/

























































































































































































































































Автор: Шалапенок Е. С Бута С.В.
Теги: invertebrata беспозвоночные систематика животных специальные зоологические науки биология зоология
ISBN: 985-6516-66-8
Год: 2002




Текст
E.C. Шалапенок
СВ. Буга
Практикум
по зоологии
беспозвоночных
Учебное пособие
УДК 592(076.5X075.8)
ББК 28.691я73
UII8
Рецензенты:
кафедра ботаники и зоологии
Гродненского государстваиюго университета;
доктор биологических наук, профессор, заведующий
лабораторией энтомологии Института зоологии МАИ Беларуси Э.И. Хотысо
ШдлавювжЕС
Ш18 Практикум по зоологии беспозвоночных: Учеб. пособие / Е.С. Шалапсиок,
СВ. Буга. — Мн.: Новое знание. 2002. — 272 с: ил.
ISBN 985-6516-66-8.
Темы практикума приведены в соответствие с накопленными за последнее
время знаниями по организации и систематике беспозюючных животных По
каждой группе объектов даны рекомендации, позволяющие обеспечить занятия
необходимым зоологическим материалом. Кратко характеризуются строение и функции
изучаемых таксонов. Книга содержит богатый иллюстративный материал.
Для студс1гтов биологических и экологических специальностей вузов.
УДК 592(076.5X075^)
ББК2&691я73
Учебное издание
Шалавою* Елена Семеновна
Буга Сергей Владимирович
Практикум по зоологии беспозвоночных
Учебное пособие
Редактор Г.И Киселе*
Корректор КА. Степанова
Художник обложки С. В. Котлеескии
Компьютерная верстка С.М.Данилюк, E.JI. Помогаем
Подписано в печать с готовых диапозитивов 28.07.i002. Формат 60^84 у**
Бумага газетная. Гарнитура Н потом. Печать офсетная. Усл. печ. л. 16.74.
Уч.-изд. л. 15.29. Тираж ЗОЮ экз. Заказ N? 1602.
Общество с ограниченной ответе гвенностыо< Новое знание*.
Л В № 310 от 14.08.2001. Минск, ул. Академическая, д. 28. к. 112.
Почтовый адрес: 220050. Минск, а/я 79. Тел./факс: (10-375-17) 211-10-33. 284-03-23.
Москва, уд. Маросейка, д. 10/1. Тел. (095) 921-67-21.
E-mail: nk&wnk.biz. http://wnx.bt7.
Рссп)<хликанское унитарное предприятие «Издательство «Белорусский Дом печати».
220013. Минск, пр. Ф. Скорины. 79.
С Шалапсиок Е.С.. Буга СВ.. 2002
ISBN 985-6516-66-8 С Оформление, ООО «Новое знание». 2002
Оглавление
От авторов 11
Технические средства юучевтшофоскотгмеких объектов 13
Микроскопическая техника 13
Микроскоп 13
Правила работы с микроскопом 16
Микроскоп бинокулярный стереоскопический (МБС) 19
Изготовление препаратов для микроскопирования 21
ПОДЦАРСТВО PROTOZOA - ПРОСТЕЙШИЕ
Тип Sarcomasrigophora - саркомястнгофоры 23
Подтип Sarcodino-езркойоъыс 23
Ндлкл&сс Rhizopoda- корненожки 23
Класс Lobosea 23
Отряд АтоеЫпа- голые амебы 23
Отряд Arctllinida (^Testacea) - пресноводные
раковинные корненожки 28
Класс Filozta 30
Отряд Foraminifera- морские раковинные корненожки 30
Налкмсс Aertnopoda 32
Класс Radiolaria - лучевикн 32
Класс Яе/ходо-солнечники 34
Подтип Mastigophora- жгутиконосцы 35
Класс Phytomastigina-растительные жгутиконосцы 36
Отряд £ц&/гло/</л1-эвгленовые 36
Отряд Dinoflasellata(mPcridinea)- панцирные жгутиконосцы . . 38
Отряд Votvocida(~Phytomonadino)-ъольвоксошс 40
Класс Zoomastigina- животные жгутиконосцы 42
Отряд Kinetoplastida 42
4 Оглавление
Подтип Opallnota 43
Отряд Opalinida- опалины 43
Тип Apicomplexa (mSporozoa) - переднекомшюксные,
или споровики 4S
Класс Gregarinina - цкгърины 45
Отряд £i<gnfjro^/j/^2-настоящие грсгарины 45
Класс Coccldiomorplta - кокинлиссбрааиые 48
Отряд Hacmosporidia - кровяные споровики 48
Тип Ciliophora (-Infusoria) - инфузории 50
Класс СШаш (Infusoria)- ресничные инфузории 50
Надотряд Oligohymenophora 50
Отряд Hymenostomata(mHolotricha) 50
Paramecium caudafum-инфузория-туфелька 50
Разнообразие инфузорий 53
ПОДЦЛРСТВО METAZOA- МНОГОКЛЕТОЧНЫЕ
Надраадел Phagocytellozoa 57
Тип Placozoa - пластинчатые 57
Класс Placozoa 57
Trichoplaxadhatrens(=n?psans) 57
Надраадел Parazoa 58
Тип Portfera(=Spongia)- губки 58
Надраадел Eumetazoa 62
Раздел Radiata - лучистые, нли Diploblastica - туслоктле ... 62
Оглавление 5
Тип Cnidaria (■ Coelenterata) - стрекающие (кишечнополостные) 62
Класс Hydrozoa-гнлроэои 62
Подкласс Hydroldea- гидроидные 62
Отряд Hydride - гидры 62
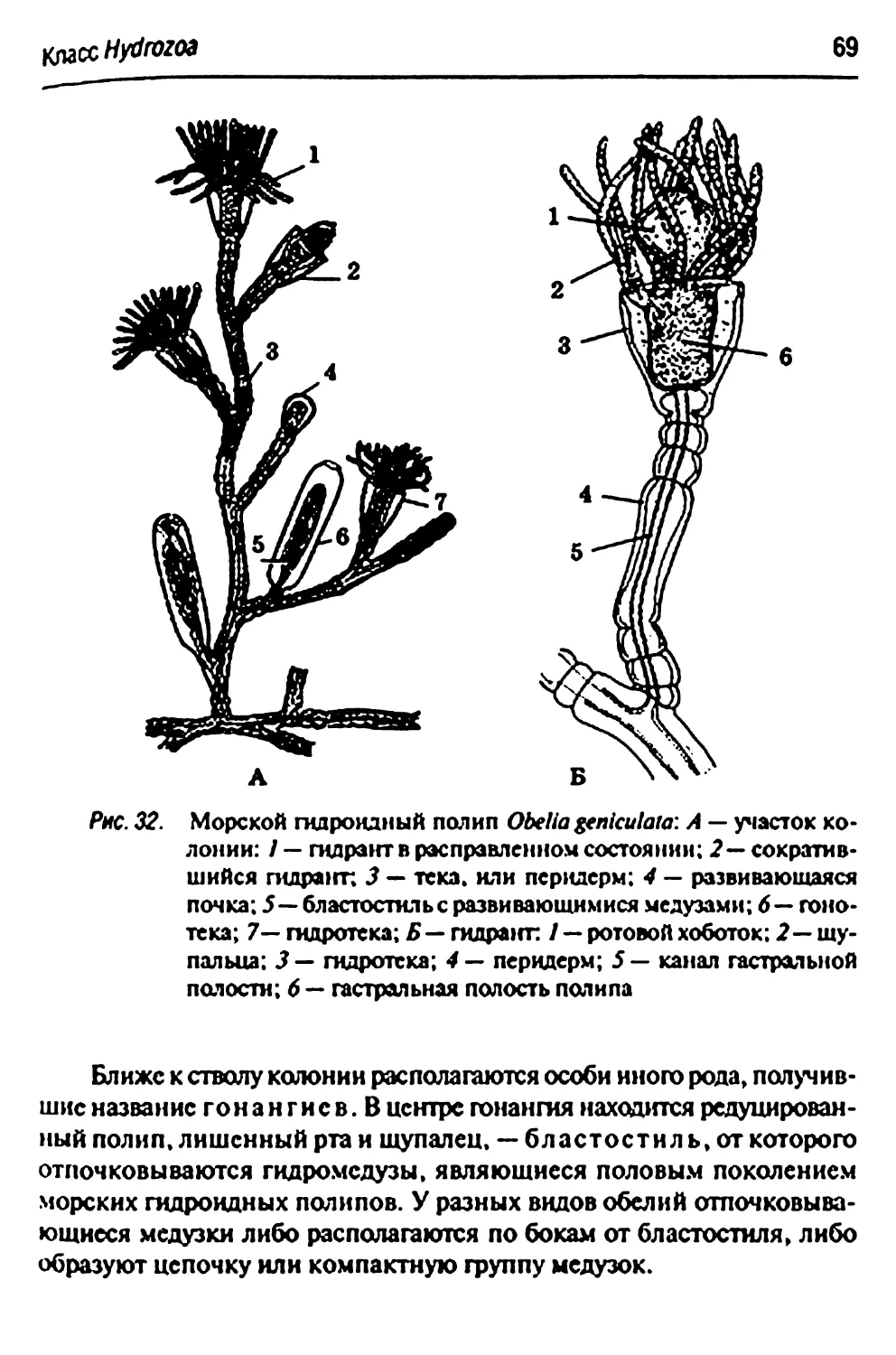
Отряд Leptdida- морские гидроидные полипы 68
Подотряд Thecaphora 68
Отряд ГлосШЛ»-трахимедузы 71
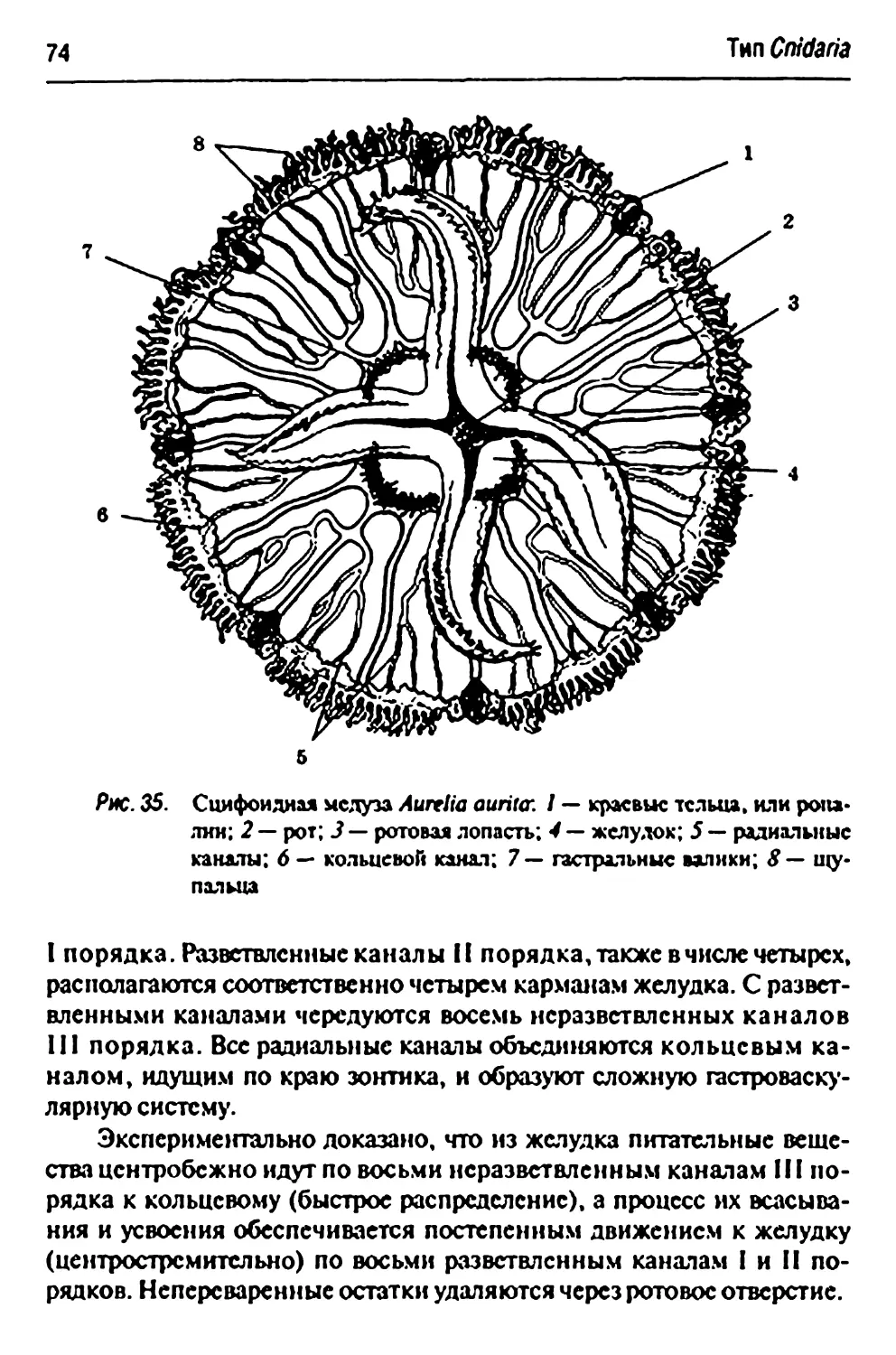
Класс Scyphozoa - сцифомсдузы 73
Отряд Scmacostomtac 73
Класс Anthozpa- коралловые полипы 77
Подкласс Hexacorollia - шсстняучсвыс кораллы 77
Отряд Лс/ш/ала- актинии 77
Раздел ЛУа/егш-двустороннесимметричные 80
Подраздел - Protostomia - первичноротые 80
Тип Plathelminthes- плоские черви 80
Надкласс TurMlaria 80
Класс Turbfllaria- ресничные черви 80
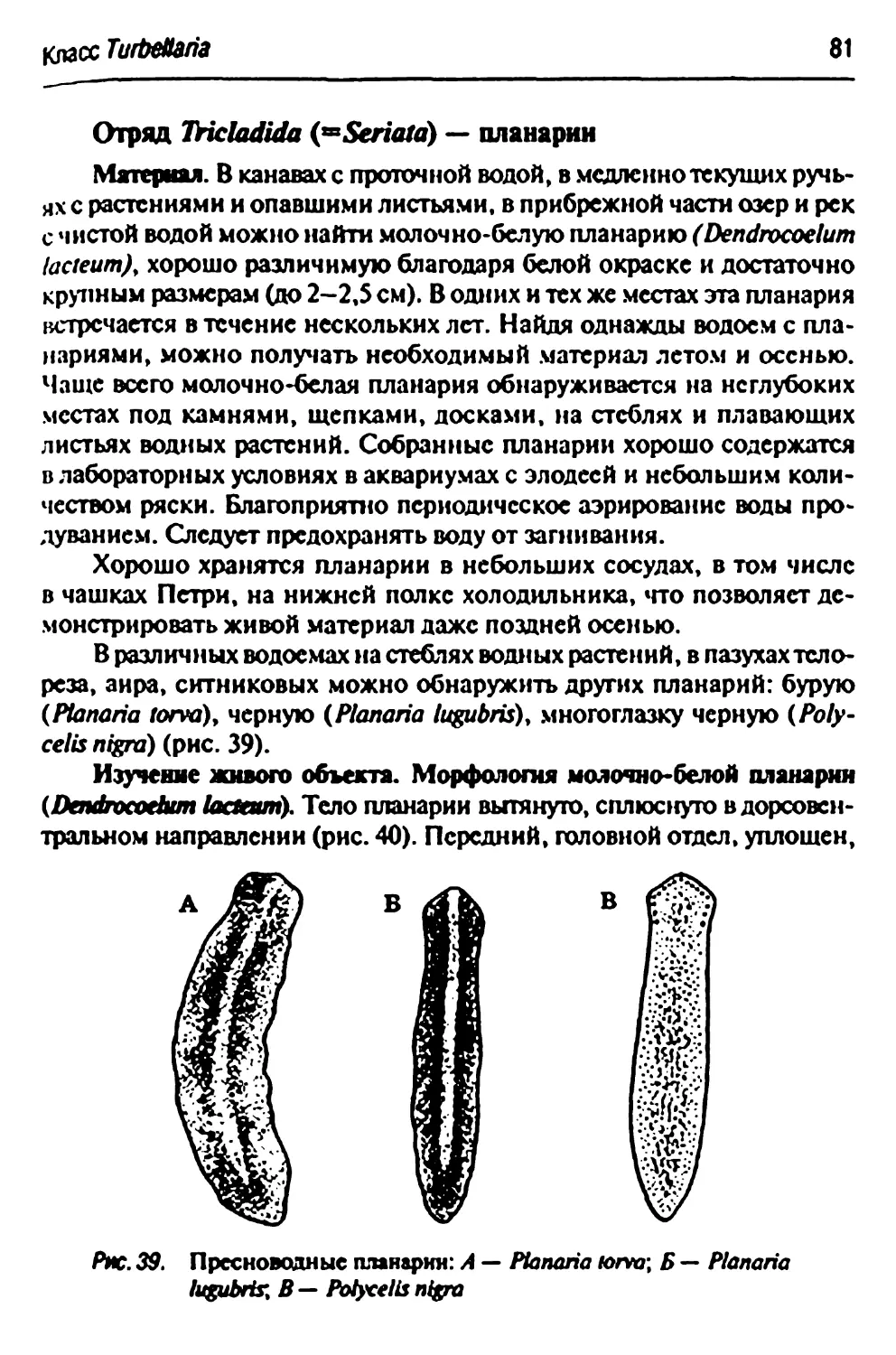
Отряд Triciadida (-&/va/0) - планарии 81
Надкласс Neoderma to 84
Класс Г/гтдЛ)^д(«0^ел^л)-дигенетичсскиесосальщики. ... 84
Dlcrocoeliumdcndritkum- ланцетовидная двуустка 84
Fasciolahepatica- печеночная двуустка 86
Изучение некоторых стадий развития печеночного сосальщика. 88
Класс Monogenoidea - моногенеи,
или моногенетические сосальщики 91
Отряд Monopisthocotyka^Polyonchoinea) 92
Potystomaintt&rrimum—лягушачий многоуст 92
Отряд Pdyopisthocoiyiea(^Oiigonchoinea) 94
Diptozoonparodoxum-сп&йпнк парадоксальный 94
Класс Cestodo- ленточные черви 94
Отряд Cyclophyilideo- цепни 95
6
От давление
Taenia solium- свиной солитер 95
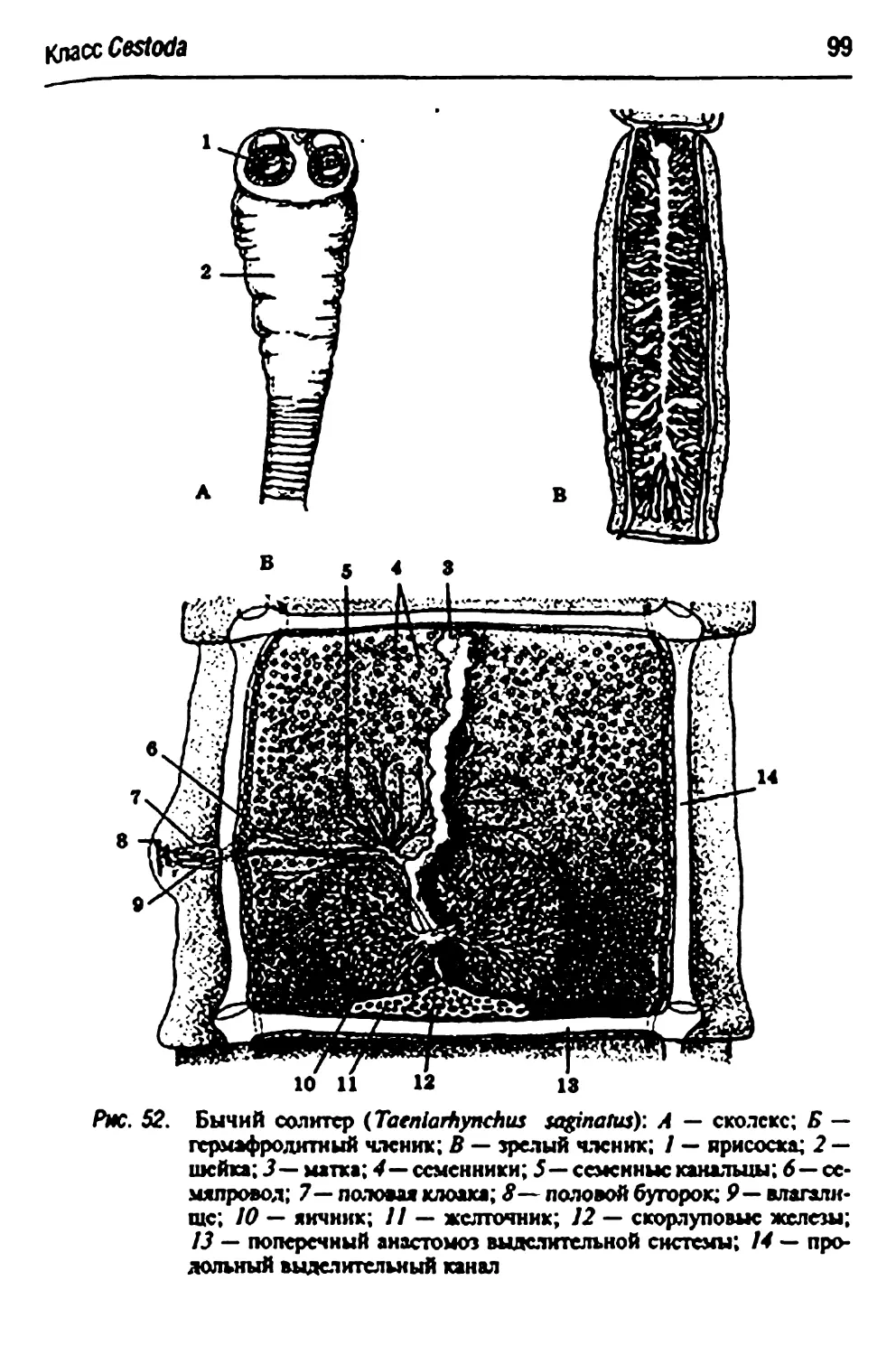
Taeniarhynchussa&inams- бычий цепень 98
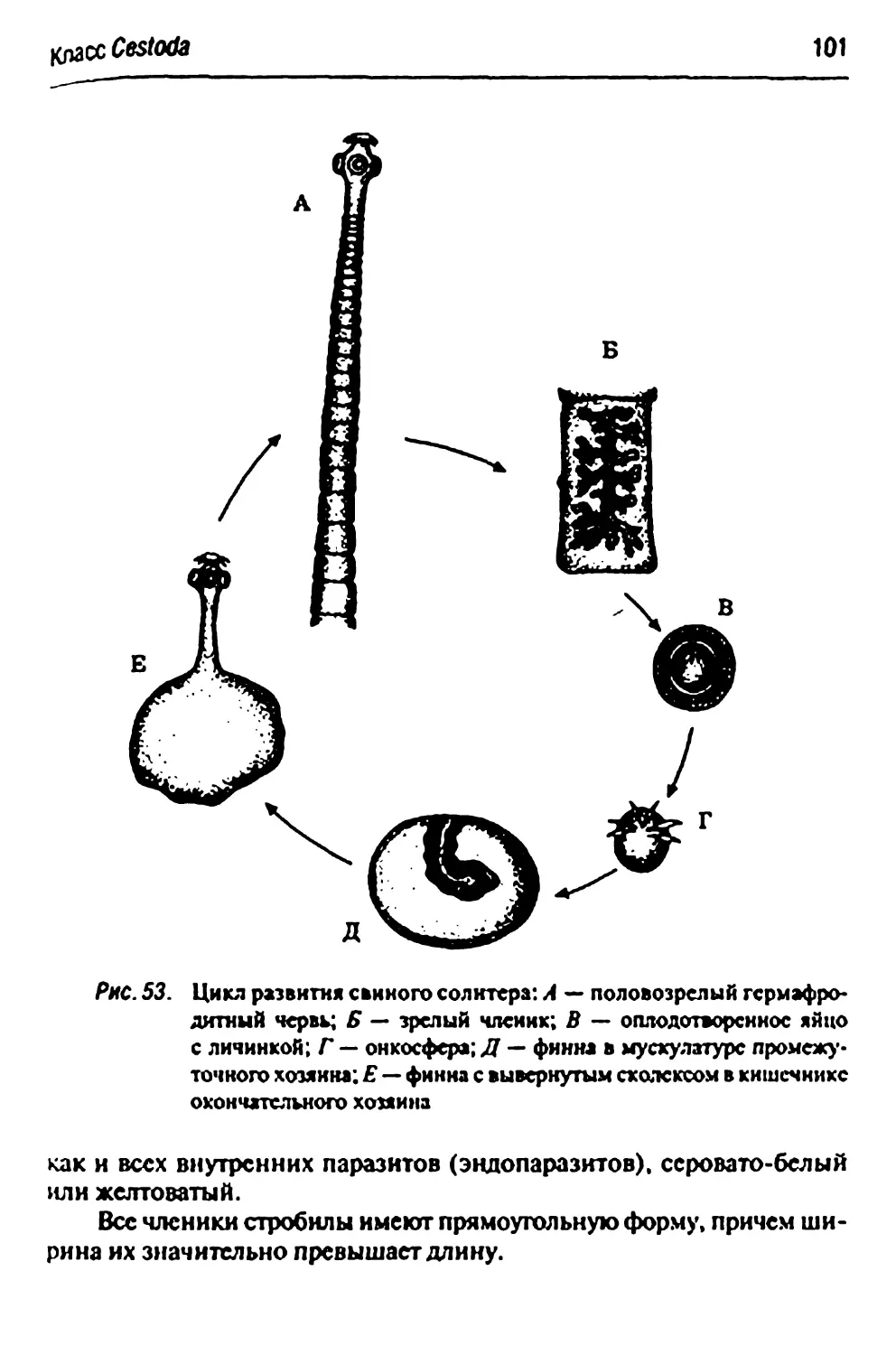
Стадии развития свиного или бычьего цепня 100
Отряд Rscudophlllidea-пссъаофнлл идеи 100
Diphylbbothrium latum-ncrntn широкий 100
Тип Nemathelminthes- круглые черви 10S
Класс Nematode—нематоды 105
Подкласс Rhabditia(mSecernentea) 105
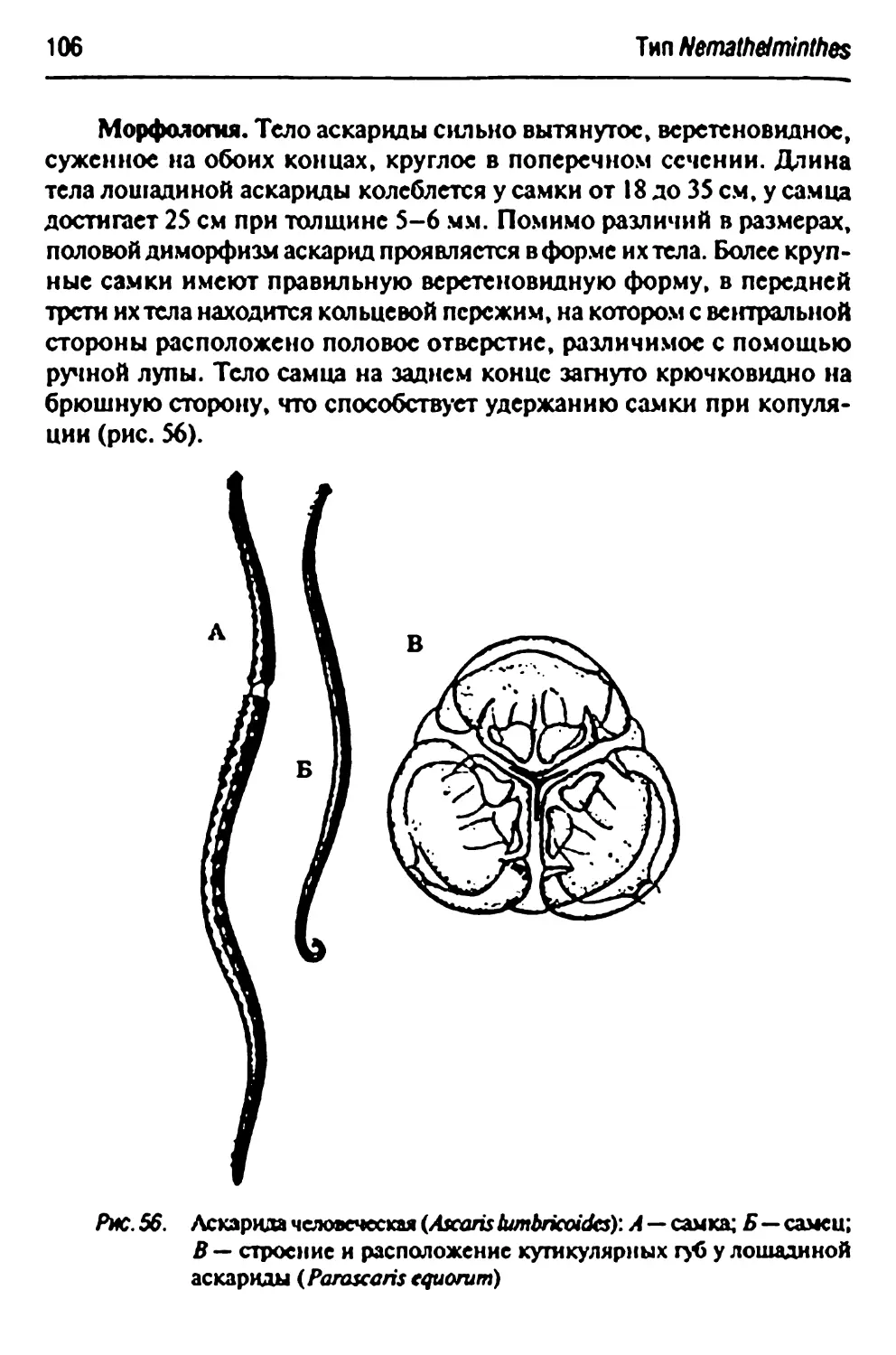
Отряд Ascaridlda- аскариды 105
Parascarisequomm- лошадиная аскарида 105
Отряд 7>^пс/гШ-шишкоиглые нематоды 112
Globoderarostochiensb- золотистая картофельная нематода ... 112
Подкласс Enoplla(**Adenophorea) 115
Отряд Trichocephalida 115
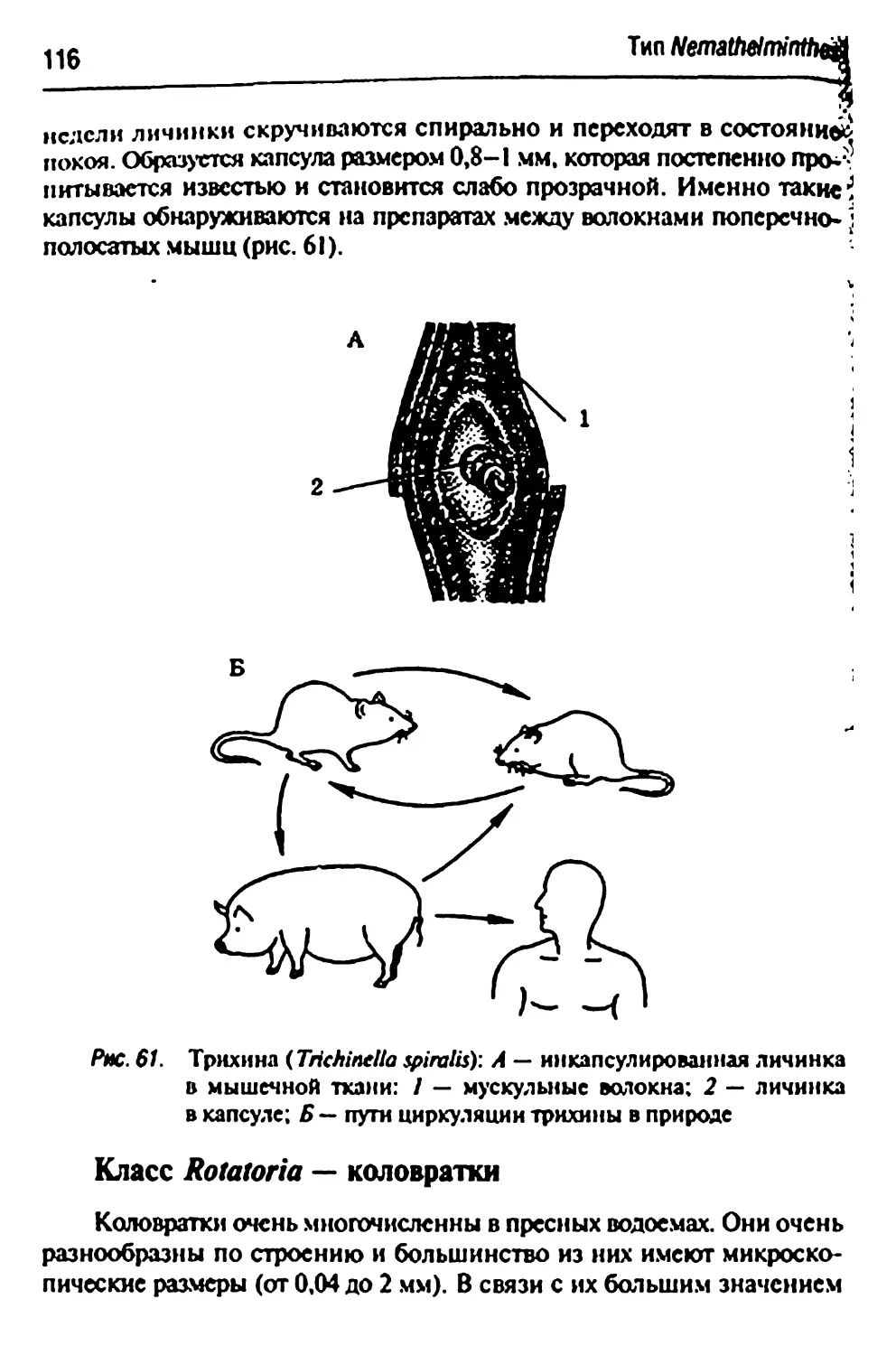
Trichlnella spiralis- трихина 115
Класс Rotatoria- коловратки 116
• Отряд Rdelhidea 117
Rotariasp 117
Ъш Annelida- кольчатые черви 120
Подпш Aclitellata - беспоясховые кольчецы 120
Класс /ОДг/юдо-многошетинковые кольчецы 120
Подкласс Errontia- бродячие 120
Отряд Nerelmorpha 120
Nereis pelagka-псосьш 120
Подкласс Sedentaria- сидячие полихеты 123
Отряд Drilomorpha 123
Arenicola marina- пескожил 123
Подтип Clitellata - поясковые кольчецы 124
Класс О/досЛде/а-малощетинковые кольчецы 124
Отряд Lumbricomorpha 124
Оглавление 7
Lumbricustcmsais- обыкновенный дождевой червь 124
Пресноводные и почвенные олигохеты отряда Naidomorpha ... 133
Класс Hirudinea- пиявки 136
Отряд Gnathobdellea-челюстные пиявки 136
llirudomcdicinalis- медицинская пиявка 136
Тип Mollusca - моллюски 142
Подткп CtWTcAi/егд- раковинные моллюски 142
Класс Gastropoda- брюхоногие моллюски 142
Подкласс Pulmonata- легочные моллюски 142
OrpnnStybmmasophom-стебельчаггоглюые 142
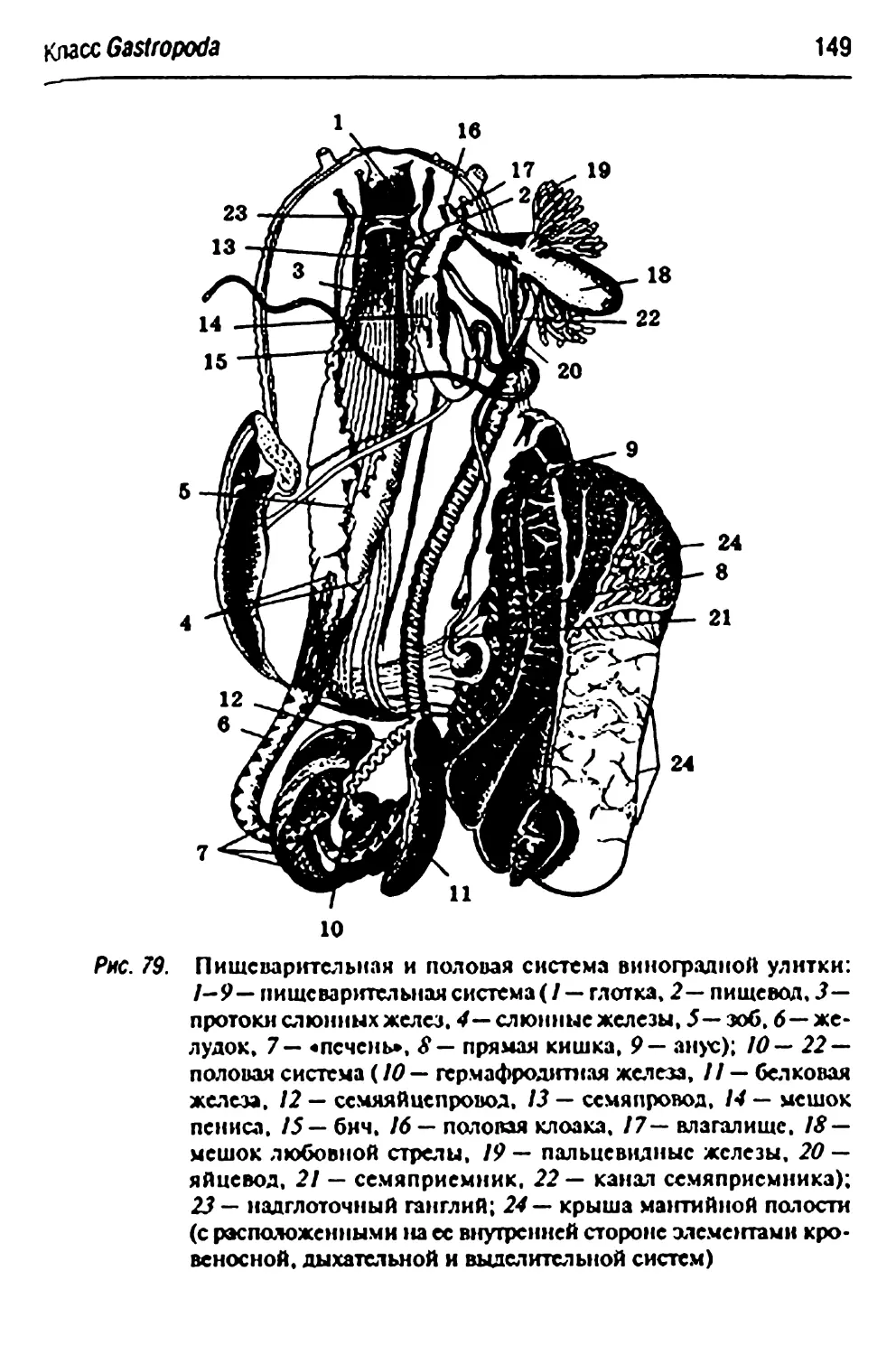
Helix ротап'а-иинотръ&пъя улитка 142
Класс BivaMa (mLamtllibranchla) - двустворчатые
(пластинчатожаберные) моллюски ISI
Отряд Eulamtllibranehia - настоящие
пластинчатожаберные моллюски ISI
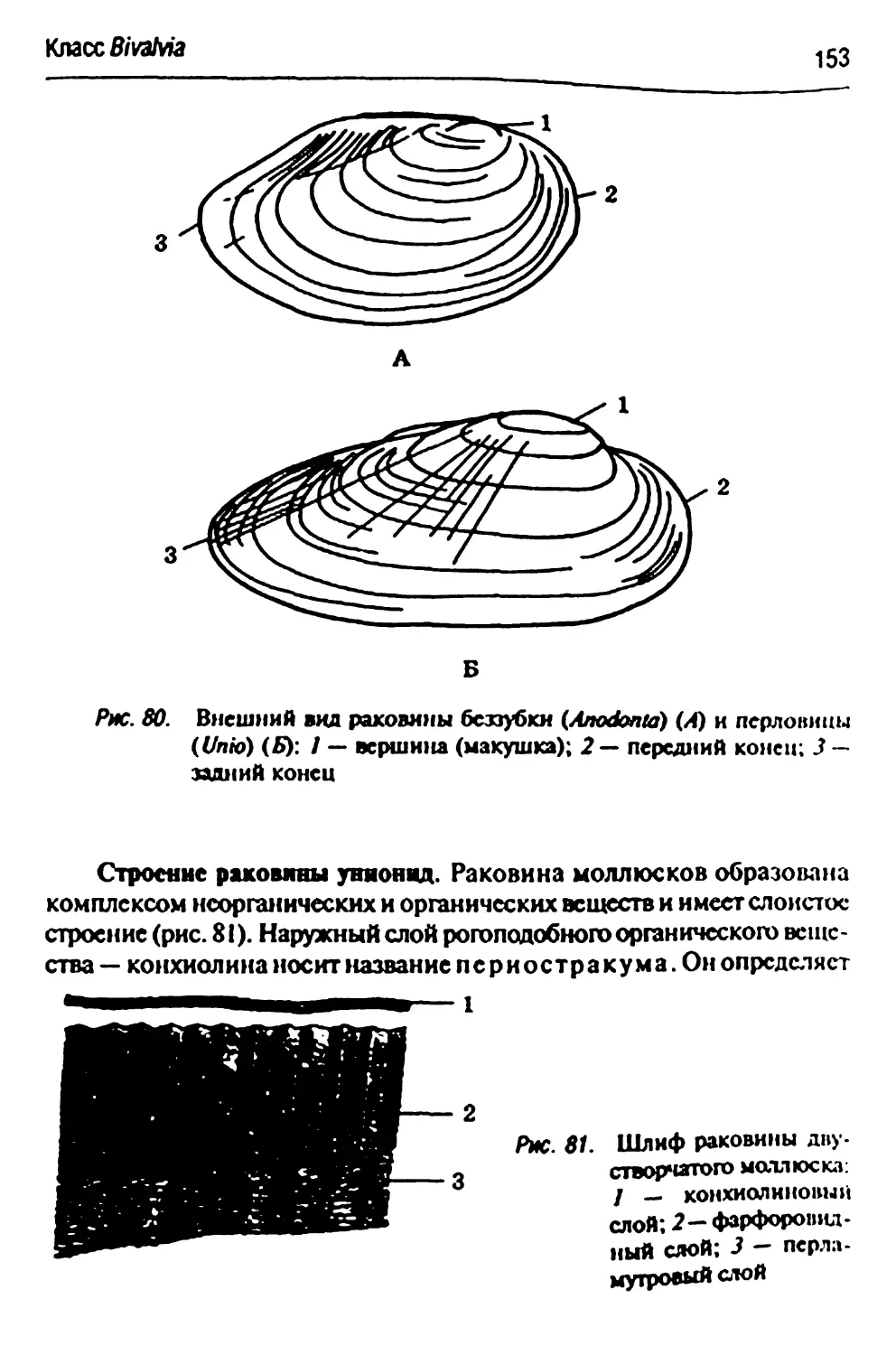
Anodontacypica- обыкновенная европейская беззубка 1S1
Тип Anhropoda - членистоногие 161
Подтип Mandibulata 161
НадклассОшаюга- ракообразные 161
Класс Branchiopoda-жабропогие раки 161
Отряд Am>mopoda(=Claabccra)-bCTbHCToychie 161
ДврЛш/вриЛгх-обыкновенная дафния (водяная блоха) 161
Класс Maxilbpoda 168
Подкласс Copcpoda- веслоногие раки 168
Отряд Cychpida-иикяопообразные 169
Cyclops spp. — циклопы 169
Класс Malacostraca - высшие раки 174
Отряд Decapoda—десятиногие раки 174
Potamobiusspp. - речные раки 174
8 Оглавление
Таксономическое разнообразие ракообразных:
основные особенности внешнего строения представителей
основных групп Crustacea 188
Класс BraiKhiopoda-жзбропогкс раки 188
Отряд Ллсзтхя—жаброноги 188
Пресноводные жаброноги 188
Artemiasalina—артсмня 190
Отряд Phylbpoda(=Notostraca)-- лисгоногиераки 191
Щитни 191
Класс MaxiOopodc 193
Подкласс Ctamcodb—ракушковые раки 193
Пресноводные остракоды 194
Подкласс ЯпаясЛ/шт»—карпосды I9S
Arjpilusfdiaccus — обыкновенный карпосд (карповая вошь) ... 196
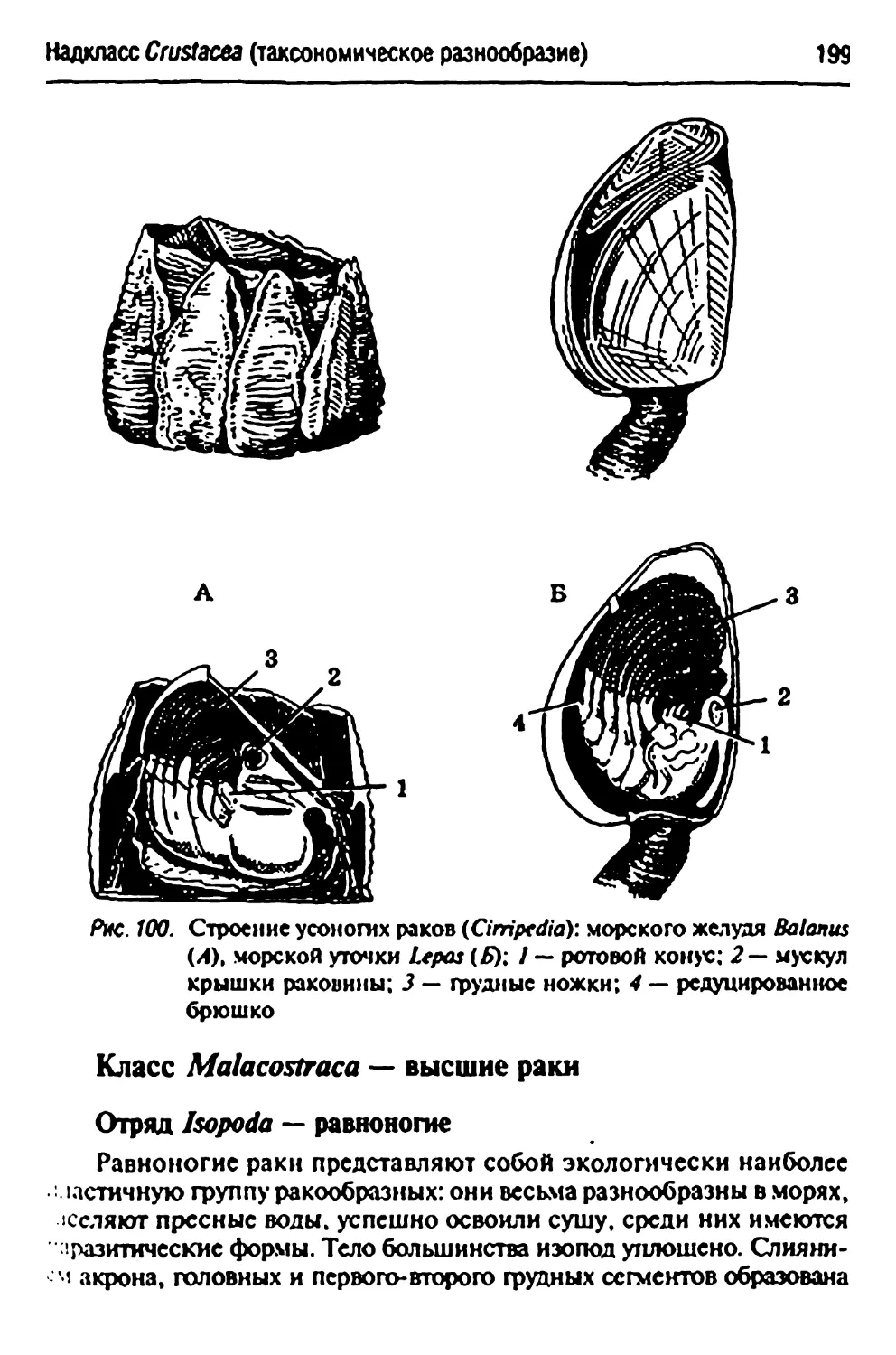
Подкласс СУ/тдо^—усоногие раки 197
Bakuutsspp,— морские желуди 198
Класс Malaantroca— высшие раки 199
Отряд Isopoda—равноногие 199
Scduria(=Mc3idoSta)entomon — uo[KKOUTbp&X2LH 200
Отряд ЛлтрЛ/ро/а—раэноиогие раки, или бокоплавы 201
Gammarusspp. — бокоплавы-гаммарусы 202
Отряд Dtcapoda—десятиногие раки 203
Palaemonspp. — травяные креветки 204
Раковинообитающис раки-отшельники 205
Carcinustcmumts—травяной краб 205
Похпяп Antennata 206
Нъдклъсс Myriapoda — многоножки 206
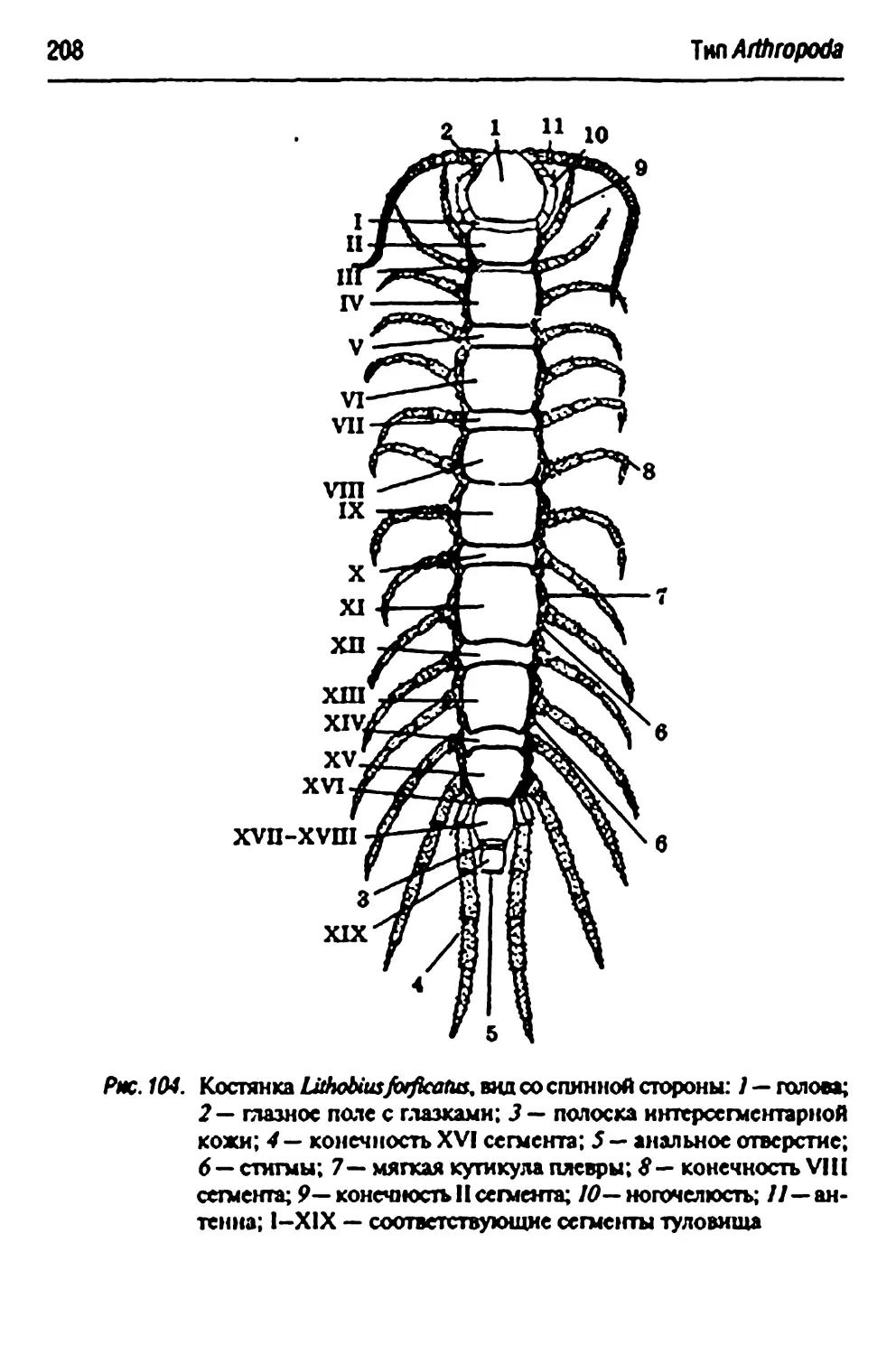
Класс Chilopoda — губоногие 207
Uthobiusfoificams — обыкновенная костянка 207
Класс Diptopoda—хвупарноногнс 210
Julidac— настоящие кивсяки 211
Ньяклъсс Hcxapoda — шестиногие 213
Класс Instcta—насекомые 213
Оглавление 9
Расчленение тела майского хруща (Mtlokmthasp.) 214
Особенности строения антенн 219
Основные типы грудных конечностей насекомых 221
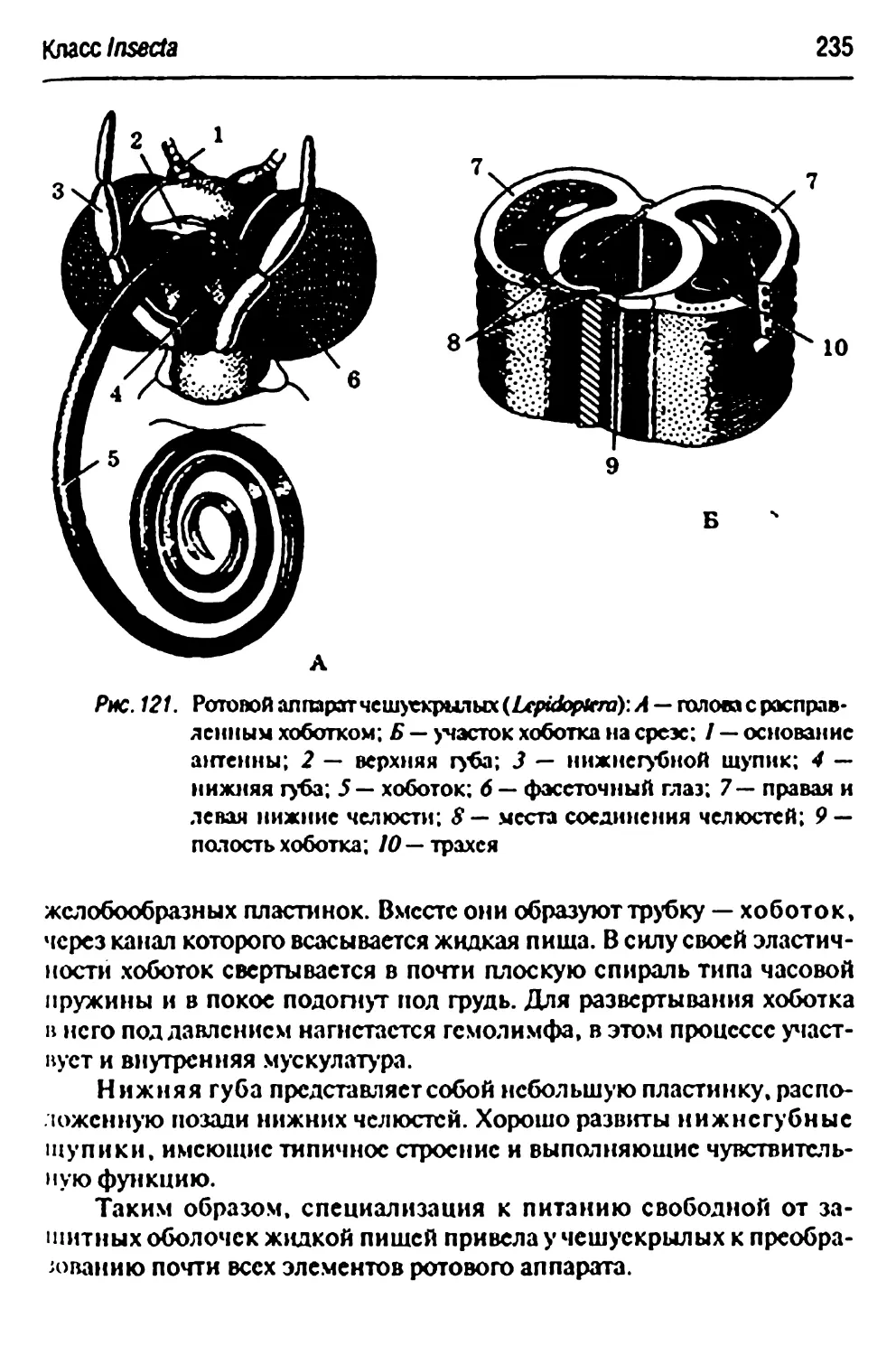
Основные типы крыльев насекомых» жилкование крыла 224
Основные функционально-морфологические типы
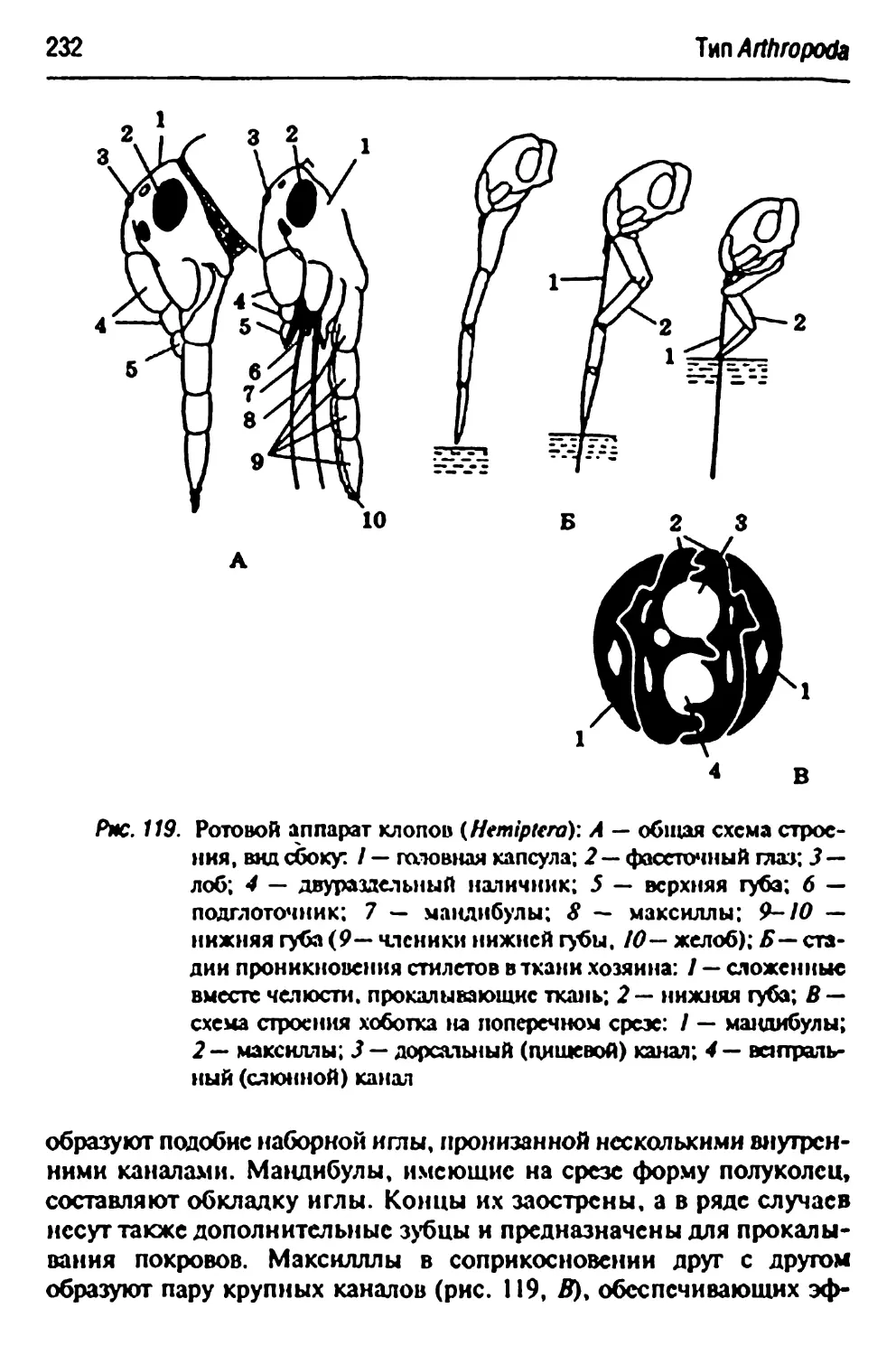
ротоных аппаратов насекомых 227
Подтип Chelicerata — хелнцеровые. . 237
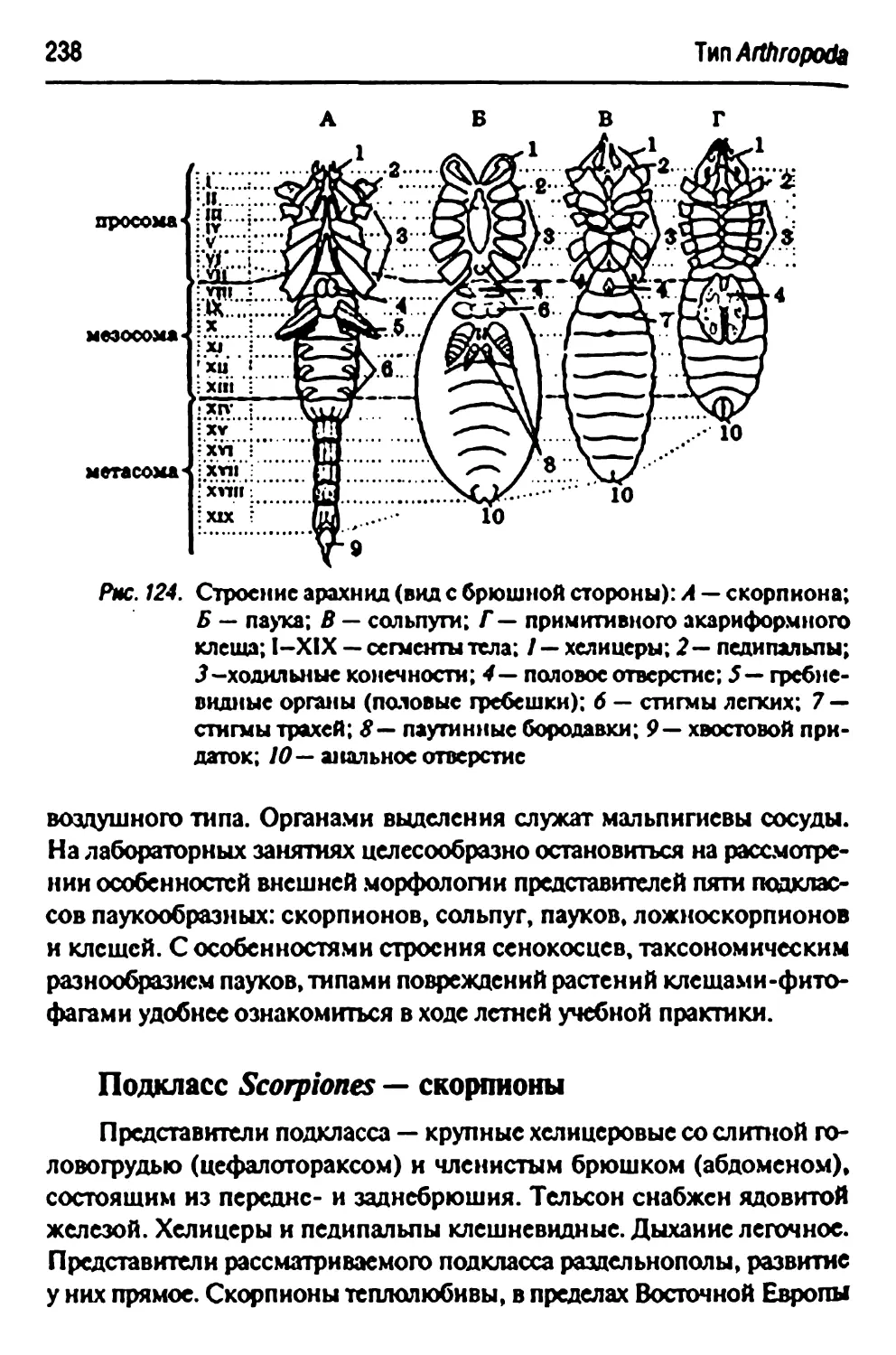
Класс Arachnida — паукообразные 237
Подкласс Scorpionts—скорпионы 238
Buthuseuptus— пестрый скорпион 239
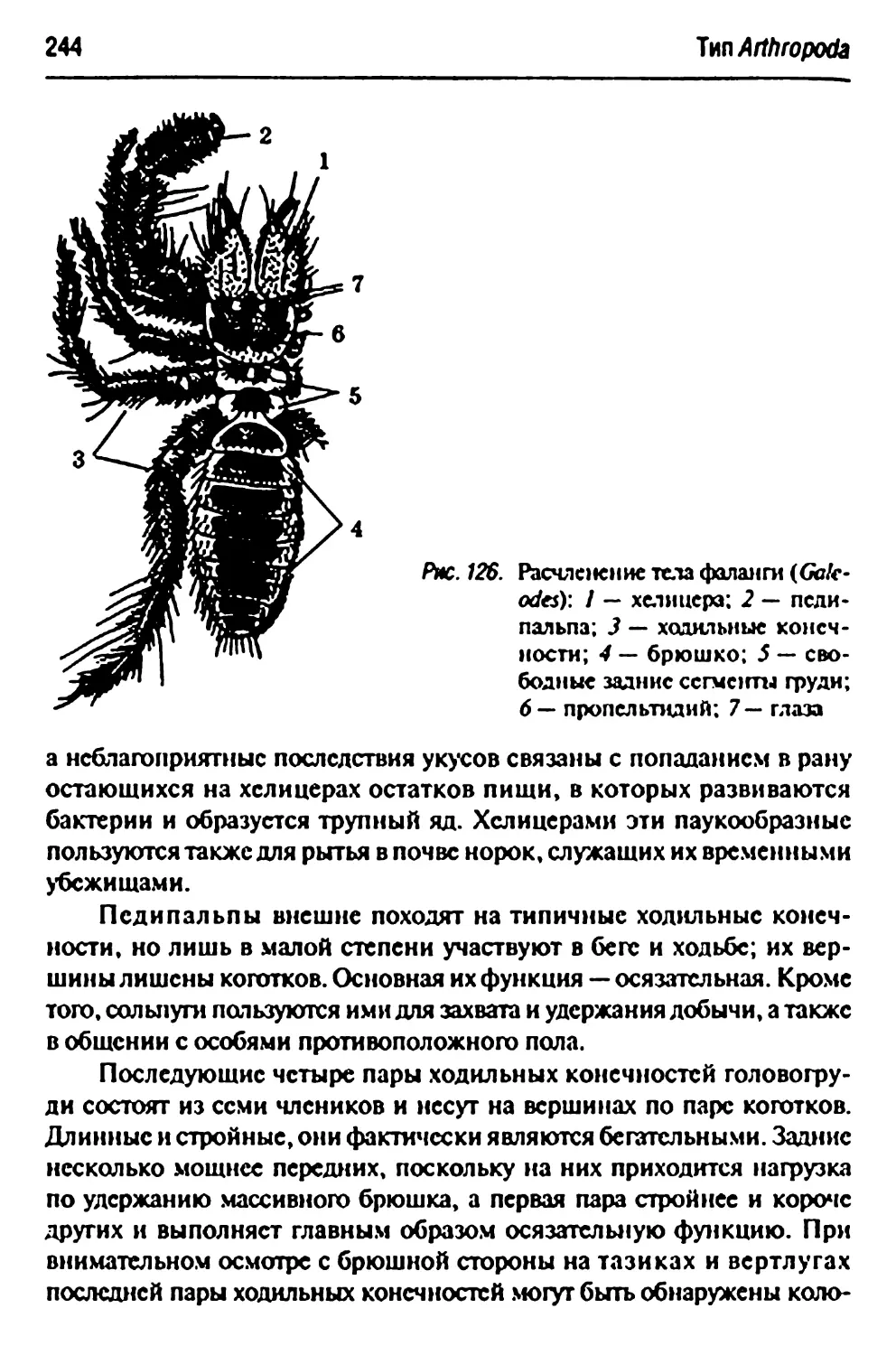
Подкласс Soipugida(mSolifidga€) — coJihnyrn 242
Galeodesaraneoides— обыкновенная фаланга 242
Поамасс Pstudoscorpionts-- ложноскорпноиы 245
Chclifcrcanavides — книжный»
или обыкновенный ложноскорпион 245
Подкласс Aronei— пауки 246
Arontus dladtmatus— паук-крестовик 247
Подкласс Acari— клещи 250
Отряд И«7/#Ь/7пю—акариформные клещи 250
Подотряд &/rxp/#brmtf—саркогпиформные клещи 251
Tyrogfyphusfaiinae — обыкновенный мучной клеш 251
Подотряд 7>т>т№//аг7Л€?-тромбидиформные клещи 253
Tetranychidae- паутинные клещи 253
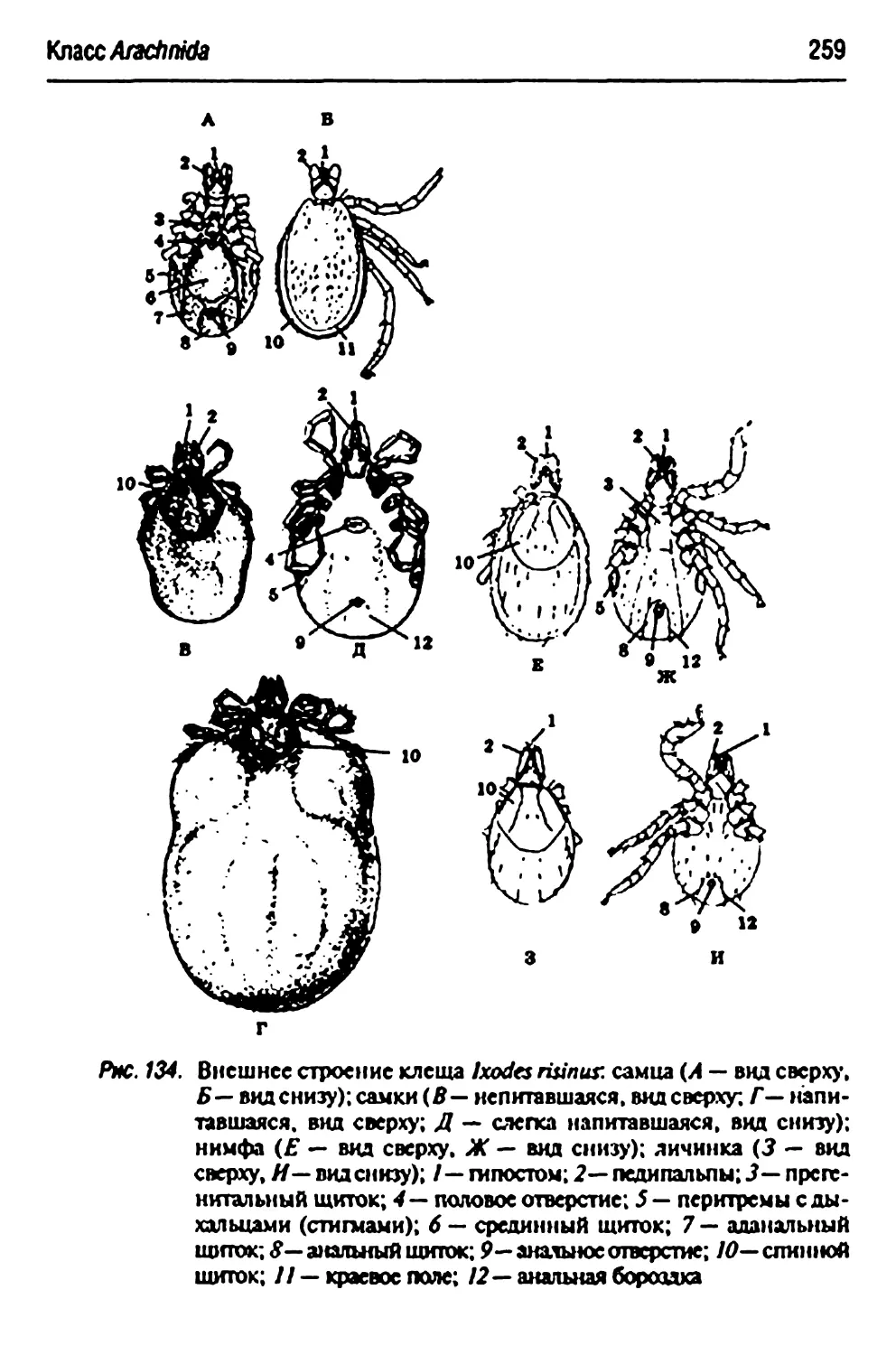
Отряд А7Аи|7^Ьппе$-паразитоформные клещи 257
Ixodes nanus- собачий клещ 257
Тип Echinodermata - иглокожие 262
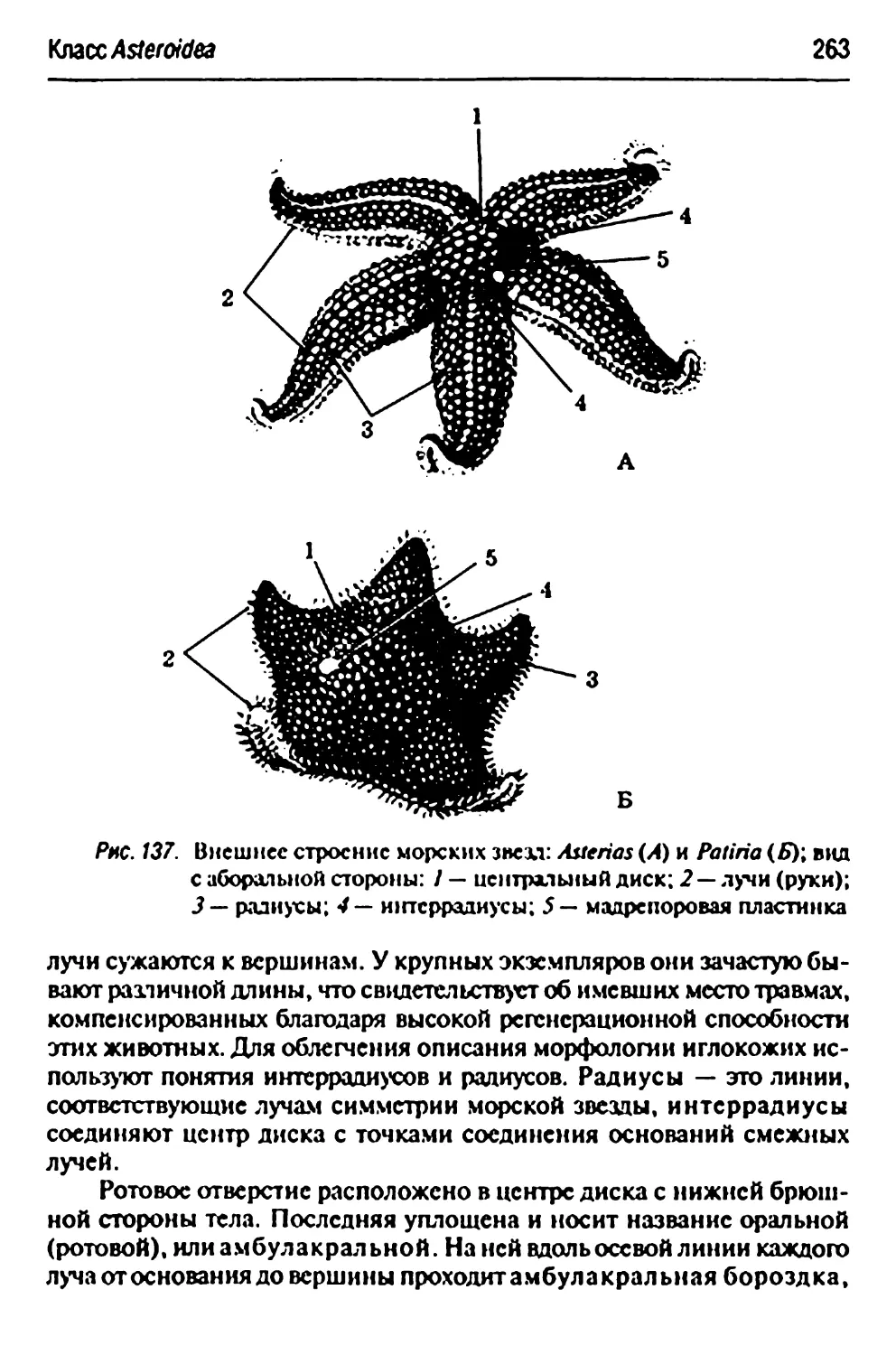
Класс Asteroidea— морские звезды 262
Класс Echinoidta — морские ежи 269
От авторов
Данный практикум по зоологии беспозвоночных предназначен для сту-
лс»гтов биологических факультетов университетов. Его создание продиктовано
рядом обстоятельств: прежде всего отсутствием, нехваткой или физическим
износом ранее изданных пособий. Так» классические лабораторные практикумы
ЕН. Павловского и СВ. Аверинцсва изданы соответственно в 1938 и в 1947 годах
и уже давно стали библиографической редкостью. Малый практикум по
зоологии беспозвоночных А.А. Зеликмана, второе издание которого вышло в свет
в 1969 году» в настоящее время в большинстве вузов имеется в ограниченном
количестве. Практикум по зоологии беспозвоночных, подготовленный ЕН.
Фроловой с соавторами (1985) для студентов педагогических институтов, рассчитан
на меньший объем курса» в нем недостаточно полно изложены некоторые
темы, рассмотрение которых представляется важным или обязательным при
подготовке биологов с базовым университетским образованием. Кроме того,
накопленный за последнее время объем зоологических представлений по организации
и систематике беспозвоночных животных требует внесения в учебные пособия
определенных изменений и дополнений.
На кафедре зоологии Белорусского государственного университета
накоплен большой опыт по организации и проведению практикума по эоаюгии
беспозвоночных. Основы его разработаны при непосредственном участии
профессора Г.Г. Винберга еше в 1950-е годы и традиционно сохранялись и
развивались его учениками. Наличие многолетнего опыта позволило нам взяться за
стольответствениый проект, как написание нового лабораторного практикума
ло эоаюгии беспозвоночных для биологических факультетов университетов
по специальности Н.04.01.00 — биология и Н.06.01.00 — экология. Объем
данного практикума рассчитан в соответствии с учебным планом на 74-78 часов.
В пособии с учетом современного состояния материального оснащения
университетских кафедр сокращен объем работ с малодоступным или
дорогостоящим материалом. Вместо этих тем предлагаются альтернативные темы и
объекты для изучения. По каждой группе модельных объектов даны
рекомендации, позволяющие обеспечить занятия необходимым зоологическим
материалом. В случаях, когда объем приводимого в пособии материала представляется
чреммерным. выделение сведений, обязательных для усвоения студентами,
может быть скорректировано преподавателями.
В настоящем практикуме отсутствует изложение материала занятий,
предусматривающих знакомство с таксономическим и эколого-морфатогическим
разнообразием некоторых трупп животных (коралловые полипы, моллюски
и др.). Такие занятия лучше всего вести на базе зоологического музея (если
таковой имеется) или на основе зоологического коллекционного материала,
которым располагает кафедра.
12
От авторов
В предлагаемом учебном пособии раздел по микроскопической технике
и темы от простейших до червей всех групп включительно написаны
доцентом Е.С. Шалалснок, темы по моллюскам, членистоногим и иглокожим —
доцентом СВ. Бутой.
В лабораторном практикуме использована современная
классификация беспозвоночных животных в соответствии со 2-м, переработанным
изданием «Жизни животных» под рсдакиией члена-корреспондента АН СССР
Ю.И. Полянского(т. 1,1987 г.), кандидата биологических наук Р.К.
Пастернак (т. И, 1988 г.) и академика АН СССР М.С. Гилярова (т. III. 1984 г.)
Учтены новые представления о системе по S. Westheide, R. Rieger.
Specielle Zoologie. I. Tell.
Gussav Fischer Vcrtog.
Stuttgart, Jena, NewJork. 1996.
Авторы весьма признательны Т. Л. Черкас за техническую помощь при
подготовке рукописи и выражают благодарность коллегам за поддержку.
Технические средства изучения
микроскопических объектов
Для изучения животных могут быть использованы различные
методы. Наибольшую информацию о внешнем строении и
функционировании организмов мы получаем, изучая животное in vivo в
обстановке естественной или максимально к ней приближенной. Это легко
удается осуществить во время летней учебной практики, но не всегда
'возможно в течение года в учебной лаборатории. Кроме того, если
достаточно легко обеспечить изучение на лабораторном практикуме
небольших водных беспозвоночных, то наземные виды могут быть
исследованы лишь после их фиксации. Многие беспозвоночные
животные, прежде всего простейшие, так малы, что изучение их возможно
только с помощью специальной микроскопической техники. С се
помощью также удастся ознакомиться с деталями строения тканей и
органов макрообъектов на препаратах срезов соответствующих структур.
Устройство оптических приборов и основные правила работы
с ними должны быть хорошо усвоены каждым студентом.
Микроскопическая техника
Любой оптический прибор, позволяющий получить увеличенное
изображение объекта, включает систему линз и устройства,
облегчающие пользование ими.
Самым простым оптическим прибором является ручная лупа (рис. I).
Основная ее часть — двояковыпуклая линза, вставленная в оправу с
ручкой. Увеличение, которое даст ручная лупа, обычно обозначено на ручке
соответствующими символами 2\ 5х, 10х.
Микроскоп
В составе микроскопа можно выделить три основных блока:
механический, осветительный и оптический (рис. 2).
Механический блок состоит из подковообразного или
прямоугольного основания (или ножки), колонки, рукоятки, предметного
столика и тубуса.
14
Технические средства изучения микроскопических объектов
Рис. 1. Ручная лупа: /-двояковыпуклая л mm; 2— оправа; J— ручка
Рис. 2. Схема устройства микроскопа: / — основание; 2 — колонка;
3 — михробинт; 4 — макровинт; 5 — окуляр; 6 — тубус; 7 —
револьвер; 8— объективы большого и малого увеличения; 9 —
зажимы; 10 — предметный столик; // — диафрагма; 12— конденсор
Аббс; 13 — зеркало; 14 — вшгт конденсора
Микроскол
15
Основание микроскола придает ему устойчивость. На колонке
закреплены предметный столик, устройства, обеспечивающие
регулирование работы оптического и осветительного блоков, и тубус. Рукоятка
позволяет удобно переносить микроскоп и перемещать его на рабочем
месте.
Предметный столик в микроскопах разных систем либо
неподвижно фиксирован на колонке, либо подвижен и положение его
может регулироваться с помощью двух крайних винтов на задней стороне
статика, обращенной к исследователю. Средний винт закрепляет
верхнюю часть предметного столика, в центре которой расположено
отверстие, обеспечивающее прохождение потока световых лучей к изучаемому
объекту.
На тубусе снизу привинчивается револьвер — подвижная
конструкция с ячейками для сменных объективов, сверху — окуляр.
Осветительный блок включает зеркало, расположенное у
основания микроскопа, и конденсор.
Зеркало имеет плоскую и вогнутую поверхности и укреплено
таким образом, что может вращаться в двух взаимно
перпендикулярных плоскостях. Это позволяет ориентировать зеркало на источник
света, находящийся в любом месте. Поток световых лучей, отражаясь
от зеркала, проходит через отверстие (прорезь) в предметном столике
и направляется на объект и в оптическую систему.
Для лучшей освещенности объекта, концентрации лучей, идущих
от зеркала, между ним и предметным столиком расположен
конденсор Аббе, представляющий собой дополнительную систему линз
и снабженный ирисовой диафрагмой и сменными
светофильтрами (матовым и синим). Все эти устройства позволяют регулировать
освещение поля зрения микроскопа в зависимости от характера
изучаемого объекта, его окраски и плотности. Перемещение конденсора
ниерх и вниз вдоль колонки осуществляется с помощью нижнего (или
переднего, расположенного ближе к зеркалу) винта.
Оптический блок состоит из окуляра и объектива, укрепленных
на концах тубуса.
Окуляр вставляется в верхнюю часть тубуса. Он представляет
собой короткую трубку с вмонтированными в нее линзами. Увеличение
окуляра в рабочем учебном микроскопе чаще всего 10- или
15-кратное (10х. 15х).
Объективы закрепляются в гнездах револьвера и легко сменяются
при микроскопировании поворотом револьвера. Установка нужного
16
Технические средства изучения микроскопических объектов
объектива наа объектом и фиксация его положения контролируется
характерным щелчком. Чаще всего используются два объектива:
объектив малого увеличения (8*)иобъектив большого
увеличения (40х). Первый — тонкий, более короткий, второй — массивный.
Для получения больших увеличений применяют 90-кратный (90х)
объектив.
Кратность увеличения объектов микроскопом при
использовании разных объективов и окуляров приведена в табл. I.
Тэблицз!
7х
10х
15х
Окуляры
Объективы
8х
56
80
120
40х
280
400
600
90х (иммерскя)
630
900
1350
Фокусировка (наведение на резкость) регулируется двумя винтами,
укрепленными на колонке. Изменением расстояние между изучаемым
объектом и объективом достигается наилучшая резкость
изображения. Для быстрого изменения этого расстояния используется
большой винт (макровинт), для более тонкой регулировки — малый,
или микровинт. Макровинт обеспечивает движение колонки с
тубусом и оптикой вдоль зубчатки (кремальеры). Движением винта
от себя (почасовой стрелке) мы опускаем всю систему, движением
на себя (против часовой стрелки) поднимаем ее.
Правила работы с микроскопом
1. Установка микроскопа на рабочем месте является важным
условием успешной работы. Микроскоп следует поставить,
ориентируя его для наблюдения левым глазом, на расстоянии около 3 см
от края стола.
2. Перед работой необходимо протереть все внешние части
микроскопа, не вынимая окуляр.
3. Для обеспечения максимального движения предметного столика
(и препарата) перед началом работы с микроскопом следует центрировать
предметный столик, т.е. движением винтов привести его в положение.
Микроскоп
17
при котором линза конденсора располагается точно посередине
отверстия предметного столика. Центрировать предметный столик нужно
и в процессе работы, и после се окончания.
4. Освещение поля зрения проводится следующим образом. При
изучении зоологических подвижных объектов лучше использовать
вогнутое зеркало, предварительно подняв конденсор вверх до упора.
Наилучшее освещение дает рассеянный дневной свет, но можно
использовать и другие источники света. Следует помнить, что
нежелательно слишком яркое и с бликах!и освещение поля зрения, беспокоящее
живые объекты и опасное для зрения исследователя. Поле зрения
должно быть освещено равномерно. При обнаружении в нем темных зон
следует проверить положение револьвера и частей конденсора. При
изучении прозрачных или бесцветных объектов поле зрения следует
притенить, прикрыв диафрагму или опустив конденсор. При
рассмотрении темных, интенсивно окрашенных объектов, диафрагму
нужно открыть.
5. Фокусировка изображения. Используя постоянный или
изготовив временный препарат, помещают его на предметный столик.
Начинать работу нужно с изучения объекта при малом
увеличении, поэтому, приступая к работе и закончив ее. следует поставить
микроскоп на малое увеличение. Понять необходимость
выполнения этого требования можно, если наблюдать изменения расстояния
между нижним концом объектива и препаратом при повороте
револьвера и смене объектива малого увеличения (10х или 15х) на большое
(40х). Исходное положение объектива малого увеличения -1 см от
нижнего его края до препарата.
Исследуя объект, основную информацию о нем можно получить
при малом его увеличении, при котором оптимальны освещенность
и резкость. Переходя на большое увеличение, мы выигрываем в
размерах объекта, но значительно проигрываем в четкости общей картины.
При малом увеличении фокусировка (наведение на резкость)
осуществляется с помощью макровинта под постоянным контролем глаза,
гс. не отнимая глаза от окуляра. Добившись необходимой резкости
О рубая наводка макровинтом), окончательную доводку фокусировки
выполняют микровинтом. При хорошей грубой наводке движение
микровинта в одну или другую сторону (от себя или к себе) не должно
превышать двух полных оборотов. В противном случае следует снова
использовать макровинт. Это особенно важно при работе с большим
увеличением, когда расстояние от края объектива до препарата очень
18
Технические средства изучения микроскопических объектов
мало. При большом увеличении следует пользоваться только
микровинтом, предварительно проведя грубую наводку при малом
увеличении, а затем поворотом револьвера установив большое увеличение.
Закончив изучение препарата при большом увеличении, микроскоп
следует сразу перевести на малое увеличение.
При работе с объективами разного увеличения нужно помнить
два важных обстоятельства.
Во-первых, микроскоп даст плоскостное изображение объекта.
Поэтому при большом увеличении мы видим четко очень тонкую
плоскость, все, что выше или ниже се, видно неясно, и нужно
постоянно работать микровинтом, чтобы рассмотреть все структуры. При
объективе малого увеличения исследуемая плоскость толще и часто
позволяет отчетливо видеть весь объект.
Вторая особенность работы при разных увеличениях связана
с освещением. Маленькое входное отверстие и свойства линз
объектива большого увеличения пропускают очень узкий пучок световых
лучей, поэтому при переходе с малого на большое увеличение мы
значительно теряем в интенсивности освещения объекта. Необходимо
открыть диафрагму конденсора.
6. Перемещение препарата на предметном столике при малом
увеличении осуществляется вручную. Определенную трудность представляет
то, что оптическая система микроскопа дает обратное изображение. Нужна
определенная сноровка, чтобы усвоить: все, «по мы видим сверху, на
самом деле расположено внизу, то, что справа, — находится слева, и
наоборот.
При переходе на большое увеличение движение препарата
должно быть очень точным и проводится винтами предметного столика.
Переходя с малого на большое увеличение, объект или его часть,
которую нужно изучить, необходимо предварительно движением
препарата разместить в центре поля зрения малого увеличения и лишь
после этого перевести на большое увеличение.
Микроскоп, как всякий точный прибор, требует бережного
обращения. Протирать его, особенно линзы окуляра и объектива,
изготовленные из мягкого, легко повреждаемого стекла, нужно осторожно,
используя мягкие, много раз странные сухие полотняные салфетки.
Нельзя использовать для очистки стекол спирт, т.к. это вызывает
растворение специальных покрытий и помутнение оптики.
Нельзя самостоятельно развинчивать окуляры и объективы. Их
повреждения может устранить только специалист.
Микроскоп бинокулярный стереоскопический (МБС)
19
Микроскоп бинокулярный стереоскопический (МБС)
Для изучения объемных организмов и наблюдения за
движением, питанием и другими формами поведения достаточно крупных
(tic микроскопических) животных, а также для их препарирования
используются бинокулярные стереоскопические микроскопы малого
увеличения (МБС). Они дают прямое изображение, имеют большое
поле зрения, широкий диапазон разрешаюших увеличений (табл. 2).
С помощью МБС можно изучать прозрачные водные объекты в
проходящем световом потоке и непрозрачные, темные — в отраженном
свете. В настоящее время используются модели МБС-9 и МБС-10
(рис. 3).
Тз6л*цз2
Кратность ушелтншя объектов бинокулярным микроскопом
при использовании разных объективов и окуляров
Объективы
о.6х
Iх
2х
4х
7х
Окуляры
3.5
6
12
24
42
6х
4.5
8
16
32
56
Sx
12.5х
7
12,5
25
50
8S
14х
8.1
14.3
2S.6
57.2
100
Оптический блок МБС включает оптическую головку и
окулярную насадку.
В оптическую головку вмонтированы все оптические детали,
включающие объектив микроскопа, выше которого установлен барабан
с галилеевскими системами. Ось барабана заканчивается
расположенными снаружи с двух сторон рукоятками, при вращении которых
происходит переключение увеличений, значения которых нанесены
на рукоятках (7х. 4х, 2х. Iх, 0,6х).
Чтобы установить нужное увеличение, необходимо, вращая
барабан, цифру на рукоятке совместить с точкой на подшипнике.
При этом персфокусировку проводить не нужно. Положение барабана
фиксируется щелчком. Фокусировка объектива оптической головки
производится винтами на направляющей, с помощью которых
поднимают или опускают головку относительно столика микроскопа.
20
Технические средства изучения микроскопических объектов
Рис. 3. Микроскоп бинокулярный стереоскопический: А — общий вил;
Б — схема устройства: / — предметный столик; 2 —кронштейн;
3 — оггтчсская головка; 4 — окулярная насадка; 5— окулярные
трубки; 6 — переключатель кратности увеличения; 7— винт
крамальеры
Изготовление препаратов для микроскопирования
21
О кул я рная насад ка состоит из двух призм, заключенных в
подвижные оправы, на которых укреплены окулярные трубки. Двигая
оправы призм, можно менять расстояние между центрами линз окулярных
тт»<х)К% адаптируя их положение к межзрачковому расстоянию глаз
исследователя.
Предметный столик съемный, устанавливается и
закрепляется винтом на специальном основании. На задней стенке основания
столика имеется гнездо для осветителя. Внутри основания
расположено зеркало с матовой и зеркальной поверхностями и рукояткой для
его вращения при регулировании освещения поля зрения.
Осветительная система, помимо зеркала, включает специальный
ос вс т и тс л ь, состоящий из конденсора и лампы накаливания,
объединенных общим корпусом.
Изучение объекта возможно в отраженном и проходящем свете.
Чтобы использовать осветитель для работы в отраженном свете, его крепят
на шарнирном кронштейне. Для наблюдения в проходящем свете
осветитель переносится в гнездо основания предметного столика.
В зависимости от структурных особенностей объекта (его
плотности, прозрачности, окраски) отверстие предметного столика может
скрываться стеклянной или металлической пластинкой, одна
сторона которой окрашена в белый, другая — в черный цвет.
Изготовление препаратов для микроскопирования
Для изучения под микроскопом используют заранее
изготовленные постоянные препараты (их изготовление требует
определенных навыков и времени) либо по ходу работы готовятся временные
препараты. Материалом для них могут служить целые
микроскопические или достаточно мелкие объекты (тотал ьн ые препараты) или
части их тела.
При работе с микроскопом любой объект помещается на
предметное стекло — стеклянную пластинку стандартного размера
(76 х 26 мм). Очень немногие объекты рассматриваются сухими, чаше
в капле воды или другой жидкости. Чтобы предохранить стекла
объективов от увлажнения, помешенный на предметное стекло в капле
жидкости материал покрывается покровным стеклом. Его обычный
размер 18 х 18 мм. Оно изготавливается из высококачественного
стекла, очень тонкое и хрупкое.
22 Технические средства изучения микроскопических объектов
Рис.4.
Накладывание
покровного стекла
на объект
Покрывать каплю жидкости
с исследуемым материалом нужно
так, чтобы на препарате не
оставались пузырьки воздуха. Для этого,
держа покровное стекло за два
уголка, противоположную его грань
ставят в каплю жидкости и постепенно
опускают стекло (рис. 4).
Оставшиеся пузырьки легко отличить: они
имеют широкий темный ободок,
а поверхность их отливает
зеркальным блеском. Крупные пузырьки
также имеют темное окаймление, а внутренняя их поверхность
напоминает запотевшее стекло.
Покрывая подготовленный временный препарат покровным
стеклом, необходимо учитывать объем изучаемого объекта, в противном
случае он можетбыть деформирован или раздавлен тяжестью стекла.
Во избежание этого на покровное стекло наносят вое ковые ножки.
Обычный чистый пчелиный воск смешивается со скипидаром при
подогреве (и тщательном соблюдении правил противопожарной
безопасности!) в пропорции 2,5:1. Такая масса может длительное время
храниться, лучше в стеклянном бюксе или коробке. Перед
использованием воск слегка разминают пальцами, чтобы он стал
пластичнее. Затем все четыре уголка покровного стекла, слегка царапая ими
по комочку, снабжают восковыми ножками желаемой высоты и
покрывают объект на препарате, ориентируя покровное стекло ножками
вниз.
Подцарство Protozoa — простейшие
Тип Sarcomastigophora — саркомастигофоры
Тип объединяет более 25 000 видов простейших. Знакомству с его
представителями в лабораторном практикуме по зоологии
беспозвоночных уделяется значительное внимание. В частности,
предусматривается ознакомление студентов с простейшими, принадлежащими
к трем наиболее значимым подтипам: саркодовых, жгутиконосцев
и опалинат.
Подтип Sarcodina — саркодовые
Изучение животных традиционно начинается с одноклеточных,
;i среди последних — с саркодовых. Наиболее удобным объектом
являемся группа пресноводных амеб. Они малоподвижны, имеют доста-
10ЧНО простое строение и легко культивируются в лабораторных
условиях. Все это облегчает их изучение.
Представители подтипа в большинстве своем характеризуются
амебоидным движением. Обитают в водной среде, почве, имеются па-
р.оитическис формы.
Надкласс Rhizopoda — корненожки
Объединяет организмы с псевдоподиями, не имеющими высоко-
дифференцированных внутренних скелетных образований.
Класс Lobosea
Амебоидные формы с лопастсвидными псевдоподиями (лобопо-
лиями), для которых не характерно слияние в сете видны с структуры.
Отряд АтоеЫпа — голые амебы
Материал. За 4-5 недель до начала работы с простейшими
подготавливаются культуры для получения нужного материала. Водой из
небольших стоячих или слабопроточных водоемов с гниющими
24
тип sarcomasagopnor^
растениями и илом заполняют сосуды объемом 0,5-1 л. Для отбора проб
можно использовать планктонный сачок, которым проводят у дна,
слегка взмучивая ил. Хороший материал может быть получен в вон
доемах с обильным развитием ряски и элодеи. После оседания ила
в пробах воды из придонного слоя можно отыскать крупных Amoeba
proteus. Для разведения амеб часто используется сенной настой. Для
этого луговое сено (2 г на 100 мл не кипяченой воды) кипятят 10-15 минут
в колбе и ставят в теплое место на 2-3 дня. На поверхности жидкости
сенная палочка образует бактериальную пленку, бактерии служат
пишейдляамеби инфузорий. Сенной настой подливают в банки
с культурами простейших.
Хорошие результаты дает разведение простейших в чашках Петри
на зернах риса (5-6 на чашку), залитых дождевой или
отфильтрованной водой из естественных водоемов (в крайнем случае,
отстоявшейся водопроводной). На появляющихся вокруг зерен риса
скоплениях бактерий, имеющих вид беловатого облачка, успешно
размножаются амебы. Пересев культуры следует производить
через 1,5-2 месяца.
Отбирать амеб следует из поверхностной пленки либо из
придонного слоя. Крупных амеб можно найти на нижней поверхности листьев
растений. Для наблюдения за живыми амебами лучше использовать
покровные стекла с восковыми ножками, которые препятствуют
гибели животных ввиду сжатия стекол при подсыхании препарата. В
последнем случае следует у края покровного стекла добавить каплю воды
из культурального сосуда.
Изучение живого объекта. Начинать рассмотрение амеб следует
при малом увеличении, предварительно притенив диафрагмой поле
зрения микроскопа.
Наиболее подходящим видом и одновременно удобным
объектом для изучения является амеба-протей (Amoebaproteus) (рис. 5). Для
нее характерны многочисленные длинные лопастевидные
псевдоподии, или ложноножки. Псевдоподин такого типа называются
л об о п од и я м и. Часто они составляют значительную часть тела
амебы. Псевдоподии непрерывно меняют свои размеры и форму, могут
исчезать или вытягиваться в направлении движения животного.
Таким образом, форма тела амебы непостоянна.
В цитоплазме амебы четко выделяется светлый тонкий наружный
однородный (гомогенный)слой —эктоплазма (лучше заметна при
медленном вращении на пару оборотов вперед-назад микровинта
Класс Lobosea
25
микроскопа). Она же образует тончайшую покровную мембрану —
плазмалемму. Клетка, то есть тело амебы не имеет еще какой-либо
оболочки, отсюда и название отряда — голые амебы. Внутренний
гетерогенный зернистый слой цитоплазмы — эндоплазма — содержит
многочисленные включения. Внутренняя часть цитоплазмы
называется илазмазол ь, периферическая — плазмагель. При
рассмотрении клетки при большом увеличении хорошо заметны токи цитоплазмы
в ее центральной части, где она менее вязкая, нежели в
периферической. Постоянные токи цитоплазмы обеспечивают образование и
изменение формы псевдоподии. При направленном движении амебы легко
наблюдать перетекание цитоплазмы и переход плазмозоля в плазмо-
гсль и обратно. Такой тип движения этих простейших носит название
амебоидного. Таким образом, псевдоподии амеб являются не посто-
ямнымн органеллами движения. Их число и форма
различаются у разных видов амеб и являются важным систематическим
признаком (рис. 5).
Питание амеб. Пищей амебам служат одноклеточные водоросли,
бактерии, частицы органики. Для изучения питания амеб при изго-
ювлении временного препарата следует перенести на предметное
стекло немного поверхностной пленки или небольшое количество
осадка со дна культуралыюго сосуда, где должно быть достаточно орга-
ники и бактерий. На препарате можно наблюдать, как амеба обтекает
пищевую частицу псевдоподиями и включает пузырек с нею в
цитоплазму. Окруженная мембраной частица представляют собой
пищевую вакуоль. В нес из цитоплазмы поступают ферменты, и вакуоль
становится пищеварительной. По завершении процесса
пищеварения и всасывания питательных веществ непереваренные остатки из
вакуоли, называемой на этой стадиидефекационной, выводятся
наружу. Как пищевые, так и дефскациониые вакуоли образуются и
опорожняются в любом месте клетки. Поэтому они рассматриваются как
непостоянные пищеварительные органеллы.
Наблюдая за амебой последовательно при малом и большом
увеличении микроскопа, можно обнаружить сократительную или
пульса и ру ющу ю вакуоль. Она имеет вид небольшого прозрачного пузырька,
который то медленно увеличивается, то резко уменьшается. При его
сокращении через временную пору в эктоплазме наружу
выбрасывается избыточная вода с растворенными в ней углекислотой и
продуктами углеводного и азотного обмена. Удаление воды обеспечивает
26
Тип SarcomastiQophora
б ^
Рис. 5. Голые амебы: А — амеба-протсй (Amoeba prottus) (по Дофлейну):
/ — псевдоподии; 2— плазмалемма; J— эктоплазма; 4 —
эндоплазма; 5— ядро; 6 — пищеварительная вакуоль; 7—
сократительная вакуоль; Б — разнообразие внешнего облика
представителей группы (по Дофлейну): / — Amoeba limax, 2— Pelomyxa
binuclcata\ 3 — Amoeba proteus;«/—A. rudiosa; 5—A. \errucasai 6 —
A. polypodia
осморегул я ци ю. поддержание в клетке на необходимом уровне
концентрации растворенных низкомолекулярных соединений, главным
образом минеральных солей. Это главная функция сократительной
вакуоли у пресноводных простейших. Вместе с водой из клетки
Класс Lobosea
27
удаляются также растворенный углекислый газ (функция
газообмена) и продукты диссимиляции (выделительная функция).
Ядро у живых амеб видно редко, так как оптическая плотность
цитоплазмы и ядерного вещества (кариоплазмы) почти одинаковы.
Лучше всего ядро различимо у Amoeba Umax. Оно выглядит как светлое
пятно, почаще маскируется включениями зернистой цитоплазмы.
Поэтому ядро следует рассматривать на окрашенном тотальном
препарате, обратив внимание на его центральное положение в клетке.
При длительном наблюдении или в период активного массового
размножения можно наблюдать процесс бесполого размножения амеб
делением (рис. 6).
размножении: / — ядро; 2 — псевдоподии;.?— эктоплазма;
4 — эндоплазма; 5 — сократительная вакуоль. 6 — митоти-
ческое деление ядра
Задание
!. Найти и рассмотреть амеб в пробах культуральной жидкости.
2. Понаблюдать за движением, образованием и изменением формы
псевдоподии.
3. Дифференцировать слои цитоплазмы.
4. Понаблюдать за процессом питания, рассмотреть
пищеварительные вакуоли.
28
Тип Sarcomastigophora
5. Найти сократительную вакуоль и последить за ее работой.
6. Рассмотреть на тотальном препарате ядро, его форму и
положение в клетке.
7. Зарисовать несколько последовательных позиций амебы при
движении, проиллюстрировав изменчивость формы тела и псевдоподии.
S. Зарисовать строение амебы.
Отряд Arcellinida (=Testacea) — пресноводные раковинные
корненожки
Материал. В культурах простейших в капле воды из
придонного слоя с небольшим количеством ила часто удастся найти
пресноводных раковинных корненожек. В илистом осадке из водоемов
с обильным развитием ряски множество корненожек из рода Arcella.
В пробах из водоемов с песчаным, слегка заиленным грунтом в
материале обычны корненожки рода Difflugia. В лабораторных культурах,
поставленных в конце лета — сентябре, можно наблюдать массовое
развитие разных видов тестацид.
Хороший материал для занятий можно получить, собрав на
верховом болоте сфагновый мох. Его высушивают и хранят в бумажном
пакете для предохранения от образования плесени. За две недели до
начала занятий мох помешают в кристаллизатор с отстойной, лучше
с дождевой водой. При комнатной температуре через 10-14 дней из
цист выходят корненожки.
Изучение живого объекта. Наиболее полное представление о
данном объекте можно получить, изучая живых корненожек под
микроскопом, используя покровные стекла с восковыми ножками.
Раковина арцеллы (Arcella sp.) дисковидная, выпуклая сверху
и вогнутая с нижней стороны, в центре которой находится круглое
устье (рис. 7). Если раковина расположена устьем кверху, оно видно
четко; при изучении раковины с противоположной стороны —
различимо слабо. Раковина образована из псевдохитина, имеет
определенную структуру и окрашена окислами железа в коричневый цвет. У
молодых арцелл раковины светлой окраски, поэтому у них достаточно
хорошо просматривается цитоплазма с включениями. Также видно, что
цитоплазма не заполняет всего пространства внутри раковины и
связана с ее стенками небольшими выростами — эпиподиями.
Наблюдая за живыми арцеллами, можно легко заметить
образование немногочисленных прозрачных лопастевидных псевдоподии
с закругленными концами.
Класс Lobosea
29
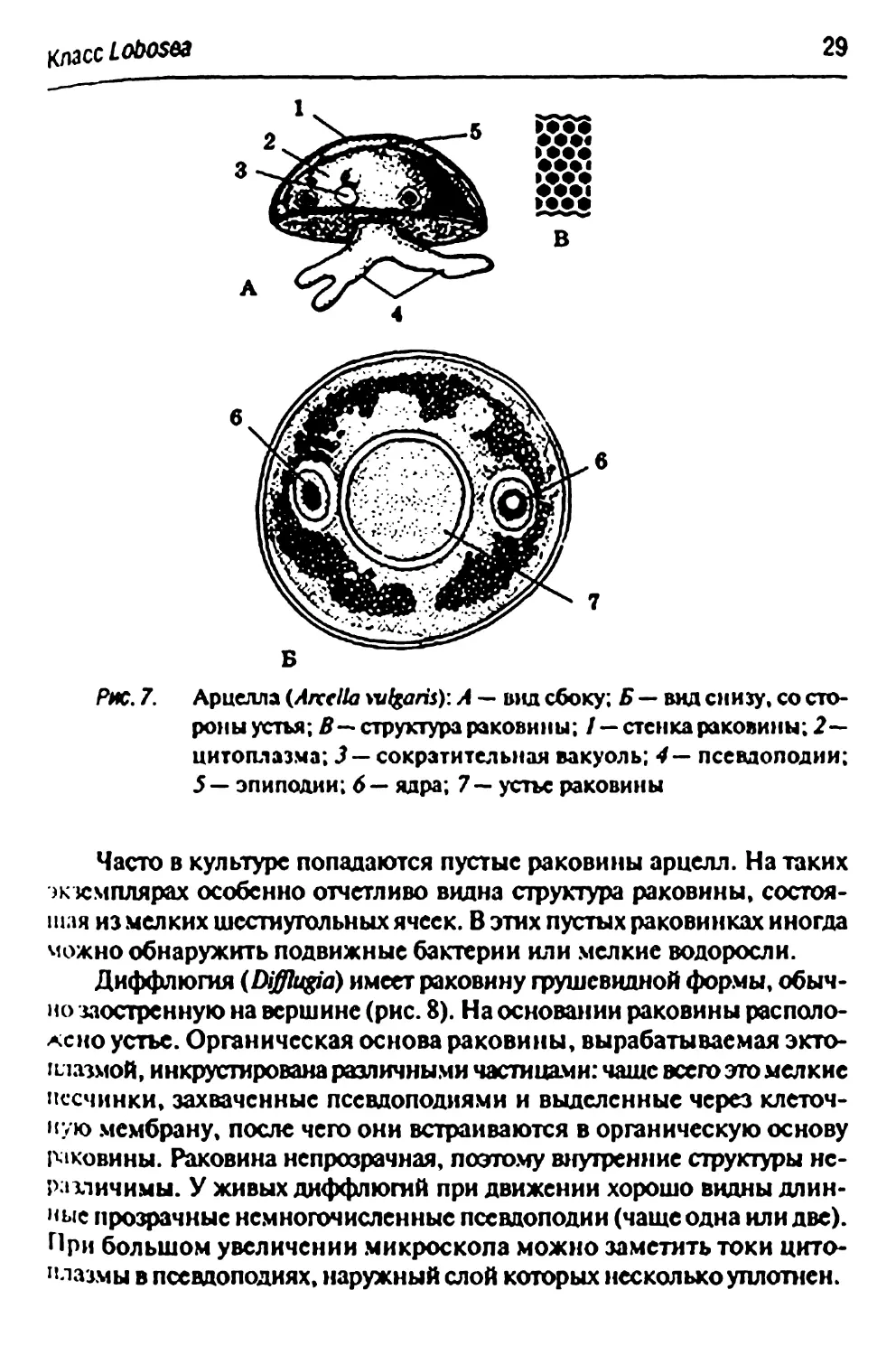
Рис. 7. Арцелла (Arcclta wfgaris): А — вид сбоку; Б — вид снизу, со
стороны устья; £— структура раковины; / — стенка раковины; 2—
цитоплазма; J — сократительная вакуоль; 4— псевдоподии;
5— эпиподии; 6— ядра; 7— устье раковины
Часто в культуре попадаются пустые раковины арцелл. На таких
ж кмплярах особенно отчетливо видна структура раковины, состоя-
шля из мелких шестиугольных ячеек. В этих пустых раковинках иногда
можно обнаружить подвижные бактерии или мелкие водоросли.
Диффлюгия (Difflugia) имеет раковину грушевидной формы,
обычно заостренную на вершине (рис. 8). На основании раковины
расположено устье. Органическая основа раковины, вырабатываемая
эктоплазмой, инкрустирована различными частицами: чаше всего это мелкие
песчинки, захваченные псевдоподиями и выделенные через
клеточную мембрану, после чего они встраиваются в органическую основу
раковины. Раковина непрозрачная, поэтому внутренние структуры
неразличимы. У живых диффлюшй при движении хорошо видны
длинные прозрачные немногочисленные псевдоподии (чаще одна или две).
При большом увеличении микроскола можно заметить токи
цитоплазмы в псешоподиях, наружный слой которых несколько уплотнен.
30
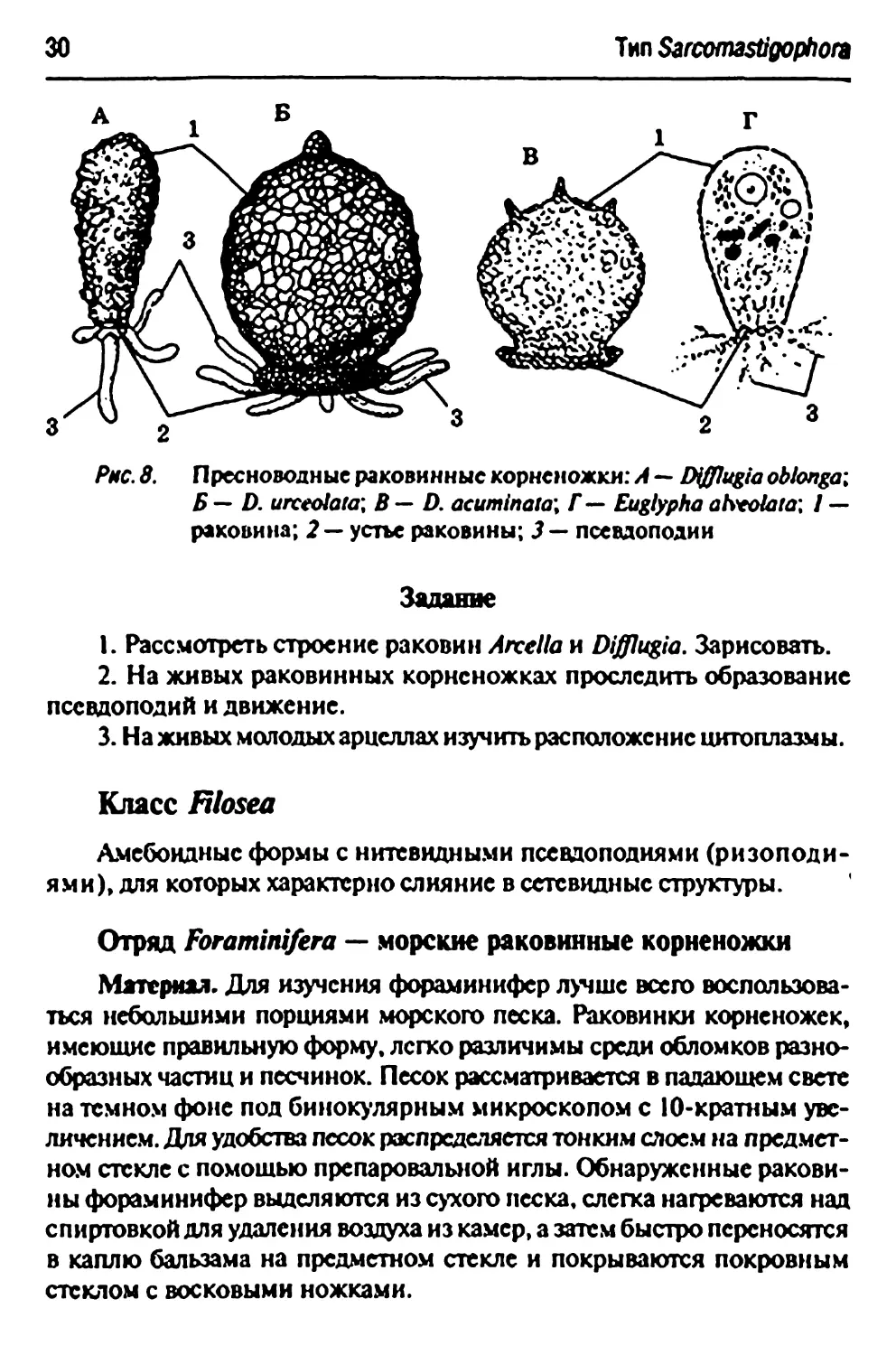
Тип SarcomastiQophora
Рас. 8. Пресноводные раковинные корненожки: А — EHfflugia obionga;
5 — D. urcedata\ В— D. acuminata'. f— Eugiyphaatvtolata\ 1 —
раковина; 2 — устье раковины; J — псевдоподии
Задание
1. Рассмотреть строение раковин Arcella и Difflugia. Зарисовать.
2. На живых раковинных корненожках проследить образование
псевдоподии и движение.
3. На живых молодых арцеллах изучить расположение цитоплазмы.
Класс filosea
Амебоидные формы с нитевидными псевдоподиями (ризоподи-
ями), для которых характерно слияние в сстсвидныс структуры.
Отряд Foraminifera — морские раковинные корненожки
Материал. Для изучения фораминифер лучше всего
воспользоваться небольшими порциями морского песка. Раковинки корненожек,
имеющие правильную форму, легко различимы среди обломков
разнообразных частиц и песчинок. Песок рассматривается в падающем свете
на темном фоне под бинокулярным микроскопом с 10-кратным
увеличением. Для удобства песок распределяется тонким слоем на
предметном стекле с помощью препаровальной иглы. Обнаруженные
раковины фораминифер выделяются из сухого песка, слегка нагреваются над
спиртовкой для удаления воздуха из камер, а затем быстро переносятся
в каплю бальзама на предметном стекле и покрываются покровным
стеклом с восковыми ножками.
Класс Filosea
31
При отсутствии морского песка изучение морских раковинных
корненожек проводится по тотальным постоянным препаратам.
Изучение раковин фораминифер. Морские раковинные
корненожки имеют разнообразные построению и форме раковины.
Однокамерные раковины встречаются редко и очень малы. Чаше можно
обнаружить многокамерные раковины. Некоторые из них инкрустированы
песчинками, большинство — пропитаны известью. Камеры у них
располагаются либо последовательно в один или два ряда, либо
закручены спирально, словно раковина моллюсков (рис. 9). Расположение
клмер, как и их форма, — основной систематический признак
фораминифер.
При изучении тотального препарата следует обратить внимание
па первую, самую маленькую, зародышевую, камеру; перегородки
между камерами, исчерченные ралиально; и поры, пронизывающие
сгенки камер и раковины. Устье раковины расположено в самой
крупной крайней камере.
Рис. 9. Морские раковинные корненожки. Раковины фораминифер:
А — Elphidium sp.: / — раковина; 2 — перегородки камер; 3 —
поры; 4— зародышевая камера; Б— Rotaliasp.\ В— Globigenna
sp.\ Г— /Ндеог&дер.'.в —видсвершины, б — виде основания;
Л— Tcxtuhhasp.
32
Тип Sarcomastigophof^
В песке средиземноморских пляжей наиболее обычны
представители родов Rotalia, Elphidium, Globigerina, Discorbis, Texiularia
(см. рис. 9).
Задание
Рассмотреть препарат фораминифер. Зарисовать строение
раковины и расположение зародышевой камеры и устья.
Надкласс Actinopoda
Класс Radiolaria — лучевики
Материал. В фиксированном формалином морском
планктоне в осадке на дне склянки можно обнаружить скелет радиолярий.
Из-за повреждений при отборе проб и не всегда бережно, как правило»
сохраняются только фрагменты центральных капсул.
В случае отсутствия морского планктона можно использовать
постоянные тотальные препараты радиолярий в бальзаме.
Изучение скелета радиолярий. На препаратах различимы
фрагменты центральных капсул и аксоподий, расположенных более или менее
строго радиально. Тонкие анастомозирующие псевдоподии,
обеспечивающие улавливание пишевых частиц и не имеющие скелетной
основы, не сохраняются и на препаратах отсутствуют. Скелет радиолярий
минеральный, состоит из окиси кремния или сернокислого стронция.
Он весьма разнообразен по форме и служит хорошим
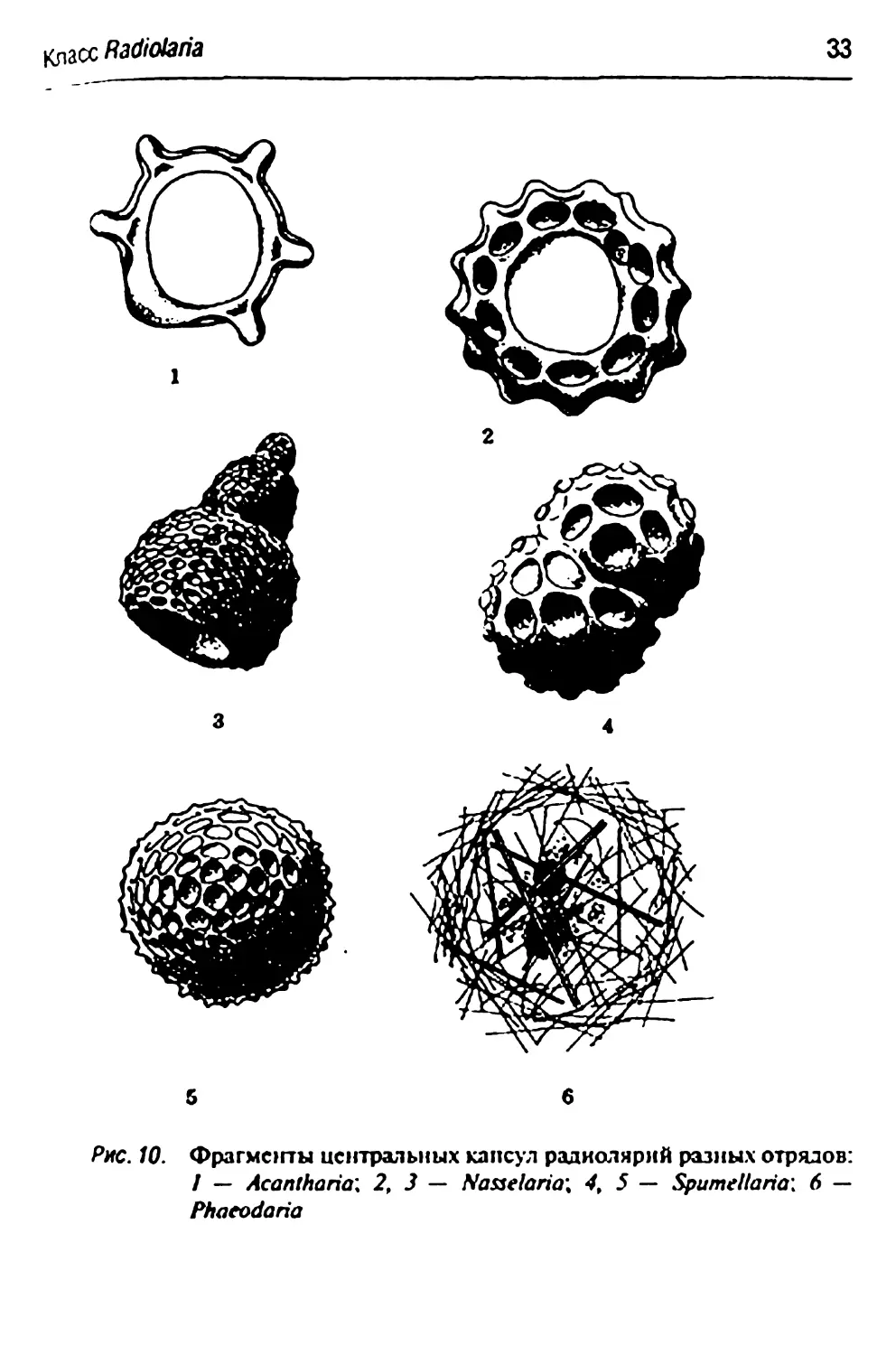
диагностическим систематическим признаком (рис. 10).
Скелет радиолярий из отряда Nasselaria также весьма разнообразен.
Основой его является чстырсхоссвая спикула, к которой
присоединяются скелетные элементы, образуя причудливые формы, напоминающие
шапочки, шлемы, шары и т.п. (рис. 10, 2,3).
Радиолярии из отряда Acantharia лишены центральной скелетной
капсулы, а иглы их образуют пять поясов по четыре иглы в каждом
(рис. 10, /). Для радиолярий из отряда Spumellaria характерны чаще
всего шаровидные капсулы, нередко вложенные друг в друга и
связанные между собой радиальными иглами. Многослойность
шаровидных капсул — результат роста радиолярий, при котором новые
скелетные капсулы большего размера образуются снаружи от
предыдущих (рис. 10, 4,5).
Класс Radiolaria
33
Рис. 10. Фрагмогты иснтральных капсул радиолярий разных отрялов:
/ — Acantharia: 2, 3 — Naaelaria\ 49 5 — Spumellaria\ 6 —
Phaeodaria
34 Тип Sarcomastigophori
Отличительной особенностью представителей отряда Phaeodartd
является наличие в капсуле одного—трех крупных отверстий вместе
многочисленных пор (рис. 10, б).
Задание
Используя тотальные постоянные препараты, рассмотреть и зари«|
совать фрагменты центральных капсул и аксоподий различных радио-]
лярий.
Класс Heliozoa — солнечники
Материал. Солнечники довольно часто попадаются в воде моч
ховых болот, а также во временных весенних водоемах и в прудах
с обильным развитием ряски в прибрежной части. В лабораторной!
культуре они сохраняются достаточно долго, иногда до следующего
сезона.
Изучение материма. Изучение живых солнечников следует
проводить л и бон а препарате с восковыми ножками, либо используя прс-!
д четные стекла с лунками. Самым удобным объектом йвляется Actil
nosphaerium eichhonti. Его величина достигает I мм, он хорошо виден
невооруженным глазом в виде белой точки. !
При изучении актиносфериума следует обратить внимание на два
четко различающихся слоя цитоплазмы: сердцевидный и корковыц
(рис. 11). В более светлом наружном (корковом) слое при длительном
наблюдении можно рассмотреть многочисленные сократительные ва^
куоли. В более темном внутреннем (сердцевидном) слое преобладаю!
пищеварительные вакуоли. Если подобные вакуоли есть и в
корковом слое, то они относятся либо к пищевым* либо к дефскационным.
При большом увеличении следует изучить псевдоподии
солнечника. В них легко обнаружить осевую нить и облекающую се текучую
цитоплазму. Такие псевдоподии носят название аксоподий.
Задание
Рассмотреть строение солнечника. Понаблюдать за движением.
mccHeJiozaa
35
Рис. 11. Акпшосфериум (Actmosphaenum cichhorni): I — корковый слой
цитоплазмы; 2 — сердцевидный слой цитоплазмы; J —
сократительные вакуоли; 4— пищеварительные вакуоли; 5— аксо-
подии
Подтип Mastigophora — жгутиконосцы
Жгутиконосцы объединяют простейших, обладающих жгутиками
как органсллами движения. Форма тела их относительно постоянная
олагодаря наличию клеточной оболочки — пелликулы. Разнообразие
типов питания (авготрофного, гетеротрофного, миксотрофного)
позволяет рассматривать жгутиконосцев как группу организмов,
пограничную между растительным и животным царствами. Изучение
растительных (эвглена зеленая, цератиум) и животных (трипаноэома, опалина)
жгутиконосцев позволяет выявить важнейшие особенности их
строения, связанные с различиями в условиях обитания и питания.
36
Тип Sarcomastigophora
Класс Phytomastigina — растительные жгутиконосцы
Отряд Euglenoidea — эвгленовые
Материал. Разнообразный материал по жгутиконосцам легко найти
весной, летом и даже ранней осенью в лужах, придорожных канавах,
небольших прулах и болотах. При массовом развитии растительных
жгутиконосцев на поверхности водоемов появляется зеленая пленка.
Такое «цветение» легко проследить по весенним лужам,
образующимся после таяния снега. Иногда фитомастигины могут развиваться
в культурах инфузорий с сенным настоем. Но лучше всего для
культивирования эвгленовых использовать специальные среды.
Ниже приводится состав такой среды. Для се подготовки
используется вытяжка из почвенной пробы (10 мл на I л дистиллированной
воды). На этот объем приготавливается раствор Дюз и, включающий:
MgS04 - 0,2 г, FcCI6 - 1 капля 1%-го раствора. КН2Р04 - 0,2 г,
пентол — 2 г, KCI — 0,2 г, вола дистиллированная — 1000 xtn.
Хорошие результаты дает среда Кнопа, в состав которой входят:
MgS04 - 0,25 г, KCI - 0,12 г, Ca(NOj)2 - 1,0 г. FcCI, - следы
(1 капля 1%-го раствора), КН2Р04 — 0,25 г, дистиллированная вода —
1000 г.
Изучение живого объекта. Эвглена зеленая имеет верстеновид-
иое тело, заостренное на конце. Длина клетки достигает 50-60 мкм.
Форма тела эвглены может изменяться благодаря эластичности
покровного слоя цитоплазмы, образующего оболочку — пелликулу. Эту
способность к быстрому изменению формы — мстаболированию
(рис. 12) — легко наблюдать под микроскопом на слегка подсыхающем
препарате. Эвглену замедляют или прекращают движение и округ-,
ляются, как перед инцистированием. j
На переднем конце тела у эвглены располагается один жгутик!
(рис. 13), основание которого погружено в резервуар, хорошо заметный
при затемнении поля зрения микроскопа. Более точное представление]
о расположении и размерах жгутика можно получить, если к капле куль-]
туры эвглены добавить немного смеси йодной тинктуры с фор мал и-]
ном (цвет смеси светло-коричневый). |
Наблюдение за живыми эвгленами позволяет изучить их посту-]
нательное винтообразное движение. Жгутиконосцы как бы ввинчи-1
вакггея в окружающую жидкую среду. Движение эвглен связано со cno-j
собностью реагировать на изменение освещения в среде их обитания.]
yaccPhytofmstigins
37
/l
F«C
Рис. 12. Изменения формы тела (мстаболирование) эвглены зеленой
(Eiftlena viridis)
Р»С. 13. Эвглена зеленая (Ещкпа viridis):
/ — жгутик; 2— резервуар
сократительной вакуоли;}—
сократительная вакуоль; 4— приводящие
каналы; 5 — хроматофоры; 6 —
ядро; 7 — парамиловые зерна;
8 — пелликула; 9 — эктоплазма;
/0— эндоплазма; // — глазок
Тип Sarcomastigophon
Эвглена зеленая отрицательно реагирует как на сильное затемнение,
так и на резкий свет. Для восприятия светового раздражения служит
светочувствительный глазок, или стигма, расположенная у
основания жгутика, и представляющая собой скопление мелкозернистого
красного пигмента.
В цитоплазме эвглены зеленой имеются овальные или слегка
удлиненные хроматофоры, содержащие зеленый пигмент — хлорофилл.
Благодаря наличию хроматофоров с хлорофиллом эвглена способна
к фотосинтезу. Такой способ питания растительных жгутиконосцев
называется автотрофным или голофитным.
Помимо хроматофоров, в цитоплазме эвглены всегда
присутствуют зерна парам ила — углевода, близкого к крахмалу. Это —
запасные вещества.
Если культуру эвглены зеленой поместить в темную камеру, в
раствор, содержащий органические частицы, эвглена утрачивает
хлорофилл, становится бесцветной и переходит к питанию
гетеротрофному (голозойному, анимальному), усваивая готовые
органические вещества из окружающей среды. Если эвглена осмотически
поглощает растворенную органику, ее способ питания называется
сапрофитным. Сочетание автотрофного питания на свету и
гетеротрофного в темноте и в среде, богатой органикой, получило
название смешанного или миксотрофного.
Около основания жгутика при большом увеличении микроскопа
хорошо различим резервуар сократительной вакуоли. Сама
сократительная вакуоль в виде прозрачного пузырька расположена
рядом с основанием жгутика и стигмой и окружена мелкими
собирательными, или приводящими, канальцами. Сначала вода
и продукты метаболизма собираются в приводящие канальцы, затем
поступают в сократительную вакуоль, а из нее в резервуар жгутика
и через выводной канал и пору выводятся наружу.
Задание
1. Понаблюдать за характером движения эвглен.
2. Рассмотреть и зарисовать строение эвглены.
Отрад DinoflogeUota (-Peridlnea) — панцирные жгутиконосцы
Мжтернал. Летом в больших озерах при помощи планктонной сетки
можно выловить массу панцирных жгутиконосцев Ceratium hirundinella.
^ccPhytomastigina 39
Р1рИ отсутствии в осенне-зимнее время живого материала можно
}(Спользовать для изучения летнюю пробу планктона,
фиксированию 10% формалином.
Изучение Ceratium hirudinella. Цератиум имеет панцирь из клст-
ч;1тки, состоящий из определенным образом расположенных
пластинок, соединенных швами (рис. 14). Скульптура панциря хорошо
видна даже при малом увеличении микроскопа. Асимметричный
и л ширь разделен почти полной бороздкой на две части, верхняя из
Коюрых имеет один вырост (апикальный), направленный вперед,
а нижняя — три (антиапикальных) выроста, направленных на-
л>1 В фиксированном материале сквозь прозрачные пластинки
[ыпциря иногда просвечивает более темная цитоплазма с ядром.
Жгутики в таком материале обычно не видны. В зависимости от периода
инпия пробы планктона длина выростов панциря Ceratium различна.
Панцирь особей летней популяции имеет длинные выросты,
облегчающие парение этих планктонных жгутиконосцев в теплой и менее
плотной воде. В осенних пробах преобладают особи Ceratium с
короткими выростами панциря. Кроме того, изменение формы тела
жгутиконосцев, образование длинных выростов, препятствует выеданию их
хищниками в летний период, наиболее благоприятный для
максимальною размножения. Это явление закономерного циклического
изменения формы тела животных в разные сезоны гола называется сезон -
ммм цикломорфозом.
Рис. 14. Цератиум (Ceratium hirudinella):
I — апикальный шип; 2—
антиапикальные шипы; 3 —
жгутиковая бороздка; 4 — пластинки
панциря
40
Тип Sarcomastigophora
Отряд Vobocida (—Phytomonadina) — волъвоксовые
Материал. Летом в небольших прудах, старицах и заводях рек удастся
найти колониальных жгутиконосцев рода Vohox. Чаще всего в пресных
водоемах встречаются два вида: Vofrox aureus и Voivoxgbbator. Первый из
них образует раздельнополые колонии диаметром 500—850 мкм, число
особей в которых достигает 500—1000. Шаровидные обоеполые
колонии V. globator круп нее — до 2 мм, с числом клеток в них до 10—20 тыс.
Собранный планктонной сеткой материал фиксируется 10%
раствором формалина. Свежий материал имеет зеленоватую окраску, но
при длительном хранении постепенно ее утрачивает.
Изучение колонии вольвокса. При изготовлении препарата
следует покрыть его покровным стеклом с восковыми ножками. Это
предотвратит деформацию и возможное раздавливание шаровидной
колонии при подсыхании препарата. Удобны для этого и стекла
с лунками.
Основную массу колонии составляет студенистое вещество —
продукт ослизнения клеточных оболочек (рис. 15). Наружный слой его
плотный, внутренний — полужидкий. Все особи колонии
располагаются по ее периферии жгутиками наружу. Хотя колония шаровидная]
в ней есть выраженная полярность. На живой колонии особи одного из
полюсов имеют более крупные стигмы — это передний полюс. Он ори-j
ентирован в направлении движения, которое обеспечивается
совместной работой жгутиков всех клеток колонии.
В колонии представлены три типа клеток. Огромное большинстве!
их способно лишь к движению и фотосинтезу. Это соматические
клетки, образующие «тело» колонии. Они соединены цитоплазмагт
ческими мостиками, хорошо различимыми при большом увеличении]
Некоторые клетки (4-10), расположенные на заднем полюсе кн
лонии, способны к активному бесполому размножению и образую!
в теле материнской колонии дочерние колонии. Эти клетки называв
ются вегетативными клетками размножения. После заверь
шения развития дочерние колонии разрывают материнскую и выходя!
в воду. Такое размножение возможно летом при активной инсоляциЦ
и благоприятном температурном режиме в водоеме. В это время нсрс-j
дко в развивающихся дочерних колониях начинается формирование кЫ
лоний следующего, «внучатого», поколения. В конце лета в планк-j
тонных пробах чаше попадаются колонии, в которых развивают^
Kmcc Phytomastigina
41
Pm. 15. Волыюкс (Volvox&obotor)'. Л — колония с вегетативными
клетками размножения; Б — общий вид летней колонии с шестью
дочерними колониями; В — участок колонии с генеративными
клетками; / — соматические клетки; 2— вегетативные клетки
размножения в разных стадиях развития молодых колоний; 3—
макропшеты; 4 — микрогаметы; 5 — цитоплазматические
мостики
i оперативные клетки, дающиежснские(макрогаметы)имуж-
кне (ми кро га меты) половые клетки. Генеративные клетки хорошо
отличимы по плотной внутренней части, окруженной светлым
ободком. Располагаются они на заднем полюсе колонии.
42
Тип Ssrcomastlgophora
Класс Zoomostigina — животные жгутиконосцы
Отряд Kinetoplastida
Материал. Традиционно с представителями данного отряда
студенты знакомятся на примере жгутиконосца Trypanosoma lewisi—
паразита крови крыс. При отсутствии живого материала изучение трипа-
нозом проводится на микроскопических препаратах мазка крови.
Вполне могут использоваться препараты (специально фиксированные
мазки крови) других трипанозом, включая виды, патогенные для
человека. На препарате, кроме жгутиконосцев, видны форменные
элементы крови: эритроциты и лейкоциты.
Изучение препарата. Трипанозомы паразитируют в плазме крови
человека и многих позвоночных животных. Размеры их очень малы
(20-25 мкм), поэтому изучать препарат следует в сочетании
объектива 90х и окуляров — 10х, 15х, т.е. при увеличении в 900 или 1350 раз,
с масляной иммерсией. Использование иммерсионного масла
уменьшает рассеивание светового потока в пространстве между препаратом
и объективом 90-кратиого увеличения и улучшает освещенность объекта.
Техника работы с иммерсией следующая: на препарат наносится
капля иммерсионного масла и пол визуальным контролем объектив
опускается до соприкосновения с ней. Между объективом и
препаратом образуется столбик масла, уменьшающий рассеивание
световых лучей. Медленным поворотом макровинта (от себя) достигается
фокусировка изображения.
Тело трипанозомы удлиненное, веретеновиднос (рис. 16). На
заднем конце от базального ядра отходит жгутик, проходящий вдоль
клетки до ее передней части, где расположен его свободный рулевой
конец. Часть жгутика, идущая шюль клетки, соединена с ней тонкой
цитоплазматической волнообразной перепонкой — у и дул ирующей
мембраной. Рядом с базальным зерном (кинетосомой)уоснования
жгутика расположено сильно окрашенное крупное тельце — кине-
т о п л а с т, обеспечивающий генерацию энергии для движения жгутика*
Цитоплазма, окрашенная в синий или бледно-фиолетовый цвет,
имеет мелкую зернистость, но лишена вакуолей. Питание и
выделение осуществляются диффузно. Ядро, занимающее почти центральное
положение в клетке, имеет овальную форму и на препарате окрашено
в красный цвет. Такую же окраску имеет краевая нить жгутика на унду-
лирующей мембране и кинстопласт.
Ютзсс Zoomastigina
43
Рис. 16. Трипаноэомы (Trypanosoma sp.) в мазке крови: А —трипаноэо-
ма среди эритроцитов (общий вид); В — строение: / — жгутик;
2— ундулирующая мембрана; J— кинетопласг, 4— кинетосо-
ма; 5—ядро
Подтип Opalinata
Представители подтипа используют для передвижения
многочисленные жгутики, организованные в параллельно идущие их ряды.
Отряд Opalinida — опалины
Материал. Живых опалин можно получить в любое время года,
если есть виварий, где содержатся лягушки. Опалины
обнаруживаются практически в каждой травяной лягушке (Rana temporaria). Извлечь
их можно при вскрытии брюшной полости лягушки, после чего
выделяются задняя кишка и клоака и помешаются в часовое стекло или
оюкс с физиологическим раствором, в котором опалины долго
остаются живыми и активными.
Для извлечения опалин из кишечника живой лягушки
стеклянную пипетку с отшлифованным концом (глазную), вводят в клоаку
и отсосанную слизь с содержащимися в ней опалинами переносят
в физиологический раствор.
При отсутствии живого материала используют постоянные
препараты, изготовленные в виде мазка из содержимого задней кишки
лягушки, фиксированного жидкостью Шаудина и окрашенного
гематоксилином.
44
Тип Sarcomastigophon
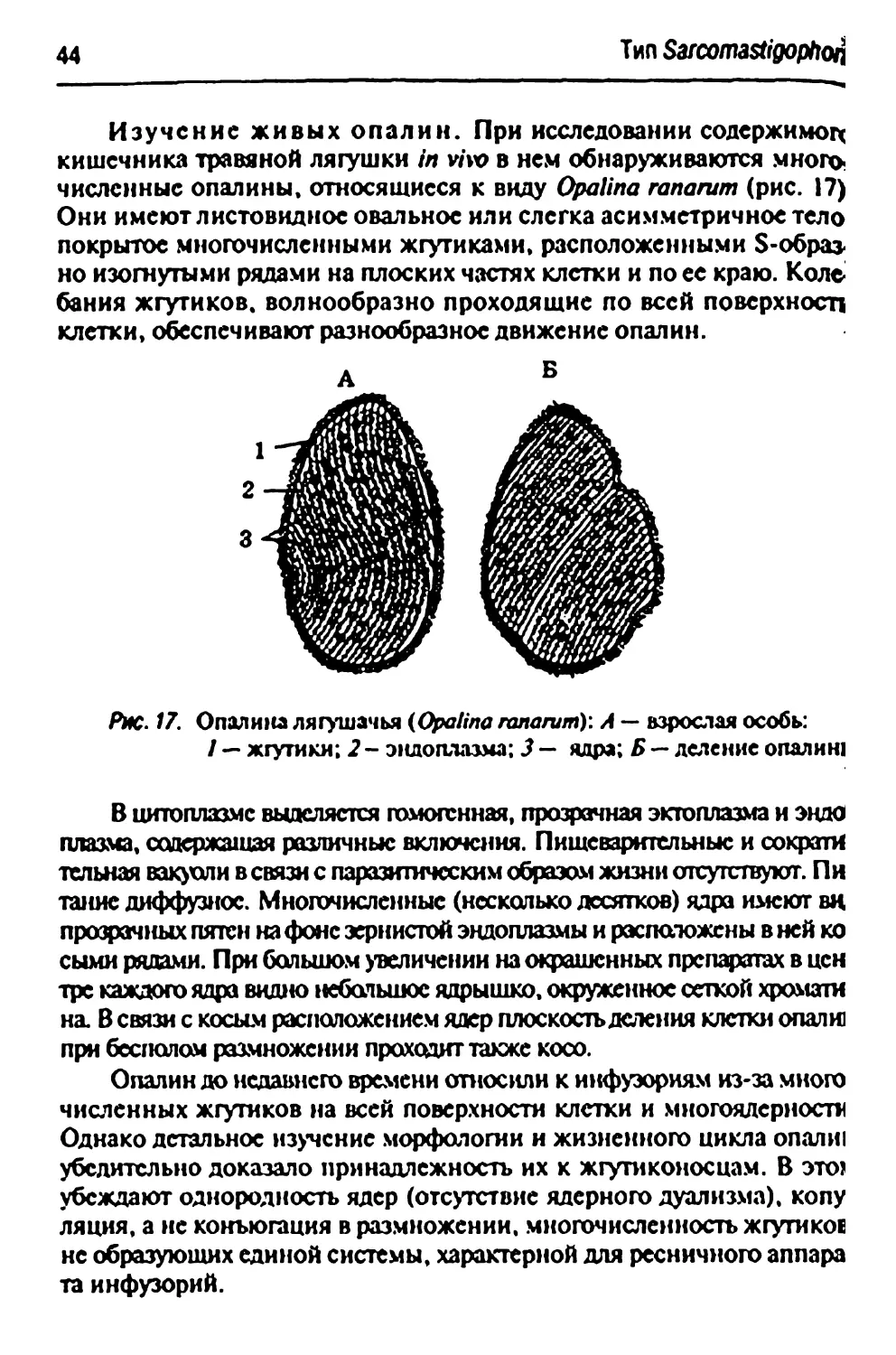
Изучение живых опалин. При исследовании содержимого
кишечника травяной лягушки in vi\x> в нем обнаруживаются
многочисленные опалины, относящиеся к виду Opalina ranarum (рис. 17)
Они имеют листовидное овальное или слегка асимметричное тело
покрытое многочисленными жгутиками, расположенными S-образ-
но изогнутыми рядами на плоских частях клетки и по ее краю. Коле
бания жгутиков, волнообразно проходящие по всей поверхносп
клетки, обеспечивают разнообразное движение опалин.
Рис. 17. Опалина лягушачья (Opalina ranarum): A — взрослая особь:
/ — жгутики; 2 - эндоплазма; J — ядра; Б — деление опалин!
В цитоплазме выделяется гомогенная, прозрачная эктоплазма и эндо
плазма, содержащая различные включения. Пищеварительные и сократи
тельная вакуоли в связи с паразитическим образом жизни отсутствуют. Пи
тайне диффузное. Многочисленные (несколько десятков) ядра имеют вн
прозрачных пятен на фоне зернистой эндоплазмы и раслаюжены в ней ко
сыми рядами. При большом увеличении на окрашенных препаратах в цен
тре каждого ядра видно небольшое ядрышко, окруженное сеткой хроматн
на. В связи с косым расположением ядер плоскость деления клетки опали]
при бесполом размножении проходит также косо.
Опалин до недавнего времени относили к инфузориям из-за много
численных жгутиков на всей поверхности клетки и многоялериосш
Однако детальное изучение морфологии и жизненного цикла опали!
убедительно доказало принадлежность их к жгутиконосцам. В это?
убеждают однородность ядер (отсутствие ядерного дуализма), копу
ляция, а не конъюгация в размножении, многочисленность жгутиков
не образующих единой системы, характерной для ресничного аплара
та инфузорий.
Тип Apicomplexa (-Sporozoa) — переднекомплексные,
или споровики
Это обширная группа простейших, ведущих исключительно
паразитический образ жизни, в связи с чем они приобрели ряд структурных
и биологических адаптации, обеспечивающих их размножение и
распространение. Паразитируют споровики у беспозвоночных и
позвоночных животных, некоторые — у человека.
Класс Gregarinlna — грегарины
Отряд Eugregarinida — настоящие грегарины
Материал. Для получения живых грегарин следует вскрыть живых
черных тараканов (Blatta orientalis). Им отделяют голову и конец
С>р:ошка и изапскают кишечник. Грегарины обитают в средней кишке.
Это г отдел кишечника легко отличим по расположению (между сле-
п!jми выростами — дивертикулами и мальпигиевыми трубочками),
прозрачности и желтовато-белой окраске. Выделенную среднюю
кишку переносят в часовое стекло с физиологическим раствором,
рл-.рывают иглой и просматривают под МБС или микроскопом при
и.сличениив-Юраз. Присутствие грегарин легко обнаружить у
зараженного таракана (даже без вскрытия кишки) по наличию беловатых
непрозрачных точек, просвечивающихся сквозь прозрачные стенки
средней кишки. Извлеченных грегарин изучают в калле воды или
фи жологического раствора. При отсутствии живого материала можно
и. пользовать окрашенные тотальные постоянные препараты.
Изучение Gregarina blattarum. При изучении живого материала
можно наблюдать медленное скольжение грегарин в растворе,
связанное с сокращением тончайших волоконец — миоием,
расположенных в эктоплазме. Легко проследить и другой способ движения, если
поместить грегарин в каплю растертой в воде туши. За движущейся гре-
' риной остается светлый след. По мнению Шевякова, это — столбик
t пин, выделяющийся через мельчайшие поры пелликулы. Механизм
и кого движения объясняется по-разному. Некоторые сравнивают его
i реактивным движением (стекающий столбик слизи словно
полеживает грегарину вперед). С другой стороны, на основании резуль-
1гов исследований с электронным сканирующим микроскопом.
46
Тип
АркххгШ
на поисрхности пелликулы обнаружена система продольно располф
женных гребней, колебания которых» вероятно, и обеспечивают
поступательное движение грегарин.
Грегарины, в связи с паразитическим образом жизни,
приобрели ряд специальных адаптации. Их тело разделено на три участка
эпимерит, протомерит и дейтомерит (рис. 18). В последнее
расположено ядро. При отделении грегарины от стенки кишечник(
на последней, обычно» остается эпимерит, обеспечивающий фиксации
паразита и препятствующий выносу его из кишечника. «
Тело грегарин покрыто плотной пелликулой (иногда ее называю]
кутикулой). Под ней располагается прозрачный тонкий слой экто^
плазм ы, а основную часть клетки составляет зернистая эндоплазма]
Включения эндоплазмы — зерна гликогена, которые легко выявлю
ются при окраске клетки жидкостью Люголя (смесь глицерина с йод!
ной тинктурой). Этим красителем зерна гликогена окрашиваются в кра|
сновато-коричневый цвет. !
Питание и все обменные процессы осуществляются диффузно
пищеварительные и сократительные вакуоли отсутствуют. !
В исследуемом материале часто можне
обнаружить попарное соединение грегарин -
сизигий, который образуется перед поло?
вым размножением грегарин, иногда на ран
них стадиях их развития.
Monocystis agilis
Материал. Грсгэрина S fonocystis agilis пара
зитируст в семенных мешках дождевых червей
Для получения материала можно использовал
живых или фиксированных червей. Практичео
ки все дождевые черви заражены разными гре
гарииами рода Monocystis.
Рис. 18. Грегарина Таракановой (Grtpmno bhnarum) -
две особи соединены в сизигий: / — прото
мерит; 2—дейтомерит; J — ядро; 4—ку
тикула; 5 — эктоплазма; б— эндоплазма
7— зерна гликогена
HXCcGregarinitu 47
Покрывают червя острым скальпелем или лезвием бритвы в
перевей трети тела, извлекают три пары беловатых семенных мешков
„помешают их в часовое стекло с небольшим количеством воды. Ткань
семенною мешка измельчают, а затем капля полученной взвеси иссле-
-летси пол микросколом. В большинстве случаев в семенных мешках
Дождевых червей встречаются небольшие (около 1 см при 80-кратном
vjic 1ИЧСИИИ микроскопа) шаровидные цисты на разных стадиях
развитии. Сами грегарины попадаются редко, имеют нсрасчлснснное
ил Л1Н-. прото- и дейтомериттело, удлиненное или округлое.
Изучение препарата. При рассмотрении препарата под
микроскопом при малом увеличении в поле зрения заметны многочисленные
к :с!ки семенных мешков червя, имеющие желтоватую окраску. Это
кодовые клетки червя на разных стадиях созревания. Среди них четко
иылеляются цисты, имеющие правильную округлую форму и серую
окраску (рис. 19). На материале можно проследить различные стадии
голового размножения грегарин. Вначале в цисте грегарины
изменяли форму, округляются, но сохраняют самостоятельность it
превращаются в гамонты. Затем в каждом гамонтс ядра многократно делятся,
каждое ядро окружается участхом цитоплазмы и образуются гаметы.
Неиспользованная цитоплазма ласт остаточное тело. При
слиянии гамет разных гамонтов образуются зиготы, позднее
покрывающиеся оболочкой и превращающиеся в ооцисты. В каждой ооцисте
Рис. 19. Моноцистис (Monocystis agilis). Цисты с ооцистами в ткани
семенного мешка дождевого червя: / — цисты с ооцистами; 2—
гамонты в цисте; J— ткань семенного мешка
48
TwApcompl&a
и процессе спорогонии ядра делятся 3 раза, образуя соответственно
восемь спорозоитов, лежащих в ооцистс своеобразными «пакетами».
Спорозоиты в ооцистс хорошо видны при большом увеличении
микроскопа.
На згой стадии развитие ооцист в цисте заканчивается. Позднее
оболочка цисты разрушается, и ооцисты со спорозоитами попадают
в семенные мешки дождевых червей и при их копуляции поступают
в семяприемники, а при оплодотворении яйцеклеток — в зиготу и
вызывают заражение нового поколения червей. Вот почему почте 100 %
дождевых червей заражены разными видами моноцистисов.
Класс Coccidiomorpha — кокцнднеобразные
Отряд Haemosporidia — кровяные споровики
Материал. Изучение малярийного плазмодия проводится по
препаратам мазков крови человека, зараженного паразитом. Это
позволяет ознакомиться с той частью жизненного цикла, которая протекает
в эритроцитах человека, т.е. с периодом эритроцитарной шизогонии
(бесполое размножение) и образованием микро- и макрогамонтов
(рис. 20).
Шюонты и шизогония в крови человека
На препарате следует рассмотреть форменные элементы крови
(лейкоциты и эритроциты). Эритроциты безъядерные, имеют форму
двояковогнутой линзы, поэтому центральная часть их окрашена слабее
за счет меньшей концентрации гемоглобина. Изучать препарат нужно
при увеличении объектива 90х с иммерсией (см. с. 42).
Развитие малярийного плазмодия в эритроците начинается с
проникновения в него мерозоита и формирования стадии «кольца».
а) Стадия «кольца» хорошо различима на препарате,
окрашенном по Романовскому-Гимза. Ядро имеет вид небольшого красного
зернышка, голубоватая цитоплазма в виде тонкого кольца окружает
центральную вакуоль. Размеры кольца составляют около уу диаметра
эритроцита (рис. 20, /).
б) Позднее цитоплазма утрачивает правильную форму, но
вакуоль еще сохраняется. Неправильная форма плазмодия на этой стадии
связана с его подвижностью (рис. 20, 2).
Класс Coctidiomorpha
49
Рис. 20. Plasmodium vivax в крови человека: / — стадия раннего кольца;
2 — амебоидная форма кольца; 3 — многоядерный растущий
шизонт; 4— шизогония; 5 — макрогамонт; 6— микрогамонт
в) Постепенно происходит рост шизонта, он принимает более
правильную округлую форму и вскоре заполняет большую часть
эритроцита. Одновременно происходит образование и накопление в ши-
зонте мелких зерен коричневого пигмента. При этом диаметр
эритроцита увеличивается почти в 1,5 раза (рис. 20, 3).
г) На следующем этапе в округлом шизонте начинается
процесс ш изогон и и. Ядро делится несколько раз, формируя до 24 ядер
(рис. 20.4). Окруженные цитоплазмой, они образуют мерозоиты. При
разрушении эритроцита мерозоиты выходят в плазму крови и
проникают в новые эритроциты, в которых описанный процесс
повторяется. Весь период развития шизонта от проникновения в эритроцит
мерозоита до конца шизогонии занимает у Plasmodium vivax48 часов.
Гамонты в крови человека, зараженного P. rivax
После завершения шизогонии в эритроцитах начинают
формироваться гамонты (гамстоциты). Молодые гамо>пы отличаются от
шиэонтов отсутствием вакуоли и правильной формой тела. Зрелые
гамонты почти полностью заполняют эритроцит, имеют сферическую
форму, одно ядро и зерна пигмента в цитоплазме.
Макрогамонты (рис. 20, 5) отличаются от микрогамонтов
(рис. 20, 6) более крупным ядром и светлой цитоплазмой.
Тип Ciliophora {-Infusoria) — инфузории
Инфузории — преимущественно свободноживущие морские и
пресноводные простейшие. Это наиболее высокоорганизованные
одноклеточные животные. Некоторые виды приспособились к жизни в
почве, в песке (псаммофилы), другие ведут паразитический образ жизни.
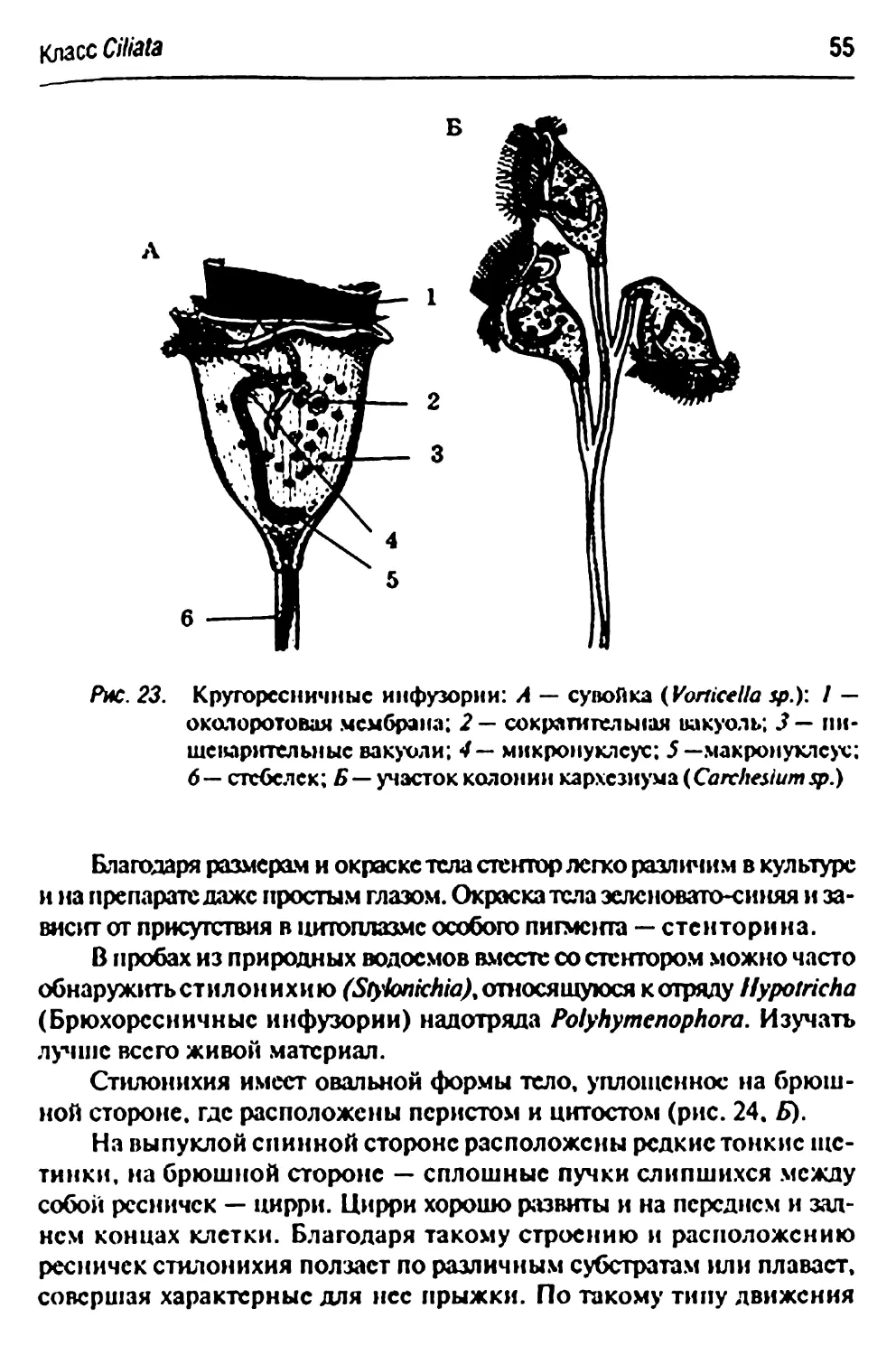
Класс Ciliata (Infusoria) — ресничные инфузории
Надотряд Oligohymenophora
Отряд Hymenoslomata (=Holotricha)
Материал. В культурах на сенных настоях (с. 23) сначала
появляются в массе мелкие ресничные инфузории рола Calpoda и рола
Glaucoma, а потом наиболее обычные Paramecium caudatum.
Размножившиеся в большом количестве, парамеции собираются в
неподвижно стоящем культуральном сосуде на его освещенной стенке в виде
узенькой белой каемки на некотором расстоянии от поверхности волы.
Для изучения небольшую каплю культуры парамеций помешают
на предметное стекло и покрывают се покровным стеклом с
маленькими восковыми ножками.
Paramecium caudatum — инфузория-туфелька
Изучение парамеций затрудняет их активное перемещение в поле
зрения микроскопа. Чтобы их остановить или замедлить движение,
можно использовать небольшой комочек гигроскопической ваты.
После того как препарат будет покрыт покровным стеклом,
парамеции застрянут между волокнами, что замедлит и ограничит их
движение. Хорошие условия для изучения объекта создаются при переносе
на препарат комочка субстрата. Инфузории охотно концентрируются
около него, привлекаемые органикой и бактериями.
Форма тела парамеции, благодаря наличию достаточно плотной
пелликулы, более стабильна, чем у эвглены зеленой, но при движении
в среде с разнообразными препятствиями может слегка изменяться
(метаболируст). В основном тело парамеции удлиненное,
закругленное на переднем конце и слегка расширенное кзади. Задний конец
заострен, на нем более длинные реснички, откуда название P.
caudatum (cauda — хвост) — парамеция хвостатая. За сходство с туфлей она
получила широко известное название инфузория-туфелька (рис. 21).
Класс Ciliata
51
Рис. 21. Инфузория Paramecium caudatum: 1 — сократительная вакуоль;
2— приводящие каналы сократительной вакуоли; 3 —
микронуклеус; 4— макронуклеус; 5— эхекреториая пора; б— реснички;
7— трнхосшеш; 8— пищеварительная вакуоль; 9— перистом; 10—
ротовая воронка; //—цитостом; /2—цнтофарникс;
13—отделяющаяся пищеварительная вакуоль; М— шгтопиг
Движение инфузории изучают, поместив ее в вязкую среду или
ограничив ее активность указанными выше способами. Если
добавить на препарат безвредного красителя (каплю растертоГ! в воде туши
или кармина) и притенить поле зрения микроскопа, удается
рассмотреть движение ресничек, покрывающих все тело инфузории. Оно
напоминает проходящие вдоль тела волны. Особенно активно работают
реснички в области околоротового углубления — п с р и с то м а, где они,
52
Тип Ciliophora
слипаясь, образуют мембраны, окружающие цитостом. Вся
совокупность ресничек, покрывающих тело инфузории, составляет
ресничный аппарат— цилнатуру.
Наблюдая за движением парамеции, можно заметить, что,
сталкиваясь с каким-либо препятствием, инфузория останавливается,
совершает движение назад (пятится), а затем продолжает двигаться вперед,
изменив направление и уклоняясь от препятствия. Доказано, что это
обусловлено деполяризацией и изменением концентрации ионов Са
в цитоплазме инфузории.
Питание парамеций, как и их движение, связано с работой
ресничного аппарата. Реснички перистомальных мембран создают ток воды
со взвешенной органикой к клеточному рту (цитостому) и дальше —
в клеточную глотку (цитофариикс). Вдоль нее также расположены
ряды слипшихся ресничек, колебание которых обеспечивает движение
органических частиц и бактерий — пищи инфузорий — ко
внутреннему концу цитофаринкса, где образуются пищеварительные вакуоли.
Процесс питания также удобнее изучать, добавив в каплю культуры
краситель (водный раствор туши или кармина).
Отрываясь от клеточной глотки, пищеварительные вакуоли
движутся сначала к заднему концу тела парамеции, затем к переднему
и снова возвращаются к заднему. Так завершается первый круг цик-
лоза. Второй круг обычно более короткий: пищеварительная вакуоль
лишь огибает макронуклеус и, завершая процесс пищеварения,
удаляет непереваренные остатки через порошицу, или цитопиг.
Движение пищеварительных вакуолей связано с круговыми движениями
цитоплазмы, циркуляция которой направляется микрофибриллами
и микротрубочками.
Сократительные вакуоли у парамеции расположены в цитоплазме
в переднем и заднем концах тела и открываются экскреторными
порами наружу на спинной стороне. Брюшной стороной считается та,
на которой находится перистом. Каждая сократительная вакуоль
состоит из центрального резервуара и системы приводящих
каналов. Вода и подлежащие удалению продукты обмена собираются
из цитоплазмы в приводящие каналы, которые постепенно
наполняются (стадия диастолы), а при их сокращении (систола) жидкость
переходит в центральный резервуар. Из него избыточная вода и продукты
обмена удаляются через выделительную пору наружу. Таким образом,
сократительные вакуоли действуют как насосы, непрерывно
откачивая воду с углекислым газом и продуктами выделения. Работают обе
Класс Ciliata
53
вакуоли в разном режиме, непрерывно обеспечивая осморегуляцию.
Частота сокращения вакуолей зависит от среды обитания, ее состава
и температуры. При 16 *С интервал между сокращениями составляет
20—25 с (т.е. 3 раза в минуту). При нарушении работы
сократительных вакуолей на поверхности клетки появляются пузыри, а затем
парамеция лопается и погибает.
Реснички и трихоцисты. Для более детального рассмотрения
ресничек и определения характера их расположения в каплю культуры
парамеций можно добавить немного смеси формалина с йодом. Пошбшис
парамеции интенсивно окрашиваются и даже при малом увеличении
хорошо видны многочисленные реснички, равномерно покрывающие
все тело простейшего.
Одновременно, особенно при переводе на большое увеличение,
можно рассмотреть и выстрелившие нити трихоцист, окружающие
клетку своеобразным «паутинным» облачком. Выстреливание
трихоцист вызвано химическим раздражителем и имеет защитное значение.
Ядра инфузорий можно изучить лишь на окрашенных тотальных
препаратах (окраска гематоксилином или борным кармином).
Макронуклеус парамеции имеет бобовидную форму и расположен в
центральной части клетки. Микронуклеус не всегда заметен, различим
при большом увеличении.
Разнообразие инфузорий
В настоящее время в классе Ciliata выделяются три надотряда,
включающие семь отрядов, представители которых могут быть
обнаружены как в пробах воды из природных водоемов, так и в
лабораторных культурах. Познакомимся с некоторыми из них.
1. К отряду Gymnostomata надотряда Kinefofragmmophora относятся
два рода хищных инфузорий — Dilepmsu Didinium. Они хорошо
развиваются в лабораторных культурах. Их жертвами часто становятся
парамеции, которых следует добавлять в культуру с хищниками. В пресных
водоемах обычно встречается Dileptus anser (рис. 22). Это крупная (до
500 мкм) инфузория, на переднем конце тела которой расположен
узкий подвижный хоботок, покрытый ресничками, составляющий
уу -у2 длины клетки. У его основания расположен рот. По средней
линии хобота проходит бороздка, лишенная ресничек. Макронуклеус
фрагментарный.
54
Тип Ciliophora
Рис. 22. Дилептус (Dilrptus anser): I — хоботок; 2 — ротовое отверстие;
3 —• ядра; 4 — сократительные вакуоли
2. Надотряд OHgphymenophora включает два отряда. К отряду Нуте-
nostomata (=Holotricha) относится рассмотренная выше Paramecium
caudatum. Отряд Periaicha (кругоресничиые) представлен в наших
водоемах обычным родом Vorticella (сувойки). Сувойки — сидячие
инфузории. Тело их располагается на длинном сократимом стебельке
(рис. 23, А). При беспокойстве стебелек скручивается спирально.
Реснички околоротовой мембраны закручены вправо (по часовой стрелке).
Они создают ток воды с органикой к цитостому и обеспечивают
удаление непереваренных остатков и продуктов обмена. Недалеко от
переднего конца тела расположена округлая сократительная вакуоль.
В цитоплазме на окрашенных препаратах хорошо различим колбасо-
видный макронуклеус. К этому же отряду относится колониальный
Corchesium sp. (рис. 23, Б).
3. Надотряд Polyhymenophora объединяет инфузорий двух
отрядов. К отряду Heterothricha (разноресничные) относится род Stentor —
трубач (рис. 24, А). Трубачи обычны в стоячих или слабопроточных
водоемах с прозрачной водой. Обнаружить их можно, зачерпнув со
дна воду с небольшими частичками органики. При изучении
трубачей следует обратить внимание на их способность быстро
сокращаться при малейшем беспокойстве и затем медленно восстанавливать
характерную форму.
Реснички околоротовой зоны закручены влево (против часовой
стрелки). На теле реснички расположены продольными рядами. Их
колебания хорошо видны на остановившемся трубаче. В связи с
крупными размерами стентора (до 1-2 мм) для изучения движения и
работы цилиатуры разных зон рассматривать его следует либо в стекле
с лункой, либо используя восковые ножки на покровном стекле. У
живого трубача и на тотальном окрашенном препарате хорошо виден че-
тковидный макронуклеус.
Класс CiHata
55
Рис. 23. Круторссничныс инфузории: А — сувой ка (Vorticella sp.)\ I —
околоротовая мембрана; 2 — сократительная вакуоль; 3—
пищеварительные вакуоли; 4 — микронуклеус; 5— макронуклеус;
6— стебелек; £— участок колонии кархезиума (Carchesiumsp.)
Благодаря размерам и окраске тела стентор легко различим в культуре
и на препарате даже простым глазом. Окраска тела зеленовато-синяя и
зависит от присутствия в цитоплазме особого пигмента — стентори на.
В пробах из природных водоемов вместе со стентором можно часто
обнаружитьстилонихию ^О'/ол/сЛ/оЛ относящуюся к отряду llypotricha
(Брюхорссничныс инфузории) надотряда Polyhymenophora. Изучать
лучше всего живой материал.
Стилонихия имеет овальной формы тело, уплощенное на
брюшной стороне, где расположены перистом и цитостом (рис. 24, Б).
На выпуклой спинной стороне расположены редкие тонкие
щетинки, на брюшной стороне — сплошные пучки слипшихся между
собой ресничек — цирри. Цирри хороню развиты и на переднем и
заднем концах клетки. Благодаря такому строению и расположению
ресничек стилонихия ползает по различным субстратам или плавает,
совершая характерные для нее прыжки. По такому типу движения
56
Тип Ciliophora
Разноресничные инфузории:
Л — трубач (Sttntor sp.)\ Б —
стилонихия (Stylonichia $р.)\
1 — перистомальное поле;
2 — мембраны
околоротовых ресничек; 3 — макро-
нуклеус; 4— микронуклеус;
5— приводящий канал
сократительной вакуоли; 6 —
сократительная вакуоль; 7 —
ундулирующая мембрана и
реснички; S— цирри
этих инфузорий легко найти на препарате. Они довольно крупные —
до 100—350 мкм. Макронуклеус стилонихии имеет гантслевидную
форму и заметен даже на живом объекте.
Таким образом, знакомство с различными представителями цилиапг
показывает, что основными систематическими признаками
простейших зпгого типа являются характер расположения ресничек и форма
макронуклеуса.
Подцарство Metazoa — многоклеточные
Надраздел PhagocyieUozoa
Тип Placozoa — пластинчатые
Класс Placozoa
Trickqplax adhaerens (=чпср(ша)
Трихоплакс — морское животное. На водорослях они образуют
скопления, хорошо заметные из-за их светлой окраски. Размеры
животного достигают 4-5 мм. Форма тела весьма изменчива благодаря
амебоидному движению.
Как показали результаты исследований, трихоплакс
распространен широко и встречается на водорослях в прибрежных водах
Атлантического океана, Средиземного и Красного морей, у берегов
Японии, в Тихом океане. Попав вместе с водорослями в морской аквариум,
трихоплакс хорошо культивируется. Такая культура трихоплакса
несколько лет успешно поддерживалась в лаборатории кафедры
зоологии беспозвоночных МГУ.
Изучение трихоплакса на окрашенных осмием тотальных
препаратах позволяет рассмотреть форму тела животного, наличие крупных
зернистых включений — пищеварительных вакуолей, содержимое
которых после завершения пищеварения выбрасывается через спинной
наружный слой уплощенных эпителиальных клеток (рис. 25).
Размножается трихоплакс бесполым способом, делением надвое и
почкованием. Иногда на препаратах можно обнаружить шаровидные,
покрытые ресничками «почки* трихоплакса после их отделения от
материнского организма. Они получили название «бродяжек» за свою
способность плавать втолше воды, обеспечивая расселение. Половое
размножение еще не полностью изучено.
Рис. 25. Трихоплакс (Trichoplax adhatrrns): изменение очертания тела
при движении
Надраздел Parazoa
Тип Porifera (=Spongia) — губки
Материал. Для изучения обшей организации губок удобнее
использовать одиночную г>6ку S>xvn raphanus, имеющую размеры 1 —2 см,
или средиземноморскую известковую губку Ascetta blanca.
Изучение скелетных образований губок следует проводить на по*
стояииых препаратах, изготовленных из элементов скелета
пресноводных губок-бадяг, а также морских губок Geodia sp. и Euspongia
officinalis.
Для приготовления препаратов скелетных образований губок
небольшие участки колонии следует поместить в пробирку с 20 %
раствором едкого калия (КОН) и, при осторожном встряхивании,
подогревать на слабом огне, не лолуская вскипания жидкости, чтобы
содержимое не выплеснулось из пробирки.
Когда все мягкие ткани растворятся, комочек губки распадается
и части скелета образуют осадок на дне пробирки. КОН сливается,
осадок осторожно промывается водой (после каждой порции воды
осадок должен осесть) и заливается 96* или абсолютным спиртом,
который, поглощая влагу из воздуха, быстро становится 96*. Осадок
кремнеземного скелета выливается в часовое стекло вместе со спиртом.
Избыток спирта нужно осторожно слить или отсосать пипеткой,
а остатки поджечь. Если весь спирт удален, спикулы скелета имеют
вид белого непрозрачного порошка. Если между иглами есть влага,
они будут бесцветными, стекловидными. Их можно подсушить,
подержав часовое стекло над слабым пламенем спиртовки.
После зтого иглой часть сухого осадка переносится на
предметное стекло в канадский бальзам и покрывается покровным стеклом
с восковыми ножками.
Роговой скелет туалетной губки из абсолютного спирта
переносится в ксилол для осветления, а затем в канадский бальзам.
Изучение скелета пресноводных б ад я г (кл. Demospongia, отр.
Cornacuspongia). Вначале с помощью МБС следует ознакомиться
с участком скелетной решетки колонии бадяг. Это позволяет
рассмотреть расположение кремнеземных спикул, спаянных
органическим роговым спонгином (рис. 26, Л).
Класс Demospongu
59
Рис. 26. Скелет г>**>к: Л — участок скелетной решетки бодяг; Б —
элементы скелета SpongUla lacustris: I — мспкклеры; 2—
микросклеры; В— мегасклераубадягр. Ephydatia\ Г— скелет Euspongia
officinalis-, Д — скелет Geodia sp.: I —шаровидные кремниевые
спикулы; 2 — четырехосевые спикулы
Отдельныеэлеме>лыскелета — спикулы
илимегасклеры,имеют всретеиовидную удлиненную форму. У SpongUla lacustris (озерная
бадяга) они слегка изогнуты, заострены с двух концов и имеют гладкую
поверхность (рис. 26, Б). Ллина их достигает 200 мкм. Кроме крупных
спикул, в скелете бадягесть мелкие шиповатые микросклеры.
60
Тип Porifera
У речной бадяги (род Ephydatio) мегасклеры изогнутые,
шиповатые в средней части, но гладкие на концах (рис. 26, В). В отличие от
озерной бадяги, у речной нет микросклер в скелетной решетке.
Таким образом, структура крупных спикул (мегасклер) и наличие или
отсутствие мелких (микросклер) — важный систематический признак
пресноводных губок-бадяг.
Скелет морской гу^ки Euspongia officinalis (туалетная, или
греческая губка), формируется внеклеточно, образован органическими,
роговыми, сильно разветвленными волокнами желтоватого цвета
благодаря содержанию в них до 14 % йода (рис. 26, Г). В старых препаратах
скелета туалетной губки цвет волокон бледнеет, поэтому
рассматривать их следует при затемненном поле зрения микроскола.
Скелет морской губки геодии (Geodiasp., отр. Tctraxonida) слагается
из элементов двух типов (рис. 26, Д). На поверхности губка покрыта
шаровидными кремниевыми спикулаяи, расположенными в
несколько слоев. Изнутри к ним прилегают чстырсхоссвысспикулы. Три оси,
ориентированные к поверхностному слою шаров, расположены так.
что угол между соседними осями составляет 120*. Четвертая ось
перпендикулярна к указанным трем. На препаратах скелета геодии обычно
видны три оси (две внешние и одна внутренняя), третья внешняя ось
при изготовлении препарата обламывается, поэтому на препарате
остается лишь место облома.
Внутренние почки (геммулы) пресноводных бадяг. Бадяги обитают
в неглубоких стоячих (пруды, озера) и проточных (речки, ручьи)
водоемах с плотным или слегка заиленным фунтом. Они образуют
неподвижные, прикрепленные к разным субстратам колонии самой
разнообразной формы. В связи с тем что подобные водоемы могут
промерзать на большую глубину, иногда до дна, бадяги живут лишь
один сезон. После отмирания колонии археоциты образуют скопления
шаровидной формы, которые покрываются двумя защитными
плотными оболочками с воздушной прослойкой между ними
(теплоизоляционный воздушный слой). Чтобы оболочки не спадали, между ними
располагаются микросклеры. Это — ге м м ул ы, ил и внутренние почки
(рис. 27). Геммулы либо остаются в скелетной решетке материнской
колонии, либо падают на дно водоема. На одном из участков
поверхности геммулы находится небольшое отверстие — микропора, через
которое весной выходят археоциты, развивающиеся в новое
поколение губок, оседающих на скелете материнской колонии или на
различном донном субстрате.
MzccDemaspongia
61
Рис. 27. Геммулы бадяг: А — озерной: / — лоровое отверстие; 2 — ме-
гасклеры; 3 — микросклеры; Б — амфидиски геммулы
речной бадягн
После отмирания колонии бадяг геммулы остаются в скелетной
решетке, что облегчает их обнаружение.
Геммулы речной бадяги (Ephydotia sp.) образуют между двумя
оболочками большое количество особых спикул — амфидисков
(рис. 27, Б). Они имеют довольно сложную форму и состоят из
срединной оси. несущей на концах звездчатые диски. Амфидиски
расположены перпендикулярно к поверхности геммулы, упираясь своими
дисками в ее оболочки, внутреннюю и наружную. Такое расположение
и форма амфидисков не только препятствуют спаданию оболочек, но
благодаря тонкой средней части оставляют значительное пространство
для воздушного слоя.
Поскольку при изготовлении препаратов внешняя оболочка
частично разрушается, на периферии геммулы при большом увеличении
можно рассмотреть внешнюю часть амфидисков.
У озерной бадяги (Spongilla lacustris) между оболочками геммулы и на
ее поверхности располагаются игольчатые микросклеры (рис. 27. А).
Таким образом, строение скелетных образований геммул. как
и других элементов скелета бадяг, является их четким
систематическим признаком.
Надраздел Eumetazoa
Раздел Radiata — лучистые, или Diploblastica —
двухслойные
Тип Cnidaria (=Coelenterata) —
стрекающие (кишечнополостные)
Класс Hydrozoa — гидрозои
Подкласс Hydroidea — гидроидные
Отряд Hydrida — гидры
Hydra oligactis — длинностебельчатая гидра
Материал. В июле - начале сентября на стеблях и листьях водяных
лилий, рдестов, телореза в прудах, медленно текущих ручьях, на
литорали озер можно отыскать гидр. В наших водоемах встречается Н. w-
ridis. зеленая окраска которой определяется присутствием в клетках ее
эктодерм ы зоохлорелл (симбиотических одноклеточных водорослей);
Н. oligactis, 6—7 щупалец которой в расправленном состоянии в 4— 5 раз
превышают длину тела самой гндры; H./usca, имеющая менее 6 щупалец
и коричневатую окраску тела Гидры могут быть обнаружены и в пробе
воды из прудов с обильным развитием ряски.
Для разведения гидр в сосуд с водой из водоема помещают
несколько растений валиснерии или элодеи. Чтобы избежать внесения
шнющих частиц и моллюсков, растения тщательно промывают. Сосуд
с гидрами ставят в светлое место, но не на прямой солнечный свет.
Каждые 1—2 дня гидр подкармливают мелкими рачками (дафниями,
циклопами и др.), а при их отсутствии — трубочниками, хирономидами,
даже дождевыми червями, разрезав их предварительно на кусочки.
Это особенно удобно зимой.
Если выполнить все указанные условия, через 1—2 недели можно
иметь большое количество гидр.
Изучение живого объекта лучше всего вест под бинокуляром в
часовом стекле или на предметном стекле с лункой. Следует рассмотреть
внешнее строение гидры, отыскав подошву, стебелек, тело,
ротовой конус с ротовым отверстием,окруженным щупальцами
(рис. 28). У расправившейся гндры легко различить прозрачные клетки
KsaccHydrozoa
63
Рис.28. Внешнее строение гидры: /—подошва; 2—стебелек; J—тело;
4 — ротовой конус с ротовым отверстием; S — щупальца; б —
зона почкования с почками
наружного слоя — эктодермы и зернистые непрозрачные клетки
внутреннего слоя — энтодермы. На щупальцах хорошо просматриваются
небольшие вздутия — скопления стрекательных клеток (см. далее).
Наблюдение за живой гидрой позволяет изучить ее движение,
питание, реакцию на раздражение, регенерацию.
Движение гидры лучше наблюдать в часовом стекле, используя МБС
Вначале пшра довольно долго остается неподвижной. Активно
двигаются, изгибаясь в разных направлениях, л ишь ее щупальца. При
достаточно продолжительном наблюдении можно видеть шагающее
перемещение гидры: закрепившись на субстрате передним ротовым
концом тела, она затем подтягивает подошву. Такие шаги повторяются
многократно. Процесс движения может быть и иным. Закрепившись
ротовым концом тела, гидра, «кувыркаясь», перебрасывает подошву
на другую сторону от ротового полюса (рис. 29). Кроме указанных форм
движения, гидры способны перемещаться по субстрату, медленно
скользя на подошве.
Питание. Если в часовое стекло поместить дафний или
циклопов, можно наблюдать захват рачков гидрой. Подвижные щупальца
64
Тип Cnktaria
Рнс. 29. Движение гидры: А — шагание; В — кувыркание
словно приклеиваются к жертве. Стрекательные клетки выделяют
секрет, парализующий добычу, а движением щупалец она
подносится корту и заглатывается. Пищеварение у гидр полостное и
внутриклеточное. Завершается пищеварение в течение 4—5 часов, после чего
непереваренные остатки выбрасываются через ротовое отверстие.
Реакция на раздражение. Для изучения действия с т р е к а тс л ь н ы х
клеток следует перенести гидру на предметное стекло с небольшим
количеством воды и покрыть препарат покровным стеклом с
восковыми ножками. Если добавить у края покровного стекла 2—3 капли
S % уксусной кислоты, можно наблюдать выбрасывание стрскател ь-
н ых н итей, которые хорошо заметны при затемненном поле зрения
микроскопа либо при большом увеличении.
Опыт по регенерации гидры можно провести в течение дня.
Для этого гидру в часовом стекле нужно разрезать острым лезвием
на несколько частей, добавить воду, небольшую веточку элодеи,
прикрыть стеклом для уменьшения испарения воды и поставить в
достаточно светлое место, но не на прямой солнечный свет.
При проведении наблюдений следует фиксировать время
регенерации и обратить внимание на этапы этого процесса.
Раэшюжошс. В летнее время в зоне почкования между телом гидры
и стебельком образуются почки, являющиеся результэтом бесполого
размножения — почкования. В почке продолжаются все части стенки
тела: эктодерма, мезоглея в виде опорной мембраны, энтодерма» а
также гастральная полость. В зависимости от стадии формирования почки
Класс Hydrozoa
65
на ней развиваются рог и щупальца. Затем она отделяется и переходит
к самостоятельному существованию. Процесс почкования идет
наиболее интенсивно при обилии пищи и благоприятном гидротермическом
режиме в водоеме и в культуралыюм сосуде в лаборатории.
В конце лета — начале осени на теле гидр можно обнаружить взду-
гия. расположенные в эктодерме верхней (семенники) и нижней
(яичники) части тела (рис. 30). Созревание в гонадах половых продуктов
предшествует половому размножению. Среди гидр есть как
раздельнополые, так и гермафродитные виды. После оплодотворения яиц в теле
материнской гидры яйца окружаются плотной оболочкой и зимуют.
Весной из них развивается новое поколение гидр.
Нервная система гцдр. Наблюдение за живыми гидрами показы-
iiacr, что малейшее беспокойство вызывает сокращение их тела. Эта
реакция — результат быстрого проведения нервного импульса диффуз-
Рис. 30. Строение гидры: А — продольный разрез; Б — поперечный разрез;
/ — рот; 2 — ротовой конус; 3 — щупальце; 4 — гастрольная
полость; 5 — мужская гонада; 6 — яйцеклетка; 7 — подошва; 8 —
почка; Р — эктодерма; /0 — опорная пластинка (мезоглея); // —
энтодерма
66
Тип Cnidaria
ной нервной системой гидры, вызывающего сокращение мышечных
волокон эпителиально-мускульных клеток эктодермы и расслабление
таких образований в энтодерме.
При отсутствии живого материала внешнее строение тела гидры
может быть рассмотрено на тотальном постоянном препарате.
Желательно иметь гидр с почками в зоне почкования.
Клеточное строение стенок тела гидры изучается на постоянных
препаратах с поперечным и продольным срезами (рис. 30, Б\ 31).
Наружный (эктодерма) и внутренний (энтодерма) слои клеток
разделены бесструктурной мезоглеей в виде опорной пластинки.
Опорная пластинка служит местом прикрепления мускульных
волокон эпителиально-мускульных клеток эктодермы и
энтодермы и обеспечивает поддержание характерной формы тела гидры.
Эктодерма образована клетками нескольких типов. Основу ее
составляют эпителиально-мускульные клетки. Каждая такая клетка
состоит из внешней цитоплазматической части с ядром и мускульного
отростка, обращенного к опорной пластинке и ориентированного
вдоль продольной оси тела животного.
Рис. 31. Участок стенки тела в разрезе при большом увеличении: / —
эктодерма; 2 — опорная пластинка; J — энтодерма; 4-9 —
клетки (4— эпителиально-мускульные энтодермальные.
выполняющие пищеварительную функцию. 5 — железистые,
6 — нервные. 7 — чувствительные, 8— иитерстициальные,
9 — стрекательные: а — пенетранты, б — вольвенты)
Класс Нубгогоа
67
Между эпителиально-мускульными располагаются
стрекательные клетки, особенно их много на щупальцах и около ротового
отверстия. Стрекательные клетки гидры различны построению,
размерам и механизму действия. Самые крупные из них грушевидные пе-
нстранты — главные орудия нападения. Каждая стрекательная
клетка состоит из цитоплазмы с ядром и стрекательной капсулы, или
книды. От стрекательной клетки наружу выдается чувствительный
волосок — кн идоцил ь. Внутри капсула заполнена жидкостью, имеет
спирально скрученную полую нить, способную выбрасываться наружу
при раздражении книдоциля. У основания нити имеется расширение —
шейка с тремя стилетами и рядами шипиков, которые пробивают тело
жертвы подобно гарпуну. Расправляющаяся нить, проникая в ранку,
вносит в нее ядовитые вещества, парализующие добычу.
Более мелкие стрекательные клетки, или вольвенты. также имеют
фушевидную форму, располагаются вокруг пенетрант и при
выстреливании не выделяют ядовитой жидкости, а лишь опутывают жертву или
части тела жертвы, затрудняя ее движение. Пснстраита с окружающими
се вольвентами образует своеобразные батареи стрекательных клеток.
У основания эпителиально-мускульных клеток эктодермы при
большом увеличении микроскопа различимы скопления мелких ин-
терстициальных недифференцированных клеток, за счет
которых после гибели выстреливших образуются новые стрекательные
клетки и обеспечивается регенерация, а в период полового
размножения формируются половые клетки.
На препаратах с мацерированными гидрами можно различить
пигментные и звездчатые нервные клетки,также расположенные
в эктодерме.
Энтодерма включает эпителиально-мускульные
пищеварительные клетки. Подобно сходным клеткам эктодермы они имеют цито-
плазматическую часть цилиндрической формы и мускульные отростки,
обращенные к опорной пластинке, но идущие поперек продольной
оси тела. Из-за такого расположения мускульные волокна
эпителиально* мускульных клеток эктодермы и энтодермы являются
антагонистами: сокращение одних всегда сопровождается расслаблением других,
что вызывает определенные изменения формы тела животного.
На стороне, обращенной в гастральную полость, энтодермальные
клетки несут чаще всего по два жгутика. Эти клетки способны к
фагоцитозу и внутриклеточному пищеварению. В них могут быть обнаружены
разного рода включения: пищевые частицы и экскреторные гранулы.
В состав энтодермы входят также железистые клетки, выделяющие
в гастральную полость пищеварительные ферменты, обеспечивающие
68
Тип Criteria
полостное пищеварение. Они содержат крупные вакуоли, но лишены
жгутиков и мускульных отростков.
Таким образом, гидра является типичным радиалыю-снммстрич-
ным двуслойным животным и служит модельным объектом при
изучении кишечнополостных стрекающих животных.
Отряд Leptolida — морские гидроидные полипы
Подотряд Thecaphora
Obelia geniculate
Материал. Изучение морских гидроидных полипов проводится
с использованием тотальных окрашенных препаратов гидромедуз и
окрашенных препаратов участка полипоидной колонии Obelia
geniculate или других видов этого рода. Гидромедузы обычны в планктоне
северных (Белого и Баренцева), дальневосточных и Черного морей.
Полипоидные колонии Obelia обитают на литорали, на небольших
глубинах, имеют древовидную форму и состоят из нескольких сотен и
даже тысяч особей. Для более полного представления о колониях
морских гидроидных полипов следует использовать влажные препараты
и музейный материал (демонстрационный).
Полипоидное поколение гидроида Obelia sp.
При изучении участка полипоидной колонии обелии хорошо
виден ствол колонии, от которого отходят многочисленные особи,
образующиеся в результате бесполого размножения почкованием. При этом
они не отделяются от материнской колонии и не переходят к
самостоятельному существованию. В результате все особи колонии имеют
общую разветвленную гастральную полость (рис. 32).
В колонии обелии образуются особи двух типов. Одни из них
имеют сходство с гидрой и получили название гидрантов (рис. 32,£).
Гидранты имеют стебелек, связывающий их со стволом колонии, и
тело, на переднем конце которого расположен вздутый хоботок с ро-
товым отверстием, окруженный венчиком щупалец. Стенки
гидранта переходят непосредственно в ствол колонии, от которой
гидрант отпочковался. Гидранты располагаются на периферии
колонии, выполняя защитную функцию и обеспечивая отлов добычи
и питание колонии. Захваченные гидрантами пищевые объекты
перевариваются самими гидрантами, а питательные вещества
распределяются между всеми особями колонии.
i^ccHydrozoa
69
Рис. 32. Морской гидроидный полип Obelia geniculate: A — участок
колонии: / — гидрант в расправленном состоянии; 2—
сократившийся гидра>гп 3 — тека, или перидерм; 4 — развивающаяся
почка; 5— бластостильс развивающимися медузами; 6— гоно-
тека; 7— гндротека; Б — гидрант. / — ротовой хоботок; 2—
щупальца; J— гндротека; ¥— перидерм; 5— канал гастрольной
полости; 6 — гастрольная полость палила
Ближе к стволу колонии располагаются особи иного рода,
получившие название гонангиев.В центре гонаигия находится
редуцированный полип, лишенный рта и щупалец, — бластостил ь, от которого
отпочковываются гидромедузы, являющиеся половым поколением
морских гидроидных полипов. У разных видов обедни
отпочковывающиеся медузки либо располагаются по бокам от бластостиля, либо
образуют цепочку или компактную группу медузок.
70
Тип Cnidaria
Вся колония обсл ии покрыта особой оболочкой, образующей свое-х
образный наружный скелет— перидерм или перисарк (иногдатека).
Оболочка эта продолжается на стебелек и тело гидранта, образуя
гидротску, и на бластостнль — гонотску. Следовательно, гонантй —
это бластостнль, окруженный го ноте кой. На стебельках всех особей
колонии перисарк разделен кольцевыми перетяжками, что
обеспечивает большую подвижность гидрантов и гонангиев.
Таким образом, в результате почкования полипоидная колония
морских гидроидных полипов формирует два типа
специализированных особей, отличающихся морфологически и функционально.
Такая колония называется диморфной (ди — два, морфа — форма).
Медузондное поколение гидроида Obelia sp.
Зонтик гидромедузы обелии в поперечнике не превышает 2-3 мм.
Эта планктонная медузка имеет плоский зонтик с редуцированным
парусом (рис. 33). По краю зонтика расположены многочисленные
щупальца и восемь статоцистов (органы равновесия). У основания
коротких щупалец находятся утолщения, образованные скоплениями
стрекаггельных клеток. Статоцисты на тотальных препаратах не всегда
видны.
Сторона зонтика, на которой располагается ротовой стебелек с
ротовым отверстием, является субумбреллой, противоположная —
Рис. 33. Гидромедуза Obelia geniculate: I — щупальиа; 2 — кольцевой
канат; J— гонада; 4— радиальные каналы; 5 — ротовой
стебелек
Класс Hydrozoa
71
эксумбреллой. Рот ведет в глотку, расположенную внутри ротового
стебелька. Эктодсрмальная глотка переходит в энтодермальный
желудок, от которого отходят четыре крестообразно расположенных
радиальных канала, впадающих по краю зонтика в кольцевой канал.
Желудок и система каналов образуют гастроваскулярную систему
гидромедуз.
На нижней стороне зонтика по ходу радиальных каналов
располагаются (соответственно в числе четырех) гонады, имеющие экто-
лермальное происхождение, характерное для всех гидрозоев. При
большом увеличении внутри гонад раздельнополых медуз можно
обнаружить крупные клетки (разные стадии роста ооцитов) или очень
мелкие зернистые образования — сперматозоиды.
Между эктодермой и энтодермой у медуз расположена прослойка
мезоглеи, не образующая, в отличие от полипов, опорной пластинки,
а имеющая вид объемной студенистой массы, на 97,5 % состоящей из
воды.
Как и у всех кишечнополостных, полипонднос и медузоидное
поколения морских гидроидных полипов — формы существования
одного вида. Основные отличия в их организации обусловлены несходством
образа жизни: активным движением одиночных, размножающихся
только половым способом медуз и прикрепленным образом жизни
колониальных полипов, размножающихся бесполым способом (табл. 3).
Отряд Trachilida — трахимедузы
Aglantha sp.
Для детального изучения строения гидромедуз следует рассмотреть
медузу Aglantha sp. У нее в цикле развития нет смены поколений,
отсутствует полипоидная стадия. Гидромедуза агланта имеет небольшие
размеры — 0,7—1 см. Зонтик куполообразный, высокий (рис. 34). Снизу
по краю зонтика расположены многочисленные короткие щупальца.
Входное отверстие в полость колокола сужается кольцевидной мускульной
перепонкой — парусом. Благодаря наличию паруса небольшие по
размеру гидромедузы способны к более активному движению.
Уменьшение выходного отверстия колокола при сокращении паруса усиливает
эффект реактивного движения при выталкивании воды из полости
зонтика. Расслабление паруса способствует заполнению полости колокола
новой порцией воды. В полость колокола свешивается длинный ротовой
стебелек с небольшими лопастями, окружающими ротовое отверстие.
72
ТипСлМа/й
Сравни гедыпв хжрбиптрмстнха
н медузоадной форм
Таблица 3
пожнпомвой
Особенности
организации
Полипоидом стадия
Мсдуэоидная стадия
Симметрия
Число слоев тс»
Стрекательные
клетки
Нервная система
Мезоглея
Пищеварительная
система
Размножение
Образ жизни
Радиальная
Двухслойные
Скопления на щупальцах
Диффузная, есть
скопления нервных клеток
на щупальцах, около рта
и на полошве
Плотная опорная
пластинка
Мешкопил1сая гастроль-
ная система у одиночных
ПОЛИПОВ
Бесполое и половое
Прикрепленные, мало-
подвижные» часто
колониальные животные
Радиальная
Двухслойные
Скопления у основания
щупалец
Двойное нервное кольцо
на краю зонтика, есть восемь
статоцист
Объемная, студенистая,
содержит 97,5 % волы
Гастроваскулярная система,
состоящая из радиальных
н кольцевых каналов. Есть
эктодермальная глотка
н энтолермальный желудок
Только половое
Подвижные, планктонные,
одиночные животные
В стенках зонтика расположены четыре радиальных какала гастроваску-
лярной системы, объединяющиеся кольцевым каналом, идущим по
краю зонтика. У половозрелых медуз хорошо
видны на с><>умбрслле колбасовидные гонады»
развивающиеся из эктодермы.
При отсутствии массового раздаточного
материала следует показать гидромедузу Agl-
antha sp. демонстрационно, используя МБС.
Рис. 34. Гидромедуза Aglantha sp.: I — гонада;
2 — радиальный какал; 3 — ротовой
стебелек; 4 — лопасти ротового
стебелька; 5 — кольцевой канал; 6 —
парус; 7— щупальца
Класс Scyphozoa
73
равноценной заменой ей может служить гидромедуза Sarsia mirabilis*
гскжс имеющая хорошо развитый парус, куполообразный зонтик
и длинный ротовой стебелек. Следует иметь в виду, что Sarsia mirabilis
является мсдузоидным поколением морского гидроидного полипа
Coryne sarsi (Lovcn) из отряда Leptolida.
Класс Scyphozoa — сцифомедузы
Отрад Semaeostomeae
Материал. Изучение сцифоидных медуз чаще всего проводится
на фиксированном материале. Объектами для изучения служат ушастая
медуза (Aurelia aurita) и ее личинка — эфира. Для исследования лучше
использовать некрупные экземпляры, на которых даже без
дополнительного окрашивания хорошо видны все особенности строения
ушастой медузы. Эфиру лучше изучать по окрашенным тотальным
препаратам.
Лиге На aurita — ушастая медуза
Строение ушастой медузы (рис. 35). Тело ушастой медузы уплощено
и несет по краю зонтика многочисленные короткие щупальца. Зона
щупалец имеет восемь неглубоких вырезов, в которых расположены
краевые тельца — ропалии. Это — органы равновесия и
светочувствительные. На внутренней стенке зонтика — субумбреллс — в центре,
на коротком стебельке располагается крестообразный рот,
окруженный четырьмя ротовыми лопастями. У плавающей медузы
ротовые лопасти свешиваются подобно заячьим ушам, откуда и название
медузы. При захватывании добычи края ротового отверстия расходятся
и рот принимает вид четырехугольного, почти квадратного отверстия.
Это позволяет медузам-хищницам захватывать довольно крупную
добычу.
Сквозь прозрачные стенки тела и мезоглею (студнеобразную,
на 97,5 % состоящую из волы) хорошо виден расположенный в центре
желудок, образующий четыре кармана, выстланные энтодермой.
В глубине этих карманов расположены гастральные нити
(валики). Это скопления железистых клеток, выделяющих
пищеварительные ферменты в полость желудка. От желудка отходит система
радиальных каналов. Четыре сильно ветвящихся канала совпадают
по направлению с ротовыми лопастями и называются каналами
74 Тип Cnidaria
Рис. 35. Сцифоидная медуза Aurtlia ourita. I — краевые тельца» или рона-
лии; 2 — рот; 3 — ротовая лопасть; 4 — желудок; 5 — радиальные
каналы; 6 — кольцевой канал; 7 — гастральние валики; 8—
щупальца
I порядка. Разветвленные каналы II порядка, также в числе четырех,
располагаются соответственно четырем карманам желудка. С
разветвленными каналами чередуются восемь неразвствлснных каналов
III порядка. Все радиальные каналы объединяются кольцевым
каналом, идущим по краю зонтика, и образуют сложную гастроваску-
лярную систему.
Экспериментально доказано, что из желудка питательные
вещества центробежно идут по восьми неразветвленным каналам III
порядка к кольцевому (быстрое распределение), а процесс их
всасывания и усвоения обеспечивается постепенным движением к желудку
(центростремительно) по восьми разветвленным каналам I и II
порядков. Непереваренные остатки удаляются через ротовое отверстие.
Класс Scyphozoa
75
При рассмотрении краевой зоны зонтика следует обратить
внимание на то, что ропалии находятся в углублениях против централь-
пых ветвей каналов I и II порядков. Здесь отсутствуют щупальца.
Половые железы, или гонады, пространственно совпадают с га-
стральными нитями карманов желудка и хорошо заметны со стороны
эксумбреллы. В отличие от половых желез гидромедуз, которые
формируются из эктодермальных клеток, гонады сцифоидных медуз
образуются в энтодерме.
Полезно провести сравнительный обзор строения гидро- и сци-
фомедуз (табл. 4).
Строение эфиры Aurelia aurita. В цикле развития ушастой медузы
происходит чередование бесполого и полового поколений —
метагенез, причем в развитии преобладает медузоидная стадия. После
формирования половых продуктов у раздельнополых медуз и
внутреннего оплодотворения яйца развиваются в особых «выводковых
камерах» на бороздках ротовых лопастей. Образовавшаяся личинка пла-
нула некоторое время свободно плавает, а осев на субстрат,
последовательно формируете ц и фи стом у, стробилу и, после отделения
горизонтальных почек от стробилы (процесс стробиляции),
последние превращаются в свободноплавающую личинку эфиру.
От взрослой медузы эфира отличается следующими
особенностями строения (рис. 36). У нее небольшие размеры, края зонтика сильно
изрезаны и образуют восемь (иногда больше) краевых лопастей. В
углублениях на вершине краевых лопастей располагаются краевые тельца,
из которых позднее развиваются ропалии. Щупальца по краю зонтика
у молодых эфир отсутствуют, но постепенно развиваются на участках
между краевыми лопастями. Крестообразный рот и желудок имеются
у самых молодых эфир. Гастроваскулярная система вначале
представлена прямыми каналами I и II порядков. По мерс роста и развития
2
Рис. 36. Эфира Aurelia aurita: I —
краевые лопасти; 2— край зонтика;
J — краевые тельца; * — рот;
5— желудок; 6— гастральные
нити
76
Тип Criteria
Тдбтщ4
Сравюггелывд характеристика строения пирондмых
н сцифоидных медуз
Признаки
Гидромедузы
Сцифоидные медузы
Размеры
Форма тела
Тело образовано
Ротовой стебелек
Гастроваскулярная
система
Движение
осуществляется
Нервная система
Половые железы
развиваются
Размножение
Мед кис (не белее 1-2
см), даже
микроскопические
Уплощенная или
куполообразная
Эктодермой,
энтодермой и прослойкой
мезоглеи
Длинный
Включает глотку,
желудок, четые радиальных
и кольцевой каналы
Сокращением м>?куль-
иых волокон зонтика
и с помощью труса
Подкожное нервное
сплетение, нервное
кольцо по краю
зонтика, органы равновесия
и светочувствительные
(в виде глазного ляпа
или глазной ямки,
иногда образуется
хрусталик)
В эктодерме
Половое
Крупные, до 1-1.5 м в
диаметре зонтика
Уплощенная или
куполообразная
Эктодермой, энтодермой
и моиоюй прослойкой
мезоглеи
Короткий
Включает желудок
с гастрольными нитями,
слошгую систему из 16
радиальных и кольцевого
каналов
Сокращением мускульных
волокон зонтика. Парус
отсутствует
Подкожное нервное
сплетение, нервное кольцо
по краю зонтика и восемь
ганглиев соответственно
местам расположения
восьми ропалий. Глаза
имеют сложное строение.
Есть органы равновесия
В энтодерме
Половое
эфиры они начинают ветвиться и заклахиывакхгся каналы III порядка.
Постепенное развитие характерно и для гастральных нитей желудка.
По мере роста выравнивается край зонтика.
Чтобы составить наиболее полное представление о процессе развитая
эфиры, желательно иметь тотальные препараты личинок разных стадий.
Класс Anthozoa
77
Класс Anthozoa — коралловые полипы
Подкласс НехасогаШа — шестилучевые кораллы
Отряд Actiniaria — актинии
Actinia equina — актиния конская
Материал. Actinia equina — широко распространенный вид
коралловых полипов. Встречается во многих морях (Белое, Баренцево,
Черное); в прибрежной части Атлантического и Тихого океанов
актинии могут образовывать значительные скопления на литорали и в зоне
отлива (в небольших углублениях грунта и скал, гас сохраняется вода).
Окраска A. equina очень разнообразна и варьирует от красной до
зеленой. Этот коралловый полип является раздельнополым,
живородящим животным. Молодые актинии развиваются в гастралыюй полости
половозрелых особей, где и могут быть обнаружены даже у
фиксированных экземпляров. При знакомстве с организацией A. equina следует
использовать окрашенные препараты поперечных срезов молодых
мелких особей, фиксированных крупных актиний, а также экземпляры,
разрезанные вдоль и поперек продольной оси тела.
Изучение внешнего строения. В расправленном состоянии тело
актинии имеет более или менее правильную цилиндрическую форму.
К субстрату актиния плотно прикрепляется подошвой. Стенки тела
часто складчатые — следствие постоянной реакции на раздражение,
когда происходит сокращение продольных и кольцевых мускульных
клеток (рис. 37).
На противоположном от подошвы конце тела находится ротовое
отверстие, имеющее форму сплющенного овала. Этот полюс слегка
выпуклый. Вокруг рта имеется широкая зона перистома, за которой
в несколько рядов располагаются щупальца. Число щупалец у
актинии разнообразно, но кратно шести в соответствии с характером ее
симметрии. При сокращении тела щупальца втягиваются, а из
ротового отверстия выбрасываются тонкие нити — аконции, несущие
многочисленные стрекательные клетки.
Внутреннее строение актиний изучается на поперечных окрашенных
срезах. Желательно рассмотреть срезы, сделанные в области глотки
и гастральной полости (рис. 38).
Под слоем эктодермы располагается хорошо развитая мезоглея.
От стенок тела внутрь гастральной полости идут септы. Число септ
кратно 6, но минимальное их количество равно 12. У актиний септы
двух типов. Длинные, глубоко вдающиеся в гастрольную полость.
78
Тип Cnidsris
Рис. 37. Внешний пил Actinia equina-. I — подошва; 2 — щупальца
расположены попарно, причем расстояние между септами одной
пары меньше расстояния между соседними парами. Это— септы I
порядка. Между парами септ I порядка расположены более короткие
Рпс. 38. Actinia aquina. Внутреннее строение. Поперечные срезы на уровне
глотки (А) и средней части тела (5): / — эктодерма; 2 —
мезоглея; J— энтодерма: 4— септы I порядка; 5—септы II порядка;
6 — продольные мышцы септ; 7— полость глотки; 8—
направительная камера; 9— сифоноглиф; /0 — меэентериальные
нити; // — внутренняя камера; 12— промежуточная камера
MzccAnthozoa
79
септы II порядка. У некоторых актиний могут образовываться септы
соответственно III и IV порядка.
Промежуток между парой септ I порядка называется внутренней
камерой, между септами соседних пар —промежуточной камерой.
Каждая септа песет на стороне, обращенной к внутренней
камере, продольную мышцу, образующую мускульный валик. На
препарате септы не прямые, имеют различную, часто сильно изогнутую
форму, что объясняется способностью животного к сокращению при
раздражении.
Энтодерма выстилает гастральную полость и стенки септ разного
порядка. Наличие септ значительно увеличивает пищеварительную
поверхность гастральной полости, обеспечивая усиленную секрецию
и активизацию пищеварения и всасывания.
Клстхи эктодермы и энтодермы разделены на два слоя:
эпителиальный и мускульный. Таким образом, у коралловых полипов впервые
образуется эктодермальный покровный эпителий, энтодермальный
железистый эпителий и два слоя мышечных клеток. Под клетками
эктодермы лежат мускульные клетки, образующие у некоторых актиний
продольные мускульные волокна. Эпителиально-мускульные клетки
энтодермы своими мускульными отростками формируют с>6энтодер-
мальный слой кольцевой мускулатуры.
Свободные края септ имеют утолщения — мезентериальные
нити, богатые железистыми и стрекательными клетками. В области
глотки септы прирастают к ее стенкам.
Глотка образуется в результате впячивания стенок ротового диска,
поэтому се передняя часть выстлана эктодермальными клетками.
Внутренний край глотки энтодермальный. Сплющенная глотка имеет
вид узкой щели. С одной ее стороны развиваются клетки с длинными
ресничками, колебание которых обеспечивает движение воды внутрь
гастральной полости. Этот край глотки получил название сифоно-
глифа. У некоторых актиний их может быть два. На краю септ,
соответствующих другой стороне глоточной щели, расположены реснички,
которые гонят воду из гастральной полости наружу. Таким образом,
реснички сифоноглифа и септ способствуют циркуляции воды в
гастральной полости, обеспечивая питание, удаление непереваренных
остатков и газообмен.
При рассмотрении поперечного и продольного срезов крупных
актиний все указанные выше особенности строения шестилучевых
кораллов могут быть изучены дополнительно.
Раздел Bilateria — двустороннесимметричные
Подраздел Protostomia — первичноротые
Тип Plathelminthes — плоские черви
Тип плоские черви по современной классификации включает
девять классов, четыре из которых объединяют исключительно
паразитических животных.
Плоские черви — двустороннесимметричные животные. Через
их тело можно провести лишь одну плоскость симметрии, которая
делит их тело на две зеркально отражающие друг друга части —
правую и левую стороны.
Соответственно, в связи с направленным движением по с><3страту,
раапичаютспиниую, или дорсальную, и брюшную, или
вентральную стороны тела, а также передний, или головной, и задний, или
хвостовой, отделы.
В отличие от ранее рассмотренных групп многоклеточных
животных у плоских червей в процессе эмбриогенеза закладываются три
зародышевых листка: эктодерма, энтодерма и мезодерма, поэтому
плоских червей относят к трехслойным животным — Triploblostka. Из
мезодермы формируются мускулатура, выделительная система,
соединительная ткань и ее производные.
Плоских червей относят к низшим червям в связи с отсутствием
у них: I) полости тела (заполнена паренхимой, отсюда название
плоских червей —паренхиматозные); 2) заднего отдела кишечника
и ануса; 3) дыхательной и кровеносной систем; 4) ганглиев в
стволовой часто нервной системы.
Вместе с тем у них впервые развивается протонсфридиальная
выделительная система и сложная половая система.
Надкласс Turbellaria
Класс Turbellaria — ресничные черви
Класс объединяет преимущественно свободноживушие формы;
покровный эпидермис имеет типичное клеточное строение.
Юйсс TurbeUaria
81
Отряд Triclodida (^Seriata) — планарий
Материал. В канавах с проточной водой, в медленно текущих
ручьях с растениями и опавшими листьями, в прибрежной части озер и рек
с чистой водой можно найти молочно-белую планарию (Dendrocoelum
lacteum), хорошо различимую благодаря белой окраске и достаточно
крупным размерам (до 2-2,5 см). В одних и тех же местах эта планария
встречается в течение нескольких лет. Найдя однажды водоем с пла-
нариями, можно получать необходимый материал летом и осенью.
Чаше всего молочно-белая планария обнаруживается на неглубоких
местах под камнями, щепками, досками, на стеблях и плавающих
листьях водных растений. Собранные планарий хорошо содержатся
в лабораторных условиях в аквариумах с элодеей и небольшим
количеством ряски. Благоприятно периодическое аэрирование воды
продуванием. Следует предохранять воду от загнивания.
Хорошо хранятся планарий в небольших сосудах, в том числе
в чашках Петри, на нижней полке холодильника, что позволяет
демонстрировать живой материал даже поздней осенью.
В различных водоемах на стеблях водных растений, в пазухах
телореза, аира, ситниковых можно обнаружить других планарий: бурую
(Planaria torva), черную (Planaria lugubris), многоглазку черную (Poly-
celis nigra) (рис. 39).
Изучение живого объекта. Морфология молочно-белой планарий
(Dendrocoelum laaetun). Тело планарий вытянуто, сплюснуто в дорсовен-
тральном направлении (рис. 40). Передний, головной отдел, уплощен,
Рис. 39. Пресноводные планария: А — Planaria torva; В — Planaria
lugybrbr, В — Pofycelis nigra
82
Тип Plalhelmifrthes]
Рнс. 40. Dcndrocoelum lacteum. Внешний *нд
плакдрии: / — присасывательная
бороздка; 2 — боковые лопасти; 3 —
глаза; 4 — рот; 5 — глотка; 6 —
глоточный карман; 7 — передняя ветвь
средней кишки; 8 — задние ветви
средней кишки
средняя часть имеет ширину около 6-7 мм.
Задний конец заострен. На головном
отделе даже при слабом увеличении хорошо
видны два черных глаза. Средняя часть
головного отдела (лобный край) слегка
выпота и утолщена, образуя
присасывательную бороздку, лучше заметную на
фиксированном материале. По бокам от
лобной бороздки располагаются короткие,
широкие, закругленные боковые лопасти,
выполняющие осязательную функцию.
Расположенные на переднем конце тела
мерцательные ямки являются хсморе-
цепторами.
В средней части брюшной стороны
тела расположен рот, за ним — половое
отверстое. Выдел кгслышс поры лежат по бокам тела со спинной стороны
и слабо различимы, как и половое отверстие.
Движение оланарин. Поместив живую планарию в чашку Петри,
можно наблюдать за ее движением. Чаше планария плавно скользит
по субстрату, благодаря движению ресничек покровного эпителия.
Колебания ресничек обеспечивают также циркуляцию воды и
улучшают кожное дыхание червей. Потревоженная планария способна к
более активному движению за счет сокращения мускулатуры
кожно-мускульного мешка, при этом края тела ее волнообразно изгибаются.
При раздражении планария может прикрепляться к субстрату
присасывательной бороздкой и резко сокращаться, подтягивая тело к
переднему концу. Затем головной отдел ее быстро выбрасывается вперед и
снова прикрепляется. Таким образом, планария способна и к
своеобразному «шагающему» движению. Движение ресничек покровного эпителия
^cc TurbeHaria 83
зу1|аЮт под микроскопом, поместив планарию в каплю воды на
предметное стекло, накрыв ее покровным стеклом с восковыми ножками
и слегка придавив стекло препаровальной иглой.
Питание. Планария — хищница. Пищей ей служат мелкие водные
беспозвоночные: водяные ослики, олигохеты, личинки насекомых,
мелкие моллюски и др. При отсутствии живого корма планарий можно
кормить кусочками мяса, дождевых червей. Потребляют они и сгустки
крови. Остатки пиши следует тщательно убирать, не допуская
загнивания воды. Кормление кровью удобно для демонстрации процесса
питания на занятиях. Перенеся животное в чашку Петри, под биноку-
ляром можно наблюдать выворачивание глотки, захват пиши и
постепенное заполнение кишечника, который при этом становится ко>прастным.
Пищеварительную систему молочно-белой планарий следует
изучать на постоянных тотальных препаратах. Препараты изготавливаются
после кормления планарий (рис. 40). На брюшной стороне в задней
трети тела расположен рот, ведущий в мускулистую глотку,
окруженную объемным глоточным карманом. Иногда глотку называют
сосательным хоботком, так как она, выдвигаясь из глоточного
кармана, способна проникать в тело жертвы, выделяя при этом секреты,
чем способствует перевариванию и последующему всасыванию
растворенных тканей добычи. Средняя кишка образует три ветви.
От глотки отходит передняя ветвь среднего отдела кишечника,
лающая вправо и влево ответвления первого, второго и даже третьего
порядка. Назад от основания передней ветви отходят две ветви средней
кишки, идущие вдоль глоточного кармана до заднего конца тела.
)ти ветви также дают незначительные боковые ответвления. В связи
с отсутствием задней кишки и анального отверстия непереваренные
остатки пиши выбрасываются через ротовое отверстие при активном
сокращении тела и кишечника.
При изучении поперечного среза планарий удается рассмотреть
элементы кожно-мускульного мешка, паренхиму и расположенные
в ней дорсовентральиые и поперечные мышцы (рис. 41).
Пол ресничным эпителием и базальной мембраной
расположены слои кольцевых, диагональных и продольных мышц.
Все промежутки между внутренними органами и стенками тела
заполнены клетками паренхимы, имеющими разнообразную
неправильную форму и рыхло соединенными друг с другом. Промежутки
между клетками паренхимы заполнены однородным промежуточным
веществом. Хорошо различимы на поперечном срезе
дорсовентральиые мускульные волокна, расположенные чаще всего вертикально.
84
Тип Plathelminthes
Рис. 41. Схематичный поперечный разрез через тело плоского червя:
/ — покровный эпителий; 2— кольцевые мышцы; 3 —
продольные мышцы; 4 — паренхима; 5 — протонефридии; 6 —
выделительная пора; 7— дорсовеитрол1»ные мышцы; S— половое
отверстие; 9— нервный тяж; /0 — половая железа; // — кишечник;
/2 — э>оодермалы1ый кишечный эпителий
Надкласс Neodermata
Объединяет паразитических плоских червей, утративших в
результате паразитического образа жизни типичный эпидермис и
выработавших новый синцитиальный покров (неодермис).
Класс Trematoda (-Digenea) — дигенстические
сосальщики
К дигенетическим сосальщикам относятся паразитические черви,
развитие которых происходит со сменой двух или более хозяев. Все
дигенстические сосальщики — эндопаразиты беспозвоночных и
позвоночных животных. Наиболее широко распространены и имеют
хозяйственное значение печеночная двуустка, паразитирующая в желчных
протоках печени овец, крупного рогатого скота,, изредка у человека,
и ланцетовидная двуустка, встречающаяся в печени тех же хозяев.
Dicrocoelium dendriticum — ланцетовидная двуустка
Морфология. Тело ланцетовидной двуустки уплощенное,
ланцетовидной формы, имеет длину 7-10 мм при ширине 1-1,5 мм. На
переднем конце расположена крупная чашевидная присоска, несколько
Класс Trematocto
85
смешенная на брюшную сторону тела. В центре ее находится ротовое
отверстие. Вторая, брюшная присоска лежит медиально на брюшной
стороне тела и отличается большими размерами и сильно развитым
мускульным валиком. Как и у печеночного сосальщика, присоски —
органы фиксации, позволяющие удерживаться в протоках печени
крупного рогатого скота и овец — основных хозяев эндопаразита
(рис. 42).
Пищеварительная система. От ротового отверстия, лежащего в центре
1*>товой присоски, отходит глотка, за ней следует короткий тонкий
пищевод, дающий две прямые ветви средней кишки,
расположенные по бокам тела и заканчивающиеся слепо. Ветви средней кишки
бывают не видны на всем протяжении, так как маскируются другими
системами органов (чаше всего желточниками).
Половая система гермафродит мая. Органы мужской половой сие-
1смы представлены двумя крупными, неправильной формы,
компактными семенниками, расположенными
не совсем симметрично ниже брюшной
присоски. От семенников вперед отходят
два семяпровода, соединяющихся вес-
мяизвергатсльный канал,
пронизывающий копулятивный орган и лежащий в
мешковидном впячивании стенки тела —
половой клоаке.
Женская половая система имеет один
яичник, расположенный позади
семенников. Короткий яйцевод открывается
в небольшой мешковидный оотип,
принимающий также протоки желточников,
Рис. 42. Ланцетовидная двуустка (Dtc/v-
coelium dendritkum): I — ротовая
присоска; 2 — глотка; 3 — ветвь
кишечника; 4 — брюшная
присоска; 5 -желточник; 6 — матка;
7 — яичник; 8 — семенник; 9 —
половая клоака; 10 — половое
отверстие
86
Тип Plathelminthes
идущих по бокам тела и имеюших гроздьсвидную форму. В оотип
впадают протоки тельца Мелиса, он также сообщается с
семяприемником и коротким лауреровым протоком (последние иногда плохо
заметны на препаратах).
От оотипа отходит сильно извитая матка. Она идет вначале
к заднему концу тела, затем поворачивает вперед и открывается
половым отверстием рядом с совокупительным органом. По мере движения
по каналу матки оплодотворенные, снабженные запасом
питательного желтка и скорлупой яйца созревают.
Fosciola hepatica — печеночная двуустка
Материал. Печеночная двуустка довольно часто обнаруживается
в протоках печени крупного рогатого скота. Зараженная печень легко
распознается, так как желчные протоки в ней вздуваются,
приподнимая местами ее поверхность. Кроме того, паразиты вызывают
сильное обызвествление стенок протоков, что определяется на ощупь.
Морфология печеночной двуустки. Тело сосальщика листовидное,
расширенное в передней части и постепенно сужающееся кзади.
Длина тела достигает 3-3,5 см при ширине около 1,5 см.
На переднем конце, на небольшом конусовидном выросте
располагается передняя, или ротовая, присоска, в центре которой
находится ротовое отверстие, а на брюшной стороне тела в конце передней
его трети — задняя, или брюшная, присоска. Ротовая присоска
имеет чашевидную форму, ограничена кольцевой мышцей и хорошо
различима под микроскопом. Брюшная присоска имеет
поверхностное положение и с внутренними органами не связана (рис. 43).
Ротовая и брюшная присоски выполняют роль органов
фиксации, обеспечивая удержание сосальщика в протоках печени.
Пищеварительная и половая системы печеночного сосальщика.
Пищеварительная система с успехом может быть изучена на
тотальных, окрашенных кармином препаратах. Ротовое отверстие ведет
в мускулистую глотку, от которой отходят две ветви кишечника,
продолжающиеся до заднего конца тела и многократно ветвящиеся.
Боковые ответвления двух основных пищеварительных каналов
достигают боковых стенок тела, что облегчает распределение продуктов
пищеварения между органами и тканями через межклеточные участки
паренхимы. Как у всех плоских червей, средняя кишка слепо
замкнута, непереваренные остатки пищи удаляются через рот.
Класс Tremstoda
87
Рис. 43. Пищеварительная система
печеночной двуустки (Fascioh hepatica):
I — ротовая присоска; 2— глотка;
3 — пищевод; 4 — брюшная
присоска; 5 — ветви кишечника; 6 —
боковые ответвления кишечника
Половая система печеночного
сосальщика, как и у остальных
паразитических плоских
червей-гермафродитов, представлена сложным
комплексом женских и мужских половых
органов (рис. 44).
В передней трети тела, ниже
брюшной присоски, справа располагается
единственный разветвленный яичник,
от него отходит я й цевод, дистальный
конец которого образ>чгт небольшое
расширение (у других сосальщиков ему
соответствует оотип). В дистальный
конец яйцевода впадает непарный проток желточииков. Сами желто-
чники имеют гроздевидную форму, занимают боковую и заднюю часта
тела. Их фолликулы образуют питательные клетхи для
формирующихся яиц. За счет особых гранул этих клеток образуется и скорлупа яиц.
Протоки жеяточников проходят вдоль тела в передней и задней его
части, а на уровне яйцевода дают поперечные ветви и соединяются
в непарный проток, расширяющийся в желточный резервуар. От
расширенной частя яйцевода отходит длинный извитый канал матки,
идущий вперед и открывающийся отверстием рядом с мужским
половым отверстием.
В расширение яйцевода рядом с резервуаром желточника впадают
протоки многочисленных скорлуповых желез, образующих в
совокупности шарообразное скопление — тельце Мелиса, и отходит лау-
рсров проток, служащий, вероятнее всего, для удаления избытка
спермы. Секрет, образуемый тельцем Мелиса, заполняет канал матки
и способствует длительному сохранению спермы после копуляции
гермафродитных червей.
88
Тип Plathelminthes
Рис. 44. Гсрмафродитная полом*
система печеночного
сосальщика: / — яичник; 2 — яйцевод
с дисталъным расширением;
J — желточник; 4 — проток
желточников; 5—
семенники; б — семяпроводы, 7 —
матка; 8— семяизьергатель»
ный канал в половой клоаке
Мужская половая система
представлена парными семенниками,
лежащими между желгочниками
в задней половине тела. От них
вперед отходят два семяпровода,
которые сливаются впереди
брюшной присоски, образуют семяизвер-
гательный канал. Расположенный
в половой клоаке, он
пронизывает совоку п игольный орган (цир-
рус) и открывается мужским
половым отверстием.
При копуляции двух герма-
фродитных особей циррус вводится
в женское половое отверстие
партнера и сперма по свободной матке
поступает в расширение яистальной части яйцевода, где и происходит
оплодотворение зрелых яйцеклеток. Желточные клетки, проходящие
по протокам желточников, обеспечивают запас питательного материала
и формирование оболочек оплодотворенных яиц, после чего яйца
поступают в матку. Здесь завершается их созревание. Зрелые яйца отличаются
более темной окраской и при изучении сосальщика под микроскопом
своеобразно преломляют свет.
Изучение некоторых стадий развития печеночного сосальщика
Материал. Живой материал для изучения стадий развития
печеночного или других видов дигенетических сосальщиков можно
получить из легочных моллюсков (прудовиков и катушек), являющихся
промежуточными хозяевами трематод. Используются два метода.
Кяасс Trematoda
89
I. Живых моллюсков (лучше взять крупных прудовиков)
помешают вбанку с водой, выставляют се на прямой солнечный свет и
наблюдают в течение 2-2,5 часа. Если моллюски заражены, из них в воду
вы идут личинки гсрмафроштюй особи, мариты, — церкарии,
имеющие вид мельчайших белых точек. При большом количестве церкарии
образуют маленькие белые облачка. Приготовив временный
препарат, церкарии можно изучить под микроскопом.
II. Если время для изучения ограничено, получить трематод на
различных стадиях развития можно при вскрытии моллюсков. Для этого
пинцетом удаляется раковина в области завитка, где находится
печень моллюска. У зараженных моллюсков печень из
зеленовато-бурой становится желтой или желто-ее рой. Выделив ее в часовое стекло
с небольшим количеством воды, печень расщепляют
препаровальными иглами, делают временный препарат и изучают под микроскопом
споро цисты, редии и церкарии.
Спдаи развития трематод. В печени моллюска из проникшей
личинки мирацидия развивается следующая стадия — спороциста
(рис. 45). Это партсногенегическая самка (партенита), способная к пар-
геногенетическому размножению. Она имеет мешковидную, слегка
вытянутую форму. Из зародышевых шаров, заполняющих тело спо-
роцисты, развивается следующее поколение партенит — редии. Редии
сосальщиков имеют вытянутую форму, на одном из концов
располагается рот, глотка и мешковидный кишечник. Все внутреннее
пространство заполнено зародышевыми шарами или сформированными
личинками мариты — церкариями на разных этапах развития. При
достаточно длительном наблюдении можно проследить выход из редий
подвижных церкарии.
Тело церкарии состоит из овального переднего отдела и
мускулистого хвоста — органа движения. На переднем конце хорошо различима
ротовая присоска, на средней части тела — брюшная.
Пищеварительная система включает рот, глотку, узкий пищевод и две ветви средней
кишки.
Церкарии после некоторого периода активного движения либо
инцистируются и переходят в стадию метацеркария (ланцетовидная
двуустка), либо оседают на прибрежной растительности,
покрываются оболочкой и превращаются в адолескарию (печеночный сосаль-
<'шк). При проглатывании адолескарии окончательным хозяином из
нес развивается марита.
При вскрытии легочных моллюсков из водоемов с
многочисленными водоплавающими птицами можно обнаружить спороцисты и
церкарии трематод сем. Schistosomatidoe. Церкарии этих трематод имеют
мускулистый, вилообразно разделенный хвост и называются фурко-
90
Тип Plethefminthes
Рис. 45. Стадии развития печеночного сосальщика: А — марита; Б —
яйио; В — мирацидий; Г— спороииста; Д — рсдия; Е —
иеркария; Ж — адолескария в капсуле; / — ротовая присоска; 2 —
брюшная присоска; 3— матка; 4—семенники; 5—желточник;
б — тельце Мелиса; 7— яичник; 8— эмбрион; 9 —
зародышевые клетки; 10— редии; //— протонефридий; 12 — иеркария;
13 — кишечник; 14— хвост; 15 — капсула
Класс Monogenoidea
91
цсркариями (рис.46). Птичьи шистосомапшы нормально завершают
развитие в теле водоплавающих птиц, но способны, проникая в покро-
пы человека, вызывать кожные поражения и аллергические реакции
разной степени тяжести. Это заболевание получило название шисто-
сомиого церкариоза, «зуда купальщиков», «водного зуда».
Рис. 46. Цсркарии птичьих шистосомзтид: А — Trickobilharzlo oceilata;
Б — Bilharzjella poionica
Класс Monogenoidea — моногенеи, или
моногенетические сосальщики
Моногенеи — исключительно паразитические животные,
преимущественно эктопаразиты. Развитие происходит без смены хозяев.
92
Тип Plathelminthes
которыми являются чаще всего рыбы, некоторые развиваются на
амфибиях. Отличительной особенностью моногеней является развитие
на заднем конце их тела прикрепительного диска с присосками,
крючьями или клапанами.
Отряд Monopisthocotytea (—Polyonchoinea)
Polystoma integerrimum — лягушачий многоуст
Материал. Лягушачий многоуст во взрослом состоянии
паразитирует в мочевом пузыре молодых лягушек. Относительно крупных
размеров (до 10 мм) паразит хорошо виден через стенки мочевого
пузыря. Выделив мочевой пузырь лягушки, его помешают в
физиологический раствор в часовом стекле, вскрывают стенки и рассматривают
червя in vivo. Хорошие результаты даст изучение тотального
препарата с брюшной стороны.
Морфология. Тело многоуста удлиненное, сужено к переднему
и расширено к заднему концу, дорсовентрально уплошено (рис. 47).
На заднем конце располагается прикрепительный диск,
отделенный от остального тела небольшим пережимом. Диск несет шесть
присосок, расположенных попарно по внешнему краю. Присоски
по краям снабжены мускульными валиками. Между присосками задней
пары хорошо различимы два крупных крючка, на диске также
находится до 16 мелких крючков, которые не всегда видны. Они
сохраняются у червя отличники и после метаморфоза не увеличиваются
в размерах.
Пищеварительная система начинается ртом на переднем конце
тела. За небольшой глоткой кишечник образует две ветви, вторично
сильно ветвящиеся, в заднем конце тела соединяющиеся и также
дающие ветви в зоне прикрепительного диска. Ветви кишечника
переплетаются, образуя сетсвидную пищеварительную систему.
Половая система гсрмафрошпная. Семенник расположен дорсаль-
но (с брюшной стороны перекрывается желточниками), семяпровод
в переднем конце тела переходит в семяизвергатсльиый канал,
открывающийся половым отверстием в половой клоаке.
Женская половая система представлена крупным непарным
яичником. Яйцевод соединяется с протоками сильно развитых желточ-
ников, скорлуповых желез и переходит в матку, половое отверстие
которой расположено в половой клоаке рядом с мужским. Имеется
два влагалища, начинающиеся на переднем конце тела червя двумя
бугорками.
цассМоподепоШеа
93
Рис. 47. Лягушачий многоуст (Potystoma intcgenimum). Общий вид червя
с брюшной стороны: / — ротовая присоска; 2— глотка; J—
наружное половое отверстие; 4 — парное влагалище; 5 —
семяпровод; 6 — матха; 7— поперечный желточный проток; 8— желто*!-
ники; 9 — кишечник; 10— присоски прикрепительного диска;
// — крючки прикрепительного диска; /2 — яйцевод; /J—
яичник
94 Тип Ptethelmiathes
Отряд Polyopistftocotylea (-Oligoncftoinea)
Diplozoon paradoxum — спайник парадоксальный (рис. 48)
Спайник паразитирует на жабрах различных карповых рыб (леща,
густеры, плотвы и др.), где и может быть обнаружен. Интересен
паразит тем, что развитие и размножение гсрмафродитных взрослых
чсрьсй возможно только при условии срастания их попарно на стадии
личинок, что обеспечивает перекрестное оплодотворение.
Морфология. Каждая особь имеет удлиненную форму, размеры
До 0,5 см. Передняя часть листовидная, сужена впереди, в месте
срастания с партнером обрззуст перехват. На заднем конце тело расширяется.
Здесь находится прикрепительный аппарат, состоящий из четырех пар
хнтнноилных прикрепительных клапанов с двумя створками в
каждом. При складывании створок между ними ущемляются жаберные
лепестки рыбы, обеспечивая фиксацию паразита.
Рис. 48. Спайник парадоксальный (Diplozoon paradoxum). Внешний вид:
/ — ротовое отверстие; 2 — ротовая присоска; 3 — глотха; 4 —
кишечник; 5— желточники; 6 — половые железы и их
протоки; 7 — прикрепительные клапаны
Класс Cestoda — ленточные черви
Ленточные черви — исключительно паразитические животные,
обитающие в кишечнике позвоночных; промежуточными хозяевами
WiccCestoda
95
}{Ч являются различные беспозвоночные и позвоночные животные.
ц связи с паразитическим образом жизни ленточные черви
изменились сильнее, чем сосальщики. Они полностью утратили
пищеварительную систему (кишечник), органы чувств, значительно
усложнились их покровы (развился тегумент) и цикл развития, в котором,
как правило, отсутствует стадия жизни во внешней среде.
Среди ленточных червей, паразитирующих у человека, наибольшее
значение имеют бычий и свиной солитеры, лентец широкий и
эхинококк, строение которых рассмотрено ниже.
Отряд Cyclophyllidea — цепни
Taenia solium — свиной солитер
Материалом для изучения строения свиного солитера являются
препараты различных участков тела червя и стааий его развития, а также
влажные демонстрационные препараты.
Морфология. Тело свиного солитера состоит из трех частей: ско-
лскса, или головки, шейки и стробилы, состоящей из большого числа
члеников — проглоттид (рис. 49).
Сколекс имеет округлую форму, в поперечнике достигает
1,5-2 мм. На нем с четырех сторон расположены присоски, а в верхней
Рис. 49. Внешний вид ленточного
червя Taenia solium: l —
сколекс; 2 — венчик
крючьев; 3 — присоски;
4 — нерасчлененная
шейка; 5— стробила; б— про-
глоттиды (а — незрелые,
б — гермафродитные, в —
зрелые)
96
Тип Phthdminthts'
части (терминально) находится хоботок, способный втягиваться
и несущий двойной венчик крючьев с маленькими зубчиками у
основания, в связи с чем свиной солитер получил название вооруженного
цепня. Присоски и система крючьев обеспечивают фиксацию червя
в кишечнике.
Шейка представляет собой нерасчлененный участок тела. Это—
зона роста. Путем почкования от шейки отшнуровываются проглот-
тиды, образуя цепочку — стробилу, включающую сотни члеников.
Стробила свиного солитера имеет длину до 2-3 м. Начальные'
членики стробилы очень малы, ширина их превышает длину. По мере
нарастания стробилы за счет отделения от шейки молодых члеников
образовавшиеся ранее проглоттиды удаляются от переднего конца тела,.
увеличиваются в размерах и становятся почти квадратными,
прямоугольными или удлиненными.
В стробиле различают три основных типа проглотгид. Самые
молодые из них не имеют сформированной половой системы и называются
незрелыми (их приблизительно около 200). Затем следуют членики,
в которых завершается развитие гермафродитной половой системы.
Они получили название гермафродитных. Концевую часть
стробилы занимают членики, содержащие зрелые яйца, поэтому и названы
они зрелыми.
Гермафродитный членик дает представление о строении
половой системы ленточных червей, так как половая система каждого
членика автономна (рис. 50).
Рис. 50. Гермафродитный членик свиного солитера (Taenia solium): I —
яичник; 2— яйисьод; J— желточиик; 4—oemm; 5—влагалище;
б — половая клооха; 7— семякзвергательный канал; £—
семенники; 9— матка; /0— выделительный какал
Класс Cestoda
97
Мужская половая система образована многочисленными
мелкими пузыревидными семенниками, рассеянными в паренхиме
членика. Отходящие от семенников семенные канальцы (часто слабо
различимые) сливаются в непарный семяпровод, переходящий все-
мяизвергательный канал, пронизывающий совокупительный
орган (циррус). Ссмяизвсргательный канал проходит через половую
клоаку и открывается на одной из боковых стенок проглоттида
половым отверстием. Обычно половая клоака и половое отверстие в
соседних члениках располагаются то слева, то справа.
Женская половая система включает один трехлопастный
яичник, боковые лопасти которого значительно больше средней. От
яичника отходит короткий яйцевод, впадающий в оотип. Желто-
чник один, распаюжен в задней части членика, имеет треугольную
форму и сстсвидную структуру. Проток желточника также открывается
и оотип. В оотип поступает секрет скорлуповых желез (тельце
Мелиса). От половой клоаки отходит прямой канал влагалища, на
внутреннем конце расширенный и образующий семяприемник,
соединяющийся с яйцеводом. Перекрестное внутреннее
оплодотворение происходит между члениками путем введения цирруса в канал
влагалища. Возможно и самооплодотворение.
От оотипа начинается канал матки. В молодых гермафролитных
члениках матка имеет вид слегка изогнутой,-слепо замкнутой трубки,
достигающей переднего края проглоттида. По мере формирования
оплодотворенные яйца поступают в матку и постепенно растягивают
се, образуя боковые ответвления. Одновременно с развитием матки
происходит редукция сначала семенников и ссмявыносящих каналов,
а затем, когда завершается образование яиц, и женской половой
системы. К этому периоду в членике остается сильно разветвленная матка,
протоки влагалища и семяпровода, циррус и половая клоака. Такой
членик и является зрелым. В его матке с каждой стороны от
основного канала есть от 7 до 12 боковых ветвей (рис. 51).
Суммарная потенциальная плодовитость свиного цепня,
живущего 10-15 лет, достигает 200-300 млн яиц.
Выделительная система цестод хорошо заметна на препаратах
гермафролитных члеников (рис. 50, 52). Продольные каналы идут вент-
рально вдоль боковых сторон от конца стробилы к сколсксу, здесь
они поворачивают назад и продолжаются дорзально до конца
стробилы. В каждом членике боковые каналы соединяются поперечными
перемычками, смешенными к заднему краю проглотгид. Таким образом.
98
Тип Plathdminthes
Р*С. 51. Зрелый членик свиного солитера:
/ — матка; 2— влагалище; J —
половая клоака
выделительная система цестод напоминает своеобразную лестницу.
При изучении препаратов обычно хорошо видны боковые каналы»
расположенные латерально от семенников, и перемычки, лежащие позади
желточника.
Отдельные элементы полового аппарата у разных ленточных
червей сильно варьируют. Однако у цестод они принципиально сходно
устроены и различаются незначительно.
Вместе с тем для диагностики заражения человека основными
видами цестод эти различия важны. Поэтому при изучении строения
бычьего цепня необходимо обратить внимание на такие
отличительные особенности.
Taeniorhynchus saginatas — бычий цепень
Сколскс бычьего цепня имеет грушевидную форму, для
фиксации на стенках кишечника он снабжен лишь четырьмя присосками
овальной формы. У него нет крючьев, поэтому бычьего солитера
называют цепнем невооруженным (рис. 52, Л).
Стробила имеет длину от4до 10 ми включает более 1000 члеников.
Гермафродитные членики содержат многочисленные (до 1000)
семенники, занимающие большую часть проглоттида. Яичник
двухлопастный, его лопасти состоят из многочисленных трубчатых долек,
которые сходятся в средней, соединительной части (рис. 52, £).
Зрелые членики сильно вытянутые, длина их составляет 16-30 мм
при ширине 5-7 мм. Число боковых ветвей матки колеблется от 17 до
35 с каждой стороны от главного канала (рис. 52, Б). Зрелые членики,
отделяющиеся от стробилы и выделяющиеся из кишечника с калом,
способны к самостоятельному движению благодаря сокращению
собственной мускулатуры.
Значительные размеры тела, мощная матка в зрелых члениках,
большая продолжительность жизни (18-20 лет) обусловили
колоссальную плодовитость бычьего цепня (до 600 млн яиц в год и около
11 млрд. за всю жизнь).
Класс Cestoda
99
Ямс. 52. Бычий солитер (Taenlarhynchus sa&natus): А — сколскс; Б —
гсрмвфродитный членик; В — зрелый членик; / — присоска; 2 —
шейка; J— матка; 4— семенники; 5— семенные канальцы;
б—семяпровод; 7— патовая клоака; 8— половой бугорок; Р—
влагалище; /0 — яичник; // — желточник; 12 — скорлуповыс железы;
13 — поперечный анастомоз выдстителъной системы; 14 —
продольный выделительный канал
100
Тип Ptithelminthes
Стадии развития свиного или бычьего цепня
Материал. Препараты яиц и финны цепней; влажные препараты
финнозного мяса и цепней демонстрационно.
Яйца. В оплодотворенном яйце до выхода члеников из кишечника
человека формируется личинка, имеющая шаровидную форму и раз*
меры до 0,03 мм в поперечнике, снабженная шестью кутикулярными
крючьями. — онкосфера, или шестикрючный зародыш. От внешних
воздействий зародыш защищен скорлупой яйца и собственной
оболочкой, которая радиально исчерчена и хорошо различима при
изучении препарата яиц цепней пол микроскопом (рис. S3, В).
Выделенные во внешнюю среду проглотгилы могут быть
заглочены подходящим хозяином (крупный рогатый скот или свинья), в
кишечнике онкосфера выхолит из яйца, оболочка которого разрушается
под действием пищеварительных ферментов хозяина. Личинка
внедряется в стенку кишечника хозяина, попадает в кровяное русло и
разносится по организму, оседая в разных органах, чаше всего в мышцах,
где образует следующую личиночную стадию — финну, или
пузырчатую глисту (рис. 53,>У).
Финна представляет собой пузырь, заполненный жидкостью,
размером с маленькую фасолину (около 7 мм). В одном из участков в стенке
финны образуется впячиванис, на котором развивается будущий ско-
лекс с присосками н крючьями (у свиного цепня) либо только с
присосками (у бычьего). В тканях тела хозяина финна сохраняется без
изменений достаточно долго, иногда несколько лет. Таким образом, для
цепней копытные и свиньи являются промежуточными хозяевами.
При потреблении финнозного мяса в кишечнике человека под
действием пищеварительных ферментов головка финны
выворачивается и закрепляется с помощью органов фиксации (присоски, крючья)
на слизистой кишечника. Пузырь финны некоторое время
удерживается на конце тела, затем отрывается и разрушается. От шейки
отделяются членики, стробила растет, в проглоттилах формируется герма-
фродитная половая система и начинается период полового
размножения червей. Человек — основной, или дефинитивный, хозяин
цепней.
Отряд Pseudophillidea — псевдофиллидеи
Diphyllobothrium latum — ленггец широкий
Лешпец широкий — один из самых крупных паразитических
плоских червей. Длина стробилы достигает 9-10 м, описаны черви длиной
до 15-20 м. Количество члеников может превышать4000. Цвет червя.
HxccCestoda
101
Рис. 53. Цикл развития свиного солитера: А — половозрелый гсрмафро-
дитный червь; Б — зрелый члеиик; В — оплодотворенное яйцо
с личинкой; Г — онкосфера; Д — финна в мускулатуре
промежуточного хозяина; £ — финна с вывернутым сколсксом в кишечнике
окончательного хозяина
как и всех внутренних паразитов (эндопаразитов), серовато-белый
или желтоватый.
Вес членики стробилы имеют прямоугольную форму, причем
ширина их значительно превышает длину.
102 TmPtathelminthts
Половая система лентеца широкого образована
многочисленными семенниками, занимающими боковые части членика по обеим
сторонам женской половой системы (рис. 54). Семенные канальцы
в задней половине членика соединяются в семяпровод, основание
его расширено в семенной пузырь, в котором собирается сперма.
Семяпровод, извиваясь, продолжается вперед и заканчивается цирру-
сом. В мешке цирруса проходитсемяизвергательный канал,
открывающийся половым отверстием в половой клоаке.
Женская половая система представлена двухлопастным яичником
в задней части членика. От средней соединительной части яичника
отходит яйцевод, впадающий в оотип, кула открываются протоки
тельца Мелиса, канала влагалища, протоки желточника.
Многочисленные (более 6000) фолликулы желточников занимают боковые
части члеников, они меньше семенников и хорошо отличимы от
последних.
Влагалище начинается шелсвидной порой в половой клоаке и
продолжается в виде прямой трубки до средней части членика, образуя
у места впадения яйцевода в оотип расширение — семяприемник.
Отоотмпа к передней части членика отходит матка. В гсрмафро-
дитных молодых члениках она имеет вид скрученного петлевилного
канала, открывающегося наружу самостоятельным отверстием
позади половой клоаки. В задних члениках стробилы матка, имеющая вид
розетки, набита яйцевыми капсулами. Таким образом, в отличие от
цепней, матха у лентецов открытая. Зрелые яйца постоянно
выделяются в просвет кишечника и выводятся наружу.
2 10 1 9 8 7.
Рис. 54. Гермафродитная полон*
система лентсдо широкого (Diphyl-
hbothrium latum): I —
совокупительный орган в половой
клоаке; 2 — семяпровод; 3 —
семенники; 4 — яичник; 5 —
оотип и скорлуповые
железы; б— желточники; 7—
проток желточника; 8 — матка;
9 — апагалище; 10 —
отверстие матки
4 5 в
Класс Cestoda
103
Рис. 55. Цикл развития лентеца широкого: / — оплодотворенное яйцо;
2 — свободноживушая личинка корацидий; 3 — лроиеркокд
в полости тела зараженного циклопа; 4 — плероиеркоид в
органах и тканях рыбы; 5 — зрелый членик червя в организме
окончательного хозяина
104
Тип Plathelminthes
Цикл развития лсегтеца широкого в связи с постоянным выделе ни -
ем яиц через отверстие матки отличается от цикла развития цепней. Из яиц
лентеца, попавших в воду, выходит свободно плавающий, покрытый
ресничками шестикрючный зародыш — корацидий. Если корацидий
проглатывается рачком-циклопом, он сбрасывает реснички и в
полости тела рачка превращается в червеобразного процеркоида. При
проглатывании зараженного циклопа рыбой в полости ее тела и
органах процеркоид превращается в плероцеркоид. Завершение
развития происходит в кишечнике человека или других рыбоядных
млекопитающих, съевших зараженных рыб (рис. SS).
Тип Nemathelminthes — круглые черви
Этот обширный тин объединяет огромное разнообразие свободно-
живущих, водных и почвенных, а также паразитических червей, раз-
питающихся в тканях растений и организме животных.
В отличие от плоских червей, они утрачивают паренхиму и между
внутренними органами и стенкой тела образуются шели.
заполненные жидкостью. Это — первичная полость, поэтому круглые черви
получили название первичиополостных. Другая отличительная черта
Nemathelminthes— редукция некоторых элементов кожно-мускулыю-
го мешка. Впервые у представителей этого типа появляется сквозной
кишечник. Все основные особенности организации собственно
круглых червей (класс Nematode) можно рассмотреть на примере широко
распространенных и хорошо изученных объектов — аскариды
лошадиной или свиной.
Класс Nematoda — нематоды
Подкласс Rhabditia (=Secernentea)
Отряд Ascaridida — аскариды
Parascaris equorum — лошадиная аскарида
Материал и методы изучения. В связи с опасностью заражения
на занятиях человеческая аскарида заменяется вышеуказанной
лошадиной или свиной {Ascaris suum). Используется материал,
фиксированный в 50% спирте или 6% растворе формалина (в нем аскариды не
сморщиваются). Если нужно получить материал в спирте, фиксацию
следует начинать с 5% спирта и постепенно, в течение одного-двух
месяцев, довести до 45-50%. За несколько часов до вскрытия аскарид,
фиксированных формалином, следует перенести из фиксатора в воду.
Вскрытие производится в препаровальной ванночке под водой.
Это правило особенно важно соблюдать, так как полостная жидкость
аскариды содержит большое количество токсичных органических
кислот, способных вызвать сильное раздражение при попадании на
слизистые оболочки глаз и дыхательных путей. Из-за упругости стенок
тела полостная жидкость находится в нем под давлением (тургор)
и при надрезе фонтанирует.
Дополнительно детальное изучение внутренних органов
аскариды проводится на поперечных срезах, окрашенных гематоксилином
или борным кармином. Лучше использовать поперечные срезы через
тело самок, имеющих более сложную половую систему.
106
Тип Nemathdminthes
Морфология. Тело аскариды сильно вытянутое, всретеновиднос,
суженное на обоих концах, круглое в поперечном сечении. Длина
тела лошадиной аскариды колеблется у самки от 18 до 3S см, у самца
достигает 25 см при толщине 5-6 мм. Помимо различий в размерах,
половой диморфизм аскарид проявляется в форме их тела. Более
крупные самки имеют правильную веретеновидную форму, в передней
трети их тела находится кольцевой пережим, на котором с вентральной
стороны расположено половое отверстие, различимое с помощью
ручной лупы. Тело самца на заднем конце загнуто крючковидно на
брюшную сторону, что способствует удержанию самки при
копуляции (рис. 56).
Рис. 56. Лскарнда человеческая (Ascaris tumbricoides): A — самка; Б—самец;
В — строение и расположение кутикулярных г><> у лошадиной
аскариды (Parascaris tquorum)
Класс Nematode
107
Ротовое отверстие расположено в переднем, или «головном», отделе
тела и окружено тремя губами, имеющими форму валиков.
Сокращение мускулатуры губ обеспечивает ущемление стенки кишечника
хозяина и удержание паразита в его просвете. Анальное отверстие
расположено несколько впереди от заднего конца тела и имеет вид
поперечной щели. Участок за анальным отверстием — хвостовой отдел.
Вдаль тела аскариды идут просвечивающие сквозь покровы и
имеющие вид водянисто-белых линий два боковых утолщения гиподермы
с выделительными каналами. Наспинной и брюшной сторонах также
проходят тонкие беловатые линии, соответствующие спинному и
брюшному утолщениям гиподермы, в которых расположены спинной и
брюшной нервные стволы.
Вскрытие аскариды. Определив брюшную сторону, нужно
прижать аскариду ко дну препаровальной ванночки, прикрепить передний
и задний конец булавками и провести по спинной стороне разрез
вдоль всего тела. Края разреза развести в стороны и с внутренней
стороны приколоть так, чтобы головки булавок были направлены в
стороны от линии надреза, а сами булавки расположены наклонно,
прижимая стенки тела аскариды не только острием (образуются разрывы),
но и частью основания. При таком положении булавки не мешают
дальнейшему вскрытию и изучению объекта.
Внутреннее строение аскариды достаточно полно изучается на
вскрытом черве (рис. S7). В передней части тела различается деление
его стенки на четыре ленты, разделенные двумя боковыми и более
тонкой брюшной линиями, соответствующими гиподермальным
валикам. В боковых валиках располагаются выделительные каналы,
в брюшном — брюшной нервный ствол, по спинному валику
проведен разрез.
К выделительным каналам впереди прилегают буроватые или
желтоватые крупные фагоцитарные клетки, имеющие звездчатую форму. Они
извлекают из полостной жидкости и накапливают в себе нерастворимые
продукты обмена. Число таких клеток колеблется от I до 4.
Пространство между стенкой тела и внутренними органами
заполнено полостной жидкостью. Это — первичная полость тела. Она
выполняет распределительную (переносит продукты обмена,
питательные вещества) и опорную функции, усиливая упругость стенок тела
(является своеобразным гидроскелетом).
Пищеварительная система. От ротового отверстия начинается
мускулистая цилиндрическая глотка длиной около 1 см. За ней идет
108
Тип Nemathelminthes
Рнс. 57. Внутреннее строение аскариды человеческой: А — вскрытая
самка; Б — половая система самца; / — ротовое отверстие; 2 —
нервное кольцо; 3— глотка; 4— средний отдел кишечника; 5—
выделительный канал; б — фагоцитарные клетки; 7 — ленты
продольных мышц; 8— половое отверстие (снаружи); 9—
влагалище; 10— правая ветвь матки; // — яичники; 12— яйцевод;
13 — задняя кишка; 14 — анальное отверстие (снизу): 15 —
семенник; 16 — семяпровод; 17 — семяиэвергательный канал;
IS— кишка; 19— отверстие клоаки
Класс Nematoda
109
средняя кишка. Передняя ее четверть сплющена дорсовситрально
и имеет поперечную исчерчен ность. Эту часть кишки иногда называют
пищеводом. Остальная часть средней кишки цилиндрическая с
глубокими продольными складками. Задняя кишка у самок не
обособлена, а у самцов образует клоаку. Заканчивается кишечник анальным
отверстием на нейтральной стороне тела. У фиксированных червей
весь кишечник имеет лентовидную форму.
Половая система аскарид трубчатая, парная у самок и непарная
у самцов. Женская патовая система образована парными яичниками,
имеющими вид тончайших нитевидных тр><ючск. Постепенно
утолщаясь, они переходятвяйцеводы. Яичники и яйцеводы, образуя
многочисленные зигзагообразные изгибы, оплетают кишечник. В задней часш
тела яйцеводы переходят в двухветвистую матку, идущую к
передней трети тела и образующую непарное, сужающееся к концу
влагалище, которое открывается наружу единственным половым
отверстием. Оплодотворение внутреннее, происходит в матке, а
сформированные яйца выделяются в кишечник хозяина и с фекалиями —
во внешнюю среду.
Мужская половая система состоит из непарного нитевидного
семенника, переходящего в семяпровод, а последний — в ссмяиз-
вергатсльный канал, открывающийся в клоаку. Семенник и
семяпровод сильно извиваются, поэтому общая длина трубчатой половой
системы самца в 5-6 раз превышает длину тела червя. Семяизверга-
тсльный канал скрыт в складке кишечника.
При копуляции из клоаки выдвигаются две кутикулярные спи-
кулы, расположенные обычно в парной совокупительной сумке.
Изучение поперечного среза. Дополнительное представление о
строении и расположении органов и тканей аскариды ласт изучение
поперечного среза (лучше всего в средней части тела) (рис. 58).
В состав кожно-мускульного мешка входит покрывающая тело
кутикула, синцитиальная гиподерма и слой продольных мышц.
Кутикула хорошо развита, толстая, многослойная. Гиподерма,
выделяющая кутикулу, образует на боковых стенках два
трапециевидных валика, в которых проходят выделительные каналы. Гиподер-
мальные валики на спинной и брюшной сторонах тела небольшие,
каплевидные. В них при микроскопировании хорошо видны нервные
стволы.
Мускулатура представлена продольными мускульными
волокнами, непосредственно прилегающими к гиподерме. Продольные ги-
110
Тип Nemthelminthes
Рмс. 58. Поперечные срезы аскариды человеческой: А — в средней части
тела; Б — через яичник; В — через яйцевод; / — куш кула; 2 —
гиподерма; J— продольные мышечные волокна; 4—
плазматические отростхи мышечных клеток; 5 — брюшной гиподермальный
валик с брюшным нервным стволом; б— мала; 7—яйцевод; £—
боковой гилсаермальный валик с выделительным каналом; 9 —
кишечник; 10 — спинной гиподермальный валик со спинным
нервным стволом; // — яичник; 12 — яйцевые клетки; 13 —
рахис; 14 — стенка яйцевода
Класс Nematoda
111
полермальные валики делят кожно-мускульный мешок на четыре
посты (две спинно-боковыс и две боково-брюшные). Каждая
мускульная клетка имеет мышечное волокно, прилегающее к гиподерме и
соединяющееся с волокнами соседних мускульных клеток. От средней
часта волокна отходит в сторону полости тела саркоплазматичес-
кий мешок с длинными плазматическими мускульными отростками.
Тела мускульных саркоплазматических мешков хорошо видны и на
некрытом черве (имеют вид беловатой бахромы на внутренней стороне
стенки тела).
Средняя кишка имеет вид щели неправильной формы,
выстлана однослойным цилиндрическим эпителием, который подстилает
плотная базальная мембрана. Микроворсинки эпителия видны при
изучении лишь свежего материала.
Половая система. На срезе, сделанном в средней части тела самки,
можно различить все части половой системы: яичники, яйцеводы,
матку. Следует иметь в виду, что трубчатая половая система образует
многочисленные петли и изгибы, поэтому на препарате можно найти
указанные органы в поперечном, продольном и косом сечениях.
В начальной части яйце водной трубки зародышевые клетки (ооци-
ты) располагаются упорядоченно, радиально и имеют пирамидальную
форму. Вершины ооцитов упираются в центральный стержень — рахис
(рис. S8, £). На дистальном конце яичника находятся мелкие, рыхло
расположенные зародышевые клетки правильной округлой формы.
Яйцевод отличается от яичника большим диаметром,
отсутствием рахиса и свободным расположением ооцитов в просвете яйцевода
(рис. 58, В). В связи с большой длиной и особым расположением трубок
яичников и яйцеводов на одном препарате обнаруживается большое
и разнообразное число срезов этих структур. Напротив, ветви матки
всегда представлены двумя крупными срезами, имеющими
неправильную округлую форму и заполненными уже сформированными
яйцами на разной стадии развития.
Яйца аскариды (рис. 59) изучаются либо на постоянном
препарате, либо при выделении их из концевого участка матки и
изготовлении временного препарата.
Для изготовления временного препарата участок матки около
влагалища переносится на предметное стекло, расщепляется
препаровальными иглами в калле глицерина и покрывается покровным стеклом.
Поскольку яйца аскариды, попав во внешнюю среду, остаются
в ней достаточно долго до проникновения в нового хозяина, они
обладают большой устойчивостью благодаря трем защитным оболочкам.
112
Тип Nemathelminthes
Рис. 59. Яйца аскариды человеческой:
А — оплодотворенные; Б —
инвазионные
А Б
Внешняя оболочка бугорчатая, выделяется эпителием половых
протоков самки и окрашена в коричневый цвет. Средняя многослойная
белковая оболочка и внутренняя липилная выделяются самим яйцом.
Внутренняя лилидная оболочка служит для защиты яйца от
химических агентов. Благодаря липидной оболочке яйца аскариды
сохраняют жизнеспособность даже в таких средах, как насыщенные растворы
сулемы (HgCI2), Cu S04, ZnS04, HCI и др.
Отряд Tylenchida — шишкоиглые нематоды
Представители отряда характеризуются кольцевидной исчерченно-
стъю кутикулы покровов и наличием выдвигающегося из ротовой полости
колющего приспособления, называемого иглой (копьем, стилетом).
Игла пронизана сквозным каналом, служащим для выведения секрета
желез пищевода. У ее основания расположено мускулистое
шишковидное расширение пищевода — бульбус. Он несет функцию насоса,
обеспечивающего всасывание соков хозяина Эти особенности строения и дали
основания для названия отряда. Наибольшее хозяйственное значение
имеют нематоды семейства Hetenxkridae, многие из которых являются
опасными вредителями культивируемых растений.
Globodera rostochiensis — золотистая картофельная нематода
Материал. Золотистая картофельная нематода, хотя и является
объектом внешнего и внутреннего карантина, однако уже имеет
достаточно широкую распространенность. Пораженные растения картофеля
выделяются своим явно угнетенным видом. На корнях таких
растений можно обнаружить характерные светлые или коричневатые
желвачки размером с маковое зерно — это прикрепившиеся самки Globodera
rostochiensis. На занятиях лучше использовать тотальные
микроскопические препараты самок в канадском бальзаме или глицерин-желатине.
Класс Nematode
113
Для демонстрации флотационного метода учета цист золотистой
картофельной нематоды осенью берут пробы почвы на участках
произрастания зараженных растений. В лаборатории почву
высушивают до воздушно-сухого состояния, просеивают через почвенное сито
с отверстиями диаметром 1 мм; прошедшую через сито фракцию в
закрытых сосудах можно хранить на протяжении многих лет.
Взрослые самки золотистой картофельной нематоды,
расположенные на поверхности мелких корней картофеля, имеют вид
молочно-белых, а затем золотисто-коричневых округлых образований (галлов)
размером с маковое зерно (рис. 60, Л). Погруженным оказывается
только передний конецтела нематоды, так что тело паразита не защищено
тканями галла. После прикрепления подвижной рассслитсльной
личинки к участку корня растения-хозяина здесь развиваются
специализированные питающие клетки, называемые гигантскими или
«нектариицами*. Они выполняют функцию посредников между
хозяином и паразитом, что определяет постоянство однажды
выбранного места питания. В результате у самок происходит постепенная
дегенерация соматической мускулатуры, и они полностью утрачивают
подвижность. Созревающие яйца остаются внутри тела материнской
особи, которая превращается в цисту. Цисты весьма устойчивы к
экстремальным условиям среды и могут переноситься с талыми водами,
на инструментах для обработки почвы (плуги, бороны, лопаты и т.п.),
колесах и гусеницах сельскохозяйственной техники, а также
посадочном материале картофеля. Выход личинок из цист растянут на
несколько лет. Основными мерами по ограничению распространения
и вредоносности золотистой картофельной нематоды являются
выращивание относительно устойчивых к этому вредителю сортов и
использование семипольных севооборотов, когда картофель выращивается
на определенном участке раз в семь лет. Невыполнение этих условий
обусловило широкое распространение глободеры на индивидуальных
приусадебных участках.
Строение тела самки. Форма ее тела близка к округлой; длина
варьирует в пределах 0,13-1,3 мм, ширина — 0,1-0.6 мм (самцы имеют
типичную для нематод форму тела, длина их составляет 0,9-1,2 мм,
ширина — 0,025-0,030 мм). Тело взрослых самок практически
полностью заполнено яйцами овальной формы (рис. 60, Б). На переднем
конце тела можно рассмотреть иглу и расположенный у ее основания
округлый бульбус, на противоположном конце тела — наружное
половое отверстие.
114
Тип Nemathelminthes
Рис. 60. Золотистая картофельная нематода (Globoderarostochfensis): А —
участок корневой системы картофеля, заселенный нематодами;
Б—молодая самка: I — ротовое отверстие; 2— стилет. J—буль-
бус; £— развивающиеся нематоды в тканях растения-хозяина:
/ — самка; 2 — самец
Класс Nematoda
115
Обнаружение в почве цист. В цилиндрический сосуд (химический
стакан, пластиковую емкость и т.п.) объемом 500-1000 см3 вставляют
полоску фильтровальной бумаги шириной 10 см и такой длины, чтобы
се концы заходили друг за друга внахлест. В сосуд вливают 250-300 мл
водопроводной воды, следя за тем, чтобы бумага плотно прилегала
к его стенкам. Затем медленно и осторожно при помешивании в сосуд
всыпают порцию (примерно спичечный коробок) воздушно-сухой
почвы. Взвеси дают отстояться около получаса, а затем добавляют
каплю обычного жидкого мокшего средства. После этого бумажную
полоску аккуратно вынимают, укладывают загрязненной стороной
вверх на пластиковую или стеклянную пластинку подходящего размера.
Поверхность полоски внимательно рассматривают под бинокуляром.
Цисты Gbbodera mstochiensis имеют округлую или округло-овальную
форму (рис. 60) и очень похожи на взрослых самок, у которых
атрофированы внутренние органы.
Подкласс Enoplia (-Adenophorea)
Отряд Trichocephatida
Trichinclla spiralis - трихина
Материалом для изучения личиночной (мускульной) стадии
развития трихины могут служить постоянные окрашенные препараты.
Большую информацию студенты получат при самостоятельном
изготовлении временных препаратов.
Для изготовления временного препарата используется
фиксированная в формалине тушка либо мышечная ткань ранее зараженного
животного.
Пинцетом с соблюдением необходимых правил
предосторожности преподаватель переносит на предметное стекло небольшой кусочек
мышечной ткани. Вышип покрывается каплей глицерина и
накрывается вторым предметным стеклом. Стекла плотно сжимаются и слегка
притираются, чтобы вышел воздух из промежутков между волокнами
мышцы. Препарат изучается под микроскопом.
Изучение мускульных трихин. После завершения развития в
кишечнике и копуляции самцы трихины погибают, а самки, проникнув
в кишечные лимфатические сосуды, отрождают огромное количество
личинок и через два месяца также погибают. Молодые личинки
разносятся током лимфы во все части тела, и внедрившись в волокна
поперечнополосатой мускулатуры, растут и разрушают волокно. Через две
116
Тип NematMm
недели личинки скручиваются спирально и переходят в состояние
покоя. Образуется капсула размером 0,8-1 мм, которая постепенно про^
пнгывастся известью и становится слабо прозрачной. Именно такие i
капсулы обнаруживаются на препаратах между волокнами поперечно-;
полосатых мышц (рис. 61).
)- -(
P$tC. 61. Трихинл (TrichincIJa spiralis): A — инкапсулированная личинка
в мышечной тхани: / — мускульные толокна; 2 — личинка
в капсуле; Б — пути циркуляции трихины в природе
Класс Rotatoria — коловратки
Коловратки очень многочисленны в пресных водоемах. Они очень
разнообразны по строению и большинство из них имеют
микроскопические размеры (от 0,04 до 2 мм). В связи с их большим значением
^cc Rotatoria
117
г1я природных водоемов (очистка воды благодаря фильтрационному
питанию, кормовые объекты для рыб) и особенностями строения и био-
;н>! ии целесообразно изучение коловраток в практикуме.
Отряд Bdelloidea
Rotaria sp.
Материал. Собрать коловраток можно в любом пресном
водоеме — от крупных озер до временных водоемов» луж и канав. Для отбора
проб планктона с этими беспозвоночными используется планктонная
сетка. Встречаются коловратки и в лабораторных сосудах с
простейшими, и в аквариумах. Часто в лабораторных культурах присутствуют
коловратки родов Rotaria и Brachionus.
Изучение живого объекта. Тело коловратки вытянутое,
червеобразное и ясно разграничено на головной, туловищный отделы и ногу
(рис. 62).
На головном отделе располагается коловрашательный
аппарат. Он образован двумя выпячиваниями, несущими по краю два
гамкиутых кольца ресничек. На брюшной стороне находится еше
один ряд ресничек, спускающийся к ротовому отверстию. Весь
коловрашательный апларатслециальными мышцами-ретракторами может
стягиваться внутрь. На спинной стороне располагается сократимый
вырост — хоботок, а позади него — длинное щупальце с чувствительными
колосками.
Туловищный отдел и нога имеют ясно выраженную наружную
кольчатость, не затрагивающую внутреннюю организацию.
Нога имеет разную длину, она способна сокращаться и
вытягиваться. На конце у нес два или три пальца. В основании пальцев
расположены «цементные* железы, протоки которых открываются на их концах.
Секрет этих желез обеспечивает закрепление коловраток на субстрате.
Тело коловраток покрыто кутикулой, под которой лежит
образующая се гиподерма.
Движение. В массовой культуре коловраток рода Rotaria можно
наблюдать разные формы движения. Закрепившись с помощью нот,
коловратка при питании совершает поисковые движения. «Шагающее»
движение (попеременное закрепление головного отдела и
подтягивание ноги) сходно с движением пиявки. Оторвавшись от субстрата,
коловратки могут «плавать» в толшс воды, используя работу ресничек
коловрашательного аппарата.
118
Тип Nemthdmlnttm
Рис. 62. Схема строения коловратки из отряда Monogononta: A —
внутреннее строение; Б— мускулатура; / — коловращательный
аппарат; 2 — слюнные железы; 3 — мастаке; 4 — пищевод;
5— желудочные железы; б — желудок; 7 — половая железа;
8 — протонефридий; 9 — кишечник; 10— мочевой пузырь;
// — клоака; 12 — анальное отверстие; 13 — цементные
железы; /4—нога; 15— пальцы ноги; 16 — туловищный отдел;
/7 — голоиа; 18— мышечные пучки
Питание Rotaria удобно наблюдать при добавлении в каплю
культуры на препарате растертого в воде кармина или водного раствора туши.
Профильтровывая воду, коловратки захватывают частицы и осветляют,
соответственно, культуральную жидкость. Коловратки-фильтраторы
в природе питаются водорослями, бактериями, взвешенной в воде
органикой. Если на временном препарате есть кусочек растения или
немного детрита, около них собираются коловратки и, активно
работая ресничками коловрашательного аппарата, создают ток воды с
пищевой взвесью ко рту.
Класс Rotatoria
119
Мускулатура тела образована несколькими пучками
продольных и кольцевых гладких и поперечнополосатых мышц, что
позволяет произвольно совершать самые разнообразные движения и
изменять форму тела, растягивая и сокращая его, втягивая головной отдел
и ногу (рис. 62, Б).
Пищеварительная система начинается ртом, расположенным
на брюшной стороне головы. Рот ведет в трубчатую глотку и ее
расширенную часть — мастаке. В нем находятся челюсти,
состоящие из кутикулярных образований — наковальни и двух молоточков,
коппорые обеспечивают измельчение пищи. Глотка переходит в короткий
пищевод, а тот — в объемный мешковидный желудок, занимающий
большую часть полости тела. В среднюю кишку на ее дистальном
конце впадают протоки выделительной и половой систем, поэтому
эта часть кишки называется клоакой (рис. 62, А).
Выделительные каналы протонефридиальной системы с
типичными терминальными клетками тянутся по бокам к головному отделу.
Половая железа, разделенная на яичник и желточник, лежит
по бокам от желудка в туловищном отделе.
Ниже приводится сравнение особенностей организации
представителей двух классов круглых червей (табл. 5).
Таблица 5
Сравнительная характеристика нематод и коловраток
Особенности
организации и биологии
Мускулатура
Клеточный состав
Ресничный эпителий
Регенерация
Терминальные клетки
протонефридий
Движения
Питание
Нервная система
Половая система,
особенности
I размножения
Коловратки
(Rctiotoric)
Пучки
поперечнополосатых мускульных волокон
Постоянный
Есть
О icy к* пуст
Есть
Разнообразные
Фнльтрзторы, хищники
Головной ганглий
и нервы
Раздельнополые,
гетерогония
Нематоды
{Stmatodo)
Продольные мышечные
волокна
Постоянный
Отсутствует 1
Отсутствует 1
Отсутствуют 1
Только змеевидные
Сапрофаги. фитофаги,
хищники
Окологлоточное кольцо
и продольные стволы
Раздельнополые,
внутреннее оплодотворение, толь-
ко половое размножение |
Тип Annelida — кольчатые черви
Кольчатые черви относятся к высшим червям, так как имеют
многие прогрессивные черты организации, отличающие их от плоских
и круглых червей. Кольчатые черви — первые сегментированные
беспозвоночные животные, в связи с чем у них развились метамерная
мстансфрилиальная выделительная система, метвмерно устроенная
нервная система и хорошо прослеживаются элементы метамерии
в строении кожио-мускульного мешка и пищеварительной системы.
Впервые у кольчатых червей появились кровеносная система и
целом — вторичная полость тела, также характеризующаяся метамер-
ным строением.
Основные особенности организации кольчатых червей,
представляющих разные классы типа, можно рассмотреть на примере много-
щетин ко во го червя нереиды, малощетинкового дождевого червя и
медицинской пиявки.
Подтип Aclitellata — беспоясковые кольчецы
Класс Polychaeta — многощетинковые кольчецы
Подкласс Errantia — бродячие
Отряд Nereimorpha
Nereis pelagica — нереида
Материал. Для изучения многощетинковых кольчецов лучше всего
использовать некрупные (длиной до 8 см) экземпляры молодых
нереид, а также препараты параподий. Благодаря плотным покровам тела
нереиды хорошо сохраняются при фиксации в формалине или (лучше)
спирте и для ознакомления с особенностями их внешнего строения
могут быть использованы многократно. Перед работой животных
следует отмыть в проточной воде.
Внешнее строение Nereis pelagica. При осмотре хорошо
различимы (рис. 63) головной отдел, однородно сегментированный
туловищный отдел и анальная лопасть (пигидиум).
Голова образована простомиумом (головная лопасть) и пе-
ристомиумом. Простомиум имеет треугольную форму и немного
сплющен в дорсовентральном направлении. На переднем суженном
его конце расположена пара щупалец, или антенн. От задней части
Класс Pdychaeta
121
Рис. 63. Передний (А) и задний (Б) концы тела Nereis pelagkcr. I —
ашгенны; 2— пальпы; J— усики; 4— глаза; 5 — обонятельные
ямки; 6 — ггараподии; 7— пигидиум; £— анальные усики
вперед по бокам отходит пара массивных пал ьп. На спинной
поверхности простомиума хорошо различимы две пары глаз, а позади них —
две обонятельные (ресничные) ямки. Все эти придатки
выполняют рецепторныс функции.
Перистом и ум образуется в результате слияния рогового и двух
последующих сегментов. На переднем крае перистомиума, который
значительно шире основания простомиума, находятся усики (цирри), —
обычно их три пары. На брюшной стороне простомиума расположено
ротовое отверстие. Иногда на нижней стороне головного отдела
может хорошо выделяться вывернутый участок ротовой полости червя —
ссбуккальный отдел с парой мощных черных хипшоидных
челюстей, серповидно изогнутых и несущих на внутренней поверхности
мелкие зубчики (рис. 64). Челюсти помогают соскребать с камней
водоросли и захватывать детрите мелкими организмами. Челюсти
можно также рассмотреть на микроскопических препаратах головного
отдела тела мелких нереид.
Туловищный отдел включает большое число (до 80-100)
однородно устроенных (т.е. гомонамных) сегментов. По бокам туловищных
сегментов расположены параподии. Их удобнее изучать на
окрашенных постоянных препаратах (рис. 6S).
122
ТипДл/Teftfe
ЯМс. 54. Передний конец тела Nereis pelagica с выпяченным бук кал ь-
ным отделом: У — челюсти; 2— буккдльный отлел; 3— просто-
миум; 4 — перистомиум; 5 — сегменты; 6 — обонятельная
ямка; 7— глаза; £— усики; 9— пальпы; /0 — антенны
Каждая параподия представляет собой мускулистый боковой
вырост стенки тела и состоит из основной часта и двух ветвей (лопастей):
верхней спинной (нотополия) и нижней брюшной (невроподия).
Каждая ветвь несет по чувствительному отростку-усику и пронизана
тонкими хитинокоными щетинками, вьшающимися за ее пределы.
Кроме того, внутри каждой ветви выделяется мощная опорная щетинка —
ацикула, выполняющая функцию внутреннего скелетного элемента.
У молодых нереид на выпуклой спинной стороне тела сквозь
тонкий слой кутикулы, гиподермы и мускулатуры иногда заметен
темный (коричнево-фиолетовый) спинной кровеносный сосуд. По
центру уплощенной нижней стороны тела тянется продольная
бороздка, над которой проходит брюшная нервная цепочка и брюшной
кровеносный сосуд.
Пигидиум имеет конусовидную форму, лишен параподий, но несет
пару анальных усиков. На его заднем крае расположено анальное
отверстие (см. рис. 63, Б).
Класс Polychaeto
123
Рис. 65. Параподия Nereispela&ka: 1 — спинной нотоподиальный усик;
2— лопасти нотоподии; J— щетинки; 4—лопасть исвроподии;
5—брюшной невропсоиальный усик; 6 — ацикулы; 7— спинная
ветвь; £— брюшная ветвь
Подкласс Sedentaria — сидячие полихеты
Отряд Drilomorpha
Arenicola marina — пескожил
Материал. Как и Nereis, этот морской червь изучается на
фиксированном материале. Пескожил является массовым видом полижет
на песчаных мелководьях северных морей, в частности Белого, где
и может быть заготовлен.
Внешнее строение. Тело пескожила вытянутое, червеобразное,
несколько сужается к переднему и заднему концам, в поперечном
сечении почти круглое (рис. 66). С помощью ручной лупы легко
рассмотреть вторичную кольчатость (на каждый истинный сегмент
приходится пять вторичных колец). В связи с роющим образом
жизни передний конец тела изменен. Маленький, слабо выраженный
простомиум лишен придатков, равно как и перистомиум. На
брюшной стороне головного отдела расположен рот, имеющий вид
поперечной щели.
Туловищный отдел подразделяется на три участка: передние
сегменты несут рудименты параподий в виде валиков, средние —
124
1m Annelida
з
Рис. 66. Внешнее строение пескожила (Artnicokt таппа): 1 — буккаль-
ный отдел: 2 — передний отдел; 3 — жабры; 4 — хвостовой
отдел; 5 — анальное отверстие
рудименты параподий и пучки нитевидных жабр, задние (хвостовые)
сегменты лишены и параподий и жабр. Пучки жаберных нитей
представляют собой видоизмененные нотоподии параподий среднего
участка туловища. Анальное отверстие располагается на заднем конце
тела почти терминально.
Отдельные экземпляры пескожилов, как и нереидыv могут иметь
выдвинутым воронковидный буккальный отдел ротовой полости.
Питаются эти черви донными илами, заглатывая при этом порции
грунта. Поэтому при легком нажатии на тело зафиксированного
пескожила можно услышать слабый характерный звук трущихся друг о
друга песчинок.
Подтип Clitellata — поисковые кольчецы
Класс Oligochaeta — малощетинковые кольчецы
Отряд Lumbricomorpha
Lumbricus terrestris — обыкновенный дождевой червь
Дождевые, или земляные, черви — обычные обитатели почв лесной
зоны Евразии. Питаясь мертвыми и разлагающимися растительными
остатками, они активно участвуют в процессах почвообразования.
Материал. Добыть дождевых червей легче всего на
окультуренных почвах: в садах, парках, на огородах и полях. Иногда они
скапливаются под камнями, досками, иными лежащими на поверхности
почвы предметами, под дерном. Наиболее эффективен сбор
дождевых червей при раскапывании навозных и компостных куч, а также
хорошо удобренных огородных грядок. Собранных червей либо фикси-
Класс Otigochaeta
руют, либо содержат в лаборатории в просторных деревянных ящиках
на 3/4 заполненных листовой землей или перегноем с измельченным
листовым опалом. Периодически почву умеренно увлажняют. Для
подкормки используют листья салата, орешника, клена, липы,
картофельные очистки, давленый отварной картофель. Яшик закрывают
мешковиной, деревянной крышкой и хранят в помещении при
температуре 12-15 *С.
Живых червей перед работой умерщвляют в парах хлороформа
или в 15-20* этаноле. Червей, предназначенных для длительного
хранения, отмывают от почвы и слизи и помещают в специальный
раствор (5 частей концентрированного формалина, 1 часть
глицерина, 94 части воды) или горячую воду, а затем переносят в 5% раствор
формалина, лучше с добавлением небольшого количества глицерина.
Перед работой такой материал нужно отмыть в воде.
Наблюдения за живыми червями. Движение дождевых червей
можно наблюдать, поместив их на лист бумаги либо в плоский стеклянный
сосуд, заполненный землей. При движении червя по бумаге хорошо
слышен шорох, вызываемый соприкосновением с ней хитиноидных
щетинок туловищных сегментов. Проведя пальцами но телу олиго-
хсты от заднего конца к переднему, можно ощутить легкое царапание
ими кожи. Щетинки облегчают передвижение червя по
горизонтальной поверхности и помогают удержаться в почвенных ходах при
попытках извлечения их хищниками.
Основной способ передвижения — изменение формы тела при
сокращении и расслаблении продольных и кольцевых мышц кож-
но-мускульного мешка. Вначале передний конец в результате
сокращения кольцевой мускулатуры утончается, вытягивается и продвигается
вперед. Сокращение продольной мускулатуры приводит к утолщению
переднего конца тела и подтягиванию заднего.
Через стеклянные стенки сосуда, заполненного землей, можно
проследить, как утончается передний конец тела, проскальзывая между
почвенными частицами. Затем они раздвигаются расширяющимися
кольцами, а задний конец тела подтягивается. Если почва плотная,
червь пропускает часть грунта через кишечник, работая как
своеобразный бур. Позднее грунт вместе с непереваренными остатками пиши
удаляется из кишечника в виде характерных комочков — копрол итов.
Внешнее строение. На живом или фиксированном метериале легко
отличить спинную (более темную и слегка выпуклую) и брюшную
(светлую и плоскую) стороны тела. Передний, головной отдел, часто
126
lunAnndkta
вздутый, интенсивно окрашенный по сравнению с тонким, несколь*
ко уплощенным и не так ярко окрашенным задним концом. На
переднем конце расположено ротовое отверстие, прикрытое
выступающей вперед маленькой складочкой — головной лопастью,
соответствующей простомиуму полихет. На заднем конце тела
находится анальная лопасть, или пигидиум, лишенный щетинок и
имеющий крупное, сжатое с боков анальное отверстие (рис. 67).
Туловищный отдел состоит из большого числа (I I0-180)
сегментов, более крупных в передней часта тела. Каждый туловищный сегмент
несет по четыре пары щетинок: две боково-брюшные и две боко-
во-спинные. Вдоль тела щетинки расположены правильными
рядами. У крупных половозрелых червей хорошо заметно в передней трети
тела, между 32-м и 37-м сегментами, утолщение кожных покровов —
Рис. 67. Передний конец тела дождевого червя (Lumbricus temstris): A —
сбоку; Б — с брюшной стороны; В — со спинной стороны; / —
головная лопасть; 2—боковые щетинки; J— брюшные
щетинки; 4— семяпроводяшая бороздка; 5— мужское половое
отверстие; б— женское половое отверстие; 7— поясок; 8— спинные
поры
Класс CHiQOChaeta
127
поясок (clirellum). На пояске многочисленны железистые клетки,
которые выделяют слизь, образующую муфточку, а затем и кокон,
в котором развивается молодь.
На спинной стороне по средней линии расположены спинные
поры, через которые полостная жидкость выделяется наружу и
смачивает покровы, облегчая скольжение по субстрату, движение муфточки,
дыхание; в задней части тела через спинные поры выводятся и
экскреты (рис. 67, В).
При рассмотрении червя с брюшной стороны с помощью лупы
можно, хоть и с трудом, различить наружные отверстия женской
(на 14-м сегменте) и мужской (на 15-м сегменте) половых систем,
имеющие вид точечных пор. В бороздках между 9-м и 10-м и 10-м
и 11-м сегментами расположены отверстия двух пар
семяприемников. От мужских половых отверстой до пояска тянутся лежащие
кнаружи от брюшных щетинок семенные бороздки, по которым
выделяемая семенная жидкость поступает к пояску (рис. 67, А, Б).
Вскрытие дождевого червя. Живых червей предварительно
необходимо умертвить, поместив на 10-15 минут в 10* этанол. Найдя у червя
передний и задний концы тела, спинную и брюшную стороны,
переносят объект в препаровальную ванночку и прикрепляют булавками,
осторожно втыкая их в головной и задний концы тела. Прикалывая
червей, их следует слегка растягивать, а чтобы не повредить
надглоточный ганглий, спереди нужно вводить булавку немного в сторону от
средней линии тела.
Разрез по спинной стороне проводят начиная от середины тела
к переднему и заднему его концам. Разрез не должен быть слишком
глубоким, он затрагивает только кожно-мускульный мешок, чтобы
не повредить кишечник. Для успешного вскрытия с боков от разреза
препаровальной иглой нужно подсечь диссспимснты. После этого
по бокам от разреза края отгибают в стороны и изнутри наискосок
прикалывают булавками.
Внутреннее строение. На вскрытом черве хорошо различимы все
части пищеварительной системы (рис. 68). Кишечник тянется ааоль
всего тела в виде довольно толстой и сильно дифференцированной
трубки. На переднем конце кишечник начинается грушевидно
вздутой мускулистой глоткой, занимающей середину первых шести
сегментов. В глотку влаааюттри пары слюнных желез. За глоткой
следует длинный тонкий трубчатый пищевод (от 7-го по 14-й сегмент),
в который открывается три пары известковых желез. Секрет известковых
желез обеспечивает нейтрализацию гуминовых кислот, содержащихся
128
lw\Anneti<b
15 16
Рис.68. Внутреннее строение дождевого червя (Lumbricusttmstris).
Передний участок вскрытого со спинной стороны червя: / —
надглоточный ганглий; 2 — глотка; 3 — пишсвол; 4 — кольцевой
сосуд («сердце*); 5 — семяприемники; 6 — семенной мешок;
7— зоб; 8— мускульный желудок; 9— спинной кровеносный
сосуд: /0— средняя кишка; // — кровеносные боковые сосуды
кишечника; 12 — диссепимент; 13 — брюшной кровеносный
сосуд; 14— нефридий; /5— брюшная нервная цепочка: /6 —
тифлозоль
Класс Oligoctoeto
129
и потребляемой пище. Пищевод впадает в тонкостенный вздутый зоб,
г* которым следует мускулистый толстостенный желудок.
Механические свойства стенок зоба и желудка можно легко определить простым
прикосновением препаровальной иглы. Глотка, пищевод, зоб и
желудок — специализированные отделы передней кишки.
Средняя кишка начинается на уровне 18— 19-го сегментов.
При ее повреждении при вскрытии червя выделяется содержимое
кишечника (частички почвы, измельченные и полупереваренные
растительные частицы), которое лучше аккуратно смыть струей воды.
На поверхности средней кишки заметны многочисленные хлораго-
тенные клетки.
Если вырезать небольшой участок средней кишки, то видно, как
из ее внутренней поверхности со спинной стороны в просвет свисает
складка — тифлозоль, увеличивающая площадь секреторной и
всасывающей поверхности и, таким образом, обеспечивающая большую
>ффективность пищеварения.
Задняя кишка короткая, внешне не отличается от средней, но
лишена тифлозоля.
Кровеносная система представлена спинным кровеносным
сосудом, расположенным сверху над кишечником. Этот сосуд
способен к сокращениям и обеспечивает движение крови от заднего
конца тела к переднему. В связи с пульсацией спинного сосуда толщина
его изменчива. В каждом сегменте к спинному сосуду подходят
боковые сосуды, хорошо различимые у крупных свежих экземпляров
дождевых червей.
В облает пищевою легко найти шесть пар мощных кольцевых
сосудов, огибающих пищевод и впадающих в продольный брюшной
кровеносный сосуд. Рассмотреть его можно, если слегка
отодвинуть в сторону кишечник. По брюшному сосуду кровь движется от
переднего конца назад, а в задней части тела по кольцевым сосудам
возвращается в спинной сосуд. Кровеносная система, таким образом,
является замкнутой. Следует обратить внимание на некоторые
особенности кровеносной системы дождевых червей. Так, огибающие
пищевод кольцевые сосуды способны к пульсации, ускоряют
циркуляцию крови и получили название «придаточных сердец».
Кровь дождевых червей красная, что обусловлено присутствием
в ней гемоглобина, растворенного в плазме крови. При
рассмотрении молодых червей обращает на себя внимание их красноватая
окраска, в значительной мерс определяемая густой сетью капилляров
130
InnAnneUda
в покровах В связи с тем что дыхание у олигохет осуществляется всей
поверхностью тела (специальные органы дыхания отсутствуют), такое
расположение капилляров обеспечивает активный газообмен.
Эффективности дыхания способствует увлажнение покровов выделениями
желез кожного эпителия и полостной жидкостью (через
упоминавшиеся выше спинные поры).
Выделительная система. По бокам от кишечника хорошо заметны
межсегментные перегородки, или диесспименты. Наличие диесспи-
ментов и даже небольших перехватов на стенках кишечника в местах
прикрепления диссепиментов позволяет судить о внутренней
сегментации дождевого червя.
Вдоль диссепиментов справа и слева от кишечника располагаются
в виде небольших петлевидно изогнутых нитевидных тр><х>чск
многочисленные метанефридии. Каждый из них открывается в полость тела
(целом) маленькой воронкой, которая переходит в трубочку,
пронизывающую стенку диссепимента и переходящую в полость другого сегмеюа.
Здесь проток метанефридия петлевидно закручивается вдоль
перегородки и затем открывается выводной порой на брюшной стороне
тела. Рассмотреть строение мстанефрндия можно, выделив его вместе с
участком диссепимента и изучив в капле воды под микроскопом.
Нервная система. Надглоточный ганглий располагается над
глоткой в передней ее части. Если препаровальной иглой оттянуть
назад мускулистую глотку, можно обнаружить ганглий в виде
беловатого двойного узелка. Если отрезать глотку и освободить кишечник
до середины тела, на брюшной его стороне четко прослеживается
расположение брюшной нервной цепочки с мстамерно
повторяющимися ганглиями, связанными продольными коннективами.
Выделив участок брюшной нервной цепочки и поместив его в часовое
стекло с водой, удастся рассмотреть отходящие от ганглиев
поперечные нервы. На переднем конце брюшной нервной цепочки находится
подглоточный ганглий.
Половая система дождевого червя устроена достаточно сложно.
Эти животные являются гермафродитами. В передней части тела
по бокам от пищевода лежат легко обнаруживаемые беловатые три пары
семенных мешков. Они представляют собой особые выросты
диссепиментов и лучше видны после удаления кишечника. Две
передние, а также задняя пара семенных мешков образуют на брюшной
стороне 10-гои II-госегме1ттовнепарные выросты — семенные капсулы,
toMCcOtiQOChaeta
131
в каждой из которых размешается по паре семенников, неразличимых
при вскрытии. Образованные в семенниках сперматозоиды
поступают в семенные мешки, где происходит их созревание. Выводятся
половые продукты через семяпроводы, открывающиеся на 15-м сегменте
выводным отверстием.
Яичники в виде двух маленьких телец находятся в 13-м сегменте,
а воронки яйцеводов, непосредственно не связанные с яичниками,
расположены в 13-14-м сегментах и открываются на последнем
выводным половым отверстием. Яичники можно обнаружить лишь
у половозрелых червей весной, в период их размножения. Яйцеводы,
семенники и семяпроводы при вскрытии не различимы. Изнутри
на брюшной стенке тела червя после удаления семенных мешков сбоку
от средней линии тела почти всегда заметны две пары семяприемников
в виде открывающихся наружу мешочков. В них после копуляции двух
гермафродитаых особей сохраняется сперма.
Изучение пооеречного среза дождевого червя (рис. 69) проводится
на окрашенных кармином постоянных препаратах (лучше, если на
одном препарате имеются серии срезов).
На препарате следует рассмотреть, начиная снаружи,
однослойный эпителий с железистыми клетками и выделяемую им тонкую
кутикулу. Глубже под базальной мембраной лежит слой кольцевых
мускульных волокон и, наконец, мощный слой продольной
мускулатуры. Волокна продольных мышц располагаются вдоль тончайших
пластинок соединительной ткани подобно иголочкам на побеге
ели. Изнутри этот слой подстилается целомическим (перитониаль-
ным) эпителием, или эндотелием, иногда разрушающимся при
изготовлении срезов.
Кожно-мускульный мешок разбивается на четыре участка:
спинной, брюшной и два боковых. Участки это разделены парными
щетинками, расположенными на брюшной и боково-спинной сторонах тела.
Даже если щетинки не попали на срез, границы участков хорошо
различимы.
Внутри вторичной полости тела (целома) центральное
положение занимает срез через кишечник, позволяющий рассмотреть слой
внутреннего энтодермалыюго железистого эпителия, кнаружи от
которого лежат кишечная мускулатура ислойхлорагогенных
клеток. На спинной стороне кишечника внутрь его паюсти вдастся складка
тифлозоля, — образования, увеличивающего секреторную и
всасывающую поверхность средней кишки. В задней кишке тнфлоэоль
отсутствует.
132 Тип Аллод
Рис. 69. Поперечный разрез из средней части тела Lumbricus terrtstrir.
1 — кутикула; 2 — цилиндрический эпителий; 3 — кольцевая
мускулатура; 4 — слой продольной мускулатуры; 5 — целом и-
ческий эпителий; б— меганефридий; 7— щетинка; £—
брюшной сосуд; 9 — субневральный сосуд; /0 — ганглий брюшной
нервной цепочки; // — нервы; /2 — слой хлорогенных клеток;
13 — полость кишечника; 14 — железистый эпителий; 15 —
тифлоэоль; 16 — спинной сосуд; 17— целом
На прогибе кишечника на спинной стороне и под кишечником
(часто в стороне от средней линии тела) видны спинной и брюшной
кровеносные сосуды. Ниже брюшного кровеносного сосуда в полости
тела располагаются элементы брюшной нервной цепочки. Если на
препарате есть срез через ганглии, хорошо видны соприкасающиеся
парные нервные узлы в общей обаючкс — неврилемме. Вдоль нервной
цепочки проходят три тонких кровеносных сосуда: два сверху и один
снизу — невральные сосуды.
Класс Oligochaeta
133
Изучение нсфридиев, трубки которых петлеобразно изгибаются»
рызывает наибольшие затруднения. В связи с таким строением
сегментарных выделительных органов на поперечных срезах они видны по
бокам от кишечника в виде круглых или продолговатых по очертанию
участков протоков. Более полную картину их строения может дать
лмализ серии последовательных срезов.
Пресноводные и почвенные олигохеты отряда Naidomorpha
Олигохеты, обитающие в пресных водоемах и в почве, и фа ют
пажную роль в природных сообществах как потребители и
разрушители мертвой, разлагающейся органики и звенья трофических цепей
и сетей. Они часто используются в качестве корма для аквариумных рыб.
Материал. В илистых канавах, на дне стоячих водоемов,
лишенных рыбного населения, очень часто можно встретить скопления
юрвей-трубочников (род Tubifex). Передним концом тела
трубочники зарываются в грунт, оставляя свободным задний конец, который
совершает колебательные движения,.активизирующие дыхание
животных в условиях низкой концс1гтрации растворенного в воде кислорода
на заиленных грунтах. Скопления червей, насчитывающие тысячи
особей, имеют вил красноватых колышащихся «подушечек». Червей,
>тмытых из грунта в мелкоячеистом сите, можно некоторое время
хранить при низкоположитсльных температурах в холодильнике. Их
содержат в широких плоских сосудах (типа кювет) под слоем воды
I -2 см, периодически (раз в сутки — двое) заменяя большую се часть.
Возможно также хранение при комнатной температуре в сосудах
с проточной, нехлорированной водой.
В водоемах среди растений, в скоплениях нитчатых водорослей
к большом количестве встречаются олигохеты рода Nais. В зарослях
йодной растительности у берегов рек и озер обычны малощетинковые
черви рода Stylaria% легко выявляемые благодаря достаточно крупным
размерам (до 2 см).
В канавах, болотах, зарастающих участках прудов, озер и рек
обитают эолосомы (Aeolosoma spp.), часто их можно обнаружить и на
стенках аквариумов. Эти микроскопические олигохеты легко
отличимы по характерным кожным железам, рассеянным по поверхности
тела и имеющим оранжевую, желтоватую или зеленоватую окраску.
Из почвенных олигохет не представляет сложности разведение
энхнтреид (Enchytraeidae), что успешно освоили аквариумисты,
называющие их «горшечными» или «молочными» червями. Так, белого
134
ТипАотмОД
энхитрея (Enchytracus albidus) можно разводить в неплотно прикрытых
ящиках или плошках с листовой землей. Кормят червей лепешками из
густого разваренного геркулеса, смоченными в молоке ломтиками
белого хлеба, кусочками вареного картофеля. Энхитреиды охотно
собираются под положенными на поверхность почвы или прикопанными
ломтиками твердого сыра, чем можно воспользоваться для
демонстрации животных или отбора материала. Червей можно от почвы
отмывать, но надо иметь в виду, что в воде они быстро гибнут и начинают
разлагаться.
Особенности строения олигохет. Трубочник (Tubifex tubifex)
относится к семейству Tubiftcidae. Длина его тела 30-60 мм (рис. 70, А). Он
чрезвычайно вынослив по отношению к загрязнению среды обитания.
Окраска тела красноватая благодаря содержащемуся в крови
гемоглобину. Наличие гемоглобина и колебательные движения задней части
тела обеспечивают необходимую интенсивность дыхания. Сквозь тонкие
покровы хорошо различимы сосуды кровеносной системы,
кишечник и внутренние межсегментные перегородки-диссепименты.
Каждый сегаеит на своей поверхности несет по два пучка щетинок,
отсутствующих на заднем конце тела.
К семейству Naididae относятся олигохсты родов Nats и Stylaria
(рис. 70, Б%В), при рассмотрении которых также удастся изучить
кровеносную и пищеварительную системы, характер сегментации и
расположение щетинок на поверхности тела. Более удобным объектом для
изучения является, благодаря своим крупным размерам (до 20 мм),
стилярия.
Мелкие (длиной 2-3 мм) эолосомы из семейства Aeohsomatidae
имеют состоящее из небольшого (от 7 до 20) числа сегментов тело,
причем сегментация выражена нечетко. На каждом сегменте имеется
по четыре пучка тонких волосовидных щетинок. Головная лопасть
расширена и несет на нижней стороне по переднему краю и около рта
реснички, способствующие скользящему движению. В покровах
множество окрашенных кожных желез. Эолосомам свойствен такой способ
бесполого размножения, как паратомия. При этом могут
образовываться цепочки из двух-пяти особей (рис. 70, Г).
Семейство Enchytraeidaeобъединяет почвенных и водных олигохет.
Размеры большинства энхитрсид составляют 10-25 мм, тело их
отчетливо сегментировано. Щетинки образуют четыре пучка на каждом
сегменте. Энхитреиды способны к бесполому размножению путем
архетомии, при котором червь распадается на части,
регенерирующие недостающие структуры.
Класс Oligochaeta
135
Рис. 70. Водные и почвенные олигохеты: А — Tubifex tubifex: I — общий
нил; 2— черви в норках на заиленном гручгте; В— Naispseudoptusa\
В — Stylaria iacustris: Г— Аеокпопш hemprichi
136 ТипЛлаа*^*
Класс Hirudinea — пиявки
Отряд Gnathobdellea — челюстные пиявки
Hirudo medicuuUis — медицинская шишка
Материал. Медицинская пиявка — эктопаразит и хищник. Образ
жизни определил ряд особенностей ее внешней и внутренней
организации, что хорошо может быть изучено на живых объектах и
фиксированном материале. Умерщвленных пиявок перед работой следует
тщательно протереть, освободив от слизи, покрывающей все их тело.
Живые медицинские пиявки используются для лечебных целей
и могут быть приобретены в аптеках. Поскольку пиявки являются
переносчиками ряда инфекционных заболеваний человека, их в настоящее
время для медицинского использования не отлавливают, как прежде,
в природе, а разводят в лабораторных условиях. По той же причине
демонстрировать питание пиявок следует на лабораторных животных,
а анатомировать либо приобретенных в аптеках еще не питавшихся,
либо заблаговременно зафиксированных пиявок. Для учебных целей
их можно собрать летом в стоячих водоемах с илистым фунтом и
обильной растительностью. Hirudo medicinalis — достаточно теплолюбивый
вид и практически не встречается севернее Белорусского Полесья,
Деснянского Полесья Украины, Центрально-Черноземного региона
России. Пиявок анастезируют добавлением в воду хлороформа, или
в 5-10% растворе хлоралгидрата, затем переводят на S минут в 5%
раствор формалина и хранят в 80* этаноле. Можно также умерщвлять
их 10* этанолом. Чтобы у анатомируемых пиявок лучше была видна
пищеварительная система, следует фиксировать напитавшихся
животных, предварительно накормив их, например, кусочками печени
либо подсаживая на лабораторных животных.
Наблюдение за движениями пиявки. При наличии живого
материала можно познакомиться с разнообразными движениями животного.
Весьма характерны «шагающие» движения пиявок. Прикрепившись
к субстрату задней присоской, червь вытягивает тело и закрепляется
на подходящем участке передней присоской. После этого, освободив
заднюю присоску и петлеобразно изгибаясь, пиявка подтягивает задний
конец тела к переднему, и фиксирует заднюю присоску рядом с
передней. Этот процесс повторяется многократно.
Отрываясь от субстрата, в большом объеме воды пиявка плывет
плавно, волнообразно, достаточно быстро изгибая тело.
Класс Hirvdinea
137
Волнообразные движения червь может совершать и оставаясь
прикрепленным задней присоской. Это свидетельствует о дефиците
и среде кислорода, а движения животного способствуют циркуляции
воды и активизации дыхания. При резком снижении содержания
растворенного кислорода пиявки выползают и присасываются к
стенкам сосуда выше уреза воды.
Ввеогаее строение. Тело медицинской пиявки слегка
уплощенное. На спинной, более выпуклой стороне достаточно четко выделяется
на темном фоне правильный рисунок в виде желтоватых продольных
цепочек (рис. 71). Брюшная сторона уплощена, имеет серо-зеленую
окраску с пятнами неправильной формы.
На переднем и заднем концах тела расположены
передняя и задняя присоски. Передняя присоска
образована слиянием головной лопасти и четырех
передних сегментов. Она имеет вид воронковидного
углубления, окруженного мускульным валиком, в
глубине его находится рот с тремя радиально
расположенными челюстями. На переднем конце тела
с помощью лупы можно рассмотреть пять пар глаз
в виде черных точек. В состав дисковидной задней
присоски входят семь сегментов. Над задней
присоской дорсально открывается анальное отверстие,
имеющее вид точки.
Остальные туловищные сегменты устроены
внешне сходно. При изучении сегментации тела пиявок
следует обратить внимание на постоянство числа
сегментов (4 + 22 + 7 = 33) и вторичную сегментацию
(на каждый истинный сегмент приходится до пяти
ложных колец).
На брюшной стороне в конце первой трети тела
на небольших бугорках едва заметны два половых
отверстия: впереди мужское, за ним — женское. Из
первого иногда выступает циррус (cirrus) — копуля-
тивный орган.
Рис. 71. Рисунок спинной стороны тела медицинской
пиявки (Hirudo medicinalb)
138
1m Annelida
Вскрытие пиявки. Прикрепив животное булавками ко дну
препаровальной ванночки, вскрытие лучше начинать со средней части тела.
Поскольку полость тела пиявки заполнена паренхимой и
пересекается дорсовентральными мышцами, вскрытие представляет
определенные трудности. После проведения неглубокого продольного разреза
необходимо подсечь препаровальной иглой диссспимснты, дорсовен-
тральныс мышцы и паренхиму, все время оттягивая стенку тела. Края
разреза разводят и сразу же закрепляют, прикалывая ко дну ванночки
булавками.
Изучение внутреннего строения. Начинают его со знакомства с
пищеварительной системой. Если перед вскрытием свежеумершвленных
пиявок освободили от потребленной ранее крови, кишечник их имеет
вил беловатой пленки. Заполненный свернувшейся кровью кишечник
фиксированных червей вздут и имеет черную окраску (рис. 72).
В переднем конце находится плотная мускулистая глотка экто-
дермалыюго происхождения, от которой к стенкам тела радиально
отходят мускульные волокна. Глотка переходит в желудок,
являющийся отделом средней кишки, имеющей энтодермальнос происхождение.
Желудок имеет 11 парных боковых выростов, из которых первая пара
часто рудиментарна, а последняя самая длинная и тянется до заднего
конца тела. Желудок служит резервуаром для всасываемой крови, за
что его часто называют зобом.
Собственно средняя кишка представляет собой короткий тонкий
участок кишечника, располагающийся между последней парой
отростков желудка, где осуществляется пищеварение и всасывание. Задняя
кишка, имеющая эктодермальное происхождение, короткая и слегка
расширенная, открывается над задней присоской анусом (рис. 72, А).
Лакунарная система у пиявок функционально замещает
кровеносную систему (примерявления субституции). При вскрытии
спинной лакунарный канал повреждается. После удаления кишечника
удастся рассмотрстьбрюшнойлакунарный канал, окружающий
брюшную нервную цепочку и образующий небольшие расширения около
ганглиев. Лучше всего выражены боковые лакуны побокам от
кишечника благодаря развитию в их стенках мускулатуры,
обеспечивающей пульсацию боковых лакун. Остальные промежутки между
внутренними органами и стенками тела заняты паренхимой.
Выделительная система представлена 17 парами нефридий,
принадлежащих 6-22-му сегментам. Они связаны с боковыми лакунами
Kn&ccHirudinea
139
Рис. 72. Внутреннее строение Himdo medkinalis: A — пищеварительная
система: / — глотка; 2 — спинной сосуд (лакуна); 3 — боковой
сосуд; 4— проток нефридия; 5— боковые карманы желудка;
б—усваивающий отдел средней кишки; 7— задняя кишка; Б—
другие системы органов (после удаления кишечника): / —
окологлоточное нервное кольцо; 2 — брюшная нервная цепочка;
. 3 — семяизвергательный канал с циррусом; 4 — семенной
пузырь; 5— яичник; б— влагалище; 7— семяпровод; £—
семенники; 9— нефридий; /0— боковой лакунарный канал
140
1m Annelida
внутренней воронкой, от которой отходит длинный, петлевидно
извивающийся канал, заканчивающийся выводной порой.
Нервная система представлена брюшной нервной цепочкой,
лежащей в брюшном лакуиариом канале. Соответственно слившимся
сегментам, образующим переднюю и заднюю присоски, п од глот ом-
н ы й га н гл и й состоит из четырех пар слившихся нервных узлов, зааняя
ганглиозная масса включает семь узлов. Над глоткой у се
основания на уровне шестого сегмента расположены надглоточные ганглии.
Половая система пиявок гсрмафродитна. Мужская половая система
представлена девятью парами семенников, от которых отходят
семенные протоки, сливающиеся в два боковых семяпровода. В передней
части тела семяпроводы образуют семенные клубочки, или
пузырьки, в которых накапливается сперма. Соединяясь, семяизверга-
тельные протоки пронизывают совокупительный орган (циррус) и
открываются половым отверстием.
° 9 ю 11 12
Рис. 73. Поперечный разрез Hirudo mtdicinolir. I — кожный эпителий и
кутикула; 2— кольцевая мускулат>ра; J—диагональная
мускулатура; 4 — продольная мускулатура; 5 — дорсовентральные
мускульные волокна; 6 — боковой лакунарный канал; 7—
боковой карман желудка; £— нефридий; 9— семенник; /0 —
семяпровод; // — брюшная нервная цепочка в брюшном
лакуиариом канале; /2— желудок; /J—спинной лакунарный канал
Класс Hirudinea
141
Женская половая система состоит из двух яичников,
располагающихся позади мужского полового отверстия. От яичников отходят
парные яйцеводы v объединяющиеся в непарное петлевидное
влагалище, открывающееся снаружи женским половым отверстием (см.
рис. 72, Б).
Строение кожно-мускульного мешка и паренхимы лучше всего
изучать на поперечном срезе тела, используя окрашенные
постоянные препараты (рис. 73).
Одновременно на поперечном срезе можно дополнительно
изучить расположение основных тканей и органов, особенности их
гистологии. Особое внимание следует обратить на развитие мускулатуры
кожно-мускульного мешка, дорсоветральных мускульных волокон
и паренхимы, заполняющей пространства между органами и тканями.
Тип Mollusca — моллюски
Моллюски, или мягкотелые, как их часто называли ранее,
представляют собой весьма многочисленную ветвь сегментированных
беспозвоночных, населяющих самые разные типы водоемов, а также
независимо от других групп животных освоивших назем но-воздушную
среду обитания. В теле взрослых моллюсков выделяются три отдела:
голова (у пластинчатожаберных моллюсков этот отдел редуцирован),
нога и туловище (висцеральный мешок). Для представителей типа
характерно наличие известковой раковины, являющейся производным
мантии — специальной складки кожи. Редукция или отсутствие
раковины — явление вторичное.
В составе типа выделяют шесть классов, с представителями двух
из них на практикуме предстоит подробно ознакомиться. Остальных
студенты изучают на занятии, посвященном ознакомлению с
биологическим разнообразием моллюсков. Содержание такого занятия
определяется возможностями вуза, его зоологического музея или
зоологического кабинета и в настоящем пособии не рассматривается.
Подтип Conchifera — раковинные моллюски
Класс Gastropoda — брюхоногие моллюски
Подкласс Pulmonata — легочные моллюски
Отряд Stylommatophora — стебельчатоглазые
Helix pomatia — виноградная улитка
Крупные размеры, особенности морфологии и относительная
доступность делают этот вид моллюсков весьма удачным объектом
для знакомства с представителями данного класса. Используемые
порой мелкие наземные моллюски (янтарка, садовая улитка), а также
крупные слизни или прудовики (последние являются
представителями отряда Basommatophora) в несравнимо меньшей степени подходят
для этих целей.
Материал. Helix pomatia является обычным обитателем тенистых
влажных парков либо схожих по условиям участков
широколиственных лесов Украины и ряда южных регионов России. В Белоруссии
в XIX веке этот моллюск выпускался в приусадебные парки. Здесь,
а также на сопредельных участках влажных лесов и кустарников
численность виноградных улиток, если она не подорвана неумеренными
Класс Gastropoda
143
заготовками, позволяет без особых сложностей собрать материал
в нужном количестве. Сбор улиток наиболее результативен в теплые
пасмурные дни при накрапывающем дожде или оседающем тумане.
Моллюсков нельзя сразу же погружать в фиксатор, так как животные
судорожно сокращают мускулатуру, сжимая туловищный мешок, и
остаются в таком малопригодном для анатомирования состоянии. Их
следует сначала поместить в сосуд, заполненный до краев охлажденной
кипяченой водой, закрыв его стеклом так, чтобы не оставалось
воздушной прослойки. Желательно анестезирование моллюсков
добавлением небольших количеств хлоралгидрата (доводя концентрацию его
раствора до 0,5%). В результате животные остаются в расправленном
состоянии, затем их тщательно отмывают и помещают в 50* этанол,
который необходимо несколько раз сменить, постепенно повышая
крепость до 80*. Это обязательное условие для успешного хранения не
извлеченных из раковин улиток. При не слишком продолжительных сроках
хранения (несколько лет) допустимо использование формалина. Перед
работой животных следует вымочить и тщательно отмыть в проточной воде.
Внешнее строение. Виноградные улитки — крупные наземные
моллюски. Известковая раковина хорошо развита и достаточно прочна.
Она цельная, спирально закрученная вправо (дексиотропная). Часто
хорошо просматривается рисунок из чередующихся светлых и темных
полос. Отверстие раковины называется устьем (рис. 74),
противоположный заостренный край раковины — вершиной, или
верхушкой. С представителями класса Gastropoda % имеющими левозакрученную
(лейотроп ную) либо редуцированную раковину, можно ознакомиться
при прохождении летней учебной практики по зоологии беспозвоночных.
Более детально рассмотреть внутреннее строение раковины лучше в
процессе вскрытия улитки.
Рис. 74. Раковина виноградной улитки (Helix
pomaria): I — устье; 2 — передний
край; J— завиток; ¥— вершина; 5 —
пупок; 6 — задний край
144
1m MoHusca
Моллюски способны целиком втягивать в раковину свое тело, но
в активном состоянии освобождают ногу и голову, тогда как
висцеральный мешок остается скрытым в ней. По краю отверстия раковины
виден более светлый край мантии (мантийный валик). Мантия
одевает весьтуловищный мешок и продуцирует саму раковину. С
правой стороны край мантии даст треугольное расширение, называемое
площадкой. Здесь открывается крупное дыхательное отверстие,
ведущее в мантийную полость, выполняющую у пульмонат
функцию легкого. Расположенное на площадке анальное отверстие
малозаметно (рис. 75).
fttc. 75. Виноградная улитка (вид сбоку, правая сторона): 1-5— голова
и ее придатки (/ — голова, 2— глазное щупальце, 3— глаз, 4—
губное щупальце, 5 — ротовое отверстие); 6 — половое
отверстие; 7 — нога; 8 — дыхательное отверстие; 9 — 13 — детали
строения раковины (9— вершина, 10— обороты спирали» // —
последний оборот, 12— край устья, 13— устье)
Наблодешя за живыми моллюсхалш. Виноградных улиток несложно
содержать в стеклянных сосудах, подкармливая ломтиками овощей.
Можно использовать также крупных прудовиков, которых перед
занятием лучше подержать некоторое время без пищи.
Для наблюдения улиток помещают на стеклянную пластинку
(прудовиков — в небольшой сосуд с водой). Спустя некоторое время
они начинают активно перемещаться. Скользящие движения
осуществляются благоюря волнообразно распространяющимся сокращениям
мускулатуры плоской ноги. За ползущими по субстрату виноградной
улиткой или слизнем остается слизистая дорожка, хорошо заметная
и после подсыхания. Аналогичным образом за скользящим по заросшим
водорослями стенкам аквариума прудовиком остается светлый след.
Класс Gastropoda
145
На переднем конце высунутой из раковины ноги различима голова
с двумя парами щупалец. Граница отдела совладает с гл><хжой
бороздой, отделяющей голову от ноги. Передняя пара более коротких
щупалец носит название губных. Глазные щупальца задней пары заметно
длиннее и несут на вершинах вздутия с черными точками глаз (у
прудовиков, принадлежащих к отряду сидячеглазых (Basommatophord),
глаза размешаются у оснований щупалец). Щупальца могут втягиваться
в соответствующие пазухи подобно пальцам перчатки. Будучи
утраченными, например в результате нападения хищника, они легко
регенерируют. Под правым глазным щупальцем расположено наружное
половое отверстие. У отдельных экземпляров из него может
высовываться наружу мужской копулятивный орган — пенис (penis).
В глубине щели между головой и передним краем ноги
расположено ротовое отверстие. Наблюдения за ползущими по обратной
стороне стекла моллюсками позволяют заметить изредка мелькающие
буроватые челюсти. Они лучше заметны, когда животные начинают
скоблить помещенный к ним в сосуд небольшой ломтик моркови или
кусочек капустного листа.
Ротовой аппарат виноградной улитки имеет в своем составе жесткую
роговую (конхиолиновую) челюсть, имеющую форму дугообразной
кутикулярной пластинки с 6-7 поперечными фебнями (рис. 76, А).
Челюсть расположена на границе ротовой полости и глотки. Для
брюхоногих моллюсков также весьма характерно наличие радулы (терки),
представляющей собой располагающийся на поверхности
мускулистого языка участок утолщенной, несушей многочисленные роговые
зубчики кутикулы (рйс. 76, Б). Радулой моллюск соскабливает
поверхностные слои растительных тканей. Такой тип повреждений растений
типичен для садовых улиток и слизней. Для ознакомления со
строением челюсти и радулы виноградной улитки лучше использовать
постоянные микроскопические препараты.
Освобождение моллюска от раковюш, ознакомление со строением
раковины. Отобранный экземпляр виноградной улитки помещают
на ударопрочную подставку, укрывают листом плохо промокающей
бумаги и легкими ударами молотка разрушают раковину. Ее осколки
имеют острые края, поэтому обращаться с ними нужно осторожно,
избегая повреждения режущими поверхностями тканей
анатомируемого животного. Особенно аккуратно следует извлекать тело
моллюска из последних оборотов раковины.
В процессе извлечения фрагментов разрушенной раковины
находят участки внутреннего столбика (колонки), которые напоминают
146
Тип McHusca
Рис. 76. Челюсть и
особенности строения
радулы виноградной
улитки: А — челюсть; Б —
участок поверхности
ранулы: / — вид
сверху, 2—вид сбоку
осевую винтовую лестницу. Столбик образуется при слиянии
соприкасающихся витков раковины. Более подробно с этим можно
ознакомиться на пустых раковинах с отверстиями. Такие раковины можно
получить выварив улиток в кипятке, удалив мягкие ткани и выполнив
надфилем ряд пропилов.
Ознакомление с мантийным комплексам органов. Очищенную от
остатков раковины и промьлую в воле улитку располагают гаювой от
вскрывающего. На треугольной площадке находят дыхательное от-
вс рсти с и вводят в него одно из лезвий тонких анатомических ножниц.
Разрез стенки мантийной полости ведут почти по краю валика вплоть
до темного пятна, соответствующего залегающему глубже сердцу.
Улитку помещают в ванночку ногой вниз и фиксируют булавками.
Стенку мантийной полости отводят пинцетом вправо, в случае
необходимости, выполняют дополнительный разрез.
В составе мантийного комплекса органов представлены
элементы кровеносной, выделительной и дыхательной систем (рис. 77).
В мантийной полости нелегочных моллюсков расположены их
органы дыхания — жабры. У легочных моллюсков, к числу которых
принадлежит винограшшя улитка, мантийная полость выполняет функции
легкого, обеспечивающего дыхание в воздушной среде. Стенки
мантийной полости пронизаны множеством кровеносных сосудов,
образующих на внутренней стороне мантийной складки рельефную
сеть (рис. 78). Среди них выделяется ствол собирающей их легоч ной
Класс Gastropoda
147
Рис. 77. Виноградная улитка с удаленной раковиной: / — «печень»; 2 -
почка; 3 — сердце; 4 — край ма»ггии
вены, впадающей в предсердие. До вскрытия сердце выглядит как
темное пятно, располагающееся рядом с крупной желтоватой областью
залегания почки. Размещается оно в мешковидном перикардии
(перикардиальной сумке), представлющем собой остаток целома.
Аккуратно вскрыв его, можно обнаружить желудочек с упругими за
счет мускулатуры стенками, а также тонкостенное предсердие.
Выделительная система у моллюсков метанефрндиального типа.
Воронка нефридия (почки) открывается в перикардиальнуюсумку,
но выявить это у анатомирумого животного затруднительно. От почки
к треугольной площадке, с которой начиналось вскрытие, идет
выводной канал выделительной системы (мочеточник). Там он
открывается наружным выделительным отверстием. Канал плохо
заметен, но его расположение легко представить, поскольку он
идет параллельно рельефно выделяющейся трубчатой задней кишке.
Остальные внутренние органы виноградной улитки залегают внутри
туловищного (висцерального) мешка и составляют так
называемый висцеральный комплекс органов. В состав этого комплекса
входят элементы главным образом пищеварительной, нервной и
Пановой систем (рис. 79). В ходе препарирования объекта необходимо
провести разрез от ротового отверстия по центральной линии головы и,
далее, по дну мантийной полости.
148
Тип Molluscs
Рис. 78. Органы дыхания и кровеносная
система виноградной улитки:
У — голова; 2— нога; 3 — край
машгии; 4 — край разреза
мантии; 5 — дно мантийной
полости; б-//— кровеносная
система (б—перикардий. 7—
желудочек ссрош. 8—
предсердие, 9— место входа легочной
вены, 10 — легочная иена;
// — легкое - сеть
кровеносных сосудов на внутренней
стороне мантии); /2—
местоположение почки; 13 —
прямая кишка; 14 — наружное
отверстие почки; /5—
анальное отверстие; /6
—дыхательное отверстие; 17 — половое
отверстие
Пищеварительная система. На границе ротовой полости и глотки
(рис. 79) расположена дугообразно изогнутая роговая челюсть,
которую легко обнаружить, зондируя ротовую полость препаровальной
иглой. Глотка отличается мощными мускулистыми стенками. Далее
она переходит в более тонкий пищевод, а тот — в длинный широкий
зоб, который также является отделом передней кишки. Дольчатые
наросты на стенках зоба представляют собой слюнные железы,
протоки которых идут вдоль пищевода и открываются в глотку. Сужаясь,
зоб переходит в среднюю кишку (желудок), которая имеет энтодер-
мальное происхождение. Желудок окружен петлями мощно развитой
«печени». В ней осуществляется переваривание поступающей из
желудка пищи. Непереваренные остатки направляются в заднюю
кишку и удаляются через анальное отверстие, расположенное около
дыхательного.
Половая система. Виноградные улитки гермафродитны, женские
и мужские половые продукты попеременно вырабатываются герма-
фродитной железой. Она располагается между лопастями
печени (рис. 79) и имеет желтоватую окраску. От железы отходит единый
гермафродитный проток, идущий к желтоватой белковой железе.
Класс Gastropoda
149
Рис. 79. Пищеварительная и половая система виноградной улитки:
1-9— пищсварител ьная система (/ — глотка, 2— пищевод, 3—
протоки слюнных желез. 4— слюнные железы. 5— эоб.
6—желудок, 7 — «печень», 8 — прямая кишка. 9 — анус); 10—22 —
половая система (10 — гермафродитиая железа, // — белковая
железа. 12 — семяяйцепровод, 13 — семяпровод, 14 — мешок
пениса. 15 — бич. 16 — полотая клоака, 17— влагалище. 18 —
мешок любовной стрелы, 19 — пальцевидные железы, 20 —
яйцевод, 21 — семяприемник. 22— канал семяприемника);
23 — надглоточный ганглий; 24— крыша мантийной полости
(с расположенными на ее внутренней стороне элементами
кровеносной, дыхательной и выделительной систем)
150
Тип MoHusca
Залегает она вблизи печени, размеры ее изменчивы и зависят от
состояния репродуктивной системы. Белковой железой вырабатываются
идущие на развитие зародыша пластические вещества. Далее следует
ссмяяйцепровод в виде широкой гофрированной трубки, он четко
разделяется на два протока: толстый трубчатый яйцевод и тонкий
семяпровод. Таким образом, протоки женской и мужской половых
систем здесь разделяются.
Яйцевод вскоре переходит в расширенное влагалище с упругими
мускулистыми стенками. Во влагалище открывается канал
семяприемника. В семяприемнике хранится семенная жидкость,
получаемая животным при копуляции (и перекрестном оплодотворении).
Он имеет шаровидную форму и красноватую (у долго хранившихся
в фиксаторе особей — буровато-серую) окраску. При
препарировании моллюска не следует извлекать семяприемник за его проток, так
как тот легко обрывается. Кроме того, во влагалище впадает проток
пальцевидных желез, выглядящих как пучок беловатых нитей.
Рядом находится продолговатый мешок любовной стрелы, имеющий
упругие мускулистые стенки. Залегающая в них любовная стрела имеет
вид толстой известковой иглы. Во время копуляции она выдвигается
из лаповых путей и вонзается в покровы тела полового партнера. Таким
образом осуществляется его стимуляция и, тем самым,
физиологическая и отологическая (поведенческая) синхронизация активности
копулирующих особей.
Что касается мужских половых протоков, то семя провод
переходит в семяизвергательный канал, пронизывающий трубчатый
копулятивный орган — пенис (penis), находящийся в сероватом
мешке пениса. От него отходит хорошо развитая нитевидная мышца —
рстрактор пениса. В функции рстрактора входит втягивание внутрь
ранее выпяченного копулятивного органа. Мешок пениса открывается
в половую клоаку. Также у основания мешка семяпровод принимает
длинный нитевидный флагеллум (бич). Это не что иное, как
придаточная железа, принимающая участие в формировании спермато-
форов — упакованных порций семенной жидкости. Именно в
пакетированном виде сперма поступает во влагалище и канал семяприемника
полового партнера. В семяприемнике оболочки сперматофоров
разрушаются, освобождая содержимое.
Наружное патовое отверстие у легочных моллюсков
располагается под правым глазным щупальцем. Им открывается наружу половая
клоака, принимающая выводные протоки как женской, так и мужской
половых систем. Через наружное половое отверстие осуществляется
Класс BivaMa 151
выведение собственного копулятивного органа, введение копулитив-
ного органа партнера, а впоследствии и откладка яиц. Расположенные
в непосредственной близости от него част половой системы
(пальцевидные железы, мешок любовной стрелы, мешок пениса и т.д.)
оказываются вследствие этого смещенными к головному отделу тела.
Нервная система моллюсков разбросанно-узловоготипа. При
описанной технике препаровки Helix pomatia можно обратить внимание
на опоясывающее начало пищевода околопищевод нос нервное
кольцо. Оно образовано идущими от церебральных ганглиев
церсбро-плевральными и церсбро-педальными коннекти-
вам и, а также под глоточной ганглиозной массой, образованной
слиянием парных париетальных и висцеральных ганглиев. К ней
прилежат парные педальные и плевральные ганглии»
дифференциация которых весьма затруднительна. Таким образом, около-
пищеводное кольцо моллюсков не гомологично окологлоточному
нервному кольцу кольчатых червей.
Класс BivaMa (-Lamellibranchia) — двустворчатые
(пластинчатожаберные) моллюски
Отряд Eulamellibranchia — настоящие
пластинчатожаберные моллюски
Anodonta cygnea — обыкновенная европейская беззубка
Бсзз><жи, равно как и другие обитающие в пресных волах
представители унионид (сем. Unionidae)% имеют крупные размеры, вполне
доступны в качестве раздаточного материала, удобны для
изучения.
Материал. Обыкновенная европейская беззубка (Anodonta cygnea)
принадлежит к числу обычных обитателей стоячих и медленногску-
щнх водоемов с песчаным или слабозаиленным фунтом Белоруссии,
Прибалтики. Украины, многих регионов Европейской части России
и Западной Сибири. Реки и многие озера большей части Евразии
населяют разные виды перловиц (род Unto), имеющие более прочные
продолговатые раковины. В реках Восточной Европы может быть
встречена узкая беззубка (Pscudanodonta complanata), которая часто
заселяет и сильно опресненные участки внутренних морей.
Предпочтительным объектом является обыкновенная европейская беззубка, но
крупные экземпляры всех перечисленных видов унионид могут быть
с успехом использованы на лабораторных занятиях.
152
Тип MoHusca
Беззубки, а тем более перловицы, очень чувствительны к
содержанию в воде растворенного кислорода, что делает содержание
большого числа экземпляров моллюсков весьма затруднительным, но
возможным при наличии бассейна или соответствующих размеров
емкости с проточной водой. Для анатомирования в учебных целях
вполне пригоден фиксированный материал. Животных умерщвляют
путем погружения на 30 минут в воду, нагретую до 35-40 #С Хранят
материал в 3-4% растворе формалина, перед вскрытием
выдерживают несколько часов в проточной воде.
Для изучения строения раковины унионид удобнее использовать
экземпляры раковин с заранее удаленными мягкими тканями.
Желательно их предварительно замочить, чтобы продемонстрировать
упругость связывающего створки лигамента, а также отшелушивание
конхиолинового слоя и нестойкость к механическим воздействиям
фарфорового слоя на макушках. Для ознакомления с устройством
замка следует иметь также раковины перловиц. Просмотр
постоянных микроскопических препаратов шлифов раковин лучше
производить, используя бинокуляры с боковой подсветкой.
Для ознакомления с особенностями строения глохидиев
беззубок могут быть использованы временные глицериновые либо
постоянные микроскопические препараты.
Внешний вид. Тело моллюска заключено в овальной формы
двустворчатую раковину. Передний конец тела несколько расширен,
задний — сужен (рис. 80). Створки раковины у беззубки
симметричны. Наиболее выпуклый участок каждой створки носит название
макушки (верхушки, пупка). Наружная окраска раковины
грязно-зеленая, серо-зеленая либо желто-бурая и определяется
поверхностным слоем органического рогоподобного вещества — конхиолина.
На поверхности створок обычно хорошо видны концентрические
линии, соответствующие периодам замедления роста моллюска. На
основе соответствующего подсчета можно сделать предположение
о возрасте рассматриваемого экземпляра. На макушке, то есть наиболее
старом участке раковины, конхиолиновый слой может стираться или
слущнватъея, обнажая имеющий белую окраску фарфоровый слой.
У отдельных экземпляров створки раковины могут быть
приоткрыты, между ними высовывается участок желтовато-бслой килевид-
ной ноги. Благодаря сокращениям ее мускулатуры моллюск движется
по дну водоема, прочерчивая в грунте неглубокую борозду. Заглянув
в щель, можно убедиться, что створки раковины подложены
складчатой беловатой пленчатой тканью — мантией.
Класс fl/va/wa
153
Б
Рис. 80. Внешний вид раковины беззубки (Anodonta) (А) и перловицы
(Unto) (£): / — вершила (макушка); 2 — передний конец; 3 —
задний конец
Строение раковины уннонид. Раковина моллюсков образована
комплексом неорганических и органических веществ и имеет слоистое
строение (рис. 81). Наружный слой рогоподобного органического
вещества—конхиолина носит название псриостракума. Он определяет
^«МНЦЩц^МШ 1
Рнс. 81. Шлиф раковины дну-
створчатого моллюска:
/ — конхиолиноиын
слой; 2- фарфороиид-
ный слой; 3 - перла-
мутровыйслой
154
Тип Molluscs
окраску раковины, которая соответствует общему тону дна водоема,
выполняя функции покровительственной. В области макушек кон-
хиолиновый слой одной створки раковины переходит на другую,
образуя узкую эластичную связку — л и гаме нт. При сведении створок
ощущается упругое сопротивление» оказываемое лигаментом. У
мертвых моллюсков створки всегда полуоткрыты. У крупных экземпляров
унионид на макушках конхиолиновый слой может быть частично
утрачен. Острием препаровальной иглы можно продемонстрировать
относительную легкость его отшелушивания и разрушения подлежащего
фарфорового слоя. Вторая важнейшая функция покровного конхи-
ол и нового слоя — защита известковой основы раковины от
воздействий абразивов (песка, например) и химически активных агентов
(главным образом, Н2С03). На препаратах шлифов раковины этот слой
имеет зеленоватую окраску и нередко выглядит расслоившимся.
Фарфоровый, илифарфоровилный, слой (остракум) является
наиболее мощным. Особенно хорошо он развит у обитателей
прибойной зоны морей. Основу его составляет карбонат кальция (СаС03).
Фарфоровклный слой образован многочисленными призмами СаС03
расположенными перпендикулярно к поверхности створки, что
можно наблюдать при рассмотрении препаратов шлифов. По этой
причине данный слой часто также называют призматическим.
Внутренний слой (гипостракум), соприкасающийся с телом
(мантией) животного, имеет гладкую (но не ровную) поверхность и
защищает нежные ткани мантии моллюска от механического
воздействия раковины. Основу его также составляет СаС03 в виде призм
или пластинок (у унионид — только пластинок), но располагающихся
параллельно поверхности раковины. Содержание конхиолина в нем
на порядок большее, нежели в фарфоровом слое. Благодаря явлению
интерференции световых волн внутренняя поверхность раковины
иризируст, а слой, обеспечивающий этот оптический эффект,
получил название перламутрового.
Узкая краевая полоска раковины образована только конхиолином.
Чуть глубже по ее внутренней поверхности проходит узкая мантийная
линия — след прикрепления мантии к раковине.
У безз><хж в составе запирающего аппарата присутствует только
лигамсит. У обитающих в текущих водах перловиц спинной край
каждой створки несет несколько выступов-з><5цов. В средней части
расположены удлиненной формы кардиальные зубцы, ближе к
переднему концу раковины—округлые латеральные зубцы. Приемы-
Класс BivaMa
155
кании створок зубцы одной из них входят в углубления — промежутки
между зубцами другой. Это образование получило название замка
оно препятствует сдвигу створок раковины относительно друг друга'
Особенности строения замка учитываются при определений'видовой
принадлежности уиионид и многих других двустворчатых моллюсков.
Вскрытие моллюска. Лезвие скальпеля аккуратно вводят между
створками, это легче сделать в задней части раковины. В
образовавшуюся шель вводят скальпель и тыльной стороной лезвия отделяют
мантийную складку от поверхности левой створки. В двух местах лезвие
наталкивается на мощные пучки соединяющих створки замыкатель-
ных мышц (аддукторов). Их аккуратно подрезают у внутренней
поверхности левой створки, после чего створки, стягиваемые лигаментом,
слегка приоткрываются. Таким образом, лигамент и
мускулы-аддукторы функционируют как антагонисты. Сводя створки, можно
ощутить упругое сопротивление лигамента. После этого аккуратно
прорезают лигамент и отделяют левую створку. На ее внутренней стороне
можно обнаружить бугристые участки — места прикрепления
мускулов-аддукторов (рис. 82). Затем моллюска укладывают в ванночку
правой створкой вниз и заливают водой.
Внутреннее строение беззубки. Тело лежащего на правой створке
моллюска укрыто левой складкой мантии (если она не была удалена
или отведена в сторону). Сзади с брюшной стороны заднебрюшной
край мантии пигментирован и несет чувствительные сосочки. Здесь
располагаются два сифональных отверстия. Нижнее соответствует
Рис. 82. Внутренняя поверхность створки раковины безз><3ки: J
редний край; 2- задний край; 3 - спинной край; 4- i
ной край; 5 — мантийная линия; б — передний мускул
катель; 7— задний мускул-замыкатель
156
Тип Mdlusca
вводному (жаберном у) сифон у, через него вода поступает к
жабрам» лежащим в мантийной полости. Верхнее — выводному
(клоакальному) сифону. Ближе к спинной стороне соответственно
спереди и сзади расположены срезы толстых мышечных пучков —
переднего и заднего мускулов-аддукторов.
Тело двустворчатых моллюсков подразделяется на два отдела: ногу
и туловище, головной отдел редуцирован. Лопастевидная нога
выдается между мантийными складками с брюшной стороны. Она
отличается особой упругостью, поскольку се внешняя часть практически
полностью состоит из мышечной ткани. После отведения в сторону
свободного края левой мантийной складки открывается мантийная
полость моллюска (рис. 83). Внимание сразу же привлекают две
мелкоисчсрчснныс желтовато-коричневые пластинки — левая
наружная и внутренняя полужабры. Приподняв их, можно обнажить
основание ноги.
Рис. 83. Беззубка с приоткрытыми створками: мантия правой стороны
тела (3) отделена от створки раковины (1)\ 2—левая створка
раковины; 4 — левая складка мантии; 5-6 — жаберный сифон;
7-10— наружные и внутренние жабры обеих сторон тела; //, 12—
губные лопасти; /J—рот; 14 — нога
Класс BivaMa
157
Ротовое отверстие располагается у переднего края основания
ноги ниже переднего мускула-аддуктора. Оно окружено двумя
парными удлиненно-треугольными листовидными ротовыми лопастями
(парусами), покрытыми мерцательным эпителием. По желобкам,
образованным лопастями каждой стороны тела, отфильтрованные
пищевые частицы направляются в ротовое отверстие.
Органами дыхания двустворчатых моллюсков являются ктс и иди -
ал ьные жабры. У настоящих пластинчатожаберных моллюсков они
состоят из нитей, соединенных мостиками и образующих пористые
решетчатые пластинки. У беззубки правая и левая жабры в своем
строении вполне симметричны, каждая состоит из двух листков
(полужабр). Обращенный к мантии листок называется наружным, или
латеральным, тогда как второй — внутренним, или медианным. Каждый
листок образован парой сросшихся по брюшному краю
перфорированных пластинок, между которыми имеется внутрижаберное
пространство. Жабры и внутренняя поверхность мантийных складок покрыты
мерцательным эпителием. Благодаря его активному
функционированию вода с растворенным в ней кислородом и взвешенными
пищевыми частицами поступает в мантийную полость и прогоняется через
жаберные ходы. Таким образом осуществляется дыхание и питание
моллюска. Далее вода поступает в наджаберную полость и, унося
экскреты (а в период размножения и половые продукты), удаляется
через выводной (клоакальный) сифон.
Кровеносная система. Жабры пронизаны сетью кровеносных
сосудов. По ним кровь поступает от жабр к предсердиям. Сердце
находится в лсрикардиальной сумке, представляющей собой участок
целомической полости и располагающейся позади переднего
мускула-аддуктора, ближе к центральной части туловища (рис. 84, 85).
Доступ к нему получают, проведя разрез по линии прикрепления
мантийной складки к спинной стороне туловища магпюска. По бокам
к продолговатому желудочку сердца примыкает пара предсердий,
имеющих тонкие пленчатые стенки. Сквозь всю перикардиальную
полость проходит кишечная трубка. При сокращении желудочка
кровь нагнетается в переднюю и заднюю аорты и далее в более мелкие
сосуды и лакуны полости тела.
Выдантельшш система представлена парными почками, или Боя-
нусовыми органами, которые залегают ниже перикардия и
выделяются своей темной окраской. Мерцательные воронки (нефростомы)
метанефридиев обращены в перикардиальную полость моллюска,
158
Тип MoHusca
Рис.84. Беззубка, извлеченная из раковины: / — нога; 2— мантия, ее
левая складка; 3— вводной (жаберный) сифон; 4— выводной
(клоакальный) сифон; 5 — брюшной край мантии; 6-10 —
мускулы (6 — передний мускул-замыкатель, 7 — задний
мускул-замыкатель. 8 — мускул-выдвигатель ноги, 9—
передний мускул-втягиватель ноги, 10— задний мускуд-втяги-
ватель); 11-13 — область расположения внутренних органов
(// — печень, 12— Кеберов орган, 13— перикардий)
кул-замыкатель раковины; 3 — мускул-втягиватель; 4 — почка;
5— правое предсердие; 6 — желудочек серша; 7— ресничная
воронка почки, открывающаяся в перикардий (целом); 8—
наружное отверстие почки; 9 — наружное половое отверстие; 10—
передняя аорта; 11 — желудок; 12 — «печень»; 13 — передний
мускул-замыкатель; 14— ротоеос отверстие; 15— левая створка
раковины; 16 — край лопасти мантии; 17 — наджаберная камера;
18— жабра; 19— половая железа; 20— кишка (частично она, как
и желудок, вскрыта); 21 — нота
Класс Bivalvia
159
выделительные отверстия открываются в мантийную полость.
Функции органов выделения выполняют также парные псрикардиаль-
ные железы, или Ксбсровы органы. Они расположены сбоку и
впереди от сердца, прилегая к внутренней стенке перикардия. У свеже-
умершоленных моллюсков перикардиальные железы имеют характерную
оранжево-красную окраску. Экскреты поступают сначала в
перикард пальну ю полость, а затем выводятся почками.
Пищеварительная система устроена достаточно просто.
Отфильтрованные частицы, а это может быть мелкодисперсный детрит (мертвая
органика), микроорганизмы (бактерии, планктонные водоросли,
простейшие) и минеральные взвеси, поступают ко рту. При повреждении
стенки пищеварительного канала видна заполняющая его зеленоватая
пастообразная масса потребленной взвеси. За ротовым отверстием
следует короткий трубчатый пищевод, переходящий вжелудок. Глотка
и ее вооружение (челюсть, радула) у двустворчатых моллюсков
редуцированы. В желудок открываются протоки «печени», которая
занимает основание ноги и некоторую часть туловища. В ней не только
вырабатываются пищеварительные ферменты, но и в значительной мере
осуществляется процесс пищеварения. Ткани печени имеют губчатое
строение, у фиксированных моллюсков они выделяются своей гор-
чично-зеленой окраской, хорошо отличаясь от буроватой полостной
паренхимы. От желудка отходит длинная и тонкая средняя кишка,
которая делает многочисленные петли в основании нош. Задняя
кишка начинается у места вхождения кишечной трубки в перикардиаль-
ную сумку, над задним мускулом-аддуктором она открывается в
клоакальную камеру.
Нервная система у пластинчатожаберных» так же как и у других
групп моллюсков, разбросанно-узлового типа. Для ознакомления
с особенностями ее строения желательно использовать свежефикси-
рованный материал, поскольку у таких экземпляров ганглии нервной
системы выделяются характерной оранжевой окраской. Парные цс-
ребро-плевральные ганглии размещаются по бокам от ротового
отверстия там, где оканчиваются ротовые лопасти,
висцеро-париетальные — внутрь от заднего мускула-аддуктора и немного вперед от
анального отверстия. Благодаря поверхностному расположению эти
ганглии легко обнаружить при вспарывании покровных тканей
препаровальной иглой. Педальные ганглии залегают в основании ноги
среди лопастей гонады и трудно различимы.
Половая система. Беззубки раздельнополы, однако половой
диморфизм не выражен. В отдельных популяциях присутствует та или
160
JmMo/lusca
иная доля гермафродитных особей. Парная гонада залегает в задней
части основания ноги и многократно пронизывается петляющей
средней кишкой (рис. 85). При вскрытии ее ткани, имеющие губчатую
структуру, выделяются желтоватой окраской, отличающей их от
тканей печени или полостной паренхимы. Трудно различимые парные
половые протоки открываются в наджаберную полость моллюска. Рс-
продуктивно активных самок беззубок (в осенне-зимний период)
и перловиц (в весенне-летний) можно распознать в ходе вскрытия
по яйцам либо эмбрионам, заполняющим внутрижаберное
пространство наружных полужабр. Начальные этапы развития унионид
проходят в межжаберных полостях материнских особей.
Строение гяоходкв. Развивающиеся из яиц личинки, глохидии,
своим строением напоминают миниатюрных беззубок. Размеры их
несколько меньше макового зерна, а форма напоминает гречневые
зерна (рис. 86). У них хорошо просматриваются расположенные на вер-
Рмс.86. Глохидии беззубки (вид с торца
раковины): / — створки
раковины; 2— краевые зубцы
створок; 3 — биссусовая нить
шинах створок острые зубцы. У некоторых экземпляров также
заметна обычно сильно скрученная темная биссусовая нить. И вершинные
зубцы, и биссусовая нить служат для прикрепления личинок к покровам
тела их хозяев — пресноводных рыб. Впоследствии они
утрачиваются, являясь, таким образом, провизорными органами.
Прикрепившиеся глохидии вызывают у рыб образование опухолей, в которых в течение
нескольких недель осуществляется их последующее развитие. Затем
стенки опухолей прорываются, молодые моллюски опускаются на дно
и переходят к характерному для взрослых особей образу жизни.
Тип Arthropoda — членистоногие
Важнейшими отличительными чертами членистоногих являются
наличие наружного скелета из хитинизированной кутикулы,
обновляющейся при линьках, и членистостъ конечностей. Тело разделено на
голову и туловище. Мозг сложный. Полость тела — миксоцсль. Дыхание
жаберное, легочное или трахейное.
Подтип Mandibulata
Надкласс Crustacea — ракообразные
Класс Branchiopoda — жаброногне раки
Отряд Anomopoda (-Cladocera) — ветвнетоусые
Вствистоусыс ракообразные — это высокоспециализированные
планктонные животные, населяющие стоячие пресные водоемы.
Обитатели морей единичны. Органами передвижения являются хорошо
развитие ветвистые антенны, — отсюда и название отряда. Тело,
исключая часть головы, защищено раковиноподобным хитннонлным кара-
паксом, 5-6 пар грудных конечностей преобразованы в фильтрующий
аппарат, обеспечивающий также и функцию дыхания. Брюшко
оканчивается фуркой (брюшной вилкой), имеющей вид когтевндных
придатков.
Daphnia pulex — обыквовеявая дафнвя (водяная блоха)
Материал. Daphnia pulex — массовый вид ракообразных во
многих мелких стоячих, обычно лишенных рыбьего населения водоемах.
Реже встречается более крупная Daphnia magna, которая также с успехом
может быть использована для учебных целей. Незначительная инюи-
дуальная продолжительность жизни этих рачков определяет
необходимость их культивирования в лабораторных условиях Питаются дафнии
бактериопланктоном либо мелкими планктонными водорослями.
Существует целый ряд методик, которые разрабатывались для
обеспечения высококачественным живым кормом молоди декоративных
и иных разводимых в искусственных условиях видов рыб.
Большинство из них технически не столь сложны, однако не всегда удастся
обеспечить стабильность культур вствистоусых рачков.
Для культивирования дафний можно использовать аквариумы
и другие емкости разных размеров, но более надежные результаты
'• fc« 16W
162
Inn Arthropods
получают при разведении их в сосудах объемом 100 л и более. Для
заселения используют несколько десятков экземпляров партсногене-
тических самок либо молоди дафний. Зимой используют
предварительно собранные зимующие яйца (эфиппии). Культуру удается
получить, если взять пробы прибрежного льда (желательно с
вмерзшими частицами берегового мусора) из водоемов, в которых
предшествующей осенью в большом количестве отмечались эти рачки. В
лаборатории пробы льда помещают в небольшие сосуды с отстоянной
водой. Один-два раза в неделю в воду добавляют несколько капель
нежирного молока, водной взвеси пекарских (не сухих!) дрожжей (из
расчета 5-7 мг на литр) либо немного прокисшего мясного бульона
(1 мл на литр). Но наиболее стабильные результаты достигаются при
использовании в качестве корма планктонных зеленых водорослей
(Chlorella, Sccnedesmus). Их культивируют на минеральных
питательных средах при искусственном освещении. Поскольку концентрация
солей в культуральной жидкости, как правило, достаточно высока, ее
нельзя доливать в сосуды с дафниями, так как те легко погибают от
осмотического шока. Лучше всего в этом случае прибегать к
центрифугированию и вносить получаемую пастообразную массу. Следует
иметь в виду, что при высокой плотности рачков накапливающиеся
в воде метаболиты замедляют размножение дафний, и это может
привести к угасанию культуры. Опасно также чрезмерное развитее бактсрио-
планктона и образование поверхностной бактериальной пленки. В этом
случае следует прекратить кормление и даже заменить часть воды на
свежую (обязательно отстойную и нехлорированную). Образующийся на дне
осадок необходимо регулярно удалять аквариумным сифоном.
Малочисленные популяции дафний можно в течение длительного времени
поддерживать в небольших сосудах с песчаным грунтом и небольшим
количеством пресноводных олигохст-трубочников (ТиЫ/ех)% которые
являются постоянным источником бактерий. Культивирование
большинства нехищных видов ветвистоусых раков осуществляется
аналогичным образом и не представляет затруднений. Для знакомства с
другими представителями отряда следует подобрать содержащие их
фиксированные пробы пресноводного планктона. В качестве
фиксатора допустимо применять 4% раствор формалина.
Наблюдение за движениями дафний удобно проводить, поместив
их в небольшой стеклянный сосуд (химический стакан, цилиндр,
пробирку). Для ветвистоусых раков характерны скачкообразные
движения, осуществляемые в результате резких взмахов разветвленных
антенн. Вслед за этим животное плавно начинает опускаться ко дну.
Класс Branchiopoda
разветвленные антенны выполняют в этом случае роль парашюта.
Подъем к поверхности обеспечивается частыми ударами антенн,
прекращение их работы ведет к погружению. Дафнии зачастую
подолгу остаются в определенном горизонте водной толщи, где
совершают маятникообразные, «толкущиеся» движения. Предугадать
траекторию движения испуганного животного весьма сложно, подобная
хаотичность уменьшает шансы хищника сделать точный бросок. Симо-
цефалюсы (Simocephalus) и шаровидные хидорусы (Chidorus) могут
прикрепляться к водной расплельности или даже к пленке поверхностного
натяжения и проводить в таком состоянии длительное время. У
представителей рода Daphnia практически вся жизнь проходит в
непрерывном движении. Дафнии обычно демонстрируют положительную
фототропическую реа> дню, собираясь в наиболее освещенной части
водоема или аквариума, где максимальна плотность планктонных зеленых
водорослей.
Строение дафнии. Длина тела взрослых дафний составляет нескол ь-
ко миллиметров. Оно полупрозрачно, поэтому для ознакомления
с особенностями строения этих рачков лучше использовать бино-
куляр, рассматривая объект в проходящем свете. Можно
воспользоваться постоянными тотальными препаратами Daphniapulex% но, имея
культуру, лучше непосредственно на занятии изготовить временные.
Отловленных стеклянной пипеткой рачков помещают в каплю воды
на часовое стекло, предметное стекло с лункой либо стандартное
предметное стекло. В последнем случае следует использовать покровное
стекло с высокими восковыми «ножками». Сохраняющих активность
животных рассматривать затруднительно, поэтому, чтобы ограничить
их движения, отбирают избыток воды полоской фильтровальной бумаги.
Тело дафнии округло-овальное (рис. 87), почти целиком
заключено в двустворчатую раковину, образованную разросшимся
головогруди ым щитом — карапаксом. На спинной стороне створки
переходят одна в другую, на брюшной имеется продольная щель, поступая
через которую, вода омывает туловище и конечности животного. Щель
также приоткрывается в момент распрямления брюшка, в обычном
состоянии подогнутого под брюшную сторону тела. Свободная от ка-
рапакса часть головного отдела тела защищена головным щитом.
Таким образом, все тело животного оказывается под прозрачным хити-
ноидным кутикулярным панцирем. Он выполняет главным образом
защитную функцию, но не в состоянии предотвратить поедание
рыбами и большинством других хищников. У представителей рода Daphnia
карапакс вытянут сзади в разной длины задний, иликаудальный,шип.
164
Тип Arthropods
Р*С. 87. Дафния (Daphnia pulex) вид сбоку. / — головной шлем; 2 —
ашеннулы; J— антенна; 4— науплиальный глаз; 5— сложный
глаз; б— мышцы сложного глаза; 7— печеночный вырост; £—
мышцы антенн; 9— средняя кишка; /0—сердце; //—яичник;
12 — выводковая камера; 13 — эмбрионы; 14 — спишюй
вырост камеры; /5—брюшко; /6—анус; /7—ф>рка; /£—третья
и четвертая пары грудных ног; 19 — жаберные мешки; 20 —
фильтрующие щетинки; 21 — брюшной край панциря (кара-
пакса); 22— шиловидный вырост карапакса
Считается, что он обеспечивает увеличение опорной поверхности,
что очень важно для планктонных форм, и осложняет заглатывание
рачков мальками. Поверхность панциря мелко скульптурирована,
что хорошо заметно даже при малом увеличении микроскопа. Контакт
с атмосферным воздухом очень опасен для этих животных:
извлеченные из воды рачки часто не в состоянии, ввиду малой смачиваемости
покровов, преодолеть пленку поверхностного натяжения и погибают,
оставаясь прикрепленными к ней.
KmccBranchiopoda
165
Тело дафнии лишено следов сегментации, его деление на отделы
неявно. Граница между головным и грудным отделами не
соответствует хорошо заметной борозде между головным шлемом и остальной
частью карапакса. Брюшко лишено конечностей, что может быть
основанием для его выделения.
Передняя часть головы клювовидно вытянута и обращена к
брюшной стороне. Из-под клювовидного отростка слегка выступают
малозаметные нсчлснистыс антсинулы, выполняющие функцию органов
химического чувства Антенны, напротив, хорошо развиты и
разветвлены. Нечленистое их основание представляет собой протоподит,
который несет две членистые ветви: экзоподит (отведен наружу) и эндо-
подит (вовнутрь), каждая из них снабжена четырьмя длинными
перистыми щетинками. На поверхности лба расположен крупный непарный
многофасеточный глаз, который закладывается как парный
орган. У живых дафний он находится в непрерывном движении
вследствие работы группы специализированных мышц, благодаря чему эти
животные, как и другие ракообразные, имеют возможность видеть
неподвижные предметы. Ниже расположен малозаметный наупли-
альный глаз, имеющий вид маленькой темной точки. Ротовые части
неразличимы.
Грудные конечности дафний листовидные. У разных
представителей Anomopoda число грудных конечностей варьирует от четырех
до шести пар, у Daphiapulex их пять, причем имеет место некоторая
функциональная дифференциация. Так, первые две пары являются
хватательными. Типичным считается строение конечностей третьей
пары, которые имеют вид пластинок, несущих многочисленные
волоски и щетинки. Здесь хорошо различимы протоподит, экзоподит
и эндоподит. Мешковидный эпиподит выполняет функцию
жаберного придатка. Ряды сидящих на эндоподитах щетинок образуют фильтр,
через который проходит нагнетаемая в полость карапакса вода.
Грудные конечности рассмотреть по отдельности весьма непросто, так как
они находятся в постоянном движении (число взмахов достигает 300
в минуту). Этим обеспечивается непрерывное поступление свежей
воды, а значит потребление пищи и жаберное дыхание. Функция
органов движения грудными конечностями утрачена.
Между раковиной и стенкой тела самки на спинной стороне
имеется полость, которая носит название выводковой камеры. В нее
поступают партеногенетические яйца, и здесь происходит развитие
эмбрионов. При подгибании брюшка готовые к самостоятельной жизни
молодые особи покидают камеру через открывающееся отверстие.
166
1m Arthropod*
На заднем конце брюшка есть пара снабженных волосками
щетинок и пара каудальных коготков, особенности строения которых
учитываются при определении таксономической принадлежности
представителей отряда.
Пищеварительная система хорошо видна на препарате, так как
изогнутая в виде скобы кишечная трубка заполнена пищевыми
частицами. Дафнии — фильтраторы. По просвечивающей окраске
можно составить представление о потребленной рачком пище: если
это частицы детрита и бактерии, преобладают красновато-бурые или
буро-серые тона, зеленые водоросли — зеленые и грязно-зеленые.
Отфильтровывая взвесь, дафнии не в состоянии отделить съедобные
частицы от несъедобных. Попавшие в проточные водоемы особи
обычно быстро погибают, так как их кишечник заполняется
непременно присутствующими в таких водах взвешенными минеральными
частицами, своим весом тянущими животных на дно.
Кровеносная система. На спинной стороне за кишечником, над
выводковой камерой расположено овальное сердце. У живых
экземпляров оно непрерывно пульсирует, совершая при комнатной
температуре до 500 сокращений в минуту. Под микроскопом иногда удается
рассмотреть крупные клетки, образующие его стенки, и даже их ядра.
При сокращениях сердечной мускулатуры кровь омывает головной
отдел, затем направляется к грудным конечностям, насыщается
кислородом и по лакунам (настоящих сосудов нет) поступает к другим
частям тела. Кровь дафний практически бесцветна, хотя и содержит
гемоглобин в растворенном состоянии. У рачков, обитающих в среде
с явным недостатком кислорода, она приобретает выраженную
окраску вследствие значительного повышения концентрации этого
дыхательного пигмента.
Выделительная система представлена парой трудно различимых
максиллярных желез и их протоков, расположенных между
перикардием и брюшной стороной тела.
Имеющаяся у самок парная половая железа (яичник) ниже
уровня сердца прилегает по бокам к кишечнику. Сформировавшиеся
яйца из нее поступают в выводковую камеру.
Знакомство с эфишшями дафний. В естественных водоемах
умеренной зоны осенью, а в лабораторных культурах при их угнетении
(ввиду перенаселенности либо по иным причинам) могут быть
обнаружены самцы, которые в несколько раз меньше самок и лишены
выводковой камеры. Оплодотворенные яйца поступают в выводковую
Кпгсс Branctiiopodd
167
камеру, где одеваются мощными зашитными оболочками и свособ-
|\иным футляром, на образование которого частично идут стенки самой
выводковой камеры. Такой футляр с покоящимися внутри яйцами
(чаще парой) называется эфиппием, или седлышком (ввиду
седлообразной формы). Продуцирующие их самки (именуемые эфиппиаль-
ными) легко выделяются среди прочих характерной темной окраской
выводковой камеры (рис. 88). Самки освобождаются от эфиппиев во
время линьки либо так и погибают с эфиппиями внутри.
Сброшенная раковина у Daphnia pulex зачастую выполняет функцию
дополнительной оболочки эфиппиев, плавающих на поверхности воды.
У Daphnia magna седлышки оседают на дно или водную растительность.
Покоящиеся яйца легко переносят высыхание, низкие температуры,
вмерзайис в лед. Описаны случаи, когда рачки нормально отрожла-
лись из эфиппиев, выделенных из кишечника рыб либо хранившихся
продолжительное время в растворе формалина. Эфиппии легко
цепляются к перьевому покрову птиц либо меху млекопитающих,
которые таким образом участвуют в расселении этих
беспозвоночных, что является примером явления зоохории. Оставшиеся на дне
высохшего водоема седлышки легко могут быть подхвачены ветром
и унесены на значительные расстояния, то есть здесь имеет место
анемохория. С внешним видом эфиппиев удобнее ознакомиться
на постоянных микроскопических препаратах либо на препаратах,
изготовленных из проб осеннего озерного планктона.
Рте. 88. Эфиппиумы дафний: Л — Daphnia pulex, Б — Daphnia торга
(внутри пустого карапакса самки)
168
T*n Arthropod*
Внешний вид босмин. Босмины (Bosmina) являются типичными
обитателями озер и крупных прудов. Во внешнем облике этих
ракообразных обращает на себя внимание короткое округлое тело и
характерный клювовидный вырост, образованный хорошо
развитыми антеннулами. Он выполняет функцию балансира и препятствует
переворачиванию животного во время гребных движений антенн
(рис. 89). Босмины составляют значительную долю в рационе молоди
озёрных рыб.
Рис. 89. Схема движений босмины (Bosmlna) в воде. Стрелками
обозначены токи воды
Класс MaxUIopoda
- Подкласс Copepoda — веслоногие раки
Веслоногие являются представителями пресноводного и морского
планктона либо придонными обитателями. Большинство Copepoda —
хищники, растительноядные виды немногочисленны. Значительную
долю составляют паразитические (главным образом эктопаразити-
ческис) формы. Они представляют серьезную опасность для молоди
многих хозяйственно ценных видов рыб и иногда заносятся вместе
с живым кормом в аквариумы. Представители рода Cyclops являются
промежуточными хозяевами некоторых гельминтов, среди которых
наиболее известны широкий лентец и ришта. С другой стороны,
веслоногие ракообразные (личинки и взрослые особи) составляют основу
рациона многих промысловых рыб.
Класс МахШородэ
169
Весьма характерным для всех Copepoda является отсутствие парных
многофассточиых глаз. У свобод ножи вущих форм в составе тела
выделяются слитный цсфалоторакс, свободные грудные сегменты
и сегментированное, лишенное конечностей брюшко. Движение
осуществляется с помощью хорошо развитых антениул и грудных
конечностей, несущих гребные поверхности (отсюда и название —
веслоногие).
Отрад Cychpida — циклооообразные
Представители отряда отличаются антеинулами, не
достигающими середины головогруди и состоящими не более чем из 19 члеников;
взрослые самки несут пару яйцевых мешков. Cychpida более
характерны для краевой и придонной зон пресных водоемов. Они не
столько флотируют (парят в толще воды), как классические планктонные
формы, сколько активно перемещаются в разных слоях водной массы.
Преобладают среди них хищники (Cycbpsspp., Mocwcydopsspp.),
некоторые питаются нитчатыми водорослями (Eucyclopsspp.) и даже
колониальными простейшими (Mesocyclopsspp.).
Cyclops spp. - цииюпы
Материи. Циклопы встречаются практически в любых стоячих
и большинстве проточных водоемов, включая временные водоемы
в дуплах деревьев, существующие несколько дней лужицы, бассейны
фонтанов и т.п. Их нетрудно содержать в аквариумах, но материал для
занятий, которые проводятся в зимний период, лучше собрать заранее
с помощью обычной планктонной сети на незамерзших участках рек.
В течение одной-двух недель циклопов можно хранить в небольшом
количестве воды без се смены и без кормления при низкоположитсль-
ных температурах, например в холодильнике. Для ознакомления с
особенностями строения фудных конечностей лучше использовать
материал, фиксированный 70* этанолом или 4% раствором формалина.
Науплиусов циклопов можно обнаружить в пробах воды из
аквариумов, в которых обитают эти ракообразные. В теплое время гола они
в массе присутствуют в планктонных пробах, отбираемых с помощью
сачка из густого мельничного газа в мелких временных водоемах. Но
на занятиях лучше использовать постоянные микроскопические
препараты окрашенных объектов либо временные препараты проб
летнего озерного планктона, фиксированных 4% раствором формалина.
170
Тип Arthropod*
Движения циклопов. Способ их передвижения заметно
отличается от свойственного вствнетоусым ракообразным. Двигаясь
характерными рывками, циклопы описывают весьма сложные траектории.
Маневренность и быстрота движений помогает этим хищникам в
поимке добычи, а также уменьшает вероятность быть съеденными более
крупными животными.
Строение циклопа. Сегментация тела веслоногих ракообразных
гетерономная (рис. 90). Слитная «сложная голова» образована в
результате объединения головных и первого грудного сегментов тела.
Далее следуют четыре свободных грудных сегмента. Брюшко
значительно уже груди и лишено конечностей. У самцов хорошо
различимы пять истинных сегментов. У самок первые два сегмента брюшка
слиты, образуя так называемый ген итальный сегмент. У взрослых
самок по бокам брюшка подвешена пара овальных яйцевых мешков.
Оканчивается брюшной отдел тела анальной лопастью, несушей
вилку (фурку) со щетинками.
На временных препаратах циклопы могут размешаться на
брюшной, спинной стороне либо на боку. Просмотрев несколько таких
особей, легче уяснить особенности строения этих животных.
Л нтс и пулы у циклопов, как и у всех свободноживущих
веслоногих, олиовствистыс и очень хорошо развиты. Основная их
функция — локомоторная. У самцов антеннулы несколько короче, слегка
заострены и имеют характерным образом закрученные вершины. Они
служат для охватывания тела самки и удержания ее во время спаривания.
Такие придатки носят название геникулирующих. На спинной
стороне сложной головы расположен небольшой науплиальный глаз.
Для дальнейшего ознакомления с придатками головного отдела
тела необходимо рассматривать животное с брюшной стороны. Антсн -
ны циклопов короче аитеннул, двувствисты и несут щетинки.
Ротовой аппарат образован четырьмя парами конечностей и ограничен
спереди и сзади складками стенки тела, которые носят название
верхней и нижней губ. Пара верхних челюстей (мандибулы)
и две пары нижних (максиллы) используются рачкамидля
обработки пиши,тогда как пара ногочслюстей (максиллопед) —для
ее захвата.
Собственно грудные сегменты» исключая последний, несут
типичные двувствистыс плавательные конечности. В
противоположность вствнетоусым ракам, у копепод грудные конечности непосред-
Класс Maxtllopoda
P*c. 90. Веслоногие (Copepoda): A — самка Cychpsstrcruuur. I — глаз; 2 -
актсннулы; J— антенны; 4— головогруаь; 5— четыре свободных
грудных сегмента; б— гснитапьныйссгментбрюшка; 7— яГшс-
вые мешки; 8— вилка (фурка); Р— кишечник; /0—
продольные мышцы груди; // — яичник; В — самец Cyclops
canthocarpoidar. 1 — глаз; 2— антеннулы; 3 — головогрудь; 4 -
свободные грудные сегменты; 5 — генктальный сспчсмт
брюшка; 6 - II-V сегменты брюшка; 7- вилка (фурка): S -.
кишечник; 9— сперматофорные карманы; /0—семяпроводы.
// — продольные мышцы груди
172
Тип Arthropods
Рис.90. Окончзние. 5 —самка Diaptomur. / —антеннула; 2— верхняя
губа; J— антенна; 4— мандибула; 5 — максилла-1; 6— макс ил-
ла-И; 7 — ногочслюсть; £ — грудные конечности; 0 — вилка
(фурка)
ственно не участвуют в добывании пиши и не отвечают за газообмен.
Строение гребною аппарата веслоногих ракообразных лучше заметно
при рассмотрении животных с боковой стороны. Для изучения
строения гребных конечностей можно использовать диагттомусов (отр. Cala-
nida) (рис. 90, В), обычно в массе присутствующих в пробах озерного
планктона.
Из внутренних органов на препаратах чаще виден только
тянущийся вдоль всего тела кишечник. По бокам от него в передней части
тела иногда просматривается крупная лопастсвидная половая
железа. Выдслитсльныя система представлена недоступными для
наблюдения парными максиллярными железами. Кровеносная система
как таковая у веслоногих отсутствует. Дыхание осуществляется всей
поверхностью тела. В отличие от дафний, циклопы малочувствительны
к низкому содержанию в воде кислорода и даже присутствию
высокотоксичных химических соединений.
Яйцевые мешки самок представляют собой конгломерат яиц,
склеенных секретом специальных кожных желез. Секрет может
обволакивать и все тело рачка, образуя подобие кокона. В таком
состоянии животное способно переносить пересыхание водоема,
вмораживание в лед и другие экстремальные условия. Это позволяет циклопам
заселять практически все пригодные для обитания водоемы.
Класс Ма»//ороЛ
173
Строение науплиусов. Развитие Copepoda осуществляется с
метаморфозом. Из яиц отрождаются личинки, называемые науплиусами.
Науплиусы циклопов проходят до пяти линек, после чего
превращаются втак называемых копеподитных л ичинок,также многократно
линяющих за время своего развития. Науплиусы имеются у
большинства ракообразных, они являются непременным компонентом
планктона любых водоемов.
Науплиусы имеют яйцевидную форму тела, внешняя сегментация
у них не выражена (рис. 91). Весьма характерен непарный простой
глазок, который сохраняется и у ряда ракообразных (например,
Anomopoda) во взрослом состоянии и носит название науплиальиого.
Науплиусы обладают парой одноветвистых придатков (антеннул)
и двумя ларами двувствистых конечностей (соответствуют
антеннам и мандибулам взрослых особей). Все они выполняют
локомоторную функцию и по необходимости используются для захвата
пищевых объектов. В отдельных случаях через прозрачные хитнноидные
покровы личинки может просматриваться пищеварительный канал,
имеющий вид широкой трубки.
Рис. 91. Науплиусы веслоногих (Copepoda): первая нлуплиальная
стадия (А), третья науплиальная стадия (£): / — антеннулы;
2 — антенны; J— мандибулы; 4— максиллы; 5— наупли-
альный глаз
174
Jm Arthropods
Класс Malacostraca — высшие раки
Отряд Decapoda — десятиногие раки
Десятиногие раки считаются эволюционно наиболее продвинутой
группой ракообразных. В то же время у них сохранилась примитивная
дифференциация головного отдела на протоцсфалон и гнатоцефалон.
Протоцсфалон (первичная голова) образован слиянием акроиа
(головной лопасти) и первого головного сегмента. Три
оставшихся головных сегмента, конечности которых вошли в состав ротового
аппарата, образуют гнатоцефалон. составляющий вместе с грудными
сегментами слитную головогрудь, одетую хорошо развитым кара-
паксом. Грудных сегментов восемь, из них три первых несут ногоче-
люсти, а пять остальных — ходильные конечности, что и стало
основанием для соответствующего названия отряда.
Классическим объектом для ознакомления с особенностями внешнего
и внутреннего строения декалод являются речные раки. Экземпляры
с вычлененными конечностями становятся малопригодными для
вскрытия, поэтому технически удобнее ознакомиться с внешним видом
и сегментацией тела, провести вскрытие и рассмотреть особенности
анатомии, а лишь затем перейти к вычленению конечностей.
Potamobius spp. — речные раки
Материал. Для анатомирования на лабораторных занятиях малого
практикума пригодны крупные экземпляры любых представителей
рода. В водоемах бассейнов Черного, Азовского и Каспийского морей
широко распространен узкопалый (длиннопалый) рак (Potamobius
(=Astacus) leptodacrylus). Он также заселяет некоторые реки и озера
Западной Сибири, прибрежные участки Азовского и Каспийского
морей, отличающиеся невысокой соленостью; в последнее время
активно проникает в водоемы бассейна Балтийского моря, которые
изначально были заселены широкопалым раком (Potamobius astacus).
Последний, ввиду существенного снижения численности популяций
и сокращения ареала, внесен в Красную книгу Международного
союза по охране природы и природных ресурсов и в Красную книгу
Республики Беларусь, что накладывает соответствующие
ограничения на его отлов. В последнее время в ряде мест успешно
осуществляется разведение узкопалого рака, и материал для занятий может
быть приобретен в этих хозяйствах. Для отлова раков в заселенных
ими водоемах используют специальные живоловки (рачни),
снаряжаемые приманкой — мертвой рыбой, лягушками либо кусочками
несвежего мяса.
Класс MaJacostraca
Хранение речных раков в живом состоянии весьма
затруднительно, для их содержания нужны большие бассейны с чистой проточной
(юлой. При фиксации животных помешают в пустые эксикаторы или
лругис герметичные сосуды и усыпляют хлороформом. Хранят
сначала в пятикратно разбавленном водой формалине, через
месяц-другой их переносят в 4% раствор формалина. Перед занятиями раков
необходимо выдержать примерно сутки в проточной или
неоднократно сменяемой воде.
Внешнее строение и сегментация речного рака. Животное помещают
в препаровальную ванночку спиной вверх. Прикосновение пинцетом
или препаровальной иглой позволяет убедиться, что покровы тела
рака обладают значительной твелостью. Отделяемая гиподермой
хитиновая в своей основе кутикула интенсивно кальцифицирована:
на долю СаС03 и других солей кальция приходится до 60 % се массы.
Это объясняет требовательность раков к жесткости воды и отсутствие
их в болотных и иных водоемах, вода в которых практически лишена
растворенных солей кальция. Покровы тела только что перелинявших
раков нежны и эластичны, так как ешс не подверглись
обызвествлению. В этот период животные отсиживаются в укрытиях. Отдельные
части тела (сегменты брюшка, членики конечностей) обладают
относительной подвижностью, благодаря мембранозным сочленениям,
где кутикула эластична ввиду отсутствия известковой пропитки.
Речные раки обычно имеют темную окраску, причем зависит она
от условий местообитания животных, варьируя от буроватой до
зеленоватой, темно-зеленой и даже голубоватой (на голубых лонных илах
и глинах). Окраска обусловлена комбинацией присутствующих в
кутикуле членистоногих меланинов и зелено-голубого пигмента циаио-
кристаллина. При высоких температурах он необратимо превращается
в красный зооэритрин, чем определяется характерная окраска
вареных раков. Хранившиеся в формалине экземпляры имеют бурую
окраску; белесые пятна и разводы на поверхности кутикулы
представляют собой конденсирующийся полимеризовавшийся
формальдегид.
Тело речного рака (рис. 92, А) подразделяется на широкоовальную
слитную головогрудь и более узкое членистое брюшко, которое в
просторечье называют также «шейкой* или «хвостом». Спереди выступает
часть первичной головы — протоцсфалона, образованного
слившимися акроном (головной лопастью) и первым головным сегментом.
Протоцсфалон несет антеннулы, антенны, а также расположенные
176
ImArttiropoda
на подвижных стебельках сложные многофасеточные глаза.
Остальные три головных сегмента вместе со всеми воссмыо грудными
сегментами образуют цельный гнатоторакс (челюстегрудь),
который со спинной стороны и боков покрыт мощным щитом — кара-
паксом, представляющим собой разросшиеся спинные пластинки
грудных сегментов тела. На поверхности карапакса различимы
неглубокие борозды. Более мощная дугообразная поперечная борозда
называется затылочной, или шейной, а пара идущих вдоль прололыюй
серединной линии спины — брахио-кардиальными (жаберно-сер-
дечными). Местоположение затылочной борозды примерно соответ-
18 17
Рис. 92. Внешний вид речного рака (Potomobius astacus): самка со
спинной стороны (А) и с брюшной стороны (Б): I — головогрудь;
2— брюшхо; 3— граница головогруди и брюшка; 4—жаберно-сер-
дечные борозды; 5 — шейный шов; 6 — рострум; 7— аитенну-
лы; 8 — аюенны; 9 — экзоподит антенны; /0 — стебельчатый
глаз; // —ногочслюсть-Ш; /2—клешня; 13— ходильные ноги;
/4—11 пара брюшных ножек; /5 — 111—V пары брюшных
ножек (плеоподы); 16 — VI пара брюшных ножек (урополы);
/7 — тельсон; 18 — анус; 19 — развивающиеся яйца,
прикрепленные к брюшным ножкам
Класс Malacostraca
177
ствует границе между последним головным и первым грудным
сегментами, а брахио-кардиальных — верхним границам жаберных
полостей и месторасположению сердца. Спереди по центру каралакса
имеется заостренный шиловидный выступ — рос тру м, по бокам есть
менее крупные шиловидные выросты, которые служат для защиты
стебельчатых глаз.
Брюшко (абдомен) состоит из шести легко различимых
сегментов и оканчивается тел ьсоном (анальной лопастью). Каждый сегмент
состоит из выпуклого сильнохитинизированиоготергита
(спинного склерита) и стернита (брюшного склерита), имеющего вид
слабохитинизированного полукольца. Боковые выросты тергитов —
эпимеры, выступают в стороны и имеют слегка заостренные края.
У самок брюшко замелю шире, чем у самцов, что связано с
необходимостью вынашивания под ним яиц и молоди. Брюшко
подвижно причленястся к гнатотораксу. Отгибая его вниз, можно увидеть
беловатую сочленовную мембрану. Аналогичным образом сочленены
между собой все шесть сегментов брюшка. Эластичность этих мембран
обеспечивает возможность изгибания и других движений брюшка.
Тельсон имеет вид плоской пластинки с расположенным на се нижней
стороне хорошо заметным анальным отверстием.
Вскрытие речного рака. Животное следует положить в ванночку
брюшной стороной вниз, а затем аккуратно согнуть тело так, чтобы
на спинной стороне обнажилась сочленовная мембрана между задним
краем гиатоторакса и первым брюшным сегментом. Ее протыкают
заостренным лезвием ножниц либо разрезают скальпелем. После
этого животное можно зафиксировать булавками. Затем лезвие ножниц
вводят под карапакс и делают, чуть отступив внутрь от
брахио-кардиальных борозд, два параллельных разреза к основаниям глаз. У
основания рострума эти разрезы соединяют. Пинцетом аккуратно снимают
вырезанную площадку. При этом важно оставить неповрежденной
тонкую пленчатую гиподерму. После ознакомления с ее строением
гиподерму удаляют и тогда открывается полость тела с внутренними
органами.
Животное разворачивают головой к препаратору и продолжают
разрезы на брюшке. Доводить их до тельсона необязательно, пройдя
несколько сегментов, продольные разрезы соединяют поперечным,
а затем аналогичным образом аккуратно, не разъединяя фрагменты
спинных пластинок отдельных сегментов, снимают соответствующий
участок покровов. Этому могут препятствовать прикрепленные к стенке
178
Тип Arthropods
тела парные межсегментариые мышечные пучки, которые идут из
одною брюшного сегмента в другой. Поскольку они расположены со
спинной стороны, в их функцию входит разгибание брюшка. После
удаления вырезанной площадки покровов и поверхностной
мускулатуры видна трубчатая задняя кишка, которую желательно не повредить.
Почти вес остальное пространство занимает мускулатура. С брюшной
стороны она намного мощнее, чем со спинной, так как при резком
подгибании брюшка совершаются мощные гребные движения
хвостовым плавником, позволяющие раку, перемещаясь скачками,
спасаться от хищников.
Используя выделенный фрагмент покровов брюшка, можно еще
раз обратить внимание на механизм подвижною сочленения его
сегментов, который характерен для всех членистоногих с гибким
членистым телом (других ракообразных, многоножек, насекомых, скорпионов
и пр.). При осторожном его растягивании обнажается
полупрозрачная сочленовная мембрана (плевра), обеспечивающая черспицеоб-
разное расположение спинных склеритов.
Внутреннее строение. Внутренности в полости гнатоторакса
располагаются в несколько ярусов, поэтому отдельные органы и их системы
приходится рассматривать в порядке, диктуемом их топографией.
Кровеносная система. После удаления гиподермальной пленки
(если таковая оставалась) открывается обширная полость, в которой
размещены внутренние органы (рис. 93). Поверхностно
располагается желтоватый пятиугольный мешочек — сердце. Кровеносная
система у ракообразных незамкнутая, нагнетаемая сердцем кровь
циркулирует по системе артерий и более мелких кровеносных
сосудов, атакже целомических синусов и лакун. Кровь бесцветна,
поэтому кровеносная система и ее содержимое не выделяются своей
окраской. Сердце располагается в перикардиальном синусе
(полости). Стенки перикардия тонкие, иногда они различимы в виде
полупрозрачной пленки. Стенки сердца, напротив, толстые и упругие,
поскольку образованы главным образом мышечной тканью.
Непосредственно от сердца отходит пять сосудов, они имеют вид тонких
полупрозрачных, слегка поблескивающих нитей. Назад, к брюшку от
сердца отходит непарная абдоминальная (брюшная) артерия.
Прямо вперед уходит непарная, или глазная, артерия, а по
сторонам — пара антеннальных артерий. Нисходящая, или грудин-
ная, артерия отходит от задней стенки сердца отвесно вниз.
Класс Malacostraca
179
Рис. ft?. Вскрытый речной рак (самец): / —
зеленая (антеннальиая) железа; 2 —
жевательный мускул мандибул; J —
печеночная (поджслудочно-печеночная)
железа; 4 — сердце (видна пара остий);
5 — задняя часть гонады (семенника);
6 — кишка; 7 — мускулы-разгибатели
брюшка, 8 — извитый семяпровод
(правый), 9 — жабры; /0— желудок
к брюшной стороне тела. Внутрь сердца кровь поступает через
снабженные клапанами щслевидные ости и, которые вполне можно
разглядеть у крупных экземпляров, зная их местоположение. Остий три
пары: дорсальные располагаются спереди на спинной поверхности
сердца, латеральные — побокам сзади, вентральные — на его
нижней стороне. Для того чтобы лучше рассмотреть оста и, следует
аккуратно вычленить сердце и, поместив в налитую в часовое стекло воду,
рассмотреть под бинокуляром.
Половая система. Ниже под сердцем расположена трехлопастная
гонада: две лопасти ее направлены вперед, а одна — назад. Нспарность
задней лопасти является результатом частичного слияния исходно
парных половых желез. Окраска и консистенция гонад определяются
полом и периодом репродуктивной активности особи. В частности,
у самок ткани железы имеют зернистую структуру и
желто-оранжевую окраску. У готовящихся к откладке яиц особей они внешне
напоминают мелкую зернистую икру. Короткие яйцеводы открываются
парными женскими наружными половыми отверстиями на
основаниях третьей пары ходильных ног. Ткани семенника губчатые.
От него берут начало семяпроводы, имеющие вид длинных
спутанных трубчатых образований. У свежеумерщвленных особей они
беловатые, у долго хранившихся в формалине — более темные,
напоминающие тонкую проволоку. Семяпроводы открываются мужскими
180
Тип Arthropods
половыми отверстиями, расположенными на основаниях пятой
пары ходильных ног. Таким образом, половой диморфизм
проявляется в месторасположении наружных половых отверстий.
Пищеварительная система. Ротовое отверстие расположено на
брюшной стороне тела. Пищевод представляет собой восходящую
к желудку короткую трубку. От желудка отходит короткая средняя
кишка, принимающая протоки массивной парной печени. Здесь
же берет начало задняя кишка, которая открывается анальным
отверстием на нижней стороне тельсона.
Изучив общую схему строения пищеварительной системы,
приступают к практическому ознакомлению с ее частями. С помощью
пинцета аккуратно удаляют яичники или семенники, при
необходимости подрезав половые протоки в местах их выхода из гонад.
Крупная парная печень занимает почти все пространство полости цефало-
торакса впереди от гонады, выделяясь характерной грязно-желтой или
горчичной окраской. У экземпляров, длительное время хранившихся
в формалине, а также при использовании фиксатора недостаточной
крепости при удалении гонад зачастую не удается избежать
повреждения печени. После успешного изъятия гонад следует отвести в
сторону расположенный спереди от печени мешотчатый желудок. При
этом становится доступным уходящий отвесно вниз короткий
трубчатый пищевод. Его аккуратно подрезают как можно ниже. Затем ближе
к заднему концу тела перерезают заднюю кишку, после чего можно
извлечь выделенный участок пищеварительного канала с
соответствующими компонентами пищеварительной системы. Его помещают в
препаровальную ванночку для детального изучения.
Положение пищевода относительно желудка определяется
расположением рта, который, как указано выше, находится на брюшной
стороне и несколько смещен назад от переднего конца тела.
Местоположение ротового отверстия легко установить, введя пинцет в
оставшийся отрезок пищевода. По короткому пищеводу измельченная пища
поступает в желудок. Пищевод и желудок представляют собой отделы
передней кишки, имеющей эктолермальное происхождение,
поэтому они снабжены внутренней хитиновой выстилкой. При линьке
рака трубчатая выстилка пищевода и фрагменты хитиновой выстилки
желудка также удаляются. Форма желудка достаточно сложна:
спереди он имеет округлые очертания, сзади — треугольный.
Поперечной бороздкой, которая хорошо выражена у крупных экземпляров,
желудок делится ha два неравных отдела: более крупный кардиаль-
ный (жевательный) и небольшой пилорический (цедильный).
Класс Malacostraca
181
Поступающая в желудок пища подвергается окончательному
измельчению в имеющем мускульные стенки кардиальном отделе. Стенки
его несут три склерита, снабженные мощными жевательными з>6цами.
Эти приспособления нетрудно обнаружить, вскрыв желудок. Сами
склериты беловатые, тогда как зубцы рыжие, бурые или совсем темные.
Прикосновением пинцетом или препаровальной иглой легко
установить степень их жесткости. В движение этот внутренний
жевательный аппарат, называемый желудочной мельницей, приводится
мощной мускулатурой. В кардиальном отделе пища также
подвергается начальной ферментативной обработке секретом слюнных желез.
Пилорический отдел желудка меньший по размерам и имеет тонкие
мембранозные стенки. Внутри хитиновая кутикула образует сложный
цедильный аппарат. Ознакомиться с его строением при
используемой технике препарирования объекта не представляется возможным.
Основной функцией данного отдела желудка является
фракционирование пищевых масс: жидкие и пюреобразные фракции поступают
в среднюю кишку, тогда как твердые частицы направляются сразу
в заднюю кишку и удаляются из организма.
Печень (поджелудочно-печеночная железа) состоит из
огромного числа трубчатых канальцев, объединенных промежуточной тканью.
В печени происходит окончательное переваривание и всасывание
пищи, в этом случае она выполняет ту же функцию, что и средняя
кишка у других животных. Помимо этого, здесь происходит
накопление резервных веществ и производится некоторое количество
пищеварительных ферментов. Таким образом, у раков этот орган не
является классической пищеварительной железой.
У давно линявших особей в специальных карманоподобных
образованиях по бокам желудка можно обнаружить линзовидные белые
«жерновки», состоящие из карбоната и фосфатов кальция. Вопреки
названию, они служат главным образом для депонирования
поступающего с пищей кальция. Сразу после линьки они рассасываются, а
запасенный кальций используется для пропитки кутикулы.
Задняя кишка трубчатая, проходит практически по дну полости
гнатоторакса, но в брюшном отделе переходит на спинную сторону,
где ее можно наблюдать в ходе начальных этапов вскрытия животного.
Этот отдел кишечника имеет эктолермапьное происхождение и
снабжен хитиновой выстилкой.
Выделительная система. Органами выделения у взрослых особей
высших раков служат парные антеннальные железы. У вскрытых
182
Inn Arthropods
животных антеннальные, или зеленые, железы обнаруживаются в
виде небольших округлых образований, залегающих в головном отделе тела
по бокам от сохранившегося отрезка пищевода. У свежеумерщвлен-
ных раков они имеют характерную зеленую окраску, что и послужило
основанием для их второго названия. У долго хранившихся
экземпляров эта окраска бывает утрачена. Препаровальной иглой или тонким
пинцетом можно подцепить и извлечь наружу белое лепешковид-
ное образование, снабженное длинным извитым канальцем. Это —
антсннальная железа, которая состоит из небольшого цсломического
мешочка и выводного протока, имеющего в дистальной части
расширение, именуемое мочевым пузырем. Выводные протоки желез
открываются выделительными порами на хорошо заметных
бугорках, расположенных на основаниях антенн.
Нервная система. Центральная нервная система представлена
надглоточным и подглоточным ганглиями (синганглиями)
и брюшной нервной цепочкой. Надглоточный ганглий (головной
мозг) иннервируетантеннулы, антенны и фасеточные глаза.
Обнаружить ею можно, удалив рострум. С подглоточным ганглием его
связывают окологлоточные коннективы (они сохранили название,
несмотря на фактическое отсутствие глотки), которые имеют вид
бело-серых тяжей, огибающих трубчатый пищевод. Можно
попытаться удалить мускулатуру и обнажить подглоточный ганглий. Он образован
слиянием трех пар головных и трех пар грудных ганглиев. Чтобы
сделать доступным грудной участок брюшной нервной цепочки,
состоящий из пяти пар пространственно разделенных ганглиев, необходимо
удалить хитиновую покрышку, образованную отростками оснований
грудных конечностей. Выделение брюшного участка нервной цепочки
осуществить проще. Удалению в этом случае подлежат
мышцы-сгибатели брюшка. При этом обнажается шесть парных ганглиев, иннервиру-
ющих соответствующие брюшные сегменты, последний иннервирует
еще и тельсон. Таким образом, концентрация ганглиев у речных раков
еще слабо выражена, а укорочение коммисур еще не привело к
слиянию парных ганглиев.
Состав и строение конечностей и придатков тела. У речных раков
сохранились конечности на всех сегментах тела, но в соответствии со
специализацией многие из них подверглись существенной
модификации. Ознакомиться с составом и строением конечностей лучше в ходе
их последовательного вычленения у ранее вскрытых экземпляров.
При этом, удерживая тело рака, пинцетом отделяют конечности
по сочленовной мембране. Практически это выглядит как
выщипывание захватываемых за основание конечностей.
Класс Maiacostracd
183
Исходным считается вариант строения двувствистой конечности,
когда ее основание (протоподит), состоящее из небольшого коксо-
и оли та и более крупного бази полита, несет две ветви — наружную
(экзополит) и внутреннюю (эндоподит). а также наружный
отросток (эпиполит). В реальности одни часта получают
преимущественное развитие или приобретают дополнительные приспособления,
а другие подвергаются той или иной степени редукции.
Выполнять вычленение конечностей технически удобнее с
заднего конца тела. Выделив очередную конечность, се рассматривают (при
необходимости — с использованием бинокуляра) и последовательно
выкладывают в препаровальную ванночку или на отдельный лист
плотной бумаги (рис. 94).
Конечности головного отдела тела. Головной отдел имеет четыре
пары модифицированных в соответствии с функциональной
специализацией конечностей и пару придатков а крона (головной лопасти).
Антеннулы считаются пр»щлткамнакрона,двуиствистостънхурсчных
раков рассматривается как вторичное явление. Обе истин антеннул
относительно коротки, имеют примерно равную длину и усеяны
множеством мелких волосков, что позволяет рассматривать их как органы
химического чувства. На верхней, спинной стороне основного
членика антеннул при внимательном рассмотрении можно обнаружить
треугольное щелевидное отверстие, закрытое густым частоколом
щетинок. Оно ведет в камеру статоциста, выполняющего функцию
органа равновесия. В качестве статолита используется песчинка или
мелкий камешек, который можно обнаружить при вскрытии статоциста.
Антенны представляют собой видоизмененные конечности.
Они двуветвисты: экзополит редуцирован до небольшой треугольной
пластинки — чешуйки. Эти образования отличаются повышенной
жесткостью, заострены на вершинах и выполняют функцию
механической защиты стебельчатых глаз (хищник, нападающий на рака
спереди, наталкивается на комбинацию заостренных шипов рострума
и лезвий чешуек). Эндополит хорошо развит и имеет вил длинного
гибкого шиловидного многочленистого отростка, называемого
жгутом (бичом) и выполняющего осязательную функцию.
Мандибулы (верхние челюсти, жвалы) используются для
предварительного измельчения пиши. В соответствии со своим
предназначением эти конечности претерпели значительные изменения:
экзополит утрачен, а эндополит подвергся редукции, превратившись
в короткий малозаметный верхнечелюстной (мандибулярный)
щупик. Протоподит преобразовался в мощную склеротизированную
184
1m Arthropods
л в в
Рис. 94. Вычлененные конечности и придатки отделов тела речного
рака: Л — головы: /—а1ггеннула; 2— антенна (вершинная часть не
показана); 3 — мандибула; 4— максилла-I; 5 —максилла-Н;
5 — груди: 6 — ногочслюсть-1; 7— ногочелюсть-Н; 8— ногоче-
люстъ-111; 9—ходилыиы нога I пары; 10-13 — ходильные ноги
ll-Vnap; Я — брюшка: /4— абдоминальные конечности! пары
(гоноподия самца, рудиментарная конечность самки); 15 —
абдоминальные конечности II пары (гоноподия самца, плсопо-
да самки); 16-18— абдоминальные конечности III, IV и V пары
(плсоподы); /9 — абдоминальная конечность VI пары (уропода)
жевательную пластинку, степень жесткости которой легко
установить с помощью пинцета или препаровальной иглы. Внутренние края
жевательных пластинок заз><)рсны, — вес это позволяет ракам
откусывать кусочки твердой пиши. Основой рациона речных раков считают
водные растения, но, по возможности, они дополняют его мягкотелыми
беспозвоночными, мертвой рыбой и другими животными. В действие
Класс Malacastmca
185
ж валы приводятся мощной мускулатурой. Обычно при препаои
ваиии мандибулы вычленяются вместе с пучками жевательных мыш
идущих к спине и прикрепляющихся там ко внутренней поверхности
карапакса.
Максиллы-I имеют вид маленьких пластинчатых образований,
в комплексе они выполняют ту же функцию, что и нижняя губа у лру-
гих членистоногих. Максиллытакже называют нижними челюстями.
Последняя, четвертая пара конечностей носит название максилл-Н,
или макс ил л ул. Они невелики, их эндолодиты миниатюрны и в
функционировании ротового аппарата играют незначительную роль. Экзо-
подит и эпиподит каждой стороны, сливаясь, образуютлопастевидный
придаток — лодочку, или скафогнатит. Именно им принадлежит
ведущая роль в обеспечении циркуляции воды в жаберных камерах.
Таким образом, основной функцией максиллул является обеспечение
процесса дыхания, участие в обработке пищевых объектов незначительно.
Ротовое отверстие располагается между мандибулами, спереди
оно окаймлено верхней губой, а сзади — нижней губой. По своему
происхождению губы представляют собой складки покровов тела.
Таким образом, нижняя губа функционально дополняет максиллы-1.
В комплексе все перечисленные конечности и придатки образуют
ротовой аппарат речного рака.
Грудные коаечносга. Передние три пары грудных конечностей
носят название ногочелюстей (максиллопед), они отличаются
наибольшей степенью расчлененности. По своим функциям эта
группа конечностей гнатоторакса является переходной. Последняя, третья
пара ногочелюстей отличается крупными размерами, внешне
походит на укороченные ходильные конечности и несет хорошо развитые
жабры. Ногочслюсги выполняют функцию захвата, удержания и
передачи ко рту пищи. Вторая пара ногочелюстей отличается от
предыдущей лишь меньшими размерами. Первая же пара самая маленькая
и лишена жаберных придатков. Ее эпиподиты имеют вид пластинок,
РИТМИЧНЫС ДВИЖеНИЯ КОТОРЫХ СПОСОбСТВуЮТ ЦИРКУЛЯЦИИ ВОДЫ Через
жаберную полость.
Десятиногие раки имеют пять пар ходил ьных ног. Они одно
ветвисты: экзоподит у взрослых особей полностью утрачен, а эшонолит
превращен в многочлениковую ходильную конечность рычажного типа
Первая пара ходильных ног вооружена мощными клешнями. Они о<:-
разованы двумя последними члениками конечности. Предпоследним
186
Тип Arthropoda
членик несет иожсвидный вырост, представляющий собой
неподвижный палец клешни. Последний членик конечности выполняет функцию
подвижного пальца, управляемого мощной мускулатурой. Именно она
является съедобной частью клешни. Внутренние края пальцев снабжены
заостренными зубцами. По их форме и расположению
дифференцируют разные виды и подвиды речных раков. Являясь мощным орудием
зашиты и нападения, клешни могут использоваться для манипуляции
какими-либо объектами. Именно с их помощью молодь
удерживается на плеоподах самки. Вторая—пятая пары имеют развитые в той или
иной степени элиподиты, представляющие собой перистые жабры.
Чтобы облегчить ознакомление с их строением, вычлененную
конечность следует поместить в воду, перистые придатки при этом окажутся
распрадоенными. Жабры размещены в парных жаберных полостях,
расположенных по бокам грудного отдела. С внутренней стороны эти
полости ограничены стенками тела, с дружной — нависающими краями
карапакса. Наружу они открываются узкими жаберными щелями.
Последние две пары ходильных конечностей исходно оканчиваются
хитиновыми коготками, а две последующие пары — миниатюрными
клешенками.
Брюшные конечности. Наличие брюшных конечностей является
отличительной чертой высших раков. Тел ьсои (анальная лопасть)
не имеет вилки (фурки) или иных придатков, что также является
особенностью представителей класса. Последний, т.е. шестой сегмент
брюшка несет пару двуветвистых сильнохитинизированных
уплощенных конечностей, называемых уроподами. Вместе с расположенным
по центру уплощенным тельсоном они образуют так называемый
брюшной плавник, используемый животным для экстренного
плавания. Интенсивное подгибание брюшка при расправленном
хвостовом плавнике обеспечивает толчкообразные движения животного
назад. Плавательные ножки (плеоподы) пятого,
четвертого,третьего, а у самок — и второго брюшного сегментов двуветвисты и в целом
наиболее близки к исходному типу строения конечностей. У самок,
помимо плавания, они используются для прикрепления яиц и
молоди. Работая плсоподами, животное медленно плывет головой вперед.
У самок первая пара брюшных конечностей рудиментарна и имеет
вил щетинкообразных придатков. У самцов первая и вторая пары
конечностей брюшного отдела преобразованы в пипеткообразные копу-
лятнвные придатки, называемые гоноподиями. Они выделяются
Класс Malacostraca
белой или желтоватой окраской и выдаются вперед под гнатоторакс.
Первая пара гонолодий утратила двуветвистость, вторая — сохранила.
Гоноподии участвуют в окончательном формировании слерматофо-
ров и их прикреплении к телу самки в области ее половых отверстий.
Таким образом, десятиногие раки располагают практически
полным набором функционально и морфологически
дифференцированных конечностей, степень гомономностн которых в целом невысока.
Таксономическое разнообразие ракообразных:
основные особенности внешнего строения
представителей основных групп Crustacea
Класс Branchiopoda — жаброногие раки
Отряд Anostraca — жаброноги
Представители отряда характеризуются следующими
особенностями строения: тело подразделяется на головной, грудной и
брюшной отделы; сегментация грудного и брюшною отделов фактически
гомономная; головофудной шит отсутствует; грудные конечности
листовидные, их эпиподиты выполняют функцию жабр; сегменты
брюшного отдела тела несут редуцированные конечности либо не имеют
таковых вообще. Большинство жаброногов являются характерными
обитателями временных (эфемерных) водоемов.
Для ознакомления с особенностями строения этих животных
лучше использовать относительно крупных пресноводных жаброногов
Bronchi pus, Chiroccphalus, а при их отсутствии — более мелких артсмий
(Artcmia salina).
Пресноводные жаброноги
Материал. В условиях большей части Восточной Европы ни один
из примерно десяти распространенных здесь видов жаброногов не
может считаться массовым либо фоновым: для обитающих во
временных водоемах представителей отряда Anostraca весьма характерна
спорадичность распространения. Это создает определенные трудности
в обеспечении учебных занятий материалом. Описываемый в
большинстве руководств Branchipus schaefferi (^stagnalis) характерен для
временных водоемов лесостепной и степной зон Восточной Европы,
в Белоруссии отмечен лишь в Полесье, где распространен также
Chirocephahis shadini. Собранных ракообразных сначала погружают
в теплую (35-40 #С) воду; затем хранят в 70-80' этаноле. Размеры тела
этих полупрозрачных животных не превышают 25 мм, для знакомства
с особенностями их строения лучше воспользоваться бинокуляром,
рассматривая объект как на черном фоне, так и в проходящем свете
с использованием полставки.
Строение жаброногов. При наличии живых ракообразных их
безусловно следует продемонстрировать на занятии. Всю свою
непродолжительную жизнь эти животные проводят в движении. Их
Надкласс Crustacea (таксономическое разнообразие)
отличительная черта — плавание спиной вниз. Находящиеся в
непрерывном движении листовидные грудные конечности одновременно
обеспечивают газообмен и выполняют функцию цедильного
аппарата, отфильтровывая мелкие частицы детрита, фито- и зоопланктон,
служащие им пищей. Живые жаброноги полупрозрачны, что делает
их малозаметными для хищников.
Тело жаброногов продолговатое, слегка сжато с боков.
Отчетливо видна его сегментация, имеющая явственно гомономный характер
(рис. 9S). Выделяются головной, грудной и брюшной отделы тела.
Головогрудиой щит (карапакс) не развит.
Головной отдел состоит из протоцсфалона, образованного
слившимися акроном и первым истинным головным сегментом, и гнато-
цсфалона, объединяющего последующие три сегмента. Протоцсфа-
лон несет пару одноветвистых нитевидных а нте н н у л. выполняющих
функцию органов химического чувства, и пару более мощных
антенн. У самок антенны одиоветвисты, у самцов они модифицированы
Рис. 95. Пресноводные жаброноги (Amstroca): самка Chiroctphalus grubei
(Л),самец Bnmchipusscha€fferi(B)\ / — науплиальный глаз; 2—
печеночный вырост кишечника; ^-Ьшгеннулы; 4-антенны; 5 —
сложный глаз; б— грудные конечности; 7— яйцевой мешок; £—
брюшко; 9 — вилка (фурка); /0 — гонада; // — средняя кишка;
/2— сердце; /J— максиллярная железа; /4— головной придаток
190
Inn Arthropods
и служат для удержания самки во время спаривания. У самцов
некоторых видов антенны снабжены многочисленными отростками
и имеют весьма причудливые очертания. Спереди протоцефалон обра-
зуетскладку стенки тела, называемую килевым или головным
придатком. Считается, что он улучшает гидродинамические
характеристики тела и экранирует ротовые части животного. На теменной
части головы расположен непарный науплиальный глаз. По бокам
головы (у некоторых видов — на специальных выростах)
размешается пара сложных фасеточных глаз. Ротовые части прикрыты спереди
складкой, носящей название верхней губы. Пара мандибул и две
пары максилл развиты слабо, рассмотреть их непросто.
Грудные сегменты несут одинакового строения листовидные
конечности. Брюшные сегменты лишены их, но на первых двух могут
быть их рудименты, выполняющие функцию наружных половых
органов. Оканчивается брюшко вилкой (фуркой). У жаброногов она
обычно покрыта многочисленными волосками. На анальной
лопасти располагается анальное отверстие.
Anemia salina — артемня
Материал. Артемии — характерные обитатели практически
лишенных животного населения мелких соленых и солоновалговодных
водоемов степной и пустынной зон Северного полушария. Так, Anemia
salina широко распространена в Прикаспии, в заливе Кара-Богаз-Гол
и озере Сиваш. Численность этих животных может быть чрезвычайно
высокой. Яйцаартемий очень легкие, и в сухом состоянии могут
переноситься ветром (явление анемохории). На берегах заселенных этими
животными водоемов часто образовывается своеобразный валик из
выброшенных с пеной яиц. В ряде мест производится их массовая
заготовка, так как выводимую из яиц молодь артемии используют для
кормления декоративных, промысловых и иных хозяйственно ценных
видов рыб. При этом обязательно производится отделение легкой
фракции яиц, которые маложизмеспособны, а также пустых яйцевых
оболочек. За некоторое время до необходимого срока порцию яиц
помещают в сосуд с водой, в которой растворено нужное количество
морской, а за неимением таковой — нефторированиой поваренной
соли (5% раствор). При более высоких температурах развитие артемий
протекает заметно быстрее, при 30 #С науплиусы отрождаются уже
менее чем через сутки.
Особенности внешнего строения артемий. Строение Anemia salina
(рис. 96) не имеет принципиальных отличий от ранее рассмотренного
Надкласс Crustacea (таксономическое разнообразие)
191
Рис. 96. Артемия (Anemia salina): самка, вид с брюшной стороны (А)%
самец, вил сбоку (£); У—непарный глаз; 2—ангеннулы; J—
парный сложный глаз; 4— антенны; 5— грудные конечности;
б— яйцевой мешок; 7— брюшко; 8— вилка(фурка)
пресноводных жаброногов. Можно лишь отметить, что тело артсмий
явно шире, а средняя скорость передвижения ниже. Кормятся
артсмий преимущественно у дна, взмучивая ил.
Отряд Phyllopoda (=Notastraca) — листоногне раки
Листоногие раки близки к жаброногам и отличаются от них
в первую очередь наличием шлемовидного карапакса и большим
количеством парных грудных конечностей. Сложные фасеточные глаза
бсссгсбельчатыс, сидячие. В настоящее время в мировой фауне
известно не более десяти современных видов Phyllopoda, любой из которых
пригоден для ознакомления с характерными особенностями их строения.
Шнтнн
Материал. Представители отряда обитают в тех же местах, что
и пресноводные виды жаброногов. Весной в большинстве регионов
Евразии в мелких временных водоемах, образующихся в результате
192
Jm Arthropods
таяния снегов, встречается весенний щктень (Lepidums opus), тогда
как обыкновенный щитень (Triops cancriformis) отмечается в самых
разнообразных мелких временных водоемах летом и осенью.
Массовое развитие Г. cancriformis нередко наблюдается в нерестовых прудах
рыбоводных хозяйств юга Европейской России и Украины. Для
отлова взрослых ракообразных утюбнее ислольэовапъ гидробиологический
скребок. При необходимости животных отмывают от ила и грязи;
хранят в 70-8(Г этаноле.
Строение пропей. Тело шишей уплощенное, что типично для
донных обитателей вообще. Овальный головогрудной щит
закрывает голову, грудь и часть брюшка животного (рис. 97). Сверху вблизи
переднего края тела размещается глазной бугорок. По бокам он
несет пару сидячих фасеточных глаз. Спереди между ними размещен
непарный науплиальный глаз, глазной бокал которого обращен
вниз. Под ним Находится оптически прозрачный столбик из
соединительной ткани, заканчивающийся на брюшной стороне тела участком
покровов, лишенным пигментации. В результате щитни могут
регистрировать освещенность находящегося под ними пространства.
1 А б
Рис. 97. Листоногис раки (Phyllopoda): весенний щктень — Lepidurusap-
us (Л), обыкновенный щитень— Triops cancriformis (Б): / —
фасеточный глаз; 2 — нитевидные придатки первой пары грудных
ножек; J — брюшко; 4 — вилка
надкласс Crustacea (таксономическое разнообразие) 193
Головной отдел тела не имеет визуально различимого деления
на прото- и гнатоцефалон. Миниатюрные антеннулы и антенны
смешены в стороны. Спереди к краю карапакса причленяется
верхняя губа ^представляющая собой складку покровов. По бокам
рогового отверстия расположены мощные, с зазубренными краями
мандибулы, за ними следуют две пары маленьких пластинчатых
максилл.
В составе грудного отдела насчитывается до 40 сегментов. Первые
10 несут по паре сходно устроенных листовидных конечностей. По
своему строению и функциям они напоминают грудные конечности
жаброногов, но имеют развитые жевательные отростки. Щит ни не
отфильтровывают пищевые частицы, а собирают пищу (частицы
детрита, мелких мягкотелых животных и даже икру рыб) со дна. Грудные
конечности используются для поиска, захвата и эстафетной передачи
пищевых частиц ко рту. Таким образом, они выполняют
трофическую, дыхательную и локомоторную функции.
Первая и вторая пара грудных конечностей несут длинные
нитевидные придатки, напоминающие антенны других членистоногих
и выполняющие осязательную функцию. На 11-м сегменте тела
расположены парные половые отверстия, конечности этого сегмента
у самок изменены и образуют яйцевую капсулу, в которой
вынашиваются яйца. У самцов они внешне не отличаются от остальных.
Начинаяс 13-го, каждый сегмент несет от четырех до шести пар л исто-
видных конечностей. Это явление получило название п о л и п од и и.
Общее число конечностей у щитней может достигать 70 пар,
размеры их постепенно уменьшаются к заднему концу тела.
Брюшные сегменты лишены каких-либо придатков. Оканчивается
брюшкофуркой. Хвостовые нити выполняют осязательные
функции и вместе с нитевидными придатками первых пар грудных ножек
образуют вокруг тела щнтия своеобразное рецепторное поле.
Класс Maxillopoda
Подкласс Ostracoda — ракушковые раки
Остракоды представляют собой очень древнюю и
характеризующуюся специфическими особенностями строения и развития группу
ракообразных, которую часто выделяют из максиллопод в самосто-
"гсльный класс. Это в большинстве своем мелкие и очень мелкие
(размеры тела большинства представителей группы не превышают
194
Тип Arthropods
I мм) животные, обладающие двустворчатой раковиной. Тело сильно
укорочено и лишено видимой сегментации. Число грудных
конечностей сокращено до одной-трсх пар либо они полностью редуцированы.
Жабры у большинства ракушковых раков отсутствуют, дыхание
осуществляется поверхностью тела. Остракоды раздельнополы, но многие
виды размножаются преимущественно партеногенстически. Развитие
с метаморфозом, раковина появляется уже на науплиальной стадии.
Остракоды имеют всесветное распространение и заселяют
буквально все типы водоемов. К самым обычным пресноводным формам
принадлежат представители родов Cypris и Heterocypris. Во время
летней учебной практики непременно следует обратить внимание
на эту фоновую группу ракообразных, провести наблюдения за
живыми остракодами.
Пресноводные остракоды
Материал. Собираютостракод в небольших прудах, других мелких
водоемах с помощью мелкоячеистого водного сачка. Ракушковых
раков можно содержать в течение длительного времени в лаборатории.
Хотя рыбы обычно отказываются их поедать, лучше для этих целей
выделить отдельный аквариум. На лабораторных занятиях при
отсутствии живого материала можно ограничиться просмотром
постоянных тотальных микроскопических препаратов пресноводных остракод.
Вследствие кальцификации раковины у многих из них непрозрачны,
и техника изготовления препаратов предусматривает процедуру дс-
кальци н ирован ия.
Строение остракод. Тело животного защищено двустворчатой
хитиноидной раковиной (рис. 98), которая зачастую пропитана
карбонатом кальция. На спинной стороне тела створки соединены
эластичной связкой, у ряда видов имеются приспособления,
аналогичные замковому механизму двустворчатых моллюсков. Смыкание
осуществляет мощный м ус кул - за м ы кате л ь, идущий от створки
к створке примерно посередине тела. Большинство представителей
группы обладают непарным науплиальным глазком, а некоторые
еще и парой сложных фасеточных глаз. Функционируют они
благодаря прозрачности раковины либо соответствующих ее участков.
Головной отдел несстантеннулы, антенны и обычный набор ротовых
конечностей. Грудных конечностей две-три пары, причем каж-.
дая из них имеет свою узкую специализацию. Последняя пара часто
преобразуется в своеобразные «чистилки» и предназначена для уда-
Надкласс Crustacea (таксономическое разнообразие)
195
Рис. 98, Ракушковые раки (Ostracoda): строение Cypridopsis vidua (Л),
движения Cypris(E); / — мускул-замыкатель раковины; 2— ан-
тсинула; J— верхняя губа; 4 — мандибула; 5 — максилла-1;
6— максилла-11; 7— наружняя ветвь максилл-I; S— задняя
грудная конечность; 9— передняя грудная конечность; /0—
ползание по растениям; // — ползание по дну; /2 — плавание
ления проникающих в раковину частиц грязи и т.п. Передние
конечности чаше ходильные, реже используются для прикрепления либо
захвата пиши. Брюшной отдел тела лишен конечностей, у некоторых
имеется фурка. Через шелевидное отверстие на брюшной стороне
тела наружу из раковины могут высовываться антенны, обычно
используемые для плавания, а у некоторых остракод — также антеннулы,
хватательные или ходильные грудные конечности и фурка. Из
систем внутренних органов на некоторых препаратах удастся разглядеть
изогнутый трубчатый кишечник.
Подкласс Branchiura — карпоеды
Подкласс объединяет около 60 пресноводных и морских видов
жтопаразитических ракообразных. Тело карпоедов листовидное;
| мзуально различаются передний, покрытый карапаксом, и задний,
196
TunArthropoda
свободный отделы. Придатки переднего отдела приспособлены для
прикрепления и питания на хозяине, заднего — для активного
плавания. Карпосды раздельнополы; из яиц отрождаются личинки с
недоразвитыми плавательными конечностями и не имеющие присосок.
Они ведут эктопаразита чес кий образ жизни и до достижения взрослой
стадии несколько раз линяют. Пищевые предпочтения у карпоедов не
выражены, кроме рыб, они могут нападать на тритонов, их личинок
и головастиков лягушек.
Argulusfoliaceus — обыкновенный карпоед (карповая вошь)
Материал. Карпосды в том или ином количестве присутствуют
в большинстве крупных пресных водоемов, имеющих рыбное
население, но осуществить массовый сбор этих ракообразных зачастую
затруднительно. Некоторое количество экземпляров можно добыть
в результате продолжительного траления сачком краевой зоны полувещной
и водной растительности на нерестилищах или в местах скопления
мальков. Иногда карпоедов удается собрать в результате внимательного
осмотра участков покровов за грудными плавниками н жаберными
крышками рыб из сетевых уловов. При ознакомлении с особенностями
строения этих ракообразных для просмотра тотальных
микроскопических препаратов удобнее воспользоваться бинокуляром.
Внешнее строение обыкновенного карооеда. Тело карповых вшей
сильно уплощенное (рис. 99), формой они слегка напоминают
некоторых скатов. Широкий пластинчатый карапакс закрывает сверху
головной и часть грудного отделов тела, ближе к его переднему краю
на спинной поверхности расположена пара сложных фасеточных
глаз, а между ними три науплиальных глазка.
При рассмотрении карпоеда с брюшной стороны хорошо видны
конечности и придатки тела. А нте н и ул ы и а нте н н ы очень
короткие, крючковидные, используются для фиксации на теле хозяина.
Пара мандибул образует острый стилетообразный хоботок. Вводя
его в ткани жертвы, карпоед сосет ее кровь. Первая пара максилл
преобразована в мощные округлые присоски. Вторая пара имеет
типичное строение, но используется также для прикрепления
эктопаразита к телу хозяина. Грудные сегменты свободные, границы между
ними хорошо различимы. Грудных конечностей четыре пары, они
хорошо развиты, двуветвисты и усажены рядами щетинок. Активно
гребя ими, карпосды оченьбыстро плавают спиной вниз. У неподвижных
экземпляров ритмичные движения грудных конечноспй не прекра-]
Надкласс Crustacea (таксономическое разнообразие)
197
ftc. 59. Карпосд Ar&dusjbBoccus. самса* /—стилет, 2— аигтсшсулац J— мак-
силла-1; 4— присоска; 5— грудные конечности: 6 — семенник;
7— печень; 8 — максилла-П; 9— антенна
щаются, и открытая поверхность тела, через которую осуществляется
газообмен, омывается водой. Брюшко сильно редуцировано,
представляя собой основание фурки. Фуркальные отростки лопастс-
видные.
Подкласс Cirripedia — усоногне раки
Группа объединяет своеобразных морских ракообразных,
ведущих прикрепленный образ жизни. Антеннулы и передняя часть
головного отдела превращены в стебельчатое или уплощенное образование,
Лепечивающее прикрепление к субстрату. Антенны и фасеточные
' лаза атрофированы. Остальное тело покрыто с боков двумя кожными
кладками, на поверхности которых формируются состоящие из карбо-
чага кальция пластики, в совокупности образующие многостворчатую
198
Тип Arthropods
раковинку. Грудные конечности вытянутые, ими животные
захватывают пишу и создают ток воды, омывающей их тело. Брюшко
недоразвито. Среди усоногих раков широко распространено явление
гермафродитизма. Развитие протекаете метаморфозом.
Balanusspp. — морские желудя
Материал. Усоногие раки чувствительны к уровню солености
воды и еше более — к содержанию в ней растворенного кислорода.
Поэтому они отсутствуют на сильно опресненных участках морей,
содержать их в морских аквариумах весьма затруднительно. Морские
желуди родов Balanusu Chthamalus в массе обитают на скальных
породах прибойной зоны Черного, Белого и дальневосточных морей. С
заселенных обрастателями мелких камней животных с осторожностью
отделяют от с>6страта острым ножом. Собрать материал можно,
отлавливая крабов; панцирь которых также служит субстратом для
поселения этих ракообразных. Желательно иметь в качестве
демонстрационного материала влажные препараты морских уточек (Lcpas)t
которые чаще поселяются на плавающих или опущенных в воду
предметах. Умерщвляют усоногих ракообразных теплой водой, иногда
достаточно оставить мелкий сосуд с животными на солнце. При этом
створки их домиков остаются несомкнутыми. В качестве фиксатора
лучше использовать 4% раствор формалина. Непосредственно перед
занятием материал переносят в водопроводную воду.
Внешний вид морских желудей. Усоногие раки, обладающие
многостворчатыми известковыми раковинами, напоминают по
внешнему виду бутоны. Их створки образованы карбонатом кальция. Белая
окраска делает их хорошо заметными на темных субстратах. У морских
желудей рода Balanus баш неподобная раковинка образована двумя
рядами известковых пластинок (рис. 100); прикрепление
обеспечивается широкой подошвой. Морские уточки (Upas) имеют слегка
уплощенную, овально-продолговатую раковину, образованную пятью-
шестью створкам и, и прикрепляются к с><хгфату с помощью
длинного мясистого стебелька. Если животное потревожить, оно прячет
венчик «щупалец» (усовидных двуветвистых конечностей) и смыкает
края створок. Грудные конечности можно рассмотреть у
фиксированных животных.
Надкласс Crustacea (таксономическое разнообразие)
19S
Рис. 100. Строение усоногих раков (Cim'ptdia): морского желудя Balanus
(Л)% морской уточки Upas (S): I — ротовой конус: 2— мускул
крышки раковины; 3 — грудные ножки; 4 — редуцированное
брюшко
Класс Malacostraca — высшие раки
Отряд Isopoda — равноногие
Равноногие раки представляют собой экологически наиболее
истинную группу ракообразных: они весьма разнообразны в морях,
«ссляют пресные воды, успешно освоили сушу, среди них имеются
'разитические формы. Тело большинства нзопод ушюшено. Слияни-
'•'. акрона, головных и первого-второго грудных сегментов образована
200
iMArthropoda
«сложная голова». Остальные грудные сегменты свободны и
несут ходильные конечности. Каралакс отсутствует. Брюшные сегменты
зачастую сливаются с анальной лопастью. Первые пять пар брюшных
конечностей выполняют дыхательную функцию, их жаберные
отростки находятся в специальной камере. Равноногие раки
раздельнополы. Начальные этапы развития яиц протекают в специальной
выводковой камере на груди самки. Для ознакомления с особенностями
внешнего строения равноногих ракообразных могут бьпъ
использованы различные мокрицы (РогсеШо, Onlscus), водяные ослики (Acellus),
но лучшими объектами следует признать морских тараканов.
Soduria (~Mesidotea) entomon — морской таражая
Материал. Это морские тараканы являются характерными
обитателями илистых отмелей Балтийского моря, Ладожского озера, средней
и южной части Каспийского моря, но ареал вида более широк. Они
фактически всеядны: питаются донными беспозвоночными,
водорослями, падалью, нападают на попавших в сети рыб. На Черном море
достаточно обычны питающиеся водорослями и имеюшие
разнообразную окраску представители рода Idotea. Собранных животных
отмывают от грязи и хранят в 4% растворе формалина. Перед занятием
отобранные экземпляры промывают в проточной воде.
Внешняя морфология. Тело Soduria entomon слабовыпуклое,
продолговатое. Длина тела крупных самцов может достигать 80 мм.
Перевернув животное спиной вверх, можно ознакомиться с
особенностями сегментации (рис. 101). Каралакс у морского таракана, как и у других
/sopoda, отсутствует. Сложная голова, которую также называют
передним (головным) щитом, компактна и образована слиянием
головных и первого грудного сегментов. По бокам ее видны
уплощенные крыловидные выросты. У их оснований располагается пара
сложных фасеточных глаз. Далее следуют семь отграниченных
друг от друга грудных сегментов, по бокам которых имеются
уплощенные и заостренные на вершинах крыловидные выросты. Первые
четыре свободных брюшных сегмента уже и короче грудных, их
боковые выросты имеют вид небольших заостренных зубцов. Слившиеся
два последних сегмента брюшка и анальная лопасть образуют мошный
ромбический задний щит.
Чтобы ознакомиться со строением конечностей и других
придатков, следует перевернуть животное брюшной стороной вверх. Антсн-
нулы несколько меньше антенн, те и другие выполняют главным
образом осязательные функции. Мощные хитинизированные манди-
Надкласс Crustacea (таксономическое разнообразие)
201
Рис. 101. Внешний вид морских тараканов: Saduria (^Aftstdotea) entomon
(Л); /dotea baltica (£); / — головной щит, 2—свободные грудные
сегменты, J— свободные брюшные сегменты, 4— задний шит
булы выделяются своей черной окраской. Далее следуют две пары мак-
силл и одна пара ногочелюстей. Первые три свободных сегмента
груди несут по паре хватательных конечностей, последующие
четыре — мощные ходильные конечности. Сегменты брюшка прикрыты
широкими створкообразными уролодами — конечностями шестого
брюшного сегмента. Отожув их, можно заглянуть в образуемую ими
камеру, где расположены листовидные конечности остальных
брюшных сегментов. Размещение их в специальной жаберной камере
позволяет этим ракообразным зарываться в ил, а также покидать на
некоторое время воду, например для обследования выброшенного на
берег мусора.
Отряд Amphipoda — разноногие раки, или бокоплавы
Отряд объединяет ракообразных, тело которых сплющено с боков,
а у многих и дугообразно изогнуто. Плавают они на боку, что и
послужило основанием для их второго названия. Видовое
разнообразие морских амфипод значительно выше, нежели пресноводных.
«Сложная голова», как и у равноногих раков, образована
слиянием акрона, головных и одного-двух передних грудных сегментов.
Большинство бокоплавов имеют пару сложных фасеточных глаз. Ка-
рапакс отсутствует. Конечности свободных грудных сегментов вы-
202
Тип Arthropods
полняют локомоторную функцию, используются для захвата и
обработки пиши и, обычно, несутлистовидныс жаберные придатки. Передние
три пары брюшных конечностей используются для плавания, тогда
как три задние являются прыгательными. Бокоплавы
раздельнополы, начальные этапы развития эмбрионов проходят в выводковой
камере, образующейся на груди самки в период размножения.
Выходящая молодь практически ничем, кроме размеров, не отличается от
взрослых.
В наших условиях наиболее доступными объектами для изучения
являются представители рода Gammarus. Gammarus lacustris и Gammarus
pulcx. В подземных волах, родниках, родниковых ручьях и озерах
Восточной Европы может быть также встречена реликтовая Synurella
ambutans.
Gammarus spp. — бокоплавы-гаммарусы
Материал. Озерный боколлав (Gammarus lacussris) широко
распространен в стоячих и медленнолпекущих водах Европы, севера Азии и
Северной Америки. Местами его численность достаточна для заготовки
раздаточного материала. Гаммарусы весьма чувствительны к
содержанию в воде кислорода, что делает затруднительным их содержание
в аквариумах. В тех же водоемах, а также на песчаных берегах
опресненных участков Балтийского моря массовым зачастую является другой
вид — Gammarus pulcx — бокоплав-прыгуи (бокоплав-блоха).
Собранных животных отмывают от песка и других загрязнений; хранят в 4%
растворе формалина либо 70* этаноле. Перед занятием отобранные
экземпляры выдерживают в проточной воде. Для изготовления
тотальных микроскопических препаратов удобнее использовать
молодых особей, обнаруженных в выводковых камерах.
Внешнее строение. Тело гаммарусов сильно сжато с боков и
дугообразно выгнуто (рис. 102). По суше они передвигаются в вертикальном
положении, но плавают на боку. Пинцетом удерживая тело
животного в нужном положении, рассматривают его спинную сторону.
. Небольшая «сложная голова» образована слиянием акроиа,
головных и первого грудного сегмента. На ней размещена пара
сложных фасеточных глаз; они сидячие, бобовидной формы. Остальные
семь грудных и шесть брюшных сегментов свободны и четко
отграничены друг от друга. Антенны длинные, двувствистые, причем одна
ветвь значительно длиннее другой; антеннулы малозаметны.
Ротовые части представлены одной парой мандибул, двумя парами мак-
Надкласс Crustacea (таксономическое разнообразие) 203
Рис. 102. Строение самки Gommamspulex. I — антеннула; 2—антенна;
3 — эпимеры первых грудных сегментов; 4 — развивающиеся
яйца; 5— задний отдел брюшка; 6— передний отдел брюшка;
7— ссмьсвободных грудных сегментов; 8— фасеточный глаз
силл, одной парой ногочелюстей (максиллопед). Грудные конечности
одноветвистые и имеют развитые жаберные придатки. У самок
вторая-пятая пары грудных конечностей несут характерные плоские
хитиновые отростки, совокупность которых образует выводковую
камеру. Специализация грудных конечностей различна: первые две пары
с изогнутыми вершинами — хватательные, остальные пять пар — пры-
гательные, из них три последние явственно отогнуты назад.
Брюшные конечности двувствисты. Из них передние три пары
приспособлены для плавания, это типичные плсоподы; задние три пары —
для прыгания, носят название уропод.Тельсои (анальная
лопасть) смешен на спинную сторону брюшка.
Отряд Decapoda — десятиногие раки
Наиболее разнообразны десятиногие раки в морях,
немногочисленные представители отряда обитают в пресных водах, а отдельные
виды крабов принадлежат к числу сухопутных форм. Среди дскапод
выделяются характерные морфо-экологическис группировки, из
которых наиболее известны креветки, раки-отшельники, крабы и
собственно раки (рис. 103). К последним принадлежат рассмотренные
204
Тип Arthropods
б в
Рмс. 103. Десятиногие раки (Dtcapoda): креветка Crangon crangon (Л),
рак-отшельник Pagurussetosus* извлеченный из
раковины-убежища (£), краб Cancer pagurus (В)
ранее речные раки рода Potamobius. Здесь же следует кратко
ознакомиться с представителями остальных.
Palaemon spp. — травяные креветки
Материал. Травяные креветки принадлежат к массовым видам
ракообразных и являются промысловыми объектами. Наиболее доступна
черноморская травяная креветка (Palaemon adspersus). Это типичный
обитатель мелководий Черного, Азовского и Балтийского морей. Она
также успешно акклиматизировалась в Каспийском и Аральском морях.
Можно использовать и других представителей этого (например,
Palaemon elegans) и иных родов (Crangon crangon и пр.). Фиксируют и
хранят креветок аналогично речным ракам.
Внешнее строение креветок. Тело креветок вытянутое и
несколько сжато с боков, что связано с приспособлением к активному
плаванию. Покровы тела эластичны вследствие слабого обызвествления;
окраска весьма изменчива и зависит от конкретных условий
обитания, являясь покровительственной, то есть адаптивной.
Протоцефалон нсссттрсхвствистыеантсннулы, а также
антенны с широкой удлиненной чешуйкой и очень длинным жгутом (би-
Надкласс Crustacea (таксономическое разнообразие)
чем). Гнатоцефалон объединен с грудными сегментами и покрыт
карапаксом, который спереди имеет длинный ростральный
отросток. Ротовые части креветок миниатюрнее, а ходильные и
плавательные конечности стройнее, чем у речных раков. У спокойно плывущих
головой вперед креветок брюшко выпрямлено, а у сидящих особей —
подогнуто под тело. В момент опасности животные расправляют
образованный уроподами и анальной лопастью хвостовой плавник
и, резко подгибая брюшко, совершают скачкообразные движения,
уплывая головой назад. Как и речные раки, самки креветок
вынашивают яйца и молодь на плавательных ножках (плеоподах) брюшка.
Раковинообитающие ракн-отшелыппси
Материал. Для простого ознакомления с представителями этой
жизненной формы ракообразных пригодны любые виды десятиногих
раков, использующих подобные укрытия. Классическим объектом
могут служить широко распространенные представители рода Pagurus.
Раков без необходимости не следует извлекать из раковины,
поскольку защищаемые ею нежные части тела легко могут быть повреждены.
Внешняя морфология раков-оттельннков. Тело
раков-отшельников спрятано в используемых ими раковинах брюхоногих моллюсков.
Наружу из домика рак высовывает только переднюю часть головы со
стебельчатыми глазами, ротовыми частями, антеннами, а также клешни
и ходильные конечности. Остальные части тела животного имеют
мягкие покровы вследствие слабого их обызвествления и надежно
защищены раковиной. При опасности рак прячется в ее глубине, закрывая
устье мощной клешней. С асимметричностью используемых раковин
связано гипертрофированное развитие той клешни, свобода
действий которой ограничена в меньшей степени. Помимо того, ввиду
спирал ьн ости раковины конец брюшка рака штопорообразно закручен.
Carcinus terraneus — травяной краб
Материал. Травяной краб принадлежит к числу обычных
обитателей галечных пляжей Черного и Балтийского морей и считается
промысловым объектом. На панцире старых животных могут
располагаться отдельные особи и колонии морских желудей. Для ознакомления
с особенностями внешнего строения крабов можно использовать
и других представителей этой морфо-экологической группы
десятиногих раков. Фиксируют и хранят крабов аналогично речным ракам.
Внешнее строение крабов. Тело крабов сильно уплощено, имеет
форму симметричного неравностороннего шестиугольника с закрут-
206
Тип Arthropods
ленными краями. Сверху голова и грудь надежно защищены мощным
и прочным карапаксом, ширина которого превышает длину. Его
механические свойства определяются повышенной степенью кальци-
фикации кутикулы. Брюшко развито слабо и подогнуто под тело.
Рассматривая краба со спинной стороны, следует найти гл азн ич -
ныс ямки, куда могут втягиваться сидящие на подвижных
стебельках крупные фасеточные глаза. Эта особенность (стсбельчатость
глаз) позволяет крабам видеть неподвижные предметы. Маленькие
щетинковидные антеннулы широко расставлены и направлены
вперед. Между ними расположены небольшие изогнутые антенны,
которые краб может прятать в специальные углубления. Из-под ка-
рапакса видны также парные клешни и четыре пары ходильных
конечностей с коготками на вершинах.
Пол крабов можно определить по форме карапакса и строению
брюшка. У самок оно значительно шире, чем у самцов. Брюшко
состоит из шести хорошо различимых сегментов. Аккуратно отогнув его,
можно рассмотреть имеющиеся на втором-пятом сегментах
брюшные ножки, которые двуветвисты и усеяны многочисленными
волосками. В этой своеобразной камере самка носит яйца, а затем молодь.
У самцов проявляется тенденция к слиянию второго-пятого
брюшных сегментов, границы между которыми едва заметны. Конечности
первых двух сегментов преобразованы в пипеткоподобные копулятив-
ные органы (гоноподии). В отличие от речных раков, самцы крабов
при копуляции вводят половые продукты непосредственно в протоки
семяприемников самок. Конечности остальных брюшных сегментов
редуцированы. Оканчивается брюшкотельсоном, несущим
анальное отверстие.
Подтип Antennata
Надкласс Myriapoda — многоножки
Надкласс объединяет почвообитающих и наземных
членистоногих с множественными гомономными туловищными сегментами,
несущими одноветвистыс членистые конечности. Дыхательная система
трахейная. В настоящее время предполагается существование двух
основных эволюционных ветвей мноюножек. Представителями одной
являются гуФоногис, другой — двуларноногие.
Класс Chilopoda
207
Класс Chilopoda — губоногие
Принадлежащие к данному таксону многоножки обладают
хорошо развитыми антеннами; фасеточных глаз нет, но могут быть стем-
моидные, образованные скоплениями простых глазков; половое
отверстие непарное и открывается на последнем туловищном сегменте.
Характерной особенностью г><х>ногих является наличие ногочелюс-
тей — хорошо развитых конечностей первого туловищного сегмента,
играющих ведущую роль в добывании пищи и обороне от врагов. Все
представители класса — хищники. Таксономическое разнообразие
Chilopoda максимально во влажных и сухих тропиках, в условиях
умеренного пояса Евразии они немногочисленны. Для ознакомления
с особенностями морфологии этой группы многоножек может быть
использована обыкновенная костянка.
Lithobiusforficatus — обыкновенная костянка
Материал. Обыкновенная костянка в условиях лесной зоны
Восточной Европы является фоновым видом, отсутствующим лишь в
некоторых полевых и болотных биотопах. Чаще всего этих многоножек
можно найти под камнями или отстающей корой деревьев. Они весьма
проворны даже при низкоположитсльных температурах, что
затрудняет их сбор. Анестезируют костянок д и этиловым эфиром или этил-
ацетатом, материал для занятий хранят в 70-80* этаноле.
Внешнее строение костянки. Тело многоножки сильно уплошено
и вытянуто в длину. Оно подразделяется на два отдела: голову и
туловище (рис. 104). Голова, а точнее головная капсула,
округло-треугольная, уплощенная, без швов и иных следов сегментации. Хорошо
заметна пара длинных нитевидных многочленистых антенн (усиков).
Они служат не только органами осязания, но и обоняния, так как
несут некоторое количество хеморецепторов. На спинной стороне
головной капсулы по бокам расположена парастеммоидных глаз,
состоящих из примерно 40 простых глазков каждый.
Туловище состоит из 19 неравной длины сегментов. Если смотреть
на костянку сверху, сразу бросается в глаза, что короткие нечетные
сегменты правильно чередуются с более длинными четными. Стигмы
(дыхальца) трахейной системы имеются только на длинных сегментах.
Таким образом, имеет место нарушение гомономности сегментации.
Первый туловищный сегмент несет ногочел юсти, функционально
относящиеся к ротовому аппарату. Последующие 15 сегментов (2-й-
16-й) снабжены однотипно устроенными х од ильным и (бегатсльными)
208
l№ Arthropod*
l и
xvn-xvm
Pm. 104. Костянка Uthobiusfafkatus, внд со спинной стороны: ) — голова;
2 — глазное пате с глазками; 3 — полоска иитерссгмснтарной
кожи; 4— конечность XVI сегмента; 5 — анальное отверстие;
6 — стигмы; 7— мягкая кутикула плевры; 8— конечность VIII
сегмента; 9— конечность II сегмента; 10>— ногочелюстъ;
/У—антенна; I—XIX — соответствующие сегменты туловища
Класс Chilopoda
209
Рис. 105. Голова и первый туловищный сегмент костянки, вид с
брюшной стороны: / — срединная вырезка верхней г><>ы; 2—
передний бугорок наличника; 3— наличник; 4-9— ногочелюстъ:
4— когтевой членик; 5— голень; б —бедро; 7—всртлуг-И;
8— вертлуг-1; 9— тазик; 10— зубцы на переднем крае тазика,
// — максилла I; 12— максилла-Н; 13— мандибула; 14— канал
ядовитой железы; 15 — верхняя губа; 16 — антенна
конечностями. Конечности 16-го сегмента имеют наибольшую
длину, направлены назад и выполняют главным образом чувствительную
функцию. Последние три сегмента брюшка отличаются малыми
размерами. Семнадцатый сегмент (прегенитальный) несет пару коротких
конечностей — половых ножек. Половое отверстие расположено на
18-м, генитальном сегменте, а Т-образнос анальное
отверстие — на последнем, 19-м, называемом анальным.
Строение ротового аппарата костянки. Рассмотреть строение
ротового аппарата можно, применяя технику последовательного
вычленения его элементов. Для этого следует зафиксировать выбранный
жземпляр в препаровальной ванночке, воткнув булавку в головную
210
Тип Arthropods
капсулу пол углом так, чтобы она была приподнята передним концом
вверх. Вычленение частей ротового аппарата удобнее начинать сзади.
Препаровальной иглой отделяют первый туловищный сегмент,
несущий ногочелюсти. Они хорошо хитинизированы, имеют
серповидно изогнутые вершины, пронизанные каналом, по которому
выделяется секрет ядовитых желез. Яд помогает губоногим справиться с
жертвой. Для человека костянки не представляют никакой опасности,
так как не могут прокусить его кожу, а яд их для него малотоксичен.
Таким образом, ногочелюсти представляют собой орган нападения
и зашиты. Далее следует пара максилл-11. Для рассмотрения их под
бинокуляром лучше изготовить временный микроскопический
препарат. Максилл ы-II одновствисты, они лишены жевательных
элементов и предназначены для ощупывания и удержания добычи. М а к -
силлы-1 также не приспособлены для измельчения пищи,
а используются лишьдля ее удержания. Мандибулы у костянок
сильно хитинизированы, особенно их зазубренные жевательные
поверхности. Вычленяются они обычно вместе с вдающимся в ротовую
полость сзади гипофар и нгсом (подглоточником),
представляющим собой складку стенки глотки. Аналогично происхождение
замыкающей спереди ротовой аппарат верхней губы, имеющей вид
дугообразной хитиновой складки. Таким образом, у губоногих имеет
место высокая степень функциональной специализации частей
ротового аппарата.
Класс Diplopoda — двупаряоногие
Представители класса обладают укороченными антеннами и
ходильными конечностями; органы зрения, если имеются, представлены
стеммальными полями; в составе ротового аппарата есть гнатохи-
лярий; намечено деление тела на три отдела: голову, грудь и
брюшко; половые отверстия парные и открываются на груди. Характерной
особенностью губоногих является незавершенное попарное слияние
собственно брюшных сегментов с образованием так называемых дип-
лосомитов, несущих по две пары конечностей и дыхалец (стигм).
Покровы тела двупарноногих обычно сильно кальцифицированы, что
придаст им особую прочность; при опасности эти многоножки
сворачиваются в кольцо, выставляя наружу панцирсподобные спинные
щитки. Большинство представителей класса являются сапрофагами
и обитают в верхнем слое почвы и подстилке. Для ознакомления
с особенностями строения многоножек этой группы может быть
Класс Diplopoda
211
использован песчаный кивсяк (Schizophytlum sabulosum) либо другие
крупные представители отрядов кивсяков (Juliformia) и многосвязов
(Pblydesmoidca).
Julidae — настоящие кнвсяки
Материал. В условиях лесной зоны Восточной Европы достаточно
обычны два вида крупных кивсяков, которые наиболее подходят для
изучения на лабораторных занятиях. Песчаный кивсяк (Schizophyllum
sabulosum) отличается характерной окраской: по бокам спины на
черном фоне идет пара ярких оранжевых продольных полос. Эти
многоножки предпочитают легкие песчаные почвы и редко появляются
на поверхности. Серый кивсяк (Sarmarojulus kessleri) является
типичным обитателем подстилки лиственных лесов. Следует остерегаться
брать этих крупных многоножек голыми руками, так как защитный
секрет спинных желез оставляет на коже долго несмываемые
фиолетово-красные пятна. При отсутствии кивсяков можно собрать
многосвязов (Polydesmus), которые обычно встречаются в не обрабатываемых
инсектицидами оранжереях, а также под отслаивающейся корой
деревьев, в старых плодовых телах шляпочных грибов. Собранных
животных анестезируют эфирами и хранят в 50—60* этаноле (в растворе
формалина материал становится хрупким).
Внешнее строение. Тело диплопол сильно вытянуто в длину;
у кивсяков оно в сечении округлое, у многосвязов — овальное с
боковыми выступами. Голова четко отграничена от туловища (рис. 106)
и несет пару коротких а итен н. Над местом их прикрепления к головной
капсуле располагаются стеммальные поля, состоящие из диффузно
размешенных простых глазков (у многосвязов они отсутствуют).
Относительно туловища голова несколько отклонена вперед и вниз.
Со стороны видно, что она причлсияется к клинообразному шейному
щитку, представляющему собой тергит (спинную пластинку)
I туловищного (грудного) сегмента. Стернит (брюшная пластинка)
этого сегмента участвует в формировании ротового аппарата.
Последующие три туловищных сегмента кивсяка несут по паре ходильных
конечностей. Вместе с модифицированным, лишенным конечностей
первым туловищным сегментом они образуют короткий грудной
отдел тела.
Последующие туловищные сегменты образуют так называемые
л и пл осом иты. Каждый диплосомит имеет единую спинную
пластинку, образованную в результате слияния двух тергитов, снабжен двумя
парами дыхалец (стигм) и двумя парами коротких ходильных ко-
212
ImA/thropoda
Рис. 106. Схема внешнего строения тела кивсяка Schlzophyltum sabuhsum:
/—голова; 2— глазное поле; J— антенна; 4— верхняя губа;
S — анальная лопасть; б — сегменты пролиферационной зоны
(зоны роста); 7— метасомит; *— просомит
нечностей. Заканчивается тело крупным цилиндрическим с округлой
вершиной тельсоном, на брюшной стороне копгорого расположено
анальное отверстие. Между ним и последним диллосомитом
расположена зона образования новых сегментов (пролиферационная
зона, зона роста). Функционирует она в течение всей жизни
животного. Зачастую здесь упастся рассмотреть несколько ешс не имеющих
конечностей колец. Таким образом, у диплопол с каждой новой
линькой число сегментов и конечностей увеличивается. Подобный тип
индивидуального развития называется анаморфозом.
У самцов конечности восьмого-девятого сегментов превращены
в копулятивные придатки — гоноподии. Они помещаются в особом
карманоподобном впячивании покровов. Морфология гоноподии
самцов двупарноногих сложна, ее признаки используются в
диагностике таксономической принадлежности этих многоножек.
Строение ротового аппарата. При изучении ротового аппарата
(рис. 107) применяют технику последовательного вычленения его
элементов. Препаровальной иглой отделяют головной отдел вместе с
шейным щитком. Ротовое отверстие снизу и сзади закрыто непарным
гнатохилярием — сложной нижней г>бой, образованной в
результате слияния второй пары максилл и элементов стернита
(брюшной пластинки) первого туловищного сегмента. Основная
его функция - удержание пищевых частиц. Первая пара максилл
полностью редуцирована. Мандибулы у двупарноногих хорошо
развиты и служат для измельчения пищи. Они снабжены несколькими
жевательными пластинками, имеющими зазубренную, бугристую
Класс Insecta
213
Рис. 107. Лицевая часть головы лвупарноногого: / — основание airren-
ны; 2— мандибула; J— гнатохилярий; 4— глазное поле
поверхность. Строение ротового аппарата диплопод в значительной
мере напоминает таковое у насекомых.
Надкласс Hexapoda — шестнногие
Класс Insecta — насекомые
Насекомые представляют собой группу беспозвоночных, наиболее
полно освоивших наземно-возлушную среду обитания. Дыхательная
система трахейного типа. У мелких форм и паразитов газообмен
может осуществляться всей поверхностью тела, а у большинства водных
личинок имеются вторичные жабры. Тело подразделяется натри
отдела (тагмы): голову, грудь и брюшко. Головной отдел насекомых
(исключая бессяжковых) несет многочлен и ковые антенны и
ротовые придатки, в большинстве своем представляющие собой
модифицированные конечности. Грудной отдел, в отличие от головного, сохра-
11яст следы сегментации. В его составе три сегмента с хорошо развитыми
конечностями. Взрослые особи (и м а го) большинства насекомых
имеют крылья, закладывающиеся как выросты стенки тела, и обладают
способностью к полету. Членистое брюшко может нести
модифицированные для выполнения специальных функций конечности.
Насекомые в большинстве своем раздельнополы; за небольшим исключени-
214
1m Arthropods
ем. развитие осуществляется с неполным либо полным превращением
(метаморфозом).
В ходе лабораторных занятий целесообразно ознакомиться с
особенностями сегментации тела насекомых, строением антенн, ротовых
аппаратов, грудных конечностей и крыльев разных типов.
Таксономическое разнообразие насекомых, особенности строения
представителей основных отрядов и экологических групп, стадии развития,
некоторые хозяйственно значимые формы насекомых (вредители
растений, опылители, гнус и другие группы эктопаразитов и тл.), типы
повреждений культивируемых растений — все эти вопросы лучше
рассмотреть в период летней учебной практики по зоологии
беспозвоночных.
Расчленение тела майского хруща (Melolontha sp.)
Материал. Лёт майских хрущей (майских жуков) на большей части
лесной зоны Европейской России, Украины и Белоруссии происходит
в мае — начале июня. В это время жуков можно собрать, отряхивая
молодые лиственные деревья рано утром либо отлавливая
энтомологическим сачком роящихся жуков в вечернее время, когда они
медленно кружат над освещенными последними лучами заходящего
солнца выступающими ветвями или вершинами деревьев и
кустарников. Усыпленных эфирами или хлороформом насекомых хранят
в 70-80* этаноле.
Майские хрущи, западный (Melolontha melolontha) и восточный
(Melolontha hippocastani), принадлежат к семейству пластннчатоусых
(Scarabaeidae) отряда жесткокрылых, или жуков (Coieoptera). Это
достаточно крупные насекомые, длина тела взрослых особей составляет
22-30 мм. Покровы тела имаго сильно склеротизированы, в чем легко
убедиться с помощью аккуратных нажатий препаровальной иглой или
пинцетом.
Тело жуков несколько уплощено в дорсовентральном
направлении и имеет овальные очертания. Со спинной стороны можно
различить небольшую слитную голову, которая подогнута подтело. Голова
имеет пару сложных фасеточных глаз (их легче рассмотреть при
увеличении) и характерной формы пару усиков (антенн), которые
могут оказаться поджатыми и вследствие этого незаметными сверху.
Далее следует хорошо развитая, сильно склеротизированная передне-
спинка, имеющая черную или темно-коричневую окраску. Почти все
остальное тело покрыто парой жестких надкрылий, или элитр (ок-
Класс Insecta
215
раска их может варьировать от светлой песочной, до почти черной но
наиболее обычна каштаново-коричневая). Надкрылья разделены швом,
проходящим по средней оси тела. Небольшая концевая часть брюшка!
имеющая вид продолговатого, направленного назад и вниз отростка!
остается не прикрытой надкрыльями и называется пигидием. У Л/е-
lolontho mclolonfha он более широкий и отличается почти
параллельными краями, у Mclolonfha hippocastani более узкий, но явственно
расширяющийся у самой вершины (рис. 108).
Рассматривая майского жука снизу, выявляют голову с
комплексом ротовых придатков (для этого лучше воспользоваться ручной
лупой или бинокуляром). Затем следует грудной отдел с тремя
парами ходильных конечностей. Брюшные сегменты имеют вид
следующих друг за другом колец все меньшего диаметра, конечностей
или придатков они лишены.
При взгляде сбоку легко установить положение продольной оси
головной капсулы относительно продольной оси тела животного.
У майского хруща, как и у многих других жуков, они составляют
прямой угол, при этом ротовые части направлены вниз. Такой тип
расположения ротового аппарата носит название гипогнатического.
У представителей других групп насекомых он может быть иным (рис. 109).
У ряда хищных личинок насекомых (жуков-плавунцов, жужелиц,
сетчатокрылых и пр.), у имаго жуков жужелиц, рогачей и многих других
Рт. 108. Майские хрущи: восточный (Mclolontha hippocastani), самка (Л)%
западный (Mclolontha mclolontha). самец (£); личинка (В)
216
l*t\ Arthropods
ротовые части направлены вперед. Такой тип расположения носит
название прогнатического. У клопов, равнокрылых, многих тараканов
ротовые части в той или иной степени подогнуты под тело и
направлены назад. В этом случае говорят об опистогнатическом типе
ротового аппарата.
Р*с. 109. Различные типы постановки ротового аппарата у насекомых:
А — гипогнатический; Б — прогнатический; В — опистогнати-
ческий
Исходно каждый сегмент тела насекомых состоит из двух хити-
низироваиных скелетных пластинок (склеритов): спинной — терги-
та и брюшной — стернита, которые имеют вид полуколец. По бокам
они соединены вкольцоплейритами,лучше развитыми в грудных
сегментах. Между собой сегменты также соединены участками тонкой
мембранозной кутикулы, — такое сочетание жестких склеротизован-
ных и эластичных мембранных участков обеспечивает необходимую
подвижность при значительном уровне прочности.
Для лучшего уяснения особенностей сегментации тела насекомых
следует вычленить отдельные сегменты или части тела майского жука
(рис. 110). Удерживая туловище, захватывают головную капсулу
пинцетом или иглой и аккуратно оттягивают ее. При этом обнажается
узкий перехват с эластичными покровами — шейка, соединяющая
голову с туловищем. Отделив головную капсулу, следует рассмотреть
ее при увеличении, воспользовавшись ручной лупой или бинокуляром.
Стенки капсулы образованы сильно хитинизированными покровами
акрона и настоящих сегментов головного отдела тела. Позади
она имеет крупное затылочное отверстие, через которое
проходят мускульные пучки, кишечная трубка, нервные
волокна, трахейные стволы, кровеносные сосуды, соединяющие головной
Класс/nsetfa
217
Рнс. 110. Расчлененный майский хрущ MehkmthameMontha: / — голова;
2— переднегруль; J— среднетрудь; 4— заднегрудь; 5— брюшко
и туловищные участки соответствующих систем. Внутри капсулы
находится защищаемый ею головной мозг.
Головная капсула несет ротовые органы, сложные фасеточные
глаза и антенны. Спереди ротовые част прикрыты наличником
в виде хитинизированной пластики, соединенной с головной
капсулой швом. По своему происхождению наличник представляет собой
вырост стенки тела. Лицевую часть головной капсулы занимает лоб,
по его бокам расположены сложные глаза и основания антенн. У
майских хрущей, как и у всех пластончатоусых, антенны пласти нчато-бу-
лавовидного типа. Размеры пластинчатой булавы различаются:
у самцов она явственно крупнее, что является проявлением полового
диморфизма.
У ряда насекомых помимо пары сложных многофасеточных глаз
имеются и простые глазки. Считается, что им принадлежит основная
роль в восприятии плоскости поляризации света и изменения
интенсивности освещения, чем широко пользуются насекомые для
ориентации в полете. Рассматривая под бинокуляром отчлененную
головную капсулу медоносной пчелы (рис. 111) или стрекозы, можно уяснить
особенности строения сложных многофасеточных глаз насекомых.
Следует также отыскать простые глазки, обратить внимание на причлененис
наличника, положение ротовых частей и оснований антенн, наличие
на головной капсуле небольшого количества швов, представляющих
собой слеша межсегментарных траниц.
218
TmArthropodd
Рис. 111. Голова рабочей особи медоносной пчелы (Apis теШ/ега): I —
антенна; 2 — верхняя челюсть; 3 — эпифариикс (нддглоточник);
4— нижняя челюсть; 5 — губной щупик; 6 — язычок; 7—
наличник; 8— фасеточный глаз; 9— глазок
Продолжая расчленение, осторожно оттягивают пинцетом или
иглой переднеспинку и отделяют первый грудной сегмент туловища.
Он представляет собой кольцо с мощной спинной частью; его
брюшное полукольцо несет первую пару ходильных конечностей. Второй
туловищный сегмент (среднегруль), имеющий вид тонкого кольца,
на спинной стороне несет пару жестких надкрылий (элитр), на
брюшной — вторую пару ходильных конечностей. После удаления
среднегруди с принадлежащей ей элитрами открывается пара
сложенных и лежащих на поверхности брюшка перепончатых задних крыльев.
Разделив брюшко и третий грудной сегмент, можно убедиться, что
заднегрудь — самый крупный грудной сегмент. Это связано с
наличием функционирующих крыльев, нуждающихся в мощной
крыловой мускулатуре. Пара ходильных конечностей, принадлежащих этому
Класс Insecto
219
сегменту, несколько крупнее прочих и служит для удержания не
имеющего собственной опоры брюшка.
Спинная поверхность брюшка отличается мягкими, легко
прогибающимися при нажатии тергитами, так как уже защищена жесткими
надкрыльями. По числу поперечных швов на более твердой нижней
(вентральной) поверхности брюшного отдела можно составить
представление о числе сегментов, вошедших в его состав. Вычленение
отдельных брюшных сегментов затруднительно, но в этом и нет
особой необходимости.
Особенности строения антенн
Антенны (усики) считаются придатками акрона (головной
лопасти) и не гомологичны конечностям. Отсутствуют они только у бес-
сяжковых (отряд Protura). Основная функция антенн —
чувствительная — обеспечивается наличием огромного числа рецепторов
(химических, тактильных и др.). В составе многочлениковых антенн принято
выделять:
- основной членик (рукоятку),
- ножку (педицелл), обычно имеющую наибольшую длину
среди всех члеников,
- жгутик (бич), объединяющий всеостальныечленики (рис. 112).
Строение антенн у насекомых разных эколого-систематических
групп весьма разнообразно. Для ознакомления с выделяемыми типами
и особенностями строения антенн используются постоянные
препараты, которые лучше просматривать под бинокуляром.
Нитевидные антенны состоят из многочисленных линейно
расположенных члеников и имеют одинаковую толщину по всей
своей длине.
Щетинковидные (шиловидные) антенны состоят из
многочисленных линейно расположенных члеников и постепенно
утончаются к вершине.
Веретеновидные антенны имеют наибольшую ширину в средней
части и постепенно сужаются как к вершине, так и к основанию.
Четковидные антенны состоят из округлых или
продолговато-овальных члеников, имеющих наименьший диаметр в местах
применения к соседним.
Пиловидные (пильчатые) усики состоят из члеников,
имеющих боковые выступы наподобие зубцов пилы.
220
JmArthropoda
ЖЗИКЛ MHO
Рис. 112. Антенны насекомых: А — общий план строения: / — жгутик:
2— ножка; J— основной членик; Б— щетинковшшая; В—
нитевидная; Г— четковндная; Д — пиловидная; £— булавовидная:
Ж— гребневидная; 3— неправильная; Я— перистая; /Г—
головчатая жука-мертвосда; Л — головчатая блохи; А/— пластинчатая;
Я— неправильная с расчлененным члеником; О—щетинконосная
Гребневидные (гребенчатые) усики состоят из члеников»
несущих с внешней стороны длинные боковые пластинчатые отростки,
которые образуют своеобразную гребенку.
Перистые антенны состоят из члеников, несущих как с
внешней, так и с внутренней стороны гибкие шетинковндные отростки,
образующие характерное оперение.
Булавовидные усики отличаются утолщающимися
вершинными члениками, образующими округлую компактную булаву.
Пластинчато-булавовидные усики иногда рассматриваются
в качестве варианта булавовидных. Вершинные членики сильно
укорочены и несут крупные пластинчатые боковые отростки.
Внутренняя мускулатура позволяет такой булаве веерообразно расправляться
и складываться.
Головчатые усики также рассматриваются как вариант
булавовидных. Их булава образована рыхло расположенными члениками,
форма которых характерна для четковидных антенн.
Класс Insecta
221
Коленчатые антенны характеризуются вытянутым основным
члеником, несушим расположенный под углом к нему относительно
мелкочленистый жгутик.
Гребенчато-коленчатые усики сочетают в своем строении черты
морфологии коленчатых и пластинчато-булавовидных антенн:
характерную для первых основную часть и типичную для вторых
пластинчатую булаву.
Неправильные антенны имеют в своем составе членики
асимметричной (неправильной) формы.
Щетинконосные антенны, как правило, состоят из трех крупных
члеников, последний из которых имеет характерную чувствительную
щетинку, гладкую или перистую.
Основные типы грудных конечностей насекомых
Грудной отдел тела насекомых несет три пары конечностей.
Исходными считаются конечности ходильного типа. При освоении
насекомыми разных жизненных сред, иных способов передвижения либо
при приобретении конечностями других функций изменилось и
строение последних, носящее адаптивный характер. В составе конечности
выделяются следующие части: тазик, вертлуг (трохантер), бедро,
голень и многочлениковая лапка (рис. 113). Члснистостьлапки
является вторичной: се членики не имеют самостоятельной мускулатуры,
обший мышечный пучок пронизывает их, как нитка в бусах или четках.
С особенностями строения конечностей основных типов следует
ознакомиться, используя постоянные микроскопические препараты.
Помимо описанных ниже, имеются и другие типы конечностей, менее
распространенные, например присасывательные или прядильные.
Строение бегательных конечностей черного таракана (Blatta orien-
talis). Бегатсльныс конечности рассматриваются как модификация
ходильных либо выделяются в отдельный самостоятельный тип. Бе-
гатсльная конечность черного таракана состоит из тазика, вертлуга,
бедра, голени и лапки (рис. 113, Л) Тазик представляет собой основной
членик, который неподвижно причленяется к грудному стерниту.
Вертлуг у таракана, как и у многих других насекомых, миниатюрен,
несколько смещен в сторону и располагается вдоль сочленения тазика
и бедра. Бедро — наиболее крупный продолговатый членик. К нему
причленяется также продолговатая, но более тонкая голень.
Членистая лапка, сравнимая по длине с голенью, на вершине несет слегка
222
Тип Arthropods
Рис.113. Типы конечностей насекомых: А — ходильная; Б— прыгалтель-
пая; Я — собирательная; Г— плавательная;Л— хватательная;
£— копательная; / — тазик; 2— вертлуг; J—бедро; 4— голень;
5 —лапка
изогнутые коготки. Бегатсльные конечности обеспечивают быстрое
перемещение по субстрату и удерживают тело высоко над его
поверхностью, что необходимо, например, при передвижении по раскаленному
солнцем песку. От ходильных конечностей бегатсльные отличаются
относительно большей длиной и стройностью, а также строением
лапки. Для конечностей ходильного типа характерны лапки с
уплощенной нижней поверхностью, несущей большое количество
волосков или щетинок и называемой подошвой.
Строение прыгательных конечностей кузнечиков (Tetrigonia spp.).
Прыгательные конечности обеспечивают передвижение прыжками,
когда значительное расстояние преодолевается практически
мгновенно. У кузнечиков и других прямокрылых прыгатсльными являются
конечности третьей пары, у которых бедренно-голенный
Класс Insects
223
сустав образует мощный рычаг. Бедра этих конечностей длинные
и толстые (рис. 113, £), с развитой мускулатурой. Голени также
удлинены пропорционально длине бедер. Сходное строение имеет третья
пара конечностей блох (Aphaniptera). У цикадовых (Cicadinea)
«рабочим» является тазобедренный сустав.
Строение плавательных конечностей жуков-плавунцов (Dytiscidae).
Плавательная (гребная) конечность характеризуется
ограниченностью свободы движений большинства ее члеников относительно друг
друга. В результате формируется единая гребная поверхность,
работающая как весло. Следует обратить внимание на размеры и форму
лапки (рис. 113. £). У плавунцов она явственно уплощена и снабжена
многочисленными длинными волосками. При выносе конечности
вперед они прижимаются к поверхности голени, а при рабочем ходе
расправляются и таким образом увеличивают гребную поверхность.
Плавательными могут быть все три пары конечностей (многие
водные личинки жуков), средняя и задняя пары (имаго плавунцов)
или только задняя (клопы-гребляки сем. Corixidac).
Строение хватательных конечностей водяного скорпиона (Nepa
cinerea). Хватательными у этого клопа являются передние конечности.
По принципу работы они напоминают складной (перочинный) нож
или капкан: удлиненные бедро и голень (рис. 113, В) при сведении
зажимают добычу. Лапка, которая могла бы затруднять
функционирование такой конечности, редуцирована, сохранив один членик.
Поскольку хватательные конечности используются хищниками для
захвата добычи, их иногда называют хищными. У обыкновенного
богомола хватательные конечности устроены аналогично, но голень
и бедро вооружены мощными заостренными хитиновыми зубцами,
которыми насекомое способно пробить даже кожу человеческой руки
(особенно, детской).
Строение копательных конечностей медведок (Gryllotalpa spp.).
Копательными у медведок являются передние конечности. Они
предназначены для рытья ходов в почве, зачастую пронизанной
многочисленными мелкими корнями растений. Все элементы такой
конечности сильно хитинизированы, укорочены и уплощены (рис. 113, Ж).
Преимущественное развитие получает голень, которая снабжена
мощными заостренными зубцами. Лапка подвергается редукции, но
сохраняется.
Копательными конечностями обладают также жуки-навозники,
некоторые клопы, жуки-усачи, короеды, то есть насекомые, которые по роду
деятельности должны разгребать с>*страт либо проделывать в нем ходы.
224
Тип Arthropods
Строение собирательных конечностей медоносной пчелы (Apis
теШ/era). Собирательными у пчел являются задние конечности.
Характерно строение голени и первого членика лапки (рис.! \\Л): внешняя
поверхность голени гладкая, но по краям ее расположены
многочисленные длинные изогнутые волоски, ограничивающие так
называемую корзинку, которая служит для накопления и переноса пыльцы.
Внутренняя поверхность очень крупного первого членика лапки
усажена многочисленными волосками, за что это образование получило
название шеточки. Пчела не занимается непосредственным сбором
пыльцы из пыльников. Посещая цветки, она накапливает на
опушенной поверхности тела пыльцевые зерна, а затем, совершая
характерные движения, вычесывает пыльцу с помощью щеточки и помещает ее
в корзиночку в виде так называемого пыльцевого комка, или обножки.
Благодаря яркой окраске пыльцевые комки хорошо заметны у
посещающих цветки пчел и шмелей.
Строение прицепных конечностей человеческой вши (Pedicalus
humanus). Прицепные конечности характерны для обитателей
волосяного покрова млекопитающих и перьевого — птиц. Для этих
эктопаразитов жизненно важно удержаться на хозяине, так как падение
с него в большинстве случаев равнозначно гибели. У человеческой
вши голень имеет специальный отросток, к которому вплотную
подходит коготок лапки (рис. 113, Г). Сама лапка одночлениковая,
сравнимый с нею по размерам коготок причленястся под углом, а просвет
между голенью и лапкой соответствует толщине волоса. В результате
формируется приспособление, напоминающее карабин в
альпинистском снаряжении. Захватываемый волос ущемляется, чем
обеспечивается надежная фиксация на нем эктопаразита. Захватывая волос за
волосом, насекомое медленно перемещается в волосяном покрове
хозяина.
Основные типы крыльев насекомых, жилкование крыла
Крылья насекомых по своему происхождению являются
нечленистыми придатками второго и третьего грудных сегментов,
образованными складками стенки тела. Форма, размеры и другие
морфологические характеристики крыльев у представителей разных отрядов
насекомых могут сильно различаться (рис. 114). Кроме основной,
локомоторной функции, крылья могут выполнять и другие, что
сопровождается соответствующими изменениями в их строении.
Класс Insecta
225
Рт. 114. Типы крыльев насекомых: А — жуков: / — жесткие (элитры);
2— перепончатые; Б— чешуекрылых: чешуйчатые; В —
стрекоз: сетчатые; Г— прямокрылых: кожистые (тегмины); Д —
полужесткокрылых: полужесткие (геыиэлнтры)
Наиболее распространенной является треугольная форма крыла.
Различают основание, которым оно причленяется к грудному
сегменту, противоположный ему апикальный (вершинный) угол и
лежащий сзади нижний, или задний угол (рис. 115). Стороны крылового
Фсугольника называются краями: костальным, или передним
(соединяет основание с апикальным углом), терминальным, или
наружным (апикальный и нижний углы), дорсальным, или задним
(основание и нижний угол).
Если крылья прозрачны, не составляет труда рассмотреть их
жилкование (расположение трахеи). Жилки представляют собой
разветвленные трахеи. Если среди них невозможно выделить крупнейшие
основные и жилкование выглядит как система мелких ячеек, такие
крылья называются сетчатыми. Они характерны для стрекоз, поденок
226
Тип Arthropods
Л00—- ^ч 3 А». 115. Общая схема строения крыльев насе-
^^ ) комых: / — основание крыла; 2— зад-
/ ний угол; J— вершина; 4— передний
/ 5 край; 5 — наружный край; 6 — виут-
_ J ренний край крыла
и сетчатокрылых. У большинства других насекомых на прозрачных
крыльях нетрудно выделить ограниченный круг радиольно расходящихся
от основания основных жилок. Эти крылья носят название
перепончатых. Таковыми являются обе пары крыльев насекомых отряда
перепончатокрылых, передняя пара — двукрылых, задняя — жесткокрылых
и полужесткокрылых. У ручейников крылья несут множество
волосков, за что отряд и получил свое название — волосокрылые. Крылья
бабочек (чешуекрылых) носят название чешуйчатых, так как их
поверхность покрыта слоем черспицеобразно расположенных чешуек.
Жилкование таких крыльев становится явным после их смачивания
авиационным бензином или аналогичным растворителем.
Воспользовавшись бинокуляром, следует рассмотреть поверхность крыльев
ручейников и бабочек. Передние крылья тараканов и прямокрылых
носят название кожистых надкрылий, или тегмин. что вполне
соответствует их механическим свойствам. У жуков передняя пара крыльев
интенсивнос)аеротизированалорогопа1обногосостояния. Такие
крылья называются жесткими, или элитрами. Основная функция
тегмин и элитр — защитная. У клопов склеротизировано лишь
основание переднего крыла, тогда как вершина его остается перепончатой
и называется мембраной. Она может сложным образом складываться,
оставаясь, наряду с задними крыльями, защищенной более жесткими
основаниями. Такие крылья получили название гемиэл итр
(полужестких). Отсюда и второе название отряда клопов — полужесткокрылые.
У двукрылых вторая пара крыльев модифицирована в лопаткообразные
жужжальца. Они выполняют функцию стабилизаторов полета и
активизируют работу крыловых мышц передней пары крыльев.
По характеру жилкования крыла часто определяют
таксономическую принадлежность насекомых. Во избежание путаницы
зоологами приняты стандартные схемы обозначения отдельных жилок
и образуемых ими ячеек. В русскоязычной специальной литературе
используется следующая генерализированная схема жилкования
(рис. 116): ближе всего к переднему (костальному) краю крыла
расположена костальная жилка. За ней слсдустсубкостальная, которая
у своей вершины подразделяется на две (первую и вторую субко-
Класс Insects
227
стальные). Далее следует ветвящаяся радиальная, а за нею — также
ветвящаяся медиальная. Следующая, кубитальная, жилка
разветвляется надвое. Последующие анальные жилки не ветвятся. Югал ь-
ная область крыла, наличие которой обеспечивает возможность
насекомому складывать крылья вдоль тела, включает целый набор нс-
ветвящихеяюгальных жил ок. Для большинства насекомых
характерны более или менее существенные отклонения от описанного
Рис. 116. Генерализированная схема жилкования крыльев насекомых:
С— костальная жилка; &,-.&>—с><жостальные жилки; /?, - Rs—
радиальные жилки; А/; - М4 — медиальные жилки; Си, - Си4 —
к>биталы(ые жилки; А, — As — анальные жилки; Ju — югальмая
жилка
варианта жилкования. Как пример можно рассмотреть особенности
жилкования крыла боярышницы. Для знакомства с разнообразием
жилкования используются препараты крыльев насекомых разных
отрядов.
Основные функционально-морфологические типы
ротовых аппаратов насекомых
Ротовые аппараты насекомых весьма разнообразны по своему
строению, что определяется в первую очередь характером пиши и
способом ее потребления. Насекомые используют в пищу самые различные
пищевые субстраты, и по этой причине ротовой аппарат претерпевает
соответствующие морфологические адаптивные изменения.
Основными функционально-морфологическими типами ротовых аппаратов
228
Тип Arthropods
насекомых считаются: грызущий» лакающий (фызушс-лижуший),
сосущий лешшогперондный. лижущий мускокдный, колюше-сосущис ге-
миптероидный и диптероидный.
Сгроевие грызущего ротового аппарата черного таракана (Blatta
orientate). Этот тип ротового аппарата характерен для наиболее древних
насекомых и считается исходным, тогда как прочие возникли в
результате его модификации. Ротовой аппарат черного таракана
отличается относительно невысокой степенью склсротизации, в связи с чем
эти насекомые могут измельчать лишь сравнительно мягкие пищевые
субстраты. Твердую пищу они предварительно должны смачивать
слюной и только после этого могут ее измельчить и съесть.
Сверху ротовые части прикрывает непарная верхняя губа (лабрум),
представляющая собой складку стенки тела в виде широкой пластинки
с прямоугольным основанием и закругленной вершиной (рис. 117). Она
подвижно причленсна к головной капсуле пол наличником. Верхняя
губа выполняет роль передней стенки предротовой полости.
Рис. 117. Ротовой аппарат черного таракана (Blotto orientalls): / —
верхняя губа; 2— жвалы (верхние челюсти); 3 — нижние челюсти;
4—основной членик; 5— стволик; 6—челюстной щупик; 7—
наружная жевательная лопасть; 8— внутренняя жевательная
лопасть; 9— нижняя губа; 10— подподбородок; //—подбородок;
12— иижнегубной щупик; 13— придаточные язычки; 14— язычки
Класс Insecta
229
Верхние челюсти (мандибулы, жвалы) представляют собой
пару коротких лопастевидных нечленистых пластинок. Они имеют
темную окраску и интенсивно склеротнзированы. Внутренние края
жвал снабжены несколькими крупными зубцами. Мандибулы не
являются строго симметричными, поскольку зубцы одной из них должны
совпадать с промежутками между зубцами другой.
Нижние челюсти (макс иллы-1) сохранили парность и член не -
тостьстроения. С головной капсулой они соединены основным
члеником, или подвеском (кардо), к которому причленяется более
массивный ствол и к (сти пес). У вершины стволика с внешней стороны
на бугорке сидит четырехчлениковый нижнечелюстной (максил-
лярный) щупик, выполняющий чувствительную функцию.
Широкая вершина стволика несет также пару жевательных лопастей.
Внутренняя жеватсльняя лопасть (лациния) сильно склеротизирована,
снабжена хитиновыми зубцами и щетинками, она принимает
непосредственное участие в измельчении пиши. Наружная
жевательная лопасть (галеа) слабо склеротизирована и покрыта
многочисленными волосками.
Нижняя губа (лабиум) непарная, но образована в результате
слияния основных члеников максилл-Н (максиллул) и сохраняет
членистое строение. В частности, лежащий в ее основании широкий
сердцевидный подподбородок (субментум) соответствует подвескам
нижних челюстей. К нему применяется более узкий подбородок
(менту м), образованный в результате слияния стволиков. Подбородок несет
щупики и язычки. Собственно язычки (глоссы), расположенные по
центру, соответствуют внутренним жевательным лопастям, а
придаточные язычки (параглоссы), занимающие промежуточное
положение между язычками и нижнегубными щупиками, — наружным
жевательным лопастям максилл-1. Нижнегубные (лабиальные)
щупики короче нижнечелюстных, состоят из трех члеников и
выполняют чувствительную функцию, причем более
специализированы к хеморсцепции. Имеющая плоскую форму нижняя губа замыкает
снизу предротовую полость и служит для слизываиия жидкой пиит.
На се внутренней стороне имеется небольшой подвижный подглоточ-
ник (гипофарингс), функционально аналогичный языку большинства
млекопитающих. По своему происхождению он представляет
непарный вырост стенки тела. Ввиду малых размеров и отсутствия склеро-
тизации гипофарингс обычно не принимается во внимание при
изготовлении демонстрационных микроскопических препаратов.
230
Тип Arthropods
Строение лакающего (грызуще-лижущего) ротового аппарата шмеля
(Bombussp.y Такой ротовой аппарат характерен для пчелиных, имаго
которых собирают нектар из цветков, и по строению (рис. 118)
напоминает рассмотренный выше грызущий, где все элементы в той или
иной степени подверглись модификации.
Рис. 118. Ротовой аппарат шмеля (Bom-
bus): / — верхняя ту6ъ\ 2 —
верхняя челюсть; 3 — язычок;
4— нижнегубной щупик; 5 —
наружная лопасть нижней
губы; б— подбородок; 7— под-
подбородок; 8 — основной
членик; 9 — стволик; 10 —
нижнечелюстной щупик; // —
нижняя челюсть
Верхняя губа представляет собой короткую, но широкую хитини-
зированиую пластинку, верхний закругленный край которой покрыт
многочисленными волосками.
Верхние челюсти внешне напоминают таковые черного
таракана, но отличаются продолговатой формой. Они не играют
существенной роли в процессе потребления пищи, но активно
используются для обработки воска, для перемещения или разгрызания травинок
и других предметов при строительстве гнезда. Известна способность
шмелей и пчел делать боковые надкусы в длинных венчиках цветков,
имеющих труднодоступные удлиненные нектарники. У медоносной
пчелы мандибулы имеют характерный желобок и используются для
сбора пыльцы.
Класс Insects
231
Нижние челюсти и нижняя губа объединены в хоботок.
Основания максилл имеют палочковидную форму. К ним причлсняются
хорошо развитые стволики. Наружные жевательные лопасти
хорошо развиты, имеют форму вытянутых клиновидных пластинок,
способных при необходимости желобоподобно изгибаться.
Внутренние жевательные лопасти, как и нижнечелюстные шупики,
рудиментарны и располагаются у оснований наружных жевательных
лопастей.
Подподбородок нижней губы имеет форму треугольника со
слегка вогнутыми сторонами и причленяется к головной капсуле.
Подбородок удлиненный, бочонкообразный. К нему причленен
единый язычок (глосса), имеющий ребристую поверхность и снабженный
многочисленными волосками. В толще язычка проходит канал,
служащий для всасывания жилкой пищи. Придаточные язычки имеют
вид коротких желобчатых пластинок. Сбоку от них отходят
нижнегубные шулики, которые претерпели морфологическую модификацию
и сменили основную функцию. Первый членик шупика развит лучше
лругих, подлине сравним с наружными жевательными лопастями
нижних челюстей, уплощен и имеет форму мелкого желоба. Остальные
три членика шупика невелики по размерам и сохранили стандартное
строение.
Хоботок является весьма эффективным приспособлением для
потребления нектара. Смыкающиеся наружные жевательные лопасти
нижних челюстей образуют дорсальную и боковые стенки, а первые
членики нижнсг>бных щупиков — вентральную стенку трубчатого
канала хоботка. Внутри размещается язычок, который при
потреблении животным жидкой пиши вибрирует. В этом отношении он
напоминает ершик для мытья узкой посуды. Мощная глоточная мускулатура
создает разрежение, что в комплексе и обеспечивает поступление
пищи. Малые количества нектара, находящиеся к тому же в наиболее
труднодоступных местах, удается достать с помощью длинного язычка
и откачать по его внутреннему каналу.
Строение гемштероишого колюще-сосущего ротового аппарата
клопов (Hemiptera). Данный тип ротовых аппаратов (рис. 119) характерен
для всех представителей отряда полужесткокрылых, или клопов (//*-
miptera). Он специализирован для потребления жидкой пищи,
находящейся под покровными тканями животных или растений.
Верхняя губа имеет вид небольшой, заостренной на вершине
пластинки, причлененной к наличнику.
Верхние и нижние челюсти преобразованы в нечленис-
тыс хитиновые стилеты. В совокупности мандибулы и максиллы
232
ImArthropoda
Рмс. 119. Ротовой аппарат клопов {Hemiptera): А — общая схема
строения, вид сбоку. / — головная капсула; 2 — фасеточный глаз; J —
лоб; 4 — двураздельный наличник; 5 — верхняя губа; б —
подглоточник; 7 — мандибулы; 8 — максиллы; 9-/0 —
нижняя губа (9— членики нижней губы, 10— желоб); Б—
стадии проникновения стилетов в ткани хозяина: / — сложенные
вместе челюсти, прокалывающие ткань; 2 — нижняя губа; В —
схема строения хоботка на поперечном срезе: / — мандибулы;
2 — максиллы; 3 — дорсальный (пищевой) канал; 4 — вапраль-
ный (слюнной) канал
образуют подобие наборной иглы, пронизанной несколькими
внутренними каналами. Мандибулы, имеющие на срезе форму полуколец,
составляют обкладку иглы. Концы их заострены, а в ряде случаев
несут также дополнительные зубцы и предназначены для
прокалывания покровов. Максилллы в соприкосновении друг с другом
образуют пару крупных каналов (рис. 119, В), обеспечивающих эф-
Класс Insecte
233
фективную транспортировку жидкости. По вектральному каналу в рану
полается слюна, содержащая пищеварительные ферменты. Дорсальный
канал предназначен для всасывания жидкой пищи.
Нижняя губа выполняет функцию кожистого чехла (ножен) для
стил сто подобных мандибул и максилл. Она представляет собой
продолговатый отросток, образованный тремя-четырьмя члениками
и снабженный спереди желобком для стилетов, которые могут
свободно скользить по нему. В состоянии покоя стилеты полностью
скрыты в канале нижней губы (рис. 119, £). При погружении их в ткани
хозяина нижнегубной чехол образует направленный назад колено-
образный или иной формы изгиб. У некоторых равнокрылых стилеты
длиннее не только нижней губы, но и всего тела своего владельца
и в покос свернуты широкой петлей или спиралью внутри головной
капсулы. Опистогнатический тип расположения ротового аппарата
позволяет этим насекомым развивать на вершине аппарата
проникновения давление, достаточное для преодоления весьма прочных
покровов их жертв.
Строение дипгтероидного колюще-сосущего ротового аппарата
кровососущих комаров (сем. Culicidae). Принадлежащие к отряду
двукрылых (Dipfera) комары указанного семейства располагают
колюще-сосущим ротовым аппаратом несколько иного строения. В отличие от
гсмигттсроидного типа, в его составе присутствует шесть стилстооб-
разных образований (рис. 120).
Верхняя губа имеет вид длинной тонкой трубки с косо
срезанным, заостренным концом. Стенки этой трубки не сплошные: вдоль
внутренней (вентральной) ее стороны идет шов с иеспаянными краями.
Кроме того, толщина стенок крайне неравномерна: спереди (с
дорсальной стороны) она больше, чем сзади (с вентральной стороны),
вокруг шва. К последнему прилегает комплекс сложенных
определенным образом остальных стнлетоподобных элементов, имеющих на
срезе слегка вогнутую форму. Мандибулы превращены в тонкие жесткие
хитиновые щетинки, заостренные на вершине и зазубренные по
краям. Нижние челюсти превратились в примерно такие же щетинки,
но сохранили длинные и тонкие членистые щупики. Ги пофари н гс
также трансформировался в жслобополобную, стилстообразную
щетинку. Внутри он пронизан тонким каналом, по которому в рану
подается слюна насекомого.
Весь описанный комплекс элементов комбинируется в стержнепо-
лобное образование, которое в нерабочем состоянии спрятано в чехле,
образованном нижней губой. В отличие от клопов, у комаров она
нечлениста и содержит больше мягких тканей. В результате можно
234
J*nArthropod3
Рис. 120. Ротовой аппарат настоящего комара (Сикх): Л — общая схема
строения; £ — проникновение стилета в ткани хозяина; 7 —
основания антенны; 2—лобный щиток; 3—основание
челюстного щупика; 4 — верхняя губа; 5 — максиллы; 6 — мандибулы;
7— подглоточник; 8— нижняя губа*. 9— сложенные и
вводимые вместе верхняя губа, мандибулы, максиллы, подглоточник
заключить, что гемиптсроилный и диптероидный варианты ротовых
аппаратов колюшс-сосушего типа устроены и функционируют
сходным образом.
Строение сосущего ротового аппарата чешуекрылых (Lepidoptera)
(рис. 121). Этот тип ротового аппарата весьма характерен для
представителей отряда чешуекрылых, или бабочек. Специализация к потреб*
лснию жидкой пищи (нектара, вытекающего из рай деревьев сока и пр.)
привела к значительным изменениям по сравнению с исходным
вариантом строения.
Верхняя губа имеет вид небольшой пластиночки или складки,
примененной к наличнику, и в функционировании ротового
аппарата участия не принимает. Верхние челюсти у большинства
представителей отряда утрачены.
Нижние челюсти в наименьшей степени подверглись редукции.
Хорошо развиты наружные жевательные лопасти в виде длинных,
Класс Insects
235
Рис. 121. Ротовой ал парат чеш)ект>ылъа(£г;>^
лепным хоботком; Б — участок хоботка на срезе; / — основание
актенны; 2 — верхняя губа; 3 — нижнегубной щупик; 4 —
нижняя губа; 5 — хоботок; 6 — фасеточный глаз; 7 — правая и
левая нижние челюсти; 8 — места соединения челюстей; 9 —
полость хоботка; 10 — трахея
жслобообразных пластинок. Вместе они образуют трубку — хоботок,
через канал которого всасывается жидкая пиша. В силу своей
эластичности хоботок свертывается в почти плоскую спираль типа часовой
пружины и в покое подогнут пол грудь. Для развертывания хоботка
» него под давлением нагнетается гемолимфа, в этом процессе
участвует и внутренняя мускулатура.
Нижняя губа представляет собой небольшую пластинку,
расположенную позади нижних челюстей. Хорошо развиты нижнегубные
щупики, имеющие типичное строение и выполняющие
чувствительную функцию.
Таким образом, специализация к питанию свободной от
защитных оболочек жидкой пишей привела у чешуекрылых к
преобразованию почти всех элементов ротового аппарата.
236
ImArthropoda
Pnc. 122. Ротовой аппарат комнатной мухи
(Afusca domestica): 1 — верхняя губа;
2— макснллярный щупик; J— рост-
рум; 4— гаустсллум; 5— лабеллумы
Строение лижущего мускоидного ротового аппарата (хоботка)
высших мух. Среди рассматриваемых типов он наиболее далек от
исходного варианта строения (рис. 122).
Основание хоботка называется рострумом. В его образовании,
помимо разросшихся краев головной капсулы, принимает участие
наличник. Спереди он несет нижнечелюстные щупики. Рострум
переходит в гаустеллум, образованный подглоточником (гипофа-
рингсом). Спереди он защищен приросшей к его поверхности верхней
губой. Далее следует парный или двухлопастный лабеллум, в
формировании которого основная роль принадлежит нижнегубным щупикам.
Рострум, гаустеллум и лабеллумы сочленены подвижно, в состоянии
покоя они складываются двумя коленами под головной капсулой
и малозаметны со стороны.
В рабочем состоянии лабеллумы имеют вид полукруглых
присосок с расположенным между ними центральным отверстием, ведущим
в глотку. К этому отверстию и идущему от него к глотке каналу
подходят мелкие канальцы, называемые псевдотрахеями. На другом
конце они открываются микропорами на вентральной поверхности ла-
беллума. Края центрального отверстия имеют хитиновые зубцы
(«зубы»)» способные дробить твердые субстраты (рис. 123). Питающаяся
на поверхности твердого субстрата муха сначала смачивает его
слюной. Если он оказывается нерастворимым, она пытается дробить его
Класс Arachnid*
237
А Б В
Рис. 123. Разворачивающиеся лабсллумы ротового аппарата высших
двукрылых (Dip/era): Л — в состоянии покоя; Б — в процессе
фильтрации жидкой пищи; В— в процессе обскребывания с>бстрота;
/ — склериты; 2— псевдотрахеи; 3— «зубы»; 4— гибкая
мембрана; 5 — мышцы
поверхность. Когда насекомое втягивает полученный раствор или
жидкую кашицу, лабедлумы функционируют наподобие поролоновой губки.
Затем животное повторяет цикл на новом месте. У осенней жигалки
(Ssomoxyscalcitrant), мух-цеце (Glossina) вооружение краев центрально-
i о отверстия представлено в виде плоских пружинящих лезвий, с
помощью которых животные способны без труда вырезать характерную
лунку даже в очень плотных кожных покровах, например крупного
рогатого скота.
Описанными типами ротовых аппаратов не исчерпывается их
разнообразие у насекомых. Общим является присутствие в их составе
соответствующим образом модифицированных конечностей и
вторичных придатков головного отдела тела.
Подтип Chelicerata — хелицеровые
Класс Arachnida — паукообразные
Класс объединяет наземных и вторичноводных членистоногих.
Первая пара головных конечностей — исходно клешневидные хел и -
не ры, занимающие предротовое местоположение. Тело
подразделяется на два отдела: просому и опистосому. последняя объединяет
^свосм составе мезосому и.мстасому(рис. 124). Просома несет шесть
"ар конечностей, включая хслицеры. Органы дыхания (если имеются)
238
lw\ Arthropod*
А Б В Г
Рпс. 124. Строение арахнид (вид с брюшной стороны): А — скорпиона;
Б — паука; В — сольпуги; Г— примитивного акариформного
клеща; I—XIX — сегменты тела; / — хелицеры; 2— педипальпы;
.?-ходильные конечности; 4— половое отверстие; 5—
гребневидные органы (половые гребешки); 6 — стигмы легких; 7 —
стигмы трахей; £— паутинные бородавки; 9— хвостовой
придаток; /0— анальное отверстие
воздушного типа. Органами выделения служат мальпигиевы сосуды.
На лабораторных занятиях целесообразно остановиться на
рассмотрении особенностей внешней морфологии представителей пяти
подклассов паукообразных: скорпионов, сольпуг, пауков, ложиоскорпионов
и клешей. С особенностями строения сенокосцев, таксономическим
разнообразием пауков, типами повреждений растений
клешами-фитофагами удобнее ознакомиться в ходе летней учебной практики.
Подкласс Scorpiones — скорпионы
Представители подкласса — крупные хслицеровые со слитной
головогрудью (цефалотораксом) и членистым брюшком (абдоменом)»
состоящим из передне- и заднебрюшия. Тельсон снабжен ядовитой
железой. Хелицеры и педипальпы клешневидные. Дыхание легочное.
Представители рассматриваемого подкласса раздельнополы, развитие
у них прямое. Скорпионы теплолюбивы, в пределах Восточной Европы
Класс ЛгасЛлйй
239
встречаются в Прикаспии, Предкавказье, Причерноморье и
Средиземноморье. Хищники.
Buthus eupeus — пестрый скорпион
Материал. Пестрый скорпион, как и другие представители
семейства Buthidae, распространен в Нижнем Поволжье, Средней Азии и
Закавказье, где местами многочислен. Для Причерноморья характерны
представители другого семейства скорпионов — Chaaidac. в Крыму
это эндемичный Euscorpius tauricus, на Черноморском побережье
Кавказа — Euscorpius italicus, в Западном Закавказье — Euscorpius
mingrclicus. Светлое время суток эти животные проводят в укрытиях.
Их можно отловить под камнями, отслаивающейся корой деревьев,
в трещинах скал и стен домов. Для сбора скорпионов используют длинный
пинцет. Из-за агрессивности и склонности к каннибализму их нельзя
держать по нескольку экземпляров в одном сосуде. Укус скорпиона не
представляет прямой опасности для взрослого человека, однако во
избежание анафилактического шока и других клинических реакций
следует проявлять максимальную осторожность. Собранных животных
немедленно помешают в заправленную морилку. Хранят материал в 70*
этаноле.
Внешнее строение. Скорпионы принадлежат к числу одних из
самых крупных (длина тела взрослых пестрых скорпионов достигает
65 мм) наземных членистоногих. Тело подразделено на головогрудь
(цефалоторакс) и брюшко (абдомен). Граница между ними, если
смотреть со спинной стороны (рис. 12S), вполне явная. Со спинной
стороны головогрудь прикрыта карапаксом, образуемым
слившимися тергитамн (спинными пластинками) шести головогрудных
сегментов и покровами акрона. Воспользовавшись ручной лупой либо бино-
куляром, можно рассмотреть располагающиеся в передней части
головогруди простые глаза: по центру размещена пара медиан ных
глаз, по переднему краю —фронтальных, по бокам —
латеральных. На нижней стороне головогруди выделяется вентральный
щит — стернум, образовавшийся в результате слияния брюшных
пластинок (стернитов) двух последних грудных сегментов.
Брюшной отдел тела состоит из внешне хорошо отграниченных
двенадцати насгоящихсегментовительсона. Различают переднебрю-
шие (мезосому). образованное семью сегментами, и заднебрюшие
(мстасому), в состав которого входят остальные пять сегментов и тель-
сон. Визуально тело скорпиона делится на широкий массивный корпус,
включающий головогрудь и перзднебрюшие, и узкое, длинное и гибкое
240
1m Arthropods
А Б
Pnc. 125. Пестрый скорпио» (Buihuseupcus): A — вид со спинной
стороны; Б— вид с брюшной стороны; /—гатовогрудь;2—хслнцсры;
3 — всртлуг педипальпы; 4 — бедро педипальпы; 5 —
подвижный палеи; 6 — ладонь; 7— голень педипальпы; 8 —
конечность II пары; 9—конечность III пары; 10-17— конечность IV
пары (10— тазик; // — иертлуг, 12— бедро; 13 — голень; 14 —
основной членик лапки; 15— I членик лапки; 16— II членик
лапки; /7—коготки); 18— ядовитая игла; 19—тедъсон;20—
заднебрюшис; 21 — переднебрюшие; 22 — анальное
отверстие; 23 — стигма; 24 — половая крышечка; 25 — стернум;
26 — челюстная лопасть ноги I пары; 27— челюстная лопасть
конечности II пары; 28— тазик конечности II пары; 29— тазик
конечности III пары; 30— гребневидный орган
Класс Arschnkto
синебрюшис, порой неправильно называемое «хвостовой часть
К последнему брюшному сегменту причленяется округлый тельсон
несущий сверху изогнутую, заостренную на вершине хитиновую иглу —
ядовитое жало скорпиона. Внутри тельсона находится парная
ядовитая железа, для выведения секрета которой служит идущий внутри
иглы канал. Скорпионы используют ядовитое жало как орудие защиты
и нападения. Потревоженное животное изгибает брюшко, удерживая
его конец над головогрудью — это типичная поза угрозы. Кроме
зашиты от врагов, яд широко используется для быстрого умерщвления
кертв: помимо различных членистоногих ими могут быть мелкие
1>сптилии и даже детеныши мелких млекопитающих. Анальное
отверстие расположено tia брюшной стороне тела фактически на сочленении
последнего сегмента заднебрюшия и тельсона. У некоторых
экземпляре оно хорошо заметно благодаря окружающим его лопастсвидным
возвышениям, представляющим собой вывернутые наружу стенки
прямой кишки.
Состав и особенности строения конечностей. Первой парой
конечностей головогруди являются короткие клешневидные хелицеры.
Расположены они фронтально (рис. 125, £)> оставляя позади ротовое
отверстие. Хелицеры трехчленисты, предпоследний их членик несет
специальный выступ, играющий роль неподвижного пальца
клешни, как у речных раков. Функцию подвижного пальца выполняет
последний членик хелицер. Основное назначение этих образований —
механическая обработка пишевых объектов.
Вторая пара конечностей, педипальпы (ногочелюсти), выделяется
среди прочих своей длиной. Они шесткчлениковые, причем послсаниедва
членика образуют мощные клешни. Помимо обороны и, главное,
захвата добычи, педипальпы выполняют осязательную функцию.
Последующие, третья - шестая, пары конечностей являются хо-
лильными и на своих вершинах несут коготки. Они лишены
каких-либо следов дву ветвистости, задние развиты несколько лучше передних.
Тазики педипальл и всех ходильных конечностей вошли в состав
брюшной стенки тела, утратив вследствие этого подвижность. Тазики
двух первых пар ходильных конечностей снабжены специальными
отростками — челюстными лопастями. Они легко различимы при
осмотре брюшной поверхности головогруди: первая пара имеет вид
греугольных, вторая — клиновидных пластинок. Лопасти окаймляют
щелсвиднос ротовое отверстие, образуя в совокупности нижнюю
губу. Скорпионы способны поглощать только жидкую и
кашицеобразную пищу, для этого им необходимо предварительно ее
измельчить и обработать слюной.
242
TmArthropoda
Брюшной отдел тела у скорпионов лишен конечностей
типичного строения, но первые его сегменты несут придатки,
рассматриваемые как модифицированные конечности. Первый брюшной сегмент
снабжен округлыми образованиями, прикрывающими расположенные
здесь половые отверстия и называемыми половыми крышечками.
У хранившихся в этаноле экземпляров они выделяются на общем фоне
своей светлой окраской. У некоторых самок под половыми
крышечками могут быть обнаружены черные пластинки. Этосперматоклей-
трумы, которыми самцы запечатывают половые отверстия после
завершения процесса спаривания. Они могут сохраняться вплоть до
времени окончательного созревания яиц. На втором брюшном
сегменте располагаются гребневидные придатки, которые несут
большое число рецепторов и участвуют в ритуале спаривания
скорпионов. На последующих четырех сегментах брюшка по бокам имеются
щелевидные отверстия — стигмы, или дыхальца. Они ведут в
легочные мешки, которые, как полагают специалисты, по своему
происхождению представляют впятившиеся внутрь дыхательные
конечности. У экземпляров, находившихся продолжительное время в воле,
края стигмальных отверстий могут быть заметно вздутыми.
Последним, седьмым, сегментом переднебрюшия, как и всеми сегментами
заднебрюшия, конечности полностью утрачены. Таким образом,
конечности брюшного отдела в большей степени, нежели головогруд-
ного, подверглись трансформации либо редукции.
Подкласс Solpugida (=Solifugae) — солытугн
Подкласс объединяет преимущественно крупных арахнид,
внешним обликом напоминающих пауков. Степень расчлененности тела
взрослых особей сольпуг выше, чем у всех остальных групп
паукообразных. Хслицеры клешневидные. Псдипальпы
ходильно-осязательного типа. Дыхание трахейное. Все сольпуги — хищники.
Для ознакомления с особенностями сегментации тела
представителей отряда можно использовать смонтированный сухой материал
либо влажные препараты взрослых особей любого вида фаланг и
других сольпуг.
Gakodes araneoides — обыкновенная фаланга
Материал. Обыкновенная фаланга характерна для степей и
предгорий Крыма, Приазовья, Предкавказья и Нижнего Поволжья. Набольшей
части территории Средней Азии вполне обычна, а местами массовым
Класс Arachnids
243
видом является закаспийская сольпуга (Galcodes caspius).
Представители рода Galeodes активны в темное время суток. Их привлекает свет,
чем пользуются для сбора этих животных. Мелкая светлоокрашенная
солнцелюбивая сольпуга (Paragaleodes hcliophilus) принадлежит к
числу дневных форм. В среднеазиатских степях и полупустынях этих
паукообразных можно наблюдать перемещающимися по
поверхности почвы даже в знойный полдень. Для учебных целей пригодны все
вышеупомянутые представители подкласса. Собранных сольпуг
анестезируют эфирами, после чего требуется специальная обработка во
избежание загнивания тканей мясистого брюшка. При хранении в
жидких фиксаторах (80* этанол, 4% раствор формалина) обязательно
с помощью шприца с тонкой иглой в брюшко делают несколько
инъекций фиксатора. При монтировании сольпуг для сухих коллекций через
небольшой разрез на нижней поверхности брюшко освобождают от
мягких тканей и набивают ватными шариками, как это рекомендуется для
крупных экземпляров пауков или жуков-маек.
Внешнее строение. Фаланги рода Galeodes принадлежат к числу
наиболее крупных: длина тела взрослых особей Galeodes araneoides
достигает 50 мм. Головогрудной отдел тела у сольпуг состоит из слитного
пропел ьтидия (образован слиянием четырех передних сегментов) и двух
свободных сегментов (рис. 126). Со спинной стороны пропельтидий
покрыт мощным склеротнзированным головным шитом. У его
переднего края расположен глазной бугорок, несущий пару выпуклых
простых глаз. Латеральные глаза у сольпуг подвержены атрофии.
Брюшко овально-цилиндрическое, состоит из десяти сегментов
с хорошо различимыми границами. Туловище, а также придатки тела
сольпуг покрыты густыми волосками и щетинками различной длины,
формы и жесткости. На нижней стороне первого брюшного сегмента
можно обнаружить половое отверстие, имеющее вид продольной
щели, прикрытой боковыми створками.
Состав и особенности строения конечностей. Первая пара
конечностей головогруди, хелицеры, получили у сольпуг преимущественное
развитие. Размерами они сравнимы с пропельтидием и представляют
собой направленные вперед, вертикально поставленные мощные
клешневидные образования. Вершины их заострены, а внутренние по-
нерхности несут многочисленные зубцы. Хелицеры являются мощным
орудием защиты и нападения, ими сольпуги захватывают, умерщвляют
и расчленяют добычу. Крупные взрослые особи могут даже прокусить
кожу человека. Бытует мнение о ядовитости сольпуг, но оно ошибочно,
244
Тип Arthropods
Рпс. 126. Расчленение тела фаланги (Gclc-
odes): I — хелицера; 2 — педи-
пальпа; 3 — хоанльные
конечности; 4 — брюшко; 5 —
свободные задние сегмогга груди;
б— пропельтидий; 7— глаза
а неблагоприятные последствия укусов связаны с попаданием в рану
остающихся на хслицерах остатков пищи, в которых развиваются
бактерии и образуется трупный яд. Хслицерами эти паукообразные
пользуются также для рытья в почве норок, служащих их временными
убежищами.
Педипальпы внешне походят на типичные ходильные
конечности, но лишь в малой степени участвуют в беге и ходьбе; их
вершины лишены коготков. Основная их функция — осязательная. Кроме
того, сол ьпуги пользуются ими для захвата и удержания добычи, а также
в общении с особями противоположного пола.
Последующие четыре пары ходильных конечностей
головогруди состоят из семи члеников и несут на вершинах по парс коготков.
Длинные и стройные, они фактически являются бе нательными. Задние
несколько мощнее передних, поскольку на них приходится нагрузка
по удержанию массивного брюшка, а первая пара стройнее и короче
других и выполняет главным образом осязательную функцию. При
внимательном осмотре с брюшной стороны на тазиках и вертлугах
последней пары ходильных конечностей могут быть обнаружены коло-
Класс Arachnids
245
кольчикоподобные малеолы (привески). Предполагается, что они
выполняют рецегтторную функцию.
Подкласс Pseudoscorpiones — ложноскорпноны
Мелкие хелицеровые по внешнему виду напоминают
скорпионов, но отличаются от последних тем, что брюшко их не имеет
подразделения на передне- и заднебрюшие. Сходство этих паукообразных со
скорпионами определяется клешневидными педипальпами. Характерно
наличие паутинных желез. Дыхание трахейное. Развитие с
метаморфозом. Хищники.
Для ознакомления с особенностями строения представителей
подкласса лучше использовать не фиксированный материал, а
постоянные тотальные препараты взрослых экземпляров книжного ложио-
скорпиона.
Cheli/er cancroides — книжный, или обыкновенный ложноскорпион
Материал. Лжескорпионы представляют собой группу широко
распространенных, но ведущих скрытый образ жизни и поэтому
малозаметных животных. Книжный ложноскорпион часто
отмечается в библиотеках, книжных складах и иных книгохранилищах,
а также чердаках, подвалах и других помещениях, где питается
главным образом клещами пыли. В природных условиях эти животные
встречаются в лесной подстилке, под отстающей корой деревьев,
в дуплах и других подобных местообитаниях. Сбор достаточного
количества экземпляров обычно затруднителен; наилучшие
результаты может дать выгонка их из проб подстилки на фотоэклекторе.
Анестезированных животных хранят в 70* этаноле, спиртовой материал
вполне пригоден для изготовления постоянных препаратов.
Внешнее строение. Тело ложноскорпиона подразделяется на голо-
вогрудной (просома) и брюшной (опистосома) отделы (рис. 127),
визуально разграниченные бороздой. Брюшко шире груди и широко
закруглено сзади, в результате чего тело имеет каплевидную форму.
Оно в значительной мерс уплощено, что позволяет этим
членистоногим использовать шелевидные укрытия, такие как полости под
отстающей корой или корешки книг. Головогрудь защищена сверху
слитным головогрулным щитом. Брюшко состоит из 11 сегментов,
границы между которыми различимы как на спинной, так и
брюшной сторонах тела.
Хслицеры уложноскорпионовнебольшие,клешневидные,силь-
но сближены между собой и направлены прямо вперед. На их вершинах
246
Тип Arthropods
Рис. 127. Книжный, или обыкновенный, ложноскорпион (Chelifer
caneroidts): I — хелицеры; 2 — годовогрудной отдел (просома);
J— головогрудной щит; 4— брюшной отдел (опистосома); 5—
ходильные конечности; 6 — педипхтьлы
открываются протоки расположенных в головогруди парных
паутинных желез. Педипальпы, вооруженные мощными клешнями,
внешне напоминают таковые скорпионов, но снабжены длинными
чувствительными волосками — трихоботриями. Педипальпами
животные захватывают добычу, но ее обработка производится хелицерами.
Остальные четыре пары головогрудных конечностей используются
для ходьбы. На вершинах ходильных конечностей расположена пара
коготков с небольшой присоской между ними. Эти приспособления
позволяют ложноскорпиоиам с легкостью перемешаться по
вертикальным поверхностям (стволам деревьев, боковым поверхностям валунов,
стенам и т.п.). Потревоженные животные пятятся либо двигаются
боком, как это свойственно крабам и паукам-бокоходам.
Расположенный на нижней стороне брюшка наружный половой аппарат имеет
достаточно сложное строение, выяснение происхождения его частей
представляет определенные затруднения.
Подкласс Aranei — пауки
Преимущественно мелкие и средних размеров хелицеровые, имеют
слитные головогрудь и брюшко, соединенные стебельчатым пере-
Класс Arachnids
247
хватом. Хелицеры и псдипальпы не клешнеобразные. Конечности
последних двух сегментов брюшка преобразованы в паутинные
бородавки. Жизнедеятельность пауков теснейшим образом связана с
прядением паутины. Покровы тела образованы главным образом
заявленными белками, а не хитином. Дыхание легочное и трахейное.
Развитие прямое. Все пауки — хищники.
На лабораторных занятиях могут быть использованы любые
крупные представители таксона, но традиционным объектом является
паук-крестовик.
Aroneus diodematus — обыкновенный тук-крестовик
Материал. Этот паук в условиях лесной зоны Восточной Европы
является одним из обычнейших представителей семейства пауков-круго-
прядов (Arancidae). Для них характерно вертикальное расположение
ловчих тенет, образованных концентрическими кругами паутины. В
сумерки либо пасмурным днем крестовики располагаются по центру
ловчей сети и их нетрудно собрать, подставляя снизу широкую пробирку
либо иной сосуд. Хранят пауков в 80* этаноле.
Внешнее строение. Тело большинства пауков сильно укорочено
и компактно. У крестовика между слитными головогрудью и
брюшком находится стебелек (рис. 128, А), образованный первым
брюшным сегментом. Головофудь уплощена с боков, ее облегают
нисходящие передний и боковые края головогрудного щита, на котором
ближе к переднему краю размещены в два ряда четыре пары простых
глаз, имеющих вил небольших черных точек.
Воспользовавшись ручной лупой или бинокуляром, можно
ознакомиться с морфологией брюшной стороны головогруди (рис. 128, Б).
По центру, между выпуклыми тазиками ходильных ног расположена
пятиугольной формы умеренно склеротпзированная пластинка — стер-
нум. Спереди от стернума лежит полукруглой формы нижняя губа,
сформировавшаяся из брюшной пластинки (стернита) II головофул-
ногосегмента. За ней следует пара челюстных лопастей,
являющихся отростками тазиков конечностей этого же сегмента. Нижняя губа
и челюстные лопасти окружают ротовое отверстие.
Хелицеры являются наиболее интенсивно склеротнзированны-
ми частями тела пауков. Они имеют фронтальное расположение; сппепень
их жесткости можно установить нажатием препаровальной иглы.
Хелицеры двучленистые; крупный основной членик имеет
цилиндрическую форму и несет на вершине крючкообразный, подвижно
прикрепленный дистальный членик. У крестовиков он выделяется своей
темной окраской. Внутри него проходит открывающийся на вершине
248
1m Arthropods
Рис. 128. Внешнее строение самки обыюювешюго паука-крестовика (st/a-
пеш diadematus):A — <&u}iu виде брюилюй стороны:
/—головогрудь; 2-3 — головные конечности (2 — хелицера, 3 — педи-
палълз); ¥— стернум; $-$— грудные ходильные конечности
(1-1Упары); 9—брюшко; 10— эпигене; // —
паутинныебородавки; /2—стигмы трахей; /J—легочные крышечки;
£—строение головогруди и расположение конечностей (хелицеры
отогнуты вперед): / — когтевндный членик хелицеры; 2 — основной
членик хелицеры; J — педипальла; 4— пучок волосков
челюстной лопасти; 5— челюстная лопасть; 6— нижняя туба;
7—тазики ног; 8— пластинчатое расширение стернума; 9— стернум;
10-16 — структурные части педипалълы (/0— тазик; // — верт-
луг, /2—бедро; 13— колено; 14— голень; 15 — лапка; 16—
коготок)
членика канал ядовитой железы. В состоянии покоя дистальный
членик поджимается к поверхности основного членика. Благодаря
заостренным вершинам и ядовитому секрету, хелицеры
представляют собой серьезное оружие нападения и зашиты. Пользуясь ими.
Класс Arachnida
249
животные захватывают, удерживают и умерщвляют добычу, а также
наносят раны, в которые затем впрыскивают содержащую
пищеварительные ферменты слюну. Всем паукам свойственно внекишечное
пищеварение.
Педипальпы выполняют главным образом осязательную
функцию, ими животное ощупывает субстрат и окружающие предметы,
фиксирует колебания нитей паутины. Как уже указывалось ранее,
педипальпы являются единственными конечностями пауков,
обладающими челюстными лопастями. У самок педипальпы по внешнему
виду напоминают ходильные конечности. У самцов они укорочены
и несут на вершинах специальные чашевидные расширения, с
помощью которых во время спаривания собирается семенная жидкость и
вводится в половые пути самки.
Ходильные конечности в числе четырех пар имеют типичное
строение, несут на вершинах хитиновые коготки и используются для
передвижения по различным с)<хгтратам, растительности или паутине.
Характерной особенностью строения ходильных конечностей пауков
является наличие дополнительного членика — колена,
расположенного между бедром и голенью.
Округло-каплевидное брюшко прикреплено к головогруди
стебельком, явственно смещенным вниз. Его дорсальная пластинка
называется лорумом, а вентральная — плагулой. Дорсальная
пластинка имеет ббльшие размеры, нежели вентральная, и вследствие этого
заворачивается на брюшную сторону. На спинной стороне брюшко
крестовика имеет светлый крестообразный рисунок, послуживший
основанием для названия живолюго. Сразу же за стебельком снизу
размещается пара щелевидных стигмальных отверстий, ведущих
в полость легочных мешков и прикрытых хитинизированнЫми
легочными крышечками. В местах расположения легочных мешков
окраска покровов более светлая. Как и у скорпионов, легкие
рассматриваются как впятившиеся вовнутрь дыхательные конечности. У самок
между стигмами открывается женское половое отверстие,
защищаемое характерной хитиновой пластинкой — эпигене. Ее форма
у самок многих групп пауков вилоспецифична и используется для их
идентификации.
На конце брюшка, снизу имеется три пары небольших конических
придатков. Это паутинные бородавки, на вершинах которых
открываются протоки паутинных желез. Бородавки первой и последней
пар доучленистые, что указывает на их происхождение в результате
преобразования брюшных конечностей. Бородавки средней пары.
250
Тип Arthropod*
на которые при осмотре из-за их мелких размеров обращают
внимание в последнюю очередь, не обладают членистостью и являются
выростами стенки тела. Между бородавками верхней лары возвышается
анальный бугорок, на котором расположеноаналыюс отверстие.
Подкласс Acari — клещи
Мелкие, иногда микроскопических размеров хелицеровыс. У
взрослых особей деление тела на отделы обычно нечеткое. Для обозначения
отдельных частей тела клешей применяется специальная терминология.
В частости, в составе тела выделяют гнатосому, подосому,
подразделяемую на проподосому и мезоподосому, а также опистосому.
Гнатосома и проподосома в совокупности образуют протеросому,
а металодосома и олистосрма — гистсросому (рис. 129). Дыхание
трахейное или кожное. Развитие с метаморфозом, личинки имеют три
пары ходильных конечностей, нимфы — четыре пары. В настоящее
время клешей подразделяют на три отряда, с представителями двух
из них целесообразно ознакомиться в ходе лабораторного практикума.
Рис. 129. Общая схема деления тела клешей на отделы: / — просом а;
2— опистосома; J— гнатосома; 4— идиосома; 5— подосома;
6— проподосома; 7— металодосома; 8— протеросома; 9— ги-
стеросома
Отряд Acariformes — акариформные клеши
Мелкие хелицеровыс, тело взрослых особей подразделяется на
протеросому (объединяет передние головогрудные сегменты) и гнете-
росому (объединяет два головогрудных и все брюшные сегменты),
Класс Arachnids
251
граница между ними обычно неявна. Ротовой аппарат устроен просто,
хелицеры грызущего или сосущего типа. Дыхание кожное или
трахейное. Для индивидуального развития характерен анаморфоз.
Эмбрионы с тремя парами зачатков конечностей. Сапрофаги, фитофаги,
хищники или паразиты. Отряд подразделяется на два подотряда и
более сотни семейств, объединяемых в серии.
Подотряд Sarcoptiformes — саркоптиформные клещи
Хелицеры фызушего типа. Почво- или растениеобитающие
членистоногие, либо паразиты животных. Расселительную функцию
выполняют взрослые особи.
Tyrogfyphusfarinie — обыкновенный мучной клещ
Материал. Мучные клещи семейства Tyroglyphidac являются
характерными вредителями запасов. При повышенной влажности
хранящегося зерна, круп, муки они в массе могут развиваться в домах,
зернохранилищах и складах. Другие тироглифокдные клещи обитают
на сухофруктах, казеиновых отходах, сыре и многих других продуктах
животного и растительного происхождения. На занятиях лучше
воспользоваться постоянными тотальными препаратами. При использовании
для их изготовления канадского бальзама клещей фиксируют 10%
раствором формалина и окрашивают. При использовании гуммиарабико-
вых заключающих сред Фора или Фора-Бсрлезс хранившихся в 70-80*
этаноле клешей нужно обязательно провести через 40* спирт.
Внешнее строение. Взрослые мучные клещи имеют
микроскопические размеры, длина тела самок не превышает 0,7 мм, самцов —
0,45 мм. Светлоокрашенное овальной формы тело Tyroglyphusfarinae
лишено видимых склеротизированных участков и одето тонкой хи-
тиноидной кутикулой. Для рассмотрения особенностей внешней
морфологии (рис. 130) лучше использовать более крупных самок,
отличающихся менее сглаженными очертаниями тела.
Рассматривая клеща со спинной стороны, следует обратить
внимание на бороздку, отделяющую треугольно-коническую протеро-
сому от продолговатой гистеросомы. Спереди хорошо различимы
направленные вперед клешневидные хелицеры. Глаз у этих
паукообразных нет. На поверхности тела определенным образом
распределены мелкие и крупные щетинки, в совокупности составляющие хетом.
252
Тип Arthropods
Рис. 130. Обыкновенный мучной клеш (Tyrogfyphusfarinoe): самка, внд
со спинной стороны (/0. самка, внд с брюшной стороны (5).
самец, внд с брюшной стороны (£). расселитсльная нимфа — ги-
попус (О; / — протсросома; 2 — ротовые органы; 3-6 — I—IV
пары ходильных конечностей; 7— гистеросома; 8— гр>эная
пластинка (стернум); 9— паювос отверстие; 10— патовые присоски;
// — анхтыюе отверстие; 12—жировые железы; 13 —
мандибулы; 14 — нижняя туба: 15 — анальные присоски; /6-2/ —
структурные части ходильной конечности (16— вертлут; /7—
бедро; 18— колено; /9— голень; 20 — лапка; 21 — коготок)
Knacc Aractinida
253
Схема размещения щетинок является важнейшим таксономическим
признаком и используется при определении клешей отряда Acariformes.
При изучении объекта с брюшной стороны (для этого выбирают
другой, лежащий на спине, экземпляр либо аккуратно
переворачивают препарат на предметном столике) на переднем конце тела видны
направленные вперед продолговатые педипальпы. Между ними
находится пластинчатая нижняя губа, несущая пару трехчлен истых
нижнегубных щупиков. Все четыре пары ходильных ног у самок
примерно одинакового строения, у самцов первая пара конечностей
отличается сильно утолщенными бедрами и коленными члениками. Все
ходильные конечности оканчиваются изогнутым коготком. На уровне
оснований задних ходильных конечностей (6-я пара) между двумя
пластинчатыми клапанами располагается половое отверстие.
Каждый клапан несет пару округлых половых присосок. В задней
части брюшной стороны тела размещено анал ьное отверстие,
имеющее виц длинной продольной щели. У самцов по бокам заднего конца
тела расположена пара анальных присосок.
Подотряд Trombidiformes — тромбиднформные клещи
Хслицеры сосущего типа. Обитатели пресных водоемов, почво-
и растснисобитающие членистоногие либо паразиты животных. Рассс-
лительную функцию выполняют личинки.
Tetranychidae — паутинные клещи
Материал. Многие паутинные клещи родов Tetranychus, Schizo-
retranychus, Oliganychus, Panonychus, как и другие представители
семейства Tetranychidae, являются массовыми вредителями культивируемых
и иных растений. Особо благоприятны для их развития условия
закрытого груша с высокими температурами и низкой относительной
влажностью воздуха. На интенсивное заселение растений паутинными
клещами указывают локальное или точечное пожелтение листовых
пластинок, других неодрсвесневших частей растений, которые при
ближайшем рассмотрении оказываются затянутыми тонкой паутиной,
что и послужило основанием для русскоязычного названия группы.
В зимне-весенний период паутинные клещи могут быть собраны на
оранжерейных или комнатных растениях. Во многих городах
Белоруссии, европейской часта России, Украины во второй половине лета
липы мелколистная (ТШа cordate), широколистная (77/ш platyphyllos)
254
Тип Arthropod*
и некоторые другие зачастую интенсивно повреждаются липовым
паутинным клешом (Schizptetranychus tiliarum). Подобно молоди пауков,
тетранихиды на паутинках легко расселяются с потоками воздуха.
Поэтому при демонстрации живых паутинных клещей следует
проявлять осторожность, чтобы они случайно не получили свободу и не
колонизировали имеющиеся в помещениях комнатные растения. Для
ознакомления с внешней морфологией клешей лучше использовать
постоянные препараты на основе заключающих сред Фора или
Фора-Бсрлсзс. В случае использования материала, хранившегося
в 70-80* этаноле, его нужно обязательно провести через 40* спирт.
Внешнее строение. Взрослые паутинные клещи имеют
микроскопические размеры, длина тела взрослых особей не превышает 0,5 мм.
Форма тела овальная или продол го взто-овальная (рис. 131). Самцы
обычно несколько мельче и стройнее самок, с коротким телом,
суживающимся к заднему концу. Спинная сторона тела выпуклая, брюшная —
умеренно выпуклая. Прижизненная окраска чаще бледно-зеленая или
бледно-желто-зеленая. У взрослых особей обыкновенного паутинного
клеща (Tetranychus urticae $./.), садового паутинного клеща (Schizo-
tetranychuspruni), некоторых других видов по бокам тела видны
характерной формы темные пигментные пятна. Летние самки красного
плодового клеща (Pononychus ulmi) имеют ярко-красную окраску, тогда
как личинки, нимфы и самцы этого вида — зеленые. Впадающие в диа-
паузу самки сливового паутинного клеша меняют бледно-желтую
окраску на лимонно-желтую.
Первичная сегментация тела утрачена, при взгляде сверху хорошо
различима лишь граница между проподосомой иметаподосомой.
На спинной стороне тела у представителей рода Tetranychus имеется
участок со складчатой поверхностью покровов, образующих
«ромбическую фигуру*. Кожные покровы тела и конечностей несут большое
количество щетинок, особенности хетома широко используются при
определении таксономической принадлежности тетранихид. На
спинной стороне по бокам проподосомы расположены простые глаза
в виде кармннно-красных точек. У большинства тетранихид их две пары.
Ротовой аппарат колюще-сосушеготипа (рис. 132, А). Гнатосо-
ма несетгипостом,стилофорсостилетами ипарупедипальп
(щупалец). Гипостом имеет вид четырехугольной пластинки,
переходящей спереди в конический ростру м, на вершине которого
располагается ротовое отверстие. Образован гипостом слившимися батальными
члениками педипальл. Стнлофор является продуктом слияния пятых
члеников (голеней) хелицер, при взгляде сверху он имеет сердцевид-
Класс Arachnid* 255
Рис. 131. Паутинные клещи (Teironychidat): Л — самка обыкновенного
паутинного клеша (Tttranychus urtieot s.1.): Б — самка содового
паутинного клеща (Schbpktranychta pnmi)\ В — самец (Г) и самка
(2) красного плодового клеща (Punonychus uimi)
ную форму. Остальная дистальная часть хелицер видоизменилась в
парные стилеты. С помощью такого ротового аппарата паутинные клещи
способны высасывать содержимое растительных клеток и даже
отдельных хлоропластов. Опустошенные клетки заполняются воздухом и при
рассмотрении при увеличении могут обусловливать характерный
256
lm Arthropods
Рис. 132. Строение ротового аппарата паутинных клешей: А — пшосома:
/ — стилет; 2— пальлы; 3—стилофор; 4— козырек; Б— пальпа
(щупальце): I — бедро; 2 — колено; J-голень; 4— лапка
серебристый опенок. Пслипальпы (щупальца) паутинных клещей
исходно шестнчлениковые, но их тазики слились в гипостом, а остальная часть
после слияния сегментов выглядит четырехчлениковой (рис. 132, Б).
Ходильные конечности паутинных клещей исходно шестичлени-
ковые. В состав конечности входит газик, неподвижно
прикрепленный к корпусу тела, вертлуг, бедро, колено, голень и лапка (рис. 133),
подвижно сочлененные друг с другом. Все они несут разнообразные
щетинки, а лапка вооружена коготками. Как и большинство клещей,
личинки тстранихнд имеют три пары ходильных конечностей, с
линькой на протонимфу появляется четвертая пара.
Рис. 133. Строение ходильной конечности паутинных клещей: / —
тазик; 2— вертлуг; J— бедро; 4— колено; 5— голень; 6—лапка
Класс Arac/Mftfo
257
Летом самки откладывают яйца прямо в местах питания, осенью —
в щелях коры, у основания почек. У ряда видов тетранихнд (например,
обыкновенного паутинного клеша) зимуют диапаузируюшие самки.
Из яиц отрождаются шестиногие личинки. Они превращаются в восьми-
ногнх протонимф, которые очень быстро линяют в дейтонимф.
Взрослые особи, прежде чем приступить к размножению, должны пройти
непродолжительный период дополнительного питания.
Отряд Parasitiformes — паразнтоформные клещи
Мелкие и средних размеров хелицеровые, тело взрослых особей
у большинства представителей лишено следов сегментации. Ротовой
аппарат сложный, сосущего типа. Дыхание преимущественно
трахейное. Эмбрионы с четырьмя парами зачатков конечностей. Хищники
или паразиты животных, растительноядные формы отсутствуют.
Переносчики возбудителей многих инфекционных заболеваний человека
и животных.
В составе отряда выделяются три основные группы: гамазовые,
иксодовые клеши и клеши-уроподы. На малом практикуме
необходимо ознакомиться с отличающимися относительно крупными размерами
и имеющими серьезное хозяйственное (эпидемиологическое, в
частности) значение иксодовыми клещами.
Ixodes acinus — собачий клещ
Материал. Клеши семейства /xodidae характеризуются высокой
степенью ландшафтно-биопгопической специализации. Так, для таежной
зоны Евразии характерен таежный клещ (Ixodes persulcatus), для зоны
мелколиственных лесов — собачий клещ (Ixodes risinus), для степей —
представители рода Dermacentor% полупустынь — Rhipicephalus,
пустынь — НуаЬтта. Сббачий клещ наиболее многочислен в тех лесных
массивах, где высока плотность копытных либо активно выпасается
скот. Взрослых клещей собирают, окашивая сачком растительность
на высоте 1,0-2,5 м либо применяя специальные экраны и иные
приспособления. Личинок и нимф коллекгируют, выполняя кошение по
травостою и кустарникам. Иксодовые клеши являются эффективными
переносчиками опасного заболевания человека — весенне-летнего
клещевого энцефалита, имеющего вирусную этиологию, а также
возбудителей ряда других болезней. Поэтому при сборе материала
необходимо предпринять все необходимые меры предосторожности для
258
Тип Arthropods
предотвращения укусов. Фиксировать клещей надо специальными
составами: эфирно-спиртовой смесью (97 частей 20* этанола, 3 части ди-
этилового (серного) эфира), жидкостью Удемса (87 частей 70* этанола,
8 частей ледяной уксусной кислоты, 5 частей глицерина) или Карнуа
(6 частей 100* этанола, 3 части хлороформа, 1 часть ледяной уксусной
кислоты). Это необходимо, чтобы предотвратить подгибание
конечностей под подосому и их повышенную хрупкость при препаровке.
На занятиях следует использовать тотальные микроскопические
препараты взрослых самцов и самок, нимф и личинок. В качестве
заключающей среды применяют канадский бальзам или жидкость
Фора-Берлсзе.
Внешнее строение. Иксодовые клещи имеют мешковидное тело,
у непитавшихся особей сильно уплощенное в дорсовентральном
направлении (рис. 134). У взрослых самок, нимф и личинок покровы
тела весьма эластичны, имеются лишь сильно склеротизированные
спинной и перитремальные щитки. Это позволяет им
многократно увеличивать объем тела при питании. Самцы потребляют мало
пищи и их тело защищено набором сильно хитинизированных щитков.
Рассматривая препараты клешей под бинокуляром. следует обратить
внимание на расположенные по бокам тела перитремальные щитки,
на которых открываются дыхальца (стигмы) трахейной системы.
Роговой аппарат колюше-сосушего типа (рис. 135). Устроен он
сложнее, чем у тстранихид. Гнатосома, которую часто называют
хоботком или головкой, несет специфически преобразованные
хслицеры и педипальпы. Хелицеры имеют подвижные вершины
(пальпы), вооружены режущими поверхностями и многочисленными
загнутыми крючьями, способны выдвигаться вперед и втягиваться
обратно. Производными педипальп являются во рот н и ч о к, г и п о с -
том и пальпы. Воротничок кольцом охватывает основание гнато-
сомы. Снизу от него отходит вырост — гипостом, вооруженный
большим числом шипиков. Сверху воротничок снабжен парой футляров
(влагалищ), в обычном состоянии скрывающих основания хелицер.
По бокам хслицеры несут четырехчлениковые щупики (пальпы).
При питании клещ пробивает покровы тела животного-хозяина
режущими поверхностями вершин хелицер и вводит ротовые части,
находящиеся в сжатом состоянии. После их расхождения он надежно зая-
коривается на хозяине. Необходимость преодоления зачастую весьма
плотных защитных покровов хозяина заставляет клещей искать
участки тела с более нежной кожей, например ушные раковины, веки,
Класс Д/эс/шй
259
Рпс. 134. Внешнее строение клеща Ixodes risinur. самца (А — вид сверху,
5— вид снизу); самки (£— иепитавшаяся. вид сверху; Г—
напитавшаяся, вид сверху; Д — слегка напитавшаяся, вид снизу);
нимфа (£ — вид сверху, Ж — вид снизу); личинка (3 — вид
сверху. Я— вид снизу); /— гапостом; 2— педипальлы; J— прспе-
нктальный щкток; 4— половое отверстие; 5 — перитремы с
дыхальцами (стигмами); б — срединный щкток; 7— аданальный
щиток; 8— анальный щкток; Р— анадыюс отверстие; /0— елкнной
щиток; II — краевое поле; /2— анальная бороздка
260
Тип Arthropods
А Б
Рис. 135. Строение ротового аппарата иксодовых клешей: А — вид со
спинной стороны; Б — вид с брюшной стороны; / — гипостом;
2 — днетальн ый членик хелицеры; 3 — пальла; 4 — футляр для
хелицер; 5 — воротничок
Рис. 136. Строение ходильных
конечностей иксодовых клешей: / —
тазик; 2— вертлуг; J — бедро;
4 — калено; 5 — голень; 6 —
лапка
подбородок, подмышечные и паховые области. Собачьи клещи в норме
питаются однократно на каждой стадии развития, после чего
приступают к подготовке линьки или откладке яиц. Питание иксодовых
клещей является сложным и длительным (многочасовым и более)
процессом и не сводится к простому поглощению пищи: в течение эпгого
периода интенсивно протекают рост и развитие организма. Попытки
механическим усилием удалтъ эктопаразита с места прикрепления
часто оканчивается разрывом тела. При этом гнатосома остается в
тканях хозяина, что может повлечь для него самые неприятные
последствия (от образования нарыва до заражения крови).
Класс Arachnids
261
Конечности иксодовых и тетраниховых клещей имеют схожее
строение (рис. 136). Личинки обладают тремя ларами ходильных
конечностей, нимфы — четырьмя. Развитие иксодид протекает
аналогичным образом, однако имеется лишь одна нимфальная стация. Не
имея глаз, клеши взбираются на возвышенные или выступающие
предметы (вершины былинок, веточек кустарников, выступающих над
открытыми участками ветвей деревьев) и неподвижно располагаются
с вытянутыми вперед передними конечностями в ожидании жертв.
Соприкосновение с предметами, фактура поверхности которых
напоминает шерсть, вызывает соответствующую реакцию фиксации этих
животных. Прокормителями личинок Ixodes ricinusобычно
становятся мышевидные грызуны, нимф — зайцы и ежи, тогда как взрослых
особей — копытные, хищные псовые (собаки, волки, лисы), а также
человек. Соответствует этому и распределение клещей по ярусам
растительности: личинки, как правило, не поднимаются выше
травяного яруса, нимфы размещаются на низкорослых кустарниках,
взрослые стерегут жертв на высоких кустарниках и ветвях нижнего яруса
крон. Таким образом, они выбирают тс места, где максимальна
возможность встречи с адекватными видами жертв.
Тип Echinodermata — иглокожие
Иглокожие — единственные представители вторичноротых,
рассматриваемые на занятиях данного практикума. Радиальная симметрия
тела у них вторична, в той или иной степени развиты внутренний
известковый скелет и амбулакральная (воднососудистая)
система. На эти особенности строения иглокожих следует обратить
особое внимание при выполнении лабораторных работ.
Иглокожие являются обитателями морей и весьма требовательны
к уровню солености воды. Содержание их в аквариумах с морской
водой весьма затруднительно. Все это осложняет знакомство с
представителями типа и обеспечение занятий необходимым материалом.
Целесообразно поэтому ограничиться непосредственным ознакомлением
с отдельными особенностями строения представителей двух классов
иглокожих: морских звезд (Asteroidea) и морских ежей (Echinoidea).
Класс Asteroidea — морские звезды
Материал. Классическим объектом для знакомства с
представителями таксона является морская звезда Asterias rubens—обычный
обитатель морских мелководий Северной Атлантики, Баренцева и Белого
морей. Ближе к берегу эти животные подходят в середине лета. Размах
лучей крупных экземпляров может достигать 30 см; окраска, вопреки
названию, разнообразна, но ограничивается красно-коричневой гаммой.
В дальневосточных морях достаточно обычны имеющие чаше всего
фиолетовую окраску Asterias amurensis, а также образующие крупные
скопления патирии (Patina). Последних можно использовать для
ознакомления с особенностями внешнего строения морских звезд, но они
малопригодны для анатомирования в учебных целях.
Морских звезд следует хранить в 70* этаноле. Хранение в
формалине ведет к разрушению скелетных элементов, плохо в нем
сохраняется и большинство внутренних органов. Собранных животных
погружают целиком в этанол указанной крепости. Затем на крупных
экземплярах следует сделать надрезы и выпустить полостную
жидкость, заменив ее этанолом. Важно проследить, чтобы в итоге
крепость спирта осталась на уровне, обеспечивающем сохранность объектов.
Внешнее строение. Тело Asterias rubens уплощено в дорсовен-
тральном направлении и очертаниями повторяет пятиконечную звезду
(рис. 137, А). Выделяются центральный диск и пять отходящих от
него лучей, называемых также руками. Более широкие у основания,
Класс Asteroidea
263
Рис. 137. Внешнее строение морских знеэп: Asterias (А) и Patina (£); вид
с аборалыюй стороны: / — центральный диск; 2 — лучи (руки);
J — радиусы; 4— интеррадиусы; 5 — мадрепоровая пластинка
лучи сужаются к вершинам. У крупных экземпляров они зачастую
бывают различной длины, что свидетельствует об имевших место травмах,
компенсированных благодаря высокой регснсрационной способности
этих животных. Для облегчения описания морфологии иглокожих
используют понятия интеррадиусов и радиусов. Радиусы — это линии,
соответствующие лучам симметрии морской звезды, интеррадиусы
соединяют центр диска с точками соединения оснований смежных
лучей.
Ротовое отверстие расположено в центре диска с нижней
брюшной стороны тела. Последняя уплощена и носит название оральной
(ротовой), или амбулакральной. На ней вдоль осевой линии каждого
луча от основания до вершины проходитамбулакральная бороздка.
264
ImEchinodermate
несущая четыре ряда амбулакральных ножек — цилиндрических, со
сочковидных выростов. С помощью ручной лупы нетрудно
рассмотреть, что их вершины снабжены небольшими дисковндными
присосками. Эта ножки отличаются от остальных участков
амбулакральной поверхности окраской, тонкими покровами и легко прогибаются
при нажатии препаровальной иглой. Именное их помощью морские
звезды передвигаются по дну и осуществляют различные
манипуляции лучами. На вершине каждой амбулакральной бороздки расположена
непарная чувствительная амбулакральная ножка, снабженная
химическими рецепторами и несущая у основания светочувствительное
пигментное пятно — глазок. Обнаружить его у фиксированных
экземпляров достаточно трудно.
Верхняя, аборальная, или антиамбулакральная сторона
тела гораздо более выпуклая, нежели оральная. В центре диска располо-
жено с трудом различимое поровидное анальное отверстие. Водном
из интеррадиусов имеется небольшая ситовидная, или мадрспо-
ровая пласти н ка, которая названа так из-за сходства в строении
с фрагментами скелета мадрепоровых кораллов. Она выполняет
функцию своеобразного керамического фильтра, через который
морская вода поступает в амбулакральную систему животного. Мадрспо-
ровую пластинку следует рассмотреть с помощью ручной лупы либо
бинокуляра, а легкое царапание препаровальной иглой дает
представление о механических свойствах этого образования.
Тело морской звезды одето мягкой тонкой кожей. На поверхности
покровов нетрудно обнаружить образования трех типов:
выступающие из кожи тупые известковые иглы и пинцстовидные либо
клювовидные педицеллярии, а также конические тонкостенные выросты —
папулы, выполняющие функции кожных жабр. Педицеллярии
имеются только у педицелляриевых звезд, к которым и принадлежит
Asterias rubens. Животное использует их главным образом для очистки
поверхности тела от загрязнения оседающими на нее частицами.
Под кожей морской звезды залегает сплошным слоем наборный
пластинчатый известковый скелет. Такое его строение обеспечивает
свободу движений лучей животного. Скелет носит название
наружного кожного, но имеет мезодермальное происхождение. В лучах Asterias
rubens с оральной стороны расположение скелетных пластинок строго
упорядочено (рис. 138), чего нельзя сказать о центральном диске либо
обо всем теле многих других представителей класса, например патирий.
Вдоль срединной оси каждой руки, образуя дно амбулакральной
бороздки, проходят два ряда амбулакральных скелетных пластинок.
KmccAsteroidea
265
Ршс. 138. Вскрытая морская звезда Asierios rubenr. I — амбулакральные
пластинки; 2 — маргинальные пластинки; 3 — печеночные
придатки (мешки); 4— гонады; 5— оральный отдел желудка;
6—лборалышй отлет желудка; 7— ректальные железы; <Р— фраг-
мент спинной стенки тела с анальным огиерстием; 9—
каменистый канал; 10 — мускулы-ретракторы желудка; II — участок
кожи с маорепоровой пластинкой; 12 — стенка осевого синуса;
13— патовой столон; 14— половой проток; 15— задняя кишка
От прочих они отличаются еще и тем, что насквозь пронизаны
тонкими поровыми канальцами, соединяющими амбулакральные
ножкисихвнутренними ампулами. По сторонам к рааам
амбулакральных примыкает по ряду адамбулакральных пластинок. Далее
266
Тип Echinodermata
следует по ряду интеррадиальных пластинок и по два ряда
нижних маргинальных (краевых) пластинок. В интеррадиусах по краям
ротового отверстия расположены так называемые зубы,
представляющие собой заостренные отростки краевых адамбулакральных
пластинок. Несмотря на подобное название, их роль в обработке пиши
незначительна. Скелет аборальной стороны луча образован либо
немногими рядами скелетных пластинок, либо многочисленными тонкими
известковыми перекладинами. Знакомиться с расположением
скелетных пластинок лучше на сухих крупных экземплярах Asterias rubens,
у которых предварительно с одного из лучей слушены
амбулакральные ножки и кожные покровы. При отсутствии такого материала
скелетные элементы целесообразно рассмотреть после вскрытия
фиксированных морских звезд.
Вскрытие морской звезды. При наличии в достаточном
количестве раздаточного материала весьма желательно, чтобы вскрытие было
выполнено каждым обучающимся. В противном случае проводят
демонстрационное вскрытие либо для знакомства с особенностями
внутреннего строения используют хранящиеся в фиксаторе уже
вскрытые крупные экземпляры.
Отыскав на аборальной стороне тела мадрепоровую пластинку,
начинают вскрытие животного с помощью анатомических ножниц.
Разрез ведут по боковой стороне тела, несколько смещая его к
аборальной. Мадрепоровую пластинку огибают по аборальной стороне,
не достигая центра диска. Центральный участок диска, несущий
анальное отверстие, также лучше обогнуть краевым разрезом. Все операции
легче выполнять, держа животное в руках. Закончив разрез, звезду
помещают в препаровальную ванночку оральной стороной вниз и
закрепляют булавками. Затем осторожно снимают кожные покровы аборальной
стороны тела, отгибая их и подсекая удерживающие связки, в
результате чего открывается полость тела морской звезды.
Внутреннее строение. Описанная техника вскрытия позволяет
вначале ознакомиться с особенностями строения пищеварительной
системы. В пределах центрального диска дорсально по
интеррадиусам залегаютгрозльевидные ректальные железы, являющиеся
выростами задней кишки. Как и часть стенок мешковидного желудка,
они зачастую удаляются вместе с кожными покровами аборальной
стороны тела. Занимающий центральное положение (рис. 138)
объемистый складчатый желудок может содержать непереваренные остатки
пищи. Однако Asterias rubens, как и другие представители семейства
Класс Asteroidea
267
Asteridac, весьма широко использует возможность выворачивать
через ротовое отверстие часть желудка и переваривать пищу, фактически
не заглатывая ее. От желудка в каждый луч отходит непарный проток,
переходящий в протоки парных бурых печеночных придатков
(мешков), которые почти целиком заполняют внутреннее
пространство луча неполовозрелых особей. Основная их функция —
продуцирование пищеварительных ферментов. При постепенном
удалении печеночных желез и желудка можно проследить, как последний
сужается к ротовому отверстию, переходя в короткий пищевод.
При этом следует подсечь мускулы-рстракторы, обеспечивающие
втягивание вывернутой через ротовое отверстие части желудка.
Половая система. Парные перистые гонады расположены в
интеррадиусах, продолжаясь в соответствующие лучи. Их размеры зависят от
степени полового созревания и могут сильно варьировать — от
малозаметных структур в пределах центрального диска до хорошо развитых
образований, заполняющих большую часть полости лучей. В
интеррадиусах на аборальной стороне тела они открываются половыми
отверстиями, практически неразличимыми на фиксированном
материале. Морские звезды раздельнополые животные, но женские
и мужские патовые железы внешне неразличимы. Половая
принадлежность особей может быть установлена только по результатам
исследования продуцируемых половых продуктов.
Амбулакральная (воднососудистая) система начинается мадрепо-
ровой пластинкой, канальцы которой ведут в S-образно извитый
каменистый канал, аходяший восевой комплекс органов.
Последний спускается с аборальной стороны центрального диска к оральной.
Друше компоненты осевого комплекса неразличимы при используемой
технике фиксации и анатомирования объекта. Стенки каменистого
канала обызвествлены, что легко выявить простым нажатием пинцета.
Каменистый канал впадает в кольцевой канал амбулакральной
системы, опоясывающий пищевод. Сам канал обычно неразличим, но
его месторасположение может быть установлено по темно-желтым или
светло-бурым размером со спичечную головкутидемановым
тельцам (железам). Они располагаются в интеррадиусах попарно и
выполняют функции лимфоцитарных органов. Общее число тндемановых
телец равно девяти, поскольку на месте расположения одного из них
в кольцевой канал амбулакральной системы впадает каменистый канал.
Вырабатываемые тидсмановыми тельцами амебоидные клетки
обеспечивают удаление поглощаемых ими продуктов метаболизма и иных
ненужных и опасных для организма соединений, т.е. выполняют
выделительную и защитную функции.
268
ImEchinodermata
Если аккуратно удалить из луча печеночные придатки (при
длительном хранении материала они могут мацерироваться и
разрушаться при прикосновении, превращаясь в кашеобразную массу), то под
ними обнажается внутренняя сторона амбулакральной бороааки. По
ней проходит радиальный канал амбулакральной системы, дающий
ответвления к ее ампулам, располагающимся здесь в четыре ряда.
Они представляют собой продолговатые мешочки с эластичными,
мускулистыми стенками. Воднососудистая система заполнена
жидкостью, основой которой является поступающая через мадрепоровую
пластинку морская вода. Сокращение ампул приводит к растяжению
ножек. Прикрепившись присосками, последние сокращаются, нагнетая
жидкость в ампулы. Одновременно животное подтягивается к месту
прикрепления присосок амбулакральных ножек. В связи с отсутствием
в лучах выраженных пучков мышечных волокон основными
функциями амбулакральной системы являются изменение формы тела
и обеспечение передвижения животных. В некоторой степени водно-
сосудистая система участвует в осуществлении дыхания и выделения.
Нервная система у морских звезд представлена тремя радиаль-
но-симметричными системами, залегающими в теле животного на
разных уровнях. Проще всего ознакомиться с залегающей
поверхностно, непосредственно под кожными покровами аборальной стороны
тела аборальной (апикальной) нервной эндоневральной
системой. Она выявляется при внимательном рассмотрении внутренней
поверхности ранее удаленной аборальной стенки тела в виде пяти
бурых радиальных тяжей, от центра диска уходящих в лучи. Эктонев-
ральная (оральная, адоральная нервная) система обнажается
после полного выщипывания амбулакральных ножек и
представляет собой пять бурых тяжей, идущих в лучи по дну амбулакральных
бороздок от околоротового нервного кольца. Залегающая в толще
тела гипоневральная система при описанной технике фиксации
и препарирования объекта выделена быть не может.
Кровеносная и перигсюльшя системы остаются за рамками
рассмотрения, поскольку требуют иной техники препаровки и анатомирования.
Специальных органов выделения у иглокожих нет. Эту функцию
выполняют амебоидные клетки полостной жидкости.
Органами дыхания служат кожные жабры — короткие
тонкостенные выпячивания стенки тела (папулы), способные к
сокращению. Они могут окружать кольцами скелетные иглы и хорошо видны
при рассмотрении под бинокуляром верхней стенки тела морской
звезды.
Класс Echinoidea
269
Класс Echinoidea — морские ежи
Материал. Наиболее доступным объектом для знакомства с
представителями данного класса являются морские ежи рода Stnwgybcentnxus,
например массовый обитатель литорали Баренцева, Белого и других
северных морей Strongylocenirotusdrvebachiensis. Животные обитают на
глубине свыше 5 м, что затрудняет их добычу. Техника фиксации морских
ежей аналогична описанной выше для морских звезд.
На лабораторных занятиях целесообразно ограничиться
знакомством с внешним видом, строением скелетного панциря и его
игловидных придатков, ротового аппарата и педицеллярий морского ежа.
Для этого используются фиксированный или высушенный материал,
смонтированные на картонках или в застекленных коробках
распиленные панцири ежей, а также постоянные микроскопические
препараты их педицеллярий.
Внешнее строение. Морской еж имеет форму сплюснутого шара
(рис. 139), тело плотно усеяно подвижно сочлененными крупными
обызвсствлснными иглами. Наиболее крупные из них носят название
первичных, более мелкие — вторичных, тогда каксамыс мелкие —
миллиарных.
Ротовой полюс располагается на более уплощенной стороне тела.
Как и морские звезды, морские ежи передвигаются по горизонтальным
поверхностям ротовым полюсом вниз. При рассмотрении
фиксированных экземпляров можно заметить, что расположенный здесь рот
окружает кожистое поле, называемое перистомальным
(ротовым), посередине которого располагаются торчащие наружу острые
Рис. 139. Морской еж Echinocidaris
с сегментом
поверхности, освобожденной от игл:
/— интерамбулакральные
ряды пластинок; 2 —
амбулакральные ряды
пластинок
270
Тип Ectiinodermata
долотообразные зубы. С противоположной стороны в окружении
коротких мелких игл размещается анальное поле (перипрокт).
Несколько в стороне от его центра открывается анальное отверстие,
выявление которого затруднительно. Среди игл расположены короткие
подвижные придатки, несущие на вершинах пинцетоподобные или
клювовидные образования — характерные для иглокожих педицел-
лярии.
Строение скелетного панциря. Скелет морских ежей имеет форму
уплощенной с полюсов сферы (рис. 140), образованной неподвижно
спаянными между собой известковыми пластинками. Выделяются
два основных их типа. Более мелкие амбулакральные пластинки
пронизаны тонкими сквозными отверстиями (эта особенность
строения выявляется при внимательном рассмотрении высушенных панцн-
Рмс. 140. Схема строения скелета морских ежей, вид с оральной
стороны: / — амбулакральные ряды пластинок; 2— интерамбулакраль-
ные ряды пластинок; J— ротовое отверстие; 4— зубы; 5—
оральная мембрана (перистом); 6 — оральные пластинки с порами
Kmcc Echinoides
271
рей); более крупные интерамбулакральные пластинки цельные.
Пластинки каждого типа образуют меридионально располагающиеся
чередующиеся парные ряды, общее их число равно десяти.
Скелетные пластинки имеют форму пятиугольников. Граница
между соседними меридиональными рядами амбулакральных и интср-
амбулакральных (т.е. разноименных) пластинок имеет вид прямой
линии, тогда как между одноименных — зигзагообразной. Принято
считать, что ряды амбулакральных пластинок соответствуют радиусам,
а интсрамбулакральных — интеррадиусам. Таким образом, здесь
выделяются все те же пять радиусов и пять интеррадиусов.
Каждая пара рядов амбулакральных пластинок заканчивается
у анального полюса генитальной пластинкой, несушей наружное
половое отверстие. Всего таких пластинок четыре, так как пятая,
имеющая несколько более крупные размеры и пронизанная
множеством мелких отверстий, представляет собой мадрепоровую
пластинку. Генитальныс пластинки чередуются с более мелкими
глазными (окулярными), каждая из которых пронизана отверстием
специализированной амбулакральной ножки, несущей глазок.
Прочие амбулакральные пластинки несут чстырс-пять пар отверстий; их
считают сложными, то есть образованными в результате слияния
соответствующего количества простых пластинок.
Наружная поверхность панциря усеяна множеством бугорков,
к которым шарнирно причленяются известковые иглы.
Строение ротового аппарата. На оральной стороне тела наружу
выступают пять радиально-симметрично расположенных острых долото-
видных зубов. Они являются частью сложно устроенного
жевательного аппарата, который называется аристотелевым фонарем. В его
состав входит 25 известковых скелетных элементов (перекладин и
пластинок), подвижно сочлененных как между собой, так и с остальным
панцирем и снабженных мускулатурой. Долотовидные з><5ы являются
видимой снаружи частью аристотелева фонаря, а просвет между
зубами — ротовым отверстием. Ежи рода Strongylocentrotus питаются
живыми обрастаниями каменистых поверхностей (водорослями и пр.),
соскребая и измельчая их с помощью жевательного аппарата.
Строение педицеллярмй морского ежа. Педицеллярии разбросаны
между иглами но всему телу морского ежа. По своему происхождению
они являются модифицированными иглами. Псдицеллярия состоит
(рис. 141) из подвижного стебелька, снабженного известковой опорной
осью, гибкой шейки и головки, образованной тремя смыкающимися
272
TmEchinodermata
Рис. 141. Псдиисллярии морского ежа Strongyhcentrotus droebochitnsis:
А — шароносная; 5— змееголовая; В— трилистная; Г—
трезубая; / — стебелек (стержень); 2 — шейка; 3 — головка; 4 —
створки
створками, работающими наподобие щипцов или пинцета. Со
строением этих образований можно ознакомиться, воспользовавшись
соответствующими постоянными микроскопическими препаратами.
Основное назначение педиисллярий — очистка поверхности тела,
промежутков между иглами от экскрементов, оседающих частиц ила
и других загрязнений. Псдиисллярии могут захватывать прикоснувшихся
к ним мелких животных, умерщвлять их и передавать, как в эстафете,
другим педицелляриям и далее к ротовому отверстию. Кроме того они,
могут травмировать мягкие ткани врагов морских ежей и отрывать
псдиисллярии хищных морских звезд. У ряда представителей класса
есть специализированные ядовитые псдиисллярии, снабженные
ядовитыми железами, которые выполняют главным образом
защитные функции.