/

























































































































































































Автор: Заренков Н.А.
Теги: invertebrata беспозвоночные систематика животных специальные зоологические науки биология
ISBN: 5-211-01395-6
Год: 1989




Текст
Н.А. Заренков
СРАВНИТЕЛЬНАЯ АНАТОМИЯ
БЕСПОЗВОНОЧНЫХ
МОЛЛЮСКИ
1989
московский государственный университет
имени М.В. Ломоносова
Н.А. Заренков
СРАВНИТЕЛЬНАЯ АНАТОМИЯ
БЕСПОЗВОНОЧНЫХ
МОЛЛЮСКИ
Издательство Московского университета
1989
ББК 28.691
3-34
УДК 592 599
Рецензенты:
доктор биологических наук А.П.Кузнецов
доктор биологических наук А.А.Шилейко
Печатается по постановлению
Редакционно-издательского совета
Московского университета
Заренков Н.А.
3-34 Сравнительная анатомия беспозвоночных.
Моллюски. - М.: Изд-во МТУ, I98& 188 с.
LSBA/ 5-2II-0I395-6.
В пособии рассматриваются типы организации моллюсков,
выделенные методом корреляции. Особое внимание уделено водо-
двигательному аппарату мантийной полости в связи с газообме-
ном и фильтрацией. Вододвижение рассматривается как компен-
сация медленному у моллюсков перемещению в пространстве.
Особенности пищедобывающего аппарата двустворчатых и брюхо-
ногих моллюсков связаны с их двигательным аппаратом и строе-
нием мацтийной полости. Строение раковины отражает особен-
ности циркуляции воды в мантийной полости.
Предназначено для студентов старших курсов, аспирантов,
преподавателей ФПК.
077(0'2) - 89 - заказное
15ВЛ/ 5-2II-0I395-6
ББК 28.691
(с) Издательство Московского
университета, 1989
ОСНОВНЫЕ ОСОБЕННОСТИ ТИПА ОРГАНИЗАЦИИ МОЛЛЮСКОВ
(M0ILUSCA)
Единство строения столь несходных на первый взгляд животных,ка-
кими представляются брюхоногие, двустворчатые, головоногие и дру-
гие моллюски, выявляется с помощью мантийного комплекса органов.
Мантийным комплексом называется система топографически и функ-
ционально взаимосвязанных органов и структур, находящихся в ман-
тийной полости или прилежащих к ней: жабры (лёгкое), сердце в пе-
рикарде, почки и мочеточник, осфрадии, гипобранхиальная железа,
задняя кишка и анус, протоки половой системы (Беклемишев, 1964 ,).
Мантия - это наружные покровы внутренностного мешка, эпителий ко-
торых выделяет раковину. При некоторых гистохимических различиях
выделение раковины протекает одинаково у брюхоногих и двустворок
{Timmermans, 1969). Мантийная полость - это пазуха между стенкой
внутренностного мешка и тем участком мантии, который свободно сви-
сает с внутренностного мешка.
Состав мантийного комплекса и его функционирмание неодинаковы
у разных моллюсков. Это объясняется особенностями других, не ме-
нее характерных для моллюсков структур, прежде вс.его внутренност-
ного мешка и ноги. Нога представляет из себя мускулистое разраста-
ние бранной поверхности между ртом и анусом и обглно служит для
передвижения. Внутренностный (висцеральный) мешок, вмещающий внут-
ренности,- это разрастание спинной стороны животн ого в направле-
нии, противоположном ноге.
Разные подтипы типа организации моллюсков представляют отдель-
ные этапы исторического развития этой группы животных. Сопоставлю
строение разных моллюсков, исследователь мысленно как бы воспроиз-
водит исторический процесс,в котором, в общем, имело место услож-
нение строения моллюсков, начиная с таких примитивных, как, нап-
ример, боконервные. Такая познавательная позиция отвечает особой
природе типа организации как становящегося во времени, развивающе-
гося объекта. Вместе с тем, важнейшие особенности типа организации
и корреляции между ними наиболее отчётливы я наглядны у высших
моллюсков - брюхоногих, двустворчатых, головоногих. Поэтому рас-
смотрение процесса становления организации удобно начать с высших
форм, т.е. в направлении, обратном историческому процессу.
Каким образом мог бы сложиться тип организации брюхоногих мол-
люсков?
Представим себе гипотетического предка моллюсков в виде немета-
мерного кольчатого червя, что в какой-то мере отвечает представле-
ниям филогенетиков (рис.1 А). У такого кольчеца, имеющего тело
червеобразной формы!, сквозной кишечник охвачен кольцевыми сосуда-
ми, наиболее крупные из которых выполняют функцию сердца, нерасч-
ленённый целом заключает пару метанефридиев, а нервная система
представлена окологлоточным нервным кольцом и парой брюшных нерв-
ных стволов. Если у такого предка произойдёт разрастание спинной
стороны, то возникнет внутренностный мешок, в который будут втя-
нуты внутренние органы (рис.1 Б). Вместе с кишечником во внутрен-
ностном мешке окажутся брюшные нервные стволы, получившие название
плевро-висцеральных, и вокругкишечное сердце.,Достаточно обширный
внутренностный мешок придаёт животному своеобразную форму: отно-
шение длины тела к её ширине таково, что перистальтика и изгиба-
ние, доступные организмам с удлинённым червеобразным телом, стано-
вятся невозможными. Вместе с тем становится невозможной энергоот-
дача через всю поверхность тела, свойственная червеобразным орга-
низмам. Поэтому исчезает кожно-мышечный мешок, своего рода непре-
рывное покрытие целома у кольчецов, через которое они осуществля-
ют энергозатраты. Вновь возникающий двигательный аппарат может
4
кожно-мышечный
мешок
целители
сердце
crb.
рог
nphp
vns
Рис.1. А - гипотетический предок моллюсков ; Б -разрастание ноги и
внутренностного мешка; В - схема строения примитивных брюхоногих.
Обозначения:crb-головной мозг,nphp.-нефридиопор,pvns(vnsj-плевро висце-
ральные (брюшные) нервные стволы, jons-ножные нервные стволы, *
мантийная полость
5
тратить энергию только через ту часть поверхности тела, которая
прилежит к субстрату. Она получила название подошвы. Скользящее дви-
жение у таких моллюсков совершается благодаря волнообразным мышеч-
ным сокращениям подошвы ноги, только отдалённо напоминающим перис-
тальтику, а также работе ресничного эпителия подошвы.
По мере разрастания внутренностного мешка возникает и усилива-
ется противоречие между массой тела и сокращающейся площадью по-
дошвы ноги, от которой зависит действенность её как двигательного
аппарата. Это противоречие отчасти устраняется разрастанием подош-
вы в стороны (уплощение) и удлинением тела позади ануса (рис.1 Б).
Последнее вызывает как бы смещение ануса вверх, к вершине внутрен-
ностного мешка. Кроме того, по мере разрастания внутренностного
мешка возникает необходимость в опорной структуре.
В отличие ст кольчецов с их мягким гидростатическим скелетом,
пригодным для изгибания и перистальтики тела, моллюски нуждаются в
твёрдом скелете, который мог бы поддерживать внутренностный мешок;
гидростатический скелет - водоносная система в ноге Natica или па-
ренхима в ноге двуотворок Undo имеет значение только для перис-
тальтики ноги. В простейшем случае поддерживающей внутренностный
мешок структурой может служить раковина, имеющая форму колпачка.
Такая раковина дополнительно может выполнять защитную функцию, в
неё при необходимости может втягиваться всё мягкое тело животного,
включая внераковинные части - ногу и голову (цефалоподиум). Вследс-
твие этого в обычном состоянии внутренностный мешок не должен______
Рис.2. А - топографические отношения почек (заштриховано), гонад (за-
чернено) и перикарда (пунктирная линия) в маЯтийной полости переднежа-
берных: 1- гипотетический предок, 2 - редукция левой послеторсионной
гоняды, 3 - редукция левой и разрастание правой гонады до состояния
Haliotis и Pleurotomaria (4 ) и Diodora (5 ), Puncturella ъТго-
chidae (6 ), Patellacea (7 ), т.е. у всех археогастропод, кроме Ne-
z'it асеа. ( 8 ) и Сосс Ипасеа, 9 - редукция правой и разрастание левой
почки как исходное состояние для Neriiacea и всех прочих Proso-
branchia (10 ). Ь строение мантийной полости у Archaeogastropoda
Zygobranchia (1 ), Patelloida Acmaeidae (2), Trochacea, Neritacea,
Valvatacea (3), PeczmibraYictila (4). (Уоп§е,1960
6
7
заполнять без остатка весь объём колпачковидной раковины, и вместе
с такой свободной,"на вырост", раковиной животное должно иметь
мышцы ретракторы, втягиватели цефалоподдума. У брюхоногих такие
ретракторы, называемые раковинной или колхнеллярной дешцей, дейст-
вительно имеются. Они протягиваются от внутренней поверхности ра-
ковины или от столбика (колюмны) к подошве ноги. Существует, одна-
ко, и другая, пожалуй, более важная причина того, что внутренност-
ный мешок не полностью заполняет внутреннее пространство раковины.
Вследствие появления раковины и прилежащей к субстрату подошвы
ноги у моллюска сокращается свободная поверхность тела, через ко-
торую изначала осуществлялся газообмен. Для возмещения сократив-
шейся поверхности тела могла бы служить мантия, выстилающая ман-
тийную полость. Пожалуй, дыхательная функция мантийной полости на-
гляднее всего проявляется у легочных моллюсков, у которых сама
складка мантии, пронизанная кровеносными сосудами, уже есть орган
дыхания (лёгкое). У некоторых лёгочных моллюсков мантийная полость
занимает до половины завитка раковины, у катушек - ещё больше
(J4u.ben.dick, 1978). У переднежаберных моллюсков дыхательная фун-
кция в основном выполняется особыми складками мантии, получившими
название ктенидиев, таким образом у них имеет место специализация
отдельных участков мантии на выполнении дыхательной функции - кон-
центрация дыхательной функции. Последнее обстоятельство объясняет
ряд других особенностей организации брюхоногих.
Прежде всего, благодаря вновь возникшим складчатым ктенидиям,
имеющим значительную величину дыхательной поверхности, появляется
возможность уменьшить объём мантийной полости. Если в простейшем
случае мантийная полость могла бы представлять из себя тор, окру-
жающий основание внутренностного мешка (циркумпедальная полость),
то вновь возникшие ктенидии позволяют уменьшить тор до сегмента.
Местоположение такого сегмента определяется необходимостью ис-
пользовать мантийную полость в качестве убежища для цефалоподиума,
У тех раковинных брюхоногих, у которых тело относительно раковины
8
Рис.З. А - схема строения ЦНС боконервных (I), переднежаберных брю-
хоногих с хиастоневрией (2) и двустворчатых (3). Б - мантийная по-
лость боконервных на поперечном срезе (I), мантийная полость двуст-
ворчатых (раковина укорочена) (2), брюхоногих (раковина укорочена)
(3), головоногих (4) и лопатоногих (5). Обозначения: * - мантийная
полость,pvn. плевро-висцеральные стволы, рп. педальные стволы,
сп- циркумпаллиальное кольцо Patellacea
9
удлинено в незначительной степени и относительная ширина подошвы
велика, например у морских блюдечек, мантийная полость сохраняет
форму тора. У них при необходимости широкий цефалоподиум сокраща-
ется и заполняет мантийную полость и его пропорции при этом меня-
ются незначительно; моллюск прижимается к субстрату. Если же голо-
ва и задний конец тела выступают из-под раковины и таким образом
цефалоподиум имеет удлинённую форму, как у большинства переднежа-
берных и раковинных брюхоногих, то мантийная полость действительно
сокращена до сегмента тора. В случае необходимости цефалоподиум
складывается вдвое, первой под защиту раковины кояомеллярная шпи-
ца втягивает голову, а затем задний отдел ноги о крышечкой (если
она имеется). Переднее положение мантийной полости определяется,
таким образом, тем, что голова моллюска находится спереди, а кры-
шечка сзади. При этом прикрепление к субстрату может не иметь мес-
та. Переднее положение сегмента торообразной мантийной полости за-
даётся 'гетерополярностью (крышечка-голова) вытянутого цефалоподиу-
ма.
Обособление дыхательной поверхности мантии, каковой можно счи-
тать жабры, имеет и другие последствия для организации моллюсков.
Теперь окисление крови осуществляется преимущественно локально, в
то время как у кольчецов оно совершалось едва ли не через всю
поверхность тела. Соответственно, разными оказываются и кровенос-
ные системы этих животных. Если у кольчецов часто не наблюдается
резко обособленного сердца и способностью сокращаться обладают
многие кровеносные сосуды ("диффузное" сердце), то у моллюсков
кроведвижущий отдел кровеносной системы обособлен достаточно чётко.
У моллюсков сердце состоит из 1-2 предсердий,которые принимают
окисленную кровь, поступающую из жабер, и мускулистого желудочка,
выталкивающего кровь в переднюю (головную) аорту. Сердце заключено
в перикард, который можно рассматривать как сократившийся туловищ-
ный целом. Сокращение целома до перикарда объясняется тем, что
вследствие появления раковины целом утратил значение опорной струк-
ю
Рис.4. А - упрощенная блок-схема распределения форм раковины у
беспозвоночных (без фораминифер); D - расстояние образующей от оси
навивания, W - скорость расширения оборотов раковины, Т - величи-
на переноса оборота. Б - схема, поясняющая параметры D,T nW.
В - раковина моллюска, у которого величина переноса оборота в ходе
роста уменыается (Каир, 1966)
11
туры (рис.1). В этом отношении моллюски напоминают членистоногих,
у которых появление наружного скелета (кутикулы) тоже вызвало рас.
пад целома, утрату им опорной функции.
У кольчецов целом сообщается с внешней средой через почки ж це«
ломодукты. У моллюсков эта связь выступает в своеобразной форме -
как связь перикарда с мантийной полостью через почку. Отверстие,
сообщающее перикард с почкой, очень характерно для моллюсков и
получило название рено-перикардиального отверстия. Гонады иногда
открываются наружу самостоятельно, но в некоторых случаях - через
мочеточник. Вообще же топографические отношения между почками и
гонадами довольно разнообразны даже в пределах одних только перед-
нежаберных брюхоногих моллюсков (рис.2). Вместе с перикардом поч-
ки и гонады обычно считаются рудиментами целома.
Помимо своеобразных топографических отношений перикарда с поч-
кой, некоторые моллюски примечательны ещё и тем, что по крайней
мере у примитивных брюхоногих желудочек сердца окружает заднюю
кишку. Эта особенность могла бы объясняться тем, что у возможного
предка брюхоногих кишечник был охвачен кольцевым сердцем (рис.1 А).
Можно считать,что обособленность сердца как кроведвижущего
отдела кровеносной системы вызвана у моллюсков обособлением орга-
нов дыхания: локальные жабры - локальный кроведвижущий отдел. Воз-
можно также, что двухтактный рабочий цикл этого органа (систола-
диастола) может считаться причиной его подразделения на предсер-
дие и желудочек: пока при сокращении желудочка кровь выталкивается
в аорту, в предсердии кровь накапливается для последующего запол-
нения желудочка. В предсердие поступает окисленная кровь из орга-
нов дыхания, и если эти органы локальны, то обособленность пред-
сердия вполне естественна. У кольчецов, с их дыханием через поверх*
ность тела, обычно нет резко обособленного сердца и вместе с этим
нет подразделения кроведвижущих "сердец" на желудочек и предсердие<
Внутренностный мешок хорошо развит у брюхоногих, головоногих и
лопатоногих, к тому же имеющих так называемый анопедиальный
2
(т.е.противоположный ноге) изгиб кишечника. Сопоставимым у этих
моллюсков остаётся и положение мантийной полости, и вместе с тем
без особых трудностей сопоставляются и планы строения этих живот-
ных в целом (рис.З А 3,4,5 ). Головоногие, образно выражаясь, - это
брюхоногие, лишённые постанального разрастания ноги (на рис.З А4
оно показано пунктиром). Лопатоногие - это брюхоногие с вентрально
направленной клиновидной ногой, их сходство усилится, если ногу
отогнуть назад (рис.З Б 5 ). У двустворчатых внутренностному меш-
ку в определённой мере отвечает направленная вентрально вниз кли-
новидная нога, вмещающая значительную часть кишки (рис.З Б2).
Двустворки, таким образом, напоминают моллюсков с развитым внут-
ренностным мешком в том смысле, что тоже имеют обширное вместили-
ще внутренних органов. Это сходство усиливается за счёт мантийной
полости, окружающей ногу. Получается, что моллюски, имеющие вмес-
тилище внутренних органов, к тому же обладают мантийной полостью.
Эта корреляция не случайна, если рассматривать мантийную полость
как восполнение сократившейся свободной поверхности тела, вызван-
ное появлением раковины и массивной ноги. Наоборот, моллюски со
слабо развитым внутренностным мешком (моноплакофоры, боконервные)
вместо глубокой мантийной полости обладают только относительно
мелким мантийным желобком, охватывающим основание ноги (рис.З Б1),
Несмотря на существенные различия, всех моллюсков объединяет
ещё одна общая черта: наличие пары ножных (педальных) стволов,
обслуживающих разросшуюся ногу. Взаиморасположение вновь возник-
ших ножных стволов и унаследованных от кольчецов внутренностных
стволов оказывается противоположным у моллюсков с анопедиальным
изгибом кишки (брюхоногие) и без оного (двустворки)(рис.З А 2,3).
Сходство между ними выражается в том, что каждый раз пара нервных
стволов размещена в двух взаимоперпендикулярных плоскостях: сагит-
тальной (в ноге у двустворок, во внутренностном мешке у брюхоно-
гих) и боковой. Этот тип ЦНС, который можно было бы назвать ани-
зопланарным, противопоставляется гомопланарному типу боконервных
13
и моноплакофор, у которых нервные стволы остаются в двух парал-
лельных боковых плоскостях (рис.З А 1). В этом факте также можно
усмотреть корреляцию между типом ЦНС, наличием вместилища внут-
ренних органов и строением мантийной полости.
Вышеописанный процесс становления организации брюхоногих, т.е.
корреляция как процесс, мог бы и не иметь филогенетического зна-
чения. В частности, гомология перикарда туловищному целому анне-
лидоподобных предков не принадлежит к общепринятым положениям.
Зато, независимо от филогении, эта корреляция может пониматься
как состояние и предлагает физиологическое объяснение фактически
наблюдаемому у брюхоногих размещению органов и структур.
При достаточно высокой абсолютной скорости движения и при условии
размещения жабер снаружи, вне мантийной полости, задача обновле-
ния воды, что необходимо для газообмена, решалась бы сама собой.
Между тем моллюски в своём большинстве, исключая прежде всего
головоногих,- медлительные животные. В этом повинен прежде всего
их двигательный аппарат - ползательная или клиновидная нога. По-
этому у моллюсков имеется специальный вододвигательный аппарат,
во вмещающий медленность ползания и не вполне удачное размещение
жабер. Обновление воды в мантийной полости у брюхоногих может
быть* весьма значительным. Известно, например, что в мантийной по-
лости некоторых Conus могут обитать колшенсальные крабы Pinno-
theridae (Wolfson, 1974).
Неглубокая желобчатая мантийная полость панцирных и моноплако-
фор служит исключительно для дыхания, тогда как функция добывания
пищи остаётся за ротовым аппаратом. Сходным образом у брюхоногих
вододвижение в мантийной полости обычно служит в основном для ды-
хания, тогда как для добывания пищи у них служат радула, челюсти,
щупальца, хобот. У двустворчатых моллюсков вследствие особенно
низкой подвижности вододвижение имеет значение не только для дыха-
ния, но и для отфильтровывания пищевых частиц. У лопатоногих пище-
вые частицы собираются ротовыми щупальцами, а мантийная полость
14
служит главным образом для дыхания. Наконец у многих головоногих
в отличие от других моллюсков вододвижение в мантийной полости
служит не только для дыхания, но и для реактивного движения. В
общем же, дыхание - наиболее обычная функция мантийной полости,
тогда как двигательная функция сохраняется за мантийной полостью
только у части головоногих.
Однонаправленное движение воды в мантййной полости наблюдается
у лопатоногих (от вершины трубчатой раковины к её устью) и боко-
нервных (спереди назад). У головоногих маятниковое движение воды
в противоположных направлениях осуществляется через отверстие ман-
тийной полости (вдох) и сифон (выдох). Поскольку мантийная полость
у головоногих в короткое время вбирает и выбрасывает большое коли-
чество воды (мгновенная энергоотдача), схема циркуляции остаётся
относительно простой. Важно подчеркнуть, что движение воды при
этом вызывается сокращениями мышц.
У прочих моллюсков токи воды в мантийной полости обычно вызыва-
ются ресничным эпителием. Вследствие этого становится возможным
распадение схемы циркуляции на относительно обособленные кругово-
роты у особенно у двустворчатых и брюхоногих. Вместе с турбулент-
ностью у двустворок и брюхоногих усложняется внутренняя поверх-
ность мантийной полости. У двустворок эта поверхность образована
2 парами жабер, лабиальными пальпами, ротовыми лопастями. Несмот-
ря на некоторое разнообразие схем циркуляции, двустворки обычно
сохраняют два основных круговорота, размещённых симметрично отно-
сительно сагиттальной плоскости животного. У брюхоногих сложная
поверхность мантийной полости, обусловленная колпачковидной или
спирально свёрнутой раковиной уже служит причиной турбулентности.
В отличие от однонаправленной (лопатоногие, панцирные) или маят-
никовой (головоногие) циркуляции, турбулентность допускает широ-
кое разнообразие формы мантийной полости, а, следовательно, и ра-
ковины, потому что наряду с поддержанием внутренностного мешка
раковина у брюхоногих поддерживает и стенку мантййной полости.
15
Больше того, надо полагать, что форма раковины в какой-то степени
определяется особенностями функционирования мантийной полости.
Спирально свёрнутая раковина брюхоногих в значительной степени
есть обызвествлённая спиральная траектория мантийной полости,
смещаемой по оси раковины по мере роста животного. Несомненно,что
у других моллюсков между строением раковины и мантийной полости
также имеют место корреляции.
Для большинства моллюсков характерны спиральные раковины, впро-
чем, свойственные и другим беспозвоночным - плеченогим и форами-
ниферам. Разнообразие спирально свёрнутых раковин упорядочено
Д.Раупом и С.Стенли (Рауп, Стенли, 1974; Каир, 1966) с помощью
четырёх параметров (рис.4).
У большинства брюхоногих раковина имеет форму полого конуса,
открытого на широком конце (устье). На одной стороне устья при-
рост идёт быстрее, чем на другом,и поэтому конусовидная трубка
сворачивается в спираль. Первый параметр, позволяющий количест-
венно изучить этот процесс, - это форма поперечного сечения оборо-
та или форма образующей кривой. На рис.4 Б образующая кривая -
окружность. У большинства брюхоногих она совпадает с формой устья.
Более точно эта кривая представляет из себя линию пересечения
расширяющейся трубки с плоскостью оси навивания. Второй параметр -
скорость расширения образующей кривой по мере обращения вокруг
оси. Он называется также скоростью расширения оборота W и пред-
ставляет из себя отношение любого линейного размера, например
диаметра,двух образующих кривых.разделённых полным оборотом спи-
рали. На рио.4 Б скорость расширения оборота W-2, т.е. при каж-
дом обороте вокруг оси линейный параметр образующей кривой удва-
ивается. Третий параметр - положение и ориентировка образующей
кривой по отношению к оси D . На рис.4 Б образующая окружность
ОТСТОИТ ОТ ОСИ на половину своего диаметра. Ориентация по отноше-
нию к оси важна в тех случаях, когда образующая кривая не являет-
ся окружностью. Наконец четвёртый параметр - перемещение образу-
16
пней кривой вдоль оси - это перенос оборота Т. Он выражается от-
ношением перемещения вдоль оси к величине перемещения от оси на
протяжении любой части оборота вокруг оси. При этом учитывается
перемещение геометрического центра образующей кривой. Если пере-
нос отсутствует, то трубка обращается вокруг оси в одной плос-
кости и образуется плоскоспиральная раковина. Обычно эти парамет-
ры остаются неизменными в течение жизни одного организма, но мо-
гут быть и исключения (рис.4 В).
В пределах класса брюхоногих скорость расширения переноса ра-
ковины изменяется от немногим более I до 5. Этим величинам отве-
чают крайние случаи - соприкосновение с осью навивания и отделе-
ние от неё пупком, конусовидной полостью внутри колюмны. Такими
раскрученными раковинами обладают 15 ныне живущих видов в
разных семействах. Функциональное значение этой особенности оста-
ётся неизвестным (Rex,Boss, 1976). Величина переноса раковины
изменяется от 0 (плоскоспиральные раковины) до очень высоких зна-
чений (башенковидные раковины). Среди современных брюхоногих нет
настоящих плоскоспиральных раковин, но в палеозое такие раковины
были довольно обычны.
На рис.4 А блок-диаграмма содержит все геометрические модели
раковины, характеризуемые четырьмя параметрами. Раковины брюхоно-
гих, как правило, занимают область медленного расширения оборота
W и вместе с тем они разнятся по величине оборота Т и расстоя-
нию образующей кривой от оси D. Спирально свёрнутые раковины голо-
воногих в большинстве своём плоские, но скорость расширения обо-
рота у них всегда невелика. В общем, основные группы раковинных
беспозвоночных занимают практически неперекрывапциеся области 3-
мерного пространства блок-диаграммы.
Малакологи уже давно выделили в разнообразии раковин брюхоно-
гих три основных морфотипа: галиотисный (Haliotis), турбиноид-
ный (Turbo,Pleurotomaria), туррителлоидный (Turritella, Per-
viacea). в этом ряду возрастает величина Т и уменьшаются W и
I) (Linsley, 1978). Более или менее средние для брюхоногих пара-
метры сохраняют раковины на рис.5 В,Г,Е, рис.6 А,Г,Д, рис.7 Д,Е.
Повышенная скорость расширения оборота наблюдается на рис.5 3,
рис.6 3,Е,Н, рис.7 Б.В.Напротив, малое значение этого параметра
свойственно раковине на рис.6 Ж. На рис.6 К изображена раковина,
имеющая значительное расстояние образующей от оси при малом зна-
чении величины переноса. Последний параметр принимает более вы-
сокое значение на рис.5 А.
Д.Рауп и С.Стенли (1974) подчёркивают, что с помощью предложен-
ных ими четырёх параметров невозможно выразить ряд других особен-
ностей раковин: декстральность и синистральность, наличие вырезок
и отверстий, столь обычных у Archaeogastropoda(psLc.7 А), а также
сифональных выростов (рис.7 Е) и других деталей скульптуры. Со-
вершенно особый морфотип представляют подверженные редукции рако-
вины заднежаберных моллюсков (рис.5 Д, рис.7 1,И)и, в частности,
крылоногих (рис.7 3), некоторых верметид и лёгочных моллюсков
(рис.6 М). Большим своеобразием отличается раковина Mungilus,
вмурованного в коралловый политик (рис.6 И). Четыре параметра не
позволяют описать строение так называемых гетерострофных раковин,
у которых первоначальная ось навивания наклонена к окончательной,
они пригодны только для обычных голострофных раковин. Наконец в
3- мерном пространстве блок диаграммы 4 не находит себе меота ра-
ковины с переменной скоростью расширения оборота (рис.6Ж).
Рис.5. Раковины переднежаберных брюхоногих: А - Сосеа ochridiana (Valva-
tidae. (HadZisceet al., 1976); Б- Notocypraea angustata (Cypraeidae), В-Лвг-
vicacea kieneri (Terebridae), T-Murexsul brazieri (Muricidae) (Macpher-
son, Gabriel, 1962). Раковины заднежаберных моллюсков Д - Spiricella
unguiculus (Hoeksema, Janssen, 1984). Раковины лёгочных моллюсков: E -
Gulella viae (Bruggen,van, 1985); I- Marisa planogyra (Pilidae); 3 - Ar-
lene megastcma (Pilidae) (Castellanoa .Fernandez, 1976)
18
19
Между тем обычные у многих раковин кили, шипы, зубцы и другие
элементы скульптуры в некоторых случаях имеют функциональное значе-
ние. Например, они увеличивают прочность раковины, когда её пыта-
ются раздавить клешнями крабы, обычные потребители брюхоногих
(Hughes, 1986). Хищный моллюск Acanthina приоткрывает створ-
ки домика балянуса с помощью шипа на передней части раковины (Sig-
nor , 1985). Так нападает моллюск на мелкие жертвы. Если же раз-
меры жертвы велики, то хищник сверлит раковины балянуса так же,
как например Natica Теребеллоидные раковины ТегеЬга с широ-
ким устьем легче раздавливаются крабами-моллюскоедами Calappi -
с1ае,чеы раковины с узким устьем, наделённые большей механической
прочностью (Signor, 1985). Зарывание в грунт вследствие медлен-
ности не спасает моллюска от хищника. Всё же следует обратить
внимание на то обстоятельство, что эта оборонительная реакция мол-
люска зависит от формы раковины. Показано (Vermej, Zipser, 1986),
что моллюски с высокой туррителлоидной раковиной зарываются быст-
рее, чем моллюски с раковинами иного типа. Желобчатые выросты края
устья у таких моллюсков, как^нога пеликана’,’участвуют в организации
токов воды. Таким выростом обладает раковина, изображённая на рис.
7 Е. Наконец, раковины различаются толщиной стенки, массивностью,
Рио.6. А - клаузилий Iphigenia plicatula (Ermann. из Лихарева, 1962).
Раковины лёгочных моллюсков: Б -Carychium tridentatum (Bank, Quit.
1985); В -Bofiliella subarcuata, у _ Ruatara киагапа, д - Соокесоп-
cha (Solem, 1976); Е- Acroloxus boetgerianus(Hu.bendick, 1969). I-
Brachypodellaangesiana, 3 - Heli carion superba (Franc, 1968a). Рако-
вины переднежаберных моллюсков: И - Mungi 1 us antiquus (Franc J968a),
К -Pseudomalaxis cornuammonis (Bieler, 1984), Л - Rhaphaulus lorra-
mi с дыхательной трубкой ( Solem, 1974a), M - Opisthostoma coro-
natizm(Solem,1964), H - Tremanotir из палеозоя (Bellerophontacea)
(Cox, i96o)
20
3
21
от которой зависит прочность раковины. В этой связи обсуждается
проблема взаимосвязи между массивностью раковин и обилием крабов-
моллюскоедов, фауна которых особенно богата в тропической зоне
океана 1978).
Раковина двустворчатых моллюсков состоит из двух створок со
слабой спиральностью: декстральной и синистральной (рис.8 А,Б).
Скорость расширения оборота, как правило, весьма велика и нередко
g
превышает 10, наоборот, перенос обычно невелик и часто имеет мес-
то смещение оборота, т.е. искривление оси раковины, показанной на
на рис.8 В прерывистой линией. В сущности такие створки турбо-
спиральны , и отдельная створка отвечает раковине брюхоногих с не-
соприкасающимися оборотами. Створки сохраняют свободным внутрен-
ний край образупцей кривой. Обычно свободный край занят замком,
который образован зубами и межзубными ямками и служит для сочлене-
ния створок. Размеры зубов и ямок увеличиваются по мере роста ра-
ковины. У некоторых двустворок спиральность особенно хорошо видна
вследствие того, что подвёрнута макушка раковины (рис.8 А). С дру-
гой стороны, некоторые раковины брюхоногих напоминают одну створку
двустворчатых (рис.7 Г).
То обстоятельство, что, несмотря на важные различия от брюхоно-
гих, створки раковин двустворчатых остаются спирально закрученны-
ми, не следует недооценивать. Можно предполагать, что спиральная
Рис.7. Раковины брюхоногих ^rchaeogastropoda: A - Monodilepas nwni-
lifera (Fissurellidae) (Dell, 1953), Б-Haliotis vulgaris (Haliotidae) (Pret-
tier, Graham, 1976) изнутри и снаружи, В - Emarginula rosea (Fissurel-
lidae) (Fretter, Graham, 1976), Г- Macroschisma producta (Fissurellidae)
(Dell, 1963), Д - Zcissurella crispata (Pleurotomariidae) (Fretter, Graham-
1976). E - Columbarium veridicum (Prosobranchia) (Dell, 1963). Раковины
Opisthobranchia К -Aglaja cylinorica (Rudman, 1963) дорзально и вент-
рально, 3 - Clio cuspidate (Tesch, 1916) (Pteropoda Thecosomata), И -
Oracle bispinosa (Tesch, 1918) (Pteropoda Thecosomata)
22
23
закрученность каким-то образом связана с турбулентной циркуляцией
воды в мантийной полости этих двух групп моллюсков.
Каждая половина тела двустворок, выделяемая сагиттальной плос-
костью, как бы отвечает целому брюхоногому моллюску. Соответствен-
но схема циркуляции воды в мантийной полости у двустворок - это
как бы удвоенная циркуляция брюхоногих. Исключение для этого со-
поставления представляют неравностворчатые двустворки, такие, нап-
ример, как устрицы. У них вследствие прикрепления к субстрату од-
на створка отличается от другой и животное несимметрично относи-
тельно сагиттальной плоскости.
Основные типы организации моллюсков более или менее отвечают
отдельным таксонам (табл. 1}
Рис. S. A-Раковина Glossus humanus, Heterodonia, со спирально свёрнутыми
макушками, вид спереди (1) и сзади (2) (Franc,1960). Б - влияние изме-
нения скорости расширения оборота на степень перекрывания, между пос-
ледовательными оборотами при фиксированной величине переноса оборота
Т и расстоянии образующей кривой от оси D (Рауп, Стенли, 1974). В -
схема роста раковины Glossus и составляющие роста: -R - радиальная ,Т-
тангенциальная, G - результирующая {Owen, 1953). Г - симметричная
створка Gfycymeris glycymeris .Taxodonta, Filibranchia,c равновеликими от-
печатками аддукторов раковины {Franc, 1960). Д - становление дисиммет-
рии у брюхоногих: 1 смещение вверх центра тяжести внутренностного
мешка у моллюска с колпачковидной раковиной (а1-а2) и переход к эк-
зогастрической планоспиральной раковине (пунктир), 2-3 моллюск
с планоспиральной экэогастрической раковиной (2 ), который претерпе-
вает поворот внутренностного мешка, вызывающий хиастоневрию,и пере-
ход к эндогастрической раковине (3), 4 - возможное смещение центра
тяжести внутренностного мешка (* ) вправо от сагиттальной плоскости
(пунктир) вследствие перехода к турбоспиральной раковине,5 - приве-
дение в сагиттальную плоскость центра .окести внутренностного мешка
вследствие регулятивного раскручивания (деторсия) и наклона турбоспи-
ральной раковины. Е строения наутилуса с экзогастрическо”
планоспиральной раковиной и задним ложением мантийной полости
24
25
Табл.I. Типы организации моллюсков
Х.Тело расчленено на более или менее развитое вместилище внутрен-
них органов (клиновидная нога, внутренностный мешок), двигатель
нус ногу и мантийную полость
II.Слабо обособленный внутренностный мешок, циркумпедальная мал
тийная полость с жабрами, гомопланарная ЦНС
III .Панцирь из подвижно сочленённых пластинок....Loricate
НО .Колпачковидная раковина..................Monoplacophorz
10.Обособленное вместилище внутренних органов, раковина, мантий
ная полость с жабрами или иными дыхательными структурами,
анизопланарная ЦНС
101.Маятниковая циркуляция воды в мантийной полости, мышечно
движение и вододвижение....................Cephalopoda
100.Однонаправленная непрерывная циркуляция воды (воздуха) в
мантийной полости
1001.Нога обычно с широкой подошвой, кишечник с анопеди-
альным изгибом, раковина обычно конусовидной формы,
турбулентная циркуляция воды (воздуха) в мантийной
полости................................Gastropoda
1000.Клиновидная нога, вододвигательная мантийная полост
I0001.Трубчатая раковина, прямоточная схема циркуля
ции воды в мантийной полости.. .Scaphopoda
10000.Двустворчатая раковина, турбулентная циркуля-
ция воды в мантийной полости...Bivalvia
0.Почти нерасчленённое округлой формы тело, раковина отсутствует
00.Червеобразные моллюски с полужёстким скелетом, желобчатой
мантийной полостью и гомопланарной URC.. ..Solenogastres
01.Червеобразное или округлой формы тело с псевдомантийной по-
лостью................Эндопаразитические Gastropoda
26
ТИП ОРГАНИЗАЦИИ БРЮХОНОГИХ МОЛЛЮСКОВ (GASTROPODA)
В.Н.Беклемишев (1064) отметил, что брюхоногие - единственная
обширная группа животных, содержание исторического развития кото-
рой состояло в выработке дисимметрии. Теория дисимметрии брюхо-
ногих неоднократно излагалась (Беклемишев, 1964; Иванов, 1940;Hy-
man, 1967; Franc,1968а; Lever, 1979), и хотя некоторые её поло-
жения нуждаются в углублённом изучении и продолжают критически
обсуждаться (Миничев, Старобогатов, 1972; Ghiseli.n, I9&6;Thomp-
son,I967; Graham, 1979), она остаётся одним из наиболее значи-
тельных обобщений сравнительной анатомии. Теория рассматривает
причины поворота внутренностного мешка, вследствие которого рако-
вина из экзогастрической сделалась эндогастрической, причины пе-
рехода от колпачковидной раковины к планоспиральной, а затем пла-
носпиральной: наконец, предлагает объяснение своеобразным топог-
рафическим корреляциям в размещении органов внутренностного меш-
ка и мантийной полости, составляющим содержание дисимметрии.
Ныне живущие брюхоногие обычно обладают спирально свёрнутой ра-
ковиной, тогда как их возможные предки имели колпачковидную рако-
вину. Последняя,симметричная сама по себе, располагалась в сагит-
тальной плоскости, подчиняясь общей двусторонней симметрии орга-
низма. Замена колпачковидной раковины на планоспиральную возможно
вызвана необходимостью сохранить низкое положение центра тяжести
внутренностного мешка, что необходимо при увеличении размеров жи-
вотного (рис.8 Д 1). Очевидно, это соображение сохраняет своё зна-
чение как для филогенеза, так и онтогенеза. Кроме того, высокая
колпачковидная раковина явно непригодна для моллюска, ползапцего
среди водорослей и на боковой поверхности камней. Поэтому если
27
у предковых форм имел место рост колпачконидной раковины, то он
должен был сопровождаться свёртыванием её в спираль. Так могла
возникнуть планоспиральная раковина. При положении завитка над
ловой раковина называется экзогастрической, если же завиток нахо-
дится сзади, то она называется эндогастрической. В теории дисим-
метрии принимается, что вновь возникшая планоспиральная ракови-
на была экзогастрической. Среди ныне живущих моллюсков такую ра-
ковину имеют наутилусы (рис.8 Е). У наутилусов раковина наполне-
на газом и служит поплавком. Мягкое тело животного занимает пос-
леднюю камеру, как бы подвешено к раковине-поплавку, и потому сох-
раняет устойчивое положение. Наутилусы не могли быть предками
брюхоногих. Теория лишь допускает, что если предки брюхоногих
имели экзогастрическую раковину, то они могли так же, как наути-
лус, обитать в пелагиали.
После того как предковые формы опустились на дно, нависающий
над головой завиток стал причинять неудобства, особенно когда
при ползании встречают^преграды в виде вертикальной поверхности.
Поэтому завиток раковины был отведён назад, причём в направлении
против часовой стрелки, и таким образом моллюски приобрели эндо-
гастрическую раковину и мантийная полость оказалась над головой
(рис.8 Д 2,3). Следовательно, согласно теории, торсион, поворот
раковины и внутренностного мешка остаётся в первую очередь след-
ствием перемены образа жизни. Оставаясь гипотезой в области фи-
логенетики, торсион является фактом в эмбриологии: личинки брю-
Рис.9. А - схема строения глаза наземного слизня Agriolimax,
Pulmonata (Р.Newell, G.Newell, 1968). Б - строение осфрадия Babylo-
nia japonica , Neogastropoda (1),часть которого, выделенная пункти-
ром, изображена отдельно (2 ) (Maeda,1983). В - эпителий осфрадия
пресноводной катушки Planorbis corneas, Puimonata (Bailey, Benja-
min, 1968) Общая схема строения желудка (Г) и пищеварительной
системы брюхоногих моллюсков (Д) {Franc, 1960; Fretter, Graham,
1968)
28
пищевод
проток пищеварительной
железы
кишечная
борозда
протостиль
цекум
титрлозоль
тяжи слизи с
пищевыми частицами
Г
секреторная клетка желудочный щит
кутикулярный I
большой
тирлозоль
сенсорное окончание
базальная клетка эпитииальная
клетка
щит
цекум
ющая
поверх-
ность
слюнная
железа
пищеводные
железы
пищеваритель-
ная железа
малый
тифлозоль
задняя
кишка
2
В
Б
стекловидное
тело
конъюнктива
сетчатка
фоторецепторная
клетка с палочкой
и терминальными
микротрубочками
трактор
линза
гемоцель
нерв
щупальца^ о
дополни- ,
тельная
сетчатка
^капсула
vj линзы
А
глазной
черв ~
29
хоногих, обитающие в планктоне, претерпевают торсион, предваряя
таким образом переход к жизни на дне. В.Фреттер (Graham, 19*^9)
указала ещё одну причину переднего положения мантийной полости,
которое установилось после торсиона. Она состоит в том, что при
опасности в мантийную полость втягивается прежде всего голова и
только во вторую очередь - нога. Однако представляется полезным
дополнительно учесть ещё одно обстоятельство, объясняющее причины
торсиона. которое как-будто бы ещё не привлекало внимания малако-
логов.
у пелагических животных, какими представляются предки брюхоно-
гих, ведущая роль в сенсорной функции могла бы принадлежать зре-
нию. У донных моллюсков значение зрения уменьшается вследствие
более слабой по сравнению с эпипелагиалью освещенности и низкой
прозрачностью придонного слоя воды, наконец, вследствие сократив-
шегося поля зрения и неровностей морского дна. К тому хе медлен-
ное ползание не всегда предполагает необходимость распознавать
удалённые от моллюска объекты, наоборот, это полезно для быстро
плавающих пелагических хищников. По всем этим причинам вместо
зрения у донных моллюсков возрастает значение хеморецепции, обо-
няния. Органы зрения у них часто сохраняются,и вследствие того,
что воздух прозрачнее воды, лучше развиты у наземных моллюсков
(рис.9 А).
Физиология обоняния донных моллюсков имеет существенные отли-
чия от физиологии зрения их возможных пелагических предков. Гла-
за даже у неподвижных или малоподвижных животных позволяют не
только мгновенно увидеть объект, отыскать направление на него или
оценить степень освещённости, но при бинокулярном зрении ещё поз-
воляют мгновенно оценить его отдалённость. Сходным образом слух
тоже позволяет мгновенно оценить градиент силы звука, воспринима-
емого парой рецепторов, а значит и мгновенно (но не обязательно
точно) определить направление на источник раздражения. Что же на*
сается обоняния, то оно позволяет решать аналогичные задачи толь-
30
go при наличии градиента запаха во времени (не мгновенно); какого-
либо аналога бинокулярности или биноринальности при этом не требу-
ется. Для проявления градиента запаха и его восприятия необходимо
время. Время уходит либо на переползание самого животного в поис-
ках градиента, либо на привлечение воды, содержащей раздражитель
(вододвижение), либо на самостоятельное перетекание воды придон-
ных течений, либо на перемещение источника запаха, либо на всё
это вместе взятое и в различных сочетаниях. Поскольку абсолютная
скорость ползания моллюска, имепцего (Уголь своеобразный движитель
как нога невелика, вододвижение приобретает исключительное значе-
ние для активного восприятия запаха.
Главным органом дистантной хеморецепции у моллюсков остаётся
осфрадий (рис.9 Б,В), хотя достаточно чувствительные хеморецеп-
торные структуры имеются в головных щупальцах {Aply&ia), мантии,
особенно на её краях {Jones, Saleuddirifl^B). Возможно, что осфра-
дий, связанный с ЦНС {Newell,Brown, 1977), выполняет не столько-
обонятельную функцию, сколько регулирует дыхание, поскольку восп-
ринимает различия в концентрации кислорода (Винников,1979). У пе-
реднежаберных насчитывается не менее 5 типов строения этого орга-
на {Maeda, 1983), что можно считать косвенным свидетельством
многозначности этого органа.
Вододвигательную функцию выполняет ресничная выстилка мантий-
ной полости. Поэтому вполне естественно, что осфрадий находится в
мантийной полости. Когда предки брюхоногих опустились на дно, зна-
чение хеморецепции в жизни животного увеличилось, сделалось целе-
сообразным переднее положение осфрадиев и необходимой для их фун-
кционирования мантийной полости. Это могло бы послужить причиной
торсиона.
Сам по себе торсион ещё не вызывает у животного наружной дис-
аометрии. Он приводит к перехлёсту плевро-висцеральных стволов
ЦНС (рис.З А 2;рис. 8 Д 3), закручиванию кишечника и двух долей
н^еварительной железы. Кроме того, правая колюмеллярная мышца
31
становится отличной от левой мышцы. После торсиона эндогастричес-
кая планоспиральная раковина остаётся в сагиттальной плоскости.
Диссимметрия вырабатывается в связи с переходом к турбоспиральной
раковине. Причины этого явления остаются, пожалуй, наиболее зага-
дочными. В теории диссимметрии принимается, что турбоспиральная
раковина выгодна тем, что представляет из себя большее приближе-
ние к сфере, нежели дисковидная планоспиральная раковина. Таким
образом сберегается строительный материал для раковины, потому
что из всех фигур с одинаковой поверхностью именно сфера имеет
наибольший объём. Указывается также, что турбоспиральная ракови-
на обладает повышенной прочностью вследствие того, что лучше за-
щищена внутренняя поверхность соприкасающихся оборотов раковины.
Однако полезно при рассмотрении этого вопроса учесть сенсорную
функцию мантийной полости, в которой находятся осфрадии.
Если бы вода, приносящая запахи, притекала к осфрадию со всех
сторон через широкое раскрытие мантийной полости, то определение
направления на источник раздражения было бы затруднено. Для того
чтобы определить искомое направление, воду к осфрадию надо подво-
дить с разных, но каждый раз с определённых направлений. Эту за-
дачу позволяет решать мантийный сифон, особенно развитый у хищ-
ников и некрофагов (рис.10 Г,Е), хотя определённое значение при
этом сохраняет подвижность всего внутренностного мешка. Подвижным
сифоном моллюск как бы обнюхивает перед собой пространство, приб-
лизительно отвечапцее четверти сферы. Совершенствование этого сво-
еобразного ощупывания пространства не предполагает биноринальности(
Рис. 10. А - строение Skeneopsisplanorbis, Trochacea, - пример умень-
шения размеров жабер вследствие миниатюризации животного (Fretter,
1948). Б - Marsenia rhombica, Lamellariacea, с мантией, покрывают
раковину (Behrens,1980). в - пара жабер в мантийной полости
Fissurella, Dlotocardia (Boutan из Hyman, 1967). Г - мантийная полость
Cfypeomorus bifusciata, Monotocardia (Houbride, 1985) с единственной
жаброй. Д - слизневидный переднежаберный моллюск Titiscania shiukishi'
nattai (Taki из Franc, 19686). E -Erato voluta, Cypraeacea , нападаю-
щий на сложную асцидию Botryllus scholsseri (Fretter, 1951)
32
A
семенной
мешок
семенник
простата
почка
card.
желудок
радула
челюсть
отверстия
желез :
передней
задней
пищеварительная
железа
и»
пищевод ганглии;
церебральный,педальный
буккальный
слюнная железа
колюмеллярный мускул
крышечка
раковина
мантия
вводной сироп
проподиальная борозда
ресничная борозда
анус
гонопора
сифон
мантия
card.
Г
семенная бурса
почка
выделительное отверстие
яйцеводная
борозда
33
имеет значение только градиент запаха во времени. Поэтому отпа^
ет необходимость в двух осфрадиях, которыми, согласно теории,
начала обладал гипотетический предок гастропод и которыми в дейс
вительности обладают некоторые Archaeogastropoda с симметрична
расположенной мантийной полостью с двойным набором мантийных орт
нов (рис.10 В).
Для совершенствования хеморецепции единственногооофрадия неое
дима однонаправленная диссимметричная схема циркуляции воды, осне
ванная на принципе "один вход-один выход". Именно по такому прщ
ципу функционирует мантийная полость с единственным приводящим
сифоном, в которой разобщены противонаправленные токи воды - прр
носящий и выносящий. Самое подходящее место для мантийного сифс
на - передний край мантийной полости возле головы, в сагиттально
плоскости моллюска. По остающимся неизвестным причинам функция
приведения воды осталась именно за левой частью мантии (левый о
фон). Приведение левой части мантии в сагиттальную плоскость оз-
начает частичную деторсию (рис.6 Д4). В действительности же
всл- пие подвижности внутренностного мешка и сифона последний
может не достигать сагиттальной плоскости и тем не менее успели:
осуществлять свои функции. Деторсию претерпело около 17% видов
брюхоногих (Graham, 1979).
После деторсии мантийная полость размещается диссимметрично
обладает собственной диссимметрией уже" потому, что содержит едш
Рио.Й. А - циркуляция воды в мантийной полости архигастропод: 1-й
dora,Fis8urellidae, с парой ктенидиев, 2- Acmaea, Patellacea, с левы»
ктенидием, 3-Patina , Patellacea, неполное кольцо мантийных жабе!
4-Patella, Patellacea, полное кольцо мантийных жабер; Б - строение:
phonaria,Р/уа51ЬгапсЬха:1-дорзально, мантия удалена, 2- удалена рано!
на, органы на потолке мантийной полости (Yonge, 1952а). Сокращен^
см,рис.10. Прерывистые стрелки обозначают токи воды, сплошные - bxoJ
щий и выходящий токи, оперённые стрелки - удаление частиц в ресни<
ных токах на полу мантийной полости. Лёгочные моллюски: В - Otina
otis (Morton, 1955), Г-Gundlachia japonica, Ancylidae (Burch, 1964) ,Л‘
Ferrissia burnupi (Brown, 1967)
34
мантия
пневмостом
анус
мантия
раковина
почка
3
колюмеллярная (раковинная) мышца
висцеральная масса
Д
кишка
мочеточник
почка
-задняя
раковинная
мышца
половое отверстие с* £ с
левая
передняя
раковинная
мышца
мантийное
отверстие
аглютинат
оз.
card.
выделитель-
ное отверстие
почка
6г.
2 вентральный ресничный
гребень, отделяющий передний
отдел мантийной полости от
заднего отдела
отверстие мантийной полости
и анус
ственный левый осфрадий. Вследствие того, что д-виждния воды сдела-
лось однонаправленным, из двух ктенидиев предка, ещё сохраняющих-
ся у некоторых ArchaeogastropodapoxpaKZxea один левый ктенидий
возле приводящего сифона. Поскольку раковину можно рассматривать
как обызвествлённую траекторию мантийной полости, в процессе рос-
та сдвигаемой вдоль оси навивания, диссимметрия мантийной полости
уже сама по себе служит причиной турбоспиральности раковины.
Турбоспиральность породила ещё одну проблему: центр тяжести ра-
ковины и внутренностного мешка вышел из сагиттальной плоскости.
В теории диссимметрии принимается, что эта проблема была решена
так: завиток раковины был отведён немного назад (деторсия) и нак-
лонён так, чтобы ось навивания раковины одним концом была направ-
лена вверх и вправо, таким образом центр тяжести раковины и внут-
ренностного мешка был приведён в сагиттальную плоскость. Выше для
деторсии было предложено дополнительное объяснение: деторсия выз-
вана тем, что приводящий сифон и единственный (левый) осфрадий
приведены в сагиттальную плоскость моллюска для успешного
осуществления сенсорной функции.
Во внутренностном мешке торсион привёл к редукции правой доли
пищеварительной железы, которая исходно была парной и располага-
лась в сагиттальной плоскости, что фактически наблюдается у личи-
нок. Впрочем, А.В.Иванов (1940) заключает, что из-за широкой из-
менчивости этого 0'ргана правильные отношения между его частями мо-
гут быть установлены не всегда. Другое следствие торсиона - скру-
чивание кишечнике, и смещение задней кишки вправо. Это вызывает из-
менения в расположении ретракторов ноги. У предка переднежаберных,
видимо, исходно имелись две такие мышцы - правая и левая. При тор-
Рис.12. А - желудок Sphyradium doliolum, Pulmonata, (Шилейко, 1984).
Б - пищеварительный тракт Akera bullata, Opisthobranchia (Morton.,
Holme, 1955) t обозначения: st- желудок, div. - место отхождения кишеч-
ного протока. В - передняя часть пищеварительного тракта Dolabella
auriculata, Opisthobranchia
зв
пищевод
ссринктер
зоб
кольцевая мышца
сфинктер
сфинктер
зоб
кишечник
жевательные
зубцы
первый
пышечный жел
желудочно - кишечный
клапан (g.v.)-
dd
тифлозоль
Б
St.
спинной
проток
второй
'желудок
отверстие задней
кишки
' отверстие протока
нижней доли
печени
проток пищеварительной
железы (d.d.)
„ Проток
в пищеварительную
железу ?
хватательные
зубцы
щетинки
кишечные клапаны
(iv.)
промежуточная камера
между первым и вторым
желудками
фильтровальная
камера .
желудочный клапан
задняя
киш КЗ
А
37
сионе задняя кишка прошла слева от левого ретрактора, который
впоследствии разросся в единственную сохранившуюся так называемую
колюмеллярную мышцу, а правый ретрактор подвергся полной редукции.
Мантийная полость и нога в их разнообразных модификациях позво-
ляют брюхоногим питаться разными способами - от фильтрации до хищ-
ничества. Поэтому строение пищеварительной системы у брюхоногих
довольно разнообразно.
При питании мелкими пищевыми частицами, добываемыми посредством
фильтрации и соскребывания, определённое значение должно сохранять
внутриклеточное пищеварение, к которому вообще предрасположены все
микрофаги (Беклемишев, 1964). Основной пищедобывающей структурой
у брюхоногих остаётся радула и поэтому пищевой материал неодноро-
ден, состоит из частиц неодинаковой пищевой ценности и нуждается
в сортировке. Последнее происходит на внутренней поверхности же-
лудка, и там же существует механизм распределения пищевых частиц
с помощью ресничного эпителия.
При макрофагии и поступлении в пищеварительный тракт крупных
кусков пищи преобладает полостное пищеварение, а распределение пи-
щевого материала в пищеварительном тракте достигается посредством
перистальтики кишки (Purchoiz, 1977). Длина кишечника в ряде
случаев скоррелирована с составом пищи. У слизней-фитофагов она
превосходит таковую хищников (Лихарев, Виктор, 1980). Предвари-
тельное размельчение пищи у макрофагов происходит в мышечном же-
лудке как у фитофагов (Fretter,Ko Вил. Шал ,1979 ), так и у плото-
ядных форм (рис.12).
Полостное и внутриклеточное пищеварение не исключают одно дру-
гого. Например, у морского блюдечка Otina (рис.II В) сначала
совершается полостное пищеварение, причём пищеварительные фермен-
ты выделяются двумя желудочными выростами. Затем пищевые частицы
перистальтическим способом поступают в желудочные выросты, которые
содержат не только секреторные, но и пищеварительные клетки, и там
фагоцитируются. В желудке непереваренные частицы пищи обволакива-
38
фея слизью, выделяемой поступающим из средней кишки протостилем,
и образуют фекальный шнур. Протостиль (рис.9 Г), хотя и не содер-
пищеварительных ферментов, однако рассматривается в качестве
предшественника обычного у двустворчатых кристаллического стебель-
gg с его широкими функциями (Morton, 1955). Сравнительно немногие
брюхоногие имеют кристаллический стебелёк, сохраняющий при этом спи-
ральную слоистость (Driscoll, 1972), но такие моллюски лишены
пищеводных желез: некоторые Cerithacea, Calyptraeacea, Rissoacea,
Strombacea (Fretter, 1962). Ч.Янг (Younge, 1930) описал эту кор-
реляцию в следующих словах: "Кристаллический стебелёк и хищничест-
во не могут нормально сосуществовать".
Отмечая широкую изменчивость строения брюхоногих моллюсков, А.
В.Иванов (1940) выделяет четыре основных типа организации.
I.Переднежаберные двупредсердные (Prosobranchia Piotocardia),
получившие впоследствии название Archaeogastropoda, облвдвхп бо-
лее или менее симметричным мантийным комплексом органов (рис.13
А,Б) и поэтому остаются наиболее примитивными среди ныне живущих
брюхоногих. Они имеют хорошо обособленный, закрученный в турбоспи-
раль внутренностный мешок, ногу с ползательной подошвой и крышеч-
кой. Впрочем, раковина иногда имеет форму колпачка и крышечка от-
сутствует. У ряда видов (Fissobranchia) раковина и мантия меди-
ально с вырезкой на переднем крае или снабжены отверстиями (рис.
7; рис.9 К). Сердце, задняя кишка и анус медиальны, с боков от
них находятся следующие органы: ктенидии, осфрадии, гипобранхи-
альные железы. Ктенидии представляют из себя аспидобранхии (дво-
якоперистые ктенидии), прикреплённые только основанием. Сердце
состоит из желудочка и двух предсердий и заключено в перикард, ко-
торый сообщается с парными почками через рено-перикардиальное от-
верстие. Левая почка обычно меньше правой, через последнюю откры-
вается единственная гонада, занимающая первые обороты внутренност-
ного мешка (рис.2 А). Кишечник с анопедиальным изгибом, задняя
окружена желудочком сердца. Имеется 2 доли пищеварительной
39
Рис. 13. Основные типы организации
А, Б - Prosobranchia Diotocardiac 2 жабрами, 2 предсердиями и 2 поя
ками; В, Г - Prosobranchia Monotocardia; Д, Е - Opisthobranchia;
Н, 3 - Pulmonata. Обозначения: ап- анус, ао. - аорта, ^.-брахиаль-
ный ганглий, Ъг.-жабры, с.-церебральный ганглий, са,- caecum,, d. -зад-
няя кишка,g.-гонады, gastr- желудок, h- печень, hbr. гипобранхи-
альная железа, т.п.~ мантийный нерв, ое.-пищевод, os-осфрадий, ov.-яй-
цевод, ра.- паллиальный ганглий, ph.- глотка,pg-педальный ганглий или
40
брюхоногих моллюсков (Иванов,1940)
педальный ствол, pl- плевральный ганглий, prt.- париетальный ганглий,
Pcrd- перикард с сердцем, ге - почка или мочеточник, r.s.- семеприем-
,1ИК» sb. - субинтестинальный ганглий, s.gl.- слюнная железа,spr- супра-
интестинальный ганглий, д-семенная борозда на пенисе, t.- семенник,
кольцевая вена, vd.-семепровод, v.g’.-висцеральный ганглий, V.m.-
мышечный желудок. Симметричная (К) и диссимметричная схема циркуля-
ции воды (ИД)в мантийной полости (Golikov, Starobogatov, 1975)
41
железы (печени), каждая из которых самостоятельно открывается в
желудок. ЦНС с хиастоневрией: левый плевро-висцеральный коннектив
проходит над кишкой, правый занимает субинтестинальное положение.
2.Переднежаберные однопредсердные (Prosobranchia Monotocardia)
тоже имеют турбоспиральную раковину, внутренностный мешок вмещает
кишечник с анопедаальным изгибом и единственную гонаду (рис.13 В,
Г). Обычно сохраняется хиастоневрия. В левой половине мантийной
полости присутствует одинарный набор органов: гребенчатый ктени-
дий, один осфрадий и единственная гипобранхиальная железа. В сво-
бодной половине мантийной полости находятся задняя кишка с анусом
и половое отверстие. Вытекающее из полового отверстия самца семя
дальше течёт по боковой ресничной борозде к пенису, который пред-
ставляет из себя боковой вырост ноги. В связи с полной редукцией
правого ктенидия отсутствует и правое предсердие, перикард сооб-
щается с единственной сохранившейся левой почкой, однако сердце
занимает боковое, не медиальное положение, и задняя кишка прохо-
дит в стороне от желудочка. Мантийная полость, таким образом, дис-
симметрична. Двупредсердные подразделяют на Mesogastropoda и
Neogastropoda . К последним относятся высокоспециализированные
в основном хищные Buccinidae, Conidae, Nassariidae, Olividae и др.
Вместе с однопредсердными двупредсердные объединены в группу
Streptoneura,T.e. имеющих хиастоневрию. Они противопоставляются
лишённым хиастоневрии лёгочным и заднежаберным моллюскам, Euthy-
пеага.
3. Заднежаберные, отличаются разнообразием внеш-
него облика и внутреннего строения (рис.13 Д,Е). У них нет крышеч-
ки, часто они лишены раковины и внутренностного мешка с мантийной
полостью, а нога не имеет подошвы. Многие из них имеют параподии,
способные загибаться на спину и закрывать раковину, ограничивая
псевдомантийную полость. Сохраняющаяся иногда мантийная полость
содержит заднюю кишку с анусом и одинарный набор левых органов:
гипобранхиальная железа, волнообразно изогнутый (пликатный) ктени-
42
сердце в перикарде, связанном с единственной почкой, ЦНС эу-
илевральног0 типа, т.е. без хиастоневрии.
4.Лёгочные, Pulmonata, (рис. 13 Н,3) иногда лишены раковины и
бособленного внутренностного мешка. Мантийная полость открывает-
ся наружу дыхательным отверстием и содержит следующие органы:
[ёгкое (система кровеносных сосудов) на месте легого ктенидия, ос-
фадяй, однопредсердное сердце в перикарде, который сообщается с
.динственной (правой) почкой , мочеточник и задняя кишка с анусом,
крышечка отсутствует. ЦНС эутиневрального типа.
Помимо системы типов организации подклассов брюхоногих моллюс-
{ов, разработанной А.В.Ивановым, существуют экологические класси-
фикации жизненных форм для отдельных подклассов. Они основаны на
трофическом принципе и учитывают особенности местообитания {Pur-
chon, 1977). Это, в сущности, классификация жизненных форм таксо-
нов. В.А.Свешников и А.Б.Станкявичус (1987) предложили систему
дорских переднежаберных моллюсков, учитывающую образ жизни и сход-
ство в пропорциях раковины; это - система фенонов жизненной формы
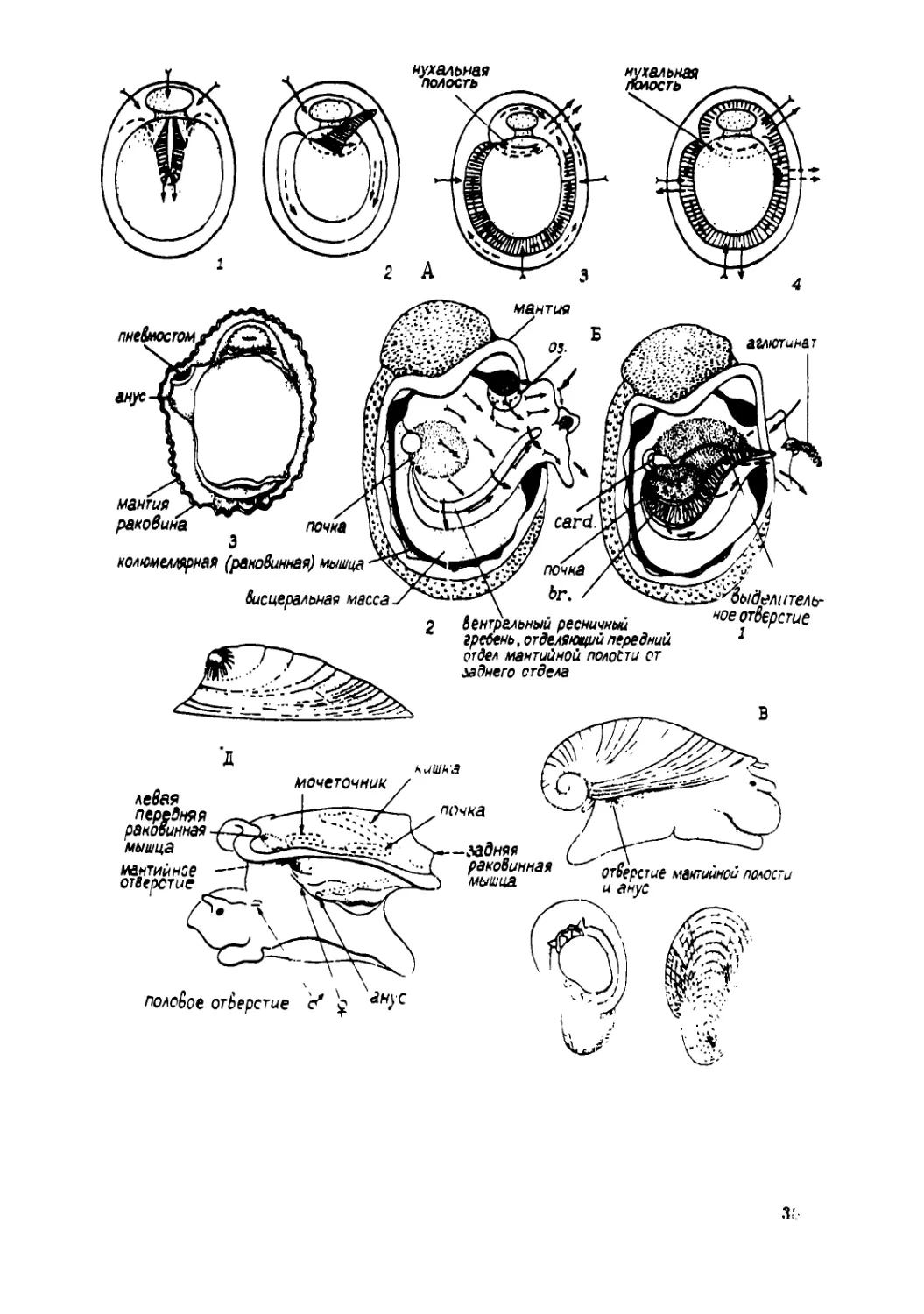
таксона переднежаберных.
Для сравнительно-анатомического изучения брюхоногих основой
остается система А.В.Иванова, усовершенствованная рядом авторов
(Миничев, Старобогатов, 1979; Golikov, Starobqgatov, 1975). Фактичес-
ки она основана на корреляциях между такими органами и структурами,
как мантийная полость, раковина, органы мантийной полости, строе-
ние ноги и пр., однако довольно часто физиологическое объяснение
таких корреляций либо остаётся неизвестным, либо носит предполо-
жительный характер. Важно подчеркнуть, что в рамках сравнительно-
анатомического исследования задачей остаётся построение системы
типов организации, фактически в той или иной степени совпадающих
с таксонами, что же касается филогении, то она полезна для истол-
кования полученных результатов, но не является целью исследования,
водным образом результаты классифицирования по сходству в гомоло-
^еских признаках могут рассматриваться с точки зрения филогене-
ТйКа» но сама филогения не есть цель систематики.
43
Дальнейшая детализация системы А.В.Иванова предполагает прив>
лечение всё более мелких признаков и частных корреляций (Табл.2),
Использование в системе таких экологических понятий, как например ।
паразитизм, несомненно есть отступление от собственно сравнителод
анатомического принципа классифицирования. Такие отступления вывь
дят исследование в несколько иную область, в сравнительную анато»
мию жизненных форм. Такие отступления трудно избежать и вместе с
тем они обогащают узко сравнительно-анатомическую классификацию.
Сравнительное изучение мантийного комплекса органов позволяет
прежде всего выявить топографические корреляции, которые выражаю!
ся во взаиморасположении органов.
У археогастропод Fissobranchia , имеющих вырезку на перед,
нем крае мантии и раковины (рис.7 К; рис. 9 К), щель или ряд от-
верстий (рис.7 В), наблюдается симметричная схема циркуляции вод
В мантийную полость вода проникает с боков, омывает ктенидии, пр
текает между жаберными листочками двух ктенидиев (рис.10 В) им
мо гипобранхиальных желез и выделительных отверстий, наконец выте>
ет наружу через вырезку (рис.2 Б 1; рис.13 К). У морского уха На-
Hotis (рис.7 Б) вода в мантийную полость притекает с обоих бокс
возле головных щупалец, а также через два крайних отверстия на
спинной стороне, а выходит через прочие три отверстия (Voltzow,
1983). Приносимые водой частицы осаждаются в слизевых потоках рео
яичного эпителия жабер и гипобранхиальной железы и выводятся на-
ружу (рис.14 I).
У брюхоногих, не имеющих вырезки края устья или отверстий, осу
ществляется диссимметричная схема циркуляции, отвечающая левой по
ловине таковой Fissobranchia (рис.2 А 3,4', рис. 13 И; рис.14 А-
Е): вода притекает слева и покидает мантийную полость с правой ст
роны. При этом органы мантийной полости размещены таким образом,
чтобы не мешать друг другу выполнять свои функции и не препятство-
вать циркуляции воды. В частности, это относится к жабрам и осфрб
дням. Они находятся возле вводного отверстия и поэтому омываются
44
Табл.2. Основные типы организации брюхоногих моллюсков
-духание совершается благодаря вододвижению и воздуходвижению.
ХХ.Ползательная нога.
III.Ресничный двигательный аппарат.
IIII.Вододвигательная мантийная полость, раковина, внут-
ренностный мешок.
IllII.Примерно симметричная схема циркуляции воды в
мантийной полости, двойной набор органов мантий-
ного комплекса...............Archaeogastropoda
НПО. Диссимметричная схема циркуляции воды в мантий-
ной полости, одинарный набор органов мантийно-
го комплекса............Раковинные Mesogastropoda,
Neogastropoda, Opisthobranchia, Pulmonata inpartium
IIIO.Мантийная полость, раковина и внутренностный мешок
редуцированы, имеются спинные жабры..Nadibranchia
ПО .Мышечный воздуходвигательный аппарат.
1101.Хорошо развитые раковина и дыхательная мантийная по-
лость с лёгкими............Раковинные Pulmonata, улитки.
ПОО.Безраковинные слизни о редуцированной мантийной по-
лостью и кожным дыханием.. .Слизни Pulmonata
10.Вододвигательная поршневидная нога в псевдомантийной полости
неподвижного моллюска.........Эктопаразитические Gastropoda
3.Специальный вододвигательный аппарат для дыхания отсутствует
01.Сохраняется способность двигаться, имеются те или иные произ-
водные ноги.
ОН.Малые размеры тела, дыхание через поверхность тела, реду-
цированные раковина и внутренностный мешок, ползание на
ресничной подошве ноги..........Opisthobranchia in partium
010.Средние размеры тела, плавание о помощью выростов ноги
мышечным способом.
0101.Плавание с помощью взмахов крыловидных выростов но-
ги, раковина имеется или отсутствует... Pteropoda
0100.Плавание посредством изгибания удлинённого тела и
непарных плавников...........................Neteropoda
ОО.Вивотиое неподвижно, многие системы органов подверглись редук-
Wui............................Эндопаразитические Gastropoda
45
свежей водой. Эти топографические корреляции сохраняются неси.
1
на то, что строение ктенидиев не остается неизменным: помимо^
коперистых аспидобранхиев, свободно лежащие в мантийной поло^
или подвешенныхк её потолку на мезентериях (рис.2 Б 3), могут
встречаться складчатые жабры на потолке мантийной полости (р^
Б 4 ).
Совершенно иной тип циркуляции устанавливается у тех архео^
тропод, у которых ктенидии полностью редуцированы и их отсутсц
восполняется многочисленными кожными складками, вторичными жц
ми. Они размещены в циркумпедальной борозде и, образуя полное
или неполное кольцо, служат поводом для названия Cyclobranchk
(Миничев, Старобогатов, 1979)(рис.12 А 3,4 ). Вследствие силь»
развития дыхательной циркумпедальной борозды и кольцевой раков
ной мышцы такие моллюски имеют развитую лакунарную циркумпедэлк
систему и циркумпедальное нервное сплетение (рис.15 А),
Нечто подобное наблюдается и у морских блюдечек Siphonariidae (рис,
Б 1,2).
Мантийная полость, лишённая жабер, но сохранившая оофрадий,
получила название нухальной полости (рис.15 Г,Д). Таким образа
у Cyclobranchia совершилось разобщение двух функций мантийн4
полости: дыхательной и сенсорной. Первая перешла к кожным жабр
циркумпедальной борозды, вторая осталась за нухальной полостью.
Рис. 14. Схема токов воды (сплошные стрелки) и ресничных токов (г
рывистые стрелки) в мантийной полости переднежаберных на поперек
срезе Pectinibranchia (А): Б - Viviparus , В - Struihiolaria, Г - Crepif
Д-SerpulorWs (Vermetldae), Е -Pyxipoma (Siliquariidae) (по разным ав^
из Owen, 1966). Ж - то же у Diodora, Diatocardia (Fretter, Gra№
1976). 3 - строение мантийной полости у слизня (Franc,1968а). Сокра^
£ - вынос наружу грубых частиц, перенос средне размерных час?
слева направо для удаления выносящим током воды, С- перенос мел5
частиц между жаберными листками и вдоль жабры для склеивания ча#1
слизью гипобранхиальной железы, еп,- эндостиль, с/. слизевой
на фронтальной поверхности жабры, fg.~ пищевой желобок,fs.- пиша*
шнур, pg. - педальная железа, г.- задняя кишка. Прочие обозначения 0
рис. 7
46
47
находящейся над головой. Показательно, что в нухальной полости
сохранилась собственная система циркуляции воды. Это обстоятель-
ство подтверждает важную роль сенсорной функции изначала единой
мантийной полости,
Дж.Мортон {Morton, 1956) проводит аналогию между мантийной
полостью моллюсков и глоткой асцидий и кишечнодышащих, пронизан-
ной жаберными щелями. Эта аналогия подразумевает не только сход-
ство в пищедобываицей и дыхательной функциях, но ещё и сходство
в сенсорной функции.
Симметричная схема циркуляции наблюдается не только у моллюс-
ков с более или менее симметричной колпачковидной раковиной
(рис.14 Н), но и у археогастропод с турбоспиральной раковиной
(рис.13 К), причём для последних характерна лёгкая диссимметрия,
которая выражается в том, что левая жабра несколько превосхо-
дит правую. С другой стороны, моллюски с симметричной колпачко-
видной раковиной могут обладать несимметричной схемой циркуляции
(рис.12 А 2,3,4 ). Эти обстоятельства ещё раз подчёркивают неод-
нозначность корреляций. Тип циркуляции в мантийной полости и
строение жаберного аппарата могут определяться и другими мало
изученными факторами (не только строением раковины). К числу та-
ких факторов относится и мантия, точнее, степень её развития.
Иногда мантия разрастается настолько, что почти скрывает ракови-
ну (рис.10 Б,Е). Обширная поверхность такой мантии не может не
иметь значения для дыхания. Вполне возможно, что она может выз-
Рис.15. Строение Docoglossa , Archaeoga&tropoda : А-ЦНС Астаеа тагси-
si (Rlghi, 1966), S- внутренностный мешок Астаеа (Walker,1968),в - попе-
речный срез Patella (Fretter, Graham, 1976). Г - жаберный листок Астаеа
(Walker. 1968). Сокращения: B.G.-буккальный ганглий,C.P.D. - церебропе-
дальный коннектив,С./?1.-цереброплевральный коннектив, L.G. - лабиаль-
ный ганглий, L.O. - левый ос^радиальный ганглий, PD-правый педальный
ганглий, RW.- правый почечный ганглий, R.O.- правый осфрадиальный ган-
глий.УЛГ.-висцеральный нерв. Прочие сокращения см.рис.7. Д - Patella
с брюшной стороны (Franc, 19686). Е - раковина Diodora, Fissurellidae
(Franc, 1968 а)
4Я
церебральная комиссура , . -о
. передний лабиальный нерв
церебральный ганглии г п о
г г щупальцевый нерв
глазной нерв
нухальная полость
педальный
ганглий
B.G.
P.D.\
педальный ствол
Б
кожные жабры
циркумпаллиальный нерв
колюмемярная (раковинная) мышца
челюсть
радула
пищевод
прямая
кишка
1_ проток
Пшцевари-
' тельной
। железы
В
49
вать уменьшение размеров ктенидия и ослабление циркуляции воды
в мантийной полости. Другой важный фактор - это размеры органу
ма моллюска (Fretter, 1948). Известно, что мелкие брюхоногие с
раковиной диаметром около 1мм вообще лишены жабер ( Omalogyr^
Rissoella) и движение воды у них вызывается только узким выно-
сящим мантийным ресничным желобком, который прослеживается от
ануса до края мантийной полости. Приносящий ток воды возникает
всего лишь как компенсация при отсутствии соответствующей nfo
структуры, прежде всего приводящего сифона. Если всё же ктени-
дий у мелких моллюсков сохраняется, то он состоит всего лишь из
9 жаберных нитей (рис.10 А), тогда как у более крупных Turrite]
lidae насчитывается 12, притом разветвлённых жаберных нитей (Moi
ton, 1953). Другой пример отсутствия жабер - это фреатические 1г
atodrobia, высота раковины которых составляет всего лишь 0,67 и
(Herschler, 1987).
Диссимметричная схема циркуляции особенно распространена ср
переднежаберных однопредсердных брюхоногих, сохранивших только
левые органы мантийной полости (рис.10 Г; рис.14 А-Е). У лёгош
так же, как у головоногих, осуществляется маятниковая циркуляр
(вдох-выдох) воздуха или воды (увторичноводных лёгочных) и ш
этом они обходятся единственным дыхательным отверстием, пневмо!
томом (рис.12 Б,В,Д; рис.13 2,3). Этим они отличаются от перед!
жаберных, имеющих сквозную однонаправленную циркуляцию с обосо
ленными вводным и выводным отверстиями мантийной полости. Так#
цикличность (вдох-выдох) объясняется тем, что вододвижение и J
духодвижение вызывается мышцами. У лёгочных и головоногих объё
воздуха (воды) при вдохе-выдохе ведёт себя как единое целое,
вдох и выдох следуют друг за другом во времени. У переднежабе?
—'
Рис. 16. A-Buccinum uxidatum с длинным ротовым хоботом и отведёнЕ^
вправо и вверх мантийным сиаоном (Сох, 1960). Б-Viviparus vivipZ
Cook, 1949) В - Struthiolaria (Morton, 1951). Г- Crepidula (Frart
1.968 6). Д- Calliostoma papillosum (Fretter, Graham, 1962). A-Gadir^
n.ivea , Pulmonata (Walsby et al., 1973)
50
51
с сквозной циркуляцией в непрерывном процессе газообмена и разд-
ражения осфрадия фазы вдоха и выдоха не выделяются, обновляемая
вода используется постепенно по мере её протекания через мантий-
ную полость. Такая непрерывность объясняется тем, что вододвиже-
ние вызывается у этих моллюсков ресничным аппаратом, обнаружива-
ющим свою действенность именно в водной среде; на воздухе он утра-
тил бы свои достоинства вследствие малой по сравнению с водой вяз-
костью воздуха. У вернувшихся в водную среду вторичноводных лё-
гочных морских блюдечков восстанавливается некоторое подобие
сквозной схемы циркуляции. У них единственный пневмостом приобре-
тает щелевидную форму, что позволяет разобщить противонаправлен-
ные токи воды (как сифоноглифы у актиний). Вода входит в мантий-
ную полость у переднего края пневмостома, омывает осфрадий, поч-
ку и вторичную жабру в передней части мантийной полости и выво-
дится наружу через обособленный задний отдел мантийной полости и
задний край пневмостома (рис.12 Б 1-3). В данном случае замеча-
тельно, что в связи с появлением вторичной жабры лёгочный мол-
люск приобрёл сложную мантийную полость и возможность сквозной
циркуляции воды.
Удаление из мантийной полости посторонних частиц - жизненно
необходимая функция у брюхоногих моллюсков. Она свойственна уже
археогастроподам. Для предотвращения загрязнения поверхности ман-
тийной полости, жабер и осфрадия посторонние частицы склеиваются
слизью в аглютинаты (псевдофекалии), выводимые наружу. Слизь вы-
деляется прежде всего гипобранхиальной железой. У Mesogastropoda
эта функция неотделима от слизевой фильтрации пищевых частиц в
Рис. 17. A- Vermetidae A- Dendropoma squmifera, поперечный срез (fonder,
1967), Б - Vermicularia spirals передняя часть с вскрытой
мантийной полостью (Morton, 1953), В - раковина Dendropoma squami-
fera (Ponder, 1967). Г - Siliquaria australis , Siliquariidae, раковина (Mac-
pherson, Gabriel, 1962). )%-Serpulorbis aureus, Vermetidae, раковина
(Hughes, 1978a).E - Crepidula fornicata, Calyptraeidae: схема поперечн
l'o среза (вверху) и спаривание (внизу) (Hoagland, 1978)
52
колюмеллярная мышца
53
противотоках слизи (Yonge, 1966). В простейшем случае, кзк у
живородки Viviparus (рис.16 В), могущей питаться с помощью раду-
лы, как большинство брюхоногих, фильтрационное питание сохраняет
подсобное значение. Взвешенные частицы обволакиваются слизью на
жабре, и получившийся таким образом аглютинат соскальзывает в пи-
щевой желобок, прикрытый нависающей складкой кожи. В пищевом же-
лобке формируется пищевой шнур, который направляется в рот рес-
ничным эпителием. Переднежаберный моллюск Bythinia может питаться
исключительно посредством фильтрации жгутиковых простейших, при-
чём аглютинаты запасаются впрок (Meier - Brook, Chong Ниап Kim.) •
Более совершенные фильтраторы - малоподвижные Crepidula (рис.
16 Г; рис.17 Е) и Struthiolaria (рис.16 В) имеют жабры особого
строения. Они образованы длинными жаберными нитями, у которых от-
ношение длины к ширине доходит до 26, обычно же у брюхоногих име-
ются относительно широкие треугольные жаберные листки (рис.15 Б).
Широкие жабры, образованные удлинёнными жаберными нитями, подраз-
деляют жаберную полость на поджаберный и наджаберный отделы
(рис.14 В,Г). Такие жабры напоминают жабры двустворок, типичных
фильтраторов, но в отличие от последних они лишены фронто-лате-
ральных ресничек. В формировании аглютината участвует слизевая
железа эндостиля (названная так по аналогии с бесчерепными), рас-
Рис . 18 . А - движение моллюска нога пеликана Aporrhais pes -peli -
cani, Strombacea (1-6), вид co стороны устья, вид со стороны осно-
вания, преодоление препятствия и положение животного в грунте (We-
ber, 1324; Уо/ge,1937/Обозначения: В,В'-основные точки опоры при дви-
жении, z'- вырост наружной губы, Z - передний вырост, сросшийся с
ситоном, «—► расстояние, которое преодолел моллюск. B-Olivella
colurnellaris,Stenoghteaip ловчими слизевыми мешками на поверхности
грунта (1) и общий вид животного (2) ( х -педальный por)(5eiiacjier,
von, 1359). в -Umbonium vestiarium, Trochacea: 1- общий вид, 2-
вид сбоку, 3 - вводной (in.siph.) выводной (ex.siph.) сифоны и щупальца
зарывшегося в грунт моллюска (Fretter, 1975). Г - передний конец
Dendropoma corallinacea , Vermetidae, во время спильтрации (1) и при
выделении ловчей слизи (2) (Hughes, 1976 6)11 -Qadinalea nivea, Pulmona-
ta, в естественном положении (Walsby et al., 1973)
54
3
2
1
55
положенная возле жабры. Аглютинаты накапливаются в пищевом желоб- '
ке, и реснички направляют их в рот. Для защиты мантийной полости
от грубых частиц Crepidula имеет слизевую сеточку на входном
отверстии мантийной полости (рис.16 Г). Зарывающиеся в грунт
Strathioiaria, по-видимому, обходятся без такого фильтра.
Фильтрационное питание свойственно по меньшей мере 6 семейст-
вам переднежаберных: Calyptraeidae, Capulidae, Siliguariidae,
Struthiolariidae, Turritellidae, Vermetidae, а также
Gadinalea, Valvata,Bythiria(Hughes,'19bS; Jorgenson, 1966; УЬп^е,1966).
Исключительно фильтрационном способом питаются прикреплённые
Siliqaariidae, Vermetidae (рис.17 А-Д). Разросшиеся жабры этих
моллюсков устроены по-разному: у первых они образованы обычными
треугольными листками, у последних они состоят из упругих нитей.
Моллюски высовывают их из раковины и, размахивая, отлавливают сео
тон. Показано, что у Vermetes alii сестон осаждается на рес-
ничном эпителии жаберных нитей (Hadfield etal., 1972). Нако-
нец, эти моллюски могут осуществлять фильтрацию с помощью нитей
слизи, выделяемой ножными щупальцами. Нити образуют ловчую .сеть,
которая вместе с уловом - прилипшими к- ней частицами, пожирается
моллюсками (рис.18 Г). Интересно, что несколько моллюсков, объе-
динившись, могут построить общую сеть (Hughes, 19786),
Сходным образом питается археогастропода Umbonitun (рис.18
Б). Вводное отверстие широкого мантийного сифона этого моллюска
образовано эпиподием, выростом ноги, и несёт радиальные выступы,
препятствующие проникновению в мантийную полость посторонних час-
тиц. Циркуляция воды в мантийной полости вызывается разросшимися
Рис. 19. Theodoxus fluviatilis, Archaeogastropoda (Whitaker, 1951)
A - вскрытые глотка и пищевод со спины, показаны ресничные токи,
Б - продольный срез глотки и пищевода с ресничными токами, В -
ротовое отверстие спереди :1- закрытое ротовое отверстие, 2-
в начале рабочего хода радула выдвинута вперёд и зубцы расправле-
ны, 3 - сближение радулярных хрящей (г. с. ) и приведение зубцов
радулы к середине в конце рабочего хода
56
Б
57
ктенидиями. Пищевые частицы обволакиваются слизью, выделяемой ги-
побранхиальной хелезой. Аглютинаты по эндостилю и окончаниям жа-
берных нитей соскальзывают к выходу из мантийной полости и там
слизываются хоботообразной мордой. Хорошо развитый сифон Umbonimn,
является производным эпиподиума. Сифон позволяет моллюску осу-
ществлять вододвижение, когда животное погружено в грунт. Впрочем,
Umbonium может отфильтровывать сестон и при обычном ползании
и к тому же не утратил способности собирать пищевые частицы со
дна. Большим своеобразием отличается фильтрационное питание
Olivella (Monotocardia) (рис.18 Б). Зарывшись в грунт, Olivella
выставляет наружу ловчие мешки, образованные выделениями роговид-
ных выростов передней части ноги. Ими улавливаются пищевые части-
цы из придонного слоя воды. Время от времени ловчие мешки втяги-
ваются, и тогда с помощью хобота моллюск пожирает их вместе с уло-
вом.
Наконец ещё один пример фильтрационного питания с помощью сли-
зевого фильтра, на этот раз выделяемого краем мантии и ноги, пред-
ставляет морское блюдечко Gadinalea (рис.16 Б; рис.18 Д). Реснич-
ные пищевые желобки у этого моллюска отсутствуют. Слизевой фильтр
схватывается околоротовыми лопастями. Показательно, что этот
моллюск длиною не больше 2 см малоподвижен, но зато обитает в
местах с сильным течением в волноприбойных вымоинах, прикрепив-
шись к их потолку подошвой, вниз вершиной раковины.
Жизненная форма фильтратора явно не представляет единства
как фенон. Она существует в разных таксонах брюхоногих независи-
мо от их положения в системе, хотя ограничена в основном Me-
sogastropoda. Она встречается и среди пелагических видов, хотя
последние в основном остаются хищниками. Фильтрация может осу-
ществляться совершенно различными морфоструктурами и э'ро обстоя-
тельство служит примером того положения, что одна и та же функ-
ция может выполняться разными органами. Это своего рода муль-
тиструктурность, понятие противоположное мультифункциональности.
58
3 общем же фильтрационное питание у брюхоногих скорее остаёт-
йСКлючением, чем правилом. Для брюхоногих в первую очередь
^ерно соскребывание пищи с помощью радулы. Этот способ пи-
требует длительного контакта животного с субстратом и при-
тгля медленно ползающих животных, неспособных к преследо-
1Нйю жертв и быстрым поисковым движениям с резкими переменами
давления движения. Сравнение с такими хищными ракообразными,
ая например ротоногие, поучительно не только в сравнительно-
натомическом, но и в этологическом плане: ротоногие в противо-
оложность брюхоногим не образуют плотных поселений.
Радулярный аппарат брюхоногих (Чухчин, 1970; Graham, 1979)
бычно включает гибкий одонтофор, который можно сравнить с язы-
:ом, и расположенную на нём радулярную мембрану, на которой раз-
мены поперечные ряды зубов радулы. В одонтофоре находятся ра-
улярные хрящи. Радула по мере истирания возобновляется в мешке
)адулы, представляющем из себя вырост вентральной стороны глотки.
г молодых животных радула возобновляется быстрее, чем у старых
Ъагапкига, Runham, 1968). Наружу радула выдвигается сокраще-
шями мышц одонтофора (рис.19). Прижимаясь к субстрату, она со-
вершает те или иные повторные движения. Обычно зубы радулы дви-
жутся поперёк одонтофора. Существует несколько основных типов ра-
дулы, их названия использованы для наименования ряда таксо-
нов (рис.20).
Рипидоглоссная радула служит для собирания свободно лежащих
пищевых частиц. Её рабочий ход - это движение назад выдвинутого
одонтофора при одновременном выдвигании вперёд радулярной мемб-
раны. При этом маргинальные зубы сметают частицы вглубь, к сере-
^не радулы. Рипидоглоссную радулу имеют многие археогастроподы,
^итаицие на мягком грунте.
Стеноглоссная и тениоглоссная радулы вооружены острыми марги-
Кздьными зубами, которые отрывают и схватывают частицы пищи при
^н^ении назад. При этом центральные и латеральные зубы подхва-
59
тывают частицы пищи. Такими радулами обладают, например, Ceri-
thacea,Cypriacea обычно обитающие на твёрдом грунте. У первч
численных трёх видов радулы одновременно функционируют зубы толк
ко одного ряда, прижатого к субстрату изогнутой поверхностью ра>
дулы,
Докоглоссная радула отличается от уже' перечисленных тем, что
благодаря совместному движению радулы и одонтофора взад-вперёд f
одновременно функционирует много поперечных рядов зубов. Таким
образом соскребают обрастания многие Patellae еа. Сакоглоссная
радула служит прежде всего для добывания сока из тканей растения,
оболочку растительной клетки моллюск прокалывает заостренным зу-
бом, остающимся единственным в каждом поперечном ряду, и высасы-
вает сок с помощью расширений и сокращений глотки. Сакоглоссная
радула встречается среди заднежаберных и у Pissoidea (Monet о-
cardia). Поперечные ряды зубов птеноглоссной и рахиглоссной ра-
дул отличаются значительной шириной и содержат очень много зубов.
Вообще же радула может содержать десятки тысяч зубов. Переднежаберни
и лёгочные моллюски имеют глоточные челюсти, выполняющие самые
разные функции: соскребывание, откусывание, прорезание. У ряда
моллюсков отдельные зубы радулы могут осуществлять самостоятель-
ные движения и таким образом у таких заднежаберных, как например
Philine , становится возможной манипуляция отдельными форами-
ниферами, которыми и питается этот моллюск. Наконец известны
радулы особого строения, которые трудно отнести к уже перечислен-
ным, а некоторые брюхоногие, в том числе паразиты и питающиеся
яйцами беспозвоночных, полностью лишены радулы (Gascoigne,1977).
Рис.20. А - радулы различных типов (Сох, 1960). Б - ядовитый зуб
и мешок зубов Gymnobela latistriata, Tu.rrid.ae, (Кантор, Сысоев,
1986), В - схема поперечных движений зубов радулы: 1,2 - рипидо-
глоссной, 3 - стеноглоссной, 4 - докоглоссной (Getting; из
Pichter, 1974). Г - зубы радулы Tarings telopia, Nudibranchia
Doridacea (Franc, 19686). Д - челюсти Piseinotecus evelinae, Nudi'
branchia Aeolidacea (Schmekel, 1960)
60
A
в|
В этом довольно широком разнообразии Ю.С.Миничев и Я.И.Ст^
богатов (1979) выделяют три основных типа строения радулы.
роглоссная радула имеет два продольных сгиба мембраны, в кар^
находится в согнутом состоянии, но распрямляется во время рабо
её зубы дифференцированы. Имеется у Pectinibranchia (передне
жаберные с гребенчатым ктенидием) и Scutibranchia (все арх^
гастроподы без Docoglossa, т.е. Zygobranchia). Мусивоглоссне,-
радула содержит многочисленные слабо дифференцированный зубы,
которые совершают движение по однообразной траектории,и имеет;
продольных сгиба (лёгочные, некоторые заднежаберники, Dextro-
branchia, Divasibranchiа).Наконец, докоглоссная радула лише-,
продольных сгибов мембраны (Patel lidae).
Длинная радула, свернутая в спираль, заполняет мешок и
лежит в головном гемоцеле. Возможно, что это обстоятель-
ство объясняет наблюдаемую у некоторых видов диссимметрию в paj
мощении зубов (рис.19 Е). Иногда строение радулы как будто бы е
связано с образом жизни. Например, среди глубоководных морских
блюдечек 9 имеют рипидоглоссную радулу, а у 5 других видов рад.^
особого строения. Однако обитающие внутри затонувших кусочков
древесины Pseudococculinidae и Cocculinidae имеют докоглосо?
радулу, a Bathysciadiidae, живущие на клювах кальмаров, им
ют радулу совсем иного типа (Rickman, 1983). Строение радул
у заднежаберников явно зависит от вида жертвы (актинии, оболов
______________________
Рис. 21. А - радулярные зубы лёгочных моллюсков: 1 - всеядной h
dosuccinea columella., 2 - преимущественно детритофага Physa vernal
(Kesler et al., 19в6).Б - тениоглоссная радула Abyssochrysos melanbides,
Prosobranchia (Houbrick, 1979) с I центральным (c.), I парой лате/
ных (I.) и 2 парами маргинальных зубцов (т.). В - радулярные 3;
цы хищной Diplomphalus seberti, Pulmonata (Solem, 1974б).Т - о ронт^
ные зубцы радулы птероподы Creseis acicula, Eutheco somata (Spoel-
1967). Д - радула соскребывающего моллюска Papuina phaeostoma (Soleflr
E - асимметричные рахидиальные зубцы радулы археогастроподы
rella. nimbosa (Hickman, 1981).Ж- радулярные зубцы .;итойага и Де1'
тофага Akera bullata, Opisthobranchia (Morton , Holme, 1955)
62
63
ники, гидроиды и пр.) и иногда зависит от возраста моллюска (Ny-
bakken, McDonald, 1961). Аналогичные примеры можно найти сре-
ди лёгочных моллюсков (рис. 19 А,В). Половой диморфизм в строении
радулы обнаружен у Drupella (Muricidae) (Fiijioka, 1982).
В отличие от соскребывателей, а также детрито- и фитофагов,хи-
щные брюхоногие вынуждены обездвиживать свои жертвы. Имеющийся
у них ротовой аппарат не позволяет им проглатывать добычу цели-
ком или крупными кусками и поэтому приём пищи у них занимает дли-
тельное время. В связи с этим у хищных брюхоногих развивается
система слюнных глоточных и хоботковых желез для наружного пере-
варивания пищи, система глоточных мышц для всасывания полураство-
рённой плоти жертвы (рис.23 ). Ядовитые зубы, пронизанные кана-
лом для впрыскивания яда, имеются у Toxoglossa, их число доходит
до 5 в каждом ряду (5АлпеА,Л'оЛп,1981)(рис.18 Б; рис. 23 Г). Сти-
лет эктопаразитических Pyramidellidae служит для впрыскива-
ния в тело хозяина выделений слюнных желез, возможно, содержащих
антикоагуляты. Яд выделяется либо железами глотки (Buccinidae),
либо гипобранхиалъной железой (Muricidae). Механический способ
обездвиживания жертв часто состоит в том, что моллюск обхватыва-
ет жертву ногой (рис.23 А). Молодые Rapana thomasianaпросверлива-
ют раковины двустворок, молодые моллюски раскрывают раковины мы-
шечным усилием ноги и затем вводят в мантийную полость выделяе-
мый гипобранхиалъной железой секрет, расслабляющий мышцы жертвы
Рис. 22. А - поперечный срез хобота и глоточной полости Volutomitra
curta, Monotocardia (Arnauxietal.,1980).B - ввернутый (1 ) и выверну-
тый хобот(2)Volutopsus,Monotocardia (сбоку)(Кантор, 1985). В -
стеноглоссная радула Buccinuin ur.da.tum на радулярных хрящах
одонтофора ( Ankel из Franc,19686^. Г - схема переднего отдела
пищеварительного тракта Toxoglossa : 1 - образование подвижной гу-
бы ринхостома, 2 - уменьшение размеров собственно хобота, 3
появление вворачивающегося псевдохобота и полная редукция собст-
венно хобота, 4 - полная редукция собственно хобота при отсутст-
вии вворачивающегося псевдохобота, 5 - появление хоботообразной
структуры из разросшейся буккальной трубки (Кантор, Сысоев, 1986)
64
65
(Чухчин, 1970). В данном случае у Я. thomasiana наблюдается кор-
реляция между способом овладения жертвы (сверление раковины -
раскрывание раковины мышечным усилием) и возрастающей как функ-
ция третьей степени массой мышц. Хищная Euglandina, нападающая
на брюхоногих, для полного пожирания жертв глубоко проникает в
полость раковины и таким образом её передняя часть уподобляется
хоботу других хищников (рис.24 А-Г; рис.25 А). Чаше же необходи-
мости в этом не возникает и передняя часть головы вытянута в мор.
ду или же имеется хобот и заменяющие его образования. К ним отно-
сится прежде всего собственно хобот (рис.22 А), ринхостом, вме-
щающий хобот (рис.22 Б 1 ,Г), псевдохобот, образованный разрас-
танием передней части головы (рис.22 Б), а также хоботообраэные
структуры, развившиеся за счёт глотки (рис.22 Б5 ). У Toxoglossa
по мере формирования этих удерживающих жертву структур постепен-
но исчезают радула, ядовитые и слюнные железы (Кантор, Сысоев,
1966). Здесь отрицательная корреляция (развитие одних структур
и редукция других) сопровождается субституцией Функции (функция
обездвиживания жертвы переходит от ядоносного аппарата к мышеч-
ным зеваниям).
Для успешного функционирования сложного ротовогоаппарата,вклг
чающего хоботообраэные структуры, радулу и сенсорные придатки,
требуется довольно сложная система мышц. Таковая действительно
существует. Она состоит из ретракторов, которые служат для выд-
вигания п втягивания хобота и радулы, причём отдельные мышцы вы-
полняют определённые частные функции и таким образом мышечная
система в целом приобретает пучковое строение (£ис.22 Г; рис.24
Б,В). Впрочем, выдвигание хобота у голожаб ерников Doridida
скорее всего совершается гидравлическим способом, он как бы вы-
-------------------------------.—-_________________________________________"
Рис. 23- А - Cassis tuberose, Топпасеа'. 1-3 - нападение на морского
ежа, 4- внутреннее строение {HughesR.,Hugh£\1981).B- прыгание Strombus
maculatus, Strombacea (Field, .1977) В - рядуляримр зубы Oenopot^
rugulata, Clavinae, Toxoglossa (Shimek, Hohn, 1981). Г- радула
Elysia. viridis, Opisthobranchia (Gascoigne, 1977)
66
•7
давливается наружу вследствие сокращения глоточной массы (Yorge,
1969).
Виды, способные просверливать раковины двустворок, встречают-
ся в 5 семействах переднежаберных брюхоногих, а также среди голо-
жаберных моллюсков. Только в семействах Naticidae, Muricidae
насчитывается 33 сверлящих вида (Carriker, 1981). Сверление в
основном осуществляется механически, радулой, поскольку установ-
лено, что покрытые воском и таким образом защищённые от воздействия
кислот створки Муа оказались всё же просверленными хищной Ро-
linices {Turder, 1953). При этом получается отверстие правиль-
ной круглой формы с ровными краями. В сверлении дополнительно
участвует находящийся в ноге железистый орган. Его выделения со-
держат кислоту, возможно ЫС1. Таким образом,сверление не следует
считать чисто механическим процессом.
Совпадение жизненной формы хищника и таксона имеет место у
Archaeogastropoda.Такие Fissurellidae., как например Diodora,
пасутся на губках (рис.14 I; рис.15 Е). Почти все Neogastropoda
тоже специализированные плотоядные формы. Mesogastropoda распре-
делены примерно поровну между фито- и детритофагами с одной сто-
роны и плотоядными с другой стороны {Morton, 1968), таким об-
разом, совпадение жизненной формы и таксона в данном случае не
имеет места. Такой же набор жизненных форм довольно полно повто-
ряется у лёгочных и заднежаберных моллюсков {Purchon,1977).
Обычный радулярный аппарат соскребывающих обрастания брюхоно-
гих, впрочем, так же как фильтрационный аппарат двустворок, поз-
воляет добывать пищу, которая нуждается в дальнейшей переработке
и прежде всего в сортировке. Она заключается в отборе пищевых час-
Рис.- 24. Philine, Opisthobranchia А - ротовой аппарат, Б - выдвигание
хобота и радулы (г.ш.- мышцы-ретракторы хобота) (Wurst, 1965). в - рото-
вой аппарат De.ndrodoris nigra,Opisthobranchia,c развитой системой ретрак-
торов глотки (Fong,1969). г - сосущий ротовой аппарат Gymnodoris oki-
navae (Vong ,1969). Д - топография органов пищеварительной системы
на контуре раковины Sphyradium doliolum, Piilmonata ('шилейко, 1984)
68
к
стенка глотки
69
Рис.25.A - нападение Euglandina rosea на Succinea(пунктировано)(1,2,3)
(Cook,1965). Б - строение Berthelinia, Sacoglossa (Kawaguti.Baba из
Myra keen,Morton, 1960). В-нападение Busyconна двустворку (Colton из
Hyman, 1967).T -Natica (Hornell из Hyman, 1967) . Natica josephinae
нападение на двустворку (E) и "галопирование" (3) (Ankel из Hyman.
1967). Д-зарывание Terebra tristis (Morton, 1964). ^"копирование" Helix.
70
тин» отделении их от постороннего материала. У соскребывателей
сортировка происходит в желудке. Посторонние частицы выводятся
в кишку, а пищевые частицы поступают в пищеварительную железу. В
этой связи уместно провести аналогию с собирающими и йильтрующи-
лл ракообразными. Их ротовой аппарат образован несколькими парами
конечностей, которые сортируют и отбирают пищевые частицы уже воз-
ле рта. Такое предротовое пространство, может быть, следует срав-
нивать с мантийной полостью двустворок. Однако и этого недостаточ-
но, и пищевой комок сохраняет свою неоднородность. Возможно, что
вследствие этого у ракообразных, так же как у моллюсков, всасываю-
щий в переваривающий отдел пищеварительного тракта обособлен от
желудка, в котором продолжается механическая переработка пищевой
массы.
Таким образом, между ползательной ногой, не способной обеспе-
чить моллюску высокую абсолютную скорость передвижения, радулой,
пригодной для растянутого во времени дпбывания сложного по соста-
ву пищевого материала,и сложным расчленением пищеварительного
тракта у большинства брюхоногих моллюсков имеет место корреляция.
Особое строение ноги, не пригодной для точных специализирован-
ных движений, когда отдельные движения как бы слиты и не отделе-
ны друг от друга, позволяет объяснить и некоторые другие особен-
ности организации брюхоногих.
В подошве ноги, пронизанной разнонаправленными в том числе
спинно-брюшными мышцами (рис.26 А), возникают волны сокращения.
Длина и высота волн не одинаковы у разных видов, число их тоже не-
постоянно - от I (Neritina) до неопределённо большого количес-
тв (Gibbula), Направление волн может быть прямым (спереди на-
зад), обратным, диагональным, латеральным, сложным (Clark, 1964)
~как у инфузорий. При попятном ползании направление волн может
Не меняться ( Olmsted,1917) .хотя у Halistis это изменение име-
ет место. Брюхоногим доступно и шагание, когда подошва ноги ка-
сается субстрата только выступающими гребнями (рис.25 1,3). Вол-
71
вы возникают ритмично в аритмично ( Olmsted., 1917). Переднежа-
берный моллюск Pomatias попеременно шагает на одной поло-
вине ноги, в то время как другая приподнята нал субстратом. В св^.
зи о относительной самостоятельностью боковых половин подошвы
они иногда отделены друг от друга продольной бороздой (рис.26 В),
Перистальтическое движение, когда форма ноги изменяется в широ-
ких пределах (рис.26 Б; рис.27 Д), тоже не представляет редкость.
Определённое значение при таком ползании имеет лакунарная систе-
ма ноги, заполненная кровью.Нога может изгибаться, следуя неров-
ностям субстрата, и складываться, когда моллюск втягивается в
мантийную полость.
Мышечное движение дополняется ресничным. Относительное значе-
ние мышечного и ресничного движения непостоянно, хотя у взрослых
организмов первое имеет большее значение, чем второе. Это соотно-
шение не связано с систематическим положением моллюска и,по-види-
мому,не зависит от размеров тела. Впрочем, у Polinices, обладаю-
щей столь же широкой подошвой, что и Natica (рис.25 Г,Е), в
связи с ростом организма наблюдается вытеснение ресничного дви-
жения мышечным, что в общем понятно: при увеличении массы живот-
ного его относительная поверхность сокращается и действенность
ресничного аппарата уменьшается. Соответственно возрастает значе-
ние мышц. В.И.Беклемишев (1964) и Кларк {Clark, 1964) удачно
сравнивают мускулистую подошву брюхоногих с телом турбеллярий, у
которых значение мышечного аппарата повышается по мере того как
увеличиваются размеры тела.
Рис.26. Prosobranchia Diotocardia\ А - подошва Neritina reclivata с
разнонаправленными мышцами, Б - поперечная волна мышечных сокращений
на подошве ноги (1-4 ) (Gainey, 1976). В -Eatoniella huttoni, ProscbrM
chia, co стороны подошвы,и её крышечка (Ponder, 1965). Prosobranchia
Moiwtocardia Г- Bullia digital!s, Stenoglossa , безглазая, с двумя ме-
таподиальными щупальцами {Cernohorsky, 1964). Д - Trichotropis borealis,
Taeniaglossa (Graham, 1954),Е-ползание Terebra gouldi, Toxoglossa
(Miller, 1975). I - безраковинный слизень Vnela odhneri, Acochlidiacea
(Delamare Deboutteville ,1960), Opisthobranchia
72
миоциты
73
Плотное прикрепление к субстрату имеет важное значение для
моллюсков, обитающих в волноприбойной зоне. Такие брюхоногие от-
вечают особой жизненной форме и в то же время представляют из
себя некоторый тип организации (мощная присасывательная подошва
ноги, обтекаемая форма раковины) и в таксономическом отношении
весьма разнородны. Прикрепление совершается согласованными сокра-
щениями мышц подошвы: раковинная мышца образует плотно прилега-
ющее к субстрату кольцо (рис.15 А), а медианные мышцы ноги сокра-
щаются таким образом, что под подошвой наступает разрежение.
В результате получается мышечная присоска. Определённое значение
при этом имеет и слизь, выделяемая железами подошвы. Она не толь-
ко улучшает плотность соединения подошвы с субстратом, но в ря-
де случаев образует своеобразные мостики, облегчающие мелким мол-
люскам преодоление нерооноотей субстрата. Вообще подошва брюхо-
ногих выделяет довольно много слизи. По-видимому, это совершенно
необходимое условие для работы ресничного аппарата подошвы,
Высокая энергетическая действенность ползания у наземных слизней
объясняется слизевыделением {Denny, 1980). 7 наземных моллюс-
ков слизь предотвращает высыхание поверхности тела. Слизь выпол-
няет и другие функции. Отделённое от субстрата морское блюдечко
Helcion(Patellidae) выделяет так много слизи, что привсплывает
и течениями уносится на новое место {Vahl, 1983). В данном
случае двигательная функция выделяющей слизь ноги выступает в
весьма своеобразной форме. После проползшего моллюска остаётся
слизевой след. Он облегчает ориентировку животного и, кроме того,
служит средой, в которой накапливаются микроскопические водорос-
PHc.27JL-BertheIinia, Sacoglossa.'. 1- Rcaribbea, пунктиром показано место
прикрепления аддуктора раковины {Edmunds, 1963), 2-В.Птах со спины
стрелками показано направление токов воды, З-Д Птах, левая створка
с протоконхом {Solem, 1974). Prosobranchia Monotocardia: Б-шагание
Lambis, Strombidae {Solem, 1974a);2 -Pomaiiopsis lapidaria, Hydrobi-
idae сбоку и подошва ноги (1-6 ).Отмечено боковое подвижное
углубление (*) между передней и задней частью ноги при неизмен-
ной общей её длине (1-3) и при её изменении (4 -6) (Davis, 1967)
74
75
ли. При повторном проползании по собственным следам моллюск пся
жирает слизь вместе с водорослями. На выделение слизи моллюск звх
трачивает до 3% получаемой им энергии (Hnghes,I986). Переднежа-
берник Litter ina irror.ata в таких слизевых следах воспринимает
направление, в котором прополз моллюск. Возможно, что при этом
имеет значение не хеморецепция (восприятие разности концентрацщ
раздражителя в слизи), а микроструктура слизевого следа (Stir-
ling', Hamilton, 1986).
Зарывание в грунт следует считать одной из форм движения, оно
совершается с помощью ноги. У Terebra нога при зарывании дей.
ствует как у двустворок: она вклинивается в грунт и образует на
конце утолщение, к которому подтягивается животное (рис.25 Д).
У hlatica нога подразделена надвое и при зарывании в грунт
главное значение имеет её передняя часть, называемая проподиумом
(рис.24 Г). В ряде случаев выявляется связь размеров и формы ра-
ковины с типом движения. Виды с высокой раковиной пользуются в
основном ресничным движением и при этом раковина волочится по
субстрату, периодически подтягиваясь вперёд. Такие моллюски осо-
бенно медлительны (Linsley, 1978), примером может служить Ceri-
thium,y которого нога составляет около трети длины раковины
(Gainey, 1976). При длине раковины 18 мм Ilyanassa ползёт со
скоростью 3,3 мм/с , но последняя связана с размерами животного
прямой зависимостью, потому что в процессе роста моллюска плошая
подошвы ноги увеличивается быстрее, чем длина раковины (алломет-
рия) {Dimock, 1985), что же касается относительной скорости
-------------------- _ _ _
Рис. 28. Prosobranchia Heteropoda: А - строение Atlanta (аг.-радулярн#
хрящи, g.ped.- педалыш:’’ ганглий, прочие ооозначения см. рис.З) (Alar
toja,Thiriot-Quievreax, 1975); L плоская раковина с продольным килем
A.plana (1,2) (Pichter, 1974); В турбоспиральная раковина A exinogyra
(Richter, 1974). Т-Jantina exigua, Ptenoglossa, прикрепившаяся подошвой
ги к собственному поплавку ( Laurse из Feininger,1972). Д распре
ленный ротовой аппарат Clione limanna, Opisthobranchia Pteropoda
Gymnosomata (Wagner ,1885). E - Acgires pnnctilucens, Opisthobranchia
Nudibranchia (Baba, 1974 a)
76
77
ползания, то она находится в обратной зависимости от размеров
животного.
Обычный для брюхоногих двигательный аппарат сохраняется у до,
вольно своеобразных двустворчатых эаднажцберников, обитающих ио,
ключительно в зарослях водоросли Caulerpa (рис.25 Б; рис.27 д)
Из двух имеющихся у них створок левая сохраняет протоконх и зак.
ладывается у личинки обычным для велигеров способом. Правая ctbqj.
ка образуется позже левой. Крышечка, имеющаяся у велигера, веко,
ре отбрасывается,и формируется, как у двустворок, раковинный ад.
дуктор и лигамент. Кроме того, имеется ретрактор ноги. Вода вхо-
дит в мантийную полость спереди и выходит сзади. Сохраняются осф.
радий и гипобранхиальная железа. ЦНС сакоглоссного типа (Kawa-
gati, 1959; Myra Keen,Morton,I3G0). Показательно, что приобре-
тя определённое сходство с двустворками, входя вместе с ними в
один фенон, эти моллюски сохранили всё же ползательную ногу и
другие структуры, свойственные брюхоногим моллюскам. Этот пример
показывает неоднозначность связи между типом двигательного аппа-
рата, строением раковины и мантийного комплекса и пр.
Освоение пелагиали переднежаберниками Heteropoda в ряде случа
ев тоже не вызвало резкой перестройки организации по сравнению
с жизненной формой донных переднежаберников (рис.26 А-Г). Сохра-
нив ногу, однако подвергнувшуюся сложному расчленению и утратив-
шую подошву, а также планоспиральную раковину, Atlanta в ряде
случаев приобрели гребень на раковине. Этот гребень, возможно
имеющий рулевое значение, имеется у некоторых личинок донных пе-
Рис.29. Prosobranchia Heteropoda: Pterotrachea hippocampus (Seapy, 1985):
А общий вид, Б глаз и головной мозг. 3 -Carinaria lamarcki ($k-s-
1976): её раковина изнутри и сбоку (Г) и внутренностный мешок (Е)
(Tesch, 1306а). й. - Prototrachea umblicata (Tesch, 1906 а). 3 - плава;®6
Pterotrachea coronata с помощью волнообразных изгибаний плав
(Morton,1964),Сокращения: g.cer.-pl. - церебро-плевральный ганглий,^/1'
глазной ганглий, rh.- риноЛор, st. - статоцист, v.s. -внутренностны-
шок, g.v.- висцеральный ганглий. Д-Firoloida kowalewskyi (Tesch,*
78
79
реднежаберннх (Fioroni, 1982). Уплощенная нога, скорее всего,
выполняет функции плавника. Причины подобного же консерватизма у
другого переднежаберника, Janthina, пожалуй, более понятны.
Ножные железы Janthiпа выделяют пористый поплавок,и животное
прикрепляется к нему как к обычному субстрату. Таким образом, ос-
воив пелагиаль, Janthi.na в определённой мере сохранила обычный
для переднежаберников образ жизни на субстрате и основные черты
организации донных представителей этой группы моллюсков. Активное
движение у них заменено пассивным дрейфом и вместо активного поис-
ка жертвы они довольствуется случайными поимками хондрофор Velel-
la, её обычных жертв, также обитающих в плейстоне.
Организация других киленогих, тоже освоивших пелагиаль -Cari-
nariidae, Pterotracheidae,напротив, подверглась весьма существен-
ной перестройке; таким образом таксон Heteropoda как жизнен-
Рис.ЗО. Pulmonata: А - соотносительные изменения в строении моллюс-
ков при переходе от улитки (1 ) к жизненной форме слизней - расти-
тельноядных полифагов (2 ) и хищников (3 ): сокращение дыхательной
поверхности мантийной полости приводит к разрастанию дыхательной по-
верхности тела вследствие обрастания раковины и складчатости кожи,
укорочение кишечника у хищника (2 ) по сравнению с фитофагом ( 3 ),
увеличение относительной массы ноги и сдвигание раковины кзади у
хищника (3 ). Обозначения:/^-часть тела, занятая внутренностями,
1 -части тела, занятые мускулатурой, -мантийная полость.(Ли-
харев, Виктор, 1979); Б - раковина и её нуклеус (справа)Candaharia
rutellum , Stylommatophora (Лихарев,Виктор, 1980)В - топография внут-
ренних органов у моллюсков, различающихся относительной величиной
раковины и внутренностного мешка :1-Aneitea simrothi, Stylommatophora,
2 - Succinea putris, Stylommatophora, 3 -Kalidos oleatus, Stylommatophora,
(Tillier, 1984). Сокращения : a - аорта, a.d- передний проток пищевари-
тельной железы, Ьт.~ буккальная масса, с. - зоб, c.d - цекум кишки, 1 •
кишечник, or. - ретрактор глазного стебелька, p.d.- задний проток
пищеварительной железы, S. желудок, s.g.- слюнная железа, задняя киа*
ка не изображена; Г - хищный слизень Testacellahaliotidea, Stylommatopho-
ra с вывернутой глоткой (Lucaz-Duthier, 2537/;Д-интерстициальный мол-
люск Hedylopsis brumbelli,Acochlidiacea, с известковыми спикулами в стеН'
ке внутренностного мешка (Swedmank, 1968). Е - Unela odhneri, Acoch-
lidiacea (Delamare-Deboutteville, 1960), Opisthobranchia
80
8!
ная форма отнюдь не представляется единством как фенон итип орга-
низации. У таких киленогих тело вытянуто, уплощено с боков. Сза-
ди оно переходит в метаподиум, а спереди продолжается в удлинён-
ную морду, кроме того, сзади иногда имеется удлинённый хвост.
Проподиум преобразован в непарный плавник. На плавнике иногда со-
храняется рудимент ноги в виде присоски, которая служит для удер-
жания жертв (рис.29). Внутренностный мешок у Pterotracheidae
несёт рудименты жабер.
Для подвижных пелагических организмов, какими являются килено-
гие, вододвижение не имеет существенного значения и для функцио-
нирования жабер не требуется мантийной полости. Внутренностный
мешок с сильно редуцированной раковиной обособлен от остального
тела и потому не стесняет изгибательных движений тела и плавни-
ка. Животное плавает внутренностным мешком вниз. Непарный плав-
ник, уплощенное тело, обособленный внутренностный мешок и руди-
ментарное состояние раковины и жабер могут считаться скоррелиро-
ванными особенностями. Следует отметить наличие у ряда киленоги
хорошо развитых глаз, что естественно для пелагических хищников,
обитающих в условиях хорошей освещённости, таким образом, хеморе-
цепторная функция, столь важная для донных брюхоногих, у килено-
гих несколько потеснена зрительной.
Жизненная форма наземных моллюсков насчитывает около 20 тысяч
видов переднежаберных и примерно 3650 видов лёгочных моллюсков
(S olem, 1974). Первые представлены исключительно раковинными
формами, улитками, тогда как вторые включают ещё и слизней. Тив
организации слизней выделяется взаимно скоррелированными особен'
костями: червеобразным телом, редукцией раковины и внутренностно'
Рис.31.Opisthobranchia, Acochlidiacea: А - Tantalum elegans , строен!1
моллюска (1 ) и его свёртывание (защитная реакция)(2 ) (Rankin,
1979), Б -Hedylopsis cornuta (Challis, 1970). Pulmonata: В-Pseudo
neitea papillata, Arthoracophoridae (Burton, 1963) ; Г слизень
Umax cinereoniger (Лихарев,Виктор, 1980) Д Crygoceras ,
Hydrobiidae (Pulmonata) (Taylor, 1974)
82
го мешка. У переднежаберных, обитающих в воде, он представлен едва
ли не единственным видом (рис.10 Д), но обычен среди переднежабер.
ных {голожаберники). Наземные переднежаберные улитки, приурочен-
ные к тропикам, утратили жабры и дышат внутренней поверхностью
мантийной полости. Для увлажнения её почки выделяют почти чистую
воду и поэтому выделительное отверстие у таких улиток открывается
в мантийную полость.У лёгочных моллюсков жабры тоже редуцированы
и не восстанавливаются даже у вторичноводных видов, вернувшихся
в водную среду. Одним из немногих исключений остаются некоторые
Basommatophora. Вместе с левой жаброй они восстановили и оагра-
рий. Место редуцированных жабер у лёгочных моллюсков занято кро-
веносными сосудами (рис.13 1,3), а иногда - особого рода губчатой
тканью (рис.14 3). Впрочем, мелкие Ancylus ограничиваются
одним только кожным дыханием, сохраняющим значение и у других
лёгочных моллюсков.
Особого рода приспособлениями к воздушному дыханию располагает
улитка Rhaphaulus (рис.6 М).0но представляет собой дыхательную
трубку в верхней части устья раковины. В засушливый сезон моллюск
втягивается внутрь раковины и закрывает устье крышечкой, впадая
в спячку. Однако при необходимости, даже не отодвигая крышечки,
он всё же может дышать. Для этого моллюск, видимо, ещё глубже
втягивается в раковину - настолько, чтобы открылось внутреннее
отверстие дыхательной трубки. Если моллюск немного выдвинется на
прежнее место, это отверстие закроется и дыхание прекратится.
Воздух - вещество значительно менее вязкое, чем вода. Поэтому
вододвижение в мантийной полости с помощью ресничного эпителия
Рис. 32 Tpisthobranchia. А - Hydatina physis, Cephalaspidea (Thompson,
1976). Б-Julia japonica, Sacoglossa, общий вид (1 ) и двустворчатая
раковина с протоконхом на левой створке ( 2 ) (Kawaguti, Уатази из
Thompson, 1976). В -Haminaea musetta, Cephalaspidea, общий вид (1,2)
и раковина (3 ). Г -Philinoglossa marcusi, Cephalaspidea, общий вид
и ротовой аппарат (2 ) (Challis, 1969а). Nudibranchia Д- Pseudover-
mis mortoni (Challis, 1969б)(£- Doto rosea(Schmekel, Kress, 1977) , E -
Aeolidia exigua (Risberc из Franc, 1966 6)
84
становится невозможным и тогда прекращается хеморецепторная функ-
ция мантийной полости. Лёгочные моллюски не имеют осфрадиев,
их сенсорную функцию принимают на себя щупальца головы, губные
лопасти, внутренняя поверхность глотки (Croll, 1983).Helisoma.
несёт рецепторные клетки на опине и на крае мантии (Jones,Sale-
uddin, 1978). Всасывание воздуха в мантийную полость (вдох) и
удаление воздуха (выдох) вызывается колебаниями стенки внутрен-
ностного мешка, т.е. мышечным и гидравлическим способом. Посколь-
ку вдох и выдох следуют во времени, лёгочные довольствуются един-
ственным пневмостомом. Схема циркуляции воздуха в мантийной по-
лости становится таким образом маятниковой.
Для обитающих в воздушной среде животных, какими в большинст-
ве своём остаются лёгочные, проблема водообмена, не существующая
для гидробионтов, приобретает исключительное значение. Её слож-
ность усугубляется тем, что она сопряжена с дыханием, поскольку
даже при воздушном дыхании потери воды неизбежны вследствие физи-
ологии механизма газообмена. Само расположение лёгкого в стеснён-
ной мантийной полости, открытой во внешнюю среду единственным
пневмостомом, величина которого поддаётся регуляции, отчасти мож-
но считать решением этой задачи. Наоборот, дыхание всей поверх-
Рис.ЗЗ. Opisthobranchia Cephalaspidea: А- плавание Akera bullate (1-4)
с помощью параподиев (р) (Morton, Holme, 1955 ; Thompson, 1976) , Б -
Akerabullata co спины и вершина раковины (Morton, 1972). Pleurob-
ranchus pumula, Notospidea: В -организм co спины (показаны входящий
и выходящий токи воды), I' - организм со спины с удалённой мантией
(пунктир) и открытым внутренностным мешком (v.sj, Д - раковина, в -
плавание с помощью изгибаний параподиев (Thompson, Slinn,1959).Сокраще-
ния: inf.р,- нижняя мантийная лопасть, р.с,- мантийный вырост. Aplysia,
Anaspidea: Л - плавание с помощью параподиев, 3 - ползание на по-
дошве ноги (1 - к субстрату прилежит расширение средней части по-
дошвы ,2 - животное опирается на расширенные переднюю и заднюю час-
ти подошвы), И - личинка в возрасте 24 часа, осевшая на водоросль
(Parker, 1917; Strenth, Blankenship, 1979)
86
87
ностью тела лишило бы животное возможности регулировать дыхание я
связанный с ним газообмен.
Более простой и действенный способ сберегания воды состоит в
том, что в сухую погоду животное прячется в укрытиях (расселины
скал, в почве, между камнями, под камнями и стволами деревьев)
или впадает в спячку. Отсюда следуют два основных способа пережи-
вания сухой погоды и два основных типа организации лёгочных.
Первый тип организации составляют раковинные улитки. Они сох-
ранили раковину и поэтому в выборе таких убежищ,как расселины, они
ограничены размерами раковины; убежищем в таком случае остаётся
прежде всего сама раковина. Зато у имеющих раковину улиток
развивается утолщение края мантии, богатое слизевыми клетками.
При необходимости слизью заклеивается устье раковины после того,
как в него втянулось мягкое тело моллюска. Засохшая слизь образу-
ет плёнку, называемую эпифрагмой, своего рода аналог крышечки
переднежаберных.У одной улитки может быть несколько эпифрагм.
С поверхности ноги слизь снимается краями утолщенной мантии, выжи-
манию слизи с краёв мантии способствуют зубы, расположенные на
краю устья раковины (рис.5 И,Е; рис.6 Б,В,Г)(Шилёйко, 1984). Сок-
ратившееся до узкой щели устье (рис.5 И) рассматривается к тому
же как приспособление против проникновения внутрь раковины хищных
насекомых. Сходную с крышечкой и эпифрагмой функцию выполняет кла-
узилий улиток Clausiliidae - известковая пластинка, подвижно
сочленённая с столбиком раковины (рис.6 А). Когда моллюск втянет-
ся в раковину, клаузилий закрывает устье. Раковинные улитки спо-
собны переносить высокую температуру засушливого сезона.
Рис. 34. Opisthobranchia,Cephalaspidea. A-Bursatella leachi: вид сбоку
(1 ), вид со спины (2 ), вскрытая мантийная полость (3 ) , вскрытый
внутренностный мешок (4 ) (.Bebbington, 1969) Б - Akera bullata.
вид со спины (1 ) и раковина (2 ) ( Thompson, 1976). в -Gastropterofl
vespertilliuni Nudibranchia Г-Harmosita sangria (Goslinger, Beh'
rens, 1986), Д - Onchidoris muricata спикулы спинных папилл в расП
равленном (1 ) и соединённом виде ( 2 ) (Millen, 1985)
88
89
Второй тип организации представлен слизнями (рис. 30 А
2, 3 ). У слизней в той или иной степени, иногда полностью, реду-
цирована раковина и вместе с ней редуцируются внутренний мешок,
колшеллярная мышца и сокращается мантийная полость. Внутреннос-
ти опускаются в цефалоподиум и таким образом складывается весьма
типичная для слизней топография внутренних органов, особенно по-
казательная в сравнении с раковинными улитками (рис.30 В 1,2,3).
Сокращение величины дыхательной поверхности, вызванное редукцией
мантийной полости, отчасти восполняется складками на поверхности
цефалоподиума. В свою очередь, кожное дыхание предполагает чис-
тую поверхность тела. Для очистки её от загрязнений служит капю-
шон, образованный выступающим передним краем мантии (рис.30 А 2).
Когда животное сокращает, втягивает передни) часть цефалоподиу-
ма, на капюшоне остаются загрязнения вместе о излишками слизи,
песчинками и пр. Дыхание поверхностью тела не поддаётся регуляции
Однако у слизней регуляция всё же имеет место, хотя принимает со-
вершенно особую форму - поведение. Благодаря червеобразной форме
тела и редуцированной раковине слизни могут проникать в
стеснённые пространства с высокой влажностью воздуха. Хищные
слизни Testacella (рис.30 Г), например, проникают даже в нор-
ки земляных червей, за которыми охотятся. Слизней-фитофагов от-
личает от хищников значительной длины кишечник и в ряде случаев
наличие слепой кишки. У фитофагов кишечник образует до 3 петель
и закручен вокруг зоба, в котором происходит измельчение и по-
лостное переваривание пищи. Более короткий,чем у фитофагов,кишеч-
ник хищников, расправлен вследствие того, что мантийная полость
вместе с рудиментом раковины смещена назад как у Testacella (рис.
30, В) (Лихарев, Виктор, 1980).
У своеобразных слизней Athorscophoridae полная редукция ра-
ковины и замена её полужёстким скелетом, образованным мелкими из-
вестковыми тельцами, привела к формированию особого, турбеллярие-
подобного облика (рис.31 В). При этом пневмостом приблизился к
90
саГитталъной плоскости. Складки, увеличивающие дыхательную повер-
хность, подверглись дихотомии, чем достигается более полное ис-
пользование поверхности тела для дыхания. Этой же цели служат мно-
гочисленные радиальные выросты мантийной полости C&rton, 1963).
ц0 непонятным причинам цефалоподиум многих слизней уплощен иным
образом, с боков, и принимает ремневидную форму (рис.31 Г).
Orygoceras, чрезвычайно своеобразные моллюски неопределённо-
го систематического положения (рис.31 Д), подверглись измельчанию
и в длину не превышают 2,5 мм.Малые размеры телА и раскрученная
раковина Orygoceras считаются приспособлением к жизни в интер-
стициали {Taylor, 1974).Вследствие малости они имеют настолько
большую относительную поверхность, что обходятся без жабер и дру-
гих дыхательных приспособлений даже при наличии раковины, за счёт
которой уменьшается величина свободной поверхности тела. Вытяну-
той формой они напоминают других довольно своеобразных моллюсков,
Acochlidiacea, также выделяющиеся своими малыми размерами тела
- до 4мм (рис.25 Н; рис.30 Д,Е; рис.31 А 1,2 ). Вследствие ма-
лости они обходятся ресничной двигательной подошвой и при отсут-
ствии жабер и раковины дышат поверхностью тела. Их удлинённый
внутренностный мешок как бы продолжает цефалоподиум, задний ко-
нец ноги укорочен и моллюск в целом приобретает червеобразный об-
лик. Такое строение моллюска пригодно для обитания в интерстици-
али, между песчинками. Необходимая при таком образе жизни гиб-
кость внутренностного мешка достигается тем, что раковина пол-
ностью редуцирована и заменена внутренним полужестким скелетом,
который состоит из многочисленных известковых спикул. Впрочем, у
Ряда видов отсутствуют и спикулы.
Вследствие растяжимости во внутренностный мешок может втяги-
ЭДгься цефалоподиум и таким образом внутренностный мешок выполня-
ет защитную функцию вместилища мягкого цефалоподиума даже при от-
сутствия мантийной полости (рис.31 А 2). Наконец, к типу органи-
зации слизней примыкают и некоторые другие заднежаберники.
91
К их числу относится крошечный (длина тела до 2 ш) червеобраз-
ный Philinoglossa (рис.32 Г), лишённый раковины и жаберного
аппарата, также червеобразный, длиною около 5 мм Pseudovermiz,
сохранивший, однако, столь характерные для Nudibranchia жабры
(рис.32 Д). В целом, однако, у Opisthobranchia организация со-
вершенствовалась не столько вследствие изменений размеров тела,
сколько прежде всего в связи с редукцией раковины.
Таксономически заднежаберные моллюски Opisthobranchia представ,
ляют из себя достаточно обособленный подкласс брюхоногих, относи-
тельно небогатый по количеству видов. Однако в сравнительно-анато-
мическом плане он неоднороден, так как объединяет весьма несход-
ные типы организации. Некоторые из них близки к переднежаберни-
кам, другие относятся к типу организации слизней, третьи прибли-
жаются к киленогим, наконец,для крылоногих Pteropoda трудно
подобрать подходящие аналогии среди других моллюсков. Свойствен-
ная заднежаберникам полная или до состояния искривлённой пластин-
ки редукция раковины (рис.5 Д; рис.7 X) и внутренностного мешка
с мантийной полостью вызывает в строении этих моллюсков изменения,
аналогичные деторсионным. Дыхательная функция переходит к боковым
выростам ноги, п арап одням, а также к спинным кожным жабрам. Эти
обстоятельства учтены в системе подкласса (Thompsort, 1976):
жабры имеются раковина наружная у взрослых моллюсков нет раковины Nudibranchia раковина внутренняя A ply si omorpha Pleurobrandiioinorpha
жабры обычно имеются Bullomorpha
жабры обычно отсутствуют Sacoglossa Thecosomata Gymnosoinata
жабры отсутствуют Acochlidiacea
92
^AS.Nudibranchia. A - плывущий Cumanotus beaumonti, Б- Fiona
Piftnataa её жабра, В - Hero Formosa, Г-Ancula cristata, Д-Qkenia
teachi (Thompson, Brown,1984)', Д -Phyllidia varicosa (Baba,Ha-
toatani, 1975), E -Eubranchus horri (Baba, 1975)
93
Становление типа организации таксона OpisthobranchiaynotiHo рассм^
ривать начиная с таких форм, как Hydatina , которые вместе с
ковиной сохранили мантийную полость с единственной жаброй и широ-
кую истончённую ногу с многочисленными складками (рис.32 А) .Вслед.,
ствие обширности такая нога имеет возможность превратиться в ор-
ган дыхания и ресничного вододвижения. Эта возможность осуществ-
ляется по мере того, как устраняется физиологическая конкуренция
со стороны дыхательной мантийной полости, а следовательно, и внут-
ренностного мешка с раковиной. Эта происходит следующим образом.
Во-первых, разрастается мантия и обволакивает раковину, что,
впрочем, наблюдается и у некоторых переднежаберников. Вследствие
этого обволакивания значение мантийной полости для дыхания умень-
шается, она погружается внутрь и постепенно редуцируется. Во-
вторых, обособляются и загибаются вверх боковые лопасти ноги, па-
раподии. Оторвавшись от субстрата, они могут участвовать в водо-
движении, необходимом для дыхания, и позволяют животному плавать.
Обрастание раковины мантией наблюдается уже у Cephalaspidea,
которые остаются наиболее примитивными среди заднежаберников
(рис.32 В; рис.33 А,В), а также у Aplysiomorpha и других форы.
Края приподнятых параподиев ограничивают у этих моллюсков щеле-
видное отверстие мантийной полости. Они могут волнообразно изги-
баться, вызывая вододвижение, и тогда мантийное отверстие изме-
няет очертания. Через переднюю часть щелевидного мантийного от-
верстия вода входит в мантийную полооть, через заднюю покидает
её, в общем, наблюдается разобщение противонаправленных токов
воды, как в глотке актиний. У Aplysia параподии разрастаются
настолько, что мантийная полость оказывается на дне довольно
глубокой выемки. Волнообразные изгибания параподиев не только
вызывают вододвижение, но позволяют животному плавать. Плавание
это, однако, несовершенно и в основном животное ползает, совмещая
перистальтику и шагание (рис.33 Н,3). Только что осевшие на суб°'
трат личинки ползают на подошве обычным для брюхоногих способом й
обладают спирально свёрнутой раковиной (рис.33 И),
м
Pio. 36. Пелагические заднежаберные моллюски: A -Phyllirrhoe,Nu-
dlbranchia (Hescheler по Hyman, 1967), Б -Thliptodon diaphanus,
tymnosomata (Meisenheimer no Tesch, 1918), В - Desirtopterus pa-
pilio, Qymnosomata (Chun, 1889), Г- раковина Cavolinia uncinata,
Thecosomata, сбоку и вентрально (5poel, 1971)
95
Среда Cephalaspidea довольно совершенными пловцами являются
Акега (рис.32 А, Б), а также Gastropteron (рис.34 В).
Pleurobranchus плавает подошвой вверх благодаря волнообразным
изгибаниям непрерывной эпиподиальной складки ноги. Она же вызыва-
ет в мантийной полости ток воды, единственную жабру .Ухо-
видная раковина остаётся под кожей и не мешает плаванию (рис.33
В,Г,Д). Несколько иной двигательный аппарат имеет Lobiger (рис.
39 Б; рис.40 3). Он состоит из двух парных придатков, являющихся,
скорее всего,производными эпиподиальной складки. Моллюск плава-
ет с помощью этих придатков и при этом изгибания тела, ограничен-
ные наличием довольно крупной раковины, по-видимому, не имеют зна-
чения.
Полная редукция раковины, колюмеллярной мышцы и внутренностного
мешка наблюдается у голожаберников. Это сделалось возможным благо-
даря двум обстоятельствам. Одно из них состоит в том, что дыхатель-
ная функция перешла к весьма характерным для этих животных кожным
жабрам (cerata), которые и послужили поводом для названия этой
группы моллюсков. Количество жабер у них непостоянно, их строение
разнообразно: нитевидные, листовидные, пластинчатые, ветвистые
и пр. (рис.32 Д,Е,1; рис.34 Г: рис.35). Возможно, что определён-
ное значение для дыхания имеют щупальцевидные выросты мантии, по-
крывающие спину моллюска или окружающие его анус (рис.35 Г,Е).
Рис. 37. Opisthobranchia Nudibranchia: А - Onchidoris bilamellata
2-движение крови в жабре, 2 - токи воды, вызываемые ресничным эпите-
лием жабер и дорзальной мантии, 3 - токи воды между папиллами дорзаль-
ной мантии, 4- собирающие (короткие стрелки) и удаляющие (длинные
стрелки) токи на главных и боковых выростах и пиннулах вокруганаль-
ных жабер, 5 - выносящие токи воды на отдельной пиннуле; Б - Archi-
doris pseudoargus, токи воды на висцеральной мантии, В - центральный
отдел кровеносной системы (Potts, 1981)', Г - книдосак Catriona
aurantia (Edmundszs Thompson, 1976); Д- Coryphella verrucosa, пожираю-
щая сцифистом Aurelia aurita (Hernroth, Grb'ndahl, 1955).
Сокращения: p.s.- мантийный синус, pd.s.-ножной синус
И
97
В связи с спинными жабрами у голожаберников сложился подкож-
ный отдел кровеносный системы, распространяющийся и в параподид
(рис.37; рис.38). В какой-то мере это означает возврат к дыха-
нию всей спинной поверхностью тела, своего рода децентрализация
дыхания. Эта децентрализация, однако, не затронула кроведвижу-
щего отдела кровеносной системы: сердце сохранилось примерно в
том виде, как у однопредсердных переднежаберников, и состоит из
желудочка и одного предсердия (рис.38 Б). Любопытно, что в ряде
случаев немногочисленные и вследствие этого с сложной поверхнос-
тью жабры, напоминающие щупальца восьмилучевых кораллов, сосре-
доточены вокруг ануса (рис.35 Г; рис.З? А 2 ), как у гипотетичес-
кого предка брюхоногих с задним положением мантийной полости.
Возможно, что эта устойчивая топографическая корреляция не слу-
чайна и свидетельствует о каких-то остающихся неизвестными осо-
бенностямидыхательной функции у голожаберников.
Возможно, что в газообмене участвуют не только жабры, но и
вся спинная поверхность тела, потому что иногда на ней выявляют-
ся токи воды, очищающие кожу от загрязнений (рис.37). Вододви-
гательную функцию, возможно, выполняет ундулирупцая мембрана на
жабрах (рис.351Разумеется, это благоприятствовало бы газообмену.
Величина дыхательной поверхности у Onchidoris регулируется
следущим образом. В коже спины у моллюска имеются спикулы. Ког-
да они расправляются, поверхность кожных папилл увеличивается,
если же спикулы складываются, папилла как бы съёживается (рис.34
Д).
Другое обстоятельство, сделавшее возможным редукцию раковины,
состоит в том, что её защитная функция перешла к весьма обычный У
голожаберникам книдосакам. Так называются удлинённые выросты спЯ'
Рис. 38. Opisth о branchia. Nudibranchia Adalaria proxima - A - крове'
носная система, В - сердце (Thompson, 1958); В -Embletonia (Marcus и3
Hyman,1967). Г-смена раковины в онтогенезе Actaeon, Cephalaspidti
(Thorsonus Hyman, 1976). Д - поперечный срез Aplysia, Anaspidea (Jfa'
les, I960)
98
продольный педальный синус
желудочек-[ ' f.
афферентный жаберный сосуд
А
стенка тела
книдосак
анус 1
почка
Личиночная
раковина
В
ринофор
предсердие
Б
гемоцель
почка
Д
перикард
кишечник
подошва ноги
дефинитивная
раковина
железа
средней кишки
.передний отдел ventriculus
правый висцеральный коннектив
гемоцель
розовая
железа
левый висцеральный*
коннектив
продольный мантийный синус
эфферентный жаберный сосуд
жабра
выделительная пора . _ „
г мантия
раковина
мантий цый ретрактор
сенсорные щетинки
гермафродитный проток
палиновая железа
ны, в которых накапливаются в пригодном для выстреливания состоя-
нии книдоцисты, ранее принадлежавшие полипам кишечнополостных,
обычной добыче голожаберных (рис.35 Д;38 В). Книдоцисты поступают
в книдосаки из кишечника, который отдаёт ответвления, заходящие
в книдосаки. Вследствие этого кишечник приобретает совершенно не-
обычную для моллюсков форму (рис.38 В). Этот случай показателен
в том отношении, что поясняет как одна и та же функция (в данном
случае защитная) может выполняться совершенно различными морфос-
труктурами (в данном случае раковиной и книдосаками) (рис.35Д).
Сенсорная функция претерпевшей редукцию мантийной полости пере-
ходит прежде всего к головным щупальцам. 7 многих заднежаберных
моллюсков, включая голожаберников, имеется 2 пары головных щупа-
лец. Первая пара обычно рассматривается в качестве боковых вы-
ростов так называемого головного щита, выполняющего хеморецептор-
ную функцию (Groll, 1983). У Cylichna cylindracea наряду с хеморе-
цепцией головной щит способствует вододвижению в мантийной по-
лости (Lemc2?e,I956). Вторая пара щупалец, называемая ринофорами,
(рис.37 А 2; 38 В; 39 А), гомологична головным щупальцам передне-
жаберных. Они служат в качестве дистантных рецепторов, обычно име-
ют складчатую поверхность и могут втягиваться в особые влагалища.
Совершенно неожиданное следствие для физиологии некоторых го-
ложаберников вызвано редукцией раковины и внутренностного мешка,
вследствие чего открылась для солнечной экспозиции спиннаясторона.
У Elysiidae (рис.38 В,Г) оно состоит в том, что хлоропласты
Рис.39. Opisthobranchia: к - голожаберник Calliopaea oophaga (Jen-
sen, 19866); Б - Lobiger souverbii, Sacoglossa (Baba, 1974), с двумя па-
рами плавательных параподиальных придатков. В - копуляция Elysia
subornata (Jensen, 1986 а), Sacoglossa ; Elysia arena (Carlson, Heft,
1377) co сведёнными на спине параподиями (Г) и с расправленными
параподиями (Д). Е - пищеварительная система Elysia viridis(Thomp'
son, 1976) (сокращения: d.gl.- пищеварительная железа, s.gl.- сланная
железа). X - строение Bulla (Actaeonidae), 3 - поперечный ср«3
через основание мантийного цекума {Franc, 1968 Ь); линией + + * *
обозначена граница мантийной полооти
100
101
растительных клеток, которыми питается моллюск, сохраняются и про-
должают функционировать в клетках пищеварительной железы среди
венозных сосудов - своего рода негативных жабер, отдапцих угле-
кислый газ (Jensen, I960). В этой связи становится понятной то-
пография пищеварительной железы, напоминающая о турбелляриях
(рис.39 Е). Другие голожаберники, Pt&raeolidia, содержат симбио-
тических зооксантелл (Hughes, 1986). Методом меченых атомов по-
казано, что органический углерод хлоропластов усваивается орга-
низмом моллюска (Greene, 1970).
Большим своеобразием отличается организация Actaeon,представи-
теля Bullomorpha (рис.39 Н,3; рис.40 Б-Е; рис.41 В-Д). Этот
крупный, длиною до 6 см, моллюск имеет проподиум (обособ-
ленная передняя часть ноги) и плоские головные щупальца, которы-
ми он способен глубоко зарываться в грунт. Как и многие другие
раковинные эаднежаберники, он имеет раковину с щелевидным устьем
и поэтому жабра у него находится не над головой, а смещена вбок.
Задняя часть мантийной полости переходит в удлинённый вырост,
мантийный цекум, достигающий вершины раковины. Внутри цекума про-
ходят два ресничных гребня и каждый из них посредине рассечён
глубокой продольной бороздой (рис.40 Г; рис.41 В). Один из них на-
зывается приносящим. Он начинается от жабры и, достигнув вершины
цекума, переходит в выносящий гребень, который заканчивается на
выводном сифоне возле ануса. Ресничные борозды вызывают вододви-
жение в мантийном цекуме и это способствует вододвижению в ман-
тийной полости. Вода проходит через жабру и фильтруется через
Рис.40. Opisthobranchia. A-Melibe, Nudibranchia (Hurst, 1968). Actaeon
tornatilis, Cephalaspidea Б - мягкое тело с раскрытой мантийной по-
лостью, В - вскрытая мантийная полость и мантийный вырост (указано
направление ресничных токов), Г - поперечный срез мантийного вырос-
та (Fretter, Graham, 1954); Д -мягкое тело без раковины, Е - ракови-
на со стороны устья (Pelseneer,1897); К - вид животного сверху
(Thompson, 1976). 3 - Lobiger serradifalci, Sacoglossa, с 2 парами па-
раподий и наружной раковиной (Gonor из Thompson, 1976) Сокращения:
inf.L- нижняя мантийная лопасть, р.с. - мантийный вырост
102
ловчий капюшон
риносрор
нога
гипобранхиальная железа
ресничный гребень правый
' Jr -
железистьи край мантии
щупальца
головное,
лабиальное
спинные
' палимы
гонада
нога
колюмеллярная
мышиа
рот
ресничный
гребень с
бороздой
мантийный
проток
пищеварительная желудок^*
железа выводной
си/рон ресничный
гребень
левый
Б
Г
ресничный гребень
мантийный
проток
рот
нога
Ж
ресничный
гребень с
бороздой
hbr.
гипобранхиаль-
ная железа
выводной си
В
р.с.
infl'.
Д
penis
вырост
проподи-
ума
пенис
ресничный гребень
3
103
слизевую завесу, образованную выделениями жабры (рис.41 В-Д).
Таким образом мантийная полость защищена от загрязнения вероят-
ность которого весьма велика при обитании животного в грунте.
Ц^кум следует рассматривать кай приращение вододвижущей фильтра-
ционной поверхности, ставшее необходимым у столь крупного и от-
носительно медлительного животного, каким является Actaeon.
Донные голожаберники только в редких случаях способны плавать,
Примером может служить Melibe (рис.40 А). Этот хищный моллюск
плавает с помощью спинных крыловидных жабер и ловит своих жертв,
рачков,околоротовыы капюшоном, края которого усажены сенсорными
папиллами. Капюшон расправляется вследствие нагнетания крови в
особые синусы. Радула у него отсутствует, и рачки проглатываются
целиком (Hurst ,1968; .РигсЛол.,1977).
Пелагические голожаберники, крылоногие Pteropoda, плавают
с помощью разросшихся параподиальных плавников, тогда как подош-
ва лишается двигательной функции. Порхающее мышечное движение
крылоногих не позволяет им строго выдерживать направление движе-
ния и развивать большую скорость. Поэтому многие из них остают-
ся микрофагами и потому нуждаются в приращении пищесобираицей по-
верхности. У части из них, Thecosomata,раковина сохраняется,
причём её форма весьма разнообразна, у других, Gymnosomata,ракови-
на отсутствует; в целом же крылоногие с их своеобразным движени-
ем представляют картину значительного разнообразия. Примером рако-
винных крылоногих может служить Limacina (рис.41 А,Б). Живот-
ное плавает, взмахивая крыловидными параподиями,и при этом поль-
зуется особым выростом тела, называемым балансиром. Пищесобираю-
щую функцию выполняет прежде всего поверхность довольно глубокой
мантийной полости, а также подошвы, освободившейся от двигатель-
ной функции, Обширность суммарной поверхности параподий и мантии
.4LLimacina retroversa (Pteropoda Thecosomata) А- плавание, Б-
строение (Morton, 1954). Actaeon tornatilis (Opisthobranchia,Cepha-
laspidea)-. В - строение мантийного выроста, Г,Д - жабра с ресничными
токами (Thompson, 1976)
104
борозды :
приносящая,
выносящая
раковина
сердце
мантийная
слизевая железа
задняя
нитка
вентральная „ „
складна
выводной
сисрон
подковообразная
складка
рот с радулой
щупальце
Б
боковая лопасть ноги
ресничное поле у основания
крылового плавника
балансир
“щевой желобок в мантийной /
полости
анус
слизевая
завеса
дорзальная
складка
медиальная
лопасть ноги с
пищевым желобком
посредине
желудка
гонада
Д
105
объясняет отсутствие у этих животных жабер. Порхающее плавание,
в отличие от Atlanta с их двусторонней симметрией, не предъявля-
ет строгих требований к симметрии животного, и во время плавания
моллюск ориентирован по-разному. Из мантийной полости и с подош-
вы ноги аглютинат поступает в ротовое отверстие, что же касается
посторонних частиц, то они удаляются ресничным эпителием основа-
ния параподий.
Крыловидные параподии имеют и многие другие раковинные крыло-
ногие. Сравнение Limacina с Creseis и с другими Thecosomata
(рис. 41 А,Б; рис.42) показывает, что относительные размеры ноги
и параподиев связаны обратной зависимостью, что и понятно, если
не упускать из виду их своего рода физиологическую конкуренцию
при собирании пищевых частиц. Наличие симметрично расположенных
параподиев возможно объясняет, почемутурбоспиральная раковина,
кстати, у крылоногих обычно синистрадьиая, сменяется двусторонне-
симметричной.Вновь сложившаяся двусторонняя симметрия не мешает
раковине выполнять защитную функцию убежища, в которое втягива-
ется мягкое тело моллюска с помощью пучковых мышц, пронизывающих
параподии и висцеральный мешок (рис.42 ЕД).
Редукция раковины прослеживается ухе у Thecosomata , например
у Gleba (рис.42 Г), у которого вместо раковины имеется псевдо-
конх, образованный плотной тканью. В полной мере раковина редуци-
рована у Gymnosomata,в частности у СИопе (рис.43). Освободи-
вшись от раковины, такие крылоногие приобретают удлинённую форму
тела и вместе с тем возможность изгибаться, хотя двигательная
функция по-прежнему сохраняется за параподиями (рис.43 Г).Возмож-
но, однако, что изгибания тела у Clionidae имеют значение при
Рис. 42, Pteropoda А - Spongiobranchia australis, Gymnosomata (Pelse-
neer из Tesch, 1913), Б-Creseis acicula, Thecosomata (Yonge, 1926),
B-Cavolinia inflexa, Thecosomata (Yonge, 1926), Г- Gleba cor data .
Thecosomata (Tesch, 1906),* её хобот (Д) (Yonge, 1926), E- пучковая
мускулатура Clio pyramidata, Thecosomata, и Cymbulia peroni (Patent
de, 1979)
IM
о^олгпище^одные мускулы
107
овладении ими жертв. Радула, пригодная в основном для соскребы^,
ния, у пелагических Pteropoda сохраняется (например,Creseh
имеет птеноглоссную радулу - рис.20 А 4), однако у хищных 4/^ъпс.
somata она дополняется другими морфоструктурами. Ротовой аппар^»
хищных Gymnosomata помимо радулы включает жёсткие крючья, голов,
ные конусы и снабжённые присосками щупальца (рис.42 А; рис.43 А,
Б,Д).Ротовые конусы служат для схватывания жертв, которыми обыч-
но служат другие крылоногие. Крючья, радула и более или менее уд.
линённый хобот используются для извлечения мягкого тела жертвы
из раковины. Поскольку приём пищи растянут во времени, приспособ,
ления для придержания иищи возле рта - щупальца с присосками -
становятся совершенно необходимыми.
Поучительно сравнение крылоногих с плавающими голожаберниками,
Melibe (рис.40 А) и Glaucus (рис.44 В). Первый в основном
обитает на дне и плавает только время от времени. Поэтому он сох-
раняет подошву ноги. Второй обитает в океанском плейстоне и ноги
не имеет. У обеих форм жабры или книдосаки (последние только у
Glaucus ) расправлены в стороны и служат для плавания и парашю-
тирования. Melibe лишён радулы и обходится ловчим капюшоном,
его жертвы - мелкие рачки, которые проглатываются целиком.
Glaucusнападает на жертв, сопоставимых с ним самим по размерам
(обычно это хондрофоры Velella ), и поэтому лишён ловчих приспо-
соблений, выгрызает куски из тела жертвы. Вместе с тем, несмотря
на указанные различия, эти два моллюска осуществили общую и иную,
нежели у большинства крылоногих, идею организации. Она состоит в
том, что животное плавает брюшной стороной вверх и продольная ось
тела, таким образом, направлена горизонтально; у большинства про-
чих крылоногих она ориентирована вертикально. Далее, у болыпинст-
____________________________________________________________________'
Рис.43.Pteropoda Gymnosomata. Схема строения Clionelimacina ; А -
голова, Б - передняя часть тела, В - висцеральный мешок, Г - плава'
ние с помощью параподий (Morton, 19586).Д - Schizobrachiuirt (Meisen'
heimer из Hyman, 1967). Сокращения: а.-анус, c.g. общее половое от-
верстие, т.- мужское половое отверстие
108
мелок крючьеВ
голодные конусы
радула
параподия
numeSoB
Д
мешки
крючьеВ
Б
радула
нога
сминая железа
простата —
проток простаты
пииевод Сумка пенш
ретракторы
голоВнык
конусоВ
Церебралы&Л
ганглий
семеприемник
cil.d.
а.
гонада
белкоВая
железа
полоВое
отберстие
герматрро
дитныа проток
слизе&ая
железа
В
пшце&аритель -
пая железа
109
ва крылоногих двигательные и парашютирующие структуры сосредото,
чены на переднем конце тела, а у рассматриваемых двух голожаб ер.
ников они рассредоточены по бокам тела. Эти различия, по-видимому
объясняются особенностями строения возможных предков этих живот,
ных.
Плавающие голожаберники в качестве исходного материала для
двигательной системы располагали жабрами и книдооаками, размещён.
ними на спине. Раковинные предки крылоногих ухе вследствие нали-
чия раковины и внутренностного мешка вынуждены обходиться параде,
днями.
Такие голожаб ерники, как Phyllirrhoe (рис. 36 А), сохраняют
сложную форму кишечника, свойственную многим донным голожаб ерни-
кам, нов остальном освоение пелагиали вызвало в их организме, по-
жалуй. более глубокие изменения, чем у Qlaucus . Тело Phyllir
гЬоё прозрачное, сплющено с боков. Сзади оно переходит в несколь-
ко обособленный хвостовой плавник и в целом сохраняет рыбообраз-
ную форму. При плавании оно изгибается в горизонтальной плоскос-
ти, так же как у рыб. Возможно, что уплощенность тела облегчает
дыхание поверхностью и поэтому у Phyllirrhoe отсутствуют специ-
альные органы дыхания,
Наблюдаемая у заднежаберников деторсия иногда доходит до 90°
Её своеобразие состоит в том, что она совершается не столько по-
воротом раковины и внутренностного мешка в направлении по яз-
вой стрелке, сколько вызвана уплощением устья раковины и её обо-
ротов в плоскости оси навивания раковины (рис.34 Б; рис.40 Е).
У безраковинных форм при этом имеет значение редукция внутреннее!'
ного мешка и втягивание внутренностей в цефалоподиум.
Рис. 44.Pteropoda Gymnosomata A - Paedoclione doliiformis, неотени-
ческая форма с ресничными кольцами. (Lain, 1972), Б - Crucibranchia
macrochira (Thompson, 1976). Nudibranchia: В -Glaucus atlanti'
cus (Thompson, 1976). Pteropoda Thecosomata: Г - Cymbulia perO'
ni (Pelseneer, 1897) \ сокращения: о. - отверстие буккальной полос-
ти , рс. - псевдоконх
110
Ill
У таких форм, как Акега и Actaeon, устье раковины остаёт-
ся примерно в сагиттальной плоскости цефалоподиума (рис.33 Б 1 ;
рис.34 Б 2; рис.40 I), и уже' одно это вызывает деформацию мантий-
ной полости, изначально находившейся с правого бока. Этим смеще-
нием объясняется оттеснение хабры из переднего положения назад,
причём ось жабры теперь направлена не вперёд, а вправо. Иными
словами,при деторсии у заднежаберных имеет место не поворот
внутренностного мешка, а его деформация. Анус, хотя и остаётся
справа, но смещается по часовой стрелке назад. Что же касается
полового отверс!ия, то оно остаётся сбоку и спереди. У Actaeon.
деторсия начинается в эмбриональном развитии, когда личиночная
экзогастрическая раковина сменяется окончательной, раскрученной
и неглубокой (рис.38 Г). Такая раковина, естественно, влияет на
размещение органов мантийной полости и внутренностного мешка.
Жизненная форма паразитов у брюхоногих ограничена только перед-
нежаберными Melanellidae и заднежаберными Pyramidellidae. Послед-
ние включают и хищников, высасывающих жертву, например, Eidima,
нападающих на офиур (Waren, 1983). Тип организации жизненной
формы эктопаразита сохраняет значительное сходство с свободноживу-
щими раковинными моллюсками. Эктопаразиты имеют раковину, полза-
тельную подбшву, обычно обладают крышечкой и мантийной полостью.
Вместе с тем для паразитического существования им понадобился хо-
бот с ротовым отверстием на конце, в глубине мышечнбй присоски,
и,кроме того,у них наметилась редукция раковины (рис.45 А,Б).
Последнее вызвано тем, что животное погружено в наружные покровы
хозяев, каковыми являются самые различные беспозвоночные. При
-—*
Рис.45. Паразитические брюхоногие моллюски. A-Pulicicochlea caloma'
ris’, Б - Р. faba 1 - раковина, 2 - общий вид (Ponder, Gooding, 1978). В-
Goodingia varicose , мягкое тело (Lutzen, 1972). Г- Stillier lincAiae,импланти-
рованные в наружные покровы морской звезды Linckia multifora (Lutsen,
1972). Д-Entocolax chiridotae (Скарлато, 1951). E - Entocolax ivanovr. 1- обДИЙ
вид, 2 - висцеральный мешок (Franc, 1968 в). Ж - Paedophorus dicoelobius
(Иванов, 1940) Сокращения: v.s.- висцеральный (внутренностный) мешо^>
d.gl.- пищеварительная железа, hbr. - гипобранхиальная железа
112
нога
113
этом головная часть паразита вытягивается, проникая в тело xogju
ина, а складки кожи вокруг основания хобота разрастаются в псеь.
домантию, обволакивающую целиком всё тело моллюска (рис.45 Г),
Таким образом возникает псевдомантийная полость (псевдопаллиум).
От настоящей мантийной полости она отличается тем, что уже не
связана, по крайней мере неявно, с дыханием, а также вододвижени^
ем и хеморецепцией. Псевдомантия, например у Styli£er, скорее
служит для того, чтобы ограничить вместилище паразита в теле хо-
зяина. Наконец, у эктопаразитов подвергается редукции радулярный
аппарат и слюнные железы.
Организация эндопаразитов претерпела значительно более глубо-
кие изменения: полностью редуцировалась раковина, мантийная по-
лость и нога, головной отдел с щупальцами и радулярным аппаратом,
Тело паразита приняло округлую, в ряде случаев червеобразную фор-
му, так что в нём невозможно признать моллюска. Таким образом,
эктопаразитические и эндопаразитические моллюски хотя и составля-
ют одну жизненную форму паразита, но не представляют из оебя
единства как фенон и тип организации.
То, что можно было бы называть рудиментом внутренностного меш-
ка, полностью обрастает псевдомантией. Специализация сопровожда-
ется у эндопаразитов полной редукцией пищеварительной системы и
обычной для паразитов гипертрофией половой системы. По аналогии
с некоторыми усоногими ракообразными у Asterophila имеет место ре-
дукция самца (рис.46 Б), который постоянно находится в псевдомаи-
тийной полости самки, точнее, в том её отделе, который по анало-
гии с семеприемником можно было бы назвать "самцеприемник".Сход-
ное явление свойственно самкам Enteroxenos (Lutzen.,1966), у которые
Рис.46. Паразитические брюхоногие моллюски. А - Enteroxenos oester-
greni (Lutzen, 1968). Б-Asiterophila japonica 1 - проксимальная часть
псевдомантийной полости (г.), в которую проникла личинка (лг.) че-
рез ресничный канал (sj , 1 - самец, 2 - самка, 3 - схема строен^
самки (vg. висцеральный ганглий)(Грузов, 1965). В - Pseudophorus
dicoelobius (Franc, 1968 в)
114
9
матка
ресничный протон
лажная мантия
А
Lk? отверстие ресничного,
^('тр—протока о центральной
полости;
самец, имплантиро-
ванный в полости
самки
Б
матка
ге.нглий
почка
левая лопасть ноги
хобот
рот
яичник
яйцевод
яйцевые
нононы
висцеральный
мешок
отверстие ломкой
.мантийной полости
яичник
кишечник
печень
суб-
интестинальный
В ганглий
нога
хобот и рот
отверстие ложной мантийной полости
ложная мантия
яйцевые иононы
яичнин
яйцевод
цеоебоо-
плевральн^я
масса
головной
кровеносным
синус
морфоструктура, поначалу считавшаяся семенником, в действитель-
ности оказалась сильно деградировавшим самцом, имплантированнну;
в стенке псевдомантийной полости (рис.46 А). В этой связи можно
предполагать, что семенник гермафродитного £ntoco2ax (рис.45 Д)
в действительности является дегенерировавшим карликовым самцом.
Псевдомантийная полость Entocolax в сущности представляет из
себя выводковую камеру, в которой скапливаются яйцевые коконы
(рис.45 Д,Е). Выводковая камера Paedophorus имеет иное происхож-
дение: она образована широкой подошвой ноги, разросшейся в ущерб
редуцированному внутренностному мешку (рис.45 X).
В онтогенезе моллюсков, так же как и у других животных, имею-
щих планктонную личинку, происходит смена ресничного двигательно-
го аппарата на мышечный вследствие увеличения размеров организма
и сокращения его относительной поверхности (рис.47 А),
Выклев личинки из яйца на стадии трохофоры наблюдается у мно-
гих низших моллюсков, а среди брюхоногих - только у некоторых ар-
хеогастропод. Обычно же из яйца выклёвывается велигер (например,
yCypraeidae - Wilson, 1985), что надо считать нормой для всего
класса брюхоногих в целом (Иванова-Казас, 1977; Fioroni, 1982).
Эмбрионизация и прямое развитие, обычные у лёгочных моллюсков,
можно считать вторичным приспособлением к жизни на суше, а у пе-
реднежаберных, обитающих в воде,- проявлением заботы о потомстве
(Purchon., 1977).
Двигательное приспособление велигера - парус (велум), представ-
ляет из себя сильно разросшийся прототрох околоротовой области
Рис.47. А - развитие. Marisa cornuarietis, Mesogastropoda, Ampullari-
Idae, стадии I, VI, VII, Ш, XIII (Demian,Yousif, 1975). Б- поворот
внутренностного мешка у велигера Patella vulgaris, Archaeogastropoda,
изображённого справа: 1-перед торсионом, возраст личинки 70 час, 2-
поворот на 90°, возраст личинки 76 час, 3- завершение торсиона, воз-
раст личинки 3,5 - 4 суток (Crofts из Сох, i960). Сокращения: an.pl,-
передняя часть ноги, d.d. - кишечный карман, ent.- энтодерма, кишеч.-
ник, l.g.- личиночный же дудок, и. с.-мантийная полость, ор.-крышечка
116
КП.
оссррадий
осфрадий
край мантии.
педальная борозда
ктенидий мочеточник
ор.
почка
мочеточник
л ампула
'Ы
^1 отдел
Ж желудка
Кишечник
оперкулярная железа
правый мышечный пучок
(ретрактор)
vl.
1
ХШ.
церебральная комиссура анальные кл
ЗаОнЯЯ
кишка
осурадиаль
висцеральный
плевро- .
педальный
глаз
педальная
комиссура
ор.
Б
V1.
2
ганглии:
церебральный,
буккальный
педальный
'СГ50/1
субинтести-
нально-
висцералытыи
коннектив
3
ор.
личиночны
ретрактор
d.d.
дефинитивный (левый)
колюмеллярный мускул
117
трохофоры. Некоторыми личинками он используется для разрыва яйце,
вых оболочек при выклеве (.Davis, 1967). Вследствие малых абсо-
лютных размеров личинки и большой относительной поверхности
велигера ресничный аппарат паруса остаётся достаточно действен-
ным приспособлением для плавания и вододвижения, и по этой же при-
чине велигер не нуждается в специальных органах дыхания. Те орга-
ны и морфоструктуры, которые понадобятся моллюску в предстоящей
жизни на дне, закладываются у велигера уже в планктонный период.
Прежде всего к ним относятся нога с полэательной подошвой, мантий-
ная полость с жаброй и внутренностный мешок в раковине. Увеличе-
ние массы тела растущей личинки вызывает сокращение величины от-
носительной поверхности. Последствия этого сокращения усугубля-
ются появлением и ростом раковины, уменьшающей свободную поверх-
ность животного. Нехватка ресничной поверхности возмещается раз-
растающимся парусом, а также того или иного рода лопастями (рис.
47 А-Г). Лопасти находятся возле рта, и ресничный покров паруса
служит не только для плавания, но и для вододвижения, привлечения
ко рту пищевых частиц {Strauthmann, Leise, 1979) (рис.46 В; рис.
48 А).
Вращение велигеров при плавании - явление того же порядка, что
и у быстро плавающих простейших, оно получает объяснение в теории
гидродинамики микроскопических объектов. Сагиттальный гребень
на раковине, вызывающий ассоциации с взрослым переднежаберником
Atlanta , мог бы свидетельствовать о прямолинейном движении, по
Рис.48. Личинки переднежаберных моллюсков. А - Nassarius reticulatus,
Neogastropoda Buccinacea : 1- общий вид сбоку, 2 - вид с брюшной сторо-
ны (Fretter, Montgomery, 1968). Б-Crucibulun spinosuin: 1 - велигер с
брюшной стороны, 2,3,4,5 рост раковины с усилением турбоспираль-
ности, 6 - поздний велигер с брюшной стороны, ползающий моллюск,
лишившийся паруса, 8- раковина позднего велигера {Fretter, 1972).
В -Tritonia hombergi, Nudibranchia: 1-только что осевшая личинка, 2-
послеличинка через несколько часов после оседания {Thompson, 1962).
118
119
Рис.47, Личинки переднежаберных моллюсков; А,Б,В,Г - велигеры с елей-
но расчленённым парусом поперечником до 16 мм (Dawydoff, 1940). Д -
Stiliger niger, Nudibranchia (Rasmussen, 1951). E - Distorsio clathrata,
Mesogastropoda (cTAsaro, 1969)
120
за-
У личинок брюхоногих такой гребень представляется редкостью (рис.
Е). Вследствие вращения спирально закрученная раковина не
^удняет плавание велигера.
Осевшая на субстрат личинка переходит к ползанию, причём,
нас-
колько это известно, поначалу ползание совершается с помощью рес-
ничного эпителия подошвы (рис.48 В). Это обстоятельство становит-
ся понятным, если принять во внимание малые размеры организма ли-
чллки. Мышечный аппарат велигера выполняет несколько иные функции:
втягивание внутрь раковины мягкого тела, втягивание паруса (Jret-
ter, 1972)(рис.49 А 2). Уже' в самом начале своей донной жизни
молодой моллюск имеет вполне сложившийся мантийный комплекс ор-
ганов в достаточно вместительной мантийной полости, а также ногу.
Торсион у разных брюхоногих имеет свои особенности. У цвупред-
сердных он совершается на стадии велигера и занимает от 2-3 мин
(Лспаеа) до 400 ч (Haliotis) (Fioroni, 1982). Торси-
он вызывается резким сокращением правого ретрактора, который впос-
ледствии сохраняется у взрослого моллюска в виде колюмеллярной
мышцы. Это сокращение производит впечатление нервно-мышечного ак-
та (Иванова-Казас, 1977). Кроме того, имеет значение неравный
рост правой и левой сторон животного, справа рост больше, чем
слева. У однопредсердных торсион начинается несколько раньше, чем
у двупредсердных,и вызывается одним только неравномерным ростом
организма. У заднежаберных мантийный комплекс закладываетсяспере-
ди. -В процессе деторсии у заднежаберных часто происходит обраста-
ние мантией и полная редукция первичной раковины и внутренност-
ного мешка. У части заднежаберных Pteropoda Thecasomata торсион
отличается особой сложностью, так как дополняется вторичным вра-
щением внутренностного мешка. В результате складывается наиболее
сложная диссимметрия среди всех первичноротых (Беклемишев, 1964).
В целом, в онтогенезе, в процессе превращения микроскопически
малой пелагической личинки в донный организм воспроизводится кар-
тина филогенетического развития, включая смену сред обитания -
пелагиали на бенталь.
121
мантийная полос
Рис. 48. Личинки брюхоногих моллюсков:А -Lacunaviveta,Mesogastropoda:
1-личинка с частично сократившимся парусом, 2- личинка, втянувшаяся
в мантийную полость (Fretter, 1972). В - krchidoris pseudoargus, Nu-
dibranchia, фильтрационные токи на брюшной поверхности (Thompson,
1959). Г - Conus omaris , Neogastropoda (Ostergaard, 1949), Д - Lacuna
с ресничным покровом возле ктенидия (Ockelmann, Nielsen, 1961). Е -
Cadlina laevis , Nudibranchia (Thompson, 1967). g _ схема, поясняпдая
топографические отношения раковины и мантии у личинки (1 ), после-
личинки (2 ) и взрослого моллюска у заднежаберных Notaspidea, Berth
ella (слева) и голожаберников Tritoniidae (справа)(3)(Thomp-
son, 1962). Обозначения!: —।—► ресничные токи, приносящие ко рту
пищевые частицы,------► вододвижение. 3 - два типа протоконхов у
заднежаберных: 1 - свёрнутый. 2 - яйцевидный (Hurst из Fioroni, 1962')
122
123
ТИП ОРГАНИЗАЦИИ ДВУСТВОРЧАТЫХ МОЛЛЮСКОВ (BIVALVIA)
Организация двустворчатых моллюсков весьма однотипна, различая
между ними в основном касаются жабер, пищеварительной системы и
раковины (Скарлато, 1981; Morton, 1983а).Таким образом, остава-
ясь таксоном, класс Bivalvia в то же время обладает значитель-
ной целостностью и как тип организации. Экологически двустворки -
прежде всего малоподвижные донные фильтраторы. Поэтому, сохраняя
относительную целостность как тип организации, таксона, они боль-
шей частью принадлежат к одной жизненной форме.
Расчленение тела двустворок своеобразно: при отсутствии морфо-
логически обособленной головы, они обладают вместительной мантий-
ной полостью, ограниченной боковыми складками мантии. Рот и анус
открываются в мантийную полость, в ней же находятся нога, предс-
тавляющая из себя клиновидный вырост брюшной стороны тела, и две
пары боковых жабер. Мягкое тело животного, включая мантию, пол-
ностью охвачено раковиной (рис.З Б 2).
Поскольку опорная функция перешла к раковине, целом как опор-
ное образование у двустворок не сохранился. Его производным, так
же как у брюхоногих, остаётся прежде всего перикард. Наличие
твёрдых скелетных частей, створок раковины, способствует сохра-
нению пучковой мускулатуры. Важнейшие в этой мускулатуре мышцы-
аддукторы раковины, ретракторы ноги и сифонов прикреплены одним
концом к внутренней поверхности раковины. Локальность пучковых
мышц, паренхимное строение большей части тела и способность ноги
и краёв мантии к перистальтическому сокращению делают возможным
существование лакунарной системы; кровеносная система, таким об-
разом, остаётся открытой.
124
friz, перемещения в пространстве обычно служит нога (рис.49 к,
Т 1;рис.50 В), значительно реже - мантийная полость (плава**
у морского гребешка Pecten. ). В связи с ногой появляются ха-
^терные для двустворок педальные ганглии. Поскольку в сравнении
с пассивным телом моллюска, включая раковину, масса мышечной
-лани ноги невелика, перемещение в пространстве обычно ограниче-
я0 и не имеет большого значения в жизни моллюска. Вследствие ря-
да обстоятельств возможности ноги как двигательного приспособле-
ния оказываются ещё более скромными.
Нога, могущая принимать червеобразную форму и выдвигаться меж-
ду складками мантии, ограничивающими ножную щель, в основном со-
вершает перистальтическое и изгибательное движение. Оно особенно
действенно, когда энергоотдача совершается через поверхность но-
ги, как у дождевых червей при ползании в почве. Однако именно
при перемещении двустворок в грунте - будет ли это зарывание или
горизонтальное ползание - сопротивление трением увлекаемому за
ногой телу моллюска остаётся значительным.
При ползании на поверхности субстрата у моллюсков возникает
развитая система мышц-ретракторов ноги (рис.49 Г 1 ),в ряде слу-
чаев появляется прикрепительная и ползательная подошва ноги (рис.
51 Б,Г,Е). Впрочем, у Nucula подошва ноги появляется в связи
с особым способом зарывания в грунт (рис.50 Г). Большим своеоб-
разием отличается ползание и плавание Lima, в котором участвуют
краевые щупальца мантии, снабжённые слизевыми клетками (рис.50 А).
Выделяемой ими слизью моллюск склеивает мертвые раковины и камни,
так что получается убежище, гнездо .Впрочем, ползанию Lima спо-
собствует и выталкивание воды из мантийной полооти и,кроме того,она
может плавать.
Значительно уступая в подвижности даже медлительным брюхоногим,
Двустворки в своей жизнедеятельности всецело зависят от вододвиже-
Дия. Оно обычно вызывается ресничным покровом мантии, хотя опре-
делённое значение при этом сохраняет мышечный аппарат аддукторов
125
(хлопание створками), а также движения жабер и слабые сокращения
тела. Для приведения и отведения воды служат два сифона - особце
участки мантии, но довольно часто для этого используется весь
свободный край мантии.
В сравнении с типичными брюхоногими обычные двустворки обла-
дают как бы удвоенной схемой циркуляции воды в мантийной полости
Две части этой единой схемы отвечают двум створкам раковины,
зобщены ногой и размещены симметрично относительно сагиттальной
плоскости животного. В целом такая удвоенная схема подчиняется
двусторонней симметрии. В свою очередь, каждая из двух её частей
диссимметрична относительно оси створки, проходящей от макушки
к брюшному краю створки (ось NX на рис.52 и рис. 53 А,Б). Дис-
симметрия циркуляции воды в каждой такой части вызвана тем, что
ротовое отверстие и некоторые пищесобирапцие структуры (губы,
околоротовые лопасти) находятся в передней части мантийной полос*
ти, тогда как вода поступает в мантийную полость обычно через си-
Рис. 49. А -обобщённая схема пучковой системы мышц двустворчатого
моллюска. Б - стадии зарывания двустворки в грунт (вид слева):
1 - створки прижаты к субстрату, сифоны открыты, нога выдвинута,
2 - сифоны замкнуты, нога продолжает выдвигаться, 3 - вследствие
сокращения аддукторов вода выталкивается из мантийной полости и
размывает грунт, кровь нагнетается в ногу,4 - нога разбухает и
заякоривается в субстрате, передний ретрактор ноги,сокращаясь,
наклоняет раковину, 5 - сокращение заднего ретрактора ноги вы-
зывает погружение моллюска в грунт, сифоны открыты, 6 - моллюск
погрузился в грунт и снова выдвигает ногу. В - то же на попереч-
ном срезе моллюска, стадия а отвечает стадиям 1,2,6, стадия
Ь отвечает стадии 3 , стадия С отвечает стадиям 4,5 (Trueman-
1968). Г - механизм прикрепления неравнобокого гетеромиарного
моллюска Mytilacea (1,4) и неравностворчатого мономиарного мол-
люска Pteriacea(2,3) на поперечном срезе и слева. Обычные стрелки
обозначают токи воды, пунктирные - удаление аглютинатов (Уоп^е,
1981). Прочие обозначения и сокращения:*-*, ► - движение крови
моллюска и воды, °-* выталкивание воды из мантийной полости,
сокращение мышц и напряжение лигамента,ат-мускул-аддуктор,ТШ'
Мускул-ретрактор, рт -протрактор ноги, tin.- поперечный педалв:гый
мускул,h.-давление в гемоцеле ноги
126
127
фон в задней части мантийной полости. Таким образом, диссимметрия
мантийной полости у двустворок - это следствие морфо-функционали
ной неодинаковости её передней и задней частей. В этой связи upejl<
ставляет определённый интерес сравнение двустворок с брюхоногими,
Если у брюхоногих диссимметрия мантийной полости устанавлива-
ется постепенно, то у двустворок она может считаться изначаль-
ной, следствием исходной неодинаковости обширной мантийной полос-
ти, вместившей всё мягкое тело моллюска, включая ротовой отдел.
Основное направление токов воды у двустворок имеет место в
парасагиттальных плоскостях, у брюхоногих токи воды пересекают
сагиттальную плоскость.
У брюхоногих турбоспиральность объясняется тем, что раковина
представляет из себя обызвествленную траекторию диссимметричной
мантийной полости, смещаемой вокруг оси навивания раковины в про-
цессе её роста (с.36). У двустворок турбоспиральность отдельной
створки объясняется тем, что она тоже может считаться обызвеств-
лённой траекторией боковой половины мантийной полости, смещаемой
в процессе роста от макушки раковины к брюшной стороне (ось NX
на рис.52).
Рис. 50. А - Lima hians (РесИпо1с1еа):1-ползанле с помощью мантийных
щупалец, 2 - поперечный срез двигательного щупальца через коль-
цо слизевых клеток, 3 - край мантии с двигательными щупялъттями
(.Gilmour, 1967). Б - топография органов мантийной полости Li~
maria orientalis, имеющей отверстие в сросшейся мантии (ау),
через которое удаляются экскременты (Grange, 1974). В - после-
довательные стадии кувыркания Scintillina zelandica (Morton.,
1957). г - закапывание в грунт Nucula nucleus (Protobnanchia) с по-
мощью боковых выростов уплощенной подошвы ноги (Stoll, 1938),
Обозначения: широкая стрелка на рис. А1 показывает нап-
равление движения моллюска, длинные стрелки показывают направле-
ние струй воды, короткие стрелки показывают направление изгиба-
ний щупалец. Сокращения: буз.^.-биссусовая железа, с.tsполоса
соединительной ткани, o.l.m.- наружные продольные мышцы, i.l.m. -
внутренние продольные мышцы, Ln..-продольный нерв
128
У брюхоногих, главным образом вымерших, помимо турбоспиральщц
имеются плоскоспиральные раковины. Сходную картину представляют
двустворки, имеющие турбо- и планоспиральные створки (рис.8 А,Г)
В этих фактах проявляется неоднозначность корреляции между стпое-
нием раковины и строением мантийной полости.
Далее, диссимметрия мантийной полости у брюхоногих есть причи-
на диссимметрии всего организма в целом. Что же касается двуство-
рок, то у них две диссимметричные части единой мантийной полости
не нарушают двусторонней симметрии всего организма в целом.
Примитивная схема циркуляции воды в мантийной полости состоит
в том, что вода притекает спереди и вытекает сзади. Эта схема осу.
ществляется у Naculidae и Lucinoidea (Haszprunar, 1985).
У подавляющего большинства двустворок вода притекает сзади (Mor-
ton , 1983а). Между формой раковины и схемой циркуляции воды име-
ет место некоторое соответствие (Tevesz, Carter, 1979).
Однообразное строение равномерно закруглённых створок у Pec-
ten и Pinctada (рис.49 Г2, рий52 ГЗ ,рис.53 В) свидетельствует
о том, что водообмен осуществляется на протяжении всего свободно-
го края мантии и что обособленные сифоны отсутствуют. Наоборот,
у Муа токи воды ограничены сильно разросшимися сифонами, тогда
как складки мантии срастаются, оставляя лишь небольшое'отверстие
Рис. 51. А - циркуляция в мантийной полости Spisula subtruncata(Mact-
ridae (1) и удаление экскрементов и аглютинатов через вентральное от-
верстие (2 ) (Monge, 1948). Б - Scintilla (Leptonidae) с раковиной,
полностью обросшей папиллярной мантией,и ползательной подошвой
(Ponder, 1967). В - схема работы жаберного насоса Parayoldiella
(Protobranchia') (Филатова, Шилейко,1985). Г - Entovalva semper!
(Montacuiidae) с широкой прикрепительной подошвой ноги (Ohshima
по Franc .I960). Д - раковина Cuspidaria cuspidate с сифональным вы-
ростом ( Franc, I960). Е -Devonie.perrieri (Montacutidae) с ракови-
ной, частично обросшей папиллярной мантией и прикрепительной подош-
вой ноги (Popham по Рцгс2юл,1977). Сокращения: o.d.-наружная п0'
лужабра, i.d -внутренняя полужабра, х - аглютинаты и экскременты.
Обозначения:----► токи воды, — ресничные токи на мантии,
токи на висцеральной массе
130
для ноги. Вследствие разрастания сифонов у Муа задние края ство-
рок расходятся, оставляя широкое зияние, и линия прирастания ман-
тии к раковине сильно изгибается, образуя мантийный синус.
Форма раковины двустворок зависит и от многих других обстоя-
тельств: положения моллюска в грунте или на грунте, прикреплённо-
го или подвижного образа жизни, особенностей строения мышц-аддук-
торов и др. В свою очередь, от формы раковины зависят энергети-
ческие затраты на зарывание в грунт,в общем возрастающее с глуби-
ной (Balakrishnan, Anzell, 1966; Eagar, 1978). Глубина зарывания
даже у одного видя зависит от возраста моллюска и его размера, от
качества грунта (Gale , 1976).
У примитивных Glycymeris (рис.52 Б 1 ), сохраняющих изомиар-
ную систему аддукторов (два мускула одинакового размера), дорзо-
вентральная ось MX и ось мантии-раковины NX совпадают и перпендику-
лярны к продольной оси АХ и оси замка БХ' две последние к тому
же параллельны между собой. Когда такой моллюск прикрепляется к
Рис. 52. А - неравно створчатая раковина Pandora inaequivalvis с ма-
кушки и сбоку (Franc, I960). Б - раковина ювенильного и взросло-
го моллюска Pseudopythina marls (Rosewater, I984).B - строение тридакны
Tridacna, правая створка (Vonge, 1953). Г - различные пути дос-
тижения мономиарного состояния у разных двустворчатых моллюсков:
1 - Glycetneris (равностворчатый, равнобокий, изомиарный),2-МуМ-
2us-(неравностворчатый, равнобокий, мономиарный), 3 *- Pecten.
('равностворчатый, равнобокий, мономиарный), 4 - Tri da спа (равност-
ворчатый, неоавнобокий, изомиарный), 5 - Bartlettia (неравност-
ворчатый, неравнобокий, гетеромиарный) ,6 - Acostaea (неравно-
створчатый, неравнобокий, мономиарный). Сокращённые названия мор-
фологических осей: ЫХ нормальная ось мантии-раковины, соединя-
ющая макушку раковины с наиболее быстро растущим свободным краем
мантии\МХ дорсо-вентральная ось тела, идущая от середины спин-
ной стороны (лигамент раковины) к середине подошвы ноги; НХ -
ось замка, вокруг которой поворачиваются створки при открывании
и закрывании раковины, АХ" продольная ось тела, проходящая
через оба мускула-аддуктора. Вращение раковины и мантии показано
пунктирной стрелкой, биссус показан на рис. 2,3,4,5. Заштрихована
передняя часть животного, пунктирована передняя часть мантии и
раковины. Все моллюски изображены с левого бока
132
субстрату как мидия, рост передней части животного уменьшается,
передний аддуктор редуцируется и наступает гетеромиарное состоя-
ние (рис.52 Б 1 ). Тогда ось НХ образует острый угол с продоль-
ной осью тела АХ и при этом дорсо-вентральная ось тела MX изги-
бается таким образом, что пересекается под прямым углом с други-
ми осями, а ось мантии-раковины NX образует со всеми прочими
осями острые углы. Поверхность раковины, прилежащая к субстрату,
становится плоской и поперечный срез животного по оси MX приб-
лижается к треугольнику (рис.49 А4).
Прикрепляющийся биссусом Pecten сохраняет способность
плавать, поэтому его раковина уплощена по оси единственного со-
хранившегося аддуктора (мономиарцре состояние) и это уплощение
раковины сказывается на способе прикрепления (рис.49 Г 2,рис.53
Г). Pinna формой раковины и биссусным прикреплением напомина-
ет мидию (рис.53 Б 3). Однако моллюск при этом не прикреплён к
субстрату, его раковина погружена в грунт примерно на 1/3. Поэ-
тому у Pinna нет уплощенности раковины в области биссусного
прикрепления и на поперечном срезе через раковину створки выгля-
лишь уплощенными. В сравнении с мидией у Pinna проис-
ходит дальнейшая редукция передней части тела и переднего аддук-
Рис. 53. А - двустворки с различным ростом раковины и размещени-
ем мягких частей тела: 1 - Glycymeris (равнобокая), Q-Siliqua
patnla (удлинённая слабо неравнобокая раковина), 3 -Ensis аг-
cuatus (сильно удлинённая неравнобокая раковина)(Уоп£е, 19526).
Б - влияние биссусного прикрепления на форму раковины: 1 -Area
(прикрепление подошвой ноги в неглубоком укрытии), 2 -Mytilxis и
3 - Pinna (прикрепление макушной вниз к вертикальной поверхности
или в грунте) (Yonge, 1953). В - схема строения Pin eta da (1 )
и Pecten (2 )(Morton Yonge , 1964). Г - биссусное прикреп-
ление Argopectenirridans (Walker, 1984). Д - топография органов
мантийной полости у детритофага Macomanasuta (1) и сестонофага
Репах gouldi (2) (Pohlo, 1973). Сокращения: Ь. биссус, U.-
макушка, add. ant.-передний аддуктор, add .post.- задний аддуктор, г,гк-
-ротовые лопасти,ex.br.-наружная полужабра, i.br. - внутренняя полу-
жабра, s.i. вводной сифон, s.ex. - выводной сифон. Прочие сокраще-
ния <?м. рис. 52
135
тора. Задний аддуктор, напротив, разрастается, и благодаря ,дОе„
му положению в центре раковины успешно схлопывает створки. Это
приводит к выталкиванию воды из мантийной полости и вызывает ре_
активное плавание.
У Cardium наблюдается только лёгкий поворот оси NX ман-
тия-раковина (рис.52 Б 5). У Tridacna ось NX изогнута так сильно
и передняя часть раковины сокращена настолько, что нога сблизи-
лась с макушкой раковины (рис.52 Б4, В). Тридакна прикрепляется
к субстрату биссусом в примакушечной области, и свободные брюш-
ные края створок обращены вверх. Так же как у Pecten, у тридакны
происходит разрастание мантии на вновь освободившейся от сокра-
тившейся части животного внутренней поверхности створки. Бё при-
крепление отмечено так называемой псевдопаллиальной линией. На
рис.52 Б 3,4,5 она показана короткими параллельными штрихами.
Смещение макушки раковины вперёд сопровождается у Ensis вытяги-
ванием задней части раковины (рис.54 А 3) и уменьшением её высо-
ты по оси МХ.Эти сопряжённые особенности позволяют моллюску быст-
ро и глубоко зарываться в грунт.
Прикрепление биссусом у Area (рис.53 Б 1 ) не вызывает иск-
ривления морфологических осей: животное лишь прижимается к субст-
рату расширенной подошвой ноги и при этом его замочный край удли-
няется.
Довольно своеобразные моллюски Etheriidae (рис.52 Б6,7; рис.54)
прикрепляются к субстрату передним выростом правой створки. Как
это видно у Bartlettia (рис.52 Б 6 ), удлинение передней части ра-
ковины приводит к искривлению створок. У Etfreria правая
створка разрастается в трубчатый "клюв", в который отчасти захо-
дит полость раковины (на рис.54 А 1 это показано пунктиром), у
Acostaea края"клюва" подворачиваются таким образом, что получа-
ется довольно обособленная камера, которая сообщается с внешней
средой только через узкую щель. Подвёрнутая поверхность клювовид-
136
Рис. 54. Двустворчатые моллюски семейства Eutheriidae: A - Euthe-
ria elliptica , створки изнутри и снаружи:1,2- правая створка, 3,4 -
левая створка. Б - Acostaea rivoli, правая створка изнутри (Уоп--
ge, 1962). В -Milneria kelseyi (Carditidae'), изнутри,с выводковой
камерой, образованной заворотом брюшного края створки (.Coan, 1974)
137
ного выроста служит для прикрепления к субстрату посредством це-
ментации. Что же касается левой створки, то она лишена клюва и
по размерам уступает правой створке. Вследствие прикреплённого
образа жизни Eutheriidae утратили ногу, педальные ганглии и ре-
тракторы ноги (Yonge , 1962).
Сверление древесины Xylbphaginae и мягких горных пород кам-
неточцами Pholadinae откладывает отпечаток на форму и строение
раковины и вызывает её редукцию. В процессе сверления камня у
Zirphaea наблюдаются примерно те же стадии, что у зарывающих-
ся в грунт обычных двустворок (рис.49 А,Б; рис.55 В,Г,Д;рис.56 Б),
Сверление осуществляется механически, острыми зубцами на перед-
ней части раковины, хотя химический способ разрушения извести
кислотой, выделяемой моллюском, по-видимому, тоже имеет место
(Yonge, 1955). Сверлящие движения створок раковины вызываются
попеременными сокращениями аддукторов. При сокращении переднего
аддуктора раковина раскрывается сзади, сокращение заднего
аддуктора вызывает раскрытие раковины спереди. Створки, смещаемые
вокруг оси МУ,при этом как бы раскачиваются. Поэтому у Pholadi-
dae и Teredinidae створки широко разведены спереди и сзади, ог-
раничивая широкое зияние, а нога в сечении круглая (.Hoagland,
Рис. 55. А - распределительная система в мантии Ostrea puelahana
(Ostreoidea'j (Harry, 1985). Б - двустворка Phlyctaenachlamys
lysioscpiillina (Leptonoidea), напоминапцая ползающих брюхоногих,
с мантией, обрастающей раковину (Popham из Franc, I960). В -
стадии сверления субстрата у Zirfaea crispata(Pholadidae'): 1-
- нога опущена, сифоны открыты, 2 -нога расширена, начинается втя-
гивание сифона, 3 - дальнейшее втягивание сифона с одновременным
сокращением вентрального аддуктора,4-сокращение заднего аддук-
тора, передняя часть расширяется, раковина скоблит субстрат, си-
фоны открыты, 5- сокращение переднего аддуктора, сифоны открыты.
Г - створки раковины Z.crispata. с брюшной стороны (.Ealahrishnar,
Nair,Ansell,1968) -Д - Pterioidea: Isognomon isognomon fl), Malleus (2)
Сокращения: add.-адцуктор, pa- передняя часть ноги.р.р. - задняя
часть ноги, л.-граница перламутрового слоя, т.п-мантийный рет-
рактор , Ьг. - передний ретрактор биссуса
138
д
Е
139
Turner, 1981). В связи с сверлением они обладают довольно развц
той системой пучковых мышц, включающей помимо обычных аддуктор^
створок,и ряд других мышц - вентральный аддуктор, передний доцОл>
нательный аддуктор и др. Для прикрепления этих мышц служат внут.
ренние скелетные структуры: апофизы, макушечно-вентральный гре-
бень, мыщелки и пр. Полость, высверливаемая камнеточцем, служит
только убежищем. Поэтому, просверлив достаточной глубины норку,
моллюск прекращает сверление. Его нога редуцируется, а переднее
зияние между створками Зарастает так называемым callum (рис,
55 Д) и края мантии почти полностью срастаются. Сифональная труб,
ка пропитывается известью, выделяемой задними краями мантии, и
получается сифоноплакс. Поскольку моллюск сохраняет связь с внеш-
ней средой только через сифоны, они развиты очень хорошо и задние
края створок расходятся широко как у Му а ; имеется и мантийный
синус.
Своеобразная форма раковины у Isognomonidae (рис.55 Е) объясня-
ется их особым способом прикрепления к субстрату и положением в
грунте. В обоих случаях макушка раковины и замок обращены вниз,
а брюшные края створок направлены вверх; в целом раковина вытя-
нута в направлении от макушки к заднему краю створок. Раковина
Pseudopythonia (рис.52 Б) изогнута вследствие обитания моллюс-
ка под элитрами полихеты Aphrodita.
Полная неподвижность, пожалуй, сильнее всего сказалась на
строении зарывающихся в грунт Brechites(pHc.56 В). Створки ра-
ковины Brechites редуцированы и неподвижно срослись с извест-
Рис.56.Teredinidae. А - Xyloredo ingolfa : 1,2-левая створка сна-
ружи и изнутри, 3,4 -правая створка, то же. Б -Xyloredo nooi: 1-
моллюск сбоку, 2- раковина со стороны вершины (Romer, 1972).
В - задний конец Bankia neztalia с палетками и их ретракторами
(p.rtr.) и сифонами (s.) (, Turner,McKoy, 1979). Г - сифоны кора-
бельного червя в обычном положении (1 ), при спаривании ( 2 ),
выводной сифон самца введён в вводной сифон самки (3 ). Д - попе-
речный срез корабельного червя (Turner, 1966). Сокращения: s.fcrv.-
наджаберная афферентная вена, su.brv.-поджаберная эфферентная вена
140
ковой трубкой, выделяемой мантией (ложная раковина). Складки
мантии срастаются весьма полно, так что остаются только отверг
тия сифонов сзади и рудимент ножной щели спереди.Вместе с рако-
виной редуцированы аддукторы (амиарное состояние), тогда как од.
фокальные и мантийные мышцы хорошо развиты ( Purchon, 1956).
Ложная раковина снизу расширяется в диск, который несёт отростки
закрепляющие животное в грунте.
Форма сильно уменьшенной в размерах раковины корабельных чер.
вей Teredinidae определяется её сверлящей функцией, тогда как
защитная функция переходит к обызвествлённым стенкам ходов, про.
сверленных в древесине (рис.56 А, рис.57).В связи с удлинённой
червеобразной формой тела у корабельных червей сильно удлинены
жабры (рис.56А) и намечается усложнение распределительной сис-
темы. Среди пищеварительных ферментов имеются целлюлазы и таким
образом им для питания доступна целлюлоза древесины, дополняю-
щая обычный рацион планктофагов. Сифоны также удлинены и своей
способностью растягиваться несколько возмещают неподвижность
моллюска (рис.57 Г 1.2.3 ).
Неравностворчатость раковин наблюдается в 51 крупном таксоне,
главным образом в семействах (Nicol, 1958). Она связана с прик-
реплением посредством цементации (Etheriidae, Ostreidae,рис.58 Б)
или биссусом (рис.58 А,В), а также объясняется особым положени-
ем животного в грунте, когда сагиттальная плоскость занимает го-
ризонтальное положение (рис.59 В,Д).
Особый случай представляет закрученная раковина Trisidos,
задняя часть которой образует горизонтальную плоскость, остаю-
щуюся на поверхности грунта (рис.59 Г). Эта закрученность не от-
ражается на размещении сифонов и строении мантийной полости и её
Рис. 57. А - строение корабельного червя Teredora vualleolus (Teredi'
nidae)(Turner, 1966). Б - моллюск-сверлильщик Pholas dactylus 1'
створка раковины изнутри, 2-мягкое тело ( Fretter, Graha/n.,1976)-
В - Brechites annulatus (Clavagellidae):1-известковая трубка, 2 -
схема строения {Franc,I960)
142
add. post,
add. ant.
сифоны:
Модной,
выводной
дополнительный вентральный
аддуктор
A
палетки
край мантии
Б
протрактор
палеток
желудочек
нога
апофиз
линия прирастания
мантии
полости :
наджаберная,
поджаберная
жабоа
еварительная add.ant
железа
гонада
иекум
раковина—j
В
2
кишка
сироональная трубка
разрез через мантию
равый ктенидий
пииеоарителеная
железа
туловищный
ганглий
анус
предсердие
почка
Иодной
си<рон
ротовая лопасть
дополнительный
вентральный аддуктор
лабиальные
пальпь!
мешок кристаллического
сгеоелька висцеральный
мешок
сращение передних частей
складок мантии
'л
ротовая лопасть
нога
мышцы передней части мантии
'периостранум
рудимент педальной апертуры
143
органов, однако сказывается на размерах внутренностей - почек,
ретракторов ноги и лр. (Morton, 19836/
Вододвижение в мантийной полости позволяет осуществлять ряд
жизненных функций, прежде всего газообмен и отфильтровывание пи-
щевых и прочих частиц. У Protobranchia (рис.60 А) жабры состо-
ят из подвешенных на мезентериях листков и их размеры относитель-
но невелики. Соответственно у них увеличены ротовые лопасти и
сильно удлинены лабиальные пальпы (рис.61 А). У Nucula послед-
ние служат для собирания пищевых частиц из толщи грунта. Таким
образом в некоторой степени осуществляется разделение функций
органов мантийной полости: дыхательная функция остаётся за жаб-
рами, нищееобирающая - за ротовыми лопастями и пальпами. Лабиаль-
ные пальпы весьма велики и у других Protobranchia.Например у
Yoldia они высовываются через задние сифоны наружу и соби-
рают частицы детрита с поверхности грунта и из его толщи (рис.
61 Б), взвешенные частицы концентрируются на жабрах (Stasek,
1965). В общем, у Protobranchia довольно чётко прослеживается
обратная корреляция между размерами ротовых лопастей и пальп с
одной стороны, и жабер - с другой стороны (рис.61 А,В). Эта кор-
реляция прослеживается и за пределами Protobranchia у разных
жизненных форы: сестонофаги имеют обширные жабры и небольшие ро-
товые лопасти, детритофаги, наоборот, обладают относительно
мелкими жабрами и обширными ротовыми лопастями (рис,53 Д).
Кроме того, существует некоторое соответствие между положени-
ем оси жабер и направлением потоков воды в мантийной полости:
Рис. 58. А - биссусная нить разных двустворок 1 - Pinna vexil-
lum (Pinnidae"), 2 - Septifer bilocularis (Mytilidae), 3 - Isogno -
mon nucleus (Isognomonidae), 4 - Pinctada mar gar it if era
(Pteriidae), 5-Modiolus thorsom (Mytilidae) (Ockelmann,1933).
Б - раковинаOstreapuelahana изнутри и снаружи (Harry, 1985).
В - биссусный комплекс Xenostrobus balani (vrg’. -висцеральный
ганглий)(0ckelmann, 1983). Г - раковина Quadrella cpadrula,
Unionidae, изнутри {Clarke, 1973). R-Adipicula tongissima (fforiko-
shi'Tsnchida, 1984)
144
145
при сквозной циркуляции спереди назад ось жабер пересекает направ-
ление преобладающего потока, если же токи воды ограничены тесно
сближенными сифонами, то ось жабры параллельна направлению токов
(рис.61 Б).
Жабры Filibranchia образованы отдельными жаберными нитями.
Они сложены вдвое таким образом, что выделяется нисходящая от мес-
та прикрепления и восходящая (к месту прикрепления жабры) части.
Нити срастаются поперечными перемычками между собой и кроме того
срастаются в той или иной степени восходящая и нисходящая части
одной нити. В итоге каждая жабра представляет из себя двойное си-
то. Выделяемая на жабрах слизь гонится ресничками покровных кле-
ток к месту перегиоа жаберных нитей, к свободному краю жабер
(рис.50 Е. рис.60 Б,В,Г), где находятся особые длинные щетинки,
ограничивающие продольную ресничную борозду. По этой борозде
частицы, облепленные слизью, гонятся вперёд к ротовому отверстию.
У части двустворок эти комочки (аглютинаты) приводятся ко рту по
Потолку мантийной полости между наружной и внутренней жабрами,
но известны случаи совмещения обоих способов приведения аглютина-
тов (рис.60 В). Сложность системы циркуляции аглютинатов в ман-
тийной полости усугубляется тем, что только часть аглютинатов ис-
пользуется моллюсками как пищевые, тогда как прочие представляют
из себя псевдофекалии и выводятся наружу как посторонние тела
(рис.51 А 1, Е ). Особенной сложностью отличается система рес-
ничных слизевых токов в околоротовой области (рис.60 Д ,рис.62 В).
Рис. 59. А - яйценосная самка Lamps!lis fascicula (Unionidae)( Kraemer,
1984). Б-Limopsis aurita (Limopsidae) (Oliver, 1981). В - Offadesma an-
gasi (Periplomatidae) (Morton, 1981). T-Trisidos yongei (Arcidaej,
искривление плоскости симметрии и закручивание раковины вдоль
продольной оси (Teves z , Carter, 1979). Д — Myadora pandoriformis
(Myochamidae) (Tevesz, 1975). E -Trapezium bublaevigatum (Trape-
ziidae) (Morton, 1979). Ж - схема вододвижения у Pisidium (Sphaerii-
dae), обитающего в собственной норке. 3 - вытянутые и сократившие-
ся сифоны одного и того же моллюска
146
147
Факт разнообразия пластинчатых хабер (рис.60 В 1 -6 ) свидетель,
ствует о том, что успешность фильтрации и газообмена в какой-то
мере связана с их строением. Это предположение относится и К дру-
гой пищесобираадей структуре - ротовым губам, отроение которых
также довольно разнообразно (рис.62 А,Б).
В организации вододвижения существенное значение принадлежат
сифонам мантийной полости. Два сифона двустворок, несмотря на bod
условность этого сравнения, напоминают глотку актинии о двумя ои-
фоноглифами. Так же как у актиний, сифоны у двуотворок служат для
разобщения противонаправленных токов воды. Особенно велико их зна-
чение у моллюсков, глубоко погруженных в грунт и не могущих поль-
зоваться ножной щелью вследствие полного или частичного её зараста-
ния. Вполне естественно, что длина сифонов неодинакова и завиоит
от глубины погружения моллюока в субстрат (рио.59 А,Б,В; рио.61 Б,'
рис.63 А,Б,В). Крошечный Pisidiuzn, обитавдий в приповерхностном
слое грунта, для отведения воды пользуется им же самим прорытым
горизонтальным ходом (рис.59 1). Размеры сифонов зависят от возрас-
та животного, его размеров. Например, молодые Abra alba ли-
шены вводного сифона и пищевые частицы поступают в мантийную по-
лость через ножную щель..У более крупных взрослых моллюсков пос-
тупление пищевых частиц имеет место в основном через вводной
сифон (Aabel, 1983).
Рис. 60. А - схема строения жабры Malletia (Protobranchia)', прерывис-
тые стрелки показывают направление биений ресничек, длинные стрел-
ки - дыхательные течения (Yonge, 1939). Б - краевая часть жабер-
ных нитей Miisculus marmoratu’s (Nfytilidae)', оперённые стрелки обозна-
чают токи воды, прочие стрелки - направление биений ресничек (At-
kins, 1938). В - поперечные срезы жабер и направление приносящих
токов 1-Nucula (Protobranchia), 2 - Pholadldea, 3-Arcopaga (Tel-
linoidea), 4-Lucinidae, 5-Lasaea (Leptonoidea)', правая жабра
является внутренней, токи, направленные ко рту, обозначены точ. -
кой (Atkins , 1937). Г - ресничные цилиндрические клетки эпите-
лия лабиальной пальпы Diplcdon delodontus (Ryriidae) (Ruga et al.
1982). Д - ротовые лопасти Modiolus metcaliei (Mytilidae) с прино-
сящими токами. Сокращения: эфферентный приносящий сосуд -eff.
148
Рис.61. Protobranchia• к-Nucinella serrei (Allen, Sanders , 1969;
Б - отношение между осью жабер и направлением водных потоков в ман-
тийной полости у имеющей сифоны Silicula (1 ) и лишённой сифонов
Pristigloma (2 ), В - кишечник Prelametila , зачернена часть
кишечника слева от желудка (st.). Г -Pristigloma nutens(Allen,San' ,
ders, 1973). Д -Fa г ay oldie Ila altraafyssalis (Филатова, ШилейкоД^
Обозначения: --—► ось жабры,pr. 1.-лабиальные палъпы.Прочие обозна-
чения см. рис.53
Рис. 62. А - схема губного аппарата Pecten maximum,состоящего из
перемекапцихся лопастей передней и задней губ (Gilmour по Риг-
chon, 1977). Б - ротовые губы двустворчатых моллюсков: 1 - простые,
2 - Cyclopecten, 3 - свободные (Limidae ), 4 - слившиеся губы с
остиями, 5 -Propeamussium, 6-Chlamys (Bernard,1972). В - рес-
ничные токи на околоротовой лопасти Phacoides (Lucinidae), вид
спереди (Allen., 1958)
151
Мантийные сифоны часто отсутствуют у относительно мелких дет-
ритофагов, например у Macula, только частично погруженных в
грунт (рис.63 А). С другой стороны, крупные Pinna (рис.64 В)
такие обходятся без сифонов, однако их жизнедеятельность протека-
ет несколько особым образом. Высоко поднимающаяся над поверх-
ностью дна часть тонкостенной раковины Pinna очень непрочна
и легко обламывается без заметного ущерба для организма. Однако
мантия в этой части раковины к створке не прикреплена и поэтому
при обламывании этой части створок мантия полностью втягивается
в мантийную полость мышцами-ретракторами. Очистку от
посторонних частиц широко открытой мантийной полости моллюска с
такой обломанной раковиной совершает особый мантийный орган -
мускулистый вырост возле задней кишки. Жабры и мантийные складки
обломанной чаоти раковины сохраняют жизнеспособность и могут вы-
двигаться наружу (Yonge, 1953).
Втягивание и выдвигание сифонов обеспечивается мышечным меха-
низмом. У одних моллюсков к нему относятся ретракторы сифона и
его радиальные мышцы. У других для этой цели служат мышцы-
аддукторы, сокращение которых вызывает сближение створок раковины,
повышение давления воды в мантийной полости и выдвигание сифонов
( Chapman., Newell, 1956; Villarroel, Stuarto, 1977; Yonge, 1971).
У Busis давление воды в мантийной полости доходит до 120 см вод*
ного столба (Trueman, 1969).
С мантией связаны и другие, частные формы двигательной актив-
ности. Пресноводные Lampsilis вынашивают глохидиев в задней час-
Рис. 63. А - топография органов мантийной полости Macula (Protob-
lanchia, находящейся в естественном положении (Fretter,Graham, 1976).
Б - Verticordia perplicata (Septibranchia) без удаленной правой
складки мантии (Bernard, 1974). В - Miatella solida (Mactridae')
с брюшной стороны (Gallardo, Osorio, 1978). Г - Temnoconcha
brasiliana (Boss, kenk, 1964); задняя часть (1 ) и брюшная сто-
рона. Д - взаиморасположение мантии и раковины у большинства дву-
створок (.1 ) и у Phlyctaenochlamys fysiosquillina (Popham ло Franc,
I960)
152
add post
(няя кишка
лериостракум
А
выводной
сисрон
щупальца
2
нога
тийный
ротовые
лопасти
вводной
сифон
щупальцевая крона
диафрагмы-------
биссусное
отверстие
раковина
мантии
add.ant.
add.post.
нога
лабиальные пальпы
сифональный
орган
задняя
кишка
выводной
сифон
вводной сифон
крестовидная
мышца
нога
ти наружных жабер (рис.59 А). Для того чтобы продлить время
бывания в воде выметанных глохидиев и таким образом повысить
ятность их прикрепления к хозяину (рыбе) и предотвратить пре^
ременное оседание тлпуидия-в и их гибель, моллюск совершает
обособленной частью заднего края мантии, взмучивая воду (Агае^
1970). Впрочем, не исключено, что эти хлопки привлекают рыб
1984).
Сенсорная функция мантии отличается значительным своеобразие
в сравнении с брюхоногими. У последних органы чувств прежде всехч
способствуют выбору направления движения и позволяют опреде-
лить направление на источник раздражения. У малоподвижных и непо
важных брюхоногих раздражение органов чувств вызывает реакцию
организма, которая выражается в схлопывании створок раковины и
замикянии мантийной полости после втягивания в неё ноги и сифо-
нов. Реакция столь общего характера обычно не вызывает значитель-
ного перемещения моллюска в пространстве. Поэтому органы чувств
у двустворок большей частью рассредоточены по краям мантии и во-
круг сифонов. Прежде всего это относится к сенсорным щупальцам
(рис. 50 А,Б; рис. 59 3; рис. 60 Б,В,Г), а также к гла-
зам (рис.65 В). Сенсорные папиллы иногда несут вторичные выросты
на боковой поверхности (рис.66 В). Статоцисты, сохраняющиеся у
Pecten, однако,находятся в ноге (рис.65 Б). Столь характерное
для двустворок рассредоточение органов чувств, очевидно, объясня-
ется прежде всего резким преобладанием вододвижения над перемеще-
Рис. 64. А - зона активности сифонов Abraninida (слева) и Abri
longicallus (справа), состоящая из переплетённых каналов, проры-
тых моллюсками в грунте. Б - сбор пищевых частиц лабиальными
пальпами у Yoldia lima.tu.la с поверхности грунта (слева) и из то-
щи грунта (справа) {Wikander, 1980). Плотной пунктировкой по»
зан слой грунта, состоящий из фекалий и аглютинатов. В - схемы
строения Pinna carnea (Pinnidae) {Yonge из Purchon, 1977).
Г - Solemya togata с брюшной стороны {Deshayes из
Franc, I960). Д - S.parckinsoni {Owen, из Purchon,1977). Е -
Calyptogena soyoae (Okutani Egava, 1975)
IM
155
нием в пространстве. Показательно также, что сенсорные щупальцу
особенно обычны на вводном айфоне (рис.59 3,Е; рис.63 Б; рис.6g
Г). Впрочем, подобным структурам приписывают помимо сенсорной и
другую функцию: они предотвращают попадание в мантийную полость
через сифон посторонних частил. Не исключено, что многочисленные
папиллы на вывернутой наружу мантии (рис.51 Б.Е) имеют значение
для газообмена. Обволакивание раковины мантией может быть полным
и приводить к редукции раковины (рис.55 Б). Об участии мантии в
газообмене свидетельствует наличие в краях мантии сосудов, час-
тично способных сокращаться и потому получивших название дополни*
тельных сердец (рис.55А). Наконец, многоклеточные железы мантии
выделяют слизь, защищапцую моллюска. Возможно, что вывернутая
наружу мантия защищает тонкостенную раковину, особенно когда к
мантии приклеиваются песчинки.
Корреляция между строением мантийной полости и пищеваритель-
ной системы неоднозначна. Это объясняется ставшим очевидностью
в последнее десятилетие фактом, что по типам питания двуст-
ворки не сохраняют единообразия и что помимо голозойного питания
свойственного большинству видов, они способны к симбиотрофии и
сапрозойному питанию.
Рис. 65. А - Рапореgenerosa (Mactridae') : 1 - общий вид, 2 - схема
строения мантийной полости, 3 -поперечный срез в области, обозна-
ченной пунктиром на 1 (Yonge , 1971). Б -Thyasira disjuncta (Lu-
С2аасе<а):органы мантийной полости (1), схема циркуляции в мантий-
ной полости.(2)(Bernard, 1972). В - Bathyarca pectunculoides
(Arcoidea), справа, выдвинута левая складка мантии (Morton-,
1982). Сокращения: ог.1.~ ротовые лопасти,ped.-нога, per. - пери-
остракум, s.in.- выводной сифон, s.ex.- вводной сифон, г. - прямая
кишка, ped.f.- ножное отверстие, mant-мантия, ped.gl.- ножная сли-
зевая железа, antped.rZr.-передний ретрактор ноги, br.rtr. - ретрак-
тор жабры, pcrd.- перикард, i.lig-внутренний лигамент, p.ex.lig.
задний наружный лигамент, S. мягкое тело моллюска, о.Ьг,-наруж-
ная полужабра, i.br. - внутренняя полужабра. Прочие сокращения см.
рис. 53
156
157
У Tridacna и Hippopus симбиотические зооксантеллы в основ-
ном содержатся в разросшихся краях мантии. Они сильно пи-
гментированы, чтобы уберечь ткани моллюска от чрезмерного света,
и могут выдвигаться наружу и расправляться, чтобы освещение для
зооксантелл сделалось оптимальным. В мантии моллюска имеются
многочисленные глазки (гиалиновые органы), которым приписыва-
ется функция концентрации и рассеяния* света в ткани мантии сре-
ди зооксантелл (рис.67 А). Зооксантеллы фагоцитируются амёбоцита-
ми. Скорее всего, необычное соотношение морфологических осей у
Tridacnidae, их своеобразное положение на грунте (рис.52 Б 5 )
является приспособлением к симбиотрофии (Yonge, 1936;Fankbo-
ner, 1981; Wilkens, 1986). Симбиотрофии у Corculum способст-
вует необычная форма раковины, уплощенной в передне-заднем нап-
равлении (рис.65 Г). Зооксантеллы, обитающие в тканях моллюска,
освещаются через особые многочисленные участки раковины, называ-
емые ’’окнами". Их волокнистая структура способствует проведению
света (рис.65 Д) (Watson, Signor, 1986). Tridacnidaeсохраня-
ют способность отфильтровывать сестон.
Симбиотрофии с окисляющими серу хемоавтотрофными бактериями
свойственна ряду животных, обитающих в гидротермали, в том чис-
ле двустворкам Calyptogena (рис.64 Е) (Cavanaugh, 1983) .
Бактерии содержатся в жабрах, вследствие этого сильно увеличен-
ных. Сходным образом увеличены размер! жабер и занимаемая ими
площадь у Lucinella divaricata, у которой хемоавтотрофные серо-
бактерии находятся в бактериоцитах(£еВеппес, 1987).
Рис.65. А - мантийный глаз Tridacna maxima (Wilkens, 1986).Б - два
статоциста и рецепторные клетки статоциста Pecten inf'lexus
(левый статоцист - слева)(Barber, 1968). В - глазное щупальце
Cardium muiicum (Franc, I960). Г - раковина Corculum cardissa
спереди (Franc, I960). Д - тонкая структура светопроводящего "окна
в раковине Corculum., включающая призматические фибриллы (ИЫ -
sen , Signor, 1986). Е - омматидий сложного глаза Area
поае (Franc, I960)
158
A
базальная
пластинка^-
глазной нерб
интеркалярная клетка
сенсорная клетка
еосрера
пигментная клетка
опорные фибриллы
кинетосома
ретинальные
клетки
ресничный пузырь
ресничные
микробилли
сетчатка
передние края створок
чувствительный
нерв
глазной
нерв
ретина
<дной
сток
статолит
рецепторные
Г клетки
:6ица
хрустальное
пигмент
щупальцевый нерв
сосудистая
оболочка
____базальная мембрана
Рио.66. A - хищничество у двустворчатых: 1, 2, 3,4
(Morton, 1987 , по разным авторам). Б - поперечный срез сифо-
нального щупальца Verticordia triangularis (Septibranchia)
(Allen, Turner, 1974). В - обычное размещение тифлозоля в же-
лудочном цекуме (схема) ( Owen по Ггапс,1960)
160
Для части Protobranchia - 5olemyidaeflucinellidae,доказано сапро-
зойное питание растворённым органическим веществом через жабры,
мантийную полость и поверхность ноги. У таких моллюсков наблюда-
ется гипертрофия жабер, частичная или полная редукция пищевари-
тельного тракта, включая желудок, а также лабиальных пальп (Куз-
нецов, Шилейко, 1984; Franc , I960; Owen, 1961; Bernard,
1980а$Что же касается хищничества, то, по-видимому, оно имеет
место в 4 семействах двустворок, хотя непосредственно наблюда-
лось только у Cuspidaria (рис.66 А, рис.51 Д). Для поимки жертв
служит особого строения сифон, расправляемый гидростатическим
давлением в мантийной полости. Втягивание сифона осуществляют
мантийные ретракторы. бифокальные щупальца выполняют сенсорную
функцию ( Morton, 1987).
Всё же, по характеру питания двустворки в основном остаются
сестонофагами, хотя детритофагия встречается довольно часто.
Иногда систематически близкие виды (например, Tellinidae -Poh-
lo, 1973) могут различаться по характеру питания, кроме того,при
использовании двустворками свободно лежащих частиц грунта детри-
тофагия не всегда чётко отличается от сестонофагии (Кузнецов,
1986). Микрофагия определяет ряд особенностей пищеварительной
системы двустворок.
Сестон, так же как и детрит, представляет из себя смесь раз-
нородных мелких частиц. Такой пищевой материал способствует сох-
ранению внутриклеточного пищеварения (Беклемишев, 1964). Фагоцито-
зу частиц пищеварительными клетками предшествует его сортировка.
Она начинается уже в мантийной полости-на жабрах, ротовых лопас-
тях и мантийных складках, где образуются псевдофекалии, выводимые
наружу. Это усложняет схему циркуляции слизевых потоков в мантий-
ной полости (рис.63 А,Б), и иногда она захватывает поверхность
мягкого тела моллюска, выдвигаемую наружу (рис.63 В).
Этих усилий по сортировке частиц не достаточно и поэтому сор-
тировка продолжается на сложной внутренней поверхности желудка
161
(рис.67, рис.68 ). Из желудка крупные частицы, не пригодные для
переваривания, выводятся в среднюю кишку благодаря деятельности
ресничных клеток. Оставшиеся частицы вовлекаются в циркуляцию,
которая в большой степени вызывается вращением кристаллического
стебелька (рис.67 Б,В). Последний образуется в особом кармане
на дне желудка и имеет слоистую структуру. Выдвигаясь в желудок,
он постепенно растворяется, освобождая пищеварительные ферменты,
и своим вращением увлекает пищевые частицы вследствие вязкости
содержимого желудка. Это способствует равномерному распределению
частиц на внутренней поверхности желудка. Строение желудка ослож-
нено наличием сортирующих полей.
Сложность строения желудка - это следствие того обстоятельст-
ва, что обработка пищевой массы в мантийной полости носит пред-
варительный характер. При отсутствии морфологически обособленной
головы двустворки вместе с тем лишены и ротового аппарата, вклю-
чая слюнные железы. Отсутствие этих структур возмещается сложнос-
тью желудка. Таким образом, пищеобрабатывающий аппарат как бы
погружен в желудок.
Существует 5 типов строения желудка. Тип отроения Gastroproteia
характеризуется отсутствием твёрдого кристаллического стебелька,
заменённого вязким секретом, а мускулистая стенка желудка изнут-
ри выстлана кутикулой (рис.68 А 1). Моллюски с желудком, устроен-
ным по типу Gastrodentia, имеют только небольшой кристаллический
стебелёк, который не может выдвигаться в желудок. Кишечные дивер-
тикулы у таких моллюсков открываются в желудок двумя узкими от-
верстиями. Значительная часть поверхности мускулистого желудка
выстлана кутикулой (рис.68 А 2). Три прочих типа строения желуд-
ка характеризуются тем, что имеется функционирующий кристалличес-
кий стебелёк и многочисленные отверстия кишечных дивертикулов.
Стенка желудка плёнчатая. Эти три типа строения различаются от-
носительными размерами неодинаково устроенного тифлозоля, разме-
щением тифлозолей и кишечной борозды (рис.68 А 3,4,5).
162
lea. g&-
Рио. 67. A - общая схема отроения желудка двустворок (Fretter,Graham,
1976). Б - вскрытый желудок Dreiaena polymorpha (Morton,1969). В -
желудок Bathyarca pectunculoides (Агсасеа) о семью желудочными ди-
вертикулами (Morton, 1982). Сокращения: с.з.-кристаллический >
белёк, g.s.-гастральный щит, 1 .р,-левый карман, 2. с. а.-левый диверти-
кул, кишечная борозда, int.-кишечник, m.t.- малый тифлозол.'
int.-пищевод, г са.-правый дивертикул, r.t. - выводной тракт же-
лудка, s.a.- сортирующая поверхность, t-большой тифлозоль,
малый тифлозоль, s.s.- мешок кристаллического стебелька
19*3
Сравнивая разные типы строения желудка, можно заметитьг что
имеет место обратная корреляция между строением стенки желудка
и кристаллическим стебельком: при наличии мышц в стенке желудка
кристаллический стебелёк либо недоразвит, либо заменён вязким се-
кретом; если же стенка желудка плёнчатая, то кристаллический сте-
белёк развит хорошо. Возможно, что в данном случае имеет место
своего рода конкуренция между двумя способами перемешивания пищи
в желудке: мышечным (Gastroproteia,Gastrodentia') и о помощью
кристаллического стебелька (прочие три типа строения). Далее,
сортирующая и распределительная структуры на внутренней поверх-
ности желудка сохраняются только у тех моллюсков, которые питают-
ся мелкими частицами, тогда как у питающихся крупноразмерными
частицами и копеподами (тип строения Gastrodentia) эти структуры
отсутствуют.
Выделяемые в пределах класса двустворчатых моллюсков три надот-
ряда примерно отвечают основным подтипам организации.
Protobranchia - это моллюски с примитивными двоякоперистыми
жабрами, состоящими из плоских жаберных листков. Они мало пригод-
ны для отфильтровывания сестона. Поэтому для сбора пищевых частиц
в первую очередь служат разросшиеся лабиальные пальпы. Пищевари-
Рис.68. А-основные! типы строения желудка у двустворчатых моллюс-
ков: 1 - Gastroproctea (Protobranchia), 2 - Gastrodenteia (Septibran-
chia), 3-Gastrotriteia (большинство Filibranchia), 4- прочие Filib-
rarchia., многие Eulamellibranchia, 5- некоторые Eulamellibranchia
(Gastropempta) (Purchon, 1959). Б - микроструктура кристалличес-
кого стебелька на поперечном срезе у Thelescopium (Alexander, Sae,
1974). В - клетки В , выделяющие кристаллический стебелёк (Judd,
1979). Сокращения: а.с.-ось кристаллического стебелька,d.h.-спин-
ной крючок, £.5.- желудочный щит, c.g- кутикулярная решётка, d d.d.-
отверстие пищеварительного дивертикула, i.g<- кишечная борозда,
mg.- средняя кишка, m.t.- малый тифлозоль, 1р. - левый карман,
1.С.- левый цекум, ое - пищевод, s.a.- сортировочная площадь, г. t.-
удаление посторонних частиц, t.y. - большой тифлозоль. Вращение
кристаллического стебелька показано стрелкой
164
тельная железа открывается в желудок 2-3 общими-протоками, внут-
реннее строение желудка относительно просто, в нём отсутствует
обычная для прочих моллюсков транспортная система - тифлозоль и
кишечная борозда (тип строения желудка Gastrqproteia).KpoMe того,
нога имеет плоскую подошву.
Autobranchia имеют жабры, состоящие из отдельных жаберных
нитей. Они функционируют как сито и вододвигательный ресничный
аппарат, и участвуют не только в дыхательной, но ив пищесобира-
ющей функции» Ротовые лопасти служат для препровождения пищевых
частиц в ротовое отверстие. В желудке функционирует хорошо раз-
витая транспортная система (типы строения Gastro trite ia
и др.). Часто моллюски прикрепляются к субстрату. Autobranchia
подразделяются на Fili branchia и Eulamellibranehiaf первых
жаберные нити обычно соединены ресничками, у вторых соседние ни-
ти и образованные ими наружная и внутренняя жаберные поверхности
соединены спайками, содержащими сосуды.
Septibranchia имеют своеобразный мембранный насос, образо-
ванный жабрами и ограничивающий субсептальную полость. Он вызыва-
ет вододвижение мышечным способом и поэтому пища может состоять
из крупных частиц и копепод. Пища поступает в рот из субсепталь-
ной полости по ротовым лопастям. Желудок приспособлен для перера-
ботки крупных частиц пищи и вследствие этого имеет кутикулярную
выстилку (тип строения Gastrodentia). Транспортная система в
желудке упрощена, а пищеварительная железа представлена только
Рис. 69. А - Planktomya henseni сбоку (1 ) и её парус снизу (2);
заштрихованы аддукторы створок раковины, пунктированы парные яд-
ретракторы паруса {Allen, Scheltema, 1972). Б - плавание Sole-
rya : 1 - нога выдвинута, аддукторы створок расслаблены, вода
втекает в раскрытую спереди мантийную полость, 2 - нога слегка
сокращена, мантийный сфинктер смыкается вокруг основания ноги,
3 - слабое сокращение ноги, створки сведены, плёнчатые края
створок загнуты внутрь, вода выталкивается через дядний вывод-
ной сифон (Reid, 1980). В - глохидий Diplodon charruanus
(Parodiz, Bonetto Г - плавание Limaria orientalis при
выталкивании воды из мантийной полости {Grange, 1974)
166
167
двумя дивертикулами. Часто наблюдается биссуоное прикрепление.
В большинстве своём Protobranchia имеют таксодонтный- замок,
и это обстоятельство можно было бы понимать как факт корреляции
данного типа замка с другими особенностями этих моллюсков, пере-
численными в диагнозе подотряда. Тем не менее, физиологическое
объяснение совместной встречаемости этих особенностей отроения
и типа замка отсутствует. Надо отметить, что вообще корреляции
между строением замка и лигамента о одной стороны и строением
раковины и мягкого тела моллюска о другой стороны, остаются про-
блематичными, хотя некоторые заключения по этому вопросу существу-
ют ( Allen, 1958; Allen, Sanders, 1973; Newell, Boyd, 1987).
В заключение надо указать на ещё одну функцию мантийной полос-
ти - вынашивание зародышей моллюсков. Это явление известно у
Sphaeriidae (Franc,I960). Мантийная полость благодаря непрес-
танному движению воды в ней особенно пригодна для такой цели.
Unionidae жпашвахп глохидиев в жабрах. Известны, однако,
моллюски, вынашивающие яйца за пределами мантийной полости, в
особых отворотах брюшного края створок раковины (рис.54 В)
и вне её (Матвеева, 1979).
у двустворок, претерпевающих метаморфоз, наблюдается три типа
личинок: I Типичный велигер, 2) видоизменённый велигер Proto-
6га nchia,3) своеобразные эктопаразитичеокие личинки Unioni-
dae ( Иванова-Казао, 1977). Велигер вследствие малости разме-
ров имеет большую относительную поверхность тела и поэтому реснич-
ный аппарат сохраняет действенность как двигательное приспособле-
ние. Показательно, что плавание личинки Pecten происходит по
винтовой траектории и при этом личинка вращается вокруг своей оси
Рис.70. А - личиночное развитие (’1,2,3 ) Ensis directus (Loosa-
noff et al., 1966). Б - развитие паразитической личинки пресновод-
ной двустворки Mntela bourguignati,Unionacea (Fryer, 1961).
Сокращения: a.f.-апикальный жгут, - зачаток ноги, b.g. биссусная
железа, о. - рот, s. - желудок, st. - статоцист, У - парус, у г. -мус-
кул-ретрактор паруса
168
169
{Cragg, 1980) как у простейших с ресничным аппаратом. Так же
как у простейших, в этом проявляются особенности гидродинамики
быстро движущихся микроскопически малых тел.
У прочих двустворок в онтогенезе размеры тела сильно увеличи-
ваются и масса тела возрастает на несколько порядков. При этом
происходит перемена пелагиали на новое местообитание - бенталь.
Вследствие сокращения относительной поверхности тела двигательная
функция переходит к мышечному аппарату. Относящиеся сюда примеры
были приведены выше (рис.49, рис.50). Дополнительно можно назвать
Limaria и Solemya (рис.68 Б,Г). Даже у глохидиев Unionidae,
хлопающих створками с помощью мышц-аддукторов (рис.69 В), мышеч-
ное сокращение служит не столько для перемещения в пространстве,
сколько для прикрепления к телу хозяина (рыба). При неподвижном
паразитическом существовании личинка не сохраняет имевшийся по-
началу ресничный покров (рис.69 Б). У такой личинки перемещение
в пространстве сменяется врастанием в тело хозяина посредством
длинных гаусториальных трубок. Они закладываются как выросты пе-
реднего края мантии и служат для питания за счёт хозяина.
Наоборот, если размеры взрослого организма по сравнению с ли-
чинкой возрастают незначительно, как у неотенической Planktomya,
то ресничный аппарат сохраняет свою двигательную функция (рис.68
А).
Однако у всех прочих двустворок размеры организма в онтогенезе
увеличиваются на несколько порядков и относительная поверхность
тела уменьшается. Поэтому у взрослых моллюсков за ресничным аппа-
ратом сохраняется только вододвигательная функция. Вследствие мед-
ленности ползания у двустворок фильтрационное питание преобладает
над другими способами питания резче, чем у более подвижных брю-
хоногих.
При возрастании веса двустворки в 10 раз скорость фильтрации
увеличивается только в 4,3 раза (Алимов, 1961). Поэтому неудиви-
тельно, что если у брюхоногих наибольшие размеры тела достигнуты
170
жизненной формой хищника (Cassis, Trigonia), то у двустворок
самыми крупными организмами остаются Tridacna, прибегшие к
симбиотрофии как к дополнению к не полностью удовлетворяющей пот-
ребности организма фильтрации.
За помощь в работе автор благодарит А.А.Львову, Л.И.Москалёва,
И.Я.Рогинскую и А.А.Шилейко.
171
ЛИТЕРАТУРА
Алимов А. Ф. Функциональная морфология пресноводных двуствор-
чатых моллюсков.//Гр.Зоол.Ин-та АН CCCP.I98I. Т.96. С.1-248.
Беклемишев В. Н. Основы сравнительной анатомии беспозвоноч-
ных. T.I.Проморфология.М. ,1964.432 о.
Богданов И. П. Морфо-функциональный анализ строения передне-
го отдела пищеварительной системы моллюсков рода Oenopoda (Gastropoda,
Turridae). //Зоол.журн.1985.1985. Т.64, Ш. С.1632-1639.
Винников Я.А. Эволюция рецепторов. Л. ,1879. 138 с.
Грузов Е. Н. Эндопаразитический моллюск Asterophila japonica
Randall et Heath (Prosobranchia: Melanellidae) и его связь с паразити-
ческими брюхоногими.//Malacologia, 1965ДЫЗ, J6 I. C.III-I8I.
Догель В. А. Класс двустворчатых моллюсков Bivalvia . /fef-
ководство по зоологии. Т.2. 1940. С.466-547.
Иванов А. В. Класс брюхоногих моллюсков Gastropoda . //Ру-
ководство по зоологии. Т.2. 1940. С.323-455.
Иванова-Казас 0. М. Сравнительная эмбриология беспозво-
ночных животных. Трохофорные, щупальцевые, щетинкочелюстные, погоно-
форы. М. ,1977. 312 с.
Кантор Ю. И. Питание и некоторые черты функциональной морфоло-
гии моллюсков подсемейства Volutopsinae (Gastropoda,Pectinibranchia).//
Зоол.журн.,1985. Т.64, й II. С.1640-1646.
Кантор Ю. И. .Сысоев А.В. Новый род и новые виды семейс-
тва Turridae (Gastropoda, Toxoglossa) северной части Тихого океана.
//Зоол.журн. ,198$.Т.65, Ki 4. С.485-498.
172
Кузнецов А. П. О детритофагии и сестонофагии в классе двуст-
ворчатых моллюсков.//Питание морских беспозвоночных в естественных
условиях.М. ,1986. С.6-10.
Кузнецов А. П.,Шилейко А. А. О беокишечных Protobran-
chia (Siva 2 via///Биол. науки. ,1964. J* 2. С.38-49.
Лихарев И. М. Клаузилииды (Clausiliidae). //Фауна СССР.
1962. М.-Л. Моллюски. Т.З, № 4. 317 с.
Лихарев И. М., Виктор А. й. Параллелизмы в отроении и
систематическое положение слизней надотряда Styl ommatophora.//
Тр.Зоол.Ин-та АН СССР.,1979. Т.80. С.70-86.
Лихарев И. М., Виктор А.й. Слизни фауны СССР и сопре-
дельных стран (Gastropoda Terrestria Nuda. Ц Фауна СССР.1980. М.-Л.
Моллюски. Т.З, № 5. 437 о.
Лихарев И.М. .РаммельмейерВ. С. Наземные моллюс-
ки фауны СССР.//Определители по фауне СССР. Л.,1952. 512 с.
Матвеева!. А. Приспособления к вынашиванию яиц у некоторых
видов двустворчатых моллюоков.//Тр.Зоол.Ин-та АН CCCP.I979.T.80.C.39.
Миничев Ю. С. Эволюция кровеносного аппарата низших Opisthob-
ranchia.// Исслед. по эвол. морф, беспозв. Л. ,1970. C.IGI-II3.
Миничев Ю. И.,СтаробогатовЯ. И. Проблема торси-
онного процесса и морфологические перестройки личинок трохофорных
животных.//Зоол.журн,,1972. T.5I, Л> 10. C.I437-J449.
Миничев Ю. С., Старобогатов Я. И. Подклассы брюхоногих мол-
люсков и их филогенетические отношения.//Зоол.журн.,1979.Т.58, № 3.
С.293-305.
Рауп Д., Стенли С. Основы палеонтологии .М.,1974 . 390 с.
Свешников В. А.,Станкявичус А. В. Система жиз-
ненных форм морских переднежаберных моллюсков.//Докл. АН СССР. Т.295,
Jj 5. С.1268-1272.
Скарлато 0. A. Entocolax chiridotaeasp,WBs!li. паразитирую-
щий в голотурии моллюск.//3оол.журн.,1951, Т.30, й 4. С.358-362.
173
Скарлато 0. А.,Старобогатов Я. И. Двустворча-
тые моллюски умеренных широт западной части Тихого океана.//0преде-
лители по фауне СССР. Л. ,1981. 479 с.
Скарлато 0. А. ,Старобогатов Я. И. Основные чер-
ты эволюции и системы класса Bivalvia. Ц Тр.Зоол.Ин-та АН CCCP.I979.
Т.80. С.5-38.
Старобогатов Я. И. Фауна моллюсков и зоогеографическое
районирование континентальных водоёмов земного шара. Л.1972. 372 с.
Татишвили К. Г., Багдасарян К. Г., К а з а х а ш-
в и л и I. Р. Справочник по экологии морских брюхоногих М. 1988.
169 о.
Филатова 3. А., Ш и л е й к о А. А. Состав, морфология и
распространение ультраабисоального рода Parayoldiella (Bivalvia). //
Тр.Зоол.Ин-та АН СССР. 1985. T.I35. С.76-94.
Ч у х ч и н В. Д. Функциональная морфология рапаны.Киев.1970. 139 с.
Ч у х ч и н В. Д. Экология брюхоногих моллюсков Чёрного моря. Киев.
1984. 176 с.
Шилейко А. А. Наземные моллюски надсемейства HelicoideaJ/
Фауна СССР. Л. ,1978.Моллюски. Т.З, М 6 . 384 с.
Шилейко А.А. Наземные моллюски подотряда Pupillina фауны
СССР (Gastropoda, Pulmonata> Geophila).// Фауна СССР.Л. ,1984.
Моллюокй. Т.З, № 3. 399 о.
А а Ъ • 1 J. Р. Morphology and function in postmetaphoral Abra al-
ba (Bivalvia: Tollinaoea.//Barsia.1985.Vol.68 no 3.P.213-22O.
Allen J,A. Observations on Coohlodesma praetunue (Pulteney) (Eula-
mellibranchia).// J.mar.biol.Ass.UK.1985. Vol.37. P.97-112.
Allen J. A. On the basio form and adaptation to habitat in the
Luoinaoea (Eulamellibranohia).//Phil.Trans.Roy.Soc.London.Ser.B.1958
No 684. Vol.241. P.421-484.
AllenJ. A., Sanders H. L. Nucinella serrei Lang (Bival-
via: Protobranchia), a monomyarian Solemyid, and possible living Ac-
tinodont.//Malacologia.1969.Vol.7t no 2-3. P.381-396.
Allen J. A.,Sanders H.L. Studies on deep-sea Protobran
chia (Bivalvia): the families Siliculidae and Lametilidae.//Bull.
Mue.Comp.Zoo1. 1975.Vol.145, no 6. Р.263-ЗЮ.
Allen J. A., Scheltema R.8. The functional morphology and geog-
174
raphical distribution of Planctomya henseni, a supposed pelagi
va^lve.//J.mar.biol.Ase.UK.l972. Vol.52. P.19*31.
Allen J. A. i Turner J. A. On the functional morphology of
the family Verticordiidae (Bivalvia) with description of new species
from the abyssal Atlantio.//Phil.Trans•Roy.Soc.London.Ser.B.1984.
No 268. P.401-536.
Alexander C. G., R a e J. C. The structure and formation
of the crystalline style of Telescopium telescopium (Linnaeus).//
The Veliger. 1974. Vol.17» no 1. P.56-60.
A t kin s D. On the ciliary mechanisms and interrelationships of
Lamellibranchia. Pt.2-7.//Quart.J.Microscop.Sci.l937.Vol.79i no 3.
P.339-373•- Types of Lamellibranchs gills and their food currents.
Pt.3//Ibdd.l937. P.375-421.-Pt.4.//Ibid.1937. P.425-445.- Pt.7//
Ibid.1938. Vol.80, no 3. P.343-436.
Arnaud P. M., Jean-Jacques , v a n, M. Anatomy, eco-
logy and distribution of the Volutidae and Volutomitridae of tne Sou-
thern Indian Ocean.//The Veliger.1980. Vo1.22, no 1. P.19-27.
Baba K. New distributional records of Aerires punctilucens (d'Or-
bigny, 1837) from Sado Island, Japan.//The Veliger.1974 a.Vol.17tno
1. P.11-12. Some comments on Lobiger sowerbii Fischer, 1856, re-
identified, of Japan.//lbid.1974 b.Vol.16.no.3.P.253-257.
Baba K. Note on the internal anatomy of a mollusc Eubranchus hor-
ii Baba, I960 (Nidibranchia, Eolidoidea: Eubranchidae).//Zoo1.Magazi-
ne. 1975. Vol.84. P.77-78.
Baba K.,Hamatani I. An illustrated list of the Phyllidi-
idae from Seto, Kii, Middle Japan.// The Veliger.1975.Vol.18.no2.
В a i 1 • у K. ,B. Anatomical and electrophysiological studies on ths
gastropod osphradium.//Symp.Zool.Soc.London,l968.No 25. P.263-268
Balakrishnan Nair N.,Ansell A. D. Cha racteris-
tics of penetration in the substratum by some marine bivalve molluscs.
//Proc.Malacol.Soc.London, 1968. Vol.38. no 3« P.177-197.
Bank R. A., Gittenberger E. Notes on Azorean and European Caryohi-
um species (Gastropoda, Basommatophora, Elobiidae).//Basteria, 1985»
No 49. P.85-100.
Barber V.C. The structure of mollusc statocysts, with particu-
lar reference to Cephalopod.//Symp.Zool.Soc.London,1968.No.23. P.37*
BebbingtonA. BurBateIla leachi guineensis aubap.nov.(Gast-
ropoda, Opisthobranchia) from Ghana.//Proc.Malacol.Soc.London, 1969.
Vol.38, no 4. P.323-341.
Behrens D.W. The Lamellariidae of the North Eastern Pacific
//The Veliger. 1980. Vol. 22, No 4. P. 324-339.
Bernard F.R. The genus Thyasira in Western Canada (Bivalvia:
Lucinacea).//Malacologia. 1972. Vol.11, no 2. P. 365-385.
175
Chun C. Bericht uber eine nach den Canarischen Inseln In Winter
ausgefuhrte Reise.//Sitzungsberg.Prouss.Akad.Wise.Berlin.1889. Bd.
29. S.519-553.
C 1 a г к R. В. Dynamics in metazoan evolution. The origin of the
coelom and segments. Oxford. 1964. 5"13 p.
Clarke A. H. The freshwater molluscs of the Canadian Interior
Bassin.//Malacologia.1973.Vol. 15, no 1-2. P.1-509.
Coan E. V. Is there double trouble in marsupial clams?//The Ve-
liger. 1974. Vol.17, no 2. P. 185-184.
Cox L. R. Gastropoda - General characteristic of Gastropoda.//
Treatise Invert.Paleont. Pt.I.Mollusca 1.1960. P.1 -168.
C r a g g S. M. Swimming behaviour of the larvae of Pecten maximus
(L.)(Bivalvia).//J.mar.biol.Ass.UK.1980. Vol .60. P.551-564.
С о о к P. M. A ciliary feeding mechanism in Viviparus viviparus (L.)
//Proc.Malacol.Soc.London.l949. Vol.27. P.265-271.
Cook A. The organisation of feeding in the carnivorous snail Eu-
glandina rosea.//Malacologia.1985. Vol.26, no 1-2. P.185-189.
Groll R. P. Gastropod chemoreception.//Biol.Rev.1985.No.58.P.293
D* A s a г о Ch. The comparative embryogenesis and early organogene-
sis of Barsa corrugate Perry and Distorsio clathrata Lamarck (Gastro-
poda, Prosobranchia).//Malacologia.1969.Vol.9, no 2. P.549-389.
Davis Ch. C. Emergence of veliger larvae from eggs in gelatnous
masses laid by some Jamaican marine Gastropods.//Malacologia.1967*
Vol.5. no 2. P.299-309.
Davis G. M. The systematic relationship of Pomatopsis lapidaria
and Oncomelania hupensos formosana (Prosobranchia: Hydrobiidae).//
Malacologia • 1967. Vol. 6. no 1-2. p.1-145.
Dawydoff C. Quelques veligeres geantes de Protobranches pro-
venant de la mer de Chine.//Bull.Biol.Franqe Beige.1940.T.74.P.497.
Delamare-Deboutteville C. Biologie des eaux
souterraines littorales et continentales.Paris. I960. 740 p.
Dell R. K. The molluscan genus Monodilepas in New Zealand.//
Trans.Roy.Soc.NZ.1953. Vol.81. Pt.1. P.145-151.
D e 1 1 R. K. Archibenthal Mollusca from northern New Zealand.//
Trans.Roy.Soc. NZ. 1965. Vol.5. no 20. P.205-216.
Demian E.S., Yousif F. Embryonic development aud ora-arofencsiE
in the snail Marisa comuarietis (Mesogastropoda: Ampullariidae.//
Malacologia.1973» Vol.12, no 1. P.125-150.
Denny M. Locomotion: the cost of gastropod crawling.//Science,
1980. Vol. 208. P.1288-1290.
Dimock R. V., Jr. Quantitative saspects of locomotion by the
mud snail, Ilyanassa obsolete.//Malacologia.1985. Vol.26 no 1-2.
P. 165-172.
177
Driscoll A. L. Structure and function of the alimentary
tract of Batillaria zonalis and Cerithidea californica, style-bear-
ing Mesogastropods.//The Veliger. 1972. Vol,14, no 4. P. 375-386..
Eagar R. M. Shape and function of the shell: a comparison of
me living and fossils bivalve molluscs.//Biol.Rev.1978.No 53.Р.16Э,
Eales N. 0. Revision of the world species of Aplysia (Gastrdpq,
da,Opisthobranchia).//Bull.Brit.Mus.(Nat.Hist.) Zool.I960. Vol.5,
no 10. P.1-404.
Edmunds M. Berthelinia caribbea.,n.sp., a bivalved gastropoda
from the West Atlantic.//J.Linn.Soc.London,1963.Vol.44 no320.P.731,
Fankboner P. V. Siphonal eyes of giant clams and their re-
lationship to adjacent Zooxanthellae.//The Veliger.1981. Vol. 23, no
3. P.245-249.
Feininger A. Traumgebilde des Heeres Muscheln und Schneckeni
Wien, Dresden.1972. 299 s.
Field L. An experimental analysis of the escape response of the I
gastropod Strombus maculatus.//Pacific Science,1977»Vol.31 no 1.P.1.
Fioroni P.,von. Allgemeine Aspecte der Mollusken-Entwick-
lung.//Zool.Jhb.Anat.1982. Bd.1O?. S.85-121.
F r a n c A. Classe des Bivalves.//Traite Zool.i960.T.5 f.2.P.1845.
F Г a n c A. Classe des Gasteropodes (Gastropoda Cuvier,1798). Gene'-
ralites et definition.//Traite' Zool.1968a. T.5 f .3«P.2-39. “ Sous-
classe des Prosobranches (Prosobranchia Milne Edwards,1848»Strepto-
neura).//Ibid.,1968b. Т.40-34.- Sous-classe des Opisthobranches.//lb.
1968c.P.608.
Frette г V. The structure and life history of some minute prosob-
ranchs of rock pools: Skeneopsis planorbis (Fabricius), Omalogyra ate
mus (Philippi), Rissoella diaphana (Alder), Rissoella opalina (Jeff-
reys).//J.mar.biol.Ass.UK, 1948. Vol23, no 3. P.597-632.
Fretter V. Some observations on the British cypraeide.//Proc.
Malacol.Soc.London,1951» Vol.29, no 1. P.14-20.
Fretter V. Metamorphic changes in the velar musculature, head
and shell of some prosobranch veliger.//J.mar.biol.Aes.UK, 1972. Vol^
52. P.161-177.
Fretter V. Umbonium veetiarium, a filter-feeding trochids.//J«
Zool.London, 1975. Vol.177. P.541-552.
Fretter V., Graham A. Observations on the opisthobranebt
molluscs Actaeon tornatilis.//J.mar.biol.Ass.UK,1954, no.3.P.565-585»
Fretter V., Graham A. British prosobranch molluscs. Lon*
don. 1962. 755 p.
Fretter V.,Graham A. A functional anatomy of Invertebfi
tes. London, NY...1976. 589 p.
Fretter V., Graham A. The prosobranch molluscs of Brit*
(78
in and Danmark. Pt.I.Pleurotomacea, Fissurellacea and Patellacea.//
J.Moll.Stud.И976a. Suppl.I.P.1 -37.
Fretter V., Ko Bun Hian. The specialization of the Aplysiid
gut.//Malacologia.1979. Vol.18 no 1-2. Р.7-11»
Fretter V., Montgomery M.C. The treatment of food by
prosobranch veliger.//J.mar.biol-.Ass.UK. 1968. No 4. P.499-520.
Frey er G. The development history of Mutela bourguignati (An>-
ceh) Bourguignat (Mollusca: Bivalvia).//Phil.Trans.Roy.Soc.London.
Ser.B. 1961. Vol.244. P.259-298.
FujiokaY. On the secondary sexual characters found in the di-
morphic radula of Drupella (Gastropoda: Muricidae) with reference to
its taxonomic revision.//Venus,1902. Vol. 4, no 4. P.203-223.
Gainey. L.F., Jr. Locomotion in the Gastropoda: functional morpho
logy of the food in Neritina reclivata and Thais rustica.//Malacolo-
gia, 1976. Vol.15, no 2. P.411-451.
Gale W. F. Vertical distribution and behavior of the fingernail
clam Sphaerium transversum.//Malacologia.1976.Vol.15» no 2. P.4o1-409.
Gallardo C.S.,Osorio C.R. Hiatella solida (Sowerby,
1834)(Mollusca, Hiatellidae) on Concholepas concholepas (Brouguiere,
1789) and other substrates.//The Veliger,1978. Vol.20 no 3 P.274-276.
Gascoigne T. Sacoglossan teeth.//Malacologia„1977. Vol.16,
no 1.. P.101-105.
Ghiselin M. T. The adaptive significance of gastropod torsion.
//Evolution,1966. Vol.2o. P.337-346.
Gilmour T. H. The defensive adaptation of Lima hians (Mollu-
sca, Bivalvia).//J.mar.biol.Ass.UK, 1967.No 47. P.209-221.
Golikov A. No, Starobogatov Y. I. Systematics of
prosobranch gastropods.//Malacologia, 1975» Vol. 15» no.1.< P. 185-232.
Gooding R. U., Lutz enJ. Studies on parasitic gastropods
from Echinoderms.Til.A description of Robillaria cemica Smith,1889,
parasitic in the Sea Urchin Echinometra meuschen, with notes on its
biology.//Dansk.Viden.aelsk.K^benhavn,1973«Biol.Skr.20, no 4.P.1-21•
Goslinger T., Behrens D. W. Two new species and ge-
nera of Aeolid Nudibranchs from the tropical Eastern Pacific ..//The
Veliger, 1986. Vol.29, no 1. P.101-11J.
Graham A. The anatomy of the Prosobranch Trichotropis borealis
Broderip et Sowerby, and the systematic position of the Capuli.de^
J.mar.biol.Ass.UK, 1954.No 33. P.129-144.
Graham A. Gastropoda.//The Systematic Ass.l979.Spec.vol.i2
Grange K. JR. The occurence of Limaria orientalis (Bivalv?vt
Limidae) in New Zealand.//The Veliger,19?4.Vol.17 no 1. P. 13-13*
Greene R.W. Symbiosis in sacoglossan opisthobranchs: trans(.ca
tion of photosynthetic products from chloroplast to host tiesu.?»//
Malacologia, 197'0. Vol.10, no 2. P.369-380.
Hadfield M.C., Kay E. A.,Gilbette M.U., Lloyd
M. C. The Vermetidae (Mollusca: Gastropoda) of the Hawaiian Island.
//Mar. Biol.1972. Vol.12. P.81-98.
Hadzisce S., Patterson С. M., Burch J. B., Lo
Verde P. T. The embryonic shell surface sculpture of Gocea and
Valvata.//Malacolog. Rev. 1976. VoL9. P.1-14.
Harry H. M. Synopsis of the supraspecific classification of li-
ving oysters (Bivalvia: Cryphaeidae and Ostraeidae).//The Veliger.
1985. Vol.28 no 2. P.121-158.
Haszprunar G. On the anatomy and fine-structure of a pecu-
liar organ in Nucula (Bivalvia» Protobranchia).//The Veliger.1985.
Vol. 28 no 1. P.52-62.
Herschler R. Phreatodrobia coronae, a new species of caves-
nail from southern Texas.//Nautilus,1987.Vol.103 no 3. P.133-139.
Hickman C. S. Evolution and function of assymetry in Archae-
ogastropod radula.//The Veliger.1981. Vol. 23» no 3. P.189-194. -
Radular patterns, systematics, diversity, and ecology of deep-sea
limpets.//Ibid.,1983.Vo1.26 no 2. P.73-92.
Hoagland К.. E. Proterandry and the evolution of environmental-
ly-mediated sex-change: a study of the Mollusca.//Malacologia. 1978
Vol.16 no 2. P.365-391.
Hoagland К. E., Turner R. D. Evolution and adaptive ra-
diation of wood-boring Bivalves (Pholadacea).//Malacologia.1981. Vol.
21. no 1-2. P.111-148.
Hoeksema D. F., Janssen A. W. Rediscovery of tne mari-
ne gastropod Spiricella unguiculus Rang, 1827 (Euthyneura, Umbraculi-
dae) in Miocene deposits of the North Sea Basin and in the recent fa-
una of South West Europe.//Basteria,1984. No 48. P.7-11.
Houbrick R. S. Classification and systematic relationship of
the Abyssochrysidae, a relict family of bathyal snails (ProsobranchiaJ
Gastropoda).//Smith.Contr.Zool.1979. No 290. P.l-21.
Houbrick R S. Genus Clypeomorus Jousse&ume (Cerithiidae: Pro-
sobranchia).//Smith.Contr.Zool.1985. No 403. P.1-103.
Hubendick B. The Baikal limpets and their phylogenetic sta-
tus. //Arch.Moll.1969. Bd.99, no 1-2. p.55-67.
Hubendick B. Systematics and comparative morphology of the
Basommatophora.//Pulmonates.l978. Vol 2a. P.1-4-7.
H u g a G. A., В r e n n e r R, R., N i v e i г о M. H. A study of
the biology of Diplodon delodontus (Lamarck,1819)(Bivalvia: Hyriidae).
I.Ecological aspects and anatomy of the digestive tract.//The Veliger.
1982. Vol.25 no 1. P.51-58.
Hughes R. N. A new species of Serpulorbis (Gastropoda: Vermeti-
dae) from South Africa.//The Veliger.1978 a. Vol.Po no 3. P.288-291.
180
Hughes R. N. The biology of Dendropoma carallinaceum and Serpu-
lorbis natalensis, two South African Vermetid gastropods.//Zool.J.
Linn.Soc.1978b. Vol.64. P.1'1'1 -127.
Hughes R. N. A functional biology of marine gastropods. Balti-
more, 1986. 24-5 p.
Hughes R. N., H u g h e s H. P. I. Morphological and behavioral
aspects of feeding in the Cassidae (Tonnacea, Mesogastropoda).// Mala-
cologia. '198'1. Vol.20, no 2. P.385-402,
Hurst A. Studies on the structure and function of the feeding ap-
paratus of Philine aperta with a comparative consideration of some
other opisthobranchs.//Malacologia.1965. Vol.2, no 3. P.28'1 -547.
Hurst A. The feeding mechanism and behaviour of the opisthobranch
Melibe leonina.//The Zool.Soc.London. Studies in the structure, physi-
ology and ecology of molluscs. 1968. P.151-166
Hyman L. H. The invertebrates.VI.Mollusca. I.Aplacophora, Polyp-
lacophora, Monoplacophora and Gastropoda.NY. 1967*
Isarunkura K., Runham N. W. Studies on the gastropod
radula.//Malacologia, 1968. Vol.7 no 1. P.71-91.
Jensen K. R. A review of Sacoglosaan diets, with comparative no-
tes on radular and buccal anatomy.//Malacol.Rev. 1980. No 13.P.55-77.
Jensen K. R. Observations on copulation in two species of Ely-
sia from Florida (Opisthobranchia; Ascoglossa.//Ophelia, 1986a, Vol.
25 no 1. P.25-32.~ Observations on feeding, copulation and spawning
of the ascoglossan opisthobranch Callipaea oophaga Lemche.//Ibid.,
1986b. Vol. 25 no 2. P.97-106.
Jones G. M., Saleuddin A. S. Ultrastructural observatio
ons on sensory cells and the peripherical nervous system in the mant-
le edge of Helisoma duryi (Mollusca; Pulmonata).//Canad.J.Zool. 1978.
Vol. 56. P.1807-1821.
Jorgenson С. B. Biology of suspension feeding. Oxford.1966.
Judd W. The secretions and fine structure if bivalve crystalline
style sac.//Ophelia, 1979. Vol. 18 no 2. P.205-254.
Kat P. W. Parasitism and the Unionacea (Bivalvia).//Biol.Rev. 1984
Vol.59. P.189-207.
Kawaguti S. Formation of the bivalve shell in a gastropod Ta-
manovalva Umax.//Proc.Jap.Acad.,1959. Vol.55. P.607-611.
Kesler D. H.,Jokinen E. H., u n n s W. R.,Jr. Trophic
preferences and feeding morphology of two pulmonate snail species.//
Canad.J.Zool.,1986. Vol.64. P.2570-2575.
Kraemer L.R. The mantle flap in three species of Lampsilis
(Pelecypoda: Unionidae).//Malacologia,1970. Vol.10 no 1. P.225-262.-
Aspects of the functional morphology of some freshwater Bivalve ner-
vous systems: effects of reproductive processes and adaptation of sen
I8I
sory mechanisms in a Sphaeriacea and Unionacea.//Malacologia, "1984.
Vol.25, no 1. P.221-259,
L a 1 1 i С. M. Food and feeding of Paedoclione doliformis Danforth.,
a neotenous Gymnosomatous Pteropod.//Biol.Bull.1972.Vol.145. P.592.
L e m c h e H. The anatomy and history of Cylichna (Gastropoda Tec-
tibranchia).//Skr.Univ.Zool.Mxis.Kohehhavn,1956.Vol.16. P.1 -278.
Le Pennec N.,Kerry A., Diouris M., M о r a g о D.,
D о n v a 1 A. Chemoautotrophic bacterienne chez le mollusque bival-
ve littoral Lucinella divaricata (Linne).//C.r.Acad.Sci.,198?.Ser.5.
T.5O5, no 1. P.1-5.
Lever J. On torsion in gastropoda.Pathways malac.1979.Utrecht.
L i n s 1 e у R. M. Locomotion rates and shell form in the Gastro-
poda. //Malacologia,1975» Vol.17, no 2. P.195-206.
Loosanoff V. L., Davis H. G., Chanley P. E.
Dimensions and shape of larvae of some marine bivalve molluscs.//
Malacologia, 1966. Vol.4, no 2. P.551-^55.
Lutzen J. Unisexuality in the parasitic family Entoconchidae
(Gastropoda: Prosobranchia).//Malacologia,1968.Vol.7 no 1. P.7-15*
Lutzen J. Studies on the parasitic gastropods from echino-
derms. II.On Stilifer Broderip, with special reference to the struc-
ture of the sexual apparatus and the reproduction.//Det.Kongl.Dans-
ke Vidensk. Selsk.1972. Biol.Skr.19, no 6. P.1-18.
Lutzen J. Studies on the life history of Enteroxenos Bonnevie,
a gastropod endoparasitic in aspidichiridotae holothurians.//Ophe-
lia, 1979. Vol.18 no 1. P.1-52.
Lucaze-Duthiere, de, H. Histoire de Testa jselle.//
Arch. Zool. Exper. Gen.,1887. 2 вег. Т.5» P.459-598.
Macpherson J. H., Gabriel C.J. Marine Mollusca of
Victoria. Handbook 2. Melbourn. 1962.
Maed a T. Types of osphradia in the Prosobranchia with special re-
ference to the relation with feeding fhabits.//Venus, 1985» Vol.41
no 4. p.264-275.
M a г t о j а И., T h i г i 0 t - Q u i e v r e u x. Donnes histo-
logiques sur 1' appareil digestif et la digestion des Atlantidae (Pro-
sobranchia Heteropoda).//Malacologia,1975» Vol. 15 no 1. P.1-27.
Meier-Brook 0. L.,Choung Kwan Kim. Note s
on ciliary feeding in two korean Bithynia species.//Malacologia,
1977. Vol.16 no 11. P.159-165.
i 1 1 e V. The Nudibranchia genera Onchidoris and Diaphoro-
doris (Mollusca, Opisthobranchia) in the Notheastern Pacific.//The
Veliger, 1985- Vol.28, no 1. P.80-95.
Miller B. A. The biology of Terebra gouldi Deechayes,1859,and
a discussion of life history similarities among other Terebrids of
182
similar proboscis type.//Pacific Science, '1975» Vol.29 no 5. P.227.
Morton J. E. Vermicilaria and the Turritellids./Proc.Malacol.
Soc.London, 1955. Vol.JO no 3» P.80-86.
Morton J. E. The biology of Limacina retroversa //J.mar.biol.
Ass.UK, 1954. Vol.35» P.297-3'12. -The functional morphology of Oti-
na otis, a primitive marine pulmonata.//Ibid. ,'1955» Vol,34. P.113-150
Morton J. E. The habits of Scintillona zelandica (Odhner) ,1924
(Lamellibranchia: Galeommatilidae).//Proc.Malacol.Soc.Londom. 1957»
Vol.32, no 5» Р.185-18Й.-Torsion and the adult snail: a re-evalua-
tion.//Ibid.,'1958a. Vol.32, no 1. P.2-10.
Morton J. E. Observations on the gymnosomatons pteropod Clio-
ne limacina (Phipps).//J.mar.biol.Ass.UK.1958b.Vol.37. P.287-289.
Morton J. E. Molluscs.Hutchinson Univ.Libr.Biol.Sci.,London,
1968. 244 p.
Morton J. E. The form and functioning of the pallial organs in
the opisthobranch Akera bullata with a discussion on the nature of
the gill in Notaspidea and other Tectibranchs.//Thr Veliger, 1972.
Vol.14, no 4. P.357-549.
Morton J. E. The ecology and digestive system of the Struthio-
lariidae species from a small New England pond, USA.//Canad.J.Zool.,
1986. Vol.64. P.2570-2575.
Morton J. E., Holme A. The occurence at Plymouth of
the opisthobranch Akera bullata, with notes on its habits and rela-
tionships. //J. mar.biol. Ass. UK, 1955» Vol.34. P.101-112.
Morton B. Locomotion. Physiology of Mollusca.1964. Vol.2.
Morton B. Studies on the biology of the Dreissena polymorphs
Pall. I.General anatomy amd morphology.//Proc.Malacol.Soc.London,
1969. Vol.38, no 4. P.301-321.
Morton B. Some aspects of the biology and functional morphol-
logy of Trapezium (Neotrapezium) sublaevigatum (Lemarck) (Bivalvia:
Arcticacea).//Pacific Science,1979. Vol35, no 2. P.177-194.
Morton B. The Anomalodesmata.//Malacologia,1981. Vol,21, no
1-2. P.35-60.
Morton B. Functional morphology of Bathyarca pectunculoifes
(Bivalvia: Arcacea) from a deep Norwegian Fjord with a discussion of
the mantle margin in the Arcoidea.//Sarsia,1982.Vo1.67. P.269-282.
Morton B. Feeding and digestion in Bivalvia.//The Mollusca.
Vol.5 Physiology. Pt.2. 1983a. P.65-14-7.
Morton B. The biology and functional morphology of the Twisted
ark Tisidos semitorta (Bivalvia: Arcacea) with a discussion on Shell
"torsion" in the genus.//Malacologia.1983 b. Vol.23, no 2.P.375-596.
Morton B. Siphon structure and prey capture as a guide to af-
183
finities in the abyssal septibranch Anomalodesmata (Bivaivia).//Bar-
ela, 1987. Vol.72, no 1. P.49-71'
Morton J.E., Y о n g e С. M. Classification and tue structu-
re of the Mollusca.//Physiol.Mollusca.1964.VO1.I.P.1-57.
Myra Keen A.,Morton J. E. Some African species of De-
ndropoma (Vermetidae: Mesogastropoda).//Proc.Malac.Soc.,1960. Vol.
54, no 1. P.56-5*1.
Newell N. D., В о у d D. W. Iteration of ligament structu-
res in Pteriomorphian Bivalves.//Amer.Mus.Novitates,1987.No 2875.P.1,
Newell P. F.,Brown A. C. The fine structure of the osph-
radium of Bulla digitalis Meuschen (Gastropoda, Prosobranchia).//
Malacologia, 1977. .Vol. 16. no 1. P.197-205.
Newell P. F., Newell G. E. The eye of the slug, Agrioli-
max reticulatus (Muller).//Symp.Zool.Soc.London,1968.No 25.Р.97-Ш.
Nicol D. A survey of inequivalve pelecypods.//J.Wash.Acad.Sci.
1958. Vol.48 no 2 Zool. P.52-62.
Nybakken J.,McDonald G. Feeding mechanisms of
West American Nudibranchs feeding on Bryozoa, Cnidaria and Ascidia-
cea, with special respects to the radula.//Malacologia,1981. Vol.20
no 2. P.459-449.
Ockelmann K. W. Descriptions of mytilid species and defini-
tion of the Dacrydiinae n.subfsm.(Mytilacea-Bivalvia).//Ophelia,1985
Vol.22, no 1. P.81-125.
Ockelmann K. W.,Nielsen C.On the biology of the
Prosobranch Lacuna parva in the Oresund.//Ophelia, 1981. Vol.20
no 1-2. P.61-95.
Oliver P.G. The functional morphology and evolution of recent
Limopsidae (Bivalvia, Arcoidea).//Malacologia, 1981.Vol.21 no 1-2.
P.61-95.
Olmsted J. M. Notes on the locomotion of certain Bermudan
mollusks.//J.exp.Zool.,1917. Vol.24.P.225-256.
Ostergaard J. M. Spawning and development of some hawaii-
an marine gastropoda.//Pacific Science, 1949. Vol.4 no 2.P.75-115»
Owen G. The shell of the Lamellibranchia.//Quart.J.Microscop.
Sci.,1955. Vol.94. P.57-70.
Owen G. Note on the habits and nutrition of Solemya parkinsoni
(Protobranchia: Bivalvia).//Quart.J.Microscop.Sci.1961. Vol.102
no 1. P.15-21.
Owen G. Feeding.//Physiol.Mollusca.1966.Vol.2. P.1-51.
Pafort van, T. I. The columellar muscle system in Clio py-
ramidata and Cymbulia peroni (Pteropoda, Thecasomata).//Malacologia,
1979. Vol.102no1-2. P.51-55.
184
Parker G. H. The pedal locomotion of sea-hare Aplyaia calilor-
nica.//J.exp.Zool.,1917. Vol.24. P.159-1*5-
Parodiz J. J.,Bonetto A. A. Taxonomy and zoogegrsphy
relationships of the South African Naiades(Pelecypoda: Unionacea
and Mutilacea).//Malacologia,1965. Vol.1 no 2. P.179-215»
Pelseneer P. Mollusques.//Traite' Zool. Fasc.16.1897. 187 Р»
Pohlo R. H. Feeding and associated functional morphology in
Tagelus califomianus and Florimetis obesa (Bivalvia: Те linacea).//
Malacologia.1975» Vol.12, no 1. P.1-11.
Ponder W. F. The family Eatonella in New Zealand.//Rec. Auck-
land Mus. 1965» Vol.6, no 2. P.47-99.
Ponder W. F. Observations on the living animal and mode of li-
fe of some New Zealand Erycinacean bivalves.//Trans.Roy.Soc.N.2.1967
Vol.10, no 5» P.21-52.- A new species of Dendropoma from New Zealand
(Mollusca: Vermetidae).//Ibid.1967a.Vol.10, no 2.P.17-20.
Ponder W. F.. Gooding R. U. Four new Eulimid Gastropods
associatea with shallow-water didematid echiniods in the Western Pa-
cific. //Pacific Science,1978. Vol.52, no.2. P.157-185»
Potts G. W. The anatomy of respiratory structures in the dori-
id nudibranchs, Onchidoris bulamellata and Archidoris pseudoargus,
with details of the epidermal glands.//J.mar.biol.Ass. UK.1981. Vol.
61. P.959-982.
Purchou D. A note of the biology of Brechites penis (L.)(la-
mellibranchia).//J.Linn.Soc.London.(Zool.),1956. Vol.45, no 289.P.45
Purchon D. Phylogenetic classification of the lamellibranchia
with special reference to the Protobranchia.//Proc.malacol.Soc.Lon-
don, 1959. Vol.55. no 5» P.224-250.
Purchon I). The biology of the Mollusca. Oxford. NY. 1977.P»560p
P u r c h о n R.D. The stomach in the Bivalvia.//Phil.Trans.Roy.Soc.
London.Ser.B.1987. Vol.516. P.185-276.
Rankin J. J. A freshwater shell-less Molluscs from the Carib-
bean: structure, biotics, and contribution to a new understanding
of the Acochlidioidea.//Life Sci.Contr.Roy.Ontario Mus.1979.no16.
Rasmussen E. Faunistic and biological notes on marine in-
verb ebrat es. I I. The eggs and larvae of some Danish marine gastropods.
//Vidensk.Medd.Dans к.Naturh.Foren.K0benhavn•1951•Vol.115•P•201-249•
R a u p D. M. Geometric analysis of shell coiling: general problems
//J.Paleont.1966. Vol.40. P.1178-1190.
Reid D. M. Aspects of a guttless species of Solemya (Bivalvia:
Protobranchia).//Canad.J.Zool.1980. Vol.58 no 5. P.586-595.
Rex M. A.,Boss K. J. Open coiling in recent gastropods.//
Malacologia. 1976. Vol.15. P.289-297.
185
Richter G. Heteropoden der "Meteor"-Expedition in den India-
chen Ozean 1964-1965.//"Meteor* Forschung-Ergebn.(D). Biol., 17.
1974. S.55-78.
R i g h i G. On the braeilian species in the Acmaea subrugosa comp,
lex (Gastropoda: Prosobranchia: Patellacea).//Malacologia,1966. Vol.
4 no 2. P.269-295.
Rosewater J.A new species of Leptonacean Bivalve from the
North-western Peru (Heterodonta: Veneroidea: Lasaeidae).//The Veli-
ger, 1984. Vol.27 no 1. P.81-89.
Rudman W.B. Three new species of the opisthobranch family Agla-
jidae from New Zealanc^’tans.Roy.Soc.NZ., 1965. Vol.10 no.2J.P.211.
Schme-kel L. Die Gattung Doto (Gastropoda: Nudibranchia) im
Mittelmeer und Armelkanal, mit Beschreibung von Doto acuta, n.sp.//
Malacologia, 1977. Vol.16 no 2. P.467-499.
S e a p у R. R. The pelagic genus Pterotrachaea (Gastropoda: Hete-
ropoda) from Hawaiian waters: a taxonomic review.//Malacologia,1985
Vol.26 no 1-2. P.125-155.
Seilacher A.,von. Schnecken in Brandungssand.//Natur u.
Volk, 1959. Bd.89. S.559-566.
Shimek R. L., К о h n A. J. Functional morphology and evoluti-
on of the toxoglossan radula.//Malacologia,1981.Vol.20,no 2.P.425.
Signor Ph. W. The role of shell geometry as a deterrent to
predation in Terebrid gastropods.//The Veliger,1985«Vol£8.no 2.Р.179»
S о 1 e m A. Malacological applications electron microscopy. III.
Radular structure and functioning.//The Veliger, 1972. Vol.14 no 4.
P.527-556.- Patterns of radular tooth structure in carnivorous land
snails.//Ibid, 1974b. Vol.17, no 2. P.81-86.
S 0 1 e m A. The shell Makers introducing mollusks. NY, London.
1974 a. 289 p.
S 0 1 e m A. Endedontold land snails from Pacific (Mollusca: Pulmo-
nata: Sigmurethra)• Fart I.Endodontidae. Chicago. 1976. 508 p.
S p о e 1 van der, S. Euthecasomata. A group with remarkable de-
velopment stages (Gastropoda, Pteropoda).//Gorinchem, Noordujin,
1967. 575 p.
S p о e 1 van der, S. New forms of Diacria quadridentata (de Bla-
inville,1821), Cavolinia Longirostris (de Blainville,1821) and Ca-
volina uncinata (Rang,1829) from the Red Sea and the East Pacific
Ocean (Mollusca, Pteropoda) .//Beaufortia,197*1 .Vol.19 no 24J.P. 1-200
S p о e 1 van der, S. Pseudothecasomata, Gymnosomata and Heteropo-
da (Gastropoda).//Zool.Meded.Rijksmus.Natuur.Hist.Leiden,1976. Deel
47. P.545-560.
S t a s e к Ch. R. Feeding and particle-sorting in Yoldia ensife-
1M
ra (Bivalvia: Protobranchia) with notes on other nuculanids.//Mala-
cologia. 1965. Vol.2 no 3. P.5*9-566.
Stirling D., Hamilton P. H. Observations on the me-
chanism of detecting mucous trail polarity in the snail Littorina
irrorata.//The Veliger, 1986. Vol.29 no 1. P.51-57.
Stoll E. Sur la mode de locomotion de quelques mollusqueв ma-
rines. //Trav,Stat.biol.Roskoff,1958. fasc.16.
Str authma nn R. B..Leise E. On feeding mechanisms and
clearance rates of molluscan veliger.//Biol.Bull.1979»Vol.157»P«52*.
Strenth N. E.,Blankenship J. E.. Laboratory cul-
ture, metamorphosis and development of Aplysia brasiliana Rang,1828
(gastropoda, Opisthobranchia).//The Veliger,1979. Vol.21 no 1.P.99.
Swedmark B. The biology of interstitial Mollusca. The Zool.
Soc.London.1968.Stud.struct.,physiol.,ecol.molluscs. P.155-1*9.
Taylor D. V. The tertiary gastropod Orygoceras found living.
//Arch.Moll.197*. Bd.104, no 1/5. P.95-96.
Tesch J. Heteropoden der Siboga-Expedition.//Siboga Exped.1906.
Livr.29 mon.51. 112 p.- The Thecasomata and Gymnosomata of the Si-
boga-Expedition.//Siboga Exped.1918.Livr.16 mon.52.
Tewesz M. J. Notes on the structure and habits of Myadora.//
The Veliger, 1975. Vol.18 no 2. P.162-165.
Tewesz M.J., Carter J.G. Form and function of Trisidos
(Bivalvia) and a comparison with other burrowing Arcoids.//Malaco-
logia, 1979. Vol.19 no 1. P.77-85.
Thompson F. G. The aquatic snails of the family Hydrobiidae
of Peninsular Florida. Gainsville.1968.298 p.
Thompson T. E. The natural history, embryology, larval bio-
logy and post-larval development of Adalaria proxima (Alder and Han-
cock Gastropoda, Opisthobranchia).//Phil.Trans.Roy.Soc.London.Ser.
B. 1958.Vol.242. P.1-58.
Thompson T. E. Feeding in nudibranch larvae.//J. mar. biol
Ass. UK. 1959.Vol.58. P.239-248.
Thompson T. E. Studies on the ontogeny of Tritonia hombergi
Cuvier(Gastropoda Opisthobranchia)•//Phil.Trans.Roy. Soc.London.Ser.
B. 1962. Vol.245. P.171-218.
Thompson T. E. Adaptive significance of gastropod Torsion.//
Malacologia, 1967. Vol.5 no 3. P.423-450.
Thompson T. E. Biology of Opisthobranch molluscs.//The Ray
Soc.ser.no 151.1976.2O7 p.
Thompson T. E.,Brown G. H. Biology of Opisthobranch
molluscs.Vol.II.//The ray.Soc.ser. no 198*.
Thompson T. E., S 1 i n n D.J.On the biology of the opis-
thobranchs Pleurobranchus membranaceus.//J.mar.biol.AS8.UK. 1959.
Vol.38. P.507-524.
187
Tullier 8. Patterns of digestive tract morphology ih the Ц,
macinasion of Helicarionid succeneid and athoracophorid snails and
slugs (Mollusca Pulmonata).//Malacologia,1984. Vol.25 no 1.P.173-193
Timmermans L. Studies on shell formation in molluscs.196g,
Trueman E. R. The burrowing activities of Bivalvia. The Zool,
Бос.London.Stud.struct.physiol.,ecol.molluscs.1968.P.167-186.
Trueman E. R. The fluid dynamics of molluscan locomotion//
Malacologia, 1969. Vol.9 no 1. P.243-248.
Turder H.J. The drilling mechanism in the Naticidae.//Eco-
logy. 1953. P.222-223.
Turner R. D. A survey and illustrated catalogue of the Teredi-
nidae (Mollusca: Bivalvia).Cambr.Mass. 1966. 265 p.
Turner R. D. Bankia neztalia n.sp.(Mollusca:Bivalvia) from Au-
stralia and New Zealand, and its relationship//.!.Roy.Soc.NZ. 1979.
Vol.9 no 4. p.465-473.
V a h 1 0. Mucus drifting in the limpet Helcion pellucidus (Prosob-
ranchia, Patellidae).//Sarsia,1983. Vol.68 no 3. P.204-212.
VermeiJ G. J.,Zipser E. Burrowing performance of some
tropical Pacific Gastropods.//The Veliger,1986.Vol.29 no2.P.200-206.
Villaroel M. ,Stuardo J. Observationes sobre la mor-
phologia general musculature у aparato digestivo an Tagelus dombei у
T.longisinuatus (Tellinacea)//Malacologia,1977.Vol.16 no 2.P.333-352
Voltzow J. Plow through and around the abalone Haliotis kam-
tscatcana.//The Veliger,1983. Vol.26 no 1. P.18-21.
IM
Ф-ка «Картолктографкя» Зак. 23^4 Тир.«5^^