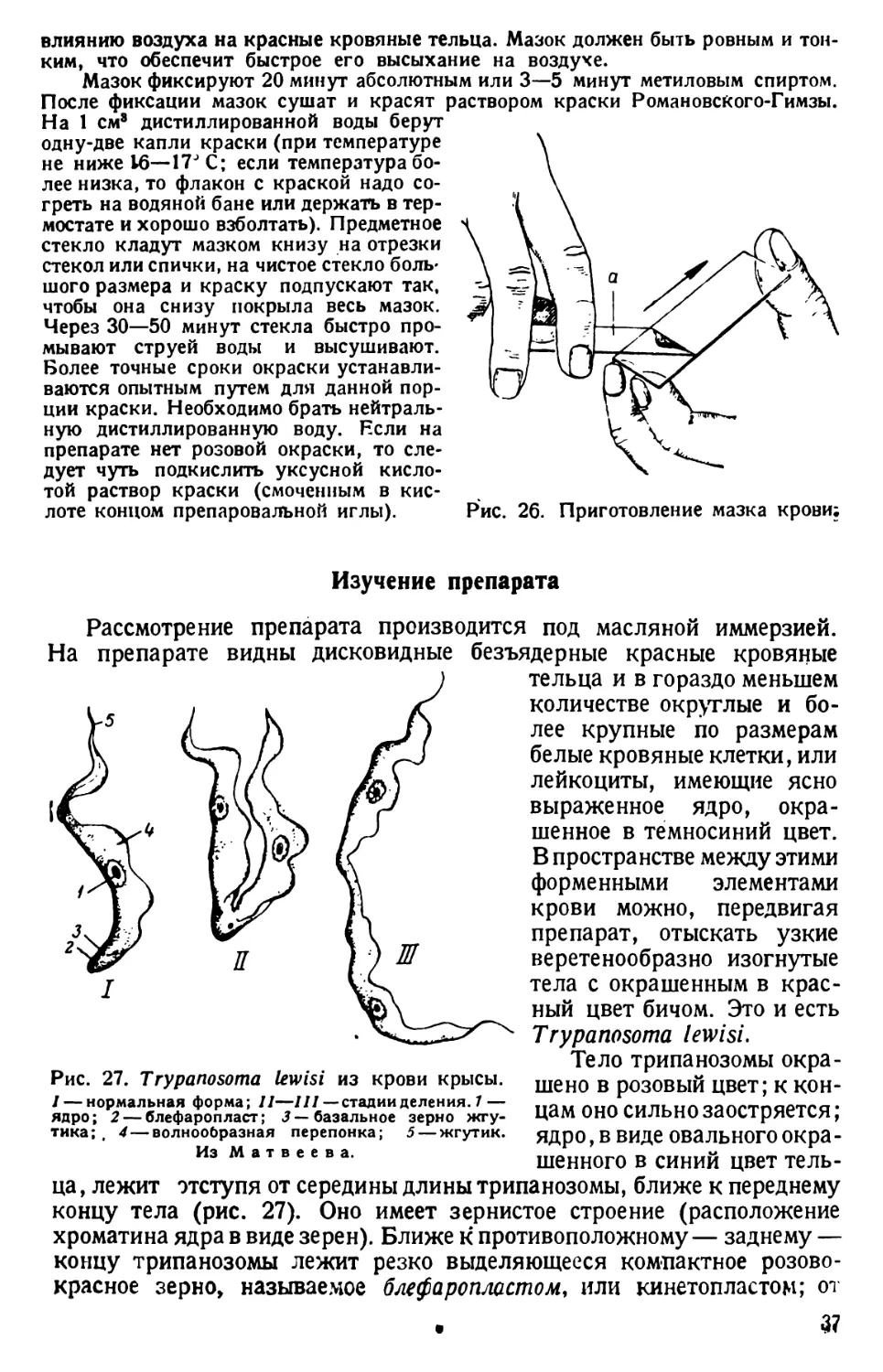
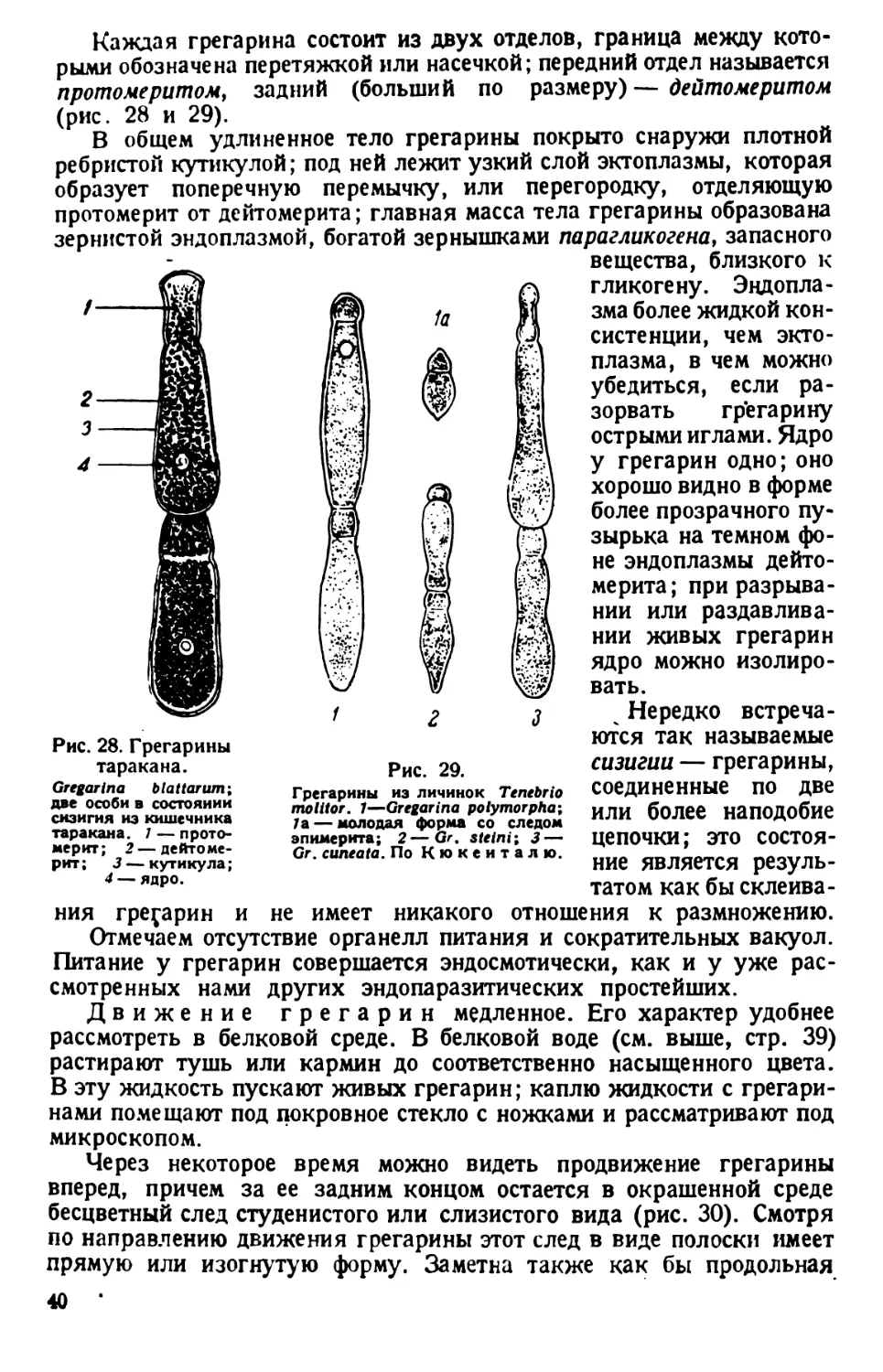
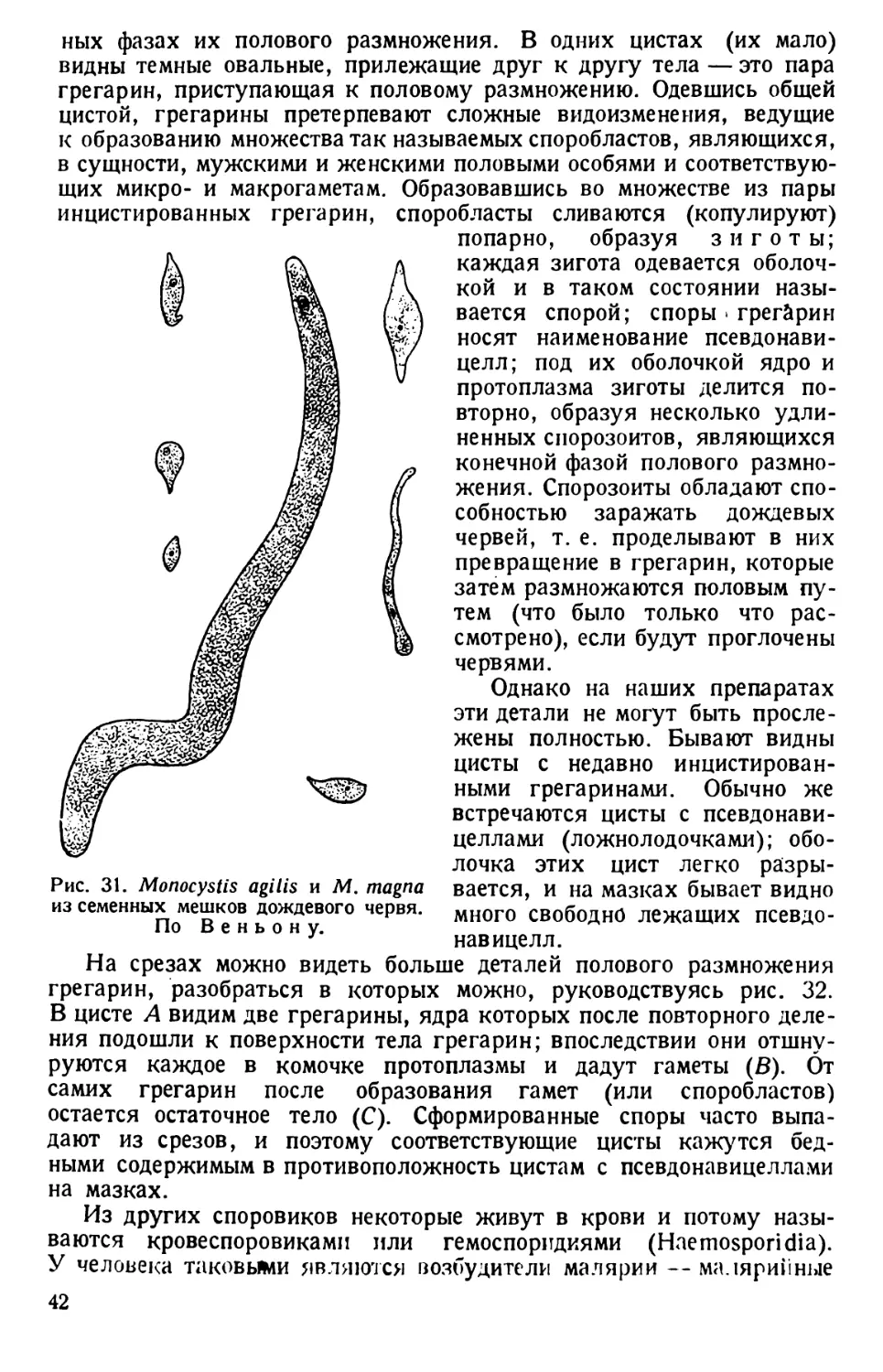
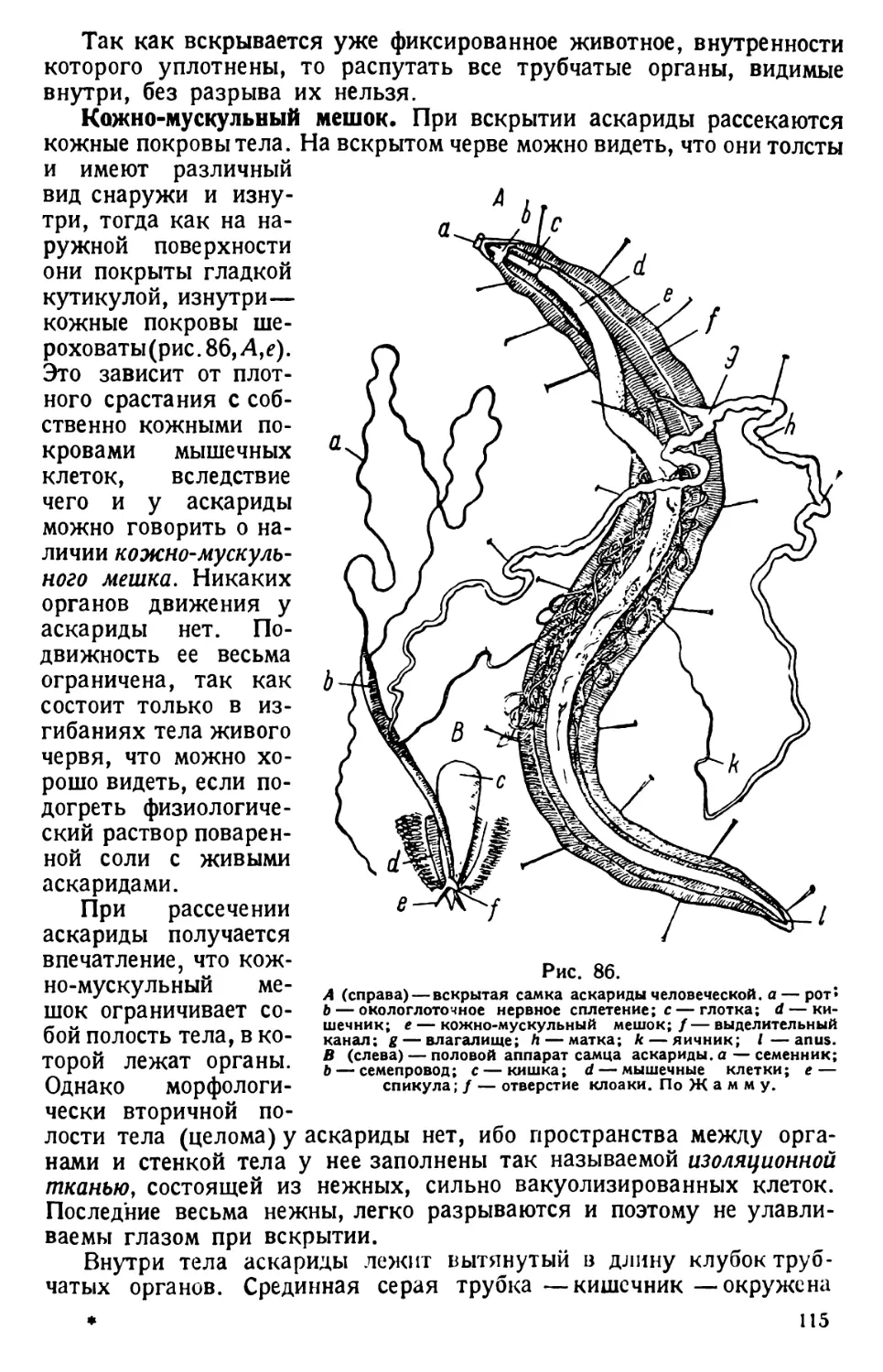
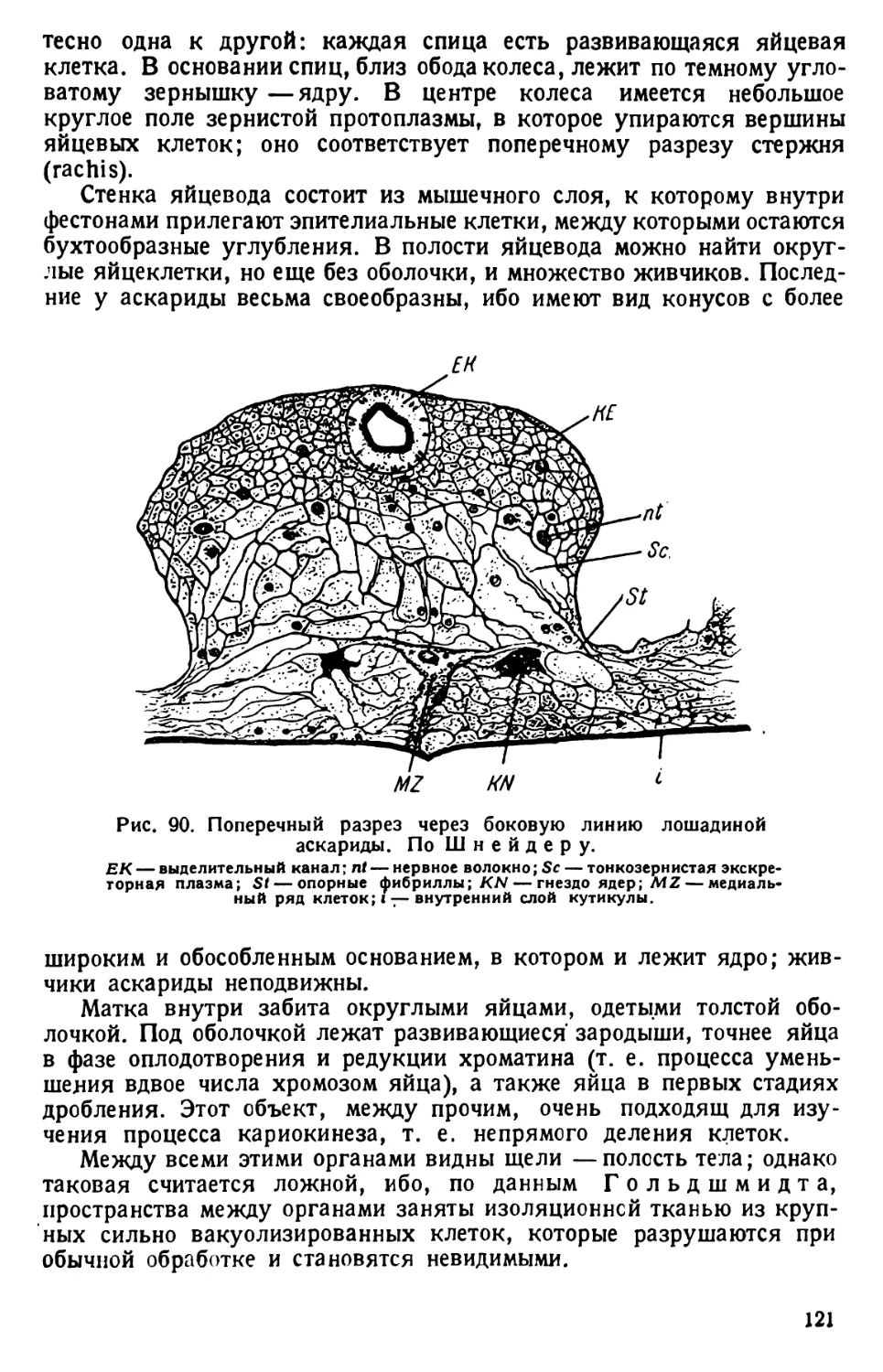
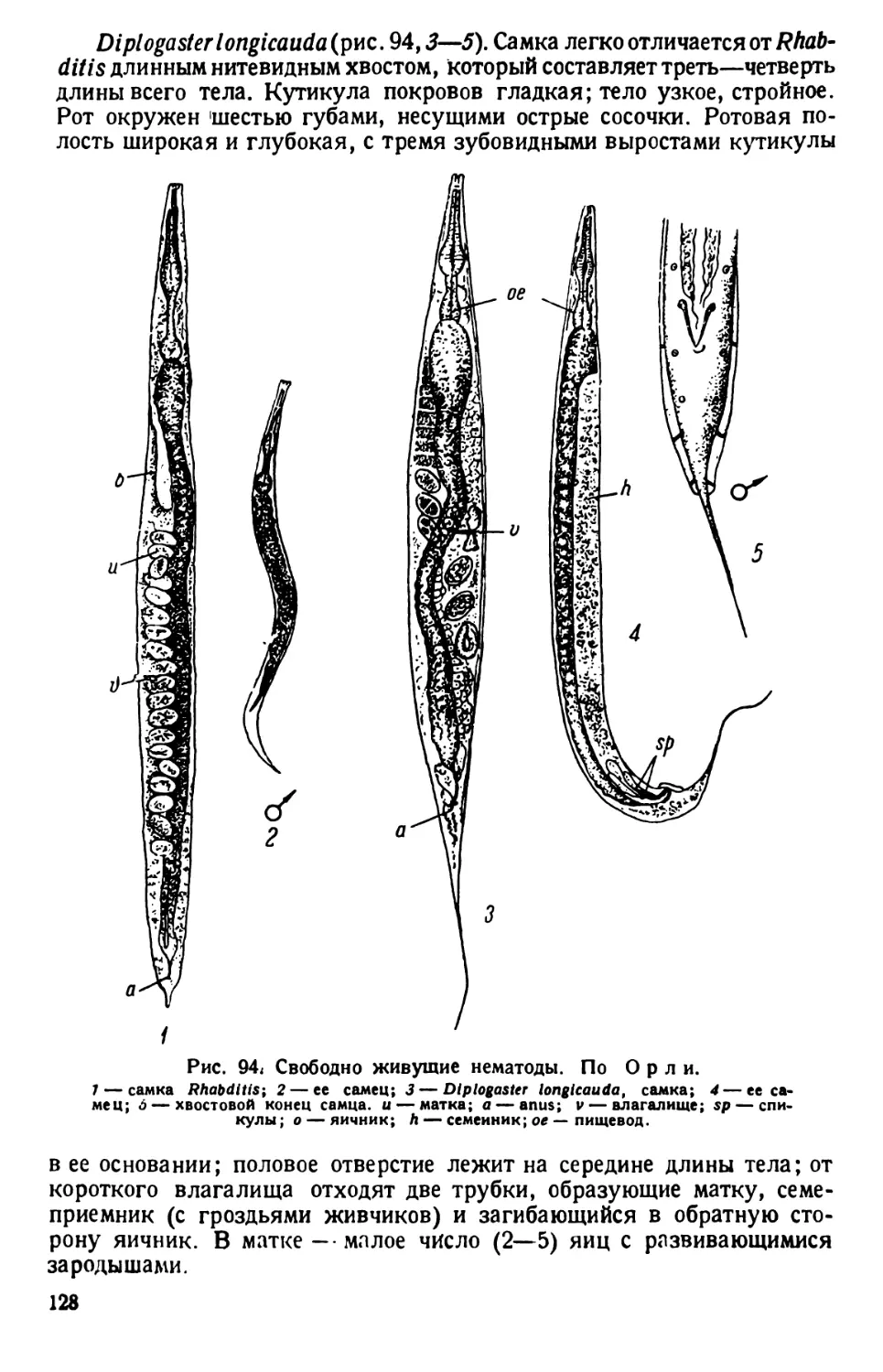
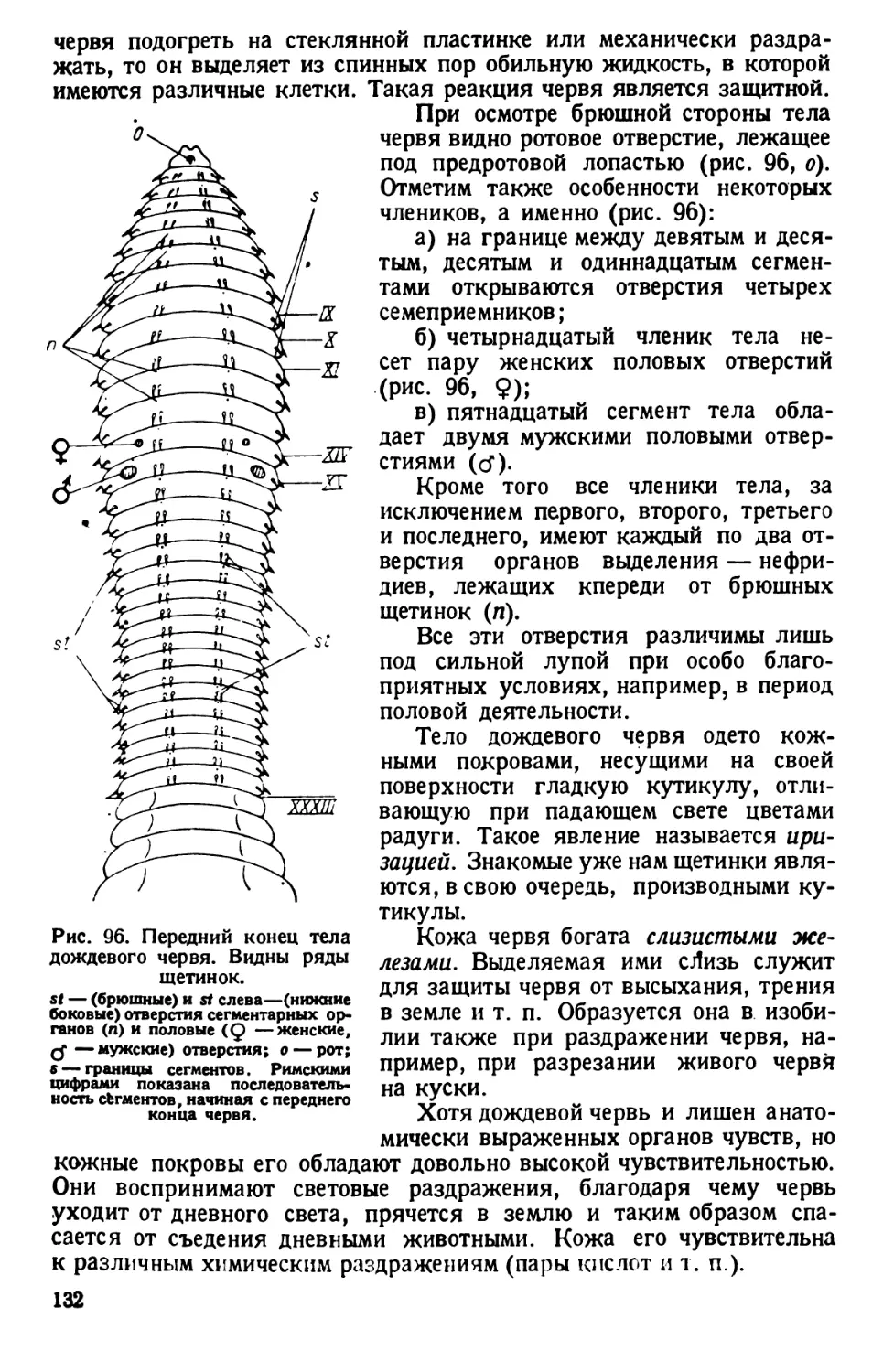
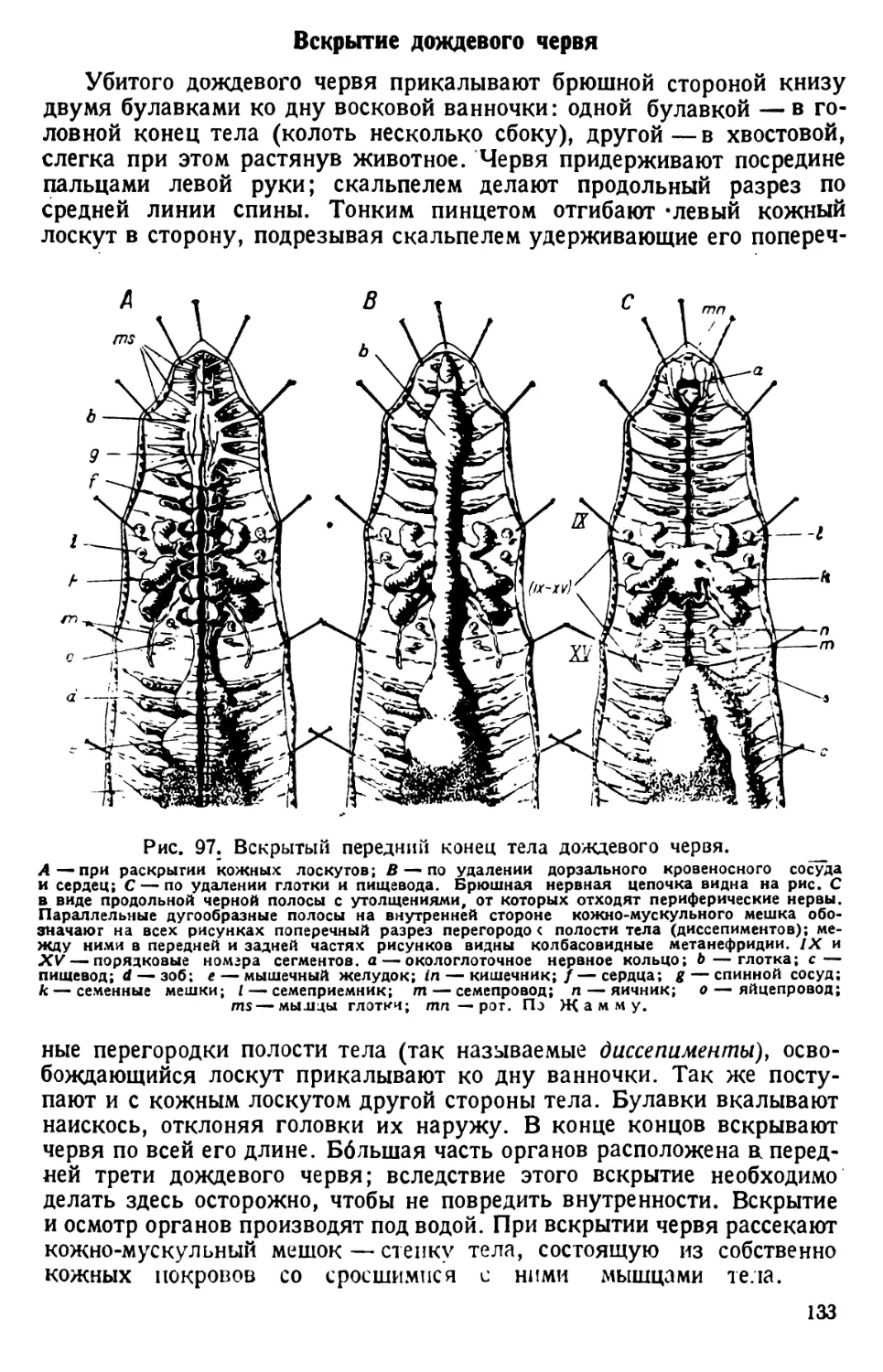
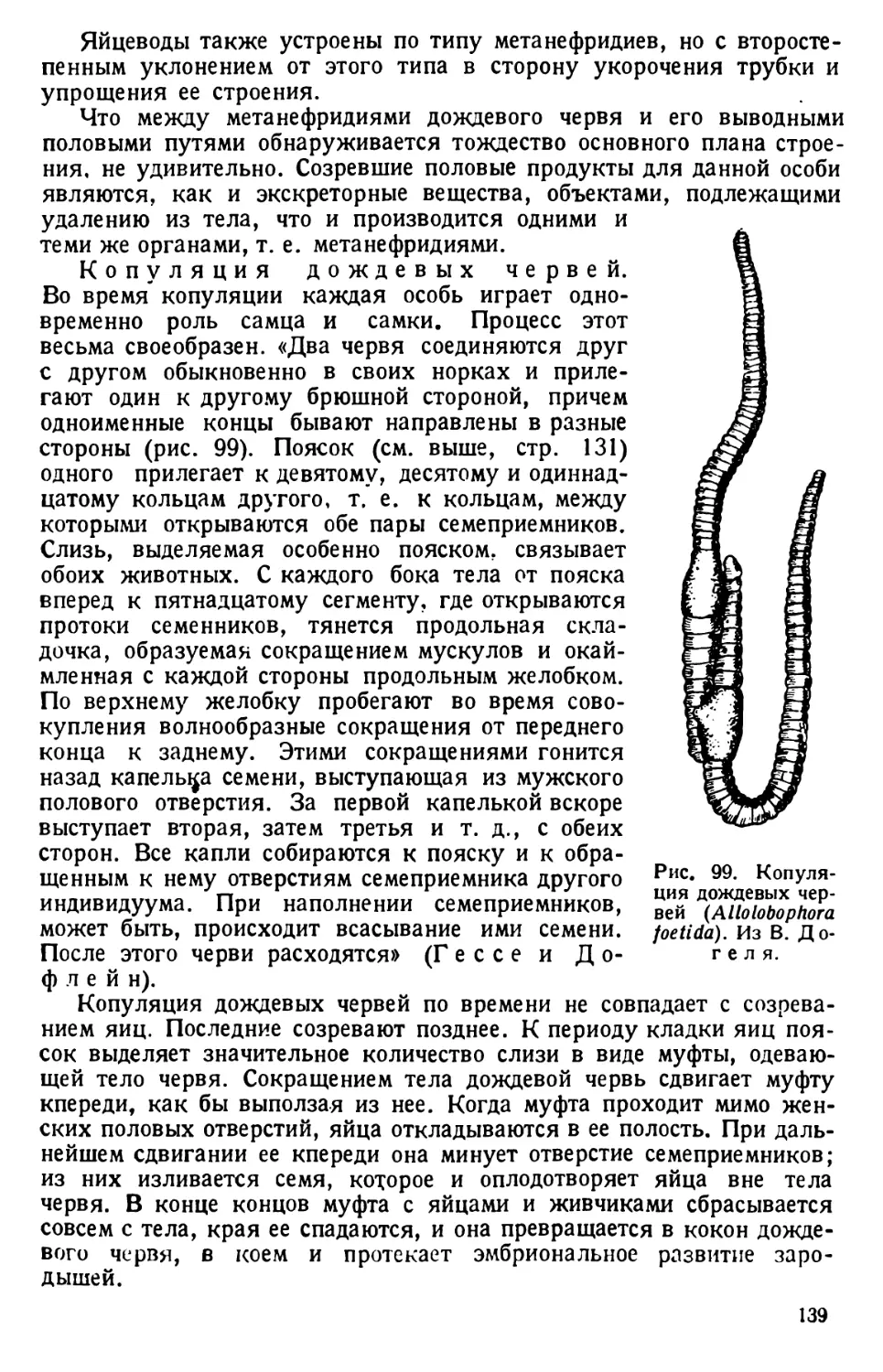
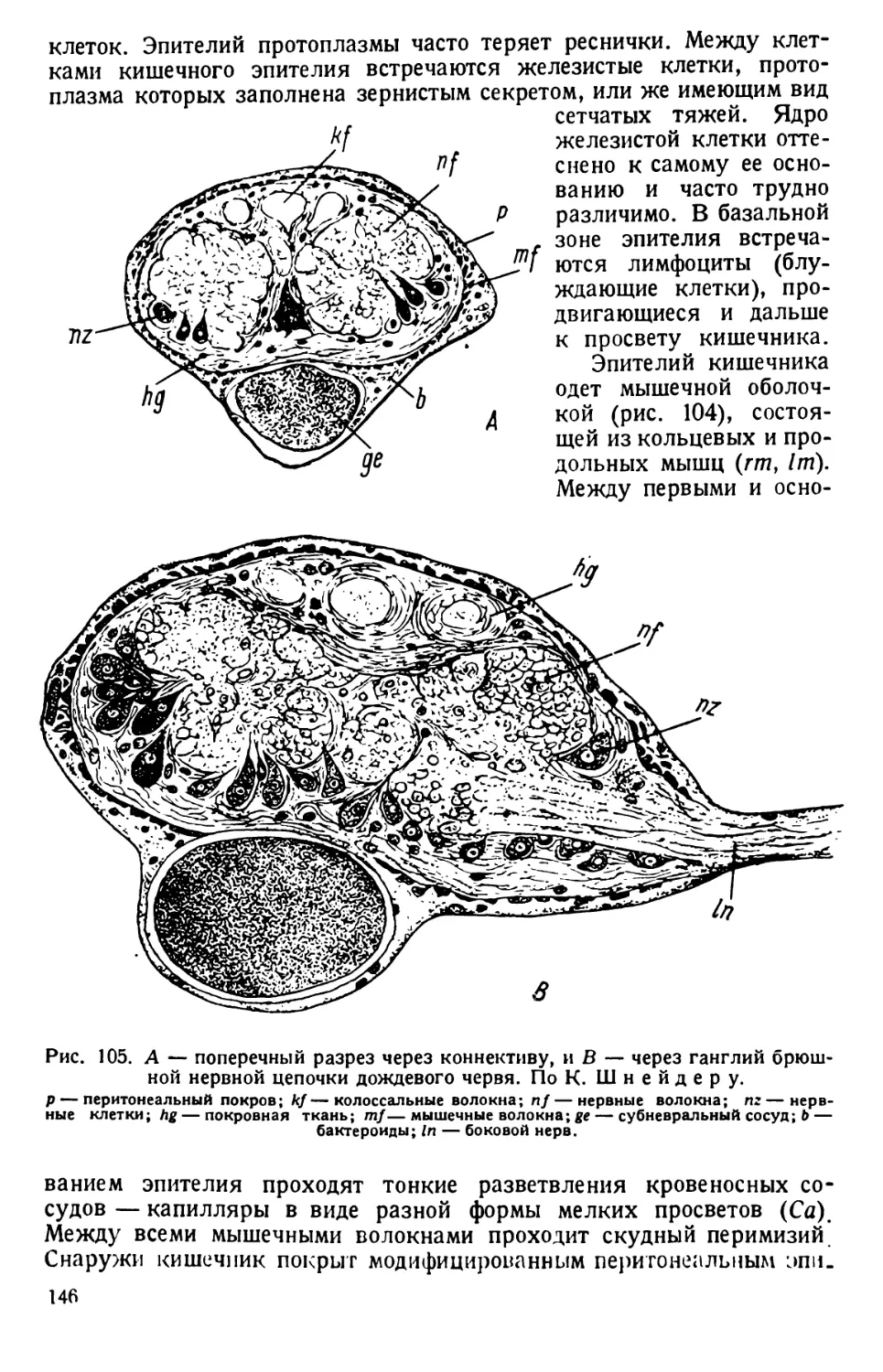
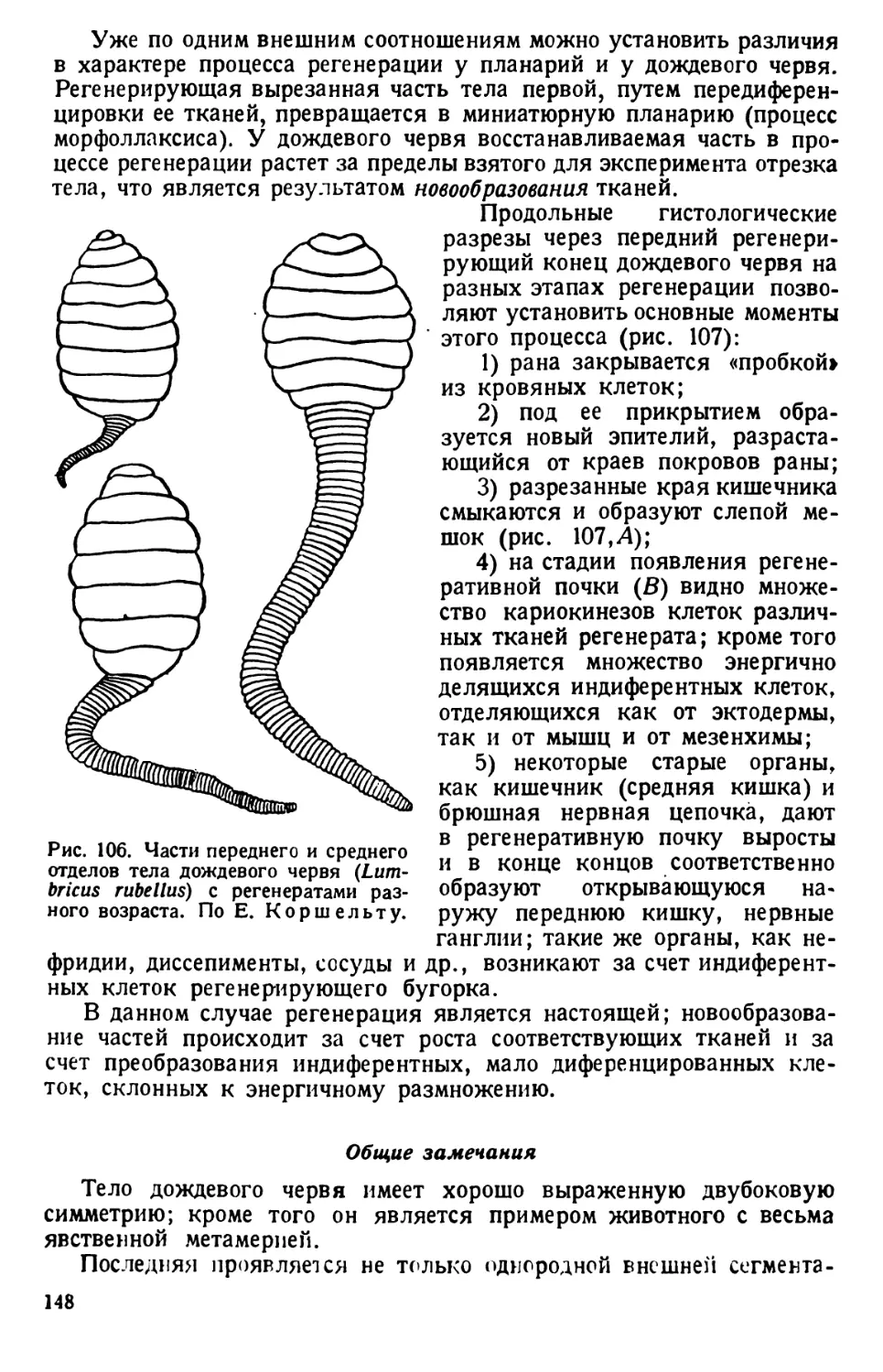
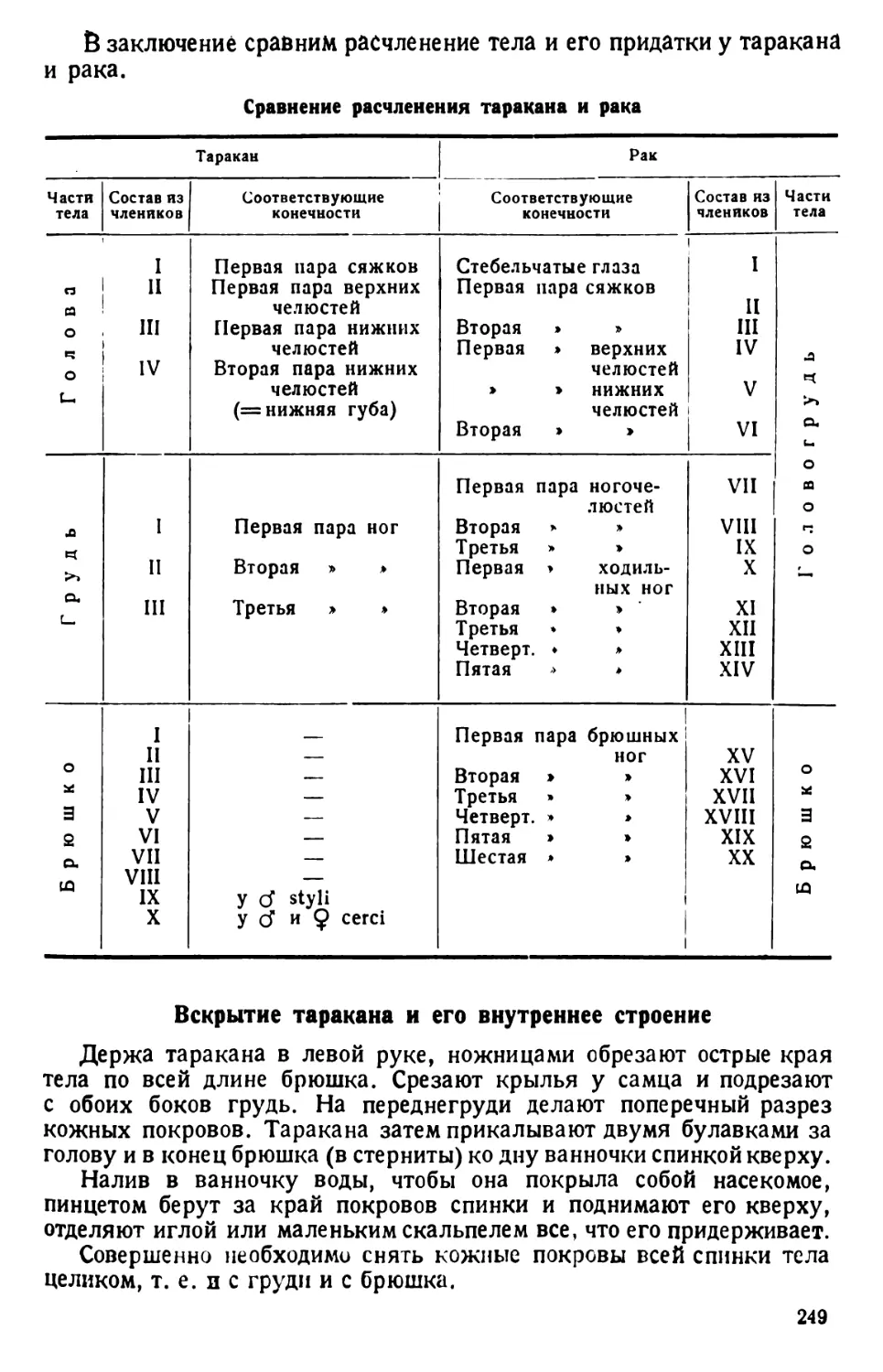
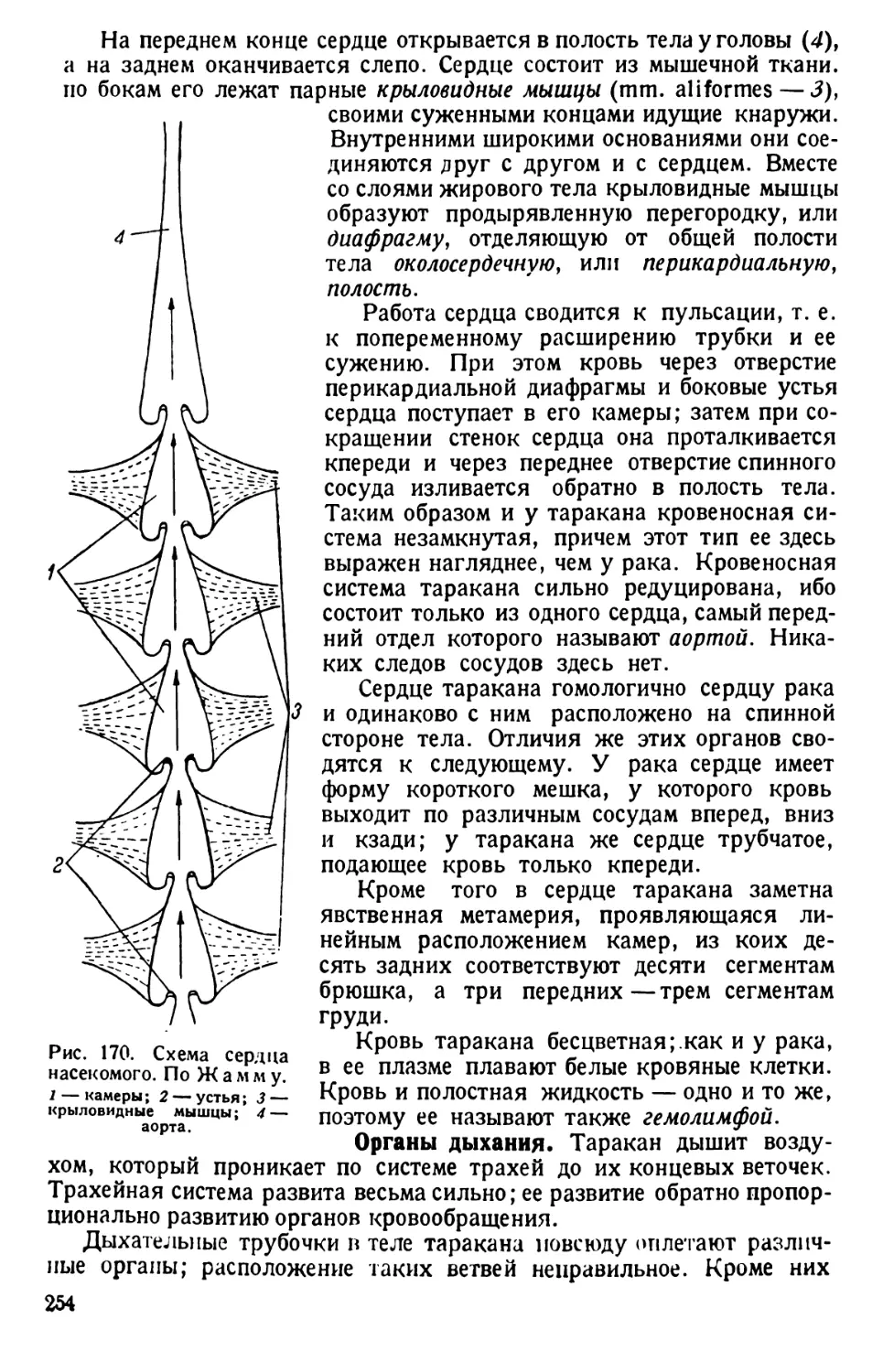
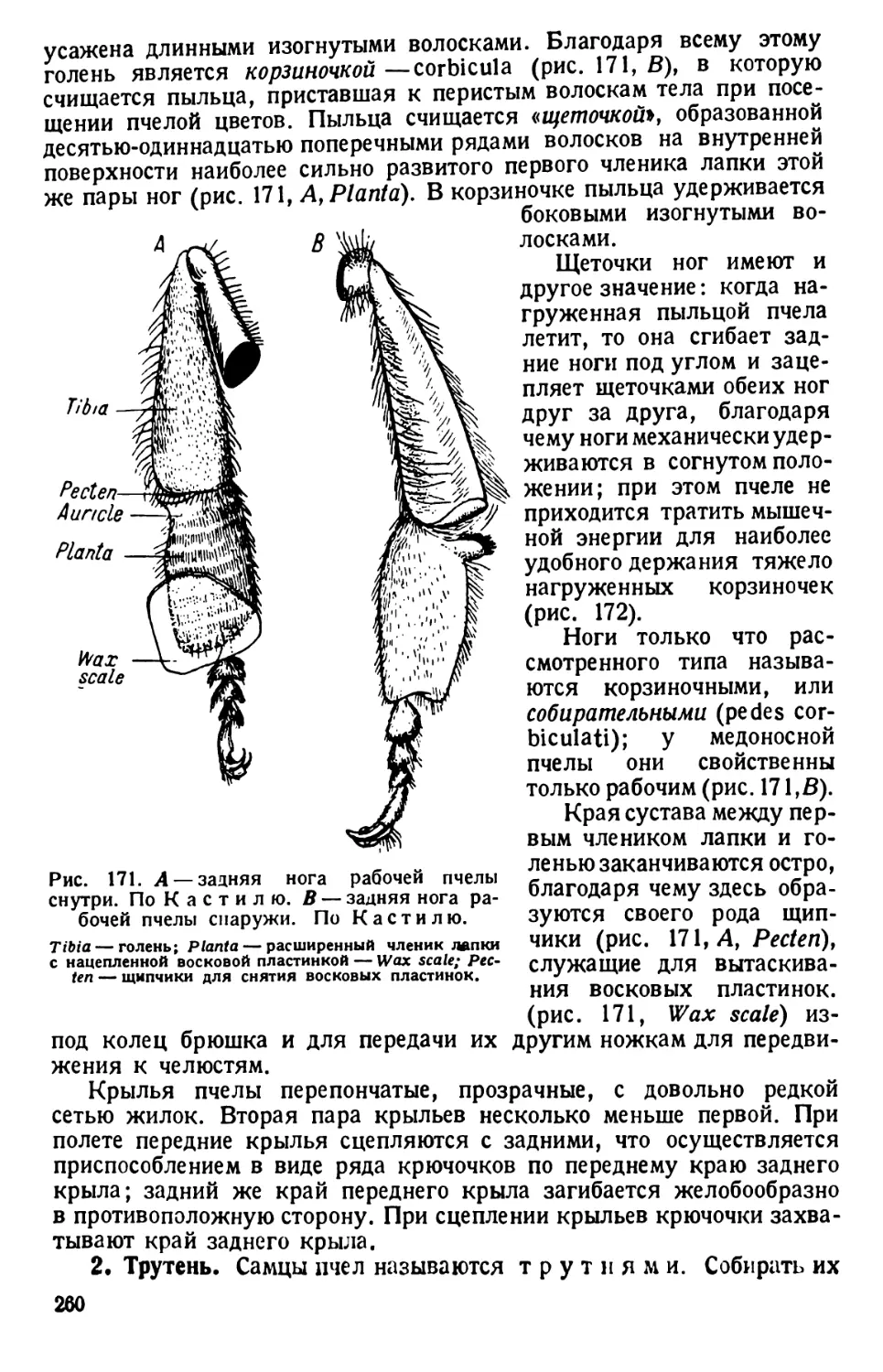
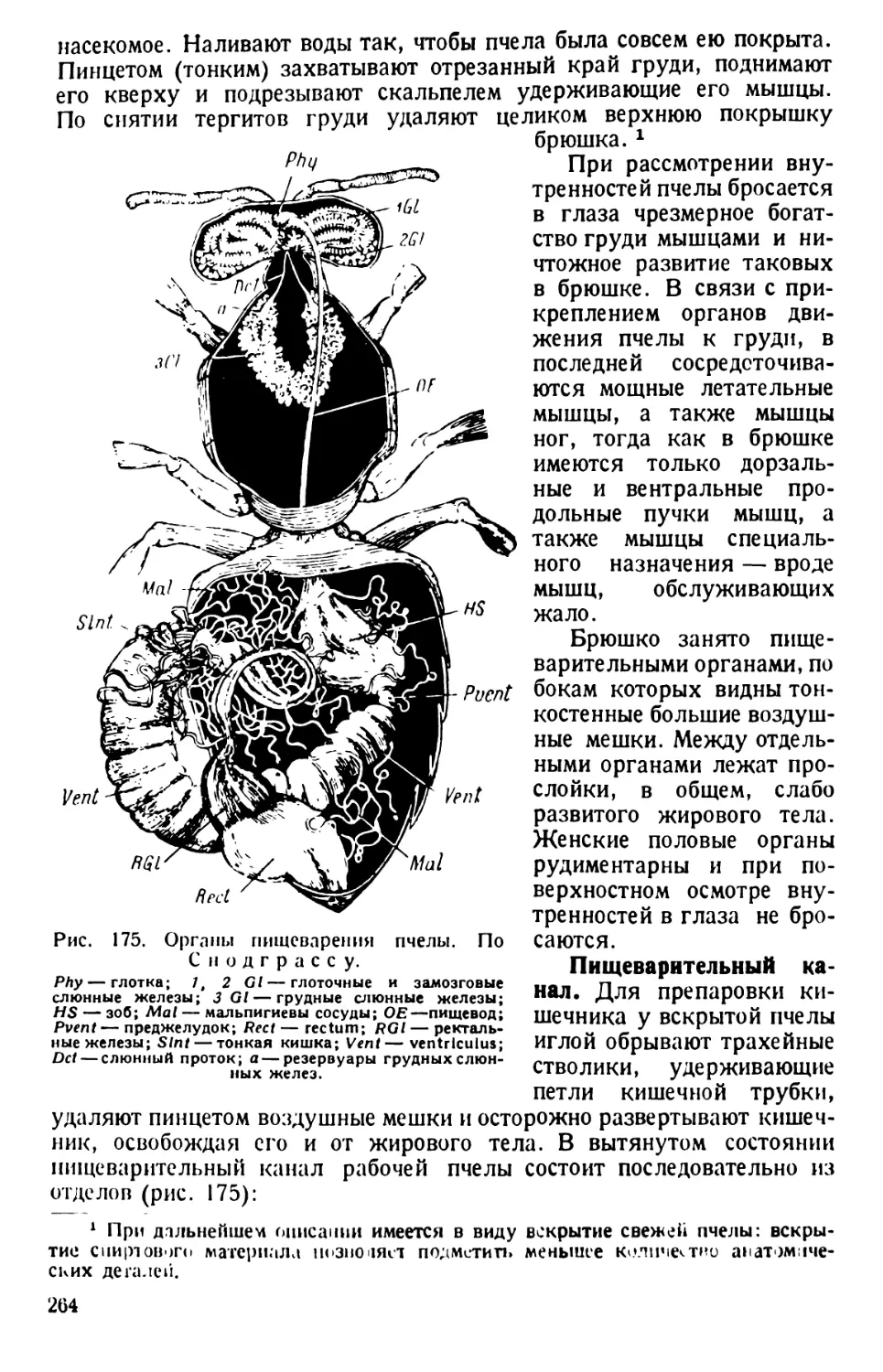
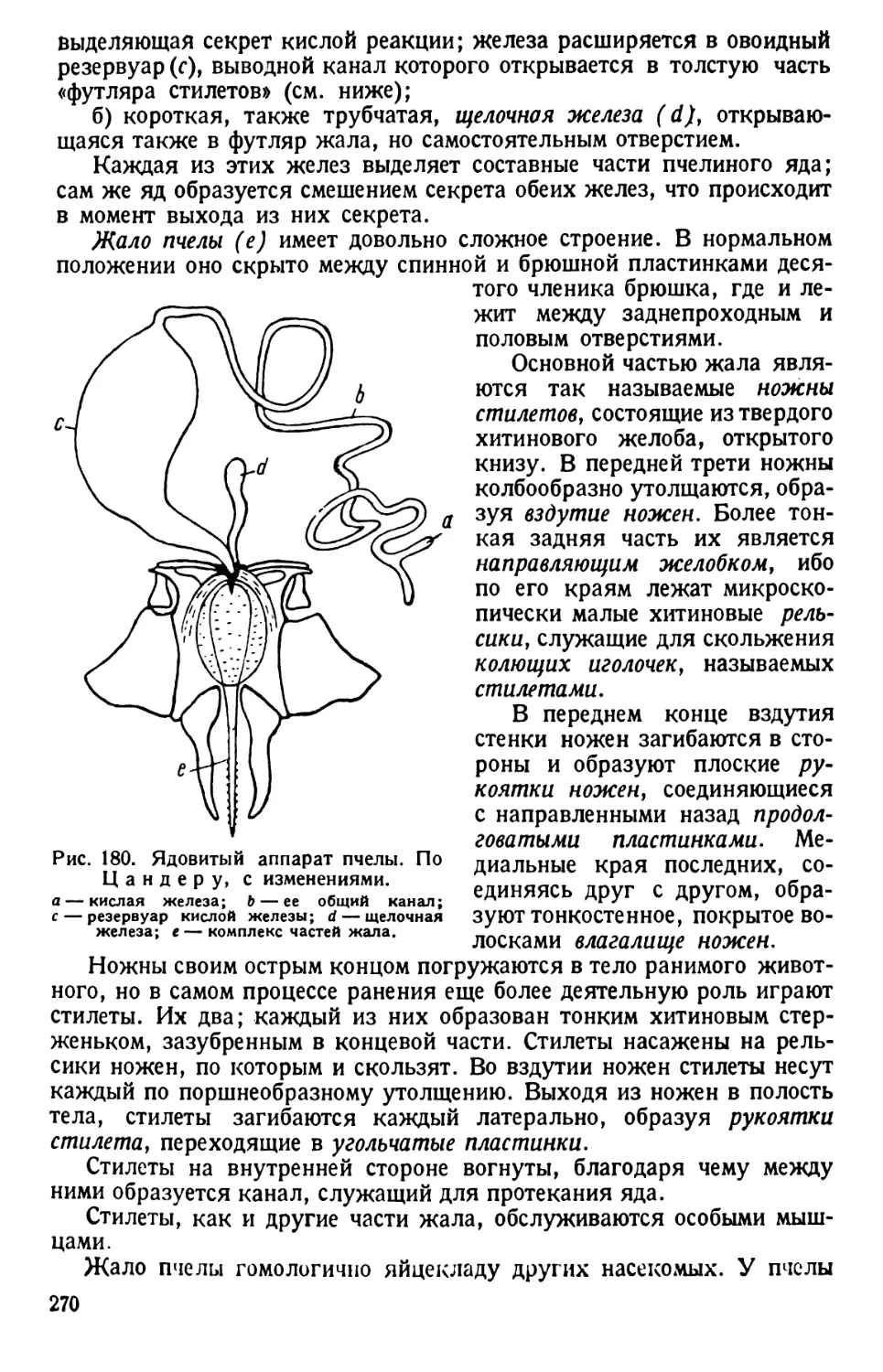
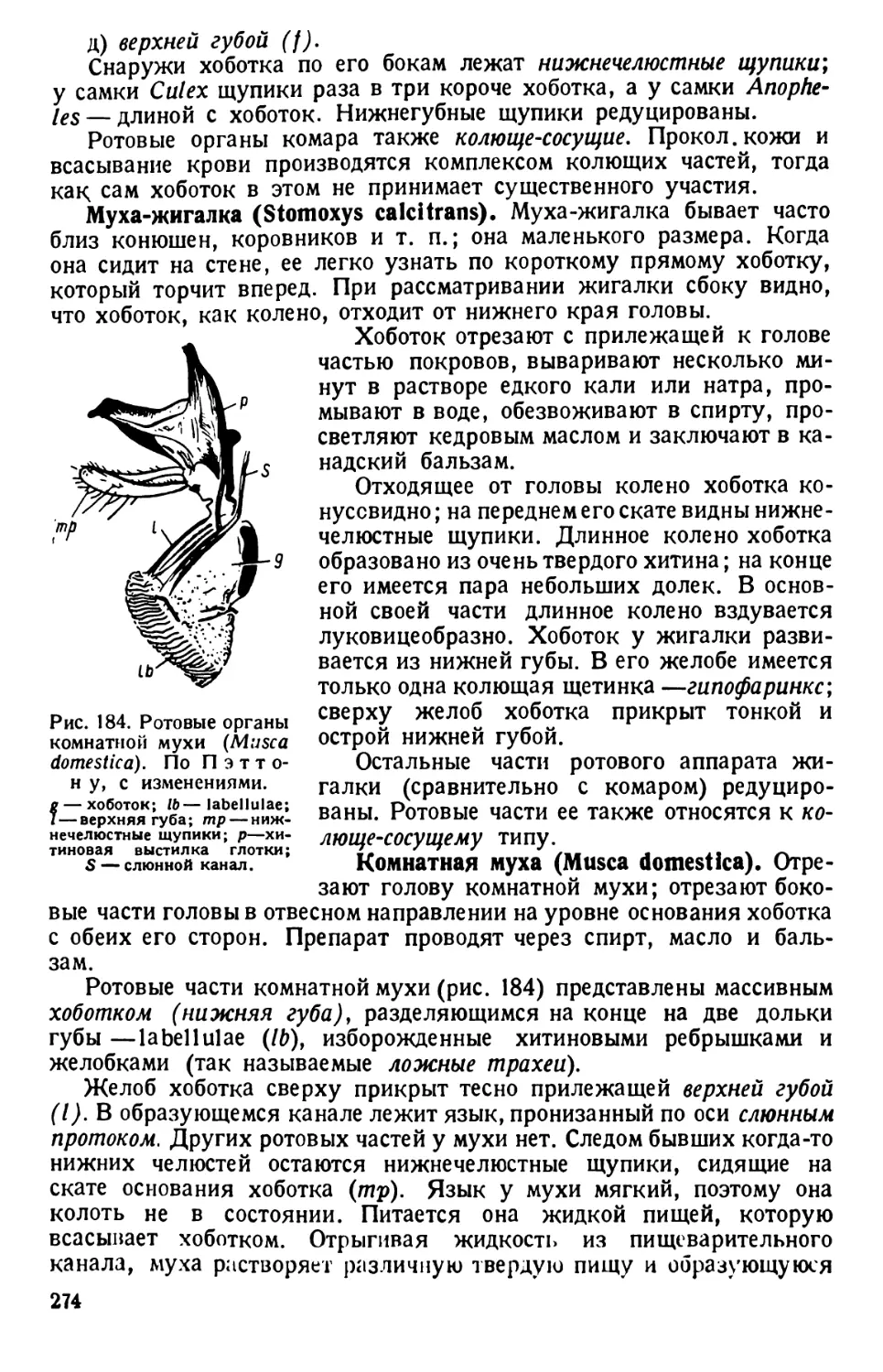
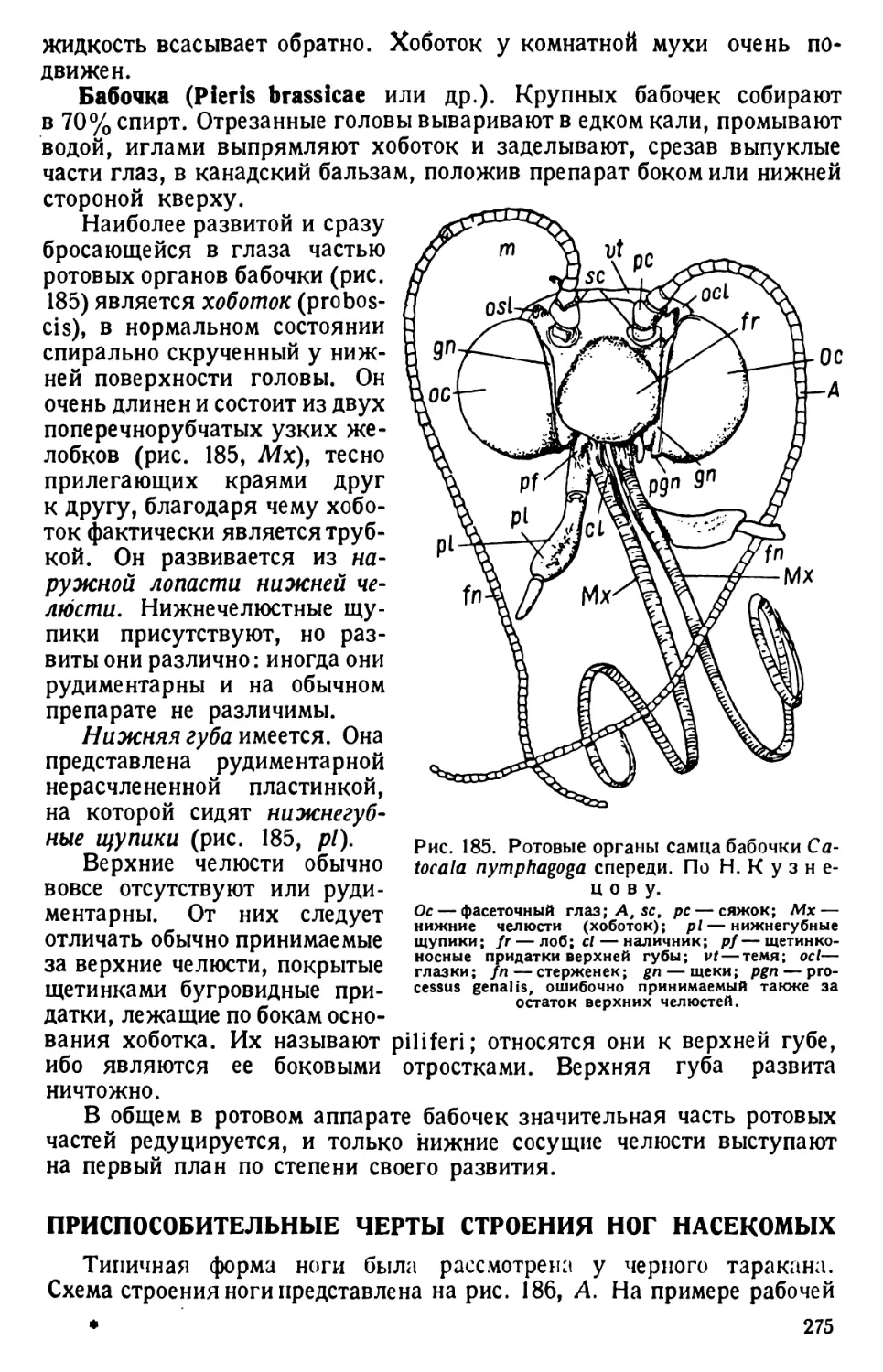
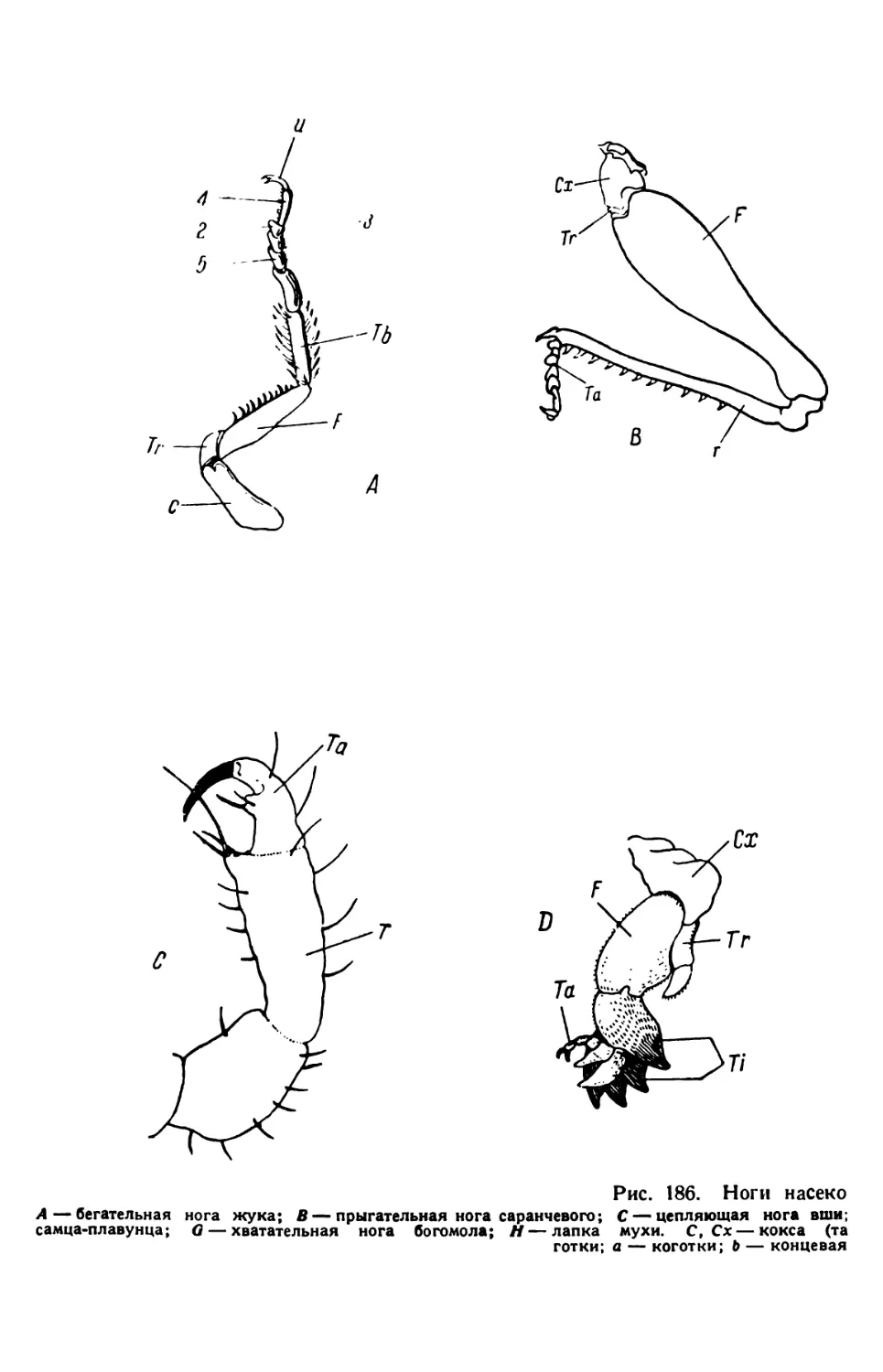
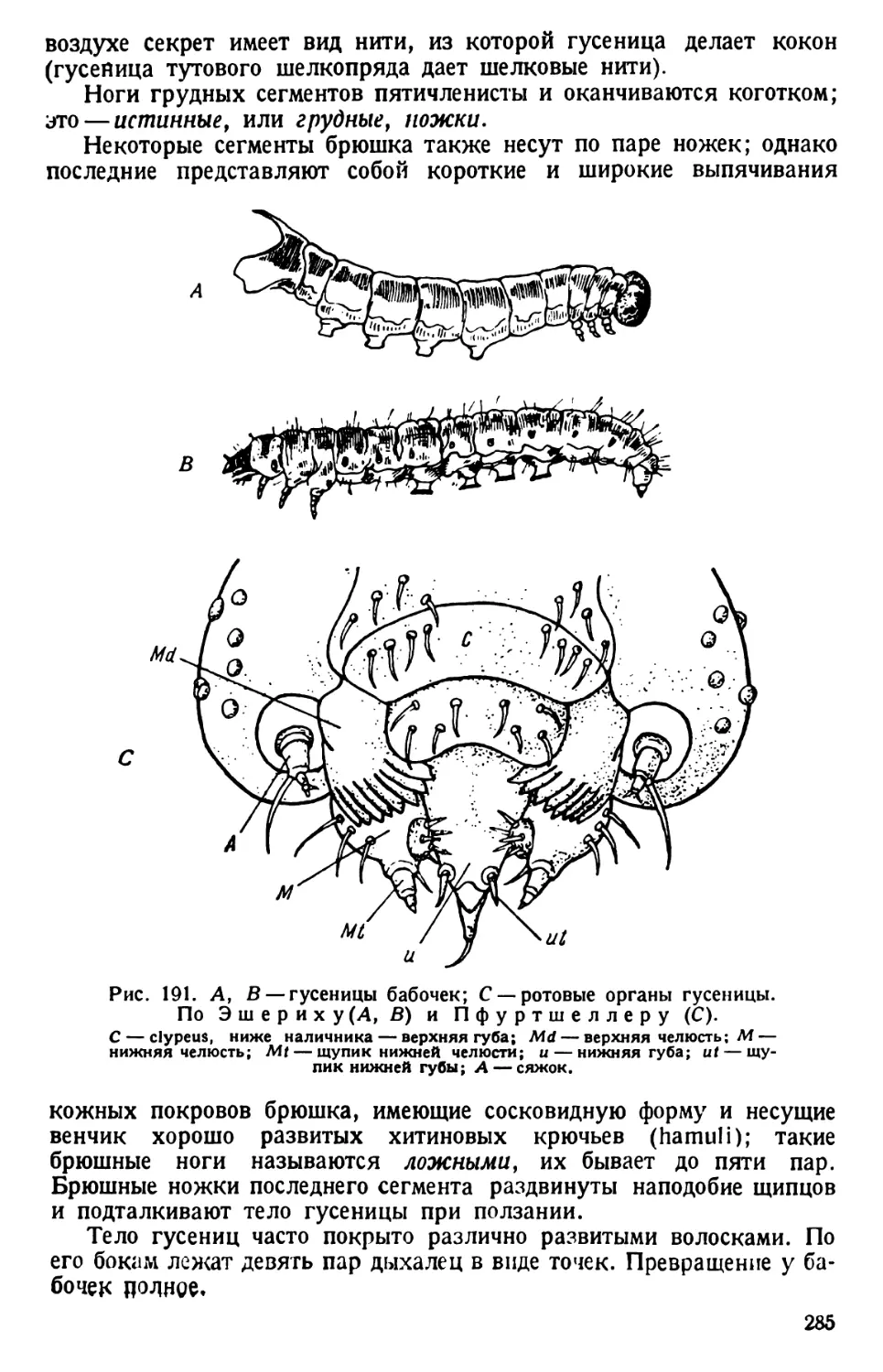
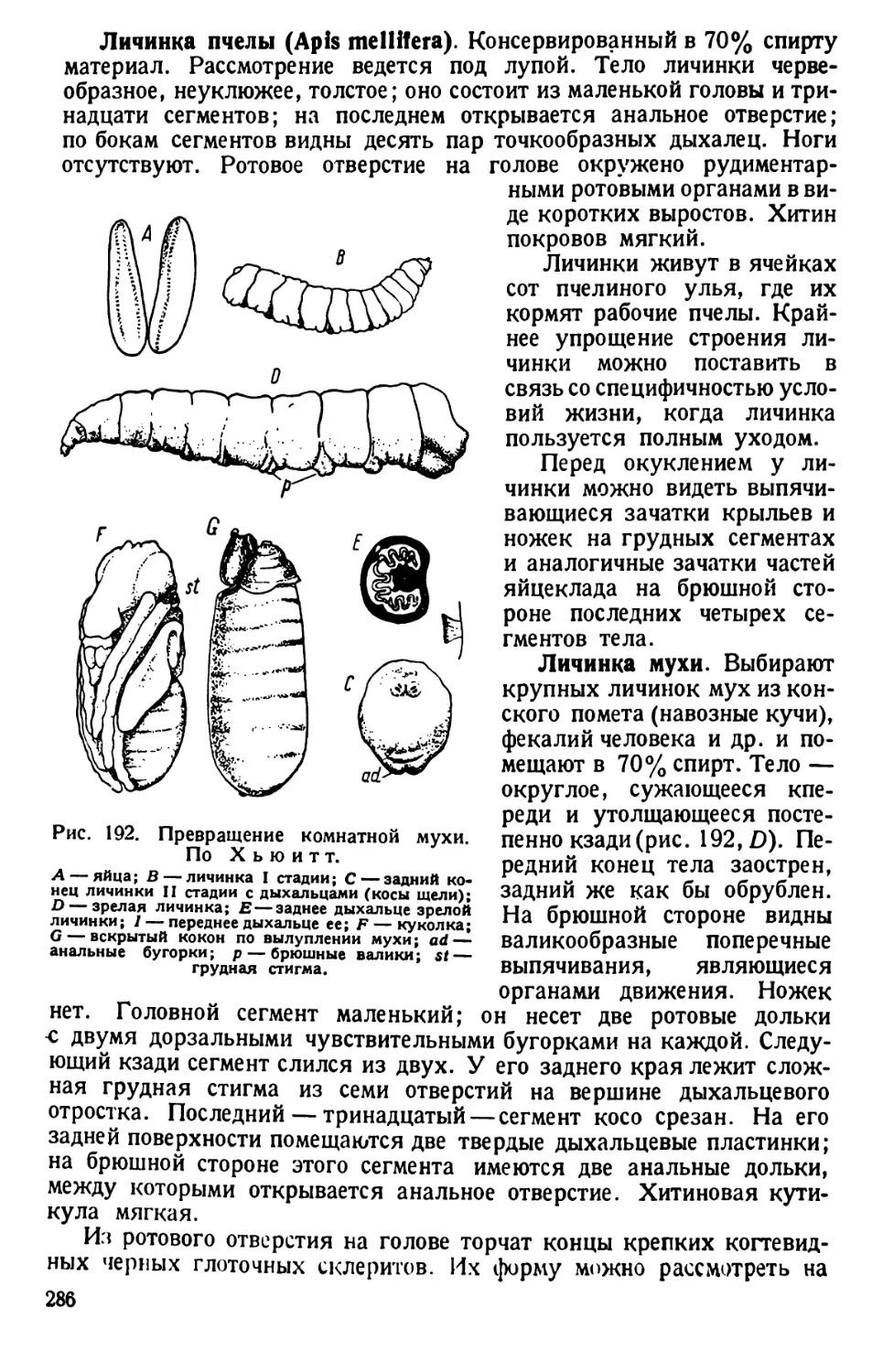
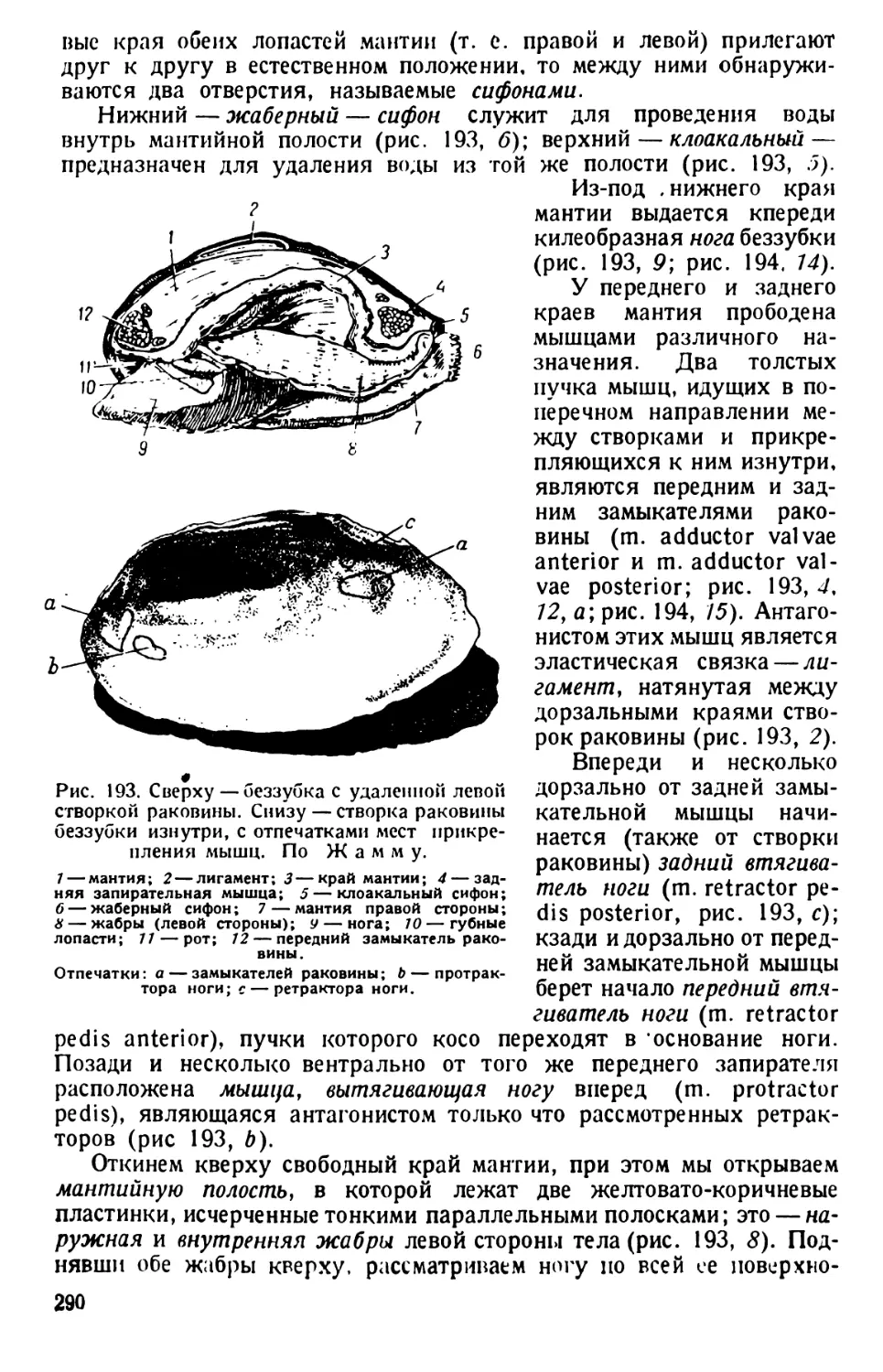
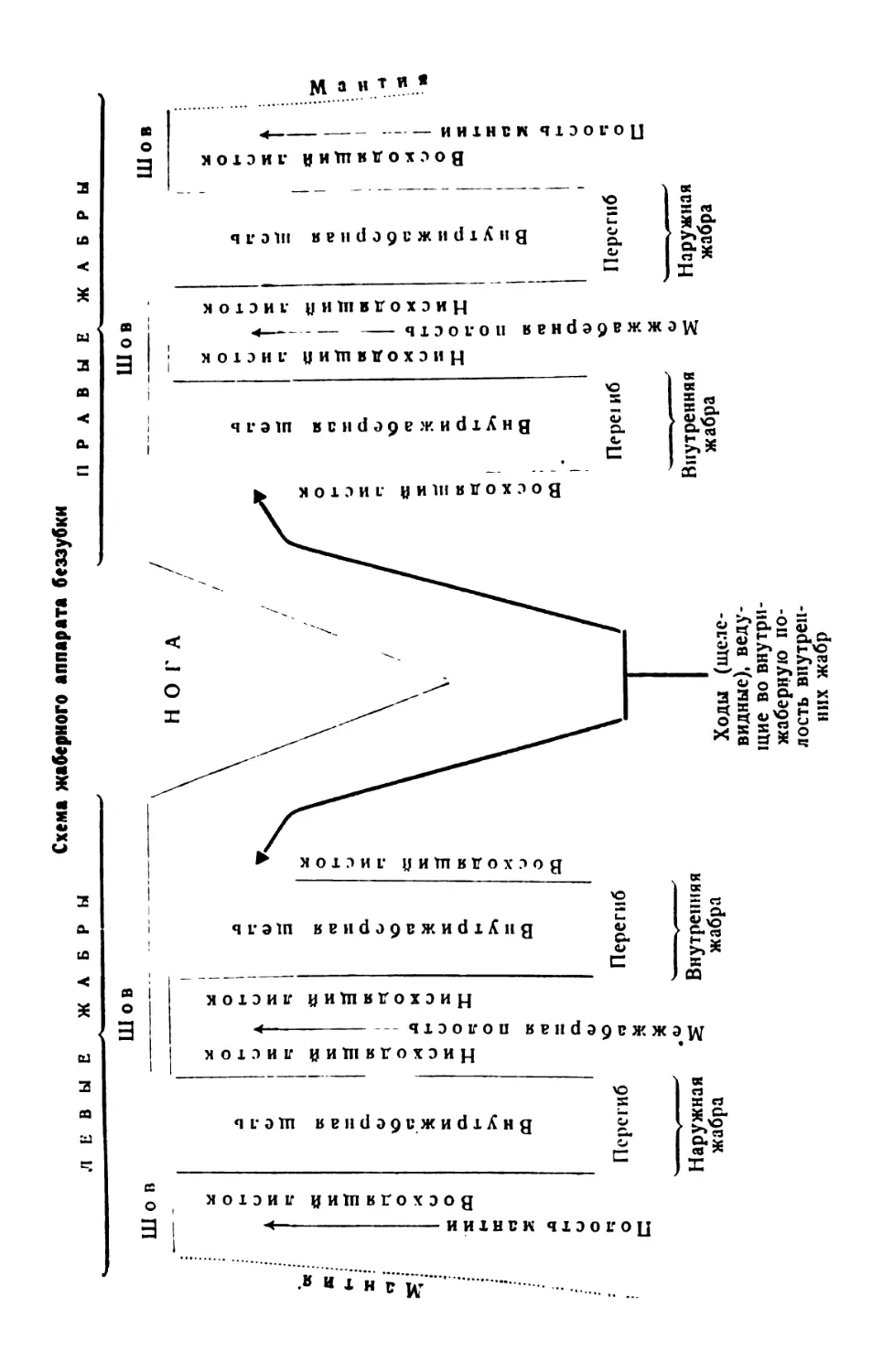
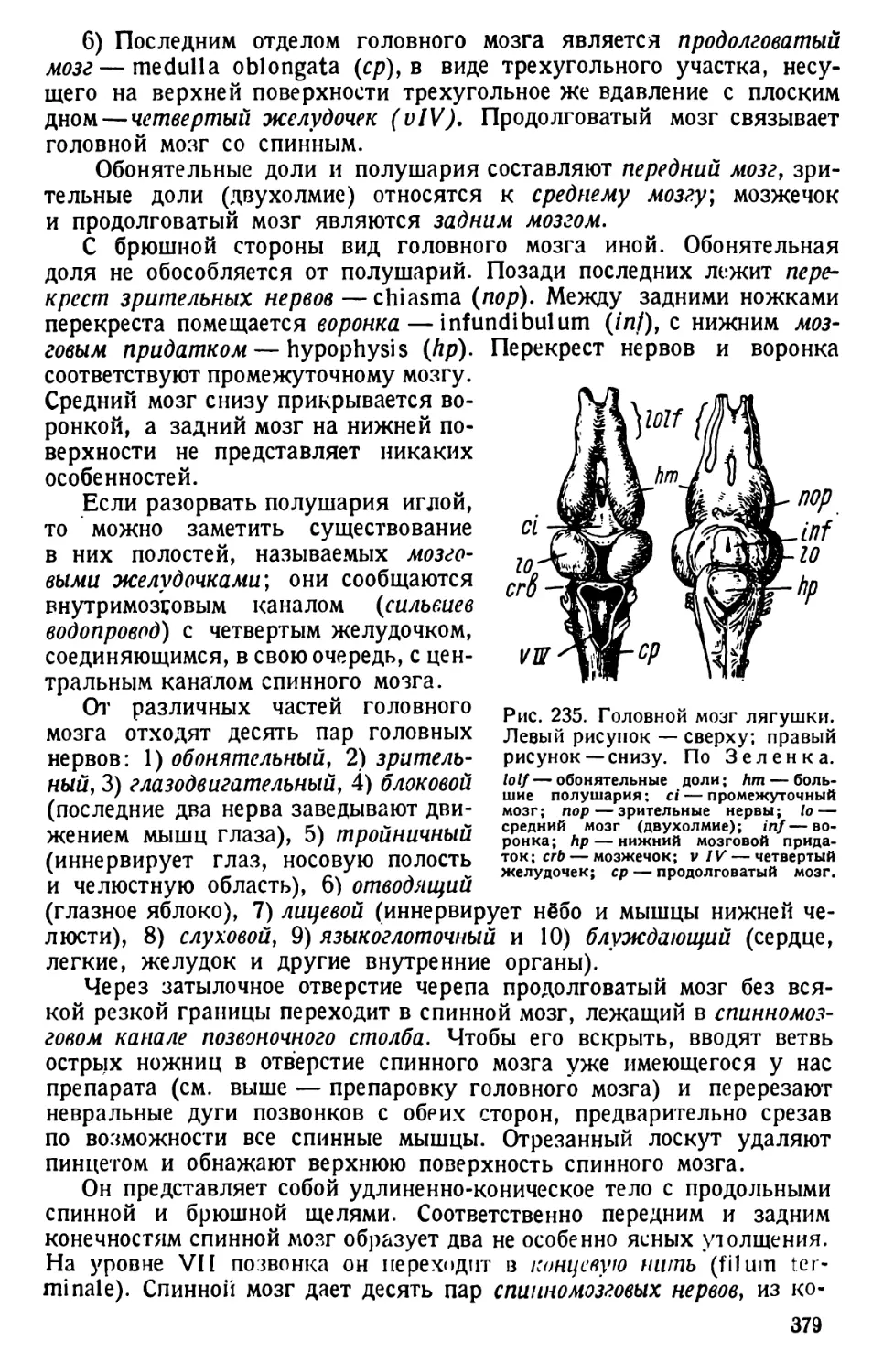
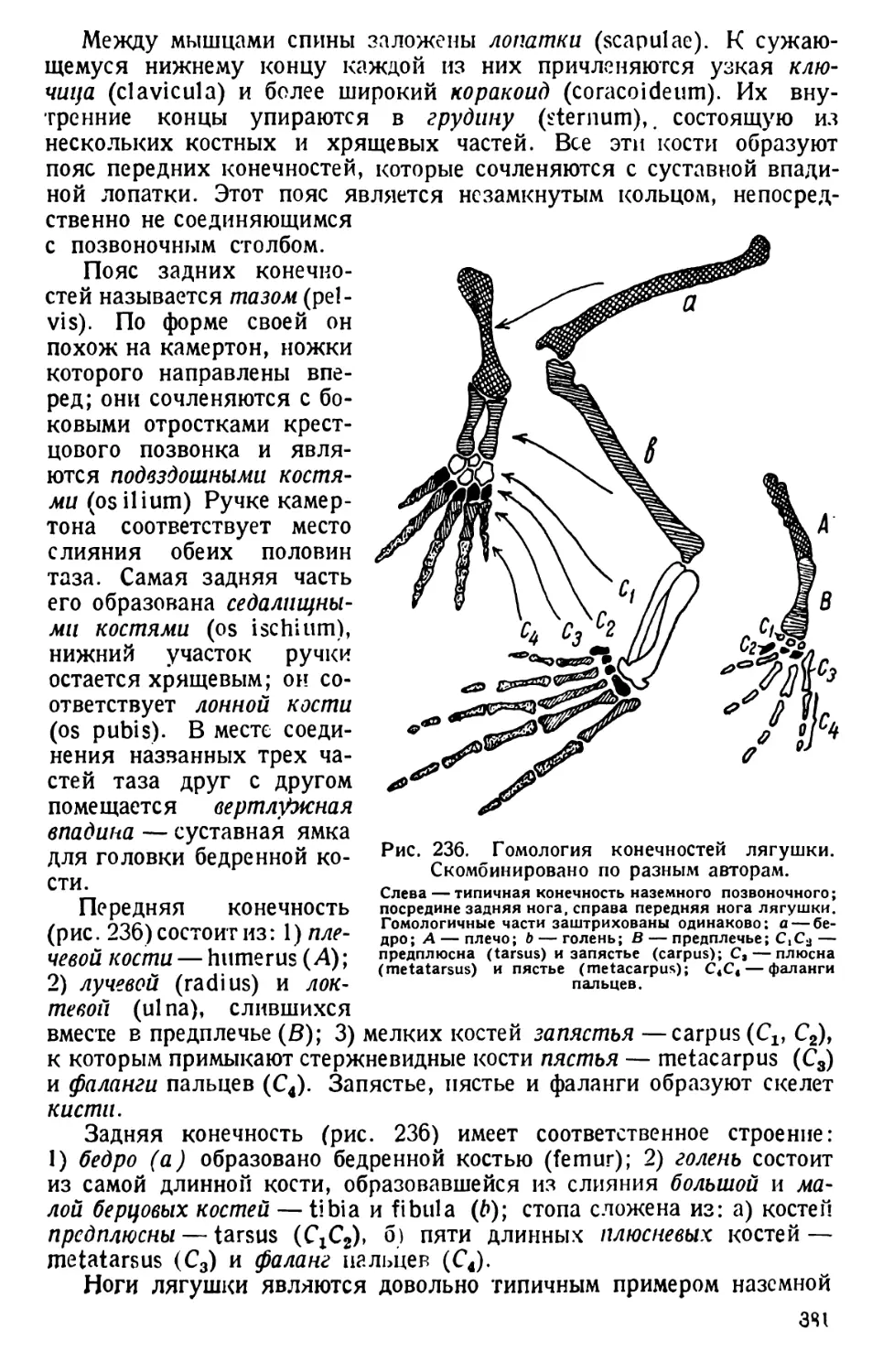
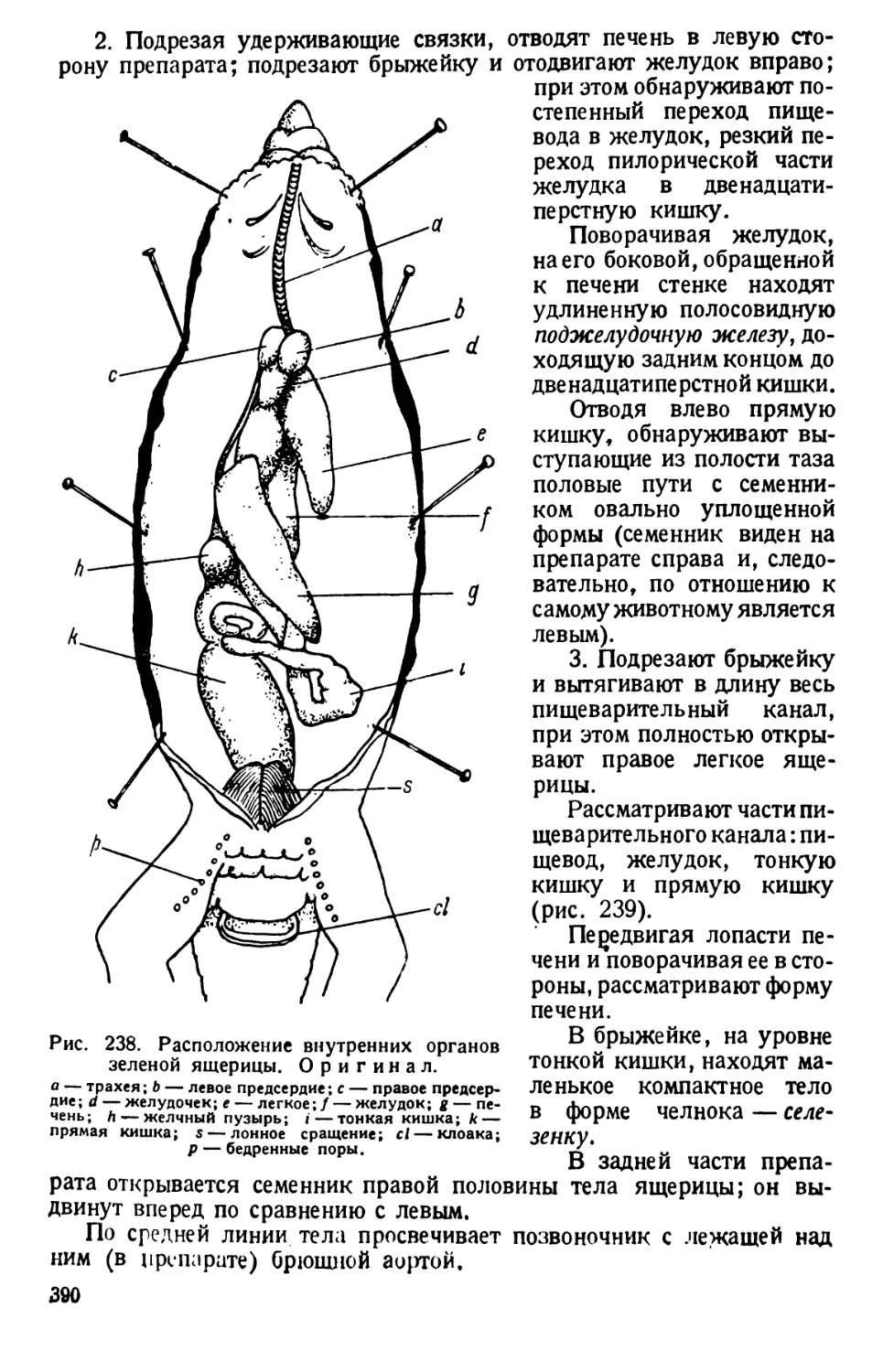
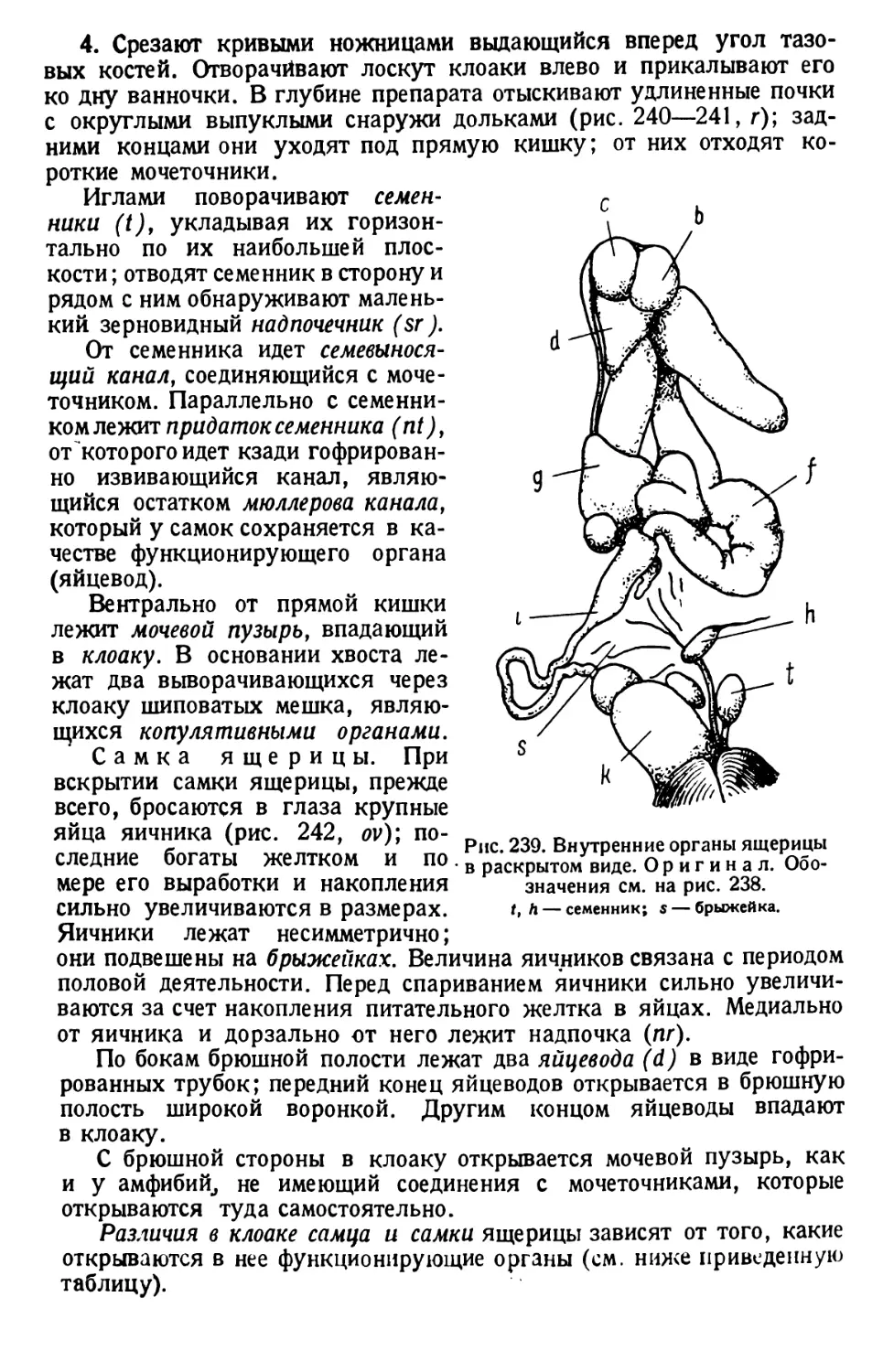
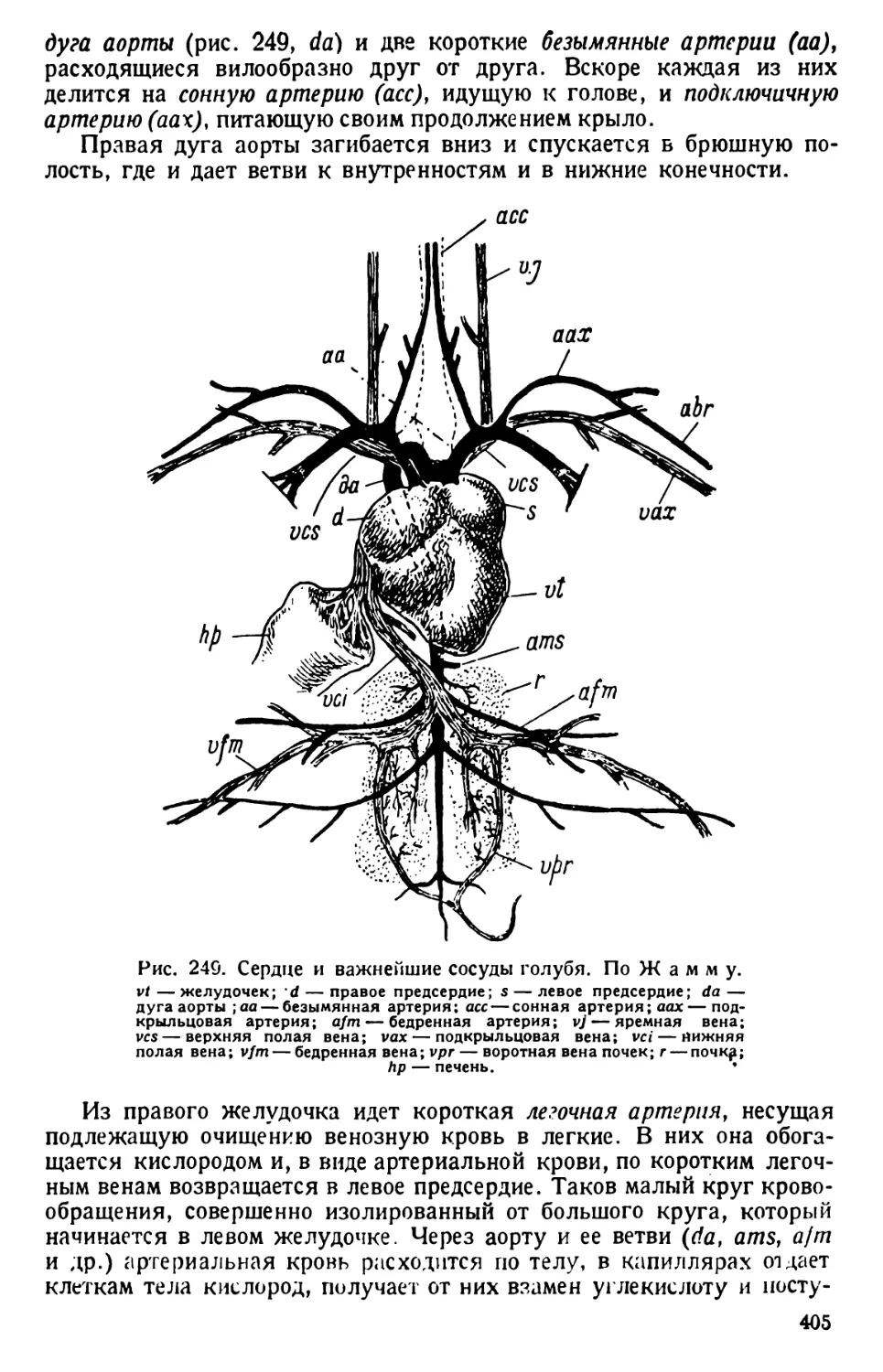
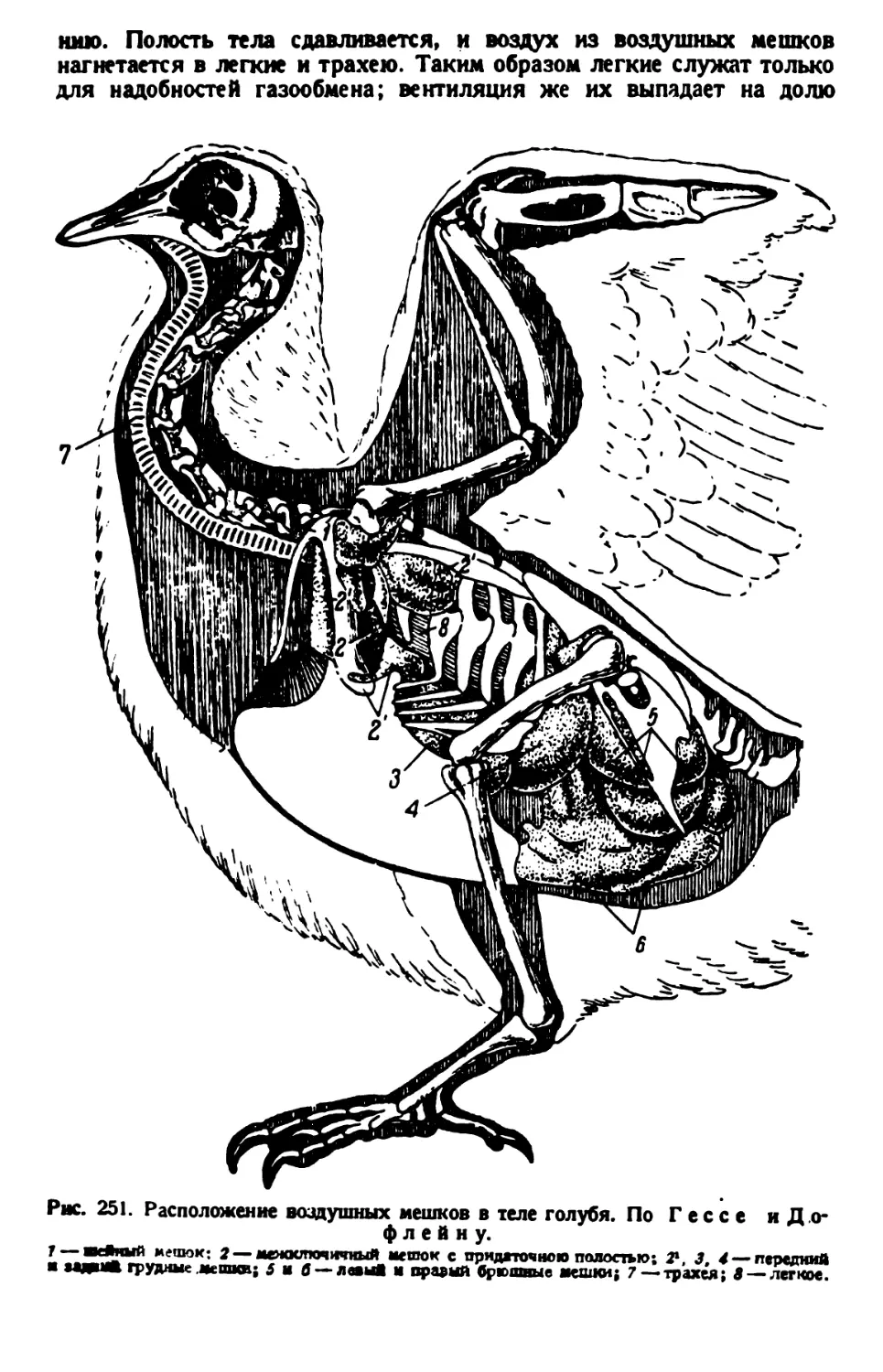
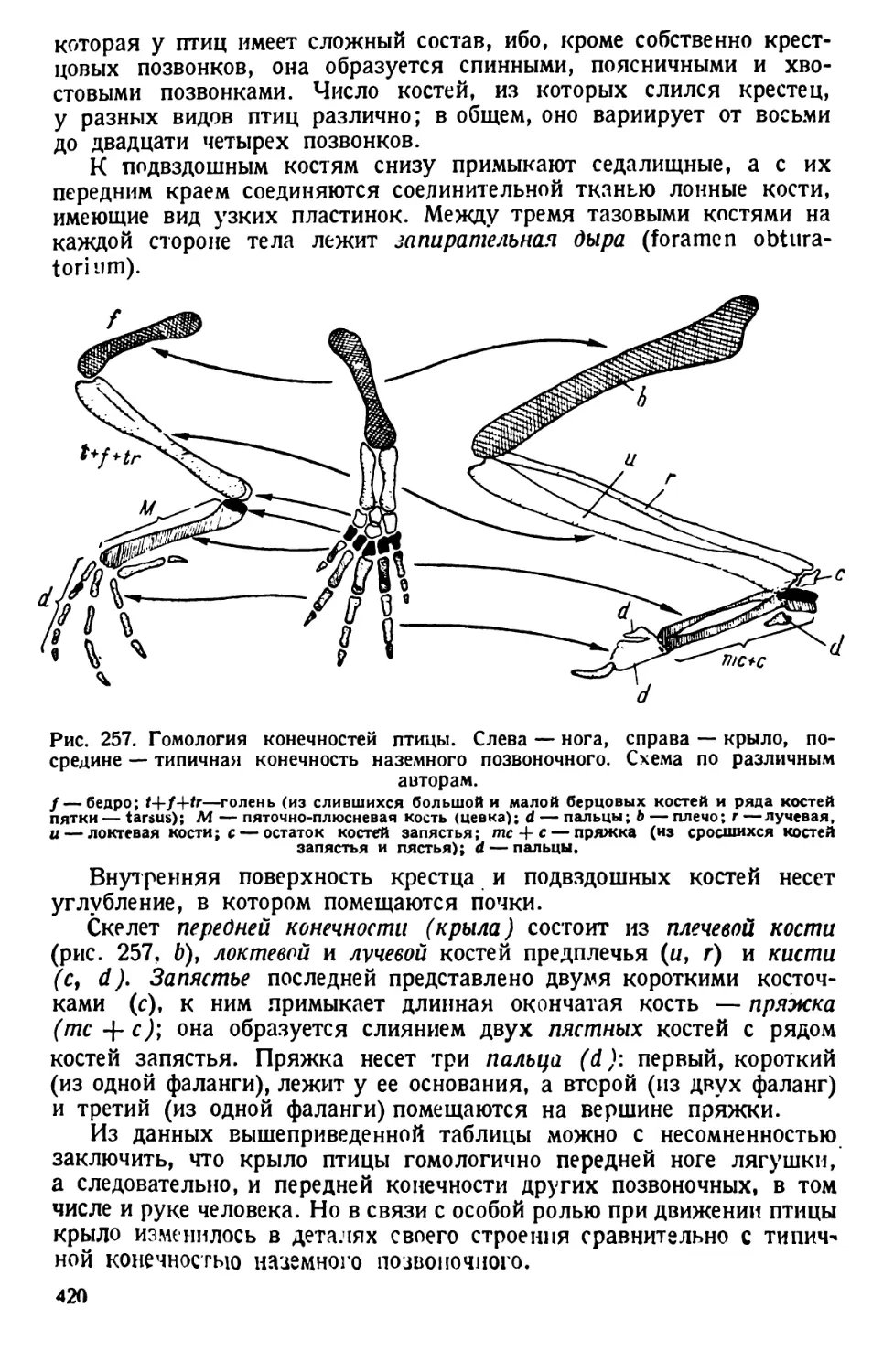
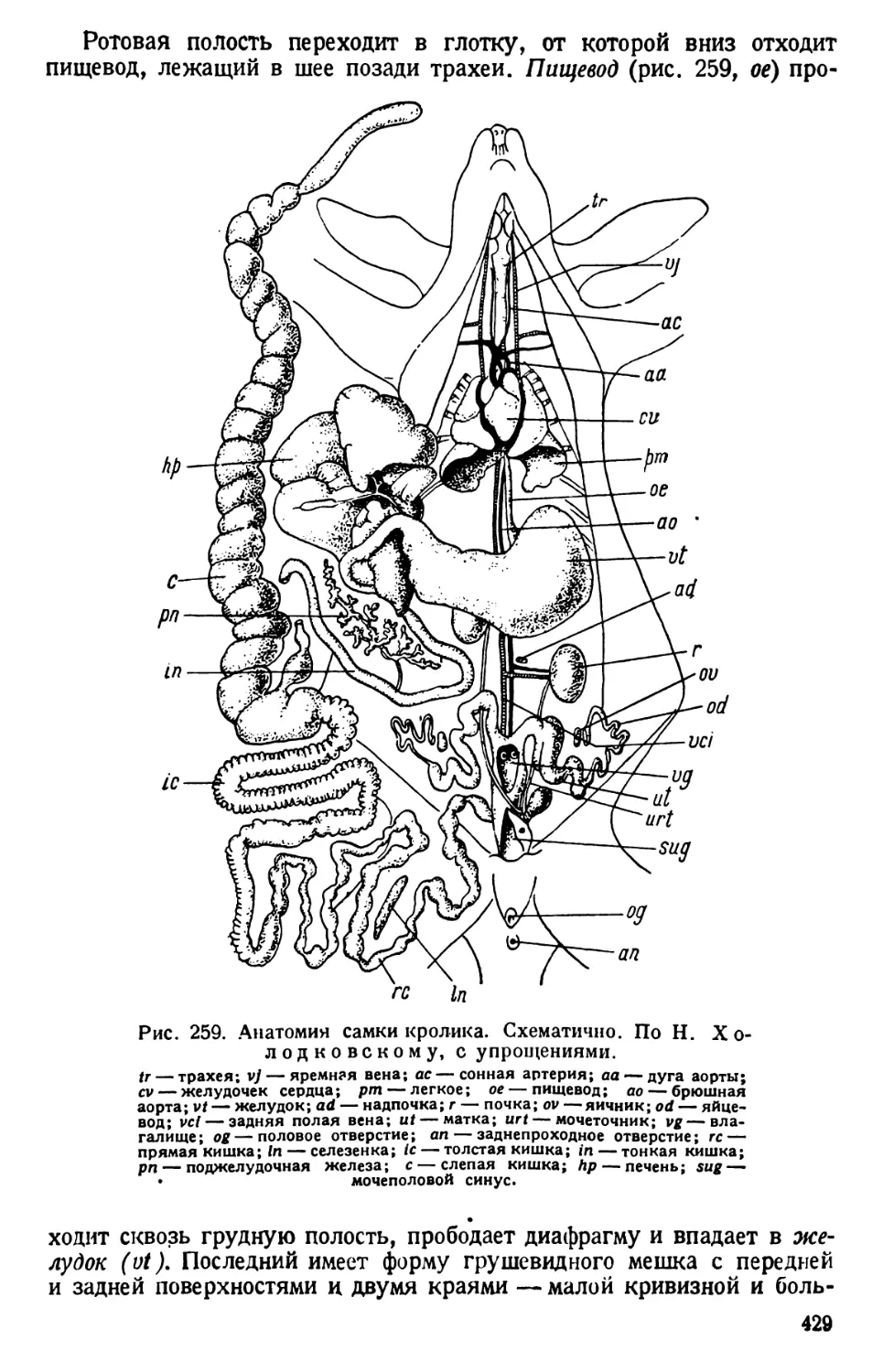
/



























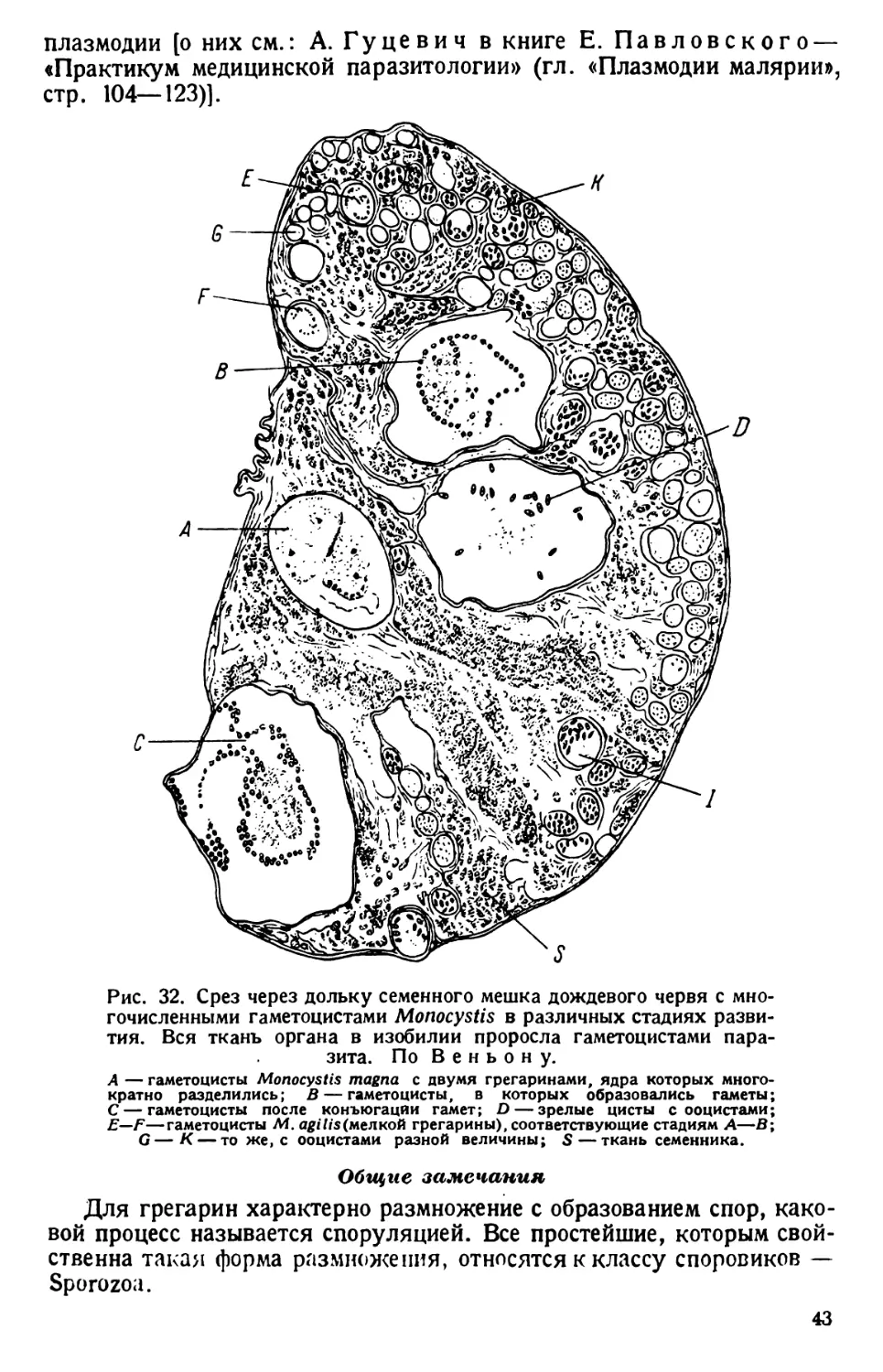


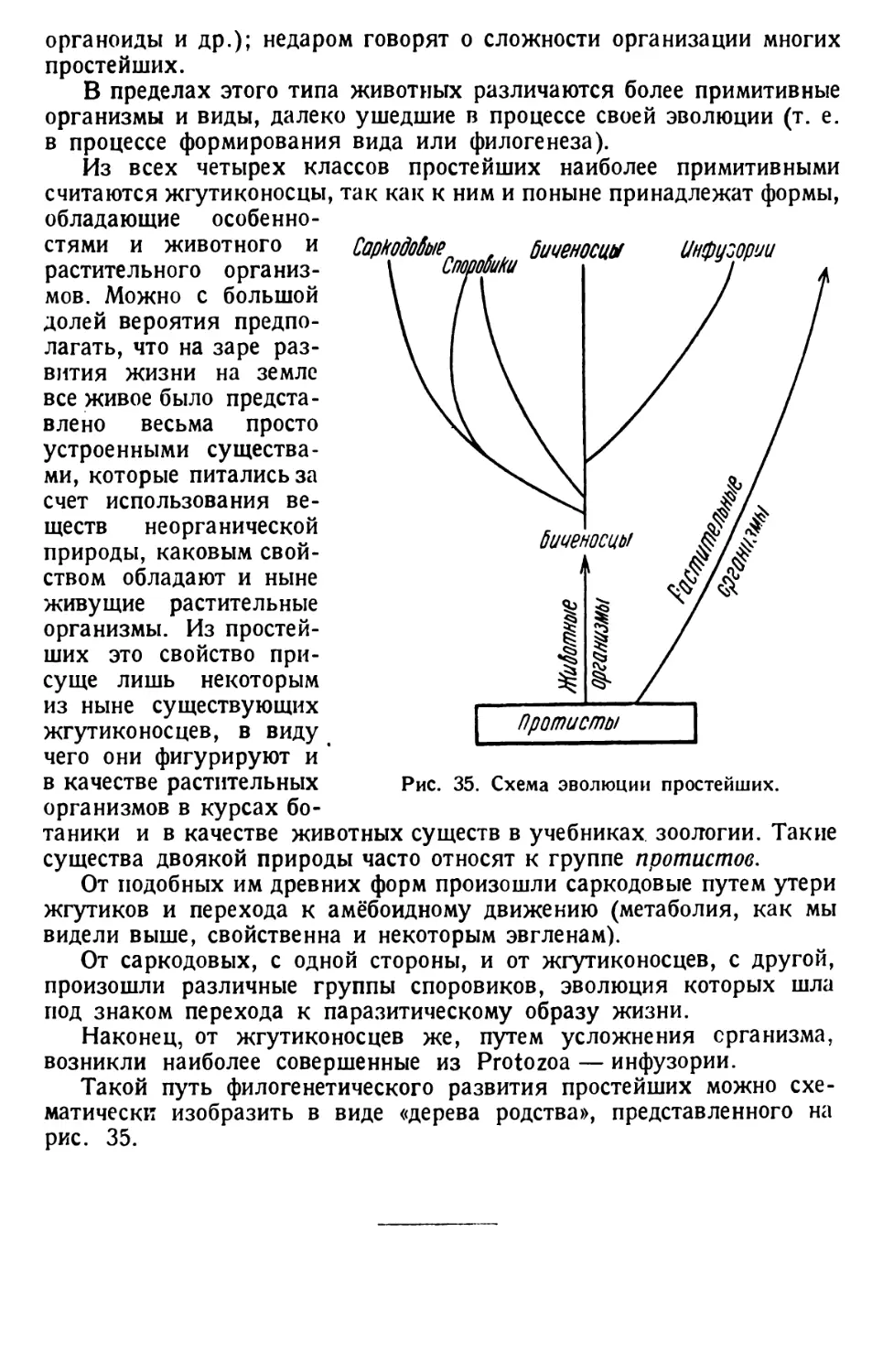


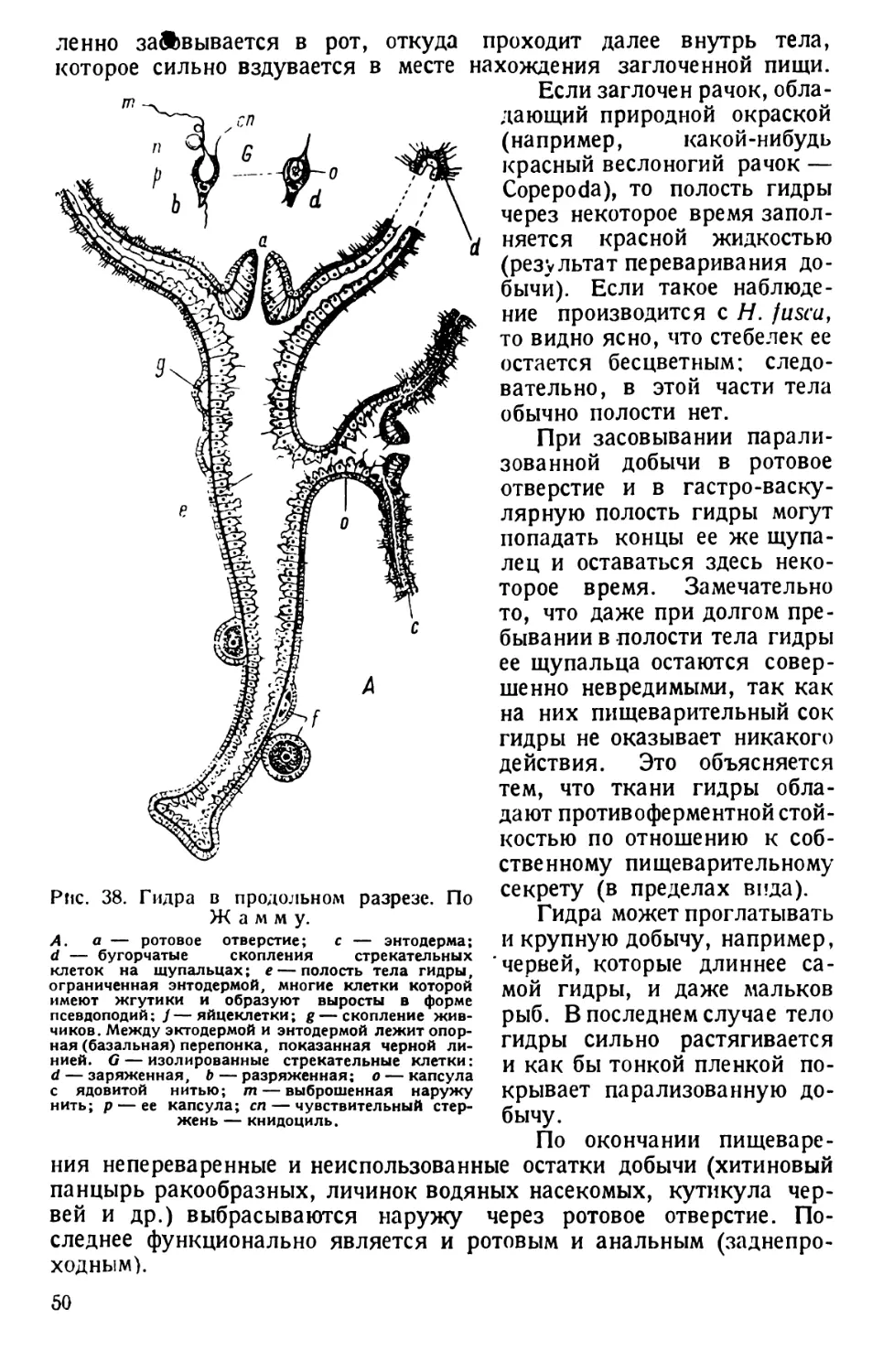








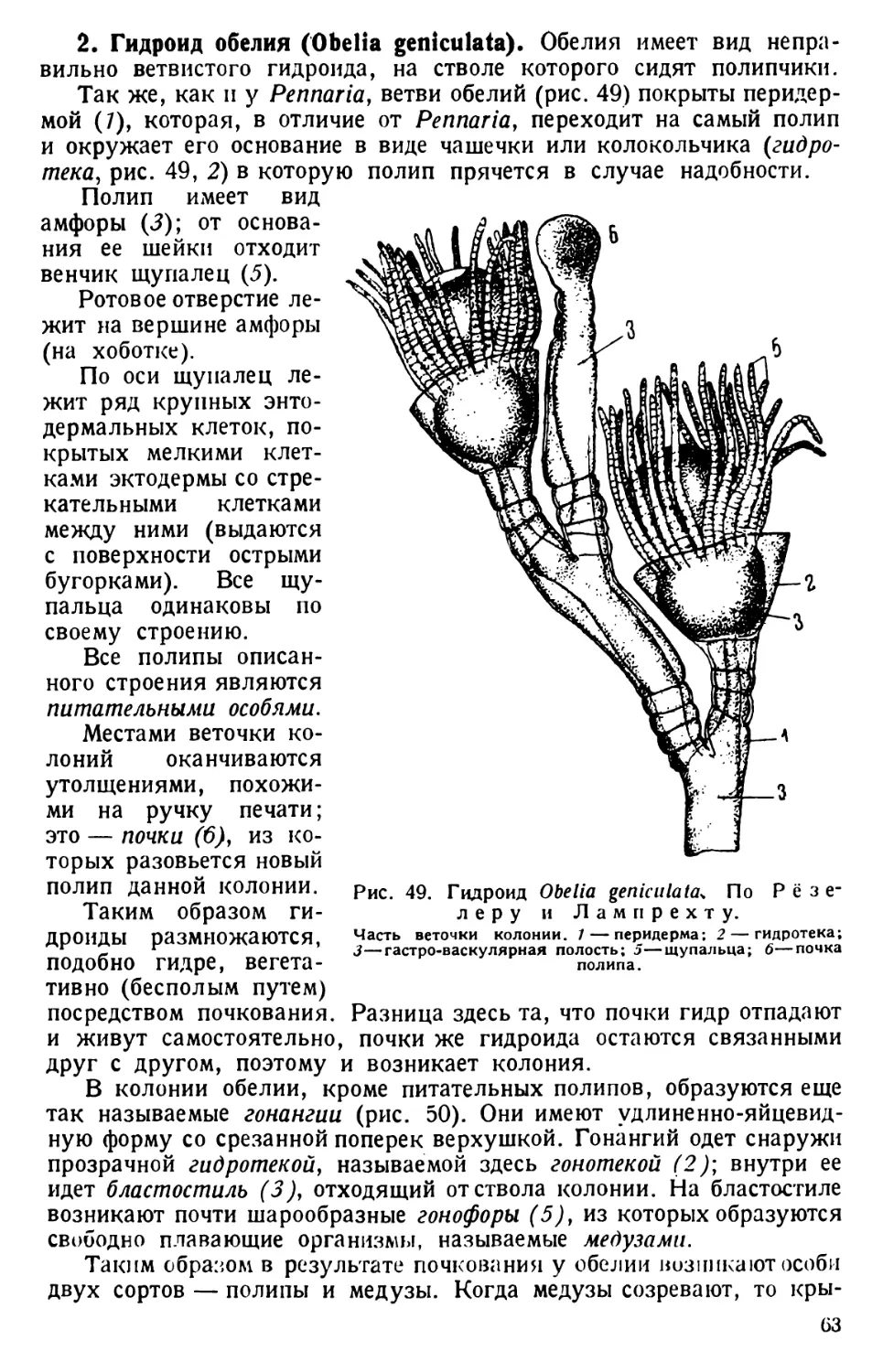









































































































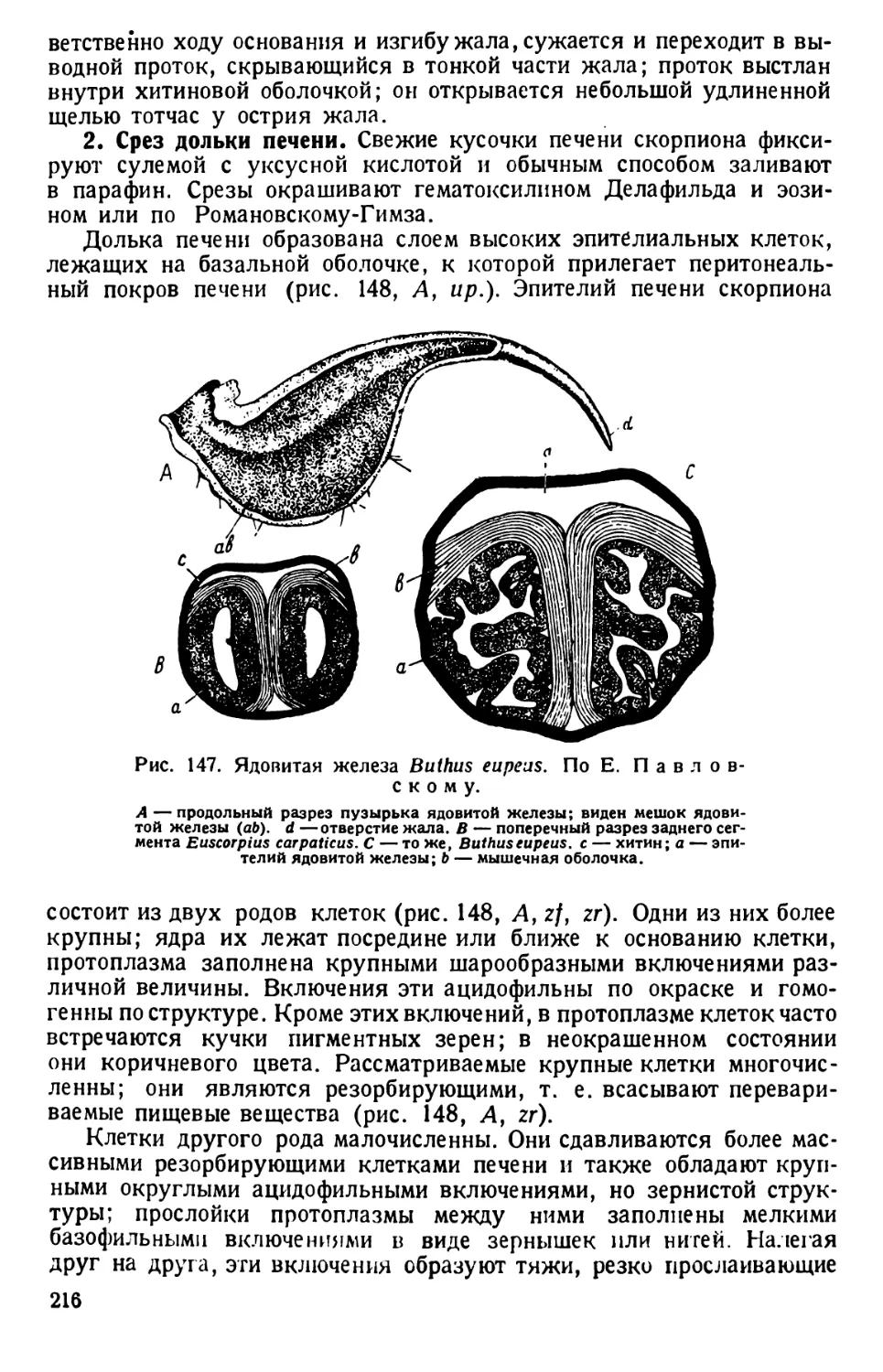
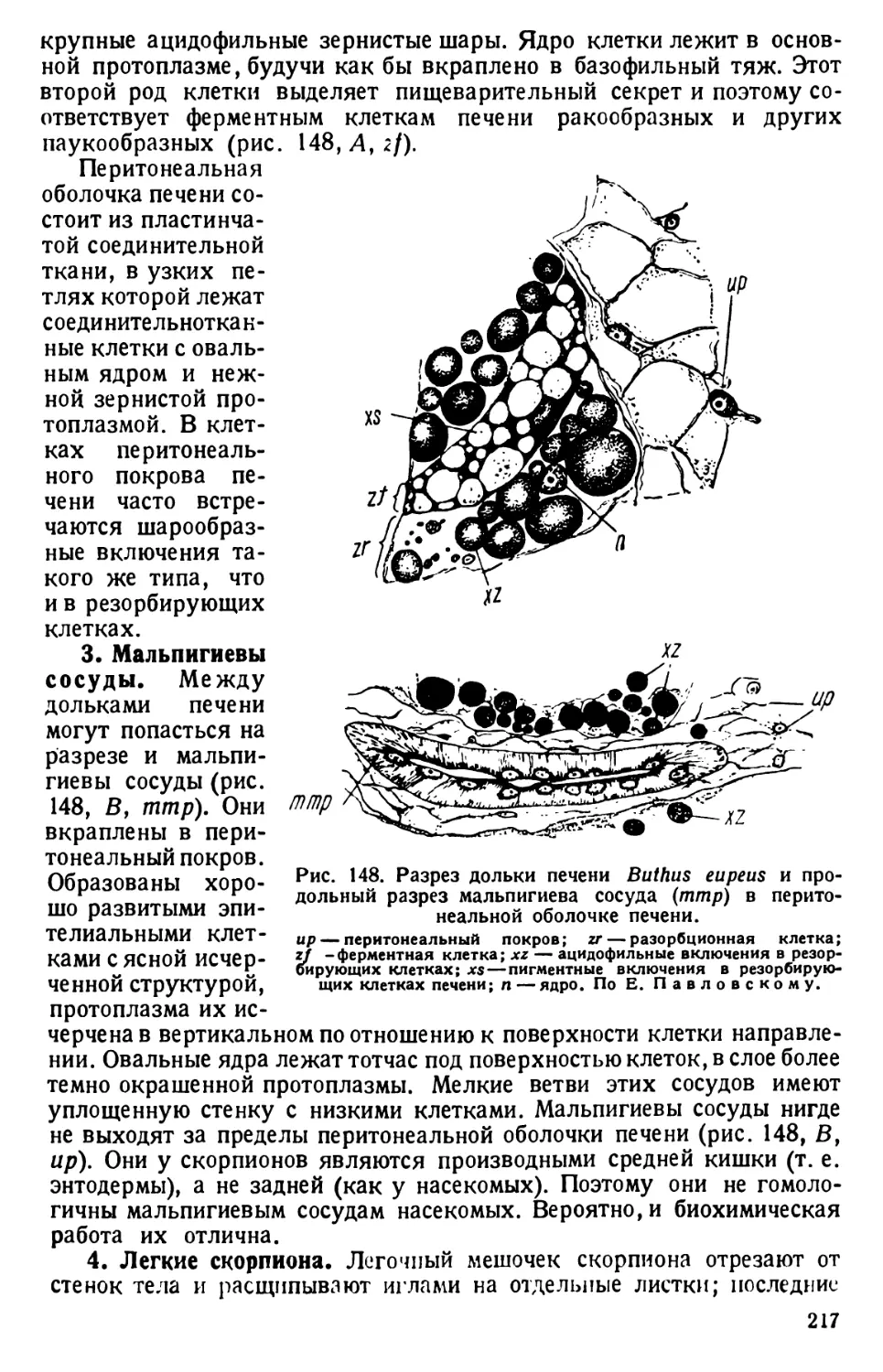






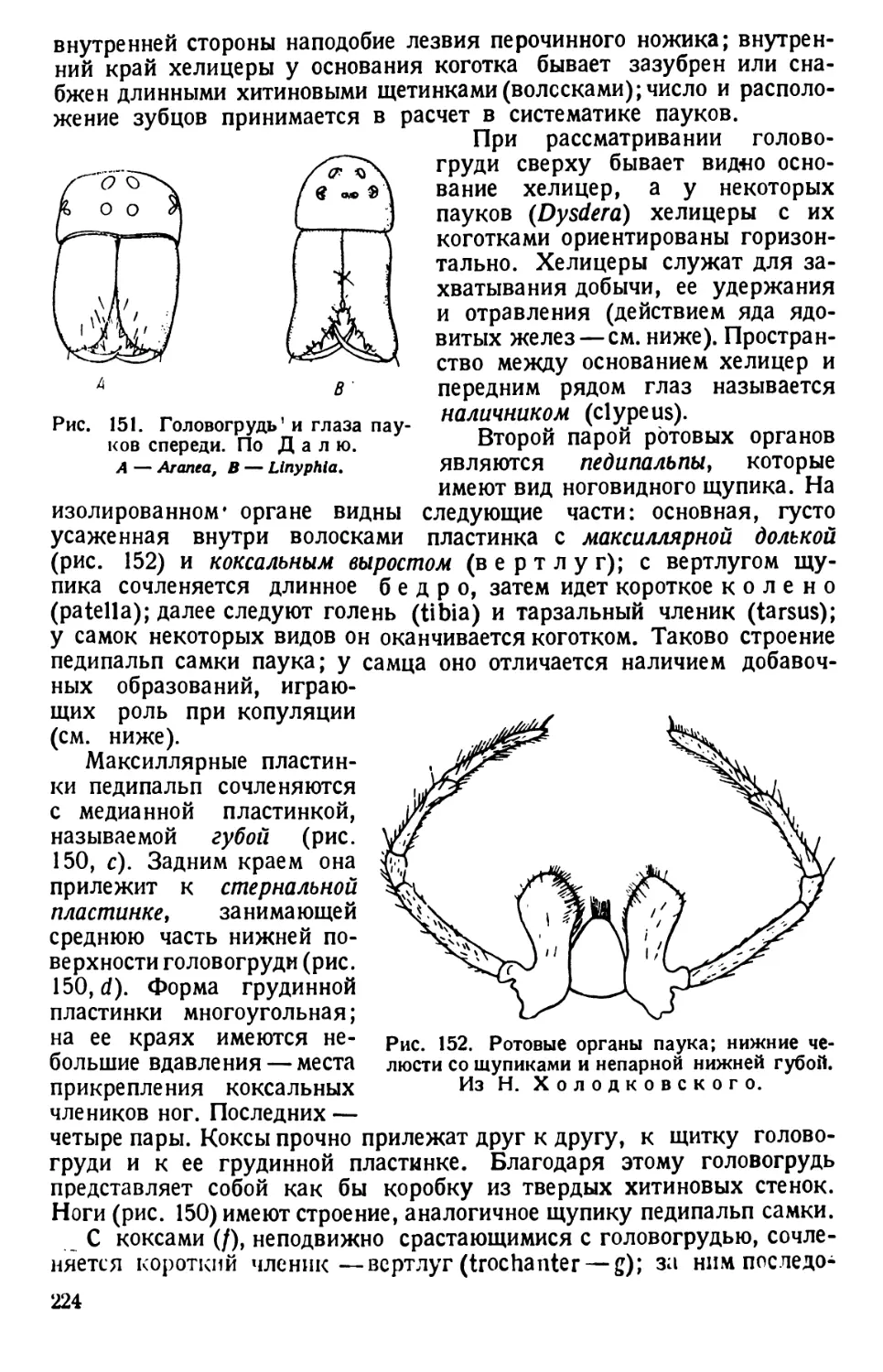
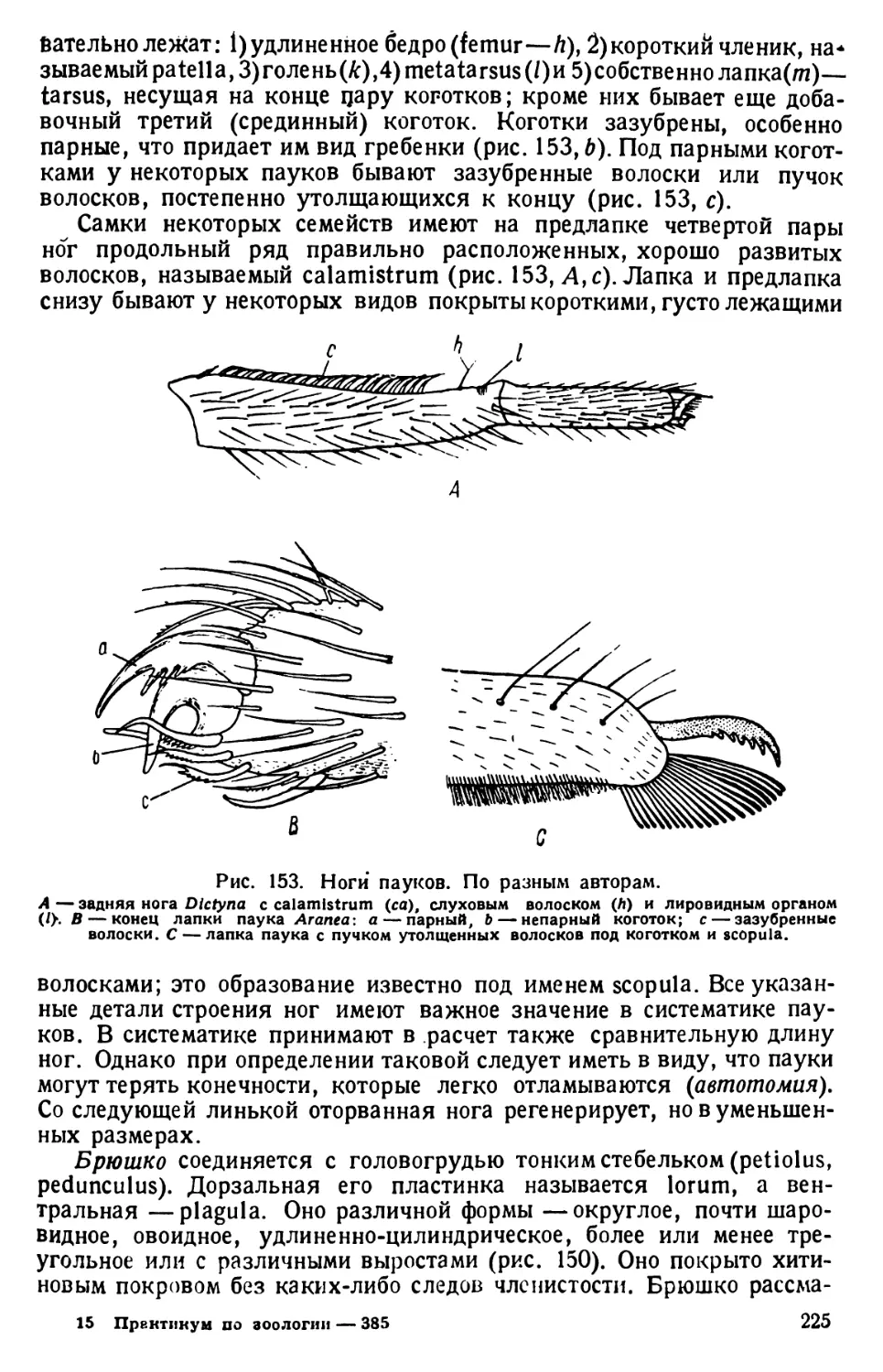



























































































































































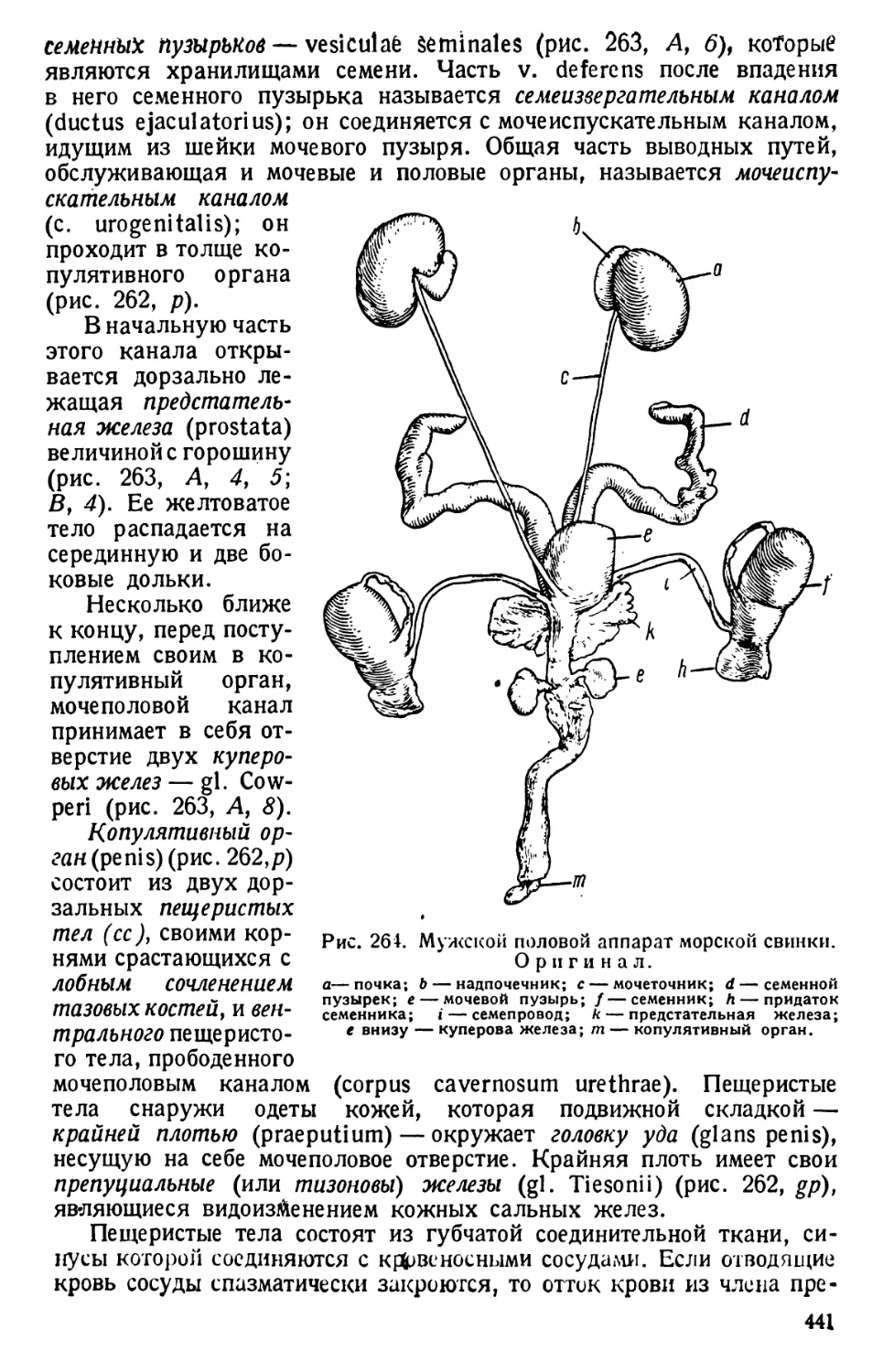
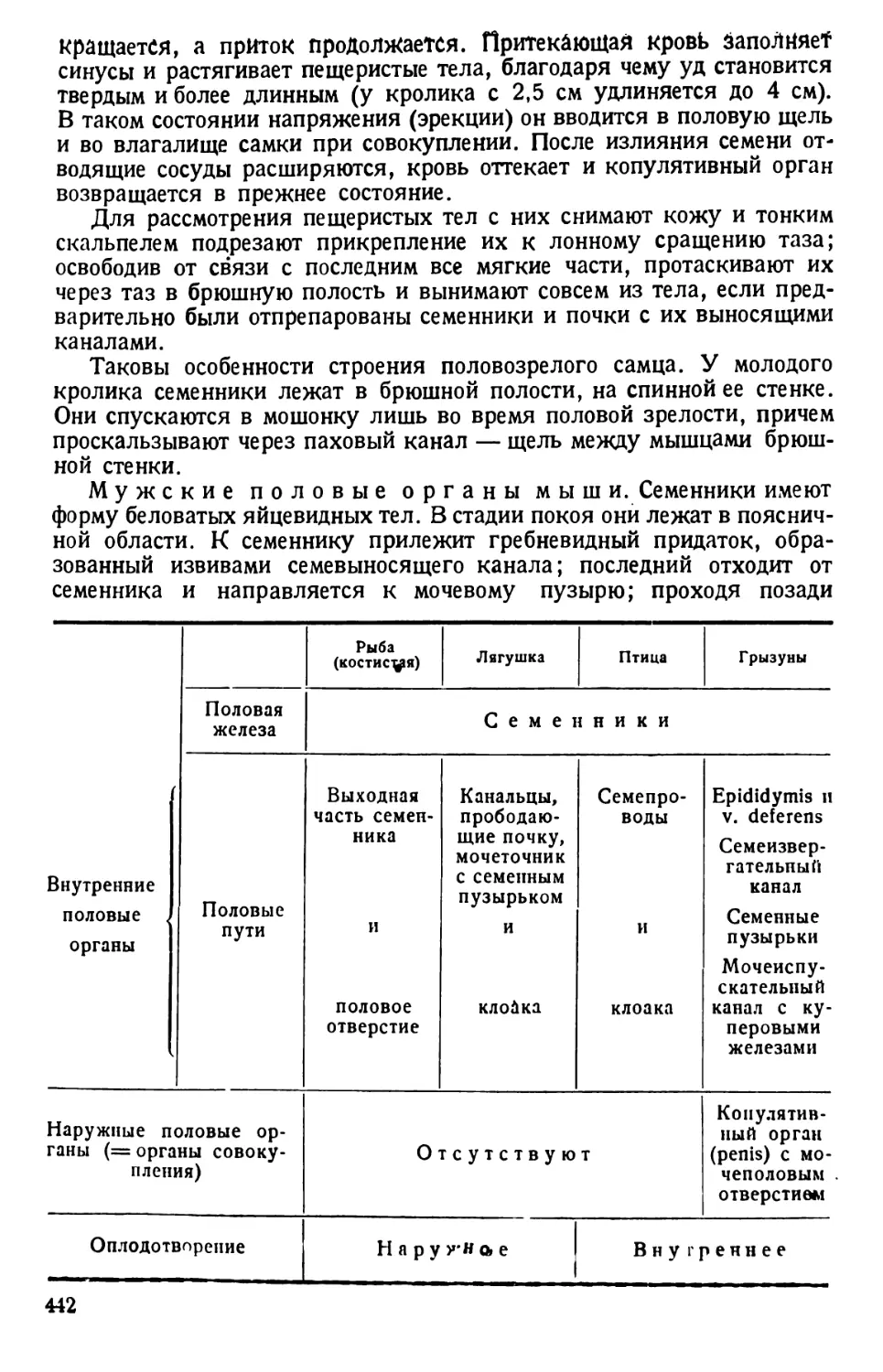



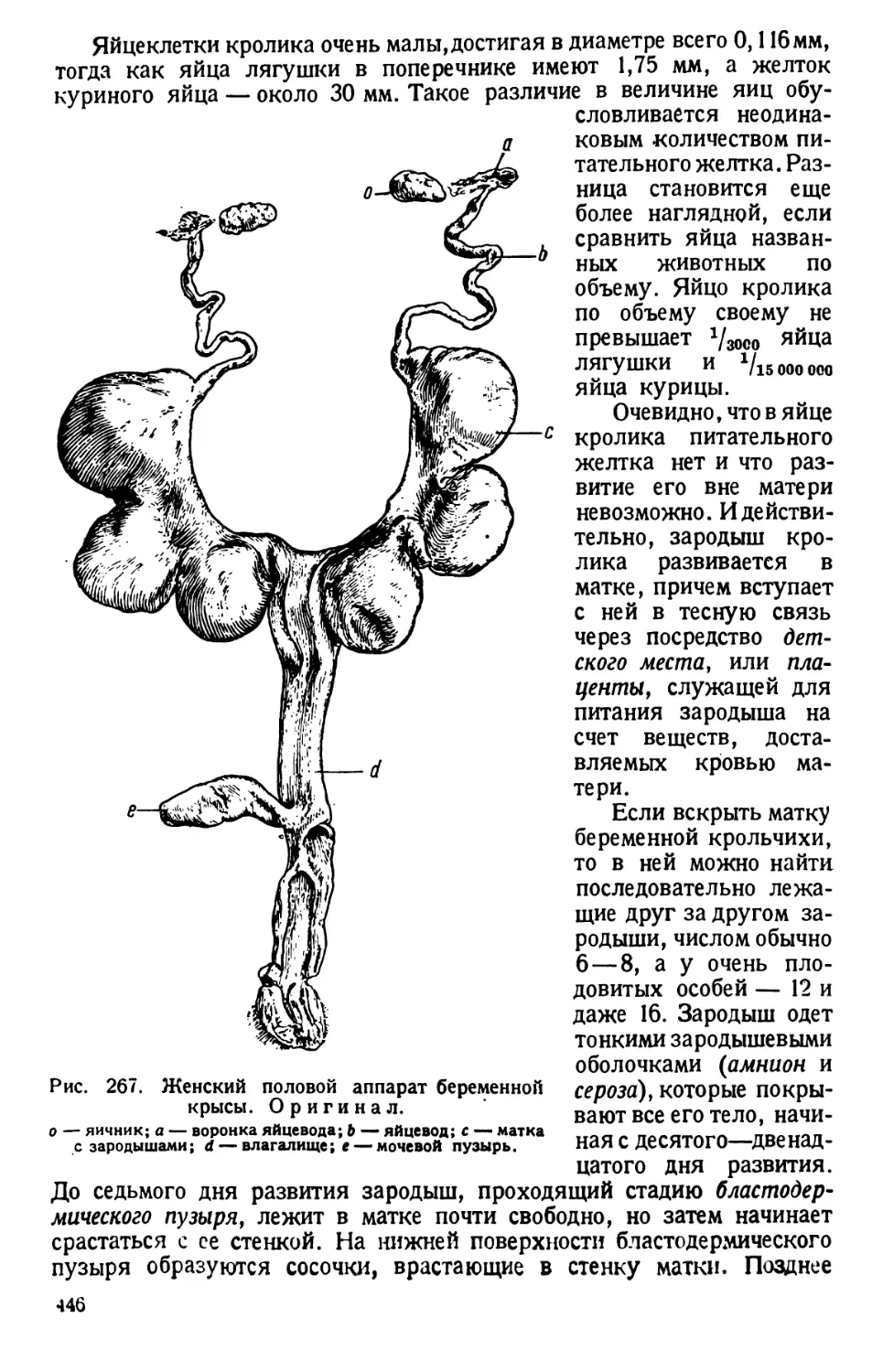


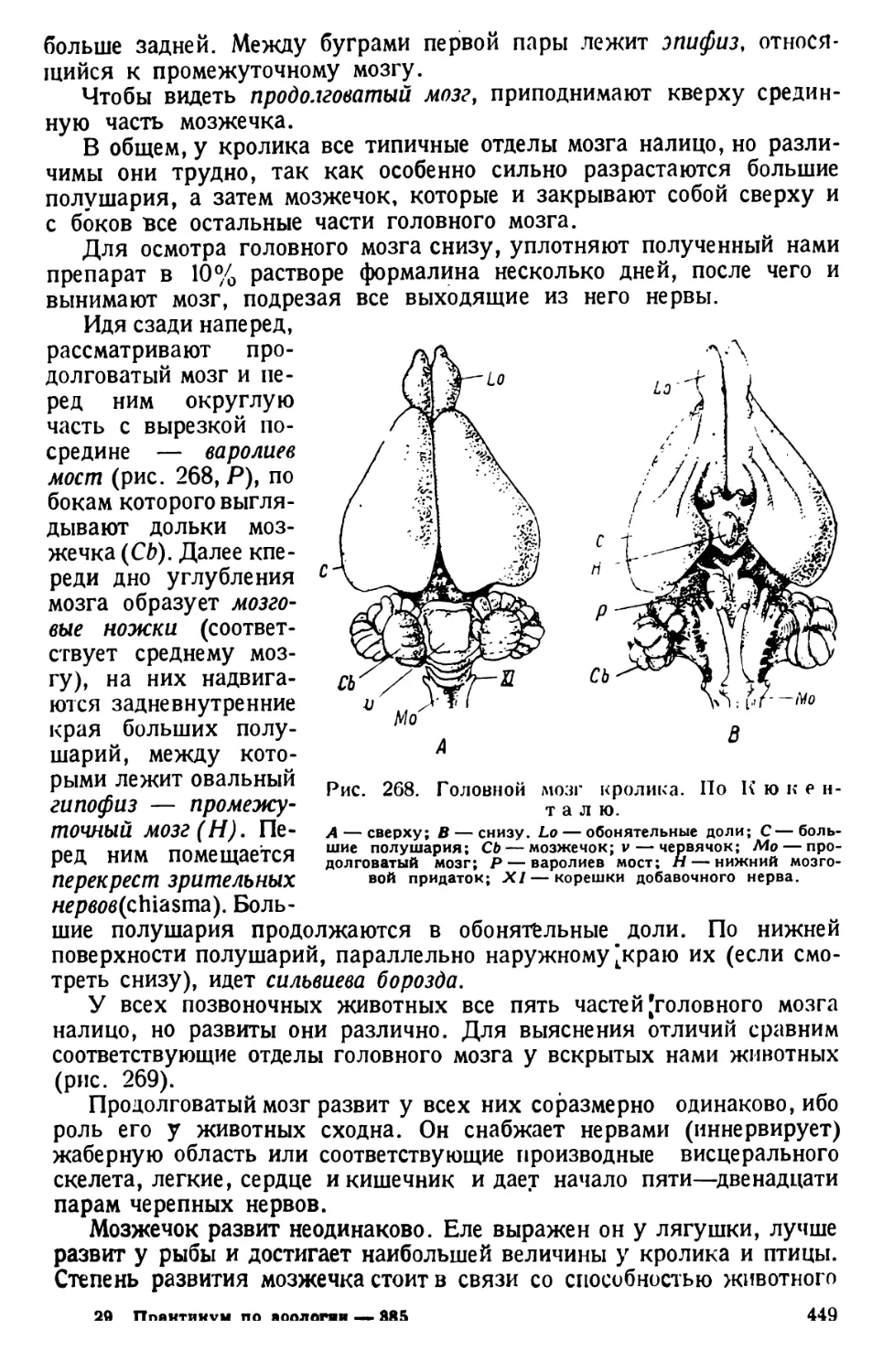






























Автор: Павловский Е.Н.
Теги: зоология экспериментальные исследования турбелярии полихеты дафнии эволюционные освещения
Год: 1938




Текст
Проф. Е. Н. ПАВЛОВСКИЙ
Заслуженный деятель науки
ПРАКТИКУМ
ПО зоологии
ИЗДАНИЕ ВТОРОЕ,
ПЕРЕРАБОТАННОЕ И РАСШИРЕННОЕ
Допущено Всесоюзным Комитетом
по делам высшей школы при СНК
СССР в качестве учебного пособия для
биологических факультетов государ-
ственных университетов и педагоги-
ческих институтов
ГОСУДАРСТВЕННОЕ
УЧЕСНО41ЕЛАГОГИЧЕСК0ВММЛХЕДЬ£1ВО НАРКОМПРОСА РСФСР
Ленинградское отделепже Ленинград
59
П— 12
Данная книга является переизданием «Руководства к прак-
тическому изучению зоологии». В настоящем виде оно сильно
расширено прибавлением новых частей и переработкой неко-
торых глав. Так, расширен отдел по простейшим, добавлены
новые объекты изучения, как-то: турбеллярии, полихеты,
дафнии, паукообразные, личинки насекомых, личинки игло-
кожих, клоп, ланцетник, пресмыкающиеся и др. Общий ха-
рактер изложения материала оставлен прежним; введены
материалы по экспериментальному исследованию животных
(например, жизненный цикл сосальщиков в моллюсках, опыты
по физиологии простейших и др.).
Книга в основе своей остается пособием к практическому
сравнительному ознакомлению с важнейшими представителями
типов животных в экологическом и эволюционном освещении.
Предназначается она для студентов-биологов.
Ответственный редактор Ф. И. Кричевская, Техн, редактор А. В. Андерсон.
Корректор С. А. Шабуневич.
Сдано в набор 1/VIII 1938 г. Подписано к печати 19/VIH 1938 г. Авт. л. 34,39 Уч.-авт. л. 34,39.
Печ. л. 30. Бум. л. 15. Тираж 10000. Бумага 60X92 Вншерского бумкомбината. Тип. зн. в
1 бум. л. 102400. Леноблгорлит № 3236. Учпедгиз № 10338. У-9. Заказ № 385.
Цена 5 р. 50 к. Переплет 2 р.
2-я типография ОГИЗа РСФСР треста .Полиграфкнига* .Печатный Двор“ им. А. М. Горького.
Ленинград, Га1чмнская, 26.
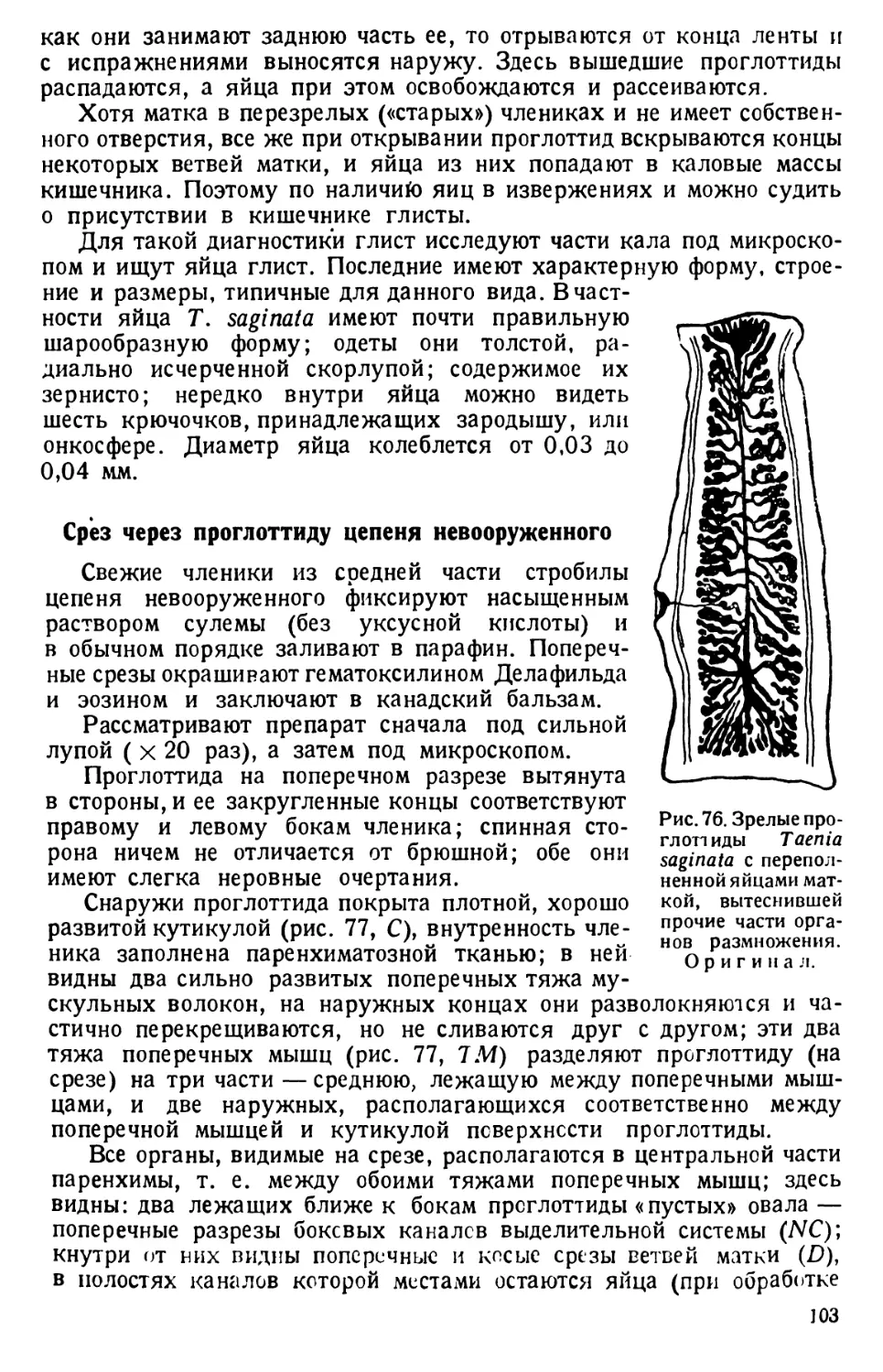
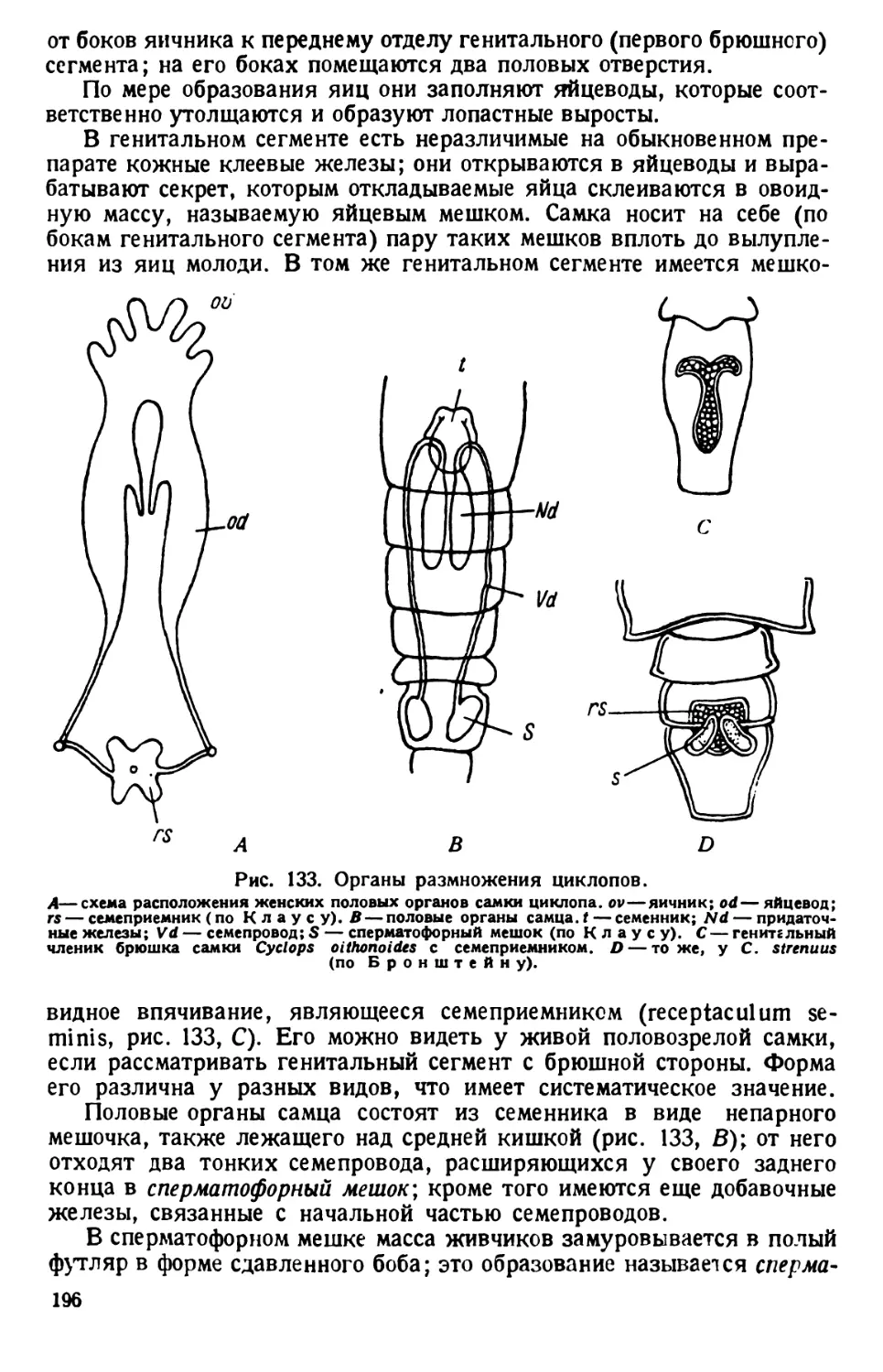
ВВЕДЕНИЕ
Для вскрытия животных необходимы следующие инструменты и
приспособления: 1) остроконечный скальпель для
разрезания тканей; достаточен скальпель средней величины; 2) пин-
цет с тонкими концами; для более крупных животных можно брать
обыкновенный «анатомический» пинцет, концы которого более толсты
и крепки; 3)2 препаровальные иглы; стальные (швей-
ные) иглы тупым концом плотно вставляются в деревянную ручку;
4) ножницы; для вскрытия мелких животных пользуются тон-
кими (малыми) остроконечными ножницами, а для крупных — более
прочными ножницами средйего размера, одна ветвь которых за-
острена, а другая на конце снабжена утолщением; 5) булавки
для прикалывания животного; годятся и обыкновенные булавки, но
удобнее и дольше служат лучшие сорта булавок со стеклянными голов-
ками.
Необходимо следить за чистотой и сохранностью инструментов и
вытирать их во время хода и по окончании работы, чтобы предохранить
их от ржавчины. Следует, в частности, тщательно вытирать замок
ножниц, особенно если они неразборные.
Препаровка небольших животных (лягушки, рыбы и т. п.) произ-
водится под водой в препаровальной ванночке. Послед-
няя представляет собой открытую сверху коробку из цинка или жести ;
бока коробки делаются покатыми или прямыми; наиболее употре-
бительны размеры ванночек: 25 х 25 х 6 см. На дно ванночки на-
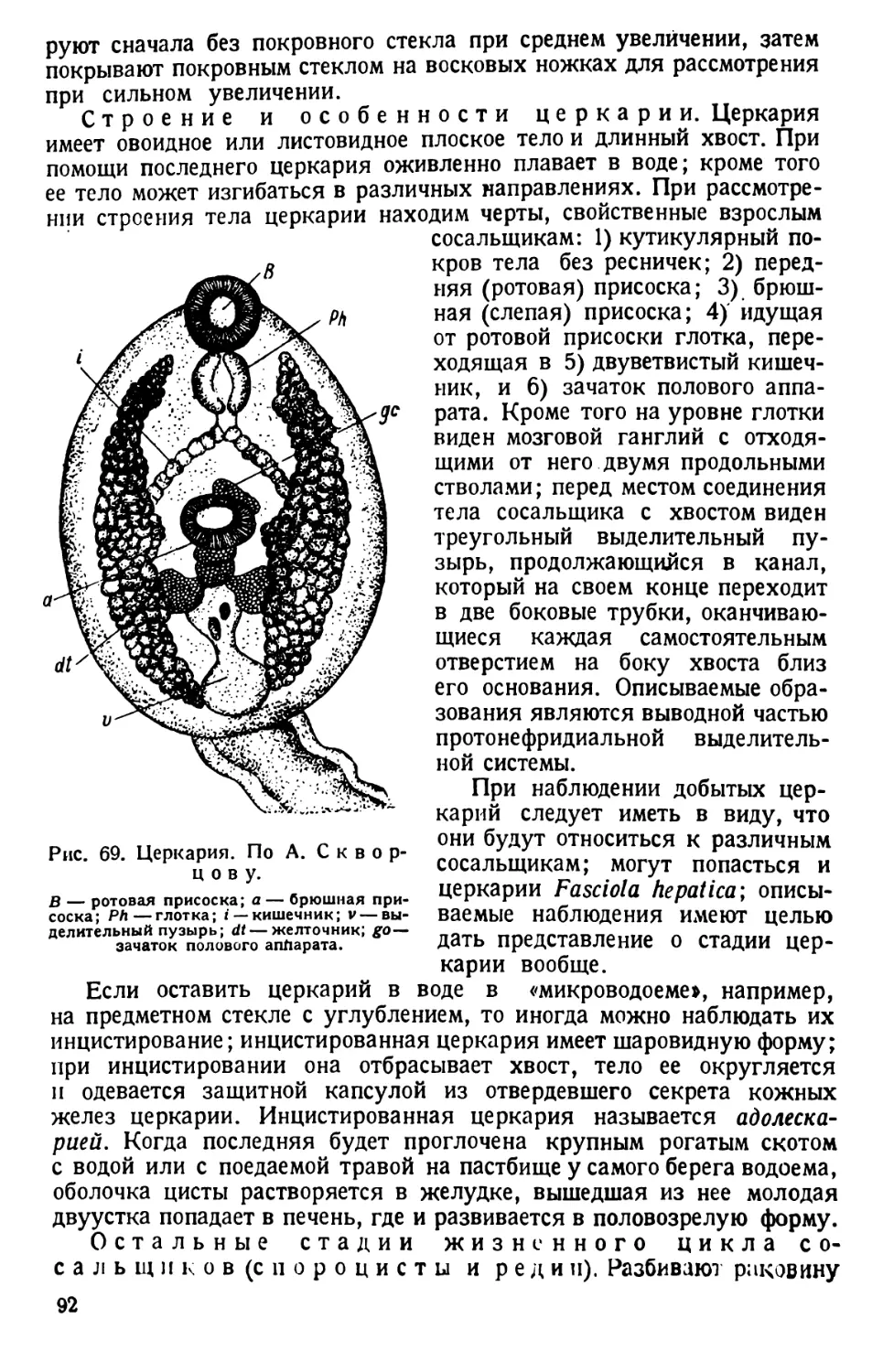
ливается расплавленная (восковая) смесь,1 которой дают застыть при
горизонтальном положении ванночки.
Чтобы образовавшееся дно не всплывало в воде, к стенкам ван ночки,
на расстоянии %—1см от дна, припаивают поперечнолежащие уголь-
ники, которые и остаются в толще воскового дна.
1 I. Церезина 200 г, воску 100 г, сала 2 ложки, голландской сажи. II. Воска
четыре части, сала одна часть, голландской сажи. III. Воска сто частей, кани-
фоли десять частей, вазелина пять частей, анилиновой черной, растворимой в масле
краски одна часть (Р а й к о в).
* 4
Вскрываемое животное прикалывают в соответствующем положе-
нии (см. текст) к дну ванночки булавками; наливают воды так, чтобы
она покрывала животное, и приступают к дальнейшей работе.
Голубя, кролика вскрывают напрепаровальныхдосках
соответствующих размеров, например 25 х 40 см. К углам досок вби-
вают гвозди, к которым за ноги привязывают вскрываемое животное.
Для препарирования мелких беспозвоночных нужны препаровальные
лупы. Необходимы также микроскопы со средним (до 400 раз) и слабым
увеличениями.
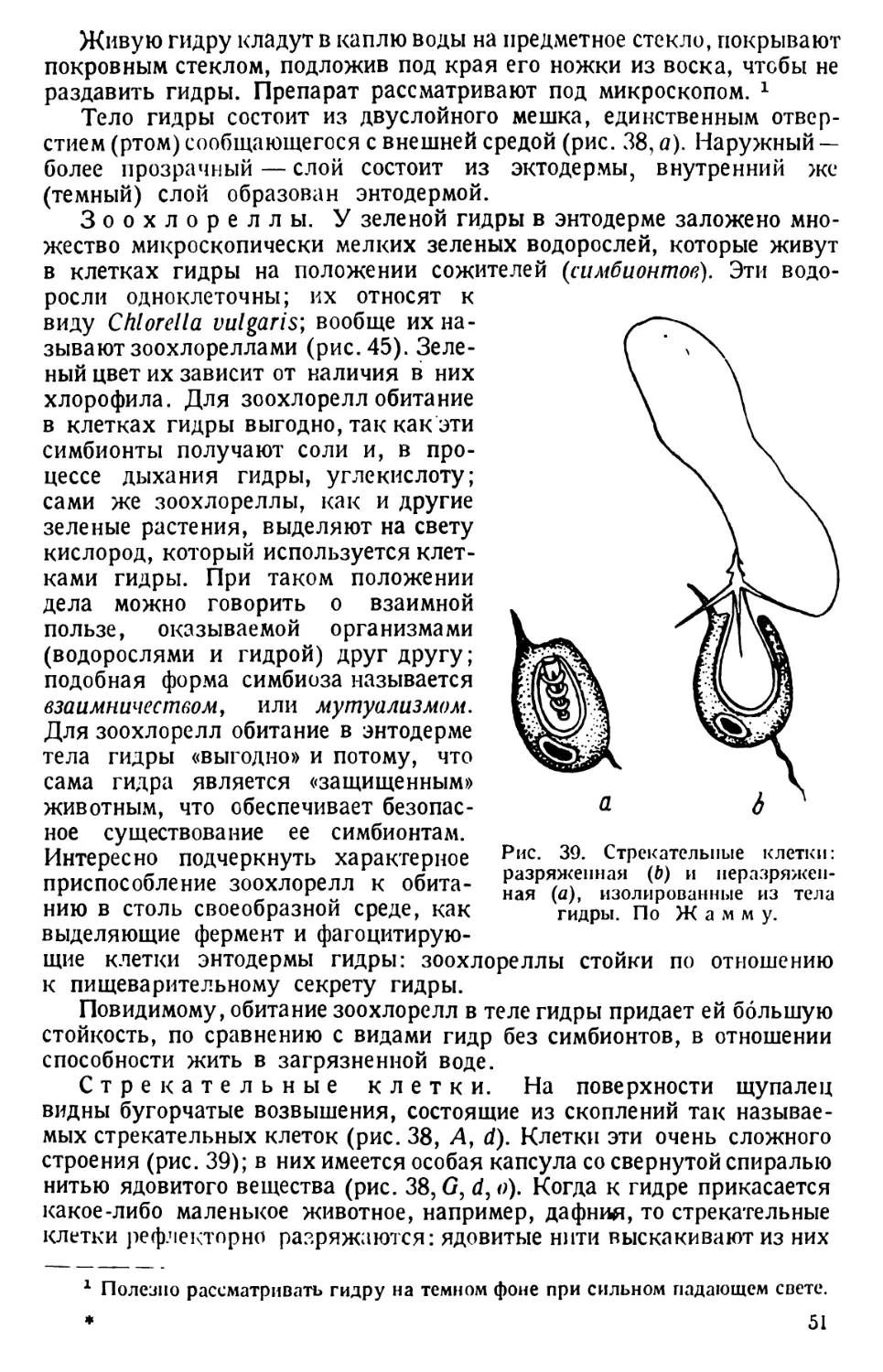
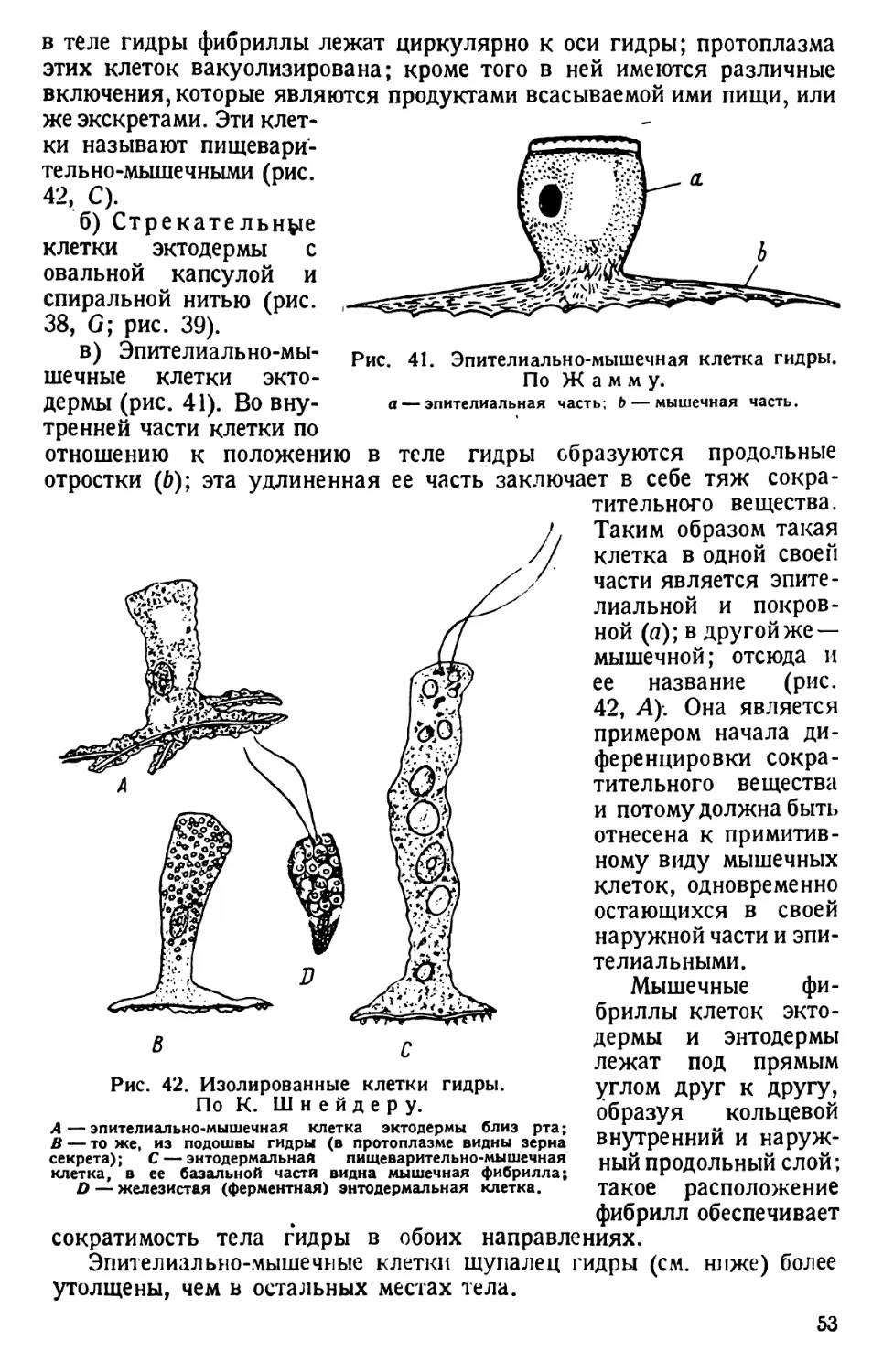
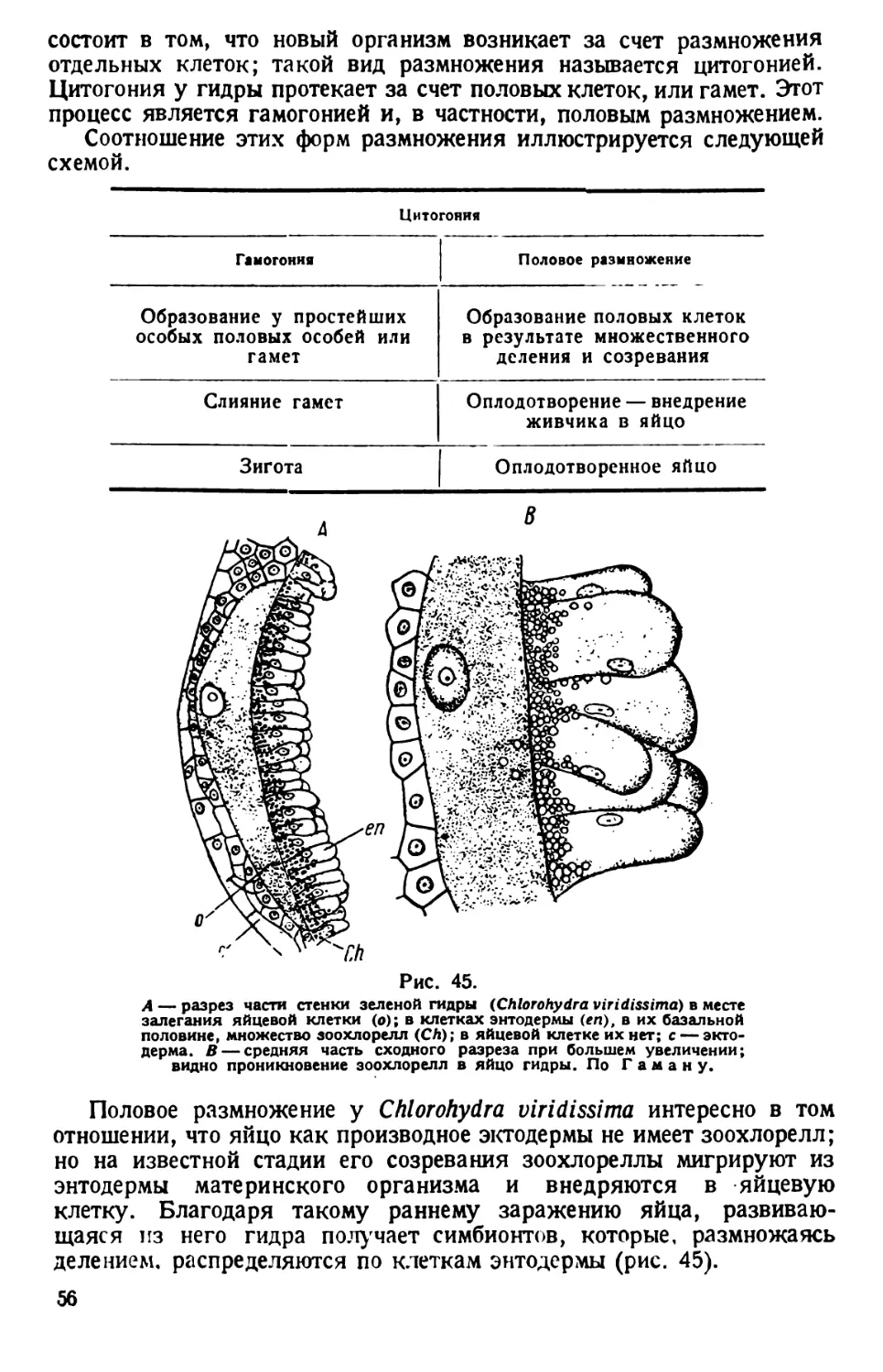
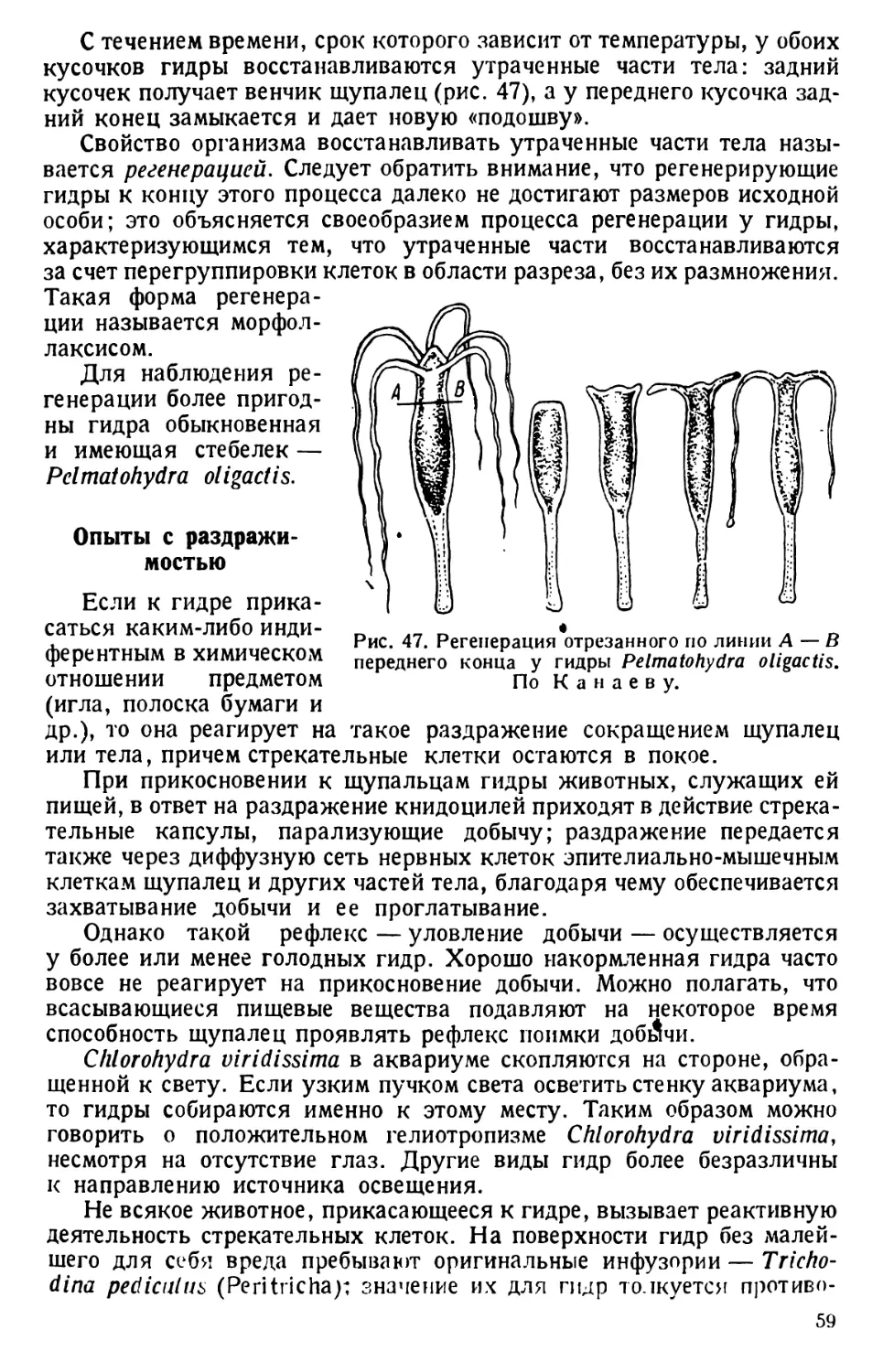
ТИП ПРОСТЕЙШИЕ (PROTOZOA)
КЛАСС РЕСНИЧНЫЕ (CILIATA s. INFUSORIA)
ТУФЕЛЬКА (PARAMAECIUM)
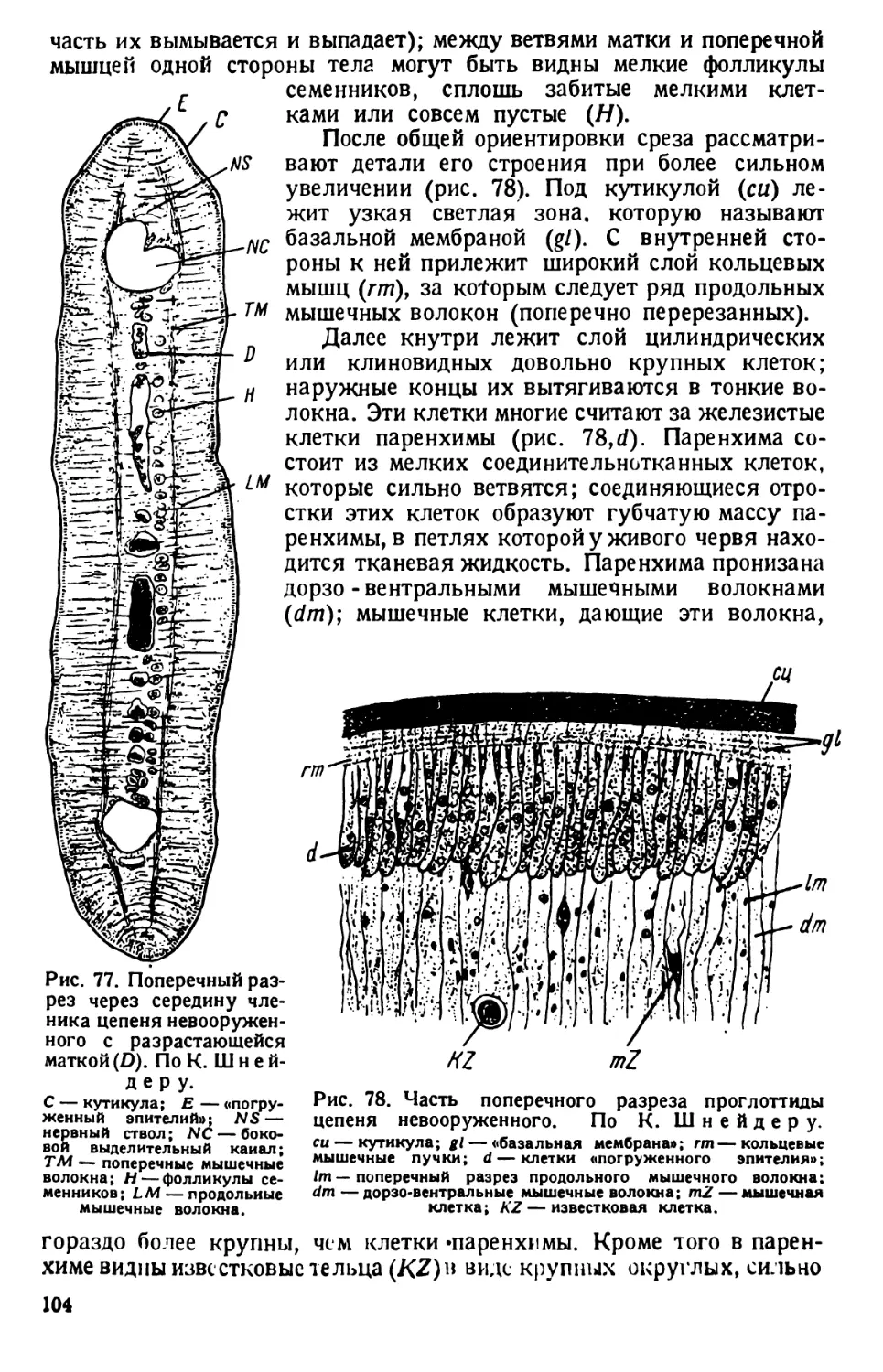
Технические замечания. 1) За 3—4 недели до начала занятий
заготовляют культуру инфузорий. Берут луговое сено, режут его и кладут в сте-
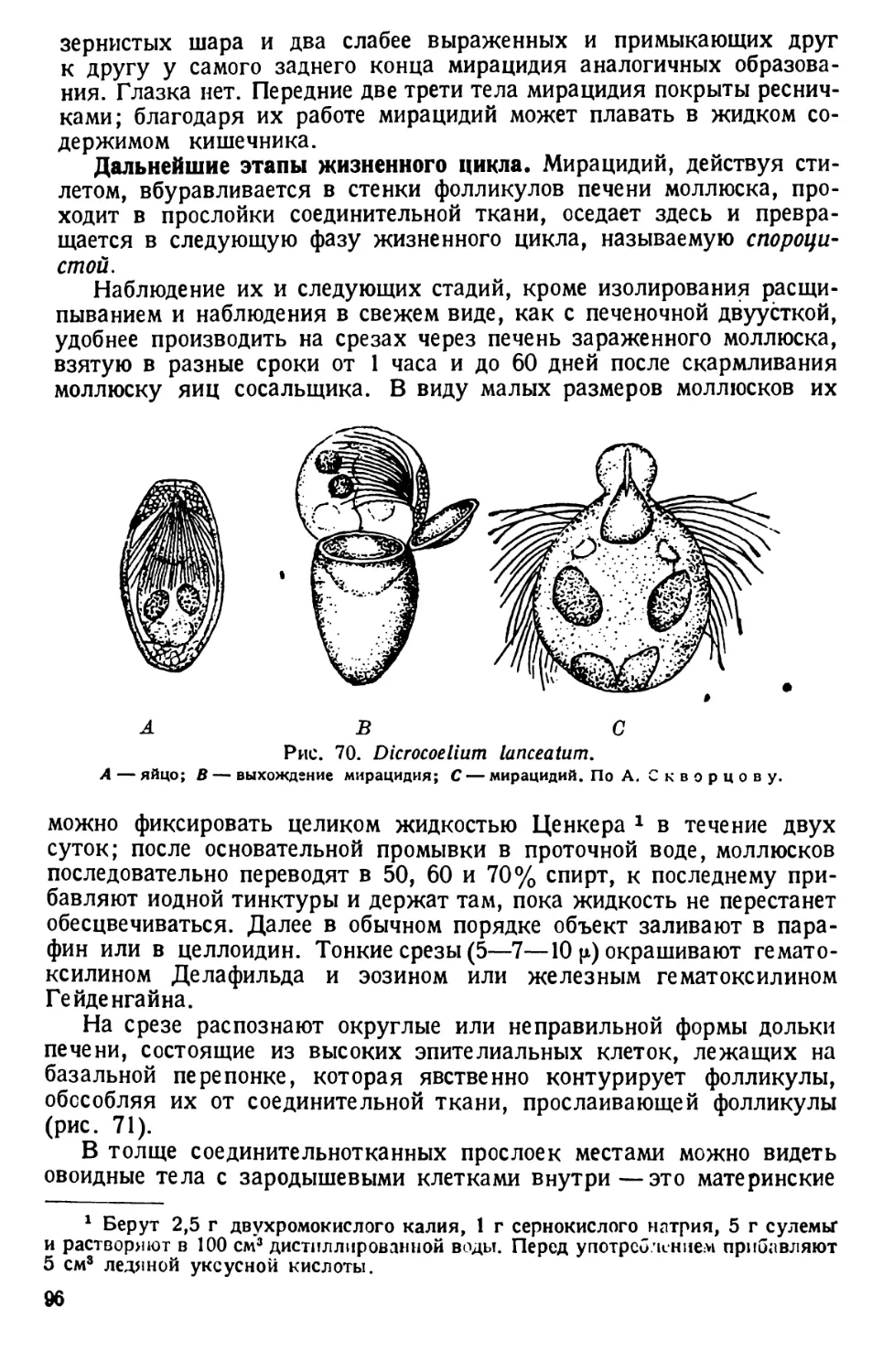
клянный сосуд с дождевой водой. Дня через три к этому сенному настою прили-
вают с полстакана воды из пруда или водоема, богатого разлагающимися органи-
ческими веществами.
С течением времени в культуре появляются различные инфузории; в конце
концов появляются и туфельки — Paramaecium. Культура оказывается наиболее
богатой недели через две после прибавления прудовой воды. На поверхности
культуры образуются точечные пленки из скопившихся бактерий, среди которых
движется много инфузорий.
2) Разнообразный материал по простейшим можно получить в пробах воды
из стоячих водоемов (пруды, старицы, канавы и др.). С водой захватывают части
грунта с разлагающимися листьями. Пробы берут из разных водоемов и сливают
в большую банку из-под варенья. Сюда же кладут несколько экземпляров ряски
или веточки водяных растений. Следят за тем, чтобы не было мелких ракообраз-
ных (дафний, циклопов и др.), присутствие которых не благоприятствует разве-
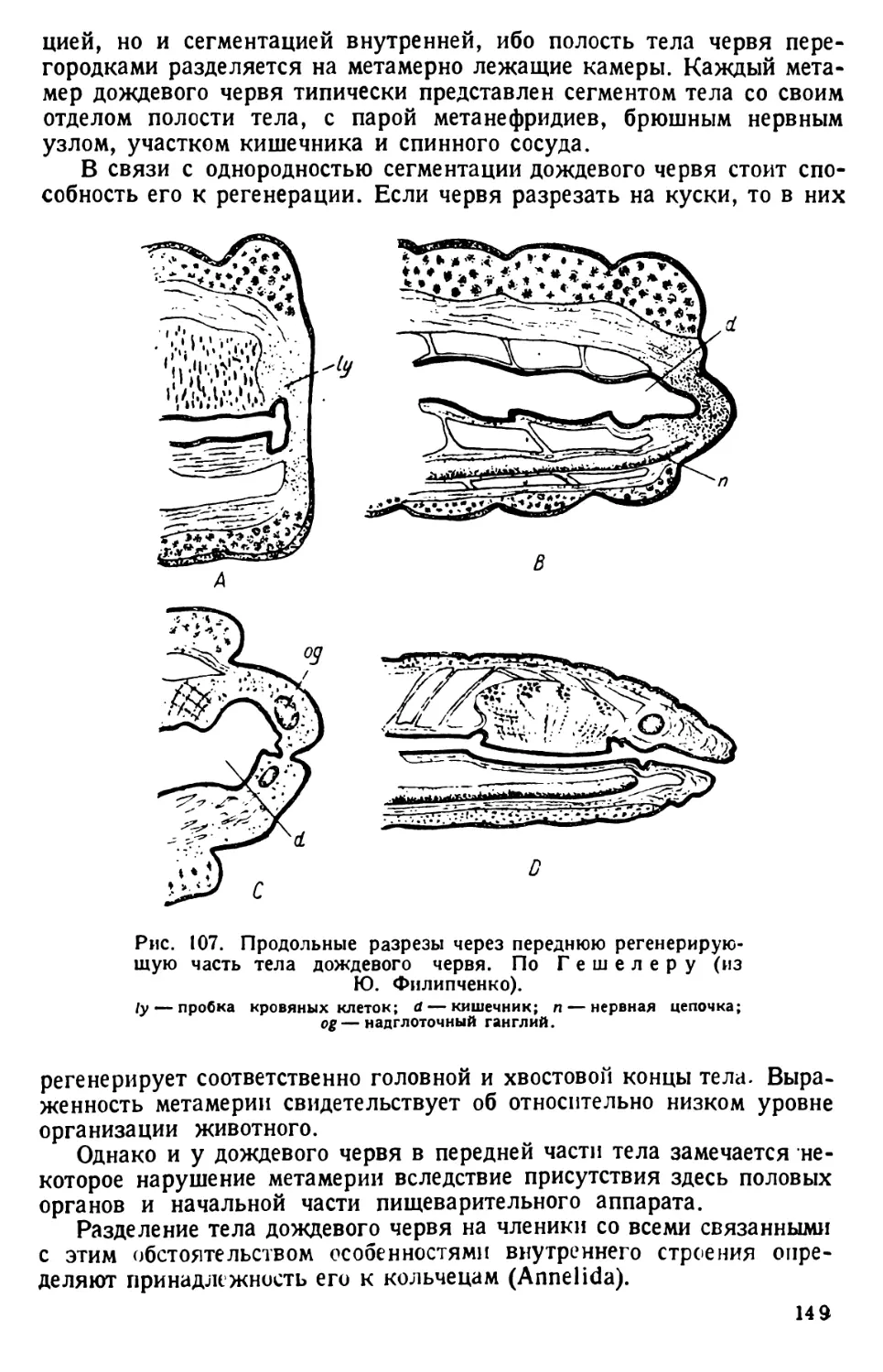
дению простейших. Банки держат у окна на рассеянном свету при комнатной
температуре под стеклянными покрышками. Протозойное население их сменяется;
поэтому необходимо время от времени просматривать пробы под микроскопом,
беря их пипеткой из разных мест периферии или середины банки из разной
глубины.
3) Массовые культуры парамеций можно получить и на другой питательной
среде. В чайные стаканы наливают кипяченой воды и опускают мешочек из «газа»
или тюля с листьями салата. Мешочек с его содержимым предварительно нужно
прокипятить. Стакан закрывают стеклянной пластинкой. В таком настое разви-
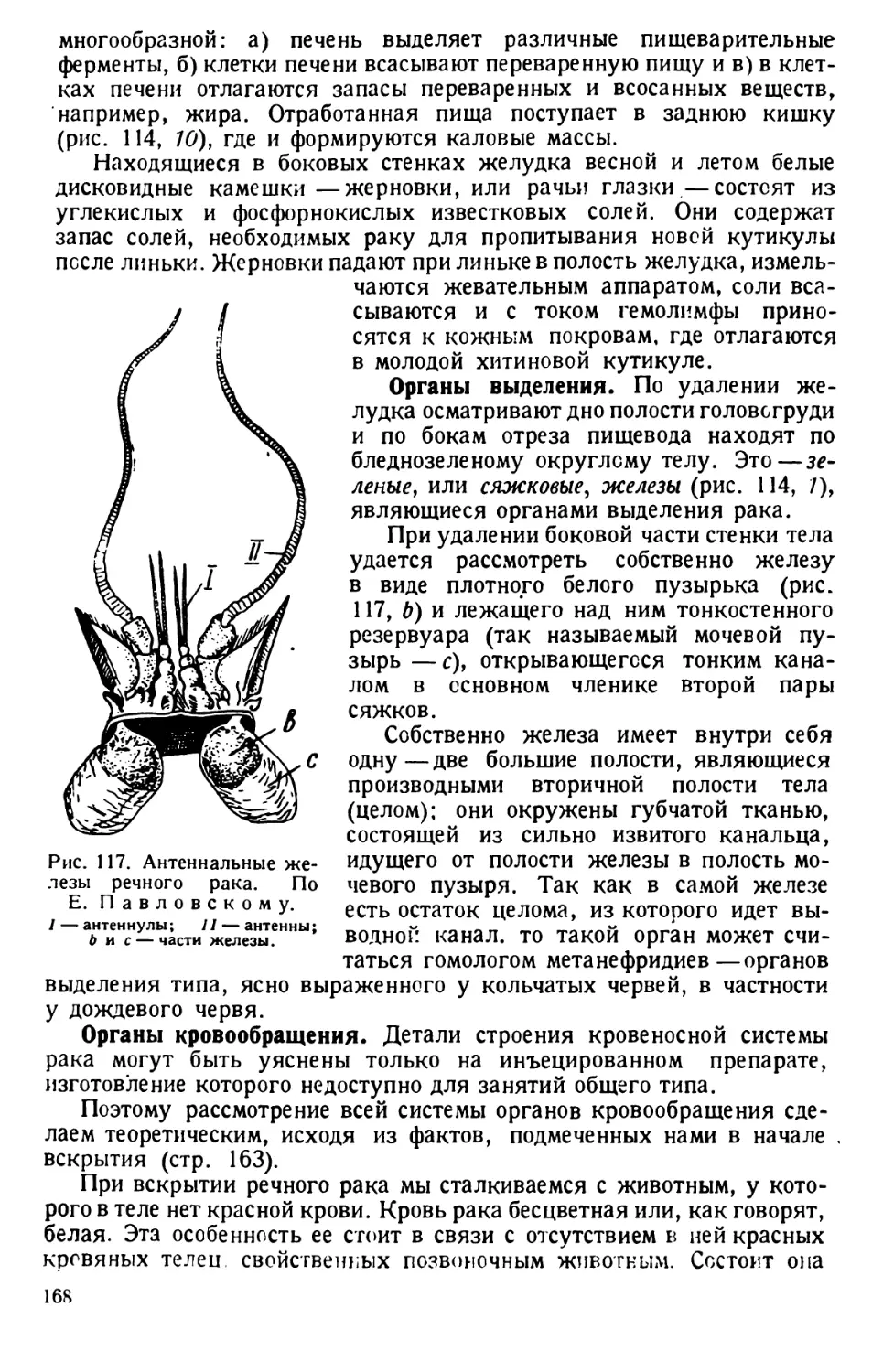
вается особенно благоприятная для питания туфелек бактериальная флора. Еще
лучше, если основной жидкостью для культуры берется 0,025% водный раствор
либихоьского экстракта.
4) Легко можно культивировать инфузорий на обыкновенном агаре, 1 * кото-
рый употребляется для культуры амёб.
Несколько инфузорий пипеткой переносят на агар-агар, где они быстро
размножаются при содержании культуры в тепле (15—20° С). Получающаяся
культура пригодна для изучения вегетативных форм и их деления.
5) Культуру инфузорий получают и на сенном отваре. 10 г сухого сена кипя-
тят в колбе с чистой водой в течение 30 минут. По остывании сенной отвар раз-
ливают в несколько стаканов или равных по величине банок для варенья или
стеклянных цилиндров. Если отвар получился крепкий, его разбавляют до цвета
жидкого чая. Отвар оставляют открытым на несколько дней, пока в нем не разо-
вьются в массе сенные бактерии (Bacillus subtilis), являющиеся прекрасной пи-
щей для инфузорий. В банки пускают по несколько штук инфузорий — туфелек,
которые быстро размножаются в питательной среде. Культура годна для исполь-
зования обычно уже через неделю.
1 Агар-агара — 0,5—1,0 г, водопроводной воцы — 90 см3, обыкновенного
щелочного питательного бульона — 10 г. После кипячения разливают в чашечки
Петри и стерилизуют.
5
Необходимо ставить несколько проб или культур, так как не всякая проба
дает непременно хорошие результаты.
Банки с культурами следует держать в покое и не встряхивать их. Дело
в том, что различные виды простейших занимают определенные места, например,
в поверхностном слое, со стороны, обращенной к свету, у дна и т. д. Из этих мест
их легко выловить пипеткой в более или менее обильном количестве; при встря-
хивании же или толчках простейшие рассеиваются по всей толще воды, что пре-
пятствует их добыванию.
6) Богатую и почти чистую культуру Paramaecium (в большинстве Р. aurelia)
Steinach получал на экстракте свиного мозга. 120 г свиного мозга разрезают
на кусочки и раздавливают в воде; смесь оставляют стоять на полдня и затем про-
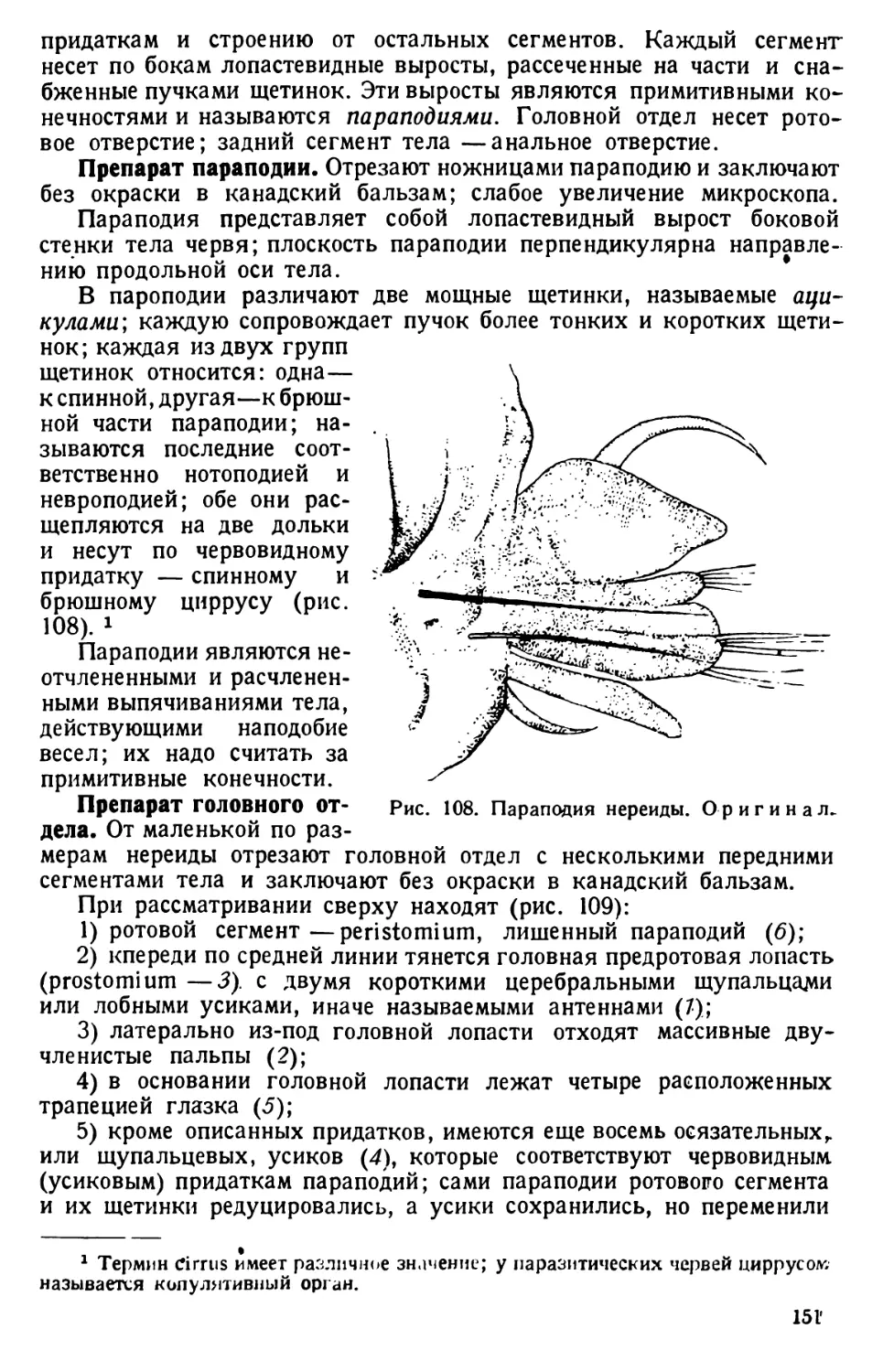
давливают ее через чистую салфетку. Полученный «мозговой сок<> разбавляют
водой до 1 л и разливают в три-четыре банки, куда прибавляют по 1 см’ сенного
настоя. Инфузории появляются дня через три. Поверхностную пленку с культуры
удаляют каждый день.
7) Для получения возможно более чистых культур 0,025% водный раствор
либиховского экстракта засевают Bacillus pro tens, который вытесняет другие
бактерии. Затем в жидкость пускают многократно отмытых в стерильной воде
парамеций, которые размножаются за счет питания бактериями.
8) Для изучения конъюгации пользуются водными культурами инфузорий,
для чего в часовое стекло помещают инфузорий из разных культур.
9) Культура держится в своей посуде известное время (недель до б, смотря
по емкости сосуда и богатству культуры). Чтобы поддерживать культуру, ее
необходимо время от времени пересевать, т. е. полстакана культуры прибавлять
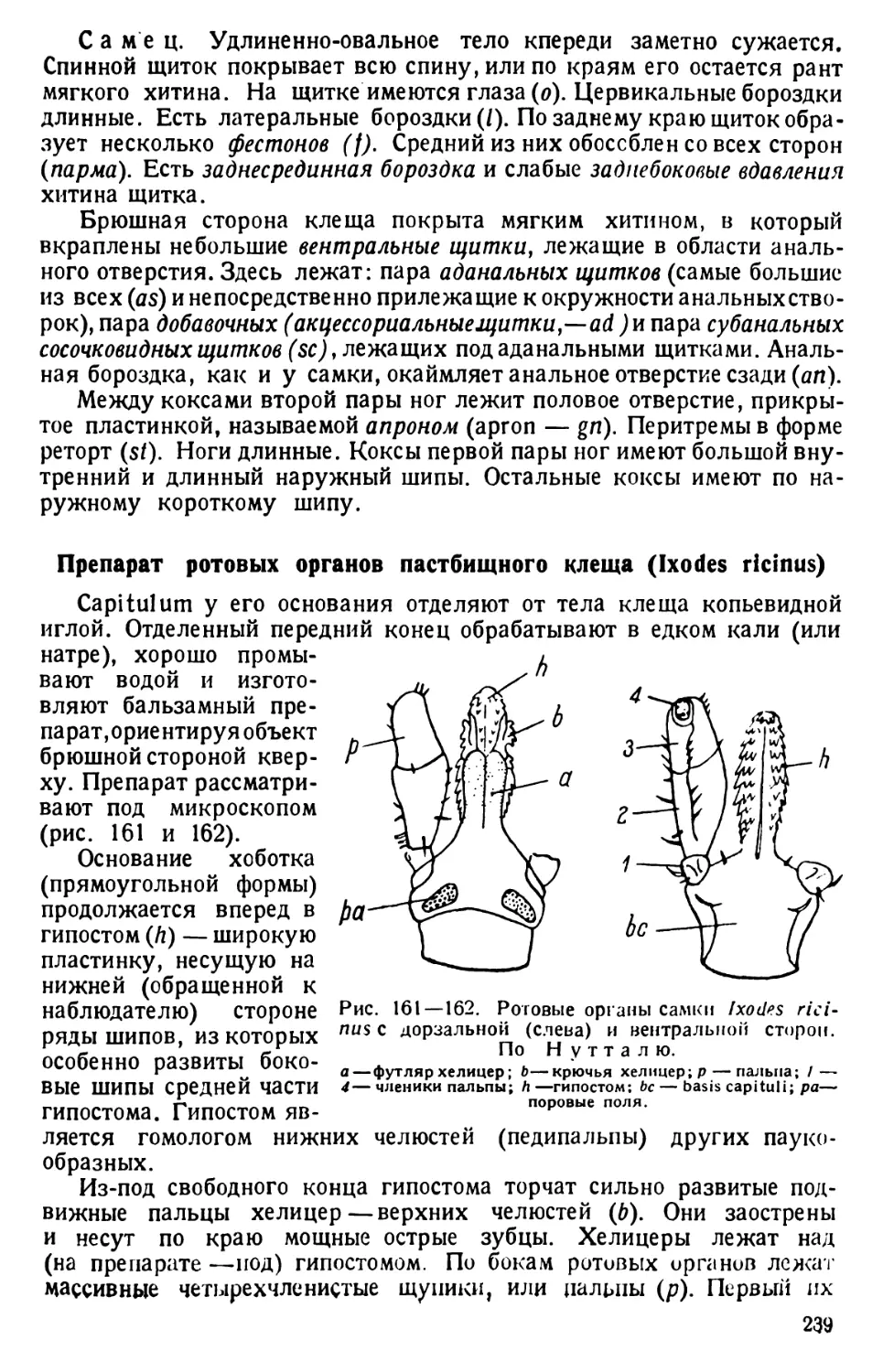
в банку со свежим сенным настоем или иной питательной жидкостью.
10) Чтобы получить достаточное количество парамеций для занятий, готовую
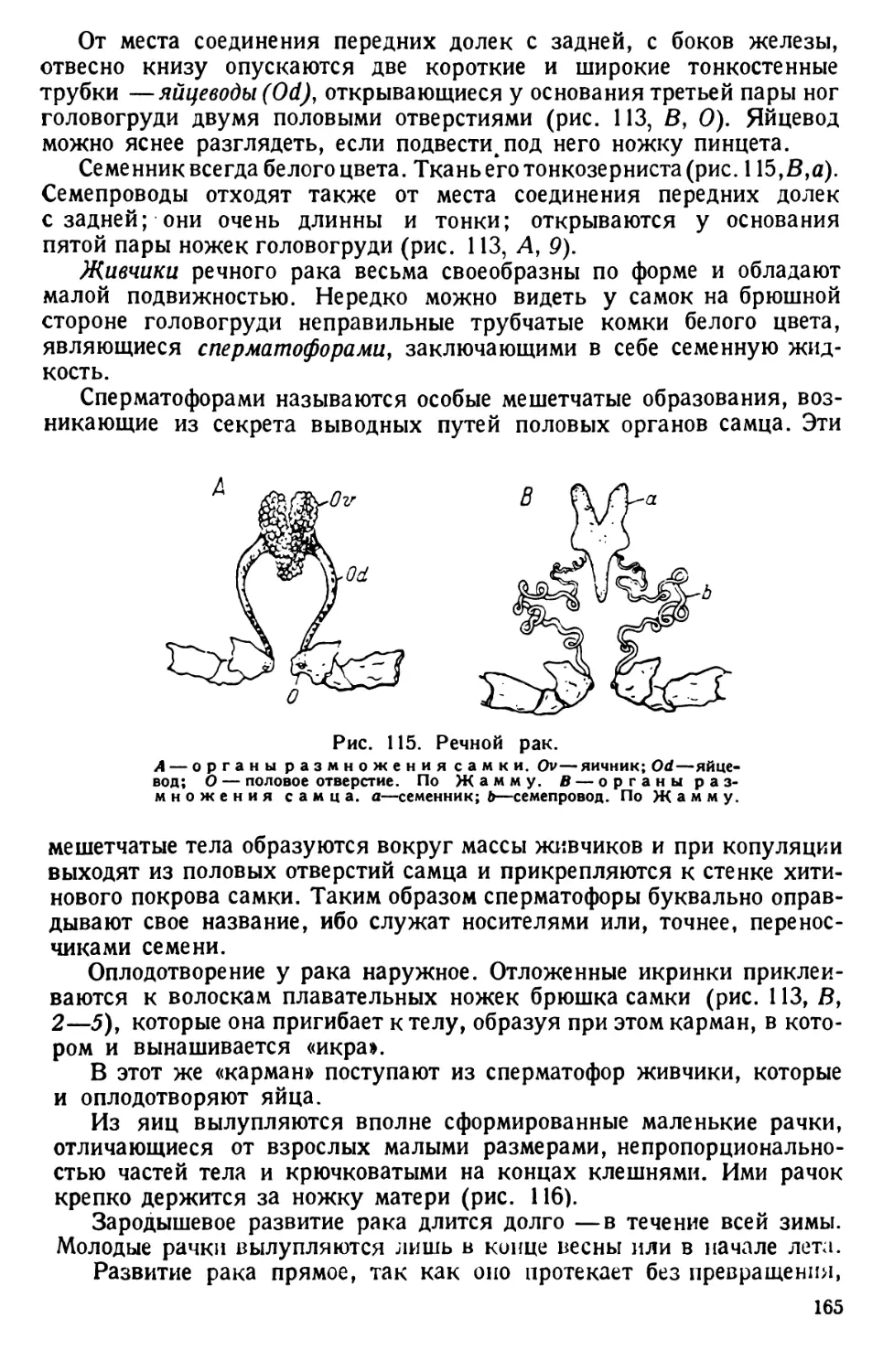
культуру с инфузориями наливают в широкую толстостенную «водомерную»
трубку с полметра длиной, заткнутую на конце пробкой. Трубку вертикально
зажимают в штатив. Через некоторое время инфузории поднимаются в верхний
слой воды, откуда их легко выбирают пипеткой. Поднятие инфузорий в верхние
слои водяного столба объясняется отрицательным геотропизмом, т. е. движением
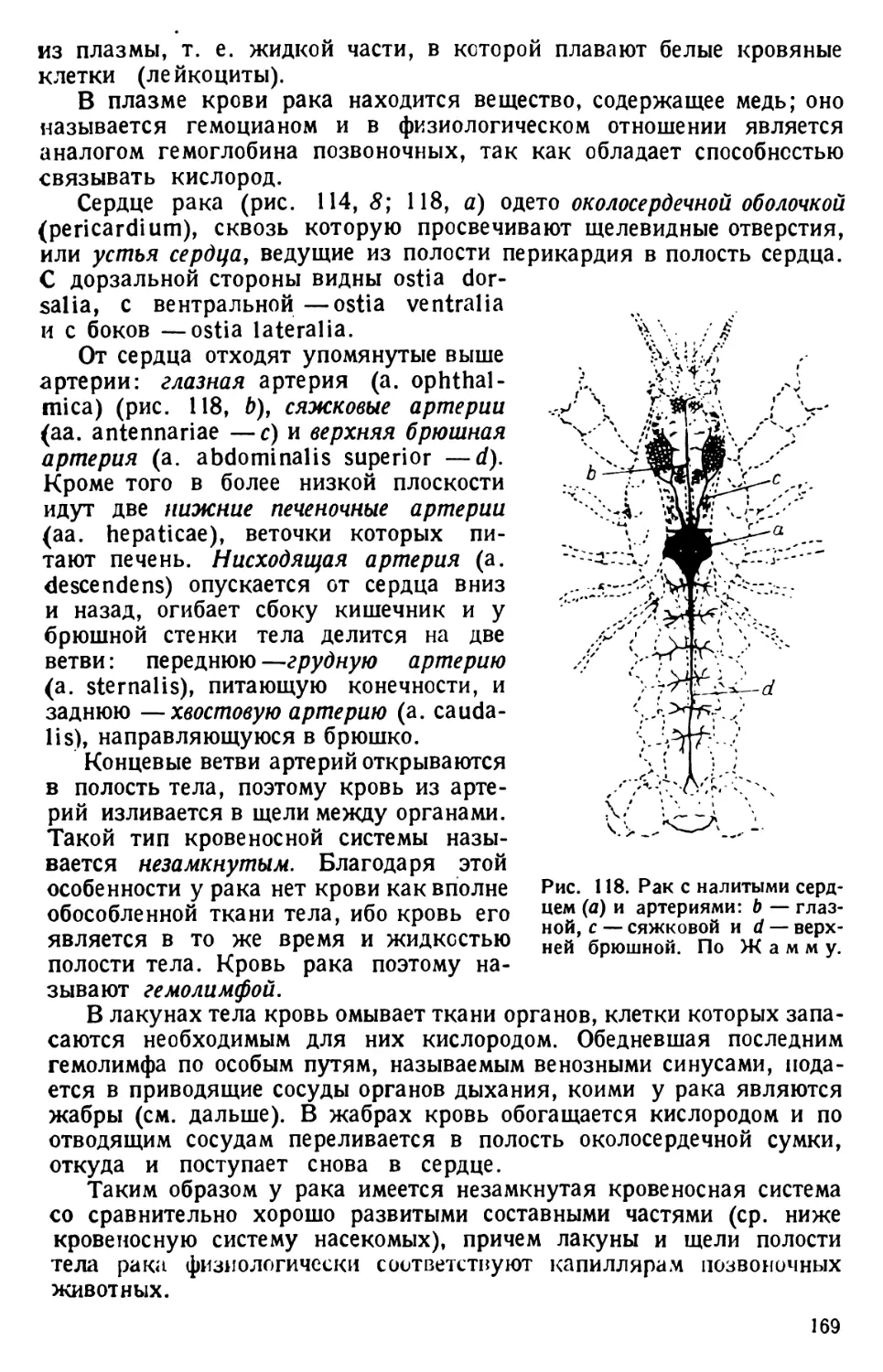
их от центра земного притяжения.
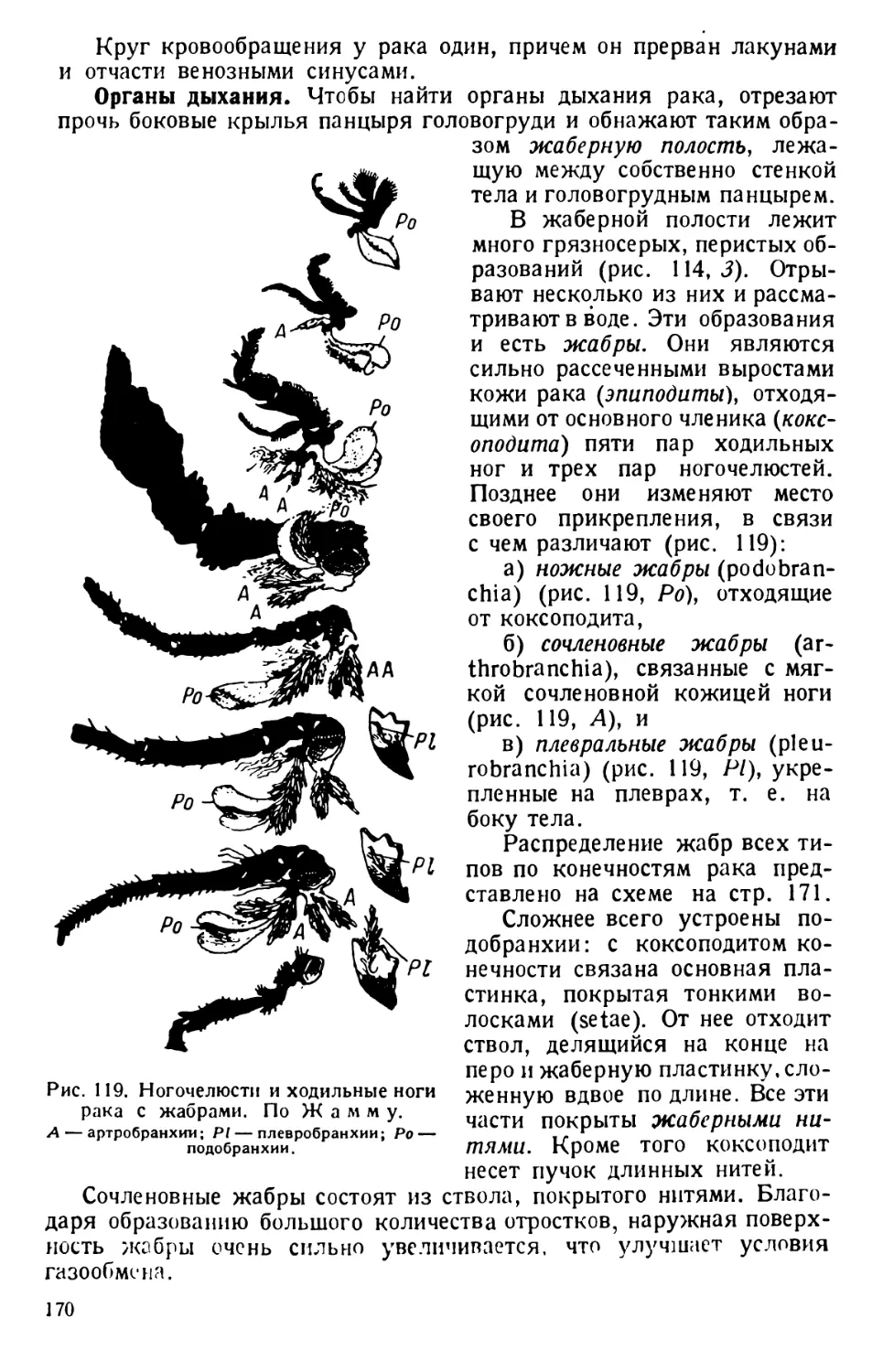
Наблюдения над живыми инфузориями
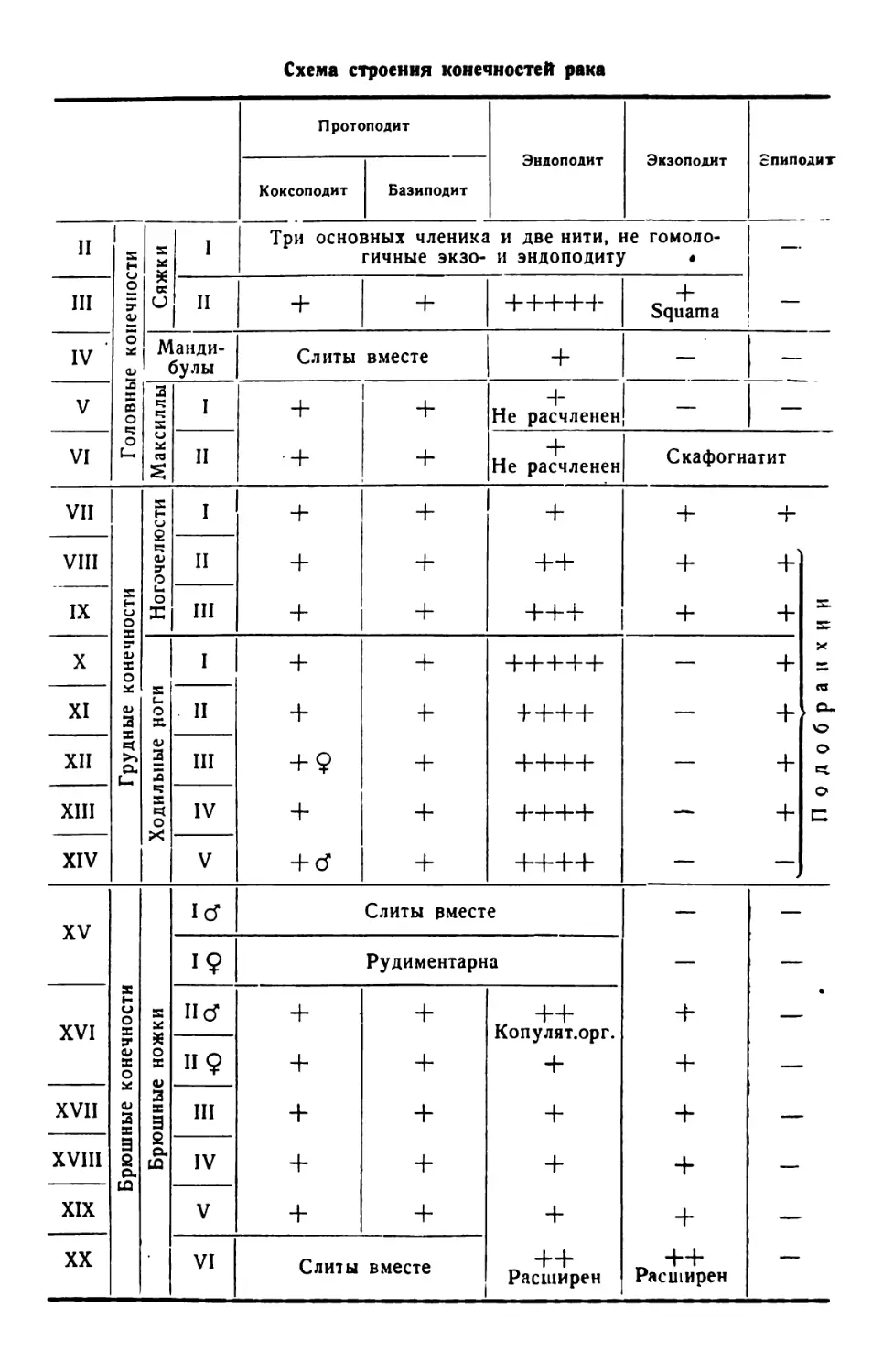
Методика наблюдений. В каплю воды на предметном стекле пи-
петкой переносят несколько туфелек, покрывают препарат покровным
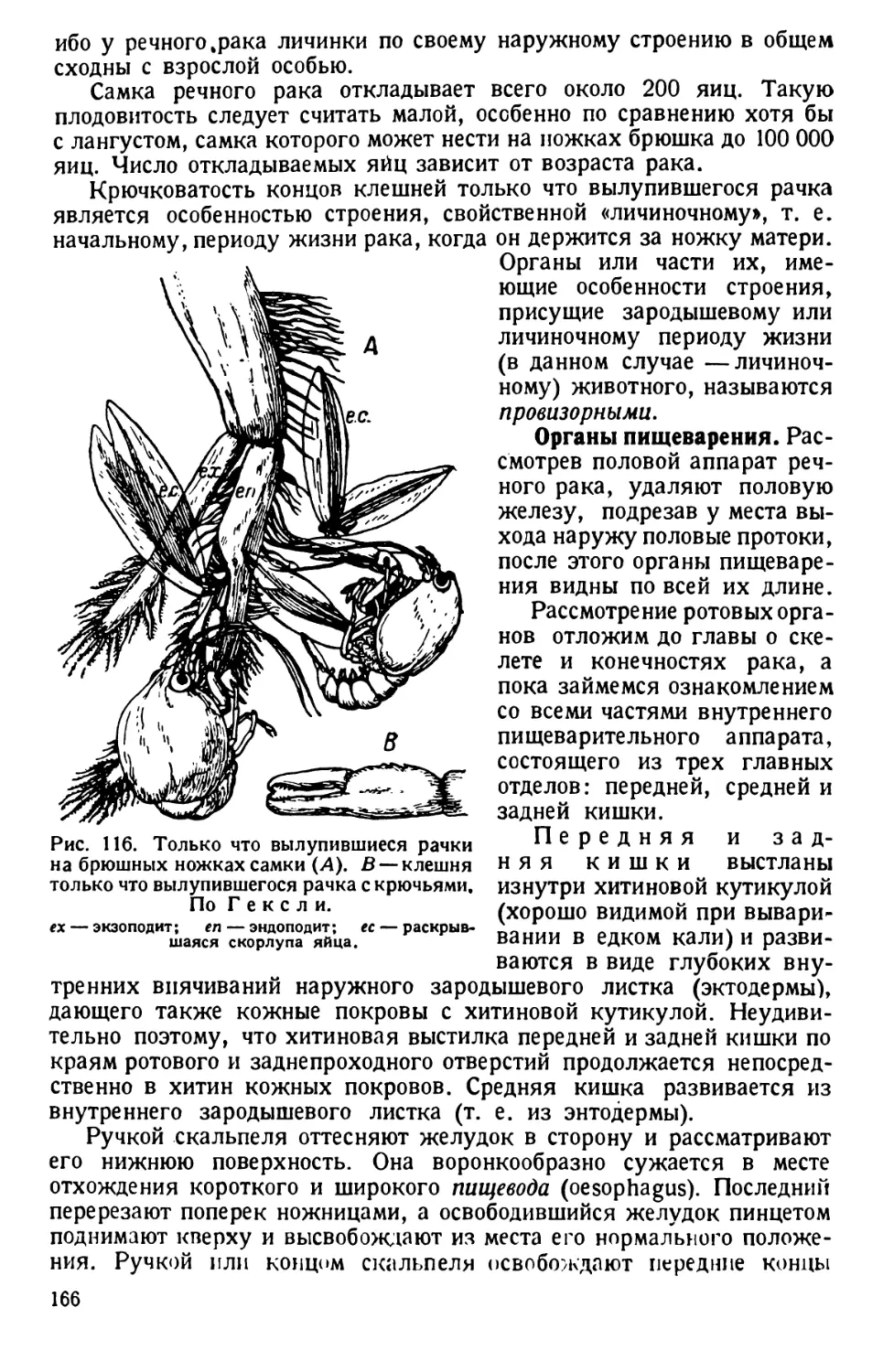
стеклом и рассматривают его под слабым увеличением микроскопа.
В поле зрения оживленно движутся маленькие организмы прибли-
зительно овальной формы. Более детально их можно рассмотреть лишь
при сильном увеличении; однако наставить препарат на фокус не
удается вследствие быстрого исчезновения инфузорий из поля зрения.
Чтобы задержать туфелек, на препарат наносят капельку раствора
гуммиарабика 1 или слабого раствора желатины, перемешивают все
осторожно иглой, покрывают препарат покровным стеклом и иссле-
дуют под микроскопом.
Еще проще можно приготовить препарат, положив на предметное
стекло несколько перепутанных волокон гигроскопической ваты; затем
на вату наносят культуру инфузорий и закрывают препарат стеклом
так, чтобы в нем не было пузырьков воздуха. Между волокнамй ваты
образуются маленькие камеры, в которых и застревают инфузории.
Из других веществ для той же цели можно применять агар-агар,
вишневый клей и даже просто слюну. Замедляющие движение инфу-
зорий вещества можно прибавлять к целой культуре или к ее более
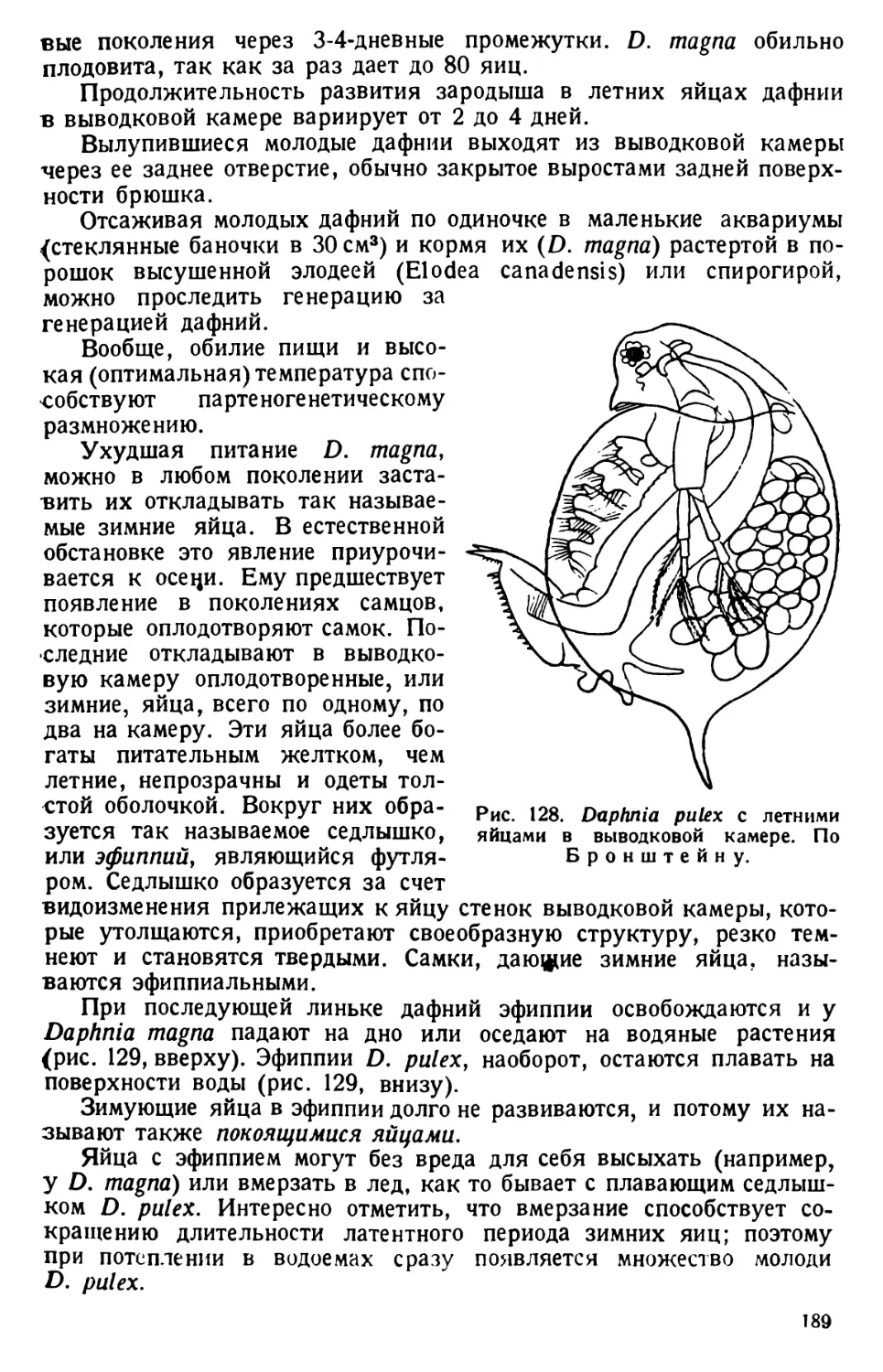
1 Гуммиарабик кристаллический (Gummiarabicutn puriss. crystallisation)
растворяют в воде в пропорции шести частей на двадцать частей воды.
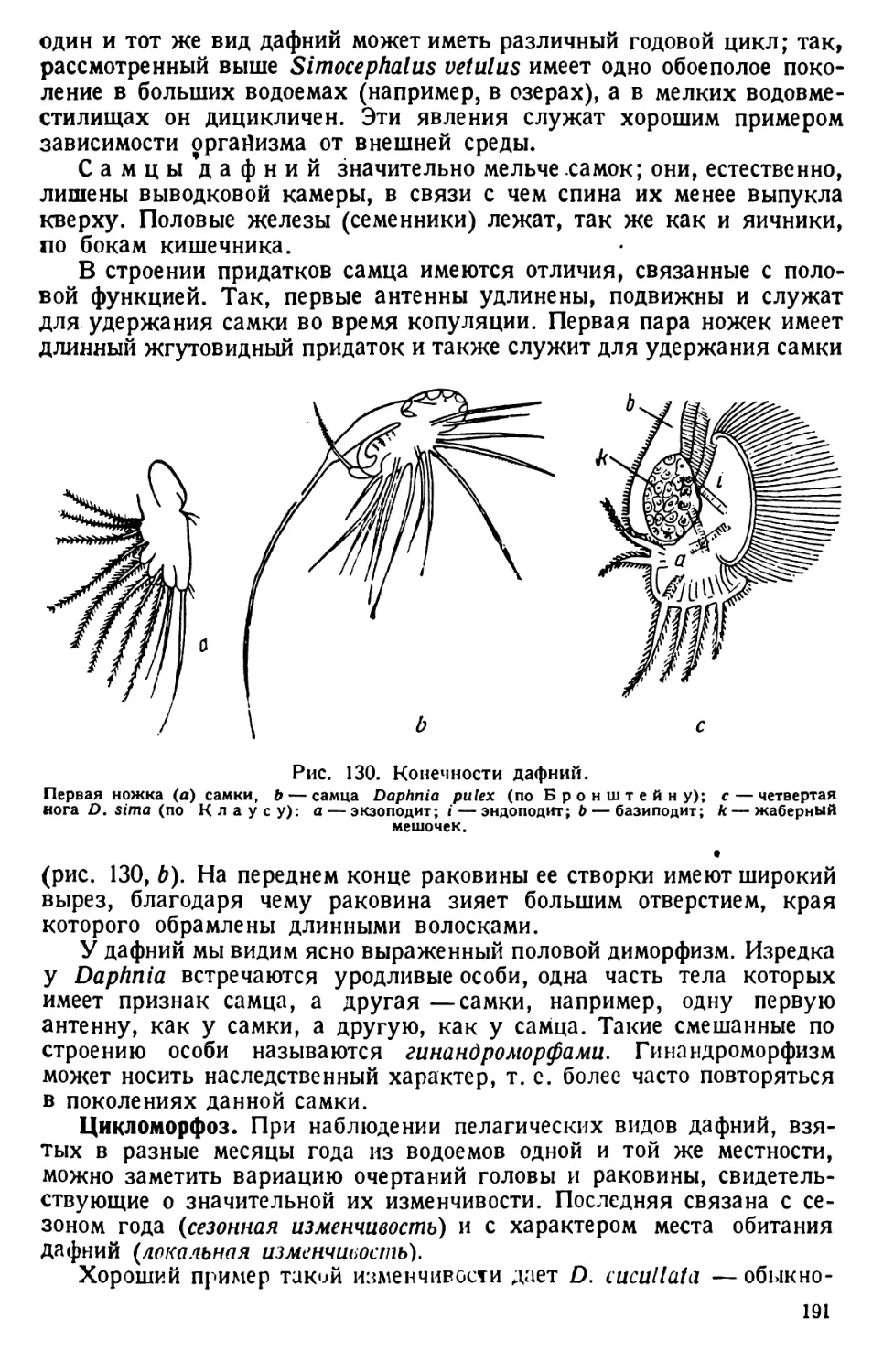
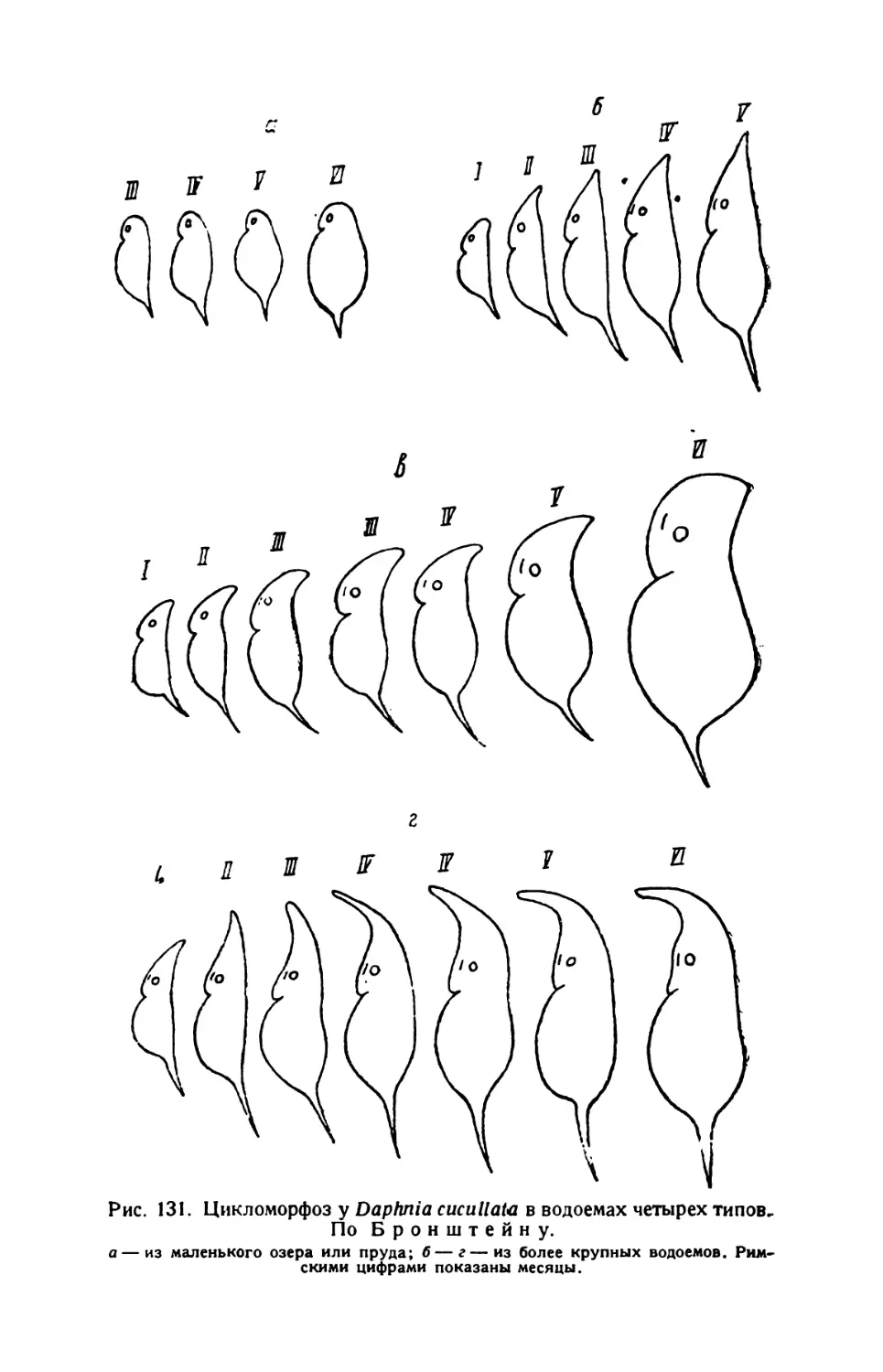
6
или менее значительной части. Для этой цели особенно удобен карра -
генсгый мех (Lichen carragaheen). Для ускорения процесса «берут
одну или две маленьких веточки каррагеновего мха, промывают
в 0,5—1% растворе соды и опускают на дно низкой и широкой про-
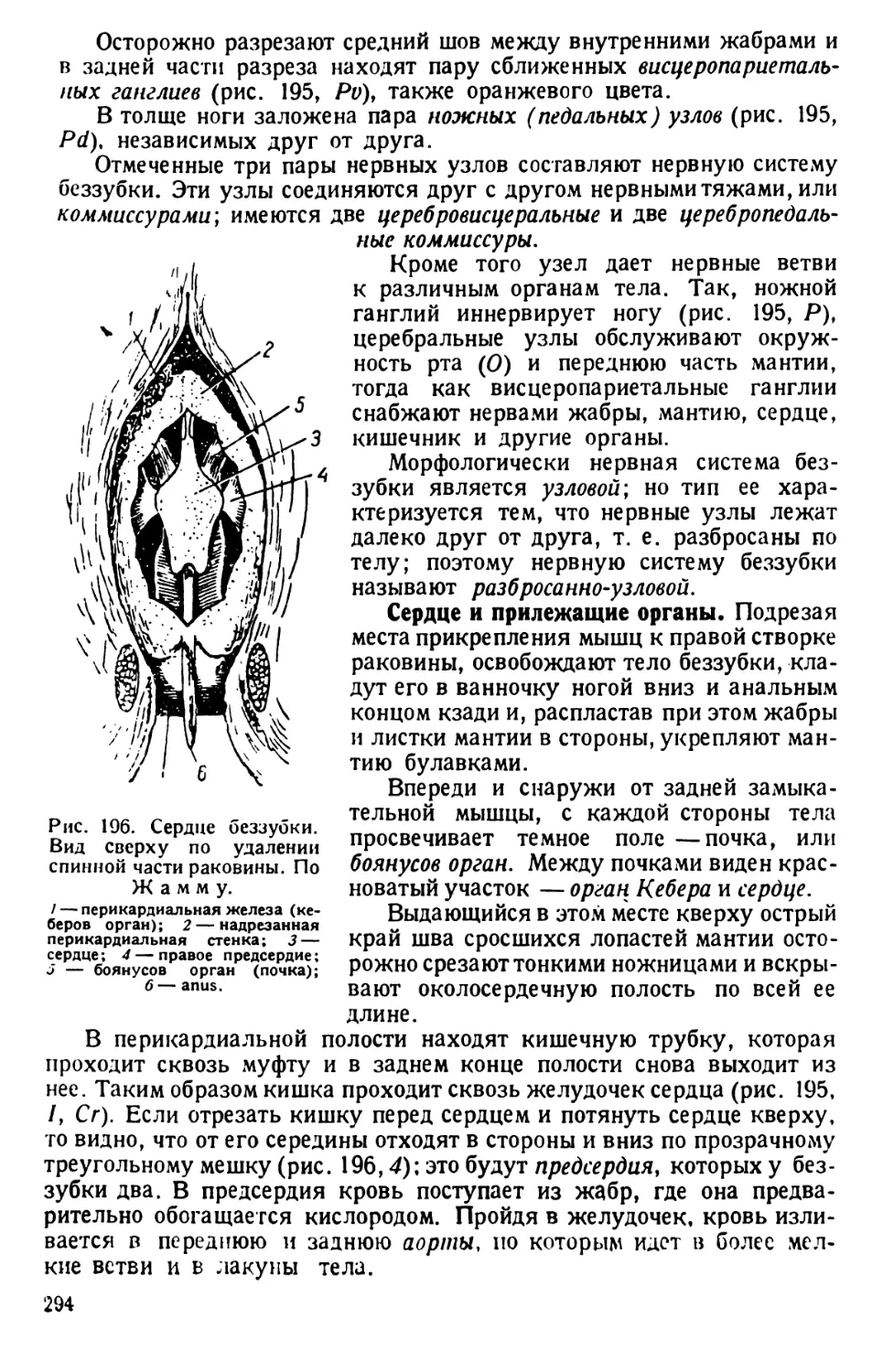
бирки. Затем в последнюю наливается 5—8 см3 обыкновенной бога-
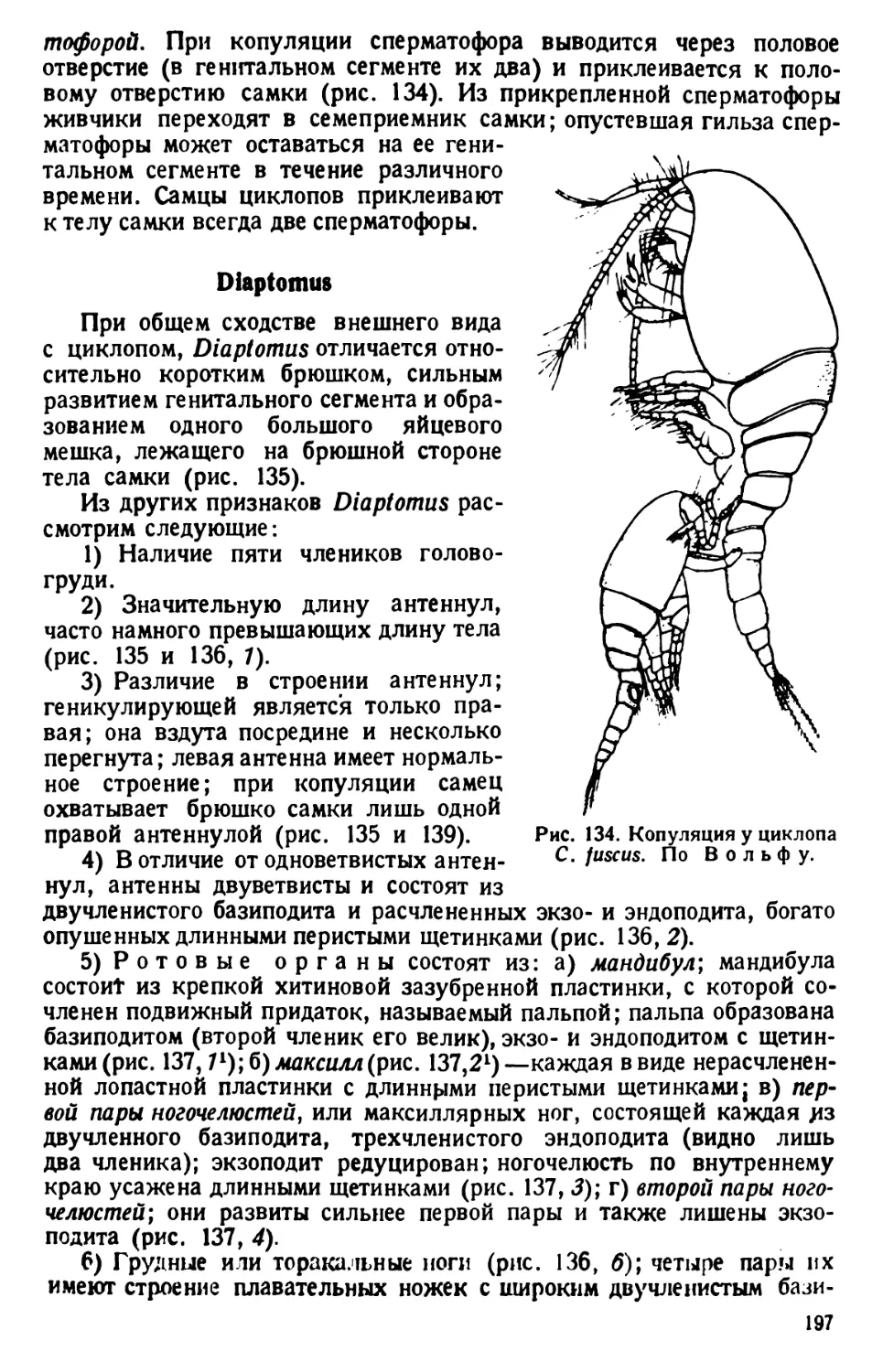
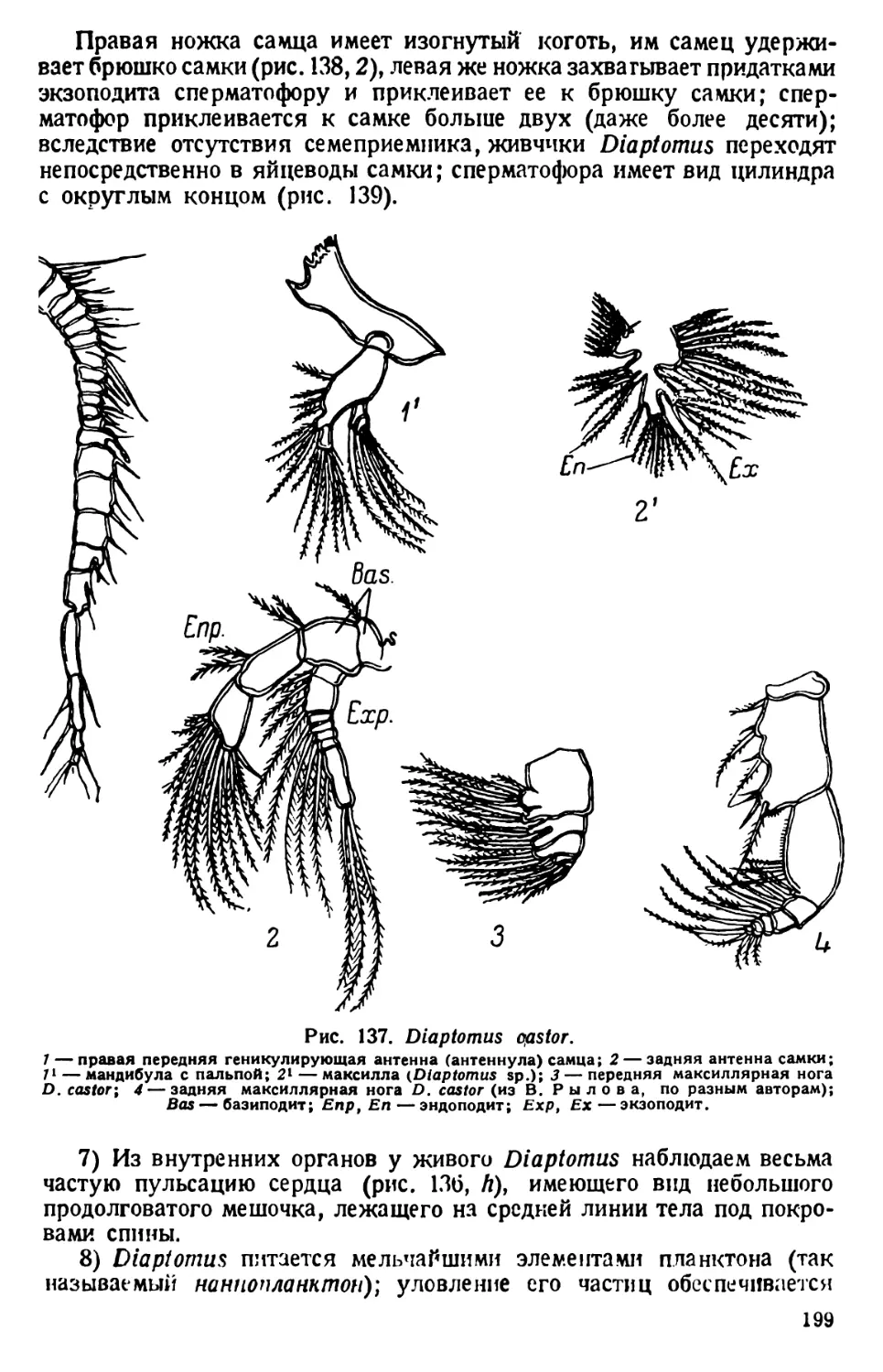
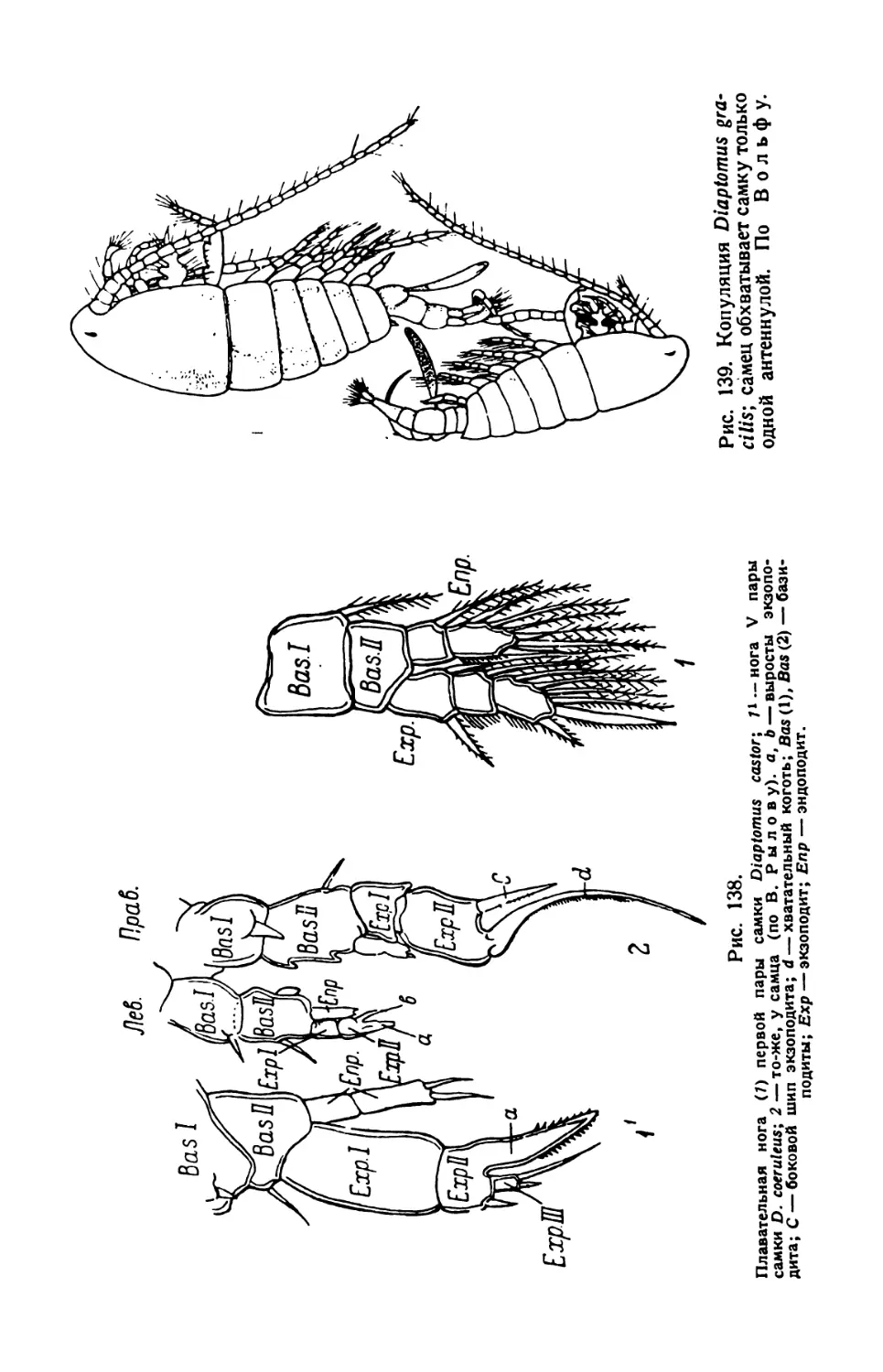
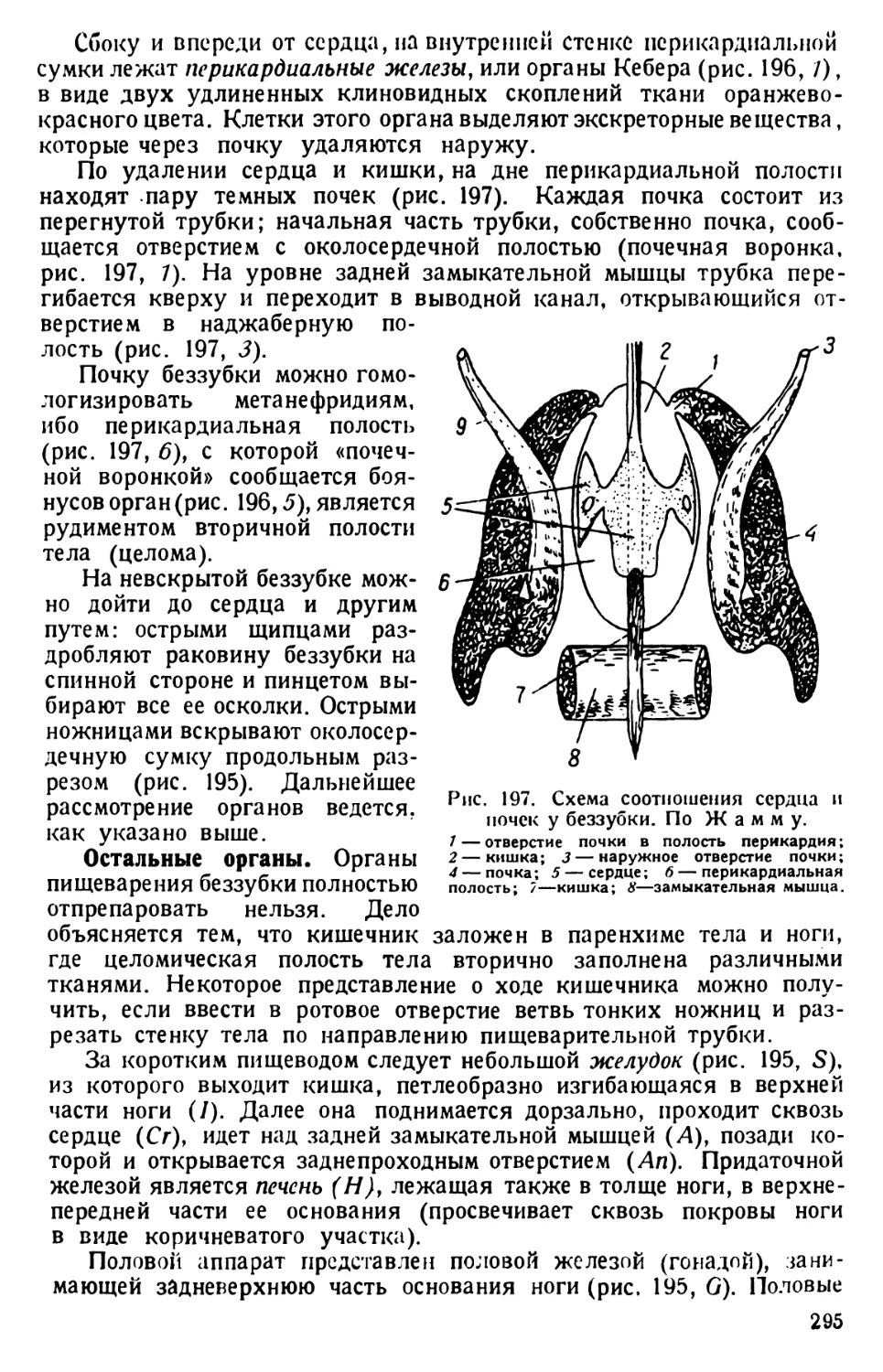
той культуры инфузорий. Уже через несколько часов, а еще лучше
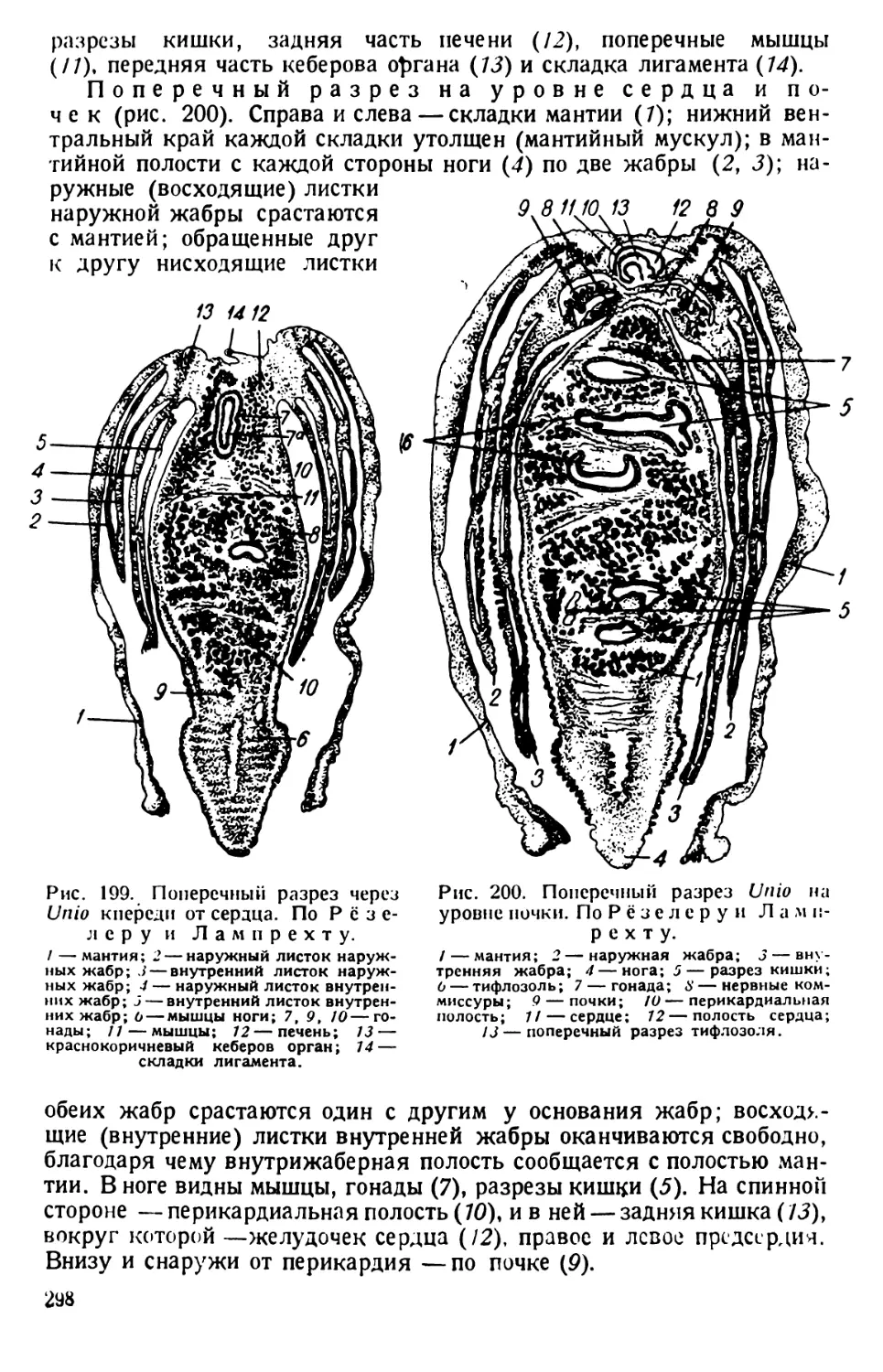
через день, образуется студенистая масса, в которой простейшие
движутся чрезвычайно медленно» (В. Догель, 1926).
Для более точного наблюдения покровное стекло ставят на высокие
ножки. Масса изготовляется сплавливанием при нагревании 2,5 частей
пчелиного воска с одной частью венецианского терпентина. Хорошо
размешанную и застывшую массу разрезают на пластинки или кубики.
Размягченную сдавливанием пальцами массу царапают углами покров-
ного стекла так, чтобы на них укрепились частицы восковой массы.
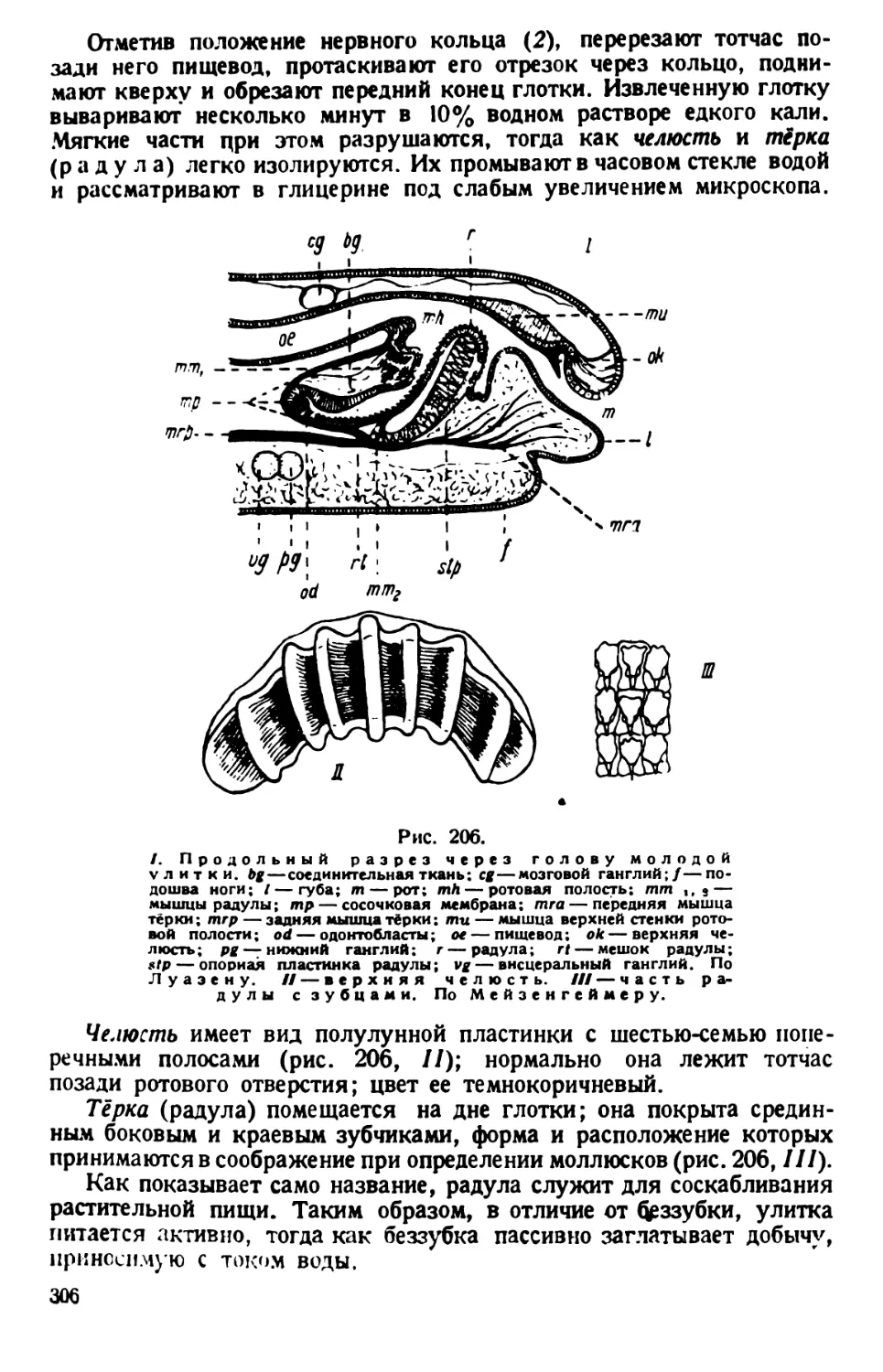
Покровное стекло держат непременно за ребра, но не за поверхность,
чтобы избежать загрязнения его отпечатком концов пальцев, мешаю-
щим последующему микроскопированию.
Покровное стекло с ножками опускают на небольшую каплю жид-
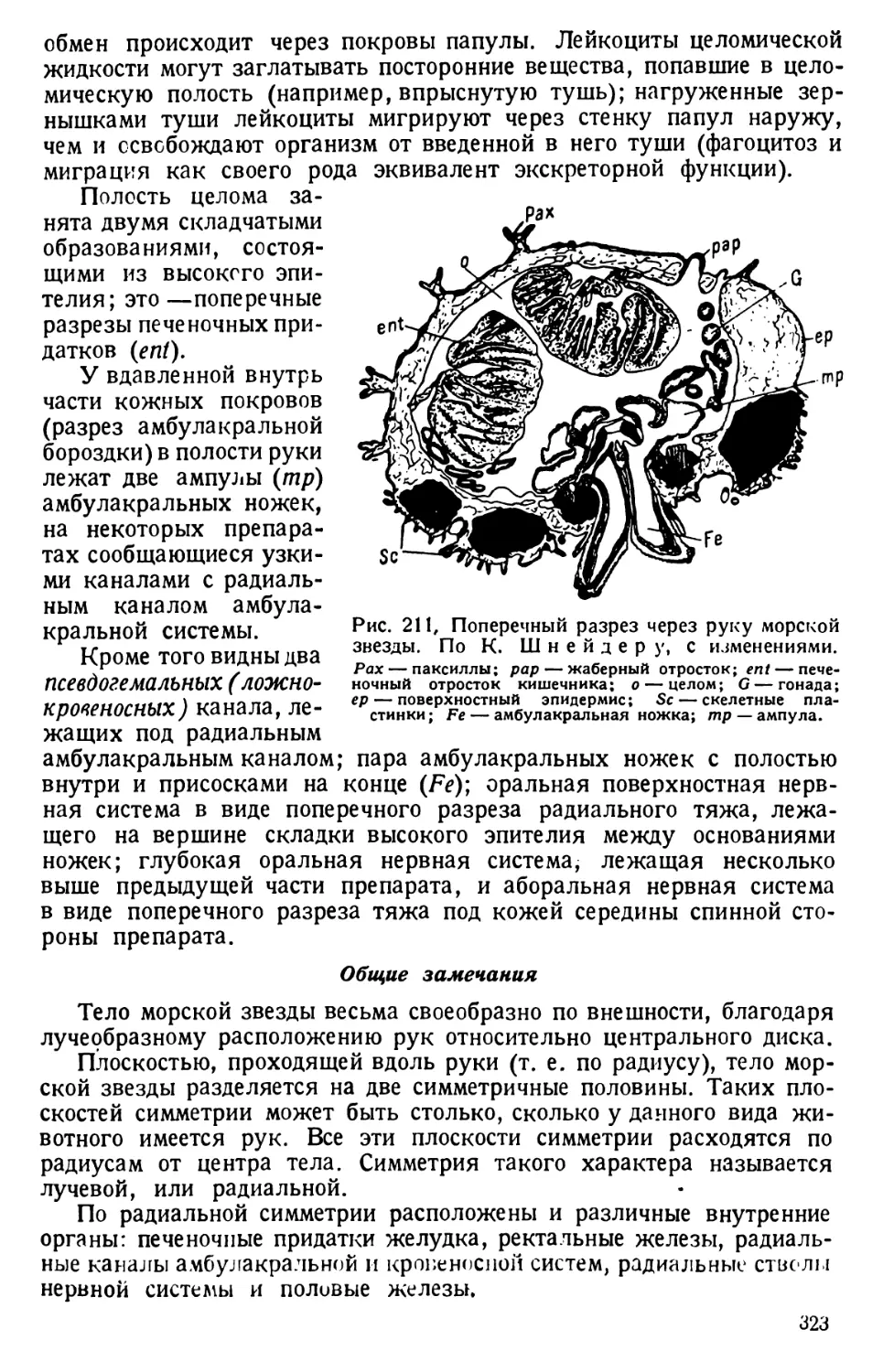
кости с парамециями и поочередно осторожно придавливают тупым
концом пинцета углы покровного стекла к предметному стеклу; при
этом находящаяся между ними восковая масса расплющивается,
а капля жидкости растекается по капиллярному пространству между
стеклами. Цель достигается лишь тогда, когда покровное стекло
придавливается к сухой поверхности предметного стекла.
Степень сдавливания контролируют под микроскопом, доводя тако-
вую до желательного замедления движения простейшего или до его
полной остановки. Испаряющуюся с краев воду возмещают добавле-
нием пипеткой жидкости культуры, из которой был взят живой ма-
териал.
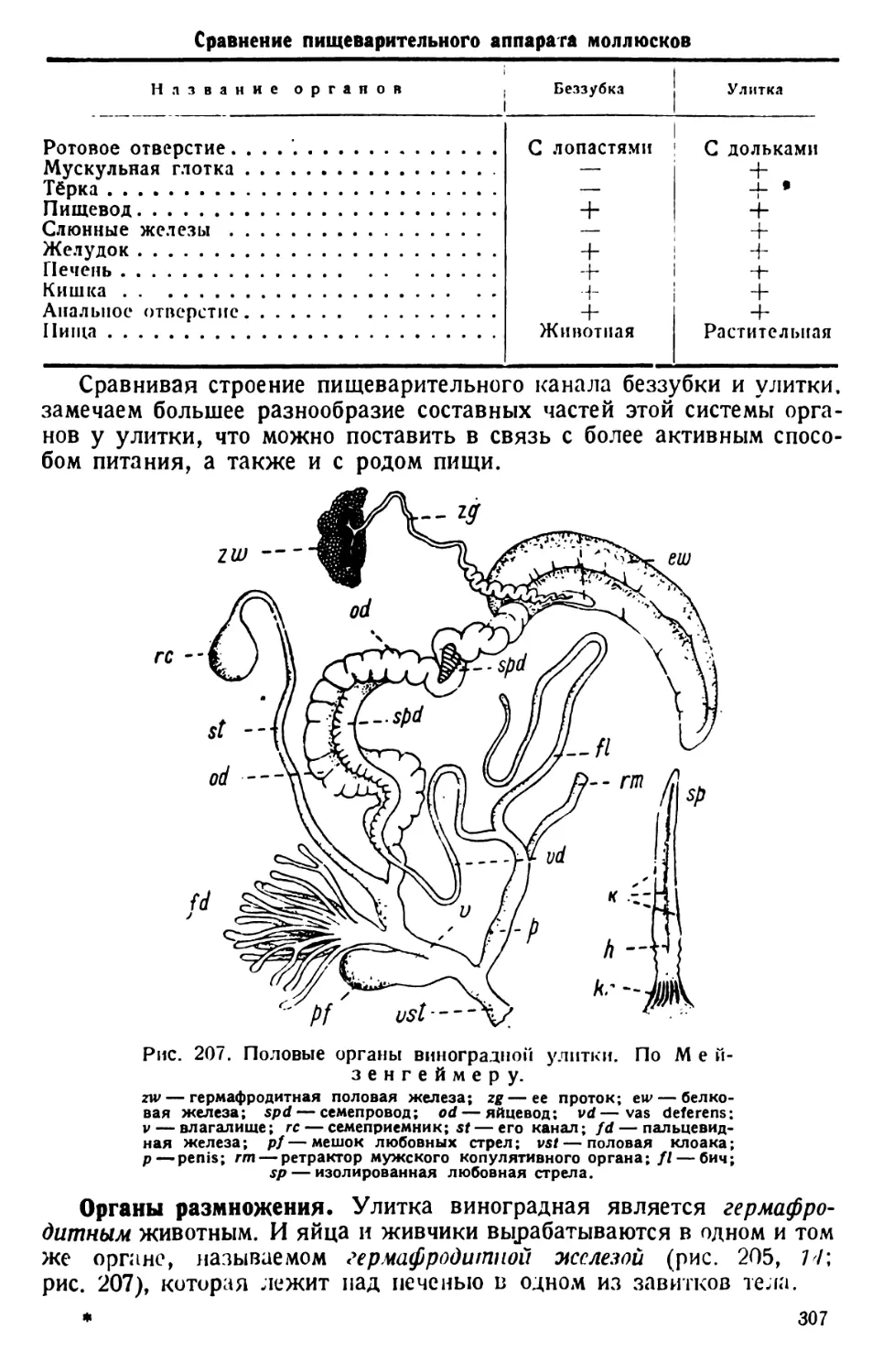
При слабом увеличении просматривают препарат инфузорий между
волокнами ваты или с замедленным движением. Найденную инфузо-
рию центрируют в поле зрения и ставят под среднее увеличение микро-
скопа, так как величина обычного вида Paramaecium caudalит коле-
блется в пределах 0,1—0,3 мм (простым глазом в часовом стекле на
черном фоне инфузории видны в виде беловатых точек). Наблюдения
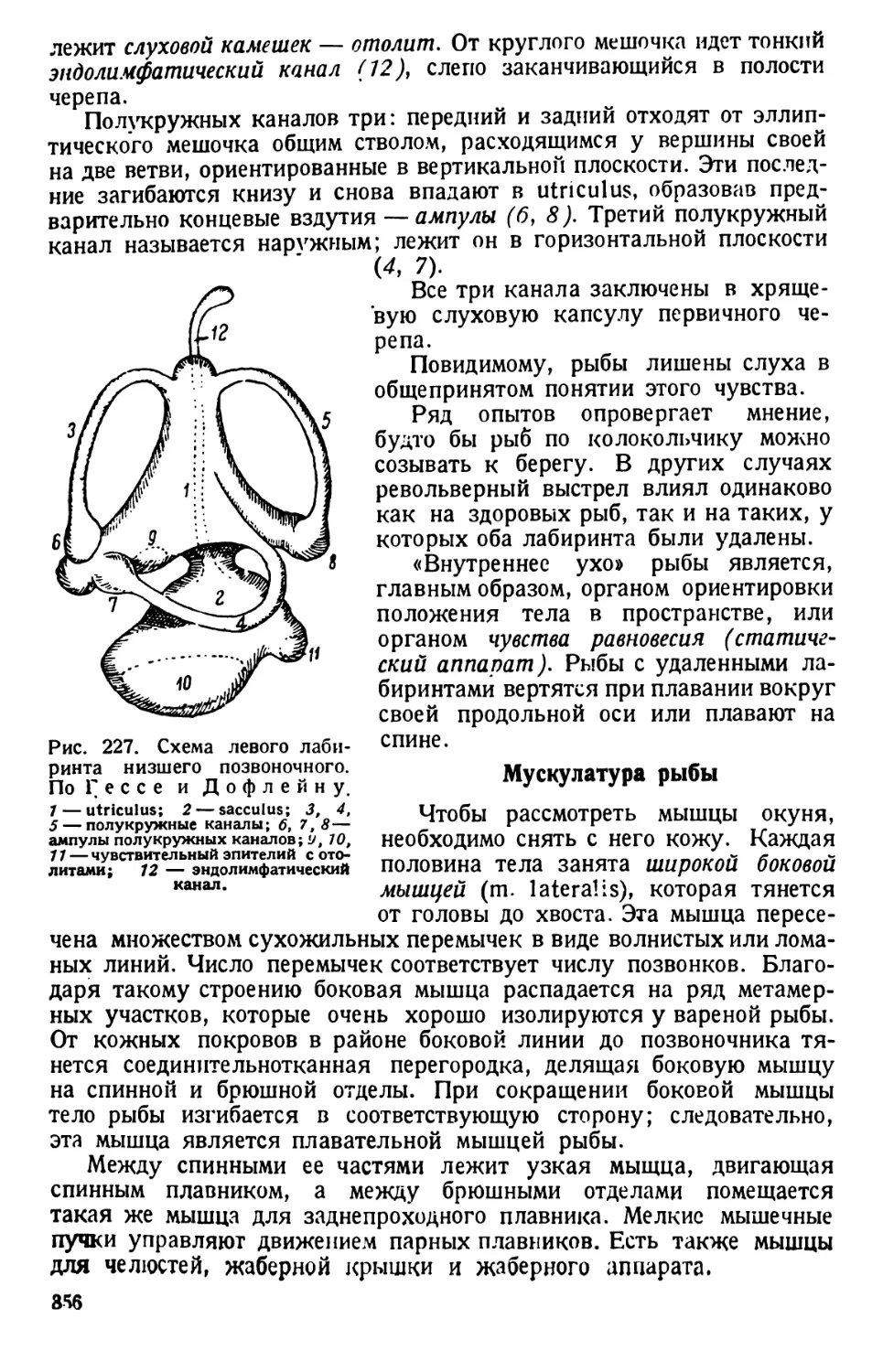
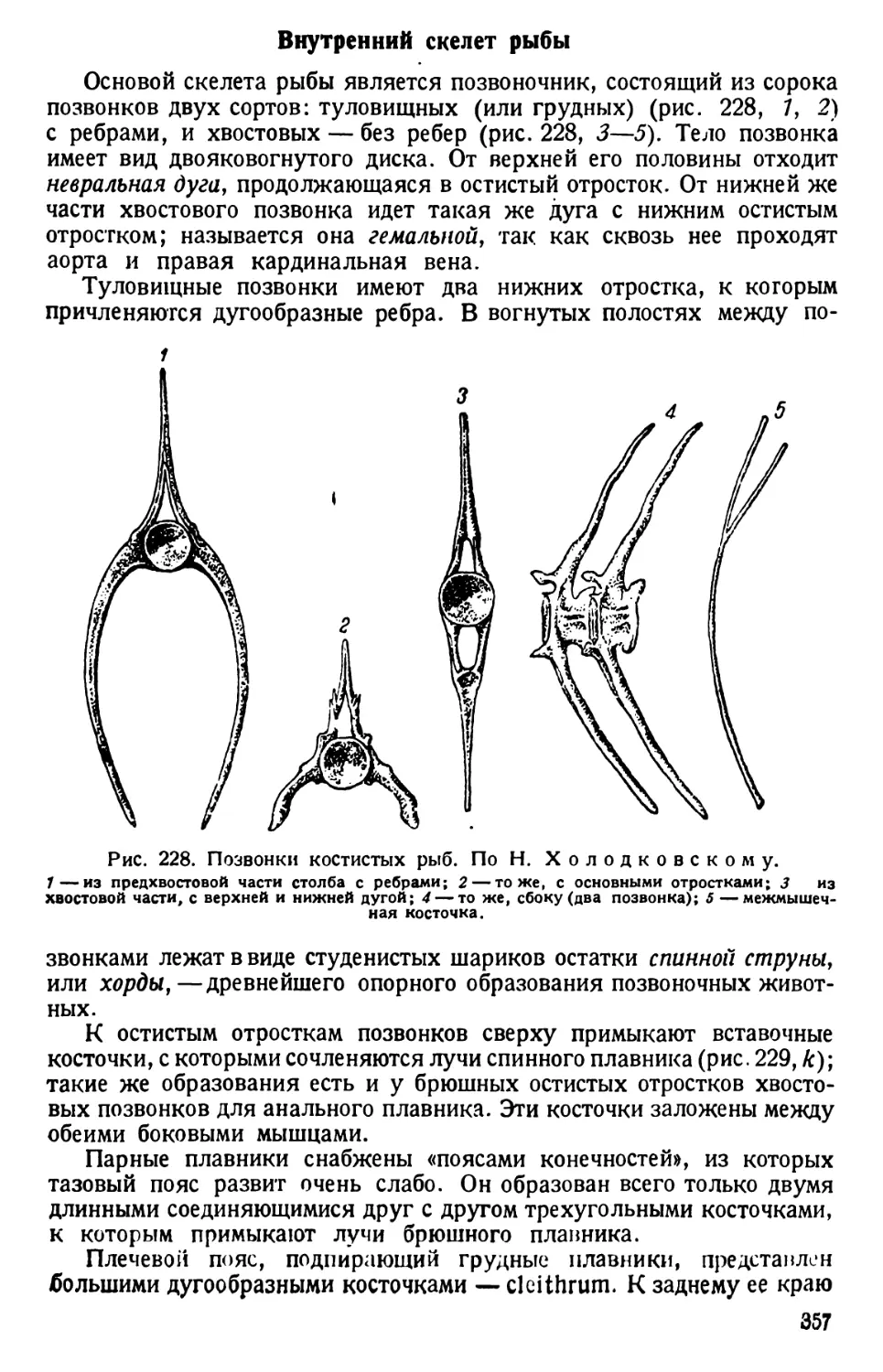
ведут при постоянном вращении микрометрического винта микро-
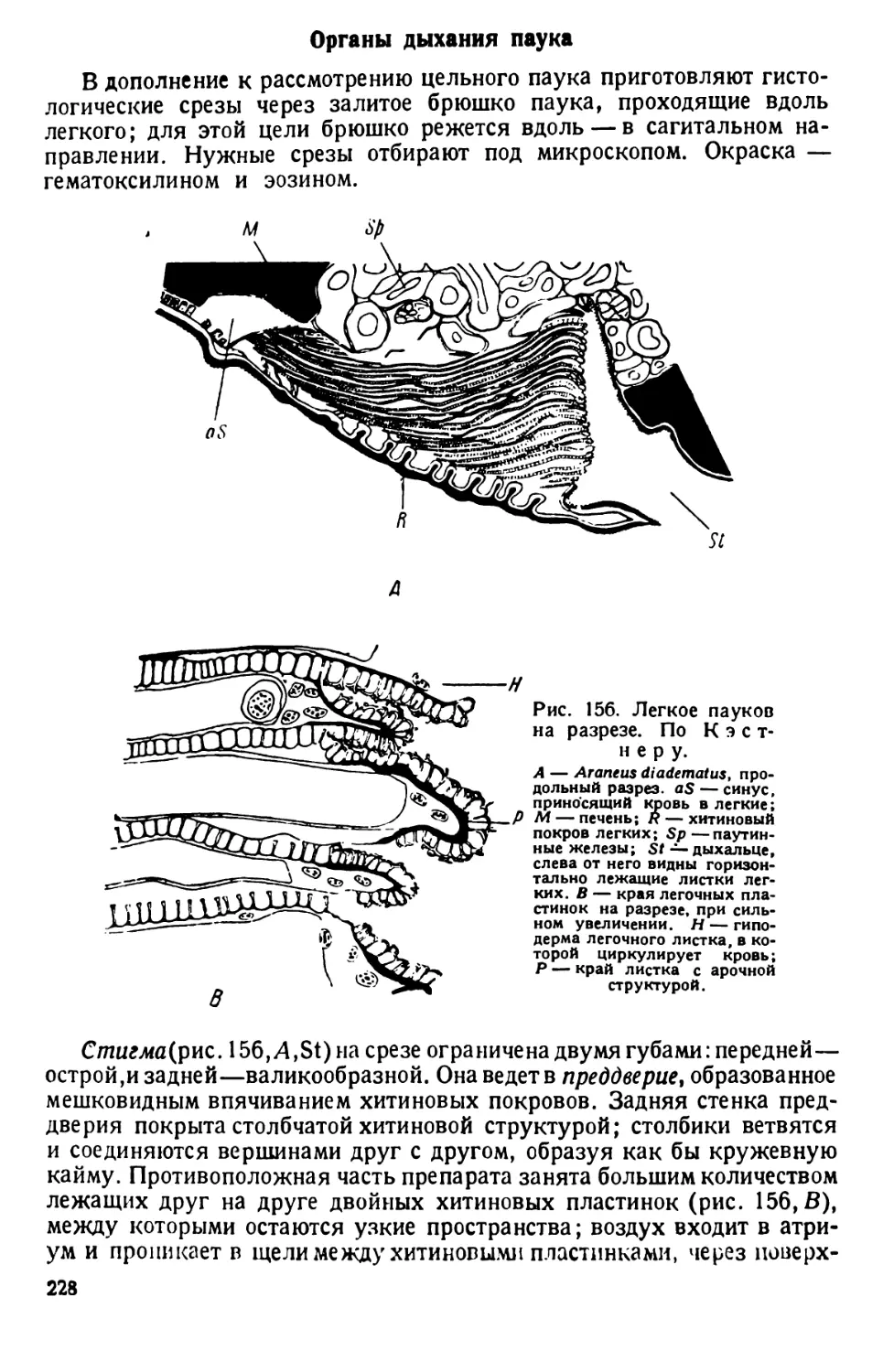
скопа.
Форма тела и общее строение. Движущиеся инфузории время от
времени вращаются вокруг оси или изгибаются и поворачиваются,
обходя препятствия в виде волоконец ваты. Наблюдая вращение инфу-
зории, отмечают вальковатую форму ее тела, которое при рассмотре-
нии его сверху (с дорзальной) или с нижней (вентральной) поверхности
имеет очертания, сходные с формой подошвы туфли, чем и обусло-
вливается наименование инфузории («туфелька»).
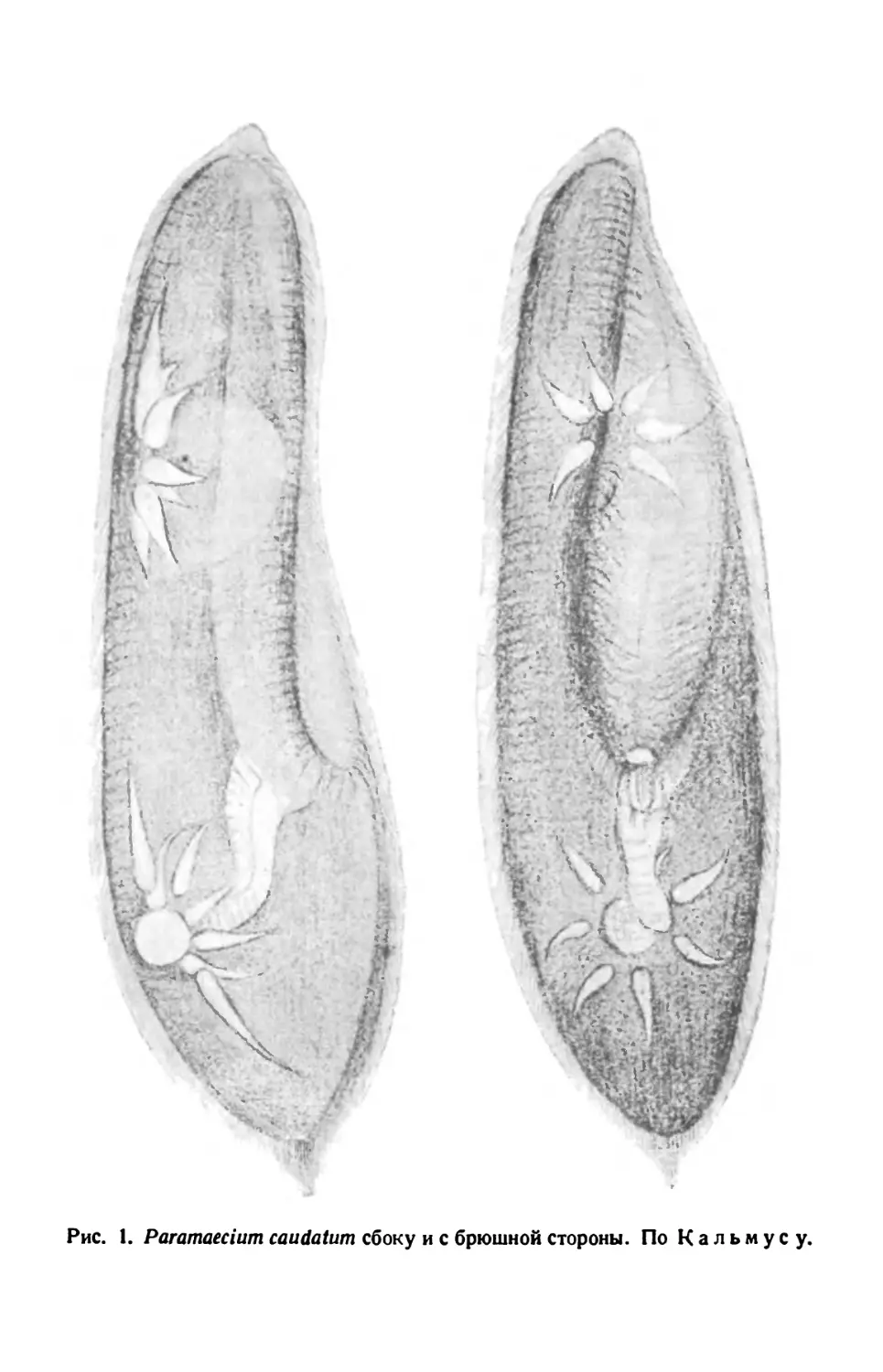
Различают передний тупо закругленный, несколько асимметричный
конец тела (при рассмотрении его со спинной или брюшной сторон) и
задний конец, который несколько более приострен (различие более
ясное при рассмотрении парамеции сбоку). Асимметрия переднего
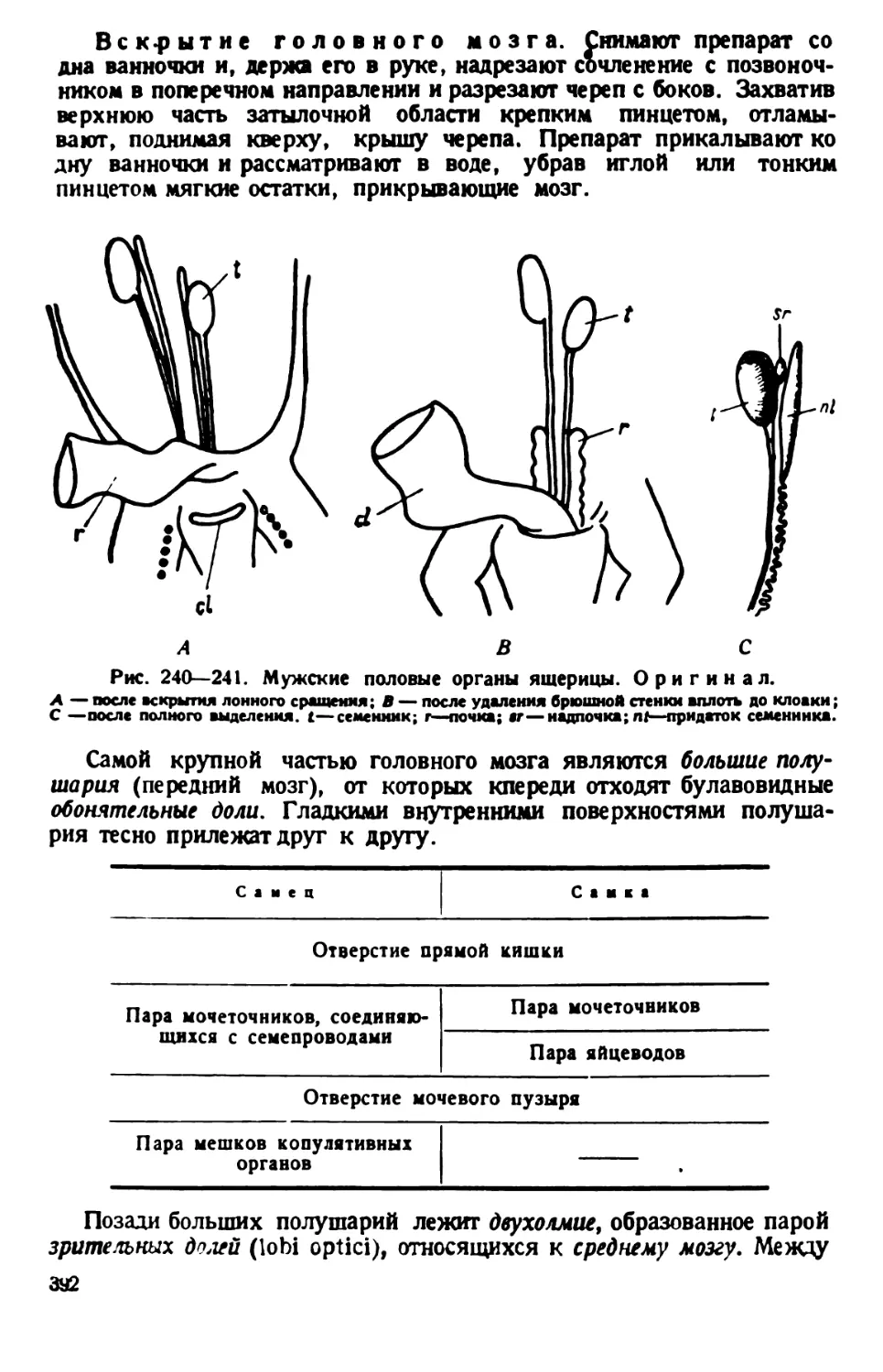
конца тела объясняется тем, что от него начинается продолговатое
«ротовое», или перистомное, углубление. Оно лежит ближе к левому
7
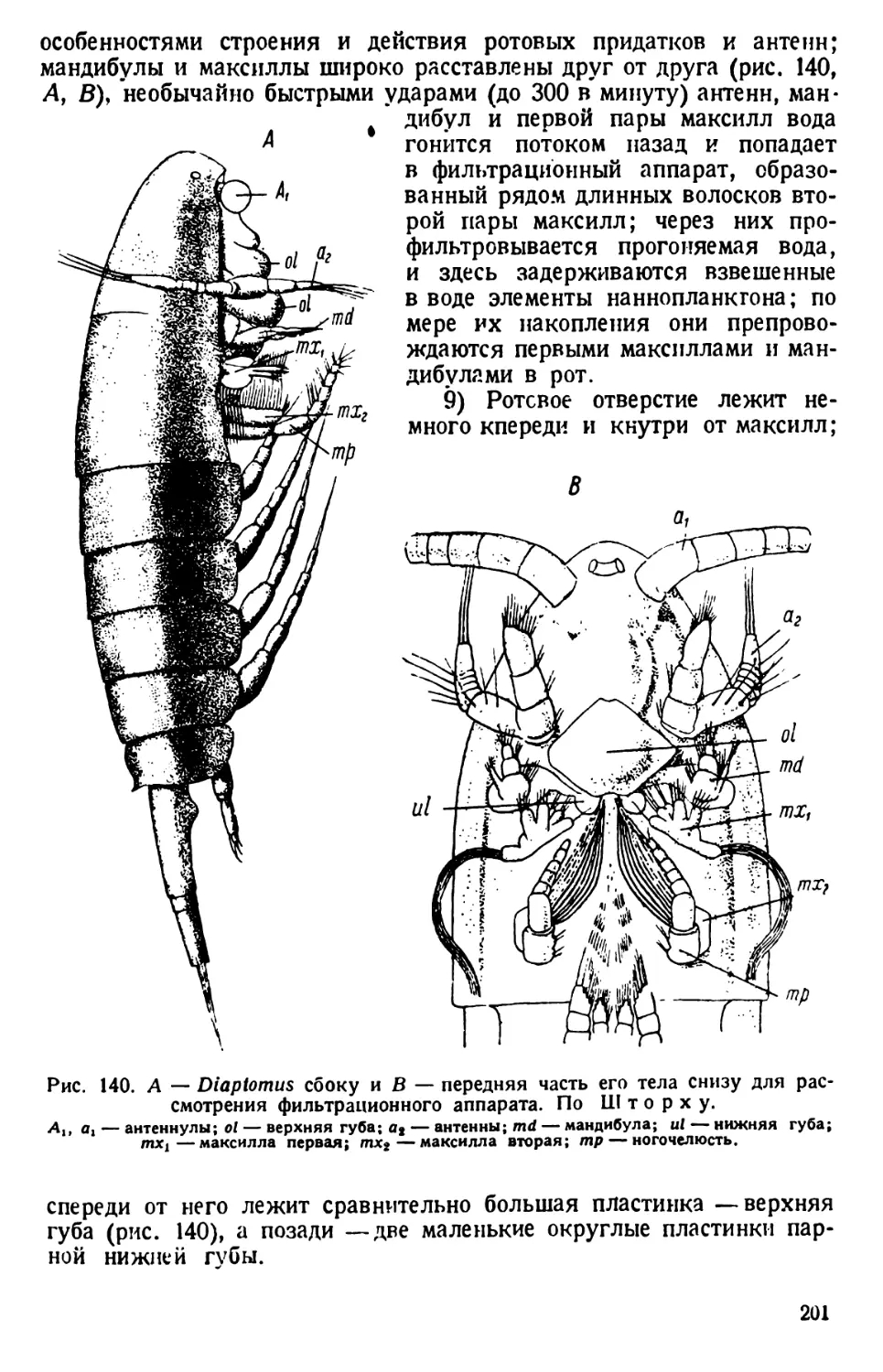
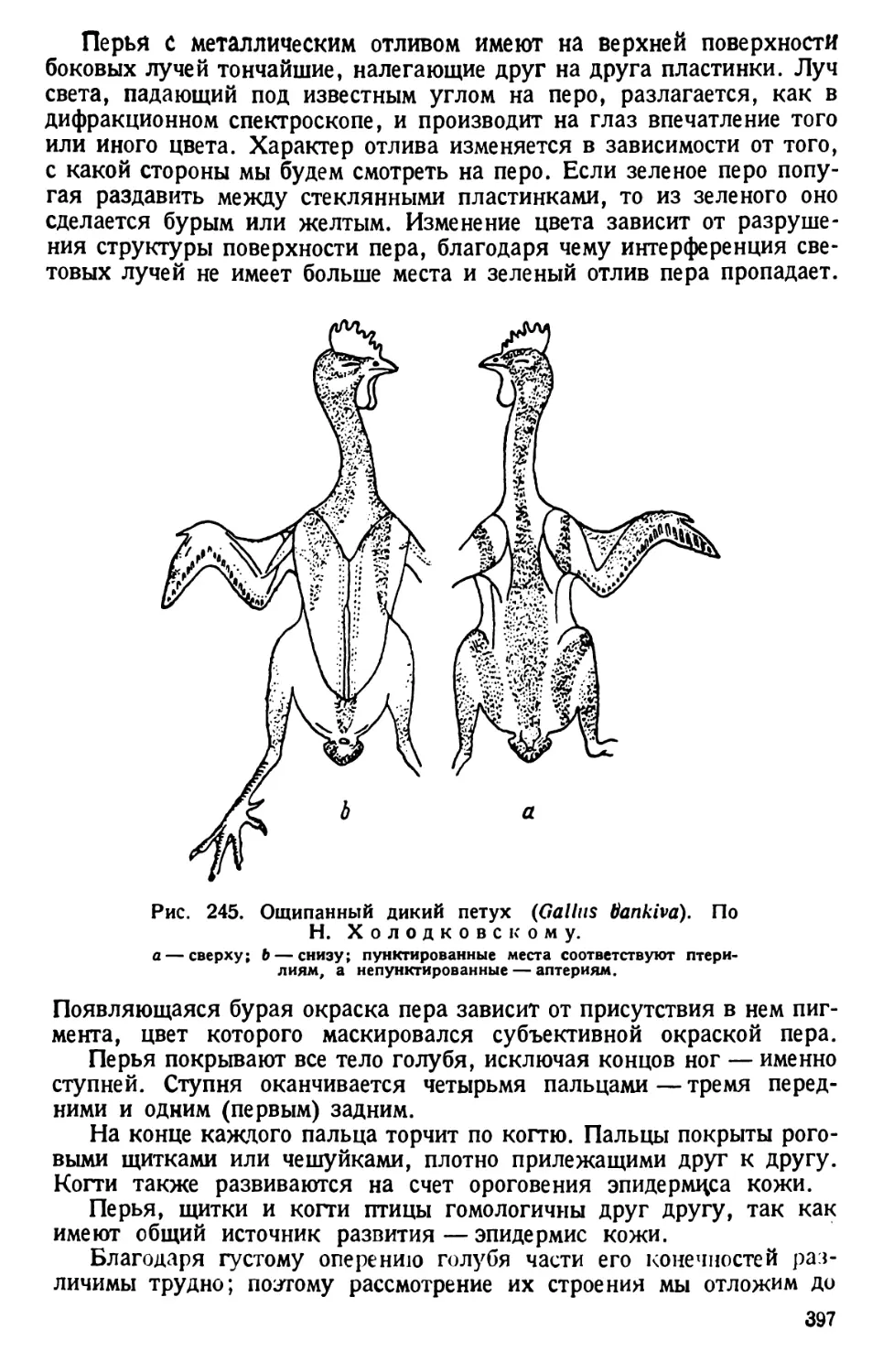
Рис. 1. Paramaecium caudatum сбоку и с брюшной стороны. По Кальмусу.
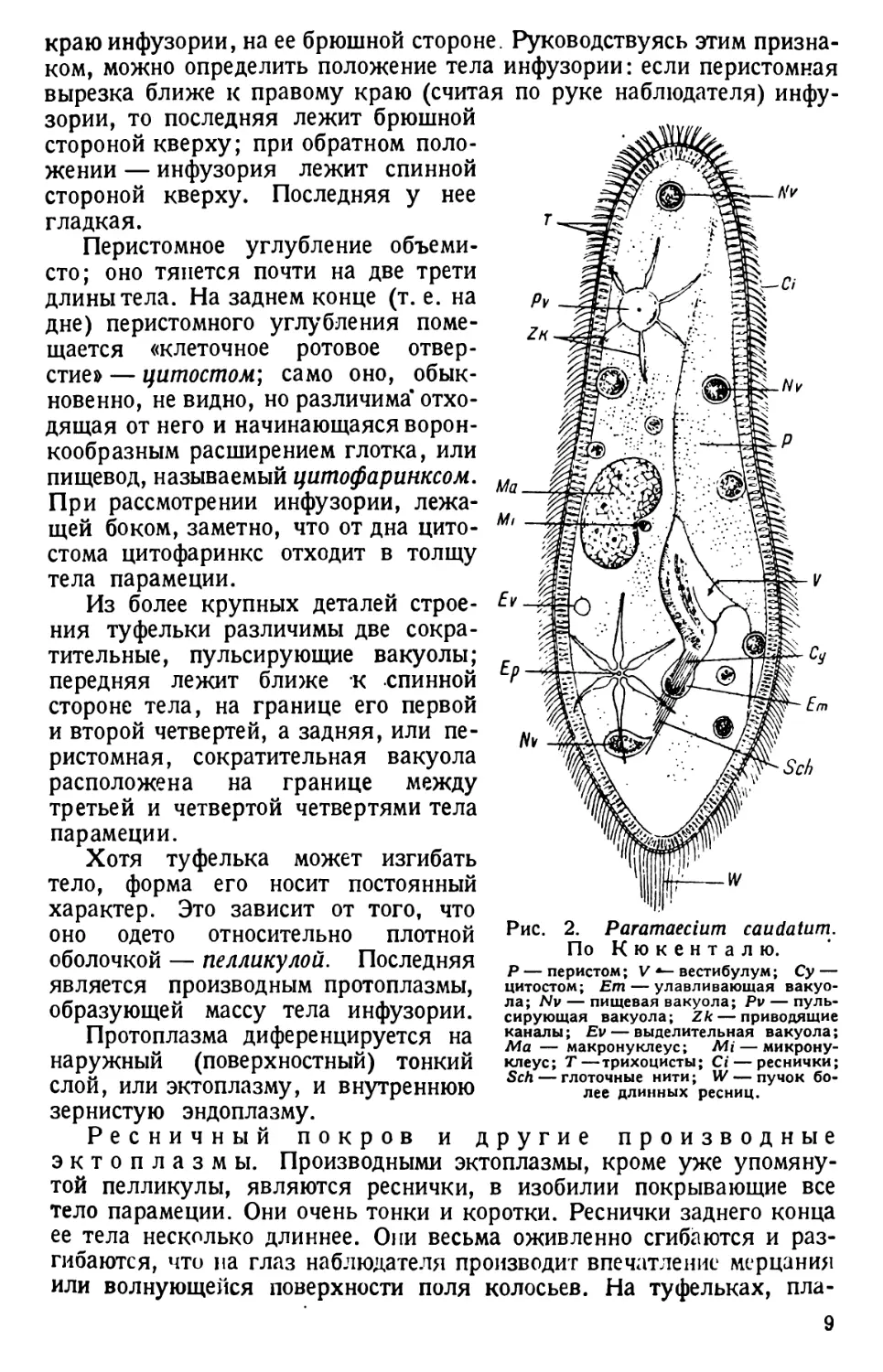
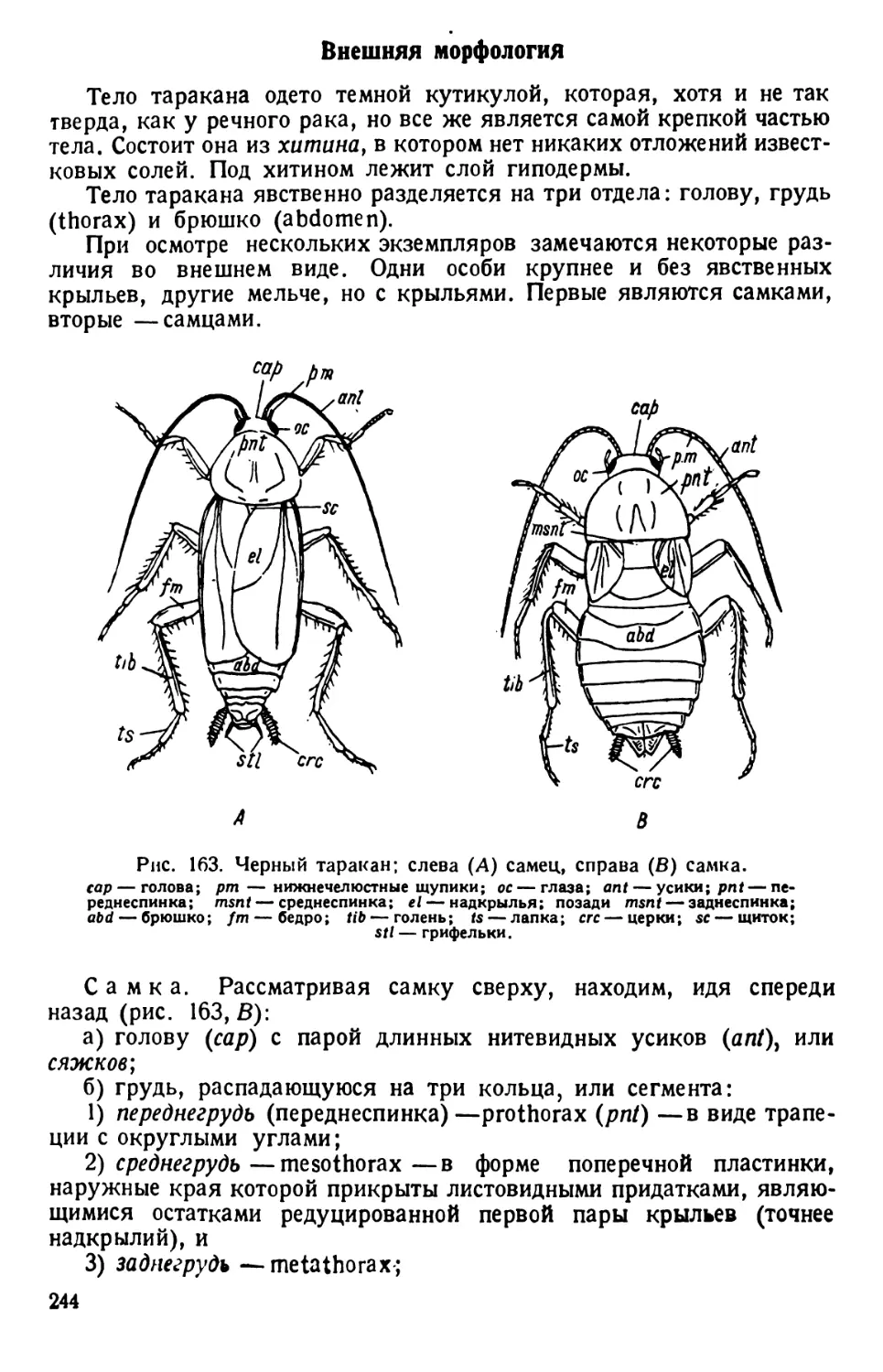
Рис. 2. Paramaecium caudatum.
По Кюкенталю.
Р — перистом; V вестибулум; Су —
цитостом; Ет — улавливающая вакуо-
ла; Nv — пищевая вакуола; Pv — пуль-
сирующая вакуола; Zk — приводящие
каналы; Ev — выделительная вакуола;
Ма — макронуклеус; Mi — микрону-
клеус; Т—трихоцисты; Ci — реснички;
Sch — глоточные нити; W — пучок бо-
лее длинных ресниц.
краю инфузории, на ее брюшной стороне. Руководствуясь этим призна-
ком, можно определить положение тела инфузории: если перистомная
вырезка ближе к правому краю (считая по руке наблюдателя) инфу-
зории, то последняя лежит брюшной
стороной кверху; при обратном поло-
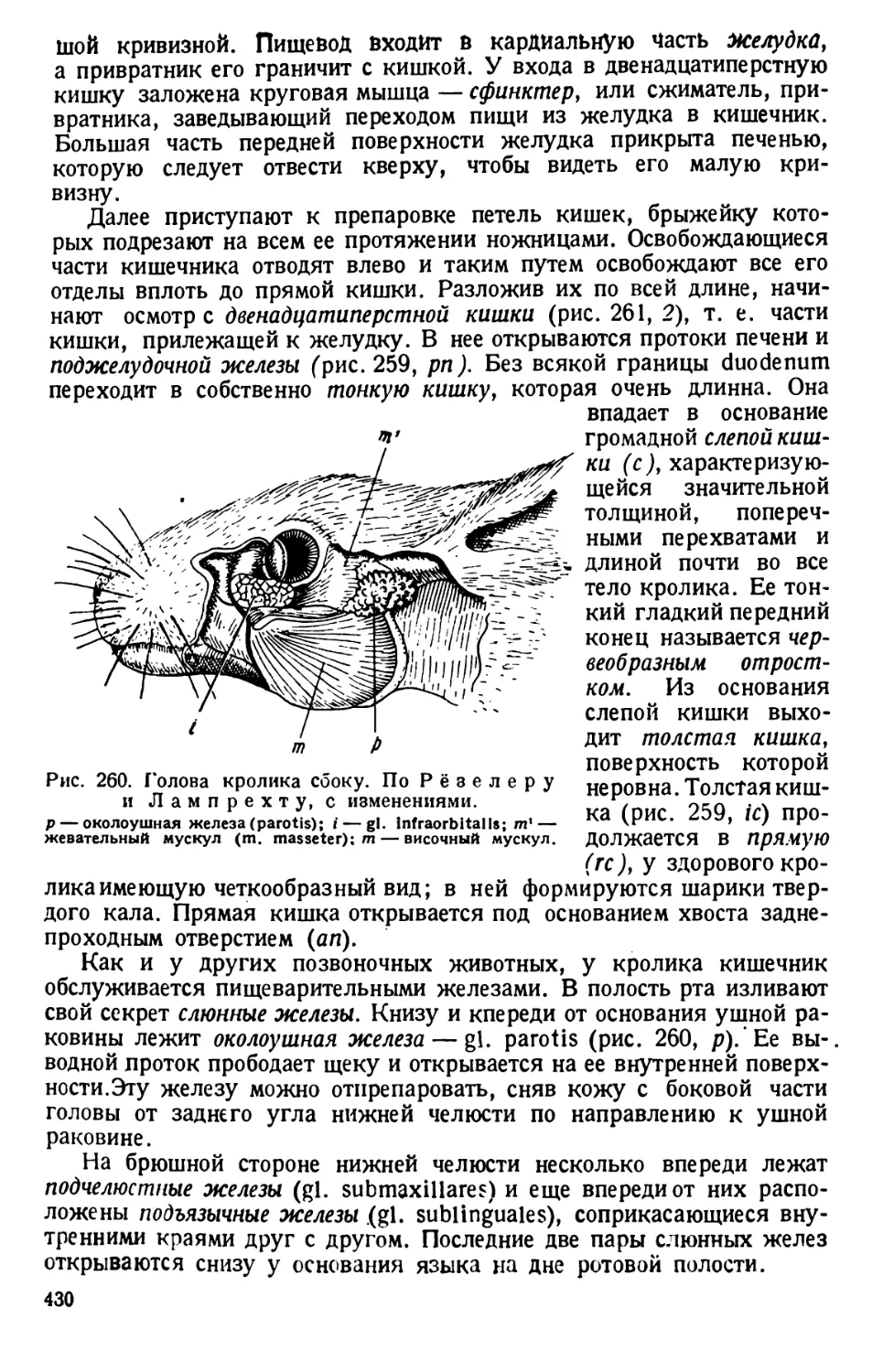
жении — инфузория лежит спинной
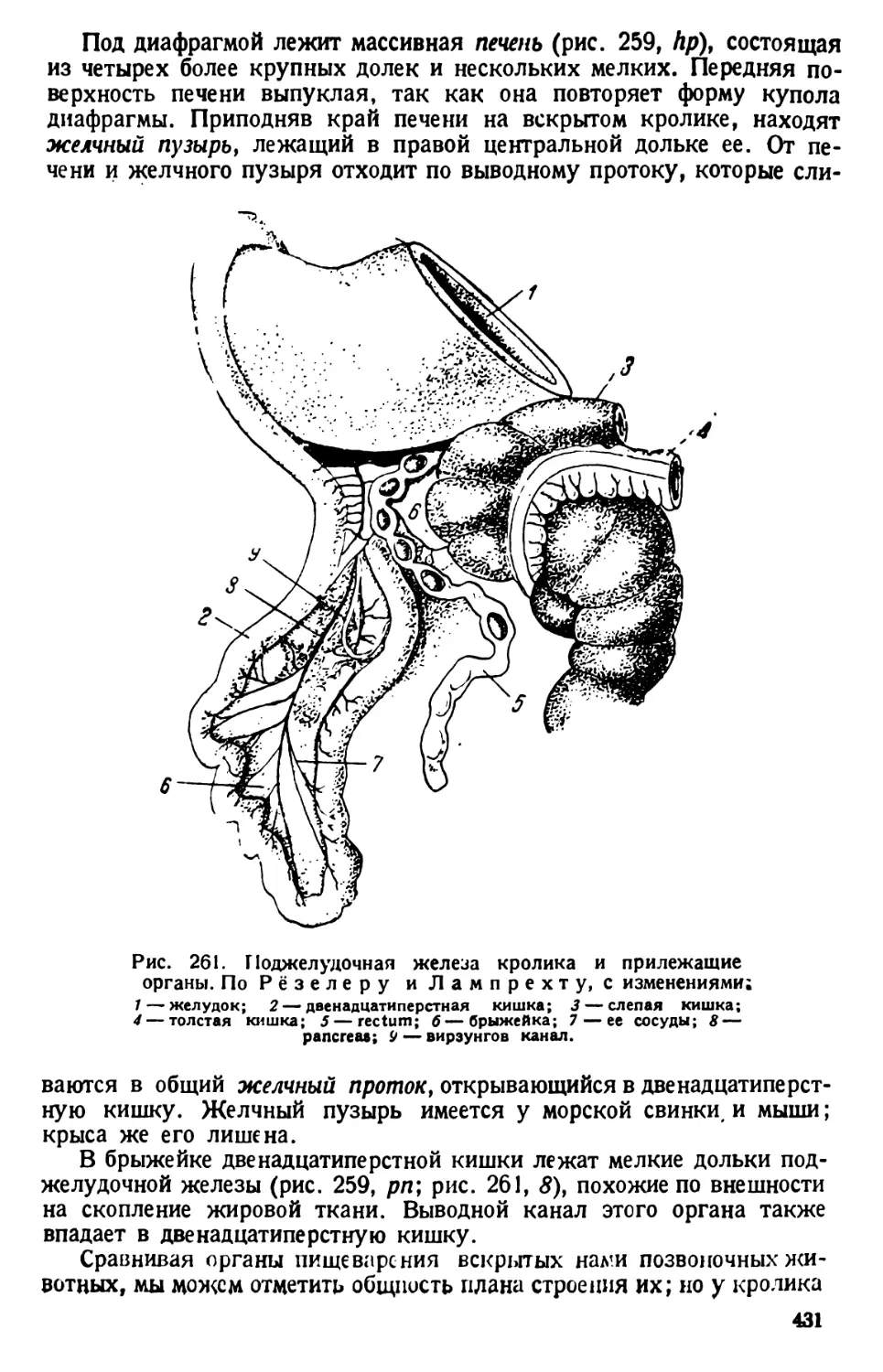
стороной кверху. Последняя у нее
гладкая.
Перистомное углубление объеми-
сто; оно тянется почти на две трети
длины тела. На заднем конце (т. е. на
дне) перистомного углубления поме-
щается «клеточное ротовое отвер-
стие» — цитостом; само оно, обык-
новенно, не видно, но различима’ отхо-
дящая от него и начинающаяся ворон-
кообразным расширением глотка, или
пищевод, называемый цитофаринксом.
При рассмотрении инфузории, лежа-
щей боком, заметно, что от дна цито-
стома цитофаринкс отходит в толщу
тела парамеции.
Из более крупных деталей строе-
ния туфельки различимы две сокра-
тительные, пульсирующие вакуолы;
передняя лежит ближе к спинной
стороне тела, на границе его первой
и второй четвертей, а задняя, или пе-
ристомная, сократительная вакуола
расположена на границе между
третьей и четвертой четвертями тела
парамеции.
Хотя туфелька может изгибать
тело, форма его носит постоянный
характер. Это зависит от того, что
оно одето относительно плотной
оболочкой — пелликулой. Последняя
является производным протоплазмы,
образующей массу тела инфузории.
Протоплазма диференцируется на
наружный (поверхностный) тонкий
слой, или эктоплазму, и внутреннюю
зернистую эндоплазму.
Ресничный покров и другие производные
эктоплазмы. Производными эктоплазмы, кроме уже упомяну-
той пелликулы, являются реснички, в изобилии покрывающие все
тело парамеции. Они очень тонки и коротки. Реснички заднего конца
ее тела несколько длиннее. Они весьма оживленно сгибаются и раз-
гибаются, что на глаз наблюдателя производит впечатление мерцания
или волнующейся поверхности поля колосьев. На туфельках, пла-
9
вающих в воде, нельзя рассмотреть детали движения ресничек; на
препаратах со студнями или слизями движение ресничек сильно
замедляется; можно хо-
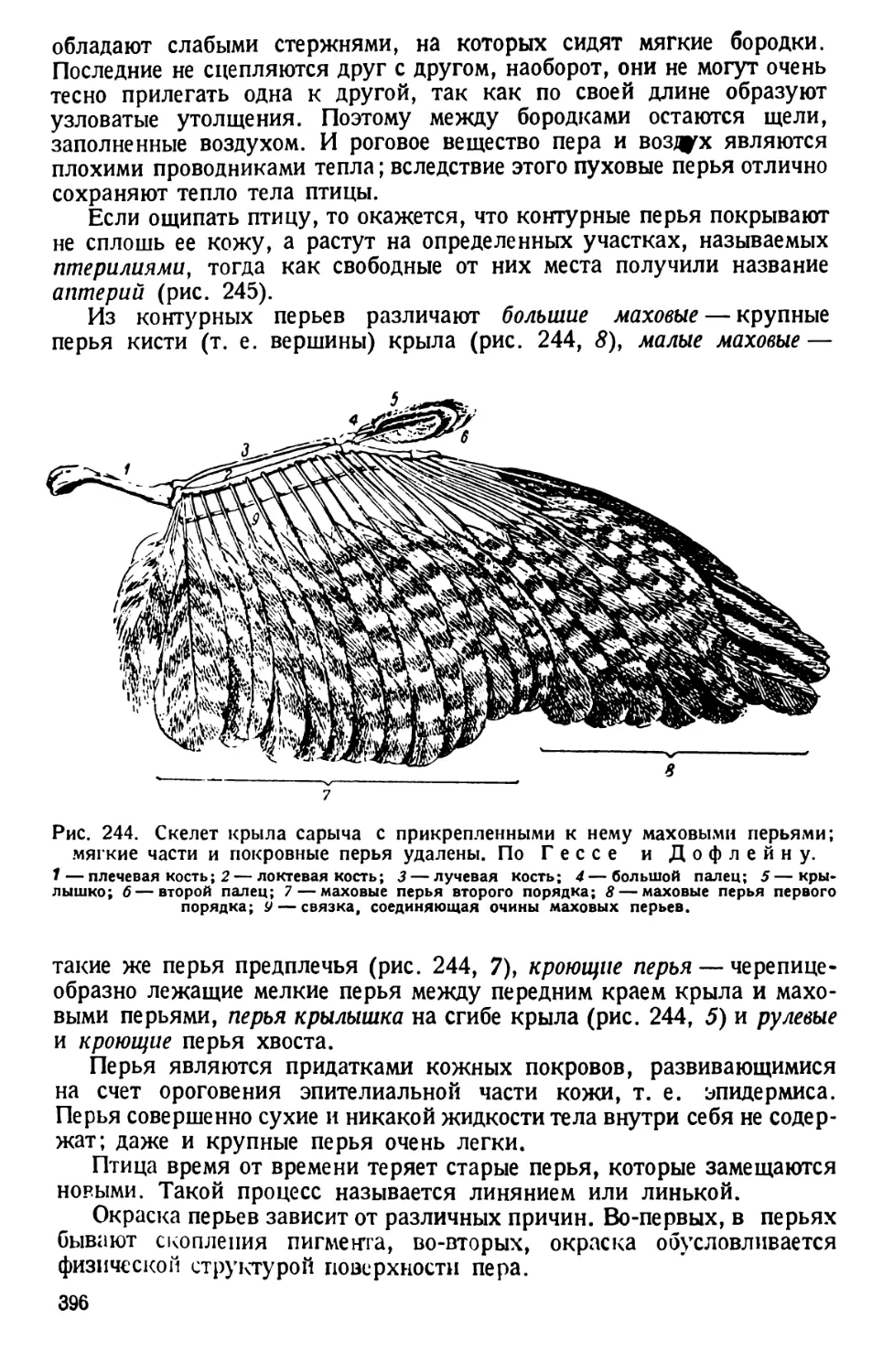
Рис. 3. Моментально фиксированное положение
ресничек Р. caudatum при движении. По Гелей.
рошо рассмотреть, что
фазы движения реснички
состоят из быстрого сги-
бания и медленного раз-
гибания. У Р. caudatum
ресничка делает 10— 11
ударов в секунду. Рес-
нички сокращаются не
синхронно (одновремен-
но), но последовательно—
метахронно, благодаря
чему по поверхности
пелликулы пробегают волны мерцания (рис. 3 и 11). Благодаря сги-
банию и разгибанию ресничек инфузория плавно передвигается вводе.
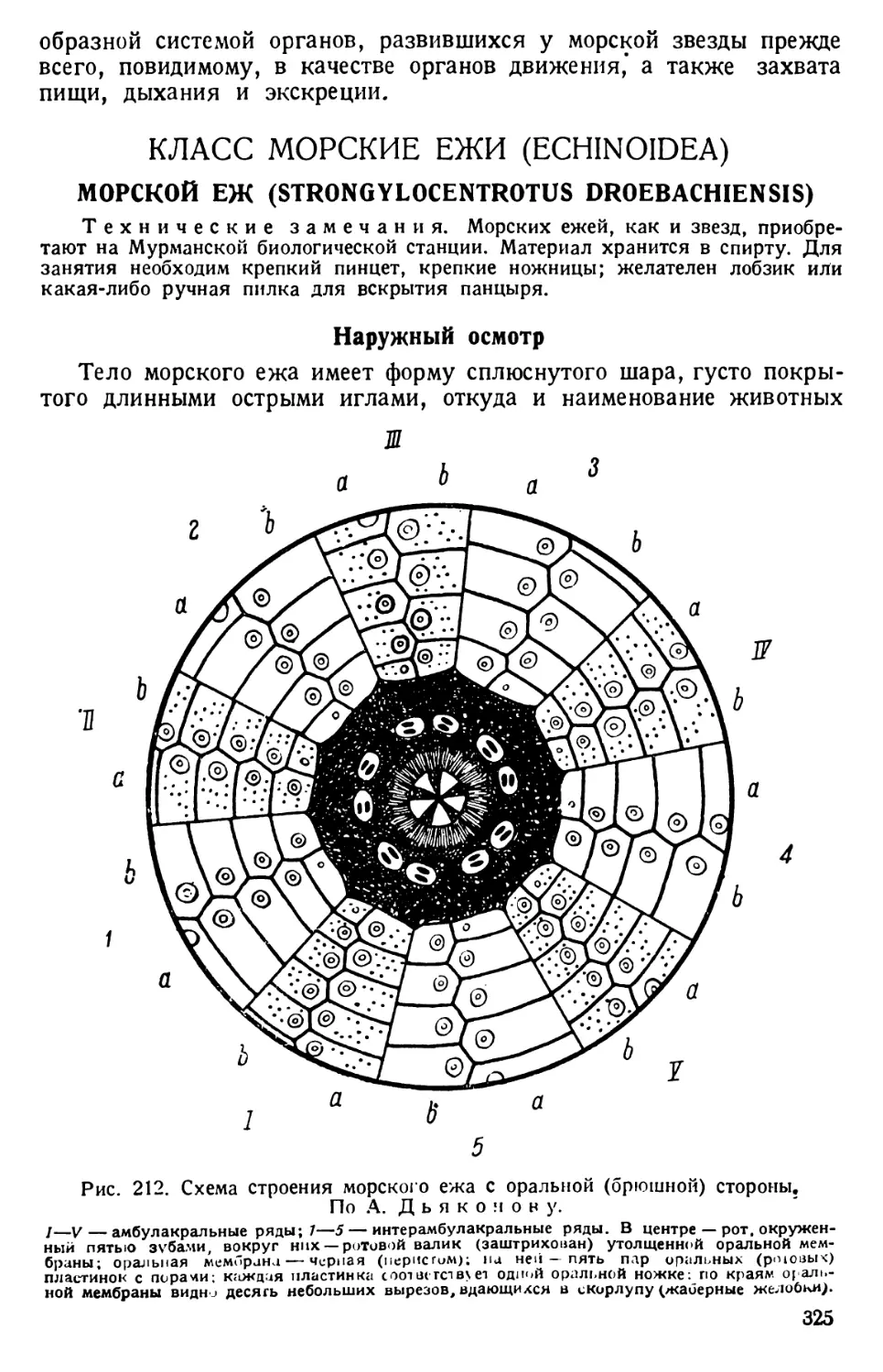
Реснички, следователь-
но, являются в качестве
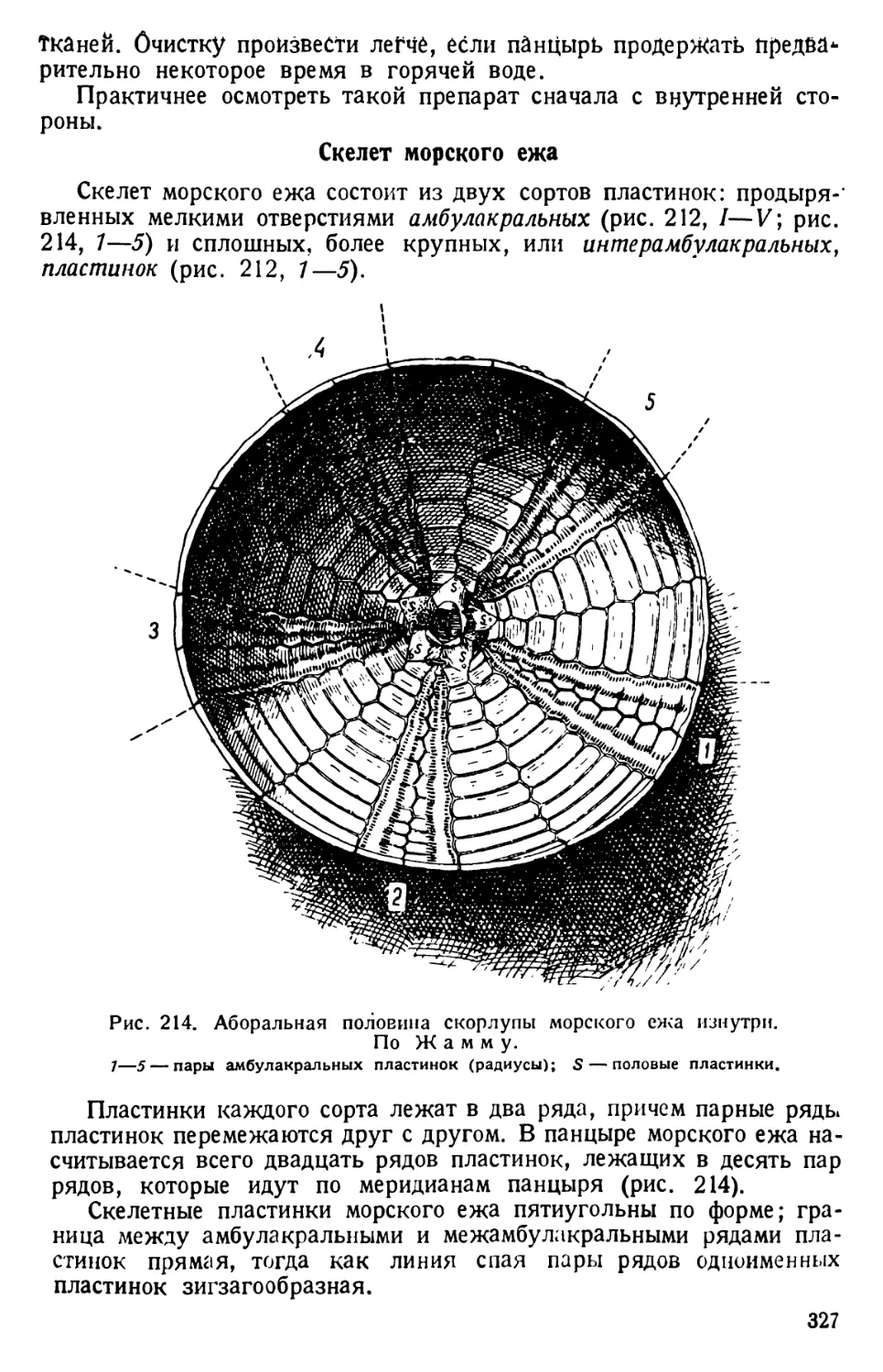
гребных ресничек орга-
ноидами движения
парамеции типа унду-
липодий.
В толще самой экто-
плазмы видны короткие,
близко лежащие друг
к другу, сильно прело-
мляющие свет палочки,
называемые трихоци-
стами, или стрекатель-
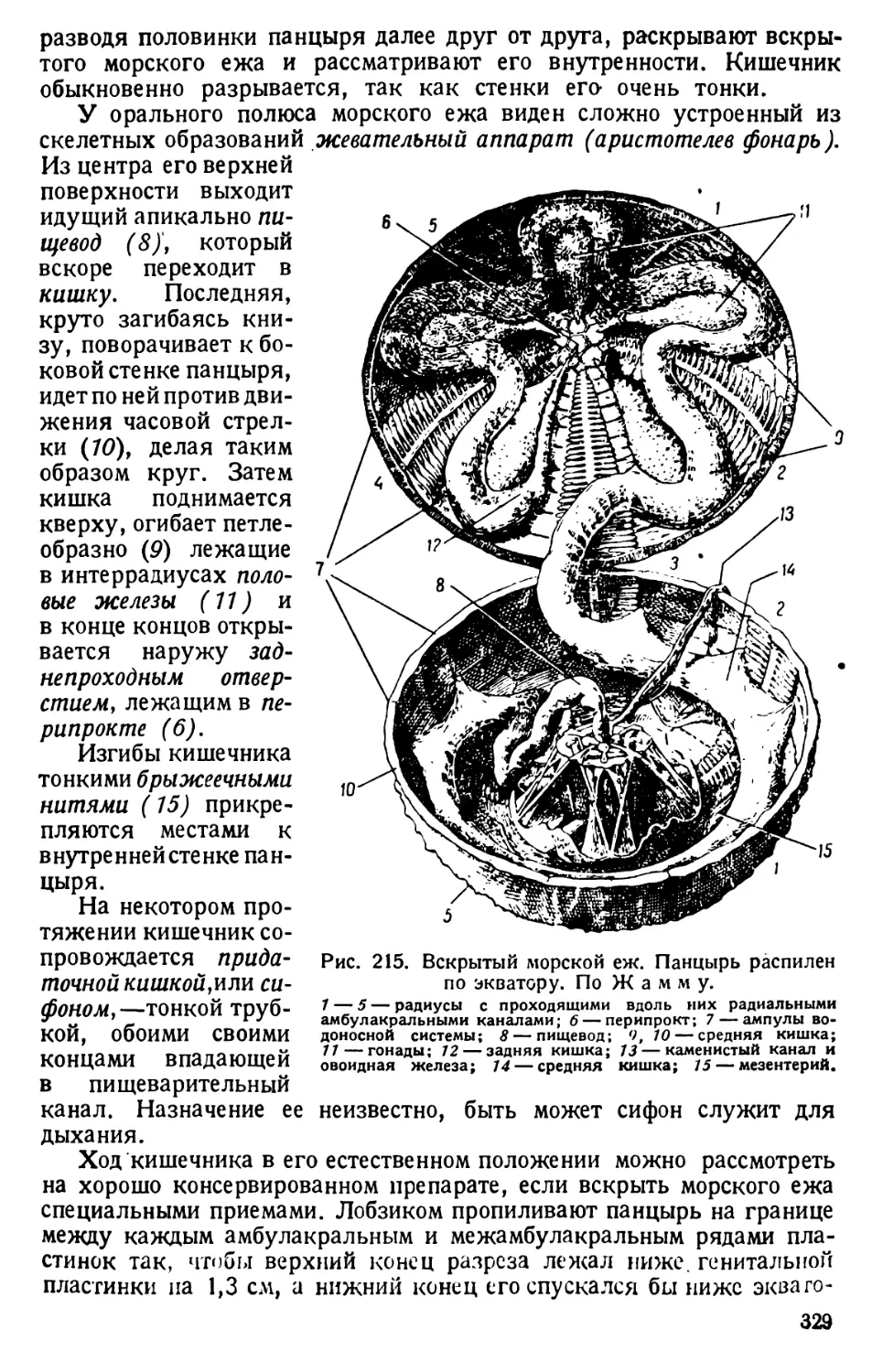
ными палочками (об их
значении см. ниже, на
стр. 18).
Сократитель-
ная вакуола. На-
конец, в эктоплазме за-
ложена уже известная
нам передняя пульсиру-
ющая вакуола. Ее ра-
боту наблюдают на жи-
вой парамеции, прида-
вленной покровным сте-
клом с восковыми нож-
ками настолько, чтобы
инфузория остановилась
на месте.
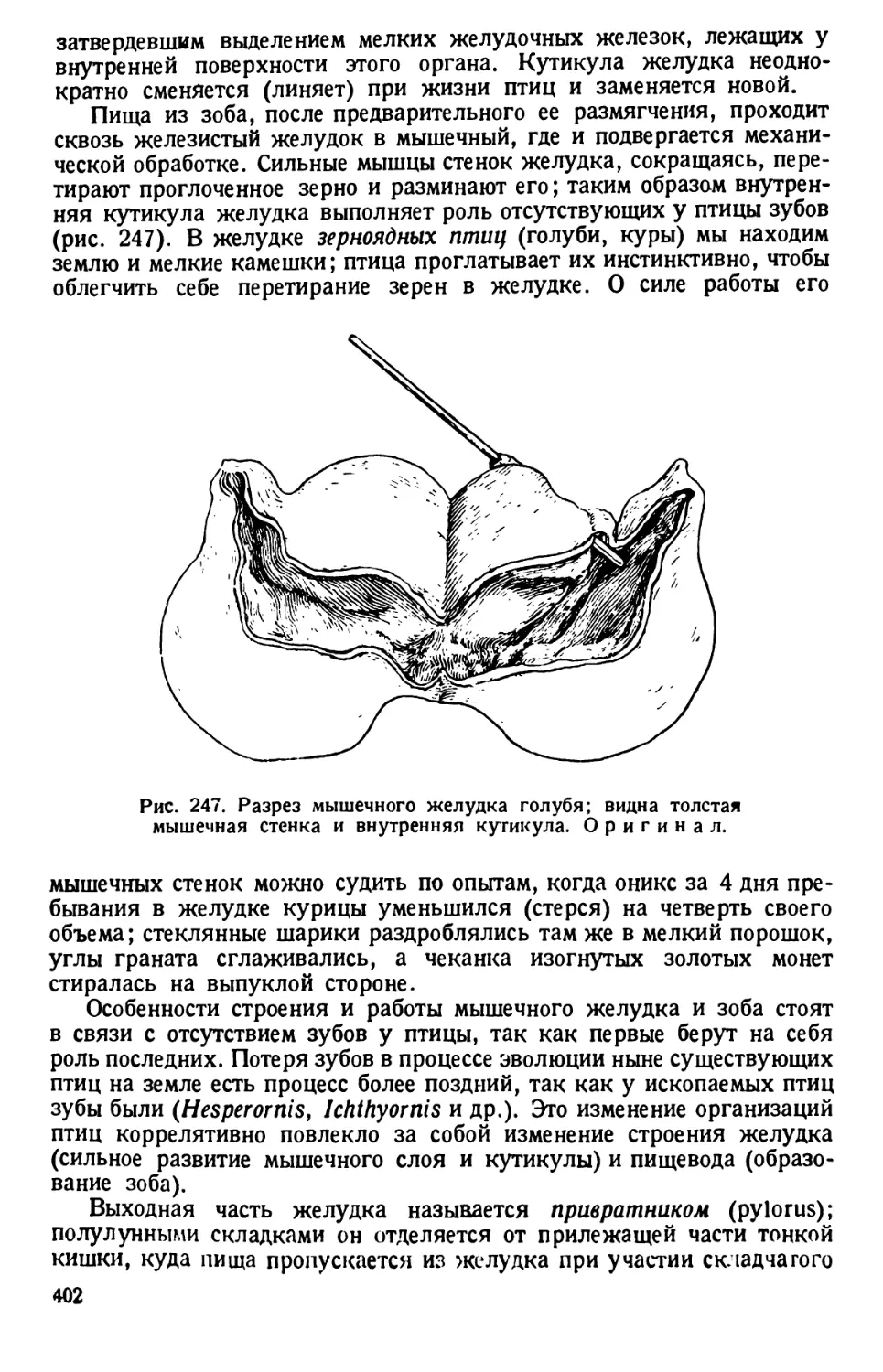
Рис. 4. Фазы пульсации сократительной вакуолы
парамеции. По Пюттеру.
7 — непосредственно перед опорожнением; 2 — тотчас
после опорожнения; 3 — образование вакуолы; 4—5 —
слияние вакуолы и начало выступания новых приводящих
каналов; 6 — начало вздутия обращенных к вакуоле кон-
цов приводящих каналов.
Наблюдая одну и ту же вакуолу некоторое время, можно озна-
комиться с характером ее пульсации. Вокруг вакуолы периодически
появляются 5—7—10 радиальных каналов. Когда вакуола при своем
сокращении (систоле) опорожняется наружу, то центральные концы
10
каналов расширяются, и скопляющаяся в них жидкость поступает
в центр звезды каналов; здесь снова появляется центральная вакуола
(диастола), тогда как сами каналы на время исчезают. Обычно обе пуль-
сирующие вакуолы работают в последовательном порядке: момент
систолы одной вакуолы совпадает с диастолой другой. Опорожнение
вакуолы происходит через выделительную пору, которая, вследствие
своих малых размеров, практически не различима.
При опорожнении пульсирующих вакуол, из тела туфельки уда-
ляется избыток воды, проникающей через эктоплазму, обладающую
свойствами полупроницаемой оболочки. Протоплазма туфельки более
богата солями, чем пресная вода; в силу неравенства осмотического
давления между протоплазмой инфузории и окружающей средой,
обеспечивается значительное поступание воды в протоплазму; под-
держание нормы концентрации ее солевого состава обеспечивается
постоянной работой пульсирующих вакуол, главная роль которых
состоит в регуляции осмотического давления. Кроме того работа этих
органоидов способствует удалению продуктов обмена веществ (фун-
кция экскреции) и дыханию; поступающая извне в тело туфельки вода
богата растворенным кислородом, а жидкость, выбрасываемая пуль-
сирующими вакуолами, наоборот, им бедна и, видимо, содержит
углекислоту.
Что опорожняющаяся вакуола действительно выбрасывает в окру-
жающую среду жидкость,— можно хорошо наблюдать на препарате
парамеций, пущенных в растертую в воде китайскую тушь или кармин.
Наблюдение ведут на препарате, покрытом покровным стеклом. При
опорожнении вакуолы видно, что в месте выхода ее жидкости наружу
зернышки туши или кармина отталкиваются от края тела инфузории.
Работа пульсирующих вакуол носит ритмичный характер и зависит
от температуры окружающей среды. Существующую взаимосвязь
можно выявить, определяя в секундах длину интервалов между двумя
опорожнениями вакуолы при данной температуре.
Соответствующие промежутки времени для парамеции составляют:
при 16° С...........21 секунду
» 20° С...........14 »
» 27° С........... 9 »
» 34° С........... 6 »
Дальнейшее повышение температуры ведет к неправильностям
в работе вакуол и к полной ее остановке; понижение же ниже 16° С
в известных пределах вызывает замедление пульсирования при сохра-
нении его ритмичности.
О величине работы пульсирующих вакуол парамеции можно судить
по сделанному расчету, согласно которому вакуолы за 46 минут
работы выделяют объем жидкости, равный объему тела инфузории
(В. Д о г е л ь).
Частота пульсирования сократительных вакуол зависит, кроме
температуры, и от степени концентрации солей в воде. Соответствен-
ная связь показана в опытах Н е г f s, выдерживавшего парамеций
в растворах поваренной соли различной концентрации. Из приводи-
мых ниже цифровых данных видно, что по мере повышения концентра-
ции солевого раствора опорожнение пульсирующей вакуолы совер-
шается все с бблыпими и ббльшими интервалами; следовательно,
требуется соответственно большее время, чтобы эти вакуолы выделили
количество жидкости, равное объему тела парамеции.
Концентрация раство- ра NaCl, в процентах Температура, в гра- дусах Цельсия Ритм сокращения ва- куолы — интервал, в секундах 1 Время, требуемое для удаления объема воды, равного объему тела парамеции, в минутах
0,00 22—23 6,2 14,7
0,25 20-22 9,3 21,3
0,50 19—20 18,4 42,7
0,75 19—20 24,8 56,9
1,00 10—20 163,0 । 379
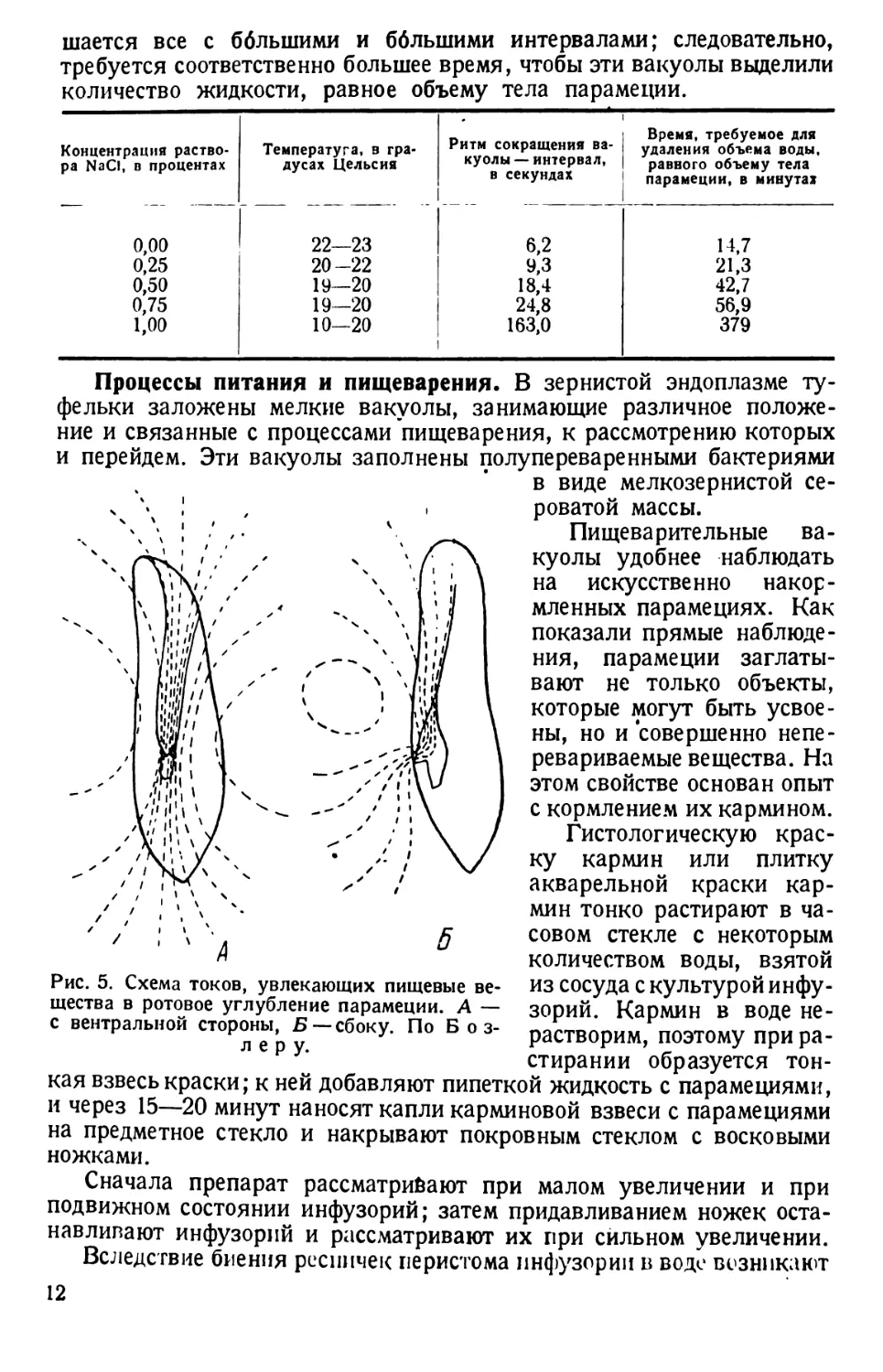
Процессы питания и пищеварения. В зернистой эндоплазме ту-
фельки заложены мелкие вакуолы, занимающие различное положе-
ние и связанные с процессами пищеварения, к рассмотрению которых
и перейдем. Эти вакуолы заполнены полупереваренными бактериями
в виде мелкозернистой се-
роватой массы.
Пищеварительные ва-
куолы удобнее наблюдать
на искусственно накор-
мленных парамециях. Как
показали прямые наблюде-
ния, парамеции заглаты-
вают не только объекты,
которые могут быть усвое-
ны, но и совершенно непе-
ревариваемые вещества. На
этом свойстве основан опыт
с кормлением их кармином.
Гистологическую крас-
ку кармин или плитку
акварельной краски кар-
мин тонко растирают в ча-
совом стекле с некоторым
количеством воды, взятой
Рис. 5. Схема токов, увлекающих пищевые ве- ИЗ сосуда С культурой инфу -
щества в ротовое углубление парамеции. А — зорий. Кармин В воде не-
с вентральной стороны Б — сбоку. По Боз- растворим, ПОЭТОМУ прира-
у' стирании образуется тон-
кая взвесь краски; к ней добавляют пипеткой жидкость с парамециями,
и через 15—20 минут наносят капли карминовой взвеси с парамециями
на предметное стекло и накрывают покровным стеклом с восковыми
ножками.
Сначала препарат рассматривают при малом увеличении и при
подвижном состоянии инфузорий; затем придавливанием ножек оста-
навливают инфузорий и рассматривают их при сильном увеличении.
Вследствие биения ресничек перистома инфузории в воде возникают
12
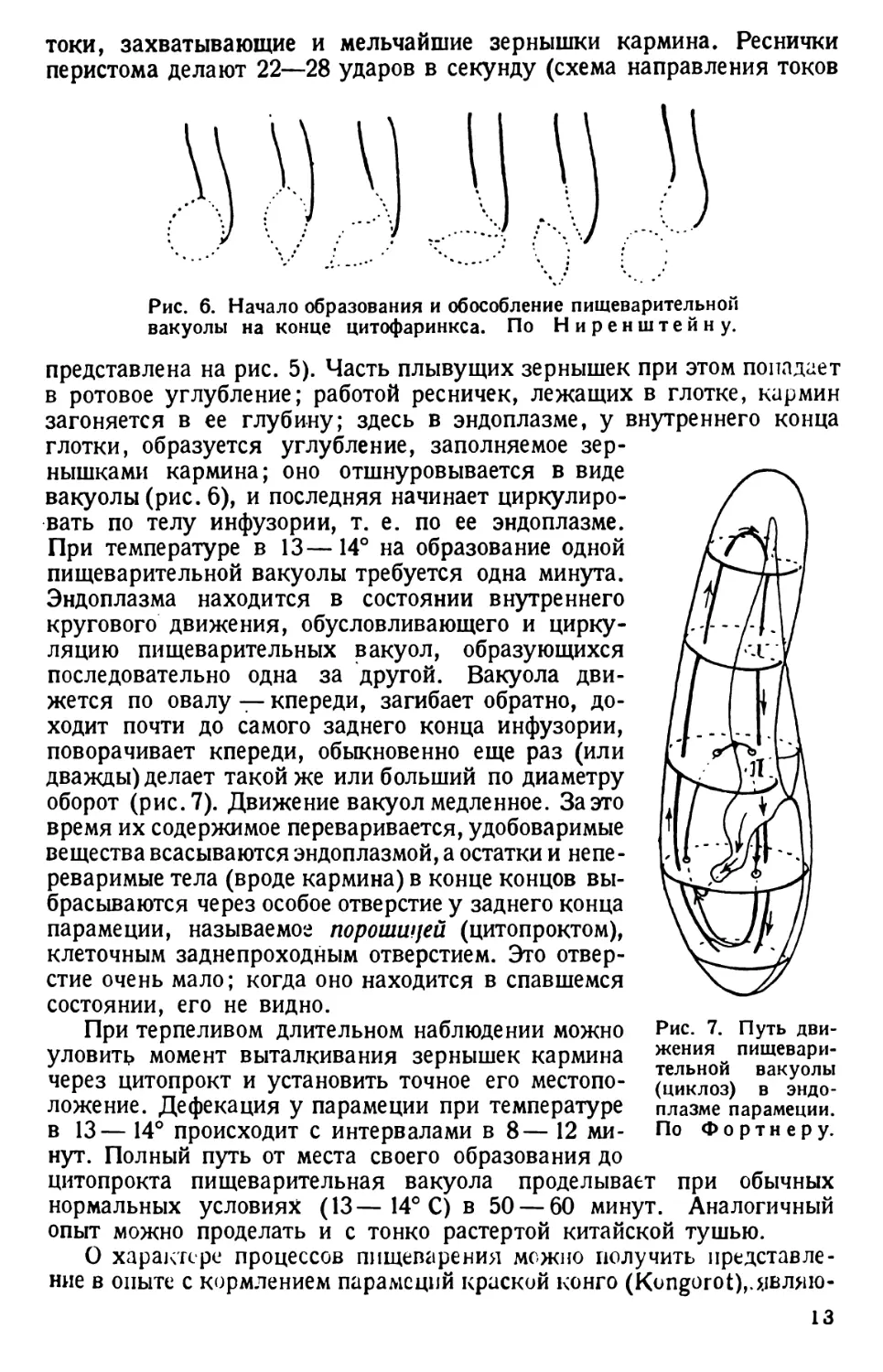
токи, захватывающие и мельчайшие зернышки кармина. Реснички
перистома делают 22—28 ударов в секунду (схема направления токов
Рис. 6. Начало образования и обособление пищеварительной
вакуолы на конце цитофаринкса. По Ниренштейну.
представлена на рис. 5). Часть плывущих зернышек при этом попадает
в ротовое углубление; работой ресничек, лежащих в глотке, кармин
загоняется в ее глубину; здесь в эндоплазме, у внутреннего конца
глотки, образуется углубление, заполняемое зер-
нышками кармина; оно отшнуровывается в виде
вакуолы (рис. 6), и последняя начинает циркулиро-
вать по телу инфузории, т. е. по ее эндоплазме.
При температуре в 13—14° на образование одной
пищеварительной вакуолы требуется одна минута.
Эндоплазма находится в состоянии внутреннего
кругового движения, обусловливающего и цирку-
ляцию пищеварительных вакуол, образующихся
последовательно одна за другой. Вакуола дви-
жется по овалу — кпереди, загибает обратно, до-
ходит почти до самого заднего конца инфузории,
поворачивает кпереди, обыкновенно еще раз (или
дважды) делает такой же или больший по диаметру
оборот (рис. 7). Движение вакуол медленное. За это
время их содержимое переваривается, удобоваримые
вещества всасываются эндоплазмой, а остатки и непе-
реваримые тела (вроде кармина) в конце концов вы-
брасываются через особое отверстие у заднего конца
парамеции, называемое порошицей (цитопроктом),
клеточным заднепроходным отверстием. Это отвер-
стие очень мало; когда оно находится в спавшемся
состоянии, его не видно.
Рис. 7. Путь дви-
жения пищевари-
тельной вакуолы
(циклоз) в эндо-
плазме парамеции.
По Фортнеру.
При терпеливом длительном наблюдении можно
уловить момент выталкивания зернышек кармина
через цитопрокт и установить точное его местопо-
ложение. Дефекация у парамеции при температуре
в 13— 14° происходит с интервалами в 8— 12 ми-
нут. Полный путь от места своего образования до
цитопрокта пищеварительная вакуола проделывает при обычных
нормальных условиях (13—14° С) в 50 — 60 минут. Аналогичный
опыт можно проделать и с тонко растертой китайской тушью.
О характере процессов пищеварения можно получить представле-
ние в опыте с кормлением парамеций краской конго (Kongorot),.5^wno-
13
Щей с я химическим индикатором на щелочность или кислотность среды.
В щелочной среде конго дает кирпично-красное окрашивание, а в кис-
лой — индигово-синее. Инфузорий помещают в насыщенный водный
раствор конго (он красного цвета). Через 10—15 минут пипеткой пере-
носят каплю смеси под покровное стекло и рассматривают, как и пре-
дыдущий препарат, при сильном увеличении.
Только что образующаяся пищеварительная вакуола окрашена
в красный цвет, как и раствор конго; через несколько минут она
делается фиолетовой и, наконец, индигово-синей; это означает, что
содержимое вакуолы приобретает кислую реакцию, pH среды вакуолы
изменяет свое значение от 5 до 3; эта фаза пищеварения аналогична
пепсинному перевариванию пищи в желудке позвоночных; позднее
содержимое вакуолы краснеет и, следовательно, становится щелочным,
что соответствует второй фазе пищеварения—трипсинному пищева-
рению кишечника. Полная смена реакций длится долго; поэтому на
занятии ограничиваются сравнением окраски различных пищевари-
тельных вакуол.
Хотя инфузории заглатывают, вообще, мелкие объекты, попадаю-
щие в ротовое углубление, все же, в известной мере, они оказывают
предпочтение веществам удобоваримым. Приготовляют две приблизи-
тельно одинаковой густоты эмульсии из мелко растертого желтка
(куриного) и кармина в двух часовых стеклах. К эмульсии прибавляют
парамеций из одной и той же культуры. Через 20—30 минут берут
из обеих эмульсий пробы и под покровным стеклом подсчитывают
под микроскопом число вакуол у 10—2Q инфузорий и определяют
среднее арифметическое для одной инфузории. Сравнение сделанных
сейчас же один за другим подсчетов покажет большее образование
вакуол с желтком, чем с кармином; следовательно, при одинаковых
условиях парамеция интенсивнее заглатывает желток, нежели непере-
варимые зернышки кармина.
Прямые наблюдения также могут подтвердить известный «выбор»
заглатываемых туфелькой веществ. Туфелька заглатывает только
мелкие зернышки кармина — не более 1—2 р. в поперечнике; при кор-
млении же ее растертым сваренным вкрутую яичным белком можно
наблюдать заглатывание гораздо более крупных кусочков (до б—10
величиной). Следовательно, ресничный цитостомальный аппарат не
является чисто механическим в своей работе, которая регулируется
в зависимости от характера воспринимаемых веществ.
Интересные опыты можно произвести над изменением хода пище-
варительных процессов при действии на парамеций различных солей.
Приготовляют молекулярные растворы необходимых солей 1 и разво-
1 Для изготовления молекулярных растворов на 1 л дистиллированной воды
берут следующее количество граммов солей:
лимоннокислого лития — CeH6O7U8 ...................... 230,64
хлористого бария — ВаС12 2Н2О....................... 244,4
магния — MgCl2........................... 95,24
лития — L1G1......................... . . . • 42,39
кальция — СаСЬ............................110,99
сернокислого магния — MgS6a или MgSO*.................1-0,39
» железа — FeSO< • 7НаО........................... 278,02
Соответствующие растворы разбавляют в несколько раз дистиллированной
водой, например тп/60, т/100, и используют для опытов.
14
дят их водой до состояния, в котором они не действуют вредно на инфу-
зорий; в соответствующий раствор пускают парамеций, накормлен-
ных тушью или кармином в течение 1% часов. Далее берут под покров-
ное стекло с восковыми ножками капельку смеси и наблюдают инфу-
зорий при большом увеличении.
Опыты с ускорением процесса дефекации производятся в слабом
растворе хлористого лития (т/60—т/100) или лимоннокислого ли-
тия, хлористого бария, хлористого магния и др. Акт дефекации зна-
чительно ускоряется, так как опорожнение отработанных пищеваритель-
ных вакуол через цитопрокт совершается
не через 8—17 минут, а гораздо скорее —
через 4—3 минуты и даже через 2— 1 ми-
нуту. Закончившие циркуляцию по телу
парамеции тушевые вакуолы скопляются
N
Со
Nv
Рис. 8. Действие некоторых
В.
Догелю и
По
солей на туфельку.
М. Исаковой-Кео.
А — туфелька, накормленная тушью, в момент дефекации
(в слабом растворе хлористого лития). Р — глотка; Cv — сократительная вакуола;
Nv—пищеварительные вакуолы, набитые тушью; N — ядро. В —туфелька в сла-
бом растворе хлористого магния; образование тушевой «кишки».
С — туфелька в слабом растворе сернокислого железа. Изры-
гание тушевой кишки через глотку обратно.
на брюшной стороне тела, в районе между задним концом глотки и
брюшной поверхностью инфузории. Вакуолы, числом 9—10, распо-
лагаются здесь под эктоплазмой в один слой, тесно соприкасаясь друг
с другом; обычно они сохраняют индивидуальность, но иногда сли-
ваются в одну большую вакуолу. Далее, поверхность тела параме-
ции над вакуолами несколько приподнимается; щель цитопрокта
раскрывается, и через нее выдавливается наружу одновременно не-
сколько вакуол. Если дефекация учащается, то одновременно выбра-
сывается по 1—2 вакуолы. Выбрасываемое содержимое имеет плот-
ную консистенцию, поэтому тушевые шарики не распадаются на
мелкие зернышки, а сохраняют свою форму (рис. 8, А).
Опыты с т/100 раствора хлористого кальция дают обратный эф-
фект, так как замедляют < кт дефекации до 30 и более минут интер
15
вала. Интересна физиологическая параллель в действии указанных
солей на парамеций и млекопитающих, несмотря на совершенно
различную структуру их органоидов и органов пищеварения: соли
магния и бария являются слабительными, тогда как соли кальция,
наоборот, действуют на кишечник закрепляюще.
Опыт с видоизменением характера поступления заглатываемых
веществ в эндоплазму производится помещением свежих (т. е. не-
кормленных) парамеций в ш/100 раствор хлористого или сернокислого
магния, в котором предварительно была растерта тушь. Наблюдение
начинают тотчас, взяв каплю смеси под микроскопом, в обычном
порядке.
Характерной особенностью является то, что заглатываемая тушь
не обособляется в отдельные пищеварительные вакуолы (как то проис-
ходит в норме), но поступает в эндоплазму парамеции сплошным колба-
совидным тяжем — «тушевой кишкой» (В. Догель); она продви-
гается сначала до заднего конца тела инфузории, затем загибается и
закручивается, делая два, три и даже четыре оборота, что наблюдается
почти у половины подопытных инфузорий (рис. 8, В).
Эти явления длятся 10—20 минут, считая с начала опыта; в даль-
нейшем «тушевая кишка» или разбивается на обычные мелкие вакуолы,
или же ее обороты спаиваются в одну большую пищеварительную
вакуолу. В обоих случаях последующее образование новых пищева-
рительных вакуол происходит в обычном порядке.
Аналогичный предыдущему опыт с ш/100 раствором сернокислого
железа приводит к иным результатам. У парамеций также образуется
более или менее извитая «тушевая кишка», но последняя отличается
значительной вязкостью и большей плотностью, благодаря чему она
долгое время (полчаса и более) может оставаться без заметных изме-
нений. У многих парамеций в растворе сернокислого железа наблю-
дается обратное медленное извергание через глотку образовавшейся
«тушевой кишки». Этот акт «рвоты» протекает уже в течение первых
10 минут опыта (рис. 8, С). Часто вышедший наружу конец «туше-
вой кишки» приклеивается к поверхности предметного стекла; ин-
фузория оживленно плавает вокруг и около точки прикрепления,
способствуя тем освобождению от заглоченной массы туши. У осво-
бодившейся парамеции в дальнейшем наблюдается обычное образова-
ние мелких пищеварительных вакуол.
Экскреторная функция осуществляется не только деятель-
ностью сократительных вакуол. При культивировании парамеций на
сенном растворе с прибавлением куска сырого мяса эндоплазма параме-
ций через несколько дней переполняется многочисленными кристалличе-
скими отложениями в виде иголочек, призм, цилиндриков и других
фигур. По Шевякову, эти образования состоят из фосфорно-
кислой извести. В особенном изобилии они скопляются вокруг сокра-
тительных вакуол. Эти кристаллы, повидимому, растворяются и
удаляются сократительными вакуолами. Временно же они являются
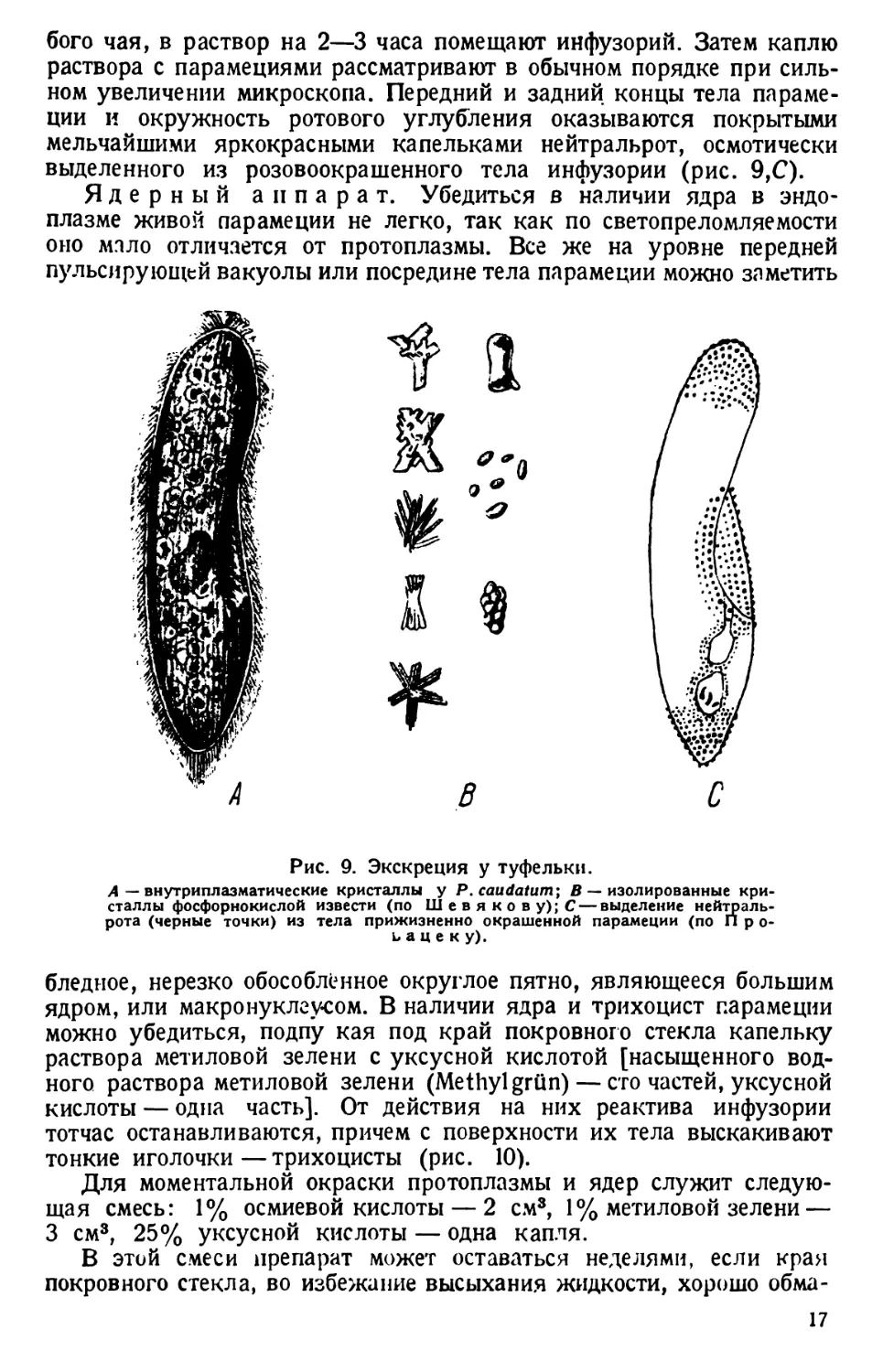
продуктами внутриклеточного выделения (рис. 9, А, В).
Своеобразная экскреторная функция присуща и некоторым частям
поверхности тела парамеции. Ее наблюдают в опыте с прижизненной
окраской нейтральротом. Эту краску растворяют в воде до цвета сла-
16
бого чая, в раствор на 2—3 часа помещают инфузорий. Затем каплю
раствора с парамециями рассматривают в обычном порядке при силь-
ном увеличении микроскопа. Передний и задний концы тела параме-
ции и окружность ротового углубления оказываются покрытыми
мельчайшими яркокрасными капельками нейтральрот, осмотически
выделенного из розовоокрашенного тела инфузории (рис. 9,С).
Я д е р н ы й аппарат. Убедиться в наличии ядра в эндо-
плазме живой парамеции не легко, так как по светопреломляемости
оно мало отличается от протоплазмы. Все же на уровне передней
пульсирующей вакуолы или посредине тела парамеции можно заметить
Рис. 9. Экскреция у туфельки.
А — внутриплазматические кристаллы у Р. caudatum; В — изолированные кри-
сталлы фосфорнокислой извести (по Шевякову); С — выделение нейтраль-
рота (черные точки) из тела прижизненно окрашенной парамеции (по Про-
ь а ц е к у).
бледное, нерезко обособленное округлое пятно, являющееся большим
ядром, или макронуклеусом. В наличии ядра и трихоцист парамеции
можно убедиться, подпу кая под край покровного стекла капельку
раствора метиловой зелени с уксусной кислотой [насыщенного вод-
ного раствора метиловой зелени (Methyl griin) — сто частей, уксусной
кислоты — одна часть]. От действия на них реактива инфузории
тотчас останавливаются, причем с поверхности их тела выскакивают
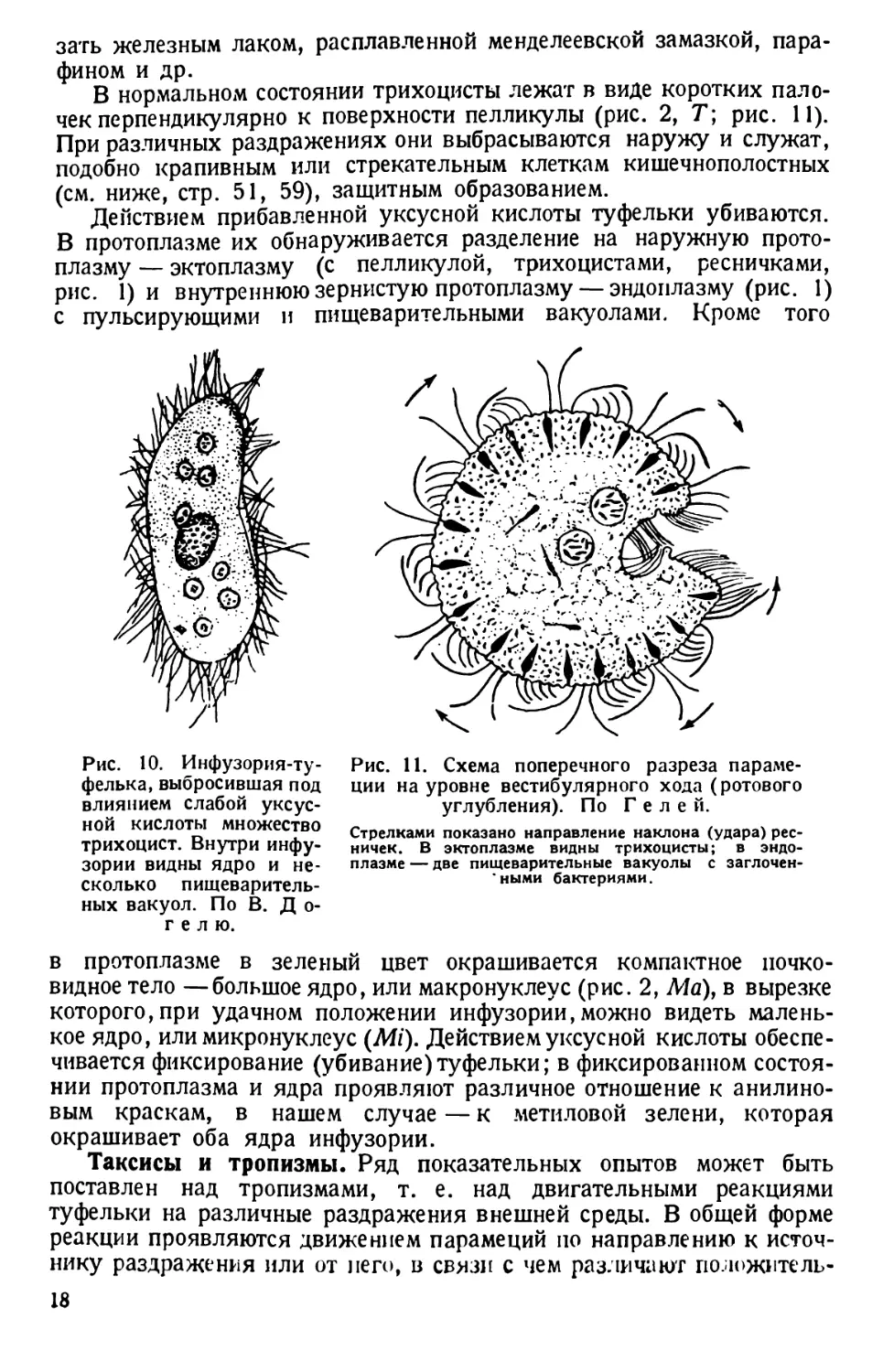
тонкие иголочки — трихоцисты (рис. 10).
Для моментальной окраски протоплазмы и ядер служит следую-
щая смесь: 1% осмиевой кислоты — 2 см3, 1 % метиловой зелени —
3 см8, 25% уксусной кислоты — одна капля.
В этий смеси препарат может оставаться неделями, если края
покровного стекла, во избежание высыхания жидкости, хорошо обма-
17
зать железным лаком, расплавленной менделеевской замазкой, пара-
фином и др.
В нормальном состоянии трихоцисты лежат в виде коротких пало-
чек перпендикулярно к поверхности пелликулы (рис. 2, Т; рис. 11).
При различных раздражениях они выбрасываются наружу и служат,
подобно крапивным или стрекательным клеткам кишечнополостных
(см. ниже, стр. 51, 59), защитным образованием.
Действием прибавленной уксусной кислоты туфельки убиваются.
В протоплазме их обнаруживается разделение на наружную прото-
плазму — эктоплазму (с пелликулой, трихоцистами, ресничками,
рис. 1) и внутреннюю зернистую протоплазму — эндоплазму (рис. 1)
с пульсирующими и пищеварительными вакуолами. Кроме того
Рис. 10. Инфузория-ту-
фелька, выбросившая под
влиянием слабой уксус-
ной кислоты множество
трихоцист. Внутри инфу-
зории видны ядро и не-
сколько пищеваритель-
ных вакуол. По В. Д о-
г е л ю.
Рис. 11. Схема поперечного разреза параме-
ции на уровне вестибулярного хода (ротового
углубления). По Геле й.
Стрелками показано направление наклона (удара) рес-
ничек. В эктоплазме видны трихоцисты; в эндо-
плазме — две пищеварительные вакуолы с заглочен-
ными бактериями.
в протоплазме в зеленый цвет окрашивается компактное почко-
видное тело —большое ядро, или макронуклеус (рис. 2, Ма), в вырезке
которого, при удачном положении инфузории, можно видеть малень-
кое ядро, или микронуклеус (Aft). Действием уксусной кислоты обеспе-
чивается фиксирование (убивание)туфельки; в фиксированном состоя-
нии протоплазма и ядра проявляют различное отношение к анилино-
вым краскам, в нашем случае — к метиловой зелени, которая
окрашивает оба ядра инфузории.
Таксисы и тропизмы. Ряд показательных опытов может быть
поставлен над тропизмами, т. е. над двигательными реакциями
туфельки на различные раздражения внешней среды. В общей форме
реакции проявляются движением парамеций по направлению к источ-
нику раздражения или от пего, в связи с чем различают положитель-
18
Рис. 12. Термо-
тропизм пара-
меции. Инфузо-
рии собрались
в месте пробир-
ки с температу-
рой 26° С. П о
В. Догелю.
ный или отрицательный тропизм. В зависимости от характера раздра-
жения говорят о фототропизме, хемотропизме, термотропизме и др.
Наблюдения над термотропизмом у туфе-
лек. Длинную (20 см) узкую, запаянную с одного конца стеклянную
трубку наполняют жидкостью с большим количеством инфузорий и
затыкают пробкой. Трубку зажимают в щипцы химического штатива.
Просматривая содержимое трубки подлупой (X Ю), устанавливают,
что инфузории распределены равномерно по всей длине трубки. Да-
лее, спиртовкой подогревают осторожно запаянный конец до 38° С.
Легко можно видеть, как парамеции быстро покидают
нагретое место и устремляются к холодному концу.
Попеременным нагреванием их можно повторно пере-
гонять от одного конца трубки к другому. Такие пере-
движения туфелек объясняются их отрицательным
термотропизмом по отношению к повышенной тем-
пературе.
Оптимальная температура, т. е. такая, при которой
все жизненные процессы протекают лучшим образом,
для Paramaecium aurelia лежит в пределах от 24 до
28° С, а для Р. bursaria — между 23—25° С (рис. 12).
Положительный хемотаксис па-
рамеций по отношению к углекислоте. На пред-
метное стекло помещают большую каплю воды с па-
рамециями и по бокам в качестве ножек для покров-
ного стекла кладут по отрезку стеклянного капил-
ляра. Препарат накрывают большим прямоугольным
покровным стеклом (26 х 32 мм) или соответствен-
ным отрезком тонкого предметного (или покровного
диапозитивного) стекла. Пускают в ход аппарат для
добывания углекислоты. Выделяемый газ отводят ре-
зиновой трубкой в тонкую капиллярную пипетку; ко-
нец последней подводят под покровное стекло и вы-
пускают из пипетки один маленький пузырек газа
так, чтобы он очутился посредине препарата. Пара-
меции, рассеянные первоначально под всем покров-
ным стеклом, вскоре сосредоточиваются каймой вокруг
пузырька углекислоты, привлекаемые, этим химиче-
ским раздражителем. Через несколько минут, по мере растворения
углекислоты в воде, химический состав раствора оказывается, вслед-
ствие своей высокой концентрации, неблагоприятным для параме-
ций; последние проявляют отрицательный хемотаксис и отходят не-
сколько к периферии от пузырька, также располагаясь кольцом в зоне
с оптимальной концентрацией раствора углекислоты. Если под стеклом
оказывается попавший одновременно пузырек воздуха (или если его
ввести преднамеренно), то можно видеть, что его присутствие ни
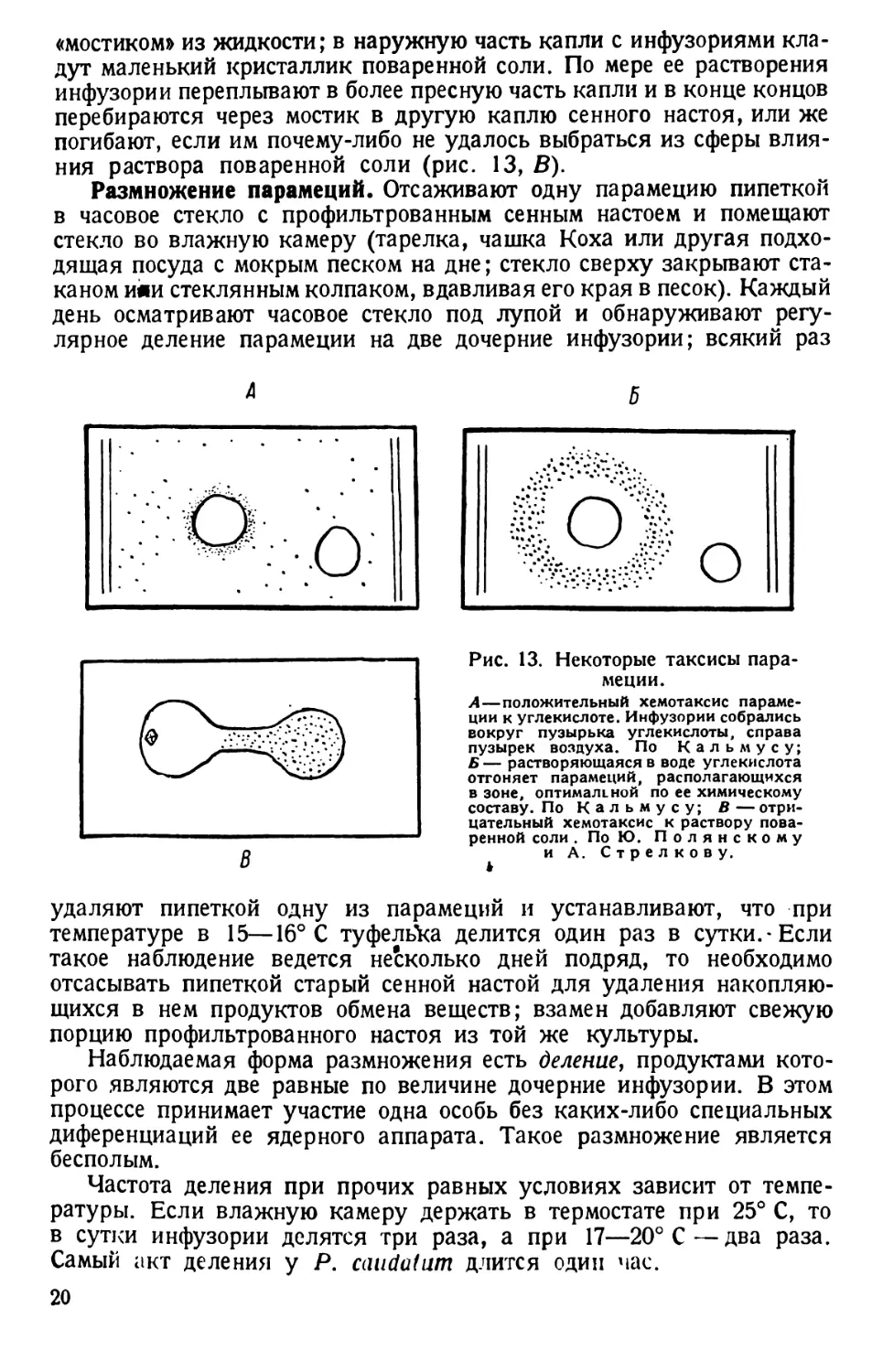
в какой мере не влияет на поведение парамеций (рис. 13, А, Б).
Хемотаксис парамеций можно наблюдать и при действии раство-
ров различных солей, например, поваренной соли. На предметное
стекло наносят недалеко друг от друга каплю культуры парамеций и
каплю стерильного сенного настоя; их соединяют друг с другом
• 19
«мостиком» из жидкости; в наружную часть капли с инфузориями кла-
дут маленький кристаллик поваренной соли. По мере ее растворения
инфузории переплывают в более пресную часть капли и в конце концов
перебираются через мостик в другую каплю сенного настоя, или же
погибают, если им почему-либо не удалось выбраться из сферы влия-
ния раствора поваренной соли (рис. 13, В).
Размножение парамеций. Отсаживают одну парамецию пипеткой
в часовое стекло с профильтрованным сенным настоем и помещают
стекло во влажную камеру (тарелка, чашка Коха или другая подхо-
дящая посуда с мокрым песком на дне; стекло сверху закрывают ста-
каном ими стеклянным колпаком, вдавливая его края в песок). Каждый
день осматривают часовое стекло под лупой и обнаруживают регу-
лярное деление парамеции на две дочерние инфузории; всякий раз
В
Рис. 13. Некоторые таксисы пара-
меции.
А—положительный хемотаксис параме-
ции к углекислоте. Инфузории собрались
вокруг пузырька углекислоты, справа
пузырек воздуха. По Каль м усу;
Б — растворяющаяся в воде углекислота
отгоняет парамеций, располагающихся
в зоне, оптимальной по ее химическому
составу. По Кальмусу; В — отри-
цательный хемотаксис к раствору пова-
ренной соли . По Ю. Полянскому
и А. Стрелкову.
удаляют пипеткой одну из парамеций и устанавливают, что при
температуре в 15—16° С туфелька делится один раз в сутки.-Если
такое наблюдение ведется несколько дней подряд, то необходимо
отсасывать пипеткой старый сенной настой для удаления накопляю-
щихся в нем продуктов обмена веществ; взамен добавляют свежую
порцию профильтрованного настоя из той же культуры.
Наблюдаемая форма размножения есть деление, продуктами кото-
рого являются две равные по величине дочерние инфузории. В этом
процессе принимает участие одна особь без каких-либо специальных
диференциаций ее ядерного аппарата. Такое размножение является
бесполым.
Частота деления при прочих равных условиях зависит от темпе-
ратуры. Если влажную камеру держать в термостате при 25° С, то
в сутки инфузории делятся три раза, а при 17—20° С—два раза.
Самый акт деления у Р. caudatum длится один час.
20
Обрабатывая большое количество парамеций метиловой зеленью
с уксусной кислотой, можно найти особей в разных стадиях деления.
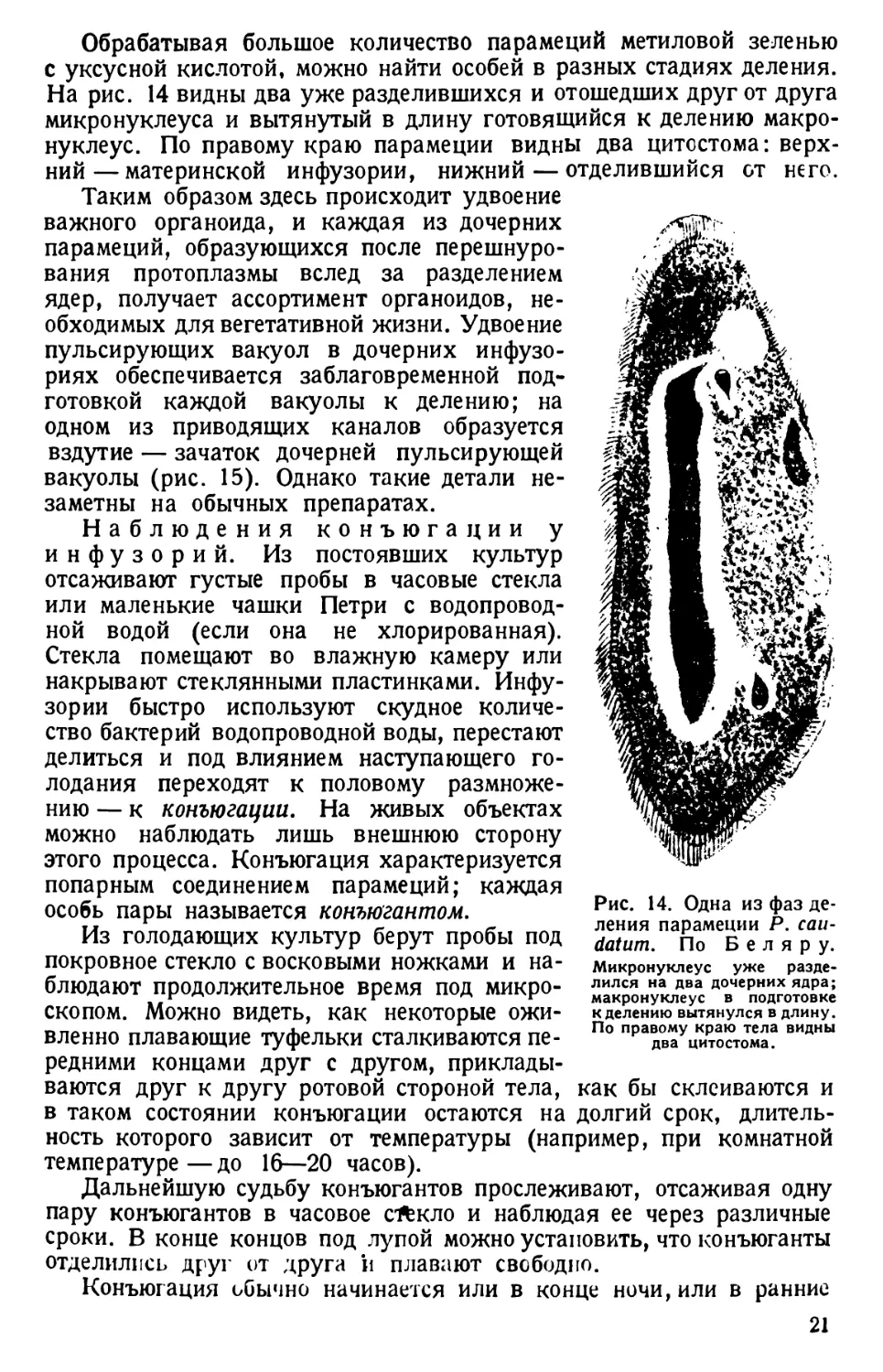
На рис. 14 видны два уже разделившихся и отошедших друг от друга
микронуклеуса и вытянутый в длину готовящийся к делению макро-
нуклеус. По правому краю парамеции видны два цитостома: верх-
ний — материнской инфузории, нижний — отделившийся от него.
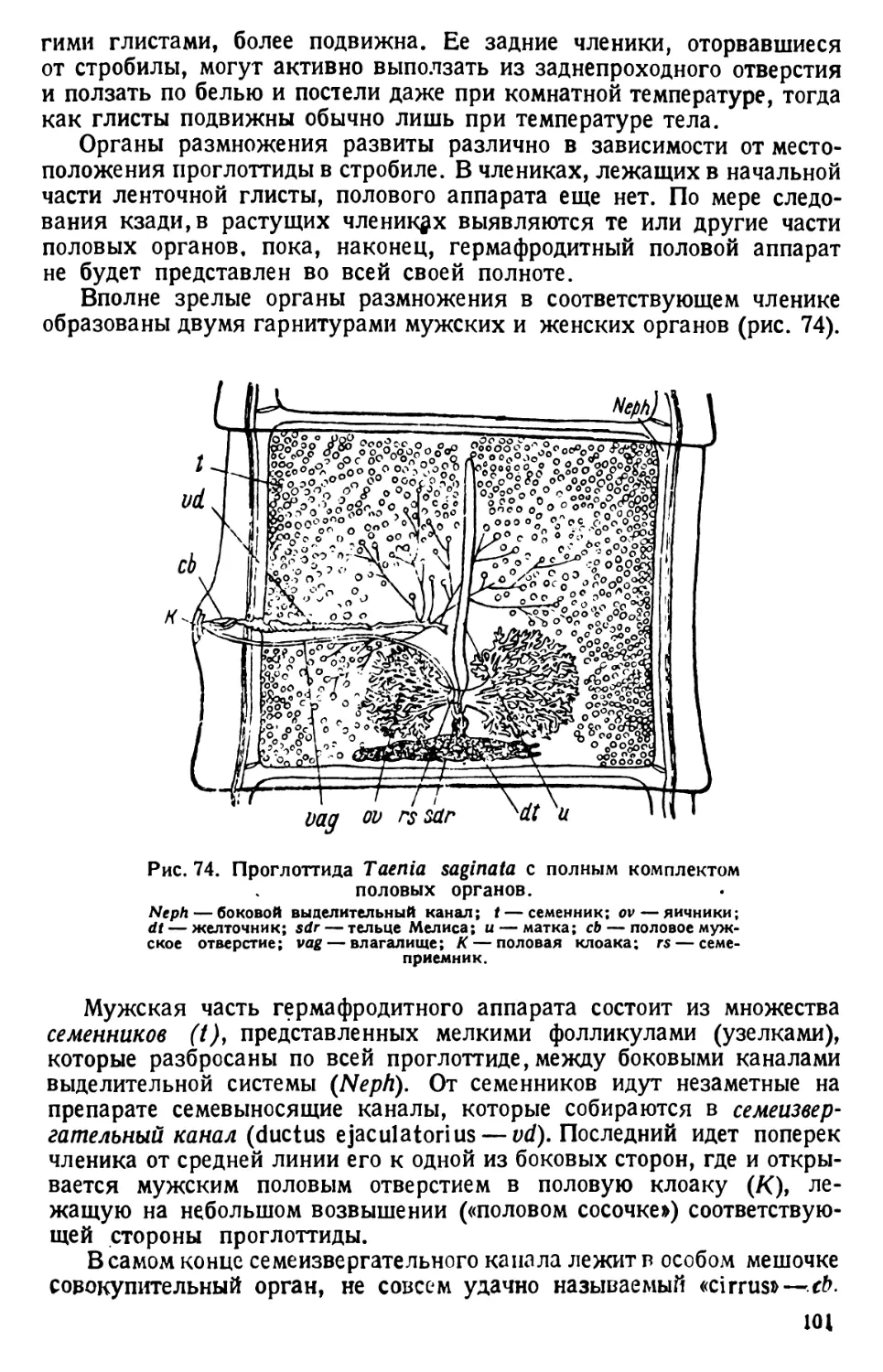
Таким образом здесь происходит удвоение
важного органоида, и каждая из дочерних
парамеций, образующихся после перешнуро-
вания протоплазмы вслед за разделением
ядер, получает ассортимент органоидов, не-
обходимых для вегетативной жизни. Удвоение
пульсирующих вакуол в дочерних инфузо-
риях обеспечивается заблаговременной под-
готовкой каждой вакуолы к делению; на
одном из приводящих каналов образуется
вздутие — зачаток дочерней пульсирующей
вакуолы (рис. 15). Однако такие детали не-
заметны на обычных препаратах.
Наблюдения конъюгации у
инфузорий. Из постоявших культур
отсаживают густые пробы в часовые стекла
или маленькие чашки Петри с водопровод-
ной водой (если она не хлорированная).
Стекла помещают во влажную камеру или
накрывают стеклянными пластинками. Инфу-
зории быстро используют скудное количе-
ство бактерий водопроводной воды, перестают
делиться и под влиянием наступающего го-
лодания переходят к половому размноже-
нию — к конъюгации. На живых объектах
можно наблюдать лишь внешнюю сторону
этого процесса. Конъюгация характеризуется
Рис. 14. Одна из фаз де-
ления парамеции Р. cau-
datum. По Б е л я р у.
Микронуклеус уже разде-
лился на два дочерних ядра;
макронуклеус в подготовке
к делению вытянулся в длину.
По правому краю тела видны
два цитостома.
попарным соединением парамеций; каждая
особь пары называется конъюгантом.
Из голодающих культур берут пробы под
покровное стекло с восковыми ножками и на-
блюдают продолжительное время под микро-
скопом. Можно видеть, как некоторые ожи-
вленно плавающие туфельки сталкиваются пе-
редкими концами друг с другом, приклады-
ваются друг к другу ротовой стороной тела, как бы склеиваются и
в таком состоянии конъюгации остаются на долгий срок, длитель-
ность которого зависит от температуры (например, при комнатной
температуре—до 16—20 часов).
Дальнейшую судьбу конъюгантов прослеживают, отсаживая одну
пару конъюгантов в часовое сЛкло и наблюдая ее через различные
сроки. В конце концов под лупой можно установить, что конъюганты
отделились друг от друга и плавают свободно.
Конъюгация обычно начинается или в конце ночи, или в ранние
21
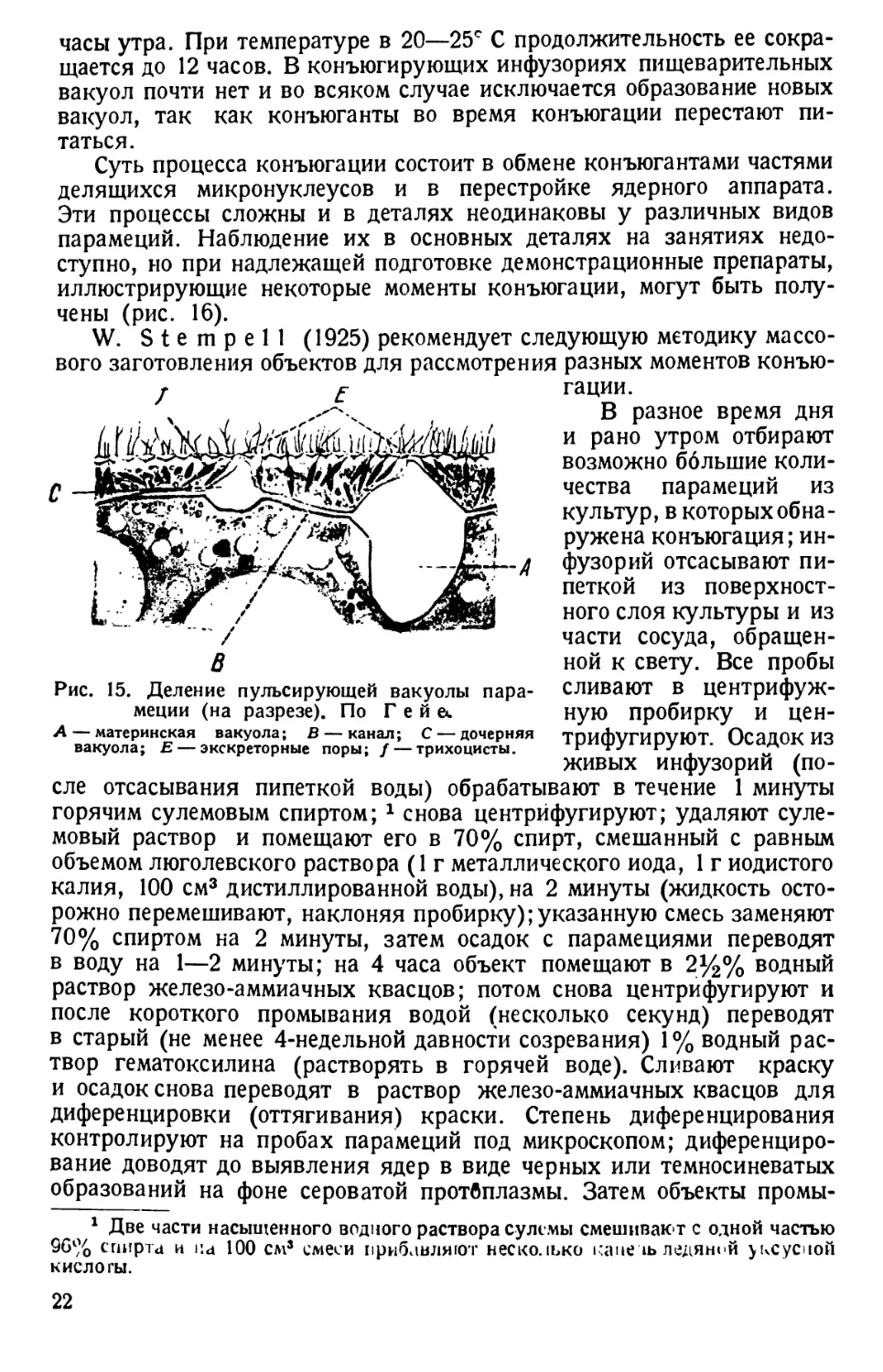
Рис. 15. Деление пульсирующей вакуолы пара-
меции (на разрезе). По Г е й &.
А — материнская вакуола; В — канал; С — дочерняя
вакуола; Е — экскреторные поры; / — трихоцисты.
с
часы утра. При температуре в 20—25е С продолжительность ее сокра-
щается до 12 часов. В конъюгирующих инфузориях пищеварительных
вакуол почти нет и во всяком случае исключается образование новых
вакуол, так как конъюганты во время конъюгации перестают пи-
таться.
Суть процесса конъюгации состоит в обмене конъюгантами частями
делящихся микронуклеусов и в перестройке ядерного аппарата.
Эти процессы сложны и в деталях неодинаковы у различных видов
парамеций. Наблюдение их в основных деталях на занятиях недо-
ступно, но при надлежащей подготовке демонстрационные препараты,
иллюстрирующие некоторые моменты конъюгации, могут быть полу-
чены (рис. 16).
W. S t е m р е 1 1 (1925) рекомендует следующую методику массо-
вого заготовления объектов для рассмотрения разных моментов конъю-
гации.
В разное время дня
и рано утром отбирают
возможно ббльшие коли-
чества парамеций из
культур, в которых обна-
ружена конъюгация; ин-
фузорий отсасывают пи-
петкой из поверхност-
ного слоя культуры и из
части сосуда, обращен-
ной к свету. Все пробы
сливают в центрифуж-
ную пробирку и цен-
трифугируют. Осадок из
живых инфузорий (по-
сле отсасывания пипеткой воды) обрабатывают в течение 1 минуты
горячим сулемовым спиртом;1 * снова центрифугируют; удаляют суле-
мовый раствор и помещают его в 70% спирт, смешанный с равным
объемом люголевского раствора (1 г металлического иода, 1 г йодистого
калия, 100 см3 дистиллированной воды), на 2 минуты (жидкость осто-
рожно перемешивают, наклоняя пробирку) указанную смесь заменяют
70% спиртом на 2 минуты, затем осадок с парамециями переводят
в воду на 1—2 минуты; на 4 часа объект помещают в 2%% водный
раствор железо-аммиачных квасцов; потом снова центрифугируют и
после короткого промывания водой (несколько секунд) переводят
в старый (не менее 4-недельной давности созревания) 1% водный рас-
твор гематоксилина (растворять в горячей воде). Сливают краску
и осадок снова переводят в раствор железо-аммиачных квасцов для
диференцировки (оттягивания) краски. Степень диференцирования
контролируют на пробах парамеций под микроскопом; диференциро-
вание доводят до выявления ядер в виде черных или темносиневатых
образований на фоне сероватой протвплазмы. Затем объекты промы-
1 Две части насыщенного водного раствора сулемы смешивают с одной частью
9G% спирта и г.а 100 см* смеси прибавляют несколько напеть ледяной уксусной
кислоты.
22
вают водой и через 70 и 96% спирт переводят (пользуясь всякий
раз ручной центрифугой) в абсолютный спирт, затем в креозот или
в *кедровое масло. Здесь окрашенные инфузории могут храниться
долгое время. Препарат приготовляют, нанося на чистое предметное
стекло каплю канадского бальзама; в ее центр помещают капельку
кедрового масла с инфузориями, взятую капиллярной пипеткой. Пре-
Рис. 16. Фазы конъюгации Paramaecium caudatum. По Штемпелю.
А — ядерный аппарат еще без изменения; В — деление микронуклеуса (л);
С — обмен микронуклеусами; D и Е — распадение макронуклеуса (гл) и деление
микронуклеусов (л).
парат покрывают покровным стеклом, предварительно перемешав
осторожно чистой иглой масло с бальзамом. Под покровное стекло
кладут вместо ножек обломки покровного же стекла или делают
восковые ножки.
На таких препаратах при массовом их изготовлении можно подо-
брать различные моменты конъюгации (рис. 16): а) начальный момент —
соединение пары конъюгантов при еще неизмененном ядерном аппарате ;
б) деление микронуклеуса; в) обмен конъюгантов частями разделив-
шихся микронуклеусов; г) распадение макронуклеуса и последующее
23
деление микронуклеусов. Детали конъюгации и последовательность
ее этапов излагаются в руководствах зоологии, протозоологии и общей
биологии. Из рассмотрения описанных препаратов можно вывести
заключение, что главную роль в конъюгации играет малое ядро пара-
меции, тогда как ее макронуклеус играет роль в процессах вегетатив-
ной жизни.
Конъюгация является процессом полового размножения, так как
она ведет к обмену ядерным веществом микронуклеусов; в резуль-
тате перестройки ядерного аппарата разошедшиеся конъюганты по-
лучают способность дальнейшего бесполого размножения делением,
каковое перед периодом конъюгации приходило в состояние депрессии.
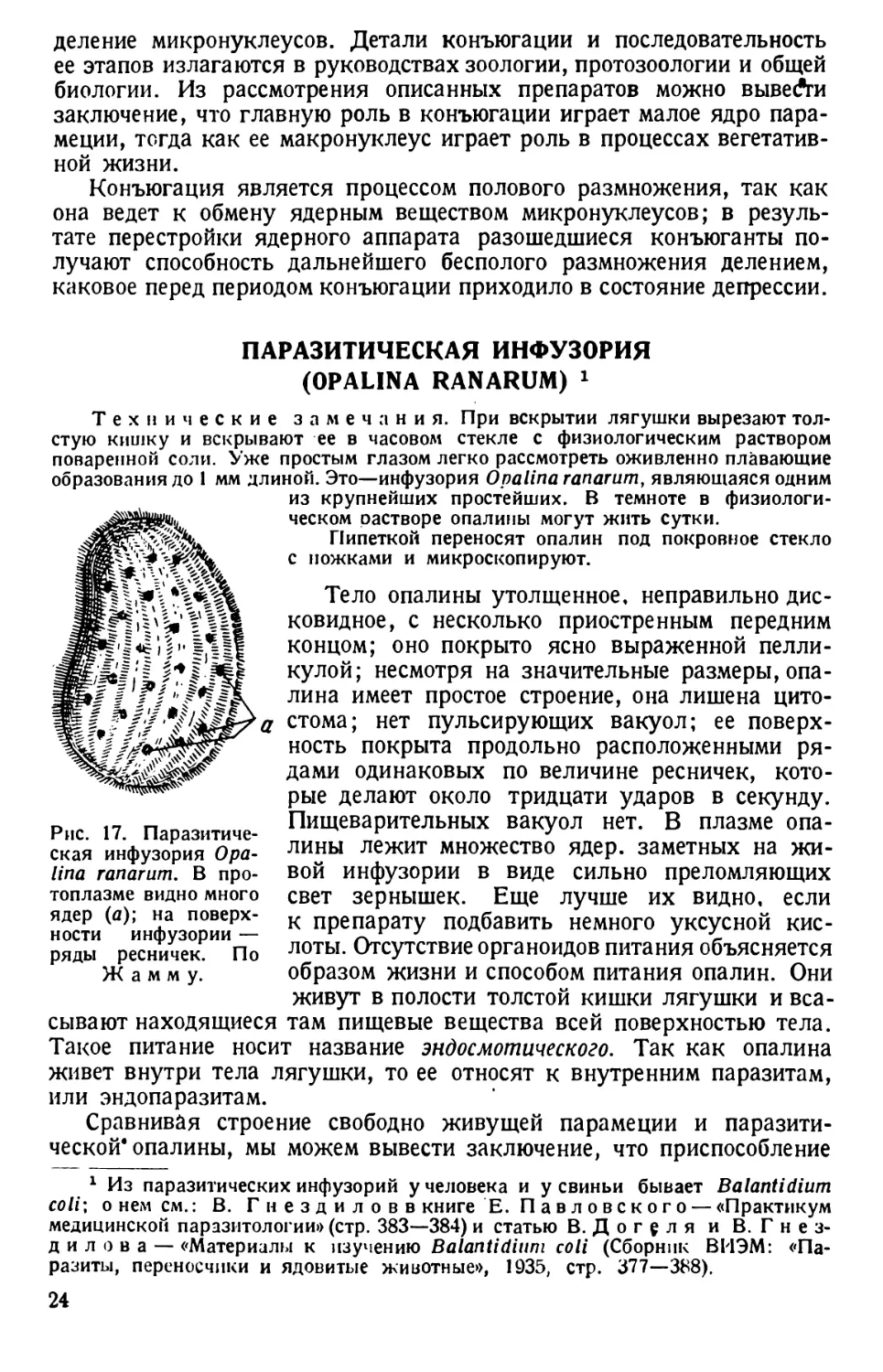
ПАРАЗИТИЧЕСКАЯ ИНФУЗОРИЯ
(OPALINA RANARUM) 1
Технические замечания. При вскрытии лягушки вырезают тол-
стую кишку и вскрывают ее в часовом стекле с физиологическим раствором
поваренной соли. Уже простым глазом легко рассмотреть оживленно плавающие
образования до 1 мм длиной. Это—инфузория Opalina ranarum, являющаяся одним
Рис. 17. Паразитиче-
ская инфузория Opa-
lina ranarum. В про-
из крупнейших простейших. В темноте в физиологи-
ческом растворе опалины могут жить сутки.
Пипеткой переносят опалин под покровное стекло
с ножками и микроскопируют.
Тело опалины утолщенное, неправильно дис-
ковидное, с несколько приостренным передним
концом; оно покрыто ясно выраженной пелли-
кулой; несмотря на значительные размеры, опа-
лина имеет простое строение, она лишена цито-
стома; нет пульсирующих вакуол; ее поверх-
ность покрыта продольно расположенными ря-
дами одинаковых по величине ресничек, кото-
рые делают около тридцати ударов в секунду.
Пищеварительных вакуол нет. В плазме опа-
лины лежит множество ядер, заметных на жи-
вой инфузории в виде сильно преломляющих
топлазме видно много
ядер (а); на поверх-
ности инфузории —
ряды ресничек. По
Ж а м м у.
свет зернышек. Еще лучше их видно, если
к препарату подбавить немного уксусной кис-
лоты. Отсутствие органоидов питания объясняется
образом жизни и способом питания опалин. Они
живут в полости толстой кишки лягушки и вса-
сывают находящиеся там пищевые вещества всей поверхностью тела.
Такое питание носит название эндосмотического. Так как опалина
живет внутри тела лягушки, то ее относят к внутренним паразитам,
или эндопаразитам.
Сравнивая строение свободно живущей парамеции и паразити-
ческой* опалины, мы можем вывести заключение, что приспособление
1 Из паразитических инфузорий у человека и у свиньи бывает Balantidium
coli-, о нем см.: В. Г н е з д и л о в в книге Е. П а в л о в с к о г о— «Практикум
медицинской паразитологии» (стр. 383—384) и статью В. Д о г { л я и В. Гнез-
дилов а — «Материалы к изучению Balantidium coli (Сборник ВИЭМ: «Па-
разиты, переносчики и ядовитые животные», 1935, стр. 377—388).
24
к паразитическому образу жизни ведет к некоторому упрощению
организации паразита, в частности к утере органоидов питания. От-
сутствие пульсирующей вакуолы, очевидно, зависит от обитания опа-
лины в среде, концентрация солей которой выше, чем в пресной воде.
Общие замечания
Инфузория «туфелька» характеризуется тем, что весь ее организм
образован из протоплазмы с заключенными в ней двумя ядрами, что
(исключая двуядерность) соответствует схеме строения животной
клетки. Поэтому инфузорий относят к одноклеточным организмам
(Protozoa), в противовес многоклеточным, или Metazoa. В то же время
одноклеточный организм не равнозначен отдельной клетке много-
клеточного животного, но может быть сравниваем лишь с целым таким
организмом.
- На основании этого признака животных вообще делят на две
группы: одноклеточных — Protozoa и многоклеточных — Meta-
zoa, причем Protozoa выделяются в качестве особого типа живот-
ных.
Туфелька отличается .микроскопически мелкими размерами.
Туфелька обладает и протоплазмой и ядром, причем ядер у нее
два — макронуклеус и микронуклеус.
Так как одноклеточному животному организму присущи все основ-
ные жизненные функции, то осуществление таковых у туфельки свя-
зывается с особыми структурами ее частей. Так, движение произво-
дится постоянными ресничками, пища заглатывается цитостомом,
неиспользованная пища выбрасывается цитопроктом, экскреция струк-
турной основой имеет сократительные вакуолы. Кроме того инфузории
свойственна способность реагировать на раздражения (например,
выбрасыванием трихоцист).
Реснички, трихоцисты, цитопрокт, цитостом и другие образования
являются «органами» инфузории, но так как сама инфузория всего
навсего — одноклеточный организм и эти «органы» ее являются обо-
собленными частями клетки, то их й называют органоидами, или орга-
неллами, в отличие от органов Metazoa.
Систематическое положение Paramaecium
Туфелька имеет тело определенной формы, покрытое постоянно
имеющимися ресничками. Эти признаки характеризуют представите-
лей класса ресничных, или инфузорий (Ciliata s. Infusoria). Так как
все реснички туфельки и опалины одинаковой величины, то этих
инфузорий относят к отряду равноресничных—Holotricha.
В отряде равноресничных туфелька занимает место в семействе
Paramaeciidae, представители которого характеризуются широким
перистомом, тянущимся от переднего конца тела до его середины;
на дне перистома лежит собственно ротовое углубление (vestibu-
lum).
К семейству Paramaeciidae принадлежит только один род Para-
maecium, распадающийся на несколько видов, отличающихся формой
25
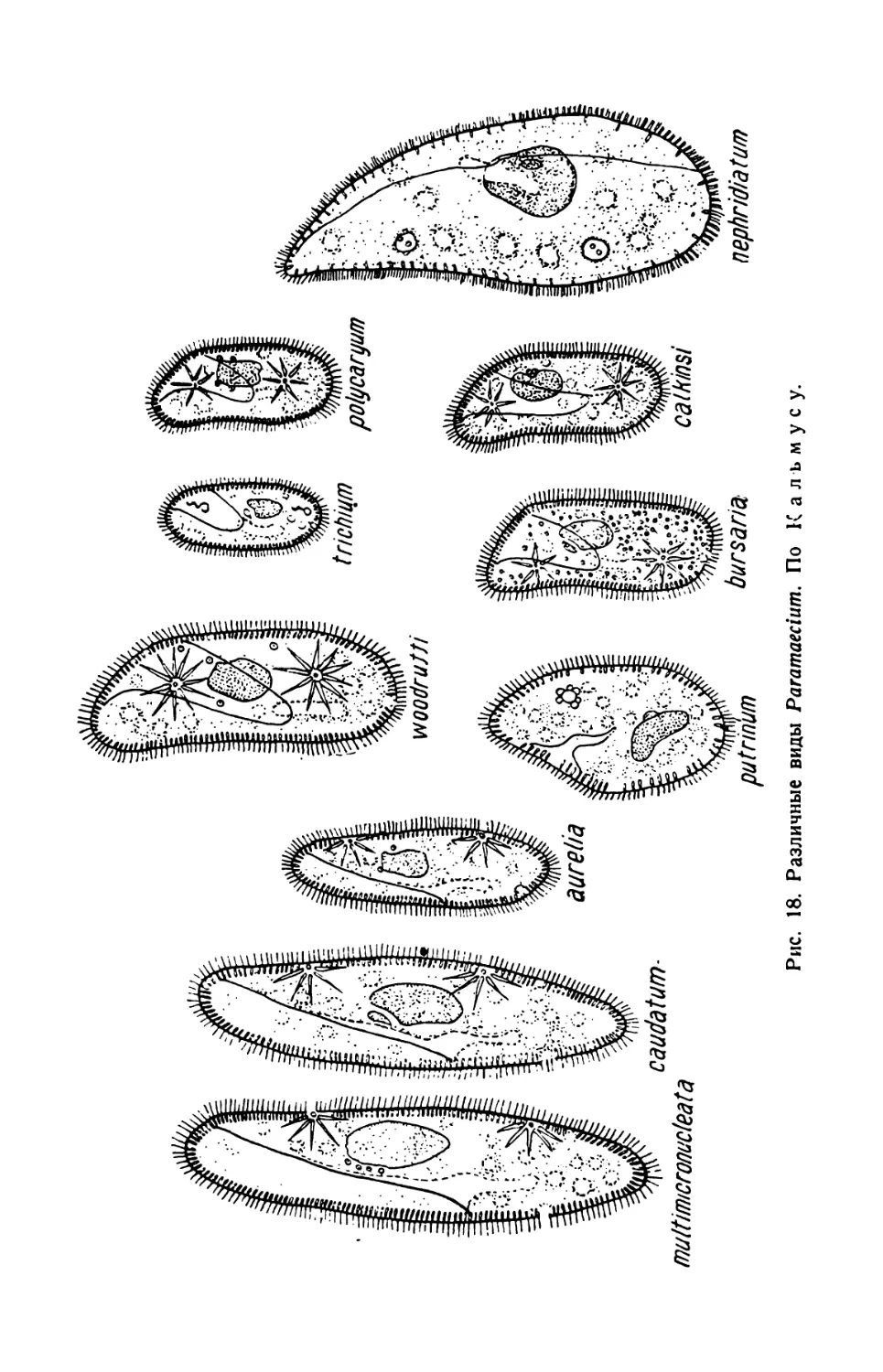
Рис. 18. Различные виды Paramaecium. По К а л ъ м у с у.
тела, количеством ядер, различными биологическими особенностями
и другими признаками (рис. 18).
Для определения видов парамеций пользуются специальной опре-
делительной таблицей.
Определитель видов рода Paramaecium
(по Н. К а 1 m и s)
1. Тело сигаровидное, не уплощенное в дорзо-вентральном направлении (группа
aurelia...............................................................2
la. Тело более короткое и широкое, ясно уплощенное в дорзо-вентральном на-
правлении (группа bursaria)...........................................3
2. В вегетативном состоянии имеется только один компактный микронуклеус . .
........................................................Р. caudiatum Ehrb.
2а. Имеются два пузыревидных микронуклеуса; задний конец тела чуть острее
переднего; обыкновенно две пульсирующие вакуолы; длина 120—180 |л . .
.......................................... . . . Р. aurelia О. F. М.
2Ь. Обыкновенно четыре маленьких микронуклеуса; часто — более двух пуль-
сирующих вакуол; длина свыше 280 ц......................................
..............................Р. multimicronucleata Powers et Mitschel
3. Только одна передняя пульсирующая вакуола с лучевидно расположенными
каналами; длина самое большее превышает ширину в два раза (80—140 ц);
трихоцист нет................................Р. pultrinum Cl. et Lachm.
За. Две пульсирующие вакуолы лежат в эндоплазме; каждая состоит из цен-
трального пузырька, двух приводящих пузырьков и одного скрученного, клуб-
ковидного выводного канала; пульсируют быстро, переменно. Один ком-
пактный микронуклеус. Длина 70—90 р...................Р. trichium Stokes
3b. Две пульсирующие вакуолы в эктоплазме с лучевидно расположенными
приводящими каналами...............................................4
4. Каждая пульсирующая вакуола с двумя выделительными отверстиями (по-
рами) .............................................•..................5
4а. Каждая пульсирующая вакуола с одной порой.........................6
5. Почти вся инфузория окрашена в зеленый цвет благодаря нахождению в ней
зоохлорелл; длина тела 90—120 ц; циклоз протекает быстро; один компакт-
ный микронуклеус.......................................Р. bursaria Ehrb.
5а. Зоохлорелл нет; длина более 150 ц..............Р. nephridiatum Gelei
6. При нормальном поступательном движении вперед парамеция вращается
(если смотреть ей вслед) по ходу часовой стрелки; два пузыревидных микро-
нуклеуса; длина 110—140 |л..........................Р. calkinsi Woodruff
ба. Парамеция вращается в направлении, обратном ходу часовой стрелки .... 7
7. Один микронуклеус...............................Р. chrysalis О. F М.
7а. Длина 70—110 ц; рот — в середине тела; 3—9 (чаще 4) мелких пузыревидных
микронуклеусов.........................Р. polycaryum Woodruff et Spencer
7b. Длина 150—210 ц; рот лежит позади середины тела; три-четыре мелкихмикро-
нуклеуса, которые часто лежат в разных частях тела . . Р. woodruffi Wenrich
К группе aurelia принадлежат сигаровидные Р. multimicronucleata,
Р. caudatum и Р. aurelia, а к группе bursaria относятся уплощенные
с широким телом Р.pultrinum, Р. nephridiatum, Р. bursaria, Р. calkinsi,
Р. polycaryum, Р. woodruffi и Р. trichium.
При определении парамеций следует считаться с возможностью
отсутствия у данной особи некоторых признаков видового значения,
в силу различных причин. Так, трихоцисты могут отсутствовать
потому, что рассматриваемая особь недавно ими «выстрелила»; при
содержании в темноте и при бедном солями корме при повышенной
температуре, у Р. bursaria зоохлореллы совершенно исчезают. Длина
тела инфузории стоит в зависимости от условий питания, температуры
и др. Все эти примеры иллюстрируют нам явления индивидуальной
изменчивости признаков.
27
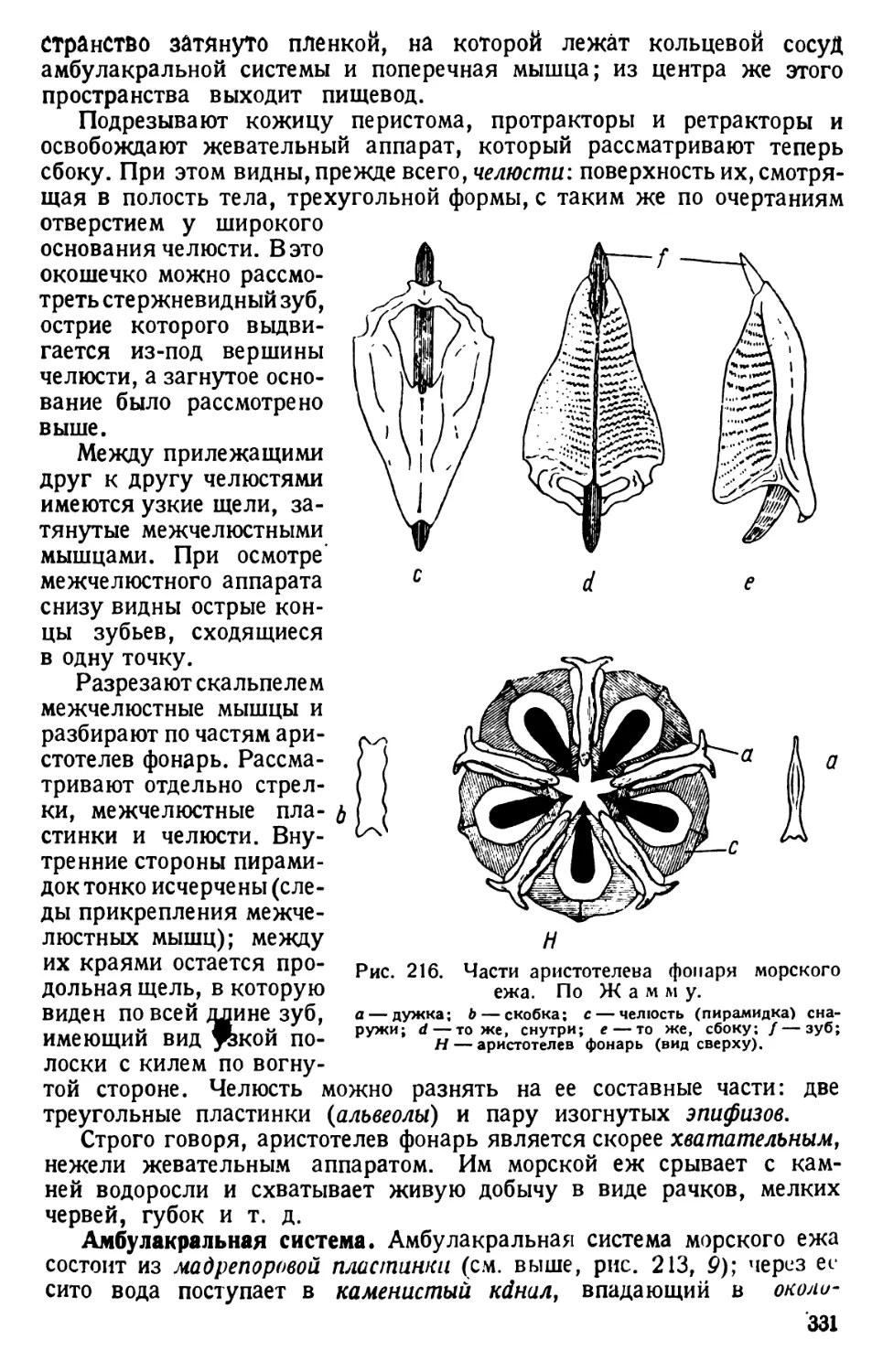
КЛАСС САРКОДОВЫЕ (SARCODINA)
ПОДКЛАСС КОРНЕНОЖКИ (RHIZOPODA)
АМЁБЫ (ОТРЯД AMOEBINA)
Технические замечания. В культурах с инфузориями обычно
разводятся и амёбы. Особенно богата ими та отливающая металлическим блеском
бактериальная пленка, которая образуется на поверхности культуры. Амёб легко
культивировать на питательной среде (см. выше, стр. 5).
Амёбы очень мелкие и не бросающиеся в глаза организмы, которых прихо-
дится отыскивать с вниманием и настойчивостью.
Неблагоприятным обстоятельством является то, что при изготовлении све-
жего препарата из живых амёб (перенесением части пленки в каплю воды под
покровное стекло) последние, вследствие механического раздражения, округ-
ляются, и требуется минут 1(Ъ—15 времени, пока они снова начнут двигаться.
Исследование надо вести при сильном увеличении микроскопа.
Наблюдения над живой амёбой
В противоположность инфузориям, амёба не имеет постоянного
очертания тела, ибо она лишена какой-либо покровной оболочки.
Тело ее состоит из комочка
Рис. 19. Движение амёбы при помощи лож-
ноножек.
7 — пищеварительная вакуола; 2 — сократительная
вакуола; 3—ядро.
слизеобразной протоплаз-
мы, в которой возможно
уловить разделение на бо-
лее прозрачную эктоплазму
и зернистую и темную эндо-
плазму (рис. 19).
Что рассматриваемый
организм есть действитель-
но амёба, можно убедиться,
наблюдая его движение.
Для этой цели время от
времени схематически зари-
совывают контуры наблю-
даемого объекта и по изме-
нению контура судят о по-
движности амёбы.
Амёба очень медленно
передвигается с места на
место, выпуская лопастные
выросты протоплазмы, на-
зываемые ложноножками
или псевдоподиями (рис. 19 и
20). Ложноножки являются
«временными органоидами»,
могущими возникать в лю-
бой точке поверхности тела амёбы. Движение при помощи ложно-
ножек называется амёбоидным; оно весьма примитивно, ибо осу-
ществляется без помощи каких-либо постоянных приспособлений или
структур.
28
Ложноножками амёба пользуется не только для движения, но и для
захватывания пищи. Встретив на пути микроскопическую водоросль,
амёба охватывает ее с боков двумя ложноножками, которые сливаются
друг с другом позади водоросли, благодаря чему последняя попадает
в эндоплазму амёбы, где и переваривается (рис. 19, 7).
Такой тип пищеварения, как у амёбы, называется внутриклеточ-
ным, или фагоцитарным. Для обслуживания этой функции у амёбы
нет никаких специальных и постоянных органоидов. Пища может
заглатываться любой частью поверхности тела, равным образом и
Рис. 20. Amoeba Птах; характер движения (направление послед-
него показано стрелкой). По Гофману.
выбрасывание непереваренных остатков происходит в любом месте,
при разрыве эктоплазмы.
В эндоплазме амёбы имеется сократительная вакуола (рис. 19, 2)
и ядро (рис. 19, 5), которое становится видимым лишь при действии
реактивов, например, уксусной кислоты, ибо светопреломляемость
живой протоплазмы и ядра одинакова.
Многие амёбы являются паразитами кишечника и некоторых
других органов различных животных и человека. [О паразитических
амёбах человека см.: В. Гнездилов в книге Е. Павлов-
с к о г о — «Практикум медицинской паразитологии» (гл. «Паразити-
ческие простейшие кишечника», стр. 365—380)].
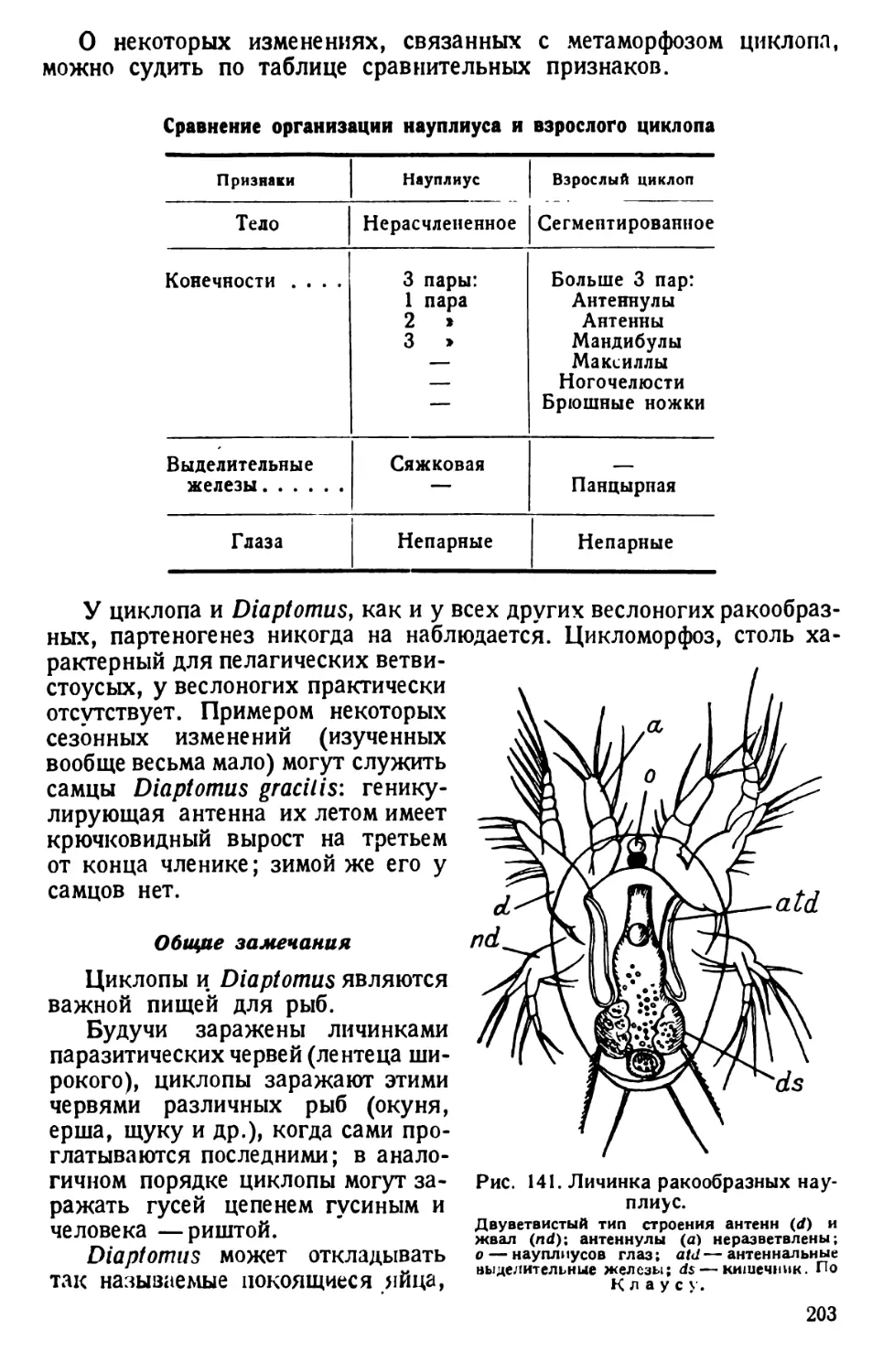
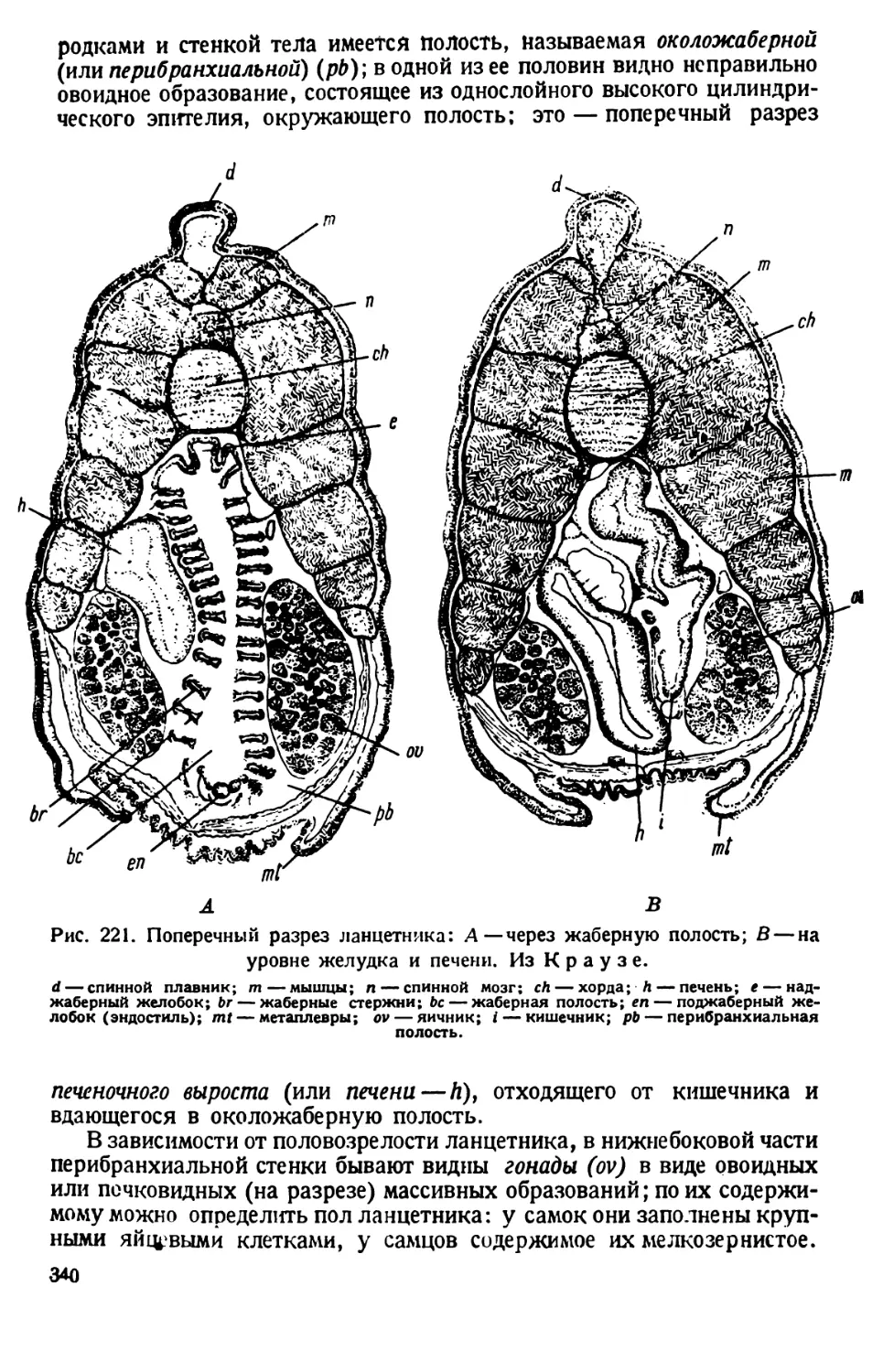
РАКОВИННЫЕ КОРНЕНОЖКИ (ОТРЯД FORAMINIFERA)
Объектом использования служит морской песок с Мурмана, из
Неаполя и др., в котором может быть много раковинок корненожек.
Если имеется сухой песок, то из него легко выделить раковинки,
смешав песок с насыщенным раствором поваренной соли. Песчинки
оседают на дно, а раковинки всплывают на поверхность. Отсюда их
выбирают шпателем, промывают водой, высушивают или помещают
в спирт.
Для непосредственного выискивания раковинок песок наносят
тонким слоем на предметное стекло и просматривают шаг за шагом при
сильном падающем свете, передвигая песчинки препаровальной иглой.
Обнаруженные раковинки переносят прикосновением влажной иглы
на чистое предметное стекло; когда будет отобрано нужное количество
29
раковинок, стекло осторожно нагревают на спиртовой лампочке и на
раковинки наносят каплю канадского бальзама; препарат закрывают
покровным стеклом, распределив раковинки так, чтобы они наподобие
ножек подпирали углы стекла. Препарат непрозрачный, и его надо
рассматривать при падающем свете. Еще лучше рассматривать сухие
раковинки без бальзама при падающем свете. Все манипуляции сле-
дует производить осторожно, чтобы не сдуть, не смахнуть отобранных
раковинок.
Совершенно естественно, что на таком препарате наблюдатель
имеет дело лишь с мертвым остатком корненожки — именно с ее скелет-
ным (опорным) органоидом, которым является наблюдаемая рако-
Рис. 21. Раковинки корненожек. По Делажу и Эруару.
1—Lagena; 2—Nodosaria; 3—Polymorphic; 4—Tetyularia; 5 — Buli-
mic; 6—Cassidulina; 7—Globigerina.
винка. Такие образования называются статическими органеллами
аллоплазматического характера, так как они являются продуктом
выделения живой протоплазмы.
Форма раковинок разнообразна, о чем можно судить по рис. 21.
Стенка их во многих случаях покрыта тончайшими отверстиями;
такие корненожки называются прободенными (Perforata); у других
раковинка имеет одно отверстие, через которое высовывается наружу
протоплазма, образующая псевдоподии.
В зависимости от строения раковинок различают однокамерные
и многокамерные формы: последние развиваются из первых путем
последовательного нарастания камер.
Раковинки раковинных корненожек имеют тонкую хитиноидную
основу, в которой отлагаются соли кремния (5Юг)или извести (СаСО8).
30
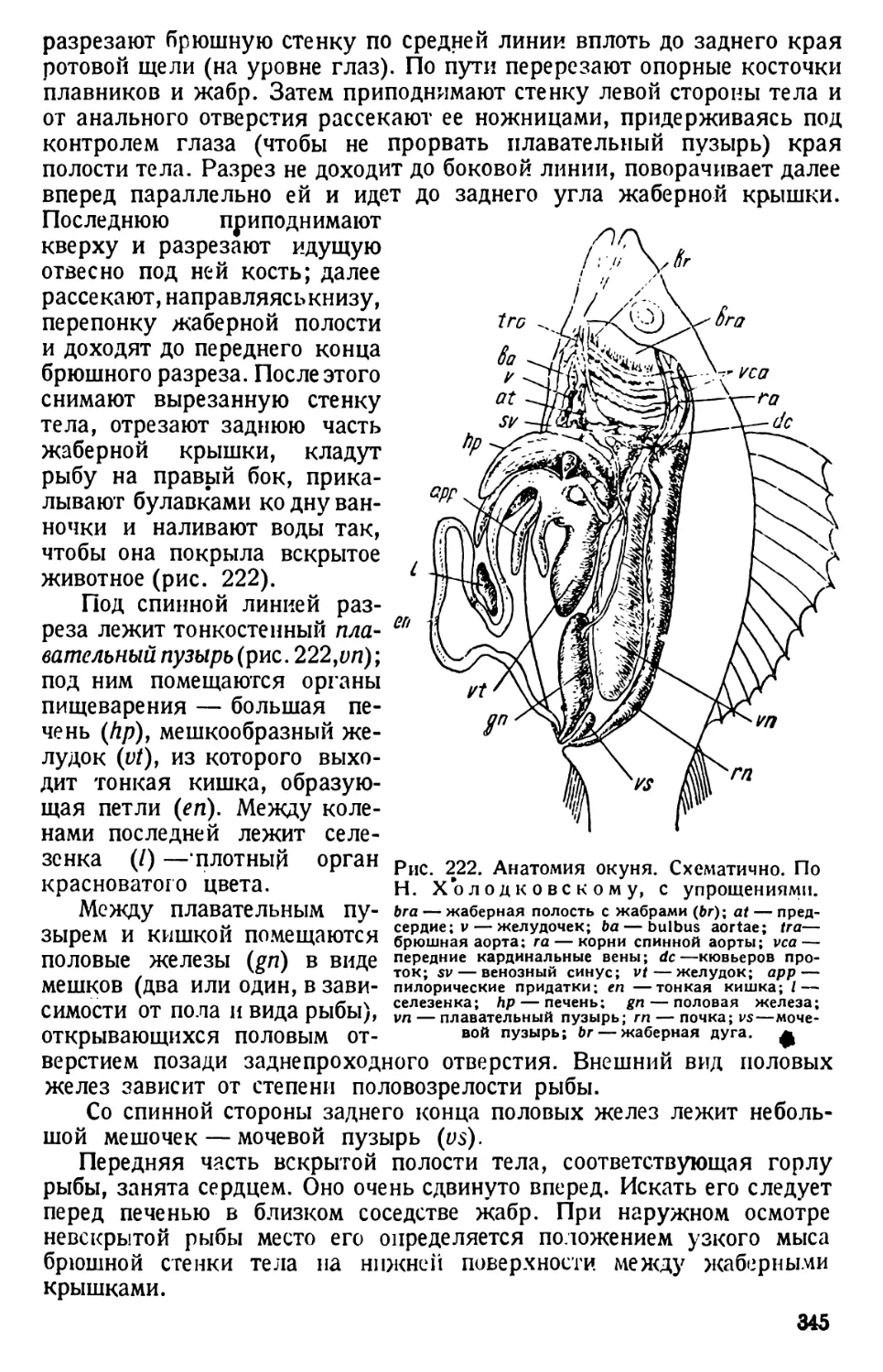
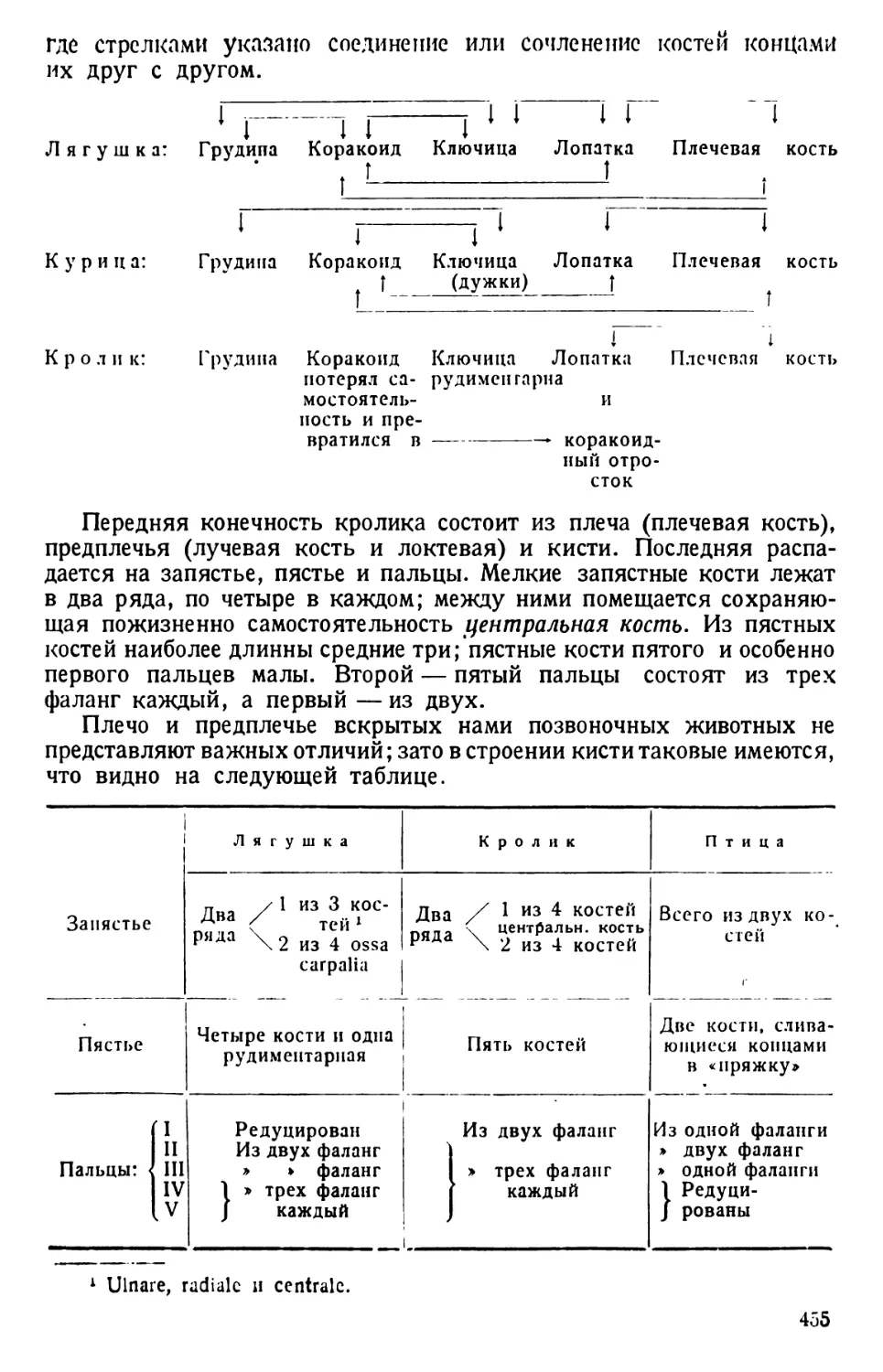
Общие замечания
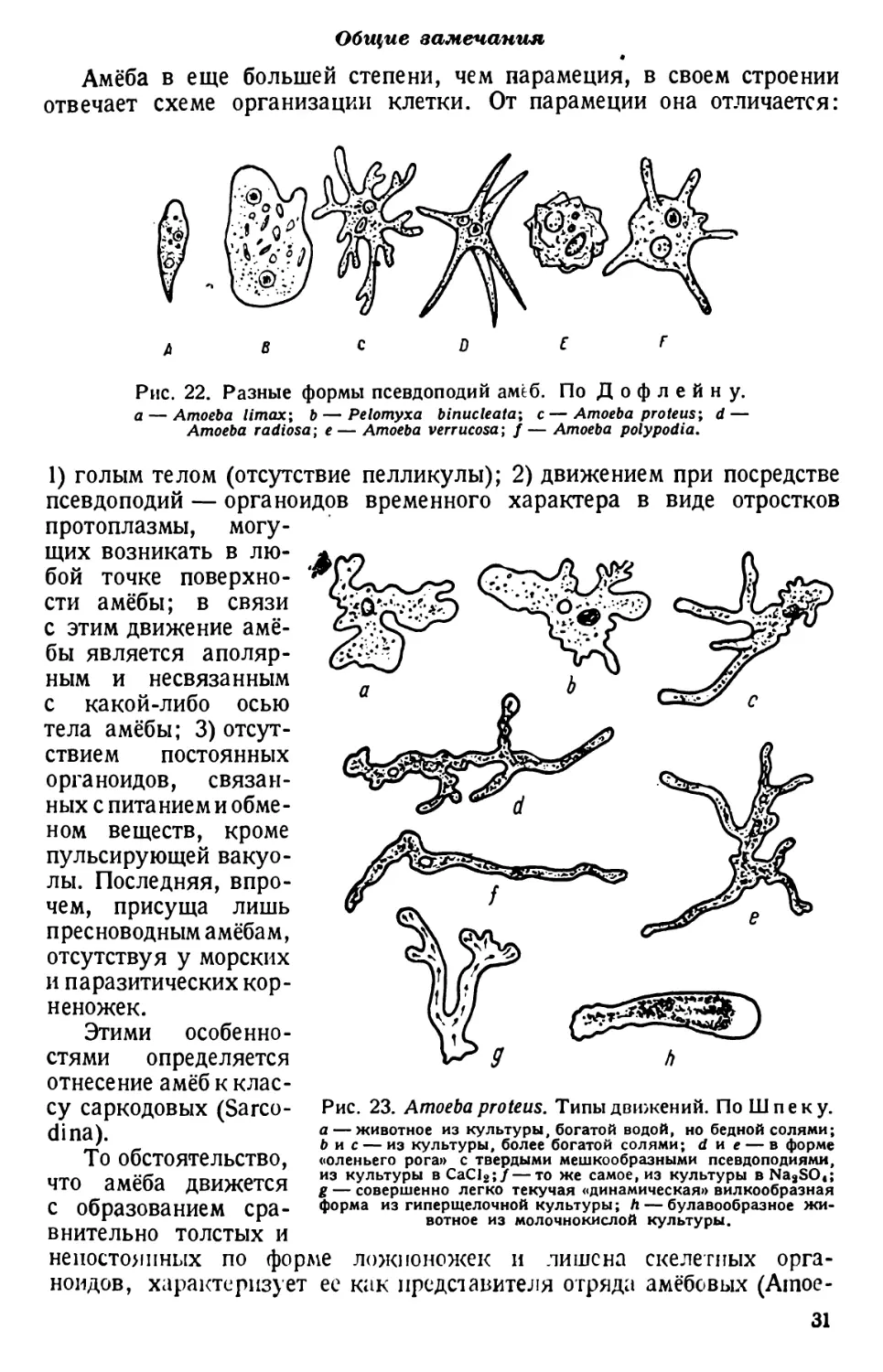
Амёба в еще большей степени, чем парамеция, в своем строении
отвечает схеме организации клетки. От парамеции она отличается:
Рис. 22. Разные формы псевдоподий амеб. По Дофлейну.
а — Amoeba Umax; b — Pelomyxa binucleata; c—Amoeba proteus; d—
Amoeba radiosa; e— Amoeba verrucosa; f— Amoeba polypodia.
1) голым телом (отсутствие пелликулы); 2) движением при посредстве
псевдоподий — органоидов временного характера в виде отростков
протоплазмы, могу-
щих возникать в лю-
бой точке поверхно-
сти амёбы; в связи
с этим движение амё-
бы является аполяр-
ным и несвязанным
с какой-либо осью
тела амёбы; 3) отсут-
ствием постоянных
органоидов, связан-
ных с питанием и обме-
ном веществ, кроме
пульсирующей вакуо-
лы. Последняя, впро-
чем, присуща лишь
пресноводным амёбам,
отсутствуя у морских
и паразитических кор-
неножек.
Этими особенно-
стями определяется
отнесение амёб к клас-
Рис. 23. Amoeba proteus. Типы движений. По Шпеку.
а — животное из культуры, богатой водой, но бедной солями;
b и с — из культуры, более богатой солями; d и е — в форме
«оленьего рога» с твердыми мешкообразными псевдоподиями,
из культуры вСаС12;/—то же самое, из культуры BNa2SO<;
g — совершенно легко текучая «динамическая» вилкообразная
форма из гиперщелочной культуры; h — булавообразное жи-
вотное из молочнокислой культуры.
су саркодовых (Sarco-
dina).
То обстоятельство,
что амёба движется
с образованием сра-
внительно толстых и
непостоянных по форме ложноножек и лишена скелетных орга-
ноидов, характеризует ее как представителя отряда амёбовых (Ашое-
31
bina). Foraminifera характеризуются, прежде всего, присутствием
раковины, чаще Ъсего многокамерной. Вследствие этого обра-
зование псевдоподий возможно у них не в любой точке поверх-
ности их тела, а через отверстие раковины и (прободенные корне-
ножки) через поры последней. В связи с этим меняется и форма
псевдоподий, которые тут чаще всего имеют вид длинных тонких
филоподий (у прободенных корненожек). Благодаря наличию рако-
винки все такие корненожки относятся к отряду Foraminifera. Дви-
жение ложноножками и преобладающее в жизни этих простейших
амёбоидное состояние являются признаками, объединяющими эти
два отряда в подкласс Rhizopoda.
Форма псевдоподий имеет систематическое значение даже для
характеристики различных видов рода Amoeba (рис. 22). Впрочем,
следует иметь в виду, что у одного и того же вида псевдоподии могут
принимать различную форму, смотря по той среде, в которой культи-
вируются амёбы. Иллюстрация такой зависимости приводится на
рис. 23.
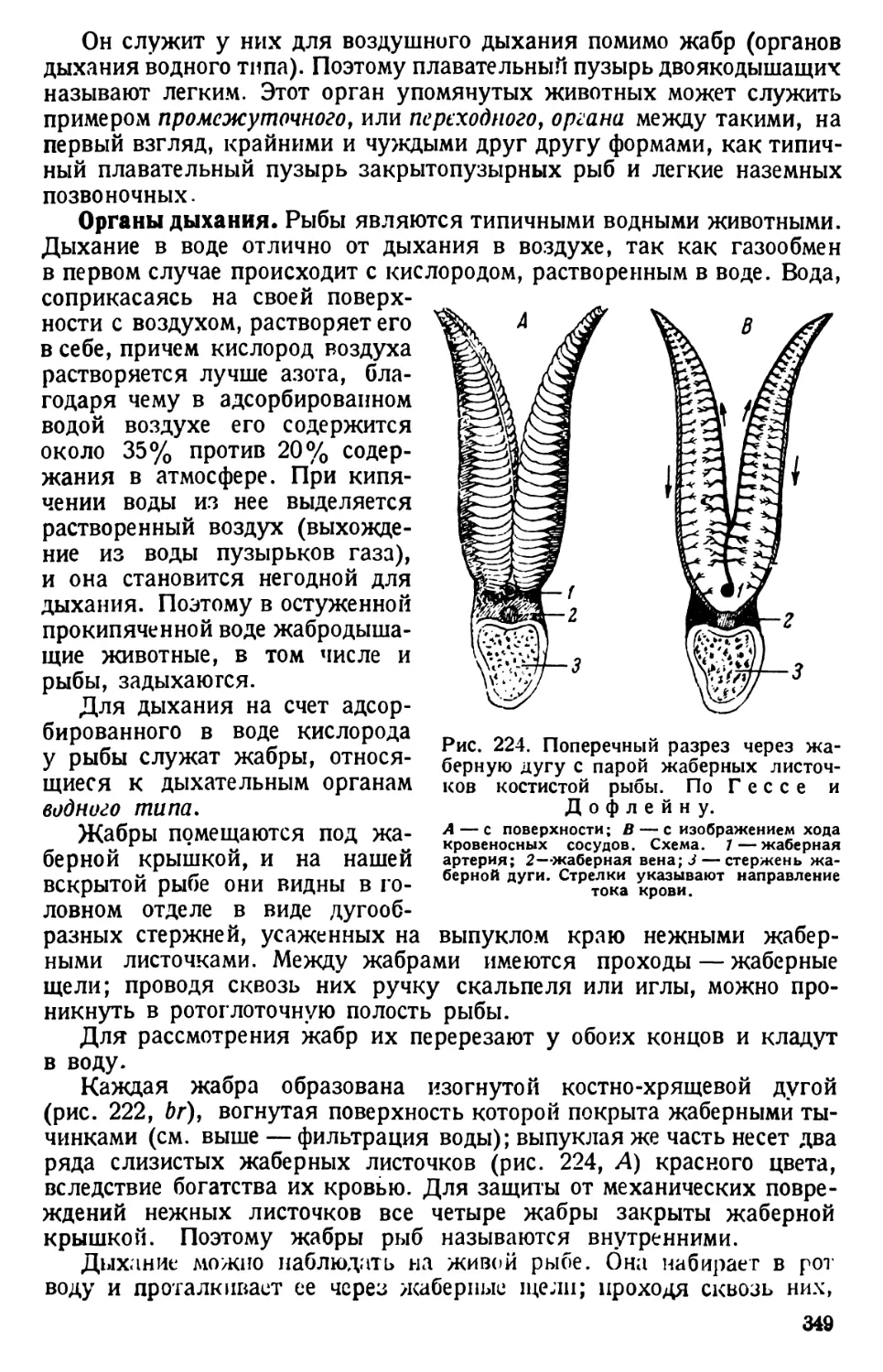
КЛАСС БИЧЕНОСЦЫ (MASTIGOPHORA)
ЭВГЛЕНА (EUGLENA)
Технические замечания. Эвглен собирают в водоемах со стоя-
чей водой, особенно обильных органическими веществами (пруды, озерки, канавы
и др.). Одни эвглены держатся у дна водоемов или на их поверхности; другие
живут в сырой земле у самого берега, образуя часто яркозеленые пленки. Мате-
риал собирают ранней весной, летом; можно его добыть и осенью. При добывании
проб захватывают поверхностный слой ила. Материал помещают в стеклянный
сосуд с водой; в нем эвглены развиваются в достаточном количестве через 1—2 не-
дели. Они положительно фототропичны и держатся поэтому у стороны сосуда,
обращенной к свету.
Для культивирования эвглен рекомендуется также питательная среда сле-
дующего состава: 0,5 г пептона, 0,5 г виноградного сахара, 0,2 г лимонной кис-
лоты, 0,2 г сернокислой магнезии, 0,05 г фосфорнокислого калия (НКаРО4) и
100 см8 воды.
Материал для занятий заготовляют в живом виде, выбирая пипеткой эвглен
из мест их обильного скопления Ъ часовые стекла, кроме того их консервируют
впрок.
Эвглен с возможно меньшим количеством ила или детрита консервируют
96% спиртом; часть их оставляют в спирту, а часть переносят в смесь равных объе-
мов 96% спирта и глицерина; на занятиях эвглен заключают в смесь глицерин-
желатины.1 *
Для заключения эвглен в глицерин-желатину на предметное стекло пипеткой
наносят капельку спирто-глицериновой смеси, вырезают небольшой кусочек
глицерин-желатины и кладут его на чистое покровное стекло, которое осторожно
подогревают снизу, держа пинцетом над спиртовкой. Следить, чтобы от нагре-
вания не появлялись пузырьки воздуха в желатине; когда она от действия тепла
расплавится, стекло быстро переворачивают каплей желатины книзу и осторожно
кладут на капельку смеси с эвгленами; желатина растекается под покровным сте-
клом и быстро застывает. В таком состоянии препарат может храниться несколько
недель; чтобы предотвратить его высыхание, края покровного стекла обводят
железным лаком.
1 21 г желатины промывают в воде и разводят в 126 см8 горячей воды, до-
бавляют 114 см8 глицерина и Зсм8 крепкой карболовой кислоты (ас. carbolicum
liquefactum); смесь перемешивают и фильтруют горячей через стеклянную пату;
разливают в широкогорлые баночки по 50—100 см3 емкостью.
32
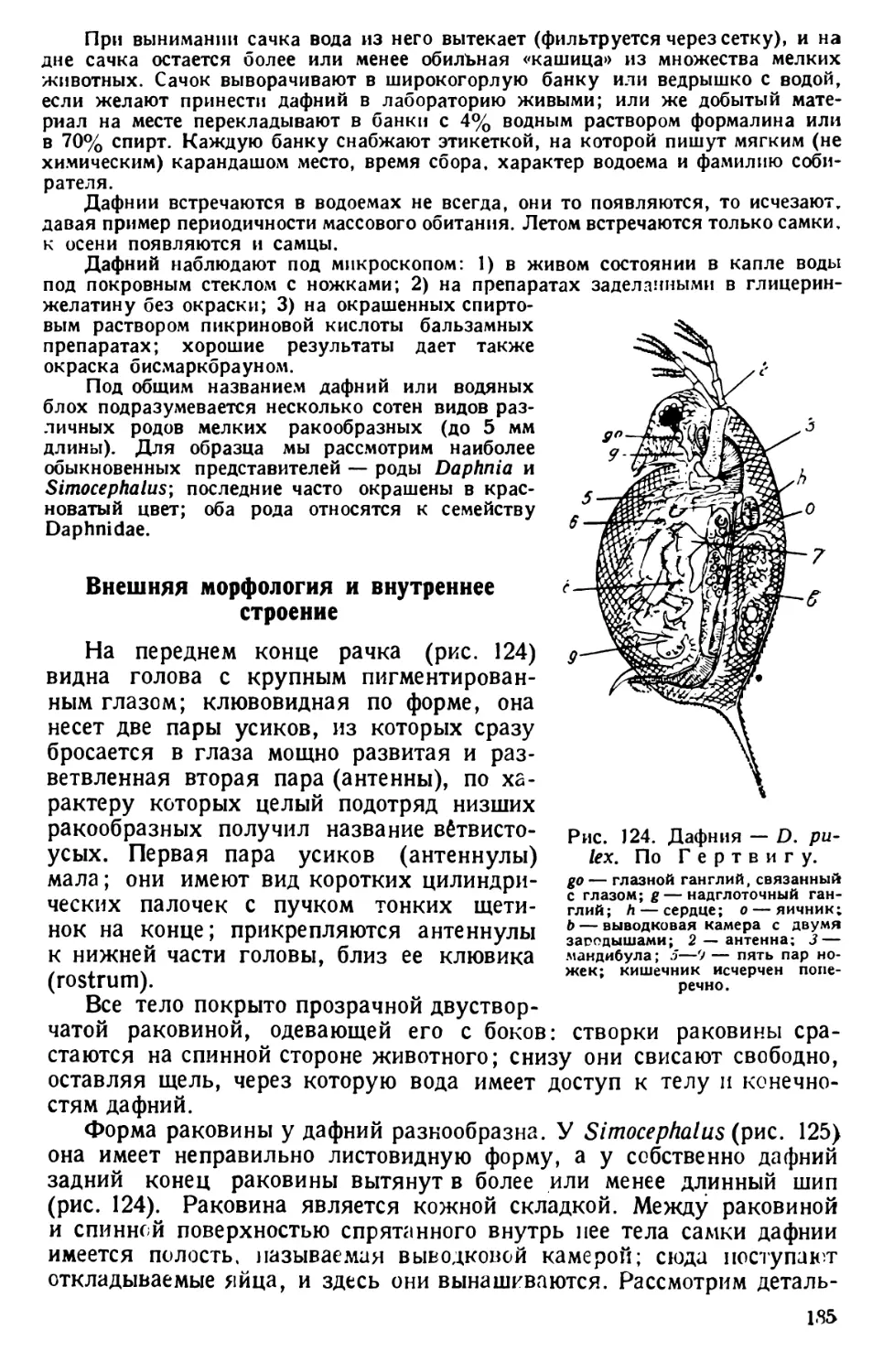
Наблюдения над эвгленами
Сухой препарат эвглены. На предметное стекло наносят пипеткой
каплю спирта с эвгленами и быстро высушивают на воздухе, покрывают
покровным стеклом или прямо рассматривают под микроскопом при
сильном увеличении.
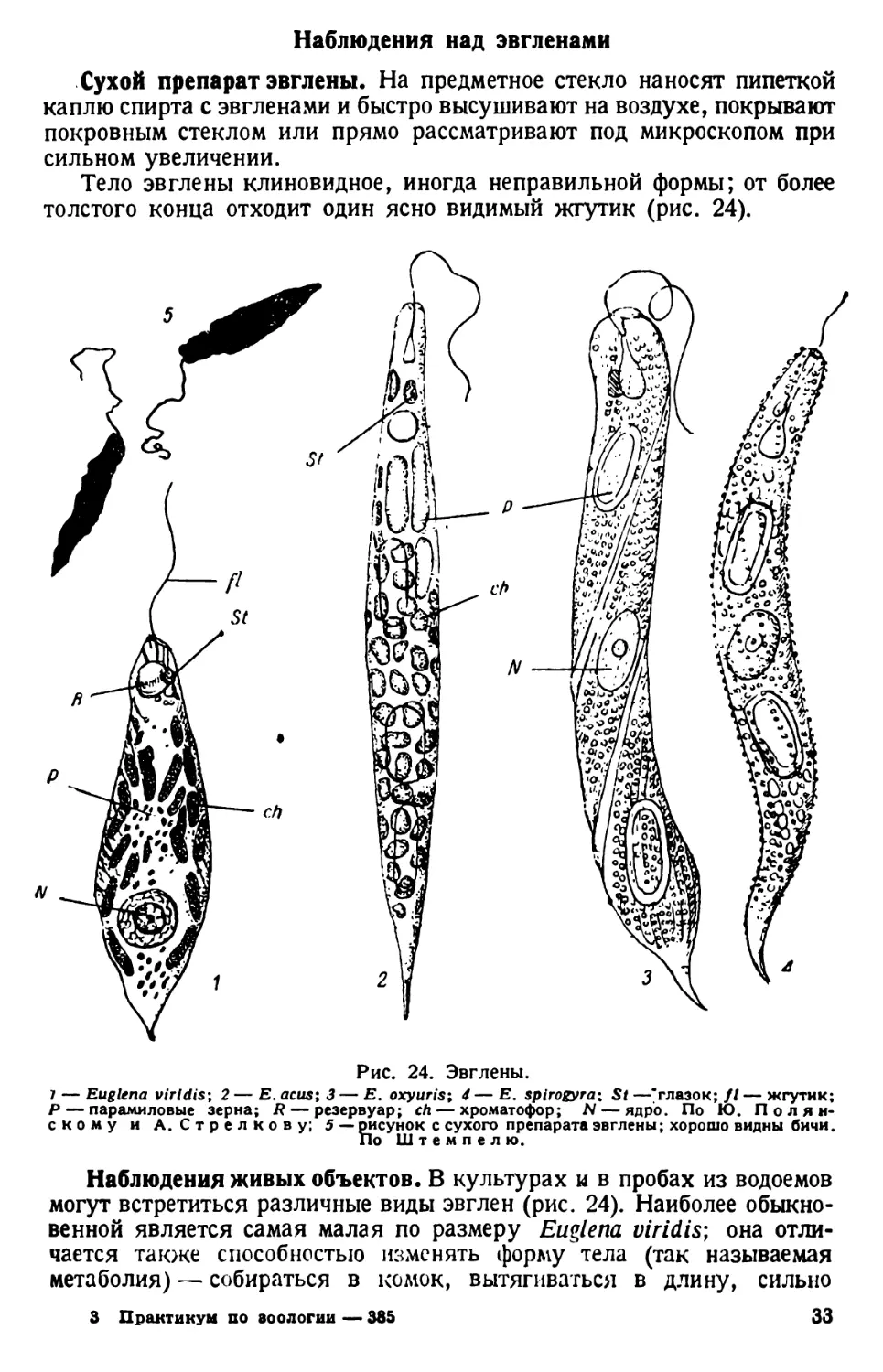
Тело эвглены клиновидное, иногда неправильной формы; от более
толстого конца отходит один ясно видимый жгутик (рис. 24).
Рис. 24. Эвглены.
7 — Euglena virldis; 2 — Е. ас us; 3— Е. oxyuris; 4— Е. spirogyra; St —"глазок; fl— жгутик;
Р — парамиловые зерна; R — резервуар; ch — хроматофор; N — ядро. По Ю. Полян-
скому и А.Стрелкову; 5 — рисунок с сухого препарата эвглены; хорошо видны бичи.
По Штемпелю.
Наблюдения живых объектов. В культурах и в пробах из водоемов
могут встретиться различные виды эвглен (рис. 24). Наиболее обыкно-
венной является самая малая по размеру Euglena viridis; она отли-
чается также способностью изменять форму тела (так называемая
метаболия) — собираться в комок, вытягиваться в длину, сильно
3 Практикум по зоологии — 385 33
так как эвглены
Рис. 25. Разрез пе-
реднего конца тела
эвглены. По В а-
г е р у.
g — бич; р — пигмент
(стигма); g внизу —
расщепившееся осно-
вание бичика; d—
уплотнение в прото-
плазме.
расширяться, принимать овальную или крестообразную форму и др.
Е. acus имеет постоянные очертания сильно удлиненного тела; еще
более велики по размерам Е. oxyuris и Е. spirogyra, но эти виды встре-
чаются реже.
Каплю культуры рассматривают под покровным стеклом при сла-
бом увеличении и наблюдают оживленное плавание, вызываемое
работой жгутика, который при слабом увеличении не виден.
При среднем (или сильном) увеличении можно видеть жгутик,
ослабляя освещение поля зрения микроскопа (суживанием диафрагмы,
боковым поворотом осветительного зеркала). Жгутики видны не всегда,
могут отбрасывать этот органоид движения.
Хотя тело эвглены и покрыто пеллйкулой,
Е. viridis, не в пример другим видам, может изме-
нять форму тела. Пелликула у Е. oxyuris и Е. spi-
rogyra образует ребристые возвышения или отли-
чается наличием мелких шипиков или бугорков.
Различают передний более тупой и задний при-
остренный концы тела. Эвглена движется жгути-
ковым концом тела вперед. Ей свойственны посту-
пательное и вращательное движения. У перед-
него конца тела лежит хорошо заметный яркокрас-
ный глазок. Он образован лежащей в вакуоле ка-
плей жирного масла, окрашенного гематохромом —
пигментом1 из группы каротиноидов. Глазок назы-
вают иначе стигмой. 2
Весьма вероятно, что глазок является свето-
преломляющим органом, направляющим световые
лучи к основанию жгутика,» чем обусловливается
передача световых раздражений, видоизменение ра-
боты жгутика и, в конечном результате, положи-
тельный или отрицательный фототаксис эвглены.
Рядом с стигмой помещается сократительная ва-
куола, окруженная часто несколькими придаточ-
ными вакуолами гораздо меньшего размера. Здесь
же лежит «резервуар» с воронкой, образованный впячиванием наруж-
ной оболочки тела эвглены и называемый «глоточным» углублением.
Со дна резервуара поднимается жгутик, начинающийся двумя кор-
нями, укрепленными в особой плазматической подушечке (рис. 25).
Тело эвглены окрашено в зеленый цвет; однако эта окраска не
диффузна; она зависит от наличия в протоплазме зеленых включений
в форме пальцевидно разветвленного хроматофора; дольки его кажутся
лежащими независимо друг от друга. Хроматофор содержит в себе
хлорофил. Форма хроматофоров у разных видов эвглен различна.
Между хроматофорами лежат сильно преломляющие свет бесцветные
зерна парамила, являющиеся отложениями запасного питательного
материала, усвоенного эвгленой в избытке против обычной потреб-
1 Пигментом называют окрашенные вещества органической природы, расти-
тельного и пн животного происхождения.
2 Не смешивать с другим шачением термина стигмы — (ыхальца \ насе-
комых.
34
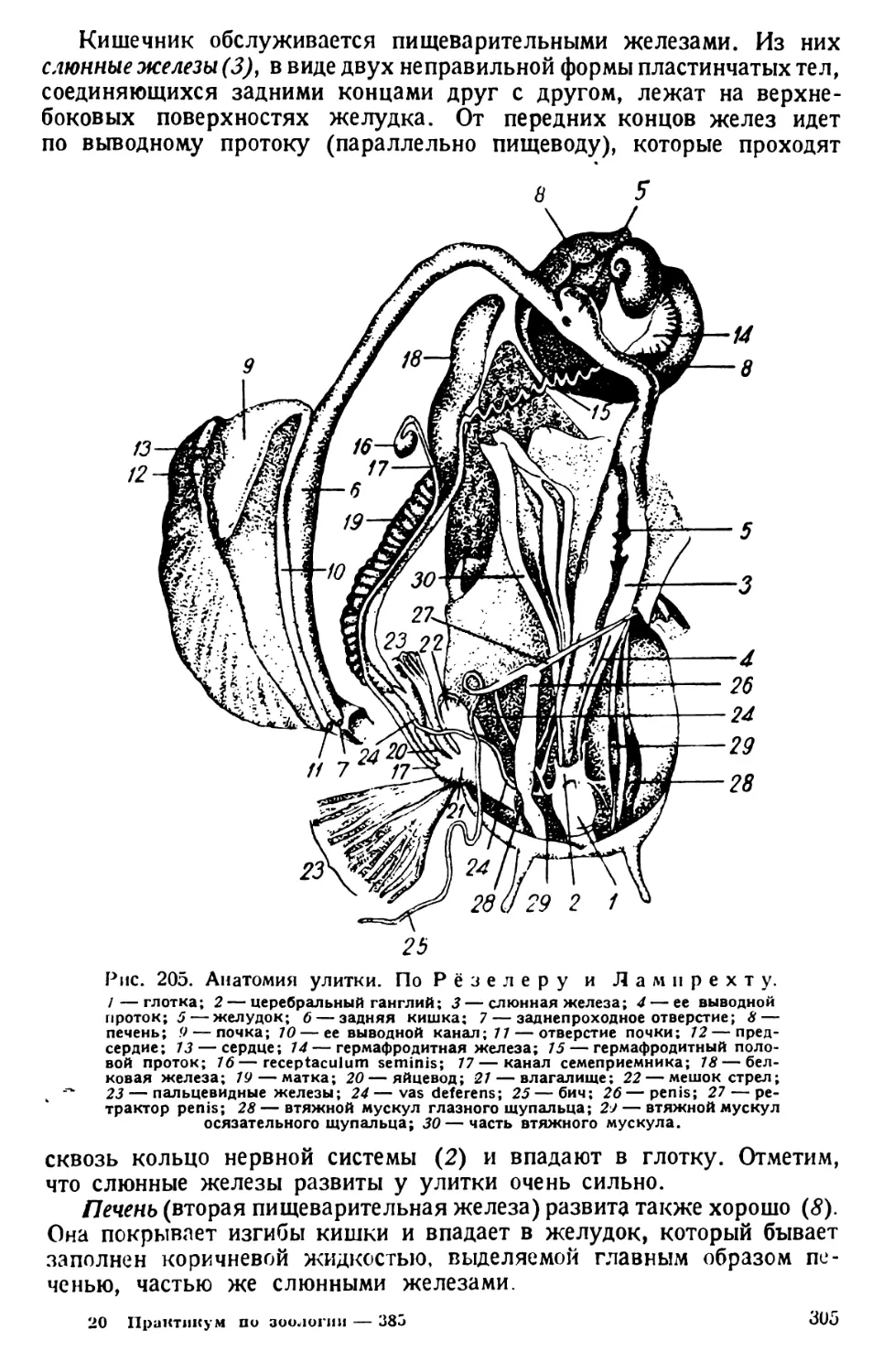
мости ее организма. Усвоение воспринятых пищевых веществ назы-
вается ассимиляцией. Парамил (равно как и капля жирного масла
глазка) является, следовательно, продуктом ассимиляции. У Е. viri-
dis его зерна мелки и многочисленны, а у Е. oxyuris и Е. spirogyra они
крупны и числом всего два. Парамил по химическому составу близок
к крахмалу. Его зерна отлагаются на поверхности хроматофора.
Ядро у живой эвглены не видно; о месте его положения можно
судить по просвечивающему участку между дольками хроматофора и
зернами парамила. Лежит оно в общем в средней части тела или не-
сколько сдвинуто кзади.
Благодаря наличию хлорофильных хроматофоров, эвглены на
свету питаются подобно зеленым растениям, т. е. за счет потребления
углекислоты и неорганических солей и синтезирования из них органи-
ческих веществ, прежде всего — углеводов (фотосинтез). Такой способ
питания называется голофитным (т. е. свойственным зеленым расте-
ниям), а соответствующие организмы — аутотрофными.
В условиях эксперимента эвглен можно перевести на питательный
режим другого порядка — на сапрофитное питание; последнее характе-
ризуется тем, что в организм через поверхность тела осмотически
поступают находящиеся в жидком состоянии в окружающей его среде
органические соединения. Поставленной цели достигают, помещая
культуру эвглен в совершенную темноту, прибавив к ней картофельного
отвара. В темноте голофитное питание не может осуществляться. Эв-
глены утрачивают хроматофоры, и тело их в конце концов стано-
вится бесцветно сероватым через 3—4 недели. Лишившиеся зеленых
хроматофоров эвглены питаются сапрофитно. Красный глазок остается
без изменения и в темноте.
Если обесцвеченных эвглен выставить снова на свет, то они позеле-
неют, так как хроматофоры у них восстановятся. Отсюда делаем заклю-
чение, что потеря и восстановление зеленых хроматофоров и, следова-
тельно, перемена форм питания у эвглены являются обратимым
процессом и сами эвглены должны быть отнесены к миксотрофным про-
стейшим, т. е. осуществляющим и голофитное и голозойное питание.
Благодаря наличию животных и растительных свойств, эвглен рас-
сматривают и в зоологии и в ботанике. Зоологи относят их к особому
отряду биченосцев — Euglenoidina, а ботаники — к простейшим орга-
низмам из групп жгутиковых отдела водорослей.
Для ясного обнаружения пелликулы, к препарату из живых эвглен
прибавляют глицерин; при начинающемся плазмолизе сжимающаяся
протоплазма отстает от пелликулы. Последняя дает реакцию на белки,
но не на растительную клетчатку, что говорит в пользу животной при-
роды эвглены.
Общие замечания
Эвглена обладает постоянным органоидом движения — жгутиком,
или бичом; в оценке постоянства наличия бича случаи отбрасывания его
отдельными особями эвглены в счет не идут. Вместе с ресничками инфу-
зорий жгутик эвглены относится к ундулиподиям, т. е. к колеблю-
щимся, сгибающимся органеллам. Бич гораздо длиннее ресничек; он
♦ •> -
является характерным признаком, определяющим принадлежность
эвглены к классу биченосцев.
Формы питания у рассматриваемых нами простейших различны.
Сопоставление их и характеристики даются на следующей схеме.
Формы питания
I
Голофитное
(как у зеленых растений)
Путем фотосинтеза за счет
углекислоты и органических
солей
I
Аутотрофные простей-
шие
I
с зелеными хроматофорами, с
осмотическим воспри-
ятием пищевых веществ по-
верхностью тела
I
Голозойное
(за счет органических веществ, вырабо-
танных другими организмами)
I
I
Гетеротрофные простейшие,
питающиеся
I I
растворенными жид-
кими веществами, на-
ходящимися в окру-
жающей среде
I
Сапрофитное - са-
прозойное питание
свободно живущих про-
стейших
I
Паразитическое
питание за счет соков
хозяина
(опалина, грегарина,
трипанозома)
оформленными твер-
дыми веществами ор-
ганической природы,
другими организмами
или их частями
I
С захватыванием пи-
щи: псевдоподиями —
амёба; привлечением
ее ресничками (па-
рамеция), бичами и др.
Осмотическое поступление пищи через поверх-
ность тела
Отсутствие органоидов питания
Организмы, питающиеся и
аутотрофно и гетеротроЛно —
миксотрофные биче-
пос ц ы (эвглена)
Обладание органоида-
ми, выполняющими
функции пищеваре-
ния
ТРИПАНОЗОМА (TRYPANOSOMA)
Технические замечания. Исследованием крови крыс отыски-
вают экземпляры с трипанозомами (Trypanosoma lewisi) в крови. Для поддержа-
ния штамма трипанозом, один раз в 5-7 дней, переливают (с соблюдением бакте-
риологических правил в отношении стерильности) кровь от зараженной крысы
белой крысе, инъецируя ей кровь под кожу.
Для приготовления препарата делают мазок крови. Каплю крови, добытую
из уха крысы или отрезанием кончика хвоста, наносят на чистое обезжиренное
предметное стекло ближе к одному из его концов. На внутренний край капли
кладут плашмя чисто вымытую препаровальную иглу или прикасаются к нему
узким краем отшлифованного предметного стекла, держа последнее под углом.
Когда кровь распределится по всей длине иглы или стекла, этот предмет влекут
по поверхности предметного стекла по направлению к противоположному концу
(рис. 26). За передвигаемым объектом увлекается кровь, распределяющаяся
тонким мазком по предметному стеклу. Вся процедура должна быть проделана
быстро, чтобы кровь не успела свернуться и подвергнуться деформирующему
36
влиянию воздуха на красные кровяные тельца. Мазок должен быть ровным и тон-
ким, что обеспечит быстрое его высыхание на воздухе.
Мазок фиксируют 20 минут абсолютным или 3—5 минут метиловым спиртом.
После фиксации мазок сушат и красят раствором краски Романовского-Гимзы.
На 1 см8 дистиллированной воды берут
одну-две капли краски (при температуре
не ниже 16—17J С; если температура бо-
лее низка, то флакон с краской надо со-
греть на водяной бане или держать в тер-
мостате и хорошо взболтать). Предметное
стекло кладут мазком книзу на отрезки
стекол или спички, на чистое стекло боль-
шого размера и краску подпускают так,
чтобы она снизу покрыла весь мазок.
Через 30—50 минут стекла быстро про-
мывают струей воды и высушивают.
Более точные сроки окраски устанавли-
ваются опытным путем для данной пор-
ции краски. Необходимо брать нейтраль-
ную дистиллированную воду. Если на
препарате нет розовой окраски, то сле-
дует чуть подкислить уксусной кисло-
той раствор краски (смоченным в кис-
лоте концом препаровальной иглы). Рис. 26. Приготовление мазка крови;
Изучение препарата
Рассмотрение препарата производится под масляной иммерзией.
На препарате видны дисковидные безъядерные красные кровяные
тельца и в гораздо меньшем
количестве округлые и бо-
лее крупные по размерам
белые кровяные клетки, или
лейкоциты, имеющие ясно
выраженное ядро, окра-
шенное в темносиний цвет.
В пространстве между этими
форменными элементами
крови можно, передвигая
препарат, отыскать узкие
веретенообразно изогнутые
тела с окрашенным в крас-
ный цвет бичом. Это и есть
Trypanosoma lewisi.
Тело трипанозомы окра-
шено в розовый цвет; к кон-
цам оно сильно заостряется;
ядро, в виде овального окра-
шенного в синий цвет тель-
Рис. 27. Trypanosoma lewisi из крови крысы.
I— нормальная форма; II—III—стадии деления. 7 —
ядро; 2 — блефаропласт; 3—базальное зерно жгу-
тика; , 4—волнообразная перепонка; 5 — жгутик.
Из Матвеева.
ца , лежит отступя от середины длины трипанозомы, ближе к переднему
концу тела (рис. 27). Оно имеет зернистое строение (расположение
хроматина ядра в виде зерен). Ближе к противоположному— заднему —
концу трипанозомы лежит резко выделяющееся компактное розово-
красное зерно, называемое блефаропластом, или кинетопластом; от
37
него отходит также окрашивающийся в красный цвет жгут; волно-
образно изгибаясь, он идет вперед, будучи связан с телом трипанозомы
прозрачной колеблющейся перепонкой, называемой ундулирующей
мембраной. От переднего конца трипанозомы жгутик свисает сво-
бодно.
При наблюдении живых трипанозом можно видеть их движение,
вызываемое работой жгутика и движениями связанной с ним ундули-
рующей мембраны. Оба эти образования являются органеллами
движения типа ундулиподий.
Для трипанозом характерно полное отсутствие каких-либо орга-
нелл питания. Живя в плазме крови хозяина, эти паразиты как бы
окружены пищей, так как в кровь поступают вещества, всасываемые
стенками пищеварительного канала. Питание трипанозом, являющихся
эндопаразитами, — эндосмотическое.
Размножение. При детальном изучении крысиных трипа-
нозом можно наблюдать фазы их размножения, происходящего в крови
хозяина. Следует иметь в виду, что период размножения делением
короток и что делящиеся формы в периферической крови после вось-
мого-девятого дня заражения встречаются редко. Рассмотренные выше
трипанозомы характерны для последних дней инфекции. Делящихся
трипанозом больше шансов найти в крови мелких сосудов внутренних
органов, в частности почек.
Подготовляющиеся к делению трипанозомы отличаются поли-
морфизмом, т. е. известным разнообразием в форме тела, по сравнению
с рассмотренными ранее типичными трипанозомами; тело первых более
широкое; ядро и блефаропласт более сближены. Вслед за разделением
ядра делится и блефаропласт; от дочернего блефаропласта отходит
зачаток нового бича, связанный с телом трипанозомы короткой мем-
браной. В дальнейшем наблюдается новое деление ядер и блефаро-
пластов, без того, чтобы протоплазматическое тело трипанозомы начало
делиться. Поэтому получаются такие многоядерные и многожгутиковые
формы, как на рис. 27, II—III. В результате полного разделения обо-
собляются мелкие округлые дочерние формы, которые разрастаются
в типичные трипанозомы.
В рассмотренном процессе нетрудно усмотреть бесполое размноже-
ние делением.
Понятие об инкубации. Если брать кровь каждый
день у только чта зараженной интраперитонеально 1 трипанозомами
крысы и искать в ней трипанозом, то в течение четырех-шести дней
в крови не находим ничего нового; первые трипанозомы появляются
лишь по истечении указанного срока. Такой скрытый период жизни
паразитов в организме хозяина, прежде чем присутствие их может быть
обнаружено в периферической крови, называется инкубационным
периодом, или инкубацией.
Последующие наблюдения длительности существования трипано-
зом в крови обнаруживают, что у различных особей трипанозомы
исчезают из организма через 3-4 и гораздо более недель.
1 То есть впрыскиванием материала в брк>шн>ю uo.iocib крысы.
38
Общие замечания
Эвглена и трипанозома, будучи представителями различных отря-
дов (Euglenoidina и Protomonadina), относятся к классу биченосцев,
так как обладают органоидом движения в форме бича.
В противоположность эвглене, трипанозомы являются исключи-
тельно гетеротрофными простейшими, т. е. питающимися растворен-
ными органическими веществами плазмы крови.
Для крысиной трипанозомы характерно обитание в замкнутой
системе кровеносных сосудов крысы. В связи с этим возникает вопрос,
как в естественных условиях осуществляется заражение свежих крыс
трипанозомами. Не касаясь деталей, отметим, что заражение крыс
происходит при непременном содействии крысиных блох (Cerato-
psyllus fasciatus). С кровью крыс они всасывают трипанозом, которые
в их организме проходят известный цикл развития. Переходя на здо-
ровых крыс, блохи передают им трипанозом и т. д. Блохи являются,
следовательно, переносчиками или передатчиками трипанозом.
КЛАСС СПОРОВИКИ (SPOROZOA)
ГРЕГАРИНЫ (GREGARINIDA) НЕКОТОРЫХ НАСЕКОМЫХ
Технические замечания. 1) Вскрывают черных тараканов (см.
ниже) и вырезают из пищеварительного канала среднюю кишку — участок
между пилорическими придатками и местом впадения мальпигиевых сосудов.
Выделенную часть вскрывают в часовом стекле в физиологическом растворе
поваренной соли, размешивают ее содержимое, дают ему осесть и в осадке под
лупой (покровное стекло с ножками) ищут грегарин. Грегарины паразитируют
далеко не в каждом таракане. Иногда при вскрытии большого числа их грегарин
обнаружить не удается, и наоборот, попадаются такие места, где тараканы во
множестве имеют этих паразитов. Поэтому для достижения успеха необходимо
брать тараканов из разных мест (из разных домов и кварталов). Паразитирующих
в них грегарин наблюдают в живом состоянии.
2) Заготовляют культуру жука — мучного хрущака (Tenebrio molitor)
семейства Tenebrionidae; в кишечнике его личинок (обычно называемых «муч-
ными червями‘>) паразитируют более мелкие грегарины — Gregarirta cuneate,
Gr. polymoprha, Gr. steini. Л/Гучных червей продают в зоологических магазинах
для корма птиц и доугих животных. Можно их достать в мучных складах. Добы-
тых личинок кладут в поместительную фаянсовую или глиняную банку, которую
на три четверти заполняют отрубями и завязывают марлей. Время от времени
этот осадок следует слегка увлажнять.
Грегарины живут в кишечнике; его извлекают пинцетом, отрезав ножницами
у захлороформированной личинки голову и задний конец тела.
Для временного содержания живых грегарин и для наблюдений над ними
приготовляют белковую воду: 20 г сырого взбитого и потом осевшего куриного
белка хорошо смешивают с 200 см8 0,5% раствора поваренной соли; фильтруют
через складчатый фильтр и прибавляют маленький кусочек камфоры; эту смесь
применяют вместо физиологического раствора.
Наблюдения над грегаринами
Микроскопируют живых грегарин черного таракана или мучного
червя (Gregarina blattarum, Gr. polymorpha, Gr. cuneatavi др.). Отмечают
хорошо оформленное удлиненное тело грегарин, молочнобелою
цвета.
39
Каждая грегарина состоит из двух отделов, граница между кото-
рыми обозначена перетяжкой или насечкой; передний отдел называется
протомеритом, задний (больший по размеру) — дейтомеритом
(рис. 28 и 29).
В общем удлиненное тело грегарины покрыто снаружи плотной
ребристой кутикулой; под ней лежит узкий слой эктоплазмы, которая
образует поперечную перемычку, или перегородку, отделяющую
протомерит от дейтомерита; главная масса тела грегарины образована
зернистой эндоплазмой,
зернышками
Рис. 28. Грегарины
таракана.
Gregarina blattarum; Гоегаои
две особи в состоянии
сизигия из кишечника ;а м‘с
таракана. 7 — прото- япмм*п»
мерит; 2 — дейтоме- Gr^?un<
рит; 3— кутикула;
4 — ядро.
Рис. 29.
ны из личинок Tenebrio
1—Gregarina potymorpha;
подая форма со следом
га; 2—Gr, steini; 3 —
2ta. По Кюкеиталю.
рагликогена, запасного
вещества, близкого к
гликогену. Эндопла-
зма более жидкой кон-
систенции, чем экто-
плазма, в чем можно
убедиться, если ра-
зорвать гр’егарину
острыми иглами. Ядро
у грегарин одно; оно
хорошо видно в форме
более прозрачного пу-
зырька на темном фо-
не эндоплазмы дейто-
мерита; при разрыва-
нии или раздавлива-
нии живых грегарин
ядро можно изолиро-
вать.
Нередко встреча-
ются так называемые
сизигии — грегарины,
соединенные по две
или более наподобие
цепочки; это состоя-
ние является резуль-
татом как бы склеива-
ния грегарин и не имеет никакого отношения к размножению.
Отмечаем отсутствие органелл питания и сократительных вакуол.
Питание у грегарин совершается эндосмотически, как и у уже рас-
смотренных нами других эндопаразитических простейших.
Движение грегарин медленное. Его характер удобнее
рассмотреть в белковой среде. В белковой воде (см. выше, стр. 39)
растирают тушь или кармин до соответственно насыщенного цвета.
В эту жидкость пускают живых грегарин; каплю жидкости с грегари-
нами помещают под покровное стекло с ножками и рассматривают под
микроскопом.
Через некоторое время можно видеть продвижение грегарины
вперед, причем за ее задним концом остается в окрашенной среде
бесцветный след студенистого или слизистого вида (рис. 30). Смотря
по направлению движения грегарины этот след в виде полоски имеет
прямую или изогнутую форму. Заметна также как бы продольная
40
исчерченность следа вследствие линейного расположения зернышек
туши между студенистыми нитями, выделяющимися наружу через
бороздки кутикулы (рис. 30). С таким обиль-
ным выделением слизи и связано скользящее
движение грегарины, являющееся особой фор-
мой движения, свойственного, кроме грегарин,
некоторым диатомеям. Хотя механизм движения
грегарин толкуется разноречиво, можно при-
нять, что движение происходит как бы вслед-
ствие «отдачи» при энергичной секреции слизи
(грубой механической моделью такого движе-
ния считают работу сегнерова колеса или дви-
жение ракеты).
У некоторых грегарин бывает еще третья
часть тела в виде эпимерита, лежащего на про-
томерите. Форма его разнообразна: то в виде ши-
поватой «головки» на вытянутой «шейке», то
в виде крючьев или, наоборот, волосовидных
придатков; он служит органом прикрепления
к стенке кишечника хозяина, в котором па-
разитирует грегарина. У некоторых грегарин
эпимерит рано редуцируется, и в окончательном
виде грегарина состоит только из протомерита
и дейтомерита.
Рис. 30.
Движение грегарины в
белковой массе с тушью;
сзади виден бесцветный
слизевой след. По Ш е в я-
к о в у.
Грегарины с подразделенным на рассмотренные нами части телом
относятся к группе Cephalina (или Polycystidea).
MONOCySTIS ДОЖДЕВЫХ ЧЕРВЕЙ
Технические замечания. Вскрывают дождевого червя со спины
(см. ниже) и осматривают дольки семенных мешков. .Вырезанные дольки рас-
щипывают на предметных стеклах, накрывают покровным стеклом (без ножек)
и микроскопируют в свежем состоянии.
Из содержимого семенного мешка приготовляют мазок на чистом покровном
стекле; взяв его пинцетом, опускают мазком книзу на поверхность 100?;» или 96%
спирта для фиксирования; через несколько минут стекло, мазком книзу, кла-
дут на каплю глицерина на предметное стекло или заключают в глицерин-жела-
тину (стр. 32).
Заливают дольки семенника в парафин; приготовляют срезы, освобождают
их от парафина, окрашивают гематоксилином Делафильда и заключают обыч-
ным путем в канадский бальзам.
Изучение препаратов
В содержимом семенных мешков лишь редко можно увидеть сво-
бодных грегарин. Чаще встречаются мелкие виды, имеющие разно-
образную форму листовидного, ленточного или иного характера
(рис. 31). Характерно, что тело Monocystis не подразделяется на
отделы и имеет строение типичной клетки; такие грегарины относятся
к группе Acephalina или Monocystidea.
Обычно на мазке встречаются шарообразные цисты — крупные
образования, одетые оболочкой и заключающие в с* бе грегарин в раз-
41
Рис. 31. Monocystis agilis и М. magna
из семенных мешков дождевого червя.
По В е н ь о н у.
ных фазах их полового размножения. В одних цистах (их мало)
видны темные овальные, прилежащие друг к другу тела — это пара
грегарин, приступающая к половому размножению. Одевшись общей
цистой, грегарины претерпевают сложные видоизменения, ведущие
к образованию множества так называемых споробластов, являющихся,
в сущности, мужскими и женскими половыми особями и соответствую-
щих микро- и макрогаметам. Образовавшись во множестве из пары
инцистированных грегарин, споробласты сливаются (копулируют)
попарно, образуя зиготы;
каждая зигота одевается оболоч-
кой и в таком состоянии назы-
вается спорой; споры • грегарин
носят наименование псевдонави-
целл; под их оболочкой ядро и
протоплазма зиготы делится по-
вторно, образуя несколько удли-
ненных спорозоитов, являющихся
конечной фазой полового размно-
жения. Спорозоиты обладают спо-
собностью заражать дождевых
червей, т. е. проделывают в них
превращение в грегарин, которые
затем размножаются половым пу-
тем (что было только что рас-
смотрено), если будут проглочены
червями.
Однако на наших препаратах
эти детали не могут быть просле-
жены полностью. Бывают видны
цисты с недавно инцистирован-
ными грегаринами. Обычно же
встречаются цисты с псевдонави-
целлами (ложнолодочками); обо-
лочка этих цист легко разры-
вается, и на мазках бывает видно
много свободно лежащих псевдо-
навицелл.
На срезах можно видеть больше деталей полового размножения
грегарин, разобраться в которых можно, руководствуясь рис. 32.
В цисте А видим две грегарины, ядра которых после повторного деле-
ния подошли к поверхности тела грегарин; впоследствии они отшну-
руются каждое в комочке протоплазмы и дадут гаметы (В). От
самих грегарин после образования гамет (или споробластов)
остается остаточное тело (С). Сформированные споры часто выпа-
дают из срезов, и поэтому соответствующие цисты кажутся бед-
ными содержимым в противоположность цистам с псевдонавицеллами
на мазках.
Из других споровиков некоторые живут в крови и потому назы-
ваются кровеспоровиками пли гемоспоридиями (Haemosporidia).
У человека таковыми являются возбудители малярии — малярийные
42
плазмодии [о них см.: А. Гуцевич в книге Е. Павловского —
«Практикум медицинской паразитологии» (гл. «Плазмодии малярии»,
стр. 104—123)].
Рис. 32. Срез через дольку семенного мешка дождевого червя с мно-
гочисленными гаметоцистами Monocystis в различных стадиях разви-
тия. Вся ткань органа в изобилии проросла гаметоцистами пара-
зита. По В е н ь о н у.
А — гаметоцисты Monocystis magna с двумя грегаринами, ядра которых много-
кратно разделились; В — гаметоцисты, в которых образовались гаметы;
С — гаметоцисты после конъюгации гамет; D — зрелые цисты с ооцистами;
Е—F—гаметоцисты М. agilis(мелкой грегарины), соответствующие стадиям А—В;
G—К — то же, с ооцистами разной величины; S — ткань семенника.
Общие замечания
Для грегарин характерно размножение с образованием спор, како-
вой процесс называется споруляцией. Все простейшие, которым свой-
ственна такая форма размножения, относятся к классу споровиков —
Sporozoa.
43
У грегарин споруляция приурочена лишь к концу их жизненного
цикла, поэтому их относят к подклассу Telosporidia.
Спора — стойкое, одетое защитной оболочкой образование,—
является стадией жизненного цикла, служащей для заражения других
особей хозяев споровиками, в нашем случае — грегаринами, что обеспе-
чивает поддержание видового существования последних.
Грегарины, как и все остальные споровики, являются внутренними
паразитами.
Экологические замечания по рассмотренным представителям
простейших
Рассмотренные нами простейшие — парамеции, амёбы, форамини-
феры и эвглены—относятся к свободно живущим организмам; средой
обитания их являются пресноводные водоемы независимо от раз-
мера последних; фораминиферы обитают в морской воде.
Опалины, трипанозомы и грегарины живут в организме других
животных и питаются за счет соков их тела; поэтому их относят к пара-
зитам. Средой обитания этих паразитов являются различные органы
хозяев: опалины и грегарины насекомых живут в полости кишечника
хозяина; трипанозомы обитают в плазме крови, заключенной в за-
мкнутую систему сосудов
млекопитающего; их назы-
вают поэтому кровепарази-
тами;. грегарины дождевого
червя обитают в семенном
мешке, т. е. являются тка-
невыми паразитами.
При всем разнообразии
сред обитания рассмотрен-
ных простейших для всех
них (т. е. сред) характерна
влажность как необходимое
условие существования Pro-
tozoa.
Рис. 33. Выхождение инфузории Stylonychia
mytilus из цисты. По Иловайскому.
Однако водоемы, в которых живут пресноводные простейшие,
легко могут пересыхать, что угрожает гибелью последним. В связи
с такой возможностью у многих простейших в процессе их эволюции
выработалась способность к инцистированию. При пересыхании
водоемов простейшие одеваются особой непроницаемой оболочкой —
цистой. Циста является затвердевшим выделением тела соответ-
ствующего простейшего.
Процесс инцистирования является защитным приспособлением
организма. Инцистированное простейшее более или менее долгое время
противостоит сухости. Оно с пылью может переноситься ветром на
далекие расстояния. Когда инцистированное простейшее попадает
снова в воду, циста растворяется или вскрывается (рис. 33), из нее
выходит простейшее и начинает активно жить.
Цисты инфузорий-туфелек встречаются редко (рис. 34, Д, В).
С инцистированием связывается у некоторых свободно живущих
простейших и размножение. Например, под оболочкой цисты Amoeba
44
proteus ее содержимое многократно делится, в результате чего обра-
зуется масса мельчайших амёб, которые освобождаются по вскрытии
цисты. Таким образом инцистирование может быть связано с за-
щитной функцией и с размножением.
Для паразитических простейших возможность высыхания в тече-
ние их индивидуальной жизни исключается, так как они обитают
в постоянно влажной среде соков тела или органов хозяина. Но особым
моментом их жизни является этап, ведущий к заражению новой особи
хозяина, другими словами — этап, обеспечивающий экстенсивность
Рис. 34.
А — Я — циста парамеций,
окрашенная гематоксилином; в левой
(А) видны еще остатки пищевых вклю-
чений. По Иваничу. С — цисты
Opalina ranarum с одним и несколь-
кими ядрами. По В е н ь о н у.
распространения паразитов (за-
ражение ими новых и новых
хозяев), что обусловливает под-
держание существования вида
последних на земле.
Такой период у многих ви-
дов паразитов связан с выходом
их из тела хозяина во внеш-
нюю среду; подготовка к этому
начинается заблаговременно еще
во время нахождения паразита в теле хозяина.
В частности, опалины, обнаруживаемые у лягушки в описанном
выше виде (стр. 24) в течение большей части года, весной начинают по-
вторно делиться, благодаря чему образуются мелкие особи с одним или
немногими ядрами. Эти дочерние особи инцистируются (30—70 н
диаметром) и в таком состоянии выпадают из кишки лягушки в воду
(рис. 34,С). Будучи проглочены головастиками, они проходят через
их пищеварительный канал и оседают в прямой кишке; здесь цисты
вскрываются и содержимое их превращается в мноюядерных крупных
опалин.
45
Инцистирование достигает еще более яркой формы у грегарин, так
как их цисты (псевдонавицеллы) падают на землю, т. е. в среду, отли-
чающуюся неизмеримо большей сухостью по сравнению с содержи-
мым кишки или с соками тканей. В то же время с инцистированием
неразрывно связано и размножение этих паразитов.
Переход простейших к паразитизму связан с новым, необычайным
моментом — с жизнью в среде, постоянно изобилующей пищевыми
веществами; это исключает из повседневных функций простейших
момент поиска пищи и упрощает функции восприятия пищевых
веществ. Последняя задача разрешается переходом паразитических
простейших к эндосмотическому питанию в смысле физиологическом
и к потере (или редукции) структурных органоидов питания, что
ведет к вторичному упрощению организации простейшего. Такие
структурные и функциональные изменения, делающие организм соот-
ветствующим условиям существования в данной среде, называется
приспособлением или адаптацией. Они присущи, конечно, и свободно
живущим животным (например: инцистирование при высыхании,
способы движения и др.).
Заключительные замечания о типе простейших
Простейшие животные являются самостоятельно живущими микрог
скопическими по величине 1 существами, которые характеризуются
одноклеточностью строения. Простейшее состоит из протоплазмы,
в которой заложено одно ядро. Многоядерное строение присуще
некоторым простейшим или в качестве обычного вегетативного суще-
ствования, или же оно возникает в период размножения.
Сложность строения одноклеточных весьма различна, начиная
от голых корненожек и кончая инфузориями. Как бы ни было просто
или сложно устроено простейшее, каждому из них присущи все
основные жизненные функции, характерные для животных организмов:
движение, обмен веществ, раздражимость (рефлекторная деятельность)
и размножение. Все эти функции организма отправляются клеткой
в целом.
В зависимости от степени усложнения строения простейшего, для
той же цели служат морфологически диференцироваиные части клетки
в виде органоидов, или органелл. Органоидов дыхания у простейших
нет, и газообмен совершается через поверхность их тела.
Понятие «простейшее», как одноклеточное животное, должно быть
трактуемо лишь в морфологическом отношении, в смысле соответствия
строения простейшего схеме строения клетки многоклеточных живот-
ных. Но в физиологическом отношении простейшее, как целостный
организм, эквивалентно всему организму многоклеточного живот-
ного.
Морфологическая равноценность простейшего клетке в известной
мере относительна, ибо диференцировка органоидов может достигать
большой сложности и тонкости (скелетные, нервные, чувствительные
1 Лишь немногие достигают величины одного или более миллиметров, и вонь
вымершие корненожки — нуммулиты— достигали 2—3 см.
46
органоиды и др.); недаром говорят о сложности организации многих
простейших.
В пределах этого типа животных различаются более примитивные
организмы и виды, далеко ушедшие в процессе своей эволюции (т. е.
в процессе формирования вида или филогенеза).
Из всех четырех классов простейших наиболее примитивными
считаются жгутиконосцы, так как к ним и поныне принадлежат формы,
обладающие особенно-
стями и животного и
растительного организ-
мов. Можно с большой
долей вероятия предпо-
лагать, что на заре раз-
вития жизни на земле
все живое было предста-
влено весьма просто
устроенными существа-
ми, которые питались за
счет использования ве-
ществ неорганической
природы, каковым свой-
ством обладают и ныне
живущие растительные
организмы. Из простей-
ших это свойство при-
суще лишь некоторым
из ныне существующих
жгутиконосцев, в виду
чего они фигурируют и
В качестве растительных Рис. 35. Схема эволюции простейших,
организмов в курсах бо-
таники и в качестве животных существ в учебниках зоологии. Такие
существа двоякой природы часто относят к группе протистов.
От подобных им древних форм произошли саркодовые путем утери
жгутиков и перехода к амебоидному движению (метаболия, как мы
видели выше, свойственна и некоторым эвгленам).
От саркодовых, с одной стороны, и от жгутиконосцев, с другой,
произошли различные группы споровиков, эволюция которых шла
под знаком перехода к паразитическому образу жизни.
Наконец, от жгутиконосцев же, путем усложнения организма,
возникли наиболее совершенные из Protozoa — инфузории.
Такой путь филогенетического развития простейших можно схе-
матически изобразить в виде «дерева родства», представленного на
рис. 35.
ТИП КИШЕЧНОПОЛОСТНЫЕ
(COELENTERATA)
КЛАСС ГИДРОИДНЫЕ (HYDROZOA)
ГИДРА (HYDRA — ОТРЯД ГИДРОПОЛИПЫ)
Технические замечания. Гидра явпяется пресноводным жи-
вотным, обитающим в стоячих водах (в канавах, прудках, озерках, заводях речек
и тому подобных местах). Живет она чаще на небольшой глубине в прибрежной
части водоема, заросшей различными водяными растениями.
Гидр собирают, вытаскивая из воды ряску, листья кувшинок и других расте-
ний и помещая их в большую стеклянную банку с водой, или же черпают прудо-
вую воду также с различными растениями, при этом избегают захватывать с во-
дой моллюсков. Когда вода отстоится, гидр можно заметить сидящими на стенках
сосуда или на растениях.
Для корма служат различные планктонные рачки, циклопы, дафнии и дру-
гие организмы. Гидр можно довольно долго содержать в неволе. Аквариум с ги-
драми засаживают растениями, чтобы не было недостатка в кислороде. Запасают
также достаточное количество корма. В случае отсутствия живого корма гидр
можно кормить мелко нарезанными кусочками дождевых червей, но после такой
кормежки необходимо выбрать из аквариума все несъеденные остатки, чтобы
избежать порчи воды (и гибели гидр) при разложении несъеденных кусочков.
При таких условиях гидры могут жить несколько месяцев.
На занятия дают живых гидр, отсаженных в часовые стекла в небольшом
количестве воды. Гидр отсаживают, пользуясь отрезком широкой (около 5—7 мм
диаметром) стеклянной трубки с гладко оплавленными краями. Закрыв конец
трубки пальцем, опускают ее в аквариум к намеченной гидре. При открывании
верхнего отверстия вода быстро входит в трубку, вовлекая с собой и гидру. Тотчас
же крепко закрывают отверстие трубки пальцем, вынимают ее из аквариума и
опорожняют ее содержимое в поместительное часовое стекло. Лишнюю воду
отсасывают пипеткой с резиновым баллоном. Наблюдения производятся под
препаровальной лупой при слабом увеличении.
Наблюдения над живой гидрой
Когда гидра расправит свое тело, то в ней можно различить трубко-
видное, сильно растяжимое тело, подошву, которой гидра прикре-
пляется к субстрату, и рот, помещающийся на противоположном конце
тела на небольшом возвышении, называемом ротовым конусом, окру-
женным длинными хорошо сократимыми щупальцами (числом 6—12).
Цвет тела гидр обыкновенно темносерый или коричне-
ватый, что свойственно представителям родов Hydra и Pelmatohydra,
в отличие от Chlorohydra viridissima, в клетках внутреннего слоя (энто-
дермы) которой имеется масса симбиотических зоохлорелл зеленого
48
Рис. 36. Pelmatohydra oli-
gactis (Hydra fusca) с очень
вытянутыми в длину щу-
пальцами. Уменьшено. По
Трамблэю.
цвета. Для микроскопирования более демонстративны Hydrehi Pelma-
tohydra.
Представители родов Hydra и Pelmatohydra отличаются тем, что
у первых тело имеет одинаковый диаметр по всей длине до подошвы
(вздутие от проглоченной пищи в расчет не
принимается); у Pelmatohydra нижние части
тела вытянуты в хорошо выраженный тон-
кий стебелек (рис. 36). ►
Размеры тела гидр колеблются у
разных видов от 1 до 3 см. При голодании тело
и, особенно, щупальца гидры вытягиваются
очень сильно в длину; например,у Pelma-
tohydra щупальца могут достигать 25 см(!)
длины (рис. 36). Если гидру обеспокоить при-
косновением иглы, то тело и щупальца ее
быстро укорачиваются, тело принимает кону-
совидную форму величиной в несколько мил-
лиметров, а щупальца с 25 см длины укора-
чиваются до 3—4 мм.
Гидра, в общем, является сидячим живот-
ным, но она все же может передвигаться
с места на место; при этом гидра вытяги-
вает тело, наклоняется в сторону, прице-
пляется щупальцами к субстрату, отрывает
подошву от старого места прикрепления,
подтягивает тело к щупальцам и прикре-
пляется подошвой к новому пункту (рис. 37).
Такое движение можно сравнить с медлен-
ным кувырканием через голову.
Питание гидры. В аквариум с го-
лодными гидрами пускают дафний или циклопов. Когда рачок слу-
чайно притронется к щупальцу гидры, то он не в состоянии больше
оторваться от него. Движениями щупалец пойманная добыча мед-
4 Практикум по зоологии — 385
49
ленно заЛвывается в рот, откуда
которое сильно вздувается в месте
Рйс. 38. Гидра в продольном разрезе. По
Ж а м м у.
А. а — ротовое отверстие; с — энтодерма;
d — бугорчатые скопления стрекательных
клеток на щупальцах; е — полость тела гидры,
ограниченная энтодермой, многие клетки которой
имеют жгутики и образуют выросты в форме
псевдоподий;/—яйцеклетки; g— скопление жив-
чиков. Между эктодермой и энтодермой лежит опор-
ная (базальная) перепонка, показанная черной ли-
нией. G — изолированные стрекательные клетки:
d — заряженная, b — разряженная; о — капсула
с ядовитой нитью; т — выброшенная наружу
нить; р — ее капсула; сп — чувствительный стер-
жень — книдоциль.
проходит далее внутрь тела,
нахождения заглоченной пищи.
Если заглочен рачок, обла-
дающий природной окраской
(например, какой-нибудь
красный веслоногий рачок —
Copepoda), то полость гидры
через некоторое время запол-
няется красной жидкостью
(результат переваривания до-
бычи). Если такое наблюде-
ние производится с Н. fusca,
то видно ясно, что стебелек ее
остается бесцветным: следо-
вательно, в этой части тела
обычно полости нет.
При засовывании парали-
зованной добычи в ротовое
отверстие и в гастро-васку-
лярную полость гидры могут
попадать концы ее же щупа-
лец и оставаться здесь неко-
торое время. Замечательно
то, что даже при долгом пре-
бывании в полости тела гидры
ее щупальца остаются совер-
шенно невредимыми, так как
на них пищеварительный сок
гидры не оказывает никакого
действия. Это объясняется
тем, что ткани гидры обла-
дают противоферментной стой-
костью по отношению к соб-
ственному пищеварительному
секрету (в пределах вида).
Гидра может проглатывать
и крупную добычу, например,
червей, которые длиннее са-
мой гидры, и даже мальков
рыб. В последнем случае тело
гидры сильно растягивается
и как бы тонкой пленкой по-
крывает парализованную до-
бычу.
По окончании пищеваре-
ния непереваренные и неиспользованные остатки добычи (хитиновый
панцырь ракообразных, личинок водяных насекомых, кутикула чер-
вей и др.) выбрасываются наружу через ротовое отверстие. По-
следнее функционально является и ротовым и анальным (заднепро-
ходным).
50
Рис. 39. Стрекательные клетки:
разряженная (Ь) и неразряжен-
ная (а), изолированные из тела
гидры. По Ж а м м у.
Живую гидру кладут в каплю воды на предметное стекло, покрывают
покровным стеклом, подложив под края его ножки из воска, чтобы не
раздавить гидры. Препарат рассматривают под микроскопом. 1
Тело гидры состоит из двуслойного мешка, единственным отвер-
стием (ртом) сообщающегося с внешней средой (рис. 38, а). Наружный —
более прозрачный — слой состоит из эктодермы, внутренний же
(темный) слой образован энтодермой.
Зоохлореллы. У зеленой гидры в энтодерме заложено мно-
жество микроскопически мелких зеленых водорослей, которые живут
в клетках гидры на положении сожителей (симбионтов). Эти водо-
росли одноклеточны; их относят к
виду Chlorella vulgaris; вообще их на-
зывают зоохлореллами (рис. 45). Зеле-
ный цвет их зависит от наличия в них
хлорофила. Для зоохлорелл обитание
в клетках гидры выгодно, так как эти
симбионты получают соли и, в про-
цессе дыхания гидры, углекислоту;
сами же зоохлореллы, как и другие
зеленые растения, выделяют на свету
кислород, который используется клет-
ками гидры. При таком положении
дела можно говорить о взаимной
пользе, оказываемой организмами
(водорослями и гидрой) друг другу;
подобная форма симбиоза называется
взаимничеством, или мутуализмом.
Для зоохлорелл обитание в энтодерме
тела гидры «выгодно» и потому, что
сама гидра является «защищенным»
животным, что обеспечивает безопас-
ное существование ее симбионтам.
Интересно подчеркнуть характерное
приспособление зоохлорелл к обита-
нию в столь своеобразной среде, как
выделяющие фермент и фагоцитирую-
щие клетки энтодермы гидры: зоохлореллы стойки по отношению
к пищеварительному секрету гидры.
Повидимому, обитание зоохлорелл в теле гидры придает ей большую
стойкость, по сравнению с видами гидр без симбионтов, в отношении
способности жить в загрязненной воде.
Стрекательные клетки. На поверхности щупалец
видны бугорчатые возвышения, состоящие из скоплений так называе-
мых стрекательных клеток (рис. 38, A, d). Клетки эти очень сложного
строения (рис. 39); в них имеется особая капсула со свернутой спиралью
нитью ядовитого вещества (рис. 38,G, d, о). Когда к гидре прикасается
какое-либо маленькое животное, например, дафния, то стрекательные
клетки рефлекторно разряжаются: ядовитые нити выскакивают из них
1 Полезно рассматривать гидру на темном фоне при сильном падающем свете.
* 51
(рис. 38, 6) и впиваются в прикоснувшийся организм (рис. 39, Ь;
рис. 40). Последний при этом парализуется, после чего и заглатывается
гидрой.
Стрекательные клетки являются примером защитных ядовитых
органов, аналогичных трихоцистам инфузорий. 1
Чтобы привести стрекательные клетки в действие, прибавляют
под покровное стекло одну-две капли 5% уксусной кислоты; под
влиянием такого резкого химического раздражителя гидра разряжает
стрекательные клетки. Пользуясь сильным объективом и суженной
диафрагмой, можно рассмотреть и самые нити, выброшенные из стре-
кательных клеток.
Рис. 40. Действие крупной стрекательной капсулы на покровы
личинки комара. По Торре.
Ch — хитиновая кутикула; D — пораженное место хитина; F — крапивная
нить; St.W— стилеты стрекательной капсулы.
Мацерирование гидры
Для того чтобы рассмотреть клетки, составляющие тело гидры,
необходимо ослабить связь между ними. Последнее достигается маце-
рированием гидры в жидкости Гертвига, состоящей из смеси равных
объемов 0,1% осмиевой кислоты и 0,5% уксусной кислоты. Маце-
рирование длится 3—10 минут, после чего гидру на несколько часов
переносят в 0,1% водный раствор уксусной кислоты, затем слегка
обмывают в воде, расщипывают иглами на предметном стекле, наносят
на препарат каплю пикрокармина и закрывают покровным стеклом
или заделывают в глицерин-желатину.
На таком препарате можно найти следующее.
а) Крупные клетки энтодермы с двумя бичами на свободной поверх-
ности (если они сохранились); эти клетки удлинены, с неровными края-
ми; у основания они несколько вытянуты; в их «подошве» заложена
сократительная фибрилла (см. ниже); при нормальном положении
1 Аналогичными называют органы или части различных организмов, имею-
щие одинаковое значение.
52
в теле гидры фибриллы лежат циркулярно к оси гидры; протоплазма
этих клеток вакуолизирована; кроме того в ней имеются различные
включения, которые являются продуктами всасываемой ими пищи, или
же экскретами. Эти клет-
ки называют пищевари-
тельно-мышечными (рис.
42, С).
б) Стрекательные
клетки эктодермы с
овальной капсулой и
спиральной нитью (рис.
38, G; рис. 39).
в) Эпителиально-мы-
шечные клетки экто-
дермы (рис. 41). Во вну-
тренней части клетки по
Рис. 41. Эпителиально-мышечная клетка гидры.
По Ж а м м у.
а — эпителиальная часть; b — мышечная часть.
отношению к положению в теле гидры образуются продольные
отростки (д); эта удлиненная ее часть заключает в себе тяж сокра-
тительного вещества.
Рис. 42. Изолированные клетки гидры.
По К. Шнейдеру.
А—эпителиально-мышечная клетка эктодермы близ рта;
В — то же, из подошвы гидры (в протоплазме видны зерна
секрета); С — эктодермальная пищеварительно-мышечная
клетка, в ее базальной частя видна мышечная фибрилла;
D — железистая (ферментная) эктодермальная клетка.
Таким образом такая
клетка в одной своей
части является эпите-
лиальной и покров-
ной (а); в другой же —
мышечной; отсюда и
ее название (рис.
42, А). Она является
примером начала ди-
ференцировки сокра-
тительного вещества
и потому должна быть
отнесена к примитив-
ному виду мышечных
клеток, одновременно
остающихся в своей
наружной части и эпи-
телиальными.
Мышечные фи-
бриллы клеток экто-
дермы и энтодермы
лежат под прямым
углом друг к другу,
образуя кольцевой
внутренний и наруж-
ный продольный слой ;
такое расположение
фибрилл обеспечивает
сократимость тела гидры в обоих направлениях.
Эпителиально-мышечные клетки щупалец гидры (см. ниже) более
утолщены, чем в остальных местах тела.
53
ками, протоплазма их вакуолиста
Рис. 43. Почкование гидры. Гидры сидят
на веточках хары в различной степени
сокращения тела. Сбоку тела некоторых
гидр видны почки. По Ж а м м у.
тельно. На теле гидры может быть
г) Железистые клетки сосредоточены преимущественно на подошве
(рис. 42. В). Они являются производными эпителиально-мышечных
клеток, от которых отличаются наличием в протоплазме эпителиальной
части множества зернышек секрета, закрывающих собой и ядро клетки.
д) Железистые клетки энтодермы мелки, приблизительно клино-
видной формы, несут один—три бича на свободном конце; выделяют
пищеварительный секрет, почему и называются ферментными клет-
рис. 42, D).
е) Нервные клетки трудно
различимы. Они полигональной
формы, с большим ядром и часто
со многими протоплазматиче-
скими отростками, идущими к мы-
шечным отросткам эпителиально-
мышечных клеток.
ж) Всеми указанными типами
не исчерпывается разнообразие
клеток гидры; кроме них есть
еще так называемые интерсти-
циальные клетки — очень мел-
кие клетки эктодермы, сохраняю-
щие эмбриональные (зародыше-
вые) свойст.ва; но их на нашем
препарате найти не удается.
Размножение гидры
Вегетативное (беспо-
лое) размножение. Нередко
можно найти почкующуюся ги-
дру. На ее теле с боку образуются
выпячивания стенки в виде почки
(рис. 43). Почка растет, на ее кон-
це развиваются щупальца,проры-
вается ротовое отверстие, и почка
принимает вид маленькой гидры.
Последняя отрывается от мате-
ринского организма, падает на
дно и начинает жить самостоя-
>дновременно и несколько почек.
Такой способ размножения называется бесполым, так как при нем
организм размножается за счет клеток тела (соматических клеток).
Точнее можно охарактеризовать размножение гидры как почкование,
при котором дочерние организмы по размерам своим в момент отделе-
ния от тела матери меньше этого последнего.
По новейшим исследованиям, почкование гидры идет за счет раз-
растания интерстициальных клеток, которые при других условиях
превращаются в половые клетки. Полость почки является продолже-
нием гастро-васкулярной полости материнской гидры. При кормлении
последней пигментированными ракообразными (см. выше), красная
54
жидкость проникает и в полость почек. Следовательно, питание их
идет за счет материнской гидры.
У Chlorohydra viridissima при почковании выпячивающаяся в почку
энтодерма заносит в себе зоохлореллы; поэтому и следующее поколение
гидр этого вида обладает симбионтами.
В случае трудности сохранения на долгий срок живых гидр, для
демонстрации почкования приготовляют тотальные препараты.
Почкующихся гидр рассаживают по часовым стеклам с малым
количеством воды. Когда они расправят щупальца и почки, их обли-
вают горячим раствором сулемы с уксусной кислотой; через 1—2 часа
гидр перекладывают в 50% спирт, через полчаса — в 70%, с при-
бавлением иода до цвета портвейна; если иод обесцветится, его доба-
вляют. Затем препарат переносят в воду для подкрашивания квасцо-
вым кармином. Далее пре-
парат заключают по общим
правилам в канадский баль-
зам, подложив под покров-
ное стекло толстые (до
0,8 мм) ножки.
Для полного наблюде-
ния почкования у живой
гидры требуется два-три
дня.
Половое размно-
жение (цитогония)-
Осенью гидра размножается
половым путем. Гидра явля-
ется гермафродитом, ибо и
живчики и яйца развива-
ются у одной и той же особи.
Семенники, в виде невы-
соких пакетов живчиков
. 38, А, е: оис. 44,
Рис. 44. Половое размножение гидры. По
Мейзепгеймеру.
Левая гидра — в стадии выработки мужских половых
клеток (S), средняя — с яйцами (О), правая — в гер-
мафродитном состоянии.
лежат ближе к венчику щупалец; яичники, содержащие всего по
одному яйцу (рис. 38, A, f; рис. 44, О), закладываются на середине
тела, ближе к его основанию.
И мужские и женские половые клетки развиваются в толще наруж-
ного слоя, причем, по новым данным, источником их образования
являются те же Y-клетки, что и при развитии почек. Развитие половых
клеток у гидры протекает не одновременно; могут встретиться экзем-
пляры только с живчиками или только с яйцами, или с обоими сортами
половых клеток.
Мужские половые клетки (живчики) по мере созревания выходят
в воду и впоследствии внедряются в яйцевые клетки гидры, остающиеся
на месте своего образования. Этот процесс называется оплодотворе-
нием. Оплодотворенное яйцо (точнее, слившиеся в одну клетку, или
зиготу, яйцо и живчик) одевается плотной оболочкой и зимует.
На следующую весну из оплодотворенного яйца развивается личинка,
превращающаяся в новую гидру.
Сущность такой формы размножения (доминирующего у животных)
55
состоит в том, что новый организм возникает за счет размножения
отдельных клеток; такой вид размножения называется цитогонией.
Цитогония у гидры протекает за счет половых клеток, или гамет. Этот
процесс является гамогонией и, в частности, половым размножением.
Соотношение этих форм размножения иллюстрируется следующей
схемой.
Цитогония
Гамогония Половое размножение
Образование у простейших особых половых особей или гамет Образование половых клеток в результате множественного деления и созревания
Слияние гамет Оплодотворение — внедрение живчика в яйцо
Зигота Оплодотворенное яйцо
Рис. 45.
Л — разрез части стенки зеленой гидры (Chlorohydra viridissima) в месте
залегания яйцевой клетки (о); в клетках энтодермы (ел), в их базальной
половине, множество зоохлорелл (Ch); в яйцевой клетке их нет; с — экто-
дерма. В — средняя часть сходного разреза при большем увеличении;
видно проникновение зоохлорелл в яйцо гидры. По Гамаку.
Половое размножение у Chlorohydra viridissima интересно в том
отношении, что яйцо как производное эктодермы не имеет зоохлорелл;
но на известной стадии его созревания зоохлореллы мигрируют из
энтодермы материнского организма и внедряются в яйцевую
клетку. Благодаря такому раннему заражению яйца, развиваю-
щаяся из него гидра получает симбионтов, которые, размножаясь
делением, распределяются по клеткам энтодермы (рис. 45).
56
Гистологическое строение гидры
Объект фиксируется, как указано выше (сулема с уксусной кисло-
той), и заливается в парафин. Тонкие срезы наклеиваются белком и,
по освобождении от парафина, окрашиваются гематоксилином Дела-
фильда с докраской эозином.
На срезе отыскивают (рис. 46, А, В, С, D, Е):
1) наружный слой тела (эктодерма) из тесно прилежащих высоких
эпителиальных клеток (покровная часть эпителиально-мышечных
клеток); между ними в эктодерме встречаются стрекательные клетки
с капсулами внутри; на
свободном конце клетки
может быть виден чувстви-
тельный штифтик, или кни-
доциль;
2) в толще эктодермы,
местами в средней зоне (счи-
тая по высоте), попадаются
стрекательные клетки в ста-
дии своего формирования;
3) в основании эктодер-
мы кое-где могут быть вид-
ны нервные клетки (при-
близительно треугольной
формы) и мелкие ядра ин-
терстициальных клеток;
4) эктодерму подстилает
базальная перепонка, резко
контурирующая границу
между экто- и энтодермой;
5) внутренний слой или
энтодерма состоит из го-
раздо более высоких кле-
А — поперечный разрез через тело гидры. Видны:
а — эктодерма, b — энтодерма и базальная перепонка
между ними. По Рёзелеру и Лампрехту.
ток, чем эктодерма; кон-
тур поверхности энтодермы
волнистый, вследствие не-
одинаковой высоты клеток; видны жгутики клеток и зернистость про-
топлазмы, связанная с секрецией пищеварительных соков;
б) если материал заготовлен осенью, то могут быть видны вздутия
эктодермы со множеством мелких ядер (мужские гонады с развиваю-
щимися живчиками) или вздутие с одной крупной клеткой в толще
эктодермы (яйцевая клетка).
Опыт с регенерацией
Гидру кладут в часовое стекло, дно которого залито воском, в пру-
довую воду и разрезают узким (глазным) скальпелем приблизительно
посредине тела. Оба кусочка кладут в чашки Коха диаметром в 7—
10 см в прудовую (не водопроводную!) воду. Воду сменяют каждый
день. Наблюдать, чтобы с водой не попадали хищные инфузории,
вредящие регенерантам.
57
Рис. 46. (Продолжение).
В— разрез экчодермы тела Pclmatohvdra uligactis. сп -книдоциль\nz - крапивная клетка;
ez— тителиально-мышечная клетка; nza — молодая крапивная клетка; п - нервная
клетка. По К. Шнейдеру. С—поперечный разрез через часть щупальца гидры.
а — эктодерма; nz— стрекательные клетки; b—клетки энтодермы, бичики их не изо-
бражены. D, £—разрез сгенки тела гидры с железистыми клетками (d— d4)
По Стрелину.
Рис. 47. Регенерация отрезанного по линии А — В
переднего конца у гидры Pelmatohydra oligactis.
По Канаеву.
С течением времени, срок которого зависит от температуры, у обоих
кусочков гидры восстанавливаются утраченные части тела: задний
кусочек получает венчик щупалец (рис. 47), а у переднего кусочка зад-
ний конец замыкается и дает новую «подошву».
Свойство организма восстанавливать утраченные части тела назы-
вается регенерацией. Следует обратить внимание, что регенерирующие
гидры к концу этого процесса далеко не достигают размеров исходной
особи; это объясняется своеобразием процесса регенерации у гидры,
характеризующимся тем, что утраченные части восстанавливаются
за счет перегруппировки клеток в области разреза, без их размножения.
Такая форма регенера-
ции называется морфол-
лаксисом.
Для наблюдения ре-
генерации более пригод-
ны гидра обыкновенная
и имеющая стебелек —
Pelmatohydra oligactis.
Опыты с раздражи-
мостью
Если к гидре прика-
саться каким-либо инди-
ферентным в химическом
отношении предметом
(игла, полоска бумаги и
др.), то она реагирует на такое раздражение сокращением щупалец
или тела, причем стрекательные клетки остаются в покое.
При прикосновении к щупальцам гидры животных, служащих ей
пищей, в ответ на раздражение книдоцилей приходят в действие стрека-
тельные капсулы, парализующие добычу; раздражение передается
также через диффузную сеть нервных клеток эпителиально-мышечным
клеткам щупалец и других частей тела, благодаря чему обеспечивается
захватывание добычи и ее проглатывание.
Однако такой рефлекс — уловление добычи — осуществляется
у более или менее голодных гидр. Хорошо накормленная гидра часто
вовсе не реагирует на прикосновение добычи. Можно полагать, что
всасывающиеся пищевые вещества подавляют на некоторое время
способность щупалец проявлять рефлекс поимки добйчи.
Chlorohydra viridissima в аквариуме скопляются на стороне, обра-
щенной к свету. Если узким пучком света осветить стенку аквариума,
то гидры собираются именно к этому месту. Таким образом можно
говорить о положительном гелиотропизме Chlorohydra viridissima,
несмотря на отсутствие глаз. Другие виды гидр более безразличны
к направлению источника освещения.
Не всякое животное, прикасающееся к гидре, вызывает реактивную
деятельность стрекательных клеток. На поверхности гидр без малей-
шего для себя вреда пребывают оригинальные инфузории — Tricho-
dina pediculus (Peritricha); значение их для гидр толкуется противо-
59
речиво; их считали паразитами, могущими объедать щупальца гидры;
данные же других авторов (R a nd) говорят скорее о пользе триходин
для гидры, так как эти инфузории поедают грибные нити, могущие
локализоваться на теле гидры; покрытие грибками является опасным
для гидры.
Общие замечания
Тело гидры, если смотреть сверху, может всякий раз быть рассечено
по длине на две симметричные половины плоскостями, идущими ради-
ально по отношению ко рту гидры. Такая симметрия называется ради-
альной, или лучевой.
Для гидры весьма характерна мешкообразная форма двуслойного
тела с единственным выходным отверстием наружу и наличие стрека-
тельных клеток. Эти признаки отличают представителей типа кишечно-
полостных животных (Coelenterata).
Внутренняя полость (рис. 38, А, е) гидры является и пищевари-
тельной и выделительной, поэтому она называется гастро-васкуляр-
ной. У плоских червей мы увидим, что органы выделения и морфоло-
гически и функционально обособлены от пищеварительного канала.
Между наружным и внутренним слоями тела у гидры заложена
тонкая опорная пластинка неклеточного характера, тогда как между
кожными покровами и эпителием кишечника плоских червей имеется
губчатая — паренхимнгы—ткань, развивающаяся из среднего за-
родышевого листка.
У гидры мы видим только обособление двух слоев тела, которые не
распадаются на отдельные органы (между тем тело плоских червей
состоит из различных систем органов: половых, пищеварения, выделе-
ния и др.).
Итак, гидра из всех изучаемых нами здесь многоклеточных живот-
ных является наиболее примитивной по строению, так как она как бы
соответствует двуслойной стадии эмбрионального развития других
животных, так называемой гаструле, приобревшей способность к само-
стоятельной жизни.
У гидры еще не произошло обособления различных органов,
и только в ее эктодерме наблюдаем диференцировку различных кле-
ток, приспособленных более или менее к определенной функции (нерв-
ные, эпителиально-мышечные, железистые и другие клетки).
Мешковидная форма тела, т. е. форма полипа, и переход эктодермы
в энтодерму по краю ротового отверстия определяют принадлежность
гидры к классу гидроидных.
Отсутствие скелета и постоянное нахождение гидры в стадии полипа
характеризуют ее как члена отряда гидрополипов. Гидра обыкновен-
ная образует иногда расу, богатую красными пигментами — каротино-
идами и липоидами.
На основании особенностей строения, гидр разделяют на роды и
виды, которые отличаются следующими признаками. Chlorohydra
viridissima (= Hydra viridis)—зеленого цвета, 1—2 см длины; щупальца
не длиннее тела, в среднем восемь щупалец, половозрелость — летом;
яйцо шарообразное, с почти гладкой оболочкой.
•60
Pelmatohydra oligadis(=Hydra fusca =H. oli gad is) — окраска измен-
чивая; длина до 3 см; ясно обособленный стебелек; щупалец в среднем
шесть; длина их может достигать 25 см; стрекательных клеток три
типа, половозрелость — осенью; яйцо шарообразное или уплощенное,
с короткими бугорками.
Hydra vulgaris (=Н. grisea) — окраска изменчивая; длина около
2 см; тело сужается к ножке, но без ясного обособления стебелька;
щупалец в среднем шесть; длина их до 5 см; четыре типа стрекательных
клеток; половозрелость — летом; яйцо шарообразное; оболочка с длин-
ными волнообразными шипиками.
Хотя гидры и могут передвигаться с места на место, но в обычном
состоянии они подолгу держатся прикрепленными в одной точке.
Поэтому гидр мы вправе отнести к животным, ведущим сидячий образ
жизни.
МОРСКИЕ ГИДРОИДЫ (ОТРЯД ГИДРОМЕДУЗЫ)
Технические замечания. Консервированных спиртом морских
гидроидов выписывают с Мурманской или Севастопольской биологических стан-
ций. От колонии гидроидов отрезают маленькие кусочки веточек, которые окра-
шивают квасцовым или борным кармином, уплотняют и обезвоживают в спиртах
возрастающей крепости и после 100% спирта переносят в гвоздичное или кедро-
вое масло для просветления, после чего заключают в канадский бальзам. Для
занятий могут быть использованы Obelia, Tubularia, Camppnularia, Gonothyrea
и другие гидроиды.
Готовый препарат рассматривают под микроскопом и зарисовывают.
Изучение препаратов
1. Гидроид пеннария (Pennaria cavolini). Гидроид пеннария по
внешнему виду имеет некоторое сходство с пером. От главного ствола
отходят поочередно в обе стороны боковые ветви, на которых через
правильные промежутки сидят короткостебельчатые грушевидные
полипы (рис. 48, А). Уже по наружному виду пеннария резко отли-
чается от рассмотренных нами животных, ибо она представляет собой
организм, состоящий из множества отдельных особей (полипов), свя-
занных вместе в одно целое; такие организмы называются колониаль-
ными.
Под микроскопом ветви колоний имеют вид трубок, покрытых
снаружи плотной хитиноподобной оболочкой (перидерма), местами
кольцеобразно-складчатой. Перидерма окружает трубку, состоящую
из двух слоев клеток — эктодермы и энтодермы, внутри которой
есть полость, сообщающая полости всех полипов друг с другом.
Полип пеннарии (рис. 48, В) имеет форму груши, обращенной тон-
ким концом вверх. От основания полипа в сторону и вверх идут длин-
ные щупальца (рис. 48, Т); они сравнительно тонки и служат для
захватывания пищи. По оси щупальца внутри заложен слой широких
эктодермальных клеток. Одеты они снаружи эктодермой, которая на
одной стороне щупальца выше, чем на другой. Полости в щупальцах
нет (рис. 48, В, внизу).
Лежащая выше часть полипа покрыта короткими булавовидными
щупальцами. Короткий стержень их также образован широкими
61
клетками энтодермы, которая покрыта эктодермой одинаковой толщины
по всей окружности щупальца. Булава последнего богата стрекатель-
ными клетками; при большом увеличении иногда удается видеть вы-
брошенные из них стрекательные нити (рис. 48, С).
Собственно тело полипа похоже на мешок, входное отверстие в кото-
рый лежит на тонком конце полипй. По краю ротового отверстия хо-
рошо различимы эктодерма в виде низких клеток поверхностного слоя
и энтодерма, состоящая из высоких темно окрашивающихся клеток.
Рис. 48.
А — часть колонии Pennaria cavoltni. По Алльману. В—отдельный полип в значительно
увеличенном виде. Sf — стебелек; 7\— длинное щупальце: Т2 — булавовидное короткое щу-
пальце; М — рот; Go 7, 2 — гонофоры; spad — осевой стержень (spadix); test — сперматозоиды;
rad — радиальный канал; Ent — энтодерма, ЕМ — эктодерма. В, внизу — часть длинного
щупальца. С — головка булавовидного щупальца со множеством стрекательных клеток, из
которых две выстрелили. По Ш л е й п у.
Полость полипа продолжается в полость ножки, ветви и ствола
колонии. Таким образом, пища, принятая данным полипом, поступает
для пользования всей колонии.
На некоторых полипах имеются боковые почкообразные придатки
(рис. 48, В, Go) различной величины. Почки эти называются гонофорами,
ибо в них развиваются половые продукты. Возникают гонофоры из
стенки полипа и являются редуцированными 1 медузами (см. ниже).
Колония пеннария прочно прирастает к субстрату, благодаря чему
переменить место (в противоположность гидре) она не может.
1 Редуцированные значит — вторично упрощенные.
62
2. Гидроид обелия (Obelia geniculata). Обелия имеет вид непра-
вильно ветвистого гидроида, на стволе которого сидят полипчики.
Так же, как и у Pennaria, ветви обелий (рис. 49) покрыты перидер-
мой (7), которая, в отличие от Pennaria, переходит на самый полип
и окружает его основание в виде чашечки или колокольчика (гидро-
тека, рис. 49, 2) в которую полип прячется в случае надобности.
Полип имеет вид
амфоры (3); от основа-
ния ее шейки отходит
венчик щупалец (5).
Ротовое отверстие ле-
жит на вершине амфоры
(на хоботке).
По оси щупалец ле-
жит ряд крупных экто-
дермальных клеток, по-
крытых мелкими клет-
ками эктодермы со стре-
кательными клетками
между ними (выдаются
с поверхности острыми
бугорками). Все щу-
пальца одинаковы по
своему строению.
Все полипы описан-
ного строения являются
питательными особями.
Местами веточки ко-
лоний оканчиваются
утолщениями, похожи-
ми на ручку печати;
это — почки (6), из ко-
торых разовьется новый
полип данной колонии.
Таким образом ги-
дроиды размножаются,
подобно гидре, вегета-
тивно (бесполым путем)
посредством почкования
Рис. 49. Гидроид Obelia geniculata^ По Рёзе-
леру и Лампрехту.
Часть веточки колонии. 1 — перидерма; 2 — гидротека;
3—гастро-васкулярная полость; 5—щупальца; 6—почка
полипа.
Разница здесь та, что почки гидр отпадают
и живут самостоятельно,
почки же гидроида остаются связанными
друг с другом, поэтому и возникает колония.
В колонии обелии, кроме питательных полипов, образуются еще
так называемые гонангии (рис. 50). Они имеют удлиненно-яйцевид-
ную форму со срезанной поперек верхушкой. Гонангий одет снаружи
прозрачной гидротекой, называемой здесь гонотекой (2); внутри ее
идет бластостиль (3), отходящий от ствола колонии. На бластостиле
возникают почти шарообразные гонофоры (5), из которых образуются
свободно плавающие организмы, называемые медузами.
Таким образом в результате почкования у обелии возникают особи
двух сортов — полипы и медузы. Когда медузы созревают, то кры-
63
шечка гонотеки открывается, медузы выплывают наружу и начинают
свободно подвижный образ жизни.
3. Медуза обелия. При рассмотрении тотального, окрашенного
квасцовым кармином препарата медузы видно, что тело ее состоит
из прозрачного студенистого колокола. Выпуклая поверхность его
(exumbrella) направлена кверху; вогну-
тая поверхность (subumbrella) обращена
книзу.
По краю медузы (рис. 51) свисает
вниз (на рисунке отходит в стороны)
множество коротких щупалец, а из
центра вогнутой поверхности медузы от-
ходит короткий стебелек (manubrium)
с ротовым отверстием на конце.
От ротового отверстия вверх идет ка-
нал, который в толще зонтика мантии
впадает в место соединения двух пере-
Рис. 50. Obelia geniculata. Го-
нангий. По Рёзелеру и
Лампрехту.
7— стебель питательного полипа;
2 — гонотека; 3 — бластостиль; 4 —
крышечка; 5 — гонофора.
Рис. 51. Obelia geniculata. Медуза снизу.
'гПо:Рёзелеру и Лампрехту.
Видны крестообразная и кольцевая пищевари-
тельная (гастро-васкулярная) полость; четыре рас-
положенные на-крест гонады (скопления половых
клеток); радиальные щупальца.
крещивающихся друг с другом трубок, идущих радиально по отно-
шению к центру зонта. По краю зонтика концы этих каналов соеди-
няются кольцевым каналом.
Система описанных полостей является гастро-васкулярной по-
лостью медузы, у которой она в значительной мере редуцирована
вследствие сильного развития богатого водой студенистого вещества,
образующего главную массу тела медузы (так называемая мезоглея).
Соответственно положению радиальных каналов, на вогнутой сто-
роне зонта медузы лежат четыре скопления половых клеток, т. е.
64
гонады. Медузы раздельнополы, причем полы различаются лишь по
характеру самих половых клеток.
Медузу можно сравнить с полипом, который подошвой повернут
кверху, причем тело его сильно раздалось в стороны, благодаря чему
и приобрело форму зонта или колокола.
Цикл развития обели и. Половые продукты, разви-
вающиеся в эктодермальных гонадах медузы, выбрасываются в воду,
где и происходит оплодотворение. Такое оплодотворение называется
наружным. Из оплодотворенного яйца образуется свободно плаваю-
щая личинка (планула), которая с течением времени оседает на дно
и превращается в полип.
Таким образом полип возникает в результате полового размноже-
ния. Полип, далее, начинает почковаться, причем новые полипы оста-
ются в связи друг с другом, вследствие чего бесполым путем возни-
кает колония. В результате того же почкования развиваются и медузы,
которые отрываются от материнской колонии и приступают к половому
размножению.
Таким образом полный жизненный цикл гидроида обелии сопря-
жен со сменой двух непохожих друг на друга поколений, которые
к тому же размножаются то цитогонией, то вегетативно. Как и у лен-
точных червей (см. ниже), мы имеем здесь пример метагенеза, т. е. раз-
множения с чередованием поколений.
Общие замечания
Полипы Pennaria и Obelia относятся к кишечнополостным живот-
ным, так как их мешковидное тело двуслойное и внутри имеет гастро-
васкулярную полость. В эктодерме их заложены стрекательные
клетки. Эктодерма переходит в энтодерму по краю ротового отверстия.
Как и гидра, они относятся к классу гидроидных, отличаясь, од-
нако, от нее колониальностью своей организации, причем в колониях
образуются особи двух сортов — полипоиды и медузоиды. Размноже-
ние их идет со сменой поколений. Эти особенности оправдывают
отнесение гидроидов к отряду, гидромедуз.
Гидра и колонии гидроидов являются сидячими пресноводными
животными, лишенными особых органов передвижения.
Питаются они мелкими планктонными 1 животными, которых ула-
вливают своими щупальцами, богатыми стрекательными клетками.
Дышат, кислородом, растворенным в воде, но не имеют особых
органов дыхания.
КЛАСС КОРАЛЛОВЫЕ ПОЛИПЫ (ANTHOZOA)
ПРОБКОВЫЙ КОРАЛЛ (ALCYONIUM PALMATUM)
Общий вид и строение. Колония пробкового коралла» (рис. 52)
имеет вид толстого, неправильно разветвленного мясистого деревца,
в ствол и ветви которого вкраплены полипы, имеющие в расправлен-
ном состоянии вид восьмилучевых звездочек. ’
1 Планктоном называют совокупность мельчайших организмов, живущих
в толще воды и не могущих противостоять ее течению.
5 Практикум по аоологии —385
65
Рис. 52. Alcyonium palmatum. Пробковый ко-
ралл-колония. По Ж а м м у.
Под препаровальной лупой рассматривают окрашенный тотальный
препарат отдельного полипа, вырезанного из ствола колонии.
Удлиненное тело полипа несет на конце венчик из восьми щупалец
(рис. 53, 77; рис. 54, А), которые начинаются от окружности ротового
отверстия широким основанием; каждое щупальце несет множество
парных боковых веточек (72).
От ротового отверстия в гастро-васкулярную полость полипа спу-
скается трубчатый пищевод, выстланный эктодермой, которая пере-
ходит в энтодерму на его внутреннем конце.
Энтодерма образует в
гастро-васкулярной поло-
сти восемь выростов — пе-
регородок (4, 5), прикре-
пляющихся к пищеводу и
оканчивающихся свободно
в полости полипа.
Свободные края перего-
родок оторочены утолще-
ниями, называемыми мезен-
териальными утолщениями
(филаменты, рис. 53, 9),
которые богаты желези-
стыми клетками. Шесть из
них коротки и толсты, два
же — нитевидного хара-
ктера. Ниже их по краю
перегородок могут быть
найдены более ограничен-
ные вздутия, образованные
скоплениями половых кле-
ток (10).
Поперечные разрезы
пробкового коралла. Изу-
чают готовые препараты
срезов окрашенного бор-
ным кармином и залитого
в парафин пробкового ко-
ралла (рис. 54, В, С).
а) С р е з на уров-
не пищевода (рис. 54, В). Снаружи препарат ограничен дву-
слойной стенкой тела; внутри видна вытянутая щель — поперечный
разрез пищевода. Щель эта изнутри выстлана эктодермой (впячивание
покровной эктодермы), а со стороны полости ограничена энтодермой.
Между пищеводом и стенкой тела натянуто восемь перегородок,
разделяющих верхнюю часть гастро-васкулярной полости на восемь
камер.
В одном из своих концов щель пищевода оканчивается округлым
жолобом (сифоноглифу, прилежащая к ней камера считается вентраль-
ной, противоположная же —дорзальной.
В толще перегородок можно видеть поперечно рассеченные мышеч-
66
Рис. 53. Продольный разрез полипа Alcyo-
nium; полусхематично. По Ж а м м у.
7—эктодермальная глотка; 2 — сифоноглиф;
3— дорзальная перегородка; 4— вентральная
перегородка; 5—перегородка; 6 — эктодерма;
7 — мезоглея; 8— энтодерма, выстилающая
каналы; У— филаменты; 10— гонады; 7 7 —
щупальца; 12— вторичные выросты щупалец.
ные пучки («мышечные гребешки»). Они вдаются с обеих сторон в брюш-
ную камеру, вовсе отсутствуют в спинной и лежат только с одной
стороны каждой из остальных камер.
Ь)Срез через собственно тело коралла
(рис. 53, С). От стенки тела отходит в гастральную полость восемь
перегородок, оставляющих центральную часть полости свободной.
В полости перегородок видны продолжения мышечных пучков. По
краю перегородок могут быть
рассмотрены разрезы мезенте-
риальных утолщений.
Скелетные элементы пробко-
вого коралла. При рассмотрении
микроскопического препарата,
который при изготовлении его
не был в соприкосновении с ка-
кой-либо кислотой, в области
глотки,втолще стенки те ла, видны
тонкие шиповатые иглы, назы-
ваемые склеродермитами, состоя-
щие из известковых соедине-
ний.
В области перехода полипа
в ствол колонии заложены скле-
родермиты более короткие и
толстые; их можно обнаружить
лишь после вываривания ку-
сочка полипняка в едком кали
(рис. 54, £>).
Размножение пробкового ко-
ралла. Пробковый коралл раз-
множается бесполым путем —
почкованием с последующим
образованием рассмотренной на-
ми колонии.
Цитогония происходит за
счет половых клеток, образу-
ющихся в энтодерме гастраль-
ной полости.
Из оплодотворенного яйца развивается личинка, дающая почко-
ванием новую колонию. Стадии медузы у Alcyonium нет.
Общие замечания
Пробковый коралл, в виде отдельного полипа, имеет по внеш-
ности тело, построенное по лучевой симметрии. Однако, вследствие
щелевидной формы пищевода, наличия сифоноглифа и особого распо-
ложения мышечных пучков *в перегородках, симметрия полипа
является только двубоковой.
От гидроидов пробковый коралл отличается следующими при-
знаками:
67
а) наличием эктодермального пищевода;
б) переходом эктодермы в энтодерму по внутреннему краю пище-
вода ;
в) присутствием энтодермальных перегородок с мезентериальными
утолщениями;
д) развитием половых клеток в энтодерме.
Рис. 54.
А — полип Alcyonium, вид сбоку на тотальном препа-
рате. t—щупальца, о — эктодермальный пищевод;
т — мезентериальные утолщения. Оригиналов —
поперечный срез полипа на уровне пищевода (О).а—экто-
дерма; b — энтодерма. По К. Шнейдеру. С — то
же, ниже конца пищевода, d — мезентериальный валик.
По К. Шнейдеру. D — спикулы Alcyonium.
Все это вместе взятое характеризует представителей класса ко-
ралловых полипов (Anthozoa), в частности восьмилучевых корал-
ловых полипов (Octocorallia).
Колония Alcyonium является мономорфной, или одноформенной,
так как все особи ее одинакового строения. Как и у гидроидов, га-
стральные полости полипов соединяются друг с другом каналами,
проходящими в толще ствола и ветвей колонии.
Как и гидроиды, коралловые полипы неподвижно прикреплены
68
к субстрату. Подвижность сохранилась только у полипоидов, кото-
рые могут втягивать свои щупальца внутрь.
Дышат коралловые полипц поверхностью тела (кожное д ы-
хание); питаются теми мелкими организмами, которые случайно
прикоснутся к щупальцам и будут парализованы ударами стрекатель-
ных нитей.
АКТИНИЯ (ACTINIA EQUINA)
Технические замечания. Actinia equina обычна в Черном море.
Получить фиксированный материал можно через Севастопольскую биологическую
станцию Академии наук. Для рассмотрения общего вида берут цельных актиний.
Для изготовления срезов через щупальца и различные участки тела, актиний
фиксируют сулемой с уксусной кислотой и заливают в парафин. Для демонстраций
срезы окрашивают железным гематоксилином или крезилэхтвиолетом.
Для рассмотрения стрекательных капсул, если есть живой материал,
отрезки щупалец или вырезанные кусочки поверхности тела актинии раздавли-
вают между предметными стеклами так, чтобы получилось подобие толстого мазка.
Мазок фиксируют жидкостью Сагпоу и окрашивают железным гематоксилином.
Препарат рассматривают при сильном увеличении; изучение стрекательных
капсул ведется под масляной иммерзией.
Общая форма и строение
Тело актинии имеет форму невысокого толстого цилиндра; широ-
кой плоской подошвой актиния прикрепляется к субстрату (камень
и т. п.) и держится крепко, так
что живую актинию нелегко ото-
рвать от места ее прикрепления;
последнее она может менять, но
подвижность ее очень мала, и
она в еще большей степени, чем
гидра, является сидячим живот-
ным.
Противоположный подошве
конец тела несет в центре рото-
вое отверстие, окруженное мно-
жеством относительно коротких
щупалец. Число их кратно
шести. При раздражении акти-
ния втягивает внутрь свой вен-
чик щупалец, ее тело сокра-
щается и принимает вид холма
с округленной гладкой верши-
ной, в центре которой видно
углубление; через последнее щу-
пальца высовываются обратно,
когда актиния снова начинает
Рис. 55. Разрез эктодермы актинии близ
ее основания. По Е. Павловскому
и А. Штейну.
Видны толстостенные (черные) овальные кап-
сулы стрекательных клеток.
расправлять свое тело. Щупальца, богатые стрекательными клет-
ками, служат для ловли добычи.
Острой бритвой разрезают фиксированную актинию поперек:
1) по середине тела и 2) ближе к ротовому отверстию. На первом срезе
видно, что в полость актинии вдается множество перегородок нерав-
69
ной величины; одни из них длинны, другие коротки; все эти перего-
родки расположены симметрично. На втором срезе можно обнару-
жить, что перегородки прирастают ц глоточной или пищеводной
трубке, свисающей от рта вглубь гастро-васкулярной полости. На по-
перечном разрезе щупалец видна полость, сообщающаяся с гастро-
васкулярной полостью тела актинии.
Препарат стрекательных клеток. На окрашенных мазках из щу-
палец преобладают тонкостенные (фуксинофильные в отношении окра-
ски) стрекательные капсулы с отлично видимой красивой спи-
ралью стрекательной нити; многие капсулы фиксируются в разряжен-
Рис. 56.
А— мазок из щупальца Actinia equina. Видно множество фуксинофильных, спиральных стрека-
тельных капсул и несколько толстостенных капсул. В — то же, мазок из стенки тела близ его
основания. Многочисленные толстостенные стрекательные клетки. По Е. Павловскому
и А. Штейну. •
ном состоянии; в таких случаях спираль на одном конце «разматы-
вается» и выпячивается в длину (рис. 56, А). В меньшем числе попа-
даются толстостенные, сплошь густо окрашивающиеся капсулы, имею-
щие вид узких длинных овалов. При разряжении у них из одного
конца выступает более или менее длинная нить. На мазках из тела
актинии преобладают именно толстостенные стрекательные капсулы
(рис. 56, В).
Срез через щупальце. Эктодерма очень высока. В ее толще во много
рядов разбросаны ядра различных клеток. Бросается в глаза мно-
жество стрекательных капсул, лежащих по всей толще эктодермы, начи-
ная от базальной перепонки и до поверхности эктодермы (рис. 55 и 57).
70
Под самой поверхностью встречающей единичные толстостенные стре-*
нательные капсулы. Попадаются крупные одноклеточные железы
с ацидофильным секретом (т. е. окрашивающиеся кислыми красками,
например, эозином). Поверхность
эктодермы густо покрыта мерца-
тельными ресничками.
Общие, замечания
Актиния является одиночным
коралловым полипом, обладающим
признаками, объединяющими ее
с пробковым кораллом в класс
Anthozoa (эктодермальный пище-
вод, мезентериальные утолщения
перегородок гастро-васкулярной
полости и др.).
В то же время актиния отли-
чается от пробкового коралла не
только тем, что она является оди-
ночным полипом, но и числом щу-
палец, которое у актинии кратно
шести. Поэтому актинию относят
к шестилучевым коралловым по-
липам (НехасогаШа).
Актинии могут медленно пере-
менять место прикрепления к суб-
страту.
Дышат коралловые полипы по-
верхностью тела (кожное дыхание)
за счет кислорода, растворенного
в воде. Питаются мелкими орга-
низмами или более крупными жи-
вотными (например, ракообраз-
ными, молодью рыб и др.), кото-
рые при плавании случайно при-
касаются к их щупальцам и па-
рализуются действием яда стре-
Рис. 57. Часть поперечного разреза
через эктодерму шупальца Actinia
equina. По Е. Павловскому и
А. Штейну.
нательных капсул.
Ядовитость многих кишечно-
полостных действительна и для
человека. Из рассмотренных нами
Видны на поверхности мерцательные во-
лоски. На разных уровнях среза— множе-
ство стрекательных спиральных капсул; под
самой поверхностью — одна толстостенная
капсула; левее — одноклеточная ацидофиль-
ная железа.
кишечнополостных животных та-
ким свойством обладают актинии; при повторных прикосновениях
руками к актиниям (при ловле губок) кожа рук воспаляется с после-
дующим омертвением покровов и их изъязвлением.
Общие замечания о типе кишечнополостных
Кишечнополостные являются самыми простыми (за исключением
губок) из всех многоклеточных животных, о которых далее будет речь.
71
Примитивными чертами их организации являются:
а) отсутствие разделения тела на органы;
б) наличие в теле единственной полости, которая обслуживает
функцию пищеварения, выделения и разнесения по телу питатель-
ных веществ (гастро-васкулярная полость);
в) двуслойность тела (в типических организмах).
Кровеносная система отсутствует.
Весьма характерной чертой кишечнополостных является облада-
ние стрекательными клетками, и этот признак часто выдвигают, на-
ряду с наличием гастро-васкулярной системы, для характеристики
типа кишечнополостных (Coelenterata), которых называют также
стрекательными животными — Cnidaria (от греческого слова knide —
крапива).
ТИП ЧЕРВИ (VERMES)
ПОДТИП ПЛОСКИЕ ЧЕРВИ
(PLATODES)
КЛАСС ТУРБЕЛЛЯРИИ, ИЛИ РЕСНИЧНЫЕ ЧЕРВИ
(TURBELLAR1A)
ПЛАНАРИЯ <DENDROCOELUM LACTEUM)
Технические замечания. Планарий ищут осенью в стоячих
или медленно текучих водоемах на водяных растениях, под камнями, обломками
веток, под упавшими на дно листьями, в илу и др. Материал черпают водяным
сачком и осматривают в лаборатории. Выкидывают также в воду труп лягушки
или какого-либо другого маленького животного и на нем собирают скопившихся
планарий. Собранных червей содержат в низких и широких стеклянных банках
(простоквашницы, большие кристаллизаторы и др.), на дно которых кладут опав-
шие листья. Планарии менее требовательны к условиям существования; поэтому
воду в аквариумах можно менять раз в три дня. Кормят их водяными осликами,
у которых*предварительно отрывают ноги; можно давать им также белый хлеб
или сгустки крови.
На севере обычными являются молочнобелая крупная, до 3 см длины,
белая планария — Dendrocoelum lacteum, бурая планария — Pl. torva, и Р1.
lugubris (последняя живет в текучих водоемах). На юге встречается Pl. gono-
cephala (рис. 58).
Наблюдают живых планарий, готовят из них тотальные препараты и срезы.
Наконец, ставят ряд опытов с их регенерацией.
Наблюдения над живыми планариями
. Рассматривают живую планарию в часовом стекле под лупой; за-
тем ее помещают в воду под покровное стекло с восковыми ножками;
придавливая их, в необходимой мере сплющивают планарию, что
позволяет видеть под микроскопом некоторые детали строения на
живом объекте.
Форма тела уплощенная, продолговатая; передний конец тупо
обрезан, задний конец несколько заострен. На боках переднего или
головного конца лежат две боковые лопасти; они подвижны и играют
роль щупалец. Несколько отступя назад лежит пара глаз в виде пи-
гментированных точек. На молочнобелом фоне тела хорошо просве-
чивают темные ветви кишечника.
Движения планарии плавные, медленные и равномерные; слизи-
стые железы планарии (см. ниже) выделяют обильную слизь; по ней,
73
как по ленте, скользит при движении планария. Dendrocoelum может
держаться брюшной стороной кверху под самой поверхностью воды
и скользить по ней в таком положении. Кроме того планария может
изгибать тело в различных направлениях, что обеспечивается сокра-
щением различных мышечных волокон в паренхиме тела (см. ниже).
Скольжение планарии зависит от работы ресничек, которые густо
покрывают тело планарии. Отсюда и название для всей группы —
ресничные черви.
Для рассмотрения ресничек осторожно сдавливают планарию и
микроскопируют край тела. Реснички покрывают свободную поверх-
ность однослойного эпителия, образующего покров тела. Эпителиаль-
1
4
Рис. 58. Планарии; внешний вид; увеличено. По Штейн-
м а н-Б реслау.
1 — Planaria torva; 2 — PL lugubris; 3 — PL gonocephala; 4 — Dendro-
coelum lac team,
ные клетки имеют на продольном разрезе форму невысоких цилин-
дров. В их протоплазме просвечивают блестящие палочковидные
тельца, называемые рабдитами; иногда их можно рассмотреть и
в глубже лежащей части тела.
Кишечник (рис. 59) *Цросвечивает в виде трех главных, слепых
на концах ветвей — одной передней и двух задних; сходятся они
в средней части животного, где просвечивает глотка, открывающаяся
ротовым отверстием. Последнее лежит на брюшной стороне тела.
Главные ветви кишечника несут множество боковых веточек, также
заканчивающихся слепо; планарии, обладающие тремя ветвями ки-
шечника, относятся к подотряду Triclada. У рассматриваемой пла-
нарии рот служит, как и у кишечнополостных животных, и для пои-
74
Рис. 59. Dendrocoelum lac-
teum. Контуры кишечника (а)
с неокрашенного препарата;
р — глотка. Оригинал.
эпителием; его реснички
йятия Пищи и для выбрасывания неиспользованных остатков пищи.
Кишечник планарий мы вправе сравнить с гастро-васкулярной по-
лостью кишечнополостных.
Глотка планарии может выпячиваться наружу. Планарии являются
хищными животными и питаются обычно мелкими водными живот-
ными.
Другие детали строения планарий
трудно различимы на живых объектах и
еще менее доступны для легкого ознако-
мления на тотальных окрашенных препара-
тах. Поэтому бегло отметим, что планарии
являются гермафродитными животными
со сложно устроенным половым аппара-
том; они имеют выделительную систему
в виде протонефридий .(см. ниже, раздел
о сосальщиках и ленточных червях); есть
у них нервная система в виде пары
ганглиев и двух продольных .стволов,
соединенных многочисленными попереч-
ными коммиссурами. Органы кровообраще-
ния и дыхания отсутствуют.
Другие обычные пресноводные плана-
рии (Triclada) отличаются меньшими раз-
мерами тела, его несколько иной формой,
темной или бурой окраской (рис. 58). У не-
которых есть несколько глаз.
Изучение среза через тело планарии
Dendrocoelum lacteum
Фиксирование сулемой с уксусной кис-
лотой; заливка в парафин; окраска попе-
речных (или фронтальных) срезов гема-
токсилином и эозином. Сначала для общей
ориентировки просматривают срез во всех
его частях при малом увеличении микро-
скопа; затем микроскопируют детали при
более сильном увеличении. Устанавливают
на поперечном срезе следующие черты ми-
кроскопического строения (рис. 60 и 61):
1) Снаружи тело покрыто однослойным
на фиксированном материале часто отпадают;
2) в эпителии видны палочковидные рабдиты (рис. 61, А, г);
3) эпителий лежит на базальной перепонке (рис. 61, Ьт);
4) все тело заполнено соединительной тканью (паренхимой), в ко-
торую вкраплены различные органы;
5) видимые в толще паренхимы просветы, ограниченные слоем
эпителия, являются разрезами ветвей кишечника и соответствуют его
полости;
6) выясняют на срезе его стороны и поверхности; приострен-
75
Рис. 60. Поперечный разрез че-
рез Dendrocoelum lacteum. По
Шнейдеру.
Ер —• эпидермис; Ent — энтодер-
ма; Но — семенные фолликулы;
Lam — продольные мышечные во-
локна; Dvm—дорзальные мышечные
волокна; Mstm — ствол центральной
нервной системы; Сот—нервная к<>м-
миссура; Drz — железистые клетки.
ные концы среза соответствуют правому
и левому краям тела; несколько вогнутая
длинная сторона среза является брюш-
ной, а противоположная ей, несколько
выпуклая — спинной стороной тела;
7) под эпителием покровов и под
базальной мембраной лежит слой коль-
цевых диагональных мышечных волокон
(рис. 61, dm, слева); далее кнутри идут
пучки продольных мышечных волокон
(1т); между спинной и брюшной поверх-
ностями в паренхиме видны местами
дорзо-вентральные мышечные волокна
(dm, справа); кольцевые и диагональ-
ные мышцы образуют кожно-мускульный
мешок;
8) паренхима представлена соедини-
тельной тканью (Ь) с ветвистыми клет-
ками; их отростки переплетаются и обра-
зуют крупнопетлистую (на разрезе) сеть;
петли, повидимому, заполнены жидко-
стью лимфоидного характера; ближе
к спинной стороне в соединительной
ткани встречаются клетки с черными
зернышками пигмента;
9) под кожно-мускульным мешком и
слоем продольных мышечных волокон
лежат крупные мешковидные клетки,
в протоплазме которых образуются раб-
диты (рис. 61, rz); отсюда рабдиты про-
никают через соединительную ткань,
мышцы, базальную мембрану и внедря-
ются в эпителий покровов, где мы уже
их наблюдали;
10) на уровне этих клеток и глубже
лежат субэпителиальные одноклеточные
железы; тело их выпячивается в узкую
длинную шейку, которая открывается
между клетками эпителия; по характеру
вырабатываемого секрета и по отноше-
нию его к краскам различают: а) базо-
фильные слизистые железы (не красятся
кислыми красками, хорошо окраши-
ваются гематоксилином) и б) ацидофиль-
ные, «белковые» железы, секрет кото-
рых на препарате имеет зернистый вид,
тогда как слизистое выделение дает
ячеистую структуру, ядра ацидофильных
желез беднее хроматином, чем ядра слизи-
стых, ядрышко в первых видно хорошо;
76
11) ближе к брюшной стороне тела находим овальный попереч-
ный разрез продольного нервного ствола; на разрезе он зернистой
структуры;
12) местами на некоторых срезах могут попасть поперечные нерв-
ные коммиссуры, сое-
диняющие оба про-
дольные ствола;
13) в нижней
(брюшной) части сре-
за^ паренхиме,видны
многочисленные мел-
кие фолликулы семен-
ников— органов, вы-
рабатывающих муж-
ские половые клетки;
14) наконец рас-
сматриваем разрезы
ветвей кишечника,
вкрапленных в парен-
химу и окаймленных
эпителием, клетки ко-
торого выделяют фер-
менты.
Рассмотрев все ука-
занные- детали препа-
рата, сделаем необхо-
димые выводы:
а) отмечаем силь-
ное развитие парен-
химы, в состав кото-
рой, кроме соедини-
тельнотканных эле-
ментов, входят мы-
шечные волокна; па-
ренхима является про-
изводным среднего
зародышевого листка,
или мезодермы; эпи-
телий покровов раз-
вивается из эктодер-
мы, а эпителий кишеч-
ника из энтодермы;
следовательно, плана-
А — Planaria gonocephala. Часть пристеночного разреза, bz —
соединительнотканная клетка; b — соединительная ткань;
Ьт — базальная мембрана; dm — диагональные мышечные
волокна; dm справа — дорзо-вентральные волокна: ed — белко-
вая железа; е — эпителий; f — реснички; 1т — продольные мы-
шечные волокна; р — пигмент; г — рабдиты; rz— клетки,
образующие рабдиты; sd—слизистая железа. По Г о ф м а н у.
В — Dendrocoelum lacteum. Часть поперечного разреза, nz —
питательные (всасывающие) клетки энтодермы; cz — желези-
стые белковые клетки энтодермы; mf—мышечные волокна;
bz — соединительнотканные клетки; pz — пигментные клетки;
dz — желточные клетки. По К. Шнейдеру.
рию мы можем хара-
ктеризовать как трехслойное животное сравнительно с двуслойным
кишечнополостным (гидра);
б) в теле планарии наблюдаем диференциацию тканей — эпите-
лиальной, мышечной, соединительной и др.; самое тело состоит из
органов, слагающихся в более или менее сложные системы, например:
органы пищеварения, размножения и др.
Опыты с регенерацией у планарий
Планарии являются прекрасным объектом для наблюдения реге-
нерации. Живую планарию разрезают в часовом стекле с воском на
дне острым узким скальпелем. Разрезы делаются в различных пло-
скостях:
а) перерезают планарию поперек на две равные части и наблюдают
восстановление соответственно передней или задней половины тела;
б) планарию разрезают поперечно на несколько частей (рис. 62, А);
каждая из них регенерирует даже и в тех случаях, когда берется
кусочек из середины тела;
в) тело планарии разрезают на две-три части косым разрезом
(рис. 62, В);
А В
Рис. 62. Опыты с регенерацией у планарий. Регенерирующие части показаны
белыми. ИзЮ. Полянского и А. Стрелкова.
Наблюдаемые кусочки планарии кладут в чашки Коха и время
от времени просматривают под лупой; воду регулярно сменяют; ре-
генерация длится долго (около месяца); сначала затягивается ранка
на месте разреза; рост регенерирующей ткани заметен с шестого —
десятого дня после начала опыта; из вырезанных кусочков тела об-
разуются соответственно маленькие планарии (рис. 62); следовательно,
у планарии, как и у гидры, налицо явление морфоллаксиса, т. е.
передиференцировка тканей червя.
Если планарию надрезать с боков или в передней части тела и не
дать надрезанным частям соприкасаться (что повело бы к их сраще-
нию), то боковые дольки регенерируют головной конец планарии,
в результате чего получается уродливое животное с несколькими
головами; такая форма регенерации, когда возникает иная часть тела,
чем та, которая была оперирована, называется гетероморфозом
(рис. 63).
78
Опыты с обратным развитием планарий
Для опытов берут Dendrocoelum lacteum и помещают в аквариум
с чистой водой; воду меняют через два-три дня и через нее постоянно
продувают воздух (нагнетательным насосом). Аквариум держат на
свету при комнатной температуре (около 17° С). При таком содержа-
нии планарии хронически голодают. Время от времени зарисовывают
контуры тела вытянувшегося червя или его измеряют. Опыт длится
до полугода и больше. Через шесть месяцев отмечается уменьшение
размеров тела планарии, в конце концов достигающей одной десятой-
----------« ---------------------- -с......., ее уменьШаетСя раз
двенадцатой нормальной величины; объем же
в триста против первоначального (рис. 64).
Наблюдаемые процессы являются далеко
идущим обратным развитием (дедиференци-
ровкой), связанным с длительным голода-
нием планарии. Из внешних процессов отме-
Рис. 64. Редук-
ция Dendrocoe-
lutn lacteum при
голодании. По
Бер нин ге ру.
Рис. 63. Гетероморфозу планарий с обра-
зованием дополнительных голов. По
Ю. Полянскому и А. Стрел-
кову.
чаем распадение глаз и последующее исчезновение пигмента. Такая
редукция глаз происходит на последних стадиях голодания.
Форма кишечника у голодающей планарии не изменяется, но он
пропорционально уменьшается, что связано с разрушением (дегене-
рацией) многих его клеток.
О внутренних процессах редукции голодающей планарии можно
судить, лишь изучая микроскопические препараты (срезы). Отме-
чается сильное растворение клеток паренхимы; нервная система,
повидимому, не изменяется, если не считать пропорционального ее
уменьшения; то же и с мышечными волокнами. Наблюдается извест-
ная последовательность в разрушении отдельных органов, из которых
исчезают, в первую очередь, менее необходимые и могущие заново
регенерировать.
79
Общие замечания
Рассмотренные нами планарии являются свободно живущими,
ползающими животными. В связи с этим и ориентировано тело пла-
нарии.
Она ползает на определенной стороне тела — на брюшной; эта
сторона определяется также и положением ротового отверстия.
Тело планарии, в общем, удлиненное и имеет передний голов-
ной, и задний хвостовой концы тела. Хотя и говорят о го-
ловном конце планарии, тем не менее обособленной головы у нее
нет, но ее передний конец имеет форму, отличную от хвостового;
на нем, несколько отступя кзади, лежат глаза; ближе к переднему
концу (а у многих планарий и почти у самого конца) помещается
ротовое отверстие; глаза лежат на спинной стороне тела; общая
форма тела уплощенная; боковые края тела несколько приострены.
Тело планарии можно разделить на симметричные половины лишь
одной плоскостью сечения, которая проходит по средней линии тела
соответственно продольной оси в дорзо-вентральном направлении.
Две боковые сходные по строению половины тела называются анти-
мерами, а соответствующая форма симметрии — двубоковой, или
билатеральной.
Двубоковая симметрия свойственна свободно по-
движным животным, а лучевая (или радиальная) — сидячим (см.
выше — кишечнополостные).
В зависимости от характера строения кишечника различают пла-
нарий ветвистокишечных (Dendrocoela) и прямокишечных (Rhabdo-
coela) — с одной ветвью кишечника или вовсе без него.
У ветвистокишечных кишечник имеет три главные ветви (Tri-
cia da — преимущественно пресноводные виды) или много ветвей
(Polycl a da — морские виды).
КЛАСС СОСАЛЬЩИКИ (TREMATODA)
ДВУУСТКА ЛАНЦЕТОВИДНАЯ (D1CROCOELIUM LANCEATUM)
Технические замечания. Двуустка ланцетовидная живет в пе-
чени овец и быков. Извлеченных из печени двуусток фиксируют формалином
(одна часть формалина на десять частей воды). Затем их сдавливают между пред-
метными стеклами, перевязав последние крепкой ниткой, и выдерживают не-
сколько часов в 95% спирту, после чего переносят для постоянного хранения
в 75% спирт. Для занятий пользуются тотальными препаратами, окрашенными
квасцовым кармином. Перед окраской двуусток кладут на один час в воду, чтобы
отмыть спирт. Препараты после окраски промывают водой, уплотняют в спиртах
возрастающей крепости (70%, 80%, 95%), обезвоживают в абсолютном спирту,
просветляют в гвоздичном масле и заключают в канадский бальзам. Препараты
изготовляют заблаговременно, чтобы канадский бальзам успел высохнуть.
Для рассматривания двуустки ланцетовидной пользуются слабым увеличе-
нием микроскопа (obj. 3, ok. 2).
Строение двуустки ланцетовидной
Двуустка ланцетовидная (рис. 65) вполне оправдывает свое наиме-
нование, так как плоское тело ее по форме действительно напоминает
80
о
и
Рис. 65. Двуустка ланце-
товидная — Dicrocoelium
lanceatum. По Б р а у н у.
i—кишечник; t—семенники;
d— желточники; и — матка;
о — яичник.
ланцет: передний конец более приострен, задний — более округлый.
На переднем конце лежит ротовая присоска, окружающая ротовое
отверстие; несколько отступя кзади, лежит брюшная присоска, имею-
щая вид кольца с радиально исчерченным толстым ободом.
Сторона тела, на которой лежит последняя присоска, считается
брюшной.
Углубление брюшной присоски оканчивается слепо, благодаря
чему этот орган служит только для прикрепления паразита к телу
хозяина. Прежде считали, что брюшная при-
соска окружает второе ротовое отверстие,
чем и объясняется неправильное по существу
название всех подобных паразитических чер-
вей двуустками.
Тело двуустки, как и у планарии, состоит
из сплошной клеточно-волокнистой массы,
называемой паренхимой, в которой и зало-
жены различные органы.
Полости тела нет, и органы лежат прямо
в паренхиме. Снаружи тело покрыто гладкой
кутикулой.
Однако в паренхиме имеются трубковид-
ные щели с резко очерченными стенками —
это и есть пищеварительный аппарат. От ро-
тового отверстия, лежащего на дне ротовой
присоски, кишечник начинается маленькой
вздутой глоткой; за ней следует короткий
пищевод, который, не доходя до брюшной
присоски, разделяется на две кишечные
ветви (/). Последние слегка извилисты; идут
они параллельно наружным краям тела, не-
сколько отступя от них.
Ветви кишечника местами прикрываются
различными органами и оканчиваются слепо,
не доходя до заднего конца тела. На этом
факте, если планарии не подвергались изуче-
нию, мы знакомимся с примером пищевари-
тельного канала, имеющего единственное
отверстие — рот, служащий одновременно и
для выведения наружу неиспользованных
частей пищи, что является признаком низ-
кой организации.
Все остальные органы, видимые в паренхиме двуустки ланцето-
видной, относятся к половому аппарату. Состоит он из следующих
частей: кзади за брюшной присоской лежат одно за другим два круп-
ных тела с округлыми очертаниями неправильной формы — это се-
менники (testes), граничащие с внутренними краями- кишечных
ветвей.
Семенники (t) сообщаются тонкими невидимыми на препарате
семевыносящпми каналами (vasa dcfcrentia) с циррусом — совоку-
пительным органом. Последний лежит между брюшной присоской и
6 Практикум по зоологии — 385
81
местом ветвления кишечника. Позади второго семенника, сбоку от
средней линии тела, виден небольшой овальный яичник (ovarium — о).
Все пространство от семенников, яичника и до заднего конца тела за-
нято толстыми изогнутыми и неправильно переплетающимися друг
с другом трубчатыми петлями матки (uterus — и). Матка набита яй-
цами.
По мере созревания яйца становятся черными, тогда как более
молодые яйца одеты прозрачной оболочкой; заключающие их в себе
изгибы матки окрашены светло. Выводная часть матки в виде узкой
трубки идет по границе между семенниками, окружает задний край
брюшной присоски и открывается наружу отверстием у переднего
края той же присоски, рядом с удлиненным мускулистым мешком со-
вокупительного органа.
К половому аппарату относятся также желточники (</), которые
лежат в виде группы мелких долек в средней части двуустки (считая
от переднего конца к заднему). Совокупность всех долек одной сто-
роны тела и образует соответствующий желточник, расположенный
между краем тела и кишечной ветвью. Желточники вырабатывают
желточные клетки, которые вместе с яйцевой клеткой одеваются плот-
ной оболочкой и образуют яйцо двуустки. Последнее поэтому отно-
сится к категории сложных яиц. Желточные клетки в них служат
для питания зародыша, развивающегося из яйца.
Яйца двуусток, как и других паразитических червей, имеют опре-
деленную для данного вида форму и размеры. С желчью они выходят
из печени и смешиваются с кишечным содержимым. В конце концов
они с испражнениями выбрасываются вон.
Обнаружение в испражнениях при микроскопическом исследова-
нии яиц двуустки позволяет установить присутствие последних у дан-
ного животного.
Двуустка ланцетовидная является гермафродитом, в котором име-
ются две полные системы и мужских и. женских органов размножения.
На рассмотренном нами препарате обыкновенно не различимы сле-
дующие части полового аппарата: так называемая скорлуповая железа,
протоки желточников, идущие из этих последних к скорлуповой
железе, и влагалище, называемое Лауреровым каналом.
Равным образом не видны выделительная и нервная системы.
Органы дыхания, кровообращения и органы чувств у двуустки лан-
цетовидной отсутствуют.
ДВУУСТКА ПЕЧЕНОЧНАЯ (FASCIOLA HEPATICA)
Технические замечания. Печеночная двуустка (в большинстве
старых учебников именуемая Distomum hepaticum) живет в желчных протоках
печени коровы. Под влиянием паразитирования этого червя стенки желчных про-
токов утолщаются, в них отлагается известь, и при резании ножом они хрустят.
Добытых из печени коровы двуусток обмывают водой и фиксируют 70% спиртом
или 10% водным раствором формалина. Часть материала запасают впрок
без дополнительной обработки для рассматривания в необработанном со-
стоянии.
Для изготовления окрашенных препаратов двуустку сжимают между двумя
предметными стеклами, перевязав их нитками, и на 1—2 минуты опускают в ки-
пяток; затем переносят в 95% спирт, снимают одно стекло и выдерживают сплю-
82
щенную двуустку часов шесть в спирте той же крепости. Через 70% спирт пере*
носят препарат в воду, затем окрашивают квасцовым кармином и заключают
препарат в канадский бальзам, как это было описано выше. На занятиях дают
готовые окрашенные препараты для рассма-
тривания под лупой и при слабом увеличе-
нии под микроскопом. Смотря по обстоятель-
ствам, можно дать и неокрашенных и несплю-
щенных двуусток в часовых стеклах в 70%
спирту для наблюдения под лупой при падаю-
щем свете.
Строение печеночной двуустки
Плоское тело печеночной двуустки
отличается большими размерами (до 3 см
длины) и своеобразной формой: широкое
листовидное тело несет спереди конусо-
видную часть, оканчивающуюся ротовой
присоской (рис. 66, т). Брюшная при-
соска лежит кзади от основания перед-
ней конусовидной части. Кишечник, так
же как и у двуустки ланцетовидной,
делится на два боковых ствола (z), иду-
щих параллельно друг другу по бокам
средней линии тела. Но эти стволы у
двуустки печеночной, в свою очередь,
сильно ветвятся, давая с наружной сто-
роны боковые ветви, концевые разве-
твления которых доходят до самых краев
тела, где и оканчиваются слепо. Такое
отличие в строении кишечника двуустки
печеночной является второстепенным;
в общем же пищеварительная система
у Fasciola и Dicrocoelium одного типа,
ибо имеет лишь единственное отверстие,
сообщающее кишечник с внешней средой.
Еще показательнее препараты инъек-
ции кишечника. Приготовляют 1% вод-
ный раствор берлинской лазури. Оття-
гивают на спиртовой или газовой го-
релке капилляр на конце тонкой сте-
клянной трубочки. В последнюю наса-
сывают раствор краски. Кончик капил-
ляра вводят в ротовое отверстие дву-
устки и осторожно вдувают краску в ки-
шечник. В удачном случае окрашенная
жидкость заполняет все разветвления
пищеварительной трубки. После инъек-
Рис. 66. Печеночная двуустка—
Fasciola hepatica.
По Стайльсу.
т — ротовой присосок; р — глотка;
I — кишечник; v — половое отвер-
стие; и — матка; о — яичник; vg —
желточник; $ — тельце Мелиса; t —
семенник.
ции двуустку опускают в 95% спирт с несколькими каплями соляной
кислоты. Часа через два двуустку слегка сдавливают между предмет-
ными стеклами и опять кладут в 95% спирт. На другой день обезвожи-
вают в 100% спирту и через ксилол заключают в канадский бальзам.
83
На обычных препаратах' (рис. 66) кишечник печеночной двуустки
закрыт различными частями половых органов. Во всех деталях ве-
твления кишечник может быть рассмотрен на живой, сдавленной
между стеклами печеночной двуустке. В таком случае просвечивает
заполняющее его содержимое буроватого цвета (изменившаяся, всо-
санная двуусткой кровь хозяина).
При рассматривании неокрашенной двуустки простым глазом можно
различить более светлое срединное поле и темные края и такую же
заднюю лопасть тела. Краевая темная оторочка занята громадным
Рис. 67. Выделительные органы сосальщиков.
А — схема строения протонефридия. Замыкающая клетка (а)
со жгутом (6); с — канал. В — схема двуустки с выделитель-
ной системой, а — отверстие; b — каналы; о — замыкающие
клетки. По Розингеру и Фурману.
количеством узелков
(фолликулов) желточ-
ников (vg), развитых
у двуустки печеноч-
ной весьма сильно.
По внутренней гра-
нице желточника идет
желточный канал, ви-
димый на окрашенном
препарате. От кана-
лов с обеих сторон
тела отходит по попе-
речному протоку, ко-
торые направляются
друг к другу и сли-
ваются вместе.
Такой поперечный
ствол желточных про-
токов отграничивает
переднюю часть тела
с частями женских
половых органов, а
именно: а) ветвистым
яичником (о), лежа-
щим в левой по отно-
шению к наблюда-
телю части тела, б) ша-
ровидным тельцем Ме-
лиса (s — прежде на-
зывалось скорлуповой железой) и в) сравнительно короткой петле-
образно изогнутой маткой (и), открывающейся наружу рядом с цир-
русом; последний помещается между брюшной присоской и углом раз-
ветвления кишечника на два главных ствола.
Кзади от поперечной ветви желточных протоков все более светло-
срединное поле двуустки занято двумя сильно ветвистыми семенни-
ками (/). Иногда на окрашенном препарате бывают видны и длинные
семепроводы, которые двумя тонкими тяжами обходят по бокам
брюшную присоску и открываются в мешок копулятивного органа.
У двуустки печеночной хорошо развита выделительная система,
но рассмотреть ее можно лишь на инъецированном препарате. Описан-
ным выше способом раствор берлинской лазури вводится или в наруж-
84
ное отверстие главного выделительного ствола (лежит на заднем конце
тела), или (что легче) в самый выделительный ствол (капилляр вкалы-
вают в паренхиму двуустки по средней линии тела). При удачной инъек-
ции выделяется главный ствол и сеть мелких боковых ветвей, дрени-
рующих всю толщу паренхимы двуустки. Ветви выделительных орга-
нов на внутренних концах замкнуты клетками со жгутом, свисающим
в полости канала (рис. 67, А, Ь). Такой тип выделительных органов
является примитивным, их называют протонефридиями.
Нервная система на обыкновенных препаратах не видна. Развита
она слабо. Начинается она по бокам глотки, над которой лежит «мозг»
в виде пары нервных узелков, соединяющихся друг с другом нервным
стволиком (коммиссурои). От «мозга» отходят три пары продольных
нервных стволов, также соединяющихся местами поперечными нерв-
ными же тяжами. Органов чувств у двуустки печеночной нет.
Особых органов движения также нет. Двуустка обладает весьма
несовершенной подвижностью, обусловленной мышечными волокнами
(гладкими) в паренхиме ее тела.
Сравнение организации печеночной и ланцетовидной двуусток
Dicrocoelium lanceatum
Fasciola hepatica
Тело..............
Присосок..........
Кишечник..........
Ветви кишечника . .
Органы размножения.
Семенники ........
Плоское
паренхиматозное
ланцетовидное I листовидное
Две I Две
В виде трубок, имеющих единственное отверстие наружу
Вторично не ветвятся | Вторично ветвятся
Гермафродитные
Яичник ...........
Желточники........
Два — в виде тел непра-
вильной лопастной формы,
лежащих в передней поло-
вине тела
Один — в виде округлого
органа
По краям средней части
тела
Два — в виде сильно ве-
твистых трубчатых тел, лежа-
щих в задней половине тела
Один — в виде ветвистого
трубчатого органа
По всей длине боков и
задней части тела, начиная
от уровня брюшной при-
Органы выделения .
Органы чувств . . . .
Вторичная полость
тела...............
Кровеносная система.
Протонефридиального типа
Отсутствуют
Общие замечания
В вышеприведенной таблице разрядкой выделены наиболее важ-
ные признаки сравниваемых сосальщиков, на основании которых
их отн<сят к известной систематической группе; остальные же при-
знаки более мелкие, дающие, в свою очередь, возможность помещать
их в отдельные роды.
85
а) У сосальщиков имеются: 1) плоское паренхиматозное тело»
лишенное вторичной полости тела, 2) органы выделения прото-
нефридиального типа, в) пищеварительный канал, лишенный задне-
проходного отверстия, и 4) отсутствует кровеносная система — все
это дает основание причислять их к подтипу плоских червей.
б) Лишенный ресничек кутикулярный покров тела, наличие ор-
ганов прикрепления в виде присосок, присутствие пищеварительного
канала и паразитический образ жизни характеризуют их как плоских
червей, принадлежащих к классу сосальщиков — Trematoda.
в) Факт паразитирования печеночной и ланцетовидной двуусток
во внутренних органах (в частности, в печени) своих хозяев опреде-
ляет принадлежность их к отряду эндопаразитных' сосальщиков
(Digena).
г) Дальнейшее систематическое положение рассматриваемых нами
червей выясняется более мелкими признаками, которые, однако,
являются признаками семейства двуусток (Distomidae), а именно:
1) положение ротовой присоски на переднем конце тела, 2) сближен-
ность с ней брюшной присоски, 3) гермафродитный половой аппарат,
4)впадение половых протоков в общую половую клоаку, которая, в свою
очередь, открывается наружу чаще всего впереди брюшной присоски.
д) Различие формы тела, ветвление кишечника, форма и положе-
ние яичника, семенников, матки и желточников — все эти признаки
дают возможность характеризовать роды сосальщиков Fasciola и
Dicrocoelium. 1
Тело двуусток построено по двубоковой симметрии, так как может
быть разделено плоскостью сечения, проходящей по длине двуустки,
на две сходные по строению половины. Симметричные половины тела
называются антимерами. Однако выдержанность двубоковой, или
билатеральной, симметрии двуусток нарушается некоторыми второ-
степенными особенностями расположения различных внутренних ор-
ганов. Так, типически симметричны присоски, ветви кишечника и
желточники обеих двуусток и семенники Fasciola hepatica. Асимме-
трично лежат яичники Fasciola и Dicrocoelium, семенники Dicrocoelium
и переплетающиеся трубки матки.
Из анатомических признаков двуусток отрицательного характера
особо должно оговорить отсутствие кровеносной системы. Отсутствие
у них органов кровообращения является особенностью первичного
значения. Этих органов у них никогда не было, нет и теперь.
Отсутствие кровеносной системы коррелятивно 2 можно связать
с отсутствием целома, т. е. вторичной полости тела (см. ниже).
Паразитическим образом жизни сосальщиков объясняются следую-
щие черты их строения:
а) узкая форма тела Dicrocoelium, приноровленных к обитанию
в тонких желчных ходах печени;
1 В новейшей систематике сосальщиков это семейство разбито на несколько
новых семейств. В частности, Fasciola относят к сем. Fasciolidae, a Dicrocoelium —
к Dicrocoeliidae.
2 Корреляцией органов называют такое соотношение их, когда изменение в од-
ном органе влечет за собой изменения в другом органе. Возникновение вторичной
полости тела коррелятивно обусловливает развитие кровеносной системы.,
86
б) присутствие пары присосок, служащих для прикрепления дву-
устки к телу хозяина в месте ее жизни;
в) отсутствие органов чувств, излишних при скрытой жизни
в условиях постоянства окружающей среды;
г) гермафродитный половой аппарат, отличающийся необычайной
производительностью. Для паразитов (особенно с таким сложным
жизненным циклом развития, как у двуусток — см. ниже) совершенно
необходимо выбрасывать бесчисленное множество яиц, так как лишь
малая часть их случайно попадает впоследствии в надлежащие усло-
вия развития; максимальная плодовитость гарантирует существо-
вание данного вида паразита на земле. Более подробную оценку ор-
ганизации паразитов отложим до следующего отдела.
Сравнение организации планарии и двуусток
Черты строения Ресничные черви (планария) Печеночная и ланцетовид- ная двуустки
Тело Плоское
Удлиненное в виде ши- рокой короткой ленты Листовидное или ланцетовидное
Мерцательные реснички по- кровов Есть Отсутствуют
Кутикула Отсутствует Есть
Присоски Отсутствуют Есть
Ротовое отверстие На брюшной стороне тела На переднем конце тела, на дне присоски
Глотка Выворачивается через ротовое отверстие Не выворачивается наружу
Пищеварительный канал .... В виде различно ветвящихся трубок, единственным (ротовым) отверстием открывающихся наружу
Органы выделения Протонефридии
Вторичная полость тела, кро- веносная и дыхательная си- стемы Отсутствуют
Центральная нервная система. Церебральные узлы с продольными анасто- мозирующими стволами
Органы размножения Гермафродитные из комплектов мужских и женских органов размножения
Глаза Есть Нет
87
По плоской форме тела, наличию пищеварительного аппарата,
гермафродитизму, обладанию протенефридиальными органами выде-
ления, паренхиматозному телу и некоторым другим признакам пла-
нарии и двуустки сходны друг с другом, что дает основание относить
их к подтипу плоских червей.
Однако приспособление сосальщиков существенно отразилось на
деталях их организации. Сосальщики филогенетически связаны с рес-
ничными червями, поэтому рассмотрение паразитических адаптаций
сосальщиков произведем, исходя из основных черт организации рес-
ничных червей.
Паразитический образ жизни сосальщиков во внутренних органах
хозяина делает ненужным мерцательный покров тела, который,
в противоположность покровам планарий, приобретает у двуусток
поверхностную кутикулу; последняя, вследствие своей прочности,
защищает внутренние части сосальщика от механического да-
вления.
Другим приобретением адаптивного характера являются при-
соски, служащие органами прикрепления к телу хозяина. С образо-
ванием ротовой присоски связано перемещение ротового отверстия
с брюшной поверхности тела ( у планарий) на передний его конец
(у двуустки).
Совершенно естественна редукция глаз, так как половозрелые
двуустки живут в полной темноте.
По степени развития гермафродитный половой аппарат доминирует
над другими органами; это ведет к огромной плодовитости, характер-
ной вообще для паразитических червей.
Таким образом переход ресничных червей к паразитическому
образу жизни в далеком прошлом привел к возникновению сосаль-
щиков; этот процесс перехода был связан: 1)с потерей некоторых черт
организации, присущих планариям, и 2)«с приобретением новых
особенностей строения, соответствующих новым своеобразным усло-
виям обитания сосальщиков во внутренних органах тела их хозяев.
НАБЛЮДЕНИЯ НАД ЖИЗНЕННЫМ ЦИКЛОМ СОСАЛЬЩИКОВ
Сосальщики имеют сложный жизненный цикл, связанный с тем,
что различные его этапы они проводят во внешней среде и в организме
хозяев, причем хозяев обычно бывает два; выше мы имели случай
познакомиться с паразитированием взрослых двуусток — печеноч-
ной и ланцетовидной —в печени крупного и мелкого рогатого скота.
Коровы и овцы являются главными, или окончательными, хозяевами
указанных сосальщиков. В печени этих животных протекает половое
размножение двуусток, которые после оплодотворения откладывают
множество яиц, по пищеварительному каналу выводящихся наружу
и с пометом хозяев попадающих на землю или в воду.
Дальнейшая судьба яиц зависит, прежде всего, от того, куда они
попадут.
Яйца печеночной двуустки могут проделывать дальнейшее разви-
тие лишь, если попадут в воду. Из яиц ланцетовидной двуустки
88
личинки выходят лишь, если последние будут проглочены сухопут*
ным моллюском Fruticicola rubiginosa, который является промежуточ-
ным хозяином для указанного паразита.
Проследить в порядке специального лабораторного практикума
полный жизненный цикл сосальщиков—дело не легкое, но рассмотре-
ние различных этапов их цикла может быть вполне осуществлено.
Печеночная двуустка
Технические замечания. На бойне необходимо достать живых
печеночных двуусток. Для получения яиц, живых двуусток кладут в физиологи-
ческий раствор в чашку Коха и ставят, закрыв чашку, в термостат при 38е С.
Через сутки выбирают пипеткой со дна отложенные яйца, хорошо промывают
водой и ставят в чашке Коха с водой в термостат при 22—24° С. Эмбриональное
развитие печеночной двуустки протекает медленно, об его ходе см.: П. П. Ив а-
нов. Общая и сравнительная эмбриология, 1937.
Начиная с двадцатого дня под оболочкой яйца можно видеть формирующегося
зародыша двуустки, который выходит из яйца в стадии личинки, называемой
мирацидием.
Яйца время от времени микроскопируют в капле воды под] покровным сте-
клом с ножками.
Влияние внешних факторов на развитие яиц. Добытые яйца раз-
деляют на несколько партий. Одну из них ставят в термостат при
25—30° С, другую оставляют при комнатной температуре, третью
ставят на ледник. Условия помещения (вода, чашки) должны быть
одинаковы.
Наблюдая пробы яиц из разных партий через 15—20 дней, уста-
навливаем, что при температуре около 0° и несколько выше в яйцах
эмбрионального развития не происходит; при комнатной температуре
(13—15° С) развитие идет очень медленно, а при 25—30° С яйца
развиваются скорее всего (optimum развития). Критерием развития
служит сравнение внутреннего содержимого развивающегося яйца
с яйцом, только что отложенным.
Вскоре после откладки яйца печеночной двуустки желточные
клетки теряют в нем свою обособленность и сливаются в общую желточ-
ную массу, в которой видны собственно яйцевая клетка (рис. 68, 7)
и ядра желточных клеток; в дальнейшем яйцеклетка (уже ранее опло-
дотворенная) проделывает полное, но неравномерное дробление; за-
родыш принимает форму компактной группы тесно лежащих клеток;
из нее постепенно формируется мирацидий, лежащий в эмбриональной
оболочке из плоских клеток; все развитие происходит за счет потре-
бления желточных клеток. Остаток желточной массы лежит сбоку
мирацидия. Последний длиннее яйца, поэтому лежит в нем изогну-
тым. Полное формирование мирацидия заканчивается при 22—24° С
на двадцатый-двадцать второй день. Созревший мирацидий выходит
из яйца, что происходит только на свету. Выход мирацидия при не-
которой усидчивости можно наблюдать под микроскопом в капле
воды без покровного стекла при среднем увеличении.
Мирацидий двигается в яйце. Затем крышечка яйца внезапно от-
скакивает, точно крышка карманных закрытых часов. Мирацидий
прорывает эмбриональную оболочку и активными движениями выби-
89
Рис. 68. Цикл развития печеночной двуустки. По С уз у к и. Из Павловского.
1—4 — развитие мирацидия в яйце; 5—мирацидий; б, 7 и 8— превращение в спороцисту;
9 — спороциста с редией; 10—13—развитие в редиях церкарий; 14—15 — церкарии; —
инцистированная церкария; 17—двуустка, вышедшая из церкарии.
рается через узкое отверстие открывшегося яйца; в яйце видна остаю-
щаяся эмбриональная оболочка (рис. 68). Через 1—минуты мира-
цидий покидает яйцо и свободно плавает в капле воды.
90
Строение мирацидия. Свежий препарат с живыми мирацидиями
в воде покрывают покровным стеклом с ножками. Придавливая по-
следние, как в работе с инфузориями, можно остановить мирацидий
и рассмотреть его при сильном увеличении.
Мирацидий имеет приблизительно клиновидное тело; широкий
конец клина является передним; на его середине видно небольшое
возвышение неправильно конусовидной формы; этот конус втяжной
(есть специальные ретракторы); все тело мирацидия равномерно по-
крыто ресничками, благодаря работе которых мирацидий свободно
плавает в воде.
На конусовидном возвышении открываются крупные одноклеточ-
ные железы, секрет которых обладает протеолитическим свойством,
растворяет живые ткани покровов промежуточного хозяина, когда
мирацидий начинает внедряться в тело моллюска (см. ниже). Позади
от конусовидного возвышения лежат расположенные крест-на-крест
пигментированные клетки глазков; под ними лежит нервный узел;
еще далее назад, ближе к бокам, лежит с каждой стороны тела по
одной клиновидной клетке с мерцательным пламенем. Это — замы-
кающие клетки протонефридиев, из которых каждая продолжается
в тонкий канал, открывающийся на боку тела ближе к его заднему
концу.
Остальное лространство первичной полости тела мирацидия за-
нято шарами, образованными из первичных половых клеток.
Рассмотрев детали строения мирацидия, охарактеризуем особен-
ности его организации.
1) Замечательно наличие ресничного покрова, который полностью
отсутствует у всех взрослых сосальщиков; по этому личиночному
признаку сосальщики сближаются с ресничными червями.
2) Ресничный покров и, следовательно, способность к плаванию
связаны со свободно подвижной жизнью мирацидия в воде; понятно
также и присутствие примитивно устроенного органа зрения в виде
пигментного глазка.
3) Плавая в воде, мирацидий отыскивает промежуточного хозяина,
в тело которого проникает благодаря работе стилета и протеолити-
ческих желез; эти оба образования являются специальными органами
мирацидия, обеспечивающими ему переход к следующей стадии жизнен-
ного цикла, которая может протекать лишь в организме промежуточного
хозяина (в нашем случае: в малом прудовике — Limnaeus minutus).
Мирацидий очень недолговечен, и в прудовой воде он живет, видимо,
не более 48 часов. Мирацидий погибает, если за это время он не про-
никает в названного моллюска.
Стадии жизненного цикла в промежуточном хозяине. Соби-
рают живых брюхоногих моллюсков (Limnaeus, Planorbis, Palu-
dina и др.) в различных водоемах. Моллюсков рассаживают
в широкие плоскодонные цилиндрики с водой и оставляют сто-
ять в комнате. Если собранные моллюски заражены сосальщи-
ками, то последние выходят из тела моллюска на стадии, на-
зываемой церкарией. Церкарии очень мелки, но все же видны
невооруженным глазом в виде подвижных белых точек. Их вы-
бирают тонкой пипеткой и переносят в каплю воды. Микроскопи-
91
нии строения тела церкарии
руют сначала без покровного стекла при среднем увеличении, затем
покрывают покровным стеклом на восковых ножках для рассмотрения
при сильном увеличении.
Строение и особенности церкарии. Церкария
имеет овоидное или листовидное плоское тело и длинный хвост. При
помощи последнего церкария оживленно плавает в воде; кроме того
ее тело может изгибаться в различных направлениях. При рассмотре-
дим черты, свойственные взрослым
сосальщикам: 1) кутикулярный по-
кров тела без ресничек; 2) перед-
няя (ротовая) присоска; 3) брюш-
ная (слепая) присоска; 4) идущая
от ротовой присоски глотка, пере-
ходящая в 5) двуветвистый кишеч-
ник, и 6) зачаток полового аппа-
рата. Кроме того на уровне глотки
виден мозговой ганглий с отходя-
щими от него двумя продольными
стволами; перед местом соединения
тела сосальщика с хвостом виден
треугольный выделительный пу-
зырь, продолжающийся в канал,
который на своем конце переходит
в две боковые трубки, оканчиваю-
щиеся каждая самостоятельным
отверстием на боку хвоста близ
его основания. Описываемые обра-
зования являются выводной частью
протонефридиальной выделитель-
ной системы.
При наблюдении добытых цер-
карий следует иметь в виду, что
они будут относиться к различным
сосальщикам; могут попасться и
церкарии Fasciola hepatica; описы-
ваемые наблюдения имеют целью
дать представление о стадии цер-
карии вообще.
Если оставить церкарий в воде в смикроводоеме», например,
на предметном стекле с углублением, то иногда можно наблюдать их
инцистирование; инцистированная церкария имеет шаровидную форму;
при инцистировании она отбрасывает хвост, тело ее округляется
п одевается защитной капсулой из отвердевшего секрета кожных
желез церкарии. Инцистированная церкария называется адолеска-
рией. Когда последняя будет проглочена крупным рогатым скотом
с водой или с поедаемой травой на пастбище у самого берега водоема,
оболочка цисты растворяется в желудке, вышедшая из нее молодая
двуустка попадает в печень, где и развивается в половозрелую форму.
Остальные стадии жизненного цикла со-
саль щи ков (спороцисты и р е д и п). Разбиваю! раковину
92
Рис. 69. Церкария. По А. Сквор-
цову.
В — ротовая присоска; а — брюшная при-
соска; Ph—глотка; z — кишечник; v — вы-
делительный пузырь; dt— желточник; go—
зачаток полового аппарата.
моллюска и вынимают печень (вскрытие—см. раздел о моллюсках,
ниже). Кусочки печени мелко разрывают иглами на предметном
стекле и выбирают под лупой беловатые или желтоватые оформленные
образования,- отличающиеся' от обрывков ткани печени. Найденный
материал рассматривают под покровным стеклом с ножками в свежем
виде или приготовляют тотальный препарат методом наклейки све-
жего объекта на предметное стекло с последующей окраской гема-
токсилином или борным кармином.
Просматривая объекты, полученные от разных моллюсков (а в удач-
ных случаях и на одном препарате), отыскивают следующие стадии
жизненного цикла сосальщиков:
1) мешковидные тела неправильной форМы; под их покровами
видны шаровидные или иной формы скопления зародышевых (поло-
вых) клеток; эти образования называются спороцистами; в спороцисту
превращается мирацидий, внедрившийся в тело моллюска; превраще-
ние носит регрессивный характер, так как мирацидий теряет рес-
ничный покров, глазок и другие органы; благодаря этому спороциста
имеет очень простое строение; 2) спороцисты вариируют по вели-
чине; кроме того, в них можно видеть развитие следующего поколения
жизненного цикла сосальщика; шары зародышевых клеток, благо-
даря размножению последних, вытягиваются в длину и диференци-
руются в образования, называемые редиями; редии выходят из спо-
роцист через особое отверстие, называемое спородуктом; редии оста-
ются, как и спороцисты, в междолечной ткани печени моллюска;
3) сформированная редия имеет удлиненное тело червеобразной формы,
часто с боковыми выростами; на переднем конце редии видна при-
соска, ведущая в зачаточный кишечник в виде короткой неветвящейся
трубки; имеется также отверстие для выхода (отрождения) следую-
щего поколения, которое развивается в полости тела редии из шаров
зачаточных клеток (по своему происхождению они связаны с аналогич-
ными образованиями спороцисты и мирацидия); 4) в редиях могут
развиваться: а) новое поколение (второе) редий же или б) поколение
церкарии, которые нам уже знакомы; в первом случае редии второго
поколения дадут в себе поколение церкарий. Церкарии (или редии
второго поколения) выходят из редии через особое («родильное»)
отверстие и покидают тело моллюска; редии второго поколения оста-
ются в междолечной ткани печени.
Общая характеристика жизненного цикла печеночной двуустки.
Печеночная двуустка в общем проделывает сложный жизненный цикл,
последовательные этапы которого сводятся к следующему:
1) попадание яйца двуустки из организма окончательного хозяина
во внешнюю среду (в воду) и вылупление из яйца свободно живущей
личинки — мирацидия;
2) короткий период свободной жизни мирацидия и активное вне-
дрение его в промежуточного хозяина (для F. hepatica — прудовик
малый — Limnaeus minutus);
3) период эндопаразитического существования в промежуточном
хозяине: а) превращение мирацидия в спороцисту; б) размножение
спороцисты с образованием поколения редий; в) размножение редий
с образованием второго поколения редий или поколения церкарий;
93
в первом случае еще размножение второго поколения редий с образо-
ванием церкарий;
4) период покидания церкариями промежуточного хозяина, ко-
роткое активное существование во внешней среде с последующим
инцистированием (стадия адолескарии);
5) пассивное попадание в окончательного хозяина (крупный ро-
гатый скот; эндопаразитическое существование); вскрытие цист, ми-
грация молодых двуусток в печень, достижение половозрелости,
размножение откладкой яиц, которые выбрасываются из тела хозяина
во внешнюю среду и дают начало нового жизненного цикла сосаль-
щика.
Половозрелые двуустки (гермафродиты) размножаются с участием
и мужских и женских половых клеток (обоеполое размножение);
редии и церкарии образуются также в процессе полового размноже-
ния (цитогония), но лишь за счет зачаточных клеток одного пола,
именно женского; такое однополое размножение называется
партеногенезом или девственным размножением.
Полный жизненный цикл печеночной двуустки, следовательно,
слагается из ряда чередующихся поколений, одни из которых (споро-
циста, (эедия) размножаются партеногенетически, тогда как полово-
зрелые двуустки размножаются с участием половых клеток обоих по-
лов. Такое чередование различных форм полового размножения назы-
вается гетерогонией. Гетерогония печеночной двуустки сопряжена
с переменой хозяев и с существованием различных стадий жизненного
цикла сосальщика во внешней среде.
Гетерогонию следует отличать от метагенеза (см. выше, стр. 65).
Метагенез характеризуется чередованием поколений, размножаю-
щихся: одно — половым путем (цитогония с оплодотворением), дру-
гое — вегетативным путем (почкование — бесполое размножение). При
гетерогонии вегетативно размножающегося поколения нет, и все
поколения организма размножаются половым путем, но одни только
за счет неоплодотворенных женских половых клеток (партеногенез),
другие же обычным путем оплодотворения.1
Ланцетовидная двуустка
Технические замечания. Необходимо иметь живых Dicrocoe-
lium lanceatum, которых добывают на бойне или в секционных ветеринарных ин-
ститутов из печени овец или крупного рогатого скота.
Живых двуусток кладут в небольшую чашку Коха с физиологическим раство-
ром, которую ставят затем в термостат на сутки при температуре 38—39° С. Дву-
1 Следует отметить, что до настоящего времени, в частности в гельминтологи-
ческой литературе, встречается изредка неправильная и уже оставленная в биоло-
гии трактовка партеногенеза как бесполого размножения; это ведет к неправиль-
ному применению понятия метагенез, которым подменяется понятие гетерогония.
Таких взглядов придерживаются К. Скрябин и Р. Шульц в ряде своих
пособий и книг. Если считать вместе с названными авторами партеногенез за бес-
полое размножение, то пришлось бы прийти к парадоксальному заключению,
что, например, самка пчелы (которая копулирует раз в жизни) размножается и
половым путем и бесполо: в первом случае она откладывает оплодотворенные
яйца, из которых развиваются рабочие пчелы и матка, во втором случае она откла-
дывает яйца при закрытом семеприемнике. благодаря чему яйцо остается неопло-
лотворенным; из таких яиц партеногенетически развиваются самцы (трутни).
94
усток вынимают, а со дна чашки пипеткой выбирают отложенные ими яйца: или
все сливают, хорошо перемешав, в центрифужную пробирку, на дно которой яйца
и оседают.
Dicrocoelium lanceatum откладывает яйца с уже вполне сформировавшейся
личинкой, как и у других сосальщиков (мирацидием). Сколько бы времени яйца
Dicrocoelium не лежали в воде, из них мирацидий наружу не выйдет.
Яйцо вскрывается тогда, когда будет проглочено и попадет в зоб наземных
брюхоногих моллюсков — Amalia marginata, Agriolimax agrestis (слизень), или
в раковинного брюхоногого моллюска —Fruticicola rubiginosa и др. Поэтому
для постановки опытов и наблюдений необходимо запасти живых моллюсков.
Слизней собирают на лугах и огородах во вторую половину лета и ближе
к осени, например: с листьев капусты, из-под камней или досок, лежащих близ
гряд. Слизни выходят для питания к вечеру. Иногда их бывает очень много,
и тогда они являются уже вредителями сельскохозяйственных культур (огороды,
поля) существенного значения. Других моллюсков ищут на лугах. Собранных мол-
люсков держат в садках следующего устройства. На дно объемистой простокваш-
ницы кладут диск проволочной сетки, края которой загибают книзу так, чтобы
сетка лежала наподобие второго дна над дном простоквашницы на расстоянии
2—3 см. На дно простоквашницы наливают воду, а на сетку кладут огородную
землю, куда и пускают одного — трех моллюсков. Садок сверху закрывают проволоч-
ной сеткой. Испаряющаяся вода поддерживает влажность земли. При кратко-
временных опытах мелких моллюсков кладут в небольшие чашки Петри, на дно
которых наносят несколько капель воды, а для более крупных берут чашки Коха
с несколько большим количеством воды.
Микроскопирование яиц Dicrocoelium lanceatum. Яйца с капель-
кой воды наносят пипеткой на предметное стекло и закрывают покров-
ным. Микроскопируют в свежем виде (рис. 70)’.
Яйца неправильно овальной формы, с ясно выраженной крышечкой
на одном полюсе; другой полюс несколько приострен. Овал яйца
слегка несимметричен в отношении продольной оси. Размеры яиц
колеблются в пределах 0,038—0,045 х 0,020—0,033 мм. Яйцо одето
толстой прочной оболочкой, у зрелых яиц — темнокоричневого
цвета.
Вследствие темной окраски оболочки у зрелых яиц трудно рас-
смотреть детали его содержимого; более ясно просвечивают два зер-
нистых шара (см. ниже).
Выход из яйца мирацидия и его строение. Зрелые яйца наносят
на кусочки моркови, которые скармливают слизню или Fruticicola.
Для упрощения моллюсков можно кормить даже кусками самих лан-
цетовидных двуусток, в которых, наряду с незрелыми яйцами, есть
и созревшие. Моллюсков вскрывают через у2—1 час после того, как
они съели яйца двуустки (вскрытие см. в главе о брюхоногих моллю-
сках). Берут содержимое зоба или начальной части кишечника и
микроскопируют его под покровным стеклом. На препарате могут быть
найдены уже пустые яйца с откинутой крышечкой, свободные мира-
цидии и мирацидии в различные моменты выхождения из яиц
(рис. 70).
В свободном состоянии мирацидий имеет округлую форму; его пе-
редняя часть обособлена в виде головной шишки. В головном отделе
в особом влагалище лежит острый стерженек — стилет. Позади
стилета видно мешковидное образование, вероятно являющееся ру-
диментарным кишечником (Л е й к а р т). Стилет с головной шишкой
может втягиваться внутрь мирацидия действием вытяжной мышцы —
ретрактора. В задней части мирацидия видны два хорошо заметных
95
зернистых шара и два слабее выраженных и примыкающих друг
к другу у самого заднего конца мирацидия аналогичных образова-
ния. Глазка нет. Передние две трети тела мирацидия покрыты реснич-
ками; благодаря их работе мирацидий может плавать в жидком со-
держимом кишечника.
Дальнейшие этапы жизненного цикла. Мирацидий, действуя сти-
летом, вбуравливается в стенки фолликулов печени моллюска, про-
ходит в прослойки соединительной ткани, оседает здесь и превра-
щается в следующую фазу жизненного цикла, называемую спороци-
стой.
Наблюдение их и следующих стадий, кроме изолирования расщи-
пыванием и наблюдения в свежем виде, как с печеночной двуусткой,
удобнее производить на срезах через печень зараженного моллюска,
взятую в разные сроки от 1 часа и до 60 дней после скармливания
моллюску яиц сосальщика. В виду малых размеров моллюсков их
ав с
Рис. 70. Dicrocoelium lanceatum.
А — яйцо; В — выхождение мирацидия; С — мирацидий. По А. Скворцову.
можно фиксировать целиком жидкостью Ценкера 1 в течение двух
суток; после основательной промывки в проточной воде, моллюсков
последовательно переводят в 50, 60 и 70% спирт, к последнему при-
бавляют иодной тинктуры и держат там, пока жидкость не перестанет
обесцвечиваться. Далее в обычном порядке объект заливают в пара-
фин или в целлоидин. Тонкие срезы (5—7—10 р) окрашивают гемато-
ксилином Делафильда и эозином или железным гематоксилином
Гейденгайна.
На срезе распознают округлые или неправильной формы дольки
печени, состоящие из высоких эпителиальных клеток, лежащих на
базальной перепонке, которая явственно контурирует фолликулы,
обособляя их от соединительной ткани, прослаивающей фолликулы
(рис. 71).
В толще соединительнотканных прослоек местами можно видеть
овоидные тела с зародышевыми клетками внутри—это материнские
1 Берут 2,5 г двухромокислого калия, 1 г сернокислого натрия, 5 г сулемы*
и растворяют в 100 см3 * 5 дистиллированной воды. Перед употреблением прибавляют
5 см8 ледяной уксусной кислоты.
96
спороцисты (рис. 71, S), в которые превратились мирацидии, сбросив
ресничный покров. Многочисленные ядра клеток, заполняющие мешок
спороцисты, повидимому, образуются за счет «зернистых шаров»
мирацидия.
Рис. 71. Срезы .через печень моллюска Helicella candidate.
s — спороциста Dicrocoelium lanceatum; sp—дочерние спороцисты; sc—спороциста сцеркариями;
р — клетки печени. По А. Скворцову.
Другие спороцисты образуют внутри себя дочерние спороцисты
(рис. 71, sp), а в этих последних развиваются стадии, называемые
церкариями (рис. 71, SC).
Другие материалы по практическому изучению трематод см.:
Е. Н. Павловский. Практикум медицинской паразитологии,
1935, стр. 269—284.
Сравнение жизненных циклов двуусток печеночной и ланцетовидной
При общем сходстве жизненных циклов печеночной и ланцетовид-
ной двуусток, протекающих со сменой поколений, отмечаем разли-
чия, свойственные каждому из этих видов сосальщиков (см. таблицу
на стр. 98).
КЛАСС ЛЕНТОЧНЫЕ ЧЕРВИ (CESTODA)
Технические замечания. Для занятия запасают ленточных
червей — лентеца широкого (Diphyllobothrium latum) и цепеня невооруженного
(Taenia saginata), изгнанных из кишечника человека. Taenia solium — солитер —
встречается редко. Отмытые от испражнений глисты распутываются в ванночке
с водой i>наматываются аккуратными рядами на стеклянную пластинку с тупыми
краями или на толстую пробирку, или цилиндрическое ламповое стекло. Концы
глисты закрепляют нитками и в таком виде опускают в цилиндр с 70% спиртом
или с раствором формалина (одна часть на десять частей воды по объему).
Для изготовления микроскопических препаратов, членики, взятые из сред-
ней части глисты, сдавливают между двумя стеклами, накладывая соответствую-
щий груз или перевязывая стекла нитками, и опускают в 75% спирт. Время от
времени стекла разнимают, чтобы дать приток свежей жидкости к препарату.
7 Практикум по «оологии — 385 97
Стадии жизненного цикла Печеночная двуустка Двуустка ланцетовидная
Яйцо Выходит в неразвитом со- стоянии; личинка развивается в течение долгого времени Откладывается с уже развив- шейся личинкой (мирацидием)
Мирацидий 1 Выходит из вскрывающегося яйца в воде и некоторое время живет свободно Яйцо вскрывается только в пищеварительном канале про- межуточного хозяина, и ми- рацидий свободно не живет
Спороциста Дает поколение редий Дает второе поколение споро- цист
Редия 1 1 Дает или второе поколение редий,или поколение церка- рий Стадии редии нет; церкарии появляются от второго поко- ления спороцист’
Церкария Инцистируется в воде (адо- лескария — инвазирующая стадия) ?
Промежуточный । хозяин Моллюск прудовик малый — Limnaeus minutus Слизни и Fruticicola
Когда утончение члеников сделается стойким, материал перекладывают в свежую
порцию спирта.
Для занятий можно брать:
1) отдельные куски ленты червя для осмотра снаружи под лупой в воде в ча-
совом стекле;
2) обезвоженные и просветленные кедровым (или гвоздичным) маслом или
ксилолом и заключенные в канадский бальзам неокрашенные перезрелые зад-
ние членики червей для рассмотрения характера ветвистости матки;
3) для той же цели можно брать выдержанные в глицерине и заключенные
в глицерин-желатину членики;
4) для анализа гермафродитного полового аппарата Taeniarhynchus saginatus
фиксированные в сдавленном состоянии средние членики окрашивают квасцовым
или борным кармином, проводят через спирты возрастающей крепости, обезвожи-
вают в абсолютном алкоголе, просветляют в гвоздичном масле и, сполоснув кси-
лолом, заключают в канадский бальзам.
Головки ленточных глист рассматривают на ином материале. При вскрытии
кишечника собак можно натолкнуться на множество ленточных глист, зрелые
членики которых по форме очень похожи на огуречное зерно. Такие лентецы на-
зываются цепенем тыквовидным (Dipylidium caninum). Головки с тонкой «шей-
кой» глист снимают со слизистой оболочки кишечника, отмывают от слизи и кла-
дут в небольшое количество теплого (+37° С) физиологического раствора. Через
некоторое время на стекло сразу приливают горячий насыщенный раствор сулемы,
который моментально убивает головки с вытянутым хоботком. Через час-два жид-
кость сливают, а головки переносят в 50% спирт; через полчаса материал перекла-
дывают в 70% спирт, к которому прибавляют иодной тинктуры до цвета мадеры.
По мере обесцвечивания спирта подбавляют иода, пока окраску спирта
не перестанет изменяться; это значит, что сулема совершенно удалена из головок.
Материал переносят в чистый 75% спирт, где и хранят. Для занятий заготовляют
окрашенные бальзамные препараты.
В целях изучения пузырчатых форм глист берут с бойни финнозное свиное
мясо. На выдавленных пальцами финках можно хорошо ознакомиться с морфоло-
гией головки ленточной глисты. Занятия сопровождаются демонстрацией музей-
ных препаратов ленточных червей.
98
ЦЕПЕНЬ НЕВООРУЖЕННЫЙ (TAENIARHYNCHUS SAG1NATUS)
Общий осмотр
Цепень невооруженный имеет очень длинное (до 12 м) и узкое
(до 1 см ширины) лентовидное тело, состоящее из множества (свыше
1000) последовательно расположенных члеников, называемых про-
глоттидами. Все тело червя, или стробила, сужается кпереди и за-
канчивается маленькой головкой — сколексом, которой глиста при-
крепляется к стенке кишечника человека.
Таким образом цепень слагается из частей двоякого рода: а) един-
ственной головки и б) множества проглоттид. Форма последних
в общем четырехугольная. Ле-
жащая за головкой узкая часть
глисты состоит из очень корот-
ких и узких члеников. Размеры
их постепенно увеличиваются,
и совершенно зрелые членики,
отделяющиеся от заднего конца
стробилы, достигают 2 см длины
при 4—7 мм ширины. Такие чле-
ники часто принимают форму
семени тыквы.
Головка. Головка Т. sagina-
tus снабжена четырьмя присо-
сками и лишена крючьев, чем
и объясняется название этой
глисты — цепень невооружен-
ный, в противоположность СО- Рис. 72. Двойной венчик крючьев на го-
литеру, ИЛИ Т. Solium, который, ловке Taenia serrata собаки. По Ж ам му.
кроме присосок, обладает еще
двойным венчиком крючьев на коротком хоботке.
Рассмотрим подробнее головку цепе ня тыквовидного — Dipylidium
caninum. Форма ее меняется в зависимости от степени сжатия самого
сколекса или от степени расширения шейки. Типически головка —
•ромбоидальной формы; может она быть и трехугольной. Сколекс
несет четыре круглых присоски и длинный хоботок, который бывает
виден лишь тогда, когда выпячивается кпереди. В таком случае на
хоботке бывает три—четыре пояса кутикулярных крючьев, похожих
по форме на шипы розы. Всего крючьев бывает 48—60. Часто хоботок
втягивается внутрь головки, и тогда он ложится по оси ее между при-
сосками. На спрятанном хоботке крючья не различимы.
Функция присосок определяется их формой. Каждая присоска
имеет вид полого полушарообразного тела, в виде ямки залегающего
на поверхности головки. Край присоски имеет вид широкого валика
или ленты, тонко исчерченной в радиальном направлении. В стенке
присоски заложено множество мышечных волокон. При сокращении
их, дно присоски уплощается. В таком состоянии она прикладывается
к стенке кишки. Когда при последующем расслаблении мышц присоска
стремится принять прежнюю форму, то в ее полости развивается
* 99
лочку. Таким
Рис. 73. Налитые
каналы выдели-
тельной системы
Taenia saginata.
Оригинал.
а — продольные ка-
налы; b — попереч-
ные каналы; с—по-
ловое отверстие.
отрицательное давление, под влиянием которого слизистая оболочка
кишечника вдавливается в углубление присоски. Таким способом го-
ловка (а с нею и сама глиста) настолько прочно держится за стенку
кишки, что нормальное изгоняющее действие кишечной перисталь-
тики не оказывает никакого действия на глисту.
Хоботок и венчики крючьев способствуют еще более надежному
укреплению паразита, ибо острые крючья впиваются в слизистую обо-
образом головка снабжена, главным образом, орга-
нами прикрепления паразита. Кроме них,в плотной
паренхиматозной головке начинаются каналы выде-
лительной системы и залегает нервная система в виде
кольцеобразных стволов, но эти образования, в ча-
стности же нервная ткань, на обычных тотальных
препаратах не видны.
Отметим здесь же, что головка лишена каких-либо
пищеварительных органов.
За головкой следует «шейка», сильно растяжимая
часть стробилы, еще не подразделившаяся на чле-
ники. В зависимости от степени растяжения шейка
бывает то уже, то шире головки.
Стробила. Стробила состоит из отдельных чле-
ников. У свежей глисты стробила имеет белый цвет
и блестящую поверхность. Снаружи членики ее по-
крыты кутикулой, под которой лежит паренхима
червя. Ни в одном членике мы не'находим следа
органов пищеварения; как и у сосальщиков, у лен-
точных глист нет также дыхательной и кровеносной
систем.
Выделительная система выражена ясно. Она со-
стоит из двух продольных боковых стволов, идущих
вдоль всей стробилы и переходящих, следовательно,
из членика в членик. Эти стволы видны на препаратах
в виде более светлых тяжей.
Если инъецировать свежую глисту берлинской
лазурью (см. выше, стр. 83), то выступают ясно не
только боковые стволы, но и поперечные каналы
(коммиссуры) между ними, идущие по переднему и заднему краям
членика, так что в общем выделительная система расположением
своих главных выводных стволов похожа на веревочную лестницу
(рис. 73). Морфологически же она, как и у сосальщиков, относится
к протонефридиальному типу.
Нервная система также представлена продольными стволами,
идущими параллельно выделительным каналам, но на тотальных пре-
паратах она не видна. В толще паренхимы заложены мышцы. Они
представлены обычно гладкими волокнами, идущими как в продоль-
ном, так и в поперечном направлениях, что можно рассмотреть только
на срезах.
Движения, которые может совершать глиста, очень ограничены.
Благодаря сокращению мышц членики (а следовательно, и вся стро-
била) укорачиваются. Taenia saginata, по сравнению со многими дру-
100
гими глистами, более подвижна. Ее задние членики, оторвавшиеся
от стробилы, могут активно выползать из заднепроходного отверстия
и ползать по белью и постели даже при комнатной температуре, тогда
как глисты подвижны обычно лишь при температуре тела.
Органы размножения развиты различно в зависимости от место-
положения проглоттиды в стробиле. В члениках, лежащих в начальной
части ленточной глисты, полового аппарата еще нет. По мере следо-
вания кзади, в растущих члениках выявляются те или другие части
половых органов, пока, наконец, гермафродитный половой аппарат
не будет представлен во всей своей полноте.
Вполне зрелые органы размножения в соответствующем членике
образованы двумя гарнитурами мужских и женских органов (рис. 74).
Рис. 74. Проглоттида Taenia saginata с полным комплектом
половых органов.
Neph — боковой выделительный канал; t — семенник; ov — яичники;
dt — желточник; sdr — тельце Мелиса; и — матка; cb — половое муж-
ское отверстие; vag— влагалище; К — половая клоака; rs — семе-
приемник.
Мужская часть гермафродитного аппарата состоит из множества
семенников (I), представленных мелкими фолликулами (узелками),
которые разбросаны по всей проглоттиде, между боковыми каналами
выделительной системы (Neph). От семенников идут незаметные на
препарате семевыносящие каналы, которые собираются в семеизвер-
гательный канал (ductus ejaculatorius — vd). Последний идет поперек
членика от средней линии его к одной из боковых сторон, где и откры-
вается мужским половым отверстием в половую клоаку (К), ле-
жащую на небольшом возвышении («половом сосочке») соответствую-
щей стороны проглоттиды.
В самом конце семеизвергательного капала лежит в особом мешочке
совокупительный орган, не совсем удачно называемый «cirrus» — cb.
101
Рис. 75. Вытеснение маткой
остальных частей полового
аппарата в стробиле Taenia
serrata. По Ж а м м у.
Т— семенники; О — яичники; р—
половая клоака.
В половую клоаку открывается также и влагалище (vag), имею-
щее вид тонкой трубки, идущей параллельно семеизвергательному
каналу позади (или книзу) от него. У своего внутреннего конца вла-
галище образует расширение, являющееся семеприемником (rs).
В задней половине проглоттиды лежат два яичника (ov), каждый
в виде густого пучка трубочек, занимающих округлое поле. Между
задним краем членика и яичниками в поперечном направлении вы-
тянута узкая полоска — желточниц (dt), значение которого точно
такое же, что и у сосальщиков.
Между обоими яичниками и желточником лежит маленькое шаро-
образное тело — железа Мелиса, прежде называвшаяся скорлуповой
железой (sdr). От этой последней кпе-
реди идет слегка изгибающаяся довольно
толстая трубка — матка (и), куда по-
ступают оплодотворенные, снабженные
желточными клетками и одетые скор-
луповой оболочкой яйца.
Все названные части женских поло-
вых органов соединяются друг с другом,
хотя детали их связи на обычных пре-
паратах заметны не всегда и не везде.
Копуляция у Т. saginata совершается
различно. Если в кишечнике имеется не-
сколько экземпляров невооруженного
цепеня, то копулятивный орган членика
одной стробилы может войти во влага-
лище какой-либо проглоттиды другой
стробилы.
Если же в пищеварительном аппарате
лежит только одна глиста, сложенная
несколько раз по длине, то копуляция
происходит между различными члени-
ками одной и той же стробилы.
Наконец, возможно и самооплодотво-
рение, когда cirrus, загибаясь, входит во
влагалище своей же проглоттиды.
По мере формирования яиц послед-
ние поступают в матку, которую и пе-
реполняют. Так как плодовитость ленточных глист необычайно ве-
лика, то неудивительно, что матка начинает давать боковые вы-
росты, также заполняемые яйцами. В конце концов ветви матки
вытесняют все остальные (уже ненужные) части полового аппарата
(рис. 75), и от описанной выше сложной системы органов остается
только средний ствол матки, дающий на каждой своей стороне от 17
до 35 вторично ветвящихся отпрысков (рис. 76).
Таков характерный вид «перезрелого» членика. (Детали —см. «Учеб-
ник паразитологии».) Из остальных органов в нем сохраняются два
продольных выделительных ствола; остается заметным и половой
сосочек.
Перезрелые членики теряют прочную связь со стробилой. и так
102
как они занимают заднюю часть ее, то отрываются от конца ленты и
с испражнениями выносятся наружу. Здесь вышедшие проглоттиды
распадаются, а яйца при этом освобождаются и рассеиваются.
Хотя матка в перезрелых («старых») члениках и не имеет собствен-
ного отверстия, все же при открывании проглоттид вскрываются концы
некоторых ветвей матки, и яйца из них попадают в каловые массы
кишечника. Поэтому по наличию яиц в извержениях и можно судить
о присутствии в кишечнике глисты.
Для такой диагностики глист исследуют части кала под микроско-
пом и ищут яйца глист. Последние имеют характерную форму, строе-
ние и размеры, типичные для данного вида. В част-
ности яйца Т. saginata имеют почти правильную
шарообразную форму; одеты они толстой, ра-
диально исчерченной скорлупой; содержимое их
зернисто; нередко внутри яйца можно видеть
шесть крючочков, принадлежащих зародышу, или
онкосфере. Диаметр яйца колеблется от 0,03 до
0,04 мм.
Срез через проглоттиду цепеня невооруженного
Свежие членики из средней части стробилы
цепеня невооруженного фиксируют насыщенным
раствором сулемы (без уксусной кислоты) и
в обычном порядке заливают в парафин. Попереч-
ные срезы окрашивают гематоксилином Делафильда
и эозином и заключают в канадский бальзам.
Рассматривают препарат сначала под сильной
лупой ( х 20 раз), а затем под микроскопом.
Проглоттида на поперечном разрезе вытянута
Рис. 76. Зрелые про-
глопиды Taenia
saginata с перепол-
ненной яйцами мат-
кой, вытеснившей
прочие части орга-
нов размножения.
Оригинал.
в стороны, и ее закругленные концы соответствуют
правому и левому бокам членика; спинная сто-
рона ничем не отличается от брюшной; обе они
имеют слегка неровные очертания.
Снаружи проглоттида покрыта плотной, хорошо
развитой кутикулой (рис. 77, С), внутренность чле-
ника заполнена паренхиматозной тканью; в ней
видны два сильно развитых поперечных тяжа му-
скульных волокон, на наружных концах они разволокняклся и ча-
стично перекрещиваются, но не сливаются друг с другом; эти два
тяжа поперечных мышц (рис. 77, 7Af) разделяют проглоттиду (на
срезе) на три части —среднюю, лежащую между поперечными мыш-
цами, и две наружных, располагающихся соответственно между
поперечной мышцей и кутикулой псверхнссти проглоттиды.
Все органы, видимые на срезе, располагаются в центральной части
паренхимы, т. е. между обоими тяжами поперечных мышц; здесь
видны: два лежащих ближе к бокам проглоттиды «пустых» овала —
поперечные разрезы боковых каналов выделительной системы (NC);
кнутри от них видны поперечные и косые срезы ветвей матки (D),
в полостях каналов которой местами остаются яйца (при обработке
103
часть их вымывается и выпадает); между ветвями матки и поперечной
мышцей одной стороны тела могут быть видны мелкие фолликулы
семенников, сплошь забитые мелкими клет-
ками или совсем пустые (Н).
После общей ориентировки среза рассматри-
вают детали его строения при более сильном
увеличении (рис. 78). Под кутикулой (си) ле-
жит узкая светлая зона, которую называют
базальной мембраной (gl). С внутренней сто-
роны к ней прилежит широкий слой кольцевых
мышц (гт), за которым следует ряд продольных
мышечных волокон (поперечно перерезанных).
Далее кнутри лежит слой цилиндрических
или клиновидных довольно крупных клеток;
наружные концы их вытягиваются в тонкие во-
локна. Эти клетки многие считают за железистые
клетки паренхимы (рис. 78,d). Паренхима со-
стоит из мелких соединительнотканных клеток,
которые сильно ветвятся; соединяющиеся отро-
стки этих клеток образуют губчатую массу па-
ренхимы, в петлях которой у живого червя нахо-
дится тканевая жидкость. Паренхима пронизана
дорзо-вентральными мышечными
(dm); мышечные
NS
NC
TM
D
H
LM
СЦ
волокнами
клетки, дающие эти волокна,
Im
dm
HZ
Рис. 77. Поперечный раз-
рез через середину чле-
ника цепеня невооружен-
ного с разрастающейся
маткой (D). По К. Ш н е й-
Д е р у.
С — кутикула; Е — «погру-
женный эпителий»; NS —
нервный ствол; NC — боко-
вой выделительный канал;
ТМ — поперечные мышечные
волокна; Н — фолликулы се-
менников; LM — продольные
мышечные волокна.
mZ
разреза проглоттиды
По К. Ш н е й д е р у.
Рис. 78. Часть поперечного
цепеня невооруженного.
си — кутикула; gl— «базальная мембрана»; гт—кольцевые
мышечные пучки; d— клетки «погруженного эпителия»;
1т — поперечный разрез продольного мышечного волокна;
dm — дорзо-вентральные мышечные волокна; mZ — мышечная
клетка; KZ — известковая клетка.
гораздо более крупны, чем клетки «паренхимы. Кроме того в парен-
химе видны известковые тельца (KZ) в виде крупных округлых, сильно
104
окрашивающихся гематоксилином образований, одетых тонким
слоем плазмы с уплощенным ядром. На препаратах, обработанных
кислотами, они исчезают.
Наконец, местами в паренхиме можно видеть концевые (замыкаю-
щие) клетки протонефридиев с узким отходящим от них канальцем.
На удачно прошедших срезах иногда можно видеть пучок ресниц
«мерцающего пламени». Выделительный канал хорошо контурирован.
Но тонкая стенка его бесструктурна.
Кнаружи от выделительного канала можно найти поперечный раз-
рез главного продольного нервного ствола, состоящего из пучка
нервных волокон и лежащих местами и неразличимых на обычном
препарате ганглиозных клеток и клеток глии (рис. 77, TVS).
Стенки матки тонки; в ней заложены ядра без каких-либо границ
между клетками (синцитий).
ЛЕНТЕЦ ШИРОКИЙ (DIPHYLLOBOTHRIUM LATUM)
Общий осмотр
По общему виду лентец широкий похож на цепеня невооружен-
ного. Стробила его достигает длины нескольких метров при четырех
и более тысячах члеников. Головка в форме удлиненного овала, 2—
3 мм. Проглоттиды большею частью короткие и широкие (отсюда
и название лентец широкий). В задней части глисты членики стано-
вятся квадратными и даже удлиненными. Цвет глисты сероватый.
Посредине зрелых члеников просвечивает темная розетка матки. Края
стробилы зубчатые, соответственно границам члеников. Половых со-
сочков на боках стробилы нет.
Головка. Головка лентеца широкого лишена настоящих присосок
взамен которых есть две присасывательные борозды (ботридии), иду-
щие одна на спинной, другая на брюшной стороне головки (рис. 79).
Каждая ботридия ограничена двумя губами, которыми и защемляются
ворсинки кишечника в целях удерживания глисты за стенку кишки.
Ботридии являются более примитивными органами, нежели настоя-
щие присоски. Ботридии на тотальном препарате головки различимы
с трудом. Их можно хорошо рассмотреть лишь на поперечном
срезе головки. За головкой следует тонкая и длинная в несократив-
шемся состоянии шейка.
Стробила. Как и Т. saginata, лентец широкий лишен органов пище-
варения, дыхания, кровообращения и органов чувств. Однако функ-
ции пищеварения ему свойственны, так же как и другим ленточным
глистам.
Лентецы, обитающие в кишечнике человека (и животных), нахо-
дятся в особо благоприятных условиях питания, ибо они окружены
пищеварительной кашицей, проходящей через кишечник и подвер-
гающейся здесь же перевариванию ферментами хозяина. Лентецы
питаются такой переваренной пищей, которую и всасывают всей
поверхностью тела. Такой способ питания называется эндосмоти-
ческим. При нем специальные органы пищеварения являются из.тиш
ними, чем и обусловливается их исчезновение у ленточных червей,
105
происшедших от плоских червей, обладающих пищеварительным
аппаратом.
Хотя лентецы питаются только пищей, находящейся в кишечнике
хозяина, и потому могут считаться нахлебниками по отношению
к своим хозяевам, однако они приносят последним известный ущерб.
В самом деле, хозяин тратит энергию на химическую переработку
пищи в своем кишечнике, и при наличии глистов он не может исполь-
зовать полностью плоды работы своего кишечника, ибо часть перера-
ботанной пищи захватывается паразитами. Таким образом волей-
неволей хозяин работает и на своих обитателей.
Кроме того глисты, живущие в пищеварительном канале, выде-
ляют продукты обмена веществ, которые всасываются кишечником,
Рис. 79. Головка лентена
малого — Diphyllobothrium
minus. По Талызину.
поступают в кровь и вызывают различ-
ной степени отравления.
Условия дыхания в кишечнике весьма
своеобразны. В этом отношении все пара-
зиты кишечника находятся в одинаковых
условиях. Поэтому для уяснения особен-
ностей дыхания эндопаразитических лен-
точных червей можно обратиться к отделу
о дыхании аскарид (стр. 117).
Органы размножения лентеца широ-
кого также гермафродитны; однако они
весьма заметно отличаются от схемы, с ко-
торой мы познакомились у цепеня нево-
оруженного.
В перезрелом членике бросается в глаза
матка, представленная толстой петлеоб-
разно изогнутой трубкой, просвечивающей
в средней продольной полосе членика.
Матка не ветвится и имеет свое собствен-
ное выводное отверстие, помещающееся на
средней линии членика, приблизительно на границе его передней
трети. Вследствие такого строения, зрелые яйца постоянно выхо-
дят из матки наружу и всегда могут быть найдены в испражнениях
хозяина.
Боковые поля проглоттиды заполнены зернистыми образованиями;
здесь, ближе к свободному краю членика, лежат диффузно рассеянные
желточники (рис. 80, dt), более же кнутри расположены семенники
(/). На препарате обыкновенно ясно различить их друг от друга не
удается.
В зрелом членике (рис. 80) кзади от матки (и) залегает яичник
(ov), по форме похожий до некоторой степени на переметную суму.
В его вырезке помещается «скорлуповая железа» (sdr). Кпереди по
средней линии тянется влагалище (vag), открывающееся, как и семе-
извергательный канал (vd), общим половым отверстием ($<5), поме-
щающимся также на плоскости членика кпереди от отверстия матки.
Яйца лентеца овальной формы, одеты тонкой двуконтурной скор-
лупой, с явственной крышечкой на одном из полюсов яйца и бугор-
ком— на другом; длина яйца около 0,07 мм. цвет его буроватый.
106
Хотя ленточные глисты время от времени теряют свои задние пе-
резрелые членики, однако величина их от этого не страдает, ибо вза-
мен утраченных проглоттид появляются новые членики.
Такой рост стробилы обусловливается деятельностью головки, ко-
торая на своем заднем конце отделяет новые молодые членики. Этот
процесс является как бы почкованием, причем продукты его остаются
в соединении друг с другом, благодаря чему и образуется многослож-
ная стробила. Последняя возникает в порядке бесполого размноже-
ния головки. Если при неудачном изгнании лентеца из кишечника
стробила выйдет вся, а головка останется на месте, то в два-три месяца
она снова отпочкует новые членики и даст новую стробилу прежнего
размера.
Рис. 80. Проглоттида с половым аппаратом лентеца широкого.
QcJ —общее половое отверстие; о—отверстие матки;ог—яичник; dt—жел-
точники; sdr — скорлуповая железа; vag — влагалище; и — матка; t — семен-
ники; vd— семеизвергательный канал; neph— выделительный канал.
ПУЗЫРЧАТЫЕ ГЛИСТЫ
В свином мясе иногда можно находить среди мышц овальные бе-
ловатые полупрозрачные тела различных размеров, от горошины до
лесного ореха и даже более (рис. 81).
При рассматривании извлеченного образования, в полупрозрачном
тонкостенном пузырьке можно видеть, у одной из его стенок, беловатый
плотный узелок. Пузырь внутри заполнен жидкостью. Если осторожно
надавливать пальцем на заполненную жидкостью часть пузырька, то
беловатый пузырек выворачивается нар.ужу. На его конце в таком
случае хорошо различима головка с четырьмя полушаровидными при-
сосками и венчиком крючьев. Головка эта весьма напоминает голову
солитера (Taenia solium).
107
Такое сходство не случайного характера, а основана на родствен-
ных отношениях между ленточной и пузырчатой глистами. Суть этих
отношений сводится к следующему.
Солитер (Т. solium), живущий в кишке человека, размножается
половым путем и дает множество яиц; я й ца в оторвавшихся члени-
ках с испражнениями хозяина выносятся из кишечника и попадают
наружу, например па зе-
млю, где случайно прогла-
тываются различными жи-
вотными, например свинья-
ми, собаками и другими
животными, роющимися на
свалках; только в кишеч-
нике свиньи (и собаки) яйца
солитера могут развиваться
дальше; тут оболочка яйца
вскрывается, и освобо-
ждается шестикрючечный
зародыш (онкосфера). Он-
косфера своими крючьями
продирается сквозь слизи-
стую оболочку стенки пи-
щеварительной трубки и
попадает в лимфатические
сосуды; с током лимфы он-
косфера заносится в мышцы
и в другие места тела, где
оседает и превращается в
вышеописанную пузырча-
тую глисту, или финку.
Образованием финки закан-
чивается развитие глисты
в теле свиньи.
Дальнейшее превраще-
ние финки возможно лишь
в том случае, если она
будет проглочена в живом
состоянии с мясом свиньи
человеком. Тогда в желу-
дочно-кишечном канале по-
следнего пузырь финки рас-
Рис. 81. Финки солитера Taenia solium. Справа
верхний — с вскрытой стенкой пузыря; ниж-
ние — со вскрытой ввернутой головкой. По
Пфуртшеллеру.
творяется; головка выворачивается наружу, прикрепляется к стенке
кишки и начинает почковать членики, в результате чего в кишке чело-
века вырастает стробила солитера.
Итак, полный жизненный цикл солитера связан с двумя хозяевами:
а) с человеком, в кишке которого живет глиста, размножающаяся
половым путем, и
б) со свиньей, в мышцах которой обитает пузырчатая форма со-
литера (т. е. финка, или цистицерк), которая способна лишь к беспо-
лому размножению (почкование стробилы).
108
На жизненном цикле солитера мы знакомимся с явлением перемены
хозяев, сопряженным со сменой поколения паразита, размножающегося
последовательно то половым, то бесполым путем. Такой тип размно-
жения, как это было у кишечнополостных (стр. 65), называется мета-
генезом.
Итак, человек заражается солитером от свиньи, а свинья—от
человека. Вне этих взаимоотношений солитер в природе не встре-
чается нигде. Условия развития Т. solium зависят в большой степени
от случайности. Случайно свинья проглатывает яйца солитера; слу-
чайно человек с полусырым мясом свиньи может проглотить живых
финок. Такое благоприятное для глисты стечение обстоятельств бы-
вает сравнительно очень редко. Из бесчисленного множества яиц,
даваемых глистой, конечно, бблыпая часть погибнет, но, по теории
вероятности, известная малая доля их попадет в благоприятные усло-
вия и даст новое поколение, которое в силу чрезвычайной плодовитости
сможет, в свою очередь, поддержать существование соответствующего
вида на земле.
Хозяев, в которых живет половозрелая форма паразита, называют
главными; хозяев, у которых обитает форма глисты, размножающаяся
лишь бесполым путем, именуют промежуточными.
Для Т. solium и Т. saginatus смена хозяев сводится к следующему.
Пузырчатая форма (финка) живет
в свинье
Пузырчатая форма (финка) живет
в быке (корове)
Ленточная форма живет в человеке
Ленточная форма живет в человеке
Промежуточные хозяева —
свинья или корова
Главный хозяин — человек
Развитие лентеца широкого еще более сложно. Пузырчатую форму
лентеца широкого ищут под кожей и в мышцах различных пресно-
водных рыб —окуня, ерша, щуки.
У ершей можно видеть просвечивающими через покровы беловатые
узелки или удлиненные образования, которые по их освобождении
оказываются подвижными «червячками». Аналогичные образования
можно находить в толще мышц (их срезают пластами), под брюшиной
различных органов, в яичниках, иногда в толще стенки желудка и др.
Освобождаемых червячков кладут в часовое стекло с водой и
наблюдают под-лупой.
Наблюдаемые существа являются плероцеркоидами лентеца широ-
кого; они хорошо движутся, изменяя ширину и длину своего тела,
по которому время от времени пробегают волны сокращения, подоб-
ные перистальтике кишечника. Плероцеркоиды движутся всегда од-
ним концом тела вперед; он является головным; на нем, так же как
ина головке взрослого лентеца, видны две продольные присасыватель-
ные бороздки, ограниченные губами. Тело плероцеркоида нерасчле-
ненное; поперечные складки тела в расчет не принимаются, так как
никакого отношения к сегментации они не имеют.
109
Все тело плероцеркоида паренхиматозное, коматозное; никакой
полости с жидкостью в нем нет; поэтому среди типичных пузырчатых
форм плероцеркоид занимает особое место.
Плероцеркоиды могут жить в воде до 8—10 дней. С ними можно
поставить ряд интересных и показательных опытов, в частности
над их способностью к миграции.
Живых плероцеркоидов пускают пипеткой в вырезанный, промытый
внутри водой и перевязанный с одного конца желудок рыбы (напри-
мер ерша); перевязывают крепко и другой конец желудка; желудок
кладут в чашку Петри с жидкостью. Через сутки можно видеть, что
некоторые плероцеркоиды уже вышли из желудка, другие же выхо-
дят из толщи его стенки.
Рис. 82. Впрыскивание плероцеркоидов рыбе в желудок. По Гнездилову
и Талызину.
Живые плероцеркоиды выдерживаются около 10 минут в водном
растворе нейтральрот (1: 1000) для прижизненной окраски; окрашен-
ных плероцеркоидов шариковой пипеткой вводят через рот в желудок
нескольким лягушкам; лягушек вскрывают через определенные
сроки.
Можно видеть выхождение плероцеркоидов через стенку желудка
и даже внедрение их в мышцы стенки тела, хотя лягушка и не является
хозяином для этих паразитов (Г н е з д и л о в и Талызин).
Окрашенных живых плероцеркоидов вводят пипеткой в желудок
живой щуке, которую пускают в акварий (рис. 82); через 1 — 2 суток
вскрывают щуку и по нахождению в ее брюшной полости розоватых
(т. е. окрашенных нейтральротом) плероцеркоидов убеждаются в спо-
собности их мигрировать через стенку желудка в различные органы
рыбы (рис. 83)..
Живых плероцеркоидов скармливают собаке, у которой предва-
рительным микроскопированием помета доказано отсутствие ленточ-
110
ных глист. Через две недели вскрывают собаку и в ее тонких кишках
находят широких лентецов, развившихся из скормленных плеро-
церкоидов.
Рис. 83. Вскрытая щука с мигриро-
вавшими из желудка плероцеркои-
дами. По Гнездилову и Т.а-
л ы з и и у.
Общие замечания
Ленточные черви являются животными со строго выдержанной ме-
тамерией: за головкой следует длинная цепь члеников, сходных по
строению друг с другом и следую-
щих один за другим. Такая сим-
метрия называется метамерной,
а самые симметричные части мета-
мерами. В тонких деталях мета-
мерия может быть проведена лишь
в различных отделах стробилы,
ибо по строению своему незрелый
членик тонкой части глисты за-
метно отличается от проглоттиды,
снабженной полным комплектом
половых органов. Но если при-
нять в расчет, что каждый членик
глисты в течение своей жизни в
разное время проходит все стадии
формирования органов, то такое
различие теряет принципиальное
значение.
Двубоковая симметрия хорошо
выражена в головке и в перезре-
лых члениках, в остальных же она
нарушается асимметрией некоторых
органов, например, положением
половых отверстий только на одном
боку проглоттиды у глист семей-
ства Taeniidae.
Длинное лентовидное тело, со-
стоящее из головки и цепи мета-
мерных члеников, а также отсут-
ствие пищеварительного канала
в паренхиматозном теле, лишенном
вторичной полости и дренирован-
ном каналами протонефридиев,—
все эти признаки определяют при-
надлежность рассмотренных нами
лентецов к подтипу плоских червей
и классу ленточных червей (Се-
stoda), собственно к расчлененным
ленточным глистам.
Форма головки и положение половых отверстий характеризуют
принадлежность лентеца широкого к семейству Diphyl lobothriidae
(головка с двумя ботридиями, половое отверстие па плоскости
членика), а Т. solium и Т. saginatus —к семейству Taeniidae (четыре
Ш
присоски и положение половых отверстий, обычно, на одной стороне
членика).
Сравнение организации ленточных червей
* ---------------------------------------------
Taenia solium, Т. sa- Diphyllobothrium
ginatus latum
Форма тела..............
Вторичная полость тела . .
Органы дыхания..........
> кровообращения . .
> чувств ............
> выделения.........
Дыхание.................
Головка.................
Пищеварительная система .
Пищеварение.............
Нервная система ........
Половые отверстия.......
Яичники ..........
Желточники.........
Матка..............
Плоская
Отсутствуют
признаки,
общие с
Отверстие матки . . .
Яйца...............
Протонефридиального типа
Интрамолекулярное
С 4 присосками | С 2 ботридиями
Отсутствует
Эндосмотическое
Продольные стволы
Сбоку членика I На поверхности
| членика
В задней половине проглоттиды, но
сосальщи-
ками
(стр. 85).
разной
По заднему краю
членика
В виде ветвяще-
гося продольного
ствола
Отсутствует
Круглые, с толстой,
радиально исчер-
ченной скорлупой
формы
В боковых частях
членика
В виде петлистой
неветвящейся
трубки
Есть
Овальные, с тонкой
скорлупой и кры-
шечкой
Соотношение сосальщиков и ленточных червей, как представителей
различных классов плоских червей, уясняется следующей таблицей:
Сосальщики
Ленточные черви
Тело
Головка .....................
Присасывательные аппараты . . .
Органы пищеварения...........
» выделения...............
» дыхания.................
> кровообращения..........
Вторичная полость тела ......
Нечленистос и
короткое ।
Отсутствуе!
Непосредственно на
теле
Есть ।
Протоне
| О т с у т
У собственно ленточ-
ных червей чле-
нистое и лентовидное
Есть
На головке
Отсутствуют
фридии
с т в у ю т
Таким образом главные отличия между сосальщиками и собственно
ленточными червями сводятся к исчезновению, т. е. к редукции,
у последних органов пищеварения, к обособлению головкй с органами
прикрепления на ней, удлинению тела и подразделению его на мно-
жество члеников
112
КЛАСС КРУГЛЫЕ ЧЕРВИ (NEMATODA)
АСКАРИДА ЛОШАДИНАЯ (ASCARIS MEGALOCEPHALA)
Технические замечания. Аскарид заказывают на лошадиной
бойне. Летом запасают материал для зимних занятий. Живых аскарид собирают
в банку с 6% формалином. При собирании материала в спирт пользуются для на-
чала очень слабым спиртом (5%) и долго и постепенно переводят аскарид в спирты
большей крепости, пока не достигнут 45—50%. За несколько часов до занятия по-
требное число аскарид переносят в воду.
Кроме лошадиной аскариды, для вскрытия при случае могут быть использо-
ваны аскариды человеческие (Ascaris lumbricoides) и аскарида свиная (A. suilla).
Все аскариды живут в толстых кишках своих хозяев.
Наружный осмотр
Аскарида имеет гладкое веретеновидное тело, на поперечном раз-
резе правильной круглой формы. Если у осматриваемого экземпляра
один из концов тела загнут крючком, то это —самец, который, кроме
того, отличается меньшими размерами тела, чем у самки (рис. 84).
При рассматривании загнутого конца тела в профиль видны малень-
кие кутикулярные иголочки, торчащие из отверстия, лежащего на
заднем (закрученном) конце самца аскариды. Эти иголочки назы-
ваются спикулами; они служат копулятивными органами
(рис. 84, cf).
У самки задний конец тела отличается от такового у самца по
нахождению несколько кпереди от конца тела заднепроходного от-
верстия (anus), лежащего в виде поперечной щели (рис. 85, а). Про-
тивоположный конец тела является передним. В центре его помещается
ротовое отверстие, окруженное тремя припухлостями, называемыми
губами; из них одна дорзальная, а две —вентральные. Передний
конец тела у обоих полов устроен одинаково.
Физиологически у аскарид не различают ни брюшной, ни спинной
сторон тела. Морфологически брюшной стороной самки считается та,
на которой лежат заднепроходное и половое (см. ниже) отверстия,
у самца же на брюшную сторону загибается задний конец тела.
Тело аскариды одето плотной блестящей белой кутикулой, иногда
слегка морщинистой; никаких следов членистости тела нет.
При внимательном рассматривании тела можно заметить четыре
продольные линии, расположенные по квадрантам поперечного сече-
ния аскариды. Две из них по своему положению являются боковыми;
из остальных же одна идет по средней линии спины, а другая —по
брюшной стороне. Боковые линии заметны лучше. Они имеют
вид темных, ровных узких полос, образованных утолщением кожи,
направленным внутрь тела. В них заложены выделительные каналы
аскариды. Соответственно спинюй и брюшной линиям тела идут нервные
стволы.
При следовании вдоль брюшной линии, приблизительно на границе
передней и средней третей длины тела самки, можно найти точкообраз-
ное женское половое отверстие. Самец же не имеет самостоятельного
полового отверстия; выводные половые пути его впадают вместе с ки-
8 Практикум по зоологии — 385 113
шечником в клоаку, которая открывается наружу на заднем конце
тела. Из нее-то и торчат спикулы.
о
Рис. 84.
Аскарида человеческая (Ascaris
lumbrco des).
Слева —самка. О — ротовое отвер-
стие; b—отверстие выделительных ор-
ганов; с — кольцевидная перетяжка тела
в области полового отверстия — d; е —
анальное отверстие. Справа— самец.
п — рот; Ъ —экскреторное отверстие; с —
спикулы; а — отверстие клоаки. По
Ж а м м у.
Вскрытие аскариды
Для правильного вскрытия ориен-
тируют аскариду брюшной стороной
вниз. Передний и задний концы тела
червя прикалывают двумя булавками
ко дну ванночки, в которую налита
вода.
Придерживая пальцами под водой
аскариду в наиболее толстой части
ее тела, скальпелем разрезают на
спинной стороне стенку тела вдоль
по средней линии на несколько сан-
тиметров. Необходимо следить, чтобы
А В
Рис. 85. Задний конец тела самки (9) и
самца (сГ) аскариды человеческой в про-
филь. По Ж а м м у.
У самца видны спикулы — о — отверстие
клоаки; а — анальное отверстие.
аскарида была покрыта водой, и не
наклоняться лицом близко к ван-
ночке, так как при первом моменте
вскрытия червя из его тела часто
выбрызгивается полостная жидкость,
могущая попасть в глаза вскрываю-
щего.
Края разреза отводят в стороны и
булавками прикалывают ко дну ван-
ночки. Разрез продолжают далее кпе-
реди скальпелем же или тонкими
ножницами; укрепляют края разреза.
Вскрытие затем продолжают, повер-
нув ванночку, до заднего конца тела,
все время старательно держась сред-
ней линии тела и отводя в сторону
лоскуты кожных покровов.
114
Рис. 86.
А (справа) — вскрытая самка аскариды человеческой, а — рот»
b — окологлоточное нервное сплетение; с — глотка; d — ки-
шечник; е — кожно-мускульный мешок;/—выделительный
канал; g— влагалище; Л — матка; к — яичник; I—anus.
В (слева) — половой аппарат самца аскариды, а — семенник;
Ъ — семепровод; с — кишка; d— мышечные клетки; е —
спикула; / — отверстие клоаки. По Ж а м м у.
Так как вскрывается уже фиксированное животное, внутренности
которого уплотнены, то распутать все трубчатые органы, видимые
внутри, без разрыва их нельзя.
Кожно-мускульный мешок. При вскрытии аскариды рассекаются
кожные покровы тела. На вскрытом черве можно видеть, что они толсты
и имеют различный
вид снаружи и изну-
три, тогда как на на-
ружной поверхности
они покрыты гладкой
кутикулой, изнутри—
кожные покровы ше-
роховаты (рис. 86, А,е).
Это зависит от плот-
ного срастания с соб-
ственно кожными по-
кровами мышечных
клеток, вследствие
чего и у аскариды
можно говорить о на-
личии кожно-мускуль-
ного мешка. Никаких
органов движения у
аскариды нет. По-
движность ее весьма
ограничена, так как
состоит только в из-
гибаниях тела живого
червя, что можно хо-
рошо видеть, если по-
догреть физиологиче-
ский раствор поварен-
ной соли с живыми
аскаридами.
При рассечении
аскариды получается
впечатление, что кож-
но-мускульный ме-
шок ограничивает со-
бой полость тела, в ко-
торой лежат органы.
Однако морфологи-
чески вторичной по-
лости тела (целома) у аскариды нет, ибо пространства между орга-
нами и стенкой тела у нее заполнены так называемой изоляционной
тканью, состоящей из нежных, сильно вакуолизированных клеток.
Последние весьма нежны, легко разрываются и поэтому не улавли-
ваемы глазом при вскрытии.
Внутри тела аскариды лежит вытянутый в длину клубок труб-
чатых органов. Срединная серая трубка —кишечник —окружена
* 115
тонкими яркобелыми разнокалиберными трубочками половых ор-
ганов.
Органы пищеварения. Пищеварительный аппарат начинается ртом,
окруженным тремя губами. За ртом следует короткая (около 1 см)
сильно мускулистая, плотная глотка; она резко граничит с собственно
кишкой —тонкостенной трубкой серого цвета (рис. 86,А, </). Стенки
ее образуют продольные складки. Нередко при неосторожном вскрытии
тела разрезается и кишка; в таком случае, чтобы увидеть остальные
органы, необходимо снять пинцетом серую пленку, т. е. стенку кишки,
закрывающую препарат. Никаких подразделений
кишечника у аскариды видеть не удается. Отме-
тим, во-первых, факт наличия пищеварительного
канала у животного, ведущего паразитический
образ жизни, т. е. в данном случае обитающего
в толстой кишке лошади, где вокруг паразита
всегда бывает изобилие пищи. Во-вторых, следует
отметить крайнюю простоту строения органов пи-
щеварения у аскариды, что особенно становится
наглядным при сравнении ее с другими животными,
в частности с дождевым червем (см. ниже).
Органы выделения. На шероховатой внутрен-
ней поверхности кожно-мускульного мешка аска-
риды ясно видны двуконтурные боковые линии,
являющиеся валикообразными утолщениями соб-
ственно кожного слоя. В этих утолщениях зало-
жены выделительные каналы. По оси каждой боко-
вой линии (рис. 86,А, /) идет канал. На переднем
конце тела оба канала соединяются друг с другом
и общим отверстием открываются на нижней сто-
роне тела, несколько отступя кзади от губ. Зад-
ний конец каждого канала заканчивается слепо.
Стенки канала образованы немногими клетками,
вытянутыми в длину и образующими в своей сово-
купности цилиндрическую трубку.
Такой тип органов выделения, как у аскариды, является своеоб-
разным. Органы выделения аскарид можно бы сравнить с протоне-
фридиями, т.е. с примитивной выделительной системой замкнутых на
своих внутренних концах трубок бесполостных животных, однако
в типичных протонефридиях внутренние концы каналов замкнуты
клетками с подвижным жгутом или с длинными ресничками, спускаю-
щимися в начало выделительного канала. У аскариды (как и у других
круглых червей) мерцательных или жгутиковых клеток нет вообще.
Органы выделения их приходится относить к образованиям sui generis,
происшедшим, быть может, из видоизмененных кожных желез.
В передней части тела, на выделительных каналах, при осматри-
вании их также изнутри, можно видеть четыре более или менее симме-
трично лежащие довольно большие кучки серого вещества, по две на
каждом канале, одна позади другой. Эти образования являются гигант-
скими по своим размерам фагоцитарными клетками (рис. 87).
Если живой аскариде впрыснуть в тело тушь и кармин в водной
116
взвеси, поместить червей в физиологическом растворе в термостат при
температуре тела и потом через несколько часов вскрыть, то можно
видеть, что фагоцитарные клетки окрасятся в цвет впрыснутого веще-
ства. Это явление зависит от того, что названные клетки захватывают
(фагоцитируют) зернышки туши или кармина и отлагают их в своей
протоплазме, что, по существу, является процессом внутренней
экскреции, или внутриклеточным выделением; при нем подлежащее
удалению из организма вещество изъемлется фагоцитарными клетками
и отлагается в протоплазме этих последних.
Нервная система состоит из двух продольных нервных стволов:
брюшного и спинного. Последний часто повреждается при вскрытии
аскариды, если разрез пройдет по средней линии спины. Чтобы найти
брюшной нервный ствол, царапают иглой шероховатую внутрен-
нюю поверхность мышечного слоя по средней линии брюшной стороны
тела и обнаруживают белый шнурочек без каких-либо следов узло-
ватости.
Оба нервных ствола соединяются с окологлоточным кольцевым
скоплением нервной ткани, которое можно видеть лишь на микро-
скопическом срезе.
Нервная система по своему типу также является своеобразной;
чего-либо подобного ей по плану строения у других животных мы
еще не видели.
Органы чувств. Анатомически выраженных органов чувств у аска-
риды нет. Это вполне вяжется с паразитическим образом ее жизни.
Живет она в толстых кишках лошади, всегда в темноте, при постоян-
ной температуре, в среде определенного химического состава, изба-
вленная от необходимости разыскивать себе пищу или укрываться от
вражеского нападения. Такие постоянные (константные) условия
жизни делают для паразита излишними различные органы чувств,
как, например, глаза, органы слуха, осязание и т. п.
Аскарида сохраняет известную долю чувствительности; но это ее
свойство связывается с наличием периферической нервной системы,
в существовании которой можно удостовериться лишь при пользова-
нии очень сложными и специальными методами исследования;
Дыхание у аскариды. Органы кровообращения и дыхания у аска-
риды отсутствуют. Дыхание совершается своеобразно уже потому, что
аскарида живет в толстых кишках, где свободного кислорода практи-
чески нет. При происходящем здесь брожении пищи выделяются раз-
личные газы, например, сероводород и пахнущие продукты распада
белков. Все это делает газовую среду полости толстых кишек анаэроб-
ной, т. е. лишенной кислорода. Обитание аскарид в анаэробной среде,
по первому взгляду, может показаться исключением из общей для
всех животных необходимости пользоваться кислородом для дыхания.
Однако принципиального отклонения здесь нет. Аскарида также дышит
кислородом (т. е. клетки ее тела получают кислород), но кислород
берется не из внешней 'среды, как у свободно живущих животных
(аэробов) При процессе разложения пищевых веществ в теле аска-
риды (в частности гликогена) освобождается кислород, который и
потребляется клетками тела паразита. Такой своеобразный способ
дыхания называют интрамолекулярным.
117
Органы размножения. Как уже упоминалось выше, аскарида раз-
дельнопола. Внутренние органы размножения самца и самки сходны
по внешности в том отношении, что они трубчатой формы.
Женские органы размножения. Подрезав концы
кишечника, пинцетом осторожно удаляют его прочь и рассматривают
женские половые органы (рис. 86,А).
Из клубка белых нитей и шнуров, окружающих кишечник, выби-
рают самые тонкие —это яичники (к). Их два. Каждый яичник пере-
ходят постепенно в более толстый яйцевод, который, в свою очередь,
продолжается в матку. Последняя представлена толстой извилистой
трубкой, лежащей, как и вторая маточная трубка, вдоль тела, парал-
лельно кишечнику. Своими передними концами обе матки (Л) соеди-
няются в тонкий общий канал —влагалище (g), открывающееся поло-
вым отверстием наружу на брюшной стороне тела и обходящее с боку
кишечник.
Если прорвать стенку матки и растереть пинцетом ее содержимое,
то оно дает в воде муть, вследствие разъединения яиц. Последние ми-
кроскопически мелки. По этим отношениям можно себе представить,
сколь плодовита аскарида. Если перевести способность ее к половой
продукции на человеческий масштаб, то, чтобы сравняться с ней
в плодовитости, женщина должна рожать каждый день по 70 человек
детей!
Оплодотворение у аскариды внутреннее; поэтому в матке, в ее
выводных частях, имеются уже развивающиеся зародыши; они ле-
жат в оболочках яйца, весьма надежно защищающих свое содержимое
от внешних воздействий. Достаточно указать, что яйца аскариды
остаются живыми даже при пятилетием хранении в растворе форма-
лина.
Яйца лошадиной аскариды являются классическим объектом для
изучения кариокинеза, процессов созревания яйца, оплодотворения и
дробления.
Мужские органы размножения. Мужской половой
аппарат устроен проще, чем женский. Он состоит всего только из
одной трубки, петли которой занимают две трети тела (рис. 86,В).
Тонкая часть трубки является семенником (а), толстая, лежащая
параллельно кишечнику, служит семенным пузырьком (vesicula semi-
nalis —b); выходная его часть, открывающаяся в клоаку, функциони-
рует как ductus ejaculatorius.
Напомним про наличие у самца совокупительных органов, коими
являются спикулы (е), лежащие в особом мешочке, открывающемся
в клоаку.
Срез через аскариду
Для рассмотрения берется поперечный срез через середину тела
самки аскариды, окрашенный гематоксилином и эозином. Для общей
ориентировки рассматривают срез под лупой или под самым слабым
увеличением микроскопа.
При общей ориентировке находя! следующие части тела (рис. 88):
а) Снаружи срез окаймлен стенкой тела, гладкой снаружи и
складчатой изнутри (</), состоящей из кожно-мускульного мешка.
118
б) Внутри препарата находят поперечный разрез кишки (g), иногда
в виде буквы Н или иных очертаний. Стенка кишки состоит из высокого
цилиндрического эпителия; в просвете кишки бывает зернистое содер-
жимое.
в) По бокам кишки бросаются в глаза два больших круга или
овала (/), заполненные мелкими шариками с точками внутри. Это — раз-
резы двух маток с развивающимися в яйцах аскариды зародышами.
г) Между матками и стенкой тела имеется много небольших круж-
ков, иногда же и длинных овалов, построенных из равномерно зерни-
Рис. 88. Поперечный разрез через тело самки аскариды. По
Ж а м м у.
а — кутикула; b — субкутмкулярный слой; с — боковой канал; d — мы-
шечные клетки; е — яичник; /—матка; g—кишечник; h — нервный
ствол.
стой ткани, без какого-либо просвета внутри —это разрезы яични-
ков (е).
д) Среди яичников попадается пять-шесть разрезов также округ-
лой формы, с гладким наружным и рифленым внутренним краями,
полость которых неравномерно заполнена разнокалиберной зернистой
массой; это—сечение яйцеводов с живчиками и формирующимися
яйцами.
После общего осмотра изучают отдельные части препарата более
детально, пользуясь более сильными объективами.
Рассматривают сначала кожно-мускульный мешок; он состоит:
из кутикулы (о), распадающейся на несколько слоев: 1) наружного —
коркового (распадающегося на вторичные слои), 2) среднего —гомо-
генного по структуре, и 3) внутренней* —волокнистого; отличаются
119
они друг от друга по окраске и строению. По толщине наружный и
внутренний слои одинаковы, тогда как в отдельности каждый из них
уступает среднему.
Под кутикулой лежит более темноокрашенный слой эпидермальных
клеток.
К эпидермальным клеткам изнутри прилегают разрезанные попе-
речно-мышечные клетки. Мышечная часть их имеет вид как бы вор-
РС
синок с более темными краями и светлой се-
рединой (d). В некоторых местах видна про-
топлазматическая часть мышечных клеток,
образованная тяжем протоплазмы с ядром,
лежащим между протоплазматической и мы-
шечной частью клетки.
Если рассматривать под микроскопом всю
окружность препарата, то можно видеть, что
мышечные клетки лежат не сплошным кру-
гом, а прерваны в четырех диаметрально про-
тивоположных местах (Л, с), вследствие чего
мышечные клетки окружности тела разби-
» ваются на четыре квадранта.
В двух местах перерыва ряда мышечных
клеток от эпителия кожных покровов отходит
стебельчатый на разрезе отросток, в вершине
которого проходит продольный ствол нерв-
ной системы. Эти места соответствуют поло-
жению спинной и брюшной линий аска-
риды. В вершине этих отростков сходятся
протоплазматические отростки мышечных кле-
ток правой и левой сторон (рис. 89).
Два других места перерыва приходятся на
середину правого и левого боков тела. От
эпителиальных клеток кожных покровов п
здесь отходит вырост, но иной формы —в виде
низкой неправильной трапеции (рис. 88,с).
У середины внутреннего края выроста виден
щелевидный овальный просвет — поперечный
разрез выделительного канала боковой линии
(рис. 90).
Стенка кишки состоит из толстой базальной
бесструктурной основной перепонки (mem-
brane propria), на внутренней стороне которой
Рис. 89. Поперечный раз-
рез через вентральную
линию лошадиной аска-
риды. По Гофману.
ЕМ — мышечные клетки; i —
внутренний слой кутикулы;
КЕ — ядра; Sf — опорные
фибриллы; N—нервные во-
локна ; РЕ — плазматические
отростки мышечных клеток.
лежит ряд очень узких и непропорционально высоких призматиче-
ских клеток эпителия с ядрами, помещающимися ровным рядом у са-
мого основания клеток; поверхность последних покрыта кутикулой
из тончайших столбиков, точно густой щеткой обрамляющих клетки.
В эпителии можно видеть различные зоны, характеризующиеся разно-
образной степенью зернистости протоплазмы, каковая связана с секре-
цией клетками пищеварительного сока и со всасыванием ими пищевых
веществ.
Яичник на разрезе сравним с колесом, спицы которого прилегают
120
тесно одна к другой: каждая спица есть развивающаяся яйцевая
клетка. В основании спиц, близ обода колеса, лежит по темному угло-
ватому зернышку —ядру. В центре колеса имеется небольшое
круглое поле зернистой протоплазмы, в которое упираются вершины
яйцевых клеток; оно соответствует поперечному разрезу стержня
(rachis).
Стенка яйцевода состоит из мышечного слоя, к которому внутри
фестонами прилегают эпителиальные клетки, между которыми остаются
бухтообразные углубления. В полости яйцевода можно найти округ-
лые яйцеклетки, но еще без оболочки, и множество живчиков. Послед-
ние у аскариды весьма своеобразны, ибо имеют вид конусов с более
Рис. 90. Поперечный разрез через боковую линию лошадиной
аскариды. По Шнейдеру.
ЕК — выделительный канал; nt — нервное волокно; Sc — тонкозернистая экскре-
торная плазма; St— опорные фибриллы; KN — гнездо ядер; MZ — медиаль-
ный ряд клеток; i 7- внутренний слой кутикулы.
широким и обособленным основанием, в котором и лежит ядро; жив-
чики аскариды неподвижны.
Матка внутри забита округлыми яйцами, одетыми толстой обо-
лочкой. Под оболочкой лежат развивающиеся' зародыши, точнее яйца
в фазе оплодотворения и редукции хроматина (т. е. процесса умень-
шения вдвое числа хромозом яйца), а также яйца в первых стадиях
дробления. Этот объект, между прочим, очень подходящ для изу-
чения процесса кариокинеза, т. е. непрямого деления клеток.
Между всеми этими органами видны щели —полость тела; однако
таковая считается ложной, ибо, по данным Г ольдшмидта,
пространства между органами заняты изоляционной тканью из круп-
ных сильно вакуолизированных клеток, которые разрушаются при
обычной обработке и становятся невидимыми.
121
Изолирование мышечных клеток аскариды
Лошадиные или человеческие аскариды фиксируются в 10% водном
растворе формалина и перед занятиями разрезаются бритвой на попе-
речные куски, длиной около 1 см, из передней и средней частей тела.
Занимающемуся дают кусочек аскариды в гли-
церине; ножницами его разрезают по длине
и двумя препаровальными иглами отделяют
под глицерином кутикулу; затем иглами рас-
щепляют клетки мышечного ряда по длине
и иглами же выделяют отдельные клетки.
Работа ведется под препаровальной лупой.
Удачно изолированные клетки заключают
в глицерин-желатину или мацерируют отрезки
стенки тела аскариды, оставшиеся после вскры-
тия в 20% едком кали в течение нескольких
часов. Затем материал сильно встряхивают
Рис. 91. Изолированная мышечная клетка
лошадиной аскариды. По Штемпелю.
в хорошо закупоренной пробирке; при этом
клетки отделяются от кутикулы, которой под-
дается действию едкой щелочи; едкий калий
сливают, дав предварительно клеткам осесть
на дно пробирки. Материал хорошо промы-
вают несколько раз водой. Окрашивают ге-
малауном, 1 промывают водой и обычным пу-
тем заключают в канадский бальзам.
Препарат рассматривают под микроскопом.
Мышечная клетка аскариды (рис. 91) имеет
при рассматривании ее сбоку протоплазмати-
ческую узкую часть, которая расширяется
по направлению к кутикуле покровов. В уз-
кой протоплазматической части лежит ядро,
Ке
Рис. 92. Попереч-
ный разрез мышеч-
ной клетки лоша-
диной аскариды.
ПоК.Ш нейдеру.
Sf — опорные фибрил-
лы; К — сократитель-
ные элементы; Ке —
ядро.
а в расширенной части (лежащей по длине тела аскариды) заложены
сократительные волокна (ссбственно мышечная часть клетки). Иногда
виден отходящий от протоплазматической части клетки протоплазма-
тический «нервный» отросток, идущий к срединному валику, заклю-
чающему в себе нервный ствол (рис. 92).
1 1 г кристаллического гематоксилина растворяют в 1 л дистиллированной
воды; к раствору прибавляют 50 г квасцов и 0,2 г йодистого натрия.
122
Строение изолированной мышечной клетки сравнивают с ее строе-
нием на поперечном срезе (см. выше). На поперечном срезе протоплаз-
матическая часть ее наиболее широка; сократительные волокна зале-
гают в поверхностных слоях —мышечной (узкой) части клетки; между
обоими корковыми слоями мышечных волокон, переходящих друг
в друга в дистальной 1 части клетки, залегает протоплазма такого же
строения, что и в толстой части клетки (рис. 92).
Строение губ аскариды
Отрезают поперечно бритвой или острым скальпелем передний
конец тела с губами и кладут кусочек разрезом вниз на предметное
Рис. 93. Губы аскариды человеческой; вид спе-
реди. По К. Скрябину и Р. Шульцу.
стекло. Наносят каплю молочной кислоты и закрывают препарат
покровным стеклом, подложив кусочки стекла соответствующей
толщины вместо ножек. Молочная кислота постепенно просветляет
препарат, причем последовательно проясняются детали строения губ.
Препарат рассматривают с апикальной поверхности (apex —вер-
шина, в данном случае—с переднего конца тела аскариды). Видны
три губы, между которыми имеются три линейные щели, сходящиеся
радиально в центре объекта. Губы имеют форму долек. Одна губа круп-
нее остальных двух; она лежит соответственно спинной стороне
1 Дистальный — лежащий далее к периферии (кнаружи); проксимальный —
лежащий ближе к телу или к его центральной части.
123
тела и потому называется дорзальной; остальные губы занимают
латеро-вентральное положение, лежат сбоку и вентрально (т. е. соот-
ветственно брюшной стороне тела). По мере просветления препарата
в периферической части губ начинают просвечивать сосочки, доходя-
щие до поверхностной кутикулы губ и являющиеся органами чувств
аскариды (рис. 93). В толще губ просвечивает пульпа, или мякоть,
состоящая из элементов субкутикулярных тканей; она имеет свое-
образные очертания.
Общие замечания
Тело аскариды построено по типу двубоковой симметрии; мета-
мерии в нем нет и следа.
Благодаря наличию веретеновидного, круглого на разрезе тела,
кишечника, обладающего и ротовым и заднепроходным отверстиями,
выделительных органов и отсутствию органов кровообращения, орга-
нов дыхания и ресничных или жгутиковых клеток где-либо в теле,
аскариды причисляются к подтипу круглых червей.
Многочисленные члены этого подтипа ведут одни свободный
образ жизни, другие живут в различных органах животных (реже
в тканях растений), т. е. являются паразитами. Аскариды принадле-
жат к числу последних.
От громадного большинства паразитических нематод аскарида ло-
шадиная отличается присутствием трех губ (кутикулярных вздутий) —
одной дорзальной и двух нижнебоковых, и веретеновидным телом.
Эти признаки характеризуют семейство аскарид—Ascaridae. Так как
аскарида лошадиная лишена крыловидных боковых придатков на
переднем конце тела и имеет две спикулы, то ее вместе с аскаридой
человеческой, аскаридой свиной и др. относят к роду Ascaris.
Совершенно своеобразные условия паразитической жизни —гото-
вая пища и относительное постоянство окружающей среды — накла-
дывают неизменный отпечаток на строение паразита в смысле упро-
щения его. В этом отношении нематоды не являются особенно хорошим
объектом, гак как у них менее, чем у других животных в аналогич-
ных условиях, имеется различий в организации свободно живущих и
паразитических форм. Все же можно отметить и здесь некоторые
поучительные моменты.
Чрезвычайная простота строения пищеварительного аппарата, от-
сутствие волосков и щетинок на кожных покровах (имеющихся у сво-
бодно живущих нематод), полное исчезновение глаз —все это связано
с переходом аскарид к паразитическому существованию.
Но паразитизм влияет на паразитов и положительно. Прогрессив-
ными особенностями организации аскарид являются:
а) сильное развитие половых органов и необычайная плодовитость,
в) выработка способности удерживаться в теле хозяина.
Паразит, обитающий в кишечнике своего хозяина, постоянно на-
ходится под влиянием изгоняющего действия перистальтики; для
паразита является вопросом жизни способность противостоять такому
действию. Если аскарида будет выброшена с калом из кишечника, она
неминуемо погибает, так как в нового хозяина она ни в каком случае
попасть не может.
124
У аскариды нет особых органов прикрепления, если не считать
слабо подвижных губ, которыми она, быть может, и защемляет сли-
зистую оболочку кишки. Но эта нематода ice же хорошо держится
в кишечнике, благодаря значительной ригидности ее длинного вере-
теновидного тела. Она точно налита и раздута жидкими соками; под
влиянием давления их изнутри на кожные покровы аскарида отли-
чается мало податливым телом. Концами его она крепко упирается
в стенки кишки, почему и не выталкивается проходящими отработан-
ными пищевыми массами. Более того, при нахождении большого числа
аскарид в кишке (их бывало иногда до 1000!) они могут образовывать
клубок, закупоривающий кишечник и обусловливающий его полную
непроходимость.
Благодаря той же ригидности и острым концам тела аскариды
могут прорывать измененную патологически стенку кишки (например,
у человека при брюшном тифе) и проникать в брюшную полость; в та-
ком случае они вызывают смертельное осложнение основной болезни
(так называемое прободение кишек и воспаление брюшины).
ТРИХИНА (TRICHINELLA SPIRALIS)
Технические замечания. 1) Добывают с бойни свиное трихи-
нозное мясо. Если возможно, берут его для занятий в свежем виде, предвари-
тельно расщипав на мелкие кусочки, параллельно ходу мышечных волокон. Ра-
ботающему выдается два толстых прямоугольных («двойное» стекло) куска стекла,
размером 5 х 10 см. Между ними крепко сжимают маленький кусочек свиной
мышцы и рассматривают под микроскопом при слабом увеличении.
2) Впрок заготовляют трихинозное мясо, консервируя его в 70% спирту.
К занятиям препараты переводят через воду в глицерин, разбавленный пополам
с водой. Разрезанные мелко и расщипанные кусочки исследуют под микроскопом
в глицерине, придавив препарат стеклом.
3) Для изготовления запаса постоянных бальзамных препаратов, кусочки
мелко расщипанной свиной мышцы проводят через 70%, 90%, 95% спирты, обез-
воживают в 100% спирту (сутки) в диализаторе с прокаленным химически чистым
медным купоросом (для надежности обезвоживания), просветляют в кедровом
или гвоздичном масле, споласкивают ксилолом (не обязательно) и заключают
в канадский бальзам или в дамар-лак. Такие неокрашенные препараты так сильно
просветляются, что трихинозные капсулы, а особенно только капсулирующиеся
трихины, различаются неопытным глазом с большим трудом.
Более показательны неокрашенные препараты, заключенные в глицерин-
желатину, но такие препараты менее прочны, чем бальзамные.
Демонстративнее — постоянные окрашенные препараты. Расщипанные мышцы
из 70% спирта на 1—2 суток переносят в борный кармин, затем диференцируют
в подкисленном спирте, сменяя его два-три раза; далее обычным путем через
спирты переводят в канадский бальзам.
4) При наличии благоустроенной лаборатории, в которой можно содержать
животных, трихин содержат в живом состоянии. Для этой цели скармливают
свежее трихинозное мясо мышам, белым крысам и даже кроликам. У таких живот-
ных можно получить кишечных половозрелых трихин, зародышей трихин в крови
и разные стадии капсуляции или личинок.
Инкапсулированные трихины
При рассматривании препарата, прежде всего, обращают внимание
на ряды параллельно идущих пучков поперечнополосатых мышечных
волокон. Их рубчатость лучше заметна при вращении микрометриче-
ского винта и сильном увеличении микроскопа. Местами мышечные
волокна разделяются прослойками волокнистой соединительной ткани,
125
а также группами сильно преломляющих свет (на глицериновых или
свежих препаратах) шаровидных телец (жировые клетки).
Рассматривая препарат, можно видеть, что кое-где мышечные
волокна оттесняются друг от друга вклиненными образованиями,
напоминающими по форме лимон. Это и будут трихинозные кап-
сулы.
В каждой капсуле можно различить стенку, полюсы и содержимое
в виде спирально скрученного или неправильно изогнутого червячка.
Иногда в капсуле бывает два таких червячка, являющихся личинками
трихины.
При долгой жизни (около Г—1% года) трихин в мышцах, в стенках
капсул отлагаются известковые соли, вследствие чего содержимое
становится невидимым. Если подбавить к препарату соляной кислоты,
то известковые соли растворяются, и капсула приобретает снова про-
зрачность. Наоборот, при начале капсулирования трихин вокруг их
личинок, внедрившихся в поперечнополосатые мышечные волокна,
капсулы еще нет. Различные стадии капсулирования обыкновенно
могут быть находимы на препаратах из мышц различных свиней,
давность заражения трихинами которых различна. При подогревании
до 40° С свежего препарата капсулированных трихин можно видеть
движение личинок внутри капсулы.
Естественного выхода наружу из капсул личинки трихин не имеют.
Живут они несколько лет и, вследствие обызвествления капсулы, по-
гибают или же погибают после смерти своего хозяина. Если же свиное
мясо будет съедено с живыми капсулированными личинками (не види-
мыми, как понятно из предыдущего, простым глазом) различными
животными —свиньями, мышами, крысами, а также и человеком, то
проглоченное мясо в желудке переваривается, капсулы вскрываются,
личинки же трихин освобождаются и начинают оживленно дви-
гаться.
С переходом пищи из желудка в кишки сюда же перебираются и
трихины. В тонких кишках они растут и достигают половой зрелости.
Самцы копулируют с самками (2—4 мм длины), после чего погибают
или уносятся с испражнениями. Самки же зарываются в слизистую
оболочку, где отрождают множество — 10 000 каждая—зародышей
(0,1 мм длины), которые попадают в кровеносные сосуды кишечника.
С током крови они разносятся по телу и оседают в мышцах; здесь
зародыши внедряются в поперечнополосатые мышечные волокна и
вырастают в уже знакомые нам личинки. Капсулы вокруг них возни-
кают в качестве реактивного образования со стороны тканей хозяина,
раздражаемых присутствием такого инородного тела, как личинка
трихины.
Таким образом один хозяин заражается непосредственно от другого,
поедая его мясо с живыми инкапсулированными трихинами.
Весьма демонстративен выход личинок трихин из капсул, что можно
видеть под микроскопом и даже показать на экране. Для проекции
пользуются обыкновенным микроскопом. Берут сильно зараженное
трихинами мясо, выделяют несколько пучков мышечных волокон,
расщипывают их и кладут в натуральный желудочный сок собаки
(добывается из фистулы желудка) или в раствор пепсина с соляной
126
кислотой. Препарат подогревают до 37° С на согревательном столике
микроскопа. Минут через 10 капсулы растворяются, а личинки начи-
нают плавать в жидкости, что очень эффектно проецируется на экране.
ПОЧВЕННЫЕ, ИЛИ САПРОФИТНЫЕ, НЕМАТОДЫ
Разрезанного на куски и вскрытого дождевого червя кладут
в чашку Коха с влажной садовой землей и закрывают плотно стеклом.
Чашку помещают в темное теплое место. Через несколько дней на
стекле можно видеть маленьких белых червячков. Их снимают тонкой
кисточкой или пинцетом и рассматривают в капле воды под микроско-
пом под покровным стеклом с ножками (свежий препарат).
Для приготовления тотальных препаратов живой материал фи-
ксируют горячим 70% спиртом с глицерином (100 см3 спирта и 5 см3
глицерина). В часовом стекле материал оставляют открытым в тер-
мостате при 40—50° С. После испарения спирта объекты заключают
в глицерин-желатину.
Вследствие прозрачности кутикулы лучшие результаты дает на-
блюдение свежих объектов.
Появившиеся червячки являются нематодами обычно двух родов —
Rhabditis и Diplogaster. Чаще встречаются виды Diplogaster longi-
cauda и Rhabditis teres. Это — очень мелкие нематоды, едва достигаю-
щие 1—1,5 мм длины. Самцы мельче самок и отличаются от них на-
личием близ заднего конца тела кутикулярных копулятивных орга-
нов в виде иголочек, стержней разной формы, называемых спику-
лами.
Rhabditis teres. Длина самки—до 1,45 мм, а самца — до 1,1 мм. Тело
самки (рис. 94, 7—2)узкое, к концам равномерно заостряющееся; одето
гладкой кутикулой; рот окружен шестью маленькими губами; рото-
вая полость длинная, узкая, с хорошо развитой кутикулярной вы-
стилкой, образующей у ее основания два кольцевидных утолщения.
Пищевод с двумя вздутиями; в заднем имеются кутикулярные зубы.
Длина пищевода равняется одной пятой длины всего тела. Кишеч-
ник тянется в виде прямой трубки; анальное отверстие лежит на
границе последней двадцать пятой части длины тела; за ним тело
сужается в короткий хвост (одной двадцать пятой длины тела).
Между вздутиями пищевода видно хорошо развитое волокнистое
нервное кольцо. Позади заднего вздутия пищевода можно рассмотреть
отверстие экскреторных органов (выделительная пора).
Органы размножения занимают все пространство, оставляемое
пищеварительным аппаратом, начиная от пищевода и до заднепро-
ходного отверстия. В матке обычно находится много мелких зароды-
шей. Края полового отверстия заметно выпячиваются наружу.
На заднем конце тела самца кутикула образует прозрачные вы-
росты в виде бурсы, или копулятивной сумки, служащей для обхваты-
вания тела самки при копуляции. Бурса подперта крупными кутику-
лярными сосочками, принимающими вид ребрышек; она окружает
хвост самца до конца.
Спикула —в виде трубочки, с ланцетовидной придаточной частью.
127
Diplogaster longicaudafjpw:. 94,3—5). Самка легко отличается от Rhab-
ditis длинным нитевидным хвостом, который составляет треть—четверть
длины всего тела. Кутикула покровов гладкая; тело узкое, стройное.
Рот окружен шестью губами, несущими острые сосочки. Ротовая по-
лость широкая и глубокая, с тремя зубовидными выростами кутикулы
Рис. 94. Свободно живущие нематоды. По Орли.
1 — самка Rhabditis; 2 — ее самец; 3 — Diplogaster longicauda, самка; 4—ее са-
мец; 6 — хвостовой конец самца, и — матка; а — anus; v — влагалище; sp — спи-
кулы ; о — яичник; Л — семенник; ое — пищевод.
в ее основании; половое отверстие лежит на середине длины тела; от
короткого влагалища отходят две трубки, образующие матку, семе-
приемник (с гроздьями живчиков) и загибающийся в обратную сто-
рону яичник. В матке — малое число (2—5) яиц с развивающимися
зародышами.
128
У самца копулятивная сумка узкая, охватывающая лишь перед-
нюю половину длины хвоста, с девятью парами сосочков. Спикулы
две, очень тонкие; их добавочная часть (так называемый рулек)
S-образной формы.
Экологические замечания. Рассмотренные нами
Rhabditis и Diplogaster являются свободно живущими нематодами,
которые питаются (особенно Diplogaster) разлагающимися раститель-
ными и животными остатками. Такой образ питания называется
сапрофитным (бт греческого sapros — гниющий). Обитают они в почве
и с землей могут заглатываться дождевыми червями. Особенно инте-
ресно то, что Rhabditis могут заползать и в метанефридии дождевых
червей и быть обнаружены в их трубках (в частности, Rh. pellia; на-
блюдать под микроскопом в капле воды вырезанные трубочки мета-
нефридиев). Рассматриваемые нематоды живут в черве, не причиняя
ему никакого вреда. После смерти дождевого червя они начинают
питаться и размножаться, что по времени совпадает с разложением
трупа.
Такую форму жизни, когда нематода живет в черве, но питается
и размножается лишь после его смерти, называют факультативным
паразитизмом.
Общие замечания
Сравнивая основные части строения паразитических и свободно
живущих нематод, мы устанавливаем в нем большое сходство; нема-
тоды отличаются от других беспозвоночных тем, что паразитизм
не налагает особенно резких отличий на паразитических круглых
червей сравнительно с их свободно живущими (в частности, сапро-
фитными) собратьями. Впрочем, следует отметить, что нет резкой гра-
ницы также между сапрофитным и паразитическим образом жизни.
Для нематод характерно сильное развитие кутикулы, которая
является, прежде всего, образованием защитным по отношению
к механическим и химическим воздействиям внешней среды (грубые
частицы почвы, воздействие стенок пищеварительного канала хозяина,
влияние ферментов и других химически действующих веществ и др.).
Интересно отметить, что у нематод способность к регенерации
практически отсутствует и в лучшем случае сводится лишь к зажи-
влению слабых ранений. Тем более при таких условиях необходима
для организма прочность кутикулы, на долю которой выпадает от-
ветственная задача надежной механической защиты нематоды от
повреждений. С кутикулой связаны и мышечные клетки, находящие
в ней опору при своих сокращениях, что увеличивает ее функциональ*
ную нагрузку. Тело нематод отличается ригидностью; оно как бы
переполнено внутренней жидкостью; под влиянием внутреннего
тургора -кутикула напряжена.
Несмотря на сильное развитие мускулатуры, движения нематод
весьма ограничены. Так как мышечные клетки лежат лишь вдоль
тела нематод, то при сокращении этих клеток спинной или брюшной
половины тела нематода изгибается то в ту, то в другую сторону,
совершая как бы качающиеся движения или волнообразные изгиба-
ния тела на спинную или брюшную сторону. Если псе мышцы сокра-
9 Практикум по зоологии — 385 129
щакугся одновременно, то соответствующий участок тела несколько
утончается, а общая длина тела возрастает.
Происхождение нематод неясно; в связи с этим 'стоит, очевидно,
и большая обособленность этой группы среди других групп типа червей.
Исследование ряда мелких нематод показало, что у них наблю-
дается постоянство числа клеток, из которых состоят те или другие
части тела особей данного вида. Например, для кроличьей острицы
(Oxyuris curvula) Е. Martini показал следующие данные:
Вся глотка................................. 80 клеток
Пищевод................................... 5 я
Эпителий средней кишки .................... Неопределенно
большое число
клеток
Мышечная оболочка средней кишки............ 9 клеток
Задняя кишка................................. 15
Крупные покровы без эпителиального синцития. 124 ,
Мезенхима..................................... 15 ,
Нервная система............................... 158 .
Половые органы.............................. 2 клетки и не-
оп ределенно боль-
шое число поло-
________________________вых клеток
Всего, не считая эпителия средней кишки, 412 постоян-
синцития покровов и половых клеток. ных клеток
Состав органов тела из малого и в то же время постоянного числа
клеток свидетельствует о весьма большой «специализации» клеток;
с этим можно связать и отсутствие способности нематод к регенера-
ции, так как специализованные клетки теряют способность к пере-
диференцировке, к замене их одна другой в функциональном отно-
шении и др.
Построение организма из определенного и относительно малого
числа строго диференцированных клеток следует считать явлением
более поздним или вторичным по отношению к составу тела живот-
ного из неопределенно большого числа клеток. Явление постоянства
числа клеток наблюдается у некоторых других животных (коловратки,
личинки иглоглавов и др.).
ПОДТИП КОЛЬЧАТЫЕ ЧЕРВИ
(ANNELIDES)
КЛАСС МАЛОЩЕТИНКОВЫЕ ЧЕРВИ (OLIGOCHAETA)
ДОЖДЕВОЙ ЧЕРВЬ (LUMBRICUS)
Технические замечания. Дождевых червей копают в огород-
ной земле, в компостных кучах; на свалках растительного мусора, подстилки из
хлевов или конюшен, одним словом в земле, богатой органическими веществами.
Целесообразнее заблаговременно запасать живой материал на зиму. Собранных
осенью червей кладут в ящики с землей или в большие цветочные горшки, кото-
рые и содержат в прохладном помещении, время от времени опрыскивая землю
водой для поддержания влажности. Чтобы животные не погибли от скученности,
рекомендуют на каждую сотню червей брать не менее трех — пяти ведер земли. Со-
бранные запасы охранять от нападения крыс, которые могут повытаскать червей.
130
Дождевых червей можно добыть и зимой, если осенью приметить места, где
их достаточно, например, в компостных огородных кучах. Перед занятием отби-
рают необходимое количество червей, обмывают их водой и кладут в банку со
слабым спиртом (около 10%). Через четверть часа они становятся совершенно
неподвижными и Годными для вскрытия. Для успеха занятий следует брать более
крупные экземпляры.
Для демонстрации движений червя оставляют несколько штук живыми.
Наружный осмотр
Вальковатое тело дождевого червя (рис. 95) состоит из большого
числа члеников, или сегментов, отделенных друг от друга кольцевыми
перетяжками. При осмотре живого животного
видно, что оно ползает на одной и той же стороне
тела, являющейся брюшной. Если червя поло-
жить на противоположную сторону тела, то он
принимает нормальное положение, переворачи-
ваясь на брюшную сторону. Спина дождевого
червя более выпукла, брюшная же поверхность
тела уплощенная.
Когда червь ползает по бумаге, то раздается
шорох, зависящий от царапания бумаги малень-
кими щетинками, которых имеется, в общем, по во-
семь на каждом сегменте (отсюда и название
малощетинковые черви — Oligochaeta). В их нали-
чии можно убедиться и другим путем, проводя
пальцем по телу червя; при этом легко ощутить,
как они цепляются за кожу пальцев. Та сторона
тела, на которой торчат щетинки, является брюш-
ной; она покрыта более светлой кожей, чем спин-
ная.
Толщина дождевого червя неодинакова. Го-
ловной конец его толще, чем задняя часть тела,
и обыкновенно окрашен темнее.
Тело половозрелых особей в области с тридцать
второго по тридцать седьмой членик заметно утол-
щено, вследствие образования пояска (clitellum).
Железы его вырабатывают секрет,- из которого
строится яйцевой кокон червя (рис. 95, с).
Над ротовым отверстием, которое поэтому не
видно со спинной стороны, нависает предротовая
лопасть (prostomium, или acron). Заднепроходное
отверстие лежит, наоборот, терминально, т. е. на
самом заднем конце тела (а).
По спинной стороне червя, начиная с десятого
сегмента, по средней линии тела, открываются так
называемые спинные поры, т. е. отверстия, веду-
Рис. 95. Дождевой
червь со спинной
стороны. ПоЖам-
м у.
Между тридцатым и
сороковым кольцами
виден clitellum(с),или
поясок; о—рот, а —
anus.
щие в полость тела. Лежат они в межсегментных перетяжках и обна-
руживаются в сильную лупу, если червя намотать, немного натя-
нув, на палец.
Если обсушенного фильтровальной бумагой живого дождевого
131
Рис. 96. Передний конец тела
дождевого червя. Видны ряды
щетинок.
st — (брюшные) и st слева—(нижние
боковые) отверстия сегментарных ор-
ганов (п) и половые (Q —женские,
—мужские) отверстия; о — рот;
е —границы сегментов. Римскими
цифрами показана последователь-
ность сегментов, начиная с переднего
конца червя.
червя подогреть на стеклянной пластинке или механически раздра-
жать, то он выделяет из спинных пор обильную жидкость, в которой
имеются различные клетки. Такая реакция червя является защитной.
При осмотре брюшной стороны тела
червя видно ротовое отверстие, лежащее
под предротовой лопастью (рис. 96, о).
Отметим также особенности некоторых
члеников, а именно (рис. 96):
а) на границе между девятым и деся-
тым, десятым и одиннадцатым сегмен-
тами открываются отверстия четырех
семеприемников;
б) четырнадцатый членик тела не-
сет пару женских половых отверстий
(рис. 96, $);
в) пятнадцатый сегмент тела обла-
дает двумя мужскими половыми отвер-
стиями (сГ).
Кроме того все членики тела, за
исключением первого, второго, третьего
и последнего, имеют каждый по два от-
верстия органов выделения — нефри-
диев, лежащих кпереди от брюшных
щетинок (л).
Все эти отверстия различимы лишь
под сильной лупой при особо благо-
приятных условиях, например, в период
половой деятельности.
Тело дождевого червя одето кож-
ными покровами, несущими на своей
поверхности гладкую кутикулу, отли-
вающую при падающем свете цветами
радуги. Такое явление называется ири-
зацией. Знакомые уже нам щетинки явля-
ются, в свою очередь, производными ку-
тикулы.
Кожа червя богата слизистыми же-
лезами. Выделяемая ими сЯизь служит
для защиты червя от высыхания, трения
в земле и т. п. Образуется она в изоби-
лии также при раздражении червя, на-
пример, при разрезании живого червя
на куски.
Хотя дождевой червь и лишен анато-
мически выраженных органов чувств, но
кожные покровы его обладают довольно высокой чувствительностью.
Они воспринимают световые раздражения, благодаря чему червь
уходит от дневного света, прячется в землю и таким образом спа-
сается от съедения дневными животными. Кожа его чувствительна
к различным химическим раздражениям (пары кислот и т. п ).
132
Вскрытие дождевого червя
Убитого дождевого червя прикалывают брюшной стороной книзу
двумя булавками ко дну восковой ванночки: одной булавкой —в го-
ловной конец тела (колоть несколько сбоку), другой—в хвостовой,
слегка при этом растянув животное. Червя придерживают посредине
пальцами левой руки; скальпелем делают продольный разрез по
средней линии спины. Тонким пинцетом отгибают *левый кожный
лоскут в сторону, подрезывая скальпелем удерживающие его попереч-
Рис. 97. Вскрытый передний конец тела дождевого червя. ________
Д—при раскрытии кожных лоскутов; В — по удалении дорзального кровеносного сосуда
и сердец; С — по удалении глотки и пищевода. Брюшная нервная цепочка видна на рис. С
в виде продольной черной полосы с утолщениями, от которых отходят периферические нервы.
Параллельные дугообразные полосы на внутренней стороне кожно-мускульного мешка обо-
значают на всех рисунках поперечный разрез перегородо < полости тела (диссепиментов); ме-
жду ними в передней и задней частях рисунков видны колбасовидные метанефридии. IX и
XV — порядковые номера сегментов, а — окологлоточное нервное кольцо; b — глотка; с —
пищевод; d зоб; е — мышечный желудок; in — кишечник;/—сердца; g — спинной сосуд;
к — семенные мешки; I — семеприемник; т — семепровод; п — яичник; о — яйцепровод;
ms — мышцы глотки; тп — рот. Пэ Ж а м м у.
ные перегородки полости тела (так называемые диссепименты), осво-
бождающийся лоскут прикалывают ко дну ванночки. Так же посту-
пают и с кожным лоскутом другой стороны тела. Булавки вкалывают
наискось, отклоняя головки их наружу. В конце концов вскрывают
червя по всей его длине. Ббльшая часть органов расположена а перед-
ней трети дождевого червя; вследствие этого вскрытие необходимо
делать здесь осторожно, чтобы не повредить внутренности. Вскрытие
и осмотр органов производят под водой. При вскрытии червя рассекают
кожно-мускульный мешок — стенку тела, состоящую из собственно
кожных покровов со сросшимися с ними мышцами тела.
133
Кожно-мускульный мешок ограничивает хорошо выраженную
вторичную полость тела (целом), разгороженную на камеры фрон-
тально стоящими перегородками (диссепименты), которые лежат
соответственно наружным границам члеников тела.
Большая часть полости тела дождевого червя занята кишечни-
ком— толстой трубкой снаружи желто-бурого цвета. По средней ее
линии идет тонкий красный шнур — дорзальный кровеносный сосуд,
тянущийся до переднего конца тела, в области которого скопляются
различные органы.
Идя спереди, находим (рис. 97,А):
а) овально или грушевидно вздутую глотку (Ь), богатую мыш-
цами (ms)', лежит она в первых шести сегментах; мышцы отходят
от нее радиально и прикрепляются изнутри к стенке тела;
б) в седьмом членике глотка переходит в тонкий и длинный пи-
щевод-,
в) по бокам пищевода видны огибающие его с боков полукруж-
ные кровеносные сосуды (так называемые сердца) (рис. 97,А, /), начи-
нающиеся от спинного сосуда;
г) в десятом, одиннадцатом и двенадцатом члениках лежат три
пары беловато-желтых лопастных тел, сливающихся по средней ли-
нии тела друг с другом; это — семенные мешки (vesiculae seminales —
А, С, к), помещаются они под и по бокам пищевода;
д) в тринадцатом сегменте пищевод переходит в вишневокрасный
зоб (proventriculus), занимающий четырнадцатый — пятнадцатый чле-
ники (Д, В, С, d);
е) за зобом следует мышечный желудок, на границе восемнад-
цатого и девятнадцатого члеников (А, С, е), открывающийся в собственно
кишечник. От этого пункта и до самого заднего конца тела строение
дождевого червя остается совершенно одинаковым;
ж) по бокам кишечника видны поперечно идущие полоски — от-
резки диссепиментов. Если качать ванночку, то можно заметить,
что между диссепиментами колеблются нитчатые образования, являю-
щиеся органами выделения метанефридиального типа.
Органы пищеварения. Дождевой червь питается перегнившими
растениями, которые он проглатывает вместе с землей. Через муску-
листую глотку пища поступает в пищевод, в который с боков впа-
дают три пары белых известковых железок (морреновские железы),
выделяющих углекислую известь; эта последняя служит для нейтра-
лизации гумусовой кислоты, которой богат растительный перегной.
Морреновские железы ищут, отодвигая друг от друга семенные мешки.
Если это не удается, то вырезают семенные мешки с одной какой-
либо стороны пищевода. Известковые железы лежат в десятом, один-
надцатом и двенадцатом сегментах в виде боковых выпячиваний
пищевода.
Далее пища переходит в зоб и мышечный желудок, отличающиеся
друг от друга различной толщиной стенок, которые в мышечном же-
лудке значительно толще, нежели в зобу.
Собственно кишка морфологически является средней кишкой, она
толста и длинна (A, in). При вскрытии стенка ее легко повреждается,
и из прорванных мест выступает черное, землистое и вязкое содер-
134
жимое кишки. Где-нибудь в задней части перерезают кишку поперек
и рассматривают плоскость разреза. От дорзальной стенки кишки
в глубь кишечника отходит складка, вдающаяся гребнем в его про-
свет. Называется она тифлозолем и служит для увеличения пищевари-
тельной поверхности кишки.
Края кишки неровны, благодаря чему она принимает до известной
степени четковидную форму, углубления на поверхности кишечника
соответствуют местам прикрепления диссепиментов. Задняя кишка
дождевого червя очень коротка и практически не отличима от средней.
Со стороны полости тела средняя кишка желто-бурого цвета. На
этом фоне ясно вырисовывается как продольный спинной сосуд, так
и мелкие боковые его веточки, отходящие от него перпендикулярно.
Если пинцетом поцарапать наружную поверхность кишки, то от
нее отходит тонкая муть. Дело в том, что кишка покрыта особыми
так называемыми хлорагогенными клетками, очень рыхло связанными
как друг с другом, так и с поверхностью кишки. При трении они
легко отделяются и дают в воде муть. Им-то и обязана кишка своей
наружной окраской. Хлорагогенные клетки относятся к выделитель-
ной системе, где и рассмотрим их подробнее.
Пищеварительный аппарат дождевого червя устроен сложнее, чем
у аскариды, что видно из сопоставления на таблице:
Аскарида Дождевой червь
Ротовое отверстие с губами Глотка Собственно кишка без каких-ли- бо придаточных частей Заднепроходное отверстие у самки) Отверстие клоаки у самца j Ротовое отверстие под предро- товой лопастью Глотка Пищевод с известковыми желе- зами Зоб Мышечный желудок Собственно кишка с тифлозолем Заднепроходное отверстие
Органы выделения. Хлорагогенные клетки происходят из эпи-
телия, выстилающего вторичную полость тела (целом) дождевого
червя. Лежат они собственно на сети нежных кровеносных сосудов,
оплетающих наружную поверхность кишки. Встречаются они и на
других органах, например, на мышечном желудке и даже, местами,
на половом аппарате.
Хлорагогенные клетки являются местом отложения экскреторных
продуктов, главным образом гуанина, от которого и зависит их жел-
тая окраска. Экскреторные вещества отлагаются в них в нераство-
римом состоянии, поэтому-то хлорагогенные клетки и могут быть от-
несены к органам внутриклеточного выделения.
Так как связь их друг с другом слабая, то они легко отрываются
и попадают в жидкость, заполняющую полость тела. Через спинные
поры или нефридиальные трубки они выходят из тела червя и, следо-
вательно, освобождают его от продуктов экскреции.
135
Собственно органы выделения у дождевого червя представлены
так называемыми метанефридиями; с их положением в теле мы уже
знакомы. Нефридии лежат по паре в каждом сегменте червя, кроме
трех первых и самого последнего; их поэтому называют также се-
гментарными органами.
Чтобы рассмотреть их строение, вырезают ножницами диссепи-
мент с прилежащей к нему трубочкой и рассматривают под микроско-
пом или сильной лупой в капле физиологического раствора поварен-
ной соли.
Нефридий (рис. 98) начинается маленькой воронкой (а), от-
крывающейся своим отверстием в полость тела. Воронка плотно
вставлена в диссепимент и по своему краю усажена венчиками рес-
ничек, работу которых можно наблюдать под микроскопом на живом
препарате. Узкой своей частью (&) воронка проходит в лежащий
позади сегме'нт, где загибается раза три (е), причем изгибы трубки
вытянуты в поперечном направлении. Далее она расширяется в кон-
Рис. 98. Схема строения метанефридия дожде-
вого червя.
а — воронка; b, с, d— части выводной трубки;
е — наружное отверстие. По Жамму,
цевой пузырек (</), откры-
вающийся наружу отвер-
стием (е) над боковой ще-
тинкой.
Когда реснички отвер-
стия воронки колеблются,
то они приводят в движе-
ние полостную жидкость,
которая, равно как и раз-
личные взвешенные в ней
клетки (например, хлора-
гогенные), загоняется в про-
свет нефридия и в конце
концов удаляется из тела.
Для органов выделения такого типа, как у дождевого червя, осо-
бенно характерно наличие мерцательной воронки, открывающейся
в полость тела. Такой тип органов и называют метанефрвдиями, в от-
личие от протонефридиев, с которыми мы познакомились у плоских
червей (стр. 85). Метанефридии свойственны лишь червям с раз-
витой вторичной полостью тела.
Органы кровообращения. С частью кровеносной системы дожде-
вого червя мы уже знакомы. Главным сосудом является спинной
сосуд, в котором кровь течет сзади наперед. В седьмом — одиннадцатом
сегментах от спинного сосуда отходят шесть пар более толстых полу-
кружных сосудов (рис. 97,A,f), огибающих с боков пищевод и впадаю-
щих под пищеводом в брюшной сосуд. Стенки полукружных сосу-
дов пульсируют, почему эти сосуды и называются сердцами. Они
перекачивают кровь из спинного ствола в брюшной. Задние пары
сердец прикрыты семенными мешками и лучше всего видны по удале-
нии последних. Затем удаляют кишечник от того места, где был про-
изведен разрез для обнаружения тифлозоля, и над нервной цепочкой
рассматривают брюшной сосуд, идущий также по средней линии
тела. Кровь течет в нем в обратном направлении, чем в спинном
сосуде, г. е. спереди назад.
136
По бокам нервной цепочки идет по тонкому околонервному также
продольному сосуду. Если отпрепаровать часть нервной цепочки и
сдвинуть ее в сторону, то под ней обнаруживается еще продоль-
ный поднервный сосуд.
Чтобы исчерпать систему продольных сосудов, остается упомянуть
о сосуде, лежащем в толще тифлозоля.
Все эти сосуды дают боковые ветви, которые своими концевыми
частями соединяются друг с другом и нигде не открываются в по-
лость тела. Следовательно, у дождевого червя имеется замкнутая
кровеносная система. О наличии такого типа органов кровообра-
щения можно заключить также по цвету крови. У дождевого червя
кровь красная, а полостная жидкость бесцветная. Очевидно, что
эти две жидкости нигде не соприкасаются друг с другом; в против-
ном случае рано или поздно они бы смешались и приобрели одинако-
вую окраску; следовательно, между кровеносной системой сосудов
и полостью тела сообщения нет нигде.
Совершенно неожиданно на примере дождевого червя мы наталки-
ваемся на некоторые черты сходства с позвоночными животными
(см. ниже), так как у последних кровь тоже красная и кровеносная
система замкнутая. Однако под этим чисто внешним сходством крове-
носной системы кроются важные различия.
Причина окраски крови червя другая, нежели у позвоночных:
красящее вещество крови —гемоглобин — у червя растворено
в плазме крови, а у позвоночных сосредоточено в красных кровяных
тельцах (эритроцитах), которые у беспозвоночных животных отсут-
ствуют.
Общность типа системы кровообращения в смысле ее замкнутости
не является признаком принципиального значения, так как крове-
носная система в обоих случаях очень различна как по существу, так
и по ориентировке в организме.
У дождевого червя, в связи с отсутствием особых органов дыха-
ния, нет разделения кровеносных сосудов на артерии и вены.
Дыхание. Особых органов дыхания у дождевого червя нет. Дышит
он поверхностью тела, в толще кожно-мускульного мешка которого
и разветвляются кровеносные сосуды. При условии влажности кожи
газообмен происходит через ее поверхность.
Возможно, что углекислота, скопляющаяся в крови червя, идет на
образование углекислой извести, выделяемой морреновыми железами
пищевода.
На примере дождевого червя мы видим животное, у которого
имеется только одно кожное дыхание.
Органы размножения. Гермафродитный половой аппарат дожде-
вого червя построен из мелких частей, которые не все доступны для
рассмотрения невооруженным глазом.
Три пары семенных мешков нам уже знакомы. Для полного рас-
смотрения их перерезают ножницами начало пищевода и, поднимая
его пинцетом, отпрепаровывают осторожно вплоть до зоба, где снова
делают поперечный разрез ножницами или отгибают здесь пищевод
назад.
Идя последовательно спереди назад, отыскивают (рис. 97, С):
137
а) первую пару семенных мешков, лежащих в девятом и десятом
сегментах;
б) первую пару семеприемников (receptaculum seminis) в виде
симметрично лежащих кнаружи от семенных мешков в девятом чле-
нике маленьких белых мешечков, являющихся выпячиванием кож-
ных покровов (/); в семеприемники поступает семя при копуляции
от другой особи и здесь хранится, пока у данного индивидуума не
созреют яйца и он не приступит к яйцекладке;
в) вторую пару семеприемников такого же строения, но лежащих
в десятом членике тела;
г) вторую пару семенных мешков в одиннадцатом сегменте;
д) третью пару семенных мешков в двенадцатом сегменте;
е) все эти три пары семенных мешков своими медиальными кон-
цами сливаются в срединный непарный мешок, в нормальном поло-
жении закрываемый сверху пищеводом (с).
Семенные мешки служат собственно для дозревания и хранения
живчиков, которые вырабатываются в двух парах семенников, лежа-
щих внутри непарной части семенных мешечков. При обычных заня-
тиях рассмотреть их не удается. Семенники лежат попарно —одна
пара позади другой. Латерально от них стенка семенного мешка про-
бодается двумя воронкообразными отверстиями. Каждое отверстие
(своей стороны тела) ведет в семевыносящий канадец (vas efferens);
оба канала, идя кзади, сливаются в семепровод (vas deferens). Семе-
проводы обеих сторон тела направляются назад и открываются двумя
самостоятельными мужскими половыми отверстиями в пятнадцатом
членике тела на брюшной его стороне (рис. 96, J)-
В строении семевыносящих путей нетрудно видеть сходство с мета-
нефридиями, благодаря присутствию воронкообразных начальных
отверстий. То, что воронки семевыносящих каналов открываются
внутрь семенных мешков, также является чертой принципиального
значения, ибо семенные мешки морфологически есть не что иное, как
разрастания диссепиментов соответствующих сегментов, и следова-
тельно, их полость является производным полости тела.
От типичных метанефридиев семепроводы дождевого червя отли-
чаются тем. что на одном конечном стволе сидят две воронки и самый
ствол проходит через четыре членика; но эти отличия второстепен-
ного значения.
Семевыносящие пути дождевого червя, равно как и женская часть
гермафродитного полового аппарата этого животного, недоступны
для обыкновенного наблюдения вследствие своих малых размеров.
Все же весной, в период кладки яиц, под лупой можно рассмотреть
пару яичников (ovaria) грушевидной формы, лежащих на брюшной
стенке тринадцатого’ сегмента.
Яйцеводы (oviducti) —в виде двух коротких трубочек с воронко-
образными отверстиями, пронизывающими диссепимент между три-
надцатым и четырнадцатым сегментами. В противоположность семе-
проводам, яйцеводы очень коротки; открываются они двумя женскими
половыми отверстиями на брюшной стороне четырнадцатого сегмента,
т. е. на членик ближе кпереди от мужских половых отверстий. Обычно
яйцевод рассмотреть нельзя.
138
подлежащими
Рис. 99. Копуля-
ция дождевых чер-
вей (Allolobophora
foetida). Из В. До-
геля.
Яйцеводы также устроены по типу метанефридиев, но с второсте-
пенным уклонением от этого типа в сторону укорочения трубки и
упрощения ее строения.
Что между метанефридиями дождевого червя и его выводными
половыми путями обнаруживается тождество основного плана строе-
ния. не удивительно. Созревшие половые продукты для данной особи
являются, как и экскреторные вещества, объектами,
удалению из тела, что и производится одними и
теми же органами, т. е. метанефридиями.
Копуляция дождевых червей.
Во время копуляции каждая особь играет одно-
временно роль самца и самки. Процесс этот
весьма своеобразен. «Два червя соединяются друг
с другом обыкновенно в своих норках и приле-
гают один к другому брюшной стороной, причем
одноименные концы бывают направлены в разные
стороны (рис. 99). Поясок (см. выше, стр. 131)
одного прилегает к девятому, десятому и одиннад-
цатому кольцам другого, т. е. к кольцам, между
которыми открываются обе пары семеприемников.
Слизь, выделяемая особенно пояском, связывает
обоих животных. С каждого бока тела от пояска
вперед к пятнадцатому сегменту, где открываются
протоки семенников, тянется продольная скла-
дочка, образуемая сокращением мускулов и окай-
мленная с каждой стороны продольным желобком.
По верхнему желобку пробегают во время сово-
купления волнообразные сокращения от переднего
конца к заднему. Этими сокращениями гонится
назад капелька семени, выступающая из мужского
полового отверстия. За первой капелькой вскоре
выступает вторая, затем третья и т. д., с обеих
сторон. Все капли собираются к пояску и к обра-
щенным к нему отверстиям семеприемника другого
индивидуума. При наполнении семеприемников,
может быть, происходит всасывание ими семени.
После этого черви расходятся» (Г е с с е и Д о-
ф л е й н).
Копуляция дождевых червей по времени не совпадает с созрева-
нием яиц. Последние созревают позднее. К периоду кладки яиц поя-
сок выделяет значительное количество слизи в виде муфты, одеваю-
щей тело червя. Сокращением тела дождевой червь сдвигает муфту
кпереди, как бы выползая из нее. Когда муфта проходит мимо жен-
ских половых отверстий, яйца откладываются в ее полость. При даль-
нейшем сдвигании ее кпереди она минует отверстие семеприемников;
из них изливается семя, которое и оплодотворяет яйца вне тела
червя. В конце концов муфта с яйцами и живчиками сбрасывается
совсем с тела, края ее спадаются, и она превращается в кокон дожде-
вого червя, в коем и протекает эмбриональное развитие заро-
дышей.
139
Таким образом у дождевого червя копуляция сопряжена с после-
дующим наружным оплодотворением.
Центральная нервная система. Чтобы рассмотреть начало нервной
системы, спинной кожный разрез доводят до самого переднего конца
тела, а лоскуты отгибают в сторону. Над глоткой видим два белых
сближенных узелка, образующих надглоточный узел (рис. 97,С, а); от
него в стороны и вниз, огибая с боков глотку, отходят глоточные
коммиссуры, сливающиеся в подглоточный нервный узел. Последний
становится заметным по отрезании глотки ножницами тотчас позади
надглоточного узла.
Надглоточный узел, боковые коммиссуры и подглоточный узел
в своей совокупности образуют окологлоточное нервное кольцо, сквозь
которое проходит глотка.
По удалении семенных мешков, зоба, мышечного желудка и кишки
обнаруживается брюшная нервная цепочка, состоящая из нервных узел-
ков, лежащих по одному в каждом членике тела (рис. 97,С). В связи
с однородностью сегментации дождевого червя и с отсутствием конеч-
ностей, все нервные узлы цепочки имеют одинаковую величину.
Таким образом нервная система дождевого червя построена по це-
почечному типу.
Органы чувств. У дождевого червя нет органов чувств, которые
были бы видны простым глазом; однако кожные покровы его обладают
чувствительностью как к световым, так и к иным раздражителям.
Поперечный разрез дождевого червя в области кишечника 1
Общая ориентировка. На поперечном разрезе дождевого червя
(рис. 100) снаружи видны: бесструктурная кутикула, покрывающая
эпидермис, состоящий из индиферентных клеток (ЕР), между кото-
рыми лежат слизистые одноклеточные железы, секрет которых
хорошо красится кислыми красками; кроме того местами лежат чув-
ствительные почки, состоящие из скопления тонких клеток с чув-
ствительными столбиками, пронизывающими кутикулу.
Под эпителием кожи лежит слой круговых (кольцевых) мышц (RM),
кнутри от которых расположен, более толстый слой продольных мышц
(dFLM, vF, IF). Эпителий и мышцы образуют вместе кожно-мускуль-
ный мешок; изнутри последний выстлан перитонеальным эпителием
(эндотелием—Per), ограничивающим вторичную полость тела, или
целом (DPI).
Местами в стенке тела видны перерезанные основания щетинок
(Во) и мышц, приводящих их в движение.
1 Так как дождевой червь проглатывает землю, то для изготовления срезов
предварительно необходимо ее удалить. Червя сажают в банку с картофельной
мукой или смоченной бумагой и выдерживают, пока его кишечник не освободится
от земли и не заполнится крахмалом. После этого червя убивают хлороформом
и фиксируют горячим насыщенным водным раствором сулемы, смешанным с вдвое
меньшим количеством спирта. Фиксирование разрезанного на куски червя длится
2—3 часа. После препарат проводят через 50%, 75%, 80%, 95% спирт, абсолют-
ный алкоголь, ксилол, ксилол с парафином и парафин (в термостате). Срезы окра-
шивают гематоксилином с эозином
140
В полости тела концентрически залегает кишечник (Ent); его
наружная поверхность покрыта хлорагогеиными клетками, под кото-
рыми лежит мышечная оболочка кишки; изнутри на ней покоится
эпителиальная выстилка кишечника; дорзальная стенка последнего
образует языкообразное впячивание внутрь, или тифлозоль (Ту).
В тифлозоле и по спинной поверхности кишки можно рассмотреть
поперечное сечение спинного сосуда и сосуда тифлозоля; между
кишкой и брюшной стенкой тела находится брюшная нервная це-
почка (Вт), которая может быть рассечена или в области самого
узла, или через коммиссуры; над и под цепочкой проходят кровенос-
ные сосуды (брюшной сосуд — vG). В боковых частях целома
Рис. 100. Поперечный разрез дождевого червя на гуровне
кишки. По К. Шнейдеру.,
ЕР — эпителий; Во — щетинка; Вт — нервная цепочка; RM — коль-
цевая мускулатура; dFLM, IF, vF, acF, Zw, BoF — спинное, боковое,
брюшное, добавочное и междущетинковое мускульные поля (продоль-
ные мускулы); Ent — эпителий кишечника; Ту—тифлозоль; ВоМ —
мускулатура щетинки; Per—перитонеальная выстилка; Ne — нефридий;
ЕВ—расширение трубки нефридия; dG. vG, snG, si (/—-спинной, брюш-
ной субневральные кровеносные сосуды и сосуд тифлозоля; DPI — вто-
ричная полость тела; DL — просвет кишечника. По К. Ш[н е й д е р у.
видны различные части метанефридиев — ЕВ (чаще извитые канальцы)
иногда попадаются на разрезе и части диссепиментов.
Детали микроскопического строения. После общей ориентировки
в препарате поперечного разреза дождевого червя рассмотрим детали
его микроскопического строения. Для этой цели используют также
срезы, приготовляемые из различных частей тела (кожа, семеприем-
ники, стенки кишечника и др.). Кроме того приготовляют временные
или постоянные препараты.
1. Эпителий покровов (эпидермис) образован узкими
и высокими индиферентными клетками, лежащими на тонкой базаль-
ной перепонке (рис. 101, Ь). Каждая клетка имеет овальное, бедное
хроматином ядро. &
141
Одноклеточные слизистые железы (рис. 101, Sd) значительно
превышают по размерам индиферентные клетки (Z) и сдавливают
прилежащие к ним клетки эпидермиса. В базальной части протоплазмы
слизистой клетки лежит ядро; остальная протоплазма занята
секретом и имеет как бы губчатую структуру. Секрет внутри клетки
окрашивается основными красками (например, гематоксилином)
в темносине-фиолетовый цвет и потому называется базофильным. Секрет
изливается наружу через тонкое отверстие (пору) в толще кутикулы
(С). В зависимости от интенсивности секреции протоплазма слизистых
клеток имеет то более зернистый, то губчатый вид.
Другие железистые клетки с зернистым секретом отличаются
иным отношением к краскам; они окрашиваются кислыми красками
(эозин и др.) и потому называются ацидофильными (их также называют
Рис. 101. Эпидермис дождевого червя, разрез. По Г о ф-
м а н у.
С — кутикула; Sd и Sa— слизистые железы; Ed — «белковая железа»;
Z—эпидермальная клетка; I—лимфоцит; b — базальная мембрана;
гт — кольцевые мышцы.
«белковыми», рис. 101, Ed). При сильном развитии одноклеточных желез
они могут доминировать над индиферентными клетками эпидермиса,
которые при этом сильно сдавливаются, ядра же их оттесняются на
разные уровни толщи эпидермиса.
В базальной части эпидермиса можно видеть мелкие округлые
клетки с крупным ядром; это — блуждающие клетки, или лимфоциты
(рис. 101, /), проникающие через базальную мембрану в толщу эпидер-
миса.
В некоторых местах тела, например, в пояске, железистые клетки
достигают колоссального развития, вследствие чего индиферентные
клетки (сильно сдавленные) обнаруживаются с трудом. Железистые
клетки не умещаются в толще эпидермы и разрастаются через базаль-
ную мембрану, принимая субэпителиальное положение. Мало того,
железистые клетки располагаются друг под другом в несколько ря-
дов (рис. 102, слева). Здесь различают три сорта одноклеточных желез:
а) «губчатые» железы (лежат в периферийной части толщи пояска — IV),
б) грубозернисты* железы (также занимающие поверхностную
142
часть толщи пояска — К) и в) тонкозернистые (самые многочислен-
ные) железы, лежащие во много рядов (Kd).
2. Щетинки своим основанием сидят в мешковидных впячи-
ваниях (сумках) эпителия покровов. Впячиваясь в мешок, эпителий
быстро уплощается; в мешке нет никаких железистых клеток. На дне
мешка молодой щетинки можно видеть щетинкообразовательную
клетку. Дно мешка образует как бы фолликулярную головку (С), со-
Рис. 102.
Слева — дождевой червь, разрез через дорзальную часть пояска в период зрелости. W—
«губчатые» железы; К — грубозернистые железы; Kd— тонкозернистые железы. По Гоф-
ман у. Справа— разрез через мешок щетинки. В — щетинка; С — фолликулярная головка;
Р — протракторы щетинки; R — ретрактор ее; Рг — брюшинный эпителий. Посредине — раз-
рез через покровы с почкой чувствительных клеток (sz). с — кутикула; sc — то же, с чувстви-
тельными щетинками; е — эпидермальные клетки; п — нервные волокна чувствительной почки;
mf— мышечные волокна; гт — кольцевая мышца; 1т — продольные мышцы; гп — круговой
нерв. По Плате.
стоящую из мышечных клеток; сюда (и к нижней трети сумки) при-
крепляются мышечные пучки протрактора щетинки (рис. 102, Р);
при их сокращении дно сумки подтягивается кверху, вследствие
чего щетинка выдвигается наружу; антагонистом этой мышцы является
ретрактор сумки (рис. 102, R).
3. На некоторых срезах, прошедших через определенную зону
сегмента, в эпителии покровов можно найти группы чувствитель-
на
ных клеток, образующих чувствительный сосочек, или почку. Они
узки и длинны (рис. 102); ядра их лежат на разных уровнях; чув-
ствительная почка несколько выдается над поверхностью покровов;
она покрыта более тонкой кутикулой с очень тонкими порами, через
которые выступают наружу короткие чувствительные щетинки —
по одной от каждой клетки (sc). В своем основании чувствительные
клетки продолжаются в нервные отростки, пучок которых пронизы-
вает мышечный слой кожно-мышечного мешка и вливается в круго-
вой нерв сегмента (рис. 102, гл). Эти почки являются, видимо, орга-
Рис. 103. Дождевой дорвь — поперечный раз-
рез через кожно-мускульный мешок. По
К. Шнейдеру.
с — кутикула; е— эпидермис; s — одноклеточные
слизистые железы; гт— кольцевые мышцы; bg—соеди-
нительная ткань; sp — соединительнотканные перего-
родки; к — «камеры» мышц; р — перитонеальный
эпителий.
нами осязания и рецепто-
рами химических и терми-
ческих раздражений.
4. Мускулатуру
кожно - мышечного мешка
рассматривают на попереч-
ном срезе сегмента (рис.
103). Под эпидермисом (е)
залегают кольцевые мышеч-
ные волокна (гт), прослоен-
ные соединительной тканью;
соединительная ткань, окру-
жающая мышеные волок-
на, называется перимизием
(bg). Мышечные волокна до-
вольно длинны и на концах
несколько заострены. По
ходу мышечных волокон
местами видны их удлинен-
ные ядра, лежащие в сред-
ней части волокна сбоку,
в некотором утолщении сар-
коплазмы. Ядра клеток пе-
римизия меньше по разме-
рам и более округлы.
В протоплазме этих клеток
бывают скопления мель-
чайших блестящих пало-
чек, которые считают сим-
биотическими бактероида-
ми (т. е. образованиями, по-
хожими на бактерий). Однако, повидимому, более правильно видеть
в них пордукты внутриклеточного выделения.
К солям кольцевых мышц непосредственно примыкают продоль-
ные мышцы. Они лежат как бы в камерах, образованных соедини-
тельной тканью (рис. ЮЗ, к), которая проходит между мышечными
элементами в виде септ, на поперечном разрезе представленных тон-
кими линиями (sp). Эти септы подходят к перитонеальному (брю-
шинному) эпителию (р), выстилающему вторичную полость тела
дождевого червя. Перитонеальный эпителий, покрывающий изнутри
кожно-мускульный мешок, называется париетальным (т. е. присте-
144
ночным) листком брюшинной выстилки. Перитонеальный эпителий
развивается из среднего зародышевого листка, т. е. из мезодермы.
Между соединительнотканными септами лежат продольные мышцы
дождевого червя; они состоят из множества мышечных волокон,
имеющих на поперечном разрезе вид коротких уплощенных образо-
ваний или более правильную округлую форму. В своей совокупности
они образуют в каждой камере фигуру как бы елочки (рис. 103),
лежа двумя боковыми, соединяющимися на концах друг с другом
рядами; между последними лежит клеточная соединительная ткань
Рис. 104. Поперечный разрез через тифлозоль кишечника дождевого
червя. По К. Шнейдеру.
Е—эпителий; "ch— хлорагогенные клетки; 1т — продольные мышечные во-
локна; гт — круговые мышечные волокна; b — соединительная ткань; g — кро-
. веносный сосуд тифлозоля; Са — капилляры; rg — спинной сосуд.
с темноокрашивающимися ядрами. Мышечные волокна также имеют
ядра, по одному на каждое волокно; вследствие значительной длины
последних, на поперечных срезах они видны редко. Соединительно-
тканные септы также имеют ядра, которые сильно уплощены (рис. 103).
5. Кишечник рассматриваем на срезе, прошедшем на уровне
тифлозоля. Изнутри кишечник выстлан высокими, узкими эпите-
лиальными клетками (рис. 104), покрытыми на свободной поверх-
ности мерцательными ресничками. Такой эпителий называют мерца-
тельным. Ядра его клеток овальной формы; лежат в средней зоне
10 Практикум по зоологии—385 145
плазма которых заполнена зернистым
71Z
клеток. Эпителий протоплазмы часто теряет реснички. Между клет-
ками кишечного эпителия встречаются железистые клетки, прото-
секретом, или же имеющим вид
сетчатых тяжей. Ядро
железистой клетки отте-
снено к самому ее осно-
ванию и часто трудно
различимо. В базальной
зоне эпителия встреча-
ются лимфоциты (блу-
ждающие клетки), про-
двигающиеся и дальше
к просвету кишечника.
Эпителий кишечника
одет мышечной оболоч-
кой (рис. 104), состоя-
щей из кольцевых и про-
дольных мышц (гт, 1т).
Между первыми и осно-
Рис. 105. А — поперечный разрез через коннективу, и В — через ганглий брюш-
ной нервной цепочки дождевого червя. По К. Шнейдеру.
р — перитонеальный покров; kf — колоссальные волокна; nf — нервные волокна; nz — нерв-
ные клетки; hg — покровная ткань; mf— мышечные волокна; ge — субневральный сосуд; b —
бактероиды; In — боковой нерв.
ванием эпителия проходят тонкие разветвления кровеносных со-
судов — капилляры в виде разной формы мелких просветов (Са).
Между всеми мышечными волокнами проходит скудный перимизий
Снаружи кишечник покрыт модифицированным перитонеальным эпи.
146
телием (его внутренностным, или висцеральным, листком), образован-
ным уже известными нам хлорагогенными клетками (ch). На попереч-
ном разрезе они имеют клиновидную, булавовидную или иную форму.
Их круглые ядра лежат на различных уровнях. Протоплазма этих
клеток имеет различный вид, смотря по состоянию деятельности кле-
ток (вакуолизованная, зернистая и др.).
Тифлозоль имеет вид складки, внутренняя часть которой имеет
полость; в ней среди хлорагогенных клеток лежит продольный крове-
носный сосуд тифлозоля (g). Над основанием тифлозоля лежит
мощный слой хлорагогенных клеток, в толще которого проходит
дорзальный спинной сосуд (rg).
6. На различных срезах можно видеть поперечное сечение нервного
узла брюшной цепочки или же соединяющего нервные узлы тяжа
(коннектива). На нашем препарате можно рассмотреть лишь общие
черты строения этих частей нервной системы (рис. 105). Нервный
узел отличается крупными размерами (В); на поперечном разрезе он
имеет форму неправильной чечевицы, от боков которой отходят боко-
вые нервы (In —видны не на всяком срезе). Под ганглием лежит
овальный в поперечном сечении поднервный кровеносный сосуд; он
одет снаружи соединительной тканью, общей с покровом ганглия.
Волокнистая соединительная ткань как бы капсулой одевает нерв-
ные элементы узла; в дорзальной его части эта ткань резко обособляет
друг от друга три гигантских нервных тяжа (рис. 105, kf), или
неврохорда, имеющих на поперечном разрезе форму овалов с тонко-
зернистым содержимым. Центральная часть препарата занята нерв-
ными волокнами (л/—нейрофибриллы); далее к периферии лежат
нервные клетки (nz); последние отличаются громадными размерами,
по сравнению с другими клетками тела; форма их часто клиновидная;
сужающаяся часть клетки является началом нервного отростка (ней-
рита), отходящего от нервной клетки; его можно проследить лишь
на коротком протяжении.
Центральная часть препарата, занятая нервными волокнами
и трудно различимыми клетками глии, подразделяется как бы
на два боковых узла, соединенных по средней части друг с другом.
Филогенетически нервная цепочка дождевого червя развилась из
двойной нервной цепочки; следом ее слияния и является описанная
особенность строения нервного ганглия; то же еще нагляднее видно
на поперечном сечении коннективы; под соединительнотканной обо-
лочкой видны разрезы двух раздельно лежащих нервных тяжей с не-
большим количеством нервных клеток (рис. 105,А).
Регенерация дождевого червя
Несмотря на сравнительно высокую организацию, дождевой червь
способен к регенерации. Даже несколько сегментов, вырезанных из
передней или средней части тела дождевого червя, восстанавливают
утраченное. Регенерат появляется в виде маленького придатка (почка),
который увеличивается в размерах, особенно в длину, вследствие
увеличения количества сегментов (рис. 106).
147
Уже по одним внешним соотношениям можно установить различия
в характере процесса регенерации у планарий и у дождевого червя.
Регенерирующая вырезанная часть тела первой, путем передиферен-
цировки ее тканей, превращается в миниатюрную планарию (процесс
морфоллаксиса). У дождевого червя восстанавливаемая часть в про-
цессе регенерации растет за пределы взятого для эксперимента отрезка
тела, что является результатом новообразования тканей.
Рис. 106. Части переднего и среднего
отделов тела дождевого червя (Lum-
bricus rubellus) с регенератами раз-
ного возраста. По Е. Коршельту.
Продольные гистологические
разрезы через передний регенери-
рующий конец дождевого червя на
разных этапах регенерации позво-
ляют установить основные моменты
этого процесса (рис. 107):
1) рана закрывается «пробкой»
из кровяных клеток;
2) под ее прикрытием обра-
зуется новый эпителий, разраста-
ющийся от краев покровов раны;
3) разрезанные края кишечника
смыкаются и образуют слепой ме-
шок (рис. 107, А);
4) на стадии появления регене-
ративной почки (В) видно множе-
ство кариокинезов клеток различ-
ных тканей регенерата; кроме того
появляется множество энергично
делящихся индиферентных клеток,
отделяющихся как от эктодермы,
так и от мышц и от мезенхимы;
5) некоторые старые органы,
как кишечник (средняя кишка) и
брюшная нервная цепочка, дают
в регенеративную почку выросты
и в конце концов соответственно
образуют открывающуюся на-
ружу переднюю кишку, нервные
ганглии; такие же органы, как не-
фридии, диссепименты, сосуды и др., возникают за счет индиферент-
ных клеток регенерирующего бугорка.
В данном случае регенерация является настоящей; новообразова-
ние частей происходит за счет роста соответствующих тканей и за
счет преобразования индиферентных, мало диференцированных кле-
ток, склонных к энергичному размножению.
Общие замечания
Тело дождевого червя имеет хорошо выраженную двубоковую
симметрию; кроме того он является примером животного с весьма
явственной метамерией.
Последняя проявляется не только однородной внешней сегмента-
148
цией, но и сегментацией внутренней, ибо полость тела червя пере-
городками разделяется на метамерно лежащие камеры. Каждый мета-
мер дождевого червя типически представлен сегментом тела со своим
отделом полости тела, с парой метанефридиев, брюшным нервным
узлом, участком кишечника и спинного сосуда.
В связи с однородностью сегментации дождевого червя стоит спо-
собность его к регенерации. Если червя разрезать на куски, то в них
Рис. 107. Продольные разрезы через переднюю регенерирую-
щую часть тела дождевого червя. По Гешелеру (из
Ю. Филипченко).
1у— пробка кровяных клеток; а— кишечник; п — нервная цепочка;
og—надглоточный ганглий.
регенерирует соответственно головной и хвостовой концы тела. Выра-
женность метамерии свидетельствует об относительно низком уровне
организации животного.
Однако и у дождевого червя в передней части тела замечается не-
которое нарушение метамерии вследствие присутствия здесь половых
органов и начальной части пищеварительного аппарата.
Разделение тела дождевого червя на членики со всеми связанными
с этим обстоятельством особенностями внутреннего строения опре-
деляют принадлежность его к кольчецам (Annelida).
14 9
Наличие у него слабо развитых и малочисленных щетинок (кож-
ных), а также гермафродитность полового аппарата характеризуют
принадлежность его к классу малощетинковых кольчецов Oligochaeta.
Экология. Дождевой червь живет в земле, где выкапывает
себе норки. Днем он держится в почве, ночью же выходит на поверх-
ность земли. У дождевого червя отсутствуют глаза, однако он ощущает
не только дневной, но и искусственный свет. Если ночью направить
свет фонаря на выползшего из своей норы дождевого червя, то он
сразу же сокращается.
Не вся поверхность тела одинаково чувствительна к свету. Более
всего чувствителен передний конец, менее —задний и еще менее —
средняя часть тела. Указанная только что способность червя объ-
ясняется присутствием в толще эпителия кожи и под ним чувствитель-
ных «зрительных» клеток, к которым подходят нервные волокна.
Дождевой червь способен воспринимать и химические раздражения.
Так, он избегает почвы с кислой реакцией, ощущает присутствие пищи,
зарытой на некоторую глубину в землю и т. п. Органами «химического
чувства» дождевого червя являются также чувствительные клетки,
рассеянные в его кожных покровах. И в этом случае наиболее раз-
дражимы концы тела (особенно передний).
Таким образом в отношении развития органов чувств дождевой
червь является низко организованным животным, ибо анатомически
выраженных органов чувств у него нет; восприятие же раздражений
извне выпадает на долю чувствительных кожных клеток.
Дождевой червь передвигается путем попеременного сокращения
и вытягивания тела. При сокращении тела передний конец прикре-
пляется к земле, а задний подтягивается к нему; при выпячивании же,
наоборот, тело упирается в землю задним концом, тогда как передний
выпячивается вперед. Такое движение тела зависит от попеременного
сокращения кольцевых и продольных мышц и напряжения тела под
давлением полостной жидкости.
Несмотря на отсутствие конечностей, дождевой червь роет ходы
в почве, работая головным концом тела. Он может также втягивать
различные предметы в норы, например, листья, куски бумаги, перья
птиц и т. п. Орудует он при этом своей глоткой, которая выворачи-
вается через ротовое отверстие и захватывает данный предмет.
Земля, прошедшая с растительными остатками через кишечник
дождевого червя, делается весьма плодородной.
КЛАСС МНОГОЩЕТИНКОВЫЕ ЧЕРВИ (POLYCHAETA)
НЕРЕИДА (NEREIS sp.)
Запасают спиртовый материал, получаемый на морских биологи-
ческих станциях (Севастополь, Мурман и др.).
Наружный осмотр
Тело нереиды вальковатое, несколько сплющенное в дорзовен-
тралыюм направлении, подразделенное на однородные сегменты. На
переднем конце тела лежит головной отдел, отличающийся по своим
150
Рис. 108. Параподия нереиды. Оригинал.
придаткам и строению от остальных сегментов. Каждый сегмент
несет по бокам лопастевидные выросты, рассеченные на части и сна-
бженные пучками щетинок. Эти выросты являются примитивными ко-
нечностями и называются параподиями. Головной отдел несет рото-
вое отверстие; задний сегмент тела —анальное отверстие.
Препарат параподии. Отрезают ножницами параподию и заключают
без окраски в канадский бальзам; слабое увеличение микроскопа.
Параподия представляет собой лопастевидный вырост боковой
стенки тела червя; плоскость параподии перпендикулярна направле-
нию продольной оси тела.
В пароподии различают две мощные щетинки, называемые аци-
кулами; каждую сопровождает пучок более тонких и коротких щети-
нок; каждая из двух групп
щетинок относится: одна—
к спинной, другая—к брюш-
ной части параподии; на-
зываются последние соот-
ветственно нотоподией и
невроподией; обе они рас-
щепляются на две дольки
и несут по червовидному
придатку — спинному и
брюшному циррусу (рис.
108) . 1
Параподии являются не-
отчлененными и расчленен-
ными выпячиваниями тела,
действующими наподобие
весел; их надо считать за
примитивные конечности.
Препарат головного от-
дела. От маленькой по раз-
мерам нереиды отрезают головной отдел с несколькими передними
сегментами тела и заключают без окраски в канадский бальзам.
При рассматривании сверху находят (рис. 109):
1) ротовой сегмент—peristomium, лишенный параподий (6);
2) кпереди по средней линии тянется головная предротовая лопасть
(prostomium —3). с двумя короткими церебральными щупальцами
или лобными усиками, иначе называемыми антеннами (Z);
3) латерально из-под головной лопасти отходят массивные дву-
членистые пальпы (2);
4) в основании головной лопасти лежат четыре расположенных
трапецией глазка (5);
5) кроме описанных придатков, имеются еще восемь осязательных,,
или щупальцевых, усиков (4), которые соответствуют червовидным
(усиковым) придаткам параподий; сами параподии ротового сегмента
и их щетинки редуцировались, а усики сохранились, но переменили
1 Термин cirrus имеет различное значение; у паразитических червей циррусом
называется копулятивный орган.
151’
•положение и функцию, превратившись из придатков параподий в при-
датки головы; отметим
ротовой сегмент (peristomium)
образовался слиянием
двух первых сегментов
тела, и вместе с предро-
товым отделом он обра-
зует голову нереиды.
На брюшной стороне
ротового сегмента лежит
ротовое отверстие; из
него может выпячиваться
наружу глотка, воору-
женная кутикулярными
челюстями.
также, что
In.
b
b
Ьг
пг
vd
1.т
tm
bm
ПО. Поперечный разрез многощетинко-
червя. ПоН. Холодковскому.
о?’ гт vv
Поперечный разрез
тела нереиды
Острой бритвой раз-
резают фиксированную
спиртом нереиду на по-
Рис. 109. Голова нереиды (Nereis). Оригинал, перечне ориентирован-
ные тонкие ломтики, вы-
бирают те, в которых имеются параподии и без окраски заключают
в канадский бальзам (ножки!); можно также фиксировать их суле-
мой. Кусочки средней части
тела свежей нереиды окра-
шивают борным кармином
и пикриновой кислотой, за-
ливают в парафин и изго-
товляют толстые микротом-
ные срезы до 20р.. Выби-
рают срезы, прошедшие че-
рез параподии и между
ними. Для более деталь-
ного изучения микроскопи-
ческого строения срезы из
неокрашенного кусочка
окрашивают гематоксили-
ном и эозином. Препараты
пригодны для общей ориен-
тировки в строении нереи-
ды. На них рассматриваем
(рис. 110):
1) поверхностную ку-
тикулу, под которой ле-
жит эпидермис, соедини-
тельнотканной подстилкой
ный мешок;
2) в центре препарата
Рис.
ВОГО
Ь, ас — щетинки; 1т — продольны^ мышцы; vd — спин-
ной сосуд жабры; de и vc — спинные и брюшные
усики; dp и vp— спинные и брюшные ветви парапо-
дий; гт — кольцевые мышцы; Ьт — брюшной нервный
ствол; vi —брюшной кровеносный сосуд; tm — косые
мышцы; ov — гонады; иг — нефридии; Ьг — воронка
нефридия; в центре — кишечник.
связанный с мышцами в кожно-мускуль-
виден поперечный разрез кишечника
152
3) между кожно-мускульным мешком и наружной оболочкой,
кишечника лежит вторичная полость тела (целом);
4) под кишечником на средней линии тела виден поперечный разрез,
брюшной нервной цепочки;
5) диаметрально противоположно на спинной стороне виден раз-
рез дорзального кровеносного сосуда;
6) из мышц различают: а) поперечные разрезы вентральных (сбоку
брюшной нервной цепочки) и дорзальных продольных мышц; б) косые
поперечные мышцы, идущие из-под нервной цепочки кнаружи и кверху
над брюшными продольными мышцами; в) слабо развитые кольцевые
мышцы, лежащие непосредственно под эпидермисом; г) мышцы, упра-
вляющие движениями ацикул, мешки которых глубоко погружены
в полость тела;
7) сбоку разреза тела может быть видна параподия; в зависи-
мости от уровня и ориентировки разреза видны щетинки ацикулы и
управляющие ими мышцы. Весьма характерно то, что.полость тела
(целом) продолжается в полость параподии, которая является не-
отчлененным от тела выростом кожно-мускульного мешка;
8) в полости тела можно заметить скопления свободно лежащих
клеток или кучки их; это —половые клетки, которые образуются
в некоторых местах перитонеального эпителия и по созревании своем
падают в полость тела, откуда и выносятся через мета нефридии;
последние видны в виде различно перерезанных трубок; благодаря
своей двойной функции —выведению экскреторных продуктов и поло-
вых клеток —их называют нефромиксиями.
Тотальный препарат
нефромиксиев
При вскрытии не-
реиды удаляют кишеч-
ник и вырезают ткани
из нижнебоковой ча-
сти стенки нереиды.
Препарат кладут в раз-
бавленный глицерин
(рис. 111, А).
Нефридий имеет
вид длинной тонкой
извитой трубки, от-
крывающейся ворон-
кой в полость тела; за
воронкой идет шейка,
которая далее обра-
зует клубок причуд-
Рис. 111. Нефридии. По Г ар.меру и Ш и пл ей.
А— Nereis, В — Arenicola. f—воронка; п — шейка' не-
фридия; t—извитой каналец; с — соединительная тканью
d— выводная часть; х— наружное отверстие нефридия £
т— пузыревидное расширение нефридия.
ливых изгибов; клубок одет соединительной тканью; значительная
часть внутренней поверхности трубки нефридии несет реснички; из
массива соединительной ткани выходит выводной проток нефридия,
открывающийся наружу у основания параподии. В каждом сегменте
тела нереиды, исключая несколько передних и задних, лежит по
паре митанефридиев, шейка которых проходит через диссепимент.
153
Общие замечания
Явственная гомономная метамерия нереиды нарушается обособле-
нием головного отдела, являющегося специально диференцированной
•сложной частью тела, на которой сосредоточены органы чувств (глаза,
усики, щупальца) и где открывается ротовое отверстие.
Кефализация, т. е. процесс образования головы за счет передних
сегментов тела характерна для многощетинковых червей, и в про-
цессе эволюции червей является важным шагом вперед в смысле подня-
тия высоты организации. Обособленная на переднем конце тела голова,
оснащенная разнообразными органами чувств, обеспечивает червю
лучшую ориентировку в окружающей среде.
Вторая характерная особенность —это образование параподий,
которые следует рассматривать как примитивные конечности; пара-
подии несут по много щетинок, отсюда и название червей —много-
щетинковые (Polychaeta); пароподии движутся спереди назад и цепляют
щетинками за субстрат дна, облегчая червю движение вперед.
Кроме того параподии снабжены органами чувств —усиками,
а у нереиды играют роль жабр, так как в их лопастях обильно ветвятся
кровеносные сосуды. В отличие от типичных пароподий, у нереиды
таковые лишены особых жабр (см. ниже —пескожил).
ПЕСКОЖИЛ (ARENICOLA MARINA)
Материал можно получить с побережья Мурмана. Пескожил —
крупная полихета длиной до двадцати пяти и более сантиметров;
живет в песке морского дна в неглубоких местах,
особенно если они лишаются воды при отливе. О место-
нахождении червей в песке можно судить по наличию
кучек колбасовидно скученных песчинок (экскременты
пескожила). Если их сгрести, то в толще песка можно
найти самого червя, который лежит изогнуто в виде
буквы U. Песчинки, прилежащие непосредственно
к телу червя, склеены в трубку, в которой и лежит
пескожил.
Рис. 112. Песко-
жил.
ках переднего
парные пучки
Наружный осмотр
При рассмотрении фиксированного спиртом жи-
вотного видно, что его кольчатое тело разделяется
на три отдела: передний, средний и задний. Средний
состоит из тринадцати сегментов, несущих по бокам
ветвистые жабры; передний —из семи сегментов,
вторично подразделенных кольцевидными перетяж-
ками по четыре—пять в каждом сегменте. Первый
сегмент является головным; из него высовывается
хобот (глотка); его бока покрыты сосочками, а перед-
няя часть имеет ротовое отверстие. На всех члени-
и среднего отделов тела, кроме головного, имеются
щетинок, торчащих на небольших возвышениях по-
154
кровов, 'они являются нотоподиями. В среднем отделе тела каждая
нотоподия несет жабру в виде пучка древовидно разветвленных вы-
ростов кожных покровов.
На брюшной стороне сегментов имеются валикообразные, попе-
речно идущие выпячивания покровов, являющиеся невроподиями.
В каждой из них спрятан ряд коротких крючковидно изогнутых щети-
нок числом 18. Таким образом у пескожила имеются оба компонента
параподий в лице сильно модифицированных ното- и невроподий.
Задний отдел тела пескожила более тонок; он лишен каких-либо»
придатков.
Вскрытие пескожила
Разрез по общим правилам вскрытия (см. дождевой червь, стр. 133)'
через толщу кожно-мускульного мешка по средней линии спины.
Отмечаем очень малое число диссепиментов —всего ясно распо-
знаем три в передней части тела.
В передней части препарата лежит способная выворачиваться
глотка (хобот) с мышцами-ретракторами; далее идет пищевод, впадаю-
щий в очень короткий желудок, имеющий два слепых мешка.
Кишка подвешена на мезентериальных нитях, являющихся остат-
ками редуцировавшихся диссепиментов; она местами покрыта хлораго-
геиными клетками.
Находим спинной сосуд; по удалении его и кишечника, по средней
линии тела отыскиваем брюшной нервный ствол; по бокам от него идут
поперечные мышцы. Срезаем ножницами поперечные мышцы на одной
стороне, вырезаем кусочек стенки тела и рассматриваем в разбавлен-
ном глицерине под микроскопом (при слабом увеличении) нефридии
пескожила (рис. 111, В).
Открывающаяся в полость тела широкая воронка имеет дорзальную
и вентральную губы с пальцевидными придатками. Воронка переходит
в широкий канал нефридия, служащий для выведения и половых и
экскреторных продуктов. Коричневоокрашенный канал расширяется
в концевой пузырек, открывающийся наружу порой. В стенке нефри-
дия видны кровеносные сосуды. Как и у нереиды, у пескожила нефри-
дии несут двойную функцию.
Общие замечания
Нереида и пескожил относятся к многощетинковым червям. Для
понимания некоторых различий в их организации (при ее однотип-
ности) следует помнить, что нереида — свободно живущее животное,
тогда как пескожил ведет сидячий образ жизни, зарываясь в песок
морского дна.
Метамерия у нереиды выражена в классической форме, тогда как
у пескожила ее однообразие нарушается разделением тела на три отдела,
имеющие различный внешний вид.
Обособления головы у пескожила, в противоположность нереиде,
нет; это можно поставить в связь с более однообразными условиями
существования пескожила в песке морского дна и упрощенным добыва-
155-
нием пищи, тогда как нереиды отличаются активностью и подвиж-
ностью.
Следует отметить полную редукцию параподий в задней области
тела и своеобразное развитие их в остальных частях; бросается в глаза
большое различие в строении ното- и невроподий, причем последние
соединяются друг с другом в общий валик на брюшной стороне соответ-
ствующего сегмента. Соотношения в степени развития ното- и невро-
подий у пескожила различны; в передней части тела они вообще слабо
развиты и совершенно разобщены друг от друга.
Типичным моментом является присутствие жабр, связанных с ното-
подиями среднего отдела тела. В то же время сильное развитие жабр
в виде кустиков веткообразных выростов покровов связано с обита-
нием пескожила в узкой трубке в толще песка, где омывание жабр
притоком свежей воды более затруднено, по сравнению с омыванием
параподий свободно живущих полихет.
Не касаясь чрезвычайного разнообразия особенностей организации
и жизни полихет, даже на двух рассмотренных примерах ясно видим,
что отдельные виды многощетинковых червей могут деградировать
с общего высокого уровня организации, достигнутого этой группой
животных в процессе эволюции типа червей. Частичная деградация
связана с последовавшим приспособлением различных видов к специ-
альным условиям обитания. Пластичность их организации обеспечила
в одних случаях утерю ранее бывших органов (параподий, придатков
головы и др.), в других — более сильное развитие некоторых частей
с соответствующим усилением или видоизменением их функций (невро-
подии, жабры).
Общие замечания по рассмотренным группам червей
Вместе с Oligochaeta, Polychaeta относятся к щетинковым коль-
чатым червям (Chaetopoda).
По сравнению с Polychaeta, Oligochaeta имеют упрощения в своей
организации, как-то: отсутствие параподий, жабр, диференцирован-
ной головы, органов чувств. Эти особенности следует считать вторич-
ными, возникшими в связи с переходом малощетинковых червей (в лице
дождевых червей) к наземному образу жизни (в толще почвы). В связи
с этим вполне обосновано считать Polychaeta предками Oligochaeta.
Сравнение важнейших особенностей их организации дано в
таблице на стр. 157.
Знаком «(+)» показана последовавшая в филогенезе редукция
органа, ранее существовавшего у предков данного животного.
Мы закончили ознакомление со строением некоторых представите-
лей важнейших подтипов червей и получили достаточно данных для
общего сравнения их с целью конечной характеристики этого типа
в целом.
Из сопоставления наиболее важных признаков можно видеть,
что тип червей в целом характеризуется лишь немногими признаками
а именно —двубоковой симметрией, более или менее удлиненным
телом, отсутствием обособленных конечностей, нервной системой,
состоящей из головных ганглиев и нервных стволов. Более точное
156
* Название органов Кольчатые червя Круглые черви Плоские червя
многоше нереида тинковые пескожил малощетинковые дождевой червь ленточные сосальщики планарии
( Тело Более или менее удлиненное
Вальковатое Округлое Плоское
Расчлененное Нерасчлепенное Членистое Нечленистое
Метамерия Двубоковая симметрия Вторичная полость тела .... Конечности Голова Органы кровообращения.... Органы дыхания Органы пищеварения Анальное отверстие Органы выделения Нервная система 4- + Вы Пара! + Замкну Жа1 i Мет ц е 1 + + )ажена очень ЮДИН (+)- тая кровенос бры 1 i а н е ф 1 ! н о ч е + + хорошо От параподий остались толь- ко одни ще- тинки (+)- :ная система Кожа + + р и д и и 1 ч н а я + Обособление < головногс 1 Интрамол + Своеобразные Шнуровая без метамерии + + ГГОЛОВКИ»— 1 конца — екулярное дь .-£+) г Узловая с + Обособление КОНЦс ixaime + ротонефриди продольным! + е переднего i тела Покровы + и 1 I стволами
Анатомически выраженные ор- ганы чувств + (+)- (+)- — — — +
Щупальца + 1 - — — — | — —
обоснование для подразделения типа червей на подтипы дается эмбрио-
логией.
Кишечнополостные и плоские черви стоят ближе друг к другу по
своим морфологическим признакам, нежели те и другие к коль-
чатым червям.
Эти соотношения вытекают из нижеприводимой таблицы.
Сравнение признаков некоторых типов животных
Кишечнополостные Плоские черви Кольчатые черви
Эктодерма + + +
Энтодерма + -J +
Мезодерма — + +
Ротовое отверстие + (+)- +
Anus — — +
Гастро-васкулярная система . . + (+)- —
Вторичная полость — — +
Кровеносная система — — +
Органы выделения • . — + +
Нервная система Диффузная Узловая Цепочечная
Половые органы • . . . Только гонады Сложно устро- Сложно ^стро-
ены ены
Стрекательные клетки + — —
Симметрия Радиальная Двубоковая Двубоковая
В сущности, тип червей является более всех других сборным типом.
ТИП ЧЛЕНИСТОНОГИЕ (ARTHROPODA)
КЛАСС РАКООБРАЗНЫЕ (CRUSTACEA)
РЕЧНОЙ РАК (POTAMOBIUS ASTACUS)
Технические замечания. Для вскрытия пользуются живыми
раками. Сохранять раков живыми очень трудно. Их можно держать лишь три-
четыре дня (редко больше) в плетенках, переложенных слоями умеренно увлаж-
ненного мха.
За */»—1 час до занятия потребное количество раков усыпляют хлороформом
в плотно закрываемой банке. Вскрытие производится в ванночке под водой. Для
ознакомления лишь с расчленением рака можно пользоваться материалом, сохра-
ненным в формалине (одна часть формалина на десять частей воды) или спирте
(70%).
Раки живут в пресной воде и держатся в норах, вырытых в берегах ниже
уровня воды. Они предпочитают водоемы с более или менее жесткой водой.
Наружный осмотр
Тело рака одето толстой и твердой скорлупой, по латинскому наиме-
нованию которой —crusta —все животные класса ракообразных
получили название Crustacea. Кожные покровы рака состоят из слоя
эпителиальных клеток, т. е. гиподермы и выделяемой ею наружной
кутикулы, образованной так называемым хитином, который к тому же
пропитывается известковыми солями. Последнее обстоятельство и
обусловливает значительную крепость скорлупы рака, 1 являющейся
в то же время его наружным кожным скелетом. При внешнем осмотре
рака видна только хитиновая кутикула, темногрязно-зеленого или
коричневого цвета. Окраска рака зависит от присутствия в кутикуле
особых пигментов, которые диффузно пропитывают хитиновую кути-
кулу. Бывают случаи нахождения раков, лишенных какой бы то ни было
пигментации, такое явление носит общее название альбинизма.
При варке скорлупа рака принимает красный цвет вследствие перехода
голубого пигмента покровов, называемого цианокристаллином, в крас-
ный зооэритрин.
Различные части тела могут двигаться одна относительно другой,
благодаря нахождению между ними мягкой сочленовной перепонки,
также покрытой снаружи хитиновой (более тонкой) кутикулой, в кото-
рой не отлагаются известковые соли.
В теле рака при осмотре его спины ясно различают два отдела:
головогрудь, покрытую сплошным панцырем, и членистое брюшко.
Головогрудь (cephalothorax) спереди оканчивается острым, в виде
‘ 40% всего веса покровов приходится на их органическое вешество, а 60% —
на углекислый кальций и другие известковые соли.
159
шипа, лобным выступом, называемым rostrum. Боковые края
последнего, в свою очередь, могут иметь зубцы (в зависимости от вида
рака).
Из-под переднего края rostrum торчит первая пара сяжков, или
усиков (antennulae). Кнаружи от них лежат длинные усики второй
пары (antennae)/ Под боковыми краями rostrum прячутся сильно
выпуклые, сидящие на подвижных ножках стебельчатые глаза.
Приблизительно посредине длины головогрудного щита, или пан-
цыря (сагарах), идет обращенный выпуклостью кзади дугообразный
желобок, называемый затылочной бороздкой (sutura cervicalis). От
Рис. 113. Речной рак. По Н. Холодковскому.
А— самец и В — самка с брюшной стороны. Ап — первая, А — вторая пара сяжков;
7— верхняя губа; 2 — верхняя челюсть; 3 — ногочелюсть; 4 — клешня;
5 — вторая, 8 — пятая нога; 9 — мужское половое отверстие; 10—13— брюшные
ножки; 14— тельсон; 15— anus; Е — отверстие зеленой железы; О— отверстие
яйцевода. У самки на ножках брюшка (7—6) икра.
выпуклой части последней идут кзади две параллельные короткие
бороздки (suturae branchiocardiacae), ограничивающие поле, под
которым лежит сердце; поэтому и называются они сердечными борозд-
ками. Кнаружи от них панцырь головогруди загибается латерально
и вниз боковыми лопастями, покрывающими жабры. Из-под краев
головогруди выступают пять пар ходильных ног (pereiopodia, рис. 113,
A, 4-8).
160
Брюшко образовано шестью сегментами и концевым отделом, назы-
ваемым telson. По бокам шестого членика лежит по паре плоских
пластинок—uropodia, являющихся видоизмененными конечностями,
которые, подобно вееру, могут складываться (рис. 113, В, 6).
При осмотре нескольких экземпляров раков со спины можно за-
метить некоторые отличия в относительной ширине брюшка. У самца
оно по ширине почти равняется наиболее широкой части головогруди,
у самки же заметно шире. Более надежные наружные половые при-
знаки могут быть обнаружены на нижней поверхности рака.
Сегменты брюшка состоит каждый из широкого спинного полу-
кольца — тергита, и узкого брюшного полукольца —стернитпа.
Боковые части тергита свисают свободно в виде эпимер. Между основа-
нием грудных ног видны узкие треугольные пластинки брюшной
стороны панцыря тела (sternum).
При осмотре брюшной стороны рака, идя спереди назад, находим
следующее:
а) Пару коротких и пару длинных сяжков; у основания последних
виден бугорок, на котором открывается сяжковая железа (рис. ИЗ,
В, Е) соответствующей стороны тела (орган выделения).
б) Группу ротовых органов, окружающих ротовое отверстие (раз-
двинуть их пинцетом и ввести осторожно конец пинцета в ротовое
отверстие, рис. 113, А, 7, 2).
в) Пять пар ходильных ног; основания их лежат между нижними
свободными краями боковых лопастей головогрудного панцыря,
плотно прилегающего к спинке тела.
Первая пара ног развита гораздо сильнее прочих; она несет на своих
концах мощные клешни (chelae), служащие для захвата пищи и для
защиты. Клешня имеет два пальца —один неподвижный, другой
подвижный.
Вторая и третья пары имеют концевые клешни, но очень маленьких
размеров; четвертая и пятая пары оканчиваются короткими когте-
видными члениками.
Если у рассматриваемого рака имеется по отверстию у основания
третьей пары ног, значит перед нами — самка. Самец отличается
расположением своих двух половых отверстий у основания пятой
пары ног (рис. 113, А, 9).
г) Брюшко рака на нижней поверхности также несет ножки по
паре в каждом членике (pleopodia). Всего их шесть пар. У самки они
являются плавательными; кроме того в период кладки и вынашивания
яиц они служат для прикрепления последних.
Брюшные ножки самца не одинаковы по форме; передние две пары
изогнуты под прямым углом и несут длинные концевые членики в виде
желобчатых палочек белого цвета; служат они в качестве копулятив-
ных органов (рис. 113, А, 10).
Все конечности рака состоят из различного числа подвижных
члеников; отсюда и название членистоногие (Arthropoda) для всех
животных, отвечающих этому признаку.
д) На нижней поверхности telson имеется продольно лежащее
щелевидное анальное отверстие (рис. 113, А, 15). Края telson и
его боковых лопастей, называемых уроподиями, рулевыми или
11 Практикум по зоологии—385. 161
прыгательными ножками, обрамлены густой каймой хитиновых во-
лосков.
В заключение наружного осмотра суммируем наружные половые
отличия рака:
Название частей Самец Самка
Брюшко Подовые отверстия . . . Ножки брюшка Не шире головогруди У основания пятой пары ног головогруди Две передние пары ножек развиты сильнее и в об- щей сложности образуют копулятивный аппарат Шире головогруди У основания третьей пары ног головогруди Одинаковые, только одна пара их рудиментарна
Иногда встречаются раки с неодинаковыми по величине клешнями,
а у иных клешня вовсе может отсутствовать. Дело объясняется легкой
способностью рака терять конечности. Если рака держать за ногу и
раздражать, то он в конце концов отламывает ногу в строго опреде-
ленном месте. Такая способность его называется автотомией, или
самокалечением. Отломанная конечность восстанавливается заново,
т. е. регенерирует; маленькие клешни на первой паре ходильных ног
и являются такими регенератами.
Так как тело рака одето крепким хитиновым панцырем, рост
животного протекает своеобразно, именно периодически. Рост рака
связан с линькой, т. е. со сбрасыванием старой хитиновой кутикулы
и с заменой ее новой. Это происходит следующим образом. У готовя-
щегося к линьке рака гиподерма вырабатывает молодую кутикулу,
отлагающуюся под старым панцырем тела. В конце концов последний
теряет связь с кожными покровами и лопается на границе брюшка и
головогруди. К этому времени рак вытаскивает ноги из их старых
футляров. В образовавшуюся щель постепенно выдвигается тело рака,
покрытое молодой кожицей. В конце концов рак совсем освобо-
ждается от старого хитинового панцыря.
Будучи одет мягкой хитиновой кожицей, рак расправляет части
тела, которые под напором внутренних органов и полостной жидкости
несколько растягиваются и увеличиваются в размерах. Дня три новая
кожица рака остается мягкой, затем в ней отлагаются известковые
соли, и новый панцырь рака затвердевает и остается в своих неизмен-
ных размерах до новой линьки и т. д.
За свою 15—20-летнюю жизнь рак линяет много раз. На первом
году жизни рак линяет восемь раз, на втором — пять раз, на третьем—
два раза; в последующие годы самка — по одному, а самец — по два
раза в год. Линька приходится на теплое время года.
Для отвердения хитиновой кутикулы требуются известковые соли;
в период перед линькой рак избирает пищу, более богатую известко-
выми солями; омары же, например, съедают сброшенный ими хити-
новый панцырь.
162
Вскрытие рака и его внутреннее строение
Для вскрытия рака прикалывают булавками спиной кверху ко дну
ванночки за боковые придатки telson и за первую пару ног, растянув
при этом животное по мере возможности.
Перерезают мягкую сочленовную перепонку между головогрудью
и брюшком скальпелем. Вводят в разрез ветвь ножниц и рассекают
кпереди и дугообразно головогрудный щит по направлению к глазу
с каждой стороны тела. Передние концы разрезов соединяют попереч-
ным разрезом позади глаз. Поднимают пальцем или пинцетом задний
край очерченного разрезами куска панцыря и скальпелем подрезают
все, что Прикрепляется к нему изнутри (при препаровке скальпелем
скрести все время по самЛ скорлупе изнутри); в конце концов пан-
цирь снимается прочь.
Поворачивают ванночку головным концом к себе и ножницами
делают два продольных боковых разреза до telson. Приподнимают
передний конец первого членика брюшка, скальпелем подрезают
удерживающие его мышцы и снимают прочь отрезанные части всех
тергитов брюшка, после чего ванночку ставят в прежнее положение.
Расположение внутренностей. При снятии панцыря головогруди
на внутренних органах часто остается пленка фиолетово-красного
цвета — это гиподерма, покрытая у готовящихся к линьке раков ново-
образованной мягкой кутикулой. Ее осторожно снимают ножницами
и пинцетом.
Из всех внутренних органов наиболее поверхностно лежит сердце.
Оно имеет вид студенистого беловатого мешочка пятиугольной формы
(рис. 118, а; 114, 8), помещающегося сейчас же кпереди от начала
брюшка.
Из-под сердца выглядывают концы трехлопастной половой железы—
яичника или семенника, величина, окраска и структура которой
изменяются в зависимости от периода половой деятельности. Две
лопасти половой железы направлены вперед, а одна —назад
(рис. 114, 4).
По бокам от сердца у самца видны многочисленные извивы тонкой
трубки яркобелого цвета —это семепроводы (рис. 114, 6).
Снаружи и спереди от половой железы и сердца вплоть до боковых
стенок тела помещается парная печень (9), образованная множеством
тесно прилегающих друг к другу трубочек грязножелтого цвета.
В переднем конце препарата лежит большой желудок (7), трех-
угольный в его задней половине. Впереди от его дорзальной стенки
отходят плоские мышцы. Стенка желудка неоднородна. В ней заложены
очень твердые пластинки и стержни, просвечивающие насквозь (скле-
риты — жевательный желудочный аппарат).
С боков трехугольной части желудка видны овальные тела с шеро-
ховатой поверхностью —поперечные разрезы челюстных мышц (2).
По бокам же переднего отдела желудка, в глубине препарата, помеща-
ются сяжковые железы (1).
Из-под сердца назад по средней линии брюшка идет прямая трубка,
часто с темным содержимым —это задняя кишка (10); проходит она
в толще белесоватых мышц брюшка.
163
Органы размножения. Поднимают пинцетом сердце и замечают,
что от него вперед, в стороны и назад тянутся тонкие, почти прозрач-
ные нити —артерии. Сердце удаляют и рассматривают лежащую под
ним половую железу.
Рис. 114. Вскрытый речной рак (самец). По Ж а м м у.
1—почка (зеленая железа); 2 — жевательная (мандибулярная) мышца; 3 — жабры (боковая
стенка панцыря снята); 4 — семенник; 5 — брюшная нервная цепочка; 6 — перерезанный
семепровод; 7 — желудок; 8 — сердце; М — печень; 10 — задняя кишка; 11 — часть разрезан-
ного семепровода; 12 — задняя кишка (перерезана). Мышцы брюшка удалены.
Яичник обычно большего размера, чем семенник, и состоит из
крупнозернистой ткани коричневатого цвета в период половозрелости
и белого во время созревания яйца (рис. 115, Ои).
164
От места соединения передних долек с задней, с боков железы,
отвесно книзу опускаются две короткие и широкие тонкостенные
трубки —яйцеводы (Od), открывающиеся у основания третьей пары ног
головогруди двумя половыми отверстиями (рис. 113, В, О). Яйцевод
можно яснее разглядеть, если подвести.под него ножку пинцета.
Семенник всегда белого цвета. Ткань его тонкозерниста (рис. 115,В,а).
Семепроводы отходят также от места соединения передних долек
с задней; они очень длинны и тонки; открываются у основания
пятой пары ножек головогруди (рис. 113, А, 9).
Живчики речного рака весьма своеобразны по форме и обладают
малой подвижностью. Нередко можно видеть у самок на брюшной
стороне головогруди неправильные трубчатые комки белого цвета,
являющиеся сперматофорами, заключающими в себе семенную жид-
кость.
Сперматофорами называются особые мешетчатые образования, воз-
никающие из секрета выводных путей половых органов самца. Эти
Рис. 115. Речной рак.
А — органы размножения самки. Ov— яичник; Od—яйце-
вод; О — половое отверстие. По Ж а м м у. В — органы раз-
множения самца, а—семенник; b—семепровод. По Ж а м м у.
мешетчатые тела образуются вокруг массы живчиков и при копуляции
выходят из половых отверстий самца и прикрепляются к стенке хити-
нового покрова самки. Таким образом сперматофоры буквально оправ-
дывают свое название, ибо служат носителями или, точнее, перенос-
чиками семени.
Оплодотворение у рака наружное. Отложенные икринки приклеи-
ваются к волоскам плавательных ножек брюшка самки (рис. 113, В,
2—5), которые она пригибает к телу, образуя при этом карман, в кото-
ром и вынашивается «икра».
В этот же «карман» поступают из сперматофор живчики, которые
и оплодотворяют яйца.
Из яиц вылупляются вполне сформированные маленькие рачки,
отличающиеся от взрослых малыми размерами, непропорционально-
стью частей тела и крючковатыми на концах клешнями. Ими рачок
крепко держится за ножку матери (рис. 116).
Зародышевое развитие рака длится долго —в течение всей зимы.
Молодые рачки вылупляются лишь в конце весны или в начале лета.
Развитие рака прямое, так как оно протекает без превращения,
165
ибо у речного .рака личинки по своему наружному строению в общем
сходны с взрослой особью.
Самка речного рака откладывает всего около 200 яиц. Такую
плодовитость следует считать малой, особенно по сравнению хотя бы
с лангустом, самка которого может нести на ножках брюшка до 100 000
яиц. Число откладываемых яйц зависит от возраста рака.
Крючковатость концов клешней только что вылупившегося рачка
является особенностью строения, свойственной «личиночному», т. е.
начальному,периоду
Рис. 116. Только что вылупившиеся рачки
на брюшных ножках самки (А). В — клешня
только что вылупившегося рачка с крючьями.
По Гексли.
ех — экзоподит; еп — эндоподит; ес — раскрыв-
шаяся скорлупа яйца.
жизни рака, когда он держится за ножку матери.
Органы или части их, име-
ющие особенности строения,
присущие зародышевому или
личиночному периоду жизни
(в данном случае —личиноч-
ному) животного, называются
провизорными.
Органы пищеварения. Рас-
смотрев половой аппарат реч-
ного рака, удаляют половую
железу, подрезав у места вы-
хода наружу половые протоки,
после этого органы пищеваре-
ния видны по всей их длине.
Рассмотрение ротовых орга-
нов отложим до главы о ске-
лете и конечностях рака, а
пока займемся ознакомлением
со всеми частями внутреннего
пищеварительного аппарата,
состоящего из трех главных
отделов: передней, средней и
задней кишки.
Передняя и зад-
няя кишки выстланы
изнутри хитиновой кутикулой
(хорошо видимой при вывари-
вании в едком кали) и разви-
ваются в виде глубоких вну-
тренних впячиваний наружного зародышевого листка (эктодермы),
дающего также кожные покровы с хитиновой кутикулой. Неудиви-
тельно поэтому, что хитиновая выстилка передней и задней кишки по
краям ротового и заднепроходного отверстий продолжается непосред-
ственно в хитин кожных покровов. Средняя кишка развивается из
внутреннего зародышевого листка (т. е. из энтодермы).
Ручкой скальпеля оттесняют желудок в сторону и рассматривают
его нижнюю поверхность. Она воронкообразно сужается в месте
отхождения короткого и широкого пищевода (oesophagus). Последний
перерезают поперек ножницами, а освободившийся желудок пинцетом
поднимают кверху и высвобождают из места его нормального положе-
ния. Ручкой или концом скальпеля освобождают передние концы
166
боковых лопастей печени и, поддерживая ее снизу, поднимают кишеч-
ную трубку на всем ее протяжении вплоть до анального отверстия,
где и перерезают ножницами.
Отпрепарованные пищеварительные органы кладут в нормальном
положении на дно ванночки и рассматривают.
Желудок рака (ventriculus) (рис. 114, 7) поперечной борозд-
кой делится на два неравных отдела: передний больший (pars
cardiaca), или жевательный желудок, и задний меньший (pars pylo-
rica), или пилорический желудок. В первом склериты перемалывают
пищу; во втором пища процеживается через своеобразную цедилку
для удаления грубых непереваримых частиц.
Пищевод и желудок (рис. 114, 7) образуют вместе переднюю кишку
(stomadaeum).
За желудком следует очень короткая средняя кишка с маленьким
дорзальным отростком (слепая кишка). С боков средняя кишка прини-
мает в себя протоки парной печени (рис. 114, 9)к которая функцио-
нально аналогична печени, поджелудочной железе и тонкой кишке
позвоночных. Разрывают пинцетом кусочек печени и рассматривают
составляющие ее мелкие, замкнутые на одном конце трубочки. Весьма
малая длина средней кишки рака компенсируется громадным коли-
чеством печеночных трубочек, развивающихся в виде выпячиваний
средней кишки.
Задняя кишка (рис. 114, 10) на всем своем протяжении идет прямо.
В ней просвечивают каловые массы, и она в тринадцать раз длиннее
средней кишки. Анальное отверстие лежит на нижней поверхности.
Речной рак является всеядным животным: он поедает побеги раз-
личных водяных растений —рдеста, хвощей, элодей и др.; пожирает
некоторых живых животных (головастиков, рыб, моллюсков, личинок
водяных насекомых), проявляет иногда каннибализм и пожирает
также трупы.
Пища захватывается клешнями и подается ими же ко рту, где
пережевывается ротовыми придатками (см. ниже). Рот легко обнару-
жить, если войти ножками сложенного пинцета в оставшийся на пре-
парате отрезок пищевода и их раздвинуть.
Пережеванная пища поступает через пищевод в желудок, где
в кардиальной его,части снова подвергается перетиранию склеритами.
Чтобы ознакомиться детальнее с его устройством, отрезают желудок
от средней кишки и удаляют мягкие части кардиального отдела нож-
ницами На оставшейся части желудка изнутри рассматривают белые
склериты с темными зубцами. Различают ossiculum cardiacum — по-
перечный дорзальный склерит, к которому прикрепляются передние
желудочные мышцы; с боков с ним сочленяются ossicula pterocardiaca
с боковыми зубцами; сзади oss. cardiacum лежит также подвижно
с ним сочлене и;ыь мъ' urccardiacum со срединным зубцом.
Нижнебоковые части выходного отдела желудка несут две пла-
стинки с волосками по свободному краю, служащими для процежи-
вания пищи, поступающей дальше в кишечник.
Обработанная и процеженная через волоски пилорической части
желудка пища поступает в среднюю кишку и в печень. Здесь происхо-
дит переваривание нищи. В связи с этим функция печени рака является
167
Рис. 117. Антеннальные же-
лезы речного рака. По
Е. Павловскому.
/ — антеннулы; II — антенны;
b и с — части железы.
многообразной: а) печень выделяет различные пищеварительные
ферменты, б) клетки печени всасывают переваренную пищу и в) в клет-
ках печени отлагаются запасы переваренных и всосанных веществ,
например, жира. Отработанная пища поступает в заднюю кишку
(рис. 114, 10), где и формируются каловые массы.
Находящиеся в боковых стенках желудка весной и летом белые
дисковидные камешки—жерновки, или рачьи глазки—состоят из
углекислых и фосфорнокислых известковых солей. Они содержат
запас солей, необходимых раку для пропитывания новей кутикулы
после линьки. Жерновки падают при линьке в полость желудка, измель-
чаются жевательным аппаратом, соли вса-
сываются и с током гемолимфы прино-
сятся к кожным покровам, где отлагаются
в молодой хитиновой кутикуле.
Органы выделения. По удалении же-
лудка осматривают дно полости головогруди
и по бокам отреза пищевода находят по
бледнозеленому округлому телу. Это—зе-
леные, или сяжковые, железы (рис. 114, 7),
являющиеся органами выделения рака.
При удалении боковой части стенки тела
удается рассмотреть собственно железу
в виде плотного белого пузырька (рис.
117, Ь) и лежащего над ним тонкостенного
резервуара (так называемый мочевой пу-
зырь — с), открывающегося тонким кана-
лом в основном членике второй пары
сяжков.
Собственно железа имеет внутри себя
одну—две большие полости, являющиеся
производными вторичной полости тела
(целом); они окружены губчатой тканью,
состоящей из сильно извитого канальца,
идущего от полости железы в полость мо-
чевого пузыря. Так как в самой железе
есть остаток целома, из которого идет вы-
водной канал, то такой орган может счи-
таться гомологом метанефридиев—органов
выделения типа, ясно выраженного у кольчатых червей, в частности
у дождевого червя.
Органы кровообращения. Детали строения кровеносной системы
рака могут быть уяснены только на инъецированном препарате,
изготовление которого недоступно для занятий общего типа.
Поэтому рассмотрение всей системы органов кровообращения сде-
лаем теоретическим, исходя из фактов, подмеченных нами в начале .
вскрытия (стр. 163).
При вскрытии речного рака мы сталкиваемся с животным, у кото-
рого в теле нет красной крови. Кровь рака бесцветная или, как говорят,
белая. Эта особенность ее стоит в связи с отсутствием в ней красных
кровяных телец свойственных позвоночным животным. Состоит она
168
из плазмы, т. е. жидкой части, в которой плавают белые кровяные
клетки (лейкоциты).
В плазме крови рака находится вещество, содержащее медь; оно
называется гемоцианом и в физиологическом отношении является
аналогом гемоглобина позвоночных, так как обладает способностью
связывать кислород.
Сердце рака (рис. 114, 8’, 118, а) о дето околосердечной оболочкой
(pericardium), сквозь которую просвечивают щелевидные отверстия,
или устья сердца, ведущие из полости перикардия в полость сердца.
С дорзальной стороны видны ostia dor-
salia, с вентральной —ostia ventralia
и с боков —ostia lateralia.
От сердца отходят упомянутые выше
артерии: глазная артерия (a. ophthal-
mica) (рис. 118, b), сяжковые артерии
(аа. antennariae —с) и верхняя брюшная
артерия (a. abdominalis superior —d).
Кроме того в более низкой плоскости
идут две нижние печеночные артерии
(аа. hepaticae), веточки которых пи-
тают печень. Нисходящая артерия (а.
descendens) опускается от сердца вниз
и назад, огибает сбоку кишечник и у
брюшной стенки тела делится на две
ветви: переднюю—грудную артерию
(a. sternalis), питающую конечности, и
заднюю —хвостовую артерию (a. cauda-
lis), направляющуюся в брюшко.
Концевые ветви артерий открываются
в полость тела, поэтому кровь из арте-
рий изливается в щели между органами.
Такой тип кровеносной системы назы-
вается незамкнутым. Благодаря этой
особенности у рака нет крови как вполне
обособленной ткани тела, ибо кровь его
является в то же время и жидкостью
полости тела. Кровь рака поэтому на-
Рис. 118. Рак с налитыми серд-
цем (а) и артериями: b — глаз-
ной, с — сяжковой и d — верх-
ней брюшной. По Ж а м м у.
зывают гемолимфой.
В лакунах тела кровь омывает ткани органов, клетки которых запа-
саются необходимым для них кислородом. Обедневшая последним
гемолимфа по особым путям, называемым венозными синусами, пода-
ется в приводящие сосуды органов дыхания, коими у рака являются
жабры (см. дальше). В жабрах кровь обогащается кислородом и по
отводящим сосудам переливается в полость околосердечной сумки,
откуда и поступает снова в сердце.
Таким образом у рака имеется незамкнутая кровеносная система
со сравнительно хорошо развитыми составными частями (ср. ниже
кровеносную систему насекомых), причем лакуны и щели полости
тела рака физиологически соответствуют капиллярам позвоночных
животных.
169
Круг кровообращения у рака один, причем он прерван лакунами
и отчасти венозными синусами.
Органы дыхания. Чтобы найти органы дыхания рака, отрезают
панцыря головогруди и обнажают таким обра-
прочь боковые крылья
Рис. 119. Ногочелюсти и ходильные ноги
рака с жабрами. По Ж а м м у.
А — артробранхии; Р1 — плевробранхии; Ро —
подобранхии.
зом жаберную полость, лежа-
щую между собственно стенкой
тела и головогрудным панцырем.
В жаберной полости лежит
много грязносерых, перистых об-
разований (рис. 114, 3). Отры-
вают несколько из них и рассма-
тривают в воде. Эти образования
и есть жабры. Они являются
сильно рассеченными выростами
кожи рака (эпиподиты), отходя-
щими от основного членика (кокс-
оподита) пяти пар ходильных
ног и трех пар ногочелюстей.
Позднее они изменяют место
своего прикрепления, в связи
с чем различают (рис. 119):
а) ножные жабры (podobran-
chia) (рис. 119, Ро), отходящие
от коксоподита,
б) сочленовные жабры (аг-
throbranchia), связанные с мяг-
кой сочленовной кожицей ноги
(рис. 119, А), и
в) плевральные жабры (pleu-
robranchia) (рис. 119, Pl), укре-
пленные на плеврах, т. е. на
боку тела.
Распределение жабр всех ти-
пов по конечностям рака пред-
ставлено на схеме на стр. 171.
Сложнее всего устроены по-
добранхии: с коксоподитом ко-
нечности связана основная пла-
стинка, покрытая тонкими во-
лосками (setae). От нее отходит
ствол, делящийся на конце на
перо и жаберную пластинку, сло-
женную вдвое по длине. Все эти
части покрыты жаберными ни-
тями. Кроме того коксоподит
несет пучок длинных нитей.
Сочленовные жабры состоят из ствола, покрытого нитями. Благо-
даря образованию большого количества отростков, наружная поверх-
ность жабры очень сильно увеличивается, что улучшает условия
газообмена.
170
Порядковый номер сегмента Название конечности Подобранхии Артробранхии передние | задние Плевробранхии
VII Ногоче- люсти I Эпиподит — — —
VIII II + 4- — —
IX III + + 4- —
X Ходильные ноги I 4- 4- + —
XI II + + 4- Рудимент
XII III + + 4“ Рудимент
XIII IV + 4- 4- Рудимент
XIV V — — — +
Всего • . 6 + эпипо- дит 6 1 5 i 1 4-3 руди- мента
Все жабры лежат в узкой щели —жаберной полости, которая
сообщается с внешней средой двумя отверстиями: а) передним —выход-
ным, лежащим у переднего конца поперечной бороздки головогруди,
где борозда упирается в край последней, и б) задне-нижним, помещаю-
щимся между краем панцыря головогруди и основаниями ножек.
Через жаберную полость прогоняется вода, вступающая через
отверстие и выходящая наружу из переднего отверстия. Вода приво-
дится в движение особой пластинкой, называемой лодочкой (скафо-
гнатит), являющейся придатком второй пары нижних челюстей.
Чтобы наблюдать воочию работу лодочки, вырезают у живого
рака кривыми ножницами край головогрудного щита в области попе-
речной бороздки и помещают рака в сосуд с водой. Направление тока
воды определяется на живом раке, если пустить каплю жидкой туши
около заднего или переднего отверстия жаберной полости.
Вода омывает жабры. Кислород, растворенный в воде, диффунди-
рует сквозь покровы жаберных листков в их полость, где и захваты-
вается кровью. Накопившаяся же в крови углекислота идет в обратном
направлении. В жабрах кровь, следовательно, обогащается кислоро-
дом и отсюда подается в полость околосердечной сумки и в сердце.
Жабры являются органами дыхания водного типа. По существу
они не что иное, как выросты кожи; следовательно, и жаберное дыха-
ние есть дыхание кожное, но приуроченное по преимуществу к опре-
деленным, особо измененным местам кожных покровов, т. е. к жабрам.
Рак может жить некоторое время и вне воды во влажной среде.
Он замыкает плотно жаберную полость, задерживает в ней воду и
тем препятствует высыханию жабр.
Центральная нервная система. Удалив rostrum, на скате нижней
стенки тела, в самом переднем ее отделе, разыскивают парный нервный
. 171
надглоточный узел рака, называемый так по положению своему впереди
его пищевода. От него отходят нервы, иннервирующие органы чувств,
а именно глаза, сяжки и слуховой аппарат.
Огибая с боков пищевод, от надглоточного узла назад отходят
окологлоточные коммиссуры к подглоточному нервному узлу. В общем
образуется, как и у дождевого червя, окологлоточное нервное кольцо,
сквозь которое проходит пищевод.
Подглоточный нервный узел является передним узлом брюшной
нервной цепочки, которую легче всего для начала найти в брюшке
рака. Ножницами срезают мышцы брюшка и на внутренней стороне
брюшных покровов тела замечают тонкий беловатый шнурок с мест-
ными узловатыми утолщениями по его ходу (рис. 114, 5). Если припод-
нять пинцетом этот шнурок, лежащий по средней линии тела, то можно
видеть, что от каждого узла в стороны отходят тонкие нервные стволики.
Число нервных узлов соответствует числу сегментов брюшка. Сово-
купность узлов, соединяющихся друг с другом коннективами, пред-
ставляет собой действительно цепочку, почему нервную систему такого
типа и называют цепочечной (рис. 114, 5).
Нервные узлы в брюшке рака развиты слабее, чем в головогрудном
отделе. В последнем нервная цепочка не нидна даже по удалении
всех органов, так как она лежит под выростами наружного скелета,
которые необходимо предварительно удалить. Скелетные выросты
видны в виде поперечных балок белого цвета, лежащих на дне пре-
парата.
Нащупывают точно края поперечных скелетных выростов и перере-
зают их возможно более латерально, держа ножницы отвесно. Так
поступают со всеми выростами на обеих сторонах тела. После этого
пинцетом удаляют отрезанные с обоих боков скелетные выросты,
убирают продольно идущую мышцу — сгибатель брюшка (m. flexor
abdominis), и на дне открывшегося таким образом грудного канала
(canalis sternalis) отыскивают нервную цепочку. Ее можно легче заме-
тить, если осторожно потягивать пинцетом за нервную цепочку в начале
брюшка; она тянет за собой и головогрудный отделх что делает послед-
ний хорошо заметным.
Грудная часть нервной системы состоит из более массивных нервных
узлов, причем парность их выражена лучше, нежели в брюшке. Всего
здесь пять пар узлов, из коих задние две пары сближены друг с другом.
Узлы соединяются друг с другом продольными коннективами, которые
также парны, но подобно узлам тесно прилежат друг к другу. Между
второй и третьей парами головогрудных ганглиев (считая сзади)
коннективы расходятся одна от другой и образуют щель для прохожде-
ния нисходящей артерии.
От головогрудных узлов отходят нервы к мышцам конечностей.
Подглоточный нервный узел по своему составу очень сложен, ибо
он возникает вследствие слияния шести пар узлов, иннервирующих
ротовые конечности рака.
Чтобы получить более цельное представление о центральной
нервной системе рака, ее выпрепаровывают целиком из тела. Лучше
выдержать предварительно препарат в жидкости следующего состава:
воды 60 см3, азотной кислоты 20 см8, глицерина 20 см8. Эта жидкость
172
Рис. 120. Речной рак.
Основание первой па-
ры сяжков сверху.
По Рёзелеру и
Лампрехту.
S — чешуя (squama);
книзу и внутри от нее —
глаз; вход в слуховой
орган, в виде трехуголь-
ного образования, лежит
в основном членике пер-
вой пары сяжков (А).
разрыхляет (или, как говорят, мацерирует) ткани, нервную же цепочку
уплотняет, что значительно облегчает высвобождение ее из тела.
В общем во всей своей совокупности нервная цепочка состоит
из: а) надглоточного узла, б) сложного подглоточного узла, равного
шести парам нервных узлов, в) грудных узлов, числом пять пар, и
г) шести брюшных узлов.
Всего, следовательно, в нервной цепочке имеется восемнадцать
пар нервных ганглиев.
При рассматривании отпрепарованной цепочки в воде на темном
фоне можно заметить начало симпатической нервной системы. Между
окологлоточными коммиссурами, тотчас позади пищевода, идет попе-
речная спайка; кпереди от нее на каждой ком-
миссуре лежит по небольшому вздутию —гло-
точному узелку. От каждого узелка отходит
по нервному стволику, образующему слиянием
так называемый внутренностный, или висцераль-
ный, нерв (п. stomachogastricus), который иннер-
вирует желудск и, вероятно, также сердце.
Нервная цепочка является для рака централь-
ной нервной системой, которая гораздо более
массивна, чем нервная система дождевого червя.
По сравнению с головным и спинным мозгом
позвоночных она неизмеримо менее массивна
и слабее концентрирована; последнее связано
с членистостью тела рака, в котором (как и у
кольчецов) каждому сегменту тела по плану
строения соответствует пара более или менее
слившихся нервных узелков.
Органы чувств. Длинные сяжки (вторая пара
сяжков) являются органами осязания рака,
которыми он может ощупывать различные пред-
меты.
Короткие сяжки (первая пара антенн) в пер-
вом оснобном членике имеют щель (рис. 120 и
122,2, St), ведущую в полость органа равновесия. Последний состоит
из кожного мешкообразного впячивания, усаженного внутри мно-
жеством волосков, у основания которых оканчиваются веточки слу-
хового нерва, идущего от надглоточного ганглия. В полости мешочка
лежат мелкие песчинки, попадающие туда извне. При перемене
положения тела песчинки в силу тяжести давят на те или дру-
гие волоски, чем и производят специфическое раздражение их,
благодаря которому рак ориентирует положение тела в пространстве.
Функция органов равновесия рака тесно связана и со слухом.
Звуковые колебания водной среды приводят в колебание песчинки
пузырька, которые своими движениями также раздражают волоски.
Поэтому песчинки пузырька называют также отолитами, а сам пузы-
рек— слуховым пузырьком. При линьке хитиновая выстилка слухового
пузырька сбрасывается, и с нею вместе удаляются отолиты. Замечено,
что после линьки раки активно заносят песчинки в слуховые пузырьки
ногами.
173
Слуховые пузырьки только что описанного типа весьма характерны
для членистоногих животных, обитающих в воде.
Органами зрения рака являются глаза; их выпуклое тело сидит на
коротком стебельке, благодаря чему глаза хорошо подвижны. Они
могут быть спрятаны под основание rostrum и, наоборот, выставлены
наружу.
В глазу различают выпуклую' роговицу (cornea), являющуюся видо-
измененной хитиновой кутикулой. Под лупой можно рассмотреть
отдельные площадки роговицы {фасетки). Глаза рака являются слож-
ными. Число фасеток глаза увеличивается с возрастом рака; при
длине тела в 4,5 см в глазу насчитываются 1930 фасеток, а при длине
в 13 см число их увеличивается до 3050.
Филогенетически глаза рака связаны, вероятно, с конечностями.
Если отрезать глаз и одновременно удалить глазной ганглий, то вместо
глаза вырастает усик. Такой тип регенерации называют гетеромор-
фозом.
Рак обладает также и чувством обоняния. Он по запаху находит
в воде приманку (например, дохлую лягушку, рыбу и т. п.).
В заключение сравним рака и дождевого червя, как животных, из
которых одно имеет специальные органы чувств, а другое лишено
анатомически выраженных органов восприятия раздражений.
Названия органов Рак Дождевой червь Примечания
Органы зрения Глаза Отсутствуют » Но червь ощущает свет особыми «зрительными клетками», рассеянными в коже Но в коже есть «чув- ствительные луювицы», воспринимающие хими- ческие раздражения Но червь способен при- нимать нормальное поло- жение тела, если вывести его из такового
Органы химического чувства Сяжки
Органы равновесия . . . Отоцисты в ос- новании I пары сяжков >
Поперечные разрезы рака
Брюшко. У другого экземпляра рака отрезают ножницами
брюшко; ножницами же подрезают кругом сочленовную перепонку
между I и 11 или II и III сегментами. Острым ножом рассекают вну-
тренние мягкие части и освобожденные сегменты рассматривают под
водой с плоскости сечения.
Препарат сверху ограничен тергитом, или спинным полуколь-
цом наружного скелета; справа и слева тергит продолжается в боко-
вые отростки —эпимеры. Нижний край сегмента ограничен скелетной
пластинкой —стернитом, с которой сочленяются брюшные ножки;
между основанием последних и эпимерой помещается соединительная
часть — плевра.
174
Внутри этого скелетного кольца лежат следующие органы: а) дор-
зально помещается мышца —разгибатель брюшка; б) вентрально
лежит мышца —сгибатель брюшка; в) между ними видна задняя
кишка (в виде кружка); г) над серединой стернита ищут нервную
цепочку.
Головогрудь. У того же экземпляра рака разрезают кру-
гом ножницами панцырь головогруди и хитиновую коробку тела
между второй и третьей парами ходильных ног. Острым ножом вле-
кущим движением разрезают внутренние органы и рассматривают
задний отрезок рака под водой на разрезе:
а) панцырь головогруди окружает препарат сверху и с боков;
б) под боковыми отростками панцыря (жаберная крышка) по-
мещается жаберная полость с жабрами;
в) изнутри жаберная полость ограничена собственно стенкой тела,
к нижнему краю которой причленяфтся ноги;
г) снизу тело ограничивается узкой пластинкой — sternum, лежа-
щей между основаниями ног;
д) от нижней стенки тела в полость тела отходят хитиновые от-
ростки (эндоскелет) —аподемы, служащие для прикрепления мышц;
е) между аподемами лежит грудной канал, в котором проходит
брюшная нервная цепочка;
ж) над аподемами ббльшую часть полости тела занимают печеноч-
ные трубочки с поперечно рассеченной кишкой;
з) над этой последней лежит половая железа и еще выше
и) сердце.
Конечности рака
Перевернув препарат брюшной стороной кверху, рассматривают
конечности рака сначала на месте, потом отделив их ножницами или
скальпелем по тонкой сочленовной кожице.
Наиболее типично устроены ножки средних брюшных сегментов
самки. Состоят они из основного стержня, называемого протоподи-
том (protopodit), который образован прилежащим к телу маленьким
члеником (coxopodit) и следующим за ним более крупным члени-
ком (basipodit). Базиподит несет на своем свободном конце две чле-
нистые веточки —внутреннюю (endopodit) и наружную (exopodit).
Эндоподит крупнее, чемэкзоподит, и состоит из более широкого основ-
ного членика и концевой веточки.
В такой конечности рака типична двуветвистость благодаря на-
личию эндо- и экзоподита.
Схема расчленения конечностей рака (рис. 121):
х экзоподит
Протоподит S
I \ эндоподит
(Коксоподит 4- базиподит)
В различных конечностях эти составные части могут частью реду-
цироваться, частью же, наоборот, разрастаться, благодаря чему ко-
нечность в целом может значительно отклоняться по своему виду от
исходной типичной формы.
175
сильной ноги рака. По
Бронштейну.
ер — эпиподит - жабра; на
basis — рудиментарный экзо-
подит — ех, и пятичлени-
стый эндоподит (собственно
нога) — еп; р — протоподит.
Памятуя последнюю, рассмотрим все конечности рака, идя спереди
назад (рис. 119 и 122).
1 пара. Стебельчатые глаза. Весьма отличны по
строению от обыкновенной конечности (рис. 122, 7), связь с кото-
рой устанавливается лишь особым экспериментом (см. выше, стр. 174,
гетероморфоз).
II пара. Малые сяжки (antennulae) состоят из: основ-
ного трехчленистого стержня и двух жгутиков —наружного и вну-
треннего. В основном членике помещается слуховой пузырек (рис. 122,
2, St). Точное гомологизирование малого
сяжка с типичной двуветвистой конечностью
затруднительно, ибо протоподит типически
двучленист, а не трехчленист. Жгуты малых
сяжков толкуются как главная и побочная
ветви (рис. 122, 2).
III пара. Длинные сяжки (anten-
nae) (рис. 122, 5). Коксоподит широк. На
нижней поверхности несет бугорок с отвер-
стием сяжковой железы. За ним следует ко-
роткий базиподит, латерально оканчиваю-
щийся острым шипом. Эндоподит развит
очень сильно; его два массивные основные
членика несут очень длинную концевую нить.
Экзоподит есть; он превращен в чешуевид-
ную заостряющуюся на конце пластинку
(squama). Таким образом двуветвистссть ко-
нечности в данном случае налицо, но с не-
которыми вторичными видоизменениями.
IV пара. Мандибулы, или верх-
ние челюсти (рис. 123, 4). Очень ко-
роткие, широкие и тонкие конечности, между
которыми лежит ротовое отверстие. Внутрен-
ний край мандибул зубчат. Мандибулы пред-
ставлены укороченным слитным протоподи-
том, к которому прикрепляется короткий
искривленный щупик (palpus), являющийся
эндоподитом. Экзоподит редуцирован.
Мандибулы, по сравнению с типичной
двуветвистой конечностью, упрощены в строе-
нии, что объясняется особой функцией мандибул —откусывать куски
пищи. Работа их весьма значительна. Приводятся в движение верх-
ние челюсти особым аддуктором—мышцей, широкое основание ко-
торой мы видели выше сбоку от задней половины желудка. При
передвигании мышцы пинцетом приходят в движение и челюсти.
V пара. Первая пара нижних челюстей (I maxil-
lae) представлена особыми пластинчатыми образованиями, в которых
различают: протоподит, состоящий из листовидных коксо- и базипс-
дита, и нерасчлененный эндоподит; экзоподит отсутствует (рис. 122,5/
VI пара. Вторая пара верхних челюстей (Il maxil-
lae) видоизменена сильнее. Коксоподит и базиподит еще более лксто-
176
л
Рис. 122. Конечности речного рака. По Г е к с л и (из Ланга).
F — стебельчатый глаз; 2 — антеннула; 3 — антенна; 4 — мандибула; 5 — первая максилла;
6—вторая максилла; 7—9— первая, вторая, третья ногочелюсти; 10— ходильная (тора-
кальная) нога; 77—12 — первая, вторая брюшные ножки самца (плеоподии, копулятивные ор-
ганы); 13 — плеоподия самки; 14—уроподия; Ва —базиподит. Со — коксоподит; Еп — эндо-
подит; Ef —эндит; Ех— экзоподит; F— чувствительная ветвь; Fp— добавочная ветвь;
Ра — щупик; St — статоцист; Da — палец; Ebr — жабра; Ер — эпиподит; Рг — протоподит;
р — проподит.
12 Практикум по зоологии — 385
видны и по наружному своему краю рассечены на лопасти. Эндоподит
очень мал и также не расчленен. Внутри челюсть несет особый при-
даток, называемый лодочкой (scaphognatit), образовавшийся за счет
слияния экзоподита с эпиподитом (см. следующий абзац) (рис. 122, 6).
Лодочка функционирует при дыхании (см. выше, стр. 171).
VII пара. Первая пара ногочелюстей (I maxilli-
pedes) и последующие ротовые придатки постепенно увеличиваются
в размерах (рис. 122, 7).
Коксоподит и базиподит срастаются в двураздельную пластинку.
Последняя несет: а) двучленистый короткий эндоподит, б) более длин-
ный экзоподит и в) направленный кзади пластинчатый придаток,
называемый эпиподитом (cpipodit). Последний принимает участие
вместе со скафогнатитом в проталкивании воды в жаберную по-
лость.
Двуветвистость конечности в данном случае имеется, но с при-
бавлением новой части, отсутствующей в типичной конечности (epi—
podit).
VIII пара. Вторая пара ногочелюстей (II maxilli-
pedes, рис. 122, 8) состоит из широкого коксоподита, от которого
в жаберную полость отходит эпиподит с нитевидными придатками.
Базиподит мал; он несет, с одной стороны, экзоподит, с другой —бо-
лее широкий эндоподит, который разделяется на пять члеников,
наименование коих рассмотрим ниже.
IX пара. Третья пара н о г о ч е л ю с т е й (III maxilli—
pedes); общий характер расчленения тот же, что и у второй пары.
Коксоподит соединяется с эпиподитом, несущим нитевидные придатки.
Базиподит несет экзоподит, состоящий из длинного основного чле-
ника и мелких концевых члеников. Эндоподит развит очень сильно;
он распадается на следующие части: а) ишиоподит (ischiopodit),
срастающийся с базиподитом, б) мероподит (meropodit), в) карпопо-
дит (carpopodit), г)проподит (propodit) и д) дактилоподит (dactylo-
podit). Эндоподит является в общем самой крупной частью третьей
пары ногочелюстей (рис. 122, 9).
IV—IX пары конечностей являются ротовыми придатками. Лежат
они скученно около ротового отверстия, одни покрывая других. Ного-
челюсти служат для подачи пищи ко рту.
X пара. П е р в а я пара ходильных ног (рис. 113, А, 4;
рис. 121 и 122, Ю) — самая крупная конечность рака с массивной
клешней на конце. Состоит последователь^ из коксоподита, базипо-
дита, сросшегося с ишиоподитом; все£ остальные части эндоподита
налицо. Дактилоподит играет роль подвижного пальца клешни, основ-
ная часть которой образована проподитом. Экзоподит редуцирован.
Эпиподит есть; он лежит в жаберной полости.*
Эта пара конечностей служит для ползанья, для захватывания
пищи и для защиты. лЯя
XI и XII пары. Вторая и третья пары х и«д ь н ы х
ног устроены сходно. В коксоподите третьей пары ног
самки лежит отверстие яйцевода (рис. 113, В, О). За ним следуют
базиподит, ишиоподит, мероподит, карпоподит. Проподит и дактилопо-
дит образуют маленькую клешню. Эпиподит функционирует в каче-
178
стве жабры (подобранхия) и снабжен нитевидными придатками. Экзо-
подит отсутствует. Служат для передвижения рака.
XIII пара. Четвертая пара ходильных ног от-
личается от второй и третьей лишь отсут-
ствием клешней. В остальном они сходны.
XIV пара. Пятая пара ходильных ног может
считаться типичной ходильной ногой. Коксо-
подит ее у самца пронизан половым отверстием (рис. 113, А, 9). Эндо-
подит развйт хорошо, тогда как экзоподит редуцирован. Клешней
нет.
XV пара. Первая пара брюшных ножек у самки
сильно атрофирована и превращена в корот-
кие стерженьки (рис. ИЗ, В, 7) или даже вовсе
отсутствует. Первая пара брюшных ножек у самца значительно
видоизменена. Протоподит срастается, видимо, с эндоподитом и обра-
зует желобоватый стержень белого цвета. Экзоподит отсутствует.
Эта пара ножек вместе со следующей играет роль копулятивного аппа-
рата (рис. 113, А, 10).
XVI пара. Вторая пара брюшных ног у самки
построена типично: протоподит (состоящий из коксо-
подита и базиподита) несет на себе почти одинаково
развитые эндо- и экзоподит.
У самца же эндоподит развит гораздо больше, чем экзоподит, при-
чем концевой членик первого загнут вперед под углом. Загнутые
части ножек вместе с первой парой ножек брюшка образуют же-
лоб для подачи сперматофор к половому отверстию самки для ко-
пуляции.
XVII и XVIII пары. Третья и четвертая пары брюш-
ных ножек (рис. 113, А, 11—13).
XIX пара. Пятая пара брюшных ножек имеет
вид типичной двуветвистой конечности.
Брюшные ножки рака могут служить и для плавания.
XX пара. Шестая пара брюшных ножек видо-
изменена очень сильно, протоподит их не рас-
членен, короток и широк. Эндоподит и экзоподит расши-
рены в виде лопастей (рис. 113, А, 14); экзоподит подразделен на две
части (рис. 122, 14). Вместе с тельсоном расширенные пластинчатые
ветви последней пары конечностей рака являются главным плаватель-
ным аппаратом. Когда рак быстро сгибает брюшко с раскрытым на
конце веером плавательных пластинок, то он дает телу сильный толчок
назад.
Для наглядности сопоставим на прилагаемой таблице схемы разви-
тия составных частей конечностей рака.
Из рассмотрения плана строения конечностей рака видим, что
основной план может изменяться при редукции или, наоборот, при
развитии той или другой части двуветвистой конечности. Так, ходиль-
ные ноги характеризуются преобладающим развитием эндоподита,
образующего главный ствол конечности; экзоподит же бесследно
исчезает. Добавочной частью их является эпиподит, присутствие кото-
рого связано с развитием жабр.
179
Схема строения конечностей рака
Протоподит Эндоподит Экзоподит сПИПОДИТ
Коксоподит Баэиподит
II Головные конечности Сяжки I I Три основных членика и две нити, не гомоло- гичные экзо- и эндоподиту « —
III II + + +++++ + Squama
IV Манди- булы Слиты вместе + — —
V Максиллы! I + + + + + Не расчленен — —
VI II + Не расчленен Скафогнатит
VII Грудные конечности Ногочелюсти I + + + + + + + +++ + +
VIII IX II + +' + + Подобранхии
III
X Ходильные ноги | I + + + 9 + + d + -и + + + +++-Ы- ++++ ++++ ++++ ++++ - + - + - + - +
XI II
XII III
XIII IV
XIV V
XV 1 Брюшные конечности 1 Брюшные ножки Id Слиты вместе + + + + + Расширен 1 1 1 1 1 1 1 1
19 Рудиментарна
XVI и d + + + + + + + + + + Копулят.орг. + + + + Расширен
П9
XVII ш
XVIII
IV
XIX V
XX VI Слиты вместе
Ногочелюсти же являются наиболее богатыми по составным ча-
стям конечностями, ибо в них, помимо эндо- и экзоподита, есть и эпи-
подит.
В связи с такой пластичностью конечностей рака стоит способность
их к перемене функции, ибо видоизмененные ноги у рака, помимо
движения, служат и челюстями (ногочелюсти) и принимают участие
в половых отправлениях (копулятивные органы) и т. д.
Распределим далее конечности рака по соответствующим сегмен-
там тела и отметим попутно разделение его на отделы в связи с иннер-
вацией конечностей из узлов нервной цепочки.
Схема сегментации речного рака и иннервации его конечностей
Номер
сегмента
Соответствующая конечность
Часть тела
Узлы нервной цепочки
I
II
III
Глаза
I пара сяжков
II »
I Надглоточный нерв-
• ный узел („мозг*4) с
тремя парами нервов
IV
V
VI
Верхние челюсти
I пара нижних челюстей
II » » »
VII VIII IX I пара ногочелюстей грудь
II III » > » »
о
X I пара ходильных ног X
XI II > > » X
XII III > » >
XIII IV > > > о
XIV V > » > о
Подглоточный нерв-
ный узел с шестью
парами нервных ган-
глиев
I торак.* ганглий
II >
III »
IV >
V * »
XV I пара брюшных ножек о I брюшной ганглий
XVI II » > > X II » >
XVII III > > » 3 III > >
XVIII IV » » > 2 IV »
XIX V > > > сх г Л V » >
XX VI » » > VI » »
Мускулатура
Чтобы рассмотреть мышцы рака, берут новое животное и разре-
зают его продольно по средней линии тела. Пинцетом снимают осто-
рожно все внутренние органы.
Кожные покровы рака представляют собой футляр, в котором спря-
таны все внутренние органы рака. Мышцы прикрепляются к внутрен-
ней поверхности кожного панцыря, более твердые части которого
соединены между собой мягкой сочленовной перепонкой. В общем
движения рака очень ограничены; они обусловливаются формой
сочленовных частей его скелета. Более всего подвижны ноги. Брюшко
181
может совершать движения только в вертикальной плоскости, т. е. i
разгибаться или сгибаться. Движение в сторону сгиба брюшка гораздо 1
большее, чем в сторону разгибания, что зависит от узости стернитов !
и ширины тергитов сегментов брюшка. В головогруди подвижных ча-
стей нет, если не считать конечностей.
В связи с большей подвижностью брюшка в нем больше всего мышц.
Разгибатель брюшка начинается в головогруди от боковой ее поверх-
ности и своими пучками прикрепляется к тергитам брюшка. Сгибатель
брюшка лежит на брюшной стороне тела.
Эти две системы мышц делают взаимно ।
противоположную работу, вследствие чего
и являются, как говорят, антагонистами
друг к другу.
В головогруди хорошо видна приводя-
щая мышца верхней челюсти, идущая к ман-
дибуле от спинной поверхности головогруд-
ного панцыря.
С клешни первой ходильной ноги сре-
зают хитиновый покров; в ее полости раз-
личают две системы пучков мышц, приводя-
щих в движение подвижный палец клешни:
приводящую мышцу (m. adductor, рис. 123, а),
служащую для захватывания клешни, и
отводящую мышцу (m. abductor —&), рас-
крывающую клешни. В связи с различием
функции первая мышца гораздо сильнее
второй. Начинаются они от внутренней по-
верхности коробки клешни и прикрепля-
ются к так называемым сухожилиям —
отросткам хитинового покрова клешни.
Сухожилие приводящей мышцы отходит от
внутреннего края основания подвижного
пальца клешни; сухожилие же отводящей
Рис. 123. Схема мышц
клешни рака с приводящими
(я—аддуктор) и отводящими
(Ь — абдуктор) мускулами.
По Ж а м м у.
мышцы связано с наружным краем основа-
ния пальца.
Мышцы у рака все поперечнополо-
сатые.
Сравнивая отношение мышц к скелету у позвоночных и у рака,
видим полную противоположность.
Позвоночные Рак
Скелет Внутренний Наружный
Мышцы и органы .... Облекают скелет снаружи Спрятаны в скелетной коробке
В связи с этим движения у наземных позвоночных разнообразны,
у рака же очень ограничены.
182
I Наблюдения над живым раком
А. На столе:
1) ползание при помощи расчлененных конечностей;
2) ориентировка тела в пространстве —перевертывание в нор-
мальное положение рака, положенного на спину;
3) замыкание клешни;
4) приведение рака в состояние гипноза. Подгибают первую пару
ног к телу, ставят рака на rostrum и согнутые ноги на стол, придер-
живают рака в этом положении, подогнув брюшко, левой рукой,
а правой поглаживают вдоль по панцырю головогруди по середине.
Осторожно отнимают обе руки, рак остается стоять в приданном ему
неестественном положении; стукнуть по столу, и рак снова становится
активно подвижным.
В. В аквариуме:
1) плавание рака при помощи брюшных ножек;
2) толчкообразное плавание при сгибании брюшка;
3) раздражимость сяжков’и глаз при прикосновении к ним;
4) кормление рака; работа ротовых конечностей;
5) работа лодочки (см. выше —органы дыхания).
Общие замечания
Тело рака имеет хорошо выраженную двубоковую симметрию.
Кроме того у него наблюдается метамерия различных частей тела и
органов, а именно: а) придатков тела, лежащих попарно, б) сегментов
брюшка и в) узлов нервной цепочки.
Метамерия у рака выражена явственно, ибо тело его состоит из
сегментов, несущих по паре конечностей и по парному нервному узлу.
Однако правильность метамерии нарушается слиянием сегментов го-
ловы и груди в общее целое —головогрудь, покрытую сплошным
панцырем. Следом такого слияния являются парные метамерные
конечности, помещающиеся на брюшной стороне головогруди. В связи
с переменой функции типичные двуветвистые конечности рака так
или иначе преобразуются, что также несколько нарушает чистоту
метамерии.
Все же все конечности рака гомологичны друг другу. Так как
гомология распространяется здесь на метамерные образования, то
можно говорить о гомодинамии конечностей. Даже глаза рака гомоди-
намны сяжкам, что подтверждается явлением гетероморфоза (стр. 174).
Метамерность нервных узлов вполне явственна; однако и от нее
есть некоторые вторичные отклонения, обусловливаемые судьбой чле-
ников, коим принадлежат соответствующие нервные узлы. В частности,
сложен и велик подглоточный нервный узел, составившийся из шести
пар нервных узлов.
Такая местная концентрация нервных ганглиев зависит от тесного
слияния соответствующих сегментов тела, следами которых остались
три пары челюстей и три пары ногочелюстей рака. Эти конечности
тесно сближены друг с другом; сблизились до степени почти полного
слияния и принадлежащие им нервные узлы (см. таблицу иннерва-
ции конечностей рака на стр. 181).
183
Теперь определим положение рака в системе животных, исходя
из оценки важности и характерности различных черт его строения:
а) присутствие наружного хитинового скелета;
б) членистость тела;
в) наличие (по типу строения) в каждом сегменте пары расчленен-
ных конечностей;
г) строение нервной системы в виде головного узла, связанного
с брюшной цепочкой ганглиев;
д) положение сердца на спинной стороне тела;
е) разделение желудочно-кишечного канала на переднюю, среднюю
и заднюю кишку.
Совокупность всех этих признаков определяет принадлежность
речного рака к типу членистоногих.
Наличие двух пар сяжков й двуветвистость типических конеч-
ностей дают основание отнести рака к классу ракообразных.
Состав тела рака из определенного числа члеников и обладание
жевательным желудком характеризуют его как высшего рака.
Наконец, слияние пяти грудных члеников с головой в голово-
грудь; сильное развитие панцыря, одевающего головогрудь сверху
и с боков; присутствие трех пар ногочелюстей и пяти пар грудных
ног с жабрами у их основания и обладание стебельчатыми глазами —
все это дает возможность отнести его к отряду десятиногих раков —
Decapoda, среди которых он занимает место в ряду длиннохвостых
раков —Macrura, отличающихся длинным брюшком с развитыми
брюшными ножками.
Экология. Речной рак является водным животным, однако
он имеет ноги наземного типа, т. е. членистые конечности, работаю-
щие как рычаги для упора о твердый субстрат.
Действительно, рак ходит по дну и прячется в норы, пользуясь
ходильными (грудными) ногами.
Собственно в воде рак передвигается различно: он плавает при
помощи быстрых ударов брюшком или огребаясь брюшными ножками,
действующими подобно веслам.
Следующей особенностью водного образа жизни является дыхание
рака жабрами, являющимися органами водного типа.
Наконец также характерным для водного животного органом
являются статоцисты, или слуховые мешочки, рака.
Зависимость рака от водной среды не абсолютная, но значитель-
ная. При благоприятных условиях (влажность среды) рак может жить
некоторое время вне воды.
ДАФНИЯ. ВЕТВИСТОУСЫЕ РАКИ (CLADOCERA)
Технические замечания. Дафний вылавливают из различных
водоемов сеткой (сачком) из мельничного газа или из другой материи, но с соот-
ветственно узкими ячеями. Дафнии живут в стоячих водоемах с пресной водой,
начиная с кратковременно существующих и кончая прудами и озерками. В про-
точных водоемах дафнии держатся в местах с очень медленным течением между
водяными растениями. Дафний иногда бывает так много, что своим присутствием
они изменяют цвет воды, поскольку о нем судят по зрительному восприятию с бе-
рега (так называемое явление «цветения воды»). При добывании дафний следует
облапливать толщу воды, не затрагивая дна.
184
124. Дафния — D. ри-
Рис.
lex. По Гёртвигу.
go — глазной ганглий, связанный
с глазом; g— надглоточный ган-
глий; h — сердце; о — яичник;
b — выводковая камера с двумя
зародышами; 2 — антенна; 3—
мандибула; 5—9 — пять пар но-
жек; кишечник исчерчен попе-
речно.
При вынимании сачка вода из него вытекает (фильтруется через сетку), и на
дне сачка остается более или менее обильная «кашица» из множества мелких
животных. Сачок выворачивают в широкогорлую банку или ведрышко с водой,
если желают принести дафний в лабораторию живыми; или же добытый мате-
риал на месте перекладывают в банки с 4% водным раствором формалина или
в 70% спирт. Каждую банку снабжают этикеткой, на которой пишут мягким (не
химическим) карандашом место, время сбора, характер водоема и фамилию соби-
рателя.
Дафнии встречаются в водоемах не всегда, они то появляются, то исчезают,
давая пример периодичности массового обитания. Летом встречаются только самки,
к осени появляются и самцы.
Дафний наблюдают под микроскопом: 1) в живом состоянии в капле воды
под покровным стеклом с ножками; 2) на препаратах заделанными в глицерин-
желатину без окраски; 3) на окрашенных спирто-
вым раствором пикриновой кислоты бальзамных
препаратах; хорошие результаты дает также
окраска бисмаркбрауном.
Под общим названием дафний или водяных
блох подразумевается несколько сотен видов раз-
личных родов мелких ракообразных (до 5 мм
длины). Для образца мы рассмотрим наиболее
обыкновенных представителей — роды Daphnia и
Simocephalus; последние часто окрашены в крас-
новатый цвет; оба рода относятся к семейству
Daphnidae.
Внешняя морфология и внутреннее
строение
На переднем конце рачка (рис. 124)
видна голова с крупным пигментирован-
ным глазом; клювовидная по форме, она
несет две пары усиков, из которых сразу
бросается в глаза мощно развитая и раз-
ветвленная вторая пара (антенны), по ха-
рактеру которых целый подотряд низших
ракообразных получил название вётвисто-
усых. Первая пара усиков (антеннулы)
мала; они имеют вид коротких цилиндри-
ческих палочек с пучком тонких щети-
нок на конце; прикрепляются антеннулы
к нижней части головы, близ ее клювика
(rostrum).
Все тело покрыто прозрачной двуствор-
чатой раковиной, одевающей его с боков:
стаются на спинной стороне животного; снизу они свисают свободно,
оставляя щель, через которую вода имеет доступ к телу и конечно-
стям дафний.
Форма раковины у дафний разнообразна. У Simocephalus (рис. 125)
она имеет неправильно листовидную форму, а у собственно дафний
задний конец раковины вытянут в более или менее длинный шип
(рис. 124). Раковина является кожной складкой. Между раковиной
и спинной поверхностью спрятанного внутрь нее тела самки дафнии
имеется полость, называемая выводковой камерой; сюда поступают
откладываемые яйца, и здесь они вынашиваются. Рассмотрим деталь-
сра-
185
нее конечности и другие придатки. Вторая (т. е. задняя) пара антенн
имеет длинный основной членик, с которым сочленены две ветви,
состоящие каждая из нескольких (до четырех) члеников, несущих
длинные перистые плавательные щетинки. У Simocephalus одна ветвь
построена из трех, а другая —из четырех члеников (рис. 125).
Ротовые органы малы, и на обычном препарате они не различимы.
Зато сквозь прозрачную сторону раковины видны ножки числом пять —
шесть пар (рис. 124, 5—9); они прикрепляются к гуювищу и у жи-
вого рачка находятся в постоянном движении, прогоняя воду через
створки раковины. Две передние пары ножек дафний узки- эни
Рис. 125. Simocephalus vefulus. По Кюкенталю.
1 — кишка; 2 — стенка тела; 3 — яйцо; 4 — сердце: 5 — щелевидное отверстие;
6 — яичник; 7 — мускулы; 8 — мандибула (верхняя челюсть); м — печеночный
отросток; 10— скорлуповая железа; 11 — головной ганглий; 12—глазной му-
скул; 13—ganglion opticum (глазной ганглий); 14 — хрустальный конус;
15—глаз; 16— добавочный глазок; 17— первая антенна; 1ч— анальное отвер-
стие; 20, 21, 22—грудные ножки пятой, четвертой, третьей пар; 23 — жаберный
мешочек одной из ножек второй пары; 24—первая грудная ножка; 25—рако-
винка; 26 — выводковая камера.
являются хватательными; ножки имеют пластинчатое строение и
снабжены щетинками или волосками; в основной части ножек видны
овоидные мешковидные вздутия, являющиеся жаберными придат-
ками. Последние гомологичны эпиподиту. При движении ножек жабры
омываются свежей водой, что обеспечивает возможность дыхания
и отфильтровывания пищевых частиц (см. ниже).
Тело дафнии оканчивается брюшком, которое загибается круто
•книзу; конечностей на брюшке нет; задний край брюшка несет выросты
и две оперенные щетинки.
Задний конец брюшка называется постабдоменом или каудой;
он несет два изогнутых копя (каудальные когти) и различные шипы
186
у их основания или на заднем своем крае. Все эти детали строения
имеют важное‘значение для систематики (рис. 126).
Внутреннее строение. Пищеварительная трубка начинается ртом
у основания головы (рис. 124): от рта идет вверх и вперед тонкий
пищевод, впадающий в длинную среднюю кишку (желудок), тяну-
щуюся вдоль по длине тела. Соответственно изгибу брюшка изги-
бается и средняя кишка, переходящая в короткую заднюю кишку,
которая открывается наружу анальным отверстием на конце кауды,
у основания ее коготков. У дафний от начальной части средней кишки
отходят вперед два слепых выроста, гомологичных печеночным при-
даткам (рис. 125, 9). У живой дафнии можно наблюдать перисталь-
тическое движение кишечника.
Рис. 126. Форма кауды у Daphnia magna, D.pulex, D. longi-
spina и D. cucullata (расположены сверху вниз). По Везен-
б е р г-Л у н д у.
Кпереди от передних ножек или над ними помещается трубчатая
раковинная (скорлуповая, или панцырная) железа, являющаяся выде-
лительным органом (рис. 125, 10). В отличие от высших ракообразных
(см. речной рак, стр. 168), выделительные органы дафнии (т. е. низ-
ших ракообразных) открываются позади максилл (третьей пары челю-
стей). Эти железы парны.
Из остальных органов —в голове находим непарный пигменти-
рованный сложный глаз, состоящий из разного числа хрусталиков
(прозрачных линз), окружающих пигментное поле (рис. 127). Глаза
имеют мышцы и благодаря их работе у живой дафнии находятся
в состоянии вибрации. Позади глаза лежит надглоточный нервный
узел, от которого идут две коммиссуры к брюшной нервной цепочке.
Кишечник дафнии часто набит пищей в виде зернистого детрита.
Добывание пищи обеспечивается фильтрацией воды через ножки
187
дафнии в процессе дыхания. Главным фильтром являются гребневид-
ные эндоподиты третьей и четвертой пар ножек. На сетке их щетинок
и волосков задерживаются инфузории, одноклеточные водоросли г
другие объекты; из них формируется пищевой комок, который дей-
ствием второй пары ног передается к ротовому отверстию. Для пол-
ного заполнения кишечника дафнии пищей требуется приблизительно
получасовая работа ее ножек.
По бокам кишечника лежит по трубковидному яичнику; выводная
часть каждого яичника —яйцевод — открывается в заднюю часть
выводковой камеры.
Кпереди от выводковой камеры лежит сердце в виде овального
мешочка (рис. 125, 4); у живой дафнии оно бросается в глаза благо-
даря ритмическим сокращениям; сердце лежит в околосердечной
Рис. 127. Сложный глаз Daphnia magna. I — сбоку, Il — сверху.
По Бронштейну.
сумке. Кровеносных сосудов нет. Кровь — бесцветная; перекачиваемая
сердцем, она омывает органы, циркулируя в пространствах между
ними. Несмотря на отсутствие кровеносных сосудов, кровь цирку-
лирует по определенным направлениям.
Наблюдения над жизненным циклом дафний
Летом (у моноциклических видов, см. ниже) встречаются только
самки дафний. Они размножаются, откладывая в выводковую камеру
яйца, которые развиваются и дают молодь без оплодотворения, т. е.
партеногенетически; отсюда и название партеногенетические самки
и яйца дафний. Партеногенез является цитогонией, но с участием
только одного сорта половых клеток, т. е. яиц.
Число откладываемых дафниями яиц партеногенетических (летних,
или субитакных) различно (рис. 128). Одна и та же самка может дать
повторные партеногенетические поколения. Из снабженных питатель-
ным желтком яиц выходит сразу молодая дафния; следовательно,
у этих ракообразных метаморфоза нет. Daphnia может давать первое
в своей жизни потомство спустя 8—10 дней после выхода своего из
яйца; за это время она проделывает четыре-пять линек. При обиль.
ной нище и подходящей температуре та же самка может давать но.
188
вые поколения через 3-4-дневные промежутки. D. magna обильно
плодовита, так как за раз дает до 80 яиц.
Продолжительность развития зародыша в летних яйцах дафнии
в выводковой камере вариирует от 2 до 4 дней.
Вылупившиеся молодые дафнии выходят из выводковой камеры
через ее заднее отверстие, обычно закрытое выростами задней поверх-
ности брюшка.
Отсаживая молодых дафний по одиночке в маленькие аквариумы
(стеклянные баночки в 30 см3) и кормя их (D. magna) растертой в по-
рошок высушенной элодеей (Elodea canadensis) или спирогирой,
можно проследить генерацию за
генерацией дафний.
Вообще, обилие пищи и высо-
кая (оптимальная) температура спо-
собствуют партеногенетическому
размножению.
Ухудшая питание D. magna,
можно в любом поколении заста-
вить их откладывать так называе-
мые зимние яйца. В естественной
обстановке это явление приурочи-
вается к осеци. Ему предшествует
появление в поколениях самцов,
которые оплодотворяют самок. По-
следние откладывают в выводко-
вую камеру оплодотворенные, или
зимние, яйца, всего по одному, по
два на камеру. Эти яйца более бо-
гаты питательным желтком, чем
летние, непрозрачны и одеты тол-
стой оболочкой. Вокруг них обра-
зуется так называемое седлышко,
или эфиппии, являющийся футля-
ром. Седлышко образуется за счет
Рис. 128. Daphnia pulex с летними
яйцами в выводковой камере. По
Бронштейну.
видоизменения прилежащих к яйцу стенок выводковой камеры, кото-
рые утолщаются, приобретают своеобразную структуру, резко тем-
неют и становятся твердыми. Самки, дающие зимние яйца, назы-
ваются эфиппиальными.
При последующей линьке дафний эфиппии освобождаются и у
Daphnia magna падают на дно или оседают на водяные растения
(рис. 129, вверху). Эфиппии D. pulex, наоборот, остаются плавать на
поверхности воды (рис. 129, внизу).
Зимующие яйца в эфиппии долго не развиваются, и потому их на-
зывают также покоящимися яйцами.
Яйца с эфиппием могут без вреда для себя высыхать (например,
у D. magna) или вмерзать в лед, как то бывает с плавающим седлыш-
ком D. pulex. Интересно отметить, что вмерзание способствует со-
кращению длительности латентного периода зимних яиц; поэтому
при потеплении в водоемах сразу появляется множество молоди
D. pulex.
189
При высыхании водоемов эфиппии дафний также высыхают без
потери покоящимися яйцами жизнеспособности. Они могу! приста-
вать как в сухом виде, так и из воды, к перьям и ногам водоплавающих
птиц и быть переносимыми ими в другие водоемы. Аналогичную роль
для сухих эфиппиев может играть ветер.
Яйца в эфиппиях защищены и от воздействия некоторых хими-
ческих факторов. Рыбы могут проглатывать их, но последние прохо-
дят через кишечник без вреда от воздействия пищеварительных фер-
ментов. Водный раствор формалина, применяемый для консервирова-
ния рыб, не влияет на жизнедеятельность яиц в заглоченных рыбой
эфиппиях.
Рис. 129. Эфиппии даф-
ний. Вверху — тонущий
эфиппий D. magna; вни-
зу — плавающий зфип-
пий D. pulex. По Брон-
штейну.
Следует отметить, что возможно сформирование седлышка без
того, чтобы в нем были покоящиеся яйца.
Если вынуть из седлйшка зимние яйца дафнии и их подсушить,
то они развиваются так же, как и неподсушенные.
Из покоящихся яиц выходят самки, начинающие класть партено-
генетические яйца.
Годичная последовательность генераций дафний характеризуется
количеством поколений самцов в году. Различают моноциклические
формы дафний, т. е. имеющие в году только один период обоеполого
размножения; бициклические формы соответственно с двумя такими
периодами и полициклические формы с несколькими половыми перио-
дами в году.
Указанные различия не всегда являются видовыми особенностями
дафнии. Отмечено, что мопоцикличность свойственна северу, а ближе
к югу наблюдается полицикличность; но даже в одном и том же месте
190
один и тот же вид дафний может иметь различный годовой цикл; так,
рассмотренный выше Simocephalus vetulus имеет одно обоеполое поко-
ление в больших водоемах (например, в озерах), а в мелких водовме-
стилищах он дицикличен. Эти явления служат хорошим примером
зависимости оргайизма от внешней среды.
Самцы дафний значительно мельче самок; они, естественно,
лишены выводковой камеры, в связи с чем спина их менее выпукла
кверху. Половые железы (семенники) лежат, так же как и яичники,
по бокам кишечника.
В строении придатков самца имеются отличия, связанные с поло-
вой функцией. Так, первые антенны удлинены, подвижны и служат
для удержания самки во время копуляции. Первая пара ножек имеет
длинный жгутовидный придаток и также служит для удержания самки
Рис. 130. Конечности дафний.
Первая ножка (а) самки, b — самца Daphnia pulex (по Бронштейну); с — четвертая
нога D. sima (по Клаусу): а — экзоподит; i — эндоподит; b — базиподит; к — жаберный
мешочек.
(рис. 130, Ь). На переднем конце раковины ее створки имеют широкий
вырез, благодаря чему раковина зияет большим отверстием, края
которого обрамлены длинными волосками.
У дафний мы видим ясно выраженный половой диморфизм. Изредка
у Daphnia встречаются уродливые особи, одна часть тела которых
имеет признак самца, а другая —самки, например, одну первую
антенну, как у самки, а другую, как у самца. Такие смешанные по
строению особи называются гинандроморфами. Гинандроморфизм
может носить наследственный характер, т. е. более часто повторяться
в поколениях данной самки.
Цикломорфоз. При наблюдении пелагических видов дафний, взя-
тых в разные месяцы года из водоемов одной и той же местности,
можно заметить вариацию очертаний головы и раковины, свидетель-
ствующие о значительной их изменчивости. Последняя связана с се-
зоном года (сезонная изменчивость) и с характером места обитания
дафний (локальная изменчивость).
Хороший пример такой изменчивости дает D. сиси!lata —обыкно-
191
б
г
4 1 И IF Г ! в
Рис. 131. Цикломорфоз у Daphnia cucullata в водоемах четырех типов.
По Бронштейну.
а—из маленького озера или пруда; б—г—из более крупных водоемов. Рим-
скими цифрами показаны месяцы.
венный озерный вид (рис. 131). Зимой эта дафния имеет округлую
голову; весной передний конец головы вытягивается и заостряется
(голова в форме шлема), что достигает максимального выражения ле-
том и в начале осени. С понижением температуры вырост головы уко-
рачивается, и зимой дафния имеет уже знакомую нам округлую форму
головы. Такие изменения связаны с температурным режимом водоема
и обилием пищи. Если содержать дафний со шлемовидной головой
при более низкой температуре и при недостаточном корме, то можно
получить от них потомство с округлой головой и обратно.
Говоря о такой изменчивости дафний, следует иметь в виду, что
под влиянием указанных факторов изменяется не сама дафния, а ее
потомство; поэтому можно встретить круглоголовых дафний, в вывод-
ковой камере которых находится молодь со шлемовидной головой.
Рассмотренная изменчивость характеризуется цикличностью, свя-
занной со сменой времен года.
Изменение морфологических признаков дафний в течение годовых
циклов ее развития называется цикломорфозом, а вариации habitus’a
(т. е. общего внешнего вида) дафний, зависящие только от влияния
известных факторов внешней среды, называются морфологическими;
формы же дафний —летняя, зимняя и др. —носят общее название
морф.
Общие замечания
Наличие двух пар антенн и расчлененных ножек позволяет отнести
дафний к классу ракообразных. Непарный глаз, наличие спинного
панцыря, отсутствие жевательного желудка, панцырная выделитель-
ная железа, значительная длина средней кишки —все это дает осно-
вание для признания дафнии за низшее ракообразное (подкласс Ento-
mostraca).
Дафния относится к отряду листоногих ракообразных — Phyl-
lopoda (так как обладает уплощенными ножками) и к подотряду С1а-
docera —ветвистоусые; кроме характерного развития и расщепле-
ния второй пары антенн, для Cladocera характерны: неясная сегмен-
тация тела, двустворчатая раковина, выводковая камера, четыре —
шесть пар туловищных конечностей и прямое развитие.
Экологические замечания. Дафнии являются ком-
понентами планктона или бентоса различных водоемов. Питаясь
микроскопическими по величине’объектами, дафнии, в свою очередь,
служат пищей рыбам разного возраста и особенно малькам. В этом
отношении роль их значительна, так как обилие дафний (и других
элементов планктона) обеспечивает прокорм большому числу рыб,
хороший их рост, что в общей сложности определяет рыбную про-
дуктивность водоема.
ВЕСЛОНОГИЕ РАКИ (COPEPODA)
Технические замечания. Для ознакомления с веслоногими поль-
зуемся самым обыкновенным рачком циклопом, а также Diaptomus. Их выла-
вливают п составе планктона из различных водоемов. В аквариумах с элодеей
циклопы могут хорошо жить подолгу и беспрепятственно размножаться.
13 Практикум по воологии — 385
193
Циклопов наблюдают живыми под покровным стеклом с ножками или в фикси-
рованном 70% спиртом или 4% формалином состоянии. Постоянные препараты
приготовляют по тому же способу, что и из дафний.
Для более детального ознакомления с наружной морфологией и для опре-
деления циклопов необходима препаровка иглами для отделения их конечностей,
усиков и др., что производится под бинокулярным микроскопом или под сильной
препаровальной лупой.
Рис. 132. Циклоп; вид сверху;
справа—геникулирующий усик
самца. Из Бронштейна.
Циклоп (Cyclops)
На препарате циклоп ложится чаще на брюшную сторону (дафния
же — на бок), поэтому рассмотрение его начинают со спинной стороны
тела. Тело циклопа расширено в перед-
ней области, где оно имеет более или
менее овоидную форму. Передний конец
тела (рис. 132) округлен, на нем виден
непарный глаз (отсюда и название ци-
клоп); справа и слева торчат в стороны
антенны.
Тело делится на овальную голово-
грудь и узкое брюшко; головогрудь со-
стоит из пяти сегментов; самый круп-
ный из них передний; он имеет форму
полуовала; собственно именно эта часть
тела циклопа является головогрудью,
так как она образовалась из слияния го-
ловы с первым члеником груди; сама
голова, в свою очередь, является слож-
ным образованием, слитым из пяти
члеников; следом этого является нали-
чие нескольких пар придатков головы
(усики, ротовые органы); следующие за
головогрудью четыре членика корот-
кие, вытянутые в поперечном направле-
нии; кзади они уменьшаются в размерах;
в своей совокупности они образуют часть
туловищем. Брюшко самки образовано
тела, называемую многими
четырьмя члениками; первый членц^ брюшка по размерам больше
других; он несет отверстия половых органов, почему и называется
половым, или генитальным, сегментом. Брюшко несет на конце две
каудальные ветви, образующие так называемую фурку, или вилку, и
несущие каждая по четыре щетинки разной длины.
По бокам основания брюшка самок часто бывают подвешены яйце-
вые мешки (рис. 132).
Другим признаком, отличающим самок от самцов, является строе-
ние усиков. Первая пара их (антеннулы) одноветвиста; у обоих полов
она развита гораздо сильнее, чем вторая (отношения — обратные по
сравнению с дафниями и другими Cladocera). У самки антеннулы
имеют вид длинных, равномерно утончающихся и дугообразно изгиба-
ющихся жгутов, членики которых несут короткие щетинки; первая
пара усиков является орудиями плавания и поддержания тела ци-
194
клопа в толще воды (своего рода «парение» в воде); при работе ножек
антеннулы функционируют также в качестве рулей; кроме того
они несут на себе различные чувствительные придатки. Антеннулы
самцов отличаются от таких же придатков самок своей формой,
связанной со специальной функцией антеннул служить для охва-
тывания брюшка самки во время копуляции. В связи с этим
антеннулызакручены у своего конца; их называютгеникулирующими
(рис. 132).
Для дальнейшего ознакомления с морфологией циклопа рассма-
триваем его с брюшной стороны. Позади уже известных нам антеннул
лежит вторая пара усиков — антенны, в форме двуветвистых обра-
зований, состоящих из четырех члеников, несущих щетинки.
Далее кзади, вокруг ротового отверстия, скученно лежат рото-
вые органы: пара верхних челюстей (мандибулы), две пары нижних
челюстей (максиллы) и пара ногочелюстей. Последние служат для
захватывания пищи, состоящей из различных микроорганизмов (одно-
клеточные водоросли, простейшие); обработка пищи производится
собственно челюстями. Циклопы могут проглатывать и живых орга-
низмов, которые затем проделывают в них известные стадии жизнен-
ного цикла; таковы, например, личинки (онкосферы) лентеца широ-
кого (Diphyllobothrium latum), личинки цепеня гусиного (Нутепо-
lepis lanceolatus) и даже таких относительных гигантов, как личинки
ришты (Dracunculus persarum). По отношению к названным парази-
там циклоп является их промежуточным хозяином. Позади ротовых
придатков на всех члениках головогруди, кроме последнего, лежит
по паре двуветвистых плавательных ножек; последние функциони-
руют в качестве весел, вследствие чего и весь отряд этих ракообраз-
ных получил наименование веслоногих.
Если отогнуть или отщепить препаровальными иглами плаватель-
ные ножки циклопа, то на брюшной стороне пятого членика голово-
груди, на границе его с брюшком, увидим рудиментарную маленькую
ножку; строение и характер вооружения ее шипиками и щетинками
важны для систематики циклопов. И у самцов и у самок циклопов
эти рудиментарные ножки устроены одинаково. Брюшко циклопа
лишено конечностей.
Из внутренних органов циклопа со спины просвечивает относи-
тельно широкий желудок; он лишен каких бы то ни было печеночных
придатков; спереди, через глотку, он соединяется с ротовым отвер-
стием, а сзади переходит в узкую заднюю кишку, лежащую в брюшке
и открывающуюся анальным отверстием между основаниями ветвей
фурки.
У циклопов сердце отсутствует, что следует считать вторич-
ной особенностью их организации. Нет также и специальных
органов дыхания; газообмен осуществляется всей поверхностью
покровов.
Органом выделения у взрослых циклопов являются максиллярные
(раковинные, панцырные) трубчатые железы, не заметные на обычном
препарате.
Половые органы самки (рис. 133, А) представлены поперечным
яичником, лежащим над желудком, и двумя яйцеводами, идущими
• 195
от боков яичника к переднему отделу генитального (первого брюшного)
сегмента; на его боках помещаются два половых отверстия.
По мере образования яиц они заполняют яйцеводы, которые соот-
ветственно утолщаются и образуют лопастные выросты.
В генитальном сегменте есть неразличимые на обыкновенном пре-
парате кожные клеевые железы; они открываются в яйцеводы и выра-
батывают секрет, которым откладываемые яйца склеиваются в овоид-
ную массу, называемую яйцевым мешком. Самка носит на себе (по
бокам генитального сегмента) пару таких мешков вплоть до вылупле-
ния из яиц молоди. В том же генитальном сегменте имеется мешко-
Рис. 133. Органы размножения циклопов.
А—схема расположения женских половых органов самки циклопа, ov—яичник; od— яйцевод;
rs — семеприемник (по Клаусу). В— половые органы самца.t — семенник; Nd — придаточ-
ные железы; Vd — семепровод; S — сперматофорный мешок (по Клаусу). С — гените льный
членик брюшка самки Cyclops oithonoides с семеприемником. D— то же, у С. strenuus
(по Бронштейну).
видное впячивание, являющееся семеприемником (receptaculum se-
minis, рис. 133, С). Его можно видеть у живой половозрелой самки,
если рассматривать генитальный сегмент с брюшной стороны. Форма
его различна у разных видов, что имеет систематическое значение.
Половые органы самца состоят из семенника в виде непарного
мешочка, также лежащего над средней кишкой (рис. 133, В); от него
отходят два тонких семепровода, расширяющихся у своего заднего
конца в сперматофорный мешок; кроме того имеются еще добавочные
железы, связанные с начальной частью семепроводов.
В сперматофорном мешке масса живчиков замуровывается в полый
футляр в форме сдавленного боба; это образование называется сперма-
196
Рис. 134. Копуляция у циклопа
С. fuscus. По Вольфу.
тофорой. При копуляции сперматофора выводится через половое
отверстие (в генитальном сегменте их два) и приклеивается к поло-
вому отверстию самки (рис. 134). Из прикрепленной сперматофоры
живчики переходят в семеприемник самки; опустевшая гильза спер-
матофоры может оставаться на ее гени-
тальном сегменте в течение различного
времени. Самцы циклопов приклеивают
к телу самки всегда две сперматофоры.
Diaptomus
При общем сходстве внешнего вида
с циклопом, Diaptomus отличается отно-
сительно коротким брюшком, сильным
развитием генитального сегмента и обра-
зованием одного большого яйцевого
мешка, лежащего на брюшной стороне
тела самки (рис. 135).
Из других признаков Diaptomus рас-
смотрим следующие:
1) Наличие пяти члеников голово-
груди.
2) Значительную длину антеннул,
часто намного превышающих длину тела
(рис. 135 и 136, 7).
3) Различие в строении антеннул;
геникулирующей является только пра-
вая; она вздута посредине и несколько
перегнута; левая антенна имеет нормаль-
ное строение; при копуляции самец
охватывает брюшко самки лишь одной
правой антеннулой (рис. 135 и 139).
4) В отличие от одноветвистых антен-
нул, антенны двуветвисты и состоят из
двучленистого базиподита и расчлененных экзо- и эндоподита, богато
опушенных длинными перистыми щетинками (рис. 136, 2).
5) Ротовые органы состоят из: а) мандибул; мандибула
состоит из крепкой хитиновой зазубренной пластинки, с которой со-
членен подвижный придаток, называемый пальпой; пальпа образована
базиподитом (второй членик его велик), экзо- и эндоподитом с щетин-
ками (рис. 137,1 *); б) максилл (рис. 137,21)—каждая в виде нерасчленен-
ной лопастной пластинки с длинными перистыми щетинками} в) пер-
вой пары ногочелюстей, или максиллярных ног, состоящей каждая из
двучленного базиподита, трехчленистого эндоподита (видно лишь
два членика); экзоподит редуцирован; ногочелюсть по внутреннему
краю усажена длинными щетинками (рис. 137,3); г) второй пары ного-
челюстей; они развиты сильнее первой пары и также лишены экзо-
подита (рис. 137, 4).
6) Грудные или торакальные ноги (рис. 136, 6); четыре пары их
имеют строение плавательных ножек с широким двучленистым бази-
197
подитом; эндоподит их короче экзоподита; оба густо усажены пери-
стыми щетинками (рис. 138). Пятая пара торакальных ног у самки
симметрична (рис. 138), у самца же асимметрична; у обоих полов они
Рис. 135. Diaptomus сверху; самка с яйцевым мешком; справа — правая
геникулирующая антеннула самца. По Бронштейну.
Рис. 136. Самка Diaptomus castor сбоку. По Г е рт в и г у.
— надглоточный ганглий и циклопический глаз; Ъ— брюшная нервная цепочка; h— сердце,
под ним яичник и желудок; Sp — сперматозоиды в сумке, прикрепленные у полового отвер-
стия; 1— антеннулы; 2 — антенны; 3 — мандибулы; 4—макси ллы; 5—ногочелюсть;
6—10—плавательные ножки.
утеряли плавательную функцию, соответственно чему изменился и их
облик как плавательных ножек, в то же время двуветвистость их
сохранилась.
198
Правая ножка самца имеет изогнутый коготь, им самец удержи-
вает брюшко самки (рис. 138,2), левая же ножка захватывает придатками
экзоподита сперматофору и приклеивает ее к брюшку самки; спер-
матофор приклеивается к самке больше двух (даже более десяти);
вследствие отсутствия семе приемника, живчики Diaptomus переходят
непосредственно в яйцеводы самки; сперматофора имеет вид цилиндра
с округлым концом (рис. 139).
Рис. 137. Diaptomus pastor.
7 — правая передняя геникулирующая антенна (антеннула) самца; 2 — задняя антенна самки;
71 — мандибула с пальпой; 21 —максилла (Diaptomus sp.); 3 — передняя максиллярная нога
D. castor; 4 — задняя максиллярная нога D. castor (из В, Рылова, по разным авторам);
Bas — базиподит; Enpt Еп—эндоподит; Exp, Ех—экзоподит.
7) Из внутренних органов у живого Diaptomus наблюдаем весьма
частую пульсацию сердца (рис. 136, Л), имеющего вид небольшого
продолговатого мешочка, лежащего на средней линии тела под покро-
вами спины.
8) Diaptomus питается мельчайшими элементами планктона (так
называемый наннопланктон); уловление его частиц обеспечивается
199
Леб. fl раб.
I
Рис. 139. Копуляция Diaptomus gra-
cilis; самец обхватывает самку только
одной антеннулой. По Вольфу.
Рис. 138.
Плавательная нога (7) первой пары самки Diaptomus castor; 71—-нога V пары
самки D. coeruleus; 2 — то-же, у самца (по В. Рылову). at Ъ — выросты экзопо-
дита; С — боковой шип экзоподита; d— хватательный коготь; Bas (1), Bas (2) — бази-
подиты; Ехр — экзоподит; Епр — эндоподит.
особенностями строения и действия ротовых придатков и антенн;
мандибулы и максиллы широко расставлены друг от друга (рис. 140,
А, В), необычайно быстрыми ударами (до 300 в минуту) антенн, ман-
дибул и первой пары максилл вода
гонится потоком назад и попадает
в фильтрационный аппарат, образо-
ванный рядом длинных волосков вто-
рой пары максилл; через них про-
фильтровывается прогоняемая вода,
и здесь задерживаются взвешенные
в воде элементы наннопланктона; по
мере их накопления они препрово-
ждаются первыми максиллами и ман-
дибулами в рот.
9) Ротсвое отверстие лежит не-
много кпереди и кнутри от максилл;
тр
ol .
та
1 Zx
В
а,
аг
тх,
тх}
тр
ol
md
Рис. 140. А — Diaptomus сбоку и В — передняя часть его тела снизу для рас-
смотрения фильтрационного аппарата. По Ш т о р х у.
AIf ах — антеннулы; ol — верхняя губа; as — антенны; md — мандибула; ul — нижняя губа;
тх1—максилла первая; тх^—максилла вторая; тр — ногочелюсть.
спереди от него лежит сравнительно большая пластинка —верхняя
губа (рис. 140), а позади —две маленькие округлые пластинки пар-
ной нижней губы.
201
Расчленение тела Diaptomus
Номера члеников по
порядку
Название конеч-
ностей
Ветви-
стость
Наименование частей тела
I [(1 + 2 + 3 + 4) + 1]
II
III
IV
V(+VI)‘
грудные-
тора-
кальные 1
ножки
Антеннулы
Антенны
Мандибулы с
пальпами
Максиллы
Ногочелюсти I
Ногочелюсти II
Плавательные
ножки I
Плавательные
ножки II
Плавательные
ножки III
Плавательные
ножки IV
Модифицирован-
ные, у d* генику-
лирующие ножки
I
I—II
I
I- II
I
I
I—II
I-II
I—II
Собственно
головогрудь
Туловище
(торакс)
Голово-
грудь в
более об-
щем по-
нимании
1 (генитальный)......
2....................
3 с фуркой* .........
(4 с фуркой).........
(5 с фуркой).........
Ножек нет
> Брюшко
Жизненный цикл Copepoda
При наблюдении под микроскопом живых самок циклопов и Dic-
ptomus можно иногда поймать момент вылупления из яиц яйцевого
мешка молоди. Сразу же бросается в глаза резкое различие в строе-
нии молоди и взрослых рачков.
Вылупляются личинки, носящие общее название науплиуса
(= пловца). Личинку вылавливают тонкой пипеткой и помещают
под покровное стекло (с ножками) в капле воды.
Науплиус циклопа (рис. 141) имеет овоидное тело с непарным
пигментированным глазом; он оживленно плавает благодаря ра-
боте трех пар снабженных щетинками конечностей, соответствующих
первым и вторым антеннам и мандибулам взрослого рачка; на задней
окружности тела торчат две пары щетинок; сквозь прозрачные хити-
новые покровы просвечивают: а) пищеварительный канал с двумя бо-
ковыми выростами и б) две сяжковые железы, открывающиеся у осно-
вания второй пары антенн науплиуса.
Науплиус несколько раз линяет, приобретает новые части и
постепенно принимает свойственное взрослому рачку строение.
Аналогичные строение личинки и ход метаморфоза наблюдаются
у Diaptomus.
1 У некоторых видов V членик торакса сливается с VI.
8 Брюшко самки состоит из трех или четырех члеников; фурка всегда на по-
следнем. Число сегментов брюшка самца на единицу больше, чем у самки.
202
О некоторых изменениях, связанных с метаморфозом циклопа,
можно судить по таблице сравнительных признаков.
Сравнение организации науплиуса и взрослого циклопа
Признаки Науплиус | Взрослый циклоп
Тело Нерасчлененное | Сегментированное
Конечности .... 3 пары: 1 пара 2 > 3 > Больше 3 пар: Антеннулы Антенны Мандибулы Максиллы Ногочелюсти Брюшные ножки
Выделительные Сяжковая
железы — Панцырная
Глаза Непарные Непарные
У циклопа и Diaptomus, как и у всех других веслоногих ракообраз
ных, партеногенез никогда на наблюдается. Цикломорфоз, столь ха
рактерный для пелагических ветви-
стоусых, у веслоногих практически
отсутствует. Примером некоторых
сезонных изменений (изученных
вообще весьма мало) могут служить
самцы Diaptomus gracilis: генику-
лирующая антенна их летом имеет
крючковидный вырост на третьем
от конца членике; зимой же его у
самцов нет.
Общие замечания
Циклопы и Diaptomus являются
важной пищей для рыб.
Будучи заражены личинками
паразитических червей (лентеца ши-
рокого), циклопы заражают этими
червями различных рыб (окуня,
ерша, щуку и др.), когда сами про-
глатываются последними; в анало-
Рис. 141. Личинка ракообразных нау-
плиус.
Двуветвистый тип строения антенн (cf) и
жвал (nd); антеннулы (а) неразветвлены;
о — науплиусов глаз; atd—антеннальные
выделительные железы; ds — кишечник. По
Клаусу.
гичном порядке циклопы могут за-
ражать гусей цепенем гусиным и
человека —риштой.
Diaptomus может откладывать
так называемые покоящиеся яйца,
203
одетые стойкой защитной оболочкой; они могут высыхать и быть пере-
носимыми ветром или различными животными в другие водоемы.
Относясь к отряду веслоногих, циклоп и Diaptomus принадлежат
к различным семействам Cyclopidae и Centropagidae.
Основные различия зтих ракообразных приведены на сравнитель-
ной таблице.
Сравнение основных признаков циклопа и диаптомуса
Признаки Cyclops Diaptomus
Геникулирующие усики . . Граница головогруди и брюшка Первые антенны Рудиментарная V пара но- же к Яйцевых мешков Сердце Семеприемник Метаморфоз Обе антеннулы По заднему краю IV чле- ника головогруди Короче длины тела; одно- ветвистые, У обоих полов одина- кова; обе ножки симме- тричны Два Отсутствует Есть » Лишь правая антеннула По заднему краю V чле- ника головогруди Значительно превышают длину тела У самцов геникулирую- щая асимметричная Один Есть Нет Есть
Сравнение основных отличий Cladocera (дафнии) и Copepoda
(циклопы, Diaptomus)
Признаки Cladocera Copepcda
Тело Неясно расчлененное Есть Двуветвистые Развиты слабо Развиты сильно, превра- щены в органы плавания Жаберные придатки брюшных ножек Есть Папцырная железа Нет Есть > Нет > Ясно расчлененное Нет Двуветвистые Развиты сильно, у (J* геникулирующие Развиты слабее • • Нет Нет (есть) У личинок сяжковая, у взрослых папцырная Железа Есть Нет Нет (есть) Нет Есть »
Выросты покровов в виде раковины
Конечности
Антеннулы
Антенны
Органы дыхания
Сердце
Органы выделения
Сперматофорное оплодотво- рение
Партеногеноз
Цикломорфоз
Гетерогония
Метаморфоз
Науплиус
204
Сравнение высших (речной рак) и низших (дафния* циклоп) ракообразных
Признаки Malacostraca Entomostraca
Сегментация тела 20 сегментов Число сегментов неопределенное
Деление на головогрудь и брюшко. . . Ясное Не всегда ясное
Глаза Парные Непарные
Средняя кишка Короткая Длинная
Печеночные трубки Развиты очень сильно; их мно- жество Два отростка или их вовсе нет
Кровеносная система Развита хорошо Сердце есть или его нет
Гетерогония Отсутствует + -
Метаморфоз -(+) + -
Антеннулы + +
Антенны + +
Далее нам остается сравнить особенности организации Cladocera
и Copepoda, признаки которых сопоставлены на соответствующей та-
блице.
Дафния, циклопы и Diaptomus, как представители Cladocera и
Copepoda, относятся к подклассу низших ракообразных, Entomo-
straca. Последние, в противоположность высшим ракообразным (см.
речной рак), характеризуются неопределенным составом тела в отно-
шении числа сегментов, малыми размерами тела, наличием фурки,
большой длиной средней кишки при слабом развитии или отсутствии
печеночных выростов и другими признаками.
В то же время наличие двух пар антенн, двуветвистость как типи-
ческий план строения конечностей, образование головогруди из
слияния нескольких сегментов, жабры как орган дыхания —все эти
признаки объединяют и низших и высших ракообразных в класс рако-
образных, или Crustacea.
КЛАСС ПАУКООБРАЗНЫХ (ARACHNOIDEA)
СКОРПИОНЫ (SCORPIO NES)
Технические замечания. Скорпионы являются обитателями юга;
в СССР они встречаются в Крыму и особенно на Черноморском побережье
Кавказа, в Закавказье и в Средней Азии.
Скорпионов отыскивают под камнями, в трещинах старых каменных и гли-
нобитных стен, в развалинах. Для ловли нужен длинный пинцет, которым скор-
пиона схватывают за хвост и пересаживают в ящик с отделениями или в баночки.
Скорпионов надо держать порознь, так как они поедают друг друга. Скорпионы
семейства Buthidae живучи и по нескольку месяцев могут голодать. Поэтому их
можно сохранять живыми, кормя мухами, личинками мучного хрущака, другими
насекомыми и пауками.
Впрок скорпионов консервируют 70% спиртом. Для рассмотрения наружной
морфологии пользуются спиртовым материалом.
205
В пределах СССР водятся скорпионы двух семейств — Buthidae (Средняя
Азия, Закавказье) и Chactidae (род Euscorpius — Черноморское побережье, юж-
ный берег Крыма). Скорпионы рода Euscorpius влаголюбивы, и их гораздо труд-
нее содержать в неволе, чем скорпионов рода Buthus.
Наиболее обыкновенными являются Buthus eupeus и Euscorpius italicus.
Внешняя морфология
Рассматривают спиртовый экземпляр скорпиона простым глазом
и с помощью простой или бинокулярной лупы, обсушив скорпиона от
спирта.
Со спинной стороны (рис. 142, А) видно разделение тела
скорпиона на передний, широкий, и задний, узкий, отделы. Перед-
ний отдел, начиная от переднего конца сверху, покрыт щиткам голо-
вогруди (с ph), имеющим в общем трапециевидную форму. Сзади идет
брюшко,широкая часть которого называется переднебрюшием(1—V11),
а задняя, узкая, часть — заднебрюшием или «хвостом» (VIII—XII).
Переднебрюшие сверху покрыто семью поперечными пластинками
твердого хитина, называемыми тергитами; бока переднебрюшия
покрыты мягкой хитиновой перепонкой —плеврами', низ передне-
брюшия имеет пять ясно различимых стернитов и два модифициро-
ванных стернита; заднебрюшие состоит из шести хитиновых удлинен-
ных члеников, одинаково твердых по всей окружности; пятый членик
гораздо длиннее остальных. С ним сочленяется концевой хитиновый
пузырек —ядовитая ампула —с острым, крепким, изогнутым жалом,
являющийся гомологом тельсона речного рака.
На рассмотренных частях наружного хитинового скелета разли-
чаем различные скульптурные элементы в виде бугорков, килей,
гребешков и др. На головогруди видим также боковые глаза, двумя
группами лежащие на ее переднебоковых углах; ближе к продоль-
ной срединной линии головогруди, на некотором расстоянии от перед-
него края последней, лежит пара главных глаз на приподнятом хити-
новом глазном бугорке.
Брюшная сторона (рис. 142, Б; рис. 143). Более сложно
устроена головогрудь, потому что к ней причленяются две пары
ротовых придатков и четыре пары ног.
Сяжков у скорпионов нет. Первой парой придатков являются
верхние челюсти —хелицеры, или жвалы (chelicerae), их удобнее рас-
сматривать со спинной стороны, вытянув вперед пинцетом из-под
переднего края головогрудного щитка.
Хелицера трехчлениста. Ее первый короткий членик спрятан под
передним краем головогруди; с ним сочленяется толстый, приблизи-
тельно яйцевидный, членик, вытягивающийся на своем передне-
внутреннем краю в крепкий изогнутый и зазубренный отросток, назы-
ваемый неподвижным пальцем (digitus immobilis); его партнером
является третий сегмент, образующий сильнее развитый подвижный
палец (digitus mobilis), сочленовной мягкой перепонкой соединяю-
щийся со вторым члеником хелицер.
Под основанием хелицер (считая по нормальному положению тела)
лежат основные членики, или коксы, второй пары ротовых при-
датков, называемых педипальпами, или ногощупальцами; их коксы
206
Рис. 142. Терминология частей тела скорпиона Buthus eupeus eupeus (С. Koch):
А — сверху, Б — снизу. По А. Б и р у л е.
1 — головогрудь (prosoma s. cephalothorax): br — предплечье (brachium); cal — шпора
(calcar); a — жвалы (mandibulae, chelicera); cph — головогрудный щиток (scutum cephalotho-
racis); ex, p— ляшки ногощупалец (coxae palporum); ex. 1, ex. 2, ex. 3 и ex. 4— ляшки (co-
xae) четырех пар ног; d. im — неподвижный палец (digitus immobilis); d. m— подвижной
палец (digitus mobilis); fr — бедро (femur); hm — плечо (humerus); /. m 1 — первая пара
челюстных люпастей (lobi maxillares primi); /. m 2 — вторая пара челюстных лопастей (lobi
maxillares secundi); m. p—тыльная сторона клешни (manus postica); mtr— плюсна (metatar-
sus); plm — клешня (manus s. palma); ptr — предстопье (protarsus); sc — вертлуг ногощупа-
лец (trochanter palporum); sp. ug — междукоготковый шип (spina pedalis); st— грудная пла-
стинка (sternum); tb — голень (tibia); tr — стопа (tarsus); trch — вертлуг (trochanter); tr. s —
шипы стопы (spinae tarsales); ung — коготки (ungues). 2 — переднебрюшье, или
брюхо (mesosoma s. abdomen): gn — половые створки (valvulae); mtst — заднегрудь, или
основная пластинка гребневидных придатков (metasternum); pct — гребневидные придатки
(pectines); spr — дыхальца (spiracula); I — Vll— верхние полукольца (terga s. scuta dorsa-
lia); Ill—Vll — нижние полукольца (sterna s. scuta abdominalia). 3 — заднебрюшье,
или хвост (metasoma s. cauda): ac — жало (aculeus); an—заднепроходное отверстие
(anus); sc. p—заднепроходный щиток (scutum anale s. pygidium); tel — ядоносный пузырек
(telson s. vesica venenifera); VIII — XII — пять колец заднебрюшья, или хвоста.
имеют вид коротких и широких косо лежащих члеников; педипальпы
(pedes maxi Hares) развиты сильно
Рис. 143. Скорпион мингрельский снизу.
Оригинал.
а — хелицеры; b — клешня педипальп; с— ко-
кса: d, е—челюстные лопасти (нижняя губа); $—
грудной щиток, позади которого лежат две поло-
вые створки; о — гребневые придатки; / спра-
ва — дыхальце; к — вертлуг (трохантер); I —
бедро; т — голень; п —лапка; о снизу — ядо-
витый пузырек; 1—IV—кольца эаднебрюшия.
и бросаются в глаза своей вели-
чиной и крупной концевой клеш-
ней (рис 142, А,Ь), они состоят
из пяти члеников, считая от кокс:
короткого вертлуга (scapula s.
trochanter), плеча (humerus), пред-
плечья (tibia palporum), клешни
(chela, manus) с неподвижным
пальцем (digitus immobilis),
образующим вместе с подвиж-
ным пальцем клещи для схваты-
вания и удержания добычи.
Коксы педипальп не сопри-
касаются друг с другом, и между
ними остается некоторое про-
странство. Тотчас кзади от каж-
дой коксы педипальп лежат коксы
первой пары ног, имеющие в об-
щем форму маленького треуголь-
ника; позади них лежат коксы
второй пары ног — в форме по-
перечных пластинок, почти
вплотную подходящих друг к
другу по средней линии тела
(рис. 142, Б, сх 1, сх 2). Про-
странство между коксами педи-
пальп первой и второй пар ног
занято двумя парами хитиновых
пластинок, называемых челюст-
ными лопастями (lobi maxilla-
res, рис. 142, Б, 1 т 1, 1 т 2;
рис. 143 d, е); они прикреплены
соответственно к коксам пер-
вой пары ног (узкие клиновид-
ные пластинки) и второй пары
ног (удлиненные, сужающиеся
кпереди пластинки, образующие
вместе нижнюю губу). Коксы
третьей и четвертой пар ног пред-
ставлены клиновидными, удли-
ненными образованиями, косо
лежащими и направленными су-
жающимися частями внутрь и
вперед.
Между их внутренними кон-
цами и коксами третьей пары
ног лежит пятиугольная или тре-
угольная срединная пластинка—
грудной щиток (sternum, рис.
208
143, s); он является производным последнего сгернита головогруди;
позади него лежит половое отверстие, прикрытое двумя округленно-
треугольными половыми створками (valvulae genitales), являющи-
мися гомологами первого стернита переднебрюшия.
За ними кзади лежит узкий поперечный валик, к боковым краям
которого прикрепляются подвижные гребневидные придатки (pecti-
nes, рис. 143, о); сам валик, являющийся основной пластинкой
гребневидных придатков (metasternum), рассматривается как видо-
измененный второй стернит переднебрюшия.
Далее кзади отмечается более однообразное строение тела, так
как позади metasternum лежат довольно длинные и широкие попе-
речные пластинки —стерниты переднебрюшия. Первый и последний
стерниты имеют форму трапеции. Первые четыре стернита несут каждый
по паре боковых, косо расположенных дыхательных отверстий (ды-
хальца, стигмы, stigmata, spiracula, рис. 142, Б,врг; рис. 143, / справа),
ведущих каждое в соответственно расположенное легкое. По сравнению
стергитами, стерниты переднебрюшия бедны скульптурными образова-
ниями; следы последних имеются на первом и пятом стернитах;
поверхность остальных гладкая.
Заднебрюшие, при рассмотрении с брюшной стороны, характе-
ризуется (у Buthus) наличием продольных килей, между которыми
лежат междукилевые площадки. Детали этих структур важны в си-
стематике. Задний край последнего (пятого) сегмента заднебрюшия
выдается кзади боковыми анальными зубчиками, или лопастями (lobi
anales).
Между пятым и последним члениками заднебрюшия в сочленов-
ной перепонке видно анальное отверстие (рис. 142, Б, ап); отверстие
anus может выворачиваться наружу, и тогда видны четыре полу-
шарообразных возвышения —выпячивания стенки задней кишки.
Анальное отверстие в нормальном положении прикрыто заднепро-
ходным щитком (pygidium или scutellum anale), связанным с задним
краем шестого сегмента.
Последний —постанальный (шестой) —сегмент образован ядови-
тым пузырьком; форму его надо рассматривать сбоку; тогда хорошо
видны узкая шейка, собственно пузырек и изогнутое жало.
Ноги скорпиона сочленены с коксами головогруди. Их четыре
пары. Каждая нога, начиная от соответствующей коксы, разделяется
на семь члеников (рис. 142 и 143), подвижно соединенных друг с дру-
гом мягкой сочленовной перепонкой: 1) вертлуг (trochanter) —
короткий членик; 2—3) бедро (femur) и голень (tibia) —самые длин-
ные членики; 4—7) лапка (tarsus), в свою очередь распадающаяся на
плюсну (metatarsus), предстопье (protarsus), стопу (tarsus) и когте-
вой членик (epitarsus) с парой подвижных коготков (ungues). Ноги
у скорпиона бегательные, но у некоторых специализированных видов
(например песчаного Liobuthus) первая пара ног приспособлена для
рытья (отбрасывания) песка.
Членики ног покрыты волосками, щетинками или имеют крепкие
выросты в форме шпоры (calcar. рис. 142, A, cal) на плюсне и шипы
у основания члеников ноги и коготков (spina pedalis). Эти образова-
ния также имеют систематическое значение.
14 Прахткжум по аоологвв — 385
209
Сегментация тела скорпиона. Головогрудь, или prosoma, образована
из шести сегментов, соответственно числу парных придатков (хели-
церы, педипальпы и четыре пары ног). Седьмой —предполовой — сег-
мент закладывается у зародыша скорпиона, но у сформированного
взрослого отсутствует; поэтому переднебрюшие —mesosoma, или abdo-
men, — состоит из семи сегментов (VIII—XIV); придатками восьмого
сегмента являются половые створки; девятому сегменту принадлежат
гребневидные придатки, которые рассматриваются как видоизменен-
ная пара брюшных конечностей; этому же членику принадлежит
metasternum. Десятому и тринадцатому сегментам принадлежат лег-
кие — по паре каждому; легкие рассматриваются как производные
жабр брюшных ножек водных предков скорпионов. Четырнадцатый
сегмент лишен каких-либо придатков и является переходным к з а д-
небрюшию (metasoma, или cauda), состоящему из пятнадцатого —
двадцатого сегментов; последний из них, лежащий позади анального
отверстия (гомолог тельсона ракообразных), видоизменен в специаль-
ный ядовитый аппарат.
Сегментация тела скорпиона
Сег- менты Спинная сторона Брюшная сторона (придатки) Отделы тела
I Хелицеры
II Педипальпы
III IV * Щиток головогруди Нога I > II S р,
V » III U о
VI » IV ю о EJ
о и,
VII Закладывается у зародыша, у взрослого отсутствует
VIII Половое отверстие и V X
половые створки а
IX Гребневидные придатки и 2 Q*
X XI ► Тергиты легкое Легкое О X
XII > Еч О
XIII » Q* О
XIV Переходный сегмент С
XV
XVI —
XVII * Хитиновые кольца заднебрюшия — X
XVIII — а л
XIX Анальное отверстие о* ю
о
। i
XX Ядовитый аппарат Парное отверстие ядо- 1 00
витых желез ।
210
Заднебрюшие чрезвычайно подвижно. Его скорпион загибает над
телом кпереди и обращенным вперед жалом ударяет врага или схва-
ченную клешнями педипальп добычу.
Вскрытие скорпиона
Предпочтительнее пользоваться живым материалом, но возможно
разобраться в строении скорпиона и на спиртовом материале.
Живых скорпионов хлороформируют или убивают серным эфиром.
Вскрытие производят в ванночке в физиологическом растворе пова-
ренной соли (если будут нужны свежие органы для каких-либо специаль-
ных целей) или в воде; к последней полезно добавить немного спирта,
если препаровка преследует цели ознакомления лишь с анатомией
скорпиона и приготовления тотальных препаратов. Сохранявшихся
в спирту скорпионов вскрывают в воде.
Острыми ножницами разрезают боковые плевры переднебрюшия,
начиная с заднего правого края; разрез ведут до переднего конца голо-
вогруди; огибают ее спереди поперечным разрезом под щитком голо-
вогруди и над ротовыми органами; далее разрез ведут спереди назад
по левому боку скорпиона до начала заднебрюшия.
Пинцетом захватывают передний конец щитка головогруди, оття-
гивают его кверху, подрезая под ним небольшим скальпелем мышцы;
чтобы не поранить желудка и печени, скальпель держат возможно
более высоко и скребут им по внутренней поверхности щитка голо-
вогруди.
Перерезая места прикрепления дорзовентральных мышц, подни-
мают кверху кожные покровы и отрезают их поперек у первого чле-
ника заднебрюшия.
Повернув ванночку в обратную сторону, разрезают с боков кольца
заднебрюшия и снимают его дорзальную часть вплоть до ядовитой
ампулы.
Поворачивают ванночку в нормальное положение и рассматривают
препарат. Идя спереди назад, находим желудочную железу (рис. 144, V)
в виде компактного довольно нежного органа, имеющего пятилопаст-
ное строение; одна лопасть идет вперед,и по две —с каждой стороны
тела; эта железа открывается в головогрудную часть средней кишки,
называемую желудком; задним краем она касается диафрагмы; у ее
средней линии видны разрезы двух дорзо-вентральных мышц; по своему
цвету и консистенции желудочная железа сходна с печенью и раньше
неправильно принималась за головогрудные отростки печени.
Печень (рис. 144, Н) занимает всю полость переднебрюшия; по сред-
ней линии печени имеется дорзальное вдавление, в котором лежит сердце
с перикардиальной сумкой; по бокам сердца в печени лежат метамерно
шесть пар отверстий, через которые проходят насквозь дорзо-вентраль-
ные мышцы; задним концом печень проходит в первый или во второй
членики переднебрюшия; печень у Buthidae сплошная, если ее рас-
сматривать сверху, тогда как у Chactidae боковая часть печени с каждой
стороны рассечена на поперечные дольки; печень коричневатого цвета;
она нежной консистенции и легко разрывается; при препаровке в фи-
зиологическом растворе разрывающаяся печень мутит воду; приба-
* 211
вление спирта облегчает препаровку, так как препятствует замутне-
нию жидкости; печень открывается в среднюю кишку (в ее часть, про-
ходящую через переднебрюшие) пятью парами про-
Рис. 144. Пище-
варительныйка-
нал скорпиона
и желудочная
железа. По
токов; средняя кишка прикрыта печенью сверху и
становится видной сверху лишь в заднебрюшии.
Средняя кишка проходит через все членики задне-
брюшия и в пятом сегменте переходит в очень корот-
кую заднюю кишку (рис. 144, /?); средняя кишка
часто бывает заполнена молочнобелым тонкозерни-
стым содержимым —гуанином (продуктом экскреции,
весьма характерным для паукообразных).
Уже отмеченное нами сердце представляет собой
сравнительно широкую трубку, образующую между
дорзо-веигральными мышцами переднебрюшия расши-
рение; в них дорзально лежит по паре отверстий, на-
зываемых устьями; последние ведут из околосер-
дечной полости в полость сердца, и через них в ука-
занном направлении идет кровь в процессе кровооб-
ращения; позади последней пары дорзо-веигральных
мышц сердце сужается и резко переходит в тон-
кий надкишечный сосуд (спинную аорту), который
идет до заднего конца тела: кровь по нему идет
в направлении спереди назад.
По рассмотрении всех этих деталей осторожно
снимаем дольки желудочной железы и печени, отщи-
пывая их тонким пинцетом и откалывая конц(А1 тон-
кого скальпеля; ванночку надо покачивать, чтобы
удалить с поверхности препарата кусочки органов.
Если материал спиртовый, то такое откалывание
кусочков печени делать осторожно и малыми пор-
циями, чтобы не порвать половые органы, лежащие
между дольками печени.
При вскрытии живого скорпиона можно удалить
печень целиком; острыми ножницами отвесно разре-
зают среднюю кишку тотчас позади диафрагмы; за-
хватывают пинцетом спереди всю массу печени и,
осторожно подергивая, поднимают постепенно пе-
чень кверху, освобождая ее от петель половых орга-
нов. В том и другом случаях следует рассмотреть
следующие детали: в головогруди, под желудочной
железой, лежит начальная часть средней кишки, над-
глоточный комплекс нервных узлов и по бокам меш-
Е. Павлов-
скому.
Н — печень; D —
дорзо-вентральные
мышцы; F—про-
токи печени; I —
тонкая кишка;R—
з< Дняя кишка ;М х—
средняя кишка;
м* — мальпигиевы
сосуды; V — же-
лудочные железы.
ковидные коксальные железы (органы вы-
деления).
Под печенью лежит средняя кишка; можно ви-
деть симметрично лежащие парные протоки печени,
отличающиеся широким диаметром; печень снизу
сильно дольчата. В конце переднебрюшия в сред-
нюю кишку открываются две пары тонких «мальпи-
гиевых сосудов) (рис. 144, Л7#); одна пара их идет
212
Рис. 145. Женские ор-
ганы размножения скор-
пиона. Оригинал.
над средней кишкой и дает боковые ветки, другая пара откло-
няется латерально; эти трубки также являются экскреторными
органами.
По удалении печени рассматриваем брюшную нервную цепочку,
на которой у Buthidae лежат или раздельно расположенные, или
неправильно скученные узлы лимфатической железы; у Euscorpius
эта железа имеет вид сплошного наднервного тяжа (рис. 149, о), кроме
того, помимо нее, имеется пара лимфоидных мешков (рис. 149, Ь),
отходящих от задней поверхности головогрудной диафрагмы по одному
с каждой стороны.
На внутренней поверхности брюшных по-
кровов, в области переднебрюшия, видны че-
тыре пары мешетчатых образований, лежащих
ближе к бокам тела и непосредственно свя-
занных с покровами —это легкие скор-
пиона.
Половые органы самки имеют различный
вид, смотря по ее зрелости и по степени раз-
вития зародышей. Яичник небеременной самки
представлен сетью трубок, которые распола-
гаются правильно и образуют как бы лест-
ничную систему из двух боковых продоль-
ных трубок (спереди сливающихся перед
половым отверстием друг с другом), сред-
ней более короткой трубки и пяти пар попе-
речных трубок (Buthus), соединяющих средин-
ную и боковые трубки друг с другом; эти
трубки являются маточными, и на них сидят
фолликулы с одной яйцевой клеткой в каждом
фолликуле (рис. 145). Не доходя до полового
отверстия боковые трубки яичника образуют
по расширению, называемому семеприемником.
У Euscorpius поперечных трубок яичника че-
тыре пары.
У беременной самки зародыши разви-
ваются в трубках яичника, куда яйца вы-
скальзывают из яйцевых фолликулов; зародыши располагаются друг
за другом, благодаря чему трубки принимают четковидную форму;
одетые зародышевыми оболочками, зародыши скорпионов разви-
ваются на месте и значительно увеличиваются в размерах; поэтому
яичники могут иметь различный вид в зависимости от стадии созре-
вания зародышей.
У скорпионов зародыши развиваются в просвете трубок яичника,
которые называют также маточными каналами. Вздутые части маточ-
ных каналов отделены друг от друга глубокими перетяжками. На взду-
тиях видны фолликулы с незрелыми яйцами, которые дадут потомство
в следующий сезон раз ножения.
По мере развития и роста зародышей, круглые четковидные утол-
щения переходят в овальные, и сами трубки яичника весьма удли-
няются.
213
Buthus и Euscorpius являются живородящими скорпионами; после
отрождения молоди растянутые трубки яичника спадаются, но неко-
торое время остаются дряблыми.
Половые органы самцов скорпионов представлены семенниками и
добавочными органами. Семенников у Buthus два; они образованы
тонкими трубочками, образующими с каждой стороны тела по три
следующих в линейном порядке друг за другом, замкнутых петли
(рис. 146); нередко семенники соединяются (анастомозируют) друг
с другом.
От каждого семенника отходит кпереди семепровод (рис. 146)
в виде трубки, идущей от передней петли до боковых придаточных
Рис. 146. Мужской половой аппарат Buthus eupeus. Оригинал.
t — семенник; р — параксиальный орган; v — концевое расширение vas
deferens; d — семенной пузырек; о — овальная железа; с — цилиндрическая
железа; е — передняя железа. Другой семенник не изображен.
органов. Последние имеют вид удлиненных пластинок или стержней
своеобразной формы; их называют параксиальными органами (р); зад-
ний конец их у Buthus утончается в придаток, называемый flagel-
lum (рис. 146). Форма этих органов своеобразна и различна в деталях
у представителей разных семейств и родов скорпионов. Передними
частями параксиальные органы соединяются друг с другом и откры-
ваются в половое отверстие. В передней части параксиальных органов
лежат передние придаточные железы (рис. 146, е).
У Buthus vas deferens перед впадением и параксиальный орган
своей стороны тела расширяется в пузырек, с которым соединяется
семенной пузырек (vesicula seminalis), имеющий форму удлиненного
мешочка, величина которого зависит от степени наполнения жив-
чиками (рис. 146, (1); кпереди от концевого пузырька лежит овоид-
214
ная или овальная железа (рис. 146, о); параллельно им* обоим
несколько кзади идет цилиндрическая железа (с). Эти детали могут
быть рассмотрены под лупой на извлеченном параксиальном органе.
Иногда у самок скорпионов (Bufhus, Euscorpius) можно, видеть
черную пластинку, выходящую краем из-под створки половой кры-
шечки. Это — сперматоклейтрум, временно запечатывающий половое
отверстие самки после копуляции и препятствующий обратному выте-
канию спермы. Сперматоклейтрум образуется, повидимому, за счет
секрета, выделяемого самцом в конце копуляции.
При вскрытии самок одного и того же вида скорпиона в разные
стадии беременности можно заметить изменения в состоянии печени:
у самок с развитыми зародышами дольки печени сильно уменьшаются
и сама печень далеко уступает объему аналогичного органа у небере-
менной самки.
Рассмотрение микроскопических препаратов
1. Ядовитые железы. Обыкновенной бритвой разрезают ядовитый
пузырек скорпиона (положив его на пробку) в поперечном направлении
на тонкие ломтики. Их обезвоживают и без окраски заключают в ка-
надский бальзам, подложив соответствующие по толщине восковые
ножки.
Объект снаружи окаймлен хитиновой кутикулой; внутри хити-
нового обода видны два овальных органа, прилежащих каждый на своей
стороне тела непосредственно к нижнебоковой поверхности покровов
ядовитого пузырька. Каждый овал является поперечным разрезом
ядовитой железы. У Euscorpius она выстлана высоким эпителием, ко-
торый не образует складок; ядовитая железа на всем протяжении,
где она не прилежит к покровам, одета толстой мышечной оболочкой;
волокна последней прикрепляются к внутренней поверхности покро-
вов ампулы. Таким образом ядовитая железа плотно вложена в ка-
меру, образованную частью хитиновых покровов (неподатливая часть
камеры) и прирастающей к ним мышечной оболочки. При сокращении
последней яд с силой выдавливается из железы, точно из шприца.
Мышечные оболочки обеих желез соприкасаются друг с другом по
средней линии ампулы (рис. 147, В).
У Bufhus ядовитая железа устроена по тому же принципу, но
отличается более сильным развитием железистой части, так как эпи-
телиальный слой железы с подстилающей его соединительной тканью
образует более или менее глубоко вдающиеся в полость железы складки.
Благодаря этому увеличивается секреторная поверхность железы.
В связи с указанными отличиями ядовитых желез говорят о двух
формах их —простой и складчатой (рис. 147, С).
Бритвой разрезают ядовитый пузырек вдоль по длине —в отвес-
ном направлении. Жало отходит к одной из половинок пузырька.
Приготовляют бальзамный тотальный препарат, положив половинки
ампулы разрезом кверху.
На препарате видна вся железа со стороны ее мышечной оболочки.
Орган занимает всю длину ампулы; свободным остается небольшое
пространство с дорзальной стороны. Кзади ядовитая железа, соог-
215
ветственно ходу основания и изгибу жала, сужается и переходит в вы-
водной проток, скрывающийся в тонкой части жала; проток выстлан
внутри хитиновой оболочкой; он открывается небольшой удлиненной
щелью тотчас у острия жала.
2. Срез дольки печени. Свежие кусочки печени скорпиона фикси-
руют сулемой с уксусной кислотой и обычным способом заливают
в парафин. Срезы окрашивают гематоксилином Делафильда и эози-
ном или по Романовскому-Гимза.
Долька печени образована слоем высоких эпителиальных клеток,
лежащих на базальной оболочке, к которой прилегает перитонеаль-
ный покров печени (рис. 148, А, up.). Эпителий печени скорпиона
Рис. 147. Ядовитая железа Buthus eupeus. По Е. Павлов-
скому.
А — продольный разрез пузырька ядовитой железы; виден мешок ядови-
той железы (ab). d —отверстие жала. В — поперечный разрез заднего сег-
мента Euscorpius carpaticus. С — то же, Buthuseupeus. с — хитин; а — эпи-
телий ядовитой железы; b — мышечная оболочка.
состоит из двух родов клеток (рис. 148, A, zf, zr). Одни из них более
крупны; ядра их лежат посредине или ближе к основанию клетки,
протоплазма заполнена крупными шарообразными включениями раз-
личной величины. Включения эти ацидофильны по окраске и гомо-
генны по структуре. Кроме этих включений, в протоплазме клеток часто
встречаются кучки пигментных зерен; в неокрашенном состоянии
они коричневого цвета. Рассматриваемые крупные клетки многочис-
ленны; они являются резорбирующими, т. е. всасывают перевари-
ваемые пищевые вещества (рис. 148, A, zr).
Клетки другого рода малочисленны. Они сдавливаются более мас-
сивными резорбирующими клетками печени и также обладают круп-
ными округлыми ацидофильными включениями, но зернистой струк-
туры; прослойки протоплазмы между ними заполнены мелкими
базофильными включениями в виде зернышек пли нитей. Налегая
друг на друга, эти включения образуют тяжи, резко прослаивающие
216
и других
ракообразных
UP
XS
zt
up
Рис. 148. Разрез дольки печени Buthus eupeus и про-
дольный разрез мальпигиева сосуда (ттр) в перито-
неальной оболочке печени.
up — перитонеальный покров; zr— разорбционная клетка;
zf -ферментная клетка; xz — ацидофильные включения в резор-
бирующих клетках; xs — пигментные включения в резорбирую-
щих клетках печени; п — ядро. По Е. Павловскому.
крупные ацидофильные зернистые шары. Ядро клетки лежит в основ-
ной протоплазме, будучи как бы вкраплено в базофильный тяж. Этот
второй род клетки выделяет пищеварительный секрет и поэтому со-
ответствует ферментным клеткам печени
паукообразных (рис. 148,4,;/).
Перитонеальная
оболочка печени со-
стоит из пластинча-
той соединительной
ткани, в узких пе-
тлях которой лежат
соединительноткан-
ные клетки с оваль-
ным ядром и неж-
ной зернистой про-
топлазмой. В клет-
ках перитонеаль-
ного покрова пе-
чени часто встре-
чаются шарообраз-
ные включения та-
кого же типа, что
и в резорбирующих
клетках.
3. Мальпигиевы
сосуды. Между
дольками печени
могут попасться на
разрезе и мальпи-
гиевы сосуды (рис.
148, В, ттр). Они
вкраплены в пери-
тонеальный покров.
Образованы хоро-
шо развитыми эпи-
телиальными клет-
ками с ясной исчер-
ченной структурой,
протоплазма их ис-
ттр
черчена в вертикальном по отношению к поверхности клетки направле-
нии. Овальные ядра лежат тотчас под поверхностью клеток, в слое более
темно окрашенной протоплазмы. Мелкие ветви этих сосудов имеют
уплощенную стенку с низкими клетками. Мальпигиевы сосуды нигде
не выходят за пределы перитонеальной оболочки печени (рис. 148, В,
up). Они у скорпионов являются производными средней кишки (т. е.
энтодермы), а не задней (как у насекомых). Поэтому они не гомоло-
гичны мальпигиевым сосудам насекомых. Вероятно, и биохимическая
работа их отлична.
4. Легкие скорпиона. Легочный мешочек скорпиона отрезают от
стенок тела и расщипывают иглами на отдельные листки; последние
217
рассматривают с плоскости, положив их в капле воды под покровное
стекло или заключив в глицерин-желатину или гуммиарабиковую
смесь.
Легочный мешочек заливают в парафин и режут в продольном
направлении, так чтобы легочные пластинки были бы рассечены по-
перек. Окраска—гематоксилином и эозином.
Легкое скорпиона представляет собой мешкообразное впячивание
хитиновых покровов переднебрюшия внутрь тела по краям щелевид-
ного дыхальца (стигма). Стенка впячивания тонкая. В полость мешочка,
в свою очередь, впячивается множество хитиновых дупликатур в виде
тонких листков, лежащих параллельно друг к другу; в толще каждого
листка имеется щелевидная полость, куда затекает кровь (гемолимфа)
скорпиона; между листками остаются щелевидные пространства, по
которым циркулирует воздух, поступающий в легкое. В общем легоч-
ные листки похожи на листы книги.
Такой орган дыхания называется легкими, потому что через его
посредство осуществляется дыхание атмосферным воздухом.
На тотальных препаратах листков можно видеть хитиновую струк-
туру их поверхности и характер обрамления свободного края.
Наблюдения над фагоцитарными органами
Методика. Необходимы живые скорпионы, которым инъеци-
руют в полость тела взвесь туши, бактерий и других веществ. Для
инъекции скорпиона иммобилизуют на пробковой пластинке, обка-
лывая его острыми дужками изогнутой проволоки. Дужками обхва-
тывают брюшко в его основании и у конца и тонкие части педипальп
ближе к их основанию.
Инъекцию производят тонкой стерильной пастеровской пипеткой;
отломав кончик, в нее набирают необходимую жидкость и вращатель-
ным движением вкалывают конец пипетки в мягкую сочленовную
перепонку ноги, оттянув и иммобилизовав последнюю пинцетом.
Когда конец пипетки вколот в ногу, ртом осторожно дуют в пипетку,
следя за движением столбика находящейся в ней жидкости; если
жидкость из пипетки не выходит, то пипетку подают несколько назад
и вращают вокруг оси. Когда впрыснуто нужное количество жидкости,
пипетку извлекают из ноги и через минуту-две скорпиона освобождают
и пересаживают в садок. При хорошо проведенной инъекции кровь
из ранки не выступает.
Записывают время постановки опыта. Скорпионов выдерживают
разные сроки и затем вскрывают, предварительно захлороформировав.
При опытах с экскреторными органами и с фагоцитозом туши ре-
зультат опыта можно видеть уже невооруженным глазом, а фагоцитоз
бактерий можно наблюдать лишь на препаратах срезов органов.
Тотальное наблюдение. Если инъецировать скор-
пиону семейства Buthidae растертые в воде китайскую тушь или по-
рошок кармина и вскрыть скорпиона через сутки, то можно видеть
почернение или покраснение узлов лимфатической железы, лежащей
над брюшной нервной цепочкой. Отыскать ее ле1*ко, удалив печень.
Скорпионы рода Bulhus имеют простую систему фагоцитарных органов.
218
Рис. 149. Лим-
фатическая же-
леза (а) и лим-
фоидный ме-
шок (Ь) скор-
пиона после
инъекции туши;
ns — брюшная
нервная цепоч-
ка. По Е. Па-
вловскому.
Если такой же эксперимент сделать с Euscorpius, то его лимфати-
ческая железа выявится в форме сплошного шнура, лежащего надбрюш-
ной нервной цепочкой в переднебрюшии. Кроме лимфатической железы,
Euscorpius имеет два лимфоидных мешка, лежащих в виде коротких
слепых выростов на заднем конце трубок; лежат они в переднем от-
деле переднегруди, несколько отступя по бокам нервной цепочки;
лимфоидные мешки отходят от диафрагмы, отделяющей переднегрудь
от переднебрюшия. Лимфоидные железы выделяют, например, ferrum
ammonito saccharatum, если его водный раствор впрыснуть скор-
пиону. Фагоцитарные функции свойственны и этим железам; поэтому
фагоцитарный аппарат Euscorpius является сложным.
Рассмотрение гистологического
среза через лимфатический узел
Buthus. Выпрепаровывают брюшную нервную це-
почку с лимфатическими узлами; вырезают из нее
кусок, фиксируют спиртом с формалином или насы-
щенным водным раствором сулемы и по общим пра-
вилам заливают в парафин. Режут на микротоме по-
перек. Окраска — гематоксилином, эозином или крас-
кой Романовского-Гимза (для неэксперименталь-
ного материала).
На препарате виден разрез или одного из нерв-
ных ганглиев, или соединяющих их нервных стволов
брюшной цепочки, тесно прилежащих друг к другу.
Над нервной цепочкой лежит наднервная артерия,
связанная с оболочками нервной цепочки. В верх-
ней стенке сосуда под иммерзией можно заметить
поперечнополосатые мышечные волокна, между вну-
тренней (intima) и наружной (adventitia) соедини-
тельнотканными оболочками артерии.
Стенка сосуда местами образует дорзальные вы-
пячивания, переходящие в лимфатическую железу.
Строма ее состоит из соединительнотканных перекла-
дин, являющихся продолжением оболочек сосуда.
В петлях стромы лежат основные клетки (лимфо-
циты или незернистые лейкоциты); бывают также
ацидофилы (зернистые клетки с оксифильной зернистостью) или другие
зернистые клетки. Основные клетки железы иногда делятся карио-
кинетически, что подтверждает кроветворную функцию лимфати-
ческой железы.
Экологические замечания
Скорпионы являются наземными обитателями, и вся их органи-
зация приспособлена к жизни в воздушной среде. Потребность их
в атмосферной влажности различна. Различают сухолюбивые ксеро-
фильные формы скорпионов (Buthus) и гигрофилов (Euscorpius), ну-
ждающихся в большой влажности воздуха.
Скорпионы хорошо бегают и благодаря наличию коготков на
ногах могут взбираться по неровным вертикальным поверх-
ностям.
219
Скорпионы являются хищными животными; они питаются другими
членистоногими, кдторых убивают ядовитым жалом. Каннибализм
у скорпионов развит сильно. Они обитают в жарких и теплых странах;
способны долго голодать. Наличие ядовитого аппарата, смертоносного
для многих животных, тем не менее не является абсолютной гаран-
тией в отношении защищенности самого скорпиона. Так, скорпионы
могут делаться добычей кара-куртов (ядовитый паук, устраивающий
из паутины гнездо); скорпионов поедают также змеи. Скорпионы
держатся днем под камнями, в развалинах, в трещинах стен, коры де-
ревьев и др. Они являются ночными животными.
Скорпионы появляются на свет в беспомощном состоянии; тело
их одето нежной хитиновой кожицей. Первое время отродившиеся
скорпионы держатся на теле матери, облепляя его со спины. Скор-
пионы живут по несколько лет.
Уплощенная форма тела скорпионов является особенностью адап-
тивного характера, так как благодаря этому скорпион легко запол-
зает в глубокие трещины, щели, под камни и др.
Общие замечания
Скорпионы отличаются от ракообразных отсутствием сяжков, на-
личием всего двух пар ротовых придатков и четырех пар ходильных
ног. При общем сходстве в разделении тела на головогрудь и брюшко,
скорпионы отличаются дальнейшим подразделением брюшка на ши-
рокое переднебрюшие и узкое заднебрюшие. Если у рака доминирую-
щей по развитию частью тела является головогрудь, то у скорпионов,
наоборот, отмечаем малую величину головогруди по сравнению с пе-
редне- и заднебрюшием.
Весьма характерно у скорпионов отсутствие брюшных ножек.
Производными одной пары их считаются гребневидные придатки;
легкие скорпионов многими рассматриваются как модифицированные
в филогенезе брюшные жаберные конечности предков скорпионов. При
втягивании внутрь пластинки жабр превратились в листки легких.
Такая перемена функции, очевидно, связана с переходом от водной
жизни к наземной и к дыханию атмосферным воздухом. От остальных
брюшных ножек не осталось никаких следов.
В отношении внутреннего строения отмечаем особенности пище-
варительного аппарата. Скорпион всасывает жидкие части добычи,
которую переминает жевательными придатками педипальп и ног.
Рак пережевывает пищу и проглатывает ее в желудок, где происходит
добавочное перетирание ее жевательными хитиновыми пластинками.
В связи с этим функция желудка рака и скорпиона не равнозначны;
к тому же эти отделы и не гомологичны друг другу, так как желудок
рака развивается из передней (эктодермальной) кишки, а у скорпиона
он является производным средней кишки (энтодермальной).
При сильном развитии печени и у скорпиона и у рака отмечается
громадное различие в соотношении средней и задней кишки. Средняя
кишка коротка, а задняя очень длинна у рака, тогда как у скорпиона
отношения обратные. Ксксальные выделительные железы скорпиона,
сяжковые (зеленые) железы рака являются производными метане-
220
фридиев. И у скорпиона и у рака видим хорошее развитие незамкну-
той кровеносной системы (сердце, сосуды, синусы), что связано с про-
странственно ограниченной локализацией дыхательных органов (лег-
ких у скорпионов и жабр у рака).
Отсутствие сяжков, наличие двух пар ротовых придатков и четырех
пар ног, подразделение тела на головогрудь и брюшко и присутствие
органов дыхания воздушного типа (в данном сулучае легкие) —все
это признаки класса паукообразных (Arachnoidea), к представителям
которых принадлежат скорпионы. В то же время членистость тела,
наличие расчлененных ножек и хитинового покрова объединяет и
ракообразных и паукообразных в один тип членистоногих.
Маленькие клешневидные хелицеры, большие клешневидные, при-
датки педипальп, модифицированная пара брюшных ножек в виде
гребневидных придатков, разделение тела на головогрудь, передне-
брюшие и заднебрюшие, оканчивающееся ампулой с жалом и ядови-
тыми железами, обладание четырьмя парами легких —все это хара-
ктерно для отряда скорпионов — Scorpiones.
Buthus и Euscorpius относятся к разным семействам—Buthidae
и Chactidae, отличающимся: первое —узкой почти цилиндрической
Сравнение организации рака и скорпиона
Название органов Рак Скорпион
Головогрудь Антеннулы Антенны Глаза Ротовые органы Ногочелюсти. . . Ходильные ноги . Есть > Стебельчатые подвиж- ные, пара Три пары (мандибулы и две . пары максилл) Три пары Пять пар (с жабрами) Нет » По краю и по середине щитка головогруди Две пары (хелицеры и педипальпы) Нет Четыре пары (жабр нет)
I Брюшко Из пяти однообразных сегментов с двуве- твистыми рудиментар- ными ножками 1 Тельсон Половые створки,гребне- видные придатки (видо- измененные конечности), ножек нет, сегменты X — XIII имеют по паре косых стигм, ведущих в легкие Переднебрюшне
Нет никаких придатков; сегменты из сплошных колец хитина Задне- брюшис
Ампула с ядовитой железой
221
клешней педипальп, тогда как у второго клешня широкая, более
или менее уплощенная. На головогрудном щитке у Chactidae с каждой
стороны по два глаза, а у Buthidae —по три или более.
ОТРЯД ПАУКИ (ARANEINA)
ДВУЛЕГОЧНЫЕ ПАУКИ (DIPNEUMONES)
Технические замечания. Пауков собирают в домах, сараях,
помещениях для скота, в отхожих местах и др. с паутины. Под сидящего паука
подводят широкогорлую баночку с 70% спиртом; при прикосновении к паутине
паук часто падает вертикально вниз, повисая на паутиновой нити. Нужно баночку
подставить так, чтобы паук упал прямо в спирт.
В природе пауков собирают стряхиванием деревьев или кустарников на
подставленный раскрытый зонт. Упавших пауков берут пинцетом. Собирают
также пауков с различных растений. Тарантулов добывают из нор заливанием их
водой. Обычная консервировка материала в 70% спирту.
Для срезов пауков фиксируют в жидкости Петрункевича: раствор А —
азотной кислоты 12 см3, медного купороса 4 г, дистиллированной воды 100 см3;
раствор Б — кристаллической карболовой кислоты 4 г, эфира 6 г, 80%
спирта 100 см3; перед употреблением смешивают одну часть раствора А с фемя
частями раствора Б. Паука опускают в жидкость и отрезают ноги у основания.
Фиксирование продолжается 1—2 дня, после чего объект переносится в 70% спирт,
сменяемый три-четыре раза. Фиксирующая смесь приготовляется всякий раз
заново из основных растворов.
А. Петрункевичем дан также рецепт другой стойкой фиксирующей
смеси в составе: 60% спирта 100 см3, азотной кислоты 3 см3, эфира 5 см3, мед-
ного купороса 2 г и кристаллического паранитрофенола 5 г. Фиксирование, как
указано выше. Заливка в парафин и окраска срезов, как обычно.
Паук-крестовик откладывает яйца в коконы, которые можно найти под кам-
нями, под корой пней; коконы одеты желтоватой мягкой паутиной. Вылупившиеся
паучки остаются сидеть в коконе продолжительное время. На юге можно найти
шаровидные твердостенные коконы кара-курта в полынной степи. Из коконов
выходит колоссальное количество молоди.
Вылупившуюся молодь фиксируют горячим раствором сулемы со спиртом,
выдерживают материал в 70% спирту с иодной тинктурой (впредь до прекращения
обесцвечивания жидкости), через 96% и абсолютный алкоголь проводят материал
в кедровое масло и заключают в канадский бальзам одних паучков спиной кверху,
а других — кверху брюшной стороной тела.
Внешняя морфология
Рассматривают под слабой лупой или бинокуляром спиртовый
экземпляр паука. Затем микроскопируют бальзамные препараты
молодого паучка и отдельных частей взрослого паука. Отдельные
части тела паука (ротовые органы, ноги, паутинные бородавки) заклю-
чают в канадский бальзам после предварительного кипячения в 20%
водном растворе едкого натра. Далее следует отмывание в воде, обез-
воживание и заключение в канадский бальзам. Для рассмотрения
полового отверстия, влагалища и семеприемников вырезают острыми
ножницами стенку брюшка на нижнепередней стороне и обрабаты-
вают, как указано выше. При лабораторном изучении препарат до-
водят до кедрового масла и рассматривают объект в часовом стекле
в масле, поворачивая препарат, как нужно.
Тело паука (рис. 150) явственно разделяется на головогрудь (А ) и
брюшко (В), соединенные друг с другом тонким и коротким стебельком.
Обычно брюшко гораздо крупнее головогруди. Последняя сверху
222
покрыта щитком, имеющим форму кубышки. Щиток часто имеет
срединное вдавление в виде маленькой ямки или продольной узкой
бороздки. Иногда бывают видны радиальные вдавления соответственно
местам прикрепления к их внутренней поверхности мышц сосатель-
ного аппарата и конечностей; радиусы идут по направлению к ямке
или бороздке.
По переднему краю щитка лежат глаза (рис. 151), числом восемь
(редко —шесть); глаза у многих видов лежат на бугорках; форма
и расположение глаз различны, что имеет существенное системати-
Рис. 150. Наружный вид паука снизу.
Справа — самка. А — головогрудь; В — брюшко; а — верхняя челюсть;
Ь — нижняя челюсть; с — губа; d — грудной щиток; е — щупик нижней челюсти;
/ — тазик; g — вертлуг; h — бедро; / — колено; к— голень; / — предлапка;
т — лапка; N — дыхальце; о — эпигина; р — отверстие трахей; q — паутинные
бородавки. По Спасскому. Слева — самец сверху, на переднем
конце головогруди видны два ряда глаз. Щупик нижней челюсти утолщен.
По Далю.
ческое значение. Форма их полушарообразная, точечная, в виде чече-
виц и др. Глаза лежат в два (реже в три или четыре) поперечных ряда.
В зависимости от расположения различают переднесредние, заднесред-
ние, переднебоковые и заднебоковые глаза.
Головогрудь, как и у скорпионов, лишена усиков, или сяжков,
что характерно вообще для всех паукообразных. При рассматривании
головогруди паука спереди видна пара сильных двучленистых хели-
цер (рис. 151); основной членик их велик, он цилиндрической или
иной формы; на его нижнем конце имеется подвижный острый кого-
ток с отверстием у самого конца, где открывается проток ядовитой
железы. Коготь прикладывается к основному членику хелицеры с ее
223
внутренней стороны наподобие лезвия перочинного ножика; внутрен-
ний край хелицеры у основания коготка бывает зазубрен или сна-
бжен длинными хитиновыми щетинками (волссками); число и располо-
:чет в систематике пауков.
При рассматривании голово-
груди сверху бывает видно осно-
вание хелицер, а у некоторых
пауков (Dysdera) хелицеры с их
коготками ориентированы горизон-
тально. Хелицеры служат для за-
хватывания добычи, ее удержания
и отравления (действием яда ядо-
витых желез — см. ниже). Простран-
ство между основанием хелицер и
передним рядом глаз называется
наличником (clypeus).
Второй парой ротовых органов
являются педипальпы, которые
имеют вид ноговидного щупика. На
жение зубцов принимается в
Рис. 151. Головогрудь’ и глаза пау-
ков спереди. По Далю.
А — Агапеа, В — Linyphia.
изолированном* органе видны следующие части: основная, густо
усаженная внутри волосками пластинка с максиллярной долькой
(рис. 152) и коксальным выростом (вертлуг); с вертлугом щу-
пика сочленяется длинное бедро, затем идет короткое колено
(patella); далее следуют голень (tibia) и тарзальный членик (tarsus);
у самок некоторых видов он оканчивается коготком. Таково строение
педипальп самки паука; у самца оно отличается наличием добавоч-
ных образований, играю-
щих роль при копуляции
(см. ниже).
Максиллярные пластин-
ки педипальп сочленяются
с медианной пластинкой,
называемой губой (рис.
150, с). Задним краем она
прилежит к стернальной
пластинке, занимающей
среднюю часть нижней по-
верхности головогруди(рис.
150, d). Форма грудинной
пластинки многоугольная;
на ее краях имеются не-
большие вдавления — места
прикрепления коксальных
члеников ног. Последних —
Рис. 152. Ротовые органы паука; нижние че-
люсти со щупиками и непарной нижней губой.
Из Н. X о л одк овс к о г о.
четыре пары. Коксы прочно прилежат друг к другу, к щитку голово-
груди и к ее грудинной пластинке. Благодаря этому головогрудь
представляет собой как бы коробку из твердых хитиновых стенок.
Ноги (рис. 150) имеют строение, аналогичное щупику педипальп самки.
С коксами (/), неподвижно срастающимися с головогрудью, сочле-
няется короткий членик—вертлуг (trochanter — g); за ним последов
224
ЬателЬно лежат: 1)удлиненное бедро (femur—Л), 2)короткий членик, на*
зываемый patella, З)голень(Л),4) metatarsus (/)и 5)собственнолапка(/п)—
tarsus, несущая на конце цару короткое; кроме них бывает еще доба-
вочный третий (срединный) коготок. Коготки зазубрены, особенно
парные, что придает им вид гребенки (рис. 153, Ь). Под парными когот-
ками у некоторых пауков бывают зазубренные волоски или пучок
волосков, постепенно утолщающихся к концу (рис. 153, с).
Самки некоторых семейств имеют на предлапке четвертой пары
ног продольный ряд правильно расположенных, хорошо развитых
волосков, называемый calamistrum (рис. 153, А, с). Лапка и предлапка
снизу бывают у некоторых видов покрыты короткими, густо лежащими
4
Рис. 153. Ноги пауков. По разным авторам.
А — задняя нога Dictyna с calamistrum (са), слуховым волоском (Л) и лировидным органом
(/>. В — конец лапки паука Агапеа: а — парный, Ъ — непарный коготок; с — зазубренные
волоски. С — лапка паука с пучком утолщенных волосков под коготком и scopula.
волосками; это образование известно под именем scopula. Все указан-
ные детали строения ног имеют важное значение в систематике пау-
ков. В систематике принимают в расчет также сравнительную длину
ног. Однако при определении таковой следует иметь в виду, что пауки
могут терять конечности, которые легко отламываются (автотомия).
Со следующей линькой оторванная нога регенерирует, но в уменьшен-
ных размерах.
Брюшко соединяется с головогрудью тонким стебельком (petiolus,
pedunculus). Дорзальная его пластинка называется lorum, а вен-
тральная — plagula. Оно различной формы —округлое, почти шаро-
видное, овоидное, удлиненно-цилиндрическое, более или менее тре-
угольное или с различными выростами (рис. 150). Оно покрыто хити-
новым покровом без каких-либо следов члснистости. Брюшко рассма-
15 Пректикум по зоологии — 385 225
'гривают снизу. Тотчас позади plaguia стебелька, по бокам средней
линии брюшка видны два узких поперечно лежащих щелевидных отвер-
стия _ стигмы, или дыхальца (рис. 150, N); они ведут каждое в полость
легкого. Покровы, одевающие легкие, окрашены бледнее, чем на
остальной поверхности. Между стигмами обычно расположено половое
отверстие. Нижняя поверхность брюшка, прилежащая к стебельку,
часто бывает несколько вогнута, образуя так называемый epiga-
strium; от остальной части брюшка этот участок бывает отделен эпига-
стральной бороздкой. В ее средней части и открывается половое от-
верстие. У большинства самок перед половым отверстием лежит видо-
измененная хитиновая пластинка, называемая эпигиной (epigyna,
рис. 150, о); она как бы чешуйкой прикрывает половое отверстие.
Форма и строение эпигины различны у разных видов пауков; нередко
вид самки пауков может быть опре-
Рис. 154. Брюшко паука снизу.
Из Аверинцева:
Р — pygidium; с — colu!us; h.sp, т.»р,
v.sp — задние, средние, передние па-
утинные бородавки.
делен лишь по строению эпи-
гины.
Половое отверстие является отвер-
стием влагалища; у большинства са-
мок пауков перед половым отверстием,
на дне общего углубления, открыва-
ются два отверстия каналов двух
семеприемников. У Epeira, Theridium
и у других родов пауков каждый се-
меприемник имеет два канала, откры-
вающихся: один —близ полового от-
верстия в atrium (наружное отвер-
стие семеприемника), и другой —во
влагалище (внутреннее отверстие). Че-
рез наружное отверстие сперма по-
ступает в семеприемник при копуля-
ции, а через внутреннее живчики
проникают во влагалище (= матка) и
оплодотворяют откладываемые яйца.
На заднем конце брюшка видны
три пары небольших конических хорошо подвижных придатков,
называемых паутинными бородавками (рис. 150, q; рис. 154). Верхние
и нижние бородавки двучленисты; они развиты сильнее одночлени-
стых срединных бородавочек и часто их прикрывают. На конце
бородавок лежат тонкие паутинные трубочки.
Верхние паутинные бородавки раздвинуты друг от друга; между
ними возвышается членистый анальный бугорок с заднепроходным
отверстием. Между нижними паутинными бородавками лежит малень-
кий конический выступ, называемый colulus.
Самки пауков, обладающие на четвертой паре ног calamistrum,
имеют перед основанием нижних паутинных бородавок поперечную
ситовидную пластинку —cribellum.
Перед паутинными бородавками лежит непарное дыхательное от-
верстие, которым открываются наружу трахеи. У пауков семейства
Dysderidae позади легочных стигм сейчас же лежит пара отверстий
трахей.
226
Половые отличия пауков. Самцы пауков меньше самок н бывают
иначе окрашены. Однако важнейшим отличительным признаком
являются особенности строения щупика нижних челюстей (педипальп),
видоизмененного в орган, играющий важную роль при копуляции.
Щупик преобразован в копулятивный аппарат, так как он служит
для введения семени в половые пути самки (рис. 150, е).
Последний членик щупика максиллы самца (лапка) шире, чем соот-
ветствующая часть у самки тсго же вида паука; называется он сут-
bium. С нижней стороны он несет углубление (alveolus); с его дном
соединяется часть, называемая гематодохой (haematodocha); под влия-
Рис. 155. Щупик педи
пальпы самцов пауког.
А — схема; В — Pholcus pha
tango ides; С — Araneus dia-
demat us; D — Pachygnatha.
c — cymblum; a — alveolus;
b — bulbus; h — haematodo-
cha; e — embolus; rs — семе-
приемник. В—D по В. В аг*
неру.
D
с
нием прилива крови она вздувается пузырьком; другая часть гемато-
дохи называется bulbus; конец bulbus выпячивается длинным носиком
или трубкой (embolus); он пронизан хитиновым каналом семенного
пузырька, лежащего в bulbus; у основания embolus лежит хитиновый
склерит, являющийся направителем embolus (conductor) (рис. 155, В.
С, D). Нами рассмотрена лишь схема строения щупика самцов пауков;
оно весьма отлично в деталях у разных видов, что имеет очень боль-
шое значение в систематике.
Копуляция состоит в том, что самец набирает сперму в семенные
пузырьки bulbus щупиков, вводит щупики в половые пути самки,
причем сперма перетекает в семеприемники самки.
227
Органы дыхания паука
В дополнение к рассмотрению цельного паука приготовляют гисто-
логические срезы через залитое брюшко паука, проходящие вдоль
легкого; для этой цели брюшко режется вдоль — в сагитальном на-
правлении. Нужные срезы отбирают под микроскопом. Окраска —
гематоксилином и эозином.
Рис. 156. Легкое пауков
на разрезе. По К э с т-
н е р у.
А — Araneus diadematus, про-
дольный разрез. aS — синус,
приносящий кровь в легкие;
М — печень; R — хитиновый
покров легких; Sp—паутин-
ные железы; St — дыхальце,
слева от него видны горизон-
тально лежащие листки лег-
ких. В — края легочных пла-
стинок на разрезе, при силь-
ном увеличении. Н — гипо-
дерма легочного листка,в ко-
торой циркулирует кровь;
Р — край листка с арочной
структурой.
Стигма(рис. 156, Д,St) на срезе ограничена двумя губами: передней—
острой,и задней—валикообразной. Она ведете преддверие, образованное
мешковидным впячиванием хитиновых покровов. Задняя стенка пред-
дверия покрыта столбчатой хитиновой структурой; столбики ветвятся
и соединяются вершинами друг с другом, образуя как бы кружевную
кайму. Противоположная часть препарата занята большим количеством
лежащих друг на друге двойных хитиновых пластинок (рис. 156, В),
между которыми остаются узкие пространства; воздух входит в атри-
ум и проникает в щели между хитиновыми пластинками, через поверх-
228
ность которых и происходит газообмен. Легочные пластинки напоми-
нают листы книги.
При большом увеличении рассматривают свободные края легоч-
ных пластинок. Каждая из них представляет собой двойную складку
тонкого хитина; по свободному краю листка хитин несколько утол-
щается, и под ним видны гиподермальные клетки; на поверхности
хитина здесь имеются столбики, образующие на разрезе кружевную кай-
му (рис. 156,В,Р). Поверхность самих листков на одной их плоскости по-
крыта прямыми столбиками хитина, которые не позволяют пластинкам
спадаться, благодаря чему между ними остается узкое пространство,
доступное для циркуляции воздуха. В толще каждой легочной пла-
стинки видна щелевидная полость, в которой циркулирует гемолимфа;
ее клетки могут быть обнаружены в различных частях препарата.
Расширенные органы дыхания соответствуют легким скорпионов.
У большинства пауков таких легких пара, отсюда и наименование —
двулегочные пауки.
Для рассмотрения других органов дыхания вываривают в растворе
едкого кали брюшко паука (например, Araneus), срезав его спинные
покровы. По растворении мелких частей препарат отмывают в воде
и рассматривают в часовом стекле в слабом спирте.
На препарате видны две пары прозрачных трубок; средняя пара
своими задними концами сливается в короткую непарную часть;
последняя принимает в себя отверстие боковых трубок и общей стигмой
(или дыхальцем) открывается наружу на нижней поверхности брюшка,
тотчас впереди от паутинных бородавок. Обнаруженные на препарате
трубки являются дыхательными и называются трахеями. Они пред-
ставляют собой впячивания гиподермы внутрь тела.
Таким образом у рассмотренного паука имеется двойная дыхатель-
ная система: пара кожных легких и две пары трахей. Последний тип
органов дыхания мы встречаем впервые из всех рассмотренных ранее
животных. Воздух насасывается в трахеи благодаря движениям
брюшка. Через стенку трахей происходит газообмен между воздухом
и гемолимфой, омывающей трахеи снаружи (т. е. в полости тела
паука).
Экологические замечания
Пауки являются воздуходышащими членистоногими; это свойство
они сохраняют даже при вторичном приспособлении к жизни в воде,
как, например, у водяного паука (Argyronet а), устраивающего под
водой колокол из паутины и приносящего туда пузырьки воздуха на
волосках тела при нырянии в воду.
Пауки отлично бегают; некоторые хорошо двигаются в боковом
направлении или прыгают. Многие из них ткут тенета из паутины;
нити ее являются затвердевшим на воздухе секретом желез паутин-
ных бородавок. Паутина липкая и к ней легко пристают случайно
соприкоснувшиеся с тенетами насекомые. Другие пауки роют верти-
кальные норы (например, тарантул) и захватывают падающую на них
добычу. Третьи сидят на цветах; часто они незаметны благодаря
сходной (подражающей) окраске. Такие пауки непосредственно схва-
тывают насекомых, садящихся на цветы.
229
Пауки являются хищниками. Они убивают ядом своих когтистых
хелицер добычу и высасывают ее соки. У них нередок каннибализм,
и самка иногда пожирает самца тотчас после копуляции.
Паукам свойственна способность есамокалечения» (автотомия),
в частности отрывания ног при опасности, когда, например, паук
бывает схвачен за одну ногу. Компенсацией такой потери является
регенерация утерянной конечности, что происходит при линьке паука.
Обжие ал .кечами»
Пауки происходят от предков с ясным расчленением брюшка.
У некоторых форм сегментация брюшка сохранилась и поныне (Liphi-
stiidae, Не pl at el a kimurai с явственными тергитами брюшка и др.),
но у громадного большинства пауков исчезли всякие следы сегмента-
ции брюшка, одетого сплошным хитиновым покровом. Впрочем,
у только что вылупившейся молоди Meta menardi, Drassodes, Philo-
Рис. 157. Следы метамерии брюшка молодых' паучков. По Г и л ь т а у.
dromus и др. заметна исчезающая с возрастом метамеризация задней
части верхней поверхности брюшка (рис. 157); этот признак имеет
несомненное филогенетическое значение.
Несмотря на отсутствие сохранившейся сегментации, можно рас-
считать, из скольких сегментов (или сомитов) построено тело паука
(по G i 1 t а и).
1. Головогрудь
А. Головные сегменты
I Сегмент с инвертированными глазами
П » с глазами обычного строения
III •> предчелюстной (с rostrum)
IV » хелицеральный (с хелицерами)
V » пальпарный (с щупиками)
Б. Грудные сегменты
VI Сегмент первой пары ног
VII » второй •>
VIII » третьей »
IX « четвертой *
230
В. Ё рюшные сегментЫ
X Первый брюшной предполовой сегмент, видоизмененный в стебелек брюшка
(petiolus, pedunculus)
XI Второй брюшной, половой, сегмент с половым отверстием и парой отвер-
стий легких или трахей
XII Третий брюшной сегмент с одним или двумя отверстиями трахей (реже
легких)
XIII Четвертый брюшной сегмент с парой плейопод, превращенных в перед-
ние бородавки
XIV Пятый брюшной сегмент с парой плейопод, превращенных в задние паутин-
ные бородавки
XV Шестой 1
XVI Седьмой I
XVII Восьмой > брюшные сегменты
XVIII Девятый
XIX Десятый J
XX Одиннадцатый брюшной сегмент с редуцированной брюшной частью
XXI Двенадцатый брюшной сегмент, или анальный бугорок, с анальным отвер-
стием.
Подразделение тела на головогрудь и брюшко у пауков выражено
в самой яркой форме. Таковое подразделение связано со слиянием
соответствующих сегментов и с потерей сегментации. Этим пауки
отличаются от скорпионов. Как у тех, так и у других нет сяжков;
ротовые органы, одинаковые по количеству, разно устроены в деталях.
Находящиеся на брюшке паутинные бородавки являются видоизме-
ненными рудиментами брюшных ножек.
Все эти особенности позволяют хорошо различать представителей
отряда пауков (Araneina) от скорпионов.
ОТРЯД КЛЕЩИ (ACARINA)
МУЧНЫЕ КЛЕЩИ (СЕМЕЙСТВО TyROGLYPHIDAE)
Технические замечания. Мучные клещи являются амбарными
вредителями; они чаще всего встречаются в складах муки и зерна; с купленными
продуктами могут быть заносимы и в жилье. Известны случаи, когда эти клещи
в несметном количестве размножались в квартирах, покрывая обои и мебель.
Клещей собирают с пораженного зерна, которое порциями просеивают через
сито; просеянный материал ссыпают в большое часовое стекло, которое рассматри-
вают под лупой. Образцы поврежденной муки насыпают конусом. Если клещей
в ней довольно много, то вследствие их движения острие конуса на глазах наблю-
дателя сглаживается.
Живых клещей помещают в каплю смеси Фора следующего состава: дистил-
лированной воды пятьдесят частей, хлоралгидрата 200 ч., глицерина 20 ч., гум-
миарабика чистого 30 ч. Смесь отстаивают несколько дней, фильтруют и разливают
по баночкам темного стекла.
Препарат покрывают покровным стеклом.
Консервирующими жидкостями для сохранения мучных клещей являются
80% спирт или глицерин, разведенный пополам дистиллированной водой.
Объекты можно заключать также в глицерин-желатину по общим правилам;
но эти препараты менее прочны для долгого хранения, чем заключенные в гум-
миарабиковую смесь Фора.
Внешняя морфология мучного клеща
Рассматривают под микроскопом живых клещей, положенных под
покровное стекло, или постоянные препараты.
231
Мучной клещ имеет микроскопические размеры: длина тела самки
до 0,7 мм, самца —до 0,45 мм. Тело одето нежной хитиновой кути-
кулой без каких-либо пластинчатых утолщений; тело мягкое, бело-
ватое; на живом объекте характерен его стеклянный отблеск при
падающем свете. Форма тела в общем овоидная; со спинки и брюшка
оно сильно выпуклое.
Рассматривают со спины самку (рис. 158,7): тело делится поперечной
бороздой на передний отдел конической или треугольной формы,
несущий на переднем конце комплекс ротовых органов (из них со
спинки видны верхние челюсти, или хелицеры, клешневидной формы),
и задний отдел с параллельными краями в средней части тела и задним
концом почти правильной полукруглой формы. Переднюю часть назы-
вают головогрудью, заднюю — брюшком: к морфологическому толко-
ванию их обратимся ниже, при рассмотрении тела клеща с брюшной
стороны.
На поверхности тела имеется несколько пар коротких или длинных
тонких щетинок; различают: а) пару волосков тотчас кзади от основа-
ния верхних челюстей; б) две пары волосков, лежащих в один ряд
впереди от поперечной борозды спинных покровов; в) на передних
углах брюшка, называемых плечевыми буграми, имеется две—три пары
плечевых щетинок; г) на спинной поверхности брюшка имеется
несколько пар коротких щетинок; д) самые длинные щетинки имеются
на заднем конце тела.
С брюшной стороны на препарате самки рассматриваем (рис. 158, 2):
а) ротовые органы (g); снизу видны нижние челюсти с лежащей между
ними пластинчатой нижней губой и с парой коротких трехчленистых
щупиков; б) анальное отверстие, лежащее в виде длинной продольной
щели перед задним концом тела (ат); в) четыре пары ног; г) половое отвер-
стие, окаймленное двумя пластинчатыми клапанами, лежащими на уров-
не основания четвертой пары ног (ago); д) пару кольцевидных половых
присосок на каждом клапане полового отверстия (pg) ;е) мелкие щетинки,
лежащие по бокам анального и полового отверстий и в других местах.
Рассматриваем детально строение ног. У самки все четыре пары
ног однородны по своему строению. Основной членик ног —кокса —
представлен сильно хитинизированной узкой тазиковой пластинкой;
тазиковые пластинки первой пары ног сливаются вместе в так называе-
мую грудную пластинку (sternum), имеющую форму буквы V(st); тазико-
вые пластинки остальных ног лежат свободно и имеют слегка дуго-
образную форму, они совершенно неподвижны; подвижная часть ног со-
стоит из пяти члеников: почти треугольного вертлуга, бедра (/), колена
(ge), голени (ti) и лапки (ta). Последняя длиннее, чем каждый из осталь-
ных члеников, которые, вообще, коротки, ибо их длина не превышает бо- •
лее чем вдвое их толщины. Лапка оканчивается одним крючковидным
коготком. Нога несет несколько щетинок. Типичными придатками ног
являются: 1) длинная вершинная щетинка голени и 2) осязательная
палочка на спинной стороне основания лапки первой и второй пар
ног. Характерно отсутствие глаз и дыхательной системы (трахеи и
дыхальца, свойственные многим другим клещам). Самец легко отли-
чается от са мки по следующим признакам (рис. 158,3): 1) меньшие раз-
меры тела; 2) наличие пары круглых анальных присосок, по одной
232
2
Рис. 158. Мучной клещ Tyroglyphus farinae. По А. Захваткину.
Z — самка сверху; 2 — то же,снизу; 3 — самец снизу; 4 — странствующая гипопальная нимфа
сверху; рг — головогрудь (propodosoma); g — ротовые органы (gnathosoma); Р 1—IV — первые-
четвертые ноги; Л — брюшко (hysterosoma); с 1—IV— тазиковые пластинки ног; st — грудная
пластинка (sternum); ago — половое отверстие; pg—половые присоски; ат — анальное отвер-
стие; gl — жировые железы; man — мандибулы; рт — щупики нижних челюстей; I — нижняя
губа; da — анальные присоски; tr— вертлуг;/—бедро; ge — коленный членик; Ц— голень;
ta — ланка; и — коготок.
Рис. 159. Схема сегментации
мучного клеша. По Фитц-
туму.
/ — gnathosoma; 2 — propodosoma;
3—metapodosoma; 4—opisthosoma.
С каждой стороны анального отверстия (da); 3) сильное утолщение пер-
вой пары ног, особенно бедра и колена, причем первое имеет снизу
конический зубцевидный вырост; на колене есть два тесно приле-
жащих друг к другу бугорка; 4) четвертая пара ног имеет каждая
по две присоски.
Вопрос о сегментации тела мучного клеща. Следы метамерии тела
клеща проявляются в расположении четырех пар ног; сегментации
тела нет и следа, если не считать наличия поперечной борозды, отде-
ляющей «головогрудь» клеща от его «брюшка». Эта борозда заходит
частично и на брюшную поверхность тела, огибая его бока позади
второй пары ног; таким образом на «головогрудь» клеща приходятся
две первые пары ног; остальные отходят к «брюшку». Уже эти факты
указывают на несоответствие указанных
частей тела клеща головогруди и брюшку
уже рассмотренных нами паукообразных,
ибо головогрудь несет на себе все налич-
ные четыре пары ног, брюшко же их
вовсе лишено или обладает лишь руди-
ментами брюшных ножек. В связи с этим
для обозначения частей тела мучного
клеща (и всех близких к нему форм) при-
меняется иная терминология (рис. 159).
Сегмент с комплексом ротовых орга-
нов вокруг ротового отверстия назы-
вается гнатозомой (gnathosoma);
«головогрудь» клеща, соответствующая
сегментам первой и второй пар ног,
именуется propodosoma; часть тела, со-
ответствующая сегментам третьей и
четвертой пар ног, называется metapo-
dosoma; задний отдел тела, соответству-
ющий прочим сегментам, образует opi-
sthosoma.
В процессе эмбрионального развития
закладываются тринадцать сегментов и
головные дольки, но сохраняются лишь границы между сегментами
максилл и первой пары ног и уже известная нам борозда между вто-
рой и третьей парами ног. В связи с таким разделением различают:
proterosoma =«головогруди»клеща, и hysterosoma = «брюшку»клеща:
Сегменты ротовых органов — gnathosoma 1
» первой н второй пар ног — propodosoma J proteroso а <головогрудь>
» третьей и четвертой .
пар ног — metapodosoma I hysterosoma — «брюшко»
Остальные сегменты тела — opisthosoma f
Развитие мучного клеща
В зараженной клещами муке можно найти все стадии развития
мучного клеща, который проделывает явное превращение, или мета-
морфоз, так как из яйца выходит дичинка, отличающаяся по своему
234
строению от взрослого клеща. Метаморфоз характеризуется прохожде-
нием следующих фаз:
1)яйца, откладываемые мучными клещами, очень мелки —
0,12—0,18X0,06—0,10 мм, беловатого цвета, правильной овоидной
формы;
2) личинка характеризуется лишь тремя парами ног и соот-
ветственно малыми размерами: после линьки она превращается в
3) первую нимфу (pronymph а) с четырьмя парами
ног, но без полового отверстия; первая нимфа при неблагоприятных
условиях (недостаток пищи и др.) линяет и превращается в
4) гипопальную фазу, или гипопус (от слова Hypopus, под
каковым именем была прежде описана эта фаза метаморфоза клеща как
самостоятельный вид); строение гипопуса мучного клеща характе-
ризуется следующими особенностями (рис. 158, 4):
а) уплощенное снизу, щитовидное, выпуклое сверху тело с более
сильно хитинизированными покровами, разделяющимися поперечной
спинной бороздкой на передний, «головной», и большой задний,
«брюшной», щитки;
б) сильно развитые ноги; при рассматривании сверху видны только
первая и вторая направленные кпереди пары ног; третья и четвертая
пары ног сверху не видны, но из-под заднего конца .тела выступает
длинная волосовидная щетинка последней пары ног; у мучного клеща
длина этой щетинки равна длине тела, достигающей 0,25 мм;
в) на брюшной стороне тела, позади четвертой пары ног, имеется
«присасывательный диск» с несколькими парами округлых присосок;
г) анального отверстия нет; ротовые органы недоразвиты.
Гипопус мучного клеща активен; ведя свободный образ жизни,
гипопус прикрепляется присосками к какому-нибудь насекомому
(например, мухе)или другому животному (например,мыши) ;при посред-
стве последних гипопус попадаете другое место, которое может оказаться
пригодным для дальнейшего завершения метаморфоза; здесь гипопус
отпадает. Поэтому такой тип гипопуса называют странствующим,
миграционным или расселительным. Гипопус обладает очень большой
стойкостью к перенесению неблагоприятных условий, гибельных для
других фаз метаморфоза клеща. Пищи гипопус не принимает. Попав
в благоприятную среду, гипопус линяет и превращается в
5) третью н и мфу (тритонимфа, второй нимфой является
сам гипопус), которая после линьки дает
6) самцов или самок, т. е. взрослые формы.
Экологические замечания
Нападая на зерно (например, пшеницы), мучной клещ выедает заро-
дыш и мучнистое содержимое зерна. В последнем образуется полость,
где клещ откладывает яйца; здесь же протекает и метаморфоз. Поэтому
в зерне можно найти клещей в разных фазах их развития. Клещ
живет также в муке, загрязняя ее своими экскрементами. Сильно пора-
женная мука негодна для пищи. Мучной клещ бывает ив других пище-
вых продуктах (например, в сыре, наряду с сырным клещем), в су-
шеных фруктах и др., бывает и в сене и остается иногда в стеклянной
посуде, которая была упакована в сене.
235
По главному объекту питания является важным амбарным вре-
дителем, который может легко распространяться с зараженными про-
дуктами, тарой (мешки) и др.
ПАСТБИЩНЫЕ КЛЕЩИ (СЕМЕЙСТВО IXODIDAE)
Технические замечания. Пастбищных клещей собирают с до-
машних животных, к коже которых они присасываются для питания кровью.
В зависимости от степени упитанности, размеры клеща сильно вариируют; это,
впрочем, касается только самок, так как размеры самца не изменяются. Собирают
клещей разных размеров, особенно отыскивая клещей среднего и малого размера,
чтобы добыть малоупитанных самок и самцов. Если имеется в виду выведение кле-
щей в лаборатории, то, наоборот, выбирают крупных самок. Клещей выбирают из
шерсти животного (крупный и мелкий рогатый скот, собаки и др.) пинцетом, осто-
рожно оттягивая клеща, чтобы его не изуродовать и не оторвать хоботка. Клещей
можно найти на животных лишь при пастбищном их содержании.
Собранных клещей консервируют 70% спиртом. Если желательно иметь
клещей с вытянутыми ногами, то живых клещей убивают горячей водой (около
75—80° С) и минут через десять перекладывают в спирт, в котором и хранят не-
определенно долгое время
Клещей изучают на цельных объектах, обтерев их от спирта и поло-
жив в сухое часовое стекло при сильном падающем свете под бино-
кулярным микроскопом, бинокулярной лупой, при слабом увеличении
микроскопа, под стативной или сильной ручной лупой.
Внешняя морфология взрослого клеща
1. Ixodes ricinus. Отбирают клещей, которые имеют половое отвер-
стие (оно лежит на брюшной поверхности тела, на уровне кокс третьей
и четвертой пар ног).
Далее клещей разбирают на две группы: у одних вся спинка
покрыта твердым хитиновым щитком; такие клещи мало вариируют
по величине; это —самцы; у других клещей щиток покрывает лишь
часть спинки, непосредственно прилегая к основанию ротовых органов;
этим отличаются самки.
Самка. Тело не имеет никаких следов членистости или под-
разделения на отделы; оно сплошное, или слитное. Со спинки рассматри-
ваем общую форму, которая вариирует в зависимости от степени насы-
щения клеща кровью, исходя в общемот овального контура. На перед-
нем конце тела виден комплекс ротовых частей, неправильно называе-
мых иногда головкой: называется он capitulum. Сверху лежит пара
верхних челюстей, или хелицер, пальцевидной формы с крючковатым
подвижным концом. По бокам хелицер лежат четырехчленистые
пальпы, или щупики (рис. 160, pb); сверху видны футляры хелицер
(рис. 160, а), переходящие своим задним концом в поперечно лежа-
щий хитиновый воротник, называемый basis capituli (рис. 160, с).
Последний является хитиновым сдавленным кольцом, на дорзальной
поверхности которого видны два участка, несущие мелкие поры;
это —поровые поля, являющиеся органом чувств. Кзади лежит плот-
ная пластинка твердого хитина — щиток (scutum); на нем имеются
слабо выраженные цервикальные бороздки или углубления.
Форма щитка коротко-овальная; в его передней вырезке сидит
основание capitulum; передние углы щитка выдаются вперед в виде
236
хорошо выраженных выступов, называемых плечами или скапулами
(scapula) (рис. 160, d); поверхность щитка слабо пунктирована; она
несет редкие светлые волоски. Глаз нет.
Остальная часть тела покрыта мягким хитином; она называется
аллоскутум. На ней разбрасаны светлые волоски. Увеличение размеров
тела самки, связанное со степенью наполнения желудка кровью,
зависит от податливости мягкого хитина. Соотношение величин щитка
и аллоскутум связано с насыщением клеща кровью. У голодной самки
задний край щитка доходит до середины тела. Цвет аллоскутум варии-
рует от беловато-желтого до коричнево-черного.
Рис. 160. Самец Hyalomma с спинной (слева) и брюшной (справа) стороны,
а — футляр хелицер; pb — пальпы; с — основание хоботка (basis capituli); d— скапула; t —
лапка; Л — орган Галлера; о — глаз; I — латеральная бороздка; f — фестоны; hp — гипостом;
сх — кокса; fm — бедро; f — голень; М — предлапка; gn — половое отверстие; tr — вертлуг;
3f — генитальная бороздка; st — перитрема; as — аданальный щиток; ad — добавочный,
аксессориальный щиток; sa — субанальный щиток; ап — анальное отверстие.
С брюшной стороны самки в комплексе частей capitulum видна
срединная непарная, покрытая мелкими зубцами пластинка, называе-
мая гипостомом (рис. 160, hp). От его основания по бокам отходят
вперед пальпы, или щупики, гипостома, состоящие из четырех члеников.
Capitulum непосредственно соединяется в одно целое с лежащей
базально хитиновой капсулой, называемой основанием или basis capi-
tuli.
Сбоку и сзади basis capituli лежат вытянутые в острый угол коксы
(рис. 160, сх) первой пары ног; за ними последовательно расположены
коксы второй — четвертой пар ног, расходящиеся друг от друга в сте-
пени, зависящей от растяжения мелкого хитина брюшной поверхности
тела. Эти коксы имеют вид лопастей, несущих каждая по одному ма-
ленькому наружному зубцу.
237
На уровне кокс четвертой пары ног лежит половое отверстие (рис. 160,
gn)в форме полулунной щели; с боков и позади него идут генитальные
бороздки, по одной с каждой стороны тела (рис. 160, з/).
Ближе к заднему концу (но не на самом конце) тела расположено
анальное (рис. 160, ап) отверстие с двумя боковыми створками или
клапанами. Спереди его огибает анальная бороздка, боковые ветви
которой идут, несколько расходясь друг от друга, назад до края тела.
Позади четвертых кокс, сбоку тела, лежит по дыхательному отвер-
стию (дыхальце, стигма) в особых пластинках твердого хитина, называе-
мых перитремами (рис. 160, St); форма и детали ее структуры имеют
значение в систематике иксодовых клещей.
Самец. Овальное тело со спины почти сплошь покрыто корич-
нево-бурым щитком, по краям которого остается узкий краевой рант
мягкого хитина. На поверхности щитка видны мелкая пунктировка и
редкие светлые волоски. Основание щитка вдавлено в его передний
край, по бокам которого выдаются скапулы. Глаз нет.
Вентральная поверхность покрыта тесно прилежащими друг
к другу плотными хитиновыми пластинками (склеритами, или щитка-
ми). Различают: предполовой (прегенитальный ) щиток, лежащий между
basis capituli и половым отверстием, которое помещается на уровне
третьей пары кокс; между половым и анальным отверстиями лежит
самый крупный—срединный(медиальный)—щиток в форме вытянутого
в длину пятиугольника; латерально от него лежит по эпимеральному
щитку, с которым связаны коксы ног (^ютветствующей стороны тела;
на этих же щитках лежит по перитреме в форме больших овалов.
Анальное отверстие лежит в передней части анального щитка, занимаю-
щего срединное положение перед задним концом тела; по его бокам
вставлено с каждой стороны по аданальному щитку.
Коксы ног несут по небольшому латеральному зубцу. Ноги состоят
из вертлуга (trochanter), бедра, голени, предлапки (protarsus) и лапки
(tarsus); на конце последних имеются по присоске (в виде подушечки)
и по паре изогнутых коготков. Лапка первой пары ног несет на дор-
зальном ребре обонятельную ямку, называемую органом Галлера.
2. Hyalomma sp. Преимущественно южный клещ, заходящий и
в зону лесостепи (рис. 160).
Самка. Удлиненно-овальное тело с щитком неправильной (но
симметричной) формы. От краев основания capitulum на щитке отходят
глубокие, изогнутые цервикальные углубления; на поверхности щитка
несколько редко разбросанных больших точек. На боковых частях
щитка с каждой стороны лежит по хорошо выраженному глазу. Хобо-
ток длинный. Basis capituli сверху в форме трапеции, с поровыми
полями овально-грушевидной формы; между полями проходит средин-
ное продольное килеобразное возвышение. На брюшной стороне тела,
между коксами второй пары ног, видно дуговидное половое отверстие.
От его краев назад до конца тела идут генитальные бороздки. Позади
анального отверстия лежит огибающая его анальная бороздка. Пери-
тремы лежат на боках тела, позади последней пары ног. Форма пери-
тремы —овал со спинным отростком.
В насыщенном состоянии гиаломмы достигают самых больших
размеров из всех иксодовых клещей —до 2% см.
238
С а м е ц. Удлиненно-овальное тело кпереди заметно сужается.
Спинной щиток покрывает всю спину, или по краям его остается рант
мягкого хитина. На щитке имеются глаза (о). Цервикальные бороздки
длинные. Есть латеральные бороздки (/). По заднему краю щиток обра-
зует несколько фестонов ((). Средний из них обособлен со всех сторон
(парма). Есть заднесрединная бороздка и слабые заднебоковые вдавления
хитина щитка.
Брюшная сторона клеща покрыта мягким хитином, в который
вкраплены небольшие вентральные щитки, лежащие в области аналь-
ного отверстия. Здесь лежат: пара аданальных щитков (самые большие
из всех (as) и не посредственно прилежащие к окружности анальных ство-
рок), пара добавочных (акцессориальныещитки,—ad ) и пара субанальных
сосочковидных щитков (sc), лежащих под аданальными щитками. Аналь-
ная бороздка, как и у самки, окаймляет анальное отверстие сзади (an).
Между коксами второй пары ног лежит половое отверстие, прикры-
тое пластинкой, называемой апроном (apron — gn). Перитремы в форме
реторт (st). Ноги длинные. Коксы первой пары ног имеют большой вну-
тренний и длинный наружный шипы. Остальные коксы имеют по на-
ружному короткому шипу.
Препарат ротовых органов пастбищного клеща (Ixodes ricinus)
Capitulum у его основания отделяют от тела клеща копьевидной
иглой. Отделенный передний конец обрабатывают в едком кали (или
натре), хорошо промы-
вают водой и изгото-
вляют бальзамный пре-
парат, ориентиру я объект
брюшной стороной квер-
ху. Препарат рассматри-
вают под микроскопом
(рис. 161 и 162).
Основание хоботка
(прямоугольной формы)
продолжается вперед в
гипостом (й) — широкую
пластинку, несущую на
нижней (обращенной к
наблюдателю) стороне
ряды шипов, из которых
особенно развиты боко-
вые шипы средней части
гипостома. Гипостом яв-
Рис. 161—162. Роговые органы самки Ixodes rici-
nus с дорзальной (слева) и вентральной сторон.
По Н у т т а л ю.
а — футляр хелицер; b—крючья хелицер; р — пальпа; / —
4— членики пальпы; h —гипостом; Ьс — basis capituli; pa—
поровые поля.
ляется гомологом нижних челюстей (педипальпы) других пауко-
образных.
Из-под свободного конца гипостома торчат сильно развитые под-
вижные пальцы хелицер—верхних челюстей (й). Они заострены
и несут по краю мощные острые зубцы. Хелицеры лежат над
(на препарате —под) гипостомом. По бокам ротовых органов лежат
массивные четырехчленистые щупики, или пальпы (р). Первый их
239
членик очень мал, второй и третий — широкие и большие, четвертый —
маленький, загнут вентрально (обращен к наблюдателю—1—4).
Подвижными пальцами хелицер клещ раздирает кожные покровы
хозяина. В образующуюся ранку погружается гипостом, направлен-
ные вершиной назад шипы которого прочно удерживают клеща на
хозяине.
Схема строения ротовых частей одинакова у всех иксодовых
клещей.
Общие замечания
Самки яйцекладущи. Из яйца выходит шестиногая личинка; после
питания на хозяине она линяет и превращается в восьминогую
нимфу, лишенную полового отверстия, которая после питания на
хозяине линяет и превращается в самца или самку, обладающих
хорошо выраженным половым диморфизмом.
Пастбищные клещи живут в открытой природе; для питания они
нападают на млекопитающих (или рептилий, или птиц). Насасывание
кровью длится несколько дней. Отвалившийся клещ проводит довольно
долгое время под травой, в трещинах почвы и в других подобных
местах, где происходит линька. В следующей фазе метаморфоза клещ
снова нападает на хозяина и т. д., пока не будет достигнуто насы-
щение самки. Ixodes ricinus в цикле метаморфоза питается последова-
тельно на трех животных, почему его и относят к треххозяинным
клещам. Hyalomma нуждается в двух или одном хозяине. Схема
соответствующих отношений дана на таблице.
Метаморфоз некоторых иксодовых клещей
Фазы метаморфоза Треххозяинный тип I Двуххозяин- ный тип Однохозяинный тип
Яйцо Личинка Насытившаяся личинка Разви Вылупивши Отпадает на землю и здесь дает нимфу вается в природе сь, нападает на хозяина Не отпадает и на самом хозяине дает нимфу Кормится на том же хозяине, что и личинка
Голодная нимфа . Нападает на второго хозяина
Сытая нимфа . . . I Отпадает па землю и даст самца или самку Остается на том же хозяине и дает на нем самца или самку
Половозрелые голодные клещи Нападают на хозяина Питаются на том же хозяине и копули- руют на нем
третьего | второго
Питаются на нем и копулируют
Насытившаяся оплодотворенная самка Падает на землю и откладывает яйца
240
Иксодовые клещи являются, следовательно, паразитами, так как
единственной пищей их является кровь хозяина. Они относятся
к наружным временным паразитам, ибо лишь на некоторые сроки
присасываются к покровам хозяина («сцепленное* с хозяином состоя-
ние); остальное же время клещи проводят свободно в природе.
Общие замечания
Мучные и пастбищные клещи сильно отличаются друг от друга
следующими признаками:
Признаки Мучные клещи Пастбищные клещн
Размеры тела Микроскопически малые Крупице
Покровы Из нежного хитина без щитков Из грубого хитина со щитками
Подразделение тела . . . На гнатозому, propodosom’y и остальную часть тела На гнатозому (capitulum с basis capituli) и остальное слитное тело
Органы дыхания Отсутствуют Есть (трахеи и дыхальца)
Половой диморфизм. . . В ы р а ж е н
Питание Растительное Животное (кровь)
Питающаяся самка.... В размерах тела не уве- личивается Увеличивается очень сильно
Метаморфоз С гипопальной фазой Без гипопальной фазы
Вся жизнь протекает . . В месте наличия пищи (зерно, мука) С чередованием пребыва- ния во внешней среде и на хозяине (для питания)
Семейство клещей.... Tyroglyphidac Ixodidae
Общие замечания по рассмотренным
представителям класса паукообразных
Все рассмотренные нами паукообразные, в лице скорпиона, паука и
клещей, при всех резких внешних отличиях имеют некоторые общие
признаки, достаточные для отнесения их к одному классу Arachnoi-
dea. Для всех паукообразных характерно: 1) отсутствие усиков, или
сяжков; 2) обладание двумя парами ротовых придатков, из которых
вторая пара (или их производное —педипальпы) несет расчленен-
ные щупики; 3) наличие в имагинальной фазе четырех пар ног; 4) вы-
двинутое вперед положение полового отверстия (кпереди от анального);
5) воздушное дыхание через легкие, трахеи или через кожные покровы
(мучной клещ). Детальное сравнение организации рассмотренных
паукообразных приведено на таблице,
J6 Практикум цо зоологии — 385 241
Сравнение организации паукообразных
Скорпион | Паук 1 Иксодовый клещ
Головогрудь . . . Есть Есть
Бпюшко Переднсбрюшие Нсрасчлененное Все тело слитное
Заднебрюшие брюшко
Расчленение па сегменты. • . . . Есть Нет Нет
Сяжки Нет Нет Нет
Хелицеры Клешневидные Когтевилпые С крючьями па концах
Педипальпы. . . . С большой клеш- ней С придатком в виде щупика Гипостом и пара пальп
Ноги Четыре пары Четыре пары Четыре пары (у imago и нимф)
Рудименты брюш- ных ножек . . . Гребневидные при- датки Паутинные боро- давки Нет
Желудочная же- леза Есть Пальцевидные выросты Слепые выросты
в виде более или
Печень ' Есть Есть менее длинных трубок
„Мальпигиевы со- суды" Две пары 1 Нет 1 1 Одна пара
Ректальный ме- шок Нет Есть "Есть
Легкие Четыре пары Одна пара Нет
Трахеи Нет Две пары Одна пара пучков трахей
242
(Продолжение)
Скорпион 1 Паук Иксодовый клещ
Коксальныс же- лезы Глаза ! Есть Есть Есть
Есть Есть Есть у некоторых видов
Ядовитые органы . Кожные железы в концевой ам- пуле с ядовитым жалом I Железы хелицер с концевым ког- тем Слюнные железы некоторых клещей
Размножение . . . Живородящи Яйцекладущи Яйцекладущи
Метаморфоз.... — (Следы) — (Следы) Есть
Центральная нерв- ная система . . . Концентрирован- ные узлы в голо- вогруди и раз- дельные в брюшке Узлы все концен- трированы в го- ловогруди, обра- зуя одну массу Весьма компактна, в виде маленького округлого тела, образовавшегося слиянием узлов
Кровеносная си- стема Не замкнутая В виде маленького мешочка, развита очень слабо
Сердце трубковидное
Развита хорошо
Лимфатические и лимфоидные ор- ганы Есть । Нет Нет
КЛАСС НАСЕКОМЫЕ (INSECTA)
ЧЕРНЫЙ ТАРАКАН (PERIPLANETA ORIENTALIS)
Технические замечания. Черный таракан является домашним
насекомым, обитающим на кухнях, чаще в пекарнях и булочных, где его можно
добыть и зимой. Для вскрытия необходимо брать живых насекомых, убивая их
серным эфиром или хлороформом. Для расчленения пригоден и спиртовый мате-
риал (из 75% спирта). В запас тараканов можно держать в поместительных садках —
деревянных ящиках с плотной крышкой и сеткой вместо одной стенки. Внутрь
кладут мятую бумагу. Кормят тараканов мокрым хлебом. Для питья ставят воду
в-низких чашках. Садок держат в тепле (близ печки).
*
243
Внешняя морфология
Тело таракана одето темной кутикулой, которая, хотя и не так
тверда, как у речного рака, но все же является самой крепкой частью
тела. Состоит она из хитина, в котором нет никаких отложений извест-
ковых солей. Под хитином лежит слой гиподермы.
Тело таракана явственно разделяется на три отдела: голову, грудь
(thorax) и брюшко (abdomen).
При осмотре нескольких экземпляров замечаются некоторые раз-
личия во внешнем виде. Одни особи крупнее и без явственных
крыльев, другие мельче, но с крыльями. Первые являются самками,
вторые —самцами.
4 В
Рис. 163. Черный таракан; слева (А) самец, справа (В) самка.
cap — голова; рт — нижнечелюстные щупики; ос — глаза; ant — усики; pnt — пе-
реднеспинка; msnt— среднеспинка; el — надкрылья; позади msnt— эаднеспинка;
abd — брюшко; fm — бедро; tib — голень; ts — лапка; сгс — церки; sc — щиток;
stl — грифельки.
С а м к а. Рассматривая самку сверху, находим, идя спереди
назад (рис. 163, В):
а) голову (cap) с парой длинных нитевидных усиков (ant), или
сяжков;
б) грудь, распадающуюся на три кольца, или сегмента:
1) переднегрудь (переднеспинка)—prothorax (pnt) —в виде трапе-
ции с округлыми углами;
2) среднегрудь—mesothorax—в форме поперечной пластинки,
наружные края которой прикрыты листовидными придатками, являю-
щимися остатками редуцированной первой пары крыльев (точнее
надкрылий), и
3) заднегрудь —metathorax;
244
в) брюшко— abdomen (abd), состоящее из десяти сегментов. В нем
менее заметны первый, восьмой и девятый членики. Последний членик
у взрослого таракана слегка рассечен (анальное отверстие). Он несет
пару мелкочленистых придатков, называемых церками—cercianales (ere).
В груди и брюшке при осмотре насекомого сверху видны спинные
полукольца члеников, называемые тергитами.
При осмотре с брюшной стороны на голове замечают ротовые
части; к стернитам груди прикрепляются три пары ног, по паре на
каждый сегмент. Брюшко гладкое и никаких придатков, кроме упомя-
нутых церков, не несет. Первый брюшной стернит рудиментарен;
следующие пять особых отличий не представляют. Седьмой же стернит
у взрослого насекомого посредине задней стороны расщеплен и снабжен
ямкой, в которую открываются женские половые органы.
Самец. У самца (рис. 163, А) со спины голова не всегда бывает
видна. За ней следует большая пластинка переднегруди, называемая
переднеспинкой (pnt); от нее кзади тянутся сложенные крылья, дости-
гающие задним концом шестого сегмента брюшка. Раздвигают крылья
в стороны. Крыльев оказывается две пары; первая называется над-
крыльями или элитрсми (-el)-, сочленяются они со среднегрудью;
вторые —собственно крылья, сочленяющиеся с заднегрудью.
Крылья являются кожистыми (хитиновыми) выростами тела; в них
можно видеть ребристые утолщения, называемые жилками (nervi,
costae). Брюшко самца отличается от брюшка самки присутствием
пары грифелевидных придатков—styli (stl), сочленяющихся с де-
вятым сегментом брюшка. Церки есть и у самца. Грифелевидные при-
датки и церки являются рудиментами брюшных ножек таракана.
Рассматривают ногу таракана в ее естественном положении,
идя от сочленения ее с грудью. Она состоит из:
а) основного конического сильно развитого членика, тазика (соха);
б) подвижно сочлененного с тазиком маленького членика —верт-
луга (trochanter);
в) стержневидного бедра (femur — /ш) с мелкими шипиками по краю;
г)'несколько более узкой голени (tibia — tib), покрытой длинными
шипиками, и
д) пятичлениковой лапки (tarsus — Is). Первый членик ее наиболее
длинен; вторые —четвертые членики короткие; пятый членик несет на
конце пару коготков (unguiculae).
Нога таракана является типичной бегательной. Она состоит, по-
добно ходильным ногам рака, из одного только нерасчлененного
стержня. Ноги таракана и рака можно гомологизировать по следующей
схеме:
Таракан Рак
Тазик (соха)..........
Вертлуг (trochanter)
Бедро (femur)....
Голень (tibia) ....
Ланка (tarsus) . . .
= CbtXI0ntfbrotopodit
= basipodit J r r
= endopodit
245
Таким образом в конечности таракана нет гомологов экзоподиту
типичной двуветвистой конечности ракообразных. Выше мы видели,
что экзоподиты редуцируются и у этих последних (стр. 179).
В наружном расчленении таракана и рака можно усмотреть черты
и сходства и различия.
Общим является состав тела из члеников, одетых хитиновой кути-
кулой, но группировка члеников в отделы тела является различной.
Количественный состав последних:
Таракан Рак
Голова по Грудь из Брюшко Головогрудь из Брюшко
крайней мере трех чле- из десяти четырнадцати чле- из шести
из шести ников члеников ников члеников
члеников 1 1
Теперь рассмотрим подробнее строение головы таракана. При
видны по бо-
осмотре головы сверху в ее нормальном положении
кам два сложных глаза; выпуклая
часть между ними называется те-
менем (vertex). Кзади голова спу-
скается покато и переходит в заты-
лок (occiput), который шейкой (col-
lum) соединяется с переднегрудью.
Спереди между основаниями сяж-
ков лежит лоб (frons); книзу и
сбоку глаз помещаются щеки(genae).
Внизу, лоб переходит в наличник
Рис. 164. Верхние челюсти (т) и
верхняя губа (ol) черного тара-
кан ’ (спичу). О р и г и и а л.
Рис. 165. Нижняя челюсть черного
таракана. О р и г и н а л.
Ог — cardc; S — stipes L — 'nbus inter-
nus; Ci—lobuo externiu; сСоку—щупик
246
(clypeus), с которым сочленяется верхняя губа (labrum, рис. 164, ol).
С боков верхней губы лежат верхние челюсти, или мандибулы (man-
dibulae, рис. 164, т) в виде широких нерасчлененных пластин с
нижне-внутренним зубчатым (режущим) краем, сочленяющихся со
щеками. Сбоку головы видны щупики нижних челюстей и нижней губы.
Чтобы рассмотреть ротовые части, отрезают голову таракана и
исследуют ее сзади. Кроме уже упомянутого выше затылка и глаз,
видны под затылочным отверстием—подподбородок (submentum),
подбородок (mentum) с членистыми придатками на свободном крае —
это все части нижней губы (рис. 166); с боков от нее лежат ниж-
ние челюсти сложного строения.
Рис. 166. Нижняя губа черного таракана. О р и г и и а л.
т — подбородок; sm—подподбородок; /—язычок (glossa); е— при-
даточные язычки (paraglossae); Т—щупик.
Отрезают осторожно ножницами нижнюю губу и откладывают
в сторону. В открывшемся отверстии виден hypopharynx (рис. 167,
Нур)—языкообразный вырост, по бокам которого расположены челюст-
ные мышцы.
Пинцетом и ножницами (или иглой) отделяют все ротовые конеч-
ности аккуратно у мест их сочленения с головой и рассматривают
под лупой в воде- под покровным стеклышком.
Верхняя губа (рис. 164 и 167, ol) и верхние челюсти (т) достаточно
знакомы по наружному осмотру. Нижняя челюсть (точнее I пара
нижних челюстей — I maxillae, рис. 165 и 167) состоит из двух
основных члеников —короткого cardo (рис. 165, Or) и более длинного
stipes (S). На свободном конце последний несет две жевательные
247
дольки—внутреннюю (lobus internus — lacinia, L) и наружную
(lobus externus — galea, Cx). Кроме того к наружному углу нижней
челюсти причленяется пятичленистый нижнечелюстной щупик (pal-
pus maxillaris, рис 167, plm).
Вторая пара нижних челюстей срастается вместе (рис. 167, plm)
основными члениками, образуя нижнюю губу (рис. 166 и 167), два
stipes образуют подбородок (т), два cardo дают подподбородок
(sm), с первым сочленяются три пары придатков:
Рис. 167. Ротовые органы таракана в искусственно раз-
вернутом виде. По Ин аза.
Epi — эпифаринкс; g — galea; gl — glossa; Hyp — гипофаринкс;
Ic — lacinia; Ma — верхняя челюсть; ol — верхняя губа; par —
paraglossa; pit —в нижнегубные -щупики; plm — нижнечелюстные
щупики.
а) лежащие на середине glossae, или язычки (lobi internes maxil
lae —z),
б) помещающиеся латерально от них придаточные язычки (para-
glossa е — е),
в) трехчленистые нижнегубные щупики (palpi labiales—Т).
Нижние челюсти и нижнюю губу можно гомологизировать дву-
членистой конечности ракообразных по следующей схеме:-
Ракообразные Таракан
Двуветвистая конечность Нижняя челюсть Нижняя губа
н £ ( Коксоподит . . . °* О | С = ( Базиподит . . . .| Придатки I । . п I Эндоподит ! Экзоподит 1 Эпиподит ! 1 Cardo Stipes Lacinia Galea Palpus maxillaris Submentum (= два слив- шиеся cardo) Mentum (=2 stipes) ! Glossa Paraglossa Palpus labialis
248
В заключение сравним расчленение тела и его придатки у таракана
и рака.
Сравнение расчленения таракана и рака
Таракан Рак
Части тела Состав из члеников Соответствующие конечности Соответствующие конечности Состав из члеников Части тела
Голова 1 11 III IV Первая пара сяжков Первая пара верхних челюстей Первая пара нижних челюстей Вторая пара нижних челюстей (= нижняя губа) Стебельчатые глаза Первая пара сяжков Вторая » > Первая » верхних челюстей > > нижних челюстей Вторая > > 1 I 11 III IV V j V, грудь
Грудь I II III Первая пара ног Вторая » > Третья > » Первая пара ногоче- люстей Вторая > » Третья » > Первая > ходиль- ных ног Вторая » > Третья ♦ ♦ Четверт. ♦ * Пятая > ♦ VII VIII IX X XI XII XIII XIV о 0Q О г; О
Брюшко I II III IV V VI VII VIII IX X 1 У d styli у d и ф cerci 1 Первая пара брюшных ног Вторая > » Третья » » Четверт. » ♦ Пятая > » Шестая > » 1 XV XVI XVII XVIII XIX XX Брюшко
Вскрытие таракана и его внутреннее строение
Держа таракана в левой руке, ножницами обрезают острые края
тела по всей длине брюшка. Срезают крылья у самца и подрезают
с обоих боков грудь. На переднегруди делают поперечный разрез
кожных покровов. Таракана затем прикалывают двумя булавками за
голову и в конец брюшка (в стерниты) ко дну ванночки спинкой кверху.
Налив в ванночку воды, чтобы она покрыла собой насекомое,
пинцетом берут за край покровов спинки и поднимают его кверху,
отделяют иглой или маленьким скальпелем все, что его придерживает.
Совершенно необходимо снять кожные покровы всей спинки тела
целиком, т. е. и с груди и с брюшка.
249
Отрезанные покровы повертывают внутренней стороной наружу
и прикалывают ко дну ванночки. На белом фоне препарата по средней
линии тела видна полупрозрачная трубочка — сердце, по бокам которого
лежат треугольные крыловидные мышцы. Рыхлая зернистая белая
ткань препарата состоит из так называемого жирового тела.
Рис. 168. Внутренние органы самца таракана. По Н. Холодков-
с к о м у, с изменениями.
pv — симпатическая нервная система; ос — глаза; ое — пищевод; т—резервуар слюн-
ных желез; D — слюнная железа; ingl — зоб; v — мускулистый желудок; ки — желу-
док; рр — пилорические отростки желудка; Malp — мальпигиевы сосуды; G— узлы
нервной цепочки; tr — трахеальные стволы; st — стигмы; test— семенники; г—се-
мевыносящий канал; vd— семеизвергательный канал; g—придаточная железа;
t — толстая кишка; С — церки; st — грифелевидные придатки.
• Обратимся теперь к основному препарату. Во вскрытом таракане
видим клубок внутренностей, целиком заполняющий полость тела
насекомого. Ббльшую часть его составляет пищеварительный канал.
Между извивами его находятся скопления жирового тела, которое
лучше всего выявить и частью удалить, направляя многократно пипет-
кой сильную струю воды на препарат. Петли кишечника при этом не
250
сдвигаются с места, ибо они укреплены тонкими ветвистыми серебри-
стыми трубочками, оплетающими различные части кишечника. Это —
дыхательные трубочки, или трахеи.
Короткими ударами иглы разрывают трахеи и освобождают петли
кишечника, который и вытягивают во всю длину, что дает возмож-
ность рассмотреть пищеварительный аппарат таракана во всех его
деталях.
Органы пищеварения. Откушенная челюстями пища поступает
в пищевод —oesophagus (рис. 168, ое). который постепенно расши-
ряется кзади и образует зоб —ingluvies (ingl). Стенки его растяжимы,
благодаря чему он может вместить значительное количество пищи,
которая поступает отсюда в мускульный желудок (у). Последний имеет
грушевидную форму; он очень короток; внутри его хитиновая выстилка
несет темнокоричневые зубцы, служащие для перетирания пищи.
Пищевод, зоб и мышечный желудок относятся к передней кишке
(stomodaeum): по всей внутренней поверхности они выстланы хитино-
вой кутикулой. В выходную часть мышечного желудка открывается
несколько пилорических придатков—изогнутых трубочек различной
величины, в общем довольно коротких (рр).
Далее следует собственно желудок (ки), имеющий вид тонкой
трубки, тянущейся до места впадения в кишечник множества мальпи-
гиевых сосудов, являющихся органами выделения таракана.
Пилорические придатки и собственно желудок относятся к средней
кишке (mesodaeum) и как производные внутреннего зародышевого
листка (энтодермы) лишены хитиновой выстилки. Мальпигиевы сосуды
(Malp) впадают в начало задней кишки (proctodaevm), неясно распа-
дающейся на части: толстую кишку (colon —I) и прямую кишку
(rectum), открывающуюся наружу заднепроходным отверстием.
Задняя кишка, как и передняя, развивается из впячивания внутрь
кожных покровов; поэтому она выстлана изнутри хитиновой кути-
кулой.
Сравнение органов пищеварения таракана и рака
Таракан Рак
к = S ( Пищевод со слюнными железами 5 а < Зоб о § 1 Мышечный желудок Пищевод Желудок
® S ( Пилорические придатки 5 i 1 Собственно желудок Очень короткая средняя кишка со множеством печеночных тру- бочек, образующих парную печень
S | Толстая кишка, образующая, как
3 ' и желудок, изгибы
$ м ( Прямая кишка
Задняя кишка, очень длинная
и совершенно прямая
251
Дополнением к пищеварительной трубке являются слюнные железы
(D), лежащие в груди таракана. Каждая железа состоит из двух
листовидных дольчатых тел и резервуара (т). Выводные протоки желез
сливаются вместе и открываются в слившиеся вместе же протоки
резервуаров, а общий слюнной проток открывается наружу под
языком —кожистым выростом внутренней поверхности нижней губы.
Листовидные тела состоят из множества мелких пузырьков (acini),
клетки которых выделяют секрет, скопляющийся в резервуаре железы.
В слюне содержится фермент амилаза, перерабатывающая крахмал
в сахар.
В общем, пищеварительный аппарат таракана развит хорошо, и
части его выражены соразмерно. Он отличается от аналогичных орга-
нов рака отсутствием печени, относительно большей длиной (извивы
кишки) и иным подразделением на части.
Общий же характер развития кишечника в обоих’случаях сходен,
ибо закладывается он из передней, средней и задней кишки.
Рис. 169. Симбионты в клетках жирового тела черного таракана.
Органы выделения. Замеченные уже нами мальпигиевы сосуды
(рис. 168, Malp) представляют собой пучок волосовидных трубочек,
открывающихся в начало задней кишки. Концы их, обращенные
в полость тела, замкнуты слепо. Мальпигиевы сосуды омываются
полостной жидкостью, в которой скопляются продукты обмена веществ.
Воспринимая их своими клетками, сосуды эти выделяют экскреторные
продукты (мочекислые соли) в свою полость, по которой последние
и стекают в заднюю кишку. Таким образом через заднепроходное
отверстие удаляются наружу и каловые массы и мочекислые соли.
Мальпигиевы сосуды являются органами выделения своеобразного
типа. Получили они свое название по имени Мальпиги, открыв-
шего их при изучении анатомии насекомых.
Из рассмотренных ранее членистоногих трубчатые органы выделения
были наблюдаемы у паукообразных (скорпион, стр. 212, 217). Их
также называют мальпигиевыми сосудами, но лишь по аналогии с соот-
ветствующими органами насекомых. Существенная разница кроется
в том, что у паукообразных они являются производными средней
кишки (т. е. эктодермального происхождения), тогда как мальпигиевы
252
сосуды насекомых развиваются из задней кишки (т. е. эктодер-
мального происхождения).
Кроме мальпигиевых сосудов, в процессах экскреции принимают
участие и некоторые клетки жирового тела. Экскреторные продукты
отлагаются в их протоплазме в виде нерастворимых соединений, что
делает их безвредными для организма. Эти соединения остаются
в клетках на все последующее время и из тела уже не удаляются.
Клетки-приемники экскреторных продуктов называются нефроцитами,
а самый способ экскреции — внутриклеточным выделением.
Таким образом экскреция достигается двумя путями:
Экскреторные продукты
Выделяются клетками органов выделения
в их полость и выбрасываются вон из
организма
Выделение внеклеточное
Почки позвоноч- Зеленая железа Мальпигиевы
Отлагаются в протоплазме за-
хватывающих их клеток в виде
нерастворимых соединений,
благодаря чему и остаются в
теле без вреда для организма
Выделение внутриклеточное
I
Нефроциты
ных рака сосуды
Аналоги
Симбионты в жировом теле таракана. Клетки
жирового тела содержат в своей протоплазме массу капелек жира; он
окрашивается спиртовым раствором Sudan III в оранжевый цвет.
По продольной оси долек жирового тела черного таракана про-
ходят ряды клеток, лишенных жировых включений; зато протоплазма
их переполнена бесчисленным количеством коротких прямых или
слегка изогнутых палочек с округленными концами (рис. 169). Длина
этих включений в нормальных условиях от 4 до 8 р., а у голодных тара-
канов она падает до 3—4 р. Описываемые образования считаются
с полным основанием за бактерий (Bacillus cuenoti), находящихся
в состоянии симбиоза с клетками жирового тела таракана (ср. выше:
симбионты у гидры); переполненные бактериями клетки называются
бактериоцитами. Симбионты проникают в яйцевые клетки, развиваю-
щиеся в яичниках таракана; поэтому выходящая из яиц молодь ока-
зывается уже «зараженной» соответствующими бактериями, и так
происходит с каждым последующим поколением.
Органы кровообращения. Кровеносная система таракана весьма
проста. Она состоит только из сердца (или спинного сосуда) в виде
длинной трубки, лежащей тотчас под кожными покровами спинки
по средней линии тела. Сердце состоит из тринадцати следующих
друг за другом отделов, или камер (рис. 170,7). Камеры отделяются одна
от другой створками, или клапанами, открывающимися только по
направлению кпереди. Кроме этого камеры имеют еще боковые отвер-
стия — устья сердца (2).
2t&
Рис. 170. Схема сердца
насекомого. По Ж а м м у.
1 — камеры; 2 — устья; 3 —
крыловидные мышцы; 4 —
аорта.
На переднем конце сердце открывается в полость тела у головы (4),
а на заднем оканчивается слепо. Сердце состоит из мышечной ткани,
по бокам его лежат парные крыловидные мышцы (mm. aliformes — 3),
своими суженными концами идущие кнаружи.
Внутренними широкими основаниями они сое-
диняются друг с другом и с сердцем. Вместе
со слоями жирового тела крыловидные мышцы
образуют продырявленную перегородку, или
диафрагму, отделяющую от общей полости
тела околосердечную, или перикардиальную,
полость.
Работа сердца сводится к пульсации, т. е.
к попеременному расширению трубки и ее
сужению. При этом кровь через отверстие
перикардиальной диафрагмы и боковые устья
сердца поступает в его камеры; затем при со-
кращении стенок сердца она проталкивается
кпереди и через переднее отверстие спинного
сосуда изливается обратно в полость тела.
Таким образом и у таракана кровеносная си-
стема незамкнутая, причем этот тип ее здесь
выражен нагляднее, чем у рака. Кровеносная
система таракана сильно редуцирована, ибо
состоит только из одного сердца, самый перед-
ний отдел которого называют аортой. Ника-
ких следов сосудов здесь нет.
Сердце таракана гомологично сердцу рака
и одинаково с ним расположено на спинной
стороне тела. Отличия же этих органов сво-
дятся к следующему. У рака сердце имеет
форму короткого мешка, у которого кровь
выходит по различным сосудам вперед, вниз
и кзади; у таракана же сердце трубчатое,
подающее кровь только кпереди.
Кроме того в сердце таракана заметна
явственная метамерия, проявляющаяся ли-
нейным расположением камер, из коих де-
сять задних соответствуют десяти сегментам
брюшка, а три передних—трем сегментам
груди.
Кровь таракана бесцветная;.как и у рака,
в ее плазме плавают белые кровяные клетки.
Кровь и полостная жидкость — одно и то же,
поэтому ее называют также гемолимфой.
Органы дыхания. Таракан дышит возду-
хом, который проникает по системе трахей до их концевых веточек.
Трахейная система развита весьма сильно; ее развитие обратно пропор-
ционально развитию органов кровообращения.
Дыхательные трубочки в теле таракана повсюду оплетают различ-
ные органы; расположение таких ветвей неправильное. Кроме них
254
можно заметить продольно идущие трахейные стволы (рис. 168, tr) —
два по сторонам брюшной нервной цепочки, два сбоку от сердца и
два боковых ствола. Эти главные продольные стволы в брюшке сое-
диняются поперечными стволиками. Сеть основных трахей в груди
более сложная, она закрывается мышцами.
От главных стволов отходят ветви к различным органам тела.
Концевые веточки трахей проникают в толщу ткани и оканчиваются
внутриклеточно.
Воздух для дыхания поступает извне через особые дыхальца, или
стигмы (st), — отверстия, лежащие по бокам тела. Из стигм воздух про-
ходит по коротким трубочкам в боковые трахейные стволы, откуда
и разносится по веточкам вплоть до их слепых концов.
Стигмы при обычной препаровке незаметны. Чтобы их рас-
смотреть, вырезают у таракана кусочек бока брюшка так, чтобы захва-
тить край тергитов и стернитов двух соседних сегментов. Несколько
минут вываривают в 20% растворе едкого кали, промывают в воде и
рассматривают в капле глицерина под покровным стеклом. В тонкой
кожице, соединяющей стернит с тергитом, между двумя члениками
лежит дыхательное отверстие на вершине маленького возвышения.
Всего у таракана десять пар стигм — семь пар в брюшке и три
пары в груди.
Рассматривают кусочек трахеи в капле воды под покровным стеклом
под микроскопом. При падающем свете на темном поле зрения трахеи
имеют вид серебристых трубочек (отражение лучей света от заключен-
ного в них воздуха). В проходящем же свете трахеи выглядят темными.
Дыхательные трубочки образуются из сильно разветвившихся впячи-
ваний внутрь тела кожных покровов. Поэтому трахеи выстланы внутри
хитиновой кутикулой, подкрепленной хитиновыми же спиральными
утолщениями, называемыми тенидиями, которые придают трахее
в целом эластичность. Снаружи (т. е. со стороны полости тела) трахея
покрыта слоем плоских эпителиальных клеток (видимых на окрашен-
ном препарате), являющихся продолжением гиподермы кожных по-
кровов.
При дыхании таракан то сжимает, то разжимает брюшко. При
сжатии брюшка внутрибрюшное давление увеличивается, трахеи
сдавливаются и бывший в них воздух выталкивается через дыхальца
наружу. При расширении брюшка трахеи, благодаря упругости тени-
дий, снова раскрываются, при этом в них развивается отрицательное
давление, в силу которого свежий воздух пассивно входит в трахейную
систему.
Заметим, что стигмы имеют особый запирательный аппарат, регули-
рующий поступление воздуха внутрь трахеи. И легкие позвоночных
и трахеи насекомых являются органами дыхания воздушного типа,
аналогичными, но не гомологичными друг другу.
Мужские органы размножения. Мужской половой аппарат (рис.
168) таракана на обычных занятиях не может быть отпрепарован
целиком. Семенники его трудно отличимы от жирового тела; кроме
того они в конце концов редуцируются; их легче найти у молодых
животных. Для обнаружения половых органов самца отрезают заднюю
кишку тотчас у заднепроходного отверстия и удаляют пинцетом
255
жировое тело из задней части препарата. Полезно также промывать
препарат пипеткой.
В полном виде органы размножения самца таракана состоят из:
а) пары дольчатых семенников (testes, test), занимающих боковые
части седьмого — восьмого члеников брюшка; лежат они в жировом
теле. От них отходит по
б) семевыносящему протоку — vas deferens (г); семепроводы соеди-
няются с
в) двумя трубковидными семенными пузырьками, у взрослых сам-
цов-переполненными живчиками; эти части открываются в
г) семеизвергательный канал—ductus ejaculatorius (vd), откры-
вающийся наружу под заднепроходным отверстием. В начальную
часть этого канала впадает множество слепых коротких трубочек при-
даточной железы, каковая часчь мужских органов всегда может быть
рассмотрена на препарате.
Женские органы размножения. При последующей препаровке
удаляют кишечник и пипеткой промывают несколько раз содержимое
брюшка. Отделяющиеся клочки жирового тела освобождают препаро-
вальной иглой или пинцетом и так шаг за шагом очищают от него
препарат. В жировом теле лежат спрятанные:
а) правый и левый яичники (ovarium), состоящий каждый из
восьми яйцевых трубочек, имеющих четковидную форму; трубочка
перетяжками разделяется на камеры,* увеличивающиеся в размерах
кзади; в камерах развиваются яйцеклетки.
Все яйцевые трубочки данной стороны тела впадают в
б) короткий яйцевод (оvi ductus). Оба яйцевода сливаются по сред-
ней линии тела в непарный рукав —
в) влагалище (vagina), открывающееся отверстием в половой кар-
ман, лежащий между седьмым и восьмым члениками брюшка.
Задняя часть брюшка занята множеством белых трубочек, обра-
зующих.
г) придаточные железы, состоящие собственно из двух желез —
левой большей и. правой меньшей; открываются они наружу близ
заднепроходного отверстия и дают материал для образования кокона;
кроме всего этого имеется еще
д) семеприемник (receptaculum seminis), состоящий из мешка и
трубки, открывающихся раздельными отверстиями в девятом стер-
пите. При копуляции в них поступает семенная жидкость самца,
хранящаяся здесь до созревания и откладки яиц.
Из всех поименованных частей лучше всего видны яйцевые тру-
бочки и придаточные железы.
Откладка яиц. Черный таракан приступает к размножению лет
пяти от роду. После копуляции брюшко самки начинает расширяться
в основании. Из-под седьмого сегмента постепенно выступает беловатый
сначала, буреющий позднее мешочек — яйцевой кокон (ootheca).
В конце концов кокон отпадает от тела самки.
Из яиц вылупляются белые личинки без крыльев. Они несколько
раз линяют, всякий раз увеличиваясь в размерах. При линьке сбрасы-
вается не только хитиновая кутикула тела, но и хитиновая выстилка
передней и задней кишки и трахей вплоть до их мельчайших раз-
256
ветвлений. На всех стадиях превращения таракан ведет подвижный
образ жизни и питается, поэтому его относят к насекомым с неполным
превращением (Hemimetabola).
Центральная нервная система. Отрезают пищевод у шейки таракана
и удаляют пищеварительные органы. Вскрывают тонкими ножницами
осторожно шейку и срезают верхнюю покрышку головы с наличником.
Удаляют мышцы груди и иглой подчищают ткани вдоль по средин-
ным брюшным трахейным стволам.
Центральная нервная система состоит из:
а) надглоточного узла (ganglion supraoesophageale s. cerebrale),
являющегося мозгом, от которого отходят зрительные нервы (пп.
орйа)кглазами сяжковые нервы (nn. antennarii) к сяжкам;
б) подглоточного узла (g. infraoesophageale), связанного с надгло-
точным ганглием боковыми коммиссурами; на препарате он трудно
различим; иннервирует он ротовые придатки;
в) брюшной нервной цепочки, составленной из девяти ганглиев
(рис. 168, О):трех крупных грудных и шести мелких брюшных. Все
эти узлы соединяются друг с другом двумя продольными коннективами,
лежащими в брюшке тесно друг около друга, а в груди более раз-
двинутыми.
Грудные узлы более массивны, ибо они иннервируют органы дви-
жения таракана. Из ганглиев брюшка выделяется ббльшими размерами
самый последний; он образуется слиянием узлов последних члеников
брюшка, чем и объясняется его величина. Нервные системы таракана
и рака сходны по плану строения и отличаются лишь второстепенными
особенностями количественного характера.
Органы чувств. Г лаза таракана (рис. 168, ос) занимают боко-
вую поверхность головы. Поверхность их испещрена микроскопиче-
скими мелкими шестигранными фасетками, почему их и относят
к сложным глазам. Кроме них есть еще пара простых глазков (ocelli),
лежащих кнутри и кверху от основания сяжков. Глазки очень малы и
заметны только в лупу.
Органами обоняния являются сяжки и церки. Волоски же, покры-
вающие тело и конечности, служат, вероятно, осязательными органами.
Общие замечания
Двусторонне симметричное тело таракана также и метамерно.
Метамерия сказывается в членистости тела, в разделении сердца на
камеры и в нервной цепочке, состоящей из узлов. Конечности также
метамерны, но они имеются только на груди, тогда как брюшко лишено
их и обладает лишь придатками, филогенетически связанными с конеч-
ностями (церки и styli).
Таракан обладает всеми главнейшими признаками, которые свой-
ственны типу членистоногих (Arthropoda) вообще (ср. стр. 287).
Однако он отличается от речного рака существенными особенно-
стями, а именно: а) разделением тела на голову, грудь и брюшко,
б) наличием лишь одной пары сяжков (против двух пар сяжков
рака) и в) присутствием только трех пар ног на груди.
Все эти признаки отличают представителей класса насекомых —
17 Практикум по зоологии — 385 257
Insecta, или Hexapoda (т. e. шестиногие), к которым и относится
таракан.
Наличие крыльев определяет принадлежность таракана к высшим
насекомым (Pterygota), среди которых он занимает место между насе-
комыми с неполным превращением (без стадии покоящейся куколки).
Такие признаки, как: а) грызущие ротовые органы, б) обладание
двумя парами крыльев, из которых передние — прямые и кожистые,
приводят таракана в надотряд прямокрылых (Orthopteroidea).
В нем таракан должен быть поставлен среди бегающих — Cursoria,
которые характеризуются: а) редукцией крыльев у самок; б) превра-
щением передней пары крыльев самца в кожистые пластинки; в) отклад-
кой яиц в коконы, которые самки носят некоторое время на себе;
г) длинными бегательными ногами и д) уплощенным овальным телом.
Таракан днем прячется от света, забираясь в различные щели
(уплощенное тело). Его можно считать поэтому ночным животным.
Черный таракан перешел к обитанию в жилье человека и в природе
не встречается.
Он питается различной животной и растительной пищей, которую
находит в доме. Таракан обладает специфическим запахом благодаря
выделению кожными железами пахучего секрета. Эта железа, в виде
булавовидного двураздельного мешочка, лежит в брюшке вентрально
от нервной цепочки, в промежутке между двумя последними ганглиями,
и открывается на брюшной стороне тела.
МЕДОНОСНАЯ ПЧЕЛА (APIS MELLIFERA)
Технические замечания. Пчел, рабочих и трутней, запасают
в 70% спирте. На спиртовом материале вскрытие внутренних органов дает часто
неудовлетворительные результаты. Дальнейшее изложение будет вестись приме-
нительно к нормальному вскрытию свежего насекомого.
Внешняя морфология
1. Рабочая пчела. Тело пчелы-работницы хорошо разделяется
на голову, грудь и брюшко.
Голова пчелы густо волосиста. На верхней выпуклой поверхности,
называемой теменем (vertex), она несет три простых глазка (ocelli);
бока головы заняты очень большими сложными (фасеточными)
глазами. К передней поверхности головы прикрепляются коленчатые
сяжки, изогнутые под углом, причем основное колено сяжка
состоит из одного длинного стержня, другое колено образовано один-
надцатью маленькими колечками. Под основанием сяжков помещается
трапециевидный лобный щиток, ниже видны ротовые придатки
пчелы.
Грудь пчелы также трехчлениста, но подразделение ее на сегменты
не так явственно, как у таракана. Более всего развита сильная выпук-
лая среднегрудь, задняя часть которой выдается в форме подково-
видного валика, называемого щитком. Передне- и заднегрудь выра-
жены слабо в виде узких хитиновых полос, не видных сверху.
Твердые хитиновые пластинки имеются и на нижней поверхности
груди; здесь также сильно развито соответствующее полукольцо
258
(episternum) среднегруди, загибающееся своими краями на ее
бока.
Брюшко пчелы соединяется твердым тонким стебельком с грудью,
причем сам стебелек и прилегающий скат груди состоят из видоиз-
мененного первого брюшного членика, который по внешнему виду
вошел в состав груди, образуя в ней четвертый спинной щиток. Таким
образом стебельчатое сочленение брюшка, по существу, лежит между
первым и вторым члениками последнего.
Собственно брюшко построено из шести более или менее явственных
сегментов. Спереди оно тупо обрублено, кзади же утончается. Спин-
ные полукольца (тергшпы) сегментов брюшка загибаются на бока и
только на нижней стороне соприкасаются с брюшными полукольцами.
На боковых частях стернитов брюшка помещаются дыхальца, по
одному на каждой стороне членика.
Из заднего конца брюшка торчит острие жала — колющего ядо-
витого прибора пчелы.
В противоположность густо волосистой груди брюшко пчелы
слабо покрыто волосками. На нижней поверхности оно несет восковые
железы, которые рассмотрим ниже.
Грудь пчелы снабжена органами движения — четырьмя перепон-
чатыми крыльями и тремя парами ног. Общий план строения ножек
пчелы такой же, что и у таракана, но характер развития отдельных
частей сильно видоизменяется в связи с особенностями функции дан-
ной конечности. Тазик, бедро, голень и лапки пчелы обильно покрыты
волосками. Ноги увеличиваются в размерах кзади (т. е. на средне-
и заднегруди). У всех ног пчелы особенно сильно выражена пяти-
членистая лапка, главным образом за счет развития ее первого чле-
ника, имеющего вид удлиненной пластинки. Каждая лапка оканчи-
вается парой коготков с липкой подушечкой между ними. Благодаря
коготкам пчела может подвешиваться и висеть на стенках улья и на
сотах. Коготками же она пользуется для передвижения по шеро-
ховатой поверхности, тогда как на гладкой поверхности она подгибает
их и ползет при помощи подушечки, которой пчела как бы присасы-
вается или даже приклеивается к гладкому субстрату.
Вторая пара ног особых отличий в своем строении не имеет. Первая
же пара ног рабочей пчелы отличается присутствием так называемой
гребенки. В основании первого уленика лапки, по его краю, имеется
правильной полукружной формы вырезка, усаженная рядом мелких
жестких волосков. На соответственном краю голени торчит несколько
пластинчатых придатков, которые при сгибании ножки ложатся на
край упомянутой вырезки и закрывают ее, играя роль клапана. Все
это приспособление служит для очистки сяжков от приставших к ним
посторонних частиц. Когда пчела намеревается совершить туалет,
то она вкладывает сяжек в вырезку лапки, прикрывает вырезку кла-
паном и через образовавшееся отверстие протаскивает сяжек взад и
вперед, причем гребенчатые волоски вырезки наподобие щетки очи-
щают сяжек.
Третья пара ног пчелы также имеет своеобразные черты строения,
объясняемые тою ролью, которую эти ноги играют при собирании
пыльцы. Голень их расширена, вдавлена ложкообразно и по краям
* 259
усажена длинными изогнутыми волосками. Благодаря всему этому
голень является корзиночкой—corbicula (рис. 171, В), в которую
счищается пыльца, приставшая к перистым волоскам тела при посе-
щении пчелой цветов. Пыльца счищается «щеточкой*, образованной
десятью-одиннадцатью поперечными рядами волосков на внутренней
поверхности наиболее сильно развитого первого членика лапки этой
же пары ног (рис. 171, A, Planta). В корзиночке пыльца удерживается
боковыми изогнутыми во-
Tibia.
Planta
Pecten
Auricle
Wax
scale
лосками.
Щеточки ног имеют и
другое значение: когда на-
груженная пыльцой пчела
летит, то она сгибает зад-
ние ноги под углом и заце-
пляет щеточками обеих ног
друг за друга, благодаря
чему ноги механически удер-
живаются в согнутом поло-
жении; при этом пчеле не
приходится тратить мышеч-
ной энергии для наиболее
удобного держания тяжело
нагруженных корзиночек
(рис. 172).
Ноги только что рас-
смотренного типа называ-
ются корзиночными, или
собирательными (pedes сог-
biculati); у медоносной
пчелы они свойственны
только рабочим (рис. 171,В).
Края сустава между пер-
вым члеником лапки и го-
Рис. 171. А — задняя нога рабочей пчелы
снутри. По Кастил ю. В — задняя нога ра-
бочей пчелы снаружи. По К а с т и л ю.
Tibia — голень; Planta — расширенный членик лапки
с нацепленной восковой пластинкой — Wax scale; Pec-
ten — щипчики для снятия восковых пластинок.
под колец брюшка и для передачи их
жения к челюстям.
ленью заканчиваются остро,
благодаря чему здесь обра-
зуются своего рода щип-
чики (рис. 171, A, Pecten),
служащие для вытаскива-
ния восковых пластинок,
(рис. 171, Wax scale) из-
другим ножкам для передви-
Крылья пчелы перепончатые, прозрачные, с довольно редкой
сетью жилок. Вторая пара крыльев несколько меньше первой. При
полете передние крылья сцепляются с задними, что осуществляется
приспособлением в виде ряда крючочков по переднему краю заднего
крыла; задний же край переднего крыла загибается желобообразно
в противоположную сторону. При сцеплении крыльев крючочки захва-
тывают край заднего крыла.
2. Трутень. Самцы пчел называются трутнями. Собирать их
260
можно только среди лета. При значительном внешнем сходстве с пче-
лой-работницей трутень отличается следующими признаками:
а) большими размерами тела;
б) очень сильно развитыми глазами, которые на темени сопри-
касаются друг с другом, благодаря чему простые глазки с темени
сдвигаются на переднюю поверхность головы, где и лежат в углу,
образуемом краями сходящихся сложных глаз;
в) более длинными тринадцатичлениковыми сяжками;
г) сравнительно короткими ротовыми органами;
д) более длинным брюшком, которое лишено воскоотделительных
желез и ядовитого аппарата;
е) отсутствием «корзиночки» в третьей паре ног;
ж) более густым опушением тела волосками, и
Рис. 172. Рабочая пчела с корзиночками ног, заполненными
собранной пыльцой, во время полета. Комки пыльцы придер-
живаются при полете второй парой ног. По К а с т и л ю.
з) сравнительно большими крыльями: наибольшие размеры перед-
него крыла трутня 11,5 мм х 3,85 мм, а рабочей пчелы—5,25 мм х 3,1 мм.
Ротовые органы пчелы. У пчелы отрезают ножницами голову
и рассматривают ее под лупой в часовом стекле. Находят части, отме-
ченные выше (см. наружный осмотр), и обращают особое внимание на
ротовые органы, которые свисают вниз. При положении головы заты-
лочным отверстием книзу, на верхней ее поверхности с основанием
головного щитка—наличника,или clypeus (рис. 173, 174, 0, —сочле-
няется узкая поперечно лежащая верхняя губа (L).
Снизу и с боков из-под нее видны верхние челюсти — mandibulae
(рис. 173 и 174, М), сочленяющиеся со «щеками», т. е. с боковыми
частями панцыря головы, обнимающими с боков головной щиток.
Верхние челюсти рабочей пчелы имеют сравнительно широкую и глад-
кую жевательную поверхность; служат они не для откусывания пищи,
а для обработки воска.
261
Раздвигают иглой или пинцетом челюсти и рассматривают осталь-
ные части ротового аппарата пчелы. Под ними лежат нижние челюсти—
maxillae (рис. 173, Мх), состоящие на каждой стороне из широкого пла-
стинчатого стволика (stipes), у сужающегося нижнего конца которого
торчит наружу рудиментарный челюстной щупик (t). Со стволиком
сочленяется покрытая волосками изогнутая серповидная пластинка,
гомологичная наружной жевательной доле (lobus externus) типичной
нижней челюсти.
Рис. 173. Ротовые органы рабочей
пчелы спереди. По Цандеру.
С — наличник; L— верхняя губа; М—
верхняя челюсть; Мх—нижняя челюсть;
G — язычок; Lt — нижнегубные щупики;
t — нижнечелюстные щупики; F — усик.
Рис. 174. Ротовые органы рабочей
пчелы сбоку. По Ц а н д е р у.
С — наличник; L — верхняя губа; М—
верхняя челюсть; Мх—нижние челюсти;
G — язычок; Lt — нижнегубные щупики;
Pg—параглоссы (добавочный язычок).
Отодвигают части нижних челюстей в сторону и рассматривают
длинный «язычок» пчелы, являющийся производным нижней губы.
Из-под краев верхних челюстей выступают наружные дольки нижней
губы, развитые очень слабо —придаточные язычки — paraglossae
(рис. 174, Pg). Внутренние дольки ее, наоборот, хорошо развиты;
срастаясь вместе, они образуют длинный волосистый хоботообразиый
язычок (рис. 173 и 174, G); по бокам его лежат членистые нижне-
губные щупики (рис. 173 и 174, Lt).
262
Повертывают голову пчелы затылочным отверстием кверху. Книзу
от последнего в головном панцыре пчелы имеется вдавление, в котором
и лежит основание ротовых частей. Здесь видны: а) маленький тре-
угольный подподбородок (submentum), помещающийся сейчас же под
затылочным отверстием; за ним следует б) удлиненный подбородок
(mentum), с концевой частью которого соединяются язык (glossa),
придаточные язычки и нижнегубные щупики. Для усвоения соотно-
шения всех этих частей рассматривают голову пчелы также и сбоку,
раздвигая эти части иглами (рис. 174).
Язык пчелы имеет форму открытого кзади желоба, который окан-
чивается маленькой ложечкой, повернутой своей вогнутостью кпереди
(при языке, опущенном вертикально вниз). Ротовой аппарат пчелы,
как то явствует из вышеизложенного, имеет своеобразное устройство,
в котором все же можно усмотреть видоизменение того характерного
плана строения, который свойствен жующему ротовому аппарату
насекомых.
Детальное сравнение соответствующих образований проведем на
таблице:
Сравнение ротовых органов таракана и пчелы
Таракан Пчела
Верхние челюсти.............
Нижние челюсти.............
Cardo.................
Stipes...................
Lobus internus...........
> externus............
Palpi maxillares.........
Нижняя губа....................
Submentum................
Mentum...................
Lobi externi.............
Lobi interni.............
Palpi labiales...........
Верхние челюсти (жующие)
Нижние челюсти
Cardo в виде тонкого поперечного стержня,
лежащего в мягкой сочленовной перепонке,
окружающей основание ротовых органов
В виде коричневой длинной пластинки
Редуцирована
Развита сильно
Рудиментарны
Нижняя губа
Submentum — подподбородок
Mentum — подбородок
Paraglossae — добавочные язычки (рудимен-
тарны)
Язык (glossa) с «ложечкой» (сильно развит)
Нижнегубные щупики
В отношении работы ротовые органы пчелы своеобразны. Питается
она собственно нектаром цветов, каковая жидкость всасывается языч-
ком; кроме того пчела обладает парой грызущих челюстей. Поэтому
ротовые органы ее называют грызуще-лижущими.
Вскрытие пчелы и ее внутреннее строение
Держа пчелу в левой руке, острыми ножницами разрезают с боков
брюшко вдоль по его длине, а также и грудь. Затем прикалывают
пчелу спиной вверх ко дну ванночки, втыкая одну булавку сбоку
в стерниты груди, а другую — у заднего конца тела также в брюшную
половину панцыря кожных покровов; при этом немного растягивают
263
По смятии тергитов груди удаляют
Phq
Slnt^
Vent
iGL
2GI
OF
HS
Puent
Vent
Mai
PSI
Sect
По
пчелы.
и замозговые
Рис. 175. Органы пищеварения
Снодграссу.
Phy — глотка; 7, 2 G! — глоточные
слюнные железы; 3 GI — грудные слюнные железы;
HS — зоб; Mai — мальпигиевы сосуды; ОЕ—пищевод;
Pvent— преджелудок; Rect — rectum; RGl — ректаль-
ные железы; Sint— тонкая кишка; Vent— ventriculus;
Det — слюнный проток; а — резервуары грудных слюн-
ных желез.
мышцы специаль-
назначения — вроде
обслуживающих
насекомое. Наливают воды так, чтобы пчела была совсем ею покрыта.
Пинцетом (тонким) захватывают отрезанный край груди, поднимают
его кверху и подрезывают скальпелем удерживающие его мышцы,
целиком верхнюю покрышку
брюшка.1
При рассмотрении вну-
тренностей пчелы бросается
в глаза чрезмерное богат-
ство груди мышцами и ни-
чтожное развитие таковых
в брюшке. В связи с при-
креплением органов дви-
жения пчелы к груди, в
последней сосредоточива-
ются мощные летательные
мышцы, а также мышцы
ног, тогда как в брюшке
имеются только дорзаль-
ные и вентральные про-
дольные пучки мышц, а
также
ного
мышц,
жало.
Брюшко занято пище-
варительными органами, по
бокам которых видны тон-
костенные большие воздуш-
ные мешки. Между отдель-
ными органами лежат про-
слойки, в общем, слабо
развитого жирового тела.
Женские половые органы
рудиментарны и при по-
верхностном осмотре вну-
тренностей в глаза не бро-
саются.
Пищеварительный ка-
нал. Для препаровки ки-
шечника у вскрытой пчелы
иглой обрывают трахейные
стволики, удерживающие
петли кишечной трубки,
удаляют пинцетом воздушные мешки и осторожно развертывают кишеч-
ник, освобождая его и от жирового тела. В вытянутом состоянии
пищеварительный канал рабочей пчелы состоит последовательно из
отделов (рис. 175):
1 При дальнейшем описании имеется в виду вскрытие свежей пчелы: вскры-
тие спиртового материала позволяет подметить меньшее количество анатомиче-
ских деталей.
264
а) пищевод (OE), идущий от головы чрез грудь и расширяющийся
в брюшке;
б) зоб (HS); величина зоба зависит от степени наполнения его
нектаром; содержимое его прозрачно; здесь проглоченная пища выдер-
живается некоторое время, подвергается известным химическим про-
цессам и отрыгивается пчелой в ячейки сот в виде меда; для питания
самой пчелы сладкое содержимое зоба время от времени поступает в
в) собственно желудок (Vent), имеющий вид сравнительно толстой
трубки с непрозрачными стенками; желудок отделяется от зоба особым
клапаном (Pvent), снабженным сильными мышцами; клапан (предже-
лудок) вдается в зоб головчатым утолщением с крестообразным про-
светом; в полость желудка он свисает трубкообразной заслонкой; все
это приспособление регулирует поступание пищи из зоба в желудок
и препятствует возвращению желудочного содержимого обратно в зоб;
желудок на своей наружной поверхности несет кольцевые перетяжки;
в нем выделяются различные ферменты, которые и переваривают
крахмальную пищу; желудок резко переходит в
г) тонкую кишку (Sint); в ее начальную часть впадает множество
сравнительно коротких
д) мальпигиевых сосудов (Mai), являющихся, как и у таракана,
органами выделения; топкая кишка расширяется в толстую пузыре-
видную
е) заднюю кишку, которая заполняется каловыми массами. В стенке
задней кишки просвечивают шесть палочковидных ректальных желез
(RGI). Задняя кишка (Red), наконец, открывается наружу задне-
проходным отверстием.
По происхождению, своему части кишечника группируются в три
отдела: пищевод, зоб и преджелудок относятся к передней кишке,
желудок — к средней, а остальные части — к задней кишке.
Сравним теперь подразделение кишечника пчелы и таракана на
части в связи с их развитием:
Пчела Таракан
Пищевод Пищевод Зоб Мышечный желудок Пилорические придатки Собственно желудок Мальпигиевы сосуды Тонкая кишка (очень короткая) Толстая кишка Прямая кишка
Зоб •
Кардиальный клапан Желудок
Мальпигиевы сосуды Тонкая кишка
Задняя кишка (ректальный пузырь) с ректаль- ными железами
Пищеварительный канал пчелы обслуживается сложной системой
слюнных желез, но отпрепаровать их в условиях обычной работы
не удается.
Органы пищеварения трутня отличаются от органов рабочей пчелы
большей длиной всего кишечника и большей длиной собственно
желудка.
265
TraSc
Рис. 176. Органы дыхания. Трахейная си-
стема пчелы-работницы по удалении спин-
ных покровов. По Снодграссу.
Видны трахейные стволы (Тга) и воздушные тра-
хейные мешки — TraSc — головы (1, 2), груди (7,
8) и брюшка (70), 1—VII Sp — стигмы, или ды-
хальца.
Органы кровообращения. Чтобы увидеть сердце пчелы, осматривают
изнутри снятые покровы спинной поверхности брюшка. На спиртовом
препарате, при удалении по отдельным частям колец брюшка, можно
высвободить все сердце в виде тонкой трубки.
Кровеносная система пчелы
состоит из трубчатого сердца,
лежащего по средней линии
тела под покровами спинки
в брюшке. Оно образовано
пятью камерами. Передняя
часть продолжается в глад-
костенную трубку — аорту;
последняя делает в стебельке
(соединяющем грудь с брюш-
ком) несколько тесно лежащих
изгибов, пронизывает тол-
щу мышц груди и в голове
открывается в полость тела.
Поэтому кровеносная система
пчелы, так же как и у тара-
кана, незамкнутая. По числу
камер имеется пять пар
крыловидных мышц.
Органы дыхания. Пчела,
как и таракан, дышит тра-
хеями, которые сильно отли-
чаются от трахей таракана
в анатомическом отношении.
Два главных продольных
ствола в брюшке пчелы очень
сильно расширяются и обра-
зуют воздушные мешки (рис.
176, TraSc). Они соединяются
друг с другом поперечными
перемычками, дают ветви в
различные органы и корот-
кими трубочками сообщаются
со стигмами брюшка. Ды-
халец в брюшке с каждой
стороны по семь, а в груди —
по три. Трахеи образуют меш-
ковидные расширения также
в груди и даже в голове.
Благодаря наличию воздуш-
ных мешков, значительная часть объема тела пчелы приходится не
на тяжелые по весу ткани, а на воздух, что значительно уменьшает
относительный вес тела. Благодаря этому пчеле существенно облег-
чается полет. Таракан — насекомое не летающее, лишен трахейных
мешков. В дальнейшем увидим, что и у птиц имеется своеобразная
особенность строения органов дыхания в виде воздушных мешков
266
Рис. 177. Женский поло-
вой аппарат рабочей пче-
лы. По Г. Кожевни-
кову, с изменениями.
о — яйцевые трубочки; g —
яйцеводы; f — влагалище;
е — семеприемник.
являющихся выростами окончания бронхов и также имеющих непо-
средственное отношение к полету.
Воздушные мешки птиц и трахейные мешки пчелы аналогичны
друг другу, так как играют сходную роль, но происхождение их
различно, и поэтому они не гомологичны друг другу.
Органы размножения. Рабочая пчела является по суще-
ству недоразвитой самкой, поэтому органы размножения у нее руди-
ментарны (рис. 177) и весьма изменчивы.
В общем имеется два яичника, иногда настолько тонких, что рас-
смотреть их очень трудно. Каждый яичник состоит из различного
числа яйцевых трубочек (о), от одной до двенадцати. Оставляя в сто-
роне детали строения, не всегда обнаруживаемые у пчел-работниц,
отметим, что яичники соединяются с яйцеводами (g); оба яйцевода
сливаются в общий яйцевод, который продол-
жается во влагалище (/). Во влагалище от-
крывается также и семеприемник — гесер-
taculum seminis (е) — маленький шарик, си-
дящий на тонкой ножке, имеющий два труб-
чатых железистых придатка. Семеприемник у
рабочей пчелы является рудиментарным и не-
функционирующим органом, который также
значительно вариирует у различных осо-
бей.
Иногда и рабочие пчелы откладывают яйца,
вероятно под влиянием особо питательной
пищи, богатой азотом. Яйца эти не оплодотво-
ряются, и из них развиваются только самцы
(партеногенез, или девственное размножение)
Подобные пчелы называются трутовками.
Трутень. В передневерхней части
брюшка лежат два бобовидных семенника
(рис. 178, Tes), длиной 4—5 мм каждый. По-
ловые протоки изгибаются, между ними про-
ходят петли пищеварительного канала, бла-
годаря чему половой аппарат можно рассмо-
треть лишь по удалении кишечника, расправлении и освобождении
половых трубок и отделении от заднего конца тела области полового
отверстия.
В изолированном и расправленном состоянии половой аппарат
составлен из следующих отделов:
а) пары семенников (testiculi), под оболочкой которых скрыто
множество семенных трубочек; из каждого семенника выходит
б) семевыносящий канал (vas deferens, V Def), более тонкий в началь-
ной своей части и толстый в выходной (Ves); vasa deferentia сливаются
друг с другом, причем в место слияния с каждой стороны впадает
довольно широкая
в) придаточная железа (AcGl);
г) непарный выводной канал, или ductus ejaculatorius (EjD), про-
должается в систему сложных образований, отличающихся большим
калибром и разнообразной внешностью (В); все последующие части
267
могут считаться копулятивными органами, ибо они выворачиваются
наружу и вдвигаются в половые пути самки.
Ductus ejaculatorius изгибается у своего конца, расширяется и
переходит в
д) совокупительную луковицу (В), на внутренней поверхности кото- •
рой лежат две треугольные хитиновые пластинки, усеянные шипиками
и загнутыми назад; половой канал за «луковицей» сужается и прини-
мает в себя два
е) перистых придаточных мешочка (/); позади от них в стенке
полового канала лежит хитиновая треугольная пластинка, густо
покрытая волосками; против нее расположен ряд поперечных замыка-
тельных пластинок; еще далее на той же
зоне полового канала
лежит вентральная
хитиновая пластинка;
в выходную часть по-
лового канала откры-
ваются
ж) два придатка
в виде рожков (Ь).
При копуляции все
части полового кана-
ла от b до g, т. е. на-
чиная с луковицы и
кончая придатками
в форме рожков, вы-
ворачиваются нару-
жу. Этот эффект мо-
жет быть достигнут
сдавливанием брюшка
только что убитого
эфиром трутня.
По окончании ко-
пуляции, совершаю-
щейся на лету («брач-
ный полет» матки),
трутень стремится вы-
Рис. 178. Половые органы трутня в естественном
расположении частей. По Снодграссу.
Tes — семенник; VDef — семепровод; Ves —семенной пузырек;
AcGl — придаточная железа; EJD — семеизвергательный ка-
нал; Реп—'выворачивающийся penis с составными частями
f,g)i & — рожковидные придатки; а — половое отверстие.
свободить свои половые части, но они так плотно сидят во влагалище
матки, что при этих попытках вырываются совсем из тела трутня, ко-
торый вскоре же и околевает. Отрывки половых органов трутня торчат
в виде белых ниточек из полового отверстия матки; по этому при-
знаку пчеловод убеждается, что матка оплодотворена.
Центральная нервная система. По удалении всех органов из брюшка
на нижней его стенке изнутри находят нижнюю диафрагму — пластин-
чатое образование, состоящее из жирового тела. Глубже лежит брюш-
ная нервная цепочка (рис. 179). В брюшке она состоит из пяти узлов,
соединяющихся парными коммиссурами; последние два узла сбли-
жены друг с другом (3—7 Gng).
В груди под мышцами и под внутренними хитиновыми выростами
эндоскелета (FuM) лежат всего только два крупных ганглия, из кото-
рых задний (2Gng) гораздо крупнее переднего (iCmg); он соответствует
268
слившимся вместе узлам средне- и заднегруди. Коммиссуры между
ними также более толстые.
В голове нервная цепочка переходит в подглоточный узел, который
соединяется с сильно развитым мозгом, т. е. надглоточным узлом
Вг).
Средняя часть мозга несет
особые грибовидные, или стебель-
чатые, тела (corpus peduncula-
tus); снизу и спереди соединя-
ются сяжковые доли, а с боков—
большие глазные доли (Ор1).
В связи с совершенной инду-
стрией пчелы и ее общественной
жизнью надглоточный узел ее
развит сильно. В частности, осо-
бое развитие мозга усматривают
в наличии именно стебельчатых
тел, образующих своего рода
мозговые извилины.
Органы чувств. Органами
чувств пчелы являются: а) три
простых глазка, б) пара слож-
ных—фасеточных—глаз, в) сяжки
как органы обоняния, на кото-
рых находятся микроскопически
мелкие чувствительные волоски,
конусы, порозные участки и дру-
гие образования, и г) органы
вкуса в виде порозных участков
в основании языка пчелы (видны
только под микроскопом).
Ядовитый аппарат пчелы.
У пчелы - работницы в заднем
конце тела скрыто легко подвиж-
ное жало, являющееся колющей
частью ядовитого аппарата. Если
держать живую пчелу пинцетом
за стебелек брюшка, то можно
видеть движение жала, высовы-
вающегося в виде изогнутой иго-
Л/Ми
Рис. 179. Центральная нервная система
пчелы. По Снодграссу.
Ор1— оптическая доля надглоточного ганглия;
Е — фасеточные глаза; 1, 2 Gng— грудные
ганглии; 3—7 Gng—брюшные ганглии; I—VII—
сегменты брюшка; Fu\, ^2+3 “ эндоскелетные
лочки из заднего конца брюшка
и прячущегося обратно.
Если только что убитую пчелу взять тонким пинцетом за жало
образования; AntNv— сяжковый нерв; W2/Vv,
W^Nv — нервы мышц крыльев.
и осторожно вытягивать его кзади, то его можно вырвать из тела, при-
чем с ним вместе будет извлечена и железистая часть ядовитого аппа-
рата.
У вскрытой пчелы, по удалении кишечника из тела, жало вынимают
с помощью пинцета и острой иглы и рассматривают под сильной лупой.
У' пчелы имеется две ядовитые железы (рис. 180);
а) тонкая трубчатая, на конце вилообразная кислая железа (а/,
269
а
Рис. 180. Ядовитый
аппарат пчелы. По
Цандеру, с изменениями.
а — кислая железа; b — ее общий канал;
с — резервуар кислой железы; d— щелочная
железа; е— комплекс частей жала.
выделяющая секрет кислой реакции; железа расширяется в овоидный
резервуар (с), выводной канал которого открывается в толстую часть
«футляра стилетов» (см. ниже);
б) короткая, также трубчатая, щелочная железа (dj, открываю-
щаяся также в футляр жала, но самостоятельным отверстием.
Каждая из этих желез выделяет составные части пчелиного яда;
сам же яд образуется смешением секрета обеих желез, что происходит
в момент выхода из них секрета.
Жало пчелы (е) имеет довольно сложное строение. В нормальном
положении оно скрыто между спинной и брюшной пластинками деся-
того членика брюшка, где и ле-
жит между заднепроходным и
половым отверстиями.
Основной частью жала явля-
ются так называемые ножны
стилетов, состоящие из твердого
хитинового желоба, открытого
книзу. В передней трети ножны
колбообразно утолщаются, обра-
зуя вздутие ножен. Более тон-
кая задняя часть их является
направляющим желобком, ибо
по его краям лежат микроско-
пически малые хитиновые рель-
сики, служащие для скольжения
колющих иголочек, называемых
стилетами.
В переднем конце вздутия
стенки ножен загибаются в сто-
роны и образуют плоские ру-
коятки ножен, соединяющиеся
с направленными назад продол-
говатыми пластинками. Ме-
диальные края последних, со-
единяясь друг с другом, обра-
зуют тонкостенное, покрытое во-
лосками влагалище ножен.
Ножны своим острым концом погружаются в тело ранимого живот-
ного, но в самом процессе ранения еще более деятельную роль играют
стилеты. Их два; каждый из них образован тонким хитиновым стер-
женьком, зазубренным в концевой части. Стилеты насажены на рель-
сики ножен, по которым и скользят. Во вздутии ножен стилеты несут
каждый по поршнеобразному утолщению. Выходя из ножен в полость
тела, стилеты загибаются каждый латерально, образуя рукоятки
стилета, переходящие в угольчатые пластинки.
Стилеты на внутренней стороне вогнуты, благодаря чему между
ними образуется канал, служащий для протекания яда.
Стилеты, как и другие части жала, обслуживаются особыми мыш-
цами.
Жало пчелы гомологично яйцекладу других насекомых. У пчелы
270
оно развивается за счет придатков рудиментарных члеников брюшка.
Ядовитые же железы пчелы являются, собственно говоря, придаточ-
ными железами женского полового аппарата. Само собой разумеется,
что у трутня, как у самца, ядовитый аппарат отсутствует.
Восковые железы. Перевертывают оставшуюся часть препарата
брюшной стороной кверху и отделяют друг от друга брюшные пла-
стинки сегментов брюшка, налегающие черепицеобразно одна на дру-
гую (рис. 181). Пластинки(стерниты) III—VI сегментов состоят каждая
из двух отделов — заднего темного и переднего, в котором вставлены
два пятигранные прозрачные зеркальца (а). В нормальном положении
зеркальца прикрыты краем впереди лежащего сегмента. Под хитино-
вой кутикулой зеркальца лежит слой высоких гипо-
дермальных клеток, являющихся восковой железой.
Выделяемый ими воск пропотевает* сквозь кути-
кулу и в виде восковой пластинки отлагается на зер-
кальце. Пчела забирает пластинки (рис. 171, А,
Wax scale) ногами, передает их челюстям, которыми
воск и разминается при постройке сот.
Восковые железы присущи только рабочим пче-
лам; они являются кожными железами пчел.
Общие замечания
Пчела, как и таракан, обладает всеми призна-
ками, характеризующими ее как животное типа
членистоногих и класса насекомых (стр. 257).
Относясь к высшим насекомым (Pterygota), пчела
Рис. 181. Восковые
железы на стерни-
тах брюшка рабо-
чей пчелы; а— «зер-
кальца». По К ft-
стилю.
вместе с тем принадлежит к насекомым с полным
превращением (Holometabola), так как послезаро-
дышевое развитие ее слагается из фаз червовидной
личинки (larva), неподвижной и не питающейся
куколки (pupa), из которой только и выходит взрос-
лое крылатое насекомое.
Такие признаки, как: а) наличие четырех перепончатых крыльев,
пронизанных редкой сетью жилок, б) грызущие (хотя бы в одной только
части) ротовые органы и в) полное превращение, определяют при-
надлежность пчелы к отряду перепончатокрылых (Hymenoptera).
Благодаря а) соединению брюшка с грудью при посредстве стебелька
и б) присутствию у самки (и рабочих) жала (видоизмененного яйце-
клада) пчелу относят к группе стебельчатобрюхих (Petiolata).
Сжатый, густо усаженный волосками первый членик лапки задних
ног, грызуще-лижущие ротовые органы и наличие особого собиратель-
ного аппарата для цветочной пыльцы — отличают группу пчел,
в которой пчела медоносная принадлежит к семейству благородных
пчел (Apidae).
Представители этого семейства ведут общественную жизнь. В пче-
линой семье имеется три сорта особей: а) одна матка, б) множество
рабочих и в) трутни. Матка и рабочие являются постоянными членами
общины, тогда как трутни терпятся в ней только в брачный период
и некоторое время спустя, после чего они изгоняются вон и погибают.
271
Община пчел существует много лет благодаря перезимовыванию
матки и рабочих.
Из особенностей строения пчелы отметим нижеследующие признаки,
связанные с образом ее жизни и с окружающей средой:
а) корзиночка на задних ногах, служащая для собирания пыльцы;
б) лижущая часть ротовых органов (язык), употребляемая для
всасывания нектара из цветов;
в) восковые железы («зеркальца») брюшка, дающие воск, который
используется для постройки сот; работа по постройке сот производится
главным образом верхними челюстями;
г) зоб («медовый желудочек») — расширение пищевода, в котором
из проглоченного нектара формируется мед, отрыгиваемый в ячейки
сот, где он и запечатывается восковыми пластинками.
Пчелы зимуют в улье, причем на зиму запасают пищу. Зимой из
улья пчелы не выходят. В это время у них прекращается акт дефе-
кации, иначе они загадили бы весь улей, что повело бы их к гибели.
Пчелы зимой едят меньше, чем летом; неиспользованные остатки пищи
и продукты экскреции мальпигиевых сосудов скопляются в ректаль-
ном пузыре, который соответственно растягивается все больше и
больше, занимая собой громадную часть полости брюшка. Указанные
соотношения становятся особенно наглядными, если отпрепаровать
кишечник летней и зимней рабочей пчелы. Весной при первом же
вылете пчела испражняется и очищает кишечник («очистительный»
вылет).
ТИПЫ РОТОВЫХ ОРГАНОВ НАСЕКОМЫХ
Мы ознакомились с ротовыми органами грызущими (таракан) и
грызуще-лижущими (пчела); этим не исчерпывается разнообразие
видоизменений ротовых частей в связи с их функцией. Далее перейдем
к ознакомлению с другими типами ротовых органов насекомых.
Постельный клоп (Cimex lectularius). От переднего конца головы
по’стельного клопа назад на брюшную сторону загибается расчленен-
ный хоботок. Его отгибают иглой вперед и отрезают ножницами
голову с переднегрудью. Острыми иглами вынимают из желоба хоботка
лежащие в нем колющие щетинки. Препарат обезвоживают в абсо-
лютном спирту, просветляют в кедровом масле и заключают в канад-
ский бальзам.
На занятия даются готовые препараты для рассматривания их под
микроскопом.
На препарате находим (рис. 182): а) два пигментированных глаза
с мелкими выпуклостями на поверхности (ос);
б) кпереди от них прикрепляются четырехчленистые сяжки ( Ant),
между основаниями которых выдается вперед
в) полукруглая верхняя губа (Lbr); из-под нее выходит
г) трехчленистый желобоватый хоботок (Lb), являющийся видо-
измененной нижней губой, которая играет роль футляра для колющих
частей ротовых органов;
Д) пары щетинковидных колющих верхних челюстей (Mnd) и
е) пары таких же нижних челюстей (Мх).
Нижнечелюстные и нижнегубные щупики отсутствуют.
272
Постельный клоп питается кровью человека. При этом он прока-
лывает кожу острыми удлиненными челюстями, по щели между кото-
рыми кровь всасывается в пищеварительный канал клопа.
У постельного клопа ротовые органы колюще-сосущего типа.
Комар (Сиlex или Anopheles). Полный комплект ротовых частей
имеется только у самок. Самцов отличают от самок по сяжкам, густо
опушенным длинными волосками, тогда как у самок сяжки усажены
мутовками коротких волосков.
В нормальном состоянии из ротовых частей виден только длинный
хоботок и нижнечелюстные щупики. Все остальные части спрятаны
внутри хоботка. Чтобы их извлечь,
хоботок сгибают иглой, благодаря
чему колющие щетинки выскаки-
вают наружу. Отрезанную голову
комара обезвоживают, просветляют
в кедровом масле и
в канадский бальзам.
Рис. 182. Ротовой аппарат постель-
ного клопа (Cimex lectularius).
По Холодковскому.
Ос — глаза: Ant — сяжки; Lbr — верх-
няя губа; Lb— нижняя губа (хоботок);
Mnd— верхние челюсти; Мх— нижние
челюсти.
Ь с
Рис. 183. Ротовые органы самки
Culex pipiens. По Пэттону,
с изменениями.
а — сяжок; b — хоботок (нижняя губа);
с—нижние челюсти; d—язык; е—верх-
няя челюсть; /—верхняя губа; g—ниж-
нечелюстные щупики; л— наличник;
I — сложный глаз.
На готовом препарате под микроскопом рассматривают (рис. 183):
а) толстую, длинную, желобоватую нижнюю губу, оканчиваю-
щуюся маленькими концевыми дольками—labellulae (b); в хоботке,
образованном нерасчлененной нижней губой, лежат:
б) пара колющих верхних челюстей (е);
в) непарная колющая щетинка, называемая языком — гипофаринкс
((/Дне смешивать с языком пчелы); она пронизана слюнным каналом,
открывающимся на самом конце гипофаринкса;
г) пара колющих нижних челюстей (с).
Все эти пять колющих щетинок лежат в трубке, образованной
глубоким желобом хоботка, прикрытым сверху длинной и узкой
18 Практикум по аоологии — 385 273
9
Рис. 184. Ротовые органы
комнатной мухи (Musca
domestica). По Пэтто-
ну^ изменениями.
— хоботок; lb—labellulae;
— верхняя губа; тр — ниж-
нечелюстные щупики; р—хи-
тиновая выстилка глотки;
$ — слюнной канал.
д) верхней губой (f).
Снаружи хоботка по его бокам лежат нижнечелюстные щупики;
у самки Culex щупики раза в три короче хоботка, а у самки Anophe-
les _ длиной с хоботок. Нижнегубные щупики редуцированы.
Ротовые органы комара также колюще-сосущие. Прокол, кожи и
всасывание крови производятся комплексом колющих частей, тогда
как сам хоботок в этом не принимает существенного участия.
Муха-жигалка (Stomoxys са lei trans). Муха-жигалка бывает часто
близ конюшен, коровников и т. п.; она маленького размера. Когда
она сидит на стене, ее легко узнать по короткому прямому хоботку,
который торчит вперед. При рассматривании жигалки сбоку видно,
что хоботок, как колено, отходит от нижнего края головы.
Хоботок отрезают с прилежащей к голове
частью покровов, вываривают несколько ми-
нут в растворе едкого кали или натра, про-
мывают в воде, обезвоживают в спирту, про-
светляют кедровым маслом и заключают в ка-
надский бальзам.
Отходящее от головы колено хоботка ко-
нусовидно ; на переднем его скате видны нижне-
челюстные щупики. Длинное колено хоботка
образовано из очень твердого хитина; на конце
его имеется пара небольших долек. В основ-
ной своей части длинное колено вздувается
луковицеобразно. Хоботок у жигалки разви-
вается из нижней губы. В его желобе имеется
только одна колющая щетинка —гипофаринкс;
сверху желоб хоботка прикрыт тонкой и
острой нижней губой.
Остальные части ротового аппарата жи-
галки (сравнительно с комаром) редуциро-
ваны. Ротовые части ее также относятся к ко-
люще-сосущему типу.
Комнатная муха (Musca domestica). Отре-
зают голову комнатной мухи; отрезают боко-
вые части головы в отвесном направлении на уровне основания хоботка
с обеих его сторон. Препарат проводят через спирт, масло и баль-
зам.
Ротовые части комнатной мухи (рис. 184) представлены массивным
хоботком (нижняя губа), разделяющимся на конце на две дольки
губы —labellulae (lb), изборожденные хитиновыми ребрышками и
желобками (так называемые ложные трахеи).
Желоб хоботка сверху прикрыт тесно прилежащей верхней губой
(I). В образующемся канале лежит язык, пронизанный по оси слюнным
протоком. Других ротовых частей у мухи нет. Следом бывших когда-то
нижних челюстей остаются нижнечелюстные щупики, сидящие на
скате основания хоботка (тр). Язык у мухи мягкий, поэтому она
колоть не в состоянии. Питается она жидкой пищей, которую
всасывает хоботком. Отрыгивая жидкость из пищеварительного
канала, муха растворяет различную твердую пищу и образующуюся
274
жидкость всасывает обратно. Хоботок у комнатной мухи очень по-
движен.
Бабочка (Pieris brassicae или др.). Крупных бабочек собирают
в 70% спирт. Отрезанные головы вываривают в едком кали, промывают
водой, иглами выпрямляют хоботок и заделывают, срезав выпуклые
части глаз, в канадский бальзам, положив препарат боком или нижней
стороной кверху.
Наиболее развитой и сразу
бросающейся в глаза частью
ротовых органов бабочки (рис.
185) является хоботок (probos-
cis), в нормальном состоянии
спирально скрученный у ниж-
ней поверхности головы. Он
очень длинен и состоит из двух
поперечнорубчатых узких же-
лобков (рис. 185, Мх), тесно
прилегающих краями друг
к другу, благодаря чему хобо-
ток фактически является труб-
кой. Он развивается из на-
ружной лопасти нижней че-
люсти. Нижнечелюстные щу-
пики присутствуют, но раз-
виты они различно: иногда они
рудиментарны и на обычном
препарате не различимы.
Нижняя губа имеется. Она
представлена рудиментарной
нерасчлененной пластинкой,
на которой сидят нижнегуб-
ные щупики (рис. 185, pl).
Верхние челюсти обычно
вовсе отсутствуют или руди-
ментарны. От них следует
отличать обычно принимаемые
за верхние челюсти, покрытые
щетинками бугровидные при-
датки, лежащие по бокам осно-
Рис. 185. Ротовые органы самца бабочки Са-
tocala nymphagoga спереди. По Н. К у з н е-
ц о в у.
Ос — фасеточный глаз; A, sc, рс — сяжок; Мх —
нижние челюсти (хоботок); pl—нижнегубные
щупики; fr — лоб; с/ — наличник; pf—щетинко-
носные придатки верхней губы; vt — темя; ocl—
глазки; fn — стерженек; gn — щеки; pgn — pro-
cessus genalis, ошибочно принимаемый также за
остаток верхних челюстей.
вания хоботка. Их называют piliferi; относятся они к верхней губе,
ибо являются ее боковыми отростками. Верхняя губа развита
ничтожно.
В общем в ротовом аппарате бабочек значительная часть ротовых
частей редуцируется, и только нижние сосущие челюсти выступают
на первый план по степени своего развития.
ПРИСПОСОБИТЕЛЬНЫЕ ЧЕРТЫ СТРОЕНИЯ НОГ НАСЕКОМЫХ
Типичная форма ноги была рассмотрена у черного таракана.
Схема строения ноги представлена на рис. 186, А. На примере рабочей
• 275
Рис. 186. Ноги насеко
А— бегательная нога жука; В — прыгательная нога саранчевого; С — цепляющая нога вши;
самца-плавунца; G — хватательная нога богомола; Н — лапка мухи. С, Сх — кокса (та
готки; а — коготки; b — концевая
мых. По разным авторам.
D — роющая нога медведки; Е— плавательная нога вертячки; F—присасывательная нога
зик); Тг — вертлуг; F — бедро; Т — голень; Та, Tz — лапка; 1—4 — членики лапки; и — ко-
подушечка; с — непарная щетинка.
пчелы было изучено видоизменение в строении ног в связи с приспо-
соблением их к специальным функциям — сбору пыльцы, очищению
усиков от загрязнения, работе с воском и др. Адаптивные (т. е. при-
способительные) черты строения ног у насекомых весьма разнообразны,
в связи с биологическими особенностями разных видов. В дальней-
шем рассмотрим примеры таких адаптаций.
Прыгательная нога прямокрылого. Объекты для рассмотрения:
саранча, кобылка, кузнечик, сверчок и др. Спиртовые или наколо-
тые на булавку экземпляры. Наблюдение простым глазом и под лу-
пой (рис. 186, В).
Задняя пара ног развита гораздо сильнее остальных двух пар. Осо-
бенно удлинены бедро (F) и голень (г); бедро, кроме того, значительно
расширено в проксимальной части, что является отражением мощ-
ного развития лежащих внутри хитиновой капсулы мышц-разгиба-
телей голени. Голень длинная, узкая. Хитиновый скелет прочен.
Насекомое быстрым сокращением мышц разгибает сустав голени
и бедра, с силой отталкивается при этом от субстрата, благодаря чему
и совершает длинный прыжок.
Хватательная нога. Наилучшим объектом является богомол (юж-
ное прямокрылое насекомое). Пригодна также верблюдка (Rhaphi-
dia, сетчатокрылое насекомое — Neuroptera). Рассматривается пер-
вая пара ног (рис. 186, G). Она удлинена и широка. Обычно короткий
тазик здесь вытянут в длину и утолщен (Сх). Бедро (F) по длине
равняется или чуть превышает тазик; по нижнему краю несет ряд
острых и крепких хитиновых зубцов. Голень укорочена, но также
вооружена зубцами (Г). Лапка тонкая, длинная (Tz). При сгиба-
нии ноги голень с силой прикладывается к бедру наподобие лезвия
перочинного ножа. Пёрвая пара ног является смертоносным орудием
ловли и захватывания добычи, состоящей из различных • насекомых
и других членистоногих. Зажатая пильчатыми краями голени и бедра
добыча пожирается богомолом. Хватательные ноги рафидии развиты
гораздо слабее, чем у богомола.
Цепляющиеся ноги. Для наружных паразитов, постоянно живу-
щих на хозяине (вши, пероеды, пухоеды и др.), важно не быть сбро-
шенными; поэтому у них выработались адаптивные приспособления,
благодаря которым эти эктопаразиты хорошо цепляются за шерсть
хозяина, в которой они живут. Эти приспособительные изменения
сказываются, например, на строении ног вшей, которые рассматри-
вают на микроскопическом препарате (рис. 186, С).
Роющая нога. Первая пара ног медведки (Gryllotalpd). Медведка роет
норы и ходы в почве садов, полей. Эту работу она осуществляет первой
парой ног, которая сильно отличается от остальных ног (рис. 186, D).
Тазик расширен и искривлен (Сх). С ним и с вертлугом (Тг), несущим
шпоровидный придаток, сочленяется короткое, широкое, коренастое
бедро (F); голень резко видоизменена в расширяющийся дистально
(треугольный снаружи) членик, вершина которого несет четыре круп-
ных зубца (Ti); внутренняя поверхность голени изогнута; к ее середине
причленяется сильно измененная лапка (Та); ее основание несет два
сильных зубца. При рытье медведка разводит передние ноги в стороны.
Для роющих ног медведки характерны — укорочение конечности,
278
расширение бедра и голени, образование зубцов на голени и лапке
и общий «коренастый» habitus конечности, сходный с таковым у крота.
О силе этих ног можно получить реальное представление, если боль-
шим и указательным пальцами сдавливать первую пару ног у живого
насекомого. Насекомые, роющие норы в сыпучем грунте, имеют
сравнительно мало измененные ноги (например, у осы Ammophila).
Присасывательная нога. Первая пара ног самца жука-плавунца
(186, F) играет важную роль при спаривании, так как при помощи
видоизмененной лапки самец прочно прикрепляется (по принципу
прикрепления присоски) к хитиновому покрову переднегруди самки.
Бедро первой пары ног искривлено. Особенно сильно изменены пер-
вые три членика лапки; они расширены, уплощены и все вместе об-
разуют большой округлый диск, усаженный по краю хитиновыми
волосками. На поверхности диска можно различить швы — следы
границ члеников лапки, вошедших в состав диска. Его поверхность
покрыта приблизительно сорока мелкими и двумя крупными хити-
новыми присосками.
Некоторые насекомые, как, например, различные виды мух, могут
ползать или сидеть на такой гладкой поверхности, как стекло. Эта
способность зависит от наличия на последнем членике ног пары по-
душечек (186, Н, Ь), усаженных мелкими железистыми волосками, бла-
годаря секрету которых лапка прилипает к гладкой поверхности
настолько прочно, что тело мухи удерживаетс>?на весу; в то же время
муха может легко отнять лапку от поверхности стекла. Кроме
подушечек, лапка несет на конце два сильно изогнутых крепких и
острых на конце коготка (а). Ими муха цепляется за микроскопические
неровности, что позволяет ей сидеть и ползать по вертикальной
плоскости. Если же коготкам уцепиться не за что (как на стекле)
и они соскальзывают, тогда вступают в действие подушечки (микро-
скопический препарат ноги комнатной мухи или другого вида,*
рис. 186, Н).
Плавательная нога. Насекомые являются типичными наземными
обитателями; однако многие виды их в процессе эволюции приспо-
собились к обитанию в воде и стали вторично водными обитателями.
Условия передвижения по твердому субстрату и плавание в воде
весьма различны, что и отразилось на адаптивных чертах строения
ног многих водяных насекомых.
Задние ноги жука-плавунца. Их коксы прочно сро-
слись с заднегрудью. Бедро уплощено и расширено, особенно в ди-
стальной части. Голень веслообразно расширена и усажена волосками
и щетинками. Лапка удлинена, ее членики постепенно сужаются
к вершине; по внутреннему краю они несут тесно лежащие длинные
плавательные волоски. В целом нога прекрасно работает,как весло;
но на суше плавунец в отношении движения беспомощен.
Жук-вертячка (Gyrinus) совершает необычайно быстрые
круговые движения на поверхности воды. Рассматривают вторую
или третью пару ног, отделив их от тела и заключив по общим пра-
вилам в канадский бальзам или в гуммиарабиковую смесь (рис. 186, Е).
Все части ноги, начиная с трохантера и кончая лапкой, листовидно
уплощены. Голень и лапка усажены по краям широкими волосками,
279
очень сильно увеличивающими плавательную поверхность ноги, что
делает ее весьма совершенным органом плавания.
Общие замечания
Сравнивая изученные формы ног насекомых, мы видим чрезвы-
чайное разнообразие их применения; в связи с разнообразием их
функций стоят и адаптивные черты их строения; но как бы ни были
изменены ноги насекомого в связи со специализацией их примене-
ния, всегда ясен основной план их строения. От основной, морфоло-
гически типичной, формы ноги возникли все виды видоизмененных
конечностей, что связано с изменением строения их отдельных частей;
общее число члеников конечности остается одинаковым; редко лишь
наблюдается редукция некоторых члеников лапки.
ЛИЧИНКИ НАСЕКОМЫХ
Личинки стрекоз. Личинок стрекоз вылавливают сачком из водое-
мов с медленно проточной или стоячей водой. Личинки часто держатся
на водяных растениях или на дне. Материал консервируют 70% спир-
том; часть сохраняют живыми для наблюдений. Спиртовый или уби-
тый хлороформом свежий экземпляр рассматриваюг под препароваль-
ной лупой. •
Личинка Aeschna (стрекоза-коромысло) характерна крупными раз-
мерами толстого тела (рис. 187, А).
Голова несет пару коротких тонких сяжков. Боковые части ее
заняты сильно развитыми фасеточными глазами. За головой лежат
три сегмента груди; средне- и заднегрудь несут по паре листовидных
зачатков крыльев. Брюшко равномерно расширяется к средней части
й круто сужается к заднему концу тела, на котором видны твердые
хитиновые шипы, окружающие заднепроходное отверстие. К груди
прикрепляются три пары ног типичного строения.
При рассмотрении головы снизу (В) бросается в глаза широкая,
длинная и толстая пластинка (mentum), к передним концам которой
причленена пара подвижных долек, несущих каждая по крепкому и
острому коготку. Рассматриваемое образование снизу прикрывает
голову личинки, почему и получило название мае к и. Оттягивая
пинцетом пластинку маски, видим, что она сочленяется с основной
длинной пластинкой (submentum), другим концом соединяющейся
с головой. Маска является, следовательно, двуколенчатым рычагом,
который может быстро выбрасываться вперед и схватывать добычу.
Она является органом схватывания добычи; морфологически она
соответствует видоизмененной нижней губе.
Над и по бокам основания submentum маски лежат нижние че-
люсти; между ними выдается «низу гипофаринкс; если удалить ниж-
ние челюсти, то хорошо видны крепкие жвалы с зубчатым свободным
краем. Над ними лежит поперечным валиком верхняя губа.
Личинка Adrion — стрелки (рис. 187, С) — отличается тонким строй-
ным телом с брюшком одинаковой толщины. Характерно присутствие
на заднем конце тела трех листовидных жабр. В толще жаберной
280
пластинки, т. е. в полости между хитиновыми покровами обеих по-
верхностей жабр, проходят ветви трахей, оканчивающиеся слепо.
Поэтому рассматриваемые жабры называются трахейными. Если
схватить живую личинку пинцетом за жабру, то личинка отрывает
ее; автотомия происходит всегда в одном и том же месте, обозначен-
ном наличием хитинового кольца видоизмененных покровов (что
видно на гистологическом срезе через жабру), к внутренней части ко-
торого прикрепляются пучки поперечно лежащих мышц. При сокра-
щении мышц крупное хитиновое кольцо ломается и жабра отпадает.1
Рис. 187. Личинки стрекоз.
А — личинка коромысла; В — откинутая маска коромысла; С — личинка
стрелки с жабрами на конце брюшка.
Отбрасывание жабр происходит, если живую личинку подвергнуть
действию паров серного эфира.
Из личинок вылупляются взрослые стрекозы без того чтобы ли-
чинка превращалась в непитающуюся покоящуюся фазу метаморфоза,
называемую куколкой (pupa/ Такой тип превращения называется
неполным метаморфозом.
Личинки поденок (Ephemeridae). Ловят сачком, как и личинок
стрекоз. Тело делится явственно на голову, грудь и брюшко. Голова
несет пару тонких сяжков, большие фасеточные глаза, три простых
глазка и ротовые органы.
1 Подробнее см.: П. П е р ф и ;i ь е в. О механизме автотомии жчбр у личинок
некоторых стрекоз. Русск. гидробиол. журн., т. II», 1923, № 3—4.
281
От заднего края среднегруди отходят крыловые зачатки в виде
пластинок, достигающих второго тергита брюшка. Брюшко оканчи-
вается тремя длинными хвостовыми нитями (церки), более или менее
густо опушенными хитиновыми волосками. Первые семь члеников
брюшка несут каждый по паре листовидных или узких трахейных
жабр (рис. 188).
по-
На
Рис. 188. Личинка
денки (Heptagenia).
груди слева удалены
конечности и зачатки
крыльев; на брюшке
справа удалены жабры.
По И м м с у.
Ими личинка ударяет по воде, точно колыхая кры-
лышками. Ротовые органы жующие. Отпре-
паровав иглами их части, можно рассмотреть:
1) лопастевидную верхнюю губу; 2) пару манди-
бул с зубцами и жевательным краем; 3) пару
максилл; их жевательная пластинка несет более
нежные зубцы; щупик максилл обычно трехчле-
никовый; 4) нижнюю губу, несущую по паре
внутренних и наружных жевательных долек
(губные щупики трех-двухчлениковые), и 5) язык
(гипофаринкс) — на внутренней стороне нижней
губы.
Личинка жука-плавунца (Dytiscus). Круп-
ная хищная личинка (рис. 189), живущая в воде.
Округлая большая голова шейкой соединяется
с удлиненной переднегрудью. Остальные членики
груди по форме сходны с члениками брюшка.
Восьмичленистое брюшко кзади заостряется и
несет две подвижные дольки — церки. Все три
пары ног густо покрыты волосками и являются
плавательными. Волосками покрыты также церки
и боковые края седьмого и восьмого члеников
брюшка. Членики со спины покрыты более твер-
дыми хитиновыми тергитами. При рассматрива-
нии личинки сбоку видны точкообразные дыха-
тельные отверстия — стигмы, лежащие на боко-
вых краях тергитов брюшка. Стигма восьмого
членика брюшка лежит на его заднем конце.
Стигма заднегруди очень мала; на среднегруди
она лежит в стерните кпереди от основания вто-
рой пары ног. Коксы передних ног более сбли-
жены, чем у остальных ног; впереди от осно-
вания ног переднегруди лежит округлая с вы-
резом хитиновая плотная пластинка — акро-
стернит. Ноги имеют длинную одночленистую
лапку.
При рассмотрении головы сверху отмечаем
(рис. 190, Д):
1) разделение хитиновых покровов головы тремя швами на предлоб-
ную (clypeus — р) и две залобные (теменные — ps) части;
2) на выпуклых боковых частях головы имеются глазные поля (а);
3) из-под переднебоковых углов головы отходят серповидные мощ-
ные верхние челюсти (7И);
4) тотчас позади и над их основанием прикрепляются длинные,
тонкие сяжки из чередующихся коротких и длинных члеников (F).
282
При рассмотрении головы снизу (рис. 190, В) находим пару сильно
видоизмененных максилл (Мх1); их cardo очень укорочен; stipes —
длинный, несет еле заметный рудимент (lobus externus) и длин-
ный щупик. Максиллы далеко отодвинуты друг от друга; между их
основаниями лежит нижняя губа
в форме выдающейся вперед пла-
стинки, несущей в переднебоковых ча-
стях по длинному щупику (Мх2). Верх-
ние челюсти снабжены глубоким же-
лобом, идущим почти от самой вер-
шины к основанию серпа челюсти;
рот посредине зарастает, а задние
отверстия желоба верхних челюстей
открываются в боковые части рото-
вой полости.
Личинка схватывает добычу манди-
булами и через их желоб впрыскивает
внутрь нее пищеварительные соки;
ферменты последних разжижают ткани
добычи, жидкие соки которой личинка
всасывает, не выпуская добычи из
мандибул. Максиллы и нижняя губа
не принимают никакого участия в про-
цессе обработки пищи. Налицо при-
мер экстраинтестинального, внекишеч-
ного, пищеварения. Впрыскиваемая
жидкость обладает ядовитыми, пара-
лизующими добычу свойствами.
Личинка плавунца превращается
в неподвижную и непитающуюся ку-
колку, из которой со временем вылу-
пляется жук. Такой метаморфоз на-
зывается полным.
Гусеница бабочки. Запасают в 70%
спирте гусениц капустницы (Pieris
brassicae), крапивницы (Vanessa urti-
сае) или других видов. Рассматривают
гусеницу под лупой при сильном па-
дающем свете (рис. 191).
Форма тела в общем цилиндриче-
ская, уплощенная на брюшной сто-
роне. Голова хорошо обособлена; сле-
дующие за ней три сегмента являются
Рис. 189. Личинка плавунца свер-
ху. По Коршельту.
А — голова; В — грудь; С — брюшко;
st — дыхальце; сс — церки; prth — пе-
реднегрудь; msth — среднегрудь; mtth—
заднегрудь; 2(1) — 9(8) — членики
брюшка.
грудными и несут расчлененные ноги;
остальные сегменты соответствуют брюшку; последний, анальный
сегмент слился из двух; в нем лежит anus. Голова покрыта крепким
хитиновым покровом, подразделенным продольным швом на боковые
отделы; спереди и снизу шов раздваивается; между его ветвями поме-
щаются лоб и наличник (clypeus).
В нижнепередней части боковых отделов головы на каждой сто-
283
роне лежат несколько простых глазков (ocelli). Под ними находится
основание сяжка данной стороны тела. Сяжки короткие, трехчле-
нистые.
Ротовые органы гусеницы грызущие (рис. 191). Ниже наличника (С)
лежит поперечно вытянутая верхняя губа; верхние челюсти (Md) млеют
Рис. 190. Личинка плавунца. Голова сверху (А) и ее перед-
ний край снизу (В). По Коршельту.
h — шея; Z — шейная борозда; р — praefrons; рз—pottfront; а — глаза;
М — мандибула; Мх< — первая максилла; Мха — щупики нижней губы
(вторых максилл); F — сяжок.
хорошо выраженный режущий, или жующий, зазубренный край; ниж-
няя челюсть несет четырехчленистый щупик (Af/). Нижняя губа сильно
видоизменена; ее двучленистые щупики очень малы (и/); они сочле-
нены с subinentum (и), несущим срединный стерженек — fusulus, на
вершине которого открывается отверстие парной паутиноюй железы,
являющейся личиночным органом гусеницы. Ее затвердевающий на
284
воздухе секрет имеет вид нити, из которой гусеница делает кокон
(гусеница тутового шелкопряда дает шелковые нити).
Ноги грудных сегментов пятичленисты и оканчиваются коготком;
это — истинные, или грудные, ножки.
Некоторые сегменты брюшка также несут по паре ножек; однако
последние представляют собой короткие и широкие выпячивания
Рис. 191. А, В — гусеницы бабочек; С — ротовые органы гусеницы.
По Эшериху(А, В) и Пфуртшеллеру (С).
С — clypeus, ниже наличника — верхняя губа; Md — верхняя челюсть; М —
нижняя челюсть; Mt — щупик нижней челюсти; и — нижняя губа; ut — щу-
пик нижней губы; А — сяжок.
кожных покровов брюшка, имеющие сосковидную форму и несущие
венчик хорошо развитых хитиновых крючьев (hamuli); такие
брюшные ноги называются ложными, их бывает до пяти пар.
Брюшные ножки последнего сегмента раздвинуты наподобие щипцов
и подталкивают тело гусеницы при ползании.
Тело гусениц часто покрыто различно развитыми волосками. По
его бокам лежат девять пар дыхалец в виде точек. Превращение у ба-
бочек родное.
286
отсутствуют, готовое отверстие на
Рис. 192. Превращение комнатной мухи.
По Хьюитт.
А — яйца; В — личинка I стадии; С — задний ко-
нец личинки II стадии с дыхальцами (косы щели);
D — зрелая личинка; Е— заднее дыхальце зрелой
личинки; 1 — переднее дыхальце ее; F — куколка;
G — вскрытый кокон по вылуплении мухи; ad —
анальные бугорки; р — брюшные валики; st —
грудная стигма.
нет. Головной сегмент маленький;
Личинка пчелы (Apis mellifera). Консервированный в 70% спирту
материал. Рассмотрение ведется под лупой. Тело личинки черве-
образное, неуклюжее, толстое; оно состоит из маленькой головы и три-
надцати сегментов; на последнем открывается анальное отверстие;
по бокам сегментов видны десять пар точкообразных дыхалец. Ноги
голове окружено рудиментар-
ными ротовыми органами в ви-
де коротких выростов. Хитин
покровов мягкий.
Личинки живут в ячейках
сот пчелиного улья, где их
кормят рабочие пчелы. Край-
нее упрощение строения ли-
чинки можно поставить в
связь со специфичностью усло-
вий жизни, когда личинка
пользуется полным уходом.
Перед окуклением у ли-
чинки можно видеть выпячи-
вающиеся зачатки крыльев и
ножек на грудных сегментах
и аналогичные зачатки частей
яйцеклада на брюшной сто-
роне последних четырех се-
гментов тела.
Личинка мухи. Выбирают
крупных личинок мух из кон-
ского помета (навозные кучи),
фекалий человека и др. и по-
мещают в 70% спирт. Тело —
округлое, сужающееся кпе-
реди и утолщающееся посте-
пенно кзади (рис. 192, D). Пе-
редний конец тела заострен,
задний же как бы обрублен.
На брюшной стороне видны
валикообразные поперечные
выпячивания, являющиеся
органами движения. Ножек
он несет две ротовые дольки
€ двумя дорзальными чувствительными бугорками на каждой. Следу-
ющий кзади сегмент слился из двух. У его заднего края лежит слож-
ная грудная стигма из семи отверстий на вершине дыхальцевого
отростка. Последний — тринадцатый — сегмент косо срезан. На его
задней поверхности помещаются две твердые дыхальцевые пластинки;
на брюшной стороне этого сегмента имеются две анальные дольки,
между которыми открывается анальное отверстие. Хитиновая кути-
кула мягкая.
Из ротового отверстия на голове торчат концы крепких когтевид-
ных черных глоточных склеритов. Их форму можно рассмотреть на
286
бальзамном микроскопическом препарате положенной на бок перед-
ней части личинки.
На фронтальном препарате отрезанного поперек заднего членика
(бальзамный препарат) можно видеть форму дыхальцевых пластинок,
прорезанных каждая тремя дыхательными щелями (рис. 192, С, Е).
Общие замечания по типу членистоногих
Рассмотренные нами представители типа членистоногих в лице
ракообразных (рак, дафния, циклоп), паукообразных (скорпион,
паук, клещ) и насекомых (таракан, пчела) имеют следующие черты
строения, которые можно считать характерными для типа членисто-
ногих:
1) расчлененное на сегменты тело (вторично может происходить
потеря сегментации вследствие слияния сегментов);
2) хитиновую кутикулу покровов, являющуюся наружным ске-
летом ;
3) наличие расчлененных парных ног, типически свойственных
каждому сегменту (при слиянии отдельных сегментов тела ноги могут
видоизменяться в строении и в отношении функции и вовсе исчезать);
4) вторичную полость тела, закладывающуюся в виде парных
целомических пузырбй, стенки которых впоследствии распадаются,
и образуется смешанная полость тела—миксоцель;
5) кишечник, который делится на переднюю, среднюю и.заднюю
кишки;
6) незамкнутую кровеносную систему; сердце лежит на спинной
стороне (оно может вовсе редуцироваться);
7) центральную нервную систему цепочечного типа, состоящую
из надглоточных узлов и брюшной нервной цепочки, лежащей на вен-
тральной стороне тела под кишечником; типически каждому сегменту
свойствен (двойной) нервный узел; при слиянии сегментов узлы
нервной системы могут сливаться, и в крайних формах видоизмене-
ния нервная система может терять внешний цепочечный вид.
ТИП МЯГКОТЕЛЫЕ (MOLLUSCA)
КЛАСС ПЛАСТИНЧАТОЖАБЕРНЫЕ
(LAMELLIBRANCHIA)
ОТРЯД НАСТОЯЩИЕ ПЛАСТИНЧАТОЖАБЕРНЫЕ
(EULAMELLIBRANCHIA)
БЕЗЗУБКА (ANODONTA)
Технические замечания. Беззубки живут в медленно текущих
речках и в водоемах со стоячей водой и песчаным дном. На зиму запасают живых
беззубок, которых содержат в аквариумах с проточной водой или в больших со-
судах, в которых периодически меняют воду. Дохлых беззубок немедленно уда-
ляют (их узнают по полураскрытой раковине и дряблой ноге).
Животных убивают, опуская на 20—30 минут в воду, нагретую до 35—40°,
или помещая их на 15 часов в 2% раствор хлоралгидрата, предварительно вдвинув
деревянный клинышек в щель между створками.
Наружный осмотр
Беззубка снаружи одета раковиной, состоящей из двух створок —
правой и левой. При осмотре беззубки сбоку различают верх-
ний (дорзальный) более ровный край, где обе створки соединяются
друг с другом, нижний округлый край, переходящий в передний
широкий и более притупленный и в задний более острый концы ра-
ковины.
На поверхности створки видны кривые линии, идущие, в общем,
параллельно наружным очертаниям раковины; лежат они более или
менее концентрически относительно выпуклой раковины, расположен-
ной в ее верхней части ближе к переднему концу. Эта часть назы-
вается пупком — umbo. Указанные линии считаются линиями при-
роста раковины. Раковина снаружи грязновато-зеленоватого цвета.
Поверхностный слой ее состоит из рогового вещества конхиолина
и называется periostracum.
При осмотре беззубки со спинной стороны видны правая и левая
створки раковины, соединенные друг с другом эластическим тяжем —
лигаментом.
Моллюск имеет неодинаковую толщину, наибольшая толщина
соответствует пупку. Оба пупка образуют вершину (apex). Кпереди
и кзади беззубка утончается и заканчивается острыми краями.
Снизу, спереди и сзади между створками беззубки видна щель,
из которой торчит нога. В щели видно, что створки изнутри покрыты
складками mhi кой ткани, образующей лопасти мантии.
288
Наблюдения над живым животным
Если взять живую беззубку в руки, то она плотно смыкает створки
раковины, и тогда, кроме наружной поверхности последней, ничего
увидеть нельзя.
В банке с водой, будучи оставлена в покое, беззубка приоткрывает
створки раковины и из образующейся щели выпускает мягкую жел-
товатую лопасть клиновидной формы. Эта лопасть служит органом
передвижения животного и называется ногой. Чтобы видеть самый
способ движения беззубки, ее кладут в аквариум с песчаным дном.
Кроме ноги, никаких других частей тела беззубки вне ее раковины
видеть не удается.
В пипетку набирают воду, в которой растерт порошок кармина;
в сосуд, где содержится беззубка, выпускают две капли этой жидкости
у заднего (более острого) конца моллюска и наблюдают по движе-
нию окрашенной струи токи воды. Вода поступает внутрь раковины
через вентральное отверстие на заднем конце; край этого отверстия
усажен сосочками. Из раковины вода выходит наружу через лежащее
вблизи и вверху другое отверстие, называемое клоакальным сифоном.
Если затем вместо кармина пипеткой пустить немного слабой кис-
лоты, то створки раковины в ответ на испытываемое раздражение
захлопываются.
Вскрытие раковины
Ручкой скальпеля раскрывают возможно шире створки раковины
и тупым концом его отделяют мантию от внутренней поверхности
левой раковины. При этой работе будут ощущаться препятствия,
когда скальпель натолкнется на замыкательные мышцы раковины.
Их отделяют у самого места прикрепления к раковине острым
ножом, ведя последний непосредственно по внутренней поверх-
ности створки. Когда все мышцы будут перерезаны, моЛлюска кла-
дут нетронутой створкой раковины книзу на стол и рассматривают,
как поднялась освобожденная от мышц другая створка. Ее поднятие
обусловливается стягиванием лигамента, которому нет больше про-
тиводействия со стороны замыкательных мышц. Сдавливая раковину
руками, можно ее замкнуть, причем ощущается некоторое противо-
действие лигамента, который снова автоматически раскрывает ра-
ковину, если ее опять положить на стол.
Разрезают ножом лигамент и удаляют прочь левую створку рако-
вины. Животное кладут на правую створку, приводят в нормальное
положение мантию и рассматривают препарат под водой.
Внутреннее строение беззубки
По удалении левой створки раковины, при нормальном положении
мантии различают верхний (дорзальный), передний, нижний (вен-
тральный) и задний края мантии (рис. 193, 7; рис. 194, 3, 4), т. е.
складки кожных покровов, или завесы, покрывающие с боков соб-
ственно тело беззубки. Задненижний конец мантии пигментирован
и покрыт чувствительными сосочками (рис. 194, 5, 6); когда сосочко-
19 Практикум по зоологии — 383 289
выс края обеих лопастей мантии (т. е. правой и левой) прилегают
друг к другу в естественном положении, то между ними обнаружи-
ваются два отверстия, называемые сифонами.
Нижний — жаберный — сифон служит для проведения воды
внутрь мантийной полости (рис. 193, б); верхний—клоакальный —
предназначен для удаления воды из той
a
Ъ
193. Сверху — беззубка с удаленной левой
Рис.
створкой раковины. Снизу — створка раковины
беззубки изнутри, с отпечатками мест прикре-
пления мышц. По Ж а м м у.
7 — мантия; 2 — лигамент; 3—край мантии; 4 — зад-
няя запирательная мышца; 5 — клоакальный сифон;
6 — жаберный сифон; 7 — мантия правой стороны;
8— жабры (левой стороны); у— нога; 10—губные
лопасти; 77 — рот; 12— передний замыкатель рако-
вины.
Отпечатки: а — замыкателей раковины; b— протрак-
тора ноги; с — ретрактора ноги.
же полости (рис. 193, 5).
Из-под . нижнего края
мантии выдается кпереди
килеобразная нога беззубки
(рис. 193, 9; рис. 194, 74).
У переднего и заднего
краев мантия прободена
мышцами различного на-
значения. Два толстых
пучка мышц, идущих в по-
перечном направлении ме-
жду створками и прикре-
пляющихся к ним изнутри,
являются передним и зад-
ним замыкателями рако-
вины (m. adductor valvae
anterior и m. adductor val-
vae posterior; рис. 193, 4,
12, а; рис. 194, 15). Антаго-
нистом этих мышц является
эластическая связка — ли-
гамент, натянутая между
дорзальными краями ство-
рок раковины (рис. 193, 2).
Впереди и несколько
дорзально от задней замы-
кательной мышцы начи-
нается (также от створки
раковины) задний втягива-
телъ ноги (m. retractor pe-
dis posterior, рис. 193, с);
кзади и дорзально от перед-
ней замыкательной мышцы
берет начало передний втя-
гиватель ноги (m. retractor
pedis anterior), пучки которого косо переходят в основание ноги.
Позади и несколько вентрально от того же переднего запирателя
расположена мышца, вытягивающая ногу вперед (m. protractor
pedis), являющаяся антагонистом только что рассмотренных ретрак-
торов (рис 193, Ь).
Откинем кверху свободный край мантии, при этом мы открываем
мантийную полость, в которой лежат две желтовато-коричневые
пластинки, исчерченные тонкими параллельными полосками; это — на-
ружная и внутренняя жабры левой стороны тела (рис. 193, 8). Под-
нявши обе жабры кверху, рассматриваем ногу по всей ее иоверхно-
290
сти (9). У переднего края основания ноги, между ней и передней за-
пирательной мышцей, помещается ротовое отверстие (рис. 193, 77;
рис. 194, 75), по бокам которого спускается по паре небольших тре-
угольных листков, называемых губными или ротовыми лопастями
или парусами (vela) (рис. 193, 10; рис. 194, 77, 72). Между задней
половиной основания ноги и не доходящей до нее внутренней жаброй
просвечивает коричневатая полоса — почка, или боянусов орган.
Позади от ноги обе внутренние
жабры (правая и левая) срастаются
друг с другом по средней линии
тела (рис. 194, 8, 10). Между нача-
лом этого срединного шва и осно-
ванием ноги имеется отверстие.
Войдя в него зондом, например го-
ловкой булавки, можно проникнуть
в наджаберную полость, через ко-
торую вода идет к выходному от-
верстию. т. е. клоакальному сифону.
По дорзальному краю внутренней
поверхности внутреннего жаберного
листка, там, где он огибает ногу,
имеется узкая щель, через которую
можно проникнуть зондом во вну-
трижаберное пространство. Дело
в том, что каждая жабра состоит из
двух листков — наружного и вну-
треннего, которые переходят один
в другой по нижнему краю жабры.
Такая же полость есть и в наруж-
ной жабре.
Прилежащие друг к другу ли-
стки обеих жабр срастаются вместе
у основания жабры; наружный же
(восходящий) листок наружной
жабры срастается, опять-таки у
основания жабры, с мантией. Схе-
матически жаберный аппарат без-
зубки представлен на стр. 292.
Каждый жаберный листок со-
стоит из густого переплета тесно
прилежащих друг к другу стержней, перекрещивающихся друг
с другом и анастомозирующих со стержнями другого листка той же
жабры.. С поверхности жаберная пластинка имеет вид решетки
с тонкими порами. Жабры покрыты мерцательным эпителием, ко-
торый имеется также на внутренней поверхности мантии и на губ-
ных дольках.
Благодаря непрестанной работе ресничек вода вгоняется в мантий-
ную полость и проникает через поры жаберных листков во внутрен-
нее пространство; отсюда вода поступает дальше в наджаберную по-
лость и через клоакальный сифон выбрасывается наружу. При таком
* 291
Рис. 194. Беззубка с приоткрытыми
створками; вид снизу. По Рёзелеру
иЛампрехту.
Мантия правой стороны тела (J) отделена
от створки раковины (7); 2 — левая створка
раковины; 4 — левая складка мантии;
5—6—жаберный сифон; 7, 8, О, 10 —
наружные и внутренние жабры обеих сто-
рон тела; 11, 12—губные лопасти; 13 —
рот; 14 — нога; 15 — передний замыкатель
раковины.
ЛЕВЫЕ ЖАБРЫ
Схема жаберного аппарата беззубки
ПРАВЫЕ ЖАБРЫ
Шов
Я
СЗ
*
S
CL
X
0Q
Наружная
жабра
Перегиб
щие во внутри-
жаберную по-
лость внутрен-
них жабр
Наружная
жабра
Шов
промывании жабр током свежей воды осуществляется жаберное ды-
хание беззубки.
Одновременно обеспечивается и возможность принятия пищи,
С током воды пассивно втягивается множество мелких планктонных
организмов (инфузорий, рачков, диатомей и т. п.), которые задержи-
ваются на жаберных пластинках, как на сите. Ударами ресничек
мерцательного эпителия они перегоняются со слизью к ротовому от-
верстию, где и поступают в пищеварительную трубку.
Чтобы покончить с наружным осмотром беззубки, остается отме-
тить положение выводных отверстий различных органов. Под задней
замыкательной мышцей лежит заднепроходное отверстие (рис. 195,
Рис. 195. Анатомия беззубки. По Ж а м м у (из Н. Холодковского).
Раковина удалена; левая лопасть мантии и жабры также; часть органов вскрыта.
А — замыкательный мускул; О —рот; С— головной нервный узел; Р1 — около-
ротовые дольки; Р — нога; Pd— ножной нервный узел; Ср — мантия;
Sb— жаберный сифон; В—жабра; Ап — анальное отверстие; Pv — внутрен-
ностный нервный узел; Sc—клоакальный сифон; Od— спинное входное отвер-
стие; R — почки; Рг — околосердечная сумка; Сг—сердце; dr—выводной канал
почки; / — кишечник; Н — печень; S — желудок (вскрыт); G—половая же-
леза. Стрелками показано направление тока воды в мантийную полость и
наружу.
Ал). Вскрыв продольным разрезом наджаберное пространство, между
основанием ноги и внутренней жаброй можно видеть лежащие одно
над другим отверстия почки (рис. 195, dr) и под ними половое отверстие.
Так как наджаберная полость сообщается с клоакальным сифоном,
то последний служит также для выведения наружу экскретов и поло-
вых продуктов.
Нервная система. В промежутке между вытягивателем ноги и ее
передним ретрактором слегка соскабливают поверхностные покровы
и обнаруживают нервный узел оранжевого цвета (ganglion cere brale,
рис. 195, С), который сливается из собственно церебрального и пле-
врального узлов. Лежит он сбоку ротового отверстия и соединяется
с аналогичным узлом другой стороны тела церебральной коммиссуро'],
огибающей спереди начало пищеварительной трубки.
293
Рис. 196. Сердце беззубки.
Вид сверху по удалении
спинной части раковины. По
Ж а м м у.
/ — перикардиальная железа (ке-
беров орган); 2 — надрезанная
перикардиальная стенка; 3 —
сердце; 4 — правое предсердие;
5 — боянусов орган (почка);
6 — anus.
Осторожно разрезают средний шов между внутренними жабрами и
в задней части разреза находят пару сближенных висцеропариеталь-
пых ганглиев (рис. 195, Pv), также оранжевого цвета.
В толще ноги заложена пара ножных (педальных) узлов (рис. 195,
Pd), независимых друг от друга.
Отмеченные три пары нервных узлов составляют нервную систему
беззубки. Эти узлы соединяются друг с другом нервными тяжами, или
коммиссурами; имеются две це ребровисцеральные и две церебропедаль-
ные коммиссуры.
Кроме того узел дает нервные ветви
к различным органам тела. Так, ножной
ганглий иннервирует ногу (рис. 195, Р),
церебральные узлы обслуживают окруж-
ность рта (О) и переднюю часть мантии,
тогда как висцеропариетальные ганглии
снабжают нервами жабры, мантию, сердце,
кишечник и другие органы.
Морфологически нервная система без-
зубки является узловой; но тип ее хара-
ктеризуется тем, что нервные узлы лежат
далеко друг от друга, т. е. разбросаны по
телу; поэтому нервную систему беззубки
называют разбросанно-узловой.
Сердце и прилежащие органы. Подрезая
места прикрепления мышц к правой створке
раковины, освобождают тело беззубки, кла-
дут его в ванночку ногой вниз и анальным
концом кзади и, распластав при этом жабры
и листки мантии в стороны, укрепляют ман-
тию булавками.
Впереди и снаружи от задней замыка-
тельной мышцы, с каждой стороны тела
просвечивает темное поле —почка, или
боянусов орган. Между почками виден крас-
новатый участок — орган Кебера и сердце.
Выдающийся в этом месте кверху острый
край шва сросшихся лопастей мантии осто-
рожно срезают тонкими ножницами и вскры-
вают околосердечную полость по всей ее
длине.
В перикардиальной полости находят кишечную трубку, которая
проходит сквозь муфту и в заднем конце полости снова выходит из
нее. Таким образом кишка проходит сквозь желудочек сердца (рис. 195,
I, Сг). Если отрезать кишку перед сердцем и потянуть сердце кверху,
то видно, что от его середины отходят в стороны и вниз по прозрачному
треугольному мешку (рис. 196,4); это будут предсердия, которых у без-
зубки два. В предсердия кровь поступает из жабр, где она предва-
рительно обогащается кислородом. Пройдя в желудочек, кровь изли-
вается в переднюю и заднюю аорты, по которым идет в более мел-
кие ветви и в лакуны тела.
294
Рис. 197. Схема соотношения сердца и
почек у беззубки. По Ж а м м у.
7— отверстие почки в полость перикардия;
2—кишка; 3—наружное отверстие почки;
4 — почка; 5 — сердце; 6 — перикардиальная
полость; 7—кишка; 8—замыкательная мышца.
заложен в паренхиме тела и ноги,
Сбоку и впереди от сердца, на внутренней стенке перикардиальной
сумки лежат перикардиальные железы, или органы Кебера (рис. 196, 7),
в виде двух удлиненных клиновидных скоплений ткани оранжево-
красного цвета. Клетки этого органа выделяют экскреторные вещества,
которые через почку удаляются наружу.
По удалении сердца и кишки, на дне перикардиальной полости
находят пару темных почек (рис. 197). Каждая почка состоит из
перегнутой трубки; начальная часть трубки, собственно почка, сооб-
щается отверстием с околосердечной полостью (почечная воронка,
рис. 197, 7). На уровне задней замыкательной мышцы трубка пере-
гибается кверху и переходит в выводной канал, открывающийся от-
верстием в наджаберную по-
лость (рис. 197, 3).
Почку беззубки можно гомо-
логизировать метанефридиям,
ибо перикардиальная полость
(рис. 197, б), с которой «почеч-
ной воронкой» сообщается боя-
нусов орган (рис. 196,5), является
рудиментом вторичной полости
тела (целома).
На невскрытой беззубке мож-
но дойти до сердца и другим
путем: острыми щипцами раз-
дробляют раковину беззубки на
спинной стороне и пинцетом вы-
бирают все ее осколки. Острыми
ножницами вскрывают околосер-
дечную сумку продольным раз-
резом (рис. 195). Дальнейшее
рассмотрение органов ведется,
как указано выше.
Остальные органы. Органы
пищеварения беззубки полностью
отпрепаровать нельзя. Дело
объясняется тем, что кишечник
где целомическая полость тела вторично заполнена различными
тканями. Некоторое представление о ходе кишечника можно полу-
чить, если ввести в ротовое отверстие ветвь тонких ножниц и раз-
резать стенку тела по направлению пищеварительной трубки.
За коротким пищеводом следует небольшой желудок (рис. 195, S).
из которого выходит кишка, петлеобразно изгибающаяся в верхней
части ноги (/). Далее она поднимается дорзально, проходит сквозь
сердце (Сг), идет над задней замыкательной мышцей (Д), позади ко-
торой и открывается заднепроходным отверстием (Ап). Придаточной
железой является печень (Н), лежащая также в толще ноги, в верхне-
передней части ее основания (просвечивает сквозь покровы ноги
в виде коричневатого участка).
Половой аппарат представлен половой железой (гонадой), зани-
мающей задневерхнюю часть основания ноги (рис. 195, G). Половые
295
клетки через половое отверстие выбрасываются в наджаберную
полость.
Беззубки раздельнополы; однако гонады их очень сходны и отли-
чаются только по характеру вырабатываемых ими половых продуктов.
Размножение беззубки
У некоторых особей беззубок наружная жабра бывает вздута: в ее
толще оказывается множество яиц, в которых развиваются личинки,
называемые глохидиями.
Глохидию в капле глицерина рассматривают под микроскопом.
Личинка беззубки одета двустворчатой раковиной. От нижнего края
створки внутрь отходит зубец, зазубренный по своему краю. Между
створками лежит сильно развитый замыкательный мускул. Если есть
случай наблюдать под микроскопом живую глохидию, то можно ви-
деть, как она захлопывает время от времени створки раковин.
От тела глохидии свисает вниз длинная клейкая нить, которая
служит (как и зубцы раковины) для прикрепления к коже рыб. Дело
в том, что глохидии падают в конце концов из жабр беззубки на дно.
При случайном приближении рыбы они прицепляются захлопываю-
щимися створками раковины й клейкой нитью к ее коже. Здесь гло-
хидии обрастают кожными покровами и в образовавшейся цисте
развиваются в молодую беззубку, которая по прорыве стенки цисты
падает на дно и становится свободноживущим животным.
Таким образом беззубка в части своего жизненного цикла является
временным паразитом рыбы.
Раковина беззубки
Для осмотра берут раковину после удаления из нее тела беззубки.
Внутренняя поверхность раковины отливает различными цветами,
что зависит от состава перламутрового слоя из тончайших пластинок;
падающий на них свет разлагается вследствие интерференции свето-
вых лучей, благодаря чему нашим глазом воспринимаются перла-
мутровые отливы раковины.
Такой род окраски не зависит от наличия каких-либо пигментов,
но объясняется особенностями структуры поверхности внутреннего
слоя раковины, почему и называется структурной окраской.
Снаружи раковина грязно-темнозеленого цвета (пигментная окрас-
ка periostracum), который в данном случае присущ конхиолиновому
слою раковины. Средний слой раковины фарфоровидный; он построен
из мельчайших призмочек углекислой извести.
Раковина в целом есть выделение клеток мантии. Если у живой
беззубки выбить кусочек раковины из ее середины, то с течением вре-
мени этот дефект будет восполнен только перламутровым слоем,
потому что вся поверхность мантии выделяет именно этот слой, тогда
как конхиолиновый и фарфоровидный слои вырабатываются клет-
ками края мантии.
Конхиолиновый слой, состоящий из органического вещества,
защищает фарфоровидный и перламутровый слои от растворяющего
действия углекислоты, могущей накопляться в воде.
296
Края раковины окаймлены мягкой оторочкой из конхиолинового
вещества, благодаря присутствию которой беззубка в случае надоб-
ности герметически замыкает раковину.
На внутренней поверхности раковины видны места прикрепления
к раковине мышц беззубки (рис. 193 снизу).
Рассмотрение разрезов
беззубки
Убитую и вынутую из
раковины беззубку уплот-
няют в водном растворе
формалина (одна часть фор-
малина на восемь частей
по объему воды) и острой
бритвой рассекают в попе-
речном направлении.
Поперечный раз-
рез на уровне ро-
товых д о л е к. На пре-
парате ориентируются при
помощи рис. 198; находят
справа и слева складки
мантии (7); посредине внизу
нога (5); между основанием
ноги и мантией с каждой
стороны по две ротовые
дольки (2, 3); сверху и сна-
ружи в щелях могут быть
захвачены разрезом листки
внутренней жабры (4).
В толще ноги —попе-
речные пучки мышц (6);
в месте перехода мышечной
части ноги в основание ле-
жат отдельные дольки го-
нады (7); над ними видны
дольки печени (77), между
которыми находятся попе-
речные разрезы желудка (9)
и кишки (70); дорзально и
поперечно лежит задняя
кишка со складкой на вен-
тральной стороне; над киш-
кой идет передняя аорта;
полость.
Рис. 198. Поперечный разрез через Unio на
уровне печени и желудка. По Рёзелеру
и Лампрехту.
7 — край мантии; 2 — наружный, 3 — внутренний па-
рус; 4 — жабра; 5 — нога; 6 — мышцы; 7 — гонады;
8— поперечный разрез нервных коммиссур; 9—тоже,
желудка; 10 — поперечный разрез ветви кишки;
11 — печень.
между мантией и ногой —мантийная
Поперечный разрез перед сердцем (рис. 199).
Находят справа и слева складки мантии (7); в мантийной полости
с каждой стороны по две жабры, в которых различают наружный
(2, 4) и внутренний (3, 5) листки. В толще ноги (6) видны гонады (9, 10),
297
разрезы кишки, задняя часть печени (/2), поперечные мышцы
(/ 7). передняя часть кеберова органа (13) и складка лигамента (14).
Поперечный разрез на уровне сердца и по-
чек (рис. 200). Справа и слева — складки мантии (7); нижний вен-
тральный край каждой складки утолщен (мантийный мускул); в ман-
тийной полости с каждой стороны ноги
ружные (восходящие) листки
наружной жабры срастаются
с мантией; обращенные друг
к другу нисходящие листки
13 U12
(4) по две жабры (2, 3); на-
Рис. 200. Поперечный разрез Unio на
уровне ночки. По Рёзелеру и Ла м и-
рехту.
/ — мантия; 2—наружная жабра; 3— вну-
тренняя жабра; 4—нога; 5 — разрез кишки;
0 — тифлозоль; 7 — гонада; 8 — нервные ком-
миссуры; 9— почки; 10—перикардиальная
полость; 11 — сердце; 12—полость сердца;
13 — поперечный разрез тифлозоля.
Рис. 199. Поперечный разрез через
Unio кпереди от сердца. По Рёзе-
леру и Л амп рехту.
/ — мантия; 2— наружный листок наруж-
ных жабр; 3— внутренний листок наруж-
ных жабр; 4 — наружный листок внутрен-
них жабр; J — внутренний листок внутрен-
них жабр; О — мышцы ноги; 7, 9, 10—го-
нады; 11—мышцы; 12—печень; 13—
краснокоричневый кеберов орган; 14—
складки лигамента.
обеих жабр срастаются один с другим у основания жабр; восходя-
щие (внутренние) листки внутренней жабры оканчиваются свободно,
благодаря чему внутрижаберная полость сообщается с полостью ман-
тии. В ноге видны мышцы, гонады (7), разрезы кишки (5). На спинной
стороне — перикардиальная полость (10), и в ней — задняя кишка (13),
вокруг которой —желудочек сердца (12), правое и левое предсердия.
Внизу и снаружи от перикардия —по почке (9).
298
Поперечный разрез через задний замыка-
тель раковины. Видны: задний замыкатель раковины; над
ним — задняя аорта и задняя кишка; в мантийной полости — четыре
жабры; над основанием жабр — наджаберные полости; снаружи —
складки мантии.
Общие замечания
Тело Anodonta мягкое, лишенное внутренних опорных частей
и покрытое твердой двустворчатой раковиной, являющейся неотъемле-
мой частью животного. Собственно тело покрыто с §оков свисаю-
щими вниз разросшимися складками кожных покровов, образующих
мантию, наружная поверхность и свободный край которой выделяют
раковину.
Из внутренних признаков особенно характерна нервная система,
построенная по разбросанно-узловому типу.
Все отмеченные выше признаки характеризуют беззубку как пред-
ставителя типа моллюсков.
Тело беззубки построено по двубоковой симметрии, выраженной
весьма ясно. Оно лишено каких-либо следов расчленения, и в нем нет
обособления головного конца.
Хорошо развитая лопастная мантия, двустворчатая раковина с за-
пирательными мышцами, пластинчатые жабры и клиновидная нога
позволяют относить беззубку к классу пластинчатожаберных (La-
mellibranchia),
В этом классе Anodonta занимает место в отряде настоящих пла-
стинчатожаберных (Eulamellibranchia), так как жабры ее имеют вид
порозных пластинок.
Замечательно отсутствие целомической полости тела у беззубки, от
которой остается только рудимент в виде полости околосердечной
сумки. Такая особенность строения является вторичной, ибо цело-
мическая полость тут вторично вытесняется сильно развивающейся
паренхимой.
Отметим, что из рассмотренных нами ранее животных бесполо-
стными были плоские черви (Platodes). У плоских червей полости
тела еще не образовалось, тогда как у беззубки полость тела уже
вытеснена паренхимой и различными органами.
Беззубка является пресноводным моллюском, живущим на дне
водоемов. Типичные придонные обитатели в совокупности своей обра-
зуют бентос.
Для движения беззубке служит нога, которая в виде клина высо-
вывается из створок раковины и упирается в дно. Движения ее
очень медленные. После себя она оставляет след в виде борозды.
При различных раздражениях беззубка быстро втягивает ногу и
захлопывает створки раковины.
Для питания ей служат планктонные организмы. Дышит она жаб-
рами на счет кислорода, растворенного в воде. В общем беззубка
тесно связана с водой как со средой обитания и вне ее скоро погибает.
299
КЛАСС БРЮХОНОГИЕ (GASTROPODA)
ОТРЯД ЛЕГОЧНЫЕ (PULMONATA)
УЛИТКА ВИНОГРАДНАЯ (HELIX POMATIA)
Технические замечания. Виноградная улитка живет в сред-
ней Европе, во влажных лесах, произрастающих на известковой почве. В случае
отсутствия этого вида можно пользоваться другими крупными улитками рода
Helix, живущими в СССР. Материал запасают впрок, положив живых улиток в боль-
шую банку, доверху наполненную 0,1% раствором хлоралгидрата в воде. Банку
закрывают пробкой так, чтобы в ней не оставалось воздуха. Через сутки улитки
дохнут обыкновенно в растянутом состоянии. Признаком смерти служит потеря
улиткой способности реагировать на укол пинцетом их тела. Для последующего
сохранения улрГгок помешают в 70% спирт. Можно также убить их, поместив в те-
плую (30—40° С) прокипяченную воду в плотно закрытом, наполненном доверху
сосуде. Делают это за час-полтора до начала занятий.
Наружный осмотр
Тело улитки одето спирально закрученной раковиной, завитки
которой уменьшаются к вершине, благодаря чему раковина имеет
вид башни. Раковина несет широкое отверстие, откуда высовывается
мягкое тело моллюска, в котором видны следующие части (рис. 201):
а) Широкая и пло-
ская нога (нижняя по-
верхность выпячен-
ного тела —pd).
б) Спинная выпук-
лая поверхность тела
шагреневого вида.
в) Мало обособлен-
ная голова, кото-
рая хорошо отделяется
от нижней поверхно-
сти ноги глубокой бо-
роздой.
г) Две пары щупа-
лец на голове; они
втяжные. Если щу-
пальца втянуты, то на их месте виднеются углубления. Первая пара
щупалец более короткая (/), вторая пара щупалец более длинная (о).
Когда они совсем высунуты наружу, то на их конце видно по черной
точке, являющейся глазом моллюска. Щупальце может впячиваться
внутрь тела, подобно вворачиванию пальца перчатки; в таком случае
прячутся внутрь тела и глаза.
д) В глубине щели между головой и передним концом ноги поме-
щается ротовое отверстие (Ь); над ним лежат две ротовые дольки.
е) Под основанием правого заднего щупальца помещается точко-
образное половое отверстие (искать внимательно! —g); иногда из
него может торчать выпяченный в различной степени копулятивный
аппарат.
ж) Противоположный задний конец тела вытянут и заострен; он
является частью собственно ноги.
300
Рис. 201. Садовая улитка. По Ж а м м у.
о — глаз на вершине щупальца; t — осязательные щупальца;
b — рот; g — половое отверстие; т— край мантии; р — ды-
хательное отверстие; ра — нога.
з) На верхней поверхности тела, почти посредине или несколько
более кзади, лежит раковина. Если ее осторожно оттянуть кверху, то
видно, что выпяченная часть тела улитки продолжается в полость
раковины, которую и заполняет. На границе входа в раковину на
мягком теле улитки находят мантийный валик (т). Под краем ман-
тии справа находим отверстие полости мантии, или дыхательное
отверстие (р).
Все перечисленные части ясно различимы или при наблюдении
живого ползущего моллюска или улитки, хорошо выпятившей на-
ружу свое тело при консервировке. В неудачно сохраненных экзем-
плярах нога бывает скрючена, голова впячена и части ее неразличимы
Кожные покровы ноги богаты слизистыми железами. Когда улитка
ползет, то оставляет за собой след в виде полосы, покрытой слизью.
Рис. 202. Раковина садовой улитки. По Мейзенгеймеру.
Ъ — основание раковины; ах — ее вершина; а—вход; и — пупок; аг — наруж-
ный край; or — передний край; иг — задний край. Справа — уродливая, скаларид-
ная, форма раковины.
На зиму, а также в жаркое и сухое время, улитка прячется глу-
боко в раковину, отверстие которой закрывается пленкой, образо-
вавшейся из затвердевшего секрета желез.
Раковина улитки. Чтобы хорошо рассмотреть раковину улитки,
кладут животное на несколько минут в кипяток, после чего пинце-
том можно легко вынуть улитку из раковины.
В раковине различают (рис. 202) широкое основание (basis—b) со
входом в раковину (apertiira — а), вершину (apex — ах) и завитки,
уменьшающиеся к вершине; в основании раковины имеется вдавле-
ние —пупок (umbo, и), внутренние стенки завитков срастаются
в осевой стержень, называемый столбиком (columella).
Отверстие раковины имеет четыре края: передний и наружный,
незаметно переходящие друг в друга (оба они правильно округлы),
301
и задний и внутренний, сходящиеся под углом друг к другу; вершина
угла схождения обращена к пупку раковины.
Раковина бывает закручена вправо или влево. Чтобы определить
характер закручивания, поворачивают раковину отверстием к наблю-
дателю, а верхушкой кверху. Если отверстие лежит справа от смотря-
щего, то раковина закручена вправо и наоборот.
Раковина наощупь крепка; прочность ее зависит от присутствия
в ней известковых солей. Часть раковины кладут в 10% водный рас-
твор соляной кислоты; наблюдающееся выделение пузырьков газа
свидетельствует о выделении углекислоты из углекислого кальция,
находящегося в раковине. Когда эта реакция заканчивается (около
получаса или часа), то остается коричневатая кожица —органическая
строма раковины, состоящая из конхина (о происхождении раковины
см. ниже).
Удаление раковины и продолжение наружного осмотра. Лёгкими
ударами молотка раздробляют раковину и пинцетом удаляют ее
Рис. 203. Садовая улитка с удаленной раковиной, вид слева.
По Рёзелеру и Лампрехту.
1—линия разреза при вскрытии; 2—просвечивающее сердце;
3 — просвечивающая почка.
куски. Особенно осторожно необходимо снимать завитки вершины
раковины; надежнее сдвигать их винтообразно в направлении, проти-
воположном ходу завитков; в противном случае при удалении рако-
вины легко оторвать тонкую часть тела.
Тело улитки совершенно точно повторяет форму раковины, ибо
везде тесно прилегает к ее внутренней поверхности (рис. 203).
В дыхательное отверстие вводят тонкий зонд, который при этом
проникает в полость мантии. Двигая зондом, определяют границы
распространения полости. Ее верхняя тонкая покрышка является
мантией, которая на всем протяжении своих краев срастается с по-
верхностью тела (валик мантии), оставляя единственную щель —
мантийное отверстие.
В толще мантии просвечивают ветви легочной вены, отходящие
от главного сосуда, направляющегося вдоль по длине завитка тела.
Кзади от легочной вены просвечивает участок, где лежит сердце
(рис. 203, 2); справа от последнего 1 видно под покровами лопасте-
видное тело коричневатого цвета —почка (рис. 203, 3).
1 При положении улитки ногой книзу и головой вперед.
302
Справа от почки, по правому краю завитка, идет задняя кишка,
открывающаяся наружу заднепроходным отверстием, которое поме-
щается под дыхательным отверстием. Три следующих кверху завитка
тела постепенно, но быстро уменьшаются в размерах. Они коричне-
ватого цвета благодаря просвечиванию печени.
У верхнего края второго завитка на темном фоне выделяется жел-
товатый участок, так называемая белковая железа (часть полового
аппарата).
Вскрытие улитки и ее внутреннее строение
Вскрытие мантийной полости. Вводят конец тон-
ких ножниц в дыхательное отверстие и перерезают поперек мантий-
ный валик. Затем разрез ведут параллельно валику мантии и над ним,
Рис. 204. Садовая улитка со вскрытой легочной полостью и
отброшенной мантией. По Рёзелеру и Лампрехту.
/—дыхательное отверстие; 2 — мантийный валик; 3—плоскость
разреза; 4 — мантия, отведенная вправо; 5 — дно мантийной полости;
6 — задняя кишка; 7 — anus; 8 — отверстие почки; 9 — ветви легочной
вены; 10— линия разреза стенки тела.
разрезают стенку тела под сердцем и под нижним краем почки и закан-
чивают его, не доходя до верхнего края первого завитка тела (рис. 203).
Захватывают пинцетом свободный край лоскута мантии и отводят
всю мантию вправо (рис. 204, 4), выворачивая при этом ее внутрен-
нюю поверхность наружу. Благодаря таким манипуляциям откры-
ваются (рис. 204): собственно стенка тела (5). гладкая на своей по-
верхности, и внутренняя поверхность мантии с лежащими в ней орга-
нами.
Пространство между этими двумя поверхностями и является поло-
стью мантии.
Внутренняя поверхность мантии покрыта сетью перекладин,
в толще которых проходят кровеносные сосуды (легочная вена с ее
303
ветвями — 9). Эта часть мантии является легким, ибо именно здесь про-
исходит обогащение крови кислородом и очистка ее от углекислоты.
Понятно теперь, почему отверстие полости мантии называется дыха-
тельным.
В толще дна мантийной полости заложены продольные и попереч-
ные мышцы. При сокращении их, выпуклая поверхность дна полости
уплощается, благодаря чему воздух насасывается в полость; при
расслаблении мышц возвращающаяся в прежнее положение стенка
полости выталкивает отработанный воздух наружу.
Позади главного продольного валика легкого лежит сердце. Оно
помещается в перикардиальной полости. Последнюю прорывают тон-
ким пинцетом и из нее извлекают сердце, состоящее из тонкостенного
предсердия (лежащего впереди) и толстостенного желудочка.
Предсердие получает богатую кислородом кровь из легкого и про-
талкивает ее в желудочек, откуда кровь направляется через аорту
(разрываемую при препаровке) в тело.
Отметим здесь же, что никаких других сосудов при последующей
работе мы не найдем. Поэтому, чтобы покончить с органами крово-
обращения улитки, упомянем, что кровеносная система ее незамкну-
тая, ибо сосуды в конце концов открываются в лакуны.
Слева от сердца (на вывернутой мантии) лежит почка, сообщаю-
щаяся маленьким отверстием (8) с перикардиальной полостью.
Выводной канал почки идет по приросшему краю мантии рядом
с задней кишкой (6) и открывается наружу рядом с анальным отвер-
стием (7).
Вскрытие полости тела улитки. Тонкими ножни-
цами рассекают стенки тела вдоль по средней линии спины, начиная
от головы, перерезают валик мантии, разрезают дно мантийной поло-
сти (рис. 204, 10) и переходят на завитки тела, придерживаясь и
здесь экватора завитков. Разрез продолжают, сколько возможно,
к вершине тела.
Далее улитку прикалывают ко дну ванночки ногой книзу и отре-
зают боковые кожные лоскуты, идя от головы по боковому краю
ноги. Удаляют перемычки между различными органами, снимают
осторожно кожные покровы с завитков тела и рассматривают.
Если препаруемый экземпляр хорошо консервирован, то рассма-
тривают его под водой. Если же приходится вскрывать сильно скрю-
ченный экземпляр, то вскрытие и рассмотрение органов лучше произ-
водить на руках. Во вскрытой полости тела лежат органы пище-
варения, размножения, мышцы и нервная система (рис. 205).
Органы пищеварения. Уже рассмотренное нами ротовое отверстие
ведет в глотку (pharynx)—овоидное мускулистое тело, лежащее
в передней части препарата (7). За глоткой последовательно идет
более узкий пищевод, постепенно переходящий в удлиненный мешко-
видный желудок (5), который также без резкой границы продолжается
в тонкую кишку, образующую петлеобразный изгиб; кишка лежит
в завитках тела, откуда она в виде задней кишки (6) идет по краю
мантийной полости и открывается наружу заднепроходным отвер-
стием (7), лежащим несимметрично справа от средней линии
тела.
304
Кишечник обслуживается пищеварительными железами. Из них
слюнные железы (3), в виде двух неправильной формы пластинчатых тел,
соединяющихся задними концами друг с другом, лежат на верхне-
боковых поверхностях желудка. От передних концов желез идет
по выводному протоку (параллельно пищеводу), которые проходят
8 5
25
Рис. 205. Анатомия улитки. По Рёзелеру и Ламп рехту.
/ — глотка; 2— церебральный ганглий; 3 — слюнная железа; 4 — ее выводной
проток; 5 — желудок; 6 — задняя кишка; 7 — заднепроходное отверстие; 8—
печень; 9— почка; 10— ее выводной канал; 77 — отверстие почки; 12—пред-
сердие; 13 — сердце; 14 — гермафродитная железа; 15 — гермафродитный поло-
вой проток; 16—receptaculum seminis; 17 — канал семеприемника; 18— бел-
ковая железа; 19— матка; 20—яйцевод; 21 — влагалище; 22 — мешок стрел;
л 23 — пальцевидные железы; 24—vas deferens; 25 — бич; 26—penis; 27 — ре-
трактор penis; 28—втяжной мускул глазного щупальца; 29— втяжной мускул
осязательного щупальца; 30—часть втяжного мускула.
сквозь кольцо нервной системы (2) и впадают в глотку. Отметим,
что слюнные железы развиты у улитки очень сильно.
Печень (вторая пищеварительная железа) развита также хорошо (5).
Она покрывает изгибы кишки и впадает в желудок, который бывает
заполнен коричневой жидкостью, выделяемой главным образом пе-
ченью, частью же слюнными железами.
20 Практикум по зоологии — 383 305
Отметив положение нервного кольца (2), перерезают тотчас по-
зади него пищевод, протаскивают его отрезок через кольцо, подни-
мают кверху и обрезают передний конец глотки. Извлеченную глотку
вываривают несколько минут в 10% водном растворе едкого кали.
Мягкие части цри этом разрушаются, тогда как челюсть и тёрка
(радула) легко изолируются. Их промывают в часовом стекле водой
и рассматривают в глицерине под слабым увеличением микроскопа.
Рис. 206.
/. Продольный разрез через голову молодой
улитки, bg—соединительная ткань; eg—мозговой ганглий;/—по-
дошва ноги; I—губа; т — рот; mh — ротовая полость; тт >, з—
мышцы радулы; тр— сосочковая мембрана; тга — передняя мышца
терки; тгр — задняя мышца тёрки; ти — мышца верхней стенки рото-
вой полости; od — одонтобласты; ое— пищевод; ок—верхняя че-
люсть; pg—; нижний ганглий: г — радула; rt — мешок радулы;
stp — опорная пластинка радулы; vg — висцеральный ганглий. По
Луаэену. // — верхняя челюсть. ///—часть ра-
дулы с зубцами. По Мейзенгеймеру.
Челюсть имеет вид полулунной пластинки с шестью-семью попе-
речными полосами (рис. 206, 11); нормально она лежит тотчас
позади ротового отверстия; цвет ее темнокоричневый.
Тёрка (радула) помещается на дне глотки; она покрыта средин-
ным боковым и краевым зубчиками, форма и расположение которых
принимаются в соображение при определении моллюсков (рис. 206,111).
Как показывает само название, радула служит для соскабливания
растительной пищи. Таким образом, в отличие от беззубки, улитка
питается активно, тогда как беззубка пассивно заглатывает добыч}',
приносимую с током воды,
306
Сравнение пищеварительного аппарата моллюсков
Название органов
। Беззубка Улитка
I I
Ротовое отверстие......................
Мускульная глотка......................
Тёрка .................................
Пищевод................................
Слюнные железы........................
Желудок................................
Печень ................................
Кишка..................................
Анальное отверстие.....................
Пища...................................
С лопастями ;
С дольками
+
+
+
Н~
+
Животная
+
+
+
+
-h
Растительная
Сравнивая строение пищеварительного канала беззубки и улитки,
замечаем большее разнообразие составных частей этой системы орга-
нов у улитки, что можно поставить в связь с более активным спосо-
бом питания, а также и с родом пищи.
Рис. 207. Половые органы виноградной улитки. По М е и-
зенгеймеру.
zw — гермафродитная половая железа; zg—ее проток; еш — белко-
вая железа; spd— семепровод; od— яйцевод; vd—vas deferens;
v — влагалище; rc — семеприемник; st — его канал; fd — пальцевид-
ная железа; р/—мешок любовных стрел; vst — половая клоака;
р—penis; rm — ретрактор мужского копулятивного органа; ft — бич;
sp — изолированная любовная стрела.
Органы размножения. Улитка виноградная является гермафро-
дитным животным. И яйца и живчики вырабатываются в одном и том
же органе, называемом гермафродитной железой (рис. 205, 71;
рис. 207), которая лежит над печенью в одном из завитков тела.
*
307
Схема гермафродитного полового аппарата улитки
Общая (выходная)
часть гермафродит*
ного полового
аппарата
о
и
Половые клетки из железы поступают в общий гермафродитный
проток (рис. 205, 15), открывающийся в малообособленную оплодо-
творителъную камеру, которую покрывает массивная белковая железа
(18); размеры последней зависят от стадии ее деятельности.
Гермафродитный проток далее разветвляется на два тесно при-
легающие друг к другу канала, служащие для раздельного выведения
яиц и живчиков.
Яйцевод (иначе называемый маткой) образован сильно складчатой
трубкой (19), рядом с которой идет простатическая железа, или се-
менной канал. Впереди эти каналы расходятся друг от друга.
Мужские выводные пути последовательно образуют семепровод (24),
продолжающийся в трубчатый копулятивный орган (26), который
обладает втяжной мышцей—m. retractor penis (27) и бичевидным
придатком —flagellum (25).
Копулятивный орган тоже открывается в половую клоаку.
Женские половые пути, отойдя от мужских, распадаются на яйце-
вод (20) и влагалище—vagina (21), впадающее вместе с копуля-
тивным органом в половую клоаку, открывающуюся наружу поло-
вым отверстием.
Сравнение гермафродитизма червей и мягкотелых
Признаки
Плоские черви ! Улитка I Беззубка
Половые железы
одной и той же
особи
Яичники и семен-
ники
Выводные пути
одной и той же
особи
Гермафродитная
половая железа,
Гермафродитный
канал
Половая железа, в
разное время года
дающая живчики
I или яйца
! I
I
Половые отверстия
одной и той же
особи
Мужское Женское
Общая половая
клоака с единствен-
ным (общим) поло-
вым отверстием
Короткое выгод-
ное отверстие
[Форма гермафро-
дитного аппарата
Половые органы
полностью раздель-
ные (анатомически)
Половые органы
раздельные частью
анатомически, ча-
стью функцио-
нально
Половые органы
раздельные только
функционально
(сезонный герма-
фродитизм)
Женские половые пути имеют ряд добавочных частей. Во влагалище
открывается длинный канал (17) семеприемника receptaculum semi-
nis (16) имеющего форму шарика; туда же впадают проток разве-
твленных пальцевидных желез (23) и плотное овоидное тело, так на-
зываемый мешок любовных стрел (22), в котором образуется известко-
вое тело, выбрасываемое при копуляции наружу, причем оно впивается
309
в кожные покровы другой копулирующей особи. Секрет пальцевид-
ных желез служит, вероятно, для облегчения выхода «стрелы» наружу
(рис. 207, sp).
Соотношения сложных частей гермафродитного полового аппарата
улитки виноградной можно уложить в схему (стр. 308). При ее
рассмотрении отметим, что собственно гермафродитной является на-
чальная часть аппарата (половая железа, ее проток и оплодотвори-
тельная камера), а также выходной отдел —половая клоака и поло-
вое отверстие; между тем средняя часть полового аппарата разде-
ляется на обособленные пути, обслуживающие порознь яйца и жив-
чики. Различие форм гермафродитизма у рассмотренных нами жи-
вотных уясняется вышеприводимой таблицей (стр. 309).
Копуляция и оплодотворение. При копуляции под действием по-
лостной жидкости копулятивный орган выворачивается через половую
клоаку и половое отверстие наружу и вводится во влагалище и канал
семеприемника другой особи.
Flagellum выделяет студневидную массу, которая поступает в се-
меприемник и обволакивает порцию живчиков, выходящую из семе-
провода; таким образом формируется сперматофора.
При копуляции сперматофора из копулятивного органа попадает
в канал семеприемника и продвигается в его резервуар. Здесь обо-
лочка сперматофоры растворяется, благодаря чему живчики осво-
бождаются.
Через несколько дней сперматозоиды спускаются по каналу семе-
приемника во влагалище, проходят в яйцевод и достигают оплодо-
творительной камеры. Когда в гермафродитной железе данной особи
созревают яйца, они по тому же самому гермафродитному протоку
поступают в оплодотворительную камеру, где их ждут живчики.
Здесь и происходит оплодотворение. Сюда же изливается секрет
белковой железы, обволакивающий яйца. По яйцеводу они выходят
во влагалище и через половую клоаку и ее отверстие откладываются
в землю.
По окончании копуляции растянутая при выворачивании копуля-
тивного органа мышца сокращается и втягивает penis обратно.
Мышцы и движения улитки. Как и беззубка, улитка движется
медленно, но все же быстрее ее. Органом движения служит широкая
и плоская нога.
Мышцы у улитки гладкие. Большая часть их заложена в толще
ноги, но, кроме того, есть несколько лентовидных мышц специального
назначения; с одной из них —m. retractor penis (рис. 205, 27) мы
уже познакомились.
Разыщем еще мышечные пучки, идущие от стенки тела к щупаль-
цам. Потягивая за эти мышцы пинцетом, можно получить предста-
вление о характере действия их в естественных условиях.
В полости тела мы найдем также несколько хорошо развитых
мышечных лент (30), переходящих в толщу ноги; другим концом
они прикрепляются к осевому столбику раковины. При своем сокра-
щении они втягивают тело улитки в раковину.
Центральная нервная система. Центральная нервная система
улитки состоит из поясовидной массы, окружающей начало пнше
ЗЮ
вода (рис. 205, 2). От этого нервного кольца отходят в разные стороны
нервные тяжи. Кольцо отпрепаровывают от соседних органов; нерв-
ные тяжи отрезают подальше от центральной массы. Освобожденное
таким приемом нервное кольцо переносят в часовое стекло с водой,
где иглами освобождают его от соединительной ткани (работа об-
легчается, если объект предварительно выдержать в мацерирующей
жидкости: воды —60 см3, азотной кислоты—20 см3 и глицерина —
20 см3).
В полном своем составе центральная нервная система представлена
правым и левым церебральными ганглиями, соединенными друг с дру-
гом соединительной тканью. От мозговых узлов идут нервы к глазам,
щупальцам, копулятивному органу, ротовым лопастям и к другим
частям. Глазные нервы отходят от церебральных ганглиев вздутым
основанием; от последних идут также два тяжа к маленьким буккаль-
ным ганглиям (це ребробуккальные коннсктивы), соединенным друг
с другом буккальной коммиссурой.
Под пищеводом лежит подглоточная ганглиозная масса, состоящая
из внутренностных узлов, образованных слиянием парных парие-
тальных и висцеральных ганглиев; к этой массе прилежит пара ножных
(педальных) и пара плевральных ганглиев.
Внутренностный комплекс узлов и ножные ганглии соединяются
с церебральными узлами самостоятельными цереброплевральными
и церебропедальными коммиссурами, благодаря чему образуется около-
пищеводное нервное кольцо.
От внутренностных узлов отходит шесть пар нервов, а от ножных
ганглиев —девять пар.
Околопищеводное нервное кольцо улитки негомологично около-
глоточному кольцу нервной цепочки кольчатых червей и членисто-
ногих, ибо морфологический состав этих органов различен, что можно
видеть на следующей схеме (см. стр. 312).
Центральная нервная система улитки отличается скученностью
ее частей. Также как и у беззубки, она состоит из немногих нервных
узлов, которые, однако, весьма сближены друг с другом, вследствие
чего нервные тяжи между ними весьма укорачиваются, и в своей со-
вокупности нервная система принимает форму широкого пояса.
Несмотря на такие внешние отличия, у улитки и у беззубки нерв-
ная система относится к разбросанно-узловому типу, характерные
особенности которого у улитки вторично сглаживаются.
Органы чувств. Глаза помещаются на вершине второй пары втяж-
ных щупалец. Органами чувств являются также первая пара щупалец
(осязание) и ротовые дольки (вкус?).
Кроме них у улитки имеется еще пара отоцистов, лежащих в виде
замкнутых пузырьков на верхних боковых поверхностях ножных
узлов в толще соединительной ткани.
В слуховых пузырьках имеются мельчайшие крупинки —ото-
литы. При обычных условиях работы отоцисты не могут быть рас-
смотрены.
Наконец, чувствительностью обладает и вся поверхность тела
улитки, равно как и выстилка мантийной полости, в чем легко убе-
диться опытами на живом животном.
3U
Сравнение нервной системы мягкотелых и членистоногих
Беззубка
Улитка виноградная
Членистоногие н кольчатые черви
Церсбро- | Церебро-
плевральный Коммиссура плевральный
узел I узел
Церебральные узлы
о
н
S
с:
ГЗ
СП
Оч
о
tr
Г5
СО
с
г-;
СО
и*"»
X
а
о
Оч
о
го
сЗ
сх
Ножной Ножной
ганглий ганглий
CTJ
Оч
X
55
55
О
X
Педальный
(парный)
ганглий
I
Внутренностный
ганглий, состоящий
из плевральных,
париетальных и
висцеральных
ганглиев
55
* 56
Примечание. Черным круж- ।
ком обозначено поперечное сече- i
ние пищевода
Висцсропариетальный
(парный) ганглий
Улитка более богата органами чувств, нежели беззубка, так как
имеет глаза и осязательные щупальца. Это объясняется тем, что
улитка может высовывать значительную часть тела из раковины и
своим телом непосредственно касаться окружающей среды, тогда
как тело беззубки всегда скрыто между створками раковины и наружу
высовывается только нога.
Общие замечания
Такие признаки, как мягкое нерасчлененное тело и наличие ра-
ковины, определяют принадлежность улитки к типу моллюсков.
Однако улитка обладает рядом особенностей строения тела, кото-
рые значительно отличают ее от уже знакомой нам беззубки.
Прежде всего остановимся на особенностях архитектуры тела
и раковины улитки, ибо здесь мы впервые встречаемся с нарушением
двубоковой симметрии. Действительно, как бы мы ни рассекали ра-
ковину и втянутое в нее тело улитки, мы никогда не получим двух
симметричных антимер. Это обстоятельство зависит от спирального
закручивания раковины, а следовательно, и спрятанного в ней тела.
У улитки имеется значительное число непарных органов, как то:
почка, легкое и все части гермафродитного полового аппарата. Все
они, как и кишечник, лежат асимметрично перекрученными и частью
сдвинутыми в сторону от середины продольной плоскости, считая
по ходу завитков тела.
Мало того, половое и дыхательное отверстия и anus с отверстием
почки сдвинуты на правую сторону тела, т. е. лежат также асимме-
трично.
Таким образом характерной особенностью плана строения тела
улитки является асимметрия. Это —вторичная особенность улитки,
ибо подобные моллюски произошли от двусторонне симметричных
предков.
Теории происхождения асимметричных моллюсков излагаются
в специальных курсах зоологии беспозвоночных. Поэтому здесь мы
отметим только, что асимметрия улитки связана с закручиванием
спиралью (во время филогенетического развития) ее раковины, что
повлекло за собой редукцию некоторых органов (например, одной
почки из прежней пары их) и смещение их.
Равным образом такое необычное положение анального, полового и
почечного отверстий, сдвинутых на правую сторону тела и лежащих
ближе к головному концу тела, объясняется наличием спирально
закрученной раковины.
Раковина улитки —производное мантии (секрет мантийных же-
лезок); спиральной трубкой, слепой на одном конце, она покрывает
значительную часть тела моллюска. Так как раковина не проницаема,
а тело моллюска совершенно выполняет ее полость, то всякие выводные
отверстия сдвинулись в те места тела, которые при выпячивании его
наружу лежали бы вне раковины.
Асимметрия закрученного тела и раковины, присутствие на го-
лове щупалец и наличие расширенной подошвообразной логи, лежащей
313
па брюшной стороне тела, определяют принадлежность улитки вино-
градной к классу брюхоногих, или Gastropoda.
Такие признаки, как присутствие легкого, лежащего на пра-
вой стороне тела, впереди сердца, строение последнего из одного
желудочка и одного предсердия, концентрация нервных узлов во-
круг пищевода и гермафродитизм, характеризуют улитку виноградную
как представителя отряда легочных моллюсков —Pulmonata.
Следующими особенностями строения, имеющими систематическое
значение, будут наличие втяжных щупалец с глазами на вершине
задних щупалец (подотряд стебельчатоглазые — Stylommatophora) и
присутствие единственного полового отверстия (группа Monogono-
рога). В пределах группы Monogonopora улитки классифицируются
по наличию и форме раковины, характеру тёрки и некоторым другим
признакам.
В легких улитки мы имеем орган дыхания воздушного типа. Они
аналогичны легким позвоночных животных и трахеям насекомых.
По существу легкое улитки является видоизмененным участком кож-
йых покровов, так как оно развивается из внутренней поверхности
мантии, которая сама по себе есть не что иное, как двойная складка
кожных покровов, нарастающая на тело моллюска и покрывающая
его на некотором протяжении. Следовательно, и легочное дыхание
улитки, по сути дела, является производным дыхания кожного.
Для передвижения улитки служит «нога» —расширенная и упло-
щенная брюшная поверхность тела. Движение улитки облегчается
выделением кожными железами слизи, которая в виде ленты остается
позади ползущего моллюска.
От различных внешних воздействий улитку защищает раковина,
являющаяся в то же время опорой для мягкого тела этого животного,
ибо никаких скелетных образований у улитки нет.
При различных раздражениях ползущая улитка прячется глубоко
в свою раковину, которая является нераздельной частью ее тела (дей-
ствие «столбикового мускула»).
Равным образом при большой сухости и жаре улитка втягивает
тело в раковину, отверстие которой закупоривается пергаментной
пленкой затвердевшего секрета кожных желез.
Питается улитка растительной пищей, для соскабливания которой
в глотке имеется особый орган в виде тёрки (радула).
Твердость раковины зависит от отложения в ее органической основе
неорганических известковых солей. С этим связана потребность улитки
принимать соответствующие вещества (в составе ее пищи) и, следова-
тельно, жить на почвах с известковыми соединениями.
СЛИЗНЯК (LIMAX • AGRESTIS)
Технические замечания. Слизняков собирают в садах, на лугу,
в лесу и других местах. Убивают их, как улиток (см. выше), и сохраняют в 70%
спирту. Слизняков вскрывать гораздо легче, чем улиток, потому что голое тело их
не закручено. В смысле внутреннего строения это животное равнозначно улитке,
но благодаря редукции раковины Lirnax не является типичным брюхоногим мол-
люском; поэтому мы рассматриваем его здесь в качестве дополнительного объ-
екта.
314
Наружный осмотр
Тело Limax вальковатое, удлиненнее (до 6 см длины); передний
(головной) конец округлый, задний (хвостовой) —заостренный; спин-
ная поверхность выпуклая, брюшная —плоская. Кожные покровы
на спине шагреневого характера. От середины спины и до заднего
конца тела выдается срединный киль. Ближе к головному концу ле-
жит гладкий овальный диск, переднебоковые края которого свободно
нависают над кожными покровами. Справа у края диска, несколько
отступя от его середины, имеется дыхательное отверстие; самый же
диск является рудиментарной мантией.
На голове помещаются две пары щупалец —пара передних (губ-
ных) и пара задних (глазных); они также втягиваются внутрь тела, как
и у улитки. На переднем конце тела имеется ротовое отверстие. С пра-
вой стороны головы расположено половсе отверстие. Брюшная сторона
тела вытянута в обособленную с боков узкую ленту с желтоватой
срединной полосой и двумя боковыми грязно-темнезелеными каймами.
Вскрытие мантийной полости
Скальпелем или тонкими ножницами разрезают мантию вдоль по
средней линии и пинцетом извлекают из заднего конца резреза оваль-
ную пластинку с концентрической по отношению к переднему полюсу
пластинки слоистостью. Извлеченный объект является рудиментар-
ной раковиной, заложенной в толще стенки мантийной полости.
Внутренние органы. Слизняка берут в левую руку зад-
ним концом к себе и тонкими ножницами разрезают стенки тела, идя
от заднего конца до головы и придерживаясь все время границы
между ногой и телом моллюска. У головы разрез поворачивают под
прямым углом вправо и рассекают кожные покровы спины, оканчивая
разрез у самой ноги сейчас же кпереди от полового отверстия.
Слизняка прикалывают ко дну ванночки ногой книзу, отворачи-
вают кожный лоскут вправо и укрепляют его также булавками. На
заднем конце комплекса внутренних органов лежит гермафродитная
половая железа. Кпереди от нее между бурыми лопастями печени идут
петли кишечника грязносерого цвета. Еще далее кпереди видны желто-
ватая белковая железа и остальные части полового аппарата.
Ориентировавшись в общем расположении органов, распутывают
тонким пинцетом клубок внутренностей, следя за тем, чтобы не отор-
вать каких-либо частей. В конце концов можно обособить пищевари-
тельный канал и половые органы.
По строению пищеварительные аппараты слизняка и улитки сходны;
поэтому повторяться здесь не будем (стр. 304). Отметим только, что
заднепроходное отверстие слизняка открывается в мантийную полость.
Центральная нервная система Limax также не требует повторного
описания, тогда как в половом аппарате есть некоторые отличия.
Гермафродитная половая железа, гермафродитный половой про-
ток, белковая железа, матка, влагалище, семенной канал, семепровод,
копулятивный орган и половая клоака —все эти органы Limax
сходны с таковыми же частями тела улитки. В остальном необхо-
димо отметить, что у Li max отсутствуют flagellum, мешок любовных
315
стрел и пальцевидные железы; кроме того отличается и семеприемник,
имеющий вид небольшого пузырька, открывающегося в клоаку.
В связи с редукцией раковины у Limax стоит исчезновение ленто-
видных мышц, втягивающих тело моллюска в раковину.
Из прочих органов остается найти органы, топографически связан-
ные с мантией. При осмотре внутренней поверхности отвернутого
лоскута спинных покровов отыскивают округлое поле, соответствую-
щее мантийному диску спины. На уровне слюнных желез обнаружи-
вают тонкую перепонку, покрывающую сердце, состоящее из одного
предсердия и одного желудочка. Указанная перепонка является стен-
кой околосердечной сумки. Далее под сердцем на препарате лежит
почка, открывающаяся в полость околосердечной сумки и в полость
мантии. Убрав сердце и почку, можно обнажить внутреннюю поверх-
ность мантийной полости, играющую роль легкого. (
Общие замечания
Limax, в общем, обладает признаками, указывающими на принад-
лежность его к типу моллюсков и к классу брюхоногих.
Однако столь характерная для брюхоногих асимметрия тела по
внешности у него проявлена не очень резко, ибо асимметрично лежат
только отверстия полости мантии и половое.
Такой кажущийся возврат к двубоковой симметрии зависит от
редукции раковины слизняка, от которой остался ничтожный рудимент
в виде пластинки. В связи с исчезновением закрученной раковины на-
ходившееся в ней также закрученное тело Limax вытянулось в длину
и вторично стало двусторонне-симметричным.
Однако двубоковая симметрия в данном случае относительна,
ибо расположение внутренних органов, в частности полового аппа-
рата и выводных отверстий легкого, кишки и половой клоаки, более
соответствует тому, что имеется у улитки, если не считаться со
спиральным закручиванием ее тела.
Филогенез Limax более многообразен в своих этапах:
1) отдаленными предками слизняка (как и вообще моллюсков) были
двусторонне-симметричные животные;
2) от них произошли типичные брюхоногие моллюски со спираль-
ной закрученной раковиной и резкой асимметрией организма (боль-
шинство нынешних Gastropoda);
3) Limax в своей эволюции пошел дальше, ибо вследствие редукции
раковины он начинает внешне снова приближаться к двусторонней
симметрии тела.
В отношении обладания признаками отряда и подотряда Helix и
Limax совершенно сходны друг с другом (стр. 313—314).
Вследствие отсутствия раковины слизняк нуждается во влажной
среде обитания; он еще в большей мере, нежели улитка, способен
выделять из кожных желез слизь, почему и получил название слиз-
няка. Для питания ему служат различные растения, в частности и
разводимые человеком, которым он может причинять вред. Расти-
тельная пища, как и у Helix, соскабливается тёркой.
316
ТИП ИГЛОКОЖИЕ (ECHINODERMATA)
КЛАСС МОРСКИЕ ЗВЕЗДЫ (ASTEROIDEA)
МОРСКАЯ ЗВЕЗДА (ASTERIAS RUBENS)
Технические замечания. Морские звезды хранятся неограни-
ченное время в 70% спирту. За несколько часов перед занятием материал перекла-
дывают в воду.
Наружный осмотр
Морская звезда имеет своеобразное по форме тело, состоящее из
центрального диска и отходящих от него пяти рук, или лучей. Руки
морской звезды относительно ее тела идут по радиусам, а места соеди-
нения оснований соседних рук лежат по интеррадиусам.
У морской звезды различают верхнюю, спинную, или аборальную,
поверхность тела, в общем выпуклую кверху, и нижнюю, брюшную,
ротовую, или амбулакральную, поверхность. Последняя характери-
зуется присутствием в каждой руке продольного желоба, называемого
амбулакральной бороздкой; ее легко обнаружить, раздвинув руками
края одного из лучей. Амбулакральная бороздка обычно заполнена
четырьмя рядами сосочкообразных выростов —амбулакральных но-
жек, служащих для передвижения морской звезды. Каждая амбула-
кральная ножка имеет на конце присоску в виде диска. На самом
конце амбулакрального желоба имеется трудно различимая на пре-
парате непарная амбулакральная ножка с пигментным пятнышком
(глазком).
В центре диска на брюшной стороне лежит довольно широкое
ротовое отверстие, из которого зачастую торчит выпяченный наружу
желудок. По краям ротового отверстия, соответственно интеррадиусам,
лежат «зубы» —отростки известковых пластинок кожного скелета
морской звезды. Анальное отверстие лежит почти в центре спинной
поверхности тела; найти его трудно.
Тело морской звезды одето более или менее мягкой кожей, в
которой заложены различные известковые образования в виде пла-
стинок, столбиков, игол и т. п. В своей совокупности они образуют
наружный кожный скелет, отличающийся, однако, от аналогичного
образования членистоногих иным происхождением. Кожный скелет мор-
ской .звезды образуется в соединительнотканной части кожи, т. е.
развивается за счет мезодермы, тогда как хитиновая кутикула члени-
стоногих является произведением наружного зародышевого листка
(эктодермы).
317
Скелетные части морской звезды подвижно соединены друг с дру-
гом, благодаря чему руки ее могут изгибаться в различных напра-
влениях.
дутся экземпляры с неодинаково
о
\ р
Рис. 208. Схема расположения пла-
стинок брюшной стороны морской
звезды Phanerozonia. По Лангу.
А — амбулакральные пластинки; В — адам-
булакральные, М — нижнемаргинальные,
N — интеррадиальные, О — ротовые пла-
стинки; Р— отверстие амбулакральных но-
жек; Т — терминальная пластинка.
Из скелетных пластинок при наружном осмотре может быть най-
дена только так называемая мадрепоровая пластинка, лежащая на
спинной стороне диска морской звезды в одном из интеррадиусов, в виде
чечевицы с мелкоточечной поверхностью.
При осмотре большого количества морских звезд наверно попа-
развитыми лучами или с лучами не-
правильного строения. Во всех этих
случаях имеет место регенерация
утерянных рук или их частей у
морской звезды. Способность этих
животных к регенерации весьма
значительна. Не только тело мор-
ской звезды может восстановить
утраченную руку, но и отпавшая
рука восстановляет заново диск и
остальные четыре луча.
Скелет морской звезды
Дно амбулакральной борозды
(рис. 208) образовано двумя рядами
амбулакральных пластинок (А);
между ними лежат отверстия, через
которые выпячиваются амбулакраль-
ные ножки. Игол на этих пла-
стинках нет. Кнаружи от амбула-
крального ряда пластинок с каждой
стороны тесно прилежит ряд адам-
булакральных (В); последние по-
крыты иглами.
В окружности ротового отвер-
стия, т. е. на самом диске морской
звезды, ряды адамбулакральных
пластинок соседних радиусов под-
ходят друг к другу и соприкаса-
ются первыми амбулакральными
пластинками, лежащими уже в ин-
террадиусе. Обе сходящиеся пластинки выступают углом к рото-
вому отверстию, образуя челюсть; поэтому их называют также рото-
выми пластинками (О); на них бывают иглы. Служат они для захва-
тывания пищи. На конце луча лежит довольно большая непарная
терминальная пластинка (Т) с пигментным глазком. На дорзальной
стороне луча лежит срединный каринальный ряд пластинок; по его
бокам идет по ряду верхних краевых (маргинальных) пластинок;
ниже лежат два нижних маргинальных ряда пластинок (М). Между
ними и адамбулакральными пластинками лежат два ряда интер-
радиальных пластинок (N). Скелет спинной стороны диска состоит
318
из неправильной сети пластинок, лежащих широкими петлями. Иглы
спинной стороны тела конической формы, довольно тонкие и редкие.
На спинной стороне те-
ла имеются многочислен-
ные мелкие щипцевидные
придатки, называемые пе-
дииелляриями; подвижные
створки их имеют форму
узкого ланцета. Имеются
также и крестообразные
педицеллярии (рис. 209)во-
круг игол маргинальных
пластинок и на других ме-
стах покровов. С педицел-
ляриями удобнее ознако-
миться у морских ежей, у
которых они крупнее. Ме-
жду спинными пластинками
скелета выступают много-
Рис. 209. Крестообразные педицеллярии мор-
ских звезд. Слева — Pedicellaster typiciis: спра-
ва — Icasterias panonla. По А. Д ь я к о и о в у.
численные папулы, или жа-
берные пузырьки (см. ниже, стр. 322 —срез руки морской звезды),
являющиеся кожными жабрами.
Вскрытие морской звезды и ее внутреннее строение
Держа морскую звезду в левой руке спинной стороной кверху,
тонкими ножницами разрезают сравнительно мягкую кожу тотчас над
твердым боковым краем каждого луча, идя от вершины его к основа-
нию. Как у вершины луча, так и у основания, разрезы соединяются
друг с другом, причем в интеррадиусе с мадрепоровой пластинкой
последнюю обходят дугообразным разрезом, огибающим внутренний
край пластинки (рис. 210, 2). Затем животное прикалывают амбула-
кральной стороной книзу ко дну ванночки за нижнюю поверхность
концов лучей. Пинцетом приподнимают кожу верхней поверхности
руки у ее вершины и подрезают тяжи, прикрепляющие изнутри пече-
ночные придатки, и удаляют таким образом всю спинную покрышку
тела целиком, причем амбулакральная пластинка остается на препа-
рате.
В диске морской звезды находим (рис. 210): слепые придатки
задней кишки, лежащие интеррадиально. Они обыкновенно прида-
влены к стенке желудка и могут оторваться при снятии спинной
покрышки тела. Эти придатки считаются ректальными железами. Под
ними лежит объемистый желудок (3) в форме большого мешка, зани-
мающего полость диска. Если оттеснить с какого-либо края стенку
желудка, подрезав придерживающие его соединительнотканные тяжи,
то можно видеть переход желудка в короткий пищевод, открываю-
щийся наружу уже знакомым нам ротовым отверстием.
В общем пищеварительный канал морской звезды весьма укоро-
чен. У некоторых видов морских звезд заднепроходного отверстия
нет (например, Astropecteri).
319
В луче морской звезды видим буроватые дольки печеночных при-
датков (5 ), впадающих в два протока. Печеночные придатки занимают
всю полость руки. Протоки их в основании луча сливаются в корот-
кий канал, открывающийся в желудок. Последний, следовательно,
имеет пять пар групп печеночных придатков, лежащих радиально,
которые ничем не соединяются с оральной стенкой тела и поэтому
легко вынимаются из соответствующего луча.
Удалив из какой-либо руки печеночные придатки, осматривают
боковые стенки двух прилежащих друг к другу лучей у места их
соединения. В желобке дна препарата находим здесь мелкодольчатую
половую железу. В каждой руке имеется по две гонады, с короткими
Рис. 210. Морская звезда вскрытая. По Рёз.елеру и
Лампрехту.
1 —наружное очертание тела; 2 — местоположение мадрепоровой пла-
стинки; 3 — 4 — слепой мешок; 5 — печеночные придатки.
канальцами в интеррадиусах на спинной стороне тела которые от-
крываются обычно незаметными половыми отверстиями. Последних,
по числу гонад, всего десять.
По удалении печеночных придатков'на дне луча видим продольно
идущее килеобразное возвышение, состоящее из мелких, тесно соеди-
няющихся друг с другом скелетных парных амбулакральных пла-
стинок; сообразно этому возвышению на нижней поверхности луча
идет амбулакральная бороздка.
Каждая пара амбулакральных пластинок образует так называе-
мый позвонок.
На боковых скатах возвышения видны лежащие'попарно малень-
кие округлые мешочки —ампулы амбулакральных ножек. Соскоблив
ножкой пинцета несколько штук ампул, находим под каждым ме-
320
шочком точкообразное отверстие, ведущее в соответствующую амбу-
лакральную ножку.
Удаляют, подрезав ножницами пищевод, весь желудок и остальные
печеночные придатки.
От мадрепоровой пластинки книзу тянется осевой синус, в котором
проходит S-образно изгибающийся каменистый канал, названный так
потому, что в его стенке имеются известковые отложения (прощупать
их твердость пинцетом). Рядом с ним идет так называемый аксиальный,
или осевой, орган, при препаровке обычно не заметный.
Каменистый канал впадает в кольцевой околопищеводный канал,
не видимый на спиртовом препарате. Местоположение канала хорошо
определяется темножелтыми тидемановыми железами, лежащими по-
парно в каждом интеррадиусе в виде бугорков величиной с була-
вочную головку. И тидемановские железы и аксиальный орган
являются своего рода кроветворными органами (лимфатическими орга-
нами ), вырабатывающими белые кровяные клетки.
От околопищеводного канала идет в каждую руку по радиальному
каналу, на препарате не видимому; эти каналы сообщаются с ампулами
и с амбулакральными ножками.
Ознакомившись со всеми составными частями амбулакральной
системы^ рассмотрим ее работу.
Через ситовидную мадрепоровую пластинку морская вода проходит
в каменистый канал и в околопищеводное кольцо, проникает дальше
в радиальные каналы и заходит в ампулы. При сокращении стенок
последних вода нагнетается в соответствующие амбулакральные ножки
(рис. 211, Fe), которые при этом напрягаются. На конце их имеются
присоски, которыми морская звезда прикрепляется к субстрату.
Напрягая и расслабляя ножки, животное медленно передвигается
с места на место.
Амбулакральная система называется также и водоносной.
Питание. Прием пищи у морской звезды весьма своеобразен.
Это животное питается, между прочим, и моллюсками, тело которых
спрятано в крепко замыкающуюся раковину. Найдя моллюска, мор-
ская звезда становится над ним, согнув свои руки книзу. Выпустив
амбулакральные ножки, она присасывается к обеим створкам рако-
вины, которые начинает медленно растягивать в стороны. Моллюск
захлопывает раковину сокращением замыкательных мышц, но под
влиянием длительного растягивающего воздействия эти сильные мышцы
в конце концов утомляются и створки раковины раскрываются.
Морская звезда выворачивает через ротовое отверстие желудок и
накрывает его внутренней поверхностью мягкое тело моллюска, кото-
рое и переваривается действием сока, выделяемого желудком.
Морские звезды питаются также кольчатыми червями, {Ачками,
другими иглокожими, падалью и др. Они могут наносить существен-
ный вред при разведении устриц, так как иногда массами нападают
на устричные банки.
Нервная система. Морская звезда имеет три системы нервных тя-
жей, лежащих в различных отделах ее тела.
Берут покровы, снятые со спинной стороны тела, поворачивают их
внутренней поверхностью кверху и прикалывают булавками за концы.
21 Практикум по зоологии — 385 321
Осматривают сетевидную поверхность кожи и находят пять радиаль-
ных буроватых тяжей, идущих вдоль по радиусам. Все они соеди-
няются вместе в центре диска. Это —апикальная (или аборальная)
нервная система.
Диаметрально противоположно лежит адоральная нервная система.
Чтобы ее найти, выщипывают все амбулакральные ножки и на дне
амбулакральных бороздок находят по продольному бурому тяжу.
Радиальные тяжи соединяются с околоротовым кольцом, лежащим
также поверхностно.
Третья система нервных тяжей заключена в толще ткани и отпре-
парована быть не может.
Прочие системы органов. Обособленной дыхательной системы
у морской звезды нет; частично роль ее выполняет амбулакральная
система, которая имеет отношение также и к экскреции.
Кровеносная система у морской звезды есть, но она развита слабо
в виде околоротового кольца и радиальных стволов, лежащих между
нервными стволами и радиальными каналами амбулакральной системы.
Рассмотреть это можно только на срезах.
Морские звезды раздельнополы. Полы различаются только по ха-
рактеру вырабатываемых половых клеток (т. е. под микроскопом).
Оплодотворение происходит в морской воде, где и протекает даль-
нейшее развитие и метаморфоз этого животного.
Поперечный разрез через руку морской звезды
Кусочки лучей морской звезды, фиксированные горячим водным
насыщенным раствором сулемы с прибавлением 5% ледяной уксус-
ной кислоты, приобретают на биологической станции. Препарат пере-
носят из спирта в воду и освобождают от извести (декальцинируют)
в 5% водном растворе азотной или соляной кислоты, пока ткани его
не сделаются мягкими. Затем препарат переносят в 5% водный раствор
калийных квасцов для нейтрализации кислоты (на 1 —2 суток). Далее
следует промывка препарата в проточной воде и проведение через
спирты. Заливают объект в парафин. Срезы (10—15 ц толщиной) на-
кладывают на стекло, смазанное белком, освобождают от парафина
ксилолом, обычным образом окрашивают гематоксилином и эозином и
заключают препарат в канадский бальзам.
Для упрощения дела препараты можно целиком окрасить борным
кармином (из спирта после декальцинации) в течение нескольких ча-
сов. Диференцируют в 70% спирту с соляной кислотой. Выдержи-
вают в спиртах и заливают в парафин.
На препарате (рис. 211) видно: кожные покровы (ер) окружают
снаружи’полость руки (целом — о). В толще кожи заметны разрезы
известковых скелетных частей (Sc) и щели ложной полости тела
(псевдоцель). В некоторых местах на поверхности покровов выдаются
разрезы игол (Рах) и выпячивания мелких жаберных отростков
(рар).
В жаберные выпячивания или папулы вдается целом; они запол-
нены бесцветной, целомической жидкостью, в которой плавают амебо-
идные клетки; снаружи папулы омываются морской водой. Газо-
322
обмен происходит через покровы папулы. Лейкоциты целомической
жидкости могут заглатывать посторонние вещества, попавшие в цело-
мическую полость (например, впрыснутую тушь); нагруженные зер-
нышками туши лейкоциты мигрируют через стенку папул наружу,
чем и освобождают организм от введенной в него туши (фагоцитоз и
миграция как своего рода эквивалент экскреторной функции).
Полость целома за-
нята двумя складчатыми
образованиями, состоя-
щими из высокого эпи-
телия; это—поперечные
разрезы печеночных при-
датков (ent).
У вдавленной внутрь
части кожных покровов
(разрез амбулакральной
бороздки) в полости руки
лежат две ампулы (тр)
амбулакральных ножек,
на некоторых препара-
тах сообщающиеся узки-
ми каналами с радиаль-
ным каналом амбула-
кральной системы.
Кроме того видны два
псевдогемальных (ложно-
кровеносных ) канала, ле-
жащих под радиальным
Рис. 211, Поперечный разрез через руку морской
звезды. По К. Шнейдеру, с изменениями.
Рах — паксиллы; рар — жаберный отросток; ent — пече-
ночный отросток кишечника: о — целом; G — гонада;
ер — поверхностный эпидермис; Sc — скелетные пла-
стинки ; Fe — амбулакральная ножка; тр — ампула.
амбулакральным каналом; пара амбулакральных ножек с полостью
внутри и присосками на конце (Fe); оральная поверхностная нерв-
ная система в виде поперечного разреза радиального тяжа, лежа-
щего на вершине складки высокого эпителия между основаниями
ножек; глубокая оральная нервная система, лежащая несколько
выше предыдущей части препарата, и аборальная нервная система
в виде поперечного разреза тяжа под кожей середины спинной сто-
роны препарата.
Общие замечания
Тело морской звезды весьма своеобразно по внешности, благодаря
лучеобразному расположению рук относительно центрального диска.
Плоскостью, проходящей вдоль руки (т. е. по радиусу), тело мор-
ской звезды разделяется на две симметричные половины. Таких пло-
скостей симметрии может быть столько, сколько у данного вида жи-
вотного имеется рук. Все эти плоскости симметрии расходятся по
радиусам от центра тела. Симметрия такого характера называется
лучевой, или радиальной.
По радиальной симметрии расположены и различные внутренние
органы: печеночные придатки желудка, ректальные железы, радиаль-
ные каналы амбулакральной и кровеносной систем, радиальные стволы
нервной системы и поливые железы.
323
Однако, если придерживаться точных отношений, то оказывается,
что не всякая пара антимер строго симметрична друг к другу. Это
объясняется тем, что мадрепоровая пластинка и каменистый канал
сдвинуты в один из интеррадиусов. Следовательно, тело морской
звезды может быть разбито на действительно симметричные половины
лишь одной единственной плоскостью сечения, которая проходит че-
рез интеррадиус мадрепоровой пластинки и противолежащую руку.
При всех других направлениях сечения мадрепоровая пластинка от-
ходит целиком в одну из антимер, что нарушает строгость симметрии.
Поэтому, если стремиться к точности выражений, то симметрию
тела морской звезды приходится называть билатерально-
лучевой.
При лучевой симметрии нет ни переднего, ни заднего конца тела
животного, которое ориентировано в дорзо-вентральном направлении,
благодаря чему различают верхнюю — спинную, и нижнюю — рото-
вую стороны тела.
Отсутствие заднепроходного отверстия у некоторых видов мор-
ских звезд есть явление вторичное, которое не имеет ничего общего
с отсутствием анального отверстия у плоских червей и кишечнопо-
лостных. У этих последних его еще не было, тогда как заднепроход-
ное отверстие первоначально было у всех иглокожих и вторично
у некоторых из них исчезло.
Такие признаки, как лучевая симметрия тела, наружный мезо-
дермальный скелет с иглами в коже, амбулакральная система и хо-
рошо развитая вторичная полость тела (целом), характеризуют мор-
скую звезду как животное типа иглокожих (Echinodermata).
В этом типе она занимает место среди представителей класса мор-
ских звезд (Asteroidea) соответственно наличию нижеследующих
признаков:
а) звездообразная форма тела с лучами, отходящими широкими
основаниями от диска, который сам по себе не обособлен;
б) радиальная симметрия пищеварительного аппарата, печеночные
отростки которого заходят в полость рук;
в) положение гонад в полости рук и
г) присутствие амбулакральных бороздок на нижней стороне лу-
чей морской звезды.
Наличие крестообразных педицеллярий, лежащих вокруг круп-
ных игол, является признаком, по которому Asterias относят к отряду
педицелляриевых морских звезд —Forcipulata. Из других признаков
отметим наличие папул и на оральной (брюшной)стороне тела. Амбу-
лакральные ножки обычно лежат в четыре ряда. Иглы простые
(без щетки мелких иголочек на конце стержня).
Asterias относится к семейству Asteriidae, для которого характерно,
что скелет лучей образован пятью продольными рядами пластинок
в дополнение к амбулакральным и адамбулакральным рядам.
Экологические замечания. Морская звезда является
обитательницей моря, где она ведет придонный образ жизни. В связи
с лучевой формой тела морская звезда передвигается медленно либо
изгибанием лучей, либо действием амбулакральных ножек.
Амбулакральная система является наиболее характерной и свое-
324
образной системой органов, развившихся у морской звезды прежде
всего, повидимому, в качестве органов движения,' а также захвата
пищи, дыхания и экскреции.
КЛАСС МОРСКИЕ ЕЖИ (ECHINOIDEA)
МОРСКОЙ ЕЖ (STRONGYLOCENTROTUS DROEBACHIENSIS)
Технические замечания. Морских ежей, как и звезд, приобре-
тают на Мурманской биологической станции. Материал хранится в спирту. Для
занятия необходим крепкий пинцет, крепкие ножницы; желателен лобзик или
какая-либо ручная пилка для вскрытия панцыря.
Наружный осмотр
Тело морского ежа имеет форму сплюснутого шара, густо покры-
того длинными острыми иглами, откуда и наименование животных
Рис. 212. Схема строения морского ежа с оральной (брюшной) стороны.
По А. Д ь я к о я о в у.
/—у — амбулакральные ряды; 1—5 — интерамбулакральные ряды. В центре — рот. окружен-
ный пятью зубами, вокруг них — ротовой валик (заштрихован) утолщенной оральной мем-
браны; оральная мембрана — черная (нерисгом); на нен - пять пар оральных (роювых)
пластинок с порами; каждая пластинка соответствует одной оральной ножке: по краям ораль-
ной мембраны видно десять небольших вырезов, вдающихся в скорлупу (жаберные желобки).
325
этого типа —иглокожие. Иглы подвижно сочленяются с кожными
покровами. Между иглами видны тонкие нитевидные, головчатые на
конце придатки —амбулакральные ножки.
Оба полюса тела лежат посредине каждой из уплощенных поверх-
ностей морского ежа. Более выпуклая называется спинной, апикаль-
ной, или аборальной, более плоская является ротовой, или оральной
(или брюшной). В центре последней заметен кожистый диск —пери-
стом, или ротовое поле, затянутое оральной мембраной; посредине
его торчат острия белых зубов, между которыми лежит собственно
Рис. 213. Околоанальная область мор-
ского ежа — Strongyloientrotus lividus.
По Ж а м м у, с изменениями.
7 — амбулакральный ряд: 2 — интерамбула-
кральный ряд скелетных пластинок; 3— аналь-
ное отверстие; 4 — радиальные, или глазные,
пластинки; 5 — генитальная пластинка; 6—по-
ловое отверстие; 7 — концевая пора; 8— око-
лоанальная мембрана (перипрокт); 9—мар-
репоровая пластинка.
ротовое отверстие.
Ротовое отверстие с зубами
окаймлено кольцевым ротовым
валиком, образованным утолще-
нием оральной мембраны. Далее
к периферии в ней видны пять
пар округлых оральных пласти-
нок, продырявленных порами;
каждая пластинка несет одну
оральную ножку (рис. 212).
Середина аборальной поверх-
ности также свободна от крупных
шипов. В центре ее виден окру-
женный короткими иголочками
диск —перипрокт (рис. 213, 8),
в котором несколько эксцен-
трично открывается анальное
отверстие (3).
Пинцетом снимают шипы в
окружности перипрокта (рис.
213); при этом обнажается апи-
кальное поле, состоящее из де-
сяти крупных многоугольных
пластинок. Пять из них назы-
ваются генитальными (5), ибо
каждая из них продырявлена
половым отверстием (6), половые
пластинки лежат в интеррадиу-
сах. Одна из генитальных пластинок больше других, неправильна
в своих очертаниях и продырявлена множеством тончайших отверстий;
это—мадрепоровая пластинка (9).
С половыми пластинками чередуются меньшие пластинки, назы-
ваемые глазными (4). Они также, имеют по отверстию, в которые
высовывается амбулакральная ножка с глазком на конце (на препа-
рате она не видна).
При удалении игол между ними попадаются особые подвижные
придатки в виде трехстворчатых щипчиков; называются они педицел-
ляриями.
Дальнейшее ознакомление с наружным скелетом морского ежа
удобнее производить на оставшихся от прежних занятий панцырях,
которые очищают от игол, педицеллярий, амбулакральных ножек и
326
Тканей. Очистку произвести легче, если панцырь продержать предва*-
рительно некоторое время в горячей воде.
Практичнее осмотреть такой препарат сначала с внутренней сто-
роны.
Скелет морского ежа
Скелет морского ежа состоит из двух сортов пластинок: продыря-’
вленных мелкими отверстиями амбулакральных (рис. 212, /—V; рис.
214, 1—5) и сплошных, более крупных, или интерамбулакральных,
пластинок (рис. 212, 1—5).
I
1
Рис. 214. Аборальная половина скорлупы морского ока изнутри.
По Ж а м м у.
;—5 — пары амбулакральных пластинок (радиусы); S — половые пластинки.
Пластинки каждого сорта лежат в два ряда, причем парные ряды
пластинок перемежаются друг с другом. В панцыре морского ежа на-
считывается всего двадцать рядов пластинок, лежащих в десять пар
рядов, которые идут по меридианам панцыря (рис. 214).
Скелетные пластинки морского ежа пятиугольны по форме; гра-
ница между амбулакральными и межамбулакральными рядами пла-
стинок прямая, тогда как линия спая пары рядов одноименных
пластинок зигзагообразная.
327
Принято считать, что амбулакральные пластинки идут по радиу-
сам, а межамбулакральные —по интеррадиусам. Следовательно,
у морского ежа в скелете будет пять радиусов (рис. 212,1—V; рис. 214,
1—5) и пять интеррадиусов (рис. 212, 7—5). Каждая пара рядов
амбулакральных пластинок упирается у апикального полюса в глаз-
ную (или первую радиальную) пластинку, тогда как интерамбула-
кральные пластинки граничат там же с генитальными пластинками
(и с мадрепоровой пластинкой) (рис. 214, S).
На противоположной—оральной — стороне тела амбулакральные
пластинки оканчиваются ушками (auriculae) для подвески сложного
жевательного аппарата, называемого аристотелевым фонарем (см.ниже).
Каждая амбулакральная пластинка пронизана четырьмя - пятью
парами отверстий (рис. 212, /), из коих каждая пара дырочек служит
для сообщения амбулакральной ножки с соответствующей ампулой
(см. ниже). Это свидетельствует о том, что амбулакральные пластинки
являются сложными, т. е. слившимися из нескольких простых пла-
стинок.
Оральная мембрана образует по граничащему с ней краю скорлупы
небольшие желобы, которые вдаются между началом радиусов и
интеррадиусов; в этих желобках залегают древовидные наружные
жабры.
Внутренняя поверхность панцыря морского ежа гладкая; более
или менее выдаются только швы между пластинками, прочно сраста-
ющимися друг с другом в неподвижный в своих отдельных частях
панцырь.
Снаружи пластинки скелета морского ежа несут множество окру-
глых возвышений-бугорков с маленькими пуговками на вершине;
к ним-то и причленяются кожные иглы ежа. Благодаря такой скульп-
туре поверхности панцыря, границы между отдельными пластинками
и между рядами их скрадываются, и нужно осматривать препарат
внимательно, чтобы ясно различить радиальные и интеррадиальные
ряды скелетных пластинок.
Бугорки сидят на ареолах—окаймленных площадках скелетных
пластинок; и бугорки и иглы вариируют в размерах. Самые крупные
бугорки и иглы называются первичными; помельче —вторичными, и
наиболее мелкие —миллиарными. Первичные бугорки и иглы ле-
жат относительно правильными меридиональными рядами; мелкие
же образования рассеяны по скорлупе без какого-либо порядка; на-
блюдается, впрочем, некоторая концентрация их вокруг крупных
бугорков. Вообще можно отметить, что бугорки и иглы интеррадиу-
сов крупнее, чем радиусов.
Все скелетные образования у морского ежа, так же как и у мор-
ской звезды, развиваются в тканях среднего зародышевого листка.
Вскрытие морского ежа и его внутреннее строение
Лобзиком распиливают панцырь по экватору и осторожно слегка
разнимают обе половинки панцыря (рис. 215). В щели между ними,
смотря jia свег, замечают идущий отвесно по оси животного тяж, со-
стоящий из каменистого канала (13; и осевого органа. Постепенно
328
разводя половинки панциря далее друг от друга, раскрывают вскры-
того морского ежа и рассматривают его внутренности. Кишечник
обыкновенно разрывается, так как стенки его очень тонки.
У орального полюса морского ежа виден сложно устроенный из
скелетных образований жевательный аппарат (аристотелев фонарь).
Из центра его верхней
поверхности выходит
идущий апикально пи-
щевод (8)', который
вскоре переходит в
кишку. Последняя,
круто загибаясь кни-
зу, поворачивает к бо-
ковой стенке панцыря,
идет по ней против дви-
жения часовой стрел-
ки (10), делая таким
образом круг. Затем
кишка поднимается
кверху, огибает петле-
образно (9) лежащие
в интеррадиусах поло-
вые железы (11) и
в конце концов откры-
вается наружу зад-
непроходным отвер-
стием, лежащим в пе-
рипрокте (6).
Изгибы кишечника
тонкими брыжеечными
нитями (15) прикре-
пляются местами к
внутренней стенке пан-
цыря.
На некотором про-
тяжении кишечник со-
Рис. 215. Вскрытый морской еж. Панцырь распилен
по экватору. По Ж а м м у.
1 — 5 — радиусы с проходящими вдоль них радиальными
амбулакральными каналами; 6 — перипрокт; 7 — ампулы во-
доносной системы; 8 — пищевод; 9, 10— средняя кишка;
7 7—гонады; 12— задняя кишка; 13—каменистый канал и
овоидная железа; 14— средняя кишка; 15 — мезентерий.
неизвестно, быть может сифон служит для
провождается прида-
точной кишкой,или си-
фоном, —тонкой труб-
кой, обоими своими
концами впадающей
в пищеварительный
канал. Назначение ее
дыхания.
Ход кишечника в его естественном положении можно рассмотреть
на хорошо консервированном препарате, если вскрыть морского ежа
специальными приемами. Лобзиком пропиливают панцырь на границе
между каждым амбулакральным и межамбулакральным рядами пла-
стинок так, чтобы верхний конец разреза лежал ниже, генитальной
пластинки на 1,3 см, а нижний конец его спускался бы ниже эква го-
329
риальной пластинки на 1,5 см. Крепким скальпелем удаляют ске-
летные амбулакральные пластинки между краями разреза, так что
в панцыре образуется окошечко. Межамбулакральные ряды, соот-
ветствующие мадрепоровой пластинке, также удаляются в указан-
ных выше пределах, причем предварительно изнутри подрезаются
брыжеечные тяжи прилежащей части кишечника. Препарат рас-
сматривают под водой. Смотря в большое окошко, различают: об-
ширную вторичную полость тела (целом); пищеварительный канал,
идущий в указанном выше направлении; аристотелев фонарь у ниж-
него полюса препарата; каменистый канал, идущий рядом с осевым
органом от мадрепоровой пластинки к аристотелеву фонарю; наруж-
ные и внутренние кровяные лакуны, лежащие по соответ-
ственной стороне кишечника в виде светлых и прозрачных тяжей,
более узких, чем сифон, и соединенных с кишечником брыжейками;
ампулы амбулакральных ножек в виде двойных рядов уплощенных
долек (лежащих на внутренней поверхности амбулакральных рядов)
(рис. 215, 7) и половые' железы (11) у верхнего полюса препарата.
При дальнейшей препаровке разрезают ножом оставшиеся меж-
амбулакральные ряды и разнимают морского ежа на две половины,
которые рассматривают изнутри.
Жевательный аппарат (аристотелев фонарь). На середине внутрен-
ней поверхности нижней половины морского ежа возвышается слож-
ная система скелетных частей, обслуживающая ротовое отверстие
этого животного. При осмотре сбоку видны парные мышцы в форме
.римской цифры V. Основания их лежат у нижнего конца соответ-
ствующего межамбулакрального ряда, а расходящиеся ветви при-
крепляются сбоку к широкому основанию собственно челюсти аристо-
телева фонаря.
При сокращении этих мышц, являющихся протракторами, весь
жевательный аппарат подается вниз.
Обратную работу проделывают их антагонисты —мышцы-ретрак-
торы, идущие от ушковидных пластинок амбулакральных рядов к
нижней, суженной части жевательного аппарата.
В общем аристотелев фонарь похож на пятигранную пирамиду с
выпуклыми сторонами и округлым основанием. Вершина пирамиды
торчит в центре перистома, а основание обращено в полость тела.
При осмотре основания пирамиды in situ различаем пять искри-
вленных узких стерженьков, похожих на компасную стрелку; лежат
они по радиусам (рис. 216, Н, а); их называют дужками (compas);
на одном конце они вилообразно раздвоены. Под ними лежат пять
более широких промежуточных (или межчелюстных) пластинок
(rotulae) (b); называются они также «скобками» (falces) и, как и
дужки, лежат горизонтально; боковыми поверхностями они соеди-
няются с эпифизами (см. ниже), лежащими в основании челюстей,
имеющих форму трехгранных пирамидок (с, d, е). Из полости пира-
мидки с ее основания загибается к оси фонаря конец стержневидного
зуба (f), лежащего соответственно интеррадиусу. Пирамидки и
зубцы расположены вертикально.
Межчелюстные пластинки не доходят до центра основания аристо-
телева фонаря. Остающееся св( бодным пятиугольное по форме про-
СТрйнСТВО затянуто пленкой, на которой лежат кольцевой сосуд
амбулакральной системы и поперечная мышца; из центра же этого
пространства выходит пищевод.
Подрезывают кожицу перистома, протракторы и ретракторы и
освобождают жевательный аппарат, который рассматривают теперь
сбоку. При этом видны, прежде всего, челюсти: поверхность их, смотря-
щая в полость тела, трехугольной формы, с таким же по очертаниям
а
Рис. 216.
отверстием у широкого
основания челюсти. В это
окошечко можно рассмо-
треть стержневидный зуб,
острие которого выдви-
гается из-под вершины
челюсти, а загнутое осно-
вание было рассмотрено
выше.
Между прилежащими
друг к другу челюстями
имеются узкие щели, за-
тянутые межчелюстными
мышцами. При осмотре
межчелюстного аппарата
снизу видны острые кон-
цы зубьев, сходящиеся
в одну точку.
Разрезают скальпелем
межчелюстные мышцы и
разбирают по частям ари-
стотелев фонарь. Рассма-
тривают отдельно стрел-
ки, межчелюстные пла- ь
стинки и челюсти. Вну-
тренние стороны пирами-
док тонко исчерчены (сле-
ды прикрепления межче-
люстных мышц); между
их краями остается про-
дольная щель, в которую
виден по всей мине зуб,
имеющий вид узкой по-
лоски с килем по вогну-
той стороне. Челюсть можно разнять на ее составные части: две
треугольные пластинки (альвеолы) и пару изогнутых эпифизов.
Строго говоря, аристотелев фонарь является скорее хватательным,
нежели жевательным аппаратом. Им морской еж срывает с кам-
ней водоросли и схватывает живую добычу в виде рачков, мелких
червей, губок и т. д.
Амбулакральная система. Амбулакральная система морского ежа
состоит из мадрепоровой пластинки (см. выше, рис. 213, 9); через ее
сито вода поступает в каменистый кйнал, впадающий в околи-
331
а — дужка;
ружи; d —
н
Части аристотелева фонаря морского
ежа. По Ж а м м у.
Ь — скобка; с — челюсть (пирамидка) сна-
то же, снутри; е — то же, сбоку; f—зуб;
Н — аристотелев фонарь (вид сверху).
пищеводное водоносное кольцо. Из последнего идут пять радиаль-
ных каналов, следующих по стенкам панцыря на границе каждой пары
рядов амбулакральных пластинок. От радиальных каналов в стороны
отходят короткие поперечные канальцы, впадающие в ампулы амбу-
лакральных ножек (плоские) (рис. 215, 7). Из каждой ампулы двумя
короткими канальцами начинается амбулакральная ножка.
Есть также и полиевы пузыри, впадающие в околопищеводное кольцо
амбулакральной системы в интеррадиусах.
Амбулакральные ножки на конце своем несут по присоске; сильно
вытягиваясь в длину, ножки морского ежа являются единственным
органом движения, так как части его скелета спаяны неподвижно друг
с другом. Впрочем, некоторые морские ежи также передвигаются,
упираясь подвижными иглами в грунт.
Оторванную амбулакральную ножку вываривают в едком кали;
осадок после промывания микроскопируют. В нем обнаруживаются
мелкие известковые элементы, называемые спикулами; обычна С-об-
разная форма спикул (рис. 217).
Органы размножения морского ежа со-
стоят из пяти гонад (рис. 215, 77) —половых
желез,- лежащих в интеррадиусах и откры-
вающихся отверстиями в генитальных пла-
стинках аборальной поверхности (рис. 213, 5).
Эти животные раздельнополы, как и мор-
ские звезды, причем наружных отличитель-
ных признаков для полов нет. Оплодотворе-
ние и развитие наружное.
Педицеллярии. Оторванную педицеллярию
рассматривают под микроскопом в свежем
виде или же изготовив обычным путем по-
репарат.
В педицеллярии различают стебелек (рис. 218, I) и сидящие на
его вершине створки, или щипчики, состоящие из трех частей. Щип-
чики образуют головку педицеллярии. Между головкой и стержнем
(стебельком) бывает мягкая шейка (рис. 218, А, 11).
По форме щипчиков педицеллярии разделяются на трехпалые
(«тридентные»), или хватательные (с длинными и утончающимися к вер-
шине ветвями, рис. 218, А), и лопастные, или железистые, или глоби-
ферные, педицеллярии с сильно расширенной головкой; ее короткие
створки имеют в себе особые ядовитые железы; верхняя часть створок
вытянута в зубец (рис. 218, В). Бывают у морских ежЯ^педицеллярии
и другой формы, похожие на головы у змей или на листок.
и Створки и ветви щипчиков приводятся в движение особыми мыш-
цами (рис. 218, A, MS), подвижны и самые педицеллярии, глав-
ным образом трехпалые, снабженные гибким стебельком.
Педицеллярии служат для очистки тела от приставших к нему
предметов; железистые же педицеллярии считаются своего рода ядо-
витыми органами.
Педицеллярии играют существенную роль при очистке поверх-
ности тела от экскреме шов, которые неминуемо его загрязняют, так
как анальное отверстие расположено на верхнем полюсе тела морского
332
Рис. 217. Спикулы из
амбулакральной ножки
Stronoyiocentrotus droe-
bachiensis. По Кларку.
стоянный бальзамный
вздутыми) мелкими
видоизмененными
MS
По А. Дьяконову.
Рис. 218. Педицеллярии.
А — трехпалая педицеллярии Echinus esculen-
tus; В — глобиферная педицеллярия Stron-
gylocentrotus intermedius. I — стержень; II— шейка; III —
головка; v — створка; MS—мышца створок.
ежа. Педицеллярии размельчают кусочки экскрементов, благодаря
чему они легко смываются водой.
Сферидии. Очищают околоротовую часть тела морского ежа от
игол и педицеллярий. При внимательном рассмотрении под лупой
можно заметить в рядах амбулакральных пластинок, в ближайшем
соседстве с ротовым полем, очень маленькие сферические тела, сидя-
щие на короткой ножке; называются они сферидиями. Морфологи-
чески сферидии счит
иглами. Физиологиче-
ское значение их не-
ясно — их считают то
органами равновесия
(статоцисты),то обоня-
ния. Сферидии встре-
чаются только у мор-
ских ежей, за исклю-
чением немногихформ.
Общие замечания
Морской еж обла-
дает всеми признака-
ми, которые характе-
ризуют тип иглоко-
жих (ср. выше, стр.
324). От морской зве-
зды он отличается
округлым телом, ли-
шенным каких-либо
выростов (в частности рук). Эта особенность, а также состав кожного
скелета из двадцати рядов плотно сросшихся пластинок, длинный
трубчатый кишечник без печеночных придатков и жевательный аппа-
рат 1 (аристотелев фонарь) характеризуют класс морских ежей—
Echinoidea.
Положение заднепроходного отверстия на верхнем полюсе тела,
хорошее развитие аристотелева фонаря, правильная более или менее
шарообразная форма тела и одинаковые размеры амбулакральных
рядов присущи представителям отряда правильных ежей—Regula-
ria, или точнее—отряда Diadematoidea, к которым и принадлежит
рассмотренный нами Strongylocentrotus.
В связи с изменениями формы тела стоят различные особенности
строения морского ежа. У него нет амбулакральных борозд, которые
заменены рядами поверхностно лежащих амбулакральных пластинок.
Таким образом у морского ежа находим лучевую симметрию тела,
хотя и не так явно бросающуюся в глаза, как у морской звезды.
Благодаря боковому положению мадрепоровой пластинки, лучевая
симметрия переходит в билатерально-лучевую, так как скелет мор-
ского ежа может быть разделен на действительно симметричные поло-
1 У неправильных морских ежей он редуцирован или сильно изменен.
333
вины лишь плоскостью, рассекающей мадрепоровую пластинку на
две половины. В отношении симметрии внутренних органов отметим,
что она нарушается спиральным ходом трубкообразного кишечника и
сдвигом в сторону от вертикальной оси тела каменистого канала. Все
остальные органы (амбулакральная система, нервная система^ гонады
и др.) расположены радиально-симметрично.
Форму тела морского ежа можно вывести из морской звезды, если
представить себе, что руки звезды загнулись на спинную сторону
тела и слились с последней, причем пространство между руками
(радиусами, парными рядами амбулакральных пластинок) превра-
тились в межамбулакральные ряды.
Для представителей огряда Diadematoidea, кроме указанных выше
признаков, характерны: наличие в ротовом поле пяти пар амбула-
кральных и отсутствие интерамбулакральных пластинок; наличие
наружных жабр, сферидий и жевательного аппарата; расширение
верхнего конца стержня стебелька педицеллярий; наличие первич-
ных бугорков на всех скелетных пластинках.
Следующие признаки характеризуют рассматриваемого нами мор-
ского ежа как представителя семейства Strongylocentrotidae: значи-
тельная сплюснутость тела в дорзо-вентральном направлении; наличие
на каждой амбулакральной пластинке более трех пар пор; прямая
или изогнутая палочковидная форма спикул амбулакральных ножек
с разветвлением на концах (у представителей семейства Echinidae —
спикулы в виде буквы С); наличие длинной шейки у глобиферных
педицеллярий, створки которых оканчиваются каждая одним конце-
вым крючком и лишены боковых зубцов.
У вида Str. droebachiensis каждая амбулакральная пластинка не-
сет четыре-семь пар пор (эхиноидный тип амбулакральных пластинок):
створки глобиферных педицеллярий крупные, вытягиваются в длин-
ную трубчатую часть, несущую короткий прямой концевой крючок.
Диаметр тела достигает 8—9 см.
Морской еж является придонным животным, медленно передвига-
ющимся при помощи амбулакральных ножек.
И для морского ежа возникновение амбулакральной системы
тесно связано с условиями жизни в морской воде. Амбулакральная
система является специфической принадлежностью иглокожих и
ни в каком другом типе животных более не встречается.
Благодаря прочности известкового наружного скелета и крепкой
спаянности его пластинок, морские ежи хорошо сохраняются в виде
окаменелостей, находимых в геологических отложениях различной
древности.
РАЗВИТИЕ ИГЛОКОЖИХ
Эмбриональное развитие морского ежа. Материал с одной из север-
ных биологических станций. Изучают тотальные, окрашенные бор-
ным или квасцовым кармином бальзамные препараты при среднем
увеличении микроскопа.
Яйцо морскою ежа шаровидное, бедное желтком, одето прозрач-
ной оболочкой.
334
Дробление полное и сначала равномерное, продукты деления
оплодотворенного яйца называются бластомерами. Находят
стадии: двух, четырех, восьми и более бластомер. Бластомеры сохра-
няют округлую форму. По мере увеличения их количества, между
ними обособляется полость дробления. Бластомеры в дальнейшем
отходят к периферии, своими боковыми частями плотно прилегая
друг к другу. На этой стадии зародыш морского ежа имеет вид одно-
слойного пузыря, или бластулы; стенка его образована клетками
характера цилиндрического эпителия (бластодерма). Полость бла-
стулы называется бластоцелем.
Далее бластула проходит в стадию гаструлы; на полюсе бластулы,
называемом вегетативным, стенка, благодаря усиленному делению
клеток, начинает впячиваться внутрь, врастая в полость бластулы.
Таким образом однослойный пузырь превращается путем инвагинации
в двуслойный мешок или колокол; эта стадия и называется гаструлой.
В гаструле различают: 1) наружный слой, или эктодерму;
2) отверстие, ведущее в гастральное впячивание, или в архэнтерон,
или в первичную кишку; 3) полость гастрального впячивания, или
гастроцель; 4) стенку гастрального впячивания, образуемую энто-
дермой, или внутренним зародышевым листком, и 5) остаток бласто-
целя между экто- и энтодермой гаструлы.
В дальнейшем происходит образование мезенхимы, закладка цело-
мических' мешков среднего зародышевого листка, или мезодермы,
(образование вторичной полости тела, или целома), и формирование'
личинки морского ежа: но эти процессы на тотальных препаратах
с достаточной ясностью проследить трудно.
Развитие морского ежа связано с метаморфозом, так как ли-
чинка его имеет совсем другое строение, чем взрослая форма. То же
имеет место у других иглокожих, в чем можно убедиться, рассматри-
вая их личинки.
Личинки иглокожих. Личинок иглокожих выводят в лаборатории,
если есть возможность иметь живой материал. Личинки иглокожих
плавают в составе планктона, иногда доминируя численно над дру-
гими существами. Планктонный материал более доступен для исполь-
зования. Личинок отбирают под сильной лупой или при слабом уве-
личении микроскопа и приготовляют из них тотальный бальзамный
препарат, предварительно окрасив квасцовым кармином.
Личинка морского ежа (Pluteus). Тело личинки
конусовидной формы. Широкая часть его имеет прямые выступы,
называемые руками. У молодой личинки четыре руки, а у взрослой —
восемь. По краю рук идет венчик или шнур ресничных клеток; при
помощи движения ресничек личинка плавает. Руки и тело личинки
подперты скелетными образованиями в виде известковых игол. В са-
мом теле виден кишечник и могут быть видны зачатки полости тела
(целомические пузыри). Плутеус морского ежа называют Е с h i no-
pluteus (рис. 219, А).
Личинка офиуры (змеехвостки) по общему виду соответ-
ствует типу Pluteus и, в отличие от личинки морского ежа, называется
О р h i о р 1 u t е u s. В ней сильно развиты руки, вытянутые в сто-
роны. Тело по отношению к размерам рук мало. В нем можно виде1ь
за?
кишечник, закладку целомических пузырей и водоносной системы
(рис. 219, В).
Личинка морской звезды, или бипиннария,
имеет приблизительно клиновидную форму с сильно рифлеными боко-
выми краями вследствие образования округленных выростов, или рук,
Рис. 219. Личинки иглокожих. А — С — по Делажу и Эруару. D — по
Шимкевичу.
А — личинка морского ежа (Pluteus) Paracentrotus lividus. d — скелет дорзальной
руки; v — скелет вентральной руки; а, с — скелет тела. В — личинка офиуры (Ophio-
pluteus). а — anus; b — рот; d— дорзальная рука; / — фронтальное поле; I — боковая рука;
v — вентральная рука. С — личинка морской звезды — бипиннария. d.i —
нижнедорзальный отросток; d md—срединный дорзальный отросток; d.s — верхний дорзальный
отросток; / — нижний боковой отросток; рг о — предротовой боковой отросток; pst, о — зарото-
вой отросток; v md — передротовой срединный отросток. D — аурикулярия, ли-
чинка голотурий. 7 и 6 — ресничный венчик; 2 — пора гидроцеля; 3 — гидроцель;
4 — передняя кишка; 5 — средняя кишка; 7 — задняя кишка; 8 — целомический пузырь;
9 — известковые тельца; 10 — пищевод; 11 — целомический пузырь.
которые очень коротки. При рассматривании личинки с брюшной сто-
роны различают среднюю дорзальную руку; далее кзади лежат после-
довательно пара преоральных рук, пара переднедорзальных рук,
пара заднедорзальных рук, пара посторальных рук и на задненаруж-
ных концах личинки пара заднебоковых рук.
336
Мерцательный шнур окаймляет края рук, частично заходя и на
спинную сторону; на брюшной стороне ресничный шнур распадается
на два участка — преоральный (малый) мерцательный шнур, лежащий
обособленно и замкнуто, и посторальную петлю шнура, идущего с боко-
вых рук, и пересекающую брюшную поверхность личинки поперек
(рис. 219, С).
Из внутренних органов видны: stamodaeum (рот), более узкий
пищевод, округлый желудок и задняя кишка с анальным отверстием.
По бокам пищеварительного аппарата лежат удлиненные целомиче-
ские пузыри. В дальнейшем бипиннария получает длинные отростки,
или руки; в такой стадии метаморфоза она называется брахиоля-
р и е й.
Личинка голотури и — а урикуляри я—имеет ши-
рокое тело с округлым передним концом и почти поперечно обрезанным
задним концом тела. В профиль форма тела овальная. Рот лежит на
дне брюшного впячивания. Мерцательный шнур окаймляет ротовое
вдавление и образует сложные извивы (рис. 219, D).
Личинка морской лилии имеет боченкообразную форму;
она несет несколько поперечных мерцательных поясков и пучок длин-
ных теменных ресничек. В толще тела личинки видны нежные узорча-
тые скелетные пластинки. Нет ни рта, ни анального отверстия.
Личинки иглокожих имеют двубоковую симметрию, которая нару-
шается в процессе преобразования личинки во взрослую форму. Они
являются планктонными организмами, так как живут, плавая в по-
верхностном слое воды открытого моря. Взрослые иглокожие, на-
оборот, являются бентоническими (донными) обитателями.
Лучевая симметрия иглокожих является особенностью, унаследо-
ванной от предков, ведших сидячий образ жизни. Свободно подвижной
образ жизни большинства иглокожих ведет как бы к некоторому вос-
становлению двубоковой симметрии; элементы ее были показаны выше
при оценке степени чистоты лучевой симметрии рассмотренных выше
представителей иглокожих.
Все иглокожие являются жителями морей. Иглокожие предста-
вляют замкнутый тип животных, происхождение которого и по сие
время остается неясным.
ТИП ХОРДОВЫЕ (CHORDATA)
ПОДТИП БЕСЧЕРЕПНЫЕ (ACRANIA
s. CEPHALOCHORDA)
ЛАНЦЕТНИК (BRANCHIOSTOMA LANCEOLATUM)
Технические замечания. Ланцетник встречается в некоторых
местах Черного моря, в частности вблизи Севастополя. Живет в песчаном грунте
дна, в который зарывается всем телом, оставляя снаружи только передний конец.
Материал для занятий в фиксированном виде можно добыть на Севастопольской
биологической станции.
Заспиртованных крупных ланцетников рассматривают в часовом стекле в 70%
спирту под лупой. Из ланцетников небольших размеров приготовляют тотальный
препарат после обезвоживания в абсолютном спирту и просветления в ксилоле.
Препарат заключают в канадский бальзам.
Фиксированных сулемой с уксусной кислотой ланцетников разрезают в по-
перечном направлении на кусочки, которые или прямо заливают в парафин, или
предварительно окрашивают борным кармином. В первом случае нанесенные на
стекло парафиновые поперечные срезы окрашивают гематоксилином Делафильда
и эозином.
Срезы приготовляют из жаберной части тела и из зажаберного отдела. Для
детального изучения необходимы полные серии срезов, особенно переднего конца
тела.
Изучение тотального препарата
В консервированном состоянии тело — белое; на бальзамном препа-
рате — желтовато-красноватое, полупрозрачное.
Форма тела при рассматривании в профиль имеет вид удлиненного,
сплюснутого с боков веретена (рис. 220). Один его конец равномерно
заострен — это задний конец тела. Передний конец снизу косо срезан
и несет щупальца, окружающие вход в жаберную полость (1). Брюш-
ная сторона несколько более выпукла, чем спинная. По спинной
стороне идет спинной плавник. Задний конец тела окаймлен хвосто-
вым плавником, имеющим сходство по форме с ланцетом. Снизу и
сбоку передних двух третей тела идут продольные складки, или ме-
таплевры, по одной с каждой стороны. Их можно наглядно видеть на
поперечных срезах.
При рассматривании тотального бальзамного препарата под микро-
скопом можно хорошо видеть метамерную исчерченность тела, завися-
щую от характера сегментации боковой мышцы ланцетника, состоя-
щей из надломленных под углом миомер, вершиной обращенных
кпереди (7). Ясные линии, разделяющие миомеры друг от друга, явля-
ются соединительно тканными прослойками, называемыми миосептами.
338
При просматривании всей толщи препарата можно увидеть длинный
веретеновидный тяж, идущий от переднего конца тела до заднего.
Этот тяж характерен вертикальной параллельной исчерченностью;
он является первичным внутренним скелетом, или хордой (12). Тотчас
над ним лежит спинной мозг (9), но на тотальном препарате он закрыт
толщей миомер и поэтому обычно не виден. Все же можно заметить
пигментные точки, которые являются глазками, заложенными в толще
спинного мозга (глазки Гессе).
Далее останавливаемся на косой исчерченности брюшного отдела
передней половины тела; здесь проходит жаберная часть кишечной
трубки, прорезанная косыми жаберными щелями (13). Далее кзади с
брюшной стороны тела можно рассмотреть кишечную трубку, особенно
если она набита пищевой массой более темного цвета.
Рис. 220. Ланцетник, общий вид сбоку. Из Гюнтера.
1 — ротовое отверстие; 2 и 3 — около^каберная полость; 4— брюшная пора; 5 — заднепроход*
ное отверстие; о—спинной и брюшной плавники; 7 — мышечные сегменты; 8 — спинной
кровеносный сосуд; 9— спинная нервная трубка; 10— глаз; 11 —cirri; 12 — спинная струна
(хорда); 13 — жаберные перегородки; 14— печень.
Поперечный срез на уровне жаберной полости
На поперечном срезе в области жаберной полости (рис. 221, А)
находим следующие части: спинной плавник в виде тупого кнопкооб-
разного выроста (d), иногда сдвинутого в сторону. Книзу от него
бока препарата заняты-наиболее массивными компактными образова-
ниями в виде тесно прилежащих друг к другу полей в общем четырех-
угольной формы, с выпуклыми краями; это — разрезы вдвинутых друг в
друга миокомм боковых мышц тела (т); на спинной стороне миокоммы
обеих половин тела соприкасаются друг с другом; ниже между ними
располагается спинной мозг( п), на поперечном срезе имеющий почти
округлотрехугольную форму.
Под спинным мозгом лежит хорда (ch) в виде крупного овального
тела с поперечной исчерченностью; ниже хорды миокоммы расходятся
в стороны; от хорды почти до самой брюшной стенки тела идут два
ряда образований в виде поперечно лежащих стерженьков, не соеди-
ненных друг с другом; срединное пространство между обоими рядами
их является полостью жаберного отдела кишечника, сами стерженьки—
разрезами жаберных перегородок (Ьг), а пространства между ними —
жаберными щелями.
По верхнему краю жаберной полосы идет продольный наджабер-
ный желобок (е); его антиподом является поджаберный желобок, или эндо-
стиль (еп), состоящий из высоких железистых и мерцательных клеток.
Под эндостилем лежит жаберная артерия. Между жаберными перего-
• 339
родками и стенкой тела имеется полость, называемая околожаберной
(или перибранхиалънои) (pb) ; в одной из ее половин видно неправильно
овоидное образование, состоящее из однослойного высокого цилиндри-
ческого эпителия, окружающего полость; это — поперечный разрез
Рис. 221. Поперечный разрез ланцетника: А—через жаберную полость; В—на
уровне желудка и печени. Из Краузе.
d — спинной плавник; т — мышцы; л — спинной мозг; ch — хорда; Л — печень; е — над-
жаберный желобок; Ьг — жаберные стержни; Ьс — жаберная полость; ел — поджаберный же-
лобок (эндостиль); mt — метаплевры; ov— яичник; i — кишечник; pb — перибранхиальная
полость.
печеночного выроста (или печени—h), отходящего от кишечника и
вдающегося в околожаберную полость.
В зависимости от половозрелости ланцетника, в нижнебоковой части
перибранхиальной стенки бывают видны гонады (ov) в виде овоидных
или почковидных (на разрезе) массивных образований; по их содержи-
мому можно определить пол ланцетника: у самок они заполнены круп-
ными яйцевыми клетками, у самцов содержимое их мелкозернистое.
340
Зрелые половые продукты через разрыв стенки гонад поступают
в околожаберную полость.
По бокам нижнего края препарата видны метаплевры в виде
боковых складок стенки тела ланцетника (mt).
Поперечный разрез в области кишечника
Поперечный разрез ланцетника в области кишечника отличается
отсутствием метаплевр и наличием под хордой ограниченного вторич-
ной полостью тела кишечника. По общему строению он сходен
с печенью. На препарате по своему развитию доминиоуют мышцы
(рис. 221, В).
Общие замечания
Пищеварительный аппарат начинается отверстием, ведущим в объ-
емистую переднюю часть кишечника, боковые стенки которого изре-
заны большим количеством жаберных щелей. В своем заднем конце
жаберная часть пищеварительного канала переходит в кишечник; от
последнего кпереди тянется печеночный отросток; кишечник — пря-
мой; он открывается’анальным отверстием в основании брюшной части
хвостового плавника.
Ланцетник вбирает воду в жаберную полость: здесь взвешенные
в ней мелкие организмы приклеиваются к слизи эндостиля, которая
движением ресничек гонится по направлению ко входу в кишечник.
Таким путем пища точно по конвейеру подается в пищеварительный
канал.
Вода прогоняется далее через жаберные щели в околожаберную
полость. При прохождении воды растворенный в ней кислород обо-
гащает кровь, протекающую в сосудах жаберных щелей; одно-
временно происходит отдача углекислоты. Вода из околожаберной
полости выходит через брюшную пору (лежащую перед брюшным
плавником) наружу (рис. 220, 4). У ланцетника налицо жаберное
дыхание.
Следует подчеркнуть характерную особенность, заключающуюся
в том, что жаберным аппаратом ланцетника является соответственно
видоизмененная передняя часть пищеварительной трубки, тогда как
у ракообразных, червей и моллюсков жабрами являются выросты или
складки покровов.
Весьма характерными особенностями строения ланцетника яв-
ляются :
1) спинное положение центральной нервной системы (спинной мозг);
2) наличие первичного (внутреннего) скелета в виде хорошо раз-
витой хорды;
3) отсутствие черепа, позвоночного столба и парных конеч-
ностей.
Вследствие наличия у ланцетника хорды его относят к типу хор-
довых (Chordata); в этом типе он занимает место в подтипе Acrania,
или бесчерепных. Это — весьма немногочисленный по составу своему
подтип, ибо к нему принадлежат всего два рода одного семей-
ства.
341
ПОДТИП ПОЗВОНОЧНЫЕ (VERTEBRATА)
КЛАСС РЫБЫ (PISCES)
КОСТИСТЫЕ РЫБЫ (TELEOSTEI)
КОЛЮЧЕПЕРЫЕ РЫБЫ (ОК^НЬ, ЕРШ, СУДАК ЩУКА)
Предварительные замечания. Окунь (Perea fluviatilis), ерш
(Acerina сегпиа), судак (Lucioperca sandra) и щука (Esox lucius) относятся к от-
ряду костистых рыб (Teleostei) класса Pisces. Рыбы являются водными позвоноч-
ными и в типе Vertebrata занимают низшее место.
Окунь, ерш и судак — обычные представители подотряда колючеперых
(Acanthopterygii), тогда как щука вместе с карасем, красноперкой, линем, кар-
пом и другими рыбами принадлежит к подотряду отверстопузырных (Physostomi),
у которых плавательный пузырь соединяется каналом с желудочно-кишечной
трубкой.
Общее строение этих рыб довольно сходно; поэтому в основу описания мы
положим анатомию окуня, причем, по мере надобности, сделаем замечания и
о строении других рыб.
Для занятий обыкновенно берут «снулую» рыбу. При покупке выбирают
экземпляры с розовыми жабрами и прозрачными глазами.
Наружный осмотр
Тело окуня веретенообразное, сдавленное с боков; оно состоит из
трех отделов: головы, туловища и хвоста. Границей между головой и
туловищем служит жаберное отверстие, а хвост начинается кзади
от заднепроходного отверстия.
Голова плотно срастается с туловищем, которое незаметно пере-
ходит в хвост. Крепость соединения частей тела рыбы в связи с ее
формой соответствует условиям передвижения в воде. Вода является
средой плотной, которая оказывает значительное сопротивление дви-
жущемуся в ней телу. Заостренная на конце и сплющенная с боков
голова рыбы способствует рассеканию воды. Удобству скольжения
в ней способствует сдавленное с боков тело. Голова должна быть крепко
соединена с телом и должна быть малоподвижной, в противном случае
при поворотах вода ее может свернугь на сторону.
Орудиями плавания являются конечности рыбы, называемые плав-
никами; их два типа — парные и непарные.
Непарные плавники лежат по средней линии тела: на спине поме-
щаются передний и задний спинные плавники. Сзади тело заканчивается
вертикально стоящим хвостовым плавником, а между основанием по-
следнего и анальным отверстием на брюшной стороне лежит короткий
заднепроходный плавник.
Парных плавников две пары: грудные, лежащие по бокам тела сзади
жаберной крышки, и сближенные друг с другом у брюшной линии
тела брюшные плавники.
Чтобы рассмотреть строение плавника, надо его расправить, взяв
пинцетом или рукой за край и оттянуть по мере возможности.
Плавник состоит из перепонки, натянутой между костными лучами.
Лучи образованы или одиночными острыми косточками (простые лучи
342
или твердые), или же мягкими разветвленными и членистыми стерж-
нями.
Первый спинной плавник состоит из 14—15 острых лучей; второй
спинной и анальный имеют только два колючих луча, остальные же —
мягкие. Хвостовой и грудные плавники построены из мягких стерж-
ней, а брюшные плавники имеют по одному колючему и по несколько
мягких лучей.
Число, положение и состав плавников у разных рыб неодинаковы;
это имеет важное значение в систематике рыб.
У ерша оба спинных плавника сливаются вместе, хотя граница
между ними остается заметной, так как лучи второго плавника (их
И—14 штук)* гораздо выше задних стержней (их 12—14) первого
спинного плавника.
У судака оба спинных плавника то совершенно разделены, то соеди-
няются перепонкой друг с другом.
Благодаря тому, что в плавниках всех этих рыб имеются острые
твердые лучи, названных выше рыб и объединяют в подотряд колюче-
перых.
Щука обладает лишь одним спинным плавником, а брюшные ее
плавники далеко отодвинуты от грудных, так как лежат ближе к аналь-
ному отверстию, чем к голове. В противоположность колючеперым,
все плавники щуки (как и других отверстопузырных) обладают мяг-
кими лучами.
Главнейшую роль в движении рыбы играет хвостовой плавник.
Ударяя им в стороны, рыба толкает свое тело вперед и движется по-
добно тому, как плывет лодка при работе только кормовым веслом.
Парные плавники играют роль рулей и способствуют сохранению
равновесия тела. Если у рыбы отрезать грудной и брюшной плавники
с одной стороны тела, то животное валится на оперированную сторону.
Некоторые рыбы, например, карпы, могут медленно плыть, работая
только парными плавниками.
Спинные и анальный плавники увеличивают боковую поверхность
тела и способствуют сохранению отвесного положения тела в воде,
другими словами, имеют значение киля лодки. Если отрезать у рыбы
эти плавники, то она плывет не по прямой линии, как обычно, а по
ломаной.
Основания лучей плавников снабжены мускулами, благодаря дей-
ствию которых плавники то раскрываются, то складываются.
Голова рыбы по переднему краю несет широкое ротовое отверстие,
ограниченное верхней и нижней челюстями. Над первой помещаются
две пары носовых отверстий, из коих передняя пара снабжена кожными
клапанами.
По бокам головы лежат два больших глаза, а по поверхности ее
рассеяны мелкие точечные отверстия, ведущие в каналы кожных орга-
нов чувств.
При осмотре головы бросаются в глаза лежащие сверху наклад-
ные кости черепа, образующие две боковые жаберные крышки. Нижние
и задние края их свободны. Если крышку приподнять, то обнаружи-
вается жаберная полость, в которой лежат четыре дугообразные
жабры. Между ними имеются пять жаберных отверстий, ведущих
343
в ротовую полость. Снизу к крышечке прилегает кожная складка,
подпертая костными лучами, отходящими от подъязычной кости.
Кожа рыбы слизистая, благодаря нахождению в толще ее эпители-
альных слоев множества одноклеточных слизистых железок. В соеди-
нительнотканной части кожных покровов заложены тонкие костные
пластинки — чешуйки, черепицеобразно налегающие друг на друга и
покрывающие почти все тело рыбы. Число рядов чешуй в различных
частях тела рыбы имеет систематическое значение. Наличие чешуек
в коже рыбы обусловливает большую прочность кожных покровов.
Чешуйки можно считать за остатки наружного, или кожного, скелета.
Благодаря слизистым выделениям кожа рыбы очень скользка, вслед-
ствие чего живую рыбу трудно удержать в руках. Кроме того слизь
выделяется в изобилии, если животное выбрасывается из воды на сушу.
Толстый слой слизи, покрывающий в таком случае рыбу, препятствует
(до некоторой степени) немедленному ее высыханию (и гибели).
По бокам тела от жаберного отверстия до хвоста тянется почти
параллельно линии спины так называемая боковая линия. Она образо-
вана кожным каналом, который открывается наружу правильно чере-
дующимися отверстиями. Боковая линия иннервируется боковой
ветвью блуждающего нерва, оканчивающейся в особых бугорках,
и является органом какого-то специфического чувства. Внутри она
заполнена слизью.
Опыты со щукой, у которой нервы боковой линии были перере-
заны, а продолжающиеся на голову каналы выжжены электричеством,
показали, что оперированная рыба не ощущает действия слабого тока
воды, направляющегося на ее тело.
Когда щука подплывает к какому-нибудь твердому предмету, то
от последнего идет отраженная волна воды; у нормальной рыбы она
ударяется о тело, давит на слизь боковых каналов, которая, в свою
очередь, действует на волоски чувствительных клеток.
Производимое ею раздражение воспринимается щукой, и она на него
реагирует тем, что заблаговременно обходит твердый предмет и избе-
гает столкновения с ним. Ослепленная щука, у которой цела боковая
линия, не натыкается на встречные препятствия, тогда как рыба без
глаз и без боковой линии в этом отношении совершенно беспомощна.
В общем этот специфический для рыб орган мы можем считать за орган
восприятия давления воды — орган, позволяющий рыбе ориентиро-
ваться при плавании в мутной воде или ночью. Очень возможно, что
боковая линия дает рыбе возможность определять также силу течения
и сообразовать с ней работу своих плавников.
Долгое время этот орган был известен под ничего не говорящим
названием органа шестого чувства.
Перед анальным плавником лежит заднепроходное отверстие, позади
которого помещается ямка с отверстиями органов размножения и
выделения.
Вскрытие* рыбы и ее внутреннее строение
Берут рыбу в левую руку и, держа ее брюхом кверху, перерезают
коротким поперечным разрезом брюшную стенку тела сейчас же кпе-
реди от анального отверстия. В прорез вводят тупую ветвь ножниц и
344
разрезают брюшную стенку по средней линии вплоть до заднего края
ротовой щели (на уровне глаз). По пути перерезают опорные косточки
плавников и жабр. Затем приподнимают стенку левой стороны тела и
от анального отверстия рассекают ее ножницами, придерживаясь под
контролем глаза (чтобы не прорвать плавательный пузырь) края
полости тела. Разрез не доходит до боковой линии, поворачивает далее
вперед параллельно ей и идет до заднего угла жаберной крышки.
Последнюю приподнимают
кверху и разрезают идущую
отвесно под ней кость; далее
рассекают, направляясь книзу,
перепонку жаберной полости
и доходят до переднего конца
брюшного разреза. После этого
снимают вырезанную стенку
тела, отрезают заднюю часть
жаберной крышки, кладут
рыбу на правый бок, прика-
лывают булавками ко дну ван-
ночки и наливают воды так,
чтобы она покрыла вскрытое
животное (рис. 222).
Под спинной линией раз-
реза лежит тонкостенный пла-
вательный пузырь (рис. 222, vn);
под ним помещаются органы
пищеварения — большая пе-
чень (hp), мешкообразный же-
лудок (vt), из которого выхо-
дит тонкая кишка, образую-
щая петли (еп). Между коле-
нами последней лежит селе-
Рис. 222. Анатомия окуня. Схематично. По
Н. X о л од к о вс к о м у, с упрощениями.
bra — жаберная полость с жабрами (br)- at — пред-
сердие^— желудочек; Ьа — bulbus aortae; tra—
брюшная аорта; га — корни спинной аорты; vca—
передние кардинальные вены; de—кювьеров про-
ток; sv — венозный синус; vt—желудок; арр —
пилорические придатки; еп —тонкая кишка; I —
селезенка; hp — печень; gn — половая железа;
vn — плавательный пузырь; гп — почка; vs—моче-
вой пузырь; br — жаберная дуга. д
зенка (/) — плотный орган
красноватого цвета.
Между плавательным пу-
зырем и кишкой помещаются
половые железы (gn) в виде
мешков (два или один, в зави-
симости от пола и вида рыбы),
открывающихся половым от-
верстием позади заднепроходного отверстия. Внешний вид половых
желез зависит от степени половозрелости рыбы.
Со спинной стороны заднего конца половых желез лежит неболь-
шой мешочек — мочевой пузырь (ns).
Передняя часть вскрытой полости тела, соответствующая горлу
рыбы, занята сердцем. Оно очень сдвинуто вперед. Искать его следует
перед печенью в близком соседстве жабр. При наружном осмотре
невскрытой рыбы место его определяется положением узкого мыса
брюшной стенки тела на нижней поверхности между жаберными
крышками.
345
Сердце рыбы состоит из предсердий (at), желудочка (и) и примы-
кающей к нему луковицы брюшной аорты (Ьа).
Органы пищеварения начинаются ртом, который окружен челю-
стями. При раскрывании рта челюсти, состоящие из нескольких под-
вижных костей, выдвигаются вперед в виде футляра, что способствует
захватыванию добычи, так как при этом увеличиваются размеры
пасти рыбы. Замечено, что ротовое отверстие велико у хищных рыб
и сравнительно мало у нехищных.
Почти все кости ротовой полости усеяны мелкйми коническими
зубами. Они имеются на челюстях, на нёбе и в глотке. Окунь хва-
тает добычу целиком, и зубы служат лишь для ее удержания. Верх-
ние зубы щуки соединяются с костями соединительной тканью, бла-
годаря чему они обладают подвижностью: при npoiлатывании пищи
они нагибаются назад. Возможно, что нижнеглоточные зубы карпо-
вых рыб служат для перетирания растительной пищи.
На дне ротовой полости лежит короткий язык в виде слизистой
оболочки, одевающей передний конец соединительного скелета жа-
берных дуг (os entoglossum). Благодаря этому, язык не имеет совер-
шенно самостоятельного движения. У щуки он усеян мелкими зубами.
Нижнезадняя стенка ротоглоточной полости образована прилежа-
щими друг к другу жаберными дугами, вогнутая поверхность которых
несет множество стерженьков, называемых жаберными тычинками.
Совокупность их образует «цедильный аппарат», служащий для
фильтрования воды, пропускаемой через жабры при дыхании. При
этом задерживаются всякие мелкие животные, которые и препро-
вождаются в воронкообразный пищееод. Его толстостенная складча-
тая внутри трубка впадает в объемистый желудок (рис. 222, vf), имею-
щий вид мешка, дно которого обращено назад. От нижней поверхности
желудка (ближе к переднему его концу) отходит кишка, принимаю-
щая в себя три короткие слепые трубочки, называемые пилорическими
придатками — appendices pyloricae (рис. 222, арр).
У щуки их нет, и желудок переходит в кишку в своей задней
части.
Пилорические придатки служат для увеличения пищеварительной
поверхности кишечника; у окуня один из них принимает в себя ко-
роткий проток желчного пузыря.
При нормальном положении органов пилорические придатки и
ббльшая часть желудка прикрываются печенью (hp). Ткань ее обычно
дрябла и легко разрывается. На внутренней, прилежащей к желудку
поверхности печени помещается грушевидный желчный пузырь.
’возвращаясь к ходу кишечника, отметим, что тонкая кишка (еп)
делает один дугообразный изгиб. Кишка подвешена на брыжейке,
в которой отлагаются жировые дольки. Тонкая кишка без особенно
резкой границы переходит в несколько более толстую заднюю кишку;
внутренней границей между ними является поперечная складка слизи-
стой оболочки. Задняя кишка направляется прямо к заднепроходному
отверстию.
Поджелудочная железа развита хорошо у щуки; она образована
рыхлой желто-бурой тканью, лежащей между кишечником и желуд-
ком. Выводной проток ее открывается рядом с желчным. У других
346
костистых рыб поджелудочная железа развита слабо, так как состоит
из мелких долек железистой ткани, заложенных между пластинками
брыжейки или же между участками ткани печени.
Плавательный пузырь окуня представляет собой замкнутый ме-
шок (рис. 222, vri) с плотными кожистыми стенками. У щуки же (как
и у плотвы) он соединяется с верхней (спинной) стенкой пищевода (рис.
223, 7—2). У обеих рыб он развивается из выпячивания пищевода,
обособляющегося от пищеварительной трубки у окуня и, наоборот,
пожизненно остающегося в связи с ней у щуки. Следовательно, не-
смотря на такое отличие в строении этого органа у названных рыб,
плавательные пузыри обеих являются органами гомологичными.
Плотная серебристо-блестящая стенка его построена из соедини-
тельной ткани и гладких мышц; благодаря сокращению последних,
емкость пузыря может изменяться.
Рис. 223. Плотва с плавательным пузырем. По Гессе и До-
ф л е й н у.
7—пищевод; 2 — канал плавательного пузыря; 3'—3 — плавательный пузырь.
В брюшной части переднего конца вскрытого пузыря видны так
называемые красные тела — участки стенки, обильно орошаемые
кровеносными сосудами, образующими здесь так называемую чудесную
сеть. Под этим именем известно явление, когда артерия распадается
по своему ходу на капилляроподобные трубочки, из которых снова
собирается артерия, дающая потом сеть истинных капилляров, ибо за
ними уже следуют вены. Красное тело является газовой железой,
служащей для выделения из крови в полость пузыря кислорода.
Плавательный пузырь заполнен смесью газов в иной пропорции,
чем в атмосферном воздухе. Состав смеси из кислорода, азота и угле-
кислоты непостоянен, что зависит, главным образом, от глубины,
с которой взята исследуемая рыба.
Этот орган является гидростатическим аппаратом, с помощью
которого рыба регулирует свой удельный вес. Изменения последнего
позволяют рыбе то опускаться, то подниматься, не затрачивая при
этом мышечной энергии. ♦
Объем плавательного пузыря может уменьшаться: а) при сокра-
щении его стенок, б) при выдавливании пузырьков газа через вывод-
ной канал в пищевод и через жаберные щели наружу (у отьерстоиузыр-
ных рыб) или в) вследствие всасывания кислорода в кровь через посред-
347
ство утонченного участка спинной поверхности пузыря, обильно
снабженного богатой сетью кровеносных сосудов.
При уменьшении объема пузыря уменьшается и количество воды,
вытесняемой телом рыбы. Следовательно, удельный вес рыбы увели-
чивается, и она в силу закона физики опускается на дно. При рас-
ширении пузыря (расслабление его стенок, выделение кислорода в по-
лость пузыря) объем рыбы увеличивается, увеличивается количество
вытесняемой воды*, благодаря чему удельный вес рыбы падает, и она
без всякого труда для себя поднимается на поверхность воды.
Рыба может так урегулировать удельный вес тела, что он будет
равен единице; в таком случае тело ее приходит в состояние безраз-
личного равновесия, и животное остается взвешенным в толще воды:
оно в ней не тонет и не всплывает.
У карповых рыб плавательный пузырь подразделен на передний и
задний отделы узким перехватом (рис. 223, 3, 3'). Сокращением мы-
шечных лент стенки пузыря рыба может перегонять воздух из одного
отдела в другой, благодаря чему изменяется удельный вес то перед-
ней, то задней половины тела, и рыба поднимается или опускается
головой или хвостом.
В общем плавательный пузырь к плаванию в истинном значении
этого слова отношения не имеет; это доказывается и тем, что луч-
шие пловцы из рыб, например, поперечноротые (акулы, скаты и т. п.),
вовсе лишены этого органа.
Поэтому его приходится считать гидростатическим аппаратом,
служащим как для изменения удельного веса рыбы, так и для опре-
деления перемены давления извне. В последнем случае животное
предпринимает те или другие действия, чтобы привести свой орга-
низм в соответствие с условиями давления, создавшимися в окру-
жающей среде.
Забегая вперед, отметим, что при общем сравнении органов лягушки
и окуня мы не находим у лягушки части, которая соответствовала бы
плавательному пузырю окуня. Однако, знакомясь с историей разви-
тия этих животных, мы не можем не отметить факта, что легкие лягушки
развиваются из выростов пищевода, т. е. из такого же (по происхо-
ждению) зачатка, что и плавательный пузырь окуня. На этом основа-
нии мы можем легкое лягушки считать гомологичным плавательному
пузырю рыбы, так как гомология устанавливается не только по сход-
ству строения, но и по сходству развития. Дальнейшая судьба указан-
ных зачатков различна: у лягушки они обособляются от пищевода и
открываются в дыхательное горло, ведущее в заднюю часть ротогло-
точной полости (легкие); у окуня зачаток обособляется в замкнутый
плавательный пузырь, который у щуки остается в соединении с пище-
водом. Гомология легких и плавательного пузыря подтверждается
тем, что у так называемых двоякодышащих рыб (Dipnoi) плавательный
пузырь имеет внутренние ячеистые стенки и открывается в глотку
с брющной ее стороны.1
1 В настоящее время взгляд на гомологию легких и плавательного пузыря
несколько изменился. Оба эти органа считаются гомологами последних жабер-
ных мешков рыб, путем смещения зачатков которых и образуются названные
органы.
348
Рис. 224. Поперечный разрез через жа-
берную дугу с парой жаберных листоч-
ков костистой рыбы. По Г е с с е и
Дофлейну.
А — с поверхности; В — с изображением хода
кровеносных сосудов. Схема. 1—жаберная
артерия; 2—жаберная вена; 3— стержень жа-
берной дуги. Стрелки указывают направление
тока крови.
выпуклом краю нежными жабер-
Он служит у них для воздушного дыхания помимо жабр (органов
дыхания водного типа). Поэтому плавательный пузырь двоякодышащих
называют легким. Этот орган упомянутых животных может служить
примером промежуточного, или переходного, органа между такими, на
первый взгляд, крайними и чуждыми друг другу формами, как типич-
ный плавательный пузырь закрытопузырных рыб и легкие наземных
позвоночных.
Органы дыхания. Рыбы являются типичными водными животными.
Дыхание в воде отлично от дыхания в воздухе, так как газообмен
в первом случае происходит с кислородом, растворенным в воде. Вода,
соприкасаясь на своей поверх-
ности с воздухом, растворяет его
в себе, причем кислород воздуха
растворяется лучше азота, бла-
годаря чему в адсорбированном
водой воздухе его содержится
около 35% против 20% содер-
жания в атмосфере. При кипя-
чении воды из нее выделяется
растворенный воздух (выхожде-
ние из воды пузырьков газа),
и она становится негодной для
дыхания. Поэтому в остуженной
прокипяченной воде жабродыша-
щие животные, в том числе и
рыбы, задыхаются.
Для дыхания на счет адсор-
бированного в воде кислорода
у рыбы служат жабры, относя-
щиеся к дыхательным органам
водного типа.
Жабры помещаются под жа-
берной крышкой, и на нашей
вскрытой рыбе они видны в го-
ловном отделе в виде дугооб-
разных стержней, усаженных на
ными листочками. Между жабрами имеются проходы — жаберные
щели; проводя сквозь них ручку скальпеля или иглы, можно про-
никнуть в ротоглоточную полость рыбы.
Для рассмотрения жабр их перерезают у обоих концов и кладут
в воду.
Каждая жабра образована изогнутой костно-хрящевой дугой
(рис. 222, Ьг), вогнутая поверхность которой покрыта жаберными ты-
чинками (см. выше — фильтрация воды); выпуклая же часть несет два
ряда слизистых жаберных листочков (рис. 224, А) красного цвета,
вследствие богатства их кровью. Для защиты от механических повре-
ждений нежных листочков все четыре жабры закрыты жаберной
крышкой. Поэтому жабры рыб называются внутренними.
Дыхание можно наблюдать на живой рыбе. Она набирает в рот
воду и проталкивает ее через жаберные щели; проходя сквозь них,
349
вода омывает жаберные листочки и затем через щель под жаберной
крышкой выталкивается вон. Во время такой циркуляции воды проис-
ходит обогащение венозной крови жабр кислородом, заимствованным
из протекающей сквозь них воды.
Органы кровообращения. Сердце окуня помещается в околосер-
дечной полости (pericardium), которая сзади ограничивается тонкой
перепончатой стенкой, лежащей перед печенью и передним концом
плавательного пузыря. В общем, сердце находится сзади жабр и впереди
от грудных плавников. Оно состоит из двух отделов—предсердия
(рис. 222, at) и желудочка (рис. 222, v). Тонкостенное предсердие темно-
коричневого цвета; оно лежит над розовым желудочком, стенки кото-
рого очень толсты. Между этими отделами имеется отверстие, снабжен-
ное двумя заслонками, которые пропускают кровь только из предсер-
дия в желудочек, но не обратно. Желудочек продолжается далее кпе-
реди в жаберный артериальный ствол (truncus arteriosus); основание
последнего расширяется в виде луковицы — bulbus arteriosus (рис. 222,
ba) и у выхода своего из желудочка несет пару заслонок, пропускаю-
щих кровь из сердца в артериальный ствол. Позади предсердия поме-
щается тонкостенный довольно объемистый мешок — венозный синус
(рис. 222, so), получающий кровь из вен тела.
Сердце рыбы является венозным, или жаберным, так как через
него подается венозная кровь в жабры. Артериальный ствол, назы-
ваемый иначе брюшной аортой (рис. 222, tra), идет вперед между
нижними концами жабр и дает в каждую жабру по жаберной арте-
рии, идущей по ее выпуклому краю до спинного конца дуги. От этого
ствола в каждый жаберный листочек ответвляется приносящий сосуд.
В жаберном листочке он распадается на систему капилляров, из кото-
рых собирается обогатившаяся здесь кислородом кровь в выносящий
сосуд (рис. 224, В). Из всех выносящих сосудов собираются жаберные
вены, числом четыре пары; соединяясь друг с другом, они образуют
настоящую, или спинную, аорту.
Аорта идет под позвоночным столбом и даст сосуды для питания
всех частей тела, кроме головы, которую питают кровью две головные
артерии (аа. cephalicae), идущие от первой пары жаберных вен. От
аорты отходят брюшная артерия (распадающаяся на ветви для же-
лудка, печени, кишек и половых желез), почечные артерии, ветви
для мышц и т. д.
Пройдя через брюшную полость, аорта внедряется в толщу хвоста
и впадает под именем хвостовой артерии (a. caudalis) в канал, обра-
зованный нижними дугами позвонков.
В конце концов артерии распадаются на капилляры, где кровь от-
дает кислород клеткам тела. Из капилляров собираются вены, или
идущие прямо к сердцу, или распадающиеся на воротные системы почек
и печени, причем печеночная вена впадает непосредственно в венозный
синус.
В общем, вены собираются в четыре крупных ствола—две передние
кардинальные, или яремные, вены—v. cardinales anteriores s. jugulares
(рис. 222, vca) и две задние кардинальные вены (v. cardinales posteriores).
На каждой стороне тела передняя и задняя кардинальные вены сли-
ваются вместе и образуют поперечный кювьерев претек — ductus
350
Cuvieri (рис. 222, de). Оба кювьеровых протока впадают, в свою оче-
редь, в венозный синус.
Таким образом кровь при своем круговороте делает только один
круг кровообращения, прерванный капиллярами в жабрах и в тканях
тела. Сердце лежит на пути только венозной крови и артериальной
крови никогда не содержит.
Органы выделения- Органами выделения рыбы являются парные
почки. Чтобы добраться до них, необходимо убрать плавательный,
пузырь, и тогда по сторонам позвоночника можно видеть две узкие
ленты нежной красноватой (иногда пигментированной ткани) —это и
будут почки (рис. 222, гп). Когда они видны неясно, их можно надежнее
заметить, если рассматривать вскрытую рыбу насквозь перед
лампой.
Передние концы почек образуют расширяющиеся лопасти — так:
называемые предпочки, или головные почки. Выделяемая почками моча
стекает по мочеточникам — тонким трубочкам, лежащим под оболочкой
почек. Оба мочеточника сливаются вместе, их общий канал образует
боковое расширение — небольшой мочевой пузырь (рис. 222, vs).
Отверстие мочевых органов лежит рядом с половым, позади задне-
проходного отверстия.
На почках можно рассмотреть правую и левую задние кардиналь-
ные вены, из них левая (по отношению к рыбе) тоньше и короче правой.
У рыбы замечательно присутствие головной почки. Этот орган
является зародышевым, так как он возникает и функционирует у за-
родышей. У лягушки также в свое время появляется головная
почка, но она потом замещается так называемой первичной почкой
(вольфово тело), которая в виде дефинитивного органа остается у этого
животного на всю жизнь. Вследствие такой последовательности
развития можно говорить о смене органов, или о подстановке одного
органа на место другого; этот процесс называется субституцией.
У рыбы субституция почек выражена не в полной мере, так как
головная почка остается и после появления первичной почки, правда,,
в виде нефункционирующего рудиментарного органа.
Органы размножения окуня имеют очень простое строение. Они:
состоят из мешковидных половых желез (рис. 222, gn), величина кото-
рых зависит от стадии половозрелости рыбы.
Мужские половые органы представлены двумя удлиненными семен-
никами моло^нобелого цвета. Ткань их нежная, ле! ко рвущаяся.
Семенники имеют крупно складчатую поверхность и к заднему концу
утончаются, переходя в слабо выраженный семевыносящий канал.
Последний открывается наружу самосгоятельным отверстием, лежащим
позади заднепроходного отверстия. В общем, семенники лежат в задней
половине брюшной полости. Как и у лягушки, мужская половая
железа окуня вырабатывает живчики. Если прорвать ее стенку в воде,
то выходит тончайшая белая пыль, мутящая воду. Каждая точка и
есть живчик, находящийся в той или другой стадии развития.
Семенники щуки лежат на брюшной стороне плавательного пузыря
в виде двух лент желтоватого цвета.
Никаких наружных половых органов у костистых рыб нет, равным
образом нет и внешних половых признаков у окуня.
351
Женские половые органы образованы у окуня непарным яичником,
лежащим в задней половине брюшной полости. В суженной части
яичник открывается наружу половым отверстием. Ткань яичника грубо
зерниста, так как яйцеклетки по объему гораздо больше живчиков.
Величина яичника зависит от стадии половозрелости рыбы. Период
брачной жизни ее называют нерестом, к этому времени яичник раз-
растается до наибольших размеров. Зрелые яйцеклетки выбрасываются
наружу — рыба, как говорят, «мечет икру».
Самка, как и самец, лишена наружных половых органов.
Оплодотворение и развитие рыбы. Окуни достигают половой зре-
лости на третий, реже на второй год жизни. Весной или в начале лета
они собираются стаями в тихих заросших затонах. Самки мечут икру,
выпуская ее из тела длинными лентами зеленовато-белого цвета;
отдельные икринки склеены друг с другом студенистым веществом.
Окунь весом в 200 г дает 200—300 тысяч икринок, а крупный—и того
больше. Самцы плавают во время нереста возле самок и выпускают
на кучи икры семенную жидкость. Оплодотворение у рыб — на-
ружное. Полагают, что род полового возбуждения воспринимается
нерестующими рыбами посредством их боковой линии, которая раздра-
жается резкими толчками воды вследствие оживленного движе-
ния рыб.
Никакой заботы об отложенной Икре родители не проявляют.
Из оплодотворенных яиц через 15—20 дней их развития выходят ма-
ленькие рыбки — мальки. Они имеют общий облик рыбы, от которой
отличаются присутствием большого брюшного желточного пузыря;
в последнем лежит питательный желток (остаток от запасов, бывших
в яйце); вскоре желток рассасывается, и малек ничем существенным
уже не отличается от рыбы. Развитие ее, следовательно, прямое, т. е.
без превращения.
Центральная нервная система окуня лежит на спинной стороне
тела в костно-хрящевой коробке черепа и в спинномозговом канале
позвоночника.
Чтобы добраться до головного мозга, берут окуня в левую руку и
срезают скальпелем в горизонтальном направлении верхнюю стенку
черепа между затылком и глазами, ведя нож сзади наперед. У рыбы
среднего размера эта операция удается хорошо, и перед нами откры-
вается черепная полость, заполненная капельками жира в виде серо-
ватой пены. Мозга пока не видно, так как последняя еЛ закрывает;
пенистую массу смывают водой из пипетки, подравнивают края разре-
зов, если они где-нибудь закрывают мозг, снимают кости между
глазами и перед ними, чтобы открыть обонятельные нервы, и обнажают
препаровкой заднюю часть мозга.
Головной мозг окуня (рис. 225), в направлении спереди назад,
состоит из следующих частей:
1) небольшие обонятельные доли с отходящими от них длинными и
толстыми обонятельными нервами (по), которые и оканчиваются в но-
совых капсулах (sn);
2) большие полушария (передний мозг, I), которые являются
второй по величине частью головного мозга;
3) средний мозг (1о), или двухолмие, —самый крупный отдел мозга;
352
Рис. 225. Головной
мозг окуня серху.
sn — обонятельные луко-
вицы; по — обонятельные
нервы; I — большие полу-
шария; to — средний мозг
(двухолмие); cb — мозже-
чок; то — продолговатый
мозг; ntr — тройничный
нерв; n.v. — блуждающий
нерв; п.ас—слуховой нерв.
4) мозжечок в виде непарного округлого тела (cb), налегающего
на переднюю часть продолговатого мозга;
5) продолговатый мозг (то) с широкой ромбоидальной ямкой на
верхней его поверхности в передней части. Утончаясь, он переходит
в спинной мозг.
При рассматривании мозга снизу (если предварительно уплотнить
препарат в слабом спирту и вынуть мозг из черепа) видны все его части,
кроме мозжечка.
Как раз на границе больших полушарий и
двухолмия приходится перекрест зрительных
нервов, причем один нерв лежит над другим.
Между задними ножками зрительных нервов
помещается нижний мозговой придаток, позади
которого лежат две нижние доли двухолмия; верх-
ние же доли их видны впереди и кнаружи от тех
же задних ножек зрительных нервов.
Все эти части головного мозга лежат в одной
горизонтальной плоскости и изгибов не обра-
зуют. Примечательно слабое развитие большого
мозга, который по своей величине уступает
двухолмию.
Головной мозг различных рыб устроен разно-
образно, и отдельные части его могут иметь
своеобразное развитие.
Спинной мозг лежит в спинномозговом ка-
нале, образованном спинными отростками по-
звонков. У основания своего отростки дуго-
образно расщепляются и ножками срастаются
с телом позвонка. Для вскрытия спинного мозга
вводят (после вскрытия головного мозга) в спинно-
мозговой канал острую ветвь ножниц, которыми
и рассекают сначала дуги позвонков одной сто-
роны тела, а затем — другой. При этом плос-
кость ножниц держат наискось к срединной плос-
кости тела.
От центральной нервной системы отходят
нервы. Головных нервов у рыбы десять пар (наименование их см. раз-
дел о лягушке, стр. 379).
Спинномозговые нервы выходят в промежутках между позвонками
двумя ветвями — спинной и брюшной, которые соединяются вне позво-
ночного канала и потом уже ветвятся. Брюшные нервы в области
плавников соединяются друг с другом, образуя сплетение. У костистых
рыб сплетение невелико, так как грудные плавники иннервируются
всего только тремя - четырьмя нервами.
Органы чувств служат для восприятия различных раздражений из
окружающей среды и передачи их по чувствительным нервам в го-
ловной мозг, где эти раздражения и претворяются в те или другие
• ощущения. Например, животное воспринимает органом слуха звуковые
колебания, которые раздражают окончание слухового нерва. Произ-
веденное раздражение по слуховому нерву передается в головной моз1;
23 Практикум по зоологии — 385 353
1 г а
Рис. 226. Глаз щуки в вертикальном про-
дольном разрезе. По Гессе и До-
ф л е й н у.
7—сетчатка; 2 — пигментный эпителий; 3 — сосу-
дистая оболочка; 4 — зрительный нерв; 5 — хру-
сталик; 6 — его связка; 7 —мышца, оттягивающая
хрусталик; 8 — ее сухожилие; 9— радужина;
10—роговица; 11—склеротика; 12 — хориои-
дальная железа, пунктиром показано положение
хрусталика в аккомодированном глазу.
в клетках его это раздражение перерабатывается в слуховое НпеЧат-
ление; в результате животное слышит данный звук.
Об органах чувств животных мы поневоле судим на основании того,
что сами ощущаем своими органами сходного значения. Такой антро-
поморфический метод крайне ненадежен, так как отправление анало-
гичных органов чувств у различных животных бывает далеко не
одинаковым. Например, сова видит ночью, муравьи ощущают ультра-
фиолетовые лучи и т. д. Еще труднее судить о роли специальных орга-
нов чувств, хотя бы о боко-
вой линии рыб. Ее долгое
время называли ничего не го-
ворящим именем органа ше-
стого чувства, и только спе-
циальными опытами удалось
подойти к разгадке ее зна-
чения как органа, восприни-
мающего степень давления
воды.
В боковой линии мы имеем
пример специфического органа
чувств, приноровленного спе-
циально к жизни в воде.
Равным образом и другие
органы чувств рыбы также
имеют свои особенности, свя-
занные с пребыванием этих
животных в водной среде.
Органы обоняния
представлены двумя слепыми
носовыми мешками, из кото-
рых каждый открывается на-
ружу парным носовым отвер-
стием. В стенке носового меш-
ка разветвляются окончания
обонятельного нерва.
Носовые полости рыбы не
сообщаются с ротоглоточной
полостью, так как у рыбы
хоан нет. Сущность обоняния сводится к тому, что частицы газо-
образного вещества (например, светильного газа) или легко испа-
ряющейся жидкости (формалин), или, наконец, постоянно (как бы
самопроизвольно) распыляющегося твердого вещества (например, му-
скуса) смешиваются с воздухом и при вдыхании попадают в носовую
полость; здесь они раздражают слизистую оболочку, в которой ве-
твятся обонятельные нервы; раздражение передается в обонятельные
доли, блогодаря чему животное ощущает запах данною вещества.
У наземных позвоночных обоняние, повидимому, связано именно
с раздражением обонятельной оболочки мельчайшими частичками
вещества. Это доказывается опытом с заполнением носа пахнущей
Жидкостью; в таком случае, при непосредственном соприкосновении
354
жидкости со слизистой оболочкой носа, запаха не ощущается; при
нюхании же жидкости запах ее воспринимается с ясностью.
Рыбы же ощущают запахи веществ, растворенных в воде, чем их
органы обоняния и отличаются от таких же органов наземных живот-
ных, начиная с земноводных.
Органами зрения рыбы являются глаза (рис. 226). При
осмотре их в нормальном положении, на голове можно видеть покры-
вающую глаз прозрачную кожицу — роговицу (10), сквозь нее про-
свечивает срединный темный кружок — зрачок, окаймленный кольцом
радужной оболочки (9). Глаза лежат во впадинах черепа, называемых
глазницами. Выступающая на поверхность черепа поверхность глаз
(роговица) мало выпукла. Вырезанное глазное яблоко на внутреннем
своем полюсе несет отрезок зрительного нерва (4), а на боковых
поверхностях — остатки глазодвигательных мышц. Наощупь оно очень
плотно; образующая глазное яблоко стенка называется белковой обо-
лочкой или склерой (11).
Если надрезать глаз и выпустить его содержимое на бумагу, то
можно найти плотный прозрачный хрусталик (5) почти шаровидной
формы, прозрачную студенистую массу неопределенной формы —
стекловидное тело, и лежащее поверхностно черное вещество — пи-
гментную оболочку (2).
Нормальное расположение в глазу этих частей таково. Под рого-
вицей, в отверстии радужной оболочки (т. е. в зрачке), лежит хрусга-
лик. Между его задней поверхностью и внутренней стенкой глазного
яблока помещается стекловидное тело. Стенка глаза составлена, идя
снаружи внутрь, из белковой оболочки (77), сосудистой оболочки (5),
пигментной оболочки (2) и сетчатки (7). Кроме этих общих для всех
позвоночных частей глаза, у костистых рыб есть еще так называемая
серебристая оболочка (argentea), содержащая в себе множество микро-
скопически мелких кристалликов. Лежит она кнаружи от сосудистой
оболочки.
Рыбы видят только вблизи,-так как, вследствие небольшой прозрач-
ности воды, которая к тому же легко мутится, далекие предметы не
могут быть рассмотрены вовсе. Глаза рыб установлены для ясного
видения ближайших предметов; этим и объясняется большая выпук-
лость хрусталика.
Органы слуха рыбы скрыты в толще ее черепа. Среднее ухо
у них отсутствует, и орган слуха представлен только внутренним ухом.
Чтобы добраться до этого органа, голову рыбы разрезают крепкими
ножницами или ножом по срединной линии тела на две половины и
вынимают из нее головной мозг и жировую его покрышку. На уровне
продолговатого мозга и мозжечка, в полости черепа, лежит большая
часть внутреннего уха.
Внутреннее ухо (рис. 227) образовано перепончатым лабиринтом
и полукружными каналами. Лабиринт распадается на большой круг-
лый мешочек — sacculus (2), лежащий внизу, и меньший верхний
эллиптический мешочек (utriculus—7), связанный с полукружными
каналами (5, 4, 5).
От sacculus спереди отходит маленький вырост — lagena (77).
являющийся зачатком улитки высших позвоночных. Внутри sacculus
• 355
Рис. 227. Схема левого лаби-
ринта низшего позвоночного.
По Гессе и Дофлейну.
1 — utriculus; 2 — sacculus; 3, 4,
5 — полукружные каналы; б, 7,8—
ампулы полукружных каналов; У,10,
11 — чувствительный эпителий с ото-
литами; 12 — эндолимфатический
канал.
лежит слуховой камешек — отолит. От круглого мешочка идет тонкий
эндолимфатический канал (12), слепо заканчивающийся в полости
черепа.
Полукружных каналов три: передний и задний отходят от эллип-
тического мешочка общим стволом, расходящимся у вершины своей
на две ветви, ориентированные в вертикальной плоскости. Эти послед-
ние загибаются книзу и снова впадают в utnculus, образовав пред-
варительно концевые вздутия — ампулы (б, 8). Третий полукружный
канал называется наружным; лежит он в горизонтальной плоскости
(4, 7).
Все три канала заключены в хряще-
вую слуховую капсулу первичного че-
репа.
Повидимому, рыбы лишены слуха в
общепринятом понятии этого чувства.
Ряд опытов опровергает мнение,
будто бы рыб по колокольчику можно
созывать к берегу. В других случаях
револьверный выстрел влиял одинаково
как на здоровых рыб, так и на таких, у
которых оба лабиринта были удалены.
«Внутреннее ухо» рыбы является,
главным образом, органом ориентировки
положения тела в пространстве, или
органом чувства равновесия (статиче-
ский аппарат). Рыбы с удаленными ла-
биринтами вертятся при плавании вокруг
своей продольной оси или плавают на
спине.
Мускулатура рыбы
Чтобы рассмотреть мышцы окуня,
необходимо снять с него кожу. Каждая
половина тела занята широкой боковой
мышцей (m. lateralis), которая тянется
от головы до хвоста. Эта мышца пересе-
чена множеством сухожильных перемычек в виде волнистых или лома-
ных линий. Число перемычек соответствует числу позвонков. Благо-
даря такому строению боковая мышца распадается на ряд метамер-
ных участков, которые очень хорошо изолируются у вареной рыбы.
От кожных покровов в районе боковой линии до позвоночника тя-
нется соединительнотканная перегородка, делящая боковую мышцу
на спинной и брюшной отделы. При сокращении боковой мышцы
тело рыбы изгибается в соответствующую сторону; следовательно,
эта мышца является плавательной мышцей рыбы.
Между спинными ее частями лежит узкая мыщца, двигающая
спинным плавником, а между брюшными отделами помещается
такая же мышца для заднепроходного плавника. Мелкие мышечные
пучки управляют движением парных плавников. Есть также мышцы
для челюстей, жаберной крышки и жаберного аппарата.
856
Внутренний скелет рыбы
Основой скелета рыбы является позвоночник, состоящий из сорока
позвонков двух сортов: туловищных (или грудных) (рис. 228, 7, 2)
с ребрами, и хвостовых — без ребер (рис. 228, 3—5). Тело позвонка
имеет вид двояковогнутого диска. От верхней его половины отходит
невральная дуга, продолжающаяся в остистый отросток. От нижней же
части хвостового позвонка идет такая же дуга с нижним остистым
отростком; называется она гемальной, так как сквозь нее проходят
аорта и правая кардинальная вена.
Туловищные позвонки имеют два нижних отростка, к которым
причленяются дугообразные ребра. В вогнутых полостях между по-
Рис. 228. Позвонки костистых рыб. По Н. Холодковскому.
7—из предхвостовой части столба с ребрами; 2 — тоже, с основными отростками; 3 из
хвостовой части, с верхней и нижней дугой; 4 — то же, сбоку (два позвонка); 5 — межмышеч-
ная косточка.
звонками лежат в виде студенистых шариков остатки спинной струны,
или хорды,—древнейшего опорного образования позвоночных живот-
ных.
К остистым отросткам позвонков сверху примыкают вставочные
косточки, с которыми сочленяются лучи спинного плавника (рис. 229, к);
такие же образования есть и у брюшных остистых отростков хвосто-
вых позвонков для анального плавника. Эти косточки заложены между
обеими боковыми мышцами.
Парные плавники снабжены «поясами конечностей», из которых
тазовый пояс развит очень слабо. Он образован всего только двумя
длинными соединяющимися друг с другом трехугольными косточками,
к которым примыкают лучи брюшного плавника.
Плечевой пояс, подпирающий грудные плавники, представлен
большими дугообразными косточками — cleithrum. К заднему ее краю
357
примыкают две небольшие косточки — лопатка и коракоид} на них
сидят основные косточки плавника, сочленяющиеся с его лучами.
Оба cleithrum соединяются друг с другом своими нижними концами.
Верхние же концы их соприкасаются с добавочными косточками,
примыкающими к черепу.
Череп рыбы состоит из множества костей, среди которых имеются
участки хряща, являющиеся остатками хрящевого зародышевого че-
репа (примордиальный череп). Кости черепа окуня двоякого происхо-
ждения: одни из них развиваются на счет хряща (хондральные кости),
другие же развиваются из соединительной ткани (покровные, или
накладные, кости).
Из поверхностных костей черепа отметим: шесть подглазничных
косточек (ossa infraorbitalia), носовые кости (оо. nasalia), кости верхней
и нижней челюстей, нёбную дугу, слагающуюся из семи косточек, и
жаберные крышки.
Рис. 229. Скелет окуня (Perea), k, t, т, п — лучи непарных плавников.
Из Н. Холодкове к ого.
По удалении этих частей, мы увидим собственно черепную коробку
окуня, состоящую также из многих костей: шести затылочных, двух
теменных, лобных, обонятельных и на нижней поверхности черепа —
сошника, парасфеноида, слуховых и других более мелких косточек.
Рыба удобна для ознакомления с жаберным, или висцеральным,
скелетом. Он образован шестью парами костных или хрящевых жа-
берных дуг. Нижними концами они соединяются со срединным рядом
соединительных косточек, из коих первая входит в язык рыбы (os
entoglossum). Верхние концы дуг подходят к основанию черепа. Пер-
вая дуга называется подъязычной} она составлена из пяти частей и несет
на себе по семи лучей, поддерживающих жаберную перепоночку;
вторая — пятая дуги также сложные: на внутренней вогнутой поверх-
ности они несут зубцы, образующие фильтровальный аппарат. Верхние
половины жаберных дуг состоят из верхнеглоточных костей, усажен-
ных зубами (глоточные зубы). Шестая пара дуг рудиментарна, так как
лишена жаберных листков и в своей основе несет лишь нижнегло-
точные кости.
358
Общие замечании
Тело окуня построено по двубоковой симметрии, которая ясно
выражена и в расположении его внутренних органов. Отчасти ее на-
рушает кишечник, хотя гораздо менее, чем у лягушки, и задние кар-
динальные вены, из которых левая развита гораздо слабее правой.
У окуня мы находим заметные следы метамерии. Так, мы имеем
метамеры, образованные позвонками и соответствующими им сегмен-
тами боковой мышцы тела. Из прочих частей скелета метамерны
жаберные дуги и опорные образования непарных плавников, а также и
лучи парных конечностей рыбы. Метамерия проявляется также и
в отхождении спинномозговых нервов; можно усмотреть сегментар-
ность строения и головы, но только выяснив детали ее эмбриональ-
ного развития, что не может входить в нашу задачу.
Конечности рыбы — водного типа: они не членисты и являются
пластинчатыми гибкими рычагами. Сравнивая парные рычаги, можно
установить гомологию строения их наружной части, образованной
в обоих случаях кожным веером, подпертым скелетными стержнями.
По своему строению парные плавники рыбы не имеют ничего общего
с членистыми конечностями наземных позвоночных; поэтому непо-
средственно гомологизировать их друг к другу нельзя. Однако
в сравнительной анатомии важный вопрос о происхождении конеч-
ностей позвоночных разрешается некоторыми теориями конечно-
стей, согласно которым наземные конечности происходят из плавников
рыб путем последовательного их изменения. Сами непарные плав-
ники счигаются остатками продольных боковых складок кожи, куда
врастают метамерные зачатки мышц, между которыми и развиваются
лучи плавников.
Подробное изложение теории конечности можно найти в курсах
сравнительной анатомии (Н. А. Холодковский; В. М. Шим-
к е в и ч).
Выберем теперь главнейшие анатомические признаки, общие для
окуня и лягушки.
1. Кожа их влажная и слизистая.
2. Опорой тела является внутренний скелет (позвоночник).
3. Нервная система компактного типа лежит на спинной стороне
тела.
4. Кровеносная система замкнутого типа.
5. Сердце лежит на брюшной стороне тела.
Значение этих признаков более широко, так как они характеризуют
вообще представителей классов рыб и земноводных, относящихся
к подтипу позвоночных (Vertebrata), причем признаки второй, третий,
четвертый и пятый общи для всех позвоночных.
Из особенностей строения рыбы укажем те, которые связаны с пре-
быванием ее в воде:
1) слизистая кожа, уменьшающая трение покровов тела о воду
при плавании;
2) боковая линия как специфический орган, служащий для опре-
деления степени и направления давления воды;
3) нечленистыс плавники, играющие роль" весел и рулей;
369
4) непарные плавники, из которых хвостовой является главным
двигателем рыбы, а спинной и анальный — се килями;
5) сплющенное с боков и заостренное на концах тело, благодаря
своей форме легко рассекающее воду и сводящее на-нет ее сопро-
тивление при плавании рыбы;
6) органы дыхания водного типа — внутренние жабры, косвенно
принимающие участие в добывании пищи (фильтрация воды для за-
хвата мелких животных);
7) плавательный пузырь как гидростатический аппарат рыбы.
КЛАСС ЗЕМНОВОДНЫЕ (AMPHIBIA)
БЕСХВОСТЫЕ (ANURA)
ЛЯГУШКА (RANA TEMPORARIA)
Технические замечания. Для вскрытия обыкновенно поль-
зуются травяной, или бурой, лягушкой (Rana temporaria) или зеленой съедоб-
ной лягушкой (Rana esculenta).
R. temporaria, в среднем, 5—7 см длины; по бокам головы несет по треуголь-
ному темному пятну, своим основанием упирающемуся в глаз.
R. esculenta — 9—11 (иногда даже 14) см длины. Спина изменчивого зеленого
цвета, переходящего в грязнобурый. У самца есть два голосовых мешка (резо-
наторы), выступающие во время кваканья из-под угла рта; лежат они в заднем
углу челюстей. Более южный вид, чем R. temporaria, в небольшом числе есть и
в Ленинграде (Кировские острова, Черная речка).
Лягушка принадлежит к классу амфибий, или земноводных, называемых так
потому, что в своей жизни они связаны с двумя стихиями — с водой и землей.
За полчаса до вскрытия лягушек кладут в банку, куда помещают вату, про-
питанную хлороформом или серным эфиром. Когда лягушки заснут, их промывают
водой, чтобы удалить пенистый секрет раздраженных анестезирующим веществом
кожных желез.
В случае отсутствия хлороформа или эфира лягушку можно* убить другими
способами. Придерживая животное в сидячем положении левой рукой, перерезают
острыми .ножницами, держа их концами отвесно вниз, позвоночный столб тотчас
позади головы. Затем перегибают лягушку в месте разреза, вводят в спинномоз-
говой канал иглу, которой и разрушают спинной мозг.
Еще проще убить лягушку ударом о стол. Животное берут за задние
ноги и, держа его брюхом кверху, быстро и сильно ударяют о край стола
головой.
Наружный осмотр
Тело лягушки состоит из короткого толстого туловища, трех-
угольней головы и двух пар ног; шея у лягушки не выражена.
Животное одето мягкой кожей, более темной окраски на спине,
чем на брюхе. Кожные покровы испещрены темными пятнами, которые
зависят от скопления в соединительнотканных слоях кожи отростча-
тых пигментных клеток, заключающих в себе множество мельчайших
зернышек пигмента (животного красящего вещества). Под влиянием
различных раздражений отростки клеток могут втягиваться; при этом
пигментные зернышки перегруппировываются и сами клетки иногда
меняют свое положение в кожных покровах. Все это до известной
степени отражается и на макроскопической окраске животного, ко-
торая может несколько*изменяться под влиянием различного осве-
360
щения. Способность к изменению цвета кожи у некоторых существ,
например, хамелеонов, развита очень сильно.
Кожа лягушки влажная, ибо она в нормальном состоянии всегда
покрыта слизистыми выделениями кйжных желез, имеющих форму
простых мешочков. Влажность кожи есть важное условие для жизни
этого животного, так как кожа принимает существенное участие в
процессе дыхания (кожное дыхание). Если поверхностные слои кожи
подсохнут, то кожное дыхание нарушается, что влечет за собой смерть
животного.
По мере старения поверхностный слой плоских клеток эпители-
ального отдела кожи, называемого эпидермисом, сбрасывается пла-
стами; это явление можно считать линькой.
Кожу лягушки легко захватить в складки; она вообще очень под-
вижна, так как лишь в некоторых местах спаяна с лежащими глубже
мышцами.
Передний конец головы округлен; по переднему краю ее поме-
щается широкая ротовая щель. Пинцетом можно раскрыть рот и со
дна его вытащить желтоватый язык, прикрепленный своим передним
концом к переднему углу нижней челюсти. Язык на конце несет пару
лопастей. Он покрыт очень клейкой слизью, выделяемой его желез-
ками.
Благодаря особенностям своего прикрепления, язык выбрасы-
вается лягушкой наружу, когда она нацеливается поймать какую-либо
движущуюся добычу, например, муху. Коснувшись ее своим липким
языком, лягушка втягивает насекомое в рот и проглатывает. Она,
как и жаба, безнаказанно для себя может проглатывать пчел и ос,
несмотря на то, что эти насекомые обладают ядовитым жалом. Дело
объясняется тем, что лягушка нечувствительна (невосприимчива)
к яду перепончатокрылых; подобное свойство организма называется
естественным иммунитетом.
Над передним концом головы, отступя от него немного назад,
лежит пара носовых отверстий (ноздри), прикрытых каждое кожным
клапаном. Если наблюдать живую лягушку, то можно видеть, что
эти клапаны то закрывают, то открывают носовые отверстия. Такая
работа чередуется с сокращением межчелюстной мышцы, играющей
роль главного дыхательного мускула.
Кзади от ноздрей помещаются смотрящие в стороны глаза лягушки,
снабженные парными веками — верхним и нижним.
Сзади каждого глаза можно различить участок кожи, плотно на-
тянутый на костном кольце. У R. temporaria в этом месте лежит
как раз височное темное пятно. Натянутый участок кожи является
барабанной перепонкой; если ее прорвать, то обнаруживается полость
среднего уха. Наружного уха у лягушки нет.
Между основанием задних ног, ближе к спинной стороне тела,
помещается отверстие клоака — небольшой камеры, принимающей
в себя выходные отверстия задней кишки, органов выделения и раз-
множения.
Передние ноги у лягушки короче задних и состоят из плеча, пред-
плечья и кисти с четырьмя свободными пальцами. Если все пальцы
передней кисти у лягушки приблизительно одинаковой формы, то мы
361
имеем перед собой самку. У самца же внутренний палец передней
ноги несет кожную бородавку, образованную подушкообразным утол-
щением кожных желез. Эта бородавка, служащая для обхватывания
самки при акте копуляции, является наружным половым признаком
самца.
Передние ноги играют второстепенную роль при передвижении
лягушки, когда она неуклюже переползает с места на место. Они
служат, главным образом, для поддерживания ее тела в сидячем по-
ложении.
Задние ноги лягушки длинны. Части их называются бедром,
голенью и стопой. Стопа состоит из пяти длинных пальцев,
соединенных между собой широкой складкой кожи, называемой
плавательной перепонкой. Задними ногами лягушка пользуется
для плавания в воде, о которую она упирается раскрытой вееро-
образно стопой; ими же она отталкивается от земли при передви-
жении скачками. В связи с такой значительной работой задние
конечности лягушки развиты гораздо сильнее передних.
Вскрытие кожных покровов и мускулатура тела
Рис. 230. Мышцы брюшной
стороны тела лягушки. По
Эккеру.
sm—межчелюстная мышца; рт —
большая грудная мышца; pmb —
ее брюшная часть; аг — прямая
мышца живота; it — ее сухо-
жильные перемычки; ое — на-
ружная косая мышца живота;
st — грудина.
друг от друга по ходу
Перед вскрытием лягушку прикалывают ко дну ванночки брюш-
ной стороной кверху, растянув в стороны ноги и воткнув в их концы
булавки. Пинцетом левой руки захватывают складку кожи между
основаниями задних ног и прорезают ее
ножницами. Затем под кожу вводят ветвь
ножниц и, держа их конец кверху, разре-
зают кожные покровы по средней линии
тела до нижней челюсти. Особенно осто-
рожно следует действовать в области го-
ловы, чтобы не прорезать тонкую меж-
челюстную мышцу. Второй кожный разрез
ведуг перпендикулярно первому на уровне
передних конечностей. Полученные четыре
кожных лоскуга оттягивают пинцетом в
стороны и подрезают ножницами или скаль-
пелем скудные перемычки, соединяющие
кожу с лежащими глубже мышцами.
Таким образом мы вскрываем подкож-
ные лимфатические пространства (мешки)
и обнажаем мышечную стенку тела.
Мышечная стенка тела, как и вообще
скелетные мышцы, состоит из поперечно-
полосатой мышечной ткани, сокращение
которой происходит по воле животного.
Присматриваясь к обнаженной части тела,
можно заметить, что брюшная стенка со-
стоит из нескольких мышц, различаемых
их волокон. Между ветвями нижней че-
люсти идет тонкая поперечная межчелюстная мышца — m. inter-
maxillaris (рис. 230, sm); кзади от нее от средней линии грудины
Ж
к передним конечностям идут удлиненно-трехугольные пучки грудной
мышцы — m. pectoral is (рт). От заднего конца грудины по средней
линии живота тянется к лонному сращению прямая брюшная мышца —
m. abdominis rectus (аг), по ходу которой имеются сухожильные пере-
мычки, числом пять (it). У самок со зрелыми яичниками эта мышца
очень растянута и тонка. В толще прямой мышцы просвечивает сре-
динная красная линия — брюшная вена (v. abdominalis), несущая
кровь из задних конечностей. С боков из-под брюшной части грудной
мышцы (pmb) выходит внешняя косая мышца — m. obliquus abdo-
minis externus (ое).
Если вскрыть кожу бедра, то под ней видны длинные мышцы ноги,
облегающие скелет конечности. В каждой мышце можно различать
собственно мышечную часть и сухожильные концы блестяще белого
цвета, прикрепляющиеся к костям скелета, которые они и приводят
в движение при сокращении мышцы.
Вскрытие полости тела и общее расположение органов
Пинцетом в левой руке захватывают всю толщу мышц брюшной
стенки между основаниями задних ног и прорезают их осторожно
ножницами. В полученное отверстие вводят тупую ветвь ножниц,
держа конец ее кверху, разрезают мышечную стенку тела кпереди,
держась средней линии. На этом пути встречают грудину (рис. 230,
st); разрезают и ее, что надо сделать с ббльшим усилием, чем разрез
мышц, но осторожно, чтобы не поранить лежащего глубже сердца.
Затем пинцетом и пальцами разводят в стороны костно-мышечные
лоскуты (части перерезанной грудины и облегающие ее мышцы) и
прикалывают их ко дну ванночки булавками. При отведении лоскутов
в стороны приходится подрезать придерживающие их изнутри соеди-
нительнотканные перепонки и тяжи. Если при вскрытии были слу-
чайно поранены кровеносные сосуды, то ванночку несколько раз сильно
покачивают, чтобы волна воды пробегала через лягушку и смывала
кровь. Необходимо, чтобы вода совершенно покрывала внутренности
животного, так как от этого зависит ясность различения внутренних
органов.
С общим расположением органов, лежащих в вы-
стланной брюшиной полости тела, удобнее ознакомиться сначала у
самца лягушки, так как картина их у него типичнее, нежели у поло-
возрелой самки.
Весь последующий осмотр следует производить, пользуясь только
пинцетом и тупым концом скальпеля. Ни в коем случае нельзя вы-
резывать органы по частям для их рассмотрения; следует сначала все
разобрать на месте, только передвигая и приподнимая различные органы.
В передней части препарата (у наркотизированной лягушки) бро-
сается в глаза бьющееся сердце (рис. 231, 4). По бокам его иногда бы-
вают видны различно растянутые мешки аспидного цвета — это
легкие (р). Они хорошо заметны лишь тогда, когда раздуты воздухом.
В спавшемся состоянии их вовсе не видно; чтобы найти их, следует
конец пинцета подвести иод сердце и, захватив им легкое, вытянуть
его на поверхность препарата.
363
Сзади и по бокам сердца ЛеЖит самый крупный орган теЛа ля-
гушки — дольчатая печень — hepar (hp), темно-красного цвета. Если
задний край ее отбросить кверху, то на нижней поверхности обнару-
живается пузырек грязнозеленого цвета, величиной с_ горошину,
желчный пузырь—vesica fellea
US
hp
ot
p
ur
лягушки.
hp
ut
cl
(vs), содержащий в себе желчь.
Из-под левого края
печени назад идет жел-
товато-белая, постепенно
сужающаяся трубка —
желудок (gt), на заднем
конце переходящий в
тонкуюкишку (it). Тон-
кая кишка образует не-
сколько подвешенных на
прозрачной складке брю-
шины — брыжейке —
петель, заполняющих
брюшную полость.
Перебирая пинцетом
петли кишки, можно
дойти до резко выражен-
ного перехода ее в тол-
стую кишку (гс), от-
крывающуюся в клоаку
(cl).
Если откинуть впе-
ред место перехода же-
лудка в тонкую кишку
и раздвинуть эти два
органа друг от друга,
то в брыжейке между
ними можно обнаружить
удлиненное оранжево-
желтое тело неправиль-
ной призматической фор-
мы — поджелудочную же-
лезу (pancreas — рп).
Между петлями ки-
шек лежит темнокрас-
ная селезенка (lien) в
форме маленькой горо-
шины.
поверхностно, и найти их
По
Рис. 231. Внутренности самки
Н. Холодковскому, с некоторыми упро-
щениями.
41—желудок: cl — клоака; hp — печень; vs — желчный
пузырь; vs (снизу рисунка) — мочевой пузырь; И — тон-
кая кишка; гс — толстая кишка; ov — яичник; рп — под-
желудочная железа; р — легкое; ovd — яйцевод; ut —
матка; ot — воронка яйцевода; г — почка; иг—мочеточ-
ник; 7 — сонная дуга; 2 — аортная дуга; 3 — кожно-
легочная дуга; 4 — желудочек сердца; 5 — левое пред-
сердие.
Все эти органы лежат более или менее
легко.
У половозрелой самки (рис. 231) картина брюшных органов затем-
няется сильно развитыми черными яичниками (ov), мешки ко-
торых занимают большую часть брюшной полости и оттесняют петли
кишек, которые вовсе могут быть не видны.
По бокам яичников лежат сильно извитые яйцеводы (ovd, не сме-
шивать их с петлями кишек!).
364
Чтобы найти кишечник у Самки лягушки, следует раздвинуть
яичники ручкой скальпеля друг от друга и со дна щели между ними
извлечь пинцетом петли пищеварительной трубки.
После осмотра поверхностных органов брюшной полости самца
лягушки отбрасывают пинцетом все брюшные органы на правую сто-
рону препарата (т. е. влево от занимающегося) и немного подрезают
скальпелем брыжейку у ее основания.
По средней линии спины открывается позвоночный столб, по бо-
кам которого лежат два уплощенно-веретеновидных тела — почки
(г) темнокрасного цвета. Почки являются органами выделения, и вы-
деляемая ими моча стекает в клоаку по двум тонким трубочкам —
мочеточникам (иг), идущим по наружному краю почек.
К органам выделения относится также и мочевой пузырь — тонко-
стенный сердцевидный мешок, открывающийся самостоятельно в
клоаку с ее брюшной стороны (vs).
У самца под почками (у вскрытого же и приколотого ко дну ван-
ночки — над ними) лежат половые железы — бобовидные семенники
желтого или темного цвета, в зависимости от степени пигментации;
на переднем конце их помещается лопастевидное жировое тело, имею-
щееся также и на яичниках лягушки.
Почки самки не имеют никакой связи с яичниками и легко могут
быть найдены у особей с маленькими яичниками. Чтобы добраться
до них у половозрелых животных, следует вырезать левый яичник
лягушки и отвести влево от себя петли кишечника.
После общего ознакомления с органами лягушки на их месте сле-
дует приступить к более подробному рассмотрению систем органов
ее тела.
Внутреннее строение
Органы пищеварения. Ротовое отверстие ведет в широкую ротовую
полость; верхняя стенка ее костная (твердое нёбо), нижняя — мяг-
кая (межчелюстная мышца). Края рта ограничены верхней и нижней
челюстями. Проведем пальцем по их поверхности и убедимся, что ниж-
няя челюсть совершенно гладкая, лишенная зубов, тогда как верхняя
челюсть усажена по краям мелкими коническими зубами. Близ внутрен-
них носовых отверстий (хоан), лежащих в передней части твердого
нёба, есть группы так называемых сошниковых зубов. Все эти образо-
вания служат только для удержания добычи, препровожденной в рото-
вую полость языком, но не для ее пережевывания.
Орудием для захвата добычи служит язык. Лягушка является
хищным, плотоядным животным; она охотится за движущейся добычей,
главным образом за насекомыми. Язык ее растяжим и вытягивается
в длину, когда выбрасывается лягушкой изо рта, чтобы прикоснуться
им к намеченной добыче. Он покрыт клейкой слизью, выделяемой
железками, открывающимися между мелкими поверхностными сосоч-
ками.
Задняя часть ротовой полости продолжается в глотку; границы
между ними нет, почему можно говорить об общей ротоглоточной
полости, выстланной внутри слизистой оболочкой, состоящей из
мерцательного эпителия. Цилиндрические клетки его на свободной
365
поверхности несут реснички, которые при Жизни животного накло-
няются то в одну, то в другую сторону. Если соскоблить скальпелем
слизистую оболочку рта недавно убитой лягушки и соскоб положить
в каплю физиологического раствора поваренной соли (т. е. 0,75%)
под покровное стекло препарата и рассматривать его при среднем
увеличении микроскопа, то можно наблюдать движение ресничек,
производящее на глаз впечатление мерцания пламени свечи.
Если ввести в рог ручку препаровальной иглы, то через короткий
пищевод можно проникнуть в желудок (рис. 231, gf). Определенной
границы между этими частями нет, и в физиологическом отношении
они до известной степени сходны. Железы слизистой (внутренней)
оболочки пищевода также выделяют пищеварительный фермент —
пепсин, служащий для разжижения белков и переведения их в пеп-
тоны.
Желудок на заднем своем конце утончается и на границе с тонкой
кишкой несет кольцеобразный перехват — привратник (pylorus),
запирающий выход из желудка.
Прилежащая к желудку часть кишки называется двенадцати-
перстной (duodenum); она принимает в себя протоки печени и под-
желудочной железы.
Без всякой внешней границы duodenum переходит в собственно
тонкую кишку (it), петли которой подвешены на брыжейке.
Чтобы увидеть последнюю, надо поднять кверху пинцетом любую
петлю кишки; еще лучше двумя пинцетами растянуть петлю в сто-
роны; тогда обнаруживается, что от кишки отходит сплошная про-
зрачная перепонка, в которой видны кровеносные сосуды. Эта пере-
понка и есть брыжейка; она представляет собой двойную складку
брюшины, выстилающей изнутри блестящим слоем плоского эпителия
брюшную полость и по средней линии спины переходящей на различ-
ные органы, в частности на кишечник.
Чтобы отпрепаровать пищеварительный канал, подрезают бры-
жейку по всему кишечнику и освобождающиеся петли отодвигают в
правую сторону лягушки. В вытянутом состоянии пищеварительный
канал ее не очень длинен, так как превышает длину тела всего раза
в полтора-два. Это объясняется тем, что лягушка питается животной
пищей, которая легче и скорее обрабатывается пищеварительными
соками. Связь развития кишечника с родом пищи становится нагляд-
ной, если сравнить пищеварительный аппарат лягушки и головастика.
Последний питается водяными растениями, и кишечник его в не-
сколько раз длиннее тела; в брюхе головастика он лежит закрученный
спиралью. Растительная пища труднее переваривается вследствие оби-
лия в ней клетчатки. Поэтому пища должна дольше задерживаться
в кишечнике, а это и достигается в более длинном пищеварительном
канапе. Отметим здесь же, что у головастика вокруг рта есть зубчатая
твердая терка, служащая для соскабливания растительных тканей.
Этот орган относится к группе личиночных органов, функционирующих
и существующих лишь в личиночной стадии развития животного.
Толстую кишку перерезают ножницами перед клоакой и отводят
в сторону. Теперь можно вскрыть кишечник, чтобы осмотретьего слизи-
стую оболочку. Сначала отрезают толстую кишку и в отверстие тон-
366
кой кишки вводят ветвь ножниц для рассечения ее стенки. Разрез
ведут кпереди. Смывают содержимое кишки и рассматривают ее вну-
треннюю поверхность. Она нежно шероховата благодаря присутствию
множества тонких складок; слизистой оболочке желудка присущи бо-
лее грубые складки.
На границе тонкой и толстой кищки имеется кольцевой внутрен-
ний клапан, или заслонка, препятствующий обратному поступанию
содержимого толстой кишки в тонкую.
В толстой кишке (рис. 231, гс) содержатся каловые массы, т. е.
неиспользованные остатки пищи, переваривавшейся в желудке и ки-
шечнике. Значительная часть ее всасывается слизистой оболочкой
тонкой кишки в кровь, с которой через брыжеечные сосуды всосан-
ные вещества разносятся по телу и потребляются на месте клетками
всех тканей.
Пищеварительных желез у лягушки две — печень (рис. 231, hp)
и поджелудочная железа (рп). Печень подразделяется на три лопасти,
или доли. На задней поверхности правой доли лежит желчный пузырь;
из него выходит пузырный проток (ductus cysticus), который соеди-
няется с печеночными каналами (d. hepatici) и образует общий желч-
ный проток (d. choledochus). Этот последний открывается в двена-
дцатиперстную кишку, предварительно приняв в себя выводные ка-
налы поджелудочной железы.
Железы слизистой оболочки желудка, тонких кишек, печень и под-
желудочная железа выделяют жидкость, богатую различными фермен-
тами, т. е. веществами, химически перерабатывающими пищу. Такая
обработка пищи происходит в полости пищеварительного канала,
откуда химически измененные пищевые вещества всасываются стен-
ками кишечника. Подобный способ пищеварения называется вне-
клеточным, или полостным.
Отрезанную прямую кишку (см. выше) вскрывают в часовом сте-
кле в воде, куда и выпускают ее содержимое. Обычно прямая кишка
является местом паразитирования множества простейших животных,
именно инфузорий родов Opalina и Nуdot herus. Отметим пока только
присутствие таких громадных по своим размерам одноклеточных
существ, как Opalina, ибо они хорошо различимы даже невооружен-
ным глазом.
Органы кровообращения. С характером обращения крови лучше
всего познакомиться на живой лягушке. Для этой цели ее пеленают
в сырую тряпочку, вытянувши задние ноги, и привязывают к проб-
ковой пластинке, в которой сделано небольшое четырехугольное
отверстие. Над этим последним растягивают плавательную перепонку,
укрепляя стопу ноги неподвижно булавками, вколотыми в пальцы.
Не следует особенно сильно растягивать перепонку и очень пере-
тягивать лягушку при привязывании, так как кровообращение в лапке
может приостановиться.
Приготовленный препарат помещают на столик микроскопа и рас-
сматривают плавательную перепонку в проходящем свете под слабым
увеличением.
В поле зрения микроскопа видно множество угловатых черных ко-
мочков — рто пигментные кдетки. В толще перепонки заметна сеть
367
тонких трубочек, между которыми кое-где проходят более толстые
каналы. По ним и движется кровь, бегущая сплошной красной струей
в крупных сосудах и медленно протекающая в тонких трубочках, где
можно различить даже отдельные эритроциты.
Сердце лягушки лежит над грудиной. Оно одето полупрозрачной
оболочкой, называемой околосердечной сумкой (pericardium). Чтобы
освободить сердце, поверхность его ощупывают пинцетом и таким
способом оттягивают перикардий и отрезают часть его ножницами.
В образовавшееся отверстие выскальзывает сердце, имеющее блестя-
Рис. 232. Вскрытое сердце бесхвостой амфи-
бии (жабы). Схема по Ж а м м у.
аа — аортная дуга; ас — сонная дуга; ар — легоч-
ная дуга; as — левое предсердие; ad — правое
предсердие; vt — желудочек; vd— желудочно-
предсердный клапан; pab — артериальная часть
артериального ствола; sv — венозный синус; vei—
задняя полая вена; ocs— передняя полая вена,
стрелками показан ход тока крови.
щую поверхность.
Оно состоит из кониче-
ской задней части — желу-
дочка (рис. 233, V), и двух
предсердий (рис. 233, D, S), ле-
жащих впереди от него. На-
ружной границы между пред-
сердиями нет. Из правой сто-
роны переднего края желу-
дочка выходит артериальный
ствол, вскоре делящийся на
две ветви.
Два предсердия (рис. 232)—
правое (ad) и левое (as) —
не сообщаются друг с другом
и порознь открываются в един-
ственный желудочек, стенка
которого изнутри морщи-
ниста: она гораздо толще,
чем в предсердиях. Если
сердце отвести вверх и впе-
ред, то можно видеть над ним
венозный синус (рис. 232, sv),
тонкостенный мешок, образо-
ванный слиянием верхних по-
лых вен (ocs) с нижней полой
веной (vei) и двумя печеноч-
ными венами. Сам же синус
открывается в правое пред-
сердие. В левое предсердие впадают легочные вены.
Работу сердца легко проследить на бьющемся органе. Она состоит
в попеременном сокращении и расслаблении мышечных стенок пред-
сердий и желудочка. Сокращение мышцы влечет за собой уменьше-
ние полости соответствующего отдела сердца; этот момент называют
систолой. Расслабление мышцы способствует, наоборот, расширению
полости; в таком случае говорят о диастоле.
Оба предсердия сокращаются одновременно. Систола предсердий со-
провождается диастолой желудочка. После их сокращения следует си-
стола желудочка и диастола предсердий. Во время систолы данная камера
сердца сжимается, бледнеет, и кровь из нее изгоняется; диастола же
характеризуется наполнением кровью соответствующей части сердца.
368
При биении сердца кровь перекачивается из одного отдела в дру-
гой, а так как к сердцу примыкают кровеносные сосуды, то есте-
ственно, что кровь из одних сосудов (вен) накачивается в сердце и
из него затем нагнетается в артерии.
Кровь течет всегда в одном определенном направлении, так как
между предсердиями и желудочком имеются две заслонки (рис. 232,
Рис. 233. Схема главнейших артериальных сосудов жабы.
По Ж а м м у.
D — правое предсердие; S — левое предсердие; V — желудочек; pb — лег-
кое; v — желудок; in—тонкая кишка; г — почка; ts — семенник;
cad — жировое тело; I— первая пара артериальных дуг; II — вторая
пара артериальных дуг; III — третья пара артериальных дуг; сгс —
общая сонная артерия; ас — кожная артерия; as — подключичная арте-
рия; abr — плечевая артерия; ат —брыжеечная артерия; ai — подвздош-
ная артерия; ар — легочная артерия.
vd), играющие роль клапанов. Они пропускают кровь из предсердий
в желудочек и препятствуют обратному току ее в предсердие при си-
столе желудочка.
Для уяснения хода крови по телу лягушки необходимо предвари-
тельно рассмотреть ветвление наиболее крупных кровеносных сосу-
дов. Отпрепаровать их сразу нелегко; поэтому мы и дадим некоторые
технические указания.
24 Практикум по зоологии — 385
369
Сердце лягушки захватывают пинцетом левой рукой, отгягивают
влево и к себе и вкалывают в него булавку так, чтобы конец ее про-
шел через все тело лягушки и вонзился бы в дно ванночки.
При таком положении сердца натягивается артериальный ствол и
его ветви. Чтобы яснее рассмотреть артерии, срезают тонкими ножни-
цами по ходу их соединительную ткань, удаляют мышечные пучки,
где они есть, и острой иглой освобождают сосуды от окружающих
тканей.
Отпрепаровав прилежащие к сердцу артерии, мы сможем просле-
дить, что каждая ветвь артериального ствола дает по три главные
артерии, идущие симметрично на каждой стороне тела. Поэтому и
говорят, что ветви артериального ствола дают три пары артериаль-
ных дуг (рис. 233): передняя называется сонной (I), средняя (вто-
рая) — аортной (II) и задняя (третья)—кожно-легочной дугой (III).
Сонная дуга (рис. 232, ас) коротка и тонка. Пройдя через так
называемую каротидную железу, она дает две ветви — язычную
артерию (a. lingualis) для питания языка и сонную — a. carotis
(рис. 233, сгс) для прочих органов головы.
Аортные дуги довольно толсгы; они загибаются назад и пере-
ходят в брюшную полость, где под позвоночником и сливаются друг
с другом в общий ствол — брюшную аорту, (aorta abdominalis,
рис. 233, II). Чтобы видеть яснее этот переход, необходимо все брюш-
ные органы отвести влево и подрезать корень брыжейки у позвоноч-
ника; затем удаляют мышечные тяжи в области основания передних
конечностей. Каждая аортная дуга дает затылочно-позвоночную ветвь
(a. occipi to vertebral is), орошающую мышцы лица и тела, и подключич-
ную артерию — a. subclavia (рис. 233, as), направляющуюся в перед-
нюю ногу.
От места слияния обеих аортных дуг отходит как бы в виде про-
должения левой дуги чревно-брыжеечная артерия — a. coeliaco-me-
senterica (рис. 233, ат), питающая желудок, кишечник, печень и
селезенку. Ее-то ветви и видны в брыжейке.
Далее назад аорта ложится между почками (рис. 233, г), к которым
и дает несколько пар мочеполовых артерий (a. urogenitales).
Сзади почек аорта делится вилообразно на две подвздошныг арте-
рии — a. iliacae (рис. 233, ai), снабжающие кровью задние конечности.
Кожно-легочная дуга дает две ветви: легочную артерию — a. pulmo-
nalis (рис. 233, ар), идущую в виде извилистого тяжа по краю легкого
до его верхушки, и кожную артерию — a. cutanea (рис. 233, ас),
которая проходит под кожей спины почти к заднему концу туловища.
Все эти артерии ветвятся на мелкие трубочки, впадающие в си-
стему капилляров; из последних кровь переходит в тонкие начальные
части вен, которые соединяются друг с другом в более крупные веноз-
ные стволы. Чем ближе кровь подходит к сердцу, тем по более тол-
стым венам она движется. Большинство этих сосудов идет параллельно
одноименным артериям, но ближайшие к сердцу вены имеют собствен-
ные наименования.
Из головы, передних конечностей и кожи венозная кровь течет
в сердце по двум верхним полым венам (v. cavae anteriores), из кото-
рых каждая образуется слиянием внешней яремной вены (у. jugularis
370
externa), большой коЖной венЪл (v. cutanea magna) и безымянной вены
(v. anonyma).
Из задних ног кровь идет по сосудам, образующим в брюшной
полости общие подвздошные вены (v. iliacae communes). Последние
погружаются в почки, где распадаются на систему тончайших ветвей,
из которых снова образуется сосуд — нижняя полая вена (v. cava in-
ferior), лежащая параллельно аорте с брюшной ее стороны. Благодаря
таким отношениям говорят о воротной системе почек.
Аналогичная воротная система есть и в печени, куда стекается
вся венозная кровь из пищеварительного аппарата по воротной вене
(v. portae). В печени воротная вена также распадается на систему
тонких трубочек, из которых собираются печеночные вены (v. hepati-
сае), впадающие в венозный синус.
Следует отметить, что препаровка вен очень трудна, и при усло-
виях обычной работы ею заниматься не приходится. Поэтому ограни-
чиваются рассмотрением доступно лежащих сосудов, как-то: нижней
полой вены, безымянных вен и других более или менее крупных ство-
лов.
Все эти вены несут кровь в венозный синус, через который она
и вливается в правое предсердие. Эта кровь в полном смысле слова
венозная, так как она содержит в себе углекислоту, выделенную тка-
нями тела, получившими из крови капилляров необходимый для сво-
его дыхания кислород.
В левое предсердие впадают короткие легочные вены (v. pulmona-
les), содержащие в себе богатую кислородом и очищенную от угле-
кислоты кровь.
Теперь рассмотрим, в какой последовательности кровь обращается
по телу, и для этого проследим полный ход некоторой ее частицы.
Возьмем ту порцию артериальной крови, которая при данной систоле
поступила в левое предсердие из легкого (рис. 232, as).
При систоле предсердия она переходит-в желудочек: при систоле
последнего эта кровь, благодаря особому устройству артериального
ствола (рис. 232, pab), попадает в первые две пары артериальных дуг —
сонную и аортную (рис. 232, ас, аа) и идет по их ветвям до капил-
ляров. Здесь артериальная кровь раскисляется, т. е. отдает кисло-
род тканям тела и взамен получает от них углекислоту, т. е. из арте-
риальной превращается в венозную. Далее она переливается в вены и
по ним возвращается в сердце, но уже в правое предсердие.
Таким образом заканчивается большой круг кровообращения,
конечными точками которого являются желудочек как исходный
пункт и правое предсердие как конечный пункт. На этом пути кровь
превращается в негодную для дыхания, и поэтому она должна очи-
ститься от углекислоты и обогатиться кислородом, что и происходит
в органах дыхания.
Из правого предсердия при его систоле венозная кровь переходит
в желудочек, откуда она выталкивается в третью (кожно-легочную)
дугу (рис. 232, ар) и частью во вторую пару артериальных дуг. В коже
и в легких кровь получает новый запас кислорода, отдает при этом
скопившуюся в ней углекислоту и по легочным венам возвращается
в сердце—в его левое предсердие. Кровь ушла из сердца и снова
• 371
йернулась в него, сделай по телу малый круг кровообращения, на пути
которого она из венозной превратилась в артериальную.
Здесь мы должны заметить, что в малом кругу кровообращения
артерии (третья пара артериальных дуг) несут венозную кровь, а вены
(легочные) заполнены кровью артериальной, т. е. создаются отноше-
ния, обратные тому, что есть в большом кругу.
Большой и малый круги кровообращения не вполне обособлены
друг от друга, так как они перекрещиваются в желудочке сердца.
В действительности кровообращение лягушки сложнее, чем только
что описано, так как предсердия сокращаются одновременно и вытал-
кивают заключающуюся в них кровь в одну общую камеру, т. е. в же-
лудочек. Следовательно, в него поступает и артериальная и венозная
кровь. Хотя внутренние стенки желудочка морщинисты, однако
артериальная и венозная кровь частично смешиваются в нем друг
с другом, благодаря чему в желудочке находятся: чистая артериаль-
ная, чистая венозная и смешанная кровь. При сокращении желудочка
все три сорта крови выталкиваются в артериальный ствол. Благодаря
известной последовательности в сокращении желудочка и особенно-
стям расположения отверстий артериальных дуг, сорта крови распре-
деляются по артериям в строго определенном порядке. Чистая веноз-
ная кровь попадает в кожно-легочные дуги (рис. 232, ар); смешанная
кровь, содержащая в себе некоторое количество кислорода, идет в
аортные дуги (аа) и служит для снабжения кислородом всех органов
тела, кроме головы; наконец,высший по своему качеству сорт — чи-
стая артериальная кровь — поступает целиком через сонные дуги
(ас) в «голову лягушки, попадая, в частности, в наиболее тонкие и
требовательные органы, именно — центральную нервную систему.
Если наличие двух кругов кровообращения связывается с легоч-
ным дыханием, то мы вправе ожидать, что кровеносная система моло-
дых головастиков, дышащих жабрами, должна быть устроена по
«рыбьему типу». И действительно, если опустить особенности ее строе-
ния, зависящие от ранней закладки и разрастания легких, то мы най-
дем сходство в строении кровеносной системы рыбы и головастика,
так как у последнего имеются четыре пары жаберных артериальных
дуг; по выходе из жабр они, в виде жаберных вен, образуют корни спин-
ной аорты. При переходе от жаберного дыхания к легочному капил-
лярная сеть жабр, как и сами жабры, исчезает, а артериальные дуги
переходят в соответствующие сосуды, свойственные взрослому живот-
ному, именно: первая пара дуг превращается в сонные дуги, вторая
пара их — в аортные дуги, третья пара редуцируется, а четвертая
пара дает легочные дуги.
Усложнение строения кровеносной системы лягушки зависит от
присутствия у нее легких. Ее трехкамерное сердце делает двоякую
работу — перегоняет кровь по телу для клеточного дыхания (большой
круг кровообращения) и перекачивает ее в легкие для артериализа-
ции (малый круг кровообращения).
Лимфатическая система. Ткани тела лягушки пропитаны жидко-
стью, называемой лимфой. Она образуется из жидких частей крови,
просачивающихся из капилляров в окружающие ткани, а также из
жидкостей, всасываемых из пищевых масс слизистой оболочкой кишеч-
372
ника. Лимфа содержит в себе белые кровяные клетки (лимфоциты).
Она обращается по системе лимфатических сосудов — тонкостенных
трубочек, открывающихся в вены. С лимфатическими сосудами нахо-
дятся в соединении подкожные лимфатические мешки и лимфатиче-
ские сердца, которые служат для накачивания лимфы в вены. .
Легче найти заднюю пару лимфатических сердец. Биение их можно
заметить на живой лягушке по движению кожи по бокам заднего конца
хвостовой кости. На препарате же снимают кожу с заднего конца
спины и прилежащих частей бедер и в двух трехугольных простран-
ствах между расходящимися мышцами обнаруживают два беловатых,
слабо бьющихся мешочка, которые своим передним концом впадают
в бедренную вену.
К лимфатической системе относится и крупный железистый ор-
ган — селезенка (lien), имеющая вид темнокрасной горошинки,
лежащей близ прямой кишки. Она построена из лимфоидной ткани и
содержит в себе громадное количество лимфоцитов, которые размно-
жаются в ней и поступают затем в кровеносную систему.
Органы дыхания. Чтобы рассмотреть легкие (рис. 231, р), у ля-
гушки удаляют сердце и печень и перерезают ножницами левое челюст-
ное сочленение, подрезают нижнюю стенку ротовой полости сей-
час же под пищеводом и отводят нижнюю челюсть влево от себя так,
как раскрывают книгу. Препарат прикалывают булавками ко дну
ванночки.
Мы имеем теперь возможность рассмотреть ротовую полость и
органы дыхания. Верхнюю стенку ротовой полости образует твердое
нёбо, на котором округлыми холмиками выдаются глаза. Кпереди от
них, близ края верхней челюсти, помещаются два внутренних носовых
отверстия (хоаны), внутри от которых идут сходящиеся под углом ряды
сошниковых зубов.
В задней части верхней стенки ротовой полости, внутрь от челюст-
ных сочленений, видны круглые отверстия евстахиевых труб —
каналов, соединяющих полость среднего уха с ротоглоточной полостью.
На откинутом влево дне ротовой полости, позади от заднего сво-
бодного конца языка, видно щелевидное отверстие гортани (larynx);
края ее выдаются кверху в виде овального бугорка. Ощупывая пин-
цетом этот бугорок, можно убедиться, что в нем заложены твердые
опорные части. Скелет гортани образован двумя трехугольными чер-
паловидными хрящами (cartilago arytaenoidea), которые своим осно-
ванием сидят на кольцевидном хряще овальной формы — cartilago
laryngotrachealis.
На внутренней поверхности черпаловидных хрящей находятся две
голосовые складки. Выталкиваемый при выдыхании из гортани воздух
приводит их в колебание, благодаря чему лягушка производит звуки
(кваканье). У самцов зеленой лягушки звуки усиливаются особыми
мешками-резонаторами, выпячивающимися наружу позади челюстных
сочленений.
Гортань помещается между задними рожками подъязычной кости
и принимает в себя отверстия двух мешковидных легких. Каждое
легкое представляет собой ячеистый изнутри мешок, в тонкой стенке
которого много эластической ткани. Вследствие этого легкое может
373
Пассивно то расширяться, то сокращаться. В стенках ячей легочная
артерия распадается на капилляры. Протекающая по ним венозная
кровь отдает углекислоту воздуху полости легкого, забирает от него
кислород и, сделавшись артериальной, по легочным венам напра-
вляется в левое предсердие.
Самое дыхание можно наблюдать на живой лягушке, у которой
носовые клапаны (выросты кожи у наружных носовых отверстий) то
открываются, то закрываются, причем попеременно опускается или
поднимается межчелюстная мышца (дно ротовой полости).
При открытых носовых отверстиях и закрытом рте лягушка опу-
скает дно ротоглоточной полости вниз; благодаря этому в последней
развивается отрицательное давление, под влиянием которого атмо-
сферный воздух проникает через ноздри, носовую полость и хоаны
в ротовую полость. По заполнении последней воздухом лягушка за-
крывает ноздри клапанами и, поднимая кверху межчелюстную мышцу,
нагнетает воздух в легкие, которые пассивно растягиваются под
влиянием его давления. Следовательно, дыхательной мышцей лягушки
является дно ротовой полости — межчелюстная мышца (рис. 230, sm).
Помимо легких, в процессе дыхания принимает важное участие и
кожа; растекаясь под эпидермисом по тонкостенным кровеносным
сосудам, кровь обменивается газами с воздухом через толщу эпите-
лиальных слоев кожи. Насколько велик такой газообмен, доказы-
вается опытами с вырезыванием легких у лягушки. Оперированные
таким образом животные продолжали жить на счет одного только
кожного дыхания. Последнее может производиться лишь влажной
кожей. При подсыхании кожи нарушается ее функция, и животное
гибнет, хотя легкие его и остаются нетронутыми.
Сравнивая органы дыхания рыбы и лягушки, т. е. жабры и легкие,
мы не находим между ними никакого сходства ни по строению, ни
по развитию — они не гомологичны друг другу. Однако у них есть
одна общая черта: и тот и другой органы сходны по функции, так
как оба служат для дыхания. Сходство органов по отправлению
(функции) называется аналогией; можно сказать, что легкие лягушки
аналогичны, но не гомологичны жабрам рыбы.
Припомним, что головастик лягушки дышит жабрами. Несомненна
аналогия его жабр таковым же органам рыбы, но есть ли между ними
гомология? Головастик вылупляется с наружными жабрами, являю-
щимися чисто личиночными органами, вторично возникшими у ля-
гушки. Они вскоре дегенерируют, и их роль переходит ко внутрен-
ним жабрам, развивающимся у головастиков в 9—10 мм длиной,
когда у них прорезаются наружу жаберные щели. Последние идут из
глотки наружу в полость, прикрытую жаберной крышкой. На осно-
вании сходства в положении жаберных щелей головастика и рыбы
мы можем гомологизировать их друг другу. Равным образом гомоло-
гичны как жаберные дуги, так и сами жабры, т. е. разрастания сли-
зистой оболочки дуг.
Таким образом жабры рыб не только аналогичны, но и гомоло-
гичны внутренним жабрам головастика, у которого последние явля-
ются временными органами, но в противоположность наружным
жабрам — органами первичного значения.
374
Рис. 234. Мочеполовые органы самца
лягушки. По Гексли и Мар-
шалу.
ts — семенник; ve —выносящие сосуды;
2 — почка; gad — надпочечник; cad — жи-
ровое тело; vs — семенной пузырек; г — пря-
мая кишка; vu — мочевой пузырь; cl —
клоака.
Мочепбловые органы. Ткани тела, потребляющие доставленные им
кровью пищевые вещества, выделяют в кровь же продукты обмена
веществ. Освобождение крови от них происходит в органах выделения,
состоящих из почек, мочеточников и мочевого пузыря.
Типичнее в анатомическом отношении устроены органы выделения
самки лягушки, потому что они совершенно не зависимы от ее поло-
вого аппарата.
Каждая почка (геп) имеет форму сплющенного широкого веретено-
видного тела темнокрасного цвета (рис. 231, г; рис. 234, z). Лежат почки
по бокам позвоночника в брюшной полости тела. К ним притекает кровь
по мочеполовым артериям и отдает мочевые продукты, вытекающие из
почек по выводным каналам — мо-
четочникам (ureter—иг)—в клоаку,
куда открывается также и тонко-
стенный мочевой пузырь, лежащий
на брюшной стенке тела, в задней
части брюшной полости (us).
Мочеточники самки по всему
своему протяжению одинаковой
толщины. В мочевом пузыре не-
редко паразитирует (у обоих полов
лягушки) крупный плоский червь
из сосальщиков — многоустка (Ро-
lystomum integerrimum).
> По брюшной стороне почки идет
узкая желтоватая полоса, не всегда
хорошо видимая, это — надпочеч-
ники (рис. 234, gad) — железы вну-
тренней секреции, т. е. такие орга-
ны, которые свои продукты (т. е.
секрет) выделяют не наружу и не
в полость органов, сообщающихся
с внешним миром, а внутрь тела,
в его кровеносную систему.
Мочевые органы самца тесно
связаны с органами размножения,
поэтому в данном случае приходится
говорить о мочеполовом аппарате.
Органами размножения являются половые железы (или семенники)
и семепроводы.
Семенников — testes (рис. 234, is) — у лягушки два. Каждый из
них имеет вид толстого плотного боба желтого или темного цвета.
От переднего полюса половой железы отходят лопастевидные лен-
точные жировые тела — corpora adiposa (cad), содержащие в себе
запас пищевых веществ на зиму. Семенники лежат под почками (а на
нашем препарате над ними). Если потянуть половую железу пинце-
том кверху, то обнаруживается тонкая прозрачная перепонка, связы-
вающая ее с почкой. В этой перепонке проходят тончайшие выводящие
канальцы (рис. 234, vef) для мужских половых клеток, или живчиков,
вырабатываемых семенником, Сквозь ткань почки семенная жидкость
?75
проникает в мочеточники, играющие у самца лягушки и вторую роль,
т. е. семепроводов. В связи с этим стоит также отличие в строении
их сравнительно с мочеточниками самки. Перед впадением своим в
клоаку у самца они образуют боковые расширения — семенные пу-
зырьки (vesiculae seminales, рис. 234, vs), служащие хранилищем
семенной жидкосги, которая извергается наружу через отверстие
клоаки.
Живчики, или сперматозоиды, имеют форму микроскопически ма-
лых хвостатых тел. Головка живчика в виде удлиненной палочки,
от заднего конца которой отходит тоненькая ниточка—хвост, обла-
дающая сократительной способностью. Ударяя хвостиком в разные
стороны, живчик плывет в окружающей его жидкости головкой впе-
ред.
Органы размножения самки лягушки состоят из яичников, яйце-
водов и маток; все эти части парные.
Женские половые железы, или яичники,—ovaria (рис. 231, ov)
образованы двумя замкнутыми мешетчатыми органами различного
цвета и величины, что зависит от степени половозрелости животного.
Мешок яичника всегда складчатый, снаружи он одет перитонеальной
оболочкой, на внутренней поверхности которой лежат развивающиеся
яйцевые клетки. В незрелом состоянии железа невелика и цвет ее
желтовато-коричневый или сероватый. Наоборот, у половозрелой самки
яичники очень велики; они занимают большую часть полости тела;
в них легко различимы крупные черные зерна — яйца. Яичник под-
вешен к спинной стенке брюшной полости на перитонеальной связке.
У переднего полюса его прикрепляется лентовидное, дольчатое жиро-
вое тело.
Выведение наружу яиц происходит по трубчатым яйцеводам —
oviducti (рис. 231, ovd). Они очень извилисты и петли их подвешены
на коротких брыжейках. Яйцеводы занимают боковые отделы брюш-
ной полости. У вскрытой лягушки они разбухают от действия воды.
Подрезая осторожно брыжейку, можно освободить яйцеводы по всему
их протяжению. Они очень длинны, и длина их превышает в несколько
раз длину тела лягушки. На переднем конце яйцевод несколько
сужается и открывается в полость тела воронкообразным отверстием
близ легких. У заднего конца он, наоборот, расширяется в мешетчатый
участок — матку, которая впадает в клоаку.
Зрелое яйцо выходит из яичника, прорывая его стенку. Вслед-
ствие этого оно попадает в полость тела и там передвигается благодаря
движению самой лягушки и ее кишечника. Случайно яйцо попадает
в воронкообразное отверстие яйцевода и втягивается в его просвет.
Благодаря сокращению мускульных стенок этого органа, яйцо мед-
ленно продвигается по яйцеводу, причем покрывается белковой обо-
лочкой, выделяемой внутренними железами этого органа. Через
матку и клоаку яйца выбрасываются наружу, где под влиянием воды
оболочка их разбухает и склеивается с оболочками других яиц; благо-
даря этому и образуются массы яиц, называемые в общежитии лягу-
шечьей икрой.
Сравнивая органы размножения лягушки и окуня, мы замечаем
ббльшую простоту устройства их у рыбы. Окунь имеет лишь половые
376
Железы, задняя часть которых является выносящим каналом. 6 общем
половые пути у окуня сравнительно с лягушкой развиты чрезвычайно
слабо и анатомически почти не выражены.
Мужские половые органы окуня совершенно самостоятельны и
никакого отношения к почкам не имеют, тогда как эти же органы
лягушки связаны друг с другом и образуют мочеполовые органы.
Женский половой аппарат окуня существенно отличается от ля-
гушечьего тем, что яйца окуня выводятся наружу через заднюю часть
яичника. У лягушки же они попадают сначала в полость тела и от-
туда уже проникают в воронку яйцевода.
Половое, равно как и мочевое отверстия рыбы не зависимы от от-
верстия задней кишки; следовательно, у рыбы клоаки нет, тогда как
этот орган характерен для земноводных.
У лягушки брачный период жизни приходится на раннюю весну,
как только растает лед. Самец обхватывают самку сверху первой па-
рой ног, и в таком состоянии спаривания проходит немало времени.
При копуляции и самка и самец выбрасывает через клоаку половые
клетки, т. е. яйца и живчики, в воду, где они находят сами друг друга.
Таким образом, при отсутствии особых органов совокупления, у ля-
гушки имеется внешнее оплодотворение, которое совершается вне
тела матери, в окружающей среде. Тут же протекает развитие потом-
ства, в судьбе которого родители не принимают никакого участия.
Органы дыхания Головастик Лягушка
Водные (жабры) Воздушные (легкие)
Органы пищеварения Приспособлены для ра- стительной пищи Приспособлены для жи- вотной пищи
Форма тела Хвостатая, изменяюща- яся в зависимости от степени развития ног Постоянная, типичная для взрослого животного
Среда обитания Исключительно вода Вода и воздушная среда
Движение Плаванье в воде при по- мощи хвоста и, позднее, ног Плаванье в воде при по- мощи задних ног, пры- гание на суше, ползание (всеми ногами)
Из оплодотворенного яйца лягушки в конце развития зародыша
выходит личинка, т. е. существо, нисколько не похожее на самую ля-
гушку. Личинка лягушки называется головастиком, так как тело ее
представляет как бы большую голову, снабженную хвостом. Самые
молодые головастики имеют наружные перистые жабры — органы дыха-
ния водного типа, ибо они приспособлены для восприятия кислорода,
растворенного в воде. Головастики, в противоположность взрослым
лягушкам, питаются растительной пищей, которую они соскабли-
вают околоротовой теркой. Во время дальнейшей самостоятельной
жизни головастики растут; наружные жабры их исчезают и замс-
377
йяются жабрами внутренними, спрятанными в особых жаберных
полостях позади головы. Все это время головастик лишен конечно-
стей. Позднее у него вырастает сначала задняя пара ног, потом по-
являются передние конечности, и головастик становится хвостатой
лягушкой. Затем хвост постепенно рассасывается, и животное прини-
мает типичный облик.
Такое развитие, которое сопряжено с изменением как внешности
животного, так отчасти его внутреннего строения и образа жизни,
называют превращением или метаморфозом.
На вышеприведенной таблице (стр. 377) сопоставлены важнейшие
отличия головастика и лягушки.
Жабры, хвост и ротовая терка головастика являются личиночными
органами, служащими лишь превращающемуся, но не вполне сформи-
рованному животному.
Нервная система лягушки состоит из центральной (головной
и спинной мозг) и периферической (чувствительные и двигательные
нервы); кроме того есть и симпатическая нервная система.
В условиях обычной работы можно рассмотреть только головной
мозг лягушки сверху. Предварительно срезают кожу с головы и пере-
резают позвоночник позади основания черепа острыми ножницами.
Затем берут лягушку в левую руку и перегибают ее так, чтобы место
разреза образовывало вершину угла. Тогда обнажается отверстие,
ведущее в череп.
В него осторожно вводят острую ветвь тонких ножниц и, дер-
жась в направлении глаза, рассекают крышу черепа. Ножницы вы-
нимают и в таком же направлении вводят их с другой стороны в то же
отверстие. Когда сделан второй разрез, захватывают задний край
очерченного костного лоскута пинцетом и оттягивают его кверху,
чтобы сломать у переднего основания. Если все сделано хорошо, то
на дне открывшейся ямки черепа обнаруживается верхняя поверх-
ность головного мозга. Подрезают и подчищают края препарата,
прикалывают его ко дну ванночки и рассматривают под водой.
Головной мозг состоит из нежной ткани, снаружи серого цвета.
Идя спереди назад, в головном мозгу можно различить следующие
части (рис. 235).
1) Непарная обонятельная доля круглой формы (lol/); от ее пе-
реднего конца отходят в носовую полость два коротких и толстых
обонятельных нерва (nn. olphactorii).
2) Сзади лежат два овальные, несколько расходящиеся своими
задними концами большие полушария — hemispheria (Jim) с продоль-
ной щелью между ними. Поверхность полушарий совершенно гладкая.
3) За полушариями следует промежуточный мозг (ci) в виде косо-
лежащего квадратного углубления; с ним связана маленькая сте-
бельчатая шишковидная железа (corpus pineale), являющаяся рудимен-
том (остатком) некогда существовавшего у позвоночных третьего,
непарного глаза на темени.
4) Позади помещаются зрительные доли—lobi optic! (Jo), пред-
ставленные двумя короткоовальными холмиками.
5) К их задней окружности прилежит тонкая поперечная пла-
стинка — мозжечок (cerebellum — crb).
378
Перекрест нервов и воронка
Рис. 235. Головной мозг лягушки.
Левый рисунок — сверху; правый
рисунок — снизу. По Зеленка.
lolf— обонятельные доли; hm — боль-
шие полушария; ci — промежуточный
мозг; пор—зрительные нервы; 1о —
средний мозг (двухолмие); inf—во-
ронка; hp — нижний мозговой прида-
ток; crb — мозжечок; v IV — четвертый
желудочек; ср — продолговатый мозг.
б) Последним отделом головного мозга является продолговатый
мозг — medulla oblongata (ср), в виде трехугольного участка, несу-
щего на верхней поверхности трехугольное же вдавление с плоским
дном — четвертый желудочек (vIV). Продолговатый мозг связывает
головной мозг со спинным.
Обонятельные доли и полушария составляют передний мозг, зри-
тельные доли (двухолмие) относятся к среднему мозгу; мозжечок
и продолговатый мозг являются задним мозгом.
С брюшной стороны вид головного мозга иной. Обонятельная
доля не обособляется от полушарий. Позади последних лежит пере-
крест зрительных нервов—chiasma (пор). Между задними ножками
перекреста помещается воронка — infundibulum (inf), с нижним моз-
говым придатком — hypophysis (hp).
соответствуют промежуточному мозгу.
Средний мозг снизу прикрывается во-
ронкой, а задний мозг на нижней по-
верхности не представляет никаких
особенностей.
Если разорвать полушария иглой,
то можно заметить существование
в них полостей, называемых мозго-
выми желудочками; они сообщаются
внутримозговым каналом (сильвиев
водопровод) с четвертым желудочком,
соединяющимся, в свою очередь, с цен-
тральным каналом спинного мозга.
От различных частей головного
мозга отходят десять пар головных
нервов: 1) обонятельный, 2) зритель-
ный, 3) глазодвигательный, 4) блоковой
(последние два нерва заведывают дви-
жением мышц глаза), 5) тройничный
(иннервирует глаз, носовую полость
и челюстную область), 6) отводящий
(глазное яблоко), 7) лицевой (иннервирует нёбо и мышцы нижней че-
люсти), 8) слуховой, 9) языкоглоточный и 10) блуждающий (сердце,
легкие, желудок и другие внутренние органы).
Через затылочное отверстие черепа продолговатый мозг без вся-
кой резкой границы переходит в спинной мозг, лежащий в спинномоз-
говом канале позвоночного столба. Чтобы его вскрыть, вводят ветвь
острых ножниц в отверстие спинного мозга уже имеющегося у нас
препарата (см. выше — препаровку головного мозга) и перерезают
невральные дуги позвонков с обеих сторон, предварительно срезав
по возможности все спинные мышцы. Отрезанный лоскут удаляют
пинцетом и обнажают верхнюю поверхность спинного мозга.
Он представляет собой удлиненно-коническое тело с продольными
спинной и брюшной щелями. Соответственно передним и задним
конечностям спинной мозг образует два не особенно ясных утолщения.
На уровне VII позвонка он переходит и концевую нить (filum ter-
minate). Спинной мозг дает десять пар спинномозговых нервов, из ко-
379
торых задние до выхода своего из позвоночника ложатся параллельно
концевой нити и образуют так называемый конский хвост (cauda
equi па).
Спинномозговые нервы можно видеть и при обычной препаровке
лягушки на верхней стенке тела с брюшной ее поверхности. Они
имеют вид тонких блестяще белых нитей.
Для головного мозга лягушки характерно слабое развитие перед-
него мозга и расположение всех его частей в горизонтальной плоско-
сти. Как наиболее нежная и чувствительная ткань, нервная система
спрятана в костно-хрящевых футлярах — черепе и позвоночном столбе,
защищающих ее от всяких вредных механических воздействий.
Симпатическая нервная система состоит из ряда мелких нервных
узелков, образующих собой с каждой стороны позвоночника погра-
ничный ствол (truncus sympathies). Соединяясь со спинномозговыми
нервами, ствол симпатической нервной системы иннервирует кишеч-
ник и образует нервные сплетения для различных внутренних орга-
нов (сердца, почек и др.).
Скелет лягушки
Ознакомиться со строением костяка можно только на специально
приготовленном препарате, сделать который на обычных практиче-
ских занятиях нельзя. Поэтом)' мы ограничимся лишь общим озна-
комлением со скелетом лягушки, отсылая интересующихся деталями
к более подробным курсам зоотомии и сравнительной анатомии.
Основой скелета является позвоночный столб (columna vertebra-
lis), состоящий из ряда мелких продырявленных косточек, называемых
позвонками (vertebra).Их девять; к заднему концу позвоночника при-
мыкает десятая косточка—длинная хвостцовая кость (os coccygis).
Передний, или первый, позвонок называется атлантом; он служит
для сочленения с черепом и является единственным шейным позвон-
ком. Следующие семь позвонков относятся к спинным, а девятый счи-
тается крестцовым. Хвостовые позвонки сливаются в одну кость — os
coccygis. Все девять позвонков имеют каждый отверстие между те-
лом позвонка и примыкающей к нему сверху невральной дугой. На-
ходясь друг против друга, эти отверстия образуют все вместе спинно-
мозговой канал, служащий для помещения в нем мозга.
Ребер у лягушки нет. Зачатки их целиком сливаются с попереч-
ными отростками II—IX позвонков.
Череп лягушки построен из хрящей (остатки хрящевой капсулы
головастика) и костей. Собственно черепная коробка узка и длинна;
края черепа ограничиваются широкой дугой челюстей. Глазницы
очень велики. Для черепа лягушки как земноводного животного ха-
рактерно присутствие двух бугорков (мыщелков) на затылочных ко-
стях, служащих для сочленения с атлантом. Кроме того типично
то, что последним нервом, выходящим из черепа, является десятый —
блуждающий нерв (n. vagus).
С позвоночным столбом в той или другой мере связаны так назы-
ваемые пояса конечностей, т. е. группы костей, соединяющие скелет
конечностей с основным скелетом тела.
380
Рис. 236. Гомология конечностей лягушки.
Скомбинировано по разным авторам.
Слева — типичная конечность наземного позвоночного;
посредине задняя нога, справа передняя нога лягушки.
Гомологичные части заштрихованы одинаково: а — бе-
дро; А — плечо; b — голень; В — предплечье; CjCa —
предплюсна (tarsus) и запястье (carpus); С, — плюсна
(metatarsus) и пястье (metacarpus); С4С< — фаланги
пальцев.
Между мышцами спины заложены лопатки (scapulae). К сужаю-
щемуся нижнему концу каждой из них причленяются узкая клю-
чица (clavicula) и более широкий коракоид (coracoideum). Их вну-
тренние концы упираются в грудину (sternum),, состоящую из
нескольких костных и хрящевых частей. Все эти кости образуют
пояс передних конечностей, которые сочленяются с суставной впади-
ной лопатки. Этот пояс является незамкнутым кольцом, непосред-
ственно не соединяющимся
с позвоночным столбом.
Пояс задних конечно-
стей называется тазом (pel-
vis). По форме своей он
похож на камертон, ножки
которого направлены впе-
ред; они сочленяются с бо-
ковыми отростками крест-
цового позвонка и явля-
ются подвздошными костя-
ми (os ilium) Ручке камер-
тона соответствует место
слияния обеих половин
таза. Самая задняя часть
его образована седалищны-
ми костями (os ischium),
нижний участок ручки
остается хрящевым; он со-
ответствует лонной кости
(os pubis). В месте соеди-
нения названных трех ча-
стей таза друг с другом
помещается вертлужная
впадина — суставная ямка
для головки бедренной ко-
сти.
Передняя конечность
(рис. 236) состоит из: 1) пле-
чевой кости — humerus (А);
2) лучевой (radius) и лок-
тевой (ul па), слившихся
вместе в предплечье (В); 3) мелких костей запястья—carpus (Сх, С2),
к которым примыкают стержневидные кости пястья — metacarpus (С3)
и фаланги пальцев (С4). Запястье, пястье и фаланги образуют скелет
кисти.
Задняя конечность (рис. 236) имеет соответственное строение:
1) бедро (а) образовано бедренной костью (femur); 2) голень состоит
из самой длинной кости, образовавшейся из слияния большой и ма-
лой берцовых костей — tibia и fibula (b); стопа сложена из: а) костей
предплюсны — tarsus (С\С2), б) пяти длинных плюсневых костей —
metatarsus (С3) и фаланг пальцев (С4).
Ноги лягушки являются довольно типичным примером наземной
341
конечности, расчлененной на три части, что дает возможность более
разнообразного движения, чем работа рыбьих нерасчлененных плав-
ников.
Скелет является образованием пассивным, отдельные кости ко-
торого соединяются друг с другом связками, хрящевыми прослой-
ками и суставными сумками. С костями срастаются своими конце-
выми сухожилиями скелетные мышцы. Сокращаясь под влиянием
импульсов, идущих из центральной нервной системы, мышцы приво-
дят в движение различные части скелета, благодаря чему двигается
и само животное.
Общие замечания
Оценивая с точки зрения симметрии расположение внутренних
органов лягушки, мы можем сказать, что двусторонне симметричными
являются все ее органы, кроме пищеварительного аппарата и селе-
зенки. Сравнивая внимательно форму и положение симметричных
органов, мы наверно найдем у разных лягушек, примеры неодинаковой
величины обеих почек, семенников, различие положения их на раз-
ных уровнях и т.п. Такие отклонения от симметрии не имеют никакого
принципиального значения, ибо свидетельствуют о склонности ор-
ганов к индивидуальной вариации, или изменчивости.
Что касается органов пищеварения, то они очень резко нарушают
двубоковую симметрию организма, так как желудок лежит слева от
средней линии тела, остальная же часть брюшной полости занята
налегающими друг на друга петлями кишек. Однако такое явное на-
рушение симметрии вторичное. На ранних стадиях развития пище-
варительный канал лягушки закладывается в виде трубки, симме-
трично лежащей на средней линии тела; и только позднее, когда на-
чинают обособляться и расти отдельные его части, они смещаются
в сторону и теряют свою первоначальную симметрию. Следовательно,
нарушение ее у лягушки нисколько нс умаляет общей типичности
двубоковой симметрии ее тела.
У лягушки мы не видим бросающейся в глаза метамерии, но r
структуре некоторых частей ее тела следы ее несомненно имеются.
Позвоночник образован из девяти сходных в общих чертах друг
с другом позвонков, следующих один за другим в линейном порядке.
Позвонки вполне можно считать метамерами по отношению к позво-
ночному столбу. Прямая мышца живота несет поперечные сухожиль-
ные полосы, которыми она также разбивается на несколько метамер.
В дополнение к этому заметим, что у молодых зародышей метамерия
вообще выражена резче, чем у взрослого животного.
Остановимся теперь на организации конечностей лягушки. Не-
смотря на неодинаковую величину их и отличие в форме различных
частей ног, можно заметить единство плана строения их даже только
при наружном осмотре. В самом деле, каждая конечность состоит из
трех частей: ближайшей к туловищу — плеча или бедра, следующей
за ней — предплечья или голени, и концевой части — кисти или стопы.
Сходство в общей архитектуре подтверждается и строением скелета.
Сопоставим на табличке соответственные части скелета доуг с другом
(рис. 236).
382
Передняя конечность -- . - - - - * Задняя конечность
Плечо Плечевая кость — А • Бедренная кость — а Бедро
Пред- плечье Лучевая и локтевая кости, слившиеся вместе — В Голенная кость (срастается из большой и малой бер- цовых костей) — b Голень
Кисть Два ряда коротких костей запястья — CtC2 Два ряда костей предплюс- ны — С± С% Стопа
Пять костей пястья—С8 Пять длинных плюсневых костей — С3
Фаланги четырех паль- цев — С4 Фаланги пяти пальцев С4
Общее соответствие частей этой таблицы бросается в глаза с не-
сомненностью. Нам остается только, коснуться некоторых расхожде-
ний в деталях.
Предплечье и голень типичной наземной конечности состоят ка-
ждое из двух костей. Кости предплечья у взрослой лягушки сливаются
вместе. Голенная кость имеет продольные бороздки, которые, равно
как и история развития, свидетельствуют, что эта кость образуется
слиянием большой и малой берцовых (следовательно, также двух) ко-
стей. В данном случае несоответствие плана строения сравнительно
с типом конечности только кажущееся, ибо оно позднейшего проис-
хождения.
Не больше значения имеет различие в числе пальцев и пястных
костей кисти лягушки. Пальцев — четыре, а пястных костей — пять.
Пятый палец у лягушки во время ее родового развития исчез. За исчез-
новением пальца такой же судьбе подверглась и соответствующая
пястная кость; но пока она еще не утеряна лягушкой, хотя и стоит
на этом пути, так как пястная кость пятого пальца очень мала по
сравнению с четырьмя остальными своими соседками. В данном слу-
чае мы имеем пример исчезновения органа, или его редукции.
Установив гомологию ног лягушки, объясним себе те второстепен-
ные отличия, которые замечаются между передними и задними ее
ногами. Отличия эти зависят от различия в функции, выпадающей
на долю этих органов лягушки.
Передние ноги служат для поддержания тела при сидении и для
переползания с места на место. Работа, выполняемая ими, невелика;
поэтому ноги коротки и мускулатура их развита соответственно слабо.
Наоборот, задними ногами лягушка отталкивается от земли при
прыжке. Они являются рычагами, приводящими животное в движе-
ние. Чем больше плечи рычага, тем, при прочих равных условиях,
значительнее эффект его действия. В связи с этим обстоятельством
мы отмечаем значительную длину всех частей задних ног, особенно
голени, кость которой является самой длинной из всего скелета ля-
гушки.
383
Для успеха действия рычага необходима крепость его пЛеч. Этому
условию отвечает строение голени, опорой которой является един-
ственная кость, образовавшаяся слиянием двух .костей — большой
и малой берцовых.
Помимо скакания, лягушка может и плавать, что она делает также
своими задними ногами. Отталкивающей поверхностью является пла-
вательная перепонка стопы задних ног. Чем больше ее поверхность,
тем лучше животное может плавать. И действительно, пальцы стопы
значительно удлинены (особенно, если сравнить их с короткими паль-
цами кисти передней ноги) и подвижны. Последнее также важно,
ибо натянутая между ними плавательная перепонка должна попере-
менно то складываться, то расправляться — складываться при под-
тягивании ног вперед (чтобы уменьшить сопротивление воды) и ра-
скрываться при толчке ног назад. В последнем случае необходимо
в возможно большей мере использовать сопротивление воды, так как
благодаря этому тело лягушки получает толчок вперед.
Итак, задние ноги являются главным органом движения лягушки.
Как скелет, так и мускулатура их развиты очень хорошо. Из этих
сопоставлений мы убеждаемся в существовании связи формы (в ши-
роком смысле этого понятия) органа с его функцией.
Работа органов, а следовательно, и жизнь животного в значитель-
ной мере зависят от окружающей среды. Например, условия передви-
жения и дыхания в воде совершенно иные, чем на суше, и т. п. Орга-
низм в течение своего родового развития (т. е. филогенеза) приспо-
собляется к условиям окружающей его среды, и в строении его тела
мы можем найти те или другие следы такого приспособления. Ля-
гушка живет и в воде и на воздухе, но при условии достаточной его
влажности. В связи с этим она может плавать и прыгать, пользуясь
приспособленными к тому задними ногами. Кожа ее, слизистая и
влажная, играет большую роль при дыхании. Личинки лягушки
приспособлены к жизни только в воде; у них органы дыхания вод-
ного типа, т. е. жабры, и для плавания служит другой личиночный
орган — хвост.
Из произведенного выше примерного анализа строения некоторых
частей тела лягушки можно прийти к заключению, что не все признаки
(или факты) анатомического строения имеют одинаковую цену. Одни
из них важны, другие же, наоборот, не имеют первостепенного зна-
чения. Важно, например, что передняя нога состоит из трех частей,
тогда как наличие в ее кисти только четырех пальцев вместо пяти
не имеет принципиального значения.
Из всей массы деталей организации животного надо уметь выбрать
те признаки, которые характерны в том или другом отношении. Зна-
чение признаков определяется путем сравнения их у различных жи-
вотных (сравнительная анатомия) и посредством изучения связи дан-
ного животного с окружающей средой (экология). Признаки экологи-
ческого характера для лягушки нами уже отмечены. Что же касается
крупнейших фактов морфологического характера, то лягушку как
животное позвоночное (подтип Vertebrata) характеризуют:
а) присутствие внутреннего осевого скелета в виде спинного
столба;
384
б) положение центральной нервной системы на спинной стороне
тела и
в) положение сердца на брюшной (нижней) стороне тела.
Как животное земноводное (класс Amphibia), лягушка обладает
мягкой слизистой кожей, конечностями наземного типа (некоторые
земноводные вторично лишены двух или всех ног), двумя затылочными
сочленовными бугорками на черепе и отличается отсутствием пере-
городки в желудочке сердца.
Лягушка относится к бесхвостым амфибиям (отряд Anura), так
как во взрослом состоянии она лишена хвоста, который бывает у нее
только в качестве личиночного органа. У развитого животного остат-
ком скелета хвоста является стержневидная хвостцовая кость — os
coccygis. которой оканчивается позвоночник.
У лягушки есть язык; поэтому она принадлежит к группе языч-
ных амфибий (подотряд Phaneroglossa); коракоиды ее плотно сра-
стаются с грудиной, как и у других неподвижногрудых земноводных
(Firmisternia). Среди последних она занимает место в семействе Ra-
ni dae, характеризующемся круглым или горизонтальным зрачком,
отсутствием железистых бугров по бокам головы, глубокой вырезкой
на заднем конце языка, нерасширенными на своих концах пальцами
и верхнечелюстными и сошниковыми зубами.
Род Rana можно распознать по сошниковым зубам, горизонталь-
ному зрачку, раздельным пальцам передних ног и хорошо развитой
плавательной перепонке.
К роду Rana относится около 150 различных видов, которые от-
личаются друг от друга по еще более мелким признакам.
Rana esculenta — съедобная лягушка — имеет косо лежащие
между внутренними носовыми отверстиями сошниковые зубы в виде
овальных, выпуклых групп. Передний и задний края их заходят за
уровни краев хоан. Морда слабо заостренная. На висках темного
пятна нет.
Rana temporaria, серая лягушка. Сошниковые зубы в виде
двух маленьких овальных групп, сдвинутых более назад, чем у пре-
дыдущего вида. Передний край их лежит сзади уровня переднего
края хоан. Задний край группы зубов заходит значительно кзади от
уровня заднего края хоан. Морда тупо закругленная. На висках тем-
ные трехугольные пятна.
Обратимся к отличиям в организации окуня и лягушки, сопоста-
вляя соответствующие признаки параллельно друг другу (табл, на
стр. 386).
Признаки, отмеченные в этой таблице номерами 3, 4, 6, 7 и 8,
отличают вообще рыб от амфибий. Об остальных же признаках
следует оговориться, так как наружный скелет у некоторых рыб (на-
пример, у угря) отсутствует. Конечности могут не быть у амфибий
(например, у червяги); утеря конечностей в таком случае вторичная.
Некоторые амфибии пожизненно дышат жабрами (наружными) и лег-
кими или же вовсе лишены органов дыхания, и тогда газообмен у них
совершается только через кожу (например, у низших хвостатых амфи-
бий — Plethodon и др.).
Что касается предпочки, то она остается в виде рудиментарного
25 Практикум по аоологии — 385 385
органа лишь у осетровых и некоторых костистых рыб; у остальных же,
как и у амфибий, она исчезает.
Признаки Окунь Ля г у ш к а
1) Наружный скелет Есть (чешуя) Отсутствует
2) Конечности Водного типа (плавники) Наземного типа (ноги по типу членистого рычага)
3) Сердце Двукамерное (венозное) Трехкамерное
4) Кровообращение Один круг Два круга
5) Дыхание Жаберное (внутренние жабры) Легочное (внутренние жабры у головастиков)
6) Головная почка (предпочка) Остается в виде руди- мента Исчезает
7) Клоака Отсутствует Есть
8) Орган слуха Внутреннее ухо (орган равновесия) Среднее и внутреннее ухо
КЛАСС ПРЕСМЫКАЮЩИЕСЯ (REPTILIA)
ЯЩЕРИЦЫ (SAURIA)
ЯЩЕРИЦА ЗЕЛЕНАЯ (LACERTA VIRIDIS)
(или другие виды)
Наружный осмотр
Тело ящерицы делится явственно на голову, удлиненное валько-
ватое туловище и длинный, постепенно утончающийся хвост. Голова
соединяется с туловищем хорошо выраженной шеей, что позволяет
ящерице двигать головой в разные стороны. Тотчас позади шеи туло-
вище несет пару передних ног. а на своем заднем конце — пару силь-
нее развитых задних ног. Обе пары ног пятипалые.
Голова и все тело ящерицы покрыты сильно ороговевшими кожными
покровами; благодаря ороговению эпителия кожи последняя очень
суха; этот признак характерен для всех пресмыкающихся, сухая кожа
которых резко отлична от влажных слизистых покровов земноводных.
Структура поверхности покровов ящерицы в разных местах тела
различна. Ороговевшие части образуют различные по величине че-
шуйки; они мелки на верхней стороне шеи, начиная от уровня слухо-
вых отверстий, и на спинной стороне ног и туловища вплоть до уровня
клоаки. Лежат они правильными поперечными рядами. Число их в по-
перечном ряду имеет значение в систематике ящериц. Хвост покрыт
правильно чередующимися кольцами узких и длинных чешуек.
386
По бокам туловища мелкие чешуйки резко граничат с крупными,
лежащими в шесть продольных рядов, поперечными пластинками.
В области плечевого пояска их расположение сменяется на панцырное;
соответственно изменяется и их форма. Несколько мельче панцирные
чешуйки на нижней стороне шеи; они заходят на голову до уровня
середины челюстей, вдав’аясь клином между крупными боковыми щит-
ками челюстей.
Позади основания задних ног видна ограниченная спереди круп-
ным клоакальным1 щитком поперечная щель клоаки.
Голова покрыта крупными щитками (рис. 237, А). Рассматривая
голову сверху, начиная спереди, находим: межчелюстной щиток (1),
спускающийся вперед и вниз к месту соединения верхних челюстей; за
ним лежат два носовых щитка (2),
в нижнебоковых частях которых видны
круглые носовые отверстия; далее
кзади лежит непарный лобноносовой
щиток (3); за ним — два предлобных
1
в 3
ове ящерицы. Из Чернова.
Д-щитки;голо вы Lacerta vivipira сверху: 1 —междучелюстной; 2— носовые; 3 — лоб-
ноносовой; 4—скуловые; 5 — предлобные; 6—верхнересничные; ✓—надглазничные; 8 — лоб-
ный; 9 — лобнотеменные; 10 — теменные; 11 — межтеменной; 12 — затылочный. В—голова
Lacerta vivipara сбоку. Щитки: 1 — междучелюстной; 2 — передний носовой; 3 — задний
носовой; 4 — передний скуловой; 5 — задний скуловой; 6—верхнересничные; 7 — ушной;
8 — барабанный; 9 — верхнегубные.
щитка; затем идет большой срединный лобный щиток (8); по его боками
лежат надглазничные щитки (7), образующие с каждой стороны про-
дольный ряд из трех щитков; два передние соприкасаются с лобным
щитком; позади последнего помещается пара лобно-теменных щит-
ков (9); за ними по средней линии головы узкий межтеменной
щиток (11), сдавленный с боков громадными теменными щитками;
наконец, между их задневнутренними участками лежит маленький
затылочный щиток (12).
При рассматривании головы сбоку (рис. 237, В) видна ротовая
щель, ограниченная челюстями. Верхняя челюсть покрыта с боков
рядом верхнегубных щитков; над ними позади носового отверстия
лежат последовательно: задний носовой щиток (двойной), передний и
задний скуловые щитки. Над глазной впадиной сверху виден ряд
1 Его нередко называют анальным щитком, что неверно, так как пресмыкаю*
щиеся не имеют анального отверстия, но обладают оз верстием. клоаь и.
387
мелких еерхнересничных щитков. Между глазом и закрытым бара-
банной перепонкой слуховым отверстием расположены два щитка:
передний — ушной, и задний — барабанный.
На нижней поверхности головы, на уровне заднего края слухо-
вого отверстия, видна горловая складка покрова чешуек, а задний
конец шеи оторочен рядом более крупных щитков в виде во-
ротника.
Теменной щиток несет в середине светлую точку или маленький
кружок; под ним лежит рудиментарный третий, или теменной, глаз
(рис. 237, А, 11).
Глаза имеют верхнее и нижнее века и мигательную перепонку
(membrana nictitans).
Передние ноги состоят, как обычно, из плеча, предплечья и пяти-
палой кисти; пальцы —тонкие, оканчиваются хорошо развитыми, креп-
кими и острыми на концах коготками.
Задние ноги значительно крупнее передних. Пятипалая стопа
с длинными пальцами; особенной длины достигает четвертый палец;
пятый палец резко обособлен от остальных. Коготки пальцев стопы
более короткие и крепкие, чем на кисти.
По задненижнему краю бедра виден ряд отверстий — пор, из кото-
рых торчат хитиновые стерженьки. Число этих бедренных пор (рис.
238, р) имеет значение в систематике ящериц.
Длина тела ящерицы, включая и хвост, определяется от переднего
края головы до конца хвоста. Измерению подвергаются экземпляры
с нормальным (нерегенерировавшим) хвостом. Хвост у ящерицы
чрезвычайно ломкий; вследствие этого она легко его отрывает в случае
опасности (автотомия). Оторванный хвост регенерирует; нередки слу-
чаи уродливой регенерации.
Окраска покровов ящерицы обусловливается наличием в коже
пигментных клеток. У некоторых видов, как хамелеоны, а в нашей
фауне — агамы, пигментные клетки легко сокращаются и меняют
свое положение в слоях кожных покровов, что сказывается изменением
цвета покровов («игра цветов» покровов).
Ороговевший чешуйчатый покров весьма характерен для ящериц
(как и для других пресмыкающихся), чем они очень резко отличаются
от земноводных. В связи с этим земноводных (Amphibia) называют
также голыми гадами, а пресмыкающихся — чешуйчатыми гадами.
Старые ороговевшие слои эпидермиса сбрасываются (этот процесс
является линькой) и замещаются новыми за счет ороговения поверх-
ностных слоев эпидермиса.
Осмотр ротовой полости. Раскрывают тупым пинцетом ящерице рот
возможно шире. На дне ротовой полости лежит мясистый язык, раз-
двоенный на конце. Оттягиваем пинцетом язык кпереди. У его осно-
вания сверху видно продольное валикообразное утолщение, вздутое
на переднем конце, где оно имеет входное отверстие. Это дыхательное
отверстие, гортань и начало трахеи. Внизу в глубине ротовой полости
широкий вход в пищевод, куда легко проникнуть ножками тупого
пинцета.
Нижние челюсти усажены рядом мелких зубов, присутствие кото*
рых легко обнаружить, проводя по челюсти ножкой пинцета. На верх"
388
ней стенке ротовой полости (нёбо) видны два отверстия в передней
ее части — хоаны, или внутренние носовые отверстия.
Срединная щель в задней половине ограничена с боков крыловид-
ными костями, несущими на себе зубы. Зубы имеются и на верхних
челюстях; форма зубов коническая.
Вскрытие ящерицы
Ящерицу прикалывают брюхом кверху ко дну ванночки или пре-
паровальной доски, булавки вкалывают в основания ног.
Сначала вскрывают кожу продольным разрезом от анального щитка
до нижней челюсти; лоскуты кожи отпрепаровывают и отводят в сто-
роны, прикалывая булавками ко дну ванночки.
По брюшной стенке тела ящерицы лежит парная прямая брюшная
мышца (m. rectus abdominis); сухожильными перемычками она под-
разделяется на метамерные части. Кпереди от нее под грудиной лежат
грудные мышцы (m. pectoral is). Между ветвями нижней челюсти натя-
нута мембранозная подъязычная мышца (m. mylohyoideus) с поперечно
лежащими волокнами.
Пинцетом захватывают складку брюшной стенки тела перед
основанием задних ног и ножницами разрезают ее насквозь. Затем
вводят ветвь ножниц в полость тела и рассекают его брюшную стенку
по средней линии вплоть до щитков нижних челюстей. При разрезе
испытывают некоторое сопротивление во время рассекания пояска
передних конечностей. Внутреннюю ветвь ножниц держат кверху,
чтобы не поранить сердца и сосудов.
Пинцетами разводят лоскуты разреза в стороны; в передней трети
разреза лоскуты отпрепаровывают скальпелем от внутренних орга-
нов, а на шее отделяют только кожу, чтобы не прорвать тонкой стенки
заднего отдела ротовой полости и начала пищевода.
Осмотр внутренних органов и их препаровка. 1. Идя спереди на-
зад, рассматривают узкую и длинную трахею (рис. 238, а), перед-
няя часть которой лежит между выпуклостями, образованными мыш-
цами, покрывающими снизу нижние челюсти; отмечаем выдвинутое
вперед положение гортани. Задний конец трахеи на препарате
прикрыт трехкамерным сердцем (b, с, d).
Глубже под сердцем (на препарате) видны спавшиеся легкие (е);
альвеолярность стрдения стенки легкого видна и на свежем объекте;
левое легкое выступает из-под сердца и лежит свободно; правое при-
крывается на значительном протяжении печенью.
Печень (g) обычного цвета лежит вдоль тела в виде узкого органа ;
из-под ее левого (на препарате) края выступает крупный шаровидный
темный грязнозеленый желчный пузырь (h). Между печенью и расходя-
щимися легкими виден узкий желудок (f).
Позади печени справа и слева на препарате тонкая кишка (z) делает
петлеобразные изгибы; справа они заходят далее назад, чем слева
(ориентировка на препарате). В задней части брюшной полости видна
прямая кишка (к), уходящая задним концом под тазовое сочленение.
Оттянутые вбок лопасти стенки тела выстланы изнутри черной
брюшиной. В их толще выступают тонкие ребра. От печени к правому
предсердию тянется хорошо выраженная печеночная вена.
389
рону препарата; подрезают
Рис. 238.
Расположение внутренних органов
зеленой ящерицы. Оригинал.
а — трахея; b — левое предсердие; с — правое предсер-
дие; d — желудочек; е — легкое;/ — желудок; g — пе-
чень; Л— желчный пузырь; i— тонкая кишка; к —
прямая кишка; $ — лонное сращение; cl — клоака;
р — бедренные поры.
2. Подрезая удерживающие связки, отводят печень в левую сто-
брыжейку и отодвигают желудок вправо;
при этом обнаруживают по-
степенный переход пище-
вода в желудок, резкий пе-
реход пилорической части
желудка в двенадцати-
перстную кишку.
Поворачивая желудок,
на его боковой, обращенной
к печени стенке находят
удлиненную полосовидную
поджелудочную железу, до-
ходящую задним концом до
двенадцатиперстной кишки.
Отводя влево прямую
кишку, обнаруживают вы-
ступающие из полости таза
половые пути с семенни-
ком овально уплощенной
формы (семенник виден на
препарате справа и, следо-
вательно, по отношению к
самому животному является
левым).
3. Подрезают брыжейку
и вытягивают в длину весь
пищеварительный канал,
при этом полностью откры-
вают правое легкое яще-
рицы.
Рассматривают части пи-
щеварительного канала:пи-
щевод, желудок, тонкую
кишку и прямую кишку
(рис. 239).
Передвигая лопасти пе-
чени и поворачивая ее в сто-
роны, рассматривают форму
печени.
В брыжейке, на уровне
тонкой кишки, находят ма-
ленькое компактное тело
в форме челнока — селе-
зенку.
В задней части препа-
рата открывается семенник правой половины тела ящерицы; он вы-
двинут вперед по сравнению с левым.
По средней линии тела просвечивает позвоночник с лежащей над
ним (в препарате) брюшной аортой.
390
4. Срезают кривыми ножницами выдающийся вперед угол тазо-
вых костей. Отворачивают лоскут клоаки влево и прикалывают его
ко дну ванночки. В глубине препарата отыскивают удлиненные почки
с округлыми выпуклыми снаружи дольками (рис. 240—241, г); зад-
ними концами они уходят под прямую кишку; от них отходят ко-
роткие мочеточники.
Иглами поворачивают семен-
ники (t), укладывая их горизон-
тально по их наибольшей плос-
кости ; отводят семенник в сторону и
рядом с ним обнаруживают малень-
кий зерновидный надпочечник (sr ).
От семенника идет семевынося-
щий канал, соединяющийся с моче-
точником. Параллельно с семенни-
ком лежит придаток семенника (nt),
от'которого идет кзади гофрирован-
ие извивающийся канал, являю-
щийся остатком мюллерова канала,
который у самок сохраняется в ка-
честве функционирующего органа
(яйцевод).
Вентрально от прямой кишки
лежит мочевой пузырь, впадающий
в клоаку. В основании хвоста ле-
жат два выворачивающихся через
клоаку шиповатых мешка, являю-
щихся копулятивными органами.
Самка ящерицы. При
вскрытии самки ящерицы, прежде
всего, бросаются в глаза крупные
яйца яичника (рис. 242, ov); по-
слрлнир бпгяты жрити-лм и пл гис.ло». внутренние иргаиымщерицв.
следние ООГаТЫ желтком И по . в раскрытом виде. Оригинал. Ооо-
мере его выработки и накопления значения см. на рис. 238.
СИЛЬНО увеличиваются В размерах. Г, Л —семенник; $ —брыжейка.
Яичники лежат несимметрично;
они подвешены на брыжейках. Величина яичников связана с периодом
половой деятельности. Перед спариванием яичники сильно увеличи-
ваются за счет накопления питательного желтка в яйцах. Медиально
от яичника и дорзально от него лежит надпочка (пг).
По бокам брюшной полости лежат два яйцевода (d) в виде гофри-
рованных трубок; передний конец яйцеводов открывается в брюшную
полость широкой воронкой. Другим концом яйцеводы впадают
в клоаку.
С брюшной стороны в клоаку открывается мочевой пузырь, как
и у амфибий., не имеющий соединения с мочеточниками, которые
открываются туда самостоятельно.
Различия в клоаке самца и самки ящерицы зависят от того, какие
открываются в нее функционирующие органы (см. ниже приведенную
таблицу).
Вс к-p ыт не головного мозга. Снимают препарат со
дна ванночки и, держа его в руке, надрезают сочленение с позвоноч-
ником в поперечном направлении и разрезают череп с боков. Захватив
верхнюю часть затылочной области крепким пинцетом, отламы-
вают, поднимая кверху, крышу черепа. Препарат прикалывают ко
дну ванночки и рассматривают в воде, убрав иглой или тонким
пинцетом мягкие остатки, прикрывающие мозг.
Рис. 240—241. Мужские половые органы ящерицы. Оригинал.
А — после вскрытия лонного сращения; В — после удаления брюшной стенки вплоть до клоаки;
С —после полного выделения, t—семенник; г—почка; аг—надпочка; п|—придаток семенника.
Самой крупной частью головного мозга являются большие полу-
шария (передний мозг), от которых кпереди отходят булавовидные
обонятельные доли. Гладкими внутренними поверхностями полуша-
рия тесно прилежат друг к Другу.
Самец
Самка
Отверстие прямой кишки
Пара мочеточников, соединяю- Пара мочеточников
щихся с семепроводами Пара яйцеводов
Отверстие мочевого пузыря
Пара мешков копулятивных
органов
Позади больших полушарий лежит двухолмие, образованное парой
зрительных долей (lobi optici), относящихся к среднему мозгу. JAewy
ЗУ2
большими полушариями и зрительными долями в центре бывает виден
верхний мозговой придаток,\или эпифиз, являющийся рудиментом
третьего глаза. Он относится к промежуточному мозгу.
Позади зрительных долей лежит в поперечном направлении слабо
развитый мозжечок. Далее кзади виден продолговатый мозг, постепенно
переходящий в спинной мозг.
Для головного мозга ящерицы характерно
значительное развитие больших полушарий.
$
DV
Рис. 242. Женские поло-
вые органы ящерицы.
Оригинал.
ov— яичник; d— яйцевод;
пг—надпочка; г — почка.
Общие замечания
Ящерица является типичным наземным
животным; опасность высыхания тканей тела
при жизни в воздушной среде, бедной в отно-
шении влажности, компенсирована орогове-
нием поверхностных слоев эпидермиса. Парал-
лельно исчезли и кожные железы. Благодаря
этому ящерица имеет сухие кожные покровы и
может жить (в лице различных представите-
лей) на каменистом субстрате и в пустынях.
Ящерица отличается от земноводных чрез-
вычайной подвижностью. Она не только очень
быстро бегает, но отлично карабкается по ше-
роховатым вертикальным поверхностям; цеп-
кость ее обеспечивается наличием крепких и
острых ороговений на концах пальцев ног.
Питается она насекомыми.
По сравнению с земноводными, ящерица,
как пресмыкающееся, делает следующий шаг
в эволюции позвоночных, переходя целиком
к наземной жизни.
Ящерица относится к классу пресмыкаю-
щихся (Reptilia); характерная черта строения их покровов уже была
отмечена выше.
Сравнение организации лягушки и ящерицы
Признаки Лягушка Ящерица
Кожа Влажная Сухая
Кожные железы Многочисленны Отсутствуют
Ороговение эпидермиса .... Слабое Очень сильное
Кожное дыхание Есть Отсутствует
Трахея Отсутствует Есть
Легкие Есть Более ячеисгы
393
Признаки Лягушка Ящерица
Сердце Трехкамерное Трехкамерное с нача- лом подразделения желудочка
Я й ц е к л а д у щ и е
Размножение Мечут икру Откладывают яйца; некоторые яйцеживо- родящи
Головной мозг Слабое развитие переднего мозга Более сильное развитие больших полушарий (передний мозг)
КЛАСС ПТИЦЫ (AVES)
КИЛЕГРУДЫЕ ПТИЦЫ (CARINATAE)
ГОЛУБЬ (COLUMBA LIVIA) И КУРИЦА (GALLUS DOMESTICA)
Технические замечания. За полчаса перед вскрытием птицу
убивают хлороформом в банке или ведре с крышкой. Для вскрытия вместо ван-
ночки берут препаровальную доску, на которой после наружного осмотра растя-
гивают голубя, захватив веревочными петлями основание крыльев и ног. Вере-
вочки натягивают так, чтобы птица не болталась, и концы их крепко привязывают
к гвоздям препаровальной доски.
Наружный осмотр
Плотное и компактное туловище голубя имеет голову на чрезвы-
чайно подвижной шее, две пары конечностей—пару задних ног и
пару передних, превращенных в крылья, и хвост.
Голова несет клюв, образованный двумя ороговевшими снаружи
половинками — надклювьем и подклювьем. Первое у своего основания
слегка вздуто и покрыто беловатой кожицей, богато снабженной
нервными окончаниями; называется она восковицей и служит органом
осязания у птицы. Перед восковицей помещаются два наружных носо-
вых отверстия в виде щелевидных ноздрей.
Края клюва ограничивают вход в ротовую полость. Раскрыв клюв,
мы на дне ротовой полости находим узкий и заостряющийся на конце
язык, по форме соответствующий впадине подклювья. Если потянуть
пинцетом язык вперед, то позади его расширенного основания легко
увидеть щелевидное отверстие дыхательного горла.
По бокам головы помещаются глаза. Они более выпуклы, чем у
рыбы, и сравнительно велики, что и понятно, так как зрение у голубя
является наиболее развитым чувством. Глаз закрывается двумя ве-
ками — верхним и более развитым нижним. Кроме век, голубь имеет
полупрозрачную мигательную перепонку, она надвигается на глазное
яблоко и совершенно его закрывает. В покоящемся состоянии эта
перепонка лежит во внутреннем (переднем) углу глаза сверху, откуда
394
Рис. 243. Схема строения контурного пера. По
Боасу (из Гессе).
7 — стержень; 2 — бородки; 3— лучи (бородки второгр
порядка), отходящие от бородки по направлению к вер*
шине пера, налегающие на лучи, отходящие от соседней
бородки в обратную сторону (3') и зацепляющие за них
своими крючечками.
ее МоЖНо йьГгянутЬ пинцетом. Мигательную перепонку можно считать
третьим веком. Благодаря своей тонкости она едва ли мешает птице
хорошо видеть даже при совершенно закрытом глазе. Служит она,
вероятно, для защиты глаз от очень сильного света и для очищения
роговицы 0*^ приставшей к ней пыли.
Под глазом и несколько позади его лежит слуховое отверстие,
которое можно найти, предварительно выщипав осторожно перья на
голове. Слуховое отверстие ведет в наружное ухо, с которым мы еще
не встречались.
Тело голубя одето тонкой сухой кожей, которая покрыта перьями
или ороговевшими пластинками (клюв, ноги). Перья являются харак-
тернейшим признаком
птиц, почему всех их на-
зывают пернатыми. Вы-
щипав несколько круп-
ных и мелких перьев, мы
убеждаемся, что их два
сорта: крупные упругие
перья называются кон-
турными, лежащие ме-
жду ними мелкие мяг-
кие перышки являются
пуховыми.
Каждое перо состоит
из стержня — rhachis
и опахала — vexillum
(рис. 243,7). Свободная от
опахала и полая ближе
к концу часть стержня
называется очином — ca-
lamus (ее именно и чи-
нят, чтобы приготовить
гусиное перо для писа-
ния); своим концом очин
погружен в перовой
мешочек кожи. Внутри
очина лежит сухое по-
лупрозрачное пластинчатое роговое вещество — душка пера.
Опахало пера образуется симметрично лежащими тонкими и длин-
ными роговыми пластинками — бородками — rami (рис. 243, 2); эти
бородки сцепляются друг с другом мелкими, боковыми отростками —
бородками второго порядка — radii (рис. 243, 3), в свою очередь несу-
щими на себе тончайшие крючечки (hamuli). Благодаря такой системе
сцепления опахало пера в целом является упругой пластинкой (рис.
244, 7, 8), цельность которой нигде не нарушается при ударе пера
о воздух.
Таково строение контурного пера; совокупность контурных перьев
обозначает наружные очертания тела птицы и чрезвычайно увеличивает
его поверхность.
Пуховые перья коротки. Они лежат в скрытых от света местах и
395
обладают слабыми стержнями, на которых сидят мягкие бородки.
Последние не сцепляются друг с другом, наоборот, они не могут очень
тесно прилегать одна к другой, так как по своей длине образуют
узловатые утолщения. Поэтому между бородками остаются щели,
заполненные воздухом. И роговое вещество пера и воздух являются
плохими проводниками тепла; вследствие этого пуховые перья отлично
сохраняют тепло тела птицы.
Если ощипать птицу, то окажется, что контурные перья покрывают
не сплошь ее кожу, а растут на определенных участках, называемых
птерилиями, тогда как свободные от них места получили название
аптерий (рис. 245).
Из контурных перьев различают большие маховые — крупные
перья кисти (т. е. вершины) крыла (рис. 244, 8), малые маховые —
7
Рис. 244. Скелет крыла сарыча с прикрепленными к нему маховыми перьями;
.мягкие части и покровные перья удалены. По Гессе и Дофлейну.
1 — плечевая кость; 2 — локтевая кость; 3 — лучевая кость; 4 — большой палец; 5—кры-
лышко; 6—второй палец; 7 — маховые перья второго порядка; 8 — маховые перья первого
порядка; 9 — связка, соединяющая очины маховых перьев.
такие же перья предплечья (рис. 244, 7), кроющие перья — черепице-
образно лежащие мелкие перья между передним краем крыла и махо-
выми перьями, перья крылышка на сгибе крыла (рис. 244, 5) и рулевые
и кроющие перья хвоста.
Перья являются придатками кожных покровов, развивающимися
на счет ороговения эпителиальной части кожи, т. е. эпидермиса.
Перья совершенно сухие и никакой жидкости тела внутри себя не содер-
жат; даже и крупные перья очень легки.
Птица время от времени теряет старые перья, которые замещаются
новыми. Такой процесс называется линянием или линькой.
Окраска перьев зависит от различных причин. Во-первых, в перьях
бывают скопления пигмента, во-вторых, окраска обусловливается
физической структурой поверхности пера.
396
Перья с металлическим отливом имеют на верхней поверхности
боковых лучей тончайшие, налегающие друг на друга пластинки. Луч
света, падающий под известным углом на перо, разлагается, как в
дифракционном спектроскопе, и производит на глаз впечатление того
или иного цвета. Характер отлива изменяется в зависимости от того,
с какой стороны мы будем смотреть на перо. Если зеленое перо попу-
гая раздавить между стеклянными пластинками, то из зеленого оно
сделается бурым или желтым. Изменение цвета зависит от разруше-
ния структуры поверхности пера, благодаря чему интерференция све-
товых лучей не имеет больше места и зеленый отлив пера пропадает.
Рис. 245. Ощипанный дикий петух (Gallus tiankiva). По
Н. X о л о д к о в с к о м у.
а — сверху; b — снизу; пунктированные места соответствуют птери-
лиям, а непунктированные — аптериям.
Появляющаяся бурая окраска пера зависит от присутствия в нем пиг-
мента, цвет которого маскировался субъективной окраской пера.
Перья покрывают все тело голубя, исключая концов ног — именно
ступней. Ступня оканчивается четырьмя пальцами — тремя перед-
ними и одним (первым) задним.
На конце каждого пальца торчит по когтю. Пальцы покрыты рого-
выми щитками или чешуйками, плотно прилежащими друг к другу.
Когти также развиваются на счет ороговения эпидермцса кожи.
Перья, щитки и когти птицы гомологичны друг другу, так как
имеют общий источник развития — эпидермис кожи.
Благодаря густому оперению голубя части его конечностей раз-
личимы трудно; поэтому рассмотрение их строения мы отложим до
397
главы о скелете, пока же установим зависимость сгибания пальцев
ног от сгибания самой ноги. Самым нижним суставом ноги является
пяточный; если мы согнем ногу в этом суставе, то пальцы непременно
скрючатся. Такая зависимость объясняется особым расположением
мышц и сухожилий ноги.
Для голубя она очень выгодна, так как позволяет ему механически
удерживаться на насесте даже во сне. Садясь на ветку, голубь всем
своим телом давит на согнутый пяточный сустав, что влечет за собой
крепкое обхватывание ветки пальцами.
На поверхности тела голубь имеет еще отверстие клоаки, которое
можно видеть снизу у основания хвоста.
Вскрытие кожных покровов голубя и мускулатура тела
Привязав голубя спиной книзу к препаровальной доске, выщипы-
вают у него с шеи, груди и брюха перья. Чтобы пух не разлетался,
щиплют мокрыми руками, крупные перья спускают в банку с водой,
а пух предварительно скатывают между ладонями.
На ощипанной птице прощупывают киль грудной кости и вдоль
него осторожно рассекают кожу скальпелем. Разрез кожи продолжают
по средней линии тела кзади до клоаки и кпереди на шее до головы.
Кожные лоскуты отпрепаровывают в стороны; препаровку на шее
производят тупым способом, т. е. оттесняют кожу ручкой скальпеля,
чтобы не прорезать зоба.
На изготовленном препарате обнажена мускулатура брюшной
стороны тела голубя. Самой крупной мышцей является большая
грудная мышца. Она начинается широким основанием от киля
грудной кости и направляется вперед и наружу к плечевой кости
крыла.
При своем сокращении она тянет поднятое крыло вниз и внутрь
и играет поэтому важнейшую роль при полете. Большая грудная мышца
относительно самая крупная цельная мышца во всем типе позвоночных
животных. У некоторых птиц вес ее равняется четверти или даже
трети веса всего тела. Эта мышца относится к мускулатуре грудной
клетки, которая обслуживается и другими мышцами (например, меж-
реберными).
Мускулатура брюшной области тела слаба. Здесь имеются пря-
мая мышца живота (без всяких следов метамерии) и наружная косая
мышца, остальные же или представлены скудными волокнами, или
вовсе отсутствуют.
Мышцы шеи и конечностей, в связи с их подвижностью, развиты
очень сильно. Заслуживают упоминания кожные мышцы, одним своим
концом или обоими прикрепляющиеся к коже птицы. Они служат для
ее натягивания и, следовательно, для движения перьев (например:
мышцы маховых и рулевых перьев; мышцы, натягивающие кожную
складку крула, и др.).
t Оценивая развитие мускулатуры голубя, мы должны отметить, что
на ней отразилась приноровленность птицы к жизни в воздухе; это
явствует из мощного развития больших грудных мышц, и наоборот,
из слабого развития мышц спины, так как компактное туловище го-
398
лубя неподвижно в своих отдельных частях, что способствует совер-
шенству и легкости полета.
Сравнивая мускулатуру брюшной стороны тела лягушки и голубя,
находим сходные части ее в виде больших грудных, наружной косой
и прямой мышц живота, которые имеют одинаковые точки прикре-
пления к скелету (например, большая грудная мышца идет от грудины
к плечевой кости). Эти мышцы гомологичны друг другу у названных
животных, хотя степень развития их резко различна по причинам
экологического характера (разная роль в зависимости от условий оби-
тания и от различия среды обитания — вода и воздух).
Вскрытие полости тела и общее расположение органов
Отрезают скальпелем место прикрепления больших грудных мышц
и отпрепаровывают их в стороны. Благодаря этому обнажается сильно
развитая грудина, несущая по средней линии киль, к сторонам кото-
рого и прирастает главная летательная мышца. Из рассмотрения
соотношений мышц и скелета разных животных выяснилось, что чем
сильнее данная мышца, тем более изменено место прикрепления ее
к кости. У человека, лягушки и других животных большая грудная
мышца развита соразмерно и грудина представляет собой плоскую
кость. У птицы грудные мышцы увеличились до крайних размеров:
увеличение их повлекло за собой также разрастание грудины в виде
киля, так как на плоской грудине мощным грудным мышцам негде
было бы прикрепиться. Из этого сопоставления можно заключить,
что изменение грудной мышцы (ее увеличение) у птицы вызвало парал-
лельным последствием появление киля на грудине. В этом частном
случае мы видим пример проявления закона соотношения, или кор-
реляции, органов, установленного Кювье. Этот закон обнаружи-
вается тогда, когда изменение одной системы органов влечет за
собой изменение других органов.
Очистив грудину от мышц, находят прилежащую к ее переднему
краю тонкую вилочку (furcula) и реберногрудинные кости (ossa sterno-
costalia), прикрепляющиеся к бокам грудины.
Далее вскрывают брюшную стенку тела по средней линии и обна-
жают клубок внутренностей. Прежде чем разобраться в них, убе-
ждаемся в присутствии у голубя так называемых воздушных мешкоз,
являющихся придатками легких. Для этой цели перерезают трахею
и в нижний конец ее вводят стеклянную трубку; в случае надобности
трахею перевязывают на трубке ниткой. В открытый конец трубки
вдувают воздух и замечают, что брюшные органы начинают шеве-
литься. Это зависит от того, что между ними заложены воздушные
мешки, которые мы и раздуваем воздухом, нагнетая его через трахею
и легкие. Стенки мешков тонки и прозрачны.
В брюшной полости можно видеть: печень, мускулистый желудок
и петли тонкой кишки. Отводя брюшные органы книзу, обнажаем
пластинчатую перегородку, ограничивающую снизу грудную полость.
Ее подрезают по краям и, вводя внутрь тупую ветвь ножниц, разрезают
боковую стенку грудной полости (ребра) наискось к сочленению крьпа
с одной и с другой сторон. Затем приподнимают задний край грудины
399
кверху, отрезают все, что прикрепляется к ней изнутри, и снимают
стенку грудной полости прочь, отрезая ножом или ножницами удержи-
вающие ее мягкие части.
В открытой таким способом грудной полости лежат легкие и сердце;
выше последнего виднеется трахея, а глубже нее — пищевод.
Внутреннее строение
Органы пищеварения начинаются ротовой полостью, ограниченной
снизу подклювьем, а сверху надклювьем и твердым нёбом. Надклювье
образуется сращением верхней челюсти, межчелюстной кости и носовых
костей, а подклювье соответствует сросшимся ветвям нижней челюсти.
Челюсти голубя лишены зубов, что характерно решительно для
всех птиц. Зубов нет ни у взрослых птиц, ни у их зародышей.
На дне ротовой полости лежит язык, по своей форме соответ-
ствующий впадине подклювья. Он имеет вид длинного равносторон-
него треугольника и покрыт многослойным эпителием, ороговевающим
на своей поверхности; ороговевает и острая вершина языка. Поверх-
ность его покрыта мелкими сосочками, также роговыми на своих
верхушках. Язык очень подвижен. Своим основанием он соединяется
со срединной частью подъязычной кости. Передний отдел ее образо-
ван внутриязычной костью (os entoglossum), сзади которой лежит
основная косточка (basihyale). От ее боков отходят длинные большие
рожки (thyrohyale) подъязычной кости. Эта кость со всеми ее придат-
ками является остатком висцерального (жаберного) скелета головы.
Сравнивая отношение языка к висцеральному скелету рыбы и птицы,
мы усматриваем гомологию ее у названных животных. У обоих язык
в основе своей имеет os entoglossum, которая у рыбы покрыта слизистой
оболочкой и почти неподвижна; у птицы же она одета мышечной мас-
сой, которая, наоборот, очень подвижна.
Язык птицы едва ли является органом ее вкуса. Он служит для
проглатывания пищи, которая не пережевывается, как, например,
зерно. Возможно, что язык является скорее осязательным органом,
так как он богат фатер-пачиниевыми нервными тельцами.
Задняя часть ротовой полости над и позади языка называется
глоткой. В ней дыхательные пути перекрещиваются с пищеваритель-
ными.
Внизу и сзади глотка переходит в пищевод (рис. 246, ое), лежащий
на шее позади дыхательной трубки (трахеи). Пищевод — склад-
чатая трубка, образующая в нижней части шеи, на передней своей
стенке, мешкообразный карман—зоб (ingluvies), служащий для хра-
нения проглоченной пищи и предварительной ее обработки. Здесь
зерна размягчаются.
Зоб (рис. 246, tng) голубя состоит из двух боковых мешков, эпи-
телий которых подвергается изменениям во время высиживания яиц
и вылупления птенцов. Многослойный плоский эпителий зоба в ука-
занный период разрастается в глубь соединительной ткани сосочками,
симулирующими собой трубчатые железы. Клетки этих сосочков под-
вергаются жировому перерождению и распадаются в творожистую
массу, которую голубь и отрыгивает в рот птенцам для их кормления.
400
Нижняя часть пищевода утолщается и образует так называемый
железистый желудок, стенка которого очень богата пищеваритель-
ными железами. Железистый, или передний, желудок выделяется под
особым именем, потому что простым глазом отличим от пищевода, но
по существу он яв-
ляется все же частью
последнего. Пища
здесь продолжает раз-
бухать; дальнейшая
же обработка ее про-
исходит в следующем
отделе пищеваритель-
ного канала — соб-
ственно желудке, или
в мышечном желуд-
ке (ventriculus саг-
nosus, рис. 246, vm).
Проследить связь
пищевода с мышечным
желудком можно толь-
ко по удалении сердца
и легких, так как в
грудной полости пи-
щевод скрыт под этими
органами. При рас-
смотрении дальней-
ших частей кишеч-
ника петли его выни-
мают из брюшной по-
лости, подрезая бры-
жейку, и отводят
влево; так отпрепаро-
вывают всю кишку до
клоаки.
Мышечный желу-
док имеет вид чече-
вицы с округлым
краем; лежит он в
брюшной полости, бу-
дучи прикрыт доль-
ками печени. Вход и
выход желудка очень
сближены, поэтому
его поверхность между
ними (называемая ма-
Рис. 246. Анатомия голубя. По Н. Холодков-
с к о м у, с упрощением.
tr — трахея; ое — пищевод; ing— зоб;// — певчая (нижняя)
гортань; vj — яремная вена; сг — сердце с отходящими от
него артериями (черные); рт — легкие; hp — печень; in —
тонкая кишка; рп—поджелудочная железа; гс — толстая
кишка; ic — слепая кишка; cl—клоака; bF— фабрициева
сумка; иг — мочеточники; г — почки; ts — семенник; vm —
мышечный желудок.
лой кривизной желудка) очень мала; в противоположность этому,
большая кривизна желудка развита сильно. Разрежем желудок по
краю и вывернем наизнанку. Стенка его очень толста; она состоит
из мышечных слоев, выстланных в полости желудка толстой, легко
отделяющейся кожицей, называемой кутикулой. Последняя является
2в Практикум по зоологии — 385
401
затвердевшим выделением мелких желудочных железок, лежащих у
внутренней поверхности этого органа. Кутикула желудка неодно-
кратно сменяется (линяет) при жизни птиц и заменяется новой.
Пища из зоба, после предварительного ее размягчения, проходит
сквозь железистый желудок в мышечный, где и подвергается механи-
ческой обработке. Сильные мышцы стенок желудка, сокращаясь, пере-
тирают проглоченное зерно и разминают его; таким образом внутрен-
няя кутикула желудка выполняет роль отсутствующих у птицы зубов
(рис. 247). В желудке зерноядных птиц (голуби, куры) мы находим
землю и мелкие камешки; птица проглатывает их инстинктивно, чтобы
облегчить себе перетирание зерен в желудке. О силе работы его
Рис. 247. Разрез мышечного желудка голубя; видна толстая
мышечная стенка и внутренняя кутикула. Оригинал.
мышечных стенок можно судить по опытам, когда оникс за 4 дня пре-
бывания в желудке курицы уменьшился (стерся) на четверть своего
объема; стеклянные шарики раздроблялись там же в мелкий порошок,
углы граната сглаживались, а чеканка изогнутых золотых монет
стиралась на выпуклой стороне.
Особенности строения и работы мышечного желудка и зоба стоят
в связи с отсутствием зубов у птицы, так как первые берут на себя
роль последних. Потеря зубов в процессе эволюции ныне существующих
птиц на земле есть процесс более поздний, так как у ископаемых птиц
зубы были (Hesperornis, Ichthyornis и др.). Это изменение организаций
птиц коррелятивно повлекло за собой изменение строения желудка
(сильное развитие мышечного слоя и кутикулы) и пищевода (образо-
вание зоба).
Выходная часть желудка называется привратником (pylorus);
полулунными складками он отделяется от прилежащей части тонкой
кишки, куда пища пропускается из желудка при участии складчатого
402
клапана привратника, препятствующего переходу в кишку круп*
ных тел.
От мышечного желудка отходит двенадцатиперстная кишка
(duodenum); она петлеобразно изгибается и тесно облегает удлинен-
ную поджелудочную железу — pancreas (рис. 246, рп). Duodenum
без всякой границы продолжается в собственно тонкую кишку (in),
являющуюся самой длинной частью кишечника.
Тонкая кишка переходит в короткую толстую, или заднюю, кишку
(рис. 246, гс); в начальную часть последней открываются две ничтожно
развитые (у голубя) слепые кишки в виде боковых карманчиков (1с).
Слепые кишки курицы, наоборот, длинны. Задняя кишка впадает
в клоаку (d), принимающую в себя, кроме половых и мочевых отверстий,
еще отверстие фабрициевой сумки (bF), непарного мешковидного органа
неизвестного значения.
Кишечник птицы относительно короче, чем у млекопитающих жи-
вотных. Так, у петуха он в четыре раза длиннее тела, тогда как у не-
которых травоядных млекопитающих раз в двадцать пять превышает
длину их тела. У хищных птиц кишечник еще короче, чем у петуха;
у сыча он лишь в два раза длиннее тела.
Относительная короткость кишечника птиц связана с необходи-
мостью по возможности уменьшения веса тела, что облегчает полет
в воздухе.
Между тем, укорочение кишки влечет за собой уменьшение поверх-
ности ее слизистой оболочки, что неблагоприятно отзывается на пере-
варивании и всасывании пищи, да еще растительной в виде зерен,
которыми питаются курица и голубь.
Такое изменение кишечника коррелятивно компенсируется уси-
лением действия пищеварительных соков и увеличением ворсинок
слизистой оболочки кишки. Ворсинками называются мелкие соско-
образные выросты внутренней стенки кишечника, способствующие уве-
личению ее всасывательной поверхности. У петуха в начальной части
тонкой кишки они не только относительно, но и абсолютно больше,
чем у человека, так как вдвое длиннее ворсинок последнего. Эта
разница станет особенно показательной, если принять во внимание
различие в величине и массе тела человека и петуха.
Ворсинки имеются и в задней кишке птиц, тогда как толстая
кишка и ее производные у млекопитающих лишены их.
Из органов пищеварения нам остается рассмотреть пищеваритель-
ные железы.
В ротовую полость птиц открываются слюнные железы (glandulae
salivariae). Их легче найти у курицы (рис. 248), где они лежат под
кожей нижней поверхности головы.
В брюшной полости помещаются массивная печень (рис. 246, hp)
и поджелудочная железа (рп). Темнокрасная печень голубя двудоль-
чата; частью своей верхней поверхности она соприкасается с сердцем,
где и имеет особое вдавление. Лежит .она, в общем, в брюшной по-
лости, хотя и заходит далеко под грудину. Выводной проток печени
открывается в двенадцатиперстную кишку. У голубя желчного пу-
зыря нет.
Печень подвешена к грудине особыми пластинчатыми связками,
* 403
образованными складками брюшины. Эта железа у птиц развита очень
сильно; она является самой крупной железой этих животных.
Расположение поджелудочной железы нами уже указано. Свой
секрет она, так же как и печень, изливает в двенадцатиперстную кишку.
Сравнивая органы пищеварения птицы, лягушки и рыбы, мы нахо.-
дим общность плана строения их, сказывающуюся разделением пище-
варительного канала на части и присутствием печени и поджелудочной
железы. Однако в деталях строения, развития и функций отдельных
органов этой системы имеются важные отличия, которые возникли
у птиц коррелятивно вследствие потери ими зубов и перехода к воз-
душному образу жизни.
Рис. 248. Слюнные железы курицы. ПоН. Холодковскому.
а — угловая; b, d, h — нижнечелюстные; с — тело подъязычной кости;
g— рог ее; f—мелкие добавочные железы; i — миндалик; к— скуловая кость.
Кровеносная система. Сердце лежит приблизительно в середине
вскрытой полости тела голубя (рис. 246, сг). Оно одето тонкой и про-
зрачной околосердечной сумкой и представляет собой мышечный
конус, основанием направленный кверху; верхушка его тупо округ-
лена. Оттянув пинцетом околосердечную сумку, разрезают ее стенку
и удаляют, чтобы рассмотреть поверхность сердца. На обращенной
к наблюдателю поверхности виднеется поперечная полоска жировой
ткани, намечающая границу между предсердиями и желудочками.
В сердце птицы мы находим новую весьма важную особенность:
оно четырехкамерно, так как состоит из двух предсердий и двух
желудочков. Правая половина сердца, т. е. правые желудочек и
предсердие, отделяется от левой половины сплошной перегородкой,
в которой никаких отверстий нет.
Потягивая пинцетом за сердце, убеждаемся, что с его основанием
связаны крупные кровеносные сосуды. В связи с разделением желу-
дочков друг от друга обособляются и выходящие из них кровеносные
сосуды. Из левого желудочка выходят видные на препарате правая
404
дуга аорты (рис. 249, da) и две короткие безымянные артерии (аа),
расходящиеся вилообразно друг от друга. Вскоре каждая из них
делится на сонную артерию (асе), идущую к голове, и подключичную
артерию (аах), питающую своим продолжением крыло.
Правая дуга аорты загибается вниз и спускается б брюшную по-
лость, где и дает ветви к внутренностям и в нижние конечности.
Рис. 249. Сердце и важнейшие сосуды голубя. По Ж а м м у.
vt — желудочек; — правое предсердие; з — левое предсердие; da —
дуга аорты ;аа— безымянная артерия; асе — сонная артерия; аах—под-
крыльцовая артерия; afm — бедренная артерия; vj — яремная вена;
ves — верхняя полая вена; vax — подкрыльцовая вена; vci — Нижняя
полая вена; vfm — бедренная вена;грг — воротная вена почек; г — почкр;
hp — печень.
Из правого желудочка идет короткая легочная артерия, несущая
подлежащую очищению венозную кровь в легкие. В них она обога-
щается кислородом и, в виде артериальной крови, по коротким легоч-
ным венам возвращается в левое предсердие. Таков малый круг крово-
обращения, совершенно изолированный от большого круга, который
начинается в левом желудочке. Через аорту и ее ветви (da, ams, а/m
и др.) артериальная кровь расходится по телу, в капиллярах отдает
клеткам тела кислород, получает от них взамен углекислоту и посту-
405
пает в венозную систему. Венозная кровь возвращается обратно в
сердце, в правое предсердие, по двум верхним (рис. 249, vcs)
и одной нижней полым венам (vei). Следовательно, большой круг
кровообращения начинается в левом желудочке и кончается в правом
предсердии, тогда как малый круг ограничивается правым желудоч-
ком и левым предсердием. Вследствие полного разделения правой
половины сердца от левой, артериальная кровь нигде не смешивается
с венозной, что является важным усовершенствованием организации
птицы по сравнению с лягушкой.
Отметим, что у голубя есть воротные системы печени и почек
(рис. 249, vpr).
Выяснив общий ход кровеносных сосудов голубя, рассмотрим по-
дробнее строение его сердца. Для вскрытия по всем правилам оно,
вследствие своей небольшой величины, подходит мало, поэтому мы
только перережем сердце поперек в области желудочков. Левый же-
лудочек толстостенен; его просвет внутри на разрезе округлый.
Стенки правого желудочка раза в два тоньше, чем у левого; полость
его узкая, серповидная. Из этих наблюдений можно с достоверностью
заключить, что левый желудочек производит гораздо большую работу,
чем правый; и это действительно так, ибо левый желудочек должен
нагнетать кровь в объемистый большой круг кровообращения, тогда
как на долю правого выпадает работа по малому кругу, проходящему
только через легкие.
О степени развигия сердца можно судить по его весу. Чем тяжелее
сердце, тем больше в нем мышечной ткани, тем оно, очевидно, силь-
нее. Показателен не абсолютный вес этого органа, а относительный
сравнительно с весом тела. У удода сердце в 50 раз легче тела, а у
куропатки в 122 раза; следовательно, сердце удода относительно
тяжелее, чем у куропатки. Эти цифры и другие наблюдения позволяют
думать, что чем лучше птица летает, тем более тяжелое и сильное
сердце она имеет. Быстрый полет сопряжен с сильной работой мышц,
которые развивают много энергии; поэтому они нуждаются в боль-
шем притоке пищевых веществ и в незамедлительном удалении нако-
пившихся в них продуктов их жизнедеятельности. Все это осуще-
ствляется быстрым кровообращением, зависящим от достаточной силы
сердца.
С внутренним его строением ознакомимся у курицы. Подрезают
ножницами кровеносные сосуды, выходящие из основания сердца, и
берут его указательным и большим пальцами левой руки так, чтобы
краем левого желудочка оно было обращено к наблюдателю. Зажав
в таком соложении сердце пальцами, разрезают его левую стенку
пилящим движением скальпеля, ведя разрез до тех пор, пока не
вскроется полость левого желудочка. Затем вводят ветвь ножниц через
желудочко-предсердное отверстие в левое предсердие и рассекают
его стенку в том же направлении, что и желудочек. Раскрывают обе
половинки сердца и осматривают его внутреннюю поверхность.
Стенка левого предсердия, по сравнению с желудочком, очень тонка;
это объясняется малой работой, выпадающей на ее долю — протал-
кивать кровь через широкое отверстие в желудочек. Отверстие между
этими частями сердца снабжено парусной сухожильной заслонкой.
406
Свободные (нижние) края створок этого клапана снабжены сухожиль-
ными нитями, прикрепляющимися другим концом к сосочковым от-
росткам изборожденной мышечными перекладинами внутренней по-
верхности желудочка. Створки клапана открываются при переходе
крови из предсердия в желудочек; когда же наступает сокращение
(систола) последнего, то кровь приподнимает пластинки клапана,
которые и захлопываются, соприкасаясь краями друг с другом, причем
натягиваются удерживающие их в этом положении сухожильные
нити.
При таком положении крови остается один выход — в аорту, куда
мы за ней и последуем.
Сердце снова берут в левую руку так, чтобы оно разрезом смотрело
на наблюдателя. В полость вскрытого желудочка вводят ветвь нож-
ниц и продвигают ее осторожно так, чтобы попасть в начало аорты.
Когда убедились в правильности положения ножниц, коротким уда-
ром смыкают их ветви и рассекают наискось переднюю стенку левого
желудочка и начала аорты. Раскрывают края разреза и рассматривают
внутреннюю поверхность препарата.
Полость левого желудочка продолжается в более узкий просвет
корня аорты; внутренняя ее поверхность, как и сама стенка, бело-
ватого цвета, тогда как ткань сердца темнокрасная. Это зависит от
того, что аорта построена из эластической соединительной ткани.
У основания аорты замечаем три пристеночных карманчика полулунной
формы — это полулунные клапаны сердца. Подведем под верхний сво-
бодный край клапана пинцет и оттянем его к себе. Такое положение
принимает клапан, когда он закрывается. Это происходит при расши-
рении (диастоле) желудочка, когда протолкнутая в аорту кровь стре-
мится возвратиться в желудочек: она оттесняет от стенки края полу-
лунных клапанов, которые и захлопываются.
Правая половина сердца также снабжена клапанами. Особенностью
птиц является так называемая мускульная заслонка между правым
предсердием и желудочком. Она имеет вид трехугольной мышечной
пластинки, лишенной сухожильных поддерживающих нитей.
Отличие в строении органов кровообращения у вскрытых нами
животных бросается в глаза. Сердце у рыбы двукамерное, у лягушки —
трехкамерное, а у птиц — четырехкамерное. В связи с обособлением
легочного дыхания оба круга кровообращения у птицы совершенно
обособлены, у лягушки они перекрещиваются в общем желудочке,
а у рыбы имеется всего-навсего один круг кровообращения, так как
дышит она жабрами.
Типичное ветвление жаберных дуг у животных с обособлением лег-
ких нарушается. У лягушки мы видели довольно совершенные остатки
артериальных дуг в виде трех пар артериальных дуг; редукция их
у птиц идет дальше. Остается первая пара дуг в виде безымянных арте-
рий; от аортных дуг сохраняется лишь правая половина, так как ме-
сто ее начала — правый желудочек — отделяется от левого желудочка.
Остальные пары артериальных дуг, закладывающиеся у зародыша
птицы по рыбьему типу, исчезают по мере развития зародыша.
Сравнивая конечные формы кровеносной системы рыбы, лягушки
и птицы, мы замечаем неизменно развивающееся отклонение ее от
407
первичной формы у рыб, объясняющееся решительным переходом
животных к наземному образу жизни. Это обстоятельство и налагает
на кровеносную систему особые черты усовершенствования, идущего
по двум направлениям: с одной стороны, сердце усложняется в строе-
нии, с другой — сложная система жаберных сосудов замещается мень-
шим числом артерий, ход которых изменяется в зависимости от возник-
новения для них новых задач.
Едва ли необходимо напоминать, что кровеносная система птиц
является замкнутой.
Органы дыхания птицы приспособлены к действию в воздушной
среде и при полете в воздухе; последнее обстоятельство налагает на
них особый отпечаток.
Органы дыхания состоят из воздухоносных путей, легких и воздуш-
ных мешков. Воздухоносные пути начинаются парными носовыми
отверстиями, лежащими у основания надклювья; ведут они в носовую
полость, которая двумя хоанами, открывающимися в непарную впа-
дину твердого нёба, сообщается с глоткой; в постжднюю воздух про-
ходит также и через ротовое отверстие. Из глотки воздух попадает
в дыхательную щель, которую можно видеть у основания оттянутого
кпереди языка. Отверстие дыхательного горла окружено жесткими
зубчатыми отростками окружающей ткани, препятствующими попа-
данию пищи в дыхательную трубку, называемую иначе трахеей.
Трахея (рис. 246, 1г) представляет собой трубку с твердыми стен-
ками; она лежит в шее спереди пищевода. Присматриваясь к ее поверх-
ности, можно видеть, что трахея состоит из множества твердых на-
ощупь колец, соединенных друг с другом более податливой тканью.
Эти кольца состоят из гиалинового хряща. Благодаря такому строению
просвет трахеи постоянно зияет, и стенки ее никогда не спадаются,
что очень важно для свободного прохождения воздуха.
Верхняя часть трахеи, прилежащая к дыхательной щели, превра-
щена в верхнюю гортань (larynx). Она состоит из перстневидного
хряща, к задней части которого примыкают маленькие четырехуголь-
ные черпаловидные хрящи.
Трахея, входя в грудную полость, делится на две очень короткие
ветви, называемые бронхами, которые и погружаются в легкие.
При переходе трахеи в бронхи лежит специфический для птиц
орган — нижняя гортань, или певчая гортань (syrinx), служащая
для произведения звуков (рис. 246, П).
Нижний конец трахеи вздувается; между ним и началом бронха
наружная стенка дыхательной трубки вдается внутрь в виде наружной
голосовой перепонки: изнутри, от внутренней стенки бронха, ей
противостоит аналогичная складка, внутренняя голосовая перепонка.
Прилежащие кольца трахеи и бронхов соединяются особыми певчими
мышцами; при своем сокращении они напрягают голосовые перепонки,
которые и приходят в колебание, когда воздух с силой прогоняется
в голосовые щели между ними.
Легкие голубя (рис. 246, рт; рис. 250, 7), как и вообще птиц,
малы; они светлорозового цвета и губчатого строения. Легкие плотно
прилежат к ребрам и даже соединяются соединительной тканью
с задней стенкой грудной полости, форму которой и повторяют. Бронх,
408
погрузившись в ткань легкого, дает четыре (спинных) бронха второго
порядка, которые распадаются на собственно легочные трубочки.
Судьба их различна; одни оканчиваются слепо, другие анастомо-
зируют друг с другом; все они на внутренней поверхности усажены
мельчайшими ячейками, служащими для газообмена, так как в их
стенках обильно ветвятся капилляры.
Некоторые из бронхов выходят из легкого с его брюшной стороны,
раздуваются в тонкостенные воздушные мешки (рис. 250, 2—5), из
которых главными являются два
шейных (рис. 251, 7), несколько
пар мелких трудных и два боль-
ших брюшных. Изнутри мешки
выстланы плоским эпителием, а с
поверхности покрыты брюшиной.
Воздушные мешки не только лежат
между различными органами (рис.
251); они распространяются даже
внутрь костей (рис. 250, 8),
большая часть которых у птиц пнев-
мо, тична, т. .е. вместо костного
мозга заполнена воздухом.
Доказать сообщение костей с ды-
хательными органами можно сле-
дующим образом. У захлорофор-
мированной курицы перепиливают
плечевую кость близ локтевого су-
става, очищают края разреза от
мягких частей и на освобожденный
конец кости крепко навязывают
резиновую трубку двойного бал-
лона. Накачивая им воздух внутрь
тела курицы, заставляют ее жалобно
кричать. Воздух через плечевую
кость поступает в воздушный ме-
шок, проходит в легкое, проникает
в бронх, колеблет перепонки пев-
чей гортани, которые и начинают
звучать.
Дыхание у птицы происходит
различно, смотря потому, находится
ли она в покое или же летит. В пер-
вом случае грудная клетка рас-
ширяется благодаря движению кпереди ребер и опусканию грудной
кости действием межреберных мышц (рис. 252); при этом расширяется
и полость тела, а заключающийся в воздушных мешках воздух разре-
жается. Развивающееся в них отрицательное давление компенсируется
притоком свежего воздуха через дыхательные пути в легкие, которые
одновременно и освежаются. Скопившийся в них отработанный воз-
дух проталкивается при этом в воздушные мешки. Выдыхание проис-
ходит при возвращении стенок грудной клетки к исходному положе -
Рис. 250. Схема воздушных мешков
голубя. По Гессе и Дофлейну.
2 — межключичный мешок; 3 — передние и
4 — задние грудные мешки; 5 — брюшные
мешки; 6—трахея; 7—легкие; 8 — пле-
чевая кость с заходящим в. нее воздушным
мешком.
ншо. Полость тела сдавливается, и воздух из воздушных мешков
нагнетается в легкие и трахею. Таким образом легкие служат только
для надобностей газообмена; вентиляция же их выпадает на долю
Рис. 251. Расположение воздушных мешков в теле голубя. По Гессе и Д о-
ф л е й н у.
1 — шейный мешок; 2— межхлючичный мешок с придаточною полостью; 21, 3, 4— передний
W* грудные мешки; 5 к 0 — левый и правый брюшные мешки; 7 —трахея; 8— летное.
воздушных мешков. Сами легкие вентилироваться не могут, так как
они неэластичны и объем их при движении грудной клетки изменяется
мало.
Иначе обстоит дело, когда голубь летает. При полете грудная
клетка, в частности грудина, крепко фиксируется, так как она служит
главной опорой для летательных мышц (большие грудные мышцы).
В это время стенки ее остаются неподвижными. Птица летит с плотно
закрытым ртом со скоростью от 15 до 90 м в секунду. Даже и при
тихой погоде она с силой врезывается в воздух, который попадает
в ноздри и под влиянием поступания новых порций воздуха механи-
чески проталкивается в трахеи, легкие и воздушные мешки. Таким
рбрэзом вдыхание у летящей птицы совершается пассивно. Выталки-
вание отработанного воздуха происходит, вероятно, активно вслед-
ствие сокращения брюш-
ных мышц, которые да-
вят на воздушные меш-
ки. Чем быстрее летит
птица, тем большее со-
противление оказывает
ей воздух, тем сильнее
проникает он в глубь ее
тела и тем энергичнее
приток кислорода; по-
этому птицы не зады-
хаются даже при самом
быстром полете на откры-
Рис. 252. Схема движения ребер и грудной кости
при дыхании у птиц. Положение при выдыхании
обозначено сплошными линиями: при вдыха-
нии — пунктиром. По Гессе и Дофлейну.
1 — позвоночник; 2 и 21 — ребра и грудино-реберные
кости; 3 — грудная кость; 4 — коракоид.
том воздухе.
Помимо главной своей
роли в дыхании птицы,
воздушные мешки имеют
еще второстепенное зна-
чение. Так, они способ-
ствуют сохранению тепла
тела птицы и заменяют собой в этом отношении жировую под-
кожную клетчатку, ибо воздух, подобно жиру, является плохим
проводником тепла. Во-вторых, «они, как эластичные подушки, под-
держивают члены тела и шею во время полета и таким образом сбере-
гают мускульную работу птицы, например, при падении или распро-
стирании крыльев для обсушивания их». В-третьих, воздушные
мешки, способствуя увеличению внутри брюшного давления, заменяюг
собой действие мышц брюшного пресса при выделении кала, который
с силой выбрасывается наружу через клоакальное отверстие.
Сравнивая легкие лягушки и птицы — органы гомологичные друг
другу, находим между ними ряд различий, зависящих от высоты орга-
низации животного и от приспособленности его к окружающей среде.
У лягушки собственно дыхательная трубка отсутствует, так как
короткое тело ее лишено шеи и легкие открываются непосредственно
в дыхательное горло. У птицы мы видим трахею, длина которой со-
ответствует длине шеи, а у некоторых видов трахея образует петлю
или спиралеобразные изгибы, помещающиеся в грудной кости. Легкое
.411
Рис. 253. cf — мочеполовые органы самца голубя,
ф — то же, самки. По Ж а м м у.
sr—надпочечник; t — семенник; г — почка; vd — семе-
провод; иг — мочеточник; cl — клоака; bF — фабрициева
сумка; ov (сверху) — яичник; odo — воронка яйцевода;
ov — левый яйцевод; ovd — правый редуцированный
яйцевод.
лягушки является слабо ячеистым, тонкостенным мешком, дыхательная
поверхность которого невелика. У птицы же оно хотя и не велико
по объему, но внутри пронизано множеством трубочек с концевыми
ячейками, что чрезвычайно увеличивает дыхательную поверхность.
Специальным приспособлением к воздушному образу жизни являются
воздушные мешки, зачатки которых появляются уже у пресмыкаю-
щихся.
Верхняя гортань птицы гомологична гортани лягушки: нижняя же
(певчая) гортань является органом, исключительно свойственным
птицам.
Органы размножения. После того, как органы пищеварения вынуты
из брюшной полости, можно приступить к рассмотрению половой систе-
мы голубя. Птицы все
раздельнополы, и самцы
отличаются от самок по
внешнему виду, иногда
даже очень резко. При-
мер хорошо выраженного
полового диморфизма мы
видим у домашних кур.
Петух отличается от ку-
рицы мясистыми выро-
стами кожи на голове,
сильно развитыми перья-
ми хвоста и голосом.
Органы размножения
его (как и у голубя) со-
стоят из пары овоидных
семенников, лежащих на
препарате над перед-
ним концом почек (рис.
253, /). От внутрен-
него края семенника
идет тонкий, извиваю-
щийся поперечными складками семевыносящий канал (vd), впадаю-
щий в клоаку (cl). Наружных половых органов нет, но, несмотря на
их отсутствие, птицы спариваются. Покрывая самку, самец плотно
прижимает отверстие своей клоаки к отверстию клоаки самки, куда и
выпрыскивает семенную жидкость.
Женские половые органы состоят из яичника и яйцевода. Асим-
метрия их проявлена очень резко, так как имеется всего один левый
яичник (рис. 253, 9 w) и левый же яйцевод (odo). Правый яйцевод
остается в виде рудиментарного придатка клоаки (pvd).
Яичник представляет собой гроздевидное тело, подвешенное на
брыжейке у верхней доли левой почки под позвоночником. В нем
видны различной величины шары — развивающиеся яйцеклетки, кото-
рые у птиц чрезвычайно богаты питательным желтком. По мере выра-
ботки последнего яйцо увеличивается в размерах и достигает
25 мм в диаметре. Питательный желток желтого цвета, поэтому и
яичник также желтый. Зрелое яйцо прорывает оболочку яичника ц
412
поступает в широкую воронку яйцевода, которая открывается в по-
лость тела. Однако при выходе яйца из яичника воронка очень плотно
прилегает к этому органу и временно составляет его непрерывный
выводной проток. Попав в яйцевод, яйцо передвигаеся далее, винто-
образно вращаясь, благодаря перистальтике яйцеводной стенки.
Яйцевод образован довольно широкой трубкой, которая брюшной
складкой подвешена к позвоночнику и почкам. На своем пути он об-
разует изгиб; за воронкой следует нисходящая часть; она возвра-
щается снова, кверху переходит во вторую петлю. Третья петля
яйцевода примыкает к клоаке. Яйцевод служит не только для
прохождения яйца, но и для его оплодотворения и снабжения обо-
лочками.
Семенная жидкость из клоаки птицы после копуляции проникает
в яйцевод вследствие активного движения живчиков. Со зрелой яйце-
клеткой живчик сталкивается в начальной части яйцевода; здесь-то
и происходит их слияние, т. е. совершается оплодотворение. Опло-
дотворенное яйцо идет по яйцеводу вниз и в его верхней половине оде-
вается слоем белка, вырабатываемого особыми железами внутренней
стенки яйцевода. В нижней части яйцевода покрытое белком яйцо
получает еще скорлуповую оболочку и первоначально мягкую скор-
лупу. В таком виде яйцо откладывается птицей, причем в момент
откладки скорлупа затвердевает и яйцо принимает свойственную ему
внешность. Часть яйцевода, вырабатывающая скорлупу, называется
иначе маткой.
Следует иметь в виду, что яйцеклетка курицы, при прохождении по
яйцеводу в оплодотворенном состоянии, начинает развиваться в заро-
дыш еще в материнском организме. Яйцо идет по яйцеводу всего
около 15—21 часа, и за это время оно начинает дробиться на верхнем
полюсе, т. е. там, где лежат его ядро и протоплазма («образователь-
ный желток»). В отложенном яйце развитие при температуре окру-
жающей среды останавливается. Эти факты указывают, что понятие
«яйцо курицы» (в общежитейском смысле) не соответствует понятию
«яйцо (т. е. женская половая клетка) птицы». Последнее надо искать
в яичнике, а отложенное птицей яйцо заключает в себе уже зародыш,
правда, остановившийся на ранней стадии развития. Последнее
тотчас же возобновляется, как только яйцо попадает в температуру
около Н-39°С. В естественных условиях это осуществляется насижи-
ванием яиц птицей.
Рассмотрим строение отложенного яйца курицы. Один полюс его
слегка заостренный, другой же более тупой. Под пористой и проницае-
мой для воздуха скорлупой лежит скорлуповая оболочка, расщепляю-
щаяся у тупого полюса, где между расходящимися ее слоями поме-
щается воздушная камера. Под скорлуповой оболочкой лежит слой бел-
ка, который у полюсов яйца заключает в своей толще спирально изог-
нутые шнуры, называемые градинками, или халадзами; они подвеши-
вают желток яйца, удерживая его в центре яйца. На верхнем полюсе
(при нормальном положении яйца) желтка помещается рубчик — уча-
сток раздробившихся клеток зародыша, лежащих на массе индиферент-
ного питательного желтка. Последний двух сортов — желтый и
белый; оба сорта желтка концентрически наслаиваются друг на друга;
413
их можно хорошо рассмотреть, разрезав желток сваренного вкрутую
яйца.
Насиживая яйца, птица согревает их теплом своего тела; перья
на брюшной стороне птицы выпадают и частью выщипываются ею
самой, благодаря этому образуются «высиживательные пятна» —
голые участки кожи, лучше согревающие яйца, так как плохо прово-
дящий тепло слой перьев на них отсутствует.
Через три недели из яиц вылупляются цыплята. Цыпленок прола-
мывает скорлупу яйцевым зубом — коническим бугорком ороговевшего
эпидермиса у конца надклювья. Яйцевой зуб является типичным
эмбриональным органом. Цыпленок тотчас по выходе из яйца способен
к более или менее самостоятельному питанию, он сам клюет пищу и
рано научается ее разыскивать. Кур поэтому относят к выводковым
птицам. Голубята же, вышедшие из яйца, совершенно беспомощны,
недостаточно оперены и слабы. Их должна первое время кормить мать,
которая отрыгивает им в рот содержимое своего зоба (так называемое
«молочко»). Голубь поэтому относится к птенцовым птицам.
Хотя птицы и откладывают яйца наружу подобно мечущим икру
лягушкам и рыбам, однако они, в противоположность последним,
заботятся о развитии яиц, равно как и о вышедшей из них мо-
лоди. Инстинкты птиц более сложны и многообразны, чем у земно-
водных и у рыб; поэтому можно говорить о большом совершенстве
психической жизни пернатых, придавая этому термину ограничитель-
ное толкование и не понимая его в антропоморфном смысле.
Возвращаясь к морфологии полового аппарата птиц, мы должны
подчеркнуть асимметрию (непарность) его женских органов размно-
жения. Правда, один яичник бывает и у некоторых рыб, но там он
симметричен, так как лежит вдоль средней линии тела. Непарность яич-
ника птицы иного происхождения — она зависит от недоразвития
органа другой половины тела, что частично отмечается также и для
яйцевода. Однобоковое развитие органов размножения птицы является
признаком вторичным, направленным к уменьшению веса тела птицы
в целях облегчения ей полета.
Органы выделения. Убрав из брюшной полости тела органы размно-
жения, находим на дне препарата органы выделения. Они представлены
двумя темнокрасными трехдольчатыми почками (рис. 253, г), кото-
рые плотно лежат в углублениях подвздошных костей по бокам позво-
ночника. От каждой почки отходит по мочеточнику (иг) в виде тонкой
трубочки, впадающей в клоаку (cl). Мочевого пузыря у птиц нет. Моча
их густая; выбрасывается она вместе с калом, в котором образует
белые прослойки. Отсутствие пузыря и густота мочи (выбрасываемой
птицей на лету) являются также приспособлением к полету (облегчение
веса тела, ненужность остановки животного для дефекации).
Почка птицы не гомологична почкам лягушки. У лягушки имеются
первичные почки (вольфовы тела), у птицы же первичные почки во
время развития исчезают и заменяются вторичной почкой. Таким
образом субституция (подстановка) органов выражена в данном случае
в крайней форме.
Нервная система. По рассмотрении всех внутренних органов отре-
зают голову голубя и кожу с его шеи чулком заворачивают так, чтобы
414
обнажилась черепная коробка. Голову берут в левую руку и скальпелем
снимают послойно костную стенку черепа. Работу производят осто-
рожно, чтобы не задеть нежной ткани мозга. С темени переходят на
затылок и боковые части черепа, где губчатые кости его обладают
большей толщиной. Пинцетом удаляют кости с краев вскрытой части
черепа и в конце концов обнажают головной мозг сверху и с боков.
Головной мозг птицы отличается своей массивностью (рис. 254);
он короток и широк. В свежем состоянии большой мозг и мозжечок
серо-красноватого цвета, остальные
же части белые. Части серого цвета,
покрыты клеточной нервной тканью
(кора головного мозга), белые же
характеризуются преобладанием в них
нервных волокон с миелиновой обо-
лочкой.
Наибольшей частью мозга являются
полушария с гладкой поверхностью^/).
Спереди из-под них чуть выглядывают
обонятельные доли; сзади примыкает
крупный мозжечок (сЫ), поверхность
которого исчерчена поперечно иду-
щими бороздами. Между мозжечком и
большими полушариями виднеются
с боков доли среднего мозга — двухол-
мие (/о); из-под мозжечка кзади и
книзу спускается продолговатый мозг
(то). Благодаря сильному развитию
больших полушарий и мозжечка, со-
прикасающихся друг с другом, про-
межуточный мозг сверху вовсе не ви-
ден, а доли двухолмия оттесняются
в стороны.
На нижней поверхности головного
мозга видны, считая спереди назад:
неясно обособленные обонятельные
доли; большие полушария, лежащие
по сторонам — доли двухолмия; сре-
динная мозговая масса между ними,
прикрытая перекрестом зрительных
нервов и крупным нижним мозго-
вым придатком, и сильно выпуклый продолговатый мозг.
Сравнивая головной мозг птицы, лягушки и рыбы, отмечаем у птиц
увеличение массы больших полушарий, разрастающихся по всем трем
направлениям. Внутри себя они содержат по боковому желудочку,
верхняя стенка которого (крышка) остается тонкой, а нижняя и боко-
вая стенки, наоборот, очень толсты благодаря сильному развитию
основных узловых скоплений мозговой ткани, называемых иначе
corpus striatum.
Примечательно также сильное развитие мозжечка, который у птиц,
помимо массивной средней части (vermis), имеет две боковые зачаточ-
Рис. 254. Головной мозг голубя
сверху. По Ж а м м у.
ос — глаз; cl — большие полушария;
1о— зрительные доли; сЫ — мозжечок;
то — продолговатый мозг; V — трой-
ничный нерв; VII— лицевой нерв;
VIII — слуховой нерв; IX — языкогло-
точный, X — блуждающий и XI — до-
бавочный нервы.
415
ные доли (flocculi). У лягушки, как и у амфибий, вообще мозжечок
ничтожен по размерам, у рыб же он очень вариирует в своемразвитии —
у одних видов мозжечок очень велик, у других же, напротив, мал.
Если рассматривать головной мозг птицы в сагитальном сечении, то
можно заметить, что он образует два изгиба: один — направленный
вверх в области промежуточного мозга, другой — в противоположном
направлении в области продолговатого мозга. Изгибы возникают вслед-
ствие разрастания головного мозга, частям которого становится тесно
в неподатливой черепной коробке. Вспомним попутно, что ни у рыб,
ни у земноводных головной мозг изгибов не делает, а лежит более
или менее в горизонтальной плоскости.
Из черепа птицы выходят двенадцать пар головных, нервов. Сравнивая
их последовательно с черепными нервами лягушки и рыбы, мы устана-
вливаем гомологию первых десяти пар вплоть до блуждающего нерва
(n. vagus) включительно. Следующей парой (XI) является добавочный
нерв — п. accessorius (рис. 254, XI), который начинается от передней
части спинного мозга, откуда выходят обособившиеся волокна блуждаю-
щего нерва. Поэтому у птиц этот нерв не имеет еще вполне самостоя-
тельного значения. Хотя он и начинается от спинного мозга, но выходит
наружу через отверстие черепа, почему и считается головным нервом.
XII пара головных нервов называется подъязычным нервом (п. hypo-
glossus); он берет начало несколькими корешками — частью от про-
долговатого, частью от спинного мозга. Спинномозговые корешки
его поднимаются через затылочное отверстие в черепную полость и
выходят наружу также через отверстие черепа.
Присутствие двенадцати пар головных нервов у птиц имеет важное
морфологическое значение; оно указывает на морфологически иной
состав головы этих животных по сравнению с земноводными и рыбами.
У птиц мы усматриваем тенденцию к соединению с головой при-
лежащих частей тела, которые можно считать метамерами, так как
они соответствуют метамерным спинномозговым нервам. Эти метамеры
сливаются с черепом, параллельно происходит присоединение и спин-
номозговых нервов к нервам черепным. Поэтому состав продолго-
ватого мозга птиц морфологически иной, чем у амфибий и рыб, так как
с ним слились новые элементы спинного мозга.
Спинной мозг птиц образует два значительных расширения в
области крыльев (грудное расширение) и в крестце; оканчивается он
тонкой нитью (filum terminale), которая заходит в хвостовую часть
позвоночника.
Спинномозговые нервы, иннервирующие переднюю конечность, обра-
зуют плечевое сплетение из четырех нервов; ноги же птицы обслужи-
ваются сплетениями поясничным (из трех пар нервов) и седалищным
(четыре крестцовых нерва).
Органы чувств. Из органов чувств голубя хорошо развиты глаза и
органы слуха. Эти чувства у птицы наиболее остры.
Общее строение глаза голубя соответствует тому, что мы видели
уже у рыбы, но есть и свои отличия. Их можно рассмотреть на глазе,
вынутом из глазницы, уплотненном в 96% спирту (сутки) и разрезан-
ном вдоль острым скальпелем. Форма глазного яблока оригинальна.
Роговица представляет собой сегмент шара с малым диаметром, а зад-
416
няя часть глаза — отрезок шара гораздо ббльших размеров. Эти два
сегмента соединены средним поясом, имеющим, в общем, форму усечен-
ного конуса.
Между роговицей и хрусталиком есть полость — передняя камера
глаза, заполненная водянистой влагой. Хрусталик сильно выпуклый;
радужная оболочка заключает в себе ресничный мускул, своим сокра-
щением способствующий изменению кривизны хрусталика, благодаря
чему он дает резкое изображение рассматриваемого предмета на сет-
чатке глаза (приспособление глаза к ясному видению предмета назы-
вается аккомодацией). Аккомодационная мышца птицы поперечно-
полосатая; поэтому она может быстро сокращаться. Для более быстрой
и сильной работы мышцы необходимы более надежные точки ее при-
крепления.
Таковой и является склеротикалъное кольцо из ряда костяных
пластинок, заложенных в толще белковой оболочки (склеры) на гра-
нице с роговицей. Сосудистая оболочка задней стенки глаза вдается
в его заднюю камеру в виде складчатого гребня, или веера, значение
которого с точностью не установлено.
Напомним, что глаз птицы имеет три века (мигательная перепонка).
В наружном углу глаза помещается слезная железа.
Из новых для нас особенностей строения глаза голубя отметим
присутствие складчатого гребня, склеротикального кольца и слезной
железы.
Орган слуха и равновесия птицы состоит из внутреннего, среднего
и наружного уха. Последнее представлено коротким наружным слу-
ховым проходом, на дне которого лежит барабанная перепонка. К ней
прилежит единственная слуховая косточка—columella (как у ля-
гушки), помещающаяся в полости среднего уха; последнее евстахие-
выми трубами сообщается с глоткой, причем общее отверстие труб
лежит позади хоан. Внутреннее ухо, или лабиринт, заложено в толще
пневматических костей стенки черепа. Оно состоит из трех полукруж-
ных каналов и лабиринта, мешочки которого сообщаются с широкой,
слегка перекрученной по оси улиткой. Полукружные каналы нетрудно
отпрепаровать, срезая губчатую кость по слоям.
Сравнивая органы слуха вскрытых животных, замечаем, что у
всех них есть внутреннее ухо, среднее присуще только птице и ля-
гушке, тогда как наружное ухо принадлежит лишь птице. Помимо
этого, есть отличия и в строении деталей. Внутреннее ухо более сходно
у костистой рыбы и лягушки, тогда как у птицы оно имеет ясно раз-
витую улитку. В строении среднего уха птица сходна с лягушкой,
ибо и та и другая обладают единственной слуховой косточкой.
Из прочих органов чувств укажем, что органом осязания являются
язык и восковица клюва, а обоняние приурочено к носовой полости,
в которой заложены носовые раковины.
Скелет
Скелет птицы, обладая всеми типичными чертами для наземного
позвоночного, отличается множеством особенностей, связанных с воз-
душным образом жизни.
27 Практикум по «оологии — 385
417
Рис. 255. Скелет сокола. По Н. X о-
л о д к о в с к о м у.
а — череп; b — шейная, часть позвоночника;
d — крестец и таз; е — хвост; g — ребра с их
крючковатыми отростками; h — грудино-ребер-
ные кости; । — грудина; к—коракоид;} — клю-
чицы; / — лопатки; т — плечевая, и—лок-
тевая; о — лучевая кости; р — запястье; q —
пясть; г, з, t—пальцы кисти; и — бедро;
v — голень; w — пяточно-плюсневая кость; х—
передние пальцы; у — задний палец стопы.
Основа скелета — позвоночник—разбивается на шейный (рис. 255,
Ь), туловищный и хвостовой (</, е) отделы, причем подвижностью обла-
дают только первый и последний.
Шейная часть позвоночника состоит из двенадцати позвонков:
шейный позвонок — атлант — несет впадину для сочленения с един-
ственным мыщелком черепа;
11 позвонок называется эпистро-
феем; он снабжен зубовидным
отростком, вдающимся в коль-
цевидный просвет атланта. ПI —
XII шейные позвонки несут по-
перечные отростки (в состав ко-
торых входят зачатки шейных
ребер), пронизанные отверстиями
для прохождения позвоночной
артерии. Шейные позвонки
весьма подвижно сочленяются
друг с другом, что позволяет
шее совершать разнообразные
движения.
Туловищный отдел позвоноч-
ника соответствует восьми спин-
ным, четырнадцати поясничным
и крестцовым и части хвосто-
вых позвонков. Эти позвонки
срастаются по несколько вместе,
часть же их сливается в одну
кость, принимающую участие
в образовании таза. По своей
подвижности этот отдел позво-
ночника — полная противопо-
ложность шейному, он совер-
шенно неподвижен.
Хвостовой отдел позвоноч-
ника образован шестью малень-
кими подвижными позвонками
и одним крупным концевым
позвонком—пигостилем; послед-
ний, как показывает история
развития, сливается из зачат-
ков нескольких (не менее пяти —
шести) позвонков.
Череп птицы (рис. 255, а) ха-
рактеризуется слитностью боль-
шинства своих костей; он образован сплошной костной коробкой, состав
которой из типичных черепных костей доказывается изучением эм-
брионального развития этой части скелета. В области затылка лежит
единственный мыщелок. Свободными остаются кости твердого нёба,
скуловые, квадратная и некоторые другие. Нижняя челюсть сочле-
няется квадратной костыо с височной областью и по своему составу
418
сложна, так как во время онтогенеза сливается из зачатков несколь-
ких костей (dentale, angulare, articulate и др.), которые у рыб лежат
самостоятельно.
Со спинными позвонками сочленяются ребра (рис. 255, g); из них
первая, вторая, восьмая пары оканчиваются свободно, тогда как
третья — седьмая пары соединяются через посредство грудино-реберных
костей (h ) с грудиной. Таким образом у птиц образуется скелет груд-
ной клетки. Ребра на своей задней поверхности несут крючкообраз-
ный вырост.
Грудина (рис. 255, /; рис. 256, 7) передневерхним краем сочленяется
с костями пояса передних конечностей (рис. 256, 3). Он образован
из непарной дужки, или вилки, — furcula (/, 4), двух коракоидов (к,3)
и двух лопаток (1,5). Дужка является соединением двух ключиц.
Рис. 256. Скелет грудной клетки птицы.
1 — основная пластинка грудины; 2 — киль грудины; 3—коракоид; 4—ключица; 5 — ло-
патка; 6—грудино-реберные кости.
Сросшимися вместе концами они примыкают к килю грудины, соеди-
няясь с ней пластинками соединительной ткани; свободные концы их
подходят к сочленовной ямке плечевой кости крыла (т). Суставная
ямка лежит на концах коракоида и лопатки. Последняя — сабле-
видной формы; она покоится на верхнебоковой стенке скелета груд-
ной клетки, будучи заложена в толще мышц. Коракоид нижним кон-
цом соединяется с переднебоковым отростком грудины. Последняя
очень велика и посредине нижней поверхности несет большой киль
(рис. 256, 2) для прикрепления летательной мышцы.
Тазовой пояс (рис. 255, 4) представляет собой изогнутое пла-
стинчатое образование, слившееся из парных подвздошных, лонных
и седалищных костей. В месте бывшего схождения их лежит вертлуж-
ная ямка, принимающая в себя головку бедренной кости (и). Под-
вздошные кости плотно срастаются с крестцовой частью позвоночника,
* 419
которая у птиц имеет сложный состав, ибо, кроме собственно крест-
цовых позвонков, она образуется спинными, поясничными и хво-
стовыми позвонками. Число костей, из которых слился крестец,
у разных видов птиц различно; в общем, оно вариирует от восьми
до двадцати четырех позвонков.
К подвздошным костям снизу примыкают седалищные, а с их
передним краем соединяются соединительной тканью лонные кости,
имеющие вид узких пластинок. Между тремя тазовыми костями на
каждой стороне тела лежит запирательная дыра (foramen obtura-
torinm).
Рис. 257. Г омология конечностей птицы. Слева — нога, справа — крыло, по-
средине — типичная конечность наземного позвоночного. Схема по различным
авторам.
f — бедро; —голень (из слившихся большой и малой берцовых костей и ряда костей
пятки — tarsus); М — пяточно-плюсневая кость (цевка); d — пальцы; b — плечо; г —лучевая,
и — локтевая кости; с — остаток костей запястья; тс Ч- с — пряжка (из сросшихся костей
запястья и пястья); d — пальцы.
Внутренняя поверхность крестца и подвздошных костей несет
углубление, в котором помещаются почки.
Скелет передней конечности (крыла) состоит из плечевой кости
(рис. 257, Ь), локтевой и лучевой костей предплечья (и, г) и кисти
(с, d). Запястье последней представлено двумя короткими косточ-
ками (с), к ним примыкает длинная окончатая кость — пряжка
(тс-1-е); она образуется слиянием двух пястных костей с рядом
костей запястья. Пряжка несет три пальца (d): первый, короткий
(из одной фаланги), лежит у ее основания, а второй (из двух фаланг)
и третий (из одной фаланги) помещаются на вершине пряжки.
Из данных вышеприведенной таблицы можно с несомненностью
заключить, что крыло птицы гомологично передней ноге лягушки,
а следовательно, и передней конечности других позвоночных, в том
числе и руке человека. Но в связи с особой ролью при движении птицы
крыло изменилось в деталях своего строения сравнительно с типич-
ной конечностью наземного позвоночного.
420
Сравнивая скелет передней конечности лягушки и птицы, мы мо
жем усмотреть следующие соотношения их частей:
Плечо Лягушка Птица
Плечевая кость Плечевая кость
Предплечье Лучевая и локтевая кости Лучевая и локтевая кости
Кисть запястье Из двух рядов шести ко- ротких костей Из двух коротких косточек (соответствующих одному ряду костей запястья)
пястье Из пяти пястных раздель- ных костей Из двух пястных костей, сросшихся концами в так называемую пряжку; с ними сливается второй ряд костей запястья
пальцы Четыре пальца (второй и третий из двух фаланг, четвертый и пятый из трех; первый палец редуцирован) Три сильно укороченных пальца: первый и третий — из одной фаланги; второй — из двух фаланг
Так, лучевая и локтевая кости предплечья неподвижны относи-
тельно друг друга, что делает конечность более прочной; кроме того
примечательна резкая редукция кисти, сказывающаяся в уменьшении
числа пальцев — фаланг, пястных и запястных костей. Пальцы у го-
лубя заметны только на скелете, так как они не выдаются наружу.
Уменьшение числа и подвижности составных частей кисти придает
ей большую компактность и прочность, что важно в общей статике
тела птедцы, ибо к покрывающей коже кисти прикрепляются большие
маховые перья (рис. 244, 8).
Анализ строения задних конечностей птицы возможен только бла-
годаря знанию истории ее развития. Нога птицы (рис. 257) состоит
‘из бедра (/), голени (/-{-/+ tf) и стопы (М, d). Скелетом бедра
является бедренная кость. Голень образована из сросшихся в раз-
личной степени большой и малой берцовых костей, причем к нижнему
концу их прирастает (без сохранения всякой внешней границы) пер-
вый ряд пяточных костей, лежащих у зародыша еще самостоятельно.
Следующая часть скелета ноги представлена единственной пя-
точноплюсневой костью — цевкой (Л4), которая возникла из слияния
второго ряда пяточных костей с четырьмя костями плюсны. С ее ниж-
ним концом сочленяются четыре пальца: первый — из двух фаланг,
второй — из трех, третий — из четырех и четвертый — из пяти.
Первый палец обращен назад, остальные смотрят вперед. Такое рас-
положение их способствует удобному и полному обхвату ветки дерева,
на которую садится голубь.
Сравним теперь скелет ноги голубя со скелетом типичной ноги
наземного позвоночного и с крылом птицы (табл, на стр. 422).
421
Задняя конечность Нога наземного позвоночного Нога голубя Крыло голубя Передняя конечность
I. Бедро Бедренная кость Бедренная кость Плечевая кость Плечо —I
II. Голеьь Большая | берцо- > вые Малая j кости Берцово-пяточная кость, слившаяся из большой и малой берцовых костей и костей первого ряда пятки Локте- вая Луче- вая кости Предпле- чье — II
пятка Ряд первый: tibi- ale, intermedium, fibulare 1 Две косточки запястья, со- ответствую- щие первому ряду костей запястья запястье
Стопа i ♦ Ряд второй: tar- salia I—V 1 Цевка — пяточно- плюсневая кость, об- разовавшаяся слия- * нием костей второго ряда пятки с косточ- ками плюсны Пряжка из двух пястных костей, слив- т ь е
III плюсна Metatarsalia I—V * шихся со вто- рым рядом ко- стей запястья п я с
пальцы Пальцы I—V Четыре пальца Три сильно ре- дуцирован- ных пальца пальцы
Эти данные убеждают нас в гомологии крыла и ног птицы, в де-
талях своего строения существенно уклоняющихся друг от друга,
что зависит от разной их роли в теле, ибо крыло птицы, служащее
для полета, не аналогично ее ноге, приноровленной для передвиже-
ния по твердому субстрату. Зато крыло птицы аналогично крыльям
других летающих животных, как, например, бабочки, стрекозы, хотя
по строению своему не имеет с ними абсолютно ничего общего.
Оценивая общее строение скелета птицы, мы отметим склонность
его к вариации в составе даже сравнительно постоянных его отделов.
Так, шейная часть позвоночника у всех земноводных состоит из
одного только позвонка, у птиц же, благодаря развитию шеи, число
се позвонков колеблется от восьми до двадцати трех; равным образом
крестцовые кости сливаются из восьми — двадцати четырех позвонков.
Развитие шеи, как внешне выраженной части тела, находит себе
отражение и в развитии соответствующего отдела позвоночника.
У рыбы и лягушки шеи нет, и шейная часть позвоночника вовсе или
почти вовсе отсутствует.
Благодаря превращению передних ног в крылья, передвижение
птицы по твердому субстрату выпало на долю только паре задних
1 Между первым и вторым рядами костей пятки лежит так называемая
центральная кость.
422
Hof, которые как в покое, таки при движении, должны поддерживать
компактное тело птицы. Для успеха действия необходимо прочное
соединение ног с телом; такое соотношение в порядке корреляции
повлекло за собой сильное развитие таза и весьма крепкое срастание
его с позвоночником на значительном протяжении последнего.
Так же коррелятивно развился и киль на грудине, служащий мес-
том прикрепления широкого основания грудной мышцы.
Для удобства полета важна неподвижность скелетных частей ту-
ловища, что достигается срастанием его позвонков в неподвижные
комплексы; наоборот, у рыбы позвоночник вообще легко сгибается
в стороны, что всегда совершается при плавании помощью хвостового
плавника или при змеевидном изгибании тела.
Общие замечания об организации птиц
Оценку организации птицы можно сделать, исходя из приспосо-
бления ее к полету. Последнее, прежде всего, сказалось в превращении
передних конечностей в летательный орган, т. е. в крылья, что вы-
звало ряд изменений в структуре скелета и мышц. Кисть крыла реду-
цировалась; образовалась довольно прочная связь плечевого пояса
со скелетом; на грудине возник большой киль, к которому прикре-
пляются большие грудные мышцы, опускающие крыло вниз при по-
лете. Развитие летательных мышц наглядно представляется тем, что
у голубя они составляют около половины веса всего тела.
Для успеха полета важно, чтобы тяжелые органы лежали воз-
можно ближе к центру тяжести тела и чтобы периферические части
последнего были бы, наоборот, более легкими. Поэтому туловище птицы
компактно и позвоночник его состоит из сросшихся групп позвонков.
Параллельно превращению передней конечности в крыло, у птиц
исчезли зубы, что отразилось на строении ее головы. Последняя стала
более легкой, кроме того исчезли или уменьшились в своем развитии
жевательные мышцы, так как птицы стали проглатывать пищу це-
ликом или кусками.
Крылья перестали служить для опоры тела, для хватания добычи
и производства других различных специальных движений (рытье,
работа по постройке нор, логовищ, гнезд и т. п.). Выпадение этих
функций замещается дополнительной работой клюва и ног. Клювом
птица хватает пищу, клювом она вьет себе гнездо, клювом пользуется
для защиты, передвижения (попугай хватается за ветку клювом
и переносит, держась им, свое тело с места на место) и т. п. Для произ-
водства столь разнообразных движений голова птицы и ее шея должны
быть очень подвижны, что и видим на самом деле.
Отсутствие зубов отразилось на внутренней организации птицы;
их роль взял на себя мышечный желудок, куда поступает пища после
предварительной обработки ее в зобе и в железистом желудке. Парал-
лельно наблюдается и чрезвычайная острота пищеварительных со-
ков, химический эффект действия которых велик.
В связи с полетом у птиц развились перья и преобразовалась
система органов дыхания, характеризующаяся наличием хорошо раз-
витых воздушных мешки в.
423
Для поддержания тела в воздухе необходимо уменьшение веса
тела, что достигается относительно малой длиной кишечника, недо-
развитием правой половины женских органов размножения, отсут-
ствием мочевого пузыря, концентрированностью мочи и пневматич-
ностью скелета. Благодаря заполненности костей воздухом, скелет
птицы относительно легче, чем у других наземных животных. Так,
у чижа вес скелета составляет 6,6% всего тела, а у равной по весу
домашней мыши — 8,4%. Уменьшение массивности скелета птиц, в ви-
дах достижения большей прочности, компенсируется отложением в нем
большего количества неорганических солей. Например, в костях зайца
содержится 75.15% неорганических зольных веществ, а в скелете
горлицы — 84,3%.
Птицы все без исключения яйцекладущи, причем кладут одно-
временно только по одному яйцу. Накопление нескольких яиц или
созревание живых зародышей в теле птицы увеличивало бы непроиз-
водительно ее вес и мешало бы легкости и быстроте полета.
Жировая ткань у птиц развита слабо; ее роль в деле сохранения
тепла тела берут на себя воздушные мешки и перовой покров, богатые
воздухом, который служит очень плохим проводником тепла.
То обстоятельство, что для передвижения по земле у птиц остались
только задние ноги, отразилось на развитии таза и отношении его
к позвоночнику.
Из этого краткого обзора особенностей организации птиц мы ви-
дим, что явления корреляции, т. е. соотношения степени развития
органов, у них имеются на каждом шагу. Птица в эгом отношении
является классическим примером.
КЛАСС МЛЕКОПИТАЮЩИЕ (MAMMALIA)
ОТРЯД ГРЫЗУНЫ (RODENTIA)
КРОЛИК (LEPUS CUNICULUS)
Технические замечания. Для вскрытия пользуются убитым
хлороформом кроликом, которого растягивают на препаровальной доске брюхом
кверху. В зависимости от обстоятельств могут быть взяты также белая крыса,
мышь, морская свинка. Дальнейшее описание ведется, в основном, по отношению
к кролику; важнейшие отличия в анатомическом строении других рекомендуемых
для вскрытия грызунов указываются в соответствующих местах текста.
Наружный осмотр
Тело грызуна состоит из головы, шеи, туловища и двух пар конеч-
ностей (ног). Голова распадается на два отдела: задний — черепной,
и передний—лицевой. По переднему и боковым краям последнего
помещается ротовое отверстие с верхней и нижней губами. Из него
торчат зубы — резцы. Над концом морды лежит нос, покрытый голой
кожей, в котором прорезаны два носовых отверстия (ноздри). Кзади
и кнаружи от основания носа помещаются глаза, прикрывающиеся
верхними и нижними веками. По краю век торчат длинные волоски —
ресницы. Мигательная перепонка, т. е. третье веко, не развита; остат-
ком (рудид\ентом) его является складка конъюнктивы во внутреннем
424
углу глаза. По бокам головы торчат ушные раковины с просвечиваю’
щими в них кровеносными сосудами. Ушные раковины обслуживают
наружное ухо и служат раструбами, предназначенными для уловле-
ния большего количества звуковых волн.
Шея невелика, но хорошо подвижна. Туловище несет пару передних
и пару задних ног. Каждая нога состоит из трех отделов, причем
задние ноги развиты сильнее передних и стопа их гораздо длиннее
кисти последних. На передних конечностях имеется по пяти пальцев,
а на задних —по четыре; пальцы оканчиваются невтяжными когтями.
На брюшной стороне туловища у самки есть несколько (4—5) пар
сосков молочных желез (у крысы их шесть пар). Присутствие этих орга-
нов очень характерно, и все животные, обладающие этим признаком
строения, относятся к классу млекопитающих (Mammalia) типа позво-
ночных.
На заднем конце туловища торчит хвост. У его основания поме-
щается заднепроходное отверстие (anus), а книзу и кпереди от него
лежит щелевидное мочеполовое отверстие у самки и копулятивный
орган (penis) у самца; на его конце—мочеполовое отверстие, а по бокам
основания этого органа лежит мошонка (scrotum), кожный мешочек,
вмещающий в себя семенники (testes — половые железы самца).
Таким образом у грызуна различие полов (половой диморфизм)
проявляется неодинаковым строением наружных половых органов и
присутствием у самки молочных желез.
Тело густо покрыто шерстью. Раздувая последнюю, заметим, что
волоски шерсти у кролика двух сортов: одни, более толстые, длинные
и окрашенные (грань, или ость), образуют собственно шерсть; другие,
тонкие, нежные, белые, сидящие между первыми, — подшерсток.
Шерсть временами выпадает, и ее волоски заменяются новыми; такой
процесс является линькой; он приурочивается к началу зимы. На зиму
у кролика появляется более густая шерсть, лучше удерживающая
тепло в теле животного, чем более редкая летняя шерсть.
На верхней губе торчат концами в стороны длинные грубые ще-
тинки — усы, или вибриссы, служащие для осязания.
Сравнивая кожные покровы известных нам позвоночных, мы убе-
ждаемся, что кожа кролика и ее придатки более близки к коже птиц,
нежели к слизистым покровам рыб и земноводных. Эта близость под-
тверждается ознакомлением с развитием шерсти млекопитающих.
Шерсть состоит из множества волосков, которые являются производ-
ными эпидермиса, подвергающегося ороговению. Припомним, что и
перья птиц также развиваются из эпидермиса и ороговевают. Благо-
даря развитию волос и перьев из общего зачатка, каковым является
эпидермис кожи, названные придатки ее могут считаться гомологич-
ными друг другу.-
Волосяной покров и млечные железы — вот два главнейших при-
знака, которые характеризуют млекопитающих вообще.
Если по богатству ороговевшими придатками кожа птиц и покровы
млекопитающих сближаются друг с другом, то по степени развития
желез они резко отличны. Кожа млекопитающих богата железами.
О млечных железах мы уже упоминали. Остается отметить микро-
скопически мелкие потовые железы и сальные железы, открывающиеся
425
в волосяные сумки — углубления эпидермиса, на дне которых сидит
волос. Потовые железы у кролика развиты очень плохо и имеются
только на губах и в паховой области. О наличии сальных и потовых
желез можно судить по микроскопическим срезам кожи.
Вскрытие кожных покровов
Ноги грызуна захватывают веревочными петлями, свободные концы
которых натягивают и привязывают к крючкам препаровальной доски.
Губкой смачивают шерсть по средней линии брюшной стороны тела
от челюстной дуги до мочеполового отверстия. От будущей линии
разреза шерсть расчесывают в стороны, чтобы получился пробор.
Пинцетом оттягивают кожу кверху и рассекают ее ножницами, начи-
ная от грудины, вперед до челюсти и назад до мочеполового отверстия.
Кожные лоскуты отпрепаровывают в стороны. При этом замечаем
скопления жира в подкожной клетчатке и беловатые млечные железы,
заложенные между кожей и мышцами. При снятии кожи вместе с ней
удаляется тонкий кожный мускул.
Нижняя стенка тела в области брюха состоит из следующих мышц:
а) прямая мышца живота, по средней линии тела разделенная соеди-
нительнотканной прослойкой белого цвета (linea alba); поперечными
перемычками такого же характера эта мышца разбивается на несколько
метамерных участков; б) с боков к ней прилежат косые мышцы живота;
другие составные части стенки тела с поверхности не видны; в)в области
грудины лежат грудные мышцы, направляющиеся к плечевым костям.
Вскрытие полости тела и общее расположение органов
Брюшную полость вскрывают, проводя разрез через толщу мышеч-
ной стенки от наружных половых органов до заднего края грудины.
Поднимая края полученных лоскутов кверху, разрезают ножницами
мышечную стенку по нижней границе грудной клетки, однако так,
чтобы не вскрыть последней. Разрезанные части стенки тела отгибают
в стороны и осматривают нормальное расположение органов брюш-
ной полости тела.
В передней части препарата, под массивной печенью, справа от на-
блюдателя, выглядывает дно желудка. Полость брюха занята петлями
кишек, прикрытыми спереди сальником (omentum) — складкой брю-
шины, свисающей вниз с края желудка. Большая часть петель кишек
относится к тонким кишкам; они отличаются от толстых гладкой и
ровной поверхностью, тогда как толстые кишки богаты кольцевыми
перехватами. Можно различить также и очень длинную слепую кишку
(coecum), которая сужается к переднему концу и отличается наиболь-
шей толщиной и обилием поперечных перехватов. Толстая кишка
переходит в прямую (rectum), открывающуюся заднепроходным отвер-
стием.
Близ левого края желудка помещается селезенка в виде длинного
пластинчатого тела красного цвета.
Если отбросить петли кишек в левую сторону препарата, то на
дне брюшний полости можно найти пару почек, из которых мочетиЧт
436
нйки направляются В мочевой пузырь, лежащий близ лонного сраще-
ния (в нижней части брюшной полости) в виде грушевидного мешочка
различной величины; последняя зависит от степени наполнения его
мочой. Между почками проходит брюшная аорта.
Брюшная полость отграничивается от грудной грудобрюшной пре-
градой, или диафрагмой, — широкой, пластинчатой мышцей, прира-
стающей к краям нижнего отверстия грудной клетки и вершиной своей
(куполом) вдающейся в последнюю. Диафрагму можно хорошо рас-
смотреть, отведя книзу желудок и оттеснив книзу же печень.
Перед вскрытием грудной полости прокалывают ножом диафрагму
и наблюдают, как вершина ее сейчас же опускается книзу. Это зависит
от того, что через искусственное отверстие диафрагмы воздух входит
в грудную полость, где и заполняет полость плевр, т. е. пространство
между легкими и стенками грудной клетки. В норме это простран-
ство безвоздушно, почему туда и устремляется атмосферный воздух.
В условиях нашего опыта атмосферное давление по обеим сторонам
диафрагмы уравнивается, почему ее купол и опускается книзу.
Грудную полость вскрывают по следующим правилам. От нижних
боковых углов грудной клетки ведут косые разрезы ножницами, рас-
секая и ребра и мышцы, кпереди и кнутри к нижней границе шеи,
причем перерезают также и пояс передней конечности. Захватывают
пинцетом передний конец заднего лоскута, оттягивают его кверху,
подрезая все, что изнутри к нему прикрепляется, и в конце концов
отрезают задний край этого лоскута от диафрагмы.
В грудной полости лежат: сердце с началом крупных сосудов, лег-
кие с бронхами и трахеей, которая продолжается в шею, где ее можно
проследить до гортани. Сзади и сбоку трахеи на шее лежит
пищевод.
После ознакомления с общей топографией органов перейдем к по-
дробному рассмотрению их по системам.
Внутреннее строение
Органы пищеварения. Открыв грызуну рот, рассматривают зубы.
На переднем конце челюстей торчат резцы: пара средних больших
и пара боковых маленьких на верхней челюсти и пара больших на ниж-
ней. Только боковая поверхность зубов покрыта эмалью; свободный
поперечник зуба постоянно стачивается при грызении растительной
пищи. У молодых кроликов за 7 дней резцы стачиваются на 3 мм.
Резцы постоянно растут и поэтому восполняют потерю зубной ткани
при ее стирании. Эта способность постоянного роста резцов у грызу-
нов врожденная, ибо она не связана с фактической работой их у дан-
ной особи. Если кролика кормить мягкой пищей, то резцы продолжают
расти и превращаются в уродливые образования, торчащие изо рта.
То же бывает в случае частичной поломки резца, нарушающей его
нормальное стачивание. Резец начинает расти в сторону, точно би-
вень; в конце концов он упирается в покровы головы или в глаз и вы-
зывает тяжелые изъязвления, ведущие подчас к гибели животного.
Непрерывный рост резцов при постоянном их стачивании является
весьма полезной и важной адаптацией. Нарушение же регуляции адап-
427
тивного роста резцов их стачиванием делает полезный адаптивный
процесс роста резцов вредным и даже гибельным для животного.
Позади резцов челюсти на некотором протяжении лишены зубов,
а дальше кзади помещается ряд коренных зубов. Сначала лежат три лож-
нокоренных зуба (на нижней челюсти два), лишенные корней, а за ними
три истиннокоренных зуба. Плоская поверхность их покрыта склад-
ками эмали, служащими для перетирания отгрызанной резцами пищи.
Клыки отсутствуют. Характер зубной системы млекопитающего вы-
ражают условно зубной формулой, где цифрами отмечают число зубов
одной половины каждой челюсти, считая спереди назад. Для кролика
2 • 0 • 3 • 3
зубная формула выражена так: t . 0.2.3 • Зубы у кролика различны
по форме, поэтому говорят, что его зубная система гетеро до нгная.
Рис. 258. Ненормальное разрастание нижнего резца у крысы.
Оригинал.
У мыши имеется с каждой стороны по одному резцу и по шести ко-
а 1-0-6
ре иных зубов j-TQTg- -
Для зубной системы вскрываемых животных характерен постоян-
ный рост несменяющихся резцов, служащих для обгрызания коры.
Такое строение и работа этих зубов настолько типичны, что все живот-
ные этого отряда называются грызунами (Rodentia). Зайцы и кролики
(представители семейства Leporidae) характеризуются присутствием
и пары малых (рудиментарных) верхних резцов.
На дне ротовой полости лежит мясистый язык, поверхность кото-
рого покрыта сосочками, еле видимыми простым глазом. По боковым
краям языка лежат крупные, до 5—6 мм длины, листовидные со-
сочки, у основания которых заложены концевые почки вкусовых нер-
вов (n. glossopharyngeus).
428
Ротовая полость переходит в глотку, от которой вниз отходит
пищевод, лежащий в шее позади трахеи. Пищевод (рис. 259, ое) про-
Рис. 259. Анатомия самки кролика. Схематично. По Н. X о-
лодковскому, с упрощениями.
tr — трахея*, у/ — яремная вена; ас—сонная артерия; аа — дуга аорты;
cv — желудочек сердца; рт — легкое; ое — пищевод; ао — брюшная
аорта; vt— желудок; ad — надпочка; г — почка; ov —яичник; od— яйце-
вод; vcl— задняя полая вена; ut — матка; urt — мочеточник; vg—вла-
галище; og—половое отверстие; ап — заднепроходное отверстие; гс —
прямая кишка; In — селезенка; ic — толстая кишка; in — тонкая кишка;
рп — поджелудочная железа; с — слепая кишка; hp — печень; sug —
• мочеполовой синус.
ходит сквозь грудную полость, прободает диафрагму и впадает в же-
лудок (vt). Последний имеет форму грушевидного мешка с передней
и задней поверхностями и двумя краями — малой кривизной и боль-
429
Рис. 260. Голова кролика сбоку. По Рёзелеру
и Лампрехту, с изменениями.
р — околоушная железа (parotis); i— gl. infraorbital Is; m' —
жевательный мускул (m. masseter); m — височный мускул.
шой кривизной. Пищевод входит в кардиальную часть желудка,
а привратник его граничит с кишкой. У входа в двенадцатиперстную
кишку заложена круговая мышца — сфинктер, или сжиматель, при-
вратника, заведывающий переходом пищи из желудка в кишечник.
Большая часть передней поверхности желудка прикрыта печенью,
которую следует отвести кверху, чтобы видеть его малую кри-
визну.
Далее приступают к препаровке петель кишек, брыжейку кото-
рых подрезают на всем ее протяжении ножницами. Освобождающиеся
части кишечника отводят влево и таким путем освобождают все его
отделы вплоть до прямой кишки. Разложив их по всей длине, начи-
нают осмотре двенадцатиперстной кишки (рис. 261, 2), т. е. части
кишки, прилежащей к желудку. В нее открываются протоки печени и
поджелудочной железы (рис. 259, рп). Без всякой границы duodenum
переходит в собственно тонкую кишку, которая очень длинна. Она
впадает в основание
громадной слепой киш-
ки (с), характеризую-
щейся значительной
толщиной, попереч-
ными перехватами и
длиной почти во все
тело кролика. Ее тон-
кий гладкий передний
конец называется чер-
веобразным отрост-
ком. Из основания
слепой кишки выхо-
дит толстая кишка,
поверхность которой
неровна. Толстая киш-
ка (рис. 259, ic) про-
должается в прямую
(гс), у здорового кро-
лика имеющую четкообразный вид; в ней формируются шарики твер-
дого кала. Прямая кишка открывается под основанием хвоста задне-
проходным отверстием (ап).
Как и у других позвоночных животных, у кролика кишечник
обслуживается пищеварительными железами. В полость рта изливают
свой секрет слюнные железы. Книзу и кпереди от основания ушной ра-
ковины лежит околоушная железа — gl. parotis (рис. 260, р). Ее вы-
водной проток прободает щеку и открывается на ее внутренней поверх-
ности.Эту железу можно отпрепаровать, сняв кожу с боковой части
головы от заднего угла нижней челюсти по направлению к ушной
раковине.
На брюшной стороне нижней челюсти несколько впереди лежат
подчелюстные железы (gl. submaxillares) и еще впереди от них распо-
ложены подъязычные железы (gl. sublinguales), соприкасающиеся вну-
тренними краями друг с другом. Последние две пары слюнных желез
открываются снизу у основания языка на дне ротовой полости.
430
Под диафрагмой лежит массивная печень (рис. 259, hp), состоящая
из четырех более крупных долек и нескольких мелких. Передняя по-
верхность печени выпуклая, так как она повторяет форму купола
диафрагмы. Приподняв край печени на вскрытом кролике, находят
желчный пузырь, лежащий в правой центральной дольке ее. От пе-
чени и желчного пузыря отходит по выводному протоку, которые ели-
Рис. 261. Поджелудочная железа кролика и прилежащие
органы. По Рёзелеру и Лампрехту, с изменениями;
1 — желудок; 2 — двенадцатиперстная кишка; 3 — слепая кишка;
4— толстая кишка; 5 — rectum; б—брыжейка; 7 — ее сосуды; 8—
pancreas; 9 — вирзунгов канал.
ваются в общий желчный проток, открывающийся в двенадцатиперст-
ную кишку. Желчный пузырь имеется у морской свинки, и мыши;
крыса же его лишена.
В брыжейке двенадцатиперстной кишки лежат мелкие дольки под-
желудочной железы (рис. 259, рп; рис. 261, 8), похожие по внешности
на скопление жировой ткани. Выводной канал этого органа также
впадает в двенадцатиперстную кишку.
Сравнивая органы пищеварения вскрытых нами позвоночных жи-
вотных, мы можем отметить общность плана строения их; но у кролика
431
пищеварительный аппарат развит относительно сильнее, чем у рыбы
и лягушки, так как распадается на несколько резко обособленных
друг от друга отделов. Примечательна также длина кишечника. У фран-
цузских кроликов она равняется почти 6 м, из коих на долю тонких ки-
шок приходится около 3 м. Пищеварительный канал хорошо развит
не только абсолютно, но и относительно. У домашних кроликов он
раз в девять-одиннадцать длиннее тела. Вспомним, что и слепая кишка
также по длине почти равняется их телу. Такое развитие органов пище-
варения объясняется растительноядностью кролика, как и других
грызунов. Растительная пища, в общем, груба, так как богата клет-
чаткой; последняя трудно переваривается сама по себе; кроме того
она оболочками одевает части тканей, состоящие из питательных
веществ. Оболочка из клетчатки трудно проницаема для пищевари-
тельных соков. Чтобы последние могли проявить свое действие, необ-
ходима хорошая механическая обработка пищи зубами (резцы и ко-
ренные зубы), которые перетирают и раздробляют клетчатку. Кроме
того потребно время для химической обработки пищи; длительность
обработки выигрывается во время прохождения пищи по очень длин-
ному кишечнику. В известных пределах мы можем, следовательно,
установить некоторую зависимость между растительноядностью жи-
вотного и длиной его кишечника, каковая в таких случаях относи-
тельно велика. Однако это соотношение не является абсолютным за-
коном, имеющим место в каждом случае, т. е. у каждого вида живот-
ного, питающегося растениями.
Органы кровообращения и лимфатические узлы. Центральный
орган кровообращения — сердце — лежит посредине грудной клетки
под грудиной. Сердце одето околосердечной сумкой (перикардиальная
сумка). Основание его прикрыто слегка рыхлой тканью зобной железы
(gl. thymus), которая хорошо развита у молодых животных; у взрос-
лых же кроликов она редуцируется; у мыши она покрывает почти все
сердце. Сердце (рис. 259, cv) состоит из четырех отделов. Темнокрасные
предсердия хорошо отличимы от розоватых желудочков, и при наруж-
ном осмотре граница между ними резка. Оба желудочка образуют кону-
совидную часть сердца, граница между ними намечается узкой жиро-
вой полоской. По удалении перикардиальной сумки можно видеть
выходящие из сердца главные кровеносные сосуды.
Из левого желудочка идет толстая аорта, загибающаяся дугой
влево (рис. 259, аа). От выпуклой части дуги выходят: безымянная
артерия, распадающаяся на правую подключичную артерию и правую
и левую сонные артерии (рис. 259, ас), и левая подключичная артерия.
Под дугу аорты подходит и скрывается под ней в толще легких легоч-
ная артерия, выходящая из правого желудочка. Дуга аорты, завер-
нув назад, прободает диафрагму и входит в брюшную полость (рис. 259,
ао), где и дает боковые ветви: для печени, брыжейки, почек (а. гепа-
les) и других внутренностей. В полости таза брюшная аорта распа-
дается вилообразно на общие подвздошные артерии (аа. iliacae com-
munes).
Венозная кровь возвращается в сердце по верхней и нижней полым
венам, впадающим в правое предсердие. Верхняя полая вена очень
коротка, нижняя же длинна; идет она параллельно аорте. У кролика
43?
есть воротная система печени. В последнюю поступает вся кровь из
кишечника по воротной вене; из печени же кровь выходит по печеноч-
ной вене, впадающей в нижнюю полую вену. Легочные сены впадают
в левое предсердие.
Благодаря обособленности камер сердца, большой круг кровообра-
щения совершенно отграничен, как и у птиц, от малого. Общий ход
кровообращения у этих животных одинаков.
Сравнивая ход главных артерий вскрытых нами позвоночных жи-
вотных, видим, что у них он все бол£е и более отклоняется от исходной
формы сосудов жаберных дуг рыб. Действительно, рыбы имеют пар-
ные артериальные дуги; число последних у лягушки уменьшается, но
все же остающиеся три пары артериальных дуг ветвятся симметрично.
У птиц симметрия сосудов, выходящих из сердца, нарушается, так как
желудочки отгораживаются один от другого и обособляются также
оба круга кровообращения. Такие же отношения видим и у кролика,
с той только разницей, что у него, как и у других млекопитающих,
остается левая дуга аорты, а у птиц — правая.
Для рассмотрения внутреннего строения сердца его вырезают из
груди и вскрывают по правилам, описанным в абзаце о кровеносной
системе птицы (стр. 404). Находят парусные и полулунные клапаны.
Правый парусный клапан состоит из трех створок, а левый — из двух,
причем правый клапан устроен из такой же фиброзной ткани, что и
левый, чем он и отличается от мышечного клапана правого сердца
птиц.
Лимфатическая система кролика развита хорошо; ее лимфати-
ческие узлы легко находимы при препаровке в виде бобовидных узел-
ков разной величины, лежащих в различных участках тела. Более или
менее постоянно узелки эти помещаются в подмышечной впадине
(gl. axillares), в месте деления трахеи на бронхи (gl. bronchiales),
в брыжейке и др. У корня брыжейки лежит скопление лимфоидной
массы, размером приблизительно в 3 см, образованное группой лим-
фатических верхнебрыжеечных узлов. Этот орган носит особое назва-
ние pancreas asellii и с настоящей поджелудочной железой ничего
общего не имеет.
К группе лимфатических узлов принадлежит красно-бурая селе-
зенка (рис. 259, In), лежащая на препарате справа от желудка.
К лимфатической системе относятся и лимфатические сосуды, по
ходу которых лежат лимфатические узлы. В противоположность кро-
веносным, лимфатические сосуды тонкостенны и очень мало заметны.
В случае их хорошего наполнения (что зависит от стадии пищеварения)
можно видеть в растянутой брыжейке беловатые тонкие полоски, иду-
щие параллельно артериям и венам к ее корню. Это и будут брыжееч-
ные сосуды, в которые поступает всасываемая пища. Лимфатические
сосуды вливаются в самый крупный сосуд — грудной проток (ductus
thoracicus), впадающий в место слияния левой яремной и подключич-
ной вен.
В заключение сопоставим для сравнения признаки, характеризую-
щие особенности строения кровеносной системы у представителей
типов позвоночных, членистоногих и червей, в пределах использован-
ного нами ранее материала.
2$ Практикум цо зоологии — 385
433
Особенности кровеносной системы позвоночных, членистоногих и червей
Кровеносная смете- ма Позвоночные Членистоногие Кольчецы
Речной рак Насекомое Дождевой черв л
Замкнутая Незамкнутая я Замкнутая
Сердце обособлено. . + + (Болес или менее) —
Сердце на брюшной стороне + 9 — —
Сердце или гомолог его на спинной стороне — Д- + +
Кровеносные сосу- ды + + — +
Венозная система . . + + — —
Кровь Красная Белая (гемолимфа) • Красная
Эритроциты + — — —
Плазма крови Бесцветная Красная
Белые кровяные клетки + + + +
Связь наличия кровеносной системы с образованием вторичной полости тела
Кровеносная система . . Животные без вторич- ной полости тела Животные с вторичной полостью тела
Кишечнопо- лостные Плоские черви Кольча- тые черви Мол- люски Члени- стоногие Позво- ночные
— — । + + +
Если у некоторых вторичнополостных животных кровеносная си-
стема отсутствует, то это объясняется редукцией ее и, следовательно,
должно оцениваться как явление вторичного характера.
Органы дыхания грызуна состоят из носовых отверстий, носовой
полости, хоан, глотки, гортани, трахей, бронхов, легких, диафрагмы
и других дыхательных мышц.
Ноздри лежат над ротовым отверстием; они ведут в носовые поло-
сти, которые вмещают в себе сильно искривленные носовые раковины,
покрытые слизистой оболочкой. В последней лежат концевые разве-
Сомнительная таблица соотношения наличия органов кровообращения у рас*
смотренных животных
Вторичная по- лость тела (целом) есть Крон снос идя СИ с т е м а
Замкнутая есть Незам развитая хо- рошо кну гля редуциру- ющаяся Отсутствует Характери- стика отсут- ствия кро- веносной си- стемы
Позвоноч- ные, коль- чатые чер- ви (дожде- вой червь) Членисто- ногие— ра- кообразные (речной рак), мяг- котелые Членисто- ногие— на- секомые (таракан) Некоторые мелкие на- секомые (редукция кровенос- ной систе- мы) Вторичное отсутствие кровенос- ной систе- мы
Вторичной по- лости тела (целома) нет — — —• Сосалыцики и другие плоские черви Первичное отсутствие кровенос- ной системы
твления обонятельного нерва, благодаря чему носовая полость является
также и органом обоняния. При прохождении через носовую полость
вдыхаемый воздух отчасти механически очищается, так как взвешен-
ная в нем пыль пристает к липкой слизистой оболочке; во-вторых,
воздух несколько согревается, ибо приходит в тесное соприкосновение
со стенками полости и раковинами, под поверхностью которых в изо-
билии ветвятся кровеносные сосуды.
Из носовой полости воздух проходит в глотку через хоаны, или
задние носовые отверстия. Они малы и щелевидны. Помещаются по-
зади и над задним краем твердого нёба.
В глотке перекрещиваются дыхательные пути с пищеваритель-
ными; это зависит от того, что пищевод лежит позади трахеи, а носовая
полость помещается кпереди и над ротовой полостью.
Гортань кролика развита гораздо лучше, чем верхняя гортань
птиц, которой она гомологична. Она состоит из широкого и высокого
щитовидного хряща (cartilago thyreoidea), отсутствующего у птиц.
Этот хрящ соединяется широкой пластинчатой связкой с дугообразной
подъязычной костью — остатком висцерального (жаберного) скелета
зародыша. Снизу щитовидный хрящ сочленяется с перстневидным
хрящом (с. cricoidea); на его задней широкой части лежат два черпа-
ловидных хряща (с. arytaenoideae), дающих начало голосовым
связкам.
У верхнепереднего края щитовидного хряща помещается основа-
ние надгортанника (epiglottis) — хряща, имеющего форму ложки. Его
можно видеть, разрезав трахею и гортань спереди.
Хрящи гортани соединяются друг с другом и с первым кольцом
трахеи соединительнотканными пластинками.
* 435
Трахея состоит (рис. 259, /г) из 48—50 прерванных йа Своей Задней
части хрящевых колец, также связываемых одно с другим соедини-
тельной тканью. Снаружи трахея имеет рубчатый вид. Изнутри она
выстлана мерцательным эпителием.
Кпереди на начальной части трахеи лежит двудольная красно-
бурая щитовидная железа. Она не имеет выводного протока; вслед-
ствие этого вырабатываемый ею секрет всасывается непосредственно
в кровь животного. Все железы такого сорта относят к железам вну-
тренней секреции; им противополагают органы внешней секреции, как,
например, потовые, желудочные железы, печень и т. п. Железы этого
порядка свои секреты изливают или на поверхность тела, или в поло-
стные органы, сообщающиеся так или иначе с внешней средой. Секрет
их удаляется из тканей тела.
Идя впереди пищевода, трахея минует шею и опускается в груд-
ную полость, где и ветвится на два бронха, погружающиеся в легкие
(рис. 259, ргп). Место ее ветвления можно хорошо видеть после удале-
ния сердца.
Розово-красные легкие очень мягки, так как вследствие губчатого
строения ткани богаты воздухом. Они дольчаты и с поверхности
покрыты гладкой блестящей оболочкой — легочной плеврой; аналогич-
ная выстилка есть и на внутренней поверхности грудной клетки —
плевра пристеночная, или париетальная. Плевра такого же строения,
что и брюшина. Она скользка, благодаря чему легкие не трутся о
стенку грудной клетки при дыхании. Способность легких к расшире-
нию можно доказать, надув легкое воздухом через стеклянную тру-
бочку, крепко ввязанную в отверстие перерезанной трахеи. Дольча-
тость надутых органов выступает очень ясно. Легкое на разрезе по-
хоже на мелкопористую губку. Если кусочек этого органа сдавить
под водой, то из него выступают мелкие пузырьки воздуха.
Грудная полость снизу замыкается диафрагмой, которая гермети-
чески отделяет ее от брюшной полости. Диафрагма является главной
дыхательной мышцей, прикрепляющейся своими краями к нижней
окружности грудной клетки. При своем сокращении купол ее, вдаю-
щийся в грудную клетку, опускается, благодаря чему увеличивается
передне-задний диаметр грудной полости. Так как последняя замкнута,
то в полости плевр при этом развивается отрицательное давление;
для его компенсирования атмосферный воздух устремляется через
дыхательные пути в легкие и растягивает их, пока они не заполнят
собой всей расширенной грудной полости. Таким образом происходит
вдыхание. За время вдоха протекает газообмен между свежим возду-
хом легких и газами крови. Воздух легких обогащается углекислотой
и удаляется при выдохе. Последний совершается обратным движением,
т. е. расслаблением диафрагмы; при этом купол ее, возвращаясь
в исходное положение, давит на легкое, сжимает его и выталкивает
часть находящегося в нем воздуха.
Помимо диафрагмы, в дыхании принимают участие и другие более
мелкие мышцы грудной клетки, но первенствующая роль принадле-
жит именно ей.
В заключение сравним последовательно части дыхательных орга-
нов воздухо дышащих позвоночных.
436
Верхняя гортань Лягушка Птица Кролик
— — Надгортанник
— — Щитовидный хрящ
— Перстневид- ный хрящ Перстневид- ный хрящ
Черпаловид- ный хрящ Черпаловид- ный хрящ Черпаловид- ный хрящ
Трахея — Есть Есть
Нижняя гортань — Есть —
Бронхи . . . : — Есть Есть
Легкие Мешетчатые Губчатые Губчатые
Воздушные мешки — Есть —
Диафрагма — Есть зачаток Развита хо- рошо
Из этого сопоставления видим, что птица и млекопитающее имеют
более совершенные и сходные друг с другом органы дыхания, чем
у лягушки.
Признаками сложного развития являются наличие дыхательных
трубок и губчатость легких, так как благодаря последнему их свойству
дыхательная поверхность легких необыкновенно увеличивается и не-
измеримо превышает поверхность ячеистой стенки мешковидных лег-
ких лягушки.
Оценивая развитие гортани, мы видим постепенное усложнение ее
у позвоночных, начиная с черпаловидных хрящей лягушки и кончая
сложно устроенным органом кролика. Птицы занимают в этом отно-
шении промежуточное положение; зато они отличаются особым ново-
образованием, свойственным только им — нижней, или певчей, гор-
танью.
Верхняя гортань — орган гомологичный у всех позвоночных жи-
вотных.
Легкие более сходно устроены у птиц и млекопитающих, чем у
каждой из этих групп животных и земноводных. Правда, характер
разветвления и концевых дыхательных частей легких у птиц иные,
чем у млекопитающих; кроме того легкое птиц срастается со стенкой
437
’судной полости и поэтому не обладает свободным движением, как
у млекопитающих животных. Однако эти отличительные признаки
птичьих легких от легких млекопитающих не так крупны, как разли-
чия в основном плане строения этих органов лягушки, образованных
мешками с ячеистыми стенками и большой общей полостью.
На первый взгляд очень резкой особенностью дыхательного аппа-
рата птиц являются воздушные мешки; они представляют собой
тонкостенные выросты поверхности легких, распространяющиеся ме-
жду различными органами тела. Воздушные мешки — это до край-
ности разросшиеся участки легкого, возникшие в связи с воздушным
образом жизни птиц. Гомологов их у млекопитающих нет, но у неко-
торых пресмыкающихся наблюдается обособление краев легких в виде
тонкостенных пузырей, которые можно рассматривать как исходную
форму возникновения воздушных мешков, достигающих такой край-
ней степени обособленности от легких, какую мы видим у птиц.
Зато и млекопитающие имеют некоторые новые органы в дыха-
тельной системе. Таковым является дыхательная мышца — диафрагма.
Правда, у птиц между легкими и печенью также лежит соединительно-
тканная перегородка со скудными мышечными пучками; ее считают
гомологом диафрагмы млекопитающих. Однако у птиц эта перегородка
пропускает сквозь себя и воздушные мешки и верхушку сердца; она
не аналогична диафрагме млекопитающих, ибо не принимает (и не
может принимать) участия в дыхании.
Сравнивая механизм легочного дыхания лягушки и кролика, за-
мечаем аналогию между межчелюстной мышцей первой и диафрагмой
последнего, так как обе мышцы принимают важнейшее участие в деле
проведения воздуха в легкие, хотя характер его поступания разли-
чен — у лягушки воздух нагнетается в легкие из ротовой полости,
у кролика он поступает в дыхательные органы вследствие присасы-
вающего действия грудной клетки.
Органы выделения. Чтобы видеть почки, необходимо удалить ки-
шечник из брюшной полости, подрезав корень брыжейки. Почек две;
каждая имеет бобовидную форму. Лежат они на препарате под боль-
шой кривизной желудка и под краем печени несимметрично, так что
левая почка кролика сдвинута ближе к хвостовому концу тела, чем
правая (рис. 259 и 262, г). Почки расположены по бокам и отступя
от позвоночника. Внутренний край синевато-красной почки несет
вырезку — устье, из которого воронкообразным расширением (почеч-
ной лоханкой) начинается мочеточник .(иг, urt). Почка часто бывает
покрыта жиром, который следует удалить при препаровке. Каждый
мочеточник идет вниз и впадает в мочевой пузырь (рис. 262, v), пробо-
дая наискось его стенку. Мочевой пузырь помещается в задней части
брюшной пол'ости. Из него выходит мочеиспускательный канал, устро-
енный у разных полов неодинаково. У самок он короток и собствен-
ным отверстием открывается в преддверие влагалища, тогда как у самца
шейка мочевого пузыря впадает в мочеполовой канал, служащий для
проведения не только мочи, но и семени. Его строение рассмотрим
ниже в связи с половыми органами.
У верхневнутреннего края почки, иногда отступя от него ближе
к позвоночнику, помещается круглая надпочечная железа, или надпочеч-
438
ник (рис. 259, ad; рис. 262, sr), желтого цвета. Она относится к группе
желез внутренней секреции.
Общее расположение почек у мыши сходно с кроличьим. Моче-
точники окружены жировой тканью, из которой их надо выпрепаро-
вать. Они прободают заднюю стенку шаровидного маленького моче-
вого пузыря, предварительно огибая сзади и снизу семевыносящие
каналы (см. ниже).
От верхней части мочевого
пузыря идет нитевидная связка,
прикрепляющаяся к передней
брюшной стенке в области пупка.
Эта связка является рудиментом
зародышевого мочевого протока,
или канала аллантоиса; послед-
ний является эмбриональным
органом, служащим для питания
зародыша (см. ниже).
Мочеиспускательный канал
самки мыши трудно различим;
он открывается на конце кли-
тора (см. ниже).
Рассмотрев на месте все части
мочевой системы, вырезаем одну
из почек. Она одета плотной
фиброзной капсулой. Зажав почку
пальцами левой руки, рассекаем
ее скальпелем, начиная с выпук-
лого края, на две половины. На
разрезе можно видеть, что ха-
рактер паренхимы почки не оди-
наковый. Наружный слой ее бо-
лее темный (корковый слой), вну-
тренняя же часть — розоватая,
тонко исчерченная в радиаль-
ном направлении (мозговое веще-
ство, состоящее 'из мочевых ка-
нальцев). Мозговое вещество
вдается в полость лоханки округ-
лым сосочком.
Как и у птиц, почка кролика
Рис. 262. Мочеполовой аппарат самца
кролика. По Фогту и Юнгу.
sr — надпочка; г — почка; иг — мочеточник;
v — мочевой пузырь; ts— семенник; ер — при-
даток семенника; vd— семевыносящий канал;
сс — пещеристые тела; р — penis; gp — тизо-
новы железы; lv — связка семенника.
третья по счету, считая закладку
ее у зародыша. Головная почка замещается первичной почкой, которая
также редуцируется, и животное вступает в жизнь со вторичной поч-
кой. Следовательно, и у кролика видим хороший пример подстановки
органов.
Почки кролика, как и млекопитающих вообще, очень плотны и
малы сравнительно с величиной тела. Почки птиц, наоборот, развиты
относительно сильнее. Зато млекопитающие обладают мочевым пузы-
рем, который отличается от аналогичного органа лягушки тем, что
дежцт на пути прохождения мочи, у лягушки же он лежит
439
в стороне от этого пути. Ход мочи у вскрытых нами позвоночных
таков:
Рыба: почка--------------------------------► мочевое отверстие.
Лягушка: почка—мочеточник—клоака—мочевой пузырь—клоака—отверстие
клоаки.
Птица: почка—мочеточник-------------------► клоака—отверстие клоаки.
Грызун: почка—мочеточник—мочевой пузырь—мочеиспускательный канал.
Рис. 263. А — Мочеполовые органы самца
кролика со спинной стороны.
7 — мочеточник; сбоку от него мочевой пузырь;
3 — v. deferens; 4 — контур простатического
пузырька; 5—предстательная железа; 6— ves.
seminal is; 7 — контур мочевого канала; 8 —
куперовы железы; .9 — мышцы; 10—ингвиналь-
ная железа; 11 — ингвинальный мешок; 12 — от-
верстие на вершине penis; 13 — penis.
В— мочевой п\зырь и мочеполовой канал
самца кролика вскрытые.
1 — мочеточник; 2— отверстие мочеточника; 4 —
вход в простату; 5 — colliculus seminails; 7—от-
верстия куперовых желез; 14—отверстие моче-
полового канала.
Сравнивая эти данные, за-
мечаем упрощение строения
мочевой системы птиц, что
связано с уменьшением веса
тела — обстоятельством, спо-
собствующим облегчению по-
лета.
Мужские половые органы.
У кролика и других грызунов
есть внутренние и наружные
половые органы. Половой ап-
парат самца довольно затруд-
нителен для препаровки.
Семенники (рис. 262, /х)
половозрелого кролика лежат
в кожном выпячивании, назы-
ваемом мошонкой (помещается
она кпереди и книзу от задне-
проходного отверстия); их
можно вытянуть в брюшную
полость, держа пинцетом за
пучок сосудов, обслуживаю-
щих семенник. Поверхность
последнего покрыта волокни-
стой соединительной тканью.
Далее разыскивают семепро-
вод—vas deferens (vd), на-
чальные извивы которого свя-
заны с так йазываемым при-
датком семенника—epididy-
mis (ер). Epididymis является
остатком первичной почки
(вольфово тело), ббльшая
часть которой редуцируется,
а оставшаяся часть меняет
функцию и из органа выделения превращается в проводящий семя
отдел половых путей. Мочеточник первичной почки, называемый
вольфовым каналом, при развитии зародыша превращается в семе-
провод. Этим и объясняется связь vas deferens с придатком семенника.
Семепровод из мошонки поднимается в брюшную полость через
так называемый паховый канал, перекрещивается с мочеточниками
(рис. 262, иг), обходя их сверху, и открывается в мочеполовой канал
со спинной его стороны, предварительно приняв в себя отверстие
440
Рис. 264. Мужской половой аппарат морской свинки.
Оригинал.
а—почка; Ь — надпочечник; с — мочеточник; d—семенной
пузырек; е — мочевой пузырь; f—семенник; h — придаток
семенника; i — семепровод; к — предстательная железа;
е внизу — куперова железа; т — копулятивный орган.
семенных пузырЪКоб — vesicular Seminales (рис. 263, А, 6), которые
являются хранилищами семени. Часть v. deferens после впадения
в него семенного пузырька называется семеизвергательным каналом
(ductus ejaculatorius); он соединяется с мочеиспускательным каналом,
идущим из шейки мочевого пузыря. Общая часть выводных путей,
обслуживающая и мочевые и половые органы, называется мочеиспу-
скательным каналом
(с. urogenitalis); он
проходит в толще ко-
пулятивного органа
(рис. 262, р).
В начальную часть
этого канала откры-
вается дорзально ле-
жащая предстатель-
ная железа (prostata)
величиной с горошину
(рис. 263, А, 4, 5;
В, 4). Ее желтоватое
тело распадается на
серединную и две бо-
ковые дольки.
Несколько ближе
к концу, перед посту-
плением своим в ко-
пулятивный орган,
мочеполовой канал
принимает в себя от-
верстие двух куперо-
вых желез — gl. Cow-
peri (рис. 263, А, 8).
Копулятивный ор-
ган (penis) (рис. 262, р)
состоит из двух дор-
зальных пещеристых
тел (сс), своими кор-
нями срастающихся с
лобным сочленением
тазовых костей, и вен-
трального пещеристо-
го тела, прободенного
мочеполовым каналом (corpus cavernosum urethrae). Пещеристые
тела снаружи одеты кожей, которая подвижной складкой —
крайней плотью (praeputium) — окружает головку уда (glans penis),
несущую на себе мочеполовое отверстие. Крайняя плоть имеет свои
препуциальные (или тизоновы) железы (gl. Tiesonii) (рис. 262, gp),
являющиеся видоизменением кожных сальных желез.
Пещеристые тела состоят из губчатой соединительной ткани, си-
нусы которой соединяются с кровеносными сосудами. Если отводящие
кровь сосуды спазматически закроются, то отток крови из члена пре-
441
кращается, а приток продолжается. Притекающая кровь Заполняет
синусы и растягивает пещеристые тела, благодаря чему уд становится
твердым и более длинным (у кролика с 2,5 см удлиняется до 4 см).
В таком состоянии напряжения (эрекции) он вводится в половую щель
и во влагалище самки при совокуплении. После излияния семени от-
водящие сосуды расширяются, кровь оттекает и копулятивный орган
возвращается в прежнее состояние.
Для рассмотрения пещеристых тел с них снимают кожу и тонким
скальпелем подрезают прикрепление их к лонному сращению таза;
освободив от связи с последним все мягкие части, протаскивают их
через таз в брюшную полость и вынимают совсем из тела, если пред-
варительно были отпрепарованы семенники и почки с их выносящими
каналами.
Таковы особенности строения половозрелого самца. У молодого
кролика семенники лежат в брюшной полости, на спинной ее стенке.
Они спускаются в мошонку лишь во время половой зрелости, причем
проскальзывают через паховый канал — щель между мышцами брюш-
ной стенки.
Мужские половые органы мыши. Семенники имеют
форму беловатых яйцевидных тел. В стадии покоя они лежат в пояснич-
ной области. К семеннику прилежит гребневидный придаток, обра-
зованный извивами семевыносящего канала; последний отходит от
семенника и направляется к мочевому пузырю; проходя позади
Рыба (костис^я) Лягушка Птица Грызуны
Половая железа Семенники
Внутренние половые органы Половые пути Выходная часть семен- ника и половое отверстие Канальцы, прободаю- щие почку, мочеточник с семейным пузырьком и клочка Семепро- воды и клоака Epididymis и V. deferens Семеизвер- гательный канал Семенные пузырьки Мочеиспу- скательный канал с ку- перовыми железами
Наружные половые ор- ганы (== органы совоку- пления) Отсутствуют Копулятив- ный орган (penis) с мо- чеполовым . отверстием
Оплодотворение Нару У’Нле Внутреннее
442
пузыря между мочеточниками, семевыносящие каналы соединяются
с семенными пузырьками, имеющими трубчатую спиральную форму;
у мыши они развиты слабо, у крысы — сильно. Семевыносящие про-
токи, сливаясь друг с другом, открываются в предстательную железу,
которая у крысы развита сильнее, чем у 'Мыши.
Тизоновы железы развиты сильно. У морской свинки отмечается
сильное развитие двух трубчатых пузырьков, значительное развитие
простаты; хорошо выражены и куперовы железы (рис. 264).
У молодых самцов-грызунов семенники лежат в брюшной полости
и к половой зрелости спускаются в мошонку.
Мужские половые органы грызунов устроены гораздо сложнее, чем
у вскрытых нами позвоночных животных; не говоря уже о разнообраз-
ных внутренних частях, грызуны обладают и наружными совокупи-
тельными органами. Соответствующие детали сравнения восстановим
по таблице на стр. 442. •
Примечательна также связь органов выделения с половым аппара-
том, которую напомним нижеследующей таблицей.
Головная почка (предпочка) pronephros Рыба Лягушка Птица Грызуны
Есть Редуцируется
Первичная почка (вольфово тело) mesonephros Есть Есть Остается в виде руди- мента при семеннике
В виде почки Придаток семенника
Мочеточник первичной почки (вольфов канал) Есть в виде мочеточника Является мочеточни- ком и семе- проводом Является только семе- проводом
Вторичная почка metanephros — — Есть является тс Есть ►лько почкой
Мочеточник вторичной почки — — Есть является то ТОЧН1 Есть лько моче- IKOM
Женские половые органы. Крольчиха имеет и внутренние и на-
ружные органы размножения.
Внутренний половой аппарат можно рассмотреть в задней части
брюшной полости по удалении кишечника. Внешность его различна,
что зависит от периода жизни самки. Крольчиха является живоро-
дящим млекопитающим, которое вынашивает в себе развивающихся
443
Рис. 265. Женские половые органы кролика.
По Рёзелеру и Лампрехту.
о— яичник; b — яйцевод; с — матка; d— мочевой
пузырь; е—rectum; f—мочеточник; v—нижняя
полая вена; а — аорта.
детенышей и, по достижении ими надлежащей стадии развития, от-
рождает их, т. е. выбрасывает наружу через половое отверстие. По-
ловые органы беременной самки велики, так как матка растянута за-
родышами.
Внутренние органы (рис. 259 и 265) состоят из двух яичников,
пары яйцепроводов, двойной матки и влагалища.
Яичники (о) имеют форму сплющенных яйцевидных тел, под-
вешенных у поясничной мышцы на широкой связке (двойной складке
брюшины). Поверхность яичника не всегда ровная. Иногда на ней
имеются бугорки, образуемые раздувающимися граафовыми пузырь-
ками, в которых развива-
ются яйцеклетки кролика.
Они не содержат в себе пи-
тательного желтка, почему
очень малы по сравнению
с яйцами вскрытых нами
позвоночных. В граафовом
пузырьке яйцо окружено
так называемыми фолли-
кулярными клетками, вы-
деляющими особую жид-
кость. Последняя давит на
стенки граафова пузырька,
которые подаются в точке
наименьшего сопротивле-
ния, т. е. со стороны по-
верхности яичника, по-
этому на ней и бывают
видны вздутия. Рано или
поздно под влиянием да-
вления жидкости стенка
граафова пузырька (т. е.
стенка самого яичника)
разрывается, и яйцо выхо-
дит наружу, т. е. в брюш-
ную полость.
Обычно в яичнике созревает сразу около шести граафовых пузырь-
ков, которые и опорожняются от заключающихся в них яиц последо-
вательно друг за другом в периоде овуляции. Периодическое выпадение
яиц (овуляция) совершается у крольчихи с апреля по июнь в правиль-
ные месячные промежутки; позднее оно также наблюдается, но уже
в неправильные сроки.
Яичник не соединяется непосредственно с яйцеводом, который от-
крывается воронкообразным отверстием в брюшную полость (рис. 259,
od); однако воронка близко прилежит к яичнику (рис. 265, Ь), по-
чему и захватывает в себя освобождающиеся яйца. Яйцевод тонок
и извилист; как и яичник, он подвешен на брюшной складке. На
своем пути яйцевод далее впадает в матку, которая, в сущности,
является расширенной его частью. Она представлена толстой трубкой,
подвешенной на широкой маточной связке (рис. 259, at). Матка
444
половины ее срастаются друг
Рис. 266. Мочевой пузырь (6) и влага-
лище (2) кролика вскрытые. По Р ё з е-
- леру и Л а м п ре хту.
1 — матка; 3 — рыльце матки; 4 — влага-
лище; 5 — мочеточник; 7 — отверстие его;
8—шейка пузыря; 9— мочеполовой тракт;
10—клитор; 12 — мочеполовая щель; !3 —
толстая кишка; 14 — анальные железы; 15 —
anus; 16— ингвинальная железа.
у крольчихи парная, и обе трубки ее независимо друг от друга откры-
ваются (рис. 266, 3) в непарный канал половых путей, называемый
влагалищем (рис. 259, vg; рис. 266, 4).
Такая матка, как у крольчихи, называется двойной (u. duplex) и
является наиболее примитивной формой у млекопитающих, так как
у большинства этих животных
с другом на большем или мень-
шем протяжении.
Влагалище представлено бо-
лее широкой, чем матка, труб-
кой, внутренняя поверхность
которой покрыта продольными
складками слизистой оболочки.
Влагалище продолжается в
преддверие влагалища, или моче-
вой синус (рис. 259, sag), в на-
чало которого открывается ко-
роткая шейка мочевого пузыря
(рис. 266, 8). Рядом распола-
гается поперечная складка сли-
зистой оболочки, соответствую-
щая девственной плеве (hymen).
Преддверие влагалища изнутри
гладко. Оно открывается поло-
вой щелью, отграниченной склад-
ками кожи — губами. В месте
вентрального соединения их друг
с другом лежит клитор (рис. 266,
10), соответствующий копуля-
тивному органу самца. Клитор
состоит из пещеристых тел, со-
единенных своим основанием с
лонным сращением тазовых ко-
стей; он достигает длины 3 см.
Все описанные детали строе-
ния можно рассмотреть, разрезав
вдоль мочеполовой синус и вла-
галище.
Строение женского полового
аппарата мыши, крысы или мор-
ской свинки соответствует той же схеме, что и у кролика. Матка у
небеременных очень тонка. При развитии зародышей образуются
мешковидные расширения матки (рис. 267). Обычно бывают бере-
менны обе матки.
Оплодотворение у грызунов, как у всех млекопитающих, — вну-
треннее. При копуляции самец вводит эрегированный член в преддве-
рие влагалища и влагалище, куда и изливает семенную жидкость.
Живчики пробираются в яйцеводы, где происходит оплодотворение
яиц. Последние же выходят из яичника через 8—12 часов после со-
вокупления.
445
Яйцеклетки кролика очень малы,достигая в диаметре всего 0,116мм,
тогда как яйца лягушки в поперечнике имеют 1,75 мм, а желток
куриного яйца — около 30 мм. Такое различие в величине яиц обу-
словливается не одина-
Рис. 267. Женский половой аппарат беременной
крысы. Оригинал.
о — яичник; а — воронка яйцевода; b — яйцевод; с — матка
с зародышами; d— влагалище; е — мочевой пузырь.
ковым количеством пи-
тательного желтка. Раз-
ница становится еще
более наглядной, если
сравнить яйца назван-
ных животных по
объему. Яйцо кролика
по объему своему не
превышает 1/ЗОсо яйца
ЛЯГУШКИ И Visoooooo
яйца курицы.
Очевидно, что в яйце
кролика питательного
желтка нет и что раз-
витие его вне матери
невозможно. И действи-
тельно, зародыш кро-
лика развивается в
матке, причем вступает
с ней в тесную связь
через посредство дет-
ского места, или пла-
центы, служащей для
питания зародыша на
счет веществ, доста-
вляемых кровью ма-
тери.
Если вскрыть матку
беременной крольчихи,
то в ней можно найти
последовательно лежа-
щие друг за другом за-
родыши, числом обычно
6—8, а у очень пло-
довитых особей — 12 и
даже 16. Зародыш одет
тонкими зародышевыми
оболочками (амнион и
сероза), которые покры-
вают все его тело, начи-
ная с десятого—двенад-
цатого дня развития.
До седьмого дня развития зародыш, проходящий стадию бластодер-
мического пузыря, лежит в матке почти свободно, но затем начинает
срастаться с се стенкой. На нижней поверхности бластодермического
пузыря образуются сосочки, врастающие в стенку матки. Позднее
446
в эти сосочки внедряются кровеносные сосуды; сюда они входят
из аллантоиса, т. е. мешковидного выроста кишечника зародыша.
Капилляры этих сосудов расширяются, также изменяются и сосуды
в прилежащей части матки. Благодаря описанным процессам, обра-
зуется детское место, или плацента. В нем сосуды матери и зародыша
тесно соприкасаются друг с другом; между ними нигде не бывает
непосредственного соединения, и кровь матери и зародыша совер-
шенно изолированы друг от друга. Однако через разделяющие их
в плаценте тонкие стенки кислород и пищевые вещества проникают
осмотически (дифундируют) из крови матери в кровь зародыша, и угле-
кислота, накопляющаяся в результате клеточного дыхания послед-
него, идет в обратном направлении, т. е. в кровь матери.
Благодаря такой тесной связи между материнским организмом
и его плодом последний питается за счет первого и растет гораздо
быстрее, чем у животных, развивающихся вне тела матери (лягушка,
цыпленок).
В общем зародышевое развитие кролика длится 30 дней, после
чего он рождается на свет в совершенно беспомощном состоянии —
голый, слепой и неспособный к самостоятельному питанию. С момента
родов период плацентарного питания заканчивается и сменяется пе-
риодом молочного кормления, в котором детеныш сосет молоко, вы-
деляемое к этому времени молочными железами. Перед родами самка
кролика устраивает из сена и собственного пуха гнездо для кроль-
чат и проявляет таким образом до известной степени родительский
инстинкт заботы о потомстве. В некоторых случаях наблюдается, что
мать поедает собственный приплод. У молодых самок эта ненормаль-
ность может объясняться ошибкой. Дело в том, что у крольчих от-
рождается и послед, который она съедает; при таком поедании воз-
можна роковая для детеныша ошибка. Однако у некоторых старых
самок укореняется подобное извращение родительского инстинкта,
и они регулярно пожирают всякий раз приплод, или же оставляют его
без надлежащего ухода, что также оканчивается его гибелью.
На шестой-седьмой день после рождения крольчата покрываются
шерстью, на десятый-двенадцатый день у них открываются глаза,
недели через 3 они выбегают из гнезда, а месяца через 3—4 переходят
на общий корм.
В заключение сравним женские половые органы вскрытых нами
животных (см. табл, на стр. 448).
Из этого сопоставления мы можем усмотреть важное сходство
между лягушкой, птицей и кроликом в том, что яйцевод не предста-
вляет собой непосредственного продолжения яичника, так как от-
крывается воронкообразным отверстием в полость тела. Такие же
отношения наблюдаются и у некоторых рыб (поперечноротые— акулы,
скаты и др.).
Центральная нервная система. Чтобы вскрыть головной мозг кро-
лика, отрезают голову от шеи в затылочном сочленении; на месте раз-
реза осматривают два затылочных мыщелка, сочленяющихся с атлан-
том. Скальпелем рассекают кожу головы сверху по средней линии и
образовавшиеся лоскуты оттягивают в стороны, перерезают хрящи
ушных раковин у их основания и таким образом обнажают черепную
447
Окунь Внутренние половые органы Наружные половые органы Раз- витие
Яичник Выводная часть яичника Половое отвер- стие — Вне тела матери Без заботы со стороны родителей
Лягушка Яичники Яйцеводы, открывающиеся в полость тела, клоака Отвер- стие клоаки —
Птица Яичник Яйцевод, открывающийся в полость тела, клоака Отвер- стие клоаки — С заботой со стороны родителей
Кролик Яичники Яйцеводы, открывающиеся в полость тела, матка, вла- галище, мочеполовой синус Половое отвер- стие Клитор и «губы» Внутреннее (плацента)
коробку. Дальнейшая препаровка ведется различно, смотря по воз-
расту вскрываемого кролика. У молодого животного кости черепа
срезают по частям в горизонтальном направлении крепким скальпе-
лем. Такое состругивание костей ведут сзади наперед, т. е. от затылка
ко лбу. Когда обнажится поверхность мозга и таким путем действо-
вать будет уже неудобно, то боковые части черепа отламываются по
кусочкам крепким пинцетом. У взрослых кроликов кости более крепки;
поэтому их приходится распиливать пилой. Сначала делают два про-
дольных разреза — кнутри и над глазами; концы их спереди соеди-
няют поперечным разрезом. При работе следят по звуку за моментом,
когда кость будет пропилена, иначе легко повредить мозг. Затем в за-
тылочное отверстие вводят ветвь крепких ножниц или костных щип-
цов и рассекают верхнезатылочную кость с двух сторон, стремясь
каждый раз дойти до заднего конца продольного разреза, сделанного
пилой. Когда крыша черепа будет со всех сторон обойдена разрезами,
ее снимают осторожно пинцетом кверху. Обычно с черепными костями
отходит и твердая мозговая оболочка (dura mater).
При осмотре мозга (рис. 268) сверху видны большие полушария
мозга (С), продолжающиеся спереди в маленькие обонятельные доли
(Lo). С задним краем полушарий непосредственно граничит мозже-
чок (СЬ). В противоположность гладкой поверхности больших полу-
шарий, мозжечок изборожден складками. В нем можно различить
срединную часть (червячок — vermis) (о) и две боковые доли с непра-
вильной формы выростами — flocculi (СЬ). Больше ничего нельзя
рассмотреть, не смещая частей мозга.
Если отвести друг от друга соприкасающиеся части больших полу-
шарий и мозжечка, то в глубине между ними можно заметить средний
мозг с четверохилмием на поверхности, причем передняя пара бугров
448
больше задней. Между буграми первой пары лежит эпифиз, относя-
щийся к промежуточному мозгу.
Чтобы видеть продолговатый мозг, приподнимают кверху средин-
ную часть мозжечка.
В общем, у кролика все типичные отделы мозга налицо, но разли-
чимы они трудно, так как особенно сильно разрастаются большие
полушария, а затем мозжечок, которые и закрывают собой сверху и
с боков все остальные части головного мозга.
Для осмотра головного мозга снизу, уплотняют полученный нами
препарат в 10% растворе формалина несколько дней, после чего и
вынимают мозг, подрезая все выходящие из
Идя сзади наперед,
рассматривают про-
долговатый мозг и пе-
ред ним округлую
часть с вырезкой по-
средине — варолиев
мост (рис. 268, Р), по
бокам которого выгля-
дывают дольки моз-
жечка (СЬ). Далее кпе-
реди дно углубления
мозга образует мозго-
вые ножки (соответ-
ствует среднему моз-
гу), на них надвига-
ются задневнутренние
края больших полу-
шарий, между кото-
рыми лежит овальный
гипофиз — промежу-
точный мозг(Н). Пе-
ред ним помещается
перекрест зрительных
Hepeoe(chiasma). Боль-
него нервы.
Рис. 268. Головной мозг кролика. По К ю к е н-
т а л ю.
А — сверху; В — снизу. Lo — обонятельные доли; С— боль-
шие полушария; СЬ — мозжечок; v — червячок; Мо — про-
долговатый мозг; Р— варолиев мост; Н — нижний мозго-
вой придаток; XI—корешки добавочного нерва.
шие полушария продолжаются в обонятельные доли. По нижней
поверхности полушарий, параллельно наружному [краю их (если смо-
треть снизу), идет сильвиева борозда.
У всех позвоночных животных все пять частей [головного мозга
налицо, но развиты они различно. Для выяснения отличий сравним
соответствующие отделы головного мозга у вскрытых нами животных
(рис. 269).
Продолговатый мозг развит у всех них соразмерно одинаково, ибо
роль его у животных сходна. Он снабжает нервами (иннервирует)
жаберную область или соответствующие производные висцерального
скелета, легкие, сердце и кишечник и дает начало пяти—двенадцати
парам черепных нервов.
Мозжечок развит неодинаково. Еле выражен он у лягушки, лучше
развит у рыбы и достигает наибольшей величины у кролика и птицы.
Степень развития мозжечка стоит в связи со способностью животного
2Q TTnewTnwvM по яоплагаи — ЯЯ5
449
к движению.У коротконогой лягушки, брюхо которой касается земли,
он очень мал; окунь (костистая рыба) является хорошим пловцом —
мозжечок его развит хорошо; нечего говорить, что у птицы, живот-
ного с наиболее совершенными движениями (полет), он достигает
Рис. 269. Головной .мозг позвоночных сверху. По Гессе и Дофлейну.
А — акулы; В — лосося (костистой рыбы); С — земноводного (лягушки); D — пресмыкаю-
щегося; Е — птицы (голубя); F—G — млекопитающих (кролика и собаки). Обонятельный отдел
и передний мозг — с продольными черточками; промежуточный — исчерчен накрест;средний—
исчерчен вдоль; малый мозг (мозжечок)покрыт пунктиром; продолговатый—зачерчен поперек.
наибольшей величины, как и у млекопитающего, мозжечок которого
еще более, чем у птицы, богат извилинами и бороздами.
Средний мозг устроен сравнительно однообразно и состоит из дву-
холмия или четверохолмия.
Промежуточный мозг, характеризующийся своими придатками —
эпифизом (третий глаз) и гипофизом, также развит приблизительно
одинаково; но и, как средний мозг, он может быть представлен относи*
450
тельно малой частью головного мозга, что зависит от крайнего раз*
растания других отделов последнего.
Передний мозг, как и мозжечок и даже более последнего, склонен
к развитию. У окуня он менее двухолмия, к которому примыкает
непосредственно спереди. У лягушки большие полушария развиты
больше, хотя они и не закрывают собой промежуточного мозга, как
у рыбы. Иное дело у птицы и, особенно, у кролика, большие полушария
которого разрастаются вверх и в стороны и образуют мощно развитую
мантию мозга, покрывающую обонятельную часть основания мозга
и лежащие над ней полосатые тела. Со степенью развития мантии
связывают способность животного к высшим отправлениям психи-
ческой деятельности. •
Оценивая головной мозг позвоночных животных как целое, мы
должны отметить, что наиболее типичным в смысле соразмерности
развития его пяти составных частей является головной мозг лягушки,
лежащий в горизонтальной плоскости. Несколько отклоняется от
этого типа мозг окуня со сдавленным и скрытым промежуточным мозгом.
Еще дальше в этом отношении идут птица и кролик, у которых раз-
растающиеся большие полушария и мозжечок закрывают собой все
остальные отделы мозга, которые вследствие этого кажутся относи-
тельно слабо развитыми.
Все эти сопоставления можно наглядно уяснить себе на прила-
гаемых схемах (рис. 269), где соответствующие части мозга однообразно
заштрихованы.
Органы чувств. Органом осязания служат крупные ще-
тинистые волосы на верхней губе, так называемые «усы». Чувстви-
тельность их объясняется тем, что у их основания разветвляются
окончания чувствительных нервов. При прикосновении к усам, осно-
вания волосков несколько сдвигаются и раздражают оплетающие
их нервы, которые и передают раздражение в головной мозг.
Орган слуха и равновесия — ухо — состоит из наружного,
среднего и внутреннего уха.
Наружное ухо характеризуется очень большой ушной раковиной —
складкой кожи, подпертой опорными хрящами. Ушная раковина
имеет вид вытянутого в длину желоба; она служит раструбом, соби-
рающим звуковые колебания и отражающим их в слуховой проход.
Дно его закрыто барабанной перепонкой; за которой лежит среднее
ухо. В нем помещаются три слуховых косточки: молоточек, соединяю-
щийся одной своей частью с барабанной перепонкой, а другим концом
сочленяющийся с наковальней, которая, в свою очередь, сочленяется
со стремячком. Благодаря таким соотношениям звуковые волны,
заставляющие дрожать барабанную перепонку, приводят в легкое
движение и слуховые косточки. Стремячко вставлено в отверстие,
ведущее в лабиринт. Последний состоит из трех полукружных ка-
налов, открывающихся в преддверие и улитку, обороты которой за-
виты спирально. Все эти части заложены в пирамидке височной кости
и при обычной работе отпрепарованы быть не могут.
Поучительно сравнить органы слуха позвоночных животных.
Общей частью из этих органов у всех позвоночных является ла-
биринт из трех полукружных каналов. У птицы совершенно ясно
451
Внутреннее ухо Среднее ухо Наружное ухо
Окунь Три полукружных капала и лабиринт с lagena 1 1 Нет ! ! И е т
Лягушка Три полукружных канала и лабиринт Барабанная полость с columella 3 ю о н к а Н е т
Птица Три полукружных канала и лабиринт со слегка изогну- той улиткой Барабанная полость с columella йены тру а я п с р с п Наружный слуховой проход
Г рызуны । Три канала, лаби- ринт со спираль- ной улиткой Барабанная полость с молоточ- ком, нако- вальней и стремячком Е в с т а х Б а р а б а и н Наружный слуховой проход и ушная ра- ковина
развивается улитка в виде слегка изогнутой трубки, которая у мле-
копитающего закручивается спирально. Среднее ухо появляется только
начиная с земноводных, к нему у птиц присоединяется короткий
наружный слуховой проход. У обеих этих групп животных одна
слуховая косточка—columella. У млекопитающих она преобра-
зуется в стремячко, и к нему добавляются еще две — молоточек
и наковальня, а к наружному слуховому проходу присоединяется
ушная раковина.
Органы зрения — глаза. Верхнее и нижнее веки по своим
краям усажены рядом ресниц. В носовом углу глаза имеется малень-
кая складочка, являющаяся рудиментом третьего века (мигательной
перепонки у птиц). Глазное яблоко типичного строения, без каких-
либо особенностей, наблюдаемых у птиц. За прозрачной роговицей ле-
жит небольшая передняя камера; задней ее стенкой служит хрусталик,
вставленный в отверстие радужной оболочки. Позади хрусталика
расположена большая камера глаза, являющаяся главной полостью
глазного яблока, заполненной стекловидным телом студенистой кон-
систенции. Изнутри большая камера глаза покрыта сетчаткой — обо-
лочкой, воспринимающей световые раздражения, в которой развет-
вляются окончания зрительного нерва.
При вылущении глаза из глазницы необходимо перерезать приво-
дящие его в движение мышцы и зрительный нерв.
В наружном (височном) углу глаза лежит слёзная железа, протоки
которой прободают верхнее веко.
1 См. стр. 355
452
Органом обоняния является носовая полость; у кролика
она объемиста и длинна. В ней к наружной ее стенке изнутри при-
крепляются изогнутые плоские кости, покрытые слизистой оболоч-
кой,— передние и задние носовые раковины; носовая полость от-
крывается наружу двумя ноздрями и внутрь в носоглотку двумя хоа-
нами. Носовые полости каждой стороны тела отделяются друг от друга
носовой перегородкой.
Раковины можно рассмотреть, распилив голову кролика вдоль по
средней линии; характер изгибов раковин легко уясняется на фрон-
тальных распилах передней части морды кролика.
Скелет
Как и у других позвоночных, основной частью скелета кролика
является спинной хребет, или позвоночник, состоящий из сорока шести
позвонков, разбивающихся на пять отделов. Шейная часть предста-
влена семью позвонками, грудная—двенадцатью, поясничная —
семью, крестцовая — четырьмя и хвостовая — шестнадцатью позвон-
ками. Все они хорошо окостеневают и соединяются друг с другом
связками и прослойками волокнистого хряща. I шейный позвонок
(атлант) имеет вид вытянутого кольца с двумя сочленовными впа-
динами для двух затылочных мыщелков черепа. I! шейный позвонок
(эпистрофей) обладает зубцевидным отростком, входящим в спинно-
мозговое отверстие атланта и сочленяющимся с его нижним краем.
Остальные шейные позвонки сходны по строению друг с другом и осо-
бенностей не представляют. Со спинными позвонками сочленяются
ребра; остистые отростки этих позвонков велики. Поясничные позвонки
крупнее всех прочих и характеризуются присутствием хорошо раз-
витых поперечных отростков, направленных своими концами вперед
и вниз. Четыре крестцовых позвонка срастаются в одну пирамидаль-
ную кость — крестец, сочленяющуюся с тазовыми костями. Задние
три крестцовых позвонка редуцируются по сравнению с передним
позвонком. Хвостовые позвонки видоизменяются еще больше и вовсе
теряют форму позвонков, представляя собой тонкие и короткие па-
лочковидные косточки.
Позвоночник пронизан спинномозговым каналом, вмещающим в себе
спинной мозг. Последний тянется до крестца, где и заканчивается
концевым конусом. Далее, до седьмого хвостового позвонка, продол-
жается его концевая нить, окруженная множеством нервов, образую-
щих в совокупности так называемый конский хвост (cauda equina).
В черепе кролика лицевая часть тесьма преобладает в своем раз-
витии над мозговой. Мозговая коробка образована из сложной заты-
лочной кости, состоящей из четырех отдельных частей—основной,
двух боковых и верхней части; между ними лежит затылочное отвер-
стие с двумя мыщелками по бокам.
Далее кпереди лежат две теменные кости; у большинства кро-
ликов между ними и за верхнезатылочной костью помещается малень-
кая межтеменная кость. По наличию этой кости череп кролика можно
отличить от черепа зайца, у которою парные части межтеменной кости
срастаются вскоре после ром<дения с затылочной костью. .Межтс-
менная кость имеется из всех позвоночных животных только у мле-
копитающих.
Снизу и с боков мозговая коробка ограничена височной костью,
в которой различают чешую со скуловым отростком, каменистую
часть, вдающуюся в полость черепа, и барабанную часть с наружным
слуховым проходом и пузыревидным вздутием. У основания скуло-
вого отростка лежит сочленовная ямка нижней челюсти.
Основание черепа образовано основной, или клиновидной, костью,
две часзИ которой дают две пары крыловидных придатков.
К теменным костям спереди прилежат две лобные кости, ограни-
чивающие сверху большие орбиты. Дальнейшим продолжением черепа
служат две длинные носовые кости, образующие верхнюю стенку но-
совой полости. Они, как и последующие части, относятся уже к ли-
цевому черепу.
К скуловому отростку височной кости примыкает скуловая кость
в виде узкой пластинки; ограничивая снизу глазную впадину, она
сочленяется с отростком верхнечелюстной кости; верхнечелюстные
кости не везде тесно срастаются друг с другом; в передней половине
между ними, в стенке ротовой полости, имеется щель, ведущая в но-
совую полость; середина щели по длине занята межчелюстной костью.
Нижняя челюсть — единственная подвижная кость черепа. Обе
половины ее срастаются спереди друг с другом. Различают горизон-
тальную ветвь челюсти с зубами и вертикальную пластинчатую часть,
несущую на вершине своей сочленовный бугорок для челюстного
сустава.
С грудными позвонками сочленяются двенадцать пар ребер: семь
первых пар их называются истинными, потому что они соединяются
непосредственно с грудиной, остальные пять пар называются ложными.
Из них первые три пары ребер сначала срастаются своими хрящевыми
концами друг с другом, а затем уже соединяются с грудиной; послед-
ние две пары ребер не доходят своими концами до нее. На брюшной
стороне тела, в грудной области, лежит грудная кость, или грудина,
состоящая из шести вытянутых друг за другом костных частей, со-
единенных между собой хрящевыми прослойками. Передний отдел
грудины называется рукояткой грудины; четыре следующих части
образуют тело грудины, и задним участком является так называемый
мечевидный отросток. Грудная часть позвоночника, ребра и грудина
образуют в своей совокупности грудную клетку.
Для опоры конечностей служат костные пояса. Пояс передней
пары ног состоит из пары лопаток и ничтожных рудиментов ключиц,
не играющих никакой роли. Лопатка образована длинной трехуголь-
ной плоской костью, лежащей в толще мышц сбоку на грудной клетке.
Ее нижний конец несет суставную поверхность для сочленения с го-
ловкой плечевой кости; от этого места кверху по наружной поверх-
ности ключицы идет костный гребень. Над нижним концом ключицы
возвышается отросток — acromion. С лопаткой срастается коракоид-
ная кость, теряющая свою самостоятельность и превращающаяся
в отросток лопатки. Таким образом плечевой пояс кролика реду-
цирован ни сравнению с таким же образованием лягушки и птицы.
Эти отношения наглядно представлены на нижеприведенной таблице,
454
где стрелками указано соединение или сочленение костей концами
их друг с другом.
1 i Лягушка: Грудипа ' ”1 1 1 1 1 1 1 Коракоид Ключица Лопатка Плечевая кость 1 ' —' I
1 ! 1 1 '
Кури п а: Грудипа Коракоид Ключица Лопатка Плечевая кость 1 (дужки) | f i ।
Крол и к: Грудипа Коракоид Ключица Лопатка Плечевая кость потерял са- рудимеп гарна мостоятель- и пость и пре- вратился в коракоид- ный отро-
сток
Передняя конечность кролика состоит из плеча (плечевая кость),
предплечья (лучевая кость и локтевая) и кисти. Последняя распа-
дается на запястье, пястье и пальцы. Мелкие запястные кости лежат
в два ряда, по четыре в каждом; между ними помещается сохраняю-
щая пожизненно самостоятельность центральная кость. Из пястных
костей наиболее длинны средние три; пястные кости пятого и особенно
первого пальцев малы. Второй — пятый пальцы состоят из трех
фаланг каждый, а первый —из двух.
Плечо и предплечье вскрытых нами позвоночных животных не
представляют важных отличий; зато в строении кисти таковые имеются,
что видно на следующей таблице.
1 । Лягушка Кролик Птица
Запястье Д»а / ' ряда \ 2 из 4 ossa carpalia _ I Два / 1 из 4 костей \ центральн. кость ряда \ 2 из 4 костей Всего из двух ко- стей
Пястье Четыре кости и одна рудиментарная Пять костей Две кости, слива- ющиеся концами в «пряжку»
(I 11 Пальцы: III IV IV Редуцирован Из двух фаланг » > фаланг 1 » трех фаланг J каждый Из двух фаланг » трех фаланг каждый । Из одной фаланги » двух фаланг » одной фаланги 1 Редуци- J рованы
1 Ulnare, radialc и centralc.
455
Пояс задних конечностей, или таз, у взрослого кролика образован
двумя безымянными костями, сочленяющимися друг с другом (лон-
ное сращение) и с крестцом. На боковой поверхности каждой поло-
вины таза имеется ямка — вертлужная впадина, служащая для
помещения головки бедренной кости. Книзу и кпереди от ямки в кости
есть широкая запирательная дыра. У молодого животного видны со-
ставные части будущей безымянной кости, а именно: подвздошная
кость (сочленяющаяся с крестцом), седалищная кость (седалищные
бугры), лонная кость и маленькая вставочная косточка* суставной
впадины (os acetabuli). Таз знакомых нам позвоночных животных,
исключая рыб, более однороден по своему составу и отличается только
различной конфигурацией частей.
Задние ноги кролика длиннее передних. С тазом сочленяется бе-
дренная кость, другим концом соприкасающаяся с большой берцовой
костью; к наружному краю ее прирастает рудиментарная малая
берцовая кость. Стопа состоит из предплюсны (пятки) (2 + 4 кости),
плюсны и четырех пальцев. Кости плюсны гораздо длиннее костей
пястья, первый палец на задних ногах отсутствует.
Бедренная кость Большая берцовая Малая берцовая Пятка Плюсна Пальцы
Первый ряд Второй ряд
Лягушка Есть Сливаются в общую голенную кость Две кости Две кости (сливают- ся из не- скольких костей) Пять костей I —V
Курица Есть Сливаются в общую голено- пяточную кость Пяточно-плюсне- вая коегь (цевка) 1 —IV
Кролик Есть Большая берцовая с приросшей частью малой берцовой кости Две кости Четыре кости Четыре кости I —IV
Из этой таблицы мы видим, что задняя нога позвоночных живот-
ных да£т вторичные отклонения от основного плана строения,
что объясняется требованиями функции. И лягушка и кролик
пользуются задними ногами для прыганья, в связи с этим они
обладают только одной костью в голени, что придает большую кре-
пость рычагу нижней конечности. У птицы отклонения еще резче,
что неудивительно, так как ноги ее являются единственной точкой
опоры о землю.
Общие замечания
Двубоковая симметрия тела грызуна нарушается ходом кишечника
и расположением брюшных органов пищеварения. Кроме того, асим-
метрично лежат почки, и есть существенные отклонения от первичной
симметрии в ходе крупных венозных стволов.
456
Метамерно лежат позвонки, ребра, части грудины и насечки пря
мой мышцы живота.
Первая й вторая пары ног гомодинамны друг другу; отклонения
в строении их немногочисленны. Как и конечности лягушки, они гомо-
логичны крыльям и ногам птиц.
Как и у птиц, скелет грызуна достигает наибольшей степени око-
стенения.
Грызун вместе с птицей, лягушкой и рыбой обладает всеми при-
знаками, типичными для позвоночных животных: внутренним скеле-
том, основой которого являются череп и позвоночник, вмещающий
в себе спинной мозг; сердцем, лежащим на брюшной стороне тела;
замкнутой кровеносной системой, красной кровью, благодаря нахо-
ждению в бесцветной плазме красных кровяных телец (эритроцитов)
с гемоглобином в их строме.
Грызуны в типе позвоночных животных занимают место среди
млекопитающих животных (Mammalia). Отличительные признаки их
сведены на нижеследующей таблице.
Таблица отличительных признаков позвоночных животных
Рыбы Земноводные Птицы Млекопитающие
Кожа Слизистая Сухая с ороговением поверх- ностных слоев эпидермиса
Роговые придатки кожи Камер в сердце Две Три Перья, когти, клюв Четыре Волосы, шерсть и когги (рога) Четыре
Дыхание Жаберное Кожно-легоч- ное Легочное Легочное
Затылочных мы- щелков Два Два Один 1 Два
Оплодотворение Питание заро- дыша В и е ш и с е На счет жслтк I В п у т р а я й ц а 1 е н н с е На счет мате- ри (через пла- центу)
Кормление дете- нышей Самостоятельное, общей пищей Молоком ма- тери
Животные Яйце к ладу щ и с Живородящие
457
Грызуны относятся к живородящим млекопитающим (Eutheria),
так как отрождают живых детенышей. Другими характерными при-
знаками принадлежности их к подклассу Eutheria s. Vivi para
являются наличие млечных желез с сосцами, отсутствие свободно
развитых коракоидов, редуцирующихся в отросток лопатки, и обо-
собленность заднепроходного отверстия от мочеполовых путей.
В подклассе живородящих кролик относится к многочисленному
отряду грызунов (Rodentia), для которых очень типично сильное
развитие резцов и отсутствие клыков; коренные зубы их—с попереч-
ными складками эмали. Послед дисковидный. Семейство заячьих —
Leporidae — характеризуется присутствием двух пар резцов в верх-
ней челюсти; пары более крупных передних и пары маленьких зад-
них. В этом семействе заключаются два рода — Lagomys (пищуха) и
Lepus (заяц). Вместе с зайцем кролик относится к роду Lepus; от
рода Lagomys он отличается зачаточным состоянием ключиц, длин-
ными ушами и длинными задними ногами. Кролик (Lepus cuniculus)
отличается от зайца (Lepus timidus) меньшей величиной, более
короткой головой и сохранением самостоятельной os interparietale.
С зайцем он дает плодущую помесь, называемую лепоридами.
Мышь и свинка относятся к семейству Muridae, а морская свинка
(Cavia cobaya)— к семейству Caviidae.
ЗАКЛЮЧЕНИЕ
В заключение коснемся общего сравнения всех типов с целью
установления их взаимоотношений, иллюстрирующих их эволюцион-
ное развитие, пользуясь теми выводами, которые мы делали всякий
раз при рассмотрении вскрываемых нами животных. Бесспорно, что
морфологически примитивными являются одноклеточные животные
организмы (тип Protozoa). Следующим этапом усложнения организмов
являются кишечнополостные животные как типически двуслойные
организмы, построенные из тканей.
С развитием производных среднего зародышевого листка, обо-
соблением органов и переходом к двубоковой симметрии от симметрии
радиальной можно от кишечнополостных перейти к плоским червям.
При возникновении вторичной полости тела и распадении его на
явственные метамеры — один шаг к группе кольчатых червей.
Следующими этапами усложнения организации является возникно-
вение членистых конечностей и обособление головы (животные чле-
нистоногие).
Что касается типов иглокожих и моллюсков, то эти группы живот-
ных являются довольно резко обособленными и ушедшими своими
путями эволюции. На их организации сильно отразился малоподвиж-
ный или сидячий образ жизни (иглокожие) и закручивание раковины
(мягкотелые).
Хордовых животных мы не можем связать так или иначе с беспо-
звоночными, потому что программа наших занятий не включала
рассмотрение тех животных, которые дали бы гозможность таковые
связи установить, но из учебников зоологии можно почерпнуть до-
казательства вероятного происхождения позвоночных как предста-
вителей типа хордовых от олигомерных червей.
'158
ОСНОВНЫЕ ЭЛЕМЕНТЫ МИКРОСКОПИЧЕСКОЙ ТЕХНИКИ 1
Приготовление тотальных микроскопических препаратов
Тотальные препараты без какой-либо обработки
Для сохранения мелких животных (например членистоногих и др.) и различ-
ных стадий их развития для последующего изучения изготовляют постоянные
микроскопические препараты, без какой-либо особой обработки, по схеме —
обезвоживание объекта и заключение его в канадский бальзам:
1) Объект фиксируется 70% спиртом (если он уже был законсервирован в вод-
ном растворе формалина, то его с час промывают водой и потом переносят часа
на 2—3 в 70% спирт).
Препарат прокалывают где-нибудь сбоку острой иглой, чтобы дать возмож-
ность спирту проникнуть через отверстие в хитине внутрь объекта.
2) Препарат переносят последовательно в 85°/0 и затем в 95°/0 спирт, выдерживая
в каждом из них, в зависимости от величины объекта, часов по б (в 95% спирту
можно оставить препарат на ночь).
3) Препарат переносят на сутки в абсолютный алкоголь для обезвожи-
вания.
Для заключения в канадский бальзам препарат необходимо пропитать на-
сквозь маслом; такое «просветление» производится гвоздичным или кедровым
маслом.
4) В узкую короткую пробирку с плоским дном наливают гвоздичного или
кедрового масла на 1 см, осторожно приливают сверху абсолютного спирта, так
чтобы обе жидкости не смешались друг с другом. В спирт опускают обезвоженные
препараты, которые задерживаются в зоне соприкосновения спирта с маслом.
Пробирку оставляют стоять в покое до тех пор, пока препараты не опустятся на
самое дно, что совершается медленно в связи с диффузией в пограничной зоне.
Пипеткой отсасывают спирт и перекладывают препараты на несколько часов в чистое
масло в часовое стекло.
5) По окончании описанных процедур контролируют прозрачность препарата.
Часовое стекло ставят на черную доску стола. Препарат должен быть невидимым;
если на нем видны белесоватые пятна, значит обезвоживание было неполное, и его
необходимо вернуть обратно в абсолютный спирт.
• 6) Если препарат прозрачный, то его на шпателе переносят на середину пред-
метного стекла и поливают несколькими каплями ксилола из капельницы, чтобы
смыть масло.
7) Сообразуясь с толщиной препарата вырезают две узкие полоски белого
картона или полоски из спичек или обламывают кусочки вытянутой стеклянной
трубочки и кладут их по бокам препарата так, чтобы они пришлись к самым краям
покровного стекла.
8) Не давая препарату высохнуть, наносят на него палочкой
канадский бальзам и накрывают препарат начисто вытертым покровным стеклом.
Если бальзама взято мало, то прибавляют каплю его к краю покровного стекла,
под которое он втягивается в силу капиллярности.
9) Поправляют иглами положение сдвинувшихся опорных полосок («ножек»)
и оставляют препарат на долгое время сохнуть в горизонтальном по-
ложении, подбавляя канадский бальзам по мере его усыхания с краев.
2 Частные указания по технике даны в тексте книги.
459
10) Такие заключенные в толстый слой канадского бальзама препараты целе-
сообразнее хранить в горизонтальном положении, так как при долгом держании
их на боку препарат опускается к нижнему краю покровного стекла даже спустя
несколько месяцев после его изготовления.
11) Вследствие толщины препарата его можно изучать только при слабых
увеличениях микроскопа.
12) Если препарат был в канадском бальзаме совершенно прозрачен, а через
некоторое время он частично или полностью почернел (при рассматрива-
нии в проходящем свете микроскопа), значит он не был доста-
точно проколот перед обезвоживанием (см. выше, § 1), из него испарился ксилол,
а бальзам внутрь не прошел. Ценные препараты можно попытаться исправить
по следующей схеме: I—растворить бальзам ксилолом и выдержать препарат
в чистом ксилоле; II —перенести (осторожно: препарат очень хрупок!) в 100%
спирт для обезвоживания; III —проколоть осторожно препарат иглой; после
суточного пребывания в 100% спирту повторяют последовательно § 4—9 настоя-
щей главы.
12-а) Если весь препарат потерял прозрачность, то его с успехом (и с некоторой
выгодой) можно рассматривать под микроскопом при сильном падаю-
щем свете (у светлого окна, близ сильной лампы и т. п.). В таком случае
с особенной ясностью видима поверхность препарата (волоски, чешуйки, бу-
горки и т. п.).
Стойкие, хотя и не так долго сохраняющиеся препараты можно приготовить,
заключая объект в глицери н-ж е л а т и н у.
Если объект хранится в формалине, то его хорошо промывают в сменяемой
воде (несколько часов); затем переносят в смесь глицерина пополам с водой для
пропитывания. Спиртовые препараты также предварительно переносят в воду.
В глицерине препарат держат до полного пропитывания им (несколько часов или
сутки).
Препарат далее кладут на предметное стекло и отсасывают лишнюю жидкость
фильтровальной бумагой. На покровное стекло берут кусочек глицерин-желатины
и нагревают его на стекле, чтобы желатина расплавилась. Полезно слегка нагреть
(но не высушить!) и объект на предметном стекле. Затем покровное стекло быстро
переворачивают и висящей каплей желатины опускают на объект так, чтобы масса
растеклась между предметным и покровным стеклами и покрыла собою объект.
Для прочности препарат с краев покровного стекла обводят железным лаком или
жидким канадским бальзамом, чтобы предупредить высыхание массы. Препарат
хранится долго. Его выгода — не такая большая прозрачность,
как бальзамного, что иногда бывает нужно.
Глицерин-желатину приготовляют: растворив в 100 см3 на водяной бане или
в горячей воде 10 г желатины в листах, к раствору добавляют 25,0 см3 глицерина
и 0,1 г тимола, растворив последний в воде, или несколько капель карболовой
кислоты (против плесени). Все перемешивают и сливают в баночку с пробкой.
Вываривание объектов в едком кали
В некоторых случаях желательно изготовить препарат из одних только хитш-
новых или известковых частей.
1) Отрезают соответствующую часть объекта и кладут ее в химическую про-
бирку, куда налит раствор едкого кали или натра (кусочек с горошину растворяют
в 5—6 см3 воды).
2) Препарат кипятят на спиртовой лампочке, пока не будут разрушены его
мягкие части.
3) Препарат переносят в чистую воду (можно с прибавлением уксусной кислоты)
для удаления едкого кали.
4) После основательной промывки препарат переносят последовательно
в 7С°/0, 95°/0 и в абсолютный спирт, просветляют кедровым маслом и заключают
в канадский бальзам.
5) При закрытии покровным стеклом подкладывают ножки под его углы (воск,
кусочки спички, бристольского картона), чтобы давлением покровного стекла не
изменило формы объекта.
6) Если изолируемые части многочисленны и мелки, то их центрифугируют и
отсасывают пипеткой со дна пробирки.
460
Наклейка тотальных препаратов на предметное стекло по способу
* Е. Павловского
1) Выпрепарованный oprarf (например: желудок, половой аппарат, мальпи-
гиевы сосуды и т. д.) на шпателе переносится в каплю физиологического раствора
на предметное стекло, где тщательно расправляется иглами и приводится в жела-
тельное положение.
2) Узкой полоской фильтровальной бумаги осторожно отсасывают физиологи-
ческий раствор и иглой приводят сдвинувшиеся части органов в их прежнее по-
ложение.
3) Наносят на объект капля по капле из пипетки жидкость С а г п о у, 1 следя
за тем, чтобы не сдвинуть частей органов.
4) Покачивают препарат из стороны в сторону, чтобы лучше профиксировать
ткани.
5) Через 1—5 минут (в зависимости от толщины препарата) жидкость
С а г п о у сливают; взамен нее прибавляют 95% спирта, которым промывают
препарат.
б) Сливши спирт, на препарат наносят несколько капель —7г% раствора
фотоксилина, коллоксилина, целлоидина или коллодия (все эти вещества раство-
ряют в смеси равных частей абсолютного спирта и серного эфира); препарат пока-
чивают, чтобы коллодий покрыл его сверху.
7) Излишний раствор коллодия сливают, наклонив стекло; препарат 1/4—
минуты держат на воздухе, чтобы коллодийная пленка чуть загустела (но не вы-
сохла!!).
8) Препарат кладут в 85% спирт на 1/2~^ час, затем перемещают его в 70%
спирт, в коем он может храниться неограниченно долгое время до окончательной
обработки.
Если наклеивается препарат, уже бывший в спирту, то начинают прямо
с пункта 6.
Окраска и окончательная обработка наклеенных тотальных препаратов
I) Из 70%спирта препарат, наклеенный на предметное стекло, переносят в воду
на 10—60 минут (в зависимости от толщины и проницаемости для воды объекта).
2) На 5—10—30 минут (установить опытом) препарат погружают в квасцовый
кармин 2 или в гемалаун.
3) Препарат переносят последовательно в несколько порций чистой воды и
в последней оставляют его на 1—2 часа.
4) На несколько часов стекло с препаратом погружают в 75% и в 95% спирт.
5) Обезвоживают препарат в 100% спирту часа (в зависимости от
препарата). При долгом держании спирт растворит коллодий и препарат
сползет.
б) Обтерев лишний спирт, стекло опускают в ксилол до полного просветления
препарата.
7) Канадский бальзам и покровное стекло с ножками (если нужно).
Окраска ненаклеенных тотальных препаратов
1) Из 70% спирта препарат переносят в 50% спирт и затем в дистиллирован-
ную воду.
2) Препарат кладут в квасцовый кармин, длительность окраски определяют
на опыте (от нескольких минут до нескольких часов).
3) Промывают в свежих порциях воды несколько часов.
4) Переносят на штативе в 50, 75, 85% спирт.
5) Обезвоживают в 100% спирте и заключают в канадский бальзам, как было
указано выше.
1 Рецепт см. ниже.
2 Квасцовый кармин приготовляют прибавлением к 100,0 см3 дистиллирован-
ной воды 2,0 г калийных квасцов и 1.0 г кармина. Кипятить 15 минут. Остудить,
отфильтровать и положить в раствор несколько кусочков тимола или камфары
(против плесени).
461
б) При отсутствии абсолютного спирта из 96% спирта препарат переносят
в карбол-ксилол, затем в чистый ксилол и после просветления — в канадский
бальзам.
Иногда подлежащие резанию на микротоме объекты окрашивают сразу целым
куском, затем заливают в парафин (см. ниже).
Для этой цели обычно применяется борный кармин:
1) Из 70% спирта препарат переносят в борный кармин на несколько часов,
i 2) Из краски препарат перекладывают в 70% спирт с 1% соляной кислоты.
3) К 100 см3 4% водного раствора буры при кипячении прибавляют 2—3 г
кармина. Затем добавляют 100 см3 70% спирта и оставляют на несколько
недель, потом жидкость сливают и фильтруют.
Кислый спирт вытягивает лишнюю краску из протоплазмы (диференциация
окраски); спирт сменяют по мере его окрашивания.
4) Выдерживают препарат в свежих порциях 75°/0 спирта для удаления кис-
лоты.
Далее объект или заключают в канадский бальзам, как уже указано, или
заливают в парафин (см. ниже).
Фиксирование объектов для изготовления срезов
Фиксирование различных животных для срезов производится или целиком,
или по отдельным частям тела и органам.
1) Простейшей фиксирующей жидкостью является 70% спирт, в котором объект
выдерживают несколько часов, после чего перекладывают в свежую порцию
спирта для хранения.
2) 70% спирт с прибавлением 5% продажного формалина.
3) Жидкость Oetcke: 96% спирта 60 частей, 40% формалина 45 частей, уксус-
ной кислоты 2 части.
4) Жидкость Фрейлинга: воды 120 частей, 96% спирта 60 частей, формалина
24 части, ледяной уксусной кислоты 4 части.
5) Насыщенный водный раствор сулемы: отвешивают по 8 г сулемы на 100 см3
воды и кипятят в колбе. Теплый раствор фильтруют в склянку, где он оконча-
тельно остывает. При охлаждении из раствора выпадают кристаллы, над кото-
рыми жидкость остается всегда насыщенной сулемой.
Для фиксирования необходимо прибавить ко взятой порции сулемы 1—2 капли
ледяной уксусной кислоты.
Сулема плохо проникает в глубь тканей и через хитин, поэтому необходимо
фиксировать разрезанное животное. Фиксировать не более 4—6 часов. Эффект
действия усиливается, если предварительно подогреть раствор до 60—70°/о.
Когда фиксирование закончено, отсасывают сулему пипеткой и на препарат
наливают 50% спирт на г/2 часа, после чего спирт заменяют 60°/о. К по-
следнему прибавляют иодной тинктуры до цвета мадеры. Сулема объекта соеди-
няется с иодом, вследствие чего спирт обесцвечивается. К нему снова прибавляют
иод и т. д. до тех пор, пока спирт не перестанет обесцвечиваться. После этого
препарат переносят в 70% спирт.
В раствор сулемы нельзя опускать никаких металлических инструментов,
которые покрываются матовым налетом и портятся. В случае необходимости поль-
зуются стеклянным или роговым шпателем, или же металлическими инструмен-
тами, покрытыми тонким слоем парафина.
6) Жидкость Сагпоу: 60 частей абсолютного спирта, 30 частей хлороформа
и 10 частей ледяной уксусной кислоты. Фиксируют от 5 минут до часа. После
фиксирования препарат выдерживают в 100 или 95% спирту, который сменяют
несколько раз.
7) Жидкость Duboscq’a приготовляется ex tempore смешиванием трех основ-
ных растворов: 15 частей 1% раствора пикриновой кислоты в 90% спирту, 6 частей
формола и 3 части ледяной уксусной кислоты.
Полезно добавить еще 10% хлороформа. Фиксируют 4 часа, после
чего препарат переносят в 85% спирт, который сменяют после того,- как он заметно
окрасится в желтый цвет. Такую смену спирта приходится повторять. Если пре-
параты не идут сразу в заливку, то для хранения их перекладывают в 75% спирт.
8) Жидкость Leeuwen’a: 1% пикриновой кислоты в абсолютном или 96%
спирту 12 частей, хлороформа 2 части, <|юрмалина 2 части, уксусной кислоты! часть.
462
Фиксирование производят в широких чашечках или баночках. Фиксирующая
жидкость по объему должна раз в 30—50 превышать объем объекта. При фиксиро-
вании жидкость вытягивает из объекта воду и разжижается; поэтому ее время от
времени перемешивают или заменяют новой, что приходится делать повторно
с крупными объектами. Бывшую в употреблении жидкость выбрасывают.
После жидкости Сагпоу препараты выдерживают в 95% спирте, который сме-
няют несколько раз для вытягивания уксусной кислоты. Если материал не идет
сразу в заливку, то его через 90% и 85% спирты переносят на хранение в 75%
спирт.
После жидкости Leeuwen’a и Duboscq’a препараты переносят в 85% спирт,
который сменяют несколько раз по мере его пожелтения. После жидкости Oetcke
и Freiling’a препараты переносят в 85% спирт, который сменяют несколько раз
по мере его пожелтения. После жидкости Oetcke и Freiling’a препараты промы-
вают в 90% спирте.
Изготовление срезов
Заливка препаратов в парафин
Для заливки объектов в парафин их необходимо провести через спирты воз-
растающей крепости, если фиксирование производилось водным раствором сулемы.
Схема работы при фиксировании смесями:
1 Duboscq’a * Сагпоу | Сулема с укс. кисл. 1 Oetcke. Freiling, | спирт.-формалин
1 После фиксирова- ния выдерживать в 85%; потом— 90% 4—6 часов; 95% 4—6 часов; (можно и дольше) После фиксирова- ния выдерживать в 100% или в 95% спиртах для оття- гивания кислоты из объекта i Спирт 50%—1/2 часа. 60%4-tinct. jodi до обесцвечивания 70% 6 часов (можно и дольше) 80% » 90% » 95% i 'Промывать в 90% и 95 % спиртах i ।
Далее препарат обезвоживают в 100% спирту, который держат в диализаторе
над слоем обожженного до белого цвета химически чистого медного купороса.
Обезвоживание лучше производить в течение 12—24 часов.
Обезвоженные препараты быстро переносят в смесь ксилола с абсолютным
спиртом на 3—4 часа, после чего их помещают в чистый ксилол до того времени,
пока они станут прозрачными (2—3—4 часа). После этого препараты переклады-
вают в парафин, растворенный в ксилоле, и ставят чашечку на термостат (с внутрен-
ней t°—55—56° С) на несколько часов, закрыв ее стеклом.
Затем открытую чашечку помещают в термостат на *j2—1 час, после чего пре-
параты переносят в чистый парафин (плавкости 55—56°), расплавленный в другой
чашечке. Через 1/2— 1 час переносят материал в другую порцию чистого парафина,
в котором оставляют его на такой же срок. Затем препарат перекладывают для
застуживания в третью порцию парафина, в часовое стекло. Перенос производится
всякий раз нагретым на спиртовой лампочке пинцетом, чтобы избежать затверде-
ния парафина и приставания объекта к инструменту.
Все процедуры переноса объектов при заливании в парафин должны быть произ-
водимы быстро, чтобы вынутый парафин не успевал застыть. С другой стороны,
совершенно необходимо не перегревать парафин выше точки
плавления, так как чрезмерно высокая температура портит объекты.
При застуживании парафина, чашечку или часовое стекло вынимают из термо-
стата и прижимают дном к ладони левой руки. При этом вскоре застывает слон
парафина у самого дна посудины. Нагретой иглой раздвигают препараты друг
от друга, так чтобы между ними везде было расстояние не менее 4 -5 мм, придают
объектам желательное положение (например, чтобы они лежали параллельно
463
поверхности дна) и опускают чашечку плавать в ванночку с холодной водой.
Когда пленка парафина застынет с поверхности, чашечку осторожно пото-
пляют; необходимо избегать очень холодной воды или не передерживать в воде
заливаемый материал, так как парафин от холода может дать трещины, из-за
которых потребуется перезаливка объектов.
Для снятия парафина из часового стекла, последнее быстро прогревают на
спиртовой лампочке, чтобы растаял прилегающий к дну слой парафина, после
чего пинцетом сдвигают парафиновую массу в ванночку с водой.
Наконец обрезают лишний парафин, так чтобы вокруг объекта осталась
прямоугольная кайма в 2—3 мм шириною.
По окончании остуживания препараты осматривают; масса парафина не должна
нигде — ни в самой себе, ни в препарате —содержать беловатых участков; нали-
чие последних говорит за то, что ксилол или хлороформ остался в блоке. Такие
препараты резать нельзя. Их надо перезалить, т. е. расплавить парафин
в термостате, переложить в одну или две порции чистого парафина, выдержав
р термостате 30—60 минут, и снова заключить в парафин, как только что описано.
Если белые пятна имеются под хитином, то полезно соскоблить парафин и ост-
рым скальпелем или копьевидной иглой срезать часть хитина над пятном, после
чего производят перезаливку указанным выше способом. При этой процедуре
препарат надо положить срезанным местом кверху, чтобы облегчить выход из
него ксилола или хлороформа.
Резание на микротоме и наклейка срезов
Перед резаньем на микротоме парафиновый блок прикрепляют расплавлен-
ным парафином к торцу деревянного кубика; по охлаждении блок обрезают
скальпелем, чтобы придать ему четырехугольную форму. Вокруг препарата должна
остаться кайма парафина в 2— 2х/2 мм шириной.
Препарат завинчивают в клемму микротома так, чтобы наибольшая его длина
была параллельна острию бритвы. Бритва должна быть острая; ее перед резаньем
наводят на ремне.
Перед резаньем запасают в стеклянный цилиндр с крышкой хорошо вымытые
и вытертые начисто предметные стекла.
Если срез закручивается, надо его при начале резанья придержать тонкой
кисточкой (остерегаться, чтобы волосы кисточки не попали бы между бритвой
и блоком); при следующем срезе надо бритвой ударить о блок чуть сильнее, чтобы
произошло склеивание срезов. Если срезы придержать кисточкой и далее, то лента
в конце концов пойдет.
Необходимо иметь в виду, что в холодной комнате серия резаться не будет.
В таком случае помогает, если перебраться с микротомом поближе к печке или
поставить около газовую или какую-либо другую горелку.
Если середина препарата выкрашивается, то это может зависеть от того, что
в препарате остался ксилол или хлороформ (на блоке видны беле-
соватые участки); в таком случае препарат можно попытаться исправить новой
заливкой, положив его в чашечку с расплавленным парафином в термостат.
Срезы наклеивают на предметное стекло куриным белком, смешанным пополам
с глицерином. На чистое предметное стекло наносят каплю смеси величиною с по-
ловину булавочной головки и растирают ее равномерно пальцем по стеклу. На
натертое место из пипетки капают дистиллированной водой (1—2 капли), которая
должна растекаться ровным слоем. На воду кладут в порядке парафиновые срезы;
когда вся партия их уложена, то стекло прогревают на спиртовой лампочке,
остерегаясь расплавить парафин срезов.
Стекло покачивают в направлении его ширины, чтобы волна воды пробегала
над срезами. От тепла парафин расширяется, срезы делаются гладкими, и длина
каждого ряда среза увеличивается.
Когда все срезы хорошо расправятся, отсасывают фильтровальной бумагой
воду, исправляют иглой положение сдвинувшихся рядов и стекло кладут сушиться
на теплый термостат, прикрыв стекло от пыли, или в термостат с температурой
Сушить препарат можно до следующего дня. Через час после начала сушения
стекло со срезами полезно минут на 30 положить в термостат с температурой
55—56° С, чтобы парафин расплавился. Затем препарат кладут на прежнее место
464
Окраска срезов
Перед окраской срезов из них предварительно удаляют парафин и переводят
их в воду в следующей последовательности:
а) растворяют парафин в ксилоле, причем препарат становится совсем
прозрачным, обтирают лишний ксилол мягкой тряпочкой и переносят
стекло
б) в абсолютный спирт, где срезы белеют,
в) далее переносят стекло в 96% спирт и, наконец, в воду.
Если в воде срезы отлетели, значит они были плохо наклеены. Из воды препа-
раты переносят в ту или другую краску.
К краям ванночки или чашки воском прикрепляют две стеклянные палочки
или трубочки на манер рельс железной дороги. На рельсы поперек кладут стекла
срезами вверх. На срезы отфильтровывают или прямо наливают раствор краски.
Из множества красочных комбинаций можно в качестве основных рекомендо-
вать нижеследующие способы окраски.
.Г ематоксилин-эозин
Гематоксилин Delafield’a: в 400,0 см3 насыщенного водного раствора аммиач-
ных квасцов размешивают раствор 4 г гематоксилина в 25 см3 абсолютного спирта.
Смесь оставляют на 4—5 дней открытой в широком сосуде. Затем прибавляют по
100 см3 глицерина и древесного спирта. Все хорошо перемешивают. Отстоявшуюся
смесь фильтруют и оставляют созревать в склянке, через пробку которой пропу-
щена узкая изогнутая стеклянная трубка (на 1—2 месяца).
Длительность окраски 3—5 минут. После промывки в воде хорошо препарат
оставить на ночь в профильтрованной водопроводной воде, где он синеет. При
перекрашивании лишнюю краску оттягивают кислым спиртом и хорошо промы-
вают в воде.
Гематоксилин Бемера
Растворяют 1 г кристаллического гематоксилина в 10 см3 абсолютного алко-
голя. Смешивают 20 г калийных квасцов в 200 см3 дистиллированной воды и ки-
пятят до их растворения. Смешивают далее оба раствора, фильтруют жидкость
и оставляют в незакрытой склянке стоять недели две на воздухе для созревания
краски, снова фильтруют и вдвое разбавляют дистиллированной водой.
Красят 3—5 минут (установить опытом), нафильтровав краску на препарат.
Препарат споласкивают в водопроводной воде и оставляют в чистой порции воды
до утра (чтобы срезы посинели).
После промывания препарат окрашивают водным раствором эозина (1 : 1000);
далее промывают в воде и заключают в канадский бальзам, проводя через спирт
(см. ниже).
Гематоксилин окрашивает ядра в синий цвет, а протоплазму в розовый.
Железный гематоксилин Гейденгайна
А) Основной раствор окраски: 1,0 г кристаллического гематоксилина раство-
рить в 10,0 см3 абсолютного алкоголя.
Б) 10 см3 основного раствора вылить в 90,0 см3 дистиллированной воды и оста-
вить в открытой посуде созревать недели три.
В) Протравой является 1% раствор железоаммиачных квасцов (ferrum ammo-
niacum sulf uric um); для растворения годны только голубые (нсвыветрившиеся)
кристаллы.
Способ окраски:
1) Стекло со срезами из воды переносят в протраву на 4—6—12 часов.
2) Быстро споласкивают дистиллированной водой.
3) Стекло кладут в раствор краски на 12—24 часа (срезы чернеют).
4) Объект перекладывают в сосуд со сменяемой простой водой на часа.
5) Стекло срезами кверху кладут в чашечку Петри с протравой (которую для
облегчения контроля откраски можно развести вдвое водой). Наблюдают за от-
краской препарата под микроскопом: протоплазма должна приобрести гравюрный
оттенок; на ее фоне ядра вырисовываются ясни.
3U Иракишум ио зоологии — 385
4Ь5
6) На 1 час стекло кладут в проточную воду.
Далее —спирты и ксилол, как выше.
Если препарат очень открашен, его опять кладут в краску и снова диферен-
цируют в протраве.
Очень прочная и красивая окраска, выявляющая тонкие детали строения ядра
и протоплазмы.
Краска Романовского —Гимза
Готовый раствор держат в тепле (+16,17°). Перед употреблением раство-
ряют в пропорции : 1 каплю краски на 1 см8 дистиллированной воды. Смесь нали-
вают в цилиндрик, куда стоя опускают препарат из дистиллированной воды.
Красят 12—24 часа.
Споласкивают стекло в дистиллированной воде и переносят в цилиндр с 95%
спиртом для диференцировки. Покачивают стекло, и когда краска будет вытянута
из протоплазмы (установить опытом и быстрым контролем под микроскопом),
срезы переносят в 100% спирт для обезвоживания (^—'/з минуты); затем—кси-
лол и бальзам.
Ядра — голубые, протоплазма — розовая. Краска в обыкновенном бальзаме
со временем выцветает. Возможна перекраска выцветших препаратов после раство-
рения канадского бальзама в ксилоле и проведения препарата через 100% и 95%
спирты в воду.
Крезил-эхтвиолет
Очень хорошая ядерная краска, дающая метахромазию. В качестве основного
раствора приготовляют насыщенный раствор в 95% спирту. Перед употреблением
1,5 части раствора смешивают с 10 частями анилиновой воды (в пробирку на 2/3 на-
ливают дистиллированной воды и добавляют капель 10 анилина, закрыв пальцем
сильно встряхивают много раз, затем фильтруют через влажный фильтр). Мета-
хромазия, в спирту пропадает; чтобы ее сохранить, препарат после промывки в воде
заключают в сироп из левулезы (10 г левулезы растворяют в 8,0 см3 горячей ди-
стиллированной воды); препарате левулезе обводят кантом из асфальтового лака
или менделеевской замазки. Быстро диференцированные 96% спиртом и заклю-
ченные в канадский бальзам препараты дают очень хорошую картину. Дополни-
тельной окраски для протоплазмы не требуется.
Заключение срезов в канадский бальзам
После промывки окрашенных срезов водой стекло переносят в цилиндрик
с 95% спиртом, где пинцетом двигают его вперед и назад. Вытерев со стекла лишний
спирт, препарат переносят в 100% спирт для обезвоживания (около минуты).
Вытерев лишний абсолютный спирт, стекло перекладывают в цилиндрик с ксило-
лом, где препараты должны стать прозрачными. Если это достигнуто, то, вынув
стекло, на срез наносят несколько капель чистого ксилола и, покачав препарат,
ксилол сливают, на срезы наносят каплю канадского бальзама и закрывают пред-
варительно вытертым покровным стеклом. Если в ксилоле некоторые места срезов
оказались белесоватыми (под микроскопом — черными), то это значит, что в них
осталась вода (см. выше обезвоживание). Препарат переносят снова в абсолютный
спирт, ксилол и бальзам.
При недостатке абсолютного спирта его можно опустить, перенося препараты
прямо из 96% спирта в карбол-ксилол. 1 Если препарат стал прозрачным, то его
переносят в чистый ксилол и заключают в канадский бальзам. Карбол-ксилол
годен также при окраске железным гематоксилином, но не анилиновыми красками
(краска Романовского, крезил-эхтвиолет и др.).
Срезы объекта, окрашенного борным кармином, освобождают от парафина
ксилолом и заключают в канадский бальзам.
1 100 см3 ксилола +22 г кристаллической карболовой кислоты,
ЛИТЕРАТУРА
Руководства по зоотомии и сравнительной анатомии
Аверинцев С. — Руководство к практическим занятиям по зоологии, СПб.,
1913.
Bonnet А. — Dissections de Zoologie. Precis-atlas, Paris, 1924.
Браун M. —Практическое руководство по анатомии животных, СПб., 1887.
Б ю ч л и О. — Лекции по сравнительной анатомии, ч. I, Пгр., 1917.
Wiedersheim R. — Grundriss der vergleichenden Anatomie der Wirbeltiere,
1898.
ГекслиТ. и Мартин Г. — Практические занятия по зоологии и ботанике,
М. 1902.
Гексли Т. — Начальные основания сравнительной анатомии, СПб., 1865.
Догель В. А. — Курс сравнительной анатомии беспозвоночных, ч. I, Учпедгиз,
1938.
Зенкевич Л. А. (ред.). — Руководство по зоологии, т. I, Биомедгиз, 1937.
Кашкаров Д. и Станчинский В. — Курс зоологии позвоночных
животных, Биомедгиз, 1935.
С о u р i n Н. — Atlas de dissections zoologiques, Paris, 1910.
Кингсли Д. С. — Сравнительная анатомия позвоночных животных, М. 1914.
Клер В. О. и Кацнельсон 3. С. — Руководство к практическим заня-
тиям по зоологии, Свердловск, 1928.
Кюкенталь В. — Руководство к практической зоологии, СПб.
Матвеев Б. (ред.). — Курс зоологии, ч. I и II, Учпедгиз, 1938.
NierstraszH. Н. und Н i г s с h G. Chr. — Anleitung zu makroskopisch-
zoologischen Ubungen, H. 1, Wirbellose Tiere, Jena, 1922; H. 2, Wirbeltiere,
Jena, 1924.
Огнев С. И. — Учебник зоологии позвоночных, Госмедиздат, 1934.
Павловский Е. Н. — Введение в зоотомию, ч. I, Позвоночные, Госиздат,
1923.
Павлович С. А. — Repetitorium к практическим занятиям по зоологии,
СПб., 1913.
Паркер, Т. — Курс зоотомии (позвоночные), М. 1904.
Паркер Т. иГасвель В. — Руководство к зоологии, т. I, Беспозвоночные,
1914; тт. II—III, Позвоночные, 1908.
Райков Б. —Практические занятия по зоологии для начинающих.
R б s е 1 е г Р. und Lamprecht Н. —- Handbuch fiir biologische Ubungen,
Berlin, 1914.
Stempell W. — Leitfaden fiir das mikroskopisch-zoologisches Praktikum,
3. AufL, Jena, 1925.
Холодковский H. A. — Учебник зоологии и сравнительной анатомии,
изд. 3, СПб., 1914.
Hoffmann Н.—Leitfaden fiir histologische Untersuchungen, Berlin, 1914.
H a t sc he k B. und Cori C. — Elementarkurs der Zootomie in fiinfzehn Vor-
lesungen, Jena, 1896.
Шимкевич В. M. — Курс сравнительной анатомии позвоночных живот-
ных, изд. 3, Госиздат, 1923. •
Schneider К. — Histologisches Praktikum der Tiere.
467
1IJ мальгаузен И. И. — Основы сравнительной анатомии позвоночных,
Учпедгиз, 1938.
Шмид Б. — Элементарная практическая зоотомия, СПб., 1910.
Schuberg. — Zoologisches Praktikum, 1910.
Jammes L. — Zoologie pratique bas6e sur la dissection des animaux les plus
r£pandus, Paris, 1904, 317 ориг. фигур.
Jhle, van Kampen, Nierstrasz, Versluys. — Vergleichende
Anatomie der Wirbeltiere, Berlin, 1927.
Простейшие
Аве рин не в С. В. — Rhizopoda пресных вод. Тр. СПб. общ. естествоиспыт.,
т. XXXVI, вып. 2, Отд. зоол. и физиол., 1906.
Вислоух С. М. — Биологический анализ воды. Определитель важнейших
показательных микроорганизмов, в книге С. Златогорова —«Учение о микро-
организмах» (ч. II, Пгр., 1916, стр. 225—305).
Догель В. А. — Как производить биологические наблюдения над простей-
шими, Госиздат, 1926.
D о f 1 е i n F. — Lehrbuch der Protozoenkunde, 5. Aufl., Jena, 1927—1928.
К a 1 m u s H. — Paramaecium, Jena, 1931. (Монография, 833 номера литератур-»
ного указателя.)
Калкинс Г. Н.—Протозоология, М. 1911.
Kiikenthal W. — Handbuch der Zoologie, Bd. 1, Protozoa, 1923/1925.
Ламперт К. —Жизнь пресных вод, СПб., 1900, гл. VIII, Простейшие жи-
вотные (Protozoa), стр. 531—600. (Определит, таблицы.)
Соколов Б. — Регенерация у Protozoa. Йзв. Биол. лаб. Лесгафта, вып. 13,
1912.
Шевяков В. — Infusoria Aspirotricha. Зап. Акад. Наук., сер. VIII, т. IV,
1896.
Wenyon С. — Protozoology, vol. I —II, London, 1926.
Кишечнополостные
Chun C. —Coelenterata, in Bro • n’s Klassen und Ordnungen des Tierreichs, II,
1889—1892.
Delage Y. et Нё г о u a r d E. — Les coelenteres. ТгаИё de Zoologie con-
crfcte, t. II, 2-me partie, 1901.
Исаев В. — Этюды об органических регуляциях, II. Тр. Лгр. общ. естество-
испыт., т. LV, 1925.
Л и н к о А. К. и Куделин Н. В. — Гидроиды. Фауна России и сопредель-
ных стран, т. I, 1911; т. II, вып. 1, 1912, и вып. 2, 1914. Изд. Акад. Наук.
Kuhn А. — Entwicklungsgeschichte und Verwandtschaftsbeziehungen der Hydro-
zoen, 1925.
К ii k e n t h a 1 W. — Coelenterata, in Kiikenthal-Krumbach, Handbuch
der Zoologie, Bd. I, Berlin u. Leipzig, 1923—1925. (Литература.)
Schluze P. — Bestimmungstabelle der deutschen Siisswasserhydrozoen. Zool.
Anz., Bd. LIV, 1923.
S tec he O. — Hydra und die Hydroiden. Monographien einheimischer Tiere,
Bd. 3, Leipzig, 1911. (Литература.)
Pax F. — Die Aktinien. Ergebn. u. Fortschr. Zoolog., Bd. IV, 1914.
To кин Б. и Горбунова Г. — Проблемы онтогенеза клетки, II, Как за-
ставить стебелек Hydra fusca регенерировать целую гидру. Биол. журн., т. III,
1934.
Трамблэ А. — Мемуары к истории одного рода пресноводных полипов,
Биомедгиз, 1937.
Турбеллярии
Беклемишев В. Н. — Некоторые вопросы географического распростране-
ния пресноводных триклад. Русск. гидробиол. жури., т. II, 1923.
Bresslau Е. — Turbellaria. in Kiikenthal, Handbuch der Zoologie, Bd. Il,
1528. .
Graff L. v. — Turbellaria, inBronn’s Klassen und Ordnungen des Tier-
reichs, Bd. IV, 1905.
468
За б у с о в И. П. — Исследования по морфологии и систематике планарий
озера Байкала. Тр. Общ. естествоиспыт. при Каз. унив., т. XLIII, вып. 4,
1911.
Ламперт К. — Жизнь пресных вод, СПб., 1900, гл. VI, Ресничные черви.
Л у с Я. Я. — Этюды по регенерации и трансплантации турбеллярий. Бюлл.
Моск. общ. испыт. прир., Отд. эксп. биол., № 1, 1924.
Nassonov N. — Les traits g£n£raux de la distribution geographique des Tur-
bellaria Rhabtfocoela dans la Russie d’Europe. Изв. Росс. Акад. Наук, 1924.
Steinmann Р. und Bresslau E. — Die Strudelwiirmer, Leipzig, 1913.
S t u m me r-Tra u nfe 1 s R. und Meixner J. — Polycladida, in В го n n’s
Klassen und Ordnungen des Tierreichs, Bd. IV, 1930—1933.
Сосальщики
Braun M. — Trematodes, in В г о n n’s Klassen und Ordnungen des Tierreichs,
Bd. IV, Leipzig, 1879—1893.
Fuhrmann O. — Trematoda, in Kiikenthal - Krumbach, Hand-
buch der Zoologie, Bd. II, Abt. 2, 1928. (Литература.)
Костылев H. H. — Сосальщики, в книге Е. Павловского — «Прак-
тикум медицинской паразитологии» (1935).
Павловский Е. Н. — Сосальщики (Trematoda). Руководство по зоологии,
т. I, 1937.
Скворцов А. А. — Исследования по циклу развития Dicrocoelium lanceatum.
Журн. Мед. паразитол. и паразит, болезней, т. III, 1934.
Скворцов А. А. (при участии В. Д. Смирновой и Е. Н. С и з я к о-
в о й). — Исследования по морфологии и биологии яйца и по циклу развития
Fasciola hepatica. Мед. паразитол. и паразит, болезни, т. V, вып. 2, 1936.
Скрябин К. и Шульц Р. — Фасциолвзы животных и меры борьбы с ними,
М. 1935.
Нематоды
Павловский Е. Н. — Курс паразитологии человека, с учением о перенос-
чиках инфекций и инвазий, изд. 3. (Печатается.)
Rauther М. — Morphologic und Verwandtschaftsbeziehungen der Nematoden.
Ergebn. und Fortschr. Zoolog., Bd. I, 1909.
Rauther M.—Nematodes, in К ii ke n t ha 1-K г u m ba c h, Handbuch der
Zoologie, Bd. II, Abt. 4, 1930.
Schneider A. — Monographic der Nematoden, Berlin, 1866.
Schneider K. — Lehrbuch der vergleichenden Histologie der Tiere, Jena,
1902.
Скрябин К. И. — Аскариды и их значение в медицине и в ветеринарии,
М. 1925.
Скрябин К. и Шульц Р. — Гельминтозы человека, т. II, 1931.
Смирнов Г. Г. — Круглые черви. Практикум медицинской паразитологии
под ред. Е. Павловского, 1935.
Олигохеты
Beddard F. —A Monograph of the Order Oligochaeta, Oxford, 1895.
Кулагин H. — Материалы по естественной истории дождевых червей. Изв.
Общ. любит, естествозн., т. LVIII, вып. 2, 1889.
Michaelsen W. — Oligochaeta, in Kiikenthal-Krumbach, Handbuch
der Zoologie, Bd. II, 1928.
Stephensen J. — The Oligochaeta, Oxford, 1930.
Vejdovsky F. — System und Morphologic der Oligochaeten, Prag, 1884.
Полихеты
Ashworth J. — Arenicola. Liverp. Marine biol. Comm., London, 1904.
Benham W. —Archiannelida, Polvchaeta and Myzostomaria. Cambridge Nat.
Hist., vol. II, 1896.
469
Дерюгин К. М. — Фауна Кольского залива. Зап. Акад. Наук, т. XXXIV,
1915.
Дерюгин К. М. — Фауна Белого моря. Иссл. морей СССР, вып. 7—8, 1928.
Ehlers Е. — Die Borstenwiirmer (Annelida chaetopoda), Leipzig, 1864—1868.
(Монография.)
Иванов П. П. — Регенеративные процессы у многощетинковых червей, СПб.,
1912.
Vogt С. und Yo u ng Е. - Lehrbuch der praktischen vergleichenden Anatomic,
Bd. 1, 1888.
Woltereck R. — Trochophora-Studien. Zoologica, Bd. XXIV, 1902.
Речной рак
Б e p к о с П. и И н г е н и ц к и й И. — Речной рак. Практическая зоотомия,
вып. II, Спб., 1899.
Бирштейн Я. и Виноградов Л. — Пресноводные Decapoda СССР
и их географическое распространение. Зоол. журн., т. XIII, 1934.
Гексли Т. — Рак. Введение в изучение зоологии, М. 1900.
Kiikenthal-Krumbach. — Handbuch der Zoologie, Bd. Ill, 1. Halfte,
Tardigrada-Crustacea, 1926—1927. (Литература.)
P e s ta O. — Decapoda. Biologic der Tiere Deutschlands, Lief. 17, 1926.
Ветвистоусые раки
Бронштейн 3. — К вопросу о культурах низших ракообразных. Русск.
гидробиол. журн., т. I, 1922.
Ke i 1 ha с k L. — Phyllopoda. Die Siisswasserfauna Deutschlands, H. 10, 1909.
Ламперт К. — Жизнь пресных'вод, СПб., 1900, гл. V.
Lilljeborg W. — Cladocera Sueciae. Nov. Acta Reg. Soc. Sci. Ups., ser. 3,
t. XIX, Upsala, 1900.
Nauman E. — Die Zucht der Daphnia magna Straus, in E. Abderhal-
den, Handbuch der biologischen Arbeitsmethoden, 1925.
R u h e F. — Monographic des Genus Bosmina. Zoologica, Bd. XXV, 1912.
Storch O. — Cladocera. Biologie der Tiere Deutschlands, Lief. 15, 1925.
Wesenber g-L und C. — Contributions to the Biology and Morphology of
the Genus Daphnia with Some Remarks on Heredity. Мёт. Acad. R. Sci. et
Lettr. Danemark, Sect. Sci., 8-me ser., t. XI, №2, 1926.
Циклопы
Brehm V. — Copepoda, in Kiikenthal-Krumbach, Handbuch der Zoolo-
gie, Bd. Ill, 1. Halfte, 1927. (Литература.)
Исаев Л. M. — О способе проникновения личинок ришты (Dracunculus medi-
nensis) в циклопов. Мед. паразитол. и паразит, болезни, т. III, 1934.
Lilljeborg W. — Synopsis specierum hucusque in Suecia observatarum gene-
ris Cyclopis. Kungl. Svenska Vet. Akad. Handl., t. XXXV, 1901.
Рылов В. M. — Свободно живущие веслоногие ракообразные (Eucopepoda).
Пресноводная фауна Европейской России, т. I, изд. журн. «Природа» и Глав-
рыбы, 1922.
Рылов В. М. — Пресноводные Calanoida СССР. Определители организмов
пресных вод СССР, вып. 1, Л. 1930. (Литература.)
Sa г s G. О. — An Account of the Crustacea of Norway, Bergen, vol. IV—VII,
Copepoda, 1903—1921.
S p a n d 1 H. — Copepoda. Biologie der Tiere Deutschlands, Lief. 19, 1926.
Скорпионы
Б иру л я А. А.—Скорпионы. Фауна России. Паукообразные, т. I, вып. 1,
Б и р у л я А. А. — Членистобрюхие паукообразные Кавказского края, ч. I,
Скорпионы. Зап. Кавк, муз., сер. А, №5, Тифлис, 1917.
470
К о 11 m a n n M. — Rec here he s sur les leucocytes et le tissu lymphoide des inver*
ttbrts. Ann. sci. nat., Zool., 9-mes£r., vol. VIII, 1908.
Kraepelin K — Die geographische Verbreitung der Scorpione. Zool. Jahrb.,
Bd. XXIII, 1905.
К r a e p e 11 n K. — Scorpiones. Das Tierreich, Lief. 8, 1899.
Павловский E. H. — Материалы к сравнительной анатомии и истории
развития скорпионов, Пгр., 1917.
Павловский Е. Н. — Сравнительная анатомия мужских половых органов
скорпионов. Тр. Пгр. обш. естествоиспыт., т. Lil 1, вып. 2, 1924.
Pawlowsky Е. — Zur Morphologic des weiblichen Genitalapparats und zur
Embryologie der Skorpione. Ежег. Зоол. муз. Акад. Наук, т. XXVI, 1925.
Ray - LankesterE., Benham W. and Beck E. — On the Muscular
and Endoskeletal Systems of Limulus and Scorpio. Trans. Zool. Soc. London,
vol. II, 1885.
Пауки
Berland L. — Les Arachnides. Encyclop. entomol., A., t. 16, Paris, 1932.
Petrunkevitsch P. — Systema aranearum. Trans. Conn. Acad. Arts a.
Sci., t. XXIX, 1928.
Petrunkevitsch A. — An Inquiry into the Natural Classification of Spi-
ders. Based on a Study of Their Internal Anatomy. Trans. Conn. Acad. Arts a.
Sci., vol. XXXI, 1933.
Schimkewitsch W. — Etude sur 1'anatomic de I'Epeire. Ann. Sci. Nat.,
Zool., vol. XVII, 1884.
Спасский C.•— Определитель пауков Донской области. Изв. Донск. инет,
ссльск. хоз., вып. V, Новочеркасск, 1925.
Vellard J. —Le venin des araignles, Paris, 1936. (Литература.)
Vogel H.— Ober die Spaltsinnesorgane der Radnetzspinnen. Zool. Anz., Bd. LIII,
1923.
Харитонов Д. — Каталог русских пауков. Ежег. Зоол. муз. Акад. Наук,
т. XXXII, 1932, Прил. (379 номеров литературы.)'
Мучные клещи
Захваткин А. — Краткий определитель амбарных клещей, М. 1935.
Порчинский И. А. — Важнейшие клеши, встречающиеся в зерне и муке,
и некоторые данные для обнаружения вредных насекомых в хлебных запасах.
Тр. Бюро по энтом., т. XI, №2, СПб., 1914.
Vitzthum Н. О.—Acari-Milben, in Kiikenthal, Handbuch der Zo-
ologie, Bd. Ill, 2. HSlfte, 1931. (Литература.)
Пастбищные клещи
Banks N. —The Acarina or Mites. Un. St. Departm. Agricult. Rep. № 108,
1915.
Bonnet A.— Recherches sur 1’anatomie сотрагёе et le developpement des Ixodi-
d4s. Ann. Univ. Lyon, nouv. s4r., 1. Sci. Mid., fasc. 20, Lyon — Paris, 1907.
Nutall G., Warburton C. and Robinson L.— Ticks. A Monograph of
the Ixodoidea, parts 1—IV, Cambridge, 1908—1926.
Оленев H. O. — Паразитические клещи Ixodoidea фауны СССР, изд. Акад.
Наук, 1931.
Павловский Е. Н. — Наставление к собиранию и исследованию клешей
(Ixodoidea), изд. Акад. Наук, 1928.
Насекомые
Be г I е ее А. — Gli Insetti, Milano, 1909.
Carpenter G. — Insect Transformation, London. 1921.
Folsom S
1923.
— Entomology' with Special Reference to Ecological Aspects, London,
471
Handschin E. — Praktische Einfiihrung in die Morphologic der Insekten.
Berlin, 1928.
I m m s A. — A General Textbook of Entomology Including the Anatomy, Physio-
logy, Development and Classification of Insects, London, 1930.
Ko r sc he 1 t E. — Der Gelbrand (Dytiscus marginalis L.), I—II, Leipzig, 1923—
1924.
Kiikenthal W. — Handbuch der Zoologie, Bd. IV, Progoneata, Chilopoda,
Insecta, 1925—1932.
Кузнецов H. — Курс физиологии насекомых, ч. I, Обмен веществ, 1927—
1929. (Литография.)
Кузнецов Н. — Насекомые чешуекрылые. Фауна СССР и сопредельных
стран, т. I, вып. 1, 1915; т. I, вып. 2, 1929.
Кузнецов Н., Ротман М. и Ильинский А. — Общая энтомоло-
гия, ч. II, Физиология и токсикология насекомых, Сельколхозгиз, 1931.
Насонов Н. В. — Курс энтомологии, ч. I, Кожные покровы насекомых,
Варшава, 1901.
Павловский Е. Н. — Методы изучения кровососущих комаров, изд. 2,
Акад. Наук, 1935.
Павловский Е. Н. — Наставление к собиранию и исследованию блох
(Aphaniptera), изд. Акад. Наук, 1927.
Поспелов В. (ред.) — Энтомология, Ленсельколхозгиз, 1935.
Patton W. and О г a g g F. — Textbook of Medical Entomology, 1913.
Patton W. and Eva ns’ A. - Insects, Ticks, Mites and Venemous Animals
of Medical and Veterinary Importance, part I, 1929.
Pawlowsky E. N.—Sammeln, Ziichtung und Untersuchung der Flohe, in
Abderhalden, Handbuch der biologiscHbi Arbeitsmethoden, Abt. IX,
Teil 7, 1930.
Schoenichen W. — Praktikum der Insektenkunde, 3. Aufl., Jena, 1930.
Schroder C. — Handbuch der Entomologie, Bd. I—III, Jena, 1912—1929.
Snodgrass R. — Anatomy and Physiology of the Honeybee, New-York, 1925.
Weber H. — Lehrbuch der Entomologie, Jena, 1933.
Римский-Корсаков M. (ред.). — Лесная энтомология, Гослестехиздат,
1935.
Шарп Д. — Насекомые (в обработке Н. Кузнецова), Спб., 1910.
Холодковский Н. А. — Курс энтомологии теоретической и прикладной,
изд. 4, Госиздат, 1927—1931.
Цандер Е. — Строение медоносной пчелы.
Моллюски
Беркос П. и Ингеницкий И. —Практическая зоотомия, вып. IV,
Беззубка, СПб., 1901.
Жадин В. И. — Пресноводные моллюски СССР, Ленснабтехиздат, 1933.
Ламперт К. — Жизнь пресных вод, Спб., 1900.
Фа усе к В. — Паразитизм личинок Anodonta. Зап. Акад. Наук, т. XIII,
№6, 1903.
La ng A. — Lehrbuch der vergleichenden Anatomie der wirbellosen Tiere. Mol-
lusca, 2. Aufl., bearbeitet von K. Haescheler, Jena, 1900.
Meisenheimer J. — Die Weinbergschnecke — Helix pomatia L. Mono-
graphien einheimischer Tiere, Bd. 4, Leipzig, 1912. (Обильная литература.)
Meyer W. — Tintenfische mit besnnderer Beriicksichtigung von Sepia und Octo-
pus. Monographien einheimischer Tiere, Bd. 6, Leipzig, 1913.
Simroth H. — Mollusca (Pulmonata), in Bronn’s Klassen und Ordnungen
des Tierreichs, Bd. 3, 1909.
Simroth H. — Die Nacktschneckenfauna des Russischen Reiches, изд. Акад.
Наук, 1901, 26 цв. табл, и 10 карт.
Иглокожие
Д ь^я ко н о^в А- М. — Иглокожие северных морей. Определитель, изд. Акад.
Дьяконов А. М. — Морские ’ежи. Фауна России и сопредельных стран.
Иглокожие, т. I, вып. 1, 1923.
472
D ej a g e Y. et Hdrouard E. — Traite de Zoologie concr6te, vol. HI, Les
Echinodermes, Paris, ,1903.
Koehler R. — Les Echinodermes des mers d’Europe. Encyclopedic scienti-
fique, vol. I, 1924; vol. II, 1927.
Mortensen Th. — Echinoidea. The Danish Ingolf Expedition, vol. IV, 1 —
2, 1903.
Mortensen Th. — Handbook of the Echinoderms of the British Isles, Oxford,
1927.
Schneider K. — Lehrbuch der vergleichenden Histologie der Tiere, Jena,
1902.
Ланцетник
Гофман. Г. — Руководство к практическим занятиям по сравнительной гисто-
логии, Биомедгиз, 1937, стр. 124—139.
Pietschmann V. — Acrania Cephalochorda, in Kiikenthal, Handbuch
• der Zoologie, Bd. VI, 1. Halfte, 1929.
Рыбы
Беркос П. и И игениц кий И. — Практическая зоотомия, вып. III,
Окунь и щука, СПб., 1899—1901.
Krause R. — Mikroskopische Anatomie der Wirbeltiere, IV, Teleostei, 1923.
Никольский A. — Гады и рыбы, СПб., 1902.
Земноводные
Англа Ж. — Лабораторные животные. Лягушка, 1905.
Беркос П. и Ингеницкий И. — Практическая зоотомия, вып. I,
Лягушка.
Брем А.— Жизнь животных. Под редакцией Н. М. Книповича, т. IV, СПб., 1914.
Гофман Г. — Руководство к практическим занятиям по сравнительной гисто-
логии, 1937.
G a u р р Е. — Anatomie des Frosches, Bd. I, II, 3. Aufl, 1896; Bd. Ill, 2. Aufl.,
1904.
Krause R. — Mikroskopische Anatomie der Wirbeltiere, III, Amphibien, 1923.
Ma к у ш о к М. — Лягушка, Госиздат, 1926.
М а р ш а л ь М. — Лягушка. Введение в анатомию, гистологию и эмбриологию,
М. 1906.
Мейснер В. — Руководство к практическим занятиям по зоологии позвоноч-
ных, вып. 1, Лягушка, Казань, 1905.
Никольский А. — Гады и рыбы, СПб., 1902.
Эккер А. — Анатомия лягушки, вып. 1, 1866.
Hempelmann F. — Der Frosch.
Пресмыкающиеся
Брем А.—Жизнь животных. Под редакцией Н. М. Книповича, г. IV—V,
Земноводные и пресмыкающиеся, СПб., 1914.
Никольский А. М. — Гады и рыбы, СПб., 1902.
Никольский А. М. —Chelonia и Sauria. Фауна России и сопредельных
стран. Пресмыкающиеся, т. I, 1915.
Терентьев П. и Черню в* С. — Краткий определитель земноводных
и пресмыкающихся СССР, Учпедгиз, 1936.
Паркер Т. — Курс зоотомии (позвоночные), 1904.
Hoffmann Н.— Reptilien, in Bronn’s Klassen und Ordnungen des Tierreichs.
Gadow H. — Amphibia and Reptiles. Cambridge Nat. Hist., vol. VIII,
London, 1908.
Птицы
Брем А. — Жизнь животных. Под редакцией Н. М. Книповича, тт. VI, VII,
VIII, IX, СПб., Пгр., 1911—1914.
Gadow Н. und Selenka Е. — Aves, in В го n n’s Klassen und Ordnungen
des Tierreichs, VI, 4, 1891-1893.
473
Krause R. — Mikroskopische Anatomic der Wirbeltiere, II, Vogel und Repti-
lian, 1922.
Kiikenthal W. — Handbuch der Zoologie, Bd. Vll, 2. Haifte, Sa uro psi da,
Aves, 1927.
Маршаль M. — Анатомия птиц, СПб., 1902.
Млекопитающие
А 1 е z a i s Н. — Морская свинка, т. 1, Анатомия, М. 1919.
Англа Ж. — Лабораторные животные. Мышь, СПб., 1906.
Бихнер Е. — Млекопитающие, СПб., 1902.
Бобринский Н. — Анатомия кролика, М. 1932.
Бр ем А. —Жизньживотных. Под редакцией Н. М. Книповича, т. X, Пгр., 1914.
VogtC. et Young Е. — ТгаИё d'anatomie сотрагёе pratique, Paris, 1894.
Weber M. — Die Saugetiere, Bd. I, 11, Jena, 1927, 1928.
Gervais P. und Lee he W. (fortgesetzt von G 6p pe rt). — Mammalia, in
Bronn’s Klassen und Ordnungen des Tierreichs, Leipzig, 1874—1914.
Gerhardt. — Das Kaninchen, Leipzig, 1919.
Krause K. — Die Anatomie des Kaninchens, Leipzig, 1884.
Krause R. — Mikroskopische Anatomie der Wirbeltiere, 1, Saugetiere, 1921.
Le i s e r i n g und M U I I e r. — Handbuch der vergleichenden Anatomie der
Haussaugetiere, Berlin, 1885.
Martin P. — Franks Handbuch der Anatomie der Haustiere, I—II, 5. Aufl.,
1919.
Можейко Б. Э. — Крыса, Симферополь, 1909.
ОГЛАВЛЕНИЕ
Стр.
Введение...................................................... 3
ТИП ПРОСТЕЙШИЕ (PROTOZOA)
КЛАСС РЕСНИЧНЫЕ (С ILIATAs. INFUSORIA)
Туфелька (Paramaecium)........................................ 5
Наблюдения над живыми инфузориями.......................... б
Паразитическая инфузория (Opalina ranarum).................... 24
Общие замечания......................................... 25
Систематическое положение Paramaecium ...................... —
КЛАСС САРКОДОВЫЕ (SARCODINA)
ПОДКЛАСС КОРНЕНОЖКИ (RHIZOPODA)
Амёба (отряд Amoebina)....................................... 28
Наблюдения над живой амёбой................................ —
Раковинные корненожки (отряд Foraminifera)................... 29
Общие замечания......................................... 31
КЛАСС БИЧЕНОСЦЫ (MASTIGOPHORA)
Эвглена (Euglena) ........................................... 32
Наблюдения над эвгленами.................................. 33
Общие замечания......................................... 35
Трипанозома (Trypanosoma).................................... 36
Изучение препарата ....................................... 37
Общие замечания ........................................ 39
КЛАСС СПОРОВИКИ (SPOROZOA)
Грегарины (Gregarinida) некоторых насекомых.................. 39
Наблюдения над грегаринами................................. —
Monocystis дождевых червей................................... 41
Изучение препаратов........................................ —
Общие замечания......................................... 43
Экологические замечания по рассмотренным представителям простей-
ших ................................................ 44
Заключительные замечания о типе простейших ......... 46
475
Стр.
ТИП КИШЕЧНОПОЛОСТНЫЕ (COELENTERATА)
КЛАСС ГИДРОИДНЫЕ (HYDROZOA)
Гидра (Hydra — отряд гидрополипы)......................... 48
Наблюдения над живой гидрой.............................. —
Мацерирование гидры..................................... 52
Размножение гидры....................................... 54
Гистологическое строение гидры.......................... 57
Опыт с регенерацией...................................... —
Опыты с раздражимостью.................................. 59
Общие замечания...................................... 50
Морские гидроиды (отряд гидромедузы)...................... 61
Изучение препаратов ..................................... —
Общие замечания...................................... 65
КЛАСС КОРАЛЛОВЫЕ ПОЛИПЫ (ANTHOZOA)
Пробковый коралл (Alcyonium palmatum)....................... 65
Общие замечания ....................................... 67
Актиния (Actinia equina).................................... 69
Общая форма и строение................................... -
Общие замечания ....................................... 71
Общие замечания о типе кишечнополостных...............: . . . —
ТИП ЧЕРВИ (VERMES)
ПОДТИП ПЛОСКИЕ ЧЕРВИ (PLATODES)
КЛАСС ТУРБЕЛЯРИИ, ИЛИ РЕСНИЧНЫЕ ЧЕРВИ
(TURBELLARIA)
Планарии (Dendrocoelum lacteum)................................. 73
Наблюдения над живыми планариями.............................. —
Изучение среза через тело планарии Dendrocoelum lacteum...... 75
# Опыты с регенерацией у планарий............................... 78
Опыты с обратным развитием планарий.......................... 79
Общие замечания........................................... 80
КЛАСС СОСАЛЬЩИКИ (TREMATODA)
Двуустка ланцетовидная (Dicrocoelium lanceatum)................ 8 0
Строение двуустки ланцетовидной............................. —
Двуустка печеночная (Fasciola hepatica)......................... 82
Строение печеночной двуустки................................. 83
Общие замечания .......................................... 85
Сравнение организации планарии и двуусток......................
Наблюдения над жизненным циклом сосальщиков .................... 88
Печеночная двуустка.......................................... 89
Ланцетовидная двуустка....................................... 94
Сравнение жизненных циклов двуусток печеночной и ланцетовидной. 97
КЛАСС ЛЕНТОЧНЫЕ ЧЕРВИ (CESTODA)
Цепень невооруженный (Taeniarhynchus saginatus)................. 99
Общий осмотр..............................’ ............. —
Срез через проглоттиду цепеня невооруженного................ 103
476
Стп.
Леитец широки! (Diphyllobothrium latum)...................... 105
Общий осмотр................................................. —
Пузырчатые глисты............................................. Ю7
Общие замечания.......................................... 111
КЛАСС КРУГЛЫЕ ЧЕРВИ (NEMATODA)
Аскарида лошадиная (Ascaris megalocephala)..................... ИЗ
Наружный осмотр ............................................ —
Вскрытие аскариды........................................... И4
Срез через аскариду....................................... 118
Изолирование мышечных клеток аскариды....................... 122
Строение губ аскариды....................................... 123
t Общие замечания........................................ 124
Трихина (Trichinella spiralis)............................... 125
Инкапсулированные трихины................................... —
Почвенные, или сапрофитные, нематоды......................... 127
Общие замечания........................................... 129
ПОДТИП КОЛЬЧАТЫЕ ЧЕРВИ (ANNELIDES)
КЛАСС МАЛОЩЕТИНКОВЫЕ ЧЕРВИ (OLIGOCHAETA)
Дождевой червь (Lumbricus)................................... 130
Наружный осмотр ............................................ 131
Вскрытие дождевого червя.................................... 133
Поперечный разрез дождевого черня в области кишечника ...... 140
Регенерация дождевого червя................................. 147
Общие замечания.......................................... 148
КЛАСС МНОГОЩЕТИНКОВЫЕ ЧЕРВИ (POLYCHAETA)
Нереида (Nereis sp.)........................................... 150
Наружный осмотр ............................................. —
Поперечный разрез тела нереиды.............................. 152
Тотальный препарат нефромиксиев............................. 153
Общие замечания.......................................... 154
Пескожил (Arenicola)............................................ —
Наружный осмотр ............................................ —
Вскрытие пескожила......................................... 155
Общие замечания .......................................... —
Общие замечания по рассмотренным группам червей................ 156
ТИП ЧЛЕНИСТОНОГИЕ (ARTHROPODA)
КЛАСС РАКООБРАЗНЫЕ (CRUSTACEA)
Речной рак (Potamobius astacus)................................ 159
Наружный осмотр ........................................... —
Вскрытие рака и его внутреннее строение.................... 163
Поперечные разрезы рака.................................... 174
Конечности рака............................................ 175
Мускулатура ............................................... 181
Наблюдения над живым раком................................. 183
Общие замечания.........................................
Дафния. Ветвистоусые раки (Cladocera) ... г...................Is4
Внешняя морфология и внутреннее строение................. 185
477
Crp.
Наблюдения над жизненным циклом дафний ............................. 188
Общие замечания ................................................
Веслоногие раки (Copepoda)............................................... —
Циклоп (Cyclops) ..................................................
Diaptomus.....................................................
Жизненный цикл Copepoda .......................................202
Общие замечания ............................................203
КЛАСС ПАУКООБРАЗНЫХ (ARACHNOIDEA)
Скорпионы (Scorpiones)...............................................205
Внешняя морфология................................................206
Вскрытие скорпиона .............................................. 211
Рассмотрение микроскопических препаратов..........................215
Наблюдения над фагоцитарными органами.............................218
Экологические замечания........................................219
Общие замечания ...............................................220
ОТРЯД ПАУКИ (ARANEINA)
Двулегочные пауки (Dipneumones)......................................222
Внешняя морфология................................................. —
Органы дыхания паука..............................................228
Экологические замечания........................................229
Общие замечания................................................230
ОТРЯД КЛЕЩИ (ACARINA)
Мучные клещи (семейство Tyroglyphidae)...............................231
Внешняя морфология................................................. —
Развитие мучного хруща............................................234
Экологические замечания........................................ 235
Пастбищные клещи (семейство Ixodidae) ...............................236
Внешняя морфология взрослого клеща................................. —
Препарат ротовых органов (Ixodes ricinus)......................... 239
Общие замечания................................................240
Общие замечания по рассмотренным представителям класса
паукообразных ...................................... 241
КЛАСС НАСЕКОМЫЕ (INSECT А)
Черный таракан (Periplaneta orientalis)..............................243
Внешняя морфология............................................... 244
Вскрытие таракана и его внутреннее строение ......................249
Общие замечания................................................257
Медоносная пчела (Apis mellifera)....................................258
Внешняя морфология................................................. —
Вскрытие пчелы и ее внутреннее строение...........................263
Общие замечания................................................271
Типы ротовых органов насекомых.......................................272
Приспособительные черты строения ног насекомых.......................275
Общие замечания................................................280
Личинки насекомых . . . -
Общие замечания по типу членистоногих . . ..............2 <7
478
Стр.
ТИП МЯГКОТЕЛЫЕ (MOLLUSCA)
КЛАСС ПЛАСТИНЧАТОЖАБЕРНЫЕ (LAMELLI-
BRANCHIA)
ОТРЯД НАСТОЯЩИЕ ПЛАСТИНЧАТОЖАБЕРНЫЕ (EULAMELLIBRANCHIA)
Беззубка (Anodonta)....................................... 288
Наружный осмотр ....................................... —
Наблюдения над живым животным...........................289
Вскрытие раковины....................................... —
Внутреннее строение беззубки ........................... —
Размножение беззубки....................................296
Раковина беззубки ...................................... —
Рассмотрение разрезов беззубки.........................297
Общие замечания.......................................299
КЛАСС БРЮХОНОГИЕ (GASTROPODA)
ОТРЯД ЛЕГОЧНЫЕ (PULMONATA)
Улитка виноградная (Helix pomatia).......................300
Наружный осмотр ........................................ —
Вскрытие улитки и ее внутреннее строение..........• • • • 303
Общие замечания......................................313
Слизняк (Limax agrestis).................................314
Наружный осмотр .......................................315
Вскрытие мантийной полости.............................. —
Общие замечания......................................316
ТИП ИГЛОКОЖИЕ (ECH I NODE RM AT А)
КЛАСС МОРСКИЕ ЗВЕЗДЫ (ASTEROIDEA)
Морская звезда (Asterias rubens).........................317
Наружный осмотр ........................................ —
Скелет морской звезды..................................318
Вскрытие морской звезды и ее внутреннее строение.......319
Поперечный разрез через руку морской звезды............322
Общие замечания......................................323
КЛАСС МОРСКИЕ ЕЖИ (Е CHINOIDEA)
Морской еж (Strongylocentrotus droebachiensis)...........325
Наружный осмотр ........................................ —
Скелет морского ежа....................................327
Вскрытие морского ежа и его внутреннее строение........328
Общие замечания......................................333
Развитие иглокожих.......................................334
ТИП ХОРДОВЫЕ (CHORDATA)
ПОДТИП БЕСЧЕРЕПНЫЕ (ACRANIA s. CEPHALOCHORDA)
Ланцетник (Branchiostoma lanceolatum)....................338
Изучение тотального препарата........................... —
Поперечный срез на уровне жаберной полости.............339
Общие замечания .....................................341
479
Стр.
• ПОДТИП ПОЗВОНОЧНЫЕ (VERTEBRATA)
КЛАСС РЫБЫ (PISCES)
Колючеперые рыбы (окунь, ерш, судак, щука)..................342
Наружный осмотр .......................................... —
Вскрытие рыбы и ее внутреннее строение...................344
Мускулатура рыбы.........................................356
Внутренний скелет рыбы...................................357
Общие замечания........................................359
КЛАСС ЗЕМНОВОДНЫЕ (AMPHIBIA)
БЕСХВОСТЫЕ (ANURA)
Лягушка (Rana temporaria).................................. 360
Наружный осмотр........................................... —
Вскрытие кожных покровов и мускула iура тела.............362
Вскрытие полости тела и общее расположение органов ......363
Внутреннее строение.....................................3( 5
Скелет лягушки...........................................38о
Общие замечания........................................382
КЛАСС ПРЕСМЫКАЮЩИЕСЯ (REPTILIA)
Ящерица зеленая (Lacerta viridis)...........................386
Наружный осмотр......................................... —
Вскрытие ящерицы.........................................389
Общие замечания ....................................... 393
КЛАСС ПТИЦЫ (А V Е S)
КИЛЕГРУДЫЕ ПТИЦЫ
Голубь (Columba livia) и курица (Gallus domestica)......... 394
Наружный осмотр .......................................... —
Вскрытие кожных покровов и мускулатура тела............. 398
Вскрытие полости тела и общее расположение органов.......399
Внутреннее строение......................................400
Скелет.................................................. 417
Общие замечания об организации птиц........................ 423
КЛАСС МЛЕКОПИТАЮЩИЕ (MAMMALIA)
ОТРЯД ГРЫЗУНЫ (RODENTIA)
Кролик (Lepus cuniculus)....................................424
Наружный осмотр .......................................... —
Вскрытие кожных покровов.................................426
Вскрытие полости тела и общее расположение органов ....... —
Внутреннее строение .................................... 427
Скелет...................................................453
Общие замечания........................................456
Заключение..................................................458
Основные элементы микроскопической техники..................459
Литература..................................................467