и вешенка (Pleurotus), в настоящее время опять относят к Agaricales. Р. eryngii при определенных условиях культивирования образует вместо пластинчатого гименофора неправильные трубочки.
К Agaricales относятся также роды Kuehnero-myces, Psilocybe, Armillaria. Omphalina, CHtocybe. Lepista, Strobilurus и Termitomyces.
10.3. Порядок: Boletales. К порядку относятся не только грибы с трубчатым гименофором типа белого гриба (Boletus edulis), но и определенные грибы с пластинчатым гименофором, с замкнутыми и корковидными плодовыми телами. Это связано с наличием переходных форм или следующих признаков: пигментированные, часто веретеновидные споры и / или диверги-рующие (расходящиеся) возле гимения гифы трамы гименофора, пигменты типа производных пульвиновой кислоты (например, вариегатовая кислота). Этими пигментами обусловлено при наличии оксидаз часто наблюдаемое синее окрашивание плодового тела (см. рис. 11.60). Эволюция в пределах порядка, как предполагается, шла от обитателей древесины (бурая гниль, см. бокс 11.4, с. 124) с прилегающими в виде корок к субстрату плодовыми телами с гладким гименофором (Coniophora) либо уже гименофором в виде складок или сот: серпула плачущая, или настоящий домовый гриб (Serpula lacrymans), образует быстро растущие, мягкомясистые плодовые тела размером до 1 м2. На древесине обитают также представители порядка с плодовыми телами — от имеющих форму раковины до дифференцированных на ножку и шляпку, образующих пластинчатый гименофор, как Tapinella atrotomentosa. Следующий эволюционный уровень — чаще всего образующие микоризу шляпочные грибы с гименофором в виде пластинок, трубочек или промежуточного типа. Paxillus involutes образует микоризу с разными деревьями, Р. fllamentosus — только с ольхой (см. 9.2.3;
бокс 11.4). У Phylloporus пластинчатый гименофор из-за наличия многочисленных анастомозов между пластинками подразделен на отдельные камеры. Gomphidius (мокруха), несмотря на ярко выраженный пластинчатый гименофор, имеет такие же микроскопические и химические признаки, как и представители порядка с трубчатым гименофором. Грибы этого порядка с труб чатым гименофором отличаются от имеющих сходный гименофор представителей других таксонов (например, Polyporusm Polyporales, Boletopsis из Thelephorales, Coltricia из Hymenochaetales) рядом особенностей. К отличиям относятся: недолговечность плодовых тел, не способных обрастать посторонние предметы; отделяемые от остальной части шляпки трубочки; другие микроскопические признаки и пигменты. Как приспособление к вулканическим почвам, сухим периодам года и т.д. можно рассматривать появление эволюционно наиболее молодых гастро-идных форм, которые также сплошь образуют микоризу, а часть из них очень специализирована в отношении физиологии питания. Виды рода Gastroboletus. растущие в Америке, можно расположить в ряд, где ножка становится короче, трубчатый гименофор претерпевает все большую деградацию, плодовое тело замыкается за счет соединения края шляпки с ножкой и начинает развиваться под землей. Подобные ряды можно проследить в родах Truncocolumella и Rhizopogon с исключительно подземными плодовыми телами.
Далее следуют порядки, объединяющие типичные формы с замкнутыми плодовыми телами (гастеромицеты, однако в настоящее время без Geastrales и Phallales). Это грибы, у которых плодовые тела, по крайней мере в молодости, замкнутые, гастроидные, ангиокарпные, их оболочка (перидий) вскрывается или распадается только после созревания спор (см. рис. 11.61, Е, F).
10.4. Порядок: Lycoperdales. Представители порядка известны под названием дождевики. Гастроидные плодовые тела, имеющие форму от шаровидной до булавовидной, только на ранних стадиях развития находятся пол землей. Снаружи они имеют оболочку, чаще всего двухслойную, состоящую из экзо- и эндоперидия. У зрелых плодовых тел экзоперидий разрывается; на жестком кожистом эндоперидии от него остаются зернышки, бородавочки или комочки. Эн-доперцдий часто открывается порой на вершине. Внутренняя часть плодового тела сначала подразделена на камеры, поверхность которых выстлана гимением. Базидии короткие, булавовидные. Они несут на сильно вытянутых стеригмах шаровидные споры, которые активно не отбрасываются, а освобождаются за счет разрушения базидий. Зрелое плодовое тело содержит только
11.2. Бактерии, грибы, растения
порошковидную массу — глебу, которая состоит из бесчисленных спор и волокон. Она возникает в результате разрушения гимения и стерильной ткани, так что остаются и составляют глебу только споры и волокна ткани (капиллиций) — остатки длинных гиф трамы. У представителей некоторых родов нижняя часть плодового тела стерильна (субглеба). В лесах часто встречается дождевик шиповатый (Lycoperdon periatum), из плодовых тел которого при надавливании высвобождается облачко порошащей бурой споровой массы. У гигантского дождевика (Langermamia gigantea), плодовое тело которого может достигать 50 см в диаметре, глеба содержит до 7,5 млрд спор. Если бы каждая спора развивалась в плодовое тело, то уже в 21-м поколении потомков образовалась бы такая грибная масса, которая в 800 раз превышала бы массу Земли.
Виды рода порховка (Bovista) растут преимущественно на лугах и пастбищах; их плодовые тела, как, например, у порховки гигантской, не имеют стерильной субглебы. Правда, ложнодождевик (Scleroderma: см. рис. 11.61, D) относится к Boletales (пигменты, ДНК!).
10.5. Порядок: Nidulariales. В плодовом теле заключены отдельные фрагменты глебы, которые могут распространяться как целые единицы — перидиоли. Внутри перидиолей базидии расположены в виде гимения. У Cyathus (бокальчика) перидиоли при созревании лежат как маленькие диски в бокаловидном перидии (см. рис. 11.61, Е). Сфероболюс (Sphaerobo-hts) образует одну-единственную перидиоль, которая из-за мгновенного выворачивания внутреннего слоя экзоперидия отбрасывается на расстояние до 1 м (см. рис. 11.61, F).
Родственные связи последних двух порядков с относящимися к Agaricales родами Сор-rinus и Agaricus (разделение порядков 10.4 и 10.5 с 10.2 выражено, таким образом, слабо!), а также с растущей на дубовой древесине печеночницей (Fistulina) вырисовываются достаточно ясно.
Применение. С древности до середины XIX столетия трут, изготовляемый из ткани плодового тела Fomes Jomentarius, служил для получения огня (откуда название гриба — настоящий трутовик). Так, в поясном мешке найденного в леднике трупа неолитического человека из долины Эца («Эци») был найден этот гриб, так же как и укрепленные на полосках кожи куски плодовых тел березового трутовика (Pipto-porus betulinus), возможно, для применения как лекарс твенного средства. Многочисленные виды гименомицетов собирают для ис
пользования в пищу, некоторые из них, например шампиньон (Agaricus bisporus), также культивируются. Ежегодный сбор культурного шампиньона во всем мире 670 000 т. Наряду с шампиньоном для питания выращиваются, особенно в Восточной Азии, различные другие базидиомицеты (например, сиитаке, Lentinus edodes). Пытаются «одомашнивать» и ранее не возделывавшиеся съедобные грибы. Однако некоторые очень ценные виды (например, белый гриб, лисичка обыкновенная) не дают в культуре плодовых тел. При введении грибов в культуру интересуются не только получением плодовых тел как приправы и пищи для человека и животных, но и использованием их для переработки различных отходов: помета животных, соломы, опилок и других материалов, содержащих лигнин и целлюлозу (рециклизации). Путем заражения древесины мицелием соответствующих грибов (например, летнего опенка, Kuehne-romyces mutabilis, или вешенки устричной, Pleurotus ostreatus) изменяют ее свойства; она становится легкой и как «микодревесина» используется для промышленной обработки (например, при изготовлении карандашей вместо древесины кедра). Виды Psilocybe (Agaricales) содержат галлюциногенные индольные производные, которые используются также при религиозных ритуалах, например в Мексике. Различные грибы выделяют в культуральную среду антибиотики, которые используются отчасти в фармацевтических целях (например, пенициллин), отчасти как фунгициды (стро-билурин из Strobilurus).
Некоторые грибы ядовиты. Об опасных поганках уже упоминалось; в целом известно примерно 150 видов ядовитых гименомицетов (аскомицетов — лишь около 10 видов), но среди них силы юядовитых немного. В старых плодовых телах съедобных грибов также образуются ядовитые вещества, как при порче мяса.
Обзор базидиомицетов. Родственные связи между аскомицетами и базидиоми-цетами и происхождение их от некоторой общей предковой группы (близкой к Taphrinomycetidae) подтверждаются гомологиями. Как и у аскомицетов, при половом размножении между пла гмогамией и
198 | ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Бокс 11.4. Встречаемость и образ жизни грибов (включая целлюлозные грибы)
Грибы, насчитывающие около 100000 видов (Oomycota 500, Eumycota свыше 90 000), обитают повсеместно как гетеротрофы, и в противоположность водорослям прежде всего на суше. Обитатели водной среды (менее 2 % всех видов) — это либо первично-водные организмы из числа размножающихся зооспорами Oomycetes и Chytridiomycetes, либо вторичноводные организмы — Ascomycetes и Fungi im-perfecti (Moniliales). Водные грибы живут преимущественно в пресной воде, однако в последнее время выявлено некоторое число морских видов (особенно аскомицетов. но также и несколько базидиомицетов). Часто на их спорах имеются приспособления для парения в воде; например, у Moniliales конидии нитевидные или с 3 — 4 лучами.
Ископаемые грибы сохранились весьма незначительно. Древнейшие находки — это хитридиомицеты во фрагментах панцирей морских животных; они восходят к кембрию. В девоне встречены лишенные поперечных перегородок гифы в остатках наземных растений; в карбоне на папоротниках уже имелись ржавчинные грибы (и вероятно, также аско-мицеты), а на корнях деревьев — микоризы; экземпляры хорошо сохранившегося пряжко-вого мицелия свидетельствуют о том, что уже в каменноугольном лесу имелись высшие ба-зидиомицеты (гименомицеты?).
Грибы питаются как саиротрофы. биотроф-ные или некротрофные паразиты (последние вызывают быстрое отмирание живых организмов; по образу жизни близки к сапротрофам на отмерших остатках) или как компоненты симбиотических ассоциаций (например, микоризы лишайников). Нередко грибами заселяются особые субстраты, например: насекомые, мхи, другие грибы и т.д. (часто лишь определенные виды).
Поскольку грибы могут разлагать самые разные органические субстраты и поражать самые разные живые организмы, причиняя им вред или убивая их, они наносят значительный ущерб. Гриб Amorphotheca resinae (Eurotia-les) освоил даже масла, бензин и смолы, питаясь содержащимися в них углеводородами; в авиации он наносит ущерб, вызывая закупорку системы питания двигателя и коррозию алюминия. Вызывая разрушение древесины, болезни человека, животных и растений, а также порчу продуктов питания и текстиля, грибы оказывают огромное отрицательное воздействие в хозяйственной деятельности. Вместе с
тем велика и положительная роль грибов. Об этом говорилось выше, при рассмотрении отдельных классов грибов (спиртовое брожение — см. 6.10.2.1; антибиотики — см. Eurotia-les; употребление в пищу — см.: применение базидиомицетов; стимуляция роста растений при микоризном симбиозе — см. 9.2.3). Из огромного разнообразия приспособлений к особым условиям жизни нам надо будет выбрать лишь немногие, экологически наиболее важные.
Грибы как разрушители древесины
В природе древесина отмирающих деревьев и пни разлагаются преимущественно грибами. Разложение древесины животными и бактериями, напротив, имеет лишь очень незначительный масштаб. Согласно современным исследованиям, бактерии, по-видимому, не бывают агрессивными разрушителями древесины. Из грибов в разложении древесины участвуют прежде всего базидиомицеты с гимениальными плодовыми телами (гименомицеты, например, Poriales, Hymenochaetales, Polyporales, Agaricales), отчасти аскомицеты (например, Ceratocystis) и несовершенные грибы. Некоторые из этих деревораз-рушителей поражают как паразиты древесину еще живых растений, как Phellinus pini и Hetero-basidion annosum — возбудители гнилей сосны и ели. а также Phellinus alni на яблонях и других лиственных деревьях или настоящий трутовик (Fames fomentarius) на буках и березах. Многие грибы живут сапротрофно только на отмершей древесине (например, виды Comlus. Trametes и Gloeophyllym). После паразитирования возможны сапротрофный образ жизни на отмершей древесине, как у опенка осеннего (Armillaria melted). который обитает на отмерших пнях деревьев и образует на них свои плодовые тела, но оттуда может переходить на живые деревья, прежде всего когда они (например, после длительной засухи) физиологически ослаблены («паразиты ослабленных хозяев»).
Видообразование шло по пути специализации на определенных хозяевах (например, Phellinus hartigii на пихте. Ph. robustus на дубе, Ph. hippophaecola на облепихе), особенно среди родов, представленных преимущественно паразитами. Некоторые грибы — опасные разрушители древесины на складах и в постройках: например, домовые грибы Coniophora puteana и особенно Serpula lacrymans могут наносить серьезный ущерб постройкам, начиная с сы
11.2. Бактерии, грибы, растения! -|gg
рых мест. Разрушение древесины грибами происходит в виде деструктивной (бурой) или коррозионной (белой, особая форма: красная) гнили. В первом случае гриб разлагает преимущественно целлюлозу, так что лигниновый компонент древесины сохраняется; древесина становится бурой, с поперечными трещинами и распадается на прямоугольные фрагменты (например. Coniophora, Serpula). Возбудители коррозионной гнили (например, Phellinus igniarius на иве) разлагают лигнин и целлюлозу, причем чаще всего они в противоположность грибам бурой гнили выделяют в субстрат фенол-оксидазы; гнилая древесина вследствие процессов обесцвечивания становится белой и продольно-волокнистой. Отдельные древесные породы противостоят разложению с помощью ядов, которые прежде всего содержатся в ядровой древесине (токсины ядровой древесины). Некоторые грибы, однако, все-таки способны разрушать защищенную таким путем древесину, причем в обезвреживании токсинов, возможно, участвуют фенолоксидазы. Только в недавнее время стали обращать больше внимания на Moderfaule как на третий тип разложения древесины. Этот вид разложения вызывают ми-куромицеты с мелкими плодовыми телами (например, аскомицетами) или грибы без полового спороношения (Fungi imperfecti). По характеру разложения болезнь представляет собой медленно развивающуюся бурую (реже белую) гниль. Примером могут служить различные виды Chaetomium (Ascomycetes), гифы которых поражают вторичную стенку трахеид или же сосудов древесины. При названных типах гнилей снижается, иногда очень значительно, сопротивляемость древесины сжатию и изгибу. Однако синева древесины сосны не влияет на ее статические качества, так как ее возбудители (виды Ceratocystis) разрушают только содержимое клеток паренхимы древесины. В процессе приобретения серой окраски строительной древесиной. например в горах, по-видимому, участвуют несовершенные грибы, если этот процесс обусловлен не исключительно погодными условиями. Некоторые дереворазрушаюшие грибы вызывают ночное свечение (биолюминесценцию), как, например, опенок осенний (Armillaria meUeay. у поселяющегося на старых оливковых деревьях гриба Omphalotus olearius светятся даже плодовые тела. Физиологически разложению древесины родственно разложение растительного опада (листьев) на лесной почве, в котором наряду с бактериями участвуют грибы (грибы лесной подстилки), и тем самым они вносят существенный вклад в образование гумуса.
Грибы как симбионты
Значительная часть высших растений образует с грибами симбиотическую ассоциацию — микоризу (см. 9.2.3). Различают облигатно и факультативно микотрофные растения. В Европе эктотрофная микориза наиболее характерна для хвойных пород и Fagales из покрытосеменных. Микориза (чаще всего эндотрофная) широко распространена и у культурных растений, таких, как земляника, томат, горох и хлебные злаки. У орхидных в природных условиях без микоризного гриба не могут прорастать мельчайшие семена, лишенные запасных питательных веществ. Микориза отсутствует лишь у немногих высших растений, например: Cyperales, Plumbaginales, Brassicaceae. Из грибов не образуют микоризы оомицеты и хитридиомицеты, в остальных классах настоящих грибов они распространены, особенно среди аскомицетов и базидиомицетов (к которым относится большинство шляпочных грибов, обитающих на лесной почве), а также и среди зигомицетов (Endogone).
Часто грибы образуют плодовые тела только в ассоциации с определенным микоризным партнером или могут жить вообще исключительно в симбиозе с ним. Так, среди «обабков» (род Leccinum) подберезовик (£. scabrum) и подосиновик желто-бурый (£. testaceoscabrum) связаны с березой, £. vul-pinum — с лвуиглыми соснами, дубовый обабок (£. quercinum) — с дубом, подосиновик красный (£. aurantiacum) — с осинами, £. саг-pini — с грабом, лещиной или осиной. Микоризные симбиозы могут «вырождаться» или переходить во взаимоотношения хозяин—паразит. Тогда или некоторые грибы паразитируют на растении-хозяине, или высшее растение, исходно служившее как партнер гриба, становится его паразитом (например, Neottia, см. 9.2.3). Исследования микориз находят практический выход при выращивании лесов на ранее безлесных территориях. Они обеспечивают теоретические обоснования внесения посевного материала грибов — микоризных партнеров. О симбиозе грибов с водорослями, который привел к стабильному партнерству в лишайниках, готорится в разделе, посвященном этим комплексным организмам (см.: Лишайники, 11.2; 9.2.4).
Исключительно многообразны симбиозы грибов с животными. Узкоспециализированные в отношении питания животные, такие, как кровососущие, питающиеся древесиной или растительными соками насекомые, содержат в определенных частях своего пищеваритель
200 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
ного тракта или в особых органах, так называемых мицетомах, симбионты — бактерии и дрожжи, для переноса которых от одного поколения к другому существуют различные приспособления. Грибы-симбионты отчасти дают своим партнерам питание (древесина для личинок рогохвоста большого), отчасти гриб поедается насекомым, так что оно обходным путем — через гриб — может использовать в пищу древесину (например, жуки-точильщи-ки). Во всех этих случаях симбионты обеспечивают своим хозяевам выживание таким образом, что они по-разному дополняют обмен веществ партнера. Тропические муравьи-листорезы культивируют в своих подземных постройках мицелий определенных видов грибов (относящихся к Agaricales): утолщенные, богатые питательными веществами концы грибных гиф идут им в пищу; гриб, разлагающий целлюлозу, вносится муравьями на субстрат, образованный из разжеванных кусочков листьев, и при закладке нового гнезда переносится в него. У термитов также тщательно культивируемый грибной мицелий (Termitomyces Agaricales) служит для питания только царицы и личинок. Сходное значение имеют так называемые грибы-амброзии, которые обитают в ходах короедов (Ipidae) и используются в пищу их личинками
Грибы как возбудители болезней
Из 162 важных инфекционных заболеваний возделываемых в Средней Европе культурных растений 83 % вызываются грибами. Потери ежегодно бывают огромными и уже приводили к случаям массового голода (см. Phytophthora). Среди организмов, которые являются паразитами на растениях, наряду с животными, бактериями и вирусами грибы играют доминирующую роль. Некоторые группы (среди слизевиков — плазмодиофоромицеты, среди других, например, Peronosporales, Erysi-phales, Uredinales, Ustilaginales) существуют почти исключительно как паразиты и возбудители болезней высших растений. Наряду с этим к паразитам растений относятся также многочисленные несовершенные грибы (см. 11.2. Deuteromycetes). Развитие паразитического гриба на растении-хозяине начинается с заражения. Гриб проникает через порезы, устьица или непосредственно через наружную стенку эпидермиса. Внедрение через кутикулу эпидермиса происходит при участии специальных экзоферментов (кутиназ) или механическим путем, когда кутикула прободается острым выростом инфекционной гифы. Обычно зара
жаются восприимчивые органы хозяина, например корневые волоски, органы проростков, лепестки или рыльца пестиков. Для успешного заражения часто бывает достаточно одной споры. Ведущее значение для прорастания споры наряду с температурой имеет достаточная влажность, поэтому в сырые годы наблюдается особенно сильное развитие грибных болезней.
На внедрение гриба растение-хозяин реагирует защитными реакциями механической или химической природы — утолщением клеточных стенок, образованием защитных веществ, например дубильных (которые обезвреживаются фенолоксидазами) или специфических фитоалексинов. Произойдет ли наконец заражение хозяина, зависит от вирулентности гриба и устойчивости хозяина. Устойчивость хозяина обусловлена его генетической конституцией, а также различными механическими, химическими и физиологическими факторами резистентности. С другой стороны, предрасположенность хозяина к заражению в соответствующее время зависит от внешних условий и может иметь решающее значение для заражения. Хорошее азотное питание приводит, например, к меньшей механической прочности клеток при быстром росте, из-за чего чаще всего повышается предрасположенность к заражению.
Вирулентность гриба показывает в пределах вида большие различия, обусловленные генетически. Среди 53 аскоспоровых культур даже из спор одного плодового тела у возбудителя мучнистой росы ячменя было получено 14 различных патотипов. У многих паразитных грибов виды разделяются на большое число форм, специфичных относительно определенных хозяев.
В качестве источников инфекции выступают все конидии и мейоспоры. Перезимовка осуществляется в виде стром, которые весной образуют плодовые тела (например, Venturia, Claviceps, Rhytisma), в виде прорастающих весной зигот (Peronosporales) и телейтоспор (Uredinales); грибы могут зимовать в корневищах, клубнях, зимующих почках растений-хозяев ит.д. Распространение возбудителя болезни осуществляется (часто очень быстро) такими же путями, как распространение плодов и семян высших растений: чаще всего потоками воздуха, нередко животными и в последнее время также человеком, который уже перенес многие болезни растений с одной части земного шара в другую.
Эпифитопии грибных болезней мы знаем только в монокультурах отдельных растений,
11.2. Бактерии, грибы, растения | 201
которые в ряде регионов даже были полностью уничтожены. Например, виноградарство на Тенерифе и Мадейре полностью прекратилось около 1850 г. из-за поражения виноградной лозы возбудителем мучнистой росы Uncinula necator.
При борьбе с болезнями растений (в противоположность бактериальным и грибным («грибковым») болезням человека) терапевтические меры играют лишь второстепенную роль. Важнейшими мерами здесь являются профи-лактикческие: препятствование контакту возбудителя и хозяина (соответствующие агротехнические мероприятия, севооборот, удаление промежуточных хозяев ржавчинных грибов) либо уничтожение патогенного гриба до или во время его прорастания (протравливание посевного материала, опрыскивание или опыление фунгицидами). Важную роль играет также выведение сортов, устойчивых к возбудителям болезней (селекция на устойчивость). Предпосылкой успешной борьбы является точное знание цикла развития и условий жизни паразита.
У человека и животных грибы вызывают самые различные заболевания. Косвенно с образом жизни грибов связаны микотоксикозы и микогенные аллергии. Такие аллергии могут
быть вызваны спорами грибов, попадающими в дыхательные пути из воздуха. Содержание спор в воздухе часто чрезвычайно велико: так, в одной сельскохозяйственной постройке было однажды обнаружено до 21 млн спор Aspergillus на I см3 воздуха. Вне помещений, напротив, эти показатели значительно ниже, а именно порядка 0,25 — 7 спор на I см3 воздуха. В легких только что забитых коров было насчитано до I 700 колоний грибов на I г сырой массы, и именно тех видов, споры которых, имеющие величину менее Юмкм, могут перемещаться по дыхательным путям. Aspergillus flavus растет на различных орехах и образует афлатоксины, вызывающие поражение печени. В противоположность таким микотоксикозам, которые могут вызываться также, например, Claviceps, при пищевом отравлении грибами говорят о мицетизме (см. Agaricales).
Грибы, вызывающие микозы, могут рассматриваться как паразиты теплокровных животных и человека. По картине болезни микозы разделяются на поверхностные (поражаются кожа, волосы, ноп и, перья, когти и копыта) и глубокие (развиваются внутри тела и могут приводить к смерти). Примерно 30—50
Рис. 11.62. Хищные грибы (А — по L. Nowakowsky; В — по Valkanow; С, D — по W. Zopf; Е, F — по Sommerstorff; G — no С. Drechsler):
А— Polyphagus, Р. euglenae с 10 сжавшимися эвгленами на разных стадиях их переваривания (200х); В — Arnaudovia, A. hyponeustica с пойманной особью Tylenchus (150х); С, D — Arthrobotrys, A. Oligospora: С — с ловчими петлями, D — с пойманной нематодой (150х); Е, F — Zoophagus'. Е — Z. insidians с тремя пойманными коловратками (90х), F— коловратка, пойманная особью Zoophagus, таллом которой разрастается внутри тела животного (125х); G — Zoopage, Z. tham-nospira с двумя амебами (500х)
202 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
возбудителей микозов представляют собой дрожжевидные (Candida, Torulopsis, Cryptococcus) и мицелиальные (Aspergillus, Trichophyton, Mucor) грибы. Некоторые виды поражают исключительно теплокровных хозяев, другие живут в почве, откуда и происходит заражение. «Ножной гриб» (Trichophyton rubrum) заражает человека поросшими его мицелием мельчайшими кожными чешуйками, рассеянными по почве. Глубокие микозы практически не передаются при контакте. Заражение ими происходит через желудочно-кишечный тракт и особенно через органы дыхания. Виды Histo-plasma — возбудители широко распространенного в теплых странах заболевания легких.
Хищные грибы
Хищные грибы (рис. 11.62) составляют особую группу факультативных паразитов. Эти грибы улавливают мелких почвенных животных (нематод, коловраток) или подвижные водоросли (Euglena) с помощью различных приспособлений, однако часто они могут культивироваться в отсутствие своих жертв на обыч
ных питательных средах. К Chytridiales относятся Polyphagus euglenae (рис. 11.62, А; см. также 11.2. Chytridiomycetes) и виды рода Amaudovia. Последние живут на поверхности воды (нейстон) и улавливают одноклеточные организмы с помощью шести длинных гиф с выростами в виде тонких волосков (рис. 11.62, В). Zoopagales (Zygomycetes) — все «охотники за животными»: они живут ловлей амеб и нематод. На гифах Zoopage thamnospira (рис. 11.62, G) удерживаются нематоды, в которые потом врастают служащие для питания гриба гаустории. Хищные грибы группы Arthrobotrys относятся к несовершенным грибам (Hyphomycetes). Они ловят своих жертв (нематод) посредством клейких гиф или сетей (рис. 11.62, С), клейких «пуговиц» (Dactylella) и несжимающихся и сжимающихся колец (Dactylium). У Oomycetes также возникло хищничество, а именно в роде Zoophagus. Гифы образуют здесь придатки, покрытые слизистым выделением, на которых зависают коловратки (рис. 11.72, Е, F). Среди базидиомицетов хишни-чество наблюдалось у некоторых видов Pleuro-tus. В целом описано около 80 видов грибов, улавливающих животных.
кариогамией вклинивается характерная двуядерная стадия (дикариофаза). Стадия парных ядер может возникать не непосредственно после плазмогамии. Более активное из двух ядер может еще пройти через многие клетки мицелия, воспринявшего его, пока не образует с генотипически отличающимся ядром пару ядер (например, у аскомицетов Neurospora или у базидиомицетов Typhula). Встречающееся на дика-риотичном мицелии многих базидиомицетов образование пряжек гомологично образованию крючков у аскомицетов; многие аскомицеты даже образуют уже пряжки (Tuberaceae) вместо крючков. Незначительное различие между крючком и пряжкой состоит в том, что первые закладываются терминально, а последние — латераль-но; кроме того, у аскомицетов образование крючков часто ограничено только конечными клетками аскогенных гиф. Напротив, плодовые тела базидиомицетов нельзя го-мологизировать с плодовыми телами аскомицетов, поскольку они состоят только из дикариотичных гиф, тогда как у аскомицетов — из гаплоидных и дикариотичных аскогенных (см. рис. 11.32, А; 11.51, В).
Если сплетению этих гиф в плодовые тела у аскомицетов всегда предшествует половой процесс или оба явления тесно связаны между собой, то образованию многих плодовых тел базидиомицетов может предшествовать один акт слияния гиф (сома-тогамии): однажды возникший дикариотичный мицелий у базидиомицетов может при воздействии соответствующих внешних факторов вновь и вновь образовывать плодовые тела. У аскомицетов дикариофаза ограничена некоторым небольшим числом клеток: в значительной мере жизненный цикл определяется гаплоидными гифами, которые питают дикариотичные гифы. Только у Taphrinomycetidae, которые эволюционно близки к базидиоми-цетам, дикариотичные гифы питаются независимо от гаплоидных. У базидиомицетов самостоятельная дикариофаза выходит на передний план всего жизненного цикла. Uredinales, рассматриваемые как исходные базидиомицеты, обеспечивают доминирование дикариофазы наличием нескольких типов спороношений, а более эволюционно подвинутые гименомицеты — выживанием дикариотичного мицелия в
11.2. Бактерии, грибы, растения | 203
течение многих лет и вегетативным размножением (оидии, фрагменты гиф и т.д.). В эволюции аскомицетов определенное значение имеет упрощение полового процесса. У базидиомицетов оно достигает своего наивысшего пункта: половые органы больше не закладываются, и соматогамия становится правилом (исключение — Uredinales). Также и повторное образование плодовых тел на дикариотичном мицелии базидиомицетов может рассматриваться как ограничение полового процесса.
1. Приложение к Mycobionts: несовершенные грибы (Deuteromycetes)
Естественная система грибов основывается, в частности, на ходе их развития и связанных с половым размножением структур полового спороношения (телеоморфы). Однако у многих грибов (порядка 30 000 видов) известны только особенности их бесполого размножения посредством конидий — бесполое спороношение (анаморфа). При этом следует различать два случая: или половое спороношение нам неизвестно, или у гриба утрачена способность к нему. Все эти грибы объединяются в искусственную группу несовершенных грибов, или дейте-ромицетов (Fungi imperfecti, или Deuteromycetes).
Несмотря на отсутствие половой стадии, генетическая рекомбинация может осуществляться благодаря наличию парасексуалыюго цикла (происходящее иногда слияние гиф разных ми-целиев: слияние ядер из этих разных мицелиев в диплоидные гетерозиготные ядра; митотический кроссинговер; гаплоидизация путем последовательной утраты хромосом).
Накопление знаний, касающихся общего набора признаков (типы гиф и септ, химизм и ультраструктура клеточной стенки, обнаружение соответствующих телеоморф), а также анализ ДНК позволяют все больше и больше распределять несовершенные грибы по классам и порядкам в естественной системе грибов. Подавляющее большинство их относится к аскомицетам и лишь немногие — к базидиомицетам.
Предварительное искусственное подразделение несовершенных грибов, служащее практическим целям, основывается на структурах, образующих конидии. Конидии возникают почти
всегда на конидиеносцах, которые располагаются поодиночке, в виде палисадного слоя или внутри вместилищ (пикнид).
I. Sphaeropsidalcs. Конидии образуются в напоминающих перитеции вместилищах (пикнидах) или полостях (камерах). Septaria apii вызывает септориоз («ржавчину») сельдерея. Сюда относятся также Fusicoccum (см. 7.6.4.1 и 9.3.3) и Lasiodiplodia (см.7.6.6.2).
2. Melanconiales. Конидии возникают на стро-матическом ложе. Gloeosporium fructigenum вызывает горькую гниль яблок (Sphaceloma, см. 7.6.3.1).
3. Moniliales (Hyphomycetales). Конидии не на строматических ложах; они развиваются на конидиеносцах, часто сильно разветвленных, которые располагаются поодиночке или в пучках (коремиях) либо же объединены вместе со стерильными гифами в слизистые ложа (спородохии). Примеры: Aspergillus, Penicillium с многочисленными видами; для некоторых видов известна связь с половой стадией (см. Eurotiales). Сюда также относят ся Histoplasma, Trichophyton, Arthro-botrys, Dactylella и Dactyliutn (см. бокс 11.4). Fusarium oxysporum f. lycopersici вызывает фузариозное увядание растений томата. (См. также Cercospora, 7.6.4.1.)
4. Blastomycetalcs. Дрожжевидно-почкующи-еся формы без половых стадий, например Cryptococcaceae (близкородственные аскомицетам) (Candida и Torulopsis, см. бокс 11.4).
5. Mycelia sterilia. Мицелии, для которых неизвестно никаких специальных репродуктивных структур (например, некоторые микоризные грибы, склероции, ризоморфы).
2. Приложение к Mycobiont а: лишайники (Lichenes)
В лишайниках гифы определенных видов грибов образуют с автотрофными водорослями (см. 9.2.4) симбиотическое объединение, которое представляет собой морфологическое и физиологическое единое целое. Находящиеся в лишайниках экто-симбиотические водоросли (фотобионты) — одноклеточные или нитчатые представители уже обсуждавшихся цианобактерий (например, Chroococcus, Gloeocapsa, Scytonema, Nostoc) или зеленых водорослей (например, Соссотуха, Cystococcus, Treboivda, Chlorella и Trentepohlia). Грибы (микобион-ты), участвующие в образовании лишайников. — это прежде всего аскомицеты (чаще виды, образующие апотеции, реже — виды, образующие перитеции), и лишь в очень
204 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Рис. 11.63. Лишайники (по К. Magdefrau):
А — Dictyonema pavw/a; В — Cladonia rangiferina; С — Cladonia pyxidata. слоевище с бокаловидными подециями; D — Thamnolia vermiculans; Е — Dermatocarpon miniatum; F — Graphis scripta; G — Parmelia acetabulum; H — Rhizocarpon geographicum; J — Roccella boergesenii; K— Usnea florida (A—К— 1/2x)
редких случаях базидиомицеты (например, Corticiaceae, Clavariaceae). Принадлежность лишайниковых грибов к различным классам в системе грибов явно говорит о том, что лишайниковые симбиозы неоднократно возникали в разных эволюционных линиях. Этим обусловлено появление новой формы организации таллофитов с собственными признаками. В результате совместного существования гриба и водоросли развились определенные морфологические и химические особенности. Лишайниковые грибы утратили в условиях симбиотического существования свою самостоятельность; они могут расти в природе только совместно с соответствующими водорослями. На этом основании лишайники рассматривались раньше как отдельная систематическая единица — как отдел Lichenes.
Морфология. Облик лишайника в редких случаях зависит от строения водоросли, чаще же всего — от строения гриба. Примерами первого случая могут быть нитчатые лишайники (например, Ephe be}, у которых гриб опутывает нитчатую цианобактерию, или слизистые лишайники. У подав
ляющего большинства родов именно гриб определяет облик лишайника. У медленно растущих накипных, или корковых, лишайников, которые живут на поверхности камня, почвы или коры деревьев, слоевище прочно соединено с субстратом. Чаше всего в определенной мере оно проникает в субстрат и имеет чаще всего четко выраженный облик (рис. 11.63, Н). Плоское, преимущественное рассеченное на лопасти слоевище листоватых лишайников (G) соединено с субстратом тяжами (ризинами)1. У некоторых листоватых лишайников (Е) дисковидное слоевище прикрепляется только в середине. Наконец, кустистые лишайники прикрепляются к субстрату очень небольшим основанием и ветвятся наподобие кустиков (J). Слоевища арктоальпий-ского вида Thamnolia vermiculans (D) свободно лежат на земле и лишь частично
1 Слоевище листоватых лишайников может прикрепляться к субстрату не только ризинами (представляющими собой выросты сердцевины), но и ризоидами (гифами, идущими от нижней коры) или гомфом (единственной прикрепляющей подошвой). — Примеч. пер.
11.2. Бактерии, грибы, растения | 205
могут прикрепляться немногими тисненными тяжами. У видов рода Cladonia (В, С) от обычно слаборазвитого первичного слоевища листоватого типа поднимаются выросты — подеции — в виде бокальчиков или кустиков, которые несут апотеции.
Гистология и физиология. Поперечный разрез через слизистый лишайник (рис. 11.64, А) показывает более или менее равномерное распределение водоросли и гриба в слоевище (гомсомсрное строение); в слизи колонии Nostoc здесь обильно развиваются гифы гриба. Гифы часто более плотно смыкаются между собой на верхней и нижней стороне и могут образовывать ко-
ровый слой. У кустистых и листоватых лишайников (рис. 11.64, В), а также у многочисленных накипных лишайников водоросли образуют определенный слой, расположенный параллельно поверхности слоевища (гетеромерное строение). В верхнем коровом слое гифы гриба часто смыкаются между собой в прочные сплетения. У листоватых и кустистых лишайников коровые слои чаще всего дифференцированы сильнее, чем у накипных лишайников (ср. В и С). У эндофлеоидных (живущих внутри коры деревьев) и эндолит-ных (живущих в трещинках камня) лишайников слоевище проникает внутрь
Рис. 11.64. Лишайники (А — по Н. des Abbayes; В — по J. Sachs; С — по Bioret; О, К — по К. Magdefrau; Е, F — по Bornet; G, Н — по E.Tschermak; J — по Plessl):
А—С — поперечные разрезы слоевищ: А — Collema pulposum (200х), 3 — Sticta fuliginosa (250х), С — Graphis dendritica (200>); D — соредий Parmelia sulcata (450х); Е—J — гаустории: Е — аппрес-сории, F— оплетающие гифы, G — внутриклеточный гаусторий, Н — внутримембранный гаусторий, J — окруженный отложением целлюлозы внутримембранный гаусторий (Е, F — 450х; G—J — бООх); К— Peltigera aphthosa (200х)
206 | ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
субстрата столь глубоко, что едва выступает на поверхность.
Гриб и водоросль живут в тесном симбиозе друг с другом, причем гриб опутывает клетки водоросли (рис. 11.64, Е, F) и внедряется в них. При этом часто возникают гаустории — выросты гриба внутрь клетки водоросли (G). Гриб, как правило, остается отделенным от протопластов водорослей, так как они отграничивают места внедрения стенками. У многих лишайников грибы образуют только внедряющиеся в стенки водорослей апрессории (Е), причем водоросль проявляет защитную реакцию в виде утолщенных стенок (J).
У некоторых лишайников наблюдаются также водоросли второго рода, которые существенно отличаются от первых по систематическому положению. Эта вторичная водоросль находится или в определенных местах внутри слоевища (например, у Solo-rina сгосеа), или же в маленьких головчатых структурах на его поверхности — так называемых цефалодиях (рис. 11.64. К): последние содержат азотфиксирующие синезеленые водоросли (например, Nostoc) и встречаются у лишайников, которые в остальной части слоевища содержат только зеленые водоросли (Chlorophyta). К нормальной симбиотической ассоциации водоросли и гриба может присоединиться второй гриб, который существует как «парасимбионт» или даже как настоящий паразит; известно большое количество таких «лишайниковых паразитов». Наконец, закономерно встречаются лишайники-паразиты в слоевищах других видов.
Углеродный обмен гриба (микобионта) полностью зависит от водоросли (фотобионта). Грибы получают от них чаще всего сахара или сахароспирты. Заключенные в сплетении гиф гриба водоросли нуждаются в получении от него воды и минеральных веществ. Кроме того, гриб обеспечивает зашиту от слишком интенсивного света. С симбиотическим образом жизни взаимосвязано наличие многочисленных специфических лишайниковых веществ, которые никогда не образуются изолированными друг от друга партнерами; они откладываются преимущественно на внешней стороне гиф и придают лишайникам ту или
иную характерную окраску. Это вещества самой разной природы: алифатические кислоты, депсиды, депсидоны, хиноны, производные дибензофурана и т.д.
Размножение. Водоросли в слоевище лишайников размножаются только вегетативно; их клетки здесь крупнее, чем при жизни в свободном состоянии, так как, очевидно, при симбиотическом образе жизни у них подавляется клеточное деление. Однако грибы развивают характерные плодовые тела (апотеции, перитеции, псевдотеции). Их гимений чаше всего не содержит водорослей. Таким образом, новое слоевище может образоваться при этом только тогда, когда прорастающий из споры мицелий случайно встретится с соответствующей водорослью. Уже удалось осуществить в отдельных случаях экспериментально подобный «лишайниковый синтез». Только у немногих лишайников (например, Endocarpori) в гимении располагаются водоросли, которые при выбрасывании спор могут увлекаться вместе с ними, так что начинающий развиваться гриб уже имеет в распоряжении нужную водоросль. Какую функцию выполняют пикнидии, встречающиеся у многих лишайников, еще неясно. Размножение лишайников осуществляется у листоватых и кустистых форм часто (в наших местах преимущественно) вегетативным путем. Прежде всего для этого служат соредии (рис. 11.64, D) — оплетенные гифами гриба клетки водоросли, которые часто образуются в определенных местах слоевища — соралях — и распространяются ветром, чтобы на подходящем субстрате снова вырасти в лишайник. У других видов на поверхности слоевища возникают выросты в виде палочек или кораллов (изидии), которые легко отламываются и также служат для вегетативного размножения. Наконец, у лишайников каждый отломанный кусок может снова вырасти в нормальное слоевище.
Встречаемость и образ жизни. Лишайники растут на самых разных субстратах: скалах, почве, коре лиственных и хвойных деревьев, мертвой древесине и т. д. В тропиках мелкие, так называемые эпифилль-ные, лишайники обитают также на листьях. Живущие на каменистом субстрате накипные лишайники, которые могут растворять
11.2. Бактерии, грибы, растения | 207
известняк (но не кварц), как первые поселенцы подготавливают субстрат для высших растений. Небольшое число видов лишайников обитает как на суше, так и в пресной воде, некоторые живут в море или в зоне попадания морских брызг (см. рис. 11.102). Наиболее пышно лишайники разрастаются в горных лесах умеренной зоны, в условиях высокой влажности воздуха и в туманных лесах тропических высокогорий, а также в тундрах, где почва часто на большом протяжении покрыта преимущественно лишайниками; они образуют здесь собственные растительные формации. Лишайники в целом избегают каменных пустынь больших городов, где они повреждаются газами, загрязняющими атмосферу (прежде всего SO2). Из-за своей различной чувствительности (некоторые накипные лишайники даже в значительной мере устойчивы) они могут служить как индикаторы уровня загрязнения воздуха.
Поглощение воды осуществляется гифами гриба. Особенно у крупных листоватых лишайников нередко часть гиф не увлажняется, так что даже при полном насыщении слоевища влагой сохраняется его аэрация; иногда на нижней стороне слоевища регулярно встречаются дыхательные поры (цифеллы). Обитатели незатененных поверхностей скал выносят не только большой нагрев (до 70 °C), но и полное высыхание на протяжении месяцев. Некоторые почвенные лишайники находятся в высушенном состоянии почти половину дней в году и тогда тоже метаболически неактивны; однако при увлажнении фотосинтез возобновляется уже через несколько минут (пойкилогидричные растения, см. 6.10.3.6). Лишайники с синезелеными водорослями в качестве фотобионтов нуждаются для фотосинтеза в жилкой воле, лишайники с зелеными водорослями могут довольствоваться и водяными парами.
Рост лишайников происходит по сравнению с другими слоевцовыми растениями очень медленно. Даже крупные листоватые и кустистые лишайники наших широт нарастают в год не более чем на 1 — 2 см. У растущего на скалах в альпийской зоне накипного лишайника Rhizocarpon geographic ит (см. рис. 11.63, Н) при опре
деленных условиях ежегодный прирост составлял примерно 0,5 мм. По среднему возрасту таких обитающих на скалах накипных лишайников определяли возраст послеледниковых морен. Продолжительность жизни лишайников колеблется от одного года (эпифильные лишайники в тропиках) до нескольких сот, возможно, даже тысяч лет (арктоальпийские накипные лишайники, обитающие на каменистом субстрате).
Лишайники как форпосты жизни внедряются в холодные пустыни высокогорий, а также Арктики и Антарктики; некоторые могут выдерживать охлаждение до -196 °C без повреждения и при -24 °C еще связывать СО2.
В ископаемом состоянии лишайники известны только с третичного периода (янтарь), однако уже как различные высокоразвитые виды, практически не отличающиеся от современных.
Применение. Cetraria islandica (исландский мох) — вид, распространенный в сухих лесах и пустошах от тундры до высокогорий, используется как целебное растение (отхаркивающее средство). Из нескольких лишайников недавно были выделены антибиотики. Считается, что обитающая в степях Северной Африки и Ближнего Востока «манна небесная» (Lecanora esculenta) — лишайник, имеющий слоевища от мелколопастных до клубневидных, — может использоваться в пищу. Некоторые лишайники, как, например, виды Roccella (см. рис. 11.63, J) в Северной Африке и на Канарских островах, дают краситель лакмус. Из Cladonia steHaris, чаще всего собираемой в Северной Европе, делают Dauer-kranze. Evemia prunastri (дубовый мох) дает ароматическое вещество (mousse de сйкпе). Кладония оленья (Cladonia rangiferina, В) наряду с другими кустистыми лишайниками составляет основной корм северного оленя. Letharia vulpina — желтый эпифитный кустистый лишайник, в основном распространенный в альпийской зоне, — единственный в Европе ядовитый лишайник; раньше он использовался для травли волков.
Систематика. Отдельные классы и порядки лишайников располагаются в фило
208 I ГЛАВА 11 СИСТЕМАТИКА И ФИЛОГЕНИЯ
генетической системе согласно систематическому положению микобионтов (ср., например, Lecanorales). Разграничение порядков и семейств лишайников, которых известно около 400 родов и более 20 000 видов, осуществляется по строению плодовых тел гриба, так как здесь мы имеем признаки, наиболее надежные для подразделения на основе родства. Важный признак — реакция на йод, если гифы или сумки окрашиваются в синий цвет, их называют амилоидные. Для различения видов служат и другие химические признаки.
Geosiphon pyriforme — замечательный пример осуществляющегося в настоящее время эндоцитобиоза между грибом (из класса зигомицетов) и цианобактерией типа Nostoc. В этом случае совместное существование в отличие от циклических эндоцитобиозов каждый раз устанавливается заново. Изучение этой чрезвычайно важной ассоциации проливает свет на возможный путь приобретения хлоропластов путем эндоцитобиоза и, кроме того, мсжет служить моделью формирования отдельных элементов клетки. Однако из-за эндосим-биотического образа жизни фотобионта в гифах гриба Geosiphon не следует относить к лишайникам. То же самое можно сказать о соответствующих ассоциациях между слизевиками (Myxobionta) и водорослями в условиях культуры.
I. Ascolichenes
(см. класс Ascomycetes)
Порядки лишайников очень близки к соответствующим порядкам грибов в системе аскомицетов и отчасти уже обсуждались при рассмотрении последних (например, Lecanorales). Также мы встречаем порядки с переходами между лихенизирован-ными формами и такими формами, которые образуют с водорослями морфологически высокоразвитые слоевища лишайников. Как и для аскомицетов, в основе разделения на порядки лежит строение сумки. За Caliciales с часто прототуникатными сумками следуют Ostropales и Graphidales с унитуникатными, Lecanorales — с преимущественно унитуникатными, а также Руге-nulales, Verrucariales и Arthoniales — с биту-
никатными сумками. Dothideales отличаются битуникатными сумками в плодовых телах, имеющих чисто асколокулярное развитие (см. Ascomycetes).
1. Порядок: Caliciales. В этом порядке имеются и нелихенизированные представители; ли-хенизированные формы образуют накипные, листоватые или кустистые слоевища. Апотеции чаще всего на хорошо выраженной ножке. Сумки распадаются, так что споры с растущими дальше парафизами образуют рыхлую массу (мазедий). Зрелые споры — от одно- до многоклеточных — находятся чаще всего по 8 в сумке. Виды рода Calicium встречаются прежде всего на коре, рода Sphaerophorus— преимущественно на силикатном каменистом субстрате.
2. Порядок: Ostropales. Сюда относятся накипные лишайники с апотециями. Бесцветные амилоидные или коричневые аскоспоры могут быть разделены как поперечными, так и продольными и поперечными перегородками; они находятся в количестве от 1 до 8 в неамилоидных сумках. Плодовые тела развиваются, как и у Caliciales, по гемиангиокарпному типу. Апотеции — или широко раскрывающиеся, дисковидные, или узкие, бокаловидные, на ножках — погружены в слоевище или в бородавочки на слоевище.
3. Порядок: Graphidales. Апотеции чаще всего в виде черточек, реже округлые, погруженные или сидячие. Сумки располагаются между простыми или разветвленными парафизами. Водоросли относятся к Chlorococcales или это виды рода Trentepohlia. Примерно 300 видов рода Gra-phis обитают преимущественно в тропиках и субтропиках на коре деревьев; в Германии на коре буков распространен «письменный лишайник» (Graphis scripta, см. рис. 11.63, F), апотеции которого напоминают старинные письмена.
4. Порядок: Lecanorales Этот очень разнообразный по строению слоевища обширный порядок (несколько подпорядков) отличается округлыми, часто блюдцевидными апотециями. Стенка сумки чаще всего толстая, унитуникатная (см. Ascomycetes), у Lichininae прототуникатная, у Peltigerinae двухслойная. Фотобионты — зеленые водоросли (например, Trebouxia, Соссотуха) и синезеленые водоросли (например, Nostoc).
Виды рода Ephebe (Lichininae) образуют войлочные нити слоевищ на влажных силикатных породах в горах средней высоты и на высокогорьях. Род Collema (Collematinae) отличается слизистыми, напоминающими листья слоевищами с видами Nostoc в качестве симбионтов; слизистые лишайники (см. рис. 11.64, А) обитают на камне и почве в равнинных местностях до снежного пояса.
11.2. Бактерии, грибы, растения | 209
Представители последующих родов в пределах Lecanorales имеют гетеромерные слоевища. Накипному типу слоевища соответствуют роды Pertusaria, Rhizocarpon (географический лишайник, рис. 11.63, Н) и Lecidea. Слоевища от накипно-чешуйчатых до чисто листоватых имеют виды Xanthoria, Acarospora и Lecanom, слоевища видов рода Xanthoria, часто имеющие окраску от желтой до оранжевой, встречаются в местах, богатых азотом. Acarospora sinopica растет на силикатном субстрате, богатом тяжелыми металлами (см. также Acarosporetum, 6.2.2.4). К листоватым лишайникам относятся виды Parmelia и Physcia. К Peltigerinae, часто имеющим весьма крупные слоевища, относятся: Peltigera — с расположенными по краям слоевища щитовидными апотециями и у некоторых видов с цефалодиями: Lobaria puhnonaria, растущий только в условиях чистого воздуха эпифит на старых лиственных деревьях, преимущественно в горах; Sticta с продырявленной цифеллами нижней стороной слоевища; Solorina с расположенными по всему слоевищу апотециями и часто также с синезелеными водорослями в слоевище, которые нередко могут в значительной степени вытеснять зеленые водоросли. Умбиликариевые (Umbilicariinae; Umbilicaria-. слоевище в диаметре до 15 см и более) обитают на силикатных скалах в горных местностях холодной и умеренной зон. Кустистые лишайники представлены следующими родами: Alectoria, Ramalina-, Letharia', Evemia; Ana-ptychia', Usnea (бородач, К) — co слоевищами в виде длинных (до 8 м) бород, свисающих с ветвей деревьев; Thamnolia (червячный лишайник, D) — на всех высокогорьях и в арктических областях; Cladonia и Stereocaulon (Cladoniinae; В, С) представлены многочисленными разнообразнейшими формами, распространенными по всем климатическим зонам; сюда же относится и исландский мох (Cetraria islandica).
5. Порядок: Pyrenulales. Накипные лишайники с перитециями, развивающимися по аскоги-мениальному типу. Многоклеточные споры располагаются обычно по 8 в сумках, которые находятся между нитевидными парафизами. Фотобионты — виды Trentepohlia. Многочисленные виды рода Pyrenula встречаются на коре деревьев прежде всего в тропиках и субтропиках.
6. Порядок: Verrucariaies. Лишайники с сидячими или, чаще, погруженными в слоевище апотециями, которые развиваются по аскоги-мениальному типу. Настоящие парафизы отсутствуют. Булавовидные или цилиндрические сумки содержат 1 — 8 бесцветных или коричневых спор. Водоросли — Chlorococcales. Многие виды Verrncaria могут быть от эндо- до эпилитов на известковых скалах, некоторые виды живут постоянно или временно под водой в пресных
ручьях и по морским побережьям. Слоевища видов Dennatocarpon, от чешуйчатых до листовидных, поселяются на скалах. Виды рода Endocarpon обладают муральными спорами (как на рис. 11.41, Е).
Если во всех ранее названных порядках (I — 6) плодовые тела развиваются по аскогимени-альному типу, в последующих порядках (7—8) имеются отклонения от этого распространенного варианта.
7. Порядок: Arthoniales. Лишайники самого разного облика (от накипных до кустистых) с округлыми (апотеции) до штриховидных (гис-теротеции) плодовыми телами. Сумки от булавовидных до яйцевидных, споры от двух- до многоклеточных. Водоросли: преимущественно Trentepohlia. Примерно 500 видов рода Arthonia, обитают в основном в тропиках, имеют тонкие накипные, живущие обычно на коре деревьев слоевища: апотеции, от округлых до звездчато-рассеченных на лопасти, не имеют края и эксципула. Также многочисленные накипные виды рода Opegrapha с округлыми до штриховидных черными плодовыми телами в стерильном эксципуле широко распространены по всему земному шару на коре деревьев, древесине и каменистом субстрате. Виды рода Roccella (рис. 11.63, J) образуют кустистые слоевища и заселяют преимущественно скалы по побережьям теплых морей.
8. Порядок: Dothideales. Этот порядок отличается псевдотециями, которые в своем развитии следуют асколокулярному типу. Гифы гриба рыхло перемешаны с различными водорослями.
II. Basidiolichenes
(см. класс Basidiomycetes)
Долгое время были известны лишь немногие тропические представители порядка Poriales, которые были ассоциированы с цианобактериями, например пантропический эпигенный вид Dictyonema pavonia (= Cora pavonia', см. рис. 11.63, А). К настоящему времени базидиомицетные лишайники найдены как в тропиках, так и в умеренной зоне. Их компонентами являются Clavariaceae или Agaricales (Omphalina) и зеленые водоросли (Соссотуха и др.).
Пятое подцарство: Glaucobionta
Отличительная особенность видов этого небольшого, очень обособленного, но тем не менее важного подцарства — пластиды,
210 | ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
которые в качестве дополнительных пигментов содержат фикобилипротеиды. Из хлорофиллов типичны хлорофиллы а, с и d\ хлорофилл b отсутствует. Крахмал как запасное вещество внутри пластид не откладывается. У Glaucobionta достигается эука-риотный тип организации у водорослей (см. бокс 11.5).
Единственный отдел: глаукофиты (Glaucophyta)
Для понимания эволюции фотоавтотрофных эукариот этот отдел, включающий всего несколько видов монадных водорослей, имеет большое значение. Хлоропласты во многих отношениях напоминают еще синезеленые водоросли; они уже давно трактовались как цианобактерии, ведущие эндосимбиотический образ жизни, и получили название цианеллы. Они (или же хлоропласты) еще окружены пептидоглюка-новой стенкой, хотя и тонкой, однако размер их генома составляет одну десятую генома свободноживущих цианобактерий и, таким образом, соответствует геному хлоропластов прочих эукариот. Значительное сходство между цианеллами и хлоропластами имеется и в организации генома.
Синезеленые пластиды содержат хлорофилл а (и т.д.) и как дополнительные пигменты фикоцианин и некоторые другие в фикобилисомах, которые расположены внутри хлоропластов на тилакоидах, не сгруппированных между собой.
Единственный класс отдела — Glaucophyceae — включает три рода водорослей (см. рис. 2.99) из пресных вод, каждый с одним видом. Клетки Glaucocystis несут два рудиментарных жгутика и уже не способны к активному движению. Cyanophom имеет два жгутика неравной длины.
Шестое подцарство: Rhodobionta
Представители этого подпарства имеют пластиды, которые напоминают пластиды у Glaucobionta. Однако оболочка хлоропласта уже не содержит пептидоглюкан Клетки, имеющие жгутики, полностью отсутствуют.
Единственный отдел: красные водоросли (Rhodophyta)
Красные водоросли, обитающие преимущественно в морях, бывают от светло-красного до фиолетового цвета, изредка от темно-пурпурного, коричнево-красного до почти черного или даже от сине- до оливково-зеленого. Формы или стадии, имеющие жгутики, такие, как монадные виды, зооспоры или сперматозоиды, отсутствуют. Одноклеточные представители встречаются только в достаточно обособленном подклассе Bangiophycidae. Как в этой небольшой группе, так и среди других красных водорослей преобладают представители с талломами в виде отдельных нитей и сплетения нитей; другие формы имеют псев-допаренхиматозное (ложнотканевое) строение. Настоящие ткани полностью отсутствуют. Сплетения нитей и псевдопаренхимы (= плектенхимы) красных водорослей образуются по одноосевому (центральнонитчатому) или многоосевому (фонтанному) типу (см. 5.3.1; рис. 5.6, 5.7). В клетках, почти без исключения одноядерных, находятся чаще всего многочисленные дисковидные, овальные или лопастные хромато-форы (пластиды), которые, однако, никогда не бывают бокаловидными.
Пластиды- В них хлорофилл а (хлорофиллов b и с нет; наличие хлорофилла d долгое время считалось спорным) и сопутствующие каротиноиды маскируются красными, сильно флуоресцирующими пигментами, которые находятся в так называемых фикобилисомах. Фикобилисомы (они имеются и у прокариотических цианобактерий) представляют собой дисковидные или шаровидные тельца размером 30—540 нм. Они располагаются в хрома-тофорах на тилакоидах и содержат водорастворимые фикобилипротеиды с просте-тическими фикобилинами, определяющими характер окраски. У красных водорослей она обусловлена преимущественно красным фикоэритрином; в фикобилисомах содержится также фикоцианин (см. рис. 2.89, В, С).
У обоих пигментов имеется несколько вариантов, которые различаются, в частности, абсорбцией и принадлежностью к тем
11.2. Бактерии, грибы, растения | 211
Бокс 11.5. Типы организации эукариотных водорослей
Виды Glaucophyta, Rhodobionta и примыкающих к ним групп (например, Heterokontophyta), как и примитивно устроенные представители Chlorobionta (Chlorophyta, Streptophytina), трактуются как водоросли. Эукариотные водоросли (т.е. водоросли, в узком смысле слова, без прокариотных синезеленых водорослей) — это одноклеточные или многоклеточные, по-разному окрашенные, первично фотоавтотрофные растения, чаще всего имеющие талломную организацию и в основном обитающие в воде. Их хлоропласты содержат фотосинтетические пигменты наряду с дополнительными пигментами. Пластиды всех эукариотных водорослей содержат хлорофилл а и большей частью еще и другие формы хлорофилла (табл. 11.3). Для оксигенного фотосинтеза в качестве донора электронов служит вода, при этом высвобождается кислород. Под дополнительными пигментами понимают различные каротиноиды (а также встречающиеся у немногих групп фикобилины, см. 6.4.2). Часто в хлоропластах или вне их имеются пиреноиды (см. 2.2.9.1). Хлоропласты ограничены либо двумя мембранами — простые пластиды, либо 3 — 4 мембранами — комплексные пластиды. У представителей некоторых групп водорослей (Cryptophyta, Chlorarachniophyta) комплексные пластиды содержат нуклеоморфу, которая рассматривается как редуцированное ядро эукариотного эндосимбионта. Согласно теории эндосимбиоза (см. 2.4.2) этот процесс обусловливает появление пластид в клетке хозяина. Предполагается, что простые пластиды (у Glaucophyta, Rhodophyta, Chlorophyta) произошли в результате единственного первичного эндоцитобиоза от прокариотных водорослей, роль которых отводится Cyanophyta. Первичный эндоцитобиоз был, возможно, уникальным эволюционным событием.
Комплексные пластиды (все другие отделы эукариотных водорослей), напротив, свидетельствуют о том, что они возникли в результате двух этапов эндосимбиоза (см. 2.4). После первичного эндосимбиоза происходили дальнейшие независимые события эндосимбиоза (вторичный эндоцитобиоз): уже эукариотные водоросли включались в клетки хозяина. Для водорослей с нуклеоморфой в пластидах такой вторичный эндоцитобиоз считается доказанным. Итак, эукариотные водоросли полифилетичны как в отношении клетки-хозяина, так и в отношении приобретения пластид в результате первичного или вторичного эндосимбиоза.
Родство пластид эукариотных водорослей и фотосинтетического аппарата прокариотных водорослей подтвердил анализ последовательностей малых субъединиц рРНК (SSU).
Органы, образующие гаметы и споры, не имеют многоклеточных покровов и чаще всего у них также отсутствуют постгенитальные, т.е. вырастающие позже, покровы. Спорообра-зуюшие органы (спорангии) всегда, а гаметангии чаще всего одноклеточные. Голые гаметангии водорослей в отличие от снабженных многоклеточными стенками и поэтому защищенных антеридиев и архегониев мхов и папоротников (см. бокс 11.8) носят названия спермогонии (сГ: со сперматозоидами, имеющими жгутики; см. рис. 11.73, Е: s) или же спер-матангии (о": со спермациями, лишенными жгутиков; см. рис. 11.67, D) и оогонии (<j>: с яйцеклеткой: см. рис. 11.73: о) или же карпо-гоны (^: с особым развитием после оплодотворения; см. рис. 11.67, F—Г).
Зиготы никогда не развиваются внутри, женских половых органов в многоклеточные зародыши. В большинстве групп водорослей репродуктивные клетки (гаметы, споры) имеют жгутики, хотя у некоторых более высокоразвитых групп жгутики имеются только у мужских гамет; лишь у немногих групп водорослей вообше не образуется жгутиковых стадий (Pennales, Rhodophyta, Zygnematophyceae). Жгутики имеют характерную для эукариот структуру 2 + 9 (см. рис. 2.16). Они обычно парные, причем один может быть направлен вперед по ходу движения, а второй — назад. Два жгутика могут быть одинаковой или разной длины, оба гладкие или разные: один гладкий, а другой снабжен перисто-расположенными волосками (перистый жгутик).
Водоросли в ходе эволюции прошли путь прогрессивного развития от одноклеточных форм до сложных ложпотканевых или тканевых талломов (см. гл. 5). Формы на уровне слоевцовых растений (таллофитов) еще не показывают дифференциации вегетативного тела на «настоящие» листья, стебли и корни. Несколько сходные образования у некоторых высокоразвитых водорослей не имеют проводящих структур, которые можно было бы сравнить с проводящими пучками сосудистых растений (например, у бурых водорослей есть лишь отдельные проводящие элементы, напоминающие ситовидные трубки). По большей части структуры, отдаленно напоминающие основные органы высших растений, не имеют
212 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
анатомической дифференциации, поэтому их обозначают как филлоиды, каулоцды и ризоиды (см. 5.3.2).
Можно различать следующие морфологические группы разной высоты организации (уровни организации).
• Амебоидный (= ризоподнальиый) уровень. Одноклеточные голые водоросли образуют псевдоподии, с помощью которых они захватывают твердые частицы пищи. Если эти придатки тонкие и нитевидные, их называют ри-зоподиями (см. рис. 11.74, С). Встречаются также объединения таких клеток.
• Монадный уровень. Одноклеточные водоросли со жгутиками, чаще всего снабженные глазками и пульсирующими вакуолями («флагелляты»; см. рис. 11.103). После клеточных делений они могут образовывать колонии, состоящие из нескольких (до очень многих) клеток (см. рис. 11.95, G; 11.74, F). Пальмеллоидная стадия (стадия пальмеллы) представляет пальмеллоидный (капсальный) уровень: при делении клеток не образуется новых жгутиков, а дочерние клетки погружены в общую слизь (см. рис. 11.75, G).
• Капсальный (= тетраспоральный) уровень. В рудиментарном виде еще сохранились различные признаки монадного уровня. Так, жгутики, если они еще сохранились, негнущиеся или редуцированные, в любом случае не способные к активному движению, могут быть только у репродуктивных клеток. Поскольку клетки после деления погружены в общую слизь, возникают колонии, которые могут быть даже вытянуты в нитевидные структуры (см. рис. 11.75, D). Клеточная стенка тонкая или отсутствует.
• Коккоцдный уровень. Вегетативные клетки не имеют жгутиков и покрыты клеточной стенкой. У них нет даже никаких остатков мо-надной организации. Могут быть одноклеточные формы, ценобии или колонии в более узком смысле (см. рис. 11.76; 11.97, А; 11.98, А).
• Нитчатый (= трихальный) уровень. Одноклеточные (моноэргидные) клетки образуют разветвленные или неразветвленные нити, нарастающие интеркалярно или верхушечно (см. рис. 11.89, А).
• Сифонокладальный уровень. Клетки, образующие нити, содержат по нескольку ядер; многоядерные клетки называют полиэнерги-дами.
• Сифональный уровень. Макроскопический таллом представлен одной-сдинственной крупной многоядерной клеткой — нитевидной или другого облика. Размеры таллома могут быть достаточно большими (см. рис. 11.73, D; 11.91; 11.92, D).
• Ложноткаиевой таллом. Боковые оси (нити) переплетаются друг с другом. Клетки часто слипаются или даже срастаются (см. 5.3.1; рис. 5.7, А).
• Тканевой таллом. Клетки, делящиеся в разных направлениях, остаются связанными между собой, образуя ткань (см. 5.3.2; рис. 5.4, G).
Кратко охарактеризованные здесь уровни организации достигались в разных эволюционных линиях водорослей независимо друг от друга, и их развитие происходило неодинаково. Тканевой таллом, например, появился в примитивной форме у Ulvophyceae и в особенно развитой форме — у Phaeophyceae, в ложнотканевой форме — у Phaeophyceae и Rhodophyceae.
или иным группам (к синезеленым или же красным водорослям). Фикобилисомы у синезеленых и красных водорослей улавливают свет и передают энергию возбуждения на собственно фотосинтезирующий пигмент. Расслоение фикобилинов в фикобилисомах (фикоцианины внутри, фикоэритрины снаружи) дает направление переносу энергии. У Ciyptophyta, также имеющих фикобилины, фикобилисомы отсутствуют.
В хлоропластах тилакоиды не сгруппированы, а располагаются с равными промежутками между собой (как у цианобактерий и глаукофитов). Двойная мембрана отграничивает хроматофоры от остальной части клетки; эндоплазматическая сеть в этом не участвует.
В качестве запасного вещества откладывается прежде всего багряпковый крахмал в форме округлых, не растворимых в воде, часто слоистых зернышек, дающих с йодом красноватое окрашивание. Это полисахарид, по своим свойствам занимающий промежуточное положение между гликогеном и крахмалом. Зернышки в отличие от крахмала у зеленых водорослей откладываются не внутри хлоропластов, а на их поверхности и в цитоплазме. Встречаются некоторые другие, присущие только красным водорослям вещества («флоридо-зиды» — соединения галактозы и глицерина), а также капельки масла.
Фибриллярный компонент клеточной стенки состоит преимущественно из цел
11.2. Бактерии, грибы, растения! 213
люлозы, микрофибриллы которой построены из цепочек, но не параллельно расположенных (как у высших растений и некоторых зеленых водорослей), а переплетенных наподобие войлока.
Аморфная часть часто содержит ослизненные галактаны (например, агар; карраге-нан = галактансульфат; галактаны — полимеры галактозы).
Жизненный цикл. Для красных водорослей характерна смена трех поколений, при которой за гаплоидным гаметофитом следуют диплоидный карпоспорофит и еще
одно диплоидное спорофитное поколение (преимущественно тетраспорофит).
При диплобионтном типе (три поколения, представленные двумя особями, например, Polysiphonia, рис. 11.65,'А) гаметофит представляет собой самостоятельное гаплоидное растение. Он развивает !j> гаметангий — карпогон. У многих красных водорослей (например, у всех флоридей) его расширенная нижняя часть (брюшко) переходит в трихогину — длинный, чаще всего тонкий воспринимающий орган (рис. 11.66, F: Г; 11.67, F: t). На других частях или дру
Рис. 11.65. Rhodobionta, Rhodophyceae. Смена поколений и ядерных фаз.
Трехчленный диплобионтный цикл Polysiphonia (А); трехчленный гаплобионтный цикл Batrachosper-тит (В). Светлые линии — гаплофаза; темные линии — диплофаза; R! — редукционное деление
214 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
гих особях гаметофитов в сперматангиях (= мужских гаметангиях) возникают лишенные жгутиков мужские (d") половые клетки — спермации. Они одноядерные, пассивно переносятся токами воды, затем прилипают к трихогине и отправляют в нее свое о" половое ядро, после чего это ядро мигрирует к ядру карпогона и сливается с ним (гамето-гаметангиогамия, или нетипичная оогамия).
Из оплодотворенной яйцеклетки возникает карпоспорофит в форме диплоидных нитей, разделенных на клетки, которые вырастают из карпогона, при этом, однако, оставаясь соединенными с гаплоидным гаметофитом. Соответственно происходит смена первых двух поколений, разделенных сменой ядерных фаз, на одном и том же растении. Карпоспорофит после ис
ключительно митотических делений дает диплоидные карпоспоры, которые, таким образом, являются митоспорами.
Тетраспорофит. У подавляющего большинства красных водорослей из карпоспоры возникает новое растение, чаще всего морфологически сходное с гаметофитом, однако диплоидное. На нем после редукционного деления из каждой материнской клетки спор образуется 4 гаплоидные тетрамейоспоры (см. рис. 11.65; 11.66, В); это поколение называется тетраспорофит. При переходе от карпоспорофита к тетраспорофиту происходит соответственно без изменения ядерной фазы смена 2-го и 3-го поколений жизненного цикла. Развитие трех поколений осуществляется, таким образом, на двух вегетативных телах (дипло-бионтное развитие); большинство красных
Рис. 11.66. Rhodophyceae (А, В — по G.G.Thuret; С, D — по Koch; Е — по F. Borgesen; F — по F.OItmanns; G — по Е. Borne):
А, В — Callithamnion corymbosum, образование тетраспор (ЗООх): А — закрытый, В — освободившийся спорангий с 4 мейоспорами; С, D — Bonnemaisonia hamifera, гаметофит и тетраспорофит (5х), С — гаметофит и зачатки цистокарпиев, D — спорофит, также известный как Trailiella intricate; Е — Ceramiales, Platysiphonia miniata, цистокарпий с видными в нем на просвет карпоспорофитами (100х); F, G — Cryptonemiales, Dudresnaya: F — оплодотворенный карпогон, на трихогине (t) которого приклеиваются еще несколько спермациев (s), дает начало разветвленным спорогенным нитям (sf), которые соединяются с шестью ауксилярными клетками (а1—аб), клетки а1—аб входят в состав ветвей, которые возникают от оси ha (250х), G — зрелое скопление карпоспор
11.2. Бактерии, грибы, растения | 215
водорослей (за исключением Nemalionales) относится к этому типу.
Гаметофит и тетраспорофит имеют чаще всего одинаковый внешний вид, но могут быть и непохожими (см. рис. 11.66, С, D), отчего раньше их относили не только к разным родам, но и даже к далеко отстоящим друг от друга порядкам. Также и паразитирующий на гаметофите карпоспорофит в отдельных случаях выглядит как нечто постороннее в такой степени, что его считали действительно инородным паразитом и давали ему особое название. Гаметофиты бывают однодомными и двудомными. В последнем случае иногда встречаются различия в строении сГ и $ растений. Часто карпоспорофит (= гонимокарп) обрастает специальными покровными нитями гаметофита, в результате чего возникает так называемое плодовое тело — цистокарпий (см. рис. 11.66, Е). Если покров закладывается уже перед оплодотворением карпогона, говорят о прокарпии.
Часто карпоспорофит поддерживается вспомогательными, или ауксилярными, клетками, которые, очевидно, имеют значение для физиологии питания. Это богатые цитоплазмой клетки карпо-спорофита, расположенные рядом с карпогоном, которые воспринимают ядро зиготы (или диплоидное ядро, возникшее после последующих делений) из карпогона, претерпевают последующие митотические деления и наконец продолжают образование карпоспорофита. Отходящие от карпогона соединительные нити (см. рис. 11.66, F: sf) могут достигать многих ауксилярных клеток. При этом диплоидные ядра размножаются и распространяются в гаметофите, так что вслед за одним-единственным актом оплодотворения от гаметофита возникают многочисленные карпоспорофи-ты, которые могут питаться от него.
Последовательность разных поколений на одном организме (гаплобионтный тип развития) можно проследить на встречающихся в пресной воде видах рода Batra-chospermum (см. рис. 11.65, В). Здесь также происходит трехчастная, гетероморфная и гетерофазная смена поколений, три члена которых, однако, остаются связанными между собой, а именно: 1) диплоидный «Chan-transia-спорофит» (предросток); 2) мутов-чато разветвленный гаплоидный гаметофит; 3) диплоидный карпоспорофит.
Гаметофит. Однодомный таллом состоит из гаплоидных, мутовчато разветвленных нитей (см. рис. 11.67, А). Многочисленные сперматангии возникают чаще всего по два из конечных клеток расположен
ных мутовками боковых ветвей (рис. 11.67, А). Каждый сперматангий состоит всего из одной клетки, все содержимое которой идет на образование одного-единственного округлого бесцветного спермация с большим ядром и очень нежной стенкой (D). Женские органы — карпогоны — сидят между ветвями, несущими сперматангии, также на концах ветвей и представляют собой длинную клетку, которая в нижней части бутылевидно расширена, а в верхней части вытянута в булавовидную трихогину (Е, F). Кар погон с трихогиной глубоко погружен в слизь. Спермаций, пассивно переносимый токами воды, может активно внедряться в эту слизь (механизм неизвестен); при этом он достигает трихогины, в которую выпускает все свое содержимое. Ядро спермация мигрирует в карпогон и сливается с находящимся там женским ядром. После этого базальная часть карпогона с образовавшимся диплоидным ядром отделяется от трихогины слизистой пробкой (G).
Карпоспорофит. Он состоит из разветвленных, разделенных на клетки нитей, которые вырастают из зиготы, однако при этом остаются связанными с гаметофитом (рис. 11.67, Н). Карпоспорофит образует во вздутых конечных клетках по одной округлой митоспоре, содержащей ядро и хлоропласт, — диплоидной карпоспоре. Карпоспоры высвобождаются из остающихся стенок конечных клеток (Ikl, k2) в виде округлых, лишенных жгутиков образований. Они вырастают в Chantransia-стадию (Chan-щотша-спорофит). Это разветвленные диплоидные нити, которые плотно прилегают к субстрату и представляют собой предросток для возникающих позже от них гаметофитов. СЛалГганяо-стадия, таким образом, еще диплоидна, но собственно гаметофит гаплоиден. Мейоз происходит без образования мейоспор в отдельных клетках нитей Chantransia. Гаплоидные клетки развиваются затем в мутовчато разветвленные гаметофиты (С).
У некоторых видов в качестве 2-го спорофита образуется маленькое, мутовчато разветвленное растение, которое в определенных местах (после мейоза) непосредственно дает начало гаметофитам; это сближает Chantransia-cra-дию со спорофитным поколением.
216 | ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Рис. 11.67. Rhodophyceae, Nemalionales Batrachospermum moniliforme (А—С — no Sirodot; D — по E. Strasburger; E—I — no H.Kylin):
A — внешний вид (Зх); В — часть таллома гаметофита с мутовкой ветвей (20х); С — диплоидный спорофит (Chantransia) с двумя сидящими на нем гаметофитами (10Ох); D — часть ветви гаметофита с 4 сперматангиями, слева вышедший спермаций (540х); Е — зачаток карпогона; F — зрелый карпо-гон; G — карпогон после оплодотворения спермацием (s), в основании его копуляция половых ядер (о); Н — диплоидный карпоспорофит (д) с гаплоидными покровными нитями (h); I — зрелый карпо-спорофит с карпоспорангиями (к); к1, к2 — освободившиеся карпоспорангии; t — трихогина
Распространение и образ жизни. Красные водоросли представлены примерно 4000 видами в составе более чем 500 родов. За немногими исключениями (например, Batrachospermum, Lemanea), они обитают в прибрежной зоне морей, особенно более теплых; многие виды очень чувствительны к колебаниям температуры. Часто красные водоросли растут значительно глубже (максимум до 180 м), куда доходит лишь слабый коротковолновый свет, где они не только могут жить как теневыносливые водоросли, но и благодаря наличию дополнительных пигментов (фикобилипротеи-дов) могут оптимально использовать пре
обладающий на глубине коротковолновый свет, комплементарный к собственной окраске этих пигментов (см. 6.4.3). Красные водоросли — представители бентоса. Они всегда прочно прирастают специальными нитями или присосками, чаще всего к каменистому субстрату, некоторые растут как эпифиты на более крупных водорослях. Такие эпифиты могут быть очень строго приурочены к определенному роду растения-форофита (например, виды рода Polysipho-nia к Ascophyllum). Красные водоросли живут автотрофно; часть из них — бесцветные паразиты, из которых несколько десятков представляют собой очень редуци
11.2. Бактерии, грибы, растения | 217
рованные формы, обитающие на других, близко родственных им видах красных водорослей («адельфопаразиты»).
Систематика. Единственный класс красных водорослей — Rhodophyceae. Он подразделяется на подклассы Bangiophycidae и Florideophycidae.
1. ПОДКЛАСС: BANGIOPHYCIDAE
Это весьма просто устроенные одноклеточные, нитчатые или листовидные водоросли с ин-теркалярным ростом. Поры обычно отсутствуют. Хлоропласты звездчатые, с пиреноидом.
1. Порядок: Porphyridiales. Объединяет одноклеточные, отчасти колониальные формы. Половое размножение неизвестно. У часто встречающейся наземной водоросли Porphyridium purpureum многочисленные отдельные клетки объединены в слизи.
2. Порядок: Bangiales. К этому порядку относятся нитчатые (например, Bangia) или листовидные (например, Porphyra} формы. Erythrotrichia образует неразветвленные нити, в которых могут возникать вегетативные моноспороцисты, а в них — по одной моноспоре. Сначала моноспоры голые, им свойственно амебоидное движение. Из них вырастают новые нити Erythrotrichia. У Porphyra (пурпурной водоросли) карпогоны чаще всего неотличимы от вегетативных клеток. Оплодотворенная спермациями диплоидная зигота после нескольких митозов непосредственно делится на некоторое число диплоидных карпоспор; таким образом, карпоспорофитное поколение либо отсутствует, либо сильно редуцировано. Карпоспора прорастает в нитчатое, очевидно. также диплоидное образование, которое внедряется в известковую раковину двустворчатых моллюсков и морских желудей. Начинается развитие так называемой Содс/госе/А-фазы, которая представляет собой спорофитное поколение. Оно заканчивается образованием конхо-спор (гомологичных тетраспорам), после чего, по всей вероятности, происходит редукционное деление. Conchocelis-фаза широко распространена по морским побережьям Европы. К этому же порядку, возможно, относится Cyanidium caldarium — вид, относительно которого дискутируется родственная связь с синезелеными водорослями (цианобактериями)* 1.
1 Практически во всех современных системах Cyanidium caldarium и некоторые родственные этому виду формы выделяются в отдельный класс Cyanidiophyceae, а он иногда выделяется в самостоятельный отдел Prerhodophyta. — Примеч. пер.
2. ПОДКЛАСС: FLORIDEOPHYCIDAE
Представители этого подкласса имеют более сложное строение таллома, основу которого составляют разветвленные нити с верхушечным ростом. Одноклеточные представители не встречаются. Даже простейшие Florideophycidae уже имеют гетеротрихальное строение (т.е. дифференцированы на подошву и вертикальные нити), однако даже наиболее высокоразвитые представители в противоположность бурым водорослям никогда не бывают паренхиматозными (см. 5.3.1), а образуют свой таллом по одноосевому или фонтанному типу (см. рис. 5.6; 5.7). Клетки часто соединены друг с другом «порами». Это отверстия или каналы со структурами в виде пробок внутри; их функция однозначно не выяснена.
1. Порядок: Nemalionales1. Ауксилярные клетки отсутствуют. Этот порядок представлен в Средней Европе видами рода Batrachospermum, растущими преимущественно в быстротекущих, затененных ручьях в форме слизистых масс, напоминающих икру, от буроватого до оливковозеленого цвета. Batrachospermum показывает отклоняющийся от нормального типа гаплобионт-ный цикл развития. Он также четко отличается от большинства других красных водорослей, тем что обитает в пресной воде, а именно чаще всего в быстротекущих ручьях с высокой чистотой воды.
Rhodochorton investiens— красная водоросль, живущая как эпифит на Batrachospermum, имеет, напротив, нормальный диплобионтный цикл развития; гаметофит и тетраспорофит в этом случае в значительной мере сходны между собой. Близкородственный морской вид Rhodochorton purpureum — гаплобионт с двучленной сменой поколений: карпоспорофитное поколение выпадает, а оплодотворенный карпогон непосредственно вырастает в тетраспорофит; последний остается сросшимся с гаметофитом того же облика.
Другой род пресноводных водорослей из этого порядка — Lemanea, тогда как виды родов Nemalion, Bonnemaisonia (см. рис. 1L66, С, D) и Gelidium живут в морях. Систематическое положение последнего рода спорное, так как здесь встречаются клетки, трактуемые как ауксилярные2.
1 Правильнее назвать этот порядок Nemalia-les. — Примеч. пер.
1 В настоящее время роды Batrachospermum, Lemanea и некоторые другие, отличающиеся сходным жизненным циклом (без образования мейоспор), обитанием в пресной воде и отсутствием красной окраски, обычно выделяются в отдельный порядок Batrachospermales; род Gelidium выделяется в порядок Gelidiales. — Примеч. пер.
218 I ГЛАВА 11, СИСТЕМАТИКА И ФИЛОГЕНИЯ
2. Порядок: Cryptonemiales. Ауксилярные клетки закладываются до оплодотворения на специальных пучках ветвей. У Corallinaceae1 (Corallina, Lithothamnion, Lithophyllum) клеточные стенки инкрустированы кристаллами кальцита (образование коралловых рифов); ископаемые представители имеют значение как образователи скальных пород. Сюда относится также Melobesia (см. рис. 5.7, В) с талломом, развивающимся по «фонтанному типу».
3. Порядок: Gigartinales. В ауксилярную клетку превращается нормальная интеркалярная клетка таллома. К этому порядку относятся гребневидно-перистый Plocamium, построенная по фонтанному типу Furcellaria (см. рис. 5.7, А) и плоско-вильчатый Chondrus.
4. Порядок: Rhodymeniales. Несущая клетка карпогона отшнуровывает перед оплодотворением дочернюю клетку, а та, в свою очередь, — ауксилярную клетку. Карпогон возникает от прокарпия (состоящего из несущей клетки, дочерней клетки, ауксилярной клетки и карпогонной ветви), который после оплодотворения становится цистокарпием. Сюда относится часто встречающаяся в Атлантике Rhodymenia с листовидным талломом.
5. Порядок: Ceramiales. Ауксилярная клетка обособляется после оплодотворения карпогона от несущей клетки карпогонной оси. Имеются прокарпий (здесь состоящий из несущей клетки, ауксилярной клетки и карпогонной ветви) и цистокарпий, как и у предыдущего порядка. Жизненный цикл соответствует представленной вначале основной схеме (см. рис. 11.65, А). Таллом построен по одноосевому типу и состоит из обильно разветвленных нитей, часто образующих кору.
Особенно сложно расчленен таллом водоросли Delesseria sanguinea, обитающей в Атлантическом океане. Листовидные лопасти ее таллома, отходящие от базального диска, имеют центральную и боковые жилки; осенью пластины отмирают, но основные оси остаются и развивают следующей весной новые пластины. К Сега-miales относятся Grinnellia (см. рис. 5.8, A), Platysi-phonia (см. рис. 11.66, Е), а также представленные в Северном и Балтийском морях роды Polysiphonia. Ceramium и Plumaria.
Раньше иногда предполагалось родство красных водорослей с аскомицетами (Mycobionta). Это предположение не было подтверждено данными анализа ДНК. Действительно, обращает особое внимание
1 В большинстве современных систем это отдельный порядок Corallinales. — Примеч. пер.
сходство следующих признаков; ф гаметангий (здесь — карпогон, там — аскогон, иногда с трихогиной; здесь — нити кар-поспорофита, там — аскогенные гифы); взаимосвязь гаметофитного и спорофит-ного поколения; отсутствие имеющих жгутики репродуктивных клеток; оплодотворение спермациями (как и у некоторых аскомицетов); слоевища лишайников с порами в септах и способностью к образованию анастомозов; сходство в химическом строении гликогена грибов и багрян-кового крахмала; запасной продукт трегалоза. Однако химический состав клеточной стенки различен; здесь целлюлоза, там хитин.
В ископаемом состоянии красные водоросли встречаются с перми во всех формациях; отнесение Solenopora из ордовика к красным водорослям является спорным.
• Отделы водорослей с комплексными пластидами без хлорофилла Ь. Последующие отделы водорослей имеют комплексные пластиды, которые возникли в результате вторичного эндосимбиоза. Поскольку эти пластиды не содержат хлорофилла Ь, но очень часто в качестве дополнительных пигментов присутствуют фикобилины. Предполагается их происхождение от предков, близких к предкам Rhodobionta, что подтверждается сходством состава пластидной ДНК.
1. Приложение к Rhodobionta: отдел криптофиты (Cryptophyta)
Представители этого отдела, за немногими исключениями (пальмеллоидные и нитчатые формы) — жгутиковые организмы (монадный тип организации). Род Bjombergiella отличается нитчатым талломом. Снабженные жгутиками асимметричные клетки, характерные для подавляющего большинства видов, не имеют клеточной стенки, а покрыты только пелликулой, которая состоит из прямоугольных или многоугольных белковых пластинок. На переднем конце выходят два жгутика, несколько различающиеся по длине. Оба жгутика несут перистые волоски: более длин
11.2. Бактерии, грибы, растения! 219
ный — в два ряда, более короткий — в один.
Жгутики чаще всего направлены вперед, реже — вдоль тела назад (рис. 11.68, С). Они выходят вместе над глубокой глоткой, которая выложена эджектосомами, сильно преломляющими свет, — это тельца, которые при раздражении выстреливают. По-разному окрашенные (например, синие, сине-зеленоватые, красноватые) хлоропласты содержат хлорофиллы а и с, а- и p-каротин и ксантофилл диатоксан-тин, а также фикобилины фикоэритрин и фикоцианин. В отличие от красных водорослей и цианобактерий эти пигменты здесь не локализованы в фикобили-сомах. Важный запасной продукт — крахмал, который откладывается в пиреноидах. Последние, правда, расположены в складке эндоплазматической сети (= оболочке хлоропласта), однако вне мембраны хлоропласта.
Вегетативное размножение осуществляется путем продольного деления; половое размножение с достоверностью не установлено.
Живущие в морях и пресных водах (примерно поровну) 120 видов распределены по 12 родам.
Хлоропласты можно рассматривать как сильноредуцированные эукариотные эндосимбионты (родственные красным водорослям) на основании наличия сохранившегося недоразвитого клеточного ядра — нуклсоморфы. Она располагается в единственном числе в пиреноиде или у поверхности хлоропласта.
Можно предполагать, что криптофиты исходно имели первичные пластиды типа цианелл (см. Glaucobionta), которые, однако, в ходе дальнейшей эволюции были заменены внедрившимися одноклеточными красными водорослями (путем вторичного эндоцитобиоза).
Отдел содержит единственный класс с единственным порядком — Cryptomonadalcs Многочисленные виды рода Cryptomonas (рис. 11.68, А) обитают преимущественно в мезотрофных водоемах.
Сапротрофные представители рода Chilo-monas (В) бесцветны, однако еще содержат один лейкопласт. Бесцветен и фаготроф Katablepharis (С). Один вид представляет собой сильноредуцированную форму, живущую как эндосимбионт в инфузории Mesodinium rubrum, которая таким образом получила способность к фотосин-
Рис. 11.68. Cryptophyta (А — по В. Fott; В — по
V. Uhlela; С — по H.Skuja)
А — Cryptomonas sp.; В — Chilomonas paramaecium; С — Katablepharis phoenicoston с одним жгутиком, направленным вперед, и другим жгутиком, направленным назад (А—С — 1 200х); с — хлоропласт с несколькими пиреноидами (показан точками); к — ядро; s — глотка; v — вакуоль
тезу. Эта симбиотическая водоросль содержит лишь один хлоропласт и немногочисленные митохондрии.
2. Приложение к Rhodobionta: отдел динофиты (Dinophyta, Pyrrhophyta, Dinoflagellate)
В большинстве своем одноклеточные формы, несущие два жгутика с тонкими ма-стигонемами; известны лишь немногие коккоидные и нитчатые формы. Наряду с вегетативным встречается половое размножение.
Хлоропласты содержат хлорофилл а; у некоторых видов установлено также наличие хлорофилла с. Окраска клеток от желто-бурой до красноватой благодаря наличию дополнительных пигментов, таких, как 0-каротин и различные ксантофиллы, из которых наиболее важен пери-динин. Оболочка хлоропласта состоит чаще всего из трех мембран, которые не связаны с эндоплазматической сетью ядра. Тилакоиды располагаются в ламеллах по три и не образуют периферической опоясывающей ламеллы (таким образом, картина иная, чем на рис. 11.72, G). Основной продукт ассимиляции — крахмал, который накапливается в виде зернышек вне хло-
220 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
poiнастов. Наряду с ним встречаются также жироподобные вещества. Для некоторых представителей имеются сведения, что исходно имевшаяся простая пластида значительно редуцировалась вплоть до того, что остался один глазок; она стала фотосинтетически неактивной и в результате вторичного эндосимбиоза была функционально заменена комплексной пластидой.
Динофиты, возможно, представляют собой гетерогенную группу эукариотных водорослей. Они содержат пластиды очень разного типа. Это взаимосвязано с их способностью к фаготрофии, которая сохранилась и у автотрофных видов. У некоторых видов пластиды не являются постоянными компонентами клеток— это клсптохлоропласты. Есть виды, у которых, возможно, имеется нуклео-морфа. Молекулярно-филогенетический анализ ядерного генома дает основание предполагать близкое родство динофитов и относимых к животным жгутиковых (Ciliata, Sporozoa или Apicom-plexa). У гетеротрофно живущего возбудителя малярии Plasmodium falciparum уже не выявляются активные остатки комплексных пластид, которые подтверждали бы родство с динофитами.
Клеточная стенка часто имеет тонкие поры, которые выходят в трихоцисты, имеющие форму мешочков. При раздражении трихоцисты выбрасывают белковые нити. Клеточная стенка у многих динофитов построена очень характерным образом из многоугольных целлюлозных пластинок, которые образуют панцирь с поперечной и продольной бороздами. Чешуйки панциря (как и створки у диатомовых водорос-
Рис. 11.69. Dinophyta. Зооксантеллы в радиолярии (Eucoronis challenged', 260х) (по Е. Haeckel)
лей) закладываются в плоских полостях внутри плазмалеммы; поверх панциря сохраняется плазматическая мембрана.
На пересечении продольной и поперечной борозд отходят два жгутика, каждый из которых располагается в одной из борозд (см. рис. 11.70, А). Поперечный жгутик несет один ряд несколько более длинных волосков, продольный жгутик — два ряда более коротких волосков. Боковые волоски значительно тоньше, чем у Heterokontophyta и Cryptophyta. Жгутик, колеблющийся в поперечной борозде, вызывает вращательное движение клетки вокруг продольной оси, тогда как жгутик, находящийся в продольной оси, обеспечивает движение клетки вперед. Например, клетка Peridinium, перемещаясь по винтовой линии, за секунду преодолевает расстояние, в несколько раз большее своей длины.
Хромосомы у большинства динофитов (как и у эвгленовых) различимы и в рабочем ядре, так как в интерфазе они останутся в конденсированном состоянии (ср. другие эукариоты; см. рис. 2.27).
При электронной микроскопии хромосомы выглядят как компактно сложенные фибриллы («гирлянды»). Фибриллы имеют диаметр 2,5 нм, что соответствует диаметру двойной спирали ДНК (см. рис. 1.5). Хромосомы других эукариот, напротив, обладают субмикроскопическими фибриллами, имеющими диаметр, примерно в 10 раз больший — 25 — 30 нм (см. рис. 2.21). Эти более толстые хроматиновые спирали из двойной спирали ДНК с центральным нуклеогистоновым тяжем в хромосомах динофитов отсутствуют. В этом имеется некоторое сходство с нуклеоплазмой прокариотных бактерий и синезеленых водорослей.
Вегетативное размножение осуществляется путем косого деления. У имеющих панцирь форм (например, Ceratium) покров, как правило, разрывается под углом к поперечной борозде, и у новых клеток восполняются недостающие половины панциря (см. рис. 2.83, D). Однако у видов некоторых родов (например, Peridimum) весь панцирь отбрасывается перед делением, так что каждая из возникших дочерних клеток полностью заново строит собственный панцирь После нескольких таких делений внутри панциря развиваются две клетки, сначала
11.2. Бактерии, грибы, растения | 221
Рис. 11.70. Dinophyta (Pyrrhophyceae)(A — по A. J. Schilling; В — по Stein; С — по Schutt; D — по Lauterborn; Е — по A. J. Schilling; F, G, Н — по G. Karsten; J — по Schutt):
А — Peridinium tabulatum (600х); В — Gymnodinium aeruginosum (ЗООх); С — Ornithocercus splendidus (125х); Dj, D2 — Ceratium hirundinella после деления (350x); E — Ceratium cornutum, циста (150x); F — Ceratium tripos (125x); G — C. palmatum (125x); H — C. reticulatum (65x); J —C. fuses (50x)
голые, которые покидают материнский покров и покрываются новыми панцирями. При неблагоприятных условиях внутри панциря возникают толстостенные цисты, способные длительно сохранять жизнеспособность.
Половое размножение на данный момент установлено лишь у немногих дино-фитов. У Ceratium оно анизогамное, с зиготической сменой ядерных фаз (мейоз при прорастании зиготы); у Glenodinium описаны изогаметы, которые возникают в клетках (гаметангиях), выходят наружу и сливаются друг с другом.
Распространение. Большинство динофи-тов из примерно 1 000 видов (120 родов) обитает в морях, где они совместно с диатомеями (отдел Heterokontophyta) составляют основную часть фитопланктона (важные первичные продуценты в морях). Наибольшее разнообразие форм достигается в более теплых водах, а наиболее массовое
развитие — в более холодных водах. В пресных водах обитают лишь немногие Peridi-niales, однако иногда в большом количестве; в высокогорных озерах они могут составлять до 50 % биомассы. Многие виды обладают характерными придатками, служащими для парения в воде (рис. 11.70, С, F—J). Noctiluca miliaris (гетеротрофный вид с голыми клетками!), а также виды Ceratium, Gonyaulax и Peridinium вызывают свечение морской воды. Массовое развитие динофи-тов («красные приливы») могут вызывать гибель рыб1. Округлые эндосимбионты разных морских животных известны под общим названием «зооксантеллы» (рис. 11.69). Все кораллы, образующие рифы, живут в симбиозе с такими динофитами. Без эндо
1 От «красных приливов» страдают не только рыбы, но и многие беспозвоночные; употребление всех этих животных в пищу может вызывать отравление человека. — Примеч. пер.
222 | ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
симбионтов кораллы выживают, однако теряют способность образовывать известковый скелет. Некоторые виды динофитов живут как паразиты на поверхности и внутри тела морских животных. Среди подобных гетеротрофных форм встречается фаготро-фия («заглатывание» бактерий и планктонных водорослей).
Систематика. Единственный класс Dinophy-сеае подразделяется на 4 порядка. Представители Dinophysiales (1) имеют стенку, две половинки которой (эпикон и гипокон) дополнительно подразделяются продольной бороздой; продольная и поперечная борозды часто окаймлены далеко выступающими пластинчатыми выростами (Omithocercus, рис. 11.70, С). У Peridiniales (2) клетка или имеет простой покров (Gymnodinia-сеае, В), или одета целлюлозными пластинками (Peridiniaceae с родами Peridinium и Ceratium, А, D—J). У морских видов имеются мешковидные «пузулы», которые выходят узким каналом в борозду, где расположен жгутик; их функция еще не выяснена. Коккоидному уровню организации соответствует порядок Dinococcales (3), нитчатому — Dinotrichales (4), с немногочисленными родами в каждом из них.
В ископаемом состоянии динофиты известны с юры и представлены большим разнообразием форм; в мелу встречаются многочисленные превосходно сохранившиеся представители разных таксонов. Кроме того, так называемые гистрикосферы в отложениях от докембрия до голоцена были идентифицированы как принадлежащие динофитам репродуктивные клетки. Они играют важную роль как «руководящие» микроскопические ископаемые.
3. Приложение к Rhodobionta: отдел Haptophyta
Этот отдел (называемый также Prym-nesiophyta) включает формы, представляющие монадный, пальмеллоидный, кокко-идный и нитчатый уровни организации. Преобладают одноклеточные формы мо-надного типа. Большинство видов обитает в планктоне морей. Монадные клетки имеют два жгутика, чаще всего равной длины, которые несут не жесткие волоски, а субмикроскопические чешуйки или узелки из органического материала. Вдобавок к этим жгутикам каждая клетка имеет еще дополнительный нитевидный придаток — гапто-нему. Она служит не для движения, а для
прикрепления. Ее субмикроскопическая структура четко отличается от строения жгутиков. На поперечном срезе гаптонемы можно различить 6 или 7 серповидно расположенных трубочек (структуры 2 + 9 нет!). У некоторых форм эта гаптонема редуцирована до короткой культи. Поверхность клетки снаружи имеет чешуйки разного размера или узелки, которые образуются в пузырьках Гольджи и затем откладываются наружу. Цистерны Гольджи в середине пузыревидно расширены. Желтые, желто-бурые или бурые хлоропласты содержат хлорофиллы а и с, p-каротин и ксантофиллы; в качестве запасного вещества откладываются хризоламинарин, масло и парамилон. Хлоропласты окружены складкой эндоплазматической сети; опоясывающая ла-мелла (см. рис. 11.72, G) отсутствует. Тила--коиды группируются по три. Глазок, состоящий из шариков, располагается в хлоропласте вплотную к его мембране. Базальное расширение жгутика отсутствует.
Систематика. В этом отделе известно около 250 видов, относящихся примерно к 45 родам. В пресных водах обнаружено лишь немного видов. Haptophyccae — единственный класс этого отдела.
1. Порядок: Prymnesiales. Представители этого порядка отличаются чаще всего очень длинной гаптонемой. Клетки покрыты двумя слоями полисахаридных чешуек, видимых только при электронной микроскопии (рис. 11.71, А). Чешуйки отличаются своей радиальной лучевой структурой; для чешуек наружного слоя характерен к тому же приподнимающийся край. Chrysochro-mulina имеет очень длинную гаптонему, которая примерно в 5 раз длиннее жгутиков. Наряду с автотрофным питанием возможно также фаго-трофное, при котором, например, могут поглощаться целые клетки Chlorella. Клетки могут переходить в амебоидную фазу. Размножение осуществляется путем продол),ного деления имеющих жгутики или амебоидных клеток на несколько дочерних клеток. Виды рода Prymnesium имеют короткую, виды рода Chrysochrotnulina — более длинную гаптонему. Prymnesium parvum прикрепляется своей гаптонемой к жабрам рыб и при массовом развитии вызывает в прудах с соленой водой гибель рыб вследствие (как и Chry-sochromulina polylepis) выделения токсина.
2. Порядок: Coccoiithophorales. Гаптонема короткая или полностью отсутствоует (см. рис. 11.71, В). Клетки несут поверх плазмалеммы два слоя тонких полисахаридных чешуек. Еще один внеш-
11.2. Бактерии, грибы, растения | 223
Рис. 11.71. Haptophyta (А— по М. Parke, I. Manton и В.Clarke; В — no Н.Lohmann, Н.А. von Stosch; С — по W. А. Р. Black):
А — Prymnesiales, Chrysochromulina chiton, пластинки панциря (ЮОООх); В, С— Coccolithophorales: В — Syracosphaera pulchra, редуцированная гаптонема между жгутиками (1 500х); С — ископаемый кокколит (Deflandhus sp.), построенный из кальцитовых ромбоэдров; нижний мел (700х)
ний слой имеет самые разнообразные чешуйки, пластинки или палочки («кокколиты»). Последние также закладываются в пузырьках Гольджи сначала как целлюлозные пластинки, на которые потом накладываются удивительные отложения кальцита, специфичные для каждого вида и поэтому крайне разнообразные. Выдвигающиеся наружу кокколиты образуют правильный панцирь вокруг клетки. В этом порядке известны также виды, ставшие в результате утраты жгутиков неподвижными (Coccolithus pelagicus) или же имеющие смену жгутиковой и прикрепленной стадий (Syracosphaera). У Hymenomonas carterae происходит гетероморфная смена поколений, причем более развитый по сравнению со спорофитом гаметофит имеет вид разветвленных нитей. Coccolithophorales (или же их кокколиты), известные в ископаемом состоянии с юры (рис. 11.71, С), объединяют характерные ископаемые виды микроорганизмов (128 родов). Они составляют важную долю в определенных известковых отложениях, где представлены в количестве до 800 млн кокколитов на 1 см3 породы (Применение — см. бокс 11.6).
4. Приложение к Rhodobionta: отдел Heterokontophyta
(= Chrysophyta, Chromophyta)1
Этот отдел при значительных различиях форм таллома отличается однообразием ультрамикроскопических структур (на
1 В настоящее время для этого отдела наиболее употребительно название охрофиты (Ochro-phyta). — Примеч. пер.
пример, гетероконтными подвижными клетками).
Здесь представлены все морфологические ступени — от монадной до сифоновой организации; наивысший уровень организации Heterokontophyta — морфологически расчлененные и анатомически дифференцированные паренхиматозные талломы.
Убедительно доказано родство автотрофных Heterokontophyta и гетеротрофных, грибоподобных Labyrinthulomycota и Oomycota. Структурные особенности их клеток, несмотря на наличие пластид, приобретенных путем вторичного эндосимбиоза, настолько совпадают между собой, что Heterokontophyta и Oomycota следовало бы объединять в общее подцарство Heterokontobionta (см. Oomycota). Только структура пластид оправдывает рассмотрение Heterokontophyta в приложении к Rhodobionta, поскольку пластиды Heterokontophyta и Rhodobionta имеют общее происхождение.
Пластцды. Иногда зеленые, однако чаще за счет наличия дополнительных пигментов желтые, желто-коричневые до коричневых хлоропласты содержат хлорофиллы а и с, p-каротин и различные ксантофиллы (см. табл. 11.3). В дополнение к двум мембранам хлоропласта пластиды окружены еще складкой эндоплазматической сети (рис. 11.72, G).
В хлоропластах тилакоиды сгруппированы по три в стопки (см. рис. 2.88) — расположение, наблюдаемое также, например,
224 | ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
emgt
Рис. 11.72. Xanthophyceae (А — по A. Luther; В — Е — по A. Pascher; F, G — по A. Massalski и G. F. Leedale, С. v.d. Ноек):
А—С — Mischococcales, Capitulariella radians (500х): А — зооспора, В — отделившийся зооспорангий, С — таллом с расположенными на концах нитей зачатками зооспорангиев; D — Chloramoebales, Ankylonoton pyreniger в процессе деления (1 ОООх); Е—G — Tribonematales: Е — фрагмент нити Tribonema с характерными Н-образными структурами клеточной стенки, попарно вдвигающимися одна в другую (бООх), F — гетероконтная зооспора Tribonema (2 ЗООх), G - хлоропласт Bumilleria (30 ОООх); а — глазок; с — хлоропласт; е — оболочка из складки эндоплазматической сети; g — опоясывающая ламелла из трех периферических тилакоидов; m — две собственные мембраны хлоропласта; р — пульсирующая вакуоль; t — стопка (ламелла) из трех тилакоидов
у Dinophyta и Euglenophyta. Непосредственно под оболочкой хлоропласта, параллельно ее внутренней поверхности, расположены тилакоиды, составляющие опоясывающую ламеллу, которая здесь очень характерна (ср., например, представителей Нар-tophyta и Heterokontophyta, имеющих многие сходные вещества, с Chlorophyta). Глазки, если они присутствуют, располагаются вблизи основания жгутиков, но внутри хлоропласта.
В качестве запасных полисахаридов образуется хризоламинарин, а иногда также ламинарии и сахароспирт маннит. Они находятся вне хлоропластов, но в основном вблизи пиреноидов. Часто накапливается масло, которое появляется у пиреноидов, однако преимущественно оно находится вторично в вакуолях.
Клеточные стенки очень по-разному усилены дополнительными защитными слоями.
Жгутики. Клетки, имеющие жгутики, ге-тероконтные. Они несут один длинный, на
правленный вперед перистый жгутик и один направленный назад гладкий жгутик. Перистый жгутик имеет два ряда волосков, которые образуются в цистернах эндоплазматической сети. Гладкий жгутик (иногда редуцированный) часто имеет расширение у своего основания.
Отдел подразделяется на пять классов. В первых двух классах хлоропласты, цвет которых бывает от зеленого до желто-зеленого, не содержат фукоксантина.
1. Класс: Chloromonadophyceae1
Этот класс состоит исключительно из представителей, находящихся на монадной ступени организации. Хлоропласты — от зеленых до желто-зеленых (дополнительные
1 В настоящее время, поскольку существует род Chloromonas, не относящийся к этому классу, вместо названия Chloromonadophyceae употребляется название Raphidophyceae. — Примеч. пер.
11.2. Бактерии, грибы, растения! 225
пигменты, табл. 11.2). В качестве запасного вещества установлено только масло в крупных, снабженных пелликулой клетках, достигающих размера 50—100 мкм. Пиреноиды отсутствуют. Под поверхностью клетки располагаются трихоцисты (см. также Dinophyta).
В этом классе известно лишь 10 видов, относящихся к шести родам. За одним-единственным исключением, они все обитают в пресной воде. В болотах можно встретить Goniostomum и Уаси-olana.
2. Класс: желтозеленые водоросли (Xanthophyceae)
Желтозеленые водоросли охватывают все уровни организации таллома — от амебоидного и монадного до сифонового. Зеленые хлоропласты окрашиваются НС1 в синий цвет и содержат вместо фукоксантина (табл. 11.2) ксантофиллы — гетероксантин и вошериаксантин. Жгутики гете-роконтного типа прикреплены сбоку. Желтозеленые водоросли, таким образом, несмотря на зеленую окраску, во многом сходны с остальными классами Heterokontophyta. От зеленых Chlorophyta этот таксон отличают отсутствие хлорофилла b и завершение заднего жгутика тонким волоском (как у бурых водорослей).
У некоторых форм клеточная стенка состоит из двух накладывающихся друг на друга половинок. Они в значительной мере состоят из целлюлозных микрофибрилл и часто пропитаны кремниевой кислотой (однако без панцирей из кремниевой кислоты!). Некоторые виды образуют эндогенные цисты с оболочкой, пропитанной кремниевой кислотой; цисты имеют форму коробки с крышкой.
Размножение. Большинство желтозеленых водорослей размножаются бесполым путем. Только в одном роде (Vaucheria) известно половое размножение в гаплонтном жизненном цикле (зиготическая смена ядерных фаз!); этот случай рассмотрен ниже1.
1 Согласно другим данным, жизненный цикл Vaucheria диплонтный (гаметическая смена ядерных фаз). — Примеч. пер.
При бесполом размножении УаисИепа концы ветвей вздуваются и отделяются от остального таллома перегородкой. Все многоядерное содержимое отделившейся клетки выходит, разрывая ее стенку, в виде одной яйцевидной крупной (размером около 1/10 мм) зооспоры (рис. 11.73, В). Ее поверхность покрыта многочисленными парными жгутиками несколько неравной длины, двигающимися синхронно. В бесцветной периферической части зооспоры позади каждой пары жгутиков расположены по два блефаропласта и одно грушевидно заостренное ядро; от них ближе к центру находятся хлоропласты (рис. 11.73, С); имеются пульсирующие вакуоли. Морфологически это образование соответствует совокупности всех зооспор, образующихся в одной клетке, и, таким образом, представляет собой синзооспору.
Половое размножение. Оогонии и спермогонии Vaucheria возникают на нитях таллома как боковые выросты (рис. 11.73, Е: о, s). Зачаток оогония (о) содержит сначала многочисленные ядра, которые, однако, все, кроме одного — будущего ядра яйцеклетки, вместе с частью хлоропластов мигрируют обратно в несущую нить; только тогда образуется поперечная перегородка. Оставшиеся хлоропласты, капельки масла и ядро отходят в нижнюю часть оогония, тогда как в клювовидном носике скапливается бесцветная плазма, которая при разрыве стенки оогония выступает в виде шарика. Много-ядерный спермогоний (s) вместе с несущей его ветвью загнуты в виде рога. Его вершина при созревании ослизняется. Многочисленные крошечные сперматозоиды выходят наружу, внедряются в отверстие оогония и скапливаются перед бесцветным воспринимающим шариком яйцеклетки. Сперматозоиды (рис. 11 73, F) гетероконтные.
После оплодотворения яйцеклетки одной из сГ гамет богатая маслом цистози-гота окружается многослойной стенкой, переходит в состояние покоя (гипнозигота) и прорастает позже после редукционного деления непосредственно в новую гаплоидную нить.
В классе Xanthophyceae описано примерно 400 видов из 40 родов, которые обитают в прес-
226 | ГЛАВА 11, СИСТЕМАТИКА И ФИЛОГЕНИЯ
Таблица 11.2. Некоторые химические признаки классов водорослей
Отделы / классы Хлорофиллы Фикобилины Каротины Ксантрофиллы
a b c a ₽ Диадино-ксантин (С) Диато-ксантин (С) Фукоксантин (D, В. А)
Glaucophyta * + - (•) + - + - - -
Rhodophyta * + - - + (•) + - - -
Cryptophyta о + - + + (•) - (+) -
Dinophyta A(O) + - + - - + (+) (+) (•)
Haptophyta A + - + - - + (+) (+) 4-
Heterokontophyta A + - + - - + + (+) (+)
Chloromonadophyceae A + - + - - + + (+) -
Xanthophyceae A + - + - - 4- 4- + -
Chrysophyceae A + - + - - + (+) (+) +
Bacillariophyceae A + - + - (•) + + + 4-
Phaeophyceae A + - + - - + (•) (•) +
Chlorohyta * + + - - (•) + - - -
Chlororachniophyta о + + - - - - - - -
Euglenophyta A + + - - - + + (+) -
Streptophyta * + + - - - + - - -
Примечание. Условные обозначения — по van den Hoek, обзор ксантофиллов — по Metzner: * — с простыми пластидами (возникшими путем первичного эндоцитобиоза); о — с комплексными пластидами и нуклеоморфой (возникшими путем вторичного эндоцитобиоза); А — с комплексными пластидами без нуклеоморфы (возникшими путем вторичного эндоцитобиоза); + — важный пигмент или запасной полисахарид; (+) — пигмент встречается; (•) — пигмент редок или только в незначительном количестве;-----пигмент или запасной полисахарид отсутствуют. В случае крахмала: + — вне
хлоропласта; © — откладывается внутри хлоропласта; А— 8-кето-каротин, например фукоксантин или сифоноксантин (последние только у Prasinophyceae и Bryopsidophyceae из зеленых водорослей);
ных водах, отчасти также в морях и на сырой почве.
Монадные формы объединяются в порядок Chloramoebales (1-й порядок — например, Апку-lonoton, рис. 11.72, D). Капсальный уровень организации представлен в порядке Heterogloeales (2-й порядок); к нему относится морской род Heterocapsa. Порядок Mischococcales (3-й поря
док) объединяет парящие в воде или сидячие формы с жесткой клеточной оболочкой. У разветвленных, обитающих в наземных условиях видов Capitulariella (рис. 11.73, С) спороцисты отрываются целиком и только потом высвобождают зооспоры (аналогично Trentepohlia, см. рис. 11.93, С, D). Tribonematales (4-й порядок) представлены в пресной воде и на влажной по-
11.2. Бактерии, грибы, растения I 227
Ксантофиллы Запасные вещества Тип пластид
Гетероксантин (С) Вошериаксантин (В) Аллоксантин (С) Перидинин (D, В) Лютеин Зеаксантин Хризол-аминарин Крахмал Багрянковый крахмал Парамилон
- - - - - + - + - - Цианеллы
- - - - + + - - + - Родопласты
- - + - - - - + - -
- - - + - - - + - -
- - - - - - + - - +
(+) (+) - - - - + - - +
- - - - - - - - - -
4- + - - - - + - - -
- - - - - - + - - -
- - - - - - + - - -
- - - - - (+) + - - -
- - - - + + - ф - - Хлоропласты
- - - - + - - - - ?
- - - - - - - - - +
- - - + - - - ф - -
В — аллен-каротин, например вошериаксантин и неоксантин (последний у Euglenophyta, Chlorophyta, Eustigmatophyta, у некоторых Heterokontophyta, Rhodophyta); С — алкин-каротиноиды; D — каротиноидные эфиры, т.е. ксантофиллы, которые на одной или обеих гидроксильных группах несут остатки жирных кислот. В таблице не учитываются 4-кето-каротины, например эхиненон у Euglenophyta + и Chlorophyta +, у Heterokontophyta (+). Цианеллы — фотосинтезирующие органеллы с пептидоглюкано-вой стенкой. Родопласты — пластиды Rhodophyta и производные от них фотосинтезирущие органеллы. Хлоропласты — пластиды Chlorophyta и производные от них фотосинтезирующие органеллы.
чве часто встречающимися видами рода Tribonema (рис. 11.72, Е), стенки неразветвленных нитей которых в продольном сечении построены из Н-об-разных элементов.
К Hetcrosiphonales (5-й порядок) относится пузыревидная, растущая на влажном иле водоросль Botrydium (рис. 11.73, G), клетка которой размером около 2 мм закрепляется в иле ризои
дами. Пузырь содержит в постенной плазме многочисленные ядра и дисковидные хлоропласты, стенка которых состоит из пектиноподобной фракции и целлюлозы. Когда Botrydium покрыт водой, то образуются многочисленные гетеро-контные зооспоры, которые после разрушения стенки пузыря выходят наружу и на подходящем субстрате вырастают в новый пузырь. При
228 | ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Рис. 11.73. Xanthophyceae, Heterosiphonales (А, В — по Goetz; С — по Е. Strasburger; D — по J. Sachs, с изменениями; Е — по F. Oltmanns; F — no М. Woronin; G — по J. Rostafinsky и M.Woronin; Н — по R.Kolkwitz):
A—F — Vaucheria (А—С — V. repens; D, Е— V. sessilis; F — V. synandra); А — закладка спорангия (150х), В — выходящая из спорангия синзооспора (150х), С — край синзооспоры (500х), D — возникающее из синзооспоры В растение с ризоидом и гаметангиями (70х), Е — участок нити с гаметангиями (150х), F — сперматозоид (700х); G, Н — Botrydium, B.granulatum: G — целое растение (ЗОх), Н — зооспора (1 ОООх); с — хлоропласты; о — оогоний; s — спермогоний; z — зачаток синзооспоры
высыхании возникает большое количество многоядерных цист из протопластов, отходящих в ризоид.
Сифональное строение имеют также широко распространенные виды рода Vaucheria. Они обитают в пресной воде или на влажной почве, прикрепляются к субстрату пучком ризоидов и имеют состоящий из лишенных поперечных перегородок разветвленный таллом, представляющий собой как бы систему нитей (рис. 11.73, D) с многочисленными ядрами и пластидами. Клеточная стенка содержит пектиноподобную массу и целлюлозу.
Стенки некоторых видов инкрустированы известью и могут образовывать известковые туфы.
К желтозеленым водорослям примыкает небольшая группа (рассматриваемая также как самостоятельный отдел Eustigmatophyta). которая
отличается от Xanthophyceae рядом ультраструк-турных признаков; хлоропласты без периферической опоясывающей ламеллы; пиреноиды только в хлоропластах вегетативных клеток; глазок на переднем конце клетки вне хлоропласта'. В жизненном цикле капсальных и коккоидных организмов могут иметься гетероконтные жгутиковые клетки. У Chlorobotrys несколько клеток
1 Эта группа, чаще всего трактуемая как класс, отличается от Xanthophyceae тем, что у большинства представителей фактически только один направленный вперед перистый жгутик; вместо второго жгутика имеется лишь базальное тело. Только у двух представителей задний гладкий жгутик выступает наружу. Кроме того, из пигментов преобладает виолаксантин, а не диато-ксантин. — Примеч. пер.
11.2. Бактерии, грибы, растения | 229
образуют колонию в слизистой обвертке; некоторые виды этого рода широко распространены в мочажинах болот.
Для рассматриваемых ниже классов (3 — 5) характерен фукоксантин в качестве дополнительного пигмента в хлоропластах, имеющих окраску от желтой до бурой.
Рассматриваемые далее классы 3 и 4 включают преимущественно одноклеточные или колониальные водоросли и реже — формы, образующие неразветвленные нити. Встречающиеся как исключение тканевые талломы имеют микроскопически малые размеры. Смена поколений неизвестна.
3. Класс: золотистые водоросли (Chrysophyceae)
Большинство видов этого класса — мо-надные одноклеточные формы, часть которых образует колонии. Реже встречаются представители, имеющие амебоидную (Rhi-zochrysis), капсальную (Chrysocapsa), кокко-идную (Chrysosphaerd), нитчатую (Phaeo-thamnion) и тканевую (Thallochrysis) организацию. Хлоропласты чаще всего от золотистых до золотисто-бурых (откуда название «фукоксантин»!). Поверхность клетки у представителей некоторых родов покрыта характерными кремнеземными чешуйками, которые образуются внутри клетки в пузырьках вблизи хлоропластов и затем в готовой форме откладываются на поверхности клетки; встречаются также окремнелые цисты (рис. 11.74, Е). У некоторых видов наблюдалось половое размножение (изогамия).
К этому классу относится около 1 000 видов, распределяющихся примерно по 200 родам. Они встречаются преимущественно в пресных, реже в солоноватых и соленых водах.
Пресноводные формы предпочитают незатененные водоемы с прохладной водой.
Питание чаще всего фотоавтотрофное, но может быть гетеротрофным и фагогроф-ным.
1. Порядок: Chrysomonadales. Одноклеточные формы золотисто-бурого цвета имеют жгутики (см. рис. 4; 11.74). У представителей некоторых
родов внутри клеток образуются эндогенные цисты, чаще всего с окремнелой стенкой и крышечкой (D, Е).
У часто встречающихся в пресноводном планктоне видов родов Uroglena (рис. 11.74, F) и Synura многочисленные радиально расположенные клетки образуют шаровидные колонии; у Synura клетки покрыты нежными кремнеземными пластинками (рис. 11.74, G).
У Mallomonas кремнеземные чешуйки несут длинные придатки, способствующие парению в воде. Часто встречающиеся в пресной воде и в море виды рода Dinobryon (рис. 11.74, Н) при круговых движениях образуют вокруг своих вытянутых клеток целлюлозные домики; после деления дочерние клетки прикрепляются на краю материнского домика и образуют новый домик, так что получаются кустовидно ветвящиеся колонии.
Для копуляции две отдельные клетки вместе с бокальчиками подплывают друг к другу, сливаются и образуют окремнелую цистози-готу.
Виды родов Ochromonas (рис. 11.74, А—Е) и Monas внешне сходны, однако у последних хлоропласты и автотрофный образ жизни утрачены.
Также гетеротрофны Craspedomonadaceae («хоанофлагелляты»); за счет ударов жгутиков пища (детрит, бактерии) подгоняется в плазматический воротничок, который расположен на верхнем конце клетки.
Только в морях встречаются голые силико-флагелляты, которые объединяются в один.
2. Порядок: Dictyochales. Они образуют изящный кремнеземный скелет, расположенный внутри клетки, например Distephanus на рис. 11.75, А. В ископаемом состоянии они известны со среднего мела.
3. Порядок: Chrysocapsales. У видов этой группы клетки в вегетативном состоянии неподвижны, в студенистых ценобиях (капсальный уровень организации, см. бокс 11.5). Напоминающие мох слоевища Hydrurus обитают в холодных горных ручьях, прикрепляясь к каменистому субстрату (рис. 11.75, В).
4. Порядок: Chrysotrichales. Клетки объединены в простые или разветвленные нити, как, например, у пресноводной водоросли Phaeothamnion (нитчатый уровень организации; рис. 11.75, Е). Зооспоры этих водорослей при определенных условиях утрачивают жгутики, окружаются плотной обверткой и размножаются делением («пальмеллоидная стадия»; рис. 11.75, G). Sticho-chrysis imnobilis полностью утратил способность образовывать подвижные стадии со жгутиками, как и многие представители следующего класса.
230 I ГЛАВА 11 СИСТЕМАТИКА И ФИЛОГЕНИЯ
Рис. 11.74. Chrysophyceae, Chrysomonadales (А—F по A. Pascher; G — no J. В. Hansen; Н — по G. Klebs):
А—Е — Ochromonas (1 ОООх): А—С — переход от нормальной формы с двумя жгутиками к амебоидному состоянию с псевдоподиями, D — образование цисты в амебоидном протопласте, Е — циста с отверстием и пробкой (заштрихована); F — Uroglena, U. americana (400х); G — Synura, S. glabra, кремнеземные чешуйки (7 200х); Н — Dinobryon, D. sertulana (350х); с — желто-бурый хлоропласт; g — жгутик; s — глазок; v — вакуоль
Рис. 11.75. Chrysophyceae (А — по К. Gemeinhardt; В — по J. Rostafinsky; С — по G. Klebs; D — по G. Berthold; Е — по A. Pascher):
А — Dictyochales, Distephanus speculum: хлоропласты в основном в эктоплазме, вне кремнеземного скелета; жгутики не показаны (1 ОООх); В — D— Chrysocapsales, Hydrurus foetidus: В — молодое растение (1 х), С — зооспора (1 200х), D — конечная часть ветви (450х); Е—G — Chrysotrichales, Pha-eothamnion borzianum: E — таллом, в котором происходит образование зооспор (400х), F — зооспора (75х), G — стадия пальмеллы (400х)
11.2. Бактерии, грибы, растения | 231
4. Класс: диатомовые, или кремнеземные, водоросли (Bacillariophyceae = Diatomeae)
Диатомеи, насчитывающие свыше 10 000 видов из примерно 200 родов, представляют собой группу крайне разнообразных коккоидных одноклеточных водорослей, иногда формирующих колонии в виде лент или звездочек. Бурые хлоропласта, иногда встречающиеся по одному или по два на клетку, содержат в значительной мере те же пигменты, что и золотистые водоросли. Также совпадают и запасные вещества. Продукты ассимиляции откладываются вне хлоропластов: хризола-минарин — в клеточном соке (у золотистых водорослей — в специальных вакуолях), масло — в особых вакуолях. Только мужские гаметы некоторых видов из по
рядка Centrales имеют один направленный вперед перистый жгутик; другие жгутиковые стадии неизвестны.
Особое положение в системе диатомеи занимают вследствие наличия внутри внешнего плазматического слоя двух кремнеземных половинок панциря, из которых одна (эпитека), как крышка коробки, накладывается на нижнюю (гипотеку) (рис. 11.76, В). К боковым загибам створок примыкает поясок из двух накладывающихся друг на друга ободков (поясковых ободков). Клетка выглядит по-разному в зависимости от того, рассматриваем ли мы ее со створки (А) или с пояска (В). От пояска иногда внутрь клетки могут отходить неполные перегородки — септы (G).
Силикатный (кремниевый) панцирь имеет, особенно на поверхностях створок, чрезвычай-
Рис. 11.76. Bacillariophyceae (А, В — по Е. Pfitzer; С — по R. Lauterborn; D, Е — no Smith; F — no Н. van Heurck; G — no В.Schroder; H — no J.Pantocsek; J — по A. Schmidt; К — no Smith; L — no G. Karsten):
A—G — Pennales: A—C — Pinnularia viridis, A — вид co створки, co швом (600x), 3 — вид с пояска (600x), С — поперечный разрез (200x), D — Licmophora flabellata (200x), E — Synedra gracilis (200x), F — Asterionella formosa (200x), G — Tabellaria flocculosa (400x); H — L — Centrales: H — Coscinodiscus pantocseki (200x), J — Triceratium distinctum (200x), К — Odontella (Biddulphia) aurita (400x), L — Chaetoceros castracanei (250x)
232 I ГЛАВА 11, СИСТЕМАТИКА И ФИЛОГЕНИЯ
Рис. 11.77. Bacillariophyceae, Pennales (А—D — по J. Helmcke и W. Krieger; Е — no W. R. Drum и H.S. Pankratz):
A—D — Pleurosigma angulatum, строение кремнеземной створки: А — общий вид средней части створки со швом, В — шов и поры, С — поры, D — реконструкция строения створки по электронномикроскопическому снимку; Е — Gomphonema parvulum, поперечный разрез через клетку в конце деления (10 ОООх); СМ — цитоплазматическая мембрана; D — диктиосомы; М — митохондрия; N — ядрышко; О — капля масла; Р — пиреноид в хлоропласте; R — шов
но сложно устроенные структуры, часто расположенные рядами; они нередко состоят из крошечных камер, которые могут быть как сверху, так и снизу открытыми или закрытыми и тогда пронизанными тончайшими порами или щелями (рис. 11.77). Силикат является отчасти кристаллическим, отчасти аморфным и поляризационно-оптически изотропным. У створок ископаемых диатомей кристаллическая решетка приобретает аморфное строение. Наряду с силикатом в покрове клеток выявлены также полисахариды («пектины»), белки и жироподобные вещества, однако целлюлоза не обнаружена. Детали для построения панциря образуются в плоских пузырьках под плазмалеммой. Эти пузырьки, возможно, происходят от аппарата Гольджи: очевидно, несколько пузырьков Гольджи сливаются в пузырек, образующий силикат.
Вегетативное размножение. Диатомеи размножаются вегетативно путем деления клетки надвое. При этом две половинки панциря (теки) отодвигаются друг от друга за счет увеличения протопласта. Каждая дочерняя клетка, унаследовав от материн
ской клетки только половину панциря, достраивает недостающую половину, которая всегда бывает гипотекой. Те дочерние клетки, которые наследовали исходную гипотеку (теперь ставшую эпитекой), и образуют соответствующую половину панциря (соответственно новую гипотеку), меньше, чем материнская клетка. Это приводит при дальнейших делениях к прогрессивному уменьшению размеров клеток до определенного минимального размера (примерно половины исходного размера), при котором наступает половое размножение, связанное со значительным увеличением размера зиготы (ауксозиготы). У некоторых видов за счет растяжимости пояска выравниваются размеры эпи- и гипотеки.
Половое размножение. Жизненный цикл диплонтный с гаметической сменой ядерных фаз; клетка диатомовой водоросли содержит, таким образом, диплоидное ядро (в противоположность, например, Zygnematophyceae). При мейозе из дипло
11.2. Бактерии, грибы, растения | 233
идных клеток возникают гаплоидные гаметы.
Пример — Centrales (прочие особенности рассмотрены ниже). Определение пола осуществляется модифика! щонно. В клетках, детерминированных как мужские, они чаще всего непосредственно превращаются в спермогоний — возникает 4 сперматозоида (рис. 11.78, d—f) с одним жгутиком каждый. В других клетках, чаше всего более крут ihbix, превращающихся в оогонии, образуются лишенные жгутиков женские гаметы — яйцеклетки. В деталях образование гамет происходит весьма по-разному, и их число может быть разным в зависимости от вида. Сперматозоиды с помощью своего перистого жгутика подплывают к яйцеклеткам. После оплодотворения внутри или вне оогония зигота одевается оболочкой, в которой откладываются силикатные чешуйки. Она вскоре прорастает, причем за счет растяжения стенки вырастает до 2—4-кратного размера исходной клетки и превращается в «ауксозиготу»'. Старые половинки панциря отодвигаются друг от друга, и внутри оболочки ауксоспоры образуются новые половинки. При этом возникает диплоидная «первичная клетка», из которой затем, как описывалось выше, при прогрессивном измельчании большей части потомства вегетативным путем возникают новые дочерние поколения.
В целом заложение половинок панциря связано с митозами. Даже ядро зиготы при образовании обеих первичных половинок панциря претерпевает митотическое деление, после которого одно из дочерних ядер дегенерирует.
Встречаемость. Диатомеи распространены в пресных водах и морях всех климатических зон. Особенно интенсивно они развиваются весной и осенью, а летом их численность падает. Многие формы живут во влажной почве и на каменистом субст рате. В тропиках они могут обитать совместно с синезелеными водорослями на листьях (эпифильные виды).
1 В отечественной литературе для этой структуры применяется название «ауксоспора», хотя даваемое здесь название «ауксозигота», по-видимому, удачнее. — Примеч. пер.
Систематика. По симметрии створок Bacilla-riophyceae подразделяют на два порядка: Centrales и Pennales1. У первой из них створка построена по радиальному плану, у второй — по билатеральному. Помимо строения створок эти два порядка сильно различаются и по типу полового размножения.
1. Порядок: центрические (Centrales). Контур створки представляет собой окружность или треугольник с закругленными углами (рис. 11.76, Н — L). Элементы орнамента створки имеют при этом радиальное или концентрическое расположение. В противоположность большинству Pennales, вегетативные клетки Centrales неподвижны. Однако мужские гаметы подвижны за счет единственного перистого жгутика (ср. рис. 11.20, А, слева), у которого отсутствуют обе центральные микротрубочки. Половое размножение (см. выше) было особенно детально изучено у Stephanopyxis и Melosira varians.
Распространение. Centrales обитают преимущественно в морях, где составляют существенную часть фитопланктона (первое место среди первичных продуцентов в Мировом океане; см. бокс 11.7). Многие из них имеют особые придатки, способствующие парению (см. рис. 11.76, L), другие объединены слизью в цепочки или соединения иного рода (К).
Виды рода Melosira. образующие нитевидные колонии из коротко-цилиндрических клеток (рис. 11.78). распространены как в морях, так и в пресной воде, многочисленные виды родов Cosci-nodiscus и Hemiaulus — только в морях. Для рода Triceratium, также морского (см. рис. 11.76, J), характерны створки с очертаниями от треугольных до многоугольных. Напоминающие обликом консервную банку, клетки Ethmodiscus gazellae (обитателя теплых морей) диаметром почти 2 мм представляют собой самые крупные клетки среди диатомей. Клетки Stephano-discus снабжены по краю круглых створок венцом из шипов. Род Rhizosolenia распространен преимущественно в морях.
2. Порядок: перистые (Pennales). Клетки имеют форму палочек или лодочек, реже клиновидные (см. рис. 11.76, A—G), поэтому их центр симметрии вытянут по одной линии, от которой перисто отходят элементы орнаментации силикатной стенки. У очень многих форм по продольной оси створки проходит трещина — «шов», тонкое строение которого сильно различается у вхо-
1 Согласно современным данным молекулярной систематики, подтверждаемым сведениями по ультраструктуре створок. Centrales представляют собой две эволюционные группы, одна из которых ближе к Pennales, чем к другой из этой группы. — Примеч. пер.
234 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Рис. 11.78. Bacillariophyceae, Centrales, Melosira varians. Половое размножение (схема) (по Н.А. von Stosch):
а—g — мужской, а' — д' — женский участки нити; а—е, а'—е' — мейоз; f — открывшийся; д — освободившийся спермогоний; сГ — мужское ядро, внедрившееся через специальную щель; f — оплодотворение; д' — молодая ауксозигота
Рис. 11.79. Bacillariophyceae, Pennales, Rhopalodia gibba, половое размножение (по H.KIebahn): А — две клетки, соединенные слизистым чехлом; В — деление материнских клеток (дегенерировавшие ядра уже исчезли); С — образование зиготы после слияния гамет; D — увеличение размера аук-созиготы (А—D 410х); Е — конечная стадия и образование нового панциря (240х)
дящих в этот таксон родов (см. рис. 11.76, А; 11.77). Полагают, что шов играет определенную роль в ползающем движении, встречающемся только у Pennales; у неподвижных (сидячих) форм шов отсутствует. В центре шов прерывается центральным узелком, а у концов створок заканчивается концевыми узелками (см. рис. 11.76, А).
Половое размножение Pennales отличается от «нормального типа» Centrales, поскольку никаких подвижных гамет не бывает. Здесь сливаются изогаметы в форме голых протопластов (единственное исключение — оогамия у Rhabdonemcr, причем О’ гаметы все же лишены жгутиков). Для спаривания две вегетативные клетки сползаются вместе и чаще всего обильно выделяют слизь.
Ядро каждой клетки делится мейотически на 4 гаплоидных ядра, из которых, однако, 2 дегенерируют, а 2 становятся ядрами гамет. Эпи- и гипотека несколько отходят друг от друга. Через получающийся разрыв между ними гаметы из разных клеток имеют возможность попарно копулировать, так что возникают две зиготы, которые сразу же вырастают в ауксозиготы. Каждая из них выделяет пару силикатных половинок панциря и становится первичной клеткой для последующих клеток разного размера. Первичные и родительские клетки располагаются перпендикулярно (рис. 11.79) или параллельно друг ДРУГУ- От этого обычного варианта имеется множество отклонений.
11.2. Бактерии, грибы, растения | 235
Распространение. Большинство подвижных Pennales обитают преимущественно на дне пресных, солоноватых или соленых водоемов (может наблюдаться массовое их развитие), эпифитно на водных растениях или в почве, однако имеются и планктонные формы. Примерно 500 видов рода Navicula, клетки которых напоминают лодочки, распространены в водоемах всех типов; виды рода Pinnularia (см. рис. 11.76, А—С) с похожими, но удлиненно-эллиптическими клетками предпочитают пресную воду. Виды Pleurosigma, клетки которых слегка S-образно изогнуты (см. рис. 11.77), могут служить как тест-объект для проверки объективов микроскопов. У видов родов Diatoma, Tabellaria (см. рис. 11.76, G), Frag/lariau др., обитающих преимущественно в стоячих пресных водах, клетки образуют длинные цепочки, у Asterionella (F) — звездчатые, у Meridion — вееровидные (иногда замыкающиеся в кольцо) колонии. Некоторые виды рода Synedra свободно парят в воде, другие прикрепляются слизистыми подошвами к более крупным нитям водорослей (Е). У также ведущих сидячий образ жизни видов Licmophora клетки после деления остаются соединенными друг с другом, так что возникают древовидные колонии из клеток, сидящих на ножках из слизи (D). Можно перечислить еще роды, упоминавшиеся в других разделах: Surirella (см. бокс 11.7), Nitzschia (см. 13.8), Gomphonema (см. рис. 11.77, Е) и Rhopalodia (см. рис. 11.79).
Филогения. Centrales, имея снабженные жгутиками сперматозоиды, являются более ранней группой, чем Pennales, у которых жгутики полностью утрачены. Предками диатомей были, возможно, водоросли типа Chrysomonadales, клетки которых уже могут нести кремнеземные чешуйки. Диатомеи в основном остались на коккоидном уровне организации, имея лишь первые намеки на нитчатую организацию.
Древнейшие диатомеи, и именно центрические формы, известны из юры. Большое видовое разнообразие их отмечается с мела.
В третичном и межледниковом периодах массовое развитие диатомей привело даже к образованию соответствующих пород (применение — см. бокс 11.6, с. 247).
В противоположность обоим классам, рассмотренным ранее, у следующего класса отсутствуют простые формы организации (например, нет одноклеточных форм или неразветвленных нитей). Часто образуются макроскопические тканевые талло
мы с сильно выраженной дифференциацией на органы и ткани. Встречаются различные типы смены поколений. В состав клеточной стенки входят только альгинаты и фукоидан.
* 5. Класс: бурые водоросли (Phaeophyceae)
Бурые водоросли — группа, представленная самыми разнообразными формами (рис. 11.80, А; см. рис. 5.5; 11.83; 11.85). Это могут быть и маленькие разветвленные нити, разделенные на клетки (гетеротрихальный тип), и псевдопаренхиматозные талломы, и многометровые растения с сильно выраженной дифференциацией на органы и_ ткани (тканевые талломы! — см. 5.3.2). Макрофиты из бурых водорослей часто подразделены на органы (филлоид, каулоид, ризоид), которые напоминают лист, стебель и корень высшего растения. Одноклеточные формы отсутствуют, т.е. монадные и коккоидные уровни организации здесь не представлены. Наряду с красными и некоторыми зелеными (Chara) водорослями бурые водоросли относят к наиболее высокоорганизованным формам.
Пластиды. Бурые хроматофоры содержат кроме характерных для всего отдела ассимиляционных пигментов прежде всего фукоксантин в качестве дополнительного пигмента, окраска которого перекрывает окраску всех других компонентов.
Клеточная стенка состоит из твердой и слизистой фракций; первая образована целлюлозными фибриллами и альгинатом, последняя — альгинатом и фукоиданом. Альгинаты — это соли альгиновой кислоты (полимера двух сахарокислот: p-D-ман-нуроновой и p-L-гулуроновой) с различными катионами (такими как ионы Са2+, Mg2+, Na+)- Подвижные стадии (зооспоры и гаметы) имеют форму от грушевидной до веретеновидной и несут чаще всего 2 жгутика неравной длины (рис. 11.80, А, В, I)1.
Вблизи жгутиков находится красно-коричневый глазок в буром хлоропласте (один,
1 Важной особенностью подвижных стадий бурых водорослей считается боковое прикрепление жгутиков. — Примеч. пер.
236 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Рис. 11.80. Phaeophyceae, Ectocarpales (А — по G.Thuret; В — D — по Berthold; Е, F — по H.Kylin; G— по Hygen; Н, J — по Р. Kuckuck):
А—D — Ectocarpus siliculosus: А — ветвь гаметофита с многогнездным гаметангием (380х), В — D — оплодотворение (В — 1 200х; С, D — 1 бООх); Е, F — Asperococcus bullosus, зигота и слияние ядер (2 ОООх); G — Nemacystis divaricates, прорастание (780х); Н — J — Ectocarpus: Н — Ectocarpus lucifugus, одногнездный мейо-спорангий на диплоидном спорофите (400х), J — Ectocarpus globifer, зооспора, волоски перистого жгутика не изображены
изредка несколько). Волоски перистого жгутика закладываются в пузырьках эндоплазматической сети или пузыревидных частях ядерной эндоплазматической сети. Гладкий жгутик в базальной части вздутый; это вздутие расположено вблизи глазка и, возможно, действует как фоторецептор. Жгутики заканчиваются (гладкий всегда, а перистый иногда) тонким волосовидным придатком. Эта особенность кроме бурых и желтозеленых водорослей больше нигде не встречается.
Жизненный цикл осуществляется в виде смены поколений, причем мейоспоры все
гда образуются в одногнездных спороцистах, а гаметы, — как правило, в многогнездных (= многокамерных) гаметангиях. Гетерофазная смена поколений изоморфная, гетероморфная — до крайне гетероморфной, с (почти) полной редукцией гаплоидного гаметофита. Увеличение значения в жизненном цикле диплоидного спорофита, появившееся среди Ectocarpales, рассматривается как прогрессивная тенденция.
Встречаемость и образ жизни. Большинство из примерно 1500—2000 видов бурых водорослей, распределяющихся по 250 родам, обитает в морях, причем наиболее интенсивного развития они достигают в водах умеренных и высоких широт. Это бентосные водоросли (см. бокс 11.7): они живут как литофиты, прочно прирастая к скалам, камням, бревнам и т.д. Некоторые из них при низкой воде остаются на воздухе. Часто бурые водоросли обитают как эпифиты на других водорослях. Они образуют в литоральной зоне скалистых побережий пышные заросли с характерным зональным распределением видов (см. рис. 11.102). Внушительное зрелище представляют собой на тихоокеанском побережье Америки подводные леса, которые образованы бурыми водорослями из родов Lessonia. Macrocystis и Nereocystis с многометровыми талломами. Крошечные нитевидные или дисковидные бурые водоросли хотя и менее заметны, но тоже широко распространены, в том числе на каменистом субстрате, морских желудях, улитках, двустворчатых моллюсках и эпифитно на более крупных бурых водорослях. Мелкие бурые водоросли могут до определенной степени жить эндофитно в более крупных водорослях. В пресных водах встречается лишь немного видов, относящихся примерно к пяти родам.
Класс подразделяется на 11 порядков, из которых следующие, имеющие меньшее значение, дальше рассматриваться не будут: Chordariales (с родами Chordaria и Leptonema, Elachista — эпифит на Fucus с вегетативной диплоидизацией в гаметофитах); Desmarestiales (характеризуются объединением нитей в псевдопаренхиматозную кору; гетероморфная смена поколений), Dictyo-siphonales (паренхиматозный таллом), Scytosipho-nales (псевдопаренхиматозные корковидные микроталломы (спорофиты) сменяются в жизненном цикле паренхиматозными мегаталломами —
11.2. Бактерии, грибы, растения | 237
гаметофиты?), Sporochnales и Sphacclariales (например, Halopteris; от Ectocarpales и др. отличается наличием верхушечной клетки).
1. Порядок: Ectocarpales. К порядку относится большинство бурых водорослей. Очень распространены виды рода Ectocarpus. Их кустовидные талломы, состоящие из разветвленных нитей, по облику напоминают зеленые водоросли из рода Cladophora (см. рис. 11.90), однако окраска их бурая. Это обитатели мелководий наших морей, которые ползучими нитями прикрепляются к субстрату (каменистому субстрату, более крупным водорослям). Нити растут интеркаляр-но, верхушечная клетка отсутствует, только часть клеток способна преобразовываться в органы размножения. Жизненный цикл в целом представляет собой изоморфную (или слабогетероморфную) смену поколений.
Гаметофит. Гаплоидный, кустовидно разветвленный нитчатый таллом гаметофита несет по бокам и на концах нитей многогнездные гаметангии, в которых вовсе не каждая клетка способна образовывать гамету. Гаметы освобождаются при разрушении внешних стенок гаметангия, выходя на его вершине. Несмотря на то что морфологически гаметы одинаковы, у многих видов рода Ectocarpus имеется физиологическая анизогамия: женские («-») гаметы вскоре после выхода из гаметангия останавливаются и отбрасывают жгутики, тогда как мужские («+») гаметы, которые привлекаются феромоном эктокарпеном, группируются возле них. Концами своих более длинных жгутиков мужские гаметы прикрепляются к покоящимся женским, и одна из них сливается с ней (рис. 11.80, В).
Спорофит. После оплодотворения зигота без периода покоя вырастает в диплоидный спорофит, состоящий из нескольких более жестких и менее разветвленных нитей. На нем возникают многочисленные яйцевидные одногнездные спорангии, в которых после мейоза образуется большое количество мейозооспор, дающих начало новому гаметофитному поколению. Определение пола гаплогенотипическое.
Эта обычная — изоморфная и гетерофазная — смена поколений может усложняться многочисленными отклонениями, как правило, трудно объяснимыми: сменяющиеся поколения не всегда связаны с определенной ядерной фазой. Каждая генерация может непосредственно снова воспроизводить себя. Пол четко не фиксирован; организмы могут обходиться и без него.
Если вместилища репродуктивных клеток у Ectocarpus возникают на концах боковых ветвей, то у Pylaiella они дифференцируются интерка-лярно.
У некоторых эпифитных видов родов Ectocarpus и Pylaiella гаметофиты и спорофиты встречаются не на тех же самых, а на разных растени-ях-форофитах (например, у Pylaiella litoralis — спорофит на Fucus. а гаметофит на Ascophyllum).
2. Порядок: Cutleriales. Смена поколе ний у Cutleria гетероморфная, с явно преобладающим гаметофитным поколением (рис. 11.81, А). Гаметофит — прямостоячее, вильчато разветвленное растение, состоящее из лентовидных лопастей, рассеченных на концах. У Cutleria multifida — водоросли из более теплых европейских морей — гаметофит обитает вблизи поверхности воды, имеет размер порядка 40 см и образует на мужских и женских растениях в микро- и мегагаметангиях мелкие (сГ) и более круп
Рис. 11.81. Phaeophyceae, Cutleriales, Cutleria multifida (А, В — no G. Thuret; C — no R Kuckuck): A — д’, В — <j> многогнездные гаметангии (400x); С — <j> и сГ гаметы, волоски перистых жгутиков не показаны (1 200х)
238 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
ные (<j>) гаметы, снабженные жгутиками (рис. 11.81). Мужские гаметы привлекаются женскими гаметами посредством феромонов мультифидов, после чего происходит копуляция (анизогамия). Ранее описывавшийся как представитель отдельного рода (Aglaozonoia) спорофит заметно меньше (его размер — несколько сантиметров); он плоский, рассеченный на лопасти, распростертый, корковидный; обитает на скалах и раковинах моллюсков на глубине от 8 до Юм. На верхней стороне паренхиматозного таллома находятся сорусы, состоящие из одногнездных спороцист. После мейоза они высвобождают зооспоры. Zanardinia имеет изоморфную смену поколений.
3. Порядок: Dictyotales. Плоские паренхиматозные талломы Dictyota размером с кисть руки многократно дихотомически разветвлены (рис. 11.82). В основе роста и вильчатого ветвления лежит деление крупной верхушечной клетки (см. рис. 5.12, В), которая отчленяет вниз от себя базальные сег
менты. Последние делятся дальше на множество клеток, которые образуют ткани (см. рис. 5.12, В—D). Они дифференцируются на периферические ассимиляционные и центральные запасающие клетки (см. рис. 11.82). Время от времени исходная верхушечная клетка разделяется перегородкой, идущей вдоль оси таллома, на две; расположенные рядом клетки, продолжая рост, вызывают дихотомическое ветвление таллома. Смена поколений изоморфная (см. рис. 11.87, В).
Гаметофит. Половое размножение — оогамия. Многогнездные спермогонии и оогонии расположены на разных растениях и всегда собраны в группы (сорусы) (см. рис. 11.82, А, В).
Каждый оогоний содержит крупную неподвижную коричневую яйцеклетку, которая выталкивается в воду и оплодотворяется сперматозоидом (С). Грушевидные сГ гаметы имеют один сильноредуцированный хлоропласт и только один боковой перистый жгутик; второй, редуцированный, жгутик с
D
Рис. 11.82. Phaeophyceae, Dictyotales, Dictyota dichotoma (f\. В, D — no G. G.Thuret; C — no Williams): A — поперечный разрез через сГ таллом с группой спермогониев (окруженной обверткой из покровных стерильных клеток, 200х); В — поперечный разрез <j> таллома с группой оогониев (200х); С — яйцеклетка с тремя сперматозоидами (400х); D — поперечный разрез таллома с тетраспорангиями (один из них освободился) и «волосками» (200х)
11.2. Бактерии, грибы, растения | 239
Рис. 11.83. Phaeophyceae, Laminariales (А — по К. Magdefrau; В — по Н. Schenck; С — по Postels и Ruprecht; D, Е, F — по J.D. Hooker);
А — Laminaria saccharina (1/40х); В — Laminaria hyperborea, вверху с остатками прошлогодней пластины (1/40х); С — Nereocystis luetkeana (1/200х); D — Lessonia flavicans (1/30x); E — Macrocystis pyrifera (1/250x); F — тот же организм, вершина таллома (1/20х)
собственным базальным тельцем остается внутри клетки в виде крошечной культи. Гаметангии развиваются только в летние месяцы; выход сперматозоидов находится в зависимости от положения Солнца и Луны и происходит только 2 дня в месяц, каждый раз в первый час после рассвета.
Спорофит. Гаплоидные гаметофиты по своему внешнему облику полностью совпадают с диплоидным спорофитом (рис. 11.87, В). Мейоспоры, возникающие по 4 в одногнездных тетраспороцистах спорофитов (рис. 11.82, D), имеют относительно большой размер и лишены жгутиков. Между тетраспороцистами выступают бесцветные волоски.
Часто встречающаяся в более теплых морях вееровидная Pndina растет посредством коровой меристемы. Dictyopteris — за счет группы специальных верхушечных клеток.
4. Порядок: Laminariales. Смена поколений гетероморфная, с явным преобладани
ем диплоидного спорофита (см. рис. 11.87, С). Спорофиты морфологически и гистологически очень дифференцированы и достигают иногда значительных размеров (рис. 11.83).
Гаметофиты всех Laminariales, напротив, имеют микроскопически малые размеры. Формы сГ и $ хорошо различаются по строению, т.е. имеются четкие вторичные половые признаки. Мужские гаметофиты относительно сильно разветвлены, быстро растут, состоят из многих, но мелких клеток (рис. 11.84, G) и несут на концах вето-чек одноклеточные спермогонии с единственным сперматозоидом в каждом. Женские гаметофиты (F) имеют значительно более крупные клетки, но растут медленнее, и клеток у них меньше; в крайнем варианте они состоят только из одной-един-ственной мешковидной клетки и образуют оогонии с одной яйцеклеткой в каждом. Голая яйцеклетка выходит через отверстие на вершине оогония, где она чаще
240 I ГЛАВА 11 СИСТЕМАТИКА И ФИЛОГЕНИЯ
Рис. 11.84. Phaeophyceae, Laminariales (А — по Р.Кискиск; В —Е — по H.Kylin; F, G — по Е. Schreiber):
А—Е — Chorda filum: А — мейозооспоры, (А') округлившиеся перед прорастанием (1 200х); В — Е — развитие одногнездного спорангия (1 ОООх): В — одноядерный, С —4-ядерный, D — 16-ядерный, Е — с почти зрелыми зооспорами; F, G — Laminaria (ЗООх): F — гаметофит, G — о" гаметофит; а — спермогонии (а, — освободившийся); е— яйцеклетка; St — s3 — молодые спорофиты, еще сидящие на освободившемся оогонии
всего и остается (F: е), а после оплодотворения (оогамии) вырастает в диплоидный спорофит (F: S|—s3).
Спорофитное поколение представляет собой макроскопическую фазу в жизненном цикле. На поверхности спорофита образуется кроме вытянутых стерильных клеток (парафиз) обширный слой булавовидных одногнездных спороцист (рис. 11.84, D). В каждой из них после редукционного деления и одновременно генотипического определения пола развивается большое число двухжгутиковых зооспор.
Спорофиты Macrvcystispyrifera (см. рис. 11.83, Е) в холодных морях Южного полушария достигают в длину свыше 50 м; их таллом, закрепляющийся на глубине 2—25 м с помощью когтевидной присоски, несет по одной стороне своих осей (каулоидов, см. 5.3.2) длинные свисающие лопасти (филлоиды, см. 5.3.2), каждая из которых имеет в основании большой воздушный пузырь. Эти пузыри поддерживают водоросль на плаву у поверхности моря. Антарктические виды Le-ssonia (D), которые имеют стволовидные основные оси толщиной с человеческое бедро и длиной до 5 м, развивают на боковых ветвях длинные свисающие филлодии, внешне напоминают
пальмы. У Nereocystis (побережье Тихого океана от Калифорнии до Аляски) длинный (до 25 м) каулоид, напоминающий канат, несет на конце крупный воздушный пузырь (с высоким содержанием монооксида углерода!), от которого отходит пучок филлоидов (С). Chorda filum («морская струна») имеет шнуровидный неразветв-ленный таллом, достигающий нескольких метров в длину. Alaria кроме филлоида на вершине имеет еще боковые «листочки» меньшего размера1.
Виды Laminaria (ср. рис. 11.102) распространены по побережьям Северной Атлантики и образуют целые подводные заросли («луга») в непосредственной близости от уровня низкой воды. Их талломы длиной до 5 м состоят из многолетней «ножки» с когтевидным ризоидом (см. рис. 11.83. В) и листовидной лопасти (А), сложенной очень многими слоями клеток. Эта лопасть (фил-лоид) каждый год заменяется за счет того, что располагающаяся у ее основания интеркаляр-ная зона роста дает начало новой «листовой пла-
1 Интересно разделение функций этих филлоидов: крупный центральный филлоид с хорошо выраженной срединной жилой стерилен, а сорусы зооспорангиев развиваются только на мелких боковых филлоидах, называемых поэтому также спорофиллами. — Примеч. пер.
11.2. Бактерии, грибы, растения | 241
станке»; старая пластина при этом отодвигается все дальше и постепенно отмирает (В). Филлоид у L. saccharina (А) простой, у L. digitata и близких по облику видов (В) — дланевидно-рассеченный. Спорофиты / . hyperborea могут жить 10—20 лет.
Дифференциация тканей. На поперечном разрезе через каулоид Laminariales в месте перехода от периферии к центру видна сильная дифференциация. Снаружи видна меристодерма (покровная ткань). Ее клетки, способные делиться в нескольких направлениях, образуют тангенталь-ные, радиальные и горизонтальные стенки. Более глубокие слои меристодермы ответственны прежде всего за рост в толщину. Рост происходит с периодичностью в зависимости от времени года, так что на более старых каулоидах имеются хорошо выраженные годичные кольца. Клетки коры от периферии к центру становятся все крупнее. За счет ослизнения клеточных стенок возникают идущие отчасти продольно, отчасти радиально рыхлые ряды клеток, а в более старых каулоидах — слизистые ходы с широкими просветами. Коровый слой обеспечивает механическую прочность каулоида и в своих наружных частях, состоящих из мелких клеток с хлоропластами, функционирует как ассимиляционная ткань; отчасти и здесь осуществляется рост в толщину. Сердцевина служит для накопления
и проведения питательных веществ. Она состоит из многоклеточных нитей (так называемых гиф), которые у поперечных перегородок расширены наподобие раструба. У видов некоторых родов (например, Nereocystis и Macrocystis) поперечные стенки таких нитей перфорированы в виде сита. С помощью радиоактивно меченных соединений углерода удалось установить транспортную функцию этих элементов. Подобные ситовидные трубки сходны по строению и функциям с уже обсуждавшимися ситовидными трубками высших растений (см. рис. 5.10).
5. Порядок: Fucales. Эти водоросли могут из-за крайней редукции гаметофита рассматриваться практически как чистые диплонты (см. рис. 11.87, D). Размножение осуществляется путем оогамии. Смена ядерных фаз гаметическая, т. е. мейоз происходит при образовании гамет. Диплоидный спорофит (рис. 11.85) образует единственное имеющееся в жизненном цикле вегетативное тело в форме таллома, иногда достигающего в длину 1 м и более. У живущих несколько лет видов Fucus кожистые, лентовидные, дихотомически разветвленные талломы укреплены своего рода «сре-
Рис. 11.85. Phaeophyceae, Fucales (по К. Magdefrau):
А — Sargassum bacciferunr, В — Himanthalia lorea; С — Ascophyllum nodosum} D — Fucus vesiculosus} E — Fucus serratus, вершина таллома (A—E — 1/4x)
242 | ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Рис. 11.86. Phaeophyceae, Fucales (по G.Thuret):
А — Fucus spiralis, двуполый (однодомный) концептакул с оогониями (о) различного возраста (25х); В — F — Fucus vesiculosus: В — ветвь со спермогониями (а) (200х), С— выход сперматозоидов из спермогония (250х), D — молодые оогонии, Е — содержимое оогония после выхода его из стенки разделяется на восемь яйцеклеток, F — освобождение яйцеклеток (D—F — 120х); р — парафизы
динной жилкой». Они прикрепляются присоской к каменистому субстрату. Концы ветвей таллома (верхушечная клетка — см. рис. 5.12, В) у некоторых видов Fucus несколько вздуты и несут тесно расположенные углубления в виде кувшинчиков — так называемые концептакулы1 (рис. 11.86, А), в которых между стерильными волосками (парафизами) расположены вместилища сГ и половых клеток (соответственно спермогонии и оогонии).
У некоторых видов спермогонии и оогонии расположены в одних и тех же концептакулах (однодомность, например, у Fucus spiralis, рис. 11.86, А); другие виды двудомны (например, F. serratus и F. vesiculosus).
Части таллома с концептакулами каждый год сбрасываются. Во вместилищах репродуктивных клеток происходит разное число митозов. Эти вместилища могут рассматриваться как одногнездные, дифференцированные по полу мейоспороцисты, а первичные продукты мейоза - как мейоспоры. Клетки, образующиеся в этих одногнездных вместилищах в результате митотических
' В русскоязычной литературе широко встречается также другое название концептакулов — скафидии. — Примеч. пер.
делений ядер, представляют собой в определенной степени крайне редуцированные гаметофиты, лишенные уже всякой самостоятельности; они, по-видимому, полностью интегрируются в соответствующие (оогонии или спермогонии) мейоспороцисты (вместилище репродуктивных клеток = мейо-спороциста = гаметангий). Из 4 возникших после мейоза гаплоидных клеток в оогонии после первого мейоза возникает 8 яйцеклеток, а в спермогонии после четырех митозов — 64 сперматозоида.
Оогонии (рис. 11.86, А: о, D) представляют собой крупные округлые образования, сидящие на одноклеточной ножке. Стенка оогония состоит из трех слоев. При созревании разрывается сначала только наружный слой стенки, так что 8 яйцеклеток остаются окруженными двумя внутренними слоями, когда они покидают концептакул (Е). В морской воде, наконец, разрывается и самый внутренний слой стенки, так что 8 яйцеклеток () начинают свободно парить в толще воды независимо друг от друга (F).
Спермогонии — овальные клетки, тесно прижатые к обильно разветвленным коротким нитям (рис. 11.86, А: а; В). Стенка спермогония состоит из двух слоев. Внутрен
11.2. Бактерии, грибы, растения | 243
няя стенка сохраняется и охватывает 64 сперматозоида (сГ), когда весь пакет при созревании выталкивается из концептакула за счет выделения слизи. Сперматозоиды состоят преимущественно из ядерно-го вещества и одного-единственного рудиментарного хлоропласта, содержащего глазок; они снабжены двумя жгутиками (в противоположность прочим бурым водорослям направленный вперед перистый жгутик более короткий).
Освободившиеся сперматозоиды (С) привлекаются к яйцеклеткам феромоном фукосерратеном (ср. 8.2.1.1). Первоначально голая зигота окружается содержащей целлюлозу стенкой, прикрепляется к субстрату и, претерпевая клеточные деления, вырастает в новый диплоидный спорофит (см. рис. 7.23).
Fucales — вершина эволюционного ряда, характеризующегося прогрессирующей редукцией гаметофита. Это четко видно при рассмотрении особенностей некоторых La-minariales. Мужской гаметофит может быть редуцирован у них до одной-единственной клетки, в то время как содержимое переходящей к покою мейозооспоры выходит и непосредственно превращается в яйцеклетку. Диплоидное растение Fucus может, таким образом, трактоваться как спорофит, мейоспоры которого непосредственно становятся почти полностью исчезающим гаметофитом.
У многих видов Fucales (как и у большинства более крупных Laminariales) таллом находится в воде также в вертикальном положении благодаря воздушным пузырям они позволяют ему раскачиваться на волнах, не перемещаясь по дну. Виды Fucus образуют в морях Северной Европы низкорослые заросли на мелководье; при низкой воде они временно остаются на воздухе, но защищены выделяемой растениями слизью (фукоидином) и поэтому даже сохраняют способность к фотосинтезу.
Fucus serratus имеет таллом с зубчатыми краями; на талломе F. vesiculosus находятся округлые воздушные пузыри, наполненные газом. Воздушные пузыри имеет и встречающийся в тех же местах Ascophyllum nodosum (см. рис. 11.85, С). У Himanthalia (В) таллом состоит из основания, напоминающего по форме юлу; на нем расположены от одной до нескольких ремневидных, вильчато разветвленных вертикальных ветвей. Более обильное ветвление таллома наблюдает
ся у распространенных в теплых морях представителей (250 видов!) рода Sargassum. Некоторые его виды благодаря наличию воздушных пузырей свободно плавают бесчисленными пучками в Саргассовом море Атлантического океана (от Гольфстрима и Вест-Индии до Азорских островов). Они размножаются здесь только вегетативно путем распада талломов (А). Сюда же относятся роды Cystoseira и Halidrys", Pelvetia (см. рис. 11.107); Coccophoraii Durvillea (бокс 11.7, с. 199).
У бурых водорослей можно проследить прогрессивный переход от изогамии к оогамии через анизогамию. У исходных форм {Ectocarpus, см. рис. 11.80, А) гаметангии обоих полов многоклеточные, одинакового внешнего вида. У более высокоорганизованных форм (например, Cutleria) с увеличением размера гамет происходит также увеличение размера гаметангиев с одновременным уменьшением числа гнезд. У Dictyota и Laminaria это приводит к образованию только одной яйцеклетки. Если у Dictyota спермогонии еще многоклеточные, то у Laminaria они уже, как и оогонии, одноклеточные, с одним сперматозоидом. Однако такое развитие признаков не позволяет делать какие-либо выводы о фактической филогении. Смена поколений (рис. 11.87) изоморфная (Isogeneratae) или гетероморфная (Heterogeneratae), причем сходство поколений рассматривается как исходный вариант, тогда как преобладание спорофита (впервые отмеченное у Ectocarpales) — как более подвинутый вариант.
Филогения. Бурые водоросли, по-видимому, произошли с золотистыми от общих предков и уже рано стали эволюционировать разными линиями. Несмотря на свои иногда очень значительные размеры, бурые водоросли в целом сохранились в ископаемом состоянии хуже, чем известковые формы зеленых водорослей. Вполне вероятно, что они существовали уже в силуре и девоне. Некоторые нижнедевонские и силурийские «стволы» {Nematophycus = Prototaxites) толщиной с человеческое бедро, которые состояли из сплетения трубковидных нитей и заканчивались мощными пучками лопастей, напоминающих пластины Laminaria, по-видимому, тоже были бурыми водорослями.
Обзор Heterokontophyta. В общем виде Heterokontophyta отражается все разнооб-
244 | ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Рис. 11.87. Phaeophyceae. Смена поколений и ядерных фаз у некоторых бурых водорослей (A—D) (по R. Harder, с дополнениями).
Гаплоидная фаза показана светлыми линиями, диплоидная — темными; G — гаметофит; S — спорофит; О — зигота; R! — редукционное деление
разие форм организации, имеющихся у водорослей. В этом смысле данный отдел выглядит очень гетерогенным. С другой стороны, очевидны родственные связи различных форм через ряд общих «консервативных» признаков, которые скорее всего либо не играют селективной роли и стабильны, либо абсолютно необходимы для дальнейшего существования вида. На основе структуры подвижных клеток и хлоропластов, а также состава хлорофиллов, дополнительных пигментов и запасных веществ можно охарактеризовать Heterokontophyta как эволюционно единую группу водорослей.
Имея паренхиматозные талломы, бурые водоросли относятся к наиболее высокоразвитым морским растениям. По расчленению вегетативного тела и проводящим элементам, аналогичным ситовидным трубкам, они напоминают строение высших растений. Усложнение строения вегетативного тела в связи с приспособлением к многообра
зию условий внешней среды — эволюционный принцип, реализуемый в самых разных группах растений. При этом постоянно можно констатировать: одна-единствен-ная эволюционная линия (группа родственных форм) развивается по пути многообразной дифференциации самых разных приспособительных форм, и наоборот — разные эволюционные линии в ходе приспособления к сходным условиям существования могли выработать сходные формы организации (конвергенция!). Пример первого положения — различные уровни организации Heterokontophyta. Второе положение подтверждается несколькими параллельными эволюционными линиями среди Heterokontophyta и зеленых водорослей, обсуждаемых дальше.
Отличительная особенность Heterokontophyta— частое использование кремниевой кислоты при образовании и оформлении жесткой клеточной стенки. Пластинки,
11.2. Бактерии, грибы, растения | 245
чешуйки и отложения из кремниевой кислоты не являются особенностью всех представителей. Так, они полностью отсутствуют, например, у голых клеток Chloromonado-phyceae и иногда в других классах. С другой стороны, как исключение они встречаются в других отделах водорослей (например, Pediastrum из Chlorobionta). Однако здесь прослеживается четкая тенденция к образованию этих структур. Целлюлоза явно не служит предпочтительным материалом для построения клеточной стенки. У Chloromo-nadophyceae жесткая целлюлозная стенка отсутствует, а у других классов, за исключением бурых водорослей, она широко не представлена.
Приспособление к наземной жизни. Обитателями влажной почвы являются, например, различные диатомеи, а среди желтозеленых водорослей, например, Botrydium (см. рис. 11.73, G). К тому же классу относится аэрофильная водоросль Capitulariella, которая распространяется путем рассеивания по воздуху целых спороцист (функционально они могут быть приравнены к апланоспорам) (см. рис. 11.72, С). Мужские репродуктивные клетки Heterokontophyta обладают (когда имеет место анизогамия или оогамия) хлоропластами, часто сильно редуцированными.
Прогрессивное эволюционное развитие по пути к чистым диплонтам происходит независимо в разных подвинутых эволюционных линиях. Так, все диатомеи диплонты, а кроме того, Pennales уже не имеют гамет со жгутиками. Среди бурых водорослей прослеживаются усиление в жизненном цикле спорофитного поколения и эволюция в сторону практически чистых диплонтов. У оогамных представителей женские репродуктивные клетки стали неподвижными яйцеклетками.
Седьмое подцарство: хлоробионты (Chlorobionta, «Viridiplantae»)
Chlorobionta содержат в своих чисто зеленых пластидах хлорофиллы а и Ь. Такая комбинация ассимиляционных пигментов
имеется у прохлоробактерий (см. царство Bacteria), а по новейшим сведениям — также у цианобактерий (см. царство Bacteria) и среди водорослей у хлорорахниофитов (см. приложение 1 к Chlorophyta) и эвгле-нофитов (см. приложение 2 к Chlorophyta), которые, однако, отличаются другими признаками, в частности отсутствием крахмала в качестве запасного вещества. Каротины и ксантофиллы (см. табл. 11.2) не маскируют зеленую окраску ассимиляционных пигментов. Хлоропласты ограничены лишь двойной мембраной, без эндоплазматической сети и опоясывающей ламеллы (см. рис. 11.72, G). Тилакоиды сгруппированы в стопки — граны. Пиреноиды, если имеются, расположены внутри хлоропласта.
Важнейший запасной полисахарид — крахмал, который откладывается в виде свободно расположенных зернышек возле пиреноида внутри хлоропласта. Иногда в клетках откладывается также значительное количество жира. Клеточная стенка состоит из полисахаридных фибрилл (преимущественно целлюлозы и отчасти маннана и глюкана), которые погружены в аморфную, часто слизевидную фракцию. Стенка образуется чаще всего непосредственно поверх плазмалеммы (в отличие от динофитов и диатомей). Аморфная фракция состоит обычно из различных полисахаридов — пектинов1.
У некоторых наземных водорослей среди Chlorobionta обнаружено стойкое вещество спорополленин (см. 2.2.7.6); оно встречается и у высших растений, относящихся к этому же подцарству, делая их споры и пыльцу устойчивыми к высыханию1 2.
У хлоробионтов в противоположность большинству ранее рассмотренных групп водорослей формы с сильно дифференцированным строением часто встречаются также в солоноватых и пресных водах. В этом подцарстве произошел переход к высшим наземным растениям.
1 У некоторых представителей подцарства другой состав, например пептидогликановый (как у зеленых водорослей из рода Chlamydomo-nas). — Примеч. пер.
2 Спорополленин имеется и у зеленых водорослей, живущих в воде (например, у видов рода Chlorella). — Примеч. пер.
246 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Первый отдел: зеленые водоросли I (Chlorophyta)
У зеленых водорослей встречаются почти все уровни организации. За исключением амебоидных форм (которые, правда, иногда представлены репродуктивными клетками), здесь выработались все типы, даже тканевый и ложнотканевый (соответственно, например, Ulva и Codium). Это могут быть микроскопически мелкие одноклеточные организмы, нитчатые водоросли — неразветвленные или разветвленные, часто образующие плотные пучки (см. рис. 11.90), а также сложно устроенные листовидные талломы, которые отчасти напоминают высшие растения.
Образование поперечных перегородок осуществляется преимущественно в фикопластах: здесь микротрубочки во время те-
Рис. 11.88. Chlorobionta, Chlorophyta. Микро-тубулярная корневая система жгутикового аппарата (по O'Kelly и Floyd; Mattox и Stewart: из van den Hoek и H.M. Jahns).
Вид на базальные тельца с корнями из двух и четырех микротрубочек в крестообразном расположении. Верхний ряд (А, С, Е) — двухжгутиковые; нижний ряд (В, D, F) — четырехжгутиковые клетки. А, В — тип «12 1асов — 6 часов»: гипотетический исходный жгутиковый аппарат, у которого базальные тельца (каждая пара находящихся друг против друга жгутиков) расположены на одной линии; С, D — тип «11 часов — 5 часов»: базальные тельца (по сравнению с А, В) слегка сдвинуты относительно друг друга против часовой стрелки; Е, F — тип «1 час — 7 часов»: базальные тельца (по сравнению с А, В) слегка сдвинуты относительно друг друга по часовой стрелке
лофазы собираются в экваториальной плоскости между разделяющимися дочерними ядрами. При этом возникает клеточная пластинка с порами для плазмодесм (см. 2.2.1).
Клетки, имеющие жгутики, чаще всего грушевидные, радиально-симметричные. Жгутики одинаковой длины в количестве 2 или 4 (редко их много) расположены на переднем конце клетки (терминально); эти подвижные клетки изоконтные, снабжены «гладкими» жгутиками. Они нередко содержат сократительные вакуоли (чаще всего 2), а также (в задней по ходу движения части) изогнутый или бокаловидый постенный хлоропласт1 с глазком (стигмой; см. рис. 11.94, А) или без него. Красный глазок состоит из глобул, которые содержат каротины; он не связан (как у Euglenophyta, Eustigmatophyta и Heterokontophyta) с расширением жгутика.
Понятие «гладкие жгутики» означает, что они не несут трубчатых волосков, однако могут иногда иметь очень тонкие волоски или чешуйки разного типа.
Большое значение для систематики хло-рофитов имеет ультраструктура жгутикового аппарата в области прикрепления жгутиков (рис. 11 88). Он состоит из базальных телец (т. е. нижних концов жгутиков в клетке), микротрубочковых корней вместе с ассоциированными с ними структурами, а также ризопласта (или ризопластов). Последние представляют собой соединения между базальными тельцами жгутиков и клеточным ядром. Жгутики чаще всего прикрепляются по крестообразному типу: четыре крестовидно расположенных микротрубочковых корня закрепляют базальные тельца жгутиков в клетке. При расположении «1 час — 7 часов» (сокращенно расположение 1 — 7) два базальных тельца располагаются, если смотреть на клетку сверху, как цифры 1 и 7 на циферблате часов. Аналогично следует понимать расположения «12 часов — 6 часов» и частый вариант «11 часов — 5 часов».
При половом размножении у подавляющего большинства образуются гаметы, имеющие жгутики. При этом копулируют
1 У немногих представителей хлоропласт может занимать и центральное положение. — Примеч. пер.
11.2. Бактерии, грибы, растения | 247
Бокс 11.6. Применение водсрослай
Из золы различных бурых водорослей (Phaeophyceae: Laminariales) можно получать йод; именно таким путем его получали еще до 30-х годов прошлого столетия. Пригодные для этого бурые водоросли могут накапливать в клетках йод (до 0,3 % сухой массы) из морской воды, где его концентрация 0,000005 %. Кроме того, бурые водоросли дают альгинаты, коллоидные свойства которых находят многостороннее применение в текстильной, пищевой промышленности, фотоиндустрии и производстве косметики. Мировая продукция их около 14000 т в год. Они используются, например, при приготовлении мороженого, пудинга, мазей, зубной пасты, диетических продуктов для похудения, капсул для лекарств, клея, красок и т. д. Из бурых водорослей получают также соду и маннит. В Китае и Японии бурые водоросли используются в пищу (комбу).
Из клеточных стенок нескольких видов красных водорослей (Rhodophyta) получают полисахариды для лекарственных и технических целей. Находят применение каррагенан из Chondrus crispus и Gigartina mamillosa с побережья Северного моря (в высушенном виде известен еще под названием «ирландский мох») и агар из различных флоридей Тихого океана (таких, как виды Gelidium и Graci-laria), а в последнее время также из европейских видов. Япония, добывая ежегодно 2 000 т агара, является важнейшим его продуцентом (использование для культур микроорганизмов, в пищевой и фармацевтической промышленности). Рог-phyra (культивируемая по морским побережьям Восточной Азии на плантациях, представляющих собой подвешенные у
поверхности воды решетки) широко используется в пищу .(нори) особенно в восточноазиатских странах.
Зеленые водоросли (Chlorophyta) имеют меньшее значение, чем красные и бурые. В Западной Сибири проводится массовый сбор нитчатых зеленых водорослей (около 1 000 000 т ежегодно с нескольких тысяч квадратных километров), которые используются для производства бумаги, а также изоляционного и строительного материала (альгилит). Для биотехнологии пригодны коккоидные зеленые водоросли (Chlorella, Scenedesmus) благодаря высокой активности фотосинтеза и возможности массового культивирования. Соответствующие исследования направлены на получение белков и витаминов для питания человека и животных (максимальные урожаи в культурах под открытым небом в тропиках — 5 т на гектар ежемесячно) С помощью «водорослевых реакторов» можно осуществлять биологический газообмен (СО2 на О2 при фотосинтезе). Испытаны соответствующие приспособления для использования этих водорослей в качестве источников кислорода и питательных веществ (например, для космических кораблей).
Прочие водоросли. Образуемые из диатомей (Bacillariophyceae) породы (диатомовая земля, полировочные сланцы, кизельгур) раньше использовались как материал для строительства, чистки и шлифовки. И поныне они еще используются как фильтрующая масса (например, для очистки воды) или как абсорбент и наполнитель. Порода, состоящая из кокколитов (Haptophyta), раньше находила применение как писчий мел.
2 гаметы (ср. рис. 11.89, F), которые часто очень сходны с зооспорами (спорами бесполого размножения) и возникают в одноклеточных гаметангиях. Мужские гаметы, как правило, имеют жгутики, женские (яйцеклетки) могут быть также неподвижными (например, см. рис. 11.100, Е). Простой гаплонтный жизненный цикл часто услож
няется наличием диплофазы (т. е. становится гаплодиплонтным). Некоторые представители за счет редукции гаметофита стали диплонтами (как Caulerpaceae). Таким образом, исходно зиготическая смена ядер-ных фаз становится промежуточной, или гаметической. Продукт копуляции — зигота — у пресноводных форм чаще всего
248 | ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Рис. 11.89. Ulvophyceae (А—К — по Dodel; L — по Р. Kuckuck; М — по H.Kylin):
А—К — Ulothrix, U.zonata: А — молодые нити с ризоидальной (г) клеткой (ЗООх); В — фрагмент нити с освобождающимися зооспорами, которые возникают по две в каждой клетке; С — отдельные четырехжгутиковые митозооспоры; D — образование и выход имеющих меньший размер двухжгутиковых гамет из фрагмента нити; Е — гаметы; F — их копуляция; G, Н — зигота; J — зигота, прорастающая после периода покоя; К — образование мейозооспор в зиготе (В—К — 480х); L — Ulva, U. lactuca (морской салат) на камне, клетки по кромке бесцветные из-за выхода зооспор (1/2х); М — Entero-morpha, Е. intestinalis, копуляция анизогамет и зигота (1 800х); 1 —3 — стадии до образования зиготы
представляет собой толстостенную, округлую покоящуюся клетку (цистозиготу), нередко окрашенную каротиноидами в красный цвет.
К зеленым водорослям относятся примерно 450 родов и 7 000 видов, большая часть которых (около 90%) обитает в планктоне или бентосе (см. бокс 11.7) пресных вод. Некоторые виды с более крупными талломами встречаются также в морях, обычно у берегов; в морском планктоне, напротив, зеленые водоросли составляют лишь незначительную часть. Некоторые зеленые водоросли живут вне воды: во влажной почве или на ее поверхности, как эпифиты на деревьях и т.д. Определенные виды даже выдерживают значительное высыхание и являются выраженными наземными растениями. Некоторые живут как симбионты в лишайниках или как внутриклеточные эндосимбионты в низших животных («зоохлореллы», например, в гидре). Некоторые представители утратили ассимиляционные пигменты и живут гетеротрофно. Родство их с автотрофными формами зеленых водорослей мож
но устанавливать по другим признакам. Систематическое положение отдельных родов зеленых водорослей остается пока неясным.
1. Класс: Prasinophyceae
Представители класса имеют своеобразные чешуйки на поверхности клеток, а также 2 или 4 одинаковых жгутика (изредка жгутик только один). Относящиеся сюда монадные (например, Pyramimonas, Pedino-monas, Platymonas), отчасти также капсаль-ные и коккоидные организмы обитают в основном в планктоне морей, лишь немногие виды — в пресных водах. Platymonas convolutae — эндосимбионт морского плоского червя.
У последующих классов (2—7) жгутики прикрепляются по крестообразному типу и жгутиковый аппарат показывает расположение 11 — 5 своих ультрамикроскопи-
11.2. Бактерии, грибы, растения | 249
ческих структур (у 8-го класса — Chloro-phyceae, напротив, расположение 1—7 или 12-6).
2. Класс: Ulvophyceae
Одноклеточные (коккоидные) и многоклеточные — нитчатые, паренхиматозные или сифонокладальные — представители этого класса, за исключением репродуктивных клеток, не имеют жгутиков. Жгутиковый аппарат (расположение 11 — 5) показывает четкий сдвиг базальных телец относительно друг друга. Образование клеточной стенки происходит бороздой без участия фикопласта. В поперечных перегородках плазмодесмы отсутствуют. Стенки содержат полисахариды. Большинство видов встречается в морских или солоноватых водах.
Талломы представлены преимущественно неветвящимися нитями, которые удлиняются за счет («диффузного») поперечного деления многих или всех клеток (нитчатый уровень организации). У рода Monostroma более старые нити за счет продольных делений клеток в одной плоскости становятся уплощенными. Крупный листовидный зеленый двухслойный паренхиматозный таллом образуется у обитающей в прибрежной зоне морей водоросли Ulva lactuca(piw. 11.89, L). Enteromorpha, водоросль также прибрежных вод, однако встречающаяся и в соленых внутренних водах, имеет таллом в виде кишки, в сечении округлой или уплощенной. Полярность у некоторых видов выражена лишь слабо. Так, например, у Ulothrix она определяется наличием единственной бесцветной ризои-дальной клетки, не способной к делению (рис. 11.89, А). Клетки имеют ядро и по одному постенному поясковидному хлоропласту, имеющему форму цилиндра закрытого либо с продольными отверстиями или же форму изогнутой пластинки с одним или несколькими пиреноидами. После ядерного деления для разграничения дочерних клеток сразу же втягивается клеточная стенка (ср. Chlorococcales).
Бесполое размножение происходит зооспорами, половое — путем копуляции гамет, имеющих жгутики. Цикл развития осуществляется или как чисто гаплонтный с
зиготической сменой ядерных фаз (Ulothrix)' либо гаплодиплонтный как гетерофазная смена поколений (Ulva).
1. Порядок: Codiolales (Ulotrichales). Часто встречающийся в пресных водах вид Ulothrix zo-nata (рис. 11.89, А) образует неразветвленные, интеркалярно нарастающие нити. Их короткие клетки содержат один лентовидный хлоропласт, который в виде незамкнутого с одной стороны кольца прилегает к клеточной стенке. Нити прикрепляются ризоидальной клеткой, чаще всего бесцветной, к камням и другим субстратам. Кроме ризоидальной клетки, любая другая клетка может служить для размножения; при этом в клетках ничего не остается. В жизненном цикле (см. рис. 11.101, А; 11.89, А—К) 4-жгутиковые гаплоидные митозооспоры обеспечивают бесполое размножение. Они имеют глазок и хлоропласт и образуются одновременно путем распада протопласта, становящегося перед этим четырехъядерным, на одноядерные подвижные клетки, которые выходят (В) через боковое отверстие в стенке материнской клетки (= спороцисте). После фазы активного движения они прикрепляются, выделяя слизь, жгутики и глазок исчезают, и клетки вырастают в новую гаплоидную. полярную нить. При неблагоприятных условиях сходным образом, но в значительно большем числе (D, Е) образуются изогаметы; они внешне сходны с зооспорами, но меньше и обладают только 2 жгутиками. Гаметы разных типов скрещивания (+, -) сливаются (F) попарно в зиготу (G). Зигота, сначала плавающая при помощи 4 жгутиков, втягивает жгутики, округляется (Н), одевается прочной стенкой (цистозиго-та) и окрашивается каротиноидами в красный цвет. Она представляет собой покоящуюся стадию, которая заканчивается мейозом и следующим за ним прорастанием 4—16 гаплоидными мейозооспорами (К)1 2. При этом происходит разделение на два типа скрещивания. Мейозооспо-ры прикрепляются к субстрату боком с образованием ризоида, так что их продольная ось при начинающемся делении становится поперечной осью. Теперь развиваются гаплоидные нити с ге
1 У Ulothrix зигота непосредственно перед мейозом прорастает в маленькое растеньице грушевидной формы; туда переходит диплоидное ядро, где уже претерпевает мейоз. Это растеньице часто трактуют как диплоидную стадию этой водоросли (Со<йо/ши-стадия), и тогда уже нельзя говорить о зиготической смене ядерных фаз. — Примеч. пер.
2 Структура, в которую прорастает зигота, иногда трактуется как диплоидное поколение (см. предыдущую сноску). — Примеч. пер.
250 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
нотипическим определением полового знака (А), которые кроме гамет могут давать также мито-зооспоры. Таким образом, эти растения гаплонт-ны с зиготической сменой ядерных фаз. Зигота (J), образующая палочковидный придаток, может отчасти рассматриваться как крайне недоразвитый спорофит.
Codiolales обитают как в пресной воде, так и в морях, отчасти также в почве. Талломы водных представителей часто прирастают к субстрату.
У Monostroma grevillei листовидный тканевой (мужской или женский) таллом образует гаметы. Зигота развивается в самостоятельный, внедряющийся в известковые раковины морских животных пузырек (Codiolum). У некоторых видов размер зиготы увеличивается в 20 (и более) раз от исходного диаметра.
2. Порядок: Ulvales с родами Ulva (морской салат) и Enteromorpha (кишечница). Здесь происходит гетерофазная изоморфная смена поколений: вклинивается диплоидная спорофитная фаза.
Эту ситуацию можно кратко рассмотреть на примере Ulva— морского салата. Гаметофит и спорофит имеют внешне одинаковые листовидные тканевые талломы (рис. 11.89, L). Смена поколений аналогична видам рода Cladophora (см. рис. 11.101. В). Гаметофиты генотипически (+ и -) дифференцированы. Между двухжгутиковыми гаметами противоположных типов скрещивания происходит изогамная копуляция. Образующиеся таким образом зиготы прорастают, как у многих морских водорослей (постоянно при благоприятных условиях для роста в противоположность пресноводным формам), сразу в диплоидный спорофит. Он дает четырехжгутиковые зооспоры, с образованием которых связаны как мейоз, так и генотипическое определение пола. Из зооспор вновь возникают гаплоидные гамсгофиты, различающиеся по полу. Как гаметофиту, так и спорофиту предшествует нитчатый предросток, который в результате продольных делений клеток становится листовидным тканевым талломом.
Род Enteromorpha отличается от рода Ulva анизогамией. Определение пола гаметофита генотипическое; мужские талломы дают с<" гаметы меньшего размера, женские — более крупные <j> гаметы с зеленым хлоропластом (ср. рис. 11.89, М). У Hormidium изучали движение хлоропластов (см. 8.2.2).
В родстве с рассмотренными порядками находится и порядок Acrosiphonales. Твердая фракция клеточной стенки состоит из переплетенных в виде войлока целлюлозных фибрилл (тип целлюлозы отличается от кристаллической целлюлозы рассматриваемых ниже Cladophorophy-сеае и большинства листостебельных растений).
Талломы Spongomorpha (гаметофит) могут быть переплетенными, как войлок. Смена поколений гетероморфная, со спорофитом типа Codiolum. Сюда же относится Urospora (см. рис. 11.102).
3. Класс: Trebouxiophyceae
К этому классу относятся водоросли, обитающие вне воды1, в том числе симбионты лишайников, от коккальной до разветвленно-нитчатой организации. Жгутиковый аппарат (у репродуктивных клеток) со сдвинутыми относительно друг друга базальными тельцами (расположение 11—5) показывает особенности, которые встречаются только в этом классе. Дочерние клетки, возникающие в результате схизогонии (быстро следующих друг за другом клеточных делений в материнской клетке), окружаются заново образующейся клеточной стенкой. Этим Trebouxiophyceae сходны с Chlorococcales из Chlorophyceae. однако митоз у них протекает иначе1 2. Виды родов Тге-bouxia и Chlorella, отличающиеся коккальным типом таллома (некоторые с апланоспорами, см. рис. 11.97, J), представлены симбионтами в лишайниках3, а также встречаются в плазме низших животных {Chlorella vulgaris в инфузориях, Chlorohydra тл др., ср. 9.2).
Сюда же примыкают Prasiolaceae. Prasiolasti-pitata имеет своеобразный жизненный цикл. В чисто вегетативных верхних частях листовидного спорофита происходят мейозы, за которыми следуют митозы. Развивающийся таким путем гаметофит остается на протяжении всей своей жизни сросшимся со спорофитом. Отдельные поля клеток гаметофита производят яйцеклетки, другие — мелкие двухжгутиковые d" гаметы (генотипическое определение пола, оогамия). Вышедшие на свободу репродуктивные клетки сливаются затем в зиготу.
1 Виды рода Chlorella очень широко встречаются не только вне воды, но и в пресных водах. — Примеч. пер.
2 Следует уточнить, в чем именно состоит отличие: у Trebouxiophyceae при митозе центриоли располагаются по бокам веретена (т. е. веретено метацентрическое), тогда как у других зеленых водорослей и многих других «протестов» — по полюсам веретена. Это основная особенность Trebouxiophyceae. — Примеч. пер.
3 Виды рода Chlorella в лишайниках не встречаются. — Примеч. пер.
11.2. Бактерии, грибы, растения | 251
Последующие классы стоят весьма изолированно; их родственные связи с другими Chlorophyta еще не выяснены.
4. Класс: Cladophorophyceae
Виды этого класса относятся исключительно к сифонокладальному типу организации; разветвленные, иногда неразветвлен-ные нитчатые талломы образованы многоядерными клетками. Клеточные стенки состоят из целлюлозы в виде фибрилл и с той же структурой, что и у наземных зеленых растений. Подвижные репродуктивные клетки имеют 2 или 4 жгутика; для жгутикового аппарата (расположение 11—5) характерно наложение базальных телец. Обитают преимущественно в морях, реже в пресных водах.
Часто обильно разветвленные талломы видов единственного порядка Cladophorales многоклеточные, а каждая клетка многоядерная. Многоядерные клетки встречаются также у видов других порядков (например, Hydrodictyon из Chlorococcales). Виды рода Cladophora имеют чаще всего гетеро-
Рис. 11.90. Cladophorophyceae, Cladophora (по F.OItmanns, с дополнениями):
А — внешний вид (1 /Зх); В — ветвление; С — гаметангий с гаметами (В, С — 250х)
фазную изоморфную смену поколений (см. рис. 11.101, В). При этом каждое поколение может воспроизводиться бесполым путем.
Рис. 11.91. Bryopsidophyceae (А — по Н.Schenck; В — по F.OItmanns; С, D — по К. Magdefrau; Е — по W. Schmitz; F, G, J — по Р. Kuckuck; Н — по R. Harder; К — по J.S. Davis):
A—D — Halimedales: А — Caulerpa prolifera, таллом (12х), В — Halimeda tuna, таллом (1 /2х), С, D — Codium tomentosum: С — таллом (1/2х), D — поперечный разрез таллома (15х); Е — Valoniales (ныне отнесены к Cladophorophyceae); Valonia utricularis, таллом (11/2х); F—К— Bryopsidales: F, G — Derbesia marina («Halicystis ovalis»): F — гаметофит (3x), G — d" и <j> гаметы (500x); H, J, К — Derbesia marina: H — фрагмент таллома спорофита (30x), J — спорангий (120х), К — зооспора (400х); g — гаметангий; г — кортикальный пузырь (вздутая клетка корового слоя)
252 | ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Изогаметы двухжгутиковые, тогда как мейо-зооспоры снабжены 4 жгутиками (у пресноводных видов их 2). Часто образующая в пресных водах пучки длиной до 30 см и более Cladophora glomerate размножается только бесполым путем1.
Пучки нитей видов Cladophora, часто встречающиеся на твердом субстрате в пресных водах (в том числе проточных) и в морях на твердом субстрате (рис. 11.90), прикрепляются у основания клеткой типа ризоида и показывают преимущественно верхушечный рост. Ветвление происходит путем выпячивания «стволовой» клетки (каждый раз под центрипетально образованной перегородкой); они продолжают рост при образовании перегородки под острым углом к продольной оси. Постенный хлоропласт сетевидный* 2. Он содержит пиреноиды с зернышками крахмала. Целлюлозные микрофибриллы клеточной стенки расположены слоями под разными углами, придавая ей большую прочность. Как и у Ulvophyceae. подвижные репродуктивные клетки (зооспоры и изогаметы) возникают в практически неразличимых внешне клетках, однако, как правило, на концах боковых ветвей (Siphonocladus — морская водоросль).
Примыкающий к Cladophorales порядок Valoniales (рис. 11.91, Е; раньше его относили к Bryopsidophyceae) отличается особым механизмом деления клеток. Протопласт первоначально не разделенного на клетки мешковидного таллома делится на несколько фрагментов разного размера, которые округляются часто еще внутри исходного таллома и окружаются новыми клеточными стенками. Таким путем может возникнуть многоклеточный псевдопарен-химатозный таллом. Талломы Valonia, содержащие крупную вакуоль, много ядер и многочисленные постенные хлоропласты, представляют собой удобный объект для изучения проницаемости клеточных стенок (см. рис. 2.66, В).
' Есть сведения, что у С. glomerata имеется и половой процесс. — Примеч. пер.
2 На самом деле у Cladophora не один хлоропласт, а много удлиненных хлоропластов неправильной формы, смыкающихся концами в постенную сеть, так что выглядят они как единая сетчатая пластинка. — Примеч. пер.
5. Класс: Bryopsidophyceae (= сифоновые)
Сифоновые, особенно распространенные в теплых морях, имеют чрезвычайно разнообразный внешний вид. Их таллом не имеет поперечных перегородок, может быть только система распорок. Клеточная стенка (в качестве компонентов которой наряду с целлюлозой встречаются маннан и ксилан) окружает, таким образом, один-единственный полиэргидный протопласт, содержащий многочисленные мелкие дисковидные хлоропласты. Только вместилища репродуктивных клеток отделяются перегородкой (сифональный уровень организации). Талломы некоторых видов представляют собой сложные сплетения нитевидных элементов. К набору пигментов, характерному для зеленых водорослей, у сифоновых добавляются сифоноксантин и сифонеин — отл1гчительные для данного класса дополнительные пигменты.
Половое размножение анизогамное, реже изогамное. Репродуктивные клетки имеют 2, 4 или много жгутиков. Жгутиковый аппарат (расположение 11—5) показывает наложение базальных телец. Жизненный цикл, согласно новейшим исследованиям, преимущественно гаплонтный, однако возможна и гетероморфная смена поколений с дикариотично-диплоидными спорофитами. Это можно рассмотреть на примере Derbesia- Halicystis. Гаметофит состоит из пузыревидного гаметангия размером 0,5 — 3 см, от которого отходит многолетний ризоид. Из-за незнания связи со спорофитом раньше это растение помещали в отдельный род («Halicystis» = гаметофит Derbesia', рис. 11.91, F). Раздельнополые растения «Halicystis» дают анизогаметы с двумя жгутиками равной длины (G). Из зиготы вырастает спорофит с разветвленно-нитевидным талломом — Derbesia (Н). В яйцевидных спороцистах этого растения, сначала дикариотичного, затем местами диплоидного, возникают после мейоза мейозооспоры с венцом жгутиков. Смена поколений гетероморфная со слабым преобладанием дикариотично-дип-лоидного спорофита, достигающего до 10 см в высоту (см. рис. 11.101, С). Состав
11.2. Бактерии, грибы, растения | 253
клеточной стенки у этих поколений, как и у Bryopsis (см. ниже), различен.
1. Порядок: Bryopsidales. Виды порядка не имеют амилопластов, служащих для образования крахмала. Виды типового рода Bryopsis образуют ползучие «корневища», прилегающие к субстрату, и вертикальные ветви, которые в верхней части перисто разветвлены. Жизненный цикл гап-лонтный. Клеточная стенка растеньиц, образующих гаплоидные гаметы, состоит преимущественно из ксилана, стенка зиготы — в основном из маннана. Derbesia (Halicystis) встречается по Атлантическому побережью.
Codiaceae отличаются талломами длиной до нескольких метров (например, некоторые виды рода Codium; рис. 11.91, С, D), представляющими собой сплетение разветвленных, лишенных поперечных перегородок нитей, жесткость которым придают кольца на клеточной стенке. Клеточная стенка состоит преимущественно из маннана. В ископаемом виде Codiaceae известны уже из нижнего палеозоя.
2. Порядок: Halimedales. Клетки представителей этого порядка имеют как хлоропласты так и амилопласты. Клеточные стенки не содержат маннана.
Семейство Caulerpaceae с распространенным в более теплых морях родом Caulerpa, представленным самыми разнообразными формами, отличается талломами, состоящими из бесцветных ползучих основных осей длиной до 1 м с идущими снизу в субстрат ризоидами, а вверх — лопастями различного облика, которые могут достигать размеров в несколько дециметров (А). Крупные растения состоят из одной-единствен-
ной многоядерной гигантской клетки, внешняя стенка которой поддерживается только многократно ветвящимися тяжами в виде балок. Клеточная стенка содержит преимущественно кси-лан. Происходит ли при образовании гамет (они выходят зелеными облачками, после чего опустевшее растение отмирает) мейоз, остается неясным.
У представителей распространенного в более теплых морях рода Halimeda (В) таллом состоит из дисковидных элементов, стенки которых инкрустированы известью.
6. Класс: Dasycladophyceae
Dasycladophyceae (Dasycladales) отличаются от типичных сифоновых (предыдущий класс) радиальной симметрией таллома и волосовидными выростами (см. рис. 4.1), которые отчасти могут сбрасываться, оставляя при этом рубцы Центральная ось несет мутовчато расположенные боковые ветви. Виды этого класса встречаются исключительно в морях.
Клеточная стенка состоит преимущественно из маннана. Таллом образован прикрепленной ризоидами к субстрату «стволовой клеткой» и мутовчато отходящими от нее боковыми ветвями (рис. 11.92, В). Эти ветви могут быть простыми или разветвленными и часто заканчиваются гаметангием.
В качестве объекта морфогенетических исследований особенно известна Acetabu-
Рис. 11.92. Dasycladophyceae, Dasycladales. Cymopolia barbata (А, В — по Н. Solms-Laubach; С, D — по F.OItmanns; Е—G — по A. De Вагу и Е. Strasburger):
А — верхняя часть растения (4х); В — продольный разрез через часть таллома, мелкими точками показан известковый покров (40х); С—G — Acetabularia mediterranea: С — взрослые талломы (естественная величина), D — продольный разрез через «зонтик», вверху венец из стерильных выростов, внизу рубцы из отпавших мутовок стерильных выростов (6х), Е — вскрывшаяся циста, из которой выходят гаметы (100х), F — гаметы (ЗООх), G— копуляция (ЗОх)
254 | ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
laria (рис. 11.92, С—G). Эта водоросль несет на неразделенной ножке «шляпку» в виде зонтика, которая состоит из радиально расположенных, плотно прилегающих друг к другу камер. Над зонтиком и под ним образуется по венцу из коротких клеток. Из верхнего венца возникает дополнительно мутовка тонких, разветвленных вверху ветвей, которые при созревании зонтика быстро отмирают. Таллом имеет сначала лишь одно-единственное первичное ядро, которое долгое время находится без изменений в ризоиде. После развития зонтика оно делится на многочисленные гаплоидные вторичные ядра, которые мигрируют в камеры, после чего начинается образование толстостенных цист. Цисты освобождаются после разрушения зонтика, вскрываются крышечкой и высвобождают гаметы (Е). Зигота (G), возникающая в результате копуляции двух изогамет, прикрепляется к дну и вырастает в новый диплоидный таллом. Согласно недавним исследованиям, Aceta-bularia не диплонт, так как уже первичное ядро, по-видимому, является гаплоидным. Однако согласно другой точке зрения мейоз происходит при образовании вторичных ядер; тогда жизненный цикл был бы дип-лонтным (с гаметической сменой ядерных фаз).
Внешние слои клеточной стенки у Dasycla-daceae очень сильно обызвествляются (В), так что после отмирания таллома остается пронизанная отверстиями известковая трубочка; на этом основана значительная породообразующая роль ископаемых Dasycladaceae, например в альпийском триасе. Начиная с кембрия известно в целом 120 родов Dasycladophyceae во всех формациях, тогда как в настоящее время — только 10 родов. На основании ископаемых находок мы можем проследить эволюцию от простых форм, у которых ветви отходят беспорядочно от стволовой клетки до высокодифференцированных, таких, как виды рода Acetabularia.
7. Класс: Trentepohliophyceae
Нитчатый таллом часто гетеротрихальный. т.е. разделенный на стелющиеся и восходящие нити, которые могут быть разветвленными (рис. 11.93, С). Клетки одноядерные. У некоторых форм стелющиеся нити склеиваются в плоский диск (Cephaleuros),
Рис. 11.93. Trentepohliophyceae (А, В — по R.Chodat; С — по К. J. Meyer; D — по G. Karsten; Е — по G.Gobi):
А, В — Pleurococcus, Р. паедеШ (бООх); С—Е — Trentepohlia'. С — Т. аигеа, участок стелющейся нити с вертикальными ветвями (на вершине правой вертикальной нити одна клетка с зооспорангием, от второй зооспорангий отделился; 500х), D — Т. umbrina, зооспорангий, из которого выходят зооспоры (ЗООх), Е — Т. umbrina, отпадение освободившегося спорангия (ЗООх)
как и у некоторых представителей Klebsor-midiophyceae (Coleochaete, см. рис. 11.106, А). Гетеротрихальным строением таллома Trentepohliophyceae напоминают еще не рассматривавшиеся Chaetophorales из Chloro-phyceae (Stigeocloniunr, см. рис. 11.99, А, В). Исключительно для Trentepohliophyceae характерны дополнительные структуры в виде колонок в жгутиковом аппарате (ориентация 11 — 5) с налегающими друг на друга базальными тельцами, а также с билатеральными килевидными углублениями в жгутиках.
Состоящая из полисахаридов клеточная стенка может дополнительно образовывать слой из спорополленина. При клеточном делении новые клеточные стенки закладываются во фрагмопласте. Большинство видов — наземные водоросли (например, эпифиты на коре деревьев или камне).
11.2. Бактерии, грибы, растения | 255
Trentepohlia (рис. 11.93, С) часто встречается как симбионт в лишайниках или как наземная водоросль на скалах (Т. аигеа — на известковых, а напоминающая запахом фиалку Т. iolithus— на силикатных породах) и стволах деревьев, а в тропиках и на кожистых листьях. Приспособление к наземному образу жизни проявляется также в том, что спороцисты с находящимися в них зооспорами часто отрываются целиком. Двухжгутиковые репродуктивные клетки копулируют как гаметы друг с другом или служат для бесполого размножения (факультативное определение функции). Очень распространенные зеленые налеты на коре деревьев и скалах вызываются водорослями типа «Pleurococcus» (Apatococcus и Desmococcus)'; некоторые из этих наземных водорослей больше не образуют подвижных клеток, т.е. они претерпели соответствующую редукцию.
8. Класс: Chlorophyceae
Эволюционно Chlorophyceae в противоположность Prasinophyceae (1), Ulvophyceae (2) и Trebouxiophyceae (3), несмотря на их часто примитивную организацию, следует, возможно, рассматривать как позже отделившуюся группу. К этому классу относятся одноклеточные или колониальные формы со жгутиками или без них, а также нитчатые (трихальные и сифональ-ные) виды. Клеточная стенка у монадных видов состоит из гликопротеинов, у неподвижных представителей — из полисахаридов, в том числе из целлюлозы. При клеточном делении новые поперечные перегородки возникают в фикопластах (см. введение к Chlorophyta); такие стенки часто перфорированы плазмодесмами. Расположение жгутиков относится к крестообразному типу; жгутиковый аппарат показывает расположение 1 — 7 или 12—6 (базальных телец и т.д.; см. рис. 11.88). Виды этого класса обитают преимущественно в пресных водах, незначительная часть их — в солоноватых и морских водах или даже во вневодных условиях.
1 . Порядок: Volvocales. К этому порядку относятся имеющие жгутики одноклеточные формы, которые могут соединяться в колонии (см. 5.1). В этом порядке хорошо
1 Род Desmococcus в большинстве современных систем относится к классу Trebouxiophyceae (см. выше). — Примеч. пер.
прослеживается переход от одноклеточных форм к колониям с различным уровнем дифференциации и увеличивающейся полярностью. Радиально-симметричные клетки имеют 2, 4 или 8 апикальных «гладких» жгутиков равной длины (ср. рис. 11.97, D). Они отходят по обе стороны апикального сосочка.
Размножение одноклеточных видов осуществляется бесполым путем зооспорами, которые образуются при повторных последовательных продольных делениях содержимого материнской клетки на 2—16 протопластов (рис. 11.94, В) и освобождаются при разрыве стенки материнской клетки, ставшей таким образом спороцистой. При половом размножении (у Chlamydomonas 10% видов) сливаются двухжгутиковые гаметы или яйцеклетки и сперматозоиды.
При изогамии (рис. 11.94, С) копулирующие гаметы имеют полностью одинаковые размер, облик и движение и в целом не отличаются от вегетативных клеток. Они могут при соответствующих обстоятельствах либо произвольно копулировать друг с другом, либо развиваться вегетативно (факультативное определение функции). Таким образом, здесь мы имеем дело с зачатками сексуальности. Гаметы могут при этом относиться к одному типу скрещивания (однодомность) или же при видимом сходстве различаться в генетическом отношении (двудомность с «+» или «-» гаметами, например, Chlamydomonas reihardui). Определение функции репродуктивных клеток отчасти зависит от внешних условий. Богатая азотом среда (ионы NH4!) обусловливает исключительно образование вегетативных клеток. Ионы Са способствуют определению функции гамет.
У видов с анизогамией (рис. 11.94, Н, J) 6" гаметы меньшего размера копулируют с более крупными <j> гаметами. У Chlamydomonas suboogama жгутики <j> гамет не функционируют, и это можно рассматривать как переход к следующей группе видов.
Оогамия. У Chlorogonium oogamum $ гаметы полностью лишены жгутиков. Они амебоидно выходят из материнской клетки (рис. 11.95, D) и становятся яйцеклетками. Яйцеклетка оплодотворяется сперматозоидами, которые возникают в количе-
256 I ГЛАВА 11 СИСТЕМАТИКА И ФИЛОГЕНИЯ
Рис. 11.94. Chlorophyceae, Volvocales, Chlamydomonadaceae (А, В — по О. Dill; С—G — по Strehlow;
Н, J — по N.Goroschankin; К — по Е. Reichenow):
А — Chlamydomonas angulosa (1 100х); В — то же, четыре дочерние клетки внутри материнской клетки (1 100х); С, D — Chlamydomonas botryoides, копуляция двух изогамет (250х); Е — Chlamydomonas paradoxa, зигота (500х); F — Chlamydomonas monoica, покоящаяся цистозигота (500х); G — Stepha-nosphaera pluvialis, прорастающая гипнозигота (ЗООх); Н, J — Chlamydomonas braunii, копуляция анизогамет (400х); К — Haematococcus pluvialis (клетка, окруженная толстым слизистым слоем, 330>); с — хлоропласт; g — жгутик; к — ядро; р — пиреноид; s — глазок; v — пульсирующая вакуоль
Рис. 11.95. Chlorophyceae, Volvocales (А—Е — по A. Pascher; F — по G. Hieronymus; G — по Stein;
Н — по N. Pnngsheim):
А—Е — Chlorogonium оодатит (240>): А — вегетативная клетка, В — сГ клетка со сперматозоидами, С— клетка с яйцеклеткой, D — выход яйцеклетки наружу, Е — яйцеклетка, окруженная сперматозоидами; F — Stephanosphaera pluvialis (250х); G — Pandorina morum (160х); Н — то же, образование дочерних колоний (стенки материнских клеток частично уже разрушены, 150х)
11.2. Бактерии, грибы, растения I 257
стве 64 или 128 в виде светло-зеленых двухжгутиковых игловидных образований в сГ особях путем последовательных делений (рис. 11.95, В). У Chlamydomonas coccifera размножение осуществляется как гамето-га-метангиогамия, так как вся 9 клетка, теряя жгутики, становится оогонием и оплодотворяется сперматозоидом.
Таким образом, уже у этих одноклеточных форм можно проследить прогрессивное развитие от изогамии через анизогамию и оогамию до слияния o' гамет с оогонием.
Имеющие жгутики репродуктивные клетки возникают чаще всего в большом количестве (2—64) в материнской клетке путем повторных продольных делений. При изо- и анизогамии они попарно объединяются в зиготы (рис. 11.94, С — Е), причем чаще всего у партнеров соприкасаются сначала вершины жгутиков, которые скручиваются друг с другом (С). При копуляции в качестве гамонов действуют гликопротеины (см. 8.2.1.1), которые привлекают друг к другу гаметы разных типов скрещивания и обеспечивают временное склеивание жгутиков. Зигота имеет 4 жгутика и поначалу еще подвижна (планозигота). Позже жгутики втягиваются, а становящаяся толстостенной зигота может переходить в состояние покоя (цистозигота; F). Гаметы всегда закладываются голыми, но могут также окружаться стенкой, и тогда при копуляции содержимое должно выходить через эту стенку. При прорастании зиготы (G) происходит мейоз, причем возникающие подвижные клетки распределяются в соотношении 1:1 по двум типам скрещивания (+ и -). Таким образом, подвижные клетки являются мейозооспорами, смена ядерных фаз зиготическая, а жизненный цикл гаплонтный. Для образования гамет в отдельных случаях задействованы целые особи.
Встречаемость. Volvocales — широко распространенные в пресных водах планктонные организмы. Они могут встречаться в таких больших количествах, что вода кажется совершенно зеленой; в морях Volvocales отсутствуют.
У многих видов развитию способствует поглощение органических веществ (миксотроф-
ность; см. 9.1), поэтому они встречаются отчасти в водоемах с органическим загрязнением. Немногие виды (например, Polytoma uvella) живут чисто сапротрофно. Хотя у них хлорофилл отсутствует, имевшийся у предковых форм хлоропласт еще можно различить и здесь как бесцветную пластиду. Вместо тилакоидов она содержит систему беспорядочно расположенных трубочек. Аналогичные пластиды отмечаются у полученных в результате облучения ультрафиолетом желтых, нефотосинтезирующих мутантов Chlamydomonas.
Классификация Volvocales. Исключительно голые представители имеются в небольшом, возможно, исходном семействе Polyblepharidaceae. Если Polyblepharides, насколько известно, размножается только путем продольного деления на две клетки, то у более высокоразвитых форм происходит также половое размножение с фенотипическим или генотипическим определением типа скрещивания (+ и -). Dimaliella salina относится к последней группе, живет в очень соленых водах и окрашена каротиноидами в красный цвет.
Семейство Chlamydomonadaceae отличается от Polyblepharidaceae наличием клеточной стенки. Исходно хлоропласт занимает центральное положение, у большинства видов Chlamydomonas он постенный, у более высокоорганизованных форм сете-видно-продырявленный или даже распадающийся на отдельные мелкие лиски. Половое размножение претерпевает прогрессивную эволюцию вплоть до оогамии.
Скорость движения имеющих жгутики клеток, например у Chlamydomonas, составляет при фототаксисе примерно 10-кратную длину клетки в секунду. Недалеко от выроста, где находятся жгутики, расположены две пульсирующие вакуоли, которые попеременно сокращаются и выталкивают воду. Они поддерживают постоянство осмотического давления клетки. Каждая клетка содержит один бокаловидный хлоропласт, в «донышке» которого обычно находится пиреноид, вокруг которого откладывается крахмал (см. 2.2.9.1 и рис. 11.94, А, ср. 11.96, В), а на переднем конце — глазок (стигма, см. рис. 11.94, А). Образование крахмала в хлоропластах не связано исключительно с пиреноидом. Составляющие глазок пигментные шарики (каротиновые глобулы) вместе образуют 3 — 8 рядов. В образовании клеточной стенки (если она имеется; как, например, у Chlamydomonas) участвуют гликопротеиды (в том числе гидроксипролин и арабиноза,
258 | ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Рис. 11.96. Chlorophyceae, Volvocales, Volvox (А — по L. Klein: В, С — по С. Janet; D—J — по W.Zimmermann; К, L — по F.Cohn; М — по С. Janet):
А — особь с 6 дочерними особями (50х); В — отдельная клетка с идущими от нее вбок к соседним клеткам плазмодесмами (1 ОООх); С— соединение клеток, общий вид (500х); D—J — развитие и выворачивание наизнанку дочернего шара (D — 250х, Е— F — 350х, G—J — 250х); К — часть однодомной особи с 5 яйцеклетками и 2 пластинками сперматозоидов (200х); L — яйцеклетка, окруженная сперматозоидами (265х); М — сперматозоид (1 ОООх); A—J, М — V. aureus; К, L — V. globator
связанная с галактозой) и полисахариды (но не целлюлоза!).
Некоторые виды (Haematococcuspluvialis, см. рис. 11.94, К) окрашивают лужи после дождя в красный цвет из-за наличия у них каротиноидов. Chlamydomonas nivalis вызывает «красный снег» в высокогорьях и в Арктике. Некоторые Chlamydomonadaceae (и другие жгутиковые) поселяются даже зимой на голом льду и снегу (см. бокс 11.7, с. 199). Виды рода Carteria имеют 4 жгутика.
Семейство Volvocaceae представляет собой результат дальнейшего эволюционного развития по пути образования колоний. Отдельные клетки, часто похожие на Chlamydomonas, оказываются связанными между собой слизью или также плазмодесмами. У Oltmansiella 4 клетки, у Gonium 4— 16 клеток объединены между собой в плоские таблички, причем все жгутики направлены в одну сторону. Колонии видов Ste-phanosphaera, живущих в остающихся после дождя лужах (см. рис. 11.95, F), представляют собой венец из 4, 8 или 16 клеток с жесткими придатками; хлоропласты имеют чаще всего по 2 пиреноида. У Pandorina 16 клеток, напоминающих Chlamydomonas,
а у Eudorina и Pleodorina соответственно 32 или 128 таких клеток объединены в один шар, полый внутри. У всех колоний жгутики совершают синхронные движения, что обеспечивается плазмодесмами (см. 2.2.7.3). От Pandorina через Eudorina до Pleodorina происходит полярная дифференциация признаков относительно направления движения в воде (размер глазков, размер клеток, способность к размножению и т.д.). Отдельные клетки в конце индивидуального развития не отмирают, а делятся или используются для образования репродуктивных клеток. Наибольшая степень организации, которая оценивается по числу клеток, их дифференцировке и полярности, отмечена у Volvox (рис. 11.96). Колонии Volvox представляют собой видимый невооруженным глазом полый шар размером до нескольких миллиметров, заполненный внутри слизью. Шар образован многочисленными (до нескольких тысяч) клетками: например, у К globator их насчитывается до 16 000. Каждая клетка имеет 2 жгутика, глазок и хлоропласт. Клетки соединены друг с другом широкими плазмодесмами (В; С). Способ
11.2. Бактерии, грибы, растения | 259
ны к размножению только некоторые клетки, рассеянно расположенные в задней части шара. Большинство клеток служит для фотосинтеза и движения; однако и они различаются размером стигмы, который постепенно уменьшается при увеличении размера клеток, от переднего к заднему полюсу (полярность!). Передний полюс шара, кроме того, определяется направлением его перемещения в воде. Шар Volvox можно рассматривать уже не как колонию, а как многоклеточную особь. Отдельные клетки уже не тотипотентны. Поскольку к размножению способна только часть клеток, большинство клеток шара отмирает после об
разования дочерних шаров или гамет («труп» как результат многоклеточности).
При бесполом размножении у Volvox (рис. 11.96, D —J) отдельные, относительно крупные клетки (D) у заднего полюса колонии несколько раз продольно делятся и образуют полую сферу (F), которая в конечном счете превращается в открытый полый шар (G). Возникший таким образом дочерний шар выворачивается наизнанку (Н) и опускается со своими жгутиками, теперь уже ориентированными наружу, внутрь еще заполненного слизью материнского шара. Таким путем возникает несколько дочерних шаров (А), которые освобождаются только после разрушения материнской особи.
Рис. 11.97. Chlorophyceae, Chlorococcales (A—G — по A. Pascher; Н —К — по Grintzesco; L, М — по G. Senn; N, О — по G. Klebs; Р—R — no R. A. Harper):
А—G — Chlorococcum (1 ОООх): А — вегетативная клетка с чашевидным хлоропластом (с), имеющим небольшую выемку лишь на переднем конце, с пиреноидом (р), к — слегка просвечивающее ядро, В — деление на 8 дочерних клеток, С — выход зооспор в пузыре, позже расплывающемся, из внутреннего слоя мембраны материнской клетки (m), D — свободно плавающая зооспора с апикальными жгутиками равной длины, Е — она же, перешедшая в состояние покоя; глазок и жгутики еще присутствуют, F, G — развитие до стадии А при потере глазка (а) и вакуолей (v); Н — К — Chlorella, С. vulgaris (500х): Н — вегетативная клетка, J, К — деление на 8 апланоспор; L, М — Scenedesmus, S. acutus (1 ОООх): L — 4-клеточная колония, М — деление; N—R — Hydrodictyon, Н. utriculatum: N — молодая сетка в клетке материнской сетки (15х), О — ячейка молодой сетки (80х), Р — часть более старой клетки с зооспорами, Q, R — соединение зооспор в новую сетку в постенном протопласте (Р—R — 10х)
260 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Половое размножение у Eudorina и Volvox оогамное. Внутри отдельных более крупных (генеративных) клеток возникают либо зеленые яйцеклетки (по одной в клетке, всего 6 —8), либо многочисленные мелкие, при освобождении расположенные в виде пластинки, сперматозоиды (рис. 11.96, К, М). Volvoxglobator— однодомный вид, V. aureus и V. carter! — двудомные. Развитие возникающих бесполым путем шаров в сГ или £ особи у двудомных видов индуцируется половым гормоном (гликопротеидом). Он образуется о" особями (или же их сперматозоидами) и необходим для того, чтобы молодые шары, детерминированные как сГ или , развивались дальше в особи определенного пола. Если половой гормон отсутствует, то образуются только бесполые шары Volvox. После оплодотворения яйцеклетка превращается в толстостенную покоящуюся зиготу, при прорастании которой совершается мейоз. У всех Volvocaceae, таким образом, клетки колонии всегда происходят от од-ной-единственной изначальной клетки.
К описанным выше формам примыкают неподвижные одноклеточные или образующие колонии зеленые водоросли с отчасти еще выраженными признаками монадных водорослей, такими, как пульсирующие вакуоли, глазки, жгутиковые стадии. Их иногда рассматривают как отдельный порядок Tetrasporales.
2. Порядок: Chlorococcales (= Protococca-les). Клетки с всего одним ядром и одним хлоропластом не имеют в вегетативном состоянии жгутиков и, следовательно, неподвижны. Только при размножении могут
возникать двухжгутиковые подвижные клетки (зооспоры, рис. 11.97, D; или гаметы). Последние чаще всего бывают голые и только после плавания в течение некоторого времени одеваются стенкой (инцис-тируются). У некоторых представителей образуются только лишенные жгутиков «апланоспоры» (К). Половое размножение, известное у очень немногих представителей. изогамное, с помощью имеющих жгутики гамет (например, Pediastrum и Hydro-dictyony, оогамия встречается крайне редко. Зиготы прорастают с редукционным делением, так что жизненный цикл протекает исключительно в гаплофазе. У некоторых видов на основе одноклеточных форм образуются характерного облика колонии (см. 5.1; например, Pediastrum, рис. 11.98; Scenedesmus, см. рис. 11.97, D). Химизм полисахаридной клеточной стенки в значительной мере неизвестен; у Pediastrum в ней встречается кремниевая кислота, а у некоторых видов откладывается спорополленин.
При клеточном делении часто (например, у Chlorococcum) сначала образуется некоторое число голых дочерних клеток, которые в дальнейшем одновременно одеваются клеточными стенками. Kirchneriella, более детально изученная с помощью электронного микроскопа, отклоняется от этого варианта: септы с материалом клеточной стенки возникают сразу же после делений (последовательно), однако вскоре опять ис
Рис. 11.98. Chlorophyceae, Chlorococcales (A—D — по А. Braun, с изменениями; Е — по G.Senn):
A—D — Pediastrum, Р. granulatum: А — дисковидная колония из клеток, в трех из которых происходит деление; из четвертой клетки выходит пузырь с 16 подвижными клетками, В — подвижные зооспоры в вышедшем наружу пузыре, С— спустя 4,5 ч началась группировка 16 дочерних особей в колонию, D — то же, вид сбоку (ЗООх); Е — Coelastrum, С. proboscideum (550х)
11.2. Бактерии, грибы, растения I 261
чезают. Образующиеся 4 дочерние клетки отходят друг от друга и окружаются каждая собственной новой клеточной стенкой, прежде чем они по отдельности покинут материнскую клетку.
Как и у Volvocales, здесь мы встречаем прогрессирующий ряд, но он направлен от одноклеточных видов к колониям (см. 5.1), которые сгруппированы в виде табличек или полых шаров.
Правда, онтогенез Kirchneriella (см. выше) позволяет предположить, что одноклеточные формы этого рода возникли от многоклеточных (колониальных) структур.
Одноклеточные формы, от округлых до эллипсоидных, представлены видами таких родов, как Chlorococcum (с зооспорами; рис. 11.97, D) и Oocystis. Группировки расположенных в один ряд клеток простейших форм в количестве чаще всего по 4 (или 8) представлены у широко распространенных в пресных водах видов рода Sce-nedesmus (рис. 11.97, L, М). Более сложно устроены колонии у также часто встречающегося в пресных водах рода Pediastrunr. они имеют вид изящных плоских табличек (рис. 11.98, А), свободно парящих в воде, и несколько напоминают Gonium, только без жгутиков. Взрослая колония Coelastrum трехмерная: клетки образуют полый шар (Е). У водяной сеточки (Hydrodictyon reticu-latum), свободно парящей в пресной воде, цилиндрические клетки смыкаются своими концами по 3—4 и образуют мешковидную колонию в форме вытянутой замкнутой сети из многих ячеек, полую внутри (см. рис. 11.97, N).
Половое размножение осуществляется изогаметами более мелкими, чем зооспоры. При прорастании зиготы сначала возникаю! 4 мейо-зооспоры, которые, недолго поплавав, превращаются в толстостенные «полиэдры». Только последние прорастают затем в новые колонии Hydrodictyon, сначала значительно меньшего размера, чем взрослые. При бесполом размножении у видов всех этих родов образуются подвижные зоо- или неподвижные апланоспоры, которые, однако, освобождаются не поодиночке. Очень рано за счет слипания стенок они связываются между собой, формируя характерную для данного вида (по числу клеток и внешнему виду) колонию (рис. 11.97; 11.98). Объединение может происходить вскоре после выхода из материнской клетки в слизистом пузыре (рис. 11.98, А)
или даже в самой материнской клетке, так что после ее разрушения уже имеется колония из соответствующего числа клеток, хотя сначала еше маленькая. Дальнейшие клеточные деления в колониях не происходя! (за исключением образования репродуктивных клеток). Вышеупомянутое сходство с соответствующим эволюционным рядом у Volvocales состоит, таким образом, только во внешнем виде, а не в характере развития. У Volvocales колонии возникают путем повторного продольного деления образующих их клеток, за счет чего положение каждой клетки в колонии определяется с самого начала. У Chlo-rococcales все клетки «выводка», возникшего в результате деления исходного протопласта, могут сначала (внутри клеток или же слизистых пузырей) свободно перемещаться относительно друг друга, прежде чем вторично соединяться между собой (рис. 11.98, А, В).
Встречаемость. Chlorococcales обитают преимущественно в пресноводном планктоне; некоторые формы перешли к наземной жизни. Такие виды — обитатели влажной почвы и скал. Почвенная водоросль Spongiochloris устойчива к высоким температурам. Chlorococcales наряду с другими водорослями являются также обычнейшими компонентами в зеленых налетах на коре деревьев и стенах. Некоторые Chlorococcales — симбионты в лишайниках (однако ср. Trebouxia, Trebouxiophyceae). Scenedesmus, Ankistrodesmus и Hydrodictyon часто используются в чистой культуре для физиологических экспериментов.
В ископаемом виде формы типа современного Pediastnim описаны уже из перми. Напоминающие Chlorococcales формы (Caryosphaerioi-des) относятся к древнейшим находкам эукари-отных клеток (см. 11.3.2).
3. Порядок: Chaetophorales. Таллом водорослей, относящихся к этому порядку, представлен разветвленными нитями, состоящими из одноядерных клеток с одним хлоропластом. Он обычно гетеротрихальный, т. е. состоит из двух частей: «подошвы» из разветвленных нитей, которые стелются по субстрату, и более или менее обильно разветвленных прямостоячих нитей, несущих органы размножения (рис. 11.99, А). Половое размножение у тех видов, у которых оно было установлено, — изо-, анизо- или оогамия.
У видов некоторых родов гетеротрихальное строение четко не выражено из-за того, что одна из двух частей таллома выражена слабо или вообще практически отсутствует. Некоторые формы представляют собой объединения нескольких особей в колонии, находящиеся в общей
262 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Рис. 11.99. Chlorophyceae, Chaetophorales (А— по J. Huber; В — по E.Juller; С — по R.N. Singh):
А — Stigeoclonium tenue (4х); В — Stigeoclonium subspinosum, зооспора (900х); С — Fritschiella tuberose; а — глазок; b — поверхность почвы; р — пиреноид; ра — первичная вертикальная нить; рг — подземная ползучая нить; г — ризоид; s — подошва (стелющиеся по субстрату нити); sf — вторичные вертикальные нити; v — пульсирующие вакуоли; w — расположенные в толще воды вертикальные нити
слизи. У Stigeoclonium (рис. 11.99, А) наряду с че-тырсхжгутиковыми зооспорами (В) встречаются двухжгутиковые изогаметы. У обитающей на почве водоросли Fritschiella (Индия, Африка; рис. 11.99, С) от подземных ползучих нитей поднимаются в воздушное пространство вертикальные разветвленные нити. Здесь проявляется функциональная дифференциация, сильно выраженная у высших растений: одна часть таллома выполняет преимущественно поглощающую функцию, другая — ассимиляционную.
Встречаемость. Большинство видов (часто как эпифиты на водорослях и других водных растениях) — обитатели пресных вод (например, Chae-tophora, Stigeoclonium).
4. Порядок: Oedogoniales. Это еще один порядок с нитчатой организацией таллома. Хотя нити здесь чаще всего неразветвлен-ные, но оогамное размножение, а также уникальная форма деления и растяжения клеток говорят о том, что это сильно подвинутые формы, прошедшие самостоятельный путь развития. Одноядерные клетки содер
жат по одному постенному сетчатому хлоропласту с многочисленными пиреноидами (рис. 11.100, А).
Уникальные деление и растяжение клеток связаны с образованием «колпачков» у верхнего конца клетки (рис. 11.100, J). Их возникновение сопряжено с началом деления ядра (профазой), когда у верхнего конца клетки образуется кольцевидный валик из сливающихся пузырьков (Гольджи?); он состоит в основном из аморфной, растяжимой фракции клеточной стенки. После завершения клеточного деления между дочерними ядрами внутри фикопласта (см. Chlorophyta, введение) появляется септа, дающая начало клеточной пластинке, сначала еще не фиксированной, — будущей поперечной перегородке. В области верхнего кольцевого валика внешняя стенка клетки в дальнейшем разрывается — образуется кольцевая трещина, после чего кольцевой валик растягивается в цилиндр. У места разрыва остается характерный колпачок. После повторений этого процесса у верхнего конца клетки число колпачков увеличивается и они как бы вставлены друг в друга (рис. 11.100, С).
Жизненный цикл гаплонтный. Относительно крупные зооспоры возникают по одной из всего содержимого клетки. Они имеют вблизи переднего конца, лишенного хлоропластов, характерный венец из многочисленных, не сгруппированных попарно жгутиков (рис. 11.100, С). В других местах нити отдельные клетки вздуваются наподобие бочонков в оогонии; их содержимое превращается в одну крупную яйцеклетку (Е), которая остается внутри оогония. Некоторые участки той же или другой нити (модификационное определение пола) в клетках, остающихся относительно короткими, образуют сперматозоиды, чаще всего по два. Они похожи на зооспоры, но мельче и имеют желтоватый цвет.
Иной путь переноса мужских репродуктивных клеток осуществляется через так называемые андроспоры и карликовые мужские растеньица («мужчинки»). В клетках, которые напоминают вышеописанные О' гаметангии, вместо сперматозоидов образуются несколько более крупные андроспоры. привлекаемые к оогониям хемотаксисом. Они не могут непосредственно оплодотворять яйцеклетки, а садятся на них или в непосредственной близости от них и прорастают в маленькие, состоящие из немногих клеток растеньица — так называемые «мужчин
11.2. Бактерии, грибы, растения | 263
ки» (рис. 11.100, Е, F), верхние клетки которых затем становятся гаметангиями и производят сперматозоиды, способные к оплодотворению. Происходящее в то же время созревание оогониев, очевидно, определяется гормонами, которые выделяются «мужчинками». С другой стороны, сперматозоиды по типу хемотаксиса привлекаются оогониями, которые теперь окружены слизистыми обвертками. Через отверстие в оогонии сперматозоиды проникают к яйцеклетке и один из них сливается с ней. После этого в оогонии развивается толстостенная красная гипнозигота. При прорастании (G) ее содержимое делится на 4 крупные гаплоидные мейозооспоры (зиготическая смена ядерных фаз), которые выходят наружу и образуют новые нити (D).
Обзор Chlorophyta. У зеленых водорослей наблюдается большое морфологическое разнообразие. Переход от одноклеточных форм к многоклеточным талломам идет через ряд уровней организации. Он кон-вергентно происходит в разных классах, соответственно в разных эволюционных ли
ниях. Это касается не только классов зеле ных водорослей, но и проявляется как общий эволюционный принцип в различных отделах водорослей. «Одетая» только одной (отчасти модифицированной) плазмалем-мой клетка (например, Polyblepharides) у зеленых водорослей получила дальнейшее развитие; во всех более высокоразвитых таксонах клетки имеют более или менее толстые клеточные стенки. Благодаря им некоторые формы могут даже жить вне воды в качестве почвенных или аэрофиль-ных водорослей.
Компоненты клеточной стенки еще более разнообразны: в качестве полисахаридного материала уже используется и целлюлоза. Как защитное вещество встречается иногда спорополленин (Trentepohliophyceae, Chlorococcales). Клеточное деление — разделение дочерних клеток поперечной перегородкой — проходит различные эволюционные стадии. В простых случаях вслед
Рис. 11.100. Chlorophyceae, Oedogoniales, Oedogonium (А— по W.Schmitz; В —D— по I. Hirn; E, F— no N. Pringsheim; G — no L.Juranyi; H — L — по K. Esser):
A — часть нити (600x); В — D — О. concatenatum, выход зооспоры и ее прорастание (ЗООх); Е —G — О. ciliatum (350х): Е, F— оплодотворение, G — прорастание зиготы; Н — L — образование колпачка при делении клетки (200х); а — карликовое мужское растеньице (наннандрий); с — хлоропласт; к — ядро; о — оогоний; р — пиреноид; z — зооспора с запасными веществами скрывающими ядро (см. D)
264 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
за делением ядра происходит разделение плазмы и еще внутри материнской клетки одновременное окружение всех частей клеточными стенками. Идущее от боковых стенок центростремительное разделение материнской клетки за счет врастания перегородки наподобие ирисовой диафрагмы между отодвигающимися друг от друга дочерними ядрами — еще один способ, который можно рассматривать как исходный. В более эволюционно подвинутых случаях в образовании новой клеточной стенки участвует фикопласт, очень редко — уже фрагмопласт (Trentepohliophyceae). Исходно бокаловидный хлоропласт в одних случаях разделяется в виде сети или на отдельные диски, в других случаях образуются крупные хлоропласты в виде пластин или лент.
Половой процесс эволюционирует от изогамии через анизогамию к примитив-
ной оогамии и наконец к ее наивысшему уровню, когда яйцеклетка уже не выходит наружу, а оплодотворяется в оогонии. Некоторые зеленые водоросли помимо наличия и так уже неподвижных яйцеклеток утратили и жгутики у остальных репродуктивных клеток. Так, некоторые виды Pleu-rococcus (Apatococcus, напротив, с зооспорами), приспособившись к жизни вне воды, распространяются лишенными жгутиков апланоспорами. В жизненном цикле можно проследить тенденцию к увеличению доли диплофазы (рис. 11.101). Обычно зеленые водоросли — гаплонты с зиготической сменой ядерных фаз; диплоидны в этом случае только зиготы (А). За счет смещения места мейоза (митотическое ядерное деление вместо мейоза) зигота прорастает в диплоидное вегетативное тело. За счет этого в жизненный цикл вклинивается дополнительно диплоидная фаза, которая закан
Рис. 11.101. Chlorophyta, смена поколений и ядерных фаз, схема основных типов (по R. Harder): А— Ulothrix, В — Cladophora', С — Halicystis-Derbesia. Дикариотичная фаза — темный пунктир; диплоидная фаза — темные сплошные линии; гаплоидная фаза — светлые линии; G — гаметофит; S — спорофит; О — зигота; R! — редукционное деление
11.2. Бактерии, грибы, растения | 265
чивается мейозом, сдвинутым во времени и в пространстве. Таким образом, возникает чередование гаплоидного гаметофита и диплоидного спорофита, т. е. гетерофазная смена поколений.
Смена поколений (рис. 11.101) может быть изоморфной (некоторые виды Cladophora, В) или гетероморфной (с преобладанием спорофита, Halicystis-Derbesia, С). Исключительно диплонтный цикл у зеленых водорослей пока неизвестен1. В целом смена поколений осуществляется на разных особях (диплобионтная). Гаплобионтная смена поколений на одной особи (что является закономерной ситуацией у мхов) у зеленых водорослей представляет редчайшее исключение (примеры — Prasiola stipitata, Bryopsis). Смену поколений ни в коей мере нельзя понимать как закономерную последовательность разных фаз. За счет бесполого размножения каждое поколение может воспроизводить само себя независимо от смены поколений. Зеленые водоросли с простым жизненным циклом (Ulothrix, А) размножаются, как правило, вегетативным и бесполым путем (например, зооспорами), тогда как половое размножение наступает только при определенных внешних условиях.
Chlorophyta — несомненно очень древняя группа низших растений. Однако с достоверностью могут быть идентифицированы только устойчивые вследствие обызвествления талломы морских Dasycladales, которые известны с кембрия. Поскольку Dasycladaceae уже в ордовике представлены большим разнообразием форм, они должны были возникнуть еще раньше. Из 120 родов, которые встречались на протяжении более 500 млн лет, в настоящее время сохранилось лишь 10.
• Отделы водорослей с комплексными пластидами, содержащими хлорофиллы а и Ь. Следующие два отдела, данные в соответствующих приложениях, содержат в своих клетках хлоропласты типа зеленых во-
1 В литературе обычно указывается, что чисто диплонтный цикл имеется у широко распространенного в Европе вида Cladophora glomerata, а также у многих (возможно, большинства) сифоновых зеленых водорослей. — Примеч. пер.
дорослей. Однако приобретение таких пластид осуществлялось путем вторичного эн-доцитобиоза (см. 2.4).
1. Приложение к Chlorophyta: Chlorarachniophyta
Этот очень маленький отдел, в который входит всего два рода с одним видом в каждом1, представляет интерес тем, что их пластиды, окруженные четырьмя мембранами, как и у криптофитов, содержат нуклеомор-фу (см. 2.4). Последняя может рассматриваться как редуцированное ядро фотоав-тотрофного эукариотного эндосимбионта. Виды этого отдела живут совместно с сифоновыми морскими водорослями теплых морей как голые амебоидные клетки, которые нитевидными плазматическими выростами соединены в сетевидные плазмодии. Светло-зеленые хлоропласты содержат хлорофиллы а и Ь. Тилакоиды в пластидах объединены в стопки по 2—6. Chlorarachniophyta считают произошедшими от нитчатых амеб путем приобретения пластид.
2. Приложение к Chlorophyta: отдел эвгленовые (Euglenophyta)
К этому отделу относятся одноклеточные формы, находящиеся на монадном уровне организации. При определенных условиях они иногда могут переходить в капсальное состояние. Размножение осуществляется путем продольного деления, половое размножение неизвестно. Зеленые хлороплас-гы содержат тот же набор пигментов, что и зеленые водоросли (хлорофиллы а и Ь, р-ка-ротин, следы ос-каротина), однако в них имеется один ксантофилл, неизвестный больше нигде в растительном царстве. В качестве запасного вещества наряду с фосфолипидами в пузырьках в плазме откладывается полисахарид парамилон в виде зернышек или дисков. Это глюкан с Р-1,3-связями, который окрашивается йодом в синий цвет. Клетки нередко спирально закручены и имеют почти всегда простую, со
1 В настоящее время в этом отделе насчитывается уже порядка 8 видов. — Примеч. пер
266 | ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
стоящую преимущественно из белков оболочку — пелликулу, которая непосредственно граничит с плазмалеммой (исключение представляет, например, Trachelomonas с домиком, содержащим железо). На переднем конце клетки имеется бутылковидное углубление — ампула, которая подразделяется на брюшную часть и канал. К ампуле прилегает пульсирующая вакуоль, которая окружена несколькими дополнительными пульсирующими вакуолями и служит как органелла, обеспечивающая осморегуляцию. У основания ампулы почти всегда отходят два жгутика, каждый от своего базального тела: один длинный и один короткий, не выходящий наружу из ампулы и срастающийся концом с длинным жгутиком. В месте их слияния находится светочувствительная органелла — фоторецептор. Вблизи ампулы расположен красный «глазок» (рис. 11.102, В). Он состоит из отдельных липидных глобул, каждая из которых окружена собственной элементарной мембраной (относительно роли «глазка» при фототаксисе см. 8.2.1.2). Длинный жгутик, снабженный волосками (см. рис. 11.20, С), описывает при движении поверхность конуса. При
Рис. 11.102. Euglenophyta (по G. F. Leedale):
А — Euglena gracilis (бООх); В — то же, передний конец (1 ОООх); С — Colacium mucronatum (500х); D — Phacus triqueter (бООх); с — хлоропласт; д, — двигательный жгутик; д2 — второй жгутик; gs — расширение жгутика (фоторецептор); к — ядро; р — свободный парамилон; ру — пиреноид; s — глазок; st — слизистая ножка; v — пульсирующие вакуоли
одновременном вращении вокруг продольной оси клетка, например у Euglena, каждую секунду перемешается вперед на расстояние, равное 2—3 длинам ее тела.
Ультраструктура эвгленовых показывает следующие особенности. В интерфазном ядре можно видеть хромосомы в конденсированном состоянии. Хлоропласты имеют оболочку из трех мембран, которая никогда не бывает связана эндоплазматической сетью с ядерной оболочкой. В хлоропластах тилакоиды по большей части сгруппированы в ламеллы по три.
Эвгленовые насчитывают более 800 видов, относящихся примерно к 40 родам. В большинстве своем они живут в пресных водах. Виды рода Euglena встречаются в основном в стоячих водоемах. богатых органическими веществами. Виды Phacus (рис. 11.102, D), наоборот, предпочитают воды, бедные питательными веществами. Colacium (С) прикрепляется с помощью слизистой ножки к свободно плавающим мелким организмам; только при размножении возможно его свободное движение с помощью жгутиков.
Хотя большинство видов питается фотоав-тотрофно. у них имеется тенденция к поглощению органических веществ дополнительно к продуктам фотосинтеза. Ряд бесцветных форм питается полностью гетеротрофно; у некоторых (например. Регапета) есть воронковидное углубление (цитостом), где находятся ловчий аппарат и палочковидный ловчий орган, с помощью которых они способны захватывать такие микроорганизмы, как бактерии, водоросли или клетки дрожжей. Таким образом, границы между растительной и животной организацией здесь еще размыты.
Euglena gracilis в темновых культурах теряет весь хлорофилл и все тилакоиды. Остающиеся тельца напоминают пропластиды; они сохраняют способность к делению и во время темновой фазы, чем поддерживается сохранность пласти-дома. При последующем освещении сохранившиеся бесцветные пластиды превращаются в хлоропласты с тилакоидами и возобновляется фотосинтез. Наряду с этим имеются разнообразные варианты данного вида, которые вообще не имеют хлоропластов. Такие формы, получаемые при определенных условиях (например, при очень быстрой последовательности делений), уже никогда не могут образовывать хлоропластов.
Эвгленовые были сначала гетеротрофной, вероятно, очень рано обособившейся группой, которая сравнительно поздно приобрела хлоро-пласты путем эндоцитобиоза с зелеными водорослями.
11.2. Бактерии, грибы, растения 1 267
Второй отдел: Streptophyta
В этом отделе жгутики либо отсутствуют (Zygnematophyceae), либо, если имеются, занимают боковое положение (KJebsor-midiophyceae; Charophyceae). Новые клеточные стенки при делении клеток (так же как у рассмотренных Trentepohliophyceae и сосудистых растений) закладываются во фрагмопластах (микротрубочки располагаются перпендикулярно плоскости будущей перегородки; ср. фикопласт: Chlorophyta, введение). Объем этого отдела, как и его разграничение с уже обсуждавшимися Chlorophyta, подтверждается данными анализа ДНК.
Первый подотдел; Streptophytina (зеленые водоросли II)
В этом подотделе объединены все группы Streptophyta от одноклеточных до многоклеточных водорослей, имеющие простую организацию. Наиболее развитые формы, связанные с водной средой, достигают довольно высокой функциональной дифференциации; у них формируются ткани (например, узлы Characeae) и защищенные органы размножения.
1. Класс: Mesostigmatophyceae
Это монадные одноклеточные организмы (например, Mesostigma) пресных вод. Их клеточный покров выглядит как орнаментированный мелкими точками. Он состоит из трех слоев чешуек. Чешуйки самого внешнего слоя (из органического материала) крупнее чешуек внутренних слоев и имеют вид корзиночек. Два жгутика несколько неравной длины прикреплены не терминально. Chiorokybus нередко образует правильной формы группы из нескольких (до 32) клеток. Из-за наличия чешуек представители этого класса раньше относили к Prasinophyceae (отдел Chlorophyta); принадлежность их к Streptophytina обоснована анализом ДНК. Создается впечатление, что Mesostigmatophyceae — предковая группа, от которой берет начало эволюция зеленых наземных растений.
2. Класс: конъюгаты, или сцеплянки (Zygnematophyceae, Conjugatae)
Конъюгаты не образуют жгутиковых стадий: у них нет ни зооспор, ни гамет со жгутиками. Половое размножение осуществляется путем конъюгации1: два голых протопласта, каждый из которых представляет все содержимое своей клетки, сливаются в зиготу. Последняя прорастает после периода покоя, претерпевая мейоз; смена ядерных фаз зиготическая. Следовательно, конъюгаты — чистые гаплонты, представленные коккоидными или нитчатыми формами. Нити неразветвленные и легко распадаются на отдельные клетки. Каждая клетка содержит ядро, расположенное в ее середине. К конъюгатам относится примерно 4000— 6000 видов (около 50 родов), обитающих в бентосе и отчасти в планктоне, почти исключительно в пресных водах.
Mesotaeniaceae — относительно примитивная группа. Водоросль существует либо в виде отдельных клеток, либо образует слизистые колонии (рис. 11.103, А; коккоид-ный уровень организации). Клеточная стенка сплошная, гладкая.
Desmidiaceae1 2 — как правило, одноклеточные формы (коккоидные). Чаще всего орнаментированные, нередко содержащие железо (поэтому желтоватые), клеточные стенки состоят из двух равных половинок — полуклеток, которые отделяются друг от друга швом или перетяжкой (перешеек, isthmus). Внутри каждой из двух точно симметричных полуклеток содержится по одному крупному центральному (не постенному) хлоропласту с одним или несколькими пиреноидами (см. рис. 2.87; бокс 4.1, рис. С; рис. 11.103, В, С). В середине клетки находится ядро.
Вегетативное размножение осуществляется путем деления клетки на две, причем (как у диатомей — см. Bacillariophyceae) у
1 Следует помнить, что термин «конъюгация» в биологии очень неудачно используется для самых разных явлений (для инфузорий, бактерий, хромосом и в данном случае). — Примеч. пер.
2 Выразительно немецкое название этой группы — Zieralgen (декоративные водоросли). — Примеч. пер.
268 | ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Рис. 11.103. Streptophytina, Zygnematophyceae; Mesotaeniaceae и Desmidiaceae (A, D—F, H —К — по A. De Вагу; В — по Palla; С, L — по N. Carter; G — по Н. Klebahn; М, N — по G. Senn; О, Р — по Delponte): А — Mesotaenium braunii (280х); В — Closterium moniliferum (200х); С — Closterium regulare, поперечный разрез через клетку, хлоропласт имеет звездчатый контур (200х); D, Е — Closterium parvulum, копуляция (ЗООх); F — Closterium rostratum, выход зиготы из оболочки (200х); G — Closterium sp., деление зиготы (200х); Н — Cosmarium botrytis (280х); J, К — то же, деление (280х); L — Micrasterias denticulate (125х); М, N — Oocardium stratum, вид сверху и продольный разрез (320х); О — Desmidium swartzii, часть цепочки из клеток; Р — то же, поперечный разрез через клетку (350х); dk — дегенерировавшее ядро; g — слизистая ножка; к — ядро; kh — известковая обвертка; р — пиреноид; v — вакуоль с кристалликами гипса
дочерних клеток должна быть восполнена половина клеточной стенки (рис. 11 103, J, К). При этом вновь возникают одноклеточные особи. Однако у представителей некоторых родов дочерние клетки остаются соединенными между собой, образуя цепочки.
При половом размножении две генотипически разные клетки прикладываются друг к другу (рис. 11.102, D) и окружаются слизью. Затем клеточная стенка расходится в середине, протопласты выходят в виде голых гамет в разбухающий, вскоре ослизняющийся копуляционный канал (мостик) и сливаются в зиготу (Е), стенка которой часто несет шипы. Рядом со зрелой гипнозиготой сначала находятся 4 пустые половинки клеточных стенок от обеих слившихся клеток. При прорастании зиготы из 4 гаплоидных ядер, возникших в результате мейоза, у большинства Desmidiaceae 2 погиба
ют, так что возникают только 2 гаплоидных «проростка».
Desmidiaceae, относящиеся к красивейшим водорослям, очень разнообразны по своему облику. Их клетки, например, имеют форму полумесяца {Closterium, рис. 11.103, В), бисквита (Cosmarium, Н) или звезды (Micrasterias, L). Euastrum (см. бокс 11.7, с. 199) имеет на концах полуклеток выемку, Staurastrum (см. бокс 11.7)— полуклетки с острыми углами. На обоих концах клеток Closterium расположены вакуоли с кристалликами гипса, которые находятся в интенсивном броуновском движении (В). Некоторые Desmidiaceae выделяют через поры в мембранах слизистые тяжи, с помощью которых клетки могут медленно передвигаться. Особи видов рода Oocardium, обитающие в богатых известью ручьях, сидят на слизистой ножке, которая инкрустирована извес
11.2. Бактерии, грибы, растения I 269
тью (М, N). Десмидиевые развиваются преимущественно в бедных питательными веществами водоемах с низким значением pH, например в торфяных болотах, где имеется большое разнообразие их видов; Pleurotae-nium и S/aurastrum обитают также в водах с щелочной реакцией.
Семейство Zygnemataceae представлено неразветвленными нитчатыми формами. Наиболее известен род Spirogyra (рис. 11.104, А). Его многочисленные виды часто встреча-
ются весной в водоемах со стоячей или медленно текущей водой в виде неприкрепленной желтовато-зеленой «тины». Нити растут интеркалярно за счет растяжения в длину и поперечного деления всех клеток; таким образом, все клетки равноценны; отсутствует и полярность нитей. Гладкие, лишенные пор целлюлозные клеточные стенки снаружи покрываются слизью, из-за чего нити слизистые на ощупь. При митозе ядерная мембрана по большей части сохраняется (внут
Рис. 11.104. Zygnematophyceae (А, В — по Н. Schenck; С — по R. Kolkwitz; D—Н — по A. Trondle; J — L — по Palla):
А—Н — Spirogyra: А — S. jugalis, клетка (250х), В — S. quinina, анизогамная копуляция (240х), С— Н — S. longata', С — участок хлоропласта у клеточной стенки (750х), D—Н — молодые и старые зиготы, D — два половых ядра перед копуляцией, Е — после слияния, F — деление ядра зиготы на 4 гаплоидных ядра, G — три мелких ядра дегенерируют (D — G — 250х), Н — одноядерный проросток (180х); J—L — Mougeotia: J, К — М. scalaris, хлоропласт повернут к зрителю широкой стороной («анфас») и узкой стороной («в профиль») (бООх), L — М. calospora, изогамная копуляция (450х); с, с,, с2 — хлоропласт(ы); к — ядро; р — пиреноид; s — крахмал; w — клеточная стенка; z — зигота; zw — стенка зиготы
270 I ГЛАВА 11 • СИСТЕМАТИКА И ФИЛОГЕНИЯ
риядерный митоз). Поперечная перегородка образуется центрипетально по типу ирисовой диафрагмы и дополнительно — как клеточная пластинка во фрагмопласте. Нити могут распадаться по поперечным перегородкам на одно- или многоклеточные фрагменты, которые служат для вегетативного размножения (см. 10.1.3.3).
Ядро находится посередине клетки и подвешено на протоплазматических тяжах внугри крупной вакуоли. Хорошо различимы один или несколько прилегающих к стенке хлоропластов, всегда в виде левовращающей спирали (см. рис. 2.87; 11.104, А, С: с) с пиреноидами (А: р).
При половом размножении две нити, чаще всего морфологически неразличимые, располагаются параллельно. По линии соприкосновения из клеток выдвигаются сосочки, так что нити снова несколько отодвигаются друг от друта и получается фигура, похожая на лестницу (лестничная конъюгация; см. рис. 11.104, В). Сосочки за счет лизиса стенки в месте соприкосновения превращаются в копуляционный канал между соответствующими клетками («гаметангиями»). Каждая клетка нити может стать «гаметангием». Определение пола модификационное (сГ и нити). Протопласт из сГ клетки в виде голой «мигрирующей гаметы» переходит в прилегающую клетку и сливается с ее протопластом («покоящаяся гамета»). Получающаяся зигота за счет отдачи воды уменьшается в объеме и превращается в гипнозиготу (рис. 11.104, В: z), которая окружается многослойной толстой коричневой стенкой, содержит много крахмала и масла и в таком виде переносит неблагоприятные условия. Хлоропласты (один или несколько) 0я «гамет» погибают. При прорастании зиготы, связанном с мейозом, 3 ядра (F, G) дегенерируют, так что возникает только один гаплоидный проросток, который вытягивается в длину и, претерпевая клеточные деления, образует новую нить (Н).
Некоторые виды Spirogyra однодомны. У них сливаются протопласты соседних клеток одной и той же нити через боковые копуляционные мостики (боковая конъюгация).
Zygnema и Mougeotia отличаются от Spirogyra строением хлоропластов. У Zygnema в каждой
клетке имеется два звездчатых хлоропласта, у Mougeotia (рис. 11.104, J, К) — единственный хлоропласт в виде плоской осевой пластинки, реагирующей на свет (ср. 8.2.2). В обоих родах есть виды, у которых зигота образуется в копуляци-онном канале (L); этот процесс напоминает таковой у представителей предыдущего порядка.
Zygnematophyceae — группа, четко отличающаяся по типу размножения и строению клетки от других зеленых водорослей, от которых она, очевидно, рано отделилась в ходе эволюции, полностью утратив жгутиковые стадии.
3. Класс: Klebsormidiophyceae (Coleochaetophyceae)
Клеточная стенка нитчатых (неветвя-щихся и разветвленных) представителей этого класса содержит целлюлозу (фибриллярное расположение). Жгут иковые репродуктивные клетки покрыты ромбовидными чешуйками из органического материала; этим они напоминают клетки Mesostigmatophyceae. Виды этого класса встречаются в пресных водах и во влажных местах вне воды. В порядке Coleochaetales виды представлены разветвленными нитями. Таллом их разделяется на подошву и вертикальные нити (гетеротрихальное строение; рис. 11.105; подобная дифференциация таллома у Trentepohliophyceae и Chaetophorales из Chlorophyceae).
Coleochaete имеет высокий уровень развития среди Chlorophyta (наличие дисковидной подошвы — рис. 11.105, А; по-особому дифференцированные волоски; оогамное размножение). Бутылевидный оогоний с бесцветной шейкой (С), которая открывается на вершине для проникновения бесцветных двухжгутиковых сперматозоидов. После оплодотворения округлая зигота увеличивается в размере и одновременно от несущей ее клетки и соседних клеток вырастают нити, так что в конечном счете зигота одевается однослойной плектенхимой — возникает «плодовое тело» (Е). При прорастании этого покоящегося органа не сразу возникают мейозооспоры, а сначала внугри зиготы после мейоза образуется гаплоидное тельце из 16 — 32 клеток, в каждой из которых развивается по одной гаплоидной зооспоре.
Klebsormidiophyceae названы по роду Klebsormidium, виды которого имеют кок-
11.2. Бактерии, грибы, растения | 271
Рис. 11.105. Streptophytina, Klebsormidio-phyceae (А — по M.Jost; В — по J. Huber; С— Е — по F. Oltmanns):
А — Coleochaete scutata, подошва (80х); В — Aphanochaete repens, развитие одетой воротничком щетинки (250х); С—Е — Coleochaete pulvinata; С — оогоний незадолго до вскрытия; D — он же, оплодотворенный; Е — зигота за счет обрастания нитями превращается в «плод» (500х); ек — ядро яйцеклетки; sk — ядро сперматозоида
коидное строение или представлены нераз-ветвленными нитями.
Среди зеленых водорослей Coleochaetophy-сеае вместе с еще не рассматривавшимися Charophyceae наиболее близки зеленым наземным растениям. Этот класс водорослей обладает такими общими признаками с мохообразными и папоротникообразными, как расположенные сбоку жгутики, тенденция к образованию тканей, защита зиготы (начало формирования зародыша), фрагмопласт и целлюлоза одного и того же типа. «Плодовые тела» Coleochaete защищены от высыхания и разрушения микробами спорополленином и соединениями типа лигнина.
4. Класс: харовые, или лучицы (Charophyceae)
Харовые представляют собой высокоразвитые зеленые водоросли с талломами, в которых строго последовательно чередуются тканеподобные и нитчатые участки
(соответственно узлы и междоузлия). Органы размножения достигают очень специфичной комплексности, отсутствующей у всех предыдущих групп. Харовые, представленные немногими родами, образуют в прудах и ручьях «подводные луга». Их известно около 300 видов. Обитают они в пресных и солоноватых водах; пресноводные виды часто особенно обильно развиваются в водоемах с высоким значением pH (pH 7 и более; жесткая вода). Characeae — единственное семейство класса, где еще сохранились ныне живущие формы.
Клеточные стенки часто инкрустированы известью, некоторые Characeae относятся к числу важнейших образователей известковых туфов. Они не переносят высоких концентраций фосфатов при загрязнении воды.
Строение. Главные и боковые оси имеют верхушечный рост, нарастая за счет одной апикальной клетки (см. рис. 5.9, В). Она отчленяет вниз от себя по очереди более узкие клетки (будущие узлы) и более вытянутые клетки (будущие междоузлия); последние больше не делятся и растягиваются в длину, претерпевая вакуолизацию, до нескольких сантиметров.
Харовые водоросли характеризуются, таким образом, регулярным подразделением таллома, достигающего нескольких дециметров, на узлы (nodi) и междоузлия (internodia).
Клетки узлов сохраняют способность к делению и развиваются в многоклеточные диски узлов, из которых вырастают расположенные мутовками членистые боковые оси разных порядков. Кроме того, возникают короткие, шиловидные клетки-«прили-с гники» и клетки коры. Коровые клетки (характерные для рода Chara и отсутствующие у видов рода Nitella и др.) образуют прилегающий к междоузлиям покров из нитевидных клеток.
Боковые ветви показывают сходное с основной осью подразделение. Они не покрыты корой, простые или несут на своих узлах короткие боковые ветви второго порядка со сходным подразделением на узлы и междоузлия.
В каждой мутовке из «пазухи» коротких «побегов» может возникать длинный «побег», сходный с главной осью (см. рис. 5.9).
ЧТ! I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Основаниями растения прикрепляются к незакрепленному грунту (илу, песку) посредством бесцветных разветвленных нитевидных ризоидов, возникающих из узлов. Некоторые Characeae образуют из нижних частей осей клубеньки, плотно забитые крахмалом, являющиеся органом перезимовки.
Молодые клетки непосредственно после клеточного деления одноядерны. В длинных клетках междоузлий ядро растет эндоми-тотически и распадается на многочисленные ядерные фрагменты, так что эти клетки становятся многоядерными. Цитоплазма находится чаще всего в интенсивном движении (цитоплазматическое движение, см. 8.2.2). Многочисленные хлоропласты находятся в постенной цитоплазме клеток.
Твердая часть клеточной стенки состоит из целлюлозы, по тонкому строению сходной с целлюлозой высших растений. Новые поперечные перегородки клеток возникаю! во фрагмопластах.
Размножение. Харовые — все оогамные гаплонты с зиготической сменой ядерных фаз. Расположенные вертикально оогонии
Chara окружены спирально обвивающими их коровыми нитями. Мужские гаметы возникают в шаровидных вместилищах сложного строения (спермогонии Chara). Двухжгутиковые сперматозоиды закручены в виде штопора, тогда как у всех других зеленых водорослей они радиально-симметричные. Округлые, в зрелом состоянии окрашенные каротиноидами в желто-красный цвет спермогонии и яйцевидные зеленые оогонии (и те и другие видны невооруженным глазом) образуются у узлов боковых осей.
Спермогонии (рис. 11.106, A: s, Е) возникают из материнской клетки, сначала делящейся на 8 клеток — октантов. Каждый октант делится затем тангентальными стенками на 3 клетки (Е). Так возникают в итоге 24 клетки, которые подразделяют круглый спермогоний следующим образом: 8 плоских внешних покровных клеток (щитков), которые из-за возникающих неполных радиальных перегородок приобретают вееровидную орнаментацию; 8 средних клеток (рукояток — manubria), кото-
Рис. 11.106. Streptophytina, Charophyceae (A, D — Chara fragilis; В, С. Е — Nitella flexilis) (А—С, Е — по J. Sachs; D — по Е. Strasburger):
А — вид сбоку на часть таллома со спермогонием (s) и оогонием (о), коровыми нитями и коронкой (с) (50х); В — рукоятка с головкой и сперматогенными нитями; С — клетки сперматогенной нити, содержащие каждая по одному сперматозоиду; D — сперматозоид (540х); Е — продольный разрез через молодой спермогоний; g — жгутики; gz — рукоятка; к — спирально закрученное длинное ядро; kz — головка; р — плазма; w — стенка
11.2. Бактерии, грибы, растения | 273
Бокс 11.7. Встречаемость и образ жизни водорослей
Преобладающее большинство водорослей — фотоавтотрофы. Им противостоят миксотрофные и гетеротрофные формы. Миксо-трофия позволяет фотосинтезирующим организмам дополнительно поглощать органику из богатой питательными веществами окружающей среды. Гетеротрофные водоросли утратили свои фотосинтетические пигменты и питаются готовыми органическими веществами; «фаготрофные» представители этой группы «поедают» твердые частицы пищи, которые поступают в пищеварительные вакуоли. Если фототрофные водоросли — типичные растения, то лишенные пигментов их фаготрофные представители имеют животный образ жизни. Среди монадных форм могут находиться близкородственные виды с автотрофной (растительной) и фаготрофной (животной) организацией. Границы между растениями и животными, таким образом, на этом относительно низком эволюционном уровне еще расплывчаты.
Про- и эукариотные водоросли хотя и встречаются почти во всех биотопах, но большинство видов обитает в воде, где они как «планктон» свободно парят в ее толще или как «бентос» прикрепляются к камням, песку и другому субстрату. Из-за различного содержания соли имеется два типа водного жизненного пространства: моря и пресные воды.
Морские водоросли
Растительный планктон морей представлен прежде всего диатомеями и динофитами (перидинеями), а также крошечными гапто-фитами (Coccolithophorales) и золотистыми водорослями (силикофлагеллятами). Представители обеих последних групп не удерживаются ячейками планктонной сетки, и их можно получить только путем центрифугирования («нанопланктон»).
Наибольшая плотность планктона (до 100 000 клеток в литре воды) наблюдается в хорошо освещаемом слое воды. В литре поверхностной воды Атлантики вблизи Фарерских островов было выявлено: 32 000 клеток динофитов, I 600 диатомей и 54 000 кокколи-тофорид. На глубине ниже 100 м число этих планктонных форм резко падает. Однако и на больших глубинах (4000 — 5 000 м) еще встречаются Coccolithophoraceae и «оливково-зеленые клетки», чье систематическое положение до сих пор неясно. Кроме того, наибольшая плотность планктона встречается в
более холодных морях и в области холодных морских течений; это объясняется тем, что воды здесь богаче соединениями азота и фосфатами. Данные вещества потребляются в слоях воды, расположенных ближе к поверхности, и накапливаются в более глубоких слоях вследствие опускания туда мертвых клеток. В холодных областях за счет как зимнего, так и ночного охлаждения поверхности моря вода перемешивается лучше чем в тропиках, что приводит к более интенсивному развитию планктона. Богатство планктона отмечается также там, где холодные глубинные воды, насыщенные соединениями азота и фосфатами, выходят на поверхность благодаря течениям.
Парение планктонных организмов в воде сопровождалось бы более или менее медленным их погружением, если бы не регулировалось специфическим весом и силой трения, а также активным движением жгутиков. Это объясняет многие особенности планктонных водорослей: наличие (образование и разложение) масла как запасного вещества, развитие выростов и выступающих клеточных стенок (см. рис. 11.70), группировка многих клеток в цепочки (см. рис. 11.76). Отмечено, что в теплых водоемах (где вязкость меньше) служащие для парения выросты крупнее, чем в холодных. Минеральные скелеты планктонных водорослей оседают на морское дно. Поскольку известь на глубине более 4000—5 000 м растворяется, на больших глубинах мы находим только скелеты диатомей, силикофлагеллят и из животных — радиолярий. На меньших глубинах (2 000 — 5 000 м) происходит также отложение извести (Coccolithophoraceae, из животных — глобигерины и т. д.). За 1 000 лет откладывается слой мощностью всего 1.5 см. Растительный бентос в морях состоит (не считая морской травы Zosteraceae) исключительно из водорослей, преимущественно бурых и красных. Чаше всего они с помощью присосок или выростов прикрепляются к твердому субстрату (камню) (см. рис. 11.83, 11.85). Подвижный субстрат (ил, песок) заселяется видами лишь немногих родов, например Caulerpa (см. рис. 11.91, А). Бентосные водоросли встречаются от зоны попадания морских брызг до глубин, где еще возможен фотосинтез (180 м).
В тропических морях водорослевая растительность не достигает такой пышности, как в морях умеренной и холодной зон (ср. причины, указанные для планктонных форм). Бурые водоросли представлены гораздо в меньшей степени, тогда как красные, напротив, доста
274 | ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
точно широко, как и некоторые теплолюбивые зеленые водоросли: Caulerpaceae, Dasycladaceae, Codiaceae, Valoniaceae. Богата также растительность коралловых рифов, причем водоросли (Halimeda, см. рис. 11.91, В; Dasycladaceae, см. рис. 11.92, В; Lithothamniori) принимают в образовании извести большее участие, чем сами кораллы. Единственное в своем роде явление представляет собой «Саргассово море», где бурая водоросль Sargassum (см. рис. 11.85, А), плавающая на поверхности воды, образует обильные заросли (сгоняемые к одному месту растения имеют до 5 т фитомассы на квадратную морскую милю).
В умеренно-теплых морях, например в Средиземном, бентос состоит преимущественно из красных и сравнительно мелких бурых водорослей. Вышеупомянутые Bryopsidophyceae и Dasycladophyceae представлены несколькими видами. Широко представлены виды Lithotham-nion. Поскольку интенсивность света зависит от времени года, основное время развития водорослей у поверхности воды — весна, на глубине — лето, а осенью водоросли вообще развиваются меньше.
В холодно-умеренных морях, например в Северном, явно доминируют бурые водоросли, как по размерам, так и по массе. Время года существенно влияет на многие виды. Так, Desmarestia теряет осенью ассимилирующие волоски, а красная водоросль Delesseria — нежные плоские части таллома, так что зимуют только «жилки».
Крупные ламинарии (см. рис. 11.83) ежегодно обновляют свои филлоиды. На рис. 11.107 на примере скалистого побережья Ла-Манша видна четко выраженная вертикальная зональность водорослевой растительности в зависимости от уровня воды при приливах и отливах. Виды верхних зон (Bangia, Porphyra, Fucus) выдерживают температуры до -20 °C, тогда как никогда не подвергающиеся высыханию обитатели более глубоких зон (Laminaria, Delesse-rid) вымирают уже при нескольких градусах ниже нуля. Холодные моря имеют бедный видовой состав водорослей, однако именно здесь талломы бурых водорослей достигают наибольших размеров: можно назвать хотя бы Macrocystis (см. рис. 11.83, Е), Lessonia (D) и Nereocystis (С) — все из Laminariales, а также Durvillea из Fucales. Они практически не уступают в размере крупным наземным растениям.
Распространение морских бентосных водорослей зависит от загрязнения и наличия питательных веществ. Е1апример, Ulva растет в очень богатой питательными веществами воде, Padina — в воде с умеренным их содержани
ем, a Sargassum и Fucus — при малом содержании органики. Между морскими и пресными водами промежуточное положение занимают солоноватые воды. Здесь за счет регулярных приливов и отливов или прибоя перемешиваются пресная и соленая вода. К местообитаниям такого типа, со спецификой планктонных и бентосных водорослей (например. Cha-гасеае), относятся также устья проточных вод.
Пресноводные водоросли
В пресной воде видовой состав растительного планктона в значительной степени зависит от наличия питательных веществ; в богатых ими (эвтрофных) водоемах они также поглощают органические вещества (миксотроф-ность). В районах умеренного климата различия температуры воды, освещенности, значения pH и других показателей в зависимости от времени года существенно влияют на состав планктона. В пресной воде амплитуда колебания температур значительно больше, чем в море. Их диапазон находится в области от температуры таяния (около 0°С) ледников и полярного льда — мест, где обитают определенные, часто окрашенные в красный цвет хламидомонады, Chlorococcales и Mesotaeniaceae, составляющие «криопланктон», до температур горячих источников, в которых еще могут развиваться некоторые диатомеи (до 50 °C) и прокариотные синезеленые водоросли (до 75 °C). В бентосе пресных вод явно доминируют по массе и видовому разнообразию цветковые растения, и только при определенных условиях доминируют водоросли (например, Chara-сеае).
Е1ейстон (сообщество поверхности воды) представлен прежде всего одноклеточными водорослями, например видами Euglena и Chro-mulina rosanoffii. Для последнего вида, который придает поверхности воды золотистый налет, отмечалась плотность до 40 000 клеток на 1 мм2. Следует различать обитающий над поверхностной пленкой воды эпинейстон и опускающийся от нее в воду гипонейстон.
Водоросли и качество воды
С усиливающимся эвтрофированием водоема существенно увеличивается образование биомассы, а с ним и потребление кислорода; на дне откладывается (вместо минеральных отложений олиготрофных озер) гниющий ил. Бедные питательными веществами (олиготрофные) водоемы вследствие искусствен-
11.2. Бактерии, грибы, растения | 275
14 -
13 -
12 -
11 -
ю -
э -
8 -
7 -
6 -
5 -
4 -
3 -
2 -
1 -
0 -
-1 -
-2 -
-3 -
5 -
-6 -
-7 -
8 -
9 -
-10 -
-11 -
-12 -
13 -
-14 -
-15
Корковидные водоросли и накипные лишайники
Verrucaria, Prasiola
Нитчатые водоросли Bangia, Urospora
Pelvetia
Fucus platycarpus
Fuctjs serratus
Alaria
Водоросли верхней части литорали Porphyra. Enteromorpha
водоросли
Laminaria digitata
Минимальный уровень квадратурного отлива
Сублиторальные водоросли с уплощенным талломом
Halidrys, Cystoseira
Комковидные известковые водоросли
__Минимальный уровень сизигийного отлива
— Максимальный уровень сизигийного прилива
Сублиторальные кустовидные водоросли
___Максимальный уровень квадратурного прилива
Кожистые водоросли-макрофиты
J L. hyperborea w
Литоральные водоросли с уплощенным талломом
Fucus vesiculosus Ascophyllum
Водоросли нижней части литорали: Rhodymenia
Литоральные кустовидные водоросли
Корковидные известковые водоросли
Рис. 11.107. Профиль растительности на побережье Ла-Манша (по W. Nienburg):
Chlorophyta: Prasiola, Urospora, Enteromorpha; Rhodophyta: Bangia, Porphyra, Rhodymenia, известковые корки (например, Lithothamnion); Phaeophyceae: Pelvetia, Fucus, Ascophyllum, Alaria, Laminaria, Halidrys, Cystoseira; лишайники: Verrucaria
ного удобрения садов и полей, а также общего загрязнения воды приходят в сильный упадок. По наличию видов-индикаторов (планктонных и бентосных) можно судить об уровне загрязнения и соответственно о качестве воды (уровне сапробности, обозначаемом цифрами от I до IV). Наряду с эукариотными водорос
лями для этого в качестве индикаторов привлекаются также прокариоты (синезеленые водоросли, бактерии), грибы и высшие растения.
Наиболее сильное загрязнение обозначается как IV (полисапробная зона). Здесь из-за недостатка кислорода преобладают процессы гниения. Кислородное истощение чрезвычай
276 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
но велико. При экстремальных условиях жизни в полисапробной зоне имеет место массовое развитие бактерий, в том числе Beggiatoa, а также прокариотных синезеленых водорослей из родов Spindina и Anabaena. Однако почти полностью отсутствуют эукариотные водоросли и водные высшие растения; единственное исключение составляют виды Euglena и Carteria. Наряду с этими немногими водорослями зеленого цвета встречается также бесцветная гетеротрофная Polytoma. Даже при сильном загрязнении, например при поступлении неочищенных сточных вод, может происходить в определенной степени самоочищение водотоков.
В сильно загрязненных водах III класса (а-мезосапробная зона) бурно включаются окислительные процессы. Высокая численность различных бактерий характерна и для этого состояния, но при этом наблюдается и массовое развитие водорослей — синезеленых, а также диатомовых и зеленых; начинают хорошо развиваться и некоторые высшие растения. Содержание кислорода может быть значительным и в дневное время превышать значение насыщения, однако ночью оно сильно падает. Из прокариотных водорослей здесь встречаются различные виды Oscillatoria (тогда как в водах IV класса — только О. putida и О. chlorina) и Phormidium', из диатомей — Stephanodiscus', из конъюгат — Closterium leibleinii и Cosmarium botrytis, из прочих зеленых водорослей — Chlamydomonas и Gonium. Также характерны грибы сточных вод из родов Leptomitus и Fusarium.
Сильно загрязненные воды II класса качества ф-мезосапробные) характеризуются еше более успешными окислительными процессами; соответственно кислородное истощение относительно незначительное. В этой зоне число живых бактериальных клеток становится еше меньше. Наоборот, наблюдается большое разнообразие диатомовых и зеленых водорослей. Синезеленые водоросли представлены Anabaena flos-aquae, Nostoc и некоторыми видами Oscillatoria. Из диатомей встречаются различные виды родов Melosira, Asterionella и др., из золотистых водорослей — Synura. Из зеленых водорослей можно назвать Pediastrum, Sce-nedesmus, Chaetophora и Oedogonium. Из десмидиевых особенно распространены различные виды Closterium.
Почти незагрязненные воды классифицируются как I класс качества (олигосапробные). В этой зоне вода, если не считать временного «цветения», прозрачна и богата кислородом. Если эта область расположена ниже по течению от загрязненного участка реки, то органика здесь уже разложена, интенсивность очень быстро протекающих процессов разложения снижается, а процессы окисления завершены. Число живых бактериальных клеток снижается до минимальных значений. Из синезеленых водорослей характерны, например, представители рода Hapalosiphon. Диатомеи представлены видами Surirella и Meridian, зеленые водоросли — видами Ulothrix, Cladophora (glomerate), Spirogyra (fluvial His) и самыми различными видами десмидиевых (из родов Closterium, Stau-rastrum, Euastrum, Micrasterias), желтозеленые — видами Vauchena Типично также наличие таких пресноводных водорослей, как Lemanea annulata и Batrachospermum moniliforme.
Аэрофильные водоросли, симбионты, образователи горных пород
Лишь немногие водоросли живут как аэрофильные организмы вне воды, прежде всего на затененной стороне скал и стволов деревьев (например, водоросли типа Pleurococcusii Trente-pohlia, см. рис. 11.93, А, С; «чернильные штрихи» из синезеленых водорослей). Наиболее часто они встречаются во влажных тропиках, где поселяются и на листьях. Подходящий известковый субстрат у поверхности (на глубину несколько миллиметров от края) часто пронизан водорослями. Еще шире распространены, но еще мало изучены почвенные водоросли. К «эдафону» — сообществу живых организмов почвы, относятся помимо синезеленых водорослей различные зеленые, желтозеленые и диатомовые водоросли. В 1 г почвы из самого верхнего слоя было обнаружено до 100 000 водорослевых клеток. Особым образом к наземному образу жизни приспособилась зеленая водоросль Fritschiella (см. рис. 11.99, С).
Важную роль в природе играют различные симбиотические водоросли. Также большое значение они часто имеют и как образователи горных пород (например, Dasycladophyceae).
рые отходят от центров щитков и вытягиваются в длину; 8 внутренних клеток (первичные клетки-головки), которые в конечном счете принимают округлую форму. Из-
за того что 8 щитков растут в ширину более интенсивно, чем в толщину, внутри шара возникает полость. К центру ее от центра каждого щитка направлены клетки-руко
11.2. Бактерии, грибы, растения
ятки с сидящими на них по одной первичными клетками-головками. Первичные головки дают начало 3 — 6 вторичным головкам. а из них вырастает по 3 — 5 длинных неветвящихся сперматогенных нитей из одного ряда клеток, заполняющих полость (рис. 11.106, В, С). В каждой из дисковидных клеток образуется по одному спирально изогнутому сперматозоиду (D) с 2 жгутиками и глазком, но без пластид.
Оогоний (см. рис. 11.106. А: о) содержит одну-единственную яйцеклетку, плотно забитую каплями масла и зернами крахмала. Этот оогоний выдается вперед и позже плотно обрастает 5 коровыми нитями, закрученными в виде левовращающей спирали. Их концы отчленяют (путем образования поперечной перегородки) клетки коронки (А: с), между которыми проникает сперматозоид. После оплодотворения зигота окружается прочной бесцветной стенкой. Кроме того, внутренние стенки коровых нитей утолщаются, становятся коричневыми и часто инкрустируются известью, тогда как внешние мягкие стенки вскоре после отделения «ооспоры» (покоящегося органа) исчезают. При прорастании зиготы происходит мейоз; из 4 гаплоидных ядер 3 дегенерируют, так что возникает только один проросток.
Своеобразное строение таллома, а также спермогонии и оогонии с их особыми защитными покровами, не имеющими аналогов среди других растений, и спирально закрученные сперматозоиды говорят о явно обособленном положении Charophyceae в системе живых организмов. Все эти признаки свидетельствуют об уникальности харовых, однако по составу пигментов и запасных веществ они сходны с другими Chlorophyta и Streptophyta.
Ископаемые харовые (особенно в форме их зигот) известны с силура; из 6 семейств, ранее существовавших на Земле, ныне сохранилось только одно.
Обзор Streptophytina. От исходных стреп-тофитов взяла начало большая часть ныне живущих наземных зеленых растений. От них возникли уже рассмотренные зеленые водоросли (II; Streptophytina) и пока еще не рассматривавшиеся мхи (см. Bryophyti-па) и сосудистые растения (см. Pteridophy-
tina и далее). Эволюционное родство этих групп подтверждается совпадением таких признаков, как: ультраструктура хлоропластов: пигментный состав (хлорофиллы а и />!); расположение и строение пиреноидов; крахмал в качестве запасного питательного вещества; изоконтные подвижные стадии с латерально расположенными жгутиками; целлюлоза в качестве строительного материала клеточной стенки, причем в упорядоченной кристаллической форме и в виде микрофибрилл. Поперечные перегородки внутри клеток возникают во фраг-мопластах. У зеленых водорослей (II) уже встречается своего рода «плодообразование», так что оплодотворение индуцирует развитие покровных нитей, обрастающих оогоний {Coleochaete). У Charophyceae покров оогония закладывается до оплодотворения. У некоторых представителей (например, Chara) в сГ гаметах хлоропласты полностью редуцированы. У Zygnematophyceae отсутствуют клетки, имеющие жгутики. У отдельных видов как защитные вещества встречаются спорополленин и лигниноподобные соединения {Coleochaete).
Однако мохообразные и сосудистые растения, несмотря на сходство по многим признакам с зелеными водорослями (II), претерпели существенное дальнейшее развитие генеративных и вегетативных структур, что свидетельствует о явном эволюционном прогрессе (приспособление к наземной жизни!).
Второй подотдел: мхи (Bryophytina)1
В большинстве своем мхи уже выработали весьма высокоорганизованные тканевые талломы1 2; этим они отличаются от зеленых водорослей из предыдущего отдела, которые еще приспособлены к жизни в воде.
В основе цикла развития мхов лежит четкая анизоморфная смена поколений
1 Авторы придерживаются далеко не общепринятой версии системы этой группы. — Примеч. ред.
2 Большинство мхов имеют побеговую организацию. — Примеч. ред.
278 | ГЛАВА 11, СИСТЕМАТИКА И ФИЛОГЕНИЯ
Воке 11.8. Эмбриофиты
К эмбриофитам относят растения, спорофит которых начинает существование как покоящийся многоклеточный зародыш, питающийся от материнского растения и часто долгое время остающийся в покоящемся состоянии (как у покрытосеменных). Правда, наличие зародыша в зачаточной форме наблюдается уже у некоторых живущих в воде водорослей (Coleochaete) с покоящимися и защищенными гаметофитом зиготами.
Типичные эмбриофиты (мхи и сосудистые растения) — это первично наземные растения с постепенно усложняющимися вегетативными органами, которые служат для закрепления в почве, поглощения воды и пита-тельньгх солей, фотосинтеза (см. рис. 4.8; 4.1.2 и 5.3.4). Из исходно талломного вегетативного тела в ходе приспособления к наземному образу жизни и в связи с увеличением размеров и разделением функций развились разные органы: у гаметофитов высших мхов — каулоид, филлоид и ризоид (см. 5.3.4), у спорофита сосудистых растений — стебель, лист и корень (см. 4.1.2).
Размножение происходит в ходе гетерофазной и гетероморфной смены поколений, при которой в жизненном цикле преобладает или гаметофит (мхи) или спорофит (папоротники, семенные растения). После оплодотворения зигота развивается в многоклеточный зародыш, питающийся за счет материнского растения. Гаметангии — здесь их называют антеридии (0я) и архегонии (<j>) — имеют оболочку (стенку) из стерильных клеток, защищающую гаметоциты. Спорангии эмбриофитов в отличие от спорангиев грибов и водорослей также имеют подобную оболочку из стерильных клеток. Такие защитные образования полностью отсутствуют у грибов и лишь изредка встречаются у гаметангиев водорослей.
Мейоспорангии грибов часто защищены сплетением гиф в плодовых телах, однако у них отсутствует оболочка из слоя стерильных клеток. У водорослей из родов Coleochaete и Chara оогоний (постгенитально) покрывается вырастающими нитями (см. рис. 11.105, 11.106, Е). С конгенитальными многоклеточными стенками антеридиев эмбриофитов можно сопоставить прежде всего стенку спермогония Chara.
Вегетативное тело построено из различных тканей, которые сильно дифференциро
ваны и выполняют различные функции. Транспирация снижена кутикулой и чаще всего регулируется имеющимися устьицами. Транспорт воды и питательных веществ иногда идет по простым проводящим тяжам (мхи) или же по все более структурно усложняющимся проводящим пучкам (папоротники, семенные растения). Пластиды содержат хлорофиллы а и Ь, а также каротиноиды. Продукт ассимиляции — образующийся в хлоропластах крахмал. Клеточные стенки состоят из целлюлозы1.
Эмбриофиты подразделяют на мхи (Вгуо-phytina). папоротникообразные (Pteridophy-tina) и семенные растения (Spermatophyta). У семенных растений антеридии и архегонии сильно редуцированы, так что почти не различимы как таковые1 2. Семенные растения поэтому уже не относят к архегониатам в узком смысле (= мхам и папоротникообразным) Сборное понятие кормофиты (или также Cormobionta; в противоположность Protobionta с более простой организацией как, например, у водорослей и грибов) происходит от понятия «кормус» — вегетативного тела, дифференцированного на стебель, листья и корни (см. рис. 4.8), — и включает мхи с иначе дифференцированными спорофитами поскольку их во всяком случае можно вывести из теломов.
Так, спорогоны мхов при нарушенном развитии, как исключение, могут вильчато ветвиться. Проводящие системы мхов, если они вообще имеются, построены достаточно просто. Однако они проявляют функциональное и даже определенное структурное сходство с проводящими элементами сосудистых растений. Все эмбриофиты близкородственны друг другу, так что их можно рассматривать как группу, имеющую общего предка, представители которой развивались в разных направлениях и достигли разного уровня развития.
1 Из целлюлозы состоит фибриллярный каркас клеточных стенок эмбриофитов, который погружен в «аморфный» матрикс из полисахаридов. — Примеч. ред.
2 У всех современных голосеменных растений и эфедры архегонии легко различимы. — Примеч. ред.
11.2. Бактерии, грибы, растения | 279
Рис. 11.108. Bryophytina.
А—С — оплодотворение (у Phaeoce-ros laevis, 900х) (А—С — по Yuasa; D — по R. Harder):
А — сперматозоид достигает яйцеклетки; В — проникновение сперматозоида внутрь яйцеклетки; С — сперматозоид в ядре яйцеклетки, остаток его цитоплазмы остается в цитоплазме яйцеклетки; D — развитие двудомного листостебельного мха (спора, протонема, гаметофит— G, оплодотворение, спорофит — S, редукционное деление, споры). Светлые линии — гаплоидная фаза; темные линии — диплоидная фаза; g — жгутики, к — клеточное ядро, р — пластиды, R! — редукционное деление
(рис. 11.108, D), при которой зеленый фото-автотрофный гаметофит чередуется со спорофитом. Гаметофит представлен либо морфологически слабо расчлененным лопастным талломом с ризоидами снизу (талломные мхи), иногда с высокой тканевой дифференциацией (например, на ассимилирующие и запасающие ткани), либо стебельками, варьирующими от лежачих до прямостоячих, покрытыми листиками и ризоидами (листостебельные мхи). Листики, за исключением средней жилки1, обычно однослойные (листья папоротников и семенных растений многослойные). Листостебельные мхи внешне несколько напоминают сосудистые растения, однако отличаются от последних, в частности, тем, что наивысшего морфологического и анатомического уровня дифференциации достиг гаметофит, а не спорофит. Кроме того, у мхов отсутствуют проводящие пучки, а в большинстве случаев и проводящие ткани. Ри
1 У юнгерманниевых жилки нет и лист целиком однослойный. — Примеч. ред.
зоиды у мхов— это одноклеточные или многоклеточные нитчатые структуры, которые ни в коем случае не сопоставимы с высокодифференцированными корнями сосудистых растений (скорее их можно сопоставить с корневыми волосками). Кутикула мхов чаще всего очень нежная, и они чаще всего быстро высыхают при недостатке воды (пойкилогидричные растения, см. 6.10.3.6). Почти у всех гаметофитов отсутствуют устьица (исключение; антоце-ротовые), однако для газообмена очень редко служат дыхательные отверстия (Маг-chantiales, см. рис. 11.100, А).
Архегонии мхов (см. рис. 11.111, J) представляют собой бутылевидные органы, части которых — брюшко и шейка — обычно имеют однослойную стенку. Брюшко заключает в себе крупную центральную клетку, которая перед созреванием делится на яйцеклетку и расположенную в основании шейки брюшную канальцевую клетку. К ней в шейке примыкают шейковые канальцевые клетки; таких клеток у мхов всегда несколько, располагающихся в один ряд (см. рис. 11.111, J).
280 | ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Антеридии (см. рис. 11.111, Е) представляют собой округлые или булавовидные образования на короткой ножке. Развивающиеся в них сперматогенные клетки, окруженные стенкой антеридия, делятся каждая на 2 сперматиды, которые обособляются друг от друга и превращаются каждая в 1 сперматозоид.
Сперматозоиды всегда короткие, не сколько спирально закрученные, нитевидные, основной объем в них занимает клеточное ядро. Вблизи переднего конца они несут 2 длинных гладких жгутика (F), направленные назад под острым углом от места прикрепления, а у некоторых представителей в цитоплазме имеется еще очень маленькая пластида (см. рис. 11.108, А: р). Яйцеклетки могут содержать также немного очень мелких хлоропластов.
Оплодотворение яйцеклетки даже у наземных форм может происходить только в присутствии воды (дождь, роса). Для этого архегоний открывается на вершине, канальцевые клетки ослизняются и выделяют определенные вещества, которые хемотактически привлекают сперматозоиды (см. 8.2.1.1). Из оплодотворенной яйцеклетки возникает диплоидный зародыш (см. рис. 11.111, К), который всегда без периода покоя развивается в спорофит.
Диплоидное спорофитное поколение развивается, таким образом, всегда на доминирующем гаплоидном гаметофите и остается связанным с ним на протяжении всей своей жизни. Изолированные спорофиты не могут полностью развиться, несмотря на наличие у них хлорофилла. Рост спорофита осуществляется, следовательно, в значительной мере за счет гаметофита. Часто встречающееся у растений питание одного поколения за счет другого (например, у красных водорослей) называют «гонотрофия» (питание за счет производителя). Транспорт веществ в спорофит сокращается или вовсе прекращается, когда он достигает примерно 2/3 своего окончательного размера. Устьица, впервые появляющиеся в ходе эволюции высших растений у мхов, образуются почти исключительно на спорофитах (см. рис. 11.127, G). Спорофит внедряется своей базальной частью (гаусторией, называемой также стопкой, см. рис. 11.127, D; 11.120, С)
в расположенную глубже ткань гаметофита, однако растет в основном в сторону вершины архегония, образуя округлый или овальный спорангий (коробочку, см. рис. 11.111, L, 11.123, М), на короткой либо более или менее длинной ножке. Весь спорофит называют спорогонием.
Во внутренней ткани коробочки — археспории — в результате двухкратных мейо-тических делений материнских клеток спор возникают мейоспоры в группах по четыре (в тетрадах), которые при созревании отделяются друг от друга и округляются. Распространение мейоспор осуществляется по воздуху. Стенка спор состоит из внутреннего нежного эндоспория и внешнего стойкого экзоспория; последний при прорастании споры разрывается. Споры прорастают в гаметофиты с образованием нитчатой или пластинчатой протонемы (см. рис. 11.123, А, В, С; 11.120, D; 11.108, D), которая вскоре переходит в зеленое растение мха1.
Наряду с размножением спорами у мхов очень часто встречается вегетативное размножение, например посредством выводковых телец (см. рис. 11.110, А; 11.114, С, G; см. 10.1.3.3), которые могут различным образом возникать на гаметофитах: на стеблях, листьях или протонеме. Они отрываются и вырастаю! в новое растение.
Рост тканей осуществляется посредством 2-, 3- или многосторонней апикальной клетки (см. 5.3.4, рис. 5.13), реже — посредством меристемы (например, Riella, Ri-ccia и Anthoceros).
Лигнина у мхов нет, лишь крайне редко встречаются сходные с лигнином соединения (как уже у некоторых зеленых водорослей — Coleochaete).
Систематика. К мхам относят примерно 24 000 видов, которые можно подразделить на четыре эволюционно далекие друг от друга группы: Marchatiopsida, Jungerma-nniopsida, Bryopsida и Anthocerotopsida. Bryo-psida называют также листостебельными мхами (всегда олиственные; листья с особыми признаками), тогда как представителей всех остальных классов объединяют: юд названием печеночники. Однако представители Anthocerotopsida, очевидно, эволюци
1 Это гаметофор. — Примеч. ред.
11.2. Бактерии, грибы, растения | 281
онно ближе к остальным наземным растениям, чем к любой другой группе мхов, так что они здесь приведены в Приложении к листостебельным мхам.
1. Класс: Marchantiopsida (= Hepaticae р.р., талломные печеночники)
Гаметофит Marchantiopsida представляет собой уплощенный, чаще всего более или менее вильчато разветвленный, сильно дифференцированный тканевой таллом. С нижней стороны он обычно несет гладкие и так называемые язычковые ризоиды, которые имеют вдающиеся внутрь утолщения стенок. В типичном случае антеридии и архегонии приподняты на особых веточках (гаметангиофорах, или подставках). В клетках содержится по одному или по нескольку окруженных одинарной мембраной «масляных телец» (представляющих собой слившиеся капли терпенов), которые в такой форме отсутствуют у всех других растений (см. рис. 11.110, G: ок). Спорофит длительное время полностью окружен разрастающейся стенкой архегония — эмбриотекой, которую он только незадолго до созревания прорывает на вершине. В созревающем спорогонии каждая клетка археспория делится на материнскую клетку споры и элатеру, которые, таким образом, являются синхронно возникающими сестринскими клетками.
Из материнской клетки споры после мейоза возникает по 4 споры. Соотношение 4: 1 гаплоидных спор (4) и стерильных диплоидных элатер (1) может в противоположность Anthocero-topsida сдвигаться в сторону числа спор (например, 8: I, 128: 1). Материнские клетки спор и элатер отделяются друг от друга продольными стенками, т.е. расположенными параллельно продольной оси спорогония.
Стенки элатер чаще всего со спиральными утолщениями (у Anthocerotopsida они обычно гладкие). Не исключено конвергентное развитие элатер у Anthocerotopsida, Marchantiopsida и Jun-germanniopsida1 1. Элатеры относятся к структурам.
1 Элатеры Anthocerotopsida настолько отличаются от элатер печеночников, что их конвергентное возникновение давно не вызывает сомнений. — Примеч. ред.
способствующим распространению спор (таким, как капиллиций — см., например, Trichia', волокна глебы — см., например, Lycoperdorr, гаптеры — см., например, Equisetum), которые следует рассматривать как аналогичные образования.
Антеридий (см. рис. 11.111, Е) возникает из одной клетки эпидермы1, которая вертикальными, взаимно перпендикулярными стенками делится на 4 клетки, после чего в квадрантах этой башнеобразной структуры происходит разделение тангенциальными стенками периферических покровных клеток и внутренних клеток, дающих начало сперматогенной ткани2.
При развитии архегониев (см. рис. 11.111, J) одна клетка эпидермы3, выступающая над соседними клетками, делится периклинальной стенкой на нижнюю клетку, дающую начало ножке, и верхнюю клетку — зачаток архегония. Три антиклинальные стенки разделяют его на центральную осевую клетку и на три покровные клетки, образующие тангенциальные секторы. На поперечном срезе через молодой зачаток архегония можно видеть все 4 клетки, на продольном срезе — осевую клетку и только две из трех покровных клеток. Осевая клетка сбоку окружена покровными, а сверху свободна; она расчленяется позже поперечной перегородкой на кроющую и внутреннюю клетки. Из покровных клеток без существенного участия клетки-крышечки образуется стенка шейковой и брюшной частей архегония, а из внутренней (центральной) клетки возникают 4—8 шейковых канальцевых клеток, одна брюшная канальцевая клетка и (в основании) яйцеклетка. Иное развитие архегониев у папоротников см. в соответствующем разделе.
Проводящие тяжи в гаметофитах чаще всего не формируются, а в спорофитах всегда отсутствуют полностью. Транспорт воды, например у Marchantia, происходит через перфорированные клетки в талломе.
1. Порядок: Sphaerocarpales4. Иногда порядок выделяют даже в отдельный подкласс (Sphaero-carpidae). Таллом5 имеет простое строение: это растущие на почве мелкие розетки (Sphaerocar-pos, рис. 11.109, А) или обитающие в воде вертикальные талломы, состоящие из оси (жилки) с
1 Точнее, протодермы. — Примеч. ред.
2 Это неточное описание развития антеридия в данной группе. — Примеч. ред.
3 Протодермы. — Примеч. ред.
4 Его целесообразно выделять в особый класс. — Примеч. ред.
5 Современные данные показывают, что это своеобразные побеги, а не талломы. — Примеч. ред.
282 | ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Рис. 11.109. Marchantiopsida, Sphaerocarpa-les (по К. Muller):
А —Sphaerocarpos michelii, таллом с обертками гаметангиев (5х); В — Riella helicophylla, <j> таллом (2,5х)
отходящим от нее в одну сторону «крылом» (Riella, рис. 11.109, В). Архегонии и антеридии окружены грушевидными обвертками, открывающимися наверху. Стенка спорогония состоит из единственного слоя клеток, разрушающегося при созревании. У Sphaerocarpos (растение, широко используемое в генетических экспериментах) были впервые (1917 г.) в растительном царстве выявлены половые хромосомы. У него, как и у некоторых других мхов, определение пола происходит при мейозе материнских клеток спор.
2. Порядок: Marchantiales. Представители порядка имеют высокодифференцированный таллом. Как пример можно рассмотреть широко распространенную в сырых местах маршанцию обыкновенную (Marchantia pofymorpha, семейство Marchan-
Рис. 11.110. Marchantiales, Marchantia, М. polymorpha (А — по К. Magdefrau; В—F — по L. Кпу; G — по К. Madgefrau, с изменениями; Н — по L. Кпу; J — по H.Leitgeb):
A—F— вегетативное размножение: А — разрез через выводковую корзиночку (12х), В — выводковое тельце в плане (80х), С — в поперечном разрезе; D —F — развитие выводкового тельца (ЗООх); G — поперечный разрез таллома (200х); Н — дыхательное отверстие, вид сверху (200х); J — развитие ассимиляционных камер; а — ассимиляторы; as — место отделения выводкового тельца; о — верхний эпидермис с дыхательным отверстием (устьицем); ок — масляное тельце; oz — масляная клетка; г — ризоид; г! — зачаток ризоида; s — клетка-ножка; и — нижний эпидермис; v — брюшная чешуйка; w — утолщения стенок запасающих воду клеток
11.2. Бактерии, грибы, растения
tiaceae). Она образует лентовидно-уплощенные, несколько мясистые, вильчато разветвленные талломы (см. рис. 11.111, A, G) шириной до 2 см со слабо выраженным срединным ребром. Они нарастают за счет деления группы меристематических клеток на своих концах. На нижней стороне таллома располагаются однослойные брюшные чешуйки (амфигастрии) и отрицательно фототропные одноклеточные ризоиды (рис. 11.110, G), которые закрепляют таллом на субстрате и проводят воду вдоль него (преимущественно капиллярно между ризоидами, как по фитилю; отчасти за счет поглощения ими воды)1.
Под эпидермисом верхней стороны, покрытым почти водонепроницаемой кутикулой, расположены обширные межклеточные пространства (см. рис. 11.110, G, J) — «ассимиляционные камеры». Они отделены друг от друга перегородками, состоящими из одного или двух слоев клеток. На поверхности таллома эти перегородки видны как светлые линии, разделяющие темно-зеленые ромбические или шестиугольные участки. Со дна камер поднимаются многочисленные короткие, состоящие из округлых клеток разветвленные ассимиляторы, которые содержат хлоропласты и образуют ассимиляционную ткань1 2 (см. рис. 11.110, G). Каждая камера сообщается с окружающим воздухом через бочонкообразное устьице. У Marchantia polymorpha оно состоит из четырех расположенных друг над другом колец, каждое из которых образовано 4 клетками. При недостатке воды устьице может даже немного сужаться, но для регуляции воды это еще не имеет существенного значения. Их строение препятствует проникновению воды в «устьице». Во всем растительном царстве едва ли найдется еще гаметофит с таким совершенным ассимиляционным и транспирационным аппаратом (см. рис. 11.122, А). Крупные, бедные хлорофиллом паренхимные клетки нижней стороны таллома служат как запаса-
1 Функции закрепления растения и поглощения воды выполняют простые ризоиды, а ее проведения — язычковые. — Примеч. ред.
2 Комплекс ассимиляторов неправомерно называть тканью. — Примеч. ред.
юшие (например, содержат масляные тельца, рис. 11.110, G: ок).
По срединному ребру таллом на верхней стороне, как правило, местами разрастается в бокаловидные структуры с зубчатыми краями — выводковые корзиночки (рис. 11.110, А; 11.111, А) с некоторым числом плоских выводковых телец внутри. Последние возникают, как показано на рис. 11.110, D — F, путем разрастания наружу и дальнейшего деления отдельных поверхностных клеток; они сидят на клетке-ножке (s), от которой позднее отрываются (В: as). В каждом углублении по бокам они имеют по точке роста и состоят из нескольких слоев клеток (С), из которых некоторые бесцветны и представляют собой зачатки будущих ризоидов (С: п). Выводковые тельца вырастают в новые талломы и очень успешно обеспечивают вегетативное размножение гаметофитов.
Жизненный цикл. Гаметангии расположены на особых вертикальных ветвях таллома гаметофита (гаметангиофорах, или подставках) (см. рис. 11.111, A, G). В нижней части гаметангиофоры имеют вид трубчатой ножки, а в верхней части повторно вильчато ветвятся, образуя звездчатые «зонтики». Антеридии и архегонии возникают на разных растениях (двудомность). Определение пола осуществляется, как и у многих других мохообразных, гаплогенотипи-чески через половые хромосомы (аналогично тому, как показано на рис. 10.19). В ножке подставки ризоиды, отходящие от ее брюшной стороны, оказываются при завертывании краев ножки внутри трубки (рис. 11.111, В, С); они доходят до нижней стороны таллома и капиллярно всасывают воду, как по фитилю.
Антеридиофоры заканчиваются горизонтальным «зонтиком», который в результате трехкратного дихотомического ветвления становится по краю 8-лопаст-ным (см. рис. 11.111, А). С верхней стороны на нем развиты антеридии, погруженные по одному в бутылевидную полость, которая открывается наружу узким отверстием (С). Эти полости отделены друг от друга ассимиляционными камерами. Вскрывание антеридиев и выход сперматозоидов происходят после дождя в ре-
284 | ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Рис. 11.111. Marchantiales, Marchantia, М. polymorpha, половое размножение (А, С—Е, G, Н, К— М — по L. Кпу; В — по К. Magdefrau; F — по Ikeno; J — по Duran):
А— сГ растение с выводковыми корзиночками и антеридиофором; точки на поверхности таллома — устьица (1,5х); В— поперечный разрез через ножку антеридиофора немного ниже «зонтика» (13х), справа — спинная сторона с воздушными камерами, слева — брюшная сторона с двумя желобками, где проходят ризоиды; С— продольный разрез через антеридиофор (18х); D— развитие антериди-ев (160х); Е — почти зрелый антеридий, продольный разрез (160х); F— сперматозоид (400х); G — <j> растение с архегониофорами (1,5х); Н — продольный разрез через архегониофор; позади ряда архегониев— псевдоперихеций (25х); J— развитие архегониев (160х); К— продольный разрез через молодой, еще заключенный в стенку архегония спорогон, окруженный псевдопериантием (35х); L — освободившийся из покровов спорогон, из которого выступают споры и элатеры; у основания ножки сохранился остаток стенки архегония (10х); М — споры и элатера (160х)
зультате того, что стенки антеридия ослизняются и расплываются. Сперматозоиды (рис. 11.111, F) собираются на антери-диальной поставке в каплях воды (роса или дождь) и задерживаются там несколько загнутым вверх краем подставки.
Архегониофоры (G) в самом начале своего развития очень сходны с антеридиофорами. Архегонии закладываются восемью радиальными лучами, причем расстояние между двумя рядами, находящимися против спинной стороны ножки, больше, чем между всеми остальными. Край молодого «зонтика» в процессе развития постепенно загибается вниз, так что группы архегониев оказываются на его нижней стороне (за счет чего исходно акропетальное зало
жение архегониев становится базипетальным). Наконец, находящиеся между группами архегониев ткани разрастаются во вторичные лопасти «зонтика» (общим число девять!); две из них развиваются между наиболее удаленными друг от друга группами архегониев (см. выше).
Оплодотворение осуществляется при дождливой погоде, когда брызги от дождевых капель, содержащие сперматозоиды, попадают с О’ на ф «зонтики». Эпидермальные клетки последних выступают вперед в виде сосочков1 и представляют собой по
1 Это не сосочки эпидермальных клеток, а особая однослойная структура псевдоперихеций. — Примеч. ред.
11.2. Бактерии, грибы, растения
285
верхностную капиллярную систему. По ней сперматозоиды проводятся к архегониям, к которым они затем привлекаются по механизму хемотаксиса — вероятно, определенными белками (см. 8.2.1.1).
Спорофит. Через несколько дней после оплодотворения зигота начинает развиваться в многоклеточный зародыш, который вырастает в небольшой овальный зеленеющий спорогон с очень короткой ножкой (рис. 11.111, К, L).
Из двух клеток, возникающих после первого деления зиготы, верхняя (обращенная к шейке архегония) дает начало округлой коробочке (экзоскопическое положение зародыша), тогда как нижняя образует гаусторию и ножку (L). Начальные стадии развития не совсем одинаковы у представителей разных родов и семейств. Периклинальными стенками зачаток коробочки разделяется на внутренние и наружные клетки (К); внутренние дают многоклеточную спорогенную ткань (археспорий).
Коробочка у Marchantia имеет стенку из одного слоя клеток с кольцевыми утолщениями стенок. Только на вершине стенка двухслойная. Коробочка вскрывается на верхушке несколькими зубчиками. Зрелая коробочка сначала некоторое время покрыта разрастающейся стенкой архегония (К), которая, однако, при вытягивании ножки разрывается и остается у основания в виде «воротничка». Кроме того, каждая коробочка окружена тонко-кожистой «частной обверткой» (= perichaetium)1 с четырьмя или пятью зубцами по краю, которая еще до оплодотворения начинает расти из короткой ножки архегония в виде мешка (Н, К). Наконец, каждый радиальный ряд архегониев окружен еще выростом таллома — «групповой обверткой» (involucrum) в виде изящных зубчиков (Н)1 2.
Коробочка высвобождает несколько сотен тысяч спор (L, М). Между спорами располагаются одноклеточные нитевидные элатеры с тонкими стенками со спираль-
1 В отечественной литературе «частную обвертку» называют периантий, а «общую обвертку» — перихеций. — Примеч. пер.
2 Она представляет собой бахромчатую по краю пластинку, а не зубчики. — Примеч. ред.
Рис. 11.112. Marchantiales, Riccia (А, В — по К. Magdefrau; С — по L. Кпу):
А— R. glauca, внизу растение со спорогонами (2х);В — R. fluitans, погруженная водная форма (2х); С— R. glauca, поперечный разрез через таллом с антеридием (125х)
ными утолщениями (М; процесс их возникновения — см. во введении к Marcha-tiopsida), которые после вскрывания коробочки совершают гигроскопические движения, разрыхляя и рассеивая споровую массу (L). Каждая прорастает в очень короткую хлорофиллоносную нить (протонему), которая сначала с помощью двусторонней верхушечной клетки, а позже более сложным образом вырастает в таллом.
Также растущий на скалах и сырой почве Conocephalum conicum (Conocephalaceae) сходен с Marchantia строением таллома, но имеет более простые устьица и у него отсутствуют выводковые корзиночки. Сперматозоиды из антеридиофора выбрасываются за счет тургорного давления на несколько сантиметров вверх. Lunularia (Lunulariaceae) имеет полулунные выводковые корзиночки и антеридиофоры без ножек. У самого мелкого представителя этой группы — Monocarpus sphaerocarpus (Мопосаграсеае; Австралия) — сильно редуцированный таллом несет единственный округлый спорогон, который окружен относительно хорошо развитым покровом.
Представители семейства Ricciaceae устроены более просто (рис. 11.112). Вильчатые ветвления таллома посредством двусторонних верхушечных клеток (см. рис. 5.12) чаще всего быстро следуют друг за другом, так что возникают небольшие розетки1 (рис. 11.112, А). У некоторых
1 Ветвления терминальные, но не вильчатые. — Примеч. ред.
286 | ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ видов таллом разделен на камеры и имеет простые устьица; однако у большинства видов верхняя часть таллома состоит из вертикальных рядов клеток, каждый из которых заканчивается более крупной бесцветной клеткой (С). Гаметангии, как и не имеющий гаустории и ножки спорофит, погружены в таллом1 1 (С). Большинство видов Riccia — напочвенные формы (А); лентовидная, дихотомически разветвленная R. fluitans (В) живет в толще воды, a Ricciocarpos natans подобно ряскам плавает по поверхности воды.
2. Класс: Jungermanniopsida (= Hepaticae р.р., преимущественно листостебельные печеночники)
К этому классу относят талломные и листостебельные формы, между которыми имеются переходы. Первые слабо дифференцированы морфологически и анатомически; с нижней стороны они несут только гладкие ризоиды. Гаметофиты листостебельных форм несут простые листья без средней жилки, расположенные в два ряда2.
Представителей класса Jungermanniopsida часто трактуют как прогрессивный ряд от талломных к листостебельным формам. Однако и листостебельные формы можно считать исходными и выводить из них талломные, предположив срастание в ходе эволюции налегающих друг на друга листьев и уплощение стебля.
Гаметангиофоры у гаметофитов отсутствуют. Масляные тела встречаются чаще всего в большом числе во всех клетках, тогда как у Marchantiopsida локализованы в особых масляных клетках. Коробочка спорого-на выносится на длинной ножке (приспособление к распространению спор ветром) и вскрывается чаще всего 4 створками (см. рис. 11.114, А). В предыдущем подклассе у спорогонов ножки отсутствуют или очень короткие. Элатеры сходны по строению и развитию с таковыми у предыдущего класса. Проводящих пучков в спорофите нет, в гаметофите они встречаются редко и тогда очень просты, т.е. сложены одними гидроидами (Symphyogyna, Haplomitrium).
1. Порядок: Metzgeriales. Таллом, нарастающий за счет одной верхушечной клетки (см. рис. 5.12, В, С) и чаще всего вильчато разветвленный, состоит из одного или нескольких слоев одинаковых клеток; у некоторых видов в нем имеется срединное ребро из удлиненных клеток (см. рис. 11.114, А, В), а у Symphyogyna даже есть примитивный центральный проводящий тяж. Род Fossombronia отличается двумя рядами косо прикрепленных листьев, многорядных в основании. Таким образом, мы имеем здесь переход от талломных к листостебельным формам. Архегонии развиваются под продолжающей дальнейший рост верхушечной клеткой; спорофиты, окруженные псевдоперихецием, сидят на талломе (А) или на коротких боковых ветвях («анакрогинно»), У видов некоторых родов элатеры в виде кисточки прикрепляются к верхним концам створок коробочки (Metzgeria, А) или в середине ее дна {Ре!На). Большинство из 500 видов этого порядка, например виды родов Riccardia, Pellia, Fossombronia, обитает на сырой почве. Напротив, Metzgeria растет на затененных скалах или как эпифит на коре лиственных пород. Род Blasia (см. рис. 11.114, С) занимает, согласно данным новейших исследований, изолированное систематическое положение между Jungermanniopsida и Marchantiopsida и поэтому, строго говоря, должен относиться к отдельному классу (Blasiopsida). У Blasia таллом имеет бугылевидные вместилища выводковых телец; он разделен по краям на листовидные лопасти и несет снизу мелкие чешуйки.
2. Порядок: Calobryales1. Вертикальные стебельки несут три ряда одинаковых листьев и прикрепляются к субстрату мясистыми разветвленными «корневищами», лишенными ризоидов, но содержащими эндотрофно живущие грибы («микориза»). В частности, встречающийся и в Европе Haplomitrium (см. рис. 11.113, С) имеет плоские, у основания многослойные листья и центральный тяж в каулоиде.
3. Порядок: Jungermanniales. Представители порядка — преимущественно тропические растения, чаще все мелкие, живущие на почве и на древесных стволах, а в тропиках и на листьях лесных деревьев. Насчитывая около 9000 видов (в Средней Европе 250), они составляют примерно 90% всех печеночников. Их тело четко дифференцировано на лежачий или приподнимающийся, разветвленный, дорсовентральный стебель и однослойные листья без средней жилки, которые располагаются в два ряда2 по бокам
1 Они располагаются в углублениях поверхности таллома, а не внутри него. — Примеч. ред.
1 За редкими исключениями, листорасположение Jungermanniopsida трехрядное. — Примеч. ред.
1 Эту группу с еще большим основанием, чем Blasia, следует выделять в особый класс. — Примеч. ред.
2 Листья чаше всего располагаются в три ряда. — Примеч. ред.
11.2. Бактерии, грибы, растения | 287
стебелька под углом к нему (рис. 11.114, D — J). В стебле нет никаких проводящих тканей. У Jun-germanniales нет ни воздухоносных щелей (как у Marchantiales), ни настоящих устьиц (как у Anthocerotales).
Листья, косо прикрепляющиеся к стеблю и поэтому расположенные подобно черепице, по-разному налегают друг на друга и весьма разнообразны по форме. Их называют набегающими, когда нижний край каждого листа покрыт верхним краем расположенного ниже (рис. 11.114,Е, F), и нисбегающими, если нижний край при рассмотрении сверху не закрыт (Н). Листья с закругленной или заостренной верхушкой (G), выемчатые (F), двузубчатые или многозубчатые, двухлопастные (Н) или рассеченные на нитевидные окончания (D). У эпифитной Frullania (J) одна из двух лопастей листа видоизменена в бокаловидную структуру, которая служит для удержания воды («водяной мешок»).
У большинства родов наряду с расположенными в два ряда спинными листьями есть еще расположенный на брюшной стороне ряд листьев меньшего размера и другого вида — это амфигастрии. или брюшные листья (например, у Frullania, Calypogeia, рис. 11.114, F, J). Образование трех рядов листьев обусловлено наличием трехсторонней пирамидальной верхушечной клетки побега, одна сторона которой, однако, дает лишь мелкие листья или — у видов с двумя рядами листьев — вообще не дает листьев. Боковые ветви возникают рядом с листьями.
Расположенные на конце побега (акрогинные) архегонии окружены периантием (рис. 11.114, G), который состоит из трех сросшихся между собой верхних листьев. У некоторых видов клетки гаметофита под оплодотворенным архегонием делятся, так что возникает мешковидное вместилище (марсупий) для защиты молодого спорофита, вырастающего после оплодотворения яйцеклетки. Спорогон бывает уже полностью развитым, когда он вследствие растяжения своей ножки (см 7.1.1) прорывает стенку архегония, остающегося в виде кожистой обвертки у его основания. В коробочке нет колонки. Оболочки клеток многослойной стенки коробочки имеют утолщения в виде колец или балок либо равномерно утолщены (если не считать тонких внешних клеточных стенок); разрыв происходит за счет силы поверхностного натяжения при потере клетками воды (см. 8.4).
Протонема Jungermanniales у разных родов устроена неодинаково, однако обычно состоит всего из нескольких клеток. У Metzgeriopsis pusilia она, напротив, представляет собой пластинку и является по сути собственно вегетативным телом, на котором сидят растеньица с немногими листьями, служащие лишь для формирования
Рис. 11.113. Takakia lepidozioides (А—В) и Haplomitrium hooked (С) (А, В — по R.M.Schuster; С — по К. МйПег):
А — внешний вид (6х); В — поперечный срез стебля (100х); С — Jungermanniopsida, Haplomitrium (6х)
половых органов. У Protocephalozia нитевидная протонема несет антеридии и архегонии'.
Вегетативное размножение также широко распространено у Jungermanniales: у одних оно происходит посредством легко отламывающихся выводковых ветвей и выводковых листьев особого облика (часты у тропических эпифиль-ных Lejeuniaceae), у других — посредством выводковых телец, образующихся преимущественно по краям или на конце листьев и состоящих из немногих или всего из одной клетки (см. рис. 11.114, Н).
К Jungermanniales относят также: Scapania, Lophozia и Trichocolea (см. рис. 11.114); Lophocolea (см. 7.1.1.); Cephalozia (см. рис. 8.38).
Эволюция- Положение Jungermanniopsida в системе противоречиво. Раньше на основании анализа ДНК предполагали их большую эволюционную близость к Bryopsida, чем к Marchantiopsida. Новейшие исследования подтверждают их самостоятельность как отдельного класса, однако одновременно указывают на более тесную связь с Marchantiopsida, чем с Bryopsida, так что оба
1 Как и у Metzgeriopsis, у Protocephalozia на долговечной протонеме образуются эфемерные побеги-гаметофоры, образующие половые органы. Непосредственно на протонеме половые органы никогда не образуются. — Примеч. ред.
288 | ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Рис. 11.114. Jungermanniopsida (А, С — по Schiffner; В — по S.O. Lindberg; D—G — по W. J. Hooker; Н, J — по К. Muller):
А—С — Metzgeriales: А — Metzgena conjugate (нижняя сторона) с несколькими сГ и одной <j> ветвями таллома, на 4 створках коробочки пучки элатер, ножка спорогона окружена псевдоперихецием (15х), В — М. conjugate, поперечный разрез таллома (ЗОх), С — Blasia pusilia, с бутылевидными выводковыми корзиночками и многочисленными ушками на верхней стороне таллома, заселенными Nostoc (4х); D—J — Jungermanniales: D— Trichocolea tomentella, лист и амфигастрий (а) (7х), Е, F — Calypogeia trichomanis: Е — растение, вид сверху, с марсупием и зрелым спорогоном (2х), F — фрагмент с 4 листьями и 2 амфигастриями, вид снизу (6х), G — Scapania undulata с периантием и зрелым спорогоном (2х), Н — Lophozia ventricosa, вид сверху, с группами выводковых корзиночек на концах листьев (10х), J — Frullania dilatata, вид снизу, с «водяными мешками» (25х)
класса (с Blasiopsida, но не с Anthocerotopsida) образуют таксон более высокого ранга. Согласно этой новой, здесь еще не используемой концепции мхи разделяют на три основные группы: Hepaticophytina (печеночники), Bryophytina (листостебельные мхи) и Anthocerotophytina (антоцеротовые).
3. Класс: Bryopsida (= Musci, листостебельные мхи)
Гаметофит листостебельных мхов всегда дифференцирован на стебель и листья. С помощью обычно разветвленных многоклеточных ризоидов с косыми перегородками (у ранее рассмотренных мхов они не-разветвленные и почти всегда одноклеточ
ные) он закрепляется в почве или подстилке (см. рис. 11.123, Е). В противоположность олиственным видам Jungermanniales побеги не дорсовентратьные, а радиально-симметричные, а листья чаще всего располагаются по спирали и при рассмотрении сверху выглядят трехрядными или многорядными. Боковые ветви образуются под листом (ср. рис. 5.13; отличие от семенных растений). Листья, нарастающие с помощью двусторонней верхушечной клетки, часто имеют среднюю жилку, а масляных телец у них нет (у листостебельных печеночников часто бывают капельки масла, а листья нарастают с помощью односторонней верхушечной клетки).
У листостебельных мхов с лежачими побегами листья при спиральном располо-
11.2. Бактерии, грибы, растения | 289
Е
Рис. 11.115. Bryopsida (A—F— по D. Н. Campbell, G — по R. Harder, Н—L— по К. von Goebel):
А—G — развитие антеридия у Funaria hygrometrica: А— поперечное деление зачатка антеридия, L — образование и С — деление верхушечной клетки, D — разделение на покров и зачаток сперматоген-ной ткани, Е — то же, на поперечном срезе (А— Е — 650>), F — почта зрелый антеридий (ЗООх), G — продольный срез собрания антеридиев Mnium hornum, часть антеридиев вццна с поверхности, часть дана в разрезе (100х); Н —L — развитие архегония у Mnium undulatum (250х): Н — ножка еще без зачатка архегония, J — архегоний (а) закладывается путем образования центральной клетки (отмечена точками), кроющей клетки (d) и клеток стенки, К — центральная клетка разделилась на яйцеклетку и брюшную канальцевую клетку, L — многочисленные шейковые канальцевые клетки отделяются от кроющей клетки; h — кроющие листья; р — парафизы; s — ножка
жении отвернуты в одну или в две стороны, так что возникает различие спинной и брюшной сторон, однако иначе, чем у печеночников (например, Fissidens, см. рис. 11.126, С).
Листостебельные мхи отличаются от других мхов также и спорофитом: он чаще всего имеет устьица и развивается как коробочка с колонкой обычно на длинной ножке1 1. Элатер нет.
Жизненный цикл. Споры листостебельных мхов прорастают в гаметофиты, представленные вначале обильно разветвленной, положительно фототропичной зеленой нитью — протонемой (см. рис. 11.123, А). Протонемы при массовом развитии видны невооруженным глазом как зеленый войлок. Сначала развиваются богатые хлоропластами нити с перпендикулярными к их оси поперечными перегородками, которые называют хлоронемой. Хлоропема постепенно переходит в стелющуюся по субстрату каулонему, которая беднее хлоропластами и имеет косые поперечные перегородки. При достаточном освещении на ней, а чаще
1 Как и у других мхов, он имеет еще и гаусторию. — Примеч. ред.
всего на ее коротких боковых ветвях, развиваются почки, дающие начало растеньицам мха1 (см. рис. 11.123, А). Из каулонемы возникают, кроме того, многочисленные, чаще всего направленные вверх боковые ветви типа хлоронем. Почка возникает таким образом, что после отделения одной или двух клеток-ножек вздувающаяся конечная клетка косыми стенками отделяет трехстороннюю пирамидальную верхушечную клетку (В, С), которая, откладывая сегменты, развивает олиственное растеньице мха. Там, где таких почек много, растеньица мха располагаются плотными дерновинками.
Половые органы располагаются у листостебельных мхов группами на концах главного побега или более мелких боковых ветвей. Они окружены самыми верхними листьями, которые часто видоизменены в особую «обвертку» — пе-рихециальные листья2 (рис. 11 115).
1 Протонема представляет собой ювенильную форму растения мха. На ней развиваются не растения мха, а гаметофоры. — Примеч. ред.
2 Перихециальные листья окружают только собрания архегониев. Вокруг собраний антеридиев находятся перигониальные листья. — Примеч. ред.
290 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Рис. 11.116. Bryopsida, Buxbaumiales (А — по Dening; В — по К. von Goebel; С, D — по К. Magdefrau):
А—С — Buxbaumia aphylla: А — <j>, Р — сГ гаметофиты (А, В — 35х), С — спорофит; D — Diphy-scium sessile
Рис. 11.117. Bryopsida, развитие спорогона Funaria hygrometrica (А—Е— по D.H.Campbell; F, G — по J. Sachs):
А, В — продольный разрез, первые деления зиготы; С—Е — поперечный разрез: С — деления в эндотеции (е) и амфитеции (a), D — дальнейшие деления, Е — более зрелый спорогон; в эндотеции самый наружный слой клеток — археспорий (аг) отделяется от колонки (со) (А— Е — ЗООх); F, G — поперечный разрез через археспорий (аг) и возникающие из него, еще не изолированные материнские клетки спор (sm) (250х); s — верхушечная клетка
Листостебельные мхи могут быть обоеполыми (гермафродитными), однодомными или двудомными соответственно в зависимости от того, развиваются ли антеридии и архегонии на одном или на разных
побегах того же растения или на разных растениях.
Между половыми органами обычно располагается некоторое число многоклеточных, часто имеющих округлые конечные клетки парафиз.
Антеридии и архегонии листостебельных мхов расположены на ножках и в отличие от остальных мхов (и архегониат в целом) (бокс 11.8, с. 204) их тело формируется из сегментов, откладываемых верхушечной клеткой (см. рис. 11.115).
Спорофит. После оплодотворения яйцеклетки хемотаксически (см. 8.2.1.1) привлекаемым сперматозоидом зигота сначала многократно делится поперечными перегородками, и развивается сегментированный зародыш, вытянутый в длину. При типичном развитии в самой верхней его клетке возникают косые перегородки, которые отделяют клиновидную двустороннюю верхушечную клетку (рис. 11.116; 11.117, А, В; 11.118). Она отделяет в обе стороны сегменты, которые делятся дальше. В клетках сегментов, образующих коробочку, перегородка появляется по вертикальной радиальной стенке, так что на поперечном срезе зародыша видны 4 квадранта (см. рис. 11.117, С). Из них при делении периклинальными стенками возникают внешние (амфитеций) и внутренние (эндотеций) клетки (рис. 11.117, С, D: а, е). Самый внешний слой эндотеция чаще всего дифференцируется в археспорий (Е, F: аг), который целиком делится на материнские клетки спор (G: sm). Каждая материнская клетка мейотически делится на 4 гаплоидные споры. В противоположность Marchantiopsida внутренние клетки эндотеция не участвуют в образовании археспория, а чаще всего образуют тяж из стерильной ткани — колонку (рис. 11.117, Е; со; 11.119, А: с), окруженную археспорием (рис. 11.119, A; sz).
Колонка служит для подведения питательных веществ и накопления воды для образующихся спор; питательные вещества спорам поставляют также богатые содержимым клетки вокруг археспория. Нижняя часть зародыша (рис. 11.118, А) — гаустория — закрепляется в ткани гаметофита.
11.2. Бактерии, грибы, растения | 291
Молодой спорофит (зародыш) сначала окружен оболочкой (эмбриотекой), которая образуется как из брюшка архегония, так и из ткани его ножки и даже из ткани стебля. По мере роста спорофита эмбриотека, не выдерживая натяжения, разрывается поперечно. При этом верхняя часть выносится на верхушке спорофита в виде колпачка, тогда как нижняя остается в виде влагалища (рис. 11.118, В).
На верхнем конце зрелой коробочки образуется своеобразная структура — колечко, предназначенное для вскрывания коробочки и рассеивания спор. Ножка выносит коробочку вверх, так что ветер может легко распространять споры. В молодом спорогоне снаружи от споровой массы на-
Рис. 11.118. Bryopsida (А — по D. Roth; В — по Leunis и Frank):
А — продольный срез молодого спорофита листостебельного мха Pogonatum urnigerum (150х); цифры указывают следующие друг за другом сегменты; сегменты 1 —7 образуют ножку спорофита; В — Pottiales, Pottia lanceolata (40х), верхняя часть побега, листья удалены, два архегония оплодотворены: зародыш слева вследствие вытягивания ножки (s) поднял верхнюю часть эмбриотеки в виде колпачка (к), оставив нижнюю как влагалище (v); эмбриотека справа еще целая; а — неоплодотворенный архегоний; SM — начало меристемы ножки
Рис. 11.119. Bryopsida, Bryidae (по G.Haber-landt, с изменениями К. Magdefrau):
А — продольный разрез через спорогон листостебельного мха Funaria hygrometrica (25х); В — ассимиляционная ткань с устьицем (90х); а — колечко; ад — ассимиляционная ткань; с — колонка; d — крышечка; р — перистом; s — устьице; sz — спорогенные клетки
ходится деятельная ассимиляционная ткань, которая покрыта эпидермой1.
На спорогоне встречаются устьица Mni-ши-типа (рис. 11.119, А, В), свойственного также папоротникам. В разных семействах они значительно различаются по числу (от
1 Ассимиляционная ткань находится значительно глубже эпидермы. К эпидерме она непосредственно примыкает в апофизе — части спо-рогона между коробочкой и ножкой. — Примеч. ред.
292 | ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
3 до 300 на одну коробочку), форме и размеру.
Проводящие тяжи. Некоторые виды имеют просто устроенные проводящие ткани. Они могут быть и в гаметофитах, и в спорофитах. Как и в проводящих пучках сосудистых растений, проведение веществ осу ществляет цепочка разных клеток. Для транспорта воды и солей служат гидроиды — вытянутые мертвые клетки, в полностью развитом состоянии, без ядра и цитоплазмы, с утолщенными боковыми стенками и косыми поперечными. В отли чие от трахеид сосудистых растений их клеточные стенки не бывают ни одревесневшими, ни укрепленными кольцевыми или спиральными утолщениями. Продукты ассимиляции проводят другие, также вытянутые клетки — лептоиды, по развитию и строению напоминающие ситовидные элементы сосудистых растений. Боковые стенки их часто утолщены и в меньшей степени, чем иногда косые поперечные стенки, пронизаны ситовидными отверстиями с плазмодесмами. Вплоть до отмирания лептоиды содержат в своей цитоплазме ядра и пластиды. Обычно гидроиды располагаются внутри, а лептоиды снаружи. часто они перемешаны с другими элементами в центральном тяже. Последний окружен тканями из тонкостенных (внутренняя кора) и толстостенных (внешняя кора) клеток. Стереиды — это живые, имеющие ядро и пластиды, вытянутые в длину клетки, соседние с гидроидами. Их утолщенные, но не одревесневшие стенки служат для придания механической прочности (сравнимы с клетками колленхимы). Граничащие с лептоидами паренхимные клетки при большом струкгурном и функциональном сходстве с ними отличаются отсутствием ситовидных пор в поперечных стенках. От такого типа тяжей имеются различные отклонения — от упрощения строения (например, в случае отсутствия леп-тоидов) до полной редукции тяжей.
Тяжи листовых следов (см. рис. 11.123, Н), присоединяющиеся к центральному тяжу или слепо заканчивающиеся в коровой паренхиме, продолжаются в листья как проводящая ткань их средней жилки. Только при непосредственном соединении между
гидроидами листа и центрального тяжа ка-улоида, т.е. при наличии настоящих листовых следов, возникают предпосылки для непрерывной системы проведения воды. Часто встречаются упрощения проводящей системы листьев: так, в коровой паренхиме каулоида гидроидов может быть очень мало или их нет совсем. В проводящих тяжах листьев для проведения ассимилятов, по-видимому, служат широкопросветные паренхимные клетки, тогда как типичные лептоиды здесь редки (Polyrrichales).
Для питания спорофита большое значение имеет его прикрепление к гаметофиту. Служащая как всасывающий орган гаустория внедряется в сильно разрастающуюся ткань ножки архегония, а в некоторых случаях (Polytrichum) — даже глубоко в ткань стебля до его центрального тяжа. Гидроиды ножки спорогона тесно примыкают к гидроидам стебля гаметофита. На гаустории спорогона иногда встречаются придатки типа ризоидов, внедряющиеся в ткань гаметофита, но чаще — передаточные клетки, которые примыкают к клеткам паренхимы или лептоидам и отличаются увеличивающими поверхность утолщениями стенки в виде шишек или горбов. Цитоплазматические связи (через плазмодесмы) между спорофитом и гаметофитом, напротив, отсутствуют. Часто сочленение тканей между гаметофитом и спорофитом довольно несовершенно.
Вегетативное размножение. Мхи обладают чрезвычайно высокой регенеративной способностью. Так, обломившиеся стебли и листья могут непосредственно или через стадию протонемы вырастать в новые растения. У некоторых видов в пазухах листьев и на верхушках побегов вырастают группы клеток, которые могут опадать как «выводковые тельца» (см. рис. 11.126, О).
Листостебельные мхи подразделяют на три подкласса (Sphagnidae, Andreaeidae, Вгу-idae). В первых двух подклассах коробочка спорогона выносится вверх с помощью гаплоидной ложной ножки (удлиненной ножки архегония1).
1 Ложная ножка имеет более сложную природу, включая часть тканей стебля. — Примеч. ред.
11.2. Бактерии, грибы, растения | 293
1. ПОДКЛАСС: ТОРФЯНЫЕ МХИ (SPHAGNIDAE)
Подкласс включает только одно семейство — Sphagnaceae, с единственным, но очень богатым видами (свыше 200!) родом Sphagnum. Сфагнум обитает в болотистых, преимущественно бедных известью местах, часто с низкими значениями pH, где образует большие подушки и ковры, из года в год нарастающие вверх, тогда как нижняя их часть отмирает и постепенно превращается в торф. В клеточных стенках откладываются лигниноподобные вещества.
Тетраэдрические споры прорастают в присутствии определенных микоризных грибов в протонему; сначала она нитевидная, а затем представляет собой маленький лопастной однослойный таллом с нитча
тыми ризоидами. Таллом образует чаще всего только один гаметофит с пучком ризоидов у основания (рис. 11.120, D).
Вертикальные, лишенные ризоидов стебли почти всегда находятся в плотных дернинках и несут через регулярные промежутки пучки боковых ветвей. В каждом пучке часть ветвей отходит вверх, а часть — вниз, плотно прилегая к стеблю (рис. 11.120. А). На верхушке побега ветви образуют плотную розетку. Некоторые виды Sphagnum (особенно обитатели верховых болот) благодаря пигментам клеточных стенок окрашены в коричневый или ярко-красный цвет. Каждый год одна из ветвей на верхушке побега развивается так же мощно, как и соответствующий прирост материнского побега, так что происходит ложное вильчатое ветвление (ложная дихотомия). По мере того как стебли снизу посте-
Рис. 11.120. Bryopsida (А, Е—Н— по К. Magdefrau; В, D — по W.Ph.Schimper; С— по Waldner; J — по Н. Schenck; К— по Kiihn):
А—Н — Sphagnidae, Sphagnum: А — S. acutifolium, растение со спорогонами (2/Зх), В — S. squrrosum, зрелый спорогон на конце веточки (10х), С — S. acutifolium, молодой спорогон в продольном разрезе (17х), D — S. acutifolium, протонема с молодым растеньицем (100х), Е — S. molluscum, участок ветви с бутылевидными клетками, накапливающими воду; листья удалены (10x), F — то же, на поперечном срезе (Юх), G— S. acutifolium, участок однослойного листа; видны крупные водоносные клетки с кольцевыми уголщениями и отверстиями, а между ними — мелкие хлорофиллоносные клетки (ЗООх), Н — то же, на поперечном срезе (ЗООх); J — К— Andreaeidae, Andreaea rupestris: J — целое растение (8х), К — продольный разрез через молодой спорофит (40х); ah — шейка архегония; aw — эмбриотека; с — колонка; d — крышечка; f — гаустория; к — колпачок; ка — коробочка; pb — листья пе-рихеция; ps— ложная ножка; s— споры; sf — гаустория спорогона; sg — спорогенная ткань; w — стенка коробочки
294 | ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ пенно отмирают, дочерние ветви становят-ся самостоятельными растениями.
Кора стеблей состоит из одно- или многослойной ткани из мертвых, пустых клеток, которые капиллярно всасывают воду; их боковые и поперечные стенки часто имеют округлые отверстия (рис. 11.120, Е). В листьях также находятся клетки с перфорированными стенками, усиленными кольцевыми или спиральными утолщениями (рис. 11.120, G); они располагаются поодиночке в ячейках однослойной сети из удлиненных живых клеток, содержащих хлоропласты. Эти своеобразные структуры служат для поглощения воды и минеральных веществ; растения могут таким образом удерживать воду, масса которой превышает сухую массу примерно в 20 раз. Листья не имеют средней жилки, а стебли — центрального тяжа.
Размножение. Отдельные ветви розетки имеют особый внешний вид и окраску. Они производят половые органы. Мужские ветви образуют в пазухах листьев округлые антеридии с длинными ножками (выходящие из них гаметы были первыми открытыми у растений сперматозоидами); женские ветви несут на конце архегонии, которые растуг в отличие от остальных листостебельных мхов без верхушечной клетки — как у печеночников. Спорогоны развивают лишь очень короткую ножку с расширенной гаусторией. Они долгое время заключены в эмбриотеку, которую разрывают на вершине, оставляя ее у своего основания в виде оторочки (рис. 11.120, В: aw). В шаровидной коробочке спорообразующая ткань (С: s) куполообразно покрывает по-лушаровидную колонку. Археспорий возникает не из эндотеция, а из самого внутреннего слоя амфитеция. Спорогон своей расширенной гаусторией погружен во вздутый конец несущего его небольшого побега; после формирования спо-рогона этот конец вытягивается в ложную ножку значительной длины и выносит спорогон вверх (В: ps). За счет избыточного давления воздуха в коробочке крышечка с хорошо различимым шумом сбрасывается, и споры вылетают на расстояние более 20 см.
2. ПОДКЛАСС: ANDREAEIDAE
Единственное семейство подкласса Andrea-еасеае представлено тремя родами. Виды рода Andreaea (их насчитывается 120) образуют мелкие плотные темно-коричневые дернинки на неизвестковых скалах в высокогорьях, в Аркти
ке и Антарктике. Спорогон, как и у Sphagnum, выносится вверх на ложной ножке, которая образуется из ножки архегония. Коробочка, сначала покрытая, как шапкой, колпачком, раскрывается четырьмя продольными щелями, причем получающиеся четыре створки остаются соединенными между собой на верхушке и у основания (рис. 11.120, J). Как и у Sphagnum, спорогенная ткань наподобие колокола окружает колонку (К). Протонема пластинчатая, разветвленная. У рода Andreaeobryum ложной ножки нет.
Ранее относимые к печеночникам Takakiales, вероятно, лучше трактовать как листостебельные мхи и включать в Andreaeidae1. Сюда относят азиатский род Takakia (см. рис. 11.113, А) с листьями, рассеченными до основания на 2 — 4 цилиндрических сегмента, и центральным тяжем в стебле.
3. ПОДКЛАСС: BRYIDAE
У представителей этого подкласса коробочка выносится вверх не принадлежащей гаметофиту ложной ножкой (как в обоих предыдущих подклассах), а образуемой спорофитом ножкой (диплоидной).
Гаметофит в этом подклассе достигает наибольшего разнообразия и наивысшей дифференциации среди листостебельных мхов; однако в немногих случаях он почти ограничен стадией протонемы (например, Ephemeropsis tjibodensis, Viridivelluspulchellum). Побеги или растут вертикально и несут на верхушке архегонии, а позже коробочку на ножке (акрокарпные мхи; см. рис. 11.123, Е), или они плагиотропные и одновременно чаще всего перисто-разветвленные, образующие архегонии, а позже коробочки на коротких боковых ветвях (плеврокарпные мхи; см. рис. 11.126, R). В стебле обычно проходит один центральный тяж (см. рис. 11.123, Н), который у наиболее высокоразвитых форм (Polytrichum) достигает значительной гистологической дифференциации (см.: Систематика, с. 233).
Листья состоят преимущественно из одного слоя клеток. Краевые клетки листовой пластинки часто образуют особую кайму (см. рис. 11.123, К: наружный слой) или зубчики. Клетки листа у акрокарпных мхов обычно паренхимные (изодиаметрические,
1 Еще лучше рассматривать их как класс Та-kakiopsida. — Примеч. ред.
11.2. Бактерии, грибы, растения | 295
см. рис. 11.123, К), у плеврокарпных, напротив, они в основном прозенхимные (вытянутые, как на рис. 11.120, G). Верхушечная клетка листа у акрокарпных форм дает несколько дочерних клеток, которые затем делятся более или менее перпендикулярными друг к другу перегородками, так что возникает сеть из изодиаметри-ческих клеток. У плеврокарпных видов дочерние клетки, отчленяемые верхушечной клеткой косыми перегородками, быстро делятся дальше на ромбические клетки, боковые углы которых вытягиваются, в результате возникает сеть из прозенхимных клеток. Листья (особенно с сетью прозенхимных клеток) часто имеют среднюю жилку (см. рис. 11.123, J — L).
Коробочка у Bryidae также достигает наибольшего совершенства. Спорогон состоит из тонкой упругой ножки (см. рис. 11.123, Е; 11.126) и радиальной (см. рис. 11.123, Е) или дорсовентральной (см. рис. 11.116, С) коробочки, сначала покрытой колпачком (верхней частью эмбриотеки: см. рис. 11.118, В; 11.123, Е, М), позднее отпадающим1. Шейка архегония вскоре засыхает и остается как шпиль на колпачке. Колпачок, таким образом, состоит не из диплоидной ткани спорофита, а из гаплоидной ткани гаметофита (см. рис. 11.108, D). Самую верхнюю часть ножки под коробочкой называют апофизой; здесь обильнее всего развиваются устьица. По всей длине коробочки проходит колонка, вокруг которой в виде полого цилиндра располагается спорогенная ткань (см. рис. 11.119, A: sz). Колонка и спорогенная ткань, кроме того, окружены крупными межклетниками (см. рис. 11.119), которые образуются в амфитеции и особенно сильно увеличиваются при созревании. Более или менее шаровидные мейоспоры чаще всего содержат многочисленные хлоропласты (см. рис. 11.123,0).
Верхняя часть стенки коробочки развивается как крышечка (см. рис. 11.119, A: d; 11.123, М). Под краем крышечки часто находится узкая зона в виде венчика— колечко (см. рис. 11.119, А: а; 11.124). Его клетки содержат расплывающуюся слизь и
1 В состав спорогона входит также гаустория. — Примеч. ред.
Рис. 11.121. Bryopsida, Bryidae, Leucobryum glaucum (по К. Magdefrau):
А — гаметофит со спорофитами (естественная величина); В — строение листа: два слоя лишенных цитоплазмы клеток, связанных друг с другом крупными отверстиями в стенках; между ними мелкие, вытянутые в длину клетки, содержащие хлоропласты (ЗООх)
Рис. 11.122. Bryidae, Polytrichales (А— по
К. Magdefrau; В — по Vaisey, С. Hebant):
А — строение листа Polytrichum formosuirr, на верхней стороне видны пластинчатые выросты из хлорофиллоносных клеток (250х); В — Polytrichum juniperinum, поперечный срез стебля (120х); аг — внешняя кора; h — гидроиды; i — межклетники; ir — внутренняя кора; I — лептоиды; z — центральный тяж
296 | ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Рис. 11.123. Bryidae, Bryales, Mnium punctatum (по К. Magdefrau):
А — протонема с почкой (20х); В — возникновение почки на протонеме; хлоропласты в верхних клетках не показаны (80х); С — заложение трехсторонней верхушечной клетки (85х); D— сГ растение (естественная величина); Е — <j> растение со спорофитом (естественная величина); F — собрание антеридиев в продольном разрезе (15х); G — собрание архегониев в продольном разрезе (15х); Н — поперечный срез стебля с центральным тяжом и тремя тяжами листовых следов (40х); J — лист (4х); К — кончик листа (25х); L — поперечный срез нижней части листа (50х); М — зрелая коробочка, рядом крышечка и колпачок (4х); N — перистом; слева наружный перистом удален; один из трех зубцов наружного перистома в сухом состоянии отогнут (ЗОх); О— спора (100х)
обеспечивают таким образом отбрасывание крышечки при созревании спор (колпачок отпадает еще раньше). По краю остающейся после опадения крышечки урночки у большинства листостебельных мхов находится перистом (см. рис. 11.119, А: р; 11.123. N), как правило, сложенный зубцами и изначально прикрытый крышечкой. У остальных мхов перистома нет. Строение перистома очень разнообразно.
У немногих мхов (Polytrichales, Tetraphidales, см. рис. 11.126, N) зубцы перистома состоят из рядов целых клеток. Однако у всех остальных мхов перистом образуется под крышечкой из утолщенных частей клеточных оболочек трех самых внутренних слоев амфитеция.
Возникновение такого перистома можно прот следить на поперечном (рис. 11.124, В) и продольном (рис. 11.124, А) срезах верхней части коробочки. Тангенциальные стенки между пер
вым и вторым слоем клеток утолщаются особенно сильно, а стенки между вторым и третьим слоем утолщаются слабее; радиальные, а также неутолщенные части тангенциальных стенок трех слоев клеток в конце концов разрушаются, так что сохраняются только утолщенные части тангенциальных стенок клеток. Они и представляют собой перистом, который, таким образом, является двойным (рис. 11.123, N) и состоит не из целых клеток, а только из сохраняющихся тангенциальных стенок. Наружный перистом состоит из 16 поперечно-полосатых зубцов (рис. 11.123, N), расположенных по внутреннему краю стенки урночки; внутренний перистом («реснички») плотно прилегает к наружному и сложен узкими пластинками и нитями, которые имеют поперечные утолщения на внутренней поверхности1 и в своей нижней части сливаются в общую мембрану (рис. 11.123, N; 11.124, A: ip). Между каждыми двумя зубцами внешнего перистома
1 На внешней. — Примеч. ред.
11.2. Бактерии, грибы, растения । 297
располагается по две реснички внутреннего перистома1 (группа диплолепидных мхов в противоположность гаплолепидным только с одним кольцом зубцов перистома).
Наружные зубцы перистома совершают гигроскопические движения (см. рис. 8.36; см. 8.4), закрывая или открывая коробочку (рис. 11.123, N) в зависимости от погоды (в сухую погоду чаще всего изгибаются наружу), и таким путем способствуют постепенному рассеиванию спор. Наклоненные спорогоны и спорогоны с широким отверстием урночки чаще всего имеют хорошо развитый перистом, тогда как у родов с вертикально расположенным спорогоном, имеющим узкое отверстие урночки, перистом часто редуцирован (рис. 11.125, D).
Поскольку можно заставить молодые спорофиты регенерировать протонему, то соответственно имеется и возможность получить диплоидные гаметофиты, которые в свою очередь могут образовывать тетрап-лоидные спорофиты. Неоднократно повторяя этот процесс, удалось получить гаметофиты с 16-кратным набором хромосом. Подсчеты числа хромосом у многих видов показали, что гаметофиты мхов часто содержат от двойных до четверных наборов хромосом на ядро и, таким образом, часто полиплоидны; и в этих случаях спорофит содержит вдвое больше хромосом, чем гаметофит.
Систематика. В основе систематического подразделения Bryidae, насчитывающих примерно 15 000 видов, лежат признаки как гаметофита, так и спорофита (особенно перистома).
1. Надпорядок: Polytrichanae. Перистом состоит из целых подковообразных или вытянутых волокнистых клеток. Побеги акрокарпные, с высокодифференцированной проводящей тканью (непрерывная система проведения воды). Подземные «корневища» отличаются от надземных частей гаметофита приблизительно радиальным расположением проводящих воду гидроидов и напоминают этим корни двудольных цветковых растений. Листья сверху с продольными пластинчатыми выростами из богатых хлоропластами клеток («ассимиляционные пластинки», см. рис. 11.122).
1 Взаимное расположение элементов наружного и внутреннего перистомов разнообразно. — Примеч. ред.
Рис. 11.124. Bryidae (А — по Sachs, с изменениями К. Magdefrau; В — по К. Magdefrau):
А — Funaria hygrometrica, продольный срез верхней части коробочки листостебельного мха перед вскрыванием (200х); В — Mnium puncta-tum, поперечный срез коробочки в области перистома (120х); ар — наружный перистом; ад — ассимиляционная ткань; с — колонка; d — зона разрыва; ed — эпидермис крышечки; ек — эпидермис урночки; ip — внутренний перистом; s — материнские клетки спор: w — толстостенные клетки, соединяющие зубцы наружного перистома с эпидермисом; I — III — клетки кольца; 1 — 3 — три самых внутренних слоя амфитеция
298 | ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ___________________________________
1.1. Порядок: Dawsoniales. Дерновинки, состоящие из стройных растений высотой до 70 см, встречаются в Австралии и других областях Южного полушария.
1.2. Порядок: Polytrichales. Отверстие урноч-ки в отличие от видов предыдущего порядка сначала закрыто тонкой пластинкой (эпифрагмой). Polytrichum (рис. 11.126, Т) с игловидными листьями, достигающий высотой 40 см, растет на почве в лесах и болотах. У Pogonatum протонема долговечная. Atrichum undulatum с волнистыми листьями в виде язычков — мох, часто встречающийся на почве в лесах.
2. Надпорядок: Dicrananae. Перистом, если он не полностью редуцирован, простой (гаплолепид-ный). Преимущественно акрокарпные мхи.
2.1. Порядок: Dicranales. Перистом обычно состоит из 16 попарно соединенных зубцов. Виды Dicranum, отличающиеся серповидными листьями, часто обитают на почве в лесах. У Leucobryum жилка, почти полностью занимающая весь лист, состоит их клеток двух типов: зеленых живых и мертвых, накапливающих воду (см. рис. 11.121). Обитает на кислом лесном гумусе. Сюда относят также Archidium (см. рис. 11.126, А) со споро-гоном без ножки, крышечки и перистома. Коро-
Рис. 11.125. Bryidae, Schistostegales, Schi-stostega pennata (A, B, D — no W. Ph. Schimper; C, E—G — по K. Magdefrau; H — no F. Noll):
A — несущее коробочку растеньице (103x); В — стерильное растеньице (10х); С— фрагмент предыдущего (50х); D — открытая коробочка (25х); Е— протонема («светящийся мох»), вид сбоку; стрелкой показано направление падения световых лучей (150х); F — то же, вид сверху (150х); G— выводковое тельце на протонеме (150х); Н — ход световых лучей в клетке протонемы
бочка раскрывается беспорядочно за счет сгнивания (клейстокарпия).
2.2. Порядок: Fissidentales. Листья расположены в два ряда, со спинным крылом. В Германии встречаются виды родов Fissidens (рис. 11.126, С, D) и Octodiceras.
2.3. Порядок: Pottiales. Клетки листьев с шишковидными утолщениями наружных стенок («папиллами»). Tortula с длинным спирально закрученным перистомом (рис. 11.126. F). Eucladium (см.: Встречаемость и образ жизни мхов, с. 228) образует чаше всего подушки, инкрустированные известняком.
2.4. Порядок: Grimmiales. Преимущественно подушковидные обитатели каменистых субстратов не имеют признаков двух предыдущих порядков. Листья, например у Grimmia (рис. 11.126, Н) и Rhacomitrium, часто вытянуты в бесцветные волоски или стекловидные окончания.
Последующие надпорядки отличаются двойным перистомом (диплолепидным).
3. Надпорядок: Bartramiales включает акрокарпные мхи. Коробочки, например у Bartramia и Timmia, обычно наклонные, булавовидные и почти округлые, бороздчатые.
4. Надпорядок: Funariales. Представлен широко распространенным по всему земному шару видом Funaria hygrometrica (рис. 11.126, J), часто встречающимся на пожарищах. Это растущий на почве акрокарпный мох с крупными гладкими клетками листьев1. Splachnum luteum (рис. 11.126, К), растущий, как и другие виды этого рода, на помете травоядных животных,— двудомный вид. Он отличается ярко окрашенной дисковидной апофизой спорогона. Женские растеньица (<j>) приземистые, с более крупными листьями по сравнению с мужскими (сГ); их архегонии окружены прилегающими листьями обвертки (у 6" они оттопырены), так что возникает своего рода почка. Уже протонемы показывают половой диморфизм. У Ephemerum протонема сохраняющаяся.
5. Надпорядок: Biyales. Внутренние зубцы двойного перистома высокодифференцированы (см. рис. 11.123, N). Имеются переходы от акрокарпных к плеврокарпным формам, а также между листьями из паренхимных и прозенхимных клеток.
Сюда относят Bryum, Rhodobryum (см. рис. 11.126, L) с крупным пучком листьев и частые на почве в лесах виды рода Mnium (см. рис. 11.123), а также Mittenia plumula (Австралия, Тасмания, Новая Зеландия), протонема которой отражает свет (ср. «светящийся мох»).
1 Интересно происхождение видового названия этого мха — hygrometrica (гигрометрическая): гигроскопичная ножка спорогония спирально скручивается в сухих условиях и раскручивается во влажных. — Примеч. пер.
11.2. Бактерии, грибы, растения | 299
Рис. 11.126. Bryopsida, Bryidae (по К. Magdefrau):
А— Archidium phascoides, целое растение (5х) и коробочка (20х); В — Dicranum scoparium, трехлетнее растение (естественная величина); С — Fissidens bryoides (4х); D — то же, лист (15х); Е —G — Tortula muralis (4х): F — перистом (ЗОх), G — лист со стекловидным волоском (10х); Н — Cnmmia pulvinata (в натуральную величину), J — Funaria hygrometrica (2х); К — Splachnum luteum (в натуральную величину); L— Rhodobryum roseum (в натуральную величину); М — Tetraphis pellucida (2х); N — перистом; О — выводковая корзиночка (8х); Р — Climacium dendroides (в натуральную величину); Q — Hylocomium splendens, четырехлетнее растение (1/2х); R — Cratoneuron commutatum (1/2х); S — Ра-pillaria deppei (1 /2х); Т — Polytrichum commune, справа молодой, покрытый колпачком спорогон (1 /2х)
6. Надпорядок; Пурпапае. Здесь преобладает плеврокарпия. Клетки в листьях прозенхимные, жилка (если имеется) в отличие от видов предыдущих групп гомогенная, т. е. состоит из клеток одного типа.
6.1. Порядок: Neckerales. Внутренний перистом прямой коробочки обычно недоразвит. Cli
macium (рис. 11.126, Р) разветвлен наподобие маленького деревца.
Часто встречающиеся в тропиках эпифит-ные виды Papillaria (S) и близких родов образуют «повислые формы». Fontinalis (см. с. 228) приспособился к жизни в воде. Виды Macro-mitrium образуют в коробочках более крупные
300 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
<j> и мелкие сГ споры. Сюда же можно отнести и Orthotrichum.
6.2. Порядок: Hookeriales. Перистом редуцирован; листья широкие, состоят из крупных клеток; пример — Hookeria lucens, обитающая на влажной почве в лесах.
6.3. Порядок: Hypnales. Внутренний перистом высокодифференцирован. Длинные ножки заканчиваются обычно наклоненными коробочками. Brachythecium, Hypnum, HylocomiumlyMC. 11.126, Q), Pleurozjum и Plagiothecium с многочисленными видами, часто встречающимися на почве в лесах: к роду Cratoneuron (R) относят важные об-разователи известкового туфа.
Систематическое положение следующих порядков до сих пор неясно.
7. Порядок: Buxbaumiales1. Протонема долговечная, богатая хлорофиллом, в значительной мере обеспечивающая питание гаметофита1 2 3 и спорофита. У Buxbaumia сильно редуцированный гаметофит1 (половой диморфизм; см. рис. 11.116, А—С) мало участвует в питании физиологически самостоятельного, снабженного обильной ассимиляционной тканью спорофита. У Diphysci-ит (D) лишенная ножки коробочка сидит в розетке из зеленых листьев.
8. Порядок: Tetraphidales. Четыре зубца перистома состоят каждый из нескольких продольных рядов клеток. Tetraphispellucida (см. рис. 11.126, М —О) часто встречается на гнилой древесине.
9. Порядок: Schistostegales. Единственный вид порядка— «светящийся мох» Schistostega pennata (см. рис 11.125); отличается вторично двухрядными листьями, отсутствием перистома и сохраняющейся протонемой, которая размножается многоклеточными выводковыми тельцами. Протонема растет в расселинах скал и ямах в земле. Она образует шаровидные клетки, которые улавливают падающий на них свет и частично отражают его (Е, Н). Листья гаметофита сначала прикрепляются косо и расположены по спирали, однако в ходе развития принимают вертикальное положение перпендикулярно лучам падающего на них света (А. В)
4. Класс: антоцеротовые (Anthocerotopsida)
Гаметофиты антоцеротовых талломные, однако отличаются от уже рассмотренных печеночников (классы 1 и 2) рядом при
1 Этот и следующий порядки, вероятнее всего, надо выделять в отдельные классы. — Примеч. ред.
2 Протонема — это стадия развития гаметофита, а не отдельное растение/поколение. — Примеч. ред.
3 На самом деле — гаметофор. — Примеч. ред.
знаков. В клетках гаметофитов нет масляных телец (у печеночников они встречаются часто) и имеется только по одному хлоропласту с пиреноидом. У гаметофита есть устьица (у печеночника Marchantia — напротив, только дыхательные отверстия). Спорофиты растут посредством интеркаляр-ной меристемы, поэтому их рост не ограничен. Способ прикрепления спорофита к гаметофиту (передаточная ткань) сходен с тем, что мы наблюдаем у Psilotopsida из папоротникообразных, но не известен ни у одного из остальных мхов. Коробочка также имеет устьица и вскрывается двумя створками. Элатеры антоцеротовых отличаются способом возникновения и внешним видом от элатер печеночников (у листостебельных мхов элатер нет).
Антеридии с самого начала закладываются эндогенно (у печеночников они сначала экзогенные, но могут потом обрастать тканью гаметофита); архегонии также погружены в таллом.
Антоцеротовые составляют небольшую группу из примерно 100 видов, которые объединяют в единственный доживший до настоящего времени порядок.
Порядок: Anthocerotales. Гаметофит представляет собой дисковидный лопастной очень просто устроенный таллом длиной несколько сантиметров, прикрепляющийся к почве ризоидами (рис. 11.127, А). Клетки более или менее однородной паренхимы содержат в противоположность всем другим мхам только по одному крупному блюдцевидному хлоропласту с пиреноидом1. Эпидерма нижней стороны таллома имеет устьица с двумя бобовидными замыкающими клетками. Расположенная за ними подустьичная полость заполнена слизью и чаще всего заселена синезеленой водорослью Nostoc (В). Листьев и чешуек на нижней стороне таллома (брюшных чешуек), как и масляных телец в клетках, нет. Ризоиды гладкостенные.
Антеридии и архегонии погружены в таллом на его спинной стороне. Оплодотворенная яйцеклетка делится поперечной перегородкой на две клетки, из которых
1 У некоторых видов хлоропластов в клетке несколько. — Примеч. ред.
11.2. Бактерии, грибы, растения | 301
Рис. 11.127. Anthocerotopsida, Anthocerotales {А— по К. Magdefrau; В, С, D— по H.Leitgeb; Е — по К. von Goebel; F — по К. Muller; G — по D.H. Campbell):
A — Phaeoceros laevis, таллом с молодыми и вскрывшимися спорогонами (2х); В — Anthoceros vincentianus, устьице на нижней стороне таллома, подустьичная полость заселена Nostoc (270х); С — Anthoceros punctatus, продольный срез молодого спорогона (130х); D — Dendroceros crispus, продольный срез почти зрелого спорогона (80х); Е — Anthoceros punctatus, неравные клеточные деления в археспории (100х); F — Anthoceros husnoti, поперечный срез спорогона с тетрадами спор и колонкой; стрелки — места разрыва стенки спорогона (ЮОх); G — Anthoceros pearsoni, устьице спорогона (125х); а — археспорий; с — колонка; е — элатеры; f — гаустория спорогона; s — споры; sm — материнские клетки спор; w — стенка коробочки
верхняя, т.е. обращенная к шейке архегония, после дальнейших делений становится спорогоном, а нижняя — более толстой гаусторией, внедряющейся в таллом клетками типа ризоидов (рис. 11.127, D)1.
Спорогон представляет собой лишенную ножки щетинковидную коробочку1 2 длиной от одного до нескольких сантиметров и вскрывающуюся, как стручок, двумя продольными створками (рис. 11.127. А). Она характеризуется (в отличие от коробочек Marchantiopsida) сложной тканевой дифференциацией. Вдоль оси коробочки располагается стерильная колонка (С, D: с), состоящая из нескольких рядов клеток. Колонка окружена тонким слоем спорогенных клеток (археспорием, а), которые помимо мейоспор образуют также стерильные клетки — элатеры. Диплоидные материнские клетки спор и клетки, становящиеся элатерами, являются сестринскими: на каждую материнскую клетку спор (соответственно тетраду спор после мейоза) при
1 Противопоставление спорогона и гаустории ошибочно, так как гаустория — это орган спорогона. — Примеч. ред.
2 Спорогон = коробочка + зона интеркаляр-ной меристемы + гаустория. — Примеч. ред.
ходится одна зрелая элатера или одна еще способная к делению стерильная клетка, в результате последовательных митотических делений которой окончательное число элатер может увеличиться многократно. Элатеры располагаются перпендикулярно оси спорогона (Е).
В отличие от всех других мхов коробочка спорофита созревает не одновременно, а длительное время удлиняется благодаря деятельности меристематической зоны у основания коробочки. В стенке спорогона имеются двуклеточные устьица (рис. 11.1-27, G), а кроме того, она содержит в своих клетках хлоропласты.
Представители семейства Anthocerotaceae развивают на талломе спорогоны длиной 1 — 7 см. В Германии встречаются два рода: Anthoceros (например, A. punctatus) — со слизевыми вместилищами, которые на верхней стороне рассеченного таллома просвечивают точками, и темными густошиповатыми спорами; Phaeoceros (например, Ph. laevis на бедном известью жнивье) — без слизевых вместилищ в лопастном талломе, с желтоватыми, покрытыми сосочками спорами. Таллом Notothylaceae по сравнению с представителями предыдущего семейства крайне мал (несколько миллиметров в диаметре).
Палеонтологические свидетельства эволюции Anthocerotopsida, к сожалению, пол
302 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
ностью отсутствуют. В отличие от печеночников и подобно листостебельным мхам, антоцеротовые имеют настоящие устьица. Этот признак сближает листостебельные мхи и антоцеротовые с высшими наземными растениями. Согласно данным анализа ДНК, антоцеротовые, очевидно, ближе к рассматриваемым дальше зеленым наземным растениям, чем прочие группы мхов.
Встречаемость и образ жизни мхов
Мхи завоевали сушу и населили ее множеством видов. Их приспособления к наземной жизни весьма разнообразны: например, значительная устойчивость к высыханию (пойкилогидрические растения, см. 6.10.3.6), сниженная или регулируемая транспирация (в частности, благодаря кутикуле; стерильным стенкам гаметангиев и спорогониев; устьицам; росту плотными подушечками и дерновинками), их приспособления к поглощению, накоплению и проведению воды, выработка различных жизненных форм, соответствующих обитанию на суше (например, древовидные — см. рис. 11.126, Р, листоватые — Q, повислые — S, войлоковидные, корковидные — R, подушечковидные — Н, дерновинные). которые обеспечивают заселение благоприятных ниш и приспособление к экстремальным условиям обитания.
Прочность тканей мхов обеспечивается набуханием клеточных стенок, а не тургором, как у высших наземных растений. Поэтому высушенные мхи при попадании в воду принимают свой исходный облик. Поглощение и отдача воды происходят, за немногими исключениями, всей поверхностью. Капиллярная система между стеблем, ризоидами и, если имеются, листьями обеспечивает значительное накопление воды, которое увеличивается у некоторых листостебельных печеночников благодаря развитию «водяных мешков» (см. рис. 11.114, J), черепицеобразному (набегающему или низбегающему — см. Jungermanniopsida) расположению листьев, наличию брюшных листьев и лопастей (см. рис. 11.114, F, Н, D), у листостебельных мхов — благодаря росту плотными высокими дерновинками.
Marchanlia, Sphagnum и Leucobryum накапливают воду также в водоносных клетках (см. рис. 11.110; 11.120, G; 11.121). Колонка в коробочке листостебельных мхов служит резервуаром питательных веществ и воды для образующихся спор. У определенных мхов (например, Funaria, Encalypta) накопление воды облегчает вздутый колпачок. Именно способностью мхов удерживать значительные количества воды в основном обусловлено буферное действие лесов в балансе воды в ландшафте. Обводненность верховых болот обусловлена осадками и очень большой способностью торфяных мхов (различных видов Sphagnum) накапливать воду.
Вышеупомянутая капиллярная система служит одновременно и внешней системой проведения воды, преобладающей у мхов. Мхи с центральным тяжем (к ним относится большинство акрокарпных листостебельных мхов и немногие печеночники) проводят поглощенную ризоидами воду по гидроидам. Особенно хорошо эта внутренняя проводящая система выражена у Polytrichanae, где даже имеются настоящие, связанные с центральным тяжом тяжи листовых следов, которые обеспечивают снабжение листьев водой (сомкнутая проводящая система).
Распространение сперматозоидов и спор. Если оплодотворение яйцеклетки сперматозоидами связано с наличием капельно-жидкой воды, то споры распространяются ветром. Их освобождение обеспечивают изменения влажности и специальные механизмы рассеивания (например, элатеры; механизмы вскрывания коробочек; см. 8.4). Зонтиковидная, ярко окрашенная апофиза у спо-рогонов видов Splachnum (см. рис. 11.126, К) способствует распространению склеенных в комочки спор насекомыми1.
Некоторые виды имеют хорошо развитые органы ассимиляции. Таллом Marchantia уже напоминает по анатомическому строению лист листостебельных растений, в том числе и приспособлениями к газообмену, хотя менее эффективными и имеющими
1 Существенную роль в привлечении насекомых играет также запах, выделяемый через устьица на апофизе. — Примеч. ред.
11.2. Бактерии, грибы, растения | 303
другое строение. На верхней стороне листьев Polytrichum образуются свободно поднимающиеся вверх пластины, поглощающие свет для фотосинтеза. Там, где уже встречаются настоящие устьица (у антоцеротовых на талломах гаметофитов и на спорофитах, у листостебельных мхов — на спорофитах), они часто вторично нефункционирующие. Как и у некоторых ныне живущих мхов, устьица исходно служили для облегчения газообмена и транспорта воды (путем транспирации). Чаще всего они расположены в плоскости эпидермы, но у некоторых видов погружены глубоко.
Ареалы распространения мхов в значительной мере совпадают по характеру с ареалами цветковых растений. Распространение некоторых видов, ныне встречающихся по всему земному шару (Marchantia polymorpha, Bryum argenteum, Funaria hygrometrica), возможно, обусловлено человеком. Мхи составляют в растительных сообществах с доминированием цветковых растений собственные сообщества подчиненного ранга (синузии, см. 14.3.4), нередко конкурируя с лишайниками. Собственные формации они образуют только в Арктике (тундре) и иногда также на верховых болотах, где продуктивность сплошного мохового покрова достигает своих максимальных значений: 200 — 900 г сухого вещества на 1 м2 в год; это соответствует урожаю сена на лугу среднего качества.
Наиболее распространены мхи как гигрофиты в областях с высокой влажностью: в лесах и на болотах. В целом печеночники более влаголюбивы, чем листостебельные мхи. Мхи чаще всего менее страдают от экстремальных условий, таких, как большая сухость, высокая температура и сильное излучение, и обходятся светом меньшей интенсивности, чем цветковые растения. Поэтому они проникают глубоко внутрь расселин и пещер, а также могут успешно расти на почве в лесах и в других затененных местах, особенно в форме войлока, дерновинок и корочек. Наибольшего разнообра зия форм, в том числе повислых, метровой длины (см. рис. 11.126, S), и эпифитов, мхи достигают в тропиках, особенно в туманных и горных лесах. У них развиты самые разные варианты приспособлений для капиллярного удерживания воды. Удивитель
но много видов мхов заселяет поверхность листьев других растений. Такие эпифиль-ные мхи в настоящее время рассматривают не как исключительно эпифиты, а иногда и как полупаразиты, которые своими ризоидами проникают под кутикулу несущего их листа и извлекают из него воду и соли.
Листостебельные мхи умеренной зоны (кроме эпифитов, а также мхов на почве и каменистом субстрате) часто проявляют ритмичность роста, четко связанную с временем года (см. рис. 11.126, В, Q). Реже они бывают однолетними, но чаще зеленые в течение всего года (вечнозеленые) и сохраняют, как облиственные печеночники, свои листья и зимой.
Ксерофитные мхи обладают высокой устойчивостью к высыханию, а также к высоким температурам и могут долгое время (Tortula muralis до 14 лет) находиться в воздушно-сухом состоянии, не теряя жизнеспособности. Споры, напротив, гораздо менее устойчивы. Находящиеся на открытом солнце и поэтому часто испытывающие сухость, мхи нередко растут низкими плотными дернинками и плотными подушками (см. рис. 11.126, Н). Зачастую они имеют серебристо-серый цвет, обусловленный длинными мертвыми окончаниями листьев. Такие «стекловидные волоски» (G), возможно, служат для защиты от избытка света и для снижения транспирации. Широкие однослойные края листьев у Polytrichum piliferum способны заворачиваться над многослойной средней частью, несущей ассимиляционные пластинки, и таким путем защищают ее от высыхания (как скручивающиеся листья, см. 4.3.3.2). Мхи могут выдерживать экстремальные температуры: их можно встретить, с одной стороны, на скалах нивального пояса высокогорий, а также в Арктике и Антарктике, с другой стороны — на открытых местах, где температура почвы может доходить до 70 °C. В эксперименте некоторые высушенные до воздушно-сухого состояния листостебельные мхи выдерживали нагревание в течение 30 мин до НО °C, оставаясь живыми.
Несколько видов вторично приспособились к жизни в воде (гцдрофиты), причем были утрачены как внешние, так и внутренние системы проведения воды.
304 I ГЛАВА 11 СИСТЕМАТИКА И ФИЛОГЕНИЯ
Fontinalis antipyretica и другие водные мхи очень чувствительны к продолжительному пребыванию вне воды. Живущие в богатых известью ручьях и водопадах мхи (например, Eucladium verticillatum, Bryum pseudotriquetrum, Cratoneuron commutatum) вносят наряду с различными видами цианобактерий (Oocardium) и Chara существенный вклад в образование известковых туфов. Извлекая из воды диоксид углерода, они превращают растворенный гидрокарбонат в малорастворимый карбонат кальция, выпадающий в осадок.
Немногие мхи (например, виды Portia) растут как галофиты по берегам морей и в засоленных местностях в глубине материков.
В полостях талломов Blasia (см. рис. 11.114, С) и Anthoceros (см. рис. 11.127, В) как симбионт живет синезеленая водоросль Nostoc. В ризоидах или клетках таллома либо стебля многих печеночников содержатся гифы гриба, однако в каждом отдельном случае бывает трудно решить, имеет ли здесь место паразитизм или же симбиоз по типу микоризы (см. 9.3.1). Бесхлорофилльный печеночник Cryptothallus mirabilis, обитающий под ковриками листостебельных мхов, питается как паразит на гифах гриба, тогда как ризоиды Marchantia и других мхов могут, наоборот, поражаться паразитическими грибами.
Обзор мхов. На связь бриофитов и рассматриваемых дальше сосудистых растений с предшествующими Chlorobionta указывает много общих признаков, в том числе одни и те же фотосинтетические пигменты, образование крахмала в пластидах, наличие целлюлозы в клеточных стенках, а также сходное строение подвижных репродуктивных клеток. Мхи, вероятно, произошли от предков, которые соответствовали стрептофитам из Chlorobionta. Из них мхи ближе к харовым (Charophyceae) по двухжгутиковым асимметричным сперматозоидам и ряду различных ультраструк-турных и биохимических признаков и, возможно, к KJebsormidiophyceae, чем к другим зеленым водорослям. Протонемы мхов, которые у отдельных форм могут непосредственно развивать гаметангии1 и у мно
1 Гаметангии развиваются на гаметофорах, хотя бы и сильноредуцированных. — Примеч. ред.
гих нитевидные, указывают на произошедший । гереход от нитчатого к тканевому таллому.
Эволюционное развитие мхов, вероятно, шло с рубежа силура и девона параллельно развитию первых наземных папоротникообразных. Неясны родственные связи антоцеротовых, которые, скорее всего, развивались параллельно печеночникам и листостебельным мхам. Действительно ли они наиболее близки (из мхов) к высшим наземным растениям, остается спорным. Исходные формы мхов, как, по-видимому, и сосудистые растения, унаследовали проводящие структуры для воды и ассимилятов от общих предков. Об этом говорят большая степень сходства в строении и функции гидроидов и лептои-дов мхов с трахеидами (сосудами) и ситовидными клетками папоротникообразных и наличие архегониев и устьиц в обеих группах. В ходе эволюции бриофитов гаметофит (зеленое растение мха) и, возможно, спорофит претерпели различные упрощения, но одновременно достигли биологического прогресса (прогрессивная редукция). Так, среди листостебельных мхов прямостоячие, высокорослые акрокарпные формы с хорошо развитой, напоминающей протостелу проводящей системой (Polytrichanae) рассматривают как исходные для лежачих, разветвленных плеврокарпных видов без проводящей системы и без жилки в листе. Наличие нефункционирующих устьиц можно трактовать также как результат регрессии. Прогрессивное развитие, напротив, претерпели клеточная сеть листьев (от паренхимной к прозенхимной) и структура устья коробочки (перистом).
Отдельные ископаемые мхи встречаются с верхнего девона; они лают мало информации для познания их происхождения. Sporogonites из девона, правда, можно рассматривать как сомнительное связующее звено между мхами и примыкающими к ним папоротникообразными (Psilophytopsida). Талломные илистостебельные печеночники, как и первые листостебельные мхи (Musettes), встречаются в карбоне Англии. Это свидетельствует о большом эволюционном возрасте бриофитов. Листостебельные мхи, которые можно отнести к Sphagnidae и Bryidae. были описаны из перми на территории России (Печора, Кузнецк). Листостебельные мхи нижнего карбона и перми имели жилки в листьях (в том числе и Protosphagnales), тогда как первые формы без жилок известны только из триаса и все более обычны в поздней юре. Большинство находок ископаемых мхов с возрастающей долей плеврокарпных известно из третичного периода. Они могут быть отнесены к современным родам.
11.2. Бактерии, грибы, растения | 305
Третий подотдел: папоротникообразные, птеридофиты (Pteridophytina)
Папоротникообразные, как и мхи, — это прежде всего растения, приспособившиеся к наземному образу жизни, однако их ткани и органы значительно более дифференцированы функционально.
В жизненном цикле современных папоротникообразных доминирует спорофит (рис. 11.128, В). Он представляет собой самостоятельное зеленое растение, и у плаунов, хвошей и настоящих папоротников дифференцирован на стебель, листья и корни. Вымершие первопапоротники, напротив, были устроены чаще всего из исключительно однотипных, лишенных листьев вильчатых осей (теломов; см. рис. 11.130, А, В). У них, как и у современных псилотовых. не было настоящих корней.
Гаплоидный гаметофит у папоротикообразных называют заростком (рис. 11.128, А). Он обычно живет лишь несколько недель, достигает не более нескольких сантиметров в диаметре и по облику часто напоминает простой талломный печеночник. В типичном случае — отклонения от правила очень разнообразны — он представляет собой простой зеленый таллом, прикрепляющийся к почве одноклеточными нитевидными ризоидами. На нем в большом числе возника
ют антеридии и архегонии. Оплодотворение, как и у мхов, возможно только в воде, т.е. при увлажнении заростков.
После оплодотворения из зиготы развивается диплоидное поколение — спорофит (рис. 11.128, Н: 3, 4), который у папоротников, однако, выглядит совсем иначе и гораздо более развит, чем у мхов. Пожалуй, лишь начальные стадии его развития протекают сходно со мхами. Заросток у большинства видов вскоре отмирает (при подавлении оплодотворения он может прожить в течение нескольких лет), однако проросток спорофита вырастает в самостоятельное многолетнее растение с корнями, стеблем и листьями: собственно папоротникообразное растение (см. рис. 11.128, Н: 4Б; 11.148, А; 11.150; 11.152, А). Спорофит папоротника, таким образом, — настоящее листостебельное растение.
Теломная теория. Органы, типичные для листостебельных растений, очевидно, возникли из лишенных листьев вильчатых теломов путем следующих основных процессов: перевершинивания, планации, срастания, редукции и изгибания (заворачивания).
Дифференциация и разделение функций исходно однородных теломов, из которых еще состояло тело первичных папоротников, на несущие основные и боковые оси должны были произойти в результате
Бокс 11.9. Сосудистые растения
Папоротникообразные и семенные растения имеют в стеблях, листьях и корнях настоящие проводящие пучки (а не только проводящие тяжи, см. ниже), которые служат для транспорта веществ (воды с минеральными веществами, продуктов ассимиляции). Поэтому их называют сосудистыми растениями (Тга-cheophyta) в отличие от мхов, у которых такие проводящие пучки отсутствуют'. Правда, у мхов с внутренней системой проведения воды
1 Сосудистые растения получили это название потому, что водопроводящие элементы у них трахеальные — трахеиды и сосуды, тогда как у мхов это гидроиды (или их нет совсем). — Примеч. ред.
уже встречаются проводящие тяжи, которые напоминают исходные типы проводящих пучков. Однако эти тяжи менее резко отграничены от окружающей ткани, а в клеточных стенках проводящих элементов не происходит отложения лигнина. Соответственно у проводящих воду элементов мхов нет утолщения стенок по типу трахеид и сосудов сосудистых растений. Совокупность проводящих пучков в стебле сосудистых растений называют стелой. В ходе все большего приспособления сосудистых растений к наземной жизни стелы становились все более развитыми в отношении проведения веществ и придания прочности стебелю (прото-, актино-, плекто-, поли-, си-фоно-, эвстела, атактостела; см. Стелярная теория, бокс 4.3).
306 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Рис. 11.128. Pteridophytina, Pteridopsida (А, В — по Н. Schenck; С—Е — по R.Sadebeck; F—Н — по R. Harder):
А, В — Dryopteris filix-mas: А — заросток (снизу) с архегониями (аг), антеридиями (ап) и ризоидами (г), В — заросток с молодым спорофитом (5х); С—G — развитие спорангия папоротника (С—Е — Asplenium, ЗООх; F, G — Polypodium, 200х): С — первые деления возникающего из одной эпидермальной клетки зачатка, D — разделение на поверхностный слой — стенку спорангия (wa) и центральную клетку (as) — археспорий, которая уже отделила клетку тапетума (t), Е — археспорий, дифференцировался на клетки тапетума и спорогенную ткань (sg), F — у клеток кольца стенки спорангия (wa) утолщены стенки, клетки тапетума лизировались, материнские клетки спор (sm) образовали тетрады спор (s), G — зрелый спорангий со спорами; Н — схема развития папоротника: светлые линии — гаплоидная фаза, темные линии — диплоидная фаза, 1 — спора, 2 — заросток с £и о" гаметангиями, 3 — заросток с молодым спорофитом, 4 — спорофит (сильно уменьшен) с сорусами спорангиев, 5 — незрелый одиночный спорангий (сильно увеличен) из соруса, 6 — зрелый спорангий с тетрадами спор; 7 — споры (s); Ь — первый лист; w — корень; 1 —5 на С, D — последовательно образующиеся стенки клеток; G — гаметофит; S — спорофит; R! — редукционное деление
11.2. Бактерии, грибы, растения | 3Q7
Рис. 11.129. Схема пяти элементарных процессов, которые согласно теломной теории привели к появлению побега (А—К — по G.Smith; L—N — по W.Zimmermann, с изменениями):
А, В — перевершинивание; С, D — планация; Е—G — редукция; Н—К — срастание (также D, N); L — N — изгибание
перевершинивания (рис. 11.129, А, В). При этом лидирующая основная ось растет более интенсивно, чем обгоняемые ею в росте сестринские оси, которые становятся ее боковыми придатками (В) и все больше берут на себя функцию ассимиляции. В ходе планации боковые оси стали ветвиться в горизонтальной плоскости (С, D). Путем конгенитального срастания эти теломы, расположенные уже в одной плоскости, превратились в плоские листовидные боковые органы (D, G). Так, по-видимому, появились крупные сложно-расчлененные листья с первоначально дихотомическим жилкованием (макро-, или мегафиллы). Располагающиеся в трехмерном объеме талломы могут срастаться между собой, в результате чего возникает более толстая паренхиматическая ось, в которой теперь проходит уже не единственный проводящий пучок (протостела) (рис. 11.130, С), а два или более (рис. 11.129, Н—К). Благодаря этому прочность оси значительно повышается. Путем редукции (F, G), как можно полагать, могли возникнуть мелкие, более или менее шиловидные листья с одной жилкой (микро-филлы). Однако не вполне ясно, не имеем
ли мы здесь дело с выростами осей, т. е. органами «sui generis», которые не могут быть выведены из теломов. Процесс изгибания можно проследить, например, на несущих спорангии осях хвощевидных (L, М) (см. рис. 11.141, F).
Развитие зародыша. У современных папоротникообразных вскоре после первых клеточных делений оплодотворенной яйцеклетки, как правило, возникают кроме гаустории (ножки) апексы корня, стебля1 и листа, которые у растущего и еще связанного с заростком зародыша (см. рис. 11.128, В; 11.155) развиваются соответственно в первичный корень, первичный стебель1 2 и первый лист (семядолю). Наличие корня характерно для большинства папоротникообразных. Противоположный «полюсу побега» конец оси проростка можно было бы назвать «корневым полюсом»; однако только у семенных растений из него развивается главный корень (см. рис. 3.1), тогда как у папоротникообразных первый корень возникает сбоку на оси как эндогенное
1 Правильно: апекс побега. — Примеч. ред.
2 Первичный побег. — Примеч. ред.
308 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Рис. 11.130. Psilophytopsida (А, В — по Edwards; С— Е — по R. Kidston и W.H. Lang; F, G — по W. Remy et al.; Н — по R.Krausel и H.Weyland):
А—Е — Rhyniales: А — Rhynia gwynne-vaughanii, реконструкция (1 /4х), В — Aglaophyton (Rhynia) major, реконструкция (1 /5х), С—Е — Rhynia: С — поперечный срез телома, показывающий протостелу (50х), D — спорангий, продольный шлиф (2х), Е — тетрада спор (100х); F—Н — Zosterophyllales: Zostero-phyllum rhenanum: F — гаметофит (= Sciadophyton), G, H — спорофит: G — молодой, связанный с гаметофитом, Н — взрослый со спорангиями; а — внешняя кора; е — эпидерма; i — внутренняя кора; р — флоэма; х — ксилема
придаточное образование* 1 (см. рис. 11.155, В: w). Проросток папоротникообразных, таким образом, не биполярный, как у семенных растений, а униполярный2. Зародышевый корень (см. рис. 11.128, В: w) быстро
1 Главный корень семенных растений образуется под подвеском и, строго говоря, эндогенный. — Примеч. ред.
1 Зародыш подавляющего большинства папоротникообразных такой же биполярный, как и у семенных растений, только искривлен соответственно своему боковому положению относительно питающего его заростка. — Примеч. ред.
отмирает, но в дальнейшем возникают многочисленные придаточные корни на побегах (первичная гоморизия, см. 4.4.1).
Три основных органа у большинства папоротникообразных нарастают благодаря деятельности апикальных инициалей (см. 3.1.1.1; рис. 3.2, А; 3.6, А). Обильно олиствен-ный побег ветвится вильчато или латераль-но (но никогда из пазух листьев!). Корни имеют корневой чехлик (см. рис. 3.6, А); боковые корни возникают не из перицикла, а из самого внутреннего слоя первичной коры (см. 4.4.2.2). Листья (по крайней мере
11.2. Бактерии, грибы, растения | 3Q9
у наиболее высокоразвитых папоротников) анатомически в основном подобны листьям семенных растений. Эпидерма надземных частей, как правило, покрыта кутикулой (важная предпосылка для наземного образа жизни на некотором расстоянии от уровня земли!) и имеет устьица (см. 3.2.2.1), однако эпидермальные клетки чаще всего еще содержат хлоропласты. В стеблях, корнях и листьях проходят хорошо развитые, состоящие из флоэмы и ксилемы проводящие пучки, которые в типичном виде в эволюции впервые появляются у папоротникообразных. Водопроводящие элементы у них — одревесневшие трахеиды; очень редко (например, у Pteridium) встречаются также сосуды (см. рис. 11.149: t). Особых механических элементов в проводящих пучках еще нет, тем не менее проводящие воду элементы часто укреплены кольчатыми или иными утолщениями своей оболочки (см. рис. 11.149). Преобладают концентрические проводящие пучки (и притом с ксилемой, окруженной флоэмой) в числе одного или нескольких, однако встречаются и другие типы пучков. У папоротникообразных можно проследить общий ряд эволюции проводящих пучков (см. бокс 4.2, рис. А). Одревеснение трахеид повышает их пропускную способность, а заодно и укрепляет побег. Поэтому папоротникообразные в противоположность мхам смогли развиться в обильно расчлененные, иногда даже древовидные растения. Клеточные стенки механической ткани, расположенной вне проводящих пучков, как правило, также содержат лигнин. Наличие корней обеспечивает интенсивное поглощение воды и позволяет образовать более крупные листья, которые производят продукты ассимиляции. Проведение питательных веществ осуществляют вытянутые продольно ситовидные клетки (см. 3.2.4.1). Вторичное утолщение благодаря деятельности камбия у современных папоротникообразных свойственно лишь отдельным представителям, да и то выражено слабо, однако оно было характерно для некоторых вымерших групп папоротникообразных.
Спорангии с мейоспорами (см. рис. 11.128, G, Н: 6) образуются на листьях, лишь в самых примитивных классах — на недиффе
ренцированных теломах. Строение спорангиев разное. Листья, несущие спорангии, называют спорофиллами. Они часто устроены более просто, чем ассимилирующие листья (трофофиллы), и располагаются по нескольку в стробиле: такие собрания спорофиллов можно называть «цветками»1. Для лучшего рассеивания спор спорофиллы обычно довольно высоко приподняты над субстратом.
Спорангии содержат археспорий со спорогенной тканью (см. рис. 11.128, Н: 5, Е: sg); ее клетки округляются, отделяются друг от друга и становятся материнскими клетками спор (чаще всего 16), каждая из которых после мейоза дает 4 гаплоидные мейоспоры, часто расположенные в виде тетраэдра.
Вокруг спорогенной ткани находятся, часто в несколько слоев, клетки, примыкающие к стенке спорангия. Они служат для питания спор и в совокупности представляют тапетум (Е: 1). Клетки секреторного тапетума выделяют содержимое через стенки. Стенки клеток плазмодиального тапетума лизируются, и высвободившиеся протопласты объединяются в периплазмодий. Последний внедряется затем между молодыми спорами из распадающихся 'гетрад, питает их, участвует в формировании стенки спор (периспория) и при этом расходуется (F, G).
Споры. Оболочка споры состоит из внутреннего эндоспория и стойкого внешнего экзоспория, на котором откладывается периспорий в виде орнамента различного облика. Споры, окраска которых бывает от буроватой до желтой, почти всегда лишены хлорофилла.
У большинства папоротникообразных (более примитивных) все споры однотипные. При прорастании из них развиваются заростки, на которых чаще всего возникают как антеридии, так и архегонии. Однако у более эволюционно продвинутых форм заростки бывают двудомными1 2. Это разделение полов в некоторых группах папоротникообразных привело к по
1 Так уже не делают с рубежа XIX—XX вв. — Примеч. ред.
2 Точнее, раздельнополыми. — Примеч. ред.
310 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
явлению двух форм мейоспор: богатых запасными веществами мегаспор (= макроспор), которые возникают в мегаспорангиях (= макроспорангиях) и прорастают в относительно крупные женские заростки, и микроспор, которые образуются в микроспорангиях и дают более мелкие мужские заростки (см. рис. 11.137). По этому признаку следует различать равноспоровые (изоспорические) и разноспоровые (гетероспорические) группы — различие, которое неоднократно независимо возникало в разных эволюционных линиях папоротникообразных.
1. Класс: псилофиты (Psilophytopsida)
Вымершие к настоящему времени Psilophytopsida — исходная группа папоротникообразных. Их вегетативное тело было построено из теломов, которые у представителей примитивных семейств были голые, а у продвинутых покрыты выростами. Спорангии располагались на концах дихотомически разветвленных осей или сбоку на главной или боковых ветвях. Все роды были изоспорическими. Настоящих корней еще не было; вместо них имелись ризомо-иды, покрытые ризоидами. В осях были развиты только простые прото- или актиностелы.
Псилофиты были древнейшими наземными растениями, имевшими проводящие пучки и устьица. Они появились на рубеже силура—девона (т. е. примерно 400 млн лет назад), быстро достигли значительного разнообразия форм и вымерли уже к началу верхнего девона1 1.
1. Порядок: Rhyniales. Риниевые (морфологически наиболее примитивные представители) имели вегетативное тело в виде голых вильчатых теломов, часто еще с терминальными спорангиями.
К роду Cooksonia, встречавшемуся с верхнего силура до нижнего девона, относят древнейших из найденных к настоящему времени наземных растений. Их дихотомически разветвленные теломы высотой около 10 см с проходившей внутри протостелой несли спорангии, ширина которых превосходила длину.
1 Отдельные представители существовали еще в конце верхнего девона. — Примеч. ред.
Rhynia (включая Aglaophyton' ранее относимый к Rhynia) — типовой род семейства Rhynia-сеае, представленный в нижнем девоне Шотландии двумя видами (см. рис. 11.130. А, В) — достигал 1 —2 м в высоту2. Спорофит, по крайней мере, у одного вида (В) отрастал от ползучих осей3 («корневищ») — горизонтальных до дуговидно изогнутых, местами покрытых ризоидами без поперечных перегородок. Вертикальные, округлые в сечении, вильчато разветвленные теломы были безлистными.
Оси имели кутикулу и устьица еще весьма простого строения (см. 3.2.2.1) и, очевидно, были органами ассимиляции. Таким образом, виды Rhynia были надземными растениями и образовывали заросли типа тростниковых. Проводящий пучок состоял из гидроидов у Aglaophyton (Rhynia) major и Homeophyton или из трахеид с очень простыми кольчатыми и. спиральными утолщениями стенок у Rhynia gwynne-i’aughanii. Протостела (см. бокс 4.3, рис. А; рис. 11.130, С) у некоторых видов уже содержала метаксилему; типичных ситовидных клеток с ситовидными полями во внешней ткани пучка — флоэме еше не было4. Не было и вторичного утолщения. Относительно крупные спорангии, имевшие форму от цилиндрической до булавовидной, находились на концах или по бокам осей5 6. Они имели стенку из нескольких слоев клеток и вскрывались продольной щелью. В них формировалось множество тетрад изоспор (рис. 11.130, D, Е).
У Rhynia gwynne-vaughanii вследствие пере-вершинивания были боковые ответвления; пе-ревершиниванию отчасти подвергались и спорангии (см. рис. 11.130, А).
У внешне сходного с родом Rhynia рода Ног-neophytotf продолговатые спорангии, располагавшиеся вместе плотными группами по 2—4, напоминают по своему строению спорогонии Sphagnum: спорогенная ткань, как колокол, облегала колонку, состоявшую из продольно вытянутых клеток. Спорангии вскрывались верхушечной порой.
1 Aglaophyton настолько своеобразен, что не может быть включен не только в Rhynia, но и в порядок Rhyniales. Возможно, что его вообще нельзя включать в Tracheophyta. — Примеч. ред.
2 Это сильно завышенная оценка. — Примеч. ред.
3 Эти оси были частью спорофита. — Примеч. ред.
4 Флоэма не сохраняется в ископаемом состоянии, поэтому неправомерно отрицать ее наличие у Rhyniaceae. — Примеч. ред.
5 У Rhyniaceae спорангии всегда были терминальными. — Примеч. ред.
6 Homeophyton существенно отличается как от Rhynia, так и от Sphagnum. — Примеч. ред.
11.2. Бактерии, грибы, растения | 31 “I
Из-за отсутствия трахеид в последнее время Aglaophyton (Rhynia) major и Horneophyton больше не рассматривают как сосудистые растения.
2. Порядок: Zosterophyllales (-сеае)1. Распространенные в нижнем девоне по всему земному шару виды этого порядка также имели вегетативное тело в виде голых вильчатых ветвей, но их латеральные спорангии, вскрывавшиеся пре-формированной поперечной щелью, были чаще всего сгруппированы в колосовидные собрания (см. рис. 11.130, Н). Их рассматривают как предковую группу плаунов, с которыми часто и объединяют.
Удалось показать, что гаметофит некоторых родов (например, Zosterophyllum) представлял собой маленькое, звездчато разветвленное растение (см. рис. 11.130, F), которое на дуговидно изгибающихся вверх ножках образовывало зонтиковидные гаметангиофоры с архегониями в центре и антеридиями на периферии. Возникавший в результате оплодотворения молодой спорофит отделялся от гаметангиофора и развивался затем в самостоятельное растение (см. рис. 11.130, G, Н). Соответственно неправильно рассматривать «корневища» Rhyniaceae как гаметофиты, которые на протяжении всей жизни связаны со спорофитом подобно гаметофитам мхов.
Таким образом, эти древние папоротникообразные имели гетероморфную смену поколений, однако эти поколения были развиты еще примерно одинаково. Отсюда эволюция могла идти как к мхам (преобладание гаметофита), так и к более продвинутым папоротникообразным (преобладание спорофита)1 2.
Как и у некоторых водорослей (например, Halycystis-Derbesia из Chlorophyta), разные поколения зостерофилловых также сначала описывали под разными родовыми названиями, пока только в последнее время не удалось установить их принадлежность к одному и тому же жизненному циклу. Так, название Taeniocrada соответствует стерильному, a Sciadophyton — фертильному гаметофиту, а спорофитное поколение носит название Zosterophyllum (см. рис. 11.130, F). Lyonophyton с ползучими, круглыми в сечении
1 Zosterophyllales вместе с плауновидными, по-видимому, представляют эволюционную линию, параллельную всем остальным сосудистым растениям, и нет оснований включать его в Psi-lophytopsida. — Примеч. ред.
2 Это явный анахронизм. Мохообразные, по современным данным, к водорослям ближе, чем сосудистые растения. Поэтому их следует считать либо сестринской группой всех сосудистых растений, либо даже предками сосудистых растений. — Примеч. ред.
Рис. 11.131. Psilophytopsida (А — по Hueber; В, С — по R. Kidston и W. Н. Lang):
А — Psilophyton, спороносная ветвь (3/4х); В — Lycopodiopsida, Asteroxylon mackiei, поперечный шлиф через актиностелу: темным показана ксилема, светлым флоэма (10х); С — то же, реконструкция (1/Зх)
осями и проводящим пучком, содержащим гидроиды. а также с вертикальными, блюдцевидно расширенными вверху гаметангиофорами предположительно представлял собой гаметофитное поколение Aglaophyton major (см. рис. 11.130, В). У него даже удалось выявить структуру антери-диев со сперматогенными клетками.
3. Порядок: Trimerophytales (-сеае). Виды этого порядка, более высокоорганизованны по сравнению с представителями предыдущего порядка, встречались от нижнего до среднего девона. На распростертых (симподиально-ветвящихся) основных осях здесь образовывались дихотомически или тройчатоветвяшиеся боковые оси с расположенными на их концах спорангиями, уже собранными в группы. Этот порядок рассматривают как предковую группу «Progymnospermae» (см. рис. 11.288).
Dawsonites в противоположность Trimerophy-ton имел обращенные назад спорангии на искривленных осях. Psilophyton ornatum нес на осях мелкие выросты (рис. 11.131, А) и представляет собой, таким образом, переход к Lycopodiopsida1.
1 Как структурные особенности, так и время существования Psilophyton исключают его из числа предков Lycopodiopsida. Авторы изложили давно устаревшие представления о происхождении плауновидных. — Примеч. ред.
312 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Psilophytopsida, будучи первыми наземными растениями, представляют исходную группу для других, филогенетически более продвинутых папоротникообразных1, а возможно, даже и определенных голосеменных.
2. Класс: плауны (Lycopodiopsida)
Спорофит плауновидных, часто вильчато разветвленный, имеет простые, нерасчле-ненные, мелкие листья (= микрофиллия), обычно расположенные по спирали. Спорангии находятся, кроме немногих ископаемых форм, поодиночке адаксиально или у основания листьев (спорофиллов); чаще всего они располагаются в верхушечных стробилах. Следуя теломной теории, можно полагать, что расположение спорангиев на листьях возникло согласно схеме, приведенной на рис. 11.133. Наряду с изоспорией широко распространена гетероспория. Сперматозоиды у немногих представителей многожгутиковые (Jsoetes), но чаще всего двухжгутиковые — этим плауновидные отличаются от всех остальных папоротникообразных. Предполагают, что Lycopodiopsida происходят от просто устроенных пси-лофитов, имевших боковые выросты на осях и верхушечные стробилы спорофиллов (Zo-sterophyllales)1 2.
1. Порядок: Asteroxylales (-сеае). Ветви были покрыты выростами в виде игл или шипов, рыхло либо более или менее плотно расположенных, они придавали растению сходство с современными плаунами.
В стеблях Asteroxylon mackiei, встречающегося совместно с Rhynia в нижнем девоне Шотландии, проходила стела, имевшая на срезе звездчатую форму (актиностела — см. бокс 4.3, рис. А; рис. 11.131, В). Лучи звезды представляли собой ответвляющиеся боковые тяжи, шедшие к месту прикрепления боковых выростов, в которых, однако, проводящего пучка не было. Ксилема стелы состояла из кольчатых и спиральных трахеид. Спорангии сидели непосредственно на стебле или в пазухах его выростов.
2. Порядок: Frotolepidodendralcs. Вымершие к настоящему времени представители этого по-
1 См. предыдущее примечание. — Примеч. ред.
2 У Zosterophyllales не было спорофиллов. — Примеч. ред.
рядка развивали спорангии уже на листьях или в непосредственной близости от них. Несмотря на рыхлое расположение листьев, по облику они напоминают современные плауны (Lycopodiales). Представители обоих важнейших семейств — Dre-panophycaceae1 и Protolepidodendraceae — найдены в нижне- и среднедевонских отложениях. Родственные связи Drepanophycus (рис. 11.132, А) неясны (связь с Zosterophyllaceae); спорангии располагались не на листьях, а между ними на коротких ножках, снабженных проводящим пучком. Листья Protolepidodendron (В) были на конце еше вильчато разделенными; спорангии находились на верхней стороне листовидных образований (спорофиллов; отчасти также вильчато разветвленных боковых осей).
3. Порядок: Lycopodiales. Представителей этого преимущественно современного порядка чаще объединяют в единственное семейство (Lycopodiaceae). Это травянистые вечнозеленые растения (400 видов; 9 из них встречаются в Германии), густо покрытые более или менее игловидными листьями. Вторичное утолщение стеблей, как и у выше приведенных порядков, отсутствует.
У плауна (Lycopodium) (см. рис. 11.134) вильчато ветвящийся побег из-за перевер-шинивания одного из разветвлений кажется моноподиальным2 (см. 4.2.5). Побеги длинные, ползучие2. От нижней стороны побегов отходят дихотомически разветвленные корни, имеющие несколько инициальных клеток в апексе. Листья, мелкие, шиловидные, по существу расположенные по спирали2, имеют неветвящуюся среднюю жилку, а в остальном сходны с листьями Asteroxylales.
Мезофилл L. clavatum однородный; лишь у немногих видов возможно различить палисадную и 1убчатую паренхиму. Эпидермальные клетки листа без хлоропластов. Как и всегда при дихотомии, ветвление не связано с листьями.
Проводящая система стебля представляет собой производную от актиностелы обильно расчлененную плектостелу (см. бокс 4.3, рис. А) с ситовидными клетками во флоэме, несущими ситовидные поля на
1 Drepanophycaceae, по-видимому, занимают промежуточное положение между Asteroxylales и Lycopodiales, и нет оснований включать их в Protolepidodendrales. — Примеч. ред.
2 Только у некоторых видов. — Примеч. ред.
11.2. Бактерии, грибы, растения | 313
продольных стенках, но еще не имеющими ситовидных пластинок. Плектостела окружена снаружи влагалищем из неодревесневших клеток, внешний слой которых содержит крахмал; далее следует одно- или двухслойная эндодерма с лигнином в тонких клеточных стенках. Как и у всех папоротникообразных, у плаунов эндодерма является самым внутренним слоем первичной коры. Внешняя кора состоит из сильно одревесневших склеренхимных клеток (рис. 11.134, L).
Часть ветвей обладает отрицательным геотропизмом. Их спорофиллы часто располагаются в плотных колосовидных стробилах над менее олиственным участком побега (рис. 11.134, G); при их образовании расходуется апикальная меристема, так что стробил завершает побег. Спорофиллы (Н) имеют вид широких чешуек и несут у основания на верхней стороне по одному крупному уплощенному почковидному спорангию, из которого высеиваются многочисленные мейоспоры — все одинакового размера (изоспоры) (J, К). С краев спорофиллов свисают вниз кожистые лопасти, каждая из которых как «индузий» прикрывает ближайший ниже расположенный спорангий.
Стенка спорангия состоит из нескольких слоев клеток, к которым изнутри примыкает секреторный тампетум. Спорангий вскрывается двумя створками, щель проходит по его вершине по линии, различимой по особому строению клеток. Споры остаются до созревания объединенными в тетрады; их многослойный экзоспорий покрыт сетью из выступающих ребрышек1 (см. рис. 11.134, J, К). Споры прорастают в природе только через 6—7 лет и за счет находящегося в них запаса питательных веществ образуют сначала 5-клеточный предросток (рис. 11.135, А), который после периода покоя развивается дальше только тогда, когда в его нижние клетки внедрятся гифы гриба, образуя подобие микоризы (В: р).
Заростки (см. рис. 11.134, Е, F; 11.135) живут в земле и представляют собой гетеротрофные беловатые клубневидные тель-
1 Это не особый покров, а скульптура самого экзоспория. — Примеч. ред.
Рис. 11.132. Lycopodiopsida, Protolepidoden-drales (А — по Н. J. Schweitzer В — по R. Krausel и H.Weyland):
А — Drepanophycus spinaeformis, нижний девон (1/4х); В — Protolepidodendron scharyanum, средний девон (1/4х)
Рис. 11.133. Переход (A—D) от верхушечного расположения спорангиев у Psilophytopsida (А) к эпифильному их расположению у Lycopodiopsida (D) (по W. Zimmermann)
ца величиной до 2 см с бугорчатыми лопастями и одноклеточными ризоидами, служащими для поглощения воды. В их питании, несомненно, большую роль играют микоризные грибы, живущие в клетках их периферических клеточных слоев (рис. 11.135, В, С). В естественных условиях половая зрелость наступает только через 12— 15 лет, а общая продолжительность жизни заростков может составлять примерно 20 лет. Однако в искусственной чистой без-бактериальной культуре все развитие за-
314 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Рис. 11.134. Lycopodiopsida, Lycopodiales, Lycopodium clavatum (A—F — по H. Bruchmann; G, H — по H. Schenck):
А — антеридий, еще не вскрывшийся, продольный разрез (75х); В — сперматозоиды (400х); С— молодой, еще не вскрывшийся архегонии; D — готовый к оплодотворению, вскрывшийся архегоний (75х); Е — взрослый заросток (2х); F — заросток с молодым спорофитом (3/4х); G — спорофилл в колосовидных стробилах; Н — спорофилл с вскрывшимся спорангием (8х); J, К — споры в двух позициях (400х); L — поперечный срез стебля (100х); b — основание листа с полостью h; е — эпидермис; еп — эндодерма; h — полость; р — флоэма; г — первичная кора; s — крахмалоносное влагалище; х — ксилема
нимает несколько месяцев. У некоторых видов заростки выступаю! верхней зеленеющей на свету частью над поверхностью земли. Заростки однодомные и несут многочисленные половые органы, преимущественно в верхней части (см. рис. 11.134, A—D; 11.135, С: a, ag). Антеридии (а) несколько погружены в ткань. Они многоклеточные, и каждая клетка, за исключением клеток стенки, дает начало одному овальному сперматозоиду, несущему у своей вершины только два жгутика (см. рис. 11.134, В). Архегонии (см. рис. 11.134, С, D; 11.135, С: ag), также погруженные, часто имеют многочисленные шейковые канальцевые клетки (до 20, однако может происходить их редукция вплоть до одной-един-ственной); самые верхние клетки стенки
при вскрывании сбрасываются. Из оплодотворенной яйцеклетки после ряда клеточных делений возникает зародыш, подвесок (рис. 11.135, D, Е: е) которого проталкивает его в ткань заростка. Развитие гаустории, всасывающей из заростка питательные вещества, и первых листьев (Ьа), остающихся чешуевидными, показано на рис. 11.135, Е. Первый корень возникает как придаточный на побеге.
У Lycopodium спорофиллы собраны в стробилы, завершающие короткие боковые ветви; у Huperzio на вертикальных, вильчато разветвленных побегах в течение года последовательно образуются трофофиллы и спорофиллы. Dipha-sium имеет стробилы из спорофиллов, как у Lycopodium, но побеги у него плоские, дорсовент-ральные, с чешуевидными листьями.
11.2. Бактерии, грибы, растения I 315
Виды Lycopodites из верхнего девона были уже очень сходны с современными представителями этого семейства. Таким образом, облик плауна сохранился неизменным на протяжении более 300 млн лет.
Если пока мы имели дело с изоспорией, то последующие порядки эволюционировали в сторону гетероспории. В пазухах листьев их представителей находится небольшой вырост — язычок (ligula) (рис. 11.136, С).
4. Порядок: Selaginellales. По облику травянистые Selaginella, или плаунки, несколько напоминают мхи1. Однако по строению своих генеративных органов и анатомическим признакам они однозначно являются папоротникообразными — соответственно настоящими сосудистыми растениями. В средних широтах они представлены несколькими видами, а в тропиках, напротив, обитает более 700 видов.
Облик. У видов Selaginella имеются отчасти лежачие, отчасти вертикальные, обильно вильчато разветвленные побеги; некоторые образуют дерновины, другие имеют лазающие побеги, поднимающиеся по кустарникам на несколько метров.
Стебель несет мелкие чешуевидные листья. Они расположены спирально или чаще всего в 4 ряда на дорсовентральных побегах: 2 ряда мелких так называемых спинных листьев и 2 ряда более крупных брюшных листьев (11.136, А; см. рис. 4.67, В; ани-зофиллия). Листья имеют только одну не-разветвленную среднюю жилку и лишь изредка наряду с губчатой паренхимой обладают и палисадной паренхимой; у некоторых видов клетки мезофилла содержат только по одному крупному бокаловидному хлоропласту. Листья селагинелл несут на верхней стороне у основания возникающую из эпидермы мелкую кожистую бесхлорофилльную чешуйку — язычок (см. рис. 11.136, С), который служит органом поглощения воды: он очень быстро всасывает дождевую воду, стекающую по стеблю, и у некоторых видов связан трахеидами с проводящим пучком.
В местах ветвления побегов у многих видов экзогенно возникают цилиндриче
1 Отсюда их немецкое название Moosfame (дословно «мхи-папоротники»). — Примеч. пер.
ские, вытянутые, растущие вниз, вильчато разветвленные бесцветные и безлистные оси — ризофоры (рис. 11.136, A: w), на свободных концах которых эндогенно возникают пучки корней. В определенных усло-
Рис. 11.135. Lycopodiales (по H.Bruchmann):
А, В — Lycopodium, L. annotinum, развитие заростка: А — 5-клеточный бесцветный предросток с оболочкой споры (sh), ризоидальной клеткой (г), базальной клеткой (Ь), апикальной клеткой (s) (580х); В — молодой заросток, в нижних клетках которого обитает эндофитный гриб (р); апикальная клетка разделилась на три клетки апикальной меристемы (s,, s2, s3) (470х); С — Е — Diphasium, D. complanatum: С — зрелый заросток с антеридиями (а), архегониями (ад) и клетками, содержащими гифы гриба (показаны интенсивно темным) (24х); D, Е — развитие зародыша: D — зародыш, претерпевший первые деления клеток; базальная стенка 1 отделяет зачаток подвеска (е) от зачатка тела зародыша; перпендикулярные ей стенки 2 и 3 (последняя в плоскости среза), а также параллельная ей стенка 4 дают расположенные в два этажа группы из 4 клеток, из которых клетки, расположенные между стенками 1 и 4, образуют гаусторию, а самая нижняя — побег, Е — средняя стадия развития (112х); Ьа — верхушка первого листа; ss — верхушка побега
316 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Рис. 11.136. Lycopodiopsida, Selaginellales, Selaginella (А — по С. Luerssen; В — по G.W. Bischoff; С — по Harvey-Gibson; D — по F.Oberwinkler):
А — S. helvetica, растение со стробилом (s) спорофиллов (2х); В — S. kraussiana, мегаспора с проростком растения (10х); С — S. lyallii, продольный срез через основание листа (b) (250х); D — S. se-laginoides, продольный разрез через стробил с мегаспорангиями (внизу) и микроспорангиями (вверху); У срезанных в медиальной плоскости спорофиллов над местом прикрепления спорангия виден язычок (I) (6х); е — эпидерма стебля; t— трахеиды; w — ризофор
виях ризофоры могут образовывать листья, как типичные побеги. Строение стелы варьирует от центральной протостелы и дистелы до сифоностелы. Вторичного утолщения нет. Крайне редко встречаются сосуды с лестничными утолщениями стенок. Эндодерма стебля (например, у A kraussiana) состоит из трубковидных отделенных друг от друга клеток с поясками Каспари (трабекул).
Selaginellaceae (единственное семейство) отличается гетероспорией и очень силыюредуцированными заростками.
Расположенные на концах побегов стробилы спорофиллов (рис. 11.136, A, D) бывают простыми или разветвленными, четырехгранными с радиальной симметрией или дорсовентральными. Каждый споро
филл несет только один спорангии, отходящий из его пазухи1. Спорангии содержат крупные мегаспоры или мелкие микроспоры: они развиваются всегда в разных спорангиях — соответственно в мега- или микроспорангиях (рис. 11.137, А, В). Однако оба типа спорангиев встречаются в одном и том же стробиле (рис. 11.136, D). Таким образом, определение пола осуществляется уже в диплофазе модификационным путем (дипломодификационное определение пола). В мегаспорангиях все заложившиеся материнские клетки спор отмирают, кроме одной, которая после редукционного деления дает начало 4 круп
1 Спорангий располагается на самом спорофилле, а не в его пазухе. — Примеч. ред.
11.2. Бактерии, грибы, растения | 3-17
ным мегаспорам ($) с бугристой поверхностью оболочки (рис. 11.137, А). В микроспорангиях возникают — также после редукционного деления — многочисленные мелкие микроспоры (о") (рис. 11.137, В).
Стенка спорангия состоит из 3 слоев клеток (средний в зрелом спорангии очень узкий). Внутренний слой — тапетум (рис. 11.137, A: t) питает споры, не разрушаясь при этом (секреторный та
петум). Спорангии раскрываются посредством когезионного механизма по заранее детерминированной линии, причем споры активно выбрасываются.
Гаметофит. Микроспоры начинают прорастать уже внутри спорангия. Спора при этом делится сначала на маленькую линзовидную клетку (рис. 11.137, С: р) и крупную клетку, которая последовательно де
Рис. 11.137. Lycopodiopsida, Selaginellales (А, В — по J. Sachs, с изменениями; С— Н — по W. С. Ве-lajeff; J, К — по H.Bruchmann; L — по R. Harder):
А, В — Selaginella inaequalifolia: А — мегаспорангий с одной-единственной тетрадой мегаспор и недоразвитыми материнскими клетками спор (70х), В — микроспорангий с тетрадами микроспор; С — G — S. stolonifera (640х); прорастание микроспоры, последовательные стадии; проталлиальную клетку (р) можно рассматривать как ризоидальную, С, D, F — сбоку, Е — со спинной стороны; на G протал-лиальная клетка не видна, клетки стенки расплывшиеся; Н — S. cuspidate, сперматозоиды (780х); J, К — S. martensii: J — проросшая мегаспора, заросток с 3 ризоидальными буграми и несколькими архегониями с поверхности (112х); К— продольный разрез: 2 архегония с развивающимися зародышами, подвесок (е), гаустория (h), ризофор (w), зародышевые листья (к) с язычком (150х); L — схема развития Selaginella; светлые линии: гаплоидная фаза; темные линии или затушеванное черным — диплоидная фаза; 1 — мейоспоры; 2 — то же, после образования заростка; 3 — мегаспора и заросток с проросшим спорофитом; 4 — спорофит (S); 5 — спорангии; 6 — мейоспоры после высеивания из спорангиев; а — клетки стенки антеридия; s — сперматогенная клетка; 1 — тапетум; G — гаметофит; R! — редукционное деление
318 I ГЛАВА 11, СИСТЕМАТИКА И ФИЛОГЕНИЯ
лится на 8 стерильных покровных клеток и 2 или 4 центральные клетки (С). Эти клетки представляют собой мужской заросток, который так и остается внутри споры. Только маленькую линзовидную клетку можно рассматривать как вегетативную, утратившую свою функцию ризоидальную клетку; остальные клетки считают одним-единствен-ным антеридием, из которых центральная клетка, окруженная покровными клетками (рис. 11.137, С — F: а), после дальнейших делений дает начало большому числу округляющихся сперматид (D—F: s). Затем покровные клетки вместе со своими стенками расплываются в слизистую массу, в которую погружена группа сперматид (F, G). Маленькая проталлиальная клетка (F: р), наоборот, сохраняется. Мужской заросток до этой стадии еще целиком находится в оболочке микроспоры; наконец, она разрывается, и 6" гаметы, возникшие из сперматид, высвобождаются в виде слабоизогнутых, булавовидных сперматозоидов, несущих на переднем конце два длинных жгутика (Н).
Не столь сильноредуцированный женский заросток образуется в мегаспоре (J). Его развитие несколько неодинаковое у разных видов. Ядро споры делится на много дочерних ядер. Они располагаются в постенной плазме у вершины споры. Клеточные стенки сначала образуются здесь, а позднее возникают и ниже. Так в большинстве случаев вся спора сверху вниз последовательно заполняется крупными протал-лиальными клетками. Одновременно в том же направлении начинаются дальнейшие деления этих клеток с образованием мелкоклеточной ткани. В верхней части заростка закладываются немногочисленные архегонии.
Стенка мегаспоры разрывается трехлучевой щелью (рис. 11.137, J); состоящий из мелких клеток бесцветный заросток немного выставляется наружу и образует на 3 бугорках своей ткани ризоиды, которые служат для поглощения воды. Затем происходит оплодотворение одного или нескольких архегониев'. Первой же перегородкой зигота делится на обращенный к шейке архегония подвесок (К: е) и собственно зародыш,
' Происходит оплодотворение не архегониев, а находящихся в них яйцеклеток. — Примеч. ред.
который для освобождения из заростка должен изогнуться наружу (К); сначала он остается заключенным в женском заростке, который в свою очередь находится в мегаспоре.
Большинство видов Selaginella живет на почве во влажных тропических лесах. Лишь немногие виды приспособились к сухим местообитаниям, как центральноамериканская S. lepidophylla, побеги которой, расположенные розеткой, при высыхании скручиваются (ложная «иерихонская роза»). Травянистые виды Selaginellites из карбона были уже разноспоровыми. Примерно 300 млн лет назад они выглядели как современные виды Selaginella.
5. Порядок: Lepidodcndrales (лепидофи-ты). Эти «древовидные плауны» (рис. 11.138) высотой до 40 м и диаметром ствола до 5 м достигли наибольшего расцвета в карбоне (см. рис. 11.163) и представляют существенный компонент каменного угля. Их линейные, спирально расположенные листья (типа микрофиллов, которые, однако, достигали до 1 м в длину) имели устьица в двух продольных желобках на нижней стороне. После их опадения на поверхности ствола оставались характерные рубцы и листовые подушки (рис. 11.138, В, D). Деревья прикреплялись к почве тянувшимися по ее поверхности (сырая почва!) повторно вильчато разветвленными ризофорами, обладавшими, как и ствол, вторичным утолщением (А, С); от них экзогенно отрастало множество относительно слабых корней своеобразного строения (так называемые аппендиксы), которые позже опадали, оставляя многочисленные рубцы на ризофоре, почему последние были названы стиг-мариями.
Листья имели простой, изредка вильчато разветвленный проводящий пучок, но в них еще не было палисадной ткани. Пятна, различимые попарно (рис. 11.138, В) или по две пары (D) на листовом рубце рядом с пятном от проводящего пучка, представляют собой места выхода тяжей рыхлой ткани, служивших для проведения воздуха и проходивших параллельно листовым следам в первичной коре. Стволы имели сифоностелу (рис. 11.139, А); тонкостенная флоэма была еще слабо дифференцирована. Кольцо малоактивного камбия образовывало в ходе вторичного утолщения новые ткани. При этом лестничные трахеиды (с несколько нетипичны-
11.2. Бактерии, грибы, растения | 31g
Рис. 11.138. Lycopodiopsida, Lepidodendrales (А—С, Е — по К. Magdefrau; D — по Stur):
А, В — Sigillaria-. А — реконструкция (1/80х), В — листовые подушки (2,5х); С —Е — Lepidodendron: С — реконструкция (1/200х), D — листовые подушки (натуральная величина), Е — стробил из спорофиллов (натуральная величина)
ми утолщениями) вторичной древесины имели одинаковый диаметр. Вторичная древесина с уже имевшимися у ряда форм однорядными лучами напоминала древесину современных хвойных (но поры трахеид были не точечные, а лестничные, и, кроме того, как почти у всех растений из карбона Северного полушария, не было годовых колец). Однако вся вторичная древесина, очевидно, не играла заметной роли ни в обеспечении механической прочности деревьев, ни в проведении воды. В стволах также была меристема, соответствующая феллогену; она особенно интенсивно откладывала производные внутрь, так что образовывалась чрезвычайно мощная по сравнению с древесиной кора (у Lepidodendron до 99 % площади поперечного сечения (!), почему виды этого рода назвали «коровыми деревьями»; см. рис. 11.139, А). Эта кора состояла главным образом из механической ткани; кроме того, она посредством язычков, сохранявшихся длительное время после отпа
дения листьев, по-видимому, участвовала в поглощении воды.
Стволы Sigillariaceae (рис. 11.138, А) были покрыты продольными рядами более или менее шестиугольных листовых подушек (В) (при вторичном росте в толщину они увеличивались вследствие дилатации). Их простые листья, имевшие длину до 10 м и ширину до 10 см, располагались пучком на конце неразветвленных или лишь слабо вильчато разветвленных стволов. В нижней части кроны на очень коротких боковых ветвях свисали крупные стробилы спорофиллов.
У Lepidodendraceae (рис. 11.138, С) листья, располагавшиеся по спирали, достигавшие нескольких дециметров в длину, сидели на ромбических листовых подушках (D). Их стволы были обильно дихотомически разветвлены и несли на ветвях длиной до 34 м терминальные стробилы спорофиллов, внешне сходные с шишками хвойных (С, Е). Очень многочисленные, чешуевидные, черепицеобразно-расположенные споро-
320 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Рис. 11.139. Lycopodiopsida, Lepidodendrales (А — по М. Hirmer; В — по McLean; С — по D. Н. Scott):
А — Lepidodendron, поперечное сечение ствола (схема); В — Bothrostrobus mundus, продольный шлиф мегаспоры с заростком (35х); С — Lepidocarpon lomaxi, продольный шлиф мегаспорангия (8х); ар — наружная перидерма; аг — наружная первичная кора; b — листовая подушка; h — обвертка (мегаспорофилл); ip — внутренняя перидерма; ir — внутренняя первичная кора; m — сердцевина: р — флоэма; pt — заросток; s — стенка споры; sp — стенка спорангия; х — ксилема
филлы закрывали спорангии, защищая их. Лепидодендроны были почти исключительно гетеро-споровыми, некоторые из них имели в мегаспорангии только одну-единственную мегаспору до 6 мм и более; у некоторых представителей (Le-pidostrobus major) мегаспора частично срасталась со стенкой спорангия, так что развитие заростка должно было происходить внутри спорангия. Заростки были сходны с заростками Selaginellaceae (рис. 11.139, В).
Большой интерес представляют некоторые каменноугольные формы (травянистые Miadesmia, Selaginellales, древовидные Lepidocarpon, Lepidodendrales) со структурами, напоминающими семя; их объединяют, несмотря на отсутствие близкого родства, под названием «Lepidospermae».
Мегаспорофилл располагался у этих «семенных» плаунов как обвертка вокруг спорангия (рис. 11. J 39, С: h); эта обвертка наверху была открытой и могла пропускать переносимые по воздуху микроспоры, обеспечивавшие оплодотворение (пока еще не известно, каким образом) заростка (pt), находившегося внутри единственной мегаспоры. Весь этот комплекс оставался на материнском растении и развивался здесь в семя1, в образовании покрова которого кроме стенки мегаспорангия участвовала также обвертка (мегаспорофилл). Мегаспорофиллы были сгруппированы в шишковидный стробил, сходный с таковым у современных голосеменных1 2.
1 Эта структура не гомологична семенам семенных растений. — Примеч. ред.
2 Шишка голосеменных только чисто внешне напоминает стробилы спорофиллов, ее организация принципиально иная. — Примеч. ред.
6. Порядок: Isoetales. Порядок в i настоящее время представлен семейством Isoeta-сеае с двумя родами. Примерно 60 видов рода полушник (Isoetes) (рис. 11.140) представляют собой подводные или живущие на влажной почве многолетние травы с клубневидной, укороченной, изредка дихотомически разветвленной осью, которая может достигать очень большого возраста.
От 2—3 продольных борозд внизу оси отходят ряды дихотомически разветвленных корней, а от верхней части оси — собранные в розетку длинные шиловидные листья (у некоторых видов до 1 м длиной!).
Пронизанные 4 воздушными каналами листья имеют на верхней стороне своего расширенного основания продольное углубление (ямка, «fovea»). Большинство листьев представляет собой спорофиллы, несущие по одному спорангию в ямке; только самые внутренние листья розетки стерильны, но по форме они не отличаются от остальных. За ямкой имеется язычок в виде кожистой треугольной структуры с погруженным основанием (рис. 11.140, В, С).
На внешних листьях розетки образуются мегаспораигии с многочисленными мегаспорами, на расположенных внутри более молодых листьях — микроспорангии с очень многочисленными микроспорами в каждом. Стенки спорангиев выстланы изнутри секреторным тапетумом. Заростки крайне редуцированы и образуются соответственно в микро- (6я) или мегаспорах (<j>). Мужские заростки на ранних стадиях развития весьма сходны с находящимися еще в оболочке споры заростками Lycopodium (см. рис. 11.135, А). В остальном они подобны мужским заросткам Selaginella, однако образуют лишь
11.2. Бактерии, грибы, растения | 321
4 спирально закрученных сперматозоида, которые несут на переднем конце пучок жгутиков. Женский заросток (рис. 11.140, О; показан у близкого рода Stylites) по строению сходен с заростком Selaginella и заполняет всю мегаспору. Он развивает небольшое число архегониев в том месте, где разрывается оболочка споры. У зародыша, развивающегося в женском заростке, находящемся в мегаспоре, нет подвеска.
Эволкщия. У Stylites (2 вида из Перу) — второго рода порядка, эволюционно, вероятно, более древнего, — стебли, покрытые листовыми рубцами, крупнее (15 см); имеется только одна продольная борозда с корнями; более выраженная тенденция к дихотомическому ветвлению. Особи вымерших представителей Isoetales из семейств Pleu-
romeiaceae и Natliorstianaceae были значительно крупнее, чем особи ныне живущих видов. Эго в меньшей мере относится к нижнемеловой Nathor-stiana и более явно — к Pleuromeia из пестрого песчаника, неветвящиеся стволы которой толщиной примерно с человеческую руку (с короткими листьями и верхушечными гетероспоровыми стробилами) достигали высоты 2 м. Современные полушники находятся в конце эволюционного ряда, который берет начало от сигиллярий (Lepidoden-drales), имевших относительно длинные листья и слабоветвившиеся или вообще неветвившиеся стволы, однако значительно более крупные, и идет дальше к Pleuromeia, Nathorstiana, а также современным видам Stylites, в котором происходило прогрессирующее укорочение стебля.
Рис. 11,140. Lycopodiopsida, Isoetales (А—С — по R. von Wettstein; D —М — по Liebig; N — по W. С. Belajeff; О — по W. Rauh и Falk; Р, Q — по D.H. Campbell):
А—С — Isoetes lacustris: A — целое растение (1/2x), В — базальный участок листа с язычком (li) и ямкой (f) (2х), С — его продольный разрез (4х); D—М — /. setacea', развитие мужского заростка и образование сперматозоидов (500х); N — /. malinverniana, сперматозоид (1 100х); О —Q — женский заросток: О — Stylites andicola, заросток во вскрывшейся оболочке споры с архегониями (аг), правый с брюшной канальцевой клеткой (Ь) и яйцеклеткой (о) (60х); Р, Q — Isoetes echinospora, развитие архегония из одной поверхностной клетки (250х); е — экзина, внутри нее интина; f — ямка; h — клетки стенки шейки; hk — шейковая канальцевая клетка; m — микроспоры; р — проталлиальная клетка; s — сперматогенные клетки; t — трабекулы; w — клетки стенки; z — центральная клетка; дает брюшную канальцевую клетку (Ь)1
1 При делении она образует брюшную канальцевую клетку и яйцеклетку. — Примеч. ред.
322 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Из изложенного выше следует, что современные Lycopodiopsida часто можно связать с вымершими формами. Последние были представлены в карбоне многочисленными древовидными родами; они были значительно более развиты, чем в настоящее время (см. рис. 11.163), и в некоторых эволюционных линиях (Lepidospermae) достигли уровня организации семенных растении. Вследствие того что климат в конце палеозоя становился все суше, древовидные плауны с их несовершенными структурами поглощения и транспорта воды вымерли или же были вытеснены появившимися формами с более совершенными проводящими системами (например, Cordaitidae) (см. рис. 11.163). Травянистые плауны и селагинеллы, напротив, оказались столь хорошо приспособленными, что сохранялись неизменными на протяжении примерно 300 млн лет вплоть до настоящего времени («персистентные типы»). Правда, в современных ландшафтах они больше не играют никакой роли, тогда как древовидные плауны вместе с каламитами и некоторыми древовидными папоротниками доминировали в каменноугольных лесах, определяя их облик (см. 11.3.2.2; рис. 11.295).
3. Класс: хвощевидные (Equisetopsida) (= Sphenopsida)
Этот класс отличается от предыдущего (И) по ряду признаков: спорофиллы четко отличаются от грофофиллов; у современных видов спорангии сидят по нескольку штук на зонтиковидных спорангиофо-рах, но никогда — в пазухах листьев; тапетум плазмодиальный (у Lycopodiopsida — секреторный тапетум); побег четко расчленен на узлы с мутовчато расположенными листьями и междоузлия.
Общие признаки хвощей: мелкие по сравнению со стеблем листья — так называемые микрофиллы, которые в противоположность остальным папоротникообразным расположены в мутовках. Стебель, чаще всего мутовчато разветвленный, четко дифференцирован на узлы и длинные междоузлия (см. рис. 11.143, А, В). Спорофиллы1,
отличающиеся от ассимилирующих листьев, чаще всего имеют форму щитков с центральной ножкой, с нижней стороны которых свисает большое число спорангиев, и собраны в шишковидные верхушечные стробилы.
Полную противоположность морфологическому единообразию современных хвощей (рис. 11.141, Е, К) представляет богатство форм ископаемых Equisetopsida.
I. Порядок: Sphenophyllales (клинолистные). Представители этого порядка полностью вымерли. Их ископаемые остатки из палеозоя (от верхнего девона до перми) отличаются (чаше всего шестичленными) мутовками листьев, на конце вильчатых или клиновидно расширяющихся, с вильчато ветвящимися жилками (см. рис. 11.142, А). Это были травянистые растения, достигавшие длины примерно 1 м, возможно, лазающие. По облику они напоминали современные виды Galium. В тонких членистых, слаборазветвленных
>
Рис. 11.141. Equisetopsida, Equisetaceae, Equi-setum (A, D — по K. von Goebel; В — no L. W. Sharp; C — no R.Sadebeck; E—К — по H.Schenck):
A — <j> заросток с архегониями с нижней стороны (17x); В — сперматозоид (1 250х); С — зародыш; 1,2 — стенки квадрантов; из верхней половины, расположенной над базальной перегородкой 1, развиваются стебель1 (s) и первая мутовка листьев (Ь), из нижней половины — корень (w) и гаустория (h) (165х); D — £ заросток с проростком (показан как более темный) сбоку, имеющим мутовки листьев и корень; Е — L — Equisetum arvense: Е — спороносные побеги (f), возникающие из почек подземного корневища, и начинающий расти бесплодный (вегетативный) побег (v) (1/2х); F и G — спорофиллы со спорангиями, на G вскрывшимися (6х); Н — спора с двумя спиральными лентами (гаптерами) периспория (ЗбОх); J — споры с развернувшимися в сухом состоянии гаптерами, при меньшем увеличении, чем на Н (100х); К— бесплодный, вегетативный побег (1 /2х); L — поперечный срез стебля, с черной ксилемой и каринальными каналами (ск) в пучках, со склеренхимными тяжами (ss) в гребнях и ложбинках (16х); а — архего-ний; Ь,, Ь2 — первые мутовки листьев; с — цитоплазма; eg — хлорофиллоносная ткань; е — эндодерма; g — жгутики; к — клеточное ядро; m — лизигенная сердцевинная полость; so — ряд устьиц; vk — валлекулярный канал
1 Листовая природа спорангиофоров хвощей не доказана. — Примеч. ред.
1 На самом деле апекс побега. — Примеч. ред.
11.2. Бактерии, грибы, растения | 323
324 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
стеблях проходил один триархный проводящий пучок; было вторичное утолщение (сетчатые трахеиды и трахеиды с окаймленными порами) (В). Четко выраженные стробилы были у некоторых видов равноспоровыми, а у некоторых — предположительно разноспоровыми1.
2. Порядок: Equisetales (хвощовые). Представители порядка составляют распространенную с конца девона и до настоящего времени основную группу класса и отличаются наличием в стебле сердцевинной полости, которая окружена венцом коллатеральных проводящих пучков1 2, к которому у древовидных палеозойских представителей снаружи примыкает вторичная древесина3.
Хвощи (Equisetaceae) в настоящее время представлены только одним родом — Equisetum, все виды которого (32) сходны между собой в основных чертах строения и развития.
Форма. От подземного ползучего корневища, находящегося в почве часто на значительной глубине, отрастают вертикальные надземные побеги с одной апикальной клеткой(см. 3.1.1.1; рис. 3.2, А, В),живущие чаще всего лишь один год. Они либо остаются простыми, либо ветвятся, образуя мутовки осей второго, третьего и т.д. порядков (см. рис. 11.141).
Ребристые оси состоят из удлиненных междоузлий. На узлах, отделенных друг от друга этими междоузлиями, расположены мутовки (см. 4.2.2) микрофиллов в форме заостренных зубпов с одним проводящим пучком каждый, которые срастаются основаниями во влагалище, охватывающее стебель (см. рис. 11.141, Е). Основания междоузлий, где они интеркалярно нарастают, одеты этими влагалищами. В узлах находится по одному кольцу внутренней ксилемы и флоэмы снаружи от нее (сифоностела). В междоузлиях проводящая система пред
1 Равно- и разноспоровыми были не стробилы, а сами растения. — Примеч. ред.
21 яжи проводящих тканей в стеблях у Equisetales не тождественны коллатеральным проводящим пучкам. — Примеч. ред.
3 Вторичная древесина находилась снаружи от первичной ксилемы, но внутри от флоэмы, поэтому неверно утверждать, что она снаружи примыкала к кольцу пучков. — Примеч. ред.
ставлена отдельными тяжами, расположенными в паренхиме (эвстела1; см. бокс 4.3, рис. А).
В области узла находятся состоящие из протоксилемы тяжи листовых следов, расположенные друг за другом под зачатками боковых ветвей и над ними. Расположенные над зачатками тяжи выходят в листья только в области расположенного выше узла. Проводящие пучки вместе со своими тяжами листовых следов в смежных междоузлиях чередуются (рис. 11.142, Е). Боковые ветви прорывают влагалища между листьями.
При малой величине листовых пластинок, которые вскоре теряют хлорофилл, функция ассимиляции переходит на зеленые стебли. Коллатеральные (см. 3.2.4.3) проводящие пучки очень бедны ксилемой. Самые старые части ксилемы вскоре исчезают, и на их местах образуются крупные полости, которые на поперечном срезе стебля выглядят как кольцо так называемых ка-ринальных полостей (см. рис. 11.141, L). В расширенной сердцевине возникает большое воздухоносное межклеточное пространство (сердцевинная полость), а в первичной коре — кольцо из так называемых валлс-кулярных (ложбиночных) полостей (под ложбинками на поверхности побега). С внутренней стороны первичная кора чаще всего отграничена одно- или двуслойной эндодермой с поясками Каспари на антиклинальных стенках клеток.
Наружные стенки клеток эпидермы стебля у хвощей более или менее сильно пропитаны кремниевой кислотой (поэтому раньше их использовали для чистки металлической посуды). В бороздках между ребрышками расположены устьица, всегда по два в продольных рядах. Им присущи следующие уникальные признаки; замыкающие клетки полностью покрыты около-устьичными клетками; при повышении тургора замыкающие клетки округляются, причем движение посредством утолщений смежных стенок передается на околоустьичные клетки, и устьица открываются.
Спорангии образуются на своеобразных спорангиофорах. Последние располагаются несколькими чередующимися мутовка-
1 Это не эвстела, а артростела. так как ее отдельные тяжи не являются пучками листовых следов. — Примеч. ред.
11.2. Бактерии, грибы, растения | 325
Рис. 11.142. А, В — SphenophyHales, Sphenophyllum (А, С — по М. Hirmer; В — по К. Magdefrau; D — по Knoell; Е — по D.H. Scott):
А — S. cuneifolium, участок побега с вильчатыми и неразделенными листьями (1/Зх); В — S. plurifolia-tum, поперечный шлиф стебля; внутри треугольная первичная ксилема с тремя протоксилемными группами, окруженная вторичной ксилемой (7х); С—Е — Eqisetales, Calamitaceae: С — Calamites са-rinatus, реконструкция (1/200х); D, Е — Arthropitys communis- D — поперечный шлиф через участок древесины (10х), Е — тангенциальный шлиф через молодой побег (10х); b — листовой след; с — ка-ринальный канал; i — внутриузловой канал; m — сердцевина; ms — сердцевинный луч; sx — вторичная ксилема; х — ксилема
ми, из-за сильно укороченных междоузлий собранными на концах побегов в шишковидные стробилы (см. рис. 11.141, Е). Сами спорангиофоры («спорофиллы») имеют форму столика на одной ножке, на нижней стороне которого располагается 5—10 мешковидных спорангиев (F, G); их снабжают концентрические проводящие пучки.
Спорогенная ткань в более молодом спорангии окружена многослойной стенкой. Ее самый
внутренний слой (плазмодиальный тапетум) в результате растворения клеточных стенок образует периплазмодий, который внедряется между округляющимися спорами и расходуется на образование их оболочек1. Таким образом, ко вре
1 Плазмодиальный тапетум выделяет вещества, используемые при формировании оболочек спор, но некорректно утверждать, что он «расходуется» на образование этих оболочек. — Примеч. ред.
326 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ___________________________________
мени созревания спор сохраняются только два внешних слоя клеток стенки спорангия; стенки клеток эпидермы имеют утолщения в форме колец и спиралей. Спорангии вскрываются продольной трещиной на внутренней стороне за счет сил сцепления между оболочками клеток стенки спорангия и водой, объем которой в этих клетках уменьшается вследствие испарения.
Вскрывшийся спорангий современных видов Equisetum высвобождает многочисленные зеленые споры со своеобразно устроенной оболочкой. На собственно споровой оболочке, состоящей из эндоспория и эк-зоспория, периплазмодий откладывает многослойный периспорий. Его самый внешний слой состоит из двух узких параллельных друг другу лент (гаптеры; см. рис. 11.141, Н, J), на концах расширенных в виде лопатки. Во влажном состоянии они спирально закручены вокруг споры. При высыхании спор гаптеры развертываются, однако остаются соединенными серединами друг с другом и с эндоспорием (J). При этом они вытягиваются, но при увлажнении снова закручиваются (см. 8.4) и своими гигроскопическими движениями могут служить для распространения спор, сцепленных группами; соответственно гаметофиты часто растут тесными группами. Споры сохраняют всхожесть только несколько дней.
Все споры одинаковые. Они прорастают в сильно рассеченные на лопасти, зеленые заростки (см. рис. 11.141, А).
Заростки представляют собой сильно рассеченные дорсовентральные курчавые пластинки, которые могут быть однодомными или двудомными1. Определение пола потенциально двуполых заростков осуществляется фенотипически под влиянием внешних факторов. В условиях голодания возникают преимущественно О" гаметофиты. Развитие до половой зрелости длится всего лишь от 3 до 5 недель — очевидно, столь быстро для того, чтобы как можно скорее завершить гаметофитную фазу, чувствительную к наличию воды и низко конкурентоспособную по отношению к мхам. Мужские гаметофиты в противоположность женским сильно пигментированы каротиноидами — явление, которое известно также у мхов (стенки антеридиев) и грибов (сГ гаметы Allomyces) и может иметь значение как защита от мутагенного облучения.
1 Правильно — обоеполыми или однополыми. — Примеч. ред.
Антеридии погружены в ткань заростка, ар-хегонии выступают над его поверхностью1. Спиралевидные сперматозоиды возникают в количестве примерно 250 — 1 000 на антеридий и имеют многочисленные жгутики (см. рис. 11.141, В).
Первая перегородка (базальная стенка, 1 — 1 на рис. 11.141. С) разделяет зиготу на две половины, из которых в противоположность Lycopodium (см. рис. 11.135, D) обе участвуют в образовании зародыша; подвесок не развивается. Первые листья появляются на побеговом полюсе сразу расположенные мутовкой и кольцевидно окружают апекс побега, который растет дальше посредством верхушечной клетки в виде трехгранной пирамиды (см. рис. 3.2, А). Зачаток первого корня находится сбоку от продольной оси зародыша (см. рис. 11.141, С: w); он растет вниз, прорывая ткань заростка (D).
Большинство видов распространенного от тропиков до холодных зон рода Equisetum предпочитает влажные местообитания. Южноамериканский Е. giganteum и некоторые другие тропические виды, будучи лазящими растениями, достигают до 12 м в длину, тогда как высота (£. telmateia) видов, обитающих в Германии, может быть максимум 2 м.
У Е. arvense — хвоща полевого (см. рис. 11.141), как и у других видов с однолетними надземными частями, короткие боковые веточки подземных корневищ превращаются в округлые зимующие клубни, содержащие запасные вещества; однако имеются и вечнозеленые виды (например, Е. hyemale).
У некоторых видов хвощей одни надземные побеги стерильны и обильно ветвятся, другие же несут на вершине стробилы спорангиофоров, ветвятся позже и слабо или вообще не образуют боковых ветвей (см. рис. 11.141, Е, К).
Оба последующих семейства полностью вымершие. Встречавшиеся только в нижнем карбоне виды Arcliaeocalamitaceae имели вильчатые листья (рис. 11.143, А), которые на соседних узлах располагались друг над другом соответственно прохождению в узлах проводящих пучков.
Calamitaceae отличаются от Equisetaceae следующими признаками. На репродуктивных осях чередовались мутовки щитовидных спорангио-
1 Над поверхностью заростка выступают только шейки архегониев. — Примеч. ред.
11.2. Бактерии, грибы, растения! 327
Рис. 11.143. Equisetales (А, В — по D.Stur; С — по M.Hirmer, с изменениями; D — по W.С.Williamson и D.H. Scott):
А — Archaeocalamitaceae, Archaeocalamites radiatus (1 /Зх); В—D — Calamitaceae: В — Annularia stellata (1/2х), С — Calamostachys Ыппеуапа, продольный шлиф стробила со стерильными листьями (4х), D — Calamostachys casheana, тангенциальный шлиф через спорангиофор, несущий три мегаспорангия и один микроспорангий (22х)
форов и ланцетовидных листьев («прицветников») (рис. 11.143, С). Наряду с равноспоровыми видами имелись и разноспоровые (D). Гаптер у спор не было. Виды широко распространенного в верхнем карбоне и перми рода Catamites (см. рис. 11.142, С) составляли важный компонент каменноугольных лесов и наряду с лепидодендронами и сигилляриями внесли существенный вклад в образование каменного угля. Экземпляры некоторых видов достигали высоты 30 м и благодаря мощному развитию вторичной древесины диаметр их стволов доходил до 1 м (С, D), однако, как и у Equisetum, имели большую сердцевинную полость («трубчатые деревья»). Стволы у большинства видов несли мутовки ветвей, но у некоторых не ветвились. Проводящие пучки (так же как и у Equisetum) на верхнем конце междоузлия разветвлялись на три веточки. Две боковые веточки сливались каждая с боковой веточкой соседнего пучка, давая пучок следующего междоузлия, тогда как третья веточка отходила наружу как пучок листового
следа1 (Е). Радиально идущие «внутриузловые каналы», возникавшие в результате лизиса тонкостенных клеток, служили, вероятно, для аэрации. Листья (рис. 11.143. В) были простые, ланцетные, с одной жилкой; на конце листа находилась — как на листовых зубчиках ныне живущих хвощей, гуттирующих воду, — гидатода. Соответственно чередованию первичных пучков в следующих друг за другом междоузлиях листья располагались чередующимися мутовками.
Класс Equisetopsida достиг наибольшего расцвета в палеозое и к настоящему времени весь, кроме рода Equisetum, вымер (см. рис. 11.163). Этот род объединяет остатки более высокоразвитых форм, поскольку в мезозое среди представителей рода Equise-
1 На с. 250 эту часть проводящей системы правильно описывали как трубчатый участок ар-тростелы («сифоностелу»). — Примеч. ред.
328 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ___________________________________
tiles были древовидные формы с вторич ным утолщением. Таким образом, наши современные хвощи — всего лишь реликты, которые мы, однако, не можем считать потомками разноспоровых представителей из палеозоя, так как гетероспория всегда должна быть производной от изоспории, но не наоборот. Стало быть, современные хвощи должны были произойти от более ранних, еще равноспоровых форм. Некоторые из вымерших форм (Catamites, Sphenophy-llum) были разноспоровыми; уровня образования семени, как у Lepidospermae, достиг род Calamocarpon.
4. Класс: псилотовые (Psilotopsida)
Сохранившиеся до наших дней виды Psilotum показывают определенное внешнее сходство с вымершими псилофитами (вильчатое ветвление побегов, отсутствие корней). Однако латеральные спорангии, сра
стающиеся в синангии, а также настоящие листья (микрофиллы) указывают на дальность псилотовых от Psilophytopsida и большую эволюционную продвинутость.
С другой стороны, помимо отсутствия корней наличие гаметофита, еще имеющего проводящие ткани, можно рассматривать как примитивный признак. Однако отсутствие корней интерпретируют и как вторичное упрощение (ср. лишенный корней водный папоротник Salvinia, утрату корней у эпифитов)1.
Единственный порядок Psilotales включает низкие многолетние дихотомически разветвленные травы (рис. 11.144, А) с вильчатыми листьями (С). Надземные стебли имеют актиностелу или уже намечающуюся сифоностелу с одревесневшей сердцевиной (В). Безлистные протостелические
1 Отсутствие корней у псилотовых связано с их микотрофностью. — Примеч. ред.
Рис. 11.144. Psilotopsida, Psilotaceae (А — по R. von Wettstein, Pritzel; В, С, Е, F — по Pritzel; D, G — по R. von Wettstein; Н — К — по Lawson):
А—Е — Psilotum, Р. triquetrum: А — внешний вид (12х), В — поперечный срез стебля с актиностелой (точнее сифоностелой) (40х), С — участок побега с синангием в пазухе вильчатого листа (2,5х), D — поперечный срез синангия (8х), Е — споры (250х); F, G — Tmesipteris, Т. tannensis: F — внешний вид (12х), G — спорофилл (2,5х); Н —К — Psilotum, Р. triquetrum: Н — заросток (15х), J — поперечный срез заростка (40х), К — сперматозоид (990х); ап — антеридии; аг — архегонии; с — внешний зеленый слой коры; ер — эпидерма; m — внутренняя кора; ту — микоризные клетки; р — флоэма; х — ксилема
11.2. Бактерии, грибы, растения | 329
корневища лишены корней (даже зародыш без зачатка корня), но несут нитевидные ризоиды и содержат микоризные грибы. Листья представляют собой чешуйки1 (мик-рофиллы, у Psilotum очень мелкие и без жилок), расположенные по рыхлой спирали. Спорангии имеют многослойную стенку, сращены по три1 2 в синангии (С, D) и еще не имеют настоящего тапетума (изоспоры получают питание от стерильных клеток археспория, которые не только окружают спорофиты, но и перемежаются с ними). Синангий сидит на очень короткой ножке в пазухе3 пары чешуек с вильчато разделенной верхушкой.
Гаметофиты, или заростки, достигают нескольких сантиметров в длину, радиально-симметричные, разветвленные (рис. 11.144, Н). Они бесцветны и живут под землей за счет микоризных грибов (J: ту). На поверхности они несут многоклеточные антеридии, в которых развиваются многочисленные сперматозоиды с множественными жгутиками. Мелкие архегонии (только с одной, редко с двумя шейковыми канальцевыми клетками) несколько погружены в ткань гаметофита. Особенно мощные заростки имеют проводящий пучок с одревесневшими кольчатыми трахеидами и эндодермой.
К Psilotopsida относят только два рода: Psilotum и Tniesipteris (с двумя видами в каждом, обитающими в тропиках преимущественно как эпифиты). У Tniesipteris вильчатые листья несколько крупнее. Они крыловидно сбегают по стеблю, а их плоскости параллельны стеблю. Однако их еще нельзя без оговорки приравнивать к листьям высших растений4. Иногда вильчатые надземные части трактуют как оси вай (листьев) с сидящими на них перистыми листочками, которые у Psilotum редуцированы сильнее, чем у Tniesipteris. Вильчатые разветвления лишь изредка соответствуют настоящей дихотомии (продольное деление трехгранной пирамидальной верхушечной
1 Вся надземная структура, включая чешуевидные образования и несущие их оси, может быть уподоблена вайе папоротников. — Примеч. ред.
2 У Tniesipteris— по две. — Примеч. ред.
3 Синангий располагается на адаксиальной лопасти перышка листа. — Примеч. ред.
4 Они соответствуют перышкам на рахисе вайи папоротников. — Примеч. ред.
клетки), чаще всего они происходят потому, что клетка, соседняя со старой верхушечной клеткой, становится второй верхушечной. Ризоиды гаметофита и спорофита развивают выводковые почки, с помощью которых осуществляется вегетативное размножение.
В ископаемом состоянии Psilotales не найдены. Однако их все-таки рассматривают как древние реликты, которые в то же время показывают четкие связи с рассматриваемыми далее Pteridopsida (через Gleicheniaceae с новокаледонским родом Stromatopteris и через Schizaeaceae).
5. Класс: папоротники (Pteridopsida) (= Filicopsida)
Представители этого класса папоротникообразных, собственно папоротники, отличаются крупными, часто сложно расчлененными мегафиллами, которые называют также «вайями». Спорангии, исходно располагавшиеся терминально, у более продвинутых форм находятся по краю листа или на его нижней стороне. Побеги, корни и листья растут — как и в ранее рассмотренном классе Equisetopsida — чаще всего благодаря деятельности единственной апикальной клетки, а не группы апикальных клеток, как у представителей Lycopodiopsida.
Вайи — чаще всего имеющие черешки и многочисленные жилки — в молодости обычно закручены на верхушке (исключение — Ophioglossales). Закручивание возникает из-за более быстрого роста абаксиальной (нижней) стороны молодых зачатков (соответственно листьев или теломов), и только потом происходит распрямление органа. Листья, развивающиеся таким ак-ропластическим образом (см. 4.3.1.2), несут на нижней стороне многочисленные спорангии, часто собранные в группы (сорусы). Стебель чаще всего неветвящийся или слабо разветвленный. Возникновение листовых пластинок из систем теломов и переход спорангиев на нижнюю сторону листа вследствие более сильного роста его верхней стороны можно представить себе по схеме, приведенной на рис. 11.145, А—Н. Ископаемая переходная форма изображена на рис. 11.145, J. Аналогично можно представлять себе и возникновение крупных перистых вай. Представители вымерших Primofilices и ныне живущих групп папо-
330 | ГЛАВА 11, СИСТЕМАТИКА И ФИЛОГЕНИЯ
Рис. 11.145. Pteridopsida (А—Н — по W. Zimmermann; J — по S.Mamay; К—М — по А.С. Seward, W. Gothan);
А — D — переход от фертильного телома к спорофиллу; Е — Н — перемещение спорангиев на нижнюю сторону листа; J — спорофилл Acrangiophyllum (напоминающего папоротник растения неясного систематического положения) из верхнего карбона (7х); К—М — жилкование перышек листьев растений, сходных с папоротниками; К — вееровидное жилкование (Archaeopteris, верхний девон), L — перистое жилкование (Alethopteris, представитель семенных папоротников из верхнего карбона), М — сетчатое жилкование (Linopteris, верхний карбон) (12х)
ротников Eusporangiatae. Leptosporangiatae и Hydropterides хотя и проявляют определенные родственные связи между собой, но являются не естественными таксонами, а ступенями развития, т.е. отражают ход развития различных признаков.
Ступень развития', первопапоротники1 1 (Primofilices) (= Protopteridiidae)
В качестве предков Pteridopsida рассматривают Psilophytopsida. Связующее звено между ними представляют Primofilices, которые, с одной стороны, еще сходны с пси-лофигами, а с другой стороны, имеют признаки более высокого развития. Общим для всех Primofilices было наличие терминальных спорангиев (рис. 11.146, В), а также то, что элементы перистых листьев располагались не в одной плоскости («объемные вайи»). Переход от псилофитов к Primofilices происходил столь постепенно, что относи тельно некоторых форм (Protopteridium и Pseudosporochnus, рис. 11.146, А—Е) может возникнуть сомнение, причислять ли их
1 Значительная часть этой группы в принятом здесь объеме принадлежит к Progymnosper-mae и не имеет отношения к папоротникам. — Примеч. ред.
еще к псилофитам или уже к папоротникам1. Филогенетическому положению Primofilices соответствует и их распространение во времени: они появились в среднем девоне и вымерли в нижней перми.
В пределах группы Primofilices происходит переход от расположенных почти пучками вильчатых ветвей с расположенными на их концах спорангиями (например, Pseudosporochnus', рис. 11.146, А, В) к клиновидно-уплощенным, неправильно вильчато разветвленным листьям и сходным с ними спорофиллам с расположенными по их краям спорангиями (например, Cladoxylon', рис. 11.146, М)2. Спорофиллы у некоторых видов располагались вперемежку со стерильными листьями, а у некоторых видов — группами, хотя еще и не были объединены в «цветки»3. Прогрессировавшее срастание теломов боками привело к появлению широких плоских листьев с вильчатым
1 Protopteridium можно причислять к Progym-nospermae, но никак не к папоротникам. — Примеч. ред.
2 Признать спорангии Cladoxylon расположенными по краям спорофиллов было бы слишком поспешным. — Примеч. ред.
3 Цветков нет и у всех остальных папоротников; они присущи только покрытосеменным. — Примеч. ред.
11.2. Бактерии, грибы, растения | 33
жилкованием, какие встречались уже в верхнем девоне (см. рис. 11.145, J; ср. также Adiantum из современных Leptosporangia-tae — см. рис. 4.60, А).
Позднее из вильчатого жилкования у папоротников и семенных папоротников постепенно развилось сетчатое жилкование. В верхнем девоне встречалось вееровидное жилкование с вильчатым ветвлением жилок, в нижнем карбоне впервые появилось перистое жилкование, а в верхнем карбоне — сетчатое, которое наиболее совершенным образом обеспечивало лист водой и питательными веществами (см. рис. 11.145, К—М).
Предпосылкой для появления таких плоских листовидных образований было то, что те-ломы расположились в одной плоскости. Однако у примитивных форм они еще отчасти располагались перпендикулярно друг к другу (как это имеет место и ныне, например, у Ophioglo-ssales), даже уплощение могло еще отсутствовать, так что «листья» были круглыми в сечении. И то и другое имело место у Stauropteris (рис. 11.146, G). верхний карбон; в таких цилиндрических ли-стьях-теломах уже могла быть палисадная паренхима.
Primofilices были преимущественно равноспоровыми; отдельные виды были уже разноспоровыми (Archaeopteridales)1. Они имели спорангии с многослойной стенкой, т.е. были эвспорангиатными. У отдельных представителей уже встречались особые механизмы вскрывания спорангиев. Увеличивалось также разнообразие стел — от протостелы до эвстелы.
Возник! ювение признаков, характерных для современных Pteridopsida, можно, таким образом, проследить на группе форм Primofilices, так что мегафильные Pteridopsida вместе с Equisetopsida можно вывести из Psilophytopsida как эволюционную ветвь, параллельную микрофильным Lycopodiopsida.
Систематика. Primofilices в целом составляют весьма гетерогенную группу, которую можно подразделить на 5 порядков.
1. Порядок; Pseudosporochnales1 2. Представители порядка встречаются в (нижнем) среднем
1 Archaeopteridales — это не папоротники, а праголосеменные. Progymnospermae. — Примеч. ред.
2 Вероятнее всего, этот порядок следует объединять с Cladoxylales. — Примеч. ред.
девоне; например, Pseudosporochnus (рис. 11.146) едва достигал высоты 1 м, с неразветвленным стволом и множеством одинаково развитых, слабо вильчато разветвленных боковых осей, которые несли многочисленные тонкие дихотоми-рующие веточки. В отдельных случаях концы ветвей были несколько расширены — начало уплощения и слияния (в терминах теломной теории). Боковые ветви с их расширенными ассимилирующими пластинками можно рассматривать как структуры, предшествующие крупным, многократно перистораздельным листьям (ме-гафиллам), или вайям.
2. Порядок: Protopteridales1. Представители этого порядка по строению своих «вай» также еше более или менее напоминают псилофиты. У некоторых родов был мощный ствол с лестничными трахеидами2 во вторичной древесине. Роды Hyenia и Calamophyton, рассматриваемые ранее как примитивные Equisetopsida, также относятся, поданным современных исследований, к этому кругу форм, равно как и Protopteridium (рис. 11.146, С), Aneurophyton, Tetraxylopteris, Rha-cophyton и Pertica (рис. 11.146, F)3.
3. Порядок: Cladoxylales. Виды порядка (Cla-doxylon, рис. 11.146, J) жили от среднего девона до нижнего карбона. Строение их стелы, которая состояла из многочисленных У-образных в поперечном сечении отдельных пучков (рис. 11.146, L), иное, чем у всех других сосудистых растений4.
4. Порядок: Coenoptcridales. Виды i юрядка (от верхнего девона до нижней перми, расцвет в нижнем карбоне) полностью характеризуются разветвленными еще в трехмерном пространстве вайями (Stauropteris. рис. 11.146, G; Botryopteris, ла-
1 Это не папоротники, а праголосеменные. — Примеч. ред.
2 У настоящих Protopteridales были точечные трахеиды вторичной ксилемы. — Примеч. ред.
3 Protopteridium, Aneurophyton и Tetraxylopteris — это праголосеменные, Hyenia и Calamophyton — представители Cladoxylales (= Pseudosporochnales), Rhacophyton принадлежит к примитивным Zygo-pteridales, a Pertica — к Trimerophytales. Их не-। |равомерно относить к одному порядку. — Примеч. ред.
4 К Cladoxylales относят растения со стелой кладоксилеевого типа. Растения, у которых не удается достоверно выявить строение стелы, включают в порядок Pseudosporochnales. Сохранение обоих порядков оправдано исключительно практикой палеоботаников строить отдельные классификации для разных органов и для органов разной сохранности — в противоположность практике неонтологов классифицировать целые организмы. — Примеч. ред.
332 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
11.2. Бактерии, грибы, растения | 333
<
Рис. 11.146. Primofilices (А — по W. Zimmermann; В — по S. Leclercq и Н. Р. Banks; С—Е, J — М — по R. Krausel и Н. Weyland; F — по Kasper и H.N. Andrews; G — по M.Chaphekar, с изменениями К. Magdefrau; Н — по D.H. Scott):
А—В — Pseudosporochnales: А — Pseudosporochnus, реконструкция, В — то же, конец ветви (натуральная величина); С—F— Protopteri-dales: С —Е — Protopteridium hostimense, девон, С — вайя (1 /4х), D — стерильные, Е — фертильные перышки (Зх), F — Pertica quadrifaria, девон, ветвь со спорангиями (s); G, Н — Соепо-pteridales, Stauropteris oldhamia, карбон: G — участок стерильной вайи, реконструкция (натуральная величина), Н — то же, спорангий с участком стенки, по которому он вскрывался (35х); J —М— Cladoxylales, Cladoxylon scoparium, средний девон: J — участок ветви (2/Зх), К — листочки (2х), L — поперечный срез плектосте-лы (4х), М — группа спорангиев (2х)
зящий папоротник Ankyropteris и многие другие1). В стенке спорангия некоторых видов уже можно распознать группу толстостенных клеток, обеспечивавших его вскрывание по заранее закладывавшейся линии разрыва, как в спорангии Osmunda (см. рис. 11.156, А).
5. Порядок: Archaeopteridales1 2. К этому порядку (разноспоровому) относят верхнедевонский космополитный род Archaeopteris, представленный многими видами (см. рис. 11.288. С). Это были уже стройные деревья с дваждыперисто-раздельными объемными вайями. Лопатковидные перышки имели перистое жилкование (см. рис. 11.145, J); по обе стороны от основания вай располагались два прилистника, как у Marattiales. Нижние перышки фертильных вай несли спорангии, расположенные по краю, обращенному к верхушке вайи. Микроспорангии содержали много микроспор диаметром 0,03 мм, а мегаспорангии — 8— 16 мегаспор диаметром 0.3 мм (см. рис. 11.288, С: 8). В стволах, достигавших высоты до 9 м и толщины до 1,5 м, имелась мощная вторичная ксилема из трахеид с араукариоидной поровостью стенок (см. 11.2, Araucariaceae). Archaeopteris объединяет, таким образом, признаки папоротников3 и голосеменных.
1 У Ankyropteris и Botryopteris были настоящие перистые вайи, вполне сравнимые с вайями большинства современных папоротников. — Примеч. ред.
2 Это праголосеменные растения, а не папоротники. — Примеч. ред.
3 Сходство его с папоротниками конвергентное. — Примеч. ред.
Некоторые авторы объединяют Protopterida-les — промежуточное звено между Psilophytopsida и Filicopsida, и Archaeopteridales, эволюционно находящийся между Psilophytopsida и Gymnosper-mae, в группу «Progymnospermae». По вторичному росту в толщину и ряду других признаков они представляют собой обширную переходную группу между птеридофитами и голосеменными1.
Ступень развития: эуспорангиатные папоротники (Eusporangiatae) (= Ophioglossidae)
Спорангии, имеющие многослойную стенку (рис. 11.147, F), развиваются каждый из нескольких клеток.
1. Порядок; Ophioglossales. Представители порядка (единственное семейство Ophi-oglossaceae, около 80 равноспоровых видов) имеют вайи, которые состоят из ассимилирующей (лишенной палисадной ткани) зеленой и перпендикулярно к ней расположенной фертильной желтоватой частей (рис. 11.147, A, D). Листья представляют собой трехмерные вайи и в этом отношении соответствуют исходному типу вай (ср. Primofilices). У спороносной части рост в ширину подавлен.
Продольный рост обеспечивает не одиночная крупная апикальная клетка, а несколько инициальных клеток, слагающих конус нарастания. На коротком подземном стебле ежегодно развивается чаще всего лишь одна-единственная вайя с длинным черешком и небольшим кожистым влагалищем. В молодости она не закручена. В нижних частях стебля находится протостела. которая выше разделяется на трубку проводящих пучков. Сильно редуцированные, имеющие в длину несколько миллиметров подземные заростки лишены хлорофилла и представляют собой многослойные клубеньки, часто многолетние, которые существуют благодаря симбиозу с микоризными грибами. Антеридии и архегонии погружены в ткань заростка (рис. 11.147, С). Зародыш, возникающий из оплодотворенной яйцеклетки, у некоторых видов в течение нескольких лет ведет подземный образ жизни.
У Botrychium как ассимилирующая часть вайи с вильчато расходящимися жилками, так и ее спороносная часть перистые. Округлые спорангии располагаются по кра
1 Progymnospermae представляет собой переходную группу между Trimerophytales и голосеменными. — Примеч. ред.
334 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
ям перьев и не срастаются между собой (рис. 11.147, D, Е). В подземном побеге происходит слабое вторичное утолщение (единственный случай среди всех современных папоротников).
У Ophioglossum зеленая часть листа имеет форму языка и сетчатое жилкование, а желтая спороносная часть — форму простого цилиндра. Спорангии погружены в ткань, располагаются в два ряда и срастаются между собой боками (рис. 11.147, А). Питанию растений, очевидно, способствуют микоризные грибы, всегда присутствующие в корнях. У Ophioglossum simplex лису большей частью вообще не имеет ассимиляционной ткани, а только несет спорангии. Заросток цилиндрический (рис. 11.147. С). Число хромосом необычайно большое (у О. vulgatum п = 256, у О. reticulatum п = 630).
2. Порядок: Marattiales. В ограниченный тропиками порядок включают также примитивные и древние папоротники. Их современные виды несут на коротком клубневидном стебле пучок вай, чаще всего имеющих несколько метров в длину, многократно перистораздельных, в молодости закру
ченных и у основания снабженных парными прилистниками. Жилкование открытое (ср. Ophioglossum с сетчатым жилкованием). Спорангии с одинаковыми спорами у некоторых родов срастаются боками в капсуловидные многокамерные, позже резко вскрывающиеся синангии (рис. 11.148, В, С), у других родов они свободные и сгруппированы в сорусы.
Долго живущие заростки содержат эндофитные микоризные грибы, однако обитают на поверхности почвы в виде автотрофных многослойных талломов. Внешне они напоминают печеночники. На нижней стороне у них развиваются погруженные антеридии и архегонии.
Современные Marattiales насчитывают примерно 200 видов из нескольких родов. Они населяют тропические леса: например, Angiopteris встречается в Азии (длина вай до 5 м!), Danaea — в Южной Америке, Ма-rattia — по всему тропическому поясу.
Эволюция. Первые Marattiales появились в карбоне. По-видимому, они произошли от изо-споровых Primofilices, которые почти все имели спорангии с многослойными стенками. До крас-
Рис. 11.147. Eusporangiatae, Ophioglossales (А, В, D, Е — по К. Magdefrau; С — по Н. Bruchmann; F — по К. von Goebel; G — по Н. Bruchmann):
А—С — Ophioglossum, О. vulgatum-. А — спорофит (1/2х), В — продольный срез верхушки фертильной части листа (2х), С — заросток с антеридиями (а), архегониями (ад), молодым спорофитом (s), имеющим первый корень, гифами гриба (h) в ткани заростка (10х); D — G — Botrychium, В. lunaria: D— спорофит (1/2х), Е — спорангии, вид снизу, F — продольный срез незрелого спорангия с многослойной стенкой; внутри материнские клетки спор, окруженные клетками тапетума (10х). G — срез заростка (35х); е — зародыш
11.2. Бактерии, грибы, растения | 335
Рис. 11.148. Eusporangiatae, Marattiales (А — по M.Hirmer; В, С — по В. Renault):
А — древовидный папоротник Megaphyton (семейство Marattiaceae), реконструкция (верхний карбон). На стволе в два ряда располагаются рубцы от отпавших вай. Основание ствола усилено мантией из отходящих от него корней, растущих вниз; В — Ptychocarpus unitus, верхний карбон, нижняя сторона перышек с синангиями (8х); С — то же, поперечный срез синангия (60х)
ного лежня их видовое разнообразие было больше, и они были более широко распространены, чем в настоящее время. Это были деревья со стволами, покрытыми корнями и достигавшими высоты Ю м (самым мощным и широко распространенным растением была Asterotheca arborescens). Тогда они полностью господствовали над Lepto-sporangiatae. Особенно приметным был Megaphyton с вайями, расположенными не по спирали, а в два ряда (рис. 11.148, А).
Ступень развития: лептоспорангиатные папоротники (Leptosporangiatae) (= Pterididae)
Спорангий (см. рис. 11.128) развивается из одной эпидермальной клетки и имеет тонкую стенку, которая после раннего лизиса тапетума состоит из одного слоя клеток. Относимые сюда папоротники — преимущественно тенелюбивые растения. Многочисленные их виды (90 % всех Pteridopsida, около 9000 видов) распространены по всему земному шару; наиболее широко они представлены в тропиках, где весьма разнообразны: от карликов размером в несколько миллиметров (например, виды Didymoglossum из семейства Hymeno-phyllaceae) до пальмовидных деревьев высотой до 20 м (см. рис. 11.150). Ствол древовидных папоротников, чаще всего толщиной с человеческую руку (семейство Суа-theaceac, роды Cyathea, Dicksonia, Cibotium), не ветвится и несет на вершине розетку многократно перистораздельных вай длиной до 3 м. В Германии встречаются папоротники. наоборот, в виде преимущественно травянистых растений с подземным многолетним вертикальным или приподнимающимся, слабо разветвленным корневищем, которое у Pteridium может быть до 40 м длины и жить до 70 лет1.
Стволы — у травянистых форм корневища — в молодости имеют чаще всего центральную протостелу, которая в более старых частях переходит в структуру типа си-фоно- и полистелы самых разнообразных форм (см. бокс 4.2, рис. А) чаще всего с центральной ксилемой и периферической флоэмой (рис. 11.151, А; 11.149; см. также 3.2.4)1 2. Изредка даже образуются сосуды (как у Pteridium aquilinum, рис. 11.149). Проводящий пучок окружен эндодермой (рис. 11.149). Вторичного утолщения нет, а прочность стволов достигается иначе, чем у Lycopodiopsida
1 Корневища ряда европейских видов, в том числе и Pteridium aqulinum, горизонтальные и ветвятся интенсивно. — Примеч. ред.
2 У папоротников бывает не полистела, а диктиостела, иногда полициклическая. Сифоностела и диктиостела находятся не в старых, а, наоборот, в более молодых участках ствола (корневища). — Примеч. ред.
336 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Рис. 11.149. Leptosporangiatae, Pterida-les, Pteridium aquilinum (по K. Magdefrau).
Проводящий пучок, поперечный и продольный разрезы (ЮОх); е — эндодерма; g — основная паренхима; s — ситовидные клетки; t — лестничные сосуды
и Equisetopsida: многочисленные пучки листовых следов проходят чаще всего на большом расстоянии в коре и вместе со склеренхимными пластинками (рис. 11.151, А) укрепляют стебель (см. также 3.2.3). У некоторых древовидных папоротников прочность ствола повышается также благодаря мантии из жестких придаточных корней. Иногда такой покров может быть чрезвычайно толстым (до нескольких дециметров!).
Мегафиллам, возникшим из многих теломов, свойственны разные варианты жилкования. Древние вильчатые листья (см. рис. 11.157) или вильчатые жилки (см. рис. 4.60, А; на листьях со слитными перышками) встречаются редко; как правило, они бывают только у листьев проростков и молодых растений. Полностью развитые листья весьма разнообразны. Часто вайи перисто-сложные (например, от дважды- до четы-реждыперистых у Pteridium aquilinum, орляка; дваждыперистые у Dryopterisfilix-mas, щитовника мужского; однократноперистые у Polypodium vulgare, многоножки обыкновенной). Однако встречаются и простые листья с доминирующей центральной и менее мощными боковыми жилками (Phyllitisscolopendrium, листовник сколопецдровый, рис. 11.151, С). Их
очень долгий, иногда неограниченный верхушечный рост в противоположность росту листьев семенных растений обусловлен деятельностью двусторонней апикальной клетки, которая, однако, позже часто заменяется группой инициальных клеток.
Лист часто развивается в течение многих лет. У орляка (Pteridium aquilinum), например, на каждой короткой веточке корневища ежегодно закладывается только один лист, которому требуется 3 года, чтобы полностью развиться. После отмирания листья оставляют — прежде всего у древовидных папоротников, где они сохраняются несколько лет после того, как полностью развернутся, — крупные, бросающиеся в глаза рубцы (см. рис. 11.150). По гистологическому строению (наличию палисадной и губчатой паренхимы) эти листья в значительной мере напоминают листья высших наземных растений, однако клетки эпидермы у папоротников чаще всего содержат хлоропласты.
Спорангии образуются в большом числе по краю или чаше всего на нижней стороне листьев (см. бокс 4.1, рис. А; рис. 11.152, В— D). Спорофиллы по внешнему облику, как правило, лишь немного отличаются от стерильных листьев (трофофиллов); однако у представителей некоторых родов они имеют явно иной облик— прежде всего
11.2. Бактерии, грибы, растения | 337
Рис. 11.150. Pteridopsida, Cyatheales, Суа-thea crinita Древовидный папоротник из Шри-Ланки (1/100х) (по Н. Schenck)
вследствие редукции пластинок перышек (ср. Mutieuccia, Blechnum, Osmunda).
В типичном случае (например, у Aspi-diales, к которым относится подавляющее большинство европейских папоротников) спорангии помногу объединяются в сорусы. Эти спорангии возникают на выступе ткани листа — плаценте (рис. 11.152, В; плаценту называют также рецептакулом) и у многих видов до созревания покрыты и защищены кожистым выростом поверхности листа — так называемым покрывальцем (или ицдузием; В — D). Отдельный спорангий в зрелом состоянии представляет собой маленькую коробочку на ножке, которая содержит большое число мейоспор почти всегда одинакового размера (изо-спория!)1. Очень характерно по-разному ус
1 У лептоспорангиатных папоротников чаще всего в спорангии развивается 64 споры. — Примеч. ред.
троенное кольцо, которое у Polypodiaceae в виде выступающего ряда клеток с сильно утолщенными радиальными и внутренними стенками проходит через спинную сторону и вершину спорангия до середины его брюшной стороны (см. рис. 11.156, D). Посредством механизма сцепления между стенками клеток кольца и находящейся в них водой (при участии клеток устья, см. 8.4) кольцо осуществляет вскрывание спорангия и выбрасывание спор (см. рис. 8.37).
Из прорастающей споры развивается недолговечный гаплоидный заросгок (см. рис. 11.128, А, В; 11.153), который не превышает нескольких сантиметров в длину и, как правило, образует как антеридии, так и архегонии. Определение пола осуществляется в норме модификационно. Заростки потенциально обоеполые. Только у представителей австралийского рода Platyzoma (семейство Gleicheniaceae) образуются споры двух типов, которые развиваются соответственно в однополые, т. е. сГ и <j> заростки (рис. 11.153, D, Е).
Рис. 11.151. Pteridopsida (А, С — по К. Magdefrau; В — по W. Troll):
А — Pteridium aquilinum, поперечный срез корневища (7х); В — Asplenium nidus, схема роста; С — Phyllitis scolopendrium (1/4х); al — внешний проводящий пучок; е — эпидерма; II — внутренний проводящий пучок; р — паренхима; s — склеренхимные пластинки; sr — склеренхимное кольцо
338 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Рис. 11.152. Pteridopsida, Aspidiales, Dryo-pteris (A, C, D — по H. Schenck; В — no L. Kny):
A — внешний вид (1/4x); В — разрез через сорус: показана плацента со спорангиями и зонтиковидным индузием (ЗОх); С — перышко с молодыми, еще покрытыми индузием сорусами; D — то же, на более поздней стадии, со сморщившимися индузиями (Зх)
Сначала вырастет нитевидная, снабженная ризоидами «протонема», которая, однако, лишь у немногих форм сильно развита и тогда, например у Trichomanes (Hymenophyllaceae) и Schizaea (Schizaeaceae), несет на своих ветвях антеридии, а на особых многоклеточных боковых ветвях — архегонии (рис. 11.153, С). Обычно нитевидная стадия существует очень недолго и, образовав всего несколько клеток, дает на своем конце клиновидную двустороннюю апикальную клетку; отчленяемые ею сегменты делятся дальше (А, В). Таким путем формируется чаше всего сердцевидный, распростертый по субстрату, тонкий, пластинчатый заросток (см. рис. 11.128, А).
Наконец, апикальная клетка заменяется несколькими иниииалями. Антеридии и архегонии возникают на стороне, защищенной от падающего света, т. е. обычно на стороне, обращенной к почве и влаге; к концу своего развития они не погружены или слабо погружены в ткань заростка1. Архегонии образуются чаще всего позже, чем антеридии. При очень плохом питании образование архегониев полностью подавлено.
Антеридий представляет собой округлое выступающее образование, которое без ножки сидит непосредственно на одной эпидермальной клетке; он произошел от нее как сосочковидный вырост, отделенный поперечной перегородкой (рис. 11.154). Образуемые в нем сперматозоиды изогнуты в виде штопора и имеют множество жгутиков (F). Как и вообще у всех архего-ниат, основной объем сперматозоида занимает ядро. Сначала он несет на своем заднем конце пузыревидный остаток цитоплазмы с мелкими пластидами и крахмальными зернами в качестве запасного вещества, который, однако, при проникновении в архегоний отбрасывается.
Архегонии возникают в многослойной средней части более взрослых заростков путем деления одной поверхностной клетки каждый. Внутри зачатка архегония образуются шейковая канальцевая клетка и центральная клетка; последняя снова делится на яйцеклетку и брюшную канальцевую клетку (рис. 11.154, М). У некоторых видов может быть несколько шейковых канальцевых клеток (сравнительно-эволюционные аспекты — см. Обзор). Архегоний становится готовым к оплодотворению после разрыва брюшной и шейковых канальцевых клеток, расплывания содержащейся в них слизи и раскрывания шейки на вершине. Сперматозоиды привлекаются к шейке архегония и яйцеклетке (см. 8.2.1.1) по механизму хемотаксиса.
После первых делений зиготы (рис. 11.155, А) апекс побега (s) зародыша дифференцируется эндоскопически рядом с будущей гаусторией (0, а зачатки первого листа (Ь) и корня (w) обращены к шейке архегония. Корень возникает на лишенном подвеска (суспензора) зародыше не против побега, а сбоку от продольной оси зародыша, как и у всех папоротникообразных. Поскольку архегоний находится на нижней стороне заростка, то побег и первый лист зародыша после выхода из архегония должны.
1 Архегонии погружены брюшком в ткань заростка, как и у остальных Pteridophyta (исключение составляют Hymenophyllaceae и Schizaeaceae с нитчатыми заростками). — Примеч. ред.
11.2, Бактерии, грибы, растения | 339
Рис. 11.153. Pteridopsida (А, В — по Dopp; С — по К. von Goebel; D, Е — по P.Tryon):
А, В — Aspidiales, развитие заростка Matteuccia struthiopteris из споры (70х); А— 11-дневный, В — 21-дневный, с апикальной клеткой (s) и образованными ею сегментами (I—X); С — Hymenophyllales, Trichomanes rigidum, нитчатый заросток с архегониофорами (а), из которых один с проростком спорофита; D, Е — Gleicheniales, Platyzoma microphyllum (20х), cf заросток; Е — то же, <j> заросток
Рис. 11.154. Pteridopsida, Aspidiales (А— Е — по L. Кпу, с дополнениями no Schlumberger и Schrau-dolf; F — Dracinschi; G—N — no L. Kny):
A—E — развитие антеридия у Dryopteris filix-mas (250x), объяснение в тексте; F — сперматозоид Thely-pteris palustris (3000x); G—N — развитие архегония у Dryopteris filix-mas (20x), объяснение в тексте
проявляя отрицательный геоторопизм, изгибаться вверх (В). Спорофит остается еше некоторое время связанным через гаусторию (1) с заростком, пока тот не отомрет. К первичному корню позже добавляются многочисленные придаточные корни. Положение оси зародыша не может быть изменено ни силой тяжести, ни светом; следо
вательно, уже заросток у Leptosporangiatae должен проявлять полярность, которая затем передается цитоплазме яйцеклетки.
На листьях нередко встречаются адвентивные (выводковые) почки, которые отпадают и участвуют в вегетативном размно-
340 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Рис. 11.155. Pteridopsida, Pteridales. Pteridi-ит aquilinum. Развитие зародыша (А — по W.Zimmermann; В — по W. Hofmeister):
А — после первых делений зиготы в архегонии; В — на более поздней стадии гаустория погружена в расширенное брюшко архегония (а); b — первый лист; f— гаустория; р— заросток; s — апекс побега; w — корень
женин; вегетативному размножению служит также превращение стеблей и даже листьев в ползучие столоны. У некоторых видов наблюдается отклонение от нормального хода смены поколений из-за апогамии и апоспории (см. 10.1.3.3); чаще всего это бывает у полиплоидных форм, многие из которых у Pteridopsida имеют большие хромосомные числа.
Систематика. Классификация многочисленных видов лептоспорангиатных папоротников основана на различиях в строении спорангиев и их положении на спорофилле. На основе этих признаков различают несколько порядков.
У представителей первых трех порядков (1 — 3) кольцо на спорангиях отсутствует или оно поперечное.
1. Порядок: Osmundales. Сорусы отсутствуют, а спорангии без кольца: стенка спорангия разрывается на его вершине благодаря группе толстостенных клеток (рис. 11.156, А). Индузия и чешуй нет. Заростки долговечные, часто даже многолетние. Виды этого порядка, объединяемые в одно-единственное семейство (Osmundaceae), имеют спорангии либо на особых спорофиллах (Osmunda cinnamomea), либо на определенных участках трофофиллов: у осмунды королевской (О. regalis) видоизменены верхние перья вай, в остальном имеющих нормальный облик, у О. cla-ytoniana — средние. Семейство известно с верхнего карбона, в настоящее время представлено немногими родами.
2. Порядок: Gleicheniales. Сидячие спорангии имеют кольцо, проходящее поперечно выше середины. Они собраны небольшими группами в сорусы, которые, однако, не защищены покрывальцем. В ископаемом состоянии представители этого порядка известны с верхнего карбона; в настоящее время они широко распространены в тропиках. Вайи (ложно-Дихотомически ветвящиеся, со «спящими» почками в разветвлениях (рис. 11.157). Принадлежность рода Platyzoma (с выраженной гетероспорией, см. рис. 11.153, D, Е) к этому порядку спорна.
3. Порядок: Schizaeales. Расположенные по краю листа сидячие спорангии вскрываются продольной щелью посредством находящегося у самой верхушки поперечного кольца (см. рис. 11.156, В). Этот порядок также известен в ископаемом состоянии с верхнего карбона; в настоящее время его распространение ограничено преимущественно тропиками. Листья у Schizaea(-ceae) напоминают листья злаков, но дихотомируют, у Anemia(-ceae) они перистые с фертильной нижней парой перьев, у Lygodium (Lygodiaceac) — вьющиеся.
Рис. 11.156. Leptosporangiatae. Спорангии (А, В — по C.Luerssen; С — no F.O.Bower):
А — Osmunda regalis (Osmundales, устье открыто, 40х); В — Anemia caudate (Schizaeales); С — Нуте-nophyllum dilatatum (Hymenophyllales); D — Dryopteris filix-mas (Aspidiales, устье в плане и сбоку) (В— D — 70х)
11.2. Бактерии, грибы, растения
341
Следующие три порядка (4 — 6) имеют косо проходящее кольцо, как, например, у Hymenophy-llum (см. рис. 11.156, С) из 4-го порядка.
4. Порядок: Hymenophyllales. Спорангии почти сидячие. Сорусы располагаются по краю листа, каждый на рецептакуле (продолжении листовой жилки), иногда сильно удлиненном, и защищены чашевидным или двустворчатым покрывальцем. Листья чаще всего тонкие, с однослойной пластинкой без устьиц. Достоверные ископаемые остатки известны только с третичного периода. Ныне живущих видов известно около 650. Они обитают преимущественно во влажных лесах тропи ков и субтропиков, как, например, виды родов Hymenophyllum (однако Н. tunbrigense очень редко, но встречается в Европе) и Didymoglossum.
5. Порядок: Matoniales. Сидячие спорангии собраны небольшими группами в сорусы, которые покрыты шитовидным покрывальцем. Этот порядок, широко распространенный в мезозое, ныне представлен лишь тремя видами, обитающими на Малайском архипелаге.
6. Порядок: Cyatheales (включая Dicksoniales). Расположенные на ножке спорангии собраны в сорусы на поверхности или по краям листьев. Эти растения появились в юре. В настоящее время это преимущественно древовидные папоротники (похожие на пальмы деревья до 20 м высотой) в горных лесах тропиков и субтропиков. Представленные многими видами роды: Cyathea (включая Alsophila, см. рис. 11.150), Dicksonia, Ci-bothium.
У следующих порядков спорангии, чаще всего имеющие хорошо выраженную ножку, вскрываются благодаря продольному кольцу (рис. 11.156, D). в редких случаях— вертикальному кольцу.
7. Порядок: Polypodiales. Сорусы образуются на нижней стороне листа; покрывальце отсутствует. Вайи у многолетних растений перисто-рассеченные или однажды перистые. Сюда наряду с родом Polypodium относят также роды Drynaria (см. рис. 11.162, В), Platycerium (см. рис. 11.162, А), Microsorium и Pyrrosia.
8. Порядок: Pteridales. Сорусы расположены по краям перьев вай. Обычный представитель этого порядка — орляк, Pteridium aquilinum (Dennstaedtiaceae), с дважды- и четыреждыпе-ристосложными вайями до 2 м длины и длинными ползучими корневищами. Сорусы прикрыты с одной стороны закрученным краем листа, а с другой — покрывальцем. К тому же семейству относится род Acrostichum. У Adian-thum (Adianthaceae) наблюдается примитивное вееровидное жилкование листьев (см. рис. 4.60, А), закрученный край которых защищает сорусы; индузий у них отсутствует. Ceratopteris
Рис. 11.157. Pteridopsida, Gleicheniales, Gleiche-nia circinata, Австралия (1/5x) (по K. Magdefrau)
(Parkeriaceae) — водное растение co стерильными плавающими листьями и фертильными водными листьями, которые несут не собранные в сорусы шаровидные, почти сидячие спорангии с вертикальным кольцом. Сюда относят также семейства Gymnogrammaceae с родом Anogrumma, Sinopteridaceae с родами No-tholaena и Cheilanthes и Davalliaceae с родом Da-vallia.
9. Порядок: Aspidiales. Сорусы, защищенные покрывальцем, располагаются на нижней стороне листа. Листовая пластинка изредка простая или лопастная, а чаще всего одно-, четы-реждыперистая. К этому порядку включающему несколько семейств, относится большинство видов папоротников, известных в Германии. У видов семейства Thelypteridaceae ( Thelypteris) сорусы сидят вдоль центральной жилки перышка, у Aspleniaceae (Asplenium, Ceterach) — по бокам конечных жилок перышек. К последнему семейству относят также листовик, Phyllitis, с цельными языковидными вайями (см. рис. 11.151, С). Представители Aspidiaceae часто образуют щитовидные или почковидные индузии (например, Dryopteris, см. рис. 11.12, С: Bolbitis). К ним очень близки Athyriaceae с часто удлиненными сорусами (как у кочедыжника женского, Athyrium ftlix-femina} У также относящегося к предыдущему семейству страусника, Matteuccia struthio-pteris. как и у представителей следующего порядка, спорофиллы отличаются от зеленых тро-фофиллов, которые воронковидно налегают друг на друга.
10. Порядок: Blechnales. У единственного встречающегося в Германии представителя этого порядка блехнума колосистого (Blechnum spi-cant) несколько темно-коричневых спорофиллов располагаются в розетке зеленых трофофил-лов, сорусы вытянуты на всю длину узких фертильных перышек. Кроме рода Blechnum сюда же относят род Salpichlaena.
342 | ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Ступень развития: водные папоротники (Hydropterides) (= Salviniidae)
К водным папоротникам относят всего несколько родов трав, обитающих в воде или на болотах. Они все разноспоровые. Их мега-и микроспорангии тонкостенные, не имеют
кольца и находятся внутри особых вместилищ, расположенных в основании листа (например, рис. 11.158, С). Мейоспоры покрыты своеобразным периспорием, возникающим из плазмодиального тапетума.
К водным папоротникам относят два порядка примерно со 100 видами.
Рис. 11.158. Hydropterides, Salviniales. Salvinia, S. natans В — no G.W. Bischoff; C—F — no E.Stras-burger; G—J — no W. C. Belajeff; К — no N. Pringsheim):
A — участок побега сбоку, видны округлые вместилища спорангиев (3/4х); В — то же, вид сверху (3/4х); С — вместилища мегаспорангиев (та) и микроспорангиев (mi) в продольном сечении (8х); D — микроспорангий (55х); Е — микроспоры, погруженные в пенистую массу (250х); F — мегаспорангий с мегаспорой, окруженной периспорием (р), в продольном сечении (55х); G — J — О’ заросток: G — деление микроспоры на три клетки I — III (860х), Н — зрелый заросток сбоку, J — вид с брюшной стороны. Клетка I поделилась на проталлиальные клетки pz, и pz2 (pz, — нефункционирующая ри-зоидальная клетка); клетка II — на стерильные клетки sz,, sz2 и две сперматогенные клетки sp,, каждая из которых образует 2 сперматозоида; клетка III — на стерильные клетки sz3, SZ4 и две сперматогенные клетки sp2. Клетки sp, sp, и sp2 sp2 представляют собой два антеридия (в состав антеридия входят не только сперматогенные клетки, но и клетки стенки. — Примеч. ред.); клетки sz,— sz4 — их стенки; цифры 1 — 1 и 2—2 отмечают положение первых клеточных стенок (640х); К— зародыш (е) на продольном срезе, заросток (рг) с хлоропластами, Ь, — Ь3— первые листья (100х); аг — остаток архегония; es — экзоспорий; f — гаустория; s — споровая клетка; ss — апекс побега; sw — стенка спорангия; wb — водный лист
11.2. Бактерии, грибы, растения | 343
1. Порядок: Salviniales (единственное семейство Salviniaceae). Относящиеся к этому порядку виды — свободно плавающие водные растения. Род Salvinia во флоре Германии представлен ставшим редким плавающим папоротником X. natans, который на каждом узле слабо разветвленного побега несет по три листа.
Два верхних зеленых листа, плавающих на поверхности воды, овальные (рис. 11.158, А), со множеством крупных межклетников. Нижние листья (wb), напротив, разделены на свисающие в воде многочисленные бесцветные нитевидные лопасти, покрытые волосками. Они выполняют функцию отсутствующих корней (гетерофиллия1, см. 4.3.2). В основаниях водных листьев сидят по нескольку шаровидных вместилищ спорангиев (А); они заключают в себе спорангии, возникающие на столбиковидной плаценте (С).
По своему заложению плацента соответствует модифицированной лопасти водного листа, тогда как покров вместилища можно рассматривать как двухслойное покрывальце. Оно возникает в форме кольцевого валика, который в виде чаши и, наконец, сферы обрастает плаценту с расположенным на ней сорусом спорангиев и затем плотно смыкается на вершине.
Каждое такое вместилище содержит один сорус — либо из более многочисленных микроспорангиев, либо из мегаспорангиев в меньшем числе (рис. 11.158, С: mi, ma); определение пола заростка осуществляется, таким образом, дипломодификаци-онно. Спорангии обоих типов расположены на ножках и имеют в зрелом состоянии однослойную стенку (D, F); в них после редукционного деления возникают мейоспоры.
Микроспорангии содержат 64 микроспоры, образующиеся в тетрадах. Они погружены в пенистое, твердеющее вещество (периспорий, рис. 11.158, Е).
Из каждой микроспоры развивается короткий трубковидный о* заросток, который состоит всего из немногих клеток. Он образует только 2 антеридия (рис. 11.158, Н). Каждый антеридий образует (из 2 сперма-тогенных клеток, дающих начало 4 сперма-
1 Правильно: анизофиллия. — Примеч. ред.
тидам) 4 сперматозоида; они выходят наружу через разрыв в клеточных стенках. Таким образом, заросток сильно редуцирован. Его развитие происходит внутри спорангия, который не вскрывается. Вместо этого заростки вытягиваются наподобие пыльцевой трубки, локально прободают его стенку, высвобождая сперматозоиды.
Мегаспорангии крупнее, чем микроспорангии, и также имеют однослойную стенку (рис. 11.158, F), однако содержат только одну мегаспору, так как лишь одна из 32 возникающих (из 8 материнских клеток спор) спор завершает свое развитие за счет остальных. Мегаспора плотно заполнена белковыми тельцами, капельками масла и крахмальными зернами; на ее вершине находится более плотная цитоплазма с ядром. Коричневая оболочка (экзоспорий) покрыта толстым ячеистым покровом — периспорием. Мегаспора остается внутри спорангия, отделяется вместе с ним от материнского растения и плавает по поверхности воды. На вершине микроспоры при прорастании возникают состоящий из мелких клеток заросток (рис. 11.158, К) и лежащая позади него крупная клетка (s), богатая запасными веществами: она служит для питания заростка и не делится дальше, хотя ее ядро в результате митозов дает многочисленные постенно расположенные дочерние ядра1.
Оболочка споры вскрывается тремя створками, стенка спорангия также разрывается, и заросток выставляется наружу в виде маленького дор-совентрального образования. Хотя он и содержит хлоропласты, однако использует запасные вещества в крупной клетке (s). Он развивает несколько архегониев, однако только одна яйцеклетка после оплодотворения дает зародыш, который своей гаусторией внедряется в расширяющееся и наконец разрывающееся брюшко архегония (рис 11.158, К). Если ни один из архегониев не оплодотворен, то образуются новые архегонии.
Второй род — Azolla — преимущественно тропический; эти изящные, обильно разветвленные плавающие растеньица несут тесно налегающие друг на друга листья, расположенные в два ряда, а на ниж
1 И мелкоклеточная ткань, и крупная многоядерная клетка представляют собой части одного заростка. — Примеч. ред.
344 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
ней стороне побега — длинные корешки (рис. 11.159, А).
Каждый лист разделен на две лопасти, из которых верхняя находится на плаву и ассимилирует, а нижняя погружена в воду и участвует в ее поглощении (А, В); кроме того, на отдельных боковых ветвях нижние лопасти листьев преобразованы во вместилища спорангиев и прикрыты выростом лопасти. В полостях верхней лопасти в качестве симбионта обитает цианобактерия Anabaena azollae. связывающая атмосферный азот; поэтому Azolla используют на рисовых полях как зеленое удобрение (ср. Cyanobacteriota). Azolla интересна своими приспособлениями к обеспечению надежного оплодотворения. После выхода из микроспорангия 64 микроспоры оказываются размешенными по нескольку в 5 — 8 образованных периплазмодием округлых комках, способных плавать, — так называемых мас-сулах. Каждая массула имеет на поверхности выросты в виде двусторонних крючков на ножке — глохидии (Н, J), которые также возникают
из вещества периплазмодиального тапетума. Эти крючки служат для прикрепления к мегаспоре, которая перемещается в воде с помощью особого поплавка, содержащего воздух (G, J). Он образуется из сильно вакуолизированного периплазмодия на верхушке мегаспорангия. Заросток развивается, как и у Salvinia.
2. Порядок: Marsileales. К порядку относят роды, виды которых обитают на заболоченной почве. Род Marsilea (Marsileaceae), который до недавнего времени был представлен в Германии видом М. quadrifolia, ныне исчезнувшим (рис. 11.160, А), характеризуется ползучим разветвленным корневищем с расположенными поодиночке листьями на длинных черешках, несущими очень близко сидящие пары перышек. Свойственные листьям никтинастии не встречаются у остальных папоротников. Над основанием черешка возникают попарно, а у других видов в большем числе, овальные
Рис. 11.159. Hydropterides, Salviniales. Azolla (А, Н — Azolla caroliniana, иначе A. filiculoides) (A, D— J — по Е. Strasburger; В, С — по К. von Goebel):
А — растение, вид сверху (4х); В — вершина побега, сверху (12х); С — то же, в поперечном сечении (12х); D — продольный срез верхней лопасти листа; в полости Anabaena azollae (70х), Е — <з" (вверху) и <j> (внизу) сорусы (20х); F — микроспорангий (65х); G — окруженный индузием мегаспорангий, содержащий мегаспору с поплавками (65х); Н — часть массулы с глохидиями (160х); J — мегаспора, у которой удалена верхняя половина индузия, чтобы были видны поплавки; к эписпорию прикрепились три массулы посредством глохидиев (65х)
11.2. Бактерии, грибы, растения | 345
Рис. 11.160. Hydropterides, Marsileales (А, Н — по G.W. Bischoff; В — по Johnson; С — по К. Magdefrau; D — по J.Hanstein; Е — по W.C.Belajeff; F — по L.W. Sharp; G — по D. Н. Campbell):
А — Marsilea quadrifolia, внешний вид (2/Зх); В — срез молодого спорокарпия; точками показан зачаток соруса (200х); С — зрелый спорокарпий (8х); D — вскрывшийся спорокарпий М. salvatrix (натуральная величина); Е — проросшая микроспора с двумя антеридиями (150х); F — сперматозоид (700х); G — архегоний (150х); Н — Pilularia globulifera, внешний вид (2/Зх); g — кольцо хрящевидной ткани; I — проводящий пучок; те — мегаспорангий; mi — микроспорангий; s — спорокарпий; ss — камеры с сорусами
вместилища спорангиев на ножках. В противоположность Salviniales у Marsileales оболочка каждого вместилища соответствует по своему происхождению ассимилирующему перышку, у которого вследствие усиленного роста нижней стороны зачатки сорусов смещаются внутрь (В). Поэтому эти вместилища называют спорокарпиями.
Сорусы, состоящие из мегаспорангия и многочисленных микроспорангиев, располагаются рядами, которые заключены в камеры (С). При созревании спор кольцо хрящевидной ткани (g), опоясывающее спорокарпий, расплывается и выдергивает наружу камеры с сорусами (D). Мужской заросток, который остается заключенным в оболочке микроспоры (Е), образует два антеридия, в которых возникают сперматозоиды (F), спирально закрученные подобно штопору. Женский заросток развивает один архегоний (G). Род Pilularia с встречающимся в Германии видом Р. glo
bulifera отличается от Marsilea простыми линейными листьями, у основания которых поодиночке возникают шаровидные спорокарпии, по происхождению также соответствующие ассимилирующему перышку (рис 11.160, Н). В спорокарпиях Pilularia имеется четыре камеры с сорусами. Листья видов обоих родов, как и у большинства других папоротников, в молодости закручены улиткой (А. Н; акропластический рост, см. 4.3.1.2).
В ископаемом состоянии Azolla известна с нижнего мела, Salvinia — с верхнего мела и Pilularia — с миоцена. В Северной Америке Salvinia в миоцене вымерла.
Распространение и образ жизни папоротникообразных
Папоротникообразные распространены во всех климатических зонах, однако наибольших размеров (древовидные папорот
346 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ___________________________________
ники!) и наибольшего видового разнообразия они достигают — прежде всего Pteridopsida и Lycopodiopsida — в тропиках. Как и бриофиты, папоротникообразные предпочитают более влажные места, однако отдельные виды заходят и в сухие области. Засоленных мест они избегают; только папоротник Acrostichum аигеит обитает в мангровых болотах всех тропических областей.
Папоротникообразные в ходе приспособления к различным условиям жизни на суше приобрели целесообразные морфологические и физиологические особенности и представлены теми же жизненными формами (см. рис. 4.19), что и семенные растения. По особенностям водного режима они занимают промежуточное положение между бриофитами и семенными растениями. Заростки большинства папоротникообразных столь же малоустойчивы к высыханию, как и протонемы мхов. Однако взрослые зеленые растения папоротников у большинства видов сами регулируют содержание воды в теле (гомойогидридные птери-дофиты) — в противоположность полностью зависящим от содержания воды во внешней среде и устойчивым к высыханию бриофитам. Правда, у некоторых пой-килогидридных видов Selaginella (Lycopodiopsida) и папоротников (Pteridopsida), напри-
Рис. 11.161. Leptosporangiatae, Polypodi-aceae. Polypodium vulgare, гидропота (80x) (no K. Magdefrau)
мер у Ceterach, Notholaena и Cheilanthes, вечнозеленые листья могут высыхать до воздушно-сухого состояния и после пребывания в нем в течение многих месяцев при увлажнении восстанавливать жизнедеятельность.
Система внутреннего проведения воды играет решающую роль в поддержании водного режима; некоторые папоротникообразные дополнительно используют приспособления к капиллярному поглощению и накоплению воды (язычки у Selaginellaceae и Lepidodendrales).
Очень немногие ксерофиты, например из Pteridopsida, защищены от высыхания восковым налетом, покровом из сухих чешуек и волосков или даже имеют сукку-лентные побеги (DavalHa) или листья (например, некоторые виды Polypodium). У обитателей влажных мест (гигрофитов) мы наблюдаем гуттацию, будь то через гидатоды на зубцах листовых влагалищ Equisetum или через своеобразные гидропоты у некоторых папоротников (рис. 11.161).
Наряду с вечнозелеными видами родов Lycopodium, Selaginella (Lycopodiopsida), Equisetum (E. Ityemale; Equisetopsida) и Polypodium (Pteridopsida) значительная часть папоротникообразных умеренной и холодной зон летнезеленые (ср. в этом плане бриофиты). Папоротникообразные в разных эволюционных линиях, некоторые уже в девоне и карбоне, выработали древовидные формы (фанерофиты; жизненные формы, см. рис. 4.19): например, среди Lycopodiopsida — Sigillaria, Lepidodendron, Pleu-romeia', среди Equisetopsida — Catamites', среди Pteridopsida — различные Primofilices, эуспорангиатные (Megaphyton, рис. 11.148, А) и лептоспорангиатные папоротники (Cyathea, см. рис. 11.150). Предпосылками к этому были укрепление осей (отчасти благодаря вторичному утолщению, но чаще посредством других приспособлений и эффективной конструкции проводящих тканей. К фанерофитам относятся также распространенные, особенно в тропиках, лазающие (лианы) и эпифитные папоротникообразные: например, тропические Gleicheniaceae (лазящие растения); Lygo-dium и Salpichlaena со вьющимися рахисами до 15 м длиной; отдельные виды рода Polypodium, взбирающиеся с помощью кор-
11.2. Бактерии, грибы, растения | 347
Рис. 11.162. Эпифитные папоротники с диморфными листьями (гетерофиллия: стерильные листья, образующие нишу для накопления гумуса, и споротрофофиллы) (по К. Magdefrau):
А — Platycerium alcicorne; В — Drynaria quercifolia (1/6х)
ней на стволы деревьев в тропиках; Platycerium и Dtynaria как эпифиты со стерильными листьями, образующими нишу для накопления гумуса (рис. 11.162). Есть основания предполагать, что все ныне живущие птеридофиты произошли от вечнозеленых древовидных предков с исходно автотрофными заростками. В дальнейшем возникли жизненные формы: хамефиты, некоторые из них имеют микотрофные (Lycopodium), а другие — существующие за счет запасов споры заростки (Selaginella), гемикриптофиты с автотрофными, недолговечными заростками (например, Dryopteris), геофиты с микотрофными (например, Ophioglossum) или автотрофными заростками (например, Pteridium, Equisetum) и терофиты (например, A nogram та). Если не считать заростков некоторых видов и в значительной мере микотрофного Ophioglossum simplex, то к гетеротрофному образу жизни (паразитизм) папоротникообразные, как и мхи (однако ср. Cryptothallus, Bryophytina), не перешли. Немногие представители вторично приспособились к жизни в воде (как гидрофиты): например, из Pteridopsida— Salvinia и Azolla (Salvi-
niales) как плавающие растения, а также Ceratopteris (Pteridales). Некоторые виды Ceratopteris живут как плавающие или в той или иной мере погруженные в воду, а некоторые обитают на влажной почве. Культивируемые в аквариумах Bolbitis heudelottii и Microsorium pteropus образуют под водой только стерильные вайи, тогда как сорусы развиваются исключительно на выступающих из воды листьях. Виды рода Isoetes (Lycopodiopsida) обитают частью на периодически заливаемой почве, частью на дне озер, нередко на глубине 1 — 3 м.
Папоротникообразные особенно конкурируют с особями сходных жизненных форм: древовидные папоротники, например, — с голосеменными, пальмами и древесными двудольными, заростки папоротников, а также спорофиты Hymenophyllaceae — со мхами и лишайниками; Equisetum — с Juncaceae и Cyperaceae; Salvinia — с Lemnaceae и т.д. Некоторые папоротникообразные при этом оказываются весьма конкурентоспособными и в благоприятных условиях встречаются в таких количествах, что образуют чистые заросли, как орляк (Pteridium aquilinum) по опушкам лесов или хвощ приречный (Equisetum fluviatile) по топким берегам озер. Некоторые виды распространены по всему земному
348 [ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
шару, например тот же орляк (как лазящее растение иногда до 4 м высотой) или плаун булавовидный (Lycopodium clavatum); можно привести примеры и ограниченных, разорванных и эндемических ареалов.
Обзор папоротникообразных. Папоротникообразные можно рассматривать как эволюционную ветвь, возникшую параллельно мохообразным от общей, возможно, уже обитавшей на суше предковой группы («Propsilophytopsida»), которая в свою очередь произошла от водорослевых предков. Среди водорослей в качестве предков этих ранних обитателей суши подходят только стрептофиты. Ранее обсуждавшаяся эволюция от мохообразных к папоротникообразным, например от форм типа Anthoceros, путем увеличения размеров, дифференциации и увеличивающейся самостоятельности спорофита (Sporogonites), напротив, представляется маловероятной. Если мхи уже с карбона не претерпевали заметного прогрессивного развития, т. е. уже примерно 250 млн лет назад были «готовыми», папоротникообразные достигли расцвета только после этого времени.
Если мхи завоевали сушу с помощью гаметофитов и при этом остались ограниченными особыми адаптивными зонами, то папоротникообразные и (прежде всего) семенные растения достигли доминирующей роли в формировании наземной растительности при прогрессирующем развитии своих спорофитов. Эволюционное преимущество спорофитных растений связано, вероятно, с их генетической стабильностью (буферный эффект) и повышенной частотой рекомбинаций (после каждого события сингамии возникающие диплоидные ядра воспроизводятся митотически; вследствие этого в мейоз вступают многочисленные диплоидные ядра), а также с развитием защитных приспособлений для чувствительного к наземным условиям гаметофита. Диплоидный спорофит папоротникообразных развит очень сильно и весьма разнообразен, что в противоположность мхам стало возможным благодаря прогрессирующему развитию в ходе эволюции одревесневших (соответственно эффективно работающих) проводящих пучков (проведение как воды, так и органических веществ1). Аналогичный
результат имеет также развитие настоящих корней. Поскольку к тому же эпидерма ку-тинизирована, то побег может расти вверх в освещенное воздушное пространство, развивать листья и ассимилировать диоксид углерода. Его обеспечение органическими веществами, таким образом, не зависит от гаметофита, из-за чего отпадает необходимость в дальнейшем росте последнего.
Верхушечный рост осуществляется чаще всего посредством апикальной клетки (рис. 3.2. А). Напротив, благодаря множественным инициалям растут эвспорангиатные папоротники и некоторые представители Lyco-podiopsida (Lycopodium, Selaginella — у последнего рода еше отчасти посредством апикальной клетки). Среди Psilophytopsida этим прогрессивным признаком отличался уже род Rhynia1.
Положение зародыша (см. рис. 11.155) чаще всего эндоскопическое (верхушка побега направлена от шейки архегония — Leptosporangiatae, а также Lycopodium, Selaginella), редко экзоскопическое (Eusporangia-tae, за исключением Marattiales, Psilotum, Equisetum, Isoetes).
У исходных форм все споры одинаковые (изоспория); у более высокоорганизованных уже появляется дифференциация на микро- и мегаспоры. Гетероспория неоднократно и независимо появлялась в разных классах папоротникообразных (Lyco-podiopsida; Equisetopsida — как у каламитов, так и у клинолистных, и Pteridopsida); с этим связано разделение функций между мелкими сГ заростками и более крупными <j> заростками. В виде «семенных плаунов» папоротникообразные независимо от семенных растений достигли такого уровня развития, как образование семени с крайней степенью гетероспории.
Гаплоидный гаметофит папоротникообразных остается (если еще не слишком сильно ре- 1 2
1 Автор этого раздела, по-видимому, своеобразно представляет роль разных тканей в проведении тех или иных веществ, равно как и значение одревеснения. К высказанной автором идее, по-видимому, следует относиться с большой осторожностью. — Примеч. ред.
2 Строение верхушки телома у Rhynia точно не известно. — Примеч. ред.
11.2. Бактерии, грибы, растения | 34g
дуцирован) на уровне организации таллома (заросток) и изредка развивает трахеиды (Psilotum). Он рано заканчивает развитие, образовав антеридии и архегонии, которые часто устроены проще, чем у мхов; крупные многоклеточные гаметангии считают примитивными по сравнению с
мелкими, состоящими из немногих клеток. Если листостебельные мхи (Bryopsida) имеют в архегонии многочисленные шейковые канальцевые клетки (10—30 или больше), то у печеночников (Marchantiopsida) их 4—8, у антоцеротовых (Anthocerotopsida) — 6, а у папоротникообразных
Рис. 11.163. Развитие важнейших групп растений в истории Земли (по К. Magdefrau)
1 Вышедшие из употребления названия.
350 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
часто только одна или несколько. У Anthocerotopsida эпидермальная клетка, становящаяся ини-циалью архегония, больше не образует клетки ножки, т. е. в отличие от Marchantiopsida и Bryopsida она сразу делится на осевую клетку и три клетки стенки архегония. У папоротникообразных нет и делений, дающих клетки стенки архегония1. Антеридии и архегоний Bryopsida и Marchantiopsida закладываются экзогенно и свободны, т.е. только позже окружаются тканью гаметофита. У Anthocerotopsida и папоротникообразных они уже на ранних стадиях развития заключены в ткань гаметофита (эндогенное образование)1 2.
История развития. Psilophytopsida, Lycopodiopsida и Equisetopsida достигли наибольшего расцвета как по разнообразию форм, так и по численности особей в палеозое. Представительство Pteridopsida значительно возросло еще в мезозое, и они лучше сохранились до сих пор, чем два других доживших до нашего времени класса (рис. 11.163). Их формы, господствовавшие от карбона до триаса, в настоящее время сохранились лишь в виде немногих видов, тогда как те семейства, которые доминируют в настоящее время, появились только в мезозое.
Четвертый подотдел.* семенные растения (Spermatophytina)
Семенные растения (примерно 250 000 видов) — самая большая современная группа растений. Они господствуют в наземной растительности почти во всех частях света и составляют основу всей наземной жизни. Для человека семенные растения имеют огромное значение, так как большинство культивируемых растений относятся именно к этой группе, в особенности к покрытосеменным.
ЧЕРЕДОВАНИЕ ПОКОЛЕНИЙ
Пяти современным группам семенных растений (Gymnospermae: Cycadopsida, Со-
1 Это утверждение весьма спорное действительности. — Примеч. ред.
2 У всех высших растений (кроме антеридиев Anthocerotophyta) гаметангии развиваются из поверхностной клетки, т.е. всегда экзогенны. — Примеч. ред.
niferopsida, Ginkgopsida, Gnetopsida; Angio-spermae: Magnoliopsida), как мхам и папоротникообразным, свойственно гетероморфное чередование поколений и диплогаплон-тное чередование ядерных фаз с диплоидным спорофитом и гаплоидным гаметофитом (рис. 11.165). Как и у современных папоротникообразных, спорофит семенных растений состоит из корня и побега со стеблем и листьями. Гаметофит в сравнении с большинством современных папоротникообразных сильно редуцирован. Семенные растения вместе с мхами и папоротникообразными составляют группу эмбриофитов, а вместе с папоротникообразными относятся к кормофитам (Kormophyta — растения, имеющие побеги1), или трахеофитам (Тга-cheophyta— сосудистые растения).
Лишь в 1851 г. W. Hofmeister обнаружил «тайное» чередование поколений у семенных растений и тем самым установил их тесную связь с мхами и папоротникообразными. Уже тогда в ботанике устоялись собственные названия для органов размножения семенных растений. И хотя их гомология с соответствующими органами папоротникообразных надежно установлена, до настоящего времени применяют обе группы понятий (рис. 11.164).
Как и отдельные папоротникообразные, семенные растения разноспоровые. В мегаспорангии (= нуцеллусе) чаще всего образуется только одна материнская клетка мегаспоры (= материнская клетка зародышевого мешка), из которой в результате мейоза получаются четыре мегаспоры. Обычно только одна мегаспора развивается в женский гаметофит (= зародышевый мешок). Женский гаметофит образует яйцеклетку2. В отличие от большинства папоротникообразных у семенных растений мегаспора не покидает мегаспорангий и соответственно материнское растение является спорофитом, и поэтому женский гаметофит развивается на материнском растении и за его счет. Кроме того, мегаспорангий семенных растений окружен стерильной оболочкой,
1 Группа включает все мхи и большинство печеночников. — Примеч. ред.
2 У многих семенных растений женский гаметофит образует архегоний. — Примеч. ред.
Мхи
Папоротникообразные
Семенные растения
Гаметофит | Спорофит
Спора ♦ Редукционное деление 4 Спороцит 4 Спорангий (коробочка)
Спорогон
4
Зародыш, развивающийся на гаметофите
4
Зигота
Сперматозоид 4 Антеридий
Яйцеклетка
4
Архегоний
Равноспоровые
Г“““----------
Спора 4 Редукционное деление 4 Спороцит 4 Спорангий 4 Спорофилл Олиственное взрослое растение
Зародыш, только вначале связанный с гаметофитом
4
Зигота
Сперматозоид Яйцеклетка
i I
Антеридий Архегоний
Разноспоровые
Голосеменные
Покрытосеменные
Мегаспора
Зеленое растение мха протонема 4_______________
Зеленый заросток
t________
Микроспора А
Редукционное деление
4
Микроспороцит
4
Микроспорангий
4
Микроспорофилл ^Мегаспорофилл
4
Мегаспороцит
4
Мегаспорангий
4
Олиственное взрослое растение
Зародыш, только вначале связанный с гаметофитом
4
Зигота
Сперматозоид
Антеридий
Упрощенный мужской гаметофит внутри микроспоры
Яйцеклетка
Архегоний
Упрощенный женский гаметофит внутри мегаспоры
4_____
Мегаспора
4
Микроспора
4
Редукционное деление
4
Микроспороцит
4
Пыльник
4
Тычинка
4 Мегаспороцит 4 Семяпочка 4 Плодолистик
Взрослое семенное растение
Зародыш в семени
Зигота
Сперматозоид или спермий
Антерцдальная и вегетативная клетки в пыльцевом зерне (или пыльцевой трубке) А
Яйцеклетка
4
Многоклеточный женский гаметофит (первичный эндосперм) в семяпочке
I
Рис. 11.164. Сравнение смен поколений и ядерных фаз у Embryophyta (Cormobionta).
Микроспора Мегаспора
4 4
Редукционное деление
4 4
Микроспороцит Мегаспороцит
4 4
Пыльник Семязачаток
4 4
Тычинка Плодолистик
Взрослое семенное растение ! Зародыш в семени f Зигота
Спермий Яйцеклетка
Генеративная Зародышевый
и вегетативная мешок клетки в пыльцевом зерне (пыльцевой трубке)
4 4________
Изображены схемы смен у мхов, равно- и разноспоровых папоротникообразных, а также у семенных растений. Гомологичные фазы развития, репродуктивные клетки и органы расположены на одном горизонтальном уровне (ср. также рис. 11.108; 11.128; 11.137; 11.165)
11.2. Бактерии, грибы, растения
СЛ
352 I ГЛАВА 11 СИСТЕМАТИКА И ФИЛОГЕНИЯ
интегументом, отсутствующим у папоротникообразных. Интегумент, нуцеллус и зародышевый мешок с яйцеклеткой в совокупности называют семязачатком1. В микро
1 Семязачаток— это комплекс из нуцеллуса и интегумента (интегументов). — Примеч. ред.
спорангии образуются многочисленные материнские клетки микроспор (= материнские клетки пыльцевых зерен), из которых после мейоза возникает множество микроспор. Мужской гаметофит (пыльцевое зерно) развивается внутри оболочки пыльцевого зерна и образует имеющие жгутики сперма-
11.2. Бактерии, грибы, растения | 353
Рис. 11.165. Чередование поколений и ядерных фаз у голосеменных (Gymnospermae, Pinus/ Coniferopsida, А—F) и покрытосеменных (G—К) (no F. Firbas).
Голосеменные: А — прорастающее семя с тестой, первичным эндоспермом (гаплоидным) и зародышем; В — побег со стеблем, листьями, а также сГ и соцветиями*; С— о" стробил и <j> констробил (молодая шишка); D — микроспорофилл с материнской клеткой микроспор, микроспорами и пыльцевыми зернами (воздушные мешки не показаны) как стадиями развития 6" гаметофитов’ кроющая чешуя шишки, над ней — семенная чешуя и открыто лежащий семязачаток, в котором развивается только 1 из 4 мегаспор; Е — семенная чешуя и семязачаток ко времени оплодотворения проросшим пыльцевым зерном, 6" и <j> гаметофит (д) с 2 крупными яйцеклетками; F — семенная чешуя зрелой шишки с (крылатым) семенем и зародышем в (первичном) эндосперме. Покрытосеменные: G — прорастающее семя; Н — целое растение с корнем, стеблем, листьями и бутоном обоеполого цветка; J — распустившийся цветок с околоцветником (чашелистики и лепестки), тычинками (с пыльцевыми зернами) и плодолистиками (завязь, содержащая семязачаток, столбик, рыльце) после опыления (пыльцевая трубка!) и перед самым оплодотворением яйцеклетки в зародышевом мешке; К — семя с тестой, вторичным (триплоидным) эндоспермом и зародышем, выпадающее из односемянного плода. Двойная штриховка: гаметофит (д) и первичный эндосперм; крап: триплоидный эндосперм; черный цвет: диплофаза (2л); светлый тон: гаплофаза (л); R! — редукционное деление (мейоз); s — спорофит
* Эти структуры правильно называть конст-робилами, а не соцветиями. — Примеч. ред.
<
ющаяся в зародыш. После оплодотворения семязачаток развивается в семя. Оно состоит из семенной кожуры, развивающейся из интегумента. многоклеточного зародыша и (почти всегда) питающей ткани (чаще всего эндосперма). Семя заменило одноклеточную спору папоротникообразных как средство распространения.
Мега- или микроспорангии расположены поодиночке или группами на простых или более или менее сложно разветвленных органах, которые можно назвать микроспорофиллами или мегаспорофиллами (тычинками и плодолистиками)1.
Хотя термины «микроспорофилл» и «мегаспорофилл» подразумевают их гомологию с листьями, совершенно не ясно, принимали ли участие и в какой мере листья в образовании микро- и мегаспорофиллов у разных групп семенных растений. У некоторых современных групп растений они совсем не похожи на листья. Также не ясна гомология микро- и мегаспорофиллов разных групп семенных растений. Термин «плодолистик» следовало бы использовать только для обозначения особых структур покрытосеменных1 2.
Спорофиллы у Spermatophytina почти всегда расположены на укороченных побегах с ограниченным ростом. Такие структуры, имеющиеся уже у некоторых папоротникообразных, можно называть цветками3, но чаще всего это понятие используют только для первично обоеполых собраний спорофиллов покрытосеменных и предположительно близкородственных им ископаемых семенных растений.
ВЕГЕТАТИВНЫЕ ОРГАНЫ
Проростки семенных растений биполярны и имеют конусы нарастания первичного стебля и первичного корня4. Число за-
тозоиды или безжгутиковые спермин. В процессе опыления мужской гаметофит переносится к семязачатку, где он прорастает, образуя пыльцевую трубку, которая, удлиняясь, доставляет к яйцеклетке или подвижные сперматозоиды, или безжгутиковые, неподвижные спермин. В результате оплодотворения яйцеклетки сперматозоидом или спермием получается зигота, развива-
1 Тычинки и плодолистики свойственны только покрытосеменным растениям. — Примеч. ред.
2 Это сделано в мировой ботанике еще в первой половине XX в. — Примеч. ред.
3 Такое чрезмерно расширенное применение термина «цветок» вышло из употребления на рубеже XIX—XX вв. — Примеч. ред.
4 Такую структуру имеют сформировавшиеся зародыши. У проростков есть уже главный корень и почка главного побега. — Примеч. ред.
354 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
родышевых листьев (семядолей) обычно 2, но может увеличиваться или уменьшаться до одного. Апикальные меристемы стебля и корня с несколькими инициальными клетками дифференцированы на несколько зон (см. рис. 3.4; 3.6). Боковые корни образуются эндогенно. У некоторых групп, главным образом из покрытосеменных (например, у однодольных), главный корень рано прекращает рост. Его функции частично принимают на себя снабженные корнями подземные побеги (например, корневище), но в первую очередь — придаточные корни на надземных побегах. Проводящие пучки корня чаще всего образуют актиностелу1. Побеги нарастают моно- или симподиально, они либо однотипные, либо дифференцированы на удлиненные и укороченные. Ветвление в типе боковое, точнее — пазушное (= аксилляр-ное). В стебле чаще всего имеется эвстела или сифоностела из расположенных кольцом открытых коллатеральных проводящих пучков, реже — атактостела с равномерно распределенными по поперечному сечению стебля закрытыми проводящими пучками. Большинство семенных растений способны к вторичному утолщению, при котором камбий образует внутрь новые слои древесины, а наружу — луб. Однако способность к вторичному утолщению часто бывает утрачена или заменена аномальным вторичным утолщением. Соответственно среди семенных растений имеются как древесные, так и травянистые представители1 2. Листья расположены спирально, двурядно, накрест супротивно или мутовчато.
Важные систематические признаки дает анатомия побега. В ксилеме встречаются или только трахеиды, или наряду с трахеидами еще и сосуды. У сосудов можно наблюдать разный наклон разделяющих стенок и разную степень их разрушения при редукции перекладин лестничных перфорационных пластинок, а также различные укорочение и расширение члеников сосудов. Однако разнообразие этих признаков отражает не
1 На стадии первичного строения в корне семенных растений развит единственный радиальный пучок. — Примеч. ред.
2 Большинство трав обладают типичным вторичным утолщением, хотя и незначительным. — Примеч. ред.
только родство, но и экологические особенности. Во флоэме у голосеменных находятся ситовидные клетки, а у покрытосеменных— происходящие из одной материнской клетки членики ситовидных трубок и клетки-спутники. У ситовидных трубок также можно наблюдать разнообразие степени наклона ситовидных пластинок, размеров ситовидных каналов в них, длины и диаметра самих члеников ситовидных трубок. Проводящие пучки мотуг быть не только коллатеральными. но и биколлатеральными благодаря образованию внутри от ксилемы еще одного тяжа флоэмы. Другие анатомические признаки стебля, используемые в систематике, — образование специализированных механических элементов (древесинных и лубяных волокон), взаимное расположение древесинной паренхимы и сосудов, пространственное разделение механических и проводящих тканей, а также степень однородности и радиальной вытянутости клеток лучей.
Весьма разнообразны также формы роста и жизненные формы семенных растений. У древесных форм иногда различают лептокаульную форму роста со сравнительно обильным ветвлением и тонкими ветвями (например, Со-niferopsida) и пахикаульную форму со слабым ветвлением и относительно толстым стволом (например, Cycadopsida). Из листопадных или вечнозеленых деревьев или древовидных форм многократно независимо возникали, особенно у покрытосеменных, кустарники и полукустарники, лианы, многолетние травы, геофиты и эпифиты с самым разным расположением покоящихся органов и, наконец, однолетние травы. Однако древесные формы могут возникать и вторично из травянистых. Это явление особенно убедительно показано для разных островных флор. Большая вариабельность типов роста и жизненных форм покрытосеменных позволила им освоить значительно больше разных биотопов, чем голосеменным. В этом, пожалуй, важная причина видового богатства покрытосеменных.
Наряду с расположением листьев на стебле (спирально, двурядно, накрест супротивно, мутовчато) есть много других признаков листьев, используемых в систематике. Особенности расчлененности листьев: мшуг быть или отсутствовать прилистники и черешок, листовые пластинки бывают цельные или в разной степени расчлененные. Форма листовой пластинки или листочка сложного листа (общие очертания, форма верхушки и основания и т.п.) также дают много признаков, равно как и форма края листа или листочка, который может быть, к примеру, цельнокрайным или зубчатым (см. рис. 4.2). Наконец, таксономически константным призна-
11.2. Бактерии, грибы, растения I 355
Односледовой/ Трехследовой/ Трехследовой/
однолакунный однолакунный трехлакунный
Рис. 11.166. Листовые следы и листовые прорывы (по J.E.Canright из A.Cronquist)
Многоследовой/ многолакунный
Рис. 11.167. Пластиды ситовидных трубок у покрытосеменных (СЭМ-фотографии Н. D. Behnke):
А-^ S-тип с крахмальными включениями: Nuphar (Nymphaeaceae) (20 ОООх); В — P-тип с кристаллоидными белковыми включениями: Gloriosa (Liliales: Colchicaceae); С — P-тип с филаментозными белковыми включениями: Allenrolfea (Amaranthaceae) (В, С 30 ОООх)
ком может быть и сложение листьев в почке. Форма листа варьирует и вследствие разной выраженности листового основания и пластинки (низовые и верховые листья, почечные чешуи и т.п.). Важна и анатомия листа. Так, листья бывают би- или унифациальными, значительно варьирует и характер жилкования листьев. Листья с вильчатым (дихотомическим) жилкованием есть, например, у Ginkgo, а у листьев с перистым жилкованием систематическое значение имеют ход и дальнейшее ветвление вторичных жилок, их возможное слияние друг с другом, вхождение в край листа и т.д. (см. рис. 4.60). У однодольных покрытосеменных часто встречается параллельное жилкование. Систематически значимы признаки анатомии зубцов листа (например, захождение жилок в зубцы, расположение в них механических тканей). Проводящие пучки стебля, в узлах продолжающиеся в проводящие пучки листьев, называют листовыми следами1. Над местом отхождения листовых сле
1 Листовым следом нередко называют всю совокупность пучков, отходящих в один лист, а не каждый такой пучок по отдельности. — Примеч. ред.
дов в проводящей системе стебля остаются так называемые листовые прорывы (лакуны). Число листовых следов и листовых прорывов изменчиво и таксономически значимо (рис. 11.166). Так, например, узлы бывают односледовыс и од-нолакунные, трехследовые и однолакунные или трехлакунные, а также многоследовые и много-лакунные.
Другие примеры систематически значимых комплексов признаков вегетативных органов — это строение устьичных аппаратов и, если имеется, опушение. Устьичные аппараты можно, к примеру, классифицировать по строению око-лоустьичных, или побочных, клеток. Например, у аномоцитных устьичных аппаратов побочные клетки не отличаются от остальных клеток эпидермы, а у парацитных аппаратов замыкающие клетки окружены двумя побочными, лежащими параллельно замыкающим клеткам. Устьичные аппараты различаются также по развитию замыкающих и побочных клеток. Они могуг происходить как от одной (мезогенные), так и от разных материнских клеток (перигенные). Волоски весьма разнообразны по числу слагающих клеток, их расположению (ветвистые или неветвистые волоски), относительному размеру, структуре стенок (жесткие, мягкие), окраске, наличию
356 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Бокс 11.10: Семенные растения (Spermatophyta)
Возникшие скорее всего в самом начале карбона (более 360 млн лет назад) семенные растения (Spermatophyta'» характеризуются прежде всего особенностями чередования поколений. Их исходно древовидные спорофиты разноспоровые и образуют микро- и мегаспоры, в которых эндоспорически развиваются гаметофиты1. Мегаспора с женским гаметофитом не покидает спорофит, а остается в невскрывающемся мегаспорангии, окруженном стерильным покровом (интегументом). Эти струкгуры (интегумент + мегаспорангий + Q гаметофит) вместе составляют семязачаток . Из семязачатка после оплодотворения яйцеклетки женского гаметофита развивается семя, давшее название всей этой группе растений. Семя исходно — единица распространения семенных растений, оно состоит из семенной кожуры (тесты), образовавшейся из интегумента, питающей ткани (эндосперма) разного происхождения и развившегося из зиготы зародыша (см. рис. 11.165). По сравнению с папоротникообразными (Pteridophyta) эволюционное новшество семенных растений, наряду с возникновением новой структуры — интегумента, состоит в том, что мегаспора больше не покидает спорофит и женский гаметофит сохраняет с hijm физический контакт. Кроме того, женский пМютофит (<j>) сильно редуцирован и иногда не образует архегониев. Также сильно редуцированный и не имеющий антеридиев мужской (о") гаметофит остается внутри микроспоры и переносится на семязачаток в процессе опыления. Затем происходит оплодотворение яйцеклетки либо подвижным 1 2
1 Спорофиты образуют не непосредственно споры, а спорангии. — Примеч. ред.
2 Семязачаток составляют мегаспорангий с интегументом/интегументами. — Примеч. ред.
жгутиковыми сперматозоидом, либо неподвижным безж1утиковым спермием.
Благодаря эволюции этих признаков оплодотворение у семенных растений стало совсем независимым от капельно-жидкой воды, а женский гаметофит и развивающийся на нем после оплодотворения зародыш снабжаются всем необходимым от материнского спорофита. Главным образом, именно в этих двух особенностях семенных растений следует видеть их преимущество среди современных наземных растений.
Гомология в чередовании поколений у мхов, папоротникообразных и семенных растений была установлена в 1851 г. W. Hofmeislcr. К этому времени для описания репродуктивных структур семенных растений уже разработали особую терминологию, которую использую! и поныне (см. рис. 11.164). Так, мегаспорангий называют нуцеллусом, а женский гаметофит — зародышевым мешком. Для микроспоры с заключенным в ней мужским гаметофитом используют понятие пыльцевое зерно.
Ныне существующие семенные растения включают пять отдельных групп: саговники (Cycadopsida), хвойные (Coniferopsida), гинк-говые (Ginkgopsida) и гнетовые (Gnetopsida) — эти четыре группы объединены в группу голосеменных (Gymnospermae), а также Magno-liopsida (покрытосеменные = Angiospermae). Их ближайшие родственники среди ныне живущих Pteridophyta — хвощи, псилотовые и папоротники.
Если семя в эволюции наземных растений, вероятно, возникло единственный раз. то се-мяподобные структуры возникали и у Lycopodiopsida. Так, у карбонового рода Lepidocar-роп мегаспорангий с мегаспорой и женским гаметофитом был окружен мегаспорофиллом и оставался на спорофите.
железок и т.п., что можно использовать в систематических целях (см. рис. 3.14). Систематическое значение также могут иметь идиобласты (например, кристаллоносные, масляные клетки), отличающиеся по строению окружающей однородной ткани.
В систематике используют и ультраструк-турные особенности, например строение пластид ситовидных трубок и эпикутикулярного воска.
Применение трансмиссионной электронной микроскопии пластид ситовидных трубок
позволило выделить разные их типы, иногда характерные для таксонов высокого ранга. Они различаются по наличию крахмальных и/или белковых включений, а также по форме и размеру последних (рис. 11.167). У голосеменных и покрытосеменных преобладает так называемый S-тип пластид, содержащих только включения крахмала. Особый P-тип с клиновидными белковыми кристаллами имеется преимущественно у однодольных покрытосеменных, а P-тип с белковыми филаментами ограничен Caryophyllales в узком смысле. Пластиды си-
11.2. Бактерии, грибы, растения | 357
товидных трубок P-типа могут содержать наряду с белком также и крахмал. Видимый в сканирующий электронный микроскоп эпикути-кулярный воск может иметь вид пластинок, волосков или трубочек (см. рис. 3.11) и иногда характеризует определенные систематические группы.
ВТОРИЧНЫЕ МЕТАБОЛИТЫ
В систематике семенных растений, и особенно покрытосеменных, учитывают большое разнообразие так называемых вторичных метаболитов. Вторичными эти весьма различные соединения названы потому, что они не обязательны для осуществления основного обмена веществ растения. Важные классы веществ для систематики покрытосеменных (рис. 11.168) — это структурно очень гетерогенные алкалоиды (например, тропановые, бензилизохинолиновые, пирролизидиновые и т. п.), флавоноиды, беталаины, глюкозинолаты (гликозиды горчичного масла), цианогенные гликозиды, полиацетилены, терпеноиды (например, монотерпеноиды, сесквитерпенлактоны, стероиды, иридоиды и т.п.) и т.д. Если беталаины и антоцианы играют важную роль как пигменты прежде всего цветков, а другие вещества обусловливают запах цветков, то многие из названных классов веществ важны, вероятно, в основном как средства защиты от травоядных животных и патогенных организмов. Причиной видового многообразия покрытосеменных наряду с многообразием форм роста оказалось, вероятно, и богатство вторичными метаболитами.
Упрощенно можно представить, что отчасти именно очень специфические взаимоотношения между растениями и травоядными животными или патогенными организмами привели к ускорению процесса видообразования, например, в связи с тем, что возникновение нового растительного вещества, защищающего от травоядных, могло привести к появлению нового вида в новой экологической нише (без травоядных).
Важнейшие пути синтеза вторичных метаболитов — шикиматный и путь ацетатмевалоновой кислоты, причем шикиматный путь у более продвинутых таксонов всегда заменяется на путь ацетатмевалоновой кислоты (рис. 11.169).
Беталаин (индикаксантин)
Н2С=СН- С-(С=С)г-СН2
Н2С= СН-(СН2)5—НС= СН
Цианогенный гликозид (пруназин)
Монотерпеноид (ментол)
Полиацетилен (дегидрофалькаринон)
Сесквитерпенлактон (партенин)
Иридоид (сверциамарин)
Рис. 11.168. Важные для систематики покрытосеменных классы вторичных метаболитов
358 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Рис. 11.169. Упрощенная схема преобразования спектра накапливаемых биологически активных «вторичных метаболитов» у примитивных (слева) и продвинутых (справа) покрытосеменных (по К. Kubitzki).
Производные шикиматного пути, среди них лигнин (показаны крапом), все больше заменяются дериватами пути ацетатмевалоновой кислоты (заштриховано). Участие ацетата в конденсации дубильных веществ и изофлавонов не рассмотрено
ЦВЕТКИ
Цветки — собрания спорофиллов, т.е. укороченные побеги с ограниченным ростом, несущие микро- и/или мегаспорофиллы. Хотя гомология спорангиофоров в разных группах семенных растений, как и гомология их листьев, никоим образом не доказаны, и в большинстве групп семенных растений микроспорангиофоры и ме-гаспорангиофоры не похожи на листья, их
следует называть соответственно микро-и метаспорофиллами. Среди современных семенных растений только Cicas имеет женские структуры, о которых нельзя говорить как об укороченных побегах с ограниченным ростом. Здесь после образования многочисленных мегаспорофиллов на побеге вновь развиваются нормальные вегетативные листья. Цветки могут быть либо однополыми — только с микро- или только с мегаспорофиллами, либо обоеполыми — с
11.2. Бактерии, грибы, растения | 35g
микро- и мегаспорофиллами. Однополые цветки могут располагаться как на разных индивидуумах (двудомность), так и на одном — однодомность. Бывает, что обоеполые и однополые цветки встречаются на одном индивидууме в разных соотношениях (полигамия; см. 10.1.3.1).
Цветки семенных растений служат для полового размножения: это и образование мужских и женских гаметофитов на спорофитах, и участие в переносе пыльцевых зерен к семязачаткам или плодолистикам (опыление) и в самом процессе оплодотворения, а также забота о развитии зиготы в зародыш, семязачатка — в семя и, у покрытосеменных, завязи — в плод.
Околоцветник
Только Gnetopsida и Angiospermae имеют снаружи от спорофиллов стерильные, но функционально явно связанные с цветком листовые органы1. Функция околоцветника, с одной стороны, состоит в защите остальных элементов цветка на стадии бутона. С другой стороны, на стадии цветка он может играть важную роль в привлечении опылителей и выполнять другие функции, связанные с опылением, а у покрытосеменных также участвовать в распространении плодов.
У Gnetopsida покров мужских и женских «цветков» состоит из одной или двух пар чаще всего срастающихся хотя бы у основания брактей, которые иногда могут трубковидно срастаться (см. рис. 11.211). Их гомология с околоцветником покрытосеменных не ясна1 2.
Покровы цветка покрытосеменных в целом называют околоцветником; он бывает весьма различной формы. Если все элементы околоцветника одинаковы, его называют простым, а отдельные листья — листочками простого околоцветника. Простой околоцветник может состоять из одного (монохламидный), двух или более кругов листочков или (при спиральном их распо
1 У Gnetopsida цветков нет. — Примеч. ред.
2 Gnetopsida далеки от Angiospermae в таксономическом отношении, и едва ли имеет смысл искать гомологии их покровов с околоцветником цветковых растений. — Примеч. ред.
ложении) из двух или более витков спирали (гомойохламидный). В цветках с неодинаковыми листочками околоцветника (двойной околоцветник, цветки гетерохла-мидные) наружные листочки, чаще всего зеленые, называют чашелистиками, составляющими чашечку (calyx). Внутренние, преимущественно яркоокрашенные листочки околоцветника — это лепестки, слагающие венчик. Общепринято, что листочки околоцветника могут иметь различное происхождение (рис. 11.170). С одной стороны, они могут развиваться из верховых листьев под цветком (обычно это признают за чашелистиками — см. рис. 4.6; переход от верховых листьев к листочкам околоцветника можно видеть, например, у Helleborus — см. рис. 4.5), а с другой стороны — из тычинок (обычно это приписывают лепесткам, и переходы между тычинками и лепестками можно найти, в частности, у Nymphaea).
Соответственно своей различной морфологической природе чашелистики и лепестки часто различаются и по снабжению проводящими пучками. В то время как в чашелистики входит обычно несколько, чаще всего три проводящих пучка, а сами они в целом анатомически сходны с листьями, в лепестки, как правило, входит только один проводящий пучок. В махровых цветках самых разных декоративных растений в качестве отклонения от нормы на месте тычинок образуются лепестки.
Элементы околоцветника покрытосеменных могут располагаться по спирали и/ или кругами. Простые околоцветники со спирально расположенными листочками можно с уверенностью считать исходными для покрытосеменных растений. Круговое расположение листочков околоцветника допускает их конгенитальное срастание. Пример сростнолистного простого околоцветника, по-г;идимому, Pofygonatum, сростнолистной чашечки — часть Caryophylla-сеае или Fabaceae,--a сростнолепестного венчика — большинство Asteridae. Элементы околоцветника могут «срастаться» и пост-генитально, например путем сцепления эпидерм папиллами или склеивания.
В спайнолепестном венчике можно различить базальную трубку и отгиб со свободными вершинами лепестков. Похожие части есть и в сростнолистном простом околоцветнике. В вен-
360 I ГЛАВА 11 СИСТЕМАТИКА И ФИЛОГЕНИЯ
А
Рис. 11.170. Переход от верховых листьев (А, В) к листочкам простого околоцветника (С) у Helleborus niger, переход от тычинок (К—F) к лепесткам (Е, D) у Nymphaea (по W.Troll)
Открытое
Квинкунциальное Скрученное Нисходящее Восходящее
Рис. 11.171. Варианты сложения элементов околоцветника в бутоне (по REndress).
Листочки околоцветника в бутоне могут не соприкасаться (открытое сложение), касаться друг друга краями (створчатое) или различным образом перекрываться (черепитчатое)
чике можно различать раннюю симпеталию: образование трубки до или одновременно с вершинами лепестков и позднюю симпеталию: образование трубки после формирования вершин лепестков. Срастания в области околоцветника обеспечивают лучшую защиту или же лучшую пространственную интеграцию органов цветка в связи с особенностями опыления.
В систематике часто используют особенности сложения листочков околоцветника в бутоне (рис. 11.171). Они могут либо соприкасаться краями (створчатое сложение), различным образом перекрываться краем (черепитчатое) или вообще не контактировать между собой (открытое). В черепитчатом сложении можно выделить далее еще несколько типов: квинкунциальное (например, чашелистики у Rosa), скрученное (например, венчик Malva, Genliana', в этом случае можно далее различать право- и левозакручен-ные варианты), а также кохлеарное (например, венчик Fabaceae с так называемым нисходящим сложением лепестков).
Наряду с защитной функцией на стадии бутона и привлечением животных-опылителей благодаря особенно заметной окраске на стадии цветения, околоцветник (как чашечка, так и венчик) может участвовать и в самом процессе опыления, например, продуцируя и/или накапливая нектар, создавая посадочную площадку для опылителя, как в цветках губоцветных, или осуществляя вторичное «предложение» пыльцы. Однако в связи с
измельчанием (например, при переходе к автогамии или при возникновении псевдантиев) цветков и при переходе к анеморфилии околоцветник может очень сильно упроститься или полностью редуцироваться в развитом цветке (ахламидные цветки). Остающиеся при плодах и иногда разрастающиеся чашелистики Moiyr способствовать и распространению плодов.
Микроспорофиллы
Микроспорофиллы разных групп семенных растений сильно различаются по строению и расположению. Их гомология не ясна, и вопрос об участии листьев в их возникновении, как это подразумевается самим названием (микроспорофилл), остается открытым. Лучше было бы называть их нейтрально — микроспорангиофо-ры. У Ginkgopsida микроспорофилл состоит из ножки с двумя висящими на ее кон чике микроспорангиями (см. рис. 11.202). В мужском цветке1 микроспорофиллы расположены спирально на удлиненной оси. Мужские цветки1 2 современных Conifero
1 У Ginkgopsida есть (мужские) микростробилы, а не цветки. — Примеч. ред.
2 Правильно — микростробилы. — Примеч. ред.
11.2. Бактерии, грибы, растения
psida шишковидные и состоят из обычно большого числа спирально или (реже) накрест супротивно расположенных микроспорофиллов. Отдельный микроспорофилл несет на нижней стороне от 2 до 20 часто сросшихся между собой микроспорангиев (см. рис. 11.203). Только у Taxus (см. рис. 11.210) микроспорангии расположены радиально на конце ножки микроспорофилла1. На нижней стороне чаще всего чешуевидных микроспорофиллов Cycado-psida (см. рис. 11.201) находится от 5 до 1 000 микроспорангиев, обычно расположенных отчетливыми группами по 3—5. Многочисленные микроспорофиллы в мужском цветке1 2 расположены спирально. У Gnetopsida микроспорофиллы расположены либо мутовками, либо терминально. Мутовчато расположены микроспорофиллы у Welwitschia, где 6 сросшихся между собой основаниями микроспорофиллов несут на концах по 3 сросшихся микроспорангия (см. рис. 11.211). В мужских цветках3 Gnetum имеется только один терминальный микроспорофилл с нитевидным основанием и один или два верхушечными микроспорангиями (см. рис. 11.211), а у Ephedra терминально располагается чаще всего один, на конце обычно раздваивающийся нитевидный микроспорофилл с 2 — 8 группами микроспорангиев, почти всегда сросшихся по 2 (см. рис. 11.211).
Микроспорофиллы (тычинки) покрытосеменных (рис. 11.172) чаще всего дифференцированы на тычиночную нить и обычно терминально расположенный пыльник. Пыльник состоит из двух соединенных связником половинок — тек, каждая из которых содержит по два сросшихся микроспорангия.
Тычинки покрытосеменных сравнительно редко отклоняются от этой базовой структуры. Микроспорофиллы бывают листовидно уплощенными и не дифференцированными на тычиночную нить и пыльник, но и в этом случае
1 Это щитовидные микроспорофиллы, характерные также для Cupressaceae s.l. и Taxodiaceace s.l. — Примеч. ред.
2 У Cycadopsida цветков нет. — Примеч. ред.
3 Это на самом деле микроконстробил. — Примеч. ред.
чаще всего отчетливо различимы две группы по два сросшихся микроспорангия. Пыльник может прикрепляться к тычиночной нити как основанием (базификсный), так и дорсальной (дорсификсный) или вентральной (вентрифик-сный) сторонами. Иногда число микроспорангиев в теке может бьпь уменьшено до одного. Благодаря поперечному и/или горизонтальному разделению микроспорангиев при сохранении в целом структуры теки могут возникать полиспорангиатные пыльники, как, например, у некоторых Rubiace е и Rhizophoraceae. У Raffle -siaceae и Viscaceac дифференциация пыльников на теки утрачена, но пыльники точно так же полиспорангиатны: их многочисленные микроспорангии вскрываются по отдельности или все вместе через одну общую пору.
Число тычинок в цветке покрытосеменных варьирует от одной до примерно 2 000. Совокупность всех тычинок называют андроцеем. Тычинки могут располагаться по спирали или мутовчато, реже — более сложным образом или беспорядочно (рис. 11.173). В то время как при спиральном расположении чаще всего число тычинок большое и нефиксированное (первичная полиацдрия), мутовчатое расположение тычинок обычно ведет к уменьшению и фиксации (олигомеризация) их числа. Цветок с двумя кругами тычинок — диплостемон-ный, с одним кругом — гаплостемонный. В цветках с исходно определенным числом тычинок в одном или немногих кругах могло произойти увеличение числа тычинок— вторичная полиандрия— благодаря разделению зачатков тычинок. При этом сначала ясно различимые отдельные зачатки могут вычленять многочисленные зачатки тычинок центробежно (рис. 11.173) или центростремительно, либо центробежно или центростремительно разделяется единый кольцевой зачаток. Благодаря расщеплению тычинок во время развития могут сформироваться «половинные» тычинки. Наконец, тычинки могут превращаться в стерильные стаминодии, которые служат как нектарники или уподобляются лепесткам для привлечения опылителей. Иногда тычинки в одном цветке устроены по-разному (гетерантерия). Мутовчато расположенные тычинки могут срастаться чаще своими нитями, но иногда и боками пыльников (синандрия) или постгениталь-
362 | ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Рис. 11.172. Тычинка покрытосеменных и детали ее строения (А, В — по A.F.W.Schimper; С, D — по Е. Strasburger; Е—J — по F. Firbas).
Общий вид тычинки у Hyoscyamus niger: А — спереди (адаксиально), В — сзади (абаксиально) (увеличено). Поперечные срезы пыльника Hemerocallis fulva: С — с еще не вскрывшимся, D — с уже вскрывшимся пыльником и проводящим пучком. Lilium pyrenaicum: Е— поперечный срез стенки пыльника с эпидермой, фиброзным слоем, промежуточными слоями и остатками тапетума; отдельная клетка фиброзного слоя снаружи (F) и изнутри (G) (150х); Н, J — схема клетки фиброзного слоя до и во время сжатия; е — эпидерма; f — тычиночная нить; fs — фиброзный слой; к — связник; I — проводящий пучок; р — гнездо пыльника; t — тапетум; th — тека; z — промежуточный слой
А В
Рис. 11.173. Ранние стадии развития цветков покрытосеменных (околоцветник удален) с многочисленными зачатками тычинок (СЭМ-фотографии L. Erbar и Р. Leins):
А — первичная поландрия у Magnolia denudata (Magnoliaceae), зачатки расположены спирально на конусовидном цветоложе; В — вторичная полиандрия у Stewartia pseudocamellia (Theaceae), центробежное заложение многочисленных зачатков тычинок на пяти расположенных между зачатками лепестков и плодолистиков секторах блюдцевидно вогнутого цветоложа
но склеиваются между собой. Сериальное срастание тычинок и лепестков или тычинок и плодолистиков также возможно.
Среди архаичных покрытосеменных наиболее широко распространено спиральное расположение тычинок, число которых варьирует.
Стенка пыльника, окружающая археспорий (ткань, образующую пыльцу'), всегда многослойна. Функционально особен
' Археспорий образует спороциты, в ходе мейоза которых формируются споры. — Примеч. ред.
11.2. Бактерии, грибы, растения | 353
но значимы слой клеток, ответственный за вскрывание пыльника, а также самый внутренний слой — тапетум. У Coniferopsida, Су-cadopsida и Gnetopsida вскрывание микроспорангия обеспечивает самый наружный слой стенки — экзотеций (см. рис. 11.201), а у Ginkgopsida и Angiospermae — субэпидермальный эндотеций (фиброзный слой, см. рис. 11.172), который иногда может отсутствовать, как, например, у Ericaceae. Как в экзо-, так и в эндотеции утолщены антиклинальные и внутренние периклинальные стенки клеток, а их наружные периклинальные стенки остаются тонкими. Как и в колечке спорангиев папоротников (см. рис. 8.37), потеря воды клетками экзо/эн-дотеция приводит к тангенциальному укорочению их наружных стенок. Вследствие этого стенка микроспорангия разрывается чаще всего в виде продольной щели (сто-миум) в строго определенном месте. В эндотеции покрытосеменных антиклинальные стенки клеток несут гребневидные утолщения, которые часто становятся толще к внутренней стенке, где и сливаются. У покрытосеменных два микроспорангия одной теки чаще всего вскрываются одной общей продольной щелью, возникающей после разрушения клеток перегородки, разделявшей прежде полости микроспорангиев.
В зависимости от ориентации шели относительно центра цветка можно различать пыльники, вскрывающиеся интрорзно (внутрь), экстрор-зно (наружу) и латрорзно (вбок).
Есть также пыльники, в которых каждый микроспорангий вскрывается собственной продольной щелью (например, Strelitzia), пыльники, вскрывающиеся порами при растворении ткани стенки в определенных местах — поры могут возникать, однако, и как очень короткие продольные щели (например, у Ericaceae) — или фиброзный слой развивается только в ограниченной области, которая отворачивается наружу в виде клапана (например, Lauraceae). Клапан может образовываться и благодаря раздвоению продольной щели на обоих концах. Иногда утолщения клеточных стенок фиброзного слоя имеют противоположную ориентацию, и микроспорангии при высыхании сжимаются (например, Welwitschia и Агасеае), выталкивая пыльцу наружу через отверстие стенки.
Экзотеций или эндотеций отделены от тапетума по меньшей мере одним проме
жуточным слоем, у покрытосеменных временным (рис. 11.172). Тапетум состоит чаще всего из богатых протоплазмой клеток с обычно эндополиплоидными ядрами и участвует в питании пыльцевых зерен и образовании веществ, откладываемых в стенки пыльцевых зерен1 либо на их поверхность (например, пыльцевой клей; вещества, важные для реакций самонесов-местимости). Секреторный тапетум долго остается интактным, в то время как пери-плазмодиальный тапетум после растворения стенок его клеток и слияния их протопластов может амебоидно проникать между развивающимися пыльцевыми зернами.
Хотя главная функция микроспорофиллов — образование пыльцы, они могут иметь и дополнительные функции, связанные прежде всего с опылением. Например, они могут сделать цветки более заметными или даже быть единственным их визуальным аттрактором. Микроспорофиллы могут продуцировать запахи, привлекающие опылителей, выделять нектар или определенным образом ограничивать движения опылителей в цветке, повышая таким образом эффективность процесса опыления. Наконец, они являются важными компонентами различных механизмов вторичного предложения пыльцы.
Из археспория возникает множество материнских клеток пыльцы, из которых после мейоза получается по четыре одноядерных пыльцевых зерна1 2. Если все клеточные стенки образуются одновременно, то говорят о симультанном способе образования микроспор. При сукцессивном способе образования микроспор первая клеточная стенка образуется уже после первого деления мейоза3.
Возникающие четыре микроспоры чаще всего располагаются в тетраэдрической тетраде (при симультанном типе образования). Но существуют и тетрады, в которых микроспоры располагаются в ряд (линейно), в одной плоскости (изобилатерально), в двух взаимно-перпендикулярных плоскостях (крестообразно) или Т-образно (при сукцессивном типе образования).
1 Правильно — микроспоры. — Примеч. ред.
2 Правильно — микроспор. — Примеч. ред.
3 Эти типы различают не по последовательности образования стенок, а по последовательности цитокинезов. — Примеч. ред.
364 | ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Пыльца
При переносе пыльцевых зерен по воздуху из микроспорангиев к женским репродуктивным органам они часто довольно долго подвергаются воздействию экстремальных условий, поэтому для размножения чрезвычайно важна защита их содержимого. Она осуществляется благодаря особому строению оболочки пыльцевого зерна, спородермы, состоя идей из двух комплексных слоев — наружной экзины и внутренней интины (рис. 11.174).
Интина, сплошным слоем окружающая протоплазму, чаще всего нежная и химически мало устойчивая. Обычно интина состоит из двух-трех слоев. Самый наружный из них часто содержит много пектинов, что позволяет ей легко отделяться от экзины. Основной компонент внут реннего слоя/слоев — фибриллы целлюлозы. При прорастании пыльцевых зерен только интина разрастается в пыльцевую трубку1.
Экзина образована в основном химически устойчивыми спорополленинами. Это терпены, которые, как полагают, возникают в результате оксидативной полимеризации из каротиноидов и их эфиров. Основной конструктивный элемент экзипы — гранулы размером около 6 нм.
У голосеменных экзина дифференцирована на внутреннюю эцдокзину, имеющую пластинчатую (ламеллярную) структуру, и наружную эктэкзину, которая состоит из внутреннего подстилающего слоя, компактного наружного слоя и гранулярного или альвеолярного среднего слоя. Эндэкзина покрытосеменных не ламеллярная, а гранулярная1 2. Эндэкзину вместе с самым внутренним слоем эктэкзины — плотным и гомогенным подстилающим слоем — у покрытосеменных (рис. 11.174) также называют нэкзиной. Называемая сэкзиной наружная часть эктэкзины покрытосеменных чаще всего сильно структурирована и скульпту-рирована. У атектатных пыльцевых зерен сэкзина представлена только столбиками в виде палочек, булав, конусов, бородавок
1 В пыльцевую трубку разрастается сифоногенная (вегетативная) клетка мужского гаметофита, покрытая И1ГГИНОЙ. — Примеч. ред.
2 У очень многих покрытосеменных эндэкзина ламеллярная. — Примеч. ред.
или сетчатой структурой на нэкзине. Столбики могут сливаться дистальными концами в дополнительный наружный слой, покров (тектатные пыльцевые зерна). Тек-тум может быть пронизан порами самой различной формы, сам бывает многослойным и скульптурированным снаружи. В полостях текгума могут откладываться про теины самонесовместимости, пыльцевой клей и т. п. Очень редко, к примеру у опыляемой под водой морской травы Zostera, экзины нет. УПпасеае могут возникать воздушные мешки за счет локального расхождения наружного и внутреннего слоев экзины (см. рис. 11.203).
Электронно-микроскопические исследова ния развития пыльцевых зерен показывают, что еще внутри каллёзной оболочки материнской клетки микроспор, на плазмалемме микроспоры откладывается тонкий слой фибриллярного материала — примэкзина1. Затем в нем появляются более компактные элементы, которые, удлиняясь и утолщаясь, становятся столбиками, расширяются на обоих концах и образуют таким образом тектум и подстилающий слой эктэкзины. Внутри от подстилающего слоя затем возни кает неламеллятная эндэкзина и, наконец, инти на. Таксоноспецифическое строение экзины контролируется самим пыльцевым зерном, но силь но зависит от тапетума спорофита. Это, среди прочего, можно видеть по реакции спорофит ной самонесовместимости (см. 10.1.3.1).
Пыльцевые зерна сильно различаются по форме, расположению и числу апертур (рис. 11.175). Бывают пыльцевые зерна без апертур (безапертурные). Обращенный к центру тетраэдрической тетрады полюс пыльцевого зерна называют проксимальным, направленный наружу — дистальным. Экваториальная плоскость перпендикулярна к оси, связывающей оба полюса. У пыльцевых зерен семенных растений бывают только дистальные, только экваториальные или разбросанные по всей поверхности апертуры. Встречающиеся у Pteridophyta проксимальные апертуры здесь не известны2. Продольно вытянутые апертуры называют бороздами (соогветственно дистальной — sul-
1 Примэкзина часто бывает везикулярной. — Примеч. ред.
2 У Pteridophyta есть (микро)споры, а не пыльцевые зерна. — Примеч. ред.
11.2. Бактерии, грибы, растения | 365
Рис. 11.174. Схема тонкого строения стенки пыльцевых зерен покрытосеменных разных типов, схематизированный рисунок (Н.Террпег, по G.Erdtman, K.Faegri и др.):
1 — эктэкзина, 2 — эндэкзина, 3 — интина, 4 — пыльцевой клей
1
2 Г""~1
Дистально-однобороздное
Трехбороздное
Трехборозднооровое
Безапертурное
Однопоровое
Трехпоровое
Тетрада
Рис. 11.175. Некоторые обычные типы пыльцевых зерен семенных растений Центральной Европы (по K.Faegri, I.Iversen, G.Erdtman, составлено Н.Террпег и М.Hesse)
Слева в каждом случае вид с дистального полюса, справа — вид с экватора. Монады (одиночные зерна): дистально-однобороздное (многие Magnoliopsida, Liliopsida), однопоровое (Роасеае), трехбороздное (часть Ranunculaceae, Quercus, Acer, Brassicaceae, Salix, часть Lamiaceae), трехпоровое (Betula, Corylus, Urticaceae, Onagraceae), трехборозднооровое (Fagus, часть Rosaceae, Apiaceae, Tilia, Asteraceae), стефанокольпатное (Rubiaceae, часть Lamiaceae), стефаноноратное (Alnus, Ulmus), пан-топоратное (Juglans, большая часть Caryophyllaceae, Amaranthaceae, Plantaginaceae), безапертурное (Populus, Callitriche) из таксонов в целом с экваториально-апертурными пыльцевыми зернами. Тетрады: у таксонов, где встречаются и монады (часть Orchidaceae, часть Typha, Ericaceae)
cus или экваториальной — colpus), округлые — порами (дистальной — ulcus или экваториальной — porus).
У голосеменных пыльцевые зерна чаще всего дистально-однобороздные. Такие пыльцевые зерна находят также среди покрытосеменных у большинства Magnoliidae и у однодольных, где, однако, имеются так
же однопоровые и безапертурные пыльцевые зерна. Большая группа настоящих двудольных характеризуется первично трехбороздными пыльцевыми зернами, борозды которых перпендикулярны экваториальной плоскости. Таким образом, исходны для покрытосеменных дистально-однобороздные пыльцевые зерн... Борозды могут быть за
366 I ГЛАВА 11 СИСТЕМАТИКА И ФИЛОГЕНИЯ
менены порами, например у трехпоровых пыльцевых зерен. Если в экваториальной плоскости встречается более трех борозд или пор, то говорят о стефанокольнатных или стефанопоратных пыльцевых зернах. Апертуры, рассеянные по всей поверхности пыльцевого зерна (глобально-апертурные, например пантопоратные), имеются, к примеру, у Cactaceae или Caiyophyllaceae. При этом число апертур может возрастать до 100 (например, у Amaranthaceae). Пыльцевые зерна с апертурами, состоящими из борозд и пор, называют борозднооровыми. Вследствие изменчивости края апертур или образования утолщенного оперкулума могут возникать очень сложно устроенные апертуры.
Невероятное многообразие типов пыльцевых зерен оказалось возможным классифицировать с помощью NPC-системы: по числу (Numerus), положению (Positio) и типу (Charakter) апер
тур. Кроме того, систематическое значение имеют еще многие различия в симметрии, форме и размере пыльцевых зерен, а также в тонкой структуре их экзины. Структуру пыльцевых зерен можно показать на так называемых палинограммах или на электронно-микроскопических фотографиях (рис. 11.176; см. бокс 4.1, рис. F).
Во время роста пыльцевых зерен тапетум образует особую клейкую субстанцию, содержащую липиды и каротиноиды, — пыльцевой клей. У опыляемых животными видов он откладывается прежде всего на поверхности пыльцевых зерен, что приводит к склеиванию их в комочки и способствует прикреплению к опылителю.
Функцию склеивания пыльцевых зерен выполняют и висциновые нити, которые чаще всего образуются внутри микроспорангия и содержат спорополленин, целлюлозу или протеины. Наконец, бывает и так, что пыльцевые зерна склеиваются секретами других элементов цветка.
Рис. 11.176. Пыльцевые зерна разных кактусов при сканирующей электронной микроскопии (А— С — по W. Klaus; D — по G. Erdtman):
А, В — Gymnocalycium mihanovichii (трехбороздное, внешний вид: 500х; детали строения остробородавчатого и пронизанного порами тектума: 5 000х); С— Opuntia sp. (пантопоратное: 1 ОООх); D — палинограмма пыльцевого зерна Centaurea scabiosa (трехборозднооровое): вид с экватора, оптическое сечение и детали структуры оболочки (световая микроскопия, 1 500х и ЗОООх)
11.2. Бактерии, грибы, растения | 367
С другой стороны, приспособления для склеивания пыльцевых зерен могут отсутствовать, например у ветроопыляемых семенных растений.
Пыльцевые зерна одной тетрады вовсе не всегда распространяются только в виде монад, т. е. по отдельности. Наряду со слипанием зерен благодаря пыльцевому клею или висциновым нитям микроспоры одной материнской клетки могут длительное время оставаться соединенными в тетраду и в таком виде распространяться (например, у Ericaceae, Drosera и др.). Путем прогрессирующей редукции трех клеток одной тетрады могут возникать и псевдомонады (например, у Сурегасеае). Если остаются объединенными пыльцевые зерна, образовавшиеся из нескольких материнских клеток, то получаются полиады, которые могут состоять из 8, 16 или 32 пыльцевых зерен (например, у Mimosoideae). Наконец, и все пыльцевые зерна одного микроспорангия могут объединяться в поллиний, а зерна двух или нескольких микроспорангиев — в поллинарий; обычно они покрыты общей спо-рополлениновой оболочкой (например, у некоторых Apocynaceae, Orchidaceae; см. рис. 11.274 и 11.227).
Мужской гаметофит
Мужской гаметофит семенных растений состоит из очень немногих клеток и никогда не имеет морфологически различимых антеридиев. Его развитие проходит в основном внутри оболочки пыльцевого зерна и лишь частично уже после его прорастания. У голосеменных, к примеру у Pinaceae и Ginkgo, путем двух неравных де
лений образуются две проталлиальные клетки и одна антеридиальная клетка, которая делится на сифоногенную клетку и генеративную клетку. Из генеративной клетки образуются клетка-ножка и сперматогенная клетка, а из последней — два сперматозоида или два спермия (рис. 11.177). Перед делением сперматогенной клетки мужской гаметофит, таким образом, пятиклеточный (2 проталлиальные клетки, клетка пыльцевой трубки, клетка-ножка и сперматогенная клетка; см. рис. 11.204). Есть немало отклонений от этого типа. Число проталлиальных клеток может возрастать до 8 (Podocarpaceae) или до 40 (Araucariaceae), или же она только одна (Cycadopsida; рис. 11.178), либо их совсем нет (Toms, Gnetum, IPe/iwzsc/iza, Angiospermae). Сифоногенная клетка развивается у всех семенных растений, а клетка-ножка у покрытосеменных отсутствует. У Microcycas (Cycadopsida) увеличено число половых клеток, так как клетка-ножка образует свыше 20 сперматозоидов.
Мужской гаметофит покрытосеменных, поскольку у него нет ни проталлиальных клеток, ни клетки-ножки, состоит только из сифоногенной клетки, которую здесь называют вегетативной, и второй, называемой генеративной, клетки, из которой образуются два спермия (рис. 11.179). Деление генеративной клетки может происходить перед или после прорастания пыльцевого зерна, время этого деления коррелирует с типом системы самонесовместимости (см. 10.1.3.1).
В любом случае пыльцевые зерна прорастают пыльцевой трубкой. Ее функция, с ‘ одной стороны, — доставить мужские по-
Первая
Микроспора
проталлиальная клетка Вторая
, проталлиальная / клетка
Вторая клетка
Антеридиальная клетка
Сифоногенная клетка (вегетативная клетка)
Клетка-ножка
Генеративная клетка
Сперматогенная___Два сперматозоида/
клетка спермия
Рис. 11.177. Развитие сГ гаметофитов от микроспоры до образования спермиев/сперматозоидов у Pinaceae и Ginkgo
368 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Рис. 11.178. Развитие сГ гаметофитов у Cycadopsida (А—Е— по Ch. Chamberlain; F— по О. W. Caldwell; G—J — по H. J.Weber):
A—E — прорастание пыльцевого зерна у Dioon edule (A—C — 840x; D — 667x; E — 420x); F — проросшее пыльцевое зерно Microcycas calocoma c 9 сперматогенными клетками {около 200x); G—J — пыльцевая трубка и сперматозоид Zamia floridana (G, H — 50x, J — 75x); a — антеридиальная клетка; e — экзина; i — интина; k — ядро; p — проталлиальная клетка; pz — сифоногенная клетка; s — клетка-ножка; sz — сперматогенная клетка
<
Рис. 11.179. Развитие сГ гаметофита у покрытосеменного растения Lilium martagon (по Е. Strasburger из I.L. L.Guignard).
Вегетативная клетка с ядром (к) и генеративная клетка (д) в пыльцевом зерне (А, В) и пыльцевой трубке (С). В переднем конце пыльцевой трубки (D) генеративная клетка разделилась на два спермия (s)
11.2. Бактерии, грибы, растения | 359
Рис. 11.180. Семязачаток и оплодотворение у Cycadopsida (А — по F. Rrbas; В — по Ch. Chamberlain): А— продольный разрез семязачатка Ceratozamia с мик|—пиле (т), интегументом (i), нуцеллусом (п) и пыльцевой камерой (р) с прорастающими пыльцевыти зернами; проросшая мегаспора: <j> гаметофит (= зародышевый мешок, е) со стенкой (w) и двумя архегониями (каждый из двух клеток шейки и одной яйцеклетки) (2,5х); В — верхняя часть нуцеллуса ко времени оплодотворения у Dioon edule, пыльцевые трубки заякорились в ткани нуцеллуса и проникли в архегониальную камеру (ак), часть сперматозоидов высвобождена, левый из двух архегониев (а) уже оплодотворен (около 100х)
ловые клетки к яйцеклеткам, но она служит также для заякоривания мужского гаметофита и как гаустория для поглошения питательных веществ, необходимых для развития и роста самой пыльцевой трубки (рис. 11.180). В качестве половых клеток у Cycadopsida и Ginkgopsida образуются подвижные многожгутиковые сперматозоиды (см. рис. 11.178; 11.180; 11.202), ay Coniferopsida, Gnetopsida и Angiospermae — неподвижные спермин (см. рис. 11.204; 11.179).
Мегаспорофиллы
Семязачатки семенных растений расположены в цветках1 очень разного вида. Хотя
1 Цветки есть только у покрытосеменных. Семенные органы других растений нельзя называть цветками! — Примеч. ред.
до сих пор точно не известны ни гомологии семяносных структур у разных групп семенных растений, ни участие листьев в их возникновении, за ними закрепилось название мегаспорофиллы вместо нейтрального мегаспорангиофоры. Понятие плодолистик традиционно применяют для обозначения мегаспорангиофора покрытосеменных растений.
Несомненно, на листьях располагаются семязачатки у саговниковых (Cycadopsida) (см рис. 11.200). У них чаще всего два семязачатка расположены на нижнем крае листовой пластинки черешкового, чешуевидного или щитовидного мегаспорофилла. До 8 семязачатков развивается на рахисе явно перистого (или по меньшей мере зубчатого) на верхушке мегаспорофилла у представителей рода Cycas. У современных хвой-
370 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Рис. 11.181. Схема этапов (А—С) развития типичного плодолистика покрытосеменных: вид спереди и продольный срез (светлый)
ных (Coniferopsida) бывает от одного до 20 семязачатков на верхней стороне уплощенной семенной чешуи (см. рис. II.205). Реже семязачатки кажутся расположенными на верхушках укороченных побегов (например, у Taxus; см. рис. 11.210). Исходя из положения семенной чешуи в пазухе кроющей чешуи, а также из сравнения с ископаемыми родичами (см. рис. 11.206; 11.207) современных хвойных семенные чешуи следует считать модифицированными укороченными побегами. Не ясно, находились ли исходно семязачатки на листьях этих укороченных побегов или на оси. У Ginkgo два семязачатка расположены на концах вильчато разветвленной ножки (см. рис. 11.202), а у родов гнетовых (Gnetopsida) каждый цветок имеет только один терминальный семязачаток (см. рис. 11.211).
В то время как у вышерассмотренных групп, объединяемых как голосеменные (Gymnospermae), семязачатки расположены открыто, у покрытосеменных (Angio-spermae) они заключены в плодолистики. Совокупность всех плодолистиков цветка покрытосеменных с содержащимися в них семязачатками называют гинецеем1. Структуру отдельного плодолистика можно понять по его развитию. При этом на ранних стадиях чаще всего возникает седловидный зачаток (рис. 11.181), более низкий край которого, называемый поперечной зоной, обращен к центру цветка. В течение некоторого времени края зачатка растут вверх со-
1 Наличие внутри плодолистика(ов) семязачатков не является критерием гинецея. Гинецей остается гинецеем, даже если он полностью или частично состоит из стерильных плодолистиков. — Примеч. ред.
вместно, благодаря чему возникает трубковидная (асцидиатная) зона (рис. 11.182). Если зачаток прекращает свой рост на внутренней стороне и далее растет только боками и спинной стороной развивающегося плодолистика, то образуется открытая в середину цветка брюшной щелью зона, называемая пликатной, или кондупликатной. Над этой областью расположен обычно стеблевидный столбик1, на котором располагается преимущественно покрытое сосочками рыльце для улавливания пыльцы. Столбик, как правило, не сплошной, а содержит чаще всего очень узкое продолжение полости асцидиатной и пликатной зон — так называемый трансмиссионный канал, по которому к семязачаткам различным образом попадают пыльцевые трубки. В развившемся таким образом плодолистике имеются снизу вверх: ножка плодолистика, с самого сначала, т.е. конгениталь-но, замкнутая асцидиатная зона и первоначально открытая вентральной щелью и лишь постгенитально замыкающаяся (с помощью секрета и/или благодаря сцеплению эпидермальных зубчиков) пликатная зона. Область полого плодолистика, содержащую семязачатки, называют завязью.
Относительные размеры асцидиатной и пликатной зон бывают весьма различными. Если в крайнем случае выше ножки образуется только асцидиатная зона, то плодолистик полностью асцидиатный. Если трубчатая зона отсутствует и образуется только пликатная зона, то плодолистик полностью пликатный (кондупликатный).
Плаценты — участки внутренней поверхности плодолистика, на которых находятся семязачатки. При этом плоские или выпуклые плаценты чаще всего находятся вблизи края вентральной щели — субмаргинально. Здесь они могут располагаться или Сообразно вокруг всей вентральной щели (очень редко) или быть U-образными, находиться только у боковых краев (латераль-но) или у нижнего края вентральной щели (медиально). Кроме того, семязачатки могут также располагаться по всей внутренней поверхности завязи (ламиналыю; рис. 11.183). Число семязачатков в плодолисти-
1 На самом деле — стилодий. — Примеч. ред.
11.2. Бактерии, грибы, растения | 37-]
Рис. 11.182. Схема строения гинецеев (по W. Leinfellner).
Продольные (А, В, С) и поперечные (А, С) срезы: А — апокарпного, В — гемисинкарпного, С — цено-карпного гинецея с асцидиатной, пликатной и соответственно синасцидиатной, симпликатной, геми-симпликатной и асимпликатной зонами
Рис. 11.183. Разные типы гинецеев, поперечные срезы через фертильную главную зону выросших завязей (частично по A. L.Takhtajan и Engler’s Syllabus):
А— апокрапный гинецей, ламинальная плацента (П); В— апокарпный гинецей, субмаргинальная П; С — гемисинкарпный гинеций, центрально-угловая П; D, G — синкарпный гинецей, центрально-угловая П (D — плодолистики пликатные, G — плодолистики асцидиатные); Е, F — паракарпный гинецей, париетальная П; Н, J — паракарпный гинецей, центральная П; многочисленные семязачатки или один базальный семязачаток
372 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Рис. 11.184. Строение плодолистиков (А—С) и их прогрессирующее срастание (D—F) (А—D — по W. Troll; Е, F — по О. L. Berg и L. F Schmidt):
А — внешний вид сформировавшегося одиночного и свободного плодолистика с вентральной стороны с замкнутым брюшным швом (у основания чашечка; около Зх); В, С — он же в поперечном разрезе, с одним дорсальным и двумя вентральными пучками, двураздельной плацентой и семязачатками на ней (около 10х); D— апокарпный, Е, F— ценокарпный гинецей соответственно со свободными и сросшимися стилодиями (А, В — Colutea arborescens; С, D — Delphinium elatum; E — Linum usitatissimunr, F — Nicotiana rustica)
ке варьирует от одного до нескольких миллионов.
В цветке бывает от одного до примерно 2 000 плодолистиков. Они могут располагаться спирально или мутовками. Как и в случае микроспорофиллов, при спиральном расположении плодолистики многочисленные, а число чаще всего не постоянно; мутовчатое расположение связано с уменьшением (олигомеризацией) и постоянством числа плодолистиков.
Если плодолистики не срастаются между собой, то гинецей называют апокарпным. Когда плодолистики срастаются — что, как правило, происходит при мутовчатом их расположении, — гинецей называют цено-карпным. Ценокарпный гинецей называют также пестиком*. Степень срастания плодолистиков может быть рахитичной: к примеру, только основаниями завязей, всеми завязями или также и стилодиями (рис. 11.184). Обычно число плодолистиков ценокарп-ного гинецея можно установить по числу рылец, но даже и рыльца могут срастись в единую структуру. Значительно варьирует
1 Пестиком называют как весь ценокарпный гинецей, так и каждый плодолистик апокарпного гинецея. - Примеч. ред.
и внутренняя структура ценокарпных гинецеев (см. рис. 11.183). Если у плодолистиков полностью развиты боковые и дорсальный участки и гинецей с гтветственно разделяется на камеры боками отдельных плодолистиков (септами), то это синкарпный (септированный ценокарпный) гинецей. В зависимости от того, страстаются ли плодолистики краями в центре или нет, мож но различать синкарпный и гемисинкарп-ный (не полностью септированный) гинецей. В таком гинецее выделяют снизу вверх синасцидиатную и симпликатпую или соответственно гемисимпликатную зоны. Плацентацию по краям отдельных плодолистиков называют центрально-угловой. Если развитие боковых сторон плодолистиков подавлено, то возникает паракарпный гинецей без септ. При этом плаценты располагаются париетально, т. е. там, где соседние плодолистики смыкаются друг с другом краями, или на дне завязи находится свободная центральная плацента, на которой семязачатки сидят на более или менее массивной колонке, не контактирующей со стенками завязи*. Впаракарпных гинецеях
1 Такой тип гинецея иногда называют лизи-карпным. — Приме ред.
11.2. Бактерии, грибы, растения | 373
могут быть и единичные семязачатки, располагающиеся на дне завязи или свисающие с ее крыши.
В ценокарпных гинецеях некоторые гнезда завязи могут быть стерильными или гинецеи кажутся одноплодолистиковыми (псевдомономерные). Могут, однако, образовываться и добавочные, так называемые «ложные» септы (см. рис. 11.265; 11.277; 11.282).
Функционально важной структурой ценокарпных гинецеев является компитум, под которым понимают общую для всех плодолистиков проводниковую ткань в области столбика и соответственно в симпли-катной зоне. Это позволяет, к примеру, в гинецеях с раздельными рыльцами, пыльцевым трубам пыльцевых зерен, попавших только на одно рыльце, прорастать во все плодолистики.
Плодолистики, как и листочки околоцветника, и микроспорофиллы могут участвовать в процессе опыления, к примеру, образуя нектар или как органы вторичного предложения пыльцы. Наконец, структуру гинецея следует понимать и в аспекте образования плодов.
Самые примитивные покрытосеменные чаще всего имеют апокарпный гинецей с варьирующим числом спирально расположенных плодолистиков. Замыкание вентральной щели таких плодолистиков происходит с помощью специального секрета.
Семязачатки
Семязачатки — это заключенные в стерильную оболочку мегаспорангии семенных растений (рис. 11.185). Они состоят из
семяножки, чаще всего одной или двух (редко трех) оболочек, или интегументов (семязачатки уни- или битегмальные), и заключенного в интегументы мегаспорангия, или нуцеллуса. Область перехода семяножки в нуцеллус называют халазой, а на противоположном полюсе семязачатка в интегументах остается отверстие — микропиле.
Во время развития семязачатка сначала образуется нуцеллус, а затем его снизу обрастают интегументы. У голосеменных в типе имеется только один интегумент. Если, как у многих покрытосеменных, образуется два интегумента, то внутренний возникает раньше наружного. Уни-тегмальные семязачатки у покрытосеменных образуются благодаря тому, что один из двух интегументов рано прекращает рост или интеркаляр-но разрастается общее основание обоих интегументов так, что на верхушке уже не заметна двойная природа или с самого начала имеется только один интегумент. Интегументы могут быть либо очень тонкими и состоять только из двух слоев эпидермы, либо между эпидермами могут быть слои клеток. Самый внутренний слой интегумента, особенно у унитегмальных семязачатков, дифференцирован в подобный тапетуму эндотелий. У некоторых паразитических покрытосеменных (например, у Loranthaceac) интегументов нет.
В зависимости от положения материнской клетки мегаспор в нунеллусе можно выделить следующие формы семязачатков. Если образованная у апикального полюса нуцеллуса материнская клетка мегаспор расположена под эпидермой, семязачаток называют тенуинуцеллятным. В нем мегаспора и развивающийся из нее зародышевый мешок сверху и с боков окружены
Рис. 11.185. Развитие и типы семязачатков у покрытосеменных (А—D— по W.Troll, схематизировано; Е—G— по G. Karsten):
А—D — развитие; Е — атропный; F — анатропный; G — кампилотропный тип; а — наружный, i — внутренний интегументы; с — халаза; е — зародышевый мешок (заштрихован); ет — материнская клетка мегаспор; f — семяножка; m — микропиле; п — нуцеллус; t — тетрада мегаспор
374 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Рис. 11.186. Развитие <j> гаметофитов покрытосеменных (А—F Hydrilla verticillate: Hydrocharitaceae; G — Polygonum divaricatum) (A—F — по P. Maheshwari; G — no E.Strasburger).
В развивающемся нуцеллусе семязачатка дифференцируется одна гиподермальная клетка (А), затем она отделяет кроющую клетку, делящуюся далее (В, С), разрастается в материнскую клетку мегаспор (D) и в ходе мейоза (Е, F) образует 4 мегаспоры, из которых только самая нижняя развивается далее в зародышевый мешок; G — зрелый семязачаток с микропиле (т), наружным (а) и внутренним (i) интегументами, халазой (с) и семяножкой (1); зародышевый мешок содержит синергиды (s) и выступающую из-под них яйцеклетку (е), вторичное ядро (к) зародышевого мешка и 3 антиподы (ар) (200х)
только эпидермой нуиеллуса. И напротив, если материнская клетка мегаспор отделена от эпидермы нуиеллуса хотя бы одной клеткой (париетальная, или кроющая, клетка), то семязачаток крассинуцеллятный (рис. 11.186). Объем ткани, окружающей апикально и с боков мегаспору и зародышевый мешок в крассинуцеллятных семязачатках, весьма различен. У семенных растений, оплодотворяемых подвижными сперматозоидами (Cycadopsida, Ginkgopsida), на апикальном конце нуцеллуса образуется углубление, называемое пыльцевой камерой (см. рис. 11.180).
Семязачатки можно различать и по ориентации их продольной оси (см. рис. 11.185). Если семяножка и микропиле расположены на одной линии, семязачаток атропный/ ортотропный. В анатропных семязачатках
вследствие изгибания интегументов и нуцеллуса в области халазы на 180” микропиле оказывается вблизи семяножки, но сам нуцеллус остается прямым. В разной степени изогнутый нуцеллус имеется в кампилотропных семязачатках.
У голосеменных встречаются только атропные семязачатки. Изгибание семязачатков многих покрытосеменных связано с достижением большей доступности микропиле для пыльцевой трубки. У покрытосеменных микропиле может быть связано с плацентой обтуратором, развившимся из семяножки, интегументов, нуцеллуса или самой плаценты (см. рис. 11.248).
В области апикального полюса нуцеллуса чаше всего возникает одна материнская клетка мегаспор. В результате мейоза из нее получается чаще всего линейная, реже Т-об-разная, тетрада мегаспор. Мегаспоры голосе
11.2. Бактерии, грибы, растения I 375
менных имеют клеточную стенку, в которой можно обнаружить спорополленин. Этого не бывает у мегаспор покрытосеменных.
У покрытосеменных исходно, вероятно, были анатропные, крассинуцеллятные семязачатки с двумя интегументами.
Женский гаметофит
В развитии женского гаметофита (зародышевого мешка) принимает участие чаще всего только одна мегаспора (моноспорический зародышевый мешок), реже две
(биспорический зародышевый мешок; некоторые покрытосеменные) или все четыре мегаспоры (тетраспорический зародышевый мешок; Gnetwn, Welwitschia, некоторые покрытосеменные). Моноспорический зародышевый мешок развивается чаще всего (в случае линейной тетрады) из самой внутренней мегаспоры, реже — из самой наружной. Женский гаметофит разных групп семенных растений редуцирован в разной степени. В любом случае его развитие начинается с делений ядер, а образование клеточных стенок происходит позже. У голо-
Мегаспорогенез - Мегагаметогенез
Материнская клетка мегаспор Первое деление Второе деление Третье деление Четвертое деление Пятое деление Зрелый зародышевый мешок
Моноспорический 8-ядерный Po/ygonwn-тип = нормальный тип 0 0 © © /©И I © I
Моноспорический 4-ядерный Oenofhera-тип 0 0
Биспорический 8-ядерный Д///ит-тип 0 с ©
Тетраспорический 16-ядерный Релаеа-тип 0 0 е 0 0
Тетраспорический 8-ядерный Fritillaria-iim 0 © © © 0
Тетраспорический 4-ядерный Plumbagella-wn © Q 0
Тетраспорический 8-ядерный ДДоха-тип 0 0 0 Q ( § 1
Рис. 11.187. Некоторые типы образования зародышевого мешка у покрытосеменных: мейоз диплоидной материнской клетки мегаспор (мегаспорогенез) и развитие гаплоидных мегаспор в зрелый зародышевый мешок (мегагаметогенез) (по Р. Maheshwari)
376 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Рис. 11.188. Опыление и оплодотворение у покрытосеменных (А — по Н. Schenck; В — по A. Jensen, существенно изменено):
А — завязь Fallopia (Polygonum) convolvulus с атропным семязачатком (схематизированный продольный срез, 48х); стенка завязи (fw), столбик (д), рыльце (п) с прорастающими и образовавшим пыльцевую трубку (ps) пыльцевыми зернами (р), семязачаток с семяножкой (f), халазой (с), наружным (а) и внутренним (i) интегументами, микропиле (т) и нуцеллусом (пи), а также зародышевый мешок (es) с яйцевым аппаратом (е), вторичным ядром зародышевого мешка (ек) и антиподами (ар); В — схема зародышевого мешка во время оплодотворения. При проникновении пыльцевой трубки (ps) в область нитчатого аппарата (fa) разрушается одна из двух синергид (s,, s2); ядра спермиев сливаются — одно (sk,) с ядром яйцеклетки (ez), другое (sk2) с двумя сливающимися полярными ядрами (рк, = ркг); в основании 3 антипода (ар(, ар2, ар3)
семенных могут возникать женские гаметофиты из нескольких тысяч клеток (см. рис. И. 180). У всех групп семенных растений, кроме Gnelum, Welwitschia (Gnetopsida) и покрытосеменных, на апикальном полюсе зародышевого мешка образуется разное число архегониев (см. рис. 11.180 и 11.204). У Ginkgo бывает два или три архегония, у хвойных (Coniferopsida) — до 60 и до 100 — у Microcycas. Архегоний состоят из одной, иногда очень крупной, яйцеклетки (диаметром до 6 мм у Cycadopsida; см. рис. 11.180), различного числа клеток шейки (шейковые канальцевые клетки отсутствуют) и иногда из одной брюшной канальцевой клетки или, по крайней мере, из ядра брюшной канальцевой клетки. У семенных рас
тений, оплодотворяемых подвижными сперматозоидами (Ginkgo и Cycadopsida), верхний конец зародышевого мешка отделен от окружающего нуцеллуса расположенной под пыльцевой камерой архегони-альной камерой (см. рис. 11.180).
Gnetum имеет тетраспорический зародышевый мешок, целлюларизация которого ограничена только нижним (халазальным) концом. До оплодотворения невозможно различить, какая клетка фукционирует в качестве яйцеклетки. Зародышевый мешок Welwitschia также тетраспорический. Неупорядоченное образование клеточных оболочек приводит здесь к появлению клеток с разным числом ядер, которые затем могут сливаться друг с другом. И у вельвичии распознать яйцеклетку до оплодотворения не удается.
11.2. Бактерии, грибы, растения
В самом обычном случае у покрытосеменных из одноядерной мегаспоры за три последовательных деления возникают два, четыре и наконец восемь свободных ядер (рис. 11.187; 11.188). По три из них окружаются собственной протоплазмой на верхнем и нижнем концах зародышевого мешка и образуют таким образом клетки, одетые сначала лишь мембраной, а позднее и тонкой клеточной стенкой1. Три верхние клетки называют яйцевым аппаратом. Средняя из них становится более крупной яйцеклеткой, а две других — синергидами. Возможно, что синергиды гомологичны клеткам шейки архегония. Три нижние клетки называют антиподами. Оба оставшихся в цитоплазме зародышевого мешка ядра называют полярными ядрами. До или после проникновения пыльцевой трубки в зародышевый мешок они сливаются во вторичное, диплоидное, ядро зародышевого мешка.
Есть много отклонений от этого типа развития (рис. 11.187). Так, в развитии зародышевого мешка могут участвовать не только одна, но и две или все четыре мегаспоры, в результате чего зародышевый мешок бывает би- или тетраспори-ческим. Другие отклонения затрагивают число ядерных делений, расположение названных групп клеток или особенности слияния ядер. В то вре-ms как, например, сформированный зародышевый мешок Репаеа-типа состоит из 16 клеток, в зародышевом мешке Oenothera-типа их всего 4.
Питание зародышевого мешка обеспечивают прежде всего антиподы, а также гаустории, которые могут образоваться из мегаспор, синер-гид или антипод.
Нектарники
Нектар — «плата» за посещение цветка опылителями. В то время как у опыляемых животными голосеменных эту функцию может выполнять опылительная капля, выделяемая через микропиле, у покрытосеменных имеются гыделяющие нектар железки — нектарники. Они могут быть множественными и не связанными друг с другом (см. рис. 11.264) или чаще всего собраны в кольцевидный диск — разрастание оси
1 Клеточная оболочка яйцеклетки неполная, она прервана на халазальном конце. — Примеч. ред.
цветка между андроцеем и гинецеем (см. рис. 11.267). Чашелистики могут образовывать нектар в определенных местах на внутренней или внешней стороне, а лепестки — на внутренней стороне. Тычинки, к примеру, могут нести нектарники на нитях или преобразуются в стаминодии и служат только для производства нектара (см. рис. 11.236). В гинецее функцию нектара может выполнять секрет рылец, а в ценокарпном гинецее бывают септальные нектарники — полости между краями плодолистиков, сообщающиеся протоками с внешней средой (см. рис. 11.228). Возможно также образование нектара и на поверхности завязи.
Нектарники за пределами цветков называют экстрафлоральньгми, они могут служить, в частности, для «расплаты» с охраняющими растение муравьями.
Расположение элементов цветка
Возможные варианты расположения элементов цветка — спиральное или мутовчатое (круговое), в более редких случаях беспорядочное — были рассмотрены выше. Устройство цветков у покрытосеменных разнообразно. В так называемых полных цветках почти всегда следуют друг за другом околоцветник, андроцей и гинецей. В зависимости от числа и положения элементов цветка его ось (цветоложе I бывает более или менее вытянутой или сильно укороченной. Она может даже превратиться в диск или даже в бокал или трубку (гипантий). Благодаря образованию гипантия основания элементов околоцветника и тычинок отдалены от основания гинецея. Существует также возможность удлинения междоузлий внутри цветка. Если вытянуто междоузлие между андроцеем и гинецеем (см. рис. 11.265), то имеется гинофор. При вытягивании междоузлия между околоцветником и андроцеем/гинецеем получается андрогинофор. В зависимости от локализации ростовых процессов в гинецее его относительное положение в цветке бывает разным (рис. 11.189). Если в первую очередь растут свободные спинные стороны (и бока) плодолистиков, то получается верхняя завязь. Околоцветник и тычинки при этом расположены у основания гинецея,
378 | ГЛАВА И. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Верхняя завязь Полунижняя завязь Нижняя завязь (гипогинный (перигинный (эпигинный
цветок) цветок) цветок)
Рис. 11.189. Положение завязи в цветках покрытосеменных (по Р. Leins)
поэтому такие цветки называют также ги-погинными. Рост цветоложа ниже перифе рического участка гинецея приводит к образованию нижней завязи. Из-за прикрепления околоцветника и тычинок выше завязи такие цветки называют также эпигин-ными. Промежуточный тип — полунижняя завязь. Если завязь свободно располагается в цветочном бокале, то она перигинная. В циклических цветках члены одного круга чаще всего располагаются в промежутках между членами предыдущей мутовки, так что члены соседних кругов чередуются. Как отклонение от правила чередования кругов органы следующих друг за другом мутовок располагаются на тех же радиусах («противолежат» друг другу). В пятикруго
вых цветках некоторых покрытосеменных (околоцветник с чашечкой и венчиком, два круга тычинок, гинецей) при соблюдении правила чередования кругов тычинки внешнего круга расположены на радиусах, чередующихся с лепестками, и противолежат чашелистиками (эписепальные), а тычинки внутреннего круга — на радиусах, чередующихся с наружными тычинками (эпи-петалытые).
Чередование кругов может нарушаться, если какой-либо круг выпадает. Если это случается, к примеру, с наружным из двух кругов тычинок, то тычинки единственного оставшегося круга расположены против лепестков (эпипетально). Из-за нарушения чередования кругов в гинецее в цветке с двухкруговым андроцеем тычинки внутреннего круга могут быть смещены наружу разрастающимися эпипетальными плодолистиками. В зрелом цветке при этом кажущийся внешний круг тычинок расположен против лепестков (эпипетально), а кажущийся внутренний — против чашелистиков (эписепально). Это явление называют обдиплостемонией.
В цветках покрытосеменных наиболее обычно мутовчатое (циклическое) расположение элементов. Число мутовок (кругов) в цветке бывает разным. Особенно часто встречаются пя
Рис. 11.190. Типы симметрии и диаграммы цветков (частично по A. W. Eichler, а также по G.Hegi): А — Sedum sexangulare, актиноморфный; В — Dicentra spectabilis, дисимметричный; С — Lamium album, зигоморфный
11.2. Бактерии, грибы, растения | 37g
тикруговые цветки с пятью мутовками (два круга околоцветника, к примеру, чашечка и венчик, два круга тычинок и чаще всего не подчиняющийся правилу чередования кругов круг плодолистиков) и — при выпадении одного круга тычинок— четырехкруговые цветки. Бывают, однако, и двух- или однокруговые цветки всего с одним кругом элементов.
Строение и расположение элементов зрелого цветка определяют его симметрию (рис. 11.190). Цветки со спиральным расположением элементов первично асимметричны. Среди циклических цветков различают радиально-симметричные (актиноморфные) с более чем двумя плоскостями симметрии, дисимметричные — с двумя плоскостями симметрии и зигоморфные цветки — только с одной плоскостью симметрии. Циклические цветки могут быть и вторично асимметричными.
При более тщательном рассмотрении симметрии выделяют медианную плоскость, которую можно провести через стебель материнского побега, цветоложе и кроющий лист цветка. Перпендикулярна к ней трансверсальная плоскость, а остальные плоскости косые. Соответственно можно различать, к примеру, медианно-, трансверсально- или косо-зигоморфные цветки.
Строение цветков можно лучше всего представить на диаграмме цветка (рис. 11.190). Эмпирические диаграммы представляют непосредственно наблюдаемое строение цветка, а теоретические содержат также интерпретации и указывают, к примеру, что какие-то из ожидаемых органов не развиты. Формулы цветков содержат информацию: о типе симметрии цветка (©> = спиральный; * = актиноморфный; -+- или | = дисимметричный; | = зигоморфный; ) = циклический асимметричный); об имеющихся элементах (Р = простой околоцветник; К= чашечка; С = венчик; А = андроцей; G = гинецей); о числе элементов в каждой мутовке (например, А5+5 = два круга тычинок по 5 в каждом; <*> = большое и неопределенное число); об отклонениях в строении отдельных элементов (например, АЗМ = стаминодии; 3° = выпадение элемента; 5“ = в~орич-ное увеличение числа элементов); о срастании элементов (число в скобках, например, С(5) = лепестки сросшиеся); о положении завязи (например, Ощ = верхняя завязь; G45>_ = полунижняя; G(5) = нижняя); об образовании ложных стенок завязи (например, G(^) или о переходах между разными типами (например, */& = от радиально-симметричного до спирального). Некоторые примеры формул:
Adonis: */& К5 С610 A. G„;
Sedum: * К5 С5 A5+s G3;
Dicentra: К2 С2+2 А2+2 соответственно
(расщепленные и сросшиеся тычинки!) (1/2 - I - 1/2) +(1/2 - I - 1/2)0(215
Lamium: l K5|C(S|Ar JG^;
Iris: * Pj() A3 + 3»G(j).
СОЦВЕТИЯ
Семенные растения имеют чаще всего несколько или много цветков, которые нередко собраны в соцветия (см. 4.2.5.3). Однозначного определения соцветия нет. Здесь мы понимаем его как репродуктивную систему ветвей, которая после завершения цветения и плодоношения далее не растет и обычно сбрасывается.
Листья в соцветиях либо отсутствуют, либо представлены более или менее невзрачными верховыми листьями (брактеями и предлистья-ми1). Отдельные мелкие цветки становятся более заметными при тесном расположении (например, Iberis), при наличии увеличенных, часто стерильных краевых цветков (например, Hydrangea, Viburnum opulus) или ярко окрашенных верховых листьев (например, Astrantia, Comus suecica). Так, благодаря разделению функций между отдельными цветками и при участии добавочных осевых и листовых образований в конце концов возникает функциональное биологическое единство, аналогичное отдельному цветку и называемое псевдантием. В качестве примеров можно назвать циатии Euphorbia, корзинки Dipsacaceae и Asteraceae или окруженный окрашенным покрывалом початок Arum. Впрочем, бывает и наоборот: такое функциональное единство представлено только частью цветка — мерантием (например, Iris).
ОПЫЛЕНИЕ
Опыление (англ, pollination) — процесс переноса пыльцевых зерен на микропиле семязачатков голосеменных или на рыльца плодолистиков покрытосеменных растений.
Различают самоопыление (автогамия) — опыление в пределах одного индивида, и перекрестное опыление (аллогамия) — опыление между двумя индивидами. Аллогамия — единственный тип опыления, когда мужские и женские репродуктивные структуры находятся на разных индивидах (двудомность), как у Cycado-psida, Ginkgo, многих Coniferopsida и Gnetopsida и
1 Предлистья — листья низовой формации. — Примеч. ред.
380 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
малой части покрытосеменных. Если однополые цветки находятся на одном и том же растении (однодомность) или хотя бы некоторые цветки обоеполые, то в принципе возможна и автогамия. Важнейшие механизмы, которые во многих ситуациях препятствуют вредной в эволюционно-биологической перспективе автогамии и самооплодотворению, — это самонесовместимость, гетероморфия, дихогамия и геркогамия (см. 10.1.3.1 и 10.1.3.2). Крайнюю степень автогамии представляет клейстогамия — самоопыление и самооплодотворение в нераскрывающемся бутоне. Например, у некоторых видов У/о/а и кислицы (Oxala acetosella) один и тот же индивид образует как клейстогамные, так и распускающиеся (хасмогамные) цветки. У Lamium amplexicaule клейстогамные цветки развиваются главным образом в начале и в конце вегетационного периода.
Важнейшими переносчиками пыльцы являются ветер, вода и разные животные.
Ветроопыление (анемофилия) требует образования большого количества пыльцы, быстрого и равномерного высвобождения ее в воздух, максимально долгого пребывания в воздухе1, а также открытых и крупных рылец, чтобы легко улавливать пыльцу. Ветроопыляемые цветки чаще всего невзрачные, без запаха и нектара. Обычно они однополые; и мужских, или тычиночных, цветков значительно больше, чем женских; пыльцевые зерна более или менее гладкие, а пыльца порошковидная вследствие отсутствия или раннего высыхания пыльцевого клея.
В связи с этими потребностями у ветроопыляемых видов возник комплекс (синдром) типичных признаков. Пыльцевые зерна у них не слипаются и способны долго держаться в воздухе либо из-за своей малой величины, либо благодаря особым воздухоносным мешкам (некоторые хвойные)1 2. Массовое образование пыльцы обусловлено увеличением пыльников и/или числа мужских цветков или тычинок. У Corylus, к примеру, один семязачаток приходится на 2,5 млн пыльцевых зерен. Высыпание пыльцы из пыльников облегчается подвижностью тычиночных нитей (например, Роасеае; см. рис. 11.232), цветоно
1 Пыльца ветроопыляемых растений теряет способность оплодотворять после часового пребывания в воздухе, поэтому пыльцевые зерна анемофилов относительно тяжелые, что позволяет сократить время пассивного полета. — Примеч. ред.
2 См. предыдущую сноску. — Примеч. ред.
жек (например, конопля: Cannabis', см. рис. 11.257) или оси всего соцветия (например, мужские сережки лещины: Corylus, ольхи: Alnus, дуба: Quercus', см. рис. 11.259 и 11.260). При этом пыльца часто сначала оседает на элементах цветка или соцветия, с которых потом ее сдувает ветер. В мужских цветках Urtica (см. рис. 8.34) и Pilea происходит взрывное высвобождение пыльцы при резком распрямлении тычиночных нитей. Длительность нахождения пыльцы в воздухе и определяемое этим расстояние ее распространения в значительной мере зависят от высоты расположения мужских цветков над землей, а также от структуры растительности. Пыльца ветроопыляемых растений вызывает у многих людей аллергию (например, сенную лихорадку). Столбики и рыльца женских цветков ветроопыляемых покрытосеменных чаще всего сильно увеличены, что повышает вероятность опыления. Число семязачатков в завязях обычно сильно редуцировано, а цветки расположены открыто. Околоцветник, только мешающий опылению, редуцирован или совсем отсутствует. Наконец, опыление облегчается и благодаря раннему цветению, в умеренном климате часто даже предшествующему распусканию листвы (например, дуб, ольха, вяз, тополь, ясень, лещина).
Географическое распространение ветроопыляемых покрытосеменных зависит от условий местообитания. Они часто растут массовыми зарослями в открытых ветру саваннах, степях и арктическо-альпийских биотопах или в древесном ярусе субтропических и бореальных лесов. Во влажных тропических лесах ветроопыляемые растения редки.
В некоторых группах растений, где встречаются как встроопыляемые, так и зоофильные виды (например, Thalictrum, Sanguisorba, рис. 11.191, Acer, Fraxinus, Artemisia), иногда легко понять воз никновение анемофилии. Это относится, в частности, к Tilia и Calluna, а у Castanea пыльца сначала покрыта пыльцевым клеем, и ее собираю! насекомые, но когда она высыхает, то разносится ветром.
Опыление водой (гидрофилия), т.е. перенос пыльцы или мужских цветков водой, встречается лишь у немногих покрытосеменных. Здесь можно различать транспорт над, по или под поверхностью воды.
В прямостоячих цветках дождевая вода служит самоопылению, реже — способствует перекрестному опылению. Гидрофилия встречается отнюдь не у всех водных растений. Чаще всего их цветки поднимаются над водной поверхностью и опыляются ветром или животными (например, Potamogetom, см. рис. 11.223). У Vallisneria (см. рис. 11.223) и Elodea оторвавшиеся мужские
11.2. Бактерии, грибы, растения | 381
цветки, а у Callitriche плавучая пыльца достигают рылец, находящихся в это время на поверхности воды. Под водой опыляются, к примеру, Ceratophyllum, Najas и Zostera .ем. рис. 11.223). У последнего рода пыльцевые зе; та нитевидные, длиной до 0,5 мм, без экзины. многих опыляемых водой видов наблюдается параллелизм в строении цветков с ветроопыляемыми формами.
Необыкновенное многообразие цветков покрытосеменных можно понять только в контексте приспособления к опылению животными (зоофилия). Зоофилия — важная причина видового разнообразия покрытосеменных, так как все возрастающая специализация цветков к опылению немногими или единственным опылителем ведет к репродуктивной изоляции — важному фактору видообразования.
Зоофилия может осуществляться только в том случае, если опылитель замечает цветки и посещает их регулярно и достаточно долго и если цветки устроены так, что опылитель соприкасается с пыльцой и рыльцем и при этом переносит пыльцу. Чтобы привлечь внимание потенциального опылителя, цветки обладают средствами дальней аттрактации (англ, advertisment), а регулярность посещения обычно обеспечивают приманки (англ, reward).
Средства дальней аттрактации имеют оптическую и химическую природу: окраска и запах. Окраску цветков в первую очередь обусловливают пигменты, содержащиеся в вакуолях (антоцианы: синий, фиолетовый, красный; антоксантины: желтоватый, белый, ультрафиолетовый; беталаины: красно-фиолетовый, желтый; халконы и ауроны: желтый, ультрафиолетовый) или в пластидах (каротиноиды: оранжевый каротин и желтый ксантофилл). Окраски могут быть модифицированы благодаря тому или иному расположению в органе клеток с пигментами, наложению слоев с различной окраской, частоте и величине межклетников, а также поверхностной структуре эпидермы. Так, гладкие эпид. умальные клетки делают окраску блестящей, а клетки с сосочками — матовой. Белая окраска цветков может возникать и за счет полного отражения света, прежде всего межклетниками.
Окраска цветка может изменяться в процессе его старения. Например, после
Рис. 11.191. Переход от энтомофилии (А) к вторичной анемофилии (В, С) у представителей рода Sanguisorba (Rosaceae) (по F. Knoll):
А — обоеполый цветок S. officinalis с 4 тычинками, сосочковым рыльцем и нектарником; В, С — однополые цветки S. minor без нектара, <j> с перистыми рыльцами, О" с многочисленными тычинками (около 6х)
опыления окраска пятен в цветках Aesculus меняется с желтой на красную. Для потенциальных насекомых-опылителей красные цветки менее заметны и привлекательны, так как воспринимаются ими как уже «бесполезные».
Понимание оптического воздействия цветков требует знания физиологии зрения опылителей, которая изучена пока у немногих животных, таких, как медоносная пчела, шмели, бражники, жужжало среди мух и некоторые колибри. Оказалось, что пчелы и шмели не видят чисто красный цвет, но зато воспринимают невидимый для человека ультрафиолет с длиной волны 310— 400 нм, а среди остальных окрасок цветков реагируют только на желтые (520 — 650 нм), синефиолетовые, включая пурпурные оттенки (400— 480 нм), и белый цвет, который воспринимают как сине-зеленый. Зрительное восприятие птиц подобно человеческому: для них наиболее заметен красный цвет. Опыты по дрессировке насекомых-опылителей показали, что оптическая привлекательность может зависеть также и от совместного действия таких факторов, как степень сочности, яркости цветка, контраст яркости и цвета, а также форма частей цветка. Подобным образом удалось доказать значенье рисунка и цветных пятен на лепестках цветков, показывающих дорогу к нектару, как, например, оранжево-желтый зев в лимонно-желтых цветках Linaria vulgaris. Распространено мнение, что цветовые указатели имитируют пыльники или пыльцу. Часто указатели заметны только для насекомых, воспринимающих ультрафиолетовый свет (на
382 | ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
пример, кажущиеся нам равномерно желтыми листочки околоцветника Caltha palustris). Оптическим аттрактантом могут быть и движения всего цветка или его частей.
Запах цветкам придают самые разные вещества. Чаще всего запахи, приятные для человека, обусловлены терпенами и бензе-ноидами, а также простыми спиртами, кетонами, эфирами органических кислот, фе-нилпропанами и др. Некоторые из этих ве ществ, например у Ophrys, могут имитировать феромоны самок насекомых, побуждая самцов совершать попытки копуляции. Неприятные запахи многих цветков или соцветий, опыляемых мясными или навозными мухами, обусловлены аминами, аммиаком или индолами.
Запах в принципе может исходить от любых частей цветка и локализован значительно менее упорядоченно, чем окраска. Соответственно и движение опылителей на запах обычно менее регулярное и отчетливое, чем при зрительной ат-грактации. Многие цветки обладают сходными со зрительными также и запаховыми указателями, которые иногда расположены в тех же местах. Примеры: корона Narcissus (см. рис. 11.228) и чешуи на основании отгиба лепестка у некоторых видов Silene (см. рис. 11.239).
У пчел и других перепончатокрылых (Ну-menoptera) повторное успешное посещение цветков одного вида вызывает ограниченное во времени стремление посещать цветки этого вида и далее. Это основано на стимулирующем действии видоспецифического запаха частей цветка, принесенного в улей нектара и пыльцы, а также высокоразвитой памяти этих животных и их способности обмениваться информацией.
Опылители посещают цветки ради разных первичных аттрактантов. Огромное значение для опылителей иМеет пищевая ценность пыльцы и нектара. В пыльцевых цветках образуется в изобилии пыльца, богатая белками, жиром, углеводами и витаминами. Такие цветки, доступные и примитивным насекомым с грызущим ротовым аппаратом, имеют многие Magnoliidae (например, Winteraceae, Victoria) и Ranun-culales (Anemone), а также таксоны с вторично полимерным андроцеем (например, Papaver, Rosa). Ряд пыльцевых цветков имеет немного тычинок со снабженными порами пыльниками (например, Solanum dulcamara), из которых пыльца высвобождается
при вибрации посещающих цветки насекомых (англ, buzz polination).
Нектар нектарных цветков — это в основном водный раствор сахаров (сахарозы, фруктозы, глюкозы), но обычно содержит также аминокислоты. Предложение нектара менее обременительно для растения, чем предложение богатой фосфором и азотом пыльцы. Нектар могут образовывать разные части цветка, он в разной степени доступен животным-опылителям. Он может находиться открыто, например на дне цветка у многих Rosaceae, но может быть скрыт глубоко в трубке венчика или в шпорцах (например, Viola, Linaria, Corydalis), откуда его могут достать лишь животные с длинным ротовым аппаратом.
Масличные цветки некоторых покрытосеменных (например, Lysimachia", Calceolaria", многие Malpighiaceae) образуют в особых железках масло, служащее опылителям пищей и иногда как строительный материал. Смолу для строительства гнезд образуют цветки Dalechampia (Euphorbiaceae) и Clusia (Clusiaceae) — смоляные цветки. В душистых цветках (например, Stanhopea: Orchidaceae; Gloxinia". Gesneriaceae) самцы пчел эуглоссин собираюг пахучие вещества, имитирующие феромоны опылителей и, как полагают, используют их при спаривании.
Цветки могут по-разному использовать стремление животных к размножению. Так, соцветия Ficus и цветки Zamia (Cycadopsi-da)1, Yucca или Siparuna служат насекомым местом откладки яиц.
У Ficus carica в течение всего года появляются кувшиновидные соцветия, в которых в разных сочетаниях находятся цветки трех типов (рис. 11.192). Наряду с мужскими имеются и женские цветки с длинным столбиком и коротким столбиком. Цветки с длинными столбиками образуют семена, тогда как цветки с короткими столбиками служат местом для откладывания яиц и развития личинок галловой осы Blastophagapsenes. Последовательность распускания цветков в течение года, с одной стороны, и в каждом соцветии — с другой, обеспечивает, во-первых, опыление и оплодотворение растения и, во-вторых, размножение насекомого.
В роде Ophrys (Orchidaceae) губа цветка имитирует формой, опушением и даже
1 У всех саговниковых есть стробилы, а не цветки. — Примеч. ред.
11.2. Бактерии, грибы, растения | 383
Рис. 11.192. Зоофилия у разных покрытосеменных:
А— шмель как опылитель; В— цветок Salvia pratensis (фиолетово-синий, несколько увеличено); С, D — рычажный механизм у Salvia pratensis-, каждая из двух тычинок имеет вытянутый в длинный рычаг связник (с), подвижно соединенный с короткой тычиночной нитью (f); только на переднем, удлиненном, конце рычага имеется фертильная тека; вторая, стерильная (s), тека образует более короткое плечо рычага, которое объединено в пластинку с соответствующей частью второй тычинки; Е, F — птица-медосос Arachnothera longirostris как опылитель Sanchezia nobilis (Acanthaceae, цветки желтые, брактеи пурпурные, около Зх); G — разрезанное продольно соцветие Arum maculatum на женской стадии развития со светло-зеленым покрывалом и невзрачными мужскими, женскими и стерильными цветками (h), загораживающими вход (2/Зх); Н — соцветие инжира (Ficus carica) в продольном разрезе (немного увеличено) с J — мужскими и К — длинностолбиковыми женскими фертильными цветками, а также L— с короткостолбиковыми женскими галловыми цветками (увеличено)
запахом самку определенных видов пчел и роющих ос, что побуждает самцов совершать попытки спаривания, которые могут привести к опылению. Недавно удалось показать, что запах цветков одного из видов имитирует феромоны соответствующих насекомых. Так как Ophrys ничем не вознаграждает опылителя, он — цветок-обманка. Конструкция цветка должна гарантировать, что опылитель в соответствии со строением своего тела будет контактировать с пыльцой и рыльцем. При этом пыльца может быть помещена на опылителя с помощью
специальных рычагов, зажимов, путем приклеивания или «выстреливания» пыльцой.
Протандрические цветки Salvia pratensis известны благодаря своему эффективному рычажному механизму, описанному С. К. Sprengel еще в 1793 г. (рис. 11.192). У них всего две тычинки, каждая из которых несет по вытянутому в длинный рычаг связнику, примыкающему к верхней губе цветка и подвижно соединенному с короткой тычиночной нитью. Только на переднем1, более длинном плече рычага находится фертиль
1 На самом деле это заднее плечо. — Примеч. ред.
384 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
ная тека. Другая, стерильная, гека образует заднее1, более короткое плечо, объединенное в пластинку с соответствующей частью второй тычинки. Эта пластинка закрывает доступ к нектару в основании трубки венчика. Когда шмель надавливает на пластинку, более длинный конец рычага изгибается и прижимает теку с пыльцой к спине зйивотного. В более старых цветках в том же положении оказывается уже рыльце, на которое насекомое и переносит пыльцу.
Пример особенно сложного механизма опыления представляет функционирующее как ловушка соцветие видов рода Arum (рис. 11.192). Однополые цветки собраны на нижнем конце толстого обоеполого протогинного початка, окруженного светлым покрывалом, которое внизу расширено, выше сужается, а наверху широко раскрыто. В самом низу початка расположены женские цветки, наверху — мужские, а между ними и над мужскими находятся стерильные цветки, преобразованные в толстые щетинки, загораживающие вход в полость покрывала. Вне полости ось початка булавовидно утолщена и у особенно хорошо изученного A. nigrum в первое после распускания утро издает резкий навозный запах. Выделение пахучего вещества стимулируется повышением температуры в початке, а его выход наружу осуществляется через отверстия системы межклетников («дырявая эпидерма»). Этот запах привлекает разных любителей навоза (мух и жуков), часть которых уже несет на себе пыльцу из других соцветий. Садясь на гладкую, покрытую маслом эпидерму внутренней поверхности покрывала или оси початка и пытаясь удержаться на ней, насекомые соскальзывают и падают в полость покрывала. Выбраться тем же путем невозможно, так как «горлышко» полости перегорожено торчащими внутрь щетинками стерильных цветков, а ее стенки, как и верхняя часть, гладкие и скользкие. Копошась внизу, насекомые опыляют принесенной пыльцой женские цветки. Ночью из вышерасположенных мужских цветков высыпается пыльца и прилипает к насекомым. Одновременно прекращается выделение запаха. Наконец, на следующий день после увядания загораживающих выход цветков нагруженное пыльцой насекомое может выбраться из ловушки и отправиться на поиски новых соцветий. «Ловушками» являются также цветки разных видов Aristolochia. У представите-1СЙ рода Ceropegia (Apocinaceae) в цветках-ловушках есть еще и зажимы.
Опылитель может получить пыльцу не только из пыльников. При вторичном предложении пыльцы другие части цветка принимают пыльцу из пыльников и нагружают ее на опылителя.
1 Переднее. — Примеч. ред.
Пример — протерандрические цветки Asteraceae, у которых пыльца высыпается в трубку, образованную склеившимися пыльниками, а уже из нее выталкивается удлиняющимся столбиком. На опылителя пыльца попадает с верхушки столбика.
Для удержания пыльцы на поверхности тела опылителей служат прежде всего пыльцевой клей и висциновые нити, но наверняка отчасти и снабженная шипиками или другими подобными структурами поверхность экзины (см. бокс 4.1, рис. F).
Можно выделить несколько типов цветков по их особенностям, тесно связанным со взаимодействием с опылителями, которые могут реализоваться на уровне части цветка, отдельного цветка или соцветия. В плоских дисковидных и чашевидных цветках (например, Апетопс. отдельный цветок; Matricaria-, соцветие; Tilia, Euporbia: соцветие) доступ к центру цветка и имеющемуся там вознаграждению практически беспрепятственный. Он ограничен в бокаль-чатых и колокольчатых цветках (например, Hyoscyamus, Crocus) и еще более — в трубчатых цветках и цветках с длинноноготковыми лепестками (например, Silene, Nico-tiana). В шпорцевых цветках нектар скапливается в шпорце (например, Linaria, Viola), а в трубчато-колесовидных цветках имеется несколько проходов к вместилищам нектара (например, Gentiana acaulis). Зигоморф-ны мотыльковые (например, Corydalis, Pisum, Polygala), одно- и двугубые цветки (например, Aconitum, Viola: отдельные цветки, Mimetes: соцветие). В щетковидных цветках обычно многочисленные тычинки далеко выступают из цветка (Syzygium, Acacia и Salix: соцветия) и, наконец, цветки-ловушки на время ловят насекомых (например, Asclepias: ловушки-зажимы1, Arum- скользкие ловушки).
1 В цветках Asclepias поллинии прикрепляются к телу опылителя специальными зажимами. Мелкие насекомые, не способные опылять виды этого рода, слишком слабы, чтобы извлечь прикрепившийся к ним поллинии. Они остаются в цветке и погибают. Опылители же без проблем покидают цветок, унося поллинии. Ничего общего с цветками-ловушками это не имеет, никакого временного удержания опылителя в цветке здесь нет. — Примеч. ред.
11.2. Бактерии, грибы, растения
Эту классификацию можно сделать еще более дробной, а некоторые цветки проявляют черты разных типов. Поэтому не всегда легко отнести цветок к какому-то определенному типу.
Строение тела и ротового аппарата, особенности поведения и пищевые пристрастия многих животных-опылителей создают специфические требования к устройству посещаемых ими цветков, обусловив их селективное преобразование. Таким образом, можно различать характерные опылительные синдромы цветков.
Правильность этого воззрения подтверждают экспериментальное исследование высокой способности к различению у большинства посетителей цветков и тот факт, что функционально очень сходные синдромы возникали на совершенно разной структурной основе: одиночного цветка, мерантия или соцветия. По особенностям цветка иногда предсказывали неизвестного еще опылителя, которого обнаруживали впоследствии. При анализе селективных влияний опылителей цветка на его строение следует принимать во внимание, что большинство цветков опыляется значительным числом разных видов животных и, таким образом, полифильны. Возрастающая специализация привела, однако, и к возникновению олиго- и монофильных цветков, опыляемых немногими или всего одним опылителем. Учитывая обратное селективное влияние цветков на опылителей, можно говорить о коэволюции между растениями и животными.
Среди (опыляемых насекомыми) энто-мофильных растений опыляемые жуками (кантарофилия) чаще всего имеют легкодоступные крупные дисковидные или чашевидные цветки зеленой или светлой окраски без нектара, обычно сильно пахнущие и продуцирующие много пыльцы. Это соответствует поведению жуков — сравнительно неловких насекомых с грызущим ротовым аппаратом, нередко повреждающих цветки. Жуки опыляют цветки многих Magnoliidae, а также дисковидные цветки эволюционно продвинутых таксонов (например, Comus, Viburnum', соцветия — псев-дантии). Гетерогенны цветки, опыляемые мухами (миофилия). К ним относятся, с одной стороны, мелкие, более или менее непахучие дисковидные цветки с открыто расположенным нектаром (например, Apiaceae, Rutd), а с другой стороны, цветки,
опыляемые навозными мухами (сапромио-филия), подражающие зелено-пурпурной пятнистой окраской и запахом навоза источникам пищи своих опылителей. Последние чаще всего являются цветками-обманками и/или цветками-ловушками и попросту «эксплуатируют» своих опылителей (например, Aristolochia: цветок; Arum: соц-ветие-псевдантий). Особенно разнообразны и обычны опыляемые пчелами цветки (мелиттофилия). Для них характерны зигоморфное строение мотыльковых и двугубых цветков с посадочной площадкой для опылителя, обычно желтая, фиолетовая или синяя окраска, легкий запах и чаще всего глубоко скрытый нектар (например, Salvia). Особые приспособления для опыления имеются у крупных тропических древесных пчел рода Xylocopa и у неотропических эуглоссиновых пчел. Цветки, опыляемые дневными бабочками (психофилия), выделяются прямым положением, узко-трубчатым околоцветником, обычно карминово-красной окраской и глубоко спрятанным нектаром (например, Dianthus carthusianorum, Nicotiana tabacum). В противоположность открытым днем психофильным цветкам опыляемые ночными бабочкам и молями цветки (сфингофилия и фаленофилия) раскрываются вечером. Это расположенные горизонтально или свисающие узко-трубчатые цветки с беловатой окраской, сильным ароматом и глубоко спрятанным нектаром (например, Oenothera, Silene, Lonicera periclymenum). Замечательна орхидея Angraecum sesquipedale с Мадагаскара со шпорцем длиной до 43 см. Предположили, что ее опыляет какой-то бражник, и в дальнейшем его действительно обнаружили. Отсюда происходит название этой бабочки — Xanthopan morgani praedicta.
Названные синдромы цветков — лишь отдельные примеры. Энтомофильные цветки столь же гетеро! енны, как и названные группы их опылителей (например, мухи и пчелы). Среди насекомых опылителями являются также Orthoptera, Hemiptera, Thysanoptera и представители других таксонов.
Опыляемые птицами цветки (орнитофилия) отчетливо отличаются от цветков, опыляемых насекомыми. Места для посадки в самом цветке чаще всего нет, и значи-
386 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
тельно более тяжелые птицы должны, посещая цветки, зависать в воздухе (колибри) или доставать до них с прочной опоры. Обычно это крупные цветки бокальча-того, трубчатого или щетковидного типа, преимущественно огненно-красные или с контрастным рисунком красных, а также синих, желтых и зеленых тонов («попугайные расцветки»). Запах у них отсутствует, и соответственно у птиц-опылителей плохо развито обоняние, зато обильно выделяется жидкий, как правило глубоко спрятанный, нектар, который птицы достают с помощью языка, трубковидного или снабженного щеточкой папилл. Пыльца переносится иногда на клюве, но чаще на других частях головы и реже на ногах (см. рис. 11.192). Орнитофильные цветки есть почти во всех тропических семействах с зоофильными цветками (например, Erythrina, Fuchsia, Hibiscus tiliaceus, Tropaeolum majus, Salvia splendens. Aloe, в том числе и виды, культивируемые в Центральной Европе). Важнейшие птицы-опылители — колибри в Нео-тропической области, нектарницы и медососы в Палеотропической. Тем не менее в обеих этих областях встречаются птипы-опылители и из многих других семейств.
Цветки, опыляемые рукокрылыми (хи-роптерофилия), ограничены тропиками. Их опылители — в первую очередь длинно-язычные крыланы в Старом Свете или летучие собаки и длинноязычные вампиры — в Новом. Для хироптерофильных видов характерны открыто расположенные, прочные, чаще всего бокальчатые с широким зевом или щетковидные цветки, раскрывающиеся ночью, часто светло- или темноокрашенные, с сильным фруктовым или винным запахом и очень большим количеством нектара (и пыльцы) (например, Camegiea, Adansonia, Cobaea, виды родов Musa и Agave).
Некоторые другие мелкие млекопитающие, к примеру грызуны, но прежде всего сумчатые, также могут опылять растения.
У современных голосеменных наряду с анемофилией известно опыление жуками (у Суса-dopsida), а также молями (например. Gnetum}. Опылителями первых покрытосеменных были, вероятно, преимущественно жуки и мухи, но. возможно, также осы и некоторые моли. Такие опылители, как чешуекрылые (дневные и ночные
бабочки), птицы, рукокрылые и высокоразвитые пчелы, появились лишь в позднем мелу или даже в третичном периоде и эволюционировали затем совместно с опыляемыми ими покрытосеменными растениями.
ОПЛОДОТВОРЕНИЕ
После того как пыльцевые зерна в результате опыления попали в пыльцевую камеру на верхушке нуцеллуса (Cycado-psida, Ginkgo), в микропиле (Coniferopsida, Gnetopsida) или на рыльце пестика (Ап-giospermae) начинается прорастание пыльцы, приводящее к оплодотворению. Опыление и оплодотворение могут быть разделены во времени, особенно у некоторых групп голосеменных, несколькими месяцами. Пыльцевое зерно прорастает пыльцевой трубкой из апертуры в оболочке.
У семенных растений с подвижными сперматозоидами (Cycadopsida, Ginkgo) функция пыльцевой трубки состоит в основном'в том, чтобы закрепить мужской гаметофит на нуцеллусе и в качестве гаустории всасывать из него питательные вещества (см. рис. 11.180). Впрочем, пыльцевая трубка растет также и по направлению к архегониям при разрушении нуцеллуса между пыльцевой и архегониальной камерами. В заполненной жидкостью архегониальной камере над архегониями сперматозоиды выходят из пыльцевой трубки и плывут к архегониям. После их проникновения в архегонии оплодотворение завершается слиянием гамет (плазмогамия) и ядер (кариогамия). Оплодотворение подвижными сперматозоидами у саговников и гинкго называют зооидиога-мией.
При оплодотворении спермиями, т.е. неподвижными мужскими гаметами (сифоногамия: Coniferopsida, Gnetopsida, Angio-spermae), пыльцевая трубка должна доставить спермин непосредственно к яйцеклеткам (см. рис. 11.204; 11.188). При этом она должна прорасти через нуцеллус, по пути локально растворяя его клетки. У Wel-witschia (Gnetopsida) обнаружено совершенно необычное явление, когда не только пыльцевая трубка растет к женскому
11.2. Бактерии, грибы, растения | 387
гаметофиту, но и от женского гаметофита навстречу ей растут трубковидные структуры.
У покрытосеменных пыльцевая трубка с поверхности рыльца должна прорасти сквозь ткани плодолистика, например столбика, чтобы достичь семязачатка. Рост идет по проводниковому каналу. При этом пыльцевая трубка может расти либо в секрете на поверхности этого канала, в его эпидерме, либо в более глубоко расположенных слоях клеток. В ценокарпных гинецеях существует возможность того, что объединенная в симпликатной области всех плодолистиков часть проводниковых каналов, компитум, распредели! трубки пыльцевых зерен, проросших только на одном рыльце, по всем плодолистикам. Путь пыльцевой трубки от плаценты к микропиле облегчается благодаря анатропному или кампилотропному строению семязачатков либо разнообразным выростам тканей между плацентой и микропиле (обтуратору).
В пыльцевой трубке покрытосеменных протоплазма с двумя спермиями и ядром пыльцевой трубки всегда находится в концевом участке, перетекая из более старых частей и часто отделяясь от них каллозной пробкой. Скорость роста пыльцевой трубки составляет 1 — 3 мм/ч, но может быть и значительно меньше, так что оплодотворение и у покрытосеменных может произойти спустя недели или месяцы после опыления (например, у орхидных). Однако задержка может быть связана также с запаздыванием развития самого семязачатка.
У хвойных и Wehvitschia (Gnetopsida) в оплодотворении участвует только один из двух спермиев. Двойное оплодотворение происходит у Ephedra и Gnetum, а также у покрытосеменных. У Ephedra благодаря слиянию одного спермия с яйцеклеткой и второго с брюшной канальцевой клеткой, а у Gnetum — двух спермиев с двумя клетками женского гаметофита образуются две зиготы, из которых далее развивается чаще всего только одна. Двойное оплодотворение покрытосеменных протекает иначе (см. рис. 11.188). Когда пыльцевая трубка добирается до яйцевого аппарата женского гаметофита, она изливает свое содержимое в одну из синергид, которая при этом разру
шается. Отмирает и ядро пыльцевой трубки, а оба спермия или их ядра перемещаются далее, вероятно, амебоидно1. В результате слияния одного спермия с яйцеклеткой возникает зигота. Второй спермий сливается с центральной клеткой зародышевого мешка, где его ядро объединяется с вторичным ядром зародышевого мешка или с обоими полярными ядрами, образуя триплоидное ядро эндосперма, из которого развивается типичная питательная ткань семени покрытосеменных. Эндосперм отсутствует, к примеру, у орхидей, а у Опа-graceae эндосперм диплоидный из-за наличия только одного полярного ядра.
СЕМЯ
После оплодотворения из семязачатка развивается семя, состоящее обычно из семенной кожуры, питательной ткани и зародыша. Семенная кожура (теста) образуется из одного или двух интегументов и может быть устроена весьма по-разному1 2. Обычно из наружной эпидермы наружного или единственного интегумента возникает экзотеста, из внутренней эпидермы — эвдотеста, из имеющихся в разном числе (но они могут и отсутствовать) промежуточных слоев — мезотеста. На зрелых семенах снаружи, как правило, можно заметить место обрыва семяножки — семенной рубчик, а в семенах, образующихся из ана-и кампилотропных семязачатков, — семенной шов, содержащий проводящий пучок (рис. 11.193).
У саговников и Ginkgo наружная зона единственного интегумента преврашается в мясистую и чаще всего ярко окрашенную саркотесту, а внутренняя часть— в
1 Пыльцевая трубка прорастает через разрушающуюся синергиду в щель между яйцеклеткой и центральной клеткой зародышевого мешка, где лопается на конце, изливая спермин (не ядра!) непосредственно на яйцеклетку и центральную клетку зародышевого мешка. Ни спермин, ни ядра не перемещаются. — Примеч. ред.
2 Теста формируется из единственного — наружного — интегумента. Из внутреннего интегумента развивается тегмен. — Примеч. ред.
388 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Рис. 11.193. Семя и его прорастание (Ricinus communis) (по W. Troll):
А— вид с брюшной стороны; В — медианный, С — трансверсальный продольные срезы семени (А—С 2х); D — проросток (натур, величина); с — карункула (элайосома); е — эндосперм; h — рубчик; hk— гипокотиль; к— семядоли; г — корешок; га — семенной шов; t — семенная кожура
твердую склеротесту'. И нтегумент хвойных дает жесткую семенную кожуру1 2. У Gnetopsida семенная кожура очень тонкая и в зрелом семени так сжата, что ее клеточная структура едва различима. У покрытосеменных с двумя интегументами в образовании семенной кожуры могут принимать участие оба или только один интегумент. В обоих случаях возможна дифференциация семенной кожуры на мясистую саркотесту и
1 В склеротесту дифференцируется средняя часть интегумента. Внутренняя часть становится эндотестой или паринхотестой бумажистой консистенции. — Примеч. ред.
2 В семенной кожуре хвойных также можно различить слои. — Примеч. ред.
жесткую склеротесту (например, Magnolia-ceae, Paeoniaceae, Punica granatum). При участии двух интегументов граница между сар-ко- и склеротестой отнюдь не должна совпадать с границей между интегументами. У многих покрытосеменных с семязачатками, имеющими два интегумента, семенная кожура развивается только из одного, а другой обычно внутренний интегумент редуцируется. При наличии только одного интегумента семенная кожура развивается из него.
Для семенной кожуры покрытосеменных важно еще одно различие. Если она возникает из наружного интегумента, ее обозначают как теста, а если из внутреннего — то как тегмен.
Кроме дифференциации семенной кожуры на сарко- и склеротесту можно обнаружить и другие вариации, особенно у покрытосеменных. Так, например, мясистыми могут быть только
Рис. 11.194. Семя и его развитие (А—С— по К.V.Goebel; D — по F.Firbas; Е—Н — по P.E.Du-chartre):
А, В — семязачатки разного возраста (70х); С — зрелое семя (9х) Epilobium angustifolium. Семена: D — Zanonia javanica (Cucurbitaceae. семя крылатое, 1/2х), Е— Papaver rhoeas, F — Pseu-dofumaria alba, G — Chelidonium majus с элайо-сомами (e), а также H — Nymphaea alba с мешковидным ариллусом (увеличено); f — семяножка; h — рубчик; m — микропиле; s — семязачаток
11.2. Бактерии, грибы, растения | 3g
некоторые участки в целом жесткой семенной кожуры. Если это происходит в области перехода семяножки в интегумент, то возникает ариллус, который может почти полностью окружать семя (например, Euonymus), быть лопастным (например. Myristiccr, см. рис. 11.218) или дифференцироваться в заполненный воздухом поплавок (напрймер, Nymphaea, рис. 11.194). Если семенная кожура мясистая в области микропиле, то говорят о карункуле (см. рис. 11.193), а если она мясиста в области семенного шва, — о строфи-оли. Такие дополнительные образования — богатые жиром, белком или сахаром элайосомы (рис. 11.194) способствуют распространению семян муравьями (например. Corydalis, Chelidonium). Часто семенная кожура ослизняется (например,
у разных Brassicaceae, Linum, томата, Plantago). образуя миксотесту. На сухой тесте могут вырастать волоски (например, ЕрНоЫит. хлопчатник — Gossipiutn) или крыловидные придатки (например, Zanonia) (рис. 11.194). В нераскрываюшихся плодах, где защитную функцию часто берет на себя стенка плода (например, Apiaceac, Asteraceae, Роасеае), семенная кожура обычно очень тонкая.
Семена сильно различаются размером. Так, семена сейшел>>ской пальмы (Lodoicea: Агеса-сеае) весят несколько килограммов, в то время как семена Orchidaceae — всего несколько тысячных долей миллиграмма. Объяснить особенности размера, формы семян и характер их поверхности можно только в связи с их приспо-
Рис. 11.195. Развитие зародыша и вторичного эндосперма у покрытосеменных (А—Н — по I. Hanstein и R.Soueges; J— по I.L.L.Guignard; К— по R.M.T.Dahlgren; L— no I.Н.Billings):
А — зигота; В —F — развитие подвеска (суспензора, s) и молодого зародыша с гипофизой (1э), а также зачатков гипокотиля (hk) и семядолей (k); G —Н — зародыши с гипофизой (h) зачатками семядолей (к) и апикальной меристемой побега (т) (А—G — Rosopsida: Capsella bursa-pastoris, Н —Liliopsida: Alisma plantago-aquatica) (ок. 200x); J —К — молодой зародыш (e) с суспензором в нуклеарном и целлюлярном эндосперме (es) (Lepidium sp. и Ageratum mexicanum) (увеличено); L — из эндосперма через микропиле выросла трубковидная ветвящаяся гаустория, которая местами прилегает к стенке завязи (w), местами — к семяножке (f); в зародышевом мешке виден также зародыш с суспензором (продольный срез молодого семени Globularia cordifolia, увеличено)
390 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Рис. 11.196. Развитие зародыша у Cycadopsida (Zamia floridana) (по I. М. Coulter и Ch. Chamberlain):
A— свободное деление ядер (12х); В — образование клеточных стенок и ткани в основании зародыша (18х); С — начинающаяся дифференцировка предзародыша в суспензор (с продольно вытянутыми клетками) и базальный зародыш (22х)
соблениями к распространению, прорастанию и устройством проростков.
Питательная ткань семени представлена чаще всего эндоспермом. У голосеменных гаплоидная ткань массивного женского гаметофита дает первичный эндосперм. Ткань нуцеллуса различима в зрелых семенах только в виде сильно спрессованной пленочки. У покрытосеменных возникает вторичный эндосперм, чаще всего трип-лоидный благодаря оплодотворению вто
ричного ядра зародышевого мешка одним из двух спермиев. Нуцеллус либо вообще неразличим, либо едва заметен, но в некоторых группах растений может служить и питательной тканью.
Вторичный эндосперм покрытосеменных обычно образуется по нуклеарному типу. В результате свободных делений ядер может образоваться более 2000 ядер, расположенных сначала в постенном слое цитоплазмы увеличивающегося зародышевого мешка (рис. 11.195). Лишь позднее начинается образование клеточных стенок (см. рис. 2.33). Вцеллюлярном типе образования эндосперма клеточные стенки образуются уже при первых делениях ядер, в гелобиальном типе развитие эндосперма в верхней части зародышевого мешка идет по нуклеарному, а в нижней части — по целлюлярному типу (см. рис. 11.223).
Образующийся эндосперм (и зародыш) поглощает питательные вещества. Прежде всего растущий зародышевый мешок вытесняет нуцеллус, большая часть которого или весь он расходуется на питание зародышевого мешка. Иногда из эндосперма или из суспензора зародыша в окружающие ткани внедряются трубковидные гаустории.
В некоторых семенах, например у мускатного ореха и пальмы Агеса, от нуцеллуса, а у некоторых Аппопасеае от интегумента, в эндосперм врастают складки ткани, разделяя его бороздами. Таким образом возникает руминированный эндосперм (см. рис. 11.218).
Кроме вторичного эндосперма у покрытосеменных питательной тканью может также служить нуцеллус. Подобного рода перисперм имеется в дополнение к эндосперму, к примеру у Nymphaeaceae, Pipera-сеае (см. рис. 11.217) и Zingiberales, и как единственная питательная ткань — у Са-ryophyllales. Наконец, возможно накопление питательных веществ в самом зародыше, в частности в мясистых семядолях (например, Fabaceae, Quercus, Juglans, Aesculus), бывают семена вообще без эндосперма (например, Orchidaceae)1.
Запасные вешества семени — это крахмал, белки или масла, откладываемые внутри клеток, либо резервная целлюлоза в клеточных стенках1 2. Соответственно этому эндосперм или другие запасающие ткани мучнистые, как у злаков, мас-
1 Семена орхидных вообще не содержат питательной ткани. — Примеч. ред.
2 На самом деле — гемицеллюлозы. — Примеч. ред.
11.2. Бактерии, грибы, растения
391
лянисгые, как у Cocos, или твердые, рогоподобные или каменистые, как у многих Liliales и некоторых пальм (например, Phytelephas-. «растительная слоновая кость»).
Развитие зародыша из зиготы может происходить по-разному (см. рис. 3.1; 11.195). Сформированный зародыш чаще всего состоит из прилегающего к микропиле подвеска (суспензора), благодаря удлинению которого остальная часть развивающегося зародыша (собственно зародыш) погружается в питательную ткань. Суспензор и собственно зародыш связаны между собой несколькими клетками, так называемой гипофизой. В зрелом семени суспензор и гипофиза чаще всего уже не различимы. Собственно зародыш состоит из зародышевого корешка, расположенного под семядольным узлом участка оси — гипокот иля, разного числа семядолей и меристемы побега с самыми молодыми зачатками листьев (почечки).
Во всех группах семенных растений с несколькими архегониями или яйцеклетками в одном семязачатке могут образоваться несколько и зигот, и зародышей (полизиготическая полиэмбриония). Правда, чаще всего в таких случаях полностью развивается только один из них, поэтому
зрелое семя содержит только один зародыш.
У Cycadopsida (рис. 11.196) и Ginkgo вначале происходят свободные деления ядер, так что может получиться более 1 000 ядер. Лишь затем начинается образование клеточных стенок. Зародыши в этих двух группах имеют по две семядоли. У Coniferopsida развитие зародыша может начинаться со свободных ядерных делений, а продолжаться путем нормальных делений клеток, но развитие его может и сразу идти по цел-люлярному типу. Зародыш чаще всего имеет несколько семядолей (рис. 11.197). Развитие зародыша у Gnetopsida целлюлярное, зародыши имеют по две семядоли. У Gne-tum и Welwitschia зародыш имеет боковое выпячивание, внедряющееся пальцевидно в питательную ткань и служащее, верояп ю, для всасывания питательных веществ.
Особенность многих хвойных состоит в том, что из одной зиготы может возникать несколько зародышей (монозиготическая полиэмбриония). У Pinus (рис. 11.198) в зиготе путем двух свободных ядерных делений получается четыре ядра, которые расположены в одной плоскости на обращенном к микропиле конце. В результате дальнейших делений ядра располагаются в два яру-
Рис. 11.197. Семя и его прорастание у Coniferopsida (Pinus pinea) (по I. Sachs):
A— семя (продольный разрез); В — I — прорастание семени; зародыш и проросток с семядолями (к), гипокотилем, главным и придаточными корнями (w); е — первичный эндосперм; s — семенная кожура; х — выпяченный и разорванный зародышевый мешок
392 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Рис. 11.198. Монозиготическая полиэмбриония у сосны (Pinus) (А—D— по I.M. Coulter, Ch. Chamberlain; Е— по J.Th. Buchholz):
A—D— Pinus nigra (100x); E — P. banksiana (80x); e — зародыш; p — базальная пластинка; г — розетка; s — суспензор с s' вторичными клетками подвеска
са по четыре ядра в каждом. В результате образования клеточных стенок и дальнейшего клеточного деления получается четыре яруса по четыре клетки в каждом. Из самого нижнего яруса путем отхождения его четырех клеток друг от друга могут образоваться четыре зародыша, а вышерасположенный ярус благодаря удлинению и делению своих клеток становится суспензором. В других семействах и родах хвойных монозиготическая полиэмбриония либо не встречается, либо значительно большее число зародышей может возникать несколько иным путем.
Монозиготическая полиэмбриония обнаружена также среди Gnetopsida — у Ephedra и Gnetum.
У Ephedra из зиготы в результате трех клеточных делений образуется восемь клеток, и все могут развиться в зародыши. У Gnetum из зиготы возникают два первичных суспензора. Путем удлинения и ветвления, но без дальнейшего деления клеток получается обычно разветвленная трубчатая система, на многочисленных концах которой образуются многоклеточные вторичные суспензоры. Их концы развиваются в зародыши, вследствие чего из одной зиготы может получиться очень много зародышей.
При монозиготической полиэмбрионии, как и при полизиготической, завершает развитие чаще всего только один зародыш. Таким образом, зрелое семя содержит только один зародыш.
У покрытосеменных зародыш развивается по целлюлярному типу (см. рис. 11.195). После одного или немногих клеточных делений возникает так называемый предза-родыш из одного ряда (но часто и Т-образно расположенных) клеток. Из него, с одной стороны, образуется суспензор, состоящий из клеток, расположенных всегда в один ряд, а с другой стороны — сначала шаровидный, состоящий из четырех или восьми клеток собственно зародыш, который затем развивается далее. Зрелый зародыш покрытосеменных имеет одну или две семядоли и может (например, у Роасеае; см. рис. 11.233) морфологически дифференцироваться далее на колеоптиль, колеоризу, щиток и т.п.
В зависимости от деталей хода клеточных делений и участия в развитии собственно зародыша и суспензора нижних и верхних клеток предзародыша различают разные типы эмбрионального развития. Наиболее часты Asteraceae-и Onagraceae-типы.
Сформировавшиеся зародыши покрытосеменных различаются по величине и степени дифференциации. Так, у Orchidaceae зародыши состоят из немногих клеток и совершенно недифференцированные. Наконец, у покрытосеменных возможно развитие зародыша без оплодотворения (агамоспермия; см. 10.1.3.3).
У Coniferopsida и Gnetopsida в распространении семян ветром и животными могут принимать участие структуры, не являющиеся частью семени. Например, имеющееся у семени Pinus крылышко — производное не семенной кожуры, а семенной чешуи (см. рис. 11.203). Сочные структуры у хвойных развиваются либо из почти всей
11.2. Бактерии, грибы, растения | 393
женской шишки (например, «шишкоягоды» Juniperus', см. рис. 11.209), либо мясистая семенная чешуя, так называемый эпима-тий, окружает семя, либо мясистой становится часть оси шишки (Podocarpaceae), либо семя окружает разрастание оси ниже его (например, Taxus; см. рис. 11.210). У Gnetopsida функцию семенной кожуры берет на себя пара чешуевидных листьев, окружающих семязачатки, которые становятся жесткими. Наружная пара чешуевидных листьев может становиться мясистой (Gnetum) или разрастаться в крылья (Welwitschia). У Ephedra только с одной парой брактей вокруг семязачатков другие брак-теи стробила могут стать мясистыми и участвовать в распространении семени.
ПЛОДЫ
У покрытосеменных семязачатки заключены в плодолистики. Поэтому по мере того как семязачатки после оплодотворения развиваются в семена, из плодолистиков, а иногда и из других частей цветка и цветоложа развивается плод, который, с одной стороны, заключает семена внутри себя и защищает их до созревания, а с другой стороны, участвует в распространении семян, высеивая их, либо вместе с ними отделяясь от растения. При развитии плодолистиков в плод они чаще всего изменяют форму, размер и анатомическое строение. Строение зрелого плода зависит от структуры гинецея, анатомии стенки плода, способа вскрывания плода при его созревании. При развитии плода из единственного плодолистика получается мономерный плод1. Апокарпные плоды развиваются из нескольких плодолистиков апокарпного гинецея. Из ценокарпных гинецеев развиваются разные ценокарпные плоды. Стенка зрелого плода (околоплодник) дифференцирована на наружную эпидерму (экзокарпий), внутреннюю эпидерму (эндокарпий) и разное число промежуточных слоев клеток (мезокарпий). Впрочем, разные слои стенки плода часто называют иначе. Если,
как, например, у костянок, имеется многослойная каменистая внутренняя часть околоплодника, то всю ее обычно называют эндокарпием. В зависимости от консистенции околоплодника различают сухие и сочные плоды. Среди сочных плодов есть ягоды с полностью сочным околоплодником и костянки с мясистой наружной частью и деревянистой внутренней частью стенки плода1. Эндокарпий может быть представлен мясистыми волосками, врастающими в виде пульпы в полость завязи (например, у Citrus). О вскрывающихся плодах говорят, если после созревания плод раскрывается и освобождается от семян. По особенностям вскрывания различают несколько типов плодов.
Если свободные плодолистики вскрываются по брюшному шву, они вентрицидные. Если ценокарпные плоды вскрываются по границам соседних плодолистиков, то говорят о септиццд-ных (вскрывающихся по перегородкам) плодах. Дорсицидные (локулицидные; вскрывающиеся по створкам) плоды вскрываются вдоль на спинной стороне плодолистика. У порицидных плодов образуются поры — сравнительно небольшие отверстия; при вскрывании поперечной кольцевой щелью на верхушке ценокарпного плода получается коробочка с крышечкой.
Невскрывающиеся плоды с однородной сухой стенкой — орехи. Но невскрывающиеся плоды при созревании могут и распадаться на остающиеся замкнутыми части (распадающиеся плоды). Если ценокарпные плоды распадаются на отдельные плодолистики, то говорят о дробных плодах, а части плодов называют мерикарпиями. Если же разломы возникают в стенках самих плодолистиков, то говорят о членистых плодах.
Многообразие плодов как средства распространения семян повышается благодаря участию других органов или их частей, а также путем образования соплодий. У Nuphar цветоложе обрастает плод, развивающийся из верхней завязи, а у многих Rosaceae гипантий (Rosa) или цветоложе (Fragaria) становятся мясистыми. В образовании плода могут участвовать простой околоцветник (к примеру, мясистый в сопло-
1 Мономерные плоды чаще всего рассматривают как один из вариантов апокарпных плодов. — Примеч. ред.
1 Типичные костянки — апокарпные плоды. Ценокарпные плоды с похожим околоплодником называют пиренариями. — Примеч. ред.
394 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Рис. 11.199. Сухие вскрывающиеся (А—G) и невскрывающиеся (Н — М) плоды (А— по G.Beck-Mannageta; В, D, Е, Н — М — по F. Firbas; С — no R. v.Wettstein; F, G — по A.F.W. Schimper).
Мономерные плоды: A— листовка (Consolida regalis): В — боб (Laburnum anagyroides). Ценокарпные плоды: С — стручок* (Chelidonium majus); D — септицидная коробочка (Hypericum perforatum); E — дорзицидная** коробочка (Iris sibirica); F — коробочка с крышечкой (Anagallis arvensis); G — коробочка с порами (Antirrhinum majus). Орех лещины (Corylus avellana): H — общий вид; J — продольный разрез. Дробный плод: К — двукрылатка (Acer pseudoplatanus; с двумя односеменными мерикарпиями). Членистые плоды: L — членистый боб (Ornithopus sativus; мономерный плод с односеменными члениками); М— членистый стручок (Raphanus raphanistrum; ценокарпный ломкий плод) (А— ок. 4х; В, С, Н —К — в натур, вел.; D, Е — Зх; F — 2х; G — 3/4х; L, М — 2/Зх); а — рубец плодоножки; с — семядоля; I — проводящий пучок к семязачаткам; m — дорсальная средняя линия плодолистика; г— корешок; s — изогнутый семязачаток; t — семенная кожура; w — стенка плода
* Типичный стручок имеет ложную перегородку и встречается у крестоцветных (Brassicaceae). Плод чистотела следует считать стручковидной коробочкой. — Примеч. ред.
** Локулинидная. — Примеч. ред.
дии Morus; волосистый у Eriophoruni), чашечка (например, сильно увеличенная и ярко окрашенная у Physalis alkekengi; представленная хохолком у Valerianaceae и Asteraceae), предлис-тья и кроющие листья (брактеи) (например, крыловидные у Carpinus или Humulus, трубковидные у Carex), плодоножки (например, мясистая у Anacardium occidentale), а также стеблевые и листовые органы соцветий (например, ку-пулы Fagaceae, мясистые ось соплодия и брактеи у Ananas).
Ниже будет более подробно рассмотрена классификация плодов по их функционированию (по способам вскрывания) и анатомии (структуре стенки плода) (рис. 11.199). Плод определяю! как цветок ко времени созревания семян, поэтому можно классифицировать плоды и на основе морфологии цветка и принимать, к примеру, первичное подразделение плодов на монокарпические (из одного плодолистика), апокарпные и ценокарпные.
Вскрывающиеся плоды
У листовок свободные плодолистики апокарпного гинецея при созревании чаще всего сухие и вскрываются по брюшному шву. У Consolida, например, в цветке только один плодолистик и соответственно плод — однолистовка. У многих Magnoliidae, Paeonia, Delphinium, Trollius или Spiraea (см. рис. I (.255) в цветке образуются многочисленные листовки, так что в целом плод можно назвать сборной листовкой1.
Бобы — это также чаще всего сухие плоды, которые развиваются из одного плодолистика, но вскрываются с брюшной и спинной сторон. Один боб в цветке характерен, например, для I eguminosae (см. рис. 11.249), несколько — Magnolia (см. рис. 11.218).
Коробочки чаще всего сухие, они развиваются из ценокарпных гинецеев. По числу полостей
1 Многолистовкой. — Примеч. ред.
11.2. Бактерии, грибы, растения | 395
(гнезд) (развиваются из синкарпных или соответственно паракарпных гинецеев) и способу вскрывания (к примеру, по всей длине, только апикально или базально, по перегородкам или по створкам, порами или крышечкой) можно выделить разные типы коробочек (см. рис. 11.199). К коробочкам, строго говоря, относится и стручок Brassicaceae (см. рис. 11.265). Он состоит из двух паракарпно сросшихся между собой плодолистиков и вскрывается створками, отделяющимися от несущей плаценты рамки, на которые «натянута» ложная перегородка. Коробочкой является и активно разбрасывающий семена плод Geranium' (см. рис. 11.243). Мясистые коробочки часто встречаются у тропических растений, а из центральноевропейских — у Еиопу-mus\ коробочка, разбрасывающая семена при прикосновении, имеется у Impatiens.
Невскрывающиеся плоды
Орехи с полностью одревесневшим невскры-вающимся околоплодником могут образоваться как из отдельных плодолистиков апокарпного гинецея (например, Anemone, Ranunculus), так и из ценокарпного гинецея (например, Betula, Ulmus, Fraxinus)1 2. У Clematis и Pulsatilla удлиняющийся перисто-волосистый стилодий, а у Geum завернутый крючком стилодий принимают участие в распространении орешков. Функцию распространения орехов у многих Dipsacaceae берет на себя наружная чашечка3 (см. рис. 11.285). Тесно прижаты друг к другу или срастаются семенная кожура и стенка плода в зерновках Роасеае (см. рис. 11.233) и в семянках Asteraceae (см. рис. 11.286), которые также являются орехами. В сборных орехах4 1 2 несколько орехов апокарпного гинецея могут быть объединены благодаря мясистому цветоложу (Fragaria) или мясистому гипантию (Rosa) (см. рис. 11.255).
К сухим невскрывающимся относят также и распадающиеся плоды. Образующиеся из ценокарпного гинецея дробные плоды разделяют на много (например, Malva) или только два (например, Acer. см. рис. 11.199) мерикарпия септи-цидно, т.е. по границам плодолистиков. У большинства Apiaceae (см. рис. 11.284) два мерикарпия остаются прикрепленными к центральной колонке (карпофору). Плоды с поперечно или вдоль разламывающимися плодолистиками на
1 Плод гераней представляет собой стеригму. — Примеч. ред.
2 Орехами следует называть ценокарпные плоды. Похожие на них апокарпные плоды называют орешками. — Примеч. ред.
3 Обвертка из прицветничков. — Примеч. ред.
4 Многоорешках. — Примеч. ред.
зывают ломкими плодами. Они могут развиваться из ценокарпных гинецеев (например, поперечно разламывающиеся членистые стручки Brassicaceae, см. рис. 11.199, продольно разламывающиеся и к тому же перпендикулярно к разлому расщепляющиеся ценобии многих Lamia-сеае и Boraginaceae; см. рис. 11.277, 11.282) либо из единственного или из отдельных плодолистиков апокарпного гинецея (например, поперечно разламывающийся членистый боб некоторых Leguminosae; см. рис. 11.199).
Костянки характеризуются тем, что наружная часть околоплодника мясистая, а внутренняя деревянистая. Только из одного плодолистика образуется, в частности, костянка Prunus (слива, вишня и т. п.; см. рис. 11.255). Ценокарпные костянки1 образуются, например, у Juglans, Olea или Sambucus. У Cocos мезокарпий волокнистый, содержащий воздух (см. рис. 11.230) и позволяющий плодам эффективно распространяться водой. Малина и ежевика имеют сборные костянки2.
Яблоко (например, Malus', см. рис. 11.255) развивается из нижней завязи ценокарпного гинецея. Наружная часть его околоплодника мясистая, внутренняя кожистая или бумажис-тая.
Целиком мясистый околоплодник характерен для ягод. Многие Annonaceae, Actaea или финиковая пальма имеют ягоды из одного плодолистика. Ценокарпные ягоды характерны для Ribes, Vitis, Vaccinium, Atropa или Convallaria. Плоды цитрусовых имеют мясистую пульпу. Если наружная стенка ягоды, как, например, у огурца и тыквы (Cucurbitaceae; см. рис. 11.258), довольно жесткая, то плод называют тыквиной.
Соплодия
Единицей распространения может быть целое соплодие. Примеры — шелковица и инжир (кувшинчатую форму у последнего имеют и соцветие, и соплодие), а также другие роды Могасеае (см. рис. 11.257), или ананас с мясистым околоцветником, осью соцветия и плодами. У Tilia несколько орехов соединены с служащим крылом предлистом (см. рис. 11.266), у Arctium единицей распространения служит корзинка с крючковидными листьями обвертки. Наконец, единицей распространения может стать и вся надземная система побегов растения (так называемое «перекати-поле»), которую веер обламывает у основания и гонит по степи, а в это время семена высыпаются. Движению способствует шаровидная форма растения.
1 Пиренарии. — Примеч. ред.
2 Многокостянки. — Примеч. ред.
396 | ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
РАСПРОСТРАНЕНИЕ ПЛОДОВ И СЕМЯН
Распространение расселительных единиц (диаспор) в пространстве выполняет разные функции. При колонизации новых, т.е. не заселенных материнскими индивидами местообитаний, распространение на небольшие расстояния важно для сохранения популяции, а на большие расстояния — для основания новой популяции. Благодаря удалению от материнского индивида уменьшается внутривидовая конкуренция между потомками и родителями, а также между сестринскими индивидами. Правда, множество диаспор почти всегда оказываются в непосредственной близости от материнского индивида (см. 10.1.3.4). Распространение диаспор можно рассматривать и как стратегию избегания травоядных животных и патогенов, «поведение» которых зависит от плотности популяции растений, и, наконец, распространение диаспор является компонентом потока генов внутри и между популяциями.
Следует учитывать, что специализация механизмов распространения диаспор существенно меньше, чем при опылении. Так, многие диаспоры распространяются несколькими способами, т.е. полихорны. Разные способы распространения могут быть и в случае различия диаспор одного индивида (гетероспермия, гетерокарпия). Например, в корзинках некоторых видов Leontodon имеются семянки как с паппусом, так и без пап-пуса, а у различных Asteraceae самые наружные семянки не имеют паппуса и удерживаются в созревшей корзинке окружающими листочками обвертки. Некоторые растения рассеивают многие или все свои плоды по соседству благодаря активному росту плодоножек, помещая их, например, в трещины скал (Cymbalaria muralis) или в почву (например, земляной орех, Arachis hypo-gaea\ Trifolium subterraneum). Плоды и соответственно их части у Stipa и Erodium (см.рис.8.36) сами вбурав.'иваются в почву. Приспособления к распространению на большие расстояния обычно свойственны растениям аридных областей.
Наблюдения за распространением плодов и семян чаще всего показывают, что расстояния, на которые разносится подавляющее большинство диаспор одного индивида, довольно малы и часто лежат в пределах нескольких метров. Большое эволюционное значение имеет встречающийся в исключительных случаях занос диас
пор на большие расстояния, что доказывает пример изолированных океанических островов, сравнительно быстро заселяемых многими видами. Молекулярные методы анализа родства показывают, что географически удаленные таксоны растений могут быть близкородственными, а их ареалы возникли в результате дальнего заноса (англ.: long distance dispersial).
Важнейшие способы распространения — с помощью животных, ветра и активное разбрасывание диаспор. Распространение животными (зоохория) может принимать разные формы. При эцдозоохо-рии животные съедают диаспоры и затем выделяют их с испражнениями.
Признаками эндозоохории являются наличие у диаспор первичных аттрактантов (питательных веществ, таких, как углеводы, белки, жиры и масла; витамины, органические кислоты, минеральные вещества), вторичных аттрактантов (например, окраска и запах) и образований (склеротеста, жесткие части околоплодника), защищающих семена от разрушения ротовым аппаратом или в кишечнике животных-распространителей. Эти признаки могут быть как у семян, так и у плодов. В то время как мясистые семена и плоды животные чаше всего быстро съедают, из сухих диаспор они создают запасы. Первоначально, вероятно, важными распространителями семян и плодов были рыбы и рептилии (ихтио- и заврохория), позднее к ним присоединились птицы (орнитохория) и млекопитающие. Как и при опылении, при эндозоохории часто возникают тесные связи между растением и животным. Соответственно у мясистых диаспор можно обнаружить характерный синдром признаков приспособления к главному распространителю. При распространении птицами диаспоры чаще всего ярко или контрастно окрашены (красные, желтые, блестящие черные), не имеют запаха, среднего размера или мелкие, с мягкой кожурой и не опадающие. Примеры — мясистые семена (Magnolia, Paeonia), костянки (Prunus avium, Ligustrum, Olea, Sambucus'), ягоды (Ribes, Wils, Vaccinium), сборные плоды (Rosa, Ru-bus) и соплодия (Moms). Млекопитающие как эндозоохорные распространители диаспор играют заметную роль в тропиках. Из-за иных, чем у птиц, органов чувств и ротового аппарата распространяемые ими диаспоры чаще всего окрашены менее ярко, но зато имеют сильный запах; часто они крупнее, с более жесткой кожурой и опадают с растения. Примеры — костянки Prunus
1 У последних трех родов — пиренарии. — Примеч. ред.
11.2. Бактерии, грибы, растения | 397
persica, многие плоды типа «яблоко», часто имеющие сравнительно жесткую кожуру ягоды (какао, Citrus, хурма, Cucurbitaceae, бананы1 1 2) и соплодия (Ficus, Artocarpus). Похожи плоды, распространяемые рукокрылыми, но они остаются висеть на деревьях.
Среди сухих диаспор есть более мелкие, распространяемые преимущественно зерноядными птицами, и более крупные (например, Quercus, Fagus, Corylus, Juglans), которые собирают в основном грызуны для запасов, причем находят не все, что прячут, так что часть семян всегда благополучно прорастает.
Распространение муравьями (мирмеко-хория) основано на том, что муравьи собирают и растаскивают семена и плоды, имеющие характерные придатки (элайосомы) с привлекающими и питательными веществами.
Элайосомы образуются из разных частей семени (например, Asarum, Chelidonium, Corydalis, Viola, Cyclamen purpurascens, Melampyrum, Allium ursinum, Galanthus nivalis) или плода (например, Anemone nemoralis, Hepatica, Lamium, Knautia). Мир-мекохория встречается в лесах умеренного и тропического пояса
Семена некоторых тропических Leguminosae с красно-черной контрастной окраской, по-видимому, имитируют элайосомы.
Сравнение близкородственных мирмекохор-ных и немирмекохорных видов показывает, что с этим способом распространения связаны и другие признаки. Так, у не имеющей элайосом Primula elatior семена созревают медленно в прямостоячих коробочках в высыхающих чашечках на длинном цветоносе, а у Р. vulgaris семена с элайосомами созревают быстро в коробочках в зеленых, ассимилирующих чашечках на коротких, полегающих на почву цветоносах.
Эпизоохория осуществляется благодаря тому, что диаспоры прикрепляются самым разным образом к поверхности животных.
Семена многих болотных и водных растений благодаря своим малым размерам могут прилипать вместе с илом к телу водных птиц и далеко разноситься ими. Возможность закрепления на теле животного повышается у семян или плодов, которые во влажном состоянии ослизняются и становятся липкими (например, Plantago, Juncus). Часто цепляются за животных диаспоры с железистыми волосками (например.
Salvia glutinosa) и в особенности — с прицепками. Прицепки бывают волосками или эмерген-цами плодолистиков (например. Medicago. Circaea. Galium aparine) или развиваются из столбиков (например. Geum urbanum1). чашелистиков (в том числе и из «наружной чашечки») или из листочков обвертки (например. Arctium, Xanthium). В то время как более мелкие цепляющиеся плоды распространяются преимущественно в шерсти мелких животных, крупные плоды с мощными прицепками (например, Tribulus. многие Pedaliaceae) приспособлены для закрепления и переноса на ногах крупных копытных.
Особая форма распространения с помощью животных обнаружена у растений-баллистохо-ров. Пробегающие животные цепляют их эластичные стебли, которые, распрямляясь, катапультируют семена или плоды (например, разные виды с плодами-коробочками. Lamiaceae, Dipsa-cus).
Уже в раннее историческое время человек стал важным фактором в распространении диаспор (антропохория). Многие сорняки были занесены непреднамеренно с посевным материалом, шерстью и кормом для скота, а культурные растения были сознательно расселены по всему миру. В результате в некоторых регионах (например, в некоторых районах Новой Зеландии или Калифорнии) антропохоры доминируют над местной флорой. Замечательно, что диаспоры некоторых полевых сорняков в результате отбора так сильно уподобились по величине и другим особенностям диаспорам соответствующих культурных растений, что их пракгически невозможно отделить механическими методами от семенного материала (например, СатеИпа, засоряющая лен).
Распространение с помощью ветра (анемохория) может быть косвенным, когда диаспоры при раскачивании стеблей вытряхиваются из соответствующих вместилищ (например, семена из коробочек — Papaver, семянки из корзинок — Bellis), или непосредственным, когда ветер сдувает и разносит диаспоры.
Во второй группе встречаются крошечные и легкие пылевидные семена (например, Orobanche, орхидеи), диаспоры с пузыревидными летучками (например, вздутые чашечки у Trifolium fragi-ferum), летучками из волосков (например, волоски на семенах, на столбике — Clematis1', ости — Stipa, паппус — Asteraceae), крылатые диаспоры (семена, крылатые орехи, дробные плоды-кры-
1 Плоды бананов распространяют рукокрылые. — Примеч ред.
1 У этого вида — из стилодиев. — Примеч. ред.
2 Стилодии. — Примеч. ред.
398 I ГЛАВА 11 СИСТЕМАТИКА И ФИЛОГЕНИЯ
латки — Acer; соплодия — Tilia; зонтиковидная обвертка «наружная чашечка» — Scabiosa) и «перекати-поле».
Гидрохория состоит чаще всего в простом переносе диаспор водой. У дождевых баллистохоров энергия падающих дождевых капель благодаря лопатообразной форме сидящих на пружинящих плодоножках плодов преобразуется в метательные движения, когда выбрасываются семена из стручочков (например, Iberis, Thlaspi) или мерикарпии из чашечек (например, Prunella, Scutellaria). Впрочем, дождевые капли могут выбрасывать семена и непосредственно из соответствующим образом устроенных плодов.
Влажность может приводить, к примеру, у Se-dum acre и многих Aizoaceae к гигрохастическо-му вскрыванию коробочек. При распространении потоками воды плавучесть диаспор достигается тем, что они не смачиваются или снабжены воздухоносным мешком (например, семена Nymphaea, мешочки разных видов осок, Carex) или особой воздухоносной губчатой тканью (например, Cocos, Iris preudacorus, Potamogeton, Cakile maritime).
При автохории внешние агенты не используются.
В то время как многие диаспоры просто падают на землю (барохория), некоторые активно разбрасываются. Этот механизм основан на тургоре (например, у «взрывающихся» коробочек Impatiens, разбрасывании семян у Oxalis и у выстреливающего семенами более чем на 12 м бешеного огурца, Ecballium; см. рис. 8.35) или на гигроскопических движениях (например, скручивание створок бобов у Dictamnus1; коробочки-катапульты у Geranium; разбрасывание семян в результате сжатия коробочек у разных видов Viola). Автохорны и соответственно ахорны также те виды, которые активно зарывают в почву диаспоры возле материнского растения (Cymbalaria muralis, Arachis hypogaea).
Все названные биологические особенности семян и плодов возможно лучше всего понять, учитывая биотоп видов. Это, например, отчетливо проявляется в том, что в нижнем травяном ярусе центральноевропейских лиственных лесов преобладает мирмекохория, у более высоких травянистых многолетников — эпизоохория, в кустарниковом ярусе — эндозоохория, в древесном
1 Плод Dictamnus синкарпный. — Примеч. ред.
ярусе — анемохория. Это соответствует вертикальному распределению агентов распространения (муравьев, млекопитающих, птиц, ветра).
ПРОРАСТАНИЕ СЕМЯН
Разнесенные тем или иным способом семена (голые или окруженные стенкой плода) попадают на поверхность или в самый поверхностный слой почвы. Здесь при подходящих условиях (см. гл. 7) и после иногда необходимого периода покоя по крайней мере некоторые семена прорастают. Однако часть семян, в особенности у видов ранних сукцессионных стадий, может оставаться в почве не прорастая, но сохраняя способность к прорастанию иногда весьма долгое время (доказано, что до 1 700 лет у Chenopodium album и Spergula arvensis). Так возникают очень большие банки семян в почве. При прорастании семя поглощает воду и набухает, а его внутренние ткани разрывают семенную кожуру (или вдобавок стенку плода). Одновременно начинает расти зародыш, расходуя запас питательной ткани. При этом выделяются (в особенности семядолями) ферменты. Некоторое время все это происходит внутри семени. Зародыш всегда расположен в семени так, что его корешок обращен в сторону микропиле, поэтому при прорастании из семени через микропиле всегда сначала появляется корень проростка с гипокотилем (см. рис. 11.193; 11.197). При надземном (эпигейном) прорастании удлиняющийся гипокотиль вытягивает семядоли из семенной кожуры и поднимает их над поверхностью почвы. При подземном (гипогейном) прорастании крупные, запасающие питательные вещества семядоли остаются в семени, и только эпикотиль появляется над ее поверхностью (например, Vicia faba, Pisum, Quercus, Juglans). Подобным образом прорастают семена и многих однодольных. Их единственная семядоля обычно представляет собой сосущий орган и извлекает питательные вещества из эндосперма.
Биология распространения и прорастания семян многих видов весьма специфична. Так, лишенные эндосперма семена Orchidaceae при прорастании должны вступить в контакт с ми-
11.2. Бактерии, грибы, растения | 39g
коризообразуюшим грибом. Также очень мелкие семена корневых паразитов (например, Oroban-che) должны для прорастания попасть на поверхность корня хозяина, а семенам или плодам стеблевых паразитов требуются особые приспособления, например повышенная клейкость, как у Viscum, что позволяет им удержаться на стебле хозяина. То же можно сказать и об эпифитах, которые отличаются либо мелкосемянностью (например, Bromeliaceae), либо клейкостью семян и плодов (например, Rhipsalis).
Необычно прорастание семян у живородящих представителей мангровых, например у Rhizophoraceae (см. рис. 11.247). В их обычно односемянных плодах семя прорастает уже на материнском растении, и из висящего на дереве плода вырастает корешок и мощно развитый (у Rhixpphora до 1 м в длину) конусовидный гипокотиль. При созревании гипокотиль и корешок отделяются от семядолей и плода и падают вниз1. При этом тяжелый зародыш втыкается в мягкую илистую почву или смывается водой и укореняется после того, как она отступит.
СИСТЕМА SPERMATOPHYTINA
Из многочисленных групп семенных растений, возникавших с верхнего девона, в настоящее время во флоре Земли еще можно встретить представителей пяти эволюционных линий. Это Cycadopsida (саговниковые), Ginkgopsida (Ginkgo), Coniferopsida (хвойные), Gnetopsida и Magnoliopsida (покрытосеменные, или цветковые, растения). Если от первых четырех групп, часто объединяемых общим понятием «голосеменные» (Gymnospermae), до настоящего времени сохранилось относительно мало видов (и только хвойные доминируют в некоторых типах растительности), то существующие с мела покрытосеменные (Ап-giospermae) представлены огромным множеством видов и являются важнейшим компонентом значительной части наземной растительности. По поводу родственных связей между различными группами семенных растений нет единого мнения. До относительно недавнего времени считали, что Gnetopsida являются ближайшими родичами покрытосеменных и, таким обра
1 Отделяются не только корешок и гипокотиль: зародыш, отбрасывая семядоли, выпадает из плода. — Примеч. ред.
зом, голосеменные представляют собой группу, парафилетическую по отношению к покрытосеменным. Однако самые различные работы по анализу последовательностей ДНК недавно показали, что вопреки этой концепции голосеменные все же представляют собой монофилетическую группу. Этим были возрождены более старые концепции, касающиеся филогении семенных растений. При последующем рассмотрении систематики и филогении семенных растений мы придаем современным пяти группам равные таксономические ранги.
1. Класс: саговники (Cycadopsida) (Palmfarne)
Ныне живущие Cycadopsida чаще всего имеют один кажущийся неразветвленным ствол, на котором по спирали расположены перистые листья, образующие терминальную розетку (рис. 11.200). Ствол бывает либо подземный, клубневидный, либо может достигать высоты 15 м.
Древесина имеет очень широкие лучи. Она может состоять только из одного древесинного цилиндра, однако у некоторых видов вследствие развития добавочных камбиев снаружи от флоэмы во время вторичного роста в толщину могут возникать многочисленные концентрические древесинные цилиндры. Центр ствола занимает содержащая крахмал сердцевина. Во всех частях растения находятся слизевые ходы. У многих видов к тому же встречаются наряду с подземной корневой системой и растущие над поверхностью земли так называемые «коралловидные» корни, в которых поселяются цианобактерии из родов Nostoc, Calothrix и иногда Anabaena, фиксирующие атмосферный азот. Установлено, что растение может использовать фиксированный ими азот.
Листья чешуевидные или одиножды-, изредка дваждыперистые (Bowenia) фотосинтезирующие. Они имеют явно ксеро-морфное строение: кутикула толстая, а устьица чаще всего погруженные; листья очень жесткие. Их длина может быть от 5 см до 3 м. Листья отличаются длительным верхушечным ростом и у некоторых видов во время развития полностью или в области отдельных перышек улиткообразно закручены, как у папоротников.
400 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Рис. 11.200: Cycadopsida (А — фото F.Ehrendorfer; В, D—G — по F. Firbras и др.; С — по J.Schuster): А— Cycas rumphii в Новой Гвинее, внешний вид. Мегаспорофиллы: В— Cycas revoluta; С— С. circinalis D — Dioon edule-, Е — Macrozamia sp.; F — Ceratozamia mexicana", G — Zamia skinned
Немецкое название этих растений — Palmfame (дословно «пальмы-папоротники») — связано с обликом, часто напоминающим пальму, и с иногда наблюдаемой закрученностью растущих целых листьев или отдельных перышек1.
Эти растения всегда двудомные, но существование половых хромосом достоверно не установлено. Мужские цветки1 2 (рис. 11.201) имеют ограниченный рост и состоят из многочисленных расположенных по спирали тычинок3 (микроспорофиллов), чаще всего чешуевидных. На их нижней стороне развивается от 5 до примерно 1 000 микроспорангиев. Они часто собраны в группы и вскрываются благодаря экзотецию. Мужской гаметофит, развитие которого происходит отчасти в микроспорангии, а отчасти после высеивания из него, содержит наряду со сперматогенной клеткой еще три клетки. Сперматогенная
1 В русскоязычной литературе, особенно в ненаучной. эти растения часто встречаются под названием «саговые пальмы»; интересно, что научное название класса происходит от родового названия Cycas, которое, в свою очередь, происходит от греческого слова kykas — пальма. — Примеч. пер.
2 Микростробилы. — Примеч. ред.
3 Называть микроспорофиллы саговниковых тычинками ошибочно. — Примеч. ред.
клетка делится на два сперматозоида с расположенными по спирали многочисленными жгутиками. Имея диаметр до 400 мкм, эти сперматозоиды являются самыми крупными из всех сперматозоидов и в растительном. и в животном мире.
Пыльцевая трубка имеет гаусториальную функцию и врастает в нуцеллус. Оплодотворение происходит по типу зоидогамии. У рода Microcycas образуются многочисленные дополнительные сперматозоиды в результате неоднократного деления другой — не сперматогенной — клетки.
Мужские цветки почти всегда занимают конечное положение на ветви. К концу цветения они смещаются в сторону разрастающейся боковой почкой. Так возникает внешне неразветв-ленный ствол, который на самом деле представляет собой симподий.
Женские цветки1 (рис. 11.201) состоят из мегаспорофиллов, также расположенных по спирали, и могут достигать 70 см. В роде Cycas спорофиллы состоят из ножки (рис. 11.200), по краям которой находится от двух до восьми семязачатков, и конечного участка, который может иметь вид перисто-раздельного листа, гребенчатой или пильчатой пластины. У остальных родов мегаспорофиллы имеют ножку и чешуевид-
1 Мегастробилы. — Примеч. ред.
11.2. Бактерии, грибы, растения | 4Q-]
Рис. 11.201. Cycadopsida (А — по A.Takhtajan; В — по W. Troll; С—Е— по L. С. Richard; F — по К. von Goebel):
А, В — <j> и сГ цветки Encephalartos altensteinii (уменьшено); тычинки: С — Cycas circinalis (примерно 2х); D — Zamia integrifolia (примерно 5х) с группами микроспорангиев (Е, ок. 15х); F — поперечный разрез стенки вскрывшегося микроспорангия Stangeria paradoxa с экзотецием (примерно 80х)
ную или щитовидную пластинку, на нижнем крае которой сидят всегда два семязачатка. Женские цветки чаще всего также имеют детерминированный рост; они расположены на конце ветви и после созревания семян, как и мужские цветки, отклоняются боковой ветвью. Только у Cycas конус нарастания не исчерпывается при образовании стробила и прорастает через женский стробил (пролиферация). Семязачатки (см. рис. 11.180) имеют интегумент, который при созревании семян дифференцируется на наружный слой (саркотесту) розового или оранжевого цвета и внутренний слой (склеротесту)1. Внуцеллусе формируются пыльцевая и архегониальная камеры. Женский гаметофит состоит из большого (до нескольких тысяч) числа клеток и возникает в результате свободного деления ядер, за которым следует образование клеточных стенок. Чаще всего он развивает от двух до шести архегониев, только у Microcycas их может образовываться до 100. Величина яйцеклетки — до 6 мм.
1 Из интегумента развивается также располагающаяся под склеротестой паренхимная па-ренхотеста, которая идет на питание развивающегося эндосперма. — Примеч. ред.
В ископаемом виде Cycadopsida достоверно известны с нижней перми. Ранние представители этой группы, как, например, юрский род Beania, имели женские цветки, в которых мегаспорофиллы были расположены значительно более рыхло, чем у современных саговниковых.
Пыльцу при опылении переносит ветер, однако отчасти и жуки. Для одного вида рода Zamia удалось показать, что жуки после завершения своего развития в мужских цветках летят, нагруженные пыльцой, к женским цветкам и опыляют их. Для представителей рода Encephalartos установлено опыление различными видами жуков. Пыльна, попавшая в каплю, выделяемую из микропиле, при ее высыхании втягивается в пыльцевую камеру, которая замыкается снаружи и соединяется с архегониальной камерой. Между опылением и оплодотворением может проходить до шести месяцев.
Cycadopsida включает около 140 видов из 11 родов и 4 семейств. Семейство Cycadaceae (1 род/около 40 видов, тропики Старого Света) с единственным родом Cycas характеризуется пролиферирующими женскими цветками. Некоторые виды имеют листовидные мегаспорофиллы. Stangeriaceae (1/1, Южная Африка) и Bowe-niaceae (1/2, северо-восточная Австралия) имеют прилистники с отчетливым жилкованием. Bowenia имеет дважды перистые листья. Zamiaceae (8/около 114, тропики, субтропики, чаще всего ' Южного полушария) представлены родами Lepidozamia (2 вида) и Macrozamia (около 14) в Австралии, Encephalartos (около 35) в Африке и
402 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Dioon (около 10), Ceratozamia (около 10), Zamia (около 40), Chigua (2) и Microcycas (1) в Америке, в том числе на Кубе (Microcycas). Большинство видов растет в тропических или субтропических лесах или саваннах.
2. Класс: Ginkgopsida (Ginkgo)
Ginkgo biloba — единственный ныне живущий представитель Ginkgopsida— сильно разветвленное листопадное дерево с удлиненными и укороченными побегами. Его древесина имеет узкие лучи. Вееровидные листья (рис. 11.202) с дихотомическим жилкованием расположены по спирали. Этот вид двудомный и имеет половые хромосомы. Тычинки1 состоят из ножки и двух висящих на ее конце пыльников1 2 (рис. 11.202). Пыльники вскрываются благодаря эндоте-пию. Мужской гаметофит состоит помимо
сперматогенной клетки еще из четырех клеток. Сперматогенная клетка делится на два сперматозоида с расположенными по спирали многочисленными жгутиками.
После опыления пыльцевое зерно прорастает пыльцевой трубкой. С одной стороны, она имеет функцию гаустории и врастает в нуцеллус, из которого извлекает питательные вещества для мужского гаметофита. С другой стороны, она растет к архегониям, в которые должны быть высвобождены сперматозоиды. Оплодотворение осуществляют сперматозоиды (зоидога-мия).
Многочисленные тычинки на оси образуют мужские цветки3, напоминающие сережки, располагающиеся в пазухах чешуевидных листьев на укороченных побегах.
Семязачатки чаще всего располагаются по два на концах вильчато разветвлен-
Рис. 11.202. Ginkgopsida (А— по A. Schenk; В— по L. С. Richard, A.W. Eichler; С— по Т. Shimamura, изменено и несколько схематизировано):
А— Baiera muensterana (рэт-лиас): вегетативный лист; семязачатки на оси <j> цветка (несколько уменьшено); радиально расположенные, закрытые и открытые группы микроспорангиев на осях о" цветков (примерно 2х); В, С — Ginkgo biloba (современный вид): В — укороченный побег с сГ цветком и молодыми листьями (натур, величина), редуцированные со спинной стороны, двучленные группы микроспорангиев (тычинки; увеличено), семязачатки (<j> цветки) и соответственно семена (несколько уменьшено); С — сперматозоид (примерно 200х)
1 Микроспорофиллы. — Примеч. ред.
2 Микроспорангиев, не гомологичных пыльникам покрытосеменных. — Примеч. ред.
3 Микростробилы. — Примеч. ред.
11.2. Бактерии, грибы, растения | 493
ной ножки (рис. 11.202). Как и мужские цветки, эта структура находится в пазухе чешуевидного листа на укороченной ветви. Семязачатки имеют один интегумент, который при созревании семени дифференцируется на наружную саркотесту, очень сильно пахнущую масляной кислотой, и внутреннюю жесткую склеротесту. Под склеротестой находится еще один мягкий слой семенной кожуры1. Нуцеллус имеет пыльцевую и архегониальную камеры. Женский гаметофит содержит хлорофилл и окрашен в зеленый цвет. Он состоит из нескольких сотен клеток, которые возникают после свободных ядерных делений, за которыми следует образование клеточных стенок. Он развивает чаще всего два или три архегония. Опыление у Ginkgo осуществляется ветром.
В ископаемом состоянии Ginkgopsida известны с нижней перми (например, роды Trichopitys и Sphenobaiera). Наибольшего разнообразия форм они достигли в юре и мелу. Род Ginkgo с видами, уже очень близкими к современному виду G. Ы-loba, впервые встречается в нижней юре, а ископаемые остатки, не отличимые от G. biloba, известны с начала третичного периода. Этот вид, таким образом, является «живым ископаемым». У вымерших представителей Ginkgopsida (например, Baiera) листья часто бывали рассечены сильнее. Пыльники располагались радиальными группами, а женские цветки1 2 содержали большое число семязачатков.
Родина G. biloba, очевидно, Китай. Поскольку этот вид не менее 1 000 лет культивируется преимущественно в Китае и Японии в парках, окружающих храмы, и иногда дичает, его природный ареал и экология достоверно не известны. Предполагают, что природный ареал его находится в провинции Чжэцзян, где этот вид растет во влажных смешанных лесах.
3. Класс: хвойные (Coniferopsida)
Современные представители Coniferopsida развиваются из зародышей с двумя — многими семядолями (см. рис. 17.197) в разветвленные деревья или, реже, кустарники с моноподиальным стволом, на котором боковые ветви часто располагаются ярусно. Вторичная древесина часто содер-
1 Эндотеста. — Примеч. ред.
2 Мегастробилы. — Примеч. ред.
жит смоляные ходы, а лучи не многорядные1. Нередко имеет место дифференциация на удлиненные и укороченные побеги, а у видов некоторых родов (например, Phyllocladus, Sciadopitys1'} укороченные побеги могут принимать вид более или менее уплощенных филлокладиев. Листья располагаются спирально, накрест супротивно или мутовчато, имеют параллельное жилкование или единственную среднюю жилку и обычно лентовидные, игловидные или чешуевидные. Есть как вечнозеленые, так и листопадные (летнезеленые) виды. Цветки3 однополые. Бывают как однодомные, так и двудомные виды.
Шишковидные мужские цветки располагаются поодиночке или рыхлыми группами (рис. 11.203). Тычинки4 располагаются по спирали или изредка накрест супротивно и имеют на нижней стороне от 2 до 20 микроспорангиев, часто срастающихся между собой. Изредка (Taxus) микроспорангии расположены радиально на конце ножки. Микроспорангии вскрываются благодаря экзотецию. Пыльцевые зерна могут иметь воздушные мешки, возникающие вследствие расслоения эктэкзины (рис. 11.203). Мужской гаметофит наряду со спермиогенной клеткой содержит еще 2 (Cephalotaxaceae, Cupressaceae, Sciadopityaceae, Taxaceae), 4 (Pi-naceae) или от 10 до примерно 40 (Arauca-riaceae, Podocarpaceae) клеток. Сперматозоиды не образуются; два неподвижных спермия, возникающих из спермиогенной клетки, доставляются к архегониям пыльцевой трубкой (рис. 11.204) (= сифоногамия).
Женские цветки5 чаще всего собраны в характерные для хвойных шишки
1 Основная масса лучей — однорядные («узкие лучи»). Многорядные («широкие») лучи, если есть, содержат по одному радиальному смоляному ходу. — Примеч. ред.
2 Строение укороченных побегов Sciadopitys очень своеобразно, но не имеет никакого отношения к филлокладиям. — Примеч. ред.
3 У хвойных нет цветков, а есть микростробилы и шишки. — Примеч. ред.
4 Это микроспорофиллы, которые нельзя отождествлять с тычинками. — Примеч. ред.
3 Точнее — это аксиллярные комплексы, а не цветки. — Примеч. ред.
404 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Рис. 11.203. Pinus (А, В—F— по О. С. Berg, С Г.Schmidt; G, Н, J, К— по E.Strasburger):
А—F, К — Р. sylvestris, G—I — Р. тидо:
А — цветущий и семяносящий побег, в пазухе опадающих чешуевидных листьев располагаются двух-иглые укороченные побеги; Е — d" цветки (G — продольный срез), тычинки с 2 микроспорангиями (Н — продольный, J — поперечный срезы); К — пыльцевое зерно с 2 воздухоносными мешками; В — <j> соцветие с комплексами из кроющих и семенных чешуй (С — сверху, D — снизу), из которых однолетние еще зеленые, и двухлетняя зрелая, раскрывающаяся шишка (А), несущая по 2 крылатых семени (F) на верхней стороне уже одревесневших к этому времени комплексов чешуй (А — несколько уменьшено, В—F увеличено, G — 10х, Н — 20х, J — 27х, К — 400х)
(рис. 11.205). Они состоят из оси, на которой располагаются спирально или накрест супротивно кроющие листья отдельных цветков, называемые кроющими чешуями. Женские цветки, или семенные чешуи, представляют собой плоские органы, на верхней стороне которых находится различное число семязачатков (от 1 до примерно 20). Семенные чешуи несмотря на их чаще всего листовидную форму, следует считать модифицированными укороченными побегами, а женская шишка современных Coniferopsida является, таким образом, соц
ветием1. Семенных чешуй вышеописанного типа нет у Тахасеае и Cephalotaxaceae1 2.
Интерпретация семенной чешуи как женского цветка и, следовательно, женской шишки как соцветия вытекает из сравнения с кордаитами и Voltziales, ископаемыми представителями Coniferopsida, а также,
1 Поскольку у хвойных нет цветка, то не может быть и соцветия. — Примеч. ред.
2 Весьма модифицированы также семенные чешуи Podocarpales. Cupressales s.l. и Araucaria-les. — Примеч ред
11.2. Бактерии, грибы, растения | 405
Рис. 11.204. Прорастание пыльцы и оплодотворение у Coniferopsida (А, В — Pinus nigra, С — Р. syl-vestris: Pinaceae; D, E— Torreya taxifolia: Taxaceae) (A, В— no J. M.Coulter, Ch. Chamberlain; C— no E. Strasburger; D— E — no J. M.Coulter, W. J. G. Land):
A, В — развитие d" гаметофита в пыльцевом зерне и пыльцевая трубка: проталлиальные клетки (р), ядро вегетативной клетки (клетки пыльцевой трубки) (к), антеридиальная клетка (а), дающая клетку-ножку (s) и спермиогенную клетку (sz), образующую в свою очередь 2 спермия (примерно 500х); С — сформировавшийся семязачаток с интегументом (i), пыльцевой трубкой (ps), а также архегониями с шейкой (h), брюшной канальцевой клеткой (Ь) и яйцеклеткой (е) (увеличено); D— пыльцевая трубка с 2 спермиями (sp,, sp2), а также ядрами клетки трубки (рк) и клетки-ножки (sk) возле яйцеклетки (ядро яйцеклетки ек, остаток шейковой клетки h); Е— слияние ядра яйцеклетки с ядром спермия, остальные ядра дегенерируют (367х)
учитывая в типе пазушное ветвление семенных растений, из положения семенной чешуи в пазухе кроюшей чешуи.
Кордаиты были деревьями высотой до 30 м, с развесистой кроной и пазушным ветвлением (рис. 11.206), вторичным утолщением и араука-риоидной поровостью трахеид вторичной древесины.
Лентовидные или ланцетные листья с дихо-томически-параллельным жилкованием располагались спирально. Тычинки находились в напоминающих шишки цветках1 (рис. 11.206). В нижней части этих цветков располагались по спирали чешуевидные листья, а в верхней части
1 Микроспорофиллы в микростробилах. — Примеч. ред.
находились также расположенные по спирали тычинки, несшие на вершине несколько микроспорангиев. Несколько таких мужских цветков рыхло располагались в пазухах кроющих листьев на удлиненных осях, выходивших из пазух обычных листьев. Сходно устроенные женские цветки располагались также в пазухе кроющего листа (рис. 11.206). За расположенными спирально чешуевидными листьями у них следовали также спирально расположенные мегаспорофиллы1, которые были палочковидными и на вершине несли по одному семязачатку (вследствие вильчатого ветвления изредка два или больше). Весь женский цветок интерпретируют как гомолог семенной чешуи современных Conifero-
1 Убедительных доказательств того, что это мегаспорофиллы, нет. — Примеч. ред.
406 | ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
psida. Наличие пыльцевой камеры в нуцеллусе указывает на то, что в отличие от современных Coniferopsida оплодотворение у кордаитов, вероятно, осуществлялось сперматозоидами. Корда-иты появились в карбоне как лесообразующие породы и вымерли в перми.
Древовидные Voltziales, имевшие в древесине трахеиды с араукариоидной поровостью, жили с верхнего карбона до нижней юры. Листья у них были игло- или чешуевидными и иногда на вершине вильчато-рассеченными. Это были однодомные растения. Как и у кордаитов, женские цветки (рис. 11.207) состояли из пазушной оси с расположенными спирально или в одной плоскости стерильными чешуевидными листьями, а также одним или чаще всего несколькими семязачатками на ножках. Однако в отличие от кордаитов эти цветки, как и у современных Coniferopsida, были собраны в компактные шишки, которые, таким образом, надо трактовать как
соцветия. Тычинки Voltziales состояли из ножки и уплощенного конечного участка. Многочисленные микроспорангии располагались главным образом на адаксиальной стороне ножки. Voltziales можно рассматривать как организмы, очень близкие к исходным формам современных Coniferopsida.
Семязачатки имеют один интегумент, который дифференцируется в твердую склеротесту.
Семена могут быть окружены мясистой оболочкой, которая, однако, никогда не возникает из семенной кожуры. Как и у Ginkgo, у хвойных многоклеточный женский гаметофит возникает путем свободных делений ядер, за которым следует образование клеточных стенок. Он может образовать до 60 архегониев. Насколько известно, все хвойные анемофильны. Пыльца либо попадает в специальную каплю, выделяемую из микропиле
Рис. 11.205. Pinaceae, Abies (А— A. nordmanniana. В—Е— A. alba) (А— по О. С. Berg, С. F. Schmidt; В—D— по F. Firbas; Е— по A. W. Eichler):
А— побег со зрелыми шишками, справа уже частично рассыпавшейся (несколько уменьшено); В — сГ цветок с чешуевидными листьями (Ь) и тычинками (sb) (примерно 2х); С, D — зрелый <j> цветок с кроющей чешуей (d), семенной чешуей (ss) и 2 семенами (s); Е— семя с верхней (о) и нижней (и) стороны (несколько уменьшено)
11.2. Бактерии, грибы, растения i 4Q7
Рис. 11.206. Кордаиты (карбон-пермь) (А, В — по C.Grand'Eury; С — no Th. Delevoryas; D, Е — по F. Florin):
А— внешний вид Cordaites sp. (длина ок. 10 м); В — олиственный побег с расположенными в пазухах листьев соцветиями Cordaites laevis (уменьшено); С — два О" цветка Cordaianthus concinnus (ок. 2,5х); D — тычинка С. penjonii с группой терминальных микроспорангиев (ок. 10х); Е — <,> цветок С. pseudofluitans с кроющим листом, стерильными чешуями и расположенными на длинных ножках семязачатками (1,5х)
семязачатка, при высыхании которой втягивается в микропиле, либо оказывается внутри семязачатка в результате роста интегумента. В ископаемом виде хвойные можно проследить до триаса, а большинство современных семейств известны по ископаемым остаткам уже в триасе и юре.
Современные Coniferopsida насчитывают семь семейств. Pinaceae (12/около 200. чаще всего в умеренной зоне северного полушария) имеют расположенные по спирали игловидные листья и могут быть вечнозелеными или листопадными. Тычинки состоят из ножки, на нижней стороне которой находится 2 микроспорангия1. Пыль
1 Микроспорофиллы сосновых пельтатные. Два микроспорангия, находящиеся на нижней половине щитовидной пластинки микроспорофилла, прирастают к его черешку. — Примеч. ред.
цевые зерна бывают с воздухоносными мешками. Семенные чешуи, более или менее несросшиеся с кроющими чешуями, несут’ два семязачатка, направленные микропиле к основанию чешуи. Растения всегда однодомные. Во флоре Средней Европы это семейство представлено родами пихта (Abies), лиственница (Larix), ель (Picea) и сосна (Pinus).
Если лиственница — дерево листопадное, то хвоинки пихт, елей и сосен живут от 3 до 9 лет, изредка дольше. Хвоинка дифференцирована на пластинку и основание. В отличие от пихты, которая сбрасывает весь лист, у ели отпадает только листовая пластинка. Основание листа срастается со стеблем в так называемую листовую подушку.
Представители родов Pinaceae имеют либо только удлиненные побеги (например, Abies, Picea, рис. 11.205; 11.208), либо удлиненные и укоро
408 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
ченные побеги. У вечнозеленых кедров (род Cedrus) и у лиственниц (рис. 11.208) каждый удлиненный побег в первый год несет зеленую хвою. На второй год на них вырастают укороченные побеги с пучками хвоинок; эти побеги могут нарастать несколько лет. У взрослых деревьев сосен хвоя располагается только на укороченных побегах. Молодые растения в первый или второй год дают и удлиненные побеги с зеленой хвоей, но после этого на них образуются только коричневые чешуевидные листья, в пазухах которых возникают укороченные побеги с немногочисленными чешуевидными листьями и пятью, тремя, двумя или даже только одной хвоинкой.
Когда пыльца попадает на семязачаток, женский гаметофит в нем еще не развит, а иногда даже не возникла и мегаспора. Соответственно между оплодотворением и опылением бывает продолжительный промежуток времени, в течение которого замыкается микропиле и скрывает прорастающую пылинку в семязачатке. У большинства видов Pinus между опылением и оплодотворением проходит один год, а семена из зрелых шишек высвобождаются только на третий год. В отличие от Pinus у ели от оплодотворения до созревания семени проходит один вегетационный период. При созревании семян шишка либо распадается (например, Abies), либо семена
высвобождаются, а шишка остается интактной (например, Picea, Pinus). Семена часто имеют кожистое крылышко, которое, однако, образуется не из семенной кожуры, а из отслаивающейся части семенной чешуи.
Экология среднеевропейских Pinaceae весьма различна. Если пихта белая (Abies alba) — дерево, достаточно требовательное в отношении почвы и климата, образует смешанные леса с буком, то формирующая однородные леса на северо-востоке Европы ель европейская (Picea abies) в Средней Европе в естественных условиях приурочена к среднегорному и субальпийскому поясам. Правда, из-за большой ценности для лесного хозяйства ель распространена гораздо шире в искусственных посадках. Лиственница европейская (Larix decidua) — светолюбивое дерево, которое часто встречается прежде всего в местах с континентальным климатом в Центральных Альпах вблизи верхней границы леса. Сосна обыкновенная (Pinussylvestris) — светолюбивое, однако в остальном неприхотливое дерево с низкой конкурентоспособностью, легко выносяшее как сухое и жаркое лето, так и сильные зимние морозы. Она встречается от равнин до верхней границы леса в горах и растет как на очень сухих, так и на очень сырых почвах. Чаще всего сосну обыкновенную можно встретить там, где более требовательные породы рас-
Рис. 11.207. Voltziales (по F. Florin):
А—D— Lebachia piniformis (ротлиген): А— верхушка ветви; главный побег с вильчатыми листьями (1/Зх); В — торчащая вверх <j> шишка с вильчатыми кроющими чешуями (1/2х); С — D — £ цветок сзади и спереди, кроющая чешуя (= кроющий лист, d), стерильные чешуи (s) и уплощенный прямой семязачаток (sa) с двураздельным интегументом* (5х); Е— Clyptolepis longibracteata (нижний триас): <j> цветок с кроющей чешуей (d), стерильными чешуями и двумя анатропными семязачатками (схематично; 2х)
* Давно показано, что это не семя, а уплощенная семяножка, на адаксиальной стороне которой располагался обращенный семязачаток. — Примеч. ред.
11.2. Бактерии, грибы, растения . 499
Рис. 11.208. Pinaceae (А—F— no G. Karsten; G, Н— по M.Willkomm):
А—F — Picea abies: А—С — побеги с сГ цветками, <j) соцветиями и шишками (уменьшено); D — тычинка; Е— <j> цветок с кроющей (d) и семенной (s) чешуями, а также семязачатком (sa) (увеличено); F — крылатое семя (в натур, вел ); G—Н — Larix decidua: G — удлиненный побег с несущими хвоинки короткими побегами (к) летом; Н — удлиненный побег с сГ цветками, <j> соцветиями и распускающимися почками укороченных побегов (к) весной (примерно в натур, величину)
ти не могут. Два других вида этого рода — сосна горная (Р. mugo) и сосна «кедровая» европейская (Р. cembra)' — обитают преимущественно в высокогорьях Средней Европы, а сосна черная (Р. nigra), распространенная преимущественно в Средиземноморье, доходит до Нижней Австрии.
Araucariaceae (3/около 23, Южное полушарие, кроме Африки) имеют чаще всего широкие листья, сидящие спирально или накрест супротивно, а в древесине — трахеиды с расположенными в виде пчелиных сот окаймленными порами (араукариоидная поровость). Это однодомные либо двудомные растения. Тычинки несут от 4 до 20 микроспорангиев, а прочно срастающиеся с кроющими чешуями семенные чешуи — лишь один семязачаток. Описанный только в 1995 г. род IVollemia из Австралии — явно
1 В России этот вид и близкие к нему (например, Р sibirica, Р. koraiensis) традиционно известны под названием «кедры». — Примеч. пер.
наиболее примитивный род семейства — имеет большое сходство с его представителями, вымершими уже в третичный период. Соответственно его можно рассматривать как «живое ископаемое».
Некоторые представители Podocarpaceae (18/около 130, Южное полушарие, от тропиков до умеренных широт) имеют очень крупные листья. Виды этого семейства, преимущественно двудомные, отличаются в первую очередь тем, что женские шишки часто редуцированы до нескольких или иногда только одного комплекса из кроющей и семенной чешуй, в котором семенная чешуя окружает семя в виде мясистого покрова (эпиматия). Кроме того, стерильные семенные чешуи могут срастаться с осью шишки в мясистую «ножку»1. Относящийся к этому семейству Parasitaxus ustus из Новой Каледонии — единственный паразит среди хвойных. Его хозяин
1 Рецептакулум. — Примеч. ред.
410 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
(Falcatifolium taxoides) также относится к Podo-саграсеае. В роде Phyllocladus (иногда выделяемом в самостоятельное семейство) функцию органов ассимиляции выполняют листоподобные, лопастные по краю филлокладии.
Cupressaceae (включая Taxodiaceae; 29/около 140; по всему земному шару, чаще всего в районах с умеренным климатом) имеют игольчатые или чешуевидные листья, расположенные по спирали, накрест супротивно или в мутовках. Растения эти однодомные или двудомные. Тычинки име-•ют от 2 до 6 микроспорангиев, а семенные чешуи, срастающиеся с кроющими чешуями, — от 2 (редко 1) до 20 семязачатков.
В Средней Европе Cupressaceae представлены двудомным можжевельником обыкновенным (Juniperus communis). Его женские шишки (рис. 11.209) состоят из многочисленных стериль-
Рис. 11.209. Cupressaceae, Juniperus communis (А, В — по F. Firbas; С—Е— по О. С. Berg, С. F. Schmidt):
А— побег растения с соцветиями (D— с опылительными каплями, а также одно- и двулетними «шишкоягодами»— Е); В— побег сГ растения с цветками (С) (А, В — ок. 2/Зх, С—Е увеличено)
пых чешуевидных листьев, а также трех семенных чешуй, каждая с одним семязачатком. Эти семенные чешуи при созревании шишки становятся мясистыми и образуют шаровидные можжевеловые «ягоды». Можжевельник обыкновенный — неприхотливое растение, типичное для выгонов и кустарничковых пустошей. Выше альпийского пояса в горах и за полярной границей леса он представлен стелющимся подвидом — subsp. alpina. Мамонтово дерево (Sequoiadendron califomicum) в Калифорнии имеет ствол диаметром более 8 м; его возраст свыше 3 000 лет. Sequoia sempervirens из прибрежных горных местностей в Калифорнии достигает высоты более 100 м. Metasequoia glyptostroboides, найденная в 1940 г. в Китае, ранее была известна только в ископаемом состоянии в третичных отложениях и является еще одним примером «живого ископаемого». Этот листопадный вид осенью сбрасывает целиком укороченные олиственные побеги. Также веткопадным и имеющим реликтовый ареал является болотный кипарис (Taxodium distichum), который образует обширные леса по болотам северного побережья Мексиканского залива. Он имеет выступающие из воды или ила выросты корней (пневматофоры), которые снабжают корневую систему воздухом.
Основанное преимущественно на форме и положении листьев разделение Taxodiaceae (листья чаще всего игловидные, расположены спирально) и Cupressaceae (листья чаще всего чешуевидные, расположены накрест супротивно или в мутовках) необоснованно. Cupressaceae, вероятно, — монофилетическая группа, возникшая из Taxodiacae. Так как Cupressaceae — название более раннее, оно должно быть оставлено при объединении этих двух семейств.
Sciadopityaceae (1/1, Япония) с единственным видом Sciadopitys verticillata отличаются наличием мутовчато расположенных игловидных филлокладиев1.
У Cephalotaxaceae и особенно Тахасеае строение женских шишек существенно отличается от характерного для вышеописанных семейств.
Женские шишки двудомных Cephalotaxaceae (1/около 6, районы умеренного климата Восточной Азии) состоят из немногих пар накрест супротивных чешуевидных листьев. В пазухе каждого из них находятся два прямостоячих семязачатка, из которых чаще всего развивается толь-
1 Это не филлокладии, а сросшиеся боками попарно игловидные листья укороченных побегов, развивающихся ярусами в пазухах чешуевидных листьев удлиненных побегов. — Примеч. ред.
11.2. Бактерии, грибы, растения | д-| -j
Рис. 11.210. Taxaceae, Taxus baccata (А, В, D — по F. Firbas; С, F — по R.von Wettstein; Е — по E.Strasburger):
А, В — цветущий о* и семяносящий побеги (с одним незрелым и двумя зрелыми семенами; 3/4х); С, D — сГ и £ цветоносные побеги каждый в пазухе хвоинки (2,5х); Е— цветоносный побег в продольном разрезе с микропиле (т), интегументом (i), нуцеллусом (п), зародышевым мешком (е), зачатком ариллуса (а) и конусом нарастания первичного пазушного побега (v) (9х); F — семя в продольном разрезе, с ариллусом, семенной кожурой, эндоспермом и зародышем (2х)
ко один. Зрелое семя окружено мясистой тканью, которая возникает из ножки семязачатка. Семенные чешуи у Cephalotaxaceae неразличимы.
У также двудомных Taxaceae (5/около 25, районы с умеренным климатом Северного полушария), имеющих древесину без смоляных ходов, единственный семязачаток находится на уплощенных коротких боковых веточках над одной из накрест супротивных пар чешуевидных листьев (рис. 11.210). Положение семязачатков на боковых веточках трактуют как конечное или боковое. Как и у Cephalotaxaceae, у Taxaceae
зрелое семя окружено мясистой тканью, которая возникает из ножки семязачатка.
Разделение Cephalotaxaceae и Taxaceae на два разных семейства крайне проблематично. Это можно видеть, например, из того, что род Атето-taxus относят то к Cephalotaxaceae. то к Taxaceae.
Интерпретация женских структур Taxaceae неоднозначна. Исходя главным образом из очевидного близкого родства Cephalotaxaceae и Taxaceae, а также из ближайшего родства с Cupressaceae/Sciadopityaceae, хорошо подтверждаемого данными по последовательности нуклеотидов ДНК, можно считать вполне вероятным, что женские структуры Taxaceae производны от шишки с кроющими и семенными чешуями, их строение не может служить основанием для часто практиковавшегося в прошлом выделения Taxaceae в отдельный порядок (Taxales). Тычинки Taxaceae также своеобразны. Так, у Taxus многочисленные микроспорангии располагаются по радиусам вокруг ножки тычинки, а у Pseudotaxus мутовки тычинок отделены друг от друга чешуевидными листьями.
В Средней Европе Taxaceae представлены тиссом ягодным (Taxus baccata), который встречается в местах с мягкой зимой, по осветленным участкам леса. Все части этого дерева кроме мясистого ариллуса ядовиты из-за наличия производных таксана, а древесина тисса — самая плотная из среднеевропейских деревьев. Таксол — вещество, содержащееся в Taxus brevifolia, — из-за его подавляющего митозы действия применяют при лечении рака.
Семейства Coniferopsida можно объединить в три группы. Первую составляют Pinaceae, вторую Лгаисапасеае и Podocarpaceae, а третью Cupressaceae, Sciadopityaceae и Cephalotaxaceae/Taxaceae. Эта группировка, основанная на молекулярных данных, подтверждается структурой мужских гаметофитов. Более тесное родство существует, по-види-мому, между второй и третьей группами.
4. Класс: Gnetopsida
Сохранившиеся доныне три рода Gnetopsida — Ephedra (35—45 видов, сухие области Евразии и Америки), Gnetum (около 30 видов, тропики всего земного шара) и Welwitschia (один вид, Юго-Западная Африка) — представляют собой древовидные растения, различающиеся в морфологическом плане. Древесина Gnetopsida имеет узкие лучи и у всех трех родов содержит сосуды1. * У
1 Наряду с сосудами в ней много трахеид.
У ряда видов Gnetum лучи весьма широкие. — Примеч. ред.
412 I ГЛАВА 11, СИСТЕМАТИКА И ФИЛОГЕНИЯ
Гомология этих сосудов с сосудами Mag-noliopsida спорна1. Виды рода Ephedra — ку
1 Членики сосудов Gnetopsida имеют эфедро-идные перфорационные пластинки, нигде за пределами класса более не известные. — Примеч. ред.
старники, небольшие деревья или изредка даже лианы. Чаще всего лианы, но также и кустарники, и небольшие деревья встречаются в роде Gnetum (рис. 11.211). Welwitschia своим неразвётвленным стволом, имеющим диаметр до 1 м, глубоко погружена в почву
11.2. Бактерии, грибы, растения | 413
Рис. 11.211. Gnetopsida. (А— по A.W. Eichler; В — по А. Н.Church; Е F — по G. Karsten, W. Lie-bisch, изменено; G, Н — по Е Markgraf, J — по W. Н. Pearson, изменено; N, Q — no G. Karsten; О, P — по O. Stapf; R — no R.von Wettstein; C, D, K, L, M, S, T— no P.R. Crane):
A — внешний вид молодого растения Welwitschia mirabilis с <j> соцветиями (ок. 1/20х); В — 6" цветок с кроющим листом (d) и двумя папами брак-тей (Ь), сросшимися между собой тычинками и стерильным семязачатком (ок. 7х); С, D — диаграммы О’ и $ цветков: Е—J — Gnetum (Е, F, J — G. gnemon, G — G. costatum, Н — G. monta-пит); Е — побег с 6" соцветиями (3/8х); F — мутовчатые частные соцветия, снаружи с фертильными б", внутри со стерильными цветками (1/5х); G, Н — 6" цветки с парой брактей (b); J — продольный срез <j> цветка с двумя парами брактей, удлиненным интегументом, нуцеллу-сом и зародышевым мешком (увеличено): К— М — диаграммы сГ и <j> соцветий и цветков Gnetum; N—R — Ephedra altissima, N — о" побег (2/Зх); О, Р — о" частное соцветие и о" цветок (7,5х); Q, R — <j> побег с незрелыми семенами (2/Зх) и терминальный <j> цветок (2х); S, Т — диаграммы о" и <j> цветков Ephedra
<
(рис. 11.211). Ephedra имеет очень мелкие чешуевидные листья, располагающиеся накрест-супротивно или в мутовках по три. У Gnetum листья простые, с перистым жилкованием, напоминающие листья многих двудольных, накрест супротивные. У взрослых растений Welwitschia имеется только одна пара супротивных лентовидных листьев с параллельным жилкованием (из двух пар, возникающих в процессе развития), которые в течение всей жизни растения нарастают у основания и отмирают на конце. Gnetopsida преимущественно двудомные.
Мужские цветки1 Gnetopsida имеют покров из одной или двух пар брактей и от одной до шести тычинок1 2 (рис. 11.211).
У Ephedra мужской цветок имеет покров, состоящий из двух сросшихся основаниями друг с другом чешуевидных листьев, расположенных в медианной плоскости. Из этого покрова чаще всего выходит ножка, на конце которой находится несколько сросшихся между собой микроспорангиев. Поскольку эта ножка может быть на конце вильчато разветвленной или же один цветок содержит две ножки, иногда считают, что мужской цветок Ephedra состоит из двух чаще всего сросшихся между собой тычинок. Также состоящий из двух медианных брактей, но часто сросшихся В трубку покров мужских цветков Gnetum окружает ножку с одним или двумя микроспорангиями на конце. Эта структура вполне соответствует тычинке. У Welwitschia мужской цветок имеет покров из двух накрест супротивных пар брактей и мутовки шести тычинок, сросшихся основаниями между собой. На вершине каждая тычинка несет три сросшихся между собой микроспорангия. Иногда считают, что здесь мы имеет дело не с шестью, а скорее с двумя трехчленными тычинками. Мужские цветки Welwitschia всегда содержат на вершине рудиментарный семязачаток над рудиментарной парой трансверсальных брактей.
Микроспорангии вскрываются короткими клапанами благодаря экзотецию. Вследствие того, что утолщения стенок клеток экзотеция у Welwitschia инвертированы, пыльца при высыхании стенки микроспорангия выдавливается из него.
Мужской гаметофит содержит кроме сперматогенной клетки еще либо четыре (Ephedra), либо только две (Gnetum, Welwitschia) дополнительные клетки. Деление сперматогенной клетки приводит к образованию двух спермиев, а оплодотворение осуществляется посредством пыльцевой трубки (сифоногамия).
Женские цветки содержат прямостоячий семязачаток с интегументом, вытянутым в длинное микропиле1. Из микропиле выделяется опылительная капля. Семязачаток окружен одной трансверсальной (Ephedra) или двумя — медианной и трансверсальной (Gnetum, Welwitschia) — парами брактей, чаще всего сросшихся между собой (рис. 11.211).
При созревании семени у Ephedra брактей цветка отвердевают и окутывают семена, а брак-теи соцветия могут становиться мясистыми У Gnetum наружная пара брактей становится
1 Цветков у Gnetopsida нет. — Примеч. ред.
2 Это микроспорангиофоры. — Примеч. ред
1 Микропилярная трубка. — Примеч. ред.
414 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
мясистой, а внутренняя — жесткой; у Welwitschia наружная пара брактей крыловидно разрастается, тогда как внутренняя твердеет. Женский гаметофит формируется путем свободных делений ядер, за которым следует образование клеточных стенок. Он состоит либо из одной мегаспоры (моноспорический: Ephedra), либо из всех четырех дочерних ядер, образующихся в результате мейоза в материнской клетке мегаспоры (тетра-спорический: Gnetum, Welwitschia). Женский гаметофит может насчитывать более чем 1 000 клеток. У Ephedra развиваются типичные ахегонии, у Gnetum и Welwitschia архегониев нет, а яйцеклетки у видов обоих родов не отличимы от остальных клеток женского гаметофита. У Welwitschia от женского гаметофита навстречу пыльцевым трубкам отрастают трубковидные структуры.
У Ephedra и Gnetum происходит двойное оплодотворение. Слияние одного спермия с яй
цеклеткой, а другого с брюшной канальцевой клеткой у Ephedra или же с любыми двумя клетками у Gnetum дает две зиготы. Которая из этих двух зигот разовьется в единственный зародыш семени, по-видимому, однозначно не предопределено.
Хотя некоторые виды Ephedra, вероятно, ветроопыляемые, по-видимому, у Gnetopsida в целом преобладает опыление различными насекомыми. Насекомых привлекают как запах, так и содержащие сахар опылительные капли.
Ископаемые Gnetopsida малоизвестны. Ископаемая пыльца, сходная с пыльцой современных гнетовых, известна с верхнего триаса, но принадлежность к Gnetopsida ископаемых макроостатков из нижней юры (например, Piroconites) или мела (Drewia. Eoanthus) сомнительна.
Предполагалось, что растущие только в пустыне Намиб растения Welwitschia mirabilis, от
Рис. 11.212. Bennettitales (А — по A. G. Nathorst; В, С — по Th. М. Harris; D — по F. Ehrendorfer; Е — по Н. Solms; F— по G. R.Wieland):
А — Wielandiella angustifolia (верхний триас), реконструкция с ветвями, листьями и цветками или плодами (1/5х); В — <j>/6" цветки Williamsoniella coronata (средняя юра) с листочками обвертки и тычинками, а также гинецеем (примерно натур, величина); С — лепестковидная тычинка (2х); D — клубневидный ствол, крона перистых листьев и цветочные почки Cycadeoidea sp. (реконструкция, сильно уменьшено); Е — продольный срез выступающего из женского цветка плода1 Bennettites gibsonianus (нижний мел) с листочками обвертки, межсеменными чешуями и расположенными на ножках семенами (примерно 1/Зх); F— продольный срез группы микроспорангиев с экзотецием у Cycadeoidea dacotensis (ок. 16х)
1 Плодов у Bennettitales не было. — Примеч. ред.
11.2. Бактерии, грибы, растения | 4-15
дельные экземпляры которых имеют возраст до 2 000 лет, являются реликтами с тех времен, когда климат там был более влажным.
С покрытосеменными часто сближали ископаемые Bennettitales (рис. 11.212). Стволы их были либо короткие неразветвленные и сильно узловатые (например, Williamsonia), либо относительно тонкие и разветвленные (например, Wielandiella). Одинождыперистые или простые листья располагались по спирали. Цветки были или однополыми, или — очевидно, у более продвинутых родов — обоеполыми1. У большинства видов цветки имели обертку из спирально расположенных кроюших листьев. Тычинки1 2 располагались в мутовках и были либо перисто-разветвленными и завернутыми на верхнюю сторону, либо нерасчлененными и листовидно уплощенными. Микроспорангии имели экзотеций и были по нескольку в виде синангия окружены двухлопастным покровом. Каждая тычинка несла несколько таких синангиев на адаксиальной стороне.
Вертикальные семязачатки имели стерильную обертку. В женском цветке или в верхней части обоеполого цветка чаще всего были многочисленные (до нескольких сотен) семязачатки, располагавшиеся по спирали вдоль удлиненной оси цветка и отделенные друг от друга стерильными межсемянными чешуями. Семязачатки с плюской и ножкой можно интерпретировать как сильно редуцированные мегаспорофиллы, а межсемянные чешуи — как стерильные мегаспорофиллы. Вероятно, Bennettiatales — по крайней мере, некоторые — опылялись насекомыми. Bennettiatales появились впервые в верхнем триасе и вымерли в мелу.
5. Класс: цветковые растения, покрытосеменные (Magnoliopsida;
Angiospermoe)
Как показывает сравнение разных изданий учебника Э. Страсбургера, представления о родственных связях среди покрытосеменных (естественно, и среди других групп растений), а отсюда — и сама система цветковых растений постоянно изменяются. Есть три причины такого продолжающего и, конечно, еще далекого от завершения изменения системы. В ранний период систематики, после К. Линнея, особенно резко возросло число вновь откры
1 Гомология этих структур цветкам покрытосеменных не доказана. — Примеч. ред.
2 Микроспорофиллы. — Примеч. ред.
ваемых видов вследствие интенсивной деятельности но сбору и изучению растений, прежде всего в ходе колонизации новых земель. Вместе с общим развитием технических средств постоянно увеличивались количество и характер изучаемых признаков растений. Если первые систематики имели в своем распоряжении только лупу, то развитие техники от простых микроскопов до современных электронных позволило проводить наблюдения со все большим разрешением, все более качественно и быстро характеризовать химический состав частей растений с помощью хроматографии и анализа ультраструктуры. В наше время для систематиков привычным средством стала уже разнообразная техника для анализа ДНК. Наконец, в ходе истории сильные изменения претерпели и сами подходы к интерпретации наблюдаемых данных. Если в начале систематики постулировали особую важность лишь некоторых признаков, то впоследствии возникла необходимость в тщательном пересмотре всех признаков с целью по возможности объективного выявления общего сходства и повторной кладисти-ческой обработки. При изучении последовательностей нуклеотидов участков ДНК применяю! такие различные методы анализа, как методы объединения соседей, максимальной парсимонии (максимальной экономии) и максимальной вероятности (см. 11.1).
Карл Линней основал применяющийся и сейчас способ наименования растений, включающий название рода и дополняющее его название вида (видовой эпитет)1. Его система, основанная в первую очередь на числе и расположении тычинок, была очень искусственной. После К. Линнея были опубликованы различные системы, принимающие во внимание все признаки, учитывавшиеся в то время, например системы М. Адансона, А. Л. де Жюссьё, А. П.Декандоля, Дж. Линдлея, С.Л. Эн-длихера, Г. Бентама и Дж. Д. Гукера, А. Бра-
1 К.Линней широко использовал этот принцип бинарной номенклатуры, ставший впоследствии общепринятым, но не был его основателем. — Примеч. ред.
416 | ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
уна, А. Энглера, С. Е. Бесси, Х.Халлира, Дж. Б. Хатчинсона и Р. фон Ветгштейна. До начала 90-х годов прошлого столетия были разработаны системы Артура Кронквиста (1981), Р. М.Т. Дальгрена (1975), А.Л.Тахтад-жяна(1980, 1997) и Р.Ф. Торна (1992). Примечательно, что опубликование дарвиновской теории эволюции очень мало повлияло на развитие самой систематики. То, что до Ч. Дарвина считали сходством, впоследствии интерпретировали как родственную близость.
Представляемая здесь система в общем плане следует опубликованной в 1998 г. и основанной на признаках ДНК (rbcL, atpB, 188гДНК) системе «группы филогении покрытосеменных» (APG) К. Бремера, М. В.Чейза, Р. А. Стивенса и др.
Эта система, доведенная только до уровня порядка, была модифицирована, главным образом в основании филогенетического древа, с учетом данных новых публикаций, в особенности последних, касающихся филогении в пределах порядков.
Главным основанием для представленной здесь системы являются результаты анализа ДНК. С постоянным увеличением числа проанализированных таксонов и сек-венированных участков ДНК (rbcL, atpB, 188гДНК, митохондриальные atp\ и matR, фитохромы А и С) они приводят к очевидному согласию о родственных взаимоотношениях разных таксонов. Добавим, что полученные результаты в большинстве слу чаев были подтверждены и классическими признаками. Тем не менее понятно, что представленная здесь система будет изменяться и далее.
Данная система в значительной степени основана на признании строгой монофилии широко известных таксонов. Вследствие этого некоторые семейства, несмотря на их фенетическую отграниченность, теперь признаны не более, чем формальными группировками в составе других семейств (например, Асегасеае будут включены в Sapindaceae, Lemnaceae — в Агасеае, Chenopodiaceae — в Amaranthaceae и т.д.). Признание их в качестве отдельных семейств привело бы к признанию парафилии остальных таксонов. Если бы, к примеру, считать и далее Lemnaceae самостоятель
ным семейством, то Агасеае было бы пара-филетическим (см. 11.1). При этом сходство как окончательный субъективный критерий классификации не принимают во внимание.
В значительной мере мы избегали употреблять формальные названия на уровнях выше порядка. Это обусловлено тем, что весьма непростая формализованная процедура присвоения названий должна применяться только тогда, когда достигнута существенная стабильность системы.
Признаваемые здесь порядки и неформальные группы более высокого уровня в неравной степени сходны по классическим признакам. Трудности, которые возникают при попытке найти классические признаки для определенных молекулярными методами родственных групп, могут в принципе иметь три основные причины. Так, возможно, что выделение какой-либо грун пы по молекулярным признакам окажется несостоятельным в ходе дальнейших исследований. Далее может оказаться, что общие признаки, возникшие в ходе быстрой фенотипической эволюции, становятся неразличимыми или же при дивергентной эволюции утрачиваются общие признаки в больших и старых родственных группах. Наконец, признаки многих ipynn не очень хорошо изучены. Здесь молекулярные гипотезы родства также имеют преимущество по сравнению с классической системати кой. Используемые в них признаки поряд ков и групп порядков далеко не всегда являются синапоморфиями.
Области распространения всех семейств, число их родов и видов даны в соответствии с различными обработками различных семейств и с очень различной точностью и достоверностью.
ЭВОЛЮЦИЯ И ФИЛОГЕНИЯ ПОКРЫТОСЕМЕННЫХ (ОБЗОР)
Покрытосеменные, включающие около 250 тыс. видов, в настоящее время — самая богатая видами группа растений, намного превосходящая все остальные группы. К важнейшим синапоморфиям покрытосеменных относятся: происходящие из од
11.2. Бактерии, грибы, растения | 4-17
ной материнской клетки членики ситовидных трубок и сопровождающие клетки флоэмы, тычинки с двумя латеральными парами гнезд пыльника, пыльники с гиподер-мальным эндотецием, пыльцевые зерна почти всегда без ламеллятной эндэкзины, трехклеточный мужской гаметофит, семяпочки, расположенные в замкнутых плодолистиках с рыльцем, стенки мегаспор без спорополленина, 8-ядерный женский гаметофит, а также двойное оплодотворение и образование вторичного эндосперма.
Вообще видовое богатство таксонов является результатом более высокой скорости видообразования и (или) более низкой скорости вымирания. Видовое богатство покрытосеменных имеет вероятно, различные причины. На первое место здесь можно поставить возникшее уже у первых покрытосеменных опыление насекомыми (шире — зоофилию). Определенная специализация опылителей приводит к репродуктивной изоляции, результатом которой, в свою очередь, может быть высокая скорость видообразования. Кроме того, возможно, что опыление насекомыми и другими животными вообще повышает степень перекрестного опыления и тем самым ограничивает генетическое обеднение, угрожающее мелким популяциям. В результате могла бы снизиться степень вымирания. Очень богатые видами орхидные (около 18 500 видов) — несомненный пример тесной связи ярко выраженной специализации по биологии опыления с видовыми разнообразиями.
Репродуктивной изоляции и повышению таким образом скорости видообразования, по-видимому, содействует расположение семяпочек в замкнутом плодолистике, благодаря чему возможно распознавание чужой пыльцы не в момент оплодотворения, а еще при опылении. Таким образом снижается вероятность оплодотворения чужеродной пыльцой и межвидового скрещивания. Неоднократно наблюдалось, что скорость роста пыльцевой трубки коррелирует с генетическим сходством производителей пыльцы и семяпочки. Поэтому относительное подавление роста пыльцевой трубки при опылении растений из генетически расходящихся популяций также может приводить к репродуктивной изоляции в пределах вида и этим способствовать повышению скорости видообразования.
Участие плодолистиков в образовании единиц расселения (плодов) в общем, пожалуй, повышает дальность расселения видов и таким образом способствует репродуктивной изоляции путем географической изоляции, а тем самым —
и повышению скорости видообразования. В то же время из-за увеличения дальности расселения, естественно, преодолевается пространственная изоляция популяций. Сочные плоды, распространяющиеся эндозоохорно. в эволюции покрытосеменных возникли, по-видимому, достаточно поздно. Эпизоохорно распространяющиеся плоды, напротив, известны уже у очень ранних ископаемых покрытосеменных.
Поскольку вторичный эндосперм покрытосеменных формируется только после успешного оплодотворения, их размножение эффективнее, чем у голосеменных. Данный факт, а также сокращение длительности гаметофитной фазы жизненного цикла способствовали, возможно, превосходству покрытосеменных в их конкуренции с разными группами голосеменных растений.
Чрезвычайное разнообразие вторичных метаболитов у покрытосеменных, очевидно, также влияет на скорость видообразования. Поскольку эти вещества большей частью обеспечивают защиту растений от травоядных животных и патогенных организмов, эволюция новых вторичных метаболитов может привести к возникновению относительно свободных от повреждающих факторов ниш, в которых возможны дивергентная эволюция и возникновение новых видов. Видовое разнообразие Asteraceae (ок. 22 750 видов), вероятно, можно объяснить богатством у них вторичных метаболитов.
Наконец, покрытосеменные, выработав разнообразные жизненные формы, смогли занять жизненное пространство, не освоенное древесными и кустарниковыми растениями. Так, травянистость в сочетании с укороченным жизненным циклом позволяет заселять относительно нестабильные местообитания. Многообразие морфологии и, по-видимому, анатомии вегетативных органов также может способствовать ускорению видообразования.
Первые ископаемые покрытосеменные появляются в отложениях раннего мела, около 140 млн лет назад. Ископаемые остатки их пыльцы и листьев обнаружены в низких широтах с семиаридным или сезонно аридным климатом. Преобладающими элементами наземной растительности они впервые становятся в среднем и верхнем мелу.
Датировка возраста покрытосеменных по «молекулярным часам» и исследования значительно более древних Gnetopsida, из которых до недавнего времени часто выводили покрытосеменные, допуская ближайшее родство этих таксонов, неоднократно приводили к предположе-
418 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Amborellales
Nymphaeales
llliciales
---Winterales, Piperales, Magnoliales, Laurales
— Ceratophyllales
— Chloranthales
Acorales
Alismatales
Liliales
Pandanales
Arecales
Poales
Zingiberales
Asparagales
Dioscoreales
Commelinales
Ranunculales
Proteales
Saxifragales
Fabales
Rosales
Cucurbitales
Malvales
Sapindales
Cornales
Ericales
Fagales Myrtales Brassicales
Geraniales Malpighiales ”
Oxalidales
---Garryales
---Gentianales
Lilianae
Commelinoidae
Caryophyllales/Polygonales
Santalales
---Lamiales
Ф co
Eurosidae I
Eurosidae II
Euasteridae I
Ф co
co
CL
----Solanales
Aquifoliales
—Asterales
Euasteridae II
----Dipsacales
Рис. 11.213. Предполагаемая филогения покрытосеменных по результатам анализа семи нуклеарных (18SrDNA, фитохромы: PHYA, PHYC), пластидных (rbcL, atpB) и митохондриальных (atp1, matR) последовательностей ДНК методом максимальной экономии (по К. Bremer, M.W.Chase и P.F. Stevens; P.S.Soltis, D.E.Soltis и M.W.Chase; Yin-Long Qui и др.; S.Mathews и M. J.Donoghue, с изменениями)
11.2. Бактерии, грибы, растения | 41g
нию. что возраст покрытосеменных не менее 300 млн лет, т.е. они возникли значительно раньше мела. Хотя это и не исключено, но столь раннее появление покрытосеменных не подтверждено ископаемыми остатками. Правда, недавно описанные ископаемые остатки цветков относятся к самым верхам юры. Есть все основания думать, что заметная диверсификация покрытосеменных, представляющих собой явно более древнюю линию эволюции, впервые произошла в начале мела.
В основание эволюционного древа покрытосеменных (рис. 11.213) ставят несколько семейств, часто включаемых в Ма-gnoliidae, к которым относится только 4 % всех ныне живущих видов. Представители этих семейств — главным образом древесные формы, но есть и травы, как правило, с простыми листьями и эфирными маслами в шаровидных идиобластах. Цветки спиральные или циклические из немногих или многочисленных элементов, пыльцевые зерна однобороздные (моносулькатные), плодолистики большей частью свободные или гинецей состоит только из одного плодолистика. Базальной линией современных покрытосеменных недавно признали, на основании различных молекулярных данных, семейство Amborellaceae, представленное единственным видом, произрастающим в Новой Каледонии. Внешний вид очень мелких (< 5 мм) цветков Amborellaceae. состоящих из немногих элементов, хорошо соответствует палеоботаническим данным, показывающим все большее число очень мелких древнейших цветков. Таким образом, весьма вероятно, что первые покрытосеменные были мелкоцветковыми. Поскольку в древесине Amborella нет сосудов, то возможно, что эти столь характерные для огромного большинства покрытосеменных элементы появились уже после возникновения покрытосеменных. Вплотную к основанию эволюционного древа покрытосеменных находится также семейство Nym-phaeaceae с распространенными в Центральной Европе кувшинкой {Nymphaea) и кубышкой {Nuphar). Представители более крупных семейств Magnoliidae — магнолия {Magnolia), лавр {Laurus) и перец {Piper). Относящийся к Magnoliidae и также произрастающий в Центральной Европе роголистник {Ceratophyllum), который несколько лет
назад рассматривали как представителя базальной линии покрытосеменных, вероятнее является ближайшим родственником однодольных или так называемых настоящих двудольных (Eudicotyledonae).
Magnoliidae — парафилетическая базальная группа, из которой возникли остальные покрытосеменные. Явным признаком того, что это остаток некогда более богатой видами группы, считается их фрагментированный ареал — преимущественно тропические и субтропические области всех континентов.
Однодольные (Liliidae, Monocotyledonae), первый большой блок остальных покрытосеменных, содержат около 22 % от общего числа их видов. Самые древние несомненные ископаемые остатки этой группы датируются возрастом около 90 млн лет. Благодаря своим характерным признакам — преобладанию травянистых жизненных форм, наличию только одной семядоли, большей частью рассеянно расположенными на поперечном срезе стебля закрытыми проводящими пучками, обычно параллельному жилкованию листьев, трехчленным цветкам и однобороздным пыльцевым зернам — однодольные образуют четко обособленные, издавна распознанные таксоны. Базальная линия однодольных представлена семейством Асогасеае с занесенным в Центральную Европу аиром {Acorus) и порядком Alismatales с такими растениями, как, например, сусак {Butomus), частуха {Alisma), рдест {Potamogetori), аронник {Arum) и ряска {Lemna). Преимущественно водный образ жизни организмов обеих линий часто давал основание предполагать, что все однодольные происходят от водных предков. Остальные однодольные более или менее ясно делятся на два больших блока. К «Lilianae» в центральноевропейской флоре, например, принадлежат лилия {Lilium), вороний глаз {Paris), орхидеи, спаржа {Asparagus), ландыш {Conva-llaria) и тамус (Tamus), а из тропических видов, например, панданусы {Pandanus). «Со-mmelinoidae», как вторая большая группа, представлена, например, семействами пальм (Агесасеае), злаков (Роасеае) и осоковых (Сурегасеае), а также бананами {Musa) и имбирем {Zingiber).
420 [ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Остальные 74% покрытосеменных относятся к Rosidae (высшие, или настоящие двудольные, от англ, eudicots), которые отличаются от Magnoliidae и однодольных пыльцевыми зернами с тремя или более апертурами и, вероятно, имеют примерно тот же возраст, что и однодольные.
К базальным линиям Rosidae принадлежат такие различающиеся таксоны, как лотос (Nelumbo), платан (Platanus), барбарис (Berbens), лютик (Ranunculus) или мак (Pa-paver). В то время как в этих линиях еще можно наблюдать вариации в количестве и расположении элементов цветка, при формировании основной группы Rosidae («core eudicots») закрепились пятимерия цветка и двойной околоцветник, состоящий из чашечки и венчика. К базальной линии этой главной группы относят порядок Caryophy-llales, который в центральноевропейской флоре представлен семейством гвоздичных (Caryophyllaceae) и к которому относятся также семейства кактусовых (Cactaceae) и аизооновых (Aizoaceae) с его «живыми камнями» — литопсами (Lithops). Большая часть Rosidae делится на два крупных блока: собственно «Rosidae» и «Asteridae».
К собственно «Rosidae», характеризующимся большей частью свободнолепестным венчиком, двумя кругами тычинок, крассинуцеллятной семяпочкой с двумя интегументами и нуклеарным типом развития эндосперма, в Центральной Европе относятся, например, герань (Geranium), зверобой (Hypericum), молочай (Euphorbia), ива (Salix), фиалка ( Viola), клевер (Trifolium), крапива (Urtica), роза (Rosa), переступень (Bryonia), бук (Fagus), береза (Betula), дербенник (Lythrum), пастушья сумка (Capsella), липа (ТШа) и клен (Acer). У «Asteridae» лепестки, как правило, сросшиеся, тычинки часто только в одном круге, семяпочка тенуинуцел-лятная с одним интегументом, целлюлярный тип развития эндосперма. К ним в Центральной Европе относятся, например, дерен (Corvus), вереск (Calluna), первоцвет (Primula), горечавка (Gentiana), подмаренник (Galium), мята (Mentha), коровяк (Verbascum), подорожник (Plantago), красавка (Atropa), вьюнок (Convolvulus), незабудка (Myosotis), падуб (Пех), тмин (Carum), ворсянка (Dipsacus), колокольчик (Campanula) и василек (Centaurea).
Хотя среднеевропейские представители «Rosidae» и «Asteridae» большей частью травянистые растения, в обеих больших группах Rosidae имеются и многочисленные древесные таксоны. Тем не менее травянистость среди «Rosidae» и «Asteridae» встречается чаще, чем среди Magnoliidae.
Краткий обзор эволюции покрытосеменных ясно показывает, что в их вегетативной сфере наблюдается отчетливая тенденция к травянистым жизненным формам. Эволюция цветка направлена к сокращению числа циклически расположенных органов, а в эволюции семяпочки происходили уменьшение и упрощение. Таким образом, у покрытосеменных продолжается та же тенденция, которую можно было наблюдать уже у разных голосеменных растений.
СИСТЕМА ПОКРЫТОСЕМЕННЫХ
1. ПОДКЛАСС: MAGNOLIIDAE
Семейства двудольных растений с примерно 10000 видами, которые в качестве подкласса Magnoliidae (или класса Magno-liopsida) во многих системах покрытосеменных помещают в основание эволюционного древа покрытосеменных, не составляют монофилетической группы. Тем не менее эти семейства имеют много сходного. В большинстве случаев это деревянистые растения с эфирными маслами (фе-нилпропаны, терпены) в шаровидных идиобластах и простыми листьями без прилистников. Цветки очень разнообразны. Так, многочисленные элементы цветка могут быть расположены по спирали, часто встречается их расположение трехчленными кругами, а иногда цветки очень простые и состоят только из немногих элементов. Пыльцевые зерна большей частью однобороздные (рис. 11.214), а плодолистики свободные (апокарпные). Растения этой группы часто содержат бензилизохинолиновые алкалоиды и/или биосинтетически близкие им неолигнаны.
Анализ самых различных молекулярных наборов данных (rbcL, atpR, 18SrDNA, митохондриальные atp\ и matR, фитохромы А и С) недавно позволил детально выяс-
11.2. Бактерии, грибы, растения | 421
Рис. 11.214. Два основных типа пыльцевых зерен покрытосеменных (фото H.Halbritter и М. Hesse): А — однобороздное зерно (Lilium martagon) с одной дистальной апертурой; В — трехбороздное зерно (здесь особая форма — трехбороздно-оровое (трикольпоратное): Ecballium elaterium) с тремя экваториально расположенными апертурами. Гидратированное состояние
нить родственные отношения в основании филогенетического древа покрытосеменных.
Amborellales
Aniborellaceae I род/1 вид. Новая Каледония
В качестве исходной линии, по этим данным, следует рассматривать семейство Amborellaceae (порядок Amborellales), представленное только одним новокаледонским видом — Atnborella trichopoda. Это двудомные вечнозеленые бессосудистые кустарники с очень мелкими цветками (< 5 мм) со спирально расположенными элементами и простым околоцветником из 5 —8 листочков. В мужских цветках содержится 10—14 тычинок, а в женских — несколько свободных плодолистиков, из которых развиваются костянки (рис. 11.215).
Nymphaeales
Nymphaeaceae, включая Cabombaceae 6/68, космополитное
Единственное семейство следующего за Amborellaceae порядка Nymphaeales, Nymphaeaceae, представлено болотными и водными растениями без масляных клеток и без бензилизохинолиновых алкалоидов. У Victoria amazonica наряду с подводными листьями есть плавающие листья, достигающие диаметра 2 м (см. рис. 4.61). Цветки обоеполые (рис. 11.216), элементы цветка расположены либо в трехчленных кругах, либо, по крайней мере, частично, спирально. Плодолистики с ламинально-диффуз-ной плацентацией, свободные или соединены чашевидным цветоложем. В Центральной Европе Nymphaeaceae представлены двумя видами кубышки (Nuphar luteum, N. pumila) и кувшинки (Nymphaea alba, N. Candida), 1 гроизрастающими в разных водоемах — от олиготрофных до эвтрофных.
Illiciales
Austrobaileayaceae 1/1. северо-восточная часть Авс тралии; Illiciaceae 1/42, Юго-Восточная Азия, Северная Америка; Schizandraceae 2/50, Юго-Восточная Азия, Северная Америка; Trimeniaceae 1/5, юго-западная часть Тихоокеанской области
422 | ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Рис. 11.215. Amborellales, Amborellaceae (А, С, Е — по A.Takhatajan; В — по Vorlage, Р. К. Endress, D — по Р. К. Endress):
А — ветвь с костянками и отдельная костянка; В — женский цветок со стаминодиями (s); С — продольный разрез плодолистика; D — мужской цветок; Е — тычинки
Рис. 11.216. Nymphaeales, Nymphaeaceae (А, В — по G. Karsten; С — по A.W. Eichler; D — по W.Troll): Nymphaea alba: А — плавающий лист; В — цветок и завязь со спирально расположенными рубцами (опавших) лепестков и тычинок (1/2х). Nuphar luteum: С — диаграмма цветка (нектарники показаны черным цветом, осевая ткань — точками); D — плод (осевая ткань отделена от свободных плодолистиков)
11.2. Бактерии, грибы, растения | 423
К порядку Illiciales, третьей ветви филогенетического древа покрытосеменных, относятся семейства Austrobaileayaceae, Trimeniaceae, Schizandraceae и Illiciaceae. Семейство Austrobaileayaceae представлено лианами с супротивными листьями. Семейство Schizandraceae — лианы с однополыми цветками, причем как в однополых, так и в обоеполых цветках видов Trimeniaceae имеется только один плодолистик. У Schizandraceae и Illiciaceae пыльцевые зерна от 3- до 6-бороздных, они возникли параллельно с 3-бороздной пыльцой настоящих двудольных.
К Illiciaceae относится бадьян (Illicium verum) и похожий на него, но ядовитый шикими (бадьян анисовый, I. anisatum).
У всех названных выше семейств очень обычны асцидиатные плодолистики с закрытой выделениями вентральной щелью.
Большое число семейств Magnoliidae принадлежит к порядкам Winterales, Piperales, Magnoliales и Laurales, очевидно образующим единую филогенетическую линию.
Winterales
Canellaceae 6/16, Мадагаскар, Африка, Южная Америка; Winteraceae 4/65, Южное полушарие
К Winterales относятся семейства Winteraceae и Canellaceae. У Winteraceae древесина бессосудистая; у Canellaceae тычинки срастаются в трубку и гинецей паракарпный.
Piperales
Aristolochiaceae 12/450-500, космополитное; Lactoridaceae 1/1, Хуан-Фернандес; Pi-peraceae 5/3000, пантропическое; Saururaceae 4/6, от Юго-Восточной до Восточной Азии, Северная Америка
Виды Piperales — часто травянистые растения с пальчатонервными листьями. Элементы цветка большей частью расположены в трехчленных мутовках. Среднеевропейские представители семейства Aristolochiaceae копытень (Asarum) и кирказон (Aristolochia) имеют почти всегда простой спайнолистный околоцветник и синкарп-ную нижнюю завязь. У видов этого семейства цветки опыляются мухами, с чем час-
Рис. 11.217. Piperales, Piperaceae, Piper nigrum (к — no G. Karsten; В — no H. Baillon):
A— побег с соплодием (1/Зх); В — костянка в продольном разрезе (5х), с мясистым мезокарпием (т), деревянистым эндокарпием (ек), семенной кожурой (s), зародышем (е), вторичным эндоспермом (se) и периспермом (р)
то связаны трубчатые околоцветники-ловушки тускло-красной или коричневой окраски и неприятный запах цветков.
Мелкие, лишенные околоцветника, обоеполые или однополые цветки Piperaceae собраны в колосья. Костянки содержат одно семя, сформировавшееся из прямой семяпочки и имеющее крупный перисперм (рис. 11.217). Плоды и семена Piper nigrum используют как черный перец. У семейства Saururaceae, очевидно близкородственного с Piperaceae, в основании колоса часто имеются хорошо заметные прицветники, отчего соцветие напоминает отдельный цветок. Единственный вид семейства Lactoridaceae — кустарник, произрастающий на острове Хуан-Фернандес, близ западного побережья Южной Америки. Его пыльца распространяется в тетрадах.
Magnoliales
Annonaceae 18/2300. пантропическое: De-generiaceae 1/2, о-ва Фиджи; Eupomatiaceae 1/2, восточная часть Австралии, Новая Гвинея; Himantandriaceae 1/2, Австралия, Азия; Magno-liaceae 7/182. от тропиков до умеренного пояса, Восточная — Юго-Восточная Азия, Америка: Myristucaceae 17/370, пантропическое
424 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Рис. 11.218. Magnoliales (А—F) и Laurales (G) (А—D, F— по A.Englers; Е — по G. Karsten; G— по Н. Bailion):
А — диаграмма цветка Michelia (черным цветом изображены прикрывающие цветок верховые листья, белым — околоцветник); В — сборный плод Magnolia virginiana со свешивающимися из листовок на проводящих пучках красными семенами (1х). С—F— Myristicaceae, Myristica fragrans: С— мужской цветок и D — женский цветок (4х); Е, F — мясистый, но растрескивающийся одноплодолисти-ковый плод (в разрезе; ок. 1/2х), красный ариллус («мацис»: пряность, лекарственное сырье) окружает темно-коричневое семя, внутри — руминированный вследствие разрастания семенной кожуры эндосперм и маленький зародыш (ок. 2/Зх); G — Lauraceae, Cinnamomum verum, цветок в продольном разрезе, гипантий, псевдомономерный гинецей, пыльники, вскрывающиеся клапанами (ок. 5х)
У Magnoliales очередные листья располагаются на трех- или многолакунных узлах, в многочисленных и часто крупных цветках (рис. 11.218) плодолистики, как правило, содержат более одной семяпочки. У семейства Magnoliaceae многочисленные элементы цветка часто расположены по спирали. Плоды их — кожистые листовки, а семена имеют мясистую оболочку. В качестве декоративных растений разводят магнолии (Magnolia) и тюльпанное дерево (Liriodendron tulipifera).
У видов семейства Аппопасеае околоцветник часто состоит из трехчленных кругов. Свободные плодолистики развиваются в ягоды. В качестве деликатеса из представителей этого семейства используют, например, плод анноны чешуйчатой (Annona squamosa). У семейства Degeneriaceae в цветке только один плодолистик, а у Eupomatiaceae отсутствует околоцветник. Только одна семяпочка развивается в плодолистике у Himantandraceae и Myristicaceae.
У Myristicaceae (рис. 11.218), кроме того, цветки мелкие однополые, а в женском цветке только один плодолистик.
Многочисленные тычинки мужских цветков срастаются в колонку.
К последнему семейству принадлежит мускатный орех (Myristica fragrans), у которого тег-мен (возникающая из внутреннего интегумента часть семенной кожуры) глубоко вдается в эндосперм (руминация).
У Magnoliales в процессе опыления очень часто участвуют элементы околоцветника или плоские тычинки и стаминодии. Так, у Аппопасеае тесно сближенные внутренние листочки околоцветника образуют опылительную камеру, у Degeneriaceae лежащие внутри от плоских тычинок и такие же плоские стаминодии подвижны и прикрывают гинецей по окончании женской фазы цветения, а у Eupomatiaceae сросшиеся основаниями тычинки и лежащие внутри андроцея стаминодии образуют полость (синан-дрий), куда жуки-опылители откладывают яйца. Развитие личинок жуков заканчивается после опадения синандрия.
11.2. Бактерии, грибы, растения I 425
Laurales
Atherospermataceae 7/12, тропическое, Азия, Австралия, Южная Америка; Calycanthaceae, включая Idiospermaceae 4/8, умеренные зоны Китая и Северной Америки, тропическая Азия; Gomortegaceae 1/1, юг Чили; Hemandiaceae 5/60, пантропическое; Lauraceae 50/2500—3500, пантропическое; Monimiaceae 18—25/150, пантропическое; Siparunaceae 2/72, Южная Америка, Африка
Laurales имеют обычно супротивные (или мутовчатые) листья и однолакунные узлы. В их цветках имеется преимущественно ярко выраженный гипантий, а плодолистики содержат только одну семяпочку. За исключением Calycanthaceae все семейства этого порядка имеют мелкие цветки; в основании тычинок, как правило, имеется пара железок, а пыльники часто вскрываются клапанами.
Виды семейств Atherospermataceae,Siparunaceae и Monimiaceae (ранее все включавшиеся в Monimiaceae) часто имеют однополые цветки с
многочисленными тычинками/пестиками и расширенным или кувшинчатым гипантием. Так, цветки у Siparuna, имеющие кувшинчатый гипантий, опыляются очень мелкими галловыми осами, которые откладывают свои яйца в женские цветки. Стилодии свободных плодолистиков в зеве кувшинчатого гипантия так тесно прижаты друг к другу, что пыльцевая трубка с одного стилодия может перейти на соседний. Эта функциональная синкарпия может быть обеспечена также путем обильного образования слизи в области рылец.
У Gomortegaceae и Hemandiaceae завязь нижняя.
Распространенные по всей тропической зоне, но проникающие и в Средиземноморье (лавровое дерево; Laurus nobilis) Lauraceae имеют гримерные циклические цветки (рис. 11.218). Единственный плодолистик развивается в ягоду или костянку.
Важные культурные растения семейства Lauraceae — камфарное дерево (Cinnamomum сат-phora), из древесины которого путем возгонки добывают камфару, и коричное дерево (С. verum и родственные виды), из коры которых получают корицу. Богаты жирами съедобные мясистые
Рис. 11.219. Ceratophyllales, Ceratophyllaceae (R.Spohn, по А.Тахтаджяну, 1980):
А — общий вид; В — вильчатый лист; С — мужской цветок; I — тычинка; Е — женский цветок
426 | ГЛАВА 11, СИСТЕМАТИКА И ФИЛОГЕНИЯ
плоды авокадо (Persea americand). В семействе есть также ведущий полупаразитический образ жизни род Cassytha с чешуевидными листьями на нитевидных побегах.
Ceratophyllales
Ceratophyllaceae 1/6. космополитное
Порядок Ceratophyllales содержит только одно семейство Ceratophyllaceae. Это распространенные по всему миру подводные травы, растущие в стоячих водоемах. Произрастающие и в Центральной Европе роголистники (Ceratophyllum) лишены корней, но прикрепляются к почве корневищами. Их часто можно найти свободно плавающими. Однополые цветки с многочисленными тычинками и единственным плодолистиком с одной семяпочкой находятся между мутовчато расположенными линейными или вильчатыми листьями (рис. 11.219).
Родственные отношения Ceratophyllaceae не ясны. По разным молекулярным данным, он является то ли сестринской группой однодольных, то ли относится к Rosidae. До недавнего времени это семейство рассматривали как самую базальную группу покрытосеменных.
Chlorantliales
Chloranthaceae 4/75, пантропическое
Столь же неясны и связи Chloranthaceae — единственного семейства порядка Chloranthales. Представители этого семейства обладают очень мелкими и большей частью голыми одно- или обоеполыми цветками с 1 — 5 тычинками и только одним плодолистиком. Chloranthaceae могли бы быть сестринской группой как однодольных, так и настоящих двудольных плюс линия Win-terales/Piperales/Magnoliales/Laurales.
Nymphaeales, Illiciales, Piperales и Chloranthales в недалеком прошлом иногда объединяли в «палеотравы». Представленные здесь их родственные отношения ясно показывают, что «палеотравы» не являются естественным таксоном.
К Magnoliidae принадлежат также представленные полностью паразитическими формами семейства Hydnoraceae (2/18, засушливые области Старого и Нового света) и Rafflesiaceae (9/ 55, пантропическое). Виды Rafflesiaceae — эндопаразиты, чья вегетативная ткань, подобно мицелию грибов, пронизывает хозяина. Прорываю
щие тело хозяина и выходящие на поверхность цветки у Rafflesia amoldii могут достигать 1 м в диаметре. Это самые крупные цветки среди покрытосеменных. Монофилия раффлезиевых весьма проблематична.
Описание семейств, стоящих в основании филогенетического древа цветковых растений, ясно показывает, что, несмотря на многочисленные сходные признаки, наблюдается также большое разнообразие в их морфологии и особенно в морфологии цветка. Так, с одной стороны, находим, например, у Magnoliaceae, крупные многочленные цветки, а с другой стороны, у Chloranthaceae цветки очень мелкие и чрезвычайно простые. Это многообразие морфологии цветка связано с чрезвычайным разнообразием биологии размножения. Сейчас склоняются к тому, чтобы исходным типом цветка считать мелкие малочленные цветки. Важный аргумент для этого — обнаружение все большего числа мелких ископаемых цветков из очень древних отложений. Правда, вместе с Archaefructus недавно было описано ископаемое покрытосеменное с очень большими цветками из отложений самого раннего мела (или даже самой поздней юры).
На большой возраст вышеназванных семейств часто указывает также их реликтовое и дизъюнктивное распространение. Тем не менее есть среди них также широко распространенные и богатые видами таксоны. Например, весьма вероятно, что еше в третичном периоде Lauraceae расселялись по разным континентам.
В целом правильнее всего вышеописанную группу семейств и порядков понимать как парафилетическую базисную группу цветковых растений, из которой возникли как однодольные, так и настоящие двудольные.
2. ПОДКЛАСС: LILIIDAE (MONOKOTYLEDON)
Однодольные — очень давно известный и четко очерченный таксон. Это обычно травянистые симподиально нарастающие растения. Первичный корень большей частью не развивается, и корневая система состоит из однородных придаточных корней (вторичная гоморизия; рис. 11.220). Раз
11.2. Бактерии, грибы, растения | 427
бросанные на поперечном срезе стебля побега или корня1 проводящие пучки (атактостела) не имеют камбия. Соответственно этому однодольные не способны к типичному вторичному утолщению. Листья их обычно параллельнонервные, очередные, без прилистников. На боковых побегах находится, как правило, только один обращенный к материнской оси предлист. Сосуды часто есть только в корнях. Пластиды ситовидных трубок почти всегда содержат клиновидные протеиновые кристаллы, иногда дополнительно в них содержатся зерна крахмала и/или фибриллярный белок. Цветки обычно трехмерные и пятикруговые. Эндотеций пыльников развивается непосредственно из слоя субэпидермальных клеток. Пыльцевые зерна однобороздные (см. рис. 11.214). Эндосперм обычно развивается по гелобиальному или нуклеарному типу, зародыш имеет только одну семядолю (рис. 11.221), которая занимает псевдотерминальное положение, смещая вбок почечку (плюмулу).
Ближайшие родственники однодольных среди Magnoliidae точно не установлены. По разным молекулярным данным, это либо Cerato-phyllaceae, либо Chloranthaceae или часть большого блока порядков Winterales/ Piperales/ Magnoliales/ Laurales, включая Chloranthaceae.
Кроме нескольких семейств неясного таксономического положения (Corsiaceae, Japonoliriaceae, Nartheciaceae, Petrosaviaceae, Triuridaceae) выделяют десять порядков. Это Acorales, Alismatales, Dioscoreales, Liliales, Pandanales, Arecales, Poales, Cornmelinales и Zingiberales; из них четыре последние вместе с отдельными далее указанными семействами, отнесение которых к определенному порядку проблематично, можно объединить в Cornmelinoidae.
Acorales
Acoraceae 1/2, Восточная Азия, широко распространившееся синантропное растение
Порядку Acorales как, вероятнее всего, самой базальной группе однодольных свой-
1 В корнях однодольных находится один радиальный пучок с полиархной ксилемой, а не атактостела. — Примеч. ред.
Рис. 11.220. Форма роста Liliidae (схема). Укрепление побега вследствие первичного утолщения (по W. Troll):
h — гипокотиль; к — семядоля; w — первичный корень; w' — узловые придаточные корни
ственны, как и многим семействам, находящимся в основании филогенетического древа покрытосеменных, маслосодержащие идиобласты. В отличие от всех остальных однодольных эндотеций не образуется непосредственно из субэпидермального слоя стенки пыльника. Клетки субэпидермального слоя делятся периклинально, и эндотеций формируется из внешнего из двух возникших таким способом слоев, что встречается и у многих не-однодольных растений. В противоположность большому числу других однодольных эндосперм развивается по целлюлярному типу. Единственный род Acorus (рис. 11.222) имеет унифациальные мечевидные листья. Невзрачные обоеполые цветки все без исключения трехмерные и собраны в початок. Зеленое покрывало кроющего листа продолжает уплощенный побег, поэтому кажет-
428 | ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Рис. 11.221. Проростки Liliidae (А—Е— по J.Sachs и R.von Wettstein, с изменениями; F— по Н. Bailion):
А — Paris quadrifolia', В — Allium сера', С — Clivia miniata-, D — Zea mays (В — D — продольный разрез; кожура семени (или соответственно плода) черная, эндосперм выделен течками, семядоля (к), влагалище семядоли (s), главный корень (w). Семядоля В—D частично или полностью трансформирована во всасывающий орган). Признаки «однодольное™» у двудольных растений: Е — проросток Ranunculus ficaria с одной семядолей; F — цветок Cabomba aquatica (Nymphaeaceae), Р3+3 А3 G3 (Зх)
<
Рис. 11.222. Acorales, Acoraceae, Acorus calamus (А — по G. Karsten; В, С — по J. Graf):
А — цветущее растение с соцветием (1/4х); В — отдельный цветок; С— завязь (поперечный срез)
11.2. Бактерии, грибы, растения | 429
ся, что початок занимает боковое положение.
В первую очередь из-за структуры соцветия Acorus до недавнего времени включали в Агасеае. В настоящее время ясно, что этот род по многим признакам отличается от Агасеае и, возможно, его следовало бы рассматривать как самое базальное из современных однодольных семейство. Аир (Acorus calamus) благодаря содержащимся в нем эфирных маслам — старинное лекарственное растение, занесенное в Европу из Восточной Азии, вероятно, в XVI в. В Центральной Европе Acorus calamus — стерильный трип-лоид, размножающийся только вегетативно.
Alismaiales
Alismataceae 12/80, субкосмополитное; Aponogetonaceae 1/45, Южное полушарие Старого Света; Агасеае, включая Lemnaceae 108/3300, космополитное, в основном тропики; Butomaceae 1/1, умеренные области Евразии; Cymodoceaceae 5/16, западная частьТи-хоокеанской области, Карибский бассейн, Средиземноморье, Австралия; Hydrocharitaceae, включая Najadaceae 18/120, космополитное; Juncaginaceae 4/12, субкосмополитное, умеренная зона; Limnocharitaceae 3/8, пантропическое; Posidoniaceae 1/9, Австралия, Средиземноморье; Potamogetonaceae, включая Zannichelliaceae 6/100, космополитное; Ruppiaceae 1/1 — 10, субкосмополитное, в основном умеренная часть Северного полушария; Scheuchzeriaceae 1/1, арктические и умеренные области Северного полушария; Tofieidiaceae 5/30, от Юго-Восточной Азии до Южной Америки; Zostera-сеае 3/18, умеренные области Северного и Южного полушарий
Порядок Alismatales включает в основном травянистые растения влажных местообитаний или водоемов. Тапетум пыльников периплазмодиальный с одноядерными клетками, а зародыши часто зеленые и содержат запасные вещества. У всех представителей кроме большинства наземных или эпифитных Агасеае в листовых влагалищах имеются мелкие чешуйки (внутри-влагалищные чешуйки, сквамулы), гинецей часто апокарпный и формирование эндосперма протекает по гелобиальному типу (рис. 11.223).
Семейство Butomaceae с единственным видом Butomus umbellatus (сусак зонтичный) характеризуется венчиковидным око
лоцветником и плодом листовкой. Плацентация ламинально-диффузная. Околоцветник Alismataceae дифференцирован на чашечку и венчик, число тычинок 6, но может быть редуцировано до 3 или, наоборот, увеличено, плод — мало- или односемянный орешек1. В то время, как цветки пастухи (Allsma) обоеполые, у стрелолиста (Sa-gittaria) они однополые. У растений этого, в основном водного, семейства подводные, плавающие и воздушные листья часто резко различаются морфологически. Виды Hydrocharitaceae — полностью погруженные (например, наяда Najas). растущие по поверхности воды (например, водокрас, Hydrocharis) или свободно плавающие в воде (например, телорез, Stratiotes aloides) растения, их цветки обычно однополые.
Занесенная в 1836 г. из Северной Америки в Европу элодея, или водяная чума (Elodea canadensis), двудомна. Вначале только женские растения. размножаясь вегетативно, стремительно распространились по Европе, вызвав в Великобритании полную блокаду системы каналов и тем самым ускорив развитие сети железных дорог. У субтропическо-тропической валлиснерии (Va-llisneria spiralis) (рис 11.223) женские цветки выносятся на поверхность воды на спиралевидном цветоносе. Мужские цветки под водой отрываются от растения и всплываю! . Достигнув поверхности воды, они раскрываются и пассивно переносятся ветром и течением к женским цветкам, где и происходит опыление.
У обитателей сырых, пресно- или солоноватоводных местообитаний из семейства Juncaginaceae цветки обоеполые, например у триостренника (Triglochin). К Potamogetonaceae относят рдесты (Potamogetom, рис. 11.223). Это укореняющиеся на дне водные растения с плавающими и погруженными листьями или только с погруженными листьями, четырехмерными, собранными в колосья цветками, большей частью ветроопыляемыми. К этому семейству относят и подводную занникеллию (Zannichellia) с однополыми цветками (рис. 11.223). Морская трава (Zostera) также имеет однополые цветки с единственной тычинкой и соответственно с одним плодолистиком. Пыльцевые зерна Zostera не имеют экзины и могут достигать 0,5 мм в длину (рис. 11.223).
К морскому образу жизни приспособлены виды семейства Cymodoceaceae, Ruppiaceae (со
1 Орешек, по определению, односемянный; малосемянные плоды следует называть орешковидными. — Примеч. ped.
430 | ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
лоноватые воды), Posidoniaceae и частично также Hydrocharitaceae. Разные Alismatales, по-видимому, по меньшей мере трижды переходили в морскую воду из пресноводных или влажных местообитаний, подвергшихся засолению.
Большое семейство Агасеае содержит в основном травянистые, но иногда также и деревянистые, главным образом наземные растения. Обычно мелкие, одно- или обоеполые цветки собраны в початковидное соцветие (спадикс), в основании которого развит большой и ярко окрашенный лист — покрывало, которое может окружать початок (рис. 11.224). Растения с однополыми цветками, как правило, однодомные, муж
ские цветки находятся в верхней части соцветия. Двудомность встречается редко (например. у Arisaema). Околоцветник есть или отсутствует, тычинок от 1 до 12, плодолистиков тоже может быть 1 или много, гинецей ценокарпный, завязь много- или одногнездная. Плод большей частью ягода.
Представители этого семейства, распространенного преимущественно в тропиках, играют большую роль в дождевых лесах (крупнорозеточные растения, эпифиты или цепляющиеся воздушными корнями лианы). Их листья обычно широкие, сердцевидные или стреловидные и часто сетчатонервные. У произрастающего в Европе аронника (Arum maculatum) початок и по-
Рис. 11.223. Alismatales (А — по A. W Eichler, с изменениями; В — по A. Englers «Syllabus»; С — по A. Kerner; D — по G. Karsten; Е — по J. Graf; F, G — по G. Hegi):
A — Alismataceae, диаграмма цветка Alisma plantago-aquatica, предлист (v) двухкилевой, обращен к материнской оси, тычинки расположены по две, плодолистики свободные, односемянные; В — Butomaceae, гелобиальный тип развития эндосперма у Butomus umbellatus (е — двухклеточный зародыш, mk — микропилярная часть эндосперма: свободные ядра, w — поперечная перегородка, ск — халазальная часть эндосперма, az — клетки-антиподы; ок. 600х); С — Hydrocharitaceae, женский цветок и оторвавшийся, свободно дрейфующий мужской цветок Vallisneria spiralis (5х). Potamogetonaceae: D— цветущий побег Potamogeton natans (1/4х); Е— женский цветок Zannichellia palustris с каймой околоцветника (р) и 4 созревшими плодолистиками (k) (6х). Zosteraceae, Zostera marine F — поперечный срез плоского початковидного соцветия (Ь) и покрывала (h) (голые <j> и d" цветки с одним плодолистиком (к) и соответственно одним пыльником (а); 20х); G — нитевидное пыльцевое зерно (натур. величина ок. 0,5 мм)
11.2. Бактерии, грибы, растения | 43-|
Рис. 11.224. Alismatales, Araceae (А — по W.Troll; В, Е — по J. Graf; С — по A. Englers «Syllabus»; D — по Ch. F. Hegelmaier):
A— Monstera deliciosa, лист (с вторично возникшими отверстиями и вырезами края) (ок. 1/1 Ох); В — Aglaonema marantifolium, соцветие с покрывалом (s), голыми <j> и о" цветками (ок. 8х); С — Pistia stratiotes, целое плавающее растение с 2 соцветиями (bs) и вегетативно образованным дочерним растением (1/Зх); D — Lemna gibba, плавающее растение, молодой членик побега (sg), мужской цветок (Ь) и плод (f); Е — Lemna trisulca, соцветие в продольном разрезе с покрывалом (s), одним женским и двумя мужскими цветками (сильно увеличено)
крывало в совокупности образуют временную ловушку для насекомых-опылителей. Опыление преимущественно жуками, падальными мухами и мелкими двукрылыми.
У родов Lemna, Spirodela, Wolffia и Wolffi-ella, ранее выделявшихся в самостоятельное семейство Lemnaceae, а сейчас включаемых в Araceae, вегетативная сфера, а также строение цветка чрезвычайно упрощены. Растения состоят из свободно плавающих по поверхности или в толще воды листовидных члеников с коротким корнем/ корнями (многокоренник: Spirodela, ряска:
Lemna) или без корней (карликовые ряски: Wolffia, Wolffiella). Размножение большей частью вегетативное. Соцветия без покрывала или с очень незаметным покрывалом и 1 — 3 однополыми цветками с одной тычинкой или одним плодолистиком. Wolffia arrhiza, размеры которой около 1,5 мм,— самое маленькое из известных цветковых растений. Также свободно плавающая и имеющая редуцированное соцветие Pistia stratiotes (рис. 11.224), по-види-мому, возникла параллельно с бывшими Lemnaceae.
432 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
«Lilianae»
Liliales
Alstroemeriaceae 5/160, Центральная и Южная Америка; Campynemataceae 2/2, западная часть Тихоокеанской области; Colchica-сеае 19/225, умеренно-тропические области Старого Света; Северная Америка; Liliaceae 16/650, Северное полушарие; Luzuriagaceae 2/6, Южная Америка, Австралия, Новая Зеландия; Melanthiaceae, включая Trilliaceae 16/170, Северное полушарие, редко Южная Америка; Philesiaceae 2/2, Чили; Ripogonaceae 1/6, Австралия, Новая Каледония, Новая Зеландия; Smilacaceae 2/310, умеренно-тропические области всего мира
Растения из семейств порядка Liliales обычно выделяют нектар у основания листочков околоцветника или тычинок. Запасающие органы у Liliaceae, например тюльпана (Tulipa), гусиного лука (Gaged), рябчика (Fritillaria) и лилии (Lilium), — всегда луковицы, у Colchicaceae (рис. 11.225) — клубни, у Melanthiaceae — корневища.
Безвременник (Colchicum autumnale) как представитель Colchicaceae содержит очень ядовитый колхицин. Это алкалоид трополоновой группы, который подавляет образование митотического веретена при делении клетки и может вызвать полиплоидию тканей. Из клубня безвременника осенью вырастает цветочная стрелка, от которой над землей виден только цветок с длинной трубкой околоцветника. Ранней весной формируются листья и плоды — коробочки и одновременно отрастает очередное междоузлие клубня.
Ядовитость чемерицы (Veratrum album), представителя Melanthiaceae, обусловлена стероидными алкалоидами. Также ядовитый вороний глаз (Paris quadrifolia) с 4-мерными цветками, мутовчатыми листьями с сетчатым жилкованием и
Рис. 11.225: Liliales (А— по H.Baillon; В— по F Firbas):
А— Liliaceae, раздельнолистный околоцветник Tulipa sylvestris (1х); В — Colchicaceae, Colchicum autumnale, цветущий и плодоносящий (2/5х)
плодом-ягодой ранее относили к семейству Trilliaceae, включенному теперь в Melanthiaceae. У видов семейства Alstroemeriaceae завязь нижняя, a Smilacaceae представлено лианами, черешки листьев которых часто снабжены парными усиками.
11.2. Бактерии, грибы, растения | 433
Asparagales
Низшие Asparagales: Asphodelaceae 15/780, Старый Свет; Asteliaceae 4/35, Южное полушарие; Blandfordiaceae 1 /4, Австралия; Вогуа-сеае 2/12, Австралия; Doryanthaceae 1/2, Австралия; Hemerocallidaceae 21/100, космополитное; Hypoxidaceae 9/100, тропическое, главным образом Южное полушарие; Iridaceae 70/ 1750, космополитное; Ixioliriaceae 1/3, от Египта до Центральной Азии; Lanariaceae 1/1, Южная Африка; Orchidaceae 788/18500, космополитное; Tecophilaeaceae 8/23, Африка, Мадагаскар, Чили, Калифорния; Xanthorrho-еасеае 1/30, Австралия; Xeronemataceae 1/2, Новая Зеландия, Новая Каледония.
Высшие Asparagales: Agapanthaceae 1/9, Южная Африка; Agavaceae 9/330, Новый Свет; Alliaceae 13/600, Северное полушарие и Южная Америка; Amaryllidaceae 59/850, космополитное; Anemarrhenaceae 1/1, Юго-Восточная Азия; Anthericaceae 9/200, космополитное; Aphyllanthaceae 1/1, Франция, Марокко; As-paragaceae 1/160—300, Старый Свет; Behnia-сеае 1/1, Южная Африка; Convallariaceae, включая Dracenaceae, Nolinaceae, Ruscaceae 26/390, космополитное; Herreriaceae 2/9, Южная Америка, Мадагаскар; Hesperocallidaceae 1/1, Северная Америка; Hyacinthaceae 67/900, большей частью Старый Свет; Laxmanniaceae 1/13, Австралия; Themidaceae 12/60, Северная Америка
В отличие от Liliales у Asparagales нектар выделяют большей частью септальные нектарники, а цветоножка часто с сочленением. Часть ее выше сочленения называют перикладием. Наконец, семена часто окрашены фитомеланином в черный цвет. Границы между семействами порядка Asparagales часто нечеткие.
Так называемым «низшим» Asparagales присуши обычно нижняя завязь, а также симультанный микроспорогенез. В центральноевропейской флоре они представлены только семействами Iridaceae и Orchidaceae. Iridaceae характеризуются наличием только одного крута тычинок. У рода Crocus, перезимовывающего в виде клубня, цветки радиально симметричные и все листочки околоцветника одинаковые, лепестковидные. Касатики (Iris) обычно имеют ползучее корневище и унифациальные мечевидные листья, влагалища которых седловидно охватывают стебель. Листочки наружного и внутреннего кругов околоцвет
ника радиально-симметричного цветка (рис. 11.226) различаются. В каждом цветке листочек околоцветника, тычинка и расширенная лопасть рыльца образуют вместе двугубую часть цветка (мерантий), в котором пыльник и рыльце эффективно разделены в пространстве. Шпажник (Gladiolus) имеет зигоморфные цветки. Крупное семейство Orchidaceae характеризуется зигоморфными цветками, редукцией андроцея до 3 — 1 тычинок, часто сросшихся со столбиком и рыльцем, а также пыльцевыми зернами, обычно собранными в тетрады или поллинии.
В умеренных широтах Orchidaceae — наземные травы, в тропиках и субтропиках это часто эпифиты, изредка лианы. Для них обычен тесный симбиоз с грибами по типу эндотрофной микоризы, но они могут и паразитировать на грибах и тогда утрачивают хлорофилл (например, гнездовка — Neottia nidus-avis, ладьян — Со-raliorhiza). Стебли эпифитных орхидей часто вздутые (псевдобульбы) и несут воздушные корни с веламеном. Зеленые воздушные корни могут фотосинтезировать. Цветки (рис. 11.227) зигоморфные и в ходе развития чаще всего поворачиваются на 180° (ресупинация). Благодаря этому губа, представленная средним листочком внутреннего круга околоцветника, переходит в нижнее положение. Она часто продолжена в шпорец. Очень редко бывает 3 фертильных тычинки (медианная наружного круга и боковые внутреннего круга андроцея), как правило, фертильных тычинок 2 (боковые внутреннего круга андроцея, медианная наружного круга преобразована в стаминодий), или 1 (медианная наружного круга; боковые внутреннего круга преобразованы в стаминодии); тычинки могут срастаться с пестиком и рыльцем в гиностемий. Пыльцевые зерна одиночные или в тетрадах, а чаще в поллини-ях или поллинариях. В поллиниях все пыльцевые зерна одного гнезда пыльника соединены спорополленином в общую массу. Поллинии имеют ножку которая возникает или из самого поллиния или выделена стерильной лопастью рыльца (клювик, ростеллум); на конце этой ножки есть клейкое тельце (висцидий), которое образовано ростеллумом и служит для прикрепления поллиния к опылителю. Если поллинии обоих гнезд теки сливаются друг с другом (вследствие разрушения перегородки между гнездами) и имеют общие ножку и висцидий или все четыре поллиния пыльника снабжены общим вис-цидием, то говорят о поллинарии. Нижние трехгнездные завязи большей частью паракарпные, но иногда бывают синкарпными. Из множества
434 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Рис. 11.226. Низшие Asparagales, Iridaceae (А — по Н. Baillon; В, С и Е—G — по W.Troll; D — по A.W. Eichler, немного изменено):
Crocus sativus: А — цветущее растение, продольный разрез (ок. 1х); В, С — продольный разрез и диаграмма верхней части цветка с околоцветником, тычинками и лопастями рыльца; Iris: D — диаграмма; /. pseudacorus: Е — внешний вид цветка (ок. 1 х); F, G — продольный разрез и диаграмма верхней части цветка; благодаря функциональной связи каждого из внешних листочков околоцветника тычинки и лепестковидной лопасти рыльца возникают три двугубных мериантия
семяпочек формируются крошечные семена без эндосперма с малоклеточным и недифференцированным зародышем. Семена из коробочек легко разносятся ветром. Развитие семени возможно только в том случае, если оно будет инфицировано спорой микоризообразующего гриба. Благодаря этому симбиозу, начинающемуся уже с момента прорастания, стали возможными отказ от резервных веществ и связанный с этим мелкий размер семени. Редкость наземных орхидей умеренной зоны объясняется как необхо
димостью тесной связи с микоризообразующими грибами, так и замедленным ростом.
Подсемейства орхидных различаются прежде всего по структуре андроцея и пыльцы. У Аро-stasioidaeae с их двумя родами, распространенными от Юго-Восточной Азии до Австралии, имеются три (Neuwiedid) или две (Apostasia) тычинки, которые едва срастаются со столбиком. Пыльцевые зерна одиночные, свободные, гинецей синкарпный. У Cypripedioideae, к которому относится прежде всего пять родов, распростра
11.2. Бактерии, грибы, растения | 435
ненных в Северном полушарии от бореального пояса до тропиков (например, венерин башмачок, Cypripedium calceolus), всегда имеется мешковидная нижняя губа и две тычинки, которые, как и у всех остальных подсемейств, срослись со столбиком в гиностемий. В этом подсемействе пыльцевые зерна также одиночные, но завязь па-ракарпная. У видов остальных подсемейств только одна фертильная тычинка. У Vanilloideae пыльцевые зерна собраны в тетрады, у Orchidoideae и Epidendroideae — в поллинии и поллинарии. Два последних подсемейства различаются тем, что у Epidendroideae тычинка изогнута над верхушкой гиностемия.
Разнообразие орхидей тесно связано со специализированной биологией опыления, которая часто сопряжена со специфическим запахом цветка. У Orchis (ятрышник) и других центральноевропейских родов губа снабжена шпорцем (с нектаром или без него), отверстие которого лежит непосредственно перед гимностемием. Насекомое садится на губу и, пытаясь дотянуться своим ротовым аппаратом до нектара в шпорце, касается головой или хоботком клейкого вис-цидия поллинария, выдергивает его из пыльника и уносит с собой. Пока оно летит на следую
щий цветок, быстро подсыхающая ножка пол-линия загибается вперед или назад и при посещении опылителем следующего цветка масса пыльцы попадает на клейкую поверхность фертильной лопасти рыльца (этот процесс можно легко воспроизвести острым карандашом). У видов рода Ophrys (офрис), не имеющих ни шпорца, ни нектара, опыление связано с приманиванием самцов перепончатокрылых путем имитации цветком формы и расцветки самки (см. бокс 4.1, рис. С). Тропические виды рода Catasetum сами отстреливают поллинарии навстречу опыляющим их пчелам, как только те, касаясь особых отростков, снимают напряжение в ткани тычинки. Цветки Cypripedium и Sranhopea действуют по принципу временных ловушек. Опыляющих виды Stanhope пчел из сем. Эвглоссовых привлекают ароматические масла, содержащиеся в цветках.
Из незрелых коробочек неотропической лазящей с помощью корней орхидеи Vanilla planifo-На (рис. 11.227) добывают ваниль. Многие тропические орхидеи культивируют из-за их своеобразных цветков великолепной расцветки и часто ароматных (например, Cattleya, Laelia, Vanda, Dendrobium, Stanhopea и др.). Многие декоративные формы возникли в результате гибридиза
Рис. 11.227. Низшие Asparagales, Orchidaceae (А — по A.W. Eichler, немного изменено; В —F — по О. С. Berg и С. F. Schmidt):
А — диаграмма цветка Orchidoideae (Orchis, до ресупинации), губа (I), во внешнем круге только одна фертильная тычинка, во внутреннем — два стаминодия (st); В — Orchis militaris, цветок, перевернутый вследствие ресупинации завязи (f): прицветник (t), наружный (ра) и внутренний (pi) листочки околоцветника, губа (I) со шпорцем (s) и гиностемий (д) (ок. 2,5х); С — гиностемий с лопастью рыльца (п), ростеллумом (г) с отростком (fs), фертильная тычинка со связником (к), двумя теками (th), содержащими внутри по поллинию (р), у которого различают ножку (с) и клейкие тельца (kk), и стаминодиями (st) (ок. 10х); D — поллинарий с вычлененным поллинием (р), ножкой (с) и клейким тельцем (kk) (ок. 15х); Е — коробочка в поперечном разрезе (ок. 8х); F — Vanilla planifolia, побег с цветками и цепляющимися корнями (уменьшено)
436 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
ции, котора: у орхидных часто случается и в природе.
Близкородственными Orchidaceae семействами считают распространенные почти целиком в Южном полушарии Asteliaceae, Blandfordiaceae, Boryaceae, Hypoxidaceae и Lanariaceae. В семействе Hypoxidaceae особенно четко выражены признаки, сближающие их с орхидными: наличие широких вдоль сложенных листьев, париетальной плацентации и только 3 фертильных тычинки. Среди низших Asparagales в семействе Asphodelaceae (например,Aloe,Kniphofla) и Хап-thorrhoeaceae имеются древовидные роды с аномальным вторичным утолщением.
Высшие Asparagales часто имеют верхнюю 'завязь и сукцессивный тип микроспорогенеза. В центральноевропейской флоре они представлены Alliaceae, Amaryllidaceae, Anthericaceae, Asparagaceae, Convallariaceae и Hyacinthaceae. Виды Alliaceae переживают неблагоприятный сезон в виде луковицы, соцветие у них — ложный зонтик.
Их узнают, кроме того, по характерному запаху содержащего серу лукового масла, которое, однако, встречается также и у Themidaceae. К этому семейству принадлежит большой род Allium (лук), к которому относятся лук посевной (А. сера), чеснок (A. sativum), порей (A. potrum) и шнитт-лук (A. schoenoprasum). Сем. Amaryllidaceae (рис. 11.228), к которому относятся, например, подснежник (Galanthus) и нарцисс (Narcissus), имеет нижнюю завязь и содержит характерные фенантиридиновые алкалоиды. У видов семейства Anthericaceae (например, венечника — Anthericum) есть корневище, а у Asparagaceae (к которому принадлежит двудомная спаржа с плодами-ягодами, Asparagus officinalis) побеги двух типов: удлиненные и укороченные, последние представлены игловидными филлокладиями. Ягоды образуют также частично ядовитые Convallariaceae (ландыш — Convalla-
Рис. 11.228. Высшие Asparagales (А, В — по J. Graf; С — по A. F.W. Schimper; D — по A.W. Eichler, немного изменено; Е — по A. Fahn из D. Frohne; F — по W. Troll):
Amaryllidaceae: А — продольный разрез цветка Narcissus poeticus (1 х); В — Galanthus nivalis (2х): нижняя завязь, столбик, соответственно свободный и сросшийся в трубку околоцветники (р), привенчик (n). Hyacinthaceae: С — Ornithogalum umbellatum, целое растение (уменьшено); D — диаграмма цветка; Е— Muscari racemosum, поперечный разрез завязи с септальными нектарниками (sp) (15х); F — Convallariaceae, сростнолистный околоцветник Polygonatum latifolium (* — места прикрепления тычинок; 2,5х)
11.2. Бактерии, грибы, растения | 437
ria; майник — Maianthemum; купена — Poly-gonatum; рис. 11.228).
В семейство Convallariaceae в настоящее время включают также палеотропические семейства Dracaenaceae и Ruscaceae. Представители бывшего семейства Ruscaceae, виды рода Ruscus, особенно обычны в Средиземноморской области. Они образуют листовидные филлокладии с сидящими на них соцветиями. В бывшее семейство Dracaenaceae входят древесные роды с аномальным вторичным утолщением стебля. У Канарского и североафриканского драконова дерева — Dracaena draco (рис. 11.229) — ствол может достигать в обхвате значительной величины.
Семейство Hyacinthaceae (рис. 11.228) с известными родами гадючий лук (Mus-cari) и птицемлечник (Omithogalum) имеет луковицу, прикорневые листья и соцветие кисть.
Аномальное вторичное утолщение стебля свойственно также распространенному в Америке семейству Agavaceae, содержащему моно-карпики Agave и Yucca. Хорошо изучены тесные взаимоотношения Yucca с опылителями. Юкко-вые моли (Tegetigula и Parategetigula) откладывают свои яйца в завязи. Так как личинки питаются развивающимися семяпочками, моли должны опылить цветок, чтобы гарантировать их развитие. Часто виды Yucca опыляет только определенный вид молей.
Dioscoreales
Burmanniaceae 9/90, космополитное тропического и умеренно теплого поясов; Dio-scoreaceae 3-20/600, пантропическое; Тассасеае 1/10, Юго-Восточная Азия, Полинезия, Америка; Tnismiaceae 4/40, космополитное тропического и умеренно теплого поясов; Tri-chopodaceae 2/2, палеотропическое
У Dioscoreales листья часто сетчатонервные (кроме Burmanniaceae и Thismiaceae), а проводящие пучки расположены не рассеянно, а в один или несколько кругов (кроме Тассасеае). Как правило, вьющиеся растения сем. Dioscoreaceae обычно имеют однополые цветки и нижнюю завязь.
Встречающиеся в тканях видов этого семейства стероидные сапонины — сырье для полу-синтетического изготовления многих гормонов (например, половых, гормонов надпочечников). Съедобные корневые шишки имеются у восточноазиатского ямса (Dioscorea batatas). На юго-востоке Центральной Европы семейство представ-
Рис. 11.229. Высшие Asparagales, Convallariaceae. Dracaena draco, старый экземпляр драконового дерева с Канарских островов (по С. Chun и G. Karsten)
лено тамусом ( Tamus communis). Большинство видов семейства Burmanniaceae — лишенные хлорофилла сапрофиты1.
Pandanales
Cyclanthaceae 12/230, неотропическое; Pandanaceae 3/800, палеотропическое; Stemo-naceae 4/45, Юго-Восточная Азия, Австралия, юго-восток Северной Америки; Velloziaceae 9/200, Центральная и Южная Америка, Африка, Мадагаскар, Южная Аравия, Китай
Выделение порядка Pandanales основано главным образом на результатах молекулярного анализа. Тетрамерия цветков как возможный общий признак порядка присуща семейству Cyclanthaceae и Stemonaceae, но очень редка у Pandanaceae и никогда не проявляется у Velloziaceae, имеющих трехмерные цветки. Однако у Velloziaceae, как иногда и у Pandanaceae, есть эпикутикулярные восковые выросты, которые не известны у вышеописанных порядков. У Cyclanthaceae листья часто похожи на листья пальм. К Pandanaceae относятся деревья, часто вильчато разветвленные, с ходульными корнями, или лазящие кустарники. Различные виды Pandanus —
1 Среди высших растений нет сапрофитов. Все так называемые «сапрофиты» — это микотрофные паразиты. — Примеч. ред.
438 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
весьма заметные прибрежные растения. Их называют «винтовыми пальмами», потому что их грубые и колючие по краям листья сидят на стеблях тремя винтообразными рядами. Оба семейства имеют однополые цветки, причем у Panda-пасеае они обычно голые. В мужских цветках обоих семейств тычинки многочисленные, а у Pandanaceae может быть увеличено также число плодолистиков. В очень плотных соцветиях различить отдельные цветки невозможно.
У Liliales, Asparagales, Dioscoreales и Pan-danales в тканях имеются рафиды оксалата кальция, у них широко распространена нижняя завязь. Далее, у них есть большей частью параллельно ориентированные эпи-кутикулярные восковые пластиночки. Эти особенности данных четырех порядков могли бы оправдать их объединение в надпорядок.
«Коммелиноидные» однодольные
Следующие четыре порядка (Arecales, Poales, Commelinales, Zingiberales) можно объединить вместе — «коммелиноидные» однодольные: наличие в клеточных стенках отражающей ультрафиолетовое излучение феруловой кислоты, отложение кремния в основном в клетках эпидермы, но отчасти также и в других тканях, дифференциация ризодермы на длинные и короткие клетки, а также образование эпикути-кулярных восковых столбиков.
Arecales
Агесасеае 190/2000, космополитное
Порядок Arecales с единственным семейством Агесасеае (=Palmae, пальмы) представлен преимущественно древесными растениями часто с неразветвленны-ми стволами (рис. 11.230), в которых происходит мощное первичное утолщение. Однако есть среди пальм и виды с тонкими ползучими или лазающими побегами (например, ротанги — Calamus и другие роды). Листья, обычно собранные в розетку на верхушке ствола, в почке многократно сложены и при развертывании более или менее глубоко разрываются по складкам. Они могут достигать длины более 20 м, жилкование их перистое (перистые паль
мы) или пальчатое (веерные пальмы; рис. 11.230). Цветки часто однополые, собраны в колос или метелку, окутанные верховым листом (покрывалом). Растения бывают как однодомными, так и двудомными. Некоторые пальмы — монокарпики, отмирающие после первого и единственного цветения и плодоношения. Цветки большей частью трехмерные (рис. 11.230), но число тычинок и соответственно плодолистиков бывает увеличенным. Гинецей апо- или ценокарпный, завязь верхняя, каждый плодолистик содержит только одну семяпочку. Виды семейства часто опыляются жуками. Плоды — ягоды или костянки. Ризо-дермис состоит из клеток одинакового размера.
При разделении семейства на шесть подсемейств особое значение придают признакам морфологии листа, соцветия, распределению цветков разных полов и морфологии плода. Распространенную в мангровых зарослях тропиков Восточного полушария Nypa fruticans можно рассматривать как изначальный вид пальм. Если у остальных пальм околоцветник слабо дифференцирован на чашечку и венчик, то у этого вида он состоит из одинаковых листочков. Пантропически распространенные пальмы относительно слабо представлены в Африке. Возможно, это результат относительно сухого климата в четвертичном периоде. Многие пальмы произрастают в нижних ярусах лесов, но некоторые доминируют в растительном покрове (например, Nypa fruticans). В Европе пальмы в настоящее время представлены только Phoenix theophrasti (Крит) и карликовой Chamaerops humilis, характерной для юго-западного Средиземноморья. Очень велико хозяйственное значение пальм. Они дают строительный материал и служат источником пишевых продуктов (например, сагового крахмала — Metroxyton sagu, Ин-домалайзия). Особенно ценны их плоды. Из мякоти плодов масличной пальмы (Elaeis guineensis, происходит из Африки) добывают масло. Финиковая пальма (Phoenix dactylifera), которую возделывают в оазисах от Сахары до Индии, может завязать ягоды из каждого из трех свободных плодолистиков, но развивается обычно только одна. Семя окружено сладкой мякотью и очень твердое благодаря накоплению в нем резервной целлюлозы1. Также очень твердый эндосперм американской слоновой пальмы
1 Запасным веществом является гемицеллюлоза. — Примеч. ред.
11.2. Бактерии, грибы, растения | 43g
Рис. 11.230. Arecales, Агесасеае (А, В— по W.Troll; С — по H.Baillon; D, Е — по J.Graf; F, К — по A. Englers «Syllabus»; G — по G. Karsten; H, J — no R. von Wettstein):
A — перистое строение листа; В — лист веерной пальмы (ок. 1 /20); С — Phoenix dactylifera, женский цветок в продольном разрезе, апокарпный гинецей (уменьшено); D, Е — диаграмма мужского и женского цветков; F — Cocos nucifera, общий вид растения (ок. 1/150); G — соцветие с покрывалом (s) и молодыми плодами, женскими (у) и остатками мужских (v) цветков (ок.1/20); Н — косточка (вид снизу) с тремя проростковыми отверстиями (уменьшено), J — костянковидный плод в продольном разрезе, с экзо-, мезо- и эндокарпием (ex, т, еп), эндоспермом (es) и зародышем (е) (уменьшено); К — Corypha taliera, общий вид (ок. 1/150)
(Phytelephas macrocarpa) используют в качестве «растительной слоновой кости». Изначально происходящая из западной части Тихого океана, но теперь распространенная по всем тропическим побережьям кокосовая пальма (Cocos nucifera) образует очень крупные, развивающиеся из ценокарпного гинецея костянки (рис. 11.230). У них гладкий экзокарпий, толстый, волокнистый мезокарпий и твердый эндокарпий. Наружная часть эндосперма плотная и богатая маслами (копра), а внутренняя жидкая (пригодное для питья «кокосовое молоко»). Двухлопастные семена Lodoicea callypige длиной до 50 см — самые крупные из всех известных семян.
Poales
Anarthriaceae 1/7, Австралия; Centroiepi-daceae 3/35, Австралия, Юго-Восточная Азия, Южная Америка; Сурегасеае 104/5000, космополитное; Ecdeiocoleaceae 2/2, Австралия; Eriocaulaceae 10/700—1400, Южное полушарие, тропическо-субтропическое; Flagellariaceae 1/4, палеотропическое; Hydatellaceae 2/10, Австралия, Новая Зеландия, Индия; Joinvilleaceae 1/2, Юго-Восточная Азия, Тихоокеанская область; Juncaceae 8/350, космолитное; Роасеае 670/9500, космополитное; Prioniaceae 1/1, Южная Африка; Restionaceae 55/500, Южное полушарие, в основном Юго-Западная Африка
440 | ГЛАВА 11 СИСТЕМАТИКА И ФИЛОГЕНИЯ
Рис. 11.231. Poales, Juncaceae (А—С) и Cyperaceae (D—Q) (А — по A. Englers «Syllabus»; В— по J. Graf; С, Е, Н, К, N, Р, Q — по A.W. Eichler; D — по F. Firbas; F, G — по Hoffmann; J, L — по G. Hegi, изменено; М, О — по Н.Walter):
А — соцветие Juncus bufonius с несколькими дрепанидиями (кроющие листья (t), предлистья (v)); В — цветок Luzula campestris (12х); С — диаграмма цветка Juncus; D — цветок Schoenoplectus lacustris (4х); Е — диаграмма цветка Scirpus sylvaticus. Eriophorum angustifolium; F — соплодие (1x); G — цветок (увеличено); H — диаграмма Kobresia myosuroides; J — частное соцветие с кроющим листом; К — женский и мужской цветки. Carex: L— общий вид Carexhirta с женским и мужским соцветиями (1/2х); М — женский цветок Carex; N — схема; Q — диаграмма; мешочек (и) сравним с кроющим листом женского цветка Kobresia, ось частного соцветия (а) редуцирована; О, Р — мужской цветок Carex sp. (15х) и его диаграмма
11.2. Бактерии, грибы, растения | 441
и Австралия; Spaiganiaceae 1/14, умеренный пояс Северного полушария, Юго-Восточная Азия, Австралия; Thumiaceae 1/3, Южная Америка; Typhaceae 1/10, космополитное; Xyri-daceae 5/300, пантропическое
Порядок Poales охватывает в основном ветроопыляемые семейства, более или менее напоминающие злаки. По этой причине у видов этого порядка нет нектарников, а листочки околоцветника очень редко лепестковидны (Xyridaceae). Зародыш Poales часто имеет линзовидную форму. В дополнение к этим морфолого-анатомическим признакам Poales узнают по разного размера делециям в определенной области пластидного генома. В пределах порядка можно выделить три четких группы семейств.
Объединяемые иногда в порядок Jun-cales семейства Cyperaceae, Juncaceae, Prioniaceae и Thumiaceae имеют хромосомы с так называемой диффузной центромерой (т.е. трубочки митотического веретена прикрепляются к ним в многочисленных точках) и пыльцевые зерна в тетрадах. Juncaceae, широко распространенные в холодно-умеренном поясе и обычно связанные с влажными местообитаниями, почти всегда имеют трехмерные цветки (рис. 11.231). У растущих в Центральной Европе ситников (Juncus) ценокарпные завязи многосеменные, а у ожик (Luzula), напротив, только с 3 семенами. Семейство Cyperaceae большей частью представлено многолетними травами, но также немногочисленными лианами, кустарниками и очень маленькими деревьями. Оно распространено преимущественно во влажных местообитаниях в холодном климате. Пыльцевые тетрады здесь — так называемые псевдомонады, в которых только одно из четырех пыльцевых зерен большое и фертильное. Морфология цветка разнообразна. У камышей (Schoenoplectus, Scirpus), болот-ницы (Eleocharis) и др. обоеполые цветки часто имеют шесть зазубренных щетинок (рис. 11.231), которые можно считать околоцветником. Они остаются при плоде и способствуют его распространению. У пушиц (Eriophorunr, рис. 11.231) цветки также обоеполые. Околоцветник у них, однако,
имеет вид каймы из белых волосков, которые также участвуют в распространении плода. Намного более упрощены однополые цветки осок (Carex, ок. 2000 видов). Мужские и женские цветки находятся в одних и тех же или в разных колосках, в пазухах кроющих чешуй. У мужских цветков только 3 тычинки, женские состоят из двух- или трехгранной завязи, которая заключена в дополнительную оболочку— мешочек (рис. 11.231). Как показывает тщательный морфологический анализ и сравнение с аркто-альпийским родом кобрезия (Kobresia), мешочек представляет собой разросшийся прицветник женского цветка (рис. 11.231). Под ним находится прицветник частного соцветия, которое у кобрезий состоит из одного мужского и одного женского цветков, а у осок — только из одного женского.
Часто объединяемые в одно (Typhaceae) семейства Typhaceae и Sparganiaceae характеризуются однополыми цветками и однодомностью. Женские соцветия расположены под мужскими. У ежеголовника (Spar-ganiuni) они шаровидные, а у цветков кожистый околоцветник, у рогоза (Typha) они вальковатые, а околоцветник состоит из волосков.
Для семейств Anarthriaceae, Centrolepi-daceae, Ecdeiocoleaceae, Flagellariaceae, Join-villeaceae, Poaceae и Restionaceae характерны апертуры пыльцевых зерен с выступающим краем (anulus) и крышечкой (operculum). Среди этих преимущественно злакоподобных растений (Flagellariaceae — лианы с листовыми усиками) наиболее близкородственны злакам (Poaceae = Gra-mineae) Joinvilleaceae. У обоих листовой эпидермис состоит из длинных и коротких клеток и имеется характерная инверсия в пластидном геноме. Poaceae (=Gra-mineae, злаки), наиболее значимые как полезные растения и как главный компонент растительного покрова, — большей частью многолетние травы, реже однолетники или древовидные растения. Их стебли (соломины) обычно округлые и полые, кроме утолщенных узлов. В основании междоузлия находится меристема. Двурядно расположенные листья состоят из охватывающего соломину и часто срастающегося краями влагалища и обычно узкой пластинки, до-
442 | ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Рис. 11.232. Poales, Poaceae, Festuca pratensis (А, В — по H.Schneck; С — по J.Schuster, сильно изменено; D — по F. Firbas):
А — отдельный цветок после удаления кроющих чешуй (6х); В — колосок с двумя колосковыми чешуями, двумя открывшимися и несколькими закрытыми цветками (Зх); С — гипотетическая диаграмма цветка злака (отсутствующие члены цветка даны пунктирной линией); D — схема колоска злака с тремя развитыми цветками; f— завязь; h — колосковые чешуи; d — нижняя цветковая чешуя (= кроющий лист); v — верхняя цветковая чешуя; I — лодикулы, наружный и (отсутствующий) внутренний круги тычинок
стирающей до 5 м в длину. На границе этих двух частей листа развит язычок (лигула), который может распадаться на отдельные волоски. Отдельные цветки собраны в так называемые колоски, которые, в свою очередь, собраны в колосья или метелки.
Каждый колосок (рис. 11.232), как правило, имеет в основании две колосковые чешуи. За ними следуют двухрядно расположенные цветковые чешуи — кроющие листья отдельных цветков.
У цветковых, реже у колосковых чешуй на верхушке может быть жесткая щетинка (ость)1. На оси цветка сидит двухкилевая верхняя 11детковая чешуя, за ней следуют две или реже три чешуйки, лодикулы, которые, разбухая, способствуют раскрытию цветка. Верхнюю цветковую чешую интерпретируют или как предлист или как рудимент внешнего круга листочков околоцветника, лодикулы — как элементы внешнего или внутреннего кругов околоцветника.
Открытия в области генетики развития указывают на то, что интерпретация верхней цветковой чешуи как элемента внешнего, а лодикул — как элементов внутреннего круга околоцветника, вероятно, верна.
1 Ость не всегда располагается на верхушке чешуи. — Примеч. ред.
В большинстве случаев развит только один трехчленный круг (редко их 2) тычинок. Ценокарпная и одногнездная завязь состоит из трех или двух плодолистиков и содержит единственный семязачаток. Цветки бывают как обоеполыми, так и однополыми (например, у кукурузы — Zea mays). В плоде злаков (зерновке) зародыш сбоку прилегает к богатому крахмалом эндосперму (рис. 11.233). Влагалище семядоли преобразовано в щитовидный сосущий орган (щиток), а пластинка ее окружает точку роста в виде колеоптиля1. Щиток нередко считают осью зародыша. Кроме того, точка роста корня окружена чехлом (колеоризой), возникающей из подвеска.
Роасеае можно в сущности разделить на две большие группы. Базальное положение в семействе занимают южноамериканские роды Апо-mochloa, Streptochaeta и Phams, а также африканская Puelia. Первая большая группа включает Bambusoideae, Oryzoideae и Pooideae («BOP-клада»). К полифилетическим Bambusoideae, вероятно, принадлежат многие деревянистые тропические растения, которые могут достигать высоты до 40 м. В цветках часто имеются 3 лодикулы и 2 трехчленных крута тычинок. В этом подсе-
1 Существует много интерпретаций структуры зародыша злаков. Авторы излагают не самую распространенную и не самую обоснованную. — Примеч. ред.
11.2. Бактерии, грибы, растения | 443
Рис. 11.233. Poales, Роасеае. Хлебные злаки, колосья и колоски (А, С, D, F, J, К — по G. Karsten; В, Е, G, Н — по F. Firbas, L— по E.Strasburger):
А, В — рожь (Secale cereale); С—Е — пшеница (Triticum): С — спельта (T.spelta); D, Е — мягкая (Г. aesti-vum); F, G — ячмень (Hordeum vulgare): F, G — двухрядная и шестирядная формы соответственно; Н — овес (Avena sativaY, J, К — рис (Oryza sativaY h — колосковые чешуи; d — нижние цветковые чешуи, их ости на В и G показаны частично; v — верхние цветковые чешуи; L — зерновка пшеницы, продольный разрез через нижнюю часть, поверхность продольной бороздки (f), слева внизу зародыш со щитком (s), проводящим пучком (I) и гаусториальной эпидермой (z), колеоптилем (с), конусом нарастания побега (vk), колеоризой (сг), зародышевым корешком (г) с корневым чехликом (w) и местом прорастания (а) (14х)
444 | ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
мействе нередко происходит по непонятным причинам синхронное цветение одного вида по всей области его распространения. Первое цветение растений, однако, часто случается спустя годы или десятилетия. Рис (Oryza) — представитель Oryzoideae. К Pooideae, у которых фотосинтез идет по типу С3, принадлежат в числе прочих Роа (мятлик), Lolium (плевел), Bromus (костер), Festuca (овсяница) — с короткими колосковыми чешуями и Arrhenatherum (райграс), Trisetum (трищетинник) и Avena (овес) — с длинными. Колоски у этих родов большей частью собраны в метелки. У родов хлебных злаков колоски собраны в колосья — Triticum (пшеница). Secale (рожь) и Hordeum (ячмень). Одноцветковые колоски среди центральноевропейских Pooideae есть у Agrostis (полевицы), Anthoxanthum (душистого колоска), Alopecurus (лисохвоста), Phleum (тимофеевки), а также у Stipa (ковыля). Вторая большая ipynna злаков охватывает Panicoidcae, Arundinoideae, Chloridoideae и Centothecoideae («РАСС-клада»). У большей частью тропических Panicoideae колоски часто уплощены в плоскости, перпендикулярной к плоскости, в которой двумя рядами располагаются цветки. Сюда относят разные просовидные злаки (Panicum, Pennisetum, Setaria), сахарный тростник (Saccharum officinale) с сердцевиной, служащей сырьем для производства сахара, а также хлебные злаки Sorghum и Zea (кукуруза). Arundinoideae, вероятно, полифилетич-ны. Две основные линии этой группы представляют Danthonia (дантония) и Phragmites (тростник). У представителей «РАСС-клады» фотосинтез обычно идет по С4-типу. Большинство распространенных в тропиках Chloridoideae характеризуются специфическими двухклеточными волосками на листовой эпидерме и С4-фото-синтезом. В Центральной Европе эта группа представлена, например, видами Cynodon (свинорой), Eragrostis (полевичка) и Spartina (спар-тина). Centothecoideae также распространены в основном в тропиках.
В настоящее время злаки доминируют примерно на 20 % покрытой растительностью земной поверхности (в саваннах, степях и на лугах). Широкое распространение злаковых сообществ очень тесно связано с эволюцией крупных копытных, таких, как парнокопытные и лошади. Переход человека около 10000 лет тому назад к возделыванию зерновых культур и связанному с этим рациональному производству полноценных продуктов питания сделал возможным возникновение высокоразвитой городской культуры. Культивируемые злаки отличаются от родственных диких форм в первую очередь увеличенным числом и размером зерновок, неразла-мывающимися при созревании осями колосьев и соответственно колосков и частично также без-
остностью плодов. Важнейшими культурными злаками (рис. 11.233) Средиземноморья и Передней Азии следует назвать пшеницу (Triticum; обычно 3 — 5-цветковые колоски с остистой или безостой нижней цветковой чешуей располагаются на оси колоса поодиночке) с гексаплоид-ной мягкой пшеницей (Т. aestivum) и тетрапло-идной твердой пшеницей (Т. durum).
Другие диплоидные (однозернянка — Т. то-пососсит) или тетраплоидные (полба — Т. di-соссит) виды сейчас уже не разводят. Ячмень (Hordeum vulgare)'. одноцветковые колоски располагаются на оси колоса по 3; у шестирядного ячменя все колоски плодоносящие и остистые, у двурядного — только средний из 3; все культурные сорта ячменя диплоидны.
Рожь (Secale cereale): двухцветковые колоски с узкими колосковыми чешуями и длинноостистыми нижними цветковыми, располагаются на оси колоса поодиночке; диплоид.
Овес (Avena): колоски в метелках; важное значение имеет только гексаплоидный овес посевной (A. sativa). В Юго-Восточной Азии важнейшая традиционная злаковая культура — рис (Oryza sativa): одноцветковые колоски в метелках, диплоид; ныне возделывается по всему миру (рис. 11.233). В засушливых областях Восточной Азии, Индии и Африки особенно большое значение имеют просо посевное (Panicum milia-сеит), могар (Setaria italica), африканское просо (Pennisetum spicatum) и сорго, или дурра (Sorghum bicolor). В Америке ок. 8 000 лет культивируют кукурузу (Zea mays) с чисто мужскими на концах стеблей и чисто женскими боковыми соцветиями.
Commeliiiales
Commelinaceae 41/650, по всему миру в тропиках и тепло-умеренном поясе; Наето-doraceae 13/100, Америка, Южная Африка, Новая Гвинея, Австралия; Phededraceae 4/5, Юго-Восточная Азия, Австралия: Ponteridia-сеае 9/33, тропическое, в основном Новый Свет
Пыльники Commelinales имеют секреторный тапетум, внутриклеточные кремнистые включения полностью отсутствуют. В этом порядке часто встречаются зигоморфные цветки, а число фертильных тычинок может быть редуцировано до одной (например, у Phylidraceae с двучленным околоцветником). К Commelinaceae относят обычные комнатные растения из родов Tradescanria и Zebrina с околоцветником, дифференцированным на чашечку и венчик. Сем. Pontederiaceae представлено водными растениями. Свободно плавающий благодаря вздутым
11.2. Бактерии, грибы, растения | 445
листовым черешкам водяной гиацинт (Eichhornia crassipes) — изначально неотропический водный сорняк. У него бывает три типа цветков, в которых два круга тычинок и пестик расположены в разных цветках в три яруса (триморфная гетеростилия).
Zingiberales
Саппасеае 1/10, неотропическое; Costaceae 4/100, пантропическое: Heliconiaceae 1/200, неотропическое, меланезийское; Lowiaceae 1/10, Юго-Восточная Азия; Marantaceae 31/550, пантропическое; Musaceae 2/40, палеотропическое; Strelitziaceae 3/6, Америка, Африка, Мадагаскар; Zingiberaceae 50/130, пантропическое
У цветков Zingiberales околоцветник большей частью дифференцирован на чашечку и
венчик, цветки зигоморфные или асимметричные, с нижней завязью. Пыльца в большинстве случаев безапертурная, семена обычно с крышечкой (оперкулюм) и часто также ариллусом.
Листья в почкосложении скручены. Часть тычинок часто видоизменена в лепестковидные стаминодии, столбик также может выглядеть как лепесток.
Опыляются они птицами, летучими мышами и другими мелкими млекопитающими, но часто также и пчелами.
У Heliconiaceae, Lowiaceae, Musaceae и Strelitziaceae тычинок шесть или пять. Для семейства Musaceae, к которому относятся важные полезные растения — банан (Musa со многими видами и гибридами, например, М. х paradisiaca) и манильская пенька (М. textilis), характерны ложные стебли из влагалищ листьев, достигающие в высоту 13 м. Простые листья могут достигать в длину 6 м.
Рис. 11.234. Zingiberales (А— по О.С. Berg и С. F. Schmidt; В— по J.D. Hooker; С— по A.W. Eichler; D — по Н.Schenck): >
А—С — Zingiberaceae: А — Zingiber officinale, цветущее растение с корневищем (2/Зх), В — цветок Curcuma australasica, С — диаграмма цветка Kaempferia ovalifolia, с кроющим листом (t), предлистом (v), чашелистиками (к) и лепестками (с), боковыми стаминодиями (ss), губой из стаминодиев (I), единственной фертильной тычинкой (fs), завязью (f); D — Саппасеае, асимметричный цветок Саппа iridiflora, три стаминодия (st1—st3), полуфертильная тычинка (sb), столбик (д) (1/2х)
446 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Однополые цветки Musa располагаются двойными поперечными рядами в пазухах крупных кроющих листьев. В повислых соцветиях женские цветки расположены внизу, а мужские — наверху.
Виды Costaceae и Zingiberaceae имеют только одну фертильную тычинку. У Zingiberaceae (рис. 11.234) два стаминодия внутреннего круга тычинок срастаются в губу. Представители этого семейства богаты эфирными маслами (например, имбирь: Zingiber officinale, кардамон: Elettaria cardamomum).
У видов Саппасеае (например, у канны: Саппа) и Marantaceae (например, маранты: Магата) асимметричные цветки имеют только одну фертильную теку единственной тычинки (рис. 11.234). Остальные тычинки, стерильная половина фертильной тычинки и столбик лепестковидные. У обоих семейств пыльца из пыльника перемещается на столбик и оттуда попадает на опылителя (вторичное представление пыльцы). У Marantaceae при этом происходя! резкие движения столбика.
Среди «коммелиноидных» однодольных есть несколько семейств с неясными родственными связями — Bromeliaceae (56/2600, неотропическое царство, Западная Африка), Dasypogonaceae (4/8, Австралия), Hanguanaceae (1/5, Юго-Восточная Азия, Тихоокеанская область, Австралия), Мауасасеае (1/10, неотропическое царство, Западная Африка) и Rapateaceae (10/80, неотропическое царство. Западная Африка).
У Bromeliaceae, Мауасасеае и Rapateaceae основная область распространения лежит в неотрописе, но некоторые представители встречаются также и в Западной Африке. Так как эта географическая дизъюнкция, по крайней мере у Bromeliaceae (Pitcairnia feliciana) и Мауасасеае (Мауаса baumii), проявляется в пределах одного рода, только один вид которого обитает в Африке, а все остальные — в Южной Америке, то западноафриканская часть ареала возникла, вероятно, вследствие дальнего заноса в прошлом. Bromeliaceae представлено в основном розеточными травами с обычно жесткими листьями. Редко встречаются также кустарники высотой до 3 м (Риуа). Бромелиевые часто ведут эпифитный образ жизни, а влагу поглощают при помощи чешуйчатых волосков.
Сильно редуцированная Tillandsia usneoides напоминает мох. Большей частью гермафродитные 3-мерные цветки с околоцветником, обычно дифференцированным на чашечку и венчик, и верхней или нижней завязью собраны в колосья, кисти или метелки. Плоды — ягоды или коробочки, семена бывают крылатыми или покрытыми волосками. Виды семейства часто опыляются птицами, поэтому цветки, кроющие листья
на оси соцветий и частично также верхние листья, нередко окрашены в яркие оттенки красного цвета.
Ряд семейств занимает неопределенное положение среди однодольных: лишенные хлорофилла Corsiaceae (2/28, Южная Америка, Новая Гвинея, Австралия; возможно, относятся к Dios-coreales) и также бесхлорофилльныс Triuridaceae (9/45, пантропическое), кроме того Japonoliriaceae (1/1, Япония), Nartheciaceae (4/40, Северное полушарие до Южной Америки; напр., Narthecium ossifragum) и Petrosaviaceae (1/3, Юго-Восточная Азия, Япония). Последние 3 семейства в прошлом объединяли вместе с Tofieldiaceae (Alismatales) в, по-видимому, очень полифилетичное семейство Nartheciaceae. У видов мексиканского рода Lacandonia (Triuridaceae) многочисленные свободные плодолистики располагаются снаружи от трех центральных тычинок. Возможно, это проявление гомеотической мутации.
3. ПОДКЛАСС: «НАСТОЯЩИЕ ДВУДОЛЬНЫЕ» (ROSIDAE)
Rosidae, как и Magnoliidae, — растения двудольные («настоящие двудольные», англ, eudicots). От Magnoliidae они отличаются отсутствием характерных эфирных масел в идиобластах, обычно циклическими цветками, но прежде всего — наличием трехбороздных (рис. 11.214) или производных от них пыльцевых зерен.
Ближайшие родственники Rosidae в пределах Magnoliidae до сих пор точно неизвестны. По разным наборам молекулярных данных в качестве сестринских групп можно признавать Ceratophyllaceae или группу порядков Winterales/ Piperales/Magnoliales/Laurales, или однодольные плюс Chloranthaceae.
Вероятно, в основании Rosidae стоят несколько изолированных семейств, представленных древесными формами (Вихасеае 5/60. космополитное, Didymelaceae 1/2, Мадагаскар; Sabiaceae 3/60, Юго-Восточная Азия, тропическая Америка; Trochodendraceae, включая Те-tracentraceae 2/2, Восточная Азия) с мелкими и часто однополыми цветками (цветки многих Sabiaceae и всех Trochodendraceae обоеполые). У Trochodendraceae древесина без сосудов, цветки с околоцветником или голые, андроцей 4- или многочленный и от 4 до 17 сросшихся друг с другом плодолистиков (рис. 11.235), а у Вихасеае, включающего распространенный в средиземноморско-атлантической области самшит (Buxus sempervirens), цветки часто двумерные. Тычинки у обоих семейств располагаются димерными кругами.
11.2. Бактерии, грибы, растения | 447
Ranunculales
Berberidaceae 13/570. умеренный пояс Северного полушария, Анды; Circaeasteraceae 2/2, Азия; Eupteleaceae 1/2, Восточная Азия; Lardizabalaceae 8/35, Восточная Азия, Северная Америка; Menispermaceae 71/450, пантропическое; Papaveraceae, включая Fumariaceae, Pteri-dophyllaceae 41/770, умеренный пояс Северного полушария, Южная Африка; Ranunculaceae 58/2500, космополитное умеренного пояса
Семейства порядка Ranunculales представлены древесными или травянистыми растениями, содержащими разнообразные бензил-изохинолиновые алкалоиды. Элементы цветка многочисленные, часто расположены по спирали или циклически, плодолистики часто несросшиеся. Ранее на основании этих признаков Ranunculales часто объединяли с Magnoliidae в группу «Ро-lycarpicae», рассматривая их как преимущественно травянистую линию развития этого таксона. Однако Ranunculales отличаются от древесных «Polycarpicae» отсутствием маслосодержащих клеток, часто рассеченными листьями, а также характерными для «настоящих двудольных» трехбороздными (или производными от них) пыльцевыми зернами.
Самое крупное семейство порядка, представленное в среднеевропейской флоре многочисленными видами, — Ranunculaceae (лютиковые). Семейство включает преимущественно крупные травы с очередными, часто рассеченными листьями. Цветки часто довольно крупные (рис. 11.236), обоеполые, с многочисленными тычинками и несколькими — многими, большей частью свободными плодолистиками (у живокости; Consolida только с одним плодолистиком). Плодолистики с несколькими или только с одной семяпочкой. Соответственно этому они развиваются или в многосемянные листовки или в односемянные плоды, часто это орешек. Изредка в семействе бывают даже ягоды (воронец — Actaea) и у родов со сросшимися плодолистиками — коробочки (чернушка — Nigella). В остальном морфология цветка в сем. Ranunculaceae очень разнообразна. Цветки могут быть радиально симметричными или зигоморфными (борец — Aconitum, живокости — Consolida, Delphinium), а многочисленные органы цветка могут располагаться по спирали или в пяти-, трех- или двучленных кругах. Околоцветник, например, у калужницы (Caltha), ветреницы (Anemone nemorosa) и прострелов (Pulsatilla) простой, а у купальницы (Trollius), зимов-
Рис. 11.235. Trochodendraceae, Trochodendron aralioides (по А.Л.Тахтаджяну):
А— цветущий побег; В — цветок (РоА»С4_ц); С— плодолистик, продольный разрез; D — пыльцевое зерно (трехбороздное); Е — незрелый плод; F — раскрывшийся плод
ника (Helleborus) и отчасти у весенника (Eranthis) его листочки связаны переходами с листьями1. Во многих родах из тычинок формируются стерильные нектарники (рис. 11.236). Они содержат нектар в ямках или в шпорцеобразных выростах и бывают малозаметны (например, Trollius, Helle-borus) или лепестковидны (например, у лютиков — Ranunculus, водосборов — Aquilegia).
Таким образом мог возникнуть двойной околоцветник, в котором листочки изначально простого околоцветника берут на себя функцию чашелистиков, а нектарники — лепестков. Сравнивая разные роды, можно представить себе формирование околоцветника и его дальнейшее расчленение как из верховых листьев (простой околоцветник, чашечка), так и из тычинок (венчик)1 2.
1 Авторы сильно преувеличивают, вероятно они принимают за околоцветник верховые листья, приближенные к цветку. — Примеч. ред.
2 Происхождение простого околоцветника Ranunculaceae не известно. При формировании двойного околоцветника он мог становиться венчиком (чашечка возникла из верховых листьев) или чашечкой (венчик возник из стаминодиев). — Примеч. ред.
448 | ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Рис. 11.236. Ranunculales, Ranunculaceae (A—U) и Berberidaceae (V) (А—С, V— по Н. Bailion; D — Н — по G. Karsten; J—О, Q — no F. Firbas; Р — по Е. Rassner; R—U — по A.W. Eichler);
А—С — Ranunculus sp., цветок (общий вид, продольный разрез); одиночный орешек (ок. 4х); D —Н — Aconitum napellus, цветок спереди (продольный разрез, после удаления околоцветника, оба нектарника свободные); молодой и зрелый апокарпные гинецеи (3/5х). Нектарник и: J — Trollius giganteus (2,5х), К — Ranunculus auricomus (Зх); L — Helleborus foetidus (4,5х), М — Aquilegia vulgaris (1 х). Плодолистики; Helleborus orientalis; N— продольный разрез (5х); О— поперечный разрез (18х); Р — Anemone nemorosa, продольный разрез, видны недоразвитые семяпочки (s) и Q — Ranunculus auricomus (10х). Диаграммы цветков: R — Cimicifuga racemose; S — Adonis vernalis; T — Aquilegia vulgaris; U — Aconitum napellus (листочки околоцветника или соответствующие чашелистики показаны штриховкой, нектарники или соответствующие лепестки — черным цветом); V — Berbens vulgaris, цветок (Зх)
Ranunculaceae особенно широко распространены во внетропических областях Северного полушария. Наряду с многолетними травами среди них есть и однолетники (например, Myosurus minimus. Ranunculus arvensis) и, изредка, древесные растения, такие, как супротивно облиственные и часто лианоидные ломо
носы (Clematis). Встречающиеся в семействе плоды-орешки чаще имеют опушенные и удлиненные стилодии (Clematis, Pulsatilla), крючковидные выросты (Ranunculus arvensis), кожистые крылья или губчатую ткань для плавания. Поэтому они распространяются ветром, животными или водой.
11.2. Бактерии, грибы, растения i 44g
Близкородственны Ranunculaceae деревянистые или травянистые Berberidaceae с обычно циклическими цветками. Околоцветник их большей частью двойной, часто с лепестковидными стаминодия ми-нектарниками и андроцеем, состоящим главным образом из нескольких трех- или реже двучленных кругов (рис. 11.236). Гинецей состоит из единственного плодолистика с верхней завязью, который развивается в ягоду. В Центральной Европе произрастает барбарис (Berbens vulgaris). У него есть листовые колючки (см. рис. 4.7) и чувствительные к раздражению тычинки; он является растением-хозяином хлебной ржавчины.
Семейство Lardizabalaceae и тропическое семейство Menispermaceae большей частью представлены лианами с часто однополыми, двудомными или однодомными, обычно трехмерными цветками. К роду Euptelea (Eupteleaceae) относятся ветроопыляемые деревья с цветками без околоцветника.
Сестринскую вышеназванным семействам (в т.ч. Circaeasteraceae) кладу составляет семейство Papaveraceae (включая Pte-ridophyllaceae, Fumariaceae) с дву- или, реже, трехмерным циклическим околоцветником и ценокарпным гинецеем с париетальной плацентацией. Pteridophyllum racemosum, единственный представитель подсемейства
Pteridophylloideae, произрастает только в Японии. У представителей подсемейства Fumarioideae (рис. 11.237) обычно бесцветный млечный сок содержится в идиобластах, а один или два из четырех лепестков, как правило, имеют шпорцы, благодаря чему цветки зигоморфны (например, дымянка — Fumaria, хохлатки — Corydalis, Pseudofuma-ria) или дисимметричны (Dicentra). Андроцей состоит из четырех тычинок, из которых две расщеплены и могут срастаться с нерасщепленными тычинками. Гинецей состоит их двух плодолистиков.
В подсемействе Papaveroideae (рис. 11.237) окрашенный млечный сок находится в идиобластах или млечниках, цветки радиально-симметричны, с одним кругом почти всегда опадающих чашелистиков, двумя кругами часто кажущихся смятыми лепестков, обычно многочисленными тычинками и верхней завязью из двух или более плодолистиков, из которой развивается коробочка. Вскрывающаяся отверстиями коробочка имеется, например, у мака (Papaver), и эндосперм семян содержит масла. Родственные отношения в пределах Papaveraceae указывают на то, что много-членность андроцея Papaveroideae вторична.
Важнейшее полезное растение семейства, известное в качестве лекарственного по мень-
Рис. 11.237. Papaverales, Papaveraceae (A, F, G — по J. Graf; В, Н — по A. W. Eichler; С—Е — по F. Firbas): Papaveroideae: А, В — Papaver rhoeas: А — цветок (3/4х), В — диаграмма цветка; С—Е — Р. somni-ferum: С — вскрывшаяся отверстиями коробочка с рыльцем (1/2х), D — часть поперечного среза плода с париетальной плацентой (2/Зх), Е — семя в продольном разрезе с семенной кожурой, эндоспермом и зародышем (8х). Fumarioideae: F—Н— Corydalis cava: F— продольный разрез цветка и тычинок (внутренние расщеплены, половинки их срослись с наружными тычинками: 1/2+1 + 1/2), G — завязь (1 х), Н — диаграмма цветка
450 I ГЛАВА 11 СИСТЕМАТИКА И ФИЛОГЕНИЯ
шей мере 3 500 лет, — мак снотворный (Papaver somniferum). Традиционным способом, надрезая развивающиеся коробочки (рис. 11.237), из него добывают млечный сок, содержащий опиум с морфиновыми алкалоидами тебаином, кодеином и морфином. Семена этого вида применяют для получения масла и в кулинарии.
Proteales
Nelumbonaceae 1/2, Юго-Восточная Азия, Австралия, Северная Америка; Platanaceae 1/9, Азия, Северная Америка; Proteaceae 79/1700, Австралия, Юго-Восточная Азия, Южная Африка, Южная и Центральная Америка
Семейства порядка Proteales, содержащие габитуально очень разные растения, характеризуются семенами со скудным эндоспермом или вовсе без эндосперма, часто прямой семяпочкой, апокарпным гинецеем и тычинками с заметными надсвязниками. Общность Nelumbonaceae, Platanaceae и Proteaceae — весьма неожиданный результат молекулярной систематики.
Внешне напоминающие Nymphaeaceae виды семейства Nelumbonaceae (Nelumbo, лотос) — водные травы с возвышающимися над поверхностью воды щитовидными листьями на длинных черешках. Благодаря отложению эпикути-кулярного воска в виде пустотелых трубочек попавшие на лист вода и грязь скатываются с его поверхности. Воск сходной структуры все чаще применяют для импрегнации технических поверхностей. Очень крупные цветки имеют многочисленные листочки околоцветника и тычинки, от 2 до 30 свободных плодолистиков, погруженных в сильно расширенное в виде обратного конуса цветоложе.
У древесных Platanaceae с единственным родом платан (Platanus) цветки мелкие и однополые, собранные в плотные шаровидные соцветия. Женские цветки содержат от 5 до 9 свободных плодолистиков. Р1а tanus hybrida часто высаживают в аллелях как устойчивое к загрязненному воздуху дерево.
Радиально симметричные или зигоморфные цветки кустарниковых или древесных Proteaceae имеют простой околоцветник из четырех лепестковидных листочков, четыре тычинки и только один плодолистик с одной или многими семяпочками. Многие виды этого семейства, особенно богатого видами в Австралии и Юж
ной Африке, хорошо приспособились к крайней сухости и частично — к естественным пожарам. Это проявляется в том, что виды, например Banksia, Накеа и Grevillea, образуют сильно одревесневающие и годами остающиеся на растении плоды, которые раскрываются только под воздействием огня. Proteaceae часто опыляются мелкими млекопитающими, а их семена обычно распространяют птицы. Формирование основных эволюционных линий Proteaceae, по-видимому, состоялось еще до распада материка Гондваны.
Основные «настоящие двудольные»
Все последующие семейства можно объединить в группу «основных настоящих двудольных» (англ, core eudicots). Если в вышеописанных группах Rosidae еще можно наблюдать вариации в числе, характере и расположении элементов цветка, пыльцевые зерна большей частью были трехбороздными и отсутствовала эллаговая кислота, то с формированием «основных настоящих двудольных» окончательно установились 5-членность кругов цветка, дифференциация околоцветника на чашечку и венчик; пыльцевые зерна у них триколь-поратные (трехборозднооровые), имеется эллаговая кислота.
В основании этой группы стоит несколько семейств с пока неясными родственными связями (Aextoxicaceae 1/1, Чили; Berberidopsidaceae 1/2, Чили, Восточная Австралия; Dilleniaceae 10/350, тропики и субтропики, по всему миру; Gunneraceae 1/40, Южное полушарие; Myrotham-пасеае 1/2, Южная Африка, Мадагаскар; Vitaceae 14/850, преимущественно тропики и субтропики, по всему миру), распространенных чаще в Южном полушарии.
Примечательны травы р. Gunnera (Gunneraceae) иногда с очень крупными листьями (листовая пластинка до 2 м в диаметре), растущие в основном во влажных местообитаниях на кислых почвах. Они содержат в слизевых железках, расположенных в области узлов, симбиотические азотфиксирующие цианобактерии Nostoc. Растущие преимущественно в виде лиан виды семейства Vitaceae имеют на побегах усики, расположенные супротивно листьям; их можно считать верхушками отдельных членов симпо-диальной системы ветвей (рис. 11.238). Соцветия занимают то же положение, что и усики. Цветки с 4- или 5-членными околоцветником
11.2. Бактерии, грибы, растения | 451
и андроцеем и гинецеем из двух сросшихся между собой плодолистиков (рис. 11.238), которые развиваются в ягоду. Лепестки на верхушке сцеплены посредством папилл и при распускании цветка отрываются и все вместе в виде колпачка выносятся вверх тычинками. Важнейшее экономическое значение среди Vitaceae имеет виноград (Vitis vinifera), который культивируют уже с раннего бронзового века. Исходная форма культурного растения — И vinifera subsp. sylvestris, возможная родина которого — сырые леса Малой Азии. Vitis vinifera в настоящее время культивируют на подвоях, устойчивых к виноградной филлоксере. У некоторых разводимых как «дикий виноград» видов р. Parthenocissus на кончиках усиков развиты прикрепительные диски. В большом роде Cissus есть стеблевые суккуленты.
Порядки Caryophyllales, Polygonales, а также несколько семейств с неясными свя
зями (Ancistrocladaceae 1/20, Африка, Азия; Asteropeiaceae 1/1, Мадагаскар; Frankenia-сеае 3/80, космополитное; Physenaceae 1 /2, Мадагаскар; Rhabdodendraceae 1/3, Южная Америка; Simmondsiaceae 1/1, запад Северной Америки; Tamaricaceae 4—5/100, Евразия, Африка) четко характеризуются эн-достомными семяпочками (с микропиле, образованным внутренним интегументом), формированием эндотеция непосредственно из субэпидермального слоя стенки пыльника и нуклеарным типом развития эндосперма.
Часто в этой группе таксонов встречается аномальное вторичное утолшение вследствие формирования добавочных кам-биев; для них также характерно произрастание в очень сухих или засоленных местообитаниях.
Рис. 11.238. Vitaceae (А — из R. Spohn по W. Troll; В — по О С. Berg и С. F. Schmidt; С — по A.W. Eichler): А, В — Vitis vinifera. А — побеговая система с побеговыми усиками, В — раскрытый цветок с редуцированной чашечкой (к), приподнятыми тычинками венчиком (с), диском (d), тычинками (s) и завязью (f) (увеличено); С— Partenocissus quinquefolia, диаграмма цветка
452 | ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Caryophyllales
Achatocarpaceae 2/6, Америка; Aizoaceae 127/2500, тропики и субтропики Южного полушария, в основном Южная Африка; Ата-ranthaceae, включая Chenopodiaceae 170/2400, космополитное; Basellaceae 4/20, Америка; Cactaceae 100/1500, Америка, один вид в Африке; Caryophyllaceae 86/2200, умеренный пояс Северного полушария; Didiereaceae 4/ 11, Мадагаскар; Molluginaceae 13/120, тропики и субтропики Нового и Старого Света; Nyctaginaceae 31/400, тропики и субтропики Нового и Старого Света; Phytolaccaceae 17/ 70, тропики в основном Нового Света; Portulacaceae 29/450, Южное полушарие, Северная Америка; Stegnospermataceae 1/3, Америка
Caryophyllales относятся к наиболее четко отграниченным и соответственно этому уже очень давно выявленным порядкам покрытосеменных. У них большей частью радиально-симметричный 5-мерный цветок с простым или двойным околоцветником и одним или двумя кругами тычинок или — вследствие вторичного увеличения числа тычинок — многочленным, центробежно развивающимся андроцеем. Особенно характерны для этого порядка азотсодержащие пигменты цветка бетаци-ан и бетаксантин (=беталаин), замещающие антоциан (кроме Caryophyllaceae/Mollugi-пасеае), феруловая кислота в составе клеточных стенок, пластиды ситовидных трубок с протеиновыми волокнами, обычно расположенными кольцом вокруг кристаллов протеина (и иногда также крахмальных зерен), кампилотропная семяпочка с изогнутым зародышем, семена с содержащим крахмал периспермом и трехклеточные пыльцевые зерна с многочисленными, распределенными по всей поверхности апертурами. Свободная центральная плацента имеется только у некоторых Amaran-thaceae (включая Chenopodiaceae). Относительно часто в порядке встречается сукку-лентность.
Виды семейств Molluginaceae и Caryophyllaceae вместо беталаина содержат антоцианы. Molluginaceae — частично деревянистые и слабо суккулентные растения, часто с простым околоцветником, трехбороздными пыльцевыми зернами и гинеце
ем с центрально-угловой плацентой. Caryophyllaceae — почти всегда травы. Соцветие их — обычно дихазий (рис. 11.239). У многих родов околоцветник простой (например, грыжник — Hemiaria), у других — двойной с чашечкой и венчиком (например, ясколка — Cerastium, куколь — Agrostemma githago, смолка — Silene viscaria; рис. 11.239). Расположение тычинок обдиплостемонное (наружные тычинки противолежат лепесткам), андроцей может быть редуцирован до одного круга, да и в нем бывают развиты не все тычинки (например, мокрица — Stellaria media А<,_>3; рис. 11.239). Число плодолистиков часто редуцировано до 3 (например, смолевка, звездчатка — Silene, Stellaria) или 2 (гвоздика — Dianthus). Некоторым видам (например, S. dioica и 5. latifo-На) присуща двудомность. Плоды, как правило, — многосемянные вскрывающиеся зубчиками коробочки, в которых в процессе развития из-за разрушения перегородок может сформироваться центральная плацента (рис. 11.239). У мелкоцветковых видов число семяпочек часто может сокращаться до одной, а вместо коробочки об разуется односемянный орех (например, дивала, грыжник — Scleranthus, Hemiaria). Традиционное разделение на три подсе мейства основано на наличии прилистников (Paronychioideae, например, Spergula, Herniaria) и соответственно свободных (Alsinoideae, например, Cerastium, Stellaria, Scleranthus) или сросшихся (Caryophyllo-ideae, например, Lychnis, Agrostemma. Silene. Dianthus) чашелистиков. Оба первые подсемейства, вероятно, не являются естественными группами.
Содержащие беталаин Chenopodiaceae имеют обычно обоеполые, но иногда и однополые цветки с часто простым и невзрачным 3- или 5-членным околоцветником, одним кругом тычинок и 2- или 3-членным гинецеем с одной или несколькими семяпочками и базальной плацентацией, из которого развиваются односемянные орехи или коробочки (рис. 11.239). Такой тип строения присущ, например, мари (Chenopodium) и свекле (Beta', рис. 11.239). Часто число членов околоцветника и тычинок еще более редуцировано и цветки однополые. Так, у солероса (Salicornia) обычно три
11.2. Бактерии, грибы, растения | 453
Рис. 11.239. Caryophyllales (А—С— по G. Beck-Managetta; D — no Р. Е. Duchartre; Е—Н, М, Q, С, U, V — по A. W. Eichler, несколько изменено; J — по Th.W. Engelmann; К, L — по F.Buxbaum; N— по H.Baillon; О, Р, S, Т — по J.Graf):
А—Н — Caryophyllaceae: А, В — продольные разрезы цветков Silene nutans и Herniaria glabra (ок. 4х), С — коробочка Cerastium holosteoides (снизу срезанная продольно) (ок. 4х), D — соцветие Cerastium sp.: дихазий (ср. также рис. 4.23, С) (ок. 1х). Диаграммы цветков: Е — Silene viscaria; F — Silene vulgaris-, G — Stellana media-, H — Paronychia sp.; J—M — Cactaceae: J — Echinocereus dubius (ребро побега с ареолами и цветком) (ок. 1/2х), К, L— продольные срезы цветков примитивного (Pereskia) и эволюционно продвинутого кактусов с воронковидным цветоложем и погруженным в него гинецеем, М — диаграмма цветка Opuntia sp. N—V — Amaranthaceae: N — цветок Beta trigyna (увеличено); О —R — суккулентные побеги с цветками (общий вид и продольный срез, а также диаграммы соответствующих цветков с А2 и A, Salicornia europaea); S—V — Amaranthus sp. (мужские и женские цветки (увеличено) и их диаграммы)
454 | ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
или четыре листочка околоцветника и одна или две тычинки (рис. 11.239), а у двудомной лебеды (Atriplex) встречаются цветки без околоцветника. Виды Chenopodiaceae часто растут на засоленных почвах и вдоль морского побережья, они нередко сукку-лентны и в таком случае не имеют листьев1. Виды семейства также широко распространены в рудеральных местообитаниях. Salicornia europaea agg. — обычный галофнтный стеблевой суккулент морских намывных берегов и мелководий (например, Северного моря). От Beta vulgaris subsp. maritima, растения приморских берегов и утесов, произошли важнейшие культурные формы свеклы (сахарная, кормовая, столовая, мангольд). В качестве овощного растения используют шпинат (Spinacea oleracea). Chenopodiaceae в настоящее время рассматривают как часть Amarantha-сеае, от которых они практически не отличаются по своим признакам. Семейство Amaranthaceae содержит различные декоративные, зерновые и рудеральные виды из р. Amaranthus (щирица), у которых цветки частично однополые (рис. 11.239).
Остальные семейства образуют только беталаин. Очевидно, одну линию эволюции составляют семейства Portulacaceae, Basellaceae, Didi-егеасеае и Cactaceae. У Portulacaceae с плодами-коробочками (например, монтия — Montia) и у вьющихся Basellaceae с невскрывающим ися плодами под цветком есть чашечковидная обертка из верховых листьев. Семейство Cactaceae (кактусовые) представлено в основном стеблевыми суккулентами. Их колонновидные и уплощенные (например, Opuntia), продольно-бороздчатые (например, Cereus) или шаровидные и бугорчатые (например, у Mammillaria) стебли почти всегда несут листовые колючки, а часто целые пучки колючек (ареолы) — модифицированные пазушные побеги и зачатки листьев (рис. 11.239). Нормальные листья есть только у базального в семействе рода Pereskia. Однако мелкие чешуйчатые или шиловидные листья бывают и у многих молодых растений других родов (например, у Opuntia). Сидячие цветки с многочленными спирально расположенными листочками околоцветника, наружными, подобными чашелистикам, и внутренними лепестковидными, многочисленными ты
чинками и большим числом сросшихся плодолистиков с нижней или полунижней завязью (рис. 11.239). Плод ягодообразный. В этом семействе опыление обычно осуществляется птицами, летучими мышами и ночными бабочками. Кактусы обитают почти исключительно в Америке, в основном в пустынях и полупустынях юго-запада США, в Мексике и на плоскогорьях Анд. Наряду с множеством мелких форм есть растения, достигающие высоты 15 м (Carnegia gigantea). Многие роды (например, Rhipsalis, Epiphyllum, Zygocactus) обитают в лесах как эпифиты. Одичавшая в Средиземноморье опунция (Opuntia ficus-indica) имеет съедобные плоды. Эндемичные для засушливых районов Мадагаскара Didiereaceae с однополыми цветками — также стеблевые суккуленты с листовыми колючками, но имеют и нормально развитые листья.
К Pltytolaccaceae принадлежит, например, одичавшая в Центральной Европе Phytolacca americana, из которой добывают красное красящее вещество, используемое для окрашивания вин. У близкородственных фитолакковым Nyc-taginaceae околоцветник, вероятно, трубковидно сросшийся, а плодолистик только один. К этому семейству принадлежат известный объект генетических экспериментов ночная красавица (Mirabilis jalapa) и культивируемая в субтропиках и тропиках лазящая лиана бугенвиллия (Bougainvillea), три цветка которой заключены в три больших и ярко окрашенных кроющих листа.
Виды семейства Aizoaceae — листовые суккуленты, имеющие цветки с очень большим числом тычинок и часто с многочисленными, образовавшимися из тычинок лепестками. Иногда они представляют собой более или менее погруженные в почву, похожие на гальку растения («живые камни» — Lithops). Плоды их — обычно коробочки, вскрывающиеся при увлажнении. Ме-sembryanthemim crystallinum — агрессивный сорняк теплых морских побережий.
Polygonales
Dioncophyllaceae 3/3, Африка; Droseraceae, включая Drosophyllaceae 4/100, космополитное; Nepenthaceae 1/75, тропики Старого Света; Plurnbaginaceae 27/650, космополитное, в основном Средиземноморье и Юго-Западная Азия; Polygonaceae 43/1100, космополитное; Simmondsiaceae 1/1, северо-запад Северной Америки; Tamaricaceae 4/78, Евразия, Африка
1 Многие суккуленты из этого семейства имеют хорошо развитые листья. — Примеч. ред.
Порядок: Polygonales: В новых системах часто включается в Caryophyllales, характе-
11.2. Бактерии, грибы, растения | 455
ризуется большей частью нафтахинонами (плюмбагином) и часто только одной базальной семяпочкой в завязи. У Polygona-сеае околоцветник состоит из двух внешне сходных 3-членных кругов или из одного 5-членного круга, тычинки расположены в двух или в одном кругах, а обычно невскрываюгцийся плод содержит только одно базальное семя (рис. 11.240). Листья очередные; их прилистники срастаются в почке в замкнутый конический колпачок, раструб (рис. 11.240), который при дальнейшем росте побега разрывается и охватывает основание междоузлия в виде кожистого футляра. Околоцветник обычно мелких обоеполых или однополых цветков часто невзрачный в связи с ветроопыляемостью, реже лепестковидный, как у гречихи (Fago-ругит) и у насекомоопыляемых горцев (Polygonum).
У щавелей (Rumex) внутренний круг околоцветника остается при плоде в качестве летучки, поплавка или цепляющегося придатка. Одногнездная завязь из трех (2— 4) сросшихся плодолистиков развивается в односемянный орех. Раньше, особенно на
бедных почвах, часто выращивали много гречихи из-за высокого содержания крахмала в перисперме. Из горных областей Центральной и Восточной Азии происходят виды ревеня (Rheum), известного как овощное и лекарственное растение. Для Plumbaginaceae характерны пентамерный, дифференцированный на чашечку и венчик околоцветник, 1 круг противолежащих лепесткам тычинок и паракарпный гинецей из 5 плодолистиков с базальной семяпочкой. К этому семейству принадлежат главным образом ксеро- и галофиты степей, полупустынь и морских побережий. В Центральной Европе широко распространены размножающиеся апо-миктически кермеки (Limonium) и армерия (Armeria) с весьма гетероморфными цветками.
Завязи с центрально-угловой, париетальной или базальной плацентацией имеют семейства Dioncophyllaceae, Droseraceae и Nepenthaceae. Виды этих семейств произрастают обычно в местообитаниях. бедных питательными веществами, и являются насекомоядными растениями (см. бокс 4.4). Dioncophyllaceae с преобразованным в
Рис. 11.240. Polygonales, Polygonaceae (А, В — по Н. Baillon; С, D — по A. W. Eichler: Е — no G. Karsten): А, В — Rheum officinale, общий вид и продольный разрез цветка (увеличено); С, D — диаграммы цветков Rheum и Rumex, Е — часть побега с листом и раструбом (st) Polygonum amplexicaule (1/Зх)
456 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Рис. 11.241. Santalales, Viscaceae, Viscum album (no F. Firbas):
A — побеги с мужскими и женскими цветками и плодами (1 /2х); В, С — мужской и женский трехцветковые дихазии (продольный разрез, листочки околоцветника срастаются с тычинками и соответственно с завязями и семяпочками: (Р4А4) соответствует (Р4С(г)) (ок. Зх)
усик кончиком листа и часть Droseraceae (росянки — Drosera, Drosophyllum) ловят насекомых с помощью образующихся главным образом на листьях клейких чувствительных волосков1. Другие Droseraceae (венерины мухоловки — Dionaea, Aldrovanda) имеют чувствительные к раздражению и складывающиеся вдоль средней жилки листья. У двудомных Nepenthaceae (Nepenthes) в качестве ловушки для животных выступает кувшиновидная пластинка листа с очень гладкой внутренней поверхностью.
Семена Simmondsia chinensis (Simmondsia-сеае) содержат масло жожоба, очень ценимое в современной косметике. Обычно многочисленные в засоленных местообитаниях Tamaricaceae представлены в Центральной Европе произрастающей на горных осыпях мирикарией (Myricaria germanica), а также часто культивируемыми видами тамарисков (Tamarix).
1 Ловчие структуры росянок — эмергенцы, а не волоски. — Примеч. ред.
Santalales
Olacaceae 25—30/250, пантропическое; Opiliaceae 9/50, пантропйческое; Loranthaceae 60 — 70/700, пантропическое, в основном Южное полушарие; Misodendraceae 1/10, умеренный пояс Южной Америки; Santalaceae 35/400, космополитное; Viscaceae 10/350, космополитное
В семействах порядка Santalales представлены (большей частью) полупаразит-ные древесные или, реже, травянистые растения. В их листьях имеются клетки, содержащие кремниевую кислоту. В цветках с верхней или нижней завязью чашечка часто редуцирована и имеется обычно только один круг тычинок, противолежащих лепесткам и срастающихся с ними. Семяпочки тенуинуцеллятные, с двумя или чаще только одним интегументом; часто (особенно у Loranthaceae, Viscaceae) интегумент и нуцеллус практически не выражены, а зародышевый мешок лежит в ткани плаценты. Плоды — обычно клейкие ягоды с единственным семенем, которое у видов без дифференцированной семяпочки не имеет семенной кожуры.
Благодаря наличию хлорофилла эндосперм у Viscaceae зеленый. Если Olacaceae и Opiliaceae паразитируют большей частью на корнях, a Santalaceae на корнях или ветвях, то Misodendraceae (только на Nothofagus) и Viscaceae — большей частью на ветвях.
Семейство Santalaceae, известное своим сандаловым деревом (Santalum album) с сильно пахнущей древесиной, в Центральной Европе представлено паразитирующими на корнях травянистыми лен-цами (Thesium). Как летнезеленый ремнецветник (Loranthus europaeus) с обоеполыми цветками, относящийся к семейству Loranthaceae, так и вечнозеленая омела (Viscum', рис. 11.241) с желтовато-зелеными листьями и однополыми двудомными цветками, представляющая Viscaceae, — древовидные растения, паразитирующие на ветвях своих хозяев с помощью мощных гаусториев.
Они попадают на ветви с помощью птиц, распространяющих их клейкие ягоды. Систематическая дифференциация омел связана, по-ви
11.2. Бактерии, грибы, растения
457
димому. со специфичностью к определенному хозяину (хвойные или лиственные деревья). Экстракты омел, содержащие вискотоксин, препятствующий росту опухолей, применяют в медицине.
Saxifragales
Altingiaceae 3/12, от Юго-Западной до Восточной Азии, Северная и Центральная Америка; Cercidiphyllaceae I/2,Китай,Япония; Crassulaceae 35/1500. космополитное, часто в аридных местообитаниях; Daphniphyllaceae 1/10. Восточная и Юго-Восточная Азия, Grossulariaceae 1/150, Северное полушарие, Анды; Halorhagaceae 8/100, космополитное, преимущественно Южное полушарие; Нагпа-melidaceae 25/80, космополитно-дизъюнктивное; Iteaceae 1 /15, Азия, восток Северной Америки: Paeoniaceae 1 /30, Евразия, редко Северная Америка; Penthoraceae 1/3, Восточная и Юго-Восточная Азия; Pterostemonaceae 1/2, Мексика; Saxifragaceae 30/550, большей частью Северное полушарие, от умеренного до арктического пояса; Tetracarpaeaceae 1/1, Тасмания
Порядок: Saxifragales. В этот порядок объединяют семейства на основе относительно однотипных последовательностей нуклеотидов ДНК, но они значительно различаются по морфологии. Поэтому в ста
рых системах их помещали в очень разных местах. Пожалуй, к общим морфологическим признакам этого порядка можно отнести: строение железок, находящихся на кончиках листьев, пыльцевые зерна, часто имеющие струйчатую поверхность, и частую встречаемость нижней завязи. Вероятно, что Saxifragales являются сестринской группой остальным Rosidae.
Ветроопыляемые и двудомные Altingiaceae, Cercidiphyllaceae и Daphniphyllaceae вместе с Hamamelidaceae, имеющими большей частью обоеполые и энтомофильные цветки, ранее объединяли в «Hamamelidae». К «Dilleniidae» относили Paeoniaceae с единственным родом Paeonia (пион), со спирально расположенными членами околоцветника, центробежно-развива-ющимся многотычинковым андроцеем и свободными плодолистиками, из которых формируются листовки. Paeonia раньше многие относили к Ranunculaceae.
Семейство Grossulariaceae (из которого в качестве самостоятельных семейств были выделены Iteaceae, Pterostemonaceae и Tetracarpaeaceae) с единственным родом Ribes (рис. 11.242) — кустарники, и их нижние завязи преобразуются в ягоды (крыжовник: Ribes uva-crispa, красная и черная смородина: Ribes rubrum, R. nigrum). Травянистые Crassulaceae — листовые суккулен
Рис. 11.242. Saxifragales, Saxifragaceae (А—D) и Grossulariaceae (E—G) (A—no A. W. Eichler; В—G — no F. Firbas):
A — диаграмма цветка Saxifraga granulata с прицветничками (a, 0); В — Saxifraga stellaris (2,5x); C —
S. granulata (1,5x), цветки; D — S. caespitosa, коробочка с чашечкой (Зх); E — Ribes uva-crispa (2,5x);
F, G — R. rubrum, соответственно цветок и ягода (3,5х и 2х)
458 | ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
ты, они имеют обычно от пяти до множества почти свободных плодолистиков в одном круге. В Центральной Европе к этому семейству принадлежат очиток (Sedum) и молодило (Sempervivum), из тропических представителей известно Kalanchoe (объект для изучения фотопериодизма, включая «Bryophyllum» с образующимися по краям листьев выводковыми почками). Виды семейства Saxifragaceae имеют гинецей, состоящий обычно из двух, часто только в основании сросшихся плодолистиков (рис. 11.242), которые более или менее глубоко погружены в цветоложе. Виды крупного рода Saxifrage (камнеломка) продвигаются в аркто-альпийскую область до климатической границы распространения сосудистых растений, образуя там различные жизненные формы (особенно подушковидные и розеточные).
Saxifragaceae в их традиционном понимании оказались чрезвычайно полифилетичными. Подлинное родство различных их представителей впервые удалось установить с применением молекулярных методов. К населяющему в основном пресные воды семейству Halorhagaceae в Центральной Европе относится уруть (MyriophyUum) с ее разделенными на тонкие доли мутовчатыми листьями и редуцированным околоцветником.
цветником и часто свободнолепестным венчиком, двумя кругами тычинок или нередко вторично полимерным андроцеем с развивающимися центробежно или центростремительно тычинками, обычно синкарп-ным гинецеем, крассинуцеллятными семяпочками с двумя интегументами и нуклеарным типом развития эндосперма. «Розиды» часто содержат тригидроксилированные флавоноиды и эллаговую кислоту.
Наряду с названными порядками к «рози-дам» относят также несколько семейств с неясными связями (Aphlodiaceae 1/1, Африка, Мадагаскар, Маскаренские острова; Crossosomataceae 3/10, запад Северной Америки; Ixerbaceae 1/1, Новая Зеландия; Krameriaceae 1/15, Америка, в основном аридные области; Picramniaceae 2/50, тропическая Америка; Stachyuraceae 1/5, от Гималаев до Японии; Staphyleaceae 5/50, Америка, Евразия; Zygophyllaceae 27/230, пантропическое, в основном аридные области).
Geraniales
Francoaceae 2/2, Чили; Geraniaceae 12/700. космополитное умеренного и тепло-умеренного поясов; Greyiaceae 1/3, Южная Африка; Ledocarpaceae 2/11, Южная Америка; Melian-thaceae 2/4, Африка; Vivianiaccae 4/6. Южная Америка
«Розиды»
Следующие 11 порядков (Geraniales, Mal-pighiales, Oxalidales, Fabales, Rosales, Cucurbitales, Fagales, Myrtales, Brassicales, Malvales, Sapindales) объединяют в «розиды». В этой группе цветки обычно с двойным около-
Рис. 11.243. Geraniales, Geraniaceae (А, С — по A.W. Eichler; В — по J.Graf);
А — Geranium pratense, диаграмма цветка; В — вскрывающийся плод (1,5х); С— Pelargonium zonale, диаграмма цветка
Порядок: Geraniales. Порядок в совершенно новом объеме, установленном по молекулярным данным, характеризуется рядом общих структурных признаков, главным образом анатомических (например, трех- или многолакунными узлами, железистыми зубчиками по краю листа), а также тестальными семенами (семенная кожура образуется из внешнего интегумента) и противолежащими лепесткам плодолистиками. Большая часть видов Geraniaceae имеет своеобразные плоды (рис. 11.243). Плодолистики очень длинные, но содержат у основания только по две семяпочки, из которых развивается лишь одна. Верхняя часть плодолистиков образует стерильный «носик». При созревании плода внутренняя часть сросшихся плодолистиков остается в виде центральной колонки, в то время как их внешние стенки, охватывающие внизу по одному семени, закручиваются вверх. При этом они либо остаются связанными с
11.2. Бактерии, грибы, растения | 45g
колонкой верхушками и разбрасывают семена (например, у многих видов гераней: Geranium), либо отделяются вместе с семенами, причем их стерильные гигроскопичные верхние части как буравчики «ввинчивают» семя в почву (например, у аистника — Erodium). Зигоморфные цветки с приросшим к цветоножке шпорцем на чашелистике свойственны преимущественно южноафриканским видам рода Pelargonium, часто разводимым как декоративные растения.
«Эурозиды I»
Порядки Cucurbitales, Fabales, Fagales, Mal-pighiales, Oxalidales и Rosales образуют одну из двух главных линий розид, выделенных по данным молекулярного анализа («эурозиды I»), для которых до сих пор не удается назвать убедительных синапоморфий.
Malpighiales
Achariaceae 3/3, Южная Африка; Balano-расеае 1 /9, юго-западная часть Тихоокеанской области; Сагуосагасеае 2/23, Южная Америка; Chrysobalanaceae 17/450, пантропическое, преимущественно Южная Америка; Clusiaceae 38/1100, пантропическое, редко умеренная зона; Dichapetalaceae 3/235, пантропическое; Егу-throxylaceae 4/200, пантропическое, преимущественно Южная Америка; Euphorbiaceae 303/6700. космополитное; Flacourtiaceae 85/800, пантропическое; Goupiaceae 1/3, Южная Америка; Hugoniaceae 7/60, пантропическое; Humi-riaceae 8/50, преимущественно Южная Америка: Irvingiaceae 4/10, тропики Старого Света: Ixonanthaceae 5/30, пантропическое; Laciste-mataceae 2/20. тропическая Америка; Linaceae 6/220, космополитное, от умеренной зоны до субтропиков; Malesherbiaceae 1—2/25. Южная Америка; Malpighiaceae 66/1200, пантропическое, преимущественно Южная Америка: Ме-dusagynaceae 1/1, Сейшельские о-ва; Ochnaceae 30/400, пантропическое, преимущественно Южная Америка; Pandaceae 3/25, тропики Старого Света; Passifloraceae 18/630, космополитное, от тропиков до тепло-умеренного пояса; Putranjivaceae 4/200, тропики Старого Света: Quiinaceae 4/40, Южная Америка; Rhizopho-гасеае 12/84, пантропическое; Salicaceae 3/390, космополитное, от умеренного пояса до Арктики; Scyphostegiaceae 1/1, Борнео; Trigonia-сеае 3/26, пантропическое; Tumeraceae 8/120, Америка от тропиков до тепло-умеренного пояса, Мадагаскар; Violaceae 25/900, космополитное, преимущественно умеренный пояс
В крупный и морфологически гетерогенный порядок Malpighiales, в основном приуроченный к тропикам Нового света, включают растения с 3-следовыми узлами, 3-членным гинецеем с сухим рыльцем и семенами с волокнистой внешней эпидермой внутреннего интегумента.
Почти полностью тропическое семейство Clusiaceae (=Guttiferae), характеризующееся обычно супротивными или мутовчатыми листьями со схизогенными секреторными вместилищами или ходами, цветками с центробежно развивающимися пучками тычинок (рис. 11.244) и обычно центрально-угловой плацентацией, представлено крупным родом Hypericum (зверобой, около 350 видов), виды которого встречаются и в Центральной Европе. Вероятно, к тропическим Flacourtiaceae, с трудом определимым как семейство и характеризующимся многочисленными тычинками, очень близки Salicaceae. Для этого семейства типичны почти всегда однополые, двудомные и более или менее голые цветки, собранные в сережковидные соцветия (рис. 11.245). В верхней завязи, состоящей из 2 плодолистиков, развиваются многочисленные семена без эндосперма, снабженные летучкой из длинных волосков. Основные роды Populus (тополь) и Salix (ива) представлены деревьями или кустарниками. Листья очередные, с прилистниками. Растения часто цветут до распускания листьев. Сидящие в пазухах кроющих чешуй цветки устроены очень просто: кроме бокальчатого цветоложа у ветроопыляемых тополей и одного или двух чешуевидных нектарников у большинства насекомоопыляемых ив в мужских цветках имеются только свободные тычинки (у Populus много, у Salix часто только две), а в женских — только один пестик. В коробочке развивается множество крошечных семян с пучком длинных волосков, которые созраняют всхожесть лишь несколько дней.
Многие ивы (например, .S’. viminalis, S.fragilis, S. alba) и тополя (например, осокорь — Р. nigra, серебристый тополь— Р. alba) выносят высокий уровень грунтовых вод и относятся к важнейшим древесным породам пойменных лесов и прибрежных кустарниковых зарослей. Как пио-
460 | ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Рис. 11.244. Malpighiales, Clusiaceae (А, В) и Violaceae (С—G) (А, С, G — по J. Graf; В, Е — по A. W. Eichler; D — по F. Firbas; F — по A. F.W.Schimper):
А, В — Clusiaceae, Hypericum: Н. quadrangulum — три многократно расщепленные тычинки, нектарник и завязь; Н. perforatum — диаграмма цветка; С—G — Violaceae: С — Viola alpina (фиалка альпийская), цветок спереди (1 х), D, Е — V. odorata (фиалка душистая), цветок в продольном разрезе (2,5х) и его диаграмма (обратить внимание на шпорец и тычинки, из которых две с нектарными придатками), F, G — V. tricolor, раскрытая коробочка (1,5х) и семена с элайосомами (10х)
Рис. 11.245. Malpighiales, Salicaceae (А—F— по G.Karsten; G — N — по A.F.W.Schimper):
А—F — Populus nigra: А, В — цветущий мужской и плодоносящий женский побеги (3/4х), С, D — мужской и женский цветки с кроющими чешуями, Е, F — плоды и семя (увеличено); G—N — Salix viminalis: G — цветущий мужской побег, J — женская сережка (1х), Н, К — мужской и женский цветки с кроющими чешуями, L, М — плоды, N — семя (увеличено)
11.2. Бактерии, грибы, растения i 4g-|
нерные растения зарастающих лесных прогалин и вырубок широко распространены осина (Р. tre-mula) и ива козья (S. саргеа). Многие виды и легко возникающие межвидовые гибриды ив обрезают каждые 2—3 года, формируя так называемые «безвершинные ивы». Ветви идут на плетение корзин. Низкорослые ползучие ивы (например, 5. refuse, S. herbacea) — характерные растения высокогорий и Арктики. Кора ив содержит различные производные салициловой кислоты, однако медицинское значение ивовой коры сейчас утрачено в связи с развитием производства синтетических производных салициловой кислоты (аспирина).
Виды семейства Violaceae имеют цветки от слабо- до полностью зигоморфных и 3-членный гинецей с париетальной плацентацией. В крупном роде Viola (фиалка, анютины глазки, 500—600 видов) передний лепесток образует шпорец (см. рис. 11.244), в который вдаются выделяющие нектар отростки обеих передних тычинок. Семена имеют элайосомы (см. рис. 11.244). Viola, вероятно, имеет андийское происхождение.
В семейство Passifloraceae, включающее часто культивируемый страстоцвет (Passiflora саеги-1еа), входят лианы с побеговыми усиками, часто с привенчиком, состоящим из многочисленных нитевидных придатков, андрогинофором и 3-членным гинецеем с париетальной плацентацией. Ариллус семян, например, у Passiflora edulis, очень вкусный плод гранадилла.
У следующих семейств в плодолистиках часто находится только по одной-две семяпочки (рис. 11.246). К семейству Lina-сеае принадлежит однолетний узколистный лен с голубыми цветками (Linum usitatissimum), введенный в культуру по
меньшей мере за 6000 лет до Рождества Христова. Лубяные волокна его стеблей служат для изготовления льняных тканей, а в десяти семенах, развивающихся в разделенной перегородками коробочке, содержится льняное масло.
Относящиеся к семейству Erythroxylaceae Erytroxylon coca и Е. novogranatense содержат алкалоид кокаин. У семейства Malpighiaceae на внешней стороне чашелистиков часто расположены парные масляные железки, служащие для привлечения опыляющих их пчел, собирающих масло. К Rhizophoraceae (рис. 11.248, см. 15.2.16) принадлежат важнейшие роды мангровых зарослей — Rhizophora, Bruguiera, Kandelia и Ceriops. Наличие опорных и дыхательных корней и виви-пария, вероятно, является приспособлением к особым условиям существования в этих тропических прибрежных сообществах.
Euphorbiaceae имеют однополые цветки с верхней, часто 3-гнездной завязью с одной или двумя анатропными семяпочками в каждом гнезде. Это деревянистые или травянистые растения с листьями, обычно имеющими прилистники, но иногда с редуцированными листьями и фотосинтезирующими стеблями. Многие представители крупного рода Euphorbia (около 2 000 видов) — стеблевые суккуленты африканских саванн и полупустынь (рис. 11.247). Внешне они сходны с кактусами, это типичный пример конвергентной эволюции (см. 4.1.1 и 4.2.6). Листья у них в основном редуцированы, а прилистники преобразованы в парные колючки. Цветки и соцветия в этом семействе очень разнообразны. Двойной околоцветник имеется, например, у тропического масличного растения Jatropha curcas (рис. 11.247). Цветки с простым околоцветником обычны, например, у среднеевропейских пролесников (Mercurialis). Околоцветник этих ветроопыляемых видов 3-член-ный. В мужских цветках большое число тычинок, а в женских, кроме завязи, имеется еще
Рис. 11.246. Malpighiales, Linaceae, Linum usitatissimum (A, В, D — no G. Dahlgren; C — no A. W. Eichler): А — цветок (lx); В— андроцей (ср. тычинки и стаминодии, st) и гинецей (Зх); С — диаграмма цветка; D — плод раскрывшийся (слева) и в поперечном разрезе (2х)
462 I ГЛАВА 11, СИСТЕМАТИКА И ФИЛОГЕНИЯ
Рис. 11.247. Malpighiales, Euphorbiaceae (A, D— по Е Pax; С, D— по R.von Wettstein, изменено; Е, F — по G. Karsten; G — по О. С. Berg и С. F. Schmidt; Н, J, N. О — по Н. Baillon; К — по A.W. Eichler, изменено; М — по J. Schweiger):
А, В — мужской и женский цветки Jatropha curcas и Mercurialis annua (С D) с чешуйками диска (d), андрофором (а), стаминодиями (s); Е, F — Ricinus communis, соцветие (1/2х) и молодой плод в продольном разрезе; G —N — Euphorbia-. G — Е. resinifera, цветущий суккулентный побег (1х), Н — К — циатий, общий вид, продольный разрез и диаграмма (пунктиром показана всегда отсутствующая железка), L — мужской цветок Е. platyphyllos с цветоножкой (st) и тычиночной нитью (f), М — продольный разрез гнезда завязи Е. myrsinites с семязачатком (sa), семяножкой (fu), присемянником (с) и обтуратором (о) (схематизировано), N — вскрывшаяся коробочка с остающейся центральной колонкой (m) Е. lathyris (увеличено), О — (мужской) цветок Anthostema senegalense с околоцветником (р) (увеличено, ср. L)
3 стаминодия. Цветки с 5-членным околоцветником и древовидно разветвленными тычинками имеет однодомная клещевина Ricinus communis (рис. 11.247, Е, F) — дерево тропической Африки с большими пальчатораздельными ли
стьями, которое в Центральной Европе можно культивировать как однолетнее декоративное растение. Семена его дают также ценное техническое масло (рициновое, или касторовое) и содержат ядовитый протеин.
11.2. Бактерии, грибы, растения | 463
Крайне простые цветки (рис. 11.247) у молочая (Euphorbia). Они собраны в своеобразное соцветие (псевдантий), который называют циа-тием. Каждый циатий состоит из женского цветка, у большинства видов голого, на длинной, часто согнутой цветоножке, который окружен пятью частными соцветиями завитками из также лишенных околоцветника мужских цветков на цветоножках. Каждый мужской цветок состоит только из единственной тычинки, отделенной от цветоножки перетяжкой. Все охвачено пятью напоминающими околоцветник верховыми листьями — кроющими листьями мужских частных соцветий, между которыми часто находятся эллиптические или полулунные нектарники. Эти циатии, в свою очередь, собраны в общие соцветия. То, что тычинка сочленена с цветоножкой, показывает, что циатии следует считать соцветиями. У других родов (например, Anthostema) в месте сочленения находится простой околоцветник. На примере циа-тия видно, как путем интеграции однополых цветков могут возникать псевдантии, опыляемые насекомыми подобно обоеполым цветкам.
Оплодотворение семяпочек происходит обычно при посредстве так называемого обтуратора — выроста плаценты, накрывающего микропиле и служащего для проведения и питания пыльцевой трубки. Плоды обычно коробочки, стенки которых отделяются от центральной колонки, разбрасывая семена.
Многие представители Euphorbiaceae имеют ядовитый млечный сок, часто содержащий каучук. К ним относится наиболее ценный каучуконос семейства — каучуковое дерево Elevea brasiliensis, происходящее из Амазонии и сейчас повсеместно культивируемое в тропиках, из него получают имеющий наибольшее значение в мировой торговле «пара-каучук». Бразильский Manihotglaziovii поставляет «сера-каучук». Из полезных растений, кроме того, следует назвать маниок Manihot esculenta, исходно неотропическое растение. Его крахмалоносные клубни («кассава», «юка», «тапиока», «маниока») нужно отваривать перед употреблением в пищу из-за наличия в них синильной кислоты. Некоторые виды Euphorbia содержат также рыбий и стрельный яды.
Oxalidales
Cephalotaceae 1/1, юго-западная часть Австралии; Соппагасеае ок. 20/300—400, пантропическое, преимущественно Старый Свет; Cunoniaceae 25/350, в основном Южное полушарие, Старый Свет; Elaeocarpaceae 10/400, пантропическое; Oxalidaceae 8/900, тропический и субтропический пояса, редко умеренный пояс; Tremandraceae 3/28, Австралия, Тасмания
Порядок: Oxalidales. Этот порядок, характеризующийся преимущественно молекулярными признаками, отличается наличием эпикутикулярных восковых пластиночек, сухим рыльцем и семенами с волокнистым (благодаря трахеидам) внешним эпидермисом внутреннего интегумента. К семейству Oxalidaceae с часто гетеростильными цветками относится среднеевропейская кислица, представитель очень большого (около 800 видов) рода Oxalis.
У кислипы подвижные, внешне похожие на клевер сложные листья. При созревании плодов семена или выдавливаются наружу при вскрывании коробочки, или выбрасываются вследствие резкого растрескивания коробочки и выворачивания наружу ее мясистого внешнего клеточного слоя. Съедобны плоды принадлежащей к этому семейству карамболы
Рис. 11.248. Malpighiales, Rhizophoraceae (А — по G. Karsten; В — по К. von Goebel; С— по W. Troll):
А—В — вивипарные Rhizophoraceae: А — Rhizophore mucronata, побег с цветками и плодами (1/5х); В, С — Bruguiera gymnorhiza, соответственно молодой плод в продольном разрезе и зрелый плод, внешний вид; h — гипокотиль; к — семядоли; р — околоплодник
464 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Рис. 11.249. Fabales, Fabaceae, Caesalpinioi-deae. Cassia angustifolia, цветущий побег и боб (1/2х) (по О.С.Berg и C.F.Schmidt)
(Averrhoa carambola). У насекомоядного растения Cephalotus follicularis (Cephalotaceae) некоторые розеточные листья удивительно похожи на кувшинчатые листья Nepenthaceae и Sarraceniaceae.
У растений следующих четырех порядков (Fabales, Rosales, Cucurbitales, Fagales) с очень редуцированным эндоспермом относительно часто встречается симбиоз с азотфиксирующими бактериями {Frankia, Rhizobium).
Fabales
Fabaceae 640/17200, космополитное; Poly-galaceae 12/750, космополитное; Quillajaceae 1/3, Южная Америка; Surianaceae 4/6, Австралия, Тихоокеанская область
Виды порядка Fabales часто имеют прилистники, апокарпный и обычно состоящий только из одного плодолистика гинецей, а также семена с большим и часто зеленым зародышем.
Для семейства Fabaceae (=Leguininosae) особенно характерна верхняя завязь из единственного плодолистика, из которого образуется своеобразный многосемянный вскрывающийся как по брюшному шву, так и по спинной жилке плод боб («бобовые», рис. 11.249). Семена обычно без эндосперма, зародыш богат крахмалом, белком и в меньшей мере — жирами, особенно в семядолях.
Fabaceae — деревянистые или травянистые растения с обычно очередными, различного вида перистыми листьями с весьма заметными прилистниками. На корнях имеются многочисленные клубеньки с симбиотическими, фиксирующими азот воздуха видами Rhizobium.
Подсемейство Caesalpinioideae (150/2200) имеет зигоморфные цветки с восходящим сложением лепестков (рис. 11.250): оба нижних лепестка охватывают два боковых, а боковые — верхний. Тычинки, как правило, свободные. Caesalpinioideae — субтропические и тропические древесные растения с преимущественно парно- или дваждыпарноперистыми листьями. Вероятно, исходные представители этой, явно не монофилетической, группы не обладали корневыми клубеньками. Среди них известно два средиземноморских вида: рожковое дерево (Ceratonia siliqua) с невскрываюшимися съедобными бобами и часто выращиваемое, обладающее каулифлорией иудино дерево (Cercis siliquastrum). К этой группе принадлежит также
Рис. 11.250: Fabales, Fabaceae, диаграммы цветков (по A. W. Eichler):
Mimosoideae: А — Mimosa pudica; В — Acacia lophantha. Caesalpinioideae: C — Cercis siliquastrum; D — Cassia caroliniaca. Faboideae: E— Vida faba (чашечка в основании более или менее сростнолистная); F — Laburnum anagyroides
11.2. Бактерии, грибы, растения | 465
Рис. 11.251. Fabales, Fabaceae, Mimosoideae, Acacia (A — по О. C. Berg и C. F. Schmidt; В — по H. Bail-Ion; C, D— no F. Noll):
A, В —A. catechu, цветущий побег (1/2x) и отдельный цветок (5х); С, D — A. nicoyensis из Коста-Рики. Побег (уменьшено) с полыми, выгрызенными муравьями (I) для жилища прилистниковыми колючками (d); листья с экстрафлоральными нектарниками (п), питательными тельцами, или «тельцами Бельме» (f), на кончиках листочков (D) (увеличено)
крупный род Cassia (см. рис. 11.249), некоторые виды рода используют в качестве лекарственных растений.
У Mimosoideae (50/3000) цветки радиально симметричные и число тычинок обычно вторично увеличено (рис. 11.251; см. рис. 11.250). Это тропические и субтропические древесные растения и травы с обычно дважды- или парноперистыми листьями и мелкими, собранными в головчатые или колосовидные соцветия цветками. Цветки часто 4-мерные с хорошо заметными длинными окрашенными тычиночными нитями. Пыльцевые зерна часто распространяются большими массами (полиадами).
К этому подсемейству принадлежат известная своими быстрыми движениями листьев мимоза стыдливая (Mimosa pudica) — пантропическое травянистое растение и очень крупный род Acacia, включающий многие виды с листовидными филлодиями, а также растения, сожительствующие с муравьями. Из коры некоторых видов добывают гуммиарабик и дубильные вещества.
У видов подсемейства Faboideae (440/ 12 000) цветки зигоморфные с нисходящим сложением лепестков. Их «мотыльковые» цветки, большей частью собранные в кистевидные соцветия, имеют 5-членную и обычно спайнолистную чашечку. В 5-член-ном, обычно свободнолепестном венчике адаксиальный (в бутоне) лепесток, или флаг, налегает на боковые лепестки, называемые крыльями, а они в свою очередь — на оба абаксиальных, которые образуют, часто склеиваясь внутренними краями, лодочку. Она заключает в себе большей частью сросшиеся нитями тычинки (все 10 или 9 + 1) и пестик (рис. 11.252).
Листья бывают парно- или непарноперистые, пальчатые (Lupinus), тройчатые (Trifolium) или простые. У разных родов (например, Vicia, Pisum) вместо конечного листочка, а часто и вместо верхних пар листочков развиты усики.
466 | ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Рис. 11.252. Fabales, Fabaceae, Faboideae (по RFirbas)
А — Astragalus gummifer, цветущий побег с листовыми колючками (1 /2х); В, С — Pisum sativum'. В — цветок, внешний вид (1х), С— цветок, разобранный по частям (1,2х): чашечка (к), венчик из флага (1), крыльев (fl) и лодочки (s), тычинки (sb) (9 + 1), а также пестик из одного плодолистика (fk) с рыльцем (п) и семязачатками (пунктир)
Главными органами фотосинтеза могут быть прилистники (Lathyrus aphaca) или, у некоторых слабо олиственных прутьевидных и колючих кустарников (жарновец — Cytisus scoparius; дрок — Genista', утёсник — Ulex), — стебли.
Цветы опыляют главным образом пчелы и шмели. Они имеют разнообразные приспособления, которые содействуют выходу или выбросу пыльников либо выдавливанию пыльцы, когда садящийся на цветок опылитель давит соответственно на крылья или лодочку. Бобы изредка бывают членистыми (разламывающимися на отдельные односемянные части) или односемянными орешками.
Рис. 11.253. Fabales, Polygalaceae, Polygala (А, В — по J.Graf, О. С. Berg и С. F. Schmidt; С — по A. W. Eichler):
А — Р. senega, внешний вид цветка; В — Р. ата-га, цветок в продольном разрезе (увеличено): зеленоватые (kJ и венчиковидные (к2) чашелистики, передний лепесток (с,) с бахромчатым придатком (а), в основании сросшимся с боковыми лепестками (с2), тычинки (s), завязь (д) со столбиком, диск (d); С — Р. myrtifolia, диаграмма цветка
Очень богатое видами подсемейство Faboideae распространено космополитно, причем в тропиках преобладают древесные, а во внетро-пических областях травянистые формы. Как растения, связывающие азот воздуха1, они предпочитают сухие, бедные питательными веществами или содержащие кальций почвы и особенно часто встречаются в евразиатских степях и полупустынях. Здесь обычен, например, содержащий более 2000 видов астрагал (Astragalus), особенно виды, относящиеся к секции Traga-cantha, куда входят колючие кустарники и по-душечники. Мотыльковые играют большую роль в различных растительных сообществах Центральной Европы.
Faboideae имеют большое хозяйственное значение. Некоторые ценные кормовые растения хорошо растут на бедных азотом почвах, а будучи запаханными, могут служить «зеленым удобрением» (различные виды клеверов — Trifolium pratense, Т. hybridum, Т. repens, Т incarnatuni; люцерна — Medicago sativa; эспарцет — Onobrychis vicifolia; сераделла — Omithopus sativus; люпины — Lupinus angustifolius, L. luteus). Другие, имеющие богатые белками, крахмалом и иногда жирами семена, дают пищевые продукты (бобы — Vida faba', горох — Pisum sativum', нут — Cicer arietinum; чечевица — Lens culinaris; фасоль — Phaseolus vulgaris, P. coccineus', соя — Glycine mas; земляной орех — Arachis hypogaea). У земляного ореха не-раскрывшиеся плоды зарываются в землю благодаря удлинению нижней части завязи и доз
1 Это неточно: атмосферный азот связывают симбиотические ризобии, а сами растения не способны это делать. — Примеч. ред.
11.2. Бактерии, грибы, растения | 457
ревают там. Средн деревянистых представителей важное значение имеет происходящая из восточной части Северной Америки робиния (Robinia pseudoacacia), применяющаяся для залесения засушливых районов и пустошей. В качестве декоративных растений разводят ядовитый южноевропейский золотой дождь (Laburnum апа-gyroides) и восточно-азиатскую глицинию (Wisteria chinensis).
Цветки многих видов семейства Polygalaceae имеют поверхностное сходство с цветками Faboideae: два лепесткообразных чашелистика, а также лодочковидная форма абаксиального лепестка с бахромчатым придатком на вершине (например, истод — Polygala). Тычинки (в основном их 8), могут срастаться в открытую сверху трубку, а обычно два плодолистика срастаются друг с другом, что нетипично для порядка (рис. 11.253).
Rosales
Barbeyaceae 1/1, Африка, Аравия; Саппа-Ьасеае 2/3, Северное полушарие; Cecropiaceae 6/280, иантропическое; Celtidaceae 9/150, космополитное, большей частью тропическое; Dirachmaceae 1/2, Сомали, Сокотра; Elaeag-пасеае 3/50. в основном умеренный пояс Северного полушария; Могасеае 40/1000, космополитное; Rhamnaceae 55/900, космополитное, в основном тропическо-субтропическое; Rosaceae 100/3000, космополитное, часто от умеренного пояса до субтропиков; Ulmaceae 6/30. космополитное; Urticaceae 45/700, космополитное, в основном тропическо-субтропическое
Порядок: Rosales. Характеризуется наличием гипантия (который, правда, у многих семейств редуцирован) и семенами с малым количеством эндосперма или без него. Многие представители порядка — деревянистые растения с прилистниками.
Представленное в европейской флоре многими видами, семейство Rosaceae включает деревянистые и травянистые растения с очередными, простыми или сложными листьями с прилистниками. Радиально-симметричные цветки часто имеют многочисленные тычинки в результате вторичного увеличения их числа. Разнообразие проявляется прежде всего в строении апокарпного гинецея и плода (рис. 11.254; 11.255).
Традиционное подразделение семейства на четыре подсемейства, основанное в большой
степени на этих признаках, должно быть частично пересмотрено с учетом молекулярных данных. Для Rosoideae обычны многочисленные плодолистики, развивающиеся в орешки или костянки (рис. 11.254; 11.255), основное хромосомное число большей частью п = 7, есть и некоторые химические особенности. У дриады (Dryas) и гравилата (Geum) стилодии вырастают в перистые или крючковидные придатки, служащие для распространения плодиков. Орешки могут также оставаться соединенными за счет разрастания мясистого цветоложа: так, у розы (Rosa) они находятся внутри кувшинчатого гипантия (плол цинародий), у земляники (Fragaria) конусовидная мясистая ось цветка несет на себе орешки снаружи. У малины (Rubus idaeus) и ежевики (R. fruticosus agg.) дру1 с другом соединены костянки. У Amyg-daloideae (=Prunoideae, косточковые), например у рода Prunus, плодолистик большей частью один и развивается в костянку, основное хромосомное число и = 8. Единственный, не срастающийся с чашевидным гипантием плодолистик образует снаружи мякоть плода, а внутри очень твердую, обычно односемянную косточку (рис. 11.254, 11.255), как у черешни (Prunus avium), вишни (Р. cerasus). сливы (Р. domestica). персика (Р. persica), абрикоса (Р. armeniaca) и миндаля (Р. amygdalus. с кожистым мезокарпием). Их (а также некоторых Maloideae) отличает содержание в семенах и в других частях растений гликозидов синильной кислоты (см. рис. 11.168). Семечковые Maloideae имеют 2 — 5 плодолистиков, нижнюю завязь и основное хромосомное число в большинстве случаев п = = 17). Плод большей частью — яблоко (рис. 11.254; 11.255). У боярышников (Crataegus) и мушмулы (Mespilus germanica) эндокарпий деревянистый и плод содержит несколько односемянных косточек по числу плодолистиков. У айвы (Cydonia oblonga), груш (Pyrus), яблонь (Matus) и видов рода Sorbus (например, 5. аиси-paria— рябина обыкновенная; .S', domestica — рябина домашняя) пергаментовидный эндокарпий состоит из нескольких частей и содержит несколько семян. В мякоти плода иногда есть еще и отдельные группы каменистых клеток. Роды, ранее включаемые в Spiraeoideae, имеющие многосемянные листовки (рис. 11.254; 11.255). не составляют монофилетической группы, они разбросаны по генеалогическому древу Rosaceae.
К Rosaceae принадлежат содержащие множество видов благодаря наличию агамоспермии роды Rosa, Rubus и Alchemilta (манжетка). Частичное ветроопыление отмечено у Sanguisorba.
Хозяйственное значение наряду с ягодными культурами (земляника, малина и ежевика) име-
468 | ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Рис. 11.254. Rosales, Rosaceae. Продольные разрезы цветков (А, В, D— F — по F. Firbas; С — по К. von Goebel):
А— Spiraea lanceolata, В — Fragaria vesca (1,5x); C— спиральное расположение (1—5) последовательно упрощающихся чашелистиков у Rosa (ср. рис. 4.6); D — Rosa canina (3/4х); Е — Pyrus communis (1,5х); F— Prunes avium (1,5х)
ют многочисленные фруктовые деревья. Из них яблоня, груша и черешня имеют дикие формы, распространенные в Европе, которые вместе с тёрном (Prunus spinosd), черемухой (Р. padus) и другими видами человек использовал уже в раннем каменном веке. Родина айвы, мушмулы, миндаля, вишни, слив — Передняя Азия, где также
есть свои дикие формы яблони, груши и черешни. Абрикосы происходят из Средней Азии и западного Китая, персик — из Китая. Их культурные формы были распространены в Европе уже в греко-римское время.
Цветки Rhamnaceae имеют только один эпи-петальный круг тычинок и полунижнюю или
Рис. 11.255. Rosales, Rosaceae. Продольные разрезы плодов (схематично) (по F. Firbas):
А — Spiraea; В — Potentilla; С — Fragaria; D — Rosa; Е — Rubus; F — Mespilus; G — Males; H — Prunus. Мякоть плода обозначена пунктиром, проводящие пучки — штриховой линией, твердый слой стенки плода (семенная кожура) — черным цветом
11.2. Бактерии, грибы, растения | 469
Рис. 11.256. Rosales, Rhamnaceae, Frangula alnus (A — no G. Karsten; В — по О. C. Berg и C. F. Schmidt;
C — no A.W. Eichler):
A — цветущий и плодоносящий побеги (0,5х); В — цветок в продольном разрезе (5х); С — диаграмма цветка
нижнюю завязь. В то время как цветки крушины ломкой (Frangula alnus), обычной в пойменных лесах, 5-мерные и обоеполые (рис. 11.256), у видов рода Rhamnus, например у крушины слабительной (R. catharticus), они 4-мерные и однополые. Костянки имеют от двух до четырех тонкостенных косточек. У видов этого семейства, так же как у близкородственного Elaeagnaceae, обнаружен симбиоз с Frankia, благодаря чему возможна ассимиляция N2. У деревянистых растений семейства Elaeagnaceae, покрытых чешуйчатыми волосками, от чего они часто выглядят серебристо-блестящими, чашечка лепестковидная, венчик отсутствует, а гинецей состоит из одного плодолистика с нижней завязью и единственной семяпочкой. К этому семейству относится произрастающая на дюнах и речных галечниках ветроопыляемая облепиха (Hippophae rhamnoides).
Все остальные семейства Rosales (кроме Dirachmaceae) ранее объединяли в порядок Urticales. Они отличаются содержащимися в отдельных клетках круглыми цистолитами карбоната кальция, невзрачными цветками с самое большее пятью ты
чинками, одногнездным гинецеем из двух плодолистиков с единственным семязачатком. У семейства Ulmaceae цветки обоеполые (рис. II.257). Это древесные растения без млечного сока, представленные в Европе вязами. Горный вяз (Ulmusglabra) произрастает в основном в горных смешанных лесах, полевой (U. minor) и гладкий (U. laevis) вязы — соответственно в равнинных и в пойменных лесах. У них двухрядные очередные резко асимметричные листья и собранные в зонтиковидное соцветие цветки, из которых уже ко времени разворачивания листьев формируются крылатые орехи.
В 1919 г. в Нидерландах впервые наблюдали вымирание вязов, вызванное грибом (Ceratocystis ulmi), который переносится жуком — вязовым за-болонником.
Семейство Celtidaceae, представленное, например, родом Celtis (каркас), выделяют из Ulmaceae главным образом по причине однополых цветков. И все последующие семейства также имеют однополые цветки. Семейство Могасеае
470 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
представлено в большинстве своем древесными растениями с млечным соком. Из них для добывания каучука используют главным образом мексиканскую Castilla elastica и азиатский Ficus elastica. У Могасеае часто формируются своеобразные соцветия и соплодия. Так, например, мелкие отдельные плоды каждого женского соцветия одно- или двудомного тутового дерева, или шелковицы (Morus), при созревании объединяются в съедобную «ягоду» благодаря разрастающимся мясистым листочкам околоцветника
(рис. 11.257). Листья белой шелковицы (М. alba) являются кормом для гусениц тутового шелкопряда. Крупные съедобные плоды индо-малайского хлебного дерева (Artocarpus) по строению похожи на плоды Moms. У видов Dorstenia и Castilla цветки и отдельные плоды собраны на блюдцевидном или кувшиновидном осевом образовании, а у рода Ficus (ок. 750 видов) они полностью погружены в кувшиновидное полое внутри образование из оси соцветия, которое становится мясистым, как и листочки около
Рис. 11.257. Rosales (А, В, К—М— no G. Karsten; С, Е, F — по A. W. Eichler; D, Н, J — по «Sillabus» A. Engler; G — по Р. Е. Duchartre; N — по J.Graf):
А—D — Ulmaceae, Ulmus minor: А, В — цветущий и плодоносящий побеги (1/Зх), С — диаграмма цветка, ( — отдельный обоеполый цветок (увеличено); Е—J — Могасеае: Е, F — диаграммы мужского и женского цветков Morus alba, G — соплодие М. nigra, Н — соцветие Dorstenia contrayerva, J — соцветие Castilla elastica (продольный разрез) (все ок. 1х, или несколько увеличено, ср. также рис. 11.192, Н —L); К—N— Cannabaceae, Humulus lupulus: К, L— цветущий мужской и плодоносящий женский побеги (0,5х), М — частное соцветие с кроющим листом и двумя женскими цветками с каймовидным рудиментом околоцветника (увеличено), N — мужской цветок Cannabis sativa (увеличено)
11.2. Бактерии, грибы, растения | 471
цветника. У средиземноморского фигового дерева (F. carica) эти соплодия съедобны (инжир). Многие виды фикусов — вечнозеленые деревья тропических лесов, часто очень мощные. Азиатский баньян (F. bengalensis) прорастает на ветвях дерева и сначала развивается там в виде мощного эпифита. Корни его опускаются до почвы. Они развиваются в массе подобно колоннообразным стволам, уничтожая в конце концов дерево, на котором баньян поселился. Так как от горизонтальных ветвей отрастают все новые корни, также достигающие почвы, то со временем из одного проростка формируется целый «лес». Такое поведение свойственно и другим видам фикуса. У фикусов очень тесные взаимоотношения с опыляющими их осами, жизненный цикл которых в сущности протекает внутри соцветий и соплодий.
К травянистым Cannabaceae (рис. 11.257), не содержащим млечного сока, относятся только роды хмель и конопля. Хмель (Humulus lupulus) — двудомное многолетнее вьющееся растение пойменных и заболоченных лесов с шершавыми от крючковидных волосков стеблями. Его шишковидные соплодия имеют хорошо заметные кроющие листья, на которых развиты железки, содержащие смолы и горечи, на чем основано применение растения в пивоварении и медицине. Конопля (Cannabissativa) происходит из Южной Азии. Она также двудомная, но однолетняя. Ее разводят прежде всего из-за длинных (1 — 2 м) лубяных волокон1, в меньшей степени— из-за богатых маслом семян. Высушенные верхушки побегов разных форм растения курят («марихуана»), а из смолы изготовляют «гашиш». У видов Urticaceae семяпочки прямые. В их мужских цветках тычинки в бутоне загнуты внутрь и зажаты, при распускании цветка они мгновенно разворачиваются подобно пружине и выбрасывают при этом пылевидную пыльцу. У некоторых родов (например, Piled) таким же способом стаминодии разбрасывают плоды. Некоторые роды из семейства Urticaceae, например крапива (Urtica), имеют жгучие волоски. Как текстильное растение используют Urtica dioica, но прежде всего — азиатскую Boehmeria nivea (рами).
1 Отдельные волокна микроскопического размера. Авторы, вероятно, имели в виду тяжи волокон. — Примеч. ред.
Cucurbitales
Anisophyllaceae 4/10, пантропическое, большей частью Старый Свет; Begoniaceae 1—5/1000, пантропическое; Coriariaceae 1/5, от Мексики до Чили, западная часть Средиземноморья, от Гималаев до Японии, Новая Гвинея, Новая Зеландия; Согупосаграсеае 1/5, Новая Зеландия, северо-восточная часть Австралии, Новая Гвинея; Cucurbitaceae 90/700, по всему миру от субтропиков до тропиков, редко умеренный пояс; Datiscaceae 2/3, запад Северной Америки, Азия; Tetramelaceae 1/1, Малайзия
Порядок: Cucurbitales. Включает в основном травянистые растения, обычно без эпи-кутикулярных восковых кристаллов, листья с пальчатым жилкованием и характерными зубцами. Цветки часто однополые, с гинецеем со свободными стилодиями, с нижней завязью и париетальной плацентацией. Плаценты сильно врастают в полость завязи и даже могут соприкасаться в ее центре. У Cucurbitaceae имеются спайнолепестный венчик и осевые усики (рис. 11.258). Проводящие пучки биколлатеральные. Однополые цветки могут быть одно- или двудомными, например у среднеевропейских переступней (соотв. Bryonia alba и В. dioica). В мужских цветках имеется пять тычинок, каждая часто только с одной текой (моно-тековые), большей частью срастающиеся группами (например, 2 + 2 + 1) или все сросшиеся (рис. 11.258), пыльник часто искривлен или s-образно согнут. Из обычно 3-гнездной паракарпной завязи развивается толстокожая и многосемянная ягода (рис. 11.258).
Известные представители семейства — происходящая из тропической Америки тыква (Си-curbita реро с сортами. выращиваемыми как овощные культуры и на маслосодержащие семена), изначально обитавший в тропической Азии огурец (Cucumis sativus), дыня (Cucumis melo) с желтой мякотью и арбуз (Citrullus lanatus) — с красной мякотью, разводимые в тропиках для производства разных сосудов горлянка, или посудная тыква (Lagenaria), используемая для изготовления растительных мочалок Lujfa и средиземноморский бешеный огурец (Ecballium). Виды семейства Begoniaceae с ярко выраженной асимметрией листьев часто разводят как декоративные растения.
472 | ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Рис. 11.258. Cucurbitales, Cucurbitaceae (А—Е, Н—К— по G.Hegi; F, G— по A. W. Eichler):
А—Е — Bryonia alba: А, В — цветущий и плодоносящий побеги, С — корнеплод (почти 1/4х), D, Е — мужской и женский цветки (почти 2х); F, G — Citrullus colocynthis, диаграммы мужского и женского цветков; Н — плоды Cucurbita pepo (почти 1/6х), J, К — Cucumis sativus (почти 1/Зх)
Fagales
Betulaceae 6/150, умеренный пояс Северного полушария, Анды; Casuarinaceae 4/100, Австралия, Малайзия, Меланезия; Fagaceae 7/700, от умеренного пояса до тропиков, в основном Северное полушарие; Juglandaceae 8/60, умеренный и тропический пояс Евразии и Америки; Myricaceae 3/45, космополитное; Nothofagaceae 1/35, Южное полушарие; Rhoipteleaceae 1/1, юго-западная часть Китая, Северный Вьетнам; Ticodendraceae 1/1, Центральная Америка
Порядок: Fagales. Семейства порядка — преимущественно ветроопыляемые кустарники или деревья с железистыми или звездчатыми волосками. Часто однополые, но обычно однодомно распределенные цветки имеют простой, как правило, очень редуцированный околоцветник и, как правило, нижнюю завязь с очень небольшим числом семяпочек в плодолистике. Пыльцевая трубка проникает в (незрелую) семяпочку через халазу, плод — обычно односемянный орех, семя без эндосперма.
У Fagaceae плодолистиков три (редко больше) (рис. 11.259). Женские цветки име-
>
Рис. 11.259. Fagales, Fagaceae (А, В — по A. W Eichler; С — по K.Prantl и W. Troll; D—Н — по G. Karsten; J — Р — соответственно no А. Е. W. Schimper и О. С. Berg и С. F. Schmidt; карты ареалов— по K.Rubner и др.):
А — С — диаграммы женских дихазиев: А — Castanea, В — Fagus (а, а,, Р, PJ, С— Quercus (прицветники (Ь) и предлистья — черные, плюска показана крапом, простой околоцветник — белый, редуцированные цветки и соответственно их прицветники и предлистья обозначены звездочкой или заштрихованы; ср. также схему на рис. 11.260, A); D—Н — Fagus sylvatica: D — цветущий побег, Е, F — мужской и женский цветки с простым околоцветником (р), G — плюска с двумя орехами, Н — поперечный разрез ореха со складчатыми семядолями зародыша (D G — натур, величина, Е, F, Н — увеличены); J— Р — Quercus robur: J — цветущий побег, К — мужской цветок с тычинками, L, М — женский цветок, внешний вид и продольный разрез (с рыльцем (п), столбиком (д), простым околоцветником (р), завязью (f), семяпочками (s) и плюской (с)), N — группа плодов, О — зрелая плюска, Р — семена в продольном и поперечном разрезе (К—М увеличены). Естественные ареалы европейских видов Fagus и Quercus
11.2. Бактерии, грибы, растения
473
474 | ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
ют, как правило, два 3-членных круга околоцветника, а в мужских цветках число листочков околоцветника и тычинок варьирует. Плоды окружены одревесневшей и покрытой чешуйками или колючками плюской, которая, возможно, возникает благодаря слиянию стерильных ветвей соцветия.
Древнейшие ископаемые находки семейства, по-видимому, относятся к дизъюнктивно распространенному в Юго-Восточной Азии и в Колумбии рода Ttigonobalanus, у которого в общую плюску заключено до 7 орехов. У настоящего каштана (Castanea sativa) частично наблюдается насекомоопыление, мужские цветки расположены в прямостоячих соцветиях, а плюска часто содержит 3 съедобных плода (каштана). Из-за этих плодов разводят в теплых районах Центральной Европы вид еще со времен Древнего Рима. У полностью ветроопыляемого бука (Fa-gus sylvatica) мужские цветки находятся помногу в (по-видимому, дихазиальных) головках, женские — в 2-цветковых дихазиях. Открывающаяся четырьмя створками плюска содержит, соответственно, два 3-гранных богатых маслами ореха (буковые орешки). Область распространения бука как доминирующей лесной породы охватывает прежде всего Центральную Европу. Особенно хорошо он растет на пологих склонах гор, но также и на равнинах, где он предпочитает не слишком бедные, хорошо аэрированные и не очень сухие почвы, проявляя в таких местах чрезвычайно высокую конкурентоспособность. Его континентальная граница, обусловленная температурным режимом и увлажнением, проходит от Мазурских озер до восточных предгорий Карпат. Мужские и женские дихазии дуба (Quercus) одноцветковые. Соответственно их плоды (желуди) сидят поодиночке в чашевидной чешуйчатой плюске. Из сильно гибридизирующих между собой аборигенных видов теплолюбивый дуб черешчатый (Q. robur, плоды на длинных плодоножках, листья короткочерешковые) распространен на большей части Европы — от Ирландии до русских лесостепей, особенно на равнинах и в нижнем горном поясе в различных местообитаниях. Дуб скальный (Q. petraea, листья длинночерешковые, плоды на коротких плодоножках) имеет меньший ареал. Родственный ему дуб пушистый (Q. pubescent), проникающий в теплые регионы Центральной Европы, происходит из Средиземноморья. Наряду с очень ценной и твердой столярной и строительной древесиной используют также и кору дуба в дубильном производстве. Из многих вечнозеленых видов следует отметить три средиземноморских — западно-средиземноморский пробковый дуб
(Q. suber), служащий сырьем для получения пробки, кермесовый дуб (Q. coccifera) и каменный дуб (Q. ilex).
Ранее включаемое в Fagaceae семейство Nothofagaceae с родом Nothofagus отличается морфологией пыльцы и наличием у семяпочек только одного интегумента. Nothofagus распространен на материках Южного полушария кроме Африки и известен в ископаемом состоянии в Антарктике.
Виды семейства Betulaceae имеют только 2 плодолистика. Исходно их цветки были целиком 2-мерными, но в дальнейшем были редуцированы. Тычинки часто расщепленные. Орешки голые или окружены оберткой. У березы (Betulcr, рис. 1L260) и ольхи (Alnus; рис 1Е260) плоды находятся в пазухах чешуй, которые образованы прицвет-ничками, сросшимися с кроющим листом; у березы чешуи опадают ко времени созревания плодов, а у ольхи одревесневают и остаются в шишковидном соплодии. У лещины (Corylus), граба (Carpinus, рис. 11.261) и хмелеграба (Ostryd) — эти три рода иногда выделяют в семействе Corylaceae — плоды окружены оберткой из припветничков и кроющего листа. У граба эта обертка служит крылом.
Из видов ольхи почти по всей Европе распространена черная ольха (Alnus glutinosa), важнейшая порода сырых болотистых и приречных лесов на равнинах. В горах, особенно на речных галечниках, такую же роль играет цир-кумбореальная серая ольха (A. incana, с сероопушенными с нижней стороны листьями), вблизи верхней границы леса — кустарниковая зеленая ольха (A. viridis). Виды ольхи существуют в симбиозе с азотфиксирующим актиноми-иетом Frankia.
Светолюбивые березы (береза повислая — Betula penduta; береза пушистая — В. pubescent) — нетребовательные древесные породы бедных почв, особенно песков и болот. Изредка встречающаяся в Центральной Европе только на болотах карликовая березка (В. папа) широко распространена в северотаежных лесах и Арктике.
У раннецветущей, распространенной в лесах и кустарниках на большей части Европы обыкновенной лещины (Corylis avellana) соцветия закладываются на прошлогодних ветвях. Из коротких, прикрытых почечными чешуями женских соцветий выдвигаются только красные рыльца Тяжелые, распространяемые птицами и белками лесные орехи содержат очень богатый жира-
11.2. Бактерии, грибы, растения | 475
Рис. 11.260. Fagales, Betulaceae (А — по A.W. Eichler, изменено; В—N — по G. Karsten):
А — диаграммы дихазиальных мужских (слева) и женских (справа) частных соцветий; схема вверху: в пазухе кроющего листа (Ь) цветок А, в пазухах его прицветников аир цветки В' и В с прицветнич-ками а', р' и аъ р, * — редуцированные цветки, штриховая линия — листочки их простых околоцветников; В—G — Alnus glutlnosa: В — цветущий побег и лист, С, Е — мужской и женский дихазии с кроющим листом и прицветничками, D — женская сережка, F — соплодие, G — орех (В — натур, величина, С—G увеличены); Н —N — Betula pendula: Н— цветущий побег и листья, J, L— мужской и женский дихазии, К— расщепленная тычинка, М — соплодие, N — крылатый орех (Н, М — 2/Зх, остальные увеличены)
476 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
ми зародыш. В торговле появляются также орехи некоторых южноевропейских видов (С. maxima — ломбардский орех; С. colurna— лещина древовидная).
Только на побегах текущего года (у мужских, часто еще безлистных) находятся цветки граба (Carpinus betulus, с остро-дваждымелкопильчаты-ми листьями). Граб распространен по всей Центральной Европе и играет большую роль вне области распространения бука в низинных лесах, так как он устойчивее бука к более континентальному (соответственно к более теплому) климату и почвам с близкими грунтовыми водами. В северной части Средиземноморья выше пояса каменно-дубовых лесов распространен хмелеграб (Ostrya carpinifolia) с орехами, полностью заключенными в обертку.
Вилы семейства Casuarinaceae, распространенные в очень сухих местообитаниях, а во влажных тропиках — на приморских песках, напоминают мутовчатым расположением своих чешуевидных листьев хвощи и эфедру (Ephedra). У мужских цветков один или два листочка околоцвет
ника и одна тычинка, у женских цветков околоцветник отсутствует, а пестик из двух плодолистиков. У видов этого семейства так же, как и у Мупсасеае, к которому относится распространенный на приатлантических болотах и пустошах душистый восковник (Myrica gale) с голыми цветками, известен симбиоз с Frankia.
Семейства Rhoipteleaceae и Juglan daceae в противоположность вышеназванным семействам имеют перистосложные листья. У Juglandaceae, например у часто культивируемого грецкого ореха (Juglans regia), цветки однополые (рис. 11.262). Его мужские цветки во множестве сидят в плотных сережках, которые распускаются из прошлогодних пазушных почек, а женские — в небольшом количестве на верхушках годичных побегов. В цветках обоих типов от трех до пяти листочков околоцветника срастаются с кроющим листом и двумя прицветничками. Плод грецкого ореха
Рис. 11.261. Fagales, Betulaceae, Carpinus betulus (область естественного распространения — по G.Karsten; Е — по М. Bflsgen; карта ареала — по К. Rubner и др.):
А — цветущий побег; В, D — мужской и женский дихазии; С — расщепленная тычинка; Е — женский цветок; F — плоды; G — орех с разросшимися кроющим листом и прицветниками (A, F, G — почти натур, величина, остальные увеличены)
11.2. Бактерии, грибы, растения | 477
Рис. 11.262. Fagales, Juglandaceae (А — по G. Hegi; В, С, Е — соответственно по О. von Kirchner, F. Fir-bas, A.W. Eichler; D— no W.Troll; F—G— no P. Hanelt; все слегка изменено):
A—E— Juglans regia: A— цветущий побег с мужскими и женскими соцветиями, В, С— мужские и женские цветки и их диаграммы с кроющим листом (d), прицветниками (v), листочками простого околоцветника (р) и рыльцем (n), D— костянка после отделения экзокарпия (е) (передняя часть удалена) от косточки (st), Е — костянка в поперечном и продольном сагиттальном разрезах с эндокарпием (склерокарпий, sk), ложным швом (медианной линией вскрывания (т), поперечной перегородкой (= настоящая перегородка, es) и медианной перегородкой (=ложная перегородка, fs), а также зародыш с семядолями (к) и почечкой (pl). Плоды: F — Engelhardtia sp. и G — Pterocarya sp. с кроющим листом (d) и прицветничками (v), трансформированными в крылья
(рис. 11.262) — костянка, его косточка при прорастании вскрывается вдоль заранее сформировавшейся борозды. В съедобных семенах запасные вещества отложены в богатых маслами лопастных семядолях.
Деревья грецкого ореха происходят из Средиземноморской области и севернее Альп страдают от морозов. Как и другие виды рода Juglans (например, североамериканский J. nigra), равно как и североамериканские виды Сагуа (гикори) и кавказская лапина (Pterocarya fraxinifolia), он ценится за свою древесину. У Pterocarya и тропической Engelhardtia орехи крылатые. Немногие семейства цветковых растений так же хорошо представлены в ископаемом состоянии и изучены, как Juglandaceae.
К настоящим «розидам I» относят семейства Celastraceae (50/800, пантропическое), Ниасеае (2/3, тропическая Африка), Pamassiaceae (2/50, умеренный пояс Северного полушария, от Центральной до Южной Америки) и Stack-housiaceae (3/25, Австралия, Новая Зеландия, Юго-западная тихоокеанская область), которые не удается включить ни в один из вышеназванных порядков. К Celastraceae из представителей среднеевропейской флоры относится бересклет (Euonymus, рис. 11.263) с окрашенной в розовокрасный цвет коробочкой, освобождающей четыре или пять семян, полностью заключенных каждый в оранжево-красный ариллус. Относимое ранее к Saxifragaceae семейство Pamassiaceae включает широко распространенный в умеренной зоне Северного полушария белозор (Par-
478 | ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Рис. 11.263. Celastraceae. Euonymus europaea (А — по J. Graf, В — по A. W. Eichler):
А — цветок; В — диаграмма цветка
nassia) с одним кругом тычинок, с которыми чередуются противолежащие лепесткам, большей частью глубокораздельные стаминодии (бокс 4.1. рис. D).
«Эурозиды II»
К «эурозидам II» с простой перфорацией сосудов, гинецеем со столбиком и семенами с крайне скудным эндоспермом относят наряду с семейством Tapisciaceae (2/3, Китай, Карибский бассейн, Южная Америка) с неясными связями также порядки Brassicales, Malvales, Sapindales и, вероятно, Му-rtales.
Myrtalcs
Alzateaceae 1/2, Южная Америка; Combre-taceae 20/400, пантропическое; Crypteroniaceae 1/4, от Индии до Малайзии; Heteropyxidaceae 1/3, Юго-Восточная Африка; Lythraceae 27/530, пантропическое, редко умеренный пояс: Melastomataceae 170/5000, пантропическое, редко умеренный пояс; Myrtaceae 148/3500, пантропическое; Oliniaceae 1/8, Африка, о-в Св. Елены; Onagraceae 17/675, от умеренного пояса до субтропического, часто Америка; Репаеасеае 7/20, Южная Африка; Psyloxylaceae 1/1, Маскаренские о-ва; Rhynchocalycaceae 1/1, Южная Африка; Vochysiaceae 7/200, тропическая Америка, I вид в Западной Африке
Порядок: Myrtales. Виды порядка часто имеют супротивные и цельнокрайные листья с прилистниками или без них, би-коллатеральные проводящие пучки, обычно 4-мерные цветки, как правило, с выраженным гипантием, в бутоне часто внутрь согнутые тычинки, один столбик и часто очень многочисленные семязачатки.
Семейству Myrtaceae свойственна, как правило, нижняя завязь. Это разнообразные вечнозеленые субтропические и тропические древесные растения, часто с лизигенными вместилищами выделений с эфирными маслами, вследствие чего они имеют значение как пряности и лекарственные растения. Нередко цветки заметны благодаря не околоцветнику, а многочисленным тычинкам (рис. 11.264) с ярко окрашенными тычиночными нитями. Из многих видов тропических родов Eugenia и Syzygium следует особо упомянуть распространенное от Шри Ланки до Борнео гвоздичное дерево (.S', aromaticum = £. са->yophyllata\ рис. 11.264). В Австралии в большинстве сухих лесов доминирует род Eucalyptus с почти 500 древесными и кустарниковыми видами. Молодые и взрослые растения часто имеют листья разной формы. Опыляют Eucalyptus главным образом птицы, а также летучие мыши и мелкие сумчатые. Многие виды могут достигать высоты до 100 м (например, Е. regnans) и относятся к самым высоким деревьям. В теплых областях Земли, как, например, в Средиземноморье, выращивают разные виды эвкалиптов, особенно £. globulus, из-за их быст рого роста. Там же встречается единственное европейское миртовое — широко выращиваемый в Европе мирт (Myrtus communis).
Древесные и травянистые виды семейства Melastomataceae особенно обычны в тропиках и субтропиках Нового света, они часто имеют дуговидные надсвязники и вскрывающиеся порами пыльники. Опыляющие их пчелы извлекают пыльцу из пыльника с помощью вибрации («опыление жужжанием»),
У преимущественно травянистых Onagraceae гипантий почти всегда удлиненный, в виде длинной трубки над нижней завязью (рис. 11.264). К этому семейству относятся происходящие из Америки, но сейчас широко распространенные по всему миру сорняки ослинники (Oenothera) — важный объект генетических экспериментов, а также происходящие из Южной и Центральной Америки и отчасти Новой Зеландии и широко культивируемые во многих местах виды рода Fuchsia, опыляемые преимущественно птицами; у них гипантий и чашечка ярко окрашены. В Центральной Европе растут виды кипреев (Epilobi-ит), двухлепестников (Circaea) и людви-гий (Ludwigid).
Преимущественно травянистые растения часто с 2-членным гинецеем относят к Lythraceae. Из них особо следует назвать
11.2. Бактерии, грибы, растения | 47g
Рис. 11.264. Myrtales (A, D— по G. Karsten; В, С, F— по A.W. Eichler; Е— по Е Firbas):
А—С— Myrtaceae: А, В— Syzygium aromaticum, цветущий побег (4/9х), бутон в продольном разрезе, раскрытый цветок (почти 2/Зх) и диаграмма, С — Myrtus communis, диаграмма цветка; D — Lythraceae, Punica granatum, цветок в продольном разрезе (4/5х); Е, F — Onagraceae, Oenothera biennis: Е— цветок в продольном разрезе с гипантием (Ь) и нектарником (n) (1/2х), F— диаграмма цветка
известный благодаря своей триморфной гетеростилии Lythrum salicaria.
В Lythraceae сейчас включают древесные и частью обитающие в мангровых зарослях Son-neratiaceae, а также кустарниковые Punicaceae и водные травы Тгарасеае. К бывшим Punicaceae относится единственный род Punica с происходящим с востока гранатовым деревом (Р. granatum), имеющим расположенные в два или три яруса плодолистики (рис. 11.264). К Тгара, единственному роду бывших Тгарасеае, относится однолетнее плавающее растение водяной орех (Тгара natans), чьи очень твердые орехи имеют формирующиеся из чашелистиков роговидные придатки.
Brassicales
Akaniaceae 2/2, Австралия, Китай; Bataceae 1/2, Южная Америка, Новая Гвинея, Австралия; Brassicaceae 350/3000, космополитное, в основном умеренный пояс; Саррагасеае 42/800, космополитное, в основном от тропиков до субтропиков; Caricaceae 4/30, тропическая Америка, Африка; Emblingiaceae 1/1, западная часть Австралии; Gyrostemonaceae 5/17, Австралия; Koeberliniaceae 1/1, Америка; Limnanthaceae 2/11, Северная Америка; Moringaceae 1/10, от Африки до Индии;
Pentadiplandraceae 1/2,Западная Африка; Re-
Pentadiplandraceae 1/2,Западная Африка; Re-sedaceae 6/70, Северное полушарие, в большинстве случаев Старый Свет; Salvadoraceae 3/12, тропики Старого Света; Tovariaceae 1/2, тропическая Америка; Tropaeolaceae 3/92, Америка
Порядок: Brassicales. Существенный признак порядка — наличие горчичных гликозидов и фермента мирозиназы. При нарушениях ткани содержащаяся в мешковидных идиобластах мирозиназа вступает в контакт с освобождающимися из других клеток гликозидами, преобразуя их в горчичное масло, служащее защитой от травоядных. Кроме Brassicales, горчичные гликозиды есть только у рода Drypetes (Putranji-viaceae, ранее Euphorbiaceae, порядок Malpighiales). Другие признаки порядка — париетальная плацентация и часто зеленые зародыши.
В основании порядка находятся, вероятно, семейства с 5-мерными цветками и прямым зародышем, как, например, Caricaceae, Moringaceae и Tropaeolaceae. Caricaceae известны по плодам папайи (Carica papaya), а к Tropaeolaceae с зигоморфными цветками со шпорцем относится
480 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
повсеместно разводимая настурция (Tropaeolum majus). Большая часть остальных семейств характеризуется 4-мерными цветками и согнутым или сложенным зародышем. Слабо зигоморфные цветки у Resedaceae, которые в Центральной Европе представлены видами рода Reseda (резеда). В этом семействе иногда плодолистики на верхушке срастаются не полностью, так что можно видеть семяпочки внутри завязи. Семейство Саррагасеае — преимущественно древесные растения. У Capparis spinosa, бутоны которого употребляют как каперсы, андроцей состоит из многочисленных тычинок. У видов Cleotne завязь разделена на 2 гнезда ложной перегородкой.
Важное семейство, легко узнаваемое по строению цветка, — Brassicaceae (=Cruci-ferae, крестоцветные). Это большей частью травянистые многолетние и однолетние растения с соцветиями кистями без конечного цветка и обычно без кроющих листьев. Их дисимметричные цветки обычно имеют 4-членную чашечку, 4 чередующихся с чашелистиками лепестка, 2 внеш
них коротких и 4 внутренних, более длинных, тычинки, верхнюю, часто сидящую на гинофоре завязь, образованную двумя плодолистиками, с вырастающей из плацент ложной перегородкой (рис. 11.265). Плод обычно стручок. При его вскрывании плацента и кожистая ложная перегородка с налегающими на нее семенами остаются на ножке плода в виде рамки с перегородкой. Семена формируются из кампилотропных семяпочек, зародыш содержит масла.
Для подразделения этого богатого видами семейства ранее использовали строение плода (кроме вскрывающегося стручка бывают также и разламывающиеся членистые стручки, распадающиеся дробные стручки и одно- или мало-семянные ореховидные стручочки), форму и положение зародыша и расположение нектарных железок. Результаты ДНК-анализа показали, что основанная на этих признаках система во многом очень искусственна.
Крестоцветные распространены преимущественно во внетропических областях Северного
Рис. 11.265. Brassicales, Brassicaceae (А — фото R. Greissl; В — по A.W. Eichler и J. Alexander; С, Н, J, М — по F. Firbas; D, Е—G. К, L— по H.Baillor):
А — Arabidopsis thaliana', В — диаграмма цветка; С, D — внешний вид цветка (2х) и цветок с удаленным околоцветником (в основании цветка видны нектарные железки; 4х) (Cardamine pratensis). Плоды: Е — Erysimum cheiri, стручок; F — Capsella bursa-pastoris, стручочек; G — Lunaria annua, стручочек, створки плода удалены, видна полупрозрачная перегородка (s — плодоножка, g — гинофор); Н — Isatis tinctoria, 1 —2-семянный крылатый орех; J — Biscutella laevigata, дробный плод. К—М — поперечные срезы семян, различное положение зародыша, семядолей (к), гипокотиля и зародышевого корешка (г): К — Erysimum cheiri («краекорешковое», 8х), L — Alliaria petiolata («спиннокорешковое», 7х), М — Brassica nigra («ортоплокное», 9х)
11.2. Бактерии, грибы, растения | 43-|
полушария. Они богато представлены в Арктике и в высокогорьях вплоть до границы распространения растительности. С точки зрения истории флор, интересен полиплоидный комплекс Biscutella laevigata (см. рис. 10.39). Многие виды широко расселились как синантропные и автогамные полевые сорняки и рудеральные растения (например, Capsella bursa-pastoris, виды Lepidium и Thlaspi). Из полезных растений имеют значение как овощные и кормовые культуры различные формы капусты (Brassica oleracea), репы (В. rapa subsp. гора), брюквы (В. napus subsp. rapifera), редьки и редиса (Raphanus sativus). как масличные и пряные растения — рапс (Brassica napus subsp. napus), масличная репа (В. rapa subsp. oleifera), черная и белая горчицы (Brassica nigra и Sinapis alba), хрен (Armoracia rusticana), как декоративные растения — желтофиоль (Erysimum = Cheiranthus cheiri), левкой (Matthiola), иберийка (Iberis) и др. Arabidopsis thaliana (рис. 11.265) с очень коротким жизненным циклом и маленьким геномом сейчас является важнейшим объектом исследования в области ботанической молекулярной генетики.
Поскольку Саррагасеае парафилетичны по отношению к Brassicaceae, оба эти семейства следует объединить в одно семейство Brassicaceae.
Malvales
Bixaceae 1/4, Америка; Cistaceae 8/200, космополитное, большей частью умеренный и тепло-умеренный пояса; Cochlospermaceae 2/20, пантропическое; Diegodendraceae 1/1, Мадагаскар; Dipterocarpaceae 16/600, пантропическое, в особенности Азия; Malvaceae s.l., включая Bombacaceae 75/1500, преимущественно пантропическое; Malvaceae s. s, 20— 30/200 космополитное, в основном умеренный пояс; Sterculiaceae 65/1000, космополитное, большей частью тропическое и субтропическое; Tiliaceae 50/450, космополитное, большей частью тропическое и субтропическое; Muntingiaceae 1/1, тропическая Америка; Neuradaceae 3/10, Африка до Индии; Sar-colaenaceae 10/30, Мадагаскар; Sphaerosepala-ceae 2/14, Мадагаскар. Thymelaeaceae 50/500, космополитное
Порядок: Malvales. Это хорошо отграниченный порядок благодаря слоистой флоэме (чередуются твердые и мягкие слои), наличию слизевых ходов и вместилищ, звездчатым волоскам и центробежному вторично полимерному андроцею. К Thymelaeaceae, часто имеющим венчи
ковидные окрашенные гипантий и чашечку и сильно редуцированные лепестки, принадлежит ядовитое волчье лыко (Daphne mezereum), растение лиственных лесов западной Евразии, чьи розово-фиолетовые цветки раскрываются еще до распускания листьев. Виды Cistaceae с многочисленными свободными тычинками представлены в средиземноморском маквисе многочисленными видами пахучего благодаря наличию ароматической смолы кустарникового рода Cistus с большими, пестрыми и быстро отцветающими цветками. Виды Helianthemum (солнцецвет), Fumana (румянка) и Tuberaria также встречаются в Центральной Европе. В азиатских тропиках важным компонентом лесов являются виды семейства Dipterocarpaceae (например, Dipterocarpus, Shorea), интенсивно используемые как источники смол и древесины.
Молекулярные исследования ясно показали, что трудно разграничиваемые семейства Tiliaceae, Bombacaceae и Sterculiaceae не являются монофилетическими. На современном уровне знаний эти три семейства лучше всего включить в широко понимаемое семейство Malvaceae s. 1. Это семейство в таком расширенном объеме характеризуется пальчатонервными листьями, нектарниками из железистых волосков, развивающимися, как правило, на внутренней стороне элементов околоцветника, створчатыми чашелистиками и часто скрученными в бутоне лепестками (рис. 11.266). Число тычинок нередко вторично увеличенное, андроцей развивается центробежно. Часто благодаря совместному росту в высоту пучков тычинок формируется тычиночная трубка, окружающая пестик и срастающаяся в основании с венчиком. Вследствие этого пыльники как бы приподняты на колонке, из-за чего раньше порядок называли «Colum-niferae» (рис. 11.266). Malvaceae в этом объеме можно подразделить на 9 подсемейств.
Более или менее свободные тычинки имеют представители бывшего семейства Tiliaceae, а также некоторые роды бывшего семейства Sterculiaceae. Из бывших Tiliaceae в Центральной Европе произрастают только виды липы (Tilid) — мелколистная липа сердцевидная (Т. cordata) и крупнолист-
482 | ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Рис. 11.266. Malvales, Malvaceae (А — по A.W. Eichler; В — по О. С. Berg и С. F. Schmidt; С, М, N — по R.von Wettstein; D —F — no G. Karsten; G — no F.Firbas; H — no H.Schneck; J —L — по H. Bailion):
A—C— Tilia: A—диаграмма цветка, В — соцветие (1х), цветонос (st), сросшийся с крыловидным предлистом (b), С — орех (поперечный разрез) с околоплодником (f), рудиментарным (s) и 1 зрелым семенем, содержащим внутри эндосперм (es) и зародыш (е) (4х); D—F — Theobroma cacao: D — цветущий и плодоносящий ствол (сильно уменьшен), Е — цветок, F — андроцей с длинными стаминодиями (почти 2х); G — диаграмма цветка Malva с подчашием (ak); Н — бутон (1 х); J — раскрытый цветок (продольный разрез, 1,5х); К— цветок с колонковидно сросшимися тычинками и выступающим сверху рыльцем (5х); L— дробный плод (4х) Malva sylvestris', М — цветок; N — раскрывшаяся коробочка с волосками семян соответственно Gossypium herbaceum и G. vitifolium (3/4х)
11.2. Бактерии, грибы, растения | 4
ная липа широколистная (Г. platyphyllos) (рис. 11.266). Tilia, по-видимому, представляет собой единственный род нового подсемейства Tilioideae.
Липы — насекомо- и ветроопыляемые деревья с дихазиальными соцветиями, которые срастаются с крупным крыловидно разрастающимся предлистом, позже служащим для распространения ветром всего соплодия. Пятигнездная завязь содержит по 2 семязачатка в каждом из 5 гнезд, из которых, однако, развивается только один, так что плод представляет собой односемянный орех. Липы произрастают в смешанных лесах на обогащенных почвах. Если поздно распускающаяся липа сердцевидная широко распространена в Европе и заходит на континентальные равнины, то раньше цветущая липа широколистная — скорее дерево среднего горного пояса, северная граница распространения которого лишь немного захватывает среднегорье Германии.
Из тропических и субтропических видов бывшего семейства Tiliaceae упомянем поставляющие джутовое волокно виды Corchorus и капскую спарманнию (Sparmannia africana) с чувствительными тычинками. Sparmannia сейчас включают в подсемейство Grewioideae. У остальных представителей Malvaceae s.l. тычинки более или менее сросшиеся. К бывшим Bombaca-сеае относят, например, роды Ceiba, волоски стенок плода которого дают не пригодное к прядению волокно (капок), и Adansonia, например вид A. digitate — африканское обезьянье дерево, или баобаб, запасающее воду в стволах и опыляемое летучими мышами. Adansonia сейчас включают в подсемейство Bombacoideae. Важнейший представитель бывшего семейства Sterculiaceae — происходящее из Америки, но сейчас повсюду разводимое в тропиках шоколадное дерево (Theobroma cacao) с крупными, простыми листьями и развивающимися прямо на стволе цветками (каулифлория) (рис. 11.266). Его большие нераскрываюшиеся плоды («какао-бобы») содержат многочисленные семена, из очень крупных семядолей которых, содержащих много жиров, получают масло какао, а после частичного отжима — какао-порошок, а также алкалоид теобромин. Виды западноафриканского тропического рода Cola содержат в семенах кофеин. Theobroma и Cola — представители бывшего семейства Sterculiaceae — на эволюционном древе, построенном по молекулярным данным, принадлежат к совершенно различным линиям.
В то время как одна часть бывших Bombacaceae и Sterculiaceae имеет 4-гнезд-
ные пыльники, другая часть этой группы, так же как и Malvaceae s.s., — 2-гнездные. Преимущественно травянистые Malvaceae s.s. (сейчас это подсемейство Malvoideae) характеризуются обычно многопоровыми пыльцевыми зернами с шиповатой поверхностью и 2-гнездными пыльниками, но очень слабо отграничены от Bombacoideae. Пестик содержит 3—5, но иногда до 50 плодолистиков (рис. 11.266). У более примитивных видов он развивается в многосемян-ную коробочку или распадается на односемянные мерикарпии по числу плодолистиков.
Коробочки свойственны хлопчатнику (Go-ssypium), кустарниковые или однолетние культурные формы которого получены путем скрещивания и полиплоидизации нескольких азиатских тропическо-субтропических видов с африканскими и американскими (например, G. arboreum и G. herbaceum: 2х, G. hirsutunr. 4х). Хлопок состоит из одноклеточных волосков на околоплоднике, достигающих в длину до 60 мм. Добываемое из семян масло имеет большое значение для производства маргарина. Дробные плоды имеются у европейских травянистых видов мальв (Mail’d), алтея (Althaea), среди последних — алтей лекарственный (A. officinalis), старинное га-лофильное лекарственное растение), а также декоративная шток-роза (A. rosea).
Sapindales
Anacardiaceae 75/600, пантропическое, редко умеренный пояс; Biebersteiniaceae 1/4, от Греции до Западной Сибири, запад Тибета; Burseraceae 16/600, пантропическое; Kirkiaceae 2/6, Африка и Мадагаскар; Meliaceae 51/550, пантропическое; Nitrariaceae 3/16, от Южной Европы до Азии, Северная Африка, Австралия, Северная Америка; Rutaceae 150/1500, космополитное; Sapindaceae 144/1630, пантропическое, редко умеренный пояс: Simarou-baceae 24/150, пантропическое
Порядок: Sapindales. Это преимущественно тропические и субтропические деревья часто с перистыми или рассеченными листьями, обычно без прилистников. Древесина содержит кремниевые включения. Обычно пятимерные и радиально-симметричные цветки часто имеют хорошо выраженный выделяющий нектар интрас-таминальный диск.
484 | ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Рис. 11.267. Sapindales, Rutaceae (А—С— по G. Karsten; D— по A. W. Eichler):
А— Citrus sinensis, цветущий побег (1/2х); В—D — Ruta graveolens, цветущий побег (1/2х), четырехмерный боковой цветок и диаграмма пятимерного верхушечного цветка с диском (d)
Рис. 11.268. Sapindales, Sapindaceae (А — по A.W Eichler; В, С — по G. Karsten и J.Graf):
А—С — Acer: А, В — A. pseudoplatanus, диаграмма цветка (ср. экстрастаминальный диск, d), женский (в продольном разрезе) и мужской цветки (почти 2х), С — A. negundo, мужской и женский цветки с редуцированным околоцветником и без диска (почти 2х)
Горькие тритерпеноиды и схизо-ли-зигенные вместилища выделений с эфирными маслами и смолами обнаружены у Rutaceae, Simaroubaceae и Meliaceae. Важнейший род семейства Rutaceae — Citrus (рис. 11.267). Его происходящие из Южной Азии невысокие вечнозеленые деревья сейчас культивируют в виде многочисленных культурных форм во всех теплых странах. К ним относятся Citrus sinensis (апельсин), С. maxima (помело), С. paradisi (грейпфрут), С. Птоп (лимон), С. medica (цитрон) и С. reticulata (мандарин). Плоды цитрусов — ягоды. Часто у них увеличено число плодолистиков и даже число кругов плодолистиков. Мякоть плодов образована сочными выростами на внутренней стороне стенки плода, врастающими в полости завязи. Phellodendron из Восточной Азии — дерево; полукустарники и высокие травы представляют теплолюбивые европейские виды: ясенец {Dictamnus alba) со слабо зигоморфными цветками и цветущая желтовато-зелеными цветками средиземноморская рута {Ruta graveolens, рис. 11.267).
11.2. Бактерии, грибы, растения i 435
Горькие тритерпеноиды, преобладающие у видов семейства Simaroubaceae, обусловливают фармацевтическое значение коры и древесины видов родов Quassia, Simarouba и Picrasma. Часто разводят восточноазиатский айлант (Ailanthus aldssima), натурализовавшийся в теплых областях Цензральной Европы. К этому семейству относят также ветроокыляемую и двудомную Leitneria из юго-восточной части Северной Америки. Ценную тропическую древесину (например, красное дерево) дают виды Meliaceae. Смоляные ходы есть у Anacardiaceae: например Anacardium occidentale— орехи кешью; Pistacia — фисташка, мастичное дерево; Rhus — сумах (красящие вещества и лаки, иногда контактные яды); Mangifera indica — манго и Burseraceae (например, мирра — Commifera, Boswellia).
В преимущественно тропическое семейство Sapindaceae со слабо зигоморфными и часто однополыми цветками в настоящее время включают Hippocastanaceae и Асегасеас. Один из двух родов бывшего сем. Hippocastanaceae — Aesculus с происходящим из горных районов Балканского полуострова и широко культивируемым в других местах конским каштаном (A. hippo-castanuth). Клен (Acer) включает около 110 видов; это один из двух родов бывшего семейства Асегасеае. Ему свойственна редукция нескольких тычинок и гинецей из 2 плодолистиков, развивающийся в дробный плод (рис. 11.268). Чаще всего длане-видные листья супротивные.
Явор (Acer pseudoplatanus) распространен в Центральной Европе в основном в горных лесах, а цветущий ранней весной клен остролистный (A. platanoides) и клен полевой (А. сатре-stre) — на низменных равнинах. Интересна тенденция к образованию однополых и частично сильно редуцированных цветков в связи с развитием в пределах рода ветроопыления. Североамериканский клен ясенелистный (A. negun-do), занесенный в Центральную Европу, — растение двудомное.
«Астероды»
Представители «астерид» имеют 5-мерные цветки с чашечкой и большей частью сростнолепестным венчиком и часто с одним кругом тычинок. Гинецей обычно це-нокарпный, число плодолистиков часто уменьшено. Семяпочки только с одним
интегументом, тенуинуцеллятные, эндосперм развивается по целлюлярному типу. В качестве вторичных метаболитов часто имеются иридоиды, индоловые и стероидные алкалоиды, полиацетилен и сесквитерпеновые лактоны.
Comales
Cornaceae 14/130, в основном умеренный пояс Северного полушария, редко тропики или Южное полушарие; Grubbiaceae 1/3, Южная Африка; Hydrangeaceae 17/170, в основном Северное полушарие от умеренного пояса до субтропиков; Hydrostachyaceae 1/20, Африка, Мадагаскар; Loasaceae 14/200, Северная и Южная Америка, редко Аравия, Африка.
Порядок: Cornales. Семейства порядка содержат иридоиды, их цветки обычно 4-мерные, часто с редуцированной чашечкой, нижней или полунижней завязью, в каждом плодолистике заключена одна или две висячих семяпочки, имеется инграста-минальный диск.
Семейство I lydrangeaceae с часто культивируемыми родами Hydrangea (гортензия; со стерильными краевыми цветками соцветия, имеющими разросшиеся чашелистики) и Philadelphus (чубушник) характеризуется нижней или полунижней завязью и 2-круговым или вторично полимерным андропеем. Родственные им Loasaceae (например, Loasa) имеют жгучие волоски.
К Cornaceae, обладающим четырехмерными цветками, только одним кругом тычинок, гинецеем из двух или трех плодолистиков и нижней завязью (рис. 11.269), из растений Центральной Европы относятся дёрен белыми цветками (Comus sanguinea), кустарник светлых лиственных лесов, и имеющий желтые цветки до распускания листвы теплолюбивый кизил (С. mas), со съедобными плодами-костянками. Травянистый дёрен шведский С. suecica — арктический вид с четырьмя окружающими компактное соцветие большими белыми верховыми листьями (псевдантий). Виды Hydrostachyaceae — обитатели проточных водоемов, у них сильно модифицированные вегетативные органы и редуцированные, однополые и, вероятно, вегроопыляе-мые цветки.
486 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Рис. 11.269. Cornales, Cornaceae, Cornus mas (no G. Karsten):
A, В — цветущий и плодоносящий побеги (1 /2х); С, D — цветок: С — вид сверху, D — продольный разрез (увеличено)
Рис. 11.270. Ericales, Primulaceae (А — по A. W. Eichler; В— по F. Firbas):
А — диаграмма цветка Primula vulgaris, имеется только внутренний, эпипетальный круг тычинок (внешний рудиментарный не показан); В — почти зрелый плод Р. elatior в продольном разрезе, с чашечкой (к), околоплодником (Г), центральной плацентой (р) и семенами (1,5х)
Ericales
Actinidiaceae 3/30. тропическая и субтропическая Азия; Balsaminaceae 2/ок. 800, в основном тропическая Африка и Азия, редко умеренный пояс; Clethraceae 1/65, тропики Старого и Нового Света без Африки; Cyrilla-сеае 3/14, Америка; Diapensiaceae 6/18, Северное полушарие, от Арктики до умеренного пояса; Ebenaceae 5/450, пантропическое; Ericaceae 130/2700, космополитическое; Fou-queriaceae 1/11, сухие области юго-запада Северной Америки; Halesiaceae 1/5, восток Северной Америки, Восточный Китай; Lecythi-daceae 20/400, пантропическое; Marcgraviaceae 5/100, тропическая Америка; Myrsinaceae 33/850, по всему миру от тропиков до субтропиков, редко умеренный пояс; Pellicieraceae 1/1, Центральная и Южная Америка; Podoste-maceae 35/200, пантропическое; Polemoniaceae 16/320, большей частью умеренный пояс, особенно запад Северной Америки; Primulaceae 20/1000, преимущественно умеренный пояс Северного полушария; Roridulaceae 1/2, Южная Африка; Sapotaceae 53/1100, пантропическое; Sarraceniaceae 3/15, Северная Америка до севера Южной Америки; Styracaceae 9/145, Америка, Средиземноморская область, Юго-Восточная Азия; Symplocaceae 1/250, тропики Старого и Нового Света без Африки; Тегп-stroemiaceae 10/300, пантропическое; Tetrame-ristaceae 2/2, Южная Америка, Юго-Восточная Азия; Theaceae 16/300, по всему миру от тропиков до субтропиков, редко умеренный пояс; Theophrastaceae 4/100, в основном тропики Нового Света; Tristichaceae 5/10, пантропическое
Порядок: Ericales. Очень широко понимаемый здесь порядок едва ли можно охарактеризовать морфологическими признаками. По меньшей мере некоторые представители всех изученных семейств все же имеют характерные анатомические особенности зубчиков листовой пластинки — одна из жилок входит в зубчик, а сам зубчик часто имеет опадающий колпачок. У видов семейства Ericaceae на месте колпачка располагается многоклеточный и часто железистый волосок.
Хорошо различима группа родственных семейств Theophrastaceae, Primulaceae и Myrsinaceae, которой присущи свободная, часто шарообразная центральная плацента и отсутствие эписепального (наружного) круга тычинок (рис. 11.270). Если у дре-
11.2. Бактерии, грибы, растения | 487
Рис. 11.271. Ericales, Ericaceae (А, В — по A.W. Eichler; С, D — по F. Firbas; Е—J — по О.С. Berg и С. F. Schmidt):
А, В — диаграммы цветков: А— Pyrola rotundifolia, В — Vaccinium vitis-idaea, тычинки (в естественном положении); С— Vaccinium myrtillus; D — Andromeda polifolia (Юх); E—J— Arctostaphylos uva-ursi: E — цветущий побег, F — цветок в продольном разрезе, G — пыльцевая тетрада, Н, J — костянка (внешний вид и поперечный разрез) с 5 косточками (F —J увеличены)
весных Theophrastaceae этот круг представлен стаминодиями, то у Primulaceae и Муг-sinaceae он отсутствует. Из травянистых Primulaceae, например у вербейника (Lysima-chia) и очного цвета (Anagallis arvensis), побеги удлиненные. Cyclamen (альпийская фиалка) с гипокотильным клубнем, имеющий центр ареала в средиземноморско- ' юго-западноазиатской области, широко распространенный, особенно в высокогорьях, Primula (первоцвет) с часто гетеро-стильными цветками, Soldanella (сольданел-ла), растущая у снежников в европейских горах, и Androsace (проломник), включающий подушечные растения нивального пояса гор, — розеточные растения.
К основному ядру Ericales принадлежат, в частности, семейства Actinidiaceae и Ericaceae. Эта группа отличается тем, что пыльники в процессе развития загибаются так, что их верхний конец направлен вниз (рис. 11.271), столбик чаще всего полый.
К Actinidiaceae с их обычно многочленным андроцеем, вскрывающимися отверстиями пыльниками и пыльцевыми зернами, частично остающимися в тетрадах относится киви (Actinidia chinensis).
В семейство Ericaceae (вересковые) сейчас включают семейства Epacridaceae, Pyrolaceae и Monotropaceae. Собственно Ericaceae — древесные, обычно кустарничковые растения часто с вечнозелеными, очень мелкими, чешуевидными или игловидными листьями, но нередки в семействе и большие деревья (например, некоторые виды рода Rhododendron) с нормально развитой листвой. Вересковые играют большую роль в растительности субарктических и арктических кустарничковых тундр, верховых болот и на грубогумусных почвах хвойных лесов, вблизи верхней границы леса в горах, в средиземноморском маквисе и на пустошах южноафриканской Капской провинции. Благодаря микотрофности они могут поселяться на крайне бедных минеральными веществами почвах. Венчик у них чаще всего спайнолепестный (свободнолепестный, например, у багульника, Ledum palustre'), пыльцевые зерна часто распространяются тетрадами (рис. 11.271). У большинства родов завязь верхняя и развива-
1 Венчик Ledum спайнолепестный с очень короткой трубкой. — Примеч. ред.
488 | ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Рис. 11.272. Ericales, Theaceae, Camellia sinensis (A—C — no G. Karsten):
A — цветущий побег (1 /4х); В — плод; С — семя
ется большей частью в многосемянную ло-кулицидную коробочку, как, например, у рододендрона {Rhododendron), подбела {Andromeda), у Erica (например, у атлантической Е. tetralix, или горной Е. сатеа) или у вереска {Calluna vulgaris), редко — в ягоду или, например у толокнянки {Arctostaphylos uva-ursi), в костянку1 (рис. 11.271). У некоторых ройов завязь нижняя, и из нее развивается ягода (например, Vaccinium myrtillus — черника; И vitis-idaea — брусника). Бывшее семейство Epacridaceae распространено исключительно в Южном полушарии. К бывшим Empetraceae с биполярным распространением и свободнолепестным венчиком относится водяника {Empetrum). Травянистыми формами среди Ericaceae представлено бывшее семейство Pyrolaceae (рис. 11.271)— вечнозеленые многолетние травы, например, широко распространенная в хвойных лесах грушанка {Pyrola) и полностью микотрофные и бесхлорофилльные бывшие Monotropaceae с вертляницей (Мо-notropa hypopytis).
Вероятно, близкородственно основному ядру Ericales семейство Theaceae (кроме Ternstroemia-сеае) с многочленным центробежным андроцеем. К этому семейству относят чайное дерево {Camellia sinensis', рис 11.272) и камелию (C.japo-nica).
1 Точнее, в пиренарий. — Примеч. ред.
К Lecythidaceae относятся распространенные преимущественно в неотропической области важнейшие деревья дождевых тропических лесов часто с крупными и сильно одревесневающими плодами. Примером может служить бразильский орех {Bertholletia excelsa), семена которого имеют очень твердую оболочку. К Sapota-сеае принадлежат, например, Butyrospermum, Ра-laquium и Рауепа, из млечного сока которых получают гуттаперчу, применяемую для изготовления изоляции проводов, а также в медицинских целях. Виды Diospyros (Ebenaceae) дают ценную эбеновую древесину, а также съедобные плоды (хурма, D. kaki). Американские насекомоядные Sarraceniaceae имеют листья в виде ловчих кувшинчиков, которые представляют удивительный пример конвергенции с очень похожими ловчими листьями Nepenthaceae и Cephalo-taceae. Возможно, близкородственно семейству Sarraceniaceae семейство Roridulaceae, имеющие клейкие железистые волоски и, вероятно, также насекомоядное. Семейство Fouqueriaceae — выразительные колючие кустарники и стеблевые суккуленты, иногда имеющие весьма причудливые жизненные формы, — распространено в аридных областях запада Северной Америки. Семейство Balsaminaceae с очень крупным родом Impatiens (недотрога, около 800 видов) отличается зигоморфными цветками, иногда ресупи-нированными, как у орхидей, т.е. перевернутыми на 180° вокруг продольной оси, с венчиковидной чашечкой. Один чашелистик со шпорцем. Семена выбрасываются из чувствительных к прикосновениям растрескивающихся коробочек. К семейству Polemoniaceae относят синюху {Pole-monium caeruleum), преимущественно североамериканский род флокс {Phlox) и опыляемую летучими мышами лиану Cobaea scandex.
Родственные отношения последних семейств (от Lecythidaceae до Polemoniaceae) до конца не ясны.
Представители Podostemaceae и часто включаемых в это семейство Tristichaceae населяют водотоки с быстрым течением и каменистым дном и отличаются сильно видоизмененным, часто похожим на мох или лишайник телом, без ясно выраженного расчленения на корни, стебли и листья.
«Эвастериды I»
«Эвастериды 1» не содержат эллаговой кислоты, и у них часто наблюдается спай-нолепестность так называемого позднего типа, т.е. трубка венчика образуется уже после закладки свободных верхних частей лепестков.
11.2. Бактерии, грибы, растения | 489
Garryales
Aucubaceae 1/4, Восточная Азия; Eucommi-асеае 1/1, запад Китая; Garryaceae 1/13, запад Северной Америки, Большие Антильские о-ва: Oncothecaceae 1/1, Новая Каледония
Представители деревянистых Garryales — большей частью (кроме Oncothecaceae и Aucubaceae) двудомные ветроопыляемые растения, часто с 4-мерными цветками. В обычно одногнездном гинецее находятся одна или две висячие семяпочки в каждом плодолистике. Вечнозеленую Aucuba japonica (Aucubaceae) с блестящими красными костянковидными плодами в Европе разводят в виде пестролистной разновидности, вызванной вирусным заболеванием.
Gentianales
Аросупасеае, включая Asclepiadaceae 335/3700, по всему миру, преимущественно тропики и субтропики; Gelsemiaceae 2/11, тропики Старого и Нового света; Gentianaceae 88/ 1500, космополитное; Loganiaceae 14/430, по всему миру, преимущественно тропики и субтропики; Rubiaceae 550/9000, космополитное
Виды Gentianales содержат особые иридоиды (секо-иридоиды) и производные от них индоловые алкалоиды и обычно имеют прилистники, которые могут быть редуцированы до каймы, соединяющей основания супротивных листьев, железистые волоски в пазухах листьев, а часто также на внутренней стороне чашечки, биколлате-ральные проводящие пучки, часто скрученный в бутоне венчик, развивающийся по нуклеарному типу эндосперм.
К Loganiaceae, большей частью древесным растениям с верхней завязью, относятся различные ядовитые растения, например из рода Stry-chnos (рис. 11.273). Многие виды содержат яды, используемые для стрел (например, южноамериканский яд кураре), а из семян Strychnos пих-vomica, азиатского рвотного ореха, добывают индоловый алкалоид стрихнин.
Большинство травянистых Gentianaceae (рис. 11.273), лишенных прилистников и содержащих насыщенные горечи (гентиопик-рин), представлены в высокогорьях Северного полушария богатыми видами родами
Рис. 11.273. Gentianales (А—Е— no G. Karsten; F—Н — по F. Firbas):
А—Е— Loganiaceae, Strychnos nux-vomica: А — цветущий побег, В, С — ягода (В — внешний вид, С — поперечный разрез), D, Е — семя (D — общий вид, Е — поперечный разрез) (1 /2х); F— Н — Gentianaceae, Gentiana lutea: F — бутон (co скрученным венчиком), G — цветок (1 х), Н — завязь, поперечный разрез (Зх)
490 | ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Gentiana и Gentianella. Gentianella достигает в своем распространении Анд и, вероятно, оттуда путем дальнего заноса достигла Австралии и Новой Зеландии. К другой линии умеренного пояса хорошо представленных в тропиках Gentianaceae принадлежат роды Centaurium (золототысячник) и Blackstonia (зеленка), из них последний имеет увеличенное число чашелистиков, лепестков и тычинок.
Семейство Аросупасеае, в которое сейчас включают также Asclepiadaceae, имеет нечленис
тые млечники, млечный сок и содержит многочисленные, часто ядовитые алкалоиды. Плодолистики с сильно развитой верхней и несросшейся зоной (рис. 11.274) почти апокарпные. Стилодии и рыльца у многих таксонов ко времени цветения срастаются постгенитально. Плоды у этих таксонов все же апокарпные. У части Аросупасеае (в узком смысле) пыльники свободные и пыльцевые зерна отдельные. К собственно Аросупасеае относят из древесных растений, например, средиземноморский олеандр (Nerium oleander), африканские виды Strophanthus (содержащие кардено-лиды, ценные сердечные гликозиды, используемые также для изготовления стрельных ядов).
Рис. 11.274. Gentianales, Аросупасеае (А, С, D — по A. Engler, В — по A. W. Eichler: Е, F — по К. Schumann):
А—D — Asclepias syriaca: А — цветок с чашечкой и венчиком, В — диаграмма цветка (в пазухе при-цветничка боковой побег), С — продольный разрез цветка с завязью (f), головчатым рыльцем (nk), пыльниками (а), короной (п), поллиниями (р) и корпускулой (kk), D — два поллиния, соединенные трансляторами (t) и корпускулой; Е, F — Strophanthus hispidus: Е — плод, F — семя
11.2. Бактерии, грибы, растения | 491
Рис. 11.275. Gentianales, Rubiaceae (А, В, G, J — по Н. Baillon; С— F — по G. Karsten; Н — по A. W. Eichler):
А, В — Cinchona calisaya'. А — цветок (4х), В — распадающаяся снизу вверх септицидная коробочка (1 х); С—F — Coffea arabica С — цветущий и плодоносящий побеги (3/8х), D — цветок, Е — костянка, мякоть плода частично удалена, F — семена соответственно без пергаментовидного эндокарпия и с эндокарпием (3/4х); G — цветок Galium odoratum, ясменника (7х); Н — диаграмма цветка Sherardia arvensis; J — мясистый дробный плод Rubia tinctoria (продольный разрез; 2,7х)
Rauvolfia (с понижающим кровяное давление индоловым алкалоидом резерпином) и разные каучуконосные растения (например, африканские Funtumia, Landolphia и бразильская Hancomia). Произрастающий в Центральной Европе барвинок (Vinca minor} — травянистое растение. У видов бывшего семейства Asclepiadaceae пыльники вместе с головчатым рыльцем соединены в гиносте-гий, пыльцевые зерна одной теки большей частью соединены в поллиний (рис. 11.274). Обычно два таких поллиния из соседних пыльников соединены друг с другом неклеточной структурой, образованной выделениями рылец (дуговидным «транслятором» с «корпускулами»). Опыляющие насекомые в поисках нектара попадают хоботком или ногами в бороздку, идущую между пыльниками, и, освобождаясь, вытягивают за корпус
кулу поллинии из пыльников. Придатки на спинной стороне тычинок могут образовывать венчиковидную корону. У Ceropegia описанный способ опыления сочетается с образованием цветков-ловушек со скользкой внутренней поверхностью трубки веничика. Помимо древесных растений в этой группе есть лианы (например, Mars-denia), иногда (например, Dischidia} эпифиты, тесно сожительствующие с муравьями, травы (например, среднеевропейский ластовень, Vincetoxicum hirundinaria, виды рода Asclepias), а также стеблевые суккуленты (например, стапелии с опыляемыми мухами цветками, особенно обычные в сухих районах Африки).
Семейство Rubiaceae имеет межчерешковые прилистники, коллатеральные прово
492 | ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
дящие пучки и нижнюю завязь (рис. 11.275). Плоды — коробочки с многочисленными семенами, или костянки1, либо дробные плоды (схизокарпии).
К этому семейству, особенно богатому видами в тропиках, где оно представлено преимущественно древесными формами, относят такие хозяйственно-ценные растения, как хинное дерево (Cinchona, содержащая хинин и другие противомалярийные индоловые алкалоиды), и изначально палеотропическое кофейное дерево (Coffea, особенно С. liberica и С. arabica). Содержащиеся в костянке по два семена («кофейные зерна») (рис. 11.275) имеют очень крупный эндосперм и содержат близкий к алкалоидам кофеин, производное пурина. С экологической точки зрения примечательны эпифиты Myrmecodia и Hydnophytum, в стеблевых клубнях которых живут муравьи, а также тропические виды родов Psychotria и Pavetta, у которых в маленьких клубневидных вздутиях на листьях обитают симбиотические бактерии. Эти бактерии определенно не фиксируют азот из воздуха, их функция неясна.
У произрастающего преимущественно в умеренном поясе травянистого Galium (подмаренник) прилистники подобны листовым пластинкам и составляют вместе с листьями 4- или многочленную мутовку. Алкалоидов у них нет, a Galium odoratum (ясменник, рис. 11.275) содержит кумарин. Родственна подмаренникам марена красильная (Rubia tinctoria), которую раньше возделывали в качестве красильного растения (содержит ализарин).
Lamiales
Acanthaceae 256/2770, по всему свету от тропиков до умеренного пояса; Avicenniaceae 1/8, по всему свету от тропиков до умереннотеплого пояса. Bignoniaceae 113/800, по всему свету от тропиков до субтропиков; Buddlejaceae 4/100, от тропиков до умереннотеплого пояса, преимущественно Азия; Bybli-daceae 1/6, Австралия, Новая Гвинея; Cyclo-cheilaceae 2/4, Африка; Gesneriaceae 126/2850, тропики по всему свету; Lamiaceae 258/6970, космополитное; Lentibulariaceae 3/180, космополитное; Муорогасеае 4/210, преимущественно Южное полушарие, большей частью Австралия; Oleaceae 29/600. космополитное; Ого-banchaceae 60/1700, космополитное, Paulowni-
1 Пиренарии. — Примеч. ред.
асеае 1/6, Восточная Азия; Pedaliaceae 18/90, по всему свету от тропиков до субтропиков; Phrymaceae 1/2, Восточная Азия, восток Северной Америки; Plantaginaceae 114/2100, космополитное, в основном умеренный пояс; Schlegeliaceae 1/12, тропическая Америка; Scrophulariaceae 24/1200, космополитное, особенно Африка; Stilbaceae 5/12, Южная Африка; Tetrachondraceae 1/2, Новая Зеландия, умеренный пояс Южной Америки; Verbenaceae 36/1035, космополитное
Порядок: Lamiales. Характеризуется супротивными листьями, железистыми волосками, в которых клетки головки возникают обычно только путем антиклинального деления, и редуцированным чаще всего до четырех или двух тычинок андроцея. За исключением Oleaceae (и Tetrachondraceae) Lamiales имеют в качестве запасного углевода олигосахариды (например, стахиозу) вместо крахмала и зигоморфные цветки. Далее порядок характеризуется химическими признаками, как, например, наличием определенных иридоидов.
Занимающее в порядке базальное положение семейство Oleaceae отличается радиально-симметричными цветками только с двумя тычинками (рис. 11.276). Плоды в этом семействе весьма разнообразны. У сирени обыкновенной (Syringa vulgaris) это коробочки. Средиземноморская, отличающаяся простыми, серебристо-серыми листьями маслина (Olea еигораеа), имеет костянки (оливки) с маслосодержащими мякотью плода и эндоспермом (рис. 11.276). Перистолистный ясень (Fraxinus), наконец, имеет односемянные крылатые орехи. Среди ясеней есть, например, субсредиземноморский белый ясень (F. omus), у которого цветки сильно пахучие, насекомоопыляе-мые, с глубоко раздельным белым венчиком. Среднеевропейский ясень обыкновенный (F. excelsior), напротив, ветроопыляемый, и его цветки, раскрывающиеся раньше листьев, не имеют околоцветника. Известные декоративные кустарники из этого семейства — Jasminum, Forsythia и Ligu-strum.
Молекулярные исследования особенно ясно показывают невозможность традиционного разграничения семейств Lamiaceae/
11.2. Бактерии, грибы, растения | 493
Рис. 11.276. Lamiales, Oleaceae (А, В — по F. Firbas; С, D — по G. Hegi; Е, F — по G. Karsten; G — по G. Hempel и К.Wilhelm):
А— диаграмма цветка Syringa vulgaris; В—D — Olea еигораеа: В — плодоносящий побег (1/5х), С — цветок (увеличено), D— плод в продольном разрезе, обнажена косточка (1х); Е—G — Fraxinus: Е— F — мужской цветок и крылатый орех энтомофильного F. ornus (несколько увеличено) G — обоеполый (слева) и мужской цветки анемофильного F. excelsior (увеличено)
Рис. 11.277. Lamiales, Lamiaceae (А— по R. Spohn, В— по Н. Schenck, С — по F. Firbas, D— по Н. Bailion):
А— продольный разрез цветка Lamium album; В — завязь (с искусственно раскрытой чашечкой) Galeopsis segetum (2х); С — продольный разрез основания цветка Lamium maculatum с чашечкой (к), венчиком (с), нектарником (п), гнездами завязи (эремами) с семязачатками (kl) и столбиком (д) (10х); D — зрелый эрем Lamium album, продольный разрез (увеличено)
494 | ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Verbenaceae и Scrophulariaceae в широком смысле.
Если прежде Lamiaceae (=Labiatae) по характеру прикрепления столбика между плодолистиками в основании завязи (рис. 11.277) и по наличию дробного плода отделяли от Verbenaceae, у которых столбик отходит на вершине завязи, то сейчас Lamiaceae понимают значительно шире и включают в него большую часть традиционных Verbenaceae с завязями обоих типов. Семейству Lamiaceae в современных границах свойственны положение семязачатков почти у края слегка свернутого плодолистика (из-за разделения последнего ложной перегородкой семязачатки оказываются на перегородке), соцветия с цимоз-но разветвленными боковыми осями, а также простые рыльца. Lamiaceae в новых границах представлены деревьями, кустарниками, полукустарниками и травами, часто с отчетливо четырехгранными стеблями. Листья супротивные, растения обладают сильным пряным запахом благодаря содержанию эфирных масел в железистых волосках. Цветки большей частью зигоморфные. Их 5-членная чашечка сростнолистная, радиально-симметричная или двугубая. Венчик обычно двугубый, нижняя губа 3-раз-дельная, верхняя — 2-раздельная. Из четырех тычинок (медианная тычинка отсутствует) одна пара часто длиннее другой. У шалфея (Salvia) и розмарина (Rosmarinus) развиты и соотвественно фертильны только две абаксиальные тычинки. Завязь образована двумя плодолистиками, поделенными ложной перегородкой, с двумя семяпочками в каждом плодолистике. Из нее формируется твердый плод, 4-семянный орех1 или характерный для Lamiaceae це-нобий, распадающийся на 4 ореховидных эрема.
Благодаря содержанию эфирных масел многие виды применяют как пряности в кулинарии и в качестве лекарственных растений (например, майоран — Majorana hortensis", базилик — Oci-тит basilicum", чабер — Satureja hortensis", лаванда — Lavandula angustifolia", розмарин — Rosma
1 Такой плод правильнее называть ореховидным, поскольку настоящий орех односемянный. — Примеч. ред.
rinus officinalis", шалфей — Salvia officinalis", тимьян — Thymus vulgaris", лимонная мята — Melissa officinalis", виды мяты, например мята перечная — Mentha piperita х М. spicata). Широко распространены в Центральной Европе аборигенные виды Ajuga (живучка). Galeopsis (пикульник), Glechoma (будра), Lamium (яснотка), Stachys (чис-теп) и Teucrium (дубровник).
К семейству Verbenaceae (в узком смысле), обладающему кистевидыми. колосовидными или головчатыми соцветиями, сидящими непосредственно на краях плодолистиков семяпочками и выдающимися, сидящими всегда на верхушке завязи столбиками с двухлопастным рыльцем, относят, например, вербену (Verbena officinalis). К кругу родства Lamiaceae и Verbenaceae, по-видимому, принадлежит также Bignoniaceae, широко распространенное в тропиках семейство, представленное древесными видами. Сюда относятся декоративное растение катальпа (Catalpa) и лиана Campsis, а также почти целиком тропическое, но большей частью травянистое семейство Acanthaceae.
Семейство Scrophulariaceae сейчас рассматривают совершенно иначе, чем ранее. Границы нижеперечисленных семейств еще толком не выяснены и их невозможно очертить по морфологическим признакам. В семейство Scrophulariaceae в узком смысле включены только такие роды, у которых нижний конец гнезда пыльника не стреловидный. К нему (рис. 11.278) относят, например, Verbascum (коровяк) с почти радиально-симметричными цветками и пятью тычинками и Scrophularia (норичник), у которого медианная тычинка сохранилась в виде стаминодия. Вероятно, близкородственно Scrophulariaceae в его новых границах семейство Buddlejaceae с произрастающей в Центральной Европе на развалинах и часто дичающей буддлеей (Ви-ddleja davidii). В Orobanchaceae (рис. 11.278) из Scrophulariaceae выделены все паразитирующие на корнях полупаразиты и паразиты. Зеленые полупаразиты — это, например, Pedicularis (мытник), Euphrasia (очанка), Rhinanthus (погремок) и Melampyrum (марьянник), а полные паразиты — Lathraea (петров крест) и Orobanche (заразиха). Большая часть Scrophulariaceae в его традиционном понимании (например, Veronica — вероника; Digitalis — наперстянка; Antirrhinum — львиный зев) вместе с представителями бывших семейств Callitrichaceae (водные растения рода Callitriche— водя-
11.2. Бактерии, грибы, растения | 495
Рис. 11.278. Lamiales (А, С — по H.Baillon; В — по F. Firbas; D, Е— по G. Karsten):
А, В — Scrophulariaceae: А— Verbascum thapsus, В— Scrophularia nodosa; С—Е— Orobanchaceae: С — Pedicularis palustris, D — бесхлорофилльный желтовато-коричневый паразит Orobanche minor на Trifolium repens (2/Зх), E — отдельный цветок (увеличено)
ная звездочка, с однополыми цветками, состоящими соотвественно из только одной тычинки или пестика с завязью, распадающейся в зрелом плоде на 4 мерикарпия, рис. 11.279), Hippuridacae (водно-болотная Hippuris vulgaris — водяная сосенка) и Glo-bulariaceae (Globularia — шаровница, имеющая головчатое соцветие и ореховидные плоды) объединены наряду с подорожниками (Plantago) и близкородственными родами в Plantaginaceae (рис. 11.279). У Plantaginaceae (иногда также называемых Vero-nicaceae) в этих очень широких границах основания пыльников более или менее отчетливо стреловидные. Из Scrophulariaceae в широком смысле в отдельное семейство выделяют часто разводимое декоративное дерево Paulownia и иногда — Calceolariaceae.
Семейство Gesneriaceae с видами Stre-ptocarpus, образующими единственную огромную семядолю, реликтовыми средизем-
номорско-монтанными родами Ramonda, Jankea и Haberlea, а также часто культивируемой восточноафриканской «узамбар-ской фиалкой» Saintpaulia ionantha имеет лишь частично подразделенные на гнезда завязи с париетальной плацентацией. В этом семействе много эпифитных видов, а также видов, опыляемых птицами. У насекомоядных Lentibulariaceae с наземными жирянками ( Pinguicula) и водными пузырчатками (Utricularia), которые с помощью образованных на листьях ловчих пузырьков ловят мелких водных животных, завязи со свободной центральной плацентой. У относящейся к этому же семейству Genlisea, произрастающей в тропической Африке и Южной Америке, недавно обнаружено химическое вещество для привлечения и отлавливания простейших с помощью сильно видоизмененных подземных листьев. К Pedaliaceae с обычно высо-
496 [ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Рис. 11.279. Lamiales, Plantaginaceae (А, Е—G — по F. Firbas; В — по Н. Bailion; Н, J, М — по «Syllabus» А. Энглера, К, L— по A. W. Eichler):
A— D — Digitalis purpurea: А, В — цветок, вид косо спереди и продольный разрез (почти 3/4х), С, D — поперечный разрез завязи, коробочка, раскрывающаяся по перегородкам и частично вдоль средней жилки (почти 1х); Е — цветок Veronica teucrium, вид спереди и сзади (1,5х); F — Anthirrhinum majus (1х); G — Linaria vulgaris (1,5x); H—M — Callitriche stagnalis: H, К — мужской и женский цветки с при-цветничками (увеличено), J, L — диаграммы цветков, М — плод в поперечном разрезе (увеличено)
коспециализированными цепляющимися плодами относится масличное растение Sesamum indicum, из которого получают кунжут.
Solanales
Convolvulaceae 55/1930, космополитное;
Hydroleaceae 1/11, пантропическое; Montini-aceae 2/4, Африка, Мадагаскар; Solanaceae 147/2900, космополитное; Sphenocleaceae 1/2, пантропическое
Порядок: Solanales. Виды порядка содержат алкалоиды стероидной и тропано-вой групп, имеющие фармацевтическое значение, у них очередные, цельные листья без прилистников и радиально-симметричные цветки с продольно сложенным в бутоне венчиком. Некоторым из них свойственны биколлатеральные проводящие пучки. У семейства Solanaceae, имеющего хозяйствен
ное значение, структура побегов часто трудно различима вследствие срастаний и смещений стеблей и листьев. Цветки часто располагаются в завитках, завязи большей частью из двух плодолистиков, часто косо расположенных (рис. 11.280), многочисленные семяпочки образуются на толстой плаценте.
Плоды-коробочки имеются у виргинского табака, Nicotiana tahacum — аллотетраплоида, возможно, происходящего от диплоидных дикорастущих южноамериканских видов N. sylvestris и, вероятно, N. tomentosiformis. Коробочки имеют также излюбленные южноамериканские декоративные растения Petunia и ядовитые рудеральные растения родов белена (Hyoscyamus) и дурман (Datura). Ягоды свойственны очень богатому видами роду Solanum, к которому принадлежит аллотетраплоидный картофель (.S', tuberosum). В XVI столетии он был завезен в Европу из Южной Америки (Анды). Баклажан (S. me/ongena), изначально растение Старого Света, и происхо-
11.2. Бактерии, грибы, растения । 497
Рис. 11.280. Solanales, Solanaceae (A, F—Н — по G. Karsten; В, Е — по A. W. Eichler; С, D — по G.Beck-Mannagetta):
А — Atropa bella-donna, симподиальный участок побега с терминальными цветками и ягодами (1/2х); В— D — Hyoscyamus: В — диаграмма цветка Н. albus. С — цветок, D — коробочка Н. Niger чашечка частично удалена (почти 1х); Е—Н — Datura stramonium: Е — диаграмма цветка, F — покрытая шипами коробочка (почти 1 х); Nicotiana tabacum: G — цветок (1 х), Н — молодая коробочка в поперечном и продольном разрезах (2х)
498 | ГЛАВА 11, СИСТЕМАТИКА И ФИЛОГЕНИЯ
дящий из Нового Света томат (5. lycopersicum) также принадлежат к роду Solanum. Ягоды имеют и происходящие из тропической Америки красный перец паприка (Capsicum аппиит) и ядовитая среднеевропейская красавка (Atropa belladonna). Фармацевтическое значение имеют прежде всего алкалоиды гропановой группы (гиосциамин, атропин, белладоннин, скополамин и др.).
Обычно вьющиеся (рис. 11.281) виды семейства Convolvulaceae большей частью с воронковидным, скрученным в бутон венчиком, как, например, у полевого вьюнка (Convolvulus arvensis) и повоя (Calystegia sepium). имеют обычно 4-семя иную коробочку. Важное пищевое растение, первоначально, возможно, неотропическое, — батат, или сладкий картофель (Ipomoea batatus) с богатыми крахмалом корневыми клубнями. В Convolvulaceae сейчас включают бывшее семейство Cuscutaceae с единственным родом Cuscuta (повилика). Это лишенные корней и листьев, более или менее бесхло-
рофилльные паразиты, растущие на разных сосудистых растениях.
Неясно положение среди «эвастерид-1» семейств Boraginaceae, включая Hydrophylla-сеае (117/2400, космополитное), Plocosperma-taceae (1/1, Центральная Америка) и Vahlia-сеае (1/5. Африка до Индии). Частично включаемые в Solanales виды семейства Boraginaceae с плодами-коробочками, как у включенного сейчас в него бывшего семейства Ну-drophyllaceae (например, у изначально североамериканской культивируемой в качестве медоноса Phacelia), и виды Boraginaceae (в узком смысле) с плодами-ценобиями из 4 эре-мов (рис. 11.282), — преимущественно травы с очередными, простыми и часто шершавыми от грубых волосков листьями. Их цветки располагаются обычно в специфичном двойном завитке и преимущественно радиалы ю-симметричны. Иногда (например, у Echium — синяк) они слабо зигоморфны. У собственно Boraginaceae венчик часто имеет 5 чешу-
Рис. 11.281. Solanales, Convolvulaceae (А — по F. Firbas; В — по J. Graf; С — по A.W. Eichler; D—G — по G. Dahlgren):
А— Calystegia sepium, цветущий и плодоносящий побег (1/Зх); В— Convolvulus arvensis, цветок в продольном разрезе (1,5х); С— диаграмма цветка (с прицветничками); D — G— Cuscuta europaea: D— безлистный побег с гаусториями и клубочками цветков (1,5х), Е—G — цветок и молодой плод, внешний вид и продольный разрез (20х)
11.2. Бактерии, грибы, растения | 499
Рис. 11.282. Boraginaceae (А — по A.W. Eichler; В — по Н. Bailion; С, G — по В. von Wettstein; D, Е — по A. Englers «Syllabus»: F— по F.Firbas):
A — диаграмма цветка Anchusa officinalis. Symphytum officinale-, В — цветок, продольный разрез, с чешуями в зеве (s) (почти Зх); С — соцветие двойной завиток (цифрами показана последовательность распускания цветков; ок. 1х); D — F — постепенное формирование ценобия: начальная (D — Bourreria) и последующие (Е — Anchusa, F — Onosma) стадии развития завязи; G — отдельный эрем Onosma visianii, продольный разрез (8х)
ек в зеве (рис. 11.282). прикрывающих вход в трубку венчика. Гинецей из двух плодолистиков с 4-гнездной завязью из-за развития ложных перегородок, развивающийся в 4 односемянных эрема. Они отличаются от похожих эремов Lamiaceae тем, что микропиле семяпочек, а поэтому и зародытевый корешок направлены вверх. Известные европейские представители семейства — медунш ia (Pulmonaria), незабудка (Myosotis), окопник (Symphytum), анхуза (Anchusa) и огуречник (Borago). В тропиках семейство представлено также родами, в которых имеются древесные формы.
«Эвастериды II»
«Эвастеридам П» свойственна спайно-лепестность так называемого раннего типа,
т. е. трубка венчика различима до заложения свободных верхних частей лепестков.
Aquifoliales
Aquifoliaceae 2/500, космополитное с дизъюнктивным ареалом; Helwingiaceae 1/5, от Гималаев до Японии; Phyllonomaceae 1/4, от Центральной до Южной Америки
Порядок: Aquifoliales. Древесные растения имеют очередные листья с очень мелкими прилистниками и часто зубчатым листовым краем. Из мелких в большинстве случаев цветков образуются костянки1 — семена с обильным эндоспермом и очень мелким зародышем. Семейство Aquifoliaceae с основным крупным родом Ilex (около
1 На самом деле — пиренарии. — Примеч. ред.
500 | ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
500 видов) характеризуется слабо сросшимся венчиком и 4-членным гинецеем (рис. 11.283). Среднеевропейский представитель семейства — падуб (Ilexaquifolium), вечнозеленый средиземноморско-атлантический кустарник или дерево с красными костянками. Листья южноамериканского /. paraguaiensis используют для приготовления чая матэ. У семейства Не1-wingiaceae и Phyllonomaceae цветки эпи-филльные.
Apialcs
Apiaceae 440/3500, космополитное, большей частью умеренный пояс Северного полушария; Araliaceae 47/1320, по всему миру, тропики и субтропики, редко умеренный пояс; Aralidiaceae 1/1, Юго-восточная Азия; Grise-liniaceae 1/7, умеренный пояс Юной Америки, Новая Зеландия; Melanophyllaceae 1/8, Мадагаскар; Pittosporaceae 9/300, от тропиков до умеренно-теплого пояса Старого Света, преимущественно Австралия; Torricelliaceae 1/2, Восточная Азия
Порядок: Apiales. Самые крупные семейства порядка — Apiaceae, Araliaceae и Pittosporaceae часто содержат эфирные масла в схизогенных ходах, эллаговая кислота отсутствует. Запасной углевод обычно — ге
мицеллюлоза. У всего порядка листья очередные и пальчатонервные. Цветки большей частью мелкие, радиально-симметричные и 5-мерные.
Семейство Pittosporaceae имеет простые листья и верхнюю завязь. У Araliaceae и Apiaceae, которые, пожалуй, лучше было бы рассматривать как одно семейство, листья от лопастных до перистых, соцветие зонтик, а в нижней завязи развивается обычно только одна висячая семяпочка в каждом гнезде. У преимущественно древесных Araliaceae завязь состоит из 2—5 плодолистиков (рис. 11.284). В развивающихся из них костянках нет секреторных ходов. К Araliaceae принадлежит произрастающий в Центральной Европе вечнозеленый и гетерофильный плющ (Hedera helix). Цветки его опыляются мухами или осами осенью, а плоды созревают следующей весной. Семейство Apiaceae (=Umbelliferae, зонтичные) содержит почти исключительно травянистые растения. Двухчленный гинецей развивается в дробный плод со схизоген-ными эфиромасличными ходами. У Apiaceae характерный внешний вид (рис. 11.284): их своеобразные стебли, разделенные на узлы и полые междоузлия, несут очередные листья, почти всегда раздельные и охватывающие стебли расширенными листо-
Рис. 11.283. Aquifoliales, Aquifoliaceae, Ilex aquifolium (A— no A.W. Eichler; В—D— no R. Spohn): A — диаграмма цветка; В — ветвь с плодами; С — женский цветок; D — мужской цветок
11.2. Бактерии, грибы, растения | 5Q-|
Рис. 11.284. Apiales, Araliaceae (А, С — по G. Hegi; В — по A. W. Eichler; D — по Thellung; Е — по F. Noll и H.A.Froebe, с изменениями; F,G, J — по G.Karsten; Н — по A.Tschirch и O.Oesterle; К, L — поО.С.Вегд и С. Е Schmidt, несколько изменено):
А—С — Hedera helix'. А — цветок (почти 4х), В — диаграмма цветка, С — плод (ягода, почти 2х); D— L — Apiaceae: Ь — цветок (Ammi majus- d — диск, g — стиллодий, f — завязь), Е — диаграмма цветка (Laser trilobium); F—J — Conium maculaturrr. F — побег (1/2x), G — зонтичек, H — цветок (продольный разрез, с двумя висячими семяпочками), J— плод, внешний вид (все увеличено); К, L— дробный плод Carum carvi, внешний вид, продольный (Юх) и поперечный (25х) разрезы, с карпофором (к), околоплодником (fw), главными ребрами с проводящими пучками (h), ложбинками с лежащими в них секреторными ходами (sg), семенной кожурой (s), эндоспермом (es) и зародышем (е)
выми влагалищами. Соцветия (рис. 11.284) обычно — сложные зонтики (зонтики из зонтичков). Их кроющие листья скучены в «обертки» и «оберточки». Мелкие, большей частью белые, реже розовые или желтые цветки пятичленны во всех кругах, кроме гинецея, но чашечка почти всегда сильно редуцирована. Кончики лепестков часто
загнуты внутрь, сами лепестки кажутся свободными. Вершина завязи часто преобразована в подстолбие, функционирующее как нектарник (рис. 11.284). В каждом гнезде завязи с перегородки свешивается анатропная семяпочка. Вторая семяпочка рано отмирает. Семя с обильно содержащим жиры и протеины эндоспермом и маленьким за
502 I ГЛАВА 11 СИСТЕМАТИКА И ФИЛОГЕНИЯ
родышем. Семенная кожура слита со стенкой плода; сухой дробный плод (схизокарпий) распадается вдоль общей стенки плодолистика на два односемянных мерикарпия. Они сначала висят на брюшном проводящем пучке (карпофоре)1, а в конце концов опадают (рис. 11.284).
У немногих Apiaceae вследствие соответствующего упрощения или редукции возникают цельные листья, например линейные у видов Bupleurum или щитовидные, как у щитолистника (Hydrocotyle vulgaris). Цветки некоторых родов, например у борщевика (Heracleum), зигоморф-ны из-за увеличения внешних по отношению к соцветию лепестков венчика. У некоторых родов заметность соцветий повышается благодаря окрашенным верховым листьям, например белой оберткой зонтиков у Astrantia или желтыми оберточками у Bupleurum. Важнейшие опылители почти всегда протандрических цветков — жуки, мухи и другие короткохоботные насекомые.
Зонтичные особенно широко распространены во внетропических областях Северного полушария, где большое число их видов произрастает в степях, на болотах, лугах и в лесах. В центральноазиатских степях они представлены вы-сокотравьем более метра в высоту (например, Ferula), а в антарктических областях — подушковидными растениями. Высокое содержание эфирных масел обусловливает большое число среди них пряных и лекарственных растений. Представлены в семействе и овощные растения. У разных зонтичных используют и плоды, и листья, и корни. Примеры — тмин (Carum carvi), анис (Pimpinella anisum), кориандр (кинза) (Coriandrum sativum), укроп (Anethum graveolens), любисток (Levisticum officinale), фенхель (Foeniculum vulgare), петрушка (Petroselinum crispum), морковь (Daucus carota), пастернак (Pastinaca sativa) и сельдерей (Apium graveolens). Очень ядовиты, например, болиголов крапчатый (Conium maculatum) и вех (Ci-cuta virosa)
Dipsacales
Adoxaceae 5/160, умеренный пояс Северного полушария; Caprifoliaceae 5/210, большей частью умеренный пояс Северного полушария; Diervilleaceae 2/22, Северная Америка, Восточная Азия; Dipsacaceae 11/290, Евразия, Африка; Linnaeaceae 5/36, бореальная зона Северного полушария, Азия, Мексика; Morinaceae 2/13, Евразия; Valerianaceae 12/400, Северное полушарие, Анды
1 Строение корпофора более сложное. — Примеч. ред.
Порядок: Dipsacales. В растениях этого порядка, распространенного преимущественно в умеренных областях Северного полушария, присутствуют секо-иридоиды. Листья их супротивные, часто сросшиеся основаниями, раздельные или более или менее зубчатые, соцветия цимозные, на лепестках имеются нектарники, состоящие из одноклеточных волосков, завязь в основном нижняя, с немногими семяпочками и часто стерильными гнездами, эндосперм развивается по целлюлярному типу.
Семейство Adoxaceae с радиально симметричными цветками и плодами-костянками1 в Центральной Европе представлено кустарниковыми и перистолистными видами бузины (Sambucus-, рис. 11.285), обычно кустарниковыми, но с простыми листьями видами калины (Viburnum), а также травянистой адоксой мускусной (Adoxa moschatellind).
В то время как первые два рода, ранее рассматриваемые в составе семейства Caprifoliaceae, имеют соцветия зонтиковидные тирсы — в них, например у Viburnum opulus, краевые цветки стерильны и выполняют функцию привлечения опылителей, — для Adoxa характерны от пяти до семи цветков, собранных почти в кубическую головку. Принадлежность Adoxaceae к Dipsacales спорна.
Семействам Caprifoliaceae, Valerianaceae и Dipsacaceae свойственны зигоморфные цветки, плоды — ягоды, коробочки или ореховидные. У собственно Caprifoliaceae развиваются ягоды или коробочки. К роду Lo-nicera с цветками, располагающимися по два и частично срастающимися завязями, относят кустарники и лианы, например каприфоль (Z. caprifolium)', род Symphoricarpos представлен натурализовавшимся снежноягодником (S. albus). Семейство Valerianaceae имеет слабо зигоморфные цветки большей частью с пятичленным венчиком, часто со шпорцем, только 1 —4 тычинками. Из 3-гнездной завязи только с одним фертильным гнездом развивается плод орех. Виды обитающего в Центральной Европе рода Valeriana (валериана) большей частью многолетники с 3 тычинками, у которых ко времени плодоношения чашечка преобразуется в кольцо волосков (рис. 11.285). Особое фармацевтическое значение имеет
1 Точнее, пиренариями. — Примеч. ред.
11.2. Бактерии, грибы, растения I 5Q3
Рис. 11.285. Dipsacales, Adoxaceae (А — по J. Graf; В — по G. Dunzinger; D, F, L — по A.W. Eichler; Е, G, Н — по F.Weberling; J, К — по G. Hegi):
А, С, D — Sambucus ebulus: А — цветок (почти 10х), С — костянка, продольный разрез (почти 5х), D — диаграмма цветка; В — S. nigra, цветок, продольный разрез (почти 10х); Е—Н — Valerianaceae, Valeriana officinalis: Е — цветок (почти 10х), F — диаграмма цветка; G — Valeriana tripteris, плод и паппус (почти Зх), Н — Centranthus ruber, цветок (почти 10х); J—L — Dipsacaceae, Scabiosa columbaria: J — корзинка, продольный разрез (увеличено), К — плод с «наружной чашечкой» (ак) и чашечкой (к) (увеличено), L — Dipsacus pilosus, диаграмма цветка
И officinalis как источник успокаивающих веществ (например, валериановой кислоты, валепотриата, эфирных масел). Разные виды однолетников из р. Valerianella употребляют в пищу в виде салата. У Dipsacaceae цветки собраны в корзинки (рис. 11.285), краевые цветки в которых часто увеличены. Одногнездные и односемянные завязи окружены 4-листной оберткой из прицветников — подчашием (рис. 11.285). Среднеевропейские роды семейства — Scabiosa, Knautia и Dipsacus. Сухие корзинки D.fullomim (ворсовальная шишка) с их твердыми остроконечными прицветниками употребляли для нанесения ворса на шерстяные ткани.
Asterales
Alseiiosmiaceae 3/12, юго-западная часть Тихоокеанской области; Argophyllaceae 1/11, Австралия. Новая Каледония; Asteraceae 1530/ 22750, космополитное; Calyceraceae 6/55, от Центральной до Южной Америки; Campanu-laceae 90/2200, космополитное; Carpodetaceae 3/5, Новая Гвинея, Австралия, Новая Зеландия; Donatiaceae 1/2, Южная Америка, Новая Зеландия, Тасмания; Goodeniaceae 11/400, Южное полушарие, в основном Австралия; Menyanthaceae 5/40, космополитное; Реша-phragmataceae 1/25, от Южной до Юго-Восточной Азии; Phellinaceae 1/11, Новая Каледония; Rousseaceae 1/1, о-в Маврикий; Styli-diaceae 6/170, Австралия, Новая Зеландия, Южная Америка
504 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Порядок: Asterales. Не считая космополитных Asteraceae, Campanulaceae и Мепуап-thaceae, основной ареал порядка явно тяготеет к Южному полушарию. Asterales характеризуются прежде всего (насколько известно) наличием полисахарида инулина, состоящего из фруктозы (вместо крахмала, состоящего из глюкозы). Кроме того, в пределах
порядка почти всегда встречается механизм вторичного предподнесения пыльцы с помощью столбика. Это достигается у разных видов разными способами: с одной стороны, путем смыкания или более или менее прочного склеивания тычинок пыльниками, с другой — с помощью различных структур на столбике.
11.2. Бактерии, грибы, растения | 505
Содержащие млечный сок Campanula-сеае характеризуются большей частью нижней завязью из 2 —5 плодолистиков и центрально-угловой плацентой с обычно многочисленными семяпочками (рис. 11.286). Их плоды — коробочки или ягоды. В подсемействе Campanuloideae цветки актиноморфные и протандрические. У колокольчиков {Campanula) прижатые к столбику пыльники еще до раскрытия цветка высыпают пыльцу на покрытую собирающими волосками поверхность столбика. Уже в бутоне пыльца с пестика вследствие отгибания собирающих волосков высвобождается на его базальную часть, и тычинки засыхают. Только потом раскрывается цветок, и, наконец, разворачиваются лопасти рыльца (рис. 11.286). Обычная в подсемействе 3-гнездная завязь развивается большей частью в коробочку, которая, напри-
<
Рис. 11.286. Asterales (А, Е — по A. W. Eichler, с изменениями; В—D — no F. Е. Clements и F. L. Long, несколько упрощено; F—К, М по— О С. Berg и С. F. Schmidt; L, N, О, Q— по Н. Bailion; Р— по F. Firbas):
А—D — Campanulaceae: А — диаграмма цветка Campanula sp., В—D — фазы распускания цветка Campanula rotundifolia (передняя часть венчика удалена): тычинки высвобождают пыльцу на столбик (В) и сморщиваются (С), пыльца сброшена, рыльца раскрыты (D) (1х); Е—О — Aste-raceae: Е — диаграмма трубчатого цветка с кроющим листом (s) и паппусом (р), F—G — продольный разрез корзинки Matricaria recutita (листочки обвертки однорядные, ложе корзинки полое, ложноязычковые цветки отвернуты вниз, пленчатых кроющих листьев нет) и Arctium lappa (листочки обвертки многорядные и крючковидные, цветки только трубчатые, есть пленчатые кроющие листья) (увеличено), Н, J — Arnica montana, К — Chamaemelum nobile: ложноязычковый и трубчатые цветки (соответственно внешний вид и продольный разрез, пленчатый кроющий лист) (увеличено), L — андроцей Carduus crispus (10х), М — столбик (д) и рыльце (n) Achillea millefolium (увеличено); N —Q— плоды (семянки): N — Hieracium villosum, О — Lactuca vi-rosa с паппусом из волосков, Р — Bidens tripartita с крючкообразными щетинками паппуса, Q — Helianthus annuus (внешний вид и продольный разрез, с зародышем) без паппуса
мер у Campanula, часто вскрывается порами. В цветках кольника (Phyteuma) лопасти венчика на верхушке срастаются пост-генитально. Цветки у него собраны в плотное соцветие, окруженное у основания листочками обвертки, и напоминает, как и соцветие букашника (Jasione), корзинки Asteraceae.
У большей части тропического подсемейства Lobelioideae цветки зигоморфные, гинецеи из двух плодолистиков, пыльники постгенитально срастаются в трубку, а высвобождаемую в эту трубку пыльцу выносит удлиняющийся столбик. Lobelia dortmanna — представитель видов этого подсемейства, населяющих умеренные широты; это редкое растение, встречающееся в олиготрофных водоемах Центральной Европы.
Семейство Asteraceae (=C<jmpositae), возникшее, вероятно, в Южной Америке вне бассейна Амазонки, — крупнейшее семейство цветковых растений, четко характеризующееся многими признаками (рис. 11.286). В основании соцветия корзинки располагаются многочисленные листочки обвертки. Ложе корзинки конусовидное или дисковидное. Кроющие листья отдельных цветков чешуевидные, пленчатые или отсутствуют. В корзинке находятся либо только радиально-симметричные пятимерные трубчатые цветки, либо только зигоморфные язычковые цветки обычно с тремя или пятью увеличенными лепестками1, либо
1 Авторы путают язычковые и ложноязычковые цветки. Первые возникли из трубчатых цветков и имеют исходно единственную губу венчика из 5 лепестков. Язычковые цветки никогда не развиваются вместе с цветками какого-либо иного типа. Ложноязычковые цветки возникли из двугубых (присущих некоторым современным Asteraceae, например, популярному декоративному растению гербере — Herbera), имевших верхнюю губу из 2, а нижнюю — из 3 лепестков. У ложноязычковых цветков сохранилась только нижняя губа, а верхняя полностью редуцирована. Как и исходные двугубые цветки, ложноязычковые цветки всегда женские и развиваются вместе с трубчатыми цветками, располагаясь по краю корзинки. Такое же положение занимают присущие некоторым сложноцветным (например, василькам: Centaurea, воронковидные цветки — стерильные цветки с увеличенным слегка зигоморфным воронковидным венчиком из вторично большого числа лепестков. — Примеч. ред.
506 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
цветки обоих типов вместе. Чашечка представлена чешуйками, щетинками или волосками (паппус) и служит для распространения плодов или же полностью редуцирована. Пять тычинок свободными нитями прирастают к лепесткам. Их пыльники слипаются в трубку, в которую они высыпают пыльцу. При удлинении столбика пыльца выносится из пыльниковой трубки либо опушенными кончиками лопастей рыльца, либо с помощью имеющихся на наружной стороне лопастей рыльца и на столбике «выметающих» волосков. Лишь после расхождения лопастей рыльца в стороны их воспринимающая внутренняя поверхность становится доступной для пыльцы. Нижняя завязь состоит из двух плодолистиков, одногнездная, содержит одну базальную анатропную семяпочку. Плод ореховидный — семянка с более или менее плотно прижатыми друг к другу, но не срастающимися околоплодником и семенной кожурой. Зародыш содержит белки и масла.
В настоящее время семейство подразделяют на I7 триб и три подсемейства. Подсемейство Barnadesioideae (только Bamadesieae) было установлено лишь недавно по отсутствию свойственной остальным сложноцветным инверсии в пластидном геноме. Цветки его обычно радиально-симметричные, а если зигоморфные, то четыре абаксиальных отгиба венчика противолежат одному адаксиальному. Парафилетическое и иногда разделяемое на несколько подсемейств подсемейство Cichorioideae имеет корзинки или только с язычковыми, или только с трубчатыми цветками. Корзинки только с одним типом цветков называют гомогамными. Сюда относят трибы Mutisieae, Cardueae, Lactuceae,Vemonieae, Lia-beae и Arctoteae. Lactuceae (=Cichorieae) имеют только язычковые цветки с пятью зубцами венчика и содержат млечный сок. Поедет авители Lactuceae — Cichorium (с цикорием, Cichorium in-tybus и эндивием, С. endivia), Scorzonera (козелец), Taraxacum (одуванчик), Hieracium (ястребинка), Crepis (скерда) и Lactuca (с L. sativa, салатом). Только трубчатые цветки с длинными узкими зубцами венчика представлены у Cardueae. К этой трибе относят бодяки (Cirsium), чертополохи (Carduus) и васильки (Ceutaurea) с васильком синим, С. cyanus, у которых краевые цветки увеличенные и стерильные)1. У лопухов (Arctium)
1 Краевые цветки васильков правильнее рассматривать как особые, воронковидные цветки. — Примеч. ред.
крючковидные листочки обвертки корзинки служат для эпизоохорного распространения семянок. У мордовников (Echinops) одноцветковые корзинки собраны в шаровидную корзинку второго порядка. В семействе нередко встречается агрегирование корзинок в корзинки второго, а то и третьего порядков. К Cardueae принадлежат некоторые полезные растения — средиземноморский артишок (Cynara scolymus) и сафлор (Carthamus tinctorium, красильное и масличное растение). Подсемейство Asteroideae с трибами Inu-leae, Plucheae, Gnaphalieae, Calenduleae, Astereae, Anthemideae, Senecioneae, Helenieae, Heliantheae и Eupatorieae часто имеет гетерогамные корзинки с краевыми зигоморфными ложноязычковыми цветками и центральными радиально-симметричными трубчатыми цветками, а также схизо-генные ходы с эфирными маслами. К Astereae относят, например, роды Aster (астра) и Bellis (маргаритка), а к Inuleae — девясил (Inula). Эдельвейс (Leontopodium) с бело-шерстистыми листочками обвертки, окружающей сразу несколько корзинок, а также цмин (Helichrysum) с окрашенными сухими кожистыми листочками обвертки относят к Gnaphalieae. Многие лекарственные и пряные растения с эфирными маслами и горькими сесквитерпеновыми лактонами входят в трибу Anthemideae. Примеры — аптечная ромашка (Matricaria recutita), пупавка благородная, или римская ромашка (Chamaemelum nobile), тысячелистник (Achillea millefolium) и разные виды ветроопыляемого рода Artemisia (полынь, например A. absinthium', полынь горькая, A. dracunculus'. эстрагон; рис. 11.287). К этой трибе относят также декоративные хризантемы (Dendranthemd). У Senecioneae листочки обвертки расположены в одном или в немногих рядах. Представители этой трибы — мать-и-мачеха (Tussilago farfara), большой род Senecio (около 1250 видов), а также небольшие розеточные деревья из рода Dendrose-necio, произрастающие в высокогорьях восточной Африки (рис. 11.287), и др. Ноготки (Calendula) с гетерокарпными плодами без паппуса относят к трибе Calenduleae. Обычно супротивные листья свойственны преимущественно американской трибе Heliantheae, например Bidens (череда), занесенной из Северной Америки Galin-soga (галинзога), часто культивируемым видам родов Dahlia (георгин) и Helianthus (с подсолнечником — Н. annuus. топинамбуром — Н. tu-berosus), трибе Helenieae, представленной декоративными растениями рода Tagetes (бархатцы), и Eupatorieae, к которой принадлежит среднеевропейский посконник (Eupatorium cannabinum).
Ближайшими родственниками Asteraceae считают Goodeniaceae и Calyceraceae. В одиночно расположенных цветках Goodeniaceae пыльца высыпается в пыльцевую чашу, образован-
11.2. Бактерии, грибы, растения i 5Q7
Рис. 11.287. Asterales, Asteraceae (А— по G.Hegi; В, С — по С. Puff):
А — ветроопыляемая Artemisia borealis, внешний вид (почти 3/4х), корзинка только с трубчатыми цветками, краевые цветки женские (увеличено); В, С — розеточные деревья из рода Dendrosenecio в тропическом альпийском поясе восточно-африканских высокогорий (около 4 000 м над уровнем моря): В — D. jonstonii subsp. refractisquamatus во влажном Рувензори, С — D. jonstonii subsp. jonstonii на засушливых склонах Килиманджаро
ную столбиком под рыльцем, которая потом почти полностью закрывается. Растущие рыльца выдавливают пыльцу из чаши, благодаря чему она становится доступной опылителям. У Caly-сегасеае, цветки которых собраны в корзинки, пыльца откладывается на верхушку столбика и здесь может быть «собрана» опылителями.
В близкородственном Asteraceae, Caly-сегасеае и Goodeniaceae семействе Menyan-thaceae завязь верхняя, а вторичное преподношение пыльцы отсутствует. К этому семейству относят вахту (Menyanthes trifoliata), болотное растение с тройчатыми листьями, и болотноцветник (Nymphoides pehata), мелкое плавающее водное растение с листьями, похожими на листья кувшинки.
Неясное положение среди «эвастерид II» занимают семейства: Bruniaceae (12/78, Южная Африка), Carllemanniaceae (1/6, от Южной до Восточной Азии); Columelliaceae (2/9, Анды), Eremosynaceae (1/1, Австралия), Escalloniaceae (7/90, Юго-Восточная Азия, Австралия, Южная Америка), Icacinaceae (52/300, пантропическое), Polyos-maceae (1/60, от Гималаев до Австралии, Новая Каледония), Sphenostemonaceae (1/7, Юго-Восточная Азия, Австралия, Новая Каледония) и Tribela-сеае (1/1, умеренные области Южной Америки).
В пределах настоящих двудольных имеется ряд семейств с неясным родством. Это Balano-phoraceae (18/44, тропическо-субтропическое, по всему миру), Bonnetiaceae (3/22, Юго-Восточная Азия, Карибский бассейн, Южная Америка; ра
нее включали в Theaceae), Cardiopteridaceae (1/2, Юго-Восточная Азия), Ctenolophonaceae (1/4, Западная Африка, Малайзия; ранее включали в Linaceae), Cynomoriaceae (1/2, от Канарских о-вов до Монголии), Cytinaceae (2/8, Южная Африка, Мадагаскар, Средиземноморье, Юго-Западная Азия, Центральная Америка; ранее включали в Rafflesiaceae), Dipentodontaceae (1/1, Южный Китай и Бирма), Elatinaceae (2/32, в основном тропическое), Geissolomataceae (1/1, Южная Африка), Hoplestigmataceae (1/2, Западная Африка), Kaiiphoraceae (1/1, Мадагаскар), Lepidobo-tryaceae (1/1, тропическая Африка; ранее включали в Oxalidaceae), Lissocarpaceae (1/2, Южная Америка), Lophopyxidaceae (1/1, Тихоокеанская область; ранее включали в Celastraceae), Medusan-draceae (1/2, Западная Африка), Metteniusaceae (1/7, Центральная и Южная Америка; ранее включали в Icacinaceae), Mitrastemonaceae (1/2, Тихоокеанская область; ранее относили к Rafflesiaceae), Paracryphiaceae (1/1, Новая Каледония), Pentaphylacaceae(l/1, Юго-Восточная Азия), Peridiscaceae (2/2, тропическая Америка), Plagio-pteraceae (1/2, Юго-Восточная Азия; ранее включали в Flacourtiaceae), Pottingeriaceae (1/2, Индия, Бирма, Таиланд; ранее относили к Celastraceae), Sladeniaceae (1/1, Юго-Восточная Азия; ранее относили к Theaceae), Strasburgeriaceae (1/1, Новая Каледония; ранее включали в Ochnaceae) и Tepuianthaceae (1/5, Южная Америка).
Семейства Bonnetiaceae и Elatinaceae, вероятно, близкородственны Clusiaceae, a Paracryphiaceae и Strasburgeriaceae, очевидно, относятся к «астеридам» В Strasburgeriaceae входит только
508 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
один вид (Strasburgeria robusta), произрастающий на Новой Каледонии. Balanophoraceac и Супо-moriaceae — корневые паразиты, не содержащие хлорофилла, их мясистые густые соцветия с очень мелкими цветками похожи внешне на грибы. К Cynomoriaceae относится произрастающий в Средиземноморье циноморий (Cynomorium cocci-пеит). Также паразитами представлено семейство Cytinaceae со средиземноморским родом Cytinus и Mitrastemonaceae. Трудность определения таксономического положения паразитных семейств обусловлена тем, что они морфологически сильно редуцированы и к тому же утратили многие пластидные гены, необходимые как маркеры для молекулярного анализа.
ПРОИСХОЖДЕНИЕ
И РОДСТВЕННЫЕ СВЯЗИ СЕМЕННЫХ РАСТЕНИЙ
Многие вопросы, касающиеся происхождения семенных растений, родственных отношений между их ископаемыми и современными группами, а также происхождения цветковых растений и возникновения цветка, все еще остаются без ответа. Основная причина этого заключается в том, что решение нужно искать в палеоботанических данных, а ископаемые следы развития семенных растений очень неполные, причем интерпретация ископаемых структур сама по себе доставляет много проблем. Соответственно имеются и альтернативы представленным ниже гипотезам.
Можно исходить их того, что семенные растения (Spermatophytina), папоротники (Pteridopsida, включая Psilotopsida) и членистостебельные (Equisetopsida) происходят от общих предков, которые могут быть представлены, например, нижнедевонским ролом Psilophyton (рис. 11.288). Сестринскую группу этой линии развития образуют плауновидные (Lycopodiopsida).
Изображенный на рис. 11.286 Р. dawsonii отличался тем, что его ветвление было анизотом-ным (неравновильчатым) и голые ветви располагались поочередно. Первичные проводящие ткани слагали протостелу. Стерильные боковые ветви были многократно разветвленными, но конечные разветвления не располагались в одной плоскости; на фертильных боковых ветвях парами располагались многочисленные спорангии на коротких ножках. Как стерильные, так и фертильные боковые ветви на концах были закру
чены назад. Веретеновидные спорангии вскрывались продольной щелью, растения были изо-споровыми.
В линии развития, ведущей к семенным растениям, возник биполярный камбий, образующий наружу вторичную флоэму, а внутрь — вторичную ксилему с древесинными лучами. В качестве ранних представителей этой линии развития можно рассматри-вать среднедевонские роды Tetraxylopteris (рис. 11.288) и Arehaeopteris (рис. 11.288), которые вместе с другими многочисленными родами часто объединяют в Progymno-spermae (праголосеменные).
Tetraxylopteris (порядок Aneurophytales) отличается от Psilophyton — дополнительно к наличию камбия — тем, что его оси ветвятся накрест супротивно и что группы спорангиев отчетливо обращены на адаксиальную сторону. Arehaeopteris (порядок Archaeopteridales) по строению спорангиев сходен с Tetraxylopteris, однако был гете-роспоровым. Эти растения имели стволы диаметром до 1,5 м и высотой до 20 м. Древесина (первоначально описанная как признак рода Cal-lyxylon) по наличию круглых окаймленных пор во вторичной ксилеме очень сходна с древесиной современных хвойных. Кроме того, у некоторых видов были развиты уплощенные листовые органы с дихотомическим жилкованием.
Из праголосеменных в позднем девоне (ок. 370 млн лет назад) произошли монофилетической эволюционной линией семенные растения. Интегумент образовавшихся в ходе эволюции семяпочек, вероятно, возник путем прогрессирующей стерилизации и срастания всех мегаспорангиев, окружавших центральный мегаспорангий, оставшийся фертильным (рис. 11.289).
Учитывая, что у Tetraxylopteris и Arehaeopteris спорангии расположены только на особых спороносных системах ветвей, эта новая синаиги-альная гипотеза возникновения интегумента кажется более убедительной, чем теломпая гипотеза, постулирующая срастание смешанных стерильных и фертильных систем осей вокруг единственного остающегося фертильным мегаспорангия (рис. 11.289).
Ранние представители семенных растений, которые некогда объединяли в так называемые семенные папоротники (Pteridospermae = Lygi-nopteridopsida), очевидно, представляют собой па-рафилетическую группу, из которой возникли разные линии семенных растений. Они имели
11.2. Бактерии, грибы, растения | 509
Рис. 11.288. Предки семенных растений (А, В — R.Spohn, А — по Н. Р. Banks, S. Leclercq и F. М. Hueber из P.Kenrickn P.R. Crane, 1997; В — по P.M.Bonamo и Н.Р. Banks из Р.Кеппски P.R. Crane; С — по С. В. Веек и С. A. Arnold из W. N. Stewart):
А— Psylophyton (нижний девон); В — Tetraxylopteris (средний девон); С — Archaeopteris (средний девон); 1 — общий вид: дерево около 6 м высотой; 2 — боковая ветвь с вегетативными и спороносными листовыми органами; 3—6 — прогрессирующее развитие плоской листовой пластинки у разных видов; 7 — микро- и мегаспорангии; 8 — микро- и мегаспоры Tetraxylopteris и Archaeopteris, представляющих так называемые Progymnospermae
Рис. 11.289. Возникновение семязачатка (R.Spohn, А—D— по J.Walton из H.N.Andrews; Е, F — по Р. Kenrick и Р. R. Crane):
A—D — из частично вегетативной, частично спороносной системы теломов дифференцируется фертильный нуцеллус (темный) и стерильная обвертка интегумент (светлая) (теломная теория); Е, F — интегумент семязачатка возникает благодаря прогрессирующей стерилизации и срастанию всех внешних по отношению к остающемуся фертильным мегаспорангию ветвей целиком спороносной системы теломов (новая синангинальная теория)
510 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Рис. 11.290. Представители так называемых семенных папоротников (Pteridospermae = Lyginopteri-dopsida) (A, F — по H.N. Andrews; В — no D.H. Scott; С — no H.Potonie; D, E — no M.Hirmer; G — no J.Walton):
А — внешний Bup,Tetrastichia bupatides (1 /Зх); В, С — структура древесины и листа Lyginopteris larischii; D, Е — ветвь с микроспорофиллами и продольный и поперечный разрезы микроспорофилла Crosso-theca (1,5х). Семязачаток с интегументом; F — Genomosperma kidstonii (1 1/2х), G — Calathospermum scoticum (3/4х)
облик, как, например, Tetrastichia (рис. 11.290), и структуру листьев и древесины Lyginopteris (рис. 11.290). Микроспорангии, как, например, у Crossotheca (рис. 11.290), располагались радиальными группами и иногда срастались друг с другом. Семяпочки большей частью имели лопастной на верхушке интегумент и находились по одной (например, Genomosperma) или по нескольку (например, Calathospermum) в обшей, часто также лопастной обвертке, которую называют ку-пулой(рис. 11.290).
Анализ родственных отношений между современными пятью группами семен
ных растений (Coniferoosida, Ginkgoopsida, Cycadopsida, Gnetopsida, Magnoliopsida), проведенный с использованием морфологических признаков, в том числе и многочисленных (около 10) ископаемых групп семенных растений, и основанный на использовании нуклеотидных последовательностей различных ядерных, пластидных и митохондриальных ДНК, приводит к весьма различающимся результатам.
При морфологическом анализе обычно Ginkgoopsida получается ближайшим
11.2. Бактерии, грибы, растения
511
родственником Coniferopsida, a Gnetopsida — группой, тесно связанной с Magnolio-psida. Gnetopsida, Magnoliopsida и ископаемые Bennettitales и род Pentoxylon обозначают как антофиты. Эта гипотеза предполагает, что Gymnospermae (голосеменные растения) не являются монофилетической линией. По отношению к Magnoliopsida они были бы парафилетичны, поскольку Gnetopsida считают наиболее близкой из ныне существующих форм к покрытосеменным.
Совершенно другую картину взаимоотношений этих таксонов дает анализ большого числа молекулярных последовательностей различных участков ДНК: Gymno-spermae оказываются монофилетической линией, сестринской покрытосеменным, в пределах голосеменных Ginkgopsida наиболее близки Cycadopsida, a Gnetopsida— к Coniferopsida. При этом о монофилии Gym-nospermae в целом можно говорить более уверенно, чем о детальных родственных отношениях внутри этой группы.
Расхождения между результатами морфологических и молекулярно-генетических исследований могут иметь две причины. С одной стороны, возможно, что морфологическое исследование приводит к неверным результатам потому, что важные вымершие группы не сохранились в ископаемом состоянии или что структуры современных растений интерпретируются неправильно. С другой стороны, допустимо, что анализы последовательностей ДНК только ныне живущих семенных растений могут привести к неправильным результатам из-за того, что в них не учтены очень многие вымершие группы. Хорошо известно, что исключение таксонов из фило
генетического анализа сильно влияет на конечный результат.
Учитывая противоречивость представлений о филогении семенных растений, рассмотрим их современные группы как таксоны равного ранга.
Представления о возникновении цветка Magnoliopsida в большой степени зависят от того, как понимать родство цветковых растений с ископаемыми и современными группами семенных растений в целом. В сущности были и есть две очень различающиеся гипотезы возникновения цветка. В то время как эвантовая гипотеза (А.Арбер и Дж. Паркин) исходит из того, что уже предки покрытосеменных имели обоеполые цветки и цветок Magnoliopsida является соответственно одноосной системой с латеральными микро- и мегаспорофиллами, псевдантовая теория (Р. фон Ветгштейн) предполагает, что цветок возник из целого соцветия однополых цветков путем конденсации систем главной и боковых осей (рис. 11.291).
Псевдантовая теория исходит из того, что покрытосеменные произошли от Gnetopsida. При этом постулируется, что обоеполый цветок покрытосеменных возник путем конденсации сложной системы осей с мужскими и женскими цветками. Плодолистик покрытосеменных, согласно этой гипотезе, возник из кроющего листа (брак-теи) женского цветка; кроме того, у покрытосеменных новый второй интегумент семяпочки, которого не было у Gnetopsida, сформировался из брактеол, расположенных под женским цветком. В этой форме псевдантовая гипотеза не подтверждается
Рис. 11.291. Гипотезы возникновения обоеполого цветка покрытосеменных (А — по A. Arber и J. Parkin; В — по R von Wettstein):
А— эвантовая гипотеза исходит из того, что уже предки покрытосеменных имели обоеполые цветки и цветки покрытосеменных, соответственно — это одноосная система с латеральными микро- и мегаспорофиллами; В— псевдантовая гипотеза, напротив, принимает, что цветок возник из целого соцветия однополых цветков путем конденсации системы главной и боковых осей
512 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Рис. 11.292. Caytonia как возможный предок покрытосеменных (R. Spohn, A, D— по Т.М.Harris из P.R.Crane; В, С, Е— по Т.М.Harris из WN.Stewardt и G.W.Rotwell):
А— перистый мегаспорофилл с боковыми купулами (5х); В — купула; С— продольный разрез купу-лы с несколькими семязачатками; D — перистый микроспорофилл с боковыми группами микроспорангиев (7х); Е — группа сросшихся друг с другом микроспорангиев в поперечном разрезе
ни молекулярными, ни морфологическими исследованиями. Согласно молекулярным данным, Gnetopsida и Angiospermae связаны отдаленным родством, а по морфологическим признакам Gnetopsida можно считать близкородственной покрытосеменным группой, но никак не их предками.
В то время как молекулярные данные, ввиду обнаруженной с их помощью монофилии современных Gymnospermae, не позволяют пролить свет на происхождение покрытосеменных, многие морфологические исследования показывают, что ближайшим родственником и возможным предком последних является род Caytonia. Происхождение покрытосеменных от предков, сходных с Caytonia, вероятно, даже не противоречит молекулярным данным, поскольку по меньшей мере в одном из этих морфологических исследований показано, что Caytonia и покрытосеменные более близкородственны между собой, чем покрытосеменные и Gnetopsida. Caytonia имели перистые микро- и мегаспорофиллы, по-видимому, в обоеполых цветках. Мегаспо
рофиллы состояли из рахиса с боковыми купулами с несколькими семязачатками в каждой (рис. 11.292), микроспорофиллы тоже были перистыми и несли боковые группы сросшихся между собой микроспорангиев (рис. 11.292). Плодолистики Magnoliopsida могли бы сформироваться, если рахис мегаспорофилла стал бы плоским и заключил в себе семязачатки. Второй интегумент семязачатка Magnoliopsida, по этим представлениям, происходит из купулы, в которой число семязачатков сократилось до единственного. Эволюция микроспорофиллов Magnoliopsida шла в сторону редукции до одной ножки с двумя синангиями и в каждом из них с двумя микроспорангиями.
Bennettitales, которые по их морфологическим особенностям рассматривают как ископаемую группу, связанную с Magnoliopsida, могли возникнуть путем редукции мегаспорофилла до единственного семязачатка на ножке (см. рис. 11.212).
Если бы микро- и мегаспорофиллы Caytonia располагались в обоеполых цветках, то это отвечало бы представлениям о возникновении по
11.3. Филогенез
растений и история растительности
крытосеменных в соответствии с эвантовой гипотезой.
В качестве резюме можно только отметить, что, как и ранее, возникновение цветка Magnoliopsida и происхождение тычинки, плодолистика и второго интегумента семяпочки неясно. Большинство авторов тем не менее склоняются к эвантовой гипотезе возникновения цветка.
11.3. Филогенез растений и история растительности
11.3.1. Обзор филогенеза организмов
Самые древние следы жизни можно обнаружить уже примерно через 1 млрд лет после возникновения Земли, т. е. примерно 3,5 млрд лет т. н. Атмосфера нашей планеты состояла тогда преимущественно из водяного пара, углекислого газа, азота (N2) и следов водорода (Н2), серы, хлора и фтора, метана, аммиака и т.п. Столь распространенный в настоящее время свободный кислород почти полностью отсутствовал, и озоновый слой, защищающий от жесткого космического и ультрафиолетового излучения, в первичной атмосфере еще не образовался. При возникновении жизни главную роль играли, очевидно, высокие температуры (например, вследствие вулканической деятельности) и электрические разряды.
Как многократно удалось показать экспериментально, при таких условиях могли возникать многочисленные органические соединения. К ним относятся аминокислоты, нуклеотиды сахара, жирные кислоты и спирты или их предшественники — важнейшие «кирпичики» жизни. До сих пор не ясно, в каких точно условиях возникли эти молекулы и соответственно их полимеры: был ли это их водный раствор («первобытный суп»), или электрически заряженные поверхности («первобытная пицца») как, к примеру, пирит. Решающим тогда стало возникновение системы, способной к идентичной редупликации и кодированию. Предпосылкой для этого, очевидно, были полинуклеотиды типа РНК с автокаталитической активностью, способные поэтому катализировать собственную репликацию. Две
или более подобные системы РНК могли включиться в так называемый «гиперцикл», в котором компоненты способствуют взаимной репликации. Благодаря связи такой кодирующей системы с липидными мембранами («компарт-ментализация») и протеинами смогли, наконец, возникнуть какие-то пока еще гипотетические прогеноты. Приходится постулировать, что в них произошли постепенное усовершенствование генетической информации путем передачи этой функции от информационной РНК к кодирующей системе ДНК (сначала основанной на двух, затем — на трех нуклеотидах) с транскрипцией и трансляцией, а также оптимизация системы обмена веществ на основе все более многочисленных и лучше интегрированных управляемых ферментами процессов.
Самыми древними достоверными ископаемыми живыми существами были уже имевшие клеточную организацию прокариоты.
Они могли покрывать свои потребности в веществах и энергии либо за счет сложных органических соединений (первичные гетеротрофы, анаэробное брожение), либо за счет анаэробного хемосинтеза (например, используя S и Н2 при высоких температурах, см. 6.9). Сокращение легко доступных органических источников энергии, очевидно, сделало необходимым выработку окислительно-восстановительной системы (цитохромы и т.п.) и затем постепенный переход к автотрофности. Сначала возникли скорее всего, различные формы анаэробного фотосинтеза (см. 6.4.10) на основе циклического фотофосфорилирования, особенно с фотосистемой I и чаще всего с H2S в качестве донора электронов. Аэробный фотосинтез с использованием воды как источника электронов и восстановлением СО2 возник у цианобактерий путем эволюции фотосистемы II (см. 6.4.5). Это привело к увеличению содержания в атмосфере кислорода и создало предпосылки для возникновения значительно более экономичного обмена веществ, основанного на аэробном дыхании. Диверсифицирование органической жизни создало возможность эволюции самых разных форм вторичных гетеротрофов, таких как, например, сапрофиты и паразиты в разных группах прокариотических организмов.
Сегодня некоторые ставят под сомнение пока еще общепринятое деление прокариотических организмов на эубактерий и архебактерий (Bacteria и Archaea). Возможно, что первые гетеротрофные эукариотические клетки с клеточным ядром и хромосомами, эндоплазматической сетью и другими внутриклеточными мембранными системами, микротрубочками и актиновы-
514 | ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
ми филаментами, иногда со жгутиками (типа 9 + 2) и, вероятно, также с митозом и мейозом происходят от общего предка с архебактериями. Впрочем, вполне допустимо, что первые эукариотические клетки возникли путем клеточного симбиоза ар-хебактерий и эубактерий (гидрогенная гипотеза, см. 2.4.2). Если возникновение многочисленных внутриклеточных мембранных систем эукариотических клеток можно объяснить вероятным впячиванием (инвагинацией) клеточной мембраны прокариотического предка, то возникновение хромосом и клеточного ядра остается непонятным. Также не известно точно, когда определенные бактерии (Negibacteriota) попали путем первичного эндоцитоза в формирующуюся эукариотную клетку, где они стали митохондриями. Возможно, что это произошло перед или сразу после возникновения первых эукариотических организмов (Archaezoa). Последнее часто принимают более вероятным, так как известны очень примитивные эукариоты без митохондрий. Сейчас, правда, считают, что эти организмы утратили свои митохондрии вторично, приспосабливаясь, например, к эндопаразитическому образу жизни, а следовательно, митохондрии или соответственно гидрогеносомы принадлежат к базовому компоненту всех эукариот (см. 2.4.2).
У эукариот количество ДНК увеличено, и она организована с гистонами в нуклеосомы, хромосомы и клеточные ядра. Распределение и дальнейшая передача наследственного материала стали более точными благодаря митозу, а возможности генетических рекомбинаций значительно возросли вследствие эволюции полового процесса с сингамией и мейозом.
Существует невероятное множество одноклеточных гетеротрофных эукариот с митохондриями. Различные представители такого организационного уровня стали исходным пунктом для дальнейшей эволюции. Вероятно, однократный симбиоз одной из этих линий развития с фотоавтот-рофными бактериями (Cyanobacteriota) в ходе второго первичного эндоцитоза (см. 2.4.2) привел к возникновению первых растительных клеток, в которых эндоцитоби-онты стали пластидами. От этих первых ра
стительных клеток происходят все растительные организмы с простыми пластидами с двойной мембраной. Эти растения предстают перед нами тремя линиями: Glaucobionta, Rhodobionta (красные водоросли) и Chlorobionta (включая зеленые водоросли), из которых Rhodobionta и Chlorobionta более близки друг к другу. В группе Chlorobionta зеленые водоросли (Chlorophyta) являются сестринской группой Stre-ptophyta, которые состоят преимущественно из первично наземных растений (Embry-ophyta).
Все остальные группы водорослей, имеющих пластиды с 3 или 4 мембранами, возникали многократно путем вторичного эндоцитобиоза, когда различные гетеротрофные эукариотические одноклеточные включали разные водоросли. Зеленые водоросли являются вторичными эндоцитобион-тами Chlorarachniophyta и Euglenophyta, а красные водоросли — Haptophyta, Crypto-phyta, Dinophyta и Heterokontophyta.
He совсем ясны отношения Dinophyta. Хотя вполне вероятно, что у них вторичными эндо-цитобионтами являются красные водоросли, но в этой группе известны случаи, по-видимому, третичного эндоцитобиоза после утраты вторичных эндоцитобионтов.
В дальнейшей эволюции разных полифиле-тически возникших групп водорослей неоднократно параллельно шла эволюция от голых мо-надных или амебоидных форм к преимущественно неподвижным, имеющим слизистые чехлы или клеточные стенки кокковидным формам, затем к нитчатым и многоклеточным трихаль-ным и, наконец, к ветвящимся в двух или трех измерениях, сложным талломным плектенхим-ным или паренхимным формам. Помимо этого среди водорослей есть и сифональные формы. Это развитие коррелирует с переходом от свободного с сидячему образу жизни и увеличением размеров. Одновременно формировалась смена поколений, происходил переход от изогамии к анизогамии и оогамии и часто к преобладанию диплофазы над гаплофазой.
Организмы, известные как грибы, имеют разное происхождение и представляют четыре линии развития одноклеточных гетеротрофных эукариот. Эти линии привели к возникновению Acrasiobionta (клеточные слизевики), Myxobionta (слизевики), Oomycota (целлюлозосодержащие грибы) и
11.3. Филогенез
растений и история растительности
Mycobionta (хитинсодержащие грибы). Предок Oomycota был также и предком выше названных Heterokontophyta, a Mycobionta, — пожалуй, ближайшие родственники многоклеточных животных.
У Oomycota и Mycobionta, как и у разных групп водорослей, можно наблюдать развитие от одноклеточных форм к сифональным нитчатым и сложным плектенхимным талломам. Заселение грибами суши и связанная с этим дальнейшая их эволюция, к примеру появление плодовых тел, тесно связаны с возникновением наземных растений.
Наконец, благодаря эктосимбиозу различных микобионтов с цианобатериями или зелеными водорослями многократно и независимо возникали разные лишайники (Lichenes).
Появившиеся в ордовике около 450 млн лет назад наземные растения (Embryophyta: Bryophytina, Ptendophytina, Spermatophytina1) находятся в тесном родстве с водорослями Streptophytina и вместе с ними составляют отдел Streptophyta, который характеризуется, к примеру, латерально расположенными (если имеются) жгутиками, клеточным делением обычно с образованием фрагмопласта, а также особой структурой целлюлозы. К Streptophytina относят, например, Charophyceae (харовые водоросли) с родом Chara и Coleochaetophyceae с родом Coleochaete, которых как близких родственников Embryophyta следует поместить в один подотдел. Streptophyta вместе с зелеными водорослями (Chlorophyta) составляют группу хлоробионтов (Chlorobionta).
Наземные растения отличаются прежде всего тем, что их гаметангии (антеридии и архегонии) имеют оболочку из стерильных клеток и спорофит питается вначале как эмбрион за счет гаметофита.
Эволюция наземных растений проходила, вероятно, по мелководьям пресноводных водоемов с сезонными колебаниями уровня воды. В пользу этого свидетельствуют наличие и ныне в пресных водах близких родственников наземных растений, а также умозаключения о том, что условия здесь для возникновения наземных ра-
1 Остатки семенных растений достоверно известны значительно позже — с верхнего девона. — Примеч. ред.
стений были значительно благоприятнее, чем на морских побережьях. Важными предпосылками для заселения суши были возникновение кутикулы, устьиц, проводящих и механических тканей, межклетников и органов поглощения воды. Почти все эти структуры могли возникать как приспособление к лимитированной на суше доступности воды.
Первоначальной группой наземных растений можно считать мохообразные (Bryophytina), у которых более сложным анатомически и морфологически поколением является гаметофит, а неразветвленный спорофит всегда остается связанным с гаметофитом, быстро становится гетеротрофным и питается за счет гаметофита. Самая древняя подгруппа Bryophytina, пожалуй, — талломные печеночники (Marchantiopsida).
Папоротникообразные (Pteridophytina) и семенные растения (Spermatophytina) имеют морфолого-анатомически сложный, разветвленный спорофит, дифференцированный на основные органы: корень, стебель и лист1, а также ксилему и флоэму. Соответственно можно объединить папоротникообразные и семенные растения в группу побеговых, или сосудистых, растений (Kormophyta, или Tracheophyta). Всегда талломное гаметофитное поколение у них постепенно редуцируется.
Пока недостаточно ясно, возникли ли Pteridophytina из в таком случае парафилетических по отношению к ним Bryophytina, поскольку они близкородственны антоцеротовым (Anthocerotopsida), или же наземные растения сразу развивались двумя линиями — Bryophytina и Pteridophytina
Pteridophytina очень давно разделились на две ветви — плаунообразные (Lycopo-diopsida), с одной стороны, и хвощеобразные (Equisetopsida), псилофитообразные (Psilotopsida) и папоротникообразные (Pteridopsida) — с другой; три последние группы являются сестринскими по отношению к Spermatophytina. Поэтому Pteridophytina по отношению к Spermatophytina парафилетичны и, соответственно, отличаются от семенных растений только отсутствием семян.
1 Такая дифференциация тела свойственна не всем Pteridophytina. — Примеч. ред.
516 | ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
В ныне почти исключительно травянистой группе папоротников, значительно различающихся характером облиствения, структурой спорофиллов, а также анатомией стеблей, в прошлом не раз возникали древовидные формы. Точно также многократно параллельно происходит переход от равноспоровости с чаще всего обоеполыми гаметофитами к разноспоровости с мужскими гаметофитами, образующимися из микроспор, и женскими гаметофитами, образующимися из мегаспор.
Возникшие, по-видимому, в самом конце девона, примерно 370 млн лет тому назад, разноспоровые семенные растения отличаются тем, что женский гаметофит (зародышевый мешок) остается на спорофите в невскрывающемся мегаспорангии (ну-целлюсе) со стерильной оболочкой (инте-гументном), по крайней мере, до опыления. Эта структура из интегумента, нуцеллюса и зародышевого мешка, называемая семязачатком, после оплодотворения развивается в семя с семенной кожурой, питательной тканью эндоспермом и многоклеточным зародышем.
Возникновение семязачатка позволило преодолеть зависимость оплодотворения яйцеклеток в архегониях, развивавшихся на самостоятельном женском гаметофите, от капельно-жидкой воды.
Среди семенных растений, по-видимому, монофилетической группе из хвойных (Coniferopsida), саговниковых (Cycadopsida), гинкговых (Ginkgopsida) и гнетовых (Gnetopsida) (голосеменные) можно противопоставить цветковые растения (Magnoliop-sida) (покрытосеменные). Таким образом, приходится пересмотреть недавно еще общепринятое представление, что покрытосеменные наиболее близки к гнетовым и, следовательно, голосеменные парафилетич-ны относительно покрытосеменых. Родственные отношения четырех групп голосеменных неясны.
Самая богатая видами (около 250000) современная группа растений — покрытосеменные — в ископаемом состоянии впервые появляется в раннем мелу, примерно I40 млн лет назад. Она демонстрирует удивительное многообразие в экологии, в образовании травянистых и древесных жизненных форм, в биологии опыления и распространения, а также в обилии вторичных
метаболитов. Поэтому сегодня покрытосеменные — доминирующий элемент почти всех типов растительности, они смогли заселить также различные экстремальные местообитания.
Вероятные родственные связи между различными группами растений и грибов обобщенно представлены на рис. 11.9.
11.3.2. История растительности
Как ясно показал обзор филогенеза организмов, флора и соответственно растительность Земли постоянно изменялись с момента возникновения жизни, вероятно, более 3,5 млрд лет назад и уж точно — с возникновения эукариотических автотрофных растений около 1,4 млрд лет назад. Однако вымирание организмов не было непрерывным (бокс 11.11, с. 517). Современный растительный покров Земли можно понять только как результат длительного исторического развития. Реконструкцией этого процесса занимаются исторические флористика и фитоценология. Изменение флоры и растительности Земли обусловлено эволюцией растений, изменением земной поверхности (расположения материков и океанов, горообразования), например благодаря тектонике плит; изменением общего состава атмосферы (концентрации кислорода и углекислого газа, появлении озонового слоя, поглощающего ультрафиолетовое излучение, изменении температуры, количества осадков и т. д.), например благодаря возникновению фотосинтеза и аэробного дыхания; изменением всех взаимодействий растений с другими, также изменяющимися организмами. Все эти различные факторы тесно связаны друг с другом.
Для реконструкции истории флор и растительности привлекают данные палеонтологии (соответственно палеоботаники), геологии, филогенетики и систематики и исторической биогеографии.
11.3.2.1. Методы
Фоссилизация растительных остатков происходит лишь в очень специфичных условиях, прежде всего в морских и озерных
11.3. Филогенез растений и история растительности
Бокс 11.11. Массовые вымирания
В истории Земли флора и растительность постоянно изменялись, одни растительные группы вымирали, появлялись новые (рис. А). Вымирание организмов, впрочем, не было непрерывным процессом. Возможно обнаружить периоды повышенной скорости вымирания, так называемые массовые вымирания. Очевидно, что массовые вымирания сосудистых растений, наземных позвоночных и морских беспозвоночных лишь в малой мере происходили одновременно. Некоторые авторы признают девять массовых вымираний сосудистых растений (девон — 391, 378 и 363 млн лет; карбон — 290 млн лет; триас — 241 млн лет; юра— 152—155 млн лет; мел— 132 млн лет; третичный период — 29 и 16 млн лет) Возможно, самое известное (но не совпадающее с вышеназванными 9 периодами) благодаря вымиранию динозавров массовое вымирание на границе мела и третичного периода, как полагают, произошло из-за столк-
новения с Землей астероида. Этот удар привел к выбросу в атмосферу огромного количества пыли. В результате на долгие годы сократилось количество достигающей Земли солнечной радиации, и соответственно произошло заметное похолодание климата. Недавно на континентальном шельфе у побережья Юкатана (Мексика) был найден кратер, появившийся в результате этого удара, первоначально предполагаемого на основании повсеместно распространенного слоя, обогащенного иридием (иридий редок на Земле, но обычен в астероидах и кометах). Предполагают, что в различных областях Земли при этой катастрофе должно было вымереть от 20 до 75% сосудистых растений (по крайней мере, локально). И хотя эта внеземная причина массового вымирания на границе мела и третичного периода вполне вероятна, вряд ли таким путем можно объяснить все массовые вымирания в истории Земли.
Млн лет
Рис. А. Относительное многообразие групп наземных растений с начала ордовика (по К. J. Niklas): L — Lycopodiopsida; Е — Equisetopsida; Р — Pteridopsida; G — голосеменные; А — покрытосеменные
518 | ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
осадках, в торфе и образующемся из него со временем каменном угле. Лучше всего в ископаемом состоянии сохранились некоторые группы водорослей, а также фрагменты побегов, листья, споры, пыльца, семена и плоды сосудистых растений. Ископаемые растения относят, насколько возможно, к современным таксонам или описывают как вымершие таксоны. Для определения систематического положения ископаемых чаще всего можно использовать только морфологические, а в случае окаменелостей — и анатомические признаки.
Непосредственно в ископаемом состоянии обычно сохраняются только скелетные элементы некоторых групп водорослей: кремнеземные диатомовые (Diato-meae), известковые Coccolithophorales (см. рис. 11.71), Corallinaceae, Dasycladaceae. В других случаях ископаемые возникают путем окаменения, углефикации или в виде отпечатков и соответственно отливок.
При окаменении (см. рис. 11.130; 11.131; 1.139) органический материал клеточных стенок и клеточного содержимого заменяется минеральными веществами (например, кремнеземом, карбонатом кальция), и возникают ископаемые, в значительной мере передающие прижизненную структуру. Углефикация происходит в ходе потери газов и воды при механическом сдавлении, вследствие чего возникают сильно деформированные остатки. Отпечатки и отливки сохраняют только форму поверхности соответствующей части растения. Заключенные в янтаре ископаемые являются чаще всего отпечатками или отливками полостей, но могут содержать также и органический растительный материал. Важный метод анализа ископаемых — получение тонких шлифов (см. рис. 11.142), с которых послойно снимают отпечатки на пластиковой пленке. На некоторых образцах хорошо различимы анатомические или даже электронно-микроскопические структуры (например, строение гран хлоропластов в листьях из миоцена).
Обычная проблема при реконструкции ископаемых находок — определить принадлежность отдельно найденных и описанных органов. Так, например, к всеобщему изумлению в 1960 г. было показано, что ранее описанные как Callixylon стволы, анатомически похожие на древесину голосеменных, и папоротникообразные листья рода Archaeopteris (см. 11.2, рис. 11.288) на самом деле принадлежали одному растению.
Особое значение, в первую очередь для более молодых отложений, имеет прекрасно сохраняющиеся благодаря устойчивой к всякого рода воздействиям экзине споры и пыльцевые зерна сосудистых растений. Анализом спор и пыльцы занимается палинология (см. 11.2— Микроспорофиллы).
Споры и пыльцевые зерна можно легко классифицировать на основе их глубокой структурной дифференцировки (см. 11.2 — Пыльца). Прежде всего, споры и пыльца ветроопыляемых, а также некоторых насекомоопыляемых растений рассеиваются в огромном количестве. В Центральной Европе ежегодно на квадратный сантиметр почвы оседает несколько тысяч пыльцевых зерен и спор, которые захораниваются в отложениях (к примеру, в морском известняке, торфе, грубогумусной почве и т. п.). По первому появлению споровых тетрад можно определить время возникновения мейоза и, соответственно, полового процесса, а самые древние трикольпатные пыльцевые зерна (см. рис. 11.268) позволяют датировать появление «Rosidae». Для изучения развития флоры и растительности в четвертичном периоде можно из подходящих отложений получить буровую колонку, послойно обработать и количественно проанализировать ее. Графическое представление пыльцевой диаграммы (см. рис. 11.306) показывает наличие и меняющееся соотношение спор и пыльцевых зерен разных видов в охваченном буровым профилем временном срезе. При знании современных количественных показателей осаждения пыльцы в различных типах растительности возможна даже реконструкция изменения количественного состава растительности вблизи от места бурения.
Знание возраста ископаемых растительных остатков имеет центральное значение для истории флоры и растительности. На смену сравнительной хронологии истории Земли (рис. 11.293), которая опирается на наличие в исследуемых слоях определенных животных и растительных руководящих остатков, сегодня пришли различные методы определения абсолютного возраста.
Возраст горных пород определяют с помощью радиометрических методов. При этом используют постоянство периода полураспада радиоактивных элементов в минералах. Например, период полураспада урана (236U —> 206РЬ) составляет 4,5 млрд лет. По относительной концентрации радиоактивного элемента и продукта его рас-
11.3. Филогенез растений и история растительности | 5-) g
Кайнозой Четвертичный период (2) Голоцен Плейстоцен Неофит
Третичный период (65) Плиоцен Миоцен Олигоцен Эоцен Палеоцен
Мезозой Мел (146) Маастрихт Кампан Сенон Турон Сеноман
Альб Алт Баррем Неоком Палеофит Мезофит
Юра(208) Верхняя Средняя Нижняя
Триас(245) Верхний Средний Нижний
Палеозой Пермь (290) Верхняя
Нижняя
Карбо I (363) Верхний карбон (пенсильваний) Нижний карбон (миссисипий)
Девон '409) Верхний девон Средний девон Нижний девон
Сигур (439) Протерофит
Ордовик (510)
Кембрий (570)
Докембрий Г ротерозсй (2 500)
Apxei. (> 4 000)
Рис. 11.293. Хронология истории Земли
лада можно определить возраст горной породы. Для датировки ископаемых остатков особое значение имеет радиоактивный калий (40К -» 40Са или 40Аг) как важная составная часть большинства осадочных пород. При использовании этого элемента хорошо удается определять возраст ископаемых находок более 100 000 лет. Для определения более молодых ископаемых (< 50 000 лет) чаще всего используют радиокарбоновый метод. Он основан на том, что в органических соединениях первоначальное отношение изотопов углерода l2C:l4C, равное таковому в воздухе, благодаря распаду |4С в l4N (период полураспада 5 730 + ± 40 лет) сдвигается в сторону |2С. Другие мето
ды абсолютной датировки ископаемых еще более молодого возраста базируются на различных процессах с выраженным годовым ритмом. Это — определение возраста по годовым кольцам прироста древесины у деревьев (дендрохронология), посредством чего иногда удается точно определить возраст до 8 000 лет. Так называемые ленточные глины и подобные им отложения, равно как и ледники, показывают слоистость, которую можно использовать для определения абсолютного возраста. Далеко разносящийся при извержениях вулканов пепел также может стать важной точкой отсчета для определения возраста. Относительно грубые палео-
520 | ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
магнитные датировки основаны на том, что магнитное поле Земли со временем периодически меняет полюса, причем ориентация магнитных полюсов «фоссилизируется» в железосодержащих горных породах. Правда, временные границы отрезков с различной полярностью приходится устанавливать изначально другими методами. На протяжении третичного и четвертичного периодов кайнозоя эпохи противоположной полярности имели длительность от 20 тыс. до 730 тыс. лет.
Для последнего тысячелетия можно установить возраст растительных остатков, если вместе с ними были найдены предметы до- или раннеисторической культуры известного возраста. Если таким образом установлена одновременность некоего доисторического периода и определенного «лесного времени», то можно, наоборот, определить возраст археологических находок неизвестного возраста по пыльцевому анализу, если в раскопе сохранилась подходящая ископаемая пыльца.
Не только ископаемые находки, но и родственные связи, и распространение современных таксонов позволяют сделать косвенные выводы об истории флоры и растительности определенной области. Многие таксоны имеют дизъюнктивные ареалы, промежутки между частями которых обычно очень велики и естественным образом в ходе обычного расселения непреодолимы. К примеру, существует дизъюнкция ареалов многих групп раст ений между Восточной Азией и восточной частью Северной Америки (см. рис. 14.24). Если исключить дальнее расселение в качестве объяснения т акого примера ареалов, то понять его можно только при допущении существования в прошлом сплошной области распространения таксона. Это подтверждают данные о климате и растительности прошлого в областях, расположенных между Восточной Азией и востоком Северной Америки. В настоящее время многочисленные исследования родства, основанные на последовательностях ДНК, позволяют в известной мере проводить абсолютное датирование, опираясь на концепцию молекулярных часов. Если удается датировать филогенетическое расхождение тех же восточноазиатских и североамериканских форм, то, исключив дальнее расселение, возможно утверждать, что по крайней мере до этого времени должна была
существовать единая общая область распространения.
Так как молекулярные часы приходится калибровать по ископаемым остаткам или геологическим событиям, существует опасность попасть в порочный «круг доказательств».
Успешные реконструкции истории флор и растительности Земли требуют географически и систематически репрезентативного и легко интерпретируемого материала. Добротную картину истории флоры и растительности, соответственно, получают главным образом для молодых геологических отложений и для областей с давними научными традициями (например, Европа, Северная Америка и во все большей степени Китай) и в основном для таких организмов, которые имеют структуры, хорошо сохраняющиеся при фоссилизации.
11.3.2.2. Докембрий и палеозой (4 млрд — 245 млн лет)
Со времени возникновения жизни более 3,5 млрд лет назад и до заселения суши многоклеточными растениями, вероятно, в ордовике (около 450 млн лет назад) теплые моря, по-видимому, населяли одноклеточные и многоклеточные организмы, сильно различавшиеся организацией, образом жизни и формами получения энергии.
В этот промежуток времени возникли эубактерий и архебактерии как прокариотические организмы, гетеротрофные эукариоты, вначале без митохондрий, а затем с приобретенными путем эндоцитобиоза митохондриями и, наконец, фотоавтотрофные эукариоты — благодаря эндоцитобио-зу между гетеротрофными эукариотами и фотоавтотрофными цианобактериями.
Несомненные ископаемые клетки прокариот, напоминающие различных бактерий, известны из формаций Fig-Tree Южной Африки возрастом 3,1 млрд лет Организмы, по образу жизни, вероятно, напоминавшие архебактерий, недавно были найдены в окаменелых морских глубоководных вулканических породах возрастом около 3,2 млрд лет. В слоях возрастом 3—1,6 млрд лет из Африки и Северной Америки многообразие прокариотических форм увеличивается. Так, находят, к примеру, похожие на Oscillatoria
11.3. Филогенез растений и история растительности | 521
цианобактерии, образовавшие ископаемые строматолиты. Первые эукариотические водоросли появились ранее 1,4 млрд лет назад, а в древних формациях Австралии возрастом от 1,5 до 0,9 млрд лет найдены на удивление хорошо сохранившиеся остатки одноклеточных водорослей (Caryosphaeroides, напоминающие Chlorococcales), у которых можно распознать разные стадии деления клетки и даже остатки клеточного ядра. Эти биоценозы содержали также бактерии, водные грибы и одноклеточных животных (Protozoa). Ранее ордовика дифференцировались две главные линии ранней эволюции растений, приведшие к зеленым и красным водорослям.
Изначальное отсутствие кислорода в атмосфере требовало получения энергии путем самых разных форм анаэробной гетеро- и автотрофии. Ранние прокариоты вели, вероятно, образ жизни, отчасти сходный с таковым современных крайне термофильных видов архебактерий. Аноксигенный и оксигенный фотосинтез появился, вероятно, уже более 3 млрд лет назад. С тех пор концентрация свободного кислорода в атмосфере постепенно нарастала. К периоду 2,8 — 2,4 млрд лет назад концентрация атмосферного кислорода достигла примерно 1 — 2 % от современной концентрации (21 %). При таких условиях сократилось число доступных для анаэробных организмов местообитаний и началась дифференциация аэробных организмов и эволюция более эффективного дыхания. Ко времени возникновения эукариотических водорослей концентрация кислорода в атмосфере достигла примерно 10%. С увеличением концентрации кислорода в атмосфере связано и возникновение задерживающего ультрафиолетовое излучение озонового слоя, ставшего важной предпосылкой заселения суши.
Ко времени заселения суши (приблизительно 450 млн лет назад) в ордовике (510—439 млн лет; здесь и ниже цифры означают число лет до нашей эры) центры современных материков располагались преимущественно в Южном полушарии (рис. 11.294), и климат Земли был теплым. Из этого времени происходят таксономически не определимые первые ископаемые находки до некоторой степени засухоустойчивых растений, представленные спорами, фрагментами кутикулы с отпечатками клеток и трубковидными клеточными элементами с поясковыми утолщениями стенок. В силуре (439—409 млн лет) встречаются остатки растений из вымершей группы Homeophytae (например, Caia, Tortilicaulis), а также род Cooksonia из линии развития
Lycopodiopsida. В девоне (409—363 млн лет) с обильными осадками и наводнениями, но также и с более выраженной засушливостью климата континенты занимали явно более северное положение (рис. 11.294). В этот период выявлена сильная дифференциация наземных растений, из-за чего девон рассматривают как начало палеофита, сменившего протерофит. Один из известных представителей девонских растений — род Rhynia (396 млн лет; см. рис. 11.130), относящийся к ныне вымершей линии развития. Почти все ныне существующие эволюционные линии растений впервые встречаются в девоне. Из Lycopodiopsida это, к примеру, Drepanophycus (примерно 410 млн лет) и Zosterophyllum (396 млн лет; см. рис. 11.130). Разделение Lycopodiopsida на Isoetales и Lycopodiales выявлено в верхнем девоне. Линия развития, ведущая к папоротникам, хвощам и семенным растениям, представлена, например, Psilophyton (390 млн лет; рис. 11.288) и Tetraxylopteris (381 млн лет; рис. 11.288). Ранний представитель хвощей— Ibyca (381 млн лет), a Rhacophyton (377 млн лет) считают представителем папоротников. Семенные растения встречаются также уже в верхнем девоне — Elkinsia (367 млн лет).
Ожидаемых различных ископаемых представителей Bryophyta, этой первоначальной группы ныне живущих наземных растений, в девоне очень мало, вероятно, из-за их очень плохой сохранности в ископаемом состоянии. Только Pal-lavicinites (370 млн лет) из Jungermanniopsida достоверно известен из девона. Впрочем, предполагали, что некоторые ископаемые из ордовика следует относить к печеночникам. Достоверно ископаемые антоцеротовые и лиственные мхи впервые встречаются в мелу.
С изменением систематического состава флоры в силуре и девоне началось и изменение растительности. Самые ранние наземные сообщества были разреженными, низкими (<50 см) и чаще всего представляли, по-видимому, земноводные заросли по берегам водоемов или в заболоченных понижениях рельефа. Позднее встречались уже и более высокие и полностью наземные сообщества. Эти наземные биоценозы в силуре и девоне были, очевидно, распространены по всему свету (рис. 11.294; 11.295).
522 ГЛАВА 11 СИСТЕМАТИКА И ФИЛОГЕНИЯ
Рис. 11.294. Расположение континентов с начала ордовика (числа — миллионы лет) (по C.Scotese и http://www.ucmp.berkeley.edu/geology/tectonics.html)
11.3. Филогенез растений и история растительности | 523
Наличие редуцентов (бактерий и грибов) на растениях показывает, что первые наземные экосистемы функционировали в сущности так же, как и современные Вероятно, что уже самые ранние наземные растения имели везикулярно-ар-бускулярную микоризу (см. 9.2.3: Микориза). Вполне возможно, что этот симбиоз сыграл решающую роль в заселении суши.
В карбоне (363 — 290 млн лет) материки еще дальше сдвинулись на север (см. рис. 11.294).
В этом периоде появились первые обширные леса с деревьями высотой более 30 м. Из остатков этих лесов образовался каменный уголь.
Центральная область этих лесов охватывала Европу и восточную часть Северной Америки, они известны также в Сибири и Восточной Азии. Во всех этих областях господствовал влажный теплый (суб-) тропический климат (рис. 11 296). Растения каменноугольных лесов не имели годовых колец древесины или покоящихся почек, анатомия их листьев подобна анатомии листьев растений современных дождевых тро-
Состав атмосферы почти достиг современных показателей. В таких условиях на сырых и переувлажненных торфяных почвах росли мощные болотные леса (рис. 11.295), в которых доминировали древовидные хвощи и плауны, а также кордаиты. Эти биоценозы были многовидовыми и сильно дифференцированными на ярусы и зоны.
Зональность определяла доступность воды. Заросли хвощей (Archaeocalamites, Catamites', см. рис. 11.142; 11.143), очевидно, занимали мелководья. На незалитых местах обитали древовидные плаунообразные высотой до 40 м и с диаметром стволов в 2 м (Lepidodendron, Sigillaria', см. рис. 11.138; 11.139) с богатой сопровождающей флорой из менее высоких древовидных папоротников, кустарниковых или лиановидных семенных растений из ныне вымерших таксонов и с нижним ярусом из травянистых клинолистов (см. рис. 11.142), лиственных мхов и печеночников. Более сухие местообитания были населены, очевидно, кордаитами (см. рис. 11.206). Благодаря мошной вторичной древесине они, вероятно, лучше поддерживали водный баланс, чем имевшие толстую кору и мало древесины лепи-
пических лесов.
додендроновые.
Рис. 11.295. Реконструкция каменноугольного леса (Музей естественной истории, Чикаго).
Слева вверху ветвь с листьями и колосками из спорофиллов лепидодендрона (Lepidodendron), правее стволы лепидодендрона и сигиллярии (Sigillaria), между ними вайя Neuropteris с образующимися семенами, а также тонкие побеги Lyginopteris (оба — Pteridospermae); в центре на переднем плане клинолист Sphenophyllum, сзади папоротник с гигантской древней стрекозой, а также другие древовидные плауновидные, справа каламит (Catamites)
524 | ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Рис. 11.296. Glossoptens, цельный лист с сетчатым жилкованием (1/Зх) (noW.Gothan)
В каменноугольных лесах встречались и животные, например амфибии, первые рептилии, пауки, многоножки и примитивные насекомые (стрекозы, тараканы). Можно обнаружить здесь и паразитические, симбиотические и сапрофитные грибы.
Одновременно с каменноугольными лесами современного Северного полушария в Южной Африке, Индии, Австралии, Антарктиде и на юге Южной Америки развилась совершенно другого типа, более скудная так называемая гондванская флора. Руководящими формами здесь считают кустарниковые семенные растения, например, род Glossoptens (рис. 11.296), разные папоротники и хвойные. Соответственно более прохладному умеренному климату этих регионов, составлявших южный континент Гондвану, древесина здешних растений иногда была с годовыми кольцами. Похолодание и аридизация климата в перми (290—245 млн лет) привели к исчезновению каменноугольных лесов и вымиранию древовидных плаунообразных и клинолистов. Зато можно наблюдать диверсификацию голосеменных, к примеру Voltziales (см. 11.2; рис. 11.207). Это уже начало мезофита. В карбоне и еще сильнее в перми Южный континент подвергся оледенению.
11.3.2.3. Мезозой (245—65 млн лет)
По крайней мере с начала аридного или полуаридного триаса (245 — 208 млн лет) в течение всей достаточно влажной юры (208— 146 млн лет) и до умеренного или теплого нижнего мела (146 —65 млн лет) флоры по всей Земле были однообразны вследствие однообразия климата, без каких-либо следов оледенений.
В этот огромный промежуток времени продолжалось движение континентов, и в мелу возникло морс Тетис, разделявшее Северный и Южный материки (см. рис. 11.294).
Главную роль во флорах в это время играли папоротники, хвощи и особенно разные 1руппы голосеменных вроде Ginkgoopsida, Coniferopsida и Cycadopsida, а также вымершие беннеттиты (см. рис. 11.212).
Обилие видов сосудистых растений и их биотопическое разнообразие сильно возросли по сравнению с карбоном, что можно объяснить экспансией растений в более сухие местообитания и коэволюцией с наземными группами животных (например, опыление насекомыми бен-неттитов, распространение семян репти лиями).
Мезофит северной континентальной области начался с наступившего в верхней перми (цехштейна) и продолжавшегося до среднего триаса длительного аридного периода, который привел к формированию ксероморфных групп растений вроде Voltziales (см. рис. 11.207) и унификации флор Северного и Южного континен тов. В более поздних флорах мезофита среди папоротников преобладали эвспорангиатные и первичные лептоспорангиатные. Ginkgoopsida были представлены примерно 11 разнообразными родами (например, Baiera, см. рис. 11.202), а представленный только одним современным видом род Ginkgo с нижней юры до верхнего мела распространен почти по всей Голаркти-ке (см. рис. 14.16). Ареал рода Araucaria из хвойных был в юре космополитным, но с третичного периода ограничивался Южным полушарием и сегодня сократился до разрозненных фрагментов в Западной Пацифике, Австралии и Южной Америке. Среди хвойных в юре и нижнем мелу были предшественники современных Podocarpaceae Южного полушария, а также Taxaceae, Cephalotaxaceae и Cupressaceae
11.3. Филогенез растений и история растительности | 525
и, возможно, также Pinaceae. Особенно типичны для флор мезофита Cycadopsida с существующими и поныне и уже вымершими таксонами (рис. 11.297).
Важным этапом в истории флор и растительности Земли стало возникновение покрытосеменных (примерно 140 млн лет) в нижнем мелу. Покрытосеменные появились, вероятно, как травы и кустарники нарушенных местообитаний в низких географических широтах в полуаридных или аридных условиях. Вскоре после возникновения покрытосеменных произошло разделение Северного (Лавразии) и Южного (Гондваны) материков морем Тетис.
Сравнительное единообразие мирового климата уступило место все более усиливающемуся градиенту температур от полюсов к экватору. Причину быстрой диверсификации и высокого видового разнообразия покрытосеменных следует искать не только во многих особенностях покрытосеменных, повышающих темпы их видообразования, но и в этих тектонических и климатических изменениях Земли. Уже в среднем мелу покрытосеменные стали доминирующей группой растений в большинстве типов растительности по всему миру. Это время отмечает начало неофита, ознаменовавшееся вымиранием
Рис. 11.297. Последовательность появления типов пыльцы и листьев древнейших покрытосеменных от нижнего мела (баррем, апт, альб) до начала верхнего мела (сеноман) на востоке Северной Америки (и в других местах) (по J. A. Doyle и L.J. Hickey).
Типы пыльцы: А, В — дистально-однобороздная (А — Clavatipollenites, В — Liliacidites и др.); С — трехбороздная (Tricolpites и др.); D —F— трехбороздно-оровая (Tricolporoidites). Типы листьев: G— обратнояйцевидный, с нерегулярным жилкованием, сходный с листьями однодольных (Acaciaephyllum); Н — почковидный (Proteaephyllum); J — яйцевидный зубчатый, с перистым жилкованием (Quercophyl-lum); К— узкообратно-яйцевидный цельнокрайный (Rogersia); L— широкоэллиптический (Ficophyl-lum); М — пальчатолопастной (Vitiphyllum); N — обратнояйцевидный (Celastrophyllum) О — щитовидный (Nelumbites, Menispermites); Р — яйцевидно-сердцевидный (Populophyllum); Q — эллиптический, зубчатый (Celastrophyllum); R, Т — перистый (Sapindopsis); S, U — трехлопастной, возрастание регулярности жилкования (Araliaephyllum и Araliopsoides)
526 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
или сильным сокращением таксонов, доминировавших в мезофите.
Более или менее непрерывные ряды ископаемых форм (рис. 11.297) показывают увеличение разнообразия пыльцы и листьев. Из этого можно сделать вывод о ранней диверсификации жизненных форм (от древесных до травянистых) и заселенных биотопов (от сухих до влажных).
Можно думать, что современные ареалы по крайней мере некоторых голарктических, пантропических или распространенных в Южном полушарии таксонов обусловлены положением континентов в мелу и раннем третичном периоде. Так, Северная Америка и Евразия сравнительно поздно отделились друг от друга в области Северной Атлантики; Африка, Аравия и Индия были долгое время связаны друг с другом, тогда как Африка и Южная Америка, напротив, рано разделились (примерно 90 млн лет), а сухопутная связь и возможность миграции между Южной Америкой, Антарктидой и Австралией с Новой Зеландией существовала еще примерно 40 млн лет назад. Пространственное сближение или соединение Индии с Южной Азией, Австралии и Новой Гвинеи с Юго-Восточной Азией, а также Северной и Южной Америк (примерно 3 млн лет) имело место лишь в самые поздние геологические эпохи. В следующих примерах звездочками отмечены достоверно известные уже из мела ископаемые остатки групп, демонстрирующие соответствующие типы распространения. Голарктическое: Magnoliaceae* (см.рис. 14.24),Cercidiphyllaceae*, Platanaceae*, Fagaceae* (см. рис. 14.17), Betulaceae*, Paeoniaceae. Пантропическое: Аппопасеае, Мо-nimiaceae, Chloranthaceae*, Myrtaceae*, Flacour-tiaceae, Sapotaceae*, Rubiaceae, Arecaceae*. Африканско-южноамериканское: Velloziaceae. В Южном полушарии: Winteraceae*, Proteaceae*, No-thofagaceae*.
Эти и другие примеры возможного мелового или раннего третичного происхождения ряда ареалов требуют критической перепроверки в свете накапливающихся данных геносистематических работ. Использование молекулярных часов, например, в исследованиях Gentianaceae и Melastomataceae показало, что миграции между тропическими континентами, вероятно, имели место и спустя долгое время после разделения континентов либо путем заноса, либо путем миграции через сегодняшние области с умеренным климатом. Для пантропических Lauraceae удалось показать, что пантропический ареал некоторых групп родов можно объяснить дрейфом континентов, тогда как другие группы родов с подобным же распространением слишком молоды для этого.
11.3.2.4. Кайнозой (65 млн лет — по настоящее время)
Палеоген-неогеновый период (65 — ок. 2 млн лет) — время очень глубоких климатических изменений. В то время как в первой половине периода, в палеогене (палеоцен 65 — 54 млн лет, эоцен 54—38 млн лет, олигоцен 38 — 26 млн лет), был широко распространен мягкий или даже субтропический климат, в более позднее время, в неогене (миоцен 26—12 млн лет, плиоцен 12 — около 2 млн лет; начало плиоцена иногда датируют 5,2 млн. лет), произошли похолодание и аридизация климата, которые продолжилось в оледенениях четвертичного периода, или плейстоцена.
Существенной причиной этих климатических изменений стало изменение поверхности Земли. Поднятие в миоцене и плиоцене Гималаев, Альп, Анд и т. п. привело к образованию обширных засушливых областей в «дождевой тени» этих новых горных систем, смещение огромных масс суши в более высокие географические широты способствовало образованию полярных ледяных «шапок» (рис. 11.294), а сокращение или высыхание больших морей вроде Тетиса привело к появлению континентального климата в удаленных от морей областях. Поэтому климат Земли в конце палеоген-неогена был значительно сложнее, чем в начале его. Эти климатические и геоморфологические измерения сильно повлияли на развитие флоры и растительности (рис. 11.298).
В палеогене даже в областях современного умеренного климата были широко распространены тропическо-субтропические флоры вечнозеленых дождевых лесов с Lauraceae (например, Cinnamomum), Moraceae (Artocarpus, Ficus), Juglandaceae (Engelhardtia), пальмами (Sabal. Elaeis, Nypa) и тропическими папоротниками (к примеру, Matonia). «Языки» этих флор достигали даже ныне арктических регионов, например, Аляски и Гренландии. В палеогене Голарктики значительно дальше к северу были распространены богатые видами листопадные лиственные и смешанные леса. Такие флоры встречались до
11.3. Филогенез растений и история растительности
527
Сухой лес на торфянике
(топяной) торф
Рис. 11.298. Реконструкция поясов растительности среднетретичного болота Центральной Европы (по M.Teichmueller из Р. Duvigneaud)
Шпицбергена и Канадского архипелага (8Г45' северной широты, в настоящее время среднегодовая температура там -20°С). Они состояли из видов Pinus, Picea, Platanus, Fagus, Quercus, Corylus, Betula, Alnus, Juglans, Ulmus, Acer, Vitis, Tilia, Populus, Salix, Fraxinus и др. родов, которые еще и сегодня встречаются, к примеру, в умеренном климате Европы. Другие роды в современной Европе уже давно вымерли, но встречаются в более теплых областях Северной Америки (Taxodium, Sequoia), Восточной Азии (Ginkgo, Cercidiphyllum) или в обеих областях (Tsuga, Magnolia, Liriodendron, Sassafras, Carya, Diospyros). Так как Северные материки тогда еще менее разошлись друг от друга, чем в настоящее время (см. рис. 11.294), в третичном периоде в циркумполярном регионе, вероятно, происходит более оживленный флористический обмен. В результате возникла так называемая аркто-третичная флора — ядро современной флоры Голар-ктиса.
Для Центральной Европы принимают среднегодовую температуру в эоцене 22 °C. В Северном полушарии граница леса была смещена к северу на 20—30°, а северная граница распространения пальм — на 10—15° по сравнению с современными. Ископаемые остатки этих палеогеновых флор найдены в Европе, например, в Экфельде — Эйфеле и Месселе под Дармштадтом, а в Гайзельтале — под Халле. Балтийская флора янтарей относится к этому же времени. В ней удалось найти дополнительно к вышеназванным родам представителей Cornaceae (Mastixia), Annonaceae, Theaceae (Stewartia), Stre-culiaceae, Sapotaceae, Symplocaceae, Pandanaceae
и Cyatheaceae, которые в настоящее время распространены главным образом в тропиках и являются реликтовыми таксонами Юго-Восточной Азии. Тогдашняя растительность, вероятно, была похожа на современные богатые представителями Lauraceae горные леса Юго-Восточной Азии. Три европейских рода (Ramonda, Jankea. Haberlea) пантропических Gesneriaceae являются, вероятно, реликтами этой тропической третичной флоры.
С эоцена до миоцена в Центральной Европе из органических отложений мелеющих озер и граничащих с ними заболоченных лесов формировались отложения бурого угля. Руководящими формами третичных лесов (рис. 11.298) были обитающие в настоящее время в теплоумеренном поясе Северной Америки реликтовые роды хвойных Taxodium и Sequoia, а также общий для Америки и Азии род Nyssa (Cornaceae).
В зарастающих водоемах (рис. 11.298) на глубине наряду с Nymphaea росли и другие ним-фейные, например Brasenia (современное распространение: Америка, Азия). Мелководье занимал пояс осок и тростника с Dulichium (Cyperaceae; Америка). Далее следовал пояс переувлажненного заболоченного леса с Nyssa, Taxodium и эпи-фитными Tillandsia (Bromeliaceae; Америка). Выше нормального положения водного зеркала росли болотистые леса. В более влажных местах встречались Myrica, Liquidambar (Америка, Азия), Cyrilla (Америка) и Osmunda claytoniana (Америка, Азия), а на более сухих — Sequoia (Америка), Sciadopitys (Азия). Sabal (Америка. Азия) и вьющийся пантропический папоротник Lygodium. Из названных родов сегодня в Европе еше растут только Nymphaea и Myrica. Характерные для тор
528 | ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
фяных лесов особенности в настоящее время сохранились в субтропической Юго-Восточной Америке (например, во Флориде).
Похолодание и частичная аридизация климата в неогене привели к смещению флор и растительных зон к югу Земли, вымиранию в Европе почти всех тропических и многих теплолюбивых аркготретич-ных таксонов и появлению обширных дизъюнкций в ареалах многих видов голарктических лиственных лесов в континентальных областях Азии и на западе Северной Америки.
С эоцена до плиоцена в Северном полушарии уменьшилась доля ранее доминировавших тропических таксонов, и стали господствовать арктотретичные представители. В Европе серьезным препятствием для миграций третичных (а также и четвертичных) флор были высокие горы, протянувшиеся с запада на восток, и Средиземное море. Отсюда понятно, почему сегодня Европа значительно беднее арктотре-тичными видами, чем климатически сравнимые области Восточной Азии и Северной Америки.
Характерные дизъюнкции: Восточная Азия — Европа — восточная Северная Америка (Fagus— рис. 14.17; Carpinus, Hepatica) и Восточная Азия — восточная Северная Америка (Tsuga, Magnoliaceae — рис. 14.24, Illicium, Sassafras, Hamamelis, Nyssa, Catalpa) и соответственно реликтовый эндемизм юго-востока Северной Америки (Taxodum) или Восточной Азии (Ginkgo. Metasequoia, Cercidiphyltum} следует понимать как этапы продолжающегося сокращения ареалов арктотретичных таксонов. Среднегодовые температуры в Центральной Европе снизились от миоцена до нынешнего времени примерно с 16 "С до 8 —9 °C.
Особенно важные рефугиумы арктотретичных таксонов (третичных реликтов) в Европе и Юго-Западной Азии — Балканы (например, Picea omorica, как близкородственная восточноазиатской P.jezoensis, и Aesculus hippocastanum — Aes-culus встречается еще в Гималаях, Восточной Азии, а также в Северной Америке), влажные биотопы восточного Средиземноморья (например, Platanus orientalis, этот род встречается только еще в Северной Америке, а в Средней и Восточной Азии вымер; Ытасеае — Zelkova. есть еще в Восточной Азии, так же как Liquidambar и Styrax) и лесные области по восточному побережью Черного и южному побережью Каспийского морей (например, Pterocarya из Juglandaceae есть еще в Восточной Азии, а в Северной Америке вымер; Albizia (Mimosaceae) и Diospyros (Ebena-
сеае) с субтропическими или даже тропическими связями).
Возникновение теплого континентального, с сухим летом климата в субмеридиональных и меридиональных зонах (например, в Средиземноморье, на западе Северной Америки, а также и в Южном полушарии, например в Чили) привело в середине палеоген —неогена к преобразованию тогдашних вечнозеленых дождевых лесов в жестколистные леса. В Европе этот про цесс ускорило многократное высыхание Средиземного моря в миоцене. Примеры этих изменений флор — появление Myrtis communis и Smilax aspera из преимущественно тропических семейств, а также Quercus ilex, Nerium oleander и Olea europaea.
Возникновение жестколистных лесов хорошо документировано, особенно в олигоненовых и миоценовых ископаемых флорах Юго-Восточной Европы с предками современных видов Laurus, Arbutus, Ceratonia, Pistacia, Phyllyrea и т. п., а также на западе Северной Америки. Леса из лавровишни на Канарских островах можно рассматривать как по меньшей мере частичный реликт этой фазы. Для некоторых обитателей лавровишневых лесов (например, Ixanthus vis-cosus (Gentianaceae), ближайшего родича средиземноморской Blackstoma) с помощью молекулярных методов удалось показать более молодой возраст. Впоследствии в дальнейшей эволюции средиземноморской флоры особенно важную роль сыграли геологически стабильные области Иберийского полуострова (рис. 11.299) и северо-западной Африки, а также юго-западной Азии.
С возрастанием сухости и континентальности удаленных от морей областей в неогене, очевидно, непосредственно связа но возникновение и прогрессирующее развитие ксерофильных безлесных флор саванн, степей, полупустынь и пустынь, а также их широкое распространение по Земле. С распространением саванн и степей повсюду была связана эволюция стадных травоядных животных.
Решающее значение для возникновения альпийской флоры Голарктики имело горообразование в миоцене. Расположение центров многообразия характерных высокогорных родов (например, Saxifraga, Draba, Primula, Gentiana, Pedicularis, Leontopodium, Crepis) показывает, что эволюция этих ро-
11.3. Филогенез растений и история растительности I 529
Рис. 11.299. Колебания климата в кайнозое (по RWohlstedt и Т. Nilsson из G.Lang).
Среднегодовые температуры реконструированы для Западной и Центральной Европы. Временной масштаб для плейстоцена и голоцена растянут, а число холодных и теплых периодов меньше, чем их было на самом деле
дов началась в центрально-азиатских горах (например, в восточных Гималаях, Западном Китае, на Алтае). Оггуда многие таксоны расселились, очевидно, через Берин-гийскую сушу, заселив циркумполярно-арктическую область, а часть их даже добралась через Центральную Америку до Южной Америки (например, Genlianella).
Несмотря на это, нельзя недооценивать и значение европейско-средиземноморских гор как центров образования альпийских таксонов. Это относится, например, к Sempervivum, Helian-themum, Rhodothamnus, Phyteuma, Achillea, Globularia и Sesleria. Здесь нередко и на примере современных представителей можно проследить последовательную дифференциацию из горных видов субальпийских и далее — альпийских.
Начавшееся в миоцене третичного периода похолодание (рис. 11.299) проявилось сильными колебаниями климата и связанными с ними оледенениями четвертичного периода (примерно от 2 млн лет до настоящего времени; разные авторы датируют начало четвертичного периода, или плейстоцена, 2,5—1,64 млн лет). Причинами резких колебаний климата в четвертичном периоде, с одной стороны, являются регулярные изменения вращения Земли и, с другой стороны, особенности земной поверхности.
Характер вращения Земли закономерно изменяется в связи с эксцентричностью ее эллиптической орбиты, с наклоном земной оси и с так называемой прецессией, которой определяется дата перигелия, самого близкого к Солнцу по
ложения Земли. Однако одних этих регулярных изменений вращения Земли недостаточно для объяснения климатических колебаний четвертичного периода, так как они имели место, очевидно, в течение всей истории Земли. Эти колебания могли возникнуть лишь потому, что благодаря тектонике континентальных плит массивы суши оказывались у полюса. Морские течения как важные переносчики тепловой энергии также повлияли таким образом, что стало возможным образование полярных массивов льда. Для начала четвертичного периода, очевидно, особенно важным было появление льдов в Арктике, так как Антарктида уже давно была покрыта льдом. Другим важным фактором для четвертичных колебаний климата было, по-видимому, изменение воздушных потоков и глобальных направлений ветров из-за продолжавшегося в четвертичном периоде поднятия гор.
В течение четвертичного периода было много (рис. 11.300) оледенений (криомер) и теплых межледниковий (термомер). В периоды оледенений на северо-западе Европы, в граничащей с ней Северо-Западной Сибири и в обширных областях Северной Америки (на юг примерно до 40° северной широты) образовывались мощные материковые ледники (до 3 000 м толщиной). Альпы также были почти полностью покрыты сплошным ледником (рис. 11.301), в то время как в горах Южной Европы, Азии, Аляски и тропиков были сравнительно небольшие ледники. Во время теплых межледниковий температуры иногда были выше, чем в настоящее время. Оледенениям в высоких географических широтах в более теп-
530 | ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Годы от настоящего времени
0-Поздний-г
Голоцен
- Weichsel
— = Ееш Saale
’5 = Holstein= x Elster
5
Cromer
Tegelen
2 млн —
Prategelen
Reuver
Июльские температуры
Годы от настоящего времени Июльские температуры
Рис. 11.300. Климато-стратиграфическое деление и реконструированные средние июльские температуры всего четвертичного периода (слева), а также среднего и позднего плейстоцена и голоцена (справа) на примере Нидерландов (по W.H.Zagwijn (слева) и J.De Jong (справа), взято из G.Lang, слегка изменено).
Цифры обозначают межледниковые периоды: 3 — Cromer IV, 4 — Cromer III, 5 — Cromer II, 6 — Cromer 1,7 — Leerdam, 8 — Bavel, 9 — Waal C, 10 — Waal A, 11 — Tegelen TC5, 12 — Tegelen TC3, 13 — Tegelen A
лых и сухих областях на юге (например, в Средиземноморье, Сахаре) соответствовали дождливые периоды (плювиальные времена), в то время как в межледниковьях там обострялись засухи. На тропических равнинах климат в периоды оледенений был холоднее и заметно суше, что приводило к отчетливому сокращению и фрагментации тропических дождевых лесов.
Среднегодовые температуры во время похолоданий в Центральной Европе опускались на 8—12 °C, в удаленных от ледников и тропических областях — на 4—6 °C. Снежная граница в Альпах проходила на 1 200— 1 400 м ниже современной (в Центральных Альпах уже на вы
соте I 600 — 2 000 м, в Северных Альпах — 900— 1 200 м). Альпийские ледники спускались в предгорья и приближались к северному леднику почти на 500 км. Но климат ледниковых периодов был враждебным для растительности и за пределами оледеневших областей. Здесь широкой каймой вокруг материкового ледника откладывалась принесенная ветром пыль. К северу от Средиземноморья на некоторой глубине в течение всего года сохранялась вечная мерзлота. Связывание огромных масс воды в леднике обусловило понижение уровня моря (примерно на 120 м) и расширение площади материков. Так, Британские острова и южная часть Северного моря во время последнего оледенения были частью материка.
Многочисленные климатические колебания четвертичного периода группируют в шесть лед
11.3. Филогенез
растений и история растительности
никовых периодов (см. рис. 11.300), которым в разных (см. рис 11.300) странах Европы и в Северной Америке даны разные названия. Похолодание началось на границе третичного и четвертичного периодов, было незначительным в претегелине (= брюгге; примерно 2,3 млн лет) и эбуроне (= донау; примерно 1,6 млн лет), усилилось в менапе (= гюнце; примерно 1,1 млн лет) и эльстере (= минделе; около 0,35 млн лет), достигло высшей точки в заале (= риссе; около 0,23 млн лет) и закончились в вейхзеле (= вюр-ме; около 0,11 млн лет). Между этими холодными периодами были потепления, которые в начале четвертичного периода были более продолжительными, чем похолодания, но в позднем чет
вертичном периоде стали существенно короче, чем периоды оледенений (голыптейнекое и эм-ское межледниковья продолжались всего по 15 000 лет, а вюрмское оледенение, напротив, около 100000 лет). Сами периоды оледенений прерывались короткими фазами потеплений (ин-терстадиалами; в вюрмское оледенение их было около 24).
Четвертичные холодные, влажные и соответственно сухие периоды сильно влияли на растительность Земли и приводили к характерным изменениям ареалов и смещениям растительных зон. Многие третичные таксоны вымерли, а новые возникли
Рис. 11.301. Растительность Европы около 20 00G лет назад (Вейхзельское оледенение к моменту максимального распространения ледника) (из G. Lang).
Зубчатая линия означает край ледника, белый цвет — тундру, редкий пунктир — тундростепь частый пунктир — степи с изолированными популяциями древесных пород
532 I ГЛАВА 11, СИСТЕМАТИКА И ФИЛОГЕНИЯ
как следствие географической изоляции, гибридизации и полиплоидизации вследствие постоянных изменений ареалов. Особенно интенсивному воздействию подвергались приледниковые области в Европе и Северной Америке.
Реконструкция растительности Европы во время максимального развития последнего оледенения показана на рис. 1L30L За исключением лесостепей и лесотундр с березами, соснами и другими холодостойкими древесными породами, распространенными локально, например, у относительно более теплой восточной оконечности Альп, Центральная Европа была тогда безлесной. По ее ведущему виду — аркто-аль-пийской дриаде восьмилепестной (Dryas octopetala) эти ископаемые флоры были названы дриадовыми. На территории дриа-довой флоры были широко распространены кустарничковые тундры и холодные степи, часто на лёссовых отложениях, заку-старенные луга, осоковые болота и бедные видами сообщества водных растений.
Из видов дриадовой флоры в настоящее время Dryas octopetala, Salix herbacea, Loiseleuria pro-cumbens, Saxifraga oppositifolia, Silene acaulis, Polygonum viviparum, Oxyria digyna и Eriophorum scheuchzeri имеют аркго-альпийские ареалы, Salix polaris и Ranunculus hyperboreus сегодня встречаются только в Арктике, a Potentilla аигеа и Salix retusa— только в альпийском поясе. Вместе с этими видами обитают и такие, которые в настоящее время встречаются еще и между Арктикой и высокогорьями, например в среднегорье (Betula папа, Empetrum nigrum), или широко распространены, так как проявляют меньшие требования к климату (Filipendula ulmaria, Ме-nyanthes tri/bliata. Potamogetori). В более сухих местообитаниях растут виды, характерные для холодных степей, которые сегодня распространены преимущественно восточнее (Artemisia, Heli-anthemum, Ephedra). На сырых почвах сегодня как сорняки растут, к примеру, Chenopodium album и Centaurea cyanus. Типичными животными этих холодных степей были мамонт, северный олень, овцебык сурок и лемминг (см. рис. 11.301).
На рис. 11.301 видно далекое отступление в Южной Европе более прихотливых древесных пород. Галерейные и долинные леса сохранялись в области более южных холодных степей, широко распространены были открытые лесостепи и лесотундры, а
рефугиумы летнезеленых лиственных или более прихотливых хвойных пород были небольшими, разрозненными и обычно находились вблизи побережий. Вечнозеленая растительность могла сохраниться, вероятно, только за пределами Европы — в Северо-Западной Африке и Юго-Западной Азии.
Идентификация четвертичных рефугиумов с помощью пыльцевого анализа сегодня дополнена применением методов анализа ДНК. При этом принято, что при миграции из рефугиума генетическая изменчивость уменьшается, соответственно в рефугиумах можно встретить больше генетических вариаций, чем в заселенных из них областях (см. 10.2.2). Широкое исследование изменчивости, к примеру, пластидной ДНК в разных частях ареалов некоторых видов деревьев показало. что Quercus robur и Q. petraea Abies alba и Alnus glutinosa имели рефугиумы в Юго-Восточной (Балканы), Южной (Италия) и Юго-Западной Европе (Иберийский полуостров), Fagus sylvatica— только в Юго-Восточной и Южной Европе, Picea abies— только в Юго-Восточной Европе. Эти результаты чаще всего подтверждают данные пыльцевого анализа, но иногда приводят и к выявлению новых рефугиумов.
В холодные времена многие виды отступали из гор и Арктики в более низкие или южные места. Они имели хорошие возможности для широких миграций по обширным взаимосвязанным равнинам. Следствием этого стал интенсивный обмен флорами. Это затрагивало не только флоры Альп, Пиренеев, Карпат и других европейских высокогорий. Например, исходно азиатские горные таксоны смогли попасть в Северную Америку через сухопутный Берингий-ский мост или в Альпы, а альпийские формы — в Арктику и наоборот. В теплые периоды эти виды, с одной стороны, снова заселяли горы и Арктику, с другой стороны, их сплошные во время оледенений ареалы оказывались разорванными распространяющейся лесной растительностью. Эти процессы объясняют многочисленные альпийские, аркто-альпийские и азиатско-альпийские дизъюкции в современной флоре, а также наличие аркто-альпийских и бореальных видов как ледниковых реликтов за пределами их основного ареала.
Многие виды Saxifraga, Gentiana, Androsace, Soldanella, Primula, Potentilla и т.п. имеют европейские горные дизъюнктивные ареалы с удаленны-
11.3. Филогенез растений и история растительности | 533
Рис. 11.302. Распространение Salix herbacea в Европе в настоящее время (•) и, по данным ископаемых находок, в послеледниковый период вюр-мского похолодания (о), а также в более ранние холодные периоды (х) (по H.Tralau из Н.Walter и H.Straka)
ми частными ареалами в Альпах, Абруцце, Пиренеях, Сьерра-Неваде, Карпатах и Балканах. Арк-то-альпийские виды чаще встречаются во многих изолированных более высоких местах среднегорий (например, в Шварцвальде, Судетах: Saxifraga oppositijblia, Veronica alpina, (jnaphalium supinum). Расстояния между разными частями ареалов достигают часто более I 000 км. Известны, например, очень удаленные находки чисто арктических видов Saxifraga nivalis и Pedicularis sudetica в Исполиновых горах, причем Pedicularis удален от основной части ареала примерно на 2 000 км.
Ива травянистая (Salix herbacea, рис. 11.302) в четвертичном периоде имела арктический ареал (северо-восток Северной Америки, Гренландия, Исландия, Шпицберген, Северная Европа) и во время ледниковых периодов проникла оттуда через Центральную Европу в Альпы, Карпаты, Пиренеи, Абруццу и Балканские горы. Ископаемые находки показывают сплошной ареал, который сегодня сильно разорван, причем между его арктической и альпийской главными частями еще можно обнаружить несколько очень изолированных реликтовых местонахождений (например, в Судетах). Подобное распространение свойственно также Betula папа, имеющей центр ареала в Северной Америке, но достигшей Азии Loiseleuria procumbens, Dryas octopetala и родственным видам, а также циркумполярной Eriophorum scheuchzeri. Из азиатских центров заселили Арктику и европейские высокогорья Saussurea alpina и Lloydia serotina, тогда как Pinus cembra и Leontopodium
alpinuni добрались только до европейских высокогорий. Alchetnilla alpina и Nigritella nigra — примеры альпийских таксонов, которые в ледниковые периоды смогли проникнуть в Арктику.
Сохранившиеся от ледниковых периодов реликтовые местонахождения аркто-альпийских или альпийских видов на равнине приурочены к болотам (например, Betula папа, Empetrum nigrum, Trichophorum caespitosum) или топям, оврагам (Saxifraga paniculata, Arabis alpina). В таких местообитаниях локальные микроклиматические условия компенсируют неблагоприятный для этих видов местный климат и сдерживают более сильные конкурирующие виды.
Горные или альпийские сосудистые растения смогли пережить ледниковые периоды как за пределами покровного оледенения, так и иногда в области оледенения на свободных от льда утесах или вершинах («нунатаках»). О положении этих рефугиумов до сих пор судили по сгущениям реликтовых или разорванных ареалов разных видов и соответственно по совпадению расположения ареалов и геоморфологически опознаваемых нунатаков. Часто в ископаемом состоянии не сохраняется ничего кроме пыльцы ветроопыляемых лесных деревьев, поэтому для определения положения рефугиумов все большее значение получает исследование ДНК.
534 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Рис. 11.303. Современный ареал и послеледниковое и голоценовое расселение бука (Fagus sylvatica) (из G.Lang).
Большие черные точки с цифрами означают время в тысячах лет по радиоуглеродным датировкам; маленькие точки — современный ареал; штриховая линия — приблизительные границы ареала 10 000 (жирно) и 5 000 лет назад
Можно ожидать разных пространственных проявлений генетической изменчивости современных растений в зависимости от того, пережили ли они оледенение вне области покровного оледенения или внутри нее. Таким образом можно показать, что альпийский Eritrichium папит переживал ледниковый период на трех географически изолированных нунатаках, в то время как арктичес-ки-субарктический (рис. 11.303) ареал Saxifraga caespitosa и арктическая часть ареала Saxifraga oppostlfolia сформировались в результате заселения освободившихся из-под ледника территорий из рефугиумов за пределами оледенения, а не с нунатаков. С помощью молекулярных методов было показано, что рефугиумом Papaver alpinum, вероятно, были западные, северо-западные, восточные и южные отроги Альп вне области покровного оледенения. Сходно расположенные рефугиумы предполагаются для очень многих других таксонов.
Похолодания обусловили, вероятно, и расширение ареалов голарктических видов и родов через экватор в Южное полуша
рие (например, Quercus, Fagus'. см. рис. 14.17), а также некоторые примечательные биполярные дизъюнкции (иногда с промежуточными местонахождениями в тропических высокогорьях).
Биполярно распространенные таксоны находят у лишайников (например. Cetraria isiandica), мхов (например, Sphagnum) и покрытосеменных (например, Carex, Epilobium, Empetrum, Primula farinosa agg). Эти таксоны в четвертичном периоде, продвигаясь по Кордильерам и Андам, попали из Северной Америки в Анды Южной Америки и антарктическую Южную Америку. Другие через горы Юго-Восточной Азии и Новой Гвинеи достигли Австралии и Новой Зеландии (которую заселяли также из Южной Америки в результате дальнего заноса, например, Gentianella), а третья миграция проходила через восточно-африканские горы на юг Африки.
В теплые эпохи четвертичного периода растительный покров был похож на совре
11.3. Филогенез
растений и история растительности
менный. Правда, в ранних межледниковьях в Центральной Европе еще встречались некоторые третичные виды, которые сейчас уже вымерли.
До начала вюрмского оледенения, с которого начался поздний плейстоцен, в Центральной Европе еще были широко распространены некоторые третичные виды: обитающая в настоящее время в Северной Америке и Восточной Азии Brasenia schreberi, эндемичная для Балкан Picea omorika, ограниченные сегодня Северной Африкой и Юго-Западной Азией кедры (Cedrus) и крупноцветковые рододендроны группы Rhododendron ponticum (см. рис. 11.306), которые сейчас встречаются еще только на Кавказе, в Северной Анатолии, Ливане, на Балканском полуострове и в юго-западной части Иберийского полуострова, т.е. в далеко отстоящих частных ареалах.
Начавшийся 10 тыс. лет назад последний отрезок четвертичного периода называют голоценом, или послеледниковьем. После последнего максимума оледенения около 20 тыс. лет назад климат постепенно становился все теплее, несмотря на возвратные похолодания. В последующие 10000 лет ледниковые массы большей частью растаяли. Благодаря этому многие виды лесов и других климатически более требовательных растительных сообществ заселили до того безлесные или
покрытые ледниками области Европы и Северной Америки. В горах это привело к поднятию границы леса, к соответствующему развитию растительных зон и к возврату альпийских тундр на большие высоты.
Обратная миграция лесообразующих пород (рис. 11.303) в позднем плейстоцене и голоцене происходила разновременно. Причина этого кроется в различиях их экологии, конкурентоспособности и скорости распространения, а также в различном расположении рефугиумов. Средняя скорость распространения древесных видов достигала примерно 400 м/год.
В холодных степях позднего плейстоцена преобладали злаки, Сурегасеае и Artemisia. Ephedra distachya (рис. 11.304) и родственные виды широко распространились по Европе с востока, но были позднее вытеснены развивавшимися лесами в засушливые внутриконтинентальные биотопы и на открытые побережья. Древесные породы позднего плейстоцена — Salix, Betula и Pinus. Тогдашние леса походили на леса современной Лапландии.
Голоцен начался с потепления климата в пребореальное время (около 10000— 8 500 лет), через бореальное время (примерно 8 500—7 500 лет) перешел в атлантическое (примерно 7 500—4500 лет) и суббореальное время (примерно 4 500—2 500 лет), когда был достигнут температурный мак-
Рис. 11.304. Распространение Ephedra distachya agg. в настоящее время и в период от позднего ледниковья до послеледнико-вья (по J. Iversen, из Н.Walter и H.Straka)
Современный ареал:
сплошное распространение
• отдельные находки
Ископаемые находки:
о позднее ледниковье и пребореальный период
+ послеледниковое
теплое время
536 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Юг Швеции Дания Британские острова Средняя Европа т
1 / II Субатлантикум (SA) 2300 111/IV Суббореальный (SB) 5300 V/VI Атлантикум (АТ) 8200 L VII/VIII Бореальный (ВО) 9900 IX Пребореаль-ный (РВ) 10300 X Древнейший Дриас (DR 3) 11100 XI Аллерёд (AL) 1-2000 XII Древний Дриас IX Субатлантикум 2500 VIII Суббореальный 5000 VII Атлантикум 8000 V/VI Бореальный 9000 IV Пребореаль-ный 10300 III Древнейший Дриас 11000 II Аллерёд 11-700 Ic Древний Дриас 1-2000 lb Бёллинг 1-2500 la Древнейший Дриас VIII Субатлантикум 2700 Vllb Суббореальный 5000 Vila Атлантикум 7500 V/VI Бореальный 9500 IV Пребореаль-ный 10300— III Верхний Дриас 10800 II Аллерёд 1-2000 I Нижний Дриас IX/X Субатлантикум 2800/2500 VIII Суббореальный 4500 VI/VII Атлантикум 7500 V Бореальный 8800/8500 IV Пребореалfa-ный 10100 III Поздний субарктический период 11000 II Средний субарктический период 1-2000 lb Древний субарктический период W W W к W к W Голоцен Позднее ледниковье
la Самый древний безлесный период к
Рис. 11.305. Климатостратиграфическое деление позднего ледниковья и голоцена в Северной, Западной и Средней Европе. Временные границы обозначены в годах до нашего времени по данным радиоуглеродных исследований (по G.Lang):
Т — температура; w — тепло или теплее; к — холод или холоднее
симум; в субатлантическое время (2 500 — по настоящее время) началось новое похолодание (рис. 11.305).
Этому соответствует развитие растительности (рис. 11.306). После короткого похолодания в конце пребореального времени началось бореальное потепление и новое распространение берез и сосен В атлантическое время начинает массово расселяться лещина, что привело сначала к возникновению сосново-орешниковых лесов. Затем из-за сокращения встречаемости березы и сосны (рис. 11.307), а также усиленного распространения вяза и дуба появились дубово-орешниковые смешанные леса.
С распространением более прихотливых широколиственных деревьев — липы, клена и ясеня — в атлантическое и суббореальное время возникают дубово-смешанные леса. На все сильнее заболачивающихся низменностях распространяются болотные ольшанники, а ели покрывают восточные Средние горы до Гарца, а также восточные Альпы и Карпаты. Сосновые леса, как и современные (рис. 11.307), встречались преимущественно в более сухих и теплых биотопах. В еще более сухих и непригодных для деревьев биотопах развились богатые видами ксерофильные низкотравные луга и степи.
11.3. Филогенез растений и история растительности
Рис. 11.306. Позднечетвертичная пыльцевая диаграмма с конца ледникового периода до настоящего времени (Luttersee, 160 м, восточнее Геттингена; пыльцевые зоны I—XII по F = Firbas, О = Overbeck) (по К.Steinberg и A. Bertsch из Н.Walter и H.Straka).
Доля пыльцы деревьев (без клена, Acer) показана черным цветом, пыльцы Corylus и недревесных пород (только важнейшие типы) — белым (и те и другие даны относительно суммы пыльцы деревьев принятой за 100 %)
Рис. 11.307. Расширение и сужение ареала сосны обыкновенной в постледниковой Центральной Европе: области одинакового содержания захороненной пыльцы (< 5%; 5—50%; 50 — 75%; >75%) в бореальном и суббореальном периодах (из F. Firbas)
538 I ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Остается неясным, возникли ли подходящие условия для распространения в Центральную Европу теплолюбивых видов каменистых пустошей, сухих низкотравных лугов и степей с юга и востока лишь в атлантическое и суббореальное время или они господствовали здесь еще до начала повторного развития лесов в голоцене. Исходно более или менее сплошные ареалы этих присредиземноморских или южно-иберийско-понтических видов сегодня чаще всего разорваны, так как распространение лесов очень сузило их жизненное пространство. Такое дизъюнктивное распространение правомерно рассматривать как реликтовое более теплых времен.
Дизъюнктивные ареалы теперь имеют мигрировавшие в теплые периоды голоцена из Средиземноморья в Центральную Европу Quercus pubescens, Comus mas, Buxus sempervirens, Cytisus nigricans, Anthericum liliagp и разные виды Ophrys. Вероятно, раньше, может быть в конце ледникового периода, в Европу попали южносибирские и южнорусские степные растения Stipa ser.Capi-data, Adonis vemalis, Lathyrus pannonicus, Scorzonera purpurea', богатый видами круг форм Linum flavum
и Astragalus exscapus, а также Eryngium campestre, Salvia pratensis и Odontites luteus попали в Юго-Восточную Европу из югозападно-азиатского Средиземноморья. Эти виды растут чаше всего на богатых известью почвах в редких зарослях кустарников на скалах, сухих низкотравных лугах и в степных сообществах, иногда — в светлых сосновых и дубовых лесах. Они заселяли с юга Центральную Европу по долйнам Роны и Рейна, через южные склоны Альп, вдоль восточных отрогов Альп и по долине Дуная. С востока теплолюбивые виды двигались севернее и южнее Карпат и через Венгрию проникли более или менее далеко в жаркие и сухие, удаленные от моря ландшафты Европы, такие, например, как континентальные долины Центральных Альп, Восточная Австрия, Внутренняя Богемия, Тюрингия и бассейн Майнпа.
В субатлантическое время заметно снизились температуры и увеличилось количество осадков. Впервые появляются бук европейский, граб и пихта, вытесняя дуб и лещину. Наконец, в историческое время сна
Рис. 11.308. Реконструкция естественной растительности в Центральной Европе к началу исторического времени (0 г. н.э.) на основании палинологических данных (по F.Firbas, из H.EIIenberg):
1 — сухие разреженные дубово-смешанные леса (без бука, годовые осадки около 500 мм); 2 — буково-смешанные леса глубоких долин (иногда с преобладанием дуба); 3 — буковые горные леса; 4 — область буковых лесов с редкими сосняками; 5 — горные леса с буком, пихтой и елью (▲ — субальпийское редколесье, Д — леса с доминированием ели) 6 — сосняки с дубом на песчаных почвах; 7 — грабовые смешанные леса; 8 — грабовые смешанные леса с елью (• — области с локальным доминированием сосны)
11.3. Филогенез растений и история растительности i 53g
чала на равнинах, а затем в низких северо-западных Средних горах начинает господствовать бук европейский, а на востоке — граб. Горные леса становятся все больше смешанными лесами с буком, пихтой и елью. Так возникла современная растительность, показанная на рис. 11.308.
Современный человек был современником изменений климата и растительности в Европе с периода примерно 40 тыс. лет тому назад, неандерталец — 200 тыс. лет, но влияние людей на флору и растительность стало заметным в пыльцевом профиле лишь с позднего неолита (около 3 000 лет до н.э.) вследствие перехода к оседлости и начала земледелия.
В почвенных профилях о земледелии свидетельствуют слои золы от выжигания леса и пыльца хлебных злаков и сорняков (Plantago, Rumex, Centaurea cyanus). Увеличение пыльцы хлебных злаков и трав означает растущие пастбищное хозяйство и луговодство. Сорняки как спутники человеческой культуры распространились по всему миру. Археофитами называют такие виды, как Agrostemma gjthago и Papaver rhoeas, которые попали в Центральную Европу вместе с зерновыми культурами еще в неолите. Sinapis arvensis и Anagallis arvensis как археофиты достигли Центральной Европы лишь в бронзовом веке. Неофиты — виды, вселившиеся в историческое время. К ним относятся, например, Impatiens glandulifera из Гималаев, Senecio vemalism Юго-Западной Азии, Elodea canadensis, Conyza canadensis и некоторые виды Aster и Solidago из Северной Америки, a Galinsoga parviflora и G. ciliata из Южной Америки. Из засушливых областей Нового Света происходит Opuntia ficus-indica и Agave americana, иногда определяющие ландшафты в Средиземноморье. Heracleum mantegazzianum (Кавказ) и Senecio inaequidens (Южная Африка) — также неофиты, которые натурализовались
в Европе лишь во второй половине прошлого столетия.
Влияние людей на окружающий мир достигло сегодня масштабов, далеко превосходящих любое естественное изменение в истории Земли. Так, например, в Европе, давно и все более плотно населенной, сохранились лишь последние остатки первобытного леса, небольшие участки тундровой и высокогорной растительности, сообщества неприступных скальных расселин, некоторые низинные и верховые болота, почти нет нетронутых участков водной и прибрежной растительности. Засоление лугов и появление дюн вряд ли прямо зависит от деятельности человека. Эти области (рис. 11.308) занимают сейчас очень маленькие площади и также подвержены непрямому воздействию человека, например вследствие попадания вредных веществ из воздуха. Считается, что на территории Германии около 1,6 % видов сосудистых растений вымерло, а 12,2% угрожает вымирание («существуют только маленькие, едва ли способные выжить остаточные популяции») или они сильно угнетены («сокращают численность, во многих местах уже исчезли»). Для тропических дождевых лесов, родины около 2/3 всех существующих видов грибов, животных и растений, принимают, что при нынешней скорости (около 1 млн км2 за каждые 5—10 лет) продолжающегося вырубания (без особой охраны известных центров разнообразия) в ближайшие 60 лет вымрет приблизительно 5 % всех видов. Подобную угрозу фауне и флоре со стороны человека можно признать и для других областей Земли.
Список литературы
Приведенные здесь книги и статьи позволяют более углубленно проработать изучаемый материал, используя цитируемую литературу.
Значительно ускорившийся прогресс исследований во многих областях науки документирован в многочисленных специальных журналах. Ввиду увеличения потока данных и возрастающей специализации все более важным становится периодическое издание обзоров. Уже в 1931 г. в Германии было основано издание «Die Fortschritte der Botanik», т.е. «Успехи ботаники» (современное название — «Progress in Botany», Springer, Berlin), которое ежегодно информирует о новых результатах исследований в ботанике. Регулярно выходят также различные ежегодные обзоры по разделам клеточной биологии и биологии развития; генетики; микробиологии; фитопатологии; физиологии растений и молекулярной биологии растений; ежегодные обзоры на английском языке (Annual Reviews of Cell and Developmental Biology; Genetics; Microbiology; Phytopathology; Plant Physiology and Plant Molecular Biology), Annu. Rev. Inc., Palo Alto. Можно рекомендовать также менее широко охватывающие периодические издания Advances in Botanical Research (Academic Press, San Diego) и Critical Reviews in Plant Sciences (CRC Press, Boca Raton). О новейших успехах все чаще сообщается в обзорах или миниобзорах, печатающихся в еженедельных или ежемесячных журналах. Эти обзоры предлагаются также собранными в особые серии, такие, как Trends in Biochemical Sciences, in Biotechnology, in Cell Biology, in Genetics, in Plant Sciences (Elsevier Science, London), in Current Opinion in Plant Biology (также издается Elsevier Science). Миниобзоры по биологии и медицине ежемесячно появляются в Current Biology (Current Biology, Ltd., London).
ВВЕДЕНИЕ
История ботаники
Cremer T (1985) Von der Zellenlehre zur Chromosomentheorie. Springer, Berlin
Jahn 1, Hrsg (2000) Geschichte der Biologic, 3. Aull. Spektrum Akademischer Verlag, Heidelberg
Magdefrau К (1992) Geschichte der Botanik, 2. Aufl. Gustav Fischer, Stuttgart
Mayr E (1984) Die Entwicklung der biologi-schen Gedankenwelt. Springer, Berlin
Moltmann UG, Hrsg (1994) 100 Jahre Stras-burgers Lehrbuch der Botanik fur Hochschulen. Gustav Fischer, Stuttgart
Лексика, терминология, даты, иллюстрации
Cole CH, HauBer-Siller I (1998) Worterbuch der Biologic, Englisch-Deutsch/Deutsch-Englisch (auch als CD-ROM). Spektrum Akademischer Verlag, Heidelbetg
Dietrich G, Stocker FW, Hrsg (1986) Fachlexikon ABC Btologie, 7. Aull. Harri Deutsch Verlag, Thun
Flindt R (2000) Biologic in Zahlen, 5. Aull. Spektrum Akademischer Verlag, Heidelberg
Lexikon der Biologic (seit 2000 im Erschcinen). 14 Bande und 1 Registerband; auch auf CD-ROM. Spektrum Akademischer Verlag, Heidelberg
Medawar PB, Medawar JS (1986) Von Aristo-teles bis Zufall. Ein philosophisches Wurterbuch der Biologic. Piper, Miinchen
Schubert R, Wagner G. (1993) Botanisches Worterbuch, 11. Aufl. Ulmer, Stuttgart
Vogel G., Angermann H. (1998) dtv-Atlas zur Biologic, 3 Bande, 10. Aufl. Deutscher Taschenbuch-verlag, Miinchen
Vogellehner D. (1983) Botanische Terminologie und Nomen-klatur, 2. Aufl. Gustav Fischer, Stuttgart
Wagenitz G. (1996) Worterbuch der Botanik. Gustav Fischer, Jena
Интернет
http://gened.emc.maricopa.edu/bio/biol81/ BlOBK/BioBook
TOC.html (Online Biology Book)
http://www.wisc.edu/botany/virtual.html (Pflanzenbilder, auch mikroskopisch; M. Clayton)
http://www.Biologe.de/menu.html (Schule-Studium- Beruf- Business)
Теория науки и познания
Lorenz К (1973) Die Rilckseite des Spiegels. Piper, Miinchen (als Taschenbuch 1997)
Список литературы | 541
Mohr Н (1977) Lectures on Structure and Significance of Science. Springer, New York
Popper KR (1994) Logikder Forschung, 10. Aufl. Mohr, Tubingen
Popper KR (1995) Objektive Erkenntnis, 3. Aufl. Hoffmann & Campe, Hamburg
Vollmer G (1985, 1986) Was konnen wir wis-sen? Hirzel, Stuttgart
Vollmer G (1995) Biophilosophie. Reclam, Stuttgart
Возникновение и развитие жизни
Bengtson S, ed (1993) Early Life on Earth. Nobel Symposium 84. Columbia University Press, New York de Duve C(1994) Ursprung des Lebens. Prabiotische Evolution und die Entstehung der Zelle. Spektrum Akademischer Verlag, Heidelberg
de Duve C (1995) Aus Staub geboren. Leben als kosmische Zwangslaufigkeit. Spektrum Akademischer Vcrlag, Heidelberg
Doolittle WF, Brown JR (1994) Tempo, mode, the progenote and the universal root. Proc Natl Acad Sci USA 9): 6721-6728
Ebeling W, Feistel R (1994) Chaos und Kosmos. Prinzipien der Evolution. Spektrum Akademischer Verlag, Heidelberg
Haken H, Haken-Krell M (1989) Entstehung biologischer Information und Ordnung. Wissensc-haftliche Buchgesellschafi, Darmstadt
Hayes JM (1996) The earliest memories of life on earth. Nature 384: 21—22
Lazcano A, Miller SL (1996) The origin and early evolution of life: prebiotic chemistry, the pre-RNA world, and time. Cell 85: 793—798
Rizztti M (2000) Early Evolution. Birkhauser, Bassel
SchopfJW, ed (1983) Earth’s Earliest Biosphere. Princenton University Press, Princeton
ГЛАВА 10. ЭВОЛЮЦИЯ
Общие работы
Avise JC (1994) Molecular Markers, Natural History and Evolution. Chapman & Hall, New York
Bachmann К (2001) Evolution and the genetic analysis of populations: 1950-2000. Taxon 50: 7-45
Briggs D, Walters SM (1997) Plant Variation and Evolution, 3rd ed. Cambridge University Press. Cambridge
Clausen J, Keck DD. Hiesey WM (1940. 1945, 1948, 1958)
Experimental Studies on the Nature of Species. Carnegie Institute of Washington Publications 520, 564,581,615
Cronk Q, Bateman R. Hawkins J, eds (2002) Developmental Genetics and Plant Evolution. Taylor and Francis, London
Futuyma DJ, Slatkin M, eds (1983) Coevolution. Sinauer, Sunderland
Futuyma DJ (1998) Evolutionary Biology, 3rd ed.
Sinauer, Sunderland
Givnish TJ, Sytsma KJ, eds (1997) Molecular Evolution and Adaptive Radiation. Cambridge University Press, Cambridge
Grant V( 1981) Plant Speciation. 2nd ed. Columbia
University Press, New York
Grant V (1991) The Evolutionary Process, 2nd ed. Columbia University Press, New York
Harper JL (1977) Population Biology of Plants.
Academic Press, London
Levin DA (2000) The Origin, Expansion, and Demise of Plant Species. Oxford University Press, Oxford
Margulis L, Fester R, eds (1991) Symbiosis as a Source of Evolutionary Innovation. MIT Press, Cambridge, Massachusetts
Niklas KJ (1997) The Evolutionary Biology of Plants. University of Chicago Press, Chicago
Raup DM (1991) Extinction. Oxford University Press, Oxford
Ridley M (1996) Evolution, 2nd ed. Blackwell, Cambridge, Massachusetts
Rose MR, Guider GV, eds (1996) Adaptation. Academic Press, San Diego
Schluter D. (2000) The Ecology of Adaptive Radiation. Oxford University Press, Oxford
Silvertown JW, Charlesworth D (2001) Introduction to Plant Population Biology, 4'h ed. Blackwell. Oxford
Smith JM (1993) The Theory of Evolution. Cambridge University Press, Cambridge
Smith JM, Szathmary E (1995) The Major Transitions in Evolution. Freeman/Spektrum Akademischer Verlag, Oxford/Heidelberg
Stebbins GL (1950) Variation and Evolution in Plants. Columbia University Press, New York
Stebbins GL (1974) Flowering Plants, Evolution Above the Species Level. Harvard University Press, Cambridge, Massachusetts
Thompson JN (1994) The Coevolutionary Process. University of Chicago Press, Chicago
Vuorisalo TO, Mutikainen PK (2001) Life History Evolution in Plants. Kluwer Academic Publishers, Dordrecht.
10.1. Изменчивость <
10.1.1. Онтогенетическая изменчивость и фенотипическая пластичность
См. литературу гл. 7.
Bradshaw AD (1965) Evolutionary significance of phenotypic plasticity in plants. Adv Genet 13: 115-155.
542 | СПИСОК ЛИТЕРАТУРЫ
Coleman JS, McConnaughay KDM, Ackerly DD (1994) Interpreting phenotypic variation in plants. Trends Ecol Evol 9: 187—191
Debat V, David P (2001) Mapping phenotypes: canalization, plasticity and developmental stability. Trends Ecol Evol 16: 555 — 561
Pigliucci M (1996) How organisms respond to environmental changes: from phenotypes to molecules (and vice versa). Trends Ecol Evol 11: 168-173
Scheiner SM (1993) Genetics and evolution of phenotypic plasticity. Annu Rev Ecol Syst 24: 35 — 68
Schlichting CD (1986) The evolution of phenotypic plasticity in plants. Annu Rev Ecol Syst 17: 667-693
Schlichting CD, Pigliucci M (1998) Phenotypic Evolution: a Reaction Norm Perspective. Sinauer, Sunderland
10.1.2. Генетическая изменчивость
См. литературу подраздела 2.2.8, 2.2.9.
Bennett MD, Leitch IJ (1995) Nuclear DNA amounts in an-giosperms. Ann Bot 76: 113—176
Brown AHD, Clegg MT. Kahler AL, Weir BS, eds (1989) Plant Population Genetics, Breeding, and Genetic Resources. Sinauer, Sunderland
Charlesworth В (1998) Adaptive evolution: the struggle for dominance. Curr Biol 8: R502 — R504
Ellstrand NC, Elam DR (1993) Population genetic consequences of small population size: implications for plant conservation. Annu Rev Ecol Syst 24: 217—242
Esser K, LisowskyT, Michaelis G, Pratje E(2000) Extranuclear inheritance: genetics and biogenesis of mitochondria. Progr Bot 62: 71 — 97
Falconer DS, Mackay TFC (1996) Introduction to Quantitative Genetics. 4th ed. Longman, Essex
Gitzendanner MA, Soltis PS (2000) Patterns of genetic variation in rare and widespread plant congeners. Am J Bot 87: 783-792
Hagemann R, Hagemann M, Block R (1997) Genetic extra-nuclear inheritance: plastid genetics. Progr Bot 59: 108—130
Hamrick JL, Godt MJW (1990) Allozyme diversity in plant species. In: Brown AHD, Clegg MT, Kahler AL, Weir BS, eds. Plant Population Genetics, Breeding, and Genetic Resources. Sinauer, Sunderland
Kimura M (1983) The Neutral Theory of Molecular Evolution. Cambridge University Press, Cambridge
Leach DRF (1996) Genetic Recombination. Blackwell, Oxford
Mitchell-Olds T (1995) The molecular basis of quantitative genetic variation in natural populations. Trends Ecol Evol 10: 324—328
Mogensen HL (1996) The hows and whys of cytoplasmic inheritance in seed plants. Am J Bot 83: 383-404
Nei M (1987) Molecular Evolutionary Genetics. Columbia University Press, New York
Nybom H, Bartish IV (2000) Effects of life history traits and sampling strategies on genetic diversity estimates obtained with RAPD markers in plants. Persp Plant Ecol Evol Syst 3: 93—114
Otto SP, Michaiakis Y (1998) The evolution of recombination in changing environments. Trends Ecol Evol 13: 145-151
Smith JM (1998) Evolutionary Genetics, 2nd ed. Oxford University Press, Oxford
Weising K, Nybom H, Wolff K, Meyer W (1995) DNA Fingerprinting in Plants and Fungi. CRC Press, Boca Raton
Seyffert W, Hrsg (1998) Lehrbuch der Genetik. Gustav Fischer, Stuttgart
Stebbins GL (1971) Chromosomal Evolution in Higher Plants. Arnold, London
10.1.3. Рекомбинативная система
См. литературу раздела 11.2.
Asker SE, Jerling L (1992) Apomixis in Plants. CRC Press, Boca Raton
Barrett SCH, ed (1992) Evolution and Function of Heterostyly. Springer, Berlin
Charlesworth D, Charlesworth В (1987) Inbreeding depression and its evolutionary consequences. Annu Rev Ecol Syst 18: 237 — 268
Falk DA, Holsinger KE, eds (1991) Genetics and Conservation of Rare Plants. Oxford University Press, Oxford
Hamrick JL, Godt MJW. Sherman-Broyles SL (1995) Gene flow among plant populations: evidence from genetic markers. In: Hoch PC, Stephenson AG. eds, Experimental and Molecular Approaches to Plant Biosystematics. Missouri Botanical Garden. St. Louis
Hartl DL, Clark AG (1997) Principles of Population Genetics, 3rd ed. Sinauer, Sunderland
Hay ward MD (1999) The genetics of apomixis. In: van Raams-donk LWD, den Nijs JCM, eds, Plant Evolution in Man-made Habitats. Hugo de Vries Laboratory, Amsterdam
Holsinger KE (2000) Reproductive systems and evolution in vascular plants. In: Ayala FJ, Fitch WM, Clegg MT, eds, Variation and Evolution in Plants and Microorganisms. National Academy Press, Washington
Levin DA, Kerster HW (1974) Gene flow in seed plants. Evol Biol 7: 139—220
Lewis D (1979) Sexual Incompatibility in Plants. Arnold, London
Lloyd DG, Barrett SCH, eds (1996) Floral Biology: Studies on Floral Evolution tn Animal-pollinated Plants. Chapman & Hall, London
Список литературы | 543
Smith JM (1978) The Evolution of Sex. Cambridge University Press, Cambridge
Michod RE, Levin BR, eds (1988) The Evolution of Sex. Sinauer, Sunderland
Mogie M (1992) The Evolution of Asexual Reproduction in Plants. Chapman & Hall, London
Nettancourt D de (2001) Incompatibility and Incongruity in Wild and Cultivated Plants. Springer, Berlin
Richards AJ (1997) Plant Breeding Systems, 2nd ed. Chapman & Hall, London
Smith JM (1978) The Evolution of Sex. Cambridge University Press, Cambridge
Takebayashi N, Morrell PL (2001) Is selffertilization an evolutionary dead end? Revisiting an old hypothesis with genetic theories and a macroevolutionary approach. Am J Bot 88: 1143 — 1150
Weller SG, Sakai AK (1999) Using phylogenetic approaches to the analysis of plant breeding system evolution. Annu Rev Ecol Syst 30: 167 — 200
Weller SG, Donoghue MJ, Charlesworth D (1995) The evolution of self-incompatibility in flowering plants: a phylogenetic approach. In: Hoch PC, Stephenson AG, eds, Experimental and Molecular Approaches to Plant Biosystematics. Missouri Botanical Garden, St. Louis
Wheeler MJ, Franklin-Tong VE, Franklin FCH (2001) The molecular and genetic basis of pollenpistil interactions. New Phytol 151: 565 — 584
10.2. Примеры и причины естественной изменчивости
10.3. Видообразование
См. литературу подразделов 11.1.1. Распознавание видов.
Abbott RJ (1992) Plant invasions, interspecific hybridization and the evolution of new plant taxa. Trends Ecol Evol 7: 401—405
Anderson E (1949) Introgressive Hybridization. Chapman & Hall, London; Wiley, New York
Arnold ML (1997) Natural Hybridization and Evolution. Oxford University Press, Oxford
Arnold ML, Emms SK (1998) Molecular markers, gene flow and natural selection. In: Soltis DE, Soltis PS, Doyle JJ, eds, Molecular Systematics of Plants II. DNA Sequencing. Klu-wer Academic Publishers, Boston
Arnold ML, Kentner EK, Johnston JA, Comman S, Bouck AC (2001) Natural hybridization and fitness. Taxon 50: 93—104
Avise JC (2000) Phylogeography: The History and Formation of Species. Harvard University Press, Cambridge, Massachusetts
Bayer R (1999) New perspectives into the evolution of polyploid complexes. In: van Raamsdonk
LWD, den Nijs JCM, eds, Plant Evolution in Manmade Habitats. Hugo de Vries Laboratory, Amsterdam
Brodie ED, Moore AJ, Janzen FJ (1995) Visualizing and quantifying natural selection. Trends Ecol Evol 10: 313-318
Ellstrand NC, Schierenbeck KA (2000) Hybridization as a stimulus for the evolution of invasiveness in plants? In: Ayala FJ, Fitch WM, Clegg MT, eds, Variation and Evolution in Plants and Microorganisms. National Academy Press, Washington
Ellstrand N, Prentice HC, Hancock JF (1999) Gene flow and introgression from domesticated plants into their wild relatives. Annu Rev Ecol Syst 30: 539-564
Endler JA (1986) Natural Selection in the Wild. Princeton University Press, Princeton
Grant V (1981) Plant Speciation, 2nd ed. Columbia University Press, New York
Harrison RG (1991) Molecular changes at speciation. Annu Rev Ecol Syst 22, 281 — 308
Harrison RG, ed (1993) Hybrid Zones and the Evolutionary Process. Oxford University Press, Oxford
Heslop-Harrison J (1964) Forty years of genecology. Adv Ecol Res 2: 159—247
Hey wood JS (1991) Spatial analysis of genetic variation in plant populations. Annu Rev of Ecol Syst 22: 335-355
Hill JD, Doebley JF (1999) Key genetic factors influencing morphological change during the domestication of maize. In: van Raamsdonk LWD, den Nijs JCM, eds, Plant Evolution in Man-made Habitats. Hugo de Vries Laboratory, Amsterdam
Howard DJ, Berlocher SH (1998) Endless Forms: Species and Speciation. Oxford University Press, Oxford
McDade LA (1995) Hybridization and phylogenetics. In: Hoch PC, Stephenson AG, eds, Experimental and Molecular Approaches to Plant Biosystematics. Missouri Botanical Garden, St. Louis
Kreitman M, Akashi H (1995) Molecular evidence for natural selection. Annu Rev of Ecol Syst 26: 403-422
Langlet О (1971) Two hundred years of genecology. Taxon 20: 653 — 722
Levin DA (1978) The origin of isolating mechanisms in flowering plants. Evol Biol 11: 185 — 317
Levin DA (1993) Local speciation in plants: the rule not the exception. Syst Bot 18: 197—208
Levin DA (2001) 50 years of plant speciation. Taxon 50: 69—91
Levin DA (2000) The Origin, Expansion, and Demise of Plant Species. Oxford University Press, Oxford
Lewis WH, ed (1980) Polyploidy: Biological Relevance. Plenum Press, New York
Orr HA (2001) The genetics of species differences. Trends Ecol Evol 16: 343 — 350
544 I список ЛИТЕРАТУРЫ
Otte D, Endler JA, eds (1989) Speciation and its Consequences. Sinauer, Sunderland
Orr HA, Coyne JA (1992) The genetics of adaptation: a reassessment. Am Nat 140: 725 — 742
Orr MR, Smith ТВ (1998) Ecology and speciation. Trends Ecol Evol 13:502-506
Ramsey J, Schemske DW (1998) Pathways, mechanism, and rates of polyploid formation in flowering plants. Annu Rev Ecol Syst 29: 467 — 501
Rieseberg LH (1997) Hybrid origins of plant species. Annu Rev Ecol Syst 28: 359—389
Rieseberg LH (1998) Genetic mapping as a tool for studying speciation. In: Soltis DE, Soltis PS, Doyle JJ, eds, Molecular Systematics of Plants II. DNA Sequencing. Kluwer Academic Publishers, Boston
Riesebeig LH (2001) Chromosomal rearrangements and speciation. Trends Ecol Evol 16: 351-358
Rieseberg LH, Wendel JF (1993) Introgression and its consequences in plants. In: Harrison RG, ed. Hybrid Zones and the Evolutionary Process. Oxford University Press, Oxford
Rieseberg LH, Burke JM (2001) The biological reality of species: gene flow, selection, and collective evolution. Taxon 50: 47—67
Schluter D (2000) The Ecology of Adaptive Radiation. Oxford University Press, Oxford
Schluter D (2001) Ecology and the origin of species. Trends Ecol Evol 16: 372 — 380
Soltis DE, Soltis PS (1999) Polyploidy: Origins of species and genome evolution. Trends Ecol Evol 14: 348 — 352
Soltis PS, Soltis DE (2000) The role of genetic and genomic attributes in the success of polyploids. In: Ayala FJ, Fitch WM. Clegg MT, eds, Variation and Evolution in Plants and Microorganisms. National Academy Press, Washington
Stace CA (1975) Hybridization and the Flora of the British Isles. Academic Press, I-ondon
Stebbins GL (1971) Chromosomal Evolution in Higher Plants. Arnold. London
Thompson JD, Lumaret R (1992) The evolutionary dynamics of polyploid plants: origins, establishment and persistence. Trends Ecol Evol 7: 302—307
10.4. Макроэволюция
Eldredge N (1989) Macroevoiutionaty Dynamics: Species, Niches, and Adaptive Peaks. McGraw-Hill, New York
Gould SJ, Eldredge N (1993) Punctuated equilibrium comes of age. Nature 366: 223 — 227
Grantham TA (1995) Hierarchical approaches to macroevolu-tion: recent work on species selection and the “Effect Hypothesis”. Annu Rev Ecol Syst 26: 301—321
Levinton J (2001) Genetics, Paleontology, and Macroevolution, 2nd ed. Cambridge University Press, New York
Niklas KJ (1997) The Evolutionary Biology of Plants. University of Chicago Press, Chicago
Stanley SM (1979) Macroevolution: Pattern and Process. Freeman, San Francisco
ГЛАВА 11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
11.1. Методы систематики
Avise JC (1994) Molecular Markers, Natural History and Evolution. Chapman & Hall, New York
Avise JC (2000) Phylogeography: The History and Formation of Species. Harvard University Press, Cambridge, Massachusetts
Ax P (1988) Systematik in der Biologic. Gustav Fischer, Stuttgart
Bisby FA (2000) The quiet revolution: biodiversity informatics and the internet. Science 289: 2309-2312
Crawford DJ (1990) Plant Molecular Systematics: Macromole-cular Approaches. Wiley, New York
Davis PH, Heywood VH (1973) Principles of Angiosperm Taxonomy. Krieger, Huntington
Erhardt W, Gotz E, Bodecker N, Seybold S (2000) Zander. Handworterbuch der Pflanzennamen, 16. Aufl. Ulmer, Stuttgart
Federov AA, ed (1969) Chromosome Numbers of Flowering Plants. Nauka, Leningrad
Forey PL, Humphries CJ, Kitching IL, Scotland RW, Siebert DJ, Williams DM (1992) Cladistics: A Practical Course in Systematics. Oxford University Press, Oxford
Hennig W (1982) Phylogenetische Systematik. Paul Parey, Berlin
Hillis DM, Moritz C, Mabie BK (1996) Molecular Systematics, 2nd ed. Sinauer, Sunderland
Jeffrey C (1982) An Introduction to Plant Taxonomy, 2nd ed. Cambridge University Press, Cambridge
Judd WS, Campbell CS, Kellogg EA, Stevens PF (1999) Plant Systematics: A Phylogenetic Approach Sinauer, Sunderland
Maddison WP, Maddison DR (1992) MacClade: Analysis of Phylogeny and Character Evolution. Version 3.0. Sinauer, Sunderland
Moore RJ (1970— 1977). Index to Plant Chromosome Numbers. Regnum Veg 68, 77, 84, 90,91, 96
Omduff R (1967— 1969) Index to Plant Chromosome Numbers. Regnum Veg 50, 55, 59
Ridley M (1986) Evolution and Classification: the Reformation of Cladism. Longman, London
Sneath PHA, Sokal RR (1973) Numerical Taxonomy. Freeman, San Francisco
Soltis DE, Soltis PS, eds (1989) Isozymes in Plant Biology. Dioscorides Press, Portland
Soltis DE, Soltis PS, Doyle JJ, eds (1998) Molecular Systematics of Plants II. DNA Sequencing. Kluwer Academic Publishers, Boston
Список литературы 1 545
Soltis PS, Soltis DE, Doyle JJ, eds (1992) Molecular Systematics of Plants. Chapman & Hall, New York
Spring O, Buschmann H (1998) Grundlagen und Methoden der Pflanzensystematik. Quelle & Meyer, Wiesbaden
Stace CA (1989) Plant Taxonomy and Biosystematics, 2nd ed. Arnold, London
Stace CA (2000) Cytology and cytogenetics as a fundamental taxonomic resource for the 20th and 21st centuries. Taxon 49: 451—477
Stafleu FA, Cowan RS (1976 — 2000) Taxonomic Literature, 2nd ed. Regnum Veg 94, 98, 105, ПО, 112, 115,116, 125, 130, 132, 134,135,137
Stearn WT (1992) Botanical Latin. History, Grammar, Syntax, Terminology and Vocabulary, 4th ed. David & Charles, Newton Abbott
Stevens PF (2000) Botanical systematics 1950-2000: change, progress, or both? Taxon 49: 635 — 659
Stuessy TF (1990) Plant Taxonomy: the Systematic Evaluation of Comparative Data. Columbia University Press, New York
Swoflord DL (2001) PAUP*. Phylogenetic Analysis Using Parsimony (and Other Methods), version 4.0b6. Sinauer, Sunderland
Sytsma KJ, Pires JC (2001) Plant systematics in the next 50 years — re-mapping the new frontier. Taxon 50: 713 — 732
Vogellehner D (1983) Botanische Terminologie und Nomen-klatur, 2. Aufl. Gustav Fischer, Stuttgart
Wagele J-W (2000) Grundlagen der phyloge-netischen Systematik. Pfeil, Miinchen
Wagenitz G (1996) Worterbuch der Botanik. Gustav Fischer, Stuttgart
Weberling F, Stiitzel T (1993) Biologische Systematik. Grundlagen und Methoden. Wissenschaft-liche Buchgesellschaft, Darmstadt
Wilson HD (2001) Informatics: new media and paths of data flow. Taxon 50: 381 — 387
Winston JE (1999) Describing Species. Practical Taxonomic Procedure for Biologists. Columbia University Press, New York
http://evolution.genetics.washington.edu/phy-lip/software.html (Phylogcnieprogramme, J.Felsen-stein)
11.1.1. Определение видов
См. литературу раздела ll.l Методы систематики. Видообразование.
Bachmann К (1998) Species as units of diversity: an outdated concept. Theor Biosci 117: 213 — 230
Claridgc MF, Dawah AH, Wilson MR, eds, (1997) Species. The Units of Biodiversity. Chapman & Hall, London
Ereshefcky M, ed (1992) The Units of Evolution. Essays on the Nature of Species. MIT Press, Cambridge, Massachusetts
Howard DJ, Berlocher SH (1998) Endless Forms: Species and Speciation. Oxford University Press, Oxford
Levin DA (2000) The Origin, Expansion, and Demise of Plant Species. Oxford University Press, Oxford
Luckow M (1995) Species concepts: Assumptions, methods, and applications. Syst Bot 20: 589—605
Otte D, Endler JA, eds (1989) Speciation and its Consequences. Sinauer, Sunderland
11.1.2. Монографии, флоры и определительные ключи
Askevold IS, O'Brien CW (1994) DELTA, an invaluable computer program for generation of taxonomic monographs. Ann Entomol Soc Amer 87: 1-16
Dallwitz MJ (1980) A general system for coding taxonomic descriptions.Taxon 29: 41 — 46
Edwards M, Morse DR (1995) The potential for computer-aided identification in biodiversity research. Trends Ecol Evol 10: 152—158
Frodin DG (2001) Guide to the Standard Floras of the World, 2nd ed. Cambridge University Press, Cambridge
Heywood VH (2001) Floristics and monogra-phy— an uncertain future? Taxon 50, 361 — 380
Pankhurst RJ (1978) Biological Identification: the Principlesand Practice of Identification Methods in Biology. Arnold, London
Pankhurst RJ (1991) Practical Taxonomic Computing. Cambridge University Press, Cambridge
The Kew Record of Taxonomic Literature (1974fi). Royal Botanic Gardens, Kew
Winston JE (1999) Describing Species. Practical Taxonomic Procedure for Biologists. Columbia University Press, New York
Wright JF, Morse DR, Tardivel GM (1995) An investigation into the use of hypertext as a user interface to taxonomic keys. Comput Appl Biosci 11: 19-27
http://www.rbgkew.org.uk/kr/KRHomeExt.html (The Kew Record of Taxonomic Literature)
11.1.3. Исследование родства
11.1.4. Филогения и классификация
11.1.5. Номенклатура
См. литературу раздела 11.1. Методы систематики
Brummitt RK, Powell СЕ, eds, (1992) Authors of Plant Names. Royal Botanic Gardens, Kew
Farr ER, Leussink JA, Stafleu FA, eds (1979) Index Nominum Genericorum (Plantarum). Regnum Veg 100—102
546 I СПИСОК ЛИТЕРАТУРЫ
Farr ER, Leussink JA, Zijlstra G, eds (1986) Index Nominum Genericorum (Plantarum). Supplementum I. Regnum Veg 113
Greuter W, Hoogland RD, Reveal JL, Crosby MJ, Grolle R, Zijlstra G, avid JC (1993) Family Names in Current Use for Vascular Plants, Bryophytes, and Fungi. Regnum Veg 126
Greuter W et al. (2000) International Code of Botanical Nomenclature (St. Louis Code). Regnum Veg 138
Holmgren PK, Holmgren NH, Barnett LC (1990) Index Herbariorum. Part I: The Herbaria of the World, 8th ed. Regnum Veg 120
Jeffrey C (1989) Biological Nomenclature, 3rd ed. Arnold, London
De Queiroz K, Gauthier J (1990) Phylogeny as a central principle in taxonomy: Phylogenetic definitions of taxon names. Syst Zool 39: 307 — 322
De Queiroz K, Gauthier J (1992) Phylogenetic taxonomy. Annu Rev Ecol Syst 23: 449—480
McNeill J (2000) Naming the groups: developing a stable and efficient nomenclature. Taxon 49: 705-720
http://www.bgbm.fu-berlin.de/iapt/nomenclatu-re/code/tokyo-e/ Preface.htm (International Code of Botanical Nomenclature)
http://www.phylocode.org (Phylogenetische No-menklaturre-geln)
http://www.inform.umd.edu/PBIO/fam/sgin-dex.html (Suprage-neric Names Arranged by Families)
http://rathbun.si.edu/botany/mg/ingform.cfm (Index Nominum Genericum)
http://www.nybg.org/bsci/ih/searchih.html (Index Herbario-rum)
11.2. Бактерии, грибы, растения
Прокариоты, эукариоты до папоротниковидных
См. литературу раздела 2.3. Строение клетки у прокариот, 2.4. Теория эндосимбионтов и водородная гипотеза, гл. 5. Принципы строения талломных растений (таллофитов).
Обобщающие работы
Bold НС (1988) Morphology of Plants, 5. Aufl.
Harper & Row, New York
Braune W, Leman A, Tauben H (1999) Pflanze-nanatomisches Praktikum, 4. Aufl. Spektrum Akademischer Verlag, Heidelberg
Bresinsky A, Kadereit J, (2001) Systematik-Poster: Botanik, 2. Aufl. Spektrum Akademischer Verlag, Heidelberg
Cavalier-Smith T (1998) A revised six-kingdom system of life. Biol Rev 73: 203 — 266
Esser К (1992—2000) Kryptogamen I und II, 2. und 3. Aufl. Springer, Berlin
Maigulis L (1993) Symbiosis in Cell Evolution.
Freeman, New York
Margulis L et al. (1989) Protoctista. Jones & Bartlett, Boston
Rabenhorst L (1884—1968) Kiyptogamenflora von Deutschland, Osterreich und der Schweiz. 45 Bande, 2. Aufl. Kummer, Leipzig
Schubert R. Handke HH, Pankow H, Hrsg (1994) Exkursionsflora fur die Gebiete der DDR und BRD. 1: Niedere Pflanzen, 3. Aufl. Gustav Fischer, Jena
Smith GM (1955) Ctyptogamic Botany. 2 Bande, 2. Aufl. McGraw-Hill, New York
Throm G (1997) Biologje der Kryptogamen, 2 Bande. Haag & Herchen, Frankfurt
Wartenbeig A (1979) Systematik der niederen Pflanzen, 2. Aufl. Thieme, Stuttgart
Weber H, Hrsg (1997) Worterbuch der Mikro-biologie. Gustav Fischer, Jena
Woese CR et al. (1990) Towards a natural system of organisms: Proposals for the domains Archaea, Bacteria, and Eucarya. Proc Natl Acad Sci USA 87: 4576-4579
Вирусы и прокариоты
1. Вирусы
Fields BN,Knipe DM (1991) Fundamental Virology, 2. Aufl. Raven, New York
Franckl RJB et al. Hrsg (1985-1988) The plant viruses. Plenum Press, New York
Harper D (1994) Molecular virology. BIOS Science Publishers, Oxford
Matthews REF (1992) Fundamentals of Plant Virology. Academic Press, San Diego Rowson KEK, Rees TAL, Maty BWJ (1981) A Dictionary of Virology. Blackwell, Oxford
Scott A (1990) Die Geschichte der Viren. Birkhauser, Basel Van Regenmortel M et al. (2000) Vims Taxonomy. Academic Press, San Diego
Voyles BA (1994) The Biology of Viruses. Mosby, St. Louis
2. Бактерии (кроме цианобактерий), археи
Balows A, Trupcr HG, Dwokin M, Harder W, Schleifer KH (1992) The Prokaiyotes, 2. Aufl. Springer, New York
Bergey (2001) Manual of Systematic Bacteriology, 2. Aufl. Williams & Wilkins, Baltimore
Brock TD, Hrsg (1997) Biology of Microorganisms, 8. Aufl. Prentice Hall, London
Fritsche F (2001) Mikrobiologie, 3. Aufl. Spektrum Akademischer Verlag, Heidelberg
Gottschalk G (1985) Bacterial Metabolism. Springer, New York
Список литературы | 547
Kandler О (1981) Archaebakterien und Phylo-genie der Organismen. Naturwissenschaften 68: 183-192
Koch A (1995) Bacterial Growth and Form. Chapman & Hall, New York
Neidhardt F et al. (1990) Physiology of the Bacterial Cell. Sinauer, Sunderland
Sigee DC (1993) Bacterial Plant Pathology: Cell and Molecular Aspects. Cambridge University Press, Cambridge
Singleton P (1995) Einfuhrung in die Bakte-riologie, 2. Aufl. Quelle & Meyer, Wiesbaden
Schlegel HG (1992) Allgemeine Mikrobiologie, 7. Aufl. Thieme, Stuttgart
3. Синезеленые водоросли (цианобактерии)
Bryant D (1994) The Molecular Biology of Cyanobacteria. Kluwer Academic Publishers, Dordrecht
Can NG, Whitton В A (1972) The Biology of Blue-green Algae. Blackwell, Oxford
Copley J (2001) The evolution of the atmosphere. Nature 410: 862—864
Mann HN, Carr NG (1992) Photosynthetic Prokaiyotes. Plenum, New York
Sleep N (2001) Oxygenating the atmosphere. Nature 410: 317 — 319
Zehr J et al. (2001) Unicellular cyanobacteria fix N2 in the subtropical North Pacific Ocean. Nature 412: 635-638
Эукариоты: грибы, лишайники, водоросли, мхи, папоротниковидные
1. Миксомицеты (слизевики)
Bonner JT (1967) The Cellular Slime Molds, 2. Aufl. University Press, Princeton
Gray WD, Alexopoulos CJ (1968) Biology of the Myxomycetes. Ronald Press, New York
Karting JS (1968) The Plasmodiophorales, 2. Aufl. Hafner, New York
Neubert H, Nowotny W, Baumann К (1993 — 2000) Die Myxomyceten, 3 Bande. Baumann, Gomaringen
Olive LS (1975) The Mycetozoans. Academic Press, New York
Stephenson SL, Stempen H (1994) Myxomycetes. Timber, Portland
2. Грибы
а) Общая группа, морфология, история развития
Ainsworth GC, Sussman AS (1965—1973) The Fungi, 4 Bande. Academic Press, London
AnkeT (1997) Fungal Biotechnology. Chapman & Hall, London
Arora D et al. (1991) Handbook of Applied Mycology. Marcel Dekker, New York
Arx JA (1981) The Genera of Fungi Sporulating in Pure Culture, 3. Aufl. Cramer, Vaduz
Bon M (1988) Pareys Buch der Pilze. Parey, Hamburg
Bresinsky A, Besl H (1985) Giftpilze. Wissen-schaftliche Veriags-gesellschaft, Stuttgart
Carlike M et al. (2001) The Fungi. Academic Press, San Diego
Dorfelt H (1989) Lexikon der Mykologie. Gustav Fischer, Stuttgart
Elliot CG (1994) Reproduction in Fungi. Chapman & Hall, London
Esser K, Kuenen R (1965) Genetik der Pilze. Springer, Berlin
Esser K, Lemke P, Bennet J. (1994— 2001) The Mycota, 10 Bande. Springer, Berlin
Frisvad J et al. (1998) Chemical Fungal Taxonomy. Marcel Dekker, New York
Gaumann E (1964) Die Pilze, 2. Aufl. Birkhauser, Basel
Gill M, Steglich W (1987) Pigments of fungi (Macromycetes). Progr Chem Org Nat Prod 51: 1-317
Hawksworth DL, Kirk, PM, Sutton, BC, Pegler, DN (1995)
Ainsworth & Biby's Dictionary of the Fungi, 8. Aufl. CAB International, Wallingford
Michael E, Henning B, Kreisel H (1958—1988) Handbuch fiir Pilzfreunde, 6 Bande. Gustav Fischer, Jena
Moore D (1998) Fungal Morphogenesis. Cambridge University Press, Cambridge
Muller E, LofTler W (1982) Mykologie, 4. Aufl. Thieme, Stuttgart
ReiB I (1998) Schimmelpize, 2. Aufl. Springer, Berlin
Schwantes HO (1996) Biologie der Pilze. Ulmer, Stuttgart
Vasilyeva L (1999) Systematics in mycology.
Bibliotheca Mycologica 178. Cramer, Berlin
Weber H (1993) Allgemeine Mykologie. Gustav Fischer, Jena
Webster J (1983) Pilze, eine Einfuhrung. Springer, Berlin
б) Оомицеты (Oomycota), настоящие грибы (Eumycota): хитрцдиомицеты, зигомицеты
Fitzpatrick НМ (1930) The lower fungi (Phy-comycetes), Reprint 1960. Johnson, London
Karting JS (1977) Chytridiomycetarum Icono-graphia. Cramer, Vaduz
Sparrow FK (1960) Aquatic Phycomycetes, 2. Aufl. University of Michigan Press, Ann Arbor
Zycha H, Siepmann R (1969) Mucorales. Cramer, Lehre
548 [СПИСОК ЛИТЕРАТУРЫ
в) Аскомицеты
Barnett JA, Payne RW, Yarrow D (2000) A Guide to Identifying and Classifying Yeasts, 2. Aufl. Cambridge University Press, Cambridge
Braun U (1987) A Monograph of the Erysi-phales. Beih Nova Hedwigia 89
Breitenbach J, Kranzlin F (1981) Pilze der Schweiz 1, Ascomyceten. Verlag Mykologia, Luzern
Dennis RWG (1977) British Ascomycetes, 3.
Aufl. Cramer, Lehre
Hawksworth DL (1994) Ascomycete Systematics. Problems and Perspectives in the Nineties. Plenum, New York
Kreger-van Rij NJW, Hrsg (1984) The Yeasts, 3. Aufl. Elsevier, Amsterdam
Moser M (1963) Kleine Kryptogamenflora Bd. Ha, Ascomyceten. Gustav Fischer, Stuttgart
Phaff HJ, Miller M, Mrak E (1966) The Life of Yeasts. Harvard University Press, Cambridge, Massachusetts
Reynolds DP, Hrsg (1981) Ascomycete Systematics. Springer, New York
Scheloske HW (1969) Beitrage zur Biologie der l^boulbeniales. Gustav Fischer, Jena
Tavares II (1969) Laboulbeniales. Cramer, Braunschweig
г) Базидиомицеты
Blumer S (1963) Rost- und Brandpilze auf Kul-lurpflanzen. Gustav Fischer, Jena
Breitenbach J, Kranzlin F (1986—1995) Pilze der Schweiz 2—4, Basidiomycetes. Veriag Mykologia, Luzern
Clemcncon H (1997) Anatomie der Hymeno-myceten. Fliick-Wirth, Teufen
Gaumann E (1959) Die Rostpilze Mitteleuropas.
Buehler, Bern
Julich W (1984) Kleine Kryptogamenflora Hb/ 1, Nichtblat-terpilze. Gallertpilze und Bauchpilze. Gustav Fischer, Stuttgart
Kreisel H, Hrsg (1987) Pilzflora der Dcutschen Demokratischen Republik. Gustav Fischer, Jena
Krieglsteiner G, Hrsg (2000—2001) Die Gro-Bpilze Baden-Wurt-tembergs, 3 Bande. Ulmer, Stuttgart
Moser M (1983) Kleine Kryptogamenflora. Bd. 11/2, Rohrlinge und Blatterpilze, 5. Aufl. Gustav Fischer, Stuttgart
Singer R (1986) The Agaricales in Modem Taxonomy, 4. Aufl. Koeltz, Konigstein
Vanky К (1994) European Smut Fungi. Gustav Fischer, Stuttgart
Zogg, H (1985) Die Brandpilze Mitteleuropas. Fliick-Wirth, Teufen
д) Дейтеромицеты
Barnett HL, Hunter BB (1987) Illustrated Genera of Imperfect Fungi, 4. AufJ. APS Press, St. Paul
Carmichael JW et al. (1980) Genera of Hypho-mycetes. University of Alberta Press, Edmonton
Domsch KH, Gams W, Anderson TH (1980) Compendium of Soil Fungi, 2 Bande. Academic Press, London
Raper KP, Thom C (1949) A Manual of the Penicillia. Baltimore (Nachdruck 1968). Hafner, New York
Raper KP, Fennel DI (1965) The Genus Aspergillus. (Nachdruck 1973). Krieger, Huntington
e) Экология
Agerer R (1987—1998) Colour Atlas of Ecto-mykorrhizae. Ein-hom, Schwabisch Gmiind
Arora DK et al. (1991) Handbook of Applied Mycology; Band 1: Soil and Plants. Dekker, New York
Ayres P, Boddy L, Hsg (1986) Water, Fungi and Plants. Cambridge University Press, Cambridge
Barron GL (1977) The Nematode-destroying Fungi. Canadian Biological Publication, Guelph
Buchner P (1953) Endosymbiose der Tiere mit pflanzlichen Mikroorganismen. Birkhauser, Basel
Dix N, Webster J (1995) Fungal Ecology. Chapman & Hall, London
Frankland J et al. (1955) Fungi and Environmental Change. Cambridge University Press, Cambridge
Henry SM (196— 1967) Symbiosis, 2 Bande. Academic Press, New York
Ingold C, Hudson H (1993) The Biology of Fun gi. Chapman & Hall, London
Jennings D, Lysek G (1996) Fungal Biology. BIOS Scientific Publishers, Oxford
Smith SE, Read, DJ (1997) Mycorrhizal Symbiosis, 2. Aufl. Academic Press, London
Vamia A, Hock В (1997) Mycorrhiza. Springer, Berlin
Wicklow DT, Carroll GC (1981) The fungal community. Dekker, New York
Zahner H (1965) Biologie der Antibiotica. Springer, Berlin
ж) Фитопатогепные грибы
Bavendamm W (1969) Der Hausschwamm und andere Bauhoiz-pilze. Gustav Fischer, Stuttgart
Brandenburger W (1985) Parasitische Pilze an GefaBpflanzen in Europa. Gustav Fischer, Stuttgart
Brandenburger W (1963) Vademecum zum Sa-mmeln parasitischer Pilze. Ulmer, Stuttgart
Borner H (1983) Pflanzenkrankheiten und Pflanzenschutz. Ulmer, Stuttgart
Borner H, Zunke U (1992) Praktikum der Phy-topathologie. Parey, Berlin
Butin H (1996) Krankheiten der Wald- und Park-biiume, 3. Aufl. Thieme, Stuttgart
Ellis MB, Ellis JP (1997) Microfungi on Land Plants, 2. Aufl.Croom Helm, London
Elstner EF, OBwald W, Schneider 1 (1996) Phy-topathologie. Spektrum Akademischer Verlag, Heidelberg
Список литературы | 549
Gaumann Е (1951) Pflanzliche Infektionslehre, 2. Aufl. Birkhauser, Basel
Hoffmann GM et al. (1994) Lehrbuch der Phy-tomedizin, 3. Aufl. Blackwell, Berlin
Prell H (1996) Interaktionen von Pflanzen und phytopathoge-nen Pilzen. Gustav Fischer, Stuttgart
Rypacek V (1966) Biologie holzzerstorender Pil-ze. Gustav Fischer, Jena
Schlosser E (1997) Allgemeine Phytopathologie, 2. Aufl. Thieme, Stuttgart
Sorauer P (1962ff) Handbuch der Pflanzen-krankheiten, Band 3 (Pilzliche Krankheiten), 6. Aufl. Parey, Berlin
Vidhyasekaran P (1997) Fungal Pathogenesis in Plants and Crops. Marcel Dekker, New York
Walker JC (1969) Plant Pathology, 3. Aufl. McGraw-Hill, New York
3. Лишайники
Ahmadjian V (1993) The Lichen Symbiosis. Wiley, New York Galun M (1988) Handbook of Lichenology, 3 Bande. CRC Press, Boca Raton
Hawksworth DL, Hill DJ (1984) The Lichenforming Fungi. Chapman & Hall, New York
Henssen A, Jahns HM (1974) Lichenes. Thieme, Stuttgart Herzig R, Urech M (1991) Flechten als Bioindikatoren (Bibl. Lichenologica 43). Bomtraeger, Stuttgart
Huneck S, Yoshimura I (1996) Identification of lichen substances. Springer, Berlin
Lange О (1992) Pflanzenleben unter StreB: Flechten als Pioniere der Vegetation an Extremstan-dorten der Erde. Univ. Wurzburg
Lutzoni et al. (2001) Major fungal lineages are derived from lichen symbiontic ancestors. Nature 411: 937-940
Masuch G (1993) Biologie der Flechten. Quelle & Meyer, Heidelberg
Mober R Holmason I (1992) Flechten von Nord- und Mitteleuropa. Gustav Fischer, Stuttgart
Poelt J (1969) Bestimmungsschlussel.der euro-paischen Flechten (Erganzungshefte 1977 u. 1981), 2. Aufl. Cramer, Lehre
Scholler H, Hrsg (1997) Flechten: Geschichte, Biologie, Systematik, Okologie, Naturschutz und kulturelle Bedeutung. Kleine Senckenberg-Reihe 27. Kramer, Frankfurt
Seaword MRD (1978) Lichen Ecology. Academic Press, London
Steiner M (1965) Wachstums- u. Entwicklungs-physiologie der Flechten. Handbuch der Pflanzen-pysiologie 15/1. Springer, Berlin
Wirth V (1995) Flechtenflora, 2. Aufl. Ulmer, Stuttgart
Wirth V (1995) Die Flechten Baden Wiirttem-bergs, 2 Bande, 2. Aufl. Ulmer, Stuttgart
Wirth V, Dull R (2000) Farbatlas Flechten und Moose. Ulmer, Stuttgart
4. Водоросли
а) Морфология, история развития, систематика
Akatsuka J (1994) Biology of Economic Algae. Academic Publishing, The Hague
Bourrelly P (1966—1970) Les algues d'eau douce, 3 Bande. Boubee, Paris
Bremer Ketal. (1987) On cladistic relationships in green plants. Taxon 36: 339—349
Brook AJ (1980) The Biology of Desmids. Black-
*well, Oxford
Buetow D, Hrsg (1968—1989) The Biology of Euglena, 4 Bande. Academic Press, New York
Chapman R et al. (1998) Molecular systematics of green algae. In: Soltis D et al. Hrsg, Molecular Systematics of Plants II. Kluwer Academic Publishers, Boston
Chapman VJ, Chapman DJ (1973) The Algae, 2.
Aufl. Macmillan, London
Cox ER, Hrsg (1980) Phytoflagellates. Elsevier, New York
Desikachary TV, Prema P (1996) Silicoflagellates (Dictyochophyceae), Bibl Phycologica 100. Bomtraeger, Stuttgart
Dodge JD (1973) The Fine Structure of Algal Cells. Academic Press, London
Drebes G (1974) Marines Phytoplankton. Thieme, Stuttgart
Ettl H (1980) GrundriB der allgemeinen Algo-logie. Gustav Fischer, Stuttgart
Fott H (1971) Algenkunde, 2. Aufl. Gustav Fischer, Jena
Gams H (1974) Kleine Kiyptogamenflora lb: Makroskopische Meeresalgen. Gustav Fischer, Stuttgart
Helmcke JG, Krieger W, GerloffJ (1962-1975) Diatomeenschalen im elektrotnikroskopischen Bild, 10 Bande. Cramer, Lehre
Huber-Pestalozzi G (1955-1982) Das Phytoplankton des SiiBwassers, 9 Bande. Schweizerbart, Stuttgart
Irvine DEG, Price HJ, Hrsg (1978) Modern Approaches to the Taxonomy of Red and Brown Algae. Academic Press, London
Irvine DT, John DM, Hrsg (1984) Systematics of the green Algae. Academic Press, London
Kornmann P, Sahling PH (1977) Meeresalgen von Helgoland. Biologische Anstalt Helgoland, Hamburg
Kowallik К (1997) Origin and evolution of chloroplasts: Current status and future perspectives. In: Schenk et al. Eukaryotism and Symbiosis. Springer, Berlin
Kylin H (1956) Die Gattungen der Rhodo-phyceen. Gleerups, Lund
Lobban CS, Wynne MJ, Hrsg (1981) The Biology of Seaweeds. Blackwell, Oxford
Marin B, Melkonian M (1999) Mesostigmatophyceae, a new class of Streptophyte green algae. Protist 150: 399-117
550 I СПИСОК ЛИТЕРАТУРЫ
Melkonian М et al. (1995) Phylogeny and evolution of the algae. In: Arai et al. Biodiversity and Evolution, pp 153— 176. The National Science Museum Foundation, Tokyo
Melkonian M (2001) Systematics and evolution of the algae, Progr Bot 62: 340—382
Moreira D et al. (2000) The origin of red algae and the evolution of chloroplasts. Nature 405: 69 — 72
Nakayama T et al. (1998) The basal position of scaly green flagellates among the green algae (Chlorophyta). Protist 149: 367 — 380
Oltmanns F (1922—1923) Morphologic und Biologic der Algen, 3 Bande (Neudruck 1974), 2. Aufl. Gustav Fischer, Jena
Palmer J (2000) A single birth of all plastids? Nature 405: 32-33
Pankow H (1990) Ostsee-Algenflora. Gustav Fischer, Jena
Pascher A Ettl H, GerloffJ, Heying H (1978ff) SuBwasserflora von Mitteleuropa. Gustav Fischer, Stuttgart
Pickett-Heaps JD (1975) Green Algae. Sinauer, Sunderland
Pringsheim EG (1963) Farblose Algen. Gustav Fischer, Stuttgart
Reitz M (1986) Die Alge im System der Pflan-zen. Gustav Fischer, Stuttgart
Sandgren CD, Smol J P, Kristiansen J, Hrsg. (1995) Chrysophyte Algae. Cambridge University Press, Cambridge
Saijeant WAS (1974) Fossil and Living Dinoflagellates. Academic Press, London
Schussnig В (1953, 1960) Handbuch der Pro-tophytenkunde, 2 Bande. Gustav Fischer, Jena
Streble H, Krauter D (1988) Das Leben im Wa-ssertropfen, 8. Aufl. Frankh, Stuttgart
Van den Hoek C, Jahns HM, Mann DG (1993) Algen, 3. Aufl. Thieme, Stuttgart
Werner D, Hrsg (1977) The Biology of Diatoms. Blackwell, Oxford
б) Экология
Chapman VJ (1970) Seaweeds and their Uses, 2. Aufl. Camelot Press, London
Dixon PS (1973) Biology of the Rhodophyta. Oliver & Boyd, Edinburgh
Ettl H, Gartner G (1995) Syllabus der Boden-, Luft- und Flechtenalgen. Gustav Fischer, Stuttgart
Golubic S (1967) Algenvegetation der Felsen. Nagele & Obermiller, Stuttgart
Hutchinson GE (1957-75) A Treatise on Limnology, 3 Bande. New York
Levring T. Hoppe HA, Schmid OJ (1969) Marine Algae. Cram, de Gruyter & Co., Berlin
Liining К (1985) Meeresbotanik. Verbreitung, Okophysiologie und Nutzung der marinen Mak-roalgen. Thieme, Stuttgart
Round FE (1975) Biologic der Algen, 2. Aufl. Thieme, Stuttgart
Sernow SA (1958) Allgemeine Hydrobiologie. Deutscher Verlag der Wissenschaften, Berlin
Schwoerbel J (1999) Einfiihrung in die Limno-logie, 8. Aufl. Thieme, Stuttgart
Tardent P (1979) Meeresbiologie. Thieme, Stuttgart
5. Мхи и папоротниковидные
а) Мхи и папоротники
Aichele D, Schwegler H (1993) Unsere Moos-und Fampflanzen, 10. Aufl. Frankh-Kosmos, Stuttgart
Esser К (1992) Kryptogamen II: Moose, Fame. Praktikum und Lchrbuch. Springer, Berlin
Frey W, Frahm JP, Fischer, E, Lobin W (1995) Kleine Kryptogamenflora IV: Die Moos- und Fampflanzen Europas, 6. Aufl. Gustav Fischer, Stuttgart
Kenrick P, Crane P (1997) The Origin and Early Diversification of Land Plants. A Cladistic Study. Smi-thonian Institution, Washington
Parihar NS (1963) An introduction to Embryophyta, 4. Aufl. Central Book Depot, Allahabad
Probst W (1987) Biologie der Moos- und Fampflanzen, 2. Aufl. Quelle & Meyer, Wiesbaden
6) Мхи
Bopp M (1965) Entwicklungsphysiologie der Moose. Handbuch der Pflanzenphysiologie 15/1. Springer, Berlin
Briining F (1994) Biyophyten als Akkumulati-ons- und Reak-tionsindikatoren (Berichte aus der Biologie). Shaker, Aachen
Chopra RN, Kumra PK (1988) Biology of Bryophytes. Wiley, New York
Correns C (1899) Untersuchungen uber die Vermehrung der Laubmoose durch Brutorgane und Stecklinge (Nachdruck 1976). Cramer, Vaduz
Dull R, Meinunger L (1989) Deutschlands Moose, 1. Teil. IDH, Munstereifel
Dull R (1994) Deutschlands Moose. Verbreitung in Deutsch-land, vertikale und zonale Verbreitung, 2 — 3. Teil. IDH, Munstereifel
Dull R (1993) Exkursionstaschenbuch der Moose, 4. Aufl. IDH, Munstereifel
Frahm J (1998) Mooseals Bioindikatoren. Quelle & Meyer, Wiesbaden
Frahm J (2001) Biologie der Moose. Spektrum Akademischer Verlag, Heidelberg
Frahm JP, Frey W (1992) Moosflora, 3. Aufl.
Ulmer, Stuttgart
Stammesgeschichte, Okologie. Gustav Fischer, Stuttgart
Goebel К (1930) Organographie der Pflanzen, Band 2, 3. Aufl. Gustav Fischer, Jena
Hebant C (1977) The conducting tissues of bryophytes. Cramer, Vaduz
Lorch, W (1931) Anatomic der Laubmoose. Handbuch der Pflanzenanatomie VII/1. Borntraeger, Berlin
Список литературы | 551
Maschke J, (1981) Moose als Bioindikatoren von Schwermetall-Immissionen (Biyophytorum Bibliotheca 22). Cramer, Vaduz
Miller NG (1988) Bryophyte Ultrastructure (Advances in Bryology 3), Bomtraeger, Stuttgart
Miller NG, Hrsg (1991) Bryophyte Systematics, (Advances in Bryology 4). Borntraeger, Stuttgart
Miller NG, Hrsg (1993) Biology of Sphagnum (Advances in Bryology 5), Borntraeger, Stuttgart
Muller К (1951) Lebermoose Europas. Band 1 (Nachdruck 1965). Kummer, Leipzig
Nebel M, Philippi G (2000— 2001) Die Moose Baden-Wurttem-bergs, 2 Bande. Ulmer, Stuttgart
Richardson DHS (2000—2001) The Biology of Mosses. Black-well, Oxford
Schuster RM (1983) New Manual of Bryology. Hattori Botanical Labo~atory, Nichinan
Schuster RM (1966—1974) The Hepaticae and Anthocerotae of North America, 3 Bande. Columbia. New York
Schuster RM (1977) Evolution and Early Diversification of Hepaticae. In Frey W, Hurka H, Oberwinkler F Hrsg, Bei-trage zur Biologie der Niederen Pflanzen. Gustav Fischer, Stuttgart
Shaw J, Goffinet В (2000) Bryophyte Biology. Cambridge University Press, Cambridge
Stetzka KM (1994) Die Waldbodenvegetation als Bioindikator fur Umweltbelastungen (Disserta-tiones Botanicae). Borntrae-ger, Stuttgart
Walther К (1983) Bryophytina. In: Engler A, Syllabus der Pflanzenfamilien. Bomtraeger, Stuttgart
Watson EV (1971) The Structure and Life of Bryophytes, 3. Aufl. Hutchinson University Library, London
в) Папоротниковидные
Barlow PW et al. (1998) Developmental Biology of Fem Gametophytes. Cambridge University Press, Cambridge
Beerling D et al. (2001) Evolution of leaf-form in land-plants linked to atmospheric CO2 decline in the late Paleozoic area. Nature 410: 352—354
Bierhorst DW (1971) Morphology of Vascular Plants. Macmillan, New York
Bower FO (1935) Primitive Land Plants (Reprint 1959). Hafner, New York
Bower FO (1922—1928) The Fems, 3 Bande, (Reprint 1964). Cambridge University Press, Cambridge
DostalJ, ReichsteinT, Fraser-Jenkins CR. Kramer KU (1985) Pteridophyten. In: Illustrierte Flora von Mitteleuropa (Hegi G, Hrsg), Band 1,1. Parey, Berlin
Eames AJ (1936) Morphology of Vascular Plants (Lower Groups). Me Graw Hill, New York
Foster AA, Gifford EM (1989) Comparative Morphology ofVascular Plants, 3. Aufl. Freeman, San Francisco
Goebel К (1930) Organographie der Pflanzen, Band 2, 3. Aufl. Gustav Fischer. Jena
Guttenbeig H (1966) Histogenese der Pteridophyten; Handbuch der Pflanzenanatomie VII/2. Bomtraeger, Berlin
Jermy AC, Crabbe JA. Thomas BA (1973) The phylogeny and classification of ferns; Bot J Linnean Soc 67, Suppl 1. Academic Press. London
Kenrik P (2001) Turning over a new leaf Nature 410: 309—310
Kramer KU, Schneller JJ, Wollenweber E (1995) Fame und Famverwandte. Thieme, Stuttgart
Kubitzki K, Kramer KU, Green PS (1990) The Families and Genera of Vascular Plants. Band 1 Pteridophytes and Gymnosperms. Springer, Berlin
Love A, Love D (1977) Cytotaxonomical Atlas of the Pteridophyta. Cramer, Lehre
Manton I (1950) Problems of Cytology and Evolution in the Pteridophyta. Cambridge University Press, Cambridge
Nayar BK, Kaur S (1971) Gametophytes of homosporous fems. Bot Rev 37: 295 — 396
Ogura Y (1972) Comparative anatomy of vegetative organs of the Pteridophyta. Handbuch der Pflanzenanatomie VI1/3. Borntraeger, Berlin
Rasbach K, Rasbach H, Wilmanns О (1976) Die Fampflanzen Zentraleuropas, 2. Aufl. Gustav Fischer, Stuttgart
Schneider H (1996) Vergleichende Wurzelana-tomie der Fame (Berichte aus der Biologie). Shaker, Aachen
Troll W (1937— 1943) Vergleichende Morphologic der hoheren Pflanzen. Bomtraeger, Berlin
Tutin TG et al. (1993) Flora Europaea, Band 1, 2. Aufl. Cambridge University Press, Cambridge
Tyron RM, Tyron, AF (1982) Fems and Allied plants. Springer, New York
Семенные растения
Смена поколений
См. литературу раздела 11.1. Методы систематики.
Cresti М, Blackmore S, van Went JL, eds (1992) Atlas of Sexual Reproduction in Flowering Plants. Springer, Heidelberg Davis GL (1966) Systematic Embryology of the Angiosperms. Wiley, New York
Friedman WE (1994) The evolution of embryo-geny in seed plants and the developmental origin of endosperm: implications for the evolution of flowering plants. Am J Bot 81: 1468—486
Gasser CS, Broadhvest J, Hauser BA (1998) Genetic analysis of ovule development. Ann Rev Plant Physiol Plant Mol Biol 49: 1—24
Gifford EM, Foster AS (1989) Morphology and Evolution ofVascular Plants, 3rd ed. Freeman, New York Johri BM, Ambegaokar KB, Srivastava PS (1992) Comparative Embryology of Angiosperms. Springer, Berlin Kapil RN, Bhatnagar AK (1991) Embryolo
552 | список ЛИТЕРАТУРЫ
gical evidence in angiosperm classification and phylogeny. Bot Jahrb Syst 113: 357 — 375
Sporne KR (1974) The Morphology of Gymnosperms, 2nd ed. Hutchinson, London
Sporne KR (1974) The Morphology of Angiosperms. Hutchinson, London
Вегетативные органы
См. литературу гл. 3. Ткани побеговых растений, гл. 4. Морфология и анатомия кормофитов раздела 11.1. Методы систематики.
Baas Р, Wheeler Е, Chase М (2000) Dicotyledonous wood anatomy and the APG system of angiosperm classification. Bot J Linn Soc 134: 3—17
Barthlott W, Neinhuis C, Cutler D, Ditsch F, Meusel I, Theisen I, Wilhelmi H (1998) Classification and terminology of plant epicuticular waxes. Bot J Linn Soc 126: 237 — 260
Behnke HD (1991) Distribution and evolution of forms and types of sieve-element plastids in the dicotyledons. Aliso 13: 167—182
Behnke HD (2000) Forms and sizes of sieve element plastids and evolution of the monocotyledons. In: Wilson KL, ed, Mono-cots: Systematics and Evolution. CSIRO, Melbourne
Bierhorst DW (1971) Morphology of Vascular Plants. Macmillan, New York
Carlquist S (2001) Comparative Wood Anatomy, 2nd ed. Springer, Berlin
Eames AJ (1961) Morphology of the Angiosperms. McGraw-Hill, New York
Endress PK, Baas P, Gregory M (2000) Systematic plant morphology and anatomy — 50 years of progress. Taxon 49: 401 —434
Fahn A (1990) Plant Anatomy, 4th ed. Pergamon Press, Oxford
Gifford EM, Foster AS (1989) Morphology and Evolution of Vascular Plants. Freeman, New York
Hickey J (1973) Classification of the architecture of dicotyledonous leaves. Am J Bot 60: 17 — 33
Hudson A (2000) Development of symmetry in plants. Ann Rev Plant Physiol Plant Mol Biol 51: 349-370
Kaplan DR (2001) The science of plant morphology: definition, history, and role in modern biology. Am J Bot 88: 1711 — 1741
Metcalfe CR et al. (1960П) Anatomy of the Monocotyledons. Clarendon Press, Oxford
Metcalfe CR, Chalk L (1950) Anatomy of the Dicotyledons. Clarendon Press, Oxford
Metcalfe CR, Chalk L (1979) Anatomy of the
Dicotyledons, 2nd ed, vol 1. Clarendon Press. Oxford
Metcalfe CR, Chalk L (1983) Anatomy of the
Dicotyledons. 2nd ed, vol 2. Clarendon Press, Oxford
Niklas К (1994) Plant Allometry. The Scaling of Form and Process. University of Chicago Press. Chicago
Schmitz G, Theres К (1999) Genetic control of branching in Arabidopsis and tomato. Curr Opin Plant Biol 2: 51 —55
Sinha, N (1999) Leaf development in angiosperms. Annu Rev Plant Physiol Plant Mol Biol 50: 419—146
Sporne KR (1974) The Morphology of Gymnosperms, 2nd ed. Hutchinson, London
Sporne KR (1974) The Morphology of Angiosperms. Hutchinson, London
Troll W (1937—1943) Vergleichende Morphologic der hoheren Pflanzen. Borntrager. Berlin
Вторичные метаболиты
См. литературу раздела 11.1. Методы систематики.
Frohne D, Jensen U (1998) Systematik des Pflan-zenreichs unter besonderer Berilcksichtigung che-mischer Merkmale und pflanzlicher Drogen, 5. Aufl. Wissenschaftliche Verlagsgesell-schaft, Stuttgart
Gibbs RD (1974) Chemotaxonomy of Flowering Plants. McGill-Queen's University Press, Montreal
Harborne J В (2000) Arsenal for survival: secondary plant products. Taxon 49: 435—449
Harborne JB, Turner BL (1984) Plant Chemb-systematics. Academic Press, Orlando
Hegnauer R (1962 — 2001) Chemotaxonomie der Pflanzen. Birk-hauser, Basel
Цветки, соцветия
См. литературу раздела 11.1. Методы систематики, 11.2. Семенные растения. Смена поколений
Baum DA (1998) The evolution of plant development. Curr Opin Plant Biol 1: 79 — 86
Coen ES, Meyerowitz E (1991) The war of the whorls: genetic interactions controlling flower development. Nature 353: 31 — 37
D'Arcy WG, Keating RC, eds (1996) The Anther: Form, Function and Phylogeny. Cambridge University Press, Cambridge
Eichler A (1875, 1878) Bliithendiagramme. Engelmann Leipzig
Endress PK (1990) Patterns of floral construction in ontogeny and phylogeny. Biol J Linn Soc 39: 153-175
Endress PK (1992) Evolution and floral diversity: the phylo-genetic surroundings of Arabidopsis and Antirrhinum. Int J Plant Sci 153: S106—S122
Endress PK (1994) Diversity and Evolutionary Biology of Tropical Flowers. Cambridge University Press, Cambridge
Endress PK, Friis EM, eds (1994) Early Evolution of Flowers. Plant Syst Evol Suppl 8
Erdtman G (1966) Pollen Morphology and Plant Taxonomy: Angiosperms. Almqvist & Wiksell, Stockholm
Frohlich MW, Meyerowitz E (1997) The search for flower homeotic gene homologs in basal angiosperms and Gnetales: a potential new source of data on the evolutionary origin of flowers. Int J Plant Sci 158: S131-S142
Grey son Rl (1994) The Development of Flowers. Oxford University Press, New York
Список литературы | 553
НеВ D (1990) Die BlOte. Eine Einfuhrung in Struktur und Funktion, Okologie und Evolution der Bliiten, 2. Aufl. Ulmer, Stuttgart
Irish VF, Kramer EM (1998) Genetic and molecular analysis of angiosperm flower development. Adv Bot Res 28: 197 -230
Lawton-Rauh AL, Alvarez-Buylla ER, Purugganan, MD (2000) Molecular evolution of flower development. Trends Ecol Evol 15: 144—149
Leins P (2000) Bliite und Frucht. Aspekte der Morphologic, Entwicklungsgeschichte, Phylogenie, Funktion, Okologie. Schweizcrbart, Stuttgart
Leins P, Erbar C (1991) Entwicklungsmuster in Bliiten und ihre mutmaBlichen phylogenetischen Zusammenhange. B1UZ 21: 196 — 204
Leins P, Erbar C (1997) Floral developmental studies: some old and new questions. Int J Plant Sci 158: 3-12
Philipson WR (1977) Ovular morphology and the classification of dicotyledons. Plant Syst Evol Suppl 1: 123-140
Punt W, Blackmore S, Nilsson S, Le Thomas A (1994) Glossary of Pollen and Spore Terminology. LPP Foundation, Utrecht
Stanley RG, Linskens HF (1974) Pollen Biology, Biochemistry, Management. Springer, Berlin
Theissen G, Saedler II (1999) The golden decade of molecular floral development (1990-1999): a cheerful obituary. Dev Genet 5: 1 — 13
Troll W (1928) Organisation und Gestalt im Bereich der Bliite. Springer, Berlin
Weberling F (1981) Morphologic der Bliiten und Blutenstande. Ulmer, Stuttgart
Опыление и оплодотворение
См. литературу раздела 10.1.3. Рекомбинатив-ная система.
Armbruster WS (1992) Phylogeny and the evolution of plant-animal interactions. BioScience 42: 12-20
Barth FG (1982) Biologie einer Begegnung: Die Partnerschaft der Insekten und Blumen. Deutsche Verlagsanstalt, Stuttgart
Barrett SCH. Harder LD (1996) Ecology and evolution of plant mating. Trends Ecol Evol 11: 73—79
Crepet WL. Friis EM, Nixon КС (1991) Fossil evidence for the evolution of biotic pollination. Phil Trans R Soc Lond Ser В 333: 187— 195
Endress PK (1994) Diversity and Evolutionary Biology of Tropical Flowers. Cambridge University Press, Cambridge
Faegri K.van der Pijl L (1980) The Principles of Pollination Ecology, 3rd ed. Pergamon, Oxford
Friis EM. Chaloner WG. Crane PR, eds (1987) The Origins of Angiosperms and Their Biological Consequences. Cambridge University Press, Cambridge
HeB D (1990) Die Bliite. Eine Einfuhrung in Struktur und Funktion, Okologie und Evolution der Bliiten, 2. Aufl. Ulmer, Stuttgart
Johnson SD, Steiner KE (2000) Generalization versus specialization in plant pollination systems. Trends Ecol Evol 15: 140—143
Kugler H (1970) Bliitenokologie, 2. Aufl. Gustav Fischer, Stuttgart
Leins P (2000) Blbte und Frucht. Aspekte der Morphologic, Entwicklungsgeschichte, Phylogenie, Funktion, Okologie. Schweizcrbart, Stuttgart
Linskens HF, ed (1974). Fertilization in Higher Plants. North Holland, Amsterdam
Lovett Doust J, Lovett Doust L, eds (1988) Plant Reproductive Biology. Oxford University Press, Oxford
Lloyd DG, Barrett SCH, eds (1996) Floral Biology: Studies on Floral Evolution in Animal-pollinated Plants. Chapman & Hall, London
Osche G (1983) Optische Signale in der Coevolution von Pflanzen und Tieren. Ber Dtsch Bot Ges 96: 1-27
Pellmyr О (1992) 1 he phylogeny ofa mutualism: evolution and coadaptation. Biol J Linn Soc 47: 337— 365
Proctor M, Yeo P, Lack A (1996) The Natural History of Pollination. Timber Press, Portland
Real L, ed (1983) Pollination Biology. Academic Press, New York
Richards AJ (1997) Plant Breeding Systems, 2nd ed. Chapman & Hall, London
Wyatt RE, ed (1992) Ecology and Evolution of Plant Reproduction. Chapman & Hall, New York
Yeo PF (1993) Secondary Pollen Presentation. Form, Function, and Evolution. Plant Syst Evol, Suppl 6
Семена и плоды. Распространение семян и плодов
Bonn S, Poschlod Р (1998) Ausbreitungsbiologie der Pflanzen Mitteleuropas. Quelle & Meyer, Wiesbaden
Corner EJH (1976) The Seeds of the Dicotyledons. Cambridge University Press, Cambridge
Dieckmann U, O’Hara B, Weisser W (1999) The evolutionary ecology of dispersal. Trends Ecol Evol 14: 88 — 90
Estrada A, Fleming TH, eds (1986) Frugivores and Seed Dispersal. Junk, Dordrecht
Fenner M, ed (1992) Seeds: The Ecology of Regeneration in Plant Communities. CAB International, Wallingford
Fenner M (1993) Seed Ecology. Chapman & Hall, London
Gadgil M (1971) Dispersal: Population consequences and evolution. Ecology 52: 253 — 261
Harper JL, Lovell PH, Moore KG (1970) The shapes and sizes of seeds. Annu Rev Ecol Syst 1: 327-356
Howe HF, Smallwood J (1982) Ecology of seed dispersal. Annu Rev Ecol Syst 13: 201 — 228
Janzen DH (1971) Seed predation by animals. Annu Rev Ecol Syst 2: 465 — 492
554 I СПИСОК ЛИТЕРАТУРЫ
Leins Р (2000) Blute und Frucht. Aspekte der Morphologic, Entwicklungsgeschichte, Phylogenie, Funktion, Okologie. Schweizerbart. Stuttgart
Levin DA (1990) The seed bank as a source of genetic novelty in plants. Am Nat 135: 563 — 572
Levey DJ, Benkman CW (1999) Fruit-seed disperser interactions: timely insights from a long-term perspective. Trends Ecol Evol 14: 41—43
Murray DR. ed (1986) Seed Dispersal. Academic Press, Sydney
Muller-Schneider P (1983) Verbreitungsbio-logic (Diasporolo-gie) der Bliitenpflanzen, 3. Aufl. Veroff Geobot Inst Eidg Tech Hochsch Stift Rubel Zurich 61: 1 — 226
Nathan R, Muller-Landau HC (2000) Spatial patterns of seed dispersal, their determinants and consequences for recruitment. Trends Ecol Evol 15: 278-285
Richards AJ (1997) Plant Breeding Systems, 2nd ed. Chapman & Hall, London
Ridley HN (1930) The Dispersal of Plants Throughout the World. Reeve, Ashford
Roth 1 (1978) Fruits of Angiosperms. Handbuch der Pflanzen-anatomie X/1
(Jibrich E (1928) Biologie der Friichte und Sa-men — Karpobio-logie. Springer, Berlin
Van der Pijl L (1982) Principles of Dispersal in Higher Plants, 3rd ed. Springer, Berlin
Salisbury EJ (1942) The Reproductive Capacity of Plants. Bell, London
Систематика общая
Beck CB, ed (1988) Origin and Evolution of Gymnosperms. Columbia University Press, New York
Biswas C, Johri BM (1997) The Gymnosperms. Springer, Heidelberg
Brummitt RK (1992) Vascular Plant Families and Genera. Royal Botanic Gardens, Kew
Chamberlain CJ (1935) Gymnosperms, Structure and Evolution. Chicago University Press, Chicago
Chase MW et al. (1993) Phylogenetics of seed plants: An analysis of nucleotide sequences from the plastid gene rbcL. Ann Missouri Bot Gard 80: 528 — 580
Crane PR (1985) Phylogenetic analysis of seed plants and the origin of angiosperms Ann Missouri Bot Gard 72: 716—793
Donoghue MJ (1994) Progress and prospects in reconstructing plant phylogeny. Ann Missouri Bot Gard 81: 405-418
Donoghue MJ, Doyle JA (2000) Seed plant phylogeny: Demise of the anthophyte hypothesis? Curr Biol 10: R106-R109
Doyle JA, Donoghue MJ (1986) Seed plant phylogeny and the origin of angiosperms: an experimental cladistic approach. Bot Rev 52: 321 — 431
Engler A (1900—1968) Das Pflanzenreich. Engelmann, Leipzig
Engler A, Prantl K, Hrsg (1879-1915, 1924— 1995) Die natur-lichen Pflanzenfamilien, 1. und 2. Aufl. Engelmann. Leipzig; Duncker & Humblot, Berlin
Kenrick P, Crane PR (1997) The Origin and Early Diversification of Land Plants. A Cladist ic Study. Smithsonian Institution Press, Washington
Kubitzki K, ed (1990(f) The Families and Genera of Vascular Plants. Springer, Berlin
Mabberley DJ (1997) The Plant-Book: A Portable Dictionary of the Vascular Plants, 2nd ed. Cambridge University Press, Cambridge
Stewart WN, Rothwell GW (1993) Paleobotany and the Evolution of Plants, 2nd ed. Cambridge University Press, Cambridge
Willis JC (1973) A Dictionary of the Flowering Plants and Fems, 8,h ed. Cambridge University Press, Cambridge
http://www.wisc.edu/botit/systematics/ (Plant Systematics Collection)
http://phylogeny.arizona.edu/tree/eukaryotes/ green-plants/ green-plants.html (Tree of Life)
http://herbaria.harvard.edu/treebase/ (Treebase) http://www.bgbm.fu-berlin.de/iopi/gpc (Provisional Global Plant Checklist)
http://www.sp2000.org (Species 2000)
http://www.gbif.org (Global Biodiversity Information Facility)
Саговниковые, древовцдные папоротники
Siehe auch Literaturhinweise unter Systematik allgemein Jones DL (1993) Cycads of the World. Smithsonian Institution Press, Washington, DC
Kubitzki K, ed (1990) The Families and Genera of Vascular Plants, vol 1. Springer, Berlin
Sporne KR (1974) The Morphology of Gymnosperms, 2nd ed. Hutchinson, London
Гинкговые, гинкго. Хвойные деревья
Siehe auch Literaturhinweise unter Systematik allgemein Dallimore W, Jackson AB (1966) A Handbook of Coniferae and Ginkgoaceae, 4,h ed. Arnold, London
Kubitzki K, ed (1990) The Families and Genera of Vascular Plants, vol 1. Springer, Berlin
Sporne KR (1974) The Morphology of Gymnosperms, 2l,ded. Hutchinson, London
Stefanovic S, Jager M, Deutsch J, Broutin J, Ma-sselot M (1998) Phylogenetic relationships of conifers inferred from partial 28S rRN A gene sequences. Am J Bot 85: 688-697
Гнетовые
Siehe auch Literaturhinweise unter Systematik allgemein Friedman WE, ed (1996) Biology and evolution of the Gnetales. Int J Plant Sci 157 Suppl
Kubitzki K, ed (1990) The Families and Genera of Vascular Plants, vol 1. Springer, Berlin
Sporne KR (1974) The Morphology of Gymnosperms, 2nd ed. Hutchinson, London
Список литературы | 555
Цветковые растения
См. также литературу по общей систематике.
Angiosperm Phylogeny Group (1998) An ordinal classification for the families of flowering plants. Ann Missouri Bot Gard 85: 531 — 553
Chase MW et al. (1993) Phylogenetics of seed plants: An analysis of nucleotide sequences from the plastid gene rbcL. Ann Missouri Bot Gard 80: 528 — 580
Crane PR, Friis EM, Pedersen К (1995) The origin and early diversification of angiosperms. Nature 374: 27-33
Cronquist A (1981) An Integrated System of Classification of Flowering Plants. Columbia University Press, New York
Cronquist A (1988) The Evolution and Classification of Flowering Plants, 2nd ed. New York Botanical Garden, New York
Dahlgren RMT (1975) A system of classification of the angiosperms to be used to demonstrate the distribution of characters. Bot Not 128: 119— 147
Dahlgren RMT (1983) General aspects of angiosperm evolution and macrosystematics. Nord J Bot 3: 119-149
Dahlgren RMT. Clifford HT, Yeo PF (1985) The Families of the Monocotyledons; Structure, Evolution and Taxonomy. Springer, Berlin
Engler A (1900—1968) Das Pflanzenreich. Engelmann, Leipzig
Engler A, Prantl K, Hrsg (1879—1915, 1924— 1995) Die natur-lichen Pflanzenfamilien. 1. und 2. Aufl., Engelmann, Leipzig; Duncker & Humblot, Berlin
Friis EM, Chaloner WG, Crane PR, eds (1987) The Origins of Angiosperms and Their Biological Consequences. Cambridge University Press, Cambridge
Huber H (1991) Angiospermen: Leitfaden dutch die Ordnungen und Famiiien der Bedeckt-samer. Gustav Fischer, Stuttgart
Hutchinson J (1973) The Families of Flowering Plants, 3rd ed. Clarendon Press, Oxford
Judd WS, Campbell CS, Kellogg EA, Stevens PF (1999) Plant Systematics: A Phylogenetic Approach. Sinauer, Sunderland
Mathews S, Donoghue MJ (1999) The root of angiosperm phylogeny inferred from duplicate phytochrome genes. Science 286: 947—950
Melchior H, Hrsg (1964) A. Engler's Syllabus der Pflanzenfamilien, Bd. 2, 12. Aufl. Borntrager, Berlin
Nandi O, Chase MW, Endress PK (1998) A combined cladistic analysis of angiosperms using rbcL and non-molecular data sets. Ann Missouri Bot Gard 85: 137-212
Rudall PJ, Cribb P. Cutler DF, Humphries CJ (1995) Monocotyledons: Systematics and Evolution. The Royal Botanic Gardens. Kew
Savolainen V et al. (2000) Phylogenetics of flowering plants based on combined analysis of plastid atpB and rbcL gene sequences. Syst Biol 49: 306 — 362
Savolainen V et al. (2000) Phylogeny of the eudicots: a nearly complete familial analysis based on rbcL gene sequences. Kew Bull 55: 257 — 309
Soltis DE et al. (1997) Angiosperm phylogeny inferred from 18S ribosomal DNA sequences. Ann Missouri Bot Gard 84: 1—49
Soltis DE et al. (2000) Angiosperm phylogeny inferred from a combined data set of 18S rDNA, rbcL and atpB sequences. Bot J Linn Soc 133: 381—61
Soltis PS, Soltis DE, Chase MW (1999) Angiosperm phylogeny inferred from multiple genes as a tool for comparative biology. Nature 402: 402 — 44
Takhtajan A (1980) Outline of the classification of flowering plants (Magnoliophyta). Bot Rev 46: 225-359
Takhtajan A (1997) Diversity and Classification of Flowering Plants. Columbia University Press, New York
Taylor DW, Hickey LJ (1996) Flowering Plant Origin, Evolution and Phylogeny. Chapman & Hall, New York
Thorne RF (1976) An updated phylogenetic classification of flowering plants. Aliso 13: 365 — 389
Thome RF (1992) A phylogenetic classification of flowering plants. Evol Biol 9: 35—106
Thorne RF (2000) The classification and geography of the flowering plants: dicotyledons of the class Angiospermae. Bot Rev 66: 441—647
Wilson KL, ed (2000) Monocots: Systematics and Evolution. CSIRO, Melbourne
Qiu Y-L et al. (1999) The earliest angiosperms: evidence from mitochondrial, plastid and nuclear genomes. Nature 402: 404—407
Zomlefer WB (1994) Guide to Flowering Plant Families. The University of North Carolina Press, Chapel Hill
http://www.csdl.tamu.edu/FLORA/newgate/ cronang.htm (Flowering Plant Gateway)
http://biodiversity.uno.edu/delta/angio/in-dex.htm (The Families of Flowering Plants: Descriptions, Illustrations, Identification, and Information Retreaval)
11.3. Филогенез растений и растительности
Allen К, Briggs D, eds (1990) Evolution and the Fossil Record. Smithsonian Institution Press, Washington, DC
Bennett KD (1997) Evolution and Ecology. The Pace of Life. Cambridge University Press, Cambridge
Birks HJB, Birks HH (1980) Quaternary Palaeoecology. Arnold, London
556 I СПИСОК ЛИТЕРАТУРЫ
Burga СА, Perret R (1998) Vegetation und Klima der Schweiz seit dem jilngeren Eiszeitalter. Ott, Thun
Chaloner WG, Hallam A, eds (1989) Evolution and Extinction. Cambridge University Press, Cambridge
Denton GH, Hughes TJ (1981) The Last Great Ice Sheets. Wiley, New York
Faegri K, Iversen J (1989) Textbook of Pollen Analysis, 4lh ed. Wiley, Chichester
Huntley B, Birks HJB (1983) An Atlas of Past and Present Pollen Maps for Europe: 0— 13 000 years ago. Cambridge University Press, Cambridge
Jablonski D (1986) Background and mass extinctions: the alternation of macroevolutionary regimes. Science 231: 129—133
Kenrick P, Crane PR (1997) The Origin and Early Diversification of Land Plants. A Cladistic Study. Smithsonian Institution Press, Washington
Lang G (1994) Quartare Vegetationsgeschichte Europas. Gustav Fischer, Stuttgart
Lovelock J (2000) The Ages of Gaia: A Biography of Our Living Earth. Oxford University Press, Oxford
Mai DH (1995) Tertiare Vegetationsgeschichte Europas. Gustav Fischer, Stuttgart
Niklas KJ (1997) The Evolutionary Biology of Plants. University of Chicago Press, Chicago
Stewart WN, Rothwell GW (1993) Paleobotany and the Evolution of Plants, 2"d ed. Cambridge University Press, Cambridge
Список сокращений
А/А — аденин — максимальная ассимиляция (СО2)
АВА/АБК — абсцизовая кислота
ADP/АДФ — аденозиндифосфат
АМР/АМФ — аденозинмонофосфат АТР/АТФ — аденозинтрифосфат bp/п.н. — пар нуклеотидов (п.н.) С/Ц — цитозин
САМ — кислотный метаболизм Сга-ssularaceae
сАМР/цАМФ — циклический аденозинмонофосфат
cDNA/кДНК — сору-ДНК сМ — сантиморганы CS — поясок Каспари d — 2'-дезокси(рибо)-
Da/Да — дальтон
DIC — контраст дифференциальной интерференции
DNA/ДНК — дезоксирибонуклеиновая кислота
DNase/ДНаза — дезоксирибонуклеаза (ДНаза) dNTP — дезоксинуклеозидтрифосфат
DR — темно-красный (синоним дальний красный) dsDNA — двуцепочечная ДНК
ЕМ — электронный микроскоп EMS — этилметансульфонат
ER/ЭР — эндоплазматический ретикулум (ЭР)
FAD/ФАД — флавинадениндинуклеотид (окисленный)
ЕАВНг/ФАДНз—флавинадениндинуклеотид (восстановленный)
g — диффузионная проводимость G/Г — гуанин
GA/ГК — гиббереллин А
GFP — протеин, флуоресцирующий зеленым светом
GTP/ГГФ — гуанозинтрифосфат (ГТФ) GUS — р-глюкуронидаза
hnRNA/гяРНК— гетерогенная ядерная РНК HPLC/ВЖХ — высокоэффективная жидкостная хроматография
HR — светло-красный
IAA/ИУК — индол-3-уксусная кислота
kb, kbp/т.п.н.
kDa/кДа
КТР
LAI/ИЛП
LAR
LHC
LTP М м,
mRNA/мРНК mtDNA/мтД Н К
МТОС/ЦОМТ
NAD+/HAД+
NADH/НАДН
NADP'/НАДФ4
NADPH/НАДФН
NMR/ЯМР
NOR
NPC
NPP
NTP
PAGE/ПААГ
PAR/ФАР
PCR/ПЦР
РЕР/ФЕП
PFD
PS/ФС у
килобазы, тысяч нуклеотидных пар килодальтон короткодневное растение индекс листовой поверхности
отношение листовой поверхности к общему весу растения
светособирающий комплекс
длиннодневное растение молярность относительная масса молекулы
матричная РНК митохондриальная ДНК центр организации микротрубочек никотинамидаденинди-нуклеотид (окисленный) никотинамидадениндину-клеотид (восстановленный) никотинамидаденинди-нуклеотидфосфат (окисленный)
н и коти нам идаде н и нди -нуклеотидфосфат (восстановленный)
ядерно-магнитный резонанс
область ядрышкового организатора
комплекс ядерной поры нетто-первичное производство
нуклеозидтрифосфат полиакриламидный гель-электрофорез
фотосинтетически активная радиация
полимеразная пепная реакция
фосфоенолпируват плотность потока фотонов фотосистема
водный потенциал («пси»)
55g | СПИСОК СОКРАЩЕНИЙ
ptDNA/птДНК — пластидная ДНК cpDNA/хлДНК — хлоропластная ДНК R — дыхание REM — растровый электронный микроскоп rER — гранулярный эндоплазматический ретикулум RNA/PHK — рибонуклеиновая кислота RNase/РНаза — рибонуклеаза RQ — дыхательный индекс rRNA/pPHK — рибосомальная РНК RubisCO — рибулозо-1,5-бифосфат-кар-боксилаза/оксигеназа RubP — рибулозо-1,5-бифосфат S — единица Сведберга SAM — апикальная меристема побега sER — гладкий эндоплазматический ретикулум SLA — специфическая листовая площадь snDNA/мяДНК — малая ядерная ДНК SRP — частица распознавания сигнала ssDNA — одноцепочечная ДНК Т/Т — тимин (частично также температура) ТЕМ — трансмиссионный электронный микроскоп TF — фактор транскрипции Тг — транспирация tRNA/тРНК — транспортная РНК U/У — урацил VP — точка роста
Единицы и символы
СИ—SI (Systeme International d’Unites): Основные единицы н символы
Величина Наименование Обозначение международное/ русское Величина Наименование Обозначение международное/ русское
Сила электрического тока (1) ампер А/А Количество вещества (N) МОЛЬ mol/моль
Длина (L) метр т/м Температура (Т) кельвин К/К
Сила света (J) кандела cd/кд Время (t) секунда s/c
Масса (М) килограмм kg/кг
Важные производные единицы СИ
Величина Наименование Обозначение международное/ русское Эквивалент в единицах СИ
Давление паскаль Ра/Па 1 Ра = 1 N • m 2 = 1 kg пг1 s-2 1 Па = 1 Н/м2 = 1 кг/(мс2)
Электрический заряд кулон С/Кл 1 С = 1 A-s= 1 J V"1 1 Кл = 1 А с = 1 Дж/В
Электрическое напряжение вольт V/B 1 V= 1 JA's1 = 1 WA 1 1 В = 1 Дж/А • с = 1 Вт/А
Электрическое сопротивление ом О/Ом 1 О = 1 ш2 kg s3 A 2 1 Ом = 1 м2 кг/(с3 • А2)
Энергия, работа, количество теплоты джоуль электрон-вольт J/Дж eV/эВ 1 J = 1 W s = 1 kg m2 • s“2 1 Дж = 1 Вт с = 1 кг м2/с2 1 eV = 1,602 • 10"19 J 1 эВ = 1,602 • 10” Дж
Поглощенная доза излучения (ионизирующее излучение) грэй Gy/Гр 1 Gy = 1 J • kg 1 = 1 m2- s"2 1 Гр = 1 Дж/кг = 1 м2/с2
Частота герц Hz/Гц 1 Hz = 1 s"1 1 Гц = 1/с
Каталитическая активность (энзимы) катал kat/кат 1 kat = 1 mol • sr1 1 кат= 1 моль/с
Сила ньютон N/H 1 N = 1 kg • m s 2 1 H = 1 кг • м/с2
Мощность, поток излучения ватт W/Bt 1 W = 1 kg m2 s’3 1 В = 1 кг • м2/с3
Освещенность люкс 1х/лк 1 lx= 1 Im m 2 1 лк = 1 лм/м2
Световой поток люмен 1т/лм 1 Im = 1 cd • sr1 (sr = стерадиан) 1 лм = 1 кд/ср
Радиоактивность беккерель Bq/Бк 1 Bq = 1 s' 1 Бк = 1 /с
560 I ЕДИНИЦЫ и символы
Единицы фотохимически действующего излучения
Величина Эквивалент в единицах СИ Величина Эквивалент в единицах СИ
Лучистая экспозиция J - пг2 Дж/м2 Световая энергия im s лм - с
Плотность потока излучения J • m 2-s 1 = W пг2 Дж/м2 • с = Вт/м2 Плотность потока квантов (фотонов) mol • m2 моль/м2
Количество энергии J Дж Поток квантов (фотонов) mol m 2 s'1 моль/(м2 • с)
Поток энергии Js~' Дж/с Скорость тока квантов (фотонов) mol s_| моль/с
Плотность световой энергии 1m m 2- s лм с/м2
Единицы процессов транспорта, роста и движения
Величина Эквивалент в единицах СИ
Транспортный поток Интенсивность (J) Поток (J) Рост, движение Скорость (v) mol • пг2 s_| или т3 т 2 - s 1 = т s-1 моль/(м2-с) или м3/(м2с) = м/с mol s1 или т3- s~', моль/с или м3/с т s-1 м/с
Часто используемые единицы в физиологии (не содержащиеся в СИ)
Величина Наименование Обозначение международное/ русское Эквивалент в единицах СИ
Давление (р) бар Ьаг/бар 1 bar = 105 Ра = 105 N пг2 1 бар= 105 Па = 105 Н/м2
Масса (т) грамм тонна g/r t/т н Г» -1 к II II II II О О о о
Молярная масса грамм/моль g/mol г/моль 1 g • mol 1 = 10 3 kg mop1 1 г/моль = 10-3 кг/моль
Константа седиментации сведберг S/S 1 S= 10-|3s 1 S= 10“l3c
Концентрация количества вещества
Молярность Моляльность Осмоляльность моль/литр моль/кг моль осмотически действующих частиц на килограмм раствора (воды) М/М mol l 1 = mol • dm 3 моль/л = моль/дм3 mol • kg 1 моль/кг osmol - kg-1 моль/кг
Единицы и символы
561
Окончание
Величина Наименование Обозначение международное/ русское Эквивалент в единицах СИ
(Молекулярная) масса частиц дальтон Da/Да 1 Da =1,6605 10"27kg 1 Да =1,6605 10“27 кг
Температура (Т) градус Цельсия °С/°С 0”С = 273,15 К 0 "С = 273,15 К
Объем (V) литр 1/л 1 1 = 10 3 m3 = 1 dm3 1 л = 10-3 м3 = 1 дм3
Время (t) минута час сутки год min/мин h/ч d/сут а/год 1 min = 60 s 1 мин = 60 с 1 h = 3 600 s 1 ч = 3 600 с 1 d= 86 400 s 1 сут = 86 400 с
Коэффициенты пересчета для больше не разрешенных единиц
Величина Наименование Обозначение международное/ русское Пересчет в единицы СИ
Давление атмосфера торр atm/атм torr/торр 1 atm (760 mm Hg) = 101 325 Ра = 1,013 bar 1 атм (760 мм рт. ст.) = 101 325 Па= 1,013 бар 1 torr = 101 325 Ра: 760 = 133,3 Ра 1 торр = 101 325 Па:760 = 133,3 Па
Энергия эрг калория erg/эрг cal/кал 1 erg = 10 7J 1 эрг = 10 7 Дж 1 cal = 4,1868 J 1 кал = 4,1868 Дж
Доза энергии рад rd/рад 1 rd = 0,01 J kg 1 рад = 0,01 Дж/кг
Доза ионов рентген г/Р 1 г = 2,58 10-4 С kg 1 1 Р = 2,58 10‘4 Кл/кг
Длина ангстрем А 1 А = 10 10 m = 10 1 nm 1А= 10-|0м= 10*' нм
Количество квантов (фотонов) эйнштейн Е 1 Е = 1 mol квантов (фотонов) 1 Е = 1 моль квантов (фотонов)
Радиоактивность кюри Ci/Ки 1 Ci = 3,77 - 10"’ Bq 1 Ки = 3,77 1010 Бк
Кратные доли единиц (приставки)*
пета- (Р/П) 1015 гекто- (h/r) 102 микро- (ц/мк) ю-6
тера- (Т/Т) 1012 дека-(da/да) 10' нано-(п/н) ю-9
гига- (G/T) 109 деци- (d/д) 10-' пико- (р/п) 10 12
мега- (М/М) 106 санти- (с/с) 10~2 фемто- (f/ф) 10-'5
кило-(к/к) 103 милли- (т/м) 103 атто- (а/а) 10 18
Для предотвращения ошибок при расчетах рекомендуется использование степеней
Предметный указатель
Автогамия 148, 169, 378—379
Автохория 398
Агамоспермия 56—57, 90, 392
Акинета 429-431
Акразиомииеты 137, 141
Актиномицеты 121
Актиностела 310—312, 328, 354
Аллогамия 54, 378
Амебозигота 138
Амфигастрия 283, 287—288
Амфитеций 290, 294, 296
Анаморфа 145, 167, 203
Андрогинофор 377, 460
Андроцей 361, 377—379, 433
Анемофилия 58, 380—381
Анизогаметангиогамия 152, 158
Анизогамия 135, 249, 255
Анизофиллия 315—316 Антеридий 280—281, 290, 300
Антеридиофор 283 — 284
Антипода 374, 377
Литофиты 511
Антропохория 397
Апертура 364 — 366
Апланоспора 144, 245, 260
Апогамия 57
Апоспория 57
Аппрессорий 205
Ариллус 389, 411,461
Артроспора 161
Артростела 324, 327
Архегониальная камера 376, 386
Архегониаты 278, 290
Архегоний 279, 281, 284
Архегониофор 284, 339
Археи 114, 132-134
— галофильные 134
— метаногенные 134
Археофит 539
Археспорий 285, 294, 362
Аскогон 144, 164—167
Аскомицеты 160— 176, 198, 208
Аскоспора 160, 162, 166
Атактостела 305, 354,427
Ауксилярная клетка 218
Ауксозигота 232 — 234
Бактериофаг 120
Бактерии 108 — 513
— грамотрицательные 122—127
— грамположительные 121—122
— коринеформные 121
— термофильные 125— 126
— фитопатогенные 126
Банк семян 298
Барохория 398
Бацилла 114
Бентос 216, 236, 273
Боб 394,464
Борозда 364
Брактея 359, 379, 394
Бутстреп-анализ 104
Вайя 329
Венчик 359, 378
Вибрион 114
Вид 95
Вирус 120
Висцидий 433
Влагалище 134, 313, 324
Водоросли 127— 132, 210—277
— бурые 235 — 245
— диатомовые 231 —235
— желтозеленые 225—229
— золотистые 229 — 230
— красные 210—218
— синезеленые 127—132
— стрептофитовые 277
— харовые 271—277
Вольва 194
Выводковая корзиночка 283
Выводковое тельце 283
Гамета 38
Гаметангий 143, 148, 151
Гаметангиогамия 135, 143, 146
Гаметангиофор 281, 286, 311
Гамето-гаметангиогамия 164, 214
Гаметогамия 135
Гаметофит 136, 151, 183
Гаметофор 280, 287, 289
Гаплодикарионт 151
Гаплодиплонт 136
Предметный указатель | 563
Гаплонт 135, 151, 153
Гаплофаза 135, 163, 260
Гаптера 261, 326
Гаптонема 222, 223
Гаустория 147, 159, 180
Гемикриптофит 347
Геофит 347, 354
Гербарий 95, 107
Геркогамия 54, 380
Гетерантерия 361
Гетерогамия 135
Гетерокарпия 396
Гетероморфия 52, 380
Гетероспермия 396
Гетероспория 312, 315, 316
Гетерофиллия 343
Гетероциста 127, 130, 131
Гидатода 126, 327
Гидроид 292, 295, 302
Гидрофилия 380
Гидрофит 303, 346
Гименофор 188—196
— пластинчатый 195, 196
— трубчатый 194, 195
Гинецей 370—377
— апокарпный 372 — 373, 420
— гемисинкарпный 372
— паракарпный 372, 423,455
— синкарпный 372, 434
— ценокарпный 372,430,485
Гиностемий 433 — 435
Гинофор 377, 480
Гипантий 377, 393,425
Гипнозигота 152, 157—158, 225
Гипноспора 141
Гипокотиль 398
Гипонейстон 142, 274
Гипоталлус 140
Гипотеза псевдантовая 511
— синангиальная 508
— теломная 508
— эвантовая 511, 513
Гипотека 231, 232, 234
Гипофиза 391
Гифы аскогенные 164—172
— септированные 160
Глаукофит 210
Глеба 193
Гломеромицеты 160
Глохидий 344
Гомоплазия 103
Гоморизия 308, 426
Гонотрофия 136, 280
Гормогония 129, 131
Грибы гетероталличные 145, 149
— гомоталличные 145
— настоящие 150, 199
— несовершенные 160, 163, 203
— хитиновые 143, 150—203
— целлюлозные 142—150
Двудомность 283, 359, 370
Диаспора 58, 113, 396
Дикарион 136, 160
Дикариофаза 151, 164. 175
Диктиостела 335
Дипланетизм 143
Диплонт 136, 151, 241
Диплофаза 136, 163, 514
Дихогамия 54, 380
Домен 111
Дрожжи 161 — 163
Жгутик плазматический 117
Жгутиковый аппарат 248, 250, 252
---акроконтный 146
---гетероконтный 142, 146
---опистоконтный 146
Завязь 370—379
— верхняя 377 393
— нижняя 378
— перигинная 378
— полунижняя 378, 464
Зародыш 56, 290, 350
Зародышевый мешок 57, 350, 516
— биспорический 375
— моноспорический 375
— тетраспорический 375, 377
Заросток 305, 318
Зерновка 395
Зигомицеты 156—160
Зигоспора 156, 158
Зигота 73, 135, 143
Зона гинецея 372
— гемисимпликатная 372
— симпликатная 372
— синасцидиатная 372
Зона завязи 370
— асцидиатная 370
— кондупликатная 370
— пликатная 370
Зооидиогамия 386
Зооспора 141—144
Зооспорангий 142, 157
Зоофилия 381, 417
Изидия 206
Изогамета 153, 221
Изогаметангиогамия 158
Изогамия 135, 151, 229
Изоспория 310—314
Индузий 313, 337
Интегумент 352, 373, 387
Интина 364
564 [ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ
Камбий 309. 354
Кантарофилия 385
Капиллиций 138, 197
Кариогамия 135, 152, 157
Карпогон 175,211,215
Карпоспорофит 213—215
Карпофор 395, 502
Карункул 389
Каулоид 212, 240, 278
Каулонема 289
Кладистика 101, 104
Классификация 104—107
Клептохлоропласт 220
Клетка генеративная 260, 367
— прогаллиальная 318, 367
— сифоногенная 367
— сперматогенная 280, 311, 367
Клетка-ножка 164, 283, 367
Клеточная стенка 114, 117
Ключ определительный 95 — 96
— дихотомический 96
— полигамический 96
Кокк 114, 121, 123-125
Кокколит 223, 247
Колеоптиль 392, 442
Колеориза 392, 442
Колечко 291, 295
Колонка 137, 138, 141
Колпачок 262, 291, 295
Кольцо 342
Компитум 373, 387
Конидио-гаметапгиогамия 144
Конидия 148, 150, 157
Концептакул 242
Конъюгация 40, 118, 267
Кора 292, 294,313
Корень 103, 235, 308
Кормофиты 278, 350
Коробочка 107, 280—286. 393
Костянка 393, 395
Криопланктон 274
Криптофиты 218 — 219
Крышечка 153, 157, 168
Ксилема 97, 309, 312
Кутикула 161, 279, 283
Лабульбениомицеты 163—164
Ламелла опоясывающая 219, 224, 229
Лепесток 31,96, 359
Лептоид 292, 304
Лигнин 197, 271,280
Лигула 442
Листовка 394,424,429
Лишайники 114, 132, 136
— гетеромерные 205
— гомеомерные 205
— корковые 204
— кустистые 204
— листоватые 204—206
— накипные 204
— слизистые 204
— эндолитные 205
— эндофлеоидные 205
Лодикула 442
Луб 553
Мазедий 208
Марсупий 287
Массула 344
Мегаспора 98, 310. 316
Мегаспорангий 310, 316, 320
Мегаспорофилл 320, 353, 358
Мегафилл 307, 329, 331
Междоузлие 271, 322, 325
Мезокарпий 392, 395,439
Мезосома 117
Мезофит 524, 526
Мейоз 115, 135, 137
Мейоспора 136, 151, 156
Мейоспороциста 151, 153, 160
Мелиттофилия 385
Мерантий 379, 385,433
Мерикарпий 393, 395, 398
Меристема 179, 239, 280
— апикальная 179. 313. 354
— интеркалярная 300
Меристодерма 241
Метод максимального подобия 103
Микобионт203, 206, 208
Микоз 167, 201
Микоплазма 122
Микориза 158, 196, 286
Микропиле 373, 374, 377
Микроспора 310, 316, 318
Микроспорангий 316, 320, 333
Микроспорофилл 353, 360 — 367
Микрофилл 307, 312, 318
Микрофиллия 312
Миксамеба 137 139
Миксобактерии 115, 118, 121
Миксотрофность 257, 274
Миксофлагслляты 137, 139
Миофилия 385
Митозооспора 151, 249
Митоспора 136, 157, 2)4
Митоспороциста 151, 153
Митохондрия 45,46, 117
Мицелий 114. 121, 122
— почкующийся 144, 161
— ценоцитный 156
Монада 367
Монофилия 102, 104,416
Мхи 277-304
— акрокарпные 294, 298
— антоцеротовые 288, 300—302
— гаплолепидные 298
Предметный указатель | 565
— диплолепидные 297, 298
— листостебельные 279, 280. 286—288
— плеврокарпные 294 298 304
— талломные 279, 286. 300
Нанопланктон 273
Нейстон 142, 202, 274
Нектарник 361, 377 — 379
— септальный 377, 433
— экстрафлоральный 377
Неофит 326, 539
Нуклеоид 114, 115
Нуклеоморфа 211,219, 220
Нуцеллус 350, 373 — 374. 376
Обвертка 130, 158, 285
Обдиплостемония 378
Обтуратор 374, 387,463
Однодомность 151, 175, 242
Оидий 144, 203
Окаменение 518
Околоплодник 393 — 395
Околоцветник 359 — 379
— двойной 359
— монохламидный 359
— простой 359
Оогамия 135, 151, 153
Оогоний 143, 154, 225
Оомицеты 142—150
Ооспора 143, 146, 148
Оперкулум 366
Орех 393, 395
Орнитофилия 385
Ость 442
Отдел 111
Палеофит 521
Палочка 114, 121, 122
Папоротники 305 — 350
— лептоспорангиатные 335 — 341
— эвспорангиатные 331, 333 — 335
Паппус 396, 506
Параллелизм 93, 99, 101
Парасимбионт 206
Парафилия 104
Парентосома 176, 193
Парсимония 102,415
Перевершинивание 305, 310, 312
Периантий 285, 287
Перидий 138, 166, 191
Перидиоль 197
Перисперм 390,423,452
Периспорий 188, 309, .326
Перистом 296—297, 304
Перитеций 164, 166, 171
Перихеций 285
Пестик 32, 372
Печеночники 132, 280. 281 — 286
Пиренарий 393, 395, 396
Пиреноид 217, 219. 224
Плазмалемма 116, 220
Плазмида 115
Плазмогамия 155, 164, 168
Плазмодий 137—40, 142
— агрегационный 137, 139
— настоящий 138, 139
Плазмодиофоромицеты 140—142
Планация 305
Планктон 273
Планозигота 138, 141, 257
Пластиды 209, 210, 218
— комплексные 211,218, 220
— простые 211, 220
Плацента 337, 370, 372
Плацентация 372, 421
— ламинальная 372, 421, 429
— париетальная 372, 436, 449
— субмаргинальная 370
— центральная 372
— центрально-угловая 372
Плектенхима 175, 190, 210
Плектостела 312
Плод апокарпный 393, 395
— вентрицидный 393
— вскрывающийся 393, 394
— дробный 393, 395, 397
— локулицидный 393, 394
— мономерный 393
— невскрывающийся 393, 395
— порицидный 393
— распадающийся 393, 395
— септицидный 393, 395
— сочный 393, 417
— сухой 39.3, 394, 395
— ценокарпный 393
— членистый 393
Плодовое тело 121, 124, 140
Плодолистик 353, 359, 369
Подвесок 314, 326
Подставка 281, 283, 284
Подцарство 111
Подчашие 503
Покрывальце 337, 340, 341
Полиада 367, 465
Полиандрия 361
Полигамия 50, 359
Полистела 335
Полиэмбриония 391, 392
— монозиготическая 391, 392
— полизиготическая 391, 392 Поллинарий 367, 433,435
Поллиний 367, 384,433
Полость валлекулярная 324
— каринальная 324
Пора 144, 160, 172
566 | ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ
Почечка 391,427
Признаки 101—107
— апоморфные 101, 102, 104
— плезиоморфные 101, 102, 107
Прилистник 271, 334, 354
Примэкзина 364
Проводящий пучок 97, 278, 324
----биколлатеральный 97, 354, 471
----коллатеральный 97, 324, 354
Прокариоты 114, 118, 125
Прокарпий 215, 218
Прорастание -
— гипогейное 167, 398
— эпигейное 209, 398
Проростковая трубка 143, 146, 149
Протсрофит 521
Протонема 280, 287, 289
Протостела 307, 316, 331
Протостелиом инеты 140, 141
Протоцит 111, 114
Прохлоробактерии 114, 246
Прохлорофит 132
Псевдантий 360, 379, 385
Псевдомонада 367, 441
Псевдопаренхима 130, 173,210
Псевдоплазмодий 124, 137
Псилофиты 310—312
Психофилия 385
Пыльник 361, 363
— базификсный 361
— вентрификсный 361
Пыльцевая камера 374, 386, 401
Пыльцевая трубка 52, 146, 353
Пыльцевое зерно 364, 366
---- атектатное 364
----бороздно-поровое 366
----глобальноапертурное 366
----стефанокольпатное 366
----стефанопоратное 366
---- тектатное 364
Развитие 189, 194—195
— гемиангиокарпное 189, 191, 195
— гимнокарпное 189, 194, 195
— псевдоангиокарппое 189, 195
Размножение 135—137
— бесполое 135, 136, 143
— вегетативное 135, 163, 203
— половое 135, 136, 137
Растение вечнозеленое 303, 312, 326
— голосеменное 114, 312, 320
— гомойогидридное 346
— двудольное 107, 297, 347
— летнезеленое 346, 403, 456
— однодольное 107, 354, 355
— пойкилогидридное 346
— покрытосеменное 97, 102, 114
— сосудистое 126, 136, 141
— хвойное 174, 184, 206
Реверсия 103
Редукция 92, 243, 307
— зиготическая 135, 157, 221
Ресупинация 433
Рецептакул 191, 193, 194
Рибосома 98, 116, 128
Ризина 204
Ризоид 157, 228, 235
Ризомицелий 144, 152
Ризомоид 310
Ризоморфа 144, 203
Ризопласт 246
Ризофор 315, 318
Риккетсии 124. 126
Ростеллум 433
Рыльце 370, 372, 376
Саговники 386, 399 — 402
Самонесовместимость 50, 52, 380
Самоопыление 40,46, 379
Сапробность 275
Саркотеста 387,401,403
Сарцина 114, 132
Связник 361,363
Семенная кожура 353, 387 — 388, 392
Семя 179, 320, 328
Семядоля 307, 391, 398
Семязачаток 352, 369, 373
— анатропный 374, 387
— атропный 374
— битегмальный 373
— кампилотропный 374, 387
— крассинуцеллятный 374, 375
— ортотропный 374
— тенуинуцеллятный 373
— унитегмальный 373
Семянка 395, 396, 506
Семяножка 373, 374
Септа 161, 178, 231
Сердцевина 241, 324, 399
Симбиоз 126, 132, 140
Симбионт 126, 250, 304
Сннангий 328, 334,415
Синандрия 361,424
Сингамия 40, 135, 514
Синергида 377, 387
Систематика 95—108
— нумерическая 100—101
— филогенетическая 101 — 103
Ситовидные • клетки 309, 354
— пластинки 313
— трубки 211,241, 354
Сифоногамия 386, 403,413
Сифоностела 316, 324, 328
Склеротеста 388, 396,401 Склероций 144, 172-173,203
Слоевище 204, 206, 208
Предметный указатель I 567
Смена поколений 136, 151, 183
----анизоморфная 136, 277
----изоморфная 136, 151, 236
----трехчленная 136
Соматогамия 151, 164, 169
Соплодие 393, 395
Сораль 206
Соредия 206
Сорус 238, 329, 334
Сосуд 127, 167, 176
Сперматангий 144, 164, 214
Сперматозоид 154, 210, 225
Спермации 164, 172, 180
Спермий 353, 367, 386
Спермогоний 225, 233, 239
Спирилла 114, 123, 132
Спирохеты 123
Спорангий 98, 137, 237
Спорангиофор 322, 324, 325
Спородерма 364
Спорокарпий 345
Спорополленин 245, 254, 260
Спорофилл 309, 312, 313
Спорофит 135, 151, 156
Спороциста 143, 147, 151
Спороцистоспора 157
Стаминодий 361, 377, 425
Стебель 279, 286, 305
Стереида 292
Стилодий 372, 395,425
Столбик 364, 370, 373
Стробил 309, 312, 313
Строматолит 131, 521
Строфиоль 389
Стручок 301, 395,480
Субглеба 197
Сумка 160, 161, 165
Суспензор 157, 338, 390
Сфингофилия 385
Схизокарпий 492, 502
Сэкзина 364
Таллом 129, 142, 150
— гетеротрихальный 254, 261, 270
— сифональный 143, 151, 515
Тапетум 98, 309, 317
— плазмодиальный 309, 322, 325
— секреторный 309, 317, 320
Тека232,361, 384
Тектум 364
Телеоморфа 145, 203
Телом 305. 309. 310
Теория псевдантовая 511
Терофит 347
Теста 356, 387
Тетрада 121, 280, 301
Тетраспорофит 214, 217
Тилакоид 126. 210, 218
Трахеиды 91, 304, 309
— лестничные 318, 331
— сетчатые 324
— точечные 319, 331
Трихогина 164, 172,213
Трихомицеты 159
Трофофилл 309, 314. 322
Тычинка эпипетальная 378, 468
— эписепальная 378, 486
Тычиночная нить 361, 380, 383
Углефикация 518
Урночка 296, 297
Устьице 126, 182, 270
Фаленофилия 385
Фанерофит 346
Фенограмма 100, 104
Фикобилисома 128, 210, 219
Фикопласт 246, 249, 255
Филлоид 212, 235, 240
Филогения 104—107
Фимбрия половая 118
Флора 95—96, 354, 397
Флоэма 97, 163, 309
Фотобионт 203, 206—209
Фрагмобазидия 177, 179, 192
Фрагмопласт 254, 264, 267
Халаза 373
Хамефит 347
Хвощи 305. 322— 328, 521
Хиастобазидия 195
Хироптерофилия 386
Хитридиомицеты 151 — 156
Хламидобактерии 123
Хлоронема 289
Хлоропласт 127, 134, 210
Холокарпий 150, 152
Хохолок 304
Хроматоплазма 128
Хромосома 220, 266, 283
Царство 111 — 114
Цветок актиноморфный 42, 397, 505
— гаплостемонный 361
— гипогинный 379
— двугубый 384, 385,433
— диплостемонный 361
— дисимметричный 379. 449
— зигоморфный 379, 384, 433
— клейстогамный 28, 380
— ловушка 384, 385, 423
— масличный 382
— мотыльковый 384, 385, 465
— нектарный 382
— обманка 383, 385
568 I ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ
— пыльцевой 382
— хасмогамный 380
— чашевидный 384, 385
— шпорцевый 384
— эпигинный 378
Цветоложе 377, 393,429
Цептроплазма 428
Цефалодий 206, 209
Цианелла 210, 219, 227
Цианобактерии 114, 127—132
Циатий 379,463
Циста 137, 152,221
Цистила 195
Цистозигота 225, 248, 249
Цистокарпий 215, 218
Цитоплазма 115, 134, 151
Цифелла 207
Чашелистик 488
Чашечка 393, 387, 394
Чешуи колосковые 79,442,444
— цветковые 442, 444
Щиток 276, 322,442
Эвстела 324, 331,354
Эдафон 276
Эджектосома 219
Экзина 364, 381, 384
Экзокарпий 393, 439
Экзоперидий 193, 196
Экзоспора 122, 129
Экзоспорий 309, 326, 343
Экзотеций 363, 400,403
Эктэкзина 364, 403
Элайосома 389, 397,461
Элатера 139, 281, 285
Эмбриотека 281,291, 294
Эмбриофит 278, 350
Эндодерма 313, 316, 324
Эндозоохория 396, 398
Эндокарпий 393,439
Эндомицетиды 161 — 164
Эндоперидий 193, 196
Эндосперм 353, 387, 390
— гелобиальный 390, 427,429
— нуклеарный 390, 420 427
— руминированный 390
— целлюлярный 390, 391,392
Эндоспора 118, 121, 129
Эндотеций 290, 294, 363
Эндоцитобиоз 142, 208, 219
Эпизоохория 397, 398
Эпинейстон 274
Эпитека 231
Эубактерии 114, 121, 126
Эукариоты 117, 118, 122
Эукарпий 150, 152, 155
Эуцит 111, 134
Яблоко 395
Ягода 96, 149,436
Ядро 163,215,234
Яйцеклетка 255, 280
Координаты для контактов
Профессор, доктор, почетный доктор Петер Зитте
Lerchengarten 1
79249 Merzhausen
peter.sitte@biologie.uni-freiburg.de
Профессор, доктор Эльмар В. Вайлер
Lehrstuhl fur Pflanzenphysiologie I кафедра физиологии растений
Fakultat fur Biologie I факультет биологии
Ruhr-Universitat / Университет в Руре
UniversitatsstraBe 150
44780 Bochum
elmar.weiler@ruhr-uni-bochum.de
Профессор, доктор Андреас Брсзински
Am Katzenbichel 22
93161 Sinzing (Viehhausen)
abresinsky@t-online.de
Профессор, доктор Йоахим В. Кадерайт
Institut fur Spezielle Botanik und Botanischer Garten / Институт специальной ботаники и ботанический сад
Johannes Gutenberg-Universitat Mainz / Университет Иоганна Гуттенберга в Майнце 55099 Mainz
kadereit@mail .uni-mainz.de
Профессор, доктор Кристиан Кёрнер
Institut fur Botanik der Universitat Basel / Институт ботаники Университета Базеля
SchonbeinstraBe 6
CH-4056 Basel
SCHWEIZ
ch.koerner@unibas.ch
Оглавление
Предисловие к 35-му немецкому изданию........................................7
Предисловие к 1-му немецкому изданию.........................................9
Авторы учебника «Ботаника»..................................................11
Издания на иностранных языках...............................................11
Хронология..................................................................12
Предисловие к русскому изданию..............................................16
Предисловие научного редактора..............................................18
Введение....................................................................21
10. ЭВОЛЮЦИЯ...............................................................23
10.1. Изменчивость..........................................................25
10.1.1. Онтогенетическая изменчивость и фенотипическая пластичность......27
10.1.2. Генетическая изменчивость........................................28
10.1.2.1. Генные мутации............................................29
10.1.2.2. Хромосомные мутации.......................................33
10.1.2.3. Геномные мутации..........................................37
10.1.2.4. Рекомбинация..............................................40
10.1.2.5. Внеядерное наследование...................................45
10.1.3. Рекомбинативная система..........................................46
10.1.3.1. Система оплодотворения....................................47
10.1.3.2. Опыление..................................................54
10.1.3.3. Система размножения.......................................55
10.1.3.4. Поток генов и жизненная форма...............................................................58
10.2. Типы и причины естественной изменчивости..............................60
10.2.1. Естественный отбор...............................................60
10.2.2. Дрейф генов......................................................67
10.3. Видообразование.......................................................69
10.3.1. Определения вида.................................................69
10.3.2. Дивергентное видообразование.....................................71
10.3.2.1. Аллопатрическое видообразование ..........................71
10.3.2.2. Репродуктивная изоляция...................................73
10.3.2.3. Перипатрическое, парапатрическое, симпатрическое видообразование и эффект основателя..................................74
10.3.2.4. Генетика видовых различий.................................77
10.3.3. Гибридизация и гибридное видообразование.........................80
10.3.3.1. Гибридизация..............................................80
10.3.3.2. Гомоплоидное гибридное видообразование....................82
10.3.3.3. Интрогрессивная гибридизация..............................83
10.3.3.4. Аллополиплоидия...........................................84
10.4. Макроэволюция.........................................................91
ОГЛАВЛЕНИЕ | 571
11. СИСТЕМА И ФИЛОГЕНИЯ .......................................................94
11.1. Методы систематики........................................................95
11.1.1. Распознавание видов.....................................................95
11.1.2. Монографии, флоры и определительные ключи...............................95
11.1.3. Исследование родства....................................................96
11.1.3.1 . Признаки.............................................................97
11.1.3.2 . «Конфликты» признаков................................................99
11.1.3.3 . Нумерическая систематика............................................100
11.1.3.4 . Филогенетическая систематика........................................101
11.1.3.5 . Метод максимального подобия.........................................103
11.1.3.6 . Статистический анализ гипотез о родстве.............................104
11.1.4. Филогения и классификация..............................................104
11.1.5. Номенклатура...........................................................107
11.2. Бактерии, грибы, растения................................................108
I. Царство (домен): Bacteria...................................................114
Первый отдел: грамположительные бактерии (Posibacteriota)......................121
Второй отдел: грамотрицательные бактерии (Negibacteriota)......................122
Третий отдел: синезеленые водоросли, или цианобактерии (Cyanobacteriota, Cyanoprokaryota, Cyanophyta)...................................................127
Четвертый отдел: прохлорофиты (Prochlorobacteriota, Prochlorophyta)............132
II. Царство (домен): археи (Archaea)...........................................132
Первый отдел: Crenarchaeota....................................................133
Второй отдел: Euryarchaeota....................................................133
III. Царство (домен): эукариоты (Eucarya, Eukaryoten).........................134
Размножение, смена ядерных фаз и поколений.....................................135
Первое подцарство: Acrasiobionta...............................................137
Единственный отдел: Acrasiomycota..............................................137
Второе подцарство: Myxobionta..................................................137
Первый отдел: Myxomycota.......................................................138
Второй отдел: плазмодиофоромицеты (Plasmodiophoromycota).......................140
Третье подцарство: Heterokontobionta...........................................142
Первый отдел: лабиринтуловые (Labyrinthulomycota)..............................142
Второй отдел: Оомицеты, или целлюлозные грибы (Oomycota).......................142
Третий отдел: Heterokontophyta.................................................150
Четвертое подцарство: хитиновые грибы (Mycobionta) ............................150
Единственный отдел: Eumycota...................................................150
1. Класс: хитридиомицеты (Chytridiomycetes).............................151
2. Класс: зигомицеты (Zygomycetes)......................................156
3. Класс: сумчатые грибы, или аскомицеты (Ascomycetes)..................160
1. Подкласс: тафриномицетиды (Taphrinomycetidae)......................161
2. Подкласс: эндомицетиды (Endomycetidae).............................161
3. Подкласс: лабульбениомицетиды (Laboulbeniomycetidae)................163
4. Подкласс: аскомипетиды (Ascomycetidae).............................164
4. Класс: базидиомицеты (Basidiomycetes)................................176
1. Подкласс: устомицетиды (Ustomycetidae).............................177
2. Подкласс: уредомицетиды (Uredomycetidae)...........................180
3. Подкласс: Hymenomycetidae..........................................185
1. Приложение к Mycobionta: несовершенные грибы (Deuteromycetes)...............203
2. Приложение к Mycobionta: лишайники (Lichenes).........................203
572 I ОГЛАВЛЕНИЕ
Пятое подцарство: Glaucobionta..................................................209
Шестое подцарство: Rhodobionta..................................................210
Единственный отдел: красные водоросли (Rhodophyta)..............................210
1. Подкласс: Bangiophycidae ... ...........................................217
2. Подкласс: Florideophycidae..............................................217
1. Приложение к Rhodobionta: отдел криптофиты (Cryptophyta)...................218
2. Приложение к Rhodobionta: отдел динофиты (Dinophyta, Pyrrhophyta, Dinoflagellata).................................................................219
3. Приложение к Rhodobionta: отдел Haptophyta..................................222
4. Приложение к Rhodobionta: отдел Heterokontophyta (= Chrysophyta, Chromophyta)....................................................................223
1. Класс: Chloromonadophyceae...............................................224
2. Класс: желтозеленые водоросли (Xanthophyceae) ...........................225
3. Класс: золотистые водоросли (Chrysophyceae)..............................229
4. Класс: диатомовые, или кремнеземные, водоросли (Bacillariophyceae = Diatomeae)..............................................231
5. Класс: бурые водоросли (Phaeophyceae)....................................235
Седьмое подцарство: хлоробионты (Chlorobionta, «Viridiplantae»).................245
Первый отдел: зеленые водоросли I (Chlorophyta).................................246
1. Класс: Prasinophyceae....................................................248
2. Класс: Ulvophyceae.......................................................249
3. Класс: Trebouxiophyceae.............................-....................250
4. Класс: Cladophorophyceae.................................................251
5. Класс: Bryopsidophyceae (= сифоновые)....................................252
6. Класс: Dasycladophyceae..................................................253
7. Класс: Trentepohliophyceae...............................................254
8. Класс: Chlorophyceae.....................................................255
1. Приложение к Chlorophyta: Chlorarachniophyta................................265
2. Приложение к Chlorophyta: отдел эвгленовые (Euglenophyta)...................265
Второй отдел: Streptophyta......................................................267
Первый подотдел: Streptophytina (зеленые водоросли II) .........................267
1. Класс: Mesostigmatophyceae...............................................267
2. Класс: конъюгаты, или сцеплянки (Zygnematophyceae, Conjugatae)...........267
3. Класс: Klebsormidiophyceae (Coleochaetophyceae)..........................270
4. Класс: харовые, или лучицы (Charophyceae)................................271
Второй подотдел: мхи (Bryophytina)..............................................277
1. Класс: Marchantiopsida (= Hepaticae р.р., талломные печеночники).........281
2. Класс: Jungermanniopsida (= Hepaticae р.р., преимущественно листостебельные печеночники)................................................286
3. Класс: Bryopsida (= Musci, листостебельные мхи)..........................288
1. Подкласс: торфяные мхи (Sphagnidae)....................................293
2. Подкласс: Andreaeidae..................................................294
3. Подкласс: Bryidae......................................................294
4. Класс: антоцеротовые (Anthocerotopsida)..................................300
Третий подотдел: папоротникообразные, птеридофиты (Pteridophytina)...........305
1. Класс: псилофиты (Psilophytopsida) .....................................310
2. Класс: плауны (Lycopodiopsida)...........................................312
3. Класс: хвощевидные (Equisetopsida) (= Sphenopsida).......................322
4. Класс: псилотовые (Psilotopsida).........................................328
5. Класс: папоротники (Pteridopsida) (= Filicopsida).......................329
Ступень развития: первопапоротники (Primofilices) (= Protopteridiidae)....................................................330
ОГЛАВЛЕНИЕ | 573
Ступень развития: эуспорангиатные папоротники (Eusporangiatae) (= Ophioglossidae)..............................................333
Ступень развития: лептоспорангиатные папоротники (Leptosporangiatae) (= Pterididae)..............................335
Ступень развития: водные папоротники (Hydropterides) (= Salvmiidae) ....342
Четвертый подотдел: семенные растения (Spermatophytina).........350
Чередование поколений......................................... 350
Вегетативные органы ............................................353
Вторичные метаболиты............................................357
Цветки .........................................................358
Околоцветник....................................................359
Микроспорофиллы..........................:......................360
Пыльца..........................................................364
Мужской гаметофит...............................................367
Мегаспорофиллы..................................................369
Семязачатки.....................................................373
Женский гаметофит...............................................375
Нектарники......................................................377
Расположение элементов цветка...........377
Соцветия........................................................379
Опыление...........................................................379
Оплодотворение.....................................................386
Семя............................................................. 387
Плоды............................................................ 393
Вскрывающиеся плоды................................................394
Невскрывающиеся плоды..............................................395
Соплодия...........................................................395
Распространение плодов и семян.....................................396
Прорастание семян..................................................398
Система Spermatophytina.......................................... 399
1. Класс: саговники (Cycadopsida) (Palmfarne)........................399
2. Класс: Ginkgopsida (Ginkgo).......................................402
3. Класс: хвойные (Coniferopsida)....................................403
4. Класс: Gnetopsida................................................411
5. Класс: цветковые растения, покрытосеменные (Magnoliopsida; Angiospermae)........................................................415
Эволюция и филогения покрытосеменных (обзор)....................416
Система покрытосеменных.........................................420
1. Подкласс: Magnoliidae...........................................420
2. Подкласс: Liliidae (Monokotyledon)..............................426
«Lilianae»......................................................432
«Коммелиноидные» однодольные....................................438
3. Подкласс: «настоящие двудольные» (Rosidae)......................446
Основные «настоящие двудольные».................................450
«Розиды»........................................................458
«Эурозиды I»....................................................459
«Эурозиды II»...................................................478
«Астериды»......................................................485
«Эвастериды I»..................................................488
«Эвастериды II»................................................ 499
Происхождение и родственные связи семенных растений.............508
11.3. Филогенез растений и история растительности.........................513
574 I ОГЛАВЛЕНИЕ____________
11.3.1. Обзор филогенеза организмов........................................513
11.3.2. История растительности.............................................516
11.3.2.1. Методы ......................................................516
11.3.2.2. Докембрий и палеозой (4 млрд—245 млн лет)....................520
11.3.2.3. Мезозой (245—65 млн лет).....................................524
11.3.2.4. Кайнозой (65 млн лет — по настоящее время) ..................526
Список литературы..............................................................540
Список сокращений..............................................................557
Единицы и символы..............................................................559
Предметный указатель...........................................................562
Координаты для контактов.......................................................569
Учебное издание
Зитте Петер. Вайлер Эльмар В., Кадерайт Йоахим В., Брезински Андреас, Кёрнер Кристиан
Ботаника. Учебник для вузов
В четырех томах
Том 3. Эволюция и систематика
Редактор Г.Г.Есакова
Технический редактор Н. И. Горбачева Компьютерная верстка: Т. А. Клименко Корректоры В. А. Жилкина, Г. Н. Петрова
Изд. № 101112452. Подписано в печать 06.03.2007. Формат 70 х 100/16.
Бумага офсетная № 1. Печать офсетная. Гарнитура «Таймс». Усл. печ. л. 46,8.
Тираж 3000 экз. Заказ №8128.
Издательский центр «Академия», www.academia-moscow.ru
Санитарно-эпидемиологическое заключение № 77.99.02.953.Д.007496.07.04 от 20.07.2004.
117342, Москва, ул. Бутлерова, 17-Б, к. 360. Тел./факс: (095)334-8337, 330-1092.
Отпечатано с электронных носителей издательства.
ОАО "Тверской полиграфический комбинат", 170024, г. Тверь, пр-т Ленина, 5.
Телефон: (4822) 44-52-03,44-50-34, Телефон/факс (4822) 44-42-15
Home page - www.tverpk.ru Электронная почта (E-mail) - salcs@tverpk.ru
Ж
Strasburger Ботаника
В четырех томах
Том 1 • Клеточная биология. Анатомия. Морфология
Том 2 • Физиология растений
Том 3 • Эволюция и систематика
Том 4 • Экология
Издательский центр «Академия» www. academia-moscow. ru