




























































































































































































































































































































































































































































































































































































































Автор: Каркищенко Н.Н.
Теги: размножение рост развитие математика биология биология человека основы биомоделирования межакадемическое издательство впк
ISBN: 5-902313-04-X
Год: 2004




Н.Н. КАРКИЩЕНКО
ОСНОВЫ
БИОМОДЕЛИРОВАНИЯ
Межакадемическое издательство ВПК
Москва
2004
УДК 612.6.052 + 615.214:51
К 043
К 043 Каркищенко Н.Н. Основы биомоделирования. — М.: Изд-во ВПК, 2004. —
608 с: ил.
Монография посвящена биомедицинскому, формально-логическому моделированию,
планированию, анализу, качеству и биобезопасности экспериментов на линейных
и аутбредных животных. На большом фактическом материале даются принципы вы-
бора адекватных животных-биомоделей для сравнительных исследований в управля-
ющих и исполнительных системах человека и животных. Анализируются противоре-
чивые тенденции биомоделирования первого и второго (альтернативного) порядков,
а также использование моделей более высоких уровней. Дан новый взгляд на фарма-
ко- и токсикинетическое моделирование, учитывающее явления аллометрии и меж-
видовой экстраполяции. Современные данные и их интерпретация могут представ-
лять интерес для физиологов, фармакологов, токсикологов, биологов-эксперимен-
таторов, в особенности для аспирантов и студентов, ищущих свой путь в науке.
Karkischenko N.N. The Basics of Biomodelling
This book concerns biomedical and formal logical modeling. It covers the planning, analysis,
quality and safety of experiments conducted on line and inbred animals. The guiding principles
worked out for the choice of appropriate animal biomodels for comparative investigations of
various operating and executing systems (e.g. nervous, circulatory, lymphatic) in animals and
humans are based on a considerable body of data. The controversial tendencies of the first
and second levels of biomodelling, as well as the use of superior models, are analyzed. The
book includes new aspects of pharmaco- and toxico-kinetic modeling, which take into account
allometry and interspecies extrapolation. Up-to-date materials and their interpretation might
be of practical use for physiologists, pharmacologists, toxicologists and bio-researchers and
to postgraduates and students seeking a career in the scientific field.
Книгу можно приобрести по адресу: www.ozon.ru
Дополнительную информацию о книге можно
получить по адресу: www.labanimals.ru
Available at www.ozon.ru & www.labanimals.ru
ISBN 5-902313-04-Х
© Н.Н.Каркищенко, 2004
© N.N.Karkischenko, 2004
ОГЛАВЛЕНИЕ
Введение 11
РАЗДЕЛ I. ПРИНЦИПЫ МОДЕЛИРОВАНИЯ
Глава 1. Формально-логические аспекты моделирования 19
О пределе и достаточности моделей 20
Соотношение животных и человека как биомоделей и прототипа 24
Функциональные системы и биомоделирование 30
Динамика, управление и устойчивость в живых системах 33
Глава 2. Биосистемы и биомоделирование 37
Экстраполяционные возможности биомоделирования 38
Структурная дискретность и функциональная непрерывность 43
Энтропия и необратимость жизни 47
Формальная аксиоматика в биомоделировании 52
Биомодели для оценки токсичности 53
РАЗДЕЛ II. КАЧЕСТВО ЖИВОТНЫХ-БИОМОДЕЛЕЙ
Глава 3. Вперед в прошлое 59
Кратко о нашей истории 60
Дельфин или хомяк: в поиске биомоделей 62
Глава 4. Стандартизация лабораторных животных 67
Требования к содержанию животных 70
Контроль качества животных и учет 75
Микробиологический контроль 77
Глава 5. Принципы выбора животных для исследований 79
Качество животных — основа эксперимента 79
Отбор линейных животных 85
Глава 6. Биобезопасность при работе с лабораторными животными 105
Уровни биологической безопасности 106
Правила биозащиты персонала 109
Эргономика 111
Зоонозы 112
Аллергии 114
Глава 7. Биориски и качество экспериментов 115
Крупные животные (мини-свиньи, овцы, бараны) 115
Хищники (собаки, кошки) 131
Крупные грызуны (кролики, морские свинки) 140
Мелкие грызуны (крысы, хомячки, мыши) 148
РАЗДЕЛ III. ЛИНЕЙНЫЕ ЖИВОТНЫЕ-БИОМОДЕЛИ
Глава 8. Генетические аспекты биомоделирования 163
Индивидуальность инбредных штаммов 164
Гетерозис и внутриклассовые корреляции 165
4
О генах-таймерах и прогнозируемых биомоделях 167
Нокаутные и трансгенные животные 172
Биомодели для исследования опухолей 176
Глава 9. Инбредные животные-биомодели 182
Генетический мониторинг 183
Криогенное сохранение эмбрионов 184
Инбредные линии кроликов 186
Инбредные линии морских свинок 191
Инбредные линии крыс 193
Инбредные линии хомячков 197
Глава 10. Коизогенные и инбредные линии мышей 203
Конгенно-резистентные линии 203
Инбредные и трансгенные линии 205
Глава 11. Мутантные линии и стоки 227
Глава 12. Гибриды F1, тетрагибриды и рандомбредные животные 237
Глава 13. SPF-животные-биомодели 243
Микробиологический контроль 245
Генетический контроль 246
Требования к корму 246
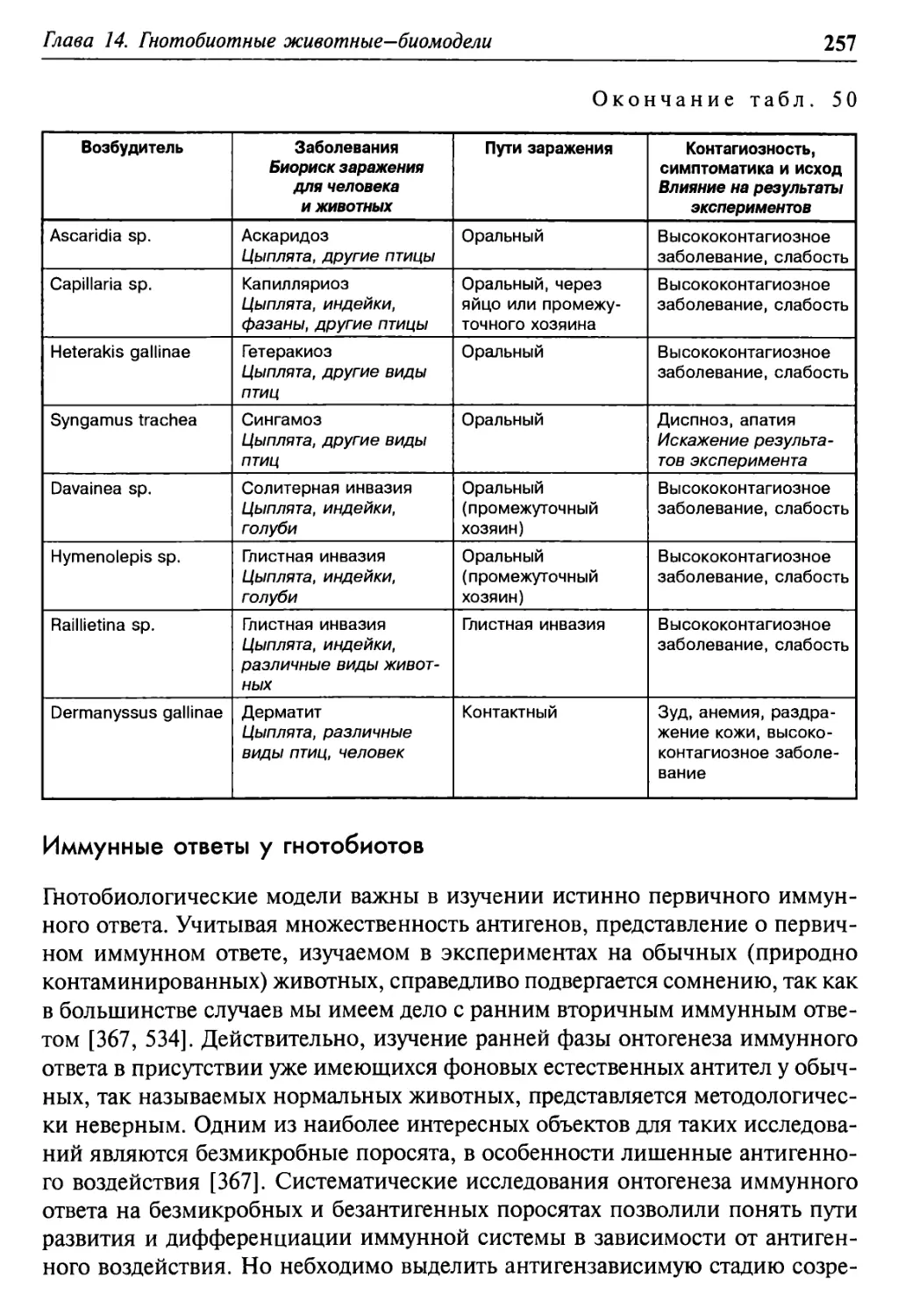
Глава 14. Гнотобиотные животные-биомодели 247
Гнотобиоты-млекопитающие 249
Птицы-гнотобиоты и биориски 253
Иммунные ответы у гнотобиотов 257
РАЗДЕЛ IV. ПЛАНИРОВАНИЕ МЕДИКО-БИОЛОГИЧЕСКИХ
ЭКСПЕРИМЕНТОВ
Глава 15. Пристрастность, репрезентативность и комбинаторика выборки 263
Глава 16. Случайный отбор и чистота эксперимента 269
Глава 17. Предел и достаточность числа наблюдений 273
Глава 18. Анализ и интерпретация результатов: разделяй и действуй 283
Последовательный анализ 284
Принятие решений 287
РАЗДЕЛ V. БИОЭТИКА В ЭКСПЕРИМЕНТАЛЬНЫХ
ИССЛЕДОВАНИЯХ
Глава 19. Гуманность при работе с животными и принципы 3R 291
Глава 20. Иммобилизация животных 297
Глава 21. Наркоз 302
Глава 22. Допустимые методы эвтаназии животных 309
РАЗДЕЛ VI. БИОМОДЕЛИРОВАНИЕ В СИСТЕМАХ УПРАВЛЕНИЯ
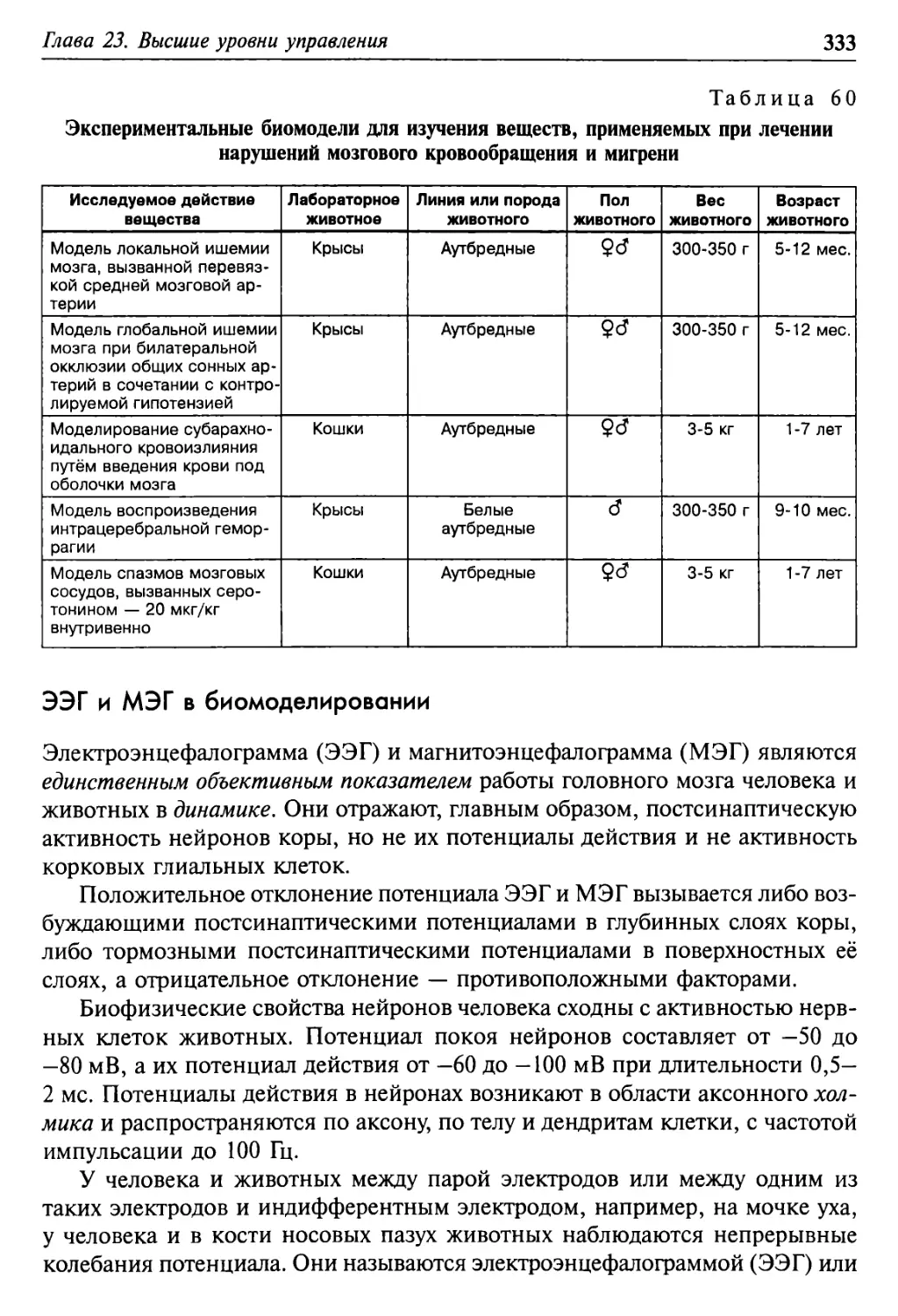
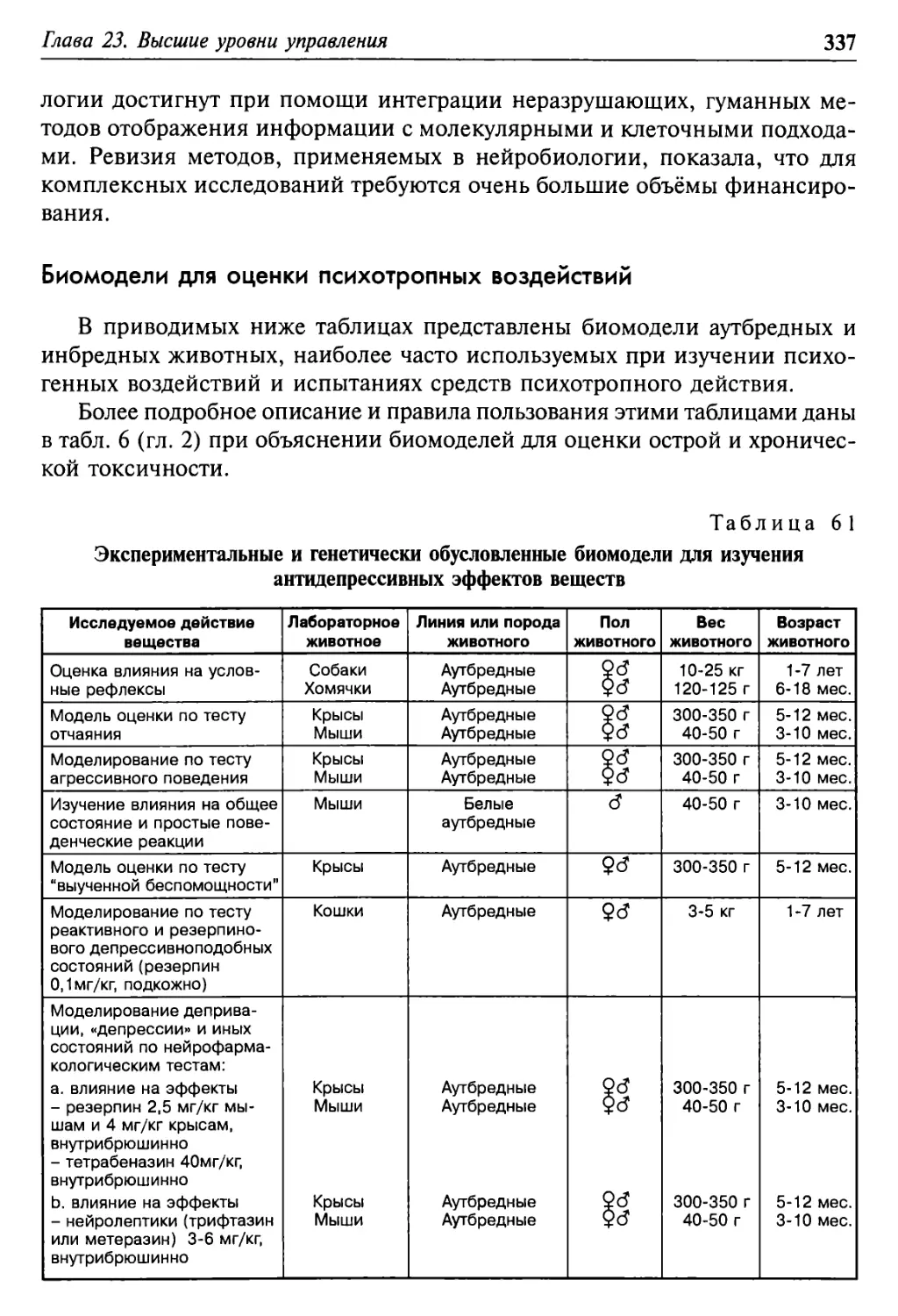
Глава 23. Высшие уровни управления 315
Головной мозг и поведение 316
5
Мозг человека и животных в сравнении 329
ЭЭГ и МЭГ в биомоделировании 333
Биомодели для оценки психотропных воздействий 337
Глава 24. Центральные уровни управления 342
Цереброспинальный уровень 343
Нейровегетативная регуляция 348
Роль N0 в нервной системе 349
Ликвор у человека и животных 352
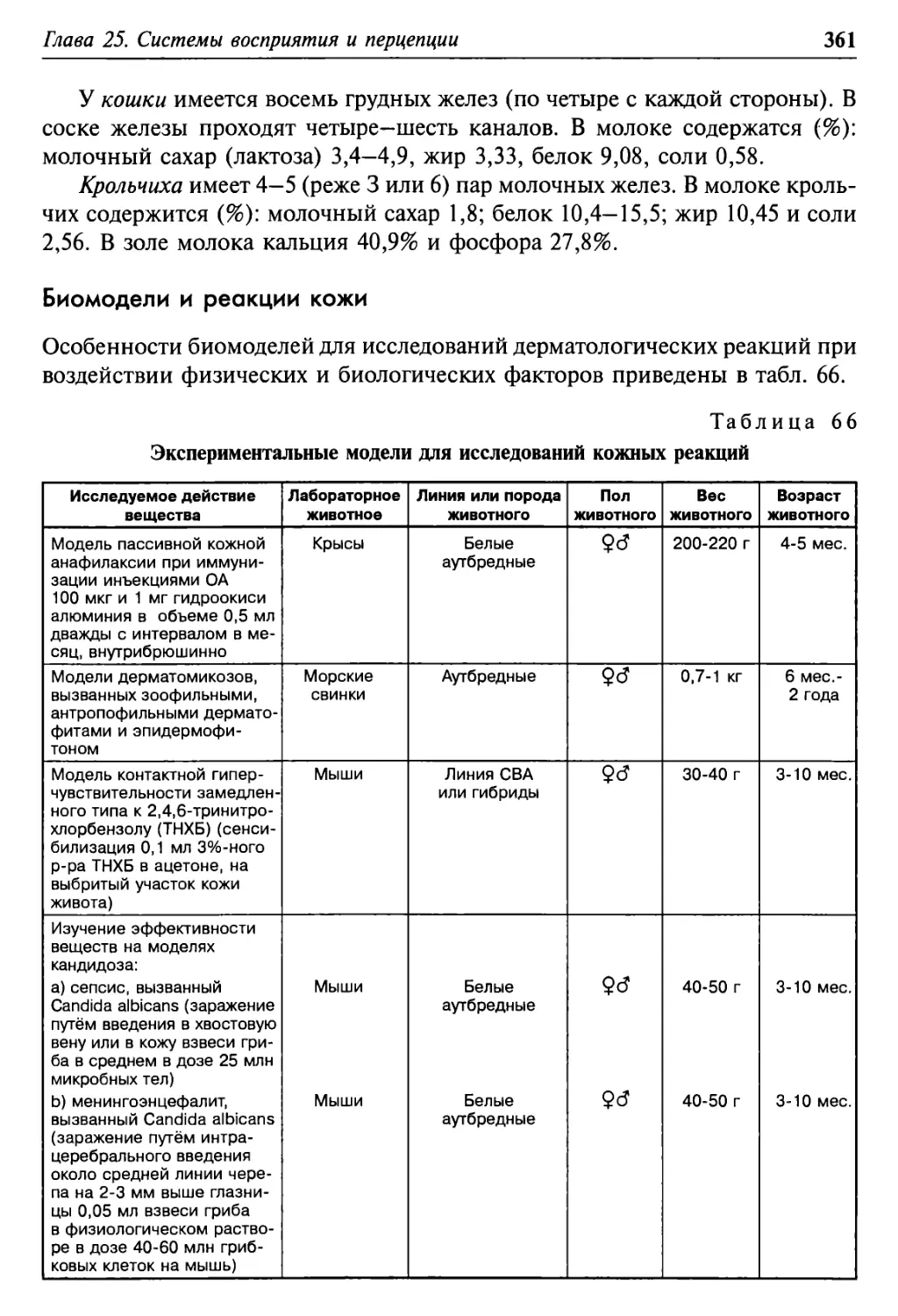
Глава 25. Системы восприятия и перцепции 354
Органы чувств в сравнении 354
Кожа и её образования 358
Биомодели и реакции кожи 361
Глава 26. Триада нейроиммуногормональных систем управления 363
Базис единства триады 364
Тимус в сравнении и биомодели 366
Гипоталамо-гипофизарный пул 368
Свет во тьме и хроноритмы 371
О железах, регулирующих основной обмен 372
Надпочечники в регуляторных процессах 375
Новые связи триады 378
РАЗДЕЛ VII. БИОМОДЕЛИРОВАНИЕ В РЕГУЛЯТОРНЫХ
СИСТЕМАХ
Глава 27. Дыхательная система 383
Сравнительная характеристика органов дыхания 383
Регистрация дыхания 386
Математическое моделирование дыхательных функций 387
Биомодели для изучения бронхо-лёгочных процессов 388
Глава 28. Кардиогемодинамика в биомоделях 391
Сердце человека и животных в сравнении 391
Биомоделирование на сердце 394
Сравнительная электрокардиография 398
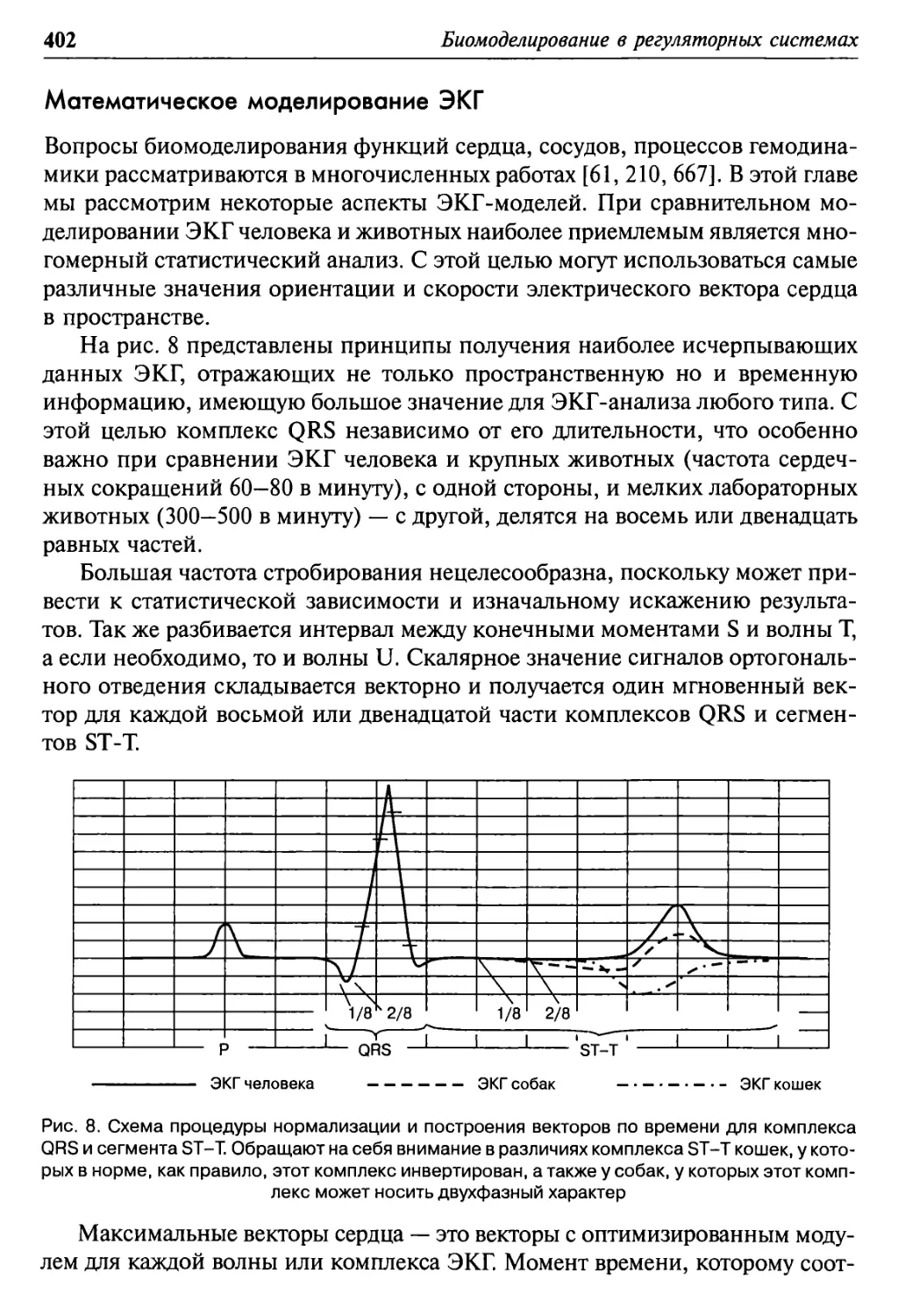
Математическое моделирование ЭКГ 402
Сосуды, гемодинамика и биомодели 404
Глава 29. Внутренняя среда организма 410
Кровь человека и животных в сравнении 410
Морфологические элементы и состав крови 414
Показатели крови инбредных животных 419
Селезенка в сравнении 421
Лимфатическая система 423
Забор крови и пункция сердца 425
Глава 30. Система интестинальных органов 430
Стартовые процессы в сравнении 431
Желудок и его сравнительные функции 434
6
Регуляторные функции печени 437
Поджелудочная железа — двуликий Янус 441
Финальные процессы в сравнении 446
Глава 31. Ренальная система и самоподобие Прокла 450
Сравнительные и холономные параметры почек 451
Моделирование почечных функций 456
Глава 32. Половое поведение и система репродукции 461
Агрессивность и половое поведение 461
Репродуктивные нюансы самок в сравнении 464
Биомодели для изучения гормональных эффектов 468
Глава 33. Остеосистема и реперы Махаланобиса 471
Скелет животных в сравнении 472
Депо или донатор: биомоделирование остеофункций 478
РАЗДЕЛ VIII. АЛЛОМЕТРИЯ И ЭКСТРАПОЛЯЦИЯ: ПРЕДЕЛЫ
И ВОЗМОЖНОСТИ
Глава 34. Критерии подобия и аллометрия 487
Многообразие подобия и его пятый вид 487
Условия переноса при экстраполяции 492
Глава 35. Физиологическое время и кинетика 496
Глава 36. Межвидовые вариации 502
Адекватность и корректировка моделей 503
Базовый план и подход Дедрика 507
Глава 37. Биологическое масштабирование 512
Токсичность и цена жизни 513
О скорости и клиренсе жизни 518
РАЗДЕЛ IX. ПЕРСПЕКТИВЫ БИОМОДЕЛИРОВАНИЯ
Глава 38. Фенотипирование и генополиморфизм 525
Поиск маркеров генов 527
Генополиморфизм и непереносимость лекарств 530
Генетический полиморфизм животных 537
Глава 39. О сейфе Творца и моделях высоких порядков 542
Аналогия отношений: новый взгляд 543
Энергия связи или энергия переноса 546
Доноры, акцепторы и электронное давление 549
Глава 40. Принцип «самовара» и нелинейные подобия 552
Неполное подобие и метаморфозы упрощения 553
Проблема измерения и безразмерности 556
Модели и модули в фармакопсихологии 559
Заключение 569
Библиография 571
TABLE OF CONTENTS
Introduction 11
SECTION I. PRINCIPLES OF MODELLING
Chapter 1. Formal logical aspects of modelling 19
Limitation and sufficiency of models 20
Correlation between animals and man as the biomodels and the prototype 24
Functional systems and biomodelling 30
Dynamics, control and stability in living systems 33
Chapter 2. Biosystems and biomodelling 37
Extrapolation capabilities of modelling 38
Structural discreteness and functional continuity 43
Entropy and irreversibility of life 47
Formal axiomatics in biomodelling 52
Biomodels for estimation of toxicity 53
SECTION II. THE QUALITY OF ANIMALS
Chapter 3. Foreward to the past 59
Our history in brief 60
Dolphin vs hamster: in searching for biomodels 62
Chapter 4. Standartization of laboratory animals 67
Keeping of animals requirements 70
The control of quality and registration of animals 75
Microbiological monitoring 77
Chapter 5. Principles of animal selection for experiments 79
The quality of animals - the basis of experiment 79
Selection of line animals 85
Chapter 6. The safe treatment of animals 105
The levels of biological safety 106
Personnel bioprotection rules 109
Ergonomics Ill
Zoonoses 112
Allergy 114
Chapter 7. Biorisks and quality of experiments 115
Large animals (mini-pigs, sheep, rams) 115
Predators (dogs, cats) 131
Large rodents (rabbits, guinea pigs) 140
Small rodents (rats, hamsters, mice) 148
SECTION III. LINE ANIMALS-BIOMODELS
Chapter 8. Genetic aspects of biomodelling 163
Specific features of inbred strains 164
Heterosis and intraclass correlations 165
8
Genes-timers and forecasted biomodels 167
Knock-out and transgenesial animals 172
Biomodels for experiments in oncology 176
Chapter 9. Inbred animals-biomodels 182
Genetic monitoring 183
Cryogenic conservation of embryos 184
Inbred lines of rabbits... 186
Inbred lines of guinea pigs 191
Inbred lines of rats 193
Inbred lines of hamsters 197
Chapter 10. Coisogenic and inbred lines of mice 203
Congenetic resistent lines 203
Inbred and transgenetic lines 205
Chapter 11. Mutation lines and stocks 227
Chapter 12. Hybrids Fl, tetrahybrids and randombred animals 237
Chapter 13. SPF-animals-biomodels 243
Microbiological monitoring 245
Genetic monitoring 246
Feeding requirements 246
Chapter 14. Gnotobiotic animals-biomodels 247
Gnotobiotic mammals 249
Gnotobiotic birds and biorisks 253
Immune responses in gnotobiotes 257
SECTION IV. PLANNING OF MEDICO-BIOLOGICAL
EXPERIMENTS
Chapter 15. Bias, representativity and combinatory of sampling 263
Chapter 16. Random sampling and pure experiment 269
Chapter 17. Limits and sufficiency of the number of observations 273
Chapter 18. Analysis and interpretation of results: divide and act 283
Successive analysis 284
Decision making 287
SECTION V. BIOETHICS IN EXPERIMENTAL STUDIES
Chapter 19. Humanity in treating the animals and the Three Rs principles 291
Chapter 20. Animals' immobilization 297
Chapter 21. Anesthesia 302
Chapter 22. Acceptable methods of animal euthanasia 309
SECTION VI. BIOMODELLING IN OPERATIONAL SYSTEMS
Chapter 23. Higher levels of the operational system 315
Cerebrum and behaviour 316
Comparison of human and animal brain 329
9
EEG and MEG in biomodeling 333
Biomodels for estimation of psychotropic influences 337
Chapter 24. Central levels of the operational system 342
Cerebrospinalis level 343
Neurovegetative regulation 348
Role of NO in nervous system 349
Liquor of man and animals 352
Chapter 25. Sensation and perception systems 354
Sense organs in comparison 354
Skin and its formations 358
Biomodels and skin responses 361
Chapter 26. Triad of neuroimmunohormonal operational systems 363
Basis of the triad unity 364
Biomodels and thymus in comparison 366
Hypothalamic-pituitary pool 368
Light in the dark and chronorhythm 371
About glands regulating basal metabolism 372
Adrenal glands in regulatory processes 375
New links of triad 378
SECTION VII. BIOMODELLING IN REGULATORY SYSTEMS
Chapter 27. Respiratory system 383
Comparative characteristics of breathing organs 383
Record of breathing 386
Mathematical modelling of respiratory functions 387
Biomodels for study of bronchi-pulmonary processes 388
Chapter 28. Cardiohemodynamics in biomodels 391
Comparison of human and animal heart 391
Heart biomodelling 394
Comparative electrocardiography 398
Mathematical modelling of ECG 402
Vessels, hemodynamics and biomodels 404
Chapter 29. Internal medium of organism 410
Comparison of human and animal blood 410
Morphological elements and blood compound 414
Inbred animals blood indices 419
Spleen in comparison 421
Lymphatic system 423
Blood sampling and puncture of heart 425
Chapter 30. System of intestinal organs 430
Starting processes in comparison 431
Gastric and its comparative functions 434
Liver regulatory functions 437
Pancreas — bifacial Janu 441
Final processes in comparison 446
10
Chapter 31. Renal system and self-similarity of Proklos 450
Comparative and cholonomic parameters of kidneies 451
Modelling of kidney functions 456
Chapter 32. Sexual behaviour and reproductive system 461
Aggressiveness and sexual behaviour 461
Reproductive features of females in comparison ^.,......„.,.. 464
Biomodels for study of hormonal effects 468
Chapter 33. Osseous system and Makchalanobis' points 471
Animal skeleton in comparison 472
Depot or donor: biomodelling of osseous function 478
SECTION VIII. ALLOMETRY AND EXTRAPOLATION: LIMITS
AND POSSIBILITIES
Chapter 34. Criteria of similarity and allometry 487
Variety of similarity and its fifth form 487
Conditions of transfer at extrapolation 492
Chapter 35. Physiological time and kinetics 496
Chapter 36. Interspecific variations 502
Adequacy and adjustment of models 503
The base plan and Dedrick's approach 507
Chapter 37. Biological scaling 512
Toxicity and the price of life 513
Velocity and clearance of life 518
SECTION IX. PROSPECTS OF BIOMODELLING
Chapter 38. Phenotyping and genopolymorphism 525
Searching of genes' markers 527
Genopolymorphism and intolerance of drugs 530
Genetic polymorphism of animals 537
Chapter 39. About the safe of Creator and models of higher orders 542
Analogy of relations: a new glance 543
Bonding energy or transfer energy 546
Donors, acceptors and electron pressure 549
Chapter 40. Principle of «samovar» and nonlinear similarities 552
Incomplete similarity and metamorphoses of simplification 553
Problem of measurement and dimensionless 556
Models and modules in pharmacopsychology 559
Conclusion 569
Bibliography 571
11
Введение
Естественная человеческая потребность находить ре-
альные прообразы абстрактных понятий в познании
биологической сущности человека и биологизации
социальных и созидательных процессов хороша и пло-
ха одновременно. Она инициирует поиск средств и
методов познания, конструкций и инструментов его
исследования, но мешает ясно видеть всю логическую
панораму. Большинство исследований по биомодели-
рованию можно разделить на две части. Первая их
часть посвящена общефилософским вопросам и логи-
ческим основам моделирования, вторая — созданию
или использованию экспериментальной базы, рас-
смотрению математического аппарата, физико-хими-
ческой и технической стороне моделирования. Мы не
будем касаться критики разных подходов, ибо, как
изрек Козьма Прутков: «Нельзя объять необъятное».
Книга, вне всякого сомнения, относится ко второй
части исследований, но, право, как хотелось бы раз-
двинуть жесткие рамки существующих пределов био-
моделирования и соединить два полюса подходов к мо-
делированию. Увы, это практически невозможно.
Метод моделей, наряду с дедукцией, использовал-
ся Аристотелем и его учениками. Их естественно-на-
учные работы, постепенно развиваясь, воплотились че-
рез тысячелетия в формально-логическое и дедуктив-
но-аксиоматическое моделирование. Биологическое
моделирование, привнося свои предметы и методы, яв-
ляется, по своей сути, лишь малой частью единых под-
ходов и принципов метода моделирования. Поэтому
мы рассмотрим формально-логические основы биомо-
делирования, его роль и место в анализе биосистем,
в управлении и устойчивости живых систем, в соотно-
12
Введение
шении дискретности и непрерывности и, конечно же, коснемся пределов и воз-
можностей экстраполяции между различными животными и человеком.
О конструкции книги
Обобщая собственный материал и данные многих авторов, мы стремились к
тому, чтобы в нашем изложении интеграция разных областей знаний не созда-
вала некий нерасчленяемый монолит, а была бы доступна в качестве отдельных
составляющих. Книга рассчитана прежде всего на тех читателей, которые пони-
мают, что если проблему удается перенести на язык формул, то она упрощается.
Математика имеет дело с упрощенными моделями явлений, а формула — это
существенный этап в построении биологической модели.
Выбор животных,
планирование
и организация
исследований
Формально-
логическое
и аксиоматическое
моделирование
Многообразие
линейных
и нелинейных
подобий
в биомоделировании
Необходимость
и достаточность
при выборе
биомоделей
Биориски, качество
экспериментов
и биобезопасность
персонала
Биомодели
для исследований
регуляторных
и исполнительных
систем
Сравнительные
количественные
параметры систем
человека и животных
Морфофункцио-
нальные
и биохимические
параметры
биомоделей
Генетически
обусловленные
биомодели и
генополиморфизм
Аллометрия
и межвидовые
вариации
в биомоделировании
Экстраполяция,
пределы
и возможности
переноса
Биомоделирование
в экспериментах,
их гуманизация
и принципы 3R
Но для тех читателей, которым это представляется не нужным или не инте-
ресным, мы даем возможность найти необходимые данные о животных-биомо-
делях, в том числе инбредных, коизогенных, рандомбредных, нокаутных и дру-
гих линий. Приводятся данные о биорисках, связанных с животными, основы
биобезопасности и гуманизации при работе с ними (см. блок-схему).
Интересными для некоторых исследователей станут сведенные в таблицы
многочисленные генетически обусловленные и экспериментальные биомоде-
Введение
13
ли лабораторных животных. Наличие в Научном центре биомедицинских тех-
нологий РАМН уникальных линий животных, имеющих врожденную патоло-
гию, аналогичную таковой у человека, например, диабет типа I и II, спонтан-
ная гипертензия, склерозирование коронарных сосудов, патология почек, пе-
чени, головного мозга и многих других органов и систем, позволяет исследо-
вателю иметь прямые, а не косвенные, экспериментальные и генетически обус-
ловленные биомодели.
Для компактности изложения мы формализовали описание разделов книги
в виде блок-схемы (см. с. 12), представляющей «срезы» материала не по главам
и разделам, а по смысловому содержанию. И хотя каждый из блоков разворачи-
вается по всей или большей части книги, читатель может легко ориентировать-
ся, поскольку в каждой главе есть ссылки на другие главы, содержащие логичес-
ки связанную информацию.
О моделях, биомоделях и биомоделировании
В биологическом моделировании можно выделить два подхода, первый из кото-
рых основан на сложных умозрительных трактовках познавательных механиз-
мов. Приверженцы этого подхода, как правило, пользуются для описания своих
теорий образным литературным языком, с присущей ему многозначностью,
риторической изящностью и эпической красотой. Это хорошо воспринимается,
но приводит к расплывчатости и запутанности всех их построений. Как прави-
ло, эти концепции и афоризмы трудно представить — можно лишь запомнить.
Существует и иной подход, но о нем чуть позже.
Огромный словарный запас русского языка дает возможность описать лю-
бые явления и объекты, но не всегда это получается компактно. Поэтому, во
избежание мультисмысловых восприятий, дадим краткие определения исполь-
зуемой в книге терминологии.
Под моделью (лат. modelus — мера, норма, образец) мы будем подразумевать
некий материальный или виртуальный объект, замещающий в процессе изуче-
ния объект-оригинал, сохраняя типичные для конкретного исследования черты.
Построение такой модели является процессом моделирования.
Биологическое моделирование или биомоделирование является процессом пред-
ставления, отображения, реализации системы, структуры или программы, в
результате которого мы получаем новую информацию об объекте. Биологичес-
кое моделирование включает использование методов самых разных областей
знания: биологии, медицины, физики, химии, математики и т.д. Пожалуй, един-
ственное, что их объединяет и на чем отражается биомоделирование, так это
триада: предмет — свойство — отношение, составляющая целевое назначение
модели. Чтобы избежать разночтения в терминах биомоделирование и биомодель,
которые зачастую у двух авторов имеют три смысла, при дальнейшем изложе-
нии материала мы будем это конкретизировать, а пока дадим определения.
Животное-биомодель — лабораторное животное, используемое в эксперименте
с целью построения демонстративных или любых других адекватных моделей
14
Введение
функционирования человека и животных для последующего описания и анали-
за изучаемых процессов.
Экспериментальная биомодель — создаваемая модель того или иного состоя-
ния, в том числе патологического, характерного или частично воспроизводяще-
го функционирование прототипа (человека или другого животного) с помощью
физических, химических или биологических факторов.
Генетически обусловленная биомодель или просто биомодель — специальные
линии животных, полученные в процессе инбридинга, нокаута, гнотобиотных
или трансгенных технологий, имеющие врожденные изменения или патологию,
характерную для заболеваний человека. Существуют прогнозируемые биомоде-
ли, когда отбор и поиск их идет на основе расчета генов-таймеров, гибридной
мощи и внутриклассовых корреляций. Также животные-биомодели незамени-
мы при поиске новых лекарств, вакцин, оценке токсинов и иных ксенобиоти-
ков, регламентации в гигиенических исследованиях, нормировании слабых хи-
мических и физических воздействий, расшифровки механизмов начала патоло-
гических процессов.
О математическом моделировании: запомнить или представить
Математическая модель — это любая совокупность элементов и связывающих
их операций, изоморфно отображающих реальные или реализуемые объекты,
процессы и явления. Любая математическая модель является несомненным
упрощением, но эта простота подчиняется жестким правилам, которые нельзя
отменить или поменять в процессе любой игры, а тем более в научной.
Когда будучи студентом четвертого курса медицинского института, я посту-
пил на вечернее отделение мехмата университета, то испытал потрясение на
первом же занятии по математической логике. Следует напомнить, что язык
математической логики является логическим воплощением языка математики,
формами которого являются абстрактные объекты и их понятия. Преподаватель
предложил представить (а не запомнить!) мячик как бесконечное множество
точек эвклидова пространства, где главным параметром был радиус, определя-
ющий расстояние от некой точки до любой другой на поверхности. При этом
надо представить, что мячик утратил свою физическую сущность: у него нет
массы, объема и т.д. На этом математическом объекте мы, используя теории
подобия, воссоздавали явления реального мира, объекты и их процессы, т.е.
осуществляли математическое моделирование.
Как ни странно, но лишь в университете я узнал (хотя, по-моему, это дол-
жно быть в курсе школьной математики), что древних мудрецов-философов
беспокоило до умопомрачения значение Л • Они хорошо знали, но не могли
понять, почему не существует дроби, квадрат которой был бы равен точно двум
( 4l = 1,414...). Например, 17/12, квадрат которой 289/144 достаточно близок к
288/144, что равнялось бы ровно двум. Можно подходить еще ближе, но мы
никогда не получим точно 2. Конечно, это можно запомнить, но понять,... По-
этому важнее научиться представлять себе это, а не запоминать механически.
Введение
15
Не потому ли простые математические истины произвели на меня, да, по-
видимому, и на любого чистого гуманитария впечатление, что сталкиваются
два полюса взглядов на обучение. Ведь на первом занятии в мединституте по
нормальной анатомии преподаватель, высыпав на стол груду костей, сказал:
«Запомните — это columna vertebralis, это — atlas, это — axis», это — vertebra
prominens». И это «запомните» — преследовало меня, как и других студентов-
медиков, все шесть лет обучения, за исключением, может быть, фармаколо-
гии, физиологии, неврологии и немногих кафедр, где говорили: «Подумайте».
Подумайте, но не представьте!
Вернемся к подходам в биомоделировании. Кроме литературно-описатель-
ного существует и другой подход, основанный на математических моделях, та-
ких, как теория алгебраических уравнений, факторный и дисперсионный ана-
лиз, теория множеств, дифференциальные и интегральные исчисления и их
уравнения, теория вероятностей и математической статистики. Наконец, суще-
ствуют методы исследования операций, принятия решений, математического
моделирования, в том числе в целенаправленной деятельности. Все эти методы
позволяют со всей строгостью, ясностью и упрощениями, свойственными мате-
матике, осуществлять биологическое моделирование. Конечно, и математика не
всесильна, особенно при описании принципов квантовой механики и взаимо-
отношений субатомных структур в живых системах. Мы коснемся вопросов био-
моделирования не с традиционной структурной, функциональной и молекуляр-
ной стороны, а, что более перспективно, со стороны энергетических межатом-
ных и субатомных отношений, обеспечивающих в конечном итоге заряды, ин-
формацию и биоэнергетику биологических систем. А адекватные математичес-
кие модели будут созданы, в этом нет сомнения. Надо лишь научиться не только
изучать и описывать эти процессы, но прежде всего их представлять.
О биорисках: животные не только кусаются
В последние годы,бурно развивается альтернативное моделирование. Мы рас-
смотрим его плюсы и минусы, а в списке литературы даны ссылки на работы в
этом направлении в качестве рекомендуемых. Тем не менее, мы считаем, что на
настоящем этапе развития науки лучшей альтернативой использованию живот-
ных является использование линейных животных, соответствующих стандартам
качества в генетическом, морфофункциональном и микробиологическом отно-
шении.
В экспериментах или иной работе с лабораторными животными следует хо-
рошо себе усвоить, что они не только кусаются и царапаются — они представ-
ляют угрозу для экспериментатора в двух аспектах. Во-первых, потому, что явля-
ются источником антропозоонозов и аллергенов, а, во-вторых, при наличии у
них инфекционной патологии дают извращенные данные в процессе экспери-
мента и могут свести на нет все усилия исследователя получить достоверные и
качественные результаты. Это столь же важный элемент любого исследования,
как и гуманное обращение с животными. Необходимо подчеркнуть, что инфи-
16
Введение
цированное или инвазированное животное не только не должно использоваться
в эксперименте, но и попадать в лабораторию из вивария. Это так же важно, как
и своевременная «зачистка» (правда, ведь очень модное словечко?) генетически
контаминированных животных в процессе инбридинга.
О чистых линиях и попутном ветре
Известные успехи в биологии, растущие требования к контролю качества ле-
карств, вакцин, пищевых продуктов, особенно трансгенных, ужесточение гиги-
енического нормирования химических, биологических и физических факторов,
развитие трансплантологии, онкологии, фармакологии стремительно сдвигают
границы используемых животных в сторону линейных. Так называемые беспо-
родные животные плавно, но безвозвратно уходят в прошлое. Это следует учи-
тывать, а для хорошего экспериментатора, имеющего дар божий, понимать и
ощущать, куда направляется поток мировой научной мысли.
По некоторым данным, в Джексоновском институте имеется более 100 000
линий инбредных животных. Естественно, что они хранятся в основном в виде
генетического материала в криобанке, но все же — это сто тысяч. В нашей стра-
не их в десятки раз меньше, однако в распоряжении исследователей имеются
все основные линии, которые составляют некий «джентльменский» научный
набор. Поэтому мы сочли важным дать описание инбредных и иных линейных
животных, литература по которым не обновлялась более 20 лет. Любой исследо-
ватель — это прежде всего оптимизатор, он ищет стержень проблемы, за кото-
рый можно ухватить. Но лишь редким творцам науки это удается, для большин-
ства из нас это лишь стержень-мираж, при попытке ухватить — он растворяется.
Остается хотя бы ловить попутный ветер, чтобы не оказаться на обочине науки.
Использование линейных животных дает лучшую воспроизводимость резуль-
татов, возможность повторения в любой другой лаборатории на этих же линиях,
позволяет раскрыть новые механизмы и укрепляет мужество экспериментатора
при заявлениях о неожиданных открытиях и находках. Поэтому пожелаем всем
исследователям и экспериментаторам попутного ветра в биомоделировании.
Об авторе
Каркищенко Николай Николаевич — директор Научного центра биомедицинских
технологий РАМН, доктор медицинских наук, профессор, академик РАРАН и Меж-
дународной академии астронавтики (Париж), член-корреспондент РАМН, лауреат
Государственных премий СССР. Автор более 360 научных работ, 12 монографий,
13 книг и учебников, 37 патентов на изобретения. Научный руководитель 12 док-
торских и более 40 кандидатских диссертаций. Основные научные интересы связа-
ны с проблемами биобезопасности, исследованиями в области оборонных и специ-
альных тематик и разработок.
Раздел! ПРИНЦИПЫ
МОДЕЛИРОВАНИЯ
Логика, которая может дать достоверность,
есть орудие доказательства.
Л.Пуанкаре
19
Глава 1 ФОРМАЛЬНО-ЛОГИЧЕСКИЕ АСПЕКТЫ
МОДЕЛИРОВАНИЯ
Модели в качестве средства познания стали упот-
ребляться уже на заре развития науки. Вместе с тем
общая теория моделирования начала создаваться
лишь с 60-х годов XX века. Употребление научных
умозаключений в виде силлогизмов зачастую дос-
таточно для определения их роли и места в системе
строго определенных рамок. Оно дает возможность
псевдологического использования термина «модель»
в различных смыслах, и наоборот, часто имеет мес-
то применение различных слов для обозначения, по
сути дела, одного и того же содержания.
Разнообразие значений термина «модель» в со-
временных биомедицинских науках бросается в гла-
за. Возникает сомнение, а можно ли говорить о мо-
делях и моделировании вообще или только о моде-
лях и моделировании в определенных разделах био-
логии и медицины. Исторически сложилось так, что
в решении многих проблем биологии и медицины
решающую роль играют экспериментальные иссле-
дования на животных, позволяющие моделировать
необходимые состояния, изучать динамику патоло-
гического процесса, метаболические сдвиги, дина-
мику и кинетику лекарств. Термин «животные мо-
дели» прочно утвердился в арсенале исследователей
в 30-х годах прошлого столетия. Boldessarins R.J. и
Fisher J.E. дали определение модели как «экспери-
ментального компромисса, где простая эксперимен-
тальная система используется для понимания гораз-
до более сложной» [76].
20
Принципы моделирования
Пока нет единства мнений по поводу употребления термина «модель»,
авторы, говоря о моделях, обычно определяют тот смысл, в котором они
употребляют этот термин [85, 372, 379, 463, 514, 596]. Это тем более важно
в том случае, когда речь идет о биомоделировании вообще, безотносительно
к той или иной области биологии, медицины, фармакологии, физиологии и
т.д. Например, существуют так называемые демонстративные модели, приме-
няемые в обучении. Они имеют много общего с мысленным эксперимен-
том, но не с моделью. Неудачным применением термина «модель» является
использование его как синонима теории, когда имеется в виду даже не изо-
морфизм разных теорий, не то обстоятельство, что данная теория обладает
одной и той же или сходной логической структурой с другой теорией, а
лишь некоторые особенности теории [16, 203, 513, 609]. Еще более неудач-
ным применением термина «модель» является его использование в качестве
синонима любого количественного описания, схемы или графического по-
строения [119, 144, 475, 488, 653, 655].
Широко распространенным, но неадекватным употреблением термина
«модель» является его использование в смысле формальной или формализо-
ванной системы, когда вместо приближения к выяснению реальной биоло-
гической сути, вводятся грубые по сути возможности исходить из посылок
подмены понятий дыхательной, сердечно-сосудистой, нервно-эндокринной
и т.д. систем, некими абстракциями типа «пневмостат», «гидростат», «гоме-
остат» [131, 168, 189, 260, 369, 518].
Поэтому при биомоделировании следует по возможности исключить из
научного языка такие значения термина «модель», для выражения которых
существуют другие прочно установившиеся термины, и сохранить этот тер-
мин для таких специфических гносеологических ситуаций, которые не по-
крываются понятиями «теория», «гипотеза», «формализм», «исследование»,
«эксперимент», «наблюдение (клиническое или экспериментальное)», «струк-
тура» и т.д. В этом случае достаточно воспользоваться возможностями фор-
мальной логики на основе приемов традиционной абстракции [327, 330].
О пределе и достаточности моделей
Приступая к процессу биомоделирования, стоит «очистить» будущую конст-
рукцию модели от несвойственных ей элементов. Для этого можно восполь-
зоваться подходом, который назовем методом исключения. Его логическую
структуру выразим следующим образом:
А(Р)(ах, ..., V -.=> (др ..., ак), (1)
где 1 < /, к < п.
Здесь А (Р) будет означать оператор, который выражает собой исключение
всего того, что не соответствует требованию Р. Символы ах, ..., ап выражают
Глава 1. Формально-логические аспекты моделирования
21
множество исходных понятий; aif ..., ак, стоящие в правой части соотноше-
ния, представляют собой те понятия, которые остались после применения к
множеству av ..., ап оператора Л(Р). Символ -. => выражает собой переход от
исходных данных к результату операции. Это некоторый аналог импликации
(=>)в научных умозаключениях, логических построениях и исследованиях,
изначально вытекающих из отрицания (—i) или исключения (см. гл. 2)
В этом случае существует одна опасность, когда исключение «неудач-
ных» значений термина «модель» доводится до того, что остается лишь одно
понятие изоморфизма законов или теорий. В таком случае сфера применения
понятия модели оказывается относящейся только к той отдельной области
науки, где было сформулировано понятие ак, и последующий логический
анализ, ценность которого всегда заключается главным образом в объе-
динении результатов различных видов, рас, систем, областей биологии, ме-
дицины, оказывается в значительной мере узким, выхолощенным и беспред-
метным.
Мы должны отчетливо себе представлять, что если в вышеизложенной
логической структуре в качестве Р вводится требование отсутствия синони-
мических терминов, таких, как «теория», «гипотеза», «структура» и т. д., то
это требование может быть обосновано лишь соображениями удобства. Зна-
чит тот, кто употребляет термин «модель» иным, «неудачным» образом, де-
лает это тоже из соображений удобства. Во многих случаях, действительно,
удобно называть теорию, гипотезу или эксперимент моделью. Но это долж-
но быть четко определено и обосновано.
Допустим, что мы откажемся от этого принципа и признаем общность
функционирования в качестве основы применения термина «модель». Тем са-
мым мы встанем на другой путь уточнения этого понятия, отойдя на время
от структурно-логического подхода в биомоделировании. Этот другой путь
предполагает анализ всех случаев употребления термина «модель» и выделе-
ние общих для всех этих случаев признаков. По сути дела, это тот путь,
который типичен для образования понятий на основе представлений, толь-
ко здесь, например, исходным материалом являются не представления о
структуре и функциях разных органов и тканей человека и животных, а
понятия и знания о фундаментальных механизмах, уже сформировавшихся
в различных областях биологии и медицины.
В противовес традиционной абстракции изберем иной тип абстрагирова-
ния, когда между исходными элементами устанавливается отношение типа
равенства. Тогда то общее, что существует между предметами, вступившими
в данное отношение, и будет тем содержанием (общим свойством), которое
нами отыскивается и извлекается в качестве элементов биомодели.
При всем различии традиционного типа абстракции и абстрагирования
с помощью отношения типа эквивалентности их результаты должны соот-
ветствовать друг другу. Здесь речь идет не о разных целях, а о разных путях
22
Принципы моделирования
их достижения, использующих корректные и доказательные элементы зна-
ний о фундаментальных основах жизнедеятельности человека и животных.
С помощью указанного метода можно выделить главные признаки биологи-
ческой модели на существующем этапе познания:
/ искомая модель является отображением, представляющим природные
или, например, трансгенные оригиналы, которые сами в свою очередь
могут быть моделью;
/ поскольку как оригинал, так и модель всегда системы, их следует опи-
сывать как упорядоченное в определенном отношении целого или мно-
жества, которое характеризуется указанием на его элементы и их свой-
ства, а также существование между этими элементами и свойствами
отношения;
/ если система состоит из одного элемента, то такую систему следует при-
знать несобственной;
/ модели никогда не охватывают все свойства оригинала, а только те,
которые существенны для того, кто применяет модель;
/ исследователь должен изначально предоставить требования и условия
последующего биомоделирования;
/ модели корректно соответствуют оригиналу, если это соответствие ус-
танавливается для определенных субъектов внутри определенных про-
межутков времени.
Подобный тип абстракции позволяет выделить в качестве общей черты
модели в любом ее понимании то, что ее исследование может в известной
мере заменить исследование прототипа, даже независимо от физической
природы того и другого. На основании метода обобщения выделяют сотни
различных употреблений термина «модель» в научной литературе. Среди них
такие значения, как описание, теория, план, абстракция, теория структуры
и т. д. Но считается, что достаточно десяти значений понятий, коррелятив-
ных понятию модели. Разумное сужение многозначности термина «модель»,
с которым не связано определенное значение, позволяет свести все его зна-
чения к единству. Такое единство достигается с помощью обобщения поня-
тия объекта моделирования, перехода к его качественному пониманию.
Иными словами, то, с чем более удобно работать, то есть то, что легче
увидеть, услышать, запомнить, записать, обработать, передать, наследовать,
с чем легче экспериментировать, и есть модель, а то, относительно чего мы
надеемся получить соответствующую информацию, работая в широком
смысле с моделью, есть объект моделирования или прототип, в нашем слу-
чае — человек.
Антиподом принципов эквивалентности и обобщения является метод ре-
дукции. В этом случае содержание обобщенного понятия накладывается на
параметры одного из исходных, к которому, таким образом, сводятся, реду-
цируются остальные. В качестве такого отношения допустимо рассматри-
Глава 1. Формально-логические аспекты моделирования
23
вать отношение аналогичности, имеющее место между различными случая-
ми употребления термина «модель». Например, скелет собаки (обезьяны,
кошки, мини-свиньи) аналогичен скелету человека, поскольку он несет иден-
тичные опорно-двигательные функции. Это верно. Однако при попытке
построения модели уже на этапе формальной логики в нее будет внесено
столько математических определителей и коэффициентов, что, возможно,
будет утрачен ее внутренний смысл. Поэтому мы должны задать более кон-
кретные условия, чтобы отношение аналогичности было рефлексивно, сим-
метрично и транзитивно, а значит, соответствовало отношениям типа экви-
валентности. Только в этом случае модель станет непосредственно иссле-
дуемым объектом, служащим для получения некоторой информации о про-
тотипе.
Выше мы говорили о методах исключения и обобщения как об отдель-
ных методах. Однако в биомоделировании чаще всего имеет место комбина-
ция обоих методов. Например, можно отбросить отдельные значения функ-
ции, резко отличающиеся от других значений, и обобщить оставшиеся. Тог-
да получим метод обобщения с элементами исключения. Если же отбрасывает-
ся не отдельное более или менее случайное значение, а целый ряд значений
и обобщаются остальные, то можно говорить о методе исключения с элемен-
тами обобщения. Разумеется, может быть и такой случай, когда исключение
и обобщение играют примерно одинаковую роль [646]. Комбинированный
метод — метод исключения с обобщением оставшегося — можно выразить
следующим образом:
A{P){av ..., яй)-1=> а,, ..., ак;
Я (а,., ..., V-1^ Kfll/> •••> fl//>)> ■■•> К1' •••» a'k)V>
C[(a\, ..., a't), ..., (alk, ..., a'k)] -. => av ..., ar
или в сокращенной форме
CBA(P)(av ...,вя)-,=><!,, ..., ar. (2)
В сущности, модель есть система, которая используется, выбирается, со-
здается, описывается, например, в терминах формальной или математичес-
кой логики, символах дифференциальных уравнений или теории вероятно-
сти третьей системой в качестве представителя сложного оригинала. Это
выполняется на основе общности с ним существенных для той или иной
определенной задачи свойств, чтобы сделать возможным или облегчить по-
нимание или овладение знанием функции оригинала, например человека,
или же чтобы его заменить. Оставляя в стороне вопрос о принципиальной
возможности дать точную количественную оценку степени правдоподобия
выводов по аналогии, отметим, что при формировании достоверной и ста-
тистически независимой выборки исследователь всегда сталкивается с опре-
деленными проблемами и трудностями. Эти трудности связаны, прежде все-
24
Принципы моделирования
го, с невозможностью при существующем уровне развития техники логичес-
кого анализа четко отграничить один признак от другого и таким образом
точно определять количество признаков. Поэтому здесь зачастую приходится
довольствоваться качественными характеристиками «много», «мало», «мень-
ше», «больше», «лучше» «хуже». Причина в том, что когда нет полной ин-
формации и точных данных, остается предполагать и строить догадки.
Было бы наивно, например, при планировании эксперимента полагать, что
все предположения сбудутся.
При планировании экспериментов с целью построения биомоделей вы-
вод по аналогии, как ни странно, будет тем более правомерен, чем менее
всеобъемлющим является переносимое качество. Но меньшее количество
свойств, входящих в состав данного качества, означает, что оно будет рас-
пространяться на большее количество объектов, будет менее специфичным.
В этом случае становится более вероятным, что произвольно выбранный
предмет будет обладать данным свойством. Чтобы быть более правдоподобны-
ми, компоненты биомодели должны заключать в себе меньшую информацию.
Это еще раз подчеркивает необходимость оптимизации предела и достаточ-
ности отбора материала при планировании эксперимента.
Соотношение животных и человека как биомоделей
и прототипа
Под биомоделью понимается такая мысленно представляемая или матери-
ально реализованная система жизнедеятельности исследуемых животных
или представителей животного мира, которая, отображая или воспроизво-
дя объект исследования, способна замещать его так, что ее изучение дает
нам новую информацию о человеке и для человека [142, 228, 282]. Матема-
тическая схема или вообще математическое описание также отображает
объект, хотя и очень односторонне [544, 567]. Но они для того и создаются,
чтобы по ним можно было в какой-то мере судить об объекте. То же самое
можно сказать обо всех формальных или формализованных системах, тем
более о тех, где изначально отсутствуют четкие данные в биомедицинских
исследованиях.
Имея в виду отмеченные трудности для уточнения понятия модели, мы
предпочтем метод обобщения. На базе полученного обобщенного понятия
модели и выделенных в процессе обобщения признаков логического ха-
рактера можно сформулировать общие подходы для биомоделей и поставить
вопрос об основах метода биомоделирования.
Совершенно очевидно, что с моделью работать более удобно, чем с прото-
типом. Но это вовсе не означает, что модель всегда замещает прототип [334].
Модель представляет интерес не сама по себе. Результат работы над нею
должен быть перенесен на прототип. Осуществляя эмпирическую интерпре-
Глава 1. Формально-логические аспекты моделирования
25
тацию теоретических терминов, модель дает возможность делать предсказа-
ния. Но это означает, что информация переносится с модели на прототип.
Можно утверждать, что конъюнкция (х)Р63 л (х)Р64 имплицирует =>(х)Р,4.
Понятие биомоделирования предполагает, таким образом, существование
двух объектов — модели и прототипа, т.е. животного и человека. Исследова-
ние одного дает возможность делать выводы о другом. Эти признаки можно
рассматривать в качестве ограничительных условий для используемых умо-
заключений. Логическими основаниями метода биомоделирования мо-
гут служить любые выводы, в которых биологические посылки относятся к
одному объекту, а научное заключение — к другому. Такие выводы представ-
ляют собой класс умозаключений, охватывающих традиционные выводы по
аналогии, позволяющие получить дополнительные знания только за счет
рационального и корректного подхода в процессе экспериментальных или
клинических исследований к построению биологических моделей.
В табл. 1 представлены основные параметры сравнительных характерис-
тик человека и крупных лабораторных животных. Эти данные имеют вполне
конкретный смысл для экспериментатора, но в чистом виде они мало при-
годны для процессов биомоделирования, поскольку, давая определенные
реперные точки для сравнения, не позволяют оценить функциональные свя-
зи модели и прототипа. Поэтому вывод о том, что, например, мозг собаки
больше мозга кошки, вовсе не означает, что функциональные способности
собаки превосходят таковые у кошки.
Таблица 1
Основные сравнительные физиологические показатели человека и крупных
лабораторных животных
Показатель
Масса тела, кг
Площадь поверхности тела, см2
Температура тела, °С
Пульс (уд./мин)
Частота дыхательных
движений
Продолжительность жизни, лет
Человек
70
18 000
37,2 (рект.)
36,6 (кожа)
60-70
20
75
Мини-свинья
37,3
8 750
39
112
15-20
15
и более
Овца
95
9 000
38,5-40
70-80
12-15
9-15
Собака
12
5 820
38,5
70-130
14-26
10-15
Кошка
3
2 466
38-39,5
110-140
10-20
14-20
Масса внутренних органов в процентах от массы тела, %
Головной мозг
Печень
Сердце
Почки
Легкие
Поджелудочная железа
Селезёнка
1,9
2,14-2,43
0,36-0,47
0,4-0,57
0,53
0,1-0,17
0,21-0,28
0,24
1,79
0,3-0,4
0,66
0,56
0,11-0,15
0,13
0,11
0,66-1
0,24-0,25
0,6-0,7
0,43-0,47
0,1-0,18
0,13-0,17
1-2,7
3,0
0,69-1,3
0,5-0,7
1,1-1,66
0,13-0,35
0,08-0,4
0,7-1,1
3,11
0,39
0,34
0,62
0,15-0,28
0,2
26
Принципы моделирования
Практически всегда, когда экспериментатор приступает к планированию
исследующих опытов, направленных на анализ исходных состояний, формиро-
вание гипотез и построение биомоделей, он просто обязан осуществить ин-
дуктивное обоснование по аналогии, с использованием математической или
формальной логики. В подтверждающих экспериментах, когда требуется лишь
ответить на поставленные вопросы, в этом нет необходимости. В этом случае
для установления правомерности вывода по аналогии нет необходимости при-
бегать к общим суждениям типа «все В с D», например «все кошки — хищни-
ки». Объектов, обладающих признаком В, может оказаться бесконечно много
(собаки, хорьки, тигры, львы и т.д.), в то время как модель и прототип могут
быть объединены значительно более узким классом.
В исследующих же экспериментах плодотворное применение вероятно-
стной логики к проблеме выводов по аналогии от одного единичного пред-
мета к другому может иметь место лишь в том случае, если удается предста-
вить единичные явления как массовые.
Когда говорится о переносе признаков с одного объекта на другой, нео-
бязательно имеются в виду два тела или его части, отграниченные в про-
странстве и времени от других тел. Например, можно переносить свойство
правильности от одного умозаключения к другому. Можно проводить ана-
логию и между свойствами, скажем, между свойством кроветворения, фаго-
цитоза и функциями стволовых клеток.
В табл. 2 представлены биохимические показатели крови человека и
животных. Хотя они представлены в виде одиночных показателей, тем не
менее, они характеризуют некую динамическую константу, а не измерен-
ную однократно величину, отражающую параметры того или иного органа.
Каждый из параметров табл. 2 значительно изменяется при воздействии
тех или иных факторов внутренней или внешней среды. Эти показатели
могут быть соотнесены в динамике с другими показателями крови, частоты
пульса или дыхательных движений. Они могут быть прокоррелированы с
изменениями продолжительности жизни (см. табл. 1). Например, при ге-
моррагическом шоке можно установить связь между динамикой изменения
гемоглобина, количеством эритроцитов, частотой пульса и дыхания. По-
этому применение принципа аналогии в этом случае будет оправданным и
корректным.
Если мы приписываем сравниваемым предметам одинаковые линейные
или многомерные свойства, то их уточнение может привести к различным
точечным свойствам [26, 47, 157, 229, 249, 517]. Если же сравниваемые свой-
ства (например, квантового выделения медиаторов) являются точечными с
самого начала, то процедура уточнения является излишней, и утверждение о
тождественности этих свойств у разных животных не будет связано ни с ка-
кими трудностями. В этом случае мы можем приступить к формулировке
правил вывода по аналогии.
Глава 1. Формально-логические аспекты моделирования
27
Таблица 2
Основные сравнительные параметры крови человека и крупных
лабораторных животных
Показатель
Гемоглобин, г/дл
Гематокрит, %
Эритроциты, млн. в мм3
Лейкоциты, тыс. мм3
Лимфоциты, %
Моноциты, %
Нейтрофилы, %
Эозинофилы, %
Базофилы, %
Билирубин, мг/дл
Глюкоза, мг/дл
Креатинин, мг/дл
Тромбоциты, тыс. мм3
Человек
11,5-14,5 (ж)
13,2-16,4(м)
36-42 (ж)
40-48 (м)
3,7-4,7 (ж)
4-5,1(м)
4-8,8
19-37
3-11
48-78
0,5-5
0-1
0,5-1,2
60-100
0,5-1,5
180-320
Мини-свинья
1,6-16,8
43,1
5-8
15-20
18,7-61,7
0,3-4
32-78,7
0,0-6,0
0-1,3
0,2-1,1
80-110
0,5-1,37
200-500
Овца
10-15
32,4
10,03
8-10
47-70
1-5
20-45
2-15
0-1
0,3-1,8
18-57
0,7-1,8
150-250
Собака
11-18
50,4
5,5-8
8-18
13-32
0-0,3
60-82
2-4
0-2
0-0,5
60-87
0,3-1,0
200-600
Кошка
12,9
35-40
7,34
15,1
23
2
65-69
5-7
0-2
2-4
39-61
0,5-1,1
285-500
В качестве общего условия правомерности вывода по аналогии является
требование того, чтобы сопоставляемые друг с другом свойства сравниваемых
предметов были точечными. В тоже время они должны быть множественны-
ми, например, биохимические или физиологические показатели человека и
животных. Для полноты выводов по аналогии целесообразно использование
нескольких видов животных, поскольку ни одно из них не может претендо-
вать на абсолютность биомодели человека.
Таким образом, вероятность индуктивного, а в конечном счете и про-
дуктивного вывода, тем больше, чем длиннее ряд исследованных случаев.
Чем больше этот ряд, тем больше оснований для отождествления вероят-
ности с наблюдаемой относительной частотой. Применительно к аналогии
это будет означать, что вывод тем правомернее, чем больше общих свойств
имеют сравниваемые предметы. Это традиционное правило логики биомо-
делирования.
Важным требованием, предъявляемым к логико-индуктивному выводу,
является требование случайности в подборе предметов, данные о которых
выражены в посылках. Большое внимание этому требованию уделяется в
статистике. Для того чтобы обеспечить случайный характер выборки, ис-
пользуются специальные приемы, например, применяются таблицы случай-
ных чисел. Применительно к аналогии получаем требование, согласно кото-
рому модель и прототип должны сравниваться по любым случайно вы-
бранным свойствам.
28
Принципы моделирования
Теория вероятностей формулирует точное определение понятий статис-
тической независимости и статистической зависимости в рядах чисел: если
переменная величина х принимает значения х 1 х2 х3 ...х, то сумма вероятно-
стей
24, = 1. (3)
Поскольку переменная х имеет определенное значение, то формула оп-
ределяет вероятность функции w, которая является пределом отношения пх,
то есть числа значений х к п. В этом случае п может бесконечно расти
. ,. п(х)
w(x) = hm —— (4)
л-»~ ft
Если в последовательности п выбрать подпоследовательность п\ то
П\Х) WJXj) _П(Х{)
„->» п w'(x2) n(x2) '
(5)
Вероятность некоторого значения х будет изменяться в том же направле-
нии и с той же частотой, что и его появление в последовательности.
Чрезвычайно важной особенностью вероятности живых систем является
абсолютная тенденция перехода от неравновесного состояния к тому равно-
весию, которое характеризуется и описывается функцией Максвелла-Боль-
цмана
N,=C ' (6)
N± = e-hv/rt
'2
где N{ и N2 — например, число электронов на верхнем и нижнем уровне
орбиталей, или распределение гибридных животных в разных поколениях,
или распределение лекарственного препарата в компартмене и его подком-
партменах (предположим, в мозге и нейронах, печени и гепатоцитах) и т.д.
Условие случайности в отборе посылок может быть улучшено. Ситуация
выглядит несколько парадоксальной, поскольку для того, чтобы добиться
действительно случайного характера отбора посылок, меньше всего можно
полагаться на волю случая. Всякое наблюдение происходит в определенных
условиях. Эти условия, не всегда осознаваемые, накладывают определенный
отпечаток на выбор предметов. В статистике широко известен поучитель-
ный пример с выбором «наугад» фамилий из справочника. Оказывается, что
здесь случайности не получится, поскольку чаще открываются именно те
страницы, которые уже открывались и бросаются в глаза, прежде всего, наи-
более длинные фамилии и т. д. Для того чтобы избежать влияния такой си-
туации, в статистике применяется так называемая расслоенная выборка, при
Глава 1. Формально-логические аспекты моделирования
29
которой разбивают исследуемые предметы на классы и берут выборку из
каждого класса.
Сущность этого приема можно понять, как стремление сделать условия,
в которых производятся наблюдения, возможно более разнообразными. Ины-
ми словами, должны быть более разнообразные объекты, данные о которых
фиксируются в посылках. Необходимо, чтобы признаки, общность которых
дана основанием, максимально отличались друг от друга и также были более
разнообразными. Например, для заявления о том, что кошка является лучшей
моделью человека в интерпретации ЭЭГ, необходимо исследовать группу
разных кошек, а также собак, крыс, обезьян и т.д., сравнить их с человеком
и лишь затем построить оптимальную биомодель. Но и это — не заверше-
ние. В гл. 40 мы представим доказательства необходимости использования в
таких случаях нелинейного моделирования.
Другим средством осуществления надлежащего отбора посылок при пла-
нировании экспериментов является исследование наиболее типичных, ха-
рактерных для соответствующего класса объектов. Если число элементов
класса, к которому относится заключение, вообще невелико и нет возмож-
ности обеспечить достаточное разнообразие и многочисленность исследуе-
мых объектов, то берутся наиболее типичные. Нетипичные, не характерные
объекты или явления при этом отбрасываются. Например, при изучении и
моделировании процессов образования и распада гликогена в печени жи-
вотных и человека можно пренебречь особенностями архитектоники пече-
ночных долей и констелляций гепатоцитов.
Существует требование, согласно которому свойства сравниваемых объек-
тов, те, общность которых дана основанием, и те, о которых говорится в
ядре вывода,— должны быть, более однородными, однотипными. Так, нельзя
устанавливать общность между человеком и животным по биологическим
признакам [295, 397, 403, 419, 636] и затем переносить на животных призна-
ки социальные или, наоборот, социальные закономерности человеческого
общества истолковывать в плане биологических законов животного мира,
как это делается в социальном дарвинизме и зоосоциологии.
Степень правдоподобия индуктивного вывода зависит также от того, ка-
ким образом образован тот класс объектов, к которому относится заключе-
ние. Мы имеем, например, класс млекопитающих, в который объединены
животные, обладающие многими существенными общими качествами, но
мы можем также формально объединить людей или животных по первой
букве их фамилий или названий. Результат исследований будет тем более
правомерен, чем в большей мере содержание понятия, охватывающего класс
исследуемых предметов, отражает их сущность.
Для свойств, о которых говорится в посылках, должно быть существенно
то, что они принадлежат именно сравниваемым явлениям. Эти свойства дол-
жны быть специфичными для сравниваемых объектов, а не такими, которые
30
Принципы моделирования
могут быть присущи чему угодно. В этом случае говорится о «силе свойства»
или, например, «генетической мощи» как противоположности его «логичес-
кой ширине». Например, при сравнении человека и животных свойства
способности к размножению, развитию и т. д. логически «уже» и потому
несравненно более важны, чем «обладают массой», «состоят из молекул»
и т. д. [183, 245, 267, 370]. Чем более специфический характер имеет данный
факт, тем менее он вероятен. Значение этого положения становится осо-
бенно ясным в связи с теорией информации. Чем менее вероятен факт, опи-
сываемый данным утверждением, тем большее количество информации оно
в себе содержит [456, 464, 525, 583]. Поэтому рассмотренное условие повы-
шения степени правдоподобия вывода по аналогии, по сути дела, экви-
валентно требованию о том, чтобы посылки содержали в себе возможно боль-
ше информации о сравниваемых объектах.
Функциональные системы и биомоделирование
В качестве примера формально-логического моделирования можно привес-
ти концепцию академика П.К.Анохина. Важным положением концепции
функциональных систем является представление об афферентном синтезе.
Прежние взгляды основывались на простой регуляторной цепи условных
рефлексов: сигнал => ответ. П.К.Анохин убедительно показал, что любому
поступку человека или животного предшествует совокупная взаимооценка
условий данного момента, основанная на прошлом опыте [24, 25].
Рассмотрим это в рамках математической комбинаторики, а именно в
задаче сочетаний с повторениями. Введем один параметр: в сочетание с по-
вторениями обязательно должны входить элементы г < п. Чтобы обеспечить
присутствие элементов r-типов, возьмем по элементу каждого типа. Тогда в
^-сочетании окажутся, заняты г мест. Остальные же к — г мест можно запол-
нить любыми элементами л-типов. Поэтому комбинаций искомого вида
столько же, сколько и сочетаний с повторениями из элементов «-типов,
содержащих по к — г элементов каждое, то есть:
В частности, если п < к и требуется, чтобы в ^-сочетания с повторениями
входил по крайней мере один элемент каждого из л-типов, то получится
C\Z\ комбинаций.
Иными словами, ответная реакция бесконечно разнообразна, вероятно-
стна и зависит от пусковых и обстановочных раздражителей, предпусковой
интеграции и всего предшествующего опыта. Интересно, что экстирпация
лобных долей нарушает процессы афферентного синтеза, о чём далее.
Важной частью этой концепции является представление о наличии кри-
териальных ориентиров сравнения исходных намерений к действию и ожи-
Глава 1. Формально-логические аспекты моделирования
31
даемых результатов на основе специального физиологического механизма —
«акцептора действия». Этот своеобразный аппарат «предвосхищения» явля-
ется конечным результатом афферентного синтеза.
В 1973 году мне посчастливилось достаточно часто и длительно общаться
с Петром Кузьмичом Анохиным, который согласился быть рецензентом моей
докторской диссертации. Это был удивительный по масштабности интел-
лекта, научному темпераменту и продуктивности логики человек. Его тео-
рия функциональных систем является блестящим примером научного миро-
созерцания и методологии науки. Использованные им теории силлогизма и
дедукции вылились в логическое построение, на основе строгих научных
фактов, системы комбинаторных взаимодействий и преобразований одно-
порядковых элементов внутри самой системы.
При фармакологическом анализе и моделировании интрацентральных
отношений головного мозга физиологов и фармакологов уже давно не удов-
летворяет трактовка результатов в рамках влияния веществ на процессы
возбуждения и торможения. Требуется методологически более совершен-
ный принцип оценки результатов. Её дает весьма продуктивная концепция
академика П.К. Анохина [23, 24, 25] и его учеников [22, 614, 617] о функ-
циональной системе и роли афферентного синтеза, в том числе мотива-
ций, обратной афферентации, акцептора результатов действия для осуще-
ствления целенаправленного поведения.
Ранее [328] нами были проведены исследования, построенные таким об-
разом, чтобы для разрешения сильной мотивации (страх, боль, голод), жи-
вотные могли в условиях конфликтных ситуаций с другими животными и
меняющихся условий опыта принимать различные решения для достижения
цели. Оценивая полученные результаты в аспекте теории функциональных
систем, следует отметить, что нейролептики (аминазин, левомепромазин и
этаперазин) и м-холиноблокаторы (атропин, амизил) нарушают мотиваци-
онные механизмы голода, страха и особенно ярости (рис. 1), снижают значи-
мость обстановочной и пусковой афферентации.
Универсальность ряда медиаторов моноаминэргической природы в про-
цессах осуществления интегративной деятельности мозга, вне зависимости
от этапа его филогенетического становления (крысы, кролики, кошки, при-
маты, человек), их способность вмешиваться в механизмы генерации и вза-
имодействия биопотенциалов различных мозговых образований, трансфор-
мацию стохастических и гармонических составляющих ЭГ и ЭКоГ, дают ос-
нование рассматривать их в качестве материального субстрата осуществле-
ния межцентральных взаимоотношений [331]. В этом аспекте центральные
эффекты психостимуляторов, психотомиметиков, антидепрессантов, транк-
вилизаторов, нейролептиков, их способность вмешиваться в синтез, метабо-
лизм и конечные эффекты моноаминов, влияние на баланс медиаторных
средств мозга, позволяют считать психотропные средства регуляторами си-
32
Принципы моделирования
стемной деятельности головного мозга и его высшего звена — коры больших
полушарий [333].
Психостимуляторы (фенамин) и психотомиметики (кокаин, LSD, BZ)
улучшают память, реакцию и пусковые стимулы, но ослабляют пищевые
мотивации. Трициклические антидепрессанты наилучшим образом устраня-
ют мотивационные механизмы страха и ослабляют агрессивное поведение.
Однако такие элементы функциональной системы, как оценка ситуации и
процесс принятия решения, в наибольшей степени подавляются нейролеп-
тиками, м-холино-и а-адреноблокаторами, в меньшей — антидепрессанта-
ми (особенно в малых дозах), но облегчаются психостимуляторами, психо-
томиметиками и антихолинэстеразными веществами (рис. 1).
Активация
Облегчение
Угнетение
Рис. 1. Фармакологический анализ функциональной системы агрессивного поведения кошек
с помощью фенотиазиновых нейролептиков (1), транквилизаторов холиноблокирующего дей-
ствия (2), трициклических антидепрессантов (3), психостимуляторов (4), психотомиметиков
(5), антихолинэстеразных средств (6): ОА — обстановочная афферентация; ПА — пусковая
афферентация; ПД — программа действия; Д — действие; ПРД — параметры результата
действия; АРД — акцептор результата действия
Интересно, что анодическая поляризация лобных зон коры, хвостатого
ядра и медиальных ядер таламуса усугубляет нарушение психотропными ве-
ществами процессов принятия решения. Подобные результаты могут быть
расценены в рамках представлений К.В. Судакова [12, 136, 137] как подтвер-
ждение особой значимости лобных долей и мозговых образований их фун-
кционального сопряжения в интегративных процессах принятия решений.
Фармакологический анализ функциональных систем, пищевого и эмо-
ционального поведения (рис. 1) также выявил исключительную заинтересо-
ванность в эффектах нейротропных и психотропных средств лобных долей
головного мозга, ответственных за интеграцию механизмов принятия реше-
ний, оценку правильности их выполнения и регуляцию интрацентральных
аппаратов их реализации. Наиболее уязвимым для различных нейро- и пси-
хотропных средств в интрацентральных системах афферентного синтеза яв-
ляется процесс принятия решения к действию. К такому выводу приходят и
Глава 1. Формально-логические аспекты моделирования
33
другие исследователи, которые объясняют этот феномен отсутствием врож-
денных или приобретенных программ принятия решения в процессе фило-
или онтогенеза организмов.
Нейролептики и центральные холиноблокаторы помимо нарушения ме-
ханизмов принятия решения, ослабляют мотивационные механизмы реак-
ций голода, страха, ярости (рис. 1). Антидепрессанты превосходят все рас-
смотренные вещества по способности тормозить мотивационные механиз-
мы страха. Особое место занимают психостимуляторы, психотомиметики
(малые дозы) и антихолинэстеразные средства, обладающие способностью
улучшать процессы принятия решения и облегчать реализацию нужного
действия [328, 332, 337].
Концепция П.К.Анохина вскрывает два прежде мало рассматривавшихся
направления исследования функций головного мозга как интегративного цен-
тра: устойчивости нелинейных регулируемых систем в целом и стабильности
вероятностно-детерминированных подсистем целостного организма челове-
ка и животных при неограниченных возмущающих воздействиях, к рассмот-
рению которых мы сейчас перейдем.
К.В. Судаков указывает [613—616], что объединение корково-подкорковых
аппаратов в доминирующее возбуждение связано с определенной функцио-
нальной системой и направлено на обеспечение результата ее деятельности.
Например, агрессивное поведение, вызванное искусственным возбуждением
гипоталамуса, можно модулировать одновременным возбуждением, времен-
ным выключением или разрушением перегородки, гиппокампа, миндалин.
Динамика, управление и устойчивость в живых системах
Любые живые организмы имеют характерную особенность приспособления
и адаптации к окружающей среде. Приспособление обеспечивает выработку
строго определенных типов контакта со средой. Все ненужные контакты ис-
ключаются.
Эволюция биосистем обеспечивает своеобразный изоляционизм, т.е. обес-
печение регулируемыми потоками энергии, информации, питания, связи,
направленными на сохранение и поддержание внутреннего баланса энергии
и энтропии организма.
Подобные свойства характерны только для динамических структур, спо-
собных обеспечивать устойчивость пространственно-кинетической стабиль-
ности. В этом заключается глубокий биологический смысл функционально-
го обеспечения распределения и управления внутри организма и с внешней
средой. У.Р. Эшби подчеркивал, что, несмотря на воздействия многочислен-
ных возмущающих факторов, организм не допускает отклонений от некоего
заданного природой оптимума. Это и есть выражение параметрической изо-
ляции и автономии организма. Оно достигается за счет использования сие-
34
Принципы моделирования
тем управления и устойчивости на разных морфо-функциональных уровнях
биосистем. Поэтому основным принципом анализа биологических объектов
является выяснение причин и условий развития динамических структур, уп-
равления и обеспечения устойчивости.
Многочисленные попытки описания принципов управления или регу-
лирования в живых системах до настоящего времени не увенчались успе-
хом. Они сводились к абстрактно-логическому описанию этих процессов,
или заканчивались попытками использования традиционных основ теории
автоматического регулирования линейных систем. Системы автоматичес-
кого регулирования представляют собой в большинстве случаев весьма слож-
ные устройства, состоящие из объекта регулирования и регулятора. Назна-
чение регулятора сводится к тому, чтобы непрерывно поддерживать в объекте
регулирования некоторое установившееся состояние или же состояние, из-
меняющееся по заданному закону. Следовательно, процесс регулирования
заключается в том, что регулятор препятствует всяким отклонениям от это-
го состояния, возникающим в объекте регулирования в результате каких-
либо нарушений его работы. С середины XX века интенсивно развиваются
исследования по развитию классических математических методов в систе-
мах автоматического регулирования в нелинейных системах на основе уче-
ния А.М.Ляпунова [437].
Одной из основных задач теории автоматического регулирования являет-
ся изучение протекания процесса регулирования во времени. Оно проводит-
ся средствами математического анализа на следующей основе. Каждой сис-
теме автоматического регулирования соответствует определенная система
дифференциальных уравнений вида:
—j*- = Xk(xl,...,xn)(k = l,...,n). (8)
Здесь х{, ..., хп — переменные, описывающие состояние системы, а Хк —
неизвестные функции этих переменных, определенные в некоторой фикси-
рованной области G пространства переменных хх, ..., хп . Это пространство
называют фазовым пространством *. Мы будем обозначать его через Е .
В пространстве Еп уравнения (8) определяют компоненты Хк вектора ско-
рости v движения некоторой точки М, называемой изображающей точкой. С
физической точки зрения уравнение (8) следует рассматривать как матема-
тическую форму записи тех законов, которым подчиняются регулируемые
системы. Свойства и особенности этих законов полностью либо приближен-
но, но с достаточной точностью отражаются характером функций Хк (xv ..., хп).
Область G определения функций Хк{хх, ..., хп) есть та часть пространства Е ,
на которую распространяется действие указанных физических законов.
Пусть величины х10, ..., хп0 обозначают начальные значения переменных
jtj, ..., хп. Они однозначно определяют исходное состояние системы регули-
Глава 1. Формально-логические аспекты моделирования
35
рования при t = 0. Каждой системе начальных значений х10, ..., хп0 соответ-
ствует решение
Xk = Xl№> *10' -' Хяо)' (9)
уравнений (8). Предполагается, что такое решение существует при всех зна-
чениях времени / > 0 и является единственным.
В дальнейшем удобно иметь дело с уравнениями, полученными из (8)
путем преобразования переменных по формулам:
Xk = X*k+Yk(k = \,...,n). (10)
По терминологии Ляпунова, уравнения, полученные после такого преоб-
разования, называются уравнениями возмущенного движения. Они имеют вид:
-± = Yk(k = l,...,n), (П)
где:
Гк<У\> •••' Уп) = Ч*\ + У» •••» х\ + Уп№ = Ь ■■•> п). (12)
Формула (10) определяет преобразование переноса начала координат в
точку с координатами х{*, ..., хп*. Вследствие этого очевидному решению
уравнений (8) соответствует очевидное решение
/,=0, ...,/„ = 0 (13)
уравнения (10). По терминологии Ляпунова, это решение следует называть
невозмущенным движением регулируемой системы.
Пусть при t = 0 переменные ylt ..., уп принимают начальные значения ylQt
..., уп0, из которых, по крайней мере, одно не равно нулю; эти значения
называются возмущениями. Каждой заданной системе таких возмущений
отвечает однозначное и непрерывное решение
Ук = Ук<У\о> -' ^о1) (* = L •••> *) (14>
Это решение называется возмущенным движением системы автоматическо-
го регулирования. Если бы мы знали все решения (14), то нам были бы
известны все возмущения движения системы. Но в общем случае получить
эти решения практически не представляется возможным, что усложняет за-
дачу о рациональном выборе параметров регулятора. Поэтому здесь нужно
качественное исследование, позволяющее обозреть все семейство возмущен-
ных движений (14) и, не прибегая к интегрированию, выяснить, стремятся
ли эти движения при / => «> к невозмущенному движению (13) независимо
от начальных значений ^10, ..., уп0.
Учение Ляпунова [437] об устойчивости движения позволяет судить об
интересующих нас свойствах возмущенных движений, не прибегая, в конеч-
ном счете, к интегрированию уравнений (13), и указывает путь к рациональ-
ному конструированию регуляторов.
36
Принципы моделирования
Если окажется, что при определенной настройке регулятора решение (13)
будет устойчивым, то это будет означать, что регулируемая система сама, без
постороннего вмешательства, изберет тот режим невозмущенного движения,
который этому решению соответствует. Если же решение (13) будет неустой-
чивым, то такой установившийся режим окажется физически невозможным.
Таким образом, определение устойчивости движения (13), данное A.M.
Ляпуновым, получает большое прикладное значение при изучении важных
вопросов современной техники [437].
Ранее нами было доказано, что нейролептики обладают способностью
повышать управляющие и сдерживающие влияния миндалевидного комп-
лекса в отношении заднего гипоталамуса и ростромедиальных ядер таламу-
са в отношении ретикулярной формации среднего мозга. Эти регулирую-
щие воздействия, в соответствии с теоремой А.М.Ляпунова, отображаются
следующим соотношением:
\/Р
Уп =
x{dyn/dt) + \
Уо- (15)
В уравнении (15) \/Р является показателем передаточной функции сис-
темы, косвенно отображающим уровень возбудимости анализируемой струк-
туры, уп — выходная характеристика системы, у0 — входной сигнал.
Экспериментальная проверка полученных соотношений позволила в даль-
нейшем подтвердить, что трифтазин, этаперазин, левомепромазин и пропа-
зин в широком диапазоне доз (0,5—10 мг/кг) обеспечивают возрастание пе-
редаточной функции в направлении миндалины —> гипоталамус и ее сниже-
ние в направлении гипоталамус —> миндалины. Антидепрессанты не изме-
няют передаточной функции системы миндалины —> гипоталамус, резко
повышая ее в направлении гипоталамус —> миндалины.
На основании многочисленных исследований в эксперименте и в клини-
ческих исследованиях нами были обнаружены эффекты психорегулирующе-
го действия в лимбической системе лекарственных препаратов, не относя-
щихся к психотропным средствам (пропранолол, оротат калия и др.) В даль-
нейшем эти препараты вошли в клинику как средства комплексной терапии
острых делириозных состояний. С другой стороны это дало импульс для
работы с химиками-синтетиками по направленному конструированию пси-
хотропных средств на основе пиримидинов. Некоторые из них нашли свое
место в клинической практике.
Подобные достаточно простые формально-логические построения позво-
ляют по-иному осмысливать получаемые экспериментальные данные и вы-
страивать их не в линеечку выводов от 1 до °°, а на основе комбинаторных
преобразований оптимизировать требования к строгости научных результа-
тов. В конечном итоге, единство содержания научных знаний, как и их раз-
личие — есть суть метода моделирования.
37
Глава 2 БИОСИСТЕМЫ
И БИОМОДЕЛИРОВАНИЕ
Биологические системы представляют собой завер-
шение определенного этапа эволюции материи. Ес-
тественно, что биосистемы вобрали в себя всю слож-
ность предбиологических форм развития материи.
В предыдущей главе мы определили, что особенно-
стью любых биосистем является устойчивость ди-
намических структур, определяемая пространствен-
но-кинетическими факторами. Исходя из этого,
можно постулировать, что функция распределения
и явления непрерывности для этой функции опре-
деляют динамичность биологических систем, начи-
ная от субмолекулярного уровня и до организма в
целом. Мы также должны осознавать, что общеби-
ологические законы роста и развития запрещают
состояние покоя. В гл. 35 мы подробнее рассмот-
рим характеристики так называемого «физиологи-
ческого времени». Здесь подчеркнем лишь, что лю-
бые отображения развития биосистем могут быть
описаны исключительно в четырехмерном про-
странстве, в котором четвертым вектором является
течение времени (т).
Биологические системы бессмысленно рассмат-
ривать в качестве закрытых или изолированных, они
по самой сути являются системами открытого типа.
Для большей убедительности рассмотрим некото-
рые свойства закрытых систем. С этой целью вве-
дем некоторые понятия, характеризующие их. Пусть
6 является эквивалентом энергетической составля-
38
Принципы моделирования
ющей (движение, тепло, увеличение температуры, информации и т.д.), т.е.
показателем процесса выделения энергии или информации и др. Символ ¥
будет означать рассеивание энергии, вещества, информации, то есть отобра-
жать явление неопределенности или энтропии. В таком случае в закрытой
системе со стабилизированными или однородными значениями 8 можно
определить такую функцию состояния ¥ , что для всех обратимых процес-
сов может быть математически обозначено как
d¥=*® , (16)
е
а для необратимых процессов и состояний эта функция выразится как
8
В этом случае соотношение можно представить как разность между ле-
вой и правой частями неравенства (17)
№-*ж, (is)
8 8
где dQ является некомпенсированным значением 8. Некомпенсированность
dQ отражает необратимость функциональных процессов внутри биосисте-
мы, a dQ характеризует энергетический обмен с окружающей средой. По-
скольку само понятие энтропии биологической системы отображает её пе-
ренос из внутренней среды во внешнюю, предьщущие уравнения могут быть
представлены как
d¥=de¥+di¥ (19)
где индекс е относится к перенесенной энтропии, а / — к энтропии, возник-
шей в самой системе.
Биологическая система имеет много уровней иерархии. На каждом этапе
иерархии можно выделить свои системы, обладающие определенной функ-
цией. Каждый элемент иерархии — это, в свою очередь, подсистема ниже-
лежащего уровня со своей функцией. На уровне современных знаний можно
условно выделить уровни иерархии с отдельными структурными элементами
и определением их функциональной роли (табл. 3).
Экстраполяционные возможности биомоделирования
Экстраполяция полученных результатов на человека — обязательный, слож-
ный и неоднозначный этап любого экспериментального моделирования. Наи-
более полно принципы экстраполяции разработаны в области оценки ост-
рой и хронической токсичности веществ для человека на моделях первого
Глава 2. Биосистемы и биомоделирование
39
порядка лабораторных животных-млеко питающих. Используется либо пря-
мой перенос данных с животных на человека, либо вводятся корректирую-
щие коэффициенты. Лабораторные животные — это классические биологи-
ческие модели, реакции которых на действие веществ во многом подобны
эффектам этих же веществ у человека [335, 337]. Критериями подобия, обус-
ловливающими адекватность моделирования и надежность экстраполяции,
являются: сходство у человека и экспериментальной модели биологических
параметров систем, реагирующих на вещество; общность характеристик ме-
таболических процессов; близость чувствительности, т.е. значении количе-
ственных показателей токсикометрии, установленных для человека и моде-
ли [388]. В настоящее время в исследованиях оценки токсичности веществ
все чаще применяются альтернативные модели второго порядка (различные
гидробионты, бактерии, ферменты, культуры клеток и др.). При этом ре-
зультаты непосредственно переносятся с тест-объектов на человека, что, как
правило, не сопровождается детальным анализом и доказательством право-
мерности такого подхода [546, 613, 627].
Следует с большой осторожностью подходить к оценке любых исследо-
ваний, но особенно острой и хронической токсичности для человека по ре-
зультатам альтернативного моделирования. Так, например, в противополож-
ность альтернативным моделям второго порядка, на лабораторных живот-
ных и, в особенности на мини-свиньях хорошо моделируется алкогольная
интоксикация. Исследования, проведенные в лаборатории НЦБМТ РАМН,
а также данные других авторов показали, что молодняк миниатюрных сви-
ней без принуждения может потреблять большое количество этанола в ком-
бинации с кормом. Оптимальной концентрацией этанола, когда около 50%
животных охотно его принимают, является 25% раствор из расчета 2,5—3,0 г
на кг массы тела. Моделируя алкоголизм на животных, следует понимать,
что идеальной модели алкоголизма практически нет. Тем не менее, мини-
свиньи дают возможность получить вполне адекватную модель для изучения
определенных аспектов алкоголизма: толерантности, симптомов отмены,
органную патологию или эндокринные и иммунные нарушения. При всех
случаях обязательным условием является попадание алкоголя в организм в
достаточном количестве для достижения фармакологически эффективных
его концентраций в крови.
Как показали исследования, выполненные на мини-свиньях [162], внут-
рижелудочное вливание этанола в дозе 1 г/кг массы тела через 2,5 ч после
его введения приводило к максимуму его содержания в плазме крови, соста-
вившему 160 мг на 100 мл плазмы. Исследования в лаборатории биомодели-
рования НЦБМТ РАМН выявили, что уже через 1,5—2 ч после потребления
свиньями 25% этанола средняя его концентрация в плазме крови составляет
100,7—111,6 мг/%, в то время как у контрольных животных, не потребляв-
ших этанол, она составляет 1,9—2,1 мг/%.
40
Принципы моделирования
Таблица 3
Место биологических и математических моделей и моделей более высокого
порядка в иерархии живых систем
а
порядк
3-го
щели
2
ские
атематиче
и «интуитивные» м
веские
Класси*
го
оких
Модели 4-
более вые
порядко
ф
2
животк
>рные
оратс
vo
и
с;
1
п
го порядк
вание 1-
моделиро
'о порядка
S
моде]
0
S
>тернативные б
1
U
ческое
1ТЭЮЩИ
2 i
о о
0 Ф
s с;
Ш 2
Иерархии
биомоделирования
Атомарный
Молекулярный
Аминокислоты
ДНК и РНК
Белок
Органеллы,
митохондрии
Рибосомы
Хромосомы
Клетка
Органы
Системы органов
и тканей организма
Анализаторные
системы
Мозг в системе ЦНС
Организм
Человек
Детерминанты
биомоделирования
Протоны, электроны
и другие элементарные
частицы
Ионы ±, в том числе их
эквивалентность
Пурины, пиримидины,
азотистые основания
А-Г, Т-Ц и А-Г, Ц-У,
основы геномики
Аминокислоты, пептиды,
основы протеомики
Ферменты, макроэрги
РНК, пептиды, белок
ДНК, белок
Органеллы (митохонд-
рии, хромосомы, рибо-
сомы, лизосомы и др.)
и их импликации
Клетки, межклеточные
субстанции в конъюнкции
и дизъюнкции
Констелляция внутренних
органов, их кванорог все-
общности и существова-
ния
Отделы анализаторов
в «автономном режиме»
Анализаторные системы
в интегративном и син-
тезирующем режиме
ЦНС, внутренняя среда,
взаимоотношения
с внешней средой, орга-
ны чувств, конечности
Разум, мышление,
подсознание, сознание,
исследование операций
и принятие глобальных
решений
Задачи, цели и функции
биомоделирования
Донорно-акцепторные отно-
шения электронов, протонов,
иных частиц, ионообразование
Химические свойства кислот,
оснований, щелочей
Способность образовывать
ДНК, РНК, пептидные связи,
;белок
Генофонд, репликация, коди-
рование и перекодирование
в белках
Ферменты, катализаторы,
энергоперенос
Накопление и перераспреде-
ление энергии
Синтез белка, пептидов
Хранение кода клетки и всех
белков
Деление, генерация потенциа
лов, действия, продукция
гормонов, транспорт кислоро-
да и др.
Выполнение механической
работы, газообмен, регуляция
эндокринных функций и др.
Стабилизация параметров
внутренней среды и межорган-
ных отношений, обеспечение
жизнедеятельности организма
Анализ и синтез частных
и обобщенных признаков
в модели образов
Управление взаимодействием
с внешней и во внутренней
среде, мышление, рассудок
Адаптация к природным
экологическим системам
Обитание в естественных
и созданных экосистемах,
проживание и управление
в общественных отношениях
и социальной сфере
Глава 2. Биосистемы и биомоделирование
41
Морфологические исследования некоторых органов этих животных (пе-
чень, сердце, половые железы), проведенные в процессе выработки алко-
гольной мотивации и изучении токсических эффектов этанола, показало,
что через 2—3 года после потребления этанола у взрослых животных и их
потомства выявляются патологические изменения во внутренних органах,
характеризующиеся изменением мембранных систем клеток с преимуще-
ственным поражением митохондрий, накоплением в клетках липидных гра-
нул, развитием внутри- и внеклеточного отека, нарушением клеточных вза-
имодействий, развитием тотального или парциального некроза клеток [453,
454, 455]. Характерной особенностью мини-свиней, потребляющих этанол,
является их выраженное ожирение. Вряд ли подобные данные могли бы быть
получены на любой другой, а тем более альтернативной модели.
В пользу использования свиньи в качестве экспериментального живот-
ного складывается мнение ученых и при изучении лекарственных веществ,
Помимо апробации антиалкогольных и антинаркотических препаратов, на
мини-свиньях, возможно проводить доклиническое испытание фармаколо-
гических препаратов [56], изучать токсичность целого ряда соединений, со-
держащих ртуть [438], свинец [58], афлатоксины [59], каменноугольные смо-
лы [60] и др. Мини-свинья может быть использована для изучения веществ,
загрязняющих воздух и отрицательно влияющих на здоровье животных или
человека [433]. Конечно могут быть использованы и другие лабораторные
животные, но трудно согласиться с тем, что реальной альтернативой могут
стать биомодели второго порядка.
В табл. 3 представлены основные уровни иерархии биомоделирования,
их детерминанты или структурные элементы, а также задачи, цели и функ-
ции объектов биомоделирования. Отметим, что ни один из этих подходов не
претендует на всеобьемлемость, не охватывает все стороны прототипа, в
нашем случае, человека. Биомодели первого порядка наиболее полно охваты-
вают собственно биологическую сферу человека, не претендуя на социальную
составляющую, то есть разум, подсознание, сознание, а также производные
этого вида деятельности homo sapiens.
Биологические модели второго порядка могут с большей или меньшей
полнотой отобразить уровни от молекулярного до клеточного (максималь-
но). Модели третьего порядка предусматривают построение математических
конструкций на основе биомоделей первого, второго порядка или аксиома-
тическое или теоретическое представление концептуальных систем, требую-
щих в дальнейшем подкрепления и четкого экспериментального обоснова-
ния тех или иных медико-биологических процессов.
Многие авторы используют такой аргумент при экстраполяции данных
биотестирования биомоделей второго порядка на человека, как высокая чув-
ствительность биотест-объектов к веществам. Однако Г.Н.Красовский счи-
тает [387], что результаты определения различий параметров токсичности
42
Принципы моделирования
веществ для моделей первого и второго порядков заставляют усомниться в
истинности утверждения о неизменно высокой чувствительности биотести-
рования [646]. Если сопоставить эффективные концентрации ЕС50, или ле-
тальные концентрации LC50 для биотест-объектов с максимально недейству-
ющими концентрациями (МНК) или ПДК веществ по токсикологическому
признаку вредности для человека, а затем определить персентили получен-
ных различий чувствительности биотест-объектов и человека к веществам,
то оказывается, что 50-й процентиль величин этих различий для светящихся
бактерий, рыб гуппи, инфузорий тетрахимена, спермы быка, дафний и т.д.,
свидетельствует о низкой чувствительности перечисленных альтернативных
моделей, по сравнению с человеком [419, 420]. Модели второго порядка часто
не реагируют именно на чрезвычайно токсичные и кумулятивные для чело-
века вещества в высоких концентрациях. Не лучшие результаты дает и тес-
тирование токсичности на клеточных культурах и водорослях: например, для
культуры клеток HeLa безопасны до 1000 МНК кадмия, водорослей сцене-
десмус до 1700 МНК бора.
Информативность биотестирования можно было бы повысить, опреде-
ляя токсичность одновременно на нескольких тест-организмах [310, 313, 416,
428]. Однако, принимая во внимание весьма низкую (по сравнению с чело-
веком) чувствительность почти всех биотест-объектов к подавляющему боль-
шинству веществ в экспресс-экспериментах, можно с уверенностью предпо-
ложить, что использование «батареи» биотестов не решит проблем надежно-
сти альтернативного моделирования токсичности веществ для человека, а
значит такая биомодель мало соответствует прототипу.
Различия в уровнях реагирования моделей первого порядка — лабора-
торных животных и альтернативных им моделей — биотест-объектов к ин-
дивидуальным веществам не позволяют получить адекватные результаты по
оценке комбинированного действия суммы этих веществ методами биотес-
тирования. Для абсолютного большинства лекарств и ксенобиотиков пря-
мая экстраполяция данных с биотестов на человека невозможна [125, 565].
Касаясь субмолекулярных, межатомарных и атомарных уровней иерар-
хии, следует, по-видимому, выделить самостоятельный четвертый уровень мо-
делирования. Этот вид моделирования не входит в прямую ни в первый, ни
во второй уровень. Теоретически его можно описать в терминах третьего
математического уровня, но интерпретация результатов требует новой сис-
темы координат и понятий. Подробнее мы рассмотрим это в гл. 39.
Таким образом, в медико-биологических и, что наиболее наглядно, в фар-
макологических и токсикологических исследованиях используются 3 вида
моделей. Биомодели первого порядка включают лабораторных животных-мле-
копитающих. Биомодели второго порядка включают альтернативные модели
(бактерии, вирусы, ферменты, различные одноклеточные организмы, куль-
туры клеток, гидробионты и др.). Биомодели третьего порядка — математи-
Глава 2. Биосистемы и биомоделирование
43
ческие модели, теоремы, аксиомы, описание процессов и функций в диффе-
ренциальных уравнениях, в понятиях теории вероятности и т. д. Все чаще
встречаются предложения об использовании моделей четвертого порядка и
более высоких порядков для описания взаимодействия неспаренных элект-
ронов, квантово-химических, микроволновых процессов, синглетных и трип-
летных отношений и т.д.
Важнейшим свойством сложных систем, к которым, вне всякого сомне-
ния, относятся биосистемы, является их структурная и функциональная нео-
днородность и многообразие. Они, в свою очередь, связаны многомерностью,
проявляющейся в большом количестве разнородных параметров, в много-
образии связей между однородными и разнородными параметрами, харак-
теризующими работу данной биосистемы. Следующей особенностью био-
систем является динамичность их взаимодействия со средой. Можно услов-
но выделить оперативную динамичность, которая проявляется в сложноор-
ганизованных реакциях на изменения и воздействия окружающей среды,
например, поведение. Существуют также онтогенетическая и филогенети-
ческая динамичность. В этих случаях мы сталкиваемся с динамикой мед-
ленных и сверхмедленных систем в структурно-функциональных измене-
ниях биосистем, например, в случае получения инбредных линий лабора-
торных животных со случайными или заданными свойствами. Достижение
высокого уровня, качества и соответствия экстраполяции в отношении че-
ловека должно строится на моделях различных порядков и использовании
инбредных моделей.
Структурная дискретность и функциональная непрерывность
Качественная неоднородность биосистем и биомоделей проявляется в тех слу-
чаях, когда работают подсистемы с различными управляющими химически-
ми, физическими, информационными и другими сигналами. Если в дости-
жении одного и того же результата подсистемы с разными постоянными
времени взаимодействуют медленные (биохимические, гормональные) или
быстрые (нервная, речевая, мыслительная) подсистемы, то создается времен-
ная неоднородность биосистем. С этими элементами непосредственно свя-
зана иерархичность биосистем, которая проявляется в постепенном услож-
нении функции на одном уровне иерархии и скачкообразном переходе к
качественно другой функции на следующем уровне иерархии. Так иерархич-
ность проявляется: в популяционных сообществах, работе нейронных ан-
самблей, взаимодействии различных органов и систем организма. Это зна-
чительно усложняет процесс биомоделирования.
Структурно-функциональная организованность и стохастичность опре-
деляют высокую устойчивость биологического вида, его формы, оптималь-
ное взаимодействие с внешней средой и, в конечном итоге, в способности
44
Принципы моделирования
биосистемы с помощью различных структурных реализаций добиваться од-
нотипного функционального результата.
Биосистемам присуще свойство нестационарности, проявляющееся в из-
менении во времени распределения реакций на один и тот же набор сигна-
лов внешней среды или смежных иерархических уровней. В то же время
сложность анализа биосистем определяется присутствием в них двух, каза-
лось бы, взаимоисключающих свойств, а именно структурная дискретность
и функциональная непрерывность.
В табл. 4 даны сравнительные морфофизиологические показатели челове-
ка и грызунов-биомоделей. Из нее можно извлечь достаточно полную инфор-
мацию об интересующих показателях, но эта таблица является и хорошей ил-
люстрацией структурной дискретности основных параметров прототипа и био-
моделей.
Таблица 4
Основные сравнительные физиологические показатели человека и лабораторных
животных — грызунов (в третьей сверху строке даны значения ректальной
температуры, у человека температура кожных покровов от 36,6 ° С и ниже)
Показатель
Масса тела, кг
Площадь поверхности
тела, см2
Температура тела, °С
Пульс, уд./мин
Частота дыхательных
движений
Продолжительность
жизни, лет
Человек
70
18 000
37,2
60-70
20
75
Кролик
4,5
3700
38,5-39,5
150-300
20-50
5-7
Морская
свинка
1,25
1500
37-39
300
100-150
4-8
Крыса
0,3
455
37-38
300-500
85
2-3
Мышь
0,05
152,5
38,5-39,3
310-840
200
2
Хомяк
0,122
366
37,5-38,5
280-412
33-127
2-3
Масса внутренних органов (в процентах от массы тела)
Головной мозг
Печень
Сердце
Почки
Легкие
Поджелудочная
железа
Селезенка
1,9
2,14-2,43
0,36-9,47
0,4-0,57
0,53
0,21-0,28
0,4-0,66
4-4,5
0,247
0,6-9,7
0,33-0,38
0,1-0,14
0,05
0,23
1,5
0,18
0,3
0,3
0,1
0,03
0,87
2,17
0,34
0,23
0,45
0,16
0,45
0,54
2,26
0,2
0,54
0,54
0,3
0,38
0,71
4,64
0,44
0,84
1,31
0,38
0,19
В табл. 5 даны параметры биохимических показателей крови человека
и животных. Как и в табл. 4, они отображают, казалось бы, точечное значе-
ние, но на самом деле в этих показателях отсутствует четвертая составляю-
щая, то есть динамика по времени. В реальной ситуации эти показатели
чрезвычайно динамичны и выражают функциональную непрерывность про-
Глава 2. Биосистемы и биомоделирование
45
цессов возникновения, развития и гибели форменных элементов крови и
исключительную подвижность изменений гемоглобина, глюкозы, креатини-
на и т.д. Иными словами, в указанных двух таблицах прослеживаются аспек-
ты неоднородности параметров биомоделей в их структурной консерватив-
ности, дискретности и лабильности функциональной непрерывности.
Таблица 5
Основные сравнительные параметры крови человека и лабораторных
животных-грызунов
Показатель
Гемоглобин, г/дл
Гематокрит, %
Эритроциты, млн/мм3
Лейкоциты, тыс./мм3
Лимфоциты, %
Моноциты, %
Нейтрофилы, %
Эозинофилы, %
Базофилы, %
Билирубин, мг/дл
Глюкоза, мг/дл
Креатинин, мг/дл
Тромбоциты, тыс./мм3
Человек
11,5-14,5 (ж)
13,2-16,4(м)
36-42 (ж)
40-48 (м)
3,7-4,7 (ж)
4-5,1(м)
4-8,8
19-37
3-11
48-78
0,5-5
0-1
0,5-1,2
60-100
0,5-1,5
180-320
Кролик
10-15
35-45
4-7
8-10
40-70
5-9
38-54
0,5-3,5
3-6
0,3
130
1,6
126-300
до 1 млн
Морская
свинка
11-15
40-50
4,5-7
5-12
45-80
8-12
20-40,4
1-5
1-2
0,24-0,3
50-120
0,4-0,7
54-100
Крыса
14,8
40-50
7,2-9,6
8-14
65-77
0-4
13-30
0-1
0
0,42
50-115
0,43
430-
до 1 млн
Мышь
12-14,9
42-44
8,7-10,5
5,1-11,6
63-75
0,7-2,6
6,7-37,2
0,9-3,8
0-1,5
0,18-0,54
108-192
0,5
100-400
Хомяк
16
40-50
7,5
7,6
75
3
30
1
1
0.5
120
0,5-0,6
336-587
Экспериментатор, работая с точечными параметрами, должен четко оп-
ределять для себя к какому объекту или процессу относятся значения диск-
ретности, ибо именно это и определяет важнейшие свойства живой материи
— различие биосистем и вариабельность количественных параметров в пре-
делах одной и той же дискретности. В этом случае, нельзя не согласиться с
определением Уолтера Р. Эшби, который считает, что сложность системы
характеризуется ее разнообразием, т.е. количеством состояний, которое мо-
жет принимать система. Число состояний сложной системы может быть
чрезвычайно велико. Поэтому для количественной оценки сложности сис-
темы используют не само число ее состояний, а логарифм этого числа. Та-
ким образом, если число состояний равно п, то сложность системы опреде-
ляется формулой
*т = lOg II. (20)
Такое обозначение сложности удобно еще и тем, что позволяет связать ее
с организацией системы. Однако оценка сложности относительно числа со-
46
Принципы моделирования
стояний системы не показывает, в каком из возможных состояний, система
находится и в какое она перейдет в следующий момент времени.
В соответствии с достаточно условным делением биосистем на детерми-
нированные и вероятностные можно различать математические модели этих
систем. Деление математических моделей по свойствам биосистем требует
использования специализированных математических методов.
Для описания детерминированных биосистем используются алгебраичес-
кие методы, которые применяются для аппроксимации экспериментальных
результатов, а также факторный и регрессионный анализы. С помощью ал-
гебраических методов можно решить задачу построения только функцио-
нальных моделей биосистем. Дифференциальные и интегральные уравнения
используются при математическом моделировании динамики биосистем, а
также для функциональных и структурно-функциональных моделей. При-
менение этих методов создает принципиальную трудность при идентифика-
ции параметров модели по экспериментальным результатам. Методы теории
управления позволяют построить более оптимизированные математические
модели, приближающиеся к теории биосистем.
Описание вероятностных биосистем можно провести с помощью теории
вероятностей, которая позволяет организовать экспериментальный материал,
соотнести элементы детерминированности и вероятности в работе изучаемой
биосистемы. Использование теории информации позволяет оценить компо-
ненты детерминизма и индетерминизма биосистем и построить математичес-
кие модели взаимодействия биосистем между собой и с окружающей средой.
На основе теорий автоматов и распознавания образов возможно построение
математических моделей адаптации биосистем, решения ими задач классифи-
кации и приспособления к изменяющейся окружающей среде.
Для выбора класса модели необходимо квалифицированное планирова-
ние, статистическая обработка данных эксперимента, определение степени
сложности и организации системы, входных и выходных переменных. Об
этом будет сказано в дальнейшем изложении. В зависимости от уровня орга-
низации объекта выбирается класс математической модели: линейная, нели-
нейная, детерминированная, вероятностная. В выбранном классе определя-
ется вид модели. Для перехода от класса модели к ее прототипу может ис-
пользоваться математический аппарат нелинейных моделей: полиномиаль-
ные, нелинейные дифференциальные уравнения и т. п.
Для определения параметров модели используются данные опытов. Кри-
терием наилучших значений параметров модели может быть минимум сред-
ней квадратичной ошибки или минимум интеграла от квадрата ошибки.
Для определения параметров модели, удовлетворяющих данному крите-
рию, составляется и решается система уравнений, получаемая из частных
производных под интегральной функции критерия по параметрам модели,
или осуществляется процедура спуска в пространстве параметров модели.
Глава 2. Биосистемы и биомоделирование
47
В настоящее время имеются готовые пакеты компьютерных программ, по-
зволяющих врачу и биологу, даже без знания математических основ моде-
лирования, рассчитать и построить корректную математическую модель
для каждого интересующего исследователя случая.
Энтропия и необратимость жизни
При неопределенности или энтропии дискретных величин р обозначим об-
щее число состояний величиной п.
Чтобы установить, в каком из состояний система находится, и в какое
состояние она перейдет в следующие моменты времени, необходимо наблю-
дать за системой длительное время. Пусть, находясь в одном из структурных
состояний, система принимает любое из п функциональных состояний. Если
достаточно долго наблюдать систему, то по частоте появления функциональ-
ных состояний можно приближенно судить о вероятностях ее пребывания в
этих состояниях. Обозначим их через рг Для общей оценки системы по ве-
роятностям К.Э.Шеннон ввел эвристическое понятие неопределенности, или
энтропии [см. формулы (16)—(19) и обозначения к ним].
Пусть измерены относительные частоты (при большом числе наблюдений
стремящиеся к вероятностям) для определения функциональных состояний
системы.
При этом выполняется соотношение
ХЛ = 1. (21)
Под неопределенностью или энтропией ¥ дискретных величин р. пони-
мают следующее соотношение
F=-5>/log/>/. (22)
Если, например, система все чаще находится в 1-м состоянии, то вероят-
ность р стремится к единице. В предельном случае, когда р. = 1, энтропия
равна нулю и система становится детерминированной. Если система безраз-
лична к своим состояниям, то вероятности равны друг другу, а неопределен-
ность, вычисленная по формулам типа (22), приобретает максимальное зна-
чение. Таким образом, неопределенность системы лежит в пределах
0<¥<¥т. (23)
Неопределенность системы характеризует ее хаотичность. Система мо-
жет считаться полностью дезорганизованной, если в любой момент она с
равной вероятностью может перейти в любое состояние. В этом случае си-
стема обладает максимальной неопределенностью, которая совпадает с ме-
рой сложности системы. Если система предпочитает одни состояния дру-
48
Принципы моделирования
гим, то закон равных вероятностей нарушается, а неопределенность систе-
мы уменьшается, т. е. ¥■<¥„. В этом случае система может принимать то же
число состояний. Уменьшение неопределенности можно связать с увеличе-
нием организации системы. Таким образом, организация системы есть реа-
лизованная в системе неопределенность:
0 = ¥т-¥, (24)
где О — абсолютная организация системы; ¥т — максимальная неопреде-
ленность системы; ¥ — текущая неопределенность системы.
Если процесс уменьшения текущей неопределенности достигает преде-
ла, то у этого процесса будет нулевое значение текущей неопределенности.
В системе будет реализована вся возможная неопределенность, и система
станет детерминированной, то есть перейдет в свою противоположность
0 = ¥т. (25)
Детерминированные системы можно сравнивать по величине реализо-
ванной в них максимальной неопределенности или логарифму числа воз-
можных состояний.
На основании введенного К. Шенноном понятия избыточности можно
оценивать организацию системы следующей мерой:
R = l~^- . (26)
Мера R характеризует величину относительной организации системы и
лежит в пределах
0<R<1. (27)
Для детерминированной системы относительная организация равна еди-
нице, а для полностью дезорганизованной — нулю.
У. Эшби, говоря о взаимодействии системы со средой, подчеркивал, что
«только разнообразие может уничтожить разнообразие». Иными словами,
для того чтобы биосистема успешно функционировала в среде, сложность и
организация ее должны быть адекватными сложности и организации среды.
Условно этот принцип можно выразить следующим образом:
(28)
где индекс е означает принадлежность к среде, a s — к системе
Ч(т)«*2(т)1
ад-ад I'
Глава 2. Биосистемы и биомоделирование
49
Из соотношений (28) и (29) следует, что разница между показателями
сложности и организации биосистемы и среды служит для биосистемы вход-
ным сигналом, заставляющим ее уменьшать исходное расхождение (рассог-
ласование) между этими параметрами. Запишем разность, соответствующую
ошибке рассогласования между системой и средой по этим показателям,
используя для общности некоторые коэффициенты пропорциональности:
§r(T) = ^(T)-M,(*)l
S0(T) = Oe(T)-*oO,(T)J"
(30)
(31)
Будем считать, что в замкнутом комплексе биосистема — среда ошибки
рассогласования являются управляющими сигналами, заставляющими не-
рвную систему так перестраивать поведение организма, чтобы уменьшить
расхождения между биосистемой и средой по показателям сложность и орга-
низация. В этом случае условия, вытекающие из принципа адекватности,
принимают вид:
f\<-°\ ■ (32)
Если в некоторый момент времени между биосистемой и средой ус-
тановилось соответствие, и только после этого, начинают изменяться слож-
ность и организация среды, то для выполнения принципа адекватности до-
статочно пропорциональности следующих величин:
d§r
dt
<0
dt
<0
(33)
В случае статической адекватности комплекс биосистема—среда аналоги-
чен системе стабилизации с обратной связью, а в случае динамической адек-
ватности — следующей системе:
d¥s
UJ m
dt
dRs
dt
dOs
dt
dK
dRe
dt
dOe
dt
(34)
50
Принципы моделирования
С точки зрения традиционной физики, термодинамики и соответствую-
щего им математического аппарата, как это не покажется странным, невоз-
можно провести четкую границу между различными биологическими форма-
ми жизни и неживой природой. Поскольку, как мы уже подчеркивали, абсо-
лютно изолированных систем в природе нет, многие авторы пытаются про-
вести эту грань через понятия антиизоморфности и энтропии. Некоторые
полагают, что живые организмы существуют в системе поглощения отрица-
тельной энтропии или негэнтропии. Этому понятию соответствует приток
свободной энергии, необходимой для компенсации её расходования в про-
цессе жизнедеятельности. В то же время любая форма жизни поддерживается
неравновесными состояниями. Чем ближе организм к состоянию равновесия,
тем скорее наступает его гибель [см. формулы (23)—(30) и их описание].
Существуют представления о том, что в процессе роста развития и стро-
ения организмов наблюдаются процессы снижения «продукции» энтропии
[302]. Эти исследования основывались на оценке производства энтропии, из-
меряемой по скорости дыхания и теплопродукции. На рис. 2 показано, что
в оогенезе формируются механизмы производства энтропии и от рождения
через развитие и динамические процессы жизни они постепенно угасают
[формула (34)].
Динамика жизни
dt
Гибель —
Рис. 2 Необратимость жизни связана со снижением продукции энтропии
Частичное обновление живых систем возможно до определенной степе-
ни за счет ауторепаративных процессов, а полное их обновление осуществ-
ляется исключительно за счет репродуктивной функции. Уже в самом рож-
дении организма заложены процессы накопления генетических ошибок и
метаболических нарушений, а явления энтропии и негэнтропии четко ха-
рактеризуют эти процессы и объясняют суть необратимости жизни. Мы вер-
Глава 2. Биосистемы и биомоделирование
51
немея к этой концепции в гл. 35, когда будем рассматривать понятия физио-
логического времени.
Резюмируя изложенные в этой главе представления, следует подчеркнуть,
что общий принцип построения модели включает в себя три основных со-
ставляющих. Во-первых, обработку информации, полученной в ходе экс-
перимента, с выбором адекватного математического аппарата исследования.
Во-вторых, определяется детерминированность или вероятностность типа
биосистемы. В-третьих, избирается оптимальный математический аппарат
для детерминированной или вероятностной модели и осуществляется ее
построение.
Эту краткую интродукцию к рассмотрению основных принципов биомо-
делирования в биосистемах различного уровня сложности оправдано допол-
нить небольшим примером. Поскольку в дальнейшем изложении нам пона-
добятся конкретные примеры, возьмём один случай, а именно регуляцию
сахара в организме мини-свиней с участием различных органов и систем.
Мы прибегнем к несложному математическому описанию процесса, полагая
что на данный момент у нас отсутствует должная полнота информации обо
всех сторонах регуляции оборота глюкозы, а главное — мы крайне мало зна-
ем о различных элементных составляющих анализируемой биосистемы.
Начнем с простейшего примера описания транспортирования и разноса
инсулина, независимо от того, каким путем он туда попал. Это может быть
определенная доза в виде инъекции или кванты эндогенного инсулина, вбро-
шенного в кровь из (3-клеток островков Лангерганса. При таких допущениях
мы можем считать, что скорость изменения содержания инсулина
в плазме крови организма пропорциональна скорости разноса инсулина кро-
веносной системой и тем меньше, чем большего значения достигло текущее
содержание инсулина:
dix , . f/(1),0<r<x/
^ + b6h=\0, />x/ ^
где /О — скорость разноса инсулина кровотоком; xi — время, в течение ко-
торого будет израсходована доза инсулина; Ь6— коэффициент пропорцио-
нальности.
Для определения параметров уравнений динамики системы сахара крови
использовались экспериментальные результаты исследования гликемичес-
ких кривых при действии на систему глюкозы и инсулина в норме. Изучение
динамических кривых сахара крови мини-свиней позволили получить дос-
таточный статистический материал для определения коэффициентов дина-
мики. Задача определения коэффициентов является задачей наилучшего
сглаживания решениями полученных дифференциальных уравнений экспе-
риментальных кривых.
52
Принципы моделирования
Учитывая инерционность работы органов и процесса распределения в
них сахара крови, а также резкие колебания его уровня вследствие быстроты
движения крови, можно пренебречь как ничтожно малой величиной его
смешиванием в сосудистом русле. Тогда простое суммирование описывает
работу общей системы управления уровнем сахара крови, включая и объект
управления:
3y=Ye (yh+y2+y, + ... +уп), (36)
где: уИ — норма, у2, ..., уп — концентрация сахара в сосудах, органах, тканях
и регулирующих системах (адреналовая, ЦНС и т.д.), которые обеспечивают
поддержание стабильного уровня сахара в организме.
Мы рассмотрели в качестве примера обоснование и процедуру синтеза
структурно-функциональной математической модели системы регуляции
уровня сахара крови, в которой взаимодействуют различные органы. Под-
черкнем еще раз, что такой метод моделирования оправдан в условиях,
когда отсутствует достоверная информация о промежуточных координатах
системы. В дальнейшем изложении, при рассмотрении функций и биомо-
делей на уровне печени, поджелудочной железы и других органов, мы вер-
немся к этому примеру и продолжим описание модели в математических
терминах.
Формальная аксиоматика в биомоделировании
Любая наука изучает некие общности объектов и присущие им свойства и
отклонения. Технологическими и инструментальными основами для этого
являются математический аппарат, информатика и ЭВМ-математика. Суще-
ствует конкретная аксиоматика, идущая от Евклида, в которой исходные
аксиомы и понятия имеют единственную систему содержательности иссле-
дуемых объектов.
Методологические основы современных математических структур, нача-
ло которым положили Д.Гильберт, А.Н.Колмогоров и Н. Бурбаки, определя-
ют не саму математику, а только объекты, которые она исследует. Это на-
правление, получившее название формализованной аксиоматики, возникло
на основе аксиоматики Евклида, но отличается точным знанием правил вы-
вода, использованием языка символов и формул, заменой содержательных рас-
суждений преобразованиями формул и счислений.
В изначале любой науки и теории находится логика, уходящая вглубь
тысячелетий. Аристотель отбросил лукавые схемы рассуждений и создал
систему силлогизмов, т. е. правил выводов. Через два тысячелетия Г. Лейб-
ниц ввел в логику математическую символику, а в XIX веке Дж. Буль реали-
зовал ее в том виде, в котором она существует во всех алгоритмах и програм-
мах современных компьютеров.
Глава 2. Биосистемы и биомоделирование
53
Здесь и далее мы используем аппарат математической логики, поскольку
ее объектами являются дискретные конечные системы, к которым относятся
и биологические, а ее задачей — структурно-функциональное и функцио-
нально-структурное моделирование таких систем на основе формальной ак-
сиоматики.
Язык математической логики хорошо приспособлен к точному описа-
нию биологических явлений, исключает двусмысленность при построении
биологических моделей. Важно, что термины «логика высказываний» и «ал-
гебра высказываний» являются синонимами. Исчисление высказываний ис-
пользует символы, обозначающие некоторые логические операции, их
смысловые значения и интерпретации:
л — конъюнкция или логическое произведение, когда р л q или р и q\
v — дизъюнкция или логическая сумма, когда р v q или р или q;
V— квантор всеобщности для каждого х со свойствами р,х), у р,х);
3 — квантор существования Зх еХ, существует элемент х множества X;
—1 — отрицание, обозначаемое как -р;
=» — импликация или логическое следствие, когда р => q, если р то q\
<=> — эквивалентность, когда р <=> q, p тогда и только тогда, когда q;
А, В, С, X — множества (обозначаются большими буквами);
а, Ь, с, х — элементы множеств (обозначаются малыми буквами);
х е X или xgl- объект х является или не является элементом множе-
ства X;
А с В — множество А содержится в множестве В, значит А является под-
множеством В.
Следует обратить внимание на возможные значения, которые в процесс
логических построений принимает объект моделирования. Если мы имеем
дело с объектами, принимающими одно из двух возможных значений (+
или —, да или нет, истина или ложь) — это двузначная логика. Если объекты
принимают значения из множества, включающего более двух элементов,
они описываются в системе многозначной логики или сводятся к двузначным
объектам.
Математическая логика и формальная аксиоматика, включающая в себя
три основные типа абстрактных структур (алгебраические структуры, струк-
туры порядка и топологические структуры), составляют математический фун-
дамент биомоделирования.
Биомодели для оценки токсичности
Выбор лабораторных животных—биомоделей для исследований, и особенно
оценки воздействия различных факторов физического, химического или
биологического генеза, является ключевой проблемой при планировании,
54
Принципы моделирования
проведении и анализе результатов. В дальнейшем изложении мы будем при-
водить необходимые материалы для исследователей, которые хотят найти
наилучшие способы и формы организации экспериментов. Мы хотели бы
обратить внимание на правильность использования этих таблиц. В колонке
«Исследуемое действие вещества» приводятся, можно сказать, классические
модели проведения экспериментов, связанных с воздействием фармаколо-
гических или иммунотропных средств, химических веществ, воздействия фи-
зических факторов. Однако наиболее важными и приемлемыми являются
биомодели для исследований, полученные с помощью генетических подхо-
дов в создании инбредных, гибридных, конгенных, мутантных и иных ли-
ний. Экспериментатор должен руководствоваться собственными целями и
задачами при выборе той или иной экспериментальной или биомодели.
В следующих колонках указывается вид, линия или порода, пол, прием-
лемые вес и возраст животных. Таблица 6
Генетически обусловленные и экспериментальные биомодели для оценки острой
и хронической токсичности фармакологических веществ, общетоксического
действия ксенобиотиков, специальных средств защиты и радиопротекторов
Исследуемое действие
вещества
Лабораторное
животное
Линия или порода
животного
Пол
животного
Вес
животного
Возраст
животного
Биомодели для оценки общетоксического действия веществ
Биомодель хлороформного
поражения (в минимальных
дозах) почек с летальным
исходом
Биомодель озоновой
интоксикации
(LD100 34-36 ppm/ч)
Биомодель высокой
резистентности к токси-
ческому действию хлоро-
форма
Гиперчувствительная био-
модель токсического дейст-
вия трихлорфторметана
и других хлор- и фтор-
содержащих интоксикантов
и ксенобиотиков
Модели для оценки
общетоксического действия
биологически активных
средств и фармпрепаратов
Модели для оценки острой
токсичности БАВ и фарм-
препаратов
Мыши
Мыши
Мыши
Мыши
Мыши
Хомячки
Собаки
Мыши
Крысы
Кролики
Мыши
Крысы
Линии A/WySnKI
CBA/CaLac
CBA/LacY
DBA/2JY
A/WySnKL
BALB/cJLac
C57BL/6J
C57L/JY
Линия: 82...2
«Английский бигл»
Гибриды
F1(CBA4C57B1/6J)
Аутбредные и/или
линии Wistar
Порода
«Шиншилла»
Гибриды
F,(CBA4C57B1/6J)
Аутбредные и/или
линии Wistar
26
$6
96
96
96
§6
96
96
96
96
96
96
96
30-40 г
30-40 г
30-40 г
120 г
до 14 кг
до 25 г
до 250 г
до 2,5 кг
18-20 г
до 250 г
3-10 мес.
3-10 мес.
3-10 мес.
6-18 мес.
8 мес-
2 года
2-3 мес.
2-3 мес.
4-6 мес.
2-2,5 мес.
2-3 мес.
Глава 2. Биосистемы и биомоделирование 55
Продолжение табл. 6
Исследуемое действие
вещества
Модели для оценки хрони-
ческой токсичности БАВ
и фармпрепаратов
Влияние на ЦНС
Лабораторное
животное
Крысы
Собаки
Мыши
Крысы
Линия или порода
животного
Аутбредные и/или
линии Wistar
«Английский бигл»
Гибриды
F,(CBA4C57B1/6J)
C57BL/6J
Аутбредные
Линии Wistar
Пол
животного
6
96
6
6
6
6
Вес
животного
150-200 г
до 14 кг
18-25 г
17-24 г
до 250 г
120-200 г
Возраст
животного
2-3 мес.
8 мес-
2 года
2-3 мес.
2-3 мес.
2-3 мес.
2-2,5 мес.
Биомодели для исследования антидотов и комплексонов, применяемых для защиты организма
от поражающего действия радиоактивных веществ
Этапы изучения:
a. Предварительный отбор
препаратов путем введения
изотопа однократно при
любой аппликации
b. Детальное исследование
эффективности
Определение зависимости
доза-эффект
Исследование с введением
фиксированной дозы токси-
канта (не менее 2 животных
обоего пола в группе)
Оценка антидотной актив-
ности при внутримышечном
(внутривенном) поступлении
токсиканта
Оценка эффективности
антидота при пероральном
поступлении токсиканта
Оценка эффективности
антидота при ингаляцион-
ном поступлении токсиканта
Оценка антидотной актив-
ности при накожном
поступлении токсиканта
Определение острой
токсичности и широты
терапевтического действия
средств и фармпрепаратов
a. Исследования на
грызунах (введение 4-5 доз
однократно, перорально
и парентерально
b. Исследования на
негрызунах (введение 2 доз,
однократно)
Крысы
Мыши
Крысы
Собаки
Мыши
Крысы
Кошки
Собаки
Мыши
Крысы
Мыши
Крысы
Крысы
Крысы
Мыши
Морские
свинки
Кролики
Мыши
Крысы
Собаки
Аутбредные
Аутбредные
Аутбредные
Аутбредные
Белые аутбредные
Аутбредные
Аутбредные
Аутбредные
Белые аутбредные
Белые аутбредные
Белые аутбредные
Белые аутбредные
Белые аутбредные
Белые аутбредные
Белые аутбредные
Аутбредные
Порода
«Шиншилла»
или альбиносы
Аутбредные
Аутбредные
Аутбредные
6
6
96
96
96
96
96
96
9<з
96
96
96
96
96
96
96
96
96
96
96
180-220 г
18-22 г
160-250 г
8-12 кг
40-50 г
300-350 г
3-5 кг
10-25 кг
18-22 г
180-220 г
18-22 г
180-220 г
180-240 г
180-220 г
18-22 г
200-300 г
2,5-3 кг
40-50 г
300-350 г
10-25 кг
2,5-3 мес.
2-2,5 мес.
2-4 мес.
1 -7 лет
3-10 мес.
5-12 мес.
1 -7 лет
1 -7 лет
2-2,5 мес.
2,5-3 мес.
2-2,5 мес.
2,5-3 мес.
2,5-3 мес.
2,5-3 мес.
2-2,5 мес.
22-27 суг.
6 мес-
3 года
3-10 мес.
5-12 мес.
1 -7 лет
56
Принципы моделирования
Окончание табл. 6
Исследуемое действие
вещества
Изучение фармакокинетики:
Исследование зависимости
концентрации препарата
в плазме крови от времени
после его введения
Изучение общей фармако-
логической активности,
влияния на основные систе-
мы жизнеобеспечения
организма
Лабораторное
животное
Собаки
Мыши
Крысы
Линия или порода
животного
Аутбредные
Аутбредные
Аутбредные
Пол
животного
в
96
96
Вес
животного
15-20 кг
40-50 г
300-350 г
Возраст
животного
1 -7 лет
3-10 мес.
5-12 мес.
Биомодели для исследования антидотов и радиопротекторов
Первичный отбор Мыши Линии C57B1, CBA б 18-24 г 2-2,5 мес.
Изучение радиопротектор-
ных свойств
Изучение фармакодина-
мики, безвредности и меха-
низма радиозащитного
действия
Высокочувствительные
биомодели для изучения
последствий действия
проникающей радиации
Биомодель чувствитель-
ности к облучению рент-
геновскими лучами:
LD50.30 менее 650
LD50.30 более 650
Мыши
Крысы
Хомячки
Собаки
Собаки
Собаки
Обезьяны
Мини-свиньи
Мыши
Крысы
Собаки
Обезьяны
Мини-свиньи
Мыши
Мыши
Мыши
Аутбредные
Аутбредные
Аутбредные
Аутбредные
Аутбредные
Аутбредные
Аутбредные
Аутбредные
Аутбредные
Аутбредные
Аутбредные
Аутбредные
Аутбредные
Линии
BALB/cLacY
DBA/2JY
Линии BALB/cJ
A/J, RF/J, SWR/J,
C57BL/6J, AHeJ
CBA/J, C3HeB/J,
SJL/J, C57BR/J,
129/J
6
6
6
6
6
6
6
6
6
6
6
6
6
96
96
96
40-50 г
300-350 г
8-12 кг
12-18 кг
18-25 кг
Ср. вес
20-30 кг
40-50 г
300-350 г
10-25 кг
Ср. вес
20-30 кг
30-40 г
30-40 г
30-40 г
3-10 мес.
5-12 мес.
6-18 мес.
1,5-5 лет
1,5-5 лет
1,5-5 лет
Взрослые
особи
Взрослые
особи
3-10 мес.
5-12 мес.
1 -7 лет
Взрослые
особи
Взрослые
особи
3-10 мес.
3-4 мес.
3-4 мес.
В этой и последующих аналогичных таблицах мы используем следующие
понятия. Под биомделью, как правило, подразумевается животное с врож-
денными, генетически предопределенными изменениями, в достаточной
степени близкими к таковым проявлениям при патологии у человека. Тер-
мин модель подразумевает создание того или иного состояния с помощью
ксенобиотиков, лекарств или физических воздействий.
Раздели КАЧЕСТВО
ЖИВОТНЫХ-БИОМОДЕЛЕЙ
Я — пёс Его Высочества в Кью.
Прошу, скажите мне, сэр, чей Вы Пёс?
Л.Поуп
{Выгравировано на ошейнике собаки,
подаренной Фредерику принцу Уэльскому)
59
Глава 3 ВПЕРЕД В ПРОШЛОЕ
Животные использовались в медицине, по крайней
мере, со времен Гиппократа [239]. В своих трудах он
пишет: «Действительно, надгортанник есть крыш-
ка, очень точно закупоривающая и не дающая про-
хода ничему, кроме питья. Вот доказательство: если
кто окрасит воду в голубой цвет или красный и даст
ее выпить очень жаждущему животному, лучше все-
го свинье, ибо это животное не отличается ни раз-
борчивостью, ни чистотою, потом перережет ей
горло в то время, как она пьет, то он найдет надгор-
танник окрашенным питьем, но эта операция не
всякому удается». Чем не прообраз биомоделирова-
ния в эксперименте?
При экстраполяции результатов исследования на
животных в отношении человека использовался
принцип аналогии, что не всегда было корректным.
Один из крупнейших врачей и философов средне-
вековья Абу Али Ибн-Сина (Авиценна) в своей «Ло-
гике» жестко критикует умозаключения по абстракт-
ной, далекой от реалий аналогии [299] в нашем по-
нимании — экстраполяции с животных на челове-
ка. Его аргументация заслуживает внимания и с со-
временной точки зрения: «это (заключение по ана-
логии) не всегда является необходимым, потому что
утверждение по сходству может противоречить ут-
верждению по другому сходству, так как есть много
вещей, которые в одном отношении схожи, а в ты-
сяче других отношений различны. В отношении
60
Качество животных-биомоделей
одного из них суждение будет правильным или может быть правильным, а в
отношении другого неправильным». Мудрые слова.
После рассмотрения в разделе I основных принципов моделирования са-
мое время напрямую обратиться к объекту биомоделирования — лаборатор-
ным животным, а затем вернуться к вопросам аналогии и экстраполяции.
Кратко о нашей истории
Проблема биомоделирования в эксперименте теснейшим образом связана с
вопросами использования и разведения лабораторных животных. Не касаясь
истории этого вопроса в других странах, отметим лишь, что в России органи-
зованный процесс выращивания лабораторных животных начался в конце XIX
века. Касаясь предыстории НЦБМТ РАМН, отметим, что накануне первой
мировой войны сельцо Коростово под Москвой купил потомственный дворя-
нин Э.Ратомский и основал здесь конный завод рысистых лошадей. После
революции он передал конный завод государству, в Коростове был организо-
ван совхоз «Светлые горы», который в конце 30-х годов был передан в ведение
Институту эпидемиологии и иммунологии им. Н. Гамалеи Наркомздрава.
12 июня 1929 года постановлением ЦИК и СНК СССР на станции Стол-
бовая Чеховского района Московской области была образована эксперимен-
тальная база Института вирусологии Наркомздрава для разведения живот-
ных генетически чистых линий с целью изучения ультравирусов. Все пред-
военные и военные годы экспериментальная база обеспечивала нужды здра-
воохранения лабораторными животными. Руководство страны всегда уделя-
ло внимание этим вопросам. Работа по разведению инбредных крыс и мы-
шей и поддержанию племенных ядер не прекращалась даже в годы Великой
Отечественной войны.
В 1957 году на базе этого учреждения был создан питомник лабораторных
животных АМН СССР, который продолжил работу по созданию новых линий
лабораторных животных. Начало строго инбредного разведения линейных жи-
вотных в питомнике «Столбовая» связано с работой сектора генетики ЛЭЖ,
руководимого М.Г. Дубиной. Все последующие специалисты в этой области,
работавшие в Столбовой и других учреждениях, были её учениками.
В 1961 году постановлением Президиума Академии медицинских наук
СССР на базе производственного отдела института была организована лабо-
ратория экспериментальных животных. Первым руководителем лаборатории
экспериментально-биологических моделей АМН СССР стал генетик, док-
тор биологических наук Н.Н. Медведев, а затем на протяжении 20 лет её
возглавлял кандидат ветеринарных наук В.А. Душкин. В 1967 году лаборато-
рия получила статус самостоятельного учреждения с названием Научно-ис-
следовательская лаборатория экспериментально-биологических моделей
АМН СССР (НИЛЭБМ).
Глава 3. Вперед в прошлое
61
К 1965 году, когда сектор генетики Лаборатории экспериментальных жи-
вотных ещё существовал на базе питомника «Столбовая», в его коллекции
имелись более 25 инбредных линий и сублиний мышей, одна линия крыс. За
этот период было налажено воспроизводство животных генетически чистых
линий для покрытия нужд научно-исследовательских учреждений. Шла ак-
тивная работа по подготовке кадров научных работников в этой области.
Так в стране появилась специализированная организация, занимающая-
ся проблемами биологии и патологии лабораторных животных. В 70-е годы
XX века в НИЛЭБМ была создана ферма мини-свиней, началась работа по
выведению линий кур, высокочувствительных и устойчивых к вирусам лей-
коза. Изучалось состояние здоровья лабораторных животных в питомниках,
отрабатывались методы микробиологического, вирусологического, парази-
тологического исследования животных.
В 1965 г. Лаборатория экспериментальных животных получила собствен-
ную экспериментальную базу в поселке «Светлые Горы» Московской обла-
сти и все племенные ядра инбредных линий мышей из питомника «Столбо-
вая» были перевезены с сектором генетики на эту базу. В питомнике оста-
лись линии крыс и мышей, имеющих производственное значение.
Центральный питомник лабораторных животных АМН СССР был со-
здан в 1959 году в соответствии с приказом Академии медицинских наук
СССР на базе четырех питомников — «Крюково», «Лыткино» Солнечногор-
ского района, «Планерная» Химкинского района и «Светлые горы» Красно-
горского района. В то время это были мелкие хозяйства с низким уровнем
материально-технической базы. С момента образования Центрального пи-
томника началась планомерная работа по совершенствованию технологии
разведения животных. За короткое время все производственные здания были
заменены новыми зданиями, пригодными конструктивно к разведению ла-
бораторных животных. Столь стремительным развитием питомник обязан
самоотверженному труду специалистов, служащих, рабочих питомника, а
главное, своему первому руководителю А.Т. Хромых, который возглавлял
питомник на протяжении многих лет.
С конца 80-х годов, т.е. в период деформации наших представлений о
«социализме с человеческим лицом» (убедительно прошу не путать с терми-
нологией об экстраполяции и интерполяции в биомоделировании и не оби-
жать братьев меньших, у которых, если и не человеческое, то, зачастую, от-
нюдь не звериное лицо) и до конца XX столетия использование лабораторных
животных в нашей стране для научных целей снизилось в сотни раз. В десят-
ки раз упало и их производство. Последовали реорганизации, реформирова-
ния, попытки акционирования и приватизации учреждений и предприятий
лабораторного производства. К чему это привело — понятно без слов.
Достаточно сказать, что лишь в 2004 году удалось достигнуть 70%-ного
уровня «доперестроечного» производства лабораторных животных в России.
62
Качество животных-биомоделей
Ряд уникальных линий утрачен. Но основной коллекционный фонд был
сохранен благодаря самоотверженности сотрудников лаборатории генетики,
которые вопреки всем правилам биобезопасности забирали домой из нео-
тапливаемых помещений самые уникальные линии животных, которых не-
чем было кормить и поить. Как врач я не могу одобрить тот биориск, но не
сказать об этом не имею права.
Так как традиционно вопросы поддержания коллекционных, инбредных
и специальных линий лабораторных животных для медицины и здравоохра-
нения решаются в рамках Российской академии медицинских наук, в 2002
году был создан Научный центр биомедицинских технологий Российской
академии медицинских наук. Это государственное учреждение образовано
на базе НИЛЭБМ, Института новых технологий РАМН, государственного
предприятия Центрального питомника лабораторных животных РАМН,
питомника лабораторных животных «Столбовая» РАМН и питомника лабо-
раторных животных «Белый мох» РАМН. В состав Центра вошли: филиал
«Андреевка», филиал «Столбовая» и филиал «Белый мох», а также Институт
новых технологий РАМН.
Сегодня коллекционный фонд Центра насчитывает более 100 линий и
стоков мышей и крыс, которые внесены в международный каталог, популя-
ции морских свинок, сирийских хомячков, кроликов, миниатюрных свиней
и безлейкозных кур. В Центре созданы и поддерживаются уникальные био-
логические модели (аналоги муковисциидоза, тетрады Фалло, сахарного
диабета, миодистрофии Дюшена, алкоголизма, трансплантации органов,
бесшовной хирургии, модели для производства вакцин, биологически ак-
тивных соединений и т.д.). Разработана особая система по технологии со-
держания лабораторных животных SPF и гнотокатегории и необходимые нор-
мативные документы.
НЦБМТ РАМН обладает 83 авторскими свидетельствами, 27 патентами
на изобретения. С 1980 года это — сотрудничающий центр Всемирной орга-
низации здравоохранения по контролируемым лабораторным животным.
Дельфин или хомяк: в поиске биомоделей
Вернемся к биомоделированию, интерес к которому в аспекте поиска, если
не идеального, то хотя бы оптимального аналога функционального воплоще-
ния человека, возникал на протяжении нескольких тысячелетий. В качестве
одного из прототипов зачастую рассматривались дельфины. Крупный дель-
фин, его отождествление с человеком, а в ряде случаев и с морским богом,
было распространено среди народов Средиземноморья еще за десятки веков
до рождества Христова. Дельфина изображали на монетах, о нем складывали
легенды, в его честь называли города и территории. Дельфинов пытались
приручить, одомашнить, а самые энергичные военачальники смотрели на них
Глава 3. Вперед в прошлое
63
как на боевые средства. Эти идеи витали задолго до Наполеона, который
считал вполне реальным форсировать Ла-Манш для завоевания Англии с
помощью дельфинов, посадив на них своих вооруженных гвардейцев.
Интерес к дельфинам вспыхнул с огромной силой в середине XX века,
когда развитие кибернетики, математической биологии, электроники и свя-
зи позволили выявить уникальные способности этих животных. Было посту-
лировано, что по своим особенностям дельфины вплотную приближаются к
человеку, оставляя за собой всех остальных животных. В дальнейшем эти
иллюзии были, конечно, развеяны, но наличие ультразвуковых сонаров,
суперуникальное строение кожи и чудесной кровеносной сети, внутривидо-
вое звуковое общение, преодоление кессонной болезни при мгновенном
погружении, а главное, выныривании китообразных с полуторакилометро-
вой глубины, не могли не привлечь внимание военных и специалистов обо-
ронного комплекса.
В некоторых странах, прежде всего в США и СССР, были созданы серь-
езные научные коллективы и разработаны программы использования дель-
финов в боевых и специальных целях. Тому были серьезные научные пред-
посылки. Несмотря на внешнее сходство с рыбами, более полусотни разных
видов дельфинов Мирового океана являются теплокровными млекопитаю-
щими, они дышат легкими, рождают живых детенышей, их хвостовой плав-
ник расположен горизонтально. В то же время они живут в среде, где при
передвижении сопротивление увеличено в 800 раз в сравнении с сухопутны-
ми животными. Упругая и гладкая кожа дельфинов лишена волос, потовых
и сальных желез. Голова дельфинов не обособлена от туловища, ноздри (ды-
хало, одно или два) открываются на темени. В соответствии со строгими
математическими расчетами и законами гидродинамики, дельфин не дол-
жен развивать скорость более 20 км/ч. Однако зафиксировано, что он в со-
стоянии в течение 20 минут двигаться со скоростью от 40 до 55 км/ч, что по
всем канонам физиологии выходит за пределы возможностей их мускульной
силы. Это означает, что у дельфинов должна быть неестественно высокая
мощность мышц, превосходящая, по меньшей мере, в 7 раз таковую у любых
других млекопитающих. В то же время у дельфинов лишь 112 основных мышц
против 170 у наземных млекопитающих. Парадокс?!
Специалисты в области гидродинамики в СССР и США установили, что
сопротивление воды при движении дельфина в 10 раз меньше, чем у любой
модели того же размера и формы. Значит, быстроходность дельфинов зави-
сит от свойств их кожи — упругой, эластичной, несмачиваемой. Было пока-
зано, что у дельфинов тонкий наружный слой кожи постоянно обновляется
и отшелушивается. Эти выводы бионики о свойствах кожи дельфинов при-
влекли внимание кораблестроителей, которые, покрывая подводные лодки
и корабли трехслойным материалом типа «ламинарфлоу» (ламинарное тече-
ние) толщиной 2—3,5 мм, получили огромный выигрыш (более 25%) в ско-
64
Качество животных-биомоделей
рости судов при той же мощности. Дальнейшее изучение кожи дельфинов и
ее моделирование позволило существенно превзойти и эти результаты.
Понятные трудности экспериментирования на высокоорганизованных
животных в морской среде, тем более находящихся в постоянном движении,
заставляют ученых искать оптимальные «сухопутные» биомодели. Первую
серьезную попытку сделал инженер ВМФ США Уолтер Рабб, который про-
вел испытание на сирийских хомячках, находившихся в камере с кремнийор-
ганическими стенами. Такой материал по закону парциальное™ давлений
непроницаем для воды, но позволяет свободно диффундировать кислороду
и углекислоте. Советские ученые проводили исследования на собаках, по-
груженных в раствор перфторуглерода на десятки минут и часы. Животные
выполняли в этот период и после извлечения все функции, свойственные
им, в том числе и условно-рефлекторные акты.
У дельфинов прекрасное обоняние, осязание и вкус. Неплохое зрение
дельфинов дополняется «акустическим разглядыванием» от инфра- до ульт-
развукового диапазона (от 1—4 Гц до 170—200 кГц). Дельфины до определен-
ной степени способны к звукоподражанию, в том числе голосу человека.
Тем не менее, 1 млн долларов, выделенный НАСА биологу Джону Лили на
обучение дельфина человеческому языку, оказался потраченным впустую.
Казалось бы, дельфин в некоторых отношениях является идеальной био-
моделью человека. Его мозг (1700 г) больше, чем человеческий (1400 г), он
имеет обилие извилин, хорошее соотношение коры больших полушарий и
мозжечка к объему мозга. Дельфин легко обучается, имитирует реакции че-
ловека, ведет семейно-стадный образ жизни. Однако непреодолимая про-
пасть от проблесков мышления дельфина до сознания человека так и не
заполнена. Дельфинам, как и другим животным, по мнению Л.В. Крушин-
ского, присуще экстраполяционное поведение и рассудочная элементарная
деятельность, в отличие от разума человека.
Тем не менее, дельфины и другие китообразные стали предметом самого
тщательного изучения и активного исследования, и прежде всего в военных
целях.
В 60-х годах XX века по решению ВПК в Казачьей бухте на Черном море,
вблизи Севастополя была создана уникальная и абсолютно секретная база
по работе с «боевыми» дельфинами. Их в обиходе так и называли. К тому
времени было известно, что за рубежом, прежде всего в США, разрабатыва-
ются потенциальные военные программы по использованию дельфинов для
боевой подводной разведки, поиска минных заграждений, гидроакустичес-
кого патрулирования портов, поиска подводных лодок и боевых пловцов-
диверсантов, доставки шпионского оборудования, съемка информации о
наличии ядерного оружия на борту кораблей и т.п.
Сообщалось об использовании дельфинов в качестве живых торпед и их
способности мгновенного уничтожения боевых акванавтов и гидронавтов (в
Глава 3. Вперед в прошлое
65
частности, во Вьетнаме), с помощью стилета, закрепленного на клюве (морде)
животного. Однако, помимо этого, дельфин является прекраснейшим объек-
том для моделирования в технике, медицинской и биологической науке.
Поэтому, когда перед нами была поставлена задача, найти приемлемый
вариант безусловного возвращения дельфинов в конкретную точку и время
после выполнения тех или иных операций, мы отнеслись к ней не только с
должной ответственностью, но и с большим научным интересом. То, что нам
в группе выдающихся ученых того времени, возглавляемых академиками
Л.Г.Ворониным, Н.П.Бехтеревой и П. Г. Костю ком, удалось внести крайне
скромный вклад и найти для афалин Tursiops truncatis Montagu и Delphinus
delphis L. — нечто аналогичное валерьянке для кошек, крайне привлекавшее
их, было осуществлено на основе биомоделирования.
Потребовалось найти биомодели и построить эксперименты на других
животных, условия работы с которыми были бы более привычными для
исследователей. Сравнительные стереотаксические, нейро- и психофизио-
логические и фармакологические исследования показали, что можно с
успехом моделировать отдельные элементы функционирования дельфинов
на собаках, кошках, даже на крысах и мышах. Но все же лучшей моделью
дельфина является другой дельфин.
Известно, что задачи повышения обороноспособности страны всегда дают
позитивный толчок развитию не только прикладной, но и фундаментальной
науке. Ведь именно эти, специальные по сути исследования, перевели дель-
фина из категории диких животных - в прекрасный экспериментальный и
лабораторный объект.
Сложно проводить экстраполяцию с любой биологической модели на про-
тотип человека, но непросто это сделать и в отношении, например, кошка —
дельфин, собака — дельфин и т.д. Было установлено, что во время сна дви-
жения хвоста дельфина синхронизированы с его дыханием, каждые 1-2 мин
он выныривает и вновь погружается. У дельфинов нет фазы парадоксально-
го или быстрого сна, присущего и обязательного для всех без исключения
млекопитающих. Это значит, что всплытие и осуществление дыхательного
акта у дельфина не требует пробуждения. Так, у дельфинов общая анестезия
(закись азота) вызывает аналогичные другим животным реакции: исчезают
некоторые рефлексы, снижается со 120—130 до 115—ПО мм Hg артериальное
давление и т.д., но реакция на некоторые психотропные средства — психо-
стимуляторы, анксиолитики — существенно отличает дельфина от других
млекопитающих.
Важнейшим для человека, но невозможным для изучения в эксперимен-
тах на сухопутных животных, является выяснение на дельфинах механизма
экскреции огромного количества NaCl, заглатываемого ими. Научившись
выводить из организма лишнюю соль, мы найдем путь лечения артериаль-
ной гипертензии. Чудесная сеть кровеносных сосудов головного мозга дель-
66
Качество животных-биомоделей
финов демпфирует колоссальные пульсовые перепады, понимание механиз-
мов работы которой и её моделирование может открыть путь к лечению
мигреней и тяжелых нарушений мозгового кровообращения. Проблемы био-
моделирования с использованием дельфинов выходят за пределы данной
книги, но интерес к ним у нас не становится меньшим.
В дальнейшем изложении мы рассмотрим различных животных, в том
числе и линейных. Селекция и инбридинг в производственных или научных
целях имеют свои рамки предела и достаточности. Тем не менее, изучение
различных животных, в том числе и дельфинов, проживающих в семейно-
стадных сообществах, где естественный, природный инбридинг неизбежен,
представляет огромный интерес для биомоделирования, расшифровки фун-
даментальных механизмов функционирования, развития и управления воз-
можными патологическими процессами.
67
Глава 4 СТАНДАРТИЗАЦИЯ ЛАБОРАТОРНЫХ
ЖИВОТНЫХ
Получение надежных и воспроизводимых результа-
тов медико-биологического эксперимента можно
достигнуть лишь при соблюдении стандартности
всех его слагаемых и условий проведения. В этом
смысле лабораторное животное является наиболее
уязвимым звеном в системе медико-биологическо-
го эксперимента. Его состояние как живого объек-
та зависит от многочисленных экзогенных и
эндогенных факторов, влияние которых далеко не
всегда бывает явным и легко регистрируемым. Сре-
ди них прежде всего следует отметить факторы ин-
фекционной и инвазионной природы. Причем раз-
личные, в том числе даже патогенные представите-
ли вирусной и бактериальной флоры, не всегда
вызывают клинически явную картину заболевания.
Часто они протекают в латентной форме или же в
виде носительства. В настоящее время актуальность
приобретает так называемая оппортунистическая,
эндогенная инфекция, активизирующаяся при им-
мунодефицитных состояниях.
Полноценность корма, сбалансированность всех
его компонентов является важнейшим условием,
обеспечивающим здоровье животного и поддержа-
ние неспецифической резистентности его организма
на высоком уровне. Велико также воздействие эко-
логических факторов на статус животного: условий
его содержания и микроклимата. Все эти моменты,
не учитываемые при проведении эксперимента, ис-
кажают и даже извращают его результаты [172, 199].
68
Качество животных-биомоделей
Достижение современного уровня медико-биологического исследования
возможно лишь при унификации всех факторов, воздействующих на орга-
низм лабораторных животных, путем строгой стандартизации как условий
содержания, так и самих животных. Введение в мировую практику требова-
ний системы GLP (Good Laboratory Practice) для проведения доклинических
испытаний лекарственных веществ предъявило еще более жесткие условия к
проблеме стандартизации животных, состоянию их здоровья.
В соответствии с международным опытом стандартность лабораторных
животных обеспечивается, с одной стороны, современной технологией их
разведения и содержания в барьерной системе, с другой — едиными крите-
риями оценки состояния здоровья. Основой этих критериев является прин-
цип недопустимости носительства ряда патогенных и условно-патогенных
агентов инфекционной и инвазионной природы: вирусов, бактерий, парази-
тов. То есть, стандартизация животных основана на исключении возможно-
сти возникновения явлений инфекционной и инвазионной патологии.
Во многих странах разработаны стандарты различных категорий качества
животных по состоянию здоровья. Они содержат перечень возбудителей, но-
сительство которых исключается. Чем выше категория качества животного,
тем больше перечень недопустимых агентов. Однако стандарты различных
стран не идентичны. Единой международной классификации лабораторных
животных по категориям качества и соответствующих стандартов не суще-
ствует. В связи с этим, животные, именуемые как SPF (Specific pathogen
free), не имеют четкой характеристики качества, и, полученные из разных
источников, могут значительно различаться по своему статусу. В последние
годы отмечается явная тенденция к унификации критериев качества жи-
вотных и создания единых стандартов. Примером могут служить разработки
группы исследователей европейских стран GV-SOLAS, FELASA.
Специалисты нашей страны совместно с зарубежными коллегами на осно-
ве мирового опыта также предприняли попытку создания требований к каче-
ству лабораторных грызунов различных категорий, которые были приняты в
1989 г. на совместном совещании в Софии. Однако опыт работы по контролю
состояния здоровья животных позволил видоизменить эти нормативы. Требо-
вания к качеству животных различных категорий максимально приближены к
общеевропейским стандартам. В качестве примера в табл. 7 приведена класси-
фикация лабораторных грызунов (мыши, крысы, морские свинки, хомячки)
по категориям.
Классификация основана на методе получения исходных данных для раз-
ведения животных, наличии и надежности барьера и глубине или уровне
контроля статуса животного по соответствующему стандарту (требованию).
Предлагается различать пять категорий качества животных:
/ категория Г. конвенциональные животные, содержащиеся в открытой си-
стеме;
Глава 4. Стандартизация лабораторных животных
69
/ категория 2\ улучшенные конвенциональные животные, находящиеся в ба-
рьерной системе неполного типа. Исходными животными этой категории
могут быть только животные более высокого класса качества (SPF). Эта
категория качества животных соответствует категории, именуемой во мно-
гих странах MD (Minimal Diseases);
/ категория 3: SPF-животные, содержащиеся в строгой барьерной системе;
/ категории 4 и 5: гнотобиоты и безмикробные или аксенные животные, со-
держащиеся в изоляторах.
Лабораторные животные высокого качества не должны иметь клиничес-
ких и доклинических заболеваний, вызванных микроорганизмами, наруше-
ниями содержания и питания, отрицательным воздействием окружающей
Таблица 7
Классификация лабораторных животных по категориям
и их использование в биомедицинских исследованиях
Категория
1
2
3
4
5а
5Ь
Характеристика
Конвенциональ-
ные (CV)
Улучшенные кон-
венциональные
(МЭили Minimal
Diseases)
SPF(CnOwin
свободные от
патогенной
флоры)
БРРилиСПФ
Безмикробные
(аксенные GF)
Гнотобиотные
Метод
получения
Случайный
От животных более
высокой категории
качества
От животных, со-
держащихся в изо-
ляторах и ассоции-
рованных с опре-
деленными микро-
организмами
Отгнотобиологи-
ческих или безмик-
робных животных,
ассоциированных
с определенными
микроорганизмами
Гистерэктомия
Свободный от лю-
бой или известной
формы жизни
низмов
Используются для
описания животных
и систем,все фор-
мы жизни в кото-
рых известны
Барьер
Отсутствует
(открытая
система)
Неполная
барьерная
система
Барьерная
система
Барьерная
система вы-
сокой степе-
ни надежно-
сти
Изолятор
Изолятор
Контроль
Стандарт
1 -й категории
Регулярный
по стандарту
2-й категории
Регулярный
по стандарту
3-й категории
Регулярный
по стандарту
3-й категории
Регулярный на
наличие ассо-
циированных
микроорга-
Использование
В целях обучения
(острые опыты)
Рутинные иссле-
дования в кратко-
временных экспе-
риментах
Изготовление
стандартных пре-
паратов, испыта-
ние на токсич-
ность препаратов.
Хронические экс-
перименты дли-
тельностью 6 мес.
Получение куль-
тур клеток при
производстве
вакцин, поддер-
жание штаммов
бактерий и виру-
сов перевиваемых
опухолей, испыта-
ние новых фарма-
кологических
средств, проведе-
ние фундаменталь-
ных исследований
иммунитета, вос-
паления и т.д.
70
Качество животных-биомоделей
среды, стрессом или врожденными патологиями. Животные определенной
генетической линии должны соответствовать характеристикам, присущим
данной линии, то есть быть генетически аутентичными.
Использование животных низкого качества может привести к необходи-
мости увеличения количества животных для проведения эксперимента и даже
полностью нивелировать результаты опыта. Например, если животные явля-
ются носителями доклинических нарушений, дополнительный, пусть даже и
незначительный стресс, причиненный в процессе содержания или в ходе
эксперимента, может привести к развитию поражения органов или тканей,
вызванного доклиническим заболеванием. В этом случае подобные наруше-
ния будут ошибочно отнесены к последствиям воздействия, оказанного на
животных в процессе эксперимента.
Выбор линии для опытов может быть обусловлен подверженностью жи-
вотных определенным заболеваниям, таким, как лейкемия или аутоиммун-
ная анемия. Если же генетическая чистота подобной линии была нарушена,
возможно, у некоторых животных данные заболевания не проявятся, и ре-
зультаты эксперимента, аналогично предыдущему примеру, окажутся недо-
стоверными.
Контроль качества лабораторных животных в процессе содержания и ходе
экспериментальных исследований должен включать в себя микробиологи-
ческий и генетический контроль, контроль внешней среды, заключающийся
в мониторинге температуры, влажности, освещения, шума и т.д., и контроле
питания.
Требования к содержанию животных
В соответствии с Европейской конвенцией по защите позвоночных живот-
ных, используемых в экспериментальных и других научных целях (Совет Ев-
ропы, Страсбург, 2004 г.), следует придерживаться нормативов содержания
лабораторных животных.
Таблица 8
Рекомендуемые температурные режимы содержания животных в клетках,
вольерах и внутренних загонах
Виды или
группы видов
Мышь
Крыса
Сирийский хомяк
Мышки-полевки
Морская свинка
Перепелиные
Кролик
Оптимальная
температура, °С
20-24
20-24
20-24
20-24
20-24
20-24
15-21
Виды или
группы видов
Кошка
Собака
Хорек
Куры
Овцы и бараны
Мини-свиньи
Лошади, жеребята
Оптимальная
температура, °С
15-21
15-21
15-21
15-21
10-24
10-24
10-24
Глава 4. Стандартизация лабораторных животных
71
В особых случаях, для молодых или голых особей может требоваться
содержание при более высоких температурах.
Все ввозимые в страну животные должны пройти карантин согласно на-
циональному законодательству. Сроки карантина в условиях лаборатории
обычно определяются соответствующим лицом исходя из обстоятельств, этим
лицом обычно является назначенный учреждением ветеринар.
Таблица 9
Рекомендуемая продолжительность карантинного периода в условиях лаборатории
Виды
Мышь
Крыса
Мышки-полевки
Морская свинка
Дни
5-15
5-15
5-15
5-15
Виды
Сирийский хомяк
Кролик
Кошка
Собака
Дни
5-15
20-30
20-30
20-30
На всех стадиях процедур необходимо обеспечить животному жизненное
пространство и учесть потенциально возможный рост животного.
Таблица 10
Размеры клеток для содержания и процедур для мелких грызунов и кроликов
Виды
Мышь
Крыса
Сирийский хомяк
Морская свинка
Кролик 1 кг
2 кг
Зкг
4 кг
5 кг
Минимальная площадь
клетки, см2
180
350
180
600
1400
2 000
2 500
3 000
3 600
Минимальная высота
клетки, см
12
14
12
18
30
30
35
40
40
Высота клетки представляет собой вертикальное расстояние между по-
лом клетки и ее верхней горизонтальной частью.
Таблица 1 1
Размеры клеток для разведения мелких грызунов
Виды
Мыши
Крысы
Сирийские хомяки
Морские свинки
Несколько самок морских
свинок с детенышами
Минимальная площадь
клетки для матери
и потомства, см2
200
800
650
1200
1 000 (на одну
взрослую особь)
Минимальная высота
клетки, см
12
14
12
18
30
72
Качество животных-биомоделей
Таблица 12
Размеры клеток для разведения кроликов
Вес самки,
кг
1
2
3
4
5
Минимальная площадь
для самки
и крольчат, м2
0,30
0,35
0,40
0,45
0,50
Минимальная высота
клетки, см
30
30
35
40
40
Минимальная
площадь гнезда,
м2
0,10
0,10
0,12
0,12
0,14
Содержание кошек в клетках должно быть строго ограничено. Их необ-
ходимо выпускать размяться хотя бы раз в день, когда это не мешает проце-
дурам. Вольеры для кошек должны быть оборудованы поддоном для туалета,
широкими полками для отдыха и предметами, подходящими для лазания и
стачивания когтей.
При подсчете минимальной площади клетки можно учитывать площадь
полок. Минимальная площадь клетки для кошки и подстилки включает в
себя площадь гнезда для котят.
Таблица 13
Размеры клеток для содержания кошек (для процедур и разведения)
Вес
кошки,
кг
0,5-1
1-3
3-4
4-5
Минимальная площадь
клетки на кошку,
м2
0,2
0,3
0,4
0,6
Минимальная
высота клетки,
см
50
50
50
50
Минимальная площадь
для кошки и подстилки,
м2
—
0,58
0,58
0,58
Минимальная площадь
вольера для кошки
и помета, м2
—
2
2
2
Собак не следует содержать в клетках дольше чем того требуют процеду-
ры. Собаки, содержащиеся в клетках, должны выпускаться из них хотя бы
раз в день для разминки, кроме случаев, когда это не совместимо с услови-
ями процедур. Должен быть установлен лимит, сверх которого животное
нельзя держать в клетке без разминки. Пространство для разминки должно
быть достаточно большим, чтобы обеспечивать собаке свободу движений.
Клетки с сетчатым полом не следует использовать для содержания собак,
кроме случаев, когда этого требует эксперимент.
Ввиду большого различия собак по высоте в холке и отсутствия стабиль-
ной зависимости между высотой и весом, минимальную высоту клетки нуж-
но определять исходя из высоты собаки в холке. Обычно минимальная вы-
сота клетки представляет высоту собаки в холке, умноженную на 2.
Глава 4. Стандартизация лабораторных животных
73
Таблица 14
Размеры клеток для содержания собак
Высота собаки
в холке, см
30
40
50
Минимальная площадь
клетки, м2
0,75
1,00
1,75
Минимальная высота
клетки, см
60
80
140
Таблица 15
Требования к вольерам для содержания и разведения собак
Вес собаки,
кг
<6
6-10
10-20
20-30
>30
Минимальная площадь
вольера на одну собаку,
м2
0,5
0,7
1.2
1.7
2,0
Минимальная площадь прилегающей зоны
для разминки на одну собаку,
м2
1-3 собаки
0,5(1,0)
1,4(2,1)
1,6(2,8)
1,9(3.6)
2,0(4,0)
больше 3 собак
0.5(1,0)
1.2(1,9)
1.4(2,6)
1.6(3,3)
1,8(3,8)
Примечание: Цифры в скобках означают общую площадь вольера на одну
собаку.
Собаки, содержащиеся вне помещений, должны иметь доступ к укрытию
от плохих погодных условий. Если собаки содержаться в вольере с решетча-
тым полом, для ночного отдыха в вольере нужно организовать зону со сплош-
ным полом. Решетчатые полы не должны использоваться, если того не тре-
бует эксперимент.
Между вольерами необходимо сохранять расстояние, не позволяющее со-
бакам нанести друг другу повреждения. Все вольеры должны быть оборудо-
ваны эффективной дренажной системой.
Таблица 16
Требования к клеткам для содержания мини-свиней
Вес мини-свиньи,
кг
5-15
15-25
25-40
Минимальная площадь
клетки, м2
0,35
0,55
0,80
Минимальная высота
клетки, см
50
60
80
Мини-свиней не следует содержать в клетках, в случае, если того не
требует эксперимент, время содержания в клетке должно быть сведено к
минимуму.
74
Качество животных-биомоделей
Таблица 17
Содержание мини-свиней, овец и баранов в загонах
Виды
и их вес,
кг
Мини-
свиньи
10-30
30-50
50-100
Овцы
<70
Бараны
<70
Минимальная
площадь
загона, м2
2
2
3
1.4
1,6
Минимальная
длина
загона, м2
1,6
1,8
2,1
1,8
1,8
Минимальная
высота
ограждения
загона, м
0,8
1,0
1,2
1,2
2,0
Минимальная
площадь загона для
групп животных,
м2/животное
0,2
0,3
0,8
0,7
0,8
Минимальная
длина кормушки
на голову, м
0,20
0,25
0,30
0,35
0,35
Стойло должно быть достаточно широким, чтобы позволить животному
удобно лежать.
Таблица 18
Содержание мини-свиней, овец и баранов в стойлах
Виды и их вес, кг
Мини-свиньи, <100
Овцы,<70
Бараны, <70
Минимальная площадь
стойла, м2
1,2
0,7
0,8
Минимальная длина
стойла, см
2,0
1,0
1,0
Минимальная
высота ограждения,
стойла, м2
0,9
0,9
0,9
Таблица 19
Требования к клеткам для содержания птиц (пространство означает объем,
полученный путем умножения внутренних длины, ширины и высоты клетки)
Виды
и их вес,
кг
Куры
100-300
300-600
600-1200
1200-1800
1800-2400
Взрослые
петухи
>2400
Перепелиные
120-140
Минимальное
пространство
на 1 птицу,
см2
250
500
1000
1200
1400
1800
350
Минимальное
пространство
для 2 птиц,
см2/птица
200
400
600
700
850
1200
250
Минимальное
пространство для
для 3 и более птиц,
см2/птица
150
300
450
550
650
1000
200
Минимальная
высота
клетки, см
25
35
45
45
45
50
15
Минимальная
длина
кормушки
на птицу, см
3
7
10
12
12
15
4
Глава 4. Стандартизация лабораторных животных
75
Сечение сетки клетки не должно превышать 10 х 10 мм для цыплят
и 25 х 25 мм для молодых и взрослых особей. Толщина проволоки клетки
должна быть не менее 2 мм. Угол наклона не должен превышать 14% (8°).
Длина поилок должна соответствовать длине кормушек. Если имеются чаш-
ки, каждая птица должна иметь доступ к двум. Клетки должны быть обору-
дованы насестами, и птицы в одиночных клетках должны иметь возмож-
ность видеть друг друга.
Контроль качества животных и учет
Контроль качества животных имеет решающее значение для исследований.
Каждое учреждение, использующее животных, должно использовать стандар-
тные процедуры обращения с животными, касающиеся размещения, питания,
физического обращения, ухода и контроля состояния здоровья. Должны учи-
тываться следующие факторы: размер клеточного оборудования, количество
животных в одной клетке, методы их удерживания и перемещения, а также
используемая система разведения. Другой важной переменной для качества
животных является питание. Основные показатели питания должны быть про-
анализированы и зафиксированы для каждого эксперимента. Дополнительно
следует проверять корм на наличие химического загрязнения.
Микробиологический статус животного также является определяющим для
результатов эксперимента. Агенты, в обычных условиях не вызывающие за-
болеваний, могут вызвать проблемы в ходе проведения эксперимента из-за
стресса, испытываемого животными. Другие агенты, как например лактат-
дегидрогеназа вируса мышей, может не вызвать заболевания в процессе эк-
сперимента, но исказить его результаты. Учитывая это, желательно исполь-
зовать как можно более «чистых» лабораторных животных с известным мик-
робиологическим статусом.
Генетический профиль лабораторных животных может быть крайне важ-
ным для успеха эксперимента. Учреждения, занимающиеся лабораторным
животноводством, должны уделять особое внимание генетической чистоте
животных. В рамках этих учреждений должны действовать программы по
предотвращению генетического загрязнения линий (по вине человека или в
результате случайного спаривания) и раннему выявлению мутаций. Каждая
линия имеет особый генетический профиль, определить который можно с
помощью биохимических, иммунологических и морфологических маркеров.
Желательно проводить предварительный генетический контроль до начала
эксперимента.
Важно вести письменный учет процедур, проводимых с животными;
обычно для этих целей заводится специальный журнал. В журнал должны
заноситься записи о ежедневных наблюдениях, проводимых процедурах и
другая текущая информация.
76
Качество животных-биомоделей
В организации должны быть приняты стандартные правила обращения
с животными, включая содержание, питание, удерживание и перемещение
и уход:
/ все новые животные, полученные из внешних источников, должны быть
изолированы и пройти процедуру карантина согласно ветеринарным тре-
бованиям;
/ на начало проведения доклинических испытаний все животные должны
быть здоровыми и не являться носителями агентов, способных повлиять
на результаты исследования. Если в ходе эксперимента животное забо-
лело или стало носителем какого-либо агента, его необходимо изолиро-
вать. Данные животные могут получать лечение, если оно не повлияет на
результаты исследования. Результаты диагностики, назначенное лечение
и его описание, а также даты проведения лечения должны быть докумен-
тированы и сохранены;
/ теплокровные животные, за исключением грызунов, еще питающихся мо-
локом матери, используемые в лабораторной практике, которых требуется
вынимать из клетки, должны быть соответствующим образом идентифи-
цированы (татуировкой, цветным кодом, сережкой, кольцом на ноге и т.д.);
/ вся информация необходимая для идентификации особи, размещенной
вместе с другими животными, должна быть доступна в местах содержания
этих животных;
/ при необходимости, животные различных видов содержат в разных поме-
щениях. Животные одного вида, но участвующие в различных экспери-
ментах, обычно также содержаться в разных помещениях во избежание ис-
кажения результатов эксперимента. Если же их смешение необходимо,
должны быть приняты меры для их адекватной дифференциации;
/ клетки, стеллажи и дополнительное оборудование должны очищаться
и стерилизоваться с достаточной периодичностью;
/ вода и корм для животных должны периодически проверяться для того,
чтобы удостовериться, что уровень их естественного загрязнения не пре-
вышает показателей, определенных в протоколе, выше которых загрязне-
ние может повлиять на результаты эксперимента. О результатах проверки
должны проводиться записи;
/ подстилки, находящиеся в клетках и загонах, не должны влиять на резуль-
таты исследования. Менять их нужно по мере необходимости для обеспе-
чения животному сухости и чистоты;
/ если используются какие-либо препараты против паразитов об этом необ-
ходимо делать запись в журнал. Препараты, способные повлиять на ре-
зультаты исследования, не используются.
Глава 4. Стандартизация лабораторных животных
11
Микробиологический контроль
Микробиологический контроль лабораторных животных посредством посе-
ва флоры на питательных средах, серологических и микроскопических ис-
следований преследует обычно две цели: диагностику заболеваний и конт-
роль микробиологического статуса. Основой диагностики заболевания явля-
ется микробиологическое обследование, но в ходе его также учитывается и
другая информация, касающаяся клинической картины, репродуктивных
функций и патологий. Образцы микробов и пробы, которые необходимо
взять у животных, определяются соответственно клиническими и патологи-
ческими отклонениями. Время проведения контроля не фиксировано и зависит
от проявления заболеваний.
Микробиологический мониторинг представляет собой процесс, призван-
ный периодически подтверждать неизменность микробиологического стату-
са животного или группы животных по сравнению с ожидаемыми показате-
лями. В этих целях у случайно выбранных животных из общей популяции
берутся пробы для выявления определенных видов микробов с целью выяс-
нения микробиологического статуса животных. Таким образом, эти исследо-
вания могут проводиться с целью обнаружения как патогенов, так и непато-
генов, то есть микробов, являющихся частью нормальной флоры.
Микробиологическое обследование чаще всего проводится с целью диаг-
ностики заболевания, тем не менее, целесообразно также проводить плано-
вый микробиологический контроль таких животных, как мыши или крысы,
используемых для точных или долгосрочных исследований. Нет необходи-
мости особо отмечать, что важно исключить вероятность наличия в организ-
ме лабораторного животного не только возбудителей инфекционных заболе-
ваний, сопровождающихся явными симптомами, но и скрытых инфекций,
которые могут под воздействием стресса, причиненном в ходе эксперимента,
вызвать симптоматические заболевания, а также исказить или извратить
результаты эксперимента. Преимуществом проведения периодического мик-
робиологического контроля является тот факт, что он позволяет выявлять
скрытые инфекции на ранних стадиях.
Несмотря на то, что виды микробов, выявляемых в ходе микробиологи-
ческого обследования, различны в зависимости от категории животных, ана-
лизы на отсутствие патогенов, опасных для человека и фатальных для живот-
ных, должны проводиться в обязательном порядке, независимо от категории
животного. Рекомендуется также проводить контроль потенциальных пато-
генов, так как их присутствие может исказить результаты экспериментов.
Исследование микробов-симбионтов и условно-патогенной флоры у мышей
и крыс может проводиться, в том числе, для оценки гигиеничности условий
содержания животных. В микробиологический контроль необходимо также
включать обследование на наличие паразитов.
78
Качество животных-биомоделей
Количество животных, у которых необходимо взять пробы для микроби-
ологического контроля, зависит от уровня инфицирования популяции. На-
пример, в случае, если показатель инфицирования превышает 50%, для об-
наружения возбудителя достаточно взять анализы у нескольких животных,
но если уровень инфицирования ниже 20%, найти возбудителя можно, толь-
ко обследовав как минимум 20 особей. В случае если искомые патогены обла-
дают средними и высокими показателями инфицирования, способными
повлиять на результаты исследования, достаточно обследовать 10 особей.
С другой стороны контроль колоний животных рекомендуется проводить
каждые 2—3 месяца, поскольку высокий уровень антител в организме живот-
ного сохраняется в течение нескольких месяцев после инфицирования. Та-
ким образом, исследование присутствующих антител является важной про-
цедурой диагностики в микробиологическом контроле.
Микробиологическое обследование и контроль, а также диагностика за-
болеваний представляют ряд трудностей ввиду ограниченного доступа к не-
обходимому оснащению лабораторий и реагентам, включая качественные
питательные среды и антигены для серологических тестов, а также по при-
чине недостаточного количества квалифицированных специалистов. Созда-
ние диагностических лабораторий во всех местах содержания лабораторных
животных представляется нецелесообразным. Вместо этого целесообразно
использовать те центры, которые проводили бы микробиологические обсле-
дования, консультировали по вопросам осуществления контроля качества
животных и осуществляли подготовку сотрудников для центров лаборатор-
ного животноводства. Обследование на присутствие паразитов может прово-
диться на месте, поскольку его осуществление проще и более экономично,
чем тесты, основанные на посевах и серологических исследованиях, хотя
возможно использование простых наборов для серологических тестов.
79
Глава 5 ПРИНЦИПЫ ВЫБОРА ЖИВОТНЫХ
ДЛЯ ИССЛЕДОВАНИЙ
В лабораторных исследованиях широко использу-
ются самые различные представители животного
мира. Однако наиболее предпочтительно в качестве
биомоделей использовать различных млекопитаю-
щих, к числу которых относятся мини-свиньи, обе-
зьяны, собаки, кошки, кролики, хомячки, мыши,
крысы и др. Существуют ли унифицированные
принципы выбора аутбредных и инбредных лабора-
торных животных для эксперимента? И да, и нет.
Качество животных — основа эксперимента
Медико-биологические эксперименты с использо-
ванием животных можно разделить на следующие
основные группы. Эксперименты, в которых лабора-
торные животные служат инструментом измерения
активности каких-либо веществ, препаратов и т. д.
(например, вакцин, сывороток или гормональных
препаратов). По Фестингу [206, 207, 208], для по-
добных опытов генотип несущественен, вопросы
экстраполяции необязательны, поэтому для «био-
пробы» можно использовать гибриды линейных
животных, нелинейных и даже аутбредных.
Для экспериментов, в которых предполагается
моделирование патологических состояний (онко-
логические исследования, изучение процессов вос-
паления, иммунитета, врожденных уродств), суще-
ственно, насколько велика роль генетических фак-
торов в развитии изучаемого состояния. Если роль
80
Качество животных-биомоделей
генетических факторов достоверно велика (как например, в возникновении
злокачественных опухолей), то их можно проводить только на определенных
линиях. Если они прямо зависят от генетических факторов (различные наслед-
ственные болезни), то для их изучения пригодны только аналогичные мутан-
тные формы мышей, крыс и т.д.
Исследования влияния различных веществ и факторов внешней среды, когда
изучается реакция живого организма на эти воздействия и в конце исследо-
вания возможна постановка вопроса об экстраполяции на человеческую попу-
ляцию, имеет свои ограничения. Для подобных исследований ограничиться
выбором одной линии или гибридной комбинации можно только в том слу-
чае, когда роль генетических факторов в исследуемом процессе очень мала.
Иногда выбор линий облегчается тем, что по изучаемому вопросу есть ин-
формация, входящая в характеристику линии.
Существуют линейные различия у животных по степени иммунного отве-
та. Часто линии, низкочувствительные к патогенному действию антигена,
отвечают на него высоким титром антител. Линия мышей BALB/c малочув-
ствительна к стафилококковой инфекции, но дает высокий иммунный ответ
на вакцину пневмококков группы А, а у мышей линии C57BL/6 отмечена
обратная зависимость [93, 143]. В то же время имеются отчетливые паралле-
ли: так, например, в случае усиления чувствительности к аудиогенным при-
падкам инбредные линии мышей располагаются в такой последовательнос-
ти: G57BL/6, BALB/c, SM, CBA, SJL, AKR. FP, 129, LP, DBA/2.
Тестирование целесообразно вести с использованием наиболее чувстви-
тельных объектов, однако линии могут проявлять совершенно неожиданную
специфичность. Необходимость экстраполяции на весь вид и далее на человека
требует проведения опытов на несколько различающихся линиях. Заранее
трудно предсказать, как будет реагировать та или иная линия животных на
новый фактор, подлежащий изучению. Для такого поискового опыта нужно
использовать набор из нескольких видов животных или их линий разного
происхождения.
Однотипный ответ будет означать малое влияние генотипа. В таком слу-
чае выбор объекта определяется лишь требованием воспроизводимости и
экономическими соображениями. Разнотипный ответ на разных линиях
будет означать существенную роль генотипа. В этом случае крайние вариан-
ты укажут на возможный размах вариаций изучаемого признака в популя-
ции. Для дальнейших опытов необходимо в зависимости от специфики ис-
следования отбирать или наиболее чувствительную, или две оппозитные ли-
нии. Если нет необходимых сведений о специфике линий, то для их выбора
можно руководствоваться данными об их родстве, отбирая для первого эк-
сперимента максимально различающиеся, неродственные линии.
При выборе для эксперимента тех или иных линий некоторое представ-
ление о степени их генетического различия дают результаты математическо-
Глава 5. Принципы выбора животных для исследований
81
го анализа скелетных измерений. Для экспериментов, результаты которых
важны для межвидовой экстраполяции, а также для экспериментов с длитель-
ным сроком наблюдения, важна устойчивость организма животных к изме-
няющимся условиям внешней среды. Подобной спецификой обладают фар-
макологические, токсикологические и гигиенические исследования. Для
таких исследований можно рекомендовать использование тетрагибридных
мышей, получаемых скрещиванием линий с разной специфичностью в отно-
шении изучаемого признака.
Поскольку в абсолютном большинстве экспериментов используются
мелкие лабораторные животные, преимущественно линейные мыши, кры-
сы, хомячки, морские свинки, полученные путем специальных приемов,
введем некоторые определения из генетики лабораторных животных.
Аллели — различные состояния гена, определяющие фенотипические раз-
личия. Для многих локусов известны только две аллели, из которых одна —
аллель «дикого типа» и вторая — мутантная. У лабораторных животных из-
вестны серии множественных аллелей, например, в локусах Agouti и Albino,
определяющих окраску шерстного покрова. В инбредных линиях лаборатор-
ных животных в гомозиготном состоянии фиксированы определенные алле-
ли, которые играют роль маркеров данной линии. В стоках неинбредных
животных наблюдается высокий уровень гетерозиготности, и слежение за
частотами разных аллелей (например, у морских свинок) позволяет контро-
лировать сохранение или изменение генофонда популяции.
Аутбридинг — скрещивание особей, состоящих между собой в менее
близком родстве, чем особи, взятые из популяции методом случайной вы-
борки.
Бэккросс — рекуррентное или возвратное скрещивание, в широком смысле
— спаривание потомков с родителем, в более узком смысле — скрещивание
гомозигот и гетерозигот типа +/+ х +/а или а/а х +/а. Бэккросс применяется
при введении какого-либо гена в инбредную линию и выведении конгенной
линии (в этом случае необходимо 10 возвратных скрещиваний).
Гибриды F1 — первое поколение животных, полученных от скрещива-
ния двух инбредных линий. Гибриды F1 генетически однородны, также как
животные инбредных линий, но гетерозиготны по тем локусам, по которым
различаются скрещиваемые родительские линии. Гибриды F1 характеризу-
ются повышенной жизнеспособностью, большей устойчивостью к измене-
ниям внешней среды и фенотипической однородностью. В биомедицинских
исследованиях широко используются мыши-гибриды F1, наиболее часто в
нашей стране — (СВА х C57BL/6)Fp за рубежом — (C57BL/6 х DBA/2)F,.
Воспроизводимость результатов экспериментов, выполненных на гибридах
F1, такая же высокая, как при использовании инбредных животных. Гибри-
ды F1 не могут использоваться для размножения, так как у их потомков
будет генетическое расщепление.
82
Качество животных-биомоделей
Инбридинг — спаривание особей, состоящих между собой в более близ-
ком родстве, чем это в среднем наблюдается при случайном спаривании в
популяции. В лабораторном животноводстве используют скрещивание бра-
тьев и сестер (в редких случаях — отца с дочерью, матери с сыном) при
выведении и размножении животных инбредных линий. При разведении не-
инбредных животных, напротив, стремятся избежать инбридинг и с этой це-
лью применяют определенные системы избегания инбридинга при спарива-
нии или рандомизации спариваний.
В настоящее время в мире выведены тысячи линий мышей, сотни линий
крыс, десятки линий морских свинок, сирийских хомячков, монгольских
песчанок, кроликов и кур.
Хотя животные достигают гомозиготности через 20 поколений братско-
сестринского инбридинга, она может нарушаться естественным мутационным
процессом, поэтому лабораторных животных размножают инбридингом по-
стоянно и систематически подвергают контролю гомозиготности. Разведение
линейных животных сопровождают постоянным отбором на жизнеспособность
и плодовитость, чтобы элиминировать возможные вредные, спонтанно воз-
никающие мутантные гены, и отбором на проявление специфического при-
знака линий (например, частоты опухолей молочных желез).
Линии коизогенные — различаются в одном локусе. Подобная ситуация
возникает в случае мутации одного гена в инбредной линии. Приближения
к коизогенности добиваются серией возвратных скрещиваний (10-12 бэк-
кроссов), которыми ген одной линии вводится в генотип другой. Ген вводит-
ся в сегмент хромосомы, содержащий, как примесь, неконтролируемые гены.
Поэтому линия, выведенная подобным способом, называется конгенной.
Существует большое количество изогенных и конгенных линий мышей, несу-
щих различные мутантные гены. Конгенные линии, различающиеся по ге-
нам тканевой совместимости, называются конгенно-резистентными (резис-
тентными к перевивке чужеродных опухолей). Эта группа линий, приобрет-
шая большое значение как инструмент исследования строения и функции
сложных полиморфных систем генов у млекопитающих, широко использу-
ется в иммуногенетических и иммунологических исследованиях.
Линии рекомбинантные — выведение начато Д. Бейли в 1971 г. в Джек-
соновской лаборатории в США. Рекомбинантные линии происходят от скре-
щивания двух неродственных инбредных линий и последующего инбридиро-
вания потомков F2 в течение не менее 20 поколений. Первая группа реком-
бинантных линий была получена от скрещивания линий BALB/c и C57BL/6.
В половине рекомбинантных линий теоретически фиксируется в гомози-
готном состоянии аллель гена одной родительской линии, во второй поло-
вине — аллель другой линии. Эта закономерность проявляется в отношении
каждого локуса. В результате набор рекомбинантных линий дает возмож-
ность исследовать корреляцию между определенными аллелями и ответом
Глава 5. Принципы выбора животных для исследований
83
на экспериментальное воздействие, плейотропные эффекты полиморфных
локусов и сцепление между ними.
Мутант — особь, у которой в результате мутации изменен какой-либо ген
или произошла хромосомная перестройка. Изменение гена приводит к из-
менению признака, развитие которого мутировавший ген детерминирует. Эти
изменения могут затрагивать любые системы и органы. Первые мутанты среди
мышей и крыс были найдены и разводились любителями до создания гене-
тики и использования в качестве лабораторных животных в экспериментах.
Это были животные с изменениями окраски шерсти (и в первую очередь —
альбиносы), структуры шерстного покрова, скелета и поведения. Наиболь-
шее количество мутантов найдено у мышей, их число постоянно возрастает
за счет выявления новых мутантов среди размножающихся инбредных жи-
вотных. Мутанты находят все большее применение в медико-биологических
исследованиях: во-первых, мутантные гены используются как генетические
маркеры для локализации новых мутаций и построения хромосомных карт
и, во-вторых, мутантные гены используются как аналоги, биологические мо-
дели наследственных патологических состояний человека.
Популяция — сообщество свободно скрещивающихся организмов с ха-
рактерными для составляющих его особей генотипом и фенотипом, обус-
ловленным отбором. Колонии (стоки) неинбредных лабораторных живот-
ных могут рассматриваться как искусственные популяции млекопитающих,
если равная возможность скрещиваний для особей обеспечивается
применением одной из систем рандомбридинга. Искусственные или синте-
тические популяции могут быть получены на основе скрещивания двух или
большего числа инбредных линий и последующего размножения потомков
F2 или путем скрещивания животных нескольких линий и гибридов F1,
полученных при скрещивании этих линий.
Рандомбридинг — система разведения, направленная на максимальное
исключение всех факторов, препятствующих и способствующих возможности
свободного спаривания каждой особи популяции с любой другой. При разве-
дении неинбредных лабораторных животных для обеспечения рандомбридин-
га применяют ротационную систему комплектования племенного поголовья,
предложенную Поили в 1960 г. Цель применения ротационной системы —
максимально возможное уменьшение влияния пространственной изоляции
животных, а также отбора; обеспечение сохранения генетической структуры
популяции с минимальной потерей гетерозиготности. Система заключается в
разделении племенного поголовья на ряд равной величины групп, субъеди-
ниц, от 4 до 8 (в зависимости от величины популяции это могут быть отдель-
ные стеллажи, группы стеллажей с клетками или целые секции). Для комп-
лектования следующего поколения обязательно используется молодняк, по-
лученный в каждой группе. Комплектование производят по одной выбранной
схеме, за счет самцов и самок, развившихся в разных группах.
84
Качество животных-биомоделей
Сток — в соответствии с рекомендациями Международного комитета по
лабораторным животным (ICLA, 1972) этим термином обозначают популя-
ции неинбредных (аутбредных) лабораторных животных, в отличие от терми-
на линия (strain), обозначающего инбредных животных. Правила разведения
инбредных животных основаны на принципах популяционной генетики, они
направлены на уменьшение генетических изменений в стоке в процессе его
разведения. Принято давать название стандартизированному стоку, если он
разводится как закрытая колония более четырех поколений. Метод разведе-
ния должен сводить к минимуму утраты гетерозиготности. Принято, что на
поколение ее потери должны быть менее 1%.
Выбор метода разведения зависит от числа производителей. Если в од-
ном поколении в стоке 10-25 размножающихся самцов, то рекомендуется
использовать систему максимального избежания инбридинга. Ее суть состо-
ит в том, чтобы каждый самец дал одного самца в следующее поколение, и
каждая самка внесла вклад в следующее поколение. При размножении избе-
гают спаривать родственников. Данная система используется чаще при раз-
ведении кошек, собак и других видов в небольших колониях.
Сублинии — инбредные линии лабораторных животных, выведенные од-
ним исследователем или в одном институте, могут передаваться для разведе-
ния в другие институты или лаборатории. В процессе разведения вследствие
остаточной гетерозиготности в момент передачи племенного ядра или вслед-
ствие мутационного процесса между разными поколениями животных од-
ной линии могут появляться генетические различия. Такие разделенные вет-
ви одной линии называют сублинией. Символ сублиния, который обознача-
ет определенного исследователя или лабораторию, включают в обозначение
линии: вслед за символом линии (СВА) пишут косую черту и за ней символ
сублинии СВА/Са (сублиния Картера линии СВА).
Сублинии могут разветвляться дополнительно на ветви (sublines), если
можно предположить генетические различия с основной группой животных
вследствие каких-либо манипуляций с частью ее членов — пересадкой яич-
ников и яйцеклеток, выкармливанием под самками-кормилицами другой
линии, выращиванием трансплантированных эмбрионов после криоконсе-
ации и т.д.
F — коэффициент инбридинга, или коэффициент возрастания гомози-
готности, предложен С.Райтом (1921) как мера степени родства между роди-
телями данной особи. Это родство (если имеется) определяется тем, насколь-
ко далеко отстает от них в родословной их общий предок (или предки). По
формуле Райта:
^=X[(l/2)"+ii1+(l + ^)], (37)
где Fx — коэффициент возрастания гомозиготности особи х; п — ряд в ро-
дословной с общим предком со стороны матери; я, — ряд в родословной
Глава 5. Принципы выбора животных для исследований
85
с общим предком со стороны отца; Fa — коэффициент инбридинга предка,
который сам инбридирован.
При разведении лабораторных животных значение F определяется мето-
дом разведения и (для неинбредных животных) величиной популяции.
В инбредных линиях теоретически F = 100% через 20 поколений братско-
сестринского инбридинга. В неинбредных популяциях F на поколение дол-
жен быть менее 1% (см. гл. 8, 11 и 12).
Отбор линейных животных
Отбор той или иной линии основывается, прежде всего, на генетических и
биологических особенностях каждой линии. Используются другие катего-
рии генетически контролируемых мышей: гибриды первого поколения (F1)
от скрещивания двух инбредных линий, конгенные и конгенно-резистент-
ные линии, мутантные линии и стоки, сложные гибриды (тетрагибриды),
рандомбредные мыши. Генетическая характеристика и специфика каждой из
перечисленных категорий мышей будет дана ниже. Выбор животных той или
иной категории определяется целью и характером самого исследования.
Ниже приводятся таблицы выбора для экспериментов и исследований,
адекватных животных-биомоделей, полученных в результате сложных гене-
тических преобразований. В левой колонке обозначены поведенческие ре-
акции, особенности метаболических процессов, уровни артериального дав-
ления, массы органов и тканей, наличие спонтанных опухолей, восприим-
чивость или резистентность к инфекциям и инвазиям, рождаемость, плодо-
витость, размеры детенышей, биохимические параметры крови, органов и
тканей, чувствительность к ксенобиотикам и фармакологическим агентам,
реакции на радиационные воздействия и различные факторы окружающей
среды. Этот список можно продолжить, но, думается, исследователь сам
найдет интересующие его феномены или проявления.
В правой колонке приводятся линии инбредных животных, у которых
обозначенные выше поведенческие, физиологические, биохимические, фар-
мако-токсические или гигиенические феномены нарастают слева неправо,
то есть усиливаются от наименьших значений к наибольшим. При необхо-
димости указаны половые особенности, возрастные показатели, характер от-
вета и иные обозначения.
Эти таблицы даны отдельно для разных видов инбредных животных и
позволяют исследователю уже в первом приближении оценить возможности
выбора оптимальных животных-биомоделей при планировании эксперимен-
тов, их проведения и определения сравнительных данных экстраполяцион-
ных процедур в процессе биомоделирования. Естественно, что эти таблицы
дают лишь некоторую интродукцию в проблеме выбора животных для экс-
перимента. По ходу изложения мы будем расширять, и комментировать эту
86
Качество животных-биомоделей
проблему. Подчеркну лишь, что более полные современные данные могли
бы составить содержание таких таблиц в значительно большем объеме, чем
предлагает читателям эта книга. Однако в перечне прилагаемой литературы
можно найти все, что к настоящему времени накоплено в мировой науке и
практике по данному вопросу.
Таблица 20
Распределение линий инбредных кроликов по характеру и выраженности
феноменов, эффектов или спонтанного поведения
Оцениваемые параметры
Гематокрит крови
Уровень гемоглобина в крови
Уровень азота мочевины
Иммунный ответ на Пли56, Лиз38, Тир6
Среднее артериальное
кровяное давление
Средний пульс
Линии животных, распределенные по выраженности
эффекта (от min к max)
вв: В, ACER С, OS, Y X, NIC, A, AXBU, IIIVO, АХ
99: ACER X В, АХ, С, OS, IIIC, AXBU, IIIVO, X, А
вв: X С, В, OS, ACER X, IIIVO, A, AX, AXBU, IIIC
99: X АХ, OS, С, X, В, AXBU, ACER NIC, IIIVO, A
вв:: A, C, X, OS, IIIVO, B, IIIC, AX, ACER Y, AXBU
99: A, C, OS, X, AX, ACER AXBU, IIIVO, IIIC, B, Y
OS, X, IIIC, AX, ACCRf, A, III, С
cJd:: OS, AC, X, C, IIIC, III, AXBU, AX, Y WH, A, ACER В
99: OS, AC, X, III, AXBU, C, AX, IIIC, WH, Y А, В, АСЕР
вв:: С, OS, AC, ACER AXBU, AX, A, III, X, Y B, IIIC, WH
99: III, AX, ACER OS, AXBU, B, AC, Y C, A, X, WH, HIE
Таблица 21
Распределение линий инбредных крыс по характеру и выраженности
феноменов, эффектов или спонтанного поведения
Оцениваемые параметры
Период выжидания до выхода
в знакомую среду
Период выжидания до выхода
в новую среду
Передвижение в открытом
пространстве
Дефекация в «открытом поле»
Линии животных, распределенные по выраженности
эффекта (от min к max)
вв: INR, A35322, TS3, WAG, A990,
MNR-a, MNR, MR, TS1, F344, ACI, IR
99: INR, A35322, WAG, TS3, A990, TSI, F344, MNR-a,
ACI, IR, MNR, MR
вв: INR.WAG, TS3, A35322, TSI, F344, A990, MNRA, IR,
MNR, MR, ACI
99: INR, TS3, WAG, A35322, IR, TSI, A990,
F344, MR, MNRA, ACI, MNR
вв: TSI, A990, ACI, A35322, F344, MNR, MR, IR, TS3,
MNRA, WAG, INR
99: TSI, A990, MNR, ACI, MR, F344, A35322,
MNRA.TS3, IR, WAG, INR
вв: MNRA, MNR, F344, TS3, WAG, TSI, INR, IR, ACI,
A35322, A990, MR
99: INR, MNRA, TS3, MNR, ACI, TSI, IR, WAG, F344,
A990, MR, A35322
Глава 5. Принципы выбора животных для исследований 87
Окончание табл. 21
Оцениваемые параметры
Активность в колесе
Содержание ГАМК в мозге
Печеночный метаболизм анилина
Печеночный метаболизм
этилморфина
Содержание гормона роста
в сыворотке
Содержание инсулина в сыворотке
Содержание тироксина в сыворотке
в возрасте 56 дней
Систолическое кровяное давление
Иммунный ответ на Гли52, Лиз33, Тир15
Иммунный ответ на синтетический
кополимер GA
Иммунный ответ на синтетический
кополимер GT
Восприимчивость
к экспериментальному
аллергическому энцефалиту
Восприимчивость
к экспериментальному
аллергическому энцефаломиелиту
Приплод: размер детенышей
Приплод: общая продуктивность
Линии животных, распределенные по выраженности
эффекта (от min к max)
3<5: А990, IR, TSI, MR, F344, ACI, TS3, INR, A35322,
MNRA, WAG, MNR
99: A990, F344, IR, INR, MR, TS3. TSI, ACI, MNRA,
A35322, WAG, MNR
WAG, RHAt, MNR, TMB, MR, TMD, RCA|, RLAf
вв: W, M520, ACI, LEW, ALB, OMt, SDt, BUF, F344, LE
99: ALB, W, OMt, M520, LEW. F344, SDt, LE, ACI. BUF
0*0*: W, SDt, BUF, LEW, LE, OMt, ALB, ACI, F344, M520
99: BUF, W, SDt, ACI, F344, ALB, LE. LEW, OMt, M520
WF, COP, F344, ACI, LEW
F344, COP, ACI, WF, LEW
ACI, COP, F344, WF, LEW
ACI, M520, WKY; WN = PETH, BUF = MNR, CAR, CAS =
LA, RLA, F344, ALB, NSD, RHA, OM, SHR
Низкий (Ag-B1): F344, LEW, S5B, CAS, CAR, NBR/1
средний (Ag-B2): WF, YO, NBR/2, OM
низкий (Ag-B3): BN, MAXX
высокий (Ag-B4): ACI, DA, COR ACP
высокий (Ag-B5): AUG, HO, PVG
низкий (Ag-B6): BUF, SD, M520, ALB
респондер: LEW, F344, WF, COP, AUG, BUF,
M520 не респондер: BN, ACI
респондер: ACI, COP, AUG
не респондер: LEW, F344, WF, BN, BUF, M520
резистентные: BN
чувствительные: LEW, DA, F344, NBR, CAS, CAR
чувствительные: AS, AGUS, LEW, AUG
резистентные: PVG/c, WAG
CAS, ACI, ALB, CAR, BUF, WN, M520, NSD, OM, MNR,
MR, F344
CAS, ALB, CAR, ACI, OM, WN, NSD, BUF, MR, MNR,
M520, F344
88
Качество животных-биомоделей
Таблица 22
Распределение линий инбредных хомяков по характеру и выраженности
феноменов, эффектов или спонтанного поведения
Оцениваемые параметры
Относительная масса сердца
взрослых хомячков
Относительная масса почки
взрослых хомячков
Относительная масса печени
взрослых хомячков
Подкожные опухоли у хомячков,
получавших бензо(а)пирен
Подкожные опухоли у хомячков,
получавших 3-метилхолантрен
Линии животных, распределенные по выраженности
эффекта (от min к max)
вв: ХЗ, 87.20, 1.5, 4.24, 54.7, 2.4, 12.14, 45.5, 86.93,
82.73, 1.26, Х68, 4.22
99: 87.20, 4.24, 12.14, ХЗ, 1.26, 1.5, 54.7, 86.93, 45.5,
2.4, 4.22, 82.73, Х68
66: ХЗ, Х68, 2.4, 54.7, 12.14, 4.24, 82.73, 1.5, 45.5,
86.93, 87.20,4.22, 1.26
99: 87.20, Х68, 4.24, 12.14, ХЗ, 45.5, 86.93, 1.26, 2.4,
1.5,82.73,4.22,54.7
63: 54.7, Х68, 12.14, 4.24, 82.73, 86.93, 1.5, 45.5, ХЗ,
1.26, 87.20,4.22, 2.4
99: Х68, 87.20, 82.73, 86.93, 54.7, 1.5, 4.24, 45.5,
1.26, 12.14, ХЗ, 2.4,4.22
66: 1.5, 12.14, 4.22, 82.73 = 54.7 = 86.93, 15.16, 45.5,
7.88, 87.20
99: 82.73, 54.7, 1.5, 86.93 = 87.20, 7.88, 4.22, 4.24,
12.14, 15.16,45.5
66: 82.73, 12.14=1.5, 54.7, 86.93, 4.22, 15.16, 87.20,
45.5, 7.88
99:86.93,4.22,82.73,1.5,12.14,54.7,45.5,15.16,
87.20, 7.88, 4.24
Таблица 23
Распределение линий инбредных мышей по характеру и выраженности
феноменов, эффектов или спонтанного поведения
Оцениваемые параметры
Приплод (количество детенышей
от самки в месяц)
Приплод (приплод/самка/неделя)
Приплод (размер детенышей
на момент отлучения от самки)
Результат разведения: размер
приплода
Линии животных, распределенные по выраженности
эффекта (от min к max)
C57BR/cd, CBAfB/Lw, C58/L.W, CBA/N, NH, NZW, STR/1,
NZB, C57L/B1, P/J, AKR/N, DBA/2, C57BL/10SC,
BRSUNT, A/He, C3Hf/He, STR/N, C57BL/Ka, AL/N, NGP,
NBL, C57BL/6, BALB/cAn
C57BL/Fa, NZB, CE, AKR, A2G-hr/+, DBA/1, 129/RrJ,
DBA/2, BI0.LP-a, A, C57L, B10.D2, NZW, C57BR/cd,
B10.A, C3H/He-mg, CBA/Ca, BALB/c, A2G, B10.BR,
СЗН/Не, СВА-Т6, C57BL/10ScSn, ICFW, NMRI
C57BL/Fa, NZB, NZW, AKR, A.B10.D2-n, DBA/1,129/RrJ,
DBA/2, BI0.LP-a, BALB/c, A2G-hr + , B10.A, A2G, CBA,
CBA-T6, C3H/He, C3H/He-mg, C57L, CE, C57BR/cd,
ICFW, B10.BR, C57BL/10, NMRI
первый: NZW, A, C3H, C3Hf, CBA/H, NZB
второй: NZW, A, NZB, C3Hf, C3H, CBA/H
третий: A, NZB, NZW, CBA/H, C3H, C3Hf
четвертый: NZB, A, NZW, CBA/H, C3H, C3Hf
Глава 5. Принципы выбора животных для исследований 89
Продолжение табл. 23
Оцениваемые параметры
Приплод (процент самок,
приносящих 4 и более детенышей)
Приплод (процент успешных
спариваний)
Размер детенышей
Размер детенышей
Процент преимплантационной
потери эмбриона
Процент выживаемости
новорожденных детенышей
Процент постимплантационных
потерь эмбрионов
Репродуктивные функции в возрасте
150 дней
Плодовитость
Генетическая идентификация
по 16 полиморфным участкам
Соотношение полов детенышей
при рождении
Процент особей, склонных
к употреблению алкоголя
Подверженность аудиогенным
припадкам в возрасте 2-6 недель
(приблизительно)
Избегание нахождения в подвижных
клетках
Стремление избежать стимуляции,
оказанной в пределах оптимальных
параметров
Подверженность электроконвульсив-
ному шоку в возрасте 2-6 недель
(приблизительно)
Эмоциональная дефекация
Исследовательская активность
в лабиринте
Прожорливость
Линии животных, распределенные по выраженности
эффекта (от min к max)
NZB, СВА/Н, NZW, А, СЗН, C3Hf
NZB, А, СВА/Н, NZW, СЗН, C3Hf
DBA/2, A, BALB/cHe, CBA, Rill, C57BL/6J, СЗН/Не,
СЗНеВ/FeJ
BALB/c, A/He, A/J, C3Hf/He, C57L, C57BL/6
BALB/c, CBA, СЗН, СО 13, C57BL, RF,
AKR/TIAId, AKR
Rill, A, DBA/2, C57BL/6, CBA, C3H/He, C3HeB/FeJ,
BALB/c
AKR, AKR^AId,C57BL,C3H,CBA,C013, RF,
BALB/c
DBA/2, Rill, A, BALB/c, CBA, C57BL/6, C3H/He
NB, DBA, A, SEA, SEC
SWR, AU/Ss, C57BL/KS, C57BL/6, C57L, C57BR/cd,
PL/J, P/J, SJL, BDR SM, DBA/2, BUB, BALB/c, C3H/HeJ,
ST/b, AKR/J, CBA/J, LP, C58, DBA/1, MA/J, 129/J, CE,
A/He, RIII/2J
At, C57BR/cdt LACAt, DBA/1, A2G, BALB/c, CBA/Ca,
C3H/He, LACGf, C57BL, CE|
DBA/2, BALB/c.A/2, C3H/2, C57BL
C57BL/6, DBA/1, C57BL/10J, DBA/2, BDR P/J
CBA, C3H/He, C57BL/6, C57BL/10, A/He, BALB/c, A/J,
C57BR/cd, DBA/2
BALB/c, 129/J, DBA/2, AKR/J, C3H/He, C57BR/cd,
C58/J, A/He, SWR
C57BL/10J, C57BL/6, DBA/1, DBA/2, BDR P/J
СЗНеВ/Fe, ND, AKR, C57BL/6, C3H, DBA/1, C57BL/10,
C57BR/a, DBA/2, A/CI, BALB/c, LP, BDR AK/e
99: A, C3H/2, DBA/2, BALB/c, I, C57BL, Rill
Qd: C3H/2, A, DBA/2, C57BL, Rill, BALB/c, I
A/CI, BDR BALB/c, C57BL/10, DBA/2, LP, C57BR/a,
AK/e, C57BL/6, ND, DBA/1, C3H, AKR, C3HeB/Fe
90 Качество животных-биомоделей
Продолжение табл. 23
Оцениваемые параметры
Внутривидовая агрессивность
Период выжидания до преодоления
преград на открытой местности
Период выжидания до выхода
из клетки
Обучаемость: количество попыток,
затраченных для определения
способа избегания шока
Обучаемость: процент ошибок
в стремлении избежать
одностороннего шока
Подвижность
Подвижность в течение 30 минут
Способность ориентироваться
в лабиринте Лешли III
Активность в открытом пространстве
Активность в исследовании
открытого пространства
Активность в «открытом поле»
Дефекация в «открытом поле»
Способность находить пищу
Хищническое поведение: период
выжидания до нападения на
насекомых
Частота опоры на задние
конечности в Y-лабиринте
Возможность подвергать условиям,
в которых необходимо избегать
шоков
Способность обучаться избеганию
шока
Линии животных, распределенные по выраженности
эффекта (от min к max)
А/Не, СВА, A/J, SWR, СЗН. RF, DBA/2, C57L, DBA/1,
C57BL/6, BALB/c, CFW, AKR, C57BR
99: C57BL, BALB/c, Rill, DBA/2, I, C3H/2, A
OO: I, C57BL, Rill, BALB/c, DBA/2, C3H/2, A
99: C3H/2, BALB/c, A, DBA/2, C57BL, I, Rill
OO: A, DBA/2, C3H/2, BALB/c, RIM, I, C57BL
C57L, C57BL/6, DBA/2, C3H/He, BALB/c, A/He, A/J
A, C3H, BALB/c, SWR, C57BL/6, CBA/J, CBA/Ca
CBA, BALB/c, DBA/2, C3H, C57BL/6
группа мышей: DBA/2, BALB/c. CBA, C3H/An, C57BL/6,
NLW
1 мышь: СВА, СЗН/Ап, DBA/2, NLW, BALB/c, C57BL/6
разница (1 группа): DBA/2. BALB/c, C57BL/6, C3H/An,
NLW, CBA
CBA, C57BL/6, C3H/He, A/He, BALB/c, C57BL/10, A/J,
DBA/2, C57BR/cd
A/He, A/J, BALB/c. CBA, C3H, DBA/1, DBA/2, SWR, AKR,
CFW, C57BL/6, C57L, RF, C57BR
A/CI, AK/e, BALB/c, C3HeB/Fe, C3H, AKR, LP, DBA/2,
BDR ND, DBA/1, C57BL/10, C57BL/6, C57BR/a
C3H, A/J, DBA/1, C57BL/10, C57BL/6, BALB/c,
C57BL/6-ala, SEC/2, SEC/1,
HRS-+ /hr, C57L, AKR, RFM
99: C3H, C57BL/10, DBA/1, BALB/c, A/J
OO: C3H, DBA/1, C57BL/10, BALB/c, A/J
A/J, C3H/He, RF, BALB/cJ, DBA/1, C57BL/10J
короткий: I, C57BL, C3H
средний: Rill, BALB/c
долгий: DBA/2, A
99: Rill, C3H/2, I, C57BL, A, BALB/c, DBA/2
OO: C3H/2, A, BALB/c, DBA/2, Rill, I, C57BL
CBA/J, C3H/He, C57BL/6, BALB/c, A/He, C57BL/10, A/J,
DBA/2, C57BR/cd
99: BALB/c, 129/J, AKR/J, C58, A/He, SWR
OO: 129/J, AKR/J, BALB/c, C58, SWR, A/He
Глава 5. Принципы выбора животных для исследований 91
Продолжение табл. 23
Оцениваемые параметры
Степень социального объединения
при встрече с агрессором
Частота случайного нажатия
на «рычажок»
Обучаемость в Т-лабиринте
Хлестание хвостом после
раздражения подушечек лап
Хлестание хвостом во время стычки
Обучаемость избегать воды
Внутривидовая агрессивность
Предпочтение алкоголя:
соотношение потребления
10% алкоголя и воды
Уровень эпинефрина надпочечников
Показатели активности арилгидро-
карбонгидроксилазы
Подверженность аудиогенным
припадкам (тесты на 21, 28, 35
и 42-й день жизни)
Показатели активности р-глюкорони-
дазы сперматозоидов
Базовый уровень сыворотки гормона
роста у самок на 78-й день
Базовый уровень сыворотки
пролактина у самок на 78-й день
Показатели специфичной активности
каталазы крови
РН крови
Артериальное кровяное давление
Уровень сахара крови
Уровень ацетилхолинэстеразы мозга
Показатели активности ацетил-
холинэстеразы мозга
Линии животных, распределенные по выраженности
эффекта (от min к max)
BALB/c, CFW, А/Не, C57BR, A/J, DBA/1, C57BL/6, АКР,,
DBA/2, SWR, C57L, СЗН, СВА, RF
SWR, CBA, A/J, C57BL/6, BALB/c, AKR, DBA/1, DBA/2,
A/He, CFW, C3H, C57L, C57BR, RF
A/He, SWR, C57BL/6, СВА, С58, C3H/He
A, DBA/2, BALB/c, C3H/2, C57BL/6
RF, CBA, C57L, C3H, SWR, C57BL/6, AKR, C57BR,
DBA/1, A/He, DBA/2, A/J, CFW, BALB/c
C3H/He, NMRI, ICFW, C57L, CBA-T6, C57BL/10ScSn
A/He, CBA, A/J, SWR, C3H/He, RF, DBA/2, C57L, DBA/1,
C57BL/6, BALB/c, CFW, AKR, C57BR
DBA/2J, DBA/2N, I, A/Crgl, A/Crgl/2, BALB/c, C3H/HeJ,
Rlll/An, C3H/Crgl/2, Rlll/Dm, C57BL/Ks, C57BL/Crgl,
C57BL/10J, C58/J, C57L, C57BR/cd, C57BL/6, C57BL
C57BL/Ka, C57BL/10, ++BALB/C, CBA, DBA/2
легкие: DBA/2, C57BL/6, AKR, SJL, NZW, NZB
печень: NZB, SJL, AKR, DBA/2, C57BL/6
подвержены: LP/J, DBA/2, AKR, SJL
среднеподвержены: LG, CBA/J, 129/J, RF, SM, BALB/c
устойчивы: C57BL/6
СЗН/Не, BALB/c, C57L, AU/Sc, DBA/2, A/J, C57BL/6J,
SM, SWR
высокий: DBA/2
низкий: C57BL/St, BALB/c, CBA/St, A/St, C3H/St
DBA/2, BALB/c, C3H/St, CBA/St, A/St, C57BL/St
XVII, C57BL, СЗН/Не, СВА, A/J, AKR,
венозной: DBA/1, SJL, SWR, DBA/2
= C3H/He = CBA/J, C57BL/6, A/He, AKR/J, 129/Re,
артериальной: C57BL/6, CBA/J = DBA/1 = 129/Re,
C3H/He, AKR/J, A/He = SJL, DBA/2, SWR
CPB-H, CPB-Q, C3H/Bi, CPB-R, CPB-W7, B10.LP, CPB-
Mo, CPB-Y NZB, CPB-P, CPB-FT, NZW, C57BL/UA, CPB-F
020, CPB-N, CP-V
AY KSA, AIM, NB, DBA/2, NBC, DDK, KSBf, C57BL/6J,
KK|, NC|
C57BL, Rill, C3H/2, DBA/2, A
C57BL/6, C57BR/cd, BALB/c, CE, CBA/Ca, LP, DBA/2
92 Качество животных-биомоделей
Продолжение табл. 23
Оцениваемые параметры
Уровень L-аминокислот
декарбоксилазы мозга
Показатели активности катехол-
О-метилтрансферазы мозга
Показатели активности
холинацетилтрасферазы мозга
Показатели активности
холинэстеразы мозга
Показатели активности декарбокси-
лазы глютаминовой кислоты мозга
Уровень декарбоксилазы
L-глютаминовой кислоты мозга
Показатели активности моноамин-
оксидазы мозга
Показатели соотношения плазмоло-
гена мозга на единицу фосфолипида
Содержание сфингозина мозга
на 1 мг протеина
Содержание стерола мозга
на 1 мг протеина
Содержание сульфатида мозга
на 1 мг протеина
Показатели активности тирозин-
гидроксилазы мозга
Показатели 45С-пробы (сердце)
Показатели 45С-пробы (скелетные
мышцы)
Циркуляция клеток, оцениваемая
по радиоактивности
Показатели активности энзима
N-метилникотинамида оксидазы
Показатели активности каталазы
эритроцитов
Показатели активности глюкозы
6-фосфатдегидрогеназы эритро-
цитов
Линии животных, распределенные по выраженности
эффекта (от min к max)
C57BL, Rill, DBA/2, A, C3H/2
LP, CBA/Ca, DBA/2, C57BR/cd, CE, C57BL/6, BALB/c
СЕ, C57BR/cd, CBA/Ca, C57BL/6, LP, DBA/2, BALB/c
DBA/2, C3H/2, C57BL, A, Rill
B10.BR, C57BL/10ScSn, C57L, ICFW, DBA/1, СЗН/Не,
A2G, CBA-T6, C3H/He-mg, NMRI
C57BL/6, C57BR/cd, CE, CBA/Ca, LP, DBA/2, BALB/c
BALB/c, LP, DBA/2, CE, C57BL/6, C57BR/cd, CBA/Ca
DBA/2, C57BL/6J-Awj, C57BL/6, SJL, BALB/c
C57BL/6J- AWJ, BALB/c, DBA/2, C57BL.6, SJL
SJL, C57BL/6J- Awj, BALB/c, C57BL/6, DBA/2
DBA/2, C57BL/6J-/Ty, BALB/c, SJL, C57BL 6
CBA, C57BL/10, DBA/2, C57BL/Ka, BALB/c
CBA/J, C57BL/6, SJL, DBA/2
CBA/J, C57BL/6, DBA/2, SJL
C57BR/cd, CBA/J, 129/J, DBA/2, SWR, BALB/c,
C57BL/10J, C57BL/6, AKR/J, C57L, SJL, C58,
RF, C3H/He, A/He, A/J, DBA/1
вв: DBA/2, A/J, BALB/c, C57BR/cd, C57BL/6, AKR,
C3HeB/FeJ
99: DBA/2, BALB/c, C57BR/cd, A/J, C57BL/6, AKR,
C3HeB/FeJ
NBL, C57BL/An, C57BL/He, SWR, C57BL/10, C57BL/6,
C57BL/Ka, C58, C57BR, DBA/2, C57L, YBR, AKR, C3Hf,
A, AL, DD, BALB/c
низкие: C57L, C57B/cdJ
средние: СЗН/Не, C57BL/6, DBA/2, SWR
высокие: A/J, A/He
Глава 5. Принципы выбора животных для исследований 93
Продолжение табл. 23
Оцениваемые параметры
Уровни эритроцитов никотинамид-
аденин динуклеотидфосфата
Показатели соотношения натрия
и калия в эритроцитах
Показатели активности глюкозы
6-фосфатдегидрогеназы
Активность печеночной бензапирен-
гидроксилазы
Активность печеночной
дельтааминолевулинатдегидратазы
Активность печеночной
аммонилазы
Активность печеночной урокиназы
Время гексобарбиталового сна
Уровень таламической гипоксантин-
гуанин фосфорибосилтрансферазы
Активность почечной
арилсульфатазы
Активность печеночной
арилсульфатазы
Активность печеночной пролин-
оксидазы
Активность печеночной
тирозинаминотрансферазы
Средний ритм сердцебиения
Скорость восстановления среднего
ритма сердцебиения в условиях
адаптации (например, адаптации
к удерживанию животных в руках)
Уровень тиреоидного гормона
Потребление кислорода в состоянии
покоя
Линии животных, распределенные по выраженности
эффекта (от min к max)
C57L, C57B/cdJ, C57BL/6, СЗН/Не, А/Не, SWR, A/J,
DBA/2
DBA/2, CBA/J, A/J, DBA/1, C57BL/6, WB, C57BL/10J,
BALB/c, 129/Sn
высокие: A/J, А/Не
средние: AKR, BALB/c, СЗН/Не, C57BL/6, С58, CBA/J,
DBA/2, RF, SEC/1 Re, SJL, SWR, 129
низкие: C57BR/cd, C57L
DBA/2, СЗН х C57BL/6F, C3H/St, A/St,
на: ICR (аутбредные), C57BL/6
C57BL/6, C57BL/KS, С58, C57BL/10Sn, C57BL/10J,
C57BL/He, DBA/2, AKR
СЗН/Mai, СЗН/HeJ, A/J, DBA/2, C57BL/6Mai, C57BL/6J
C57BL/6Mai, СЗН/Mai, C57BL/6J, A/J, C3H/He, DBA/2
66: CE, ICFW, NZW, AKR, NMRI, CBA/Ca, C57BL/Lac,
BALB/c, C57BR/cd, 129/RrJ, C3H/He, DBA/2, A2G, C57L
DBA/1
99: NZW, CE, NMRI, AKR, ICFW, BALB/c, DBA/2 = DBA/
1, C57BL/Lac, C3H/He, A2G = C57BR/cd = CBA/Ca,
C57L= 129/Rrj
LP, CBA/Ca, CE, DBA/2, C57BL/6, C57BR/cd, BALB/c
LG, A/He, C57L, MA/My, C57BR/cd, C57BL/6, AKR,
DBA/2, C3H/He, SWR, BALB/c, RF
C3H/He, MA/My, A/He, AKR, LG, DBA/2,
BALB/c, C57L, C57BR/cd, SWR, RF, C57BL/6
высокие: C57BL/6, ST/b, 129/Re, DBA/2
низкие: PRO
DBA/2, DBA/1, SWR, C57BL/10, C57BL/6, RF, C3H/He
129/J, BALB/c, LP, DBA/2, C57BL/6, C57BR/cd, SJL
LP, 129/J, BALB/c, C57BL/6, DBA/2, C57BR/cd,
BUC, YBR, C3H/Wi, BUB, C57BL/Fn
101, C3H, C57BL, CBA, A, SWR, DBA
94
Качество животных-биомоделей
Продолжение табл. 23
Оцениваемые параметры
Уровень метаболизма
Сон
Процентное соотношение
парадоксального или REM сна
Скорость проводимости
периферического нерва
Уровень холестерина в плазме
Показатели активности
холинэстеразы в плазме
Уровень тестостерона в плазме
Содержание порфирина в железах
у женских особей
Уровень простагландина
в эпидидимисе
Уровень кальция в сыворотке
Уровень холестерина в сыворотке
(у обоих полов)
Уровень гаптоглобина в сыворотке
Линии животных, распределенные по выраженности
эффекта (от min к max)
66: AKR/J, ST/b, A/J, P/J, CBA/J, СЗНеВ/Fe, BDP,
BALB/c, RR SM, DBA/1, 129/J, C57BL/6, C57BR/cd,
SWR, MA/J, SJL, DBA/2
99: AKR/J, CBA/J, СЗНеВ/Fe, P/J, RF, A/J, BDP, DBA/1,
ST/b, MA/J, BALB/c, C57BL/6, SM, SJL, SWR, DBA/2,
129/J, C57BR/cd
процент времени на сон: C57BR/cd, СЗН/Не, C57BL/6,
DBA/2, CBA, BALB/c
процент времени фазы медленного сна: C57BR/cd,
СЗН/Не, C57BL/6, DBA/2, BALB/c, CBA
процент времени на фазу парадоксального сна: BALB/
с, СВА, СЗН/Не, DBA/2, C57BL/6, C57BR/cd
соотношение дневного и ночного сна: BALB/c, СЗН/Не,
CBA, DBA/2, C57BL/6, C57BR/cd
CBA/J, А, СЗН/Не, BALB/c, C57BL/6, DBA/2, C57BR/cd
СЗН/Не, A/J, C57BL/6, DBA/1, CBA/J, BALB/cJ
66: 12-я неделя: C57BL/6, RF/J, BALB/c, CBA/J, SJL,
BDP, 129/J, LP
24-я неделя: C57BL/6, SJL, BALB/c, RF, CBA/J, 129/J,
BDP, LP
99:12-я неделя: C57BL/6, SJL, BALB/c, RF, CBA/J,
BDP, 129/J, LP
24-я неделя: SJL, C57BL/6, CBA/J, BALB/c, BDP,
129/J, LP
66: AKR, RF, SJL, LP, MA, SM, СЗНеВ/Fe, C57BR,
BALB/c, C57L, ST/b, CE, CBA/J, A, C58, DBA/2, C57BL/
6, PL, Rill, 129, SEC/1, AU
99: AKR, RF, C57BR, BALB/c, СЗНеВ/Fe, C57L, SJL,
C57BL/6, SM, ST/b, СЕ, А, С58, 129, CBA/J, SEC/1, RIM,
DBA/2, MA, PL, DBA/1, AU
B10.BR, C57BL/10ScSn, СЗН, В10.А, A
C57BL/6, BDP = C57L, C58 = C57BR/cd, SEC/1 Re,
DBA/2, RF/J, BALB/c, SJL, A/He, SWR, SM, CBA/J,
C3H/He, AKR/J
PGF. C57BL/6, DBA/2, CE, BALB/c, AKR/J, A/J
PGE: C57BL/6, DBA/2, CE, AKR/J, BALB/c, A/J
4 месяца: A/Fg, C3H/Fg, AKR, LCS, C57BL/Fg, C3H/He
7 месяцев: AKR, C3H/Fg, A/Fg, СЗН/HeJ, C57BL/Fg,
LCS
C57BL/6-ata, DBA/1, SEC/2, SEC/1, C3H
AKR, C3H, BALB/c, A, SWR, NBL, C57BL/6, C57BL/An,
C58, DD, DBA/2
Глава 5. Принципы выбора животных для исследований
95
Продолжение табл. 23
Оцениваемые параметры
Показатель активности
церулоплазмина в сыворотке
Систолическое кровяное давление
Температура
Осмолярность мочи
Чистый вес мозга
Надпочечники: частота возникнове-
ния вакуолизации в Х-зоне
Количество эритроцитов
Гематокрит крови
Гемоглобин крови (г/100 см3 крови)
Средний объем клеток крови
Процент ретикулоцитов крови
Вес мозга без полового различия
Соотношение веса мозга и тела
Линии животных, распределенные по выраженности
эффекта (от min к max)
66: BALB/c, C57BL/10J, C57BL/6, А/Не, A/J, C57L,
C57BR/cd, СЕ, C3HeB/Fe, AKR/J, ST/b, DE, SM, DBA/2,
С58, СЗН/Не, MA/J, SJL, LP/J, SWR, CBA/J, СВА/Са,
SEC/1 Re, 129/J, RF, DBA/1
99: ST/b, СЕ, С58, 129/J, SJL, DE, AKR/J, CBA/J,
C57BR/cd, A/J, BALB/c, A/He, C3HeB/Fe, C57BL/10J,
DBA/2, C57BL/6, MA/J, SM, SEC/1 Re, SWR, C57L, LP,
P/J.CBA/Ca, DBA/1, RF/J
СЗНеВ/FeJ, C57BL/10J, A/J, C57BR/cd, CE, C58/J,
129/J, DBA/1, DBA/2, AKR/J, СЗН/Не, C57BL/6J, A/He,
RF, SJL, CBA/J, C57L/J, BALB/c, SWR
ректальная: BALB/c, C57L, DBA/2, C3H, 101, C57BL,
CBA, CFW, A/He
конечностей: C57BL, C57L, A/He, DBA/2, C3H, BALB/c,
101, CFW, CBA
хвостовая: A, BALB/c, CBA, C3H, DBA/2, CFW, 101,
C57L, C57BL
STR, STR/1, C57L, AL/N, DBA/2, A/He, BALB/cAn
63: DBA/2, SJL, DBA/1, SM, BDP, ST/b, C57BR/cd, A/J,
129/J, AKR/J, C57BL/6, Rp MA/J, SWR, P/J, C3HeB/Fe,
CBA/J, BALB/c
99: DBA/1, C57BR/cd, SJL.DBA/2.A/J, STb,
129/J, C57BL/6, SM, BDP, MA/J, SWR, AKR/j, P/J, RF,
CBA/J, C3HeB/Fe, BALB/c
Rill, C57L, C57BR, SW, C3H, Af/A
СЗН/J, AKR, A/J, A/HeJ, Rill, СЗН/Sc, C57BL/6J,
C57BL/6-/7, C57L/He, ST/J, DBA/Wn, CBA/J, BALB/cAn,
l/J, DBA/2, BALB/cJ, DBA/1, C57BR/cd
СЗН/J, A/J, A/HeJ, DBA/2, СЗН/Sc, DBA/Wa, C57BL/6-p,
DBA/1, C57BL/6, ST/J, Rill, CBA/J, AKR, BALB/cAn, l/J,
BALB/cJ, C57BR/cd, C57L/He
ST/J, СЗН/J, DBA/Wa, DBA/2, A/HeJ, A/J, C57BL/6-p,
DBA/1, СЗН/Sc, C57BL/6, l/J, CBA/J, Rill, AKR,
BALB/cAn, C57BR/cd, C57L/He, BALB/cJ
DBA/2, DBA/1, DBA/Wa, ST/J, СЗН/Sc, C57BL/6-p, A/
HeJ, CBA/J, СЗН/J, A/J, C57BL/6, l/J, BALB/cJ,
BALB/cAn, Rill, C57BR/cd, AKR, C57L/He
DBA/1, ST/J, СЗН/Sc, AKR/J, C57BR/cd, l/J, C57BL/6-p,
C57BL/6, C57L/He, CBA/J, DBA/Wr, Rlll/J, СЗН/J, A/HeJ,
BALB/cJ, DBA/2, BALB/cAn, A/J
DBA/2, A/J, C57L, A/He, C57BL/6, BALB/c
RF, C3HeB/Fe, C58, CBA/J, A/J, AKR/J, C57BR/cd, ST/b,
C57L, DBA/2, SJL, LP/j, C57BL/6, Rill, MA/J, SWR, SM,
129/J, BALB/c, P/J
96
Качество животных-биомоделей
Продолжение табл. 23
Оцениваемые параметры
Вес тела на 21-й день жизни
Вес тела на 56-й день жизни
Вес тела на 112-й день жизни
Щетинки на подошвенной стороне
лапы
Типы клеток в аденогипофизе
(высокий/низкий процент каждого
типа)
Содержимое перитонеального
экссудата
Объем гиппокампа
Частота развития добавочной
селезенки
Иммунокомпетентные клетки
селезенки мышей
Объем неокортекса
Количество пейеровых бляшек
Процентное соотношение гемо-
глобинизированных и ретроидных
клеток в красной и белой пульпе
селезенки
Линии животных, распределенные по выраженности
эффекта (от min к max)
66: СЗН/Bi, AKR/Lw, C57BL/6Ka, C57BL/6, AL/N, DBA/
2, A/HeCr, STR, SM/J, C57L, СЗН/Не, BSVR, BRVS,
BALB/cAn, CBA/J, NZW, BRVR, CFW, SJL, C3Hf/He,
C57BL/10SC, NIH/PL, A/J, PRI/P1
99: СЗН/Bi, C3H/He, C57BL/6Ka, C57BL/6, AKR/Lw,
AL/N, SM/J, A/HeCr, C57L, STR, DBA/2, BRVR, NZB,
BRVS, CBA/J, C3Hf/He, BSVS, BALB/cAn, BSVR, NZW,
CFW/P1, SJL/J, NIH/P1, C57BL/10SC, A/J, PRI/P1
66: SM/J, A/HeCr, C57L, A/J, C3Hf/Hk, BALB/c, C57BL/
6, DBA/2, C57BL/6Kg, SJL/J, STR, СЗН/Bi, AKR/Lw,
BRVS, CBA/J, C57BL/10SC, BRVR, AL/N, NIH/P1, CFW/
P1, C3H/He, NZB, NZW, PRI/P1, BSVR
99: SM/J, C3Hf/He, A/HeCr, C57BL/6, C57L, SJL, A/J,
BALB/cAn, C57BL/6Ka, DBA/2, BRVR, STR, СЗН/Bi, AKR/
Lw, BRVS, NZB, CBA/J, C57BL/10Sc, CFW/P1, NIH/P1,
AL/N, BSVR, NZW, C3H/He, PRI/P1
66: SM, A/HeCr, C57L, C3Hf/He, DBA/2, BALB/c,
C57BL/6, SJL, A/J, AKR/Lw, AL/N, NZB, C3H/He, CBA/J,
NZW, PRI, C57BL/10SC
99: SM, C57L, A/HeCr, C57BL/6, BALB/c, DBA/2, C3Hf/
He, SJL, AKR/Lw, CBA/J, A/J, NZW, AL/N, NZB, C3H/He,
C57BL/10SC, PRI
CBA-T6, NMRI, A2G, C3H/He, NZW, B10.BR, BIO.LP-a,
B10.A
ацидофильные: A/Sy, C57BL/Sy, Rlll/Sy, C3H/Sy, DBA/Sv
базофильные: C57BL/Sy, A/Sy, C3H/Sy, DBA/Sy, Rlll/Sy
хромофобные: DBA/Sy, C3H/Sy, Rlll/Sy, C57BL/Sy, A/Sy
процент макрофаги: A/WySn, SJL/J, BALB/cAn, B10.A,
C57BL/10Sn
процент лимфоцитов: C57BL/10Sn, B10.A, SJL/J,
процент огранулоцитов: A/WySn, BALB/cAn, SJL/J,
B10.A, C57BL/10Sn
C57BL, DBA/2, SWR, RF, A/He, 129, CBA/J, MA,
C3H, CE, Rill, A, DBA/1, BALB/c, C58, C57BL/6, C57L
высокий: A/Ph, A.CA
средний: A.SW
низкий: CBA/J, СВАД6, C3H, NZB, C57BL/10, C57BL/6,
B10.BY B10.D2, B10.X B10.A, BIO.ARv, B10.LP, B10.ARII
DBA/2, 129, A/He, MA, SWR, C57BL/6, BALB/c,
CBA/J, RF
C57BL/6, A, C58, Rill, BALB/c, C3H, DBA/1
красные, 49 дней: PL, Af, MAf, AKR, C57BL
90 дней: Af, MAf, PL, C57BL, AKR
белые, 49 дней: MAf, PL, A, C57BL, AKR
90 дней: PL, MAf, C57BL, A, AKR
Глава 5. Принципы выбора животных для исследований
97
Продолжение табл. 23
Оцениваемые параметры
Рост детенышей с 0 до 12 дня
Относительный вес почки
Относительный вес вилочковой
железы
Вес селезенки на единицу массы
тела
Вес яичек на единицу массы тела
Соотношение веса вилочковой
железы и массы тела
Вес щитовидной железы на единицу
массы тела
Вес гипофиза на 78-й день у самок
Количество клеток перитонеального
экссудата
Дистрофическая кальцификация
сердечной мышцы у старых особей
Склонность к опухолям
Процент нерожденных мышат
с врожденными пороками развития
Линии животных, распределенные по выраженности
эффекта (от min к max)
DBA/2, C57BL/6J, Rill, A, СЗНеВ/FeJ, СВА,
СЗН/Не, BALB/cHe
C57BL/10J, А/Не, C57BL/6, Rlll/An, LP, A/J, ST/b, DE,
AKR/J, CE, SWR, C57L, C57BR/cd, СЗНеВ/Fe, СЗН/Не,
P/J, CBA/J, MA/J, BDp, BALB/c, HRS
14-й день: MA, C3Hf, C3H, Af, BALB/c, AKR, PL, C57BL
49-й день: СЗН, C3Hf, C57BL, MA, Af, BALB/c, PL, AKR
90-й день: C3Hf, C3H, Af, BALB/c, MA, C57BL, PL, AKR
на 1-й день: AKR, C3Hf, MA, PL, C3H, MAf, BALB/c,
C57BL
на 7-й день: AKR, MA, PL, C3H, C3Hf, C57BL, Af, MAf,
BALB/c
на 14-й день: MA, PL, AKR, C3Hf, C3H, Af, C57BL, MAf,
BALB/c
на 28 -й день: СЗН, MA, AKR, PL, Af, MAf, C57BL, C3Hf,
BALB/c
на 49-й день : PL, AKR, MA, Af, C3H, C57BL, BALB/c,
MAf, C3Hf
на 90-й день: PL, AKR, C3Hf, MA, Af, C57BL, C3H,
BALB/c
8 недель: AKR/J, CBA/Fa, CBA/J, C57BL/10J, C57BL/6,
DBA/A, YS
11 недель: AKR/J, C57BL/10J, CBA/KW, SJL, C57BL/6,
DBA/2, YS
A/He, C3H/He, BALB/c, C57BL, AKR
ST/J, CBA/J, DBA/2, CD-1t, C57BL/6
A/St, BALB/c, C57BL/St, DBA/2, CBA/St, C3H/St
A/WySn, SJL/J, BALB/cAn, B10.A, C57BL/10Sn
высокая вероятность: СЗН/J, СЗНеВ/Fe, DBA/1, DBA/2
средняя: A/J, BALB/c
низкая: C57BL/6, C57BL/10, C57L, Rill,
129/J, MA/J, SWR
девственницы 99: MA/J, ST/b, SM, SWR, C57BL/6,
129/J, DBA/1, P/J, BDP, BALB/c, A/J, C57BR/cd, DBA/2,
RF, Rill, СЗНеВ/Fe, LP/J, PL, SJL, CBA/J, C58, AKR
66: BALB/c = SM = MA/J, ST/b = DBA/1,
BDP, P/J, C57BL/6, Rill, 129/J, A/J, SWR, DBA/2, PL,
LP/J, C57BR/cd, SJL, СЗНеВ/Fe, RF, CBA/J, C58, AKR
ноль: BALB/cGrKt-tk, BT/Kt
0.1-0.5%: CBA/J, СЗН/J, DBA/1, 129/RrSu-oe
менее 0.5%: 129/RrSvKt-jt, A/J, C57BL/6JKt
98 Качество животных-биомоделей
Продолжение табл. 23
Оцениваемые параметры
Частота возникновения калыдифици-
рованного перикардита у особей
одного рода
Частота развития лейкоза: процент
частоты
Вероятность обнаружения вирусо-
подобных частиц в саркомах,
вызванных химическими веществами
Продолжительность жизни
в конвенционных условиях
Продолжительность жизни особей,
выращенных в SPF - условиях
Продолжительность жизни
в конвенциональных условиях
Вероятность лейкемии у самок
мышей (приблизительно, в различ-
ные периоды жизни)
Вероятность развития аденокарци-
номы молочной железы у конвенцио-
нальных особей
Дегенерация сетчатки и другие
схожие заболевания сетчатки
(диагноз на основании данных
гистологического исследования)
Вероятность возникновения аденомы
легких (среднестатистически,
за различные периоды жизни)
Вероятность случайного возникнове-
ния опухоли молочных желез
в 2-летнем возрасте
Линии животных, распределенные по выраженности
эффекта (от min к max)
CHI/St, DBA/St, C3H/St, C/St, DBA/2
0-10: A2G, BALB/c, DBA/LiA, DBA/LiAf, LIS/A, LTS/A,
020/A, RIII/ScA, RIII/ScAF, STS/A,WLL/BrA
11 -20: A/BrA, A/BrAf, AKR/FuAf, СЗН/HeA, CBA/BrA,
GRS/A, LTS/Af
21-50: AKR/FuA, BIMA/A, СЗН/Hef, C57BL/U, MAS/A,
TSI/A, TSI/Af
51-70: -71-100: AKR/FuRdA
A, BALB/c, C3Hf, C3H, AKR, C58
99: AKR/J, C58, P/J, BDP, ST/b, DBA/1, MA/J, SJL, A/J,
PL/J, CBA/J, BALB/cJ, SM, SWR, RF/J, C3HeB/Fe,
C57BL/6, 129/J, Rlll/An, C57BR/cd, DBA/2, LP
66: AKR/J, C58, SJL, PL/J, RF/J, BDP, SWR, P/J, ST/b,
CBA/J, BALB/c, MA/J, A/J, SM, 129/J, Rlll/An, СЗНеВ/
Fe, C57BL/6, C57BR/cd, DBA/2, DBA/1, LP
вв: AKR, NZB, C57L, CBA/Ca, DBA/1, CE, BALB/c, A/Gr,
C57BR/cd, C3H/He, DBA/2, A2G, C57BL, 129/RrJ, NZW
99: AKR, NZB, A/Gr, BALB/c, C57BL, C57BL, A2G,
C57BR/cd, 129/RrJ, C3H/He, DBA/1, CE, DBA/2, NZW,
CBA/Ca
66: BALB/c, A/J, DBA/2, C57BL/6
99: DBA/2, A/J, BALB/c, C57BL/6
высокая (< 3%): DBA/Li, WLL/Br, WLLf/A, LTS
средняя (3-10%): DBAf/A, Af/A, CBA/Br, 020
низкая (> 10%): C57BL/U, C3Hf/A, AKR/Fu, MaS/A
9, < 10%: C57BL/6, C57BR/cd, C57L, MA/My, P/J,
BALB/c, AKR
<5, > 10%: DBA/2, DBA/1, C3HeB/J (только инбредные),
C3H/He, A/J, A/He
анормальная: rd: BDP/J, CBA/J,
C3H/(10 подлиний), CFW, DA, FL, P/J, PL/J,
SJL, ST, SWR, WB/Re, WC/Re, WH/Re
нормальная: rd+: A (5 подлиний), A2G, AK, AKR, Bl, AU,
BALB/cGn, BALB/cJ, BSL, C/St, CBA/Ca, CBA/St, CE,
CHI, C57BL (5 подлиний), C57BR/cd, C57L, C58, DBA/1
(3 подлиний), DBA/2 (3 подлиний), DE, F, I, LG, MA/J,
RF/J, Rlll/An, SM, WK/Re, 129/J, 129/Rr
низкая: C57BL/UA, CBA/Br, C3Hf/A, DBAf/2, WLL/Br,
WLLf/A
высокая: 020/A, A/Br, Af/A, AKR/Fu, MaS/A
C57BL, TS, 020, MAS, BALB/c, C3Hf, GR
Глава 5. Принципы выбора животных для исследований
99
Продолжение табл. 23
Оцениваемые параметры
Линии животных, распределенные по выраженности
эффекта (от min к max)
Вероятность развития опухолей
у особей старых линий
все виды опухолей, 66: 129, DBA/2, LR СЗН, СВА,
C57BL/10, СЗН.К
99: 129, LP, C57BL/10, DBA/2, СВА
лимфома 66: LR 129, СВА, СЗН.К, DBA/2, СЗН,
C57BL/10
99: 129, LP, DBA/2, СВА, C57BL/10
гепатома, 66: 129, DBA/2, C57BL/10, LRC3H, СВА,
СЗН.К
99: ноль: DBA/2, СВА, C57BL/10
положительно: LR 129
гастрокишечные опухоли, 66: нуль: LP, 129, DBA/2,
СВА, C57BL/10
положительно: СЗН, СЗН.К
99: ноль: LR СВА
положительно: 129, DBA/2, C57BL/10
саркома тканей легких: 66: нуль: СВА, C57BL/10, СЗН
СЗН.К
положительно: 129, DBA/2, LP
99: ноль: C57BL/10, DBA/2
положительно: 129, СВА, LP
опухоли репродуктивных органов,
66: ноль: 129, DBA/2, СВА, C57BL/10, СЗН, СЗН.К
положительно: LP
99: ноль: C57BL/10
положительно: LR 129, СВА, DBA/2
опухоли легких, 66:129, DBA/2, СВА, СЗН.К
положительно: LR СЗН
99: ноль: DBA/2, СВА
положительно: 129, C57BL/10, LP
опухоли молочных желез, 66: нуль: 129, DBA/2, СВА,
C57BL/10, СЗН.К
положительно: LP
99: ноль: C57BL/10, 129
положительно: LR DBA/2, СВА
Канцерогенный эффект
9,10-диметил-бензантрацена
опухоли легких: C57BL, СЗН, СВА, IF, BALB/c, A
лейкемия: A, C57BL, IF, СЗН, СВА, BALB/c
опухоли печени: BALB/c, A, C57BL, IF, СВА, СЗН
Развитие амилоида после введения
казеина
C57BL, NIH (аутбредные), СЗН/Не, BALB/c, AKR, AL
Развитие амилоида после введения
эндотоксина Escherichia coli
AKR, NIH (аутбредные), BALB/c, C57BL, СЗН/Не, AL
Эозинофильная реакция на ацетат
кортизона
чувствительные: C57BL/6, C57BR/cd, C57BR/a
нечувствительные: 129/Rr, DBA/1, BALB/c
Фибросаркома: возникновение
опухоли
66: I. F, JK, CBAN, JKL, N, FC, С, СВА, С I, A/St,
C57BL, L, СЗН/St, CHI
99: F, I, JKL, N, CBAN, FC, JK, C, A, C12l, CHI, CBA,
C57BL, L, C3H/St
100 Качество животных-биомоделей
Продолжение табл. 23
Оцениваемые параметры
Время гексобарбиталового сна
Чувствительность к гистамину
Выживание в среде со 100% 02
при давлении в 3 атм.
Вероятность развития волчьей пасти
особей инбредных линий в ответ
на введение 2,5 мг кортизона
ежедневно в течение 4 дней,
начиная с 11 -го дня беременности
Индукция арилгидрокарбона
гидросилазы в ответ на ингаляцию
сигаретного дыма
Подверженность облучению: цент-
ральный нервный синдром после
больших доз рентгеновского
облучения
Опухоли легких: вероятность разви-
тия в ответ на уретан
Вероятность развития опухоли мо-
лочных желез, вызванных уретаном
3-метилхолантрен: провоцирование
развития опухолей кожи
3-метилхолантрен: вероятность
провоцирования развития подкожных
опухолей
Чувствительность к 3-метилхолант-
рену (доза Щ50, вызывающая
развитие опухоли)
Чувствительность к 3-метилхолант-
рену (провоцирование возникно-
вения опухоли)
Чувствительность к радиации
Чувствительность самок
к рентгеновскому излучению,
влияющему на репродукцию
Линии животных, распределенные по выраженности
эффекта (от min к max)
NIH, CFW/N, СЗН/Не, GP, BALB/c, STR/N,
C57BL/6, DBA/2, AL/N
C57BR/cd, C57BL, LACGf, CBA/Ca, DBA/1, BALB/c,
A2G, A/Lac, C3H/He
CBA/J, C3H/He, C3HeB/Fe, C57BL/6, RF, DBA/2, SWR,
AKR, C57BL/10, 129/J, C57L, A/He, DBA/1, SJL, A/J,
BALB/c, C58, C57BR/cd
Линии % детенышей, рожденных
СВА 12
C57BL 19
СЗН 68
DBA 92
А 100
C57BL/6, СЗН/Не, A/J, DBA/2, AKR, SWR
SWR, C57BL/6J, C57BR/cd, BALB/cJ, CBA/J
C57BL/Fa, CBA/Fa, Rill, JU, KL, A
C57BL, 020, TS, MAS, BALB/c, C3Hf, GR
I, BALB/c = C3H = Rill, C57BL
DBA/1, AKR/J, DBA/2, SJL, 129/J, SWR, C57BL/10ScSn,
BALB/cCr, C58, C3H/He, C57BL/6, B10.BR, C3H/fMai
СЗН/fMai, C3H/He, C57BL/6, BALB/cCr, BALB/c,
C57BL/6Mai, C57BL/6J, C57L
C57BL/6, СЗН/fMai, BALB/cCr, C57BR/cd, C3H/He, SWR,
C58, C57L, SJL, AKR/J, DBA/2, DBA/1
66: BALB/c, A/J, RF/J, SWR, C57BL/6, CBA/J, СЗНеВ/
Fe, SJL, C57BR/J, 129/J
99: BALB/c, SWR, A/J, C57BL/6, CBA/J, C3HeB/Fe, RF/
J, SJL, C57BR/J, 129/J
SEC, A, SEA, NB, DBA
Глава 5. Принципы выбора животных для исследований 101
Продолжение табл. 23
Оцениваемые параметры
Чувствительность к токсичности
хлороформа
Чувствительность молочной железы
к эстрадиолу и прогестерону
Чувствительность матки
к кратковременному введению
эстрогена
Чувствительность к тиреотропину
Чувствительность к общему
рентгеновскому облучению
Подверженность развитию карцино-
генеза ободочной кишки, вызванному
1,2-диметилгидразином
Подверженность возникновению
врожденной волчьей пасти в резуль-
тате введения кортизола в период
созревания
Восприимчивость к изониазиду
Чувствительность к озону
Подверженность возникновению
тератогенного эффекта в результате
применения кортикоидов
Подверженность возникновению
опухолей в результате нанесения
на кожу 4-нитрохинолина N-оксида
Тератогенный эффект ацетазоламида
Токсический эффект сальмонеллы
T.murium
Вероятность возникновения опухолей
после применения 3,4-бензопирена
Чувствительность к рентгеновскому
излучению
Количество антител, производимых
в ответ на введение антигена (АЬ)
Линии животных, распределенные по выраженности
эффекта (от min к max)
чувствительны: A, BALB/c, СЗН, HR, DBA/2
устойчивы: C57BL/6, C57BL, C57BR/cd, C57L, ST
чувствительны: Rill, I
промежуточность: DBA/2, СЗН, BALB/c
устойчивы: C57BL, А
СЗН/Не, C57BL/6, C3HeB/Fe, DAB/1, 129
C57BR/cd, C57BL/6, C57L, SWR, C58, СЗН/Не, СЗНеВ/
FeJ, BALB/c LAF C3D2F1, RF, 129, C57BL/10J, B6AF1,
AKR, A/He, AKD2F1, CAF1, DBA, A/J, CBA/J
CBA/J, BALB/cJ, СЗН/Не, Rlll/Pn, СЗНеВ/FeJ, A/HeJ,
DBA/1, A/J, AKR, DBA/2, SM/J, C57BL/6-C, C57BL/10,
C57BL/6, RF, C57BR/cd, LP, SWR, C57L, BDR MA/My,
C58, P/J, ST/J, SJL, MA/J, 129/J
чувствительны: SWR/J, P/J, ICR/Ha
устойчивы: DBA/Li, C57BL/Ha, C57BL/6, AKR/J
C57BL/10ScSn, C57BL/6, B10.A, A/J
BALB/c, C57BL/6 = 129/J, AKR/J, AJ = DBA/2, C57L,
C58 = SWR, C3H/He
NZB, CBA, AKR, RF, A, SJL, SWR, C3HeB/Fe, 129,
BALB/c, C57BR/cd, C57BL/10, C58, C57BL/6,
C57BL/6-cJ, C57BL/10cz-pd, WB/Re, DBA/2, C57e/Ha
гидрокортизон: C57BL/6, A/J, CBA, C57BL/10
кортизона ацетат: A/J, C57BL/10, CBA, C57BL/6
IF, C57BL x IFFj, A, C57BL, NZY
SWR, C3H/He, DBA/2, A/J, C57BL/6, CBA/J
устойчивы: СЗН/Не
чувствительны: NMRI, DE/J, C57BL/6, A/He, DBA/2,
CBA/J
AKR, C58, BALB/c, A, C3Hf, C3H
BALB/c, SWR, A/J, C57BL/6, C3HeB/Fe, CBA/J, RF,
C57BR/cd, SJL
B10.D2-n, C57BL, C3H, A/J, SWR, SIMPSON
102 Качество животных-биомоделей
Продолжение табл. 23
Оцениваемые параметры
Амилоидоз: чувствительность
к индукции с адьювантом Фрейда
Относительное сходство антител
Выработка антител в ответ на введе-
ние синтетической двойной скручен-
ной РНК
Выработка антител бляшкообразую-
щими клетками в ответ на пневмо-
кокковый полисахарид
Выработка антител в ответ на пневмо-
кокковый полисахарид III типа
Концентрация антител анти-DNR
после иммунизации на 11-й неделе
Эпсилон-антиген, антиген присут-
ствующий в жидкостных экстрактах
из опухолей и тканей
Восприимчивость к эксперименталь-
ному аллергическому энцефало-
миелиту
Иммунный ответ на декстран
Иммунный ответ на ферритин
Иммунный ответ на GAT
(Glu60, Ala30, Туг10)
Иммунный ответ на Про-Гли-Про-
овальбумин
Иммунный ответ на (Про66, Гли34)п
Иммунный ответ на (Про-Гли-Про)п
Иммунный ответ на синтетический
полипептид Гли57, Лиз38, Ала5
Первичный иммунный ответ
на альбумин бычьей сыворотки
Линии животных, распределенные по выраженности
эффекта (от min к max)
печень: AL/N, A/He, STR/N, C57BL/6, DBA/2.GP, AKR/N,
СЗН/HeN, NBL
лимф, узлы: А/Не, STR/N, AL/N, DBA/2, GP, AKR, C3H/
Не, NBL
сердце: AL/N, STR/N, A/He, C57BL/6, NBL, DBA/2, AKR,
GP, СЗН/Не
C57BL, B10.D2-n, CBA, SWR, СЗН, SIMPSON, A/J
BALB/c, C57BL/6, СЗН/Не, NZB, NZW, DBA, AL/N
C57BL/KS, CBA, DBA/2, 129, C57BL/6, A,
C3H/He, BALB/c
66: CBA/J, CBA/H, DBA/2, BALB/c
99: CBA/J, DBA/2, AKR/N, DBA/H, NZB, C57BL/6, CBA/
H-T6, C3H/He, BALB/c
DBA/2, BALB/c, C57BL/6, A/He, AKR, C3H/He, AL/N
эпсилон-позитивные: C58/J, DBA/1 J, DBA/2J, BALB/cJ,
MA/J, ST/J, 129/J
эпсилон-негативные: A2G, A/J, СЗН/HeJ, CBA/J, RF/J,
LG/J
резистентные: RF/J, BALB/c, DBA/2, ST/b, BRVR, BSVS
чувствительные (низко ... высоко) CBA/J, AKR/J, A/J,
129/J, C57BL/6, C57L, SWR, SJL
высокие респондеры: BALB/c, 129, C58, BAB/1
низкие респондеры: C57BL/6, SJL, CBA, AKR, A/He,
NZB
A-Thyl.l, C3H/He, AKR, I/St, DBA/2, A.SW, C57BL/6, 129,
BALB/c, A/J, B10.A, B10.BR,
C57BL/10, B10.D2, C57BL/6-7/a, B10.M
нуль-респондеры: SJL
респондеры (низкие...высокие): C57BL/6, C57L,
B10.BR, CBA, СЗН/Не, DBA/2, A/J, BALB/c, A/He
C57BL/6, DBA/2, BALB/c, SJL, DBA/1, AKR,
SWR = C3H.SW, C3H/He
SJL, C57BL/6, AKR, C3H/He, SWR, C3H.SW, DBA/2
DBA/2 = DBA/1, AKR, C3H.SW, SJL, SWR, BALB/c,
C3H/He, C57BL/6
респондеры: СЗН/Не, BALB/c, 129/J
не респондеры: C57BL/6, C57BL/10, A/J, CBA/J
C57BL, C3H, BALB/c, CBA, DBA/1, A/HeJ
Глава 5. Принципы выбора животных для исследований 103
Продолжение табл. 23
Оцениваемые параметры
Первичный гемагглютининовый
ответ на эритроциты овцы
Первичная гемолизная реакция
на эритроциты барана
Первичный иммунный ответ
на бактериофаг fd
Ответ на Vi антиген
Подверженность анафилактическому
шоку в ответ на введение
овальбумина
Чувствительность к эндотоксину
липополисахарида
Подверженность пассивной кожной
анафилаксии
Развитие лейкемии после заражения
Эктромелия: смертность при
естественном эпизоотии
Гипергликемия у мышей, инфициро-
ванных вирусом энцефаломиокардита
Индуцирование диабета вирусами
энцефаломиокардита
Летальный исход через 28 дней
после внутривенной инъекции
Mycobacterium marinum
Восприимчивость к Mycoplasma
fermentens
Возникновение случайных опухолей
молочной железы и связь с анти-
геном опухоли молочной железы
Чувствительность к Salmonella
typhimurium
Восприимчивость к Salmonella
typhimurium
Линии животных, распределенные по выраженности
эффекта (от min к max)
dose 3 х 107: А/Не, C57BL, СЗН, DBA/1, CBA, BALB/c
3 х 108: C57BL, А/Не, СЗН, BALB/c, CBA, DBA/1
3 х 109; C57BL, DBA/1, A/He, CBA, СЗН, BALB/c
Dose 3 x 107: A/He, C57BL, СЗН, CBA, DBA/1, BALB/c
3 x 108: A/He, CBA, C3H, C57BL, DBA/1, BALB/c
3 x 109: CBA, C57BL, C3H, A/He, DBA/1, BALB/c
BALB/c, STR, AKR, BALB/cHo, C3H/He, CBA/Mh, C57BL,
NMRI (аутбредные)
неспецифичные антитела: AL/N, DBA/2, C57BL
специфические (низкие...высокие титры): СЗН, BALB/c
высокая чувствительность: NC, IDT
промежуточная: CFW, DDD, ICR, DDY CF-1
низкая: КК, DBA/2, BALB/c, СЗН/Hef, C57BL/6, IVCS
Rill, СЗН/HeJ, СЗН/Crgl, C57BL/Crgl, A/Crgl
lgG1:C57BL/10J = B10A = B10.D2-n, = AKR, B10.BR =
A/J = A.By = A.SW, C3H/He, SJL/J, DBA/1 = DBA/2
IgE: C57BL/10J = DBA/1 = DBA/2, A.BY B10.BR = B10.A,
B10.D2-n, A/J, = A.SW = C3H/He = SJL, AKR
повышенная чувствительность: BALB/c, DBA/2J, DBA/
2N, BUB/BnJ, SJL
чувствительны: DBA/1, C3H/He, AKR/J, A/J
устойчивы: C57BL/6, C57L
AKR, C57BL/6, MA/Nd, MA, BALB/c, C3H, DBA/1, A
гипергликемия: DBA/2, CD-I
без гипергликемии: CBA/J, C57BL/6N, BALB/c, A/J
ненормальная толерантность к глюкозе: DBA/2, SWR,
SJL, C57L, A/He, C58, RF
нормальная толерантность к глюкозе: AKR, BALB/c,
CBA, СЕ, СЗН/Не, СЗНеВ/Fe, LP
101, C57BL, C57L, CFW, DBA/2, CBA, BALB/c, СЗН, А
C57BL/6, C58, AKR/J, BALB/c, DBA/1, C3H/He
BALB/c = 129/J = DBA/1, CBA/Jf, DBA/2, C57BL/6,
C57BR/cd, СЗНеВ/Fe, A/He, A/J, СЗН/HeJ, AKRut,
C57Lf
BALB/c, C57BL, B10.D2-n, DBA/2, C3H/He, A/J, CBA
C57BL/6, BALB/c, C3H/He, DBA/2, A/J
104 Качество животных-биомоделей
Окончание табл. 23
Оцениваемые параметры
Чувствительность к вирусу лейкемии
Friend В
Чувствительность к вирусу лейкемии
Friend S
Восприимчивость к вирусу простого
герпеса
Восприимчивость к вирусу
лимфоцитарного хориоменингита
Восприимчивость к вирусу лейкемии
Молони (процент лейкемии
у реципиентов)
Восприимчивость к вирусу гепатита
3-го типа мышей
Восприимчивость
к Plasmodium berghei
Восприимчивость к гельминтам
Opisthorchis felineus
Линии животных, распределенные по выраженности
эффекта (от min к max)
чувствительны: DBA/2, DBA/2f, NZB, NZW, SJL, CBA/J,
C3H, C3Hf, BALB/c, 1, T6, A, Af, A/J, СВА/Н, СВА-Т6
устойчивы: C57BR/cd, C58, C57BL/1, C57BL/6, C57BL/
10, C57BL/Ks, B10.D2-old, B10.D2-new
чувствительны: DBA/2, DBA/2f, NZB, NZW, SJL,
CBA/J, C3H, C3Hf, AKR, СВА/Н, СВА-Т6
устойчивы: BALB/c, I, T6, A, Af, A/J, C57BR/cd, C58,
C57BL/1, C57BL/6, C57BL/10
AKR, SWR, A, DBA, CBA, BALB/c, СЗН/Не, C57BR,
C57BL/6, C57BL/KS, C57BL/10
C3H, B10.D2-O, B10.D2-n, Websterf, SWR.
NH, C58, C57BL/Ka, C3Hf/Fg, AKR, RF, C3Hf/Bi, C3H/Lw,
BALB/c
устойчивы: A/J, A/Orl
промежуточные: CBA/Orl, C3H/He, СЗН/Umc, CBA/H-
T6, AKR, l/Umc
чувствительны: BALB/c, NZB, B10.D2-O, C57BL/6, A/Ps
DBA/1, C58, DBA/2, A/He, AKR, A/J, ST/b, C57L
DBA/2, CBA/Lac, A/He, SWR/J, CC57W/Mv, C57BL/6J
105
Глава 6 БИОБЕЗОПАСНОСТЬ ПРИ РАБОТЕ
С ЛАБОРАТОРНЫМИ ЖИВОТНЫМИ
При проектировании помещений для лаборатор-
ных животных следует предусмотреть их изоляцию
от персонала лаборатории в случае необходимос-
ти, а также возможность деконтаминации и дезин-
фекции.
Проектирование помещений для животных, так-
же как и лабораторий, должно осуществляться
с учетом степени опасности исследуемых микроор-
ганизмов; здесь также выделяют четыре уровня био-
логической безопасности (1, 2, 3, и 4). Во внимание
принимаются и иные факторы, такие, как объем
и концентрация исследуемого патогенного агента,
путь заражения животного и возможный путь выве-
дения агента из организма животного. Учитывают
особенности животного: его агрессивность или спо-
собность кусать или царапать, его природных экто-
и эндопаразитов, зоонозные заболевания, которым
подвержено данное животное, а также возможное
распространение аллергенов.
Определенные требования существуют и к ос-
нащению лаборатории, ее проектированию, причем
меры предосторожности усиливаются по мере воз-
растания уровня биологической безопасности лабо-
ратории.
Эти требования изложены и кратко представ-
лены в табл. 24.
106
Качество животных-биомоделей
Таблица 24
Лабораторные животные и уровни биологической безопасности: свод правил
работы и перечень безопасного оборудования
Группа
риска
1
II
III
IV
Уровень
безопасности
1
2
3
4
Лабораторная практика
Ограниченный доступ,
защитная одежда и перчатки
Ограниченный доступ и знаки
биологической опасности;
защитная одежда и перчатки;
деконтаминация отходов и
клеток перед уборкой
Контролируемый доступ;
специальная защитная
одежда; остальное, как для
уровней 1 и 2
Строго ограниченный доступ;
3-й уровень деятельности
плюс помещения для
переодевания и душевые; все
отходы деконтаминируются
перед удалением
из помещения
Оборудование
обеспечивающее
безопасность
Средства личной защиты
и гигиены
I и II классы боксов
биологической безопасности
(БББ) для видов работ,
сопровождающихся
образованием аэрозолей;
средства личной защиты
I и II классы БББ для всех
видов работ; средства
личной защиты
Класс III БББ или ламинар-
ные костюмы с положитель-
ным давлением
Уровни биологической безопасности
Первый уровень биологической безопасности. К этому уровню биологи-
ческой безопасности относится большинство лабораторных животных (за
исключением приматов), а также животных, зараженных инфекционными
агентами группы риска I. При работе с ними требуется хорошая микробиологи-
ческая техника (ХМТ).
Второй уровень биологической безопасности. Сюда относится работа с
лабораторными животными, зараженными инфекционными агентами груп-
пы риска II. Необходимо соблюдение следующих предосторожностей:
/ на дверях и в соответствующих местах должны вывешиваться знаки
биологической опасности;
/ проектирование должно предусматривать эффективную очистку
и уборку помещений;
/ двери должны открываться и легко закрываться;
/ отопление, вентиляция и освещение должны быть адекватными;
/ при механической вентиляции она должна быть приточной и обеспе-
Глава 6. Биобезопасность при работе с лабораторными животными
107
чиваться путем выброса воздуха в атмосферу. Воздух не должен рецир-
кулировать в остальных помещениях, т.е. используется система «тоталь-
ного выброса»;
/ доступ разрешается только ограниченному кругу лиц;
/ содержаться должны лишь животные, используемые в эксперименте;
/ должна быть программа наблюдения за членистоногими и грызунами;
/ необходимо использовать средства защиты от насекомых;
/ после использования рабочие поверхности следует деконтаминировать
соответствующими дезинфектантами;
/ боксы биологической активности уровней 1 и 2 должны быть приспо-
соблены для работы с образованием аэрозолей;
/ автоклав должен быть расположен в помещении или рядом с ним;
/ подстилки из клеток животных удаляются с минимальным образова-
нием аэрозолей или пыли;
/ перед удалением подстилки все использованные материалы подлежат
деконтаминации;
/ материалы для автоклавирования или сжигания транспортируются
в безопасных закрытых контейнерах;
/ после использования клетки животных подлежат деконтаминации;
/ трупы умерщвленных животных подлежат сожжению;
/ должны быть предусмотрены устройства для мытья рук. Персонал дол-
жен мыть руки перед уходом из помещений, где содержатся животные;
/ в помещении следует носить защитную одежду, снимаемую при уходе
из лаборатории. Следует предусмотреть наличие перчаток;
/ все травмы, даже незначительные, должны быть запротоколированы;
/ в помещениях, где содержатся животные, запрещается прием пищи
и воды, курение, использование косметики.
Третий уровень биологической безопасности. Относится к работе с лабо-
раторными животными, зараженными патогенными агентами группы риска III.
Приемлемы все рекомендации, разработанные для лабораторий с уровнем
биологической безопасности 1 и 2, а также:
/ доступ в лаборатории должен быть строго ограничен;
/ помещения, в которых содержатся животные, должны быть отделены
от остальной части лаборатории и вивария воздушным шлюзом с дву-
мя дверями;
/ при входе в помещения устанавливаются раковины для мытья рук
и душ;
/ должна быть предусмотрена механическая вентиляция с поступлением
воздуха во все комнаты. Отработанный воздух фильтруется через ВД-
фильтры перед выбросом в атмосферу («полный выброс»). При проек-
тировании системы предусматриваются устройства, препятствующие
108
Качество животных-биомоделей
случайному обратному току воздуха и его нагнетанию ко всем клеткам
с животными;
/ в помещении устанавливается автоклав;
/ сжигатель должен находиться в помещении и быть легкодоступным.
Его можно заменить иным аналогичным прибором с согласия соответ-
ствующих служб;
/ животные, инфицированные патогенными агентами группы риска 3,
помещаются в клетки, изолированные от остального помещения, или
в лаборатории перед вытяжными отверстиями вентиляции;
/ подстилки для животных при использовании должны образовывать ми-
нимальное количество пыли;
/ в помещении носится защитная одноразовая одежда. При выходе из
помещения она снимается и автоклавируется перед уничтожением;
/ следует обеспечить регулярную иммунизацию персонала.
Четвертый уровень биологической безопасности. Работа в данном поме-
щении обычно тесно связана с работой в максимально изолированной лабо-
ратории — группа риска IV. Поэтому правила, разрабатываемые наци-
ональными и местными органами здравоохранения, должны соответство-
вать обоим типам лабораторий. Применяются все правила работы с лабора-
торными животными, разработанные для уровней биологической безопас-
ности 1, 2 и 3, а также:
/ доступ строго контролируется (наличие ключей), вход возможен лишь
лицам, имеющим разрешение директора данного учреждения;
/ персоналу не разрешается работать в одиночку, используется правило
«работы вдвоем»;
/ персонал должен иметь прекрасную подготовку по микробиологии
и знать все факторы риска, сопряженные с работой, а также необходи-
мые меры предосторожности;
/ если рабочее помещение не является частью максимально изолирован-
ной лаборатории — с обозначением «уровень биологической без-
опасности», то оно должно быть расположено в отдельном здании;
/ вход в помещение осуществляется через воздушный бокс, перед входом
в который с «чистой» стороны располагаются раздевалки и душевые;
/ помещение вентилируется через ВД-фильтры вытяжной вентиляцией,
обеспечивающей отрицательное давление;
/ при проектировании вентиляционной системы должна быть ликвиди-
рована случайная возможность обратного тока воздуха, а также созда-
ния в помещении положительного давления;
/ в комнате рядом с лабораторным помещением должен находиться ав-
токлав с двумя дверцами;
/ при входе в лабораторию персонал обязан снять уличную одежду и на-
Глава 6. Биобезопасность при работе с лабораторными животными 109
деть защитную одежду одноразового использования. После окончания
работы одежда снимается и помещается в специальные емкости для ав-
токлавирования и уничтожения; персонал обязан принять душ;
/ для поступления материалов используется воздушный бокс;
/ все манипуляции с животными производятся в боксах 3-го класса био-
логической безопасности;
/ все животные должны находиться в изоляторах;
/ перед удалением из лаборатории все отходы и подстилки для живот-
ных автоклавируются;
/ иммунизация персонала и медицинский контроль осуществляются
в соответствии с указаниями и международными правилами.
Правила биозащиты персонала
При работе с лабораторными животными в качестве абсолютного посту-
лата следует принять, что здоровый персонал более ценен для учреждения, чем
самое высокое качество животных.
Содержание лабораторных животных в питомниках и эксперименталь-
но-биологических клиниках (ЭБК) создает потенциальную возможность вза-
имного инфицирования человека и животных (антропозоонозы), а также
перекрестного инфицирования животных, во избежание чего необходимо
строгое соблюдение правил личной гигиены работников.
Персонал, работающий с животными, дважды в год должен обследо-
ваться в медицинском учреждении по правилам, действующим для пище-
вой промышленности. Носители патогенной для животных флоры, вклю-
чая антропозоонозы, как и больные люди, к работе с животными не до-
пускаются.
Персонал питомника и ЭБК должен быть обеспечен требуемым количе-
ством спецодежды, санитарная одежда должна быть персонально маркиро-
Таблица 25
Перечень спецодежды для персонала в зданиях барьерного типа
Наименование предметов
1. Санитарные брюки
2. Санитарные халаты
3. Колпаки
4. Маски
5. Носки
6. Трусы
7. Нижняя рубашка (майка)
8. Полотенце
9. Тапочки
Количество на 1 чел.
в «чистой» зоне
6 пар
6 шт.
6 шт.
6 шт.
6 пар
6 пар
6 шт.
6 шт.
4 пары
Количество на 1 чел.
в «грязной» зоне
4 пары
4 шт.
4 шт.
4 шт.
4 пары
4 пары
4 шт.
4 шт.
2 пары
по
Качество животных-биомоделей
вана. Ее количество на одного работника «чистой» зоны в питомниках и
ЭБК барьерного типа должно позволять стирку не реже чем через 1 день и
ежедневное автоклавирование. Комплект спецодежды для персонала пред-
барьерной (конвенциональной) зоны имеет тот же набор одежды, но в коли-
чествах, обеспечивающих возможность стирки не реже двух раз в неделю.
Таблица 26
Перечень спецодежды для персонала при конвенциональном
содержании животных
Наименование предметов
1. Санитарные брюки
2. Санитарные халаты
3. Колпаки
4. Полотенце
5. Тапочки
Количество на 1 чел.
в «чистой» зоне
4 пары
4 шт.
4 шт.
2 шт.
2 пары
Количество на 1 чел.
в «грязной» зоне
4 пары
4 шт.
4 шт.
2 шт.
2 пары
Необходим утвержденный обязательный перечень спецодежды для работни-
ков «чистой» и «грязной» зон.
Санитарная одежда подлежит стирке 1 раз в 3 дня. При содержании
улучшенных конвенциональных животных одежда подвергается автокла-
вированию.
Резиновые перчатки (хирургические и бытовые) должны быть в доста-
точном количестве из расчета на одного человека 2 пары в месяц. Работники
дезинфекционно-моечного отделения должны обеспечиваться резиновыми
сапогами и прорезиненными фартуками. Руководители и другие сотрудни-
Таблица 27
Содержимое комплектов первой помощи
Предметы
Руководство по биобезопасности
Индивидуально упакованный стерильный
лейкопластырь
Стерильная повязка на глаза
Треугольная повязка
Стерильная повязка для наложения
на обширные раны
Безопасные булавки
Стерильная лекарственно необработанная
повязка среднего размера
Стерильная лекарственно необработанная
повязка большого размера
Стерильная лекарственно не обработанная
повязка очень большого размера
Количество сотрудников
1-5
1
10
1
1
1
6
3
1
1
6-10
1
20
2
2
2
6
6
2
2
11-50
1
40
4
4
4
12
8
4
4
Глава 6. Биобезопасность при работе с лабораторными животными
111
ки, не имеющие доступа в «чистую» зону, пользуются санитарными халата-
ми, легкой обувью и пластиковыми бахилами.
Вход персонала в «чистую» зону помещения барьерного типа произво-
дится через санпропускник, работник снимает одежду, кольца, цепочки, серь-
ги и др. и оставляет их в шкафу «грязной» раздевалки санпропускника, тща-
тельно принимает душ, используя мыло, мочалку, банную щетку, вытирается
досуха полотенцем и одевается в полный комплект стерильной одежды. Очки
протираются дезраствором. В течение рабочего дня работник из чистой зоны
не выходит. В случае экстренного выхода процедура входа в «чистую» зону
через санпропускник полностью повторяется заново. Таким же образом вхо-
дят в чистую зону сотрудники, выполняющие эксперимент на животных и
при необходимости технические работники. Рабочий инструмент подверга-
ется автоклавированию или дезинфекции и проводится через шлюзы.
Персонал обеспечивается в необходимых количествах мылом и кремом
для ухода за кожей. В помещениях с животными питомника и ЭБК необхо-
димо иметь емкости с дезинфицирующими растворами для обработки рук.
Сотрудники, работающие с лабораторными животными, должны ежегод-
но проходить медицинское обследование. Особое внимание необходимо
обращать на симптомы аллергии и нарушения опорно-двигательного аппа-
рата. Для подтверждения или исключения причин заболеваний могут потре-
боваться дополнительные лабораторные анализы. Респираторные функции
сотрудников должны контролироваться независимо от их нарушений.
Рекомендуется проводить раз в 3 года медицинское обследование персо-
нала, сходное с проводимым при приеме на работу, включающее сдачу об-
разцов сыворотки.
Работа с лабораторными животными и производство биологических об-
разцов для опытов требуют специализированной подготовки. Особое значе-
ние имеет квалификация сотрудников питомников. Работа с лабораторными
животными требует соблюдения норм биобезопасности, так как существует
опасность инфицирования человека болезнетворными бактериями и микро-
организмами. Человек может являться носителем микробов, способных пе-
редаваться животным. Животные и пыль, находящаяся в лабораториях, где
их содержат, являются сильнейшими аллергенами, опасными для людей
работающих в подобных лабораториях. Лица, работающие с животными в
условиях лаборатории, должны проходить проверку перед назначением и
обучением.
Эргономика
В процессе подготовки персонала для лабораторий, в которых содержатся
животные, необходимо уделять должное внимание наблюдению за состоя-
нием здоровья персонала и принятию превентивных мер. Работа должна
112
Качество животных-биомоделей
быть организованна таким образом, чтобы избегать излишней физической
нагрузки на конечности сотрудников, а также на плечевой пояс и спину.
Многие нарушения опорно-двигательной системы имеют тенденцию к хро-
низации.
Уход за животными подразумевает частое поднимание самих животных,
их клеток, поилок и корма. Если в клетках есть подстилки, то они должны
быть вычищены раз или два в неделю, иначе будет превышен уровень амми-
ака. Клетки с животными часто расположены на стеллажах в несколько ря-
дов. Ввиду ограниченности пространства клетки располагают как на очень
низком, так и на очень высоком уровне. Для того чтобы достать клетку с
нижнего стеллажа придется нагибаться, а с верхнего — тянуться с руками
поднятыми под углом больше 90°. Нагрузка в таких случаях существенно
превышает нормальную. Большая нагрузка возникает и при работе с изоля-
торами. Даже в гибком пластиковом изоляторе сложно достать до всех углов,
и приходиться поднимать существенный вес на вытянутых руках. Исследо-
вания показали, что при работе с изоляторами нагрузка на треть превышает
максимальное добровольное мышечное сокращение. Сотрудники лаборато-
рий в связи с этим должны уделять внимание своей физической подготовке
и состоянию здоровья.
Сотрудники лаборатории должны представить личную и семейную исто-
рию болезни, касающуюся аллергий. Уже существующая аллергия на какой-
либо вид животного обычного для лаборатории является явным противопо-
казанием. Нарушения опорно-двигательного аппарата, дыхательной систе-
мы и кровообращения также препятствуют утверждению кандидатуры, по-
скольку работа с животными предполагает высокую физическую нагрузку.
При медицинском обследовании до приема на работу рекомендуется прово-
дить тесты на аллергические реакции. Также необходимо проверять канди-
дата на носительство некоторого вида бактерий. Лицо, в организме которого
эти бактерии присутствуют, не должен работать с животными, поскольку
может передать им эти бактерии. Необходимо также вести наблюдение за
респираторными функциями, сделать флюорографию. Рекомендуется брать
образцы сывороток, для того, чтобы в случае возникновения зоонозов, мож-
но было провести анализы.
Зоонозы
ВОЗ зарегистрировала до 200 зоонозов, то есть заболеваний прямо или кос-
венно передающихся от животных человеку. При прямой передаче для воз-
никновения зоонозов необходим только контакт. Циклозоонозы вызваны
микроорганизмами, для размножения которых необходимы 2 вида позво-
ночных существ. Возбудители метазоонозов развиваются в беспозвоночных
организмах, например в насекомых. Сапрозоонозы могут сохраняться в мине-
Глава 6. Биобезопасность при работе с лабораторными животными 113
Таблица 28
Биологические опасности антропозоонозов и групповые номера (ГН) уровня
опасности для человека в классификации Консультативного комитета ВОЗ
по опасным патогенам (2002)
Виды патогенов
Возбудители и их «хозяева»-животные
Групповые номера
Кролики, крысы, хомячки, мыши и другие грызуны
Бактерии
Лептоспиры
Вирусы
Грибы
Паразиты
и гельминты
Salmonella (например, S. Typhimurium)
Yersinia pseudo-tuberculosis
Actinobacillus moniliformis (крысиная лихорадка)
Leptospira (различные виды)
Lymphocytic choriomeningitis
Sendai (вирус)
Hantaan (вирус)
Trichphyton
Micosporon
Hymenolepisnana
2
2
2
2
3
2
3
см см
2
Птицы и другие летающие виды
Бактерии
Вирусы
Грибы
Salmonella
Вирус болезни Newcastle
Пситтакозы и орнитозы
Aspergillus
Candida
2
со го
ю ю
Собаки, кошки и другие хищники
Бактерии
Лептоспиры
Вирусы
Грибы
Паразиты
и гельминты
Salmonella
Pasteurella
Возбудитель кошачьей лихорадки
Leptospira иктерогеморрагическая
Кроличьи вирусы (специфические)
Microsporum
Toxoplasma Гонди
Тохосага (собачья)
Ancylostoma
Echinicoccus granulosus
Sarcoptes scabiei
Ctenocephalides (собачья)
Ctenocephalides (кошачья)
2
2
Не классифицирован
2
3
2
3
2
2
3
Не классифицирован
Не классифицирован
Не классифицирован
Мини-свиньи, овцы и иные крупные животные
Бактерии
Вирусы
Паразиты
и гельминты
Bacillus anthracis (сибирская язва)
Brucella species
Mycobacterium tuberculosis
Staphylococci
Clostridia
Erysipelothrix
Q fever (Coxiella burnetti)
Cow Pox (чума коров)
Entamoeba histolytica
Giardia
Balantidium coli
3
3
3
2
2
2
см см
см см см
114
Качество животных-биомоделей
ральной почве в течение долгого времени. Наиболее часто встречающиеся
зоонозы: туберкулез, бруцеллез, оспа, псевдотуберкулез, сальмонеллез, рожа,
пастереллез, листереллез, бешенство, орнитоз, пситтакоз, трихофития, ток-
соплазмоз, эхинококкоз, тениоз, туляремия и ботулизм. Многие из них
вызывают у человека тяжелые заболевания.
Аллергии
Развитие аллергических реакций одна из серьезнейших проблем работы в
лаборатории с животными. У четверти сотрудников в процессе работы раз-
вивается аллергия на перхоть, мочу или кровь животных. Продолжитель-
ность нахождения в контакте с аллергеном до возникновения симптомов
может быть от полугода до двух лет.
Исследователи, находящиеся в долгом контакте с животным, предназна-
ченным для опытов, могут проявлять гиперчувствительность. Поэтому ран-
ний перевод животных в исследовательские лаборатории нежелателен. Ины-
ми словами, с точки зрения биобезопасности, чем меньше контакт с живот-
ными и ближе к началу эксперимента, тем меньше риск заболевания.
Из лиц, изначально страдавших от ринита или конъюнктивита, у поло-
вины, скорее всего, разовьется астма. Чаще всего пациенты реагируют на
аллергены различных животных. Профилактика заключается в использова-
нии перчаток и масок при контакте с животными. Животные должны содер-
жаться отдельно от рабочих мест сотрудников.
У гиперчувствительных пациентов часто обнаруживали аллергические
реакции на пыль, состоящую из эпителиальных клеток животных. Белки,
содержащиеся в выделениях животных, таких как моча или слюна, также
являются мощными антигенами.
Исследования показали, что уровень содержания животного протеина в
воздухе обратно зависим от степени вентиляции помещения. Количество
перхоти меняется в зависимости от проводимых работ. В связи с этим реко-
мендуется оснащение лаборатории локальной вентиляцией и ношение пер-
соналом индивидуальных масок. Важно также развивать направления ежед-
невной работы, нацеленные на снижение количества перхоти в воздухе.
Исследователи и техники также находятся в контакте с потенциальными
аллергенами, работая с животным или его тканями. Они должны соблюдать
те же правила, что и основные сотрудники.
Несмотря на следование правилам обращения с животными, последние
бодаются, царапаются, а иногда и кусаются. В момент укуса в ткани челове-
ка заносятся не только антигены, но болезнетворные бактерии, чего следует
не только остерегаться, но и серьезно опасаться.
115
Глава 7 БИОРИСКИ И КАЧЕСТВО
ЭКСПЕРИМЕНТОВ
НА ЛАБОРАТОРНЫХ ЖИВОТНЫХ
Крупные животные (мини-свиньи, овцы,
бараны)
Мини-свиньи все шире используются в медико-
биологических целях. Из всего разнообразия лабо-
раторных животных, пожалуй, самыми «молодыми»
можно назвать мини-свиней. Именно мини-свинь-
ям в последнее время начали уделять особое внима-
ние исследователи многих стран мира. В то же вре-
мя свиньи, как объект исследования, использова-
лись и описаны еще в трудах Гиппократа [239].
Из многих видов диких свиней, обитающих в
настоящее время, очень кратко остановимся на био-
логии кабанов, поскольку кабаны использовались
как исходный вид при создании многих линий
мини-свиней. Согласно результатам многочислен-
ных исследований, род кабанов имеет древнейшее
происхождение. К нему относится значительное
число подвидов и разновидностей. Данные архео-
логических раскопок позволяют считать, что райо-
ном первичного обитания диких свиней была юго-
восточная Азия, в последующем они расселились в
Центральную Азию, Африку и Европу. Кабаны ме-
няют окраску в зависимости от возраста. Молодые
кабанята продольно-полосатые, а осенью линяют и
становятся бурыми. У кабанов есть также зимняя,
утепленная подшерстком шуба и летняя — щетини-
стая. Во второй половине ноября у кабанов гон, а в
116
Качество животных-биомоделей
апреле появляются новые выводки поросят. Самка делает для поросят удоб-
ное, теплое и сухое гнездо, где кабанята находятся 2—3 недели, а затем она
их выводит в лес и учит всем премудростям лесной жизни. Самки
с детенышами довольно часто объединяются и вместе их воспитывают.
В случае гибели одной из них заботу о кабанятах принимает на себя другая
самка. Защищают свое потомство кабанихи яростно, нападая даже на тигра.
Использование природного генофонда диких видов свиней продолжается
и в настоящее время при выведении различных пород свиней, в том числе
и лабораторных мини-свиней.
Свиньи всех существующих пород по их происхождению подразделяют-
ся на две основные группы: азиатского и европейского корня. От европей-
ского дикого кабана произошли свиньи коренных пород Европы, а от ази-
атских диких форм — свиньи пород Азии. В дальнейшем европейские
и азиатские породы смешались и образовались среднеземноморские древ-
ние породы свиней.
Азиатские дикие свиньи отличаются от европейских значительно мень-
шими размерами, более округлыми формами туловища, повышенной ско-
роспелостью и склонностью к ожирению. Череп у азиатских свиней относи-
тельно короткий и широкий, лоб выпуклый, ряды коренных зубов располо-
жены непараллельно друг другу. У европейских свиней череп длинный
и узкий, лицевая часть развита сильно, коренные зубы расположены парал-
лельными рядами. Эти отличия свойственны только для взрослых особей, на
ранних стадиях развития различий в строении черепа у азиатских и европей-
ских диких свиней не наблюдается.
У домашних свиней произошли существенные изменения по сравнению
с дикими предками. Если дикие свиньи ведут преимущественно ночной образ
жизни, то у домашних свиней активная жизнедеятельность переместилась
на дневные часы. У домашних свиней повысилась скороспелость, они при-
обрели склонность к более раннему размножению, у них повысилось много-
плодие, исчезла сезонность в проявлении половой активности, что способ-
ствовало получению опоросов в любое время года, изменилась конституция,
мясные и сальные качества свиней культурных пород. Эти изменения про-
изошли под влиянием доместикации диких свиней, в результате направлен-
ной селекции и подбора производителей желаемого генотипа.
Дикие свиньи были одомашнены в разных географических зонах в пери-
од 5000—4000 лет до нашей эры. В одомашнивании участвовало несколько
подвидов диких кабанов. В настоящее время на территории Евразии обитает
около 25 подвидов дикого кабана, в нашей стране живут 5 подвидов, отли-
чия их довольно незначительные: оттенки окраски, размеры тела, строение
черепа, зубов. Кабанята легко приручаются и содержатся как обычные сви-
ньи в хозяйстве, а домашние свиньи, отпущенные или убежавшие на волю,
очень быстро дичают, возвращаются к привычкам своих предков и даже
Глава 7. Биориски и качество экспериментов на лабораторных животных 117
внешне становятся похожи на кабанов. Домашние свиньи и кабаны легко
скрещиваются, поскольку домашняя свинья происходит от кабана. Напри-
мер, гибрид кабана и ландрасса показывает неполное доминирование альби-
низма над диким типом окраса.
В процессе разработки проекта «Манхеттен» по созданию американской
атомной бомбы куратором проекта генералом Лесли Гровсом, по предложе-
нию ученых-ядерщиков, в 1943 году была поставлена задача перед генетика-
ми по созданию линий карликовых свиней. С этого периода начались ин-
тенсивные работы по их разведению и использованию не только в радиаци-
онной биологии, но и в других областях медицины. В последние 20 лет во
многих странах мира наметилась определенная тенденция к быстрому рас-
ширению масштабов использования свиней в качестве лабораторных живот-
ных. Главным отличием лабораторных свиней от обычных, используемых
в сельском хозяйстве, является их миниатюрность, за что их называют мини-
свиньями (до 20-30 кг) или миди-свиньями (50 кг и более).
Мини-свиньи, созданные к настоящему времени, как правило, в 3—5 раз
меньше обычных свиней. Наиболее востребованные животные, во взрослом
состоянии, т. е. в возрасте 2 лет, они весят 70—80 кг, а некоторые виды — 40—
50 кг, тогда как обычные свиньи в этом возрасте весят до 300 кг.
Небольшие размеры мини-свиней не только значительно сокращают
площади для их содержания, но и в несколько раз сокращают расходы на
кормление. Небольшие размеры мини-свиней позволяют содержать их
в обычных вивариях. Например, в ФРГ их содержат в двухярусных клетках.
В США, некоторых странах западной Европы и в Японии выведены линии
миниатюрных свиней для изучения на них различных аспектов медико-
биологических проблем. Первые генетически стабильные линии мини-сви-
ней были получены в 1949—1953 гг. в Миннесотском университете США,
где в качестве источников генов карликовости использовали мелкие формы
гвинейских и одичавших лесных свиней Калифорнии и Луизианы.
Другая линия мини-свиней была выведена в Питман-мурской лаборато-
рии (штат Флорида, США) на базе одичавших свиней. Эти питман-мурские
миниатюрные свиньи характеризуются не только темной окраской щети-
ны, но и темной кожей. Сейчас они разводятся в Японии и послужили
исходным материалом для некоторых других пород мини-свиней. Путем
скрещивания питман-мурских свиней с палоусскими в Ричмонде (штат Ва-
шингтон, США), были выведены хэнфордские мини-свиньи. Эти мини-
свиньи известны под названием «лабко».
В университете штата Небраска вывели пятнистых мини-свиней, скре-
щивая миннесотских мини-свиней с гвинейскими одичавшими хряками.
Недавно стало известно о выведении томпсоновских миниатюрных инб-
редных свиней (штат Иллинойс, США), используемых в исследованиях
специального центра по атеросклерозу при Чикагском университете. В США
118
Качество животных-биомоделей
проводится настойчивая и большая работа по созданию миниатюрных ла-
бораторных свиней с различными вариантами фенотипов и генотипов.
В странах Европы широкое распространение получили геттингенские
мини-свиньи (белая и черная линии), которых создали в Геттингенском
университете ФРГ в 1960—1963 годах. Эти мини-свиньи получены путем скре-
щивания миннесотских свиней из США с домашними свиньями двух пород:
черной вьетнамской и немецкой ландрас. В Англии выведена линия мини-
свиней на основе хэндфордских миниатюрных свиней США. Во Франции
в качестве лабораторных животных используют диких корсиканских свиней.
Работать с дикими или одичавшими свиньями довольно сложно из-за их
агрессивности или пугливости, а также грубости и пигментированности кожи
и щетины. В 1976 году в ГДР путем скрещивания вьетнамской черной, улуч-
шенного ландраса и сэттльшванской пород была получена тригибридная
линия миниатюрных свиней, названных мини-леве. Потомство их расщеп-
ляется по масти на белую, черную и пятнистую.
В нашей стране создание мини-свиней начато в 1970 году в Новосибир-
ском институте цитологии и генетики СО РАН. Была предпринята отдален-
ная гибридизация и сложное воспроизводительное скрещивание домашних
свиней породы ландрас и вьетнамской породы Й с диким кабаном европей-
ского и азиатского происхождения. Полученная линия миниатюрных сви-
ней названа мини-сибс. Скажем прямо, название не очень удачное, так как
сибсами в генетике называют родных братьев и сестер.
В лаборатории биомоделирования НЦБМТ РАМН работы по созданию
линии тетрагибридных миниатюрных свиней начались в 1974 году на основе
скрещивания белых геттингенских свиней с частично отселектированной по-
пуляцией новосибирских свиней. Тогда еще работа по созданию линии мини-
сибс не была завершена.
Таким образом, в мировой практике в настоящее время используют спе-
циально выведенные лабораторные породы: в США — хормельские, хэн-
фордские, питман-мурские, небрасские, белтсвиллские белые, в ФРГ — гет-
тингенские и мини-ЛЕВЕ, в Японии — омини, в России — минисибс (ми-
ниатюрные сибирские свиньи) и миниатюрные светлогорские свиньи MSY.
Индекс Y означает принадлежность к НЦБМТ РАМН.
Генофонд мини-свиней России позволяет совершенствовать их в виде
специализированных линий, отвечающих необходимым требованиям к ла-
бораторным свиньям для всех областей медико-биологического моделиро-
вания [642]. Их генофонд отличается широким спектром полиморфизма по
иммуногенетическим и цитогенетическим характеристикам, благодаря ис-
пользованию при их выведении домашних и диких исходных форм европей-
ского и азиатского происхождения. Использование диких кабанов при вы-
ведении отечественных мини-свиней обеспечило получение животных
с повышенной крепостью конституции, необходимой для содержания и эк-
Глава 7. Биориски и качество экспериментов на лабораторных животных 119
спериментирования. У свиней как производственных пород, так и лабора-
торных линий выделяются три особенности роста.
Первая видовая особенность заключается в низкой скорости роста эмбри-
онов до рождения и высокой после рождения. Поросята в эмбриональный
период растут медленно. По скорости роста в этот период они уступают
кроликам в 5 раз. Вторая видовая особенность роста свиней выражается
в высокой его интенсивности в период после рождения. По интенсивности
роста в этот период свиньи превосходят кроликов в 8—10 раз. Третья видовая
особенность роста свиней состоит в сочетании большой его продолжитель-
ности с высокой интенсивностью. Все эти процессы находятся под генети-
ческим контролем.
Большинство животных интенсивно растет до полового созревания, за-
тем темп роста их значительно замедляется. В отличие от животных других
видов свиньи продолжают расти и после полового созревания, которое на-
ступает в 4—5 месячном возрасте. Рост хряков и свинок продолжается до
трехлетнего возраста. Свиньи разных пород отличаются по характеру роста,
что обусловлено различиями генетических вариантов, контролирующими рост
у разных пород. Например, лабораторные мини-свиньи разных линий отли-
чаются по весовым характеристикам и по темпу роста в разные периоды
онтогенеза. Имеются существенные различия в живой массе хряков и сви-
нок. Если до 4 месяцев различий не наблюдается, то уже к 2—2,5 годам хряки
производственных пород весят от 280 до 300 кг, а свиньи от 200 до 250 кг.
У светлогорской мини-свиньи, наоборот, хряки весят значительно мень-
ше, чем свинки. Двухлетний хряк весит 28—32 кг, а свинка этого же возраста
весит 37—40 кг. Это связано с тем, что идет интенсивный отбор, направлен-
ный на уменьшение размеров и живой массы, а поскольку мужской генотип
скорее отвечает на селекцию, то, естественно, у мини-свиней вес самцов
меньше, чем самок, в то время как у производственных пород, которые от-
бирались на большую массу и большую интенсивность роста, наоборот, вес
самцов больше, чем самок. Такова уж специфика ответа на селекцию у осо-
бей мужского пола.
Кратко остановимся на особенностях развития мини-свиней MSY. Мини-
свиньи достигают половой зрелости к полугоду, но для воспроизводства обыч-
но используют 7—9 месячных животных, так как в это время развитие внут-
ренних органов, костяка и других физиологических показателей достигает
полной зрелости. Если у производственных пород продолжительность бере-
менности 113—116 дней, то у лабораторных мини-свиней светлогорской по-
пуляции она составляет 112—113 дней, то есть селекция на уменьшение мас-
сы тела коррелирует с уменьшением периода беременности, что необходимо
учитывать при проведении экспериментов, для которых необходимы эмбри-
оны точного срока развития или при проведении гнотобиологических экспе-
риментов, когда начинают операцию перед самым рождением.
120
Качество животных-биомоделей
Поросята мини-свиней растут значительно медленнее, чем поросята про-
изводственных пород, и к 6 месяцам достигают 8—12 кг, тогда как поросята
производственных пород в этом возрасте весят 70—95 кг. Уменьшение веса
поросят мини-свиней при правильном рационе кормления не уменьшает их
жизнеспособности, они так же энергичны, крепки, как и крупные поросята
производственных пород. Технология выращивания поросят мини-свиней
практически такая же, как и производственных пород. Свинья неприхотлива
к уходу и еде, условиям содержания, легко дрессируется, имеет сравнитель-
но короткую беременность, ее потомство многочисленно и скороспело,
а генофонд разнообразен. Это дает им преимущество перед многими лабо-
раторными животными.
В лаборатории биомоделирования НЦБМТ РАМН установлена корреля-
ционная связь живой массы и признаков экстерьера у светлогорских мини-
свиней. Оценка особей по экстерьеру — один из важнейших разделов селек-
ционной работы. По признакам экстерьера определяют тип конституции,
породную и линейную принадлежность, особенности телосложения, состо-
яние здоровья. Кроме того, известно, что люди в зависимости от типа тело-
сложения имеют предрасположенность к некоторым видам заболеваний, что
необходимо учитывать при моделировании ряда патологий. На 94 мини-сви-
ньях было проведено изучение коррелятивных связей живой массы особей и
ряда признаков экстерьера.
Была установлена положительная корреляция между живой массой в 1-, 3-
и 6-месячном возрасте и длиной туловища Г, = 0,8, Г3 = 0,9 и Г6 = 0,9 и
обхватом груди за лопатками Г, = 0,6, Г3 = 0,9 и Г6 = 0,9 в те же возрастные
периоды. Живая масса также положительно коррелирует с обхватом пясти
Т{ = 0,1, Г3 = 0,8 и Г6 = 0,5 и с высотой в холке Т{ = 0,7 , Г3 = 0,7 и Г6 = 0,5.
Так как обхват пясти и высота в холке связаны с развитием скелета, то
можно предположить, что у светлогорских мини-свиней скелет в меньшей
степени, чем прочие органы и ткани, имеет удельный вес в общей живой
массе особи, на что указывают более низкие коэффициенты корреляции по
сравнению с длиной туловища и обхватом груди за лопатками.
Поскольку длина туловища, обхват груди за лопатками и высота в холке,
обхват пясти не одинаково коррелируют с живой массой, можно считать,
что при интенсивном отборе по живой массе в ряде поколений будут полу-
чены изменения телосложения у светлогорских мини-свиней в соответствии
с задачами и заказом экспериментаторов.
Наряду с совершенствованием стандартных мини-свиней с живой мас-
сой во взрослом состоянии 50—60 кг ведутся работы по созданию особо мел-
ких лабораторных свиней (15—17 кг) с использованием карликовых непаль-
ских свиней. В Грузии сохранилась мелкая аборигенная сванетская порода
свиней в высокогорных районах средней и верхней Сванетии. Генофонд этих
популяций представляет ценность для выведения новых линий лаборатор-
Глава 7. Биориски и качество экспериментов на лабораторных животных 121
ных мини-свиней путем скрещивания с уже созданными, а также селекцией
путем чистопородного разведения.
Спорадическое использование в качестве модели мини-свиньи позволи-
ло ученым заключить, что она является великолепным животным для лабо-
раторных изысканий, поскольку по ряду анатомо-физиологических показа-
телей ближе всего стоит к человеку после обезьяны. С человеком ее роднит
строение и физиология сердечно-сосудистой системы [190, 441, 460], пище-
варения [80, 620], строение зубов [660], почек [496], кожи [485], глаза [536].
Мини-свинья подвержена стрессам [458], имеет сходный с человеком состав
крови и цифры артериального давления. С конца 40-х годов, когда впервые
была селекционирована более удобная для содержания в условиях вивария
миниатюрная свинья, средний вес которой не превышал 50 кг, работа про-
должалась и была селекционирована свинья с массой тела 20—28 кг. Ведутся
исследования по созданию линии еще более мелких животных с массой тела
12-15 кг [484].
Система кровообращения и сердце мини-свиньи имеют большое сход-
ство с человеческими. Многочисленные исследования показали, что в ко-
ронарных сосудах и сердце у мини-свиней могут формироваться атероскле-
ротические бляшки, сходные с таковыми у человека. Атеросклероз также
легко моделируется на мини-свиньях путем их содержания на рационе с
добавкой топленого жира или холестерина в сочетании с кокосовым маслом
и желчным экстрактом [317]. Исследования, выполненные на светлогорс-
кой популяции миниатюрных свиней, содержащейся в нашей лаборатории
биомоделирования НЦБМТ РАМН, показали, что у этих животных атерос-
клероз моделировался в течение 6—12 месцев при сочетании в рационе сли-
вочного масла или маргарина с холестерином на фоне регулярных стрессов.
Стрессы создавались путем перемещения животных из одного стада в дру-
гое. Морфологически у таких свиней в аорте и коронарных сосудах обнару-
живались атеросклеротические бляшки, а электрофизиологические иссле-
дования выявили реакции на различные раздражители и введение лекар-
ственных веществ, сходные с таковыми у человека. Изучая атеросклероз на
мини-свиньях, необходимо и достаточно легко получать информацию и о
биохимическом составе крови: содержании общих липидов, триглицеридов,
фосфолипидов и др. [9].
Иммунологи довольно активно используют свиней для решения своих
задач. Так, было показано, что механизм подавления иммунитета после ише-
мии связан с образованием иммуносупрессорных веществ в сыворотке крови
[479]. На свиньях ведутся исследования по изучению гистосовместимости,
исследуются иммунологические аспекты постнатального развития плода [65,
647]. Исследования, выполненные в НЦБМТ РАМН, свидетельствуют, что
алкогольная интоксикация приводит к уменьшению у животных содержания
Т- и В-клеток крови, заметной тенденции к усилению реакции бласттранс-
122
Качество животных-биомоделей
формации лимфоцитов [132, 133]. Есть также данные, что при этом происхо-
дит угнетение активности ферментов метаболизма ксенобиотиков [325].
На свиньях возможно моделирование вирусных заболеваний, свойствен-
ных человеку [382], а также целого ряда протозойных, паразитарных и бак-
териальных заболеваний, общих с человеком. Было, например, показано,
что свинья является лучшим объектом для изучения энцефалита, вызванно-
го вирусом простого герпеса типа 1, поскольку, наряду с прочими достоин-
ствами, позволяет получить от 5 до 10 мл спинно-мозговой жидкости, в то
время как от кролика можно получить лишь 1 мл [136].
В радиобиологии свинью можно использовать как экспериментальное
животное, заменяющее обезьяну, с целью проверки влияния на организм
различных доз облучения [120]. Было показано, что при дозах 2400—5000 рад
у животных стрессовое состояние длится до 5 мин, а при более высоких
дозах (13 000 и выше) животные надолго впадают в состояние тяжелого стрес-
са. На свиньях возможен анализ действия ионизирующего облучения и изо-
топов (стронция-90, америция-241, кобальта-60) на различные системы орга-
низма: репродуктивные органы и потомство [71], мозг [112], кости и мышцы
[181]. Миниатюрных свиней можно также использовать в целях обнаруже-
ния и определения сравнительной степени нейтрализации ряда радиоизото-
пов в живых организмах [182, 407].
Исследования на мини-свиньях включают как консервативные методы,
например, измерение систолического давления хвостовой артерии [480] или
оригинальный метод введения в рот таблеток с помощью поршня или пис-
толета [106], так и различные хирургические вмешательства. Свиньям воз-
можно вживление в мозг электрокортикографических электродов [530], уда-
ление гипофиза [171], а также различных желез: слюнных, паращитовидной,
щитовидной [161, 431, 562]. Для различных манипуляций в области грудной
клетки проводят латеральную [56] или срединную [272] торакотомии с пос-
ледующими необходимыми для эксперимента манипуляциями: удалением
легкого, пересадкой сердца, ваготомией и т. д. Экспериментальные опера-
ции на брюшной полости включают введение канюли в желудок [3], наложе-
ние фистул слепой кишки [541] и кишечника, поджелудочной железы [516].
На этом объекте можно производить биопсию печени [409], удаление над-
почечниковых желез [442], селезенки [574], конструировать перекрестное кро-
вообращение [580] и создавать портокавальные анастомозы [146].
Целый ряд хирургических вмешательств можно проводить и в области
мочеполовой системы: пересадку зародышей, гистерэктомию [219], удале-
ние яичников [120], кастрацию и удаление семявыносящего протока [107].
Можно вводить канюлю в яичниковую вену [236], проводить кесарево сече-
ние, вводить канюлю в мочеточник [434], проводить биопсию почки и мик-
ропункцию нефронов [273]. Возможно даже создавать анастомозы между
фаллопиевой трубой и подвздошной кишкой [670].
Глава 7. Биориски и качество экспериментов на лабораторных животных 123
Перечень возможностей использования в медико-биологических изыс-
каниях миниатюрных свиней свидетельствует, что этот вид лабораторного
животного может быть прекрасной моделью для решения целого ряда важ-
ных исследовательских задач, о чем мы будем говорить в дальнейшем из-
ложении.
Таблица 29
Вирусные инфекции мини-свиней, потенциально опасные
для человека и животных и искажающие результаты исследований
Возбудитель
Aujeszky virus
(herpesvirus)
Transmissible
gastroenteritis virus
(TEG) (coronavirus)
Myxovirus influenza sp.
(A.suis, A.equi)
Pox virus свиней
a. original virus
b. vaccine virus
Teschen virus
(picornavirus)
African fever virus
свиней (iridovirus)
Rabies virus
(rhabdovirus)
Vesicular exanthema
virus (calicivirus/
picornaviruses)
Enteric cytopathic
orphan (ESCO)
viruses свиней
(picornavirus)
Заболевания
Биориск заражения
для человека
и животных
Псевдобешенство,
болезнь Ауешки
Мини-свиньи и многие
другие виды животных
Передающийся гастро-
энтерит
Собаки, мини-свиньи
Оспа мини-свиней
Мыши, морские свинки,
кролики(вакцинный
вирус), свиньи (ориги-
нальный вирус оспы
мини-свиней)
Полиомиелит, болезнь
Тешена
Мини-свиньи
Африканская лихорадка
мини-свиней
Мини-свиньи, (экзоти-
ческие копытные живот-
ные)
Бешенство
Различные виды живот-
ных, человек
Полиоэнцефаломиелит
(в экспериментальной
инфекции у поросят, не
получающих молозиво)
Мини-свиньи
Пути заражения
Оральный, воздушно-
капельный
Оральный, воздушно-
капельный, внутри-
маточный
Оральный, воздушно-
капельный
Оральный, воздушно-
капельный, внутри-
маточный
Кожный
Оральный, воздушно-
капельный
Оральный, кожный,
воздушно-капельный
Через укусы бешеных
животных
Оральный
Контагиозность,
симптоматика и исход
Влияние на результаты
экспериментов
Нервные нарушения,
специфические пора-
жения органов, выки-
дыши, смерть поросят
Нарушения развития,
поражения нервов, вы-
кидыши, смерть
Ухудшение самочувст-
вия (состояния), диа-
рея, смерть поросят
Смерть.
Искажение результа-
тов эксперимента
Изменения кожи
Нервные нарушения,
в большинстве случаев
бессимптомное тече-
ние
Высокая смертность.
Искажение результа-
тов эксперимента
Смерть.
Искажение результа-
тов эксперимента
Поражения органов,
нарушение разведе-
ния (выкидыши), низ-
кая смертность
Выкидыши.
Искажение результа-
тов эксперимента
Fever virus свиней
(togavirus)
Свиная лихорадка
Мини-свиньи
Грипп мини-свиней
Мыши, хорьки, мини-
свиньи, человек
Везикулярная экзантема
Мини-свиньи
Оральный, кожный
124 Качество животных-биомоделей
Окончание табл. 29
Возбудитель
Foot and mouth
disease virus (FMV)
(rhinovirus/picorna-
viruses)
Adenovirus
Заболевания
Биориск заражения
для человека
И ЖИВОТНЫХ
Болезнь стоп и рта,
афтозная лихорадка
Мини-свиньи, жвачные
животные
Не определенные
заболевания
Мини-свиньи
Пути заражения
Оральный, воздушно-
капельный, кожный
Оральный,
назальный
Контагиозность,
симптоматика и исход
Влияние на результаты
экспериментов
Изменения кожи
Скрытое течение,
энцефалит у новорож-
денных поросят,
выкидыши
Таблица 30
Микоплазменные, бактериальные и грибковые инфекции мини-свиней,
потенциально опасные для человека и животных и искажающие
результаты исследований
Возбудитель
Mycoplasma sp.
(A.granularum, hyor-
hinis, hyopneumoniae,
suipneumoniae)
M.hyosynovia
M.axanthum (так же
у гусей) М. flocculare
Escherichia coli
(некоторые серотипы)
Erysipelothrix
rhusiopathiae
Bordetella
bronchiseptica
Pasteurella multocida
Salmonella sp.
Заболевания
Биориск заражения
для человека
и животных
Ринит, пневмония,
артрит
Мини-свиньи
Дизентерия свиней,
колиэнтеротоксемия
Различные виды живот-
ных, человек
Рожа мини-свиней
Различные виды живот-
ных, человек
Ринит, пневмония
Различные виды живот-
ных, человек
Септицемия
Различные виды живот-
ных, человек
Дизентерия свиней,
колиэнтеротоксемия
Различные виды живот-
ных, человек
Пути заражения
Воздушно-капельный
Оральный, восходя-
щая инфекция моче-
выводящих путей
Оральный, кожный
Воздушно-капельный
Оральный, воздушно-
капельный
Дизентерия мини-
свиней, колиэнтеро-
токсемия
Контагиозность,
симптоматика и исход
Влияние на результаты
экспериментов
Поражения органов,
нарушение развития,
смерть
Смерть.
Искажение результа-
тов эксперимента
Поражение органов и
суставов, нарушения
развития при хрони-
ческом течении.
Смерть при остром
течении
Смерть.
Искажение результа-
тов эксперимента
Поражения органов,
смерть.
Искажение результа-
тов эксперимента
Смерть.
Искажение результа-
тов эксперимента
Глава 7. Биориски и качество экспериментов на лабораторных животных 125
Продолжение табл. 30
Возбудитель
Erysipelotrix
rhusiopathiae
Brucella sp., esp.
В. suis
Actinomyces sp.
Pasteurella
haemolytica
Pasteurella
pneumotropica
Staphylococcus
aureus
Streptococcus
pneumoniae
Streptococcus
pyogenes
Haemophilus suis
Listeria monocyto-
genes
Mycobacterium sp.
(tuberculosis, bovis,
avium)
Bacillus anthracis
Заболевания
Биориск заражения
для человека
И ЖИВОТНЫХ
Сальмонеллез, паратиф,
инфекционный энтерит,
тиф поросят
Различные виды живот-
ных, человек
Бруцеллёз, эпидемиче-
ские выкидыши
Различные виды живот-
ных, человек
Актиномикоз
Мини-свиньи, быки
Пастереллёз,
пневмония
Мини-свиньи, овцы,
быки
Пастереллёз,
пневмония
Мини-свиньи
Абсцессы, пиодерма
Различные виды живот-
ных, человек
Пневмония, бронхо-
пневмония
Различные виды живот-
ных, человек
Стрептококковая
септицемия
Различные виды живот-
ных, человек
Фибринозный поли-
серозит
Морские свинки,
мини-свиньи
Листериоз
Различные виды живот-
ных, человек
Туберкулёз
Различные виды живот-
ных, человек
Сибирская язва
Различные виды живот-
ных, человек
Пути заражения
Оральный
Оральный, кожный,
внутриматочный
Кожный
Оральный, воздушно-
капельный
Оральный, воздушно-
капельный
Оральный, кожный
Воздушно-капельный
Воздушно-капельный,
кожный
Оральный, воздушно-
капельный
Оральный, внутри-
маточный
Оральный, воздушно-
капельный
Оральный
Контагиозность,
симптоматика и исход
Влияние на результаты
экспериментов
Скрытое течение или
энтерит и диарея
Выкидыши, орхит
Поражение молочных
желез, нарушение
рождаемости
Поражения органов,
смерть.
Искажение результа-
тов эксперимента
Поражения органов,
смерть.
Искажение результа-
тов эксперимента
Локальные поражения
тканей.
Искажение результа-
тов эксперимента
Изменения в легких,
смерть.
Искажение результа-
тов эксперимента
Поражения органов,
смерть.
Искажение результа-
тов эксперимента
Хромота, смерть.
Искажение результа-
тов эксперимента
Поражения органов,
нервные нарушения,
смерть
Нарушение развития,
поражения органов,
смерть.
Нарушение развития
при хронических фор-
мах, смерть при ост-
рых и подострых
формах
126
Качество животных-биомоделей
Окончание табл. 30
Возбудитель
Clostridium sp.
Fusobacterium
necrophorum
Spirillum suis
Eperythrozoon sp.
Leptospira sp.
Treponema
hyodysenteriae
Trichophyton sp.
Microsporum sp.
Заболевания
Биориск заражения
для человека
И ЖИВОТНЫХ
Газовая гангрена,
столбняк, ботулизм
Различные виды живот-
ных, человек
Дифтерия мини-свиней
Мыши, мини-свиньи,
жвачные животные,
человек
Дизентерия мини-
свиней
Мини-свиньи
Иктероанемия,
пневмония
Мини-свиньи, быки
Лептоспироз
Различные виды живот-
ных, человек
Диарея
Мини-свиньи
Дерматомикоз
Различные виды живот-
ных, человек
Дерматомикоз
Различные виды живот-
ных, человек
Пути заражения
Оральный, кожный
Оральный, кожный
Оральный
Трансплацентарный
Оральный, кожный
Оральный
Воздушно-капельный,
кожный
Воздушно-капельный,
кожный
Контагиозность,
симптоматика и исход
Влияние на результаты
экспериментов
Поражения органов,
смерть
Некрозы пораженных
тканей, смерть.
Искажение результа-
тов эксперимента
Смерть.
Искажение результа-
тов эксперимента
Незначительные об-
щие проявления,
анемия.
Искажение результа-
тов эксперимента
Выкидыши, снижение
жизнеспособности
поросят
Диарея.
Искажение результа-
тов эксперимента
Недомогание.
Искажение результа-
тов эксперимента
Недомогание.
Искажение результа-
тов эксперимента
Таблица 31
Инвазивные заболевания мини-свиней, потенциально опасные
для человека и животных и искажающие результаты исследований
Возбудитель
Eimeria spp.
(herpesvirus)
Isospora suis
Sarcocystis spp.
miescheriana
Заболевания
Биориск заражения
для человека
и животных
Коккцидиоз
Мини-свиньи
Саркоспоридиоз
Мини-свиньи, ягнята
Пути заражения
Оральный
Оральный
Контагиозность,
симптоматика и исход
Влияние на результаты
экспериментов
Энтерит.
Искажение результа-
тов эксперимента
Ухудшение самочувст-
вия (состояния)при
тяжелом течении ин-
фекции, тяжелая бо-
лезнь
Глава 7. Биориски и качество экспериментов на лабораторных животных 127
Продолжение табл. 31
Возбудитель
Toxoplasma gondii
Balantidium sp.
(coli, suis)
Ascaris suum
Trichuris suis
Globocephalus
urosubulatus
Hyostrongylus rubidus
Metastrongylus spp.
Oesophagostomum sp.
Strongyloides ransomi
Trichinella spiralis
Cysticercus tenuicollis
(Taenia hydatigena)
Cysticercus cellulosae
(Taenia solium)
Diphyllobothrium
latum
Заболевания
Биориск заражения
для человека
И ЖИВОТНЫХ
Токсоплазмоз
Различные виды живот-
ных, человек
Балантидиоз
Мини-свиньи, человек
Аскаридная инвазия,
аскаридоз
Мини-свиньи, человек
(другие виды: миграция
личинок)
Трихуроз
Мини-свиньи, иногда
обезьяны, человек
Нематодная инвазия,
глобоцефалез
Мини-свиньи, редко
человек
Остричная инвазия же-
лудка, гиостронгилез
Мини-свиньи
Остричная инвазия
легких, метастронгилез
Мини-свиньи
Эзофагостомоз
Мини-свиньи, редко
человек
Стронгилоидоз
Кролики,
мини-свиньи
Трихиноз
Мини-свиньи,
человек, грызуны
Цистицеркоз
Мини-свиньи, жвачные
животные
Цистицеркоз
Мини-свиньи, жвачные
животные
Дифиллоботриоз
Кошки, собаки, мини-
свиньи, человек
Пути заражения
Оральный, транспла-
центарный
Оральный
Оральный
Оральный
Оральный
Оральный
Оральный (промежу-
точный хозяин)
Оральный
Чрезкожный
Оральный
Оральный
Оральный
Оральный
Контагиозность,
симптоматика и исход
Влияние на результаты
экспериментов
Нарушение разведе-
ния, смерть поросят
Энтерит.
Искажение результа-
тов эксперимента
Истощение, измене-
ния органов.
Искажение результа-
тов эксперимента
Нарушение развития,
истощение в случае
тяжелой инвазии
Энтерит и анемия,
особенно в случае
тяжелой инвазии
Истощение, анемия
вследствие энтерита
и потери крови
Поражения легких,
нарушение развития
в случае тяжелой
инвазии
Хронический энтерит.
Искажение результа-
тов эксперимента
Заболевания кишеч-
ника, анемия, диарея
грудных поросят
Поражения кишеч-
ника (кишечный три-
хиноз). Поражения
мышц (мышечный три-
хиноз)
Необратимые пораже-
ния печени в случае
тяжелой инвазии
Слабость, истощение.
Искажение результа-
тов эксперимента
Незначительные кли-
нические симптомы
128
Качество животных-биомоделей
Окончание табл. 31
Возбудитель
Echinococcus hydati-
dosus (Echinococcus
granulosus)
Macracanthorynchus
hirudinaceus
Demodex suis
Haematopinus suis
Sarcoptes suis
Заболевания
Биориск заражения
для человека и животных
Эхинококкоз
Мини-свиньи, человек
жвачные животные
Акантоцефалоз
Собаки, свиньи,
обезьяны, человек
Демодекоз
Собаки, свиньи,
обезьяны, человек
Педикулёз
Мини-свиньи
Чесотка
Мини-свиньи
Пути заражения
Оральный
Оральный, промежу-
точный хозяин
Контактный
Контактный
Контактный
Симптоматика и исход
Влияние на результаты
экспериментов
Поражение печени.
Искажение результа-
тов эксперимента
Искажение результа-
тов эксперимента
Пустулы
Раздражение кожи
Зуд, дискератоз
Овца — парнокопытное животное рода баранов, семейства полорогих.
Происходит от диких горных баранов (муфлонов и архаров), которые были
одомашнены более 8 тыс. лет назад. Эволюция домашних овец проходила,
прежде всего, в направлении преобразования шёрстного покрова и форм
телосложения.
Рост овцы (высота в холке) 55—100 см, длина тела (от затылка до корня
хвоста) 60—110 см. Матки весят 30-100 кг, бараны (так же называют самца
домашних овец) 60—180 кг. У баранов большинства пород хорошо развиты
рога; матки безрогие или с небольшими рожками. Морда с прямым, иногда
горбоносым профилем. Нижняя часть её заострена, губы тонкие, очень под-
вижные, резцы поставлены под тупым углом к челюсти. Благодаря такому
строению головы овцы могут очень низко скусывать траву и полнее, чем
другие виды животных, использовать пастбища. Ноги крепкие, что делает
овец способными к длительным переходам. У взрослых овец 32 зуба. Смена
молочных зубов начинается с 12-18 мес. и заканчивается в 3,5—4 года. Масть
белая, чёрная, рыжая, серая. Овцы с тонкой и полутонкой шерстью, как
правило, белые. По форме хвоста овец делят на короткотощехвостых (хвост
тощий, не достигающий скакательного сустава, 10—12 позвонков), длинно-
тощехвостых (хвост тощий, достигает скакательного сустава, иногда ниже,
20—22 позвонка), короткожирнохвостых (хвост из 10—12 позвонков, с жиро-
выми отложениями), длинножирнохвостых (хвост из 20—22 позвонков, с жи-
ровыми отложениями), курдючных (хвост из 5—6 позвонков, жировые отло-
жения на ягодицах и вокруг хвоста называются курдюком).
Продолжительность жизни овцы при благоприятных условиях составля-
ет 14—15 лет. Половая зрелость наступает в 5—7 мес. К спариванию допуска-
Глава 7. Биориски и качество экспериментов на лабораторных животных 129
ют в 15—18 мес. При естественном спаривании на одного барана назначают
60—70 маток; при искусственном осеменении семенем одного барана за се-
зон (45 суток) осеменяют более 3000 маток. Беременность продолжается 145-
155 суток. Большинство овец даёт по одному ягнёнку, некоторые по два-три,
овцы романовской породы — до пяти. Новорождённые ягнята весят 3—5 кг.
Рост заканчивается к 2—4 годам. Шёрстный покров грубошёрстных овец со-
стоит из смеси грубых волокон диаметром 100—200 мкм и более тонких пу-
ховых; тонкорунных — из однородных пуховых волокон диаметром в сред-
нем 25 мкм. Длина шерсти у тонкорунных овец 5-9 см, у полутонкорунных
— до 40 см, у грубошёрстных — 10-15 см. Племенная работа с овцами на-
правлена на получение и выращивание животных желательных качеств пу-
тём отбора, подбора и скрещивания.
Основные корма для овец — трава естественных и сеяных пастбищ, сено,
преимущественно мелко-стебельчатое, яровая солома, силос и концентраты.
Пасут овец с ранней весны до поздней осени, а в районах с небольшим
снежным покровом и в районах с жарким климатом — круглый год. Содер-
жат в кошарах; на сезонных пастбищах устраивают затиши. Овцы хорошо
переносят различные температуры воздуха, успешно содержатся на высоко-
горных пастбищах. Очень чувствительны к сырости, подвержены простуд-
ным, гельминтозным и копытным заболеваниям. Разводят овец почти во
всех странах.
До использования овец на протяжении десятилетий в иммунологических
исследованиях в качестве экспериментальных моделей широко использовали
мелких инбредных грызунов, которые дали возможность производить пере-
садки тканей и органов между сингенными и аллогенными животными.
Благодаря различным манипуляциям, проводившимся с этими животными
in vivo и in vitro, наши представления об организации и функционировании
иммунной системы значительно расширились. Вместе с тем проведение
модельных экспериментов на овцах оказалось плодотворным при онто- и
филогенетических исследованиях и при изучении физиологии лимфоидной
и иммунной систем.
На овцах можно проводить разнообразные хирургические операции, что
было особенно полезным при изучении иммунного и воспалительного от-
вета на введение антигенов и пирогенных веществ; трансплантационных
реакций на пересаженные органы и ткани; влияния тимэктомии у плодов
на онтогенез иммунной системы; миграции популяций лимфоцитов между
сосудистой и лимфоидной системами у плодов, новорожденных и взрос-
лых животных; а также модулирующих эффектов антигенов на процессы
миграции.
Возможность хирургического введения поливиниловых или пластмас-
совых канюль в афферентные и эфферентные протоки лимфатических уз-
лов, в кишечный и грудной лимфатические сосуды позволяет исследовать
130
Качество животных-биомоделей
кинетику ответных клеточных реакций на подкожное или внутрилимфати-
ческое введение иммуногенов и на пересадку тканей и органов. С помощью
этих методик можно контролировать кинетику процессов регуляции лим-
фоидных и других тканей циркулирующими в крови костномозговыми лим-
фоцитами. Исследуя лимфу, оттекающую от лимфатических узлов, можно
качественно и количественно оценивать клеточные реакции лимфоидных
тканей. Именно так можно исследовать реакцию одиночного лимфати-
ческого узла на введение антигена в афферентный сосуд, если в его эффе-
рентный проток вставлена трубка, выведенная наружу. С помощью этого
же приема одновременно можно исследовать образцы из других компарт-
ментов лимфатической системы, намеренно не подвергнутых действию
иммуногена.
Такие исследования реальны и на ранних плодах овец, что позволяет
изучать временную последовательность дифференцировки лимфоидных тка-
ней в отсутствие экзогенных или получаемых от матери антигенов (эпителио-
хориальная плацента непроницаема для белков или иммуноглобулинов ма-
теринского организма). Именно в таких экспериментах и были установлены
закономерности развития первичных и вторичных лимфоидных органов и
время достижения компетентности по отношению к иммуногенам. Канюли-
рование лимфатических сосудов плода in utero и фетальная тимэктомия были
использованы для изучения возникновения пула рециркулирующих лимфо-
цитов плода, а также роли тимуса в онтогенезе лимфоидных тканей и в раз-
витии иммунологической компетентности.
Подвздошные пейеровы бляшки и тимус овцы обладают уникальными
свойствами. Показано, что пейеровы бляшки подвздошной и тощей кишки
овцы различаются морфологически и, вероятно, функционально. Пейеровы
бляшки тощей кишки содержат В-клеточные фолликулы и отчетливые Т-
зависимые зоны, в то время как пейеровы бляшки подвздошной кишки
связаны в основном с В-клеточным лимфопоэзом, и могут частично рас-
сматриваться как первичный орган лимфоидной системы. Следует отметить,
что у овец костный мозг не единственный источник лимфоцитов. Установ-
лено, что у них первичным лимфоидным органом для Т-клеток является
тимус, причем и у плодов, и у ягнят имеются цервикальный и торакальный
тимусы. В настоящее время отработаны методы канюлирования лимфати-
ческих протоков, дренирующих пейеровы бляшки подвздошной кишки и
цервикальный тимус до точки их входа в лимфатические узлы. Подобные
методы весьма полезны при исследовании популяций клеток, «покидающих»
эти лимфоидные органы.
Разработка гибридомной технологии получения моноклональных анти-
тел к поверхностным маркерам лимфоцитов овец позволяет в настоящее
время более полно охарактеризовать субпопуляции их лимфоцитов. Даль-
нейшие исследования субпопуляций лимфоцитов in vivo и in vitro должны
Глава 7. Биориски и качество экспериментов на лабораторных животных 131
прояснить вопросы их онтогенеза, путей миграции и функциональной роли
лимфоцитов в иммунитете не столько овец, сколько человека.
Хищники (собаки, кошки)
Собаки относятся к хищникам, семейству псовых (Canidae). Происходят они
от двух диких предков — шакала (в южных странах) и волка (в северных
странах). Археологические раскопки с достоверностью подтверждают, что
собака была одомашнена свыше 14 тыс. лет тому назад — раньше других
животных.
Существуют несколько сот пород собак, имеются также линейные (бигл
и др.). Наиболее пригодными являются различные помеси от скрещивания
нескольких пород собак. В ряде исследований опыты проводятся на восточ-
ноевропейских (немецких) овчарках, русских европейских лайках, средне-
азиатских овчарках, южнорусских овчарках и таксах. Собака подходит для
воспроизведения длительных хронических опытов. Линейные собаки требу-
ют специальных условий, так как хуже переносят хронические опыты. Осо-
бенности нервной, эндокринной, сердечно-сосудистой систем, органов пи-
щеварения, дыхания и выделения собаки близки к таковым у человека.
Собака породы бигль является наиболее распространенной и общеприня-
той линейной биомоделью для проведения стандартизованных исследова-
ний и оценки лекарств, ксенобиотиков, нормирования факторов окружаю-
щей среды, испытания медицинской техники и т.д. Бигль принят во всех
лабораториях мира как стандартная модель в физиологии, фармакологии,
токсикологии, в исследованиях центральной нервной системы и высшей
нервной деятельности с помощью метода условных рефлексов (выработка
секреторных и двигательных временных связей) и электроэнцефалографии,
для изучения функций сердечно-сосудистой системы, пищеварения, норми-
рования таких факторов, как ионизирующее излучение, пребывание в био-
спутниках, гипоксия и т.д.
Бигл — это небольшая, не грубая гончая компактного сложения, произ-
водящая впечатление большой жизненной силы и энергии. Высота в холке
должна быть не больше 40,5 см. Голова довольно длинная, но не грубая, лоб
выпуклый, достаточно широкий, с немного выступающим затылочным буг-
ром. Переход ото лба к морде хорошо выражен. Морда не заостренная, губы
сухие, плотно прилегающие. Мочка носа черная, широкая, с широко рас-
крытыми ноздрями. Глаза темно-коричневые или коричневые, не должны
быть запавшими или выпуклыми, со спокойным выражением. Уши длин-
ные, низко посаженные, с тонкой шерстью, висящие в красивой складке,
тесно прилегают к скулам.
Шея умеренной длины, с небольшими складками под горлом. Туловище
короткое, грудь глубокая с выпуклыми ребрами. Поясница крепкая и невы-
132
Качество животных-биомоделей
пуклая. Передние конечности и плечи мускулистые, хорошо сформирован-
ные. Передние ноги абсолютно прямые. Предплечья массивные, круглого
сечения. Задние конечности и бедра очень мускулистые, коленные и скака-
тельные суставы с хорошо выраженными углами. Плюсны короткие, мас-
сивные. Лапы округлой формы, сводистые, с толстыми подушечками. Хвост
умеренной длины, высоко поставлен, энергично держится серпом над спи-
ной. Не должны опускаться хвосты, свернутые над спиной кольцом или
крючком. У короткошерстной формы шерсть гладкая, очень густая и не слиш-
ком тонкая или короткая. Жесткошерстная разновидность имеет очень гус-
тую и проволокообразную шерсть. У лабораторных биглей возможен любой
окрас, допустимый у гончих.
Пороки, исключающие использование биглей в эксперименте: узкая или
бульдожья голова, опущенная мочка носа, уши V-образной формы, глаза
малые или близко посаженные, черноватый окрас головы, слишком тонкая
шея, провислая спина, длинная поясница, отставленные локти, плоские
бедра, прямой постав, хвост слишком длинный, слишком толстый или зак-
рученный.
В научных лабораториях лучше всего использовать выносливых,
неприхотливых к пище, физически хорошо развитых собак. Эти собаки дол-
жны пройти необходимый карантин, ветеринарный осмотр, им необходимо
сделать профилактические прививки против бешенства и дегельминтизацию.
Реакции дыхания, кровообращения и других важнейших систем организ-
ма собаки при изменении условий внешней среды, а также воздействии
многих фармакологических и токсических веществ приближаются к измене-
ниям, наблюдаемым у человека. Собак используют для наложения различ-
ных фистул внутренних органов, воспроизведения различных неинфекци-
онных заболеваний: болезни сердца и сосудов, неврозы, поражения органов
желудочно-кишечного тракта и органов выделения, эндокринных заболева-
ний и т. д. Собаки являются хорошими моделями воспроизведения ряда
инфекционных заболеваний, а щенятам легко прививают корь, коклюш,
лептоспирозы, чуму собак.
Кошки. Хищники, рода кошачьих, широко используются в разных обла-
стях биологии и медицины: для электрофизиологических и антропометри-
ческих экспериментов, для проведения острых опытов с регистрацией кро-
вяного давления и дыхания, для определения токсичности веществ, биоло-
гической стандартизации сердечных гликозидов и для других целей. Со-
временная кошка (Felidae) не имеет отношения ни к дикому лесному коту,
ни к степной кошке. Ее предок — это североафриканская аравийская бу-
ланая кошка, которая жила в пустыне, питалась ящерицами и грызунами.
Первыми, кому удалось приручить кошку, были египтяне, и произошло это
почти 4 тыс. лет назад (из Египта кошку вывезли сначала в Индию и Ки-
тай, а уж потом в Европу). В отличие от собаки, которая жила с человеком
Глава 7. Биориски и качество экспериментов на лабораторных животных 133
как компаньон, кошку изначально заводили в доме как животное декора-
тивное.
В Средние века в Европе кошек использовали отнюдь не в медико-био-
логических целях — их (особенно черных) объявили исчадиями ада и вместе
с ведьмами сжигали заживо на костре. Потом, в эпоху Возрождения, живот-
ных все же амнистировали, но в некоторых странах их по-прежнему исполь-
зовали не по назначению. К примеру, в цивилизованной Швеции вплоть до
начала XX века сохранялся обычай замуровывать живьем кошку в фунда-
менте строящегося дома, чтобы не заводились крысы, а водились деньги. Но
все это в прошлом. На нашей планете живут больше 400 миллионов кошек,
причем значительное число их обитает в домах. В Австралии на каждых 10
жителей приходится по 9 кошек. Кстати, в Австралии, как и в Новой Зелан-
дии, кошкам запрещено гулять самим по себе. Потому что они хоть и до-
машние, а все же — хищники и наносят непоправимый ущерб уникальной
фауне. Хвостатые киллеры покушаются даже на национальное достояние но-
возеландцев — птичку киви. А посему был издан закон, запрещающий вла-
дельцам кошек выпускать животных на улицу без ошейника с микрочипом
после 9 вечера. Если животное поймают в неурочное время и в неположен-
ном месте, владельцу выписывают штраф. А кошкам без ошейника грозит
уничтожение. Ничего не поделаешь, закон есть закон.
Среди кошек, представляющих исследовательский интерес, различают
длинношерстные и короткошерстные породы. К длинношерстным породам
принадлежат: персидская (ангорская, или азиатская) кошка, требующая
тщательного ухода, европейская длинношерстная кошка и бирманская кош-
ка. Для экспериментальных исследований наиболее подходят короткошер-
стные кошки, особенно разные помеси их. Кошки длинношерстных пород
мало пригодны для экспериментальных исследований. Кошки короткошер-
стных пород выносливые, мало требовательные и вполне подходящие для
экспериментальных целей.
При проведении различных физиологических и фармакологических
опытов используют изолированные органы кошки (сердце, кишечник,
матку, селезенку, почки). Чаще всего наиболее оптимальными для опытов
являются взрослые крепкие кошки (весом 2—3 кг) в возрасте одного —
четырех лет.
На взрослых кошках относительно хорошо экспериментально воспроиз-
водятся стафилококковые инфекции, сап, бешенство, венерическая лимфог-
ранулема, дерматомикозы, глистные инвазии. У котят вызывают экспери-
ментальный коклюш, амебную дизентерию, болезнь Ауэшки, туляремию,
холеру.
134 Качество животных-биомоделей
Таблица 32
Вирусные инфекции хищников (собаки, кошки), потенциально опасные
для человека и животных и искажающие результаты исследований
Возбудитель
Заболевания
Биориск заражения
для человека
И ЖИВОТНЫХ
Пути заражения
Контагиозность,
симптоматика и исход
Влияние на результаты
экспериментов
Собаки
Rabies virus
(rhabdovirus)
Distemper virus
(paramyxovirus)
Canine parvovirus
(CPV)
Contagious hepatitis
(H.c.c.) virus
(adenovirus)
Canine herpesvirus
(CHV)
Бешенство
Кошки, различные виды
животных, человек
Собачья чума, тяжелые
поражения подушечек
лап
Собаки, хорьки
Миокардит (внезапная
смерть щенков), гемор-
рагический энтерит
Собаки
Контагиозный гепатит,
болезнь Рубарта
Собаки
Обесцвечивание шерсти
Собаки
Через укусы живот-
ных
Контактный,
воздушно-капельный
Оральный (воздуш-
ный), контактный
Оральный, контакт-
ный
Воздушный, контакт-
ный, через плаценту
Слюнотечение, куса-
ние, необычная про-
жорливость, общий
паралич
Высококонтагиозное
заболевание, высокая
смертность
Диарея, рвота, иногда
повышенная заболева-
емость и смертность
Лихорадка, рвота,
диарея, смерть
Смерть молодых щен-
ков (до 2-недельного
возраста)
Кошки
Rabies virus
(rhabdovirus)
Feline panleucopenia
virus (FPV)
(parvovirus)
Feline infectious
peritonitis virus
(Coronavirus)
Feline rhinotracheitis
virus (FVRV)
(herpesvirus)
Feline calicivirus (FCV)
(picornavirus)
Бешенство (водобоязнь)
Кошки, другие виды
животных, человек
Панлейкопения, коша-
чий энтерит
Кошки, норки
Комплексное иммунное
заболевание
Кошки
Кошачий ринотрахеит,
кошачий грипп, ринит
Кошки
Кошачий грипп,
кошачья калицивирус-
ная инфекция
Кошки
Через укусы
Оральный, воздушно-
капельный, через
плаценту
Контактный
Воздушный, контакт-
ный
Воздушный, контакт-
ный
Слюнотечение, куса-
ние, повышенная про-
жорливость, общий
паралич
Лихорадка, гастро-
энтерит с высокой за-
болеваемостью и
смертностью
Острая или хроничес-
кая инфекция, гипер-
протеинемия, перито-
нит, плевральный ас-
цит, гидроторакс
(наличие жидкости в
плевральной полости)
Эндемический ринит,
конъюнктивит, лихо-
радка
Конъюнктивит, ринит,
глоссит, стоматит.
Высококонтагиозное
заболевание.
Искажение результа-
тов эксперимента
Глава 7. Биориски и качество экспериментов на лабораторных животных 135
Окончание табл. 32
Возбудитель
Feline leukaemia virus
(FLV)
Felinereovirus, cepo-
типы 1, 2, 3
Pseudorabies virus
(Herpesvirus suis)
Заболевания
Биориск заражения
для человека
И ЖИВОТНЫХ
Лейкемия, опухоли,
кошачья калицивирус-
ная инфекция
Кошки
Часто ассоциируется
с кошачьим гриппом,
кошачья реовирусная
инфекция (КРИ)
Кошки и другие тепло-
кровные животные,
человек
Болезнь Ауешки
Кошки, собаки, норки
Пути заражения
Контактный
Контактный
Оральный
Контагиозность,
симптоматика и исход
Влияние на результаты
экспериментов
Острые и латентные
инфекции, изменения
гематологических по-
казателей, различные
опухоли в результате
угнетения иммунитета
Обострение течения
первичного заболева-
ния (следствие вторич-
ного эффекта).
Искажение результа-
тов эксперимента
Агрессивность, зуд,
самоизувечивание,
смерть
Таблица 33
Микоплазменные, бактериальные и грибковые инфекции хищников
(собаки, кошки) потенциально опасные для человека и животных и искажающие
результаты исследований
Возбудитель
Mycoplasma sp.(felis,
gatae, spumans, canis,
Acholeplasma granula-
rum)
Brucella sp.
Salmonella sp.
Shigella sp.
Bordeteila
bronchiseptica
(вторичная)
Заболевания
Биориск заражения
для человека
и животных
Микоплазмоз, заболева-
ния респираторного
тракта, урогенитальные
повреждения, конъюнк-
тивиты
Кошки, собаки
Кошачий бруцеллёз
Многие виды животных,
человек
Сальмонеллёз
Многие виды животных,
человек
Бактериальная
инфекция
Собаки, обезьяны,
человек
Вторичный, бронхосеп-
тический бордетеллёз
Пути заражения
Воздушный, контакт-
ный
Оральный, контакт-
ный
Оральный
Оральный
Воздушный
Контагиозность,
симптоматика и исход
Влияние на результаты
экспериментов
Предрасположенность
к инфекциям при
стрессах.
Искажение результа-
тов эксперимента
Выкидыши, детская
смертность, стериль-
ность, орхиты,
метриты
Энтериты, смерть
молодых животных
В большинстве слу-
чаев латентно проте-
кающие энтериты
Ринит, кашель, лихо-
радка, бронхопневмо-
ния. Высококонтагиоз-
ное заболевание
136 Качество животных-биомоделей
Продолжение табл. 33
Возбудитель
Proteus sp.
Haemobartonella
felis, H. canis (host
specific)
Chlamydia psittaci
Rickettsia canis,
Coxiella burnetii
Nocardia asteroides
Listeria
monocytogenes
Pasteurella sp.
Actinomyces sp.
Dermatophilus
congolensis
Campylobacter
coli, c.jejuni
Mycobacterium sp.
(tuberculosis, bovis,
avium)
Заболевания
Биориск заражения
для человека
И ЖИВОТНЫХ
Диарея, энтериты
Многие виды животных,
человек
Гематобартонеллёз
Кошки, собаки, другие
виды животных, человек
Кошачья хламидийная
инфекция
Многие виды животных,
человек, птицы
Риккетсиоз, лихорадка-Q
Кошки, собаки, домаш-
няя птица, человек
Нокардиоз (стрептотри-
хоз)
Кошки, собаки, обезья-
ны, человек
Листериоз
Все животные и человек
Пастереллёз
Многие виды животных,
человек
Актиномикоз
Собаки, кошки, другие
виды животных,
человек
Дерматофилёз
Собаки, кошки, кролики,
обезьяны, человек
Геморрагический
энтерит
Человек, собаки, кошки,
птицы, хомячки
Туберкулёз
Многие виды животных,
человек
Пути заражения
Оральный
Через укусы насеко-
мых
Контактный, ораль-
ный, воздушный
Через укусы клещей
Воздушно-капельный,
контактный
Оральный,
пренатальный
Воздушно-капельный,
оральный
Контактный
Контактный
Оральный
Воздушно-капельный,
оральный,
контактный
Контагиозность,
симптоматика и исход
Влияние на результаты
экспериментов
Высококонтагиозное
заболевание, смерть
при стрессах
Лихорадка, анемия,
спленомегалия, смерть
щенков, после сплено-
мегалии повышенная
чувствительность
к инфекциям, парази-
тарное носительство
Ринит, конъюнктивит,
Высококонтагиозное
заболевание. -.
Искажение результа-
тов эксперимента
Лихорадка, коньюнкти-
вит, бронхопневмония,
отеки суставов.
Высококонтагиозное
заболевание
Заболевания кожи,
лихорадка, абсцессы,
плевриты
В большинстве случа-
ев скрытое течение,
выкидыши
В большинстве случа-
ев скрытое течение
Узелки на коже,
в ротовой полости,
свищи
Поражения кожи, язвы.
Искажение результа-
тов эксперимента
Геморрагическая
диарея.
Искажение результа-
тов эксперимента
В большинстве случа-
ев хроническое тече-
ние, смерть при
стрессе.
Искажение результа-
тов эксперимента
Глава 7. Биориски и качество экспериментов на лабораторных животных 137
Окончание табл. 33
Возбудитель
Francisella tularensis
Leptospira sp.
Trichophyton sp.
Microsporum sp.
Epidermophyton sp.
Заболевания
Биориск заражения
для человека
И ЖИВОТНЫХ
Туляремия
Многие виды животных,
человек
Лептоспироз
Многие виды животных,
человек
Трихофитоз
Собаки, кошки, морские
свинки, кролики, чело-
век
Микроспороз
Собаки, кошки, человек
Эпидермофитоз
Собаки, кошки, кролики,
человек
Пути заражения
Воздушно-капельный,
оральный,
контактный
Оральный (чрезкож-
ный, пренатальный)
Контактный,
воздушно-капельный
Контактный,
воздушно-капельный
Контактный
Контагиозность,
симптоматика и исход
Влияние на результаты
экспериментов
Лихорадка, спленоге-
патомегалия, бронхо-
пневмония, высоко-
контагиозное забо-
левание
В большинстве случа-
ев латентная форма,
при стрессах повы-
шенная заболевае-
мость и смертность,
поражения селезенки
печени и почек
Локальный дерматит,
единичные случаи или
энзоотии, незначитель-
ные проявления и вме-
шательство с дерма-
тологическим экспери-
ментом
Локальный дерматит,
выпадение волос,
незначительные
общие проявления
Дерматит
Искажение результа-
тов эксперимента
Таблица 34
Инвазивные заболевания хищников (собаки, кошки), потенциально опасные
для человека и животных и искажающие результаты исследований
Возбудитель
Isospora spp.
Giardia spp. (G.canis)
Toxoplasma gondii
Babesia canis
Заболевания
Биориск заражения
для человека
и животных
Кокцидиоз
Собаки, кошки, другие
виды животных
Лямблиоз
Собаки, кошки, другие
виды животных, человек
Токсоплазмоз
Многие виды животных,
человек
Пироплазмоз, бабезиоз
Собаки
Пути заражения
Оральный или через
промежуточного хо-
зяина
Оральный
Оральный
Через укусы клеща
Контагиозность,
симптоматика и исход
Влияние на результаты
экспериментов
Энтериты у щенков и
котят. Высококонта-
гиозное заболевание
Энтеропатия.
Искажение результа-
тов эксперимента
В большинстве случа-
ев симптомы не прояв-
ляются
Лихорадка, анемия,
желтуха
Искажение результа-
тов эксперимента
138
Качество животных-биомоделей
Продолжение табл. 34
Возбудитель
Leishmania donovani
Toxocara canis
Toxascaris leonina
Toxocara mystax (cati)
Ancylostoma
caninum
Capillaria sp.
Strongyloides
stercoralis
(S. canis)
Trichuris vulpis
Uncinaria
stenocephala
Aelurostrongylus
abstrusus
Заболевания
Биориск заражения
для человека
И ЖИВОТНЫХ
Лейшманиоз, «Кала
азар»
Кошки, собаки, человек
Токсокароз
Собаки, другие предста-
вители рода собак,
человек (личинки пере-
летных птиц)
Инвазивный аскаридоз
Собаки, кошки, другие
виды кошачьих, человек
(личинки перелетных
птиц)
Токсакароз
Собаки, кошки, другие
виды кошачьих, человек
(личинки перелетных
птиц)
Анкилостомоз
Собаки, другие предста-
вители рода собак,
кошки, человек (личин-
ки перелетных птиц)
Капилляриоз
Собаки, кошки
Стронгилоидоз
Кошки, собаки, другие
виды животных, человек
Трихуроз
Собаки
Унцинариоз
Айлуростронгилёз
Пути заражения
Через укусы моски-
тов
Оральный, пренаталь-
ный, через молоко
Оральный, промежу-
точные хозяева
Оральный, через
молоко, чрескожный,
какие-то хозяева
Оральный, через
молоко, какие-то
хозяева
Оральный
Чрескожный
Оральный
Оральный
Оральный
Контагиозность,
симптоматика и исход
Влияние на результаты
экспериментов
Поражения кожи,
печени и селезенки
Высококонтагиозное
заболевание, при
стрессах смерть
Высококонтагиозное
заболевание, при
стрессах смерть
Высококонтагиозное
заболевание
Искажение результа-
тов эксперимента
Высококонтагиозное
заболевание, при
стрессах смерть.
Искажение результа-
тов эксперимента
Высококонтагиозное
заболевание, при
стрессах смерть.
Искажение результа-
тов эксперимента
Высококонтагиозное
заболевание, при
стрессах смерть.
Искажение результа-
тов эксперимента
Высококонтагиозное
заболевание, при
стрессах смерть.
Искажение результа-
тов эксперимента
Высококонтагиозное
заболевание, при
стрессах смерть.
Искажение результа-
тов эксперимента
Высококонтагиозное
заболевание, при
стрессах смерть.
Искажение результа-
тов эксперимента
Глава 7. Биориски и качество экспериментов на лабораторных животных 139
Продолжение табл. 34
Возбудитель
Ollulanus tricuspis
Dirofilaria repens
Opisthorchis felineus
Echinochasmus
perfoliatus
Echinococcus
granulosus
Diphyllobothrium
latum
Taenia spp.
Multiceps multiceps
Mesocestoides
lineatus
Demodex sp.
Sarcoptes sp.
Заболевания
Биориск заражения
для человека
И ЖИВОТНЫХ
Гельминтная инвазия
желудка
Филяриатоз
Собаки, кошки, в ред-
ких случаях - человек
Инвазия печёночной
двуусткой, описторхоз
Трематодная инвазия
Собаки, кошки, другие
виды животных, человек
Эхинококкоз
Собаки (окончательный
и промежуточный хозя-
ин), различные виды
животных и человек
Дипилидоз
Собаки, кошки, другие
виды животных, человек
Солитёрная инвазия,
дипилидоз
Собаки, кошки, другие
виды животных
Солитёрная инвазия,
мультицептиоз
Собаки, другие виды
животных
Солитёрная инвазия,
мезоцистоидоз
Собаки, кошки, другие
виды животных, человек
Демодикоз
Собаки, другие виды
животных, в редких слу-
чаях - человек
Чесотка
Собаки, другие виды
животных, человек
Пути заражения
Оральный
Через укусы комаров
Оральный (промежу-
точный хозяин)
Оральный (промежу-
точный хозяин)
Оральный
Оральный (промежу-
точный хозяин)
Оральный (промежу-
точный хозяин)
Оральный (промежу-
точный хозяин)
Оральный (промежу-
точный хозяин)
Прямой контакт
в раннем возрасте
Контактный (прямой
или непрямой)
Контагиозность,
симптоматика и исход
Влияние на результаты
экспериментов
В редких случаях
гастриты.
Искажение результа-
тов эксперимента
Высококонтагиозное
заболевание, при
стрессах смерть.
Искажение результа-
тов эксперимента
Высококонтагиозное
заболевание, при
стрессах смерть.
Искажение результа-
тов эксперимента
Высококонтагиозное
заболевание, при
стрессах смерть.
Искажение результа-
тов эксперимента
В большинстве случа-
ев латентная, риск
инфицирования для
человека и многих
видов животных
Заболеваемость при
стрессе.
Искажение результа-
тов эксперимента
Заболеваемость при
стрессе.
Искажение результа-
тов эксперимента
Заболеваемость при
стрессе.
Искажение результа-
тов эксперимента
Заболеваемость при
стрессе.
Искажение результа-
тов эксперимента
Ухудшение самочувст-
вия (состояния), дер-
матит, выпадение во-
лос, смерть
Ухудшение самочувст-
вия (состояния), дер-
матит, выпадение во-
лос, чесотка
140
Качество животных-биомоделей
Окончание табл. 34
Возбудитель
Notoedres cati
(N. cuniculi)
Otodectes cynotis
Cheyletiella spp.
Ctenocephalides spp.
Ixodes ricinus
Rhipicephalus
sanguineus
Linognathus setosus
Neotrombicula
autumnalis
Odectes canis
Felicola
subrostratus
Заболевания
Биориск заражения
для человека
И ЖИВОТНЫХ
Ушная чесотка
Кошки
Заражение ушным
клещом
Кошки, другие кошачьи,
собаки
Заражение ушным
клещом
Собаки, кошки, человек
Заражение блохами
Собаки, кошки, человек
Заражение клещом
Собаки, другие виды
животных, человек
Заражение клещом
Собаки, другие виды
животных, человек
Педикулез
Собаки
Заражение клещом
Кошки, собаки, другие
виды животных, человек
Педикулёз
Собаки
Педикулёз
Кошки
Пути заражения
Контактный (прямой
или непрямой)
Контактный (прямой
или непрямой)
Контактный
Контактный
Контактный
Контактный
Контактный
Контактный
Контактный
Контактный
Контагиозность,
симптоматика и исход
Влияние на результаты
экспериментов
Ухудшение самочувст-
вия (состояния), дер-
матит, выпадение во-
лос, чесотка
Ухудшение самочувст-
вия (состояния), дер-
матит, выпадение
волос, чесотка
Дерматит, чесотка.
Искажение результа-
тов эксперимента
Чесотка, кожные ал-
лергические реакции
Раздражение кожи
вследствие укусов,
переносчик инфекции
Раздражение кожи
вследствие укусов,
переносчик инфекции
Ухудшение самочувст-
вия (состояния), дер-
матит, выпадение
волос
Сильные расчесы,
раздражение кожи
Раздражение кожи
Раздражение кожи
Крупные грызуны (кролики, морские свинки)
Кролики. В экспериментах чаще используются кролики породы шиншилла,
название которых произошло из-за сходства окраски меха с южноамери-
канским зверьком. Окраска волосяного покрова серебристо-голубовато-
серая. Венский голубой кролик отличается быстротой развития. Вес взрос-
лого кролика более 4 кг. Порода шампань имеет равномерную серебристую
окраску волосяного покрова. Живой вес крольчат при рождении около 50 г,
а взрослого кролика 2,6—3,7 кг. Венский белый по форме тела напоминает
венского голубого. Окраска волосяного покрова белоснежная, блестящая.
В отличие от альбиносов у них радужка глаз имеют голубую окраску. Гене-
тически выведено около 60 пород кроликов, среди них в нашей стране
Глава 7. Биориски и качество экспериментов на лабораторных животных 141
распространение получили фландр, белый великан, серый великан, совет-
ский мардер и др.
Кролики служат прекрасным биологическим объектом для физиологи-
ческих, фармакологических и токсико-биологических наблюдений; оценки
активности гормональных препаратов (инсулин адреналин и др.); для био-
логического контроля вакцин (сибиреязвенной, паратифа телят, дизентерии
ягнят, противочумной и др.); проверки активности агглютинирующих сыво-
роток (противостолбнячной, противосибиреязвенной и т. д.). Особенности
овуляции крольчих делает их классическим объектом для изучения функции
яичников.
Кролики служат для приготовления гемолитической сыворотки для ре-
акции связывания комплемента, незаменимы для биологической проверки
материала на бешенство, для приготовления антирабических вакцин. Кроме
этого, их используют для создания моделей таких заболеваний, как экспери-
ментальные опухоли (карцинома Броун-Пирса, дегтярный рак кожи), холе-
стериновая болезнь, стафилококковые и стрептококковые инфекции,
сибирская язва, листереллез, псевдотуберкулез, сальмонеллезы, столбняк,
газовые раневые инфекции, ботулизм, сифилис, для воспроизведения ал-
лергических реакций и т. д. У крольчат вызывают бактериальную дизенте-
рию, менингококковые инфекции.
Кроликов используют также для установления активности культур возбу-
дителей листереллеза, туляремии, анаэробов, стафилококков.
Таблица 35
Вирусные инфекции кроликов и морских свинок, потенциально опасные
для человека и животных и искажающие результаты исследований
Возбудитель
Заболевания
Биориск заражения
для человека
И ЖИВОТНЫХ
Пути заражения
Контагиозность,
симптоматика и исход
Влияние на результаты
экспериментов
Кролики
Myxomatosis virus
(pox virus)
Aujeszky virus
(herpesvirus)
Pox virus кроликов
Миксоматоз
Кролики
Псевдобешенство
Кролики, кошки, собаки,
мини-свиньи, жвачные
животные
Оспа кроликов, керато-
конъюнктивит
Кролики
Чрезкожный (через
укусы комаров),
контактный
(слизистая оболочка)
Оральный
Оральный, воздушно-
капельный
Локальные отеки,
блефароконъюнктивит,
отеки, апатия, 99% за-
болеваемости при пер-
вичном контакте
Ухудшение самочувст-
вия (состояния), спе-
цифические пораже-
ния органов, смерть
Энзоотии, высокий
уровень смертности.
Искажение результа-
тов эксперимента
142 Качество животных-биомоделей
Продолжение табл. 35
Возбудитель
Borna virus (не клас-
сифицирован)
Adenovirus
Papilloma virus кро-
ликов papilloma
shape (papovavirus)
Fibroma virus кроли-
ков. Shope fibroma
(pox virus)
Вирус кроликов
(herpesvirus cuniculi)
Kidney
vacuolating virus
кроликов (RKV)
(polyomavirus)
Заболевания
Биориск заражения
для человека
И ЖИВОТНЫХ
Инфекционное воспале-
ние структур головного
и спинного мозга
Кролики
Энтерит
Кролики
Папилломатоз
Кролики
Фиброматоз
Кролики
Неопределенные
заболевания
Кролики
Неопределенные
заболевания
Кролики
Пути заражения
Оральный
Оральный
Чрезкожный (через
укусы комаров)
Чрезкожный
Воздушно-капельный
Оральный
Контагиозность,
симптоматика и исход
Влияние на результаты
экспериментов
Нарушения ЦНС.
Искажение результа-
тов эксперимента
Субклиническая
инфекция.
Искажение результа-
тов эксперимента
Спонтанные формиро-
вания новообразова-
ний
Изолированная
инфекция.
Искажение результа-
тов эксперимента
Хроническая перси-
стирующая инфекция.
Искажение результа-
тов эксперимента
Неизвестно.
Искажение результа-
тов эксперимента
Морские свинки
Lymphocytic
choriomeningitis virus
(LCM) (arenavirus)
Pneumonia virus
мышей (PVM)
(paramyxovirus)
Sendai virus or para-
influenza 1 virus
[paramyxovirus)
Adenovirus
Reovirus тип 3
Cytomegalovirus
морских свинок
(herpesvirus)
Лимфоцитарный хорио-
менингит(пневмопатия)
Мыши, морские свинки,
хомячки, собаки,
обезьяны, человек
Пневмония
Мыши, крысы, морские
свинки, хомячки
Пневмония
Мыши, крысы, морские
свинки, хомячки, мини-
свиньи, человек
Бронхопневмония
Морские свинки
Неопределенные
заболевания
Мыши, морские свинки,
хомячки, человек
Неопределенные
заболевания
Морские свинки
Оральный, транс-
плацентарный
Воздушно-капельный,
оральный
Оральный, воздушно-
капельный
Воздушно-капельный
Оральный
Оральный
Хроническая перси-
стирующая инфекция.
Искажение результа-
тов эксперимента
Обычно субклиничес-
кая инфекция.
Искажение результа-
тов эксперимента
Обычно субклиничес-
кая инфекция.
Искажение результа-
тов эксперимента
Спорадические случаи
смерти
Обычно субклиничес-
кая инфекция
Искажение результа-
тов эксперимента
Хроническая персисти-
рующая инфекция.
Искажение результа-
тов эксперимента
Глава 7. Биориски и качество экспериментов на лабораторных животных 143
Продолжение табл. 35
Возбудитель
Н-1 virus (Toolan
virus) (parvovirus)
SV-5 (paramyxovirus)
Polioma virus
(papovavirus)
Заболевания
Биориск заражения
для человека
И ЖИВОТНЫХ
Неопределенные
заболевания
Мыши, крысы, морские
свинки, хомячки
Неопределенные
заболевания
Сирийские хомячки,
морские свинки, чело-
век, обезьяны
Онкогенные
Мыши, крысы, хомячки
Пути заражения
Оральный
Воздушно-капельный,
трансплацентарный
Оральный
Контагиозность,
симптоматика и исход
Влияние на результаты
экспериментов
Заболевание вслед-
ствие вторичной ин-
фекции
Респираторные симп-
томы и нарушения со
стороны ЦНС.
Искажение результа-
тов эксперимента
Бессимптомное.
Искажение результа-
тов эксперимента
Та блица 36
Микоплазменные, бактериальные и грибковые инфекции кроликов
и морских свинок, потенциально опасные для человека и животных
и искажающие результаты исследований
Возбудитель
Actinomyces sp.
Mycoplasma sp.
(pulmonis, caviae),
Pasteurella multocida
Bacillus piliformis
Yersinia pseudotuber-
culosis
Streptococcus
(серогруппы A, G, C)
Заболевания
Биориск заражения
для человека
и животных
Актиномикоз
Кролики, кошки, хомяч-
ки, собаки
Ринит, бронхит, пневмо-
ния, артрит
Мыши, крысы, морские
свинки, кролики
Пастереллез, энзоотич-
ный пастереллез, пнев-
мония, отсутствие ды-
хания через нос у кро-
ликов
Многие виды животных,
человек
Болезнь Тизера
Различные виды живот-
ных
Псевдотуберкулез, ро-
дентиоз
Различные виды живот-
ных
Септицемия, пневмония,
редко абсцессы
Мыши, крысы, кролики,
морские свинки, кошки
Пути заражения
Воздушно-капельный,
контактный
Воздушно-капельный
Воздушно-капельный,
оральный
Оральный
Воздушно-капельный,
оральный
Воздушно-капельный,
оральный
Контагиозность,
симптоматика и исход
Влияние на результаты
экспериментов
Искажение результа-
тов эксперимента
Часто скрытое течение
Искажение результа-
тов эксперимента
Серьёзные проблемы
с разведением кроли-
ков и с проведением
экспериментов, еди-
ничные случаи смерти,
энзоотии
Энтерит, диарея,
смерть
Недомогание, абсцес-
сы, в редких случаях
энзоотии.
Искажение результа-
тов эксперимента
От единичных до час-
тых случаев смерти.
Искажение результа-
тов эксперимента
144
Качество животных-биомоделей
Продолжение табл. 36
Возбудитель
Francisella tularensis
Fusobacterium
necrophorum
Bordetella
bronchiseptica
Klebsiella pneumoniae
Streptobacillus
moniliformis
Streptococcus
pneumoniae
Haemophilus sp.
Listeria
monocytogenes
Mycobacterium sp.
(tuberculosis, bovis,
avium)
Escherichia coli
(некоторые серотипы)
Заболевания
Биориск заражения
для человека
И ЖИВОТНЫХ
Туляремия
Кролики, мыши,
морские свинки, крысы,
человек
Некротические процес-
сы, изъязвления, под-
кожные абсцессы, сеп-
тицемия
Мыши, кролики, человек
Пневмония, бронхо-
пневмония, плевро-
пневмония
Различные виды живот-
ных
Пневмония, бронхо-
пневмония, плевро-
пневмония
Мыши, крысы, морские
свинки, кролики,
человек
Гнойный полиартрит,
анемия, диарея, конъ-
юнктивит, паралич
Мыши, крысы, кролики,
морские свинки, индей-
ки, человек
Пневмония, бронхо-
пневмония, плевро-
пневмония, менингит
Различные виды живот-
ных, человек
Энтерит, гнойные пора-
жения, пневмония, ме-
нингит
Морские свинки, кроли-
ки, мыши, кошки,
человек
Листериоз
Различные виды живот-
ных, человек
Туберкулез
Различные виды живот-
ных, человек
Колиэнтерит, инфекции
мочевыводящих путей
Морские свинки, кро-
лики, собаки, кошки,
человек
Пути заражения
Чрезкожный,
воздушно-капельный,
оральный,
контактный
Чрезкожный,
воздушно-капельный
Воздушно-капельный
Воздушно-капельный,
оральный
Оральный,
от укуса кролика
Воздушно-капельный
Аэрогенный,
оральный
Оральный, транс-
плацентарный
Воздушно-капельный,
оральный
Оральный, восходя-
щая инфекция моче-
выводящих путей
Контагиозность,
симптоматика и исход
Влияние на результаты
экспериментов
Незначительные
общие проявления,
абсцессы.
Искажение результа-
тов эксперимента
В редких случаях абс-
цессы, смерть.
Искажение результа-
тов эксперимента
Редко смертельные
исходы, недомогание.
Искажение результа-
тов эксперимента
Редко смертельные
исходы, недомогание.
Искажение результа-
тов эксперимента
Недомогание, повреж-
дения органов, абс-
цессы, смертельный
исход
Редкие случаи смерти
или энзоотии (в слу-
чае гиповитаминоза С)
Недомогание, редко
смертельные исходы.
Искажение результа-
тов эксперимента
Выкидыши, редкие
случаи смерти или
энзоотии
Искажение результа-
тов эксперимента
Недомогание, диарея,
смерть.
Искажение результа-
тов эксперимента
Глава 7. Биориски и качество экспериментов на лабораторных животных 145
Продолжение табл. 36
Возбудитель
Proteus sp.
Staphylococcus
aureus
Treponema cuniculi
Bacillus piliformis
Corynebacterium
murium (kutscheri,
pseudotuberculosis
murium)
Pasteurella
multocida
Pasteurella pneumo-
tropica (nomen
dubium)
Salmonella sp.
Clostridium
perfringens
Trichophyton sp.
(quinckeanum,
schonleinii, menta-
grophytes, gypseum,
asteroids)
Microsporum canis
Заболевания
Биориск заражения
для человека
И ЖИВОТНЫХ
Энтерит, абсцессы,
пиометра, дерматиты,
инфекция мочевыводя-
щих путей
Морские свинки, кролики,
мыши, крысы, человек
Подкожные абсцессы,
септицемия, мастит
Различные виды животных
Сифилис кроликов
Кролики
Болезнь Тизера
Различные виды животных
Абсцессы, свищи, энте-
рит, пневмония
Мыши, крысы, морские
свинки, хомячки
Пастереллёз, септице-
мия, пневмония
Различные виды животных
Бронхопневмония,
абсцессы
Мыши, крысы, хомячки,
собаки, мини-свиньи,
человек
Энтерит
Энтерит
Различные виды животных
Трихофитоз
Морские свинки,
кролики, человек
Микроспороз
Мыши, крысы, кошки,
кролики, собаки,
человек
Пути заражения
Оральный, восходя-
щая инфекция моче-
выводящих путей
Воздушно-капельный,
оральный,
чрезкожный
Чрезкожный,
оральный
Оральный
Оральный, воздушно-
капельный
Воздушно-капельный,
оральный
Аэрогенный,
оральный
Оральный
Оральный
Воздушно-капельный,
контактный
Воздушно-капельный,
контактный
Контагиозность,
симптоматика и исход
Влияние на результаты
экспериментов
Недомогание, смерть
Искажение результа-
тов эксперимента
Спорадические случаи
смерти.
Искажение результа-
тов эксперимента
Единичные случаи или
энзоотии
Тифлит, поражения
печени, смерть при
стрессах
Недомогание, связан-
ное с изменениями
в легких и гнойными
процессами.
Искажение результа-
тов эксперимента
Смерть при стрессах.
Искажение результа-
тов эксперимента
Гнойные процессы,
смерть, изменения
в легких.
Искажение результа-
тов эксперимента
Смерть при стрессах.
Искажение результа-
тов эксперимента
Редкие случаи смерти
токсемия
Искажение результа-
тов эксперимента
Локальный дерматит,
единичные случаи или
энзоотии, незначитель-
ные проявления и вме-
шательство с дермато-
логическим экспери-
ментом
Локальный дерматит,
выпадение волос, не-
значительные общие
проявления
146
Качество животных-биомоделей
Таблица 37
Инвазивные заболевания кроликов и морских свинок, потенциально опасные
для человека и животных и искажающие результаты исследований
Возбудитель
Eimeria
Eimeria stiedae
Giardia sp.
Encephalitozoon
(Nosema) cuniculi
Sarcocystis cuniculi
Toxoplasma gondii
Cryptosporidium sp.
Graphidium strigosum
Paraspidodera
uncinata
Passalurus
ambiguus
Trichostrongylus
retortaeformis
Protostrongylus sp.
Заболевания
Биориск заражения
для человека
И ЖИВОТНЫХ
Кишечный кокцидиоз
Кролики, морские
свинки
Печеночный кокцидиоз,
кокцидиоз билиарного
тракта
Кролики
Лямблиоз
Кролики, морские
свинки, другие виды
ЖИВОТНЫХ
Нозематоз
Кролики, морские свин-
ки, мыши, крысы, дру-
гие виды животных,
человек
Саркоспоридиоз
(саркоспоридический
миозит)
Кролики
Токсоплазмоз
Различные виды
ЖИВОТНЫХ
Кишечный кокцидиоз
Кролики
Гельминтная инвазия
Морские свинки,
кролики
Гельминтная инвазия
кишечника
Морские свинки
Инвазия острицами
Кролики
Гельминтная инвазия
Кролики
Легочная инвазия
Кролики
Пути заражения
Оральный
Оральный
Оральный
Оральный, транс-
плацентарный
Оральный
Оральный, транс-
плацентарный
Оральный
Оральный
Оральный
Оральный
Оральный
Оральный
Контагиозность,
симптоматика и исход
Влияние на результаты
экспериментов
От умеренной до
высокой патогенности,
диарея
Высокая патогенность,
широкое распростра-
нение заболевания,
смерть при тяжелых
формах инвазии,
возможны энзоотии
Энтеропатия.
Искажение результа-
тов эксперимента
Генерализованная
болезнь.
Искажение результа-
тов эксперимента
Повреждения органов.
Искажение результа-
тов эксперимента
Чаще латентное тече-
ние. Высококонтагиоз-
ное заболевание
Искажение результа-
тов эксперимента
Редко небольшие
геморрагические
высыпания на стенке
желудка
Выкидыши, смерть
молодых животных
Геморрагический
тифлит и колит при
тяжелой инвазии
Диарея, потеря веса,
слабость при тяжелой
инвазии
Диспноэ при тяжелой
инвазии.
Искажение результа-
тов эксперимента
Глава 7. Биориски и качество экспериментов на лабораторных животных 147
Окончание табл. 37
Возбудитель
Taenia pisiformis larva
(Cysticercus
pisiformis)
Echinococcus
granulosus larva
Fasciola hepatica
Dicrocoelium
dendriticum
(D. lanceolatum)
Cheyletiella sp.
Chirodiscoides caviae
Gliricola porcelli
Gyropus
Notoedres cuniculi
Psoroptes cuniculi
Sarcoptes scabiei
Заболевания
Биориск заражения
для человека
И ЖИВОТНЫХ
Цистицеркоз
Кролики
Однокамерный
эхинококкоз
Человек, собака (окон-
чательный и промежу-
точный хозяин)
Трематодная инвазия
печени
Морские свинки,
кролики, другие виды
животных, человек
Трематодная инвазия
печени
Морские свинки, кроли-
ки, другие виды живот-
ных, человек
Заражение клещами
Кролики, кошки, собаки,
иногда человек
Заражение клещами
Морские свинки
Педикулез
Морские свинки
Педикулез
Морские свинки
Чесотка тела
Кролики
Ушная чесотка
Кролики
Чесотка
Морские свинки,
кролики
Пути заражения
Оральный
Оральный
Оральный (промежу-
точный хозяин)
Оральный (промежу-
точный хозяин)
Контактный
Контактный
Контактный
Контактный
Контактный
Контактный
Контактный
Контагиозность,
симптоматика и исход
Влияние на результаты
экспериментов
Анемия и возможно
нарушение пищеваре-
ния при тяжелой
инвазии
Спорадические еди-
ничные случаи, серь-
езные заболевания,
даже смерть при ло-
кализации в мозге,
легких или печени
Поражения печени,
смерть.
Искажение результа-
тов эксперимента
Поражения печени.
Искажение результа-
тов эксперимента
Изменения кожи
Общее ухудшение
самочувствия
(состояния)
Зуд, ухудшение само-
чувствия (состояния)
Ухудшение самочувст-
вия (состояния)
Ухудшение самочувст-
вия (состояния)
в случае сильного
поражения
Ухудшение самочувст-
вия (состояния)
в случае сильного
поражения
Ухудшение самочувст-
вия (состояния)
в случае сильного
поражения
Морская свинка — Cavia, guinea—pig или Porcellus — принадлежит к от-
ряду грызунов. В Европу морские свинки были завезены испанцами в XVI
столетии из Перу, их родины, поэтому во Франции, Испании, Италии, Пор-
148
Качество животных-биомоделей
тугалии их называют перуанскими свинками, в Бельгии — индийскими или
горными, в Англии — гвинейскими, в Германии и у нас — морскими свин-
ками. Форма головы, похрюкивание послужили, по-видимому, причиной
того, что этих животных называют свинками. В зависимости от длины шер-
сти различают такие основные породы морских свинок. Гладкошерстная или
короткошерстная порода наиболее широко распространена в лабораториях
России. Гималайская свинка характеризуется белой окраской туловища. Гол-
ландская морская свинка выведена в Голландии. Агути золотистый (с золо-
тисто-коричневой окраской туловища) и агути серый (с серой окраской ту-
ловища). Розетчатая (японская) порода выведена в Англии. Длинношерст-
ная (ангорская) порода привезена из Перу.
Новорожденные морские свинки зрячие, их кожа покрыта шерстью,
в первый день рождения свободно передвигаются, находят и поедают корм,
т. е. приспособлены к самостоятельной жизни. На основании этого говорят,
что морские свинки рождаются взрослыми. Морские свинки являются клас-
сическим объектом для изучения аллергических реакций (анафилаксии),
а также авитаминозов С и Р.
Изолированные органы морской свинки используют для общефизиологи-
ческих и фармакологических исследований. У морских свинок можно вызвать
такие инфекционные заболевания, как туберкулез, псевдотуберкулез, дифте-
рию, чуму, лептоспироз, сап, раневые газовые инфекции, столбняк, бруцел-
лез, туляремию, холеру, листереллез, сальмонеллезы, риккетсиозы и др.
Большое количество открытий в области бактериологии сделано благо-
даря экспериментальным исследованиям на морских свинках. Важную роль
сыграла морская свинка как лабораторное животное, на котором проводи-
лось экспериментальное обоснование внедрения различных прививок у
людей, изучены методы десенсибилизации. Сыворотка морских свинок со-
держит много комплемента и используется для постановки реакций Борде
Жангу, Вассермана. Эритроциты морской свинки — хороший объект для ге-
магглютинации в диагностике вируса гриппа. Кроме того, морская свинка
служит ценным объектом для выявления возбудителей туберкулеза, сапа,
бруцеллеза, чумы и лептоспироза.
Мелкие грызуны (крысы, мыши, хомячки)
Биомодели-грызуны являются самым распространенным объектом для прове-
дения исследовательских и испытательных экспериментов во всех лаборато-
риях мира.
Крысы относятся к роду Rattus, семейству мышеобразных (Muridae). Для
экспериментальных исследований в лабораториях используют белых крыс,
которые являются альбиносами черной и серой пород. Останки крыс обна-
ружены в археологических находках 1 и II тысячелетий до н. э. Черная крыса
Глава 7. Биориски и качество экспериментов на лабораторных животных 149
появилась в Европе в XII веке. Места распространения диких крыс связаны
с жильем человека.
Важное преимущество белых крыс как лабораторных животных заключа-
ется в том, что они довольно устойчивы к инфекционным заболеваниям и
дают большой приплод. Небольшой вес белых крыс и относительно простое
содержание и разведение их в лабораторных условиях позволяют проводить
массовые опыты.
Крысы необходимы для установления токсичности лекарственных веществ
и ядов, широко используются при изучении вопросов питания, проведении
биологической стандартизации гормональных препаратов, постановке науч-
ных исследований по витаминологии, эндокринологии, биохимии. Исполь-
зуют белых крыс также для воспроизведения на них экспериментальных
опухолей (саркомы Кричевского и Синельникова) и инфекционных заболе-
ваний (бешенство, амебиаз, грипп свиней и др.).
Для лабораторных целей чаще всего используют белую мышь — Mus
musculus L., которая является альбиносом серой, домашней мыши. Принад-
лежит она к отряду грызунов (Rodentia), семейству мышиных (Muridae),
подсемейству Murinae. Как лабораторные животные, мыши нашли широкое
распространение в Советском Союзе и за рубежом. Их используют для все-
возможных научных целей в биологии, онкологии, токсикологии, фармако-
логии, физиологии, микробиологии, генетике. Белые мыши являются наи-
более подходящим объектом для определения токсичности химических и
биологических препаратов, стандартизации гормональных препаратов вак-
цин, сывороток, для изучения злокачественных опухолей, лейкозов. Исполь-
зуются мыши также для изучения патогенеза различных инфекционных за-
болеваний и для воспроизведения таких заболеваний, как сибирская язва,
пастереллез, листереллез, сальмонеллезы, ботулизм, столбняк, раневые газо-
вые инфекции, риккетсиозы, грипп, энцефалиты, пневмококковые инфек-
ции и др. Мыши являются незаменимыми лабораторными животными для
разработки проблем наследственности, диагностики вирусных и других ин-
фекционных заболеваний.
Таблица 38
Вирусные инфекции мышей и крыс, потенциально опасные
для человека и животных и искажающие результаты исследований
Возбудитель
Ectromelia virus
(pox virus)
Заболевания
Биориск заражения
для человека
И ЖИВОТНЫХ
Эктромелия, мышиный
сифилис, высыпания на
коже
Мыши, крысы, морские
свинки, кролики
Пути заражения
Чрезкожный, ораль-
ный, воздушно-
капельный
Контагиозность,
симптоматика и исход
Влияние на результаты
экспериментов
От молниеносного до
субклинического тече-
ния инфекции.
Искажение результа-
тов эксперимента
150 Качество животных-биомоделей
Продолжение табл. 38
Возбудитель
Hepatitis virus мышей
(MHV) (coronavirus)
Reovirus тип 3
(diplornavirus)
Sendai virus or para-
influenza 1 virus
(paramyxovirus)
Pneumonia virus
мышей (PVM) (para-
myxovirus)
Theiler's virus (GD VII)
(picornavirus)
Toolan's H-1 virus
(parvovirus)
Kilham virus крыс
(KRV) (parvovirus)
Coronavirus крыс
(RCV) (coronavirus)
Sialodacryoadenitis
virus (SDA)
(coronavirus)
Lethal intestinal
virus мышат
(LIVIM) (coronavirus)
Заболевания
Биориск заражения
для человека
И ЖИВОТНЫХ
Вирус гепатотэнтеро-
тропный или нейро-
тропный (в зависимос-
ти от серотипа)
Мыши, крысы
Неопределенные заболе-
вания взрослых мышей
Мыши, человек
Пневмония
Мыши, крысы, хомячки,
мини-свиньи, морские
свинки, человек
Пневмония
Мыши, крысы, хомячки,
морские свинки
Энцефаломиелиты
Мыши, хомячки
Неопределенные забо-
левания
Крысы
Неопределенные забо-
левания, смертность
и врожденные пороки
у детенышей, у KRV-не-
гативно инфицирован-
ных самок
Крысы
Пневмония, в основном
у крыс на грудном
вскармливании
Крысы, мыши
Сиалодакриоаденит,
кератоконъюнктивит
Крысы, мыши
Диарея у детенышей
мышей
Мыши
Пути заражения
Оральный, воздушно-
капельный
Оральный, воздушно-
капельный
Воздушно-капельный,
оральный
Воздушно-капельный
Оральный
Оральный
Оральный, через
молоко
Воздушно-капельный,
оральный
Воздушно-капельный,
оральный
Воздушно-капельный,
оральный
Контагиозность,
симптоматика и исход
Влияние на результаты
экспериментов
От молниеносного до
субклинического тече-
ния инфекции
Искажение результа-
тов эксперимента
Энзоотии у новорож-
денных мышей с угне-
тением роста, пораже-
ния ЦНС, диарея и по-
вышение жирности во-
лосяного покрова
Генерализованная суб-
клиническая инфекция,
появление осложнений
при стрессах или при
смешанных инфекциях
Генерализованная суб-
клиническая инфекция,
появление осложнений
при стрессах или при
смешанных инфекциях
Нейропатогенная
у молодых животных,
в других случаях бес-
симптомная кишечная
инфекция у мышей
Нейропатогенная
у подсосных крыс,
позднее - субклини-
ческая инфекция
Субклиническая ин-
фекция.
Искажение результа-
тов эксперимента
В большинстве случа-
ев субклиническая ин-
фекция.
Искажение результа-
тов эксперимента
Интерстициальная
пневмония у новорож-
денных животных
От острого до бес-
симптомного течения
инфекции
Глава 7. Биориски и качество экспериментов на лабораторных животных 151
Продолжение табл. 38
Возбудитель
Epizootic diarrhoea
мышат (EDIM)
(rotavirus)
К virus (pneumonia
virus новорожденных
мышей) (papovavirus)
Adenovirus мышей
Polyoma virus
(papovavirus)
Encephalomyocarditis
virus (EMC)
(picomavirus)
Lymphocytic chorio-
meningitis virus (LCM)
(arenavirus)
Lactic dehydrogenase-
stimulating virus (LDV)
(togavirus)
Cytomegalovirus
мышей (MCMV)
(herpesvirus)
Mammary tumor
virus (MTV)
Endogenous murine
viruses тип С (сино-
нимы: retroviruses,
oncornaviruses,
leukaemia viruses)
Некоторые примеры
этих вирусов:
Murine sarcoma
viruses (MuSV)
- для крыс:
Kirsten sarcoma virus
Заболевания
Биориск заражения
для человека
И ЖИВОТНЫХ
Диарея, чувствительны
детеныши до 18-го дня
жизни
Мыши
Пневмония у мышей
моложе 10 дней
Мыши
Неопределенные забо-
левания
Мыши, хомячки, мини-
свиньи
Онкогенные
Мыши
Энцефаломиокардит
Мыши, крысы, морские
свинки, кролики
Лимфоцитарный хорио-
менингит
Мыши, морские свинки,
хомячки, собаки,
обезьяны, человек
Неопределенные
заболевания
Мыши
Угнетение роста
Мыши
Карцинома молочной
железы
Мыши
Пути заражения
Оральный
Воздушно-капельный,
оральный
Воздушно-капельный,
оральный
Оральный
Оральный
Оральный
Оральный, через
молоко или рану
(укусы)
Оральный (через
инфицированную
слюну)
Через молоко
Контагиозность,
симптоматика и исход
Влияние на результаты
экспериментов
Высококонтагиозное
заболевание, низкая
смертность у подсос-
ных мышей
Субклиническая инфек-
ция у мышей старше
10 дней
Грудные мыши: острая
летальная инфекция.
Позднее - субклини-
ческая инфекция
Субклиническая инфек-
ция, редко спонтан-
ные новообразования
Грудные мыши:
субклиническая
инфекция
От острой до хрони-
ческой персистирую-
щей инфекции, в за-
висимости от вида
инфекции; хроничес-
кое носительство
Увеличение уровня
лактатдегидрогеназы
плазмы, отдаленные
нарушения в эндо-
кринной и иммунной
системах
Хроническая персисти-
рующая инфекция
Болезни у взрослых
животных
Искажение результа-
тов эксперимента
Диагностика только в специализированных лабораториях
Саркома
Крысы или мыши
Контактный
Часто скрытая, ново-
образования, смерть.
Искажение результа-
тов эксперимента
152 Качество животных-биомоделей
Окончание табл. 38
Возбудитель
Harvey sarcoma virus,
Sarcoma virus крыс
(in vitro)
- для мышей:
Moloney sarcoma
virus, FBJ osteosar-
coma virus, «sarcoma»
virus мышей линии
Balb/c, Gazdar sar-
coma virus, sarcoma
virus мышей (in vitro)
Murine leukaemia
viruses (MuLV)
Moloney MuLV,
Rauscher MuLV,
AKR MuLV,
Rad LV (для C57BL/
Ka),
Gross MuLV
Заболевания
Биориск заражения
для человека
И ЖИВОТНЫХ
Саркома
Крысы или мыши
То же
Лейкемия
Мыши
То же
«
«
«
«
Пути заражения
Контактный
То же
Контактный
То же
«
«
«
«
Контагиозность,
симптоматика и исход
Влияние на результаты
экспериментов
Часто скрытая, ново-
образования, смерть.
Искажение результа-
тов эксперимента
То же
Часто скрытая, лейке-
мия, смерть.
Искажение результа-
тов эксперимента
Таблица 39
Микоплазменные, бактериальные и грибковые инфекции мышей
и крыс, потенциально опасные для человека и животных
и искажающие результаты исследований
Возбудитель
Mycoplasma sp.
Pulmonis, arthritides,
neurolyticum,
Acholeplasma
Bacillus piliformis
Corynebacterium
kutschery (murium,
pseudotuberculosis)
Salmonella sp.
Заболевания
Биориск заражения
для человека
и животных
Ринит, бронхопневмо-
ния, артрит, нарушение
нервных функций, вос-
паление среднего уха
Крысы, мыши
Болезнь Тизера
Различные виды живот-
ных
Абсцессы, свищи, энте-
риты, пневмония
Мыши, крысы, хомячки,
морские свинки
Сальмонеллез
Различные виды живот-
ных, человек
Пути заражения
Воздушно-капельный,
трансплацентарный
Оральный
Оральный, воздушно-
капельный
Оральный
Контагиозность,
симптоматика и исход
Влияние на результаты
экспериментов
Часто скрытая, нару-
шение развития,
смерть
Повреждения печени,
диарея, высокая
смертность
Недомогание, соответ-
ствующее изменениям
в легких и процессы
нагноения
Энтериты, энзоотии,
часто скрытое течение,
смерть при стрессах
Глава 7. Биориски и качество экспериментов на лабораторных животных 153
Продолжение табл. 39
Возбудитель
Pasteurella pneumo-
tropica (nomen
dubium)
Streptobacillus
moniliformis
Yersinia pseudo-
tuberculosis
Pasteurella multocida
Bordetella bronchi-
septica
Francisella tularensis
Pseudomonas
aeroginosa
Streptococcus
pneumoniae
Klebsiella
pneumoniae
Заболевания
Биориск заражения
для человека
И ЖИВОТНЫХ
Бронхопневмония,
абсцессы, конъюнкти-
виты
Мыши, крысы, хомячки,
собаки, мини-свиньи,
человек
Гнойный полиартрит,
анемия, диарея,
конъюнктивит, паралич
задних конечностей
Крысы, мыши, морские
свинки, кролики, индей-
ки, человек
Псевдотуберкулёз,
Крысы, мыши, морские
свинки, кошки, кролики,
обезьяны, человек
Пастереллёз, бронхо-
пневмония, абсцессы,
отит
Различные виды живот-
ных (особенно кролики),
человек
Бронхопневмония,
пневмония, плевро-
пневмония
Различные виды живот-
ных, изредка человек
Абсцессы, туляремия
Кролики, мыши, морс-
кие свинки, крысы, че-
ловек
Септицемия, пневмония,
инфекции мочевыводя-
щих путей, абсцессы
Различные виды живот-
ных, человек
Пневмония, бронхо-
пневмония, плевро-
пневмония, менингит
Различные виды живот-
ных, человек
Пневмония, бронхо-
пневмония, плевро-
пневмония
Мыши, крысы, морские
свинки, обезьяны,
человек
Пути заражения
Оральный, воздушно-
капельный
Оральный, от укуса
крыс (содоку или
хейверхиллская лихо-
радка)
Оральный, воздушно-
капельный
Воздушно-капельный,
оральный
Воздушно-капельный
Чрезкожный, воздуш-
но-капельный, ораль-
ный, контактный
Воздушно-капельный,
оральный
Воздушно-капельный
Воздушно-капельный,
оральный
Контагиозность,
симптоматика и исход
Влияние на результаты
экспериментов
Конъюнктивит, энзо-
отии, некрозы, изме-
нения £ легких; может
быть бессимптомное
течение
Искажение результа-
тов эксперимента
Недомогание, высокая
смертность, пораже-
ние органов, абсцессы
Искажение результа-
тов эксперимента
Недомогание, абсцес-
сы (кишечника, лимфа-
тических узлов, печени,
селезенки)
Энзоотии,тетанус
мышц шеи, смерть
при стрессе
Искажение результа-
тов эксперимента
Недомогание, измене-
ния в легких, смерть
Искажение результа-
тов эксперимента
Незначительные
общие проявления
Искажение результа-
тов эксперимента
Смерть, особенно пос-
ле облучения, ожогов,
хирургических вмеша-
тельств
Недомогание, измене-
ния в лёгких, смерть
Искажение результа-
тов эксперимента
Скрытая. Смерть при
стрессах.
Искажение результа-
тов эксперимента
154
Качество животных-биомоделей
Продолжение табл. 39
Возбудитель
Staphylococcus
aureus
Streptococcus
(серогруппы А, С, G)
Haemophilus sp.
Listeria monocyto-
genes
Citrobacter freundii
Clostridium
perfringens
Actinobacillus sp.
Enterobacter sp.
Erysipelothrix
rhusiopathiae
Escherichia coli
(некоторые серотипы)
Proteus sp.
Заболевания
Биориск заражения
для человека
И ЖИВОТНЫХ
Инфекции кожи,
абсцессы, маститы
Различные виды живот-
ных, человек
Гнойные поражения
Различные виды живот-
ных, человек
Гнойные поражения,
энтерит
Кролики, мыши, кошки
Листериоз
Различные виды живот-
ных, человек
Колит, абсцессы
Мыши, крысы, человек
Энтеротоксемия
Различные виды живот-
ных, изредка человек
Гнойные поражения
Различные виды живот-
ных
Энтериты
Мыши, крысы, человек
Рожа, септицемия
мышей
Мыши, крысы, кролики,
мини-свиньи,
домашняя птица,
человек
Энтерит, инфекции
мочевыводящих путей,
абсцессы, пиометра,
дерматиты
Различные виды живот-
ных, человек
Энтериты, инфекция
мочевыводящих путей,
абсцессы, пиометра,
дерматиты
Морские свинки, кроли-
ки, мыши, крысы,
человек
Пути заражения
Чрезкожный, ораль-
ный, воздушно-
капельный
Чрезкожный, воздуш-
но-капельный
Воздушно-капельный,
оральный
Оральный, транс-
плацентарный
Оральный
Оральный
Воздушно-капельный
Оральный
Оральный,
чрезкожный
Оральный, восходя-
щая инфекция
мочевыводящих
путей
Оральный, восходя-
щая инфекция моче-
выводящих путей,
воздушно-капельный
Контагиозность,
симптоматика и исход
Влияние на результаты
экспериментов
Абсцессы (от единич-
ных случаев до энзо-
отии.
Искажение результа-
тов эксперимента
Абсцессы, маститы,
пневмония, септице-
мия
Единичные случаи
смертельных исходов
Выкидыши, моноцитоз,
изменения в печени,
поражение нервов
Незначительные
общие проявления,
смерть при стрессах
От единичных до
эндемичных случаев
заболевания
Недомогание, часто
скрытые симптомы
От единичных до час-
тых случаев смер-
тельных исходов при
стрессе
Только в эксперименте:
недомогание, смерть
Искажение результа-
тов эксперимента
Недомогание, фаталь-
ная диарея.
Искажение результа-
тов эксперимента
Недомогание, смерть.
Искажение результа-
тов эксперимента
Глава 7. Биориски и качество экспериментов на лабораторных животных 155
Окончание табл. 39
Возбудитель
Eperythrozoon
coccoides
Haemobartonella muris
Leptospira (различ-
ные серотипы)
Trichophyton
Microsporum canis
Заболевания
Биориск заражения
для человека
И ЖИВОТНЫХ
Мыши, крысы
Бартонеллез
Мыши
Лептоспироз
Различные виды живот-
ных, человек
Трихофитоз
Различные виды живот-
ных (особенно морские
свинки), человек
Микроспороз
Мыши, крысы, кошки,
кролики, собаки,
человек
Пути заражения
Через укусы
Через укусы
Чрезкожный, ораль-
ный
Воздушно-капельный,
контактный
Воздушно-капельный,
контактный
Контагиозность,
симптоматика и исход
Влияние на результаты
экспериментов
Гиперпластическое
увеличение селезенки,
гепатит
Скрытое течение, тяже-
лая анемия у животных
после спленэктомии
Часто скрытая, потеря
веса, изменение гема-
тологических показа-
телей
Истощение, в редких
случаях смерть, выпа-
дение волос, незначи-
тельные общие прояв-
ления, поражения кожи
(могут быть скрытыми)
Выпадение волос, по-
ражения кожи, незна-
чительные общие про-
явления
Искажение результа-
тов эксперимента
Таблица 40
Инвазивные заболевания мышей и крыс, потенциально опасные
для человека и животных и искажающие результаты исследований
Возбудитель
Encephalitozoon
(Nosema) cuniculi
Chilomastix
bettencourti
Spironucleus muris
(Hexamita muris)
Giardia muris
(Lamblia muris)
Заболевания
Биориск заражения
для человека
и животных
Нозематоз
Кролики, мыши, крысы,
хомячки, морские свин-
ки и другие грызуны,
собаки, кошки
Кишечная инвазия
Мыши, крысы, хомячки
Спиронуклеоз (хрони-
ческий энтерит)
Мыши, крысы, хомячки
и другие грызуны
Лямблиоз (энтерит)
Мыши, крысы, хомячки
и другие грызуны
Пути заражения
Оральный, транс-
плацентарный
Оральный
Оральный
Оральный
Контагиозность,
симптоматика и исход
Влияние на результаты
экспериментов
Энцефалит, нефрит,
часто скрытое тече-
ние
Ухудшение самочувст-
вия (состояния),
энзоотии
Часто скрытая, дис-
комфорт, диарея,
стимуляция макро-
фагов
Ухудшение самочувст-
вия (состояния),
диарея, энзоотии
156
Качество животных-биомоделей
Возбудитель
Eimeria sp.
Cryptosporidium sp.
Toxoplasma gondii
Hepatozoon sp.
(Leucocytozoon sp.)
Aspiculuris tetraptera
Syphacia muris
Syphacia obvelata
Trichosomoides
crassicauda
Strongyloides ratti
Capillaria hepatica
Multiceps serials
larva (Coenurus
serialis)
Hydatigera
taeniaeformis larva
(Strobilocercus)
Hymenolepis sp. spp.
Leptopsylla segnis
Myobia musculi
Заболевания
Биориск заражения
для человека
и животных
Кишечный кокцидиоз
Мыши, крысы
Кишечный кокцидиоз
Мыши
Токсоплазмоз
Различные виды живот-
ных, человек
Гепатозоонозы
Крысы, мыши
Инвазия острицами
Мыши, крысы
Инвазия острицами
Крысы, мыши
Инвазия цистицерка
Крысы
Мыши, крысы, грызуны
Инвазия нематод
Мыши, крысы, кроли-
ки, редко - человек
Крысы и другие виды
животных, собаки
(окончательный хозяин)
Цистерикоз
Солитерная инвазия
Мыши, хомячки, крысы
и другие грызуны,
человек
Тунгиозная инвазия
Мыши, крысы
Заражение клещами
Мыши
Продо
Пути заражения
Оральный
Оральный
Оральный, транс
плацентарный
Оральный, через
промежуточного
хозяина
Оральный
Оральный
Оральный
Оральный,
чрезкожный
Оральный
Оральный
Оральный
Оральный, промежу-
точный хозяин: раз-
личные насекомые
(Н. Nana также без
промежуточного
хозяина)
Контактный
Контактный
жение табл. 40
Контагиозность,
симптоматика и исход
Влияние на результаты
экспериментов
Энтерит, фатальный
или интермиттирую-
щий
Энтерит.
Искажение результа-
тов эксперимента
Латентные, незначи-
тельные общие про-
явления, смерть
Анемия, энтерит,
дегенеративные
изменения печени
Анемия в случае мас-
сивной инвазии, по-
теря веса, энтерит
Анемия в случае мас-
сивной инвазии, по-
теря веса, энтерит
Гранулемы легких,
повреждение эпителия
желчного пузыря
Диарея.
Искажение результа-
тов эксперимента
Изменение печени,
диарея и смерть
у мышей
Повреждения соеди-
нительной ткани
Дегенеративные
изменения печени,
вздутия
Изменения метаболиз-
ма, энтерит
Беспокойство, транс-
миссивное заболева-
ние
Воспаление кожи,
выпадение волос
Глава 7. Биориски и качество экспериментов на лабораторных животных 157
Окончание табл. 40
Возбудитель
Myobia musculinus
Nosopsyllus fasciatus
Notoedres muris
Sarcoptes scabeii
Polyplax (serrata,
spinulosa)
Psorergates simplex
Radford ia sp.
Trichoecius sp.
Hoplopleura sp.
Заболевания
Биориск заражения
для человека
И ЖИВОТНЫХ
Заражение клещами
Мыши
Тунгиозная инвазия
Крысы, мыши
Ушная чесотка
Крысы
Чесотка
Мыши, крысы
Педикулез
Мыши, крысы
Чесотка мышей
Мыши
Клещевая чесотка
Мыши, крысы
Клещевая чесотка
Мыши
Педикулез
Пути заражения
Контактный
Контактный
Контактный
Контактный
Контактный
Контактный
Контактный
Контактный
Контактный
Контагиоэность,
симптоматика и исход
Влияние на результаты
экспериментов
Воспаление кожи,
выпадение волос
Беспокойство, транс-
миссивное заболева-
ние
Изменения кожи на
рыле, ушах, хвосте:
избыточные грануля-
ции, экссудативный
дерматит
Изменения кожи
Анемия, кахексия,
экзема с расчесами,
беспокойство
Воспаление ушей,
изменения кожи
(узелки)
Изменения кожи, вос-
паление; выпадение
волос
Изменения кожи,
выпадение волос
Изменения кожи,
выпадение волос
Сирийский хомячок {Mesoczicetus auzatus) принадлежит к отряду грызунов
(Rodentia), семейству мышиных (Muridae), подсемейству хомяков (Cricetinae).
В последние годы сирийский хомячок во многих странах мира стал ис-
пользоваться как лабораторное животное. Весьма интересна история его
одомашнивания и использования в экспериментах. Сирийский хомячок
в недалеком прошлом относился к числу вымерших животных. Впервые это
животное было поймано в Сирийской пустыне и описано английским зоо-
логом Уотерхаузом. Лишь единичные музейные экземпляры этого вида гры-
зунов подтверждали открытие ученого. Однако на протяжении долгого вре-
мени никому не удавалось обнаружить сирийского хомячка в природе.
В 1930 г. во время экспедиции по Сирии проф. Ахарони удалось заметить
и выкопать из норы беременную самку золотистого (сирийского) хомячка,
потомство которой было легко одомашнено, хорошо размножалось и дало
начало новому виду лабораторных животных.
Золотистый хомячок как лабораторное животное получил распростране-
ние во многих странах мира и успешно конкурирует с такими традиционны-
158
Качество животных-биомоделей
ми экспериментальными животными, как морские свинки, мыши и крысы.
Любопытно то, что в природных условиях золотистый хомячок был обречен
на вымирание и почти вымер, в условиях одомашнивания он оказался не-
прихотливым, выносливым и весьма плодовитым.
Хомячков используют, прежде всего, для воспроизведения инфекцион-
ных заболеваний (туберкулез, лептоспироз, проказа, полиомиелит, сап, бе-
шенство, бруцеллез, коксаки-инфекция, лейшманиоз, токсоплазмоз, вирус-
ный энцефалит, амебная дизентерия, ящур, сибирская язва, столбняк и др.).
Кроме того, они используются для изучения авитаминозов, лучевых пораже-
ний, проведения разнообразных исследований в областях онкологии, физи-
ологии, патофизиологии и фармакологии.
Хомячки — миролюбивые, спокойные и чистоплотные животные, кото-
рые легко привязываются к человеку. По размеру хомячки уступают крысе.
Вес взрослого животного составляет 130—150 г. Они имеют желто-коричне-
вую окраску.
Таблица 41
Вирусные инфекции хомячков, потенциально опасные
для человека и животных и искажающие результаты исследований
Возбудитель
Lymphocytic
choriomeningitis virus
(LCM) (arenavirus)
Theiler's virus (GD VII)
(picornavirus)
Reovirus тип 3
(diplornavirus)
Sendai virus or para-
influenza 1 virus
(paramyxovirus)
Cytomegalovirus
(herpesvirus)
Adenovirus мышей
Заболевания
Биориск заражения
для человека
И ЖИВОТНЫХ
Лимфоцитарный
хориоменингит
Мыши, морские свинки,
хомячки, собаки,
обезьяны, человек
Энцефаломиелит
Мыши, морские свинки,
хомячки, собаки,
обезьяны, человек
Неопределенные
заболевания
Мыши, морские свинки,
хомячки, человек
Пневмония
Мыши, крысы, морские
свинки, хомячки, мини-
свиньи, человек
Менингит
Хомячки
Онкогенные в экспери-
ментальной инфекции
Мыши, хомячки
Пути заражения
Оральный, транс-
плацентарный
Оральный
Оральный, воздушно-
капельный
Оральный, воздушно-
капельный
Оральный
Воздушно-капельный,
оральный
Контагиозность,
симптоматика и исход
Влияние на результаты
экспериментов
Обычно хроническая
персистирующая ин-
фекция, хронический
носитель
Проявляющаяся
инфекция.
Искажение результатов
эксперимента
Бессимптомная инфек-
ция.
Искажение результатов
эксперимента
Обычно субклиничес-
кая инфекция
Искажение результатов
эксперимента
Обычно хроническая
персистирующая ин-
фекция
Бессимптомная инфек-
ция.
Искажение результатов
эксперимента
Глава 7. Биориски и качество экспериментов на лабораторных животных 159
Окончание табл. 41
Возбудитель
Leukaemia virus
complex
Некоторые примеры
этих вирусов:
a) Hamster virus С
b) Bernhard virus
c) Moloney virus
d) Mouse sarcoma
virus MSV,
(Moloney) (M)
Pseudomonas
aeruginosa
Klebsiella pneumoniae
Streptococcus sp.
Staphylococcus
aureus
Campylobacter sp.
Proteus sp.
Leptospira sp.
Cryptococcus
neoformans
Заболевания
Биориск заражения
для человека
И ЖИВОТНЫХ
Хомячки
Хомячки
Мыши, крысы, хомячки
Мыши, крысы, хомячки
Септицемия
Различные виды живот-
ных
Бронхопневмония,
пневмония
Мыши, крысы, морские
свинки, кролики,
хомячки
Абсцесс, септицемия
Различные виды живот-
ных, человек
Стафилодерма, абсцес-
сы, септицемия
Различные виды живот-
ных, человек
Илеит, тифлит, колит
Хомячки
Энтерит, инфекция моче-
выводящих путей,
абсцессы, пиометра,
дерматит
Морские свинки, кроли-
ки, мыши, крысы,
человек
Лептоспироз
Различные виды живот-
ных
Микозы
Мыши, хомячки
Пути заражения
Воздушно-капельный,
Воздушно-капельный,
оральный
Контактный, ораль-
ный, через укусы
Контактный, ораль-
ный, через укусы
Оральный
Оральный, восходя-
щая инфекция моче-
выводящих путей
Чрезкожный, ораль-
ный
Контактный, ораль-
ный
Контагиозность,
симптоматика и исход
Влияние на результаты
экспериментов
Индуцированная
лейкемия.
Искажение результатов
эксперимента
Смерть при стрессах
Искажение результатов
эксперимента
Смерть при стрессах
Искажение результатов
эксперимента
Ухудшение самочувст-
вия (состояния)
Искажение результатов
эксперимента
Ухудшение самочувст-
вия (состояния)
Искажение результатов
эксперимента
Диарея, смерть
Недомогание, смерть.
Искажение результатов
эксперимента
Нарушение развития,
поражение селезенки
и печени
Ухудшение самочувст-
вия (состояния)
Искажение результатов
эксперимента
160
Качество животных-биомоделей
Таблица 42
Инвазивные заболевания мышей и крыс, потенциально опасные
для человека и животных и искажающие результаты исследований
Возбудитель
Lymphocytic
choriomeningitis virus
(LCM) (arenavirus)
Toxoplasma gondii
Syphacia sp.
Trichosomoides
nasales
Hymenolepis sp.
Demodex sp.
Notoedres sp.
Sarcoptes scabiei
Заболевания
Биориск заражения
для человека
И ЖИВОТНЫХ
Лимфоцитарный
хориоменингит
Мыши, морские свинки,
хомячки, собаки,
обезьяны, человек
Токсоплазмоз
Различные виды живот-
ных
Инвазия острицами
Хомячки
Крысы, хомячки
Солитерная инвазия
Мыши, крысы, хомячки,
другие виды животных,
человек
Демодикоз
Хомячки
Чесотка
Мыши, крысы, хомячки,
другие виды животных
Чесотка
Мыши, крысы, хомячки,
другие виды животных
Пути заражения
Оральный, транс-
плацентарный
Оральный
Оральный
Оральный
Оральный, промежу-
точный хозяин: насе-
комые (Н. Nana так-
же без промежуточ-
ного хозяина)
Контактный
Контактный
Контактный
Контагиозность,
симптоматика и исход
Влияние на результаты
экспериментов
Обычно хроническая
персистирующая ин-
фекция, хронический
носитель
Ухудшение самочув
ствия (состояния)
Неизвестна
Ухудшение самочувст-
вия (состояния)
Ухудшение самочувст-
вия (состояния),
энтерит
Гиперкератоз
Поражения кожи,
выпадение волос
Поражения кожи,
выпадение волос
Резюмируя изложенные материалы, следует напомнить хорошую пого-
ворку: «Каков поп — таков и приход». Но если она в большей степени может
быть отнесена к экспериментатору, его опыту, эрудиции и интеллекту, то в
отношении животных уместно сказать: «Каково животное — таков и резуль-
тат». Разумеется, животное должно быть здоровым соматически. Однако
при наличии инфекционной паталогии у животных экспериментатор дол-
жен проститься с корректным результатом изначально. Более того, работа с
больными животными — это научный суицид!
Раздел III ЛИНЕЙНЫЕ
ЖИВОТНЫЕ-БИОМОДЕЛИ
Среди Ротшильдов начало инбридингу положил Джеймс,
младший сын Мейера, женившись в 1824 году на дочери
своего брата. Через шестьдесят лет после смерти основателя
династии «коэффициент инбридинга» составлял 0,07,
а он сам стал источником обеих аллелей в 6% генных
локусов всех своих праправнуков.
Р.Коннифф
163
Глава 8 ГЕНЕТИЧЕСКИЕ АСПЕКТЫ
МОДЕЛИРОВАНИЯ
Многие лаборатории могут получить хорошие ре-
зультаты в ходе регулярного, например ежемесяч-
ного, контроля тех характеристик, которые для ис-
следований института являются наиболее важными.
Резкие изменения могут быть связаны либо с вли-
янием генетики, либо окружающей среды, но в лю-
бом случае исследователи узнают об этом и можно
будет изучать причины изменения [117, 198, 425].
Плодовитость должна отслеживаться регулярно
для обеспечения эффективного управления воспро-
изводственной колонией. Внезапный и резкий рост
плодовитости должен быть тщательно исследован,
поскольку он может быть вызван гибридной мощью,
являющейся результатом спаривания, противореча-
щего обычной системе размножения данного орга-
низма. Хорошо известно, что гибриды F1 обычно
воспроизводятся гораздо лучше, чем инбреды. Так,
несмотря на то, что для повышения плодовитости
при регулярном ведении воспроизводственных ко-
лоний рекомендована селекция, к очень большим
пометам следует относиться с некоторым подозре-
нием, если существует возможность того, что гене-
тическая контаминация уже имела место.
Поведение должно изучаться экспериментатора-
ми тщательно (хотя обычно субъективно). Многие
штаммы обладают спокойным темпераментом, в то
же время гибриды, как правило, бывают гораздо бо-
лее нервными и активными. Так, если темперамент
штамма меняется, то это должно стать поводом для
164
Линейные животные-биомодели
тщательного исследования с применением способов пересадки кожи, иссле-
дования формы нижней челюсти или биохимических способов. Нет нужды
подчеркивать, что весь персонал, работающий с инбредными штаммами, дол-
жен быть хорошо подготовлен и должен обладать как можно более высокой
квалификацией.
Индивидуальность инбредных штаммов
Каждый инбредный штамм представляет собой уникальное сочетание гене-
тического материала, порождающее уникальный фенотип. Многие такие
фенотипные черты полезны в исследованиях, некоторые позволяют вывести
«модели» болезней, прочие дают полезные физиологические, анатомичес-
кие или поведенческие характеристики (примеры приведены в табл. 43). Эти
индивидуальности, хорошо отраженные в литературе, представляют одну из
самых полезных инбредных черт.
Таблица 43
Примеры моделей болезней и интересующих характеристик в инбредных
линиях мышей
Характеристика
Предпочтение спирта (10%)
Агрессия/борьба
Аудиогенные судороги
Аутоиммунная анемия
Амилоидоз
Расщепление нёба
Синдром Чедиака-Хигаши
Гипертония и/или пороки сердца
Гиперпролинемия и пролинурия
Ожирение и/или диабет
Остеоартропатия коленных суставов
Полидипсия
Устойчивость к миксовирусным
инфекциям
Опухоли:
Лейкемия
Ретикулоклеточная саркома
(болезнь Ходжкина)
Опухоль легких
Гепатома
Опухоль молочной железы
Овариальная тератома
Приобретенная плазмацитома
Тестикулярная тератома
Полное отсутствие спонтанных
опухолей
Расстройство питания
Линии
C57BL, C57BR/cd
SJL, NZW
DBA/2
NZB
YBR, SJL
CL, А
SB
BALB/c, DBA/1, DBA/2
PRO
NZO, PBB, KK, AY
STR/1
SWR, SWV
A2G
AKR, C58, PL, RF
SJL
A
C3Hf
СЗН, СЗН-4ПУ, GRS/A, Rill
LT
BALB/c, NZB
129/trSv
X/Gf
A2G
Глава 8. Генетические аспекты моделирования
165
Хотя каждый инбредный штамм обладает собственным уникальным на-
бором характеристик, для многих экспериментов нужен только хороший,
стандартный, воспроизводимый, легкодоступный экспериментальный мате-
риал. Некоторые более общие «общецелевые» штаммы, например, C57BL,
СВА, BALB/c и СЗН в случае мышей и F344, LEW и PVG в случае крыс,
идеальны для таких экспериментов, хотя всегда необходимо помнить, что
экспериментальные результаты, полученные от единичного инбредного
штамма, не могут обобщаться применительно ко всем особям. Там, где
необходима широкая применимость, следует использовать несколько штам-
мов. Хотя аутбредный материал также представляет собой уникальное со-
брание генотипов, ничто не свидетельствует о том, что единичный аутб-
редный материал представляет особи в целом с большей представительно-
стью, чем единичный инбредный штамм. Для повышения общепримени-
мости результатов исследований использовать единичный аутбредный ма-
териал невозможно.
Гетерозис и внутриклассовые корреляции
Хорошо известно, что животные из инбредных линий, как правило, облада-
ют меньшей генетической мощью и более слабой детородной функцией по
сравнению с большинством гибридов или аутбредным материалом. Это яв-
ляется результатом инбредной депрессии, общего ослабления готовности к
воспроизводству в связи с повышением гомозиготности. Примечательной
чертой такой депрессии является снижение показателей по всем парамет-
рам, связанным с общей способностью производить молодняк, доживаю-
щий до полового созревания — «готовностью». Так, у мышей повышение
коэффициента инбридинга на 10 процентов приводит в среднем к уменьше-
нию величины помета на 0,6 и снижению веса в возрасте 6 недель на 0,58 г
у самок. В качестве постулата принято считать, что противоположностью
инбредной депрессии является гибридная мощь животных. Она поддается
математическому описанию [206-208] и обозначается как гетерозис.
Когда измеряемая переменная является дискретной (мертвый/живой,
респондер/нереспондер и т.д.), возможность рассчитать /*7 сохраняется. Спо-
соб заключается в произвольном назначении величины животному 1, нахо-
дящемуся под воздействием, и животному 0, не находящемуся под воздей-
ствием. Если ni является общим количеством животных в /-м штамме, г. из
них не являются нормальными и имеются с штаммов с математическим
ожиданием количества животных, равным 7V<=> Е п{, то средний квадрат D(x)
между штаммами равен
Д*) =
V 2/ (X//)2
(с-1),
(38)
166
Линейные животные-биомодели
а средний квадрат в штаммах равен
In2
ЪП--!—
D{y) =
\N-c).
Затем можно оценить внутриклассовую корреляцию по
[D(x)-D(y)]
Г; =
где
' [D(x) + (k-l)D(y)]'
k = {N-^/N)/{c-\).
(39)
(40)
(41)
Пример приводится в табл. 44.
Одна из проблем с расчетным значением rf, полученным таким путем,
заключается в том, что оно зависит от среднего уровня наблюдаемых ано-
малий.
Таблица 44
Вариационный размах между нормальными и аномальными (после введения
ацетазоламида) линиями мышей для качественной характеристики
Штамм
CBA/J
C57BL/6
A/J
DBA/2
СЗН/Не
SWR
Нормальные
28
38
18
15
57
29
Аномальные
15
33
8
6
5
1
68
Итого
43
71
26
21
62
30
253
D(x) =
D(y) = W-
152 ЗЗ2 Г
—< + +...+ —
43 71 30
152 ЗЗ2 I2
+ +...+ —
43 71 30
68^
253
75 = 1,380
7(253-6) = 0,173
tf = {253-(432+712+...+302)/253}/5 = 40,5
1,380-0,173
гн =
= 0,147
1,380+ 39,5 0,173
С поправкой на независимую от частоты корреляцию (гвк):
р(\-р)
ГНК - ГЯ,01 2
(42)
(43)
(44)
(45)
(46)
Глава 8. Генетические аспекты моделирования
167
для р = 0,269, z = 0,385
ЛЛАП 0,269(1-0,269)
^=0Д47- 0,3852 =0Д91- <47>
Таким образом, если встречалось больше животных с аномалиями, то г7
менялось. Это можно скорректировать путем перехода на модель постоян-
ного исходного распределения ответственности, при которой отдельные лица
попадают под воздействие, если они переступают определенное пороговое
значение ответственности. Внутриклассовую корреляцию на шкале 0, 1 гн 01
можно обратить в корреляцию на непрерывной шкале следующим образом:
Р(1-Р)
гнк=гн$\ 2—' (48)
z
гдер = Е rt/Nявляется общей частотой особей с аномалией, a z — ординатой
на стандартизированной кривой нормального распределения, соответствую-
щей вероятности р. Приблизительные стандартные погрешности гн как для
непрерывных, так и для дискретных переменных вычисляются по приведен-
ным формулам, а в табл. 44 даны количественные различия между линиями.
О генах-таймерах и прогнозируемых биомоделях
Ряд качеств инбредных животных делает их уникальным инструментом био-
моделирования, а также использования в биомедицинских и биологических
исследованиях. Инбредные животные гомозиготны и генетически однород-
ны, что обеспечивает получение на них надежных и воспроизводимых ре-
зультатов. Новые направления — гибридомная технология, создание транс-
генных животных — также основаны на использовании мышей инбредных
стандартных линий. Инбредные мыши используются при тестировании ле-
карственных препаратов, химических соединений на мутагенность, канце-
рогенность и построения моделей. Подробнее этот вопрос будет рассмотрен
далее в этом разделе, а также в гл. 38.
В лаборатории генетики НЦМБ РАМН заведующим лабораторией
А.М. Малашенко и его сотрудниками Е.Л. Игнатьевой и Т.Б. Бесковой пред-
принята попытка создания модели периодически меняющегося прогнозиру-
емого ответа на мутагенные воздействия у мышей CBA/LacY в ряду поколе-
ний инбридинга [446]. Для тестирования химических мутагенов исследован
и выделен ряд линий, отобрано несколько линий с различными ха-
рактеристиками и чувствительностью к мутагенам. Среди этих линий СВА/
LacY (СВА) была выбрана как нормальная устойчивая линия по результатам
опытов с тиоТЭФ и другими мутагенами.
В ходе последующих исследований было обнаружено резкое достоверное
повышение чувствительности мышей СВА к кластогенному эффекту тиоТЭФ.
168
Линейные животные-биомодели
Обнаружены также случаи невоспроизведения результатов как в нашей, так и
в других лабораториях. Изучение этого явления привело к заключению о су-
ществовании генетически детерминированных колебаний нормы реакции
организма в ряду поколений инбридинга. Предположили, что существуют
генетические системы «генов-таймеров», которые от поколения к поколению
меняют с определенным ритмом активность генома, плодовитость и норму
реакции организма.
Этот вывод был основан на анализе корреляций между колебаниями
плодовитости в разных сублиниях СВА и корреляции между плодовитостью
и чувствительностью к мутагену тиоТЭФ (г = —0,91 + 0,21). Кроме того,
была обнаружена высокая корреляция колебаний коэффициента плодовито-
сти Q в ряду поколений инбридинга у мышей, разводимых неодновременно
— максимальный разрыв между инбредным возрастом (F) в лаборатории
генетики и филиале «Столбовая» достигал пяти поколений (это соответству-
ет 2,5—3 годам). Обе сублинии происходили от одной пары сибсов в F93 и
животных, живущих в разное время в разных условиях, объединял только
геном их общих предков. Этот разрыв возник случайно и далее он стал со-
кращаться.
Моделирование подобной ситуации было осуществлено с целью провер-
ки предложенной гипотезы. Ранее использовались данные по оценке чув-
ствительности мышей в ряду шести поколений, при этом период цикла ко-
лебаний уровня плодовитости составил четыре поколения. Поэтому было
начато создание двух колоний мышей СВА — двух сублиний (смена поколе-
ний в которых происходила с разной скоростью) при регулярном наблюде-
нии уровня ответа на мутаген тиоТЭФ мышей каждого последующего поко-
ления, предполагая достигнуть разрыва в 8—10 поколений.
Эта задача требует длительного разведения, однако в итоге возможно
создание двух колоний одновременно существующих изогенных мышей с
резко различающейся чувствительностью к мутагену. Они могли бы стать
объектами для начала исследования генетических факторов, периодически
меняющих норму реакций организма, плодовитость и, вероятно, продолжи-
тельность жизни (так же, как в линиях дрозофилы). Когда были получены
два поколения: в «быстрой» сублинии (на племя используются мышата пер-
вого помета) и «медленной» сублинии (размножение до конца репродуктив-
ного периода) — FI46 и F142 соответственно, разводимые одновременно и
достоверно различающиеся по чувствительности к тиоТЭФ, было выполне-
но исследование их чувствительности к кластогенному эффекту классичес-
кого канцерогена бензпирена.
Оценена чувствительность к тиоТЭФ мышей 19 последовательных по-
колений, размножаемых с возможно более быстрой сменой поколений («бы-
страя» сублиния). Кривая колебаний признака согласуется с ранее получен-
ными данными и предложенной гипотезой, объясняющей эти колебания.
Глава 8. Генетические аспекты моделирования
169
При среднем значении 30% клеток с индуцированными аберрациями хромо-
сом частота колебаний варьировала от 20 до 40%.
Минимальные и максимальные значения достоверно отличаются не толь-
ко друг от друга, но и от среднего уровня (при используемых выборках раз-
личия достоверны при разнице более 5%). Спектр хромосомных поврежде-
ний, индуцированных тиоТЭФ, представлен в табл. 45 по поколениям F|42
и F146. Различия между максимально различающимися по чувствительности
поколениями иллюстрируют данные по двум последовательным поколениям
- F146 (42,6% клеток с повреждением хромосом) и F147 (20,5%). При про-
смотре 500 клеток зарегистрировано: F146 — 415 делений, 103 обменные абер-
рации (кольца и транслокации) и 13 клеток с множественными аберрация-
ми. Количество разрывов хромосом на одну клетку 1,56; F147 — 150 делений,
33 обменные аберрации, 0,43 разрыва на клетку. Таким образом, у мышей
поколения F146 в клетках костного мозга возникло в 3 раза больше повреж-
дений хромосом, чем у их потомков — поколение F147.
На основе выровненной кривой колебаний признака статистическим
методом, предназначенным для анализа периодических функций (цикли-
ческие изменения численности популяции и т.д.), доказана периодичность
колебаний и полное соответствие данным, полученным при исследовании
колебаний плодовитости мышей. Циклические колебания идут с тем же
ритмом: 4,3 ± 0,3 поколения инбридинга на период цикла.
Чувствительность мышей «медленной» сублинии, прошедшей с начала
раздельного разведения двух сублиний четыре поколения инбридинга, со-
ответствовала ожидаемой на основе оценки признака у мышей «быстрой»
сублинии. В табл. 46 представлены данные по оценке эффекта тиоТЭФ
и бензпирен у мышей F142 и F146 («медленная» и «быстрая» сублиния соот-
ветственно), разводимых одновременно. Мыши поколения F]46 высокочув-
ствительны к тиоТЭФ, мыши F142 находились на среднем «нормальном»
уровне, различия между ними достоверны при Р < 0,001. Чувствительность
мышей F]42 «быстрой» сублинии, живших два года назад до проведения дан-
ной работы, была аналогичной (32,6%).
Бензпирен был выбран для данной работы для расширения характеристи-
ки различий нормы реакций двух поколений, различающихся по чувстви-
тельности к алкилирующему мутагену. Полученные данные демонстрируют,
насколько норма реакции мышей определенного поколения влияет на ре-
зультаты тестирования мутагенной активности исследуемого химического
соединения.
Бензпирен является классическим канцерогеном, представителем поли-
циклических углеводородов. Его присутствие обнаружено в загрязненном
промышленными выбросами воздухе, выхлопных газах автомобильных дви-
гателей, табачном дыме, консервированных копчением продуктах питания
и т.д. При воздействии на животных в эксперименте бензпирен оказался
170
Линейные животные-биомодели
активным локальным канцерогеном и индуктором опухолей тканей, не кон-
тактировавших с ним (опухоли легких, лейкемии). Для этого вещества уста-
новлена корреляция канцерогенных и мутагенных свойств. При этом индук-
ция генотоксичности обусловлена целым комплексом полиморфных мета-
болизирующих генов. У мышей бензпирен способен индуцировать доми-
нантные летальные мутации в постмеиотических половых клетках самцов
и хромосомные нарушения в клетках костного мозга. Поэтому он был пред-
ложен для позитивного контроля в программе ВОЗ по оценке краткосроч-
ных тестов для исследования канцерогенности химических соединений.
Таблица 45
Кластогенный эффект тиоТЭФ в дозе 2,5 мг/кг и бензпирена в дозе 400 мг/кг
в клетках костного мозга мышей CBA/LacY поколений F142 и F146,
существовавших одновременно
Поколение
F*
F,«
Мутаген
введения
ТиоТЭФ
Бензпирен
Контроль
ТиоТЭФ
Бензпирен
Контроль
Метод
введения
в/б
в/б
п/о
в/б
в/б
п/о
Число
клеток
500
400
400
500
500
500
500
500
Число хромосомных нарушений
Пробелы
3
1
1
1
13
2
1
Фраг-
менты
253
5
4
2
415
10
11
2
Кольца
16
23
Обме-
ны
70
80
1
1
Множест-
венные
аберрации
18
13
Число клеток, содержащих
структурные
аберрации
150(30,0±2,0)
5(1,3±0,6)
4(1,0+0,5)
2(0,4±0,3)
208 (41,6±2,2)
11 (2,2±0,7)
12(2,4±0,7)
2(0.4±0.3)
хромосомные
нарушения, %
150(30,0±2,0)*
6(1,5±0,6)
5(1,2±0,5)
3(0,6±)
213(42,6±2,2)*
11 (2,2±0,7)
14(2,8±0,7)*
3(0.6±0.3)
" Достоверность различия опыта и контроля Р < 0,001.
В табл. 45 представлены результаты оценки кластогенного эффекта бенз-
пирена у мышей двух поколений (F]42 и F146) при внутрибрюшинном и внут-
рижелудочном введении. При низкой частоте клеток с хромосомными нару-
шениями эти группы одного поколения не различались. Однако в реакциях
между поколениями установлены достоверные различия. Если у мышей F142
при объединении групп с разным путем введения канцерогена найдено 1,5%
клеток с повреждениями хромосом, что недостоверно отличается от контроля,
то у мышей F146 — 2,5% ± 0,7 (отличие от контроля достоверно при Р < 0,001,
от ^142 ~~ ПРИ Р < 0,01). Полученная оценка для F146 близка к найденной
ранее [448]. Совершенно очевидно, что оценка эффекта бензпирена у мышей
поколения F142 дает ложный отрицательный результат.
Проверка чувствительность к тиоТЭФ мышей четырех последовательных
пометов поколения F147 дала следующие оценки: 21,2—16,2—15,8—12,4% кле-
Глава 8. Генетические аспекты моделирования
171
ток с аберрациями хромосом. Налицо определенное направленное измене-
ние признака, в данном случае — снижение. Снижение чувствительности
свидетельствует, что оно обусловлено именно возрастом отца, так как старе-
ние матери может дать только снижение резистентности к мутагену.
На следующих этапах необходимо более полное исследование, включа-
ющее оценку чувствительности потомков мышей, находящихся на противо-
положных точках кривой колебания признака в ряду поколений. Можно
ожидать разнонаправленные изменения. Есть косвенное подтверждение это-
го предположения в опытах по индукции опухолей канцерогеном у мышей
двух родственных сублиний Стронга: у потомков Prunt по мере увеличения
номера помета предрасположенность к раку возрастала, в сублинии 5-Prunt
она падала. Эти сублинии отличались инбредным возрастом: вероятно, ро-
дители изученных потомков обладали разной чувствительностью к канцеро-
гену и находились на разных позициях по кривой колебаний признака.
Полученные данные демонстрируют, насколько важен учет колебаний
нормы реакции в поколениях инбридинга при исследовании канцерогенеза
и эффекта генетически активных соединений на инбредных мышах. Резуль-
таты исследований хорошо согласуются с ранее полученными данными и
предложенной гипотезой о генетической детерминированности наблюдаемых
колебаний анализируемых признаков (плодовитость, норма реакций). Коле-
бания чувствительности к мутагену в ряду поколений выглядят как регуляр-
ный, с определенным ритмом процесс, не зависящий от средовых воздей-
ствий и контролируемый геномом. Следовательно, существуют гены-тайме-
ры, которые меняют активность генома и реакции организма от поколения
к поколению. Вероятно, эти гены функционируют под влиянием информа-
ции об активности генома предыдущего поколения с использованием про-
цесса генетического импринтинга. Это предположение согласуется с гипоте-
зами о природе и биологической цели генетического импринтинга.
Модель существования функционирующих во времени генетических
часов предполагает наличие каких-то единиц измерения. Наиболее вероят-
но, что это — циклы деления половых клеток. Трансформированные куль-
туры соматических клеток прекращают существование после определенно-
го числа делений. Поскольку предшественники мужских половых клеток
делятся непрерывно, то норма реакции потомков должна меняться в зави-
симости от возраста родителей и номера помета. Из этого следует, что ус-
тановленный ритм колебаний плодовитости и нормы реакции (4,3 ± 0,3)
характерен для принятого в НЦБМТ РАМН и филиале «Столбовая» комп-
лектования следующего поколения в племенном ядре из мышей 2—3-го
пометов. Можно ожидать изменения ритма при ином выборе (1-го или 6-
го пометов) и построить прогностическую модель.
Полученные данные позволяют надеяться на перспективность исследо-
вания обнаруженных закономерностей. В опытах на линиях дрозофилы ус-
172
Линейные животные-биомодели
тановлено, что колебания плодовитости, радиочувствительности и продол-
жительности жизни в ряду поколений коррелированы. Изучение гипотети-
ческих генов-таймеров может дать полезную информацию для разработки
оптимизации качества и продолжительности жизни человека. Интересна
в этом отношении гипотеза о возможной регуляции скорости процесса ста-
рения видоспецифическими генами-таймерами (см. гл. 34-37). С другой сто-
роны, эти данные практически полезны для планирования и организации
экспериментов с использованием инбредных мышей, интерпретации полу-
ченных результатов и построения оптимизированных моделей.
Нокаутные и трансгенные животные
Хотя в основном все медико-биологические исследования чаще выполня-
ют на таких животных, как мыши, крысы, хомячки, кролики, однако по
своим биологическим и физиологическим особенностям ближе всех к че-
ловеку — мини-свинья. Именно ее органы и ткани все чаще используют
для трансплантологии.
Как мы уже подчеркивали, в процессе получения мини-свиней, в том числе
и светлогорской породы, были использованы шведские ландрасские свиньи
— от них белая кожа и большие уши, вьетнамские черные свиньи — от них
малый рост. Дикие кабаны наградили мини-свиней физической крепостью. В
результате у животных, весящих до 50 килограммов, размеры сердца и его
клапанов, селезенки, поджелудочной железы, печени, почек, надпочечников
и простаты аналогичны размерам соответствующих органов человека. У мини-
свиней сходный с человеком липидный обмен, что позволяет исследовать на
них ишемическую болезнь, разрабатывать методы ее ранней диагностики и
лечения. Сердечные клапаны мини-свиней идеальны для пересадки человеку.
В 90-х годах начали изучать анатомические особенности сердца и коронар-
ных сосудов мини-свиней. Удалось установить, что они очень схожи с наши-
ми. Мини-свиньи годятся для пересадки сердца и его частей.
Выращивание специальных животных-доноров снимает проблемы заго-
товки и консервации органов, отныне можно будет иметь наготове орган не-
обходимого размера от гарантированно здорового донора. Американские, а в
дальнейшем и российские ученые клонировали генетически измененных мини-
свиней, подходящих для трансплантации органов и тканей человеку. Это се-
рьезный научный прорыв, поскольку наиболее реальный путь трансплантоло-
гии — пересадка органа от одного биологического вида другому. Большие
надежды поэтому возлагаются на мини-свиней, но при трансплантации ос-
новная проблема — иммунологическая реакция организма, которая вызывает
отторжение чужой ткани или органа из-за того, что человеческие антитела
атакуют клетки чужого организма. Мишенями служат молекулы Сахаров на
поверхности клеток мини-свиней. Чтобы уничтожить мишени, ученые «нока-
Глава 8. Генетические аспекты моделирования
173
утировали» ген, который обеспечивает их образование, так называемый GGTA1
ген. Антитела не могут атаковать клетки без мишеней, поэтому реакция от-
торжения блокируется [429]. Следующим шагом стало клонирование генети-
чески измененных мини-свиней и создание новой линии этих животных,
которая предназначена специально для трансплантации.
Мини-свинья по имени Голди, появившаяся на свет в лаборатории уни-
верситета штата Миссури (США), стала научной сенсацией. Она является
клоном — генетический материал для ее создания был взят у свиньи-донора.
У Голди заблокированы два гена, т. е. они есть, но не выполняют свои функ-
ции. С ними проделали манипуляции, которые в генетике называют нокау-
том, — попросту говоря, «выбили» из колеи. Эти гены отвечают за выработ-
ку альфа-1,3-галактозилтрансферазы (GGTA1). Именно через этот полиса-
харид запускается мощная иммунная реакция отторжения. Именно эта про-
блема препятствует успеху ксенотрансплантологии, т. е. пересадки органов
животных человеку. А если у животного нет GGTA1, то антитела человека
бессильны, и отторжение не происходит.
Возможно, органы Голди, выведенной специалистом по репродуктивной
эмбриологии профессором Рэндалом Прэзером, уже сегодня не вызвали бы
реакции отторжения, если их пересадить человеку. Но до этого на самом
деле еще далеко. Во-первых, нужна не одна такая Голди, а целое стадо. Во-
вторых, требуется еще не одна серия экспериментов на животных, прежде
чем достижение найдет дорогу в клинику.
Проведенные в Каролинском институте Стокгольма клинические испы-
тания показали, что свиные панкреатические железы, введенные человеку,
способны производить инсулин и годятся для лечения диабета. Есть также
данные об успешных результатах лечения больных людей, страдающих бо-
лезнью Паркинсона, болезнью Хантингтона и эпилепсией: это может быть
осуществлено путем переноса им нервных клеток головного мозга от эмбри-
она-свиньи.
Что касается изучения онтогенеза иммунного ответа на разные антигены,
то мини-свиньи являются просто незаменимой моделью. Во-первых, невелики
по размеру и ими можно легко манипулировать в лабораторных условиях; во-
вторых, многоплодны, что позволяет в одном помете иметь несколько конт-
рольных и подопытных животных; в-третьих, возможно выращивание в кон-
тролируемых условиях с определенной антигенной экспозицией благодаря
достижениям гнотобиологии; и, в-четвертых, поросята рождаются лишенны-
ми иммуноглобулинов, обладающих активностью антител.
В Кембридже (Англия) ученые путем внедрения генов человека в геном
свиньи вывели породу трансгенных свиней, генетический код которых по-
зволяет предположить, что органы этих животных не будут отторгнуты орга-
низмом человека в случае их пересадки. Это происходит за счет генов-регу-
ляторов, которые регулируют иммунную систему организма человека-реци-
174
Линейные животные-биомодели
пиента. Первые опыты по пересадке почек трансгенных свиней обезьянам
показали обнадеживающие результаты — почки нормально функционирова-
ли в организме обезьяны более двух месяцев. Это указывает на реальную воз-
можность пересадки почек, сердца и других органов от свиней человеку.
В связи с этим британское и израильское правительства вскоре, по-видимо-
му, разрешат пересадку сердца и почек от трансгенных свиней человеку. Та-
ким образом, ксенотрансплантация, которая до недавнего времени рассмат-
ривалась как нереальная альтернатива донорству человеческих органов, сей-
час провозглашается реальной возможностью. Но есть опасность, что вместе
с печенью, почками или свиным сердцем в организм человека может проник-
нуть вирус, малоизученный на настоящий момент.
Использование более мелких животных, и в частности лабораторных
мышей, в качестве биомоделей наследственных болезней человека и в транс-
плантологии основано либо на применении инбридинга — в этом случае
ценность представляет весь генотип, который детерминирует весь комплекс
биологических характеристик, либо на применении мутантов — в этом слу-
чае генотип играет важную роль, но не основную.
Большая часть мутантов спонтанного происхождения, однако существует
ряд индуцированных радиацией хромосомных аномалий и химическими
мутагенами генных мутаций. Индуцируемые этими факторами мутации про-
исходят в любом сайте независимо от экспериментатора. В последние деся-
тилетия начат принципиально иной способ индукции мутаций. Индуциру-
ются направленные мутации конкретных генов. Активно создаются две фор-
мы направленных мутаций — трансгенные организмы и нокаутные.
Трансгенные формы несут сегмент чужой ДНК, который введен в геном
путем гомологичной рекомбинации, вставкой инфекционным агентом — рет-
ровирусным вектором или негомологичной вставкой (микроинъекцией в про-
нуклеус). В JAX более 100 трансгенных линий.
В 2004 году в НЦБМТ РАМН из Института канцерогенеза РОНЦ РАМН
получены трансгенные мыши B6-Tg N(ACT в EGFP) 10 Sb, несущие ген
green fluorescent protein (зеленое свечение в УФ).
Нокаутные формы получают микроинъекцией генетически измененных
эмбриональных стволовых клеток в бластоцисту хозяина. Разрушение, заме-
щение или удвоение гена в стволовых клетках производят путем гомологич-
ной рекомбинации между экзогенной ДНК и эндогенным геном (например,
блокированием работы целевого гена встройкой гена резистентности
к неомицину). Большая часть трансгенных линий создана для эксперимен-
тальной онкологии и иммунологии. В JAX лаборатории поддерживается более
300 нокаутных линий, в Институте канцерогенеза РОНЦ — 6 линий.
Методы направленного мутагенеза распространяются на самые различ-
ные макроорганизмы: от мышей до свиней и обезьян. Из литературы изве-
стно,что планируется инактивировать ген лактоглобулина у коров, чтобы мо-
Глава 8. Генетические аспекты моделирования
175
локо было не аллергенным. Американские компании «Гемател» и «Даунпрот
вакцин корпорейшен» пытаются вывести и затем клонировать коров с чело-
веческими генами для производства антител человека к токсину ботулизма
(в целях защиты от биотерроризма).
В России наиболее интенсивно работа с трансгенными мышами ведется
в РОНЦ РАМН Л.А.Побезинским, Е.Л.Побезинской и сотрудниками [532,
533]. В последний год начата активная работа в этом направлении в лабора-
тории генетики (зав. к.б.н. А.М.Малашенко) нашего Центра в сотрудниче-
стве с РОНЦ. Мы сошлемся на любезно предоставленные вышеуказанными
авторами данные по исследованиям на трансгенных мышах.
Как известно, аллогенные трансплантаты успешно отторгаются иммун-
ной системой реципиента даже при трансплантации между конгенными
линиями животных, различающимися только аллелями МНС. Поэтому до-
пущение возможности участия рецепторов врожденного иммунитета в алло-
генных реакциях потребовало бы также допустить и гетерогенность в их
специфичности, не только сопоставимую по широте разнообразия, но
и скорректированную с экспрессией различных аллельных форм молекул
главного комплекса гистосовместимости (МНС). Таким образом, возникла
проблема в понимании механизмов участия системы врожденного иммуни-
тета в отторжении аллогенных трансплантатов.
В работах [532, 533], наряду с традиционными линиями мышей на осно-
ве C57BL/6, использованы мыши-нокауты по молекулам (32-микроглобули-
на — C57BL/6J-B2tf2tmiUnc(H-2b), транспортеров, ассоциированных с процес-
сингом (ТАР) - С51Ъ\./Ы-ТарГт1АгР, ос-цепи ЕСК - C57BL/6J-7,C7?6tmIMom
(H-2b), CD4 - B6.\29S2-Cd4tmIMak, CD8 - B6A29S2-Cd8atmrMak, CD400-
0B6.129P2- Tnfrsf5lmIKik.
Одним из ранних проявлений иммунного ответа мышей на клетки алло-
генных опухолей является накопление клеток с повышенной гранулярно-
стью в селезенке. Их появление может быть обнаружено на следующий день
после иммунизации и достигает пика на 5—6-й день. Эти клетки не экспрес-
сируют маркеров Т- и В-лимфоцитов, макрофагов, естественных киллеров
и дендритных клеток [1, 221]. Они экспрессируют CD lib (Mac-1) и Gr-1
(Ly6G) b и по морфологии являются нейтрофилами. Отсутствие экспрессии
CD31 (РЕСАМ-1) указывает на их активированное состояние, возникшее в
процессе воспалительного ответа и проникновение в ткани. У мышей, име-
ющих дефекты развития клеток CD8+, ответ нейтрофилов на иммунизацию
клетками аллогенных опухолей существенно подавлен. Он полностью от-
сутствует у генетических нокаутов по ос-цепи Т-клеточного рецептора (TCR).
У нокаутов по CD4 и CD40 интенсивность ответа сопоставима с ответом
животных дикого типа, что указывает на его независимость от Е-хелперов
CD4+ и активации профессиональных антигенпрезентирующих клеток ре-
ципиента.
176
Линейные животные-биомодели
Использование мышей, конгенных по МНС, а также животных, нокаути-
рованных по молекулам, участвующим в презентации, и корецепторам CD4 и
CD8, показало, что появление активированных нейтрофилов в селезенке ре-
ципиентов опухолевых клеток зависит от экспрессии аллогенной молекулы
МНС класса I опухолевыми клетками и корецептора CD8+ Т-лимфоцитами
в организме реципиента. Полное отсутствие эффекта у нокаутов по а-цепи
TCR однозначно свидетельствует о том, что эффект полностью зависит от
распознавания аллоантигена Т-клетками, экспрессирующими TCRa/(3, фун-
кция которых не может быть замещена естественными киллерами и Т-клет-
ками, экспрессирующими TCRy/5. Подавление ответа у нокаутов по генам
ТАР, (32-микроглобулина и CD8 ни в одном случае не было полным, что, по
мнению авторов [532, 533], объясняется высокой способностью внутрити-
мусной селекции к компенсации нарушений, вызванных дефицитом специ-
фических лигандов (собственных молекул МНС) и костимуляторов. У нока-
утов по генам ТАР и |32-микроглобулина в периферических лимфоидных
органах обнаруживается небольшое количество клеток CD8+, тогда как у но-
каутов по корецептору CD8 внутритимусная селекция может приводить к по-
явлению клеток CD4~CD8", способных реагировать в VLR на молекулы МНС
класса I в отсутствие корецептора.
Интересен тот факт, что отсутствие корецептора CD4 и костимуляторно-
го рецептора CD40 на клетках реципиента не оказывают влияния на интен-
сивность накопления нейтрофилов в селезенке животных, иммунизирован-
ных клетками аллогенных опухолей.
Таким образом, у животных с нокаутом в генах удается выявить прин-
ципиально новые эффекты и расшифровать механизмы формирования и
течения воспалительного ответа, формирования реакций при транспланта-
ции и в целом точнее представлять себе формирование иммунных реакций
и процессов.
Биомодели для исследования опухолей
Таблица 46
Генетически обусловленные и экспериментальные биомодели для исследования
механизмов развития опухолевых процессов и оценки эффективности
противоопухолевых средств
Исследуемое
действие вещества
Модели для изуче-
ния спектра про-
тивоопухолевой
активности на
основе перевивае-
мых опухолей
Лабораторное
животное
Мыши
Линия или порода
животного
Линии DBA2 или
гибриды
BDF^DBAjXCst
BL/6J]
Пол
животного
9(5
Вес
животного
18-20 г
Возраст
животного
2-2,5 мес.
Глава 8. Генетические аспекты моделирования У11
Продолжение табл. 46
Исследуемое
действие вещества
Лимфолейкоз Р-388
(прививочная
доза-106внутри-
брюшинно, внутри-
венно или под-
кожно)
Лимфолейкоз
L-1210 (прививоч-
ная доза-105 внутри-
брюшинно, внутри-
венно или под-
кожно)
МеланомаВ-16
(прививается
взвесью опухолевых
клеток по 30-60 мг в
0,3-0,5 мл питатель-
ной среды на мышь,
подкожно или
внутривенно)
Эпидермоидная
карцинома лёгкого
Lewis (прививается
подкожно взвесью
опухолевых клеток
по 30-60 мг в
0,3-0,5 мл питатель-
ной среды на мышь)
Аденокарцинома
молочной железы
Са-755(прививается
подкожно взвесью
опухолевых клеток
по 30-60 мг в
0,3-0,5 мл питатель-
ной среды на мышь)
Лимфаденоз
Фишера L5178Y
(прививается под-
кожно взвесью
106клеток, внутри-
брюшинно)
Опухолевый плев-
рит лимфолейкоза
Р-388(прививочная
доза 5x105-1x106
кл./мышь в объеме
0,2-0,3 мл)
Лабораторное
животное
Мыши
Мыши
Мыши
Мыши
Мыши
Мыши
Мыши
Линия или порода
животного
Линии DBA2 или
гибриды BDF
Линии C57BL/6j
и гибриды BDF,
илиР,[СВАхС57В1/6Л
Линии C57BL/6j
и гибриды BDF,
или F,
Линии C57BL/6j
и гибриды BDF,
или F,
Линии DBA2
или гибриды BDF,
Линии DBA
или гибриды BDF,
Линии СВА
Пол
животного
Яд
Яд
Яд
Яд
Яд
Яд
Яд
Вес
животного
30-40 г
30-40 г
30-40 г
30-40 г
30-40 г
30-40 г
30-40 г
Возраст
животного
3-10 мес.
3-10мес.
3-10 мес.
3-10мес.
3-10 мес.
3-10мес.
3-10 мес.
178
Линейные животные-биомодели
Продолжение табл. 46
Исследуемое
действие вещества
Рак шейки матки
РШМ-5 (прививается
подкожно взвесью
опухолевых клеток
по 30-60 мг в
0,3-0,5 мл питатель-
ной среды на мышь)
Аденокарцинома
толстой кишки
АКАТОЛ (прививает-
ся подкожно
взвесью опухолевых
клеток по 30-60 мг
в 0,3-0,5 мл пита-
тельной среды на
мышь)
Гепатома 22 солид-
ная и асцитная(при-
вивается подкожно
(Г-22)иливнутри-
брюшинно (АГ-22)
суспензией асцит-
ных клеток из под-
кожно растущей
опухоли по 30-60 мг
в 0,3-0,5 мл питатель-
ной среды на мышь)
Карцинома Эрлиха
асцитная и солидная
(прививается внутри-
брюшинно (АРЭ) или
подкожно (СЭ) сус-
пензией клеток асци-
та по 107кл./мышь
или взвесью клеток
из подкожно расту-
щей опухоли по
30-60 мг на мышь
в разведении пита-
тельной среды)
Спонтанные опухоли
молочных желез:
60% случаев
75%
85-100%
20%
75%
84%
<
<
<
<
Лабораторное
животное
Мыши
Мыши
Мыши
Мыши
Мыши
«
»
«
«
«
Линия или порода
животного
Линии BALB/c
Линии Сд/НА
Линии СЭНА
Линии Balb/c,
гибриды F,,C57
BL/6J или С3НА
АНЕ J
CBA/J
СЗН/HeLac
C3Hf
DBA/ULac
DD/He
Пол
животного
9tf
9с5
9с?
9с?
Я£
9
9
У
9
Вес
животного
30-40 г
30-40 г
30-40 г
30-40 г
30-40 г
Возраст
животного
3-10 мес.
3-10 мес.
3-10 мес.
3-10 мес.
380-400 дн.
3-10мес.
До 1 года
3-10мес.
>1 года
3-10мес.
Глава 8. Генетические аспекты моделирования 179
Продолжение табл. 46
Исследуемое
действие вещества
Биомоделирование
на основе гепатом
и других спонтанных
опухолей печени:
10-12% случаев
27%
28%
26-30%
64%
Биомоделирование
на основе спонтан-
ных опухолей легких:
7% случаев
10%
12,5%
30%
50%
Биомодель спонтан-
ных опухолей (тера-
томы до 5%) с хао-
тичной дифференци-
ацией клеток, про-
исходящих от одной
стволовой клетки
Биомодель спонтан-
ной лимфоидной
лейкемии (с гибелью
к 300-му дню):
более 30% случаев
70-80%
91%
Модель натурального
лейкоза:
26% случаев
36%
72%
73%
80-88%
Биомодель ретикуло-
клеточной саркомы
типа Ходжкина
до 91%
Модель лейкемии,
вызванной эстро-
генами (через
180 дней)
Лабораторное
животное
Крысы
Мыши
«
«
«
Мыши
«
«
«
«
Мыши
Мыши
«
«
Крысы
Крысы
Мыши
Мыши
Мыши
Мыши
Мыши
Линия или порода
животного
AUG/LacSto
СЗНА/Mv
CBA/CaLac
CBA/LacY
CBA/J
CBA/CaLac
C3H/HeDiSn
AHJ
CC57BR/MV
AHeJ
129/J.4
DBA/2JY
AKR/JY
AKR/JY
F344/N
F344/N
HRS/J
C58/JY
C58/JY
SJL/J
A/WySnKL
Пол
животного
Q в
Я в
в
ев
ев
в
9г?
^
Я/
в
яв
^J,^
в
я
QJ?
в
я
в
я
яв
яв
Вес
животного
196-285 г
30-40 г
«
«
«
30-40 г
«
«
«
«
30-40 г
30-40 г
«
«
250-300 г
«
30-40 г
«
«
30-40 г
30-40 г
Возраст
животного
2,5мес.-3г.
3-10 мес.
«
«
«
3-10мес.
«
«
«
«
3-10 мес.
3-10 мес.
«
«
5-12 мес.
«
3-10мес.
«
«
3-10 мес.
3-10мес.
180 Линейные животные-биомодели
Продолжение табл. 46
Исследуемое
действие вещества
Высокочувствитель-
ная биомодель
к цитогенетическому
эффекту тиоТЭФ:
в дозе 5мг/кг внутри-
брюшинно 43% кле-
ток с хромосомными
нарушениями (спон-
танно 3,4%)
Биомодель ретикуло-
клеточной опухоли
типа В(спонтанная
в 55%)
Биомодель природ-
ной аденомы гипо-
физа в 25-36%
Биомодель натураль-
ной интерстициально-
клеточной опухоли
семенников до 85%
Биомодель карцино-
мы щитовидной же-
лезы:
25% случаев
27-40%
Биомодель высокой
чувствительности
к образованию опухо-
лей под воздействи-
ем бензопирена и
триметилхолантрена
Биомодель спонтан-
ных опухолей желудка
Биомодель спонтан-
ных опухолей тонкой
кишки
Биомодель спонтан-
ных опухолей матки
Биомодель спонтан-
ной доброкачествен-
ной аденомы коры
надпочечников
Лабораторное
животное
Мыши
Мыши
Мыши
Крысы
Крысы
Крысы
Крысы
Хомячки
Хомячки
Хомячки
Хомячки
Хомячки
Линия или порода
животного
A/WySnKL
C57L/JY
F344/N
F344/N
F344/N
WAG/GSto
Линии: 1,5
4,24
12,14
82,73
4,22
54,7
4,22
4,22
54,7
4,24
Пол
животного
$г?
9
9с?
9 с?
<3б
Q в
Я с?
Q6
%%
в в
96
Q6
9 с?
9с?
о
V
9
Вес
животного
30-40 г
30-40 г
30-40 г
300-400 г
300-400 г
300-400 г
300-400 г
120г
«
«
«
120 г
«
120 г
120 г
«
120г
Возраст
животного
3-10мес.
3-10 мес.
3-10мес.
22-31 мес.
5-20 мес.
5-20 мес.
5-20 мес.
6-18 мес.
«
«
«
6-18 мес.
«
6-18 мес.
6-18 мес.
«
6-18 мес.
Глава 8. Генетические аспекты моделирования
181
Окончание табл. 46
Исследуемое
действие вещества
Высокочувствитель-
ная естественная
биомодель к терато-
генному действию
лекарственных
средств
Биомодель карцино-
мы железистого же-
лудка под воздейст-
вием триметилхол-
антрена
Высокочувствитель-
ная биомодель фиб-
росаркомной опухо-
ли, индуцированной
диметилбензантра-
ценом
Биомодель рака
молочной железы,
индуцированного
метилхолантреном
Биомодель опухолей
желудка, индуцирован-
ных N-нитронатрий-
этиламином
и N-нитронатрий-
метиламином
Лабораторное
животное
Хомячки
Хомячки
Хомячки
Хомячки
Хомячки
Линия или порода
животного
4,24
45,5
15,16
15,16
15,16
54,7
87,20
Пол
животного
9с?
9<5
9 Q
9 9
9 с?
Вес
животного
120г
«
120 г
120 г
120 г
«
120 г
Возраст
животного
6-18 мес.
«
6-18 мес.
6-18 мес.
6-18 мес.
«
6-18 мес.
Гомозиготность и генетическая однородность животных дают возмож-
ность воспроизвести эксперимент многократно и в любой лаборатории мира
с одним и тем же результатом. Это повышает надежность медико-биологи-
ческих исследований. Но есть и сложности. Они, как ни странно, множатся
с увеличением количества новых штаммов животных по принципу «больше
штаммов — труднее выбор». Правильность и обоснованность выбора адек-
ватной биомодели может быть достигнута лишь в комплексе специалистов
по генетике лабораторных животных, с одной стороны, и экспериментато-
ров и испытателей — с другой.
182
Глава 9 ИНБРЕДНЫЕ
ЖИВОТНЫЕ-БИОМОДЕЛИ
Линейные или инбредные животные представля-
ют закрепленный инбридингом генотип. Они го-
мозиготны и генетически однородны. Линейные
животные обеспечивают полноценную воспроиз-
водимость результатов и возможность их повторе-
ния в любой лаборатории. Генетическая однород-
ность позволяет расходовать меньшее число жи-
вотных для получения доказательных результатов.
[385, 468, 469].
Не секрет, что для некоторых, а возможно и мно-
гих экспериментаторов, к которым честно отношу
и себя, или иных пользователей лабораторными жи-
вотными, результаты генетических исследований
представляются в виде продукта неких магических
и тайных игр «касты избранных». Поэтому не будем
перегружать изложение избыточными специальны-
ми терминами.
Остановимся на самых общих представлениях.
В инбредных линиях генетическая однородность,
или гомозиготность, животных сохраняется посто-
янным спариванием родных братьев и сестер в пле-
менном ядре линии. Племенное ядро линии пред-
ставляет собой группу животных одной линии, раз-
множаемую в соотношении 1:1 родных братьев и
сестер.
Если инбредная линия поддерживается в кол-
лекции для сохранения ее уникального генотипа и
нет необходимости в большом количестве живот-
ных, достаточно иметь племенное ядро и ограни-
Глава 9. Инбредные животные-биомодели
183
читься работой с ним. Если животных необходимо тиражировать, то плем-
ядро передается в питомники.
В филиалах НЦБМТ РАМН ведется работа по разведению линейных жи-
вотных для использования в экспериментах. К работе с племенными ядрами
инбредных линий допускаются лица, имеющие биологическое или зоотех-
ническое образование и прошедшие соответствующую стажировку.
Но даже при тщательном соблюдении всех правил содержания инбред-
ных животных существует опасность возникновения генетической изменчи-
вости, гетерозиготности в высокоинбредных линиях. Причиной могут быть
спонтанные мутации или случайные скрещивания [42, 67-72, 163, 179, 180,
391, 598, 599].
Если в процессе экспериментов было случайное скрещивание с животными
другого генотипа, восстановить гомозиготность линии, сохранив исходный ге-
нотип, практически невозможно. Такие животные подлежат уничтожению.
Для гарантии гомозиготности животных инбредных линий необходимо
контролировать их однородность по генетическим признакам.
Для контроля генетической однородности инбредных линий мышей и
крыс применяется реципрокная изотрансплантация кожи. Она позволяет кон-
тролировать гомозиготность по большому числу генов, так как совмести-
мость тканей — полигенный признак. Этот прием дает возможность выя-
вить очень слабые генетические различия между животными одной линии,
обусловленные остаточной гетерозиготностью или спонтанными мутация-
ми. Метод технически достаточно прост и не требует больших материальных
затрат. Мышей и крыс для контроля отбирают в возрасте 25—30 дней. Транс-
плантация осуществляется на 2—3 месячных мышах и 3—3,5-месячных кры-
сах, но не раньше 6-недельного возраста. Существуют и другие методы, но
их описание не входит в наши задачи. Это иная область, в которой работают
совершенно иные профессионалы.
Генетический мониторинг
Не вызывает сомнения тот факт, что «генетическое заражение» нарушает
чистоту линии животных и достоверность результатов эксперимента. Гене-
тический контроль не может предотвратить «генетического заражения», но,
тем не менее, является важным методом обеспечения генетической каче-
ственности лабораторных животных. Генетический контроль может быть
разделен на три категории:
характеризация — проводится для подтверждения генотипа инбредных
линий и для создания генотипа новых линий путем проверки возможно боль-
шего числа локусов;
мониторинг I — проводится с целью подтверждения генетического про-
филя;
184
Линейные животные-биомодели
мониторинг N — проводится для подтверждения критических подгрупп,
которые представляют собой минимальный набор отдельных локусов, по-
зволяющих выделять данную группу животных в обособленную линию.
В генетическом контроле локусов, позволяющем выделять животных
в отдельную лабораторную линию, называются маркерами. В соответствии
с биологическими функциями маркеры делятся на шесть групп: биохимичес-
кие, морфологические, иммуногенетические, молекулярные генетические, фар-
макогенетические и цитогенетические. Для избрания локусов в качестве мар-
керов для генетического анализа их обнаружение должно быть точным, про-
стым в исполнении, эффективным и экономичным. Наиболее подходящие
маркеры были найдены в биохимической и иммуногенетической группе,
поскольку количество локусов в них известно лучше, чем в других маркерах,
и их обнаружение проще в осуществлении. Для работы достаточно 19 мар-
керов: 15 биохимических и 4 иммуногенетических. Предложена следующая
схема генетического контроля. Каждая линия сразу после внедрения должна
пройти характеризацию в целях подтверждения генотипа. Если внедряемая
линия соответствует ожидаемым характеристикам, она периодически (раз в
год) подвергается мониторингу I. Через несколько лет после внедрения вновь
проводят процедуру характеризации. Мониторинг II проводится по необхо-
димости. Эффективным механизмом контроля является сверка критических
подгрупп. Критические подгруппы представляют собой ряд локусов, строго
отобранных из генетических профилей ограниченных инбредных линий.
Например, пять наиболее часто встречающихся инбредных линий (AKR,
СЗН/Не, DBA/2, BALB/c и C57BL/6) можно распознать, используя четыре
биохимических маркера (Hbb, Саг-2, Gpi-1, Idh-1). Количество маркеров
критических подгрупп зависит от количества проверяемых линий, но обыч-
но не больше пяти.
Криогенное сохранение эмбрионов
Из эмбрионов мышей, замороженных в жидком азоте 12-15 лет назад, были
успешно получены молодые особи животных. Несмотря на то, что были
предприняты экспериментальные попытки применить данный метод сохра-
нения к другим видам (включая человека), предстоит глубже изучить его,
прежде чем использовать на постоянной основе в отношении различных
видов живых существ [240]. Основными причинами криогенной заморозки,
например, эмбрионов мышей являются следующие:
/ гарантия того, что вся линия или популяция не будет потеряна в ре-
зультате болезни или несчастного случая;
/ криогенная заморозка — наиболее экономичный метод поддержания
значительного количества будущих особей, необходимости в которых
в данный момент нет, но может вскоре возникнуть;
Глава 9. Инбредные животные-биомодели
185
/ криогенная заморозка может использоваться для перевозки по воздуху
коллекционных или редких линий;
/ криогенное сохранение используется для предотвращения генных из-
менений в инбредных линиях, вызванных аккумуляцией мутаций;
/ криогенно замороженные эмбрионы используются для экспериментов
в области преимплантационной эмбриологии;
/ создание популяции для получения высококачественных и достовер-
ных экспериментальных данных.
Для проведения успешного криогенного сохранения эмбрионов мышей
необходимо недорогое, но качественное оборудование, реактивы высокой
степени очистки, но главное — специалисты в области преимплантацион-
ной эмбриологии, имеющие успешный опыт криогенного сохранения эмб-
рионов. Оборудование, необходимое для криогенной заморозки, включает
в себя частично опорожненные колбы, приспособления для обращения
с жидким азотом, С02-инкубаторы, термопары, регистрирующий прибор и
бинокулярные микроскопы.
Донорами эмбрионов обычно являются препубертатные мыши в воз-
расте около 21 дня. С помощью сыворотки беременной кобылы и челове-
ческого хорионического гонадотропина у самок вызывают гиперовуляцию
и затем их спаривают с племенными самцами. Доказательством успешно-
го спаривания является наличие вагинальной пробки, со дня ее обнаруже-
ния начинается отсчет дней. На третий день мышь убивают и извлекают
матку и фаллопиевы трубы. Из фаллопиевых труб с помощью соответству-
ющего инструмента вымываются эмбрионы, которые на этот момент дол-
жны находиться на стадии 8-клеточного развития. Далее, из общей массы
эмбрионов выбраковываются поврежденные или недоразвившиеся. Остав-
шиеся эмбрионы в среде с соответствующим криопротектором (диметил-
сульфоксид) помещают в пробирки или трубочки, подготовленные для
заморозки. Далее они охлаждаются до почти полной заморозки и колбы
закупориваются кристаллом льда для предотвращения переохлаждения.
После этого эмбрионы замораживают. Обычно скорость охлаждения низ-
кая (около 0,4 градуса в минуту). С момента достижения эмбрионами тем-
пературы —80° С скорость охлаждения можно увеличить, пока степень
заморозки не будет соответствовать температуре жидкого азота (—196° С).
При этой температуре эмбрионы могут сохраняться неограниченное ко-
личество времени.
Когда возникает необходимость в эмбрионах, их размораживают, исполь-
зуя принятый в лаборатории метод. Обычно это происходит при комнатной
температуре или в ванне с температурой воды 37° С. Неповрежденные эмб-
рионы отбирают, омывают и выращивают 24 часа в подходящей среде в С02-
инкубаторе. Если эмбрионы жизнеспособны, они развиваются в бластоцис-
ту, подходящую для имплантации.
186
Линейные животные-биомодели
За 2 дня до размораживания эмбрионов нескольких псевдобеременных
мышей, будущих суррогатных матерей, подготавливают к приему эмбрионов
путем спаривания со стерильным самцом (стерилизованным с помощью
вазектомии или генетически стерильным). Будущая суррогатная мать полу-
чает анестезию, ей делают один кожный надрез и далее по надрезу в верхней
брюшине по обеим сторонам от позвоночника с тем, чтобы обнажить боко-
вые стороны матки. С обеих сторон матки делается по маленькому надрезу,
через которые помещается 4—6 эмбрионов. Кожный надрез закрывается
с помощью зажима. Если операция прошла успешно, через 18—19 дней дол-
жны родиться мышата. Квалифицированный специалист может получить 30—
40% жизнеспособных молодых особей из общего количества замороженных
эмбрионов.
Инбредные линии кроликов
В поддержании и использовании в эксперименте инбредных кроликов важ-
ная роль принадлежит их ранжированию. Ранжирование зависит от того,
какие конкретно штаммы были включены в исследования, и может менять-
ся в результате взаимодействия между генотипом и окружающей средой. Были
исследованы эстеразы сыворотки преальбумина у перечисленных здесь штам-
мов. Во многих штаммах фенотипы были единообразными, как и следует
ожидать от штаммов с высокой степенью инбридинга. Отсутствие аллели
Est-3d в материале было заметно, так как она является преобладающей алле-
лью в колониях диких кроликов.
A/J
Инбридинг 95%. Альбиносный штамм, выведенный Чаи (Chai) в 1958 г.
из материала, состоящего на 50% из III, на 25% из R (R имеет то же проис-
хождение, что и штаммы В и Y) и на 25% из разных штаммов. Разделение по
ангорскому гену 1. Вес тела зрелой особи приблизительно 2500—2700 г. Гено-
тип крови HgF. Генотип гамма-глобулина Аа3/Аа3, Ab4/Ab4. Высокое артери-
альное давление крови. Высокий гематокрит крови (самцы, самки). Высо-
кий гемоглобин крови. Высокие уровни азота мочевины крови, низкий уро-
вень сывороточного сахара, низкий уровень сывороточного фосфора и низ-
кий уровень сывороточных общих белков у самок.
AC/J
Инбридинг 85%. Черный, с отступающими белыми отметинами
{aaEfiE?iduddud). Выведен Сауином (Sawin) из голландского материала Луизы
Пирс (Louise Реагсе) в Институте Рокфеллера (Rockefeller Inst). Скрещен с X
и прочими штаммами и впоследствии подвергнут инбридингу с селекцией
на ген хондродистрофии. Вес зрелой особи составляет около 2400 г. Генотип
Глава 9. Инбредные животные-биомодели
187
гамма-глобулина Ла3/Ла3, АЬ5/ЛЬ5, Ас7/Ас7. Имеет рецессивный ген хондро-
дистрофии ас. У гомозигот уменьшенный размер, непропорционально умень-
шенные конечности и большая голова. Хрящ аномален, и имеется заметный
недостаток окостенения. Животные умирают при рождении или вскоре пос-
ле рождения. Состояние напоминает изменения, встречающиеся у человека,
скота и собак. Штамм также имеет ген расщелины позвоночника (sb). Гомо-
зиготы имеют расщепленный позвоночник, переходящий из заднего свода
черепа в хвост, покрытый тонким слоем кожи. У большинства животных —
расщепленная губа, расщепленное нёбо, заметная деформация позвоночни-
ка и загибающийся к брюху конец хвоста. Также имеют место и другие ано-
малии. Приблизительно у 50% особей есть добавочные селезенки. Печеноч-
ная ариловая углеводородная гидроксилаза не индуцируется. Низкое арте-
риальное давление крови. Низкая способность связывания сывороточного
железа. Низкий уровень серопротеинсвязанного йода. Высокие уровни гор-
монов роста у самок.
ACEP/J
Инбридинг 93%. Голубоглазые белые (v/v) голландского происхожде-
ния, селектированные Сауином в 1951 г. для исследования предрасполо-
женности к аудиогенным судорогам, которые во многих отношениях похо-
жи на судороги, обнаруженные у некоторых инбредных штаммов мышей.
Судороги вызваны рецессивным геном ер. Генотип гамма-глобулина Аа3/
Аа3, Ab5/Ab5, Ac21/Ac21. Животные имеют малые размеры, отличаются гипе-
рактивностью, вес тела зрелой особи составляет приблизительно 2400 г.
Штамм также отличается генной гипогонадией (hg), в связи с чем оба пола
стерильны. У самцов сперматогенные канальцы имеют малый размер и лишь
один ряд клеток Сертоли. Аномальные яичники имеют малый размер и
лишены фолликул. Степень аномальности бывает различной. Либидо нор-
мальное. Ариловая углеводородная гидроксилаза не индуцируется. Высо-
кое артериальное давление крови у самцов и самок. Высокие уровни сыво-
роточного железа у самок. Низкий гематокрит (самцы, самки). Высокое
общее содержание сывороточного холестерина у самок. Высокий уровень
серопротеинсвязанного йода у самок. Высокие уровни гормонов роста. Вы-
сокие уровни сывороточной щелочной фосфатазы у самок, высокие уровни
сывороточного фосфора у самок и низкие уровни сывороточного общего
белка у самцов.
AX/J
Инбридинг 82%. Штамм шиншиллы (ссИЗ, cch3, AAww), выведенный Сауи-
ном в 1927 г. из материала Института Басси (Bussey Institute). Этот материал
сначала был назван «семейством ПЬ», а затем — «семейством V». Материал
был селектирован по гену атаксии после того, как этот порок наблюдался
188
Линейные животные-биомодели
впервые в 1936 г. Он был скрещен со штаммами X и III, получил название
АХ в 1943 г. Генотип гамма-глобулина Аа3/Аа3, Ab4/Ab4, Ac7/'Ас21.
Вес зрелой особи составляет 3500 г. Имеет рецессивный ген атаксии (ах).
У гомозигот наблюдается прогрессивный паралич, который начинается в
возрасте приблизительно 2 месяцев. Он начинается с задних лап, но быстро
прогрессирует и приводит к чрезвычайной возбудимости, неспособности
принимать пищу или пить, нерегулярности дыхания и смерти. Гистологи-
чески атаксия связана с регрессивным изменением в ворсистых окончаниях
коры мозжечка. Низкая частота пульса у самок. Высокая способность связы-
вания сывороточного общего железа у самцов. Высокий гематокрит у сам-
цов, низкие уровни гемоглобина крови у самок, но высокие у самцов. Вы-
сокое общее содержание холестерина в плазме. Низкий уровень серопроте-
инсвязанного йода. Низкие уровни сывороточного сахара (самцы, самки),
низкие уровни сывороточного фосфора у самок.
Таблица 47
Основные биохимические параметры крови инбредных кроликов
Параметры
Гематокрит крови
Гемоглобин крови
Сывороточный сахар
Азот мочевины крови
Общий уровень серопротеинов
Связывание сывороточного
железа
Серопротеинсвязанный йод
Уровни гормона роста
Уровень сывороточного
фосфора
Содержание сывороточного
холестерина
Сывороточная щелочная
фосфотаза
Уровни
Высокие
A/J, X/Y(9), AX/J(d)
A/J, AX/J(d), AXBU/J(0*)
X/J
AJ, B/J(9), Y/J
X/J, Y/J
ACEP/J(9), AX/J(0*), B/J(9)
Y/J, lll/J
ACEP/J(9), Y/J(0*), lll/J
AC/J(9), ACEP/J, C/J,Y/J(d)
ACEP/J(9), X/J(0*), Y/J
ACEP/J(9), AX/J, B/J(d)
ACEP/J(9), C/J, X/J*
Низкие
B/J, Y/J, ACEP/J
AX/J(9), C/J, Y/J
AJ, AX/J, B/J
C/J
AJ(9)
AC/J(9), AXBU/J(d)
AC/J, AX/J
AXBU/J(d)
A/J, ACEP/J(d), AX/J(9)
b/j(9), x/j(d)
B/J
AXBU/J
Инбридинг 82%. Альбиносный (ее) штамм, выведенный Сауином из
мутации, вызывающей буфтальм в материале АХ, и выделенный в 1967 г.
Ранее назывался AXbubu. Генотип гамма-глобулина Ab4/Ab4, Ac1/Ac21. Вес тела
зрелой особи составляет приблизительно 3500 г. Рецессивный ген bu вызы-
Глава 9. Инбредные животные-биомодели
189
вает болезнь, подобную детской глаукоме у человека. Он повышает внутри-
глазное давление, диаметр роговицы, васкулярность роговицы, способствует
помутнению роговицы и ороговению, препятствует оттоку жидкости из гла-
за и может приводить к атрофии зрительного нерва. У самцов снижается
скорость сперматогенеза. Причиной этого порока может являться дефект
обмена (3-каротина или витамина А. Уровни аскорбата во внутриглазной жид-
кости заметно снижаются у животных с тяжелой формой болезни. У самцов
из этого штамма наблюдается низкий уровень сывороточного железа. Высо-
кий уровень гемоглобина крови — у самцов. Низкий уровень гормонов ро-
ста плазмы — у самок.
B/J
Инбридинг 91% (также называется ACCR (В)). Выведен Сауином из 62,5%
голландского материала, 6,25% X, 7,8% Шс и прочего остального материала.
Коуэну на F3 и Чаи на F7. Альбинос, атласный (с/с, sa/sa, dud/dud). Генотип
крови HgF/HgF. Гистосовместимый. Генотип гамма-глобулина Аа3/Аа3, АЬ5/
АЬ5, Ас7/Ас7. Вес тела приблизительно 2000 г. Высокое артериальное давле-
ние крови (самцы, самки). Низкая способность связывания общего сыворо-
точного железа у самок. Низкий гематокрит крови (самцы, самки). Высокое
общее содержание холестерина в плазме у самцов, но низкое у самок. Высо-
кие уровни азота мочевины крови у самок, низкие уровни сывороточного
сахара и сывороточной щелочной фосфатазы у самцов и самок.
C/J
Инбридинг 85%. Штамм дикой (АА) или черной (аа) окраски, выведен-
ный Чаи в 1958 году из материала, состоящего на 50% из III, на 25% из R6
(R имеет то же происхождение, что и штаммы Y и В) и на 25% из разного
материала. Генотип гамма-глобулина Aat/Aa1, Ab4/Ab4, Ac7/Ac7. Характерис-
тики: вес тела — приблизительно 2950 г, низкая частота пульса
у самцов; интенсивный иммунный ответ на Glu56, Lys38, Туг6; низкие уровни
гемоглобина крови (самцы, самки); высокие уровни гормонов роста плазмы
(самцы, самки); низкие уровни азота мочевины крови, высокие уровни сы-
вороточной щелочной фосфатазы (как у самцов, так и самок).
X/J
Инбридинг 88%. Материал темно-желтого окраса (C/^/aaeebb), выведен-
ный в 20-х годах XX века в результате гибридизации мелкого штамма А Касла
(Castle) с прочим материалом. В 1935 г. был введен ген Dw, который впослед-
ствии был перенесен на штамм III. Штамм имеет 12 ребер и использовался
Сауином в исследованиях роста. Генотип гамма-глобулина Аа3/Аа3, Ab4/Ab4,
Ас7/Ас7. Вес тела зрелой особи составляет приблизительно 2000 г. Имеет ре-
цессивно наследуемый по аутосоме ген ha (гемолитическая анемия), вызы-
190
Линейные животные-биомодели
вающий иммунопатию, похожую на ту, что встречается у мышей NZB. По-
страдавшие животные бледные и слабые. В некоторых формах возникает
тяжелая форма желтухи. Среднее время выживания составляет приблизи-
тельно 5 месяцев. При вскрытии обнаруживается гемоглобинемия, желтуха,
бледность печени с субкапсулярным кровотечением, геморрагическая селе-
зенка и гемоглобинурия. Штамм также характеризуется тем, что около 49%
особей имеют добавочные селезенки. Печеночные ариловые углеводороды
не индуцируются. Низкое артериальное давление крови. Слабый иммунный
ответ на Glu56, Lys38, Туг6. Высокий гематокрит крови у самок. Низкое общее
содержание холестерина в плазме у самцов. Высокие уровни сывороточного
сахара, очень высокие уровни сывороточной щелочной фосфатазы, прибли-
зительно в два раза выше, чем у других штаммов, высокий уровень сыворо-
точного фосфора у самцов и высокие уровни общих серопротеинов у самцов
и самок.
Y/J
Инбридинг 84%. Также называется ACCR (Y). Альбиносный штамм {ее,
sa, sa), состоящий на 62,5% из голландского материала из Института Рок-
феллера, имеет гены ас, стере небольшим процентом X, III, IIIc и прочего
материала. Коуэну на F2 и Чаи на приблизительно F7. Тип гамма-глобулина
Аа3/Аа3, Ab5/Ab5. Вес тела приблизительно 2300 г. Гистосовместимый. Около
42% особей имеют добавочные селезенки. Высокая способность связывания
сывороточного железа (самцы, самки). Низкий гематокрит (самцы, самки),
низкий гемоглобин крови. Высокие уровни связанного серопротеинами йода
у самцов. Высокие уровни гормонов роста плазмы у самцов. Как у самок, так
и у самцов высокие уровни азота мочевины крови, высокое содержание сыво-
роточного фосфора, высокий общий уровень серопротеинов.
III/J
Инбридинг 81%. Альбиносный штамм (с/с, А/А, Е?/Е?), выведен Сауином
из новозеландского белого штамма Касла в 1932 г. Штамм ранее назывался III
или Што. Генотип гамма-глобулина Aat/Aa1, Ab4/Ab4, Ac7/Ac7. Этот очень
спокойный штамм был селектирован для пресакрального роста, имеет 13 ре-
бер и был использован в исследованиях роста и развития. Высокий продуцент
антител. Кровные родственники принимают кожные трансплантаты. Из этого
же материала выведены семь дополнительных штаммов или подштаммов. Вес
тела зрелой особи составляет приблизительно 3800 г. Печеночная ариловая
углеводородная гидроксилаза не индуцируется (в отличие от пяти прочих
штаммов). Низкая частота пульса у самок. Высокий иммунный ответ на Glu56,
Lys38, Туг6. Высокие уровни сывороточного железа. Как у самцов, так и у са-
мок — высокие уровни серопротеинсвязанного йода.
Глава 9. Инбредные животные-биомодели
191
Инбредные линии морских свинок
Линия 2 (Family 2, Strain 2, STR 2)
Инбридинг 92%. Генотип а, вР, s. Окраска шерсти трехцветная: черно-
рыже-белая. Из Министерства сельского хозяйства США (где разводились
с 1906 г.) попали к Райту на F11 и поддерживались братско-сестринским
скрещиванием на протяжении 33 поколений до 1933 г., затем размножались
рандомбредно до 1940 г., когда линия перешла к Хестону, который возобно-
вил систему братско-сестринского разведения. В 1950 г. на FI2 линия была
передана в Национальный институт здравоохранения (г. Бетесда, США); все
сублинии, отделившиеся впоследствии, происходят из этого института. Ги-
стосовместимы. Относительно резистентны к туберкулезу. В сублинии N со
средней частотой встречаются случаи кальцификации (обызвествления) в
области большой кривизны желудка, ободочной кишки, почек, полосатой
мускулатуры брюшной стенки, легких и аорты у старых животных (в 34 ме-
сяца) [206, 207, 208]. Дают ответ на PLL, имеют антигены GPL-A-B.1. Вес
тела самки 780 ± 69 г, самца — 802 ± 65 г. Относительный вес органов (грамм
на 1 кг живого веса): печень самки — 37,45, самца — 31,63; легкие самки —
6,64, самца — 6,50; сердце самки — 2,65, самца — 2,64; щитовидная железа
самки — 0,074, самца — 0,076; почки самки — 3,58, самца — 3,67; надпочеч-
ники самки — 0,505, самца — 0,501; селезенка самки — 1,32, самца — 0,97
[206, 207, 208].
Репродуктивность средняя: «деловая» плодовитость 7,1 на самку в год;
средняя величина помета при отъеме 2,48+0'03 [206, 207, 208].
Линия 13 (Family 13, Strain 13, STR 13)
Инбридинг 93%. Генотип а, ер, s. Окраска шерсти трехцветная: черно-рыже-
белая. Происхождение такое же, как у линии 2. К Райту попали в 1915 г. на
F13, инбредировались на протяжении 33 поколений (до 1933 г.), затем разво-
дились рандомбредно до 1940 г., после чего снова инбредировались. В Наци-
ональный институт здравоохранения были переданы в 1950 г. на F12. Все
последующие сублинии происходят из этой колонии, гистосовместимы, более
чувствительны к туберкулезу, чем линия 2. Не дают ответа на PLL, имеют
антиген GPL-A-B.1. Более крупные по величине, чем линия 2. Вес тела самки
составляет 940 ± 99 г, самца — 1044 ± 69 г [208]. Относительный вес органов
(грамм на 1 кг живого веса): печень самки — 37,76, самца — 32,27; легкие
самки — 7,91, самца — 6,92; сердце самки — 2,40, самца — 2,31; щитовидная
железа самки — 0,097, самца — 0,075; почки самки — 2,48, самца — 2,46;
надпочечники самки — 0,302, самца — 0,297; селезенка самки — 0,99, самца
— 0,699 [207]. Репродуктивность средняя: «деловая» плодовитость 5,5 на самку
в год; средняя величина помета при отъеме 2,69 ± 0,06 [208]. Сексуальное
поведение менее активное, чем у морских свинок линии Family 2.
192
Линейные животные-биомодели
B/Lac
Инбридинг 82%.Генотип еР, п. Выведены Лундбергом в Институте сыво-
роток (Копенгаген) селекцией на резистентность анафилаксии, вызываемой
яичным альбумином. Переданы в Национальный институт медицинских ис-
следований (Лондон) в 1966 г., а в 1976 г. — в Центр лабораторных животных
(Lac). Большинство кожных трансплантатов приживается без последующего
отторжения. Резистентны к анафилаксии, вызываемой яичным альбумином
[206]. Полиморфны по локусу PGM (фосфоглюкомутаза), несут аллели PGM1
и PGM2 [116]. Вес тела самки 1036 г, самца — 1093 г. Относительный вес
органов (грамм на 1 кг живого веса): селезенка самки — 0,6, самца 0,51;
тимус самки — 0,35, самца — 0,29; надпочечники самки — 0,50, самца —
0,24; мозг самки — 4,11, самца — 4,00. Средняя плодовитость: по числу
рожденных на одну самку в год — 10,4, по числу выкормленных — 9,3; сред-
няя величина помета — 2,5; смертность до отъема — 11%.
ОМЗ/Lac
Инбридинг 76%. Генотип d3, s, Y. Заложена Роджерсом в 1952 г. из ран-
домбредного коммерческого стока, затем передана в Эллингтон-Фарм в
1964 г., а оттуда — в Центр лабораторных животных (Lac) в 1965 г. Нет спон-
танных опухолей. Отсутствует токсемия беременности [208]. Несут аллель
PGM1 (фосфоглюкомутаза) [208]. Хорошо размножаются. Средняя плодови-
тость: по числу рожденных на одну самку в год 6,5; по числу выкормленных
— 5,6, средняя величина помета — 2,3, смертность до отъема — 14%.
R9/Lac
Инбридинг 75%. Генотип е?, s, п. Заложены Роджерсом в 1941 г. от ран-
домбредных предков. В Центр лабораторных животных (Lac) поступила в
1965 г. из Эллингтон-Фарм. Частота спонтанных опухолей в возрасте свыше
3 лет составляет 14% [208]. Несут аллель PGM2 (фосфоглюкомутаза) [116].
Показатели крови следующие: количество эритроцитов (миллионов на 1 мм3)
— самки 4,45-5,48, самцы 5,70-4,88; гемоглобин (грамм на 100 см3): самки
12,11—12,3, самцы 12,5—13,54; гематокрит — самцы 40,4—42,9%, самки 39,0—
40,4%; количество лейкоцитов (тысяч на 1 мм3) — самки 3,68-4,46, самцы
3,68—4,73 [643, 668;]. Вес тела самки 844 г, самца — 971 г. Относительный вес
органов (грамм на 1 кг живого веса): селезенка самки — 1,23, самца — 0,84;
тимус самки — 0,24, самца — 0,29; надпочечники самки — 0,44, самца —
0,42, мозг самки — 4,50, самца — 4,29 [643]. Средняя плодовитость по числу
рожденных на одну самку в год 8,3; по числу выкормленных — 7,0; средняя
величина помета — 2,5; смертность до отъема — 16%. Средняя продолжи-
тельность жизни более 4 лет.
Глава 9. Инбредные животные-биомодели
193
IMM/R
Инбридинг 81%. Генотип с, ер, z. Окраска шерсти белая, альбиносы. За-
ложена Лундбергом из колонии морских свинок-альбиносов (Копенгаген).
Отселектированы на резистентность к анафилаксии, вызываемой яичным
альбумином. Спонтанных опухолей нет. Более восприимчивы к инфекциям,
чем IMM/S. Не дают анафилактических реакций и не продуцируют антител
против яичного альбумина (в количестве, поддающемся измерению). Плохо
размножаются [206, 207, 208].
IMM/S
Инбридинг 83%. Генотип с, еР, z. Окраска шерсти белая, альбиносы.
Продукция антител такая же, как у IMM/R. Отселектированы на чувстви-
тельность к анафилаксии, вызываемой яичным альбумином. Спонтанных
опухолей нет. Крайне сильная анафилактическая реакция у 100% животных
на ингаляцию яичного альбумина в очень низких дозах. Хорошо продуци-
руют антитела. Мало восприимчивы к инфекциям. Размножаются хорошо.
Инбредные линии крыс
Как и инбредные мыши линии крыс обозначены нами в соответствии с
международной номенклатурой. Инбридинг в линиях (F), разводимых в
России, указан, как и в линиях мышей, по данным на декабрь 2004 г. Про-
исхождение и характеристики линий крыс, кроме особо оговоренных случа-
ев, даются в соответствии с общепринятыми стандартами.
AUG/LacSto Поддерживаются в Sto (August)
Инбридинг F68. Генотип И, р. Окраска шерсти: капюшон осветленный
(цвета кофе с молоком), остальная, часть белая. Выведены от одной из
сублиний. В Sto поступили в 1958 г. из Lac на F16. А990, А7332, А28807 и
A35322 — линии, родственные AUG, которые иногда также называют «Ав-
густ». Обладают сходными биологическими характеристиками, но различа-
ются по генам окраски. Встречаются гепатомы, которые могут трансплан-
тироваться. Чувствительны к экспериментальному аллергическому энце-
фалиту. Чувствительны к Haemophilus bronchosepthicum. Резистентны к ин-
дукции аутоиммунного тиреоидита [206, 207, 208]. В Sto живой вес крыс в
возрасте 1 месяца у самок составляет 43,1 г, у самцов 44,4 г, в 3 месяца
самки весят 127,2 г, самцы — 157,7 г, в 9 месяцев самки — 196,0 ± 1,29 г,
самцы — 285,8 ± 1,93 г. Обладают высокой жизнеспособностью и плодови-
тостью. В Sto величина помета при рождении составляет 6,57, при отъе-
ме — 6,37 ±0,10 крысят.
194
Линейные животные-биомодели
F344/N Поддерживаются в Sto (Fischer 344, Fischer) До 1980 г. Y
Инбридинг F141. Генотип а, с, h. Окраска шерсти белая, альбиносы. Начал
инбредировать Куртис в 1920 г. в Институте раковых исследований Колумбий-
ского университета (США). В Y в 1976 г. на F130, переданы в Sto на F132.
Небольшой размер тела. Частота опухолей молочных желез составляет 41%
у самок и 23% у самцов. Аденомы гипофиза у 36% самок и 24% самцов.
Частота интерстициально-клеточных опухолей семенников колеблется от 65
до 85% в разных сублиниях. Карциномы щитовидной железы встречаются
у 22% крыс этой линии, мононуклеарно-клеточная лейкемия у 24%. Поли-
плоидные опухоли эндометрия матки развиваются у 21% самок, подкожные
фиброаденомы у 9% самок, узелковая гиперплазия печени у 5% животных
обоих полов.
Частота опухолей в условиях SPF следующая: лейкемия у 26% самцов и
у 36% самок; опухоли молочных желез у 12% самцов и у 20% самок; опухоли
других локализаций у 9% самцов и у 5% самок. Крысы линии F344 дают рост
следующих трансплантируемых опухолей: гепатома Даннинга, гепатома LC-
18, гепатома Novikoff, карцинома молочных желез НМС и R-3230, опухоли
гипофиза MtT и MtTf, карциносаркома Walker-256, лейкемия Даннинга,
лейкемия HLF1, IRC-741 и R-3149, лимфосаркома R-3251, фиброма мо-
лочных желез F609, фибромосаркома R-3244, саркомы IPS 9802 и R13259,
саркома матки F529, а также лейкемии R-3323, 3330, 3399 и 3432.
Сублиния N обеспечивает рост трансплантируемых опухолей 1RC741,
IRS1548, L = C18, R3244, R3149, R3330, R3323, R3432, R3604, IRS9802, R3211,
R3230, R3251, R3259 и трех типов аденокарцином молочных желез. В этой
сублинии наблюдаются спонтанные опухоли передней доли гипофиза у 15%
крыс, опухоли клеток Лейдига у 30—50%, встречаются грануломатозные по-
ражения лимфатических узлов и печени. Особенности морфологии хромо-
сом: № 3 — субтелоцентрическая, Х-хромосома — телоцентрическая,
№ 4 и № 9 имеют большую С-полосу, № 5 и № 7 не обнаруживают
С-исчерченности или она слабо выражена.
Отиты обнаруживаются у 40—60% старых животных, бронхоэктазы у 61 —
70%, нефриты у 21—30%, миокардиты у 21-30%. У молодых крыс нефриты
возникают редко. Отмечен высокий метаболизм этилморфина и анилина в
печени у самцов. Резистентны к развитию солевой гипертензии. Имеют вы-
сокую специфическую активность, но низкую способность к индукции ре-
дуктазы НАДФ-цитохрома С (по сравнению с аутбредными крысами Sprague
— Dawley). Чувствительны к индукции опухолей молочных желез
2-ацетиламинофлюорином. У животных происходит быстрая абсорбция
имплантированных таблеток диэтилстилбэстрола, что приводит к смерти.
Уровень LD50 к действию раствора пентабарбитала (70 мг/кг) низкий. Чув-
ствительны к цистецеркозу. Уровень инсулина в сыворотке низкий. Низкий
Глава 9. Инбредные животные-биомодели
195
первичный и вторичный иммунный ответ на эритроциты барана [208]. Ан-
тигены Т-лимфоцитов: Ly-1+, Ly-2,1', Ly-2,2+ [676].
Средняя продолжительность жизни составляет, по данным разных авто-
ров, 24—31 месяц у самцов и 22,5—29 месяцев у самок. Хорошо размножаются,
средняя величина помета (1/12). У самцов низкий уровень дефекации в от-
крытом поле. Активность в колесе (как у самцов, так и у самок) низкая [208].
MSUBL/lcgn (Строевой)
Инбридинг F66. Генотип а. Окраска шерсти черная. Выведены О.Г. Стро-
евой в Институте биологии развития РАН от скрещивания диких крыс с MSU-
микрофтальмическими крысами, полученными от Броумана из Университета
штата Монтана. Затем были переведены в Институт цитологии и генетики СО
РАМН (Новосибирск). Высокая частота микрофтальмии и анофтальмии.
Высокая реактивность надпочечников к АКТГ.
WAG/GSto Поддерживаются Sto (Wistar albino Glaxo)
Инбридинг F109. Генотип а, с. Окраска шерсти белая, альбиносы. Вы-
ведены Бачарачем в 1924 г. от крыс Wistar. В филиал АМН СССР «Столбо-
вая» (Sto) поступили в 1966 г. (5 пар) из Glaxo Laboratory на F?. Разные
сублинии WAG имеют различные гены окраски, возможно, в прошлом было
генетическое засорение. Поэтому необходимо указывать сублинейный сим-
вол. Частоты опухолей следующие: карцинома щитовидной железы 27—40%,
феохромоцитома 20%, островковоклеточная аденома 1%. При средней про-
должительности жизни 31 месяц у самок аденомы встречаются у 69%, аде-
нома коры надпочечников — у 29%, фиброаденома молочных желез — у
21%. В группе из 290 животных этой линии были обнаружены опухоли 19
различных типов. Наблюдается высокая частота полиспермии. Резистент-
ны к экспериментальному аллергическому энцефаломиелиту. Некоторые
сублинии дают анафилактическую реакцию на декстран. Чувствительны к
индукции аутоиммунных тиреоидитов, а также к дефициту железа. Имеют
низкий вторичный ответ по антителам к полипептиду (Т, G) Pro-L [208].
Вес тела крыс в возрасте 1 месяца: самки — 37,5 ± 0,81 г, самцы — 39,9 ±
0,85 г; в 3 месяца: самки — 154,1 ± 1,36 г, самцы — 207,7 ± 2,40 г; в 9
месяцев: самки — 228,2 ± 2,15 г, самцы — 332,0 ± 3,36 г. Плодовитость в
Sto: число пометов на одну самку — 4,47 ± 0,14, величина помета при рож-
дении — 7,08 ± 0,12, при отъеме — 6,1 ± 0,15 крысенка. Средняя про-
должительность жизни превышает 31 месяц у самок и 22 месяца у самцов.
Средняя величина помета — 8,7.
BDI
Инбридинг F101. Генотип Л, С, Н, р. Окраска шерсти желтая, розовогла-
зые. Их начал выводить Друкрей в 1937 г. из колонии желтых розовоглазых
196
Линейные животные-биомодели
крыс. Инбридированы были в конце 40-х годов XX века. Линия BD была
получена скрещиванием с крысами Wistar с последующим инбридингом.
Остальные линии этой группы (от BDIII до BDX) выведены от скрещивания
первых двух линий BDI и BDII и селектированы по генам окраски. Однако
линии BD имеют четыре различных гаплотипа Н-1 (AgB, или RT1 по новому
обозначению): Н-ld, H-lw, Н-1й и Н-1е вместо двух, как следовало бы ожи-
дать для подобного скрещивания. Все линии BD имеют низкую частоту
опухолей при средней продолжительности жизни в 700—950 дней (в зависи-
мости от линии). Антигены Т-лимфоцитов BDII: Ly-1', Ly-2,1'; Ly-2,2+; BDV:
Ly-1+, Ly-2,2*, Ly-2,1-, BDVII: Ly-1+, Ly-2,1, Ly-2,2+ [676]. BDII - высокая
частота микрофтальмии; BDVII — дают низкий вторичный ответ по антите-
лам к полипептиду (TG)-Pro-L [208]. BDVIII — иногда встречается ваги-
нальная атрезия [208].
LEW (Lewis, W/Lew)
Инбридинг F97. Генотип а, И, с. Окраска шерсти белая, альбиносы. На-
чал инбридировать Льюис из колонии. Обеспечивают рост трансплантируе-
мых опухолей: лимфомы 8, саркомы почек, фибросаркомы МС-39, LM-1,
ML-7, карциномы 10 Льюиса, саркомы 3 Льюиса (FEDEX-LEW, CaM-LEW).
Чувствительны к индукции аллергического энцефаломиелита после воздей-
ствия основного белка миелина морских свинок. Высокочувствительны к
индукции аутоиммунного миокардита и аутоиммунного гломерулонефрита
(коррелирует с главным комплексом гистосовместимости). Наблюдаются адъ-
ювантиндуцированные артриты. Уровень метаболизма этилморфина у самок
в печени высокий. Имеют высокий уровень тироксина, инсулина и ростово-
го гормона в сыворотке [207, 208]. Антигены Т-лимфоцитов: Ly-1+, Ly-2,1+,
Ly-2,2r [676]. Используются как инбредный партнер к ряду КР-линий. Ста-
новятся тучными на диетах с высоким содержанием жира. Высоко плодови-
ты. Послушны.
SHR (спонтанно-гипертензивные крысы)
Инбридинг F93. Генотип с. Окраска шерсти белая, альбиносы. Выведены
в Японии. Окамото в 1963 г. скрестил спонтанно-гипертензивного самца из
колонии крыс Wistar в Киото с самкой, обладающей повышенным кровя-
ным давлением. Селекция в процессе инбридинга велась на спонтанную
гипертензию. Сублинии также селектировали на развитие цереброваскуляр-
ных поражений и параличей. Несмотря на высокую частоту гипертензии, не
наблюдается явных поражений почек и надпочечников. Гипертензия очень
тяжелая, кровяное давление часто свыше 200 мм рт. ст. Высокая частота сер-
дечно-сосудистых заболеваний. Животные реагируют на антигипертензив-
ные лекарства и являются подходящей моделью для скрининга антигипер-
тензивного действия. У молодых SHR-крыс уровень норадреналина и допа-
Глава 9. Инбредные животные-биомодели
197
минбета-гидроксилазы увеличен по сравнению с другими линиями, но об-
щее количество катехоламинов одинаково. Содержание катехоламинов в над-
почечниках уменьшено. Повышенный уровень циркулирующего тиротропи-
на. Вес щитовидной железы ниже, чем у крыс Wistar.
WKY/N (Kyoto-Wistar)
Инбридинг F15 (находится в процессе выведения). Генотип с. Окраска
шерсти белая, альбиносы. Аутбредный сток крыс Wistar из Медицинской
школы Киото (Япония) передан в Национальный институт здравоохранения
(N) (г. Бетесда, США) в 1971 г. Линия селектировалась на нормальное дав-
ление крови как контрольная к SHR (в экспериментах с гипертензивными
крысами SHR в качестве контроля по норме могут служить крысы любой
инбредной линии с нормальным давлением крови). В отечественных иссле-
дованиях по физиологии высшей нервной деятельности были использованы
крысы Крушинского-Молодкиной. Эти крысы не соответствуют стандарт-
ным требованиям, предъявляемым к животным инбредных линий, так как
степень инбридинга точно не известна, поэтому для их обозначения больше
подходит термин «сток».
Сток КМ (Крысы Крушинского-Молодкиной)
Аутбредный сток был заложен на основе популяции крыс Л.В. Крушин-
ским и Л.Н. Молодкиной в 1947 г. На протяжении ряда поколений разводи-
ли, селектируя на высокую чувствительность к звуковому раздражителю (при
ограниченном инбридинге).
Инбредные линии хомячков
Если принять во внимание историю использования лабораторных животных
в биомоделировании, то можно сказать, что сирийские хомячки были одо-
машнены сравнительно недавно. Все лабораторные штаммы происходили от
трех особей из одного помета, пойманных неподалеку от Алеппо в Сирии в
1930 году, хотя в 1971 г. были пойманы дополнительные животные. Необыч-
ная реакция сирийских хомячков на кожные аллотрансплантаты привлекла
большое внимание. Во многих случаях пересадки кожи между членами зак-
рытой (но не инбредной) колонии особи жили в течение неопределенного
срока, хотя эти трансплантаты, сделанные между животными из разных ко-
лоний, могут отторгаться с такой же силой, как и других особей. Принятие
пересаженной кожи, естественно, не означает, что штамм хомячка является
полностью инбредным, как и в случае с большинством прочих особей.
На примере сирийских хомячков был тщательно изучен химический
онкогенез. Большинство работ выполнялось на неинбредных животных.
Считается, что большинство противоречащих результатов наблюдений, по-
198
Линейные животные-биомодели
лученных в ходе исследований химического онкогенеза у случайно разве-
денных хомячков, может быть объяснено генетическими различиями. Инб-
редные штаммы должны применяться гораздо шире.
Серьезной сравнительной работой на инбредных хомячках с применени-
ем трех из четырех штаммов продолжает оставаться работа Хомбургера и его
коллег. Если имеются данные по нескольким штаммам, они ранжируются, а
верхний и нижний квартили указываются хотя бы приблизительно.
Номенклатура инбредных штаммов хомячков стандартизирована лишь
частично. Некоторые штаммы получили кодовые названия, подобные тем,
что даны мышам. Совокупность инбредных штаммов, с которыми работали
«Телако» (Telaco) и «Био-рисёч консалтентс» (Bio-Research Consultants),
Кэмбридж и Массачусетс, имеет численную номенклатуру, основанную на
генеалогических номерах самца и самки, которые были использованы для
образования инбредного штамма, разделенных точкой. Пока не наступит
ясность с разработкой стандартной системы номенклатуры, исследователям
возможно использовать приведенные здесь названия штаммов.
ALAC
Инбридинг F74. Акромеланический альбиносный штамм, выведенный
в Центре животных Лаборатории Совета по медицинским исследованиям
(Lac), Великобритания, в 1966 году, из коммерческого аутбредного матери-
ала. Размножается удовлетворительно в составе моногамных пар, хотя не
очень активно. Величина помета 5—6 детенышей, продуктивность — около
0,5 детенышей на самку в неделю.
RL
Инбридинг F71. Цвет рыжеватый. Происхождение: д-р П.Лемон
(P. Lemond), Институт Армана-Фрапье (Armand-Frapier Inst., Монреаль),
из помета, купленного у «Био-рисёч консалтентс» в 1961 г. Характеристики:
средний жизненный цикл 687 ± 12 (максимум 961) дней в выборке из 202
самцов и 531 ±8 (максимум 811) дней в выборке из 202 самок. Спонтанные
опухоли: 6,9% у самцов и 2,5% у самок. Обычные причины смерти: интер-
стициальный нефрит, тромбоз левой ушной или легочной вен, при этом оба
порока часто сопровождают друг друга, что приводит к отеку и водянке.
Гиперволемия и серозная или геморрагическая киста печени, часто сопро-
вождаемая нефритом и тромбозом. Крупный помет, но плодовитость зави-
сит от сезона: от 29% в декабре—январе до 87% в апреле—октябре. Самки
агрессивны и иногда убивают самцов при спаривании.
ХЗ
Инбридинг F79. Акромеланический альбиносный штамм, выведенный
«Био-рисёч консалтентс» из материала, поставленного Лакассом (LaCasse).
Глава 9. Инбредные животные-биомодели
199
Средний жизненный цикл — 527 дней. Чувствителен к тератогенному дей-
ствию лекарств. Относительно маленькие сердце, легкие и печень, но
у животных в возрасте относительно большой вес печени у самок, малый вес
сердца у самцов и малый вес почек. Первый помет появляется в позднем
возрасте, малая величина помета.
Х.68
Инбридинг F71. Акромеланический альбиносный штамм, выведенный из
материала Швенткера (Schwentker) — Лакасса. Средний жизненный цикл со-
ставляет 531 день. Увеличение сердца приблизительно у 30% животных в
возрасте. Амилоидоз и цирроз печени, наблюдаемые у самок в возрасте 1—2
года. У животных в возрасте низкий относительный вес печени и низкий от-
носительный вес почек, большой относительный вес сердца. Первый помет
в позднем возрасте.
1.5
Инбридинг F68. Акромеланический альбиносный штамм, выведенный
из материала NIH. Длинный жизненный цикл — 590 дней. Предрасположен
к зубному кариесу. Относительно умеренная кардиомиопатия. Первоисточ-
ник гена дистрофии, который в настоящее время перешел и на прочие штам-
мы, особенно на 14.6. Повышенный уровень сывороточной креатинфосфат-
ной киназы перед наступлением клинических признаков дистрофии. Боль-
шое сердце и легкие, но маленькие яички в возрасте 180 дней. Малый раз-
мер помета. Малый относительный вес сердца у самцов в возрасте 200 дней.
Предрасположен к образованию опухолей в результате действия бензпирена
и триметилхолантрена у самцов и самок. Малый вес тела у взрослых особей
(самцы и самки), большой размер помета, малая частота сердцебиений у
взрослых самцов и самок.
1.26
Инбридинг F86. Акромеланический альбиносный штамм, выведенный из
материала Лакасса — Швенткера. Короткий жизненный цикл — 476 дней. Ма-
лое скопление ткани бурого жира в межлопаточной области в возрасте 56 дней,
малый размер щитовидной железы в возрасте 21 день, большой помет. У жи-
вотных в возрасте большой относительный вес сердца у самцов, большой от-
носительный вес почек у самцов. Малый вес тела у взрослых самцов и самок,
большой помет, малая частота сердцебиений у взрослых животных обоего пола.
2.4 (син: LSH)
Инбридинг F79. Штамм дикой окраски, выведенный Лондонской школой
гигиены и тропической медицины. Жизненный цикл средний — 532 дня. По
данным Хомбургера и Никсона (Nixon), у этого штамма развивается доброка-
200
Линейные животные-биомодели
чественная гипертрофия предстательной железы, но не смогли подтвердить
этот факт, что означает наличие либо различий между сублиниями, либо не-
установленный компонент окружающей среды. Цирроз и амилоидоз печени
наблюдаются у самок в возрасте 1—2 лет. Относительно стоек к аденовирусу
типа 12 по сравнению с аутбредным материалом. Большой относительный вес
печени (самцы и самки) и малый относительный вес почек у самцов.
4.22
Инбридинг F94. Штамм дикой окраски, выведенный из материала Швен-
ткера. Жизненный цикл короткий — 481 день. Увеличение сердца происхо-
дит приблизительно в 30% случаев у животных в возрасте 90—100 дней. Ами-
лоидоз и цирроз печени наблюдаются у самок в возрасте 1—1,5 лет. Относи-
тельно стоек к появлению опухоли молочных желез под воздействием ме-
тилхолантрена. Малый помет. Часты случаи возникновения опухолей пред-
желудка, тонкой кишки и матки, но редки случаи опухоли молочных желез
под воздействием 20-метилхолантрена. У животных в возрасте 150 дней боль-
шой относительный вес печени (самцы и самки), большой относительный
вес сердца (самцы) и большой относительный вес почек. Самцы подверже-
ны возникновению опухолей под воздействием бензпирена, а самки чув-
ствительны к триметилхолантрену. Малый вес тела особей у самок.
4.24
Инбридинг F83+. Штамм дикой окраски, выведенный из материала
Швенткера. Жизненный цикл длинный — 589 дней. Самки тучные и тяже-
лее самцов по достижении зрелости. Часты случаи возникновения доброка-
чественных аденом коры надпочечников, особенно у самок. Цирроз и ами-
лоидоз печени встречаются у самцов в возрасте 1—2 лет. Чувствительны
к тератогенному действию лекарств. Самки обладают стойкостью к возник-
новению опухолей под действием триметилхолантрена и бензпирена. Боль-
шой помет. Малый относительный вес сердца и малый относительный вес
почек у самок. Большой вес тела у взрослых особей (самцы и самки), малый
помет, частые сердцебиения у взрослых особей.
12.14
Инбридинг F70+. Штамм кремовой окраски, выведенный из материала
Уоррена (Warren). Жизненный цикл короткий — 473 дня. Прогрессивный
паралич задних лап встречается у самцов в возрасте 6—10 месяцев, при этом
страдают только мускулы задних лап; также может возникать дегенерация
спинной колонны спинного мозга. Наследуется рецессивно по полу. Прояв-
ление у самок не всегда предсказуемое, возможно, в связи с мозаицизмом.
Самцы предрасположены к опухолям под воздействием бензпирена и три-
метилхолантрена. Малый вес тела взрослых особей у самцов, первый помет
Глава 9. Инбредные животные-биомодели
201
в раннем возрасте, низкая частота сердцебиений (самки, самцы). Низкий
относительный вес печени у самцов и низкий относительный вес сердца
у самок.
14.6
Инбридинг F76. Акромеланический альбиносный штамм, выведенный
из материала Швенткера — Лакасса, с рецессивным геном миопатии, вве-
денным от штамма 1.5. Жизненный цикл короткий — 200 дней. Мышечная
слабость появляется в возрасте 60—200 дней. Она может проявляться
у отьемышей в том, что те занимают положение, как при плавании. У 43%
животных в возрасте до 30 дней обнаружены дистрофические поражения в
сократительных мышцах щек. Болезнь обычно прогрессирует. Гистологичес-
кие поражения включают варьируемый диаметр мышечных волокон, сухой
некроз и, на более поздних стадиях, жировую инфильтрацию и пролифера-
цию соединительной ткани. Изменения напоминают дистрофическую мио-
тонию и миопатию Дюшена у человека. Уровень сывороточной креатинфос-
фатной киназы заметно повышен. Большинство животных умирают от зас-
тойной сердечной недостаточности в возрасте приблизительно 150 дней. Кар-
диомиопатия сопровождается характерными поражениями языка. Для опре-
деления наследственной предрасположенности к застойной сердечной не-
достаточности может применяться ЭКГ. Кардиомиопатия связана с измене-
ниями в сывороточных электролитах и водным балансом, но уровни селена
в ткани нормальные. Большой вес тела взрослых особей у самок.
15.16
Инбридинг F72. Штамм кремово-дымчатой окраски, выведенный из ма-
териала Уоррена. У животных, попавших под воздействие триметилхолант-
рена, наблюдается карцинома железистого отдела желудка. Высокочувстви-
тельны к индуцированию фибросаркомной опухоли диметилбензантраценом.
Чувствительны к индуцированию рака молочных желез метилхолантреном,
но обладают низкой чувствительностью к большинству опухолей прочих
видов. Самки обладают стойкостью к индуцированию опухолей бензпире-
ном. Первый помет появляется в раннем возрасте.
45.5
Инбридинг F79. Штамм кремовой окраски, выведенный из материала
Ингхама — Тулана - Галфа (Ingham - Toolan - Gulf). Жизненный цикл длин-
ный — 562 дня. У обоих полов в возрасте 1—2 лет часто встречается цирроз
и амилоидоз печени. Чувствительны к тератогенному действию талидомида.
У 17% особей встречается опухоль надпочечника. Стойкость к индуцирова-
нию подкожных опухолей бензпиреном (самцы, самки), а самцы обладают
стойкостью к триметилхолантрену. Маленький помет.
202
Линейные животные-биомодели
54.7
Инбридинг F74. Штамм янтарной окраски, выведенный из материала
Уоррена. Жизненный цикл средний — 509 дней. Гетерохромия радужки.
У животных обоих полов в возрасте 1—2 лет наблюдаются амилоидоз и цир-
роз печени. При воздействии 20-метилхолантрена часто встречаются опухо-
ли преджелудка, желудка, матки и молочной железы. Самки чувствительны
к индуцированию опухолей бензопиреном. Малый вес печени у самцов
и высокий относительный вес почек у самок. Низкий вес тела взрослых осо-
бей у самцов, большой помет.
Любая генетически разнородная популяция может быть расчленена на
ряд инбредных линий или, что чаще встречается в зарубежной литературе,
штаммов. «Чистые линии», как в 1903 г. их определил Иоганнсен, стали
использоваться сначала в ботанике и агрономии на основе самоопыляю-
щихся растений. В растительном и животном мире присутствуют генетичес-
ки различные особи, которые и переходят в гомозиготное состояние в виде
разных аллелей и генов.
Столетняя история исследований в этом направлении показала, что мать
Природа ставит существенные ограничения возможностям инбридинга, при-
чем тем более жесткие, чем более высокоорганизованны животные. Макси-
мальное разнообразие инбредных конгенно-резистентных, мутантных, но-
каутных и иных линий получено на мышах. Им и будут посвящены две сле-
дующие главы
203
Глава 10 КОИЗОГЕННЫЕ И ИНБРЕДНЫЕ
ЛИНИИ МЫШЕЙ
Коызогенными называются генетически идентичные
линии, т. е. линии, имеющие различия по одному
локусу. Истинная коизогенность возможна только
в случае единичной мутации в инбредной линии.
Такие линии, полученные специальными генетичес-
кими методами, названы конгенными. Изогенное со-
стояние достигается введением гена одной линии
на генетическую основу другой линии. Конгенные
линии, которые отличаются друг от друга по локусу
тканевой совместимости и, следовательно, взаимно
резистентны к трансплантату ткани друг друга, на-
зываются конгенно-резистентными линиями (КР),
пара таких линий — КР-парой.
Число КР-линий мышей составляет несколько
сотен и они непрерывно увеличиваются в результа-
те получения рекомбинантных гаплотипов по Н-2,
ценность которых особенно велика в изучении ге-
нетической организации сложной системы Н-2.
Вторым источником получения новых КР-линий
являются мутантные гаплотипы Н-2, индуцирован-
ные или спонтанные [180, 471]. Известны десятки
аллелей локуса Н-2К и аллеля H-2D, входящих в
комплекс Н-2.
Конгенно-резистентные линии
Конгенные линии обозначаются соединением сим-
волов двух родительских линий, разделенных точ-
кой. Первая часть содержит полный или сокращен-
204
Линейные животные-биомодели
ный символ основной линии. Вторая часть может содержать полный или
сокращенный символ донорской линии. Например, КР-линия B10.D2 про-
исходит от основной линии C57BL/10-B10 и донорской DBA/2-D2. Иногда
вторая часть содержит символ дифференциального локуса и его аллель В6.С-
Н-2Ьа или Вб-Ly-l.l. Иногда в обозначении употребляются символы субли-
ний, если есть основания полагать, что сублинии отличаются: B10.DI/Ph и
B10.DI/Y, например, выведены почти одновременно в разных лабораториях.
Области использования КР-линий мышей охватывают онкологию, транс-
плантологию, иммуногенетику, онкогенную вирусологию, фармакологию и
токсикологию.
А. СА (М506) (сокращенно М506) Поддерживается Y
Инбридинг F34. Генотип Н-2?т1. Получена в лаборатории генетики (Y) от
мыши F1 (А.СА х A/Sn) со спонтанной мутацией в Н-2^[ 180]. Мутация в Н-
2К типа замены антигена GL. Трансплантаты кожи от донора с нормальным
аллелем (Н-20 отторгаются на мутантном на 21-й день, в обратном направ-
лении — значительно медленнее (на 35—65-й день), иногда не отторгаются.
Имеются четкие серологические отличия от мышей А.СА и различия в реак-
циях клеточного иммунитета: смешанная культура лимфоцитов (mixed
lymphocyte culture, MLC) — СКЛ, цитотоксический эффект иммунных лим-
фоцитов (cell mediated lympholysis, CML) — ЦЭЛ, реакция трансплантата
против хозяина (graft versus host reaction, GVHR) — РТПХ [372].
B6.C(Hz-l) Поддерживается Y
Инбридинг F45. Генотип H-2bml. Получена Bailey от мыши Fl(C57BL/6 x
BALB/c), у которой была обнаружена спонтанная мутация в Н-2Ь путем
возвратных скрещиваний с C57BL/6, поступила в Y на NI0F18. Мутация
типа GL, локализована в Н-2К [42]. Реципрокные трансплантаты кожи меж-
ду мышами с исходным гаплотипом и мутантным отторгаются на 12—14-й
день. Различия выявляются в реакциях клеточного иммунитета: СКЛ, ЦЭЛ,
РПТХ. Серологические отличия проявляются в том, что Н-2.5 отсутствует на
эритроцитах и имеется на лимфоцитах у мутантов, кроме того, у них мень-
шее количество антигена Н-2К.ЗЗ [179, 180, 372].
B6.M505/Y Поддерживается Y и Sto
Инбридинг F34. Генотип Н-2Ьм3. Происхождение от мышей C57BL/6Sto со
спонтанной мутацией в Н-2, обнаруженной при проверке на гомозиготность
племенного ядра В6 в филиале «Столбовая» [179, 189]. Мутация типа GL в
локусе Н-2К. Трансплантаты нормального аллеля отторгаются на мутантном
животном через 19,0 ± 1,25 дня, в обратном направлении — через 13,3 ± 0,04
дня. Мутантные аллели H-2Kbml и Н~2КЬт3 не идентичны, так как при рецип-
рокном обмене трансплантаты отторгаются в среднем на 14-15-й день [28].
Глава 10. Коизогенные и инбредные линии мышей
205
Различия между нормальным аллелем и Н-2КЬт3 выявляются в реакциях СКЛ,
ЦЭЛ, РПТХ [372, 483].
B6.C(Hz-170) Поддерживается Y
Инбридинг F? + 8N2F33. Генотип Н-2Ьт4. Происхождение от Bailey на F?
в Институте общей генетики РАН, в Y поступил один самец Hz-170, для
сохранения мутации был скрещен с В6, в F2 отобраны гомозиготы Ьт4/Ьт4,
размножаются инбредно. Спонтанная мутация в Н-2КЬ типа GL. Трансплан-
таты кожи В6 отторгаются на Fj (H-2bmlx H-2bm4). Серологически исходный
и мутантный аллели не различаются.
B10.D2(M504)-M504 Поддерживается Y
Инбридинг F36. Генотип H-2dml. Происхождение: получена от мыши
B10.D2, у которой обнаружена мутация в H-2d, индуцированная диэтил-
сульфатом.
Мутация типа GL в локусе H-2D [591]. Реципрокные трансплантаты кожи
мышей с мутантным и нормальным аллелем отторгаются до 20-го дня. Му-
тация вызвала серологически определимые изменения антигенов: аллоанти-
генные специфичности Н-240,49 гаплотипа H-2d утрачены, появилась спе-
цифичность Н-2.50 [163]. Кроме того, она вызывает стимуляцию в СКЛ,
ЦЭЛ, РПТХ.
CBA.M523/Y Поддерживается Y
Инбридинг F32. Генотип H-2RM1. Происхождение от мыши CBA/LacSto
(H-2R) со спонтанной мутацией в Н-2, обнаруженной при проверке на гомо-
зиготность племенного ядра СВА в филиале «Столбовая» [67, 71]. Мутация
типа GL в области К, кожа СВА отторгается на М523 быстрее (в среднем 23—
26 дней), чем в обратном направлении (37-44 дня), у самок быстрее (первая
цифра), чем у самцов. Серологических отличий не обнаружено [67, 71], но
проявляются различия с нормальным аллелем в СКЛ, ЦЭЛ, РПТХ [372].
Кроме того, наблюдается дефектность функционирования иммунокомпетент-
ных клеток Fl (M523 х СВА) в семисингенном реципиенте, предварительно
облученном [212].
Инбредные и трансгенные линии
Инбредные линии мышей обозначают, например, так: A2G/Lac Y, где A2G
— название линии, Lac и Y — обозначение лаборатории или института, где
линия поддерживается. Ниже приводится описание инбредных линий мы-
шей, разводимых и поддерживаемых в нашем Центре в лаборатории генети-
ки (зав. к.б.н. A.M. Малашенко). Линии сублинии и сублинии обозначены
в соответствии с международными правилами. Даны полные сублинейные
206
Линейные животные-биомодели
символы, отражающие хронологическую последовательность передачи ли-
нии. Если линия выведена в НЦБМТ РАМН, она имеет один сублинейный
символ — Y. Линии, поддерживаемые в центральных лабораториях нашего
Центра, также отмечены «Поддерживаются Y»; линии, находящиеся в фили-
алах, отмечены соответствующими символами: Sto — филиал «Столбовая»
And — филиал «Андреевка», Вт — филиал «Белый мох» НЦБМТ РАМН. После
названия приведена окраска шерсти мышей данной линии, далее материал
расположен в том же порядке что и в «Стандартной номенклатуре инбред-
ных линий мышей» [604]. После инбридинга F (суммарное число поколений
братско-сестринского инбридинга за известную историю линии) указывает-
ся генотип и окраска шерсти, глаз, гаплотип Н-2, гены других полиморфных
локусов. Далее дается происхождение линии, ее передач и время поступле-
ния в Центр, затем приводится краткая характеристика.
A/HeJ Поддерживается Sto
Инбридинг F170. Генотип а,Ь,с,Н-2?. Окраска шерсти белая, альбиносы.
Получена Стронгом в 1921 г. от скрещивания белых мышей из колонии
Cold Spring albino и Bagg albino. В питомник «Столбовая» (Sto) поступила
из США в 1961 г. на F113. У 57,9% размножающихся самок в питомнике
«Столбовая» возникают опухоли молочных желез. Средний возраст возник-
новения опухолей — 413,9 дня, продолжительность жизни с опухолями —
463 дня; 12,3% самок имели первичные опухоли легких в возрасте 602 дней;
у 12,3% самок лейкозы в возрасте 555,5 дня. Первичные опухоли легких
встречаются у половины самцов. На них легко перевивается штамм гепато-
мы А [321]. Высокая частота амилоидоза, нефритов у старых животных [604]
и расщепление нёба у молодых [604]; у 20% самок и 13% самцов — откры-
тые веки. Имеют малый вес тимуса [253, 254], вес надпочечников мини-
мальный в сравнении с СВА/Са. Общая радиочувствительность самцов
LD50.30 = 518 рад. [166]. Ткань легких мышей этой линии очень чувстви-
тельна к метилхолантрену [604]. Чувствительны к действию аминазина: 62%
животных неактивных в тесте открытого поля после введения 4 мг/кг. Двух-
месячные самцы чувствительны к заражению метацеркариями Opisthorchis
felineus [689].
Устойчивы к Naturallia musculi Muratov. Наиболее чувствительны к зара-
жению вирусом мышиной оспы (экстромелии) [195], гриппом типа А, ин-
тенсивно вырабатывают интерферон после введения вируса псевдочумы кур,
слабо реагируют на эритроциты барана [234]. Вес тела самок и самцов 27 г
в возрасте 168 дней. Продолжительность жизни самок в условиях SPF — 558
дней, самцов — 512 дней [207], в конвенциональных условиях продолжи-
тельность жизни самок 464 + 9,87 дня. Плодовитость: 97,3% фертильных
самок, 4,43 помета на фертильную самку (п = 259), 4,19 мышонка в помете
при отсадке.
Глава 10. Коизогенные и инбредные линии мышей
207
A/WySnKI Поддерживается Y и Sto
Инбридинг F85. Генотип а,Ь,с, Н-2°. Окраска шерсти белая, альбиносы.
Выведена Стронгом в 1921 г. В публикациях известна как сублиния A/Sn.
Подвержены раку молочных желез, частота и возраст проявления не просле-
жены. Имеют 25 пресакральных позвонков [604]. Вес семенных пузырьков
снижен, что коррелирует с гаплотипом Н-2а [246]. Яичник содержит на-
ибольшее число ооцитов (до 15 000). Низкое число Пейеровых бляшек. Низ-
кий относительный вес гипофиза — 1,1 мг сухого веса на 100 г веса тела.
Содержат большое количество жира в надпочечниках (у самок больше, чем
у самцов). Повышена чувствительность к действию озона (летальная доза
34—36 ppm/ч), которая связана с дефицитом тирозиназы и пероксидазы
в легких и коже [235]. Минимальные концентрации хлороформа в воздухе
вызывают гибель от поражения почек, особенно у самцов.
Введение эстрогенов вызывает лейкемию через 180 дней, у самцов возни-
кают интерстициональные опухоли семенника. Надпочечники у гонадэкто-
мированных самок (30 дней после операции) не уменьшаются, обработка
эстрадиолбензоатом не приводит к их увеличению. Чувствительна к цитоге-
нетическому эффекту тиоТЭФ (доза 5 мг/кг, в/б): 42,8% клеток с хромосом-
ными нарушениями при спонтанном уровне 3,4%. Избегают алкоголя в любых
концентрациях. Высокочувствительна к капсульному и бескапсульному штам-
мам сибирской язвы [2]. Является инбредным партнером нескольких кон-
генно-резистентных (КР) линий. Потенциальная плодовитость высокая —
7,6 ± 0,3 живых эмбриона на самку (из 11 исследованных линий), эмбрио-
нальные потери относительно низкие — 21,2%.
AKR/J Поддерживается Y
Инбридинг F137. Генотип а, с, //-2е. Окраска шерсти белая, альбиносы.
Высоколейкозная линия Фэрса с 1928 г. Поступила в филиал НЦБМТ РАМН
питомник «Столбовая» в 1961 г. из Jax на F60, затем в Yb 1965 г. на F80. Мыши
обоих полов подвержены лимфоидной лейкемии. Около 91% самок сублинии
Y погибают от лейкозов в среднем к 300-му дню, в других сублиниях просле-
жена частота лейкозов у самцов (70—80%); чувствительны к агенту лейкемии
[12, 604]. Вирус мышиного лейкоза в высоком титре обнаруживается в бере-
менной матке, яичниках, в плаценте его в 10—30 раз меньше; в тканях эмбри-
онов не определяется до 18—20-го дня, у сосунков содержится во всех тканях,
в наиболее высоком титре в костной ткани, матке и селезенке. В возрасте 5—
10 месяцев содержание вируса в тканях практически одинаково, максималь-
ный титр в сыворотке, однако у больных лейкозом вирус содержится в более
высоком титре, чем в тканях здоровых сингенных мышей [555]. После рожде-
ния в высоком титре продуцируется эндогенный лейкозный вирус типа Гросса
с последующим развитием лейкоза тимусного происхождения на 7—9-м меся-
це жизни. Развитие лейкоза сопровождается увеличением числа копий в ДНК
208
Линейные животные-биомодели
только опухолевых клеток. Из мышей линии AKR выделено четыре группы
эндогенных мышиных лейковирусов, различающихся по биологическим свой-
ствам: 1) экотропный хс+ MuLV, содержащийся в больших титрах в сыворотке
и нормальных тканях мышей в течение всей жизни; 2) экотропный xc~MuLV,
который активно продуцируется лейкозными клетками; 3) ксенотропный ви-
рус, обнаруженный только у взрослых мышей; 4) политропный MuLV, выде-
ленный из тимусов взрослых мышей.
Показано, что xc~MuLV и политропный MuLV обладают высокой онко-
генной активностью [499]. Встречаются протеинурия и гломерулонефрит
у 3-месячных мышей (18% случаев), у 9-месячных (68%) и у 12-месячных
(83%) [504]. При инфекции вирусом Раушера (RLV) у мышей развивается
эритролейкоз [640, 641]. Устойчивы к тератогенному эффекту 1-этил-1-нит-
розомочевины [164, 165] и 7,12-диметилбензантрацена [408]. Относительно
устойчивы к канцерогенному действию 3-метилхолантрена [383].
Повышенная чувствительность к озону (летальная доза 34—36 ррт/ч)
вследствие дефицита тирозиназы и пероксидазы в легких и коже [235]. Крайне
чувствительны к ипрониазиду, гибель наступает от поражения печени. Отно-
сительно резистентны к токсическому воздействию препаратов декстрана
(самцы больше самок) [129]. Высокочувствительны к повреждающему дей-
ствию кадмия, который вызывает у них некрозы семенников [630, 631]. Об-
работка тестостерон-пропионатом приводит к многократному (в 95—125 раз)
увеличению концентрации р-глюкоронидазы в почках. Недостаточность
комплемента С5 [604], с чем связывают высокий процент спонтанных лей-
козов: сыворотка мышей, содержащих С5, оказывала противолейкозное дей-
ствие на AKR [356].
Резистентны к заражению салмонеллами [174], к протозойной инфекции
Besnitia jallisoni, к вирусу вакцины [427], полностью резистентны к вирусу
саркомы Молони [640], относительно резистентны к вирусу диареи инфан-
тильных мышей [592]; резистентны к инфекции Leishmania [604]; чувстви-
тельны к вирусу лимфоцитарного хориоменингита [374]; наиболее чувстви-
тельны к внутрибрюшинному введению коклюшной вакцины [294].
AKR/J вырабатывают большое число антителообразующих клеток на ди-
нитрофенол-гемоцианин [82,83]. В антителах AKR/J, как и у мышей DBA/2,
CBA/Lac преобладает лизин [662]. Не реагируют на бычий у-глобулин [501];
не вырабатывают антител к эритроцитам хомяка [498]. Низка концентрация
жира в надпочечниках [604]. Резко выражен половой диморфизм по относи-
тельному весу гипофиза: у самок — 2,5, у самцов — 1,7 мг (сухого веса на 100 г
веса тела); у самцов самый высокий относительный вес подчелюстной железы
в этих линиях. Фактор тимической активности сыворотки крови содержится
в количестве 1:64. Содержание общего кортикостерона в крови составляет 13,3
±1,16 мкг%. Высока концентрация кальция в сыворотке крови, которая близ-
ка к отмеченной у линии СЗН/Не в 4-месячном возрасте [51].
Глава 10. Коизогенные и инбредные линии мышей
209
Вес тела половозрелых самок — 21,05 г, самцов — 23,27 г [528]. Плодови-
тость низкая при конвенциональных и SPF условиях. Она возрастает при
содержании на диетах, включающих в свой состав от 19 до 23% белка и
около 6% жира [287]. В сублинии Y — 92,0% фертильных самок (п = 264), 2,9
помета на фертильную самку, средняя величина помета при отсадке — 5,2
мышонка. Средняя продолжительность жизни самок составляет 279—298
дней, самцов — 326 [604]. В условиях SPF продолжительность жизни не
меняется [206, 207]. Низкий уровень эмоциональности: 34% по дефекации в
«открытом поле».
BALB/cJLac Поддерживаются Y и Sto
Инбридинг F131. Генотип Ь, с, Н-21. Окраска шерсти белая, альбиносы.
Линия происходит от белых мышей, приобретенных Баггом в 1913 г., инбри-
динг начал Макдауэлл в 1923 г. В Sto на F70 в 1958 г., в Y на F105 в 1971 г.
Частота опухолей молочных желез низкая, но высокая при введении факто-
ра молока [604]. В сублинии Sto и Y у размножавшихся самок частота опу-
холей молочных желез — 1,8% в возрасте 473 дней, у 12,2% самок опухоли
легких в возрасте 582 дней, в других сублиниях у обоих полов опухоли лег-
ких наблюдаются с частотой 20—30%. У 5% самок в возрасте 572 дней наблю-
даются злокачественные ретикулярные новообразования. У старых живот-
ных встречаются ангиомы, опухоли яичников и надпочечников, лимфосар-
комы; почти у всех самцов к 20 месяцам развивается амилоидоз селезенки
[604]. Введение вируса Раушера увеличивает селезенку в 30—40 раз, 100%-ная
гибель к 40-му дню после инфицирования при низком содержании ин-
терферона и антител в сыворотке крови [638], 100%-ная передача лейкеми-
ческого вируса потомству [314]. Кроме того, у инфицированных мышей
в возрасте 1—2 месяца развивается гломерулонефрит [504]. Характерны очень
большие ретикулоэндотелиальные органы по сравнению с весом тела [604].
В ряду BALB имеют высокий относительный вес подчелюстной железы,
у самок самый высокий относительный вес надпочечников. Большая актив-
ность фермента фенилэтаноламин-1Ч-метилтрансферазы в сравнении с мы-
шами CBA/J и DBA/2J, причем не обнаружена связь между активностью
и весом надпочечников. Высокая активность щелочной фосфатазы
в межальвеолярных перегородках [250]. Фактор химической активности в сы-
воротке крови равен 1:32.
Высокочувствительны к радиации: LD50.30 самцов < 570 рад, LD50.30 са-
мок = (585 ± 12) рад. После рентгеновского облучения самок на 10-й день
беременности (доза 150 рад) у 5—8% эмбрионов наблюдается полидактилия
задних конечностей, что значительно ниже, чем у C57BL/6. Чувствительны
к действию озона вследствие дефицита тирозиназы и пероксидазы в легких
и коже. Летальная доза составляет 34—36 ppm/ч. Обладают слабой чувстви-
тельностью к тератогенному действию временной гипоксии на 9-м дне
210
Линейные животные-биомодели
беременности, уродства позвоночника возникают у 17% эмбрионов. Уретан
индуцирует опухоли у обоих полов, семенники резистентны к повреждающе-
му действию кадмия [630, 631]. Минеральное масло (bayol—553 и primol-
355) вызывает образование плазмоклеточных опухолей, при этом ослабляет-
ся интенсивность иммунных реакций [389].
Чувствительны к канцерогенному действию метилхолантрена [427],
к тератогенному действию 7,12-диметилбензантрацена [408]. Самцы чувстви-
тельны к токсическому действию тиоТЭФ, у них обнаружен резко вы-
раженный пороговый эффект при LD50.30 = 23,75 мг/кг: смертность от 0 до
100% колеблется в диапазоне доз от 22,5 до 25 мг/кг. Инсулин вызывает
слабый тератогенный эффект. Относительно резистентны к тератогенному
действию на задние конечности эмбрионов 5-флуороурацила.
Восприимчивы к S. typhimurium [174, 526, 527], к вирусам Раушера, сар-
комы Молони [230, 465], Фрейнда [137], коксаки [680], желтой лихорадки
[152], оспенной вакцины [427], гриппа, диареи инфантильных мышей [592].
Чувствительны к Leishmania [604]. Резистентны к вирусу ЛХМ [374], к виру-
су Граффи [137], стафилококковой инфекции. Наиболее резистентны из вось-
ми тестированных линий к Eimeria [604]. Имеют среднюю чувствительность
к вирусу кори [604], к заражению В. antracis [2]. Дают высокий иммунный
ответ на эритроциты барана [82, 83, 234], салмонелезный и пневмококковый
полисахариды [143], на стрептококки группы А [93], на липополисахариды
[557]. Характеризуются низкой выработкой интерферона на вирус опухолей
молочных желез, вирус болезни Ньюкасла, вирус лейкоза Раушера. Облада-
ют высокой активностью комплемента [604]. В костном мозге дифференци-
ровано 17 типов клеток. Количество клеток зернистого ряда возрастает на
4—5% каждый месяц с одного до четырех [307]. Наблюдаются поражения
сердца у самок и артериосклероз у старых животных обоих полов в боль-
шинстве сублиний этой линии. Поражение сердца выражается в тромбозе
левого предсердия, что приводит к 3—5-кратному его увеличению. Эта пато-
логия наблюдается у 60—66% многократно рожавших самок в возрасте от 270
до 800 дней. Она зависит от повторных беременностей, так как почти не
встречается у старых виргинных самок и самцов (5%). Патогенез и этиоло-
гия заболевания неизвестны. Нарушений клапанного аппарата сердца нет.
Однако, несмотря на частые тромбозы, мыши BALB/c страдают от серь-
езного дефицита необходимых для коагуляции крови факторов, которые
содержатся в следующих количествах: 1) фактор IX (компонент плазменного
тромбопластина, фактор Christmas) — около 40% от нормы, 2) антигемофи-
лический фактор (фактор ХШ) — около 60% от нормы, 3) Stuart-Prower фак-
тор (фактор X) — около 50% от нормы, 4) протромбин — около 33% от
нормы. При этом содержание других компонентов, таких, как Hageman-
фактор и фактор V (лабильный фактор, проакцелерин, акселератор-глобу-
лин), всегда в норме. Указанные аномалии протромбинового комплекса об-
Глава 10. Коизогенные и инбредные линии мышей
211
наруживаются у беременных самок незадолго перед родами и исчезают через
несколько дней после них. Отдача плазменного протромбина может состав-
лять на 20—25% больше нормы в течение 2 дней после родов. Фаза отдачи
является умеренной или же совсем не выражена в первой стадии протром-
бинового времени. Фибрионолитическая активность в крови тромботичес-
ких мышей настолько низка, что ее невозможно измерить. Явление про-
тромбиновой отдачи у мышей BALB/c чрезвычайно сходно с состоянием
инфарктного больного, подвергающегося лечению дикумарином и гепари-
ном. Сравнительное изучение состояния антисвертывающейся системы
(АСС) у BALB/c и C57BL/6 показало, что уровень антигемофилического
глобулина и скорость свертывания контрольной плазмы у них не отличают-
ся; по реакции на введение тромбина нет основания полагать, что причиной
тромбоза левого предсердия является угнетенное состояние АСС.
Вес половозрелых самок в возрасте 2 месяцев составляет 21,2 ± 0,10 г, сам-
цов — 24,3 ±0,12 г. Мыши этой линии опережают других в скорости морфо-
логического созревания, по времени появления первой пары сосков и откры-
тия влагалища, которое происходит на 37-й день. Им свойственны длитель-
ный период размножения, в среднем 4,96 помета на самку; 98,4% фертильных
самок, 5,4 мышонка при отсадке.
Самая высокая потенциальная плодовитость: 7,8 ± 0,4 живых эмбриона на
самку — при самом высоком уровне эмбриональных потерь 49,4%. Средняя
продолжительность жизни самок сублинии StoY — 417 ± 9,78 дня, самок суб-
линии J — 575 дней, самцов — 539 дней, существенно не отличается от про-
должительности жизни в условиях SPF Lac [207]. Сравнительно не агрессив-
ны. Высокоэмоциональны: уровень эмоциональности — 82% по дефекации в
«открытом поле». Избегают алкоголя в любых концентрациях. Сильно выра-
жен инстинкт запасания пищи.
BRSUNT/N Поддерживаются Y
Инбридинг F151. Генотип а, Ь, Н-21 (определен в Y). Окраска шерсти
коричневая. Выведена Стронгом как ветвь BRS, размножаемая без обработ-
ки метилхолантреном получена в Национальном институте здравоохране-
ния США. В Y поступила из Ог1 в 1976 г. на F134. Склонны к ожирению,
страдают от желудочных поражений. Заболевание зубов и околозубного про-
странства (Periodontal disease) у 100% животных [604].
СВА/Са Lac Поддерживаются Sto и Y
Инбридинг F97. Генотип +, Н-21. Окраска шерсти — агути. Одна из суб-
линий линии СВА, выведенной Стронгом в 1920 г. от скрещивания мышей
Bagg albino с DBA. В 1 Sto из Lac на F33 в 1958 г., в Y на F53 в 1965 г. Частота
опухолей молочных желез колеблется от 1 до 65% [604]. В сублинии Y 1,7%
размножающихся самок имеют опухоли молочных желез, 14,6% самок и 26,4%
212
Линейные животные-биомодели
самцов — опухоли печени, опухоли легких у 5,1% самок и 6,6% самцов;
указанная патология встречается в возрасте старше 700 дней. В сублинии Sto
наблюдалось 7% спонтанных опухолей яичников у девственных самок в
возрасте старше 20 месяцев, у размножавшихся и л актировавших — 58% опу-
холей, период развития опухолей сокращался у них до 15 месяцев; развитию
опухолей предшествовал период 5-7 месяцев, когда самки не беременели
(после 1-6 пометов) [306]. В селезенке на ранних стадиях канцерогенеза,
индуцируемого вирусом S47 (С8), накапливаются клеточные антитела к эм-
бриональным фибробластам, в то время как у C57BL/6, резистентных к
онкогенезу, подобных антител не обнаружено [10].
Имеют небольшие скелетные изменения. Третий моляр — самый мел-
кий, у 18% мышей отсутствует полностью. Нижние моляры видоизменены
сильнее верхних [604]. Чувствительны к хроническому облучению всего тела
[604]. Имеют самый низкий уровень спонтанных хромосомных нарушений.
Чувствительны к минимальным дозам хлороформа: гибнут от поражения
почек. Наиболее резистентны к токсическому действию тиоТЭФ среди сам-
цов четырех испытанных линий: LD50.30 = 31,73 мг/кг.
Мыши этой линии устойчивы к салмонеллезной инфекции [174, 526, 527],
к Leishmania [604]. Они восприимчивы к заражению метациркариями
Opisthorchis felineus [689]. Высокочувствительны к вирусу гриппа типа А,
восприимчивы к вирусу лимфоцитарного хориоменингита [374], к Согупе-
bacterium kitscheri [285, 286]. Резистентны к заражению эндогенным вирусом
мышиного лейкоза [49]. Устойчивы к бескапсульному и восприимчивы к
капсульному штаммам сибирской язвы В. antracis [2]. Дают высокий иммун-
ный ответ на общий антиген энтеробактерий [241]. Имеют самый высокий
титр антител против эритроцитов барана [522]. Относительный вес тимуса,
селезенки, лимфоузлов (мезентериального) — самый низкий из шести ис-
пытанных линий. Имеют наибольший вес надпочечников в сравнении с А/
Не. Уровень кортикостерона в плазме крови выше, чем у В6 [577]. Фактор
тимической активности содержится в том же количестве, что у AKR, B6
(1:64), но выше, чем у BALB/c. В сублинии N обнаружен Х-сцепленный
иммунный дефект: очень низкий уровень сывороточного IgM и неспособ-
ность к ответу на некоторые антигены; кроме того, с возрастом не образуют-
ся антинуклеарные антитела.
Вес тела половозрелых самок в возрасте 2 месяцев составляет 19,02 ±
0,07 г, самцов — 23,0 ± 0,10 г; фертильных самок — 98,9%, средняя величина
помета при отсадке — 6,0 ± 0,1. Продолжительность жизни самок составляет
705 ± 20,0 дней, самцов — 686 ± 17,6 дня, максимальная продолжительность
жизни тех и других — 1031-1071 день. В условиях SPF продолжительность
жизни самок 825 дней, самцов — 486 дней [206, 207]. Мыши CBA/CaLacStoY
гистосовместимы с СВА/СаН [70].
Глава 10. Коизогенные и инбредные линии мышей
213
CBA/J Поддерживаются Y
Инбридинг F183. Генотип +, rd, H-2?. Окраска шерсти диких мышей, агу-
ти. Выведена Стронгом в 1920 г. от скрещивания Bagg albino X DBA.
В And из Jax в 1970 г. на F149, в Y 1972 г. на F154. Частота спонтанных опу-
холей молочных желез у виргинных самок составляет 75%; у 64% самцов ге-
патомы. Относительный вес гипофиза сравнительно низок: у самцов 1,3 и у
самок 1,5 миллиграмм сухого веса на 100 г веса тела. Подчелюстная железа
самцов имеет самый низкий относительный вес среди испытанных линий.
По устойчивости к радиации занимают среднее положение: для самцов
LD50.30 = (656 ± 9) рад, для самок LD50.30 = (689 ± 8) рад, однако чувствитель-
ны к облучению всего тела [604]. Семенники самцов высокочувствительны к
повреждающему действию кадмия [630, 631], восприимчивы клистериям [52].
По числу антителообразующих клеток в селезенке дают высокий ответ на
эритроциты барана [108] и низкий на пневмококковый полисахарид Sill. При
активном поведении имеют минимальную продукцию тиреоидного гормона.
В возрасте 56 дней вес тела самцов 24,32 г, самок — 23,52 г [528]. Яичники
содержат (postpartum) малое количество ооцитов (10 000). Плодовитость в Jax:
90,6% фертильных самок, 3,9 помета на фертильную самку, 5,7 мышонка в
помете [244]. Продолжительность жизни виргинных самок — 527 ± 15,4 дня,
самцов — 527 ± 17,0 дня. При одинаковом гаплотипе Н-2 негистосовместима
с СВА/Са [71, 72].
CC57BR/Mv Поддерживаются Y
Инбридинг F109. Генотип а, Ь, Я-26. Окраска шерсти коричневая. Выведе-
на Н.Н. Медведевым (Mv) в 1943 г. от скрещивания BALB/cN x C57BL/N. В
Y получена от Mv в 1976 г. на F93. При введении фактора молока спонтанные
опухоли молочных желез развиваются у 60% животных [468]. Частота альвео-
логенных опухолей легких составляет 26,9%, спонтанные опухоли других ло-
кализаций не превышают 1—1,5%; вирус полиомы отсутствует [469]. Переви-
вается лейкоз Мазуренко. Из спонтанной лимфосаркомы выделен вирус С-
типа [439]. Устойчивы к действию радиации в сравнении с тремя другими
линиями: LD50.30 = 559 рад для самок и 538 рад для самцов. Высокочувстви-
тельны к S. typhimurium [174]. Чувствительны к вирусу диареи инфантильных
мышей [592]. Наиболее чувствительны из восьми линий к Nuttallia musculi
Muratoy. Высокорезистентны к капсульному штамму сибирской язвы В. antracis
[2]. Интенсивно вырабатывают интерферон после введения вируса псевдочу-
мы кур. Самый низкий уровень интерферона после введения вируса болезни
Ньюкасла [585]. Активность водорастворимой и липидосвязанной каталазы
печени равна соответственно 213 и 99 ед. Содержание общего кортикостерона
в периферической крови (12,7 ± 1,56) мкг%. Плодовитость — 6,3 мышонка в
помете, отход за подсосный период — 5%, главным образом в первых пометах,
продолжительность жизни — 17,48 месяца.
214
Линейные животные-биомодели
CC57W/Mv Поддерживаются Y
Инбридинг F 101. Генотип а, Ь, с, Н-2Ь. Окраска шерсти белая, альбино-
сы. Выведена Н.Н. Медведевым в 1943 г. от скрещивания BALB/cN x C57BL/
N. В Y получены от Mv в 1974 г. на F81. После введения фактора молока
опухоли возникают у 60% животных [468]. Частота опухоли легких составля-
ет 28,1%, опухоли других локализаций — 1-1,5%, свободны от вируса поли-
омы [469]. Чувствительны к заражению S. typhimurium [174]. Мало воспри-
имчивы к заражению метациркариями Opisthorchis felineus [689], восприим-
чивы к нуталиозу Наиболее чувствительны к заражению Encephalitozoon
cuniculi [323]. Чувствительны к заражению чумным микробом. Особеннос-
тью является высокое фагоцитарное число 5,01 ± 1,1 в сравнении с другими
пятью линиями [478]. Плодовитость — 7,0 мышат в помете, общий отход за
подсосный период — 5%. Продолжительность жизни — 15,62 ± 0,4 месяца
[468]. Рекомендуются как модель для экспериментального изучения инвазии
Е. cuniculi.
СЗН/HeDiSn Поддерживаются Y
Инбридинг F91. Генотип +, Н-Я. Окраска шерсти — агути. Происхожде-
ние такое же, как у СЗН/Не, в Y на F70 в 1965 г. Известна как сублиния
СЗН/Sn. Как все сублинии СЗН, сублиния Sn высокораковая. Спонтанные
опухоли легких возникают у 5—10% [384]. Инъекция уретана самцам вызы-
вает аденомы легких и повышает частоты гепатом [179,180]. Вероятно, ха-
рактеристика СЗН/Не во многом приложима к этой сублинии. Среди 10
испытанных линий дали минимальный титр антител на иммунизацию жи-
выми и убитыми лептоспирами, максимальный — на анатоксин С1.
oedematiens [522]. Используются в онкологии и иммуногенетике как инбред-
ный партнер многих конгенно-резистентных линий.
СЗНА/Mv Поддерживаются And, Y
Инбридинг F94. Генотип +, Н-2к. Окраска шерсти — агути. Выведена от
скрещивания самки СЗН с самцом А в 1944 г., в Sto на F48 в 1962 г., в Y на
F57 в 1965 г. Частота опухолей молочных желез — до 30 % За весь период
размножения в Sto и Y частота опухолей резко упала. Частота опухолей лег-
ких у самок — 10,7%, у самцов — 19,9%, у 3,3% самок и 26,3% самцов наблю-
дались гепатомы в возрасте свыше 2 лет. Чувствительны к индукции опухо-
лей печени канцерогенами. Самцы наиболее чувствительны как к летально-
му действию тотального облучения рентгеновскими лучами (LD50.30 =413
рад), так и к индукции доминантных деталей (доза 400 рад). Однако по ча-
стоте хромосомных аберраций в половых клетках (46,3%) они занимают про-
межуточное положение среди трех испытанных линий (A/HeJ, СЗНА/Mv,
C57BL/6) [166]. Высокочувствительны к вирусу диареи инфантильных мы-
шей [592], резистентны к нутталиозу. Содержание гликогена в печени —
Глава 10. Коизогенные и инбредные линии мышей
215
от 2,2% сырого веса печени в 6-месячном возрасте и до 7,9% в возрасте 11
месяцев [362]. Гемоглобин «гомогенного типа». Фертильных самок — 96,0%,
на одну фертильную самку — 3,15 помета; средняя величина помета при
отсадке — 6,52, нежизнеспособных пометов — 13,7%. Средняя продолжи-
тельность жизни самок — 683 ± 20,5 дня, самцов — 701 ± 21,2 дня.
C57BL/6J Поддерживаются Sto, Y
Инбридинг F113. Генотип a, H-2f}. Окраска шерсти черная. Линия C57BL
получена Литтлом в 1921 г. В Sto на F62 в 1961 г., в Y на F78 в 1965 г. Опухоли
молочных желез отсутствуют, изредка встречаются у размножавшихся самок
(около 1%), низкая частота опухолей других локализаций у животных даже в
возрасте свыше 18 месяцев [244]. В возрасте 785 дня у 31,1% самок наблюда-
ются лимфолейкозы, ретикулезы, лимфогрануломатозы [435]. Возникают
спонтанные аномалии развития. 12% животных имеют дефекты глаз: от слиш-
ком маленьких одного или обоих глаз до полного их отсутствия, разная сте-
пень микрофтальмии одного или обоих глаз. 13—15% животных имеют ске-
летные аномалии: уродства черепа (5%), позвоночного столба (11%), конеч-
ностей (1,5%) [604]. Самое низкое число пейеровых бляшек — 7,4 [299]. Яич-
ники старых самок содержат повышенное количество цероидного пигмента.
В селезенке иногда наблюдаются черные пятна, гистологически характеризу-
ющиеся диффузным расположением пигментированных клеток.
Чувствительны к радиации: для самцов LD50.30 = 647 рад, для самок —
670 рад. Наблюдается высокая частота гепатом после облучения [604]. Чув-
ствительны к тератогенному действию временной гипоксии на 9-м дне бе-
ременности: уродства грудины возникают у 23% эмбрионов. Относительно
резистентны к летальному действию озона. Резистентны к таким концен-
трациям хлороформа в воздухе, которые смертельны для мышей А, СЗН и
DBA. Малочувствительны к летальному действию ипрониазида. Ре-
зистентны к летальному действию тиоТЭФ (LD50.30 = 28,96 мг/кг). Облада-
ют резко выраженной индивидуальной изменчивостью по этому признаку.
Чувствительны к патогенетическому эффекту тиоТЭФ при высоком уров-
не спонтанных хромосомных нарушений. Чувствительны к индукции |3-
нафтофлавоном активности монооксидаз (например, арилгидрокарбонгид-
роксилазы и цитохрома Р—450). У 55% потомства от матерей, обработан-
ных на 18-й день беременности 1-этил-1-нитрозомочевиной, возникает
лейкоз, у 7% — гепатомы [164, 165].
Чувствительны к канцерогенному действию 3-метилхолантрена [383].
6-аминоникотинамид вызывает сращение позвонков у 58% эмбрионов и
расщепление нёба у 11% (по сравнению с 89 и 76% соответственно в ли-
нии А). Этот признак наследуется цитоплазматически. Задние конечности
эмбрионов наиболее чувствительны к тератогенному действию 5-флуороу-
рацила. Кортизон индуцирует расщепление нёба у 20% эмбрионов. Чув-
216
Линейные животные-биомодели
ствительны к тератогенному действию 5-фтор-2-дезоксицитилин, 3-метил-
холантрена и 7,12-диметилбензантрацена [408]. Кортизон вызывает эози-
нопению с максимумом эффекта при 6 мг (по сравнению с 96 мг в других
линиях). Подчелюстная железа крайне чувствительна к действию ми-
нимальных доз андрогенов и к некоторым дозам эстрогенов (по присут-
ствию амилоида в трубочках). Устойчивы к действию гистамина и серото-
нина в сравнении с SJL [648]. Чувствительны к действию аминазина: 90%
животных становятся неактивными при дозе 4 мг/кг.
Высокочувствительны к заражению салмонеллами [174]. Фенольная вак-
цина из S. typhimurium вызывает интенсивную выработку опсонизирующих
антител, но не защищает от заражения [467]. Чувствительны к стафилококко-
вой инфекции, резистентны к Corynebacterium kutscheri [285, 286]. Резистен-
тны к вирусам эктромелии [195] и диареи новорожденных [592]. При внутри-
брюшинном введении в 300 раз более резистентны к вирусу простого герпеса,
чем DBA/2 [368]; резистентны к вирусу Молони и Фрейнда, но чувствитель-
ны к вирусу Граффи [137]. Обладают слабой чувствительностью к инфекции
цуцугамуши, которая протекает у них бессимптомно [359]. Наиболее чувстви-
тельны к внутричерепному введению коклюшной вакцины [294] и к возбуди-
телю везикулезного риккетсиоза (D. murinus) [559]. Резистентность к листе-
риозу в 10 раз выше, чем у BALB/c [123]. Высокореактивны на лептоспиры
[522] и на общий антиген энтеробактерий в сравнении с DBA/2 [241].
Высокий иммунный ответ на бычий у-глобулин [501], вырабатывают ан-
титела к полимеру из глутаминовой кислоты, аланина, тирозина. У 45% мышей
в 6-месячном возрасте обнаруживаются антитела к вирусу лейкоза AKR [304].
Обладают хорошей продукцией интерферона на вирус лейкоза Раушера [593].
В 3 раза активнее, чем BALB/c, продуцируют сывороточный интерферон на
введение вируса опухоли молочной железы мышей. Образуют большее коли-
чество циркулирующего интерферона, чем BALB/c, на внутривенное введе-
ние вирусов болезни Ньюкасла, гриппа А, Сендай, лейкемии Фрейнда, поли-
инзиновой и полицитидиловой кислот. В нормальной сыворотке обнаружен
фактор тимической активности, способный увеличивать в костном мозге нор-
мальных мышей число розеткообразующих клеток с эритроцитами барана.
Имеют самый высокий относительный вес селезенки (4,71 г) из шести испы-
танных линий, самую низкую активность сывороточного лизоцима (14,4%),
высокую переваривающую способность нейтрофилов (36,9 ± 1,9).
Плодовитость увеличивается при содержании на диетах с повышенной
концентрацией белка (23% по сравнению с 19%) и жиров (11—12% по срав-
нению с 6% для других линий) [287]. Дефицит рибофлавина и избыток га-
лактофлавина вызывает увеличение числа аномалий развития по сравнению
со спонтанным уровнем: частота расщеплений нёба доходит до 13%, возра-
стает частота брахигнахии и дефектов глазной мускулатуры. Средний вес
мышей этой линии в возрасте 168 дней составляет 28 г [528]. Вагинальное
Глава 10. Коизогенные и инбредные линии мышей
111
открытие происходит позже, чем у BALB/c, на 9 дней. Фертильных самок —
99,4%, 4,32 помета на фертильную самку (п — 437), 6,22 мышонка в помете
при отсадке. Средняя продолжительность жизни самок в Y — 656 дней, в Jax
у самок — 692 дня, самцов — 676 дней. Продолжительность жизни у самцов
— до 878 дней при содержании с 6-недельного возраста в контролируемых
условиях, у самок — до 794 дня. Предельный возраст самцов — 1132 дня
[404].
C57BL/1 OScSn Поддерживаются Y
Инбридинг F122. Генотип а, Н-Я. Окраска шерсти черная. Сублиния
C57BL (см. C57BL/6) поступила к Скотту на F26 в 1940 г. В Y на F97 в 1971 г.
Частота опухолей молочных желез очень низка, не имеют фактора молока
[604]. Уретан вызывает повышение частоты лейкозов, аденом легких, геман-
гиом печени и аденом Гардериевых желез. Чувствительны к канцерогенному
действию 3-метилхолантрена [383]. Встречаются глазные дефекты и гидро-
цефалия. Вес семенных пузырьков сравнительно высок, коррелирует с гап-
лотипом Н-2f> [246]. Резистентны к летальному действию озона [235]. На
вирус Раушера отвечают высоким уровнем антител [638]. Резистентны к за-
ражению Corynebacterium kutscheri [285, 286]. Дают максимальный титр ан-
тител на иммунизацию живыми и убитыми лептоспирами. Чувствительны к
S. typhimurium и инфекции Leishmania [604]. Высокочувствительны к гис-
топлазмозу [508]. Активность печеночной каталазы снижена на 50% по срав-
нению с другими линиями и сублиниями C57BL/6He и C57BL/6An. Сред-
ний вес самок в возрасте 168 дней — 36 г, самцов — 37 г [528]. Самки и
самцы агрессивны, особенно в условиях конкуренции; агрессивное поведе-
ние самцов элиминируется через 25 дней после кастрации. Склонность к
запасанию пищи выражена слабо. Являются инбредным партнером многих
КР линий.
C57BL/KsJ - db Поддерживаются в Y
Инбридинг Fn+58. Генотип: a, db. Получены из Jax в 1978 г. Мыши несут
рецессивный ген диабета db (4-я хромосома), детерминируют развитие диа-
бета, сходного с diabetis mellitus человека. Характеризуются всем спектром
аномалий этой болезни от ожирения до полного бесплодия у гомозигот по
гену db. Дефицита инсулина нет. Используется как биомодель аналогичной
патологии человека: диабета типа II и по отдельным феноменам этиопато-
генезу, диабета типа I.
05781-/6-^ /+ Поддерживаются в Y
Линия, несущая на генотипе C57BL/6. Мутантный ген (Dominant spoiling)
— Y(WY), выявленный в 1962 г. в Sto. Современное обозначение — Kif'y. Ген
детерминирует формирование рецепторов тирозинкиназы клеточной мемб-
218
Линейные животные-биомодели
раны и определяет передачу межклеточной информации и взаимодействие
клеток в эмбриогенезе. Одна из наиболее актуальных биомоделей биологии и
генетики развития, ранее использовалась как модель наследственной анемии;
гомолог человеческой «пегости» — белой пряди на голове.
C57L/JY Поддерживаются Sto, Y
Инбридинг F137. Генотип а, Ь, In, H-2. Окраска шерсти свинцовая. Был
обнаружен Мэрреем среди коричневых мышей C57BR. В Sto на F80 в 1961 г.,
с 1965 г. поддерживается в Y.
Наблюдаются гипофизарные опухоли у старых животных. Ретикулокле-
точные опухоли типа В встречаются у 54,5% размножавшихся животных [604].
В сублинии (Y) опухоли разных локализаций встречаются с частотой 1,5—
4,4%, и только разного типа ретикулоклеточные опухоли наблюдаются у 16,4%
самок и 7,9% самцов в возрасте старше 600 дней. Кисты яичников у 10%
самок. Более резистентны к хлороформу в воздухе в сравнении с линиями
СЗН, A, DBA. При внутрибрюшинном введении 1-этил-1-нитрозомочевины
самкам на 18-й день беременности 42% потомства поражается лейкозом,
возникают аденомы легких, гепатомы [164, 165]; кроме того, обнаружен те-
ратогенный эффект: наибольшая эмбриональная смертность (57,3%) на 14-
й день беременности и наибольшая частота уродства мозга и глаз (69,6%) по
сравнению с другими четырьмя линиями [164]. Их семенники высокочув-
ствительны к повреждающему действию кадмия [630, 631]. Чувствительны к
заражению P. berghei, гиперчувствительны к инфекции Echinococcus
multilocularis [604]. Вес тела у обоих полов в возрасте 140 дней — 25 г [528].
Плодовитость следующая: 63% фертильных самок, на фертильную самку при-
ходится 2,87 помета, средняя величина помета при отсадке — 5,24. Продолжи-
тельность жизни в условиях SPF у самок — 604 дня, у самцов — 473 дня [604].
В Y средняя продолжительность жизни у самок — 537 дней, у самцов — 459
дней. Высокая подвижность в «открытом поле» (9,9) и высокая активность в
колесе (10,6) коррелирует с низким уровнем декарбоксилазы глутаминовой
кислоты в мозге. Используются в онкологических исследованиях.
C58/JY Поддерживаются Y
Инбридинг F173. Генотип а, Н-2!1. Окраска шерсти черная. Выведена
Макдауэллом. Из Jax поступила в Sto на F114 в 1961 г., в Y поддерживается
с 1965 г. Высокая частота лейкозов у мышей обоих полов: в Y — у 80,4%
самок в возрасте 317 ± 11,7 дней, в Jax — у 88% самок и 73% самцов. Аплазия
почек наблюдается у 10% животных [604]. В яичниках часто встречаются
полиовулярные фолликулы, содержащие от 2 до 5 яйцеклеток. Резистентны
к летальному действию озона [235], к заражению P. berghei [604]. Высокочув-
ствительны к заражению S. typhimurium [174], чувствительны к дерматофи-
там. BY— 76,9% фертильных самок, 2,65 помета на фертильную самку,
Глава 10. Коизогенные и инбредные линии мышей
219
средняя величина помета при рождении — 4,22 мышонка и при отсадке —
3,58. Эмбриональные потери высокие. Соотношение полов: 0,552 самцов.
Продолжительность жизни самок — 351 день, самцов — 373 дня.
DBA/U Lac Поддерживаются Y
Инбридинг F85. Генотип a, b, d, H-21. Окраска шерсти осветленная корич-
невая. Старейшая инбредная линия, заложена Литтлом в 1909 г. Из Англии в
Sto на F28 в 1958 г., с 1965 г. поддерживается в Y. Резистентны к большинству
опухолей мышей DBA/2. Штамм Р1534 растет у 50% животных DBA/1, a S91
— у обеих линий [138]. Опухоли молочных желез развиваются у 75% размно-
жавшихся мышей в возрасте старше года и у некоторых виргинных самок
старше 18 месяцев [604]. Частота опухолей молочных желез у размножавших-
ся самок в Y в возрасте 460 дней — 61,5%, погибали самки в возрасте 503 дня.
Относительно устойчивы к канцерогенному действию 3-метилхолантрена:
канцерогенный индекс < 31, у чувствительных > 64 [383].
Устойчивы к инфекции P. berghei, но чувствительны к заражению S.
typhimurium C5, чувствительны к инфекции Leishmania, Coxiella burnetii
[604]. Высокочувствительны (после А/Не) к мышиной оспе. На инфициро-
вание вирусом Раушера 8-недельные самцы отвечают 10-кратным увеличе-
нием селезенки на 20-й день после введения вируса, далее наблюдается
полная или частичная ремиссия заболевания, выживает 90% животных. Ре-
зистентность к этому вирусу, видимо, связана с высоким уровнем антител
и высоким титром сывороточного интерферона [638]. Дают низкий иммун-
ный ответ на синтетические полипептиды в сравнении с мышами SJL/J
[490]. Не вырабатывают антитела к линейному полимеру из глутаминовой
кислоты, аланина и тирозина [232]. Имеют самый низкий как абсолютный,
так и относительный вес мозга среди испытанных линий. Половой димор-
физм по этому признаку у данной линии отсутствует. Число пейеровых
бляшек в их лимфатической системе — самое высокое из семи испытанных
линий [299].
Чувствительны к содержанию минимальных доз хлороформа в воздухе,
гибель происходит от поражения почек. Семенники высокочувствительны к
повреждающему действию кадмия [630, 631]. Введение кортизона беремен-
ным самкам вызывает расщепление нёба у значительного числа эмбрионов:
от 20 до 100% в зависимости от дозы и времени обработки. Желтые тела в
яичниках сохраняются на протяжении многих эструсных циклов; с возрас-
том они гиалинизируются и обызвествляются. Тестостерон-пропионат инду-
цирует незначительное (в 4—5 раз) увеличение (З-глюкуронидазы в почках.
Яичники неполовозрелых самок чувствительны к гонадотропину человечес-
кого хориона.
Страдают фенилкетонурией, так как несут аллель гена, снижающего ак-
тивность фенилаланингидроксилазы на 50% . Чувствительны к тератогенно-
220
Линейные животные-биомодели
му действию временной гипоксии на 9-м дне беременности: аномалии гру-
дины у 74% эмбрионов. Скелетные аномалии развиваются у эмбрионов при
кормлении беременных самок рибофлавиндефицитными и галактофлавин-
содержащими диетами: дефицит рибофлавина вызывает расщепление нёба у
41% эмбрионов. В Y 92,5% фертильных самок, 3,9 помета на фертильную
самку, 4,85 мышонка в помете при отсадке. Продолжительность жизни са-
мок — 514 дней. Низкая подвижность в «открытом поле» и крайне низкая
активность в колесе коррелирует с относительно высоким уровнем декар-
боксилазы глутаминовой кислоты в мозге [266].
DBA/2J Поддерживаются Sto, Y
Инбридинг F124. Генотип a, b, d, H-21. Окраска шерсти осветленная
коричневая. Происхождение — DBA/1, из Jax в Sto в 1964 г. на F80, в Y —
с 1965 г. К большинству опухолей мышей DBA/1 резистентны. Опухоли мо-
лочных желез редко встречаются у старых размножавшихся самок, снижение
частоты произошло с 1955 г. Около 1/3 самок имеют лейкемии [244]. Пере-
вивается нейробластома С-1300, ее рост сопровождается появлением в кро-
ви фермента дофамингидроксилазы, специфического для симпатической
нервной системы [255]. Устойчивы к канцерогенному действию 3-метилхо-
лантрена [383]. Внутрибрюшинное введение 1 -этилнитрозомочевины на 16-
й день беременности вызывает лейкозы у 65% потомков, гепатомы — у 22%
[164, 165]. У самцов самый низкий абсолютный вес мозга среди испытанных
линий. Очень низкий относительный вес гипофиза (у самок 1,3, у самцов 1,2)
и у самцов — подчелюстной железы. Абсолютный вес яичников самый вы-
сокий (25—65 мг). Желтые тела сохраняются на протяжении многих эструс-
ных циклов, с возрастом они гианилизируются и обызвествляются. Абсо-
лютный вес семенников значительно больше, чем у самцов B10/J, имеет
тенденцию с возрастом увеличиваться (с 188 мг в 8-недельном возрасте до
236 мг в 16-недельном), с возрастом усиливается сперматогенез, так же, как
у В6 в отличие от B10/J [569].
Крайне чувствительны к содержанию паров хлороформа в воздухе и про-
дуктов окисления этилена [604]. Резистентны к индукции (3-нафтофлавоном
активности некоторых монооксигеназ (например, арилгидрокарбонгидрок-
силазы) [497]. Обладают слабой чувствительностью к токсическому действию
ипрониазида. Чувствительны к инсулину. После введения аминазина 67%
животных становятся неактивными в тесте «открытого поля». Чувствитель-
ны к тератогенному действию винкристина. Относительно устойчивы к те-
ратогенному эффекту 1-этил-1-нитрозомочевины [164]. Семенники высоко-
чувствительны к повреждающему действию кадмия [630, 631]. Тестостерон-
пропионат дает относительно слабое увеличение концентрации (3-глюкуро-
нидазы в почках (в 4—5 раз). Интактные мыши резистентны к индукции
лейкемии метилхолантреном, тогда как кастрированные чувствительны к ней.
Глава 10. Коизогенные и инбредные линии мышей
221
Яичники неполовозрелых самок чувствительны к гонадотропину человечес-
кого хориона.
Относительно резистентны к P. berghei [604], резистентны к подкожному
заражению S. typhimurium (инфицирующая доза — более 105 микробных тел)
[526, 527]. Наиболее восприимчивы к возбудителю везикулезного риккетси-
оза D. murinus испытанных линий [559], к заражению метацеркариями
Opistorchis ielinous из шести испытанных линий [689]. Чувствительны к ви-
русу гепатита мышей. В 300 раз чувствительнее, чем В6, к вирусу простого
герпеса [368]. Не способны продуцировать антитела к эндотоксину кишеч-
ной палочки [82, 83]. Вырабатывают меньшее количество антител в сравне-
нии с В6 на общий антиген энтеробактерий [241]. В отличие от DBA/1 вы-
рабатывают антитела к линейному полимеру из глутаминовой кислоты, ала-
нина и тирозина [232].
Дают низкий иммунный ответ на ДНК [216], на вакцину из стреп-
тококков группы А [93], против пневмококкового полисахарида III типа,
минимальный иммунный ответ на первичную иммунизацию а-анатокси-
ном CI. oedematiens [522]. По образованию антител высокореактивны к им-
мунизации полинозин-полицитидиновой кислотой, липополисахаридом Е.
coli или одним из видов Salmonella по сравнению с СЗН/Не [659]. Отмече-
на низкая концентрация липидов в надпочечниках, наблюдается кальци-
нация сердца [604].
Страдают фенилкетонурией, так как несут аллель гена, снижающего ак-
тивность фенилаланингидроксила на 85%. При активном поведении на-
блюдается минимальная продукция гормона щитовидной железы в сравне-
нии с наиболее спокойными мышами Swiss и ST/J. Самцы гибнут при де-
фиците в рационе витамина К. Плодовитость снижается при содержании
жиров в диете свыше 6% и белков свыше 19%. Вес тела самок и самцов в
возрасте 168 дней достигает 28 г [528]. Плодовитость в Y: 86,3% фертильных
самок, 4,0 помета на фертильную самку, средняя величина помета — 4,4
мышонка. Средняя продолжительность жизни виргинных самок — 714 дней,
самцов — 707 дней.
DBA/2JY Поддерживаются в Y
Инбридинг F178. Генотип: a, b, d, H-21. Окраска шерсти осветленная ко-
ричневая. Получены Jax в Y в 1964 г. У трети самок развивается лейкемия,
устойчивы к канцерогенному действию 3-метилхолантрена. Самцы крайне чув-
ствительны к парам хлороформа в воздухе. Аудиогенные припадки можно
вызвать у 100% мышей в возрасте 35 дней. Средняя продолжительность жизни
самок — 714 ± 21,4 дня, самцов — 707 ± 22,4 дня. Используется как партнер
для производства популярных гибридов C57BL/6 х DBA/F1 и BALB/c x DBA/
2F1 (для получения моноклональных антител генотипа BALB/c). Линия ис-
пользуется в онкологических исследованиях.
222
Линейные животные-биомодели
129/J.Y Поддерживаются Y
Инбридинг F127. Генотип: Aw, с, с ch, р/ср, Н-2Ь. Окраска шерсти палевая.
Получены Y из Jax в 1964 г. Мыши этой линии наиболее резистентны
к радиации. У самцов иногда развиваются тератомы (1—5%), характеризую-
щиеся хаотичной дифференциацией клеток, происходящих от одной ство-
ловой клетки. Эта особенность сделала линию 129 наиболее используемой
при производстве многочисленных трансгенных линий мышей.
HRS/J Поддерживаются Y
Инбридинг F120. Генотип hr, Ъ, с, d, H-2!1 поддерживаются скрещиванием
гетерозиготных (+/пг) самок с гомозиготными (hr/hr) самцами. Окраска шер-
сти белая, голые. Мутантный сток мышей hr (hairless — безволосые) получен
Грином в 1959 г., от него в Jax в 1964 г., оттуда в Y в 1974 г. Несут рецессивную
аутосомную мутацию hairless, которая первоначально в 1926 г. была обнаруже-
на у диких домовых мышей в Лондоне. У гомозигот нормальный шерстный
покров развивается до 10-дневного возраста. В это время они начинают терять
волосы, которые выпадают из фолликулов. Через некоторое время нарастает
очень мало тонких пуховых волос, которые вскоре тоже выпадают. Такой рост
волос повторяется с интервалом около месяца, однако животные выглядят
совершенно безволосыми. Ногти исключительно длинные и искривленные.
Гистологически выявляется гиперкератинизация многослойного эпителия и
верхней части волосяных канальцев, начиная с 14 дней. Формирование воло-
сяных луковиц аномально, и нижняя часть фолликула обычно отделяется от
верхней. В изолированной нижней части развивается киста. Иногда кисты
возникают в изолированных сальных железах. Гомозиготы обладают нормаль-
ной плодовитостью, но большинство самок плохо выкармливают мышат.
Некоторые +/hr самки теряют большую часть шерсти в возрасте старше 4 ме-
сяцев [604]. 45% гомозигот hr/hr к 8—10 месяцам имеют лейкемию, к 18 меся-
цам частота лейкемии достигает 72%. У гетерозигот +/hr частота лейкемии
составляет к 10 месяцам только 1% и 20% — к 18 месяцам [604]. У мышей
генотипа hr/hr наблюдается атрофия коры тимуса. Так как безволосые мыши
более чувствительны к развитию лейкемии, чем нормальные, полагают, что
ген hr обусловливает дефект иммунной системы [278].
1/StY Поддерживаются Y
Инбридинг F118. Генотип a, b, d, p, s, H-2J. Окраска шерсти пятнистая.
Вывел Стронг в 1926 г. из группы беспородных мышей. В Y поступили от
Клейна из Чехии в 1968 г. на F88. Устойчивы к химической индукции опухо-
лей [604]. Высокочувствительны к возбудителю туберкулеза [38]. Комплемент
не обнаруживается. Наблюдается сцепленная с полом недостаточность фос-
форилазной киназы [604]. Активность альдегидоксидазы в печени в 2 раза
ниже, чем у мышей линии C57BL/6J [59].
Глава 10. Коизогенные и инбредные линии мышей
223
NZB/BI Orl Поддерживаются Y
Инбридинг F92. Генотип а, В, +rd, H-21. Окраска шерсти черная. Выведе-
ны Белыпевским (В1) в 1948 г. из смешанной колонии университета
г. Отаго, которая была перевезена в Новую Зеландию из Англии в 1930 г.
Происходит от черной пары, отделившейся от NZO на F3.
Страдают аутоиммунной гемолитической анемией, которая обусловлена
взаимодействием нескольких генов. Наблюдаются антинуклеарные антитела
и волчанкоподобные нефриты. Характерны ретикулоцитоз, экстрамедулляр-
ный эритропоэз, нередко развиваются желтуха и спленомегалия. Компле-
мент не определяется [604]. Резистентны к инфекции Leishmania [604]. Вы-
сокореактивны к иммунизации ДНК, дают самый низкий титр интерферона
в сыворотке крови, индуцированного вирусом лейкоза Раушера [593]. Сред-
ний вес тела в возрасте 112 дней у самок — 31 г, у самцов — 34 г [528].
Средняя продолжительность жизни животных обоих полов составляет 270—
280 дней [604], в условиях SPF продолжительность жизни самок — 441 день,
самцов — 459 дней [207, 208]. Используются для изучения аутоиммунных
процессов, преимущественно в гибридной комбинации с NZW.
NZW/LacY Поддерживаются Y
Инбридинг F50. Генотип Ь, с. Окраска шерсти белая. Выведены в 1948 г. от
пары белых мышей из основного стока в Новой Зеландия. В Y получены
в 1972 г. из Англии (Lac) на F25. Мыши этой линии обладают высокой спо-
собностью к выработке аутоантител. У них не развивается толерантность к
бычьему гамма-глобулину [94]. Гибридные самцы, несущие гаплоидный на-
бор генов линии NZW, высокочувствительны к токсическому действию пре-
паратов декстрана [129]. У гибридов F (NZB x NZW) наблюдаются волчан-
коподобные нефриты с положительной реакцией на LE-пробу [604]. Для
них характерно спонтанное возникновение антител к нуклеиновым кисло-
там: против РНК — в 2,5—5,5 месяца и против ДНК — в 5,5—7,5 месяца
[610]. Чувствительны к инфекции Leishmania [604]. Средний вес тела самок
в возрасте 112 дней — 30 г, самцов — 35 г [528]. Средний размер помета в Lac
— 5,0 мышонка, средний интервал между пометами — 27,1 дня [207]. Про-
должительность жизни обоих полов и конвенциональных условиях — 630
дней [604], в условиях SPF самки живут 733 ± 42,8 дня, самцы — 702±34,0
дня [208]. Гибриды (NZB x NZW)F, служат для получения культур тканей.
SWR/JY Поддерживаются Y
Инбридинг F128. Генотип А, В, с, Н-21. Окраска шерсти белая, альбино-
сы. Выведена от неинбредных мышей Swiss, в Y с 1974 г. на F109. Частота
опухолей молочных желез в различных сублиниях очень низка: от долей
процента до 30%. Лейкемия наблюдается у 10% самок. Первичные опухоли
легких возникают у 36—50% мышей [604]. Чувствительны к действию 3-ме-
224
Линейные животные-биомодели
тилхолантрена (канцерогенный индекс > 64) [383]. После внутрибрюшинно-
го введения матерям 1-этил-1-нитрозомочевины на 18-й день беременности
лейкоз возникает у 85% потомства, на 16-й день — у 74% [165]. На гистоло-
гических срезах обнаруживаются аномалии в строении сетчатки глаз. В воз-
расте 6—10 месяцев у 314 самок развиваются полидипсия и полиурия. Арте-
риосклероз встречается у животных обоих полов [604]. Средняя резистент-
ность к радиации LD50 = 629 рад. Отмечена высокая смертность у самцов,
подвергнутых действию продуктов окисления окиси этилена [604]. Продук-
ция тиреоидного гормона в сравнении с более активными по поведению
мышами DBA/2 максимальна. Замещающая андрогенная терапия лишь ча-
стично восстанавливает агрессивное поведение у кастрированных самцов.
Доза тестостерон-пропионата в 600 мг/день вызывает у них депрессию. Ре-
зистентны к гистоплазмозу [508], к заражению метацеркариями Opisthorchis
felineus [689]. Имеют самое высокое давление крови из 19 тестированных
линий [604]. Значение рН артериальной крови сравнительно высоко (среди
16 испытанных линий): в левом желудочке рН 7,43), рН венозной крови —
7,28. Потребление кислорода в спокойном состоянии самое высокое из семи
испытанных линий. Самцы SWR чувствительны к дефициту витамина К [604].
В Y средняя продолжительность жизни виргинных самок, страдающих по-
лидипсией и полиурией, составляет 496 дней, нормальных — 655 дней, сам-
цов — 616 дней. В Y происходит 45,0% фертильных скрещиваний, на одну
фертильную самку приходится 3,5 помета, средняя величина помета — 6,8
мышат. Имеют самый низкий уровень общих эмбриональных потерь (в Y),
потенциальная плодовитость по первому помету — 6,9.
TPS/Y Поддерживаются Y
Инбридинг F27. Генотип аа. Выведена в Y при селекции на повышенную
чувствительность к цитогенетическому эффекту тиоТЭФ, проведенной сре-
ди мышей-октагибридов, полученных скрещиванием восьми неродственных
линий: A/Sn, BALB/c, CBA/Ca, C57BL/6, DBA/2, GLF, I/St и 101/Н. Наибо-
лее чувствительны к цитогенетическому эффекту тиоТЭФ среди 20 испы-
танных линий и гибридов, к цитогенетическому эффекту митомицина С,
фторофура и циклофосфамида среди четырех испытанных линий. Предназ-
начены для испытания на мутагенность химических соединений.
YT/Y Поддерживаются Y
Инбридинг F35. Генотип а, Ъ, cch, p, d, go, In. Окраска шерсти грязно-
белая, шерсть длинная. Выведена как линия-анализатор в отделении генети-
ки Y, ее родоначальники — мыши, гомозиготные по семи рецессивным ге-
нам, были отобраны среди потомков от скрещивания линий C57L/Y, DBA/
2Y, C57BL/Lac-go и 129/JY в 1967 г. Линия имеет маркеры-гены окраски
шерсти (2, 4, 7, 9, 19). Предназначена для использования в экспериментах,
Глава 10. Коизогенные и инбредные линии мышей
115
направленных на определение частоты индуцированных проникающими
излучениями или химическими агентами генных мутаций в маркерных ло-
кусах. Схема организации экспериментов следующая: самцов дикого типа
подвергают воздействию изучаемого мутагенного фактора и затем скрещи-
вают с самками линии-анализатора. Если в какой-либо гамете самца прои-
зойдет мутация в одном из семи маркерных локусов, то в первом поколении
среди потомков, имеющих дикий тип окраски, появится особь, гомозигот-
ная по мутировавшему гену. Используются для изучения мутагенного эф-
фекта диэтилсульфата [448] и генетической структуры популяции неинбред-
ных мышей [392, 393, 394].
101 /Н Поддерживаются Y
Инбридинг F30. Генотип Aw, H-2R . Окраска шерсти — агути белобрюхие.
Выведена Данном (Dunn) в Радиобиологическом центре Англии в 1948 г.
Поступили в Y в 1969 г. без данных об инбредном возрасте.
Имеют низкую частоту опухолей молочных желез. Высокочувствительны
к индукции кожных и легочных опухолей [604]. Наиболее чувствительны к
токсическому действию тиоТЭФ из четырех испытанных линий: LD50.30 =
21,53 мг/кг. Линяя чувствительна к патогенетическому эффекту тиоТЭФ, при
этом имеет самый высокий уровень спонтанных хромосомных нарушений
из девяти исследованных линий. Мыши этой линии имеют самый высокий
относительный вес мезентериального лимфоузла (0,98 ± 0,06) из шести ис-
пытанных линий. Количество эритроцитов высокое: (10,1 ± 0,40) млн/мм3,
лейкоцитов (13,8 ± 0,90) тыс/мм3. Размножаются медленно, обладают сред-
ней фертильностью [604]. Потенциальная плодовитость составляет 4,7 ± 0,31
живых эмбриона на самку. Уровень общих эмбриональных потерь высокий.
Используются главным образом в опытах по радиационному и химическому
мутагенезу, реже — в иммунологических и морфологических исследованиях.
Иногда служат для приготовления культур тканей, для изучения проблем
тканевой совместимости и защиты от радиации (гибриды 101 х СЗН). Пред-
ложена как чувствительная линия для тестирования химических мутагенов.
129/RrJ Поддерживаются Y
Инбридинг F79. Генотип Aw, cch, р/ср, Я-26. Окраска шерсти белая или
палевая. Получена Данном как производные линии 101 с генотипом с°Ир/^И р
и ср/ср. Из J поступила в Sto в 1964 г. на F39, с 1965 г. поддерживается
в Y скрещиванием компаундов c°h/c или гомозигот с/с с c°h/cch. Опухоли раз-
личных локализаций относительно редки — они встречаются у 20% виргин-
ных самок и 12% самцов. Нечувствительны к фактору молока. У самцов этой
линии иногда развиваются тератомы семенников; их частота колеблется в
разных сублиниях от 1 до 5%. Наблюдается высокая частота отложения кам-
ней в мочевом пузыре [604]. Старые самки часто страдают закупоркой вен в
226
Линейные животные-биомодели
надпочечниках и в матке [604]. Мыши 129 наиболее резистентны к радиации
из 10 испытанных линий: для самцов LD50.30 = 734 ± 10 рад; для самок LD50.30 =
= 774 ±13 рад. Высокочувствительны к действию эстрогенов в любом возра-
сте [604]. Относительно резистентны к обработке тестостерон-пропионатом:
концентрация Р-глюкуронидазы в почках увеличивается лишь в 4—5 раз. Ре-
зистентны к инфекции Leishmania и наиболее чувствительны к вирусу Sendai
из 19 испытанных линий [604]. Давление крови относительно низкое (систо-
литическое 88,7 ± 2,0 мм рт. ст.); ритм работы сердца — самый медленный из
семи испытанных линий (частота пульса 630 ± 8 уд./мин), рН венозной
крови — 7,32, артериальной — 7,37. Продолжительность жизни колеблется от
648 дней у виргинных самок и 679 дней у самцов до 819 дней для каждого
пола. BY— 66,3% фертильных самок, на фертильную самку — 2,67 помета,
нежизнеспособных пометов — 23,2%; средняя величина помета — 5,19 мы-
шонка. Используются в эмбриологических исследованиях, онкологии, фар-
макологии.
227
Глава 11 МУТАНТНЫЕ ЛИНИИ И СТОКИ
Термин сток означает закрытые колонии (популя-
ции) лабораторных животных, размножаемых лю-
бым способом, кроме инбридинга. Стоки обознача-
ют как Lac:A2G, где Lac — источник животного,
A2G — название, символ стока. В филиале «Андре-
евка» размещаются две популяции неинбредных
(аутбредных) мышей: Kv:SHK и Kv:ICR (Kv — Крю-
ково) — символ, источник развития, SHK — сим-
вол популяции. В дальнейшем аббревиатура Kv за-
менена на And.
Постоянно протекающий в природе мутацион-
ный процесс является источником новых мутантных
форм животных с наследственными изменениями
признаков организма, любых систем и органов.
Мутанты широко применяются в медико-биологи-
ческих исследованиях, являясь естественными мо-
делями наследственных заболеваний человека. Мо-
делирование наследственных болезней стало воз-
можным в связи с выявлением целого ряда генов,
обусловливающих нарушение развития, аналогич-
ное врожденным болезням человека. Среди наибо-
лее ценных мутантов следует упомянуть ген мышеч-
ной дистрофии dy (dystrophia muscularis); гены rd
(retinal degeneration), mi (microphtalmia), dw (dwarf,
карликовость), db (diabetes), гены Ww. SI, обуслов-
ливающие дефекты кроветворения и т. д. В Y боль-
шинство мутантных стоков поддерживается инбри-
дингом. Если мутация обнаружена в инбредной
линии и поддерживается инбридингом, такая му-
228
Линейные животные-биомодели
тантная линия является коизогенной (см. гл. 10). Если же мутантный ген
вводят в стандартный генотип инбредной линии серией возвратных скрещи-
ваний, то в этом случае линия является конгенной.
BALB/cY- wal Поддерживаются Y
Инбридинг F29. Генотип с, wal. Окраска шерсти белая, альбиносы. Му-
танты найдены среди мышей производственного стада линии BALB/c в Sto
в 1969 г. Wal — waved alopecia (волнистые, лысеющие), рецессивный ген не
локализован. Гомозиготы в 14-дневном возрасте покрыты короткой волни-
стой шерстью. В возрасте одного месяца, после первой смены шерстного
покрова, вырастают более короткие и редкие, чем в норме, волосы. Курча-
вость и последующее облысение развиваются от межлопаточной к задней
части спины, постепенно охватывая бока и живот.
У взрослых мутантов шерстный покров состоит из тонких извитых волос,
остевые волосы отсутствуют. Облысение более выражено у самок, чем у сам-
цов [602].
C57BL-goY Поддерживаются Y
Инбридинг F32. Генотип a, goY. Окраска шерсти черная. Спонтанная ре-
цессивная мутация (go,) — ангорская, вызывающая рост длинной шерсти,
обнаружена в 1961 г. у мышей C57BL/Lac в Sto.
Гомозиготы go/go обладают нормальной жизнеспособностью и плодови-
тостью. Шерстный покров мутантов состоит из длинных волос, которые к
моменту отъема в 2 раза длиннее, чем у нормальных мышей. Вибриссы также
очень длинные. Мутантный ген длинношерстности был идентифицирован
как повторение мутации go по результатам двух тестов. При проверке на сцеп-
ление с геном ^(XVII группа сцепления, 5-я хромосома) среди 153 потом-
ков F2 найдены 93 короткошерстные пестрые мыши, 45 черных длинношер-
стных, 6 пестрых ангорских и 9 черных короткошерстных мышей. Это рас-
щепление свидетельствует о том, что ген длинношерстности находится в 5-й
хромосоме; частота кроссинговера с геном W составляет (9,8 ± 2,5) %. Тест
на аллелизм с мышами C57BL/6-go, полученными из Джексоновской лабо-
ратории, подтвердил идентичность генов goY и go1.
C57BL/KsJ-db+/+m Поддерживаются Y
Инбридинг Fn+8. Генотип a, db+/+m. Окраска шерсти черная. Поступи-
ли в Y из JAX в 1978 г. Мыши несут рецессивный ген diabetes-^ (8-я группа
сцепления, 4-я хромосома). Ген db в гомозиготном состоянии вызывает ди-
абет, сходный с diabetes mellitus, сопровождающийся аномальным ожирением,
дегрануляцией бета-клеток в островках поджелудочной железы, гиперглике-
мией, полиурией, глюкозурией. Содержание сахара в крови очень высокое,
уровень утилизации глюкозы снижен. Дефицита инсулина нет. Диабетики
Глава 11. Мутантные линии и стоки
119
обоих полов бесплодны. Ген w-misty — рецессивный, осветляющий окраску
— маркер оппозитной хромосомы, не несущей гена db.
C57BL/6Y-Hi/+ Поддерживаются Y
Инбридинг N17F8. Генотип аа, +Hi. Hare tail («заячий хвост») — полудоми-
нантный аутосомный ген, обнаруженный в питомнике «Столбовая» в 1967 г.
среди гибридов F1(CBA/Ca х В6). Ген Hi обладает полной пенетрантностью.
Гетерозиготы имеют сильно укороченный, толстый хвост, загнутый вверх или
в сторону. У гомозигот отмечена пониженная жизнеспособность. Гомозигот-
ные самки не размножаются, хотя плодовитость самцов близка к нормальной.
Фенотипически гомозиготы характеризуются более сильной выраженностью
мутантных признаков: тело резко укорочено, позвоночник часто искривлен
на границе грудной и поясничной области, хвост сильно укорочен. Ген не
локализован.
С57В1./6У-а1ип8 Поддерживаются Y
Инбридинг N23F18. Генотип а]+/а'Ш5 (2-я хромосома). Окраска шерсти
черная, желтобрюхие. Доминантный ген, вызывающий в гетерозиготном
состоянии сильное укорочение и искривление хвоста, найден в 1964 г.
у мышей линии CC57BR в питомнике «Столбовая». Сцеплен с геном а (не-
агути, 2-я хромосома), частота кроссинговера между геном, вызывающим
укорочение хвоста, и геном а составляет (4,97 ± 1,11) %. Идентифицирован
как доминантный аллель гена ип по результатам скрещивания с мышами
B10.UW. Ген Uns детален и в гомозиготном состоянии. Введен в генотип
линии C57BL/6 серией возвратных скрещиваний.
Рецессивный мутантный ген a1 (black-and-tan) — «загорелые агути» — най-
ден у мышей C57BL/6 в 1968 г. в питомнике «Столбовая». Мутантный ген
идентифицирован как повтор мутации а' по результатам теста на аллелизм с
мышами, имеющими стандартный ген а'; поддерживается на одной хромосо-
ме с Un. Ген а' рецессивен по отношению к гену А, но доминантен по отно-
шению к гену а. Вызывает отсутствие феомеланина в шерстном покрове на
спине и боках, но усиливает желтую пигментацию на животе. Используются
гены Unsw а' как генетические маркеры, ген £/«•* — для моделирования наслед-
ственных аномалий человека. Современное обозначение гена — Kit w'y.
С57В1./6-\ЛЛ /+ Поддерживаются Y
Инбридинг F47. Генотип a, +/JVY. Окраска шерсти черная с белыми пят-
нами. Доминантная мутация пятнистости была найдена у мышей C57BL/6 в
1962 г. в Sto. С 1965 г. поддерживается в Y.
Мутантный ген WY — Dominant spotting Yurlovo — идентифицирован как
аллель серии И7 по фенотипическому проявлению: ослабленный уровень пиг-
ментации, белая проточина на голове, белое пятно на животе, иногда на
230
Линейные животные-биомодели
спине и боках. Кроме того, это было подтверждено в тесте на аллелизм с
аллелем IVе, при скрещивании мышей +/W* и +/№* получили 150 потомков,
среди которых было 54+, 44 +/Ff*, 44 +/WY и восемь черноглазых белых
компаундов JP/WY. Компаунды WA/WY обладали пониженной жизнеспособ-
ностью, из найденных восьми мышей только три дожили до 2-месячного
возраста. Доминантные мутации в этом локусе очень часты, гомозиготы и
компаунды с мутантными аллелями серии И'часто страдают опухолями яич-
ников, снижением числа половых клеток, что обусловлено снижением про-
дукции полового гормона и увеличением количества гонадотропного гормо-
на гипофиза.
Мутантные аллели варьируют от давно найденного гена W (гомозиготы
по которому погибают в течение недели после рождения от анемии) до IVе,
гомозиготы по которому жизнеспособны, хотя и бесплодны. Гетерозиготы
по гену W имеют белую пятнистость, размеры которой варьируют в зависи-
мости от генотипа. У них нормальная кровь, и они полностью жизнеспособ-
ны и плодовиты. Гомозиготы страдают макроцитарной анемией и погибают
в течение недели после рождения.
Найденный в Sto ген Неблизок по эффекту к гену W, но гомозиготы WY/
WY всегда погибают сразу после рождения. Поэтому среди потомков от
скрещивания гетерозигот наблюдается расщепление на два класса: «пест-
рых» и черных (нормальных) в отношении 2:1 (среди 1354 мышат было 927
пятнистых и 427 черных). Живых анемичных гомозигот можно обнаружить
при вскрытии беременных самок на 17—19-й день беременности. Количе-
ство красных кровяных клеток у гетерозигот + Wyhq отличается от нормы.
Количество лейкоцитов выше, чем у В6. Некоторые аллели серии Wисполь-
зуются для изучения наследственной анемии человека. Современное обо-
значение гена — Kit w~y.
C57BL/6J-W7+ Поддерживаются Y
Инбридинг F5N100. Генотип a, W°/+. Окраска шерсти черная с белыми
пятнами. Поступила в Y из Jax в 1978 г. на N97. Мыши несут мутантный ген
жизнеспособной доминантной пятнистости (Dominant spotting viable) W° (XVII
группа сцепления, 5-я хромосома). Гетерозиготны по этому аллелю имеют
белую пятнистость, несколько ослабленную пигментацию окрашенных учас-
тков тела и легкую форму макроцитарной анемии. Гомозиготы W^/W* стра-
дают анемией в тяжелой форме, но часть из них выживает и вырастает чер-
ноглазыми белыми мышами.
C57BL/6-a* Uns Поддерживаются Y
Генотип а'+/а' Uns (2-я хромосома). Окраска шерсти черная, желтобрю-
хие (эффект гена а'). Доминантный ген, вызывающий в гетерозиготном
состоянии дефект осевого скелета, найден в 1964 г. у мышей линии CC57BR
Глава 11. Мутантные линии и стоки
231
в питомнике «Столбовая». Летален в гомозиготном состоянии; анализ му-
тации показал, что эта спонтанная делеция (302 килобаз) гена Paxl. Мо-
дельная система для работы в области эмбриологии, биологии и генетики
развития.
C57BL/10 - fzY Поддерживаются Y
Инбридинг F5. Генотип a/afzY/fzY. Мутант найден сотрудниками НЦМБ
РАМН (Y) в линии C57BL/10Sn в 1980 г. Окраска шерсти черная. Гомозиго-
ты имеют тонкие извитые волосы. Остевые волосы отсутствуют. Плодови-
тость и жизнеспособность нормальны. Ген идентифицирован как повторная
мутация fuzzy по результатам скрещивания с мышами, гомозиготами по гену
fz, полученными из Jax через Igg (XIII группа сцепления, 1-я хромосома).
C57BL/10-hrrhY Поддерживаются Y
Линия несет мутантный ген Rhino в 14-й хромосоме, выявленный в Y в
1980 г. Мутантный ген (точковая мутация) детерминирует отсутствие мест-
ного покрова у гомозигот, бесплодие самок и раннюю потерю плодовитости
у самцов, раннее старение и развитие аутоиммунных процессов в организме.
Используется в экспериментальной биологии, дерматологии, иммунологии,
фармакологии.
C57BL/HeJ-pe Поддерживаются Y
Импортирована из Jax в 1988 г. Ген вызывает отсутствие феомеланина,
осветление окраски и дефект лизосом, ген нестабилен. Окраска шерсти -
осветленные агути (серые). Используется в экспериментальной генетике,
фармакологии.
C57BL/6Y-smK(smoky) Поддерживаются Y
Рецессивная мутация найдена в линии C57BL/6JY в Y. Ген не локализован,
вызывает осветление окраски и бесплодие самок (атрезия влагалища и дефект
оогенеза). По мнению А.М.Малашенко,это самые красивые мыши, которые
трепетно ждут исследователя. Окраска шерсти светло-серая. Могут быть ис-
пользованы для моделирования паталогии женской репродуктивной сферы.
C57BL/6Y-+/Hi, до Поддерживаются Y
Мутация Hare tail (Hi) — полудоминантный ген, найден в 1967 г., гетеро-
зиготы имеют дефект осевого скелета (укороченный хвост). У гомозигот
снижена жизнеспособность, самки Hi/Hi бесплодны. Ген не локализован.
Рецессивная мутация в 5-й хромосоме у andora (go) обнаружена в 1961 г.
в питомнике «Столбовая». Ген в гомозиготном состоянии влияет на шерст-
ный покров (длинная шерсть). Модельная система для биологии и генетики
развития млекопитающих, для фармакологии и токсикологии.
232
Линейные животные-биомодели
В10. CW/Y Поддерживаются Y
Инбридинг F30. Генотип аа, се. Выведена в Y путем введения гена альби-
низма с (из линии CC57W) на генетическую основу линии C57BL/10. Ис-
пользуются при производстве сложных гибридов-альбиносов, а также для
получения тетрагибридов CBWA [390—394] в интересах токсикологии и эм-
бриологии.
(СЗНх101 )F1 -nuY Поддерживаются Y
Мутация nudeY (11-я хромосома) возникла в линии мышей 101/Н в Y в
1986 г. Рецессивная точковая мутация обуславливает у гомозигот дефект
иммунной системы — отсутствие тимуса и Т-лимфоцитов, а также отсут-
ствие шерстного покрова. Используется в онкологии, иммунологии, дер-
матологии, фармакологии, токсикологии.
ВЮ.Т/Sn Поддерживаются Y
Инбридинг N51. Генотип а, Т/+, Н-2Ь. Получена от Sn в Igg, в Y на N28.
Мыши этой линии имеют доминантный маркер 17-й хромосомы — локус
Brachyury (T), расположенный ближе к центромере, слева от Н-2. Линия
конгенная, размножается путем возвратных скрещиваний с C57BL/10SnY.
Окраска шерсти — черная.
Ген N утрачен - Splotch Поддерживаются Y
Инбридинг F45. Генотип a, Sp/+. Поступила в Y в 1973 г. из J на F28.
Гетерозиготна по гену Sp. N-naked («голый»), полудоминантный ген (15, VI).
£/?-splotch («грязное» пятно), полудоминантный ген (I, XIII). Спонтанная
мутация в линии C57BL/6. Гетерозиготы имеют белую пятнистость на животе
и иногда на лапах и хвосте. Гомозиготы погибают на 13-й день эмбриональ-
ного развития от уродств, включающих расщепление позвоночника в люмбо-
сакральной области, разрастание нервной ткани, редукцию или отсутствие
спинальных ганглиев и их производных, нарушение строения хвоста. Приме-
няется в эмбриологии.
RaSd Поддерживаются Y
Генотип ++/RaSd (2-я хромосома), Ra и &/локализованы на оппозитных
концах 2-й хромосомы. Окраска темная, короткохвостые агути. Разводятся на
генотипе гибридов FI (CBA/Ca x B6), N8. Сток мышей с мутантными генами
Ra и Sd получен в Y из Института генетики животных в Эдинбурге (Шот-
ландия) в 1973 г. Ra, ragged («оборванный»). Полудоминантная спонтанная
мутация, у гетерозигот первый шерстный покров растет несколько медлен-
нее, чем у нормальных мышей. Шерстный покров почти не содержит пухо-
вых волос. Окраска агути модифицируется таким образом, что мыши +/Ra
выглядят необычно темными. Гетерозиготы обладают нормальной жизнеспо-
Глава 11. Мутантные линии и стоки
233
собностью и плодовитостью. Гомозиготы почти полностью голые. Они часто
страдают отеками при рождении и могут погибнуть до отъема. Некоторые
выживают и могут размножаться. Изучение развития мышей +/Ra показало,
что рост поздно дифференцирующихся волосяных фолликулов, которые про-
дуцируют пуховые волосы, резко заторможен или остановлен. У мышей Ra/
Ra почти все волосяные фолликулы не развиваются.
Небольшое число гетерозигот имеет белую млечную жидкость в брюш-
ной полости, начиная с момента рождения до 1—2-недельного возраста. Sd,
Danforth's short tail (короткий хвост Данфорта), спонтанная мутация, обна-
руженная Данфортом. Многие гетерозиготы и все гомозиготы погибают вско-
ре после рождения от нарушений в урогенитальной системе. Выживающие
гетерозиготы могут иметь хорошую жизнеспособность и плодовитость. Гете-
розиготы имеют искривленный короткий хвост с уменьшенным количеством
хвостовых позвонков. Хвост может отсутствовать полностью, 3-й и 4-й кре-
стцовые позвонки спаяны. Одна или обе почки могут быть редуцированы
или отсутствовать. Гомозиготы имеют сходные, но более серьезные наруше-
ния. Кроме того, у них заращенный анус и иногда отсутствуют прямая киш-
ка, уретра и мочевой пузырь. Дефекты развития могут быть прослежены,
начиная с аномального строения нотохорда.
ru ce f Поддерживаются Y
Инбридинг F20. Генотип се, f, ru. Окраска сильно осветленная, красные
глаза. Неинбредные мыши, гомозиготные по трем генам — се, /, ги, были
получены Y из Института генетики животных (Эдинбург, Шотландия). (?-
extreme dilution («сильное осветление») — один из рецессивных аллелей се-
рии albino в локусе с, 7-я хромосома. Гомозиготы се / се имеют черные глаза
с сильно осветленным шерстным покровом, что обусловлено снижением ак-
тивности тирозиназы. / — flexed tail («изогнутый хвост»), 13-я хромосома,
рецессивный ген, гомозиготы рождаются с транзиторной анемией, обуслов-
ленной нарушением гематопоэтической функции печени эмбриона. Боль-
шинство гомозигот имеет также изогнутый хвост и белое пятно на животе,
но эти признаки не постоянны. Анемия развивается с 12-го дня жизни эм-
бриона, когда печень начинает продуцировать клетки крови. Ru—ruby-eye
(«рубиновые глаза»), 19-я хромосома. Рецессивный ген, гомозиготы при рож-
дении имеют не пигментированные глаза, которые позднее становятся тем-
но-рубиновыми. Черный пигмент в шерсти ослаблен до темно-серого, жел-
тый пигмент слегка осветлен. Сильно уменьшено количество меланоцитов в
ретине, коже ушей и век.
REX Поддерживаются Y
Производное мышей ICR. Мыши несут рецессивную мутацию, вызыва-
ющую дефект шерстного покрова и вторичное отсутствие глаз. Сток ждет
исследователей.
234
Линейные животные-биомодели
T/t6 Поддерживаются Y
Генотип T/t6 (17-я хромосома). Сток, имеющий сбалансированную леталь-
ную систему, получен Y на основе скрещивания мышей В10. Т и T/t6, импор-
тированных в 1967 г. из Англии. Т, Brachyury — полудоминантный ген, найден
в 1927 г. Добровольской-Завадской. Длина хвоста у гетерозигот варьирует от
почти нормальной до полного его отсутствия. Некоторые (или почти все)
хвостовые позвонки могут отсутствовать. Изучение развития мышей показа-
ло, что нотохорд в хвостовой части аномален. У мышей генотипа Т/Т ното-
хорд никогда не развивается, нервная трубка и сомиты неправильны, и задняя
часть тела сильно редуцирована. Гомозиготы погибают на 10-й день беремен-
ности. Ген Г входит в сложную систему локуса Т, в котором известно большое
количество как доминантных, так и рецессивных мутаций. Мыши
Т/+ имеют короткий хвост, ?/+ — нормальный хвост, T/f — бесхвостые, Т/Т
и f/f1 — гомозиготы легальны. Гомозиготы по некоторым аллелям /-серии
жизнеспособны и имеют нормальные хвосты.
Компаунды по некоторым аллелям (flf1) обычно имеют нормальный хвост
и жизнеспособны, но самцы — стерильны. Аллели t сильно уменьшают часто-
ту кроссинговера в этом районе IX группы сцепления. Ген t6 также детален
в гомозиготном состоянии и соответственно модифицирует эффект гена Т
у компаундов T/t6. Для локуса Г характерны аномально высокая частота внут-
ригенной рекомбинации и появление новых аллелей (1:500), в связи с чем
были предприняты попытки использовать этот локус для оценки генетическо-
го эффекта облучения. Ген Т используют в качестве диагностического мар-
кера для поиска рецессивных аллелей /-серии в популяциях диких и лабора-
торных мышей. Расщепление в потомстве самок генотипа f/+ или Т/ f
соответствует менделевскому, но среди потомков от самцов этих генотипов
наблюдается избыток особей от гамет, несущих /-аллель.
Tabby Поддерживаются Y
Генотип Та, А'-хромосома. Окраска шерсти агути, «муаровые». Поддержи-
ваются на генотипе Fl (CBA/Ca x B6), N24. Аллель TaJ возникла спонтанно
в линии 129/Sv в Джексоновской лаборатории. Получены из Джексоновской
лаборатории в 1972 г. на N11. Впервые полудоминантная мутация Tabby про-
изошла в линии, селектируемой Фолконером на большой размер тела. Геми-
зиготные самцы и гомозиготные самки имеют одинаковый фенотип. Они
характеризуются отсутствием остевых волос по всему телу, безволосыми уча-
стками около ушей, голым хвостом, слегка искривленным на конце, умень-
шенным размером век, респираторными расстройствами и изменением ок-
раски агути. Гетерозиготные самки легко узнаются, если они имеют окраску
агути, в этом случае они имеют поперечные темные полосы. Волосы, образу-
ющие темные полосы, не имеют желтого кольца. Самцы Tabby размножаются
удовлетворительно, но гомозиготные самки часто стерильны. Гетерозиготные
Глава 11. Мутантные линии и стоки
235
самки характеризуются нормальной плодовитостью. Ген Та используется для
исследования частот потерь и нерасхождения половых хромосом.
Tufted Поддерживаются Y
Инбридинг F38. Генотип //"(17-я хромосома). Сток, гомозиготный по tufted
(«клочковатый), получен в Y в 1967 г. из Англии (Н). Ген tf, вероятно, возник
спонтанно. Гомозиготы характеризуются периодической потерей волос и но-
вым ростом шерсти, начинающимся с головы и распространяющимся вдоль
тела. Жизнеспособность и пенетрантность хорошие.
WR/Y Поддерживаются Y
Инбридинг F80. Генотип: — a, +/WY. Мыши имеют сильно осветленную
окраску, черноглазые. Выведена в Y от скрещивания линий C57BL/6 — WY и
129/J. Среди гетерозигот до 20% особей имеют черные пятна-участки с нор-
мально пигментированной шерстью; на данном генотипе ген WY (Kit-wY)
нестабилен. Высокочувствительны к мутагенам прямого действия, в том числе
к цитогенетическому эффекту тиоТЭФ, но плохо активируют промутагены,
такие, как митомицин С и циклофосфамид. Используются в эксперимен-
тальной генетике, эмбриологии и для тестирования химических мутагенов.
YT/2 Поддерживаются Y
Инбридинг F14. Генотип а, Ь, bpH, d se, go, р, s. Окраска шерсти светло-
коричневая, пятнистая. Линия-анализатор выведена в Удля изучения гене-
тической структуры популяций неинбредных лабораторных мышей и мута-
генеза. Мыши гомозиготны по восьми маркерным рецессивным генам. Ли-
ния получена на основе гибридизации линий. Данная линия более удобна
для изучения структуры популяций по генам окраски, чем линии YT и РТ,
так как она не имеет гена cch (chinchilla), который в комбинации с геном
альбинизма с затрудняет у компаундов c/c°h идентификацию фенотипа.
101 .A/Y Поддерживаются Y
Инбридинг N7F20. Генотип с. Линия выведена в Y путем введения гена
альбинизма с из линии A/Sn на генетическую основу линии 101/Н. Цитоге-
нетический анализ показал, что мыши линии 101.А и линии 101/Н обладают
повышенной чувствительностью к мутагенному эффекту тиофосфамида.
Линия предназначена для использования при производстве сложных гибри-
дов, создания искусственных популяций и проведения исследований в фар-
макологии и токсикологии.
129/ReDg-Fu/+ Поддерживаются Y
Инбридинг N31F34. Генотип Aw, cchp, Fu/+ . Окраска шерсти желтая. Из
J в Y в 1969 г. на N31F9. Fu — fused («сплавленный»), полудоминантный ген
236
Линейные животные-биомодели
(17-я хромосома), вызывающий искривление и укорочение хвоста. Мутация
произошла в Bassey Institute до 1931 г. Проявление этого мутантного гена
очень вариабельно. Как гомозиготы, так и гетерозиготы могут иметь укоро-
ченный и искривленный хвост, хотя отдельные особи могут быть нормаль-
ными. Гомозиготы обычно поражены сильнее, чем гетерозиготы. Проявле-
ние гена не зависит от остального генотипа, но у потомков матерей генотипа
Fu/+ или Fu/Fu признак проявляется слабее, чем у потомков матерей +/+.
Иногда гомозиготы и гетерозиготы страдают глухотой и обладают аномаль-
ным поведением, сходным с поведением мутантов circling (один из маркеров
17-й хромосомы). Применяются в эмбриологии.
CC57BR-bpH Поддерживаются Y
Инбридинг N1. Мутанты получены Y из Джексоновской лаборатории,
ген вводится в генотип линии CC57BR. Ьрн — brachypodism — Н («коротко-
ногие»), 2-я хромосома. Этот рецессивный ген обусловливает поражение
скелета — ахондроплазию. Применяются в эмбриологии.
237
Глава 12 ГИБРИДЫ F1, ТЕТРАГИБРИДЫ
И РАНДОМБРЕДНЫЕ ЖИВОТНЫЕ
Многие исследователи отдают предпочтение гибри-
дам F1: они широко используются в биологических
и медицинских исследованиях, особенно в экспе-
риментах по фармакологии, токсикологии, физио-
логии, транспланталогии, иммунологии, микроби-
ологии, вирусологии, онкологии, радиологии,
а гибриды КР-линий — в иммуногенетике.
Гибриды первого поколения (F1) являются потом-
ками от скрещивания двух высокоинбредных линий.
Поэтому они генетически однородны, также как
животные инбредной линии, но гетерозиготны по
тем парам генов, по которым отличаются друг от
друга две скрещиваемые линии. Огромным преиму-
ществом гибридов F1 являются их фенотипическая
однородность, большая устойчивость к изменени-
ям внешней среды, чем у животных инбредной ли-
нии, обладающих пониженной плодовитостью и
жизнеспособностью.
Гибриды F1 наследуют генетические особеннос-
ти родительских линий, но не сохраняют всех их
характеристик. Ответ их организма на эксперимен-
тальные воздействия однотипный и стабильный.
Воспроизводимость результатов исследований, вы-
полненных на гибридах F1, такая же высокая, как и
у инбредных животных. Гибриды F1 не могут быть
использованы для размножения, поскольку у по-
томков F2 будет генетическое расщепление.
Тетрагибриды — потомки животных, полученных
от скрещивания гибридов F1 инбредных линий.
238
Линейные животные-биомодели
Исходным материалом тетрагибридов являются четыре инбредные линии,
которые постоянно поддерживаются для получения тетрагибридов. Тетра-
гибриды представляют гетерогенную популяцию, которая по генетической
структуре близка к рандомбредным или неинбредным животным. Тетрагиб-
риды сложнее получить, чем рандомбредных мышей, так как для этого нуж-
но поддерживать четыре инбредные линии, однако они обладают некоторы-
ми преимуществами.
Определенная гибридная комбинация может быть получена в любой
лаборатории, располагающей исходными линиями, в то время как любая
выборка из рандомбредной колонии не может быть воспроизведена повтор-
но и, будучи использована для племенных целей, даст начало колонии, отли-
чающейся от исходной. Причем, чем меньше взята выборка, тем больше
различий можно ожидать. Другим преимуществом тетрагибридов является
возможность большего генетического контроля и достижения большей гене-
тической гетерогенности животных в эксперименте, так как для их получе-
ния могут быть использованы линии с известной специфичностью в отно-
шении изучаемого признака. В НЦБМТ РАМН предложена комбинация тет-
рагибридов мышей из четырех инбредных линий, имеющих белую окраску
шерсти [448, 449], а также светлогорская линия мини-свиней.
В качестве иллюстрации приведем пример получения в НЦБМТ РАМН,
в лаборатории биомоделирования, а территориально в пос. «Светлые горы»
Красногорского района Московской области, новой линии гибридов мини-
свиней. Процесс селекции светлогорской мини-свиньи проводился следую-
щим путем. Гибриды от скрещивания ладрасской породы и вьетнамской
вислобрюхой свиньи, полученные академиком РАСХН В.Н. Тихоновым
в Новосибирске, были завезены в НЦБМТ РАМН. В дальнейшем в НЦБМТ
РАМН для снижения инбридинга и повышения уровня изменчивости про-
водилось скрещивание дигибридов LV с геттингенской мини-свиньей /g/,
завезенной из ФРГ еще раньше. Полученные тригибриды LVg использова-
лись как для разведения в себе, так и для скрещивания с другой тригибрид-
ной формой, несущей в себе гены дикого кабана LVS. Результатом такого
воспроизводительного скрещивания явилось создание сложного тетрагиб-
рида с генами LVgS. Естественно, такое сочетание генов давало возможность
предположить, что при разведении их в себе выявится большой диапазон
изменчивости, что необходимо для проведения отбора.
Но, видимо, у первых двух поколений такого скрещивания были упуще-
ны или не проявились желательные генотипы, а последующая селекция не
внесла существенных изменений в весовые характеристики тетрагибридов,
хотя они являются вполне пригодными для лабораторных работ, так как вес
годовалых самок составил около 29 кг, а самцов — 22 кг. Вес тригибридов
LVS, несущих гены кабана, также широко варьировал, и в процессе селекции
на протяжении трех поколений существенно уменьшить этот показатель у
Глава 12. Гибриды F1, тетрагибриды и рандомбредные животные
239
мини-свиней этой сублинии не удалось. Однако интересно отметить, что
относительный прирост в подростковый период у мини-свиней этой группы
в 5-м поколении значительно меньше, чем этот показатель во 2-м поколе-
нии (133% F2 и 75% в F5). У мини-свиней LVS пятого поколения этот пока-
затель значительно ниже и в период полового созревания (4-6 месяцев).
Однако после полового созревания они продолжают расти так же, как и во
втором поколении. Это, по-видимому, обусловлено влиянием генов кабана.
Именно на второй год жизни кабаны матереют и набирают соответствую-
щий вес. Сравнение относительного прироста у мини-свиней (сублинии LVg)
в 1-м и 3-м поколении также показало существенное снижение этого пока-
зателя в подростковый период. Самцы мини-свиньи LVg в 12 месячном воз-
расте весят 24 кг, а самки — 36 кг, что является вполне приемлемым показа-
телем карликовости этих свиней.
У всех исследованных сублиний имеет место половой диморфизм по
размерам и массе тела. Интересно, что на ранних этапах постнатального
роста он, как правило, не проявляется, а если проявляется, то не резко. Если
в природных популяциях свиней самцы намного крупнее самок, что необхо-
димо для борьбы за существование и продолжение рода (имеются в виду
известные схватки между самцами в период гона), то в нашей популяции,
когда отбор направлен на уменьшение размеров веса, самцы во всех субли-
ниях намного меньше самок. Это подтверждает известную гипотезу о том,
что генотип самцов более податлив на различные воздействия внешней сре-
ды, а, следовательно, в генотипах самцов имеется тот запас генетических
вариантов, который у них быстрее проявляется, чем у самок, при изменении
давления отбора, будь этот отбор естественным или искусственным.
Воспроизводительная способность всех сублиний светлогорских мини-
свиней остается, несмотря на небольшие размеры популяции, очень хоро-
шей. В разных сублиниях плодовитость по 1—2-му опоросу колеблется от 6,3
до 8,3 поросенка на опорос, а по 3—4-му опоросу от 7,3 до 11 поросят на
опорос. Процент мертворожденных не коррелирует с процентом инбридин-
га. Светлогорские мини-свиньи остаются вполне плодовитыми и рождают
нормальное потомство даже при 25% инбридинга, в то время как мини-
свиньи зарубежных линий не выдерживают и 10% инбридинга. Это указыва-
ет на то, что в предыдущие годы прошел жесткий отбор на жизнеспособ-
ность светлогорской мини-свиньи.
Селекция на уменьшение размеров светлогорских мини-свиней привела
к незначительному уменьшению срока супоросности и в среднем у них су-
поросность равна 113 дням, тогда как у обычных свиней этот срок равен 113-
116 дням. Это важно учитывать при гнотобиологических и других экспери-
ментах, когда нужно точно знать срок супоросности.
Сведения о стандартных гематологических, иммунобиохимических по-
казателях здоровых лабораторных животных являются необходимым уело-
240
Линейные животные-биомодели
вием планирования биомедицинских экспериментов. Исследование кле-
точных элементов крови у светлогорских мини-свиней в возрастном ас-
пекте выявило большую схожесть этих показателей с таковыми у человека.
Число лейкоцитов и лимфоцитов возрастает от однонедельного до поло-
возрелого возраста животного и затем несколько снижается в период мате-
рости.
Количество розеткообразующих Т-лимфоцитов увеличивается с возрас-
том и достигает наивысшего значения у 8-месячных животных. Эти данные
хорошо согласуются с результатами исследования динамики этого показате-
ля у человека. Исследование Т-лимфоцитов в возрасте от 7 до 60 месяцев
показало, что их число немного повышается до периода матерости (22—24
месяца), а затем сохраняется практически без изменений.
Число естественных киллеров, изученных на перевиваемом штамме опу-
холевых клеток К562 в возрасте 8—60 месяцев, как показали исследования,
остается на всех сроках практически одним и тем же. Это указывает на важ-
ную роль естественных киллеров в иммунологическом статусе организма.
Определен уровень общего белка плазмы (8,0 г %) и уровень свободного
холестерина (90—188 мг %) у взрослых (24—48 месяцев) животных.
Рандомбредные животные — неинбредные, нелинейные животные зак-
рытых колоний, размножаемые по определенной, в большинстве случаев
ротационной системе, обеспечивающей рандомизацию скрещиваний. Каж-
дая такая колония характеризуется определенными частотами генов и ге-
нотипов, животные гетерозиготны по неопределенному числу генов, и
поэтому сама колония и каждая выборка из нее генетически гетерогенны.
Животные этой категории фенотипически менее однородны, чем гибриды.
Методы генетического контроля стали применяться также при разведении
и производстве нелинейных, генетически гетерогенных животных, кото-
рые все еще составляют большую часть экспериментальных биологических
объектов.
Необходимым условием сохранения биологических особенностей нели-
нейных животных и воспроизводимости результатов экспериментов являет-
ся поддержание гетерозиготности при сохранении стабильности генетичес-
кой структуры колонии. Выполнение этого условия обеспечивается при раз-
ведении животных по определенной системе, обеспечивающей рандомиза-
цию скрещиваний.
Стандартными могут считаться животные из закрытых колоний, размно-
жаемые по ротационной системе на протяжении не менее четырех поколе-
ний при потере гетерозиготности менее 1 % на поколение. Для обозначения
стандартных неинбредных колоний (их предложено обозначать термином
«сток») используют буквенные символы сток Lac: LACA обозначают коло-
нию мышей LACA, поддерживаемую в английском центре лабораторных
животных (Lac). В гл. 11 дана их более подробная характеристика.
Глава 12. Гибриды F1, тетрагибриды и рандомбредные животные
241
В медико-биологических исследованиях значительно чаще используются
менее крупные животные: кролики, морские свинки, крысы, мыши, хомяч-
ки. В их ряду имеются гибриды F1, рандомбредные и аутбредные штаммы.
Но в последние годы растет интерес к использованию мини-свиней, собак,
овец и других крупных животных
В НЦБМТ РАМН популяции неинбредных лабораторных животных от-
вечают всем международным требованиям. В частности, это колонии мор-
ских свинок, крыс и мышей в филиале «Андреевка» (And) и кроликов в
филиале «Белый мох» (Вт), крыс, сирийских хомячков и мышей в филиале
«Столбовая» (STO). Развивается производство мини-свиней в And и овец и
баранов — в Вт. Приведем лишь некоторые примеры.
SHK(And) Поддерживается And
Генотип: се, полиморфны по локусам А, В, D и Н-2, концентрация рецес-
сивных аллелей генов окраски следующая: а — 0,34, Ь — 0,50, d — 0,01.
Мыши Swiss-H, родоначальники колонии, были завезены из Чехии в 1955 г.
в Центральный питомник «Крюково» (ныне филиал «Андреевка»). Суще-
ствуют с 1961 г. как закрытая колония, а в 1972 г. введена ротационная сис-
тема разведения. Средняя величина популяции — 10 000 размножающихся
самок и самцов, потеря гетерозиготности на поколение — менее 0,01% [392,
393, 394]. Мыши негистосовместимы (выявлены следующие гаплотипы ос-
новного комплекса тканевой совместимости Н-2: a, b, d, i, h, q, s, x — не-
идентифицированные гаплотипы, составляющие более 40%) [390]; самки под-
вержены раку молочных желез (частота заболевания выше 50%), в связи с
чем средняя продолжительность жизни составляет только 12,5 ± 0,5 месяца.
Самки SHK более склонны к возрастному ожирению, чем линейные СЗНА
[103]. Высокоплодовиты: среднее число желтых тел на самку первой бере-
менности 8,7 ± 0,12, живых эмбрионов 7,3 ± 0,16; максимальные показатели
в пятую беременность — 12 желтых тел и 9,99 живого эмбриона на самку.
Эмбриональная смертность после имплантации 5%. Средний вес в возрасте
56 дней: у самок (23,7 ± 0,2) г, у самцов (28,3 ± 0,2) г.
SHR(And) Поддерживается And
Генотип: се, полиморфны по локусам А, В и D, концентрация рецессивных
аллелей генов окраски: а — 0,33, Ъ — 0,24, d— 0,07. Родоначальники завезены
из Центрального питомника «Рапполово» в 1962 г. С 1971 г. разводятся по
ротационной системе, средняя величина популяции 3000 размножающихся
самок и самцов, потеря гистерозиготности на поколение 0,02%. Имеют гап-
лотипы Н-2 (b, d, h, s, x = 35%), плодовиты, у самок первой беременности
среднее количество желтых тел 9,07 ± 0,17, живых эмбрионов 7,42 ± 0,20.
Эмбриональная смертность после имплантации 3,4% [392, 393, 394.]. Генети-
ческая структура колоний SHK и SHR по исследованным локусам (окраски
242
Линейные животные-биомодели
и Н-2) достоверно отличается, хотя в пределах колоний сохраняется ста-
бильность по частотам генов [390, 391, 392]. Это обстоятельство необходимо
учитывать экспериментаторам при использовании нелинейных мышей.
CBWA Поддерживается Y
Альбиносы-тетрагибриды, получаемые скрещиванием мышей четырех
линий: BALB/c, B10.CW, CC57W, A/Sn.
Генотип: се, полиморфны по локусам А, В и Н-2. Средний вес самок в
возрасте 40 дней составляет (19,8 ± 0,1) г, самцов — (22,0 ± 0,2) г; среднее
количество желтых тел на самку в первую беременность (10,44 ± 0,19), жи-
вых эмбрионов 9,44 ± 0,30. Эмбриональная смертность после имплантации
около 4% [392, 393,3 94]. Тетрагибриды CBWA производятся в лаборатории
генетики НЦБМТ РАМН.
243
Глава 13 SPF-ЖИВОТНЫЕ-БИОМОДЕЛИ
Классификация лабораторных животных по кате-
гориям дана в гл. 4, табл. 7, посвященной их стан-
дартизации. Об условности этой классификации
можно судить хотя бы по тому, что ни одна из пред-
ложенных терминологических систем не получила
всеобщего признания. Поэтому в целях унифика-
ции в тексте будут использованы определения из
словаря Коэйтса и Густавссона (1984) и номенкла-
туры ВОЗ (2001).
Конвенциональные животные являются микроби-
ологически неопределенными биологическими си-
стемами, то есть животными, являющимися носи-
телями различных неизвестных микроорганизмов.
Такие животные могут содержаться как в стериль-
ных, так и в обычных условиях, и являются потен-
циальными носителями патогенов. Можно доста-
точно длительное время поддерживать этих живот-
ных в хорошем состоянии здоровья, используя ме-
тоды, сходные с клиническими методами (табл. 7,
гл. 4).
Обычные лабораторные животные, используе-
мые в биомедицинских исследованиях, не гнотоби-
оты, а просто здоровые особи. Теоретически иде-
альным для исследовательских целей было бы муль-
тиассоциированное лабораторное животное-гното-
биот, то есть животное с хорошо известной «нор-
мальной» флорой, не содержащей облигатных или
факультативных патогенов, и являющееся нейтраль-
ным для исследования.
244
Линейные животные-биомодели
В реальности же использовать в исследованиях подобных микробиологи-
чески известных и лабораторно поддерживаемых животных невозможно ввиду
их чрезвычайной дороговизны, объяснимой тем, что для поддержания ги-
гиенического статуса животного-гнотобиота его необходимо содержать в
изоляторе.
Противоположностью этому являются SPF-животные. SPF-животные
являются носителями целого ряда неизвестных видов микроорганизмов. При
этом SPF-животное свободно от одного или нескольких специфических
патогенных микроорганизмов (табл. 7, гл. 4).
Таким образом, SPF-животное — это негативно определенная биологи-
ческая система, предполагающая носительство «хозяином» некоторых мик-
роорганизмов. Это — биологическая колония различных видов неизвестных
микроорганизмов, неизученных ассоциаций, в которой точно установлено
отсутствие одного или нескольких видов патогенных микроорганизмов. До тех
пор, пока SPF-статус таких животных подтверждается, они считаются кли-
нически здоровыми.
Здоровые животные, микробиологический статус которых подтвержден
(SPF-животные), являются более экономичной альтернативой гнотобиотам.
Однако гнотобиотные технологии представляют наилучший способ разви-
тия базовой линии SPF-животных. В 1975 г. Дж.Э. Рейнер рекомендовал без-
микробную технологию для предотвращения заражения колоний лаборатор-
ных животных. Это стало одним из методов выращивания колоний здоро-
вых лабораторных животных.
Производство SPF-животных выглядит следующим образом. Первона-
чально безмикробных животных с неразвитой системой иммунной защиты
ассоциируют с «коктейлем» аэробных и анаэробных микробов-симбионтов.
Эта непатогенная микрофлора способствует развитию системы иммунной
защиты, которая в свою очередь позволяет животным выжить вне изолятора
в условиях обычного содержания. Ожидается, что животное получит из ок-
ружающей среды дополнительно некоторые непатогенные организмы, что
завершит или стабилизирует кишечную флору.
Здоровью колоний лабораторных животных постоянно угрожают инфек-
ции. Невозможно ограждать колонии животных от вторжения нежелатель-
ных организмов вечно. Для того чтобы как можно дольше или, по крайней
мере, до окончания эксперимента поддерживать здоровую популяцию жи-
вотных, необходимы квалифицированные научные сотрудники и подготов-
ленный персонал.
При надлежащем оборудовании и уходе можно поддерживать удовлетво-
рительное состояние здоровья колоний SPF-животных на протяжении не-
скольких лет. Но даже в оптимальных условиях статус здоровья колонии будет
постепенно снижаться, что приведет к необходимости вновь создавать попу-
ляцию животных из свежего биоматериала. Перед тем как приступить к
Глава 13. SPF-животные—биомодели
245
воссозданию колонии, необходимо предварительно тщательно вычистить и
продезинфицировать отсек, в котором содержались животные. Хорошо ос-
нащенная и действенная шлюзовая система и навыки персонала являются
гарантией снижения риска проникновения инфекции в отсеки размноже-
ния животных и содержания особей, предназначенных для эксперимента.
Как уже упоминалось ранее, для создания здоровой группы животных
с подтвержденным микробиологическим статусом для получения племенного
потомства требуются квалифицированные специалисты, высококачественное
оборудование и гнотобиотная технология. При проведении работ по возоб-
новлению популяции лабораторных SPF-животных необходимо также нали-
чие лаборатории, осуществляющей генетический контроль (см. гл. 4, 9 и 14).
Поскольку внутреннее пространство изолятора ограничено (и в него можно
поместить лишь определенное количество пар для получения потомства), для
воссоздания колонии необходимо отбирать животных с учетом генетического
профиля имеющихся лабораторных линий и будущей колонии.
Учитывая стоимость работ и оборудования, во избежание неудачи целесо-
образно приобрести группу животных для размножения в зарекомендовавших
себя лабораториях, например, в лаборатории генетики НЦБМТ РАМН.
В ходе работ или производства определенных видов лабораторных жи-
вотных представляются необходимыми микробиологический и генетичес-
кий контроль.
Микробиологический контроль
Для проведения микробиологического контроля могут применяться различ-
ные методики. В рамках этой книги невозможно дать детальное представле-
ние о них. В общем виде методы проверки могут быть сведены к некоторым
обязательным моментам (см. также гл. 4 и 6).
Использование животных—«индикаторов» с известным микробиологическим
статусом. Животных помещают в ту же окружающую среду, в которой нахо-
дится проверяемая популяция, и периодически умерщвляют и обследуют.
Голые (атимичные) мыши являются прекрасными индикаторами, поскольку
у них присутствует иммунодефицит и они особенно подвержены воздей-
ствию патогенов. Но данные мыши не подходят для обнаружения измене-
ний в серологическом титре, который может указывать на наличие вирусов,
поскольку антигенный ответ у голых мышей отсутствует. Для определения
серологического титра необходимо использовать животных с соответствую-
щим иммунным статусом.
Выборка животных из популяции колонии для проведения лабораторных ис-
следований. Это — общее посмертное микробиологическое и гистологичес-
кое обследование, выявление паразитов, и т.д. Степень и детальность прово-
димой проверки зависят от величины расхода вида и качества оборудования.
Частота проведения проверок и количество животных, используемых в ходе
246
Линейные животные-биомодели
нее, определяются многими факторами; шансы обнаружить инфекцию зави-
сят от количества проверенных животных по отношению к общему количе-
ству особей в колонии (в процентном выражении), а также от степени рас-
пространения инфекции среди животных.
Плановая проверка животных, которые умерли или были умерщвлены, вклю-
чая особей, использовавшихся для экспериментов. Такая проверка позволяет
обнаружить инфекции или повреждения, угрожающие здоровью колонии и
качеству проводимых экспериментов.
Генетический контроль
Описать детально в рамках данной работы процесс проведения генетическо-
го контроля не представляется возможным, для получения полной инфор-
мации рекомендуем обратиться к изданиям по проблемам генетического
контроля мышей и крыс ICLAS. Регулярная проверка имеющихся животных
поможет выявить генетическое «заражение» от других линий или мутации,
которые могут отрицательно повлиять на исследования. Подробнее принци-
пы генетического контроля изложены в гл. 4, 5, 8 и 9.
Требования к корму для SPF-животных
Корм для SPF-животных не должен содержать насекомых, паразитов и па-
тогенов, перед использованием его рекомендуется стерилизовать. Если сте-
рилизация невозможна, особое внимание необходимо уделить подготовке и
хранению кормов во избежание их микробного заражения.
Ингредиенты для кормов должны быть тщательно отобраны из высоко-
качественного сырья с низким микробным показателем. Продукты живот-
ного происхождения, такие, как мясо или костная мука, могут содержать
высокое количество патогенов и поэтому должны быть исключены. Обору-
дование и помещения для подготовки корма должны тщательно очищаться,
а подготовленный корм сразу же должен быть помещен в плотно закрываю-
щиеся емкости для защиты от паразитов.
Тепловая обработка при средней температуре эффективна для уничтоже-
ния большинства насекомых и паразитов, которыми может быть заражен
корм. Подготовка при 70—80° С гранулированного и сухого корма, имеюще-
гося в продаже, является преимуществом, поскольку позволяет существенно
снизить в нем количество живых организмов.
Для современной науки требуются лабораторные животные высокого
качества, а это стоит дорого. Эксперименты на животных с известным мик-
робиологическим статусом дают лучшие результаты и их реже приходиться
проводить заново. Преимуществом SPF-животных также является то, что
они здоровее других. Производство высококачественных лабораторных жи-
вотных должно быть дополнено их содержанием в оптимальных условиях и
регулярной проверкой.
247
Глава 14 ГНОТОБИОТНЫЕ
ЖИВОТНЫЕ-БИОМОДЕЛИ
Вопрос о возможности безмикробной жизни был
поднят в XIX веке Пастером. Чтобы найти ответ на
этот вопрос, ученые стали разрабатывать безмик-
робную технологию гнотобиотехники и безмикроб-
ные гнотобиотные исследования, гнотобиологию.
С тех пор большое количество видов млекопи-
тающих, включая обычные виды лабораторных гры-
зунов, сельскохозяйственных животных и челове-
ка, были выращены в безмикробной среде. Целью
исследований было и остается выяснение взаимо-
действия между животным и его ассоциированны-
ми микробами, то есть между «хозяином» и фло-
рой, и между самими микробами, населяющими
организм животного.
Безмикробные и гнотобиотические модели не
являются альтернативой экспериментам с исполь-
зованием обычных животных, хотя значительная
часть исследователей, в том числе и автор этой кни-
ги, рассматривают безмикробных животных как
объекты, пригодные для специфических иссле-
дований, а не рутинных испытаний — для этого они
слишком дороги и не всегда надежны.
Использование лабораторных животных для
контроля качества пищевых продуктов, лекарствен-
ных препаратов, ксенобиотиков, трансформируе-
мых микрофлорой в токсические соединения, не-
прерывно растет. Одновременно повышаются и
требования потребителей и контролирующих ин-
ститутов к качеству, знанию долгосрочных и воз-
248
Линейные животные-биомодели
можных побочных эффектов, и, соответственно, безопасности их приме-
нения. С бурным развитием технологии создания генно-инженерных мик-
ро- и макроорганизмов и их продуктов решение вопроса об их выпуске в
окружающую среду и широком использовании потребует тщательных ис-
пытаний на гнотобиотических моделях.
Для описания живых существ, живущих изолированно или в ассоциа-
ции с другими полностью известными видами организмов, был предложен
термин гнотобиотика (происходящий от греческих слов gnotos — извест-
ный, и bios — жизнь), по аналогии с другими дисциплинами. В соответ-
ствии с этим гнотобиотика рассматривается как область исследований,
а гнотобиота (включая безмикробных и моно-, ди-, три-, и т. д. полиассо-
циированных живых объектов) — как объект исследований. Накопленные
данные в этой области позволили ввести более широкий термин — гното-
биология [440]. Термин гнотобиология, или точнее гнотобиотология (букваль-
но: познанная жизнь), происходит от терминов гното и биото (флора и
фауна).
Гнотобиотехнология представляет собой набор методов и технологий,
используемых в гнотобиотных исследованиях. Гнотобиология — это произ-
водство в научных целях биологических организмов (животных, растений и
людей), наличие в которых полезных или нетипичных микроорганизмов
полностью известно в пределах современной диагностики. Из вышесказан-
ного следует, что гнотобиотные животные могут быть:
безмикробными — то есть свободными ото всех видов микроорганизмов
(вирусов, бактерий, грибков и паразитов), которые можно обнаружить со-
временными методами диагностики;
моно-, ди- и полиассоциированными — в организме которых, присутствуют
1, 2 или несколько видов известных микроорганизмов (изначально безмик-
робное животное, заселенное одним или более видом известных микроорга-
низмов).
Итак, гнотобиотное животное — это позитивно определенная биологичес-
кая система, состоящая из «хозяина» и симбиотических микроорганизмов. Это
биологический конгломерат видов, каждый из которых известен.
Животные-гнотобиоты должны выращиваться в изоляторах. Изолятор
должен обладать некоторыми обязательными признаками:
/ внутренняя поверхность и содержимое безмикробного пространства,
клетки или помещения должны быть выполнены из безмикробных ма-
териалов и в них должна поддерживаться безмикробная среда;
/ к внутреннему пространству изолятора и его содержимому должен быть
обеспечен зрительный доступ;
/ необходимо наличие специальных приспособлений для манипуляций
внутри изолятора, а также для помещения в него гнотобиотного содер-
жимого извне без риска нарушения гнотобиотных условий;
Глава 14. Гнотобиотные животные—биомодели
249
/ должны быть оборудованы шлюзы для перемещения безмикробных жи-
вотных и материалов в изоляторы и из них без нарушения внутренних
гнотобиотных условий;
/ во избежание нарушения безмикробных условий должна быть обору-
дована вентиляционная система, позволяющая проветривать простран-
ство помещения и создающая внутри него положительное давление.
Производство безмикробных млекопитающих возможно благодаря тому,
что эмбрион в чреве здоровой матери является микробиологически стериль-
ным. Незадолго до окончания беременности плод может быть асептично
извлечен посредством гистерэктомии или кесарева сечения и помещен в сте-
рильный изолятор. Молодняк может быть вскормлен в изоляторе безмик-
робной приемной матерью или искусственным молоком, по составу при-
ближенным к натуральному. Данная процедура на сегодняшний день редко
используется в лабораториях для получения гнотобиотных мышей и крыс,
поскольку они есть в продаже. Популяции гнотобиотов должны включать
в себя достаточное количество особей репродуктивного возраста для поддер-
жания генетического однообразия потомства.
Производственные масштабы, необходимые для поддержания стабиль-
ного приплода безмикробных крыс или мышей требуют больших финансо-
вых вложений и редко укладываются в рамки бюджета большинства иссле-
довательских лабораторий.
Безмикробные животные иных видов, нежели мыши или крысы, должны
выращиваться самим потребителем. Не представляет особой сложности по-
лучение безмикробных морских свинок, поскольку их детеныши достаточно
развиты на момент появления на свет и не требуют длительного (а иногда и
вовсе не требуют) искусственного вскармливания. Низкие показатели вы-
живания характерны для кроликов. Кошек, собак и некоторые виды сельс-
кохозяйственных животных можно вырастить безмикробными, но из-за их
значительного размера они представляют трудности для длительного содер-
жания в изоляторах.
Гнотобиоты-млекопитающие
Существуют оперативные и консервативные способы получения гнотобио-
тических животных первой генерации. Возможно, основу этих работ соста-
вили наблюдения над особенностями развития новорожденных младенцев у
человека, а также детенышей животных: щенят, котят и мышат — у живот-
ных. Ведь новорожденные являются гнотобиотами в той степени, в какой
они контактировали с внешней средой после рождения и были контамини-
рованы микроорганизмами.
Технологически для получения гнотобиотов применяются различные
модификации гистерэктомии (сухой и влажной) и гистеротомии. В первом
250
Линейные животные-биомодели
случае извлечение матки с эмбрионами производится в асептической среде,
вне изолятора, с последующей ее перевязкой в области шейки, ампутацией
и переносом через бактерицидный гидрошлюз в стерильное пространство
камеры изолятора. В камере плоды извлекаются и освобождаются от плод-
ных оболочек.
Во втором случае операция осуществляется с использованием операци-
онного блока хирургического изолятора. Операция производится изнутри
изолятора через пленку операционного блока и склеенные с ней кожные
покровы живота беременной самки с последующим извлечением плодов из
беременной матки непосредственно в стерильное пространство камеры хи-
рургического изолятора. Гнотобиоты необходимых инбредных линий могут
быть получены путем одной из модификаций кесарева сечения с использо-
ванием лактирующей гнотобиотической самки для выкармливания новорож-
денных животных в изоляторе [534].
Из консервативных способов в литературе описаны методы получения
гнотобиотов из взрослых особей путем обработки антибиотиками и
антимикробными препаратами. Однако в настоящее время такой метод рас-
пространения не получил. В связи с коммерческой доступностью гнотобио-
тических животных различных видов и категорий, получение племядер из
коммерческих источников остается наиболее распространенным путем по-
лучения и последующего разведения гнотобиотов.
Методы получения поросят-гнотобиотов включают гистерэктомию и
основаны на факте непроницаемости здоровой интактной плаценты для
большинства микроорганизмов. В НЦБМТ РАМН разработаны методы по-
лучения и выращивания новой категории гнотобиотов — безантигенных
животных, контролируемых по антигенам микробного и животного проис-
хождения [62]. Животные-гнотобиоты применяются для изучения некото-
рых аспектов респираторных болезней человека, приготовления тканевых
культур, стандартных моноспецифических сывороток, изучения эффектов
изменения стандартной микрофлоры, а также целого ряда иммунологичес-
ких проблем.
С использованием хирургических изоляторов у свиней применялся так-
же метод гистеротомии для получения безмикробных поросят [440, 451, 452].
Безмикробные поросята и свиньи представляют особую ценность для ис-
следований в области иммунобиологии и патологии, учитывая отсутствие у
них трансплацентарной передачи иммуноглобулинов и антител материнс-
кого происхождения [452, 611]. Кроме того, благодаря морфофункциональ-
ным сходствам целого ряда систем (кровообращения, пищеварительного
тракта, кожи, сердечно-сосудистой и др.) мини-свиней с аналогичными си-
стемами и органами у человека этот вид животных находит все большее
применение в моделировании заболеваний человека. В экспериментах осо-
бую ценность представляют миниатюрные породы свиней [377]. Первые ме-
Глава 14. Гнотобиотные животные—биомодели
251
тодические приемы получения безмикробных поросят методом гистерото-
мии в России были отработаны в середине 70-х гг. в лаборатории биомоде-
лирования НЦБМТ РАМН [534].
Морские свинки были первыми животными, полученными в безмикробном
состоянии еще в 1895 г. Способность к активному образу жизни и самостоя-
тельному питанию сразу после рождения дали возможность получения и вы-
ращивания животных этого вида. Однако выращивание безмикробных морс-
ких свинок до половозрелого возраста представляет серьезную технологичес-
кую проблему, связанную с подбором адекватных диет и устранением чрезмер-
ного растяжения слепой кишки. Последний феномен, отмечаемый у безмик-
робных грызунов, в значительной степени проявлялся у этого вида гнотобио-
тов. Поэтому размножение морских свинок в безмикробных условиях удалось
получить значительно позднее, чем безмикробных крыс и мышей. В России
первые эксперименты с безмикробными животными также были начаты с
получения первой генерации безмикробных морских свинок в НЦБМТ РАМН.
Как показали дальнейшие наблюдения, для получения первой генерации
безмикробных морских свинок целесообразнее выращивать их непосредствен-
но под конкретные экспериментальные исследования, а не воспроизводить
в производственных масштабах. Получение гнотобиотических морских сви-
нок осуществляется оперативными методами. Попытки получения гнотоби-
отических морских свинок путем деконтаминации к успеху не привели. Для
оперативного получения безмикробных морских свинок используют хирур-
гические гнотобиологические изоляторы (для гистеротомии) или изолято-
ры, снабженные бактерицидным гидрошлюзом (для гистеротомии). Продол-
жительность беременности у морских свинок варьирует от 61 до 68 дней.
При отборе беременных самок для операции ориентируются на объектив-
ные признаки: размягчение лонного сочленения, расхождение лонных кос-
тей на 20—22 мм.
Первые эксперименты по получению безмикробных кроликов были начаты
в 40-х гг. XX века [542]. В России модель безмикробных кроликов была впер-
вые получена и применена в изучении патогенеза холерной инфекции и ме-
ханизмов интоксикации в конце 70-х гг. в НЦБМТ РАМН [77, 78]. Принци-
пиально методика оперативного получения первой генерации безмикроб-
ных кроликов мало отличается от таковой для морских свинок. Поскольку
беременность у крольчих длится ровно 30 дней, отбор беременных самок для
операции трудностей не представляет. Обычно животных оперируют в пос-
ледний день беременности. Для операции используются две модификации
кесарева сечения — гистеротомия и гистерэктомия, но, учитывая преимуще-
ство местной анестезии для выживания крольчат, более предпочтителен ме-
тод гистерэктомии.
Работы по получению безмикробных хомячков были начаты в 1960 г. Без-
микробных хомячков получают с помощью как оперативных методов, так и
252
Линейные животные-биомодели
методами деконтаминации. Методика кесарева сечения у беременных самок
хомячков подробно описана в литературе [106]. Возможно перекрестное
вскармливание новорожденных хомячков лактирующими самками других
видов грызунов.
Методом селективной деконтаминации у сирийских хомячков можно
освободить желудочно-кишечный тракт этих животных от кишечных
жгутиковых и Pasteurella pneumotropica. Процедура включала введение с
питьевой водой сульфата дигидрострептомицина (2 г/л), а также диметр ида-
зола (800 мг/л). Для устранения P. pneumotropica в защечные мешки живот-
ных вводили 0,5 мг пасты, состоящей из неомицина (27 мг/г) и дигидрост-
рептомицина (27 мг/г). Подобная процедура продолжается 4 недели, после
чего животных содержат в стерильных камерах с ламинарными потоками
стерильного воздуха. Методика позволяет устранить аэробные, грамотрица-
тельные бактерии, а также кишечные жгутиковые. Для более полной декон-
таминации используют купание хомячков в бактерицидной ванне с последу-
ющим переносом в стерильный гнотобиологический изолятор. Смесь анти-
биотиков для деконтаминации включает (мг/дл): амфотерицин — 10, ампи-
циллин — 50, бацитрацин — 500, хлорамфеникол — 100, колистинсульфат —
500, фузидин — 50, канамицин — 400, налидиксин — 500, неомицинсульфат
— 500, пенициллин G — 100, полимиксин В — 25, стрептомицин — 250,
тетрациклин — 50, триметоприм—сульфаметоксазол — 15, тилозинтартрат
— 50. Методика позволяет выращивать деконтаминированных хомячков в
течение ряда генераций.
Получение плотоядных лабораторных животных, к которым относятся
собаки и кошки, в безмикробных условиях представляет особый интерес
для экспериментальной медицины, и в частности таких ее разделов, как
нормальная и патологическая физиология, инфекционная и неинфекци-
онная патология, аллергология, токсикология, экспериментальная хирур-
гия и др.
С середины XX в. начаты работы по получению и применению в меди-
ко-биологических экспериментах безмикробных обезьян [322, 674]. Для по-
лучения их в безмикробном состоянии пользовались операцией гистерото-
мии у беременных самок павианов с использованием хирургического пле-
ночного изолятора Трекслера. В подобных изоляторах осуществляли и их
последующее выращивание с использованием для искусственного вскарм-
ливания коммерческой диеты SMA, обогащенной железом, разведенной
водой в соотношении 1:1,1. Через 1—2 дня обезьяны приучались к самосто-
ятельному питанию. В возрасте 2 месяцев гнотобиотические обезьяны пе-
реводились на брикетированный корм, смешанный с молоком. Опыт вы-
ращивания безмикробных обезьян в течение 10 месяцев показал хорошие
результаты [322].
Глава 14. Гнотобиотные животные—биомодели
253
Птицы-гнотобиоты и биориски
Методики получения гнотобиотов были также разработаны и адаптированы
и в отношении других сельскохозяйственных животных и птиц. Так, описа-
ны методы получения гнотобиотических ягнят, козлят, телят, свиней, а так-
же гнотобиотических птиц (цыплят, перепелок, индеек, уток). Получение
этих животных основано на тех же принципах, которые описаны для других
лабораторных животных.
Получить безмикробных птиц относительно легко, но дорого — у них
выше, чем у млекопитающих, риск случайного заражения. Процедура их
получения детально описана. Яйца нужно брать от здоровых родителей во
избежание риска инфицирования эмбриона на этой стадии. Наиболее час-
тым источником заражения являются микроорганизмы, находящиеся под
скорлупой и на ее поверхности, поэтому брать можно только яйца с абсо-
лютно чистой скорлупой. Они помещаются в обычный инкубатор, но из-
влекаются оттуда за несколько дней до предполагаемого вылупления птен-
цов. Во время инкубационного периода яйца необходимо поддерживать в
идеальной чистоте. Обращаться с яйцами можно только в перчатках, мыть
или скрести нельзя, так как это может повредить их защитный слой. Инку-
батор также должен быть тщательно вымыт и помещен в зону, очищенную
от пыли.
За 2 дня до вылупления птенцов яйца, предварительно деконтаминиро-
вав, помещают в изолятор. На этой стадии яйца еще раз проверяют и выб-
раковывают те, на которых обнаружены пятнышки, трещинки или грязь. Их
просвечивают и в изолятор помещают только оплодотворенные яйца. Обыч-
но яйца помещают в шлюз, опрыскивают 2%-ным раствором уксусной кис-
лоты и оставляют на 30 минут. Лишь после этого они могут быть помещены
в изолятор. Температура в изоляторе должна быть 37—38° С при относитель-
ной влажности воздуха 70-80%. Увеличить влажность можно с помощью
емкостей с водой или смоченных хлопковых ватных тампонов. Для контроля
условий внутри изолятора туда помещают влажный и сухой термометры.
В период вылупления необходимо поддерживать достаточный доступ возду-
ха в изолятор.
После вылупления изолятор очищают от скорлупы и дают птенцам пищу
и воду. Птенцы быстро учатся самостоятельно есть и пить. Температуру в
изоляторе снижают до 35° Сив течение двух недель постепенно доводят до
комнатной. От перьев птиц и раскрошенной еды появляется большое коли-
чество пыли. Жидкие выделения и разлитая питьевая вода могут существен-
но повысить уровень влажности. Для поддержания влажности на комфорт-
ном уровне необходимо обеспечить хорошее проветривание изолятора. Вы-
ходные фильтры могут забиваться пылью и конденсированной влагой, в этом
случае рекомендуется установить еще одну ступень механической очистки и
254
Линейные животные-биомодели
влагопоглотитель. Вместо выходных фильтров можно использовать альтер-
нативные приспособления.
Выращивать и поддерживать популяцию гнотобиотов и безмикробных
животных, как мы уже подчеркивали, — дорогостоящее занятие. Гнотобиотех-
нология требует высокой и узкопрофильной специализации, наличие квали-
фицированного персонала и многообразия специального оборудования.
Таблица 48
Вирусные инфекции цыплят, потенциально опасные
для человека и животных и искажающие результаты исследований
Возбудитель
Marek's disease
herpesvirus (MD-V)
Infectious laryngo-
tracheitis virus (ILT)
(herpesvirus)
Egg drop syndrome
virus (EDS)
(adenovirus)
Infectious bronchitis
virus (IB)
Avian encephalitis
virus (AE)
(picornavirus)
Newcastle Disease
virus (NDV)
(paramyxovirus)
Infectious bursitis
virus (non-classified
RNA virus)
Avian leucosis virus
(ALV)
Заболевания
Биориск заражения
для человека
И ЖИВОТНЫХ
Болезнь Марека
Цыплята, индейки,
фазаны, другие виды
животных
Инфекционный ларинго-
трахеит
Цыплята, фазаны
Снижение яйценосности
Цыплята
Инфекционный бронхит
Цыплята
Энцефаломиелит
Цыплята, фазаны
Болезнь Ньюкастла
Цыплята, фазаны, мно-
гие виды птиц, человек
Инфекционный бурсит
Цыплята
Комплекс лейкозов:
лимфоидный лейкоз,
миелобластоз, миело-
цитоматоз, эритроблас-
тоз, саркоматоз, остео-
петроз
Цыплята, фазаны,
(в редких случаях дру-
гие птицы)
Пути заражения
Воздушно-капельный
Воздушно-капельный
Оральный
Воздушно-капельный
Через яйца,
оральный
Воздушно-капельный,
оральный
Оральный
Воздушно-капельный
Контагиоэность,
симптоматика и исход
Влияние на результаты
экспериментов
Антитела в желтке,
высококонтагиозное
заболевание и смерть
Антитела в дейтерио-
плазме, высококонта-
гиозное заболевание
Возможны антитела
и вирусы в яйцах
Возможны вирусы
в яйцах или антитела
в желтке, высококонта-
гиозное заболевание
Вирусы в яйце, анти-
тела в желтке, высоко-
контагиозное заболе-
вание, смерть цыплят
Антитела в желтке,
возможны вирусы
в яйцах, высококонта-
гиозное заболевание
и смерть
Антитела в желтке,
вирусы в желтке,
высококонтагиозное
заболевание и смерть
цыплят
Вирусы в яйце, анти-
тела в яйце, высоко-
контагиозное заболе-
вание и смерть
Глава 14. Гнотобиотные животные—биомодели
255
Окончание табл. 48
Возбудитель
Fowl-pox virus
(pigeon-, canary-,
chicken-, pheasant-,
quail- или Agaporni-
dae-pox в зависи-
мости от вида
поражения)
Avian influenza A virus
(orthomyxovirus)
Paramyxovirus
Parainfluenza virus
2и 3
Avian reoviruses
Заболевания
Биориск заражения
для человека
И ЖИВОТНЫХ
Оспа птиц
Цыплята (другие птицы
в зависимости от
штамма вируса)
Не определенные
заболевания
Цыплята, гуси, фазаны,
индейки (другие птицы)
Не определенные
заболевания
Цыплята, индейки,
различные виды птиц
и млекопитающих,
человек
Не определенное
заболевание
Различные виды домаш-
них и диких птиц
Пути заражения
Кожный
Воздушно-капельный,
оральный
Воздушно-капельный,
оральный
Воздушно-капельный
Контагиозность,
симптоматика и исход
Влияние на результаты
экспериментов
Наличие антител и ви-
русов в яйце точно не
доказано, высококон-
тагиозное заболева-
ние
Антитела в желтке,
возможно наличие
вирусов в яйце,
высококонтагиозное
заболевание
Антитела в желтке,
возможно наличие
вирусов в яйце
Синдром пониженного
всасывания, артрит,
тендосиновиит, мио-
кардит у гусят
Таблица 49
Микоплазменные и бактериальные инфекции цыплят, потенциально опасные
для человека и животных и искажающие результаты исследований
Возбудитель
Mycoplasma
gallisepticum
Mycoplasma synoviae
M. meleagridis
Mycoplasma
gallinarium
Pasteurella multocida
Заболевания
Биориск заражения
для человека
и животных
Микоплазмоз
Цыплята, индейки,
фазаны, перепела
Инфекционные
синовииты
Цыплята, индейки,
фазаны, голуби
Не определенные
заболевания
Цыплята, индейки,
фазаны, голуби
Птичья холера
Цыплята, индейки
Пути заражения
Воздушно-капельный
Оральный
Воздушно-капельный
Оральный
Контагиозность,
симптоматика и исход
Влияние на результаты
экспериментов
Наличие возбудителя
в яйце,
высококонтагиозное
заболевание
Высококонтагиозное
заболевание
Возможно наличие
возбудителя в яйце
Высококонтагиозное
заболевание и смерть
только в случае
стресса
Линейные животные-биомодели
Окончание табл. 49
Возбудитель
Salmonella
gallinarium
Haemophilus
paragallinarum
Clostridium
perfringens
Escherichia coli
Listeria
monocytogenes
Mycobacterium avium
Staphylococcus
aureus
Streptobacillus
moniliformis
Заболевания
Биориск заражения
для человека
И ЖИВОТНЫХ
Белая бациллярная
диарея
Цыплята, почти все
птицы, млекопитающие
Контагиозный куриный
ринит
Цыплята, индейки
Некротический энтерит
Различные виды живот-
ных, человек
Колибациллез
Различные виды живот-
ных, человек
Листериоз
Различные виды живот-
ных, человек
Птичий туберкулез
Цыплята, различные
виды птиц и млекопита-
ющих, человек
Стафилококкоз
Цыплята, индейки,
другие виды птиц
Птичий стрептобациллез
Индейки, крысы, мыши,
морские свинки, кроли-
ки, человек
Пути заражения
Оральный
Воздушно-капельный
Оральный
Оральный
Оральный
Оральный, при опре-
деленных обстоятель-
ствах воздушно-
капельный
Кожный, воздушно-
капельный
Через укусы крыс
(содоку или хейвер-
хиллская лихорадка)
Контагиозность,
симптоматика и исход
Влияние на результаты
экспериментов
Возможно наличие
возбудителя в яйце,
высококонтагиозное
заболевание и смерть,
особенно у цыплят
Высококонтагиозное
заболевание
Вьсококонтагиозное
заболевание, в боль-
шинстве случаев
латентное течение,
смерть
Высококонтагиозное
заболевание, особен-
но в случае стресса,
важно как вторичное
проникновение
Низкая распростра-
ненность заболевания
Возможно наличие
возбудителя в яйце,
высококонтагиозное
заболевание и смерть
Изменения кожи,
артриты, иногда
септицемия
Искажение результа-
тов эксперимента
Таблица 50
Инвазивные заболевания цыплят, потенциально опасные
для человека и животных и искажающие результаты исследований
Возбудитель
Eimeria spp.
Histomonas
meleagridis
Заболевания
Биориск заражения
для человека
и животных
Кокцидиоз
Цыплята, индейки,
голуби, гуси
Гистомониаз
Цыплята, индейки
Пути заражения
Оральный
Оральный
Контагиозность,
симптоматика и исход
Влияние на результаты
экспериментов
Высококонтагиозное
заболевание, слабость,
смерть
Энтерит, гепатит
Искажение результа-
тов эксперимента
Глава 14. Гнотобиотные животные—биомодели
257
Окончание табл. 50
Возбудитель
Ascaridia sp.
Capillaria sp.
Heterakis gallinae
Syngamus trachea
Davainea sp.
Hymenolepis sp.
Raillietina sp.
Dermanyssus gallinae
Заболевания
Биориск заражения
для человека
И ЖИВОТНЫХ
Аскаридоз
Цыплята, другие птицы
Капилляриоз
Цыплята, индейки,
фазаны, другие птицы
Гетеракиоз
Цыплята, другие виды
птиц
Сингамоз
Цыплята, другие виды
птиц
Солитерная инвазия
Цыплята, индейки,
голуби
Глистная инвазия
Цыплята, индейки,
голуби
Глистная инвазия
Цыплята, индейки,
различные виды живот-
ных
Дерматит
Цыплята, различные
виды птиц, человек
Пути заражения
Оральный
Оральный, через
яйцо или промежу-
точного хозяина
Оральный
Оральный
Оральный
(промежуточный
хозяин)
Оральный
(промежуточный
хозяин)
Глистная инвазия
Контактный
Контагиозность,
симптоматика и исход
Влияние на результаты
экспериментов
Высококонтагиозное
заболевание, слабость
Высококонтагиозное
заболевание, слабость
Высококонтагиозное
заболевание, слабость
Диспноэ, апатия
Искажение результа-
тов эксперимента
Высококонтагиозное
заболевание, слабость
Высококонтагиозное
заболевание, слабость
Высококонтагиозное
заболевание, слабость
Зуд, анемия, раздра-
жение кожи, высоко-
контагиозное заболе-
вание
Иммунные ответы у гнотобиотов
Гнотобиологические модели важны в изучении истинно первичного иммун-
ного ответа. Учитывая множественность антигенов, представление о первич-
ном иммунном ответе, изучаемом в экспериментах на обычных (природно
контаминированных) животных, справедливо подвергается сомнению, так как
в большинстве случаев мы имеем дело с ранним вторичным иммунным отве-
том [367, 534]. Действительно, изучение ранней фазы онтогенеза иммунного
ответа в присутствии уже имеющихся фоновых естественных антител у обыч-
ных, так называемых нормальных животных, представляется методологичес-
ки неверным. Одним из наиболее интересных объектов для таких исследова-
ний являются безмикробные поросята, в особенности лишенные антигенно-
го воздействия [367]. Систематические исследования онтогенеза иммунного
ответа на безмикробных и безантигенных поросятах позволили понять пути
развития и дифференциации иммунной системы в зависимости от антиген-
ного воздействия. Но небходимо выделить антигензависимую стадию созре-
258
Линейные животные-биомодели
вания мультипотентных некоммитированных Т- и В-клеток и антигензави-
симую стадию их дифференциации и пролиферации. В антигеннезависимой
стадии индуктивная роль в стимуляции развития стволовых клеток в Т-клет-
ки (в тимусе) и В-клеток (в бурсо-эквивалентных условиях) отводится микро-
химической среде этих органов. В то же время поликлональные активаторы
В-клеток не обладают стимулирующим влиянием на иммунологически «дев-
ственные» В-клетки, полученные от безмикробных поросят [367].
Исключение антигенной стимуляции проявляется у безмикробных жи-
вотных в недостаточном развитии лимфоидной ткани, клеточных и гумораль-
ных факторов иммунитета. Лимфоидная ткань у них находится в том исход-
ном состоянии, которое отмечается у новорожденных животных и на основе
которого происходит ее дальнейшее развитие после первого контакта с мик-
робными и другими антигенами при рождении животных. Эти различия
выражены в лимфоидной ткани и лимфатических узлах, дренирующих орга-
ны и ткани, испытывающие наибольшее воздействие микроорганизмов,
в частности кишечник. Так, если вес мезентериальных лимфоузлов у обыч-
ных мышей составляет около 134 мг/100 г веса тела, то у безмикробных та-
кого же возраста этот показатель в 3 раза ниже [534].
В корковом веществе тимуса безмикробных крыс отмечается понижен-
ное содержание лимфоцитов и отсутствие плазматических клеток. Гистоло-
гическими исследованиями лимфоидной ткани показано намного меньшее
количество зародышевых центров у безмикробных животных по сравнению
с обычными. При цитологических исследованиях вторичных лимфатичес-
ких органов у безмикробных поросят наличие плазматических клеток выя-
вить не удавалось. Понижены у безмикробных животных и уровни сыворо-
точных глобулинов, что объясняется сниженной антигенной стимуляцией
по сравнению с обычными животными.
Для гнотобиотов характерно недоразвитие лимфоидной ткани в лимфо-
идных органах, дренирующих слизистые оболочки, контактирующие с мик-
робной средой. Это проявляется в гипоплазии перибронхиальной лимфоид-
ной ткани и ее пониженной лимфоцитарной инфильтрации. У безмикробных
животных отмечается также пониженная масса подчелюстных лимфатичес-
ких узлов. Соответственно установлены и отличия в содержании клеточных
элементов в лимфоидной ткани безмикробных животных. Отношение их
абсолютного числа у обычных и безмикробных мышей составляет: для плаз-
мобластов — 12,3; больших лимфоцитов — 12,3; незрелых плазматических
клеток — 14,4; зрелых плазматических клеток — 8,3; средних лимфоцитов —
3,7 и малых лимфоцитов — 3,0. Различия в лимфоидной ткани у обычных и
безмикробных животных проявляются и в содержании иммунокомпетент-
ных клеток. В лимфоузлах безмикробных животных особенно снижено ко-
личество больших и средних лимфоцитов, больших пиронинофильных
и плазматических клеток.
Глава 14. Гнотобиотные животные—биомодели
259
Пониженные уровни иммуноглобулинов у безмикробных животных не
означают снижения иммунокомпетентности лимфоидной системы и ее спо-
собности к иммунному ответу. Образование антител у безмикробных живот-
ных наблюдается уже через 48 ч после антигенной стимуляции. После ассоци-
ации с гексафлорой у гнотобиотических мышей отмечалось повышение кон-
центрации сывороточных IgGl, IgG2a и IgA, причем уровни IgA и IgGl даже
превышали соответствующие значения у обычных животных [496].
У безмикробных поросят в возрасте 1,5 месяцев установлены резко сни-
женные уровни иммуноглобулинов классов М и G, в то время как уровни
IgA были одинаково низкими в сыворотке крови как у безмикробных, так
и обычных поросят. Эти показатели у гнотобиотов повышались более чем в
2 раза после моноассоциации с непатогенной кишечной палочкой Escherichia
coli 083. Особенно высокие уровни IgG, не отмечавшиеся у безмикробных
животных, выявлялись у гнотобиотических поросят, моноассоциированных
с непатогенными микроорганизмами Bifidobacterium bifidum. Показатели
сывороточных фракций глобулинов и агглютининов у безмикробных живот-
ных в 1,5—3 раза ниже, чем у обычных животных. В лимфоидных элементах
слизистой оболочки кишечника синтеза иммуноглобулинов не выявлено,
в то время как у обычных животных, а также у гнотобиотических мышей
после конвенционализации отмечается секреция в просвет кишечника им-
муноглобулинов, в основном IgA класса.
В лаборатории лекарственной профилактики НЦБМТ РАМН исследова-
лось влияние инсулина (1 ед/100 г веса в день, в течение недели) на форми-
рование иммунного ответа на эритроциты барана (ЭБ; 50% взвесь по 0,1 мл
под подошвенный апоневроз обеих лапок) у крыс линии Wistar и крыс-гно-
тобиотов. В качестве контроля использованы крысы обеих групп (без инсу-
лина).
На 3-й и 7-й день после иммунизации ЭБ в лимфоузлах и селезенке
исследовали число РОК и АОК на 103 ядерных клеток, коэффициент плаз-
матизации лимфоузлов и уровень гемагглютининов в крови. Результаты
показали, что фоновая активность Т- и В-систем у интактных гнотобиотов
несколько ниже, чем у Wistar (по числу РОК и АОК). У контрольных гното-
биотов на 3-й день после иммунизации ЭБ число РОК в лимфоузлах
и селезенке было достоверно меньше (27,8 ± 2,9 и 37,0 ± 1,1), чем у Wistar
(соответственно 45,0 ± 1,8 и 50,1 ± 2,1; Р < 0,01). На 7-й день эти величины
несколько выравнивались. У крыс, получавших инсулин, во все сроки на-
блюдения (в обеих группах) в лимфоузлах и селезенке их число резко (вплоть
до полного исчезновения) уменьшалось.
Число АОК на 103 ядерных клеток у гнотобиотов также было меньшим на
3-й день (в лимфоузле — 44,3 ± 2,2, в селезенке — 52,3 ± 2,2), чем у Wistar
(58,0 ± 2,6 и 67,8 ±2,1; Р < 0,01). Введение инсулина значительно увеличи-
вало число АОК в лимфоузлах и селезенке крыс обеих групп: на 3-й день у
260
Линейные животные-биомодели
крыс Wistar их число в селезенке было равно 96,1 ± 7,1, в лимфоузлах —
178,0 ± 10,9 (Р < 0,01); у гнотобиотов соответственно 80,0 ± 8,2 и 71,7 ± 6,0
(Р < 0,01 в сравнении с контрольными данными). На 7-й день опыта у крыс
Wistar, получавших инсулин, сохранялось увеличение числа АОК в лимфоуз-
лах и селезенке (в сравнении с контролем Р < 0,01), тогда как у гнотобиотов
их число уменьшалось.
В лимфоузлах развивалась пролиферация клеток плазматического ряда,
а в крови выявлялись гемагглютинины. Однако их различие у гнотобиотов
и контрольных крыс было несущественным. Кроме того, у гнотобиотов, по
сравнению с контролем, на 7-й день после иммунизации под влиянием
инсулина происходит отчетливое угасание иммунного ответа, тогда как у
крыс Wistar стимулирующий эффект на гуморальное звено сохраняется от-
четливым (Р < 0,01). Полученные результаты свидетельствуют о том, что
инсулин тормозит клеточное звено иммунитета и стимулирует гуморальное,
хотя механизм такого эффекта инсулина до конца не выяснен, тем не менее,
можно констатировать, что по интенсивности формирования иммунного
ответа крысы-гнотобиоты значительно уступают таковому у интактных жи-
вотных.
В свете приведенных данных возникает естественный вопрос: нужны ли
такие сложные биомодели, как SPF-животные и гнотобиоты?
Этот вопрос, и не без оснований, волнует исследователей уже десятиле-
тия. Высокая стоимость, огромная трудоемкость, высокие технологии, с
одной стороны, и относительно скромные результаты — с другой, делают
поставленный вопрос отнюдь не праздным. Тем не менее, опыт жизни гово-
рит о том, что наука — это лучший бизнес, но дает выход не ранее чем через
10 лет. Действительно, многих результатов мы можем достигнуть если не на
конвенциональных, то на менее дорогих и менее капризных, чем гнотобио-
ты, MD-животных. Но всегда ли это удаётся в полной мере?
Думаю, что каждому вдумчивому и скрупулезному исследователю стано-
вится ясно, что понимание сложнейших фундаментальных механизмов фор-
мирования иммунной защиты, интегрированной с формированием термо-
регуляторных процессов, воспаления, лихорадочных состояний, регенрации
и ауторепарации, вряд ли достижимо без использования гнотобиотов. Ины-
ми словами, SPF-животные и гнотобиоты постепенно, но стабильно займут
свою реальную нишу в научных исследованиях и биомоделировании слож-
нейших процессов, протекающих в организме человека на уровне систем
вирус—хозяин, бактерия—хозяин и т.д., а также при поиске принципиально
новых химиотерапевтических средств.
Раздел IV ПЛАН И РОВАН И Е
МЕДИКО-БИОЛОГИЧЕСКИХ
ЭКСПЕРИМЕНТОВ
Наука — это в лучшем случае здравый смысл —
строго точный в наблюдении и милосердный
к ошибкам в логике.
Т. Гексли
263
Глава 15 ПРИСТРАСТНОСТЬ,
РЕПРЕЗЕНТАТИВНОСТЬ
И КОМБИНАТОРИКА ВЫБОРКИ
Итак, предположим, что на основании вышеизло-
женного уже сформулированы цели, определены
методы исследования и оптимальные животные.
Каковы же дальнейшие шаги на пути к получению
конкретных результатов экспериментов и биологи-
ческому моделированию? Естественным продолже-
нием этого процесса является, конечно же, плани-
рование экспериментов с лабораторными животны-
ми и оценка их результатов.
Процесс планирования экспериментов может
быть различным по объему, поскольку зависит от
конечной цели, которую ставит перед собой иссле-
дователь. Испытания новых хирургических техно-
логий представляют собой наиболее простую фор-
му экспериментов, так как они могут проводиться
с использованием лишь одного животного. Их про-
тивоположностью являются многосторонние экс-
перименты, в ходе которых исследуются тысячи
животных, получивших составляющие апробируе-
мой субстанции в различных дозах и при разных
условиях.
Использование статистики в первоначальном
процессе планирования всех экспериментов, за ис-
ключением простейших, имеет большое значение,
так как позволяет не только избежать ошибок, но и
повысить шансы успешного проведения исследова-
ния. Ведь в конечном итоге мы осознанно или не-
целенаправленно осуществляем это в целях биомо-
делирования.
264
Планирование медико-биологических экспериментов
Исследователь должен четко сформулировать вопросы, на которые он
надеется ответить в ходе эксперимента. Если для эксперимента используют-
ся животные, важно, чтобы поставленные вопросы имели значимость, но-
визну и прикладное значение. Недобросовестное планирование эксперимента
может привести к расточительному использованию жизней животных, что
является недопустимым. При планировании экспериментов с участием жи-
вотных протокол должен составляться в письменной форме и содержать
информацию о цели исследования, поставленных вопросах, проверяемых
гипотезах и о предполагаемом практическом и научном значении результа-
тов для прототипа модели, в нашем случае — человека. Важно также обес-
печить использование возможно лучших методик и технологий.
Высококачественные животные, хорошее оборудование и тщательный
статистический анализ не смогут исправить упущения, возникшие в резуль-
тате плохого планирования. Необходимо также минимизировать последствия
использования участниками любого качественного исследования или про-
екта различных методик в ходе проведения эксперимента. Все эти факторы
должны быть изучены и включены в протокол, а в дальнейшем представле-
ны в виде биологической модели. Другие факторы, такие, как вид и катего-
рия животных для эксперимента, также должны быть учтены на стадии пла-
нирования. Например, в случае, когда требуется минимизировать различия
между животными, в протокол вносится положение об использовании ис-
ключительно инбредных линий. Для целей других экспериментов могут,
напротив, понадобиться гетерогенные животные.
Для того чтобы сделать правильный выбор, исследователь должен обла-
дать высоким уровнем знаний о различных факторах, влияющих на экспе-
римент. Вид клеточного оборудования и такой на первый взгляд незначи-
тельный фактор, как его размещение, должны учитываться, поскольку мо-
жет потребоваться его разнообразить. Например, размещение всех клеток с
тестируемой группой животных на верхних стеллажах может привести к нео-
жиданным результатам, если степень освещения повлияет на ход экспери-
мента. Исследования должны планироваться таким образом, чтобы исклю-
чить все переменные, кроме тех, которые необходимы для эксперимента. На-
пример, количество, качество и разнообразие корма является важной пере-
менной, которую часто недооценивают. Квалифицированный учет всех необ-
ходимых факторов и отбор значимых параметров составляют основу планиро-
вания экспериментов.
Область математики, в которой изучаются вопросы о том, сколько раз-
личных комбинаций, подчиненных тем или иным условиям, можно соста-
вить из заданных объектов, называется комбинаторикой. Теоретическое ис-
следование вопросов комбинаторики предприняли в XVII веке французские
ученые Паскаль и Ферма. Дальнейшее развитие комбинаторики определило
её место в биомоделировании.
Глава 15. Пристрастность, репрезентативность и комбинаторика выборки 265
Предположим, в каждую серию экспериментов нами взято по 8 живот-
ных. Нам необходимо получить независимый, т.е. непристрастный ответ через
каждый час эксперимента, продолжающегося в течение 4 часов, хотя бы от
одного животного. Сколькими способами мы можем этого достигнуть?
Рассмотрим эту задачу в общем виде. Пусть даны предметы или эивотные,
относящиеся к п различным видам. Из них составляют всевозможные расста-
новки по к предметов в каждой, или, как будем в дальнейшем кратко гово-
рить, к-расстановки. При этом в расстановки могут входить и предметы одно-
го вида, а две расстановки считаются различными, если они отличаются друг
от друга или видом входящих в них предметов, или порядком этих предметов.
Расстановки описанного типа называются k-размещениями с повторения-
ми из элементов п видов, а число всех таких расстановок обозначают А„ .
Итак, мы хотим доказать, что число ^-размещений с повторениями из
элементов п видов равно
Лк=пк. (49)
Доказательство проводится с помощью математической индукции по к —
числу элементов в размещении при фиксированном значении п. При к = 1
ответ ясен — каждое размещение (с повторениями) состоит только из одно-
го элемента, и разные размещения получаются, если брать элементы различ-
ных видов. Но так как число видов равно я, то и число размещений равно п.
Итак, А{п =п в соответствии с формулой (49).
Предположим теперь, что уже доказано равенство Ак~х = пк~1 , и рас-
смотрим ^-размещения с повторениями. Все такие размещения можно по-
лучить следующим образом. Возьмем любое (к— 1)-размещение (с повторе-
ниями) (ар ..., ак j) и припишем к нему элемент ак одного из имеющихся
п видов. Мы получим некоторое ^-размещение (ар ..., ак1 ак). При этом
ясно, что из каждого (к— 1)-размещения получится столько ^-размещений,
сколько есть различных видов элементов, то есть п размещений. Очевидно,
что, действуя описанным образом, мы не пропустим ни одного £>разме-
щения и ни одного не получим дважды, если (ар ..., ак 7) Ф (Ьр ..., Ък 2),
или, если акФ Ьк , то (ар ..., ak_lf ак) Ф(Ь}, ..., Ьк_{ Ьк). Поэтому число к-
размещений с повторениями из элементов п видов в п раз больше, чем
число (к— 1)-размещений с повторениями из элементов тех же видов. Та-
ким образом, А* = пАпк~1 . Но мы считаем уже доказанным, что Ак~1 = пк~1 ,
поэтому
Ак=п-пк~1=пк . (50)
Тем самым равенство (49) доказано для всех значений к. Формула (49)
встречается в целом ряде решений биомоделирования. Мы рассмотрим те-
перь решение поставленной выше задачи. Восемь животных могут распреде-
литься между четырьмя одночасовыми отрезками 48 способами, из них
266
Планирование медико-биологических экспериментов
в З8 случаях — на любом одночасовом отрезке, 28 — на двухчасовых отрезках,
и в одном случае на трехчасовых отрезках не будет участвовать ни одно жи-
вотное. По принципу включений и исключений получаем ответ 48 — 4 х 38+6 —
— 4 = 40 824 варианта. Иными словами, всего лишь для 8 животных, взятых
в эксперимент с четырьмя выборками, мы имеем около 41 тысячи возмож-
ных комбинаций. А возможно ли эту задачку упростить? Да, возможно, если
мы перейдем в иную систему счисления
Наряду с десятичной системой счисления применяются и другие — дво-
ичная, троичная, восьмеричная и т. д. В я-ичной системе счисления исполь-
зуются я цифр. Подсчитаем, сколько в я-ичной системе натуральных чисел,
записываемых ровно к знаками. Если допустить записи, начинающиеся с
нуля, то каждое А;-значное число в я-ичной системе счисления можно рас-
сматривать как размещение с повторениями, составленное из к цифр, при-
чем цифры бывают я видов. По формуле (49) получаем, что количество чи-
сел, имеющих такую запись, равно пк.
Но для натуральных чисел не применяют записей, начинающихся с нуля.
Поэтому из полученного значения пк надо вычесть количество чисел, я-ич-
ная запись которых начинается с нуля. Если отбросить у этих чисел первую
цифру — нуль, то получим {к— 1)-значное число (быть может, также начина-
ющееся с нуля). Таких чисел по формуле (49) будет пк~К
Значит, общее количество А>значных чисел в я-ичной системе счисления
равно:
пк-пк-1 = пк~1х(п - 1). (51)
Например, в десятичной системе счисления имеем 103х9 = 9000 че-
тырехзначных чисел; из 10 000 чисел от 0 до 9999 надо отбросить тысячу
чисел, а именно числа от 0 до 999.
Полученную нами формулу можно вывести и иным способом. Ведь в
А;-значном числе, записанном по я-ичной системе счисления, первой циф-
рой может быть любая из цифр 1, 2, ..., я—1, а второй и последующими —
любые из цифр 0, 1, 2, ..., я—1. В эксперименте с 8 животными их комбина-
торика распределится следующим образом: на первый час у нас я—1 живот-
ное, а на остальные к—1 часов — по я кандидатов. Отсюда легко получаем,
что искомых чисел может быть (я — 1) пк~1.
Рассмотрим ещё один пример. Например, необходимо получить гибриды
Fp т. е. первое поколение животных, полученных от скрещивания двух ин-
бредных линий. Для этих целей отобраны 5 самок и 3 самца. Необходимо
разбить их на две группы по 4 особи, чтобы в каждой было хотя бы по од-
ному самцу. Если в первой линии один самец, а в другой — два, то самцов
можно разбить на группу 3 самок из 5. Это можно сделать С35 = 10 спосо-
бами. Всего по правилу произведения получаем 3 х 10 = 30 способов разбив-
ки на группы. Это уже более реально.
Глава 15. Пристрастность, репрезентативность и комбинаторика выборки 267
Применение методов математической статистики к обработке наблюде-
ний основано на глубокой аналогии между производством наблюдений и
отбором из некоторой генеральной совокупности. При этом в качестве ге-
неральной совокупности рассматривается чисто гипотетическая совокуп-
ность всех возможных результатов наблюдений при данном комплексе ус-
ловий испытания. Отбор данных из этой совокупности производится в про-
цессе наблюдений независимо от нашей воли. Благодаря этому основным
фактором отбора наблюдений всегда является случайность, что и позволяет
применять для обработки наблюдений основные положения теории вероят-
ностей.
Отбор данных, происходящий помимо нашей воли, можно назвать есте-
ственным отбором; именно таким является отбор данных при наблюдениях.
Однако при производстве наблюдений и при дальнейшей их обработке часто
возникает необходимость и в других, искусственных способах отбора. Напри-
мер, при анализе субстанций приходится делать пробы из разных партий,
чтобы нейтрализовать возможную неоднородность материала; отбор проб
при этом находится целиком в нашем распоряжении. Во время контроля за
производством лекарств приходится выбирать образцы из общей продук-
ции. При различных клинических и демографических исследованиях также
нужен предварительный отбор объектов для изучения. Примеры таких слу-
чаев, когда исследователь вынужден делать сознательный выбор, можно еще
долго продолжать. Отметим лишь, что даже заготовленный цифровой мате-
риал может нуждаться в дополнительном отборе в целях сокращения объе-
ма, удаления неподходящих данных, проверки правильности и добросовес-
тности полученных данных и подготовительных расчетов. Подобный отбор
также производится исследователем целиком по его воле.
Существует много способов искусственного отбора; выбор того или ино-
го способа зависит от цели отбора, от поставленной задачи. В самом общем
плане способы отбора делятся на две группы: пристрастные и репрезента-
тивные (см. гл. 16).
Пристрастными называются такие способы отбора, при которых отбира-
ются элементы по какому-либо заранее намеченному признаку, при этом
проверке подлежат все элементы совокупности, из которой делается отбор.
Например, из совокупности чисел отбираются п самых больших или отбира-
ются все числа, не достигающие нужной величины. Пристрастный отбор
применяют и для того, чтобы изъять все наблюдения с нарушенными усло-
виями испытания. Так, при идентификации линейной принадлежности
животных, вычислении функций обобщенного Мехаланобисова расстояния
(см. гл. 33) морфологических параметров животных, не соответствующих
неким заданным краниометрическим характеристикам, выделяют в отдель-
ную группу и в дальнейшем не учитывают данные по больным и смешанно-
породным особям. Пристрастный отбор является важной стадией экспери-
268
Планирование медико-биологических экспериментов
мента; его задача состоит обычно в том, чтобы устранить все заметные
нарушения условий испытания. С помощью пристрастного отбора нередко
удается ликвидировать те или иные доминирующие (несимметричные) фак-
торы, нарушающие нормальность распределения.
Рассмотрим простой случай пристрастного отбора, например, при иссле-
довании действия психотропных средств у крыс по методам УРПИ или УРАИ.
Возникает вопрос о необходимости учета важнейших параметров, в частно-
сти теплового колебания и взаимодействия молекул в теле животного. Хотя
взаимодействие исследуемых веществ, несомненно, происходит на молеку-
лярном и даже атомарном уровне, необходимость учета этого фактора ка-
жется странной. На самом деле, какова связь условно-рефлекторных реак-
ций с термодинамическими молекулярными процессами?
Атомы колеблются вокруг положения равновесия с большими скоростя-
ми, но движение это хаотическое или, скорее, псевдохаотическое. Атомы и
частицы, движущиеся в одну сторону, уравновешиваются атомами, летящи-
ми в другую. Но представим себе ситуацию, когда большинство атомов дви-
жется в одну сторону, не уравновешиваясь с другой стороны. Тогда УРПИ
и УРАИ будут извращены по непонятной экспериментатору причине. Допу-
стимо ли это? Теоретически — да, практически — сомнительно. Как же быть
с учетом такого фактора? Но оставим это на совести пристрастного отбора
исследователя.
Пристрастный отбор всегда является сознательным с ясной характерис-
тикой данных, подлежащих отбору, поэтому он редко вызывает затруднения.
Сложнее обстоит дело со второй группой случайных способов отбора,
о которой речь пойдет в следующей главе.
269
Глава 16 СЛУЧАЙНЫЙ ОТБОР И ЧИСТОТА
ЭКСПЕРИМЕНТА
Способы отбора называются репрезентативными,
если отобранная группа элементов достаточно пол-
но характеризует всю совокупность, из которой был
сделан отбор. Разумеется, как бы ни был удачен
отбор, в суждениях о всей совокупности будет эле-
мент случайности. Более того, некоторые особен-
ности всей совокупности вообще не отразятся на
отобранной группе элементов, поэтому «репре-
зентативность» отбора, вообще говоря, является
относительной и связана с конкретной числовой
характеристикой совокупности, которая изучается
с помощью отобранных элементов.
Репрезентативный отбор применяется тогда, ког-
да судить о характеристиках совокупности, исполь-
зуя все ее элементы, невозможно либо слишком
трудно из-за того, что эта совокупность чересчур
велика, а возможно, и не вся доступна анализу. Если
объем Л^этой совокупности очень велик, то его прак-
тически можно считать бесконечным. В этом слу-
чае заданную совокупность можно рассматривать
как генеральную, а отобранные элементы как вы-
борку, применяя в дальнейшем все достижения об-
щего выборочного метода. Если же N не очень ве-
лико по сравнению с количеством п отобранных
элементов (скажем, iV < 10 п), то с числом N нужно
считаться при интерпретации результатов отбора.
Количество отбираемых элементов можно опре-
делить, если известна дисперсия ст2 изучаемой ха-
рактеристики (по каким-либо предыдущим дан-
270
Планирование медико-биологических экспериментов
ным). Это количество будет зависеть от той точности, с которой мы хотим
получить значение характеристики. Если эта точность задана допустимой
дисперсией s2, то п определяется из формулы
которая при очень большом N преобразуется в известную нам уже формулу
S2=~. (53)
П
Последнюю формулу легко объяснить: а2 есть как бы генеральная дис-
персия, a s2 есть дисперсия среднего выборки из п элементов, которая всегда
в п раз меньше «одиночной» дисперсии.
Допустим, что нам известна дисперсия, связанная с неоднородностью
вещества и равная а2 = 0,20. Допустим, что нам нужно знать содержание
вещества с дисперсией s2v не превосходящей 0,40, причем на методику ана-
лиза приходится дисперсия s22 = 0,35 (ошибка воспроизводимости). Это зна-
чит, что на долю неоднородности допускается оставить дисперсию s2< 0,05.
Если в нашем распоряжении N = 50 проб вещества, то необходимое число
проб п определится из равенства
0Ж50-1Л
(54)
50-1
\ У
то есть равно 5 (точное решение уравнения равно 40/9).
Выбор того или иного способа репрезентативного отбора зависит от сте-
пени наших знаний о всей совокупности. Если, например, нам известно, что
элементы в совокупности расположены случайным образом, то можно при-
менять механический отбор — отбирать каждый пятый или каждый десятый
и так далее элемент. Если же в последовательности элементов имеется неко-
торая ритмичность, то нужно применять аритмичный отбор, например, в
первой десятке брать первый элемент, во второй десятке — второй и т. д.
Скажем, анализируя один раз в день качество продукции, мы не должны
брать пробы в одно и то же время суток.
Иногда вся совокупность заведомо разбивается на отдельные части, ко-
торые желательно равномерно учесть. В этом случае применяют типический
отбор, т. е. отдельно отбирают элементы из каждой части и лишь, потом
сводят их в общую группу.
Так, анализируя виды или линии животных, мы должны изначально рас-
членить их на группы сообразно требованиям, заложенным при формулиро-
вании задач и целей эксперимента. При этом оправдана избыточность вы-
борки каждой группы. Затем мы вправе в каждой выборке отобрать для эк-
сперимента нужное число животных. В тех случаях, когда о всей совокупно-
сти ничего не известно, единственной гарантией репрезентативности может
служить случайный отбор. Для того чтобы отбор был случайным, нужно все
Глава 16. Случайный отбор и чистота эксперимента
211
элементы совокупности пронумеровать; номера отобранных элементов дол-
жны образовывать случайную последовательность чисел. Проверить случай-
ность какой-нибудь последовательности чисел нетрудно хотя бы методом
серий. Но здесь речь идет о том, чтобы создать случайную последователь-
ность, собственной волей имитировать случай.
Имитация случая является весьма трудной задачей. Как бы мы ни стара-
лись, выписывая числа, делать их случайными, в них обязательно проявится
(особенно при больших объемах) какой-то бессознательно выбранный план.
Почти всегда можно узнать, как получена последовательность, «подделать»
случайность трудно. Тем не менее, в очень многих исследованиях случайный
отбор или случайное перемешивание или рандомизация данных настоятель-
но необходимы. Как же быть в таких ситуациях? На помощь случаю может
прийти сам случай. Например, если нужно 10 мышей для эксперимента, то
из клетки, содержащей 50 или 100 животных, можно их отобрать. Случай-
ность вытянутых мышек будет полностью обеспечена. Однако стоит предо-
стеречь экспериментатора: ведь те, которых легче поймать, менее подвижны
и могут быть больны.
Можно воспользоваться и другим способом. Если взять, например, вось-
мизначные таблицы логарифмов (или синусов, тангенсов и т. п.), то после-
дние три-четыре цифры в табличных числах будут абсолютно случайными.
Возьмем подряд нужное количество двух-, трех- или четырехзначных чисел,
составленных из этих последних цифр, и случайность выбора нам вновь будет
гарантирована. Для удобства пользования, получаемые, таким образом слу-
чайные числа выписывают в виде специальных таблиц.
Пользоваться таблицей случайных чисел очень просто. Допустим, нужно
отобрать 10 элементов из совокупности, содержащей 100 элементов. Прону-
меруем все элементы от 00 до 99. Затем, начиная с любого места таблиц,
возьмем две последние цифры десяти идущих подряд чисел. Получившиеся
номера и покажут, какие элементы надо отобрать. Так, начиная с первого
числа, мы получим номера: 82, 49, 18, 48, 09, 50, 17, 10, 37, 51... (повторяю-
щиеся цифры надо опустить).
Выбранную последовательность случайных чисел в дальнейшем никоим
образом нельзя изменять. Нужно помнить: именно случайность служит ос-
новой применения методов математической статистики, именно она обес-
печивает репрезентативность отбора. Нарушение случайности, как правило,
ведет к искажению результатов. Аналогично отбору производится случайное
перемешивание элементов. При этом нужно выписывать случайные номера
до тех пор, пока они не охватят все заданные элементы. Это и обеспечит
репрезентативность искомой выборки.
Прежде чем приступить к выбору числа наблюдений или исследований,
следует выполнить ряд процессуальных шагов. Законодательства, акты, инст-
рукции или иные документы большинства стран мира предусматривают, что
272
Планирование медико-биологических экспериментов
каждое исследование должно проводиться в соответствии с утвержденным
письменным протоколом, содержащим цель исследования и все методы его
проведения. В протоколе должна быть отражена следующая информация:
/ описательный заголовок и цель исследования;
/ название, химическая формула и код тестируемых и контрольных суб-
станций;
/ наименование заказчика и исполнителя эксперимента;
/ информация о статистических методах, которые использованы при пла-
нировании эксперимента, и применении методов рандомизации;
/ способ идентификации тестируемых животных {пристрастный или типи-
ческий отбор);
/ номер, вес, пол, вид, линия и возраст животного, а также источник его
происхождения (вносятся по необходимости), а также количество живот-
ных в эксперименте и принципы их отбора {комбинаторика, аритмичный,
типический или случайный отбор);
/ план проведения эксперимента с указанием пристрастного отбора и опи-
санием всех методов исключения оценки хода и результатов с предубежде-
нием {репрезентативность);
/ состав корма и растворителей, эмульгаторов и других материалов, исполь-
зуемых для получения растворов и суспензий для контроля субстанций.
Описание должно содержать сведения о допустимых уровнях загрязнения
корма, превышение которых способно повлиять на ход эксперимента и
исказить его результаты {метод исключения, пристрастный отбор);
/ дозировка тестируемой и контрольной субстанции, выраженная в милли-
граммах на килограмм веса или иной форме, а также метод, путь и частота
введения {в системе СИ);
/ вид и периодичность анализов и замеров, используемый математический
аппарат обработки данных {статистические, дифференциальные, теория
автоматического регулирования и другие методы);
/ требования к ведению письменного или иного учета и хранения информа-
ции;
/ дата утверждения протокола заказчиком и дата и подпись руководителя
исследований.
Пересмотр протокола и все, вносимые в него изменения, а также причи-
ны, послужившие этому, должны быть документально оформлены, датиро-
ваны, подписаны руководителем исследования и приложены к протоколу.
Количество животных, необходимых для эксперимента, зависит от целого
ряда факторов: целей эксперимента, степени требуемой точности, ожидае-
мых различий последствий введения тестируемой субстанции, структуры и
методов анализа результатов. Эти факторы также должны быть учтены в
процессе планирования эксперимента, что требует участия в планировании
специалиста в области статистики.
273
Глава 17 ПРЕДЕЛ И ДОСТАТОЧНОСТЬ
ЧИСЛА НАБЛЮДЕНИЙ
Если для выявления результатов требуется сравнить
две группы животных — подвергшихся воздействию
и контрольных, то количество животных, необхо-
димых для демонстрации действительных отличий,
будет зависеть от предполагаемой степени этих от-
личий. Степень отличий иногда можно предполо-
жить исходя из результатов предыдущих экспери-
ментов и, соответственно, определить необходимое
количество животных. В случае если требуется боль-
шая точность, количество животных можно увели-
чить. Увеличение их количества целесообразно лишь
до определенного уровня. Излишне большая выбор-
ка влечет за собой статистическую зависимость и
искажение результатов.
Увеличение числа параллельных наблюдений п
является основным способом повышения точности
статистического анализа. Действительно, среднее
выборки объема п имеет дисперсию в п раз меньше,
чем одиночные наблюдения. Поэтому для опреде-
ления необходимого числа наблюдений достаточно
знать генеральную дисперсию наблюдений а2 и до-
пустимую дисперсию результата s2; при этом
а2
» = 7- (55)
Увеличивая число параллельных наблюдений,
можно неограниченно повышать точность найден-
ного результата, лишь бы только в процессе наблю-
дений не менялись условия испытания. Именно этим
274
Планирование медико-биологических экспериментов
приемом пользуются, чтобы уменьшить доверительный интервал при неиз-
менной доверительной вероятности.
Сложнее обстоит дело с выбором числа наблюдений при проверке гипо-
тез. Это число должно одновременно служить двум целям: уменьшать веро-
ятность а ошибки первого рода (отклонение правильной гипотезы) и вероят-
ность р ошибки второго рода (принятие неверной гипотезы). Первая веро-
ятность совпадает с принятым уровнем значимости р. Вторая вероятность
зависит от многих факторов, и, в первую очередь, от того, насколько невер-
на принимаемая гипотеза. Поэтому под Р понимают обычно наибольшую
возможную вероятность ошибки второго рода при любых отклонениях от
правильной гипотезы. Контроль ошибок второго рода является трудной за-
дачей и требует специальных методов проверки гипотезы. Общий принцип
такого контроля мы поясним на примере конкретной гипотезы.
Допустим, изучается нормально распределенная случайная величина £, с
дисперсией а2. Высказывается гипотеза, что генеральное среднее этой вели-
чины а < О (односторонняя гипотеза). Требуется проверить эту гипотезу по
одному наблюдению х. Если мы хотим, чтобы вероятность ошибки первого
рода была не больше а, мы должны считать гипотезу неверной, только если
х > их_а<з, где Wj_a — квантиль стандартного нормального распределения.
Действительно, с доверительной вероятностью 1 — а справедлива односто-
ронняя оценка
a > х - ы,_аа, (56)
которая позволяет утверждать, что а > О лишь при х > их_аа.
Если окажется, что х< и{_а(5, то мы будем вынуждены рассматриваемую
гипотезу признать справедливой, хотя, возможно, допустим при этом ошиб-
ку второго рода. Вероятность неравенствах < их_а<з зависит от того, каким на
самом деле является а. Если обозначить через F(x) функцию нормального
распределения со средним а и дисперсией а2, то
P{x<ul_aG} = F(u]_aG). (57)
Используя функцию Лапласа, получим:
Р{х<и1_ао} = ±+Ф
|Ч_аа-оЛ 1
= -+Ф
2
щ_ас—
(58)
Если a > 0, то принимаемая гипотеза неверна. Однако при этом а может
быть сколько угодно близким к нулю, а это значит, что
(59)
будет сколько угодно близко к
Ф("1-а)=т-а
2 (60)
Глава 17. Предел и достаточность числа наблюдений
275
Иными словами, максимальная вероятность ошибки второго рода равна
Р=Д+Ф(и,_а) = 1-а. (61)
Например, при обычном уровне значимости а = 0,05 будет (3 = 0,95, т. е.
мы почти всегда ошибемся и примем неверную гипотезу.
Итак, желая застраховать себя от несправедливого отклонения верной
гипотезы, мы впадаем в другую крайность, объявляя правильными почти
сплошь неверные гипотезы. Точно такая же картина возникнет, если в каче-
стве отправной точки взять фиксированную малую вероятность р\ Для того
чтобы вероятность принять неверную гипотезу не превышала (3, мы должны
будем отклонять гипотезу уже при х > —иХа<5. А это значит, что большой
станет вероятность а, по-прежнему равная 1 — |3. Соотношение вероятнос-
тей а и Р выражается площадями участков под нормальной кривой. Пло-
щадь, расположенная справа от критического значения, равна а, располо-
женная слева — равна р\ Стараясь уменьшить ос, мы сдвигаем это критичес-
кое значение вправо, стараясь уменьшить (3 — влево. Ясно, что с помощью
единого критического значения невозможно даже сделать так, чтобы обе эти
вероятности были меньше 1/2. На практике обычно вероятности ошибок
обоих родов задаются заранее и являются малыми числами (не больше 0,1).
В этом случае при проверке гипотезы приходится пользоваться одновременно
двумя критическими значениями: считать гипотезу верной, если х < —м,_оО",
и неверной, если х > и{_аа. Такой подход обеспечивает заданный уровень ве-
роятностей ошибок первого и второго рода, однако при этом появляется
участок неопределенности [—и{аО, и^а]. Длина этого участка
5 = (Wj_p + «!_a) a (62)
называется неопределенностью критерия. Вопрос о том, какой вывод делать в
случае, если я: попадет на участок неопределенности, решается примени-
тельно к условиям задачи; чаще всего в этом случае гипотеза объявляется
сомнительной и подвергается повторному анализу.
Неопределенность критерия сильно ухудшает эффект статистического ана-
лиза гипотез, поэтому ее всячески пытаются, если не устранить вообще (мы
уже видели, что это невозможно), то, по крайней мере, сделать как можно
меньше. Непосредственно из формулы (62) видно, что это можно сделать и
при неизменённых заданных а и (3, если уменьшать дисперсию а2. Уменьшать
дисперсию можно, повышая точность методики. Но более универсальным и
надежным средством является увеличение числа параллельных наблюдений.
Действительно, если вместо одиночного провести п параллельных на-
блюдений и рассмотреть их среднее х, то его дисперсия будет
2 а2
о—=—
X П *
276
Планирование медико-биологических экспериментов
Проверяя гипотезу по значению х, мы должны будем принять ее, если
а а
х<-и1-р~7= , и отвергнуть, если *>-wi-p-7= .
yjn у/П
Неопределенность станет теперь равной
Ь = (их_р+щ_а)-= 5 (63)
т. е. уменьшится в V« раз.
В практических задачах допустимая неопределенность критерия задается
обычно заранее, наряду с вероятностями аир. Это позволяет сразу же оп-
ределить необходимое число параллельных наблюдений:
« = ("l-P+"l-a) "g . (64)
Как бы тщательно мы не изучали и использовали математические, и в
частности, статистические методы, мы не должны упускать конечную цель -
биологическое моделирование процесса, функции, эффекта в плане строго-
го переноса вновь полученных данных на прототип, т.е. на человека.
Обратимся в этом ключе к случаям планирования фармако-токсиколо-
гических экспериментов. Прежде чем осуществить конкретные исследова-
ния, необходимо осуществить доэкспериментальные мероприятия — от это-
го зависит корректность полученных результатов. Необходимо весьма строго
подойти к выбору конкретных биологических моделей, их соответствию для
исследования совершенно определенных субстанций, веществ или препара-
тов. Без такого подхода, выбранная биомодель может не только не соответ-
ствовать прототипу, но и заведомо извращать результаты экстраполяции.
Это особенно важно в случае использования биомоделей второго поряд-
ка т. е. при альтернативном моделировании. Удачный логический анализ
в этом направлении осуществлен Г.Н. Красовским [386]. Во избежание оши-
бок при оценке генотоксичности и канцерогенности объектов окружающей
среды, последствия которых для человека становятся негативными, должны
быть представлены данные с показателями суммарной мутагенной активно-
сти. Следует учитывать, что существенный вклад в токсичность исследуемых
субстанций могут вносить не сами исследуемые вещества, а содержащиеся
в них примеси.
Очевидно, что в таких случаях прямая экстраполяция данных с биотестов
на человека некорректна. Использование альтернативных моделей в токси-
кологических оценках должно основываться на параметрах стандартных уров-
ней реагирования модельных объектов на индивидуальные вещества и со-
держащиеся в них примеси. Только в этом случае можно говорить о допус-
тимости прямой экстраполяции полученных при биотестировании результа-
тов на прототип. Рассмотрим этот подход в рамках вышеизложенных мате-
матических дефиниций.
Глава 17. Предел и достаточность числа наблюдений
111
Проанализируем ситуацию, когда в исследуемом на токсичность веще-
стве присутствуют вредные примеси. К сожалению, мы сталкиваемся с
такой ситуацией в большинстве случаев, поскольку 100%-ный уровень очи-
стки невозможен, будь то продукты химического синтеза, генно-инже-
нерные или рекомбинантные конструкции, вещества природного проис-
хождения.
В Институте новых технологий РАМН нами был разработан принципи-
ально иной подход к созданию 0,1%-ного нитроглицерина для инъекций.
Субстанцию мы получали от крупного оборонного предприятия. Хотя все
требования фармстатьи были соблюдены и выходной контроль подтверждал
это, нас не удовлетворял желтоватый цвет (откуда ему было появиться?)
субстанции и иные нюансы, которые здесь мы обсуждать не будем.
Поскольку мы создавали принципиально новую технологию получения
нитроглицерина в глюкозе (в дальнейшем — в лактозе, галактозе и т.д.), а не
на спиртовой основе, мы провели тщательный анализ всей технологической
цепочки и поняли, что в процессе стерилизации автоклавированием, по-
видимому, происходит окисление глюкозы, дающее примеси. Потребовались
многие месяцы для перехода на фильтрационную стерилизацию, на новый
техрегламент, согласование его с технологами, на новые доклинические и
клинические испытания, согласование и утверждение в Минздраве, патен-
тование [696]. Но этот труд стоил результата. В конечном итоге, нам удалось
получить не «желтоватый», а свободный от примесей, т.е. более безопасный
для человека, с меньшими побочными проявлениями чистый препарат, ут-
вержденный в качестве новой ФСП в Российской Федерации.
На основе этого примера из нашей научно-производственной практики
попытаемся формализовать этот процесс и представить его в более общем
виде.
Зададим условие, при котором содержание побочного продукта не долж-
но превышать 0,04%. Ошибка воспроизводимости одиночного анализа
а = 0,002%. Требуется, чтобы вероятность наличия вредных примесей не
превышала (3 = 0,02. Вместе с тем мы не должны исключать из эксперимента
вещества, входящие в выборку с примесями до 0,04%. Если содержание при-
меси в субстанции «на пределе», какие-либо гарантии дать трудно. Но мы
можем, например, потребовать, чтобы вероятность исключения веществ с
содержанием примеси ниже 0,036% (считаем их соответствующими нашим
требованиям) не превышала а = 0,05. Нужно определить число параллель-
ных анализов, которое позволило бы удовлетворить всем поставленным тре-
бованиям.
Чтобы использовать проведенные ранее теоретические рассуждения, бу-
дем в качестве случайной величины £ рассматривать отклонение результата
анализа примеси от заданного предела 0,04. Генеральное среднее а величи-
ны § даст истинное значение этого отклонения. Предположение «субстан-
278
Планирование медико-биологических экспериментов
ция пригодна» совпадает с гипотезой а < 0. Вероятности аир заданы в
явном виде, допустимую неопределенность критерия легко найти:
5 = 0,04 - 0,036 = 0,004.
Отсюда
" = К95 +"o.988)2ff ] =(l,64 + 2,05)2f^l =3,42 , (65)
т. е. для выполнения всех требований задачи нужно делать четыре парал-
лельных анализа. Субстанция будет при этом признана безоговорочно год-
ной, если среднее отклонение
*< -2,05^2 «-0,002, (66)
т. е. если по результатам четырех анализов содержание примеси (в среднем)
окажется меньше 0,038%.
Если результат анализа будет больше чем 0,038%, то субстанция является
или сомнительной (в пределах участка неопределенности), или неприемле-
мой. Обычно участок неопределенности присоединяют к участку неприем-
лемой субстанции, считая вещество непригодным всегда, как только резуль-
тат анализа окажется больше 0,038%. Это связано с тем, что в процессе син-
теза и очистки субстанции никогда не работают на пределе значений приме-
сей. Вероятность получения готовой субстанции с содержанием примеси,
близким к 0,04%, достаточно мала.
Описанная схема анализа предполагает, что число параллельных наблю-
дений п выбирается до токсикологического эксперимента и в процессе ис-
следования не меняется. Однако, если бы первый же анализ примесей суб-
станции был < 0,02%, то токсикологические исследования можно было бы
осуществлять как с совершенно «чистым» веществом. Иными словами, выб-
ранное заранее п всегда достаточно для получения заданных вероятностей а
и Р, но отнюдь не всегда необходимо.
Излишних наблюдений можно избежать, поступая, например, следую-
щим образом. Вначале делают одно наблюдение х1 и сравнивают его с пре-
делами [и^пв и Wj_aa]. Если это наблюдение будет меньше [и^а], тогда
гипотезу а < 0 можно сразу же принимать; если xY > w,_aa, то гипотезу мож-
но сразу отвергнуть. И только если х, попадает на участок неопределенности
[ttjna, «!_ao"], нужно проводить второе наблюдение.
Сделав второе наблюдение х2, находим среднее:
Xt +Х7
х2=~^- (67)
_ а с
Это среднее сравниваем с пределами -Mi-p-y= и Щ-а^= .
Глава 17. Предел и достаточность числа наблюдений
279
Гипотеза может быть снова отвергнута или принята, и тогда наблюдения
кончаются. Если же х2 попадает на новый участок неопределенности
ст а
7ГМ1-аЛ
"1-р-7г>м1-а-7г
то делают третье наблюдение х3, находят среднее
х1+х2+х3
Х-> =■
(68)
и начинают новую проверку по пределам
-м,
1-р
VJ и l-«S
Процесс продолжается до тех пор, пока количество наблюдений не дос-
тигнет выбранного заранее по формуле числа п.
Ясно, что при таком способе проверки гипотезы число необходимых
наблюдений будет в большинстве случаев значительно меньше первоначаль-
но выбранного п.
Разумеется, последовательное увеличение числа наблюдений неприемле-
мо там, где наблюдения проводятся заранее намеченными сериями и добав-
ление новых наблюдений сопряжено с большими трудностями. Например,
при исследовании в гистохимии или электронной микроскопии необходи-
мое число снимков нужно определять заранее. В противном случае для каж-
дого нового снимка или соседнего участка придется повторять весь процесс.
До сих пор во всех наших рассуждениях проверялась гипотеза а < 0. Ги-
потеза а > 0 проверяется аналогично, только теперь ее нужно принимать,
если
х>и
1-Р "Г
л/л
(69)
Обе эти гипотезы односторонние и проверяются с помощью односто-
ронних критериев.
Если же проверяется двусторонняя гипотеза а = 0 (ни больше, ни мень-
ше), то для проверки нужно применить двусторонний критерий. Пусть, по-
прежнему, вероятность отклонить верную гипотезу равна а, а вероятность
принять гипотезу, в то время как на самом деле а > 5, равна (3.
Тогда число наблюдений определится формулой
п =
+ и
1-р
(70)
и гипотеза признается справедливой только, если среднее л: одновременно
280
Планирование медико-биологических экспериментов
удовлетворяет двум неравенствам:
Мы рассмотрели вопрос о выборе числа параллельных наблюдений, ког-
да дисперсия наблюдений а2 известна заранее. Если же а2 заранее неизвест-
на, то для оценок нужно применять критерий Стьюдента. Выбор числа на-
блюдений п при этом более сложен и изложен в специальной литературе по
математической статистике.
Важным открытием биологии конца XX века была разгадка генетическо-
го кода. Удалось выяснить, каким образом наследственная информация
передается потомству. Поскольку информация записана в гигантских моле-
кулах дезоксирибонуклеиновой кислоты (ДНК), то существенно, как раз-
личные молекулы ДНК отличаются друг от друга, в каком порядке идут в
них 4 азотистых основания: аденин, тимин, гуанин и цитозин. Эти осно-
вания определяют порядок построения белков организма из двух десятков
аминокислот, причем каждая аминокислота зашифрована кодом лишь из
трех азотистых оснований, а не четырех.
Но откуда взялось число 3? Почему не 2 или 4? Ведь с помощью комби-
наций двух оснований можно зашифровать лишь 42 = 16 аминокислот, а
этого недостаточно. Если же брать по 3 основания, то получим 43 = 64 ком-
бинации. А этого с избытком хватит, чтобы зашифровать два десятка. Было
бы весьма интересно узнать, как используется в природе избыточность ин-
формации — ведь число комбинаций равно 64, а число аминокислот втрое
меньше. Природа предусматривает достаточность резервирования, но отнюдь
не избыточность — она рачительная, но и экономная хозяйка.
Так, в одной хромосоме содержится несколько десятков миллионов азо-
тистых оснований. Число различных комбинаций, в которых они могут
идти друг за другом, невообразимо велико. Ничтожной доли этих комби-
наций достаточно, чтобы обеспечить все разнообразие живой природы за
время существования жизни на Земле. Разумеется, надо иметь в виду, что
лишь ничтожная доля теоретически возможных комбинаций приводит к
жизнеспособным организмам. Оно равно 4N, где N — число оснований в
хромосоме.
При решении многих комбинаторных задач целесообразно воспользовать-
ся методом сведения к аналогичной задаче для меньшего числа предметов,
или методом рекуррентных соотношений (от латинского recurrere — возвра-
щаться), который позволяет свести задачу об п предметах к задаче об п — 1
предметах, потом к задаче об п — 2 предметах и т. д. Последовательно умень-
шая число предметов, доходим до задачи, которую уже легко решить. Во многих
случаях удается получить из рекуррентного соотношения явную формулу для
решения комбинаторной задачи.
Глава 17. Предел и достаточность числа наблюдений
281
В 1202 году итальянский математик Фибоначчи среди многих других задач
привел следующую: пара кроликов приносит раз в месяц приплод из двух
крольчат (самки и самца), причем новорожденные крольчата через два ме-
сяца после рождения тоже приносят приплод. Сколько кроликов появится
через год, если в начале года была одна пара кроликов?
Из условия задачи следует, что через месяц будет две пары кроликов.
Через два месяца приплод даст только первая пара кроликов, и получится 3
пары, а еще через месяц приплод дадут и исходная пара кроликов, и пара
кроликов, появившаяся два месяца тому назад. Поэтому всего будет 5 пар
кроликов.
Обозначим через F{n) количество пар кроликов по истечении п месяцев
с начала года. Мы видим, что через п + 1 месяцев будут эти F(n) пар и еще
столько новорожденных пар кроликов, сколько было в конце месяца п — 1,
то есть еще F(n — 1) пар кроликов. Иными словами, имеет место рекуррент-
ное соотношение
F(n + 1) = F(n) + F(n - 1) (72)
Так как по условию, F(0) = 4 и F(\) = 2, то последовательно находим
F(2) = 3, F (3) = 5, F(4) = 8, и т.д.,в частности F(\2) = 377.
Числа F(n) называют числами Фибоначчи. Они обладают целым рядом
свойств. Выведем выражение этих чисел через Скт. Для этого установим связь
между числами Фибоначчи и следующей комбинаторной задачей. Найти
число «-последовательностей, состоящих из нулей и единиц, в которых
никакие две единицы не идут подряд.
Чтобы установить эту связь, возьмем любую такую последовательность и
сопоставим ей пару кроликов по следующему правилу: единицам соответ-
ствуют месяцы появления на свет одной из пар «предков» данной пары (вклю-
чая и исходную), а нулями — все остальные месяцы. Например, последова-
тельность 010010100010 устанавливает такую «генеалогию» — сама пара по-
явилась в конце 11-го месяца, ее родители — в конце 7-го месяца, «дед» —
в конце 5-го месяца и «прадед» — в конце второго месяца. Исходная пара
кроликов зашифровывается при этом последовательностью 000000000000.
Ясно, что при этом ни в одной последовательности не могут стоять две
единицы подряд — только что появившаяся пара не может, по условию,
принести приплод через месяц. Кроме того, при указанном правиле различ-
ным последовательностям отвечают различные пары кроликов, и обратно,
две различные пары кроликов всегда имеют разную «генеалогию», так как,
по условию, крольчиха дает приплод, состоящий только из одной пары
кроликов.
Установленная связь показывает, что число «-последовательностей, об-
ладающих указанным свойством, равно Fiji).
282
Планирование медико-биологических экспериментов
Докажем теперь, что
т = с°„+1 + с\ + с*„_, + - + <7_« - (73)
где р = (п + 1)/2, если п нечетно, и р = л/2 если п четно.
Иными словами, р — целая часть числа (п + 1)/2 (в дальнейшем мы будем
обозначать целую часть числа а через Е(а)), таким образом,
р = Е(п+ 1)/2. (74)
В самом деле, F(ri) — это число всех «-последовательностей из 0 и 1, в
которых никакие две единицы не стоят рядом. Число же таких последова-
тельностей, в которые входит ровно к единиц и п — к нулей, равно Ckn_k+V
Так как при этом должно выполняться неравенство к<п — к + 1, то & изме-
няется от 0 до Е(п+\)/2. Применяя правило суммы, мы вновь приходим к
соотношению (74).
Таким образом, правильное использование статистических приемов и
математических методов в планировании эксперимента, анализе результатов
и формулировании результатов научных исследований имеет наиважнейшее
значение. Статистика является базовым приемом для определения соотноше-
ния единичного экспериментального образца и определенной популяции.
Она является инструментом, позволяющим исследователю определить, ка-
кое количество изменений следует отнести на долю случая, а какое вызвано
определенным воздействием в целях эксперимента. Надлежащее использо-
вание статистических методов позволяет определить количество животных,
необходимых для эксперимента, способствуя, таким образом, их экономии.
В нашем случае статистические приемы позволяют спланировать, выпол-
нить исследования, построить модель и осуществить корректный экстрапо-
ляционный перенос новых данных на прототип, т. е. на человека
283
Глава 18 АНАЛИЗ И ИНТЕРПРЕТАЦИЯ
РЕЗУЛЬТАТОВ: РАЗДЕЛЯЙ
И ДЕЙСТВУЙ
Интерпретация результатов для некоторых экспе-
риментов может заключаться в описании, а для
других в аналитическом сравнении различных ви-
дов полученного лечения или воздействия субстан-
ции в двух или нескольких различных группах. Как
выше отмечалось, эксперименты можно разделить
на два вида: подтверждающие и исследующие. В под-
тверждающих экспериментах делается попытка от-
ветить на заранее поставленные вопросы. В иссле-
дующих экспериментах описывается исходное состо-
яние и формулируются гипотезы для дальнейшей
проверки и построения модели.
Для подтверждающих экспериментов использу-
ются различные статистические тесты. Эти тесты
основаны на гипотезе нулевого эффекта, то есть
предполагается, что оказанное воздействие не дало
никакого эффекта. Также предполагается естествен-
ное отличие тестовой и контрольной групп, но если
было найдено отличие, которое нельзя считать ес-
тественным или случайным, считается, что их выз-
вало причиненное воздействие.
В исследующих экспериментах также использу-
ется статистика, но не в целях определения значи-
мых отличий последствий воздействия. Статисти-
ческие приемы в этих экспериментах скорее исполь-
зуются для математической оценки, описания ре-
зультата воздействия на определенную популяцию
или математического моделирования.
284
Планирование медико-биологических экспериментов
Последовательный анализ
В предыдущей главе приводились соображения, показывающие, что число
наблюдений можно сократить, если по ходу анализа учитывать уже сделан-
ные наблюдения. Обобщением этой идеи служит разработанный Вальдом
метод последовательного анализа. При этом методе после каждого нового
наблюдения решают, принять гипотезу, отклонить или продолжать испыта-
ния. Последовательный анализ позволяет сокращать число необходимых
наблюдений в среднем в два-три раза по сравнению с обычными методами,
фиксирующими число наблюдений заранее.
Сейчас последовательный анализ — это уже обширная теория. Мы рас-
смотрим его только в применении к анализу генерального среднего а на-
блюдаемой случайной величины.
Допустим, что нам нужно сделать выбор между гипотезами а < ах и а > а2
(предполагается, что а, < а2). Вероятность принять гипотезу а > а2, когда в
действительности а < ар обозначим через а. Вероятность противоположной
ошибки, т. е. принятия гипотезы а < ар когда в действительности а > а2,
обозначим через (З.Числа а и (3 обычно малы и задаются заранее.
Основная идея последовательного анализа заключается в следующем.
При каждой совокупности наблюдений х{, х2, ..., хп мы можем найти веро-
ятность р ' того, что эти наблюдения получены из совокупности с генераль-
ным средним а{', и вероятность /?и" того, что они получены из совокупности
с генеральным средним ос2. Согласно принципу максимума правдоподобия
на практике осуществляются события с максимальной вероятностью. Это
значит, что при рп' > рп" нужно считать более правдоподобным значение
а = ctj (a с ним и всю гипотезу а < а,). Если же рп" > рп', то предпочтение
нужно отдать второй гипотезе а > а2. Итак, все решается отношением прав-
доподобия рп"/рп'.
Существенно важно, будет ли оно больше или меньше единицы. Ясно,
однако, что в случае, когда отношение правдоподобия лишь немного отли-
чается от единицы, предпочтение соответствующей гипотезе будет весьма
сомнительным, и лучше всего продолжить испытания. Точные показатели,
насколько должно отношение правдоподобия отличаться от единицы, чтобы
между гипотезами можно было сделать уверенный выбор, определяются за-
данными вероятностями аир. Вальд показал, что гипотезу а < а} можно
принять, если
и гипотезу а > а2 можно принять, если
Глава 18. Анализ и интерпретация результатов: разделяй и действуй 285
Если же
Р *Л 1-Р
ГГ^ ~* (77)
то испытания надо продолжать.
При каждом новом наблюдении границы для отношения правдоподобия
не меняются, меняется лишь само отношение. Это облегчает применение
последовательного анализа, позволяет свести его к простым алгоритмам.
Если наблюдаемая случайная величина имеет нормальное распределение
с заранее известной дисперсией а2, то условие продолжения испытаний мож-
но преобразовать к виду
А1 + Ьп < Jjc, < А2 + Ьп, (78)
где
а то °2 1 Р а то °2 1 1_Р и а1+а2
4=2,3 18-г~> 4=2,3 lg—-,b= ' (79)
а2-а! 1-а а2-а1 а 2 • к у'
Числа А{, А2и b подсчитываются сразу же по исходным данным, благо-
даря чему дальнейшая проверка ведется только по сумме проделанных на-
блюдений и их числу.
Последовательный анализ нормально распределенной случайной вели-
чины особенно удобно проводить геометрически. Для этого после каждого
наблюдения строят точку на координатной плоскости, откладывая по оси
абсцисс число проделанных наблюдений п, а по оси ординат их сумму X*,..
Испытания нужно продолжать, пока эти точки будут находиться в полосе
между прямыми у = Ах + Ьп и у = А2 + Ьп (рис. 3). Если же хоть одна точка
окажется ниже этой полосы, анализ нужно прекращать и принимать гипо-
тезу а < а,. Точно так же гипотеза а > а2 принимается, как только хоть одна
точка окажется выше указанной полосы.
Рассмотрим следующий пример. Необходимо построить эксперимент по
отбору животных (в нашей лаборатории это осуществляется на крысах Wistar)
с целью получения гибридов первого поколения, устойчивых к гипоксии.
Зададим условие, что число слабо устойчивых к гипоксии не должно превы-
шать 60%. В связи с естественной дисперсией и генетической неоднородно-
стью групп животных возникает средняя квадратичная ошибка результатов,
известная по большому числу предыдущих анализов и равная а = 0,24%.
Чтобы гарантировать добротность, требуется отбраковать животных, имею-
щих низкую устойчивость к гипоксии и попадающих в выборку, превышаю-
щую 60%, с вероятностью не меньшей 0,95. В то же время нельзя отбрако-
вывать животных, составляющих выборку, превышающую 40%. Поэтому
ставится еще одно требование — животные, попадающие в выборку не более
чем на 58%, принимать с вероятностью 0,90.
Покажем, как для контроля устойчивых к гипоксии животных приме-
нять последовательный анализ. В поставленных условиях otj = 58, (32= 60.
286
Планирование медико-биологических экспериментов
Допустимые вероятности ошибок: а = 1 - 0,90 = 0,10 и Р = 1 — 0,95 = 0,05.
Непосредственно вычисляем
. О10,242, 0,05 .,,
Ах =2,3 lg = -8,31 ,
1 0,20 0,90
. О10,242. 0,05 111f.
^2=2'3Wg^=13'10'
, = M±M = 5j9.
2
Область продолжения анализов будет ограничена прямыми
у = -8,31 + 5,9л и у = 13,10 + 5,9л ,
(80)
(81)
(82)
(83)
которые построены на рис. 3. Анализ
тат 42%. Пусть ему соответствует на
ординатой у = 4,2. Результаты иссле-
дований данной выборки (обозначим
её точкой Мх) находится в статисти-
ческой области «продолжения испы-
таний», поэтому берем вторую вы-
борку. Её анализ дал результат 39%,
что соответствует условной точке М2
с абсциссой п = 2 и ординатой у =
4,2+ 3,9 = 8,1. Испытания снова
нужно продолжать. Третий анализ
дал значение 36%; ему соответствует
некая точка Мъ с абсциссой п = 3 и
ординатой у = 8,1 + 3,6 = 11,7. Чет-
вертому анализу с результатом 40%
соответствовала бы точка М4 (п = 4,
у = 15,7). Наконец, пятая точка М5,
соответствующая еще одному анали-
зу с результатом 44%, получает коор-
динаты п = 5, у = 15,7 + 4,4 = 20,1,
и выходит за пределы «полосы М4
продолжения испытаний» в ожидае-
мом нами направлении. Данную вы-
борку животных можно считать со-
ответствующей требованиям устой-
чивости к гипоксии (рис. 4).
Может показаться, что устойчи-
вость к гипоксии видна уже из пер-
первой выборки животных дал резуль-
рис. 4 точка Мх с абсциссой п = 1 и
у 1
30-
20-
10-
0
к
г^
I
1
г%
I I I
2 3 4
n
Рис. З. Координатная плоскость и оси, опреде-
ляющие проведенные эксперименты, иллюс-
трирующие, что в этих исследованиях нет жи-
вотных, устойчивых к гипоксии
Рис. 4. Точка М5 выходит за пределы необхо-
димости продолжения испытаний, иллюстри-
рующие наличие животных, устойчивых к ги-
поксии
Глава 18. Анализ и интерпретация результатов: разделяй и действуй 287
вого анализа. К сожалению, такой вывод поспешен, так как из-за высокого
значения ст мы не можем гарантировать заданные вероятности ошибок а и р.
Для решения поставленной задачи с помощью метода последовательного
анализа нам понадобилось пять наблюдений. Если же мы захотим опреде-
лить число наблюдений п заранее по формулам предыдущего пункта, то мы
получим:
к2
" = («0,90+"0,95)
( „ \ ^0,24^
v<X2-<Xt,
(1,28 + 1,65)"
0,20
13, (84)
т. е. классический метод потребовал бы 13 серий экспериментов. В то время
как при планировании и использовании математического аппарата Вальда,
т.е. последовательного анализа мы получаем репрезентативную выборку и
корректный результат уже при пяти сериях. Преимущества планирования и
оптимизации эксперимента и последовательного анализа неоспоримы.
Принятие решений
Как бы тщательно не был осуществлен анализ и дана интерпретация резуль-
татов экспериментов, перед исследователем стоит задача принятия решения.
Под понятием «принятие решения» подразумевается применение математи-
ческих и количественных методов для обоснования решений. Оно включает
в себя: постановку задачи; построение математической модели; нахождение
метода решения; проверку и корректировку модели; реализацию найденно-
го решения в виде нахождения экстраполяционных параметров переноса с
биомодели на прототип (человека или животного).
Построение математической модели осуществляется на основе достаточ-
но строгой, логически непротиворечивой, содержательной постановки зада-
чи. Этот процесс называется формализацией задачи.
В том случае, когда все действующие в эксперименте факторы определены
и достоверны, а случайные воздействия отсутствуют, мы сталкиваемся с ситу-
ацией принятия решений в условиях определённости. Например, необходимо
испытать активность нового жаропонижающего средства при эксперименталь-
ной гиперэргии. Результат исследования даст абсолютно однозначный ответ о
наличии эффекта. Более того, исследователь получит однозначные количе-
ственные показатели этого действия, выраженные в градусах Цельсия. Есте-
ственно, что решение о результатах отвечает условиям определенности.
Если же мы находимся в ситуации, в которой результаты экспериментов
не дают однозначного ответа, а относятся к одному из множества возмож-
ных откликов с разными вероятностями их осуществления, а исход конкрет-
ной реализации случаен и непредсказуем, метод называют принятием реше-
ний в условиях риска. Например, эксперимент в полном развитии, а под ок-
ном рабочие затеяли рубить асфальт.
288
Планирование медико-биологических экспериментов
В тех случаях, когда результаты экспериментов зависят от непредсказуе-
мых факторов, такая задача, называется принятием решений в условиях нео-
пределённости. Например, требуется выдать заключение о радиопротектор-
ном действии препарата в условиях комплексного воздействия радиацион-
ных, акустических и электромагнитных факторов. Чтобы исключить явле-
ния неопределенности, необходимо исследовать протекторные свойства каж-
дого из факторов, их попарного воздействия и комплексного влияния. Это
требует огромного количества экспериментов и достаточно сложного мате-
матического анализа. Экспериментатор вправе, без дополнительных иссле-
дований, принять решение в условиях неопределенности. Иными словами,
если нет полной информации, то остаётся либо предполагать, либо строить
догадки. Поэтому в формулировке таких задач следует делать оговорку «по
возможности», «по-видимому», «вероятно».
В процессе принятия решений можно выделить несколько этапов:
/ определить основные и альтернативные способы действия, адекватный
набор целей и установить нормы и пределы искомых результатов;
/ условия неопределённости, связанные с альтернативными решениями,
должны быть выражены через распределение вероятностей, желательно чис-
ленно;
/ искомые результаты должны быть представлены через ранжирование
предпочтений в числовой форме;
/ рациональный синтез информации должен быть осуществлен таким
образом, чтобы можно было построить общую модель принятия решений.
Многие исследователи и специалисты в области исследования операций
и теорий принятия решений руководствуются принципом «разделяй и дей-
ствуй», иными словами, необходимо любую анализируемую ситуацию разде-
лить на части, а затем действовать в соответствии с указанными этапами, т.е.
каждую часть анализировать отдельно. Только после этого можно построить
общую модель для принятия решений.
Некоторые, не лишенные чувства юмора специалисты определяют про-
цессы принятия решений и исследования операций, как искусство давать
плохие ответы на те научные вопросы, которые при других подходах являют-
ся ещё худшими. Но если говорить серьезно, то решение, принятое в усло-
виях неопределённости, но на основании математических расчётов, будет
лучше, чем взятое наугад первое попавшееся или, которое некоторые иссле-
дователи не без юмора называют эвристическим или интуитивным.
Разделу БИОЭТИКА
В ЭКСПЕРИМЕНТАЛЬНЫХ
ИССЛЕДОВАНИЯХ
Сострадание к животным так тесно связано
с добротою характера, что можно с уверенностью
утверждать, что не может быть добрым тот,
кто жесток с животными.
А.Шопенгауэр
291
Глава 19 ГУМАННОСТЬ ПРИ РАБОТЕ
С ЖИВОТНЫМИ И ПРИНЦИПЫ 3R
Некоторые национальные и европейские законода-
тельства, регулирующее проведение экспериментов
на животных, требуют, чтобы всякий раз, когда это
возможно, применялись методы без использования
животных. В ряде стран желающие получить лицен-
зию на осуществление работ с лабораторными жи-
вотными должны представить детальную програм-
му мероприятий по сокращению, усовершенствова-
нию и замене экспериментов с использованием жи-
вотных. Это и есть предмет биоэтики. Биоэтика изу-
чает моральные, юридические и социальные про-
блемы, возникающие по мере развития биологии и
медицины. Она основывается на достижениях био-
логии, медицины, логики, философии [378]. Важ-
ное место в этой дисциплине занимают и вопросы
этики проведения научных экспериментов на жи-
вых животных [87, 373].
Необходимость сокращения, усовершенствова-
ния и замены экспериментирования на животных и
перехода к исследованиям на основании компро-
миссных походов отражена в многочисленных на-
учных и публицистических работах [149, 151, 160,
215, 366, 372, 377, 412, 481, 582].
В последние годы наблюдается жесткое проти-
востояние между теми, кто отстаивает разумное и
обоснованное сокращение, усовершенствование и
замену экспериментов на животных, и теми, кто
считает, что подобные эксперименты не могут быть
292
Биоэтика в экспериментальных исследованиях
оправданы никакими соображениями, если принять во внимание вероятные
страдания животных. В этой ситуации твердую, взвешенную позицию зани-
мает «Фонд по замене животных в медицинских экспериментах» (FRAME).
Эта международная организация последовательно выступает как против дей-
ствий экстремистских групп, стремящихся возбудить общественное мнение
требованиями запрета альтернативных методов исследований, так и против
предоставления безграничной свободы ученым, которые проводят биомеди-
цинские исследования на лабораторных животных [32, 214, 281].
Противники проведения исследований, как правило, говорят о страда-
ниях животных и соблюдении их прав, тогда как сторонники черпают свои
аргументы в необходимости продолжения исследований над животными с
тем, чтобы ускорить прогресс в медицине и биологии, найти новые средства
предотвращения и лечения болезней. Наука нуждается в общественном со-
гласии, а это как раз и предполагают принципы сокращения, усовершен-
ствования и замены экспериментирования на животных, суть которых со-
стоит в том, чтобы, обеспечив гуманное отношение к животным, сохранить
главное — проведение исследований в интересах жизни и здоровья людей.
Идея защиты животных и природы достаточно сильно, и раньше, чем в
других странах, проявилась в Великобритании, где в 1926 году в Великобри-
тании было основано Общество по охране животных Лондонского универ-
ситета (с 1938 года — Университетская федерация защиты животных, UFAW).
Эта организация сыграла и играет важнейшую роль в развитии этических
концепций экспериментирования на животных и в становлении практичес-
ких направлений, улучшающих эксперименты на животных. Группа иссле-
дователей во главе с М. Хьюмом, созданная в рамках актива UFAW, пред-
приняла попытку разработать принципы гуманной экспериментальной тех-
ники. Работа группы была продолжена У. Расселом и Р.Берчем, когда в 1959
году под их редакцией вышла книга The Principles of Humane Experimental
Technique («Принципы гуманной экспериментальной техники»). Авторы
показали четкую корреляцию между гуманностью научного эксперимента и
его научной эффективностью, подчеркнув, что «если используется набор
принципов экспериментирования, то критерии гуманизации являются луч-
шим из того, что мы можем предложить». В монографии Рассела и Берча
обоснована концепция гуманного использования животных в эксперимен-
тах, которая получила название «Концепция трех R». Концепция трех R имеет
биологическое содержание, и поэтому ее с полным правом можно имено-
вать «Биоэтическая концепция трех R», в основу которой легли три основных
положения: Replacement — замена, Reduction — сокращение, Refinement —
улучшение. Эта концепция находит широкое распространение в исследова-
ниях и публикациях последних лет [487, 455].
Replacement — замена в опыте, когда это возможно, высокоорганизо-
ванных животных менее развитыми живыми объектами или применение аль-
Глава 19. Гуманность при работе с животными и принципы 3R 293
тернативных методов исследования (эксперименты на культурах клеток и
тканей, использование изолированных органов, физико-химических и био-
химических системам, эксперименты на микроорганизмах и растительных
объектах, компьютерное и математическое моделирование). Есть неплохие
результаты в использовании культур клеток тканей (методы in vitro) с целью
определения общей токсичности химических соединений для человека и жи-
вотных, оценки иммунотоксичности, нефротоксичности, гепатотоксичнос-
ти, нейротоксичности, фототоксичности, раздражающего действия, канце-
рогенное™. Успехи в разработке биополимеров позволили получить струк-
туры эквивалентные человеческой коже.
Активные приверженцы биомоделирования второго порядка считают, что
необходимо оказать полную поддержку широкому распространению среди
научной общественности информации об альтернативных методах, имею-
щихся в стране или регулярно описываемых во многих международных жур-
налах по альтернативным методам [149, 266].
Reduction — достижение воспроизводимых результатов с использова-
нием минимального количества животных; адекватный выбор лаборатор-
ных животных; использование стандартных по микробиологическим, гене-
тическим и экологическим параметрам животных; оптимальное планиро-
вание и, что крайне важно, использование статистических методов не только
при обработке полученных данных, но и на стадии планирования. Одним
из наиболее надежных путей снижения количества животных, используе-
мых в экспериментах, является дальнейшее развитие и осуществление
стндартизации лабораторных животных по генотипу, микрофлоре и эколо-
гическим параметрам. Благодаря уменьшению количества переменных фак-
торов, стандартизация может помочь в получении более надежных резуль-
татов на меньшем количестве животных [482]. Исследователи должны вла-
деть информацией о значительном количестве моделей уже имеющихся для
исследований. Отсутствие соответствующей информации или ее недоступ-
ность для широкой научной общественности может стать одним из неиз-
бежных факторов неправильного использования лабораторных животных в
экспериментах. Вопросы оптимизации в использовании лабораторных
животных и планировании экспериментов подробно изложены в предыду-
щей главе.
Необходимо оказать полную поддержку широкому распространению
среди научной общественности информации о моделях, имеющихся или
регулярно описываемых во многих биомедицинских журналах. В качестве
примера можно привести American Journal of Pathology, который в каждом
номере в специальной рубрике публикует статьи по этому вопросу, а также
каталог Фестинга [206, 207, 208] и журналы Laboratory Animals и Laboratory
Animal Science. Широкий обмен информацией об экспериментах на жи-
вотных, позволяет избежать неоправданного дублирования, а грамотное
294
Биоэтика в экспериментальных исследованиях
планирование экспериментов и их анализ дает дополнительную и коррект-
ную информацию при гораздо меньшем количестве использованных жи-
вотных [594].
Refinement - улучшение условий содержания лабораторных животных и
использования их в экспериментах, уменьшение дистресса животных во время
экспериментов и применение обезболивающих средств, но не в ущерб цели
эксперимента. С точки зрения внешних проявлений (а также физиологии),
животные реагируют на непереносимые боль и дискомфорт так же, как и
люди. Для того, чтобы захотеть предотвратить страдания животных, нужно
поверить, что животные их испытывают [251]. Даже поверив, что животное
может испытывать страдания, и желая предотвратить их, человек может не
выполнить обезболивание, если не знает, как это делается, или занят своей
работой настолько, что забывает об этом. Для регистрации болевых реакций
у животных необходимо углубленное изучение особенностей поведения
животных и изменения этого поведения при болевых воздействиях. Концеп-
ция исследования боли на животных включает в себя представления о том,
что животное не должно подвергаться боли в большей степени, чем может
выдержать человек. Следует согласиться с И.П.Павловым, что субъективный
подход для строгой науки малоприемлем, но отчасти приемлем в биоэтике
экспериментирования на животных, если даже считать, что страдание и боль
у животных никем субъективно не переживаются и не могут быть оценены
и измерены строго объективно [214, 215, 378].
Активная пропаганда и внедрение биоэтических принципов трех Rs на-
чались с 1969 г, когда в Великобритании появилась организация по охране
животных FRAME. Она играет значительную роль в прогрессе гуманной
технологии эксперимента. В настоящее время FRAME имеет международ-
ный статус и пользуется поддержкой Европейского Союза. Выходит журнал
FRAME, который называется ATLA (Alternatives to Laboratory Animals). Это
— читаемый и цитируемый биомедицинской прессой журнал. Идеи концеп-
ции трех R используются для подготовки законов по охране эксприменталь-
ных животных. Биоэтическая концепция трех R отражена в Директивах ЕС
1986 года. В последнее десятилетие растет число симпозиумов и конферен-
ций по этой проблеме.
В последние годы под эгидой Европейского Совета в Северной Италии
возник Европейский Центр по утверждению (валидации) альтернативных
методов (ECVAM). Центр способствует разработке альтернативных (заме-
няющих эксперименты на животных) методов, проверяет и утверждает
адекватность новых или уже имеющихся методов, проводит информаци-
онную работу, организует научные конференции и семинары. Биоэтичес-
кая концепция трех Rs распростраяется все шире и находит применение в
странах Западной Европы, в США, Канаде, Австралии, Новой Зеландию,
и Японии.
Глава 19. Гуманность при работе с животными и принципы 3R 295
Замена экспериментов на животных альтернативными методами не все-
гда возможна. С целью обеспечения безопасности для человека вновь разра-
батываемых лекарственных препаратов или пищевых добавок во многих стра-
нах мира имеются законы, не только защищающие животных, но и обязы-
вающие разработчиков апробировать новые продукты на животных по сис-
теме «хорошей лабораторной практики» — GLP (Good Laboratory Practice).
Многие научные эксперименты невозможно проводить на менее сложных
системах, чем целостный организм. Возрастает потребность в эксперимен-
тах на генетически модифицированных животных. В этой связи обсуждают-
ся этические вопросы трансгенной технологии (J.W.Gordon, 1997; A.Hoog,
et al 2000, отчет МВД Англии).
Концепция гуманного эксперимента способствовала развитию альтерна-
тивных методов во многих областях: при производстве вакцин, в вирусоло-
гии, токсикологических исследованиях, при тестировании безопасности раз-
личных продуктов и лекарств, в физиологических исследованиях и санитар-
но-гигиенических работах.
Однако имеются еще более широкие перспективы в поиске альтернатив
животным в экспериментальной практике. По отдельным направлениям
биоэтической концепции трех Rs выполняются многочисленные работы
химиками, биологами, математиками, специалистами в области компью-
терных технологий с целью замены экспериментов на живых животных
альтернативными методами и снижения количества животных в экспери-
менте.
В 1991 году в г. Ростове-на-Дону на I съезде токсикологов России, пред-
седателем которого был избран автор этой книги, впервые была выделена
самостоятельная секция по альтернативным методам, на которой обсужда-
лись возможности замены экспериментов на животных альтернативными
методами при изучении механизмов токсического действия веществ.
Столетия назад были созданы многие из экспериментальных моделей,
использующих животных, а изменения на развитие альтернативы в значи-
тельной степени оставили ученым и их интуитивной прозорливости.
По данным литературных источников, общественное мнение в США,
Канаде и большинстве стран Европы одобряет подходы, исключающие ис-
пользование животных в исследованиях. Правительство теперь признает, что
такие подходы могут позволить преодолеть некоторые из ограничений, пред-
полагающих проведение экспериментов над животными. Так, рекомендации
Специального комитета Палаты лордов Великобритании предполагают со-
здание «Центра трёх направлений», который должен стать административ-
ным органом по координации деятельности исследовательских коллекти-
вов, использующих лабораторных животных и занятых в существующих на-
учных центрах высокого уровня. Правительство Великобритании и ряд госу-
дарств Евросоюза поддерживает эти идеи.
296
Биоэтика в экспериментальных исследованиях
Основной сутью концепций по замене лабораторных животных является
междисциплинарность, которая в решение биомедицинских вопросов могла
бы привнести целый набор применимых подходов от признанных исследо-
вательских групп учёных, предполагающих отказ от использования живот-
ных в экспериментах. Они предусматривают финансирование и стимулиро-
вание высококачественных исследований с целью разработки методов заме-
ны животных в исследованиях, обучение исследователей и распространение
знаний об альтернативных подходах и методах
297
Глава 2 0 ИММОБИЛИЗАЦИЯ
ЖИВОТНЫХ
Обездвиживание животных представляет собой ком-
плекс мер физического или фармакологического
воздействия, направленный на сдерживание есте-
ственной подвижности животного в целях проведе-
ния необходимых действий по уходу, обследованию
или проведению экспериментальных действий,
включая анестезию. Качество полученных образцов
напрямую зависит от обращения с животным во
время эксперимента. Правильное обращение и обез-
движивание может дать животному ощущение бе-
зопасности и таким образом уменьшить его боль и
испуг. Это не только позволяет в некоторых случаях
провести эксперимент без анестезии, но и удовлет-
воряет принципу гуманного обращения с животны-
ми. Одновременно с этим, правильное обращение с
животными обеспечивает также безопасность пер-
сонала и исследователей, поскольку успокаивает
животное.
Персонал должен быть обучен правильному об-
ращению с животными, поскольку физический
контакт с ними является частью их ежедневной
работы по уходу за животными и их размноже-
нию. Исследователи также должны иметь опыт об-
ращения с животными, так как именно они про-
водят большинство экспериментов. В противном
случае желательно, чтобы обученный сотрудник
ассистировал исследователю при проведении экс-
перимента.
298
Биоэтика в экспериментальных исследованиях
Для того чтобы не испугать животное, не причинить ему вред или боль,
движения при обращении с ним должны быть осторожными и мягкими. Во
многих случаях целесообразно использовать сети и другие специальные при-
способления для поимки и обездвиживания животного.
В исследованиях на животных возникает вопрос надежной фиксации
отдельных частей тела или всего животного в целом. Без фиксации у собак
или других крупных лабораторных животных трудно или невозможно про-
водить длительные внутривенные введения. Для иммобилизации животных
их прикрепляют к столу или специальному устройству в положении на спи-
не. В таком положении трудно проводить манипуляции на животном. Пре-
бывание животных в таком неестественном положении является стрессор-
ным фактором. Процесс фиксации животного в положении на спине явля-
ется трудным и вызывает агрессивную реакцию у животных.
Существуют устройства фиксации крупных лабораторных животных,
которые позволяют сохранить естественное положение животных, обеспе-
чить надежную иммобилизацию тела всего животного или отдельных конеч-
ностей, на которых предполагается определенная манипуляция. Принцип
этого устройства заключается в том, что и лоток и все стойки для конечно-
стей и для морды легко трансформируются. Детали такого устройства можно
легко подгонять под любые размеры животных по длине и высоте. В задней
части устройства должен быть установлен пластиковый моче- и калоприем-
ник. Под приемник подставляют посуду для сбора мочи или кала.
Мини—свиньи фиксируются матерчатым подбрюшником и тканевыми или
синтетическими бинтами, надежно закрепляя их в станке в положении на
животе.
Первое, с чем приходится сталкиваться исследователям при эксперимен-
тировании на свиньях, — это с необходимостью ограничить подвижность и
провести анестезию. Механически частичную подвижность, необходимую,
например, при взятии крови, можно ограничить путем подвешивания на
ремнях, подводимых под передние и задние конечности и закрепляющихся
на спине животного, как это делается в нашей лаборатории. Panepinto с
соавторами [84] предлагают устройство для иммобилизации в виде гамака из
грубого хлопчато-бумажного волокна типа парусины, снабженного рукава-
ми для конечностей, укрепленным на стальных рамах. Есть и другие спосо-
бы фиксации. Однако, каковы бы они ни были, важно соблюдать одно усло-
вие — свести к минимуму стрессовые факторы.
Подбрюшник размером 60 х 20 см изготавливают из ткани (лучше син-
тетической или полусинтетической). По углам и в середине к подбрюшнику
пришивают четыре пары тесемок. Подбрюшником охватывают живот, грудь,
затем его выводят между передними ногами животного на шейную часть,
после чего фиксируют тесемками к продольным рейкам станка, и животное
лишается возможности двигать корпусом в направлении сверху вниз и впе-
Глава 20. Иммобилизация животных
299
ред. Кроме того, такая фиксация уменьшает нагрузки на передние конечно-
сти, а опираясь на подбрюшник мини-свинья лишается возможности дви-
гать ногами и наносить удары. Бинтом животное может быть дополнительно
фиксировано к продольным рейкам. Мини—свиньи зачастую бывают крайне
возбудимы и до начала основных экспериментов их следует приучить к ус-
ловиям иммобилизации.
Собаки столь неоднозначны по своему поведению, что их во всех случаях,
до начала иммобилизационных мероприятий, необходимо предварительно
приучить к пребыванию в станке. Этому способствует кормление после того,
как животное постоит привязанным в станке. После двух-трех таких трени-
ровок собаку и других животных удается легко фиксировать и проводить
необходимые процедуры без помощи посторонних лиц. Для полного обезд-
вижения собаки прибегают к наркозу как рауш—ингаляционному, так и не-
ингаляционному.
Кошку без особых усилий можно фиксировать на короткое время. Креп-
ким захватом берут кошку за кожу в области затылка, а другой рукой — за
кожу в области поясницы и легко прижимает ее к столу. Фиксированному
таким образом животному без затруднений проводят ряд несложных мани-
пуляций (оральное, подкожное или внутримышечное введение). Кошки ста-
новятся очень неспокойными и агрессивными при попытке ограничить их
движения на более длительное время или при ряде насильственных вмеша-
тельств. Поэтому фиксировать животное нужно обдуманно, быстро и надеж-
но. Обездвижить кошку можно, если запеленать голову и конечности кус-
ком крепкой ткани.
Кролики отличаются от других животных своим спокойствием и относи-
тельно слабым сопротивлением при фиксации. Привязывают их к специаль-
ным станкам или вивисекционным столам таким же образом, как кошек в
собак. Следует иметь в виду, что при сильном запрокидывании головы у
кроликов легко возникает смерть от удушья. Удобен способ фиксации кро-
ликов в боксе с круглой прорезью в передней стенке. При этом голову жи-
вотного выводят сквозь отверстие передней стенки наружу, а туловище раз-
мещают внутри. Для вживления электродов в головной мозг кроликов (это
относится и к другим животным) используют специальный стереотаксичес-
кий аппарат, надежно фиксирующий голову животного.
Фиксация морских свинок не представляет затруднения. Помощник удер-
живает животное за спину и под грудь так, чтобы большой и указательный
пальцы охватывали шею, а другие пальцы обездвиживали передние конеч-
ности и ограничивали движения головы. Снизу он удерживает заднюю
часть тела и задние конечности. Фиксированной таким образом морской
свинке придают нужное положение. Если нужно фиксировать морскую
свинку на длительное время, то ее привязывают к станку или операцион-
ному столику.
300
Биоэтика в экспериментальных исследованиях
Белых лабораторных крыс берут за спину или хвост. Для фиксации жи-
вотного вполне достаточно бывает взять его за кожу в области спины, а
большим и указательным пальцами, удерживая крысу с боков, выдвинуть
передние конечности кпереди. После этого животному придают нужное
положение. Во избежание укусов нужно работать в резиновых или кожаных
перчатках. Для фиксации крыс предложен ряд других специальных приспо-
соблений — гильзы из проволочной сетки, цилиндры. Удобно фиксировать
крысу, завернув ее в салфетку или в сетку из мягкой проволоки. Для дли-
тельного обездвиживания белых крыс фиксируют к операционному столу
или к специальному станку.
Чтобы фиксировать белую мышь, ее ловят рукой или корнцангом за хвост,
ставят на стол и большим и указательным пальцами левой руки захватывают
кожу в области спины, а остальными пальцами удерживают задние конечно-
сти и хвост. Фиксированному животному придают нужное положение. Мож-
но фиксировать при помощи корнцанга, захватив им кожу в области затыл-
ка, для более длительной фиксации применяют специальные гильзы, окуты-
вают тело салфеткой или мягкой проволочной сеткой или привязывают к
операционному столику. Для введения жидкостей в вену мышей можно при-
менять специальные методы фиксации.
Сирийского хомячка для длительного обездвиживания следует укутать в
салфетку или привязать к операционному столу, стараясь не причинять ему
боль. При необходимости вызвать длительную иммобилизацию прибегают к
наркозу, для чего можно использовать эфир, хлороформ, хлоралгидрат дру-
гие препараты.
Все группы перечисленных животных могут с успехом использоваться
для изучения механизмов действия и эффективности местных анестетиков.
Таблица 5 1
Генетически обусловленные и экспериментальные биомодели для изучения
местноанестезирующего действия веществ
Исследуемое действие
вещества
Модель для оценки
терминальной анестезии
Модель для оценки
инфильтрационной
анестезии
Модель для оценки
проводниковой
анестезии
Лабораторное
животное
Кролики
Кошки
Крысы
Морские
свинки
Кролики
Мыши
Лягушки
Мыши
Крысы
Крысы
Кролики
Кошки
Линия или порода
животного
Аутбредные
«
Аутбредные
Rana temporaria
Аутбредные
«
Пол
животного
в
б
6
в
6
в
в
в
6
6
Вес
животного
2-2,5 кг
3,5-4 кг
200-250 г
200-250 г
2-2,5 кг
18-20 г
200-250 г
18-20 г
180-220 г
200-250 г
2-2,5 кг
3,5-4 кг
Возраст
животного
7-10 мес.
1 -7 лет
5-12 мес.
2-30 сут.
5-6 мес.
2-2,5 мес.
2-30 сут.
2-2,5 мес.
3-4 мес.
4-5 мес.
5-6 мес.
1 -7 лет
Глава 20. Иммобилизация животных 301
Окончание табл. 51
Исследуемое действие
вещества
Модель для оценки
эпидуральной анестезии
Модель для оценки
спинномозговой
анестезии
Модель для оценки
спинальной анестезии
Модель для оценки
местнораздражающего
действия веществ по
методу Норре, Alexander
и Miller, Kollzer и Wehr
Лабораторное
животное
Кролики
Крысы
Крысы
Крысы
Линия или порода
животного
Аутбредные
Аутбредные
Аутбредные
Белые
аутбредные
Пол
животного
6
9<J
6
6
Вес
животного
2,8-3,6 кг
200-250 г
200-250 г
Не менее
120 г
Возраст
животного
7мес.4 года
3-4 мес.
3-4 мес.
С 2 мес,
Хотелось бы ещё раз подчеркнуть, что использование фармакологичес-
ких средств, если они не влияют на результат эксперимента, целесообразно
и с точки зрения гуманности необходимо при выполнении вивисекционных
и иных травмирующих животное исследований.
302
Глава 2 1 НАРКОЗ
Наркоз у животных традиционно осуществлялся раз-
личными фармакологическими средствами (хлоро-
форм, эфир, морфин, хлоралгидрат, уретан, гексе-
нал, тиопентал натрия, хлоралоза и др.) и различны-
ми путями (ингаляционный, интратрахеальный,
внутривенный, внутримышечный, ректальный).
Чаще всего пользуются комбинированным морфин-
но-эфиро-хлороформным наркозом. Для этого мор-
фин в виде 1-2 %-ного раствора вводят под кожу из
расчета 5-8 или 10 мг на 1 кг веса животного. Лучше
пользоваться раствором морфина с атропином. Че-
рез 10—20 мин после инъекции у собак может быть
рвота, дефекация (не всегда), дремотное состояние
и заметная адинамия. Спустя 30—35 мин после вве-
дения морфина собака почти не оказывает сопро-
тивления и ее легко привязывать.
Ингаляционный наркоз. После фиксации круп-
ных лабораторных животных (мини-свиньи, овцы,
бараны, собаки, кошки) на морду надевают маску и
дают эфиро-хлороформную смесь (одна часть чис-
того хлороформа и две части этилового эфира для
наркоза). Чтобы ускорить наступление наркоза,
первоначально в течение 1—3 мин можно давать
один чистый хлороформ. Не следует пользоваться
хлороформом у кошек, к которому они весьма чув-
ствительны. Необходимо внимательно и регулярно
следить задыханием, состоянием корнеальных реф-
лексов, равномерно добавлять наркотическую
смесь.
Глава 21. Наркоз
303
При необходимости проведения оперативных вмешательств у лаборатор-
ных животных {мини-свиней, овец, собак, кошек, кроликов) следует применять
внутритрахеальный наркоз. Вначале дают эфироморфинный или хлорофор-
мно-эфиро-морфинный наркоз. Под визуальным контролем интубируют
через рот широкую трубку. Глотку вокруг трубки рекомендуется тампониро-
вать влажной марлей. Интубационную трубку присоединяют к наркозному
аппарату. Чаще всего пользуются эфирнокислородной смесью.
Энтеральный наркоз включает в себя ректальный и оральный. За сутки
до начала наркоза животным дают слабительное, лучше сульфат магния или
натрия, а перед наркозом делают очистительную клизму. Под кожу вводят
морфин с атропином и спустя 10—20 мин приступают к ректальному введе-
нию наркотика. Нельзя вводить морфин кошкам, так как у них он вызывает
не угнетение, а возбуждение нервной системы с явлениями галлюцинаций и
даже судорог. Для ректального наркоза чаще всего используют 10%-ный
раствор хлоралгидрата, приготовленный со слизью крахмала, салепа или
настоя алтейного корня. Наркотической дозой хлоралгидрата является 0,3—
0,5 г/кг. Хлоралгидрат можно использовать для орального введения в дозах
0,4—0,6 г/кг, а также внутривенного и внутрибрюшинного введения из рас-
чета соответственно 0,1—0,15 и 0,3—0,4 г на 1 кг веса. Хорошим наркотичес-
ким веществом является уретан. Дозы для внутривенного введения собакам —
0,7—1 г/кг, для внутрибрюшинного и внутримышечного — 1—1,2 г/кг. Хлора-
лозу (глюкохлоралозу) вводят собакам и другим лабораторным животным
внутривенно в виде теплого, приготовленного на физиологическом растворе
(0,8%), хлористого натрия. Доза 40 мг/кг у собак вызывает лишь возбужде-
ние. Наркотические дозы 100—120 мг/кг, а дозы в 150 мг/кг уже токсичны.
Таким образом, хлоралоза обладает небольшой наркотической широтой.
Весьма чувствительны к оральному введению хлоралозы кошки. Смерть у
них наступает от введения 65 мг/кг, в то время как собаки погибают от дозы
650 мг/кг.
Парентеральный наркоз следует проводить в сочетании с морфином.
Тиопентал натрия (пентотал натрия) в виде свежеприготовленного 2%-ного
раствора для внутрибрюшинного введения берется из расчета 1,5 мл на 1 кг
веса, для внутримышечного — по 2 мл на 1 кг веса собаки, т. е. 30—40 мг/
кг. Гексенал в виде 10%-ного свежеприготовленного раствора медленно
вводят внутривенно из расчета 30—40—50 мг/кг. Маленьким собакам, кош-
кам и кроликам гексенал в виде 1—2%-ного раствора можно вводить внут-
рибрюшинно. Для этого животное фиксируют следующим образом: зад-
нюю часть туловища приподнимают и каудальнее пупка, несколько отсту-
пив от средней линии, делают прокол брюшной стенки. Для крупных жи-
вотных раствор гексенала при внутрибрюшинном введении должен быть
более концентрированным (2—5%-ный), доза наркотика составляет при этом
от 50 до 70 мг/кг.
304
Биоэтика в экспериментальных исследованиях
Спинномозговое обезболивание проводят введением новокаина или
совкаина в спинномозговой канал между 1-м и I хвостовыми позвонками
(сакральное эпидуральное обезболивание) или на уровне верхних углов под-
вздошной кости. Для этого животное фиксируют в боковом положении или
спиной кверху, подложив ему под живот, находящийся у края стола, подуш-
ку и дугообразно выгнув его спину. Выстригают шерсть, дезинфицируют кожу
в области первых хвостовых позвонков и в этом месте делают прокол. В
зависимости от величины собаки ей вводят от 2 до 10 мл 2%-ного новокаина
или 0,1%-ного совкаина. У кошек можно проводить люмбальную спинно-
мозговую анестезию, для чего делают прокол между последним поясничным
и 1-м хвостовым позвонком с последующим введением 1%-ного раствора
новокаина или 0,1%-ного совкаина
Местная анестезия слизистых оболочек достигается 2—5—10%-ными ра-
створами кокаина или дикаина.
Комбинированный наркоз является наиболее распространенным и об-
щепринятым видом обездвиживания, выключения сознания и обезболива-
ния у лабораторных животных. Этот вид наркоза является наиболее оптими-
зированным подходом в клинической анестезиологии, что, в свою очередь,
делает его приемлемым в необходимых случаях биомоделирования лабора-
торных животных.
Так, например, при комбинированном наркозе у средних и мелких жи-
вотных (кошки, кролики) лучше начинать с этилового эфира. Животное по-
мещают под стеклянный колпак, куда кладут кусок ваты, обильно смо-
ченный эфиром. Для достижения наркоза необходимо 20—30 мл эфира (в за-
висимости от веса животного). Внимательно следят за дыханием и тонусом
животного, так как в этих условиях легко передозировать наркотик. В состо-
янии наркоза фиксируют животное к операционному столу и в дальнейшем,
по мере надобности, поддерживают глубину наркоза нанесением эфира на
маску. Гексеналовый наркоз в дальнейшем следует проводить с осторожно-
стью, поскольку барбитураты потенцируют эффекты морфина в отношении
дыхательного центра. Лучше применять этаминал натрия (нембутал) внутри-
брюшинно или подкожно в виде 5%-ного раствора по 40-60 мг/кг. Наркоз
наступает через 10—15 мин после введения. Если же наркотическое состоя-
ние при этом не развилось, необходимо ввести дополнительную дозу, рав-
ную 1/4 предыдущей дозы. Естественно, что для парентерального введения
растворы должны быть стерильными.
Для анестезии при небольших хирургических процедурах можно внутри-
мышечно вводить различные нейролептики: фентанил [111], диазепам [86],
смесь пентобарбитала натрия и скополамина гидрохлорида [283]. Некоторые
авторы [293] для длительной анестезии предлагают свой способ, заключаю-
щийся в последовательном внутримышечном введении вначале смеси аналь-
гетика мередина гидрохлорида и транквилизатора азаперона, а через 20 мин —
Глава 21. Наркоз
305
смеси наркотиков: кетамина гидрохлорида и сульфата морфия. Недостатком
внутримышечного введения обезболивающих препаратов является невоз-
можность регулирования уровня иммобилизации. Этого можно добиться
путем внутривенного введения обезболивающих препаратов, например, че-
рез ушную вену [621]. Многие исследователи считают наиболее эффектив-
ным ингаляционный наркоз, в том числе и с предварительным внутривен-
ным введением барбитуратов [175, 660].
Собакам и другим крупным животным {мини-свиньи, овцы) можно вво-
дить сернокислую магнезию в виде 25%-ного раствора внутривенно по 1,0—
1,5 г/кг или внутримышечно по 1,25 г/кг. Лучшие результаты этот препарат
дает в комбинации с морфином. Наркоз можно вызывать и иными наркоти-
ческими средствами, которые указаны в дальнейшем изложении.
Следует особо подчеркнуть, что перед дачей наркоза, а во многих случаях
и перед началом эксперимента, абсолютно необходима премедикация, или
лекарственная подготовка.
У кроликов и кошек лекарственная подготовка, или премедикация, вклю-
чают последовательное, комбинированное или раздельное применение сле-
дующих препаратов: атропин 0,2 мг/кг подкожно (п/к) за 30 мин.; фента-
нил-флюанизон 0,1—0,5 мл/кг внутримышечно (в/м); диазепам или мидазо-
лам 2 мг/кг внутривенно (в/в), интраперитонеально (и/п) или в/м; кетамин
25 мг/кг + ксилазин 4 мг/кг в/м.
После премедикации можно перейти к общей анестезии. Для этого неболь-
шими концентрациями этилового эфира, но не хлороформа (кролики и кошки
весьма чувствительны к хлороформу и быстро от него погибают), можно вы-
зывать неглубокий наркоз. Для получения глубокого наркоза эфир нужно до-
бавлять небольшими порциями, осторожно. Необходимо помнить, что при од-
новременно даваемой большой дозе может наступить остановка дыхания и
смерть. Кролики хорошо переносят уретановый наркоз. Уретан вводят по 0,6—
1 г на 1 кг веса в/б и в/м (20—40%-ный раствор). Хлоралгидрат вводят в виде
10—12%-ного раствора в/в в дозе 100—150 мг/кг или ректально в дозе 300—
500 мг/кг. В смеси с морфином (100—250 мг/кг хлоралгидрата в 0,5—1 мг/кг
морфина) его можно вводить в/б. Для наркоза можно пользоваться также бар-
битуратами кратковременного (гексенал, тиопентал) и среднего (этаминал, бар-
бамил) действия. Наряду с классическими схемами наркоза в настоящее время
рекомендуются следующие средства инъекционной анестезии.
/ фентанил-флюанизон 0,3 мг/кг в/м, диазепам или мидазолам 2 мг/кг
в/в или и/п спустя 5 мин.;
/ кетамин 10мг/кг в/в + ксилазин 3 мг/кг в/в;
/ медетомидин 0,3 мг/кг в/в + кетамин 20мг/кг + диазепам 1 мг/кг п/к.
При желании, экспериментатор может прибегнуть к ингаляционной ане-
стезии:
/ галотан 1,5-2,0%;
306
Биоэтика в экспериментальных исследованиях
/ энфлюран или изофлюран;
/ метоксифлюран 0,4— 1,0%.
Для получения более глубокого обезболивания можно ввести бупренор-
фин 0,02 мг/кг в/м или в/в 12 р/ч.
У морских свинок премедикация осуществляется следующими вещества-
ми и в следующей последовательности: атропин 0,05 мг/кг и/к/, фентанил-
флюанизон 1,0 мл/кг в/м; диазепам 5 мг/кг и/п (не оказывает обезболиваю-
щего действия!); кетамин 100 мг/кг и/п.
Следует помнить, что наряду с отсутствием обезболивающего действия
кетамина, применение эфира или хлороформа может дать негативные по-
бочные эффекты со стороны сердечно-сосудистой и центральной нервной
систем морских свинок. Современные средства для общей ингаляционной
анестезии, хорошо переносимые морскими свинками, включают метоксиф-
люран и галотан. Для проведения наркоза ингаляционными средствами
животное помещают под колпак, куда кладут ватку, смоченную одним из
этих наркотиков. Вызвав таким образом неглубокий наркоз, животное при-
вязывают и через маску продолжают вводить наркотики ингаляционным
путем. Применяется также и хлоралгидрат. Хлоралгидрат лучше вводить
вместе с морфином. Для наркоза могут быть использованы также другие
неингаляционные наркотики: гексенал, тиопентал, уретан. В качестве неин-
галяционных средств общей анестезии используются:
/ фентанил-флюанизон 1 мл/кг в/в + диазепам 2,5 мг/кг и/п 5 мин
спустя;
/ кетамин 40 мг/кг и/п + ксилазин 5 мг/кг и/п;
/ медетомидин 0,5 мг/кг + кетамин 40 мг/кг и/п или в/м;
/ фентобарбитон 25 мг/кг и/п.
В отличие от крупных животных морским свинкам для усиления обезбо-
ливающего эффекта рекомендуется вводить:
/ аспирин 100 мг/кг п/о 4 р/ч;
/ бупренорфин 0,05 мг/кг п/к 12р/ч.
У белых крыс премедикация осуществляется следующими веществами в
нижеизложенной последовательности:
/ атропин 0,05 мг/кг п/к, в/м за 30 мин.;
/ фентанил-флюанизон 0,2-0,5 мл/кг в/м или 0,3—0,6 мл/кг и/п;
/ диазепам или мидазолам 2 мг/кг в/в или и/п (не оказывает обезболива-
ющего действия!); кетамин 25 мг/кг в/м дает слабый расслабляющий и
обезболивающий эффект;
/ ксилазин 1-3 мг/кг в/м.
Ингаляционный наркоз у белых крыс с осторожностью можно проводить
при помощи этилового эфира, а также галотана и метоксифлюрана. Живот-
ное помещают под небольшой колпак, куда кладут ватку, смоченную эфи-
ром, и следят за наступлением наркоза. Неингаляционный наркоз вызывают
Глава 21. Наркоз
307
подкожным или внутрибрюшинным введением уретана (10%), этаминала (в/
б 40—50 мг/кг), барбамила (п/о 50-80 мг/кг), хлоралгидрата (200—250 мг/кг)
и других наркотиков, например:
/ фентанил-флюанизон 0,4 мг/кг и/п;
/ фентанил-флюанизон 0,3 мг/кг + диазепам 2,5 мг/кг и/п;
/ медетомидин 0,5 мг/кг + кетамин 75 мг/кг и/п или п/к;
/ фентобарбитон 40 мг/кг и/п — без обезболивания!
Так же, как у морских свинок, для усиления обезболивающего эффекта
вводят: аспирин 100 мг/кг п/о 4 р/ч; бупренорфин 0,1-0,5 мг/кг п/к 12 р/ч.
Для премедикации мышей используют следующие вещества:
/ атропин 0,04 мг/кг п/к, в/м за 30 мин.;
/ фентанил-флюанизон (гипнорм) (1:10 в 0,9%-ном растворе хлорида на-
трия) 0,01-0,03 мл/10 г; ксилазин 0,1 мг/10 г и/п на мышах дает слабый
обезболивающий эффект.
Полное обездвиживание мыши можно получить под наркозом. Наркоти-
зирование мышей проводят с осторожностью под этиловым эфиром, меток-
сифлюраном или хлороформом. Животное помешают под небольшой кол-
пак, куда кладут ватку, смоченную средством для ингаляционной анестезии,
и внимательно следят за дыханием и тонусом мышц.
Неингаляционный наркоз можно вызывать введением гексенала, барба-
мила, уретана а также других средств для неингаляционной анестезии:
/ фентанил-флюанизона (гипнорма) (1:10 в 0,9%-ном растворе хлорида
натрия) 0,03 мл/10 г + диазепам 0,05 мг/10 г и/п.;
/ налоксона в качестве антагониста 0,00001—0,001 мл/10 г и/п или в/в;
фентобарбитона 40 мг/кг и/п — без обезболивания!
Для усиления обезболивания вводят:
/ аспирин 1,2 мг/10 г п/о 6 р/ч;
/ бупренорфин 0,01—0,025 мг/10 г и/п или п/к 12 р/ч.
Таблица 52
Экспериментальные биомодели для изучения снотворных, наркотических
и антинаркотических эффектов веществ
Исследуемое действие
вещества
Моделирование пролонги-
рования снотворного дей-
ствия барбитуратов (вве-
дение гексенала в дозе
50 мг/кг, внутрибрюшинно)
Моделирование гипноти-
ческого эффекта по боково-
му положению животных
(введение гексенала в дозе
30-60 мг/кг, внутри-
брюшинно)
Лабораторное
животное
Мыши
Крысы
Мыши
Крысы
Линия или порода
животного
Аутбредные
Аутбредные
Аутбредные
Аутбредные
Пол
животного
9с?
9сГ
Яд
9д
Вес
животного
40-50 г
300-400 г
30-50 г
300-350 г
Возраст
животного
3-10 мес.
5-12 мес.
3-10 мес.
5-12 мес.
308 Биоэтика в экспериментальных исследованиях
Окончание табл. 52
Исследуемое действие
вещества
Моделирование снотвор-
ного действия с электро-
физиологических экспери-
ментах:
a. модель для оценки
влияния в-в на циклы
«сон-бодрствование»
b. моделирование депри-
вации парадоксальной
фазы сна по методу Жуве
(модель нарушений сна
при невротических рас-
стройствах)
c. моделирование нару-
шений структуры «сон-
бодрствование» при сдви-
гах дофаминхолинерги-
ческих взаимодействий
(при парксиническом синд-
роме и депрессивном со-
стоянии)
Биомодель для исследова-
ния антинаркотического
действия
Лабораторное
животное
Крысы
Крысы
Крысы
Крысы
Линия или порода
животного
Белые
аутбредные
Белые
аутбредные
Белые
аутбредные
Аутбредные
Пол
животного
б
б
б
96
Вес
животного
300-350 г
300-350 г
300-350 г
300-350 г
Возраст
животного
5-12 мес.
5-12 мес.
4-5 мес.
Взрослые
особи
Пояснения к таблице можно найти в табл. 6, гл. 2.
309
Глава 2 2 ДОПУСТИМЫЕ МЕТОДЫ
ЭВТАНАЗИИ ЖИВОТНЫХ
Эвтаназия мышей осуществляется углекислым газом
в течение 10 мин, шейной дислокацией, фентобар-
битоном (5%-ный раствор 0,5 мл и/п). Умерщвле-
ние мышей производят также эфиром, хлороформом,
пропусканием через головной мозг электрического
тока.
Эвтаназия крыс осуществляется углекислым га-
зом в течение 15-20 мин, шейной дислокацией, фен-
тобарбитоном (5%-ный раствор 0,5 мл и/п), гильо-
тиной. Крыс умерщвляют также хлороформом или
эфиром, помещая их в небольшую закрытую посу-
ду, или пропусканием электрического тока через
головной и спинной мозг.
Эвтаназия морских свинок осуществляется угле-
кислым газом, фентобарбитоном 90 мг/кг и/п. Мор-
ских свинок умерщвляют также хлороформом, эфи-
ром, вводя их ингаляционно, внутрибрюшинно и в
толщу легких, а также пропусканием электрическо-
го тока от городской сети через головной и спин-
ной мозг.
Эвтаназия кроликов осуществляется фентобарби-
тоном 100 мг/кг в/в или и/п. Кроликов можно
умерщвлять нанесением удара по основанию чере-
па (при этом держат их за задние конечности вниз
головой), внутривенным введением хлороформа,
эфира или воздуха, а также пропусканием электри-
ческого тока из городской сети через спинной и
головной мозг (один электрод в виде зажима Пеана
310
Биоэтика в экспериментальных исследованиях
накладывают на угол рта, другой в виде иглы вводят под кожу в области
крестца). Время воздействия тока 3—5 с.
Эвтаназия крупных лабораторных животных (мини-свиньи, собаки, кошки
и др.) осуществляется введением в толщу легких или кровь хлороформа
(5-7 мл), эфира (15-20 мл) или пропусканием электрического тока из го-
родской сети через центральную нервную систему (один электрод в виде
иглы вводят в области поясничных позвонков, а другой в виде зажима
Пеана накладывают на угол рта). Более приемлемые методы изложены в
табл. 53.
Таблица 53
Характеристика приемлемых методов эвтаназии экспериментальных
животных [378]
Виды
животных
Скорость
1. ХИМИЧЕСКИЕ ME
А) Препараты для v
1. Натрия пентобар
Грызуны
Кролики
Плотоядные
Птицы
Крупные
млекопитаю-
щие
++
++
++
++
++
2. Препарат Т-61
Грызуны
Кролики
Плотоядные
Птицы
Крупные
млекопитаю-
щие
++
++
++
++
++
3. Секобарбитал /1
Плотоядные
++
4. Хиналбарбитон /
Крупные
млекопитаю-
щие
++
Эффективность
тодь
нъеки
битон
++
++
++
++
++
++
++
++
++
++
*ибука
++
нупер
++
Удобство
ИИ
ат (не
+
+
+
+
+
ин
—
каин
—
Риск
мбута
+
+
+
+
+
+
+
+
+
+
+
+
Гуманность
л, пег
++
++
++
++
++
++
++
+
++
++
++
++
Оценка в баллах
nbutal
5
5
5
5
5
4
4
4
4
4
4
5
Вещества и лекарственные препараты,
физические методы выбора, дозы, формы
и особенности применения для эвтаназии
)
Обычно используется 18%-ный раствор
(200 мг/мл) препарата в дозе 200 мг/кг в/в
или в/б; для крупных птиц в/б или внутри-
черепная инъекция (Foramen magnum), для
грызунов в/в и в/б инъекции; для кроликов
и крупных млекопитающих в/в
Т-61 является комбинацией местного анесте-
тика, снотворного и курареподобного веще-
ства. Средство предназначено только для
медленного в/в введения, иначе оно вызыва-
ет боль у животных; птицам массой < 250 г
можно вводить в грудную мышцу
Приемлемо в/в введение
Приемлемо в/в введение лошадям
Глава 22. Допустимые методы эвтаназии животных 311
Продолжение табл. 53
Виды
животных
Л
1-
о
О
а
о
и
Л
1-
о
о
X
а
s
ф
■е-
■е-
о
со
1-
о
\0
О
5
1
О
X
X
л
X
а
с;
с;
а
VD
а
(9
X
Ф
Я1
О
Вещества и лекарственные препараты,
физические методы выбора, дозы, формы
и особенности применения для эвтаназии
Б) Ингаляционные средства
5. Двуокись углерода
Грызуны
Птицы
Кролики
Крупные
млекопитаю-
щие
++
++
+
+
++
++
+
+
++
++
++
+
++
++
++
++
+
+
+
4
4
1
1
С02 в концентрации > 70% приемлем для
грызунов, кроликов и свиней; для морских
свинок и для цыплят в возрасте 72 ч —
100% С02. С02 не следует использовать для
животных, способных нырять
6. Окись углерода
Грызуны
Птицы
Кролики
+
+
+
+
+
+
+
++
++
—
++
+
++
2
2
1
Опасен для оператора
7. Летучие анестетики — галлотан, энфлуран, изофлуран
Грызуны
Плотоядные
Птицы
Кролики
Крупные
млекопитаю-
щие
++
++
++
++
+
++
++
++
++
+
++
+
++
++
+
+
+
+
+
+
++
++
++
+
5
4
4
2
2
Приемлемы; смерть необходимо подтвер-
дить; у кроликов возникают признаки
дистресса; для рыб используется только
галлотан; из крупных млекопитающих
применяются у ягнят и козлят
II. ФИЗИЧЕСКИЕ МЕТОДЫ Методы использования физических
факторов
8. Церквильная дислокация
Грызуны
Кролики
Птицы
++
++
++
++
++
++
+
++
++
++
—
4
4
4
Приемлем для грызунов массой < 150 г,
для кроликов < 1 кг.
9. Декапитация
Грызуны
Кролики
+
+
+
+
+
+
++
++
—
2
2
Можно использовать у грызунов и кроликов
массой < 1 кг, но предпочтительнее другие
методы
10. Сотрясение мозга (оглушение)
Грызуны
Кролики
Птицы
Плотоядные
Крупные
млекопитаю-
щие
++
++
++
++
++
+
+
+
++
++
+
+
4
2
2
Приемлем для грызунов массой < 1 кг,
для птиц < 250 г и для новорожденных
плотоядных; для крупных млекопитающих
необходимо немедленное обескровливание
312
Биоэтика в экспериментальных исследованиях
Окончание табл. 53
Виды
животных
о
о
кор
о
зность
екти
-е-
■е-
го
о
1-
о
ю
о
£
II. ФИЗИЧЕСКИЕ МЕТОДЫ
11. Оглушение электротоком
Крупные
млекопитаю-
щие
Кролики
Плотоядные
Птицы
++
++
++
+
++
+
++
+
+
++
—
+
12. Мацерация
Птицы
++
++
++
13. Микроволновое облучение
Грызуны
Кролики
Птицы
++
++
++
++
++
++
—
—
—
14. Быстрое замораживание
Грызуны
Кролики
—
+
+
+
++
++
15. Привязной гарпун
Крупные
млекопитаю-
щие
Кролики
Плотоядные
++
++
++
++
+
++
++
—
—
16. Отстрел
Пресмыкаю-
щиеся
Крупные
млекопитаю-
щие
Плотоядные
++
++
++
++
++
++
++
—
—
иск
о.
—
—
—
—
++
++
++
++
++
++
+
+
++
—
—
—
J3
о
о
i
со
2
i^
—
+
—
—
—
+
+
+
+
+
+
+
++
+
—
—
баллах
со
со
цен
О
4
3
3
1
4
3
3
3
1
1
5
4
3
4
2
1
Вещества и лекарственные препараты,
физические методы выбора, дозы, формы
и особенности применения для эвтаназии
Методы использования физических
факторов
Смерть следует подтвердить. Не следует
применять у кошек, так как их шерсть отли-
чается высокой электропроводностью,
и у рогатых животных, если рога мешают
наложению электродов
Приемлем для цыплят в возрасте до 72 ч
Применяется нейробиологами для фиксации
метаболизма мозга; для мышей приемлемо
облучение всего тела
Приемлем, если это предусмотрено планом
эксперимента. Приемлем для новорожденных
грызунов и плодов кроликов с массой < 4 г
Приемлем для крупных млекопитающих;
гарпун не всегда эффективен для умерщвле-
ния крупных свиней и взрослых быков из-за
толщины и плотности их черепов
Приемлем только в полевых условиях; пред-
почтителен для лошадей
Примечание. Приемлемость метода оценивается следующими показателями:
/ скорость действия агента: «++» - очень быстро, «+» - быстро «—» - медленно;
/ эффективность метода: «++» - высокоэффективен,«+» - эффективен,«—» - неэффективен;
/ удобство метода:«++» - прост в применении, «+» - требует опыта, «—» - требует специаль-
ного обучения экспериментатора;
/ риск для оператора: «++» - безопасен, «+» - опасность минимальная, «—» - опасно;
/ гуманность: «++» - приемлем для всех экспертов, «+» - приемлем для большинства экспер-
тов, «—» - неприемлем для всех экспертов и экспериментаторов,
оценка в баллах «1-5»: 1 - не рекомендуемый метод, 5 - наиболее рекомендуемый метод.
Разделе БИОМОДЕЛИРОВАНИЕ
В СИСТЕМАХ УПРАВЛЕНИЯ
Между разумом и рассудком такая же граница,
как между книгой кулинарных рецептов и пирогом.
К.Берне
315
Глава 2 3 ВЫСШИЕ УРОВНИ
УПРАВЛЕНИЯ
Высший пик эволюции живой материи — головной
мозг человека имеет студнеобразную консистенцию,
массой -1,5 кг. С механической точки зрения он
построен очень ненадежно, но в то же время гаран-
тирует его носителю хорошую устойчивость в не-
стабильной внешней среде. Само по себе наличие
гигантского количества нервных путей и миллиар-
дов нервных клеток еще не обеспечивает устойчи-
вость системы. Однако способность головного моз-
га преобразовывать химические, электрические и
магнитные воздействия в универсальные коды, обес-
печивать их хранение и направлять в исполнитель-
ные органы и системы по принципу «надо полу-
чить такой то результат» (а не «расслабься, сокра-
тись, выдели») делает его высшим законодателем не
только для человека и животных, но, в определен-
ной степени, и для природы (см. гл. 2).
Возмущения в неравновесных функциях распре-
деления строго ограничены, что иллюстрируется
процессами распространения нервного импульса с
последующим восстановлением распределения по-
тенциалов. Это одно из проявлений универсальнос-
ти нервного кода. Эволюция создала и четко разде-
лила кодовую и параметрическую части.
В основе параметрической части функций не-
рвной системы лежит рефлекторный процесс. Не-
рвная система разделяется на центральную и пе-
риферическую. Центральная нервная система
316
Биомоделирование в системах управления
включает головной и спинной мозг вместе со спинномозговыми ганглия-
ми. Вместе с периферической нервной системой ЦНС, благодаря принципу
обратных связей, защищает от внешних и внутренних разрушающих воз-
действий и обеспечивает устойчивость и согласованность всех систем орга-
низма, моделируемых (см. гл. 1) на основе теории управления.
Головной мозг и поведение
Головной мозг состоит из большого и ромбовидного мозга. Большой мозг
включает в себя средний, межуточный и конечный мозг. Последний состоит
из плаща и обонятельного мозга. Плащ имеет на поверхности борозды и
извилины, степень развития которых, а также их направление не одинаковы
у различных животных, но очень обеднены в сравнении с человеком. У собак
и кошек относительно мало борозд и извилин, подобно другим хищным они
концентрически расположены вокруг сильвиевой борозды. У мини-свиней мозг
непропорционально маленький в соотношении с массой тела. У кроликов,
крыс, хомячков и мышей поверхность плаща сглажена и лишена дифференци-
ровки на явные и соответствующие человеку борозды и извилины. Обоня-
тельные извилины в области основания черепа сливаются, образуя у всех
животных тракт.
Для получения различных экспериментальных моделей поведения, регу-
лируемого высшими отделами ЦНС широко используются методы прямого
электрического раздражения корковых и глубинных структур головного мозга.
Эти работы начались с классических исследований Хосе Дельгадо и нашли
свое применение в самых различных направлениях нейрофизиологии, нейро-
и психофармакологии, неврологии, нейрохирургии и психиатрии. Мы в своих
работах широко пользовались этим приемом для оценки эффективности пси-
хотропных средств и ксенобиотиков [326, 328, 344]. Это исключительно на-
дежные информативные и легко воспроизводимые методы исследования на
самых различных животных.
Измерения порогов возникновения тех или иных поведенческих реак-
ций можно осуществлять через хронически имплантированные электроды у
самых разных животных (табл. 54, 56, 57, 58). Подчеркнем, что во многих
клинических исследованиях при электрическом раздражении мозга человека
развиваются такие же реакции как и у животных, что дает хороший инстру-
мент для исследователей и расширяет экстраполяционные возможности при
биомоделировании.
При микроскопическом изучении кору головного мозга разделяют на
шесть слоев: молекулярный, наружный зернистый, слой малых пирамидных
клеток, внутренний зернистый, слой больших пирамидных клеток и слой
полиморфных клеток [75]. В затылочной области коры содержатся девять
слоев, а в обонятельной области — пять слоев.
Глава 23. Высшие уровни управления
317
Таблица 54
Сложноорганизованные реакции у мини-свиней, собак, кошек и кроликов
при электрической стимуляции нео- и архипалеокортекса
Электростимуляция
мозговых структур
Прореальная изви-
лина, преоптичес-
кое поле, передний
гиппокамп,уздечка
Височная часть вер-
хушки и орбиталь-
ная кора
Пириформная кора,
гиппокамп в ростро-
базальной части
Передняя поясная
извилина коры
Каудо-вентральный
и задний отделы
гиппокампа
Характер
поведенческих
реакций
Страх, переходящий
в ярость (при увели-
чении порогов раз-
дражений)
Возбуждение без
ярости
Реакция страха
и ярости
Реакция ярости.
Поведенческие
эквиваленты гнева
Реакции,подобные
страху; облегчение
обучения методом
условных рефлек-
сов
Вегето-
эмоциональные
реакции
Расширение зрачков,
взъерошивание, тре-
мор, остановка дыха-
ния вслед за гипер-
пноэ, прижатые уши,
уменьшение реактив-
ности
Оглядывание, насто-
раживание
Пищевой автоматизм
и автономные реак-
ции, животное испуга-
но или взволновано
Ориентировочные
проявления
Расширение зрачков,
взъерошивание, при-
жимание ушей, пока-
зывание зубов, шипе-
ние, фыркание, хрюка-
ние, дефекация и мо-
чеиспускание
Атака
Ненаправ-
ленное
кусание
Попытки
атаки,
кусание
Без агрессии
Атаки нет
Направлен-
ные удары
и кусание
Избегание
Бегство в укры-
тие
Иногда побег
Ненаправлен-
ное бегство
Попытки избе-
гания
Направленный
побег
На поверхности полушарий головного мозга человека различают следу-
ющие доли: лобную, теменную, затылочную, височную и характерную для
животных обонятельную. Кора головного мозга у млекопитающих пред-
ставлена неокортексом, архикортексом, палеокортексом и межуточной
корой.
Лобные доли коры у животных представляет прореальная извилина,
а у человека это префронтальная кора (в узком смысле слова включает поля
9-12 по Броку), а также поля 13 и 14 на их орбитальной поверхности. Аффе-
рентные сигналы поступают от дорсомедиального ядра таламуса. У лобной
коры обширные двусторонние связи с отделами лимбической системы, вклю-
чая поясную извилину, гиппокамп, миндалину и гипоталамус. Лобные доли
можно рассматривать как неокортикальную часть лимбической системы.
Лимбическая система играет особую роль в видоспецифичном поведении,
эмоциях, влечениях, мотивациях и т. д. Важнейшей функцией лобной коры
является управление врожденными формами поведения при помощи накоп-
ленного опыта и принятых решений.
318 Биомоделирование в системах управления
Таблица 55
Экспериментальные и генетически обусловленные биомодели для изучения
ноотропного действия веществ*
Исследуемое действие
вещества
Модель амнезии по УРПИ,
вызванной электрошоком
15-20 мА, 200-500 мс, на
роговицу глаз; 10-100 мА,
200-500 мс на поверхность
ушных раковин
Модель амнезии, вызванной
скополамином 0,5-1,5 мг/кг,
(внутрибрюшинно)
Модель для транскалло-
зальных вызванных потен-
циалов 20-40 В, 0,2-1 мс
Модель для изучения про-
тивогипоксических свойств:
введение нитрита натрия
в дозе 150-250 мг/кг, под-
кожно
Моделирование влияния на
физическую работоспособ-
ность - «бегущая дорожка»,
«беличье колесо», плава-
ние с дополнительной
нагрузкой
Моделирование амнести-
ческих нарушений при
перинатальных поврежде-
ниях - введение 25%
этанола в дозе 4000 мг/кг
в течение всей беремен-
ности (21 день), через
желудочный зонд
Моделирование амнести-
ческих нарушений при
введении:
a. морфина в дозе 10 мг/кг
2 раза в день, внутри-
брюшинно
b. феназепама 10 мг/кг
внутрь
c. фенобарбитала 30 мг/кг
внутрь (во время беремен-
ности)
d. ингибитора белкового
синтеза циклогексамида
0,6 мг/кг, подкожно
e. нитрита натрия 40 мг/кг,
внутрибрюшинно (между
10-м и 19-м днями бере-
менности)
Лабораторное
животное
Крысы
Мыши
Крысы
Мыши
Крысы
Кролики
Кошки
Мыши
Крысы
Мыши
Крысы
Крысы
Крысы
Крысы
Крысы
Крысы
Линия или порода
животного
Аутбредные
Аутбредные
Аутбредные
Аутбредные
Аутбредные
Аутбредные
Аутбредные
Аутбредные
Аутбредные
Аутбредные
Аутбредные
Аутбредные
Аутбредные
Аутбредные
Аутбредные
Аутбредные
Пол
животного
96
96
96
96
96
96
9^
Q6
§6
9
9
9
9
96
9
Вес
животного
300-350 г
40-50 г
300-350 г
40-50 г
300-350 г
2,5-4 кг
3-5 кг
40-50 г
300-350 г
40-50 г
300-350 г
300-350 г
300-350 г
4-6 г
300-350 г
4-6 г
Возраст
животного
5-10 мес.
3-10 мес.
5-10 мес.
3-10 мес.
5-12 мес.
7 мес. -
4 года
1 -7 лет
3-10 мес.
5-12 мес.
3-10 мес.
5-12 мес.
в период
беремен-
ности
5-12 мес.
5-12 мес.
7-14 дн.
5-12 мес.
7-14 дн.
Глава 23. Высшие уровни управления
319
Окончание табл. 55
Исследуемое действие
вещества
Биомодели для изучения
памяти при естественном
старении
Нарушение памяти в про-
гериатрических моделях:
применение пищи, содер-
жащей 2% холестерина в
течение 2-3 месяцев
введение этанола в течение
20-36 недель (в виде 15%
водного раствора), средне-
суточная доза 0,58-0,63 мл
введение скополамина
в дозе 1 мг/кг, внутрибрю-
шинно, в течение 20 дней
Моделирование когнитив-
ных нарушений, вызванных
фронтальной лобэктомией
Моделирование когнитив-
ных нарушений, вызванных
ишемическими воздейст-
виями:
a. фотоиндуцированный
тромбоз - введение фото-
чувствительной краски бен-
гальского розового 40 мг/кг
в подключичную вену
и облучение светом ксено-
новой лампы 250 Вт
b. двух- или четырехсосуди-
стая перевязка артерий
Влияние на ССС, на дыха-
ние и на периферические
отделы нервной системы
(на интактных или наркоти-
зированных животных)
Лабораторное
животное
Мыши
Крысы
Крысы
Мыши
Мыши
Крысы
Крысы
Крысы
Крысы
Мыши
Крысы
Песчанки
Кошки
Кошки
Крысы
Линия или порода
животного
Аутбредные
Аутбредные
Аутбредные
Аутбредные
Аутбредные
Аутбредные
Аутбредные
Аутбредные
Аутбредные
Линии ddY
Аутбредные
Аутбредные
Аутбредные
Аутбредные
Аутбредные
Пол
животного
Qc?
9%
9с?
9
9с?
9с?
9с?
Q6
§<?
§с?
§с?
9с?
Вес
животного
40-50 г
300-350 г
300-350 г
40-50 г
40-50 г
300-350 г
300-350 г
300-350 г
300-350 г
30-40 г
300-350 г
средний
вес
3-5 кг
2,5-4 кг
300-350 г
Возраст
животного
14-19 мес.
16-22-24
мес.
5-12 мес.
3-10 мес.
3-10 мес.
5-12 мес.
5-12 мес.
5-12 мес.
5-12 мес.
3-10 мес.
5-12 мес.
Взрослые
особи
1 -7 лет
7 мес-
4 года
5-12 мес.
* См. табл. 6 в главе 2
Образования новой, старой и древней коры и глубокие структуры лимби-
ческой системы участвуют в формировании и реализации сложноорганизо-
ванных поведенческих актов. Электрическим раздражением через вживлен-
ные электроды можно вызвать оборонительные и пищевые эквиваленты по-
ведения. Обычно используют импульсный ток с частотами от 5 до 100 имп/с
и напряжением от долей до нескольких вольт. Определив порог исходной
реакции, можно по его снижению или повышению количественно оценить
уровень возбудимости структур мозга при введении психотропных средств или
любых ксенобиотиков, действием факторов окружающей среды.
320
Биомоделирование в системах управления
В табл. 54 и 56 представлены реакции разных животных при электрости-
муляции нео- и архипалеокортикальных и лимбических структур мозга.
Табл ица 5 6
Активно- и пассивно-оборонительные реакции у мини-свиней, собак, кошек
и кроликов при электрической стимуляции миндалевидного комплекса
Электростимуляция
мозговых структур
Медиальные и цент-
ральные ядра мин-
далины
Базальные и лате-
ральные ядра мин-
далины
Латеральные и цен-
тральные ядра мин-
далины, вентраль-
ная часть внутрен-
ней капсулы
Вентро-медиальные
и каудальные участ-
ки латерального и
базального ядер
Околожелудочковое
серое вещество
около миндалин
Характер
поведенческих
реакций
Ярость
Съеживание, прояв-
ление страха
Реакция страха
(временами сме-
шанная)
Реакции гнева
Поиски соперника
Вегето-
эмоциональные
реакции
Мочеиспускание,
дефекация
Рычание, слюноотде-
ление, взъерошива-
ние
Поиски, испуганные
взгляды, расширен-
ные зрачки, пресмы-
кание, беспокойство
Рычание, шипение,
припадание к земле,
прижимание ушей,
удары лапой или копы-
том
Расширение зрачков,
взъерошивание, при-
жимание ушей
Атака
Пассивно-
оборони-
тельные
реакции
Кусание и
царапание
Отсутствует
Направлен-
ная атака
(редко)
Нанесение
направлен-
ных ударов
Избегание
Избегание
Стремление
спрятаться
Подпрыгивает,
убегает, пря-
чется
Редко
Слабое
Если у человека на новую кору приходится 95,6% всей площади полуша-
рия, то у собаки она составляет 84,2%, у кошек 83%, у мини-свиней около
80%, у грызунов значительно меньше. В цитоархитектоническом и функци-
ональном отношении кора головного мозга животных также разделена на
зоны, которые в свою очередь разбиваются на области, подобласти, поля и
подполя.
Мы не будем здесь останавливаться на хорошо известных холинэргичес-
ких, моноаминергических и пептидергических механизмах регуляции инт-
рацентральных отношений головного мозга (подробнее в гл. 24). В после-
дние годы резко возрастает интерес к роли оксидазотных механизмов в обес-
печении функций головного мозга. Следует сразу отметить, что в осуществ-
лении многочисленных функций мозга принимает участие оксид азота (N0).
Содержащие NO-синтазу (фермент, отвечающий за продукцию и оборот ок-
сида азота) нейроны локализованы в различных частях ЦНС. Максимальное
содержание NO-синтазы обнаружено в нейронах мозжечка и обонятельной
Глава 23. Высшие уровни управления
321
луковице. В мозжечке мРНК NO-синтазы находится в основном в грануляр-
ном слое, а иммунореактивная NO-синтаза — в молекулярном и грануляр-
ном слоях. В высокой концентрации фермент содержится в корзинчатых
клетках мозжечка. Абсолютно лишены NO-синтазы клетки Пуркинье.
В нервных клетках содержится только конститутивная NO-синтаза. Ак-
тивация этого фермента происходит под влиянием ионов Са, возбуждающих
аминокислот, ацетилхолина (АЦХ) и некоторых других факторов. Бычий мозг
(цитозоль) содержит NO-синтазу, которая конкурентно ингибируется раз-
личными блокаторами синтеза N0. Результатом повторного внутрибрюшин-
ного введения крысам одного из блокаторов, N-нитро-Ь-аргинина (L-NA),
вызывает необратимое ингибирование NO-синтазы мозга. После 90%-го
ингибирования ферментной активности с помощью L-NA, активность NO-
синтазы не возвращалась к норме по меньшей мере в течение 5 дней[358].
Мы только начинаем понимать роль оксида азота в механизмах деятельно-
сти ЦНС, но это несомненно перспективный путь познания мультимедиа-
торных систем мозга в сравнительном аспекте.
Белое вещество полушарий построено из проекционных, комиссураль-
ных и ассоциационных мякотных волокон. В полушариях большого мозга
серое вещество группируется в подкорковые узлы. Ядра полушарий объеди-
нены под названием полосатого тела, которое состоит из: хвостатого ядра, в
котором различают головку, тело и хвост, плохо выраженный у собак; чече-
видного ядра, разделенного на скорлупу и бледный шар с внутренним и
наружным члениками; миндалевидного ядра. Недалеко от чечевидного ядра
расположена ограда. В табл. 57 показаны реакции животных при электро-
стимуляции разных участков и ядер миндалин.
Хвостатое ядро и скорлупа составляют филогенетически более новый
стриатум, а бледный шар чечевидного ядра — более древний паллидум. По-
вреждение центров стриапаллидарной системы приводит к характерным
нарушениям движения, возникновению гиперкинезов, развитию паркинсо-
низма. В полушариях головного мозга находятся боковые желудочки.
У основания обоих полушарий плаща расположен обонятельный мозг.
Его размеры и функции тем значительнее, чем менее организован головной
мозг. Обонятельный мозг относительно более развит у грызунов, чем у хищ-
ников, а тем более у человека. Он состоит из обонятельных луковиц, обоня-
тельных трактов, обонятельных треугольников, грушевидных долей, хвоста-
тых ядер и аммонового рога.
В луковице животных находится желудочек, стенки которого покрыты
мерцательным эпителием. В луковицу входят волокна п. olfactorius. Груше-
видные доли животных лишены извилин. Двигательные анализаторы распо-
лагаются в передней центральной извилине. В верхней части моторных зон
размещены центры регуляции движениями конечностей, а в нижней — уп-
равляющие мышцами головы.
322
Биомоделирование в системах управления
Таблица 57
Оборонительные и пищевые реакции у мини-свиней, собак, кошек и кроликов
при электрической стимуляции гипоталамуса
Электростимуляция
мозговых структур
Ростральный отдел
гипоталамуса
Передний гипота-
ламус, околосводо-
вое и преоптиче-
ское поле
Средний отдел
гипоталамуса
Латеральный гипо-
таламус, супра-
оптическое ядро
Периферический
или латеральный
гипоталамус
Каудо-латеральный
отдел гипоталамуса
Каудо-медиальный
отдел гипоталамуса
Характер
поведенческих
реакций
Реакция побега
Истинная ярость,
агрессивная атака
Агрессивная
реакция
Аффективная
атака
Атака с подкрады-
ванием
Псевдоаффектив-
ные реакции, без
изменении в спон-
танном поведении
в ответ на ласку
Тревога, бегство,
реакции,харак-
терные для страха
Вегето-эмоциальные
реакции
Расширение зрач-
ков, рычание, шипе-
ние, хрюкание
и визжание
Фыркание, шипение,
хрюкание, рычание,
дефекация, моче-
испускание струей
Расширение зрач-
ков, прижимание
ушей, выгибание
спины дугой, вытя-
гивание конечно-
стей, остановка
спонтанного поведе-
ния
Тревога, расшире-
ние зрачков, шипе-
ние, рычание, выги-
бание дугой спины,
мочеиспускание,
выпущенные когти,
слюнотечение,
гиперпноэ
Скрытая тревога,
расширение зрач-
ков, незначительные
проявления подкра-
дывания
Расширение зрач-
ков, взъерошивание,
прижимание ушей,
слюнотечение,
гиперпноэ, припаде-
ние к земле, рыча-
ние, хлестание хво-
стом
Специфическая
локомоторная актив-
ность, попытки к бег-
ству, реакции,
подобные страху
и боли
Атака
Удаление от
эксперимен-
татора
Движение
атаки
Яростная
целенаправ-
ленная атака
Хватание
крысы и дру-
гих мелких
животных
(не у кроли-
ков)
Злобные
укусы
Не направ-
ленные ку-
сания и ца-
рапание
Пассивно-
оборони-
тельная
реакция
Избегание
Хорошо
ориентиро-
ванное бег-
ство, п ры-
гание наверх
клетки
Агрессия
может
сменяться
бегством
Отсутствует
Отсутствует
Редко
Слепые
попытки
к бегству
Бегство и
облегчение
обучения
избегания
Глава 23. Высшие уровни управления
323
Экстирпация затылочной доли нарушает зрительные условные рефлек-
сы; височной доли — нарушает слух; теменных областей — кожную чувстви-
тельность; аммонова рога — обонятельные восприятия.
Промежуточный, средний и задний отделы головного мозга образуют ство-
ловую часть, или ствол мозга. Задний и продолговатый мозг объединяются
под названием ромбовидный мозг. Нижнюю, базальную поверхность головно-
го мозга, от которой отходят 12 пар черепно-мозговых нервов, принято назы-
вать основанием мозга, а её верхние слои — конвекситальной корой.
Межуточный мозг состоит из зрительных бугров, надбугровой, субтала-
мической, подбугровой и забугровой областей. В таламусе имеются пере-
дние, внутренние, средние, наружные и вентральные отделы, а также клетки
ретикулярной формации. К таламусу прилегают наружное коленчатое тело,
вливающееся в зрительный тракт и внутреннее коленчатое тело, переходя-
щее в задние ножки четверохолмия. Эпиталамус состоит из эпифиза. Ядро
уздечки связано с обонятельным анализатором.
Гипоталамус состоит из 20 ядер. К воронке подвешен нижний придаток
мозга или гипофиз. Он защищен от повреждения турецким седлом и состоит
из трех частей. Задняя доля гипофиза, или нейрогипофиз, образована окон-
чаниями аксонов нервных клеток, тела которых находятся в гипоталамусе и
его функции регулируются нейромедиаторами. ЦНС функционирует как эн-
докринная железа. В ней вырабатываются нейромедиаторы и гормоны. Их про-
дукция генетически предопределена. Так, например, уровень норадреналина в
гипоталамусе линий BALB/cJLac — 0,499 мг/г, что в 3 раза выше, чем у мышей
C57BL/6 [184]. У линий C57BL/6J уровень гипоталамического норадреналина
(0,165-0,152 мг/г сырого веса) в 3 раза ниже, чем у BALB/c [184].
Передняя доля гипофиза, или аденогипофиз — это скопление клеток,
секретирующих гормоны. Передняя и задняя доли гипофиза разделены ме-
жуточной долей, которая иннервируется нервами, идущими из гипоталаму-
са. Промежуточная доля имеет большое значение у низших позвоночных и
значительно меньше у млекопитающих. Её функции у человека неясны.
Ядра гипоталамуса более дифференцированы у мини-свиней, обезьян, со-
бак, кошек и менее у грызунов. В табл. 58 представлены сложноорганизован-
ные поведенческие реакции животных, вызванные электрическим раздраже-
нием ядер гипоталамуса (100 имп/с; 5 мс, 2—5 В).
Поведенческие реакции животных при раздражении гипоталамуса имеют
такое количество отдельных компонентов и нюансов, что их описание заня-
ло бы слишком много места. По собственным наблюдениям можно сказать,
что в сравнении с другими участками мозга, дающими как бы монохромати-
ческое освещение поведения, электростимуляция гипоталамуса вызывает
гамму цветов и оттенков, двигательных, эмоциональных и вегетативных
компонентов. Гипоталамус, как призма, преобразует серию электрических
импульсов в яркий спектр цветов психовегетативного поведения.
324
Биомоделирование в системах управления
Таблица 58
Поведенческие реакции у мини-свиней, собак, кошек и кроликов
при электрической стимуляции таламуса и среднего мозга
Электростимуляция
мозговых структур
Латеральный отдел
таламуса
Латеральный отдел
покрышки, рост-
ральная часть цент-
рального серого ве-
щества, задневент-
ральное ядро тала-
муса
Средний мозг, цент-
ральное серое ве-
щество (медиальная
часть)
Серое вещество,
окружающее силь-
виев водопровод
(каудальная часть)
Характер
поведенческих
реакций
Реакция страха, об-
легчение условно-
рефлекторной дея-
тельности
Реакции, подобные
страху, закрепление
условных рефлек-
сов
Аффективная обо-
рона, угроза,атака,
иногда убегание,
поведение бегства
Реакции,показыва-
ющие боль, поворо-
ты головы, движе-
ния глазных яблок
Вегето-
эмоциональные
реакции
Расширение зрачков,
прижимание ушей,
шипение, фыркание,
хрюкание, дефекация
и мочеиспускание
Расширение зрачков,
взъерошивание, «ис-
пуганное» выражение
морды, пронзитель-
ные голосовые реак-
ции, активно-оборо-
нительные движения,
мочеиспускание, де-
фекация, тахикардия,
учащение дыхания
Рычание, шипение,
прижатые уши, расши-
рение зрачков, взъе-
рошивание, сгорблен-
ная спина
Расширение зрачков,
борьба, гиперпноэ,
мочеиспускание, де-
фекация
Атака
Защитные
реакции в
виде ударов
и кусания
Хватание и
кусание на-
ходящихся
в пределах
досягаемо-
сти предме-
тов
Направлен-
ная атака
Нет
Избегание
Направленные
или хаотичные
реакции избе-
гания
Большие прыж-
ки, защитные
телодвижения,
бегство
Иногда
Бегство или
оборона
Нагревание или охлаждение определенного участка переднего гипотала-
муса вызывает усиление теплоотдачи или термогенеза. Регистрацией актив-
ности одиночных нейронов идентифицированы температурно-чувствитель-
ные пулы, в которых при локальном нагревании увеличивается частота раз-
рядов. Усиление активности этих нейронов предшествует запуску противо-
действующего нагревания механизма.
Преоптическая область и передний гипоталамус содержат не только теп-
лочувствительные клетки, но и нечувствительные к температуре нейроны, а
также холодочувствительные клетки, которых значительно меньше, чем теп-
лочувствительных. Термочувствительные структуры в заднем гипоталамусе
не встречаются.
Между зрительными буграми находится третий желудочек мозга, покры-
тый сосудистой пластинкой. Передняя стенка её образована концевой пла-
стинкой, а задняя — задней комиссурой. В состав межуточного мозга входит
Глава 23. Высшие уровни управления
325
area postrema через которую в мозг моноамины и ксенобиотики проникают
лучше по сравнению с другими участками ствола мозга.
Удаление переднего мозга у собак и кошек сохраняет регуляцию мышеч-
ного тонуса при движениях головы, в то время как у человека, обезьяны и
мини-свиней утрачивается рефлекс выпрямления.
Средний мозг содержит четверохолмие, черное вещество, проводящие
пути от коры полушарий большого мозга к продолговатому мозгу, мосту и
спинному мозгу, ножки, красное ядро, скопление нервных клеток, от кото-
рых отходят III,IV и отчасти волокна V пары черепно-мозговых нервов, а
также проводящие пути из спинного мозга в четверохолмие и таламус.
При локальном нагревании среднего и продолговатого мозга можно вы-
зывать терморегуляторные реакции, но их термочувствительность ниже, чем
в преоптической области и переднем гипоталамусе.
Четверохолмие у человека и животных состоит из зрительных и каудаль-
ных слуховых холмов. Зрительные холмы обычно большой величины, чем
слуховые, но у хищников, в сравнении с грызунами, слуховые холмы разви-
ты лучше и по размерам превышают ростральные.
Клетки четверохолмия принимают участие в координации слуховых (пе-
редние четверохолмия) и зрительных (задние четверохолмия) рефлексов.
Искусственное раздражение в области четверохолмия у животных вызывает
движение глазных яблок и шевеление ушами.
В средний мозг входят ножки мозжечка. Через средний мозг проходит
сильвиев водопровод, соединяющий третий и четвертый мозговые желудоч-
ки. Нейрональные констелляции вокруг сильвиева водопровода отвечают за
очень многие реакции организма, в том числе поведенческие (табл. 58). Они
участвуют в регуляции эндорфиновой системы и центрального управления им-
мунитетом.
Существует некая джентльменская договоренность ученых, согласно ко-
торой полосатое тело, черную субстанцию, красное ядро, люисово тело и
мозжечок включают в экстрапирамидную систему, которая является комп-
лексом двигательных подкорковых образований среднего и промежуточного
мозга и, через нее кора связана с двигательными эффекторными нейронами.
Церебротомия на уровне межуточного и среднего мозга не вызывает ги-
пертонии мышц-разгибателей и животное сохраняет способность регулиро-
вать тонус. При изменении положения головы эти животные способны воз-
вращать ее в исходное состояние. Нервные клетки, красное ядро регулируют
тонус скелетной мускулатуры и обеспечивают через моноаминэргические
системы [63, 277, 538, 678] выполнение координированных движений и со-
хранение позы и равновесия (см. табл. 59).
Мозжечок разделяется на три доли. Передняя доля состоит из язычка,
центральной доли (которая у высших животных имеет крылья, но у собак
они отсутствуют), передней части, которая у мини-свиней слабо выражена,
326 Биомоделирование в системах управления
четырехугольных долек. Средняя доля состоит из ската, бугра, задней части
четырехугольной дольки, петлевидной (полулунной) дольки и парамедиан-
ной дольки или миндалины. Задняя доля состоит из пирамиды, язычка, узел-
ка, клочка и парафлокуса.
Таблица 59
Генетически обусловленные и экспериментальные биомодели для изучения
антипаркинсонического действия веществ
Исследуемое действие
вещества
Модель оценки антипаркин-
сонических эффектов по
методике каталепсии,
вызванной галоперидолом
- 1 мг/кг, внутрибрюшинно
Модель каталепсии,
вызванной трифтазином -
2 мг/кг, внутрибрюшинно
Модель экстрапирамидных
нарушений, вызванных
резерпином - 2,5 мг/кг,
внутрибрюшинно
Модель ПС, вызванного
системным введением
нейротоксина МФТП:
30 мг/кг
40 мг/кг
30 мг/кг
30 мг/кг
Модель ПС, вызванного
6-гидроксидофамином -
100-250 мкгб-ГОДА в
10-25 мкл растворителя,
в боковые желудочки
Модель оценки потенциро-
вания эффектов апомор-
фина - 0,3-1 мг/кг, под-
кожно
Модель оценки потенциро-
вания эффектов амфета-
мина:
2,5-5 мг/кг, подкожно
2,5-10 мг/кг, подкожно
Модель тремора, вызван-
ного оксотреморином -
р-р 1 мкл оксотреморина
в 20 мл дистиллированной
воды, 0,2 мл на 100 г мас-
сы тела, внутрибрюшинно
Модель тремора, вызван-
ного ареколином - 25 мг/кг
подкожно
Лабораторное
животное
Крысы
Крысы
Мыши
Мыши
Крысы
Крысы
Крысы
Крысы
Крысы
Крысы
Крысы
Мыши
Крысы
Мыши
Линия или порода
животного
Аутбредные
Белые
аутбредные
Белые
аутбредные
Линии
С57В 1/6
Белые
аутбредные
Белые
аутбредные
Белые
аутбредные
Аутбредные
Белые
аутбредные
Белые
аутбредные
Аутбредные
Аутбредные
Аутбредные
Аутбредные
Пол
животного
в
с?
9с?
9с?
9с?
9с?
9(5
9с?
9с?
9с?
°с?
§с?
9с?
9с5
Вес
животного
180-220 г
300-350 г
40-50 г
30-40 г
200-250 г
200-250 г
200-250 г
300-350 г
300-350 г
300-350 г
300-350 г
40-50 г
300-350 г
40-50 г
Возраст
животного
3-4 мес.
5-12 мес.
3-10 мес.
3-10 мес.
4-5 мес.
7-8 мес
12 мес
и старше
5-12 мес.
5-12 мес.
5-12 мес.
5-12 мес.
3-10 мес.
5-12 мес.
3-10 мес.
Глава 23. Высшие уровни управления
327
Окончание табл. 59
Исследуемое действие
вещества
Модель ПС, вызванного
внугримозговым введением
1 -метил-4-фенилпиридиния
(МФП+) - 10 мкг в 2 мкл
физраствора, в каждое
ядро ЧС
Модель ПС, создаваемого
введением каиновой
кислоты в ХЯ мозга - 5 мкл
(0,1-0,15 мкг), в каждое
ядро
Лабораторное
животное
Крысы
Крысы
Линия или порода
животного
Белые
аутбредные
Аутбредные
Пол
животного
6
9<J
Вес
животного
300-350 г
300-350 г
Возраст
животного
9-10 мес.
5-10 мес.
Кора мозжечка состоит из трех основных слоев: наружного молекуляр-
ного, ганглионарного, образующегося в основном клетками Пуркинье, и
зернистого слоя. Функции мозжечка достаточно изучены и описаны в лю-
бом вузовском учебнике. Но его роль в системной деятельности мозга мало-
понятна до сих пор.
В последние годы интенсивно развивается исследования по выяснению
оксидазотных механизмов функционирования и метаболических процессов
мозжечка. Установлено, что NO-синтаза, выделенная из крысиного мозжеч-
ка, как и эндотелиальный фермент, нуждается в кальмодулине. Она стехио-
метрически фосфорилируется цАМФ-зависимой протеинкиназой и Са-каль-
модулинзависимой протеинкиназой. Очищенный энзим мигрирует в элект-
рофорезе как одиночная субъединица массой 150 кД и, скорее всего, явля-
ется мономером. Используя антитела, созданные к NO-синтазе, выделенной
из мозжечка, локализация этого энзима в мозге была картирована иммуно-
гистохимически. Кросс-реактивность антител с эндотелиальной NO-синта-
зой сделала возможной визуализацию фермента в сосудистом эндотелии и
периферических нейронах [579]. Клонирование комплементарного мозго-
вой NO-синтазе ДНК показало, что в данном энзиме есть сайты для узнава-
ния NADPH, FAD, FMN и кальмодулина, которые являются сайтами фос-
форилирования. Таким образом, NO-синтаза потенциально может регули-
роваться всеми названными факторами. Эти данные дают новые представле-
ния об интегративных процессах в мозге и роли в них мозжечка.
Под корой мозжечка находится белое вещество в виде сложного рисунка.
В белом веществе находятся скопления серого вещества в виде трех ядер.
При удалении мозжечка нарушается мышечный тонус (атония); понижается
обмен веществ, быстро развивается утомление (астения); возникают дрожа-
тельные движения головы и конечностей (астазия). Раздражение коры моз-
жечка может вызвать движение конечностей, как и при раздражении коры
больших полушарий. Мозжечок регулирует обмен веществ, соматическую и
вегетативную систему у животных и человека. Показано наличие серотони-
328
Биомоделирование в системах управления
нэргических связей с ядрами шва и норадренэргических - с голубым пят-
ном. В коре мозжечка действуют ГАМК-эргические тормозные и аспартат-
и глютамат- возбуждающие клетки. У разных видов животных, в особенно-
сти у инбредных мышей и крыс обнаружены большие различия в содержании
серотонина и норадреналина в коре мозжечка.
Воролиев мост животных в сравнении с человеком развит слабо. Со сто-
роны продолговатого мозга в варолиев мост проникают участки серого ве-
щества, которые являются частью ядер тройничного и вестибулярного не-
рвов.
Продолговатый мозг состоит из ядер черепно-мозговых нервов (V-XII) и
промежуточных ядер, расположенных по ходу проводящих путей. У хищни-
ков, в сравнении с грызунами, хорошо выражено наружное тело продолгова-
того мозга, осуществляющее связи со стволом, мозжечком и спинным моз-
гом. По-видимому, следует отказаться от взглядов на продолговатый мозг,
как некий триггер-проводник. Это — крупный нейровегетативный центр.
Так, обнаружена четкая корреляция между активным поведением мышей
В6 и уровнем норэпинефрина в продолговатом мозге [451, 452]. У этих живот-
ных наблюдается сильно выраженная склонность к дракам и потреблению
алкоголя в высоких концентрациях (12,5%). Содержание алкогольдегидроге-
назы в печени значительно увеличивается после принятия 10%-ного этанола,
тогда как количество других печеночных ферментов остается без изменения.
Между мозжечком и продолговатым мозгом находится четвертый желу-
дочек, с помощью которого центральный канал спинного мозга соединяется
с подпаутиновым пространством и полостью третьего мозгового желудочка.
Желудочки мозга это не только емкость для ликвора. Как и в венозном си-
нусе, в их стенках обнаружены миофибриллы, чувствительные к кофеину и
другим психостимулирующим средствам. Не потому ли у женщин, страдав-
ших гипотонией в молодости, гипертонические кризы прекрасного возраста
купируются именно кофеином, стимулирующим ставшие слабенькими ми-
офибриллы, уменьшая переполнение мозговых емкостей и давление на со-
судорегулирующие центры. Но это уже специальная биологическая модель,
ждущая своего исследования.
Ретикулярная формация — это группа диффузно рассеянных ганглиоз-
ных клеток, которые берут начало в спинном мозгу и входят в состав ство-
ловой части мозга животных и человека, клетки, которой контактируют ден-
дритами и аксонами с нейронами продолговатого, среднего и межуточного
мозга. Ретикулярная формация у низших позвоночных животных выполняет
роль главного центра координации разнообразных реакций и поведения, но
в процессе филогенеза эти функции берут на себя новые отделы мозга и его
коры. По современным данным её роль заключается в следующем:
/ активация коры, обеспечение уровня осознания стимулов и реакций,
ритма сон/бодрствование;
Глава 23. Высшие уровни управления
329
/ передача аффективно-эмоциональной афферентной информации в
лимбическую систему;
/ регуляция жизненно важных рефлексов (кровообращения, дыхания,
глотания, кашля и чихания), требующих интеграции нескольких аф-
ферентных и эфферентных потоков импульсов;
/ регуляция позы и целенаправленных движений.
Барьерная система мозга обеспечивается огромным и до конца не рас-
шифрованным комплексом, включающим гемато-энцефалические, астроци-
тарные и мембранные механизмы защиты [8, 35, 312, 461, 505, 515, 688, 690].
В целом эта система защиты обозначается как гемато-энцефалический барь-
ер. Она обеспечивает высший уровень защиты мозга от целого ряда эндоген-
ных и экзогенных веществ и токсикантов [436, 462 ,637] и является излюб-
ленным объектом для моделирования.
Мозг человека и животных в сравнении
Головной мозг человека в среднем весит 1350—1450 г, однако вариации объе-
ма мозга человека достаточно велики. Так, в литературе приводятся данные
сравнения массы мозга у двух великих писателей Анатоля Франса (1050 г) и
И.С.Тургенева (2012 г). Хотя массы их мозга разнятся почти в два раза, ин-
теллектуальные способности вряд ли могут иметь подобные различия. По-
этому прямые экстраполяции по массе мозга между людьми, а тем более
между человеком и животными, вряд ли могут быть признаны корректными.
Тем не менее абсолютные значения массы мозга животных некоторые авто-
ры рассматривают в качестве критериальных значений в биомоделировании.
Относительные показатели массы мозга к массе тела человека и животных в
процентном отношении приведены в табл. 1 и 3 (гл. 1).
У мини-свиней размеры головного мозга не превышают 75—90 г. Некото-
рые особенности высшей нервной деятельности у мини-свиней также необ-
ходимо знать при работе со свиньями. Результаты изучения особенностей
высшей нервной деятельности у мини-свиней показывают, что уже у ново-
рожденных поросят легко вырабатываются рефлексы на определенный со-
сок матери. У них вырабатываются рефлексы, связанные с кормлением. В
этот период хорошо выражены рефлексы подражания и стадности. Напри-
мер, рефлекс кормления от одной свиноматки распространяется в течение
нескольких минут на всех свиноматок и поросят в секции. Рефлекс стадно-
сти проявляется в том, что поросята на прогулке всегда держатся группой, и
при испуге убегают всегда группой в одном направлении. Условно-рефлек-
торная деятельность у молодняка мини-свиней нормализуется после начала
полового созревания. Мини-свиньи отличаются широким индивидуальным
разнообразием в проявлении условных рефлексов. У мини-свиней, по
А.В.Квасницкому, наблюдается 4 типа высшей нервной деятельности (ВНД):
330
Биомоделирование в системах управления
/ сильный неуравновешенный тип, характеризующийся хорошей активно-
стью нервной деятельности, но из-за преобладания возбудительного процес-
са неуступчивы и драчливы;
/ слабый тип, с плохой тормозной реакцией, пугливостью;
/ сильный уравновешенный тип, со сдержанностью как возбудительного,
так и тормозного процессов;
/ сильный уравновешенный подвижный тип, характеризуется примерно оди-
наковой силой раздражительных и тормозных процессов и хорошей под-
вижностью.
Мини-свиньи с уравновешенным типом нервной системы для экспери-
ментальных целей, безусловно, предпочтительнее, так как работать с этими
животными гораздо удобнее, легче строить экстраполяционные переносы на
человека. Но у людей те же типы ВНД. Возможно, мой пример покажется
неуместным, но он реален и заключается в том, что поведение толпы при
катастрофах, террористических актах и в экстремальных ситуациях весьма
напоминает реакции испуганного стада животных. Моделирование подоб-
ных ситуаций на мини-свиньях, на наш взгляд, не только оправдано, но и
необходимо.
У собак различных пород абсолютный вес мозга не одинаков и колеблет-
ся от 54 до 150 г, что составляет у мелких собак 1:37, а у крупных 1:100 часть
веса тела. Отношение веса спинного мозга к весу головного у разных живот-
ных составляет 1:4,5 и даже 1:9. На серое вещество мозга у собак приходится
61,1% мозговой ткани. Подробнее анатомо-физиологические особенности
мозга собак будут приведены также в гл. 24.
Вес головного мозга кошки составляет 21—34 г (в среднем 30 г), или 0,7—
1,1% веса тела. У кошек головной мозг тяжелее спинного в три-четыре раза,
а вес спинного мозга у животных средней величины — 7,5 г. Полушария
головного мозга несколько округленной формы. Обонятельные луковицы
развиты в меньшей степени, чем у собак. Борозд и извилин мало, идут они
преимущественно продольно. Лобная доля занимает 6,9% всей поверхности
головного мозга.
Центральная нервная система кролика характеризуется примитивностью
строения, так как слабо развита кора полушарий головного мозга. Полушария
небольших размеров, сужены кпереди, не имеют борозд и извилин. Вес цен-
тральной нервной системы по отношению к весу тела составляет 0,6-1%, т. е.
около 15—17 г. На долю спинного мозга приходится 1/3 веса всей центральной
нервной системы. Спереди большого мозга выделяются значительные по объе-
му обонятельные луковицы. Нечетко выражен варолиев мост. Мозжечок не
имеет компактной формы, уплощен спереди назад, имеет небольшие боковые
полушария (клочки). Морфологическое созревание коры у кролика происхо-
дит к 10—15-му дню со дня рождения (цитоархитектоника коры к этому вре-
мени приобретает вид, свойственный взрослому животному). Новорожден-
Глава 23. Высшие уровни управления
331
ный кролик не приспособлен к самостоятельной жизни. Из черепномозговых
нервов глазодвигательные, языко-глоточные и блуждающие включают в себя
парасимпатические волокна.
Вес полушарий головного мозга морских свинок 2,5—6,5 г, а размеры сле-
дующие: длина 20-25, ширина 21-13, высота 13-16 мм. Вес головного мозга
у морской свинки весом 200 г составляет 2,8 г, а у свинки весом 500 г — 3,2 г.
Вес спинного мозга соответственно 1—1,3 г. Наружная поверхность мозговых
полушарий разделяется на четыре доли: лобную, теменную, височную и заты-
лочную. Кора больших полушарий головного мозга морских свинок не имеет
выраженных извилин, а лишь неровности, поэтому эти животные относятся
к группе гладкомозговых. Имеются четыре продольные борозды примитив-
ного типа: ринальные передняя и задняя, гипокампова и борозда мозолисто-
го тела. Кроме них, имеются еще сильвиева, носовая и каудальная борозды.
Поверхность большого мозга разделена сильвиевой бороздой на переднюю
(лобно-теменную) и заднюю (височно-затылочную) области. Морская свин-
ка, по сравнению с другими животными, рождается с наиболее развитым
головным мозгом. К моменту рождения у нее заканчивается морфологичес-
кое и биохимическое развитие коры головного мозга. Так, на 42—45-й день
эмбриональной жизни в коре больших полушарий головного мозга выраже-
ны микроскопические пять слоев, характерные для взрослых животных.
Головной мозг взрослой крысы весит в среднем 2,4—2,8 г, что составляет
0,9—1% веса тела. Варолиев мост развит очень слабо. Полушария переднего
мозга гладкие. Обонятельные доли большие. Строение головного и спинно-
го мозга, отхождение черепно-мозговых и спинных нервов принципиально
такие же, как и у других млекопитающих. Сравнительно близки функции и
адаптационные механизмы.
Так, в стоке крыс КМ первоначально 98—99% животных проявляли рез-
кое возбуждение с судорожными припадками в ответ на резкий звук, впос-
ледствии частота снизилась до 80—90% [204]. Для сравнения: вес щитовид-
ной железы 622 ± 30 мг (выше, чем у крыс Wistar). Абсолютный вес надпо-
чечников 23,4 ± 1,7 мг. Активность как щелочной, так и кислой РНКазы в
коре, подкорке и стволовой части мозга выше, чем у Wistar. Уровень актив-
ности (фермента) АХЭ в продолговатом мозге 713 ± 22, а в подкорковых
образованиях 841 ± 19 (выше, чем у Wistar). Коэффициенты долгосрочной
памяти 13,4 ± 2,2 и 1,14 ± 0,02; краткосрочной памяти 0,044 ± 0,002. Крысы
высоковозбудимы, со слабым тормозным процессом [204] — удобные био-
модели в нейрофизиологии, фармакологии, гигиене.
У линий CC57BR-H рецессивный ген, жизнеспособность гомозигот по-
нижена, аномалии поведения, обусловленные поражением лабиринта и
центральной нервной системы, микрофтальмии.
У крыс WAG/GSto поведение эмоциональное, высокий уровень амбу-
ляции в открытом поле и активности в колесе. Хорошо ориентируются
332
Биомоделирование в системах управления
в лабиринте. Быстро адаптируются к изменениям окружающей среды [206,
207, 208].
Строение и функции головного и спинного мозга мыши принципиально
такие же, как и у других млекопитающих. Головной мозг взрослой мыши
весит 0,27 г. Животные линии BALB/cJLac не имеют мозолистого тела в
мозге (этот признак наследуется полигенно с эпистатическим взаимодей-
ствием генов). Нервный пучок свода часто сталкивается с передней комис-
сурой, которая у некоторых мышей BALB/c перемещена и проходит позади
или через нервный пучок свода, дефект обусловлен полигенным аддитив-
ным наследованием с пороговым проявлением. Самки имеют самый высо-
кий вес мозга как абсолютный, так и относительный к весу тела из всех
испытанных линий; относительный вес мозга довольно высок и у самцов.
У линий DBA/1J Lac защитные дозы некоторых противосудорожных
лекарств как и для мышей DBA/1 в целом следующие: хлорпромазин PD50 =
= 4,8 мг/кг; мепробамат PD50 = 4,5 мг/кг; резерпин PD50 = 5,0 мг/кг. Ули-
ний DBA/2J аудиогенные припадки можно вызвать у 100% мышей в возра-
сте 35 дней и у 5% — старше 55 дней [604]. Ранее предполагалось, что аудио-
генный шок связан с метаболическим дефектом, который детерминируется
генетическим локусом, расположенным вблизи локуса dilution на 9-й хромо-
соме. Теперь доказано, что эти локусы не сцеплены, так как рекомбинация
между ними происходит с частотой 0,51 ± 0,013 [138]. Относительно резис-
тентны к летальному действию озона [235].
Линии DBA/2J обладают хорошей способностью к «запоминанию» экс-
периментальных задач с избеганием электрического тока в сравнении с В6,
слабой склонностью к потреблению алкоголя [604]. Симптомы хронической
недостаточности магния в рационе более выражены, чем у В6 [58].
В заключение этой части стоит напомнить, что у всех млекопитающих
головной и спинной мозг покрыт тремя оболочками. Твердая мозговая обо-
лочка состоит из листков, выстилающих полости черепа. Между листками
расположены пространства, называемые венозными пазухами, или веноз-
ными синусами (sinus venosi), в них собирается кровь из вен головного моз-
га, области глазницы и черепных костей. Из синусов кровь поступает во
внутреннюю яремную вену.
В табл. 60 представлены биомодели для исследования и оценки мозгово-
го кровообращения и средств его коррекции.
Между твердой мозговой оболочкой спинного мозга и надкостницей
позвоночного канала имеется эпидуральное пространство, которое предох-
раняет спинной мозг от повреждений и способствует его смещению во вре-
мя изменений положения позвоночника.
Кроме твердой, имеются еще паутинная и мягкая мозговые оболочки
головного и спинного мозга.
Глава 23. Высшие уровни управления
333
Таблица 60
Экспериментальные биомодели для изучения веществ, применяемых при лечении
нарушений мозгового кровообращения и мигрени
Исследуемое действие
вещества
Модель локальной ишемии
мозга, вызванной перевяз-
кой средней мозговой ар-
терии
Модель глобальной ишемии
мозга при билатеральной
окклюзии общих сонных ар-
терий в сочетании с контро-
лируемой гипотензией
Моделирование субарахно-
идального кровоизлияния
путём введения крови под
оболочки мозга
Модель воспроизведения
интрацеребральной гемор-
рагии
Модель спазмов мозговых
сосудов, вызванных серо-
тонином — 20 мкг/кг
внутривенно
Лабораторное
животное
Крысы
Крысы
Кошки
Крысы
Кошки
Линия или порода
животного
Аутбредные
Аутбредные
Аутбредные
Белые
аутбредные
Аутбредные
Пол
животного
9с?
9с?
93
в
9с?
Вес
животного
300-350 г
300-350 г
3-5 кг
300-350 г
3-5 кг
Возраст
животного
5-12 мес.
5-12 мес.
1 -7 лет
9-10 мес.
1 -7 лет
ЭЭГ и МЭГ в биомоделировании
Электроэнцефалограмма (ЭЭГ) и магнитоэнцефалограмма (МЭГ) являются
единственным объективным показателем работы головного мозга человека и
животных в динамике. Они отражают, главным образом, постсинаптическую
активность нейронов коры, но не их потенциалы действия и не активность
корковых глиальных клеток.
Положительное отклонение потенциала ЭЭГ и МЭГ вызывается либо воз-
буждающими постсинаптическими потенциалами в глубинных слоях коры,
либо тормозными постсинаптическими потенциалами в поверхностных её
слоях, а отрицательное отклонение — противоположными факторами.
Биофизические свойства нейронов человека сходны с активностью нерв-
ных клеток животных. Потенциал покоя нейронов составляет от —50 до
-80 мВ, а их потенциал действия от -60 до —100 мВ при длительности 0,5-
2 мс. Потенциалы действия в нейронах возникают в области аксонного хол-
мика и распространяются по аксону, по телу и дендритам клетки, с частотой
импульсации до 100 Гц.
У человека и животных между парой электродов или между одним из
таких электродов и индифферентным электродом, например, на мочке уха,
у человека и в кости носовых пазух животных наблюдаются непрерывные
колебания потенциала. Они называются электроэнцефалограммой (ЭЭГ) или
334
Биомоделирование в системах управления
электрограммой (ЭГ). Частота регистрируемых колебаний составляет от 1 до
50 Гц, а их амплитуда до 90—120 мкВ.
Частота и амплитуда волн ЭГ зависит от вида животного, расположения
электродов и степени бодрствования. Впервые электрические изменения в
мозгу в ответ на стимуляцию сенсорных органов зарегистрировал в 1875 г.
Ричард Катон. В 1929 г. Г.Бергер подтвердил данные Р.Катона уже без стиму-
ляции органов и зафиксировал спонтанные биопотенциалы на коже головы
человека, а в 1935 г. Эдриан Мэтьюз классифицировал эти ритмы.
Ритмы ЭГ индуцируются главным образом активностью подкорковых
структур, особенно таламуса. При декортикации ритмичная активность тала-
муса не изменяется. При изучении глубинных структур зрительных бугров
обнаружено множество таламических пейсмекеров, генерирующих и поддер-
живающих ритмичную активность. Выраженным синхронизирующим и де-
синхронизирующим действием на таламус обладает ретикулярная формация.
У кроликов к 10—15-му дню со дня рождения цитоархитектоника коры
приобретает вид, свойственный взрослому животному. К этому же времени
устанавливается биохимическое и электроэнцефалографическое созревание
коры. Спонтанные электрические колебания коры головного мозга впервые
появляются у крольчонка старше пяти дней. Электрическая активность коры
головного мозга становится сформированной к 10—15-му дню постнаталь-
ной жизни кролика. Новорожденный кролик не приспособлен к самостоя-
тельной жизни.
Спонтанные электрические потенциалы коры головного мозга у морской
свинки регистрируются на 46—49-й день эмбриональной жизни и становятся
еще более выраженными у новорожденных. Морские свинки и куры рожда-
ются с хорошо сформированной (морфологически, биохимически и элект-
роэнцефалографически) центральной нервной системой, выполняющей
многообразные сложные реакции.
У крыс слабо выраженные электрические потенциалы коры головного
мозга регистрируются на пятый день после рождения, и нормальная ЭЭГ
отмечается с 15-го дня, хотя морфологическое развитие коры заканчивается
раньше, к 10-му дню.
Спонтанные электрические потенциалы коры головного мозга у морской
свинки регистрируются на 46—49-й день эмбриональной жизни и становятся
еще более выраженными у новорожденных. Морские свинки и куры рожда-
ются с хорошо сформированной (морфологически, биохимически и элект-
роэнцефалографически) центральной нервной системой, выполняющей
многообразные сложные реакции.
Как и волны ЭЭГ, вызванные потенциалы (ВП) отражают не импульса-
цию, а прежде всего синаптическую активность нервных клеток. Если ре-
гистрировать ВП в разных слоях коры, то форма ВП будет меняться: первона-
чальное положительное отклонение (верхние слои) исчезнет и вместо него
Глава 23. Высшие уровни управления
335
появится первичная отрицательная волна с коротким латентным периодом.
Нижние слои, иными словами, афферентные импульсы, вызывают деполя-
ризацию нейронов IV — V слоев коры.
Биопотенциалы записываются как с поверхности обнаженной коры
(ЭКоГ), так и с кожи головы. В последнем случае получают интегральную
электроэнцефалограмму (ЭЭГ). Записи ЭЭГ в принципе те же, что и в слу-
чае ЭКоГ, но в связи с большим электрическим сопротивлением амплитуда
зубцов и частота волн ЭЭГ меньше.
У животных и человека при закрытых глазах регистрируется основной ос-
ритм (ос-волны с частотой 8—12 Гц, в среднем 10 Гц) или синхронизирован-
ная ЭЭГ При открытых глазах или поступлении сигналов от других органов
чувств сс-волны исчезают (блокада ct-ритма) и сменяются (3-волнами с боль-
шей частотой (14—30 Гц, в среднем 20 Гц) и меньшей амплитудой. Это де-
синхронизированная ЭЭГ. Более медленные и высокоамплитудные колеба-
ния — 0-волны (тета-ритм 4—7 Гц, в среднем 6 Гц) и 5-волны (дельта-ритм
0,5—3,5 Гц, в среднем 3 Гц), но в норме у людей в отличие от животных они
выявляются только во сне. В ЭЭГ детей характерны более медленные и
нерегулярные ритмы даже в бодрствующем состоянии. «Созревание» четко-
го сс-ритма у детей происходит к 5—7 годам, у животных же {мини-свиньи,
собаки, кошки, кролики, крысы и т.д.) низкочастотные ритмы в бодрствую-
щем состоянии присутствуют на протяжении всей жизни. При разложении
ЭЭГ быстрым преобразованием Фурье выявляется, что спектральные мощ-
ности плотности животных значительно обеднены в высокочастотной обла-
сти. Кластерный анализ, проведенный нами, показал, что единой соотноси-
мой частотой у человека и разных животных является частота 9 ± 0,75 Гц,
причем 0- и 8—волны у животных несут гораздо большую информационную
содержательность, чем у человека. У человека же основные информативные
характеристики лежат между верхними значениями 9-ритма и нижними и
средними 6-ритма. Это следует строго учитывать при биомоделировании про-
цессов деятельности головного мозга на основе анализа ЭЭГ и ЭКоГ.
При движении электрических зарядов возникает также магнитное поле.
Мозг генерирует как электрические, так и магнитные волны. Магнитные поля
можно зарегистрировать с помощью высокочувствительных датчиков на
основе сверхпроводящих квантовых интерференционных устройств. Пре-
имущество этой методики перед ЭЭГ заключается в повышенной точности
локализации очага корковой активности, поскольку сигналы от соседних
участков не накладываются друг на друга.
В конце 80-х годов мы провели исследования сравнительной информа-
тивности ЭЭГ и МЭГ на базе Института атомной энергии им. И.В. Курча-
това. Не вдаваясь в детали, следует подчеркнуть, что ЭКГ и МКГ (магнито-
кардиограмма) являются практически близнецами и разнятся лишь в более
мощной амплитуде МКГ. В то же время ЭЭГ и МЭГ абсолютно несопоста-
336
Биомоделирование в системах управления
вимы по своим амплитудно-частотным характеристикам и информативно-
сти. МЭГ имеет несомненные преимущества перед ЭЭГ, поскольку несет
большую, а главное, новую информацию о функционировании мозга, дей-
ствии психотропных средств и т.д. В МЭГ гораздо больше сопоставимых
феноменов между человеком и животным, в частности, в гораздо большем
расширении эквивалентного частотного диапазона. Мы не сомневаемся в
исключительной перспективности МЭГ для биомоделирования. Проблема
лишь в цене вопроса. Методики МЭГ на основе жидкогелиевых сверхпро-
водящих материалов дороги и сложны в эксплуатации ввиду огромных зат-
рат по экранированию от магнитных возмущений Земли и окружающих
территорий.
Нейрофизиологи и нейрофармакологи, использующие методы визуали-
зации для замены экспериментов над животными, как правило, испытывают
трудности с привлечением средств для своей работы. Так, требуется не менее
3—5 лет, чтобы обеспечить финансирование и развертывание мощностей по
МЭГ мозга во вновь создаваемых центрах. МЭГ используется в единичных
лабораториях США и стран Европы. Для сравнения, Япония имеет 20 лабо-
раторий магнитной энцелофалографии (МЭГ).
Применяемые стратегии финансирования исследований имеют тенден-
цию не поощрять замену, отказ от использования животных, а находить аль-
тернативы в виде оптимальных подходов в биомоделировании. Развитие новых
методов по замене вивисекционных экспериментов над животными рассмат-
ривается как пограничная деятельность. Внедрение принципиально новых
технологий определяет прогресс в биологии и медицине. Например, аппарат
отображения при помощи магнитного резонанса (MRI) имеет всё увеличива-
ющееся поле применений, и может в настоящее время использоваться для
изучения количественных изменений гематоэнцефалического барьера паци-
ентов с рассеянным склерозом или с опухолями. Изучение человека при по-
мощи спектроскопии магнитного резонанса может ускорить разработку ле-
карств, сокращая затраты и использование животных.
Методы, подобные магнитному резонансу или магнитной энцефалогра-
фии, не являются альтернативами сами по себе, но они станут таковыми,
если их использование позволит сократить эксперименты на животных, в то
же время приблизив их к человеку.
Многие исследователи хотели бы использовать сложное визуальное обо-
рудование отображения, подобное аппарату отображения при помощи маг-
нитного резонанса, которое позволит им изучать как людей, так и живот-
ных, но они не имеют доступа к этим современным технологиям.
Психические расстройства стали теперь существенной причиной недее-
способности в мире, превращая исследования мозга и поведения в страте-
гически важное направление. Методы нейровизуализации представляют со-
бой ценную замену экспериментов над животными. Прогресс в нейробио-
Глава 23. Высшие уровни управления
337
логии достигнут при помощи интеграции неразрушающих, гуманных ме-
тодов отображения информации с молекулярными и клеточными подхода-
ми. Ревизия методов, применяемых в нейробиологии, показала, что для
комплексных исследований требуются очень большие объёмы финансиро-
вания.
Биомодели для оценки психотропных воздействий
В приводимых ниже таблицах представлены биомодели аутбредных и
инбредных животных, наиболее часто используемых при изучении психо-
генных воздействий и испытаниях средств психотропного действия.
Более подробное описание и правила пользования этими таблицами даны
в табл. 6 (гл. 2) при объяснении биомоделей для оценки острой и хроничес-
кой токсичности.
Таблица 6 1
Экспериментальные и генетически обусловленные биомодели для изучения
антидепрессивных эффектов веществ
Исследуемое действие
вещества
Оценка влияния на услов-
ные рефлексы
Модель оценки по тесту
отчаяния
Моделирование по тесту
агрессивного поведения
Изучение влияния на общее
состояние и простые пове-
денческие реакции
Модель оценки по тесту
"выученной беспомощности"
Моделирование по тесту
реактивного и резерпино-
вого депрессивноподобных
состояний (резерпин
0,1 мг/кг, подкожно)
Моделирование деприва-
ции, «депрессии» и иных
состояний по нейрофарма-
кологическим тестам:
a. влияние на эффекты
- резерпин 2,5 мг/кг мы-
шам и 4 мг/кг крысам,
внутрибрюшинно
- тетрабеназин 40мг/кг,
внутрибрюшинно
b. влияние на эффекты
- нейролептики (трифтазин
или метеразин) 3-6 мг/кг,
внутрибрюшинно
Лабораторное
животное
Собаки
Хомячки
Крысы
Мыши
Крысы
Мыши
Мыши
Крысы
Кошки
Крысы
Мыши
Крысы
Мыши
Линия или порода
животного
Аутбредные
Аутбредные
Аутбредные
Аутбредные
Аутбредные
Аутбредные
Белые
аутбредные
Аутбредные
Аутбредные
Аутбредные
Аутбредные
Аутбредные
Аутбредные
Пол
животного
Ос?
§с5
$с?
96
6
9с?
9с?
§с?
Вес
животного
10-25 кг
120-125 г
300-350 г
40-50 г
300-350 г
40-50 г
40-50 г
300-350 г
3-5 кг
300-350 г
40-50 г
300-350 г
40-50 г
Возраст
животного
1 -7 лет
6-18 мес.
5-12 мес.
3-10 мес.
5-12 мес.
3-10 мес.
3-10 мес.
5-12 мес.
1 -7 лет
5-12 мес.
3-10 мес.
5-12 мес.
3-10 мес.
338
Биомоделирование в системах управления
Окончание табл. 61
Исследуемое действие
вещества
Лабораторное
животное
Линия или порода
животного
Пол
животного
Вес
животного
c. влияние на эффекты
- L-дофа 100-200 мг/кг и
500 мг/кг, внутрибрюшинно
d. влияние на эффекты
- фенилэтиламин 100 мг/кг
и 10-20-40 мг/кг, подкожно
или внутрибрюшинно
e. влияние на эффекты
- апоморфин 10-25 мг/кг,
подкожно
f. влияние на эффекты
- триптамин 5 мг/кг кры-
сам и 25 мг/кг мышам, в/в
д. влияние на эффекты
- клофелин 0,1-0,5 мг/кг,
в/в или внутрибрюшинно
Крысы
Мыши
Крысы
Мыши
Крысы
Мыши
Крысы
Мыши
Мыши
Крысы
Кошки
Кролики
Аутбредные
Аутбредные
Аутбредные
Аутбредные
Аутбредные
Аутбредные
Аутбредные
Аутбредные
Аутбредные
Аутбредные
Аутбредные
Аутбредные
300-350 г
40-50 г
300-350 г
40-50 г
300-350 г
40-50 г
300-350 г
40-50 г
40-50 г
300-350 г
3-5 кг
2,5-4 кг
Моделирование деприва-
ции, «депрессии» и иных
состояний по нейро-
фармакологическим
тестам:
h. влияние на эффекты
- фенамин 2,5-5-7,5-
10 мг/кг мышам, подкожно,
0,5-1 мг/кг, в/в-кошкам
и кроликам
i. влияние на эффекты
- 5-окситриптофан
50 и 300 мг/кг, внутри-
брюшинно
j. влияние на действие
- ареколин 0,25-0,5 мг/кг,
- галантамин 1-2мг/кг,
- физостигмин 0,1 -0,2 мг/кг
внутривенно,
- оксотреморин
0,25-0,5 мг/кг, внутри-
брюшинно
Мыши
Кошки
Кролики
Мыши
Кролики
Кролики
Кошки
Мыши
Аутбредные
Аутбредные
Аутбредные
Аутбредные
Аутбредные
Аутбредные
Аутбредные
Аутбредные
40-50 г
3-5 кг
2,5-4 кг
40-50 г
2,5-4 кг
2,5-4 кг
3-5 кг
40-50 г
Исследование антидепрес-
сантов биохимическими
методами
Крысы
Кошки
Хомячки
Аутбредные
Аутбредные
Аутбредные
300-350 г
3-5 кг
120-125 г
Модель для тестов низкой
активности «в колесе» (5,8)
и слабой активности в «от-
крытом поле» (6,0), связан-
ными с очень высоким
уровнем декарбоксилазы
глутаминовой кислоты
в мозге
Мыши
Линия
СВА/Н-Т6
30-40 г
Глава 23. Высшие уровни управления
339
Таблица 62
Экспериментальные и генетически обусловленные биомодели для изучения
седативных и транквилизирующих эффектов веществ и физических факторов
Исследуемое действие
вещества
Моделирование конфликт-
ной ситуации (вариант
Vogel-a)
Моделирование по методу
четырех пластин
Модель условного закапы-
вания пищи
Модель для изучения пове-
дения в «открытом поле»
мышей с активным и пас-
сивным типами эмоцио-
нально-стрессовых реакций
Исследования по расши-
ренному изучению анксио-
литической активности
Лабораторное
животное
Крысы
Крысы
Мыши
Мыши
Крысы
Мыши
Хомячки
Линия или порода
животного
Аутбредные
Аутбредные
Аутбредные
Линии С57В1/6
и BALB/C
Аутбредные
Аутбредные
Аутбредные
Пол
животного
96
96
96
96
§6
96
Вес
животного
300-350 г
300-350 г
40-50 г
30-40 г
300-350 г
40-50 г
120-125 г
Возраст
животного
5-12 мес.
5-12 мес.
3-10 мес.
3-10 мес.
5-12 мес.
3-10 мес.
6-18 мес.
Таблица 63
Экспериментальные и генетически обусловленные биомодели для изучения
нейролептических эффектов веществ и физических факторов
Исследуемое действие
вещества
Поведенческие методы:
влияние на модель для
оценки ориентировочно-
двигательной реакции и
локомоторной активности
Модель для оценки влияния
нейролептиков на условные
рефлексы
Модель для изучения
каталептогенных свойств
веществ
Модель для изучения
искусственной гипотермии
по влиянию на ректальную
температуру
Модель оценки влияния на
угнетение "стартл-реакции"
и ее препульсовое ингиби-
рование
Модель для изучения
влияния на спровоцирован-
ную агрессивность
Лабораторное
животное
Мыши
Крысы
Мыши
Крысы
Мыши
Крысы
Мыши
Крысы
Крысы
Мыши
Крысы
Линия или порода
животного
Аутбредные
Аутбредные
Аутбредные
Аутбредные
Аутбредные
Аутбредные
Аутбредные
Аутбредные
Аутбредные
Аутбредные
Аутбредные
Пол
животного
$6
96
9^
96
$6
96
$6
96
6
6
96
Вес
животного
40-50 г
300-400 г
40-50 г
300-400 г
40-50 г
300-400 г
40-50 г
300-400 г
250-300 г
40-50 г
300-350 г
Возраст
животного
3-10 мес.
5-12 мес.
3-10 мес.
5-12 мес.
3-10 мес.
5-12 мес.
3-10 мес.
5-12 мес.
4-5 мес.
3-10 мес.
5-12 мес.
340 Биомоделирование в системах управления
Продолжение табл. 63
Исследуемое действие
вещества
Моделирование стерео-
типного поведения,
вызванного:
a. введением апоморфина
0,3-1 мг/кг, подкожно
мышам и 4 мг/кг крысам,
b. введением малых (пре-
синаптических) доз апо-
морфина (0,01-0,15 мг/кг,
подкожно)
c. формированием амфета-
миновой стереотипии
(2,5 мг/кг, подкожно)
Моделирование феномена
"вертикализации", вызван-
ного введением апоморфи-
на (2-5 мг/кг, подкожно)
Моделирование амфетами-
новой гиперактивности
(2,5-10 мг/кг, подкожно)
Модель 5-окситриптофано-
вого гиперкинеза (300 мг/кг,
внутрибрюшинно)
Моделирование ареколино-
вого тремора (25 мг/кг,
подкожно)
Моделирование эметичес-
ких проявлений, вызванных
апоморфином
(0,1 мг/кг, в/в)
Модели для определения
нейрорецепторного про-
филя нового соединения
Животные для биохимичес-
ких и других дополнитель-
ных методов, используе-
мых для определения
дискинетического потен-
циала нейролептиков
и для более углублённого
анализа их действия на
моноаминергические
системы
Биомодель агрессивного
(драчливого) поведения
для оценки нейролептиков
и кальмативов
Биомодель для оценки и
дифференцирования психо-
тропных веществ и инкапа-
ситантов по уровню
эмоциональности 52-58%
Лабораторное
животное
Крысы
Мыши
Крысы
Крысы
Мыши
Мыши
Мыши
Мыши
Собаки
Крысы
Крысы
Мыши
Мыши
Линия или порода
животного
Аутбредные
Аутбредные
Аутбредные
Аутбредные
Аутбредные
Аутбредные
Аутбредные
Аутбредные
Аутбредные
Аутбредные
Аутбредные
Линия C57BL/BJ
Линия СЗН/HeLac
Пол
животного
°<5
§<5
9(5
9(5
9с?
9(5
9с5
96
в
9(5
9с5
66
в
Вес
животного
300-350 г
40-50 г
300-350 г
300-350 г
18-25 г
30-40 г
30-40 г
30-40 г
10-25 кг
300-350 г
300-350 г
30-40 г
30-40 г
Возраст
животного
5-12 мес.
3-10 мес.
5-12 мес.
5-12 мес.
2-3 мес.
3-10 мес.
3-10 мес.
3-10 мес.
1 -7 лет
5-12 мес.
5-12 мес.
3-10 мес.
3-10 мес.
Глава 23. Высшие уровни управления
341
Окончание табл. 63
Исследуемое действие
вещества
Биомодель 70%-ного
блокирования активности
аминазином (4 мг/кг)
в тесте «открытого поля»
Высокочувствительная био-
модель УРАИ (условная
реакция активного избега-
ния) для фармакологии,
токсикологии, физиологии
Биомодель чувствитель-
ности к аминазину (62%
животных неактивны в тесте
«открытого поля» после
введения 4мг/кг)
Лабораторное
животное
Мыши
Мыши
Мыши
Линия или порода
животного
Линия DBA/2J
Линия DBA/2J
Линия AHeJ
Пол
животного
9cJ
9с?
96
Вес
животного
30-40 г
30-40 г
30-40 г
Возраст
животного
3-10 мес.
3-10 мес.
3-10 мес.
Каждый исследователь, занимающийся исследованием эффектов и по-
иском психотропных средств, понимает, что не существует раз и навсегда
признанных и надежных методик. Несмотря на то, что нами была проведена
большая работа по систематизации методик для оценки психотропных
средств, они весьма условны для оценки и экспертизы действия психоген-
ных физических факторов, например, инфразвука или иных упругих акусти-
ческих волн. Но многочисленность и относительная ограниченность этих
методов связана со сложностью моделируемой системы — психики. При ин-
терпретации результатов в этом случае мы уходим из предметной области
нейрофизиологии и психофармакологии в философские сферы чувственно-
сти общественного человека. А в этой сфере проблемы биомоделирования
становятся до непреодолимости сложными, что в гл. 40 и будет проиллюс-
трировано.
342
Глава 24 ЦЕНТРАЛЬНЫЕ УРОВНИ
УПРАВЛЕНИЯ
Слово nervus древние римляне использовали для
обозначения любого волокна в теле человека или
животного — связки, сухожилия, а также нервов в на-
шем сегодняшнем понимании. Со временем это
слово стало использоваться только в последнем
смысле.
В 1891 г. немецкий анатом X. Вальдейер счел
нужным заменить слово «нерв», потому что оно
означало только длинные отростки нервных клеток,
видимые под микроскопом. В своих поисках назва-
ния всей нервной клетки в целом он обратился к
греческому эквиваленту этого латинского слова —
neuron и назвал её нейроном.
Небольшие по размеру сильно ветвящиеся
отростки нервных клеток называются дендритами
(от греч. dendron) — дерево, которое эти отростки
напоминают по форме. Длинный неветвящийся от-
росток получил название аксон (от греч. axon — ось).
Аксон одного нейрона обычно разветвляется на
конце, соединяясь с дендритами другой клетки или
ее телом. Но истинного соединения и слияния не
происходит — между мембраной аксона и мембра-
ной другой клетки остается промежуток около 200
ангстрем. Такое соединение называется синапсом.
По-гречески synapsis — соединение, связь; это в дей-
ствительности область соприкосновения нервных
клеток друг с другом.
Глава 24. Центральные уровни управления
343
В этой главе мы рассмотрим биомодели на спинальном уровне ЦНС,
периферической нервной системы и иные сравнительные аспекты функци-
онирования нервной системы.
Цереброспинальный уровень
В биомоделировании на уровне центральной и периферической нервной
системы часто прибегают к основам теории автоматического регулирования
(см. гл. 1), используя показатель уровня демпфирования системы, который
обеспечивает стабильность регуляции. Рассмотрим пример терморегуляции,
в котором участвуют периферические нервы, рецепторы и высшие уровни
ЦНС. Естественно, что в нервном волокне должен быть оптимальный сиг-
нал, он не должен затухать, но и не должен нейрохимически «взорвать»
волокно по типу флаттера. Кожные рецепторы — это наружные датчики.
Тонус мышц, их дрожание, изменение кровотока, закрытие пор для тепло-
изоляции и многие другие проявления — это исполнительный уровень, а ЦНС
— это система регуляции. Тогда: х* = (ос — $у)х описывает скорость реагиро-
вания системы в регуляции теплопродукции, а у* = (ух — &) у отражает ре-
акцию системы по теплоотдаче.
При этом: х, у — системы соответственно теплопродукции и теплоотда-
чи. Скорость х* пропорциональна х, но коэффициент пропорциональности
а — $у тем меньше, чем больше теплопродукция. И наоборот, коэффициент
ух — 5 тем больше, чем больше теплоотдача.
Решения уравнения показывают, что система имеет колебательный ха-
рактер, но постоянно стремится к срединной точке равновесия, когда х ~ у,
х*~ у* (рис. 5).
На рис. 5 изображена диаграмма авторегулирования процесса, когда по-
вышение температуры среды (ab) включает механизмы теплопередачи (be),
падение температуры (cd) усиливает теплопродукцию (da). Этот процесс плав-
ного регулирования терморегуляции происходит до тех пор, пока система
придет к некоему уровню равновесия (ё).
В реальности этого не может быть у теп-
локровных, у которых система регули-
рования стремится к (е), но никогда ее
не достигает, оставаясь близкой к (е) (но
не равной).
На рис. 6 представлены другие состо-
яния уровня терморегуляции для экст-
ремальной ситуации, например при бру-
тальной инфекционной гипертермии. В
этих случаях система перепрыгивает из _ „
Рис. 5. Динамика терморегуляции при
ТОЧКИ (а) В ТОЧКУ (Ь) И Дальнейшее ДВИ- градуальном изменении температуры
У *
344
Биомоделирование в системах управления
У ♦
жение может переходить по эллипсу (bf). Если на уровне (bf) система будет
достаточно оптимизирована, она перейдет в режим градуального регулирова-
ния (рис. 5), если нет — то произойдет скачкообразное перемещение на уро-
вень (cd). В ином случае, например при падении человека в ледяную воду, мы
можем столкнуться со скачкообразным смещением с уровня (bf) на уровень
(ag), но следующие действия будут аналогичны вышеописанным.
Для поддержания уровня равновесия
должны включаться механизмы опере-
жающего управления, когда сигналы идут
не через десятки релейных станций, в
которых оцениваются, интегрируются,
анализируются многочисленные сигна-
лы, а непосредственно к периферичес-
ким волокнам с командами для испол-
нительных органов. Поэтому мы соглас-
ны с теми авторами, которые считают
периферические нервы намного более
сложными объектами, нежели простое
проводящее волокно. В экстренных си-
туациях ожидания команд «сверху» чревато негативными последствиями. Это
потом все будет оценено и сбалансировано, но в экстренных ситуациях не-
обходимо мгновенное, автономное реагирование и внутреннее биомоделиро-
вание ситуации. Мы еще приведем сравнительные данные терморегуляции у
различных животных, а теперь рассмотрим некоторые другие аспекты био-
моделирования.
Ниже представлены биомодели (табл. 64) для изучения формирования и
развития судорожных реакций и исследования средств антиконвульсивного
действия на уровне ЦНС и периферической нервной системы.
Таблица 64
Генетически обусловленные и экспериментальные биомодели для изучения
противосудорожного действия веществ
Рис. 6. Динамика терморегуляции при
скачкообразном изменении температуры
Исследуемое действие
вещества
Модели для изучения
противосудорожного дейст-
вия веществ:
а. проба с коразолом
50-120 мг/кг, подкожно
Ь. проба с пиротоксином
2,5 мг/кг, подкожно
с. проба с бикукуллином
2,7 мг/кг, подкожно
d. проба со стрихнином
1,2 мг/кг, подкожно
Лабораторное
животное
Мыши
Линия или порода
животного
Белые
аутбредные
Пол
животного
в
Вес
животного
19-29 г
Возраст
животного
2-4 мес.
Глава 24. Центральные уровни управления 345
Продолжение табл. 64
Исследуемое действие
вещества
e. проба с NMDA или
квискалатом 0,2 мг/5 мл
для клонических судорог и
3 мг/5 мл для тонических
судорог, в желудочки мозга
f. проба с аппликацией
пенициллина, раствор в
концентрации 20000 МЕ/мл,
на кору головного мозга
Модель для оценки анти-
эпилептического действия
веществ на линиях живот-
ных с генерализованными
эпилептическими
абсансами
Модель аудиогенных
судорог - 110 дБ( 12 кГц)
в течение 60 с
Модель эпилептического
статуса при введении НСТ
в дозе 5,5 ммоль/кг, разве-
денной в 3,5 мл/кг физио-
логического раствора, в/м
Модель для оценки про-
тивосудорожного действия
веществ (проба с камфо-
рой - 1 г/кг, подкожно)
Модель для оценки про-
тивосудорожных эффектов
по методике киндлинга
Моделирование эпилепто-
генного хронического очага
коры мозга, вызванного
аппликацией кобальта у
крыс - на кору левого полу-
шария: 1,5 мм вперёд от
брегмы и 1,5-2 мм лате-
ральнее сагитального шва
Биомодель гиперактивных
животных для изучения ме-
ханизмов предрасположен-
ности к аудиогенным судо-
рогам, с высоким уровнем
серопротеинсвязанного
йода (преимущественно
у самок)
Модель парциальных судо-
рог, вызванных апплика-
цией оксида алюминия -
на дорзальный гиппокамп
или миндалину
Модель инсулинзависимых
судорог
Лабораторное
животное
Мыши
Крысы
Крысы
Мыши
Крысы
Мыши
Крысы
Крысы
Кролики
Кошки
Мыши
Линия или порода
животного
CrhNMRI BR
Белые
аутбредные
или линии
Wistar
Линия
Линии
DBA/2J
Белые
аутбредные
Белые
аутбредные
Белые
аутбредные
или линии
Wistar
Белые
аутбредные
Линия ACEP/J
Аутбредные
A2G/Lac
Пол
животного
в
9с5
9<?
в
6
9<5
6
9с?
9с5
9с5
Вес
животного
19-29 г
180-250 г
300-350 г
6-10 г
220-250 г
20-24 г
250-300 г
180-220 г
2,4 кг
3-7 кг
30-40 г
Возраст
животного
2-4 мес.
2,5-3 мес.
5-12 мес.
21-22 дня
3-5 мес.
2-3 мес.
5-12 мес.
3-4 мес.
7 мес-
4 года
1 -7 лет
3-10 мес.
346
Биомоделирование в системах управления
Окончание табл. 64
Исследуемое действие
вещества
Биомодель аутогенной
эпилепсии
Биомодель аудиогенных
судорог (в 100%) и аудио-
генного шока
Биомодель аудиогенных
судорог у 80-95% живот-
ных
Лабораторное
животное
Крысы
Мыши
Крысы
Линия или порода
животного
Линия
Крушинского-
Молодкиной
Линии
DBA/2J
DBA/2JY
Сток КМ
Пол
животного
9с5
96
9с?
Вес
животного
250-300 г
15-30 г
300-350 г
Возраст
животного
5-12 мес.
35 дней
5-12 мес.
Спинной мозг является важнейшим регуляторным и исполнительным
уровнем ЦНС одновременно. Он заключен в позвоночном канале и по-
крыт тремя оболочками, между которыми имеются эпидуральное, субду-
ральное и подпаутинное пространства. В шейной и поясничной частях
спинного мозга имеются утолщения. Средней бороздой и срединной ще-
лью спинной мозг разделяется на правую и левую половины. В разрезе
спинной мозг округлой формы, в центре его находится спинномозговой
канал.
Важнейшей функцией ЦНС является обеспечение локомоции. Считает-
ся, что они координируются высшими отделами головного мозга, но запрог-
раммированы на уровне спинного. По-видимому, у человека есть спиналь-
ные локомоторные центры, что может быть смоделировано на спинальном,
курарезированном животном. Чередующиеся залпы мотонейронов сгибате-
лей и разгибателей напоминают их разряды при ходьбе (см. табл. 65).
Серое вещество спинного мозга концентрируется вокруг центрального
канала и имеет форму крыльев бабочки. Боковые рога, находятся в грудных
сегментах спинного мозга, в которых заложено наружное симпатическое ядро
Якобсона. В верхние рога сегментов спинного мозга входят чувствительные
нервные пучки, а из нижних рогов соответствующего сегмента отходят ко-
решки двигательных нервов, которые объединяются в один смешанный спин-
номозговой нерв [402]. От спинного мозга отходят с правой и левой стороны
2-4 десятка спинномозговых нервов, что соответствует числу позвонков. В
конечном отделе спинной мозг суживается, образуя мозговой конус, и окан-
чивается на уровне поясничных позвонков.
С помощью гистохимических методов показано, что ЦНС имеет ряд
интегративных, нисходящих трактов. Так норадренергическая и дофаминер-
гическая системы простираются от голубого пятна к спинальным нейронам.
Иммунофлуоресцентными методами установлено наличие нисходящих се-
ротонинэргических трактов от ядер шва до спинальных нейронов. Полагают,
что они участвуют в системной регуляции не только локомоции, но и сома-
тосенсорной регуляции и вегетативных функциях.
Глава 24. Центральные уровни управления
347
Спинной мозг структурно является как бы негативом головного, посколь-
ку его серое вещество покрыто белым веществом спинного мозга. Белое ве-
щество состоит из нервных волокон, которые создают чувствительные (аф-
ферентные, восходящие, центростремительные) и двигательные (нисходя-
щие, центробежные) проводящие пути. На этом уровне обеспечиваются слож-
ные нейрохимические процессы центральной и периферической нервной
системы [101, 237, 228, 629]
Спинной мозг достаточно термочувствителен. У собак, кошек и крыс при
повышении температуры спинного мозга по всей его длине лишь на не-
сколько десятых долей градуса наблюдаются одышка, расширение кровенос-
ных сосудов и угнетение термогенеза. Охлаждение спинного мозга вызывает
дрожь, но для этого требуется значительное изменение температуры.
У морской свинки для запуска терморегуляторных реакций достаточно ло-
кального изменения температуры в области шейного отдела спинного мозга.
Термочувствительность клеток спинного мозга уступает клеткам переднего
гипоталамуса, однако, роль каждой из них в локальном изменении темпера-
туры до конца не изучена. Мало известно о локализации и морфологии струк-
тур, обусловливающих термочувствительность спинного мозга и его взаимо-
отношения с температурочувствительными структурами дорсальной стенки
брюшной полости и в мускулатуре.
Чем более организовано животное, тем большее количество двигатель-
ных волокон входит в состав белого вещества; так, у собак двигательные
волокна составляют 10% общей массы белого вещества спинного мозга, у
мини-свиней — 15, у обезьян — 20, а у человека — 30%.
Таблица 65
Генетически обусловленные и экспериментальные биомодели для изучения
механизмов атаксии и прогрессивного паралича
Исследуемое действие
вещества
Биомодель атаксии и про-
грессивного паралича с ре-
грессивными явлениями де-
генерации коры мозжечка,
развивающейся с 2-х меся-
цев
Модель прогрессивного па-
ралича задних конечностей
(в возрасте 6-10 мес.)
с явлениями миопатии и
дегенерации колонны спин-
ного мозга
Лабораторное
животное
Кролики
Хомячки
Линия или порода
животного
Линия AX/J
(семейство V)
Линия 12.14
Пол
животного
9с?
в
Вес
животного
3500 г
120 г
Возраст
животного
7 мес-
4 года
6-18 мес.
Спинной мозг выполняет проводниковую функцию. В нем проходят
нервные импульсы с периферии в головной мозг и из центров на перифе-
рию. Центры спинного мозга обеспечивают большое количество двигатель-
348
Биомоделирование в системах управления
ных рефлекторных актов, поддерживают тонус поперечнополосатой муску-
латуры, обеспечивают дефекацию и регуляцию функций мочеполовой и ве-
гетативной нервной системы. Разрушение спинного мозга выше III—V шей-
ных сегментов спинного мозга ведет к параличу дыхания и смерти.
Центральный спинномозговой канал, субарахноидальное пространство
и полости желудочков мозга заполняет лыквор (цереброспинальная жид-
кость).
Нейровегетативная регуляция
Периферические нервы образуются тысячами длинных волокон. Нервы,
отходящие от спинного мозга, называются спинномозговыми, а от голов-
ного мозга — черепно-мозговыми. Судя по тому, какой характер раздраже-
ний проводят нервные волокна и в каком направлении идут нервные им-
пульсы, нервы разделяются на афферентные (чувствительные) и эфферен-
тные (двигательные, или секреторные). Нервы состоят из пучка нервных
волокон, которые могут быть мякотными (миелиновыми, т. е. покрытыми
миелиновой оболочкой) и безмякотными (немиелиновыми). Миелиновая
оболочка образована швановскими клетками и разделена перехватами Ран-
вье на сегменты длиной от 0,1 до 1 мм. Все нервные волокна имеют соеди-
нительную оболочку, состоящую из эпиневрия, периневрия и эндоневрия.
Эпиневрий покрывает весь нервный ствол, периневрий — лишь группы
нервных волокон, а эндоневрий одевает каждое нервное волокно и выпол-
няет не только защитные, но и специфические функции [15, 276, 311]. В
толще нервного ствола имеются нервы нервов и собственные кровеносные
и лимфатические сосуды. Кроме мякотных волокон, в каждом нерве нахо-
дятся и безмякотные нервные волокна, относящиеся к вегетативной не-
рвной системе.
Число спинномозговых нервов у животных соответствует количеству сег-
ментов спинного мозга. К каждому спинномозговому нерву от погранично-
го симпатического ствола присоединяются соединительные ветви.
В отличие от человека, у животных обычно имеется пять-шесть хвосто-
вых нервов, которые разделяются на дорсальные и вентральные ветви.
У млекопитающих животных, как и у человека, имеется 12 пар черепно-
мозговых нервов, из них только два нерва имеют одни афферентные волокна
(слуховой и зрительный), остальные, кроме афферентных, содержат и эффе-
рентные волокна и являются смешанными нервами. В связи с этим в цент-
рах смешанных черепно-мозговых нервов выделяются ядра чувствительные
и двигательные (секреторные).
Вегетативные нервы существенно отличаются от двигательных (сомати-
ческих) нервов, так как состоят из преганглионарных и постганглионарных
волокон. Преганглионарные волокна в отличие от постганглионарных по-
Глава 24. Центральные уровни управления
349
крыты мякотной оболочкой. Центры вегетативной нервной системы разме-
щены во всех отделах коры головного мозга, главным образом в лобных и
теменных долях, подкорковых образованиях ствола мозга (nucleus caudatus,
putanem, globus pallidus, tuber cinereum, corpus mammilare, thalamus opticus),
стенках третьего и четвертого мозговых желудочков, в гипоталамусе и рети-
кулярной формации.
Вегетативную нервную систему разделяют на симпатический и парасим-
патический отделы. Выделяется третий отдел вегетативной нервной системы
- энтеральный, к которому относятся интрамуральные автономные нейро-
ны интестинальных органов, функционирующие независимо от команд ЦНС.
Центры парасимпатического отдела нервной системы заложены в среднем,
продолговатом мозгу и в клетках крестцовых сегментов спинного мозга. Цен-
тры симпатических нервов заложены в грудино-поясничном отделе спинно-
го мозга.
Симпатический и парасимпатический отделы вегетативной нервной си-
стемы находятся в сложной функциональной синергии. Возбуждение сим-
патического нерва вызывает учащение ритма и усиление сердечного сокра-
щения (положительное хронотропное и инотропное действия), расширение
зрачка, бронхиол, замедление перистальтики кишечника, скудную секрецию
пищеварительных желез, сужение кожных и мышечных сосудов, тогда как
коронарные, мозговые, легочные сосуды расширяются. Возбуждение пара-
симпатических нервов сопровождается отрицательным хроно- и инотроп-
ным действием на сердце, сужением зрачка и бронхиол, значительным уси-
лением перистальтики желудочно-кишечного тракта, спазмами сфинктеров,
расширением сосудов тазовых органов и сужением коронарных сосудов.
Через парасимпатические волокна можно усиливать симпатический эффект
и наоборот. Некоторые органы не имеют двойной иннервации, так скелет-
ные мышцы, потовые железы, мышцы, поднимающие волосы, и радиальные
волокна радужки глаза иннервируются только симпатическими нервами, а
ресничная мышца глаза — только парасимпатическим нервом.
В тех случаях, когда органы иннервируются симпатической и парасимпа-
тической системой, доминируют регуляторные влияния последней. Но регу-
ляция гликогенолиза в печени и липолиза в жировых клетках, концентрации
свободных жирных кислот и глюкозы в крови, как и иные метаболические
процессы находятся в сфере исключительно симпатической системы.
Роль NO в нервной системе
Практически все нейромедиаторы являются производными аминокислот или
самими аминокислотами. В последние годы особое внимание привлекла
аминокислота аргинин (L-Arg) после того как в 1987 г стала известна её роль
как эндогенного источника N0 или оксида азота. Полагают, что N0 образу-
350
Биомоделирование в системах управления
ется из терминального атома гуанидиновой группы L-Arg, при этом дезам и -
нирование ведет к появлению NH, с последующим окислением до N0. Этот
процесс регулируется NO-синтазой. Супероксиддисмутаза активирует N0-
синтазу, а каталазы её блокируют. Ввиду чрезвычайной краткости жизни N0
(от 5 до 20 с) о его наличии судят по локализации NO-синтазы. Одна из трех
форм NO-синтазы, являющаяся мозговым энзимом, относится к цитозоль-
ным изоферментам.
N0 является внутриклеточным регулятором и соответствует критериям
нейромедиатора по наличию ферментов биосинтеза, воспроизводимости
эффекта при стимуляции нервов или введения агонистов, блокирование
эффектов ингибиторами биосинтеза. Но в отличие от других медиаторов это
архипростая молекула, диффундирующая в соседние клетки, обеспечивая
межклеточную интеграцию во всех тканях, находится в цитоплазме клеток, а
не в синапсах.
По-видимому, роль N0 в нервной системе значительно важнее, чем сей-
час нам представляется. Помимо максимальных количеств NO-синтазы в
мозжечке и обонятельной луковице, она наблюдается в 2—5% нейронов гип-
покампа, стрио-паллидума и неокортекса. NO-синтаза обнаружена в ней-
ральных пулах других отделов мозга и астроцитах. Индуцибельная NO-син-
таза астроцитов ближе к таковой у макрофагов, нейтрофилов, микроглии,
клеток эндотелия (гл. 23 и 26) и не зависит от метаболизма Са2+. Для ней-
ронов характерна цитозольная её форма. Синтез индуцибельной NO-синта-
зы происходит во всех клетках при кислородном взрыве, характерном для про-
оксидантного стресса. Регуляция NO-синтазы зависит от эндогенного или
экзогенного N0, например, нитроглицерином.
N0 участвует двояко в синаптической передаче, вызывая длительную
активацию в неокортексе, стриатуме, гипоталамусе и относительно дли-
тельную депрессию в гиппокампе. Введение L-Arg в желудочки мозга при-
водит к сомногенному эффекту, а введение в этих условиях ингибиторов
NO-синтазы препятствует развитию сна и оказывает пробуждающий эф-
фект.
Пересматриваются взгляды на регуляцию психомоторных механизмов в
том числе участия N0 в воспроизведении ранее выработанных навыков у
мышей и крыс. Показана зависимость глюкокортикоидных и кг-опиоидных
медиаторных систем от N0 в регуляции этих эффектов. N0 регулирует про-
цессы привыкания и склонности к употреблению алкоголя у животных, а так-
же пищевую и питьевую мотивации. Этанол подавляет активность NO-син-
тазы и снижает образование N0 в нейронах неокортекса. Стрессогенные
нарушения питания, по-видимому, также регулируются N0.
Хотелось бы особо подчеркнуть, что N0 участвует как в обеспечении
психомоторной активации, так и регуляции страха и депрессий. Ингибито-
ры NO-синтазы обладают анксиолитическим действием у крыс по стандар-
Глава 24. Центральные уровни управления
351
тным тестам, а также по продолжительности в тесте времени зоосоциально-
го взаимодействия. По-видимому, механизмы N0 и NO-синтазы в дальней-
шем будут рассматриваться через призму использования в качестве экспери-
ментальной биомодели тревожности и страха и оценки на её основе перс-
пективных и принципиально новых психотропных средств.
Однако понимание роли и места N0 в регуляторных процессах ЦНС
далеко от завершенности. Так в работах А.А. Каменского и соавторов [357]
показано, что L-Arg в дозе 100мг/кг L-Arg не изменял уровень тревожности,
а в дозе 250 мг/кг он усиливал проявления пассивно-оборонительных реак-
ций. При выработке навыков с отрицательным подкреплением по тесту УРПИ
L-Arg в дозе 250 мг/кг улучшал обучение крыс.
В тех же исследованиях [357] показано, что N0, ухудшая обучение крыс
с положительным подкреплением, улучшает показатели УРПИ при болевых
раздражениях. Оксид азота в больших количествах вызывает нейродегенера-
тивные изменения вплоть до гибели нейронов, что говорит о цитотоксичес-
ком действии. Однако многие авторы склоняются к тому, что нейротоксич-
ность N0 обусловлена механизмами, не связанными с NO-синтазой. Иссле-
дование роли N0 в развитии расстройств деятельности мозга говорит о пре-
имущественно нейрпопротекторном его действии.
NO играет роль не только в ЦНС, но и в периферической нервной сис-
теме. Результаты некоторых экспериментов позволяют предположить, что
путь L-Arg-NO участвует в сенсорной передаче. N0 может сам выполнять
функции медиатора в неадренергических и нехолинергических (non-adrenergic
non-cholinergic, NANC) нервах. С другой стороны, N0 может быть модуля-
тором активности других медиаторных систем.
N0, высвобождающийся не только из нейронов, но и при участии спе-
циальных NANC нервов, представляющих чрезвычайно разветвленную сеть,
участвует как важнейший регулятор основных органов и систем организма.
NANC — нервы принимают участие в иннервации, наряду с адрено-, холи-
но- и пептидергическими системами. Роль N0 в системной регуляции орга-
низма ещё предстоит изучать и особенно у инбредных и иных чистых линий.
Это даст новые, перспективные биомодели для исследований.
Следует подчеркнуть, что основные медиаторы нервного возбуждения
(ацетилхолин, дофамин, адреналин, норадреналин, серотонин, ГАМК и т.д.),
выделяясь на окончаниях нервных волокон иннервируемых органов, тканей
или ганглиев, взаимодействуют со специальными рецепторами, осуществля-
ют передачу нервного возбуждения. Рецепторы являются точкой приложе-
ния действия многих лекарственных препаратов и ксенобиотиков. Посколь-
ку NO отвечает всем критериям, предъявляемым к нейромедиаторам, можно
полагать, что расшифровка оксидазотных механизмов будет также способ-
ствовать прогрессу в биомоделировании.
352
Биомоделирование в системах управления
Ликвор у человека и животных
Спинномозговая жидкость или ликвор у человека продуцируется главным
образом сосудистыми сплетениями. Из боковых желудочков она через мон-
роевы отверстия переходит в III желудочек, откуда по сильвиеву водопрово-
ду попадает в IV желудочек. Из IV желудочка она идёт в центральный канал
спинного мозга, а через отверстия сосудистой оболочки — Мажанди и Луш-
ка — в большую цистерну и субарахноидальное пространство. В субарахно-
идальном пространстве спинного мозга ток ликвора направлен и вниз, и
вверх — к головному мозгу.
Из субарахноидального пространства жидкость по периневральным ще-
лям черепных нервов (главным образом обонятельного, зрительного и слу-
хового) и по венам оболочек удаляется в лимфатическую и венозную систе-
му. Жидкость образуется медленно, по 2-3 мл в час. При ускорении эваку-
ации жидкости усиливается и её продукция.
В организме взрослого человека содержится 150—200 мл спинномозговой
жидкости, причём на долю цистерн субарахноидального пространства го-
ловного мозга приходится 50 мл, на долю желудочков 40—60 мл, остальное
количество приходится на долю субарахноидального пространства спинного
мозга.
Спинномозговая жидкость, покрывая со всех сторон головной и спин-
ной мозг, играет роль жидкого буфера, защищающего мозг от механических
травм. Кроме того, она принимает участие в питании центральной нервной
системы и выведении продуктов обмена веществ. Отсюда понятно, что все
патологические процессы в нервной системе должны в той или другой сте-
пени отражаться на свойствах и составе спинномозговой жидкости. В спин-
номозговой жидкости человека содержится 0,18—0,33%о белка, а также 1—2
лимфоцита в 1 мм3. Нормой считается наличие от 3 до 6 лейкоцитов в счёт-
ной камере.
У животных ликвор также образуется из крови в сосудистых сплетениях
желудочков мозга. Оттекает ликвор у животных главным образом через лим-
фатические сосуды и вены паутинной оболочки. Следует ещё раз под-
черкнуть, что в отличие от животных у человека отток происходит через
пахионовы грануляции. Лишь у собак и некоторых крупных животных име-
ются внутрисинусные пахионовы грануляции.
Давление ликвора у собаки в среднем составляет 145 мм вод. ст. (с коле-
баниями от 30 до 230 мм). Цереброспинальная жидкость собак бесцветная,
прозрачная и имеет следующий состав: форменных элементов в 1 мм 3 от 5
до 8; воды 99% и 1% сухого остатка; удельный вес 1,006-1,007; рН 7,4-7,5;
щелочной резерв в % СО 42-50; белка 15—20 мг %, молочной кислоты 15-
25, сахара 45-77, хлористого натрия 602-683, кальция 5,13-7,44, магния 2,58-
3,87, хлора 365—475 и неорганического фосфора 2,82—3,47 мг%. При различ-
Глава 24. Центральные уровни управления
353
ных заболеваниях состав цереброспинальной жидкости меняется, что может
служить диагностическим показателем.
Спинномозговая жидкость у кошек бесцветная, прозрачная, содержит от
О до 5 форменных элементов в 1 мм3. Общее количество белка в спинномоз-
говой жидкости 8—16 мг%, количество хлоридов (NaCl) — 670—723, сахара —
57-67 мг%.
Спинномозговая жидкость кролика прозрачная, бесцветная, содержит у
здоровых животных 5-10 лимфоцитов в 1 мм3, гликогена 0,65—1 (0,91) мг%,
сахара 45—79 (65) мг%, молочной кислоты 20—40 мг%; удельный вес — 1,005.
354
Глава 2 5 СИСТЕМЫ ВОСПРИЯТИЯ
И ПЕРЦЕПЦИИ
Органы чувств в сравнении
Орган зрения. Якоб Фриз в 1818 г. писал: «Там, где
речь идет о познании Природы, люди руководству-
ются тем, что они видят». Человек и животное ви-
дят не глазом, а мозгом, — глазом мы лишь воспри-
нимаем. Быстрые скачки глаз длительностью от 10
до 80 мс и амплитудой от долей градуса до 90° на-
зываются саккадами. Человек воспринимает в виде
света электромагнитное излучение в диапазоне от
400 до 700 нм.
Глаз состоит из глазного яблока, защитных и
вспомогательных приспособлений глаза (веки, слез-
ная железа, глазные мышцы и фасции). В глазном
яблоке имеется три оболочки: наружная фиброзная
оболочка, в которой различают прозрачную рого-
вицу и белочную оболочку; средняя сосудистая обо-
лочка, в которой различают собственно сосудистую
оболочку, ресничное тело и радужную оболочку;
внутренняя оболочка, или сетчатка.
Сетчатка состоит из светочувствительных эле-
ментов — колбочек, воспринимающих дневной свет,
и палочек, воспринимающих световое раздражение
в сумерках. В палочках находится зрительный пур-
пур (родопсин), который является 11-цис-ретина-
лем, т. е. альдегидом витамина А. Процесс транс-
дукции в фоторецепторе начинается с поглощения
фотона л—электронами сопряженных двойных свя-
Глава 25. Системы восприятия и перцепции
355
зей ретинала. Чувствительность палочек к свету в 1000 раз превышает свето-
чувствительность колбочек. К глазному яблоку относят хрусталик (lens
crystallina) и стекловидное тело (corpus vitreum).
Различают бинокулярное зрение, когда поле зрения одного глаза насла-
ивается на поле зрения другого глаза, и монокулярное зрение, когда поля
зрения правого и левого глаза не совпадают. У собаки зрение бинокулярное.
Угол между обеими зрительными осями (угол зрения) у различных живот-
ных значительно колеблется. Так, у собак он равен 92,5°, у кошек 77°. Амп-
литуда и частота саккад у большинства животных превосходят таковые у
человека, а диапазон воспринимаемых электромагнитных излучений лежит
ниже 300 нм для «ночных» и выше 850 нм для «дневных» животных.
Большинству млекопитающих животных присуще восприятие цветового
ощущения, и эту функцию выполняют колбочки. Причем наибольшей чув-
ствительностью к цветовому ощущению обладает область так называемого
желтого пятна сетчатки, которое у собак отсутствует.
Зрение у кошек хорошо развито. На сетчатке глаза много палочек, осо-
бенно большое количество их сконцентрировано в центральной ямке. Ост-
рота ночного зрения у кошки в четыре раза больше, чем у человека, но днев-
ное зрение у кошек в пять раз хуже дневного зрения людей. Восприятие
кошками цветов подтверждается электрофизическими исследованиями.
Зрение у кролика монокулярное. Глаза кролика позволяют осматривать
все вокруг, поскольку поле зрения правого и левого глаза наслаивается спе-
реди на 27° и сзади — на 9°. Несмотря на то, что у кроликов удается выра-
ботать с помощью метода условных рефлексов дифференцировки на некото-
рые цвета, многие авторы склонны считать, что у кроликов цветное зрение
отсутствует. Объясняют это тем, что дифференцировки вырабатываются не
на цвет, а на разности интенсивности источников положительных и диффе-
ренцировочных раздражителей.
Глазное яблоко у кролика больших размеров и имеет такое же строение,
как и у других животных. Глаз кролика имеет три века — верхнее, нижнее и
третье, расположенное во внутреннем углу глаза. Под третьим веком нахо-
дится железа третьего века, которая по характеру секрета относится к саль-
ным железам. Слезный аппарат представлен слезной железой, расположен-
ной в височном углу глаза. Глазодвигательные мышцы: четыре прямых, две
косых и оттягиватель глазного яблока.
У животных с цветовым зрением установлена, как и у человека, цвето-
специфинная организация колбочек в сетчатке. С помощью микроспектрофо-
тометрии получены различия в горизонтальных и биполярных клетках, одни
из которых гиперполяризуются монохроматическим светом одного диапазо-
на и деполяризуются другим. С помощью этого метода возможно с большим
или меньшим приближением оценить цветочувствительность животных-био-
моделей.
356
Биомоделирование в системах управления
Орган слуха и равновесия. Звук - это колебания молекул упругих волн
со скоростью в воздухе около 335 м/с. Уровень звукового давления измеря-
ется в децибелах (дБ), частота в герцах (Гц). Функцию органа слуха и равно-
весия выполняет ухо (auris) которое разделяется на наружное (auris externa),
среднее (auris media) и внутреннее (auris interna).
Органы слуха расположены в улитке. Участок стенки перепончатого ка-
нала улитки, обращенный к барабанной лестнице, называется основной
мембраной. Основная мембрана состоит из поперечных соединительноткан-
ных волокон («струн»), длина которых от 0,05 до 0,5 мм. На основной мем-
бране расположен спиральный, или кортиев, орган (organon spiralis Corti),
в нем оканчиваются ветки слухового нерва. Кортиев орган имеет рецептор-
ные внутренние (у человека около 3 500) и наружные (у человека 12 000)
волосковые клетки. Чувствительные клетки на своей поверхности содержат
немерцающие волоски и играют роль слуховых рецепторов.
Звуковые колебания через барабанную перепонку, слуховые косточки и
овальное окно передаются перилимфе и эндолимфе улитки, вызывая резо-
нанс у струн основной мембраны, что улавливается волосковыми рецептор-
ными клетками.
Вестибулярный аппарат состоит из преддверья и полукружных каналь-
цев, в которых находятся рецепторные клетки, содержащие субмикроскопи-
ческие стереоцилии (до 80 на клетку) и киноцилии ( по одной). Он воспри-
нимает положения тела и головы и способствует сохранению равновесия
тела. При изменениях положения головы происходит рефлекторная переда-
ча импульсов с вестибулярного аппарата на мышцы глаз, шеи, туловища и
конечностей.
У человека верхняя граница слухового восприятия достигает 20 000 Гц, а
собаки воспринимают такие высокие тоны, как 40 000—90 000 Гц, которые
человек не слышит. Чувствительность слухового анализатора у собаки высо-
кая. Собаки различают 1/8 тона, т. е. способны отличить 800 Гц от 812 Гц.
Зрение у кабанов не очень острое, но зато слух и чутье превосходные. Челове-
ка кабан слышит за 600 м. Столь же острый слух и у кошек. Барабанная по-
лость у кролика относительно большая и вмещает слуховые косточки: молото-
чек, наковальню, чечевицеобразную косточку и стремечко. Внутреннее ухо,
имеющее строение, близкое к человеческому, удобно для биомоделирования.
Орган обоняния представлен участком слизистой оболочки обонятель-
ной части носовой полости, которая выстлана эпителием. У человека в обо-
нятельной области 107 рецепторов на 10 см2 площади. Обонятельный рецеп-
тор — это биполярная клетка, с дендритом, содержащим специальный кино-
цилии. Нейроэпителиальные клетки посредством нерва соединены с обоня-
тельными центрами.
Обонятельная область покрывается секретом обонятельных железок, ко-
торый растворяет пахучие вещества и этим способствует их воздействию на
Глава 25. Системы восприятия и перцепции
357
обонятельные клетки и, кроме того, предохраняет слизистую от высыхания.
Собаки очень чувствительны к запаху предельных кислот, они оказывают на
собаку такое же действие, как запах корня валерианы на кошек.
Функцию этого органа у кроликов и других грызунов выполняет обоня-
тельная часть носовой полости. Слизистая оболочка в области верхней но-
совой раковины и носовой перегородки выстлана специальным однослой-
ным обонятельным эпителием, имеющим пигмент, и богата альвеолярными
трубчатыми железами.
До последнего времени не были ясны первичные механизмы восприятия
запахов. Открытие механизмов медиации оксида азота в ЦНС и перифери-
ческой нервной системе позволяют по иному взглянуть на эти процессы.
Система NO-цГМФ играет существенную роль в регуляции обоняния: силь-
ные запахи вызывают стойкое повышение концентрации цГМФ в обоня-
тельной луковице, сопровождающееся увеличением синтеза N0. Полагают,
что взаимодействие NO и цГМФ имеет значение при адаптации к сильным
запахам [358]. Высокие концентрации NO-синтазы в обонятельной системе
как позвоночных, так и беспозвоночных позволяют предположить, что N0
участвует в передаче хемосенсорной информации. На насекомых показано,
что система NO-цГМФ является компонентом молекулярного механизма,
вовлеченного в адаптивные и интегративные процессы во время передачи
хемосенсорной информации. За расшифровку интимных механизмов ощу-
щения запахов группа американских исследователей удостоена в 2004 году
Нобелевской премии
Орган вкуса. Вкусовые ощущения воспринимаются при раздражении
заложенных в слизистой оболочке ротовой полости вкусовых почек (луко-
виц). У человека они расположены на поверхности языка и состоят из 40—
60 сенсорных вкусовых и опорных клеток каждая. Рецепторные белки вку-
совых рецепторов у человека и животных достаточно близки. Вкусовые поч-
ки имеют вкусовые неироэпителиальные клетки, выходящие в специальные
канальцы — вкусовые поры. Посредством вкусовых пор они сообщаются с
ротовой полостью. К вкусовым почкам подходят разветвления вкусовых не-
рвов (барабанная струна, иннервирующая 2/3 передней части языка, язы-
коглоточный нерв, иннервирующий заднюю часть языка, и блуждающий
нерв, который подходит к вкусовым почкам в области глотки). Локализация
вкусовых почек у млекопитающих преимущественно в стенках углублений
валиковидных сосочков, в других сосочках (листовидных, грибовидных и
нитевидных), а также частично в зеве, мягком нёбе и надгортаннике. Для
человека и животных считают основными вкусовыми ощущениями кислое,
сладкое, горькое и соленое. Имеются данные, указывающие, что у собак
восприятие вкусовых раздражителей происходит менее дифференцирован-
но, а у кошек найдены «Н20-волокна», реагирующие только на воду или
пищу, содержащую значительные количества воды.
358
Биомоделирование в системах управления
Орган осязания. Осязание воспринимается у животных кожным покро-
вом и слизистой губ, языка, ротовой полости и т. д. Участки тела, принима-
ющие участие в осязании, богаты окончаниями чувствительных нервов, кон-
центрирующихся или вокруг волосяных влагалищ, или в эпителии кожи.
У животных особой чувствительностью отличаются губы, кончик носа, по-
душечки на лапах и концы пальцев. У некоторых животных (кошка, кролик)
хорошо развиты осязательные волосы (вибриссы), корни которых богаты
нервными окончаниями или связаны с осязательными тельцами Меркеля.
Весьма чувствительны и хорошо развиты вибриссы на верхней губе. Если
удалить вибриссы, то у животных наблюдаются затруднения в ориентировке,
особенно в темноте. У крыс хорошо выражены вибриссы, которые в виде
длинных волос размещены над глазами и на нижней губе, а основная их
масса сконцентрирована на верхней губе. Вибриссы служат органами осяза-
ния и воспринимают не только соприкосновение с предметами, но и улав-
ливают колебания воздуха. В носовой полости грызунов очень много обоня-
тельного эпителия.
Вулси и Ван-дер-Лоос наблюдали в мозге мыши хорошо выраженные
морфологические сдвиги после удаления вибрисс. В соматосенсорной зоне
были отмечены скопления клеток, имеющие характерную форму. Если смот-
реть со стороны поверхности, то эти скопления имели вид колец. На сре-
зах через толщу коры они напоминали бочонки, или цилиндры с менее
плотной центральной частью. «Бочонки» выстраивались правильными ря-
дами. Поначалу это казалось странным, потому что такие скопления не
обнаруживаются в других областях коры мыши. При подсчете цилиндров и
изучении их распределения в сенсорной коре была найдена корреляция
между ними и числом вибрисс на морде животного: каждый «бочонок»
соответствовал одной вибриссе на противоположной стороне морды. Так
же, как в зрительной системе, морфология скоплений существенно и ста-
бильно менялась в зависимости от условий, в которых протекал ранний
онтогенез. Если у новорожденной мыши удаляли ту или иную вибриссу, то
у взрослого животного в коре отсутствовал соответствующий «бочонок».
Вопросы о том, имеются ли «бочонки» у новорожденных мышей, есть ли
критический период и каковы последствия сенсорного опыта и неупотреб-
ления, еще не изучались. Тем не менее, это наблюдение служит еще одним
примером пластичности нейрональной архитектуры в самых разнообразных
системах.
Кожа и её образования
Кожный или общий покров представляет собой внешнюю оболочку живот-
ного. Такое положение делает его посредником между внешним миром и
скрытыми частями тела и защищает последние. Кожа человека и животных
Глава 25. Системы восприятия и перцепции
359
является, по-видимому, самым крупным иммунным органом. Наряду с этим
кожа является мощным рецепторным полем и осуществляет функции осяза-
ния, выделения, терморегуляции, а также функции защиты организма [40,
98, 298, 301, 316, 675, 686].
Кожа (cutis) имеет три слоя: надкожицу, или эпидермис, состоящую из
многослойного плоского эпителия, глубокую часть которого составляют
живые клетки, постоянно размножающиеся путем деления, а поверхност-
ную образуют ороговевшие, постоянно отмирающие и слущивающиеся клет-
ки; собственно кожу, в которой находится множество чувствительных не-
рвных окончаний, капилляров, корни волос и железы; подкожную клетчат-
ку, или подкожный слой, толщина которого зависит от скопления в нем
жира и у собак достаточно выражена лишь в области когтевого валика.
Мини-свиньи являются прекрасной моделью человека и в дерматологии,
так как кожа свиньи во многом схожа с кожей человека [486]. Свидетель-
ством этому, в частности, является тот факт, что меланомы, спонтанно воз-
никающие у свиней, имеют сходство с некоторыми типами пигментных
опухолей у человека [481]. Свинья является удобной моделью и при иссле-
довании кожных трансплантатов, так как позволяет изучать различные их
типы, изменять характер трансплантатов в зависимости от специфики
экспериментальных условий [361].
К производным кожи относят волос. На морде у хищников и грызунов
имеются осязательные волосы (усы, вибриссы); ресницы плохо развиты или
отсутствуют.
Характерными образованиями кожи у собак являются мякиши и когти.
Мякиши находятся в области лап (шесть штук на передних и пять на зад-
них), снабжены большим количеством нервных окончаний, служат для опо-
ры и играют важную роль в осязании.
Волосяное одеяние у некоторых млекопитающих представлено довольно
причудливыми рисунками, хотя и не так разнообразно в оттенках цветов,
как оперение птиц. Сами волосы бывают то очень тонкие и нежные, то тол-
стые (осязательные), а иногда и очень грубые, до твёрдости игол ежей и
дикобразов, то очень короткие, то длинные (более метра).
Покров млекопитающих, помимо заметных простым глазом волос, со-
держит в своей толще ещё и углубления, выстланные клетками, т. е. железы.
Железистые образования оказываются также разнохарактерными по стро-
ению, а следовательно, и по значению: одни выделяют пот, другие — кожное
сало, третьи — специальную пахучую жидкость. Даже такой продукт пита-
ния, как молоко, вырабатывается кожными железами.
Через отверстия тела — рот, ноздри, задний проход, а также через мо-
чеполовые отверстия — кожный покров заходит то на большую, то на мень-
шую глубину внутрь, вступая в соединение со слизистыми оболочками. Осо-
бенно глубоко покров проникает во время развития зародыша в полость рта,
360
Биомоделирование в системах управления
так что приспособления для выработки слюны, слизи и даже зубы происходят
из кожного покрова.
Физиологическое значение кожи состоит в том, что она является важ-
нейшим органом иммунитета, защитным покровом и барьером для микроорга-
низмов, играет роль выделительного и осязательного органа и принимает уча-
стие в терморегуляции.
Характерной особенностью кожи кролика как органа выделения является
то, что потовые железы выражены слабо и локализованы преимущественно
в области морды. Сальные железы особенно хорошо развиты на наружном
ухе. Кожа кролика обладает большей проницаемостью для ядов, чем кожа
человека.
Когти — роговые образования последних фаланг пальцев. Они во мно-
гом помогают хищникам и грызунам при еде, обороне, нападении и т. д.
Потовые железы у собак на коже сконцентрированы на мякишах и кон-
чике носа. Служат для выделения пота (чем способствуют удалению из орга-
низма некоторых веществ (гормоны, промежуточные продукты обмена ве-
ществ, яды и токсины).
Сальные железы связаны с волосяным влагалищем, вырабатывают кож-
ное сало, которое покрывает кожу, волосы и предохраняет их от действия
многих вредоносных факторов. У самцов сальные железы хорошо развиты в
коже мошонки.
К кожным образованиям относятся молочные железы, выполняющие
специализированную функцию. Молочные железы функционируют в связи
с беременностью и родами. Собакам, как и мини-свиньям, свойственно мно-
жественное вымя с пятью, реже четырьмя холмами с каждой стороны. На
коже сосков имеются редкие нежные волоски, сальные и потовые желёзки.
На вершине соска открываются (варьирующие в количестве 6—12) сосковые
каналы. Ходы в соске во время лактации слегка расширяются, формируя
подобие цистерны.
В первую неделю после родов выделяется молозиво. Молоко собак со-
держит (%): жира 9,26-9,57, сахара 3,05-3,11, казеина (казеиногена) 3,5-
4,15, альбумина и других белковых веществ 5,57—11,17, солей 0,73—0,9. Кро-
ме казеиногена и альбумина, в молоке содержится глобулин (а-, р- и у-
глобулин). Причем 60% белков сыворотки молока приходится на долю |3-
лактоглобулина, который больше других белков способствует росту молод-
няка. Альфа- и гамма-глобулины сыворотки молока играют роль антител.
Минеральный остаток молока суки включает 25,4% фосфора и 36,5% каль-
ция, а остальная часть приходится на натрий, калий, железо, магний, серу и
другие химические элементы.
Молоко собак отличается от коровьего большим содержанием жира и
белка, меньшим содержанием углеводов. На 1 л молока собаки приходится
4,4 мг железа, что в семь-восемь раз больше, чем в молоке коровы.
Глава 25. Системы восприятия и перцепции
361
У кошки имеется восемь грудных желез (по четыре с каждой стороны). В
соске железы проходят четыре—шесть каналов. В молоке содержатся (%):
молочный сахар (лактоза) 3,4—4,9, жир 3,33, белок 9,08, соли 0,58.
Крольчиха имеет 4—5 (реже 3 или 6) пар молочных желез. В молоке кроль-
чих содержится (%): молочный сахар 1,8; белок 10,4—15,5; жир 10,45 и соли
2,56. В золе молока кальция 40,9% и фосфора 27,8%.
Биомодели и реакции кожи
Особенности биомоделей для исследований дерматологических реакций при
воздействии физических и биологических факторов приведены в табл. 66.
Таблица 66
Экспериментальные модели для исследований кожных реакций
Исследуемое действие
вещества
Модель пассивной кожной
анафилаксии при иммуни-
зации инъекциями ОА
100 мкг и 1 мг гидроокиси
алюминия в объеме 0,5 мл
дважды с интервалом в ме-
сяц, внутрибрюшинно
Модели дерматомикозов,
вызванных зоофильными,
антропофильными дермато-
фитами и эпидермофи-
тоном
Модель контактной гипер-
чувствительности замедлен-
ного типа к 2,4,6-тринитро-
хлорбензолу (ТНХБ) (сенси-
билизация 0,1 мл 3%-ного
р-ра ТНХБ в ацетоне, на
выбритый участок кожи
живота)
Изучение эффективности
веществ на моделях
кандидоза:
a) сепсис, вызванный
Candida albicans (заражение
путём введения в хвостовую
вену или в кожу взвеси гри-
ба в среднем в дозе 25 млн
микробных тел)
b) менингоэнцефалит,
вызванный Candida albicans
(заражение путём интра-
церебрального введения
около средней линии чере-
па на 2-3 мм выше глазни-
цы 0,05 мл взвеси гриба
в физиологическом раство-
ре в дозе 40-60 млн гриб-
ковых клеток на мышь)
Лабораторное
животное
Крысы
Морские
свинки
Мыши
Мыши
Мыши
Линия или порода
животного
Белые
аутбредные
Аутбредные
Линия СВА
или гибриды
Белые
аутбредные
Белые
аутбредные
Пол
животного
9<5
9<5
9<5
96
96
Вес
животного
200-220 г
0,7-1 кг
30-40 г
40-50 г
40-50 г
Возраст
животного
4-5 мес.
6 мес-
2 года
3-10 мес.
3-10 мес.
3-10 мес.
362
Биомоделирование в системах управления
Окончание табл. 66
Исследуемое действие
вещества
Изучение эффективности
на моделях глубоких
микозов:
a) кожный спиротрихоз
(введение суспензии дрож-
жевой фазы гриба Sporo-
trichum в объёме 1 мл,
подкожно)
b) криптококкоз (интрацере-
бральное заражение через
«родничок» - 3 млн клеток
гриба в 1-2 каплях физио-
логического раствора)
Ультрафиолетовая эритема
Лабораторное
животное
Мыши
Мыши
Морские
свинки
Линия или порода
животного
Белые
аутбредные
Белые
аутбредные
Альбиносы
Пол
животного
96
96
96
Вес
животного
40-50 г
40-50 г
250-500 г
Возраст
животного
3-Ю мес.
3-10 мес.
2-4 мес.
Основные пояснения и особенности пользования таблицей можно найти
в главе 2 (табл. 6). Тем не менее, следует ещё раз подчеркнуть, что кожа и её
образования у человека и животных является самым большим рецепторным
полем восприятия и перцепции, а также крупнейшим органом иммунной сис-
темы. Яоэтому мы сочли оправданным привести здесь модели для изучения
эффектов лекарств и ксенобиотиков как непосредственно на кожу, так и
через ее участие в системных проявлениях и реакциях на воздействие физи-
ческих, грибковых и микробных факторов.
363
Глава 2 6 ТРИАДА
НЕЙРОИММУНОГОРМОНАЛЬНЫХ
СИСТЕМ УПРАВЛЕНИЯ
Древние римляне в соответствии с незыблемыми
правилами совершали пожертвования своему горо-
ду или несли повинность (munis). Это могли быть
денежные взносы, постройка храмов, а также несе-
ние гражданской или военной службы. Те же из них,
кто был освобожден от таких действий, входили в
категорию immunis (im- означает «не»). Их называ-
ли «иммунисом» — свободным от чего-либо, от
какой-либо повинности.
Этот термин в дальнейшем вошел в биологию и
медицину. Тот, кто переболел определенным инфек-
ционным заболеванием, не заболевает им вновь, ста-
новясь иммунным — невосприимчивым к этому за-
болеванию. В крови образуются белковые молеку-
лы, которые, соединяясь с болезнетворным агентом,
обезвреживают его.
Молекулы могут соединяться с ксенобиотиками
и с ядами, которые выделяются микроорганизма-
ми. Такие яды иначе называются токсинами, от
греческого слова toxikon — яд. Его происхождение
восходит к тем временам, когда наконечники стрел
смазывали ядом. Греки называли эту операцию
toxikon pharmakon. По большому счету вся фунда-
ментальная суть борьбы за выживание может быть
сведена к обеспечению защиты организма от экзо-
и эндотоксинов
В системе регуляции организма особая роль
принадлежит согласованному действию иммунной
364
Биомоделирование в системах управления
и гормональной систем. По большому счету эта единая система обеспечения
гомеостатических функций, которая только ради удобства поделена и рас-
сматривается как бы независимо друг от друга и от функций ЦНС. Однако
природа не знает о нашем делении на науки, она едина. Одни и те же струк-
турные элементы и функциональные системы осуществляют свои регуля-
торные воздействия на нейромедиаторные, гормональные, иммунные и ис-
полнительные звенья организма.
Базис единства триады
Соединения, синтезируемые в небольших железках, всегда привлекали при-
стальное внимание врачей. Железами стали называть все органы тела незави-
симо от их размера, которые выделяли различные соки. Печень, например,
тоже является железой, недаром в русском языке ее название связано со
словом «печь» — готовить, обрабатывать пищу, ее называют также пищева-
рительной железой.
Большинство гормонов образуются в аппарате Гольджи из самых различ-
ных клеток (нервных, железистых, паренхиматозных) путем трансляции ге-
нетической информации, закодированной в РНК, в аминокислотную после-
довательность. Как и в других белках существуют гормоны, производные
аминокислот и липидов.
Гормональным веществом считается лишь то, для которого в клетках-
мишенях есть специфические рецепторы, позволяющие считывать информа-
цию, закодированную, в гормоне. Гормон-рецепторный комплекс может
индуцировать или подавлять синтез белка за счет экспрессии генетической
информации.
Гормон взаимодействует с рецептором, локализованным на плазматичес-
кой мембране, через второго посредника внутри клетки. Такими посредни-
ками служат циклический аденозинмонофосфат (цАМФ) из аденозинтри-
фосфата (АТФ), циклический гуанозинмонофосфат (цГМФ), кальмодулин и
фосфоинозитол. Гормон-рецепторный комплекс активирует аденилатцикла-
зу или другие ферменты, которые катализирует дефосфорилирование АТФ и
превращение его в цАМФ. На последующих этапах цАМФ влияет на функ-
циональную активность клетки.
Гормоны вырабатываются секреторными клетками желез (щитовидная
железа, гипофиз и др.), либо разбросаны на а- и (3- клетки островков Лан-
герганса. Гормоны хранятся в гранулах — внутриклеточных органеллах, от-
деленных от цитоплазмы мембраной. В гранулах содержится большое число
молекул гормона, погруженных в белковый матрикс. Когда гормон высво-
бождается, мембрана гранулы сливается с плазматической мембраной и в
месте слияния образуется отверстие, через которое молекулы гормона выб-
расываются в межклеточное пространство путем экзоцитоза.
Глава 26. Триада нейроиммуногормональных систем управления
365
Характеризуя деятельность эндокринной системы, следует коснуться па-
ракринного действия гормонов. Если гормоны — это носители химической
информации, вырабатываемые секреторными клетками и выделяемые в
кровь, которая доставляет их к органу-мишени, то характерным признаком
органа-мишени является способность считывать информацию, закодирован-
ную в гормоне. В организме человека и животных есть органы-мишени, на-
ходящиеся в непосредственной близости от гормональной клетки. Гормоны
диффундируют через межклеточное пространство к клетке-мишени. В этом
случае их называют паракринными гормонами местного действия или тка-
невыми гормонами. Простагландины, простоциклины, тромбоксаны действу-
ют именно таким образом. Некоторые классические гормоны могут действо-
вать паракринным путем. Наряду с классическими гормонами, паракрин-
ным действием обладают и классические нейромедиаторы, выделяемые нерв-
ными клетками. Нейромедиаторы диффундируют через синаптическую щель
и на постсинаптической мембране, как и гормон, связываются со специфи-
ческим рецептором. При кажущейся общности проявлений мы сталкиваем-
ся с совершенно различными аспектами биомоделирования процессов, кото-
рые рассмотрим в дальнейшем изложении.
Высокочувствительные гибридомные иммунологические методы позво-
ляют получать моноклональные антитела, которые связываются только с
конкретным гормоном. Они неуязвимы для системы деградации, а комплекс
гормона с антителом можно осадить и определить количественно, напри-
мер, в тестах на беременность, тиреотоксикоз, инсулин.
Методы радиоиммунологического анализа основаны на использовании
специфических антител, которые инкубируют со смесью радиоактивно ме-
ченных и интактных молекул гормона. Общее число молекул гормона должно
превышать связывающую способность антител, благодаря чему происходит
конкуренция за места связывания. После отмывки молекулы разделяют, изме-
ряют радиоактивность связанной фракции и по калибровочной кривой опре-
деляют количество гормона в биологическом образце. Эти методы и позволя-
ют измерять ничтожно малые количества гормона в нано- и пикограммах.
Открытие многогранных функций оксида азота определяет новый взгляд
на процессы гормональной регуляции. Так, содержащие NO-синтазу нейро-
ны обнаружены в гипоталамусе, преимущественно в паравентрикулярном и
супраоптическом ядрах: аксоны этих нейронов проецируются в нейрогипо-
физ. NO-синтаза обнаружена также в срединном возвышении гипоталамуса,
которое связано с аденогипофизом посредством гематоневральной контакт-
ной зоны. Показано, что N0 участвует в регуляции репродуктивных функ-
ций. N0 способствует высвобождению гонадотропных гормонов и их рили-
зинг-факторов. Окситоцин стимулирует NO-синтазу через норадреналин, в ре-
зультате чего образуется N0. N0 вызывает выделение люлиберина из гипо-
таламуса. В свою очередь люлиберин не только индуцирует высвобождение
366
Биомоделирование в системах управления
лютеинизирующего гормона, но и провоцирует половое поведение (подроб-
нее в гл. 32). Более того, NO может действовать по механизму обратной связи
на окситоцинергические терминали и подавлять секрецию окситоцина.
Тимус в сравнении и биомодели
Вопросам биомоделирования в иммунной системе посвящено большое ко-
личество публикаций. Исследование иммунной системы представляет собой
самостоятельный раздел медицинской науки. В данном случае мы сошлемся
на последние работы в этой области [21, 176, 418, 396, 489], которые имеют
непосредственное отношение к вопросам альтернативного биомоделирова-
ния. Поскольку иммунокомпетентные органы, в том числе лимфатическая
система, будут представлены в последующих главах, здесь мы остановимся
на характеристике тимуса.
Тимус у человека и млекопитающих — это дольчатый орган, сильно раз-
витый лишь у эмбрионов и в первые годы жизни; затем он постепенно ис-
чезает и у взрослых особей большей частью совсем отсутствует. Развитый
тимус находится в области шеи и трахеи, а главным образом в краниальном
отделе грудной полости. Этот расположенный за грудиной орган вырабаты-
вает ряд пептидов. Считается, что эти пептиды участвуют в механизмах
иммунологической защиты.
Поскольку тимус телят является излюбленной моделью в иммунологи-
ческих и эндокринологических экспериментах, следует напомнить, что у них
он относительно большой и следы его часто остаются надолго в грудной
полости. У молодых телят он состоит из большой непарной грудной доли и
парных шейных частей, выступающих справа и слева из грудной полости по
трахее вплоть до гортани. Эта железа расположена в переднем отделе груд-
ной полости, между I и VI ребрами на грудине. Два вилкообразных выроста
зобной железы размещены по бокам трахеи. Построена железа из долек,
состоящих из коркового и мозгового вещества. Наряду с известными функ-
циями по регуляции иммунологического статуса, деятельность тимуса связа-
на с половыми железами, начало функционирования, которых совпадает с
обратным развитием тимуса. Размеры тимуса также значительно уменьша-
ются при беременности и лактации.
У мини-свиней тимус очень сильно развит; в нём также различают груд-
ную непарную и шейную парную часть, простирающуюся до гортани и даже
до глотки.
У овец также имеется непарная грудная часть; она лежит слева и вент-
рал ьно от трахеи, пищевода и передней полой вены, простираясь даже на
левую поверхность сердечной сорочки. Выступающий между первыми рёб-
рами передний конец делится приблизительно в средней трети шеи на две
парные доли, достигающие краниально гортани.
Глава 26. Триада нейроиммуногормональных систем управления
367
У собак железа относительно мала и лежит в грудной полости на грудной
кости в пределах от 1-го по 6-е ребро. Парные, выступающие в отдел шеи,
доли незначительны, и левая из них несколько больше правой. К 2—3 годам
в грудной полости от неё сохраняются лишь незначительные остатки. Ви-
лочковая железа щенков в возрасте до двух недель составляет 0,58%, а в
возрасте двух-трех месяцев вес ее равняется уже 0,06—0,08% веса всего тела.
Тимус у молодых кроликов хорошо развит. У взрослого животного парен-
хима железы заменяется жировой и соединительной тканью.
Тимус у морских свинок заходит на шею.
Вилочковая железа у крыс, хомячков и мышей довольно больших разме-
ров. Расположена она под трахеей и состоит из двух долей.
В табл. 67 представлены некоторые биомодели, используемые в иммуно-
логических и гормонально-биохимических исследованиях.
Таблица 67
Линейные животные и экспериментальные биомодели в иммунологии
Исследуемое действие
вещества
Моделирование иммуно-
модулирующей активности
(1000, 100, 10 мкг на мышь)
Моделирование для изуче-
ния влияния препарата на
неспецифическую рези-
стентность организма
(1000, 100, 10 мкг в объёме
0,5 мл физиологического
раствора, внутрибрюшинно
или подкожно)
Модель для оценки влияния
препарата на гуморальный
иммунный ответ при имму-
низации эритроцитами ба-
рана
Модель для оценки про-
аллергического/противо-
аллергического действия:
a) Системная анафилаксии
(подкожная сенсибилиза-
ция чужеродным белком)
b) Продукция аллергенспе-
цифических IgE и IgG анти-
тел (антиген - трижды
перекристаллизованный
овальбумин куриных яиц)
Моделирование пассивной
кожной анафилаксии
Моделирование анафилак-
сии изолированных органов
Модель для оценки анафи-
лактогенной активности
Лабораторное
животное
Мыши
Мыши
Мыши
Морские
свинки
Мыши
Крысы
Морские
свинки
Морские
свинки
Линия или порода
животного
СВА,
BALB/c,
СВ57/В1
СВА или
(CBA/C57BL)
Линии СВА
и C57/BL
Аутобредные
СВА х C57/BL
F1
Аутбредные
Аутбредные
Аутбредные
Пол
животного
9с?
9с?
9с?
9с?
9с?
9с?
9с?
9с?
Вес
животного
18-20 г
18-20 г
18-20 г
10,7-1 кг
18-20 г
180-250 г
0,7-1 кг
0,7-1 кг
Возраст
животного
2-2,5 мес.
2-2,5 мес.
2-2,5 мес.
6 мес-
2 года
2-2,5 мес.
3-4 мес.
6 мес-
2 года
6 мес-
2 года
368
Биомоделирование в системах управления
Окончание табл. 67
Исследуемое действие
вещества
Моделирование активной
кожной анафилаксии
Биомодель первичного
иммунодефицита у мышей-
нюдов с глубокими наруше-
ниями иммунной системы,
отсутствием тимуса,
Т-лимфоцитов, шерстного
покрова
Основные продуценты
моноклональных антител
Биомодель высокоинтенсив-
ного иммунного ответа на
аминокислоты: глютамин,
лизин, тирозин и др.
Биомодель со слабым
иммунным ответом на
аминокислоты: глютамин,
лизин, тирозин и др.
Высокоактивный продуцент
антител как биомодель
в иммунологии
Лабораторное
животное
Морские
свинки
Мыши
Мыши
Мыши
Кролики
Кролики
Кролики
Линия или порода
животного
Аутбредные
Аутбредные
Мутантный
сток
Линии
BALB/cLacY
DBA/2JY
Линии C/J
III/J
Линия X/Y
Линия III/J
Пол
животного
9с?
9с?
9с?
9с?
9с?
9с?
9с?
Вес
животного
0,7-1 кг
40-50 г
30-40 г
30-40 г
2,9 кг
3,8 кг
2,0 кг
3,8 кг
Возраст
животного
6 мес-
2 года
3-10 мес.
3-10 мес.
3-10 мес.
7 мес-
4 года
7 мес-
4 года
7 мес-
4 года
7 мес-
4 года
Гипоталамо-гипофизарный пул
Древние греки и римляне полагали, что мозг служит охлаждению воздуха и
крови. Позже долгое время существовало представление, будто в головном
мозге образуется флегма (phlegma) — слизь, мокрота, которая покрывает внут-
реннюю поверхность носовых раковин и особенно докучает нам при про-
студных заболеваниях. «Мукус» (mucus) — латинский перевод слова «флег-
ма». Ученые и медики долго не понимали функции мозга и лишь в середине
XIX века Б. Уальдер, описывая строение мозга, дал одной из желез название
гипофиз (от греч. hipo — под, внизу wphyo — расти, что означает буквально
«расту под»). Так ее ныне и называют. Другое название у этого органа питу-
ита сохранилось в медицинской терминологии, например в слове апитуи-
таризм, что означает отсутствие гипофиза.
Гипофиз, или нижний придаток (hypophysis cerebri, s. glandula pituitaria), —
железа внутренней секреции, расположена в ямке турецкого седла под осно-
ванием головного мозга. В гипофизе различают переднюю, среднюю и зад-
нюю доли. На верхней части передней доли имеется туберальная часть, ко-
торая охватывает гипофизарную ножку.
Паренхиму передней доли гипофиза составляют переплетающиеся меж-
ду собой трабекулы (тяжи), состоящие из главных, хромофобных недеятель-
Глава 26. Триада нейроиммуногормональных систем управления
369
ных клеток. Из последних образуются хромофильные клетки, которые раз-
деляются на ацидофильные и базофильные. Туберальная часть передней доли
гипофиза образуется эпителиальными тяжами, у собак — эпителиальными
трубочками. Передняя доля гипофиза (аденогипофиз) вырабатывает шесть
гормонов (табл. 68).
Таблица 68
Гормоны аденогипофиза человека и животных
Сокращенное название
АКТГ
ТТГ
ФСГ
ЛГ
ГР
пл
Название
Адренокортикотропный
гормон (кортикотропин)
Тиреотропный гормон
(тиреотропин)
Фолликулостимулирующий
гормон
Лютеинизирующий
гормон
Гормон роста
(соматотропный гормон)
Пролактин
Орган-мишень
Кора надпочечников, продукция
кортикоидов и андрогенов
Щитовидная железа, органы
регуляции основного обмена
Гонады, активация созревания
в яичниках
Гонады, овуляция, образование
желтого тела
Все клетки тела, более точная
мишень не известна
Многие клетки тела, в том числе
молочные железы, гонады
АКТГ, ТТГ, ФСГ и ЛГ называют гонадотропными, а ГР и ПЛ — эффек-
торными гормонами (ФСГ и ЛГ — это два гонадотропина).
ФСГ и ЛГ вызывают разрыв фолликула, овуляцию и образование желто-
го тела. Оба этих гормона, получившие свои названия по функциям, кото-
рые выполняют у женщин, имеются и у мужчин. ФСГ играет у мужчин важ-
ную роль в созревании спермы, а ЛГ стимулирует синтез тестостерона в ин-
терстициальных клетках Лейдига.
Клетки аденогипофиза со стороны центральной нервной системы регу-
лируются гипоталамусом. В гипофизе они вызывают либо высвобождение
гипофизарного гормона и тогда называются рилизинг-факторами (РФ) или
рилизинг-гормонами, либо угнетение его секреции и тогда называются ин-
гибирующими факторами или гормонами. Иногда рилизинг-факторы назы-
вают либеринами, а ингибирующие факторы — статинами (табл. 69).
Аминокислотная последовательность предшественника АКТГ включает
39 аминокислотных остатков, составляющих АКТГ, а также аминокислот-
ные последовательности эндогенного опиоидного пептида (3-эндорфина и
а-меланоцит-стимулирующего гормона (сс-МСГ). Генетически запрограмми-
рованные на синтез этого предшественника клетки являются проопиомела-
нокортин-синтезирующими клетками (ПОМК-клетки). В них из одного
предшественника образуются Р-эндорфин, сс-МСГ и АКТГ. Основные клет-
ки передней доли гипофиза это на самом деле ПОМК-клетки, а АКТГ - это
важный продукт секреции этих клеток.
370
Биомоделирование в системах управления
Таблица 69
Гипофизотропные гормоны
Сокращенное
название
Рилизинг- гормоны
ТРГ
ЛГ-РГ (ЛГ/ФСГ-РГ)
КРГ
ГР-РГ
ПРЛ-РГ
Полное название
(описательное)
Тиреотропин-рилизинг-
гормон
Рилизинг-гормон
лютеинизирующего гормона
Кортикотропин-рилизинг-
гормон
Рилизинг-гормон гормона
роста
Рилизинг-гормон пролактина
Ингибирующие гормоны
ГР-ИГ
ПЛ-ИГ(ПИФ)
Ингибирующий гормон
гормона роста
Ингибирующий гормон
пролактина
Другое название
Тиреолиберин
Люлебирин
(гонадолиберин)
Кортиколиберин
Соматолиберин
Пролактолиберин
Соматостатин
Пролактостатин
Действует на
ттг
ЛГиФСГ
АКТГ
ГР
ПРЛ
ГР
ПЛ
Средняя доля гипофиза состоит из многослойного эпителия. Задняя, кро-
ме невроглии, имеет небольшие клетки с отростками. Эти клетки называются
питуицитами, они являются производными эпендимы. В средней доле гипо-
физа выделен интермедии (меланоформный, или пигментный гормон), кото-
рому придают важную роль в регуляции распределения пигмента в организме.
Задняя доля гипофиза продуцирует гормоны: окситоцин, усиливающий
сокращения мускулатуры матки, вазопресин, который суживает сосуды и по-
вышает кровяное давление, и антидиуретический гормон (АДГ). Под влия-
нием АДГ увеличивается реабсорбция воды в канальцах почек и уменьшает-
ся диурез. Недостаток этого гормона вызывает несахарный диабет.
Вокруг сильвиева водопровода находятся мощные образования нервных
пулов, имеющее название центральное серое вещество. В нем и в других
нервных образованиях обнаружены новые функции, в том числе пептид-
ные и белковые гормоны, которые выделяются в кровь, т. е. являются
нейрогормонами. Большие участки ЦНС способны выполнять эндокрин-
ную функцию. Об общности и различиях между гормоном и нейромедиа-
тором мы уже говорили. Терминали и коллатерали аксона нейронов окан-
чиваются на других клетках и высвобождают вещества в виде нейромедиа-
торов, а также нейрогормонов, нейропептидов, которые также следует счи-
тать нейромедиаторами.
Нейрофармакологически установлено, что нейроны, выделяющие тот или
иной нейромедиатор, могут быть подразделены на субпопуляции, высво-
Глава 26. Триада нейроиммуногормоналъных систем управления
371
бождающие разные нейропептиды. Представляется вероятным исследова-
ния этих механизмов в аспекте биомоделирования на разных уровнях.
Приведем некоторые сравнительные данные. У мини-свиней и крупных
собак гипофиз весит 60—70 мг. У кошек гипофиз очень маленький, весит 10—
15 мг. В середине задней доли гипофиза сохраняется продолжение полости
воронки, выстланной эпендимой. Гипофиз у кролика имеет размеры 5x3 мм
и составляет 0,0016% веса тела. Как и у других животных, состоит он из трех
долей. Гипофиз крысы состоит из задней, передней и слабо развитой проме-
жуточной долей. Вес гипофиза у взрослых самок значительно больше, чем у
взрослых самцов. У мышей линии C57BL/6J относительный вес гипофиза
сравнительно высок, половой диморфизм по этому признаку резко выра-
жен: у самок 2 мг сухого веса на 100 г веса тела, у самцов — 1,4 мг/100 г. У
самцов В6 самый высокий относительный вес надпочечников из восьми
испытанных линий. Средний темп суточной секреции L-тироксина (4,19 мг
на 100 г веса тела) значительно выше, чем у BALB/c. Имеют низкий уровень
кортикостерона в плазме крови при сравнении с СВА/Н [577].
Свет во тьме и хроноритмы
Эпифиз, или верхний мозговой придаток (шишковидная железа) возникает
из стенки третьего желудочка и представляет собой маленькое овальной
формы образование. У человека и животных он выполняет множество фун-
кций. У низших животных эпифиз обладает светочувствительностью и опос-
редует влияние света на половые и хроноритмы. Но и у млекопитающих
фотоны могут проникать в железу через кожу, череп и ткани мозга. Нейро-
медиатором в эпифизе служит норадреналин. Клетки эпифиза получают ин-
формацию о ритмичном чередовании света и темноты через риногипотала-
мический тракт.
Синтез и высвобождение мелатонина в эпифизе уменьшаются на свету и
увеличиваются в темноте. Суточные колебания можно выявить при опреде-
лении концентрации мелатонина в крови у людей. У животных функции
мелатонина связаны с размножением. У человека его роль или более скром-
на, или пока не изучена.
Сравнительно недавно стало известно, что эпифиз играет важную роль
не только в регуляции суточных ритмов, но и в механизмах, обеспечиваю-
щих приспособление организма к влиянию неблагоприятных факторов ок-
ружающей среды. Эту функцию эпифиз выполняет также посредством сек-
реции мелатонина, но не исключается участие и других мессенджеров или
гормонов, структура которых пока неизвестна.
Способность шишковидной железы вырабатывать мелатонин существен-
но изменяется на протяжении жизни человека. У новорожденных гормон
образуется в ничтожно малых количествах и в крови практически не опре-
372
Биомоделирование в системах управления
деляется. В дальнейшем функциональная активность железы повышается и
максимального уровня в плазме крови мелатонин достигает в возрасте 3-5
лет. В период полового созревания уровень гормона резко снижается в свя-
зи с уменьшением его удельной секреции на фоне увеличения массы тела.
Менее значительно концентрация мелатонина в плазме уменьшается в зре-
лом и пожилом возрасте.
В последние годы изменились представления о механизмах возрастного
снижения мелатонинобразующей функции эпифиза. Если раньше снижение
связывали с кальцификацией железы и уменьшением количества пинеало-
цитов, то в настоящее время все большее число исследователей указывают
на функциональное угасание с возрастом активности эпифиза. С возрастом
уменьшается и плотность (3-адренорецепторов на поверхности пинеалоци-
тов, снижается образование цАМФ в клетках железы и при адренергической
стимуляции нарушается их взаимодействие с норадреналином.
Меняются и наши представления о возможных путях моделирования и
коррекции функциональных состояний эпифиза. При кальцификации же-
лезы и потере пинеалоцитов ранее восполняли проведением заместительной
терапии мелатонином. Новые подходы к биомоделированию функциональ-
ных изменений активности эпифиза, открывают перспективу для воздей-
ствий, направленных на восстановление способности железы продуцировать
собственный мелатонин. К ним относятся облучение ярким светом в днев-
ное время суток, введение экстрактов эпифиза или их синтетических анало-
гов, например эпиталамина или эпиталона, отказ от приема лекарственных
препаратов, подавляющих выработку мелатонина ((3-адреноблокаторы, не-
стероидные анальгетики, пурины, бензодиазепины, нейролептики), умень-
шение или устранение факторов, действие которых подавляют функциональ-
ную активность пинеальной железы, поиск агонистов мелатонина. Учиты-
вая последние данные о влиянии L-Arg и N0 на продукцию мелатонина,
редукцию кальцификации и снижения числа пинеалоцитов, можно наме-
тить один из путей биомоделирования функций эпифиза.
Микроскопически эпифиз состоит из клеток двух видов — мелких, базо-
фильных с отростками и крупных клеток, слабо воспринимающих окраску.
По мере старения уменьшается количество железистых клеток за счет разра-
стания соединительной ткани и появления известковых отложений (конкре-
менты, мозговой песок). Эпифиз крысы и других мелких грызунов расположен
между большими полушариями головного мозга в виде маленького пузырька.
О железах, регулирующих основной обмен
Тиреоидные гормоны щитовидной железы стимулируют метаболизм во всем
организме а механизмы, лежащие в основе разнообразных эффектов тире-
оидных гормонов, представляют исключительный интерес в плане биомоде-
Глава 26. Триада нейроиммуногормональных систем управления
373
лирования, поскольку они далеки от понимания. Зачастую вместо конкрет-
ных представлений о процессах мы сталкиваемся с догадками разных авто-
ров, которые преподносятся на основании отрывочных фактов, плохо согла-
сующихся друг с другом. Безусловным является то, что тиреоидные и пара-
тиреоидные гормоны являются регуляторами основного обмена организма.
Гормон щитовидной железы трийодтиронин связывается с рецепторами
в ядре клетки и воздействует на геном, вызывая усиление транскрипции и
трансляции, и стимулирует синтез белка во всех клетках тела. Тиреоидные
гормоны влияют на выход из клеток Na+ и поступление в них К+, но в силу
чего это происходит — не понятно. Повышая, активность многих ферментов
тиреоидные гормоны, усиливают интенсивность метаболизма углеводов,
особенно в митохондриях, которые увеличиваются в размерах. Существует
прямая зависимость между концентрацией тиреоидных гормонов в крови и
интенсивностью основного обмена.
У детей и детенышей животных тиреоидные гормоны способствуют нор-
мальному развитию мозга в постнатальный период. Плод получает достаточ-
ное количество тиреоидных гормонов от матери, но после рождения дефи-
цит собственных тиреоидных гормонов становится фатальным для развития
мозга. Так, например, средний темп секреции железы у C57BL составляет
2,44 мг L—тироксина на 10 г веса, что в 2 раза выше, чем у детёнышей не-
которых линий инбредных мышей (см. гл. 9), которые погибают из-за не-
хватки тиреоидных гормонов.
Щитовидная железа у млекопитающих почти всегда двудольная, соеди-
ненная перешейком, причём у одних из них этот признак резко выражен, у
других — слабо. Она расположена у гортани в пределах первых (2—3) колец
трахеи и распадается на правую и левую приблизительно симметричные
боковые доли. Нередко у животных наблюдаются добавочные щитовидные
железы.
У мини-свиней железа тёмно-красного цвета, не разделена на доли и ле-
жит вентрально на трахее у щитовидного хряща, так что перешеек, за редки-
ми вариациями, не выражен. Этот сравнительно плоский орган достигает
4,0—4,5 см в длину, 2,0—2,5 см в ширину и 1,0-1,5 см в толщину, причем от
медианного участка к бокам он утончается и имеет здесь нечто вроде отро-
стков, напоминающих доли. Форма железы сильно изменчива.
Делаются попытки исследовать возможность пересадки некоторых видов
эндокринной ткани (например, щитовидной железы) и высказывается мне-
ние, что лучшая сохранность достигается при их имплантации в большой
сальник, ямку желчного пузыря или капсулу почки свиньи [564, 595].
У овец боковые доли железы вытянуты (3—4 см длиной, 1,25—1,5 см ши-
риной и 0,5-0,75 см толщиной) и располагаются на трахее между пищево-
дом и грудинно-щитовидным мускулом. Перешеек в большинстве случаев
слабо выражен. У баранов щитовидная железа значительно варьирует в вели-
374
Биомоделирование в системах управления
чине, и полная симметрия между правой и левой долями сравнительно ред-
ка. Лежит она на трахее от гортани до 3-го и даже 7-го кольца. Размеры её:
2,5-5,0 см в длину, 1,0—1,5 см в ширину и 0,5 см в толщину. Перешеек с же-
лезистыми частями встречается очень редко, а чаще обнаруживается в виде
слабого соединительнотканного тяжика, но иногда и такового не имеется.
У собак железа имеет вид миндалины. Доли соединены тонким перешей-
ком, но последний нередко отсутствует. В исключительных случаях он мо-
жет быть вытянут в длинную дугу до входа в грудную полость. Ввиду разли-
чия величины собак установить средние размеры органа трудно. Железа у
собак средней величины весит около 2 г. У собак паращитовидные железы
размещены у переднего края щитовидной железы, на ее боковой поверхно-
сти, а также могут находиться под ее капсулой. У крупных собак паращито-
видные железы могут быть до 7 мм в диаметре.
У кошек вес щитовидной железы колеблется в пределах 0,5—2,8 г. По форме
и расположению она во многом соответствует щитовидной железе собак.
Паращитовидные железы относительно большие и размещены так же, как у
собак.
У кролика щитовидная железа весом в 2 кг составляет в среднем 0,2 г.
Состоит из двух долей и перешейка. У самок железа больших размеров, чем
у самцов. Околощитовидные железы представлены у кролика в виде двух пар.
Передняя пара находится в паренхиме щитовидной железы, а задняя распо-
ложена самостоятельно у каудальной части щитовидной железы. В зобной
железе имеется ткань добавочных эпителиальных телец, поэтому у кроликов
при удалении обеих пар околощитовидных желез отсутствует тетания.
Добавочные щитовидные железы встречаются и у других животных, ко-
торые по положению, величине, числу и частоте обнаружения значительно
варьируют и рельефнее выступают лишь после оперативного удаления щи-
товидной железы. В таких случаях они увеличиваются, компенсируя отсут-
ствие главной железы. Чаще добавочные железки располагаются неподалеку
от щитовидной железы, но могут встречаться на всем протяжении трахеи,
особенно у овец и собак.
Щитовидная железа у морских свинок небольшая, без перешейка. Вес ее
0,06-0,1 г.
Щитовидная железа у крыс парная и расположена у основания трахеи на
ее латеральной поверхности. Обе доли уплощенной формы и верхней час-
тью соединены между собой еле заметным перешейком (на уровне 2-3-го
трахеальных колец). Вес щитовидной железы 13—60 мг, у крысы весом
200 г - 23-28 мг.
У крысы имеются две паращитовидных железы. Они находятся на передне-
боковой поверхности правой и левой долей щитовидной железы, в дорсаль-
ной их части, и выделяются в виде беловатого пятнышка округлой формы. У
отдельных животных встречаются и дополнительные паращитовидные желе-
Глава 26. Триада нейроиммуногормональных систем управления
375
зы. Крысы довольно устойчивы к недостатку гормона паращитовидной желе-
зы и погибают от судорог лишь на четвертый-пятый день после операции.
Говоря о потребности йода для организма различных животных, ряд ис-
следователей предлагают пересчитывать его в отношении к человеку, для
которого суточная потребность составляет около 150 мкг. Исходя из массы
тела животных, можно пересчитать потребность, хотя, учитывая различия в
метаболизме (см. гл. 35), это заявление кажется излишне смелым.
Надпочечники в регуляторных процессах
Напомним еще раз, что латинское слово glans означает желудь. По ассоци-
ации с плодами дуба гландами называли небольшие плотные комочки ткани
в организме человека. Такие комочки находили, к примеру, над почками, их
называли супраренальными (буквально надпочечниковыми) или адреналовы-
ми. По-латыни renalis — почечный, a ad означает при. Адреналовые гланды со
временем стали переводить как надпочечниковые железы или просто надпо-
чечники.
В 1895 г. биохимики Джордж Оливер и Эдуард Л. Шарпи-Шэфер обна-
ружили, что в надпочечниках содержится какое-то вещество, которое вызы-
вает сокращение стенок артерий, что приводит к повышению кровяного
давления. Выделить это вещество удалось в 1901 г. японскому ученому Иокихе
Такамине. Это был первый случай получения вещества, которое железа вы-
деляет в небольших количествах в кровь, вызывая определенную реакцию
какого-то органа. В 1902 г. английские физиологи Уильям Бейлис и Э. Стар-
линг предложили называть такие вещества, выделяемые железами внутрен-
ней секреции, гормонами — от греческого слова hormao, что значит «приво-
жу в движение, побуждаю». Первый гормон назвали адреналином: от латин-
ских ad— при, над и renalis — почечный. Его греческий эквивалент epinephrine:
по-гречески epi — над, сверху, a nephros — почка.
Надпочечники, закладываясь из целотелия, сливаются в сплошной пар-
ный или даже непарный орган. У млекопитающих и те и другие нераздельно
слиты в два (правый и левый) компактных органа, которые и называются
надпочечными железами. Они лежат в брюшной полости рядом с почками
и связаны с последними жировой капсулой и сосудами. На разрезе этих
желез ясно выделяются три слоя: корковый (интерреналовая ткань или клу-
бочковая зона), мозговой или пучковый, состоящий из хромаффинной тка-
ни, и внутренний или сетчатая зона. Эти зоны вырабатывают разные гормо-
ны. В сетчатой зоне образуются в основном андрогены, а также глюкокор-
тикоиды, в пучковой зоне — глюкокортикоиды, а в клубочковой — минера-
локортикоиды.
Минералокортикоиды синтезируются в клубочковой зоне надпочечни-
ков. Альдостерон, основной гормон, участвует в регуляции обмена эритро-
376
Биомоделирование в системах управления
цитов и водного баланса, повышая путем осмоса реабсорбцию натрия и воды
в почечных канальцах, но способствует выделению калия и протонов. Транс-
порт ионов и воды в кишечнике, а также в слюнных и потовых железах
также регулируется альдостероном.
У человека и животных в сетчатой зоне коры надпочечников вырабаты-
вается большое количество андрогенов. У мужчин и у самцов животных
андрогенов большую часть секретируют все же семенники и только лишь
менее трети — надпочечники. Высокие концентрации АКТГ стимулируют
андрогенез. Наибольшей активностью среди андрогенов обладает тестосте-
рон [657], однако в коре надпочечников он образуется в незначительном
количестве. С19—стероиды уступают по активности тестостерону. У женщин
и у самок животных дегидроэпиандростерон выделяется надпочечниками,
но его секретируют и яичники при нарушении функций этой железы.
Глюкокортикоиды стимулируют глюконеогенез в печени, а под влияни-
ем кортизола аминокислоты метаболизируются с образованием глюкозы в
крови. Пополнение аминокислот в организме происходит в результате рас-
щепления мышечных белков, а значит, глюкокортикоиды оказывают ката-
болическое действие на белки. Под действием глюкокортикоидов синтез
мышечных белков сильно снижается, поскольку глюкокортикоиды угнетают
транспорт аминокислот в мышечные клетки. Кортизол подавляет использо-
вание глюкозы клетками тела и противодействует эффекту инсулина, вызы-
вая накопление глюкозы в крови и тканях. Кортизол обладает антианаболи-
ческим действием, усиливает поглощение аминокислот, которые использу-
ются клетками печени для глюкознеогенеза.
Глюкокортикоиды участвуют в расщеплении триглицеридов. В крови
повышается, а в жировых клетках уменьшается соответственно жирных кис-
лот и триглицеридов
Мозговой слой надпочечников вырабатывает адреналин и норадреналин.
Адреналин участвует в углеводном обмене, регуляции функций ЦНС и ис-
полнительных органов через вегетативную нервную систему. Мозговой слой
надпочечников содержит клетки, которые в процессе эмбриогенеза мигри-
ровали из нервных складок. Несмотря на свое «дворянское» происхождение,
эти клетки дифференцируются не в нейроны, а в клетки, лишенные дендри-
тов и аксонов. Хотя они вырабатывают норадреналин или адреналин, но
иннервируются холинергическими волокнами. Такое вот хитросплетение.
Вернемся к математическому моделированию регуляции углеводного об-
мена теперь уже с участием гормонов и медиаторов, вырабатываемых надпо-
чечниками. Будем исходить из посылки, что понижение уровня сахара кро-
ви относительно нормы способствует выработке корой надпочечников адре-
налина, поэтому будем считать, что скорость выработки адреналина пропор-
циональна разности у — уп и тем меньше, чем большего значения достигло
текущее содержание адреналина:
Глава 26. Триада нейроиммуногормональных систем управления
377
— + с2х = а6(у-уп) (85)
где а6, с2 — коэффициенты пропорциональности при у < уп.
Это математическое описание достаточно корректно описывает систему
нейроэндокринной регуляции по принципу отрицательной обратной связи
и наглядно демонстрирует один из фундаментальных принципов регулятор-
ных процессов в живых системах. Подобный подход может быть приложим
и к другим метаболическим процессам в организме человека и животных.
Универсальность математического аппарата позволяет осуществлять наибо-
лее оптимальное биомоделирование, чем простое экспериментальное иссле-
дование.
Клетки мозгового слоя надпочечников вырабатывают также пептиды,
выполняющие регуляторную функцию в ЦНС, желудочно-кишечном тракте
и многих других органах и тканях. К настоящему времени идентифицирова-
ны вещество Р, интестинальный вазоактивный полипептид, соматостатин,
(3-энкефалин, холецистокинин и др.
У мелких жвачных надпочечные железы по форме и величине уподобля-
ются большой фасоли. У мини-свиней они продолговаты и на поверхности
изрезаны бороздами. У собак надпочечные железы также имеют вытянутую
овальную форму. Надпочечники у кошек бобовидной или шаровидной фор-
мы, расположены в передней части околопочечной жировой ткани на зна-
чительном расстоянии от почек.
Обособленные части надпочечных органов в качестве маленьких доба-
вочных желёзок встречаются и в других местах тела. Они состоят из межпо-
чечной или околопочечной ткани или, наконец, из той и другой вместе:
/ желёзки из межпочечной ткани можно иногда встретить около основных
надпочечных желёз, под капсулой почек и на прилежащих сосудах, а так-
же на сосудах и нервах, расположенных вне брюшинной выстилки (экст-
раперитонеально);
/ желёзки из межпочечной и околопочечной ткани находят вокруг основ-
ного органа, в солнечном сплетении, в семенном канатике;
/ желёзки из хромаффинной (околопочечной) ткани или параганглии более
распространены. Их можно встретить около основной железы и по пути
следования шейной и грудной частей симпатического нерва, в окружнос-
ти корня крупных брюшных сосудов, в широкой маточной связке. Боль-
шой параганглий находится впереди места деления сонной артерии и на-
зывается paraganglion intercaroticum s. glomus caroticum.
Надпочечники у кроликов располагаются асимметрично, правый нахо-
дится на уровне X грудного позвонка у внутреннего края правой почки, а
левый — на уровне II поясничного позвонка. Вес одного надпочечника со-
ставляет 0,021—0,026% веса тела. На разрезе хорошо выражен корковый слой
378
Биомоделирование в системах управления
(интерреналовая ткань) и мозговой слой (хромаффинная ткань). Для кроли-
ков характерно, что интерреналовая ткань обнаруживается и в других учас-
тках организма (в придатке семенников, в придатках яичников, по ходу полых
вен), чем можно объяснить выносливость кроликов к экстирпации надпо-
чечников. Надпочечники прилегают кпереди и медиально к почкам глини-
сто-желтото цвета, довольно объемистым, весом 0,1—0,2 г.
Надпочечники крыс имеют желтоватый цвет и расположены кпереди и
медиальнее почек. Вес надпочечников 13—38 мг, причем у взрослых самок
надпочечники более тяжелые, чем у самцов. Абсолютный вес надпочечни-
ков в стоке КМ = 23,4 мг. Встречаются и добавочные надпочечники.
В главах 30—32 будут рассмотрены регуляторные функции других гормо-
нальных систем и приведены некоторые данные их биомоделирования.
Новые связи триады
Существует достаточно большое и убедительное число работ, посвященных
поиску связей и взаимовлияний между нервной, иммунной и эндокринной
системами.
Открытие значения оксида азота в регуляторных процессах вызвало взрыв-
ной интерес во всех областях биологии и медицины, заставило пересмотреть
взгляды на многие вопросы развития патологии у человека, а значит осуще-
ствить ревизию подходов к биомоделированию. Попытаемся проанализиро-
вать в этом плане принципы взаимоотношений внутри триады в новых по-
нятиях и терминах.
Оксид азота вырабатывается эндотелиальными клетками сосудов, макро-
фагами и нейтрофилами. Молекула N0 может существовать в разных элек-
тронных состояниях:
/ относительно стабильный N0° нейтрально заряженный радикал;
/ присоединение электрона переводит его в анион N0 (нитроксил);
/ потеря электрона трансформирует его в ион NO+ (нитрозоний).
Эти формы N0 имеют клеточные мишени. Главная мишень NO — раство-
римая гуанилатциклаза, содержащая 4 гема, которые являются рецепторами
для NO. NO- имеет лишний электрон, что повышает его химическую актив-
ность (в три раза выше, чем у кислорода). Связывание NO с группой гема
индуцирует растворимую гуанилатциклазу. Это сопровождается усилением син-
теза циклической ГМФ. В клетках гладкой мускулатуры цГМФ снижает концен-
трацию внутриклеточного кальция, что приводит к расслаблению клетки и
вазодилатации. Кроме того, цГМФ в крови совместно с предсердным натрий-
уретическим гормоном, вырабатываемым в эндокринных клетках предсердия,
взаимодействует со специфическими рецепторами почечного эпителия, глад-
кой мускулатуры артерий, коры надпочечников и гипофиза, легких, печени,
тонкого кишечника, вызывая разнообразные биологические эффекты.
Глава 26. Триада нейроиммуногормональных систем управления
379
Точкой приложения N0 является матричная РНК. Особый интерес пред-
ставляет способность N0 экспрессировать ряд важнейших белков и фермен-
тов, причем как на уровне транскрипции, так и трансляции (стресс-белков,
ферритина, белков антиоксидантной защиты, белков-рецепторов трансфер-
ритина, ядерного белка с53, ответственного за блокаду злокачественных но-
вообразований и других белков). В разных электронных состояниях молеку-
ла N0 может активировать или подавлять активность таких ферментов и
белков как гуанилатциклазы, РНК-редуктазы, компонентов дыхательной
цепи и гликолиза, ядерных факторов транскрипции, белков ионных каналов
и многих других.
Роль оксида азота в функционировании различных систем организма
можно представить следующим образом:
/ в ЦНС и периферической НС наблюдаются нейромодулирующая активность,
определяющая долговременное потенцирование, формирование памяти,
восприятие боли, и активация зрительного анализа;
/ в иммунной системе развиваются антипатогенные реакции, неспецифичес-
кая цитотоксичность, противоопухолевая защита, токсемии, отторжение
трансплантата, изменения регуляции взаимодействия лейкоцитов со стен-
ками сосудов, активности тромбоцитов;
/ в эндокринной системе оксид азота обеспечивает регуляцию синтеза и секре-
ции гормонов: инсулина, пролактина, тиреоидного гормона, паратиреоид-
ного гормона, гормонов надпочечников, гормонов репродуктивного цикла.
Помимо этого N0 обеспечивает релаксацию кровеносных сосудов
мозга, сетчатки глаза, сердца, легких, почек, кишечника, кавернозной тка-
ни, мышцы сердца и расслабление гладкомышечной ткани трахеи, желудка,
кишечника, мочевого пузыря, матки.
NO стимулирует синтез цитопротекторных простагландинов групп Е и 12
за счет активации циклогеназ. Гиперпродукция N0 в иммунокомпетентных
клетках — макрофагах и нейтрофилах — может иметь положительное значе-
ние для организма. Именно в них N0 действует как один из основных эффек-
торов системы клеточного иммунитета, уничтожающих патогенные микро-
бы и злокачественные клетки. При стрессовых язвенных поражениях желуд-
ка уменьшение продукции N0 является причиной адренергических и ише-
мических повреждений слизистой оболочки.
Гиперпродукция N0 путем активации NO-синтазы, ключевого фермен-
та, приводящего к гиперпродукции N0, наблюдается при увеличении концен-
траций свободных жирных кислот и внутриклеточного Са2+, а также появле-
нии свободных радикалов. При этом активируются конститутивная и инду-
цибельная NO-синтазы, что может привести к гипотензии, недостаточности
кровообращения и кардиогенному шоку.
Следует подчеркнуть, что высокие концентрации N0 не реализуют свой
эффект через цГМФ. Напротив, они оказывают прямое цитотоксическое и
380
Биомоделирование в системах управления
иммуногенное действие, связываясь супероксидным радикалом, и образуют
пероксинитрит, который индуцирует повреждения ДНК и мутации, ингиби-
рует функцию ферментов. Таким образом, в высоких концентрациях N0
принадлежит исключительно важная роль в течении и исходе критических
состояний, что делает его фактором эндогенной интоксикации.
Многие исследователи считают, что цитотоксическое и цитостатическое
действие N0 обусловлено его способностью продуцировать пероксинитрит
в реакции с супероксидным радикалом. Пероксинитрит сравнительно стаби-
лен при физиологических значениях рН, но при патологических состояниях
в течение секунд распадается, оказывая окислительное действие на различ-
ные внутриклеточные мишени. Оно и определяет цитотоксическое действие,
вызывающее гибель клеток и тканей по механизмам апоптоза и некроза.
N0 подавляет действие таких вазоконстрикторов, как тромбоксан А2
и серотонин, выделяемых из тромбоцитов. Поскольку NO является мощ-
ным эндогенным вазодилататором и играет важную роль в предупрежде-
нии избыточной вазоконстрикции, недостаточный синтез этого соедине-
ния может явиться индуцирующим фактором в патогенезе гипертензии
и атеросклероза.
Снижение продукции или ускоренный распад N0 приводят к развитию
тяжелых сердечно-сосудистых заболеваний, связанных с нарушением функ-
ции эндотелия, патологическим повышением сосудистого тонуса и арте-
риального давления. К их числу относятся артериальная гипертензия, стено-
кардия, атеросклероз, диабетическая ангиопатия и др.
При диабете, когда накапливающиеся продукты гликозилирования
усиленно разрушают N0, дефект вазодилатации можно снизить ингибиро-
ванием гликозилирования аминогуанидином. Коррекция эссенциальной
и вторичной гипертензии, гиперхолестеринемии и посттрансплантационных
состояний может быть достигнута L-аргинином.
Мы ещё вернемся к свойствам и эффектам N0 в следующих главах. Здесь
же, вместо резюме, можно сказать, что NO-система, сопряженная с L-Arg,
в свою очередь, являющимся предшественником орнитина, цитрулина, глю-
тамата, глутамина, глутатиона, ГАМК, спермина, спермидина, агматина,
пролина, путресцина и многих других регуляторных веществ, играет исклю-
чительно важную роль в процессах саморегуляции в организме. Помимо того,
что это новые знания о жизнедеятельности и развитии патологии, дающие
важные направления биомоделированию, это принципиально иной взгляд
на интегративные механизмы триады нервной, иммунной и гормональной
систем управления.
Раздел VII БИОМОДЕЛИРОВАНИЕ
В РЕГУЛЯТОРНЫХ
СИСТЕМАХ
Телефон для Арманда — как гидрант для собаки. Он не
может пройти мимо, не воспользовавшись им.
Разница только в том, что вместо лапы он поднимает
трубку.
Миссис Арманд Хаммер о своем муже
383
Глава 2 7 ДЫХАТЕЛЬНАЯ СИСТЕМА
В этой главе мы рассмотрим сравнительные данные
дыхательной системы человека и животных, не ка-
саясь многогранных функций и физиологических
особенностей органов дыхания. Мы рекомендуем
читателю обратиться к специальной литературе и
последним исследованиям, которые являются прин-
ципиальными в аспекте построения биомоделей и
экстраполяции экспериментов на животных в от-
ношении человека [11, 37, 53, 357, 558, 681].
Сравнительная характеристика органов
дыхания
Носовая полость человека и животных делится но-
совой перегородкой на две половины. От носовой
перегородки в правую и левую половины носовой
полости отходят по две завитые раковины, образуя
при этом узкие проходы. Слизистая оболочка, по-
крывающая носовую полость, богата кровеносны-
ми сосудами и имеет сложные складки, чем спо-
собствует очищению поступающего воздуха от пыли,
а также увлажняет и обогревает его. Внутри гортани
(larynx) имеются связки, участвующие в формиро-
вании голоса.
Трахея овец и баранов состоит из 48—50 колец.
В грудной полости до бифуркации трахея отдаёт в
верхушку правого лёгкого особый эпартериальный
бронх.
384
Биомоделирование в регуляторных системах
Трахея мини-свиней цилиндрическая и состоит из 32—36 колец. Концы
колец на дорзальной стороне или заходят друг за друга, или же едва касают-
ся. У мини-свиней, как и у жвачных, трахея отдает для верхушечной доли
правого лёгкого эпартериальный бронх.
Трахея собак содержит более 40 колец (42—46) и формой напоминает ци-
линдр, лишь слегка уплощённый в дорзо-вентральном направлении. Дор-
зальные тонкие и гибкие концы колец трахеи не сходятся друг с другом, а
скрепляются соединительнотканной мембраной. Поперечный мускул трахеи
лежит снаружи, дорзально на кольцах трахеи. Слизистая оболочка довольно
плотно примыкает к мембране с внутренней стороны.
У кошек трахея состоит из 38—43 хрящевых колец. В гортани кошек нет
клиновидных хрящей, слизистая оболочка гортани не образует карманов,
позади надгортанника имеются боковые выросты.
У морских свинок трахея около 3,5 см в длину, диаметром 3—5 мм. Хряще-
вые кольца сзади не замкнуты.
У крысы трахея состоит из 30 хрящевых полуколец и выстлана двухслой-
ным эпителием.
Лёгкие мини-свиней ясно дольчатые: на правом лёгком выделяется верху-
шечная доля, отграниченная широкой вырезкой от сердечной доли, которая
узкой и глубокой щелью отъединена от большой диафрагмальной доли. До-
бавочная доля имеется. Левое лёгкое состоит из долей того же числа и наи-
менования, за исключением, разумеется, добавочной доли. На поверхности
лёгкого заметен ячеистый рисунок, отражающий характер структуры органа
из мелких долек, разделённых прослойками соединительной ткани; однако,
рисунок не так рельефен, как у рогатого скота.
Лёгкое жвачных ясно разделено на доли значительными надрезами. В
левом лёгком насчитывают три доли: верхушечную, сердечную и диафраг-
мальную. В правом лёгком, помимо указанных трёх долей, существует лежа-
щая медиально-добавочная доля. Весьма характерно для жвачных, что бронх
для правой верхушечной доли отделяется от трахеи до её бифуркации; сама
верхушечная доля подразделяется широкой вырезкой на две лопасти, в ре-
зультате чего в правом легком оказывается пять долей.
Дольчатость лёгких у собак выступает особенно ясно, так как междоле-
вые вырезки (за исключением передней левой) непосредственно достигают
основных бронхов. Левое лёгкое имеет три обычные доли: верхушечную,
сердечную и диафрагмальную, причём сердечная вырезка в промежутке между
третьим и седьмым межрёберными пространствами оставляет вентральный
участок сердца (сердечной сорочки) неприкрытым. В правом лёгком насчи-
тывают четыре доли благодаря наличию добавочной доли. Верхушечная доля
этого лёгкого нередко раздвоена.
Общая площадь альвеол у собак составляет 100 м2. Весят легкие прибли-
зительно 1/60—1/90 часть тела собаки и характеризуются хорошо выражен-
Глава 27. Дыхательная система
385
ной дольчатостью. Левое легкое имеет три доли (верхушечную, сердечную и
диафрагмальную), а правое — четыре (верхушечную, которая может быть
раздвоена, сердечную, диафрагмальную и дорсальную добавочную). Жизнен-
ная емкость легких собаки весом около 10 кг составляет 500—550 мл, вели-
чина выдоха 40—60 мл.
Суженные куполы плеврального мешка у собаки углубляются в шейную
область за пределы I ребра. Правая и левая плевральные полости сообщают-
ся между собой, имея в нижнем участке средостения отверстие. Это под-
тверждается не всеми авторами, особенно в отношении щенков. Но осторож-
ность при пункции плевры следует соблюдать.
Легкие кошек составляют всего лишь 0,62% веса тела, причем на правое
легкое приходится 41,5% (7,9 г), а на левое — 58,5% (11,1 г). Частота дыхания
20—30 в минуту. Минутный объем легких равен 1000 см3. Верхушечный ку-
пол плеврального мешка, как и у собак, справа и слева вступает в шейную
область за передний край первого ребра. Особенностью строения органов
грудной полости кошек является то, что у взрослых животных правая и ле-
вая плевральные полости сообщаются между собой в области заднего средо-
стения. Поэтому при вскрытии одной из полостей наступает двусторонний
пневмоторакс, как и у собак.
У кролика легкие и дыхательная часть носовой полости по сравнению с
другими животными развиты недостаточно. Вес дыхательного аппарата по
отношению к весу тела составляет около 1,28%, причем вес лёгких 0,33—
0,38% веса тела. Частота дыхания от 50 до 100 в минуту. Выдох на 1/3 длин-
нее вдоха, дыхательный коэффициент при 20° С равен 0,83. Правое легкое
разделено на четыре доли (верхушечную, сердечную, диафрагмальную и до-
бавочную), левое — на три (редуцированную верхушечную, сердечную и ди-
афрагмальную). Правое легкое тяжелее левого и весит 12 г, а левое — 11 г.
Плевральные полости правого и левого легкого у кроликов между собой не
сообщаются.
У морских свинок правое легкое разделено на четыре доли: верхушечную,
сердечную, диафрагмальную и добавочную; левое на три: верхушечную, сер-
дечную и добавочную. Вес обоих легких у морских свинок равен около 3,8 г.
Правое легкое весит на 10—15% больше левого. Частота дыхательных движе-
ний 80—130 в минуту. Гладкая мускулатура дыхательных путей весьма чув-
ствительна к воздействию гистамина и претерпевает выраженные изменения
при анафилактическом шоке. Реактивность лимфатической системы легких
морской свинки настолько высока, что даже при незначительных механи-
ческих и бактериальных раздражениях быстро возникает лимфаденит, ост-
рые аллергические реакции, отек Квинке, развивается туберкулёз (табл. 70).
У крыс левое легкое состоит из одной, а правое из четырех долей: верху-
шечной, сердечной, диафрагмальной и добавочной. У молодых белых крыс
весом 130—150 г правое легкое весит 0,45 г, а левое — 0,4 г. У взрослых крыс
386
Биомоделирование в регуляторных системах
(200-250 г) правое легкое весит 1,05 г, а левое — 0,8 г. Частота дыхания 110—
150 в минуту.
Левое легкое у хомячка состоит из одной доли, а правое имеет верхушеч-
ную, сердечную и диафрагмальную и добавочную доли.
Аналогично строение лёгких и у мышей. Интересно, что при прочих рав-
ных условиях потребление кислорода у разных линий одного вида может
разниться. У линий C57BL/6J самки В6 потребляют несколько меньше кис-
лорода, чем самцы; так, например, в возрасте 35—42 дня при температуре
29° С самки потребляют 1,41 мг/г кислорода в час, а самцы — 1,47 мг/г (у мы-
шей DBA/2 наблюдается обратная картина) [63]. Мыши — удобная биомо-
дель для изучения туберкулеза (табл. 70)
Регистрация дыхания
В сравнительных исследованиях дыхательной функции исследователи стара-
ются избегать хирургического прокола грудной стенки, поэтому давление
можно измерить с помощью небольшого баллончика с тензодатчиками. Бал-
лончик фиксируется в пищеводе, в месте прилегания к задней стенке груд-
ной полости. Величина давления колеблется в диапазоне от 0 до 20 см. во-
дяного столба. Необходимо надежная цифровая фильтрация низкочастот-
ных сердечных помех (1—10 Гц), дыхательных и мышечных наводок и шумов
свыше 10 Гц. Дыхание регистрируют также на кимографе при помощи нало-
жения на грудную клетку датчиков пневмографа, введения и фиксации ка-
нюли в трахее, а также введением канюли в один из носовых ходов, в кото-
ром предварительно производят анестезию слизистой.
Дыхательные колебания у мини-свиней, овец и собак регистрируют специ-
альным пневмографом или полиграфом. Датчики закрепляются на грудной
клетке животного. В остром опыте колебания выдыхаемого воздуха можно
регистрировать при помощи канюли, вставленной в одну из ноздрей. Часто-
та дыхания у здоровых собак в покое составляет 10—30 периодов в минуту.
Дыхание у кошек регистрируется при помощи полиграфа или пневмогра-
фа, который привязывают к грудной клетке, а также путем введения в тра-
хею специальной канюли с двумя отводами или введением канюли в один из
носовых ходов (слизистую носового хода перед введением канюли анестези-
руют раствором дикаина или кокаина).
Полиграф позволяет очень точно регистрировать глубину дыхания, дли-
тельность и характер вдоха, выдоха и паузы между ними. Частота дыхания у
здоровых кошек 20—30 периодов в минуту.
У морской свинки вставляют в трахею стеклянную канюлю с тройником,
наружный конец которого соединяют с капсулой Марея, и записывают ды-
хание полиграфом. Канюлю можно вставлять, в один из носовых ходов у
ненаркотизированного животного, но слизистую следует анестезировать раст-
Глава 27. Дыхательная система
387
вором кокаина или дикаина. Регистрация дыхания возможна также пневмо-
графом. Частота дыхания у морских свинок 80—130 периодов в минуту.
В острых опытах на крысах дыхание регистрируют из трахеи, куда встав-
ляют канюлю. В хронических опытах удобнее всего пользоваться полигра-
фом, что позволяет точно учитывать глубину дыхания, длительность вдоха и
выдоха, длительность паузы между ними не только у взрослых, но и у ново-
рожденных. Регистрировать дыхание у крыс (и мышей) можно также при
помощи широкой полоски марли, одетой на грудную клетку в виде петли и
соединенной с рекодером. Частота дыхания у здоровых крыс 100—150 пери-
одов в минуту.
В хронических опытах на мышах лучше пользоваться полиграфом. Часто-
та дыхания у здоровых мышей от 140 до 210 периодов в минуту.
Математическое моделирование дыхательных функций
В качестве одной из простейших модельных математических систем, исполь-
зуемых в исследованиях на человеке и животных, а также для экстраполя-
ции, могут быть взяты уравнения первого порядка. Методы измерения и
регистрации дыхания приведены в этой главе выше.
Полное давление Р{, развиваемое в дыхательной системе человека и жи-
вотных, должно преодолеть упругое сопротивление легких и вязкое сопро-
тивление потоку воздуха. Давление Ре/,требуемое для преодоления упругого
сопротивления, равно эффективной жесткости легких Е, умноженной на
обьем легких V:
Pei = EV. (86)
Поскольку абсолютно достоверные методы определения объема легких как
в покое V0, так и в заполненном воздухом состоянии неизвестны и в экспе-
рименте трудно определимы, принимается упрощенная формула (86). В этом
случае упругие свойства легких выражают через их растяжимость, т. е. величи-
ну, обратную модулю объемной упругости и выражаемой в см3/см. водяного ст.
Растяжимость С вычисляется по следующей формуле:
r-Vi~Ve
С"7~р-' (87)
где Vj — объем в конце вдоха, Ve — объем в конце выдоха, Р. — давление в
конце вдоха, а?е- давление в конце вдоха.
Вязкое сопротивление будем считать постоянным, а требуемые для его
преодоления давление Рг равным сопротивлению, умноженному на поток Я:
р dv d
Pr=-jfR> (88)
388
Биомоделирование в регуляторных системах
откуда
Wf- <89)
Теперь можно объединить упругость и вязкость и получить расчетное
выражение для модели в целом:
п п п У ndV Р-Ре ndV
P,=Pel+P= — + R— = -* - + R—. (Qf)\
' el r С dt Vt-Ve dt W
Поскольку все физиологические процессы являются динамическими,
следовательно, объем также является функцией времени. Поэтому с помо-
щью определенного интеграла можно вычислить изменения объема легких
VT во времени:
Vx = )(dv/dt)dt, (91)
о
а следовательно, получить величину Ре1, и, зная величину Р0, получить Рг.Через
известный дифференциал (dV/dt)Moymo определить значение R и, зная эти
параметры и величину мощности, затрачиваемой в каждый момент времени,
равную произведению давления на поток, мы вычислим величину работы W,
затрачиваемой на дыхание:
(92)
где fj — момент начала вдоха, t2 — момент окончания выдоха.
Путем подобных несложных вычислений можно довольно полно описать
соотношение дыхательных функций у человека и различных животных и
установить наиболее оптимальные экстраполяционные коэффициенты. Это
позволит получить соотносительные модели растяжимости легких, вязкого
сопротивления и в целом работы дыхательного аппарата. В свою очередь эти
математические модели позволят наиболее полно оценить действие различ-
ных лекарственных препаратов, ксенобиотиков и факторов окружающей сре-
ды на периферические и исполнительные звенья дыхательной системы че-
ловека и животных.
Биомодели для изучения бронхо-легочных процессов
Физиология дыхания является с количественной точки зрения, наиболее
изученной областью. Стремительное внедрение компьютерных технологий
еще более обогащает этот процесс. Однако существующий разрыв между эк-
спериментальными, генетически детерминированными и математическими
моделями не заполняется. Нет единства и в системе измерений. Например,
'2
h
(n-4>i£
Глава 27. Дыхательная система
389
если вместо измерения потока воздуха применяются плетизмограмма или
УЗИ, некоторые их преимущества сводятся на нет неудобствами пересчета и
верификации.
Да и в случае приведенных нами в табл. 70 и 71 биомоделей требуется
творческий подход и обоснованный отбор оптимального подхода. Иными
словами, во всех случаях должна действовать важнейшая формула достиже-
ния успеха (У): У = БМ + Ж + И, где, БМ — оптимальная биомодель, Ж —
качественное животное, И — интеллект исследователя.
Таблица 70
Экспериментальные и генетически обусловленные биомодели
для изучения противотуберкулёзного действия веществ *
Исследуемое действие
вещества
Модель туберкулёзной
инфекции:
a. заражение 0,1 мг культу-
ры в 0,5 мл физиологичес-
кого раствора для «острого»
опыта
или 0,025 мг в 0,5 мл, под-
кожно в правую паховую
область)
b. заражение 0,5 мг культу-
ры в 0,5 мл физиологичес-
кого раствора, в вену хво-
ста или 0,1 мг в 0,2 мл
раствора, в венозный си-
нус глаза
Высокочувствительные
биомодели воспроизведе-
ния туберкулёза при зара-
жении микобактериями
Лабораторное
животное
Морские
свинки
Мыши
Мыши
Мыши
Морские
свинки
Морские
свинки
Линия или порода
животного
Аутбредные
Линии BALB/c
или СВА
l/stY
l/st
STR2
STR13 (наиболее
чувствительны)
Пол
животного
9с?
9с?
9<3
9с?
Вес
животного
250-300 г
18-25 г
30-40 г
30-40 г
780-800 г
750-810 г
Возраст
животного
1-2 мес.
2-3 мес.
3-10 мес.
3-10 мес.
5-12 мес.
5-12 мес.
* См. пояснения к табл. 6 (глава 2).
Таблица 71
Линейные животные и экспериментальные биомодели для изучения бронхиальной
астмы и других заболеваний дыхательных путей и действия веществ,
применяемых при их лечении
Исследуемое действие
вещества
Модель для первичной
оценки бронхолитической
активности ХС на экспери-
ментальных моделях in vitro
Модель гистамин-и ацетил-
холининдуцированного
бронхоспазма (гистамин
5-10 мкг/кг или ацетилхо-
лин 40 мкг/кг, внутривенно)
Лабораторное
животное
Морские
свинки
Морские
свинки
Линия или порода
животного
Аутбредные
Аутбредные
Пол
животного
9с?
9с?
Вес
животного
500-600 г
350-400 г
Возраст
животного
4-5 мес.
1-3 мес.
390 Биомоделирование в регуляторных системах
Окончание табл. 71
Исследуемое действие
вещества
Модель бронхоспазма у не-
наркотизированной мор-
ской свинки с аэрозольным
воздействием бронхоконст-
риктора (1%-ный раствор
гистамина или 0,5%-ный
раствор ацетилхолина)
Модель анафилактического
бронхоспазма при аэро-
зольном воздействии анти-
гена
Модель лёгочной анафилак-
сии с увеличением сосуди-
стой проницаемости дыха-
тельных путей
Модель антигениндуциро-
ванного бронхоспазма у
активно сенсибилизирован-
ных животных (сенсибили-
зация 0,5 мл раствора, со-
держащего ОА в дозе
20 мкг и гидрокись алюми-
ния в количестве 100 мг на
свинку по методу Andersson,
внутримышечно)
Модель гиперчувствитель-
ности замедленного типа
к эритроцитам барана при
сенсибилизации введением
2-107 суспензии эритроци-
тов барана в 0,2 мл фос-
фатного буфера, подкожно
Модель бронхоальвеолита
на основе ингаляционного
введения сефадекса А-25
Модель индуцированного
отёка лапы при введении
ПАФ (полный адъювант
Фрейнда) - (0,1 мл ПАФ,
субплантарно в правую
лапу)
Модель кашля, индуциро-
ванного аэрозольным вве-
дением лимонной кислоты
Модель для оценки муко-
литической активности ис-
следуемых в-в (вводят
внутрибрюшинно или
внутрижелудочно в объёме
1 мл (доза 1,2 мг/кг)
Лабораторное
животное
Морские
свинки
Морские
свинки
Морские
свинки
Морские
свинки
Мыши
Крысы
Крысы
Морские
свинки
Морские
свинки
Линия или порода
животного
Аутбредные
Аутбредные
Аутбредные
Аутбредные
Линии BALB/c
или
(CBAxC57B1)F1
Линии Wistar
Аутбредные
Аутбредные
Аутбредные
Пол
животного
9с?
9с?
9с?
9с?
с?
в
с?
9с?
9с?
Вес
животного
500-600 г
300 г
300-400 г
350-400 г
30 г
200-240 г
200-300 г
0,7-1 кг
0,7-1 кг
Возраст
животного
4-5 мес.
1 мес.
1-3 мес.
1-3 мес.
3-10 мес.
4-5 мес.
4-6 мес.
6 мес-
2 года
6 мес-
2 года
391
Глава 2 8 КАРДИ О ГЕМОДИНАМИКА
В БИОМОДЕЛЯХ
Сердце всех млекопитающих представляет собой
четырехкамерный мышечный орган, который состо-
ит из двух предсердий и двух желудочков. Полости
желудочков и предсердий выстланы тонкой внут-
ренней оболочкой (эндокард) Между висцеральным
листком перикарда и внутренней оболочкой сердца
находится мышечная оболочка (миокард).
Прежде чем рассмотреть сравнительные особен-
ности строения сердца человека и животных, а так-
же их отличия у каждого вида, дадим предельно
краткую характеристику общих морфо-функцио-
нальных черт. Итак, у человека и животных сердце
— это центральный орган кровообращения.
Сердце человека и животных в сравнении
Сердце покрыто серозным мешком, который имеет
париетальный и висцеральный листки. Эпикард
срастается с мышцей сердца и является его наруж-
ной оболочкой. Наружный листок перикарда срас-
тается с перикардиальной плеврой и образует сер-
дечную сорочку, от которой в области верхушки
сердца отходит грудино-перикардиальная связка.
Эта связка фиксирует сердечную сорочку к грудной
кости. Таким образом, серозный мешок образует
околосердечную полость, в которой в норме нахо-
дится небольшое количество серозной жидкости.
От желудочков предсердия отделены венечной
бороздой и имеют ушки сердца. У хищных живот-
392
Биомоделирование в регуляторных системах
ных левое ушко меньше, чем правое. У собак в 87% случаев ушки одинако-
вые, а в 9% — левое ушко больше правого. Стенки предсердий тонкие, оди-
наковой толщины справа и слева.
В правое предсердие впадают задняя полая и передняя полая вены. В пе-
реднюю полую вену вливается кровь из правой непарной вены. Расширен-
ное устье передней полой вены называют венозным синусом. В левое пред-
сердие впадают четыре легочные вены, образуя при этом три муфтообразных
выступа со сложными переплетениями сердечных волокон в виде сфинкте-
ров. Эти выступы называются лакунами легочных вен. Между правым и левым
предсердиями находится предсердная перегородка, в которой на месте оваль-
ного отверстия в эмбриональный период имеется овальная ямка (как у чело-
века, так и у животных).
Предсердия сообщаются с желудочками посредством атриовентрикуляр-
ных отверстий. Правый атриовентрикулярный (трехстворчатый) клапан,
кроме основных трехстворок, имеет еще слабую четвертую. Левый атриовен-
трикулярный клапан состоит из двух основных створок (у собак находится
еще одна добавочная) и называется двустворчатым или митральным.
Стенки правого желудочка сердца тоньше стенок левого желудочка.
Межжелудочковая перегородка имеет выпуклость в сторону правого желу-
дочка. В желудочках находятся мускульные перекладины и поперечные мус-
кулы сердца. Левый желудочек имеет более развитую мускулатуру, но мус-
кульные перекладины выражены слабее. В устьях легочной артерии и аорты
находятся клапанные устройства. Клапаны состоят из трех полулунных ство-
рок. На середине свободных краев клапанов имеются утолщения — узелки
полулунных створок, которые обеспечивают плотное соприкосновение кра-
ев створок. Аранциевы узелки всегда встречаются в аорте, а в легочной ар-
терии — лишь в 30% случаев.
Проводящая система сердца осуществляет передачу нервных импульсов
и поддерживает автоматизм сердца. Состоит из специальных мышечных
волокон Пуркинье, образующих: сино-предсердный узел Кейт-Флюка, ко-
торый размещается в правом предсердии между правым ушком и передней
полой веной; предсердно-желудочковый узел Ашоф-Тавара, размещенный в
нижней части перегородки между предсердиями и отчасти переходящий в
желудочковую перегородку; пучок Гиса, который спускается к желудочковой
перегородке и играет роль соединительного звена между предсердиями и
желудочками. Пучок Гиса распадается на две ножки, одна из которых идет
по внутренней поверхности правого, другая — по внутренней поверхности
левого желудочка. В дальнейшем обе ножки у всех животных и человека
разветвляются на волокна Пуркинье.
Симпатические нервы сердца отходят от звездчатого ганглия. От блужда-
ющего нерва к сердцу отходит депрессорный нерв, который вместе с симпа-
тическими нервами образует на сердце шесть сплетений: два у основания
Глава 28. Кардиогемодинамика в биомоделях
393
легочной артерии, одно между полыми венами в области правого предсер-
дия, два в области легочных вен и одно сплетение на передней стенке пред-
сердий.
В кровоснабжении сердца принимают участие две венечные артерии, устья
которых находятся у начала аорты, в области ее каналов. Левая венечная
артерия, крупнее правой, отдает нисходящую левую ветвь. Правая венечная
артерия сама переходит в нисходящую правую ветвь.
Снабжение сердца кровью осуществляется главным образом во время
систолы. Причем в коронарные сосуды поступает до 10% крови, нагнетае-
мой в аорту. Три четверти этой массы крови проходит через левую венечную
артерию (у всех животных и у человека).
Венозная кровь оттекает по большой сердечной вене, в которую впадает
средняя сердечная вена. Кроме них, имеются малые сердечные вены. Вены
сердца несут кровь в правое предсердие.
Лимфатические сосуды сердца направляют лимфу в передние средостен-
ные и бронхиальные лимфатические узлы.
Сосудистая система состоит из большого и малого кругов кровообраще-
ния, которые начинаются и оканчиваются в сердце, а также из лимфатичес-
кой системы.
В стенках артерий и вен различают три оболочки: внутреннюю, среднюю
и наружную. В артериальных сосудах наиболее развита средняя оболочка,
которая имеет эластические и круговые мышечные волокна. Внутренняя
оболочка содержит слой коллагеновых эластических волокон и эндотели-
альных клеток. Артериальные сосуды характеризуются большой прочнос-
тью, эластичностью и способностью сокращаться. Например, сонные арте-
рии мини-свиней, собак и кошек способны выдерживать давление, в 20 раз
превышающее нормальные цифры, а просвет аорты может возрастать на 30%.
Стенки вены отличаются хорошо развитой адвентицией (в то время как ос-
тальные стенки развиты слабо), большим просветом и наличием клапанов.
В организме животных находится обширная сеть капилляров, которые
связывают артерии с венами. На один квадратный миллиметр мышцы серд-
ца приходится свыше 2600 капилляров.
Легочной ствол выходит из конуса правого желудочка и несет венозную
кровь к легким. У корня легких он разделяется на два сосуда, один из кото-
рых идет в левое, а второй — в правое легкое. Каждый из них в свою очередь
многократно разветвляется, и в результате образуется густая сеть капилля-
ров, которые охватывают альвеолы.
Сердечная деятельность человека и животных состоит из трех строго пос-
ледовательных фаз: систолы, диастолы и паузы. Такая закономерность рабо-
ты сердца обеспечивает постоянное движение крови по сосудистому руслу в
одном направлении: из предсердий в желудочки; из левого желудочка в аор-
ту и далее в большой круг кровообращения.
394
Биомоделирование в регуляторных системах
Венозная кровь из большого круга кровообращения поступает в правое
предсердие, в то время как в левое предсердие собирается артериальная кровь
из малого круга кровообращения. Клапанный аппарат сердца препятствует
обратному току крови.
Биомоделирование на сердце
По наружной форме сердце мини-свиньи занимает промежуточное положе-
ние между сердцем крупных и средних лабораторных животных. В правое
предсердие, близ каудальной полой вены, впадает левая непарная вена. Лё-
гочных вен только две. Добавочной продольной борозды нет или она едва
намечена. Гребешковые мускулы в предсердиях, особенно левом, сильно
развиты. Foramen ovale часто (до 20% случаев) не зарастает. Трёхстворчатый
клапан, помимо трёх больших створок, имеет слабо развитую четвёртую.
В двухстворчатом клапане находят два маленьких добавочных клапана. Сухо-
жильных струн в правом желудочке пять и они более слабые, чем в левом,
где их одиннадцать. Сосочковые мускулы длинные, из них в правом желу-
дочке сильно развит пристеночный. Аранциевы узелки на полулунных кла-
панах аорты встречаются в 96% случаев, в лёгочной артерии — редко (в 26%
случаев). Сердечный хрящ один. Сердечные артерии распределены, как у
лошадей. Вес сердца достигает 0,28% от живого веса. Сердце лежит несколь-
ко больше (около 3/6) в левой половине грудной полости, чем в правой, но
с большим наклоном по отношению к грудной кости. Краниально оно до-
стигает середины 3-го ребра, каудально — середины 6-го. Верхушка сердца
располагается в области соединения 7-го рёберного хряща с грудной костью.
Сердечная сорочка прикрепляется к грудной кости в области от 3-го до 7-го
рёберного хряща.
На мини-свинье возможно изучение как нормальных физиологических
процессов при нагрузке (тренировке), так и патологических изменений в
миокарде при ишемии и инфаркте, моделируемых, например, путем посте-
пенного перевязывания коронарной артерии [665].
Особо следует сказать о крайней чувствительности свиней к стрессовым
факторам. Многочисленные железы, расположенные у свиней в области
спины и шеи, выделяют особые вещества липидной природы — феромоны,
образующие сложную гамму, индивидуальную для каждого животного [151],
вследствие чего свиньи способны различать животных, принадлежащих к
различным группам. При изменении окружающей среды и возникновении
стрессовых ситуаций свиньи отвечают изменением поведенческих и пище-
вых реакций, гормонального статуса, рН крови [4]. Сопоставление данных,
полученных на человеке, с экспериментальными исследованиями на свиньях
показало идентичность некоторых изменений минерального обмена, мета-
болизма и структуры костной ткани, состава белка, возникающих при стрес-
Глава 28. Кардиогемодинамика в биомоделях
395
совых нагрузках [20]. Все эти факты свидетельствуют о несомненной при-
годности свиньи для моделирования стресса.
Мини-свинья широко используется в экспериментальной хирургии. Име-
ются данные о проведении операций по трансплантации сердца [19] и поис-
ке возможностей для использования сердечных клапанов свиньи в качестве
гетеропластического материала в хирургии сердечных клапанов [432]. Экс-
периментирование на свиньях при изучении различных вопросов трансплан-
тологии неоценимо. На свиньях можно моделировать пересадку самых раз-
личных органов: печени, селезенки, поджелудочной железы, костной и эн-
докринной тканей [24, 360, 443, 492,].
У овец форма сердца встречается трёх типов. В области венечной и про-
дольных борозд обычно находится много субэпикардиальной жировой клет-
чатки. На каудальной поверхности сердца резко выступает третья продоль-
ная добавочная борозда. Гребешковые мускулы хорошо развиты. Овальное
отверстие у 22% взрослых животных остаётся незаросшим. Аранциевы узел-
ки на полулунных клапанах аорты встречаются в 65% случаев (у овец в 83%),
а в лёгочной артерии — в 47% (у овец в 70%). Сердце на 5/7 отодвинуто
влево от средней сагиттальной плоскости и между 3-ми 4-м ребром непос-
редственно прилежит к левой грудной стенке. Передний контур сердца до-
ходит до 3-го ребра, а задний — до 6-го. Верхушка сердца лежит против 5-
го рёберного хряща или позади него, не достигая грудной кости на 2 см, а
впереди от диафрагмы — на 2—5 см, в зависимости от фазы дыхания. Сер-
дечная сорочка прикрепляется двумя связками к грудной кости у 6-го рёбер-
ного хряща. Левая венечная артерия развита значительно сильнее правой и
отдаёт нисходящие ветви для левой, правой и добавочной продольных бо-
розд. Правая венечная артерия питает только правый желудочек. Величина
сердца различна и зависит от породы, пола и возраста животного. Вес со-
ставляет около 0,4% от живого веса.
У собаки различают четыре типа сердца: эллипсовидное (42,6%), конусо-
видно-эллипсовидное (24%), эллипсовидно-шаровидное (26%) и шаровид-
ное (7,4%). Вес сердца колеблется в зависимости от породы и возраста собак
и по отношению к весу тела составляет от 0,76 до 1,2%. Сердце собаки по
внешней форме более округлое, чем у других животных, и с более притуп-
лённой верхушкой, на которой ясно выступает завиток, образованный пуч-
ками мускульных волокон. Аранциевы узелки на аортальных клапанах встре-
чаются всегда, а в лёгочной артерии редко (в 30% случаев). В фиброзное
кольцо аорты между клапанами заложены три маленьких хряща. У старых
животных они обизвествляются. В грудной полости сердце занимает почти
горизонтальное положение от 3-го до 7-го ребра. Верхушка сердца обращена
каудально, с уклоном влево и вентрально, и находится в области 6—7-го и
даже 8-го рёберного хряща. Справа сердце прилежит к грудной стенке в
области 5-го ребра, а слева — в области 4—6-го ребра. Вес сердца колеблется
396
Биомоделирование в регуляторных системах
в очень широких пределах (от 0,85 до 1,4% к живому весу) в связи с разно-
образием пород собак. Артерии сердца идут, как и у жвачных. Частота пульса
у здоровой собаки в среднем 90—130 ударов в минуту, у новорожденного
щенка — 140—160 и больше. Минутный объем сердца у здоровой собаки
весом 10 кг составляет 1450 мл. Максимальное кровяное давление в общей
сонной и бедренной артериях находится у здоровых животных в пределах
120-160, а минимальное — 30—60 мм рт. ст. Полный кругооборот крови про-
исходит за 15—18 с.
Сердце кошки весит 10-15 г (около 0,39% веса тела). Оно овальной фор-
мы, верхушка сердца выражена нечетко. Частота пульса составляет 110—180
ударов в минуту. Максимальное артериальное давление крови в среднем
составляет 150 мм рт. ст. Скорость циркуляции крови в среднем — 6,7 с.
Сердце кролика имеет следующие размеры: длина 3,5—3,8 см, ширина в
спинно-брюшном направлении 2,2—2,5 см. Вес сердца, лишенного крови,
у взрослого кролика составляет 0,274% веса тела. Правый желудочек серд-
ца большой, тонкостенный, левый несколько длиннее, имеет толстую стенку
и образует верхушку сердца. Правое предсердие имеет хорошо развитое
ушко и синусовую область, в которую впадают передняя и задняя полые
вены. В левое предсердие впадают центральный, левопередний и правопе-
редний коллекторные стволы легочных вен. Для кролика характерно то,
что лакуны легочных вен отсутствуют, а мышечные волокна левого пред-
сердия по стенкам легочных вен проникают вглубь легких. Такое внутри-
легочное предсердие во многом благоприятствует кровообращению у жи-
вотных с частым сердцебиением. Частота пульса у здорового кролика 150—
140 и реже 320—360 ударов в минуту. У кролика весом 2 кг минутный объем
сердца составляет 440 мл.
Сердце морской свинки четырехкамерное, весит 2—2,5 г (в среднем 2,3 г).
Сердечный толчок слабый, разлитой. Дыхательных изменений сердечного
ритма нет. Большая часть сердца покрыта долями легких. Покрывающий
сердце перикад с помощью ligamentum sternopericardiacum соединен с гру-
диной. Кровяное давление в сонной артерии 70—80 мм рт. ст. Средняя час-
тота сердечных сокращений у морской свинки 250—355 в минуту. Кровь в
зависимости от места взятия дает неодинаковые показатели состава формен-
ных элементов.
Сердце крысы 1,3 см длиной, в диаметре в среднем 0,79-0,95 см, а окруж-
ность у основания 2,5—3 см. Вес его составляет в среднем 1,3 г (у белой
крысы весом 130-150 г сердце весит 0,55 г, а у крысы весом 200—250 г —
1,5 г). Сердце почти полностью окружено легкими и свободно лишь в пере-
дне-нижней части. Кровоснабжение сердца осуществляется за счет левой и
правой венечных артерий. Кровяное давление в сонной артерии составляет
100—130 мм рт. ст. Минутный объем крови у крысы весом 200 г равен 122 мл.
Скорость кровотока в аорте в момент систолы достигает 255 мм/с. Частота
Глава 28. Кардиогемодинамика в биомоделях
397
сердечных сокращений у белой крысы 286—500 в минуту. От дуги аорты от-
ходят безымянная, левая общая сонная и левая подключичная артерии. Реже
безымянная артерия отсутствует, и тогда от дуги аорты вместо нее отходят
правая общая сонная и правая подключичная артерии. В правое предсердие
впадает задняя полая вена, парные безымянные и сердечные вены. Пере-
дней полой вены, которая у других животных возникает от слияния безы-
мянных вен, у крыс нет.
Сердце сирийского хомячка имеет форму усеченного конуса и большей
своей частью располагается в левой части грудной клетки.
Сердце мыши покрыто перикардом, с помощью которого фиксируется к
грудине. Верхушка сердца расположена в I межреберье. На поперечном разре-
зе сердце слегка овальное. Ушки сердца довольно большие. Частота сердцеби-
ения 520—780 ударов в минуту. У взрослой мыши сердце весит в среднем 0,1 г.
У мини-свиней средняя частота сердечных сокращений составляет 70 со-
кращений в минуту. Верхушечный толчок — 3—5-е-межреберное простран-
ство, абсолютная сердечная тупость — слева внизу во 2—3-межреберном про-
странстве.
У овец частота сердечных сокращений в минуту зависит от возраста жи-
вотных: ягнята — 115, молодняк — 85-95, взрослые овцы — 70-80, старые
овцы — 55—60, бараны — 68—80. Верхушечный толчок — 3-5-е межреберное
пространство.
У собак частота сердечных сокращений зависит от размера. Маленькие
собаки — в среднем 115, большие собаки — 85 сокращений в минуту. Вер-
хушечный толчок — 4-6-е межреберное пространство в нижней трети груди
слева рядом с грудиной; сердечная тупость — слева в 4-6-м межреберье; верх-
няя граница — реберная дуга, в 6-м межреберье находит на перкуторную
область печени. Ударный объем 14 мл (на 10 кг массы тела), минутный объем
145 мл (при частоте 100 уд/мин).
У кошек пульс взрослого животного в среднем 125 уд./мин, пульс котят
180—200 уд./мин. Частота пульса кролика в среднем составляеи 175, а у мор-
ских свинок — 300 уд./мин.Для сирийских хомячков характерны большие раз-
личия в частоте сердечных сокращений: 280—420 уд./мин. У крыс средние
значения пульса 400 уд./мин. У крыс WAG/GSto при эндокардите (у 4% крыс
в возрасте 3 месяца) частота пульса до 550 уд./мин. У мышей линии DBA/1J
Lac с обызвествлением сердца (обнаруживается почти в 100% случаев) час-
тота пульса превышает 850 уд./мин. У мышей BALB/cJLac наблюдаются элек-
трокардиографические нарушения в деятельности сердца, изменена кривая
левожелудочкового давления: фаза расслабления меньше, чем у нормальных
мышей (0,04 с и 0,07 с), а длительность фазы покоя почти вдвое больше.
Другие данные и биомодели см. в табл. 72.
398
Биомоделирование в регуляторных системах
Таблица 72
Генетически обусловленные биомодели для изучения действия
кардиотропных веществ
Исследуемое действие
вещества
Биомодель спонтанной
гипертрофии миокарда
у 30% животных
Биомодель натуральной
дистрофии миокарда с по-
вышенным уровнем сыворо-
точной креатинфосфатной
киназы (первопричина
в гене дистрофии) перед
клиническим проявлением
дистрофии
Биомодель кардиомиопатии
и миопатии, сопровождае-
мых гиперкальцификацией
сердца и скелетных мышц
и последующей некротиза-
цией этих тканей
Биомодель натуральной
генно-детерминированной
брадикардии с частотой
сердечных сокращений
до 50% от нормы (при
норме 280-412)
Биомодель брадикардии
с высоким содержанием
холестерина и низким
содержанием йода в плаз-
ме крови
Лабораторное
животное
Хомячки
Хомячки
Хомячки
Хомячки
Кролики
Кролики
Линия или порода
животного
Линия X3
Линия 1,5
Линия 82 ... 2
Линия 82.73
Линия AX/J
C/J
Пол
животного
96
96
96
96
96
6
Вес
животного
120 г
120 г
120 г
120 г
3,5 кг
2,9 кг
Возраст
животного
6-18 мес.
6-18 мес.
6-18 мес.
6-18 мес.
7 мес-
4 года
7 мес-
4 года
Сравнительная электрокардиография
ЭКГ записывают чаще всего в трех и реже в четырех отведениях. Первое
отведение — электроды накладывают на пясти правой и левой конечностей.
Второе отведение — электроды накладывают на пясть правой передней ко-
нечности и плюсну левой задней конечности. Третье отведение — ток отво-
дят от пясти левой передней конечности и плюсны левой задней конечнос-
ти. Четвертое отведение — грудное.
Зубцы электрокардиограмм животных, как и человека, обозначаются бук-
вами Р, Q, R, S, Т. Наибольшая величина зубцов отмечается во втором отве-
дении (рис. 7 А и В).
На ЭКГ у собак рельефно выступает дыхательная аритмия. Принципи-
ально у здоровых собак разных пород характер ЭКГ одинаковый, лишь для
восточноевропейских овчарок свойственно увеличение зубцов. Для собак
характерным является отрицательный зубец Т чаще всего во всех трех отве-
дениях (рис. 7В) .
Глава 28. Кардиогемодинамика в биомоделях
399
Величина интервала P-Q равна 0,11 с, а величина интервала QRS 0,04-
0,05 с. Электрическая ось комплекса QRS составляет 30—70° у нелинейных
и 30—75° у собак чистых линий. Интервал S—Т у большинства здоровых со-
бак размещается на изоэлектрической линии, а в 1/3 случаев пересекает ее
снизу вверх (от +0,2 до —0,2 с). Величина интервала S—Т у собак колеблется
в пределах 0,04—0,1, а интервала QRST — в пределах 0,16-0,24 с.
В систолическом периоде электрокардиограммы (от начала зубца Р до
конца зубца Т) различают:
/ предсердный комплекс — от начала зубца Р до начала зубца Q . Зубец Р
отображает возбуждение предсердий, его вершина соответствует началу си-
столы предсердий. Интервал Р—Q характеризует время, затраченное на пе-
редачу возбуждения из предсердий в желудочки;
/ начальную фазу желудочкового комплекса — QRS;
/ конечную фазу желудочкового комплекса — от начала зубца Т до его конца.
0,05"
r Ч Сердечный период ► r a.
р -
л
А
Л
/\
/1
/
V/
о
<У
S
— -
" Т "
'\
j i
"V"
F
/
\
(
А
л
/\
/\
/ \
J
1
_^~
s
■^
\j'
т
P-Q Предсердия Т-Р Сердечная пауза
Q-T Желудочки
Рис. 7. Сравнительные особенности электрокардиограммы человека (А) и кошек и собак (В)
в норме. Следует обратить внимание на зубец Т, который у многих животных инвертирован,
что следует учитывать в процессах биомоделирования. Инверсия зубца Т у человека харак-
теризует грубую патологию, например, инфаркт миокарда
Зубец Q отвечает возбуждению внутренней поверхности мускулатуры же-
лудочков, правой сосочковой мышцы, перегородки, верхушки левого желу-
дочка и основания правого желудочка. Зубец R возникает в результате по-
степенного охвата возбуждением наружной мускулатуры правого и левого
желудочков и основания левого желудочка. Его верхушка соответствует на-
чалу систолы желудочков. Высокий зубец R свидетельствует о высоком воль-
таже токов сердца, что свойственно хорошему функциональному состоя-
нию мускулатуры сердца или повышенной возбудимости симпатической не-
рвной системы. Низкий зубец R указывает на дистрофические изменения
в миокарде.
400
Биомоделирование в регуляторных системах
Отрицательный зубец R в первом и третьем отведениях часто свидетель-
ствует о нарушениях проводящей системы.
Изменение формы комплекса QRS в виде расщепления или притупления
зубцов, а также расширение интервала QRS могут указывать на поражение
конечных ветвей проводниковой системы и поражение миокарда.
Зубец S совпадает с максимальным возбуждением желудочков. Отрезок
S-T находится на изопотенциальной линии ЭКГ и указывает на то, что ле-
вый и правый желудочки полностью охвачены возбуждением и в это время
происходит их сокращение (рис. 7В).
Зубец Т соответствует конечной фазе возбуждения желудочков и в опре-
деленной мере отражает состояние биохимических процессов в миокарде.
Вторым периодом ЭКГ является диастолический, охватывающий интер-
вал — от зубца Т до начала зубца Р. Регуляция работы сердца осуществля-
ется экстра- и интракардиальными нервными воздействиями, передавае-
мыми волокнами симпатических и блуждающих нервов, а также гумораль-
ным путем.
Электрокардиограмма кошек имеет следующие особенности: зубец Р все-
гда положительный, его высота 0,1—0,15 мВ и длительность 0,03—0,04 с. Зубец
Q встречается редко или он очень маленький. Зубец R имеет амплитуду 0,3—
0,5 мВ. Зубец S маленький. Интервал QRS составляет в среднем 0,04 с. Зубец
Т в 25% случаев отрицательный, а в остальных — положительный. Положи-
тельный зубец Т достигает 0,1—0,15 мВ, а отрицательные зубцы — меньше.
Интервал QT в среднем равен 0,2 с, а промежуток между зубцами Р и Q
колеблется в пределах от 0,06 до 0,09 с (рис. 7В).
Электрокардиограмма кролика характеризуется тем, что сегмент RST в
большинстве случаев лежит на изолинии. Высота зубца R в третьем отведе-
нии несколько большая, чем во втором, и составляет: R2 — 0,07...0,25 (чаще
0,1...0,15) мВ, a R3 - 0,08...0,35 (чаще 0,15-0,2) мВ.
Зубец Т у кроликов очень высокий, особенно во втором отведении (вы-
сота его в два раза больше комплекса QRS). Зубец Q встречается не всегда,
во втором отведении всего лишь в 4,8%, а в третьем — в 6,3% случаев.
Зубец Р в первом отведении очень маленький или отрицательный, а во
втором и третьем отведениях всегда положительный, высота его 0,1-0,15 мВ
и продолжительность 0,03—0,04 с. Интервалы между зубцами составляют:
PQ - 0,07, QRS - 0,04 и QT - 0,14 с.
Запись электрокардиограммы у морских свинок производят под наркозом,
потому что у не наркотизированных животных возникают помехи вслед-
ствие дрожания мышц.
Зубец Р во всех отведениях положительный, особенно выраженный во
втором отведении, а в первом и третьем отведениях бывает сглаженным или
может отсутствовать. Зубец Pj имеет амплитуду 0—0,25 мВ. Зубец Q во всех
отведениях отсутствует. Интервал от зубца Р до точки Q составляет 0,06— 0,08 с.
Глава 28. Кардиогемодинамика в биомоделях
401
Колебания величин зубца: Rj — 0,3...1,1 мВ, R2 — 0,4...1,5 мВ, R3 — 0,2...0,7
мВ. Зубец S во втором отведении отсутствует, а в первом и третьем отведениях
регистрируется в 20% случаев. Длительность интервала QRS у морских свинок
составляет 0,02—0,04 с.
Зубцы Т обычно отрицательные (в первом отведении этот зубец доволь-
но часто отсутствует), их величины следующие: Tj — 0...0,2, Т2 — 0,05...0,35,
Т3 — 0,05...0,3 мВ. Длительность интервала S-T составляет 0,03—0,12 с. Дли-
тельность интервала QRST 0,11—0,2 с.
Запись электрокардиограмм у крыс чаще всего проводится под наркозом
(у ненаркотизированных животных из-за дрожания мышц возникают неус-
транимые помехи). В первом отведении зубец R очень низкий, а остальные
зубцы отсутствуют или с трудом различимы, в связи с этим первое отведение
у белых крыс не имеет никакого значения. Во втором и третьем отведениях
регистрируются отчетливые электрокардиограммы.
Зубец Р почти всегда положительный, хотя в редких случаях может быть
и отрицательным как во втором, так и в третьем отведениях. Величина Р2
колеблется в пределах 0,1—0,35, а Р3 — в пределах 0,1-0,3 мВ; их длитель-
ность 0,01-0,02 с.
Зубец Q почти всегда отсутствует во всех отведениях. Интервал Р—Q (до
точки Q) равен 0,04-0,05 с. Высота зубца R2 составляет 0,3-0,85, a R3 —
0,35-0,7 мВ.
Зубец S во втором отведении встречается в 10,2%, а в третьем отведе-
нии — в 37,5% случаев. Интервал S—Т у белых крыс отсутствует, и если на
электрокардиограмме отмечается зубец S, то он сразу же переходит в зубец
Т, образуя соединение S—Т. В подавляющем большинстве случаев зубец S
отсутствует, и нисходящее колено зубца R непосредственно переходит в зу-
бец Т. Длительность интервала QRS 0,01—0,025 с.
Зубец Т всегда положительный, величина его колеблется в пределах: Т2 —
0,3...0,7 мВ, Т3 — 0,35...0,65 мв, т. е. почти не уступает по величине зубцу R.
Интервал QRST составляет 0,07-0,1 с.
Интервал Т—Р, указывающий на величину диастолической паузы, неред-
ко может вовсе отсутствовать — зубец Т сразу же переходит в зубец Р — или
составляет 0,01—0,05 с.
Электрокардиограмма мыши. Зубец Р во всех отведениях положительный,
но небольшой величины. Интервал PR равен 0,016-0,045 с. Зубец R во всех
отведениях направлен кверху, но в первом отведении он очень низкий, а
наивысшая высота его во втором и третьем отведениях достигает 0,25—0,3 мВ.
Интервал QRS составляет 0,02—0,04 с. Зубец S довольно часто отсутствует
или слабо выражен. При отсутствии зубца S нисходящее колено зубца R
переходит в мелкий отрицательный зубец Т, который в своей конечной ча-
сти может переходить в положительную фазу. Амплитуда зубца Т составляет
0,07-0,1 мВ.
402
Биомоделирование в регуляторных системах
Математическое моделирование ЭКГ
Вопросы биомоделирования функций сердца, сосудов, процессов гемодина-
мики рассматриваются в многочисленных работах [61, 210, 667]. В этой главе
мы рассмотрим некоторые аспекты ЭКГ-моделей. При сравнительном мо-
делировании ЭКГ человека и животных наиболее приемлемым является мно-
гомерный статистический анализ. С этой целью могут использоваться самые
различные значения ориентации и скорости электрического вектора сердца
в пространстве.
На рис. 8 представлены принципы получения наиболее исчерпывающих
данных ЭКГ, отражающих не только пространственную но и временную
информацию, имеющую большое значение для ЭКГ-анализа любого типа. С
этой целью комплекс QRS независимо от его длительности, что особенно
важно при сравнении ЭКГ человека и крупных животных (частота сердеч-
ных сокращений 60—80 в минуту), с одной стороны, и мелких лабораторных
животных (300—500 в минуту) — с другой, делятся на восемь или двенадцать
равных частей.
Большая частота стробирования нецелесообразна, поскольку может при-
вести к статистической зависимости и изначальному искажению результа-
тов. Так же разбивается интервал между конечными моментами S и волны Т,
а если необходимо, то и волны U. Скалярное значение сигналов ортогональ-
ного отведения складывается векторно и получается один мгновенный век-
тор для каждой восьмой или двенадцатой части комплексов QRS и сегмен-
тов ST-T.
/
У
\
\
1— р —1
■
1
/
/
f
/
V
У\
\ v
А
Д-
/Т
L\
1
\
\
\
\
■V
v^
•> /-./о
^*
\ "
\
\
V —
\
4
— к -
\
I/O <£/0 I/O £/0
L- OF
is -J
ST-T
/
/..
А
/
. «-*
\
■\
ч.
•* " """
, — • —
^
1 1
ЭКГ человека
ЭКГ собак
ЭКГ кошек
Рис. 8. Схема процедуры нормализации и построения векторов по времени для комплекса
QRS и сегмента ST-T. Обращают на себя внимание в различиях комплекса ST-T кошек, у кото-
рых в норме, как правило, этот комплекс инвертирован, а также у собак, у которых этот комп-
лекс может носить двухфазный характер
Максимальные векторы сердца — это векторы с оптимизированным моду-
лем для каждой волны или комплекса ЭКГ. Момент времени, которому соот-
Глава 28. Кардиогемодинамика в биомоделях
403
ветствует такой вектор, определяется пиковым положением кривой модуля.
Ориентации вектора определяют пространственные и временные характери-
стики параметров комплекса QRS, волны Т и других параметров ЭКГ.
Предполагается, что совокупность измеренных параметров, например, пос-
ледовательность мгновенных векторов, распределена по многомерному нор-
мальному закону. Для каждой группы животных или людей из выборки вы-
числяются параметры соответствующего нормального распределения, т. е.
оценки величин \xv £р ц2, £2, ..., где ^ij — среднее значение измерений пер-
вой группы, а Ер Е2 — соответствующие матрицы ковариаций и т.д.
Для электрокардиографической записи, которую необходимо отнести
к какому-либо классу, вычисляются следующие величины:
^ =(-ЯГ-ц1)2;г1(Аг-ц1) + С1, (93)
d2={X-\i2)J^{X-\i2) + C2, (94)
где X— вектор электрокардиографических изменений и С — константа, най-
денная эмпирическим путем. Величины dl и d2 представляют собой вектор-
ные разности между вектором сердца исследуемого и векторами средних
значений двух сравниваемых групп животных или людей. Если величина dx
оказывается меньше, чем d2, то анализируемую запись относят к группе 1,
в противном случае — к группе 2. В дальнейшем анализ можно продолжить
в отношении любого количества групп данных для людей или животных.
Векторы средних значений \х{ вычисляются по формуле
M^-IX, t (95)
где Xk — наблюдаемые значения у'-й измеряемой величины для категории
записей /. Элемент с индексом ij матрицы ковариаций Е£ вычисляются по
формуле:
. 1 n
^г^Ж-^К^-^-) , (96)
где Хи и Х.( — наблюдаемые значения в /-м и у'-м измерениях величин для
категории к. Постоянные Сх и С2 определяются методом линейного про-
граммирования из условия минимизации количества ложных решений для
исходных электрокардиографических записей.
Процедура классификации схематически показана на рис. 9. Следует об-
ратить внимание, что области рассеивания пространственных векторов яв-
ляются трехмерными и имеют форму эллипсоида. Плотность распределе-
ния уменьшается с увеличением расстояния от среднего значения находяще-
гося в центре области рассеивания. Это уменьшение плотности изображено
изменением плотности окраски. Граница между областями, изображенная
жирной линией, получена на основе рассмотрения функций плотности рас-
404
Биомоделирование в регуляторных системах
пределения. На этом рисунке видны отличия плотности распределения и
рассеивания векторов сердца у кошек, для которых характерен инвертиро-
ванный характер зубца Т и у мини-свиней, характер рассеивания вектора
сердца которых ближе к таковому у человека.
Рис. 9. Распределение пересекающихся областей рассеивания вектора сердца у кошек (А),
человека (Б) и мини-свиней (В) в норме. Границы между областями изображены жирными ли-
ниями, полученными на основе функций плотности распределений
Расстояние некоторого исследуемого вектора от данной группы можно
определить через плотность распределения вероятности. Чем меньше плот-
ность вероятности, или концентрация сравнительных параметров в окрестно-
сти исследуемого случая, тем больше его расстояние от среднего значения
данной группы. Задача экстраполяции таким образом сводится к вычислению
расстояний исследуемой электрокардиографической записи в различных ис-
следуемых группах животных, равно как и между животными и человеком.
Сосуды, гемодинамика и биомодели
Древние анатомы не находили в венах крови, а потому полагали, что по
этим сосудам к различным частям тела доставляется воздух. Они думали, что
эти сосуды являются продолжением трахеи. Однако они видели и другие
сосуды с более толстыми, чем у вен, стенками. По-гречески полая трубка
Глава 28. Кардиогемодинамика в биомоделях
405
называлась arteria, что происходит от слов аег — воздух и tereo — содержать.
Поэтому артерия может переводиться как сосуд, содержащий воздух. Это
название было перенесено и на пустые, похожие на вены сосуды, которые
мы сегодня называем артериями.
Древнеримский врач Гален обнаружил, что артерии, как и вены, перено-
сят кровь. Но еще долгие века так никто и не мог понять, для чего же нужны
два типа кровеносных сосудов. Гарвей, давший наиболее полное описание
сосудистой системы, не установил звено, где артерии соединяются с венами.
В 1661 г. биолог и врач М.Мальпиги обнаружил недостающее звено— ка-
пилляры. Именно по ним кровь перетекала из артерий в вены, омывая на
своем пути каждую клетку. Эти мельчайшие сосудики были названы капил-
лярами (от лат. capillaries — волосяной).
Аорта является главной магистралью большого круга кровообращения.
Ее разделяют на дугу аорты, грудную и брюшную части. При выходе из ле-
вого желудочка сердца на ней отмечается расширение — луковица аорты
является местом отхождения ряда важных артерий. Дуга аорты является
местом отхождения ряда важных артерий. У различных животных эти сосу-
ды отходят по-разному.
От подключичных артерий отходят: позвоночная артерия, реберно-шей-
ный ствол, от которого отходят поперечная шейная и глубокая шейная ар-
терии; плече-шейный ствол, начинающийся в области I ребра; внутренняя
грудная артерия довольно значительных размеров; небольшая наружная груд-
ная артерия. Подключичные артерии направляются в передние конечности
уже как подмышечные артерии.
Общая сонная артерия разветвляется на наружную и внутреннюю сон-
ные артерии. В месте бифуркации имеется очень много нервных оконча-
ний — хемо- и барорецепторов, которые называют рефлексогенной зоной
sinus caroticus. Они играют большую роль в регуляции гемодинамики, дыха-
ния и кровяного давления [399]. Открытие роли оксида азота в регуляции
сосудистых реакций дало основание думать не только о его роли как меди-
атора, но и как регулятора сосудистых реакций. Впервые N0 был охаракте-
ризован как эндотелиальный фактор релаксации сосудов (endothelium-derived
relaxation factor, EDRF), природа которого оставалась неясной до 1987 г.,
когда установили, что EDRF представляет собой N0. Как вскоре оказалось,
N0 синтезируется не только в эндотелиальных, но и во многих других, если
не всех, клетках, тканях, органах, в том числе в сердце и стенках сосудов.
Концепция об изменении сосудистой реактивности в зависимости от ба-
зального NO-тонуса может также объяснить некоторые аспекты толерантно-
сти к нитровазодилататорам, которые вводят в течение длительного времени
(десенситизация рецепторов к N0).
Соединительные ветви правой и левой внутренних сонных артерий, сли-
ваясь между собой, формируют вокруг гипофиза артериальное (виллизиево)
406
Биомоделирование в регуляторных системах
кольцо. В передней части кольца берет начало назальная мозговая артерия,
через которую кровь поступает к мозолистому телу, передним отделам полу-
шарий и обонятельным луковицам.
Каудальные соединительные артерии внутренней сонной артерии обра-
зуют каудальную мозговую артерию, которая осуществляет кровоснабжение
четверохолмия, задних отделов полушарий, и отдает артерию для заднего
сосудистого сплетения боковых желудочков.
Мозг, как и сердце, тоже очень чувствителен к нарушению кровоснабже-
ния, но проявляется это иначе. Если уменьшить приток крови к мозгу, то
наступит смерть. Но прежде человек «заснет». Древнегреческие фокусники
развлекали публику «усыплением» животных, особенно коз. У коз длинная
шея, и фокус осуществить очень легко, нажимая по обе стороны шеи на
артерии, несущие кровь к мозгу и голове. В результате этого коза «засыпа-
ла», на самом же деле она теряла сознание (если можно так выразиться о
животном). Как только давить на артерию прекращали, кровь снова устрем-
лялась к мозгу, неся с собой кислород и питание, и коза «просыпалась».
Артерии эти получили название сонных.
В формировании задней части виллизиевого кольца у собак принимают
участие ветви наружных сонных артерий — затылочные артерии и ветви под-
ключичных артерий — позвоночные артерии. Позвоночные артерии анасто-
мозируют с затылочными соответствующих сторон и вскоре после этого де-
лятся на передние и задние ветви. Задние ветви образуют непарную вентраль-
ную спинномозговую артерию. Правые и левые передние ветви, соединив-
шись вместе, образуют основную мозговую артерию, которая впадает в задний
конец виллизиевого кольца. От основной мозговой артерии к мозжечку отхо-
дят передние и задние артерии мозжечка и внутренняя слуховая артерия.
От брюшной аорты отходят: чревная артерия, от которой отходят пече-
ночная, левая желудочная и селезеночная артерии; передняя брыжеечная
артерия; парные диафрагмально-брюшные артерии; почечные артерии; над-
почечниковые артерии; внутренняя семенная артерия; задняя брыжеечная
артерия, которая делится на левую ободочную и переднюю прямокишечную
артерии; глубокая окружная подвздошная артерия; семь поясничных арте-
рий. Брюшная аорта на уровне поясничных позвонков дает правую и левую
наружные подвздошные артерии, после чего она переходит в общий ствол
внутренних подвздошных артерий.
Движению крови по венам способствуют такие факторы, как сокраще-
ние скелетной мускулатуры и вследствие этого сдавливание вен, наличие
клапанов, насасывающая способность отрицательного давления грудной
полости и отрицательного давления в полостях предсердий во время диасто-
лы. Краниальная полая вена образуется в области первых ребер слиянием
правой и левой безымянных вен. В краниальную полую вену впадают общий
ствол внутренних грудных вен, парный позвоночно-реберный ствол и пра-
Глава 28. Кардиогемодинамика в биомоделях
407
вая непарная вена. Безымянная вена объединяет подмышечную и общую
яремную вены.
Общая яремная вена образуется в результате слияния внутренней и на-
ружной яремных вен. Наружная яремная вена собирает кровь от наружной
и внутренней челюстных вен. От передней конечности кровь оттекает по
подкожной и глубокой венозным магистралям. От парных органов брюш-
ной полости, тазовых органов, диафрагмы, мышц задних конечностей и т. д.
кровь оттекает по системе сосудов каудальной полой вены, которая образу-
ется на уровне последнего поясничного позвонка путем слияния правой и
левой общих подвздошных вен. Общие подвздошные вены начинаются пос-
ле слияния наружных и внутренних подвздошных вен.
Из вен задних конечностей следует указать на латеральную подкожную
вену голени и стопы, более известную под названием малой подкожной вены.
Она образуется из вен, собирающих кровь из дорсальной и плантарной по-
верхностей лапы. Эта вена у мини-свиней, кошек и собак легко доступна
для внутривенного введения лекарственных веществ и взятия крови.
Медиальная подкожная вена голени и стопы (большая подкожная вена)
начинается в проксимальной части голени дорсальной и плантарной ветвя-
ми. По сравнению с малой сафеной у собак она развита слабее. Обе вены
сафены впадают в бедренную вену, которая продолжается в наружную под-
вздошную вену.
Кровь из желудка, селезенки, поджелудочной железы и кишечника по
воротной вене поступает в печень. В печени воротная вена рассеивается,
образуя междольковые вены и внутридольковые венозные капилляры. Из
венозных капилляров кровь переходит в печеночные вены и попадает в зад-
нюю полую вену.
Скорость течения крови в аорте при поперечнике аорты в 0,1 см состав-
ляет 184 см/с. Скорость течения крови в сонной артерии составляет от 10 до
34 см/с. Полный кругооборот кровь завершает в среднем за 7,8 (4,71—10,4) с.
Кровяное давление в сонной и бедренной артериях — 80—130 мм рт. ст.
Особенностями аорты у кроликов являются резкая изогнутость дуги и ее
низкое расположение и некоторое смещение влево. От дуги аорты сосуды
отходят по рассыпному типу.
Общие сонные артерии идут в нервно-сосудистом пучке шеи вдоль тра-
хеи. Внутренняя сонная артерия через канал височной кости проходит в по-
лость черепа и снабжает кровью головной мозг, глаза и носовую полость.
Как и в предыдущих главах, мы приводим варианты выбора биологи-
ческих моделей для физиологических, фармакологических и гигиеничес-
ких исследований, ориентированных в аспекте экстраполяции на человека
(см. табл. 73).
408
Биомоделирование в регуляторных системах
Таблица 73
Генетически обусловленные и экспериментальные биомодели для изучения
гипотензивного действия веществ и физических факторов
Исследуемое действие
вещества
1 этап - скрининг
(50 мг/кг ФС, внутрь)
(5 мг/кг ФС, внутривенно)
введение мезатона, тира-
мина, ангиотензина-1
и ангиотензина-2
Биомодель гипертоничес-
кой болезни на спонтанно-
гипертензивных крысах с
постоянным АД = 200 мм Нд
Изучение влияния ФС на
основные показатели цент-
ральной гемодинамики
Изучение эффективности
ФС при курсовом введении
(на животных с эксперимен-
тальной гипертонией)
Модель для оценки мио-
тропных свойств гипотен-
зивных лекарств
Биомодель спонтанной
гипертензии
(АД до 200 мм Нд) (у самок
с низким уровнем сыворо-
точных глюкозы, фосфора
и общих белков)
Биомодель для изучения
гипотензивных состояний
и гипотонии, сопровождае-
мых низкими уровнями
сывороточного железа
и йода
Биомодель спонтанной ги-
пертензии (АД > 180мм Нд)
с высоким общим уровнем
сывороточного холестерина
Лабораторное
животное
Крысы (с экс-
перимен-
тальной на-
следственной
гипертонией
Крысы
(нормотен-
эивные)
Кошки
Собаки
Крысы
Крысы
Кошки
Кролики
Собаки
Крысы
Кролики
Собаки
Кролики
Кролики
Кролики
Кролики
Линия или порода
животного
Аутбредные
Аутбредные
Аутбредные
Аутбредные
Линия SHR
Аутбредные
Аутбредные
Аутбредные
Аутбредные
Аутбредные
Аутбредные
Аутбредные
Аутбредные
Линии: A/J
B/J
Линия AC/J
Линии: ACEP/J
ACCR(B)
Пол
животного
9с?
9с?
§с?
яв
§с?
§с?
9с?
9с?
9
9
9
с?
Вес
животного
300-350 г
300-350 г
3-5 кг
12-20 кг
300-400 г
180-250 г
2-4 кг
2,5-5 кг
10-25 кг
300-350 г
2,5-4 кг
10-25 кг
2,5-4 кг
2,6 кг
2,0 кг
2-3 кг
2,4 кг
2,0 кг
Возраст
животного
5-12 мес.
5-12 мес.
1 -7 лет
1 -7 лет
5-18 мес.
5-12 мес.
1 -7 лет
1 -4 года
1 -7 лет
5-12 мес.
1 -4 года
1 -7 лет
7 мес-
4 года
7 мес-
4 года
7 мес-
4 года
7 мес-
4 года
7 мес-
4 года
7 мес-
4 года
В табл. 73 даны лишь некоторые моменты выбора оптимальных живот-
ных — биомоделей для изучения гипотензивных эффектов фармакологичес-
ких, токсикологических и иных средств. Не случайно говорят, что медицина
и кардиология стремительно меняется, а нитроглицерины неизменны. Речь,
конечно, идет о большой группе центральных и периферических вазодиля-
таторов. А в этом случае изменения наших взглядов меняются стремительно.
Глава 28. Кардиогемодинамика в биомоделях
409
Мы хотели бы ещё раз подчеркнуть роль оксида азота в эффектах лекар-
ственных средств. Несмотря на то, что действие N0 и участвующих в его
метаболизме веществ на кровообращение в норме и патологии достаточно
хорошо изучено, лишь в единичных работах исследуется взаимовлияние эф-
фектов N0 на сердечно-сосудистую систему и на другие физиологические
показатели. Относительно малые дозы веществ, оказывающие благоприят-
ное действие на сердечно-сосудистую систему, могут негативно влиять на
другие параметры, и наоборот. Данный аспект очень важен с точки зрения
возможного терапевтического применения средств, влияющих на синтез N0.
Предложенные выше биомодели могут стать основой для изучения этих про-
цессов. Иные биомодели, удобные для оценки системного действия веществ
или физических факторов, можно найти в таблицах других глав.
410
Глава 2 9 ВНУТРЕННЯЯ СРЕДА ОРГАНИЗМА
Греческое название красных кровяных клеток —
эритроциты. Греческое erytros означает красный,
a kytos — полость, кисту, клетку. Белые клетки на-
зываются лейкоцитами, от греческих leukos — бе-
лый, бесцветный, и того же — kytos.
Левенгук называл красные тельца еще глобула-
ми, шариками, но он не мог видеть в свой микро-
скоп, что на самом деле эритроциты имеют диско-
образную форму с углублением в центре каждой
стороны. В 1805 г. Йене Берцелиус получил из крас-
ных телец бесцветный белок и назвал его глобули-
ном, потому что считал, что получил этот белок из
глобул, шариков. Внутри телец глобулин соединен
с красящим веществом, которое тогда называли ге-
матином, от греческого haima — кровь. Всё вместе
называли поэтому гематоглобином, что со време-
нем сократилось до гемоглобина, как мы сегодня и
называем этот важнейший красный белок крови.
Кровь человека и животных в сравнении
Кровь и лимфа являются внутренней средой организ-
ма. Кровь человека и животных состоит из плазмы
и форменных элементов (эритроциты, лейкоциты и
тромбоциты), которые чрезвычайно широко исполь-
зуются в сравнительных исследованиях и биомоде-
лировании [477, 576].
Продуцентом форменных элементов крови яв-
ляется красный костный мозг, который по мере ста-
Глава 29. Внутренняя среда организма
411
рения организма заменяется жировой тканью и превращается в желтый
костный мозг. У большинства животных красный костный мозг находится
лишь в плоских костях, но у кроликов, крыс, белых мышей и морских свинок
он имеется и в трубчатых костях. В красном костном мозгу образуются
эритроциты, зернистые лейкоциты и тромбоциты. Эритропоэз у млекопи-
тающих происходит интраваскулярно в венозных синусах костного мозга
из эритробластов. Созревая, эритроцит проходит стадии нормобласта, ре-
тикулоцита, нормоцита, а затем поступает в кровоток. В небольшом коли-
честве в периферическую кровь поступают и ретикулоциты. В костном мозгу
экстраваскулярно образуются также лейкоциты в ретикулярных клетках
стромы костного мозга. Регуляция гемопоэза осуществляется нейрогумо-
ральным путем. Костный мозг имеет богатую сеть интерорецепторов. Важ-
ную роль играют факторы гуморальной регуляции (витамин В12, витамин
В6, фолиевая кислота, витамин С, микроэлементы — железо, кобальт, медь,
цинк).
Желтый костный мозг участвует в жировом обмене. Эндотелиальные
клетки венозных синусов костного мозга входят в состав ретикуло-эндоте-
лиальной системы.
Продолжительность жизни эритроцитов составляет в среднем 80-120 дней,
лейкоцитов — всего несколько дней. Разрушение старых эритроцитов в нор-
ме происходит главным образом в селезенке, где ежедневно гибнет 0,6—3%
всего количества красных кровяных телец. Пополнение эритроцитов идет за
счет молодых клеток, которые образуются и созревают в костном мозгу. Лей-
копоэз осуществляется в селезенке, лимфатических узлах и костном мозгу.
Моноциты образуются клетками ретикуло-эндотелиальной системы.
На долю крови у взрослого человека приходится примерно 6—8% общей
массы тела, а у детей, в связи с более высоким содержанием в организме
воды, — 8—9%. У взрослого это соответствует 4—6 л крови (нормоволемия).
Повышение общего объема крови называют гиперволемией, а снижение —
гиповолемией.
Реакция крови слабощелочная и в нормальных условиях поддерживается
буферными системами на одинаковом уровне (рН 7,35—7,50). Удельный вес
цельной крови 1,05—1,06, а удельный вес плазмы крови 1,029—1,034. Фор-
менных элементов в крови здоровых животных 40—45, а плазмы 55—60%. Вес
общего количества крови лабораторных животных по отношению к весу тела
составляет 5—8%.
В состав плазмы крови входят различные соли (электролиты), белки,
углеводы, липиды, ферменты, гормоны, витамины, промежуточные продук-
ты обмена веществ и газы. Плазма крови состоит из 90—92% воды и 8—10%
сухого вещества. Основную массу сухого вещества плазмы крови составляют
органические вещества и лишь 0,8—0,9% неорганические вещества. В 1 л
плазмы человека содержится 900—910 г воды, 65—80 г белка и 20 г низкомо-
412
Биомоделирование в регуляторных системах
лекулярных соединений. Удельная масса плазмы равна 1,025-1,029, а ее рН
незначительно колеблется в пределах 7,37—7,43, составляя в среднем (в ар-
териальной крови) около 7,4.
Плазма крови животных, как и человека, соломенно-желтого цвета и
содержит следующие фракции белков: сывороточный альбумин (4—5%), сы-
вороточный глобулин (около 2,5—3%) и фибриноген (0,4—0,5%). Сыворо-
точные глобулины путем «высаливания» разделяются на две группы: эугло-
булины и псевдоглобулины. Путем электрофореза глобулины расщепляются
на три фракции: а-, (3- и у- глобулины. В образовании антител важное зна-
чение придают у-глобулину.
В крови животных постоянно содержатся кислород, углекислый газ и
азот. Азот находится в плазме лишь в растворенном состоянии (около 1 об.%).
Кислород в небольшом количестве растворен в плазме, в основном же он
находится в связанном с гемоглобином состоянии в виде оксигемоглобина.
Его содержание в артериальной крови 17,8—20,58 об.%, в венозной крови —
6,97-14,88 об.%. Процент насыщения кислородом крови у собаки равен 93,6.
Углекислый газ находится в крови в трех состояниях: небольшая часть в виде
раствора в плазме; в виде двууглекислых щелочей; в виде непрочных хими-
ческих соединений с гемоглобином и белками плазмы крови. Углекислоты в
артериальной цельной крови содержится 39,21—49,47 об.%, в венозной —
47,18-50,02 об.%.
Белковые фракции плазмы крови у собак следующие (в % по отношению
к общему количеству белка): альбумин — 39,6, (Xj-глобулин — 16,9, ^-гло-
булин — 8, Р-глобулин — 13, у-глобулин — 9,3.
Электрофоретические фракции сыворотки здоровых собак (при общем
белке, равном 6,49%) составляют (%): альбумин — 50,3, oCj-глобулин — 5,3,
сс2-глобулин — 11,4, Pj-глобулин — 9, Р2-глобулин — 12,2, у-глобулин — 11,6.
Фибриноген относится к глобулинам, он имеет большое значение в свер-
тывании крови. Количество фибриногена в крови собаки 0,42—0,64% (в сред-
нем 0,52%).
Если принять вязкость воды за 1, то средняя относительная вязкость крови
у здорового взрослого человека составит 4,5 (3,5-5,4), а вязкость плазмы 2,2
(1,9—2,6). Увеличение гематокрита сопровождается более быстрым, чем при
линейной зависимости, возрастанием вязкости крови (т. е. ее внутреннего
трения).
Роль белков плазмы заключается в поддержании коллоидоосмотического
давления в сосудистом русле, транспортировке некоторых веществ (липоп-
ротеиды, витамины и т. д.) и выполнении защитных функций.
Пропердин является эуглобулином с молекулярным весом, в восемь раз
превышающим вес у-глобулина. В сыворотке содержится 0,02—0,03% про-
пердина. Пропердин инактивирует бактерии, простейшие, вирусы в присут-
ствии комплемента и ионов магния. Количество пропердина у разных видов
Глава 29. Внутренняя среда организма
413
животных неодинаково. Больше всего его содержится в сыворотке мышей
(титр равен 10—20 ед/мл) и особенно у крыс (20—50 ед/мл) — вот почему эти
животные обладают высокой степенью естественного иммунитета. У кролика
титр пропердина примерно равен содержанию его в сыворотке человека (4-
8 ед/мл). Наименьшее количество пропердина в сыворотке морской свинки
(1-2 ед/мл), с чем связывают большую восприимчивость этих животных к
инфекциям.
Таблица 74
Экспериментальные и генетически обусловленные биомодели для изучения
действия препаратов, влияющих на гемостаз
Исследуемое действие
вещества
Модель для оценки специ-
фической активности анти-
агрегантов
Модель для оценки воздей-
ствия антикоагулянтов на
плазменный и тромбоци-
тарный гемостаз с разви-
тием экспериментального
тромбоза
Модель для оценки специ-
фической активности анти-
коагулянтов прямого дейст-
вия, влияющих на тромбин
без помощи ATIII
Модель для оценки эффек-
тов новых гепариноидов,
а также антитромботической
активности антикоагулянтов
Модель для оценки гемор-
рагической активности
Модель для оценки актив-
ности антикоагулянтов не-
прямого типа действия
Модель для оценки специ-
фической активности тром-
болитических средств
Модель для оценки специ-
фической активности гемо-
статических средств мест-
ного и резорбтивного дей-
ствия
Лабораторное
животное
Кролики
Кролики
Крысы
Морские
свинки
Мини-свиньи
Кролики
Крысы
Кролики
Крысы
Морские
свинки (реже)
Крысы
Кролики
Крысы
Морские
свинки
Кролики
Собаки
Кролики
Крысы
Собаки
Линия или порода
животного
Аутбредные
Аутбредные
Аутбредные
Аутбредные
Аутбредные
Аутбредные
Аутбредные
Аутбредные
Аутбредные
Аутбредные
Аутбредные
«Шиншилла»
Линия Wistar
Аутбредные
Аутбредные
Аутбредные
«Шиншилла»
Аутбредные
Аутбредные
Пол
животного
9с?
9с?
9с?
9с?
9с?
9с?
§с5
9с5
9с?
9с5
9с5
9с?
9с?
§с?
Вес
животного
2,5-4 кг
2,5-4 кг
300-350 г
0,7-1 кг
35-55 кг
2,5-4 кг
300-350 г
2,5-4 кг
300-350 г
0,7-1 кг
300-350 г
3-4 кг
250-350 г
0,7-1 кг
2,5-4 кг
10-25 кг
2-4 кг
300-350 г
10-25 кг
Возраст
животного
7 мес-
4 года
7 мес-
4 года
5-12 мес.
6 мес-
2 года
1,5 года
7 мес-
4 года
5-12 мес.
7 мес-
4 года
5-12 мес.
6 мес-
2 года
5-12 мес.
7 мес-
4 года
5-12 мес.
6 мес-
2 года
7 мес-
4 года
1 -7 лет
7 мес-
4 года
5-12 мес.
1-7 лет
Кроме белков, имеются и другие азотсодержащие органические плазмен-
ные вещества (мочевина, мочевая кислота, аминокислоты, аммиак, креатин,
414
Биомоделирование в регуляторных системах
креатинин, пуриновые основания, гиппуровая кислота и индикан), которые
известны под объединенным названием остаточный азот и составляют в
норме 0,02-0,04%.
Характерной особенностью крови является способность свертываться.
Согласно современным представлениям, свертывание крови является слож-
ным ферментативным физиологическим процессом, в осуществлении кото-
рого участвует большое количество факторов. Одни из этих факторов (акце-
лераторы) ускоряют, другие (ингибиторы) подавляют внутрисосудистое свер-
тывание крови. Действие акцелераторов и ингибиторов взаимосвязано.
В настоящее время известны следующие факторы, участвующие в про-
цессе свертывания крови человека (многие из этих факторов присущи и
животным): фактор I — фибриноген, II — протромбин, III — тромбоплас-
тин, IV — кальций, V — проакцелерин, VI — акцелерин, VII — проконвер-
тин, VIII — антигемофилический глобулин, IX — Кристмас-фактор, X —
фактор Коллера.
Кроме указанных факторов, для свертывания крови необходимы пред-
шественник тромбопластина плазмы (ПТП) и играющие роль ускорителей
реакции I, II и 111 факторы пластинок. Процесс свертывания крови осуще-
ствляется трехфазно: образование тромбопластина, образование тромбина,
образование фибрина, не растворимого в плазме. Свертывание завершается
выпадением фибрина. Отсутствие или недостаток одного из перечисленных
факторов, а также задержка его активности приводят к нарушениям гемос-
таза, в то время как избыточное образование или чрезмерная активность
одного или нескольких компонентов свертывающей системы может способ-
ствовать ускорению свертывания крови. В здоровом организме человека и
животных внутрисосудистому свертыванию крови препятствует наличие
антикоагуляционной системы (гепарин, антитромбин) и установление дина-
мического равновесия между коагуляционными и антикоагуляционными
системами. Например, у собаки свертывание крови происходит на протяже-
нии 2,5—5 мин, но может затягиваться до 8 мин.
Морфологические элементы и состав крови
Количество эритроцитов, лейкоцитов и тромбоцитов зависит от различных
влияний внешней среды, вида и пород животных, способа и момента взятия
крови, пищевого режима и других факторов.
У собак диаметр эритроцитов в среднем составляет 7,19 мкм, минималь-
ная резистентность эритроцитов 0,54-0,58, максимальная — 0,33-0,41. Форма
тромбоцитов лабораторных животных и человека почти одинакова — это
овальные пластинки, но у собаки тромбоциты значительно крупнее. У соба-
ки средняя величина кровяных пластинок составляет 8,93 ± 0,25 мкм, а у
человека — 3,4 ± 0,04 мкм. Удобная модель?!
Глава 29. Внутренняя среда организма
415
Таблица 75
Генетически обусловленные биомодели для изучения анемий
Исследуемое действие
вещества
Биомодель аутоиммунной
гемолитической анемии
Биомодель для изучения
патогенеза макроцитарной
анемии
Биомодель транзиторной
анемии, обусловленной на-
рушением гемопоэтической
функции печени эмбриона
Биомодель гемолитической
анемии (аутосомный ген ha)
с явлениями иммунопатии
Спонтанно высокие уровни
гематокрита и гемоглобина
крови, как биомодель для
гематологии человека
Лабораторное
животное
Мыши
Мыши
Мыши
Кролики
Кролики
Линия или порода
животного
Линия NZB/BL
Линии:
C57BL/6-WV+.
C57BL/6J-WV/+
Линия ru ce f
Линия X/J
Линия A/J
Пол
животного
Яб
Яд
9<5
9<?
9
Вес
животного
30-40 г
30-40 г
25-30 г
2,5-4 кг
2,5-3 кг
Возраст
животного
3-10 мес.
3-10 мес.
3-10 мес.
7 мес-
4 года
7 мес-
4 года
Кровь животных и человека имеет весьма существенные различия по
количеству эритроцитов и содержанию гемоглобина, поэтому нельзя пользо-
ваться формулой вычисления цветного показателя крови, указанной для че-
ловека, при определении его у лабораторных животных. Многие экспери-
ментаторы допускали подобную ошибку, в связи с чем можно встретить за-
ниженные или завышенные цифры цветного индекса крови для здоровых
животных. Определять цветной показатель следует по формуле
_ ЕпхНЬ
°=НЬ^Ё' (97)
где G — цветной показатель, Еп — количество эритроцитов в 1 мм3 крови для
данного животного, НЬп — количество гемоглобина у исследуемого живот-
ного, Е и НЬ — количество эритроцитов и гемоглобина в норме.
У здоровых лабораторных животных, в том числе и у собак, цветной
показатель крови колеблется от 0,82 до 1,23 (в среднем 1,02). Количество
эритроцитов в 1 мм3 крови колеблется от 4,2 до 7,413 млн.
У собак обнаружено шесть факторов крови: А, В, С, D, Е, Е Некоторые
из этих факторов имеют характер естественных антител, но для определения
несовместимости практическое значение имеет лишь фактор А. Антитело
фактора А представляет собой гемолизин в агглютинин. Однако у собак
имеется только одна группа крови, аналогичная Ооф у человека, т. е. в сыво-
ротке крови собак имеются оба агглютинина, реагирующие с эритроцитами
крови человека групп А и В. Эритроциты собак не имеют агглютиногенов.
В связи с этим для первичной гемотрансфузии кровь можно брать от любой
416
Биомоделирование в регуляторных системах
другой здоровой собаки. При повторных гемотрансфузиях у собак могут на-
блюдаться анафилактоидные реакции. Возникают они ввиду наличия у до-
нора внегруппового изоантигена, на который организм реципиента после
первого переливания крови вырабатывает изоиммунные антитела, обуслов-
ливающие изоантигенную несовместимость. Выявляемые в сыворотке реци-
пиентов изоиммунные антитела сходны с Rh-антителами в изоиммунных
сыворотках человека. Можно использовать изоантигенную несовместимость
крови собак в качестве метода для изучения лекарственной десенсибилиза-
ции и изоиммунизации. Но немного истории.
В лесах индийской Бенгалии живет обезьяна-макак, которую Жан Оде-
бер в 1797 г. назвал резусом. В 1900 г. Карл Ландштейнер открыл, что чело-
веческая кровь может содержать одно из двух веществ или не содержать их
вовсе. Два года спустя было обнаружено, что человеческий организм может
содержать и оба вещества. Эти вещества были названы А и В. Таким обра-
зом, возможны четыре типа, или группы, крови: А, В, 0 и АВ. Люди с одной
группой крови отличаются от людей с другой группой крови наличием или
отсутствием у них определенных антигенов в эритроцитах и антител в плаз-
ме крови. Определяют группу крови по реакции гемагглютинации — склеи-
вания эритроцитов. Открытие Ландштейнера сделало возможным перелива-
ние крови.
В 1940 г. К.Ландштейнер и А.Винер обнаружили в крови макака-резуса
вещество, которое обозначили начальными буквами его латинского назва-
ния — Rh-фактор. Это — антиген, содержащийся в эритроцитах макаков-
резусов и людей. По наличию или отсутствию резус-фактора выделяют ре-
зус-положительные (около 85 % людей) и резус-отрицательные (около 15 %
людей) организмы. При переливании резус-положительной крови резус-от-
рицательным лицам или при беременности резус-отрицательной женщины
резус-положительным плодом возможны осложнения. А как у животных?
Кровь различных кошек совместима, поскольку изогемоагглютинация
встречается крайне редко. У кошек описано две группы крови с факторами
А (85% случаев) и В (15% случаев) с соответствующими им антителами —
агглютининами. Общее количество крови составляет в среднем 1/20 часть
веса тела, или 5%.
Количество эритроцитов в 1 мм3 крови может колебаться от 6,6 до
10 млн/мкл. Диаметр эритроцитов составляет 5—7 мкм (в среднем 6 мкм).
Количество ретикулоцитов составляет 0,2% общего числа эритроцитов.
Кровяные пластинки у кошек больших размеров — 7,39 мкм, что делает их
удобными для изучения тромбоцитов, поскольку их величина у других
животных намного меньше. Общее количество тромбоцитов у кошек со-
ставляет в среднем 285 тыс. в 1 мм3. Гемоглобина в крови кошек содержит-
ся 60-90% (в среднем 12 г/дл); общее количество лейкоцитов — 17 тыс./
мкл (в среднем).
Глава 29. Внутренняя среда организма
417
Лейкоцитарная формула следующая (в %): нейтрофилы 61-75 (65); эози-
нофилы 6-8 (7); базофилы 0-2; лимфоциты (10-65 (30)) 30-22 (26); моно-
циты 1—3. РОЭ по Вестергрену: за 1 ч — 4 мм, за 2 ч — 10 мм.
Кровь можно взять пункцией большой подкожной вены нижней конеч-
ности (вены сафены).
Состав крови у мини-свиней
Количество эритроцитов, млн/мм3
Размер эритроцитов, мкм
- у поросят
Содержание гемоглобина в 100 мл крови
Количество тромбоцитов в 1 мм3
Размер тромбоцитов, мкм
Количество лейкоцитов 1 мм3
- у взрослых животных
- у молодняка
Размер лейкоцитов, мкм
- юные лимфоциты
- взрослые лимфоциты
- моноциты
- нейтрофилы
- эозинофилы
- базофилы
Лейкоцитарная формула (%)
- лимфоциты
- нейтрофилы
- базофилы
- моноциты
Минеральные вещества сыворотки, мг%
- кальций
- фосфор
- калий
Удельный вес крови (г/см3)
Показатель рН
Свертывание крови, мин
Объем крови от массы тела
Гематокрит, %
Резистентность эритроцитов
к раствору NaCI, %
Содержание сахара в крови, %
5-8(6,5)
6-6,2
5,8-6,0
1,6-16,8 г/мл
200 000-500 000 (300 000)
2
15 000-20000
10 000-15 000
5,5-11,0(9,3)
12,1-17,6(14,3)
11,0-22,0(14,4)
6,6-15,4(10,9)
9,9-16,5(10,7)
1,0-15,4(13,2)
18,7-61,7(36,8)
32,0-78,7(58,0)
0,0-1,3(0,3)
0,3-4,0(1,9)
9,04(5,37-12,7)
7,93(4,66-10,1)
24,15(19,30-24,25)
1,050-1,060
7,3-7,4
10-15
1/22(4,6%)
43,1
Удельный вес крови, г/см3
Показатель рН
Время свертывания крови, мин
Объем крови от массы тела
или 8 мл крови на 100 г массы тела
Гематокрит, %
Резистентность эритроцитов
к раствору NaCI, % (минимум )-(максимум
0,42-0,62
0,08-0,10
Состав крови у овец
1,051(1,041-1,061)
7,3-7,4
4-8
1/12-1/13
32.4
0,60-0,45
418
Биомоделирование в регуляторных системах
Состав
Количество эритроцитов, млн/мм3
Размер эритроцитов, мкм
Плотность эритроцитов
Содержание гемоглобина, г/Hg в 100 мл
Количество тромбоцитов в 1 мм3
Размер тромбоцитов, мкм
Количество лейкоцитов в 1 мм3
Лейкоцитарная формула, %
- базофилы
- эозинофилы
- нейтрофилы
- лимфоциты
- моноциты
Минеральные вещества сыворотки (мг%)
- кальций
«
- натрий
- калий
Состав крови у кошек
Эритроциты, млн/мкл
Гемоглобин, г/дл
Лейкоциты, тыс./мкл
Зрелые нейтрофилы, %
Лимфоциты, %
Моноциты, %
Эозинофилы, %
Тромбоциты (х 109/л)
6,0-7,34
9,9-12,9
14,8-19,9
60-71
22-30
1-2
5-8
285-500
Состав крови у морских свинок
Объем крови, мл/100 г
массы тела
Гематокрит, %
Гемоглобин, г/дл
Эритроциты, млн/мкл
Лейкоциты, тыс./мкл
Лимфоциты, %
Моноциты, %
Нейтрофилы, %
Эозинофилы, %
Базофилы, %
Билирубин, мг/мл
Глюкоза, мг/дл
5-7
40-50
11-15
4,5-7
5-12
45-80
8-12
20-40
1-5
1-2
0,24-0,30
50-120
у собак
5,5-8,0
7,2
2,4
11-18
200 000-600 000
2-3,5
8-18 000
1 (0-2)
3(2-4)
74(60-82)
22(13-32)
0,1 (0-0,3)
(крупные породы) 12,28 (12,02-12,54)
(мелкие породы) 8.37(7,6-9,9)
331,3(295,0-360,0)
20,32(16,7-27,87)
Состав крови у сирийских хомячков
Гемоглобин, г/дл
Гематокрит, %
Эритроциты, млн/мкл
Лейкоциты, тыс./мкл
Лимфоциты, %
Моноциты, %
Нейтрофилы, %
Эозинофилы, %
Базофилы, %
Билирубин, мг/дл
Глюкоза, мг/дл
Креатинин, мг/дл
16
40-50
7,5
7,6
75
3
30
1
1
0,5
120
0,5-0,1
Состав крови у кроликов
Объем крови, мл/100 г
массы тела
Гемоглобин, г/дл
Гематокрит, %
Эритроциты, млн/мкл
Лейкоциты, тыс./мкл
Лимфоциты, %
Моноциты, %
Нейтрофилы, %
Эозинофилы, %
Базофилы, %
Билирубин, мг/дл
Глюкоза, мг/дл
Креатинин, мг/дл
5-7
10-15
35-45
4-7
8-10
40-70
5-9
38-54
0,5-3,5
3-6
0,3
130
1,6
Глава 29. Внутренняя среда организма
419
Состав крови у крыс Состав крови у мышей
Гемоглобин, г/дл
Гематокрит, %
Эритроциты, млн/мкл
Лейкоциты, тыс./мкл
Лимфоциты, %
Моноциты, %
Нейтрофилы, %
Эозинофилы, %
Базофилы, %
Билирубин, мг/дл
Глюкоза, мг/дл
Креатинин, мг/дл
14,8
40-50
7,2-9,6
8-14
65-77
0-4
13-30
0-1
0
0,42
50-115
0,43
Гемоглобин, г/дл
Гематокрит, %
Эритроциты, млн/мкл
Лейкоциты, тыс./мкл
Лимфоциты, %
Моноциты, %
Нейтрофилы, %
Эозинофилы (%)
Базофилы, %
Билирубин, мг/дл
Глюкоза, мг/дл
Креатинин, мг/дл
12-14,9
42-44
8,7-10,5
5,1-11,6
63-75
0,7-2,6
6,7-37,2
0,9-3,8
0-1,5
0,18-0,54
108-132
0,5
Показатели крови инбредных животных
У линий мышей A/HeJ рН венозной крови из правого желудочка составляет
7,30 ±0,0011, рН артериальной крови из левого желудочка — 7,40 ± 0,009
[63]. Количество эритроцитов 9,48 ±0,18 млн/мм3, гематокрит 42,5 ± 0,5 %,
гемоглобин 12,7 ± 0,2 г/100 см3, лейкоцитов 6,14+0,68 тыс/мм3, ретикуло-
цитов 2,9%, гранулоцитов у самок 19,8 ± 2,3 %, у самцов 37,2 ± 7,5 % [63].
Систолическое давление крови 82 мм рт. ст., самое низкое содержание
ос-антитрипсина в сыворотке из 34 линий [604].
У инбредных мышей линии AKR/J рН венозной крови из правого желу-
дочка составляет 7,32 ± 0,008, рН артериальной крови из левого желудочка
7,39 ± 0,007 [63], эритроцитов 9,38 ± 0,24 млн/мм3, гематокрит 45,6 ± 1,0 %,
гемоглобин 13,9 ± 0,2 г/100 см3, ретикулоцитов 2,3%, лейкоцитов 6,80 ± 0,65
тыс/мм3, гранулоцитов у самок 21,6 ± 4,5 %, у самцов 23,8 ± 5,2 % [63]. Име-
ют высокоактивную каталазу крови [604].
У линий BALB/cJLac систолическое давление крови у 9-месячных сам-
цов составляет 105 ± 2,0 мм рт. ст., частота пульса 494 ± 11 уд./мин [63, 570],
эритроцитов 10,51 ± 0,16 млн/мм3, гематокрит 48,0 ± 0,7 %, гемоглобин 15,0
± 0,2 г/100 см3, ретикулоцитов 8,54 ± 1,04 тыс/мм3; гранулоцитов у самок
14,9 ± 2,1 %, у самцов 15,3 ± 3,8 % [63].
У линий CBA/J кровяное давление (систолическое) 97,4 ± 1,9 мм рт. ст.,
частота пульса 657 ± 8 уд./мин [63]. Эритроцитов 10,04 ± 0,27 млн/мм3,
гематокрит 45,0 ± 1,3 %, гемоглобин 13,5 ± 0,2 г/100 см3, ретикулоцитов
2,6%, лейкоцитов 6,44 ± 0,33 тыс/мм3, гранулоцитов 26,8 ± 4,5 % у самок,
24,2 ± 2,5 % у самцов [63], уровень лейкоцитов наименьший, уровень гра-
нулоцитов наивысший из 18 исследованных линий, рН венозной крови из
правого желудочка — 7,29 ± 0,009, артериальной крови из левого желудоч-
ка - 7,37 ± 0,012.
У линий CC57BR/Mv эритроцитов 6 млн в 1 мл, гемоглобина 15,4 г%,
лейкоцитов 10,7 тыс. в 1 мл. Отличаются большим содержанием гранулоци-
420
Биомоделирование в регуляторных системах
тов и базофилов в крови и клеток гранулоцитарного ростка в костном мозге
и селезенке. Гемоглобин «гомогенного» типа, аллель Hbbs [612].
У линий CC57W/Mv эритроцитов в крови 8,2 ± 0,4 млн/мм3, лейкоцитов
15,2 ± 1,3 тыс./мм3, самое низкое число нейтрофилов 1,8 ± 0,3. Диффузный
тип гемоглобина, аллель Heed [612].
У линий СЗН/HeLac соотношение белковых фракций (в % от общего
белка сыворотки) следующее: альбумины 3,38, ос-глобулины 0,092, |3-глобу-
лин 0,463, у-глобулин 0,058, рН венозной крови 7,27 ± 0,010, артериаль-
ной — 7,38 ± 0,014. Давление крови самое высокое из девяти испытанных
линий в любом возрасте. Среднее систолическое давление (в мм рт. ст.) в
возрасте 2-5 месяцев — 111 (колебания 95-138), 13-14 месяцев — 136 (114—
165), 31-32 месяца - 151 (138-164) [63].
У линий C57BL/6J давление крови среднее. В возрасте 9 месяцев у сам-
цов систолическое давление крови 93,3 ± 2,2 мм рт. ст., а частота пульса
633 ± 11 уд./мин. Значение рН артериальной крови самое низкое из 16 ис-
пытанных линий (рН артериальной крови в левом желудочке 7,36 ± 0,010,
венозной — 7,30) [63]. У 12-дневных эмбрионов мышей этой линии общее
число клеток красной крови (в сотнях тысяч на 1 мм3) и доля ядерных
предшественников эритроцитов (в %) на разных сроках развития 4,6 ± 0,4
(97%), у 13-дневных 5,4 ± 0,2 (61%), у 14-дневных 9,8 ± 0,4 (32%), у 15-
дневных 16,7 ± 1,0 (7%), у 16-дневных 26,6 ± 0,6. Число эритроцитов среди
18 испытанных линий у взрослых мышей среднее — 9,66 ± 0,09 млн./мм3,
гематокрит 44,0 ± 0,4 %, гемоглобин 13,3 ± 0,2 г/100 см3, число лейкоцитов
высокое — 11,43 ± 1,04 тыс/мм3, гранулоцитов низкое: у самцов 10,4 ±
3,5%; у самок 8,2 ± 1,2 % [63].
У линий C57L/JY гематокрит крайне высок, систолическое давление крови
97 мм рт. ст. [604]. Наиболее низкая активность глюкозо-6-фосфат-дегидро-
геназы в эритроцитах (из восьми испытанных линий) коррелирует с низким
содержанием в них никотин-аденин-динуклеотидфосфата [194].
У линий DBA/1J Lac имеют самую низкую рН венозной крови из 16
испытанных линий: в орбитальном синусе 7,23 ± 0,020, правом желудочке
7,26 ± 0,014; рН артериальной крови в левом желудочке 7,37 ± 0,011 [62];
высокое количество эритроцитов — 10,52 ± 0,27 млн/мм3; самое низкое ко-
личество ретикулоцитов — 1,5-10%о, лейкоцитов — 8,60 ± 1,60 тыс/мм3.
У линий DBA/2J самки в возрасте 35-42 дня при температуре 29° С по-
требляют кислорода больше, чем самцы. Систолическое давление крови
относительно низкое — 89,1 ± 2,0 мм рт. ст., частота пульса 614 ± 11 уд./мин
[63]. Количество эритроцитов относительно высокое: 10,3 ± 0,25 млн/мм3,
лейкоцитов 9,28 ± 0,29 тыс/мм3. рН венозной крови из правого желудочка
7,30 ± 0,007, артериальной крови из левого желудочка — 7,41 ± 0,009 [63].
У линий SJL/J отмечены: у{- и у2-парапротеинемия и низкое содержание
эритроцитов у самцов, среднее кровяное давление, самая высокая частота
Глава 29. Внутренняя среда организма
421
пульса из семи испытанных линий, низкая частота аудиогенных припадков,
амилоидоз печени и селезенки после 6 месяцев.
В стоке B/Lac показатели крови следующие: количество эритроцитов (в
миллионах на 1 мм3) — самки 4,28-4,95, самцы 5,98-5,99; гемоглобин (грамм
на 100 см3) — самки 12,3-14,4, самцы 13,7-15,36; гематокрит — самки 39,2—
44,4%, самцы 43,3-47,6%; количество лейкоцитов (в тыс. на 1 мм3) — самцы
4,50-6,39, самки 5,07-5,50 [643, 668].
В стоке ОМЗ/Lac показатели крови следующие: количество эритроцитов
(в млн. на 1 мм3) — самки 4,77—4,83, самцы 5,39—5,60; гемоглобин (грамм на
100 см3) — самки 12,44—12,9, самцы 14,3—14,31; гематокрит — самки 40,7-
42,1%, самцы 45,0—46,0%; количество лейкоцитов (в тыс. на мм3) — самки
5,74-6,02, самцы 7,70-10,79 [643, 668]. Вес тела: самки 848 г, самцы 943 г.
Относительный вес органов (граммы на 1 кг живого веса): селезенка— самки
1,22, самцы 0,77; тимус — самки 0,19, самцы 0,26; надпочечники — самки
0,34, самцы 0,30; мозг — самки 5,11, самцы 5,03 [643].
Селезенка в сравнении
У мини-свиньи селезёнка вытянутая, длинная, с суженными концами, на
поперечном разрезе треугольная; париетальная поверхность её плоская, а
висцеральная имеет вдоль всего органа гребень; по краю его лежат ворота.
Цвет селезёнки яркокрасный, консистенция непостоянная, но довольно
плотная; на разрезе ясно выступают фолликулы. Длина селезёнки 38—45 см,
ширина 5—8 см, вес 0,1—0,3% от общего веса тела. Лежит она дорзо-вент-
рально слева от желудка, дорзально граничит с левой почкой, вентрально—
с печенью и выступает несколько за последнее левое ребро.
В лаборатории биомоделирования НЦБМТ РАМН осуществлено изуче-
ние иммунологических показателей у светлогорских мини-свиней, которые
являются анатомически и физиологически нормальными животными, обла-
дающими небольшими размерами и массой тела. Работа проводилась на хря-
ках (7-дневные—60-месячные животные). Исследовали число лейкоцитов и
лимфоцитов в камере Горяева и пересчитывали на 1 мм3 крови у мини-сви-
ней в возрасте от 7 дней до 22 месяцев, а также число розеткообразующих Т-
лимфоцитов по методу Зальберга у животных в возрасте 7—22 месяца. Опре-
деляли число естественных киллеров на перевиваемом штамме опухолевых
клеток К562 у животных в возрасте 7—60 месяцев. Весь материал обрабаты-
вали статистически по методу Фишера — Стьюдента. Всего было исследова-
но 335 животных.
Исследование клеточных элементов иммунной системы у светлогорских
мини-свиней в возрастном аспекте показало, что число лейкоцитов и лимфо-
цитов возрастает у животных до 4-месячного возраста, оставаясь в дальней-
шем без изменений у животных до 14-месячного возраста, и затем несколько
422
Биомоделирование в регуляторных системах
понижается. Так, число лейкоцитов (тысяч в 1 мм3 крови) у 7-дневных живот-
ных равно 12,1 ± 1,12, у 4-месячных — 15,3 ± 1,24, у 12-месячных — 14,0 ±
0,86, у 22-месячных — 12,0 ± 0,80). Число лимфоцитов (тысяч в 1 мм3 крови)
у 7-дневных животных равно 8,2 ± 0,63, у 4-месячных — 12,6 ± 0,96, у 12-
месячных — 10,3 ± 0,85, у 22-месячных — 7,6 ± 0,62. Количество розеткооб-
разующих Т-лимфоцитов увеличивается с возрастом и достигает наивысшего
значения у 8-месячных животных, достигнув своего пика, оно практически
остается без изменений до 22-месячного возраста и несколько понижается у
50—60-месячных животных. Число естественных киллеров в возрасте 7—60
месяцев остается практически одним и тем же. Это указывает на особую роль
естественных киллеров в иммунологическом статусе организма.
У овцы селезёнка плоская, округло-треугольной формы, красно-корич-
невого цвета и довольно мягкой консистенции. Дорзо-краниальный угол её
на висцеральной поверхности несёт ворота. Лежит она слева на рубце, со-
единяясь с ним в большей своей части рыхлой клетчаткой.
У собаки селезёнка плоская, неправильной треугольной, очень непосто-
янной формы, вытянутая дорзо-вентрально. Вентральный конец её расши-
рен, дорзальный сужен. Передний край — с вырезкой, задний — более пря-
мой. В середине селезёнка несколько сужена и более тонкая. Цвет её крас-
ный, с несколько синеватым отливом, консистенция плотная, на разрезе
трабекулы и фолликулы незаметны. Селезёнка лежит косо каудо-вентраль-
но, вентральным концом выходит за край последнего левого ребра, достигая
сегментальной плоскости 2-го и даже 4-го поясничного позвонка и горизон-
тальной плоскости, проведённой через 7-е и 10-е рёберно-хрящевые соеди-
нения. Вес и величина селезёнки у собак в связи с многообразием их пород
ещё более изменчивы, чем у других животных; вес колеблется в пределах
0,08—0,4% от веса тела.
Селезенка покрыта соединительной капсулой. Кроме капсулы, имеется
строма, которая состоит из трабекул, густо переплетающихся между собой.
Паренхимой селезенки является пульпа, образованная ретикулярной тканью,
кровеносными сосудами и форменными элементами крови (красная пуль-
па). Кроме того, имеется и белая пульпа, состоящая из лимфоидных узелков
(мальпигиевы тельца).
В селезенке образуются лимфоциты и моноциты. Селезенка является
органом ретикуло-эндотелиальной системы, играет роль депо крови, уча-
ствует в обмене железа. У собак, как и у человека, селезенка имеет венозные
синусы. Селезенка, являясь продуцентом моноцитов, а они составляют до
8% от всех лейкоцитов крови, обеспечивают через них поставку в организм
цитотоксинов, лейкотриенов, интерлейкинов, интерферонов, факторов ро-
ста клеток и регуляторов воспаления
Основными кроветворными органами кролика являются костный мозг,
селезенка, лимфатические узлы, а также лимфатические образования ки-
Глава 29. Внутренняя среда организма
МЪ
шечника. Селезенка у кролика маленькая, темно-красного или темно-зеле-
ного цвета, удлиненной формы. Длина её 5 см, ширина около 1,5—2 см, вес
составляет 0,05% веса тела, причем с возрастом относительный вес селезен-
ки уменьшается. Костный мозг кролика, как и других грызунов, является
деятельным не только в плоских, но и в трубчатых костях.
Лимфатическая система
У самых мелких кровеносных сосудов стенки настольно тонки, что водная
фракция крови легко проходит сквозь них, омывая каждую клетку и образуя
так называемую интерстициальную жидкость. Это название она получила
потому, что заполняет межклеточные пространства. Латинское interstitium
переводится как «заполняющий между, промежуточный».
Интерстициальная жидкость по химическому составу близка к плазме
крови, но содержит меньше белков. Клетки, которые окрашивают кровь в
красный цвет, не могут пройти сквозь стенки сосудов, поэтому интерстици-
альная жидкость бесцветна, что и нашло отражение в ее названии: limpha
означает чистую воду, влагу.
Из межклеточных пространств лимфа попадает в мелкие сосуды, кото-
рые соединяются в более крупные, а те в свою очередь в еще более круп-
ные, и так до тех пор, пока наконец все они не образуют два протока,
идущие через грудную полость к шее. Здесь они впадают в вены, и таким
образом лимфа возвращается в ток крови. Тот из двух протоков, что по-
больше и располагается слева, называется торакальным или грудным прото-
ком. Сосуды и узлы, содержащие лимфу и отводящие ее из тканей и орга-
нов в венозную систему, называются лимфатическими и образуют лимфа-
тическую систему.
Лимфатические узлы располагаются по ходу лимфатических сосудов,
особенно на шее, под челюстью, в подмышечных впадинах. Они представ-
ляют собой небольшие уплотнения ткани. Древние анатомы сравнивали их
с желудями, поэтому и назвали гландами.
В лимфатических узлах (а также селезенке и костном мозге) образуются
клетки, которые называют лимфоцитами. Лимфоциты участвуют в борьбе с
болезнетворными бактериями, поэтому нередко при заболеваниях лимфа-
тические узлы увеличиваются в размерах, разбухают, становятся болезнен-
ными.
Лимфоидные органы у млекопитающих представлены солитарными фол-
ликулами; скоплениями фолликул, называемыми лимфатическими (пейеро-
выми) бляшками; миндалинами и лимфатическими узлами. В лимфатичес-
ких узлах происходит образование лимфоцитов, обезвреживание токсинов и
патогенных микроорганизмов.
Лимфатические сосуды напоминают вены, но мышечный слой стенок
424
Биомоделирование в регуляторных системах
значительно тоньше, хотя они и осуществляют сократительную функцию.
Лимфатические сосуды имеют клапаны. В грудном протоке на уровне пояс-
ничных позвонков, между ножками диафрагмы, находится поясничная ци-
стерна. Грудной проток вливается в краниальную полую вену, или в начале
левой яремной вены. В месте впадения лимфатического грудного протока в
венозную систему имеется один или два полулунных клапана.
У млекопитающих лимфа передвигается благодаря сокращению мышц
тела, присасывающей способности грудной клетки, а также сокращению
стенок самих лимфатических сосудов. Лимфатическая система включает в
себя начальные сети лимфатических капилляров, внутриорганные сплете-
ния лимфатических сосудов, внеорганные лимфатические сосуды с лимфа-
тическими узлами и главные лимфатические протоки, которые соединяются
с венозной системой.
Среднее количество лейкоцитов лимфы грудного протока собаки состав-
ляет 10 511 в 1 мм3, из них лимфоцитов — 86%, больших мононуклеаров —
5,1%, эозинофилов — 2,5%, полиморфно-ядерных нейтрофилов — 1,2%.
В грудном протоке лимфа содержит 94—96% воды, от 2 до 4,5% белка (из них
0,05% фибриногена), 0,1% сахара, 0,2-0,9% жира (количество жира резко
возрастает при поедании жирной пищи).
В периферической лимфе, кроме лейкоцитов, имеются и эритроциты. В
периферической лимфе конечности собаки в среднем содержится около 550
лейкоцитов в 1 мм3, и из них более 50% лимфоцитов.
В лимфе содержится значительно меньше белка (0,3—3,3%), чем в плазме
крови, что объясняется слабой проницаемостью стенок капилляров для бел-
ка. Лимфа, хотя и содержит фибриноген и протромбин, свертывается значи-
тельно медленнее, чем кровь.
• По количественному содержанию небелкового азота и сахара химичес-
кий состав лимфы мало отличается от состава плазмы крови. Но количество
хлоридов в лимфе несколько большее, а по содержанию кальция и особенно
органического фосфора лимфа беднее плазмы крови. Лимфа содержит маг-
ний, железо и ферменты (диастазу, липазу и гликолитический фермент).
Реакция лимфы щелочная, удельный вес 1,010-1,018 г/см3. С возрастом
интенсивность лимфообразования уменьшается. Так, у молодых собак из
грудного протока вьщеляется лимфы значительно больше, чем у старых. Через
трудной лимфатический проток за сутки вьщеляется в среднем (63—64 мл
лимфы на 1 кг веса собаки).
У собак насчитывается около 60 лимфатических узлов, паренхима кото-
рых состоит из ретикулярной ткани, группирующей фолликулы.
Важнейшие лимфатические узлы мыши: lymphocentrum subiliacum,
lymphonodulus axillaries, lymphocentrum mandibulare.
Глава 29. Внутренняя среда организма
425
Забор крови и пункция сердца
Очень важным этапом в исследованиях на мини-свиньях является взятие крови.
Основной недостаток свиньи как лабораторного животного — это отсутствие
поверхностных кровеносных сосудов, пригодных для взятия достаточно боль-
ших объемов крови (10-15 мл). В связи с этим разработан ряд методов для
получения образцов крови. Среди наиболее доступных, используемых, в
частности, в нашей практике, является способ получения крови путем пун-
кции орбитального синуса, находящегося у внутреннего угла глаза животно-
го. Для этого хорошо использовать специальные заточенные иглы длиной
10—12 см с наружным и внутренним диаметрами соответственно 3 и 1,5 мм
[222, 297]. Другие исследователи предлагают взятие крови из подхвостового
синуса введением иглы между 4-м и 5-м или 5-м и 6-м копчиковыми по-
звонками [233].
Все более широкое использование свиней в качестве модели часто требу-
ет многократного взятия крови, а также раздельного получения образцов
артериальной и венозной крови из различных органов, необходимых для
биохимических исследований, анализа колебания гормонов, скорости вса-
сывания различных лекарственных препаратов и многого другого. С этой
целью применяют метод введения катетеров в магистральные сосуды (аорту,
полые вены и т. д.) или в крупные сосуды различных органов. Исследователи
предлагают способы введения катетеров, изготовленных из полиэтилена [549],
тефлона [663], полиуретана [100], силиконизированной резины [671] и дру-
гих материалов, в яремную вену, печеночную, воротную, нижнюю полую и
подкожную брюшную вены [43, 202, 411, 494, 673]. Катетеризировать можно
и артериальные сосуды, в частности, аорту и сонную артерию [268]. Возмож-
на катетеризация не кровеносных сосудов и протоков, например, печеноч-
ного лимфатического и общежелчного протоков [186].
Пункцию сердца у овец и баранов лучше производить под наркозом.
Место предполагаемого укола освобождают от шерсти и дезинфицируют.
Указательным пальцем, смазанным настойкой йода, определяют сердеч-
ный толчок и прокалывают грудную клетку, отступив 2—3 мм от левого
края грудины. Прокол обычно производят в 11-м межреберье. Иглу вводят
на глубину 2—2,5 см. При попадании иглы в полость желудочка в шприце
появляется кровь, которая самотеком или при постепенном вытягивании
поршня заполняет его полость. Пункцией сердца удается получить 25—30 мл
крови.
Пункция сердца у собаки осуществляется после фиксирования животом
кверху. Обрабатывают место укола (выстригают шерсть и кожу смазывают
спиртом или раствором йода). Для пункции берут иглу длиной около 8—
10 см. Чтобы взять кровь из правого желудочка, укол производят в третьем
межреберном промежутке справа по парастернальной линии.
426
Биомоделирование в регуляторных системах
Иглу следует при этом держать перпендикулярно кожной поверхности.
Для получения крови из левого желудочка пункцию производят в третьем
межреберном промежутке по левой парастернальной линии. Можно прово-
дить пункцию сердца в месте наиболее сильного сердечного толчка, ощуща-
емого при пальпации. Если при первой попытке кровь в шприце не появля-
ется даже при легком вытягивании поршня, то следует иглу немного вытянуть
и затем, медленно продвигая ее внутрь, нащупать пульсацию сердца кончи-
ком иглы и проколоть его. У собаки средней величины можно взять без ущер-
ба для ее здоровья до 150—250 мл крови. После взятия крови нужно ввести
подкожно физиологический раствор в количестве, в два раза большем объема
взятой крови, или внутривенно ввести какой-либо кровезаменитель.
Капли крови у собаки можно брать из края уха или из мочки уха после
насечки, а также из мягкой части ступни после укола иглой. Перед взятием
крови выстригают шерсть и место укола дезинфицируют раствором йода или
спиртом.
Большие количества крови у собак берут из малой подкожной вены го-
лени, из подкожной вены предплечья или из наружной яремной вены. Тех-
ника забора отличается от внутривенных введений тем, что наложенный на
конечности жгут снимается лишь после окончания взятия крови. При пун-
кции одного из бедренных сосудов собаку фиксируют на столе животом
кверху. На одной из конечностей участок ниже пупартовой связки (место
укола) выстригают и дезинфицируют. Определяют пульсацию бедренной
артерии. Отступив на 2—5 мм медиальнее от пульсирующей артерии и дер-
жа шприц перпендикулярно сосудам, прокалывают вену. Постепенно про-
двигают иглу вглубь тканей и оттягивают поршень шприца до появления
темной венозной крови.
Таким же образом берут кровь из бедренной артерии, в 4—5 см от пупар-
товой связки. Появление алой крови и быстрое самостоятельное наполне-
ние полости шприца демонстрирует успех процедуры.
Из подкожных вен голени, предплечья и наружной яремной вены без
затруднения у собак можно взять 10—20 мл крови. При необходимости по-
лучить максимальное количество крови животное наркотизируют или под
местным обезболиванием отделяют наружную яремную вену, вставляют в
неё канюлю и производят кровопускание. Подобным образом кровопуска-
ние можно проводить и из общей сонной артерии.
У кошки кровь можно получить после надсечек ушных вен, а также про-
колом или надрезом ступни при соблюдении правил асептики.
Брать кровь у кошек можно из поверхностной вены голени и поверхно-
стной вены предплечья. Для этой цели животное следует надежно фикси-
ровать. Пережимают вену наложением жгута на соответствующую конеч-
ность и производят пункцию вены. Венопункцией можно получить 2—5 мл
крови.
Глава 29. Внутренняя среда организма
All
Максимальное количество крови можно получить под наркозом, отпре-
парировав и вскрыв наружную яремную или бедренную вены, а также арте-
рии. После взятия крови останавливают кровотечение и зашивают рану. Кровь
можно получить пункцией сердца.
У кролика кровь можно получить путем надреза или прокола краевой
вены уха. Проколы следует делать на мелких ветвях ушной вены, начиная с
верхушки уха. Для получения 2—5-10 мл крови наружную поверхность уха
покрывают тонким слоем жидкого парафина, и ухо с внутренней стороны
протирают ксилолом, после чего прокалывают или надрезают вену и выте-
кающую кровь собирают в пробирку. Под наркозом кровь у кролика можно
брать после отсепарирования и вскрытия бедренной или наружной яремной
вены. Максимальное количество крови получают после вскрытия общей
сонной артерии, причем перерезанный сердечный конец сосуда вставляют в
колбочку, в которую вытекает кровь, а на мозговой конец накладывают ли-
гатуру. Этим путем у взрослого кролика можно получить 50—70 мл крови и
сохранить ему жизнь, для чего перевязывают конец артерии, из которой
вытекала кровь, зашивают рану и сразу же под кожу или внутрибрюшинно
вводят физиологический раствор, подогретый до 37° С.
У морских свинок кровь можно брать из кавернозного синуса Пети. Для
этого во внутренний угол глаза, между орбитой и глазным яблоком, прово-
дят иглу вдоль кости в горизонтальном направлении и шприцем насасывают
кровь. Пункцию сердца у морских свинок проводят под наркозом, и кровь
получают вакуумным способом. Перед пункцией выстригают шерсть и де-
зинфицируют кожу. Местом пункции служит II межреберье слева на 1—1,5 см
краниальнее от конца procesus xyphoideus. От левого края грудины отступают
на 2 мм и вертикальным уколом прокалывают грудную клетку. Если игла
находится в желудочке сердца, то при потягивании поршня кровь поступает
в полость шприца. Если в шприц поступают пузырьки воздуха или серозной
жидкости, это указывает на то, что игла находится в легких. В таких случаях
иглу нужно вытянуть несколько назад. Пункцией сердца у крупных морских
свинок можно получить до 10—12 мл крови. Забор следует производить не
чаще одного раза в неделю.
У крысы кровь можно получить после нанесения насечек на ушах или
хвосте. Для получения больших количеств крови у взрослых крыс прибегают
к пункции хвостовой вены. Хвост обогревают теплой водой, дезинфициру-
ют; вену сдавливают у корня хвоста, вводят в сосуд иглу и шприцем отсасы-
вают кровь. Нередко для этого обрезают кончик хвоста, после чего собирают
кровь, вытекающую из раны. Из кончика хвоста удается получить значи-
тельное количество крови, вакуумно отсасывая ее.
Кровь из бедренной вены берут под наркозом. Вену отпрепарируют и
вскрывают. Из вскрытого сосуда кровь накопляется в ране. После взятия
крови рану тампонируют и зашивают.
428
Биомоделирование в регуляторных системах
Весьма удобно брать кровь у крыс, а также у других мелких лабораторных
животных из ретробульбарного венозного сплетения при помощи пастеров-
ской микропипетки (кончик пипетки должен быть слегка заточенным и иметь
в диаметре не более 1 мм). Для этого крысу захватывают за кожу шеи боль-
шим и указательным пальцами, а другими пальцами надежно удерживают за
кожу спины. Концом пипетки пробуравливающими движениями прокалы-
вают конъюнктиву внутреннего угла глаза и проводят ее на глубину 1-2 мм
за глазное яблоко, где находится венозное сплетение. При правильном вве-
дении в капилляр микропипетки из ретроорбитального сплетения самоте-
ком поступает кровь. При необходимости взять большое количество крови
следует натянуть кожу в области шеи, чтобы сдавить яремные вены и создать
венозный застой, т. е. повысить венозное давление в ретроорбитальном ве-
нозном сплетении. Описанный способ прост, позволяет брать кровь при
хронических наблюдениях. Редко возникают осложнения в виде поврежде-
ния глаза и его слепоты.
Пункция сердца у крыс. У обездвиженных животных выстригают шерсть
в области предполагаемого укола и дезинфицируют кожу. Пальпаторно оп-
ределяют место конечного толчка сердца. На 1 см краниальнее от установ-
ленной точки, отступив на 1—2 мм от левого края грудины, делают укол,
держа иглу вертикально. Пункцией сердца у крупных крыс удается получить
до 6—8 мл крови. При пункции сердца лучше пользоваться вакуумным мето-
дом отсасывания крови. Пункцию проводят не чаще одного раза в неделю.
После взятия крови подкожно вводят 0,9%-ный раствор хлористого натрия.
У мыши кровь (две-три капли) удается получить после прокола лапы или
ампутации кончика хвоста (перед обрезанием хвост подогревают водой с
температурой 40—45° С и дезинфицируют). После взятия крови рану на хво-
сте прижигают раствором йода. Можно получать кровь у мышей из хвосто-
вых сосудов вакуумным способом. Кончик хвоста смазывают ксилолом и
отрезают его. На рану наносят каплю 5%-ного раствора лимоннокислого
натрия, после чего хвост вставляют в одно из отверстий специального бал-
лончика, стенки которого парафинированы. Сквозь второе отверстие ваку-
умным насосом отсасывают воздух и при давлении 20—40 мм рт. ст. в про-
бирку насасывают кровь. Указанным способом получают 0,2—0,5 мл крови,
сохраняя жизнь животного. Повторное взятие крови производят через два—
пять дней.
Весьма удобным является взятие крови у мышей из ретроорбитального
венозного сплетения. Техника взятия крови такая же, как и у крыс. Кончик
микропипетки должен быть заточен под углом 45°. Микропипеткой у мы-
шей можно взять кровь и из подъязычного венозного сплетения. Для этого
левой рукой следует удерживать мышь за кожу шеи, слегка натягивая ее и
сдавливая вены шея. Пинцетом открывают рот, поднимают язык, прокалы-
вают кончиком микропипетки слизистую в области подъязычного венозно-
Глава 29. Внутренняя среда организма
429
го сплетения, которое отчетливо выступает. В капилляр пипетки кровь по-
ступает самотеком.
У мышей перед пункцией сердца выстригают шерсть и дезинфицируют
место укола. Животное фиксируют животом кверху. Пальпаторно определя-
ют место наилучшего ощущения сердечного толчка и на 4-5 мм краниаль-
нее от этого пункта у левого края грудины делают прокол. Пункцией сердца
удается получить до 0,5 мл крови. Путем декапитации у мыши получают
примерно 0,5—0,6 мл крови.
У сирийских хомячков небольшое количество крови можно получить из
хвоста, отсекая его кончик. Эту манипуляцию лучше производить под лег-
ким эфирным наркозом. Применяется вакуумный способ отсасывания кро-
ви при кардиопункции у кроликов и других мелких лабораторных живот-
ных, этим способом можно получить от взрослого кролика 60-70 мл крови
и сохранить ему жизнь. После взятия крови парентерально вводят теплый
0,9%-ный раствор поваренной соли в двойном объеме от взятой крови.
Техника пункции сердца хомячков и взятия из него крови такая же, как и
для других мелких лабораторных грызунов. Повторную пункцию сердца сле-
дует проводить не ранее чем через две недели. Под наркозом можно брать
кровь из v. jugularis и v. saphena anterior оперативным путем, освободив их от
окружающих тканей. Довольно удобно брать кровь у золотистых хомячков
из ретроорбитального сплетения при помощи тоненькой пастеровской пи-
петки. Принцип взятия такой же, как у крыс и мышей. Подобным образом
пастеровской пипеткой можно брать кровь из сублингвального венозного
сплетения.
430
Глава 3 0 СИСТЕМА ИНТЕСТИНАЛЬНЫХ
ОРГАНОВ
Несмотря на то, что процессы пищеварения счита-
лись важнейшими ещё в древние времена, только в
начале XIX в. возникло предположение, что в орга-
низме образуются особые вещества, которые осуще-
ствляют определенные химические реакции. Стало
понятно, что желудочный сок способствовал пере-
вариванию пищи и усвоению ее организмом.
Сначала эту способность приписывали только
действию соляной кислоты, содержащейся в желу-
дочном соке. Но в 1835 г. Теодор Шванн сообщил,
что в желудочном соке помимо соляной кислоты
содержится вещество, также обладающее перевари-
вающей способностью. Он назвал это вещество пеп-
сином от греч. pepsis — варить, растворять.
По-латыни «подниматься», «заставлять подни-
маться» —fermentare, это слово происходит от fervere
— гореть, кипятить. Отсюда название, данное впос-
ледствии веществам, — ферменты — и явлению —
ферментация (от лат. fermentum — закваска).
Поэтому в 1878 г. Вильгельм Кюне стал назы-
вать неорганические ферменты энзимами, что зна-
чит «содержащийся в закваске», чтобы показать, что
по поведению они похожи на ферменты в дрожже-
вых клетках.
В 1897 г. Эдуард Бухнер обнаружил, что дрожже-
вой фильтрат обладает ферментативной способнос-
тью. Это свидетельствовало о том, что для проведе-
ния реакции важно присутствие клеточного сока, а
не самих клеток. Так было открыто бесклеточное
Глава 30. Система жтестинальных органов
431
брожение и доказано, что все ферменты сходны в своей основе независимо
от того, где они находятся — внутри или вне клетки. А термины «фермент»
и «энзим», образованные соответственно от латинского и греческого кор-
ней, сохранились оба, и ими пользуются в равной мере.
Стартовые процессы в сравнении
Рот служит началом пищеварительного тракта. Пища через пищевод попада-
ет в желудок. Из желудка она поступает в кишечник.
В нижней части кишечника кишка толще, чем в верхней части, и она
называется толстой кишкой. Тонкую кишку делят на три части. Первая часть,
выходящая непосредственно из желудка, называется двенадцатиперстной
кишкой.
Следующая часть тонкой кишки носит название тощая кишка. Римский
ученый и писатель Авл Корнелий Цельс считал, что в ней пища не перева-
ривается, а проходит не задерживаясь. Последняя часть тонкой кишки назы-
вается подвздошной кишкой (лат. botulus — колбаса). От этого слова произош-
ло название тяжелого токсинного заболевания — ботулизма.
Как у человека, так и у животных моторная и секреторная активность
желудочно-кишечного тракта регулируется вегетативной нервной системой.
ЖКТ обильно снабжен парасимпатическими и симпатическими нервными
волокнами. Преганглионарные волокна блуждающего нерва, выходящие из
продолговатого мозга, иннервируют пищевод, желудок, тонкий кишечник,
проксимальный отдел толстого кишечника, печень, желчный пузырь и под-
желудочную железу, а волокна, выходящие из крестцового отдела спинного
мозга, иннервируют сигмовидную кишку, прямую кишку и анус. Парасим-
патические волокна, иннервирующие ЖКТ, оканчиваются в ганглиях интра-
муральных сплетений или в ганглиях, расположенных в стенках слюнных
желез и печени. Нейромедиатором в преганглионарных нервах служит аце-
тилхолин, который взаимодействует с н-холинорецепторами, локализован-
ными на мембранах ганглионарных клеток. Ацетилхолин служит медиато-
ром и в постганглионарных нервных окончаниях, но в этом случае он вза-
имодействует с м-холинорецепторами эффекторных клеток. Серотонин и ряд
пептидов (вазоактивный интестинальный полипептид, энкефалины, веще-
ство Р и серотонин) выполняют роль постганглионарных медиаторов.
У человека слюна образуется и продуцируется со скоростью приблизи-
тельно 1 л в сутки. Она смачивает и очищает полость рта и пережеванную
пищу, и обладает бактерицидным действием лизоцима и ионов тиоцианата.
Под действием слюны начинается переваривание углеводов. При уменьше-
нии слюноотделения возникает чувство жажды.
Скорость кровотока через слюнные железы человека в покое составляет
0,1-0,6 мл/мин/г ткани, но может возрастать в 5 раз.
432
Биомоделирование в регуляторных системах
Удельный вес слюны близок к 1,0. Минеральными компонентами слюны
являются Na+, K+, С1~ и HCOj . Во время прохождения через протоки,
обладающие низкой проницаемостью для воды, слюна из изотонической ста-
новится гипотонической. Конечная осмолярность слюны составляет 2/3 ве-
личины осмолярности плазмы. Состав электролитов слюны зависит от ско-
рости секреции: при увеличении объема слюны концентрация Na+ и С1~
повышается, а концентрация К+ падает. рН слюны составляет 5,45—6,06, а
при стимуляции повышается до 7,8.
Белки являются самыми важными соединениями, содержащимися в пище.
Собаки, в рационе которых были только жиры и углеводы, умирали пример-
но через месяц. Герард Мульдер в 1839 г. назвал альбуминозные субстанции
протеинами (от греч. protos — первый, самый важный). По-русски протеины
— это простые белки. Некоторые белки, в том числе белок яйца, называют
альбуминами. Альбумин же самого куриного яйца называется яичным или
овальбумином (от лат. ovum — яйцо).
Кроме простых белков, протеинов, есть еще сложные белки, протеиды. В
их состав кроме аминокислот входят и небелковые вещества. Однако слово
протеиды употребляют не всегда, и тогда все белки без исключения называ-
ют протеинами.
К числу протеидов относятся, например, липопротеиды (липопротеины)
— протеины, соединенные с липидами, то есть жироподобными вещества-
ми, а также нуклеопротеиды (нуклеопротеины), имеющие в своем составе
нуклеиновые кислоты.
Ферменты слюнных желез включают амилазу, гликопротеины, мукопо-
лисахариды, лизоцим, иммуноглобулины и вещества, определяющие груп-
пу крови, ое-амилаза секретируется околоушными железами, а мукопроте-
ины — подчелюстными и подъязычными железами, сс-амилаза стабильна
при рН 4-11 и максимально активна при рН 6,9. Она гидролизует а-1,4-
гликозидную связь в полисахаридах, расщепляя крахмал на мальтозу и
мальтотриозу.
Состав слюны у мини-свиней
Количество слюны Различно в зависимости от вида кормления,
около 500-800 мл ежедневно
Удельный вес, г/см3 1,002-1,009
Содержание воды, % 98,0-99,0
Показатель рН 7,32
Неорганические составляющие Хлориды, фосфаты, бикарбонаты калия,
натрия и магния, следы сульфатов, нитратов,
аммония и солей железа
Органические составляющие Альбумины и глобулины, муцин,
ферменты: диастаза, следы мальтазы
Состав слюны у овцы
Количество слюны, от общего Обильное, 2/3 - в период приема корма,
количества 1/3-в жвачный период
Глава 30. Система интестинальных органов
433
Удельный вес, г/смэ
Содержание воды, %
Показатель рН
1,002-1,009
98,0-99,0
7,99-8,27(8,10)
Состав слюны у собаки
Количество слюны, мл
Удельный вес, г/см3
Содержание воды, %
Показатель рН
Неорганические составляющие:
Органические составляющие:
700-1000 ежедневно
1/3 - в жвачный период
1,001-1,008
98,0-99,0
7,56
Хлориды, фосфаты, бикарбонаты калия„натрия,
кальция и магния, следы сульфатов, нитратов
и аммиака
Альбумины, глобулины, муцин,
ферменты: диастаза
В последние годы предложено немало формально-логических, математи-
ческих и биологических моделей. Каждая из них отражает те или иные сто-
роны деятельности пищеварительной системы, сложных метаболических
процессов и важнейшей функции живых систем — функции питания. В ка-
честве примера мы остановимся на одной из простых математических моде-
лей, отражающих систему регуляции и обмена углеводов, а если более кон-
кретно, то Сахаров. Медико-биологические исследования были выполнены
на кроликах-шиншилах. В процессе исследования регистрировались глике-
мические кривые при действии инсулина на систему обмена глюкозы. Ма-
тематические модели построены на основе современных эндокринологичес-
ких концепций.
При построении модели будем исходить из следующих посылок. Кровь с
содержанием сахара в мг% обозначим у. Она снабжает желудок и кишечник,
печень, клетки нервной системы, поджелудочную железу и ткани организма.
В нашем построении мы исключим фармакокинетическую составляющую,
хотя, справедливости ради, следует отметить, что в этом случае мы имеем
многокомпартментную модель. Упростим нашу модель, выделив в качестве
основных параметров лишь те, которые касаются регулирования уровня са-
хара в организме. В этом случае наша система имеет следующие посылки.
Обозначим символом Sl глюкозу, поступающую энтеральным путем в желу-
док и из него через систему воротной вены в печень. Символом S обозначим
глюкозу, которая может быть введена парэнтерально. Символом s{ обозна-
чим препараты инсулина, вводимые внутримышечно, а символом s — эндо-
генный инсулин; Ьх — коэффициент пропорциональности и размерности.
\Slt 0<t<Ts
>*=)s,
0>t>x
*i
(98)
Мы полагаем, что концентрация сахара в воротной вене превышает теку-
щее значение сахара крови на величину yv В силу определенного регулятор-
434
Биомоделирование в регуляторных системах
ного механизма концентрация сахара на выходе печени (в печеночной вене)
будет также отличаться от текущего значения сахара крови на величину у2.
Ткани организма потребляют глюкозу и поэтому содержание сахара в крови,
оттекающей от тканей, отличается от концентрации сахара в притекающей
крови на величину у3, которая характеризует безвозвратное потребление са-
хара при расходе энергии. При превышении неких параметров сахара в кро-
ви и тканях поджелудочная железа выделяет инсулин s, влияющий на по-
требление глюкозы тканями. При снижении уровня сахара в крови и тканях
адреналовая система (кора надпочечников) активизирует процессы выработ-
ки и выброса глюкагона, адреналина и других факторов, способствующих
повышению уровня сахара крови. Гипоталамо-гипофизарный комплекс че-
рез релизинг — факторы и исполнительные органы — в зависимости от те-
кущего содержания сахара крови и затрат, необходимых организму для ре-
шения той или иной задачи, осуществляет управление сахаром крови. В даль-
нейшем мы рассмотрим элементы математической модели в отношении
разных систем организма и, в первую очередь, на уровне поступления глю-
козы в кровь и ткани из желудка.
Желудок и его сравнительные функции
Кардиальный отдел желудка содержит трубчатые железы с многочисленными
извитыми ответвлениями. Дно и тело желудка человека, составляющие 3/4
всего желудка, содержат прямые или слегка изогнутые железы, в стенках ко-
торых помимо аргентофинных и выделяющих слизь клеток присутствуют па-
риетальные или обкладочные клетки, секретирующие НС1, и зимогенные клет-
ки, секретирующие пепсиноген I и П. Пилорический отдел желудка человека
имеет разветвленные трубчатые железы, секретирующие слизь, а также
G-клетки, вырабатывающие гастрин. Слизь покрывает внутреннюю поверх-
ность желудка, образуя слой толщиной около 0,6 мм, защищая ее от механи-
ческого и химического повреждения. Слизистые клетки желудка человека сек-
ретируют Na+, K+, С1~, НСОз , а обкладочные клетки выделяют Н+, К+ и С1~
. При увеличении секреции активность обкладочных клеток в общем секрете
повышается; содержание Na+ снижается и полностью исчезает HCOj .
Как мы уже указывали, одними из первых были открыты ферменты, или
энзимы, пищеварительных соков, которые растворяют или переваривают
мясо. Это их свойство отражено в названии пепсина, который обнаружили
в желудочном соке. Вскоре были открыты новые ферменты, названия кото-
рых образовывали по аналогии с названием пепсина.
Пепсиноген представляет собой смесь предшественников протеаз. Под
действием НС1 он превращается в протеолитический фермент пепсин. Су-
ществуют 8 протеолитических ферментов. Пять быстро мигрирующих пеп-
синогенов, составляющих группу I, обнаруживаются только в области глав-
Глава 30. Система интестинальных органов
435
ных и обкладочных клеток. Остальные пепсиногены, составляющие группу
II, распределены по всему желудку и, кроме того, присутствуют в брунне-
ровых железах тонкого кишечника. Пепсин действует только в диапазоне
рН 1,8-3,5.
Внутренний фактор желудочного сока — это гликопротеин с молеку-
лярной массой 42000, секретируемый обкладочным клетками. Внутренний
фактор и белок, связывающий витамин В12, (R-белок, от англ. rapid —
быстрый, названный так в связи с высокой подвижностью при электрофо-
резе), способствуют всасыванию витамина В12. В кислом желудочном соке
витамин В12 связывается главным образом с R-белком. Высвобождающий-
ся витамин В12 взаимодействует с внутренним фактором. В12 всасывается в
кровь воротной вены. Часть поглощенного витамина В12 депонируется в
печени, а остаток в виде комплекса с белком транскобаламином II цирку-
лирует с кровью.
Остановимся на некоторых особенностях физиологии пищеварения мини-
свиней. Это — истинно всеядные животные. В природных условиях свиньи
питаются корешками растений, травой, желудями, орехами, листьями, хво-
ей, корой деревьев, корнеплодами, грибами, ягодами. Кроме того они по-
едают насекомых и их личинки, червей, некрупных змей, яйца птиц, гнездя-
щихся на земле, яйца рептилий, некрупных животных, таких, как мыши,
бурундуки, землеройки, кроты, не брезгают и падалью. В домашних услови-
ях или на фермах свиней кормят концентратно-картофельными и концент-
ратно-корнеплодными рационами в количестве 2—2,5% от веса тела для взрос-
лых животных.
В первые недели жизни у поросят анатомически и функционально не-
доразвит и пищеварительный аппарат. Установлено, что пищеварительные
органы (желудок, тонкие и толстые кишки) у них после рождения малы по
размерам, но отличаются очень интенсивным ростом. Рост желудка и его
вместимость у свиней имеет определенную специфику в разные периоды
постнатального онтогенеза. Поросята рождаются с относительно слабо раз-
витым желудком. К 10-дневному возрасту, когда живая масса поросят по-
чти удваивается, масса желудка увеличивается в три раза. К месячному
возрасту развитие желудка значительно опережает рост всего организма:
если живая масса поросенка увеличивается почти в три-четыре раза, то масса
желудка возрастает в 6 раз, а его емкость возрастает почти в 16 раз по срав-
нению с новорожденным. В целом за два месяца жизни масса желудка уве-
личивается в 22 раза, а его емкость в 46 раз, тогда как вес поросенка за этот
период увеличивается в 10 раз. Следовательно, в первые два месяца жизни
интенсивность роста желудка у свиней в 2 раза, а его вместимость почти в
5 раз превышают скорость роста всего организма. В последующем интен-
сивность роста желудка несколько снижается, однако продолжает опере-
жать рост всего организма. В подсосный период желудок поросят составля-
436
Биомоделирование в регуляторных системах
ет около 12% массы всего желудочно-кишечного тракта, а у взрослых сви-
ней до 18%.
В первые три недели жизни поросят в их желудочном соке нет соляной
кислоты, без которой ферменты желудочного сока (пепсин и химозин, пе-
реваривающие белки, и липаза, расщепляющая жиры) не могут нормально
функционировать. Если поросята в этот период питаются только молоком
матери, оно переваривается в тонких кишках под воздействием секрета
поджелудочной железы, сока кишок и желчи. Кишечное пищеварение в
это время компенсирует неполноценность желудка. Период, когда в желу-
дочном соке поросят нет соляной кислоты, и до возраста, когда она у них
появляется, называют периодом возрастной неполноценности желудка.
Изучение особенностей желудочного пищеварения у поросят на ранних
стадиях постэмбрионального развития показало, что возрастная неполно-
ценность желудка, выражающаяся в отсутствии свободной соляной кисло-
ты, имеет биологический смысл и является важным приспособительным
механизмом только в период молозивного питания.
В первые два дня жизни поросят всасываются интактные иммунные бел-
ки матери, находящиеся в молозиве, и тем самым создается пассивный
иммунитет новорожденных. Это возможно потому, что в желудке нет соля-
ной кислоты, осуществляющей гидролиз белков, а в молозиве в большом
количестве содержатся ингибиторы трипсина. Уже на 4-5-й день жизни по-
росят рН желудочного сока равен 4,5, т. е. достаточно кислая реакция, чтобы
створаживать молоко. Однако свободной соляной кислоты в этот период
пока еще не обнаруживается.
Попутно заметим, что состав молозива у свиней и человека практически
идентичны, и пассивный иммунитет у младенцев также создается через мо-
лозиво матери, поскольку в этом возрасте у младенцев нет соляной кислоты
в желудке. К 6—7-месячному возрасту пищеварительные органы у свиней
достигают размеров, достаточных для переваривания больших количеств кор-
ма и становятся вполне зрелыми как в анатомическом, так и в физиологи-
ческом отношении.
Специфической особенностью в деятельности желудочных желез у сви-
ней является непрерывное отделение секрета, не прекращающееся и при го-
лодании.
Пищеварение у свиней наиболее изучено и ему постоянно уделяется
внимание исследователей, так как свиньи на протяжении всей своей исто-
рии откармливались для получения высоко питательных пищевых продук-
тов, необходимых человеку.
В предыдущих главах мы рассмотрели некоторые посылки математичес-
кого моделирования регуляции уровня сахара в крови и тканях. Уместно
описать этот процесс на этапе всасывания Сахаров из желудка. Введение
глюкозы в желудок приводит за счет процессов всасывания в кишечнике
Глава 30. Система интестинальных органов
437
к повышению концентрации сахара в крови воротной вены. Учитывая спе-
цифику физиологического процесса, естественно предположить, что ско-
рость изменения концентрации сахара в крови воротной вены пропорцио-
нальна скорости всасывания и тем меньше, чем большего значения достига-
ет концентрация сахара в воротной вене:
^ + ад=(о, ,>*, ' (99)
где (7j(1) — удельная скорость всасывания глюкозы, т — время всасывания,
ах — коэффициент пропорциональности и размерности.
Естественно, что полученные соотношения отражают лишь одну сторону
процесса всасывания, а именно пассивное поступление его из желудка в
систему крови. Но мы специально огрубили систему математического опи-
сания, чтобы сопоставить его в единой системе счисления с процессами,
проходящими в других органах и, в первую очередь, в печени.
Регуляторные функции печени
Печень (hepar) представляет собой самую большую железу в организме, по-
строенную из колоссального количества работающих клеток. В настоящее
время значительное число экспериментальных исследований по пересадке
печени выполняется на свиньях. На карликовых свиньях изучены различные
способы консервации печени и их возможные последствия. Кроме того, было
показано, что эффективным средством профилактики одного из серьезней-
ших осложнений, возникающих при трансплантации печени — гиперкали-
емии, связанной с потерей гепатоцитами К+ и выходом его в кровь, является
диализ [457]. Стало возможным с большей степенью вероятности экстрапо-
лировать на человека данные, полученные при изучении биохимических
сдвигов, возникающих в крови реципиента после пересадки печени [33]. На
свиньях с успехом разрабатывается техника операций по орто- и гетеротопи-
ческой пересадке печени с анализом различных этапов операции и проблем,
связанных с функционированием трансплантата [443, 679].
Значение печени довольно разнообразно, так как она служит промежу-
точным органом (барьером) в кровообращении между желудочно-кишечным
трактом и всеми остальными частями организма, что дает возможность на-
ходить самые различные направления для биомоделирования и экстраполя-
ции [128, 238, 269, 315, 381, 682].
Одной из важнейших функций печени является обеспечение процессов
терминального оксиления. Это достигается участием системы цитохромов Р-
450. В последнее время стала известна роль в этом процессе оксида азота и
его ферментов. Так NO-синтаза на 58% гомологична цитохром-Р-450-редук-
тазе позвоночных и бактериальной сульфитредуктазе по аминокислотной пос-
438
Биомоделирование в регуляторных системах
ледовательности С-конца (641 аминокислота). Все эти энзимы уникальны
тем, что содержат сайты для FMN, FAD и NADPH в одном полипептиде.
Печень на своём вентральном крае имеет у большинства животных (со-
баки, кошки, мини-свиньи, овцы) то более, то менее глубокие надрезы, отде-
ляющие различной величины участки, называемые уже не дольками, а доля-
ми печени.
У млекопитающих печень — единственный (кроме лёгких) орган в теле,
в котором поступающая (по воротной вене) венозная кровь вновь растека-
ется по капиллярным сосудам и вступает в очень близкое соприкосновение
с печёночными клетками, изменяя свой состав. В печени происходит пред-
варительная обработка венозной крови с извлечением из её состава углево-
дов, откладывающихся в печени в виде гликогена, совершается синтез моче-
вины и нейтрализация весьма многих ядовитых веществ.
Относительная величина печени не одинакова: среди позвоночных она
больше у низших классов (рыбы, амфибии), среди млекопитающих она от-
носительно массивнее у собак по сравнению с травоядными. Пользуясь тем
же критерием в отношении печени лабораторных животных, можно выявить
большое сходство в строении этого органа у жвачных и у человека. У собак,
помимо средней доли с её подразделениями и отростками, особенность
присуща и правой доле, так как она в свою очередь из-за надреза распада-
ется на две части, из которых ближайшую к жёлчному пузырю принято на-
зывать правой медиальной долей, а правую краевую —правой латеральной
долей. То же самое имеет место и на левой доле, в результате чего и разли-
чают левую медиальную долю и левую латеральную долю. На печени мини-
свиней существуют те же соотношения, что и у собак, но квадратная доля
имеет вид треугольника, так как нижний конец жёлчного пузыря и круглая
связка сходятся друг с другом.
Печень у собак сравнительно большая, с отчётливыми долями, отделён-
ными друг от друга глубокими вырезками. Особенно сильно выступают че-
тыре доли: левые латеральная и медиальная, а также правые латеральная и
медиальная. Из них наиболее объёмистая левая латеральная доля.
Печень у мини-свиней относительно больше, чем у растительноядных, но
меньше, чем у хищных. Она обладает тремя вырезками и ясно распадается
на четыре доли: левые латеральную и медиальную доли и правые латераль-
ную и медиальную доли. Правая медиальная доля жёлчным пузырём на вис-
церальной поверхности отграничивается под воротами печени от небольшой
клинообразной квадратной доли, не достигающей своим вентральным зао-
стрённым концом края соседних долей. На поверхности печени заметны
очень маленькие, различной формы дольки, придающие органу рябоватый
вид, что характерно для печени.
Поскольку в ряде экспериментов в иммунологии, вакцинологии и ради-
ационной медицине используются лошади и жеребята, следует подчеркнуть
Глава 30. Система интестинальных органов
439
Таблица 76
Генетически обусловленные и экспериментальные биомодели
для изучения гепатопротекторов
Исследуемое действие
вещества
Модели острой патологии
печени, вызванной гепато-
токсикантами
Биомодель аутосомной тя-
жёлой формы желтухи, со-
провождаемой гемоглоби-
немией, гемоглобинурией,
субкапсулярными геморра-
гиями в печени и селезёнке
Биомодель врождённого
цирроза и амилоидоза
печени
Лабораторное
животное
Крысы
Мыши
Мыши
Хомячки
Линия или порода
животного
Аутбредные
или линейные
Аутбредные
или линейные
Линия ХД
Линии: 2.4
(LSH)
4.22
4.24
54.7
Пол
животного
9с5
9<3
9с*
9с?
96
Вес
животного
300-350 г
40-50 г
2,5-4 кг
120-125 г
120-125 г
120-125 г
120-125 г
Возраст
животного
5-12 мес.
3-10 мес.
7 мес-
4 года
6-18 мес.
6-18 мес.
6-18 мес.
6-18 мес.
существенные отличия в структуре печени по сравнению с другими живот-
ными. Печень лошадей лишена жёлчного пузыря. В ней, благодаря надрезу
на вентральном крае, можно выделить большую правую долю, а по локали-
зации круглой связки — левую долю, распадающуюся на левую медиальную
и левую латеральную доли в средней доле отношения остаются обычные.
Следует обратить внимание на характерное положение жёлчного пузыря:
у жвачных он сильно свешивается за пределы вентрального края, у всеядных
этого не отмечается; у собак он также не опускается, однако ввиду наличия
глубоких разрезов печени жёлчный пузырь виден не только с висцеральной,
но и с диафрагмальной поверхности, чего у многих животных не бывает.
Дольки печени по своим размерам обнаруживают значительные колеба-
ния, причём дольки самой малой величины свойственны собакам (955—
968 мкм в поперечнике), затем идут мелкие жвачные, за ними лошади
(1326 мкм), крупный рогатый скот (1380 мкм) и, наконец, свиньи (1570—
1700 мкм). Цвет печени в зависимости от кровенаполнения меняется в от-
тенках от бурого до красно-бурого. Лишь у молодых сосущих животных при
наличии жира оттенок оказывается желто-бурым.
Печень всегда занимает определённое положение, а именно тотчас поза-
ди диафрагмы в подреберьях. У одних собак она лежит в левом и несколько
больше в правом подреберье, тогда как желудок, наоборот, помещается не-
сколько больше в левом подреберье. У других животных печень сильно сдви-
нута в правое подреберье (лошади), а желудок большее место занимает в
левом подреберье. Наконец, у рогатого скота печень всецело лежит в правом
подреберье, а огромный желудок передними участками занимает почти всё
левое подреберье.
440
Биомоделирование в регуляторных системах
Гепатоциты печени человека и животных образуют пластинки толщиной
в одну клетку, разделенные узкими щелями (пространство ДиссеЛ представ-
ляющими собой заполненные кровью синусоиды, эквивалентные капилля-
рам. В стенках синусоидов имеются поры, через которые могут проходить
такие крупные макромолекулы, как альбумин и липопротеины. По мельчай-
шим канальцам желчь собирается в более крупные канальцы Геринга. Ка-
нальцы внутри долек печени и между ними сливаются, образуя печеночный
проток, который отходит к желчному пузырю. Печеночный и пузырный
протоки образуют общий желчный проток, открывающийся в двенадцати-
перстную кишку на вершине фатерова сосочка позади протока поджелудоч-
ной железы или рядом с ним.
Вырабатываемый в печени сок, или секрет, носит название желчи. Если
обтурация мешает выходу желчи из желчного пузыря, она начинает поступать
в кровь, и в крови появляются желчные пигменты. Желчные пигменты имеют
темно-зеленый или желто-красный цвет. Желто-красный пигмент желчи на-
зывается билирубин. Желчные пигменты, попадая в кровь, накапливаются в
подкожной клетчатке и придают коже нездоровую желтушную окраску.
С желчью выводятся конечные продукты обмена: билирубин, ксенобио-
тики, лекарства, токсины и др. Вьщеляемый с желчью холестерол регулирует
его баланс, а желчные кислоты способствуют эмульгированию и всасыва-
нию жиров.
При выборе биомоделей следует учитывать, что у мышей у линий C57BL/
6J активность печеночной каталазы снижена на 50% по сравнению с други-
ми линиями, тогда как в остальных тканях она не отличается от обычной.
Для этих мышей характерен низкий уровень холестерола в сыворотке [324]
и низкая концентрация жира в надпочечниках [604].
Желчь содержит мине-
ральные соли и слизь. В сут-
ки у человека выделяется
около 600 мл желчи.
Емкость желчного пузы-
ря человека составляет всего
50—60 мл, хотя печень сек-
ретирует желчь со скоростью
500-700 мл/сут.
Многогранность функ-
ций печени и регуляторных
механизмов, протекающих в
ней процессов отражена в
многочисленных публикаци-
ях и монографиях. Вряд ли
возможно в пределах совре-
Таблица 77
Состав печеночной и пузырной желчи человека
Компоненты
Na +
К +
Са2+
С1-
нсо3-
Желчные
кислоты
Лецитин
Желчные
пигменты
Холестерол
РН
Печеночная желчь,
ммоль/л
165
5
2,5
90
45
35
1
0,8
3
8,2
Пузырная желчь,
ммоль/л
280
10
12
15
8
310
8
3,2
25
6,5
Глава 30. Система интестинальных органов
441
менных возможностей математики дать всеобъемлющую модель ее функци-
онирования. Мы заведомо упрощаем задачу и рассмотрим только один из
аспектов работы печени, касающийся преобразования глюкозы в гликоген и
его депонирование.
Отмечена способность печени откладывать гликоген из сахара, притека-
ющего с кровью. Этот процесс осуществляется через систему контроля уров-
ня инсулина, его синтеза и либерации. Если текущее содержание сахара в
крови мало, то вследствие распада гликогена печень может повышать содер-
жание сахара в крови, увеличивая концентрацию^ в печеночной вене. Про-
цессу распада гликогена способствуют ряд регуляторных факторов, в том
числе адреналин. Повышенное содержание сахара в крови воротной вены в
силу инерционности процесса образования гликогена тоже способствует по-
вышению концентрации у2. Эти функции печени формально можно запи-
сать в виде следующего уравнения:
-^ + a4y2=a3yl+a2(y-yn)-b[i + clx-bAiu (ЮО)
где уп — регуляторные воздействия нейроэндокринной системы; а2, а3, а4, Ъ1У
Ь4, с, — коэффициенты пропорциональности. Член айу2 характеризует огра-
ниченную возможность участия печени в процессе образования и распада
гликогена.
Представленный элемент модели, описывая одну из функций печени,
дает возможность адаптивного моделирования. При желании это математи-
ческое описание может быть существенно расширено и интегрировано с
другими элементами регуляторной модели, в том числе с описанием процес-
сов выработки инсулина в поджелудочной железе.
Поджелудочная железа — двуликий Янус
Поджелудочная железа — pancreas — первоначально в филогенезе предста-
вляла собой серию небольших трубкообразных выпячиваний по всему тон-
кому кишечнику, и лишь впоследствии эти отпрыски, уменьшаясь в числе и
увеличиваясь в объёме, сконцентрировались у начала кишечника. Впрочем,
и в настоящее время у некоторых млекопитающих имеют место редкие слу-
чаи появления аномальных придатков панкреатических желез на двенадцати-
перстной, тощей и подвздошной кишках. В тонком кишечнике пища пере-
варивается за счет секретов поджелудочной железы, желчного пузыря и тон-
кого кишечника. Секрецию сока поджелудочной железы регулируют гормо-
ны секретин и холецистокинин, и блуждающий нерв.
Так, в 1874 г. Вильгельм Кюне открыл энзим в соке поджелудочной же-
лезы — трипсин, фермент, расщепляющий белки и второй фермент со сход-
ными свойствами — химотрипсин.
442
Биомоделирование в регуляторных системах
Существуют железы, не имеющие выводных протоков называющиеся
беспроточными или эндокринными. Вырабатываемые ими специфические
вещества — гормоны — выделяются непосредственно в кровь или лимфу.
Эндокринные железы называют иначе железами внутренней секреции.
В 1889 г. было установлено, что после удаления поджелудочной железы
подопытная собака жила не более нескольких недель, при этом у нее разви-
вались симптомы, сходные с симптомами, которые наблюдались у людей,
страдающих сахарным диабетом. Слово «диабет» означает «что-то, прошед-
шее сквозь». Его применяют для названия заболеваний, при которых моча
не задерживается в организме, а вьщеляется в избытке — жидкость проходит
«насквозь». При сахарном диабете в организме нарушается правильное рас-
пределение сахара в тканях, поэтому он накапливается в крови и выделяется
затем в повышенных количествах с мочой.
В 1916 г. английский физиолог Эдуард А. Шарпи-Шэфер предположил,
что островки Лангерганса являются эндокринной частью поджелудочной
железы — скоплениями клеток, которые производят гормон, регулирующий
уровень сахара в крови. А поскольку этот гормон продуцируется островка-
ми, или инсулами, то гормон этот Шарпи-Шэфер предложил назвать инсу-
лином. А когда этот имеющий пептидную природу гормон был выделен,
название «инсулин» так и осталось, как осталось в истории науки и имя
давшего его ученого.
Поджелудочная железа человека, имеющая массу около ПО г, способна
выделять в сутки 1,5 л секрета. Главный проток поджелудочной железы (вир-
зунгиев проток) проходит через всю железу и открывается в двенадцатипер-
стную кишку позади общего желчного протока, а у 30—40% людей вместе с
ним на верхушке большого дуоденального (фатерова) сосочка.
Панкреатический сок человека содержит основные анионы С1~ и НСОз ,
катионы Na+ и К+. Панкреатический сок изотоничен плазме крови. Панкре-
атический сок содержит Са2+, Mg2+ и Zn2+, а также сульфат и фосфат.
Концентрация катионов при стимуляции остается постоянной, а кон-
центрации С1~ и HCOJ меняются в противоположных направлениях. При
максимальной секреции концентрация HCOJ составляет 130-140 ммоль/л,
а рН равен 8,2.
Гидролазы белков панкреатического сока составляют почти 90%. Среди
них основными являются протеолитические ферменты: пептидазы и фосфо-
липаза А. Они секретируются в виде зимогенов, т. е. предшественников, под-
лежащих активации, тогда как липаза, амилаза и рибонуклеаза — в активной
форме. Активацию катализирует энтерокиназа, выделяемая слизистой две-
надцатиперстной кишки. Энтерокиназа катализирует превращение трипси-
ногена в трипсин. Трипсин активирует и другие протеазы. Ингибитор трип-
сина блокирует действие трипсина и препятствует перевариванию собствен-
ных тканей.
Глава 30. Система интестинальных органов
443
Активаторами экзокринного отдела поджелудочной железы являются
гормоны секретин и холецистокинин. Секретин стимулирует клетки, секре-
тирующие бикарбонат. Холецистокинин стимулирует ацинозные клетки,
секретирующие ферменты. Секретин действует на ацинусы, а холецистоки-
нин — на эпителий протоков. Вазоактивный интестинальный полипептид
по строению близок к секретину, а гастрин — к холецистокинину. Оба они
обладают значительно более слабым действием и конкурируют с гормонами
за рецепторы. Слабой стимулирующей активностью обладают также веще-
ство Р и нейротензин. Соматостатин и глюкагон угнетают секрецию панкре-
атического сока человека.
Значение для пищеварения поджелудочной железы, как поставщика в про-
свет кишечника различных ферментов, огромно. Она заключает в своей толще
панкреатические островки, играющие роль железы с внутренней секрецией.
Поджелудочная железа желтоватого цвета и обычно пронизана жировой
тканью. Как у человека, так и у животных она состоит из головки, левой и
правой (хвоста железы) долей. Железа в целом располагается приблизитель-
но в пределах двух последних грудных и двух первых поясничных позвонков.
Ее единственный выводной проток открывается на расстоянии 15—25 см от
пилоруса.
Интересные данные получены в опытах, выполненных на обычных и
карликовых свиньях с трансплантацией поджелудочной железы. При этом
было показано, что наиболее успешная пересадка наблюдается между сибса-
ми, когда даже через 4—6 месяцев после операции в ткани поджелудочной
железы сохранялось сравнительно много Р-клеток [492]. Удалось также уста-
новить, что в качестве материала для трансплантации островковых клеток
наиболее подходящим является поджелудочная железа 45-50-дневных пло-
дов поросят, поскольку у карликовых свиней эндокринные клетки подже-
лудочной железы созревают раньше, чем экзокринные [567]. Проводятся ис-
следования по созданию модели диабета и показано, что приемлемой явля-
ется модель «толерированного» экспериментального диабета с частичной пан-
креатэктомией и применением стрептозотоцина [672] (см. также табл. 78).
Поджелудочная железа у собак представляет сравнительно узкий, удли-
нённый орган. Правая доля железы расположена вдоль двенадцатиперстной
кишки, а в сторону желудка по большому сальнику от неё ответвляется левая
доля. Выводных протоков бывает один или два, а иногда даже три. Когда их
оказывается два, один из них, а именно поджелудочный проток, открывает-
ся вместе с жёлчным протоком на сосочке двенадцатиперстной кишки, а
другой добавочный проток — на расстоянии 3—5 см позади первого. У ко-
шек иногда наблюдается образование в виде поджелудочного пузыря (напо-
минает отношение жёлчного пузыря к печени).
Установить хотя бы приблизительно общую форму поджелудочной желе-
зы невозможно, так как у различных животных она значительно разнится:
444
Биомоделирование в регуляторных системах
она бывает то в виде неправильного треугольника (мини-свиньи), то в виде
вытянутой пластинки (овцы, козы), то в виде перегнутой почти по перпенди-
куляру ленты (собаки). Поджелудочная железа, как двуликий Янус, является
экзокринным и эндокринным органом. Экзокринные функции можно оце-
нить с помощью перорального введения флуоресцеина или парааминобен-
зойной кислоты, а также секретина — холецистокинина. По изменению их
концентрации оценивают эффективность панкреатической секреции
На заре химической науки, когда не существовало никаких приборов, рас-
познавание веществ осуществлялось на вкус. Именно так идентифицировали
класс кислых соединений. Таким же образом были открыты и сладкие ве-
щества. По-гречески сладкий — glykys, поэтому это слово применили для на-
звания одного из Сахаров, а именно глюкозы. В небольших количествах глю-
коза присутствует в крови, являясь источником энергии. Поэтому её ещё на-
зывают сахаром крови. В 1857 г. французский физиолог Клод Бернар открыл
в печени соединение, удивительно похожее на крахмал и при необходимости
образующее глюкозу. Он назвал его гликоген, то есть глюкозообразующий.
Наряду с экзокринными функциями, поджелудочная железа участвует
в системных процессах гормональной регуляции в организме человека и жи-
вотных. В поджелудочной железе островки Лангерганса состоят из а- и |3-
клеток. (3-клетки продуцируют гормон инсулин, влияющий на углеводный
обмен. Инсулин снижает содержание сахара в крови, способствует переходу
глюкозы в гликоген, активирует гексокиназу, препятствует образованию уг-
леводов из белка и жира, участвует в активизации синтеза белков из амино-
кислот и др. а-клетки островков Лангерганса вырабатывают гормон глюка-
гон, который проявляет антагонистическое действие по отношению к инсу-
лину. Активизируя фермент печени — фосфорилазу, глюкагон способствует
переходу гликогена в глюкозу и этим не допускает резкого снижения сахара
в крови при гиперинсулинемии. Кроме того, глюкагон посредством активи-
зации инсулиназы разрушает избыточное количество инсулина.
Клетки выводных протоков поджелудочной железы выделяют гормон
липокаин, продуцируемый поджелудочной железой, который предохраняет
печень от жировой инфильтрации, способствует выходу фосфолипидов из
печени в кровь, активизирует окисление жирных кислот в печени. Гормо-
ны, участвующие в поддержании постоянного уровня сахара в крови —
инсулин, глюкагон и соматостатин, секретируются островками Лангерганса
в поджелудочной железе. Эти структуры представляют собой скопления эн-
докринных клеток, так называемых а-, (3- и А-клеток, разбросанных в виде
групп по несколько тысяч клеток среди экзокринной ткани, составляющей
основную массу поджелудочной железы. Около 60% островковых клеток
приходится на (З-клетки, вырабатывающие инсулин, 25% — на а-клетки,
секретирующие глюкагон, и остальные 15% — на А-клетки, выделяющие
соматостатин.
Глава 30. Система интестинальных органов
445
Таблица 78
Генетически обусловленные биомодели
для изучения механизмов нарушения обмена веществ
Исследуемое действие
вещества
Биомодель для изучения
механизмов ожирения
Биомодель для изучения
нарушений пуринового об-
мена и подагры с высокими
уровнями общего азота
и мочевины крови
Биомодель для изучения
диабета типа II и стандарти-
зации антидиабетических
препаратов (дефицита
инсулина нет)
Лабораторное
животное
Мыши
Кролики
Мыши
Линия или порода
животного
Линии:
BRSUNT/NY
BRSUNT/NY
Линии:
A/J
B/J
Линии:
C57BL/KS-
db+/+m и
C57BL7KsJ-db
Пол
животного
яа
Яд
я
Яд
Вес
животного
30-40 г
2,6 кг
2,0 кг
30-40 г
Возраст
животного
3-10 мес.
7 мес-
4 года
7 мес-
4 года
3-10 мес.
Инсулин, образующийся в (3-клетках поджелудочной железы, представ-
ляет собой полипептид, состоящий из двух пептидных цепей. А-цепь, состо-
ящая из 21 аминокислотного остатка, соединена дисульфидным мостиком с
В-цепью, состоящей из 30 аминокислотных остатков. Инсулины животных
разных видов лишь слегка различаются по аминокислотной последователь-
ности и обладают одинаковым биологическим действием. Достаточно ска-
зать, что инсулин свиней отличается от человеческого лишь одной амино-
кислотой.
Суммарный результат различных метаболических эффектов инсулина
сводится к понижению концентрации глюкозы в крови, составляющей в
норме 0,8—1,0 г/л. При повышении уровня сахара в крови после приема
углеводов происходит выброс инсулина. Под влиянием инсулина увеличи-
вается поглощение глюкозы почти всеми клетками тела, и ее концентрация
в крови опять снижается. Считается, что превышение уровня сахара крови
над нормой включает механизм выработки инсулина островковой тканью
поджелудочной железы, поэтому можно считать, что скорость выработки
инсулина / пропорциональна превышению у над уп (где уп — норма) и тем
меньше, чем больше количество выработанного инсулина:
-^ + b5i = a2(y-yi), Приу>уп, (101)
где а2и Ь5 — коэффициенты пропорциональности.
Эта несложная математическая модель отражает регуляторные механиз-
мы выработки инсулина. При всей кажущейся простоте, ее достоинствами
является возможность соотнесения к вышеизложенным описаниям процес-
сов поступления глюкозы из желудка в систему кровообращения и утилиза-
446
Биомоделирование в регуляторных системах
ции глюкозы в гликоген в печени, а также ее интеграции с моделями глю-
кокортикоидной регуляции и утилизации глюкозы тканями, о чем речь пой-
дет ниже.
Глюкагон, образующийся, подобно инсулину, в альфа-клетках островков
Лангерганса, представляет собой полипептид. Он состоит из 29 аминокис-
лотных остатков. По своим функциям глюкагон является антагонистом ин-
сулина. Он стимулирует расщепление гликогена в печени (гликогенолиз),
обеспечивая таким образом быстрое повышение концентрации глюкозы в
крови при чрезмерном ее падении (гипогликемии).
Глюкагон, образуя комплекс с рецептором, расположенным на поверхно-
сти плазматической мембраны, активирует аденилатциклазу, которая пре-
вращает АТФ в цАМФ.
Соматостатин — это пептид, состоящий из 14 аминокислотных остатков.
Впервые он был обнаружен в гипоталамусе. Образующийся здесь соматоста-
тин угнетает синтез в гипофизе гормона роста, чем и обусловлено его назва-
ние. Впоследствии соматостатин был найден во многих тканях, где он вы-
полняет роль преимущественно ингибитора. В островках Лангерганса он
образуется в А-клетках и, действуя паракринным путем, угнетает секрецию
инсулина и глюкагона. Кроме того, он угнетает перистальтику желудочно-
кишечного тракта и желчного пузыря и уменьшает секрецию пищева-
рительных соков, вследствие чего замедляется всасывание пищи. Таким об-
разом, действие соматостатина направлено в целом на подавление пищевари-
тельной активности и, следовательно, на предотвращение слишком больших
колебаний уровня сахара в крови.
Финальные процессы в сравнении
Поскольку процесс пищеварения у человека и животных имеет определен-
ные различия, напомним о принципах утилизации и всасывания у людей
углеводов, белков, липидов, микроэлементов и воды.
Через тонкий кишечник человека за сутки проходит около 9 л жидко-
сти. Приблизительно 2 л поступают из крови и 7 л с эндогенными секре-
тами желез и слизистой кишечника. Более 80% этой жидкости всасывается
обратно в тонком кишечнике: около 60% в двенадцатиперстной кишке и
20% в подвздошной кишке. Остальная жидкость всасывается в толстом
кишечнике и только 1%, или 100 мл, выделяется из кишечника с каловыми
массами.
Особенности питания людей, находящихся в разных климатических зо-
нах, регионах, относящихся к различным этносам, столь многогранны, что
можно говорить лишь о некоем абстрактном рационе. При биомоделирова-
нии следует очень строго подходить к выбору лабораторных животных, ибо
конечная модель может быть существенно искажена!
Глава 30. Система интестинальных органов
447
Очень приблизительно можно сказать, что суточный рацион человека
включает 250—280 г углеводов, что эквивалентно 4,3—13,7 кДж/г. Если чело-
век не является чистым вегетарианцем, то он потребляет с пищей 70—90 г
белков в день.
Гидролиз белков осуществляется эндопептидазами (трипсин, химотрип-
син, эластаза) и экзопептидазами (карбопептидазы А и В). Эндопептидазы
расщепляют белки до олигопептидов, а экзопептидазы отщепляют амино-
кислоты от пептидов. Около 30% конечных продуктов гидролиза составляют
нейтральные и основные аминокислоты и 70% — олигопептиды, состоящие
из 2—6 аминокислот.
80—90% экзогенных и эндогенных белков всасывается в тонком кишечни-
ке. Около 50—60% белков пищи всасывается в двенадцатиперстной кишке и
около 30% — по мере прохождения химуса до подвздошной кишки. Пример-
но 10% белков достигает толстого кишечника, и расщепляются бактериями.
Некий «усредненный человек» потребляет в день около 60—100 г жиров.
До 90% жиров пищи составляют триглицериды, содержащие жирные кис-
лоты с цепью из 16 (пальмитиновая кислота) или 18 (стеариновая, олеи-
новая, линолевая кислоты) атомов углерода. Триглицериды, содержащие
жирные кислоты с цепью в 2—4 углеродных атома или 6-8 атомов, состав-
ляют небольшую часть жиров пищи. Примерно 10% жиров пищи приходят-
ся на фосфолипиды (лецитин), эфиры холестерола и жирорастворимые
витамины.
В слепую кишку человека поступает 1—1,5 л жидкости в день. Около 90%
этого объема всасывается в толстом кишечнике и только около 100 мл вы-
деляется. При постепенном поступлении жидкости всасывается до 5 л. Ионы
Na+ поступают в клетки кишечника путем простой диффузии за счет боль-
шого градиента концентраций (130 ммоль/л) и разности потенциалов (30 мВ).
Выведение Na+ из клеток осуществляется в базолатеральной области клеточ-
ной мембраны насосом, активируемым (Ыа+-К+)-АТФазой. Ионы К+ выхо-
дят по электрохимическому градиенту из межклеточного пространства в
просвет кишечника через относительно проницаемые для них плотные кон-
такты. Несмотря на высокую концентрацию ионов К+ в клетке (около
80 ммоль), лишь небольшая их часть диффундирует в просвет кишечника
непосредственно через клеточную мембрану. Ежедневно в толстый кишеч-
ник поступает 5—10 ммоль К+, а выводится с каловыми массами 10—15 ммоль,
при этом концентрация в последних К+ составляет примерно 90 ммоль/л.
Сорбция Na+ и экскреция К+ регулируются минералокортикоидами и
глюкокортикоидами. Эти гормоны стимулируют (Na+-K+)-ATOa3y, а альдо-
стерон повышает также проницаемость для Na+ апикальной области мемб-
раны клеток кишечника.
В толстом кишечнике человека активно всасываются хлорид и бикарбо-
нат Na+. Ежедневно в толстый кишечник поступает около 60 ммоль ионов
448
Биомоделирование в регуляторных системах
С1~ (и его концентрация здесь составляет около 60 ммоль), а выводится только
около 2 ммоль ионов. Поглощение С1~ сопряжено с выделением НСОз в
мембранных клетках. Накопление ионов НСОз в просвете кишечника обес-
печивает слабощелочную реакцию каловых масс как у человека, так и у ряда
животных.
Сравнительно-анатомические исследования позволили установить опре-
деленные различия кишечника человека и животных. Так средний, или тон-
кий, отдел кишечника мини-свиней состоит из весьма многочисленных мел-
ких мотков, вследствие чего длина его достигает 20 м. В этом отношении
свиньи, как всеядные, занимают промежуточное положение между хищны-
ми животными с коротким кишечником и растительноядными с длинным
кишечником.
Свинья вообще активно используется при изучении ожирения и, види-
мо, является идеальной моделью в применении к человеку [262]. Склон-
ность к ожирению у свиней объясняется значительно большей эффективно-
стью превращения энергии корма в жир, чем у других животных. В настоя-
щее время липогенез и обмен липидов у свиньи интенсивно изучается в ряде
лабораторий [474, 608, 667, 668]. Отмечено, что число жировых клеток у сви-
ней достигает максимума к 5-месячному возрасту [19]. Новорожденный по-
росенок по минимальному содержанию жира (1%) сходен с младенцем ма-
лой массы, что позволяет использовать поросенка в качестве модели при
изучении питания преждевременно родившихся детей. Такое использование
оправдано еще и тем, что физиология пищеварения и потребность в пита-
тельных веществах у поросят и новорожденных детей также имеет большое
сходство и дает возможность экспериментальной проверки рецептов детско-
го питания [531].
Особенно ценны поросята для изучения протеиново-углеводного голода-
ния, так как оно сопровождается такими же морфологическими и биохими-
ческими сдвигами, как и у детей с общим истощением, получавших рацион
с протеиново-углеводным дефицитом [41, 50].
С проблемой кормления тесно связано применение свиней для изучения
язв желудка и 12-перстной кишки, которые экспериментально легко моде-
лируются у этих животных введением, например, гистамина [296] или скар-
мливанием соответствующего рациона (желатинизированная кукуруза, яч-
менная солома в количестве 5-10% от рациона). Язвы индуцируются с помо-
щью потерь крови [371] и физического воздействия [48]. Немаловажным
обстоятельством является и то, что язвы, спорадически обнаруживаемые у
свиней, сходны с таковыми у человека [519]. В настоящее время многими
исследователями высказывается мысль, что свинья более пригодна для гас-
троэнтерологических исследований, чем собака [247, 317, 473, 548, 650].
Двенадцатиперстная кишка мини-свиней 40—90 см длиной, от пилоруса
желудка в правом подреберье тянется по печени и правой дорзальной части
Глава 30. Система интестинальных органов
449
брюшной стенки к тазу до заднего конца правой почки. Лимфатические
фолликулы и пейеровы бляшки у мини-свиней отчётливо выделяются. Пей-
еровы бляшки в количестве от 16 до 38 по направлению кзади заметно удли-
няются, растягиваясь на 50 см, а одна из них к концу подвздошной кишки
имеет длину от 1,15 до 3,20 м и даже простирается на толстый отдел.
Двенадцатиперстная кишка у собак сравнительно короткая, подвешена на
длинной брыжейке и по ширине просвета не уступает толстому отделу. Она
отходит от пилоруса в правом подреберье, направляется вдоль печени вправо,
дорзально и назад по правой стенке брюшной полости. Подслизистые железы
имеют чрезвычайно короткую область распространения от 1,5 до 2 см.
Лимфатические фолликулы очень маленькие. Пейеровы бляшки обнару-
живаются в различном числе, причем у молодых собак они несколько мно-
гочисленнее (14—25). Почти все они короткие — от 7 до 8,5 мм, и лишь одна
в конце подвздошной кишки у молодых собак растягивается на 10—40 см.
Резюмируя изложенные в этой главе сравнительные данные, следует под-
черкнуть, что каждая из медико-биологических наук своими методами пы-
тается наблюдать и описывать свойства сложных систем, в частности, обес-
печивающих пищеварение. Необходимо привлечение методов математичес-
кого моделирования. Однако классическая математика не в состоянии спра-
виться с огромным числом взаимодействий и переплетением переменных.
Это особенно заметно в интестинальных органах, когда кусочек пищи не
только разлагается на тысячи метаболитов, но дает и ещё нечто неосязаемое
— энергию. В главах 37 и 39 мы рассмотрим эти аспекты подробнее.
Конечно, можно использовать чисто литературные подходы, красиво и
афористично описав процессы пищеварения. Можно использовать сухие ма-
тематические термины, упростив и примитивизировав биопроцессы, но дав
им соответствие в общих системах. Осознавая ограниченность математичес-
кого моделирования, мы, тем не менее, отдаем им предпочтение хотя бы из-
за возможности проводить сравнительные и экстраполяционные построения.
450
Глава 3 1 РЕНАЛЬНАЯ СИСТЕМА
И САМОПОДОБИЕ ПРОКЛА
Явление самоповторения целого в собственных ча-
стях, многократного включения своих копий пре-
вратилось в общефилософскую категорию за 400 лет
до Рождества Христова, когда неоплатоник Прокл
начал провозглашать: «Все во всем». Вряд ли он имел
в виду строение почек, но последние, как ничто
лучше, подтверждают его парадигму. Дело в том, что
почки не просто парный орган, а орган, в котором
его субъединица — нефрон — является автономно
функционирующий частью. Поэтому биомоделиро-
вание функций нефрона является вполне благопри-
стойным процессом построения модели почки. Но
в то же время имеются значительные межвидовые
особенности строения почек, которые следует учи-
тывать при биомоделировании.
Итак, главными органами мочевыделительного
аппарата человека и животных являются почки.
Скорость клубочковой фильтрации обусловлена
интенсивным почечным кровотоком. Масса обеих
почек у человека составляет 300 г, т. е. всего 0,4%
средней массы его тела (70 кг). Через них проходит
1,2 л крови в минуту, это около 25% общего сердеч-
ного выброса, составляющего в покое 5 л/мин.
У некоторых видов млекопитающих в отличие
от человека эмбриональная дольчатость проявлена
так сильно, что у них и во взрослом состоянии об-
наруживают большое количество полностью отде-
лённых друг от друга маленьких почек, в силу чего
Глава 31. Ренальная система и самоподобие Прокла
451
весь орган представляет конгломерат одинаковых мелких образований—по-
чечник. Такую почку имеют медведи и китообразные млекопитающие. Такую
структуру относят к типу множественных почек.
Другой тип бороздчатой многососочковой почки свойствен, в частности,
рогатому скоту. У него же существует ещё небольшая особенность, заключа-
ющаяся в том, что стебельки от почечных чашек короткие и впадают обычно
в два крупных ствола, сливающихся в мочеточник.
К типу гладких многососочковых почек относят такие, у которых пери-
ферическая мочеотделительная зона слита в одно компактное образование,
так что с поверхности орган бывает гладкий, но при разрезе его ясно видны
сосочки, как, например, у свиней. У сосочков имеются почечные чашки, но
нет уже стебельков к мочеточнику.
Тип гладких однососочковых почек характеризуется слиянием в одно
компактное целое не только мочеотделительной зоны, но и отводящей; пос-
ледняя представляет у овец, собак и кошек сплошной, в виде гребня, продол-
говатый общий сосочек.
Опыты по пересадке почек между различными линиями гибридных кар-
ликовых свиней с известным фенотипом АГ класса I и II позволили вьывить
целый ряд антигенных факторов, влияющих на функции трансплантата и
высказать предположение, что отторжение трансплантата генетически опос-
редовано [520] не только у свиней, но, очевидно, и у других животных.
Сравнительные и холономные параметры почек
Почка человека содержит около 1,2 млн нефронов. Нефрон состоит из
клубочка с боуменовой капсулой, проксимального извитого канальца, петли
Генле и дистального извитого канальца. Дистальные извитые канальцы не-
скольких нефронов открываются в собирательную трубочку. Каждый нефрон
способен самостоятельно обеспечивать специфические виды транспорта и
экскреторные функции.
Почки человека и животных, как мы понимаем, не идентичны. Что же
их объединяет, и что не совпадает? В середине XX века крупнейший мате-
матик Дэвид Бом экстраполировал идеи Прокла целиком на Вселенную и
предложил холономную парадигму. Её суть в том, что наш мир — это некая
голограмма, каждая частичка которой содержит всю информацию о целом.
Поэтому различия в структуре почки человека и разных животных практи-
чески нивелируется самоподобием нефронов у них. Рассмотрим это несколь-
ко подробнее.
Почки имеют хорошо выраженные ворота. На разрезе почки различают
следующие зоны: корковую, или мочеотделительную, содержащую большое
количество кровеносных сосудов; мозговую, которая является мочеотводя-
щей частью почки, и пограничную.
452
Биомоделирование в регуляторных системах
Высокая скорость клубочковой фильтрации обеспечивает мощный кро-
воток через корковое вещество почек, где отфильтровывается около 1/5 объе-
ма почечного плазмотока. За сутки у человека образуется 170 л фильтрата,
но через почки за сутки проходит воды почти в 10 раз больше, чем содер-
жится во всем организме. Это достигается тем, что сосудистый клубочек и
капсула Шумлянского — Боумена формируют почечное (мальпигиево) тель-
це. Мальпигиево тельце вместе с канальцами составляет нефрон. Обычно
лишь 25—50% всех нефронов принимают участие в работе, а остальные на-
ходятся в «резерве». В сосудистых клубочках почек происходит фильтрация
и образование первичной мочи, которая по составу весьма близка к плазме
крови.
Проходя по извитым канальцам, первичная моча подвергается значитель-
ным изменениям, а именно: происходит реабсорбция воды, хлорида натрия,
аминокислот, сахара. В почках образуются и выводятся с мочой гиппуровая
кислота, аммонийные и фосфорнокислые соли.
Восходящее колено петли Генле контактирует с сосудами. В месте этого
контакта находится группа специальных эпителиальных клеток, которые
содержат многочисленные гранулы с ферментом ренином. Он представляет
собой пептидазу, расщепляющую ангиотензиноген-синтезируемый в печени
белок плазмы-с образованием декапептида - ангиотензина I. Под действием
превращающего фермента, он преобразуется в октапептид ангиотензин, ко-
торый стимулирует секрецию альдостерона и, участвует в регуляции тонуса
почечных артериол. Несомненно, ангиотензин II самое мощное сосудосу-
живающее вещество, образующееся в организме человека и животных.
Работа почек контролируется центральной нервной системой, вегетатив-
ным отделом нервной системы, гормонами задней доли гипофиза (антиди-
уретический гормон) и надпочечников — альдостерона и адреналина.
При угрозе избыточной потери NaCl из плотного пятна юкстагломерул-
лярного аппарата выделяется ренин. Ренин через ангиотензин II стимулиру-
ет продукцию альдостерона, реабсорбцию Na+ в дистальных отделах нефро-
на, образование мРНК, индуцирующей синтез мембранных белков. Поступ-
ление Na+ в клетку стимулирует натриево-калиевую АТФазу. Отношение
выводимых из нее ионов Na+ и входящих ионов К+ равно 3:2.
Однако вернемся к сравнениям функционирования почек у человека
и животных. Как известно, люди делятся на предпочитающих или исключи-
тельно употребляющих растительную пищу, и предпочитающих мясо. При
биомоделировании следует это четко учитывать. И вот почему.
При активации Ыа+-Н+-обмена в канальцевую жидкость поступает боль-
ше ионов Н+. Повышенная альдостероном Н+секреция приводит к внутри-
клеточному алкалозу, увеличивающему калиевую проводимость апикальной
мембраны. Ионы К+ накапливаются в канальцевой жидкости и выводятся
с мочой. При диете, состоящей преимущественно из мяса, в клетки поступает
Глава 31. Ренальная система и самоподобие Прокла
453
много К+ и мало Na+; кроме того, катаболизм белков дает избыток кислотных
ионов. Альдостерон у человека и животных препятствует нарушению тонкого
баланса электролитов, стимулируя К+ и Н+ секрецию и 1Ча+реабсорбцию.
Поэтому кошки и собаки более оптимальные животные для биомоделирова-
ния этого типа людей.
У людей, предпочитающих в системе питания молоко и его продукты,
субпродукты и растительную пищу, экскреция натрия должна усилиться, а
К+ и Н+ — ограничиться, что обеспечивается пониженным выделением аль-
достерона, а дополнительная экскреция натрия требует самостоятельного гор-
монального регулирования. Более оптимальными биомоделями для этого
типа людей будут мини-свиньи.
Известно, что в предсердиях образуется пептидный гормон, получивший
название предсердного натрийуретического фактора. Он повышает скорость
фильтрации и замедляет реабсорбцию Na+ в дистальном отделе нефрона.
Предсердный натрийуретический фактор накаливается в пузырьках в ткани
предсердий и высвобождается при нервной стимуляции, или при растяже-
нии. Объем внеклеточной жидкости зависит от содержания в организме
натрия.
На транспорт Na+ и С1~ влияют такие гормоны, как АДГ, ангиотензин II,
брадикинин и простагландины. Механизм их участия в поддержании гоме-
остаза до конца не понятен.
Прежде чем перейти к сравнительным характеристикам, следует напом-
нить, что почки являются эндокринным органом, поскольку они секретиру-
ют ренин и витамин D-гормон, а также эритропоэтин, играющий заметную
роль в эритропоэзе.
Почки у свиней по своему типу гладкие многососочковые. Как и у мелких
жвачных, они имеют почти правильную бобовидную форму, но бывают бо-
лее значительной длины и более плоски. Они одеты фиброзной капсулой и
часто полностью окружены жировой тканью (жировая капсула).
Почки мелких жвачных (овцы и козы) сильно отличаются от таковых
крупного рогатого скота. Они принадлежат к гладкому однососочковому типу
с ярко выраженной бобовидной формой. Продольный разрез показывает
наличие общего сосочка. Количество почечных пирамид дает основание
говорить о слиянии 10-16 долек.
Почки у хищников по строению относятся к гладкому однососочковому
типу, форма их, как и у большинства млекопитающих, близко подходит к
бобовидной, однако, особенно у семейства кошек, они короче и толще, чем
у мелких жвачных и свиней. Почки у хищников расположены под первыми
тремя поясничными позвонками. Правая почка несколько заходит в правое
подреберье, достигая печени. Они имеют бобовидную форму, покрыты фиб-
розной капсулой, окружены околопочечным жиром и почечной фасцией.
Снизу прилегает брюшина так, что они лежат забрюшинно.
454
Биомоделирование в регуляторных системах
Почки у собаки однососочковые с гладкой поверхностью. Они составля-
ют 0,5-0,71% веса всего тела. Особенностью внутреннего строения почек
собак и кошек являются очень длинные петли Генле, чем объясняется выра-
ботка у этих животных концентрированной мочи. Из почечной лоханки моча
через мочеточники попадает в мочевой пузырь, который легко растягивается
и может вмещать большое количество мочи. Мышечные волокна стенки
мочевого пузыря образуют опорожнитель, а циркулярные волокна в области
мочеиспускательного канала составляют замыкатель пузыря. За сутки круп-
ные собаки выделяют 1—2 л, а мелкие — 40—250 мл мочи. Правда, количе-
ство выделяемой мочи зависит от количества принятой воды и пищи. Удель-
ный вес мочи собаки колеблется в пределах 1,016—1,060 (в среднем 1,025).
У собак, питающихся мясом, реакция мочи кислая, а у находящихся на без
мясной диете — щелочная.
Моча собак и других животных содержит: общий азот — 0,26%; мочевую
кислоту — 0,006; мочевину — 0,46 мг%; креатинин — 0,009 мг%; золу —
2,00 мг%; СаО - 0,035 мг%; Р205 - 0,05 мг%; MgO - 0,05 мг%; H2S04 -
0,02 мг%; С1 - 0,3261; Si02 - 0,1 мг%.
Из физических свойств мочи собаки следует отметить высокую величину
депрессии (А), равную 3,29, и электропроводность — 26,1.
Почки у кошки короткие, толстые и округлые, имеют один сосок кони-
ческой формы, а признаки деления на доли отсутствуют. Вес почек состав-
ляет 0,34% веса тела. На их поверхности наблюдаются борозды от вен. По-
крыты почки очень плотной фиброзной капсулой. За сутки кошки выделяют
75—200 мл мочи. Физико-химический состав мочи кошек следующий: рН —
7,5; удельный вес — 1,055; величина депрессии — А4,73; электропроводность
при 18° С (10"3) 22,37; общий азот — 0,9878%; мочевая кислота — 0,008;
мочевина — 0,14; креатинин — 0,02; минеральные вещества — 2,47, в том
числе Р205 - 0,19; СаО - 0,098; MgO - 0,034; H2S04 - 0,103; Cl - 1,007;
Si02 - 0,008%.
Вес обеих почек кролика составляет 0,6—0,7% веса тела. Лежат они забрю-
шинно. Размеры почек взрослого кролика следующие (см): длина — 3, шири-
на — 2, толщина — 1,5. Правая почка весит больше, чем левая. Почки кроли-
ков характеризуются наличием одного соска. Мочеиспускательный канал у
самцов кроликов отличается от других животных прямолинейным расположе-
нием. У крольчих мочеиспускательный канал довольно длинный, оканчивает-
ся на нижней стенке влагалища и его отверстие прикрывается специальной
складкой. Почки кролика за сутки выделяют 180—440 мл мочи желтого цвета
с удельным весом 1,010—1,015. Величина депрессии (А) колеблется в пределах
0,55-1,22. Реакция мочи щелочная (рН 8,0). В состав мочи входят (%): общий
азот — 0,7322, мочевая кислота — 0,009, мочевина — 0,2069, креатинин —
0,001, зола - 1,19, СаО - 0,206, Р205 - 0,09, хлор - 0,26.
Состав мочи и ее реакция могут изменяться в зависимости от диеты.
Глава 31. Ренальная система и самоподобие Прокла
455
Почки у морской свинки обычной формы; правая расположена краниаль-
нее левой. Вес одной почки у молодых свинок — 2,6, а у взрослых — 5 г.
Взрослые морские свинки за сутки выделяют около 50 см3 мочи. Удельный
вес мочи 1,033—1,036. В моче содержится 3,5% мочевой кислоты.
Таблица 79
Генетически обусловленные и экспериментальные биомодели
для исследования ренальной системы
Исследуемое действие
вещества
Биомодель патологии почек,
в 25% случаев сопровожда-
ющейся гидронефрозом
Модель генетически детер-
минированного интерсти-
циального нефрита, со-
провождающегося тромбо-
зом лёгочных и перифери-
ческих вен в 63% случаев
Модель гипертрофии пред-
стательной железы (добро-
качественная, без призна-
ков малигнизации)
Лабораторное
животное
Мыши
Хомячки
Хомячки
Хомячки
Линия или порода
животного
Линия
Т(9;17)138Са
Линия RL
Линия LSH
87.20
Пол
животного
Яв
Яв
ев
ев
Вес
животного
30-40 г
30-40 г
120 г
120 г
Возраст
животного
3-10 мес.
6-18 мес.
6-18 мес.
6-18 мес.
Почки у крысы бобовидной формы, длина их около 16—19 мм. Правая
почка лежит в правом подреберье и по отношению к левой отклонена не-
сколько вперед. У крыс весом 130—150 г правая почка весит 0,63 г, левая — 0,6
г, у крыс весом 200—250 г правая почка весит 2,05 г, левая — 2 г. Левая почка
прилегает к тазу. Почка у крысы однососочковая. Мочевой пузырь такой же,
как и у других млекопитающих, но его стенка толще, чем у кролика.
Почки у мыши имеют типичные для этого органа формы, их размеры
0,9 х 0,5 х 0,4 см. Левая почка размещена каудальнее правой. Вес правой
почки у взрослых мышей в среднем 0,14, а левой — 0,13 г.
Надпочечники величиной с маковое зерно (весом по 2—3 мг) отличаются
от окружающей жировой ткани оранжевым цветом. Топографически распола-
гаются они кпереди и кнутри от почек, причем правый надпочечник лежит на
соответствующей почке, а левый от почки несколько отдален. Кортизон у
линий BAL/cJL/6 вызывает эозинопению, максимальный эффект наблюдает-
ся при введении 96 мг (доза высокая по сравнению с другими линиями).
Почки у золотистых хомячков лежат ретроперитонеально. Они бобовид-
ной формы и с гладкой поверхностью. Между почками размещены лимфа-
тические узлы, покрытые жировой клетчаткой. Правая почка немного выде-
ляется кпереди. Мочевой пузырь в наполненном состоянии круглой формы,
величиной с горошину. Надпочечники маленькие, яйцевидной формы обра-
зования, которые расположены на передне-внутренней поверхности почек и
покрыты жировой клетчаткой.
456
Биомоделирование в регуляторных системах
Моделирование почечных функций
Сравнительные особенности функционирования ренальной системы чело-
века и животных можно установить с помощью прямых и опосредованных
методов исследования. Результаты этих экспериментов могут быть положе-
ны в основу оптимизированного моделирования [130, 566].
Измерение объемов жидкостных компартментов, определение распреде-
ления жидкостей крайне важно для фармако- и токсикокинетического мо-
делирования. В табл. 80 приведены значения относительного содержания
воды в различных органах и тканях человека и животных. Процентное со-
держание воды имеет значительные колебания в зависимости от возраста
человека и его конституции. Более значительный разброс у животных. По-
этому при любых экстраполяционных переносах следует вводить коэффици-
енты размерности и пропорциональности. В табл. 80 они обозначены как (3.
Для таких животных как овцы или мини-свиньи, у которых запасы жировой
ткани претерпевают существенные колебания необходимо использовать спе-
циальные поправочные коэффициенты (3*, указанные в табл. 80.
В организме и в различных органах и тканях содержание воды можно
измерить методом разведения соответствующего индикатора. Если индика-
тор ввести в компартмент, т.е. исследуемые органы или системы и дать из-
вестному количеству вещества равномерно раствориться, то этот объем мож-
но будет рассчитать по конечной концентрации индикатора. Оно определя-
ется как количество вещества S в некотором объеме V:
S S
к = у> откуда V = — . (102)
Общий объем воды в организме определяют с помощью тяжелой или
меченой тритием воды, а также веществ, легко проходящих через клеточные
мембраны (антипирин). Объем внеклеточной жидкости измеряют с помо-
щью инсулина или тиосульфата, т. е. веществ, не способных диффундиро-
вать через клеточную мембрану. Объем плазмы определяют с помощью ра-
диоактивного 1311, красителя Эванса и других веществ, остающихся в про-
свете кровеносных сосудов. Объем внутриклеточной жидкости определяется
разностью между общим и внеклеточным объемами, а межклеточной — раз-
ностью между внеклеточным и плазматическим. Используемые индикаторы
попадают в плазму, их концентрацию определяют при анализе крови. По-
скольку за время распределения часть их выводится из организма, то ис-
пользуют несколько образцов крови, и путем экстраполяции определяют
равновесную концентрацию.
Общее количество воды в жидкостных компартментах организма опреде-
ляют методом разведения индикаторов. Ее содержание в различных органах
и тканях неодинаково. В жировой ткани ее 10%. У человека и у мле-
Глава 31. Ренальная система и самоподобие Прокла
457
Таблица 80
Расчет примерного содержания воды в различных компартментах человека
и животных-млекопитающих (в крайней правой колонке даны цифры
для человека массой 70 кг)
Ткань или орган
Кровь
Почки
Сердце
Легкие
Селезенка
Мышцы
Мозг
Кишечник
Кожа
Скелет
Жировая ткань
Содержание
воды, %
83
83
79
79
76
75
74
74
72
22
10
Процент
от веса тела
8,0
0,4
0,5
0,7
0,2
42
2,0
1.8
18
16
10-55
Количество воды (л)
в организме
4,7 хр*
0,3 хр
0,3 хр
0,4 хр
0,1 хр
22 хр
1,5хр
0,9 хр
9,1 хр
2,5 хр
0,7 хр*
копитающих на долю воды в среднем приходится 73% массы всех тканей.
Эту величину используют для определения относительного содержания ли-
пидов в организме:
Gv = (0,73 xk)/Hn (103)
где: Gv — количество внеклеточных липидов (%), Нп — количество несвя-
занной воды {%), к — коэффициент размерности для различных животных.
При общем содержании воды в организме, равном 65% массы тела, око-
ло 10% его приходится на жировые отложения. Масса жировой ткани может
достигать до 50% при содержании воды в организме около 37%.
Скорость почечной фильтрации измеряется по принципу Фика с помо-
щью индикаторного вещества. Оно не должно связываться с белками плаз-
мы, задерживаться при прохождении через клубочковую мембрану, реабсор-
бироваться и секретироваться в почечных канальцах, расщепляться и синте-
зироваться в почках, быть токсичным. Наиболее оптимальным индикатором
является инулин, полифруктозид, состоящий из 20 остатков фруктозы, ко-
торый не образуется в человеческом организме.
Расчет скорости почечной фильтрации Vp основывается на соотноше-
нии, при котором количество выделенного с мочёй индикатора определяет-
ся его концентрацией умноженный на объем выделенной жидкости. В этом
случае:
v>=i*st-
(104)
458
Биомоделирование в регуляторных системах
где Cg — концентрация индикатора в моче, Pg — концентрация индикатора
в плазме, St — объем жидкости, выделяемой за единицу времени.
Скорость фильтрации в нефроне измеряется тем же способом. В обыч-
ных условиях у здорового человека она составляет 50 нл/мин. Для оценки
скорости фильтрации в нефроне животных осуществляется микропункция
петли канальца, и за определенный период времени производится забор
жидкости. Расчет скорости фильтрации нефрона Vn, например для инулина
(i), можно получить из следующего уравнения:
GfxVr
где Cif — концентрация инулина в фильтрате мочи, Cip — концентрация
инулина в плазме крови, Vx — объем фильтрата, поступающего в каналец за
период времени т.
Клиренс реабсорбирующихся веществ, таких, как электролиты, сахара, а
также некоторые ксенобиотики, в отличие от «чисто» фильтрующихся, на-
пример инулина, ниже, поскольку они не только фильтруются в клубочках,
но и активно извлекаются канальцевыми клетками. Расчет клиренса может
быть осуществлен по следующей формуле:
О х Vt
К = , (105)
Ср
где О — концентрация индикаторного вещества в моче, Ср — концентрация
индикатора в плазме, Vt — объем мочи за единицу времени.
В реальных условиях для оценки клиренса часто используется оценка кли-
ренса эндогенного креатинина. Его преимущество в том, что не нужно вво-
дить индикаторные вещества. Но точность этого метода несравнимо меньше,
нежели инулиновая проба. Креатинин образуется при обмене веществ в мыш-
цах и его суточная продукция у человека составляет 9 мг/л (80 мкмоль/л). Он
экскретируется путем клубочковой фильтрации. Расчет клиренса креатинина
производится по формуле, близкой к формуле (105):
Ckt х Vkt
Kk = х р ,
Ckp
где Ckt — концентрация креатинина в моче, Ckp — концентрация креатини-
на в плазме, Vkt — объем фильтрации за единицу времени, Р — коэффициент
пропорциональности и размерности в аллометрических пересчетах для жи-
вотных разных видов.
При биомоделировании на животных широко используются методы мик-
ропункции, а также микроперфузионные исследования. Они дополняются
электрофизиологическими методами, оценивающими перенос заряженных
частиц через поверхности плазматических мембран. Эти методы оценива-
Глава 31. Ренальная система и самоподобие Прокла
459
ют активность ионов Н + , Na+, K+, С1" или Са2+ и позволяют измерить их
перенос через мембрану. Информативным подходом в фармако- и токси-
кокинетике является «пэтч-кламп-метод» локальной фиксации потенциа-
ла. С помощью высокочувствительной компьютерной аппаратуры можно
на изолированном одиночном канале зарегистрировать прохождение по
нему ионов или его чередующиеся открытые и закрытые состояния. В на-
стоящее время, возможно, культивировать нефроны и проводить модели-
рование воздействия на них различных физических, химических и биоло-
гических факторов.
Как мы уже подчеркивали, наиболее универсальным принципом моде-
лирования является математическое моделирование. Обратимся к уже при-
водимым примерам математического моделирования регуляции и поддер-
жания уровня сахара в крови и тканях человека и животных. В данном слу-
чае рассмотрим процессы утилизации Сахаров тканями организма, в том числе
в почках. Известно, что вследствие асимметричности строения канальцевых
клеток в почках осуществляется активный, сопряженный транспорт и под-
держивается клубочково-канальцевое равновесие. В канальцах осуществля-
ется не только секреция воды, ионов мочевины, глюкозы, многих ксеноби-
отиков, но и реабсорбция. Реабсорбция глюкозы осуществляется столь ин-
тенсивно, что с точки зрения моделирования этого процесса её можно рас-
сматривать наряду с продукцией или иньекцией. Ткани почек поглощают
глюкозу вследствие процесса диффузии, и скорость поглощения зависит от
разности у — уъ при у > уу В то же время способность тканей поглощать
глюкозу ограничена, а увеличению количества поглощенного сахара способ-
ствует инсулин. При учете этих явлений мы приходим к следующему диффе-
ренциальному уравнению:
-^- + asy3=-b3i-b5i-a7(y-y3), (Ю6)
где а7, as, bv b5 — коэффициенты размерности и пропорциональности.
Системой дифференциальных уравнений мы можем описать упрощен-
ные регуляторные процессы поддержания уровня сахара в крови и тканях. В
свою очередь, это позволяет нам определить также текущее содержание са-
хара в крови. Учитывая, что инерционность системы смешивания различных
составляющих уровня сахара крови, которая вследствие быстроты действия
кровеносной системы пренебрежимо мала по сравнению с инерционностью
работы органов, для получения текущего содержания уровня сахара в крови
можем провести простое суммирование:
У = Уп + У2 + Уз- 0°7)
Это равенство, наряду с вышеизложенными дифференциальными урав-
нениями, уже можно использовать для расчетных счислений, хотя его дос-
460
Биомоделирование в регуляторных системах
тоинство заключается в том, что мы получаем возможность дать корректное
описание работы системы управления уровнем сахара крови, включая и
объект (объекты) управления этими процессами.
Таким образом, особенности автономного функционирования нефрона,
ни в коем случае не отменяющие системное функционирование и регулиру-
ющие влияния ренальных органов на гомеостаз организма, позволяют осу-
ществлять хорошее экстраполяционное межвидовое моделирование, и под-
тверждают холономную парадигму Прокла-Бома. Действительно как нигде в
организме, почечная система построена на принципах самоповторения и
самоподобия.
461
Глава 3 2 ПОЛОВОЕ ПОВЕДЕНИЕ
И СИСТЕМА РЕПРОДУКЦИИ
Репродуктивные органы у разных видов животных
имеют существенные различия. Также большие раз-
личия имеются между органами размножения жи-
вотных и человека. Поэтому, несмотря на общность
функций, следует достаточно осторожно относить-
ся к вопросам экстраполяции полученных на жи-
вотных данных в отношении человека. Последние
работы в этом направлении указывают на трудно-
сти в биомоделировании [64, 683] и оценки токсич-
ности и тератогенности[150, 201] различных веществ
у различных животных и в альтернативных биомо-
делях. Мы полагаем, что выбор адекватной биомо-
дели должен быть сделан исследователем в отноше-
нии каждого конкретного случая и задач, которые
он формулирует для себя. Поэтому мы дадим срав-
нительные характеристики органов размножения у
различных животных и их особенности, которые
могут быть использованы при построении биомо-
делей репродуктивных функций.
Агрессивность и половое поведение
Регуляция репродуктивных функций находится под
контролем ЦНС. В этом процессе участвуют слож-
ные механизмы управления со стороны гипотала-
мо-гипофизарной системы, которые были рассмот-
рены в гл. 26. Главное предназначение половых
функций заключается, естественно, в воспроизвод-
462
Биомоделирование в регуляторных системах
стве и поддержании вида. Однако не меньшая роль принадлежит половым
гормонам и в регуляции поведения, причем это поведение также ориентиро-
вано на сохранение как индивидуума, так и популяции.
Речь идет прежде всего об агрессивном поведении. Андрогенные гормо-
ны, важнейшим из которых является тестостерон, концентрация которого
в крови мужчин не должна быть менее 1—2 нг/мл, обеспечивают агрессивное
половое поведение. У животных это проявляется в еще более яркой форме,
например, в тестах оценки нейролептических и транквилизирующих средств
используется так называемый «бой самцов». При этом наиболее выражен-
ные проявления агрессивности и результативности наблюдаются у самцов
с повышенным уровнем тестостерона в крови.
Хотя превышение уровня тестостерона над границами естественной нор-
мы не коррелирует с возрастанием агрессивности, блокаторы эффектов ан-
дрогенов или кастрация животных нивелируют либидо и агрессивность. Ге-
нетически созданы инбредные и другие специальные линии животных с раз-
ным уровнем агрессивности, имеющих существенные различия уровня тесто-
стерона в крови (см. раздел III). Эти животные могут служить хорошими
биомоделями не только с точки зрения исследования репродуктивных функ-
ций, но и анализа сложноорганизованных форм поведения.
У животных-самцов половой аппарат состоит из семенников с придатка-
ми, семяпроводов, мочеполового канала, полового члена и придаточных по-
ловых желез — предстательной, луковичных и пузырьковых. Семенники име-
ют яйцевидную форму и находятся в мошонке. В дольках семенника нахо-
дится соединительная ткань, в которой проходят кровеносные, лимфатичес-
кие сосуды и нервы. В этой ткани размещены группы железистых клеток,
которые называются интерстициальными. В интерстициальной мужской
железе вырабатываются гормоны внутренней секреции. У верхнего края се-
менника находится придаток. Гормональная деятельность семенников осу-
ществляется преимущественно интерстициальными клетками Лейдига, ко-
торые иначе называются мужской железой зрелости, или пубертатной желе-
зой. Интерстициальные клетки продуцируют мужской половой гормон тес-
тостерон.
Семенники мини-свиней очень большие и бывают почти правильной эл-
липсоидной формы. Предстательная железа, как и у крупных жвачных, со-
стоит из тела и рассеянной части. Тело достигает в ширину 2,0—2,5 см, при-
крыто пузырьковидными железами и снаружи совершенно незаметно. Рас-
сеянная часть формирует слой вокруг слизистой оболочки тазовой части
мочеполового канала и снаружи окружена, за исключением дорзальной сто-
роны, мускулом мочеполового канала. Половой член, длиной до 45—50 см,
имеет большое сходство со жвачными животными. Наружное отверстие мо-
чеполового канала лежит на вентролатеральной стороне, почти рядом с за-
острённым концом члена.
Глава 32. Половое поведение и система репродукции
463
Семенники жвачных относительно крупные. Вес семенников достигает у
барана 200-300 г, у козла 145-150 г.
У козлов и баранов выражена только одна рассеянная часть предстатель-
ной железы, а тело отсутствует. Половой член имеет приблизительно 8 см
в длину. Общая длина его достигает у баранов 30—35 см.
Семенники собак относительно небольшие и имеют округло-овальную
форму. Вес семенника с придатком у собак средней величины около 30 г, т. е.
0,23% веса тела. Семенной канатик длинный и широкий. Хорошо выражена
луковица мочеполового канала. Пузырьковые и луковичные железы отсут-
ствуют. Предстательная железа большая, состоит из двух долей. В каудаль-
ном участке полового члена находятся два хорошо выраженных пещеристых
тела. Половой член имеет два пещеристых тела, а в головке его заложена
кость полового члена.
У самцов кошек половой член короткий, в нем размещена косточка (дли-
ной 3—4 см), а на слабо выраженной головке находятся небольшие шипы.
Семенники почти круглые, у взрослых животных их вес составляет 4—5 г.
Луковичные железы очень маленькие, величиной с горошину. Пузырьковые
железы отсутствуют, а предстательная железа слабо развита.
Вес семенников у кроликов составляет 0,2—0,3% веса тела. Придаток се-
менника значительно утолщен на заднем конце, и от этой утолщенной части
отходит семявыносящий проток. Семяпроводы в своем заднем конце обра-
зуют расширение, в котором находятся слизистые железы, после чего впада-
ют в мочеиспускательный канал. К мужскому половому аппарату кролика
относятся:
/ предстательная железа, состоящая из пяти долей;
/ мужская матка — железа, соответствующая семенным пузырькам дру-
гих млекопитающих, она открывается в мочеиспускательном канале
вблизи впадения семяпроводов;
/ редуцированные пузырьковидные железы;
/ железы луковицы мочеиспускательного канала — парные крупные же-
лезы, расположенные на границе с пещеристыми телами мочеполово-
го канала. Половой член имеет пещеристые тела и в стадии эрекции
достигает 4 см в длину.
Вес семенников у морских свинок до 2,3 г. В большинстве случаев левый
семенник больше правого. Благодаря наличию сильно развитого наружного
эректора яичка семенники могут втягиваться в брюшную полость. Из при-
даточных желез мужских половых органов сильно развиты пузырьковые
железы, которые имеют вид длинных выростов. У самцов морских свинок на
конце полового члена имеются два роговых образования — эпидермальные
рожки. Кастрация новорожденных самцов приводит к тому, что эпидермаль-
ные рожки не развиваются в процессе роста. У некастрированных животных
ампутированные рожки способны регенерировать.
464
Биомоделирование в регуляторных системах
Вес обоих семенников у молодых крыс (весом 130—150 г) составляет 0,7 г,
а у взрослых (весом 200-250 г) — 2,5 г. В семенниках мало интерстициаль-
ных клеток. Семенники чаще располагаются в мошонке и могут втягивать-
ся в паховые ходы. В теле полового члена имеется косточка. Как и у кроли-
ка, у крыс хорошо выражена мужская матка, которая открывается в моче-
половой проток. Пузырьковые железы очень развиты. У основания мужс-
кой матки находятся две пары ампулярных желез. Предстательная железа
развита хорошо.
Семенники у самцов мыши относительно большие, располагаются чаще
всего в мошонке, но могут находиться и в паховом канале и брюшной поло-
сти. Придатки несколько отделены от яичка. Пузырьковая железа двойная,
хорошо развита. От семенных придатков отходит семяпровод, который впа-
дает в общий мочеполовой синус. Мочеполовой синус закрывается прилега-
ющими к нему предстательной железой и железами Купера.
Репродуктивные нюансы самок в сравнении
В отличие от мужчин и самцов животных, у которых семенники функциони-
руют на протяжении всей жизни и, несмотря на возрастное снижение уровня
тестостерона, сперматогенез не угасает, у женщин и у самок животных гормо-
нальная регуляция идет волнообразно с затуханием во второй трети жизни.
Помимо известных со студенческой скамьи механизмов полового пове-
дения и репродукции с участием рилизинг-факторов, половых гормонов, в
том числе октапептида окситоцина, в этих процессах участвуют и другие
пептиды. Так гонады секретируют вазоактивный кишечный пептид и ряд
эндогенных опиоидных пептидов, а также значительную группу пептидов с
пока неизвестными функциями. Возможно, некоторые из этих пептидов, а
точнее изменения их активности, определяют глубокие вазодистонические,
кардиальные и психосоматические изменения в климактерическом периоде.
Наличие специальных линий животных-биомоделей и исследования на них
могут дать ответы и по этим вопросам
У самок органы размножения состоят из яичников, яйцеводов, или ина-
че называемых маточных (фаллопиевых) труб, и матки. На разрезе яичника
различают две его зоны: корковую (фолликулярную), в которой среди соеди-
нительной ткани заметны фолликулы (граафовы пузырьки), и мякотную
(сосудистую). В граафовых пузырьках созревают яйцеклетки. При созрева-
нии граафовы пузырьки лопаются и происходит выход яйцеклетки — овуля-
ция. Гормональная деятельность яичников осуществляется за счет функци-
онирования фолликулярного аппарата, желтого тела и интерстициальной
ткани корковой части яичников. Эпителиальные гранулезные клетки созре-
вающих фолликулов выделяют женские половые гормоны — эстрогены (эс-
трон, эстриол и эстрадиол). Желтое тело возникает из гранулезных клеток
Глава 32. Половое поведение и система репродукции
465
лопнувшего фолликула. В нем вырабатывается гормон прогестерон.
С наступлением половой зрелости в половых органах самок происходят
циклические изменения. Половой цикл у животных подразделяется на сек-
суальный период и период покоя. Сексуальный период наступает с появле-
нием течки и возникновением у животных охоты. Происходящие в сексу-
альном периоде изменения в половых органах самок представлены следую-
щими фазами:
/ предтечка — в это время происходит увеличение размеров фолликулов;
/ течка — фолликулы увеличиваются до предельных размеров, созрева-
ют, лопаются, т. е. возникает выход яйцеклеток в область верхних учас-
тков яйцеводов (овуляция). В это время под влиянием фолликулярно-
го гормона у самок отмечается гиперемия и опухание матки и влагали-
ща, шейка матки приоткрывается, железы шейки и преддверья проду-
цируют секрет, который выделяется наружу. В период течки самки боль-
шинства животных проявляют половое возбуждение. У многих живот-
ных кожные железы самок выделяют в это время секрет со специфи-
ческим запахом, привлекающим самцов;
/ послетечка — на месте лопнувших фолликулов развиваются желтые
тела. Уменьшается и исчезает гиперемия половых органов. Самка пе-
рестает быть беспокойной и возбужденной;
/ фаза покоя — если не наступила беременность, стенки матки резко
уменьшаются. В яичнике хорошо выражено желтое тело, в связи с чем
задерживается развитие фолликулов. Наступает пролиферация желез
матки. С рассасыванием ложного тела вновь наступает фаза предтеч-
ки. У сук и кошек течка обычно наступает весной и осенью (два цик-
ла), у грызунов наблюдается несколько циклов.
Если наступает оплодотворение яйцеклетки, развивается беременность.
У некоторых самок (крольчихи, кошки) овуляция наступает лишь после
совершения коитуса, т. е. рефлекторно. У других животных наступление
овуляции и ее продолжительность зависят от длительности течки. Так, у сук
течка затягивается на семь—девять дней, овуляция же происходит в середине
или конце течки и продолжается три—четыре дня. На месте лопнувшего фол-
ликула образуется желтое тело, которое при наступлении оплодотворения
функционирует на протяжении всей беременности (истинное желтое тело).
Если же беременность не наступила, то желтое тело вскоре рассасывается
(ложное желтое тело).
В корковой части яичников имеется интерстициальная ткань, которая
выполняет роль женской железы зрелости (женская пубертатная железа). Во
время беременности интерстициальная ткань разрастается. Например, яич-
ники неполовозрелых самок линий BALD/cJLac чувствительны к гонадот-
ропину человеческого хориона: в ответ на гонадотропин они дают высокую
добавочную овуляцию.
466
Биомоделирование в регуляторных системах
Таблица 8 1
Генетически обусловленные биомодели
для изучения репродуктивных функций и возрастных особенностей организма
Исследуемое действие
вещества
Биомодель на линиях мы-
шей-долгожителей для ис-
следований в геронтологии
Спонтанная биомодель
артериосклероза и тромбо-
за левого предсердия
Биомодель аутоиммунной
гемолитической анемии ,
а также красной волчанки
у человека
Оптимальные биомодели
для получения механизмов
передачи межклеточной
информации, взаимодейст-
вия клеток в эмбриогенезе,
генетики развития
Биомодель для изучения
процессов раннего старе-
ния и развития аутоиммун-
ных процессов в организме
Биомодель для изучения
причин бесплодия, наруше-
ний оогенеза (у мышек
обнаруживаются атрезии
влагалища)
Биомодель детской глауко-
мы человека с нарушением
оттока внутриглазной жид-
кости с низким уровнем
аскорбата и атрофией зри-
тельного нерва
Биомодель замедления
сперматогенеза, сопровож-
даемого нарушением обме-
на р-каротина и витамина А
Биомодель для эндокрино-
логии на животных с пре-
дельно высокими уровнями
гормона роста
Лабораторное
животное
Мыши
Мыши
Мыши
Мыши
Мыши
Мыши
Кролики
Кролики
Кролики
Линия или порода
животного
Линия CBA/LacY
Линия CBA/LacY
Линия NSB/Orl
Линии: C57BL/6-W
T/t6 (сток)
C57BL/6-a4lns
C57BL/HeJ-pe
C57BL/6Y-+Hi,go
C57/BL/10-hzzhY
C57BL/16Y-smk
(smoky)
Линия AXBU/J
Линия AXBU/J
Линии: AC/J
ACEP/J
Пол
животного
9c?
9c?
9c?
9c?
9c?
9c?
9c?
9c?
9c?
9c?
Bee
животного
19-23 г
25-30 г
25-30 г
25-30 г
25-30 г
25-30 г
3,5 кг
3,5 кг
2,4 кг
2,4 кг
Возраст
животного
Старше
3 лет
5-20 мес.
5-20 мес.
5-20 мес.
5-20 мес.
5-20 мес.
7 мес-
4 года
7 мес-
4 года
7 мес-
4 года
7 мес-
4 года
Яйцеводы (фаллопиевые трубы) начинаются в виде воронки с бахромой
и открываются в перитонеальную полость отверстиями. У собак они очень
извилистые, тонкие, длиной 5—11см, покрыты жировой тканью. Стенка
яйцевода имеет мускульный слой. Проксимальные части правой и левой
фаллопиевых труб сливаются между собой и образуют шейку матки. В ней
выделяют шейку, тело и рога матки.
Яичник мини-свиней относительно большой (до 5 см длиной), округлой,
слегка вытянутой формы, бугристый — на его поверхность выступают фолли-
Глава 32. Половое поведение и система репродукции
467
кулы в месте прилежащей оболочкой яичника. Нередко большое число таких
бугорков придает яичнику свиней вид ягоды ежевики. Яичник скрыт в сильно
развитой яичниковой бурзе.
Яичник мелкого рогатого скота по сравнению с таковым у крупных жи-
вотных значительно меньше и имеет эллипсоидную форму. Его овуляцион-
ная поверхность обширна и выпукла. У взрослых животных правый яичник
обычно несколько больше левого (у новорождённых он меньше). У овец и
коз яичник более округлый и относительно крупнее. Матка относится к
двурогому типу. Тело матки снаружи довольно длинное (10—15 см), однако
внутри большая часть матки разделена перегородкой надвое (так называемое
«ложное тело»). Неразделённой остаётся лишь небольшая её часть, длиной
5—6 см, переходящая в шейку матки.
Яичник у собак имеет удлинённую и слегка уплощённую эллипсоидную
форму. У средней величины собак яичник достигает в длину 0,5—2 см. На его
поверхности нередко можно заметить возвышения созревающих фолликулов
(граафовых пузырьков). Брыжеечный край яичника обращен дорзо-меди-
ально. Матка собак относится к типу двурогих. Она почти всецело разме-
щается в брюшной полости и отличается длинными, тонкими и прямыми
рогами, расходящимися друг от друга в виде римской цифры V. Тело матки,
длиной около 3 см, в передней части имеет небольшую перегородку. Длины
рогов имеют тонкие стенки и небольшую (0,5—1,5 см) срединную перегородку
в краниальном участке, что напоминает до некоторой степени отношения,
свойственные телу матки рогатого скота.
У самок кошек яичники малых размеров, до 1 см длиной. Матка двуро-
гая, плацента отпадающая.
У крольчих в каждом фолликуле яичников находится одно или очень редко
два яйца. Матка у крольчих двойная. Длина ее рогов 7 см, в полость влага-
лища они открываются отдельными отверстиями. Мускулатура матки хоро-
шо развита. Влагалище у взрослой крольчихи 7—8 см в длину. В нижнюю
стенку влагалища впадает мочеиспускательный канал, а в передней части
находится устье матки. В стенках мочеполового канала имеются малые и
большие железы преддверья. У крольчих клитор состоит из двух пещеристых
тел и достигает в длину до 2—3 см. Овуляция осуществляется спустя 10—15 ч
после спаривания. Поэтому покрытие крольчих необходимо для стимуляции
выхода яйцеклеток из фолликулов. Начавшаяся течка у крольчих продолжа-
ется до тех пор, пока не будет коитуса, а при отсутствии его длится, пока не
кончится сезон размножения (май—август и в теплые осень и зиму). Таким
образом, не допуская крольчиху к спариванию, можно изучать гормональ-
ную функцию яичников, в которых не содержится желтых тел. Искусствен-
ным образом овуляцию у крольчих вызывают введением гонадотропного
гормона. Наступает овуляция спустя 10—13 ч после введения гормона, вслед-
ствие чего возникает ложная беременность с образованием желтых тел. Из
468
Биомоделирование в регуляторных системах
этих соображений крольчиха может служить хорошей биомоделью в фарма-
кологии, экспериментальном акушерстве, биомедицинских технологиях.
Яичники морских свинок размещены позади почек. Они овальной формы,
их длина в среднем составляет 3 мм. Фолликулы заметны невооруженным
глазом.
Матка мини-свиней принадлежит к двурогому типу. Рога матки весьма
длинные (до 200 см) и у своего начала узкие, так что граница их с яйцепро-
водами не выражена. На своём протяжении рога описывают многочислен-
ные петли наподобие петель тонкого отдела кишечника. Они целиком лежат
в брюшной полости, подвешенные на маточной брыжейке.
Яичники у взрослой самки мыши круглой формы, величиной с просяное
зерно. Правый яичник располагается немного кпереди левого. Оба яичника
окутаны жировой тканью. Матка двурогая. Рога до 3—4 см длины. Продол-
жительность полового цикла зависит от климатических условий (в холодные
месяцы он удлиняется). В среднем он равен трем—девяти дням, длитель-
ность различных стадий полового цикла белой мыши следующая: стадия
покоя — около 50—60 ч; предтечка — 12 ч; течка — 8—10—18 ч; послетечка —
24-30 ч.
О функции яичников грызунов и о стадии полового цикла можно судить
по данным, полученным при исследовании влагалищных мазков. В среднем
продолжительность полового цикла у самок, отсаженных от самцов, состав-
ляет шесть—семь дней. В каждой фазе полового (эстрального) цикла наблю-
дается свой клеточный состав мазка, взятого из половой щели крыс.
После кастрации самок половые циклы прекращаются, а после введения
женских синтетических половых гормонов эстральные циклы возобновля-
ются. Этот факт используют для выявления веществ гормонального действия
и проверки их активности.
Матка у крыс двураздельная. В месте перехода непарной части матки во
влагалище имеется достаточно хорошо развитый сфинктер, образующий
шейку матки. У девственных самок при входе во влагалище имеется hymen.
Длина рогов матки около 5 см, а диаметр 2—3 мм.
Матка у хомячков двурогая. Рога S-образной формы оканчиваются на
уровне нижнего полюса почек. Мочеотделительный канал у самок открыва-
ется самостоятельным отверстием вентральнее влагалища. В настоящее вре-
мя благодаря искусственному отбору при разведении хомячков выведено
несколько пород этих лабораторных животных: альбиносы, частично альби-
носы, желтые, бежевые, пегие, черные, агути.
Биомодели для изучения гормональных эффектов
В последние годы в исследованиях эмбиотоксичности, тератогенности и реп-
родуктивной токсикологии все шире используются специальные животные-
Глава 32. Половое поведение и система репродукции 469
Таблица 82
Моделирование гормональных эффектов на гормонозависимых штаммах
опухолей репродуктивных органов и рекомендуемые штаммы
Исследуемое действие
вещества
1. Для веществ с эстроген-
ной активностью:
a. высокометастазирующий
рак ВМР-П (прививание
50 мг опухолевой массы
подкожно в область правой
подмышечной впадины)
b. перевиваемая адено-
карцинома предстательной
железы Dunning (R3327-H)
c. рак яичников, индуциро-
ванный по методу Бискин-
ДОВ
2. Для веществ с антиэстро-
генной активностью:
a. эпидермоидная карцино-
ма лёгкого Льюис (LLC, 3LL)
(введение 50 г опухолевой
массы подкожно в область
правой подмышечной
впадины)
b. ДМБА-индуцированные
опухоли молочной железы
крыс
3. Для веществ с прогести-
новой активностью:
a. рак шейки матки РШМ-5
при ортотопической пере-
вивке
b. аденокарцинома молоч-
ной железы Са-755 (введе-
ние 50 мг опухолевой массы
подкожно в область правой
подмышечной впадины
c. меланома В-16
d. аденокарцинома толстой
кишки АКАТОЛ
4. Для агонистов и аналогов
гормонов гипоталамуса
(RH-LH и соматостатина)
a. ДМБА
b. перевиваемая аденокар-
цинома предстательной
железы Dunning
Лабораторное
животное
Мыши
Крысы
Мыши
Крысы
Мыши
Крысы
Мыши
Мыши
Мыши
Мыши
Крысы
Крысы
Линия или порода
животного
Линии A/snell
Copenhagen
Линейные или
аутбредные
Линейные или
аутбредные
Линии C57BL6
или гибриды
BDF1 [DBAxC57BL6]
Линии Wistar
Линии СВА
Линии C57BL6
или гибриды
BDF1 [DBAxC57BL6]
Линии C57BL6
или гибриды
BDF1 [DBAxC57BL6]
Линии BALB/c
Линии Wistar
Copenhagen
Пол
животного
9
6
96
9с?
в
9
в
9
9с?
6
9
в
Вес
животного
18-20 г
200-250 г
20-25 г
150-200 г
18-20 г
От 100 г
18-20 г
18-20 г
18-20 г
18-20 г
От 100 г
200-250 г
Возраст
животного
2-2,5 мес.
3-4 мес.
2-3 мес.
2-3 мес.
2-2,5 мес.
С 55 дней
2-2,5 мес
2-2,5 мес.
2-2,5 мес.
2-2,5 мес.
С 55 дней
3-4 мес.
биомодели с генетически обусловленными опухолями или животные, на ко-
торых подобные явления легко воспроизводятся. Этот путь является доста-
470
Биомоделирование в регуляторных системах
точно перспективным, результаты легко воспроизводимы, а методы — адек-
ватными.
Следует подчеркнуть, что предлагаемые биомодели достаточно хорошо
коррелируют по результатам исследований с данными альтернативного мо-
делирования. В то же время не во всех случаях достигается простой перенос
результатов с животных на человека [150]. По-видимому, следует использо-
вать дополнительные методы математического моделирования и сглажива-
ния функций в процессе экстраполяции.
Резюмируя сравнительный обзор репродуктивных органов у животных,
следует упомянуть о процессе формирования гонад, т.е. семенников и яич-
ников. После завершения миграции половых клеток у эмбрионов генетичес-
ки женского пола, гонады дифференцированы относительно слабо. В них
продолжается митотическое деление примордиальных половых клеток и раз-
витие гранулезных, а затем образуется примордиальный фолликул.
У эмбрионов мужского пола, имеющих Y-хромосому, интенсивная про-
лиферация ткани гонад начинается раньше и приводит к образованию го-
над, из которых и образуются сперматозоиды. Экспериментально показано,
что после ранней кастрации у плода мужского пола дифференцировка гени-
талий идет по женскому типу. Для развития женских гениталий половые
гормоны не нужны, тогда как у плодов — самцов в этом процессе участвует
фактор, вырабатываемый клетками Сертоли, а также андрогены, секретиру-
емые клетками Лейдига.
Эти моменты нелишне помнить при проведении эмбриологических ис-
следований, в особенности с использованием ксенобиотиков эмбриотокси-
ческого действия, а также гормонов, памятуя, что, например, стероиды, лег-
ко превращаются в андрогены. Это немаловажный фактор при биомодели-
ровании, в том числе агрессивного поведения
471
Глава 3 3 ОСТЕОСИСТЕМА
И РЕПЕРЫ МАХАЛАНОБИСА
Согласно древним описаниям, начиная с папируса
Эберса, человеческий скелет представлялся высох-
шим человеком, отсюда и происхождение термина
скелет: от греч. skeletos — высохшее тело. Форми-
рование скелета у человека проходит несколько
стадий. Окончательное развитие скелета заканчи-
вается лишь спустя многие годы после рождения
человека.
Постепенное развитие скелета происходит не
только у человека, но и у всех млекопитающих,
птиц, рептилий, амфибий и костных рыб: сначала
возникает хрящевой скелет, который затем замеща-
ется костной тканью. Только у акул и их родствен-
ников скелет остается хрящевым на всю жизнь. На-
звания костей давались исходя из их формы, так
clavicula по-латыни «маленький ключ», то есть клю-
чица. Лопаточная кость называется по-латыни
scapula от греческого слова skaptein — копать, рыть,
лопатить.
Г.А.Илизаров доказал, что кость способна не
только отлично восстанавливаться, но и расти в
любом возрасте. Его операции и эксперименты оп-
ровергли мнение о костях как инертных, слабо ре-
генерирующих тканях.
Мне он рассказывал, как скупал сотни черенков
для лопат и, распиливая их вдоль и поперёк, искал
варианты фиксации в разных шинах, дугах и, нако-
нец, кольцах.
472
Биомоделирование в регуляторных системах
Этот пример я привел с целью показать один из видов физического, а
также механического подхода к моделированию. Сразу подчеркнём, что
модель, по сути, является чисто биологической, а зародилась она в голове
Г.А.Илизарова как абстрактно-логическая. Просто на первом этапе она на-
шла своё физическое воплощение в виде механической модели. И лишь в
дальнейшем она по итерационному принципу воплотилась в моделирование
на животных и в клинических исследованиях [73, 86, 95].
Скелет животных в сравнении
Рассмотрим некоторые особенности скелета животных. Это делается по мно-
гим причинам. Во-первых, отдельные элементы скелета нам понадобятся при
обозначении реперных точек в процессе манипуляций на животных, в том
числе при осуществлении стереотаксических операций. Во-вторых, иссле-
дование количественных параметров нижней челюсти или мандибулярный
метод является важнейшим приемом генетического контроля [656]. В-тре-
тьих, а это одна из главных задач, необходимо соотнести точки опорного
аппарата человека и тех животных, которые понадобятся при биомоделиро-
вании. В-четвертых, мы хотели бы выделить те особенности скелета живот-
ных, которые определяют главные задачи и цели борьбы за выживание [84].
Оси вращения имеются в организме человека и животных. Примерами
являются позвоночный столб и многочисленные суставы, благодаря кото-
рым, например, рука может вращаться вокруг своей оси. Но все же наиболее
яркий пример — это наш позвоночный столб. Его латинское название spina
— хребет. Первоначально «спина» означало длинную, тонкую колючку тер-
на. А наш позвоночный столб получил название спина за острые остистые
отростки позвонков, которые выступают на спине. Отдельные кости
позвоночного столба называют позвонками, или по-латыни vertebra от лат.
vertere — вертеть, крутить, так как сочленения отдельных позвонков позво-
ляют поворачивать голову и туловище.
У человека позвоночный столб насчитывает 33 позвонка и подразделяется
на отделы. Первые 7 позвонков образуют шейный, или цервикальный, отдел
позвоночника. Затем идет грудной, или торакальный, отдел, состоящий из 12
позвонков. Ниже пояса находится поясничный отдел позвоночника, содержа-
щий 5 массивных позвонков, который называется люмбальным.
Крестцовый отдел состоит из 5 позвонков. Позвонки здесь вместе с та-
зовыми костями образуют некоторое подобие креста. Древние называли эти
позвонки сакральными, потому что у животных именно эту часть обычно
использовали для жертвоприношений.
Имеется 4 копчиковых, или каудальных, позвонка, от cauda — по-латыни
хвост. Это рудимент хвоста, а позвонки слились воедино, и называются од-
ним словом — копчик.
Глава 33. Остеосистема и реперы Махаланобиса
473
Скелет различных животных имеет разную плотность и крепость. Так,
скелет кошки очень легкий и крепкий, подвижный и эластичный. Хребет уд-
линенный, поясничные и крестцовые позвонки сросшиеся, а остальные —
подвижные. Грудная клетка в начальной части сужена больше, чем у собак.
Ключица у кошек развита сильнее, чем у собак. Конечности прямые. На кон-
цах пальцев имеются подвижные когти, которые кошка может втягивать и
выпускать в случаях необходимости. Мышцы и сухожилия хорошо развиты.
Скелет головы млекопитающих (череп) имеет мозговой и лицевой отделы.
Мозг человека и животных защищен плотной костной коробкой. Греки
называли череп kranion. От этого слова происходит краниология — раздел
антропологии, изучающий черепа, краниометрия — измерение черепа, метод
исследования в антропологии; краниальный — черепной или расположенный
ближе к голове.
Лицевой отдел черепа располагается впереди и снизу (оровентрально) по
отношению к мозговому отделу. Такое взаиморасположение может зна-
чительно варьировать в зависимости от степени развития того или другого.
Для приблизительного уяснения их взаимоотношения измеряют так назы-
ваемый лицевой угол. Линии, образующие этот угол, проходят от середины
наружного слухового прохода до тела резцовых костей и отсюда по касатель-
ной к выдающейся точке лба. Например, лицевой угол скелета головы чело-
века и некоторых обезьян приближается к прямому, а соответствующий угол
у мини-свиней, овец, кошек, собак и других животных — острый (34—41°).
Это показывает, что у человека сильно развит мозговой отдел черепа и уко-
рочен лицевой, а у большинства животных их можно признать развитыми
более или менее равномерно.
Мозговой отдел мини-свиней, как и у других животных, включает заты-
лочную кость, имеющую уплощённое тело. У длинноголовых рас животных
череп сильно вытянут по сагиттальной оси. Лицевой отдел скелета головы
мини-свиней имеет форму четырёхгранной пирамиды вследствие сильного
сужения лицевого черепа к резцовой области. У самцов старше 3 лет изо рта
торчат клыки сверху и снизу — чем старше зверь, тем больше клыки. Как
всякий дикий вид, кабан имеет широкие вариации изменчивости, вот поче-
му его можно использовать при выведении различных пород мини-свиней и
использовать в биомоделировании.
На свиньях успешно изучаются различные аспекты трансплантации ко-
стной ткани. Большинство исследователей сходятся во мнении, что наибо-
лее успешно происходит приживление денатурированной кости, поскольку
это стимулирует дифференцировку костной ткани трансплантационного ложа
и формирование соединительно-тканных мостиков [360, 572].
Имеются экспериментальные данные, правда, пока малочисленные, о воз-
можной коррекции болезни Виллибранда путем трансплантации костного
мозга от здоровых свиней к больным [33].
474
Биомоделирование в регуляторных системах
У мелких жвачных особенно резко отличается устройство лобных костей
вследствие неодинакового расположения, по сравнению с собаками и кош-
ками, роговых отростков. В силу этого лобные кости сильно выпуклы в
области основания роговых отростков, причём последние в местах отхода от
лобных костей сближены друг с другом.
Скелет головы собак по своей форме чрезвычайно разнообразен. Суще-
ствуют длинноголовые (дог, борзые и др.), короткоголовые (мопс, бульдог) и
с головой умеренной длины (пинчер, английский бигл) собаки, причём ко-
лебания в длине падают на лицевой отдел черепа. У длинноголовых живот-
ных мозговой отдел черепа на уровне скуловых дуг уже, чем у короткоголо-
вых, наружный саггитальный гребень сильнее развит, а профиль лба и носа
очень слабо вогнут. Черепная коробка кошек удлиненная, с большими глаз-
ными впадинами и хорошо выраженным сводом. Череп у кошек укорочен,
хорошо развита задняя часть. Лицевая (передняя) часть небольшая. Скелет
головы кролика состоит из 25 костей. Высота и ширина черепа уступает дли-
не, так как у кроликов хорошо развит лицевой отдел черепа. Череп крысы
состоит из 11 парных и 6 непарных костей, из них 15 костей приходится на
лицевую и 13 на мозговую часть черепа.
Позвоночный столб животных, как и у человека, состоит из позвонков, в
которых различают тело, дугу, позвоночное отверстие и отростки: остистый
и два поперечных. Кроме того, в каудальной части грудного и поясничного
отделов у собаки выражены добавочные отростки. Позвоночные отверстия
всех позвонков составляют позвоночный канал, в котором расположен спин-
ной мозг.
Сравнивая шейный отдел осевого скелета у лабораторных животных сле-
дует остановить внимание на следующих различиях, связанных с особен-
ностями образа жизни, т. е. со способами питания и поведением животных.
Шейный отдел у мини-свиней. Пять последних позвонков имеют очень
короткие тела, без вентральных гребней, с плоскими головками и пологими
ямками. Поперечно-рёберные отростки разделены на меньший, направлен-
ный в боковую сторону и назад, отпрыск и больший, направленный вниз
пластинчатый, увеличивающийся от 3 до 6-го позвонка, отпрыск. Эти от-
прыски налегают черепицеобразно один на другой. Остистые отростки срав-
нительно сильно развиты и удлиняются спереди назад, так что самый высо-
кий из них находится на 7-м позвонке. У собак шейный костный рычаг
умеренной длины, (с прочно скреплёнными сегментами, на которых рель-
ефно выступают как поперечно-рёберные отростки с отпрысками вниз, так
и остистые отростки, повышающиеся от 3 к 7. Позвоночный столб собаки
имеет следующее количество позвонков по различным его отделам: в шей-
ном — 7, грудном — 13, поясничном — 7, крестцовом — 3 сросшихся по-
звонка, которые составляют крестцовую кость, в хвостовом отделе число
позвонков не постоянно — от 18 до 22.
Глава 33. Остеосистема и реперы Махаланобиса
475
Грудной отдел у мини—свиней состоит из 14 грудных сегментов, но бывают
нередко вариации в сторону увеличения до 15, реже 16, а в исключительных
случаях до 17 сегментов. Грудной отдел собак обладает 13 сегментами. На телах
позвонков вентральных гребней нет. Остистые отростки более цилиндричны,
чем у других животных. Из них первые шесть мало разнятся по своей длине
и до 10-го наклонены назад, 11-й принято считать диафрагматическим.
Сравним общие и отличительные параметры скелета у разных животных
и определим реперные точки. Форма поперечного сечения грудной клетки
значительно разнится у различных млекопитающих, особенно в передней
половине. Это сечение может иметь: форму поперёк поставленного овала,
т. е. с короткой высотой и значительной шириной, как, например, у челове-
ка; приблизительную форму круга, как, например, у хищников, и форму
отвесного овала, или точнее, треугольника, т. е. с высотой, более значитель-
ной, чем ширина, как, например, у домашних животных. Шейный отдел
собак относительно короче, чем у крупных животных, но длиннее, чем у
овец и свиней. На 4, 5, 6 и 7-м позвонках имеются ясно выраженные ости-
стые отростки, напоминающие таковые у овец и другого рогатого скота.
Мини-свиньи обладают длинной поясницей (7 позвонков), но соединения
суставных отростков имеют у них устройство с уклоном в сторону проч-
ности, так как передние суставные отростки вправлены в задние, как ось во
втулку колеса, в силу чего получаются очень крепкие замки. Обширность
поясницы и прочность связи соответствуют здесь большой ёмкости брюш-
ной полости. Количество длинных поясничных позвонков варьирует: обыч-
но их бывает 7, иногда 6 или, редко, 5. Хвостовый отдел содержит 20—23
позвонка.
У мелкого рогатого скота (овцы, бараны) налицо все признаки обширно-
го и прочного поясничного отдела, причём сравнительно большая протяжён-
ность поясницы (6 позвонков) обусловлена удлинением тел позвонков, а
ширина — хорошо развитыми поперечно-рёберными отростками.
Поясничный отдел собак с 7 (6) позвонками, довольно длинный. Нерв-
ные дужки сильно отростчатые. Суставные отростки напоминают таковые у
крупных животных.
Хвостовый отдел собак включает 20—22 позвонка, из которых первые пять
хорошо развиты и имеют даже суставные отростки с суставными поверх-
ностями. Начиная с 4-го (5-го), тела позвонков снабжены гемальными отро-
стками; постепенно уменьшаясь, они сохраняются до 16-го позвонка. Дру-
гие хищные, особенно из семейства кошек, обладают длинной поясницей
(7 позвонков); устройство соединений суставных отростков позволяет раз-
нообразить движения. Между грядой остистых и поперечнорёберных отро-
стков сравнительно сильно развита ещё гряда сосцевидных и гряда добавоч-
ных отростков. Таким образом, у хищных все оттенки построения пояснич-
ного отдела соответствуют облегчённой и диференцированной подвижности.
476
Биомоделирование в регуляторных системах
Позвоночник кролика имеет 46 позвонков, из них шейных — 7, грудных
— 12 или 13, поясничных — 7 или редко 6, крестцовых — 4 и хвостовых —
16 или реже 15. Крестцовые позвонки сливаются в одну крестцовую кость.
Позвоночный столб крысы состоит из пяти отделов: шейного — 7 по-
звонков; грудного — 13 позвонков, от которых отходят 13 пар ребер; пояс-
ничного — 6 позвонков; крестцового — 4 сросшихся позвонка; хвостового —
27 или 30 позвонков.
Позвоночник морских свинок состоит из 36—37 позвонков, из них 7 шей-
ных, 13 грудных, 6 поясничных, 4 крестцовых и 6-7 хвостовых. В грудной
части имеется 13 ребер, из них 6 истинных и 7 ложных. Грудина состоит из
5 поздноспаивающихся частей, причем рукоятка и мечевидный отросток
срастаются лишь у старых животных.
Крестцовый отдел образовался вследствие слияния 3—5 позвонков, (у овец
и коз 4, а нередко 3, у мини-свиней 4, у собак и кошек 3) и представляет
неподвижный отдел позвоночного столба. Крестцовая кость прочно вклине-
на между крыльями подвздошных костей таза, который через её посредство
упирается в позвоночный столб. Благодаря таким взаимоотношениям тазо-
вые конечности могут легко толкать туловище вперёд.
Значение хвоста у животных довольно разнообразно. Водные позвоноч-
ные используют его как главное орудие передвижения. У наземных хвост
играет роль руля (белка, лиса), дополнительного орудия хватания (обезьяна),
отпугивания насекомых (большинство животных) или, наконец, за ненадоб-
ностью совершенно исчезает как таковой, оставляя лишь небольшие остат-
ки сросшихся в одну хвостовую косточку позвонков {человек). У животных
количество позвонков хвоста довольно сильно варьирует даже у одного и
того же вида {у лошадей 15—49, у рогатого скота 18—20, у овец 3—24, у коз 12—
16, у мини-свиней и собак 20—23). Лишь у хищных хвост до некоторой сте-
пени может играть роль руля (особенно у кошачих).
Следует остановиться на особенностях соединения костей конечностей у
животных. Локтевой сустав лишь у собак и кошек построен так, что позволя-
ет не только разгибание и сгибание, но и небольшое вращение; у остальных
домашних животных этот сустав может производить движения, как у лоша-
ди, т. е. вокруг одной оси. На связках незначительное различие выражается
в том, что боковая латеральная связка, а у собак и кошек и медиальная, рас-
щеплена в дистальном направлении на две ветви, закрепляющиеся одна на
лучевой, а другая—на локтевой кости.
Движение между костями предплечья имеется только у собак и кошек, и
то очень ограниченное, допускающее слабое вращение лучевой кости около
локтевой. Эта функция ограничена как боковыми связками, так и особой
кольцевой связкой луча, которая начинается от связочного бугра лучевой
кости и оканчивается на дорзо-медиальном крае локтевой кости. Запястный
сустав в смысле движений имеет некоторые особенности у собак и, особенно
Глава 33. Остеосистема и реперы Махаяанобиса
477
у кошек в том отношении, что позволяет движения отведения и приведения,
а также слабо выраженного вращения по продольной оси. В области соеди-
нения дистального ряда костей запястья с пястными костями движения у
кошек и собак также возможны, тогда как у крупных животных они сведены
к нулю.
Особенность связочного аппарата животных заключается в том, что, в
зависимости от количества костей запястья, межкостных связок бывает то
больше, то меньше: у жвачных — меньше, у всеядных —больше и ещё больше
у собак. Связочные волокна между пястными костями у всеядных (3—4) и у
собак (2—5) создают подвижность одной кости около другой. У жвачных
существуют связки, соединяющие рудиментарную пястную 5-ю кость с за-
пястной 4-й и с основной пястной костью (3 + 4). Биомодели паталогии
представлены в табл. 83.
Таблица 83
Генетически обусловленные биомодели для исследования
костной и мышечной систем
Исследуемое действие
вещества
Биомодель для изучения
врожденного нарушения
костной и опорно-двигатель-
ной систем и для исследо-
ваний в эмбриологии
Биомодель хондромуко-
идальной патологии чело-
века, обезьян, скота, собак
и других животных при
рецессивном гене хондро-
дистрофии, определяющем
аномалии хрящей и сла-
бость окостенения
Генетически Sb предопреде-
лённая модель расщеплен-
ных губы и нёба, а также
деформаций позвоночника
Биомодель для исследова-
ния мышечной дистрофии
с пролиферацией ядер
сарколеммы и увеличением
количества интерстициаль-
ной ткани
Модель миопатии Дюшенна
человека с дистрофической
миотонией со 100%-ной
врождённой миопатией
в возрасте 60-200 дней
(у 43% - до 30 дней), с су-
хим некрозом, жировой ин-
фильтрацией и пролифера-
цией мышц соединительной
тканью
Лабораторное
животное
Мыши
Кролики
Кролики
Мыши
Хомячки
Линия или порода
животного
Линии:
B10.UW/Sn
C57BL/6Y-HJ/+
Naked-Splotch
RaSd, rucet,
129/ReDg-Fu/+
Линия AC/J
Линия AC/J
Линия
129/ReJ-+dy
Линия 14.6
Пол
животного
9с?
9
9
9(5
9с?
Вес
животного
30-40 г
2,4 кг
2,5-4 кг
30-40 г
120 г
Возраст
животного
До 2 мес.
7 мес-
4 года
7 мес-
4 года
До 6 мес.
До 200
дней
478
Биомоделирование в регуляторных системах
Депо или донатор: биомоделирование остеофункций
Костная ткань является резервуаром и донатором жизненно важных веществ,
которые избыточно расходуются в процессе борьбы организма за выжива-
ние. Остеопенический синдром неизбежно развивается при многих тяжелых
заболеваниях с разным патогенезом и с возрастом, особенно у женщин пос-
ле менопаузы.
Все известные этиологические факторы, приводящие к возникновению
остеопороза, объединены общим патогенетическим ауторегуляторным меха-
низмом воспроизведения костного матрикса. Он служит основой развития
остеопенического или остеопоротического синдрома. Внеклеточный матрикс,
будучи продуктом биосинтетической деятельности клеток, не остается пас-
сивным и активно влияет на их функции, тем более что сам он не постоя-
нен. Важнейшим условием ауторегуляторного остеогенеза служит процесс
взаимодействия остеоцитов, остеобластов и внеклеточного матрикса, что
можно хорошо моделировать на животных (табл. 84).
Следует также иметь в виду, что стойкая гиперкальциемия может обусло-
вить ряд тяжелых расстройств сердечной деятельности. Кальциево-фосфор-
ный гомеостаз поддерживается комплексным механизмом его регуляции.
Гормональный уровень обеспечивается паратгормоном кальцитонином, 1,25-
диоксикальциферолом, являющимся метаболитом витамина Д с гормональ-
ной активностью, кортизолом, половыми стероидами и другими гормонами.
Экскреторный уровень обеспечивается кишечником, почками и скелетом,
как резервуаром Са и Р, откуда они могут при необходимости извлекаться,
а при избытке — откладываться. Концентрация Са2+ в крови может повы-
шаться при гиперпаратиреоидите.
Эти гормональные изменения могут служить одной конечной цели —
сохранению минеральной составляющей костного матрикса. В биологи-
ческом отношении это целесообразная адаптивная и компенсаторная ре-
акция гормональной системы, регулирующей Са-Р-гомеостаз в ответ на
структурные изменения в костных тканях (повышение элиминации из них
фосфатных анионов, снижение их минерализации, увеличение пирофос-
фатов в моче), которые являются проявлением системного остеопеничес-
кого синдрома вследствие разрушения полифосфатных соединений гидро-
ксиапатита.
Методом инфракрасной спектроскопии показана связь развития лейке-
мии с увеличением содержания в костях кислых форм ортофосфата каль-
ция — Са(Н2Р04)2 который обладает наибольшей растворимостью среди
других фосфатов — СаНР04 и Са3(Р04)2. Этот переход среднего ортофосфа-
та в кислые формы обусловливает повышенную элиминацию фосфатных
анионов из кристаллической компоненты костей, что, в свою очередь, при-
водит к нарушению минерализации костной ткани. Вынос кальциевых и фос-
Глава 33. Остеосистема и реперы Махаланобиса
479
фатных соединений из костей способствуют прогрессивному развитию осте-
опороза и изменению минерального состава мочи. Практически весь фос-
фор в моче находится в виде ортофосфата.
Органическая матрица вследствие разрыва С—О связей деформируется и
теряет способность накапливать и удерживать ионы Са, что также усиливает
процесс деминерализации в твердых тканях. Ускорение элиминации фос-
фатных анионов из кости вызывает повышение уровня пирофосфатов в моче,
что является дополнительным подтверждением наличия генерализованного
или системного остеопороза.
Как известно, митохондриальная ДНК (мДНК) обеспечивает автоном-
ный биосинтез белков, в том числе ферментов, снабжающих организм энер-
гией за счет окислительного фосфорилирования — синтеза АТФ. Митохон-
дриальный геном отличается выраженной нестабильностью — скорость эво-
люции мДНК превышает таковую ядерной ДНК (яДНК) в 10—20 раз. Высо-
кий уровень мутагенеза мДНК объясняется отсутствием эффективных сис-
тем репарации, измененным генетическим кодом и протекающими в мито-
хондриях окислительно-восстановительными процессами. мДНК подверга-
ется атаке свободными радикалами, поскольку митохондрии являются ос-
новным источником таких соединений в организме, а их ДНК особенно
чувствительна к окислению. Скорость окисления ДНК в митохондриях зна-
чительно выше, чем в ядре. Уязвимость мДНК обусловлена тем, что она не
содержит гистоновых белков, которые связаны с ядерной ДНК и защищают
её. Кроме того, митохондральные гены только в минимальной степени об-
служиваются ферментами, заменяющими окисленные участки ДНК в кле-
точном ядре.
В экспериментальной стоматологии для изучения особенностей форми-
рования дентина, периодов кальцинации, тканевых перестроек в процессе
развития зубов, а также изыскания возможностей их трансплантации [547] в
качестве биомодели функционирования костной ткани активно использу-
ются мини-свиньи [39, 104, 320].
Парность зубов позволяет строить числовую формулу лишь по одной по-
ловине дорзальной и вентральной аркад. При биомоделировании в стомато-
логии следует учитывать некоторые особенности животных. Взяв половину
зубов каждой аркады, составляют формулу, где цифры над чертой означают
половину количества зубов дорзальной аркады, а под чертой вентральной
аркады. У морской свинки 20 зубов. Жевательная поверхность коренных зу-
бов у свинок снабжена невысокими тупыми бугорками. У крыс нет клыков
и премоляров, всего зубов 16. Эмаль покрывает резцы лишь на передней
поверхности, в связи, с чем эти зубы имеют острый край и легко затачива-
ются. У большинства млекопитающих аркады верхних и нижних зубов, сле-
довательно, и челюсти, строго противолежат друг другу и совпадают. Че-
люсти с такими зубами называются изогнутыми, или совпадающими {хищ-
480
Биомоделирование в регуляторных системах
ные, всеядные). У других млекопитающих верхняя аркада имеет более широ-
кие коренные зубы и верхние челюсти шире расставлены, так что только
часть каждого нижнего коренного зуба совпадает с частью противолежащего
верхнего. Такие челюсти называются анизогнатными, или несовпадающими
(у жвачных).
Другим приемом биомоделирования является контроль генетической
стандартности лабораторных мышей, осуществляемый методом мандибуляр-
ного анализа. Этот метод был предложен сотрудником Британского центра
лабораторных животных М. Фестингом в начале 70-х годов XX века [207].
Мандибулярный метод генетического контроля лабораторных мышей, заклю-
чающийся в многофакторном анализе с помощью компьютеров ряда пара-
метров нижней челюсти. Этот метод является разновидностью остеометричес-
кого или краниометрического
анализа. В основу метода по-
ложен один из генетических
принципов согласно которому
форма мандибулы детермини-
руется большим числом генов,
локализованных в разных хро-
мосомах (рис. 10).
Этот метод основан на ис-
пользовании и вычислении
функции обобщенного Махала-
нобисова расстояния, характери-
зующего вероятность принад-
лежности особи к одной из
групп. Данный метод оценива-
ет не гомозиготность, а степень
соответствия отдельных индивидов краниометрическому стандарту линии. Он
позволяет определять, к какой линии или колонии относится данное животное
или группа животных и идентифицировать линейную принадлежность. Можно
выяснить, насколько данная выборка животных соответствует стандартным об-
разцам той линии, к которой она предположительно относится [588, 589]. Дан-
ный метод позволяет обнаруживать расхождение сублиний, происходящее за
счет накопления невидимых мутаций и оценивать степень генетического сход-
ства или отличия соответствующих линий и популяций.
В табл. 84 представлены данные нашей сотрудницы Е.Ф. Шмидт по клас-
сификации линейных мышей. Аббревиатурой МС обозначен мандибуляр-
ный стандарт линий или тестируемой выборки. Следует подчеркнуть, что
уже далеко не новый мандибулярный метод позволяет надежно идентифи-
цировать линейную принадлежность и аутентичность исследуемых живот-
ных генетическому стандарту линии или стока. Это определяется тем, что
Рис. 10. Основные реперные точки, снимаемые с пра-
вой ветви мандибулы: 1 -6 - расстояние по вертикали,
7-11 - расстояние по горизонтали
Глава 33. Остеосистема и реперы Махаланобиса
481
Таблица 84
Результаты идентификации линейной принадлежности мышей путём
вычисления реперов Махаланобиса и расстояний (MP) между ними
№
п/п
1
2
3
4
5
6
7
8
9
10
Мер
Сигма
MP относительно МС линии
A/Sn
77,16
31,48
72,02
59,64
32,64
86,18
57,34
77,41
97,81
85,17
67,72
21,13
AKR
14,02
11,67
15,08
16,21
14,84
42,89
8,45
23,25
105,52
28,63
28,06
27,53
BALB/c
127,3
81,27
171,15
153,05
91,76
92,18
63,31
41,91
125,69
70,95
101,86
39,10
CC57W
53,12
54,28
78,78
99,01
71,61
38,59
84,88
88,87
171,77
46,55
78,75
36,28
NZW
30,54
38,62
26,13
20,84
24,02
42,72
19,51
29,72
76,28
56,53
36,49
17,05
SWR
82,60
57,62
68,92
61,32
30,89
58,20
21,95
27,55
134,19
84,71
62,80
31,60
Отнесены по
минимальному
MP
AKR
AKR
AKR
AKR
AKR
CC57W
AKR
AKR
—
AKR
AKR
—
Причина
ошибки
идентификации
—
—
—
—
—
?
—
—
Калыдификация
—
—
—
для составления морфометрических стандартов выбрана нижнечелюстная
кость (мандибула), поскольку: во-первых, ее форма, описываемая 13 пара-
метрами, зависит от большого числа генов, т. е. является полигенно-обуслов-
ленным признаком; во-вторых, в отличие от большинства количественных
показателей она представляет собой высоконаследуемый признак;
в-третьих, условия содержания и особенности кормления животных могут
изменить размеры челюсти, но не влияют на ее форму.
Модифицированный метод мандибулярного анализа используется в
НЦБМТ РАМН с 1976 г. [588, 589].
Костная ткань является резервуаром и донатором жизненно-важных ве-
ществ, которые избыточно расходуются в процессе борьбы организма за
выживание. Остеопенический синдром неизбежно развивается при многих
тяжелых заболеваниях с разным патогенезом и с возрастом, особенно у жен-
щин после менопаузы.
Все известные этиологические факторы, приводящие к возникновению
остеопороза, объединены общим патогенетическим ауторегуляторным меха-
низмом воспроизведения костного матрикса. Он служит основой развития
остеопенического или остеопоротического синдрома. Внеклеточный матрикс,
будучи продуктом биосинтетической деятельности клеток, не остается пас-
сивным и активно влияет на их функции, тем более, что сам он не постоя-
нен. Важнейшим условием ауторегуляторного остеогенеза служит процесс
взаимодействия остеоцитов, остеобластов и внеклеточного матрикса, что
можно хорошо моделировать на животных (табл. 83).
482
Биомоделирование в регуляторных системах
Следует также иметь в виду, что стойкая гиперкальциемия может обусло-
вить ряд тяжелых расстройств сердечной деятельности. Кальциево-фосфор-
ный гомеостаз поддерживается комплексным механизмом его регуляции. Гор-
мональный уровень обеспечивается паратгормоном кальцитонином,: 1,25^
диоксикальциферолом, являющимся метаболитом витамина Д с гормональ-
ной активностью, кортизолом, половыми стероидами и другими гормонами.
Экскреторный уровень обеспечивается кишечником, почками и скелетом,
как резервуаром Са и Р, откуда они могут при необходимости извлекаться, а
при избытке — откладываться. Концентрация Са2+ в крови может повышать-
ся при гиперпаратиреоидите.
Эти гормональные изменения могут служить одной конечной цели —
сохранению минеральной составляющей костного матрикса. В биологи-
ческом отношении это целесообразная адаптивная и компенсаторная ре-
акция гормональной системы, регулирующей Са- Р- гомеостаз, в ответ на
структурные изменения в костных тканях ( повышение элиминации из них
фосфатных анионов, снижение их минерализации, увеличение пирофос-
фатов в моче), которые являются проявлением системного остеопеничес-;
кого синдрома вследствие разрушения полифосфатных соединений гидро-
ксиапатита.
Методом инфракрасной спектроскопии показана связь развития лейке-
мии с увеличением содержания в костях кислых форм ортофосфата каль-
ция — Са(Н2Р04)2 который обладает наибольшей растворимостью среди
других фосфатов — СаНР04 и Са3(Р04)2. Этот переход среднего ортофосфа-
та в кислые формы обусловливает повышенную элиминацию фосфатных
анионов из кристаллической компоненты костей, что в свою очередь, при-
водит к нарушению минерализации костной ткани. Вынос кальциевых и
фосфатных соединений из костей способствуют прогрессивному развитию
остеопороза и изменению минерального состава мочи. Практически весь
фосфор в моче находится в виде ортофосфата.
Органическая матрица вследствие разрыва С-0 связей деформируется и
теряет способность накапливать и удерживать ионы Са, что также усиливает
процесс деминерализации в твердых тканях. Ускорение элиминации фос-
фатных анионов из кости вызывает повышение уровня пирофосфатов в моче,
что является дополнительным подтверждением наличия генерализованного
или системного остеопороза.
Как известно, митохондриальная ДНК (мДНК) обеспечивает автономный
биосинтез белков, в том числе ферментов, снабжающих организм энергией за
счет окислительного фосфорилирования — синтеза АТФ. Митохондриальный
геном отличается выраженной нестабильностью — скорость эволюции мДНК
превышает таковую ядерной ДНК (яДНК) в 10-20 раз. Высокий уровень му-
тагенеза мДНК объясняется отсутствием эффективных систем репарации,
измененным генетическим кодом и протекающими в митохондриях окисли-
Глава 33. Остеосистема и реперы Махаланобиса
483
тельно-восстановительными процессами. мДНК подвергается атаке свобод-
ными радикалами, поскольку митохондрии являются основным источником
таких соединений в организме, а их ДНК особенно чувствительна к окисле-
нию. Скорость окисления ДНК в митохондриях значительно выше, чем в
ядре. Уязвимость мДНК обусловлена тем, что она не содержит гистоновых
белков, которые связаны с ядерной ДНК и защищают её. Кроме того, мито-
хондральные гены только в минимальной степени обслуживаются фермента-
ми, заменяющими окисленные участки ДНК в клеточном ядре.
Другим аспектом биомоделирования функций костной системы является
физиологический механизм устранения избыточных и аномальных клеток,
необходимый для поддержания тканевого гомеостаза, получивший название
апоптоз. Появление нового класса антирезорбентов костной ткани — бис-
фосфонатов, способных индуцировать апоптоз остеокластов, повысило ин-
терес к проблемам биомоделирования процессов остеопороза с целью повы-
шения эффективности его терапии.
Этому способствовало, наряду с позитронно-эмиссионной, магнитно-
резонансной томографией и спектроскопией, внедрение однофотонной эмис-
сионной компьютерной томографии (ОФЭКТ) для прижизненной визуали-
зации апоптоза в костной ткани. Эти методы важны и при исследовании и
биомоделировании процессов деградации хрящей и костей при ревматоид-
ном артрите. С помощью ОФЭКТ на мышах C57BL/6Y-Hi/+ и кроликах
FC/J показано, что 14C-DG — дигоксин (радиоактивный зонд) резко накап-
ливается в области воспаления и костно-хрящевой деструкции. Противовос-
палительные, иммуносупрессивные и гормональные препараты, в частности
преднизолон (в течение 4 суток после введения животным бычьего коллаге-
на II типа), снижали содержание количества радиозонда по сравнению с
животными контрольной группы.
При использовании однофотонной и ультразвуковой остеоденситомет-
рии, рентгеноморфометрии и других методов констатируется факт сниже-
ния минеральной плотности и прочностных характеристик костной ткани,
её ускоренное старение у лиц, подверхшихся воздействию малых доз внеш-
него облучения и других экологически неблагоприятных факторов. В луче-
вой модели остеопороза отмечены преостеобластический «провал», деструк-
ция костного матрикса, диспластический фиброз, кумуляция радионукли-
дов в кристаллах гидроксиапатита, нарушение энергетического метаболиз-
ма. При радиационном поражении страдают делящиеся клетки, прежде все-
го преостеобласты, в которых тормозится митоз. Его восстановление, как и
явления ликвидации радиационного блока митозов в других клеточных си-
стемах, требует дополнительных источников энергии, которые вовлекаются
в восполнение тканевого ущерба в виде фосфатных ресурсов, АТФ, парат-
гормона, кальцитонина и неспецифических гормонов (инсулина, картизола,
сомототротина, половых гармонов и т.д).
484
Биомоделирование в регуляторных системах
В последние годы признается общепринятым использование ОФЭКТ и
других методов анализа для выявления нарушений остеообразования на ран-
них стадиях, а также для мониторинга больных остеопорозом, что делает
этот метод наиболее оптимальным приемом в биомоделировании на совре-
менном этапе развития науки.
Если остеопению и остеопороз рассматривать как общебиологическое
явление, то все изменения биоэнергетических процессов, которые просле-
живаются при системном остеопорозе, можно перенести на другие клини-
ческие формы, так как конечный результат один и тот же. В последние
годы формируется мнение о том, что состояние костной ткани является
более вероятным предиктором продолжительности жизни, нежели уровень
артериального давления или холестерина. В гл. 35 и 37 мы вернемся к этим
вопросам. Но в данный момент можно с уверенностью сказать, что итера-
ционные математические модели могли бы сделать эту версию более прав-
доподобной.
Раздал VIII АЛЛОМЕТРИЯ
И ЭКСТРАПОЛЯЦИЯ:
ПРЕДЕЛЫ И ВОЗМОЖНОСТИ
Исследуй всё,
пусть для тебя на первом месте будет разум;
представь ему руководить тобой
Пифагор
487
Глава 3 4 КРИТЕРИИ ПОДОБИЯ
И АЛЛОМЕТРИЯ
Эмпирическая аналогия отношений между челове-
ком и животными лежит в основе медико-биологи-
ческого моделирования. Развитие практики моде-
лирования привело к необходимости решения ряда
логических проблем этих умозаключений. Посколь-
ку традиционная логика игнорировала эти пробле-
мы, как и саму форму вывода, врачи и биологи взя-
лись за их решение сами. Важнейшими из них яв-
ляются проблемы условий правомерности исполь-
зования таких аналогий.
Многообразие подобия и его пятый вид
Исходным понятием теории подобия, как указыва-
ет само название теории, является понятие подобия.
Простейшей формой подобия является простран-
ственное подобие. В биомедицинских моделях под
ними понимается сходство морфо-функциональных
параметров.
В медико-биологических исследованиях, как мы
уже неоднократно подчеркивали, в основном ис-
пользуются три вида моделей: лабораторные живот-
ные, альтернативные тест-объекты и математичес-
кие модели. Для того чтобы перенос эксперимен-
тальных данных с модели на человека был правоме-
рен, надежен и в принципе допустим, необходимо
соблюдение основного условия моделирования, а
именно принципа подобия экспериментальной
модели оригиналу, отклик которого на воздействие
488
Аллометрия и экстраполяция: пределы и возможности
любого внешнего фактора окружающей среды она должна воспроизвести.
Условия, понятия и принцип подобия реализуются через критерии и кон-
станты подобия.
Прежде всего остановимся на необходимых условиях подобия и призна-
ках, входящих в определение понятия подобия и делающих последние подоб-
ными. Но всегда ли эти признаки совместимы друг с другом и не противоре-
чит ли в некоторых случаях требование тождественности основного уравне-
ния, описывающего явления, требованию наличия преобразований подобия?
Уравнение должно описывать подобные явления, иметь одну и ту же форму
для целой группы явлений и отвечать условиям гомогенности функций.
Проблема выбора констант подобия связана с определением конкретной
формы ограничений, накладываемых на этот выбор. Рассмотрим условия
инвариантности функции по отношению к масштабным преобразованиям.
Пусть функция имеет вид F(uv ..., ип) = 0. Такое уравнение описывает
целый класс явлений. Выберем из них два явления, для которых имеют место
масштабные преобразования в условиях квантора всеобщности Vw :
Vh F(u\, ..., и'п) => 0; (108)
\/uF(u'\, ..., и"п) => 0. (109)
Здесь и',, ..., и'п; и'\, ..., и"п — различные значения величин «,, ... ип.
Если два выбранных нами явления подобны, то имеют место масштаб-
ные преобразования:
и", =V„; (ПО)
""„ = *У„- (I")
Согласно определению подобия, для того чтобы два явления были по-
добны, требуется сосуществование уравнений:
VuF(uv ..., «Л)=>0; (112)
\/uF{kxuv ..., *йий) => 0. (113)
Для этого необходимо, чтобы F(k{uv ..., knun) тождественно обращалось в
нуль при значениях переменных, удовлетворяющих F{u{, ..., ип) = 0. Это
возможно лишь при
Vw F{k{uv ..., knun) => ф(£,, ..., kn) F{uv ..., un), (114)
когда равенство нулю правой части неминуемо влечет за собой равенство
нулю левой.
Таким образом, в том случае, если F описывает подобные явления при
умножении всех величин, стоящих под знаком функции, на постоянные мно-
жители, все постоянные выносятся из-под знака функции, образуя некоторый
множитель для функции в целом. Масштабное преобразование отдельных
Глава 34. Критерии подобия и аллометрия
489
переменных биомодели приводит к масштабному преобразованию всей функ-
ции в целом. Функции, обладающие таким свойством, носят название гомо-
генных. Уравнение же, которое при переносе всех его членов в левую часть
принимает вид #= 0, где Я— гомогенная функция, называется гомогенным
уравнением.
Итак, чтобы функция могла описывать подобные явления, необходимо,
чтобы она, так же, как и соответствующее уравнение, была гомогенной в
биомодели.
Существует класс степенных функций, которые являются гомогенными
независимо от выбора множителей кг Возьмем уравнение, в левой части
которого находится степенное выражение
W F{ux, ..., ип) => и, «», ..., и™ => 0. (115)
Подставляя и( = кр^ получим:
W ЖМр •••> кпип> =* (Vi)al - (W" => °» С116)
или
Vi/ Л^Ир ..., *„«>» (А:^1, ..., ^«Хи,»1, ..., и„а") = 0, (117)
то есть, полагая кхаХ... к™ => к, получим
Vw W,«p ..., *„ия) =» Щ«р ..., ип) => 0. (118)
Такие функции можно назвать безусловно гомогенными. Они остаются
инвариантными по отношению к любым масштабным преобразованиям
вследствие самой структуры функции. Остальные функции являются инва-
риантными лишь по отношению к определенным типам масштабных
преобразований, удовлетворяющих тем или иным условиям. Эти функции
называются условно гомогенными. К ним относится подавляющее большин-
ство функций, с которыми приходится встречаться при решении задач био-
медицинского моделирования. В медицинской биологии, как и в целом в
формально-логическом моделировании, общепринятыми являются пять кри-
териев подобия.
Остановимся на первых четырех:
/ первый критерий подобия отображает пространственное сходство или
общность морфо-функциональных характеристик органов и систем у
прототипа человека и его биологической модели;
/ второй критерий подобия отображает единство или сходство метаболи-
ческих, нейрорегуляторных, двигательных, эндокринных и экзокрин-
ных функций;
/ третий критерий подобия отображает единство или максимальное сход-
ство эффектов критических систем и органов по их реагированию на
выбранное или исследуемое воздействие;
/ четвертый критерий подобия выражает сопоставимость констант или
490
Аллометрия и экстраполяция: пределы и возможности
иных параметров, количественно оцениваемых и математически опи-
сываемых в системе гомогенных функций.
Соблюдение в модели этих критериев подобия позволяет осуществлять
эффективную экстраполяцию с модели на прототип и обратно. Некоторые
авторы a priori полагают, что лишь при соблюдении этих критериев можно
достигнуть идеальной однозначности в биомоделировании. Это не так, а точ-
нее — далеко не так.
Существуют формулировки достаточных условий подобия, в которых ис-
ключено понятие о критериях. Так как условия однозначности относятся
тоже к некоторой системе, составляющей часть рассматриваемой системы,
то понятие подобия условий однозначности уже должно содержать в себе и
условие инвариантности их критериев. Поэтому можно сказать: системы
подобны, если их условия однозначности подобны. При этом все время
предполагается, что уравнения, описывающие оба явления, имеют одну и
ту же форму. Таким образом, понятие подобия можно сформулировать как
подобие условий однозначности при тождественности основной системы
уравнений. Это дает достаточное основание для утверждения подобия явле-
ний. То есть для подобия явлений, кроме тождественности уравнений, до-
статочно геометрического подобия, временного подобия, подобия биоло-
гических констант и подобия начальных и граничных условий, функций и
эффектов.
Итак, мы столкнулись с пятым, временным, видом подобия, не входящим
в одно из подмножеств пространственного подобия. Определим его место в
биомоделировании. Временное подобие определяется аналогично геометри-
ческому. Момент времени численно определяется величиной интервала между
ним и другим моментом, принятым за начало отсчета. Отрезок времени, в
течение которого происходит явление, состоит из бесконечного множества
таких моментов — временных «точек».
Два биологических явления будут подобными во временном отношении
в том случае, если все временные точки одного из них можно получить при
помощи умножения в одно и то же А: число раз чисел, выражающих соответ-
ственные временные точки другого биологического же явления.
1" = к?. (119)
Например, такое преобразование во времени аналогично однородной кон-
формации биомолекул в пространстве, при помощи которой определяется
пространственное подобие. Его можно рассматривать как масштабное пре-
образование. Тогда, как и в случае пространственного подобия, соответствен-
ные временные точки подобных явлений будут эквивалентны в числовом
отношении, отличаясь друг от друга лишь масштабом.
Поскольку для любых двух пар соответственных точек
ъ" = kz{ и т2" = Лт2', то т/ - т/Д/' - т2" = к, (120)
Глава 34. Критерии подобия и аллометрия
491
то есть отношение любого временного отрезка явления к соответственному
отрезку подобного ему явления есть величина постоянная. Она, как и в
случае пространственного подобия, носит название множителя преобразо-
вания.
Группы подобных во временном отношении явлений отличаются друг от
друга лишь абсолютной величиной множителя преобразования. Временное
подобие носит название гомохронии. В частном случае, при к = 1, оно пере-
ходит в синхронию, так же, как пространственное подобие при к = 1 превра-
щается в равенство.
О биологическом подобии систем, то есть о подобии их параметров, оп-
ределяющих физиологические свойства, можно говорить лишь в том случае,
если эти параметры не являются величинами постоянными. В противном слу-
чае любые две биосистемы были бы подобны, так как отношение двух по-
стоянных есть величина постоянная. Иными словами, мы имели бы в
качестве модели кошки не другую кошку, а ее саму. Это, конечно, идеальная
модель, но вряд ли она нам интересна с точки зрения медицинской биоло-
гии и биомоделирования.
При моделировании любых характеристик, изменяющихся во времени и
пространстве, будь то скорость развития лекарственных эффектов или по-
бочных проявлений параметров фармакокинетики, динамики процессов
воспаления и репарации при воздействии микробных факторов и т. д. [345,
551, 552, 553, 690], необходимо определение степени подобия используемой
модели по временному критерию обязательно и требует внесения соответ-
ствующих констант.
Математические уравнения, связывающие протекающие во времени био-
логические реакции организма с массой тела, дают возможность рассчитать
коэффициенты или константы подобия относительных скоростей этих реак-
ций для лабораторных животных и человека. Константы подобия показыва-
ют, во сколько раз быстрее (или медленнее) может развиться интоксикация
у животного по сравнению с человекам из-за большей (или меньшей) ин-
тенсивности биологических процессов, и составляют, по нашим расчетам,
если человека принять за единицу: для мыши — 9,7 , крысы — 5,2, морской
свинки — 4,3, кролика — 2,5, собаки — 1,7, овцы — 1,4, свиньи — 0,83 и
лошади — 0,75. При переходе на реальное время это означает, что, напри-
мер, интоксикация или побочная реакция лекарств, развивающаяся у крысы
за 3 месяца, проявится у мыши уже через 1,5 месяца, у кролика — через 6
месяцев, собаки — 9 месяцев, человека — 16 месяцев, а у лошади — только
через 22 месяца воздействия вещества. Существование аллометрических за-
висимостей времени развития эффектов интоксикации необходимо прини-
мать во внимание, оценивая достаточность длительности фармакологичес-
ких и токсикологических опытов при экстраполяции данных с животных на
человека [81, 185, 380, 400, 401].
492
Аллометрия и экстраполяция: пределы и возможности
Однако даже при соблюдении всех условий результаты единичного меди-
ко-биологического опыта, выраженные в форме зависимостей между раз-
мерными величинами, не могут быть распространены на другие явления.
Такие результаты характеризуют лишь единичное. Например, можно пере-
нести некоторые критерии моделирования гидродинамики на рыбках гуппи
в отношении дельфина или тюленя, но не в отношении слона или человека.
Иначе будет обстоять дело в том случае, если мы будем выражать результаты
опытов не в форме зависимостей между размерными величинами, а пользу-
ясь критериями подобия. Тогда записанный в таком виде результат будет
характеризовать уже не единичное явление, а целую группу явлений, то есть
все явления, подобные друг другу. Пусть удастся показать, что п = к, где п —
критерий подобия, к — определенное число. Данное равенство соответствует
различным условиям опыта со всевозможными численными результатами.
Оно утверждает, что при всем разнообразии числовых данных определенные
их комбинации, составляющие критерии подобия, подчинены соотношению,
выраженному в данном равенстве. Поскольку таких комбинаций множество,
устанавливается связь между разнообразными явлениями, которые, однако,
все должны принадлежать к одной группе, то есть быть подобными друг
другу.
Условия переноса при экстраполяции
Установив эмпирически зависимость п = к, мы получим право переносить
результаты опытов с одними явлениями на другие явления, существенно
отличающиеся от первых. Например, результаты, полученные при наблю-
дении движения крови в артерии определенного размера, могут быть пе-
ренесены на движение жидкости с другими физическими свойствами при
других определяющих размерах системы. Таким образом мы можем исполь-
зовать не биологическую, а физическую или физико-химическую модель
сосуда, насытить ее датчиками и поэтапно исследовать все интересующие
нас параметры крови: скорость движения, вязкость, изменения агрегатно-
го состояния и т. д. Это может оказаться весьма полезным устройством для
изучения влияния лекарств и ксенобиотиков. Последующие математичес-
кие описания процесса дадут нам право использовать принципы подобия
систем для биомоделирования на животных и экстраполяции в отношении
человека.
Корректность экстраполяции особенно важна при использовании в ис-
следованиях альтернативных биологических моделей 2-го порядка. Такие
наиболее часто используемые биологические модели 2-го порядка, как даф-
нии, рыбы, инфузории, светящиеся бактерии, сперматозоиды крупного ро-
гатого скота, клеточные культуры, по всем или нескольким критериям не
подобны человеку [2]. В силу особенностей таксономических механизмов
Глава 34. Критерии подобия и аллометрия
493
организации живых структур имеет место резкое и теоретически почти не
предсказуемое варьирование различий концентраций реагирования биотест-
объектов на отдельные вещества значимых, т. е. близких к предельно допу-
стимым концентрациям этих соединений для человека [197, 205, 414, 420].
Независимо от того, какой метод биотестирования используется, почти во
всех случаях для большинства тестируемых веществ концентрации реагиро-
вания биотест-объектов в сотни и тысячи раз превышают ПДК для человека
[159, 170, 173, 178, 187, 363, 388, 459].
Меганечувствительность биотестов к концентрациям для ряда веществ,
значимым для человека, помимо биологического неподобия моделей, имеет
и другие причины одна из которых — экстраполяция данных кратко-
временных, большей частью острых, опытов на биотест-объектах на условия
хронического воздействия на человека, когда летальные (или близкие к ним)
концентрации, полученные при биотестировании, приравниваются к уров-
ням гигиенических ПДК. Кроме того, такой фактор, как кумуляция, весьма
мало значим для биотест-объектов по сравнению с человеком. В частности,
различия в оценках кумулятивное™ веществ для человека и дафний дости-
гают 4—6 порядков. Поэтому биотестирование не пригодно для оценки ку-
мулятивных свойств веществ для человека. При прямой экстраполяции дан-
ных к безопасной для населения может быть отнесена вода, содержащая вы-
сокие, заведомо токсичные для человека концентрации загрязнений. Поэто-
му для избежания или минимизации вероятности ошибок экстраполяции
необходимо всегда приводить концентрации реагирования биотест-объек-
тов на тестируемые соединения и применять биотестирование в строго рег-
ламентированных рамках. [55, 139, 472].
Хотя математические модели уже давно и успешно используются в био-
логии и медицине, проблемы оптимизации и систематизации таких моде-
лей, оценки их эвристической значимости и определения граничных усло-
вий применения в конкретных условиях нуждаются в дальнейших исследо-
ваниях и интерпретациях.
Важнейшее значение для интерпретации экспериментов, проведенных
на животных, с учетом размера тела и основных физиологических показате-
лей различных видов млекопитающих позволяет делать некоторые заключе-
ния относительно характеристик организма человека и его функций.
Важными в этом отношении являются такие показатели, как интенсив-
ность метаболизма, анатомия, частота сердечных сокращений и химия орга-
низма. Существуют впечатляющие корреляции этих показателей с размера-
ми организма [90, 91, 406, 605]. Наиболее наглядным явлением и порази-
тельным примером является постоянство удельной поверхности тела.
Как видно из рис. 9, внешняя удельная поверхность тела млекопитающих
и других позвоночных остается равной удвоенной удельной поверхности сфе-
ры с таким же объемом при изменении массы тела в 107 раз. Более высокие
494
Аллометрия и экстраполяция: пределы и возможности
Ю5
104
103
102
1П1
I
#п
.
Л/^
•V. уг
1 I I I I I
10° 101
102 10э 104
Масса тела, г
105 106 107
значения на графике относятся к буковым деревьям. Отсюда следует, что эта
удивительная закономерность распространяется даже на растения.
Сплошной линией показана зависимость между поверхностью и массой
для шара плотностью 1,0. Точки в правом верхнем углу относятся к буковым
деревьям.
Относительный объем, занимаемый отдельными органами, по существу,
не зависит от размеров тела, как показано на рис. 10 на примере объема
легких млекопитающих. Это
справедливо даже для таких
крупных обитателей океана,
как дельфины и киты. Отсут-
ствие зависимости характер-
но и для других органов. Од-
нако иногда встречаются не-
которые видовые особеннос-
ти, и они, как правило, не
являются неожиданными.
Так, например, обычно серд-
це составляет 0,5—0,6% от
массы тела почти всех млеко-
питающих, кроме скаковых
лошадей и борзых собак, у ко-
торых оно крупнее.
Однако эта анатомическая
закономерность не может
распространяться на все уров-
ни, поскольку живые орга-
низмы существуют в услови-
ях некоторых ограничений,
зависящих от их размеров.
Одним из таких ограничений
для наземных животных яв-
ляется необходимость в меха-
нической опоре для противо-
действия силе тяжести, в свя-
зи с чем наблюдается утяже-
ление скелета у более круп-
ных животных. Однако утя-
желение скелета поразитель-
но мало. Было найдено, что
масса скелета возрастает про-
порционально массе тела в
Рис. 9. Зависимость между площадью поверхности
и массой тела у позвоночных
100
10
1,0
0,1
0,01 -
0,001
.Кит
Дюгонь т
Ламантин ^
Медведь У м Корова
Свинья ?/* Дельфин
Коза ••^Человек
Собака •
Енот*
Кошка • у'
-. <»ХОбезьяна
Сурок. >«Кролик
Броненосец
Крыса -/^Морская свинка
Мышь
Землеройка
Летучая мышь
J
0,01 0,1
1 10
Масса тела, кг
100
1000
Рис. 10. Зависимость между объемом легких и мас-
сой тела у млекопитающих. Наклон линии указывает
на простую пропорциональную зависимость
Глава 34. Критерии подобия и аллометрия
495
степени 1,13 (т1,13); для крупных животных многие авторы рассматривают
как большое техническое достижение природы.
Галилео Галилей считал [227], что «...если деревянный брус выдерживает
тяжесть, скажем, десяти равных ему брусьев, то подобная ему, но больших
размеров балка не сможет выдержать веса десяти одинаковых с нею балок.
Кто не знает, что лошадь, упав с высоты трех-четырех локтей, ломает себе
ноги, тогда как собака при этом не страдает, а кошка остается невредимой,
будучи брошеной с высоты восьми-десяти локтей, точно так же, как свер-
чок, упавший с верхушки башни, или муравей, упавший на Землю хотя бы
из лунной сферы...».
Понятно, что лошадь не спит на деревьях и ей не нужно взбираться туда
или спрыгивать. Тем не менее, при моделировании аллометрических харак-
теристик и использовании их для экстраполяции следует основываться на
критериях подобия, в данном случае, безусловно, на третьем и четвертом.
В то же время трудно даже представить какое-либо построение в виде био-
логической модели, в котором мы смогли бы проигнорировать и пятое по-
добие, т. е. временной вектор модели. Именно этот аспект и будет нами рас-
смотрен в гл. 35, где надеемся показать необходимость учета в аллометри-
ческих построениях такого фактора, а точнее параметра, как физиологичес-
кое время.
496
Глава 3 5 ФИЗИОЛОГИЧЕСКОЕ ВРЕМЯ
И КИНЕТИКА
Тема межвидовых вариаций в фармакокинетике и
токсикокинетике до последнего времени являлась
эмпирической и описательной, за исключением
нескольких работ [90, 156]. Типичным является при-
мер, когда препарат вводится двум или более видам
млекопитающих, затем измеряются его уровни в
крови и моче, вычисляются параметры, ищутся
корреляции и результаты представляются в таблич-
ной или графической форме. Этот подход привёл к
заключению, что, в общем, другие виды млекопита-
ющих элиминируют большинство препаратов более
быстро, чем человек.
Затем было установлено, что элиминирующая
эффективность организма или общий клиренс орга-
низма, когда они выражаются отношением объёма
ко времени на единицу веса тела, обычно выше у
животных, чем у человека. Поэтому исследователи
традиционно полагали, что два вида с различными
величинами клиренсов, выраженных таким спосо-
бом, имеют различные величины фармакокинети-
ческих параметров. Но так ли это на самом деле?
Возьмём, например, данные для мыши и коровы,
приведенные в табл. 85. Мышь (0,03 кг) имеет печё-
ночный клиренс препарата 52,3 мл/мин/кг веса тела,
а корова (760 кг) имеет печёночный клиренс 14,1 мл/
мин/кг веса тела. Вследствие того, что вес печени
мыши составляет 5,83% от веса тела, а у коровы
1,57%, соответствующие величины клиренса стано-
вятся идентичными, когда вес тела заменяется на вес
Глава 35. Физиологическое время и кинетика
497
печени. А поскольку скорость перфузии печени у обоих видов является при-
мерно одинаковой на каждый кг веса печени [90], то и среднее печёночное
экстракционное отношение у обоих видов является приблизительно одинако-
вым (около 0,60).
Более важным для нас, однако, является повышение интенсивности мета-
болизма с уменьшением размерой тела. Как видно из рис. 11, скорость мета-
болизма изменяется пропор-
ционально массе тела в степе-
ни 3/4. Не существует никаких
объяснений этой поистине
замечательной закономернос-
ти, справедливой даже для
очень малых организмов. На-
личие такой закономерности
имеет важные последствия
для анатомического строения
и химии крови. В то же время
эффективность энергетичес-
ких превращений у млеко-
питающих не зависит от раз-
меров тела.
С помощью согласования
параметров интенсивности
метаболизма и величин клиренса с индивидуальной анатомией и физиоло-
гией организма можно устранить кажущиеся различия. Однако существует и
другой способ оценки этих данных, а именно с помощью введения концеп-
ции «физиологического времени» [96, 97, 115].
Boxenbaum вводит понятие физиологического времени [90], определяя его
как единицу измерения хронологического времени, зависящую от вида, не-
обходимую для завершения некоторого физиологического события, не зави-
симого от вида. Если говорить более простым языком, то физиологическое
время — это хронологическое время, нормированное на вес тела в некото-
рой степени (что и будет показано ниже). Возьмём, например, процесс ста-
рения двух видов: собаки (с продолжительностью жизни 14 лет) и человека
(с продолжительностью жизни 14 и 98 лет).
Собака старится со скоростью 7,14% её жизни в год, а человек — со
скоростью 7,14% его жизни за 7 лет. Поэтому 1 год для собаки и 7 лет для
человека являются эквивалентными физиологическими временами необхо-
димыми, чтобы произошло независимое от вида физиологическое событие,
т. е. проживание 7,14% от продолжительности жизни.
Применяя это понятие к вышеупомянутому примеру, мы наблюдаем, что
в среднем каждый микролитр крови у мышей проходит через печень один раз
ю-12 ю-9 ю3 ю6
Масса тела, г
Рис. 11. Связь между интенсивностью метаболизма и
массой тела у различных организмов: I - гомойтерм-
ные (39° С), II - пойкилотермные (20° С), III - однокле-
точные организмы
498
Аллометрия и экстраполяция: пределы и возможности
Таблица 85
Теоретические данные для гипотетического печёночного метаболизма
препарата у мыши и коровы
Вес тела, кг
Объём крови, л
Вес печени, кг
Печёночный кровоток, л/мин
Печёночный клиренс, мл/мин/кг веса печени
Печёночный кровоток, мл/мин/кг веса тела
Печёночный клиренс, л/мин
Среднее печёночное экстракционное отношение
Печёночный клиренс, мл/мин/кг веса тела
Время полного обращения печёночного кровотока, мин
Доля объёма крови очищения препарата на время обращения
Мышь
0,030
0,0021
0,00175
0,00262
898
87,3
0,00157
0,60
52,3
0,802
0,60
Корова
760
53,2
11,9
17,8
898
23,4
10,7
0,60
14,1
2,99
0,60
за каждые 0,802 мин. У коровы каждый микролитр крови в среднем проходит
через печень один раз за каждые 2,99 мин. Мы распознаем эти величины как
времена обращения, равные объему крови, делённому на печёночный крово-
ток. Эти времена, или более правильно «физиологические времена», также
равны хронологическому времени, которое необходимо для того, чтобы пол-
ный объём крови прошел через печень в среднем один раз. То есть, проще
говоря, каждые 0,802 мин у мыши эквивалентны 2,99 мин у коровы (табл. 85).
Рассматривая элиминацию препаратов у каждого вида, видим, что 60%
(печёночное экстракционное отношение, в процентах) препарата в крови
будут элиминироваться за время одного обращения. С другой точки зрения,
отношение времени обращения просто является величиной обратной отно-
шению клиренсов, если последнее выражено как мл/мин/кг веса тела.
Физиологическое время оценивается с помощью темпа изменений в
живых организмах, а хронологическое время обычно измеряется при помо-
щи движения стрелок по циферблату, которые в свою очередь синхронизи-
рованы с некоторым другим движением, например вращением Земли вокруг
оси. Хорошие примеры физиологического времени даны в работах [242, 243,
256, 605]. Как время дыханий, так и сердечных сокращений значительно
различаются у разных видов млекопитающих, варьируя как показательная
функция от веса тела с показателем равным 0,28.
Время дыханий (с) = 0,169 х В0'28 (121)
Время сердечных сокращений (с) = 0,0428 х 2?0>28 (122)
где В — вес тела в граммах.
Площадь альвеолярной или диффузионной зоны легких изменяется про-
порционально скорости потребления кислорода, а не площади наружной
поверхности тела, что показано на рис. 12.
Глава 35. Физиологическое время и кинетика
499
1000
2 100
3
3
g 10
с
со
1,0
0,1
0,01
Мини-свинья i
Медведь
Кролик 9
Броненосец
Овца #
Человек—j
Собака •.
Енот
Кит
1 Корова
i Дельфин
Ламантин
Дюгонь
Кошка
Обезьяна
Крыса
.Мышь
• Землеройка
• Летучая мышь
• Морская свинка
/т
0,1
10
Масса тела, кг
100
1000
10000
Рис. 12. Связь между площадью альвеолярной поверхности и потреблением кислорода
у млекопитающих
Деление времени дыханий (пневматохрон) на время сердечных сокраще-
ний (кардиохрон) даёт величину, равную 4,0. Другими словами, у всех мле-
копитающих 4 сердечных сокращения соответствуют 1 дыхательному циклу.
Кроме того, у многих видов млекопитающих, исключая долгоживущих при-
матов (см. ниже), продолжительность жизни также пропорциональна весу
тела в степени 0,28. Это показывает, что большинство млекопитающих име-
ет одинаковое число дыханий и сердечных сокращений в течение их жизни.
Быстроживущая мышь имеет частые сердечные сокращения и короткую
продолжительность жизни, в то время как медлительный гиппопотам имеет
редкие сердечные сокращения и относительно длительную жизнь. Аналогия
с фармакокинетикой ясна: мелкие короткоживущие животные обычно бо-
лее быстро удаляют препарат из организма (если использовать хронологи-
ческое время) на единицу веса тела, чем более долгоживущие животные. Но
измеренные с помощью их собственных внутренних часов, млекопитающие
удаляют препараты с одинаковой скоростью.
Физиологическое время становится синонимом фармакокинетического вре-
мени, когда оно применяется к фармакологическим событиям.
Эффективность массопереноса через альвеолярную поверхность не зави-
сит от размеров. У мелких животных плотность капилляров выше, однако,
согласно [571], она недостаточна, чтобы удовлетворить запросы повышен-
ного метаболизма. В связи со сказанным химия крови зависит от размеров
организма.
500
Аллометрия и экстраполяция: пределы и возможности
Влияние размеров млекопитающих на химическое поведение крови выг-
лядит следующим образом (рис. 13):
/ при данном рН гемоглобин мелких животных отдает кислород легче, чем
гемоглобин крупных животных;
/ гемоглобин мелких животных освобождает больше кислорода в ответ на
снижение рН или увеличение давления С02 (эффект Бора).
Отмеченные интересные закономерности могут быть полезны для коли-
чественных предсказаний и, кроме того, позволяют понять некоторые ас-
пекты проблем метаболизма.
100
CD
3
о
га
I
50 -
60 80
мм рт. ст.
Рис. 13. Кривые диссоциации оксигемоглобина различных млекопитающих: 1 - слон, 2 - ло-
шадь, 3 - человек, 4 - овца, 5 - лиса, 6 - кошка, 7 - крыса, 8 - мышь. Видно, что кровь мелких
животных отдает кислород при более высоких напряжениях кислорода, что обеспечивает до-
ставку кислорода в количестве, достаточном для поддержания высокого уровня метаболизма
у этих животных. Пунктирной кривой показано влияние подкисления (эффект Бора) на кровь
мыши (кривые 8 и 8Б)
Важной характеристикой физиологического или фармакокинетического
времени является то, что оно обычно зависит от возраста. Возьмём наиболее
часто цитируемый пример фармакокинетического времени — диспозицион-
ный период полувыведения. Для многих препаратов этот параметр возраста-
ет вместе с хронологическим возрастом. Понятие о диспозиционном перио-
де полувыведения можно так же использовать для иллюстрации связи физи-
ологического времени с хронологическим временем. В общем, процессы, оп-
ределяемые скоростью, в фармакокинетике описываются с помощью хроно-
логического времени, которое является независимой переменной в знамена-
теле, например клиренс является объёмом в единицу (хронологического)
Глава 35. Физиологическое время и кинетика
501
времени. То есть физиологическое событие, в этом случае очищение опреде-
лённого объёма крови, происходит как функция времени. Но в фармакоки-
нетическом времени, например диспозиционный период полувыведения,
50%-ное уменьшение содержания препарата становится независимой пере-
менной, а время становится зависимой переменной, т. е. время становится
«процессом» [220]. И в этом заключается методология подхода ко многим
проблемам межвидовых вариаций в фармакокинетике: нормирование хроноло-
гического времени на приемлемый внутренний барометр или часы. Как будет
обсуждаться ниже, один методический подход состоит в поиске связи между
фармакокинетическими параметрами и весом тела и/или возможной про-
должительностью жизни.
502
Глава 3 6 МЕЖВИДОВЫЕ ВАРИАЦИИ
Наиболее важное различие между видами животных,
обычно используемых в кинетических исследовани-
ях, — их размеры, но исследователи, за исключением
Дедрика [156], молчаливо игнорируют этот факт.
К сожалению, это говорит о том, что изучение раз-
меров и их последствий (аллометрия), вероятно, вы-
ражает основной аспект базового плана для кине-
тических процессов. Экстраполяция полученных ре-
зультатов на человека — обязательный, сложный и
неоднозначный этап любого экспериментального
моделирования в кинетике. На моделях 1-го порядка
(лабораторных животных-млекопитающих) исполь-
зуются либо прямой перенос данных животных на
человека, либо вводятся коэффициенты запаса. Ре-
акции лабораторных животных на действие ксено-
биотиков во многом подобны таковым эффектам у
человека.
Критериями подобия, обусловливающими адек-
ватность моделирования и надежность экстраполя-
ции, являются: сходство у человека и эксперимен-
тальной модели биологических параметров систем,
реагирующих на ксенобиотик; общность характери-
стик метаболических процессов; близость чувстви-
тельности в единых количественных показателях для
человека и модели.
В настоящее время в исследованиях все чаще
применяются альтернативные модели 2-го порядка
(различные гидробионты, бактерии, ферменты,
культуры клеток и др.). При этом результаты непо-
Глава 36. Межвидовые вариации
503
средственно переносятся с тест-объектов на человека, что, как правило, не
сопровождается детальным анализом и доказательством правомерности та-
кого подхода.
Адекватность и корректировка моделей
В сложных системах, к которым относятся и все биологические системы,
даже самая красивая модель отражает реальный процесс лишь частично.
Поэтому мы должны честно оценить пределы ее допустимости в отношении
прототипа, осуществить проверку степени соответствия и адекватности. Рас-
смотрим это в приложении к биомоделям первого и второго порядков.
Если ошибка моделирования превышает некоторый допустимый предел,
то это свидетельствует о том, что в модели не учтены важные взаимосвязи и
факторы, и, следовательно, ее нельзя считать адекватной изучаемому объек-
ту в требуемой степени. Потребуются корректировки, дополнительные ис-
следования, выбор новой математической модели, изменения ее перемен-
ных коэффициентов — и этот процесс может повторяться вновь и вновь до
достижения удовлетворительного приближения к параметрам оригинала.
Основной аргумент, используемый при обосновании возможности экст-
раполяции данных биотестирования на человека, — это высокая чувствитель-
ность биотест-объектов к токсичным веществам [122, 424, 509, 512]. Однако
результаты определения различий параметров токсичности веществ для мо-
делей 1 -го и 2-го порядков заставляют усомниться в истинности утверждения
о неизменно высокой чувствительности биотестирования. Если сопоставить
эффективные концентрации ЕС50 или летальные концентрации LC50 для био-
тест-объектов с максимально недействующими концентрациями (МНК) или
ПДК веществ по токсикологическому признаку вредности для человека, а
затем определить персентили полученных различий чувствительности био-
тест-объектов и человека к веществам, то оказывается, что 50-я персентиль
для светящихся бактерий равна 1609 (т. е. на половину тестируемых веществ
они начинают реагировать лишь с концентраций, в 1609 раз превышающих
ПДК), для рыб гуппи — 337, инфузорий тетрахимена — 275, спермы быка —
4516 и лишь для дафний относительно меньше — 44, в то время как 95-я
персентиль составляет от 13 500 для дафний до 467 660 для спермы быка.
Это свидетельствует о низкой чувствительности перечисленных альтер-
нативных моделей к веществам по сравнению с человеком. Проблема усу-
губляется еще и тем, что модели 2-го порядка часто не реагируют именно на
чрезвычайно токсичные и кумулятивные для человека вещества в высоких
концентрациях. Так, дафнии не чувствительны к концентрациям до 130 000
МНК дихлорэтилена, 13 500 — дихлорэтана, 20000 — таллия; светящиеся
бактерии — к присутствию в воде до 6400 МНК бензола, 3500 — ДДТ, 6000 —
гептанола; рыбы гуппи выживают при более, чем 400 000 МНК дихлорэти-
504
Аллометрия и экстраполяция: пределы и возможности
лена, 8000 — тетрахлорметана и 24 000 — хлорбутана; сперматозоиды быка
сохраняют жизнеспособность при концентрациях до 560 000 МНК акрила-
мида и 350 000 — метилметакрилата; инфузории тетрахимена переносят до
90 000 МНК дихлорэтана [387].
Не лучшие результаты дает и тестирование токсичности на клеточных
культурах и водорослях: например, для культуры клеток HeLa безопасны до
1000 МНК кадмия, водорослей сценедесмус до 1700 МНК бора. Правильнее
было бы сопоставлять эффективные концентрации для тест-объектов не с
МНК, а с пороговыми концентрациями (ПК) хронического опыта, хотя ве-
личины МНК более доступны. Следует учитывать, однако, что они отлича-
ются от пороговых иногда на порядок [386].
Различия в уровнях реагирования моделей 1-го порядка — лабораторных
животных и альтернативных им моделей — биотест-объектов к индивиду-
альным веществам не позволяют получить адекватные результаты по оценке
комбинированного действия суммы этих веществ методами биотестирова-
ния. Суммарный эффект ксенобиотиков при альтернативном моделирова-
нии будет определяться наиболее токсичными для тест-организмов веще-
ствами, в то время как для лабораторных животных и человека этот эффект
будет обусловлен совершенно другим набором веществ из этой суммы. Ре-
зультаты оценки комбинированного действия вещества или смеси веществ
на моделях 2-го порядка могли бы быть применимы к человеку в одном
малореальном случае — при четком параллелизме в чувствительности моде-
лей 1-го и 2-го порядка, т. е. при одинаковых коэффициентах различия мо-
делей ко всем действующим веществам в изучаемой смеси [388].
Информативность биотестирования стремятся повысить, определяя ток-
сичность одновременно на нескольких тест-организмах. Однако, принимая
во внимание весьма низкую (по сравнению с человеком) чувствительность
почти всех биотест-объектов к подавляющему большинству веществ в эксп-
ресс-экспериментах, можно с уверенностью сказать, что использование «ба-
тареи» биотестов не решает проблему надежности альтернативного модели-
рования действия веществ для человека.
Отождествление функций на основании тождества отдельных, вырван-
ных из системы признаков или структур, то есть структурно-функциональ-
ная аналогия как форма умозаключения, довольно часто рассматривается
как форма вывода. Она порочна сама по себе. Так, французский физиолог
П. Косса, опровергая вывод математика Куффиньяля, протестует против
использования в процессе этого вывода структурно-функциональной ана-
логии. Он считал, что такая манера рассуждать вызывает разве что удивле-
ние: из морфологического сходства делать заключение о функциональном
тождестве означает перейти границы экстраполяции. Доводя до карикатуры
этот способ рассуждения, вполне возможно сказать: «Клетка Пуркинье
мозжечка с ее специфическими древовидными отростками походит на се-
Глава 36. Межвидовые вариации
505
мисвечник древнееврейской литургии. Следовательно, клетка Пуркинье
является органом, в котором помещаются религиозные наклонности чело-
века». Иронию Косса можно распространить и на функционально-струк-
турную аналогию, то есть на отождествление структур на основе тождества
функций. С ее помощью, наряду с ценными результатами, также было по-
лучено много ошибочных выводов. Однако это не означает, что следует
отбросить саму форму вывода. Нужно выяснить условия, снижающие веро-
ятность ошибок или даже исключающие их. По крайней мере, в простей-
ших случаях определение таких условий допустимо, но именно в самых
простейших моделях при установлении свойств отношений.
Логические свойство отношений обнаруживаются, как и вообще всякие
свойства, лишь в отношениях к другим вещам, в данном случае к другим
отношениям. Это предполагает, что наряду с множеством элементов модели
ар ..., ап и прототипа bv ..., bn мы должны также иметь множество отноше-
ний на них: ар ..., аки соответственно (3,,..., {37. Сказанное означает возмож-
ность представления R в модели а,,..., алкак совокупность отношений мно-
жества ар ..., ак и отношения R' в прототипе bv ..., bn как совокупности
отношений множества (Зр ..., |3Л
Изложенные соображения дают основание для включения в число усло-
вий правомерности аналогии биомоделей 1-го и 2-го порядков. Требование,
согласно которому отношения в модели и прототипе должны быть разбиты
на такое количество отношений между подмножествами элементов сравни-
ваемых систем, чтобы оказалось возможным выявить логические свойства
этих отношений, будет дано нами чуть ниже.
Непосредственно заданные отношения в сравниваемых системах явля-
ются интегральными, то есть относятся к системам в целом. В связи с этим
при анализе отношений их разложение на множество элементарных отно-
шений является непременным условием.
Далеко не каждые равномощные множества критериев подобия биомо-
делей оказываются изоморфными. Изоморфными могут быть лишь такие
системы, у которых равномощны не только множества элементов, но и
множества отношений на них, которые сами в свою очередь рассматривают-
ся как самостоятельные элементы. Десять биотест-систем (дафнии, инфу-
зории тетрахимена, водоросли, рыбки гуппи и т.п.) и десять функций ЦНС
человека и животных могут быть с точки зрения альтернативного моделиро-
вания признаны изоморфными системами, поскольку налицо соответствие
элементов и пространственных соотношений. Но согласно приведенным
выше уточнениям, у нас нет оснований считать их изоморфными, поскольку
неизвестно, соответствуют ли метаболические процессы, критические ре-
цепторные образования или воспроизводимость биологического отклика
и его порог для человека и альтернативной модели. Если же два множества
изоморфны в рассмотренном выше смысле, то собственные для этих мно-
506
Аллометрия и экстраполяция: пределы и возможности
жеств логические свойства определенных на них отношений будут одинако-
выми и приемлемыми в качестве биомодели.
Пусть множество А состоит из элементов ар ..., ап, на которых определены
отношения а,, ..., ак. На другом множестве В из элементов Ьх,..., Ък определе-
ны отношения (Зр ..., РЛ. Возьмем произвольное отношение аД1 < / < к). Внут-
реннее свойство выразится в виде зависимости высказывания o.n(aiV ..., ос,,),
(где а.р ..., аа — совокупность элементов, входящих в поле отношения а,) от
множества высказываний, содержащих другие отношения данной системы:
аДсс,,, ..., аи) =>f[a{ fa,, ..., аь), ..., а^,(а(/_1)Р ..., а(/_1)й)),
•••> <Wa(i+i)i' -' Vnv)» •••» а*Кр ■••» акЛ (123>
Согласно условию изоморфизма, в системе В этому будет соответство-
вать выражение
РАр •■•» *й) => Ф[?1 (*р •••> *i,)> •••> P/-i(Vdp -' Vu»>>' -'
1)1'-' "(/+l)v)' •"' P*(V -. М- (124)
Поскольку мы рассматриваем логические свойства а, и |3,, то, согласно
определению логических свойств Б. Расселом, мы можем заменить все по-
стоянные в выражениях (125) и (126) переменными. Всегда можно сделать
так, чтобы соответствующие друг другу элементы и отношения множеств
охватывались областью изменения одной и той же переменной. Таким обра-
зом, выражения (125) и (126) превращаются в
ri(X il, •••' -V =>/[г1 (*р "•' x\s'> "•' r/-l(X(/-l)l» •■•' *(/_!)«))'"•'
..., ',/+1Ц;+|)1,-, x(/+i)v)' "•' rkSxkl> •"> xkr>\-> (125)
'"Ail, •"' Xlt' => V 1Г1 (*1> •"' *1j)> •"' r/-l(*(/-l)l> "•' X(i-\)n)''•"'
•••' r/+l'*(/+l)p —> X(/+l)v)> •"' Г*'*ЛР -•' XAr)l- (126)
Видимо, что аргументы и значения логических функций ц/ и / в обоих
полученных выражениях совпадают. Поскольку в этих выражениях нет вне-
логических констант, отмеченное совпадение будет иметь место при любых
конкретных значениях переменных, то есть /<=> у.
Мы далеко не абсолютизируем роль лабораторных животных в биомоде-
лировании функций человека. Модели любого порядка имеют право на жизнь
при условии учета их места и времени. Однако мы не можем согласиться с
безапелляционностью утверждения о том, что альтернативные модели реша-
ют абсолютно все проблемы биомоделирования в отношении человека.
Так, одно из направлений альтернативного моделирования связано с оцен-
кой суммарной мутагенной активности в бактериальном тесте Эймса. Зача-
стую значимость теста абсолютизируется и отрицательный результат тести-
рования непосредственно экстраполируется на человека: вещества не мута-
генные в тесте Эймса, считаются не генотоксичными и не канцерогенными
Глава 36. Межвидовые вариации
507
для человека. Однако, ряд веществ, для которых тест Эймса отрицателен,
канцерогенны для лабораторных животных и потенциально для человека,
например: акриламид, атразин, ДДТ, 2,4-Д, линдан, 2,4,6-трихлорфенол, 1,4-
дихлорбензол, бериллий, нитрилотриуксусная кислота (группы 2Б и 2В по
классификации МАИР). Для мышьяка и бензола, канцерогенность которых
доказана для человека (1 группа по классификации МАИР), реакция теста
Эймса также отрицательная.
Неадекватность реакций теста Эймса на канцерогенные для человека
вещества приобретает особое значение при его использовании для оценки
опасности галоформных соединений, образующихся при хлорировании пи-
тьевой воды, в частности таких, как тетрахлорметан (четыреххлористый уг-
лерод), дихлорметан, тетрахлорэтилен (группы 2Б и 2В по классификации
МАИР), для которых тест Эймса отрицателен [388]. Даже наиболее харак-
терный представитель этих соединений — хлороформ (группа 2Б по класси-
фикации МАИР) — при изучении на генотоксичность давал как положи-
тельные, так и отрицательные результаты. Очевидно, во избежание ошибок
при оценке генотоксичности и канцерогенности в тесте Эймса, последствия
которых для населения могут быть весьма тяжелыми, так как сопряжены с
ростом онкологической заболеваемости, необходимо честно говорить о сум-
марной мутагенной активности канцерогенных веществ, не выявляемых с
помощью этого метода.
Для основной же массы веществ и лекарств прямая экстраполяция дан-
ных с биотестов на человека попросту невозможна. В случаях использования
альтернативных моделей необходимо приводить стандартные уровни реаги-
рования тест-объектов на индивидуальные вещества для того, чтобы иссле-
дователи могли ориентироваться в допустимости прямой экстраполяции на
человека полученных при биотестировании результатов. Это тем более важ-
но, что даже у лабораторных животных-биомоделей имеются большие раз-
личия, например, в относительном потреблении воды (рис. 14), а значит и
делюции лекарственных препаратов и ксенобиотиков.
Базовый план и подход Дедрика
В контексте этого обсуждения базовый план для фармакокинетики относится
к логической простоте характеристик кинетических событий. Как часть это-
го базового плана более мелкие млекопитающие имеют относительно боль-
шие размеры органов, элиминирующих препараты (печень, почки), чем у
больших млекопитающих [7, 156, 220,]. А вследствие того, что скорость пер-
фузии этих органов кровью грубо пропорциональна их размерам [6, 7], более
мелкие животные, за счёт одной лишь анатомии и физиологии, имеют боль-
ше возможностей избавляться от молекул препарата. Никто в фармакокине-
тике, по-видимому, не оценивал тонкостей, связанных с размерами, больше,
508
Аллометрия и экстраполяция: пределы и возможности
чем Дедрик [156]. В своём убедительном анализе авторы [290, 606] приводят
основные принципы межвидовых вариаций в фармакокинетике, а именно:
небольшие млекопитающие могут рассматриваться как истинные физиологи-
ческие модели больших животных в инженерном смысле этого термина.
100
юоо
10000
Вес, кг
Рис. 14. Относительные показатели потребления воды млекопитающими в переводе на кило-
грамм массы тела по сравнению с человеком, показатели которого условно взяты за единицу
(остальные обозначения см. на рис. 15)
С этой целью авторы взяли плазменные уровни метотрексата (Мтк) у пяти
видов млекопитающих (мышь, крыса, обезьяна, собака, человек), которые раз-
личались в 17 000 раз, нормировали и масштабировали их так, чтобы они
приближались к единственной кривой. Мтк элиминируется преимуществен-
но почечной экскрецией. Процесс нормализации, или масштабирования, пред-
ставляется следующим образом: на обычном графике логарифм уровня препа-
рата в плазме изображается как функция хронологического времени. Для пра-
вильного масштабирования этого графика необходимо сделать две вещи.
Во-первых, требуют внимания различия уровней в плазме, обусловленные
различиями в дозах (в мг/кг). Это легко устраняется делением уровня в плаз-
ме на дозе мг/кг.
Во-вторых, различия в значениях клиренса (мл/мин на кг веса тела) по-
требовали разработки приёмов определённого типа, чтобы согласовать хро-
Глава 36. Межвидовые вариации
509
нологическое время с фармакокинетическим. Вследствие того, что более мел-
кие виды имеют большие значения клиренсов, то хронологическое время
у этих видов ускорялось бы (сокращение времени) по отношению к более
крупным видам животных.
Авторы проводили нормализацию клиренсов, деля время на оси X на вес
тела В в степени 0,25. Тогда как первая методология установления дозы ка-
жется довольно логичной, то эта, последняя, может представляться на пер-
вый взгляд абсурдной — и в этом заключается камень преткновения. То, что
исследователи понимали и совершенно правильно использовали, заключа-
ется в осознании, что любой процесс, который включает хронологическое
время как измерение, зависит от размеров [156]. Всё, что они сделали, зак-
лючалось в устранении зависимости от размеров, т. е. они масштабирова-
ли^ хронологическое время на фармакокинетическое время.
Для мыши массой 0,022 кг (быстрого элиминатора метотрексата Мтк)
В = 2,89. Взяв отношение 0,385/2,89 мы приходим к сотношению 0,13 к 1.
Поэтому одна минута человеческого хронологического времени эквивален-
тна 0,13 мин хронологического времени мыши. Переводя это в более точные
фармакокинетические термины: мышь будет очищать такой же объём пре-
парата на 1 кг веса тела за 0,13 мин, какой человек очищает за 1 мин.
Подыскивая физиологическую основу степенного преобразования веса
тела, Dedrick et al. использовали эмпирическую связь из работы Adolph [10]:
CLcr = 4,2 х Я0-69 (127)
CLcr/B = 4,2xB-W (128)
где CLcr — клиренс креатинина (мл/ч), а В — масса тела (г). Поэтому у раз-
личных видов млекопитающих клиренс креатинина, выраженный в виде от-
ношения к весу тела, изменяется пропорционально весу тела в отрицательной
степени 0,31. Поскольку Мтк элиминируется главным образом почками, впол-
не логично, что межвидовые вариации клиренса Мтк коррелируют с меж-
видовым клиренсом креатинина, а показатель степени 0,31 близок к 0,25.
Признавая основной вклад Дедрика, рассмотрим несколько дополнитель-
ных соотношений. Для этого используем первичные данные для Мтк (из
работы [9]), а значения плазматического клиренса (CL) будем вычислять на
основании дозы и площади под кривой [156]. Отметим, что CL (мл/мин)
изменяется в зависимости от В (кг) в степени 0,69:
"«„,,= 10,9 х Я0'69 (129)
Уравнения (129) и (131) можно легко скомбинировать. Однако, уравне-
ние (129) сначала необходимо переписать так, чтобы установить единицы:
a,cr=8,2-i?069 (130)
где CL — клиренс креатинина (мл/мин), а В — вес тела (кг).
510
Аллометрия и экстраполяция: пределы и возможности
Разделив уравнение (129) на уравнение (130), получим:
Cij^/CL,,-1,33. (131)
Поэтому межвидовое отношение CLMmK и CLcr является константой, не-
зависимой от вида и видовых размеров. В известном смысле, все виды сход-
ны, экскретируя Мтк из своих организмов со скоростями, коррелирующи-
ми с внутренними факторами, вносящими вклад в их индивидуальную экс-
крецию креатинина.
Клиренс Мтк также можно отнести к общему количеству нефронов.
Поэтому в соответствии с Adolph [10]:
N= 188 000 х В °'62, (132)
где N — общее число нефронов у видов млекопитающих, а В — вес тела (кг).
Разделив уравнение (131) на уравнение (134), получим:
CLMmK/N=5,Sx\0-5-B^ <133>
Показатель степени экспоненты 0,07 в уравнении (133) называется экс-
понентой остаточной массы или ЭОМ (6): чем ближе ЭОМ к 0, тем ближе
дэом к J Уравнение (134) показывает, что для каждых 100 000 нефронов,
клиренс Мтк равен приблизительно 5,8 мл/мин. Эта связь подтверждает,
что гломерулы у различных млекопитающих сходны анатомически и физио-
логически. Подтверждение этого довода получено исследователями, кото-
рые продемонстрировали, что средняя длина гломерулярного капилляра
постоянна (733 мкм) и не зависит от размера млекопитающего [290]. Кроме
того, средняя скорость прохождения через капилляры гломерул также по-
стоянна, так как определяется значением времени нахождения в гломеру-
лярном капилляре (1,88 с). Другие процессы в почке также являются отно-
сительно инвариантными. Поэтому Edwards [177] показал, что безотноси-
тельно к виду млекопитающего почечный кровоток составляет 25,9% от сер-
дечного выброса. Аллометрический анализ, сходный с тем, который был
использован для получения уравнения (135), был применён и здесь, и зна-
чение ЭОМ было равно 0,02.
Возвращаясь к данным о Мтк, следует добавить предостережение. У более
мелких животных, например у мышей и крыс, значительная часть неизменён-
ного препарата появляется в фекалиях (экскреция с желчью после паренте-
рального введения [281]). Это количество составляет примерно 30% дозы у
мышей и 50% у крыс. Может быть, как утверждал Дедрик [156], Мтк, как
фармакокинетическое время, более тесно соотносится со средним временем
удержания в сосудистой системе, чем с клиренсом креатинина (см. ниже).
Имея значительные межвидовые вариации клиренса, давайте теперь вер-
нёмся к вариациям объёмов распределения (К„), используя данные, приве-
Глава 36. Межвидовые вариации
511
денные в [9]. зависимость Кр от В для Мтк, получаем:
Кр = 0,859 хВ°>ш, (135)
где ^ — объём распределения (л), а В — вес тела (кг).
С другой стороны, известно, что общее количество воды в организме
{ОKB) в литрах задаётся выражением:
ОКВ = 0,703 х50-963. (136)
Комбинируя уравнения (135) и (136), имеем:
Кр / ОКВ = 1,22 х Я"0-045, (137)
Кр/ ОКВ= 1,22. (138)
Снова межвидовой фармакокинетический параметр становится относи-
тельно инвариантным, когда масштабируется по отношению к биологичес-
кому параметру внутри организма. Эти последние две зависимости пара-
метра от веса тела являются специфическим примером аллометрии или ге-
терогонии.
Вследствие того, что CL и Кр для Мтк могут быть соотнесены с весом
тела, используя простое аллометрическое уравнение (127), кажется интерес-
ным начать исследование моделирования природы связи между аллометри-
ей и эпюрой Дедрика.
Из вышеизложенного следует, что важно с большой осторожностью под-
ходить к оценке экстраполяции на человека результатов альтернативного мо-
делирования. Необходимо придерживаться тактики взвешенного и крити-
ческого отношения к возможностям и ограничениям экстраполяции для всех
без исключения методов биотестирования и альтернативного биомоделиро-
вания, принимая во внимание, что при оценке безопасности лекарств или
любых иных ксенобиотиков применение таких моделей крайне ненадежно и
во многих случаях может нанести вред здоровью человека.
512
Глава 3 7 БИОЛОГИЧЕСКОЕ
МАСШТАБИРОВАНИЕ
Биологическое масштабирование трудно считать
новой дисциплиной. Уже в 1637 г. Галилей [227]
обсуждал связь между размерами скелета и массой
организма. Но по отношению к физиологическим
и фармакокинетическим проблемам работа Adolph
[6, 7], вероятно, оказала наиболее мощное влия-
ние на дальнейшее развитие этой области иссле-
дований.
В своей, сегодня уже классической статье, опуб-
ликованной в журнале Science в 1949 г., Adolph сде-
лал следующее замечание, касающееся использова-
ния простого аллометрического уравнения, также
именуемого гетерогоническим: «Следует сознавать,
что нет никаких ограничений, определяемых чем-
либо, кроме времени существования организма и того
массива свойств, которые рассматриваются как след-
ствие этих ограничений» [6, 7].
Размеры органов человека и животных, также
как биохимические и физиологические процессы,
являются масштабированными по отношению к весу
тела, и вследствие того, что эти органы и процессы
влияют на распределение препарата, мы можем с
уверенностью ожидать наличия корреляции между
фармакокинетическими процессами и нормальны-
ми повседневными эндогенными процессами. С
этой точки зрения, однако, связь межвидовых ас-
пектов анатомии и физиологии с особенностями
фармакокинетики требует дальнейшей разработки
[46, 54, 92, 217, 264].
Глава 37. Биологическое масштабирование
513
Фактически концепция Adolph'a [6] предполагает выполнение следую-
щих теоретических предпосылок, которые названы теориями:
/ теория взаимозависимости: ни одна компонента не регулируется неза-
висимо от всех других;
/ теория требований: скорости оборота любой одной компоненты зави-
сят от содержания и скоростей обмена других компонент;
/ теория сохранений: скорости оборота обычно лежат вблизи минимума,
совместимого с независимым продолжением функций;
/ теория масштабирования времени: одна компонента или группы ком-
понент имеют тенденцию среди многих видов к разгрузке (избавлению)
в относительно сходные времена.
Автор этой книги наверняка согласился бы с этим и охотно поддержал
эту концепцию и вытекающие из неё теории, если бы «времена», на которые
ссылаются, являлись физиологическими временами.
Токсичность и цена жизни
Поскольку как физиологические, так и фармакокинетические процессы ре-
гулируются тем или иным способом с помощью аллометрии, представляется
интересным исследовать, масштабируются ли эндогенные процессы (свя-
занные со скоростями) тем же способом, что наблюдается для распределе-
ния препаратов. По словам Drabkin, «метаболическая машина изнашивает-
ся». В таком случае этот процесс может быть назван «токсичностью жизни»
или «платой за жизнь» [169]. Это действительно является «токсичностью жиз-
ни», которая вызывает уменьшение скорости распределения препаратов,
обычно наблюдаемой у пожилых.
При анализе литературы вскоре стало ясно, что большинство доступных
данных о скоростях при межвидовых вариациях в физиологических и биохи-
мических процессах представляют собой данные, относящиеся либо к вре-
менам оборота, либо к терминальным периодам полувыведения в экспонен-
тах. Поэтому мы попытались выяснить, масштабируются ли времена оборо-
та и периоды полувыведения для эндогенных процессов способом, анало-
гичным тому, которым масштабируются на вес тела периоды полуэлимина-
ции лекарственных препаратов.
Для эндогенных веществ в стационарном состоянии время оборота в
организме равно среднему времени удержания, т. е. средней продолжи-
тельности жизни молекулы или частицы в организме. Для процессов перво-
го порядка в стационарном состоянии время оборота, /* задаётся следую-
щим выражением:
t*p =(\,44xt]/2)(Vss/Vtp)=\y22, (139)
где ty2 — период полувыведения терминальной tp — фазы, Vss— объём рас-
514
Аллометрия и экстраполяция: пределы и возможности
пределения в стационарном состоянии, и V — объём распределения в тече-
ние терминальной /р-фазы.
В случае исследования метотрексата Мтк, описанного в гл. 36, вслед-
ствие того, что как У(, так и CL могут быть выражены в терминах алломет-
рических уравнений, t,n может так же быть выражена:
1/2
\/2Мтк
= (0,693) х (1000)(0,859 х Я0-918)/(Ю,9 х В °-69),
или Л
(НО)
W™ = 54,6x*°^, (141)
где В — вес тела {кг), a tl/2MmK — период полувыведения Мтк (мин).
В табл. 86 суммированы данные по межвидовым вариациям периодов
полувыведения антипирина, дигоксина, гексобарбитола, фенилбутазона, ани-
лина и диазепама. Эта группа представляет различные соединения с преиму-
щественными путями элиминации, которыми являются почечная экскреция
для метотрексата и дигоксина, и метаболизм для других препаратов. Хотя
все регрессии достигают удовлетворительной степени статистической зна-
чимости, существует вполне определённый тренд. Экспоненты алломет-
рических уравнений имеют тенденцию группироваться около величины 0,25,
а не 0, 0,5, 0,75 или 1,0.
Таблица 86
Аллометрические параметры, описывающие межвидовые вариации
периодов полувыведения препаратов*
Препарат
Метотрексат
Циклофосфамид
Антипирин
Дигоксин
Гексобарбитал
Фенилбутазон
Анилин
Диазепам
Число видов
животных
5
6
10
5
5
7
5
4
Коэффициент
корреляции
0,994
(Р<0,01)
0,752
(Р<0,01)
0,503
(Р<0,01)
0,941
(Р<0,05)
0,912
(Р < 0,05)
0,335
(Р<0,5)
0,650
(Р<0,3)
0,989
(Р<0,05)
Аллометрический
коэффициент
54,6
36,6
74,5
98,3
80,0
34,0
62,2
122
Среднее:
Аллометрическая
экспонента
0,228
0,236
0,269
0,234
0,348
0,0613
0,176
0,428
0,222
* Примечания:
/ Периоды полувыведения использованы в минутах, вес тела — в килограммах.
/ Взяты только те данные, которые соответствуют не менее чем 4 видам.
/ Данные по человеку были исключены из анализа.
Глава 37. Биологическое масштабирование
515
Это соответствует основному плану энергетических затрат у млекопита-
ющих, а распределение препаратов является энергетическим процессом по-
требления. Таким образом, вес тела является системой отсчёта для ритми-
ческих явлений [7, 256], и теория сохранения химической энергии требует,
чтобы время оборота энергии было пропорционально В °-25 [366]. Соответ-
ственно масштабирование остаточного потребления кислорода на вес тела
(пропорционального основной скорости метаболического обмена) для пой-
килотермных и гомойотермных животных приводит к идентичным алломет-
рическим показателям с экспонентой равной 0,75 [280, 537].
Поэтому энергический метаболизм у пойкилотермных одинакового веса
идентичен [257]. Энергетические траты являются исключительно важными
для понимания межвидовых вариаций в фармакокинетике вследствие того,
что они тем или иным образом влияют на количество энергии, расходуемой
на элиминацию ксенобиотиков и препаратов. Это в свою очередь отражает-
ся количественными показателями фармакокинетики. В значительной сте-
пени это определяется параметрами относительного веса внутренних орга-
нов животных (рис. 15).
I j 1 L_! 1 1—1_1 l_i 1 L_U !_М—I Bec кг
0.1 1 10 100 1000 оес-кг
Рис. 15. Кривые регрессии некоторых внутренних органов млекопитающих. По оси ординат —
относительный вес органов (в граммах) к массе тела (в килограммах). • - печень, А - почки,
■ - надпочечники
516
Аллометрия и экстраполяция: пределы и возможности
Анализируя скорости метаболизма и время энергического оборота у мле-
копитающих, Gunther утверждал, что увеличение размеров тела в общем свя-
зано с прогрессивным снижением скорости метаболизма на единицу массы
тела, которое, в свою очередь, влияет на скорость оборота на клеточном
уровне и потому может изменять время протекания биологических процес-
сов [256, 257, 258, 259]. В этом отношении Kleiber [366] показал, что ско-
рость метаболизма (М) организма пропорциональна весу тела в степени 0,75.
М=ахВ°<75, (142)
где а — константа в соответствующих единицах.
В симилометрически организованных пулах животных, т. е. взятых про-
порционально весу тела, содержание энергии или термодинамической потен-
циональной энергии прямо пропорционально массе организма или весу тела.
Поэтому время оборота (ВО) пропорционально В0'25 [366]:
химическая энергия, содержащаяся в организме
время оборота =
метаболическая скорость
или
ВО= (щхВ^)/(а2хВ^75) = В°>25 , (143)
где aj и а2 — константы.
Поэтому время оборота энергии в организме пропорционально В0'25.
Поскольку целое является просто суммой частей, то следует сказать, что в
среднем индивидуальное время оборота для процессов потребления симило-
метрической энергии относится к весу сходным образом. Это продемонст-
рировано в табл. 87, где можно наблюдать, что аллометрические выражения
для времён оборота эндогенных процессов имеют показатели экспонент,
группирующиеся около величины 0,25. Обращаясь к распределению препа-
ратов в стационарном состоянии, когда удовлетворены описанные выше
условия, видим, что значения времени оборота препаратов приблизительно
пропорциональны периодам полувыведения соответствующих видов.
Другими словами, межвидовые вариации параметров распределения пре-
паратов у млекопитающих в большой степени регулируются теми же факто-
рами, которые регулируют другие процессы в организме, например энерге-
тические траты. Такая связь даже более очевидна, когда мы считаем, что вся
живая материя фактически является системой для захвата, преобразования и
высвобождения энергии [542]. И, как обсуждалось выше, энергетические
траты у млекопитающих просто являются функцией размеров тела, с време-
нем оборота пропорциональным В в степени 0,25.
Gunther сформулировал это следующим образом: «...на всех уровнях био-
логической организации мы можем наблюдать, что онтогенетическое и
филогенетическое увеличение размеров непосредственно связано с возрас-
танием продолжительности всех периодических явлений, общая особенность
Глава 37. Биологическое масштабирование
517
Таблица 87
Аллометрические параметры, описывающие времена оборота
или соответствующие оценки для эндогенных субстратов или процессов
Субстанция
Сывороточный
альбумин
Общее содержа-
ние воды в орга-
низме
Эритроциты
Сердечное
кровообращение
Количество
видов
6
8
11
11-12
Коэффициент
корреляции
0,993
(Р<0,01)
0,940
(Р<0,01)
0,778
(Р<0,01)
Аллометрическии
коэффициент
5,68
6,01
68,4
0,422
Среднее:
Аллометрическая
экспонента
0,296
0,161
0,102
0,21
0,192
* Примечания:
/ Параметр В в аллометрических уравнениях имеет размеренность — килограммы.
/ Время оборота в днях, вес тела в килограммах.
/ Время оборота аппроксимировалось как 1,44 г1/2.
/ Общее содержание воды в организме делили на величину суточной скорости экскреции
воды с мочой, чтобы получить оценку времени оборота.
/ Среднее время прохождения крови по сосудистой системе оценивается из двух алломет-
рических уравнений, объём крови как функция веса тела, делённая на сердечный выброс
(как функцию веса тела). Время оборота (или время прохождения) — в минутах.
которых может быть сформулирована с помощью степенной функции для
биологического времени: Tb = a IY0*27» [256, 257, 258, 259].
Практически, хронологическое время нормируется на биологическое делени-
ем на В в степени 0,25, и в этом возможно содержится основа преобразова-
ния времен, первоначально использованная в фармакокинетической модели
[156].
Процессы фармакокинетической элиминации зависят главным образом
от размеров и функции печени и почек. Такие органы, как печень и почки,
отвечая метаболическим потребностям, увеличивают свои относительные
размеры так, что аллометрическии показатель экспоненты равен примерно
0,85 [242, 243]. Это может объяснить, почему некоторые фармакокинетичес-
кие параметры элиминации в аллометрическом уравнении ближе к 0,85, чем
к 0,75. Например, Boxenbaum показал, что внутренний клиренс (несвязан-
ного препарата) для антипирина у 10 видов млекопитающих был функцией
веса тела в степени 0,885 (см. ниже) [90, 91]. В этом случае фармакокинети-
ческое время масштабируется на В в степени 0,115.
Существует ещё один способ рационализировать фактор масштабирова-
ния В в степени 0,25. Дедрик с соавторами отмечали, что ткани более мел-
ких животных получают больший кровоток на единицу объёма тканей, чем
ткани более крупных животных [156]. Среднее время нахождения (хроно-
518
Аллометрия и экстраполяция: пределы и возможности
логическое) крови в сосудистой системе (объём крови/сердечный выброс)
масштабируется на вес тела в степени 0,21(см.табл. 87). Поэтому более мел-
кие животные могут инактивировать препараты быстрее, чем более круп-
ные животные, благодаря кровотоку в некоторых или всех элиминирующих
органах.
В своей книге «Математический подход к физиологическим проблемам»
Riggs отмечал преобладание экспоненционального роста и задержки и по-
давляющего превосходства логарифмически нормальных распределений в
природе [545]. Последовательно, вслед за более общим подходом Reiser [542],
Riggs рекомендовал принять логарифмический взгляд на скорости биологи-
ческих процессов.
Польза аллометрии в фармакокинетике уже была хорошо описана [91,
440]. Так, например, Boxenbaum [90, 91] продемонстрировал, что у 10 видов
млекопитающих (исключая человека), внутренний клиренс несвязанного пре-
парата антипирина соотносится с весом тела следующим образом:
С/,и1т=0,00816хЯ0'885, (144)
где В — вес тела (кг), СЬиЫ — внутренний клиренс несвязанного препарата
(л/мин).
Было также продемонстрировано, что печёночный кровоток QH (л/мин)
также связан с весом тела (кг):
QH= 0,0554 х В °'894. (145)
О скорости и клиренсе жизни
Как обсуждалось выше, аллометрическое уравнение, по-видимому, имеет
большую пользу при анализе межвидовых фармакокинетических данных.
Однако не было проведено тщательного рассмотрения при выборе парамет-
ров, коррелирующих с весом тела. Как только что было показано, периоды
полувыведения могут коррелировать как следствие математического «упроще-
ния». Величины клиренсов могут коррелировать, по крайней мере, по двум
причинам. У тех препаратов, которые метаболизируются только в печени и
имеют высокое экстракционное отношение, клиренсы достигают по вели-
чине значений скорости печёночного кровотока. Эти аллометрические зави-
симости могут быть не более чем вторичными корреляциями, возникающи-
ми вследствие первичной аллометрической связи между печёночным или
легочным кровотоком и весом тела. Они также зависят от показателей ды-
хательного объема (рис. 16).
Продолжительность жизни видов контролируется генетически определен-
ной скоростью старения и является основным биологическим свойством орга-
низма [147, 148]. Другими словами, каждый вид имеет характерную продол-
жительность жизни, что коррелирует с такими специфическими функциями
Глава 37. Биологическое масштабирование
519
3 я
^ *
й *
&s
о m
5 и
(0
i
« к *
5 л s
* m с;
3 Ф О
О Ю Q.
* о *
я
*
а я
VO J
о m
о о
ИНЬЯ
а) и
ш '
о 5
£ i
ш i
з- 5
И
Q.3
о о
* с;
I I I I I |_| I I
0.1 1 10 100 1000 Вес, кг
Рис. 16. Сравнительные показатели дыхательного объема млекопитающих (см3 кг/мин) и че-
ловека (дыхательный объем человека принят за единицу). По оси ординат — коэффициент
экстраполяции в отношении человека (остальные обозначения см. на рис. 15)
видов, как длительность беременности, продолжительность полового созрева-
ния, частота сердечных сокращений и скорость метаболизма [167]. Большин-
ство диких животных никогда не достигают своей полной продолжительности
жизни, поскольку их убивают хищники или же они гибнут от несчастных
случаев до того, как их жизнедеятельность серьёзно ухудшится за счёт старе-
ния [148]. Но, исключая несчастные случаи, генетические болезни, гибель от
хищников и т. д., млекопитающие обычно умирают от болезней, которые сами
вызваны недостаточностью функций кровообращения или иммунной систе-
мы. Существует максимум возможной продолжительности жизни (МПЖ), свя-
занной с этой формой «естественной смерти», который частично реализуется
с помощью генетически контролируемых биологических часов. Эти биологи-
ческие часы регулируют скорость, с которой невероятно низкая энтропийная
организация организма уступает силам природы, т. е. умирает [575]. Биологи-
ческие часы могут также влиять и на скорость метаболизма препарата.
Максимальное потребление калорий (МПК) за период возможной продол-
жительности жизни можно определить как произведение МПЖ на специ-
фическую скорость метаболизма (ССМ) [147, 561]. Считается, что МПК пред-
ставляет общую «жизненную ёмкость» организма и скорость, с которой ра-
520
Аллометрия и экстраполяция: пределы и возможности
страчивается определённая жизненная ёмкость, определяется величиной,
обратной МПЖ. Другими словами, скорость жизни (скорость энергетичес-
ких трат) влияет на её продолжительность. Следовательно, быстро живущая
крыса не может прожить столько же, сколько медлительный гиппопотам.
Из предшествующего обсуждения ясно, что в терминах повышения про-
должительности жизни организм экономит на метаболической активности,
включая метаболизм ксенобиотиков и экскрецию. Процесс этой экономии
таков, что количества ферментов, включая ферменты метаболизма лекар-
ственных препаратов, обычно существуют на уровнях, примерно в 2 раза
превышающих минимум, требуемый организму [265, 586]. Это обеспечива-
ет генетически удовлетворительный «фактор надёжности». Можно возра-
зить, что количество энергии, расходуемое на распределение препаратов,
пренебрежительно мало и что препараты обычно даже не присутствуют в
организме.
Однако в данном случае достаточно сказать, что распределение препа-
рата — это просто «модель» для распределения ксенобиотиков и что повсе-
местное присутствие ксенобиотиков в природе таково, что организмы мле-
копитающих почти непрерывно «детоксицируют» ксенобиотики со значи-
тельными метаболическими затратами. Как единичный пример, анализ ме-
таболизма кофе и шоколада [270] показал наличие более чем 700 соедине-
ний, с которыми взаимодействуют отдельные ферменты. Вот цена, которую
платят, чтобы насладиться чашечкой кофе мокко. Следовательно, ксеноби-
отики из этих двух препаратов сами по себе могут оправдывать большую
часть качественного механизма лекарственного метаболизма. Даже такой,
казалось бы, безвредный напиток, как натуральный апельсиновый сок, со-
держит, по крайней мере, 217 ксенобиотиков в летучей фракции. Эти ксе-
нобиотики включают кислоты, спирты, альдегиды, эфиры, углеводы, кето-
ны и т. д.[14].
Во всех случаях мы сталкиваемся с изнашиваемостью метаболических
систем, с тем, что мы называем отравлением рецепторных структур, а более
образно это явление действительно может быть отнесено к понятию токсич-
ности жизни.
Из предшествующего обсуждения следует, что скорость некоторых
процессов потребления энергии может быть обратно пропорциональна МПЖ
(продолжительность жизни), и тому есть примеры.
Boxenbaum [90, 91] утверждает, что параметр, который лучше всего опре-
деляет способность организма отражать химическое вторжение ксенобиоти-
ков (или препаратов), — это внутренний клиренс не связанного с белками
препарата CLuint. В этом отношении метаболизм растворимых в липидах ве-
ществ, выведение которых осуществляется печенью в режимах, не достига-
ющих насыщения, могут обычно описываться моделями с хорошим переме-
шиванием, обсуждавшимся Pang [511].
Глава 37. Биологическое масштабирование
521
Boxenbaum [90, 91] успешно использовал параметр CLuint. при анализе
межвидовых вариаций в фармакокинетике. У 10 видов млекопитающих, но
не у человека, этот параметр был пропорционален весу печени; однако, у че-
ловека эта величина составила 1/7 предсказанной им величины.
В то же время межвидовой метаболизм липофильных препаратов имеет
тенденцию распадаться на две категории. В первой категории, для которой
прототипом является антипирин, другие млекопитающие склонны иметь
величины CL на единицу веса печени примерно в 5-20 раз выше, чем у
человека.
Вследствие того, что человек обычно является уникальным как в отно-
шении наличия низких величин CXuint, так и повышенной продолжительно-
сти жизни, представляется интересным определить, нет ли некоторых кор-
реляций, способных возникать из аномалий. Существует достаточное осно-
вание для таких корреляций. Продолжительность жизни коррелирует с ана-
томическими, связанными со временем развития организма, специфически-
ми метаболическими процессами, такими, как температура тела, вес тела,
размеры мозга, продолжительность беременности, частота сердечных сокра-
щений, скорость базального метаболизма, уровень психического развития,
скорость постнатального развития, возраст полового созревания.
Как первый шаг в установлении любых таких взаимосвязей, необходимо
получение оценок МПЖ. В этом контексте МПЖ определяется как макси-
мальная документированная продолжительность жизни вида, исключая че-
ловека; в общем, подавляющее большинство данных о МПЖ получается на
домашних животных.
Используя методы множественной регрессии, Sacher [560, 561] вывел сле-
дующее уравнение:
МПЖ = 10,839 X (Bw) 0.28.^0,225 (146)
где МПЖ — максимально возможная продолжительность жизни (годы); Bw —
вес мозга (г); В — вес тела (г).
Табл. 88 обобщает данные, на основе которых были получены оценки
максимально возможной продолжительности жизни.
Таблица 88
Данные о весе мозга, весе тела и максимально возможной
продолжительности жизни
Животные
и человек
Мышь
Крыса
Морская свинка
Кролик
Собака
Вес врослого
организма, г
23
250
270
2 550
14 200
Вес врослого
мозга, г
0,334
1,88
3,42
9,97
75,4
Вес мозга
(% от веса тела)
1,45
0,751
1,27
1,391
0,531
Вычисленная МПЖ
(годы)
2,67
4,68
6,72
8,01
19,7
522
Аллометрия и экстраполяция: пределы и возможности
Окончание табл. 88
Свинья
Овца
Коза
Крупный рогатый
скот
Макака-резус
Человек
Вес врослого
организма
77 200
57 600
31300
310 000
4700
70 000
Вес врослого
мозга
58,2
110
130
252
62
1530
Вес мозга
(% от веса тела)
0,072
0,191
0,416
0,081
1,32
2,19
Вычисленная МПЖ
(годы)
11.4
18,3
23,3
21,2
22,3
93,4
Первоначально анализу был подвергнут самый типичный препарат —
антипирин, затем подобные данные были получены для фенитоина и клоно-
зепама, соответственно.
Интересно, что регрессионные уравнения, могут быть преобразованы
таким образом, что время (хронологическая МПЖ) становится тем, что Frank
называет «процессом», т. е. физиологическое время становится зависимой
переменной [220].
Таким образом, скорость метаболизма любого препарата или ксенобио-
тика, сама по себе, имеет заметное влияние на МПЖ, но скорее то, что
скорость метаболизма ксенобиотика или препарата, в частности, и его фар-
макокинетика, в общем, является показателем более общего основного биоло-
гического плана жизнедеятельности организма млекопитающих в общем, и
человека — в частности.
Раздела ПЕРСПЕКТИВЫ
БИОМОДЕЛИРОВАНИЯ
Удовольствие исследователя:
задирать юбки природе.
Ж.Ростан
525
Глава 3 8 ФЕНОТИПИРОВАНИЕ
И ГЕНОПОЛИМОРФИЗМ
Целый ряд направлений биомоделирования осно-
вывается на исследованиях генетического полимор-
физма фенотипов человека, связанных вопросами
медико-биологического характера при установлении
генетической компоненты в предрасположенности
индивидов к определенным заболеваниям [213, 292,
539] и чувствительности к лекарствам.
При воздействии на клетку различными краси-
телями её органеллы реагировали на них по-разно-
му. Так было открыто ядро клетки. Внутри его об-
наружились участки, которые прочно связывали
определенные краски, становясь хорошо видимы-
ми. В 1871 г. Вальтер Флемминг назвал внутрикле-
точное вещество, образующее такие участки, хро-
матином (от греч. chroma — цвет).
Когда метод окрашивания применили для ис-
следования стадий деления клетки, оказалось, что
на определенной стадии деления окрашенный ма-
териал собирается в нитевидные окрашенные струк-
туры, или тельца, которые назвали хромосомами, то
есть окрашенными тельцами (греч. «хрома»+soma —
тело). Сам же процесс деления с образованием ни-
тевидных структур назвали митозом (от греч. mitos
— нить). Митоз — простое деление ядра без выяв-
ления нитевидных хромосом. Деление клетки, при
котором происходит уменьшение числа хромосом
вдвое, называется мейозом. Было установлено, что
клетки тела человека содержат 46 хромосом, а каж-
дая хромосома состоит из тысяч отдельных единиц,
526
Перспективы биомоделирования
ответственных за развитие определенного признака. Эти единицы были на-
званы генами (от греч. genes — рождающийся, рожденный).
Термин генетика предложил в 1907 г. У. Бейтсон. А термин ген был введен
в научный обиход В. Иогансеном. В 1909 г. он же предложил термины гено-
тип — наследственная основа организма, совокупность всех генов, локали-
зованных в его хромосомах, и фенотип (от греч. phaino — являю) — совокуп-
ность всех признаков и свойств организма, сформировавшихся в процессе
его индивидуального развития.
В последние годы сложилось более оптимальное определение этих про-
цессов — «генополиморфизм» или генетический полиморфизм. Оно отно-
сится в большей степени к описанию явлений лекарственного метаболизма
в различных этнических группах и популяциях. Более традиционное назва-
ние «биохимический полиморфизм» закрепилось в исследованиях, касаю-
щихся медико-генетических и этногеографических аспектов. В нашей рабо-
те мы будем использовать оба терминологических определения.
В течение многих лет осуществлялся настойчивый поиск биохимичес-
ких маркеров для фенотипирования [74]. В качестве примера можно обра-
тить внимание на обнаруженную достоверную зависимость некоторых фе-
нотипов ингибитора протеаз — otj-антитрипсина и генетических разновид-
ностей системы HLiA, с одной стороны, и рядом болезней — с другой.
Результаты таких исследований, проведенных в разных странах, нашли свое
отражение в общей сводке ВОЗ (2001). Что касается ферментных систем,
то имеются данные об ассоциации изоферментов сывороточной амилазы,
вариант D плацентарной щелочной фосфатазы с заболеваниями. Значение
изучения изоферментов для медицины подчеркивается в публикациях о
многих заболеваниях и в многочисленных направлениях медицинской
биологии.
В основе метаболизма лекарственных препаратов и ксенобиотиков в орга-
низме человека лежит преобразование их в водорастворимые фракции, ко-
торые вьщеляются с мочой. Преобразования медикаментов и ксенобиотиков
осуществляется в две фазы, а ферменты, ускоряющие их метаболизм подвер-
жены генетическому полиморфизму. Активность этих ферментов варьирует
в широком диапазоне. Генетический полиморфизм обеспечивает индивиду-
альные особенности метаболизма ксенобиотиков и лекарств.
Перспективным направлением, непосредственно связанным с проблемой
изучения генетического полиморфизма, является применение большого спек-
тра радиоактивных лигандов. Это позволило идентифицировать генетичес-
кие разновидности белков сыворотки крови, специфически связывающих и
транспортирующих биологически активные вещества — ферменты, витами-
ны, гормоны. Более перспективным подходом к исследованию генетическо-
го полиморфизма является оценка спектральных характеристик и резонан-
сных спектров интересующих нас веществ. Собственно говоря, явления «спек-
Глава 38. Фенотипирование и генополиморфизм
527
троскопии» представляют собой огромный набор методических приемов,
основанных на анализе сигналов, идущих из молекул и атомов.
С помощью спектрометров — устройств, регистрирующих отклик моле-
кул на действия излучений, измеряют инфракрасный (ИК) и ультрафиоле-
товый (УФ) спектры. Значительно более сложным методом является масс-
спектроскопия. Она позволяет взвесить молекулы и точно установить коли-
чество тех или иных атомов в молекуле. Все названные методы не позволяют
установить, как атомы связаны друг с другом и какую форму имеет образу-
емая ими молекула. Привлечение для исследований в биохимии и генетике
СВЧ-техники, электронного парамагнитного резонанса или ЭПР-спектро-
скопии позволили неразрушающими белки методами анализировать фено-
типические особенности организмов в динамике, сверхточно оценивая ха-
рактер кинетических процессов интра— и межмолекулярых отношений.
Спектроскопическая информация может быть получена на основе ядер-
ного магнитного резонанса, т. е. ЯМР-спектрометром. С помощью этого
прибора измеряется магнитное поле, создаваемое каждым атомом, напри-
мер водорода, находящегося в различных окружениях микрочастиц. Суще-
ствуют и другие, к сожалению, чрезвычайно дорогостоящие принципы ис-
следования резонансных спектров.
Такие разработки способствуют установлению функциональных разли-
чий генетических разновидностей в пределах каждого биохимического локу-
са. Практическая ценность этих исследований для фармакологии, терапии и
в целом для клинической медицины очевидна, так как открывает перспек-
тиву дифференцированного подхода к выбору и дозировке лекарственных
препаратов, индивидуализированной и дифференцированной фармакотера-
пии в зависимости от фенотипических, этнических и расовых особенностей.
Поиск маркеров генов
Большой толчок генетическим исследованиям, наряду с классической физи-
кой, дали ядерная физика и радиационная медицина. Эти два направления
позволили по-иному взглянуть на процессы физической химии в биологии
и медицине [213, 415]. К началу 70-х годов XX века стала понятна роль не-
спаренных электронов и свободных радикалов в кинетике ферментативных
реакций и иных каталитических процессах, в изучении структуры молекул,
механизмов повреждения и защитных механизмов тканей.
Световые лучи, как и любые другие волны, имеют собственную длину
волны, которая измеряется в нанометрах (нм), единицах длины, равных од-
ной миллиардной доле метра (10~9 м). Свет с длиной волны, равной 700 нм,
воспринимается нами как красный, а с длиной волны 420 нм — как фиоле-
товый. Между этими крайними точками лежат оранжевый, желтый, зеленый
и синий цвета — все это составляет спектр видимого света.
528
Перспективы биомоделирования
Но видимый свет — это лишь небольшое окошко в спектре других излу-
чений, используемых для исследований, причем сдвинутое в сторону малых
энергий. Если «осветить» фотопластинку лучами, длина волны которых ле-
жит за фиолетовым концом спектра, она засветится еще сильнее, чем на
видимом свету, хотя мы этого излучения и не видим, что свидетельствует о
существовании какой-то радиации. Эти лучи назвали ультрафиолетовыми
(лат. ultra — далее, более, сверх). Длины волн ультрафиолетового света лежат
в пределах от 400 до 10 нм. Еще короче длины волн рентгеновского и гамма-
излучений.
Уильям Гершель в 1800 г. обнаружил, что в области, лежащей за красным
светом, термометр отмечал более сильное повышение температуры, чем в
видимой части спектра. Это указывало на присутствие там излучения. Излу-
чение, непосредственно примыкающее к красному концу видимого спектра,
назвали инфракрасным, или тепловым. «Инфра» по-латыни означает «под,
снизу». Инфракрасное излучение располагается в самом низу энергетичес-
кой шкалы. Длина его волны достигает от 1—2 мм до 760 нм. Только радио-
волны имеют большие длины волн. Как инфракрасные, так и ультрафиоле-
товые диапазоны исследования стали использоваться для идентификации
маркеров при оценке генетического полиморфизма.
В последние годы в арсенал исследователей вошли методы электронного
парамагнитного резонанса (ЭПР), двойного электронно-ядерного резонанса
(ДЭЯР), спектроскопии Мёссбауэра или ядерного у-резонанса (ЯГР). Суть
этих методов заключена в способности неспаренных электронов, обладаю-
щих собственным моментом количества движений или угловым моментом,
называемым спином, под воздействием магнитного или электронного поля
приобретать общую координационную ось.
В отсутствие такого воздействия спины неспаренных электронов в фер-
ментах, свободных радикалах и других молекулах ориентированы случайным
образом. При наложении же внешнего магнитного, электрического или у-
поля неспаренные электроны разделятся на две группы: у одних спины будут
ориентированы параллельно, а у других антипараллельно направлению поля.
Все вещества, содержащие неспаренные электроны, подразделяются на две
группы. В первой группе неспаренные электроны связаны со всей молеку-
лой или большей её частью. Во второй группе неспаренные электроны обычно
связаны с атомами переходной группы — железом, кобальтом, медью, нике-
лем, и их число в атоме, как энергия, изменяется с изменением валентности
этих атомов.
Исследование генетического полиморфизма человека и животных и их
фенотипических особенностей производится на основе глубокого изучения
особенности ферментативных функций. Естественно, что используются са-
мые различные биохимические методики. Но поскольку в состав большин-
ства ферментов входят ионы металлов — железо, медь, марганец, молибден,
Глава 38. Фенотипирование и генополиморфизм
529
кобальт, ванадий, никель и др., ЭПР является наиболее оптимальным при-
емам для их изучения (рис. 17, 18).
о2 со2
Рис. 17. Классическая рентгеноструктурная модель гема
С этой точки зрения всесторонне изучены свойства ферментативных ре-
акций ксантиноксидаз, альдегидоксидаз, дегидрогеназ, каталаз, пероксидаз,
цитохромоксидаз и др. Исследованы гем и его окружение. Достаточно на-
помнить, что гем входит в состав пероксидазы, каталазы, цитохромов и
многих других биологических молекул. На рис. 17 изображен гем, т. е. пор-
фириновое кольцо с атомом железа в центре; через пятую координационную
связь к гену присоединяется глобин, через шестую — группа X (кислород,
двуокись углерода и т.д.).
gi~ g = 2Vl + 8sin29
Рис. 18. Структура гема на основе ЭПР-спектроскопии. g-фактор отражает электронно-маг-
нитный резонанс для свободного неспаренного электрона или неспаренного электрона, свя-
занного с атомом или молекулой. Зависимость значения g-фактора у высокоспинового про-
изводного гемоглобина составляет в плоскости гема g= 6,0 во всех направлениях
530
Перспективы биомоделирования
На рис. 17 приведена полученная с помощью ЭПР структура гема.
В гемомоглобине содержатся четыре гема, а значит, и четыре атома железа
на молекулу, а в миоглобине — только один гем.
Стоит подчеркнуть, что перечисленные выше ферменты и многие другие
белки являются естественными биохимическими маркерами генов и широ-
ко используются для фенотипирования и определения расовой и этничес-
кой принадлежности человека к той или иной популяции.
По своей сути положение точки парамагнитного резонанса характеризу-
ет синглетную линию поглощения. Это означает, что значение поля, при
котором имеет место резонанс, определяет величину g-фактора для данного
свободного неспаренного электрона или неспаренного электрона, связанно-
го с атомом или молекулой (рис. 18).
Генополиморфизм и непереносимость лекарств
Генетический полиморфизм играет исключительно важную роль в адаптаци-
онных механизмах защиты от ксенобиотиков. Одним из маркеров является
гликопротеин Р, который принадлежит к семейству плазменно-мембранных
белков, кодируемых MDR генами, и действует как трансмембранная помпа,
удаляющая лекарственные средства из клеточной мембраны и цитоплазмы.
Субстратами гликопротеина Р являются дигоксин, нифедипин, ловастатин,
фексофенадин и другие, а веществами, снижающими активность — верапа-
мил, хинидин, кетоконазол. Географические, этнические и расовые различия
в частоте полиморфных аллелей являются базисом, по крайней мере, для
нескольких выявленных различий в фармакокинетике и/или лекарственного
эффекта или токсичности между популяциями. Полиморфизм в 26 экзоне, в
позиции 3435(С3435Т), не вызывает изменения в аминокислотах, но влияет
на уровень экспрессии гликопротеин Р в тонком кишечнике и, соответствен-
но, на кишечную абсорбцию субстратов гликопротеин Р. Так, выделяют два
генотипа, С и Т, по наличию С или Т аллелей соответственно, где Т аллель
ассоциируется с редукцией экспрессии гликопротеин Р, а С аллель — с индук-
цией гликопротеин Р.
Генетический полиморфизм ферментов, участвующих в метаболизме ле-
карств, является ответственным за измененный уровень и активность эн-
зимов, а также определяет риск неблагоприятных реакций на лекарствен-
ные препараты и возникновение некоторых видов рака. В работе [118] ис-
следовано 325 человек русского происхождения, которые были здоровыми
людьми или пациентами, не имеющими злокачественных болезней. Работа
включала полное исследование двух ферментов I фазы, CYP1A1 и CYP2D6,
одного фермента II фазы, NAT 2. Проверены наиболее общие изменения
NAT 2: 191 G>A, 2820T, 341 Т>С, 590ОА, 803A>G, 857G>A и новое из-
менение 111Т>С.
Глава 38. Фенотипирование и генополиморфизм
531
Определение генотипа восьми различных единичных нуклеотидных по-
лиморфизмов гена NAT 2 предусматривает генотип, определяющий медлен-
ное ацетилирование 59,7% индивидов; 34,7% (29,6—40,2%) участников экс-
перимента оказались носителями, по меньшей мере, одной аллели, в кото-
рой кодируется способность к быстрому ацетилированию, и 5,6% (3,3—8,6%)
обладают гомозиготными аллелями быстрого ацетилирования (неизменен-
ная аллель*4 или измененная аллель *12А). Частотность изменений аллелей
и генотипов фермента NAT 2 у русской группы были схожими с такими же
показателями кавказской группы. Оказалось, что, как и в других европейс-
ких группах, преобладание медленных ацетиляторов у русских людей яви-
лось результатом высокой частотности *5 аллелей. Изменение 191 G>A, ха-
рактерное для темнокожего населения, не было зафиксировано в русской
популяции.
Остановимся на характеристике некоторых значимых ферментных сис-
тем, обеспечивающих генетический полиморфизм человека.
Гаптоглобин (Нр-Hb) является а2-гликопротеином, содержание которо-
го в норме составляет 25—30% всей ос2-глобулиновой фракции и 1—2% бел-
ков сыворотки; однако концентрация его в обычных условиях может сильно
варьировать.
Наиболее характерным свойством гаптоглобина является способность
образовывать с гемоглобином трудно диссоциирующий комплекс. Гемогло-
бинсвязывающий белок содержит около 83% чистого белка, остальные 17%
приходятся на углеводные компоненты. Образование комплекса предотвра-
щает выведение из организма находящегося в крови свободного гемоглоби-
на через почки.
Мировое распределение факторов Нр представляет существенный инте-
рес в антропологическом отношении. Так, различные по происхождению
популяции, живущие в экваториальной и тропических зонах Африки, Аме-
рики и Океании, характеризуются весьма высокой концентрацией аллеля
Hpl. Население Индийского субконтинента и Австралии отличается самы-
ми высокими частотами Нр2 в общемировом отношении. Народы Европы
имеют промежуточные частоты аллелей Нр, где в среднем концентрация
первой аллели колеблется в пределах 0,38—0,40. Монголоидам свойственны
относительно низкие величины
Система трансферрина (Tf). Молекулярная масса трансферрина состав-
ляет 88 000—90 000. Трансферрин, или сидерофилин, в соответствии с фено-
типическими особенностями относят к |3,-глобулинам сыворотки крови.
Основными функциями трансферрина являются: трансформация доста-
точно токсичных для организма ионов Fe++ и Fe+++ в деионизированную
форму; перенос железа к органам кроветворения (рис. 55); подавление бак-
териального роста в плазме лишением патогенных микроорганизмов железа;
угнетение вирусного размножения.
532
Перспективы биомоделирования
Хотя большинство вариантов Tf, по-видимому, случайным образом рас-
пределяется в популяциях, тем не менее, при геногеографическом анализе
обнаруживается определенная закономерность в локализации типов этого
белка, главным образом в распределении В01, В2, Dchi и D,. Первый из них
наиболее характерен для североазиатских монголоидов и американских ин-
дейцев, второй — для европеоидов, третий — для азиатских монголоидов, а
последний — для африканских и меланезийцев. Так, наибольшая частота
гена Tfcl выявлена у негроидных групп, наименьшая — у монголоидов; по-
пуляции Европы в этом отношении занимают промежуточное положение.
Столь же интересно распределение гена Tfc2, частота которого увеличивает-
ся в ряду: монголоиды — европеоиды — негроиды.
Система альбумина (AI). Альбумин человека характеризуется наличием
единственной полипептидной цепи с молекулярной массой 66 000—69 000,
содержащей 33—37 половинных остатков цистеина. Он состоит из высоко-
организованного компактного ядра, окруженного достаточно диффузной
оболочкой со складчатой поверхностью. Для альбумина, по всей видимости,
свойственно множество молекулярных вариантов, что частично обусловлено
различной локализацией внутримолекулярных дисульфидных связей.
Локус альбумина принадлежит к серии генетических локусов, наиболее
изменчивых в популяции человека. На основании ЭПР-спектроскопии сы-
вороточных белков обнаружено около 98 вариантов этого белка, а также не
менее 35 уникальных аллоальбуминов.
Наконец, отдельные формы альбумина в определенных этнических груп-
пах достигают полиморфных частот и поэтому получили соответствующее
наименование.
Церулоплазмин (Ср). Альфа-2-гликопротеин сыворотки крови, облада-
ющий оксидазной активностью (атакует р-фенилендиамин и некоторые фе-
нолы), является основным белком, который связывает и транспортирует ионы
меди (более 90% меди в плазме). Эта фракция содержит около 7% карбогид-
ратов, а концентрация ее в плазме клинически здоровых взрослых составля-
ет 35—38 мг/100 мл. Отмечена низкая оксидазная активность церулоплазми-
на у новорожденных; у женщин при беременности она увеличивается; отме-
чены этнические различия в иммунохимических характеристиках церулоп-
лазмина.
Показано, что Ср является мономерным белком с молекулярной массой
135 000, очень чувствительным к протеолитическому расщеплению
Система иммуноглобулинов (Gm). Молекула иммуноглобулина (Ig) со-
стоит из одной пары соответственно идентичных тяжелых (Н) и легких (L)
цепей, соединяющихся между собой посредством дисульфидных связей. L-
цепи имеют молекулярную массу около 23 000 дальтон; половина из них
имеет постоянную первичную структуру (С-часть), а другая половина — ва-
риабельную (V-часть). Сходным образом построены и Н-цепи, но в отличие
Глава 38. Фенотипирование и генополиморфизм
533
от L-цепей они почти в два раза длиннее и тяжелее, около 3/4 длины ее
принадлежит к постоянной части. Под действием протеолитических фер-
ментов молекула Ig расщепляется на Fc- и РаЬ-фрагменты; фрагмент Fab, раз-
деляющий Н-цепь, называется Fd-частью.
У человека и высших позвоночных известны два типа L-цепей (% и X) и
5 классов Н-цепей (у, ос, г\, 5, е). Последние характеризуют 5 классов моле-
кул (IgG, IgA, IgM, IgD и IgE), различных по иммунологическим свойствам.
IgG-класс составляет основную массу иммуноглобулина нормальной сыво-
ротки.
Открытие наследственного полиморфизма IgG принадлежит Граббу, об-
наружившему, что сыворотка крови больных ревматоидным артритом обла-
дает способностью агглютинировать эритроциты, сенсибилизированные
неполными «анти-О-антителами». Эта реакция, однако, в норме подавляет-
ся примерно у 60% людей европейского происхождения.
Система псевдохолинэстеразы сыворотки крови (Ph). Фермент псев-
дохолинэстераза (КФ 3.1.1.8), синтезируемый, по всей вероятности, в пече-
ни, легко гидролизует различные эфиры холина, такие, как ацетилохолин,
бутирилхолин, бензоилхолин, а также некоторые другие, например эфиры
ацетилсалициловой кислоты.
Система эритроцитарной фосфоглюкомутазы (PZM1). Фермент фосфог-
люкомутаза (КФ 2.7.5.1) является фосфотрансферазой, катализирующей
перенос фосфатной группы из положения 1 в положение 6. Фермент широ-
ко распространен в живой природе и играет важную роль в углеводном об-
мене. Его активность проявляется не только в эритроцитах, но и в различ-
ных органах и тканях, а также в некоторых продуктах выделения.
После электрофоретического разделения в крахмальном геле гемолиза-
тора различных людей наблюдаются три характерных типа изоферментных
спектров, что дало основание для предположения о генетической природе
подобной изменчивости. Эти данные пордтверждены и дополнены ЭПР-
спектроскопией.
Система супероксидцисмутазы (SOD-A) — фермент, катализирующий
окисление дихлорфенолиндофенола. Этот белок обнаруживается во всех
тканях, и был отождествлен с супероксидцисмутазой, которая воздействует
на весьма токсичные для клеток радикалы супероксида с образованием пе-
рекиси водорода и кислорода.
Существуют два варианта супероксидцисмутазы: митоходриальная фор-
ма SOD представлена в клетках всех тканей, за исключением эритроцитов;
растворимая форма фермента наблюдается также и в эритроцитах. Первая
разновидность супероксидцисмутазы относится к SOD-В и находится под
контролем аллелей одного локуса, картированного в хромосоме 6. Вторая
форма получила название SOD-A; локус, ответственный за ее детермина-
цию, находится на хромосоме 21.
534
Перспективы биомоделирования
Система экскреции b-аминоизомасляной кислоты (BAIB). Среди ами-
нокислот, выделяемых из организма с мочой, наибольшее значение в антро-
пологии имеет Р-аминоизомасляная кислота (ВАШ), полиморфизм экскре-
ции которой в популяциях человека исследован пока недостаточно. При ис-
следовании при помощи хроматографии аминокислотного состава мочи сре-
ди веществ, окрашивающихся нингидрином, выделяется пятно, обозначаемое
как «Т». В дальнейшем это вещество идентифицировано как Р-аминоизомас-
ляная кислота — продукт деградации пиримидинового основания тимина.
Интерес со стороны врачей и биологов к вариабельности экскреции BAIB
обусловлен тем обстоятельством, что частота встречаемости выделителей
большого количества BAIB колеблется у разных народов в широких преде-
лах: у большинства европейцев она незначительна, у негроидов составляет в
среднем 20—30%, у монголоидов и ориенталов еще выше и отличается высо-
кой дисперсией.
Щелочная сывороточная фосфатаза. К настоящему времени стало из-
вестно, что кроме локусов АВО и Se, существуют, по-видимому, и другие
генетические факторы, играющие роль в детерминации фенотипов щелоч-
ной сывороточной фосфатазы. Тот факт, что группы крови J и R некоторых
домашних животных, аналогичные группам крови АВО человека, также
имеют сходную с фенотипами Рр природу ассоциации, наводит на мысль о
возможной существенной роли этой связи.
При рассмотрении характера пространственного распространения типов
щелочной фосфатазы среди населения Северной Азии следует обратить вни-
мание, с одной стороны, на высокую частоту встречаемости среди северных
монголоидов фенотипа Рр2, концентрация которого во многих случаях дости-
гает и превышает 40%; с другой стороны — на обнаруживающуюся высокую
дисперсию распределения типов Рр, подобную распределению факторов АВО.
В сводке, опубликованной в 1975 г., указывается, что PGM,6 достаточно
широко распространен в популяциях Тихоокеанского бассейна, в частности
среди китайцев и северных таиландцев, в Индии, у тибетцев Непала и авст-
ралийских аборигенов Квинсленда, а также в Японии. Аллель PGM,7 с не-
значительной частотой представлен в нескольких префектурах Японии, у
таиландцев, аборигенов Западной Малайзии, в некоторых популяциях Ин-
дии. У китайцев Индонезии этот ген обнаружен с частотой 2,4%.
Система альбумина (AI). Одновременно с типированием разновиднос-
тей трансферрина, группоспецифического компонента посттрансферрина
учитывалась фенотипическая картина области альбумина.
Исследование альбумина представляет специальный интерес после того,
как стали известны полные сводки о полиморфизме по этой системе среди
аборигенов Северной Америки. Оказалось, что альбумин «Наскапи» (AINa)
является распространенным вариантом среди восточных субарктических
племен американских индейцев. В племени наскапи он встретился с доста-
Глава 38. Фенотипирование и генополиморфизм
535
точно высокой полиморфной частотой — 0,031, у аппачей резервации Сан-
Карл ос в Аризоне — с меньшей концентрацией, однако и здесь этот фактор
оказался полиморфным (0,016). Тем не менее, у эскимосов Северной Амери-
ки и Гренландии не обнаружено генетических разновидностей этого белка.
Именно поэтому идентификация редких типов альбумина у чукчей и других
народностей Северной Азии имеет важное значение для обнаружения гене-
тических связей аборигенов Нового и Старого Света.
Сравнительно небольшая пропорция экскреторов отмечена среди рус-
ских и украинцев, что соответствует литературным данным о низкой их ча-
стоте среди населения Европы. Относительная доля выделителей в группе
негров Западной Африки оказалась несколько выше. Наибольшая частота
фенотипа высокого уровня экскреции Р-аминоизомасляной кислоты уста-
новлена в индийской и одной из эвенкийских групп.
Частота встречаемости лиц с высоким уровнем экскреции ВАШ и оценка
генных частот в некоторых этнорасовых группах: русские — 14,08% (0,375);
украинцы - 12,50% (0,354); татары - 16,66% (0,408); казахи - 18,18% (0,426);
арабы — 19,44% (0,441); латиноамериканцы — 19,05% (0,437); негры Запад-
ной Африки — 24,34% (0,493); индийцы — 36,36% (0,603); эвенки (Эконда,
ЭНО) - 38,57% (0,612).
Изучение генетического полиморфизма значимо даже не столько для
биохимии и генетики, сколько для фармакологии и токсикологии. По своей
сути генетический полиморфизм предопределяет индивидуальную чувстви-
тельность человека к лекарствам и ксенобиотикам (табл. 89). Поиск опти-
мальных биомоделей, выбор животных для исследований имеет решающее
значение в предупреждении многих проявлений побочных реакций для че-
ловека.
Таблица 89
Непереносимость некоторых лекарственных средств в связи с особенностями
генетического полиморфизма
Дефектный
фермент
Глюкозо-6-
фосфат-
дегидрогеназа
Распространение
среди населения
Арабы, американс-
кие негры, греки,
евреи,индонезийцы,
курды, малайцы,
новогвинейцы, паки-
станцы, персы,ру-
мыны, сардинцы,си-
цилианцы, таи, фил-
липинцы, жители
Южного Китая
Непереносимые
лекарственные
препараты
Более 120 лекарств,
из них: акрихин и ана-
логи, аспирин и анало-
ги, сульфапиридазин
и др. сульфаниламиды,
фурадонин и др. фура-
ны, дифенилсульфон,
викасол, производные
анилина, ПАСК. Лево-
мицетин, новарсенол,
толбутамид, БАЛ,
фенилгидразин
Характер
реакции
Тяжелая гемо-
литическая
анемия, резкие
боли в поясни-
це и животе,
обмороки,
лихорадка
Тип
наследования
Сцепление с
Х-хромосомой
536 Перспективы биомоделирования
Продолжение табл. 89
Дефектный
фермент
Ацетилтранс-
фераза
Каталаза
Псевдохолин-
эстераза
Глютатион-
редуктаза, мет-
гемоглобин-
редуктаза
В стадии
изучения
Неизвестно
Данные
недостоверны
Неизвестно
P-N-ацетил-гек-
созаминидаза
Распространение
среди населения
4,6% эскимосов,
13,3% японцев,
21,5% американских
индейцев, 30% насе-
ления Кавказа,
32,8% латиноамери-
канцев, 50% населе-
ния США
В Корее, Японии,
Южном Китае
и Швейцарии
1 случай на 2000
человек
1% гетерозиготных
5% населения США
В 2 больших семьях
31,5% европейцев,
10,6% китайцев,
2,7% африканцев
1 случай на 20 000
человек
1/зо среди американ-
цев еврейского про-
исхождения и 1/зоо
среди прочих амери-
канцев
Непереносимые
лекарственные
препараты
Изониазид, фенилзин,
гидралазин, дапсон,
сульфадимезин
Перекись водорода,
этиловый и метиловый
спирт
Дитилин, отдельные
миорекласанты
Нитроглицерин и др.
нитраты и нитриты,
сульфаниламиды,
левомицетин, ПАСК,
фурадонин,хинин,
ацетанилид,параце-
тамол
Кортикостероиды
в глазной практике
Варфарин
Вещества, содержа-
щие группу N-C-S,
фенилтиомочевина,
метил- и пропилтио-
урацил, фенилтио-
карбамид
Галотан, эфир, цикло-
пропан, метоксифлу-
ран
Сердечные гликозиды,
препараты, содержа-
щие ганглиозиды и
активирующие сахара
Характер
реакции
Медленное
ацетилирова-
ние препаратов
вызывает повы-
шение их токси-
чности
Медленное
разрушение
препаратов
усиливает их
токсичность
Усиление
токсичности
из-за замедле-
ния метабо-
лизма
Метгемоглоби-
немия,цианоз
Глаукома
Устойчивость
к препарату
Отсутствие
ощущения
вкуса этих
веществ
Повышение
температуры
тела с развити-
ем мышечной
ригидности
и смерти
Болезнь Тея-
Сакса. Побоч-
ные реакции
вследствие
медленного
разрушения
ганглиозидов
Тип
наследования
Аутосомно-
рецессивный
Аутосомно-
рецессивный
Аутосомно-
рецессивный
Аутосомно-
рецессивный
Аутосомно-
рецессивный
Аутосомно-
рецессивный
Аутосомно-
рецессивный
Аутосомно-
рецессивный
Аутосомно-
рецессивный
Глава 38. Фенотипирование и генополиморфизм
537
Окончание табл. 89
Дефектный
фермент
Амило-1,6-
глюкозидаза
Киназа фосфо-
рилазы
Распространение
среди населения
Жители Средиземно-
морья (евреи,ливан-
цы, сирийцы и др.)
Афроамериканцы,
гавайцы, таи, евреи,
арабы Ближнего
Востока
Непереносимые
лекарственные
препараты
Сульфаниламиды,
нитриты, стрептоми-
цин, неомицин, не-
наркотические аналь-
гетики
Барбитураты,транк-
вилизаторы, нейро-
лептики, нитриты
и нитраты
Характер
реакции
Резкое увели-
чение печени,
гипогликемия,
гиперурикемия,
гипергликемия
Небольшое уве-
личение печени,
гипогликемия,
обмороки,
лихорадки
Тип
наследования
Аутосомно-
рецессивный
Связана с по-
лом, сцепле-
ние с Х-хромо-
сомой
Аллельная частотность генов, в которой кодируется информация о фер-
ментах I и II фаз, описана в отношении целого ряда национальностей, вы-
явлена их широкая этническая вариантность, однако, отсутствуют данные о
полиморфизме ферментов, участвующих в метаболизме лекарств для рус-
ской нации, которая является самой многочисленной среди славянских на-
родов. Это определило направление исследования генотипов некоторых ге-
нов, в которых закодирована информация о ферментах I и II фаз, преобра-
зующих лекарства, проведено сравнение полученных результатов с соответ-
ствующими данными исследований у других наций, а также предпринята
попытка выяснить, являются ли русские люди генотипически схожими с
другими кавказскими группами или существует влияние Востока [118].
Генетический полиморфизм животных.
Ферменты I фазы, принадлежащие к семейству цитохромов Р-450 (CYP450),
являются самыми важными в метаболизме лекарств и ксенобиотиков. Среди
изоформ CYP тип CYP2D6 является наиболее активным. Он катализирует
метаболизм около 25% всех наиболее клинически важных лекарственных пре-
паратов, таких, как антиаритмические препараты, трициклические антидеп-
рессанты, (3-адреноблокаторы и другие. Фермент CYP1A1 отвечает за окисли-
тельный метаболизм токсичных препаратов: диоксина и бензпирена.
Такой фермент II фазы, как ариламин N-ацетилтрансфераза 2 (NAT 2),
играет важную роль в биотрансформации ряда ароматических и гетероцик-
лических аминов и гидразинов, которые содержатся в лекарствах, а также в
окружающей среде [118]. В предыдущих главах мы приводили данные иссле-
дования активности цитохрома Р-450 у мини-свиней и указывали на важную
роль этой ферментной системы в обеспечении генетического полиморфизма
у животных. Поскольку нестероидные противовоспалительные лекарствен-
ные препараты в своих эффектах в значительной степени зависят от особен-
538
Перспективы биомоделирования
ностей генополиморфизма мы сочли необходимым дать таблицу биомоде-
лей для их изучения.
Таблица 90
Природно обусловленные и экспериментальные биомодели для изучения
генетического полиморфизма нестероидных противовоспалительных веществ
Исследуемое действие
вещества
Моделирование воспали-
тельного процесса:
a. керрагенинового отёка
лапы (0,1 мл 1%-ного
раствора керрагенина,
субплантарное введение)
b. перитонита (1%-ный
раствор уксусной кислоты
- 1 мл на 10Ог массы тела,
внутрибрюшинно)
c. фетровой гранулёмы
(имплантация под кожу
живота 4 простерилизован-
ных фетровых листков
массой 10 г)
d. адьювантного артрита
(0,1 мл адьюванта Фрейнда,
субплантарное введение
в правую заднюю лапу)
e. моделирование субвос-
палительных проявлений
по тесту ультрафиолетовой
эритемы
Модель для оценки аналь-
гетического действия
веществ:
a. механическое раздраже-
ние воспалённой лапы
b. корчи, вызванные уксус-
ной кислотой (0,75%-ная
кислота, 0,1 мл/10 г -
внутрибрюшинно и 1%-ная
кислота, 0,5 мл/100 г,
внутрибрюшинно)
Модель для оценки жаро-
понижающих эффектов
веществ по тесту дрожже-
вой лихорадки (20% сус-
пензия пекарских дрожжей,
подкожно)
Модель ульцерогенного
действия НПВС
Биомодель для изучения
механизмов спонтанных
эрозивных поражений
желудка
Лабораторное
животное
Крысы
Крысы
Крысы
Крысы
Морские
свинки
Крысы
Мыши
Крысы
Крысы
Крысы
Мыши
Линия или порода
животного
Линейные
Линейные
Линейные
Линейные
Альбиносы
Линейные
Линейные
Линейные
Линейные
Линейные
Линии: BRSUNT/N
и BRSUNT/NY
Пол
животного
Яд
Яд
Яд
Яд
Яд
Яд
Яд
Яд
Яд
Яд
Яд
Вес
животного
130-180 г
130-180 г
130-180 г
130-180 г
250-500 г
130-180 г
20-26 г
130-180 г
130-180 г
130-180 г
30-40 г
Возраст
животного
3-5 мес.
3-5 мес.
3-5 мес.
3-5 мес.
2-4 мес.
3-5 мес.
2-2,5 мес.
3-5 мес.
3-5 мес.
3-5 мес.
3-10 мес.
Глава 38. Фенотипирование и генополиморфизм
539
С целью поиска оптимальной биомодели генетического полиморфизма
человека в лаборатории биомоделирования НЦБМТ РАМН под руковод-
ством В.В.Осипова были проведены исследования на мини-свиньях. С этой
целью определяли генетический полиморфизм трансферрина, церулоплаз-
мина, ферментов метаболизма и их фенотипическую гетерогенность.
Трансферрин. Генетически детерминированный полиморфизм трансфер-
рина (основного белка плазмы крови, переносящего железо) проявляется в
различной электрофоретической подвижности алломерных форм.
Трансферрины при анализе генетических вариантов дают нечеткие дан-
ные, так как различные типы трансферринов меняют электрофоретическую
подвижность при взаимодействиях с другими белками крови, как у челове-
ка, так и у животных. Фенотипы трансферрина определяли с помощью элек-
трофореза в крахмальном геле по Смитису. В норме трансферрин сыворотки
не насыщен железом, но для определения фенотипа его необходимо насы-
тить ионами Fe3+, поскольку свободный белок и комплекс его с железом
имеют различные изоэлектрические точки. Различные формы трансферрина
у мини-свиней слабо различаются по электрофоретической подвижности,
поэтому определение фенотипа следует проводить путем тщательного срав-
нения картины разделения данного белка с электрофореграммами извест-
ных фенотипов трансферрина. У мини-свиней каждый из аллелей трасфер-
рина обнаруживается в виде трех основных полос.
Исследование трансферринов сыворотки крови у 180 взрослых (9—12-
месячных) животных показало, что у светлогорских мини-свиней встреча-
ются 4 кодоминантных аллеля, контролирующих полипептидную цепь трас-
феррина. Наиболее часто встречается генотип ВВ (0,97), остальные гомози-
готные генотипы АА, СС и ДЦ встречаются приблизительно с одинаковой
частотой (около 0,005). Гетерозиготные фенотипы трансферрина у мини-сви-
ней очень трудно различимы из-за сильной вариабельности полос. Более
того, разные варианты электрофореграмм, сделанные из одной и той же
пробы, имеют неодинаковые наборы полос.
Фенотипическую гетерогенность внутри одного варианта приписывают
различному содержанию нейраминовой кислоты. Присоединение к транс-
феррину нейраминовой кислоты контролируется отдельным геном. Такой
вид вариабельности не зависит от основных вариаций фенотипа, контроли-
руемых структурным локусом, кодирующим полипептидную цепь трансфер-
рина. Наличие различных остатков нейраминовой кислоты размывает точ-
ность определения фенотипа трансферрина. Такая гетерогенность не соот-
ветствует истинному генотипу трансферрина и дает нечеткие данные. Это
мешает использованию трансферрина как генетического маркера в селекции
мини-свиней.
Церулоплазмины. Показано, что содержащий медь гликопротеид плаз-
мы — церулоплазмин, существует в трех фенотипических вариантах, облада-
540
Перспективы биомоделирования
ющих различной электрофоретической подвижностью. Церулоплазмины де-
терминируют двумя кодоминантными аллелями. Частота встречаемости в 150
обследованных сыворотках мини-свиней такова: Ср1/ Ср1 — 0,01; Ср2/ Ср2
— 0,97; Ср1/ Ср2 — 0,02. Это дает основание считать, что наличие двух генов,
контролирующих формирование полипептидных цепей церулоплазмина,
может в некоторых случаях быть использовано при селекции светлогорских
мини-свиней. В качестве генетического маркера наиболее пригоден доволь-
но редко встречающийся ген Ср1, так как наличие его в гомозиготном состо-
янии у производителей дает дополнительную возможность проследить гени-
алогию всего семейства, установить степень родства и кровность.
Ферменты метаболизма. У взрослых 7—60-месячных мини-свиней оп-
ределяли активность ферментов метаболизма; цитохром-с-редуктазу, цитох-
ром Р-450 и глютатион-Б-трансферазу. Эти ферменты играют большую роль
в тканевом дыхании.
Показано, что активность цитохром-с-редуктазы у взрослых мини-сви-
ней на всех изученных сроках остается одинаковой. Активность цитохром
Р-450 повышается с 7 до 18 месяцев, а затем остается без изменений. У мини-
свиней 7—22 месяцев отмечается повышение активности глютатион-Б-транс-
феразы.
По-видимому, период от 7 до 22 месяцев является у мини-свиней очень
важным, так как в этот период происходит окончательное формирование
животного. Это подтверждают данные по относительному приросту, актив-
ности иммунологических компонентов и некоторых ферментов метаболиз-
ма. Иными словами, этот возраст является наиболее оптимальным для мо-
делирования метаболических процессов в аспекте экстраполяции на взрос-
лого человека.
Генополиморфизм у грызунов. Многие общие инбредные штаммы мы-
шей отличаются наличием двадцати или более полиморфных очагов. Неко-
торые очаги характеризуются иммунологически, но многие могут быть иден-
тифицированы электрофорезом в крахмальном геле.
Биохимические методы обладают исключительной чувствительностью и
могут показывать изменение штаммов и/или сублиний в результате мутации
или генетического контаминирования. Так, изучен ряд европейских коло-
ний и показано, что некоторые сублинии СЗН отличались друг от друга в
различных очагах, возможно, ввиду генетической контаминации в какой-
либо момент в прошлом. Таким же образом изучены совокупности 69 евро-
пейских и японских штаммов в пяти очагах (Hbb, Mod-1, Id-1, Es-1 и Es-2),
в которых обнаружены случайные сублинии, отличавшиеся от ранее опубли-
кованных стандартов.
Основной недостаток биохимических методов — это сроки и необходи-
мое «ноу-хау». В будущем эта проблема может быть решена путем автомати-
зации. В настоящее время биохимические методы чрезвычайно ценны в
Глава 38. Фенотипирование и генополиморфизм
541
качестве средства периодической проверки характеристик штаммов, напри-
мер раз в 1 — 5 лет, но для опытов, проводимых с более высокой периодич-
ностью, потребуются иные, более экономичные, способы.
Можно сослаться на упрощенный способ с использованием полосок
Целлогеля, для выполнения которого требуется лишь 55 минут. Данный спо-
соб применим по отношению к очагам гемоглобина, трансферрина, эстера-
зы-1 и других биохимических субстратов.
Поиск надёжных корреляций генетического полиморфизма человека и
животных — весьма перспективный путь биомоделирования в интересах здо-
ровых людей.
Вне зависимости от того, какой метод будет разработан или предпочтен
автором для исследования фенотипических особенностей и генетического
полиморфизма, метод ЭПР—спектроскопии будет оставаться неким тесто-
вым приемом, позволяющим соотносить в единой системе разные данные.
Однако в поиске новых принципов биомоделирования и экстраполяции с
животных на человека следует помнить, что генетические маркеры-фермен-
ты являются прежде всего катализаторами, функционально увеличивающи-
ми скорости биохимических реакций.
Эти ферменты-маркеры не только высокоэффективны, но и строго спе-
цифичны по отношению к субстрату катализа. Существуют абсолютно спе-
цифичные ферменты, например, уреаза, гидролизирующая мочевину до С02
и аммиака, группоспецифичные, например, пепсин, действующий на пептиды
с ароматическими группами, а также ферменты, специфичные к определен-
ному типу реакций или химической связи. Существует и группа ферментов,
специфичных к стереоизомерам субстратов, например, лактатдегидрогеназа,
катализирующая окисление только L-формы молочной кислоты.
Поэтому при планировании экспериментов с целью поиска генетических
маркеров, свойственных во всей полноте биохимическим и физиологичес-
ким процессам, следует не только констатировать наличие того или иного
фермента у человека и животного, а соотносить его по возможности наибо-
лее широко со всей гаммой эффектов, вызываемых им. С этой точки зрения
инбредные животные могут, с одной стороны, идеально моделировать дей-
ствие лекарства на китайца или монгола, но, с другой — не отражать тяжелое
побочное действие в отношении кавказоида. Именно поэтому моделирова-
ние генетического полиморфизма представляет одну из сложных областей
биомоделирования. Не случайно ведь говорят: «что для одного благо, для
другого — смерть».
542
Глава 3 9 О СЕЙФЕ ТВОРЦА И МОДЕЛЯХ
ВЫСОКИХ ПОРЯДКОВ
Древнегреческие философы Левкипп и Демокрит в
V в. до н. э. пришли к выводу о невозможности бес-
конечного деления вещества на все более мелкие
части. На определенном уровне их дальнейшее де-
ление невозможно. Большое разнообразие подоб-
ных частиц позволяет создавать разные вещества.
Частички, которые нельзя больше разделить, назва-
ли неделимыми (по-греч. неделимый — atomos).
Дальтон назвал свои неделимые частички атомами,
и на этот раз атомная теория получила признание.
В 1896 г. было обнаружено, что некоторые атомы
могут самопроизвольно распадаться, испуская при
этом частицы намного меньших размеров. Это яв-
ление, открытое Анри Беккерелем, было названо ра-
диоактивностью .
Некоторые молекулы газов состоят из одного
или нескольких атомов (например, аргон и инерт-
ные газы). Известны молекулы, состоящие из мил-
лионов атомов — ДНК, РНК, белки. Теория о мо-
лекулярном строении определяет молекулу как
мельчайшую частицу вещества, обладающую всеми
его химическими свойствами. Латинское слово moles
означает массу, а суффикс cula имеет уменьшитель-
ное значение, поэтому слово «молекула» дословно
переводится как «маленькая масса».
В качестве единицы массы вещества принимают
единицу, называемую моль. Моль — это масса ве-
щества, выраженная в граммах, число которых рав-
Глава 39. О сейфе Творца и моделях высоких порядков
543
но его молекулярной массе. Например, 1 моль этилового спирта — это 46 г
спирта, так как его молекулярная масса равна 46.
В силу закона действующих масс удобнее оперировать именно молями,
потому что вещества реагируют между собой в количествах, пропорциональ-
ных молям. Дело в том, что моль любого вещества содержит вполне опреде-
ленное, постоянное для всех веществ количество молекул. Число молекул
в моле равно 6,02-1023. Это так называемое число Авогадро, потому что его
рассчитали, исходя из закона Авогадро.
Как атомы, так и молекулы стали объектами моделирования различных
ситуаций, а также биомоделирования в живых системах. Казалось бы, это
наиболее простой раздел моделирования, при котором можно легко дать
любое математическое описание протекающих процессов, однако, на прак-
тике все оказалось намного сложнее.
Аналогия отношений: новый взгляд
Классики-естествоиспытатели выстроили для нас картину мира, в фунда-
мент и стены которой положены принципы структурной организации и со-
пряженные с ними функциональные отношения. Эти модели мироздания до-
статочно хорошо описываются в рамках математических моделей третьего
порядка с очевидностью 1 + 1 = 2.
Но, увы, эта очевидность стала совсем не очевидной уже с первой поло-
вины XIX столетия, когда наш соотечественник Н.И. Лобачевский в 1826 г.
опроверг незыблемый с III века до Рождества Христова пятый постулат Ев-
клида, согласно которому через точку, лежащую вне прямой, проходит един-
ственная прямая, не пересекающая исходную.
Возможно в тот момент, когда начал свои работы Н. Лобачевский, крайне
секретный сейф Творца чуть приоткрылся, и он увидел существование неев-
клидовой геометрии. Н. Лобачевский с легкостью доказал, что сумма углов
треугольника может быть меньше п, а всякое число равно нулю и 1 = 2 или,
по крайней мере, 1 + 1 > 2. В те годы школяров пороли розгами и за меньшие
издевательства над математикой. Но когда Н. Лобачевский предложил заме-
нить пятый постулат Евклида аксиомой о том, что через точку, лежащую вне
прямой, проходят, по крайней мере, две прямые, не пересекающие исходную,
возмущению приверженцев классической математики уже не было предела.
Аксиома Н.Лобачевского казалась виртуальной до 1868 г., когда через
40 лет после его первой работы Бельтрами показал, что новая геометрия вы-
полняется на поверхностях с постоянной отрицательной кривизной. А уже
в начале XX века Ф. Клейн предложил свою модель: в качестве плоскости —
внутренность круга с хордами в виде прямых. Модель строилась как элемент
классической геометрии, что определяло геометрию Лобачевского непроти-
воречащей классике Евклида.
544
Перспективы биомоделирования
Построение биологических моделей первого и второго порядка — это
область Евклидовой геометрии, коль скоро мы стараемся представить мате-
риал в графиках, диаграммах, гистограммах и таблицах. Моделирование же
на уровне атомов и элементарных частиц потребует иных представлений, а
главное — понимания процессов в рамках геометрии Н.Лобачевского. В этом
случае в силу вступают и иные принципы аналогий.
Как мы уже подчеркивали в гл. 1 и 2, в современной науке основной
принцип моделирования основан на аналогиях, которые можно назвать струк-
турно-функциональными и функционально-структурными. Первый вариант мы
рассмотрели в приложении биомоделей первого и второго порядков. Второй
вариант, т. е. функционально-структурный, можно охарактеризовать как ана-
логию отношений. Если с модели на прототип переносится любое отношение
переменных единиц, то мы имеем дело уже с чистой аналогией отношений,
которые в некоторых случаях возможно описать математическими моделями
третьего порядка. А почему не во всех случаях?
Дело в том, что при использовании аналогий отношений само понятие
изоморфизма структур модели и прототипа отодвигается на второй план.
Аналогия установления изоморфизма становится не предпосылкой, а резуль-
татом вывода. В табл. 3 (гл. 2) показаны разные уровни биосистем и возмож-
ные варианты выбора принципов биомоделирования. Мы специально выде-
лили аспект моделирования четвертого порядка. Кому-то это покажется спор-
ным, но, тем не менее, нельзя не согласиться, что на этом уровне вступают
в силу такие представления, которые отрицают законы неадекватности, прин-
ципы изоморфизма. К ним мало приложимы постулаты и теоремы классичес-
кой физики и математики.
Перед тем как перейти к их рассмотрению, вспомним историю класси-
ческой физики и химии, т. е. те моменты, которые сложились в понятные
для нас представления о строении материи и законы природы.
К концу XX века было известно более 30 элементарных частиц. Тщатель-
ное изучение позволило выявить их общие свойства. Так основными харак-
теристиками элементарных частиц считают их массу покоя и элементарный
заряд.
Суммарная масса всех частиц, образующих молекулу, атом или ядро,
является массой данного микрообъекта, если её уменьшить на величину
дефекта массы. Дефект массы прямо пропорционален энергии, которую
нужно затратить, чтобы расщепить микрообъект на элементарные частицы.
Электрический заряд атома или его ядра равен сумме зарядов составляющих
его частиц.
Спин является очень важной характеристикой элементарной частицы (см.
гл. 38). Спин элементарной частицы — квантовая величина, которая не име-
ет аналогов в классической механике и электродинамике. Это такое же
неотъемлемое свойство элементарной частицы, как заряд или масса. Его
Глава 39. О сейфе Творца и моделях высоких порядков
545
можно объяснить как момент импульса элементарной частицы, который не
связан с её движением и не зависит от внешних условий.
Для элементарных частиц выполняется закон сохранения электрическо-
го заряда: при любом взаимном превращении частиц алгебраические суммы
электрических зарядов исходных и конечных частиц равны. Все точки в
пространстве физически равноправны, что позволяет считать однородным
пространство. Следствием является закон сохранения момента импульса,
который определяется изотропностью пространства, то есть физического
равноправия всех направлений. В микромире осуществляется четыре типа
фундаментальных взаимодействий:
/ гравитационное взаимодействие для процессов микромира настолько
мало, что обычно им пренебрегают. Это связанно главным образом с
тем, что массы элементарных частиц очень малы. Однако оно присуще
всем частицам без исключения;
/ электромагнитное взаимодействие основано на процессах, происходя-
щих с частицей в электромагнитном поле. Для электронейтральных
частиц (нейтрино, антинейтрино, фотон) оно не осуществляется. Элек-
тромагнитное взаимодействие обуславливает существование атомов и
молекул, их связь между ядром и электронами осуществляется благо-
даря их разноимённым зарядам;
/ слабое взаимодействие охватывает процессы, происходящие с участием
нейтрино или антинейтрино. Это самое медленное из всех взаимодей-
ствий микромира;
/ сильное взаимодействие проявляет себя в связи протонов и нейтронов в
ядре, хотя ядерные силы — это только частный случай сильного взаимо-
действия, это силы притяжения. Они обеспечивают связь между нук-
лонами и одноименно заряженными протонами. Это не электричес-
кие силы, потому что они действуют не только между заряженными
протонами, но и нейтронами, не имеющими заряда. Они также не яв-
ляются гравитационными силами, которые ничтожно малы для объяс-
нения ядерных эффектов.
Таким образом, физика элементарных частиц наглядно доказала, что
далеко не все физические процессы укладываются в рамки классической
электромагнитной модели. Две частицы (нейтрон и позитрон) выявили не-
возможность объяснения физических явлений, исключительно электромаг-
нитными и гравитационными взаимодействиями. Стабильность электронов,
протонов и фотонов — это исключение в природе элементарных частиц.
Остальные элементарные частицы произвольно или в результате столкнове-
ний превращаются в другие частицы.
Интуитивно понимая, что все биологические процессы — это взаимодействие
элементарных частиц, что именно с помощью электронов молекулы объеди-
няются в высокоорганизованные структуры, мы никогда не учитываем эти
546
Перспективы биомоделирования
процессы в моделировании системных функций. Но надо ли это и, главное,
возможно ли учесть такие аспекты обеспечения жизнедеятельности?
Но именно этот вопрос определяет необходимость построения биологи-
ческих моделей сложнейших и совершенных биологических реакций в терми-
нах энергетики элементарных частиц и их связи и взаимодействий. Существу-
ют три типа превращений энергии: превращение энергии электрона в энер-
гию связи, реакции переноса групп (обмен энергетических связей) и выпол-
нение работы. При этом выполнение работы, т. е. исполнение функций, яв-
ляется переходом энергии связи в энергию электрона. По-видимому, мы многое
поймем, когда сможем представить эти процессы на уровне элементарных
частиц, энергетики АТФ, изменения скользящих нитей миофибрилл, сокра-
щения мышцы, выполнения работы в соответствии с психофизиологически-
ми установками в виде модели четвертого, пятого или иного порядка.
Энергия связи или энергия переноса
Мы были вынуждены напомнить о характеристиках, превращениях и взаи-
модействии элементарных частиц потому, что в биомоделях четвертого по-
рядка на первое место выдвигается не структурная или функциональная
компоненты, а энергетическая составляющая, которая с ними не коррелиру-
ет, а зачастую и не связана впрямую. Электронное окружение молекул игра-
ет в биоэнергетике особое значение, пока мало понятное современной на-
уке. В любом научном описании стыдливо опускается один нюанс — пере-
ход от принципов квантовой механики субатомных единиц к межмолеку-
лярным взаимоотношениям в биологических процессах. И это не лукавство
ученых. Попросту отсутствует адекватная научная база для исследования и
логико-математического (реального, а не спекулятивного) аппарата описа-
ния этих процессов и оптимального моделирования.
В то же время все прекрасно понимают, что атом — это система электро-
нов, стабилизируемая ядром, а молекулы — это атомы, удерживаемые вместе
поделенными электронами. Биологические реакции представляют собой реак-
ции электронов, т. е. вся энергетика живых систем — это всего лишь энерге-
тика субатомных частиц. И это не просто подмена понятий. Горячая крышка
на самоваре выглядит так же, как холодная. На глаз это так, но на ощупь...
Не советую. Говоря о моделях четвертого и более высоких порядков, мы рас-
сматриваем энергетическую составляющую объекта, т.е. объектом становится
не крышка самовара, а ее теплоемкость или количество тепла, или... все что
угодно, но отражающее внутренние процессы энергопреобразования.
В биомоделях четвертого порядка отсутствует, как уже нами подчерки-
валось, один из основополагающих законов жизни — закон неаддитивности.
Одна частица плюс еще одна частица, соединенные наугад, — это всего лишь
две частицы: 1 + 1 = 2; система аддитивна. Но если две частицы объединить
Глава 39. О сейфе Творца и моделях высоких порядков
547
каким-то определенным образом, то возникает нечто новое, большее чем их
простая сумма: 1 + 1 > 2. Это важнейшее уравнение биологии. Оно спра-
ведливо для всех объектов — от самых простых до самых сложных. Если
один электрон и одно ядро объединить соответствующим образом, то воз-
никнет атом водорода, который являет собой нечто большее чем просто ядро
и просто электрон. Когда атомы объединяются в молекулу, эта молекула будет
чем-то качественно новым, что уже нельзя описать только лишь на языке
атомов.
В качестве иллюстрации к биомоделированию четвертого порядка обра-
тимся к оригинальным суждениям лауреата Нобелевской премии Альберта
Сент-Дьёрди [623, 624, 625], рассматривающим соотношения ионизацион-
ного потенциала с реакционной способностью электронов. Всем исследова-
телям известен парадокс, что энергия и реакционная способность электрона
обратно пропорциональны его ионизационному потенциалу, т. е. чем ниже
ионизационный потенциал, тем выше энергия и реакционная способность и
наоборот.
В начале третьего тысячелетия нет предпосылок выделять неделимые или
самые элементарные кирпичики природных процессов. Любой, самый ма-
лый объект исследования бесконечно сложен и может иметь бесконечное
число параметров. Главное при построении модели — цель исследования,
учет важнейших факторов и отбрасывание несущественных. Поэтому любая
модель неполна, поскольку не тождественна абсолютно объекту-оригиналу.
Заложенная изначально неполнота модели исключает её идеальную адек-
ватность оригиналу. Для чего же нужна модель четвертого порядка? В об-
щем, требования к ней те же, что и к моделям иных порядков, но как бы
более заострены на выяснении понятий, лежащих за горизонтом наших взгля-
дов на механизмы функционирования нашего организма. Итак, модель чет-
вертого порядка необходима для понимания:
/ энергетической структуры конкретного объекта, законов развития, вза-
имодействия элементарных частиц при выполнении тех или иных функ-
ций, а также в процессе саморазвития и взаимодействия с внешней сре-
дой;
/ принципов управления, диагностики и лечения в живых объектах и про-
цессах на уровне ядерных сил, взаимодействия и превращения элемен-
тарных частиц;
/ путей прогнозирования прямых и косвенных последствий реализации
различных способов и форм воздействия на сильные и слабые кванто-
во-механические связи и их взаимодействие с гравитационными и элек-
тромагнитными процессами в организме человека и животных.
Парадоксальность ситуации заключается в том, что все биологические ре-
акции, обеспечивающие существование жизни на планете Земля, укладыва-
ются в диапазон от 11,3 до 12,6 электронвольт. Для этого диапазона пред-
548
Перспективы биомоделирования
лагается ввести иное масштабирование, вьщелив некую энергетическую шкалу
биопотенциалов, выраженных в электронвольтах (рис. 19).
В живых системах электроны имеют минимальную энергию, если они
связаны с кислородом в молекуле воды. Вода обладает наименьшей энерги-
ей и самым высоким ионизационным потенциалом (12,56 эВ).
По-видимому, А. Сент-Дьёрди удалось загля-
нуть в сейф Творца, когда в качестве рабочей ги-
потезы он предложил четко разграничить поня-
тия энергии связи и энергии электрона и различия
между ними. Энергия электрона — это высокий
биопотенциал отдельного электрона, энергия свя-
зи — это общая энергия электронов, образующих
эту связь. Обозначим высокоэнергетическую связь
символом г), а энергию электрона обозначим сим-
волом е. В таком случае процесс окислительного
и фотосинтетического фосфолирирования мож-
но было бы кратко записать в виде:
0 -
1 -
2 -
3 -
4 -
5 -
6 -
7 -
8 -
9 _
10 _
11 _
12 _
13 _
14 _
оо
>s
S
J
X
ш
ш
Q.
X
о
ф
Т
L.
о
с;
о
S
ю
X
си
о
m
л
о
га
ю
О
I
\
t
I
н2о
л
е.
(147)
Поскольку жизнь — это превращение энер-
гии, например химической в механическую, элек-
трическую или осмотическую, а её поставляет
АТФ, то энергия высокоэнергетической связи
должна перейти в энергию электрона е\
х\ ~ ее
(148)
Рис. 19. Диапазон всего жи-
вого на Земле (выраженный в
электронвольтах) не велик, но
в него укладываются все био-
логические процессы [625]
причем эта реакция обратима и способна обеспе-
чивать системы, производящие ту или иную ра-
боту Р:
eenh~P (149)
Таким образом, можно описать превращение энергии электрона в энер-
гию связи реакции переноса групп, в которых одна высокоэнергетическая связь
обменивается на другие эндергонические связи и выполнение работы.
Реакция переноса заряда идентифицируется и если это а-электрон, то
мы говорим о доноре или акцепторе а-электронов. В п -электронах донором
является неподеленная электронная пара атома азота или кислорода. Вакан-
тная орбиталь в этом случае называется v-акцептором. Перенос заряда осо-
бенно легко протекает между молекулами, обладающими развитой системой
71-электронов и плоской конфигурацией, допускающей максимальное сбли-
жение молекул. В присутствии таких развитых систем 71-электронов перенос
заряда может осуществляться и с одного атома, электронная плотность ко-
Глава 39. О сейфе Творца и моделях высоких порядков
549
торого особенно высока, а дырку затем может занять электрон из общего
фонда 71-электронов.
Одна и та же молекула может быть поочередно или одновременно и до-
нором и акцептором, если она содержит одновременно занятую орбиталь.
Акцептор и донор — это не два различных типа молекул, а лишь различный
характер действия, хотя доминирующими в реакциях данной молекулы мо-
гут быть либо высшая занятая, либо низшая незанятая орбитали, в результа-
те чего молекула будет как хорошим акцептором, так и неплохим донором.
Реакции переноса заряда можно записать в виде образования комплекса с
переносом заряда и диссоциацией:
R + S = RS = R°SE = R° + SE. (150)
И всё же для описания электронных процессов язык классического мо-
делирования недостаточен и неточен. Более точно процесс образования
комплекса с переносом заряда записывается на языке волновой механики
следующим образом [623, 624]:
\j/N - го|/ (RS) + 8\|/ (R° SE) , (151)
где N — означает основное состояние, \|/ — волновую функцию, а коэффи-
циенты а и 8 — долю электрона, принимающую участие в реакции.
Для возбужденного состояния * (слабый перенос заряда) можно записать
уравнение:
\|/Е « 5* (RS) - a* (R° SE) . (152)
Донорно-акцепторные отношения трактуются исследователями по-раз-
ному. Так если для биолога кислота — это Н+—ионы, а основание ОН —
ионы, то для химика кислотой является соединение, способное принимать
электроны, а основанием — способное отдавать их. В этом случае кислотно-
щелочное равновесие не что иное, как перенос заряда. Тогда доноры пред-
ставляются основаниями, а акцепторы — кислотами.
Доноры, акцепторы и электронное давление
Попробуем проанализировать в рамках этих представлений взаимоотноше-
ния в биосистемах. Начнем, казалось бы, с простейших. Так, каждая пептид-
ная связь содержит две электронно-активные группы — СО и NH. Сама
пептидная связь существует в нескольких резонансных формах. Акцептор-
но-донорные свойства этих форм до конца не изучены. Можно ожидать, что
свойства этих форм неодинаковы и поэтому другие доноры и акцепторы
должны оказывать существенное влияние на стабильность изомерных форм
пептидной связи. В то же время сами эти формы должны активно влиять на
акцепторно-донорные свойства. Атом серы может быть не только акцепто-
550
Перспективы биомоделирования
ром или донором электронов, но и катализатором — переносчиком неподе-
ленной пары электронов азота и кислорода к группам SH группы S—S. В бел-
ках S-атом цистеина отделен от полипептидной цепи «насыщенным» угле-
родным атомом (всего четыре связи одинарные). Электроны атома серы изо-
лированы от цепи. Однако SH-группа может присоединяться к пептидной
цепи, образуя тиозольное кольцо, в результате чего электронная структура
SH-группы цистеина сливается с электронной структурой пептидной цепи.
Уже из этих, далеко не оригинальных данных, становится понятно, что
подход к моделированию на основе стерических представлений о природе
белков, а пептидных связей как их статистических элементов, отражает лишь
поверхностные свойства и признаки, а не их сущность. К сожалению, мы
ещё очень мало знаем о квантовых механизмах биологических процессов.
Существующие модели также далеки от аналогий типа изоморфизма, как
изоморфные изображения человека в прямом и кривом зеркале.
Безусловным является лишь то, что донорно-акцепторные взаимоотно-
шения в живой клетке протекают иначе, чем в иных химических системах.
Как установлено, клетка избыточно богата донорами [623].
Основные химические элементы, из которых построена клетка — это С,
N, Н, О и S. Число способов, при помощи которых эти атомы могут быть
соединены, ограничено. Атом азота NH2-rpynnbi аминокислот может отда-
вать электроны своей неподеленной пары хлоранилу. Эти же электроны мо-
гут принимать кислород и рибофлавин. Первичные амины — более сильные
доноры по сравнению со вторичными, и в ряду этиламин—диметиламин-
триметиламин донорная способность убывает. Но в то же время, сс-амино-
кислоты или амиды кислот — более сильные доноры, чем даже этиламин.
У кислорода тоже есть пара неподеленных электронов, и поэтому он может
быть сильным донором [624].
Атом S имеет неподеленную пару электронов и поэтому он является хо-
рошим донором даже в составе дисульфидной группы S—S. Донорной актив-
ности этого атома благоприятствует его способность к конформации. Итак,
в клетке имеется богатый источник лабильных электронов.
Благодаря своей системе л-электронов СО-группа может вести себя и
как акцептор кетоидного 71-электрона. Альдегидный карбонил — более сла-
бый акцептор, чем кетонный; в водном растворе альдегид R—НСО суще-
ствует, главным образом, в виде гидрата. Поскольку СО-группа способна
быть и акцептором я-электронов, и донором 71-электронов, её роль в уста-
новлении донорно-акцепторного равновесия в клетке, вероятно, очень ве-
лика. То же относится и к CS-группе. Таким образом, живая клетка богата
донорами, но бедна акцепторами электронов [625].
Резюмируя изложенное, можно сказать, что энергоресурсом для жизни
служат электроны или, точнее, энергия, полученная ими в процессе фото-
синтеза от фотонов. Источниками лабильных электронов является неподе-
Глава 39. О сейфе Творца и моделях высоких порядков
551
ленные электронные пары азота, кислорода и серы; такими электронами
могут быть и электроны, активированные в процессах обмена. Главным ак-
цептором является карбонил, активность которого может повыситься за счет
увеличения фонда я-электронов.
Процессы жизнедеятельности в большей степени предопределены изме-
нениями донорно-акцепторного равновесия, т. е. определенного соотноше-
ния между высшими заполненными и низшими незаполненными орбиталя-
ми, иными словами — сдвигом в соотношении потенциалов ионизации и
величин электронного сродства. Сдвиг равновесия в пользу доноров означа-
ет увеличение концентрации высокоактивных электронов, своего рода элек-
тронное давление. Электронное сопряжение, которое возникает в результате
перекрывания орбиталей, участвующих в образовании межмолекулярных и
внутримолекулярных связей, должно, очевидно, сообщать подвижность ак-
тивным электронам, что способствует вовлечению воды в процессы белко-
вых преобразований.
Попытки математического описания и моделирования триплет-синглет-
ных связей в живых организмах потерпели фиаско. Биомодели четвертого
порядка скорее относятся к будущему медико-биологической науки, способ-
ной объединить квантовую механику, биологию и новую математику. Пока
же эти построения целесообразно осуществлять в рамках биолого-математи-
ческих описаний и математических моделей третьего порядка и в гибридных
моделях.
При написании книги я долго размышлял, а нужна ли в ней такая глава,
как эта. Взвесив все pro и contra, я решил, что если её прочтут хотя бы три
любящих науку студента, то начнут задавать вопросы своим профессорам.
Ответы их вряд ли удовлетворят. Вот тогда-то студенты превратятся в истин-
ных естествоиспытателей и, без сомнения, попытаются заглянуть в сейф
Творца. Дай им Бог удачи, чтобы дверца чуть приоткрылась, и одной тайной
Природы стало меньше.
552
Глава 4 0 «ПРИНЦИП САМОВАРА»
И НЕЛИНЕЙНЫЕ ПОДОБИЯ
С точки зрения классической математики построе-
ние любых биологических моделей — это процесс
сведения тех или иных явлений или функций к стро-
гим формально-логическим, интегральным, диффе-
ренциальным или вероятностно-статистическим
счислениям. С точки зрения биолога или медика —
это процесс построения принципиально новых кон-
струкций у различных животных и человека.
И немаловажно при выборе модели найти наи-
более простое подобие. Нильс Бор часто повторял,
что если человек не понимает проблемы, он пишет
много формул. В связи с этим мне вспомнилась одна
ситуация из жизни.
В один из промозглых вечеров периода ускоре-
ния и перестройки мы обсуждали, как на нищенс-
кую зарплату инженера привлечь в Волгодонск наи-
более талантливую научную молодежь страны. Мы
- это академик Е.П.Велихов, профессор А.А.Коко-
шин и автор этой книги. И тут Евгений Павлович
сказал, что дело не только в деньгах, надо популя-
ризировать ядерную энергетику. Он говорил, что
многие даже не представляют себе ядерный реак-
тор, который, по его словам, является не чем иным,
как аналогом самовара. Ну, истинный самовар, при-
чем не какой-то электрический, а «бабушкин», на
дровишках. Дровишки он уподобил ядерному топ-
ливу, воду — теплоносителю. Разве что стоит одеть
ещё одну рубашку для второго контура воды. Мо-
дель реактора готова.
Глава 40. Принцип самовара и нелинейные подобия
553
Мне это сравнение показалось поразительным. Ведь атомный реактор
представляет собой невообразимо сложное инженерное сооружение, пик
конструкторской мысли. Процессы расщепления ядра идут в соответствии
со сложнейшими законами природы и описываются нетривиальными ма-
тематическими формулами. И вдруг — самовар! Парадокс? И, да и нет?
В сущности, самовар представляет собой простую, физически осязаемую
модель, иными словами, неполное приближение, возможно, даже мульти-
пликативное подобие реактора. В дальнейшем такое чрезвычайное упро-
щение в наших обсуждениях так и стало звучать: «принцип самовара». Те,
кому по душе обозначение и представление сложных систем и процессов в
простых аналогиях и подобиях, на основании таких «моделей» могут кон-
струировать более сложные, в том числе и биомодели. Моделирование в
медицине и биологии несомненно требует разработки нового математичес-
кого аппарата, сопоставимого со сложностью жизни биологических существ
и их адаптации к окружающей среде. Поскольку сама среда требует реаги-
рования в нелинейных и нестандартных условиях, человек и животные
«запрограммированы» самой природой и Господом Богом к существова-
нию в многомерных, со многими степенями свободы, стохастических си-
стемах.
Это, естественно, определяет условия моделирования, подразумевающие
поиск более сложных подходов и принципов подобия, хотя изначально мы
можем отталкиваться от «принципа самовара».
Неполное подобие и метаморфозы упрощения
Некоторые известные математики считают, что наступивший XXI век будет
веком нелинейной математики, поскольку все линейные процессы изучены,
а многообразие нелинейностей бесконечно. Вряд ли это так, хотя в том, что
касается биомоделирования, они несомненно правы. Действительно, требу-
ются новые взгляды и решения.
В предыдущих главах мы рассмотрели принципы подобия и соответству-
ющего им анализа размерностей к мультипликативным отношениям между
величинами. В этом случае отношение одной величины к другой означает,
во сколько одна из них больше другой.
В медицинской биологии все более активно разрабатываются методы,
позволяющие анализировать неполное, приближенное подобие. Создание
новой, «неклассической» теории подобия позволяет показать, как перемен-
ные, характеризующие элементы сравниваемых систем, преобразуются друг
в друга иным, более сложным образом в биомоделях.
В отличие от обычного, линейного подобия, основанного на мультипли-
кативных отношениях между величинами, предлагается использование не-
линейного подобия биомодели и прототипа.
554
Перспективы биомоделирования
Например, рассматриваются п переменных величин хи, х21, ..., хи1, кото-
рые находятся в функциональных зависимостях с переменными х[2, ..., хп2
Х\2 ~f\2X\VX22 =J22X2V '••» Хп2 ~~ *п2ХпУ U-^)
Заданные функции/j2(l < i < п) представляются как функции перемен-
ных хи, х2х, ..., хп{. На них накладывается ряд ограничений. Они должны
быть непрерывными и дифференцируемыми, однозначными или конечно
многозначными. В этом случае предполагается особое условие о выборе
однозначной ветви соответствующей функции. Еще одним ограничивающим
условием являются непрерывные и дифференцируемые частные производ-
ные. Функции/,2, ...,/и2, удовлетворяющие приведенным условиям, называ-
ются функциями нелинейного преобразования переменных хп, ..., хп1.
Затем формулируются положения, аналогичные обычным положениям
классической теории подобия. При этом, если два явления нелинейно по-
добны, то искомые нелинейно-относительные величины явлений удовлет-
воряют тождественным замкнутым системам уравнений, выраженных в не-
линейно-относительной форме [646].
С проблемой нелинейного моделирования мы столкнулись в начале те-
перь уже далеких 70-х годов XX столетия, когда возник вопрос о необходи-
мости экстраполяции результатов психофармакологических исследований на
разных видах животных и пациентах неврологической и психиатрической
клиник с повреждением лобных долей. Попытки описать процессы в поня-
тиях и терминах мультипликативных, а тем более аддитивных отношений
классического линейного подобия оказались обреченными на неуспех. Уп-
рощения же свелись к таким метаморфозам, к которым вполне применим
афоризм: «Если в задаче одна переменная — это не задача, если больше двух
— она неразрешима».
Невропатологи и нейробиологи справедливо называют лобные доли «не-
мыми» зонами. Их разрушение не дает столь явных неврологических симп-
томов, как это можно видеть в отношении других участков конвекситальнои
коры. Поражения лобных долей не позволяет больным справляться со слож-
ными тестами на интеллект. У них наблюдаются отсутствие мотиваций, твер-
дых намерений и планов, основанных на прогнозировании. В жизни они
ненадежны, грубы, нетактичны, легкомысленны, вспыльчивы. У таких боль-
ных обнаружена тенденция продолжать начатое действие, когда жизненные
ситуации или условия эксперимента требуют иных реакций, а главное реше-
ний [661].
При повреждении лобных долей в поведении млекопитающих, как и у че-
ловека, наблюдается выраженная тенденция к персеверации, т. е. патологи-
ческая настойчивость в выполнении начатого действия, когда требуется уже
совершенно иное. Животные гораздо хуже выполняют задания с отсрочен-
ным подкреплением, хотя легче справляются с задачей, если в период экс-
Глава 40. «Принцип самовара» и нелинейные подобия
555
перимента их держать в темноте. Повышенная отвлекаемость животных,
подвергнутых лобэктомии, подтверждается их гиперактивностью и гиперре-
активностью.
Мы апплицировали на прореальные извилины кошек и кроликов хрони-
чески вживленные охлаждающие термосистемы. Интересно, что локальное
охлаждение до 20° С вызывало те же эффекты, что и удаление лобных долей.
Восстановление температуры приводило к возвращению всех утраченных
функций, в том числе условных рефлексов.
Диазепам в дозе 0,5—1 мг/кг и другие седативные средства аналогично
темноте улучшают результаты выполнения заданий у лобэктомированных
кошек. Животные, перенесшие префронтальную лобэктомию, особенно
плохо справляются с тестами, в которых им предъявляют много стимулов
или вариантов выбора. Аналогичные изменения мы наблюдали и у испыту-
емых людей с разрушениями лобных долей.
Таким образом, префронтальная область коры человека и прореальная
извилина мозга животных играют главную роль в выработке стратегии пове-
дения. Нарушение этой функции приводит к задержке или исключению
координации между постановкой задачи и началом ее решения. В этот пе-
риод успевают накопиться раздражители, требующие правильного включе-
ния в целостную поведенческую реакцию [13, 22, 395]. При биомоделирова-
нии таких сложных ситуаций трудно обойтись без использования принци-
пов нелинейного подобия, и никакие упрощения не облегчают решение
задачи.
Переход от обычного подобия к нелинейному представляет собой суще-
ственный шаг в развитии математического аппарата. Это развитие вполне
естественно, потому что нет оснований для предпочтения мультипликатив-
ных отношений всем остальным.
Органы чувств человека являют тождественные мультипликативные от-
ношения, например, мы воспринимаем как одну и ту же форму мультипли-
кативно подобных фигур. Если же фигуры будут нелинейно подобны друг
другу, то их форма будет восприниматься как разная. Но нет никакой уве-
ренности в том, что это имеет место у всех живых существ [466]. Если чело-
век различает вариации геометрических фигур как разные, то для многих
животных важен общий облик, а не разные формы начертания многоуголь-
ника — это ещё одна метаморфоза упрощения.
Как ни странно «сила» или «мощь» упрощения, как его именуют сами
авторы, становится зачастую самодовлеющей и даже философской катего-
рией. С важностью великих они доказывают, что это необходимо в просве-
тительных и педагогических целях, а вот на следующем этапе... Но такой
будущий этап не только не нужен, но и вреден, поскольку создает, простите
за выражение, только заморочку. Поэтому следует иметь в виду, что такие
популистские упрощения не имеют ничего общего с понятиями приближен-
556
Перспективы биомоделирования
ного или неполного подобия и создают нереальные метаморфозы в биоло-
гическом моделировании.
Проблема измерения и безразмерности
Мы должны отдавать себе отчет в том, что при построении моделей сложных
психических процессов лукавство является почти обязательным элементом.
По мере развития математики и техники мозг эволюционировал в умах уче-
ных от аппарата Морзе, релейной станции, компьютера до некоей гологра-
фической конструкции по Прибраму. Эти «модели» являются временной
уступкой обстоятельствам на определенном уровне развития науки.
При построении модели на основе нелинейной теории подобия необхо-
димо иметь в виду трудность, связанную с проблемой измерения. Обычно
измерение связано с определением мультипликативного отношения между
измеряемой величиной и некоторой другой, принятой за единицу измере-
ния, на которой строится теория подобия.
В биологии и медицине между мультипликативным и аддитивным физи-
ческим подобием имеется существенное различие. В одном случае рассмат-
ривается совокупность разнородных величин, характеризующих один и тот
же объект. Тогда константы подобия различны для разных величин и требу-
ется уравнение связи для переноса отношений с модели на оригинал. Сам по
себе этот факт не вытекает из идеи биологического подобия. В самом деле,
если нам известно, что один объект подобен другому, то мы можем перено-
сить соотношения между размерами одного на другой, не интересуясь урав-
нением. В другом случае, если сравниваются однотипные величины, напри-
мер размеры, отношения будут иметь безразмерный характер, и мы можем
их корректно математически описать.
Аддитивное биологическое подобие ближе к пространственному. Посколь-
ку соотношения подобия аддитивны, имеет смысл сопоставлять в сравнива-
емых системах также аддитивные, а не мультипликативные соотношения.
Тогда величины хх', ..., хп' и соответственно х", ..., хй" нужно складывать и
вычитать, а не множить и делить. Это означает, что они должны быть од-
нотипными. Бессмысленно складывать и вычитать, например, длину и мас-
су. В то время как в обычной теории подобия модель, как и прототип, пред-
ставляет собой группы свойств одного объекта, в аддитивной теории, наобо-
рот, модель и образец должны представлять одно свойство, распределенное
на множество различных объектов. Например, измеряя артериальное и ве-
нозное давление, скорость пульсовой волны и ДР между ними у разных
животных и человека, мы имеем дело с равными значениями одной величи-
ны, характеризующей многие биообъекты.
Однородность сопоставляемых величин дает возможность делать выво-
ды от модели к человеку без анализа уравнений, связывающих все харак-
Глава 40. «Принцип самовара» и нелинейные подобия
557
теризующие их величины. Очевидно, что в случае аддитивного подобия
речь идет о переносе аддитивных соотношений. Эти соотношения должны
быть аддитивно безразмерными и при делении друг на друга давать величи-
ны одинаковой размерности. Мультипликативное отношение, не завися-
щее от абсолютных значений сравниваемых величин, определит нелиней-
ное подобие [646].
Можно определить аддитивное отношение, если из одной величины
вычесть другую. Полученная разность будет выражать отношения при самых
различных значениях соотносящихся величин. Если мы из одной длины
вычтем другую, то получим величину той же размерности, но связанную с
мультипликативным отношением к единице измерения. В качестве такой
единицы можно взять одну из соотносящихся величин. В аддитивном случае
роль единицы играет нуль. В качестве нуля берется значение одной из срав-
ниваемых величин. Полученная разность показывает, на сколько другая ве-
личина превышает этот нуль. Давайте вернёмся в гл. 39 и вспомним гениаль-
ного Н.Лобачевского, который, декларируя «1 = 2», выводил следствия из
разряда «всякое число равно нулю». Маститые ученые считали это чудаче-
ством, а автора определили в безграмотные шутники. Поистине, никогда
нельзя торопиться с преждевременными заключениями. Стоит посоветовать-
ся с Историей, она достаточно беспристрастная Дама.
Если все константы нелинейного аддитивного подобия одинаковы, то любое
аддитивно безразмерное отношение, найденное в модели, можно переносить
на прототип.
Совокупность всех аддитивно безразмерных отношений между отдель-
ными величинами, характеризующими модель и образец, можно рассматри-
вать как одно отношение — R1. Если константы подобия сх, ..., сп различны
для разных величин, то отношения между отдельно взятыми величинами jcj',
..., х ' и соответственно х", ..., хп" не будут одинаковыми. Но при условии
тождественности формы уравнений, связывающихх/,..., хп' и соответствен-
но х", ..., дс ", можно приравнивать друг к другу соответствующие отноше-
ния отдельных элементов по всем остальным, вместе взятым. Поэтому тож-
дественность уравнений означает вместе с тем тождественность отношений,
понимаемых в указанном смысле.
Наличие констант подобия и тождественность уравнений связи необходи-
мо совместить для того, чтобы явления могли считаться подобными и рав-
няться единице. Приведем рассуждение, соответствующее тому, которое име-
ет место в обычной теории подобия, но уже для случая нелинейного подобия.
Пусть одна из сравниваемых систем описывается уравнением
Ф/(х/, ...,х/) = 0. (154)
Для второй будет верно
срДх,", ..., */') = 0- (155)
558
Перспективы биомоделирования
Подставляя в (157) равенство (156), получим
ф[(с, + *,'), .... (сп + */)]. (156)
Параллельное существование уравнений (155) и (156) будет возможно
только в том случае, когда все константы аддитивного подобия выйдут из-
под знака функций ф,. в виде одного
Ф/ [(с, + V),...,(c„ + */)] = ^(с,, ..., сп) + уДх,', ..., */) = 0. (157)
При условии у{(с{, ..., сп) = 0 уравнения ф,= 0 будут инвариантными по
отношению к произведенному преобразованию.
Функции ф/? обладающие указанным свойством, и отвечающие им урав-
нения можно назвать аддитивно гомогенными [646].
В теории мультипликативности свойством гомогенности обладают фун-
кции типа степенных комплексов XjOti, ..., хпап, в которых степени ар ..., ап
— отвлеченные числа. В аддитивном случае аналогичную роль будут играть
функции типа а{х{ + ... + ос^, где ар ..., ап — отвлеченные числа. Если
ос^/ + ... + ос^х/ = 0 и otjXj" + ... + сс^" = 0, то, пользуясь соотношениями
подобия, получим
(аЛ+...+ а^п) + (а1х1'+...+ а„хп") = 0. (158)
Такое равенство (158) можно записать в виде
(а^ +...+ апсп) + (<х1х1'+...+ a„x/') = 0. (159)
Отсюда очевидно, что условием инвариантности уравнения по отноше-
нию к аддитивным преобразованиям подобия будет следующее соотноше-
ние: (XjC, +...+ апсп = 0. Левую часть этого уравнения можно назвать подоби-
ем аддитивности.
В этом случае все члены уравнения соединены знаком плюс. Очевидно,
что некоторые из членов должны иметь отрицательное значение. Ноль пра-
вой части означает, что сумма положительных членов равна сумме отрица-
тельных: Еос^. — Еос^ = 0.
Вместо ct и ck подставим их значения из уравнений (154) и получим
ЕосДх," - х!) = Ъа.к{хк" - хк'). (160)
Эти соотношения преобразуем так, чтобы каждая часть уравнения содер-
жала величины, относящиеся к одной системе:
Yapc" - lafa" = Yap! - lafa'. (161)
Безусловно, аддитивно безразмерные комбинации величин можно рас-
сматривать в качестве аналогов критериев подобия. Это уже аддитивные кри-
терии нелинейного подобия. Как и в обычном случае, такие критерии оди-
наковы у обеих сравниваемых систем и могут стать базой для расчетов в
Глава 40. «Принцип самовара» и нелинейные подобия
559
реальных условиях биологического моделирования и экстраполяции в наи-
более трудных для трактовки случаях переноса, например в построении
моделей психофармакологического воздействия на мнестические и сложные
психические функции, но статистический принцип организации нейрональ-
ных механизмов мозга обусловливает сложность извлечения информации из
интегральных электрограмм. Это требует привлечения современных методов
анализа случайных процессов и использования компьютеров.
Модели и модули в фармакопсихологии
Теперь, как и обещали, вернёмся к рассмотрению лобных долей и попыток
анализа их функций в плане моделирования, основанного на принципах не-
линейного подобия. Покажем это на примере из наших исследований.
Известно, что электрография мозга (ЭГ) является одним из широко ис-
пользуемых физиологами и клиницистами методических подходов, позво-
ляющих выявить некоторые моменты функционального состояния или па-
тологии мозга. Нами на моделях психоподобных нарушений у животных,
а также в условиях клинического исследования ЭЭГ у больных с психичес-
кими нарушениями отрабатывались принципы анализа электрокортикограмм
(ЭКоГ) и электросубкортикограмм (см. также гл. 23).
Мы начали эти работы со студентом факультета прикладной математики
ТРТИ, а ныне профессором В.П.Омельченко. Было установлено, что элек-
трограммы мозга являются аддитивным процессом, включающим в себя как
квазипериодические, так и стохастические компоненты. Для исследования
деятельности мозга по ЭГ нами был использован математический аппарат
анализа временных рядов, который был представлен в виде комплексной
компьютерной программы. В биологическом аспекте задача формулирова-
лась следующим образом:
/ выяснение нейрохимической организации системной деятельности
мозга и определение роли нейромедиаторов в генезе различных час-
тотных составляющих электрограмм;
/ выявление изменений многомерных вероятностно-статистических ха-
рактеристик ЭГ мозга животных и психически здоровых людей в усло-
виях действия психоактивных агентов или физических факторов;
/ установление различий реагирования здоровых и больных людей (по
ЭГ) на введение психоактивных веществ;
/ построение стохастических и нелинейных моделей, отображающих па-
тологические изменения в ритмической деятельности мозга человека
и животных до и после воздействия психотропных факторов и веществ.
Решение данных задач было необходимо для получения банка эталонов
электрограмм мозга, характеризующих определенные психические наруше-
ния человека. Второй целью, преследуемой нами, являлось нахождение об-
560
Перспективы биомоделирования
щих признаков изменения ЭЭГ мозга человека и ЭГ животных, необходи-
мых для последующего скрининг-анализа при изыскании новых психоак-
тивных агентов с целью фармакопсихологического моделирования.
В дальнейшем эти исследования расширились, и новая работа, но на
этих же принциах, была выполнена как на животных (мини-свиньи, соба-
ки, кролики и кошки) с хронически имплантированными электродами кор-
ковые и субкортикальные образования, так и на людях с экстракорпораль-
ной регистрацией биопотенциалов. Запись ЭГ мозговых структур живот-
ных и ЭЭГ человека осуществлялась с помощью компьютерных электроэн-
цефалографов.
Касаясь принципов анализа, контроля и прогноза ЭГ и ЭКоГ, следует
отметить, что они являются отражением вероятностных процессов деятель-
ности мозга, т. е. будущие изменения электрограммы не могут быть заранее
точно предсказаны. Анализ электрограмм мозга как стохастического про-
цесса осуществлялся по комплексной программе, в которую входили следу-
ющие этапы:
/ проверка реализации на стационарность и выделение стационарных
участков;
/ вычисление автокорреляционных функций и установление интервала
корреляции;
/ определение плотности распределения мгновенных значений ЭГ и
ЭКоГ и проверка полученных распределений на нормальность, нахож-
дение основных моментов распределения;
/ получение автоспектральных характеристик;
/ вычисление взаимной корреляционной функции;
/ определение функций когерентности и фазовых характеристик;
/ определение частотных характеристик;
/ анализ определенного вида нестационарных и переходных процессов.
Наиболее разработанной областью теории случайных функций являются
стационарные случайные процессы. Часто при анализе ЭГ делается допуще-
ние, что процесс стационарен и удовлетворяет условию эргодичности. Одна-
ко, как показывает практика, необходимы дополнительные исследования для
установления стационарности процесса. Наименьшая длина реализации,
необходимая для спектрального анализа ЭГ, выбирается согласно следую-
щей зависимости:
Т=\0//н, (162)
где^ — наименьшая частота, присутствующая в электрограмме.
Нами использовался следующий подход к исследованию стационарности
ЭГ. Если принять, что отдельные участки мозга (а именно: сравнительно
небольшие констелляции нейронов, ядра, циклические организованные си-
стемы типа лимбической, стриопалидума и т. п.) являются регуляторами,
Глава 40. «Принцип самовара» и нелинейные подобия
561
обеспечивающими наиболее оптимальный режим функционирования, а био-
электрическая активность этих мозговых образований есть прямое отображе-
ние их регулирующих функций, то, очевидно, стационарность ЭГ будет ото-
бражать работу регулятора в привычном режиме, когда возмущающие воз-
действия не превышают допустимых пределов.
В случае возникновения закритических возмущений регулятор перево-
дит систему организма на новый режим работы, т. е. возникает переходной
процесс, параметры которого зависят как от особенностей самой системы и
ее регулятора, так и от параметров возмущающего воздействия. Чем быстрее
система перейдет к стационарному состоянию (при одинаковых возмущаю-
щих воздействиях), тем выше ее устойчивость. Исходя из этих соображений
можно произвести классификацию ЭГ мозга и в соответствии с характером
этого процесса использовать адекватные методы его анализа. Акцентируем
внимание на той части настоящей работы, которая посвящена анализу ста-
ционарных участков ЭГ и созданию алгоритмов оценки спектральной плот-
ности нестационарных процессов определенного вида, удобных для нелиней-
ного моделирования.
Исходя из допущения, что различным установившимся состояниям че-
ловека и животных соответствуют стационарные участки ЭГ мозга, нами
применялся метод выбора и комплектования стационарных участков задан-
ной длины.
Отбор стационарных участков ЭГ мозга для одно- и многомерного кор-
реляционного и спектрального анализа проводился нами в два этапа. На
первом этапе в результате экспресс-анализа отбрасывались участки с явно
выраженными артефактами, мышечными наводками, переходными процес-
сами и т. д. Второй этап предусматривал компьютерную проверку отобран-
ных реализаций на внутреннюю стационарность в широком смысле. Извест-
ны несколько показателей стационарности процесса, основанных на изме-
нении математического ожидания (или математического ожидания и дис-
персии) и корреляционных функций. Нами исследовались изменения мате-
матического ожидания, дисперсии и автокорреляционных функций (АКФ)
односекундных сегментов на протяжении 10—15 секунд. Индексы стацио-
нарности определялись по следующим формулам:
£м,. (163)
562
Перспективы биомоделирования
\ k-\ \ k \ \т .
h^ij1- х -^тъю-т? , (165)
k-lMk-j i=j+iM\x=i
где 1м — индекс стационарности по математическому ожиданию; ID — ин-
декс стационарности по дисперсии; IR — индекс стационарности по АКФ;
М — среднее значение процесса на i-u участке; Dt — значение дисперсии на
/-м участке; к — число участков; R{i) —значение АКФ на шаге т; М — мак-
симальное число запаздывания АКФ.
Выделение сегментов, отличающихся по АКФ, производилось по формуле
1 к-\ 1 \М
k-\i=\ M yT=i
Л2
(166)
Кроме того, применялись критерии серий и критерии тренда для выяснения
независимости выборочных оценок и выявления монотонных колебатель-
ных трендов.
Если полученные критерии и индикаторы стационарности реализации
превышали допустимые пределы, ожидаемые за счет обычной выборочной
статистической изменчивости, то применялась цифровая фильтрация иссле-
дуемых временных рядов на основе быстрого преобразования Фурье. Циф-
ровые фильтры снижали мощность частот, лежащих до 1—2 Гц, что приводи-
ло к значительному улучшению показателей стационарности.
В результате экспериментов было установлено, что изменения индикато-
ров стационарности зависят от функциональной организации конкретной
мозговой структуры, состояния организма, но особенно, от воздействия раз-
личных психотропных средств или физических факторов.
Выделенные стационарные реализации служили для определения авто-
корреляционных и спектральных функций. Кроме самостоятельной харак-
теристики ЭГ мозга, указывающей на связь между отсчетами, отстоящими
на время т, АКФ служит для установления интервала корреляции.
Определение некоррелированных отсчетов необходимо для проведения
амплитудного анализа. Как известно, определению плотностей распределе-
ния мешает рисунок мгновенных значений амплитуд вероятностных про-
цессов необходимо использовать независимые отсчеты. Однако в некоторых
случаях достаточно условия некоррелированности рядом стоящих отсчетов,
что и определяется по интервалу корреляции.
Вероятность попадания мгновенных значений амплитуд ЭГ в коридор
[X; X + Щ определялась согласно формуле:
Pl ~ w ~Ш- <167>
где xt — значение стационарного случайного процесса; W — ширина диффе-
Глава 40. «Принцип самовара» и нелинейные подобия
563
КРОЛИК
КОШКА
СОБАКА
МИНИ-СВИНЬЯ
ЧЕЛОВЕК
Фон
0,2 -
0.1 -
0,05 -
0,02 -
0,01 -
10 20 30 40
i \gW(J)
II
1111
5,0-
4,0-
3,0-
2.0-
1,0-
0 5 10
5.0-
4,0-
3,0-
2,0-
1,0-
0 5 10
Н2(/)
Hz
0.2-
0,1-
0,05-
0,02-
0,01-
10 20 30 40
il
li
ll
м л
т 'ч
'1л^ х-^-
1 1 Г "i—Y-
5,0 ■
4.0 -
3.0 -
2,0 -
1,0 -
0 5 10 15
Н2(/)
/fl
Л
Hllli
ill
5,0-
4,0-
3,0-
2,0-
1,0-
0 5 10 15
H2W
Щф
Hz
— Анафранил (доза для человека 25 мп, для животных 1 -3 мг/кг)
Рисполепт (доза для человека 2-3 мг, для животных 0,5 мг/кг)
Рис. 20. Биологическое моделирование по показателям спектральной плотности мощности (С)
электрограмм лобных долей мозга человека и животных и переходных процессов на основании
расчета модулей частотных характеристик под влиянием антидепрессанта анафранила (А) и
нейролептика рисполепта (Р). МЧХ выявляют и дают четкие геометрические отображения срав-
нительной эффективности действия различных психотропных средств и общности их проявле-
ния в электрограммах животных, стоящих на разных уровнях филогенеза
ренциального окна; nt — число отсчетов, когда выполняется событие
xt<xt<x+ W\ N — общее число некоррелированных отсчетов; / — номер
канала.
564
Перспективы биомоделирования
Для характеристики формы кривой распределения определялись цент-
ральные моменты: математическое ожидание, дисперсия, асимметрия. Кро-
ме того, нами проводилось исследование с целью проверки кривой распре-
деления на нормальность. Применение критерия Пирсона показало, что
кривые ЭГ мозга не удовлетворяют нормальному распределению. В резуль-
тате экспериментов были установлены значимые различия в кривых плотно-
стей распределения и их характеристик при действии различных психофар-
макологических агентов [329].
Определение сглаженной выборочной оценки нормированной спектраль-
ной плотности производилось по формуле Дженкинса и Ваттса:
G(f) = 2h
М-1 (wtk\
1 + 2 X D(t)*(t)coJ^
т=1 \М J.
(168)
где к = 0, 1, ..., М\ Дт) — весовая сглаживающая функция Тьюки, которая
определяется как
Дх) =
1 + cos— |;т = 0,1,2,...,М
V М) (169)
1/2
0; %>М
Для полученных значений 80%-ный доверительный интервал равен
0,85G(/); \,\SG(f), т. е. получается 15%-ная относительная ошибка.
Для анализа реализаций, не отвечающих требованиям стационарности,
применялся метод определения нормированных спектральных оценок 1- и
2-секундных сегментов, трехмерные графики которых отображали динамику
спектральных составляющих во времени. Выборочная оценка спектральной
функции всей реализации определялась по формуле
G{f) = \iG]{f), (170)
где Gf.(/) — спектральная плотность i-ro сегмента.
Для определения изменения связей между различными мозговыми обра-
зованиями производился многомерный спектрально-амплитудный ана-
лиз. В результате анализа определялись кросскорреляционные функции,
квадрат спектра когерентности и фазовые характеристики между попарны-
ми отведениями.
С Помощью компьютера производилось определение текущих оценок ЭГ
мозга, а также введение управляемого эксперимента с выдачей стимулирую-
щих воздействий при достижении исследуемым сигналом пороговых значе-
ний. Были получены текущие значения математического ожидания, диспер-
сии, асимметрии и эксцесса, кроме того, определялись значения ковариаци-
Глава 40. «Принцип самовара» и нелинейные подобия
565
онных и корреляционных коэффициентов за короткие промежутки времени
и информативность оценок.
Для выяснения некоторых нейрохимических механизмов системной орга-
низации мозга нами производилась аппликация 70—250 мкг ацетилхолина,
норадреналина, серотонина или имипрамина, рогипнола и аминазина на про-
реальную и переднюю сигмовидную извилины. При аппликации нейролеп-
тиков и антидепрессантов сохранялись характерные черты их системного
действия не только в самой коре, но и в сопряженных образованиях.
Регистрация электрограмм хвостатого ядра и гипоталамуса и их последу-
ющий спектральный анализ позволили выяснить, что аппликация имипра-
мина на прореальную извилину вызывала перераспределение мощностей
между 0,5 и 1 Гц в сторону возрастания первой составляющей до 0,45, т. е. в
2 раза, в хвостатом ядре и относительное возрастание 1 и 2 Гц (до 0,25 и 0,24)
в заднем гипоталамусе. При аппликации на прореальную извилину рогип-
нола значительно уменьшалась мощность 1—4-герцевых составляющих (0,19—
0,04), но имел место выброс в области 6 Гц (0,1) в хвостатом ядре и наблю-
далось увеличение мощности компонента в 2 Гц (0,12) в заднегипоталами-
ческом ядре. Следует также подчеркнуть, что аппликация галоперидола и
рогипнола на прореальную извилину приводила к резкому снижению мощ-
ностей высокочастотных составляющих (10—60 Гц) в хвостатом ядре. Эти
данные можно расценивать как дополнительное подтверждение участия нео-
кортекса в пространственной синхронизации биопотенциалов не только
корковых формаций, но и определенных отделов субкортекса.
Знание спектральных мощностей ЭГ мозговых структур до и после вве-
дения фармакологических агентов позволяет определить абсолютное значе-
ние частотной характеристики воздействия вещества. Поскольку нами были
установлены изменения частотной структуры важнейших мозговых образо-
ваний при введении либераторов или прекурсоров эндогенных моноаминов
и психотропных средств, мы имели возможность приступить к построению
частотных моделей эффектов исследуемых веществ, которые строились на
основании следующего соотношения:
Н =W)' (171)
где Ifiif) — модуль частотной характеристики (МЧХ) определенной мозговой
структуры под влиянием изучаемого вещества; G, — спектральная плотность
ЭГ или ЭКоГ интактной структуры; G*B — спектральная плотность ЭГ или
ЭКоГ структуры, находящейся под воздействием вещества;/= 0,5; 1; 2;...; 60 Гц.
Мы ввели новое математическое понятие tPij), обозначив его как МЧХ
или модуль частотных характеристик по ЭЭГ, ЭКоГ или ЭГ. По своей сути
он отражает модельные элементы переходных процессов мозговых структур.
Иными словами, это нормированная функция, которая как бы «очищает»
566
Перспективы биомоделирования
все иные показатели, кроме тех, которые характеризуют эффект психотроп-
ного препарата. Это «чистый портрет» препарата уже не в психофармаколо-
гическом, а в истинном фармакопсихологтеском проявлении.
Будь это линейная модель, мы бы вычли из исходных значений спектра
ЭЭГ его же значение после введения препарата и могли бы интерпретиро-
вать в терминах «лучше-хуже», «больше-меньше». Мы же работаем в области
моделирования на основе нелинейного подобия и используем адекватный
математический аппарат, а значит, как бы отбрасываем такие понятия, как
«механизм», «эффект» психотропных средств. Мы получаем очищенные элек-
трографические эквиваленты изменений психики, происходящих под или
вне влияния внешних факторов.
Изменения МЧХ могут служить хорошим дифференциальным показате-
лем действия новых и классификации уже известных нейротропных средств.
Так, рисполепт, в противоположность анафранилу, подавляет большую часть
высокочастотного спектра, что выражается в уменьшении значений МЧХ в
диапазоне 14—60 Гц (рис. 20).
Несмотря на высокую информативность и динамичность мгновенных
значений спектров (рис. 20) видовые различия прослеживаются недостаточ-
но наглядно. Так, у грызунов различия в спектральных характеристиках не-
велики. Большее приближение к спектральным характеристикам человека
наблюдается у хищников (кошка, собака) и мини-свиней. Антидепрессант
анафранил существенно увеличивает мощность спектральных характерис-
тик в диапазоне 8—11 Гц. У человека этот пик всегда приходится на 9 Гц-
составляющую. Эти изменения могут служить прогностическим признаком
при оценке антидепрессантов и поисках новых средств антидепрессивного
действия, поскольку ранее [327, 328] мы обнаруживали и у других антидеп-
рессантов, таких, как имипрамин и амитриптилин. У грызунов и мини-сви-
ней, как и у человека, антидепрессанты повышают мощность спектра в вы-
сокочастотном диапазоне, вплоть до 35 Гц.
Нейролептик рисполепт заметно смещает значение мощности спектра в
диапазон, лежащий ниже 9 Гц, с доминирующим ритмом 2—4 Гц (рис. 20С).
Мы хотели бы еще раз подчеркнуть, что эти изменения регистрировались
в прореальных извилинах животных и соответственно в лобных долях челове-
ка, которые можно определить в качестве информативных зон мозга для оценки
психотропных средств. При поиске информативных параметров мы исходили
из принципов теории и подобия для нелинейных систем [(см. уравнения (156-
163)]. Полученные значения МЧХ оказались достаточно информативными и
наглядными как в сравнительном аспекте (кролик, кошка, собака, мини-сви-
нья, человек), так и для разграничения эффектов антидепрессанта анафрани-
ла (рис. 20А) и нейролептика рисполепта (рис.20Р).
МЧХ для анафранила дают весьма характерную геометрическую картину
нелинейного подобия. Значения fPif) существенно снижались в диапазоне
Глава 40. «Принцип самовара» и нелинейные подобия
567
1-5 Гц и возрастали в диапазоне 10-15 Гц. Столь же наглядные изменения
прослеживались в значениях FP-if) при действии рисполепта. Они характери-
зовались возрастанием значений Ff-if) в диапазоне 1—4 Гц и снижением в
диапазоне 6—9 Гц.
Мы полагаем, что по значениям МЧХ можно не только разграничивать
эффекты психотропных средств в отношении животных разного филогене-
тического уровня, но и осуществлять экстраполяции с максимальным при-
ближением в отношении человека. Кроме того, это весьма удобная модель
для поиска перспективных психотропных средств и выявления их психофи-
зиологических эффектов.
С целью выявления изменений в спектральных характеристиках ЭГ при
психических нарушениях, а также при психофармакологических воздействиях
на здоровых и больных людей нами исследовались психически здоровые
добровольцы и больные с лобными повреждениями.
Следует отметить, что у здоровых людей с визуально обнаруживаемым
(при прикрывании глаз) в затылочных долях ос-ритмом спектральный анализ
ЭГГ позволил нам выявить отчетливое доминирование 11—12-герцевых со-
ставляющих во фронтальных, лобных сочетательных, затылочных областях,
зонах Брока и Вернике. Фенамин (10 мг), анафранил (25 мг) и рисполепт
(2 мг) не устраняли доминирования этих составляющих. У больных с апати-
ко-абулическими синдромами, связанными с повреждениями лобных долей
мозга, анафранил вызывал расширение эффективной полосы частот с 0,5—
18 Гц до 1—26 Гц. Но наряду с относительно равномерным распределением
мощностей по всей эффективной спектральной полосе анафранил значимо
уменьшал мощность 4—5-герцевых составляющих, в исходном состоянии яв-
ляющихся доминирующими. Аналогичное уменьшение мощности 4—5 Гц мы
отмечали и у здоровых людей. Рисполепт чрезвычайно резко суживал эф-
фективную полосу частот (с 1—9 Гц до 1—11 Гц) с отчетливым всплеском
мощности 3-герцевой составляющей.
Не вдаваясь в подробное описание реагирования здоровых и больных людей
на введение средств, мы лишь подчеркнем существенную общность измене-
ний спектральных составляющих электрограмм мозга животных (мини-сви-
ней, собак, кроликов и особенно кошек), с одной стороны, и человека — с
другой, при действии психоактивных агентов. Нами были обнаружены опре-
деленные закономерности в реагировании как здоровых, так и больных лю-
дей на введение психотропных веществ, что в свою очередь дает возможность
использовать эти средства в качестве диагностических фармакологических
индикаторов или оценки физических факторов [327, 349].
Вне всякого сомнения, процессы моделирования психики являются слож-
нейшими процессами, а исследования действия психотропных препаратов
попросту невозможно представить в виде какого-то подобия или аналогии.
Например, нельзя измерить и оценить улучшения настроения после приме-
568
Перспективы биомоделирования
нения антидепрессантов. Поэтому мы прибегаем либо к описательным мо-
делям, либо используем простенькие альтернативы. Один из авторов экспе-
риментальной биомодели для оценки эффективности антидепрессантов
А. Винтер, предложивший метод взбегания мыши по скользящей веревочке,
реально оценивая свою идею, сказал: «Мышка может сколько угодно бегать
по этой веревочке, но никогда не накинет её на шею». В этом высказывании
заключен печальный результат завершения депрессии в виде суицида.
Мы не случайно рассматриваем вопросы моделирования психических
функций и роли в них лобных долей в заключительной главе, ибо это и есть
самый сложный аспект биологического моделирования, требующий исполь-
зования нетривиальных биологических, психофармакологических и матема-
тических подходов. К сожалению, на сегодняшний день не существует еди-
ной концепции и единого взгляда на моделирование психики. Нет даже
согласия в том, думают ли животные и как соотнести психические процессы
у них в отношении человека.
Различия в подходе к построению таких моделей и взглядов на эти про-
цессы хорошо отражены в следующей шутке. Однажды математик спросил у
биолога: «Перед Вами пустой самовар без топлива и воды. Как вскипятить
воду?» Биолог ответил: «Наполнить самовар водой, наполнить топливом и
зажечь его». Математик согласился с этой версией задачи, но усложнил ус-
ловие: «А теперь решите вторую задачу: самовар заполнен водой и дровами.
Как вскипятит воду?» «Это ещё проще, — ответил биолог, — надо разжечь
самовар». Такой ответ привел математика в крайнее возбуждение: «Ничего
подобного! Надо не разжигать самовар, а вылить из него воду извлечь дрова
и мы придем к первой задаче, которую уже умеем решать]» Поэтому при
моделировании, в тех случаях, когда задачу сводят к уже решенной, шутя
говорят, что применяют «принцип самовара», т. е. предельно её упрощают.
Резюмируя вышеизложенное, можно сказать, что применение комплекс-
ных методов оценки позволяет как экстенсифицировать, так и интенсифи-
цировать экспериментальные и клинические нейрофизиологические иссле-
дования, а основываясь на принципах подобия, можно выстроить оптималь-
ную биомодель изучения патологии человека. В этих исследованиях «прин-
цип самовара» является неподходящим методом. В этом случае более опти-
мальным является использование итерационного моделирования, а не све-
дение модели к решаемым или уже решенным уравнениям.
Конечно, это не простые подходы к биомоделированию. Это, скорее даже,
— гибридное моделирование. Именно оно дает блестящий результат на со-
временном уровне знаний по биомоделированию высших функций челове-
ка, да ещё и в самой пикантной области — в лобных долях головного мозга,
интегративных центрах формирования психики.
569
Заключение
Величайший драматург, Природа создала суперсце-
нарий самой увлекательной пьесы, которая называ-
ется «жизнь». В соответствии с законами жанра, в
ней все было дуально разделено на хорошее и пло-
хое, черное и белое, доброе и злое. Однако в этот
сценарий, по-видимому, кто-то проник, все, сме-
шав и разметав. По сути, это не мешало развитию
пьесы, но что-то в ней стало непонятно, что-то не
сходилось. Вот тогда-то появились исследователи-
драматурги и экспериментаторы-сценаристы. Одна-
ко Природа слишком велика и необъятна, чтобы
уложить весь сценарий в одну пьесу и потребова-
лись понятия дедукции, аналогии, подобия и, на-
конец, моделирования, которые создавали интригу
процесса и кульминацию его развития.
Принцип дуальности присутствует в моделиро-
вании, как и во всех иных областях, знаний. Здесь
также сталкиваются структура и функция в их пред-
ставлении, а в целом — единство и противоречие
недостижимого идеала оригинала и реальности и
ограниченности модели. Биомоделирование сродни
иконописи, поскольку мы хотим на вполне матери-
альном холсте, вполне осязаемой кистью и ограни-
ченным спектром красок нарисовать то, что мы
никогда не видели, не слышали, не осязали. Мы
моделируем не только то, что ощущаем, а стремим-
ся материализовать наши абстрактные понятия и
представления. Всегда хочется построить зримую
модель того, что никогда не удается увидеть, на-
пример, атомные или молекулярные взаимоотноше-
ния, внутри- или межклеточные процессы.
570
Заключение
Мозг, по крайней мере человеческий, так устроен, что способен проник-
нуть в суть вещей и смоделировать их математическими, физическими, хи-
мическими или биологическими методами. Сколь совершенной ни была бы
модель, как бы мы не пытались отразить в ней динамику, в конечном итоге
это будет лишь квазистатическое отображение гипердинамических процес-
сов Природы. Поэтому, представив на суд читателей свою книгу, в которой
попытались дать новые подходы и взгляды на биомоделирование, мы отдаем
себе отчет в тщетности объять необъятное.
Жесткие и бескомпромиссные споры сторонников альтернативного мо-
делирования и биомоделирования на лабораторных животных представля-
ются мне беспредметными. Крестовые походы за или против использования
животных-млекопитающих в эксперименте должны рассматриваться как
попытки к реанимации мракобесия в науке, когда приверженцы или про-
тивники того или иного учения в лучшем случае подвергались остракизму, а
в худшем — попадали в ГУЛАГ. И призывы к «гуманизму» здесь не при
чем — надо быть гуманным в сути своей, а не по отношению только к хо-
мячкам или приматам. Инфузории или гуппи — тоже живые организмы.
Поэтому деление на модели разных порядков — весьма относительная и
далеко не идеальная форма познания. Формы моделирования должны не
нумероваться, а быть адекватными, оптимальными и максимально анало-
гичными прототипу. Это мы и хотели донести до читателя.
Автор благодарен всем тем, кто помог в получении малодоступных изда-
ний и первичных данных (проф. В.В.Хоронько, проф. Ю.С.Макляков, к.б.н.
А.М.Малашенко, к.м.н.Э.Х.Абдрашитова), в подготовке и переработке фак-
тического материала (к.м.н. Е.В.Брайцева, Е.В.Иванова, А.В.Шевцов), в
научном и техническом редактировании (доц. Р.ГКостогрызова, Б.М.Боро-
денков).
Любой автор стремится продемонстрировать свои несомненные данные,
а в идеале — абсолютные истины. То же, что не может объяснить — вольно
или невольно пытается скрыть. В этом также проявляется дуальность чело-
веческой психики. Конечно, это значительно проще сделать в публичном
выступлении, когда автору помогает страстность риторики. Беспристраст-
ное изложение в книге, во-первых, эмоционально обедняет аргументацию
автора, а во-вторых, лишает его возможности парировать критические заме-
чания. Поэтому автор заранее принимает критику за возможные издержки и
опечатки (увы, не застрахованы!) и готов участвовать в дискуссии по всем
замечаниям, исходящим из научных принципов и истин. Наш электронный
адрес: niknik @ atlan-tv.ru.
571
БИБЛИОГРАФИЯ
1. Aasen Т., Hodgins M.B., Edward M. et al. The relationship between connexins, gap
junctions, tissue architecture and tumour invasion, as studied in a novel in vitro model
of HPV-16-associated cervical cancer progression. Oncogene, 22, 6025-6036, 2003.
2. Абалакин В.А., Черкасский Б.Л. Использование мышей инбредных линий как
модели для индикации штаммов В. antracis. ЖМЭИ, 2, 146-147, 1978.
3. Abel М., Buck W.B. A technic for gastric cannulation of swine. Cornell Vet., 57,383,
1967.
4. Aberle E.D., Merkel R.A., Forrest J.С Physiological responses of stress susceptible and
stress resistant pigs to heat stress. J. Anim. Scl, 38, 954, 1974.
5. Abramov V.M., Vasiliev A.M., Vasilenko R.N. et al. Structurial base of polyfunctional
role of СаП protein and Caf 1M chaperone in providing Yersinia pestis circulation in
ecological systems of natural plague foci. Protein structures: Kaleidoscope of Structural
and Functions, ISBN: 81-7736-177-5, 2003.
6. Adolph E.F. Physiological Relations. The Jaques Cattell Press, Lancaster, Pa., 1943.
7. Adolph E.F. Quantitative relations in the physiological constitutions of mammals. Science,
109, 579-585, 1943.
8. Агаджанян Н.А. Развитие идей А.П. Авцына в области экологической физиоло-
гии. Вестник РАМН, № 9, 50-53, 2002.
9. AgarvalO.P, Arora R.B. Blood lipid levels in pigs and their possible role in development
of aortic atherosclerosis. Indian J. Exp. Biol., 11, 343, 1973.
10. Агеенко А.И., Epxoe B.C. Аутореактивность спленоцитов мышей чувствительных
линий в латентном периоде канцерогенеза, индуцированного вирусом S47 (С8).
Вопр. вирусологии, 6, 734-737, 1976.
11. Agopyan N, Bhatti Т., Yu S. et al. Vanilloid receptor activation by 2- and 10- particles
induces responses leading to apoptosis in human airway epithelial cells. Toxicol, and
Appl. Pharmacol, 192, 21-35, 2003.
12. Акимова Л.А., Бландова З.К., Кременская Л.В. Генетическая однородность мы-
шей и крыс инбредных линий питомника «Столбовая» АМН СССР. В кн.: Лабо-
раторные животные в медицинских исследованиях. М., 11-12, 1974.
13. Акмаев И.Г., Волкова О.В., Гриневич В.В., Ресненко А.Б. Эволюционные аспекты
стрессорной реакции. Вестник РАМН, № 6, 24-27, 2002.
14. Alberola J. and Izquierdo I.J. The volatile fraction of orange juice. Methods for extraction
anil study of composition. Chemistry and Technology, Academic Press, N. Y, 283-304,
1978.
15. Alexander P.S., Smith S., Wood A. Transcriptional profiling of neuronal differentiation
by human embryonal carcinoma stem cells. In vitro Stem Cells, 21, 459-471, 2003.
16. Altenburger R., Nendza M., Schuurmann G. Mixture toxicity and its modeling by
quantitative structure-activity relationships. Envir. Toxicol, and Chemistry, 22, 1900-
1915, 2003.
17. Altman PL., Ditlmer D.S. Biology Data Book. 2nd ed., v I, Fed. of American Soc.for
Exper. Biol, Bethesda, Md., 100, 416-422, 1972.
18. Аляутдин Р.Н., Кройтер Й., ХаркевичД.А. Доставка лекарственных препаратов в
мозг с помощью наночастиц. Экспер. и клин, фармакол., 2, 65, 2003.
572
Библиография
19. Anderson D.B., Kaufmann R.G. Cellular and enzymatic changes in porcine adipose tissue
during growth. J. Lipid Res., 160, 1973-14.
20. Anderson J.J.B., Milin L, Crackel W.C. Effect of exercise on mineral and organic bone
turnover in swine. /. Appl. Physiol, 30, 810, 1971.
21. Andersson M., Agurell E., Vaghef H. et al. Extended-term cultures of human T-
lymphocytes and the comet assay: a useful combination when testing for genotoxicity in
vitro? Mutation Research, 540, 43-55, 2003.
22. Анохина И.П., Проскурякова Т.В. Участие нейропептида холецистокинина в ме-
ханизмах регуляции эмоций и влечений. Вестник РАМН, № 6, 36-40, 2002.
23. Анохин П.К. Биология и нейрофизиология условного рефлекса. М., Медицина,
1968.
24. Анохин П. К. Теория функциональной системы. Усп. физиол. наук, 1,1,19-54,1970.
25. Анохин П. К. Принципиальные вопросы общей теории функциональных систем.
В кн.: Принципы системной организации функций. М., Наука, 5-48, 1973.
26. Anon. Dentistry Today. ATLA, 21, 30-31, 2002.
27. Антипов И.Г., Карпова Г.В., Новицкий В.В. и др. Характеристика показателей кро-
ветворной системы здоровых линейных мышей. В кн.: Использование лаборатор-
ных животных в разработке, производстве и контроле биологических медицинских
препаратов. М., 207-208, 1976.
28. Apt A. S., Blandova Z., Dishkant I. et al. Study of H-2 mutations in mice. A comparison
of the mutants M505 and Hz-1 by skin grafting and serological techniques.
Immunogenetics, v. 1, 444-451, 1975.
29. Arbab A.S., Bashaw L.A., Miller B.R. et al. Intracytoplasmic tagging of cells with
ferumoxides and transfection agent for cellular magnetic resonance imaging after cell
transplantation: methods and techniques. Transplantation, 76, 1123-1130, 2003.
30. Арчаков A.M. Геномика, протеомика и биоинформатика - науки XXI столетия.
Медицинская кафедра, 3, 6-13, 2002.
31. Armstrong, M. Т., Armstrong, P.B. Growth factor modulation of the extracellular matrix.
Exper. Cell. Research, 288, 235-245, 2003.
32. ArpornmaeklongP, KochelM., Depprich R. etal. Influence of platelet-rich plasma (PRP)
on osteogenic differentiation of rat bone marrow stromal cells: an in vitro study. Int. J. of
Oral and Maxillofacial Surgery, 33, 60-70, 2004.
33. Arrias P.J., Alter R.M.A., Estebanez S.J. Alteraciones bioquimicas peroperatorias an
el transplante ortotopico de Rigato en el cerdo. Rev. esp. enferm. apar. digest, 68, 1-9,
1985.
34. Арзамасцев А. П., Северина И. С., Григорьев Н.Б. и др. Экзогенные доноры оксида азо-
та и ингибиторы NO-синтаз (химический аспект). Вестник РАМН, 12, 88-95, 2003.
35. Asashima Т., Iizasa H, Terasaki Т. et al. Rat brain pericyte cell lines expressing cc2-
adrenergic receptor, angiotensin II receptor type 1A, klotho, and CXCR4 mRNAs despite
having endothelial cell markers. /. of Cell. Physiol., 197, 69-76, 2003.
36. Ашмарин И.П., Каразеева Е.П. Нейропептиды. В кн.: Биохимия мозга. СПб., 232-
266, 1999.
37. Aufderheide M., Knebel J.W., Ritter D. An improved in vitro model for testing the
pulmonary toxicity of complex mixtures such as cigarette smoke. Exper. and Toxicol.
Pathol., 55, 51-57, 2003.
Библиография
573
38. Авербах М.М., Мороз A.M., Литвинов В.И. и др. Естественная резистентность к
туберкулезу и некоторые вопросы иммуногенетики. В кн.: Иммунология и имму-
нопатология туберкулеза. М., Медицина, 106-116, 1976.
39. Avery J.К. New concepts concerning the development of the teeth of domestic pig and
man. Thesis Univ. of Rochester, N.Y., 324, 1952.
40. Backvall H, Wassberg C, Berne B. et al. Similar UV responses are seen in a skin organ
culture as in human skin in vivo. Exper. Dermatol., 11, 349-356, 2002.
41. Badger T.M., Tumbleson M.E., Hutcheson D.P. Proteincalorie malnutrition in young
Sinclair (S-l) miniature swine. Growth, 36, 235, 1972.
42. Bailey D. W., Snell G D., Cherry M. Complementation on serological analysis. In:
Immunogenetics of H-2 system. Basel: Karger, 155-162, 1971.
43. Bailie M.B., Wixson S.K., Land M.S. Vascular-access postimplantation for serial blood
sampling in conscious swine. Lab. Anim. Sci., 36, №4, 431-433, 1986.
AA. Bak Ung-Bok, Yamamoto H, Azuma R. Experimental actinomicotic abscess in mice of
several strains. Nat. Inst. Anim. Health Quart., v. 12, 232-233, 1972.
45. Bakshi D.K., Sharma P. Genotoxicity of textile dyes evaluated with Ames test and rec-
assay. /. of Envir. Pathol. Toxicol, and Oncol, 22, 101-109, 2003.
46. Balcarcel R.R., Clark L.M. Metabolic screening of mammalian cell cultures using well-
plates. Biotechnologe Progress, 19, 98-108, 2003.
Al. Ballard P. Leahy D.E., Rowland M. Prediction of in vivo tissue distribution from in
vitro data. 3. Correlation between in vitro and in vivo tissue distribution of a homologous
series of nine 5-n-alkyl-5-ethyl barbituric acids. Pharmaceutical Research, 20, 864-
872, 2003.
48. BarletJ.P. The influence of porcine calcitionin given intracastrically on restraint-induced
gastric ulcers in swine. Horm. Metab. Res., 6, 517, 1974.
49. Barnes R.D., Tuffrey M., Wills E.J. etal. The innate resistance of CBA mice to endogenous
murine leukaemia virus infection. Brit. J. Cancer, v. 34, 35-38, 1976.
50. Barnes R.H., Moore A.U., Pond W.G. Behaviorale abnormalities in young adult pigs
caused by malnutrition in early life. /. Nutr., 100, 49, 1970.
51. Barrett Ch. P., Donati E. J., Volz J. E. et al. Variations in serum calcium between strains
of inbred mice. Lab. Anim. Sci., v. 25, 638-640, 1975.
52. Baughn R.E., Bonventre P.F. Nonspecific resistance to Listeria monocytogenes in mice
infected and elicited with Staphylococcus aureus. Med. Microbiol, and Immunol, v. 161,
243-252, 1975.
53. Barton G.M., Medzhiton R. Control of adaptive immune responses by Toll-like receptors.
Curr. Opin. Immunol, v. 14, 380-383, 2002.
54. Bearer C.F., Jacobson J.L., Jacobson S. W. et al. Validation of a new biomarker of fetal
exposure to alcohol. Journal of Pediatrics, 143, 463-469, 2003.
55. Beck СМ., Geyh A., Srinivasan A. et al. The impact of a building implosion on airborne
particulate matter in an urban community. Jour, of the Air and Waste Management Ass.,
53, 1256-1264,2003.
56. Becker P.M., Lord L., Dobell A.R.C. Techniques and pitfalls of anestesia and thoracic
surgery in the pig. /. Surg. Res., 13, 215, 1972.
57. Beeken W. L. W., VolwilerP. D., Goldsworthy L. E. etal. Studies of I-albumin catabolism
and distribution in normal young male adults. /. Clin. Invest., 41, 1312-1333, 1962.
574
Библиография
58. Belknap J.К, BergJ.H., Cocke R. etal. Induction and reversal of the magnesium deficiency
syndrome in inbred mice. Exp. Neurol., v. 57, 506-515, 1977.
59. Bell R.R., Blanchard Ch.A., Haskell B.E. Metabolism of vitamin B6 in the I-strain
mouse. II. Oxidation of pyridoxal. Arch. Biochem. and Biophys., v. 147, 602-611,
1971.
60. Березов Т. Т., Яглова Т. Т., Дмитриева Т.Е., Жирков Ю.А. и др. Направленный транс-
порт лекарственных средств с помощью липосом. Вестник РАМН №5,42, 2004.
61. Berglund J. D., Mohseni M. M., Nerem R. M. et al. A biological hybrid model for collagen-
based tissue engineered vascular constructs. Biomaterials, 24, 1241-1254, 2003.
62. Berlin N.I., Waldmann T.A., Weissman S.M. Life span of red blood cell. Physiol. Rev.,
39, 577-616, 1959.
63. Bernstein B. W. Dissection and culturing of chick ciliary ganglion neurons: a system well
suited to synaptic study. Methods in Cell Biology, 71, 37-50, 2003.
64. Bianca S., Li Volti G., Caruso-Nicoletti M. et al. Elevated incidence of hypospadias in
two Sicilian towns where exposure to industrial and agricultural pollutants is high.
Reproductive Toxicol, 17, 539-545, 2003.
65. Billingham R.E., Silvers W.K. Free skin grafting in mammals. In: Transplantation of
tissues and cells. Philadelphia: Wistar Inst. Anat. and Biol., 1-23, 1966.
66. Rubinstein E. Bioterrorism: role of antimicrobial agents. KMAX3, №4, 2001.
67. Blandova Z., Mnatsakanyan Y.A., Egorov I.K. Study of H-2 mutations in mice.
VI M523, a new К end mutant. Immunogenetics, v. 2, 291-295, 1975.
68. Бландова З.К., Душкин В.А., Малашенко A.M. и др. Линии лабораторных живот-
ных для медико-биологических исследований. М., Наука, 1983.
69. Бландова З.К., Гребенюк В.А., Кременская Л.В. Гомозиготность линии August/Sto
в опытах с трансплантацией кожи. В кн.: Биология лабораторных животных. М.,
3, 41-42, 1971.
70. Бландова З.К. Гомозиготность мышей инбредных линий коллекционного фонда
научно-исследовательской лаборатории экспериментально-биологических мо-
делей АМН СССР в опытах с трансплантацией кожи. В кн.: Использование лабо-
раторных животных в разработке, производстве и контроле биологических ме-
дицинских препаратов. М., 194-196, 1976.
71. Бландова З.К. Идентификация гаплотипов Н-2 трех сублиний СВА. В кн.: Лабо-
раторные животные в медицинских исследованиях. М., 21-23, 1974.
72. Бландова З.К., Кременская Л.В., Гребенюк В.А. и др. Генетическая однородность
мышей инбредных линий питомника «Столбовая» в опытах с трансплантацией
кожи. В кн.: Генетика лабораторных животных и эксперимент. М., Медицина, 28-
29, 1974.
1Ъ. Bobacz К, Gruber R., Soleiman A. et al. Expression of bone morphogenetic protein 6 in
healthy and osteoarthritic human articular chondrocytes and stimulation of matrix
synthesis in vitro. Arthritis and Rheumatism, 48, 2501-2508, 2003.
74. Бочков Н.П. Генетика в современной кардиологии. Вестник РАМН, 5, 7, 2004.
75. Боголепов Н.Н. Ультраструктура синапсов коры большого мозга человека в воз-
растном аспекте. Вестник РАМН, 6, 27-31, 2002.
76. Boldessarins R.J., Fisher J. E. Model systems in Biological Psychiatry. Cambrige 5 MFT
Press, 251, 1975.
Библиография 575
77. БолотскихЛ.А., Малашенко A.M., Лушникова З.С. и др. Создание коллекционно-
го фонда лабораторных животных SPF статуса. Ланимология, 1, 43-44, 1993.
78. БолотскихЛА. Опыт получения и выращивания безантигенных миниатюрных по-
росят для медико-биологических исследований. Вестник АМН СССР, 5, 42-43,1981.
79. Boogerd W., Boudewyn P.A.C. A simple method for obtaining cerebrospinal fluid from a
pig model of Herpes encephalitis. Lab. Anim. Sci., 4, 386-388, 1986- 36.
80. Book S.A., Bustad L.K. The fetal and neonatal pig in biomedical research. Anim. Sci.,
997, 1974.
81. Boraldi F., Croce M.A., Quaglino D. et al. Cell-matrix interactions of in vitro human
skin fibroblasts upon addition of hyaluronan. Tissue and Cell, 35,37-45, 2003
82. Borel Y, Kilham L. Carrier-determined tolerance in various strains of mice. The role of
isogenic IgG in the induction of hap specific tolerance. Froc. Soc. Exp. Biol, and Med.,
v 146, 470-474, 1974.
83. Borel J.F. Comparison of the immune response to sheep erythrocytes, tetanus toxoid
and endotoxin in different strains of mice. Agents and Actions, v. 4, 277-285, 1974.
84. BorschelG.H., Dennis R.G., Kuzon W.M. Contractile skeletal muscle tissue-engineered
on an acellular scaffold. /. Plastic and Reconstr. Surg., 113, 595-604, 2004.
85. Bostrom J., Bohm M., Gundertofte K. et al. A 3D QSAR study on a set of dopamine D4
receptor antagonists./. of Chemical Inform, and Сотр. Sci., 43, 1020-1027, 2003.
86. Botchwey E.A., Pollack S.R., El-Amin S. et al. Human osteoblast-like cells in three-
dimensional culture with fluid flow. Biorheology, 40, 299-306, 2003.
87. Bottermann H. Animal welfare aspects in case of therapeutical emergency. Deutsche
Tierarztliche Wochenschrift, 110, 205-206, 2003.
88. Boulos B.M., Jenkins W.L., Davis L. Pharmacokinetics of certain drugs in the
domesticated goat. Am. J. Vet. Res., 33, 943-952, 1972.
89. Bowie J. W., Solberg L.A., Fass D.N. Transplantation of normal bone marrow into a pig
with severe von Willebrand's disease. J. Clin. Invest, 26-30, 1986-76.
90. Boxenbaum H. Interspecies scaling, allometry. Physiological time, and the ground plan
of pharmacokinetics. J.Pharmacokin. and Biopharm., v. 10, №2, 201-227, 1982.
91. Boxenbaum H. Interspecies variation in liver weight, hepatic blood flow, and antipyrine
intrinsic clearance: extrapolation of data to benzodiazepines and phenytoin. J. Phar-
macokin. and Biopharm, 8, 165-176, 1980.
92. Bradbury S.P., RussomC.L.,AnkleyG.T. etal. Overview of data and conceptual approaches
for derivation of quantitative structure-activity relationships for ecotoxicological effects
of organic chemicals. Envir. Toxicol, and Chemistry, 22, 1789-1798, 2003.
93. Braun D.G., Kindred В., Jocobson E. B. Streptococcal group A carbohydrate antibodies
in mice: Evidence for strain differences in magnitude and restriction of the response,
and for thymus dependence. Europ. J. Immunol., v. 2, 138-143, 1972.
94. Braverman L. M., Slesinski I. Study of autoimmune disease in New Zealand mice. IV.
Immunologic reactivity and reticuloendotheli-al function. /. Invest. Dermatol., v. 55,
317, 1970.
95. Brittberg M., Peterson L., Sjogren-Jansson E. et al. Articular cartilage engineering with
autologous chondrocyte transplantation. A review of recent developments. /. of Bone
and Joint Surgery — American, v. 85-A, 109-115, 2003.
96. Brody S. Bioenergetics and Growth. Hafner, N. Y, 592, 1964.
576
Библиография
97. Brody S. Relativity of physiologic time and physiologic weight. Growth 1, 60-67, 1937.
98. Broekmans W.M.R., Vink А.Л., Boelsma E. et al. Determinants of skin sensitivity to
solar irradiation. Eur. Jour, of Clin. Nutrition, 57, 1222-1229, 2003.
99. Брондз Б.Д., Рохлин О.В. Молекулярные и клеточные основы иммунологическо-
го распознавания. М., Наука, 336, 1978.
100. Brown D. Е., King G.J., Hacker R.R. Polyurethan indwelling catheters in piglets. /. Anim.
Sci., 37, 303, 1973.
101. Brown A. Live-cell imaging of slow axonal transport in cultured neurons. Methods in
Cell Biol., 71, 305-323, 2003.
102. BrubakerR.R. Yersinia pestis and bubonic plague. The prokaryotes, an evolving electronic
resource for the microbiological community. V Springer Verlag, N.Y., 2000.
103. Бунто Т.В., Обухова JI.K. Частота злокачественных новообразований и продол-
жительность жизни нелинейных мышей SHK. В мышей линии СЗНА. В кн.: Ис-
пользование лабораторных животных в разработке, производстве и контроле био-
логических медицинских препаратов. М., 209-210, 1976.
104. Buch D.L., Jump E.B., Weaver M.E. Orthodontic tooth movement in miniature swine.
L.A.D.R. 43rd General Meeting Abs., 80, 1965.
105. БулдаковЛ.А., Калистратова B.C. Радиоактивное излучение и здоровье. М., Ин-
форм-Атом, 165, 2003.
106. Burke J., van HosierG, Trentin J. Caesarian derivation and foster nursing of strain LSN
inbred hamsters. Lab. Animal Care., v. 20, №2, 238, 1970.
107. Burns J.J. Species differences and individual variations in drug metabolism. In
B.B. Brodie and E.G. Erdos (Eds.). Proc. Int.Pharmacol. Meeting, Pergamon Press, N. Y,
v. 6, 277-287, 1962.
108. Buschmann H., Krausslich H., Meyer I. et al. Variation of the immune response to sheep
erythrocytes in several strains of mice and their crosses. Med. Microbiol, and Immunol.,
v. 158, 71-82, 1972.
109. Быков В.А., Денисов-Никольский Ю.И., Ребров Л.Б. Использование биотест-сис-
тем в обеспечении биобезопасности. Вестник РАМН, №10, 42, 2002.
ПО. Calder W.A. Scaling of physiological processes in homeothermic animals. Ann. Rev.
Physiol, 43, 301-322, 1981.
111. Callear J.F.F., Y.F.F., van Gestel. An analysis of field experiments in pigs in the U.K.
and freland with sedative neuroleptic Azaperone. Vet. Rec, 89, 453, 1971.
112. Carlyle JR. The use of swine in brain interstitial radiation studies. Lab. Anim. Sci., 36,
No. 4, 381-385, 1986.
113. Camevalli, C. M., Soares, С P., Zangaro, R.A. et al. Laser light prevents apoptosis in Cho
K-l cell line. J.of Clin.Laser Med. and Surg., 21, 193-196, 2003.
114. Carrasco R.A., Stamm N.B., PatelB.K. One-step cellular caspase-3/7 assay. Biotechniques,
34, 1064-1067, 2003.
115. Carrel A. Physiological time. Science, 74, 618-621, 1931.
116. Carter N, Festing M. Erythrocyte enzyme and protein variation in three guinea-pig
strains. Guinea-Pig News Lett., 6, 12-16, 1972.
117. Chan К. Т., Hsieh D.P. H, Lung M.L. In vitro aflatoxin Bl-induced 53 mutations. Cancer
Letters, 199, 1-7,2003.
118. Чернов Ю.Н., Гайкович Е.А., Heap Poomc. Генотипирование ферментов лекар-
Библиография 577
ственного метаболизма в русской популяции. Клиническая фармакология в Рос-
сии. М., 289-290, 2004.
119. Chang C.J., Нои К.Н. High-resolution optical Doppler tomography for in vitro and in
vivo fluid flow dynamics. Chang Gung Medical Journ., 26, 403-411, 2003.
120. Chaput R.L., Wise D. Miniature pig incapacitation and performance decrement after mixed
gamma-neutron irridiation. Armed Forces, Radiobiol. Res. Inst. SR, Md.24, 69-12, 1969.
121. Chassagnole C, Quentin E., Fell D.A. et al. Dynamic simulation of pollutant effects on the
threonine pathway in Escherichia coli. Comptes Rendus Biologies, 326, 501-508, 2003.
122. Cheema U., Yang S.Y., Mudera V. et al. 3-D in vitro model of early skeletal muscle
development. Cell Motility and the Cytoskeleton, 54, 226-236, 2003.
123. Cheers C, Mc.Kenzie I., Pavlov H. etal. Resistance and susceptibility of mice to bacterial
infection: course of listeriosis in resistant or susceptible mice. Infect, and Immun., v. 19,
763- 770, 1978.
124. Черкасский Б.Л., Бакулов И.А., Пчелинцев СЮ. Зоонозы и биобезопасность. Ве-
стник РАМН, 10, 30, 2002.
125. ChiariniA., PetriniP, BozziniS. etal. Silk fibroin/poly(carbonate)-urethane as a substrate
for cell growth: in vitro interactions with human cells. Biomaterials, 24, 789- 799, 2003.
126. Cho M.S., Yee H., Chan S. Establishment of a human somatic hybrid cell line for
recombinant protein production. J.of Biomedical Sci., 9, 631-638, 2002.
127. Cho M.S., Yee H., Brown C, Mei B. et al. Versatile expression system for rapid and stable
production of recombinant proteins. Biotechnol. Progress, 19, 229-232, 2003.
128. Choi J., Oris J. T. Assessment of the toxicity of anthracene photo-modification products
using the top minnow (Poeciliopsis lucida) hepatoma cell line (PLHC-1). Aquatic
Toxicol., 65, 243-251, 2003.
129. Cholnoky E., Fischer J., Varga M., Gyorffy G. Aspects of genetically defined populations
in toxicity testing. II. Phenotypic differences in sensitivity to a toxic dextran preparation.
Ztschr. Versuchs tierk. Bd., 16, 43-48, 1974.
130. Choudhary N., Sharma M., Verma P. etal. Hepato and nephrotoxicity in rat exposed to
endosulfan. /. ofEnvir. Biol., 24, 305-308, 2003.
131. Chun C, Heineken K., Szeto D. et al. Application of factorial design to accelerate
identification of CHO growth factor requirements. Biotechnol. Progress, 19, 52-57,2003.
132. Чуич H.A., Рябчиков О.П., Марданова Г.В., Осипов В.В. Реакция бласттрансфор-
мации лимфоцитов плода мини-свиньи на конковалин А при хронической ал-
когольной интоксикации. Актуальные вопросы современной гистопатологии, М.,
107-108, 1990.
133. Чуич Н.А., Токарева О.И., Марданова Г.В. и др. Иммунологическая характерис-
тика печени мини-свиньи при хронической алкогольной интоксикации. Акту-
альные вопросы современной гистопатологии, М.,109-110, 1990.
134. Ciftci M., Demir Y, Ozmen I. et al. In vivo and in vitro effects of some plant hormones on
rat erythrocyte carbonic anhydrase and glucose-6-phosphate dehydrogenase activities.
J. of Enzyme Inhibition and Medicinal Chemistry, 18, 71- 76, 2003.
135. Claranello R.D., Barchas R., Kassler S. et al. Catecholamines: strain differences in
biosynthetic enzyme activity in mice. Life Sci., v. 11, pt I, 565-572, 1972.
136. Clark W.J., Palmer R.F., Howard E.B. et al. Stroncium 90: effects of chronic injestion
on farrowing performance of miniature swine. Science, 169, 3945, 598-600, 1970.
578
Библиография
137. Claude J., Leclerc L., Gomard E. Characteristics of cytolytic T-cells from resistant and
sensitive strains in murine leukemia. Proc. Amer. Assoc. Cancer Res. andAmer. Soc. Clin.
Oncol., v. 16, 202, 1975.
138. Collins R.L. Maltese dilution, chromosome 9, and audiogenic seizures in DBA/2 mice:
experimental evaluation. Brain Res. v. 70, 541-546, 1974.
139. Comber M.H.I, Walker J.D, Watts C. et al. Quantitative structure-activity relationships
for predicting potential ecological hazard of organic chemicals for use in regulatory risk
assessments. J. Envir. Toxicol, and Chemistry, 22, 1882-1828, 2003.
140. Condon R.G., Schaefer E.J., Santoro M. et al. Development of a Chinese hamster ovary
cell line for recombinant adenovirus-mediated gene expression. Biotechnology Progress,
19, 137-143, 2003.
141. Cook J.C, Jacobson C.F., Gao F. et al. Analysis of the nonsteroidal anti-inflammatory
drug literature for potential developmental toxicity in rats and rabbits. Developmental
and Reproductive Toxicol., 5-26, 2003.
142. Cox M. Progress on regulations for human-derived therapeutic products. Med. Device
Technol, 14, 32-34, 2003.
143. Cramer M., Braun D.G. Genetics of restricted antibodies to streptococcal group
polysaccharides in mice. /. Exp. Med., v. 139, 1513-1528, 1974.
144. Cui S., Wang X., Liu S. et al. Predicting toxicity of benzene derivatives by molecular
hologram derived quantitative structure activity relationships (QSARS). SAR andQSAR
in Envir. Res., 14, 223-231, 2003.
145. CuillouxL., DzvigaC, Gourgaud-Massias С etal. Flow cytometry versus histamine release
analysis of in vitro basophil degranulation in allergy to Hymenoptera venom Lambert.
Cytometry, 52B, 13-19, 2003.
146. Cuschieri A., Baker P.P., Holley M.P. et al. Portacaval shunt in the pig. I. Effect on
survival, behavior, nutrition and liver function. /. Surg. Res., 17, 387, 1974.
147. Cutler R.G. Evolution of human longevity: a critical overview. Mech. Ageing Dec, 9,
337-354, 1979.
148. Cutler R. G. Evolution of longevity in primates. Hum., v. 5, 169-202, 1976.
149. Cyranoski D. Are China's bioethics under control? Nature, 424, 239, 2003.
150. CzeizelA.E., Petik D., Vargha P. Validation studies of drug exposures in pregnant women.
Pharmacoepidemiology and Drug Safety, 12, 409-416, 2003.
151. Dantzer R. The pig as a model for behavioral research. Lab. Anim. Sci., 36, 362, 1986.
152. Darnell M.B., Koprowski H, Lagerspetz K. Genetically determined resistance to infection
with group В arboviruses. J. Infect. Diseases, v. 129, 240-247, 1974.
153. De Cabo R., Furer-Galban S., Anson R.M. et al. An in vitro model of caloric restriction.
Exper.Gerontology, 38, 631-639, 2003.
154. De Maeyer E., De Maeyer-Guignard I., Hall W. et al. A locus affecting circulating
interferon levels induced by mouse mammary tumor virus. /. Gen. Virol., v. 23, 209-
211, 1974.
155. De Maeyer E., De Mayer-Guignard J. A gene with quantitative effect on circulating
interferon induction — further studies. Ann N. Y., Acad. Sci., v. 173, 228-238, 1970.
156. Dedrick R.L., Bischoff K.B., Zaharko D.Z. Interspecies correlation of plasma
concentration history of methotrexate (NSC-740). Cancer Chemother., Rep: Part 154,
95-101, 1970.
Библиография
579
157. Demarteau О., Jakob М., Schafer D. et al. Development and validation of a bioreactor
for physical stimulation of engineered cartilage. Biorheology, 40, 331-336, 2003.
158. Denny H.R., MesservyA. Surgical technique for the extirpation of the mandibular salivary
glands and the collection of salivary secretions in the pig. Vet. Rec, 90, 650, 1972.
159. Desai T.R., Tyrrell G.J., Finlay W.H. In vitro evaluation of nebulization properties,
antimicrobial activity, and regional airway surface liquid concentration of liposomal
polymyxin В sulfate. Pharmaceutical Res., 20, 442-447, 2003.
160. Dey G., ForsterS. Veterinary authorities and animal welfare organizations — is cooperation
possible? Deutsche Tierarztliche Wochenschrift, 110, 199-205, 2003.
161. Di Pauli R. Genetics of the immune response. Immunology, v. 109, 394-400, 1972.
162. Dienhart G.B., Tubbleson M., Hicklin K.W. Plasma lactic acid and pyruvic acid
concentrations following intragastric infusion of ethanol in adult miniature swine.
Internal. J. Biohem., 6, 211, 1975.
163. Дишкант И.П., Ведерников А.А., Егоров И.К. Изучение мутаций Н-2 мышей. Ге-
нетика, т. 9, №10, 83-90, 1973.
164. Diwan B.A. Strain-dependent teratogenic effects of 1-ethyl-1-nitro-sourea in inbred
strains of mice. Cancer Res., v. 34, 151-157, 1974.
165. Diwan D.A., Meier H. Strain- and age-dependent transplacental carcinogenesis by 1-
ethyl-1-nitrosourea in inbred strains of mice. Cancer Res., v. 34, 764-770, 1974.
166. Домарева О.П. Генетическая радиочувствительность мышей разных линий. Ин-
форм. бюл. Науч. совета по пробл. радиобиологии АН СССР, 15, 117-119, 1973.
167. Donckla W.D. A time to die. Life Sci., 16, 31-44, 1975.
168. Doucette W.J. Quantitative structure-activity relationships for predicting soil-sediment
sorption coefficients for organic chemicals. Envir. Toxicol, and Chemistry, 22, 1771-
1788, 2003.
169. Drabkin D.L. Imperfection: biochemical phobias and metabolic ambivalence. Persp.
Biol. Med., 1, №2, 473-517, 1959.
170. Drexler H.G., Quentmeier H., Mac Leod R.A. Malignant hematopoietic cell lines: in
vitro models for the study of MLL gene alterations. Leukemia, 18, 227-232, 2004.
171. Du Mesnildu Buisson F., Leglise PC, Anderson L.L. Hypophysectomy in pigs. /. Anim.
Sci., 23, 1226, 1964.
172. DucattiA., Vargas V.M. Mutagenic activity of airborne particulate matters an indicative
measure of atmospheric pollution. Mutation Res., 540, 67-77, 2003.
173. Dumont L.J., Vanden Broeke T. Seven-day storage of apheresis platelets: report of an in
vitro study. Transfusion, 43, 143-150, 2003.
174. Душкин В.А. Характеристика чувствительности мышей инбредных линий к S.
typhimurium. В кн.: Мат. конф. по биологии лабораторных животных. М., 27-29, 1967.
175. Dziuk P.J., Phillips T.N., GraberJ. W. Halothane closedcircuit anesthesia in the pig. An.
J. Vet. Res., 25, 1773, 1964.
176. Earnhart С G., Kaattari S. L. The humoral response to in vitro generated parasite antigens
is enhanced by the removal of a defined media component prior to immunization. J.of
Immunol Methods, 278, 67-78, 2003.
177. Edwards N.A. Scaling of renal function in mammals. Сотр. Biochem. Pliysiol., 52A, 63-
66, 1975.
580
Библиография
178. Eggen R.I.L., Segner H. The potential of mechanism-based bioanalytical tools in
ecotoxicological exposure and effect assessment. Analytical and Bioanalytical Chemistry,
377, 386-396, 2003.
179. Egorov I.K., Mnatsakanyan Y.A., Pospelov L.E. Do histocompatibility antigens recognize
themselves? Immunogenetics, v. 5, 65-74, 1977.
180. Егоров И. К, Бландова 3. К. Генетическая однородность мышей инбредных линий
питомника «Столбовая». Генетика, т. 4, № 12, 63- 69, 1968.
181. Eisele G.R., Bernard S.R., Nestor С. W. Gastrointestinal absorbtion of Americium-241
by orally exposed swine: comparison of experimental results with prediction of metabolic
models. Radiat. Res., 112, 62-73, 1987.
182. Ekman L. Distribution and excretion of radiocesium in goats, pigs and hens. Acta Vet.
Scand, 2, 4-15, 1961.
183. El-Amin S.F., Lu H.H., Khan Y. et al. Extracellular matrix production by human
osteoblasts cultured on biodegradable polymers applicable for tissue engineering.
Biomaterials, 24, 1213-1221, 2003.
184. Eleftheriou B.E. A gene influencing hypothalamic norepinephrine levels in mice. Brain
Res., v. 70, 538-540, 1974.
185. Elegbede J.A., Flores R., Wang R.C. Perillyl alcohol and perillaldehyde induced cell cycle
arrest and cell death in BroTo and A549 cells cultured in vitro. Life Sciences, 73, 2831-
2840, 2003.
186. Elliott M.H., Furneaux R. W. Surgical preparation of the pig for in vivo hepatic function
studies. Res. Vet. Sci., 12, 594, 1971.
187. Ellis R.J., van den Heuvel M.R., Bandelj E. et al. In vivo and in vitro assessment of the
androgenic potential of a pulp and paper mill effluent. Envir. Toxicol, and Chemistry, 22,
1448-1456,2003.
188. Emanuel L.A. In vitro testing for allergy diagnosis. Otolaryng. Clinics of North America,
36, 879-893, 2003.
189. Enache M., Dearden J.C., Walker J.D. QSAR analysis of metal ion toxicity data in
sunflower callus cultures (Helianthus annuus "Sunspot"). QSAR and Combinatorial
Science, 22, 234-240, 2003.
190. Englehardt W. V. Swine cardiovascular physiology, Swine in Biomedical Research. Pacific
Northwest Lab., 317, 1966.
191. Ermens J., Comber M. Quantitative structure-activity relationships. Envir. Toxicol, and
Chemistry, 22, 1822-1828, 2003.
192. Eller B.C., Bobber J.S. The use of toxicity bioassays to monitor the recovery of oiled
wetland sediments. Envir. Toxicol, and Chemistry, 22, 1945-1955, 2003.
193. Quilichini P.P., Diabira D., Chiron С et al. Effects of antiepileptic drags. Gozlan, H.
Epilepsia, 44, 1365-1374, 2003.
194. Erickson R.P. Erythrocyte nicotinamide - adenine dinucleotide phosphate levels and
the genetic regulation of erythrocyte glucose 6-phosphate dehydrogenase activity in the
inbred mouse. Biochem. Genet., v. 11, 33-40, 1974.
195. Ермолаева С.Н., Бландова З.К., Душкин В.А. Генетическое исследование чувстви-
тельности мышей разных линий к вирусу эктромелии. Генетика, т. 8, № 5, 161 -
163, 1972.
Библиография
581
196. Ершов Ф.И. Медицинская значимость интерферонов и их индукторов. Вестник
РАМН, 2, 9-11, 2004.
197. Espuelas M.S., Legrand P., Campanero M.A. et al. Polymeric carriers for amphotericin.
B: in vitro activity, toxicity and therapeutic efficacy against systemic candidiasis in
neutropenic mice. /. of Antimicrobial Chemotherapy, 52, 419-427, 2003.
198. Evandri M.G., Mastrangelo S., Costa L.G. et al. In vitro assessment of mutagenicity and
clastogenicity of BDE-99, a pentabrominated diphenyl ether flame retardant. Envir.
and Molecular Mutagenesis, 42, 85-90, 2003.
199. Evseeva T.I., Geras'kin S.A., Shuktomova I.I. Genotoxicity and toxicity assay of water
sampled from a radium production industry storage cell territory by means of Allium-
test. J. of Envir. Radioactivity, 68, 235-248, 2003.
200. Fabricant J. D., Dunn G., Schneider E. L. Maternal age related pre- and postimplantation
fetal mortality: a strain survey. Mech. Ageing and Develop., v. 8, 227-231, 1978.
201. Falcon M., Vinas P., Osuna E. et al. Environmental exposures to lead and cadmium
measured in human placenta. Arch, of Envir. Health, 57, 598-602, 2002.
202. Farrias L., Woodle E.S., Frey Ch.F. A simple technique for experimental hepatic vein
catheterization in swine. Lab. Anim. Sci., 36, №4, 406-407, 1986.
203. Faulon J. L., Visco D.P. Jr., Pophale R.S. The signature molecular descriptor. /. ofChem.
Inf. and Computer Sci., 43, 707-720, 2003.
204. Федоров В.К., Еремеев НС, Ивонин А.А. К вопросу об установлении и сохране-
нии линейных особенностей у животных. В кн.: Биология лабораторных живот-
ных. М., 3, 42-45, 1971.
205. Feierman D.E., Melinkov Z., NanjiA.A. Induction of CYP3A by ethanol in multiple in
vitro and in vivo models. Alcoholism, Clin, and Exp. Res. 27, 981-988, 2003.
206. Festing M.F. W. International index of laboratory animals. Carshalton: MRC Lab. Anim.
Centre, 141, 1980.
207. Festing M.F.W., Blackmore D. Life span of specified-pathogen-free MRC category 4
mice and rats. Lab. Anim., 5, 179-192, 1971.
208. Festing M.F. W. Inbred strains in biomedical research. ATLA, 26, 283-301, 2002.
209. Firoved A.M., Deretic V. Microarray analysis of global gene expression in mucoid
Pseudomonas aeruginosa./. ofBacterioi, 185, 1071-1081, 2003.
210. Fischer D., Li. Y, Ahlemeyer В., Krieglstein J. et al. In vitro cytotoxicity testing of
polycations: influence of polymer structure on cell viability and hemolysis. Biomaterials,
24, 1121-1131, 2003.
211. Fiske R.A., Klein P.A. Effect of immunosupression on the genetic resistance of A2G
mice to neurovirulent influenza virus. Infect, and Immun., v. 11, 576-587, 1975.
212. Fontalin L.N., Kondratyeva T.K, Novikova Т.К. et al. Genetic resistance of CBA and A
mice to transplanted lyinphoid and hemopoietic cells of CBA, M523 mutants and their
F hybryds. Immunogenetics, v. 11, 323-340, 1980.
213. Forbes Z.G., Yellen B.B., Barbee K.A. et al. An approach to targeted drug delivery based
on uniform magnetic fields. IEEE Transactions on Magnetics, 39, 3372-3377, 2003.
214. Forster S. Veterinary authorities and animal welfare organizations — is cooperation
possible? Deutsche Tierarztliche Wochenschrift, 110, 199-205, 2003.
215. Forster S., Henderson P. Wants stand on animal-friendly farming. J. of the Amer. Veter.
Med. Assoc, 222, 1504, 2003.
582
Библиография
216. Fournie G.J., Labert P.H., Miesher Р.Л. Features of the immune response to DNA in
mice. I. Genetic control. Clin, and Exp. Immunol., v. 26, 46-51, 1976.
217. FoyJ.W., Bombick B. R., Bombick D. W. et al. A comparison of in vitro toxicities of cigarette
smoke condensate from Eclipse cigarettes and four commercially available ultra low-
tar cigarettes. Food and Chem. Toxicol., 42, 237-243, 2004.
218. Franek F., Eckschlager T., Katinger H. Enhancement of monoclonal antibody production
by lysine-containing peptides. Biotechnol, Progress, 19, 169-174, 2003.
219. Frank E.R. Veterinary surgery. Minneapolis, Burgess Publ. Co., 1964.
220. Frank L.K. Structure, function and growth. Phil. Sci., 2, 210-235, 1935.
221. Friedrich M.J. Studying cancer in 3 dimensions: 3-D models foster new insights into
tumorigenesis. J. oftheAmer. Med. Assoc, 290, 1977-1979, 2003.
222. Friend D. W., Brown R.G. Blood sampling from suckling piglets. Can. J. Anim. Sci., 51,
547, 1971.
223. Fuchs J.R., Hannouche D., Terada S. et al. Fetal tracheal augmentation with cartilage
engineered from bone marrow-derived mesenchymal progenitor cells. /. of Pediatric
Surgery, 38, 984-987, 2003.
224. Fujino Т., Sato Y, Une M. et al. In vitro farnesoid X receptor ligand sensor assay using
surface plasmon resonance and based on ligand-induced coactivator association. /. of
Steroid Biochem. and Molec. Biol, 87, 247-252, 2003.
225. Gabridge M.G., Cohen L.J. Development of an animal model for mycoplasma-related
reproductive failure. Lab. Anim. Sci., v. 26, 206-210, 1976.
226. Gaitonde M. K., Festing M. F. W. Brain glutamic acid decarboxylase and open field activity
in ten inbred strain of mice. Brain Res., 103, 617-621, 1976.
227'. Galilei G. Dialogues concerning two new sciences, 1637 translated by H. Crew and A.
De Salvio. Macmillan, N. Y, 1914.
228. Gallagher J .A. Human osteoblast culture. Methods in Molec. Med., 80, 3-18, 2003.
229. Garcia M., Forster V., Hicks D. et al. In vivo expression of neurotrophic and neurotrophin
receptors is conserved in adult porcine retina in vitro. Invest. Ophthalm. and Visual Sci.,
44, 4532-4541, 2003.
230. Gaidar A.F., RusselE., Harverman R.B. Mouse strain-related differences in the biologic
and immunologic responses to a murine sarcoma virus. /. Nat. Cancer Inst, v. 50, 971 -
978, 1973.
231. George R.E., ChaputRL., VerrelliD.M. etal. The relative effectiveness of fission neutrons
for miniature pig performance decrement. Radiac. Res., 48, 332, 1971.
232. Gershon R.K., Maurer PH., Merryman C.F. A cellular basis for genetically controlled
immunologic unresponsiveness in mice: tolerance induction in T-cells. Proc. Nat. Acad.
Sci., v. 70, 250-254, 1973.
233. Getty R., Ghoshal N.G. Applied anatomy of sacrococcygeal region of the pig as related to
tailbleeding. Vet. Med. Small. Anim. Clin., 62, 361, 1967.
234. GhaffarA., James K. The effect of antilymphocytic antibody on the humoral immune
response in different strains of mice. Immunology, v. 24, 455-475, 1973.
235. Goldstein B.D., Lai L. Y., Ross S.R. etal. Susceptibility of inbred mouse strains to ozone.
Arch. Envir. Health, v. 27, 412-413, 1973.
236. Gomes W.R., Herchler R.C., Erb R.E. Progesterone levels in ovarian venous effluent of
the non-pregnant sows. /. Anim. Sci., 24, 723, 1965.
Библиография
583
237. Gomez T.M., Harrigan D., Henley J. et al. Working with Xenopus spinal neurons in live
cell culture. Methods in Cell Biol., 71, 129-156, 2003.
238. Gomez-Lechon M.J., Ponsoda X., O'Connor E. et al. Diclofenac induces apoptosis in
hepatocytes. Toxicology in Vitro, 17, 675-680, 2003.
239. Гиппократ Избранные книги (Перевод с греческого проф. В.И.Руднева.). М.,
Сварог, 1994.
240. Gorti G.K., Lo J., Falsafi S. et al. Cartilage tissue engineering using cryogenic
chondrocytes. Arch. ofOtolaryng. — Head and Neck Surg., 129, 889-893, 2003.
241. Gorzynski E., Neter E., Ambrus I. L. Differences in antibody responses of mouse strains
to enterobacterial common antigen. Proc. Soc. Exp. Biol, and Med., v. 134, 776-779,
1970.
242. Gould S.J. Allometry and size in ontogeny and phytogeny. Biol, Rev. 41, 587-640, 1966.
243. Gould S.J. One standard lifespan. New Scientist, 81, 388-389, 1979.
244. Green M.C. Genetic nomenclature for the immunoglobulin loci of the mouse.
lmmunogenetics, v. 8, 89-97, 1979.
245. Greenfeder S., Gilchrest H., Cheewatrakoolpong B. et al. Real-time assay of tryptase release
from human umbilical cord blood-derived mast cells. Biotechniques, 34, 910-912, 2003.
246. Gregorova S., Ivanyi P., Mickova M. et al. The influence of H-2 haplotypes on vesicular
gland weight. Folia biol., v. 22, 44-46, 1976.
247. Gregory R.A., Tracey H.Y. The preparation and properties of gastrin. /. Phisioi, 156,
523, 1961.
248. Григорьев A.M., Богомолов В.В., Моргун В.В. Медицинская реабилитация космо-
навтов после длительных космических полетов. Руководство по реабилитации
лиц, подвергшихся стрессорным нагрузкам. М., Медицина, 400, 53-81, 2004.
249. Grikscheit Т., Ochoa E.R., Srinivasan A. etal. Tissue-engineered esophagus: experimental
substitution by onlay patch or interposition. /. of Thoracic and Cardiovasc. Surg., 126,
537-544, 2003.
250. Грицюте Л.А. К вопросу об активности щелочной фосфатазы в легких мышей
разных инбредных линий. В кн.: Вопросы медицинской генетики и генетики чело-
века. Минск, 214-220, 1971.
251. Gross D.R., Tranquilli W.J., Greene S.A. et al. Critical anthropomorphic evaluation and
treatment of postoperative pain in rats and mice. J. oftheAmer. Veter. Med. Assoc, 222,
1505-1510, 2003.
252. GrunbergJ., KnoglerK., Waibel R. et al. High-yield production of recombinant antibody
fragments in HEK-293 cells using sodium butyrate Biotechniques, 34, 968-972, 2003.
253. Грунтенко Е.В., Беляев Д.К. Возрастная изменчивость тимуса у мышей высоко-
и низкораковых линий. В кн.: Проблемы генетики развития. М., Наука, 58-61,
1972.
254. Грунтенко Е.В. Иммунитет и возникновение злокачественных опухолей. Ново-
сибирск, Наука, 184-194, 1977.
255. Guerinot P., Bonnay M., Bohuon С. Dopamine P-hydroxylase (ECl. 14.2.1) dans le sang
de souris porteuses d'un neuroblastoma (C 1300). Relation avec la croissance tumorale.
Acad. Sci, v 272, 544-546, 1971.
256. GuntherB. Physiological time and its evolution. In A. Locker (Ed.). Biogenesis Evolution
Homeostasis, Springer-Verlag, Heidelberg, 127-133, 1973.
584
Библиография
257. Gunther В., Guerra Е. Theory of biological similarity applied to some data of comparative
physiology. Acta Physiol. Latinamerica, 7, 95-103, 1957.
258. Gunther В., Lebn B. de la Barra On the space-time continuum in biology. Acta Physiol.
Latinamerica, 16, 221-231, 1966.
259. Gunther E.C., Stone D.J., Gerwien R.W. et al. Prediction of clinical drug efficacy by
classification of drug-induced genomic expression profiles in vitro. Proceedings of the
National Academy of Sci., USA, 100, 9608-9613, 2003.
260. Gupta S.P., Maheswaran V., Pande V. et al. A comparative QSAR study on carbonic
anhydrase and matrix metalloproteinase inhibition by sulfonylated amino acid
hydroxamates. /. of Enzyme Inhibition and Med. Chem., 18, 7-13, 2003.
261. Gupta U., Cook J.C., Tassinari M.S. et al. Comparison of developmental toxicology of
aspirin (acetylsalicylic acid) in rats using selected dosing paradigms. Birth Defects
Research Part В — Devel. and Reprod. Toxicol., 68, 27-37, 2003.
262. Gurr M.I., Kirtland J., Philip M. Adipose tissue development in the pig. Proceedings of
the 1st International Congress on Obesity, ed. A.Howard, London, Newman Publishens
Limited, 109, 1974.
263. Hahn B. H., Stevens M. B., Remington M. et al. Heightened circulating antibody responses
in New Zealand mice. J. Lab. and Clin. Med., v. 77, 558-562, 1971.
264. Haines ТА., May T.W., Finlayson R.T. et al. Factors affecting food chain transfer of
mercury in the vicinity of the Nyanza site, Sudbury River, Massachusetts. Envir.
Monitoring and Assessment, 86, 211-232, 2003.
265. Haldane J.B.S. The theory of the evolution of dominance. Genet., 37, 365-374, 1939.
266. Halila R. The role of national ethics commissions in Finland. Bioethics, 17, 357-368,
2003.
267. Hamard P., Blondin C, Debbasch С et al. In vitro effects of preserved and unpreserved
antiglaucoma drugs on apoptotic marker expression by human trabecular cells. Graefes
Archive for Clin, and Exper. Ophthalm., 241, 1037-1043, 2003.
268. Hand M.S., Phillips R. W., Miller Ch. W. et al. A method for quantitation of hepatic
pancreatic and intestinal function in conscious Yucatan miniature swine. Lab. Anim.
Sci., 31, №6, 728-731, 1981.
269. Hanumegowda U.M., Copple B.L., Shibuya M. et al. Basement membrane and matrix
metalloproteinases in monocrotaline-induced liver injury. Toxicol. Sci., 76, 237-246,
2003.
270. Harborne J. B. Introduction to Ecological Biochemistry. Academic Press, N. Y., 143,
1977.
271. Hare D. Time to think outside the box. Canadian Veter. Journ., 44, 949-951, 2003.
272. Harvey R.C., Jones E.E. A technique for bioinstrumentation of the thorax of miniature
swine. Lab. Anim. Sci., 32, Ml, 94-95, 1982.
273. Hatfield P.J., Cameron J.S., CadenheadA. Renal biopsy in the pig. Res. Vet. Sci., 19, 88,
1975.
21 A. Hattan D., Cerilli G. J. Spontaneous reticulum cell sarcomas developing in C3H/HeJ
mice on prolonged immunosuppressive therapy. Transplantation, v. 11, 580-581, 1971.
275. HePrevostC, Levy-LeblondE., VirelizierJ. L. etal. Immunopathology of mouse hepatitis
virus type 3 infection. I. Role of humoral and cell-mediated immunity in resistance
mechanisms. /. Immunol., v. 114, 221-225, 1975.
Библиография
585
276. Не Y, Baas P. W. Growing and working with peripheral neurons. Methods in Cell. Biol,
71, 17-35, 2003.
277. Heidemann S.R., Reynolds M., Ngo K. et al. The culture of chick forebrain neurons.
Methods in Cell. Biol, 71, 51-65, 2003.
278. Heiniger H. I., Meier H., Kaliss N. et al. Hereditary immunodeficiency and
leukemogenesis in HRS/J Mice. Cancer Res., v. 34, 201-211, 1974.
279. Hemmingsen A. M. Energy metabolism as related to body size and respiratory surface
and its evolution. Rep. Steno Mem. Hosp. Nord. Insulin Lab., 9, 1, 1960.
280. Henderson E.S., Adamson R.H., Denham C. et al. The metabolic fate of tritiated
methotrexate. Cancer Res., 25, 1008-1017, 1965.
281. Henon P. R. Human embryonic or adult stem cells: an overview on ethics and perspectives
for tissue engineering. Advances in Exp. Med. and Biol, 534, 27-45, 2003.
282. Herd J.A., BargerA.C. Simplified technique for chronic catherization of blood vessels.
/. Appl. Physiol, 19, 791, 1964.
283. Hintz H.F., Booth A.N., CuculluA.F. etal Aflatoxin toxity in swine. Proc. Soc. Exp. Biol.
Med., 124, 266, 1967
284. Hirst R.G., Wallace M.E. Inherited resistance to Corynebacterium kutscheri in mice.
Infect, andlmmun., v. 14, 475-482, 1976.
285. Hirst R.G., Olds R.J. Corynebacterium kutscheri and its alleged avirulent variant in mice.
J. Hyg., v. 80, 349-356, 1978.
286. Hoag W.G. Spontaneous cancer in mice. Ann. N. Y. Acad. Scl, v. 108, 805-831, 1963.
287. Hollenbeck P.J., Bamburg J.R. Comparing the properties of neuronal culture systems: a
shopping guide for the cell biologist. Methods in Cell. Biol, 71, 1-16, 2003.
288. Holme A., Daniels M., Sassoon J. et al. A novel method of generating neuronal cell lines
from gene-knockout mice to study prion protein membrane orientation. Eur. Jour, of
Neuroscience, 18, 571-579, 2003.
289. Holt J. P., Rhode E.A. Similarity of renal glomerular hemodynamics in mammals. Am.
Heart J., 92, 465-472, 1976.
290. Horan I., Clotworthy M., Fokunang C.N. et al. The development of an in vitro screening
strategy for topically applied products. J. of Ethnopharmacology, 89, 81-90, 2003.
291. Hui K., Lipkind G., Fozzard HA. et al. Electrostatic and steric contributions to bloc of the
skeletal muscle sodium channel by m-conotoxin. J.Gen. Physiol, 119(1), 107-111,2002.
292. Hoyt R.F., Hayre M.D., Dodd К. Т. et al. Long acting intramuscular anesthetic regimen
for swine. Lab. Anim. Sci., 36, 4, 413-416, 1986.
293. Хромачева Р. П. К вопросу об использовании инбредных мышей при стандарти-
зации коклюшной вакцины. В кн.: Генетика лабораторных животных и экспери-
мент. М., 48-49, 1974.
294. Huard J., Li Y., Peng H. et al. Gene therapy and tissue engineering for sports medicine.
/. of Gene Med., 5, 93-108, 2003.
295. Huber W.G., Wallin R.F. Experimental production of porcine gastric ulcers. Vet. Med.,
60, 551, 1965.
296. Huhn R.G., OsweilerG.D., Switzer W.P. Application of the orbital sinus bleeding to swine.
Lab. Anim. Care, 19, 403, 1969.
297. Huhtala A., Nurmi S.K., Tahti H et al. The immunohistochemical characterisation of
an SV40-immortalised human corneal epithelial cell line. ATLA, 31, 409-417, 2003.
586
Библиография
298. Hummel K.P., Richardson F.G., Fekete E. Anatomy. In: Biology of the laboratory mouse.
McGraw-Hill, 247-308, 1966.
299. Ибн-Сина. Даниш-наме. Сталинабад, 116-117, 1957.
300. Igarashi M., Irwin C.R., Locke M. et al. Construction of large area organotypical cultures
of oral mucosa and skin. /. of Oral Pathol, and Med., 32, 422-430, 2003.
301. Зотин А.И., Зотина PC. Скорость дыхания, теплопродукция, производствен-
ная энтропия. Общая биология, XXX, 1, 94, 1969.
302. Игнатов Ю.Д., Скоромец А.А., Амелин А.В. Современные представления о мигре-
ни и механизмах действия средств для её лечения. Вестник РАМН, 10, 13-19,
2003.
303. Ihle J.N., DomotorJ., Bengali K.M. Strain-dependent development of an autogenous
immune response in mice to endogenous С type viruses. Bibl. haematol, 43 177-179,
1976.
304. lautu J.H. The eradication of feral pigs. Australian Veterinary Journ., 81, 704, 2003.
305. Ирд Е.А. Опухоли яичников у мышей линии СВА. Вопр. онкологии, т. 21, 12, 72-
74, 1975.
306. hoard P., Danchaud J.P., Abrigeone E. et al. Etude du medullogramme de la souris
BALB/c. Soc. biol., v. 166, 1448-1456, 1972.
307. Ивани П., Егоров И.К. Иммуногенетика совместимости тканей (HL-A и Н-2).
М., Наука, 102-218, 1975.
308. Jacoby R.O., Bhatt P.N. Genetic resistance to lethal flavivirus encephalitis. /. Infect.
Diseases, v. 134, 158-165, 1976.
309. Janeway C.A. Jr., Medzhitov R. Innate immune recognition. Ann. Rev. Immunol., v.20,
197-216, 2002.
310. Jeffrey PL., Balcar V.J., Tolhurst O. et al. Avian Purkinje neuronal cultures: extrinsic
control of morphology by cell type and glutamate. Methods in Cell Biol., 71, 89-109,
2003.
311. Jeliazkova-Mecheva V. V, Bobilya D.J. A porcine astrocyte/endothelial cell co-culture
model of the blood-brain barrier. Brain Research Protocols, 12, 91-98, 2003.
312. Jenkins S.M., Barone S. The neurotoxicant trimethyltin induces apoptosis via caspase
activation, p38 protein kinase, and oxidative stress in PC 12 cells. Toxicol. Letters, 147,
63-72, 2004.
313. Jenson A.B., GroffD.E., Mc Conahey P.J. et al. Transmission of murine leukemia virus
(Scripps) from parent to progeny mice as determined by p30 antigenemia. Cancer Res.,
v. 36, 1228-1232, 1976.
314. Jiang G, Qiu W., Deluca P.P. Preparation and in vitro/in vivo evaluation of insulin-
loaded poly (acryloyl-hydroxyethyl starch)-PLGA composite microspheres. Pharma-
ceutical Res., 20, 452-459, 2003.
315. Jones P.A., King A. V High throughput screening (HTS) for phototoxicity hazard using
the in vitro 3T3 neutral red uptake assay. Toxicol, in Vitro, 17, 703-708, 2003.
316. Jonsson A.C. Occurence of metenkephalin, metenkefalin-Arg6-Phe7 and meten-
kephaIin-Arg6 -Gly7 -Leu8 in gastrin cells of hod antral mucosa. Cell and Tissue Res.,
2, 361, 1985.
317. Jakshi D.K. Sharma, P. Genotoxicity of Ames test and rec-assay. /. of Envir. Pathol.
Toxicol, and Oncol, 22, 101-109, 2003.
Библиография
587
318. June М., Sato Y. In vitro farnesoid X receptor ligans sensor. /. of Steroid Biochem. and
Molec. Biol, 87, 247-252, 2003.
319. Jump E.B., Weaver M.E. The miniature pig in dental research. In: Swine in biomedical
research.Pacific North-West Lab., 543-557, 1966.
320. Каледин В.И., Поляченко В.М. Новый перевиваемый штамм гепатомы мышей
линии А/Не («гепатома А»). В кн.: Лабораторные животные в медицинских иссле-
дованиях. М., 50-52, 1974.
321. KalterS., EichbergJ., HeberlingR. Raisinggermfreebaboons (papio cynocephalus). In:
Germfree research. Academic Press., N.Y. & London, 611-613, 1973.
322. Калякин В.Н. О различиях в инвазионной чувствительности мышей некоторых
инбредных линий к Encephalitozoon cunicule. В кн.: Биология лабораторных жи-
вотных. М., 3, 160-162, 1971.
323. KandutschA.A., Coleman D.L. Inherited metabolic variations. In: Biology of the laboratory
mouse, 377-386, 1966.
324. Караулов А.В., Огарков В.И., Осипов В.В. Клеточный иммунитет и активность
ферментов метаболизма ксенобиотиков в лимфоцитах у мини-свиней в норме
и при действии алкоголя и анабола. Антибиотики и медицинская биотехнология,
32, 11, 862-864, 1987.
325. Каркищенко Н.Н. Попытка автоматизации работы экспериментатора по изме-
рению уровня возбудимости мозговых структур. Сб.: Некоторые вопросы экспе-
риментальной и клинической медицины. Ростов н/Д, 14, 1966.
326. Каркищенко Н.Н. Математическая модель оценки психофармакологического
эффекта. Веб.: Проблемы моделирования психической деятельности. Новосибирск,
270, 1968.
327. Каркищенко Н.Н. Фармакология системной деятельности мозга. Росиздат, 152,
1975.
328. Каркищенко Н.Н, Омельченко В.П. Об одном комплексном методе анализа био-
потенциалов мозга на ЦВМ и АВМ. Вопросы кибернетики, №25, Биотехнические
системы, М., 78, 1975.
329. Каркищенко Н.Н. Биологическое и математическое моделирование в нейрофар-
макологии эмоций. В кн.: Фармакология здравоохранению. Л., 95, 1976.
330. Каркищенко Н.Н. Нейрохимические основы агрессивного поведения. Изв. СКНЦ
ВШ, Естеств. науки, № 3, 94, 1976.
331. Каркищенко Н.Н. Количественный анализ электрограмм мозга при действии
биогенных аминов. Фармакол. и токсикол., № 6, 666, 1978.
332. Каркищенко Н.Н. Катехоламинергическая регуляция экспрессивного поведения.
В кн.: Катехоламинергические нейроны. М., Наука, 75, 1979.
333. Каркищенко Н.Н. О возможности экстраполяции преклинических психофарма-
кологических исследований на человека. Vсимпозиум по клинич. фармакологии,
Венгрия, 93, 1980.
334. Каркищенко Н.Н. Экстраполяция экспериментальных данных на методику ис-
пытания лекарственных средств в клинике. Фармакол. и токсикол., № 3, 22,
1982.
335. Каркищенко Н.Н. Пиримидины — эндогенные анксиолитики? Известия СКНЦ
ВШ, Естеств. науки, №4, 81, 1983.
588
Библиография
336. Каркищенко Н.Н, Тараканов А.В. О роли трициклических антидепрессантов в
центральной регуляции гипералгезии и стрессаналгезии. Бюл. эксп. биол. и мед.,
8, 193, 1985.
337. Karkischenko NN. Uridine: possible endogenus anxiolytic. Abstr. of the 6th General Meet.
oftheEurop. Soc. For neurochemistry: Molecular basis of neural function, Prague, 112,1986.
338. Каркищенко Н.Н. К сравнительной оценке эффективности новых методов фар-
макологической профилактики болезни движения. Изв. СКНЦ ВШ, Естеств.
науки, 4, 122, 1986.
339. Каркищенко Н.Н., Хайтин М.И. Количественная оценка анксиолитического и
ноотропного эффектов калия оротата в широком диапазоне доз. Фармакол. и
токсикол., №1, 14, 1986.
340. Karkischenko N. N. Psychounitropism in the drug action. Proc. of 7th Int. Conf. on Clinical
Pharm. ofCMEA countries, Plzen, 103, 1987.
341. Karkischenko N. N. Clinical pharmacology of pyrimidines. Pharmacologia Clinica Polona,
IV. Abstr. of 12th Int. Symp. on Clin. Pharm., Wroclaw, Poland., 5, 1988.
342. Karkischenko NN. Clinical pharmacology problems in the field of preparation for the
work in the emergency situations. Abstr. of 14th Symp. on Din. Pharm., Berlin 48, 1989.
343. Каркищенко Н.Н. О требовании к «идеальному» лекарственному средству про-
филактики космической болезни движения. Космич. биол. и авиакосм, мед., 6,
33, 1989.
344. Каркищенко Н.Н. Производные пиримидина: психотропные свойства и моле-
кулярные механизмы центрального действия. Фармакол. и токсикол., 4, т. 53,
67, 1990.
345. Каркищенко Н.Н. Психоунитропизм лекарственных средств. М., Медицина, 208,
1993.
346. Каркищенко Н.Н. Фармакологические основы терапии. М., IMP-Медицина, 560,
1996.
ЪА1. Каркищенко Н.Н, Пчелинцев С.Ю., Каркищенко В.Н. Психотропное, антистрес-
сорное и антиноцицептивное действие интерферона. Вестник РАМН, 10,18,1999.
348. Karkischenko N.N. Designing of the drugs based on cytokines with determined
pharmacokynetical properties. Eur. Cytokine Netw., 11, 156, 2000.
349. Каркищенко Н.Н. Лекарственная профилактика. М., Воентехлит, 752, 2001.
350. Karkischenko N.N., Pchelinctcev S. Y, Denisov L.A. Perspectives of biological defense in
local conflicts and bioterrorism. 7th Intern. Symp. of Protection Against, Stockholm,
Sweden, Chemical and biological warfare agents, 213, 2001.
351. Каркищенко Н.Н, Хоронъко В.В., Сергеева С.А. Фармакокинетика. Ростов-на-Дону,
Феникс, 2001.
352. Каркищенко Н.Н. Лекарственная профилактика в повышении популяционной
резистентности к биологическим факторам. Вестник РАМН, 10, 19-24, 2002.
353. Каркищенко Н.Н. Лекарственная профилактика - новый путь защиты при био-
терроризме и биокатастрофах. Медицинская кафедра, 4, 4-9, 113-120, 2002.
354. Каркищенко Н.Н (ред.) Лабораторные животные. Положение и руководство. М.,
ВПК, 138, 2003.
355. Kassel R.L., Hardy W.D., Day N.K. Complement in cancer. In: Biol. Ainplif. Syst.
Immunol., N. Y, 277-294, 1977.
Библиография
589
356. Кулаков В.И.,Барашнев А.Г., Антонов А.Г. Перинатальный стресс и постстрессо-
вая реабилитация. Руководство по реабилитации лиц, подвергшихся стрессорным
нагрузкам. М., Медицина, 400, 53-81, 2004.
357. Каменский А.А., Савельева КВ. Оксид азота и поведение. М., Изд. НЦССХ им.
А.Н.Бакулева, РАМН, 156, 2002.
358. Кекчеева Н.Г., Кокорин И.Н. О различиях в чувствительности некоторых линий
мышей к экспериментальной инфекции цуцугамуши. В кн.: Вопросы инфекци-
онной патологии и иммунологии. М., Медицина, 5, 208-213, 1976.
359. Keller F., Wolff W., Bocher R. Morphologischstrukturelle differenziernung
denaturierter orthotoper osteoimplantate in reaktion mit dem hagerknochen. ///
Aussagenzumkollagen-polymorphismus, Wiss.Z.-Friedrich-Schiller-Univ. Jena, 35, 345-
347, 1986.
360. Kerrigan C.L., Zelt G., Thomson J.G. The pig as an experimental animal in plastic surgery
for the study of skin flaps, myocutaneus flaps and fasciocutaneus flaps. Lab. Anim. Sci.,
36, №4, 408-412, 1986.
361. Keca Л.Ю. Возрастные изменения количества гликогена в печени мышей инб-
редной линии СЗНА. В кн.: Экспериментальная и клиническая онкология. Таллин,
Валгус, 66-68, 1972.
362. KestnerS.N. Metabolism and size of organs. J. Physiol, 87, 39-41, 1936.
363. Хаитов P.M. Физиология иммунной системы. М., ВИНИТИ РАН, 224, 2001.
364. ХаркевичД.А. Успехи и перспективы фармакологии. Фармакол. и токсикол., Т52,
6, 106-120, 1989.
365. Kleiber M. Metabolic turnover rate: a physiological meaning of the metabolic rate per
unit body weight. /. Theor. Biol., 53, 199-204, 1975.
366. Kim G., Okumura M., Bosnakovski D. et al. Biological properties of allogenic articular
chondrocytes on the surface of bovine cartilage explants in vitro. J. ofVeter. Med. Series,
A50, 418-423, 2003.
367. Kirchner H, Kochen M., Hirt H.M. et al. Immunological studies of HSV-infection of
resistant and susceptible inbred strains of mice. Ztschr. Immunitatsforsch., Bd. 154, 1978.
368. Kirkwood T.B., Proctor C.J. Somatic mutations and ageing in silico. Mechanisms of Ageing
and Devel., 124, 85-92, 2003.
369. Kirsch- Volders M., Sofuni Т., Aardema M. et al. Report from the in vitro micronucleus
assay working group. Mutation Res., 540, 153-163, 2003.
370. Kivilaakso E., Kalima TV., Lempinen M. Gastric ulceration in the pig subjected to
raemor-ragicshock. Scand. J. Gastroenterol., 9, 685, 1974.
У11. Klein C, Kaiser D., Kopp S. et al. Similarity based SAR (SIBAR) as tool for early ADME
profiling. /. of Computer-Aided Molecular Design, 16, 785- 793, 2002.
372. Koch V. Reporting noncompliance in the animal facility. Lab Animal, 32, 27, 2003.
373. Kristen U., Use of higher plants as screen for toxicity assessment. Toxicol, in Vitro, 11,
181-191, 1997.
374. Кочеровская М.Ю. Чувствительность мышей различных линий к хронической
инфекции вирусом лимфоцитарного хориоменингита. В кн.: Арбовирусы. М., 9,
109-112, 1974.
375. Kohn H.I., Klein J., Melvold R. W. The first H-2 mutant workshop. Immunogenetics, v.
7, 279-294, 1978.
590
Библиография
Ъ1в. Kohn H.I., Melvold R. W. Spontaneous histocompatibility mutations detected by dermal
grafts: significant changes in rate over 10-year period in the mouse H-system. Mut. Res.,
v. 24, 163-169, 1974.
Ъ11. Кольчик Ю.А., Душкин В.А., Горбунова В.А. Миниатюрная свинья в медико-био-
логических исследованиях. Лабораторные животные в медико-биологических ис-
следованиях, М., 188-190, 1974.
378. Копаладзе Р.А. Биоэтика. Эксперименты наживотных — история, состояние воп-
роса, перспективы. М., Спутник +, 66, 2003.
379. Kort E.J., Jones A., Daumbach М. et al. Quantifying cell scattering: the blob algorithm
revisited. Cytometry, 51 A, 119-126, 2003.
380. Koshimune M., Takamatsu K, Nakatsuka H. et al. Creating bioabsorbable Schwann cell
coated conduits through tissue engineering. Bio-Med. Mater, and Eng., 13, 223-229,
2003.
381. Kostrubsky V.E., Strom S.C, Hanson J. et al. Evaluation of hepatotoxic potential of drugs
by inhibition of bile-acid transport in cultured primary human hepatocytes and intact
rats. Toxicol. Sci., 76, 220-228, 2003.
382. Koukai M. I., Trebichavsky I, Smolova M. et al. Pig as a model for Ruman viral diseases.
Z. Versuchsterk, 30, №12, 53, 1987.
383. Kouri R.E., Salerno R.A., Whitmire C.E. Relationships between aryl hydrocarbon
hydroxylase inducibility and ensitivity to chemically induced sufcutaneous sarcomas in
various strains of mice. J. Nat. Cancer Inst., v. 50, 363-368, 1973.
384. Красковский Г.В., Порубова Г.М. Роль генетических факторов в канцерогенезе
легких у мышей. В кн.: Вопросы медицинской генетики и генетики человека. Минск,
203-213, 1971.
385. Красникова НО. О преимуществах использования линейных животных при ис-
пытании стимуляторов заживления ран кожи. В кн.: Генетика лабораторных
животных и эксперимент. 46-48, 1974.
386. Красовский Г.Н. Прикладные аспекты использования аллометрии в экологии
человека. Вестник РАМН, 7, 39-42, 2000.
387. Красовский Г.Н, Егорова НА., Антонова М.Г. Проблема экстраполяции резуль-
татов биотестирования на человека. Токсикол. вестник, 3, 12-17, 2002.
388. Krasovskii G.N. Extrapolation of experimental data from animals to man. Envir. Health
Perspectives, v. 13, 51-58, 1976.
389. Kripke M. L., Weiss D. W. Studies on the immune responses of BALB/c mice during
tumor induction by mineral oil. Intern. J. Cancer, v. 6, 422-430, 1970.
390. Крышкина В.П., Бландова З.К. Стабильность генетической структуры рандомб-
редных мышей колонии SHK по комплексу генов Н-2. Генетика, т. 13, 11, 1955-
1959, 1977.
391. Крышкина В.П., Климанова Е.А. Исследование генетической однородности ин-
бредных линий AKR/Rap, СЗН/Rap, СЗНА/Rap, C57BL/6Rap питомника «Рап-
полово» АМН СССР. В кн.: Генетика лабораторных животных и эксперимент.
М., Медицина, 30-31, 1974.
392. Крышкина В.П., МалашенкоА.М. Генетическая изменчивость белых нелинейных
мышей. Генетика, т.8, 11, 76- 82, 1976.
393. Крышкина В.П., Малашенко A.M. Исследование генетической изменчивости не-
Библиография
591
линейных лабораторных мышей с помощью линий-анализаторов. Генетика, т. 9,
5, 52-55, 1973.
394. Крышкина В.П., Малашенко A.M. Сложные гибриды мышей CBWA — новая мо-
дель для медико-биологических исследований. Бюл. эксп. биол. и мед., 11, 118-
119, 1975.
395. Крыжановский Т.Н. Патологические системы в деятельности ЦНС. Вестник
РАМН, 6, 18-23, 2002.
396. Krouse J.H., Stachler R.J., Shah A. Current in vivo and in vitro screens for inhalant
allergy. Otolaryng. Clinics of North America, 36, 855-868, 2003.
397. Kuai X.L., Cong X.Q., Li X.L. et al. Generation of hepatocytes from cultured mouse
embryonic stem cells [see comment]. Liver Transplantation, 9, 1094-1099, 2003.
398. Кубатиев А.А., Федосова Н.Ф., Алисиевич СВ. и др. Механизмы опосредованной
регуляции функций нейтрофиллов у больных сахарным диабетом второго типа.
Бюл. эксп. биол. и мед., 2, 164, 2004.
399. Kubo К, Kuroyanagi Y. Effects of vascular endothelial growth factor released from
cultured dermal substitute on proliferation of vascular endothelial cells in vitro. J. of
Artificial Organs, 6, 267-272, 2003.
400. Kuhn J., Molle K, Brinkmann T. et al. High-density tissue-like cultivation of JAR
choriocarcinoma cells for the in vitro production of human xylosyltransferase. J. of
Biotechnology, 103, 191-196, 2003.
401. Kuhn R., Pagano A., Stoehr N. et al. In vitro and in vivo characterization of MPEP, an
allosteric modulator of the metabotropic glutamate receptor subtype 5: review article.
Amino Acids, 23, 207-211, 2002.
402. Kuhn T.B. Growing and working with spinal motor neurons. Methods in Cell Biol, 71,
67-87, 2003.
403. Kunick C, Lauenroth K, Wieking K. et al. Evaluation and comparison of 3D-QSAR
CoMSIA models for CDK1, CDK5, and GSK-3 inhibition by paullones. /. of Medicinal
Chemistry, 47, 22-36, 2004.
404. В.Г. Кукес. Метаболизм лекарственных средств: клинико-фармакологические
аспекты. М., Реафарм, 2004.
405. Lagerspetz К. V. Koprowski Н, Darnell M. et al. Thermoregulation in group В arbovirus-
resistant and group В arbovirus-sesceptible mice. Amer. /. Physiol., v. 225, 532-537,1973.
406. Лайтфут Э. Явления переноса в живых системах. Биомедицинские аспекты
переноса количества движения и массы. М., МИР, 520, 1977.
407. Lake S. Biological availability of radionuclides produced by Plowshare Event Schooner.
Health. Phys., 20, 225-317,577, 1971.
408. Lambert G.H., Nebert D.W. Genetically mediated induction of drug-metabolizing
enzymes associated with congenital defects in the mouse. Teratology, v. 16, 147-153,
1977.
409. Lamberth J.L. Liver biopsy of weaner pigs using open circuit halothane anesthesia.
/. Austral. Vet., 43, 272, 1967.
410. LandyJ., Sandberg R. Delivery of germfree pig. Fed. Proc, v. 20, 369, 1961.
411. Langner P.M., Benson G.J., Neff-Devis C. et al. A nonrestrictive method for maintaining
grouped swine for blood sampling applications to physiologic assessment of stress.Lab.
Anim. Sci., 35, №2, 182-185, 1985.
592
Библиография
412. Lanicault G, Merritt A., Rosato E. Comparative relationship between serum gastrin
concentration and gastric acid output. Am. J. Dis., 17, 523, 1972.
413. Лапин Б.А., Джакидзе Э.К., Крылова P.M. и др.Инфекционная патология обезь-
ян: краткая характеристика, возможности экспериментального изучения, био-
безопасность для человека при содержании обезьян в приматологических пи-
томниках и лабораториях. Вестник РАМН, 4, 43, 2004.
414. Laquerriere P., Grandjean-Laquerriere A., Jallot E. et al. Importance of hydroxyapatite
particles characteristics on cytokines production by human monocytes in vitro.
Biomaterials, 24, 2739-2747, 2003.
415. Lark B.S., Mahajan R.K., Walia T.P. Determination of metals of toxicological
significance in sewage irrigated vegetables by using atomic absorption spectrometry and
anodic stripping voltammetry. Ind. J. ofEnvir. Health, 44, 164-167, 2002.
416. Lazarova M., Slamenova D. Genotoxic effects of a complex mixture adsorbed onto
ambient air particles on human cells in vitro: the effects of vitamins E and C. Mutation
Research, 557, 167-175, 2004.
417. LedbetterJ.A., Herzenberg L.A. Xenogeneic monoclonal antibodies to mouse lymphoid
differentiation antigen. Immunol. Rev., v. 47, 63-90, 1979.
418. Lee J. E., Dietert R. R. Developmental immunotoxicity of lead: impact on thymic function.
Birth Defects Research Blood, 67, 861-867, 2003.
419. Lee J.H., Park T.G., Park H.S. et al. Thermal and mechanical characteristics of poly
(L-lactic acid) nanocomposite scaffold. Biomaterials, 24, 2773-2778, 2003.
420. Lee R.F., Steinert S. Use of the single cell gel electrophoresis/comet assay for detecting
DNA damage in aquatic (marine and freshwater) animals. Mutation Research, 544,
43-64, 2003.
421. Leigh-Browne G, HarpurP.P. Intestinal intubation in the unanesthetized miniature pig:
technique for sampling and measurement of gut length per fistulumff Rev. Vet. Sci., 18,
6, 1975.
All. Лепахин В.К., Астахова А.В. Безопасность фармакотерапии — одна из важней-
ших проблем здравоохранения. Клин, исслед. лек. средств в России, 2, 326, 2001.
423. Leparc-Goffart I., Poirier В., El Zaouk A. et al. New generation of cell culture assay for
smallpox vaccine potency. /. of Clin. Microbiol., 41, 3687-3689, 2003.
424. LiX.Q., BjorkmanA., Andersson T.B. etal. Identification of human cytochrome P(450)s
that metabolise anti-parasitic drugs and predictions of in vivo drug hepatic clearance
from in vitro data. Eur. J. of Clin. Pharmacol., 59, 429-442, 2003.
425. Lichtenberg-Frate H, SchmittM., GellertG. etal. Ayeast-based method for the detection
ofcytoandgenotoxicity. Toxicol, in Vitro, 17, 709-716, 2003.
416. Sasaki Y, Suto Т., Ambo A. et al. Biological properties of opioid peptides replacing Tyr
at position 1 by 2,6-dimethyl-Tyr. Chem. Pharm. Bull., 47(10), 1506-09, 1998.
All. Lilly F., Duran-Reynals M.L. Combined neoplastic effects of vaccina-virus and
3-methylcholanthrene. /. Nat. Cancer Inst., v. 48, 105-112, 1972.
428. Linares A.F., Loikkanen J., Jorge M.F. et al. Antioxidant and neuroprotective activity of
the extract from the seaweed, Halimeda incrassata (Ellis) Lamouroux, against in vitro
and in vivo toxicity induced by methylmercury. Veter. and Human Toxicol., 46,
1-5, 2004.
Библиография
593
429. Ling W.L., Deng L., Lepore J. et al. Improvement of monoclonal antibody production
in hybridoma cells by dimethyl sulfoxide. Biotechnol. Progress, 19, 158-162, 2003.
430. Link R.P, Pensinger R.R. Lead toxicosis in swine. Am. J. Vet. Res., 27, 759, 1966.
431. Littledike E.T., St.Clair L.E., Notzold R.A. Effects of parathyzoidectomy of the pig.
/. Vet. Res., 29, 635, 1968.
432. Litzke L.F., Berg R. Quantitativ-morphologische unter-suchungen am herzen das
miniaturschweines MINI-LEWE. Exper. Vet. Med., Leipzig, 4, 547- 556, 1977.
433. Lillie R. Y. Air pollutants affecting the performance of domestic animals — a literature
review. f/S Agricultural Handbook, 380, 109, 1972.
434. Lloyd W.E., Buck W.B. Technique for semipermanent cannulation of ureters in bovine,
ovine, porcine and canine species. /. Am. Vet. Res., 32, 817, 1971.
435. Лобанова З.И., Малашенко A.M., Шмидт Е.Ф. Наследственная патология почек
мышей линии T138/V. В кн.: Биологическая характеристика лабораторных жи-
вотных и экстраполяция на человека полученных данных. М., 69-70, 1980.
436. Lockman PR., Koziara J, Roder K.E. et al. In vivo and in vitro assessment of baseline
blood-brain barrier parameters in the presence of novel nanoparticles. Pharmaceutical
Research, 20, 705-713, 2003.
437. Ляпунов A.M. Общая задача об устойчивости движения. М., Гостехиздат, 1950.
438. Loosemore R.M., Harding J.D.J., Lewis G Mercury poisoning in pigs. Vet. Res., 81,
268, 1967.
439. Ловенецкий А.П., Киселев Ф.Л., Зарецкий И.З. и др. Биохимические и физико-
химические характеристики вируса типа С, выделенного из спонтанной лим-
фосаркомы мышей линии CC57BR. Вопр. вирусологии, 1, 14-20, 1975.
440. Luckey T.D. Germfree Life and Gnotobiology. N.Y., London: Academic press, 512,
1963.
441. Luginbuhe H. Spontaneous atherosclerosis in swine, swine in biomedical research. Pacific
Northwest Lab., 347, 1966.
442. Lustgarten C, Bottoms G.D., ShaskasJ.R. Experimental adrenalectomy of pigs. Am. J. Vet.
Res., 34, 279, 1973.
443. Lygidakis N.J. Segmental auxiliare liver transplantation: a new approach to an old
problem. /. Surg. Res., 38, 246-251, 1985.
AAA. Maclean С W. Observation on coaltar poisoning in pigs. Vet. Rec, 84, 594, 1969.
445. Madhusudhan Т., Richhariya A., Majumdar S.S. et al. An in vitro model for grafting of
hematopoietic stem cells predicts bone marrow reconstitution of myeloablative mice.
/. of Hematotherapy and Stem Cell Res., 12, 243-252, 2003.
446. Maes L., Vanden B.D., Germonprez N. et al. In vitro and in vivo activities of a triterpenoid
saponin extract (PX-6518) from the plant Maesa balansae against visceral leishmania
species. Antimicrobial Agents and Chemotherapy, 48, 130-136, 2004.
447'. Малашенко A.M., Чернушенко В.К., Гребешок В.А. и др. Выявление гетерозигот-
ности по рецессивным генам у мышей инбредных линий. В кн.: Лабораторные
животные в медицинских исследованиях. М., 7-8, 1974.
448. Малашенко A.M. Исследование мутагенного эффекта малых доз диэтилсульфата
у лабораторных мышей методом определенных локусов. Генетика, т. 12, №3,
163-165, 1976.
594
Библиография
449. Малашенко A.M. Мутация типа Brachyury у мышей линии РТ. Генетика, т. 11,
№1, 146-147,1975.
450. Малашенко A.M., Игнатьева Е.Л., Бескова Т.Б. Моделирование периодически
меняющегося прогнозируемого ответа на мутагенные воздействия у мышей СВА/
LacY в ряду поколений инбридинга. Генетика, т. 37, №10, 1353-1357, 2001.
451. Mandel L., Travnicek J., Sterzl J. Our experiences in rearing germfree piglets: history,
contemporary status and outlooks. Acta Vet. Brno., v. 46, Suppl. 4, 3-11, 1977.
452. Mandel P. Genetic control of biogenic amines and the possibilities offered by new
microanalytical techniques for the investigation of human cerebrotical techniques for
the investigation of human cerebrospinal fluid. In: Catecholamines and schizophrenia.
Oxford, 23-294, 1975.
453. Марданова Г.В. Ультраструктурные особенности некоторых внутренних органов
миниатюрных поросят в норме и при антенатальном воздействии этанола. Сб.
трудов научн. конфер. Вопросы экспериментальной и клинической медицин, Куйбы-
шев, 18-222, 1990.
454. Марданова Г.В., Осипов В.В., Кострюков Г.И. Ультраструктурные особенности
миокарда новорожденных мини-свиней при длительной алкоголизации роди-
тельской пары. Бюл. эксп. биол. и мед., 4, 398-400, 1990.
455. Марданова Г.В. Миниатюрная свинья в экспериментальной биологии и меди-
цине. М., 99, 2004.
456. Marin L., Minguela A., Torio A. et al. Flow cytometric quantification of apoptosis and
proliferation in mixed lymphocyte culture. Cytometry, 51A, 107-118, 2003.
457. Marino I.R., Dehuca G. Orthotopic liver transplantation in pigs. An evaluation of different
methods of avoiding the revascularization syndrome. Transplantation, 40, 494-498, 1985.
458. Marple D.N., Cassens R.G. A mechanism for stress susceptibility in swine. J. Anim. Sci.,
37, 546, 1973.
459. Marques P.F., Oliveira M.E., Franca A.S. et al. Modeling and simulation of pulsatile
blood flow with a physiologic wave pattern. Artificial Organs, 27, 478-485, 2003.
460. Martin J.E. Physiology diseases of swine. Iowa State University Press, 186, 1964.
461. Martin Т.Е., Grishanin R.N. PC 12 cells as a model for studies of regulated secretion in
neuronal and endocrine cells. Methods in Cell Biol., 71, 267-286, 2003.
462. Martinez, C. Y., Hollenbeck, P.J. Transfection of primary central and peripheral nervous
system neurons by electroporation. Methods in Cell Biol., 71, 339-351, 2003.
463. Mattioni B.E., Kauffman G. W., Jurs P.C. et al. Predicting the genotoxicity of secondary
and aromatic amines using data subsetting to generate a model ensemble. /. ofChem.
Inform, and Computer Sci., 43, 949-963, 2003.
464. MayorS. Research bodies disappointed by decision to cancel primate research laboratory
British Medical Journal, 328, 306, 2004.
465. McKenzie EC, Morgan J.M., Melvold R.W. et al. Serological and complementation
studies in four C57BL/6 H-2 mutantes. Immunogenetics, v. 3, 241-256, 1976.
466. Meberg P. J., Miller M. W. Culturing hippocampal and cortical neurons. Methods in Cell
Biology, 71, 111-127, 2003.
467. Medina S., Van S.I., Robson E.G. Effect of nonspecific stimulation on the defense
mechanisms of inbred mice. J. Immunol., v. 114, 1720-1725, 1975.
Библиография
595
468. Медведев Н.Н. Об инбридинге, плодовитости и жизнеспособности. Генетика, т.
5,№3, 132-147, 1969.
469. Медведев Н.Н., Ольховская И.Г. К онкологической характеристике мышей
CC57BR и CC57W. Вопр. онкологии, т. 20, № 5, 84-87, 1974.
470. Mellet L.B. Comparative drug metabolism. Proc. Drug. Res., 13, 136-169, 1969.
All. Melvold R. W., Kohn H.J. Eight new histocompatibility mutations associated with the
H-2 complex. Immunogenetics, v. 3, 185-191, 1976.
All. Medzhitov R., Janeway C.A. Decoding the patterns of self and non-self by the innate
immune system. Science, v. 296, 298-300, 2002.
A13. MerrittA.M., Brooks F.P. Basal and histamineinduced gastric acid and pepsin secretion
in the conscions miniature pig. Gastroenterology, 58, 801, 1970.
A1A. Mersmann H.J., Phinneg G., Brown L. Y. Factors influencing the lipolytic respons in
swine (sus domesticus) adipose tissue. Gen. Pharmac, 6, 193, 1975.
475. Meylan W.M., Howard P.H. A comparison of model performance for six quantitative
structure-activity relationship packages that predict acute toxicity to fish. Envir. Toxicol.
and Chemistry, 22, 1799-1809, 2003.
A16. Meylan W.M., Howard P.H. A review of quantitative structure-activity relationship
methods for the prediction of atmospheric oxidation of organic chemicals. Envir. Toxicol.
and Chemistry, 22, 1724-1732, 2003.
All. MiceliM., AlbertiL., Bennardini F. etal. Effect of low doses of ethanol on platelet function
in long-life abstainers and moderate-wine drinkers. Life Sci., 73, 1557-1566, 2003.
478. Михайлова Т.П., Зингер ТВ., Шубина ТС. и др. Характеристика некоторых нор-
мативных показателей линейных мышей. В кн.: Использование лабораторных
животных в разработке, производстве и контроле биологических медицинских пре-
паратов. М., 208-209, 1976.
479. Miller C.L., Lim R.C. Postishemia immunosuppression in miniature swine model. Hab.
Anim. Sci., 36, №4, 375-380, 1986.
480. Miller K.N., TwohillS.A. Method for measuring systolic blood pressure in the conscious
swine (us scrofa). Lab. Anim., 12, №6, 51-52, 1983.
481. Millikan L. W., Molon J.L., Hook R. R. Melanoma in Sinclair swine: a new animal model.
J. Invest. Dermatol, 92, 20, 1974.
482. Mirkes R. The wrongs of animal rights. The National Catholic Bioethics Quarterly, 3,
287-307, 2003.
483. Schulte-Frohlinde E., Reindl W. et al. Effects of oral casokefamide on plasma levels,
tolerance, and intestinal transit in man. Peptides, v. 21(3), 439-442, 2000.
484. MollerJA. Frontiars of pigdom. Sci. News, 26, 129, 1986.
485. Montagna W., Yun I.S. The skin of the domestic pig. /. Invest. Derm., 11, 1964.
486. Meltz M.L. Radiofrequency exposure and mammalian cell toxicity, genotoxicity and
transformation. Bioelectromagnetics, 24, Suppl.6, 196-213, 2003.
487. Montague P. Research ethics and the precautionary principle: marching toward
environmental decay. Cambridge Quarterly of Healthcare Ethics, 12, 466-467, 2003.
488. Moody R.P., MacPherson H. Determination of dermal absorption QSAR/QSPRs by
brute force regression: multiparameter model development with molsuite 2000. J. Toxicol,
and Envir. Health, Part A 66, 1927-1942, 2003.
596
Библиография
489. Moon Y, Uzarski R., Pestka J.J. Relationship of trichothecene structure to COX-2
induction in the macrophage: selective action of type В (8-keto) trichothecenes. J. of
Toxicol. andEnvir. Health , Part A 66, 1967-1983, 2003.
490. Mozes E., Shearer G.M., Seta M. et al. Genetic control of immune responses to synthetic
polypeptides in mice: Cellular analysis of the phenotypic correction of the Ir-3 gene
defect by polyadenylic - polyuridylic acid. J. Immunol., v. 3, 439- 447, 1973.
491. SpivakC.E., BeglanC.L., SeidleckB.K. etal. Naloxone activation of mu-opioid receptors
mutated at a histidine residue lining the opioid binding cavity. Mol. Pharmacol., v. 52
(6), 983-992, 1997.
492. Mullen Y., Yoneda K., Fujiva H. Survival of cultured fetal pancreas transplants in
miniature pigs. Transplan Proa, 1, 17, 417-419, 1986.
493. Munro H.N., Downie E.D. Relationship of liver composition to intensity of protein
metabolism in different mammals. Nature, 203, 603-604, 1964.
494. Mustard R.A., Lipohar Ch., Finley R.J. et al. Asimlified method for metabolic studies in
conscious swine. Lab. Anim. Sci., 36, №4, 393-395, 1986.
495. Neumann Т., Nicholson B.S., Sanders J.E. Tissue engineering of perfused microvessels.
Microvascular Res., 66, 59-67, 2003.
496. Nielsen T.W., Maaske C.A., Booth N.H. Some comparative aspects of porcine renal
function. Swine in Biomedical Research. Pacific Northwest Lab., 259, 1966.
497. Niwa A., Kumaki K, Nebert D. W. et al. Genetic expression of aryl hydrocarbon
hydroxylase activity in the mouse. Distinction between the "responsive" homozygote
and heterozygote at the Ah locus. Arch. Biochem. and Biophys., v. 166, 559-564, 1975.
498. Nomoto K, Harada Т., Koyanagi P. et al. Immune response against hamster erythrocytes
in the low-responder mouse strains. Jap. J. Microbiol., v. 20, 375-384, 1976.
499. Nowinski R.C, Esther F. Oncogenicity of AKR endogenous leukemia viruses. J. Virol,
v. 27, 13-18, 1978.
500. Numata H, Lizuka H, Shemada A. et al. Studies on pharmacological characteristics on
the Gottingen miniature pig. Jap. J. Pharmacol. Suppl., 30, 241, 1980.
501. Ohertman M., Sulica A. Genetic control of humoral immune response to bovine gamma
globulin. Rev. roum. biochim., v. 12, 35-41, 1975.
502. Okita G. T. Species difference in duration of action of cardiac glycosides. Fed. Proc, 26,
1125-1130, 1967.
503. Okinewick J.P, Meredith R.F., Brozovich B. et al. Stimulation of immune response in
hybrid foloowing Raucher virus infection. Proc. Soc. Exp. Biol, and Med., v. 157, 449-
452, 1978.
504. Oldstone M.B.A., Tishon A., Tonietti G. et al. Immune complex disease associated with
spontaneous murine leukemia: Incidence and pathogensis of glomerulonephritis. Clin.
Immunol, and immunopathol., v. 1, 6-14, 1972.
505. OteyCA., Boukhelifa M., Maness P. B35 neuroblastoma cells: an easily transfected, cultured
cell model of central nervous system neurons. Methods in Cell Biol., 71, 287-304, 2003.
506. OverlandG, Morath S., YndestadA. etal. Lipoteichoic acid is apotent inducer of cytokine
production in rat and human Kupffer cells in vitro. Surg. Infections, 4, 181-191, 2003.
507. Palm J.E. The laboratory rat, Rattus norvegicus. In: Handbook of Genetics, v. 4, 243-
254, 1975.
508. Patton R.M., RiggA.R., Chick E. W. et al. Histoplasmosis in purebred mice: influence of
Библиография
597
genetic susceptibility and immune depression on treatment. Mycopathologia, v. 60, 39-
43, 1976.
509. Pan G, Shawer M., Oie S. et al. In vitro gene transfection in human glioma cells using a
novel and less cytotoxic artificial lipoprotein delivery system. Pharmaceutical Res., 20,
738-744, 2003.
510. Panepinto L.M., Phillips R.W., NordenS. etal. Acomfortable, minimum stress method
of restraint for Joukatan Miniature Swine. Lab. Anim. Sci., 33, Ml, 95-97, 1983.
511. Pang K.S. Hepatic clearances of drugs and metabolites. Trends Pharmacol. Sci., 19, 42-
53, 1980.
512. Papini S., Cecchetti D., Campani D. et al. Isolation and clonal analysis of human epidermal
keratinocyte stem cells in long-term culture. Stem Cells, 21, 481-494, 2003
513. Patlewicz G, Rodford R., Walker J.D. Quantitative structure-activity relationships for
predicting mutagenicity and carcinogenicity. Envir. Toxicol. andChem., 22, 1885-1893,
2003.
514. Paula S., Tabet M.R., FarrC.D. et al. Three-dimensional quantitative structure-activity
relationship modeling of cocaine binding by a novel human monoclonal antibody. /. of
Med. Chem., 47, 133-142, 2004.
515. PavoneA., Cardile T. An in vitro study of new antiepileptic drugs and astrocytes. Epilepsia,
44, 34-39, 2003.
516. Pekas J. С Permanent physiological fistula of the pancreas and other digestive glands.
/. Appl. Physiol., 20, 1082, 1965.
517. Perez G, Tabares В., Jover R. Semi-automatic quantitative RT-PCR to measure CYP
induction by drugs in human hepatocytes. Toxicology in vitro, 17, 643-649, 2003.
518. Perkins R., Fang H., Tong W. et al. Quantitative structure-activity relationship methods:
perspectives on drug discovery and toxicology. Envir. Toxicol, and Chemistry, 22, 1666-
1679, 2003.
519. Perry T W., JimenezA.A., ShivelyJ.E. Incident of gastric ulcers in swine. Science, 139,
349, 1963.
520. Pescovitz M. D., Thistlethwaite J. R., Sharp T. Class 11 major histocompatibility complex-
matched renal allografts in swine: summary of current results and continuing studies.
Transplant., 686-688, 1985.
521. Петров В.И. Коррекция проявлений постстрессорного синдрома у участни-
ков ликвидации последствий Чернобыльской катастрофы. Руководство по ре-
абилитации лиц, подвергшихся стрессорным нагрузкам. М., Медицина, 400, 160-
180, 2004.
522. Петров Р.В., Пантелеев Э.И., Машиашвили И.Я. и др. Иммунный ответ мышей
разных инбредных линий на анатоксин Clostridium oedemations. Бюл. эксп. биол.
и мед., 2, 198-200, 1977.
523. Петров Р.В. Формы взаимодействия генетически различающихся клеток лим-
фоидных тканей (трехклеточная система система иммуногенеза). Успехи соврем,
биол., т. 69, №2, 261-271, 1970.
524. Петров Р.В., Хаитов P.M. Искусственные антигены и вакцины. М., Медицина
288, 1988.
525. Pitsillides СМ., Joe E.K., Wei X. et al. Selective cell targeting with light-absorbing
microparticles and nanoparticles. /. Biophysical, 84, 4023-4032, 2003.
598
Библиография
526. Plant I., Glynn А.Л. Natural resistance to Salmonella infection, delayed hypersensitivity
and Ir genes in different strain of mice. Nature, v. 248, 345-347, 1974.
527. Plant J., Glynn A.A. Genetics of resistance to infection with Salmonella typhimurium in
mice. /. Infect. Diseases, v. 133, 72-78, 1976.
528. Poiley S.M. Growth tables for 66 strains and stocks of laboratory animals. Lab. Anim.
Sci.,2, 759-779, 1972.
529. Покровский В.И. Медицинские проблемы биобезопасности. Вестник РАМН, 10,
6, 2002.
530. Понд У.Дж., Xayn T.K.A. Биология свиньи. М., Колос 334, 1983.
531. Pond W.G., Snyder W., Walker E.F. Comparative utilization of casein, fish protein
concentrate and isolated soybean protein in liquid diets for growth on baby pigs.
/. Anim.Sci., 33, 587, 1971.
532. Побезинский Л.А., Побезинская Е.Л., Петрищев В.Н. и dp. Накопление нейтро-
филов в селезенке мышей, иммунизированных клетками аллогенных опухолей.
Вестник РАМН, 7, 121, 2004
533. Побезинская Е.Л., Побезинский Л.Ни др. Кросс-реактивность Т-клеточного ре-
цептора клона клеток памяти CD8+, полученного в ответ на иммунизацию клет-
ками аллогенной опухоли. Бюл. эксп. биол. и мед., 137, №5, 563, 2004.
534. Подопригора Г.И. Медицинская гнотобиология. Мед. информ. агентство (МИА),
М., 2003.
535. Prichard Th.J., Madara J.L., TapperD. W. Failure of cyclosporine to prevent small bowel
allograft rejection in pigs. J. Surg. Res., 553-558, 1986.
536. Prince J.H., Diesim CD., Eglitis J. et al. Anatomy and histology of the eye and orbit in
domestic animals. Thomas, Springfield, 111, 1960.
537. Prothero I.W. Maximal oxygen consumption in various animals and plants. Сотр.
Biochem. Physiol., 64A, 463-466, 1979.
538. Quilichini P.P., Diabira D., Chiron С et al. Effects of antiepileptic drugs on refractory
seizures in the intact immature corticohippocampal formation in vitro. Epilepsia, 44,
1365-1374, 2003.
539. Quinn G.P., AxelrodJ., and Brodie B.B. Species, strain and sex differences in metabolism
of hexoharbitone, amidopyrine, antipyrine and aniline. Bioch. Pharmacol., 1,152-159,
1958.
540. Ramoshebi L.N., Matsaba T.N., Teare J. Renton L. et al. Tissue engineering: TGF-p
superfamily members and delivery systems in bone regeneration. Expert Rev. in Molec.
Med., 4, 1-11, 2002.
541. Redman D. R., Teaque H. S., Hehderickx H. K. et al. Cecal fistulation of the pig using two
forms of indwelling cannulas. /. Anim. Sci., 23, 1032, 1964.
542. ReiserO.L. Philosphyandthe Concepts of Modern Science. Macmillan, N. Y., 43-78,239-
263, 1935.
543. Reyniers J.A. Introduction to the general problems of isolation and elimination of
contamination. Illinois: Charles С Thomas Springfield., 95, 1943.
544. Rhim J.S. Generation of immortal human prostate cell lines for the study of prostate
cancer. Methods in Molec. Med., 81, 69-77, 2003.
545. Riggs D.S. Mathematical Approach to Physiological Problems. Williams and Wilkins,
Baltimore, 295-296, 1963.
Библиография
599
546. Rivedal Е., Myhre О., Sanner Т., Eide I. Supplemental role of the Ames mutation assay
and gap junction intercellular communication in studies of possible carcinogenic
compounds from diesel exhaust particles. Archives of Toxicol., 77, 533-542, 2003.
547. Roberta Т., Federico M., Federica B. et al. Study of the potential cytotoxicity of dental
impression materials. Toxicology in Vitro, 17, 657-662, 2003.
548. Roberts M.E., Stewart P.S. Modeling antibiotic tolerance in biofilms by accounting for
nutrient limitation. Antimicrob. Agents and Chemother, 48, 48-52, 2004.
549. Robinson C.A., Hengeveld C.A., Verster F.B. Improved polyethylene cannulation
technique. Physiol. Behav., 4, .123, 1969.
550. Roehrig S., TabbertA., Ferrando-May E. In vitro measurement of nuclear permeability
changes in apoptosis. Analytical Biochemistry, 318, 244-253, 2003.
551. Rollin B.E. An ethicist's commentary on whether veterinarians should support activist
groups. Canad. Veter. Journ., 44, 955, 2003.
552. Романов Ю.А. Общие положения теории пространственно-временной органи-
зации биологических систем. Вестник РАМН, 6, 13-18, 2002.
553. Романов Ю.А., Антохин А.И., Козлова А.Ю. Пространственно-временная орга-
низация пролиферативной системы в эпителии крипты тонкой кишки интакт-
ных мышей. Бюл. эксп. биол. и мед., 12, 678, 2003.
554. Романов Ю.А., Маркина В.В. О пространственно-временных закономерностях
содержания гликогена в дольке печени крыс. Бюл. эксп. биол. и мед., 11, 563,
2003.
555. Rowe W.P., Pincus T. Quantitative studies of naturally occurring murine leukemia virus
infection of AKR mice. /. Exp. Med., v. 135, 429-436, 1972.
556. RowlandM., Blaschke Т.Е., Mefrin P.J. etal. Pharmacokinetic states modifying hepatic
and metabolic function. Am. Pharm. Assoc. Acad. Phann. Sci., Washington, D.C., Chap.,
4, 53-75, 1976.
557. Rudbach J.A., Reed N. Immunological responses of mice to lipopoly-saccharide: lack of
secondary responsiveness by СЗН/HeJ mice. Infect, andlmmun., v. 16, 513-517, 1977.
558. Ruiz V., Ordonez R.M., Berumen J. et al. Unbalanced collagenases/TIMIP-1 expression
and epithelial apoptosis in experimental lung fibrosis. Amer. Journ. of Physiol. - Lung
Cell. andMolec. Physiol., 285, 1026-1036, 2003.
559. Rybkina N.N., Kabanova E.A., Truongdinh K. Interlineal susceptibility of mice to the
causative agent of vesiculous rickettsiosis. Folia microbiol., v. 21, 501, 1976.
560. SacherG.A. Maturation and longevity in relation to cranial capacity in hommed evolution.
Primate Functional Morphology and Evolution. Mouton, The Hague, 417-441, 1975.
561. Sacher G.A. Relationship of lifespan to brain weight and body weight in mammals. Ciba
Found. Colioq. Aging, 5, 115-133, 1959.
562. Sage M.J., Pond W.G., Kroop L. et al. Bone metabolism in thyroidectomized young
pigs. Cornell. Vet., 59, 547, 1969.
563. Sakai Y., Fukuda O., Choi S. H et al Development of a biohybrid simulator for absorption
and biotransformation processes in humans based on in vitro models of small intestine
and liver tissues. /. of Artificial Organs, 6, 273-281, 2003.
564. Saks P., Bendix Kn., Luring N.I. Autologons splenic implantation in pigs. A comparison
of two methods. ActaChir. Scand., 151, 409-411, 1985.
565. Sanderson H, Johnson D.J., Wilson C.J. et al. Probabilistic hazard assessment of
600
Библиография
environmentally occurring pharmaceuticals toxicity to fish, daphnids and algae by
ECOSAR screening. Toxicol Letters, 144, 383-395, 2003.
566. Santoso J. Т., LucciJ.A., Coleman R.L. etal. Saline, mannitol, and furosemide hydration
in acute cisplatin nephrotoxicity: a randomized trial. Cancer Chemother, and Pharmacol.,
52, 13-18, 2003.
567. Sasaki-Nobuo, Yoneda Kumiko, Bigger Ch. Fetal pancreas transplantation in miniature
swine. I. Developmental characteristics of fetal pig pancreas. Transplantation, 38, 335-
340, 1984.
568. Saxen I. Effects of hydrocortisone on the development in vitro of the secondary palate
in two inbred strains of mice. Arch. Oral. Biol., v. 18, 1469-1479, 1973.
569. Schire J.G.M., BartkeA. Strain differences in testicular weight and spermatogenesis with
special reference to C57BL/10J and DBA/2J mice. /. Endocrinol, v. 55,163-171,1972.
570. Schlager G., Dickie M. M. Natural mutation rates the house mouse. Estimates for five
specific loci and dominant mutation. Mutat. Res., v. 11, 89-96, 1971.
571. Schmidf-Nielsen K. Energy metabolism, body size, and problems of scaling. Fed. Proc,
29, № 4, 1524, 1970.
572. Schmidt W., Bocher R., Wolff W. Morphologisch — strukturelle differenzierung
denaturierter orthotoper osteoimplantate in reaktion mit dem lagerknochen. Wiss. Z.
Friedrich-Schiller-Univ., Jena, Naturwiss., 35, 340-341, 1986.
573. Schmidt-Nielsen K. Scaling in biology: the consequences of size. /. Exp. Zool, 194, 297-
308, 1975.
51 A. SeamerJ., Walker R.G. Splenectomy and the spleen weight of young pigs. Res. Vet. Sci.,
1, 125, 1960.
575. Seely S. The evolution of human longevity. Mod. Hypotheses, 6, 873-882, 1980.
576. Seitzer U., Gerdes J. Generation and characterization of multicellular heterospheroids
formed by human peripheral blood mononuclear cells. Cells Tissues Organs, 174, 110-
116, 2003.
577. Семенков В.Ф., Молотков О.В. Уровень кортикостерона у инбредных мышей и
проявление иммунологической реакции на слабые трансплантационные анти-
гены. Докл. АН СССР, т. 214, №6, 1437-1439, 1974.
578. Семенов Х.Х., МалашенкоА. М. Цитологическое исследование ранней эмбриональ-
ной смертности у лабораторных мышей. Бюл. эксп. биол. и мед., 10, 107-110, 1975.
579. Savelieva К. V., Tarasova O.S., Mikoyan V.D. et al. Elevation of brain nitric oxide impairs
learning in rats. Behav. Pharmacol, v. 7, suppl. 1, 100, 1996.
580. Serge E., Westhues M., Siller W.G. etal Veterinary operative surgery. Copenhagen, Medical
Book Co, 1966.
581. Сергеев П.В., Шимановский Н.Л., Петров В.И. Рецепторы. Волгоград, 640, 1999.
582. Стенина М.А., Савчук В.И., Ярыгин В.Н. Изучение наследственной мышечной
дистрофии MDX мышей как гомологичной модели для внедрения клеточных
технологий в практику лечения прогрессирующих мышечных дистрофий у че-
ловека. Бюл. эксп. биол. и мед., 5, 2004.
583. Shalev M. European Commission proposes radical overhaul of animal transport rules.
Lab. Animal, 32, 16, 2003.
584. Shand D.G., Kornhauser G., Wilkinson R. Effects of ration and blood flow on hepatic
drug elimination./. Pharmacol Exp. Ther., 195, 424-432, 1975.
Библиография
601
585. Щегловитова О.Н., Миткевич Л.М. Продукция интерферона — тест для характе-
ристики биологических особенностей организма. В кн.: Использование лабора-
торных животных в разработке, производстве и контроле биологических медицин-
ских препаратов. М., 147-148, 1976.
586. SheppardRM. Natural Selection and Heredity. Harper & Row, N. Y., 129-145, I960.
587. Shire J.G.M., BartkeA. Strain differences in testicular to C57BL/10J and DBA/2J mice.
J. Endocrinol., v. 55, 163-171, 1972.
588. Шмидт Е.Ф. Мандибулярный анализ как экспресс метод генетического конт-
роля лабораторных животных. В кн.: Основные принципы организации биомеди-
цинских экспериментов на животных. М., 27-32, 1979.
589. Шмит К.К., Мейсик К.С., О' Браэн АД. Бактериальные токсины: друзья или вра-
ги? КМАХ, т.2, №1, 2000.
590. Shreffler D., David С, Gotz D. et al. Genetic nomenclature for new lymphocyte antigens
controlled by the I region on the H-2 complex. Immunogenetics, v. 1, 189-190, 1974.
591. Шумова Т.Е., Крышкина В.П., Егоров И.К. Изучение мутаций Н-2 мышей. Ана-
лиз мутации 504 методом гибридов Р1. Генетика, т. 8, № 12, 171-174, 1972.
592. Шустрова И.Е., Ермолаева С.Н. Вирусная диаррея лабораторных мышей. В кн.:
Биология лабораторных животных. М., 3, 109-113, 1971.
593. Siegel В. V., Brown-McKay, Morton J. Interferon induction in New Zealand black mice
by murine leukemia virus. J. Immunol, v. 2, 644-646, 1973.
594. Silverman J., Willan P.C., Depinto A. A. et al. The case of the (over) zealous IACUC
member. Lab Animal, 32, 18-19, 2003.
595. Simenovic ChJ., Dhall D.P., Wilson J.D. A comparative study of transplan sites for
endocrine tissue transplantation in the pig. Austral., J. Exp. Biol, and Med. Sci., 64, №1,
37-41, 1986.
596. Sippl W. Development of biologically active compounds by combining 3D QSAR and
structure-based design methods. /. of Сотр.-Aided Molec. Design, 16, 825-830, 2002.
597. Смирнова Л.А., Ольхова Н.В., Нерадовский В.А. Изучение чувствительности мы-
шей различных линий к возбудителю чумы. В кн.: Лабораторные животные в
медицинских исследованиях. М., 54-60, 1974.
598. Snell G.D. The H-2 locus of the mouse: Observations and speculations concerning its
comparative genetics and its polymorphism. Folia biol. (CSSR), v. 4, 335-358, 1968.
599. Снелл Г., Доссе Ж., Нэтенсон С. Совместимость тканей. М., Мир, 498, 1979.
600. Софронов Г.А., Румак B.C., Епифанцев А.В. Экотоксиканты и здоровье населе-
ния. Вестник РАМН, 11, 24-28, 2002.
601. Sorg R.V., Ozcan Z., Brefort T. et al. Clinical-scale generation of dendritic cells in a
closed system. /. of Immunotherapy, 26, 374-383, 2003.
602. Сорокина Ю.Д., Бландова З.К. Наследственное изменение кожного и шерстного
покрова у мышей BALB/c-walY. В кн.: Биологическая характеристика лабора-
торных животных и экстраполяция на человека экспериментальных данных. М.,
79-80, 1980.
603. Spector W.S. et al. Handbook of Biological Data, W.B. Saunders. Philadelphia, 163-
164,339, 1956.
604. Staats J. Standardized nomenclature for inbred strains of mice: seventh listing. Cancer
Res., v. 40, 2083-2128, 1980.
602
Библиография
605. Stahl W.R. Physiological design criteria. Prof. 17th Ann. Con. Ens. in Med. Biol., 60,
1964.
606. Stahl W.R. Physiological similarity and modelling. In: The Application of Dimensional
Analysis and Physical Similarity Theory to Mammalian Physiology, Appleton-Century-
Crofts, 1970.
607. Stahl W.R. The analysis of biological similarity. Adv. Biol. Med. Phys., 9,355-464, 1963.
608. Steel N.C., Frobish L. Т., Кеепеу М. Lipogenesis and cellularity of adipose tissue from
genetically lean and olese swine. J. Anim. Sci., 39, 712, 1974.
609. Stefanovski D., Moate P.J., Boston R.C. WinSAAM: a windows-based compartmental
modeling system. Metabolism: Clinical and Experimental, 52, 1153-1166, 2003.
610. Steinberg A. D., Pincus Т., Fatal N. The pathogenesis of autoimmunity in New Zealand
mice III. Factors influencing the formation of antinucleic acid antibodies. Immunology,
v. 20, 523-531, 1971.
611. StezU. Gnotobiological models and methods in immunology. Folia Microbiologica, v.
24. №1, 58-69, 1979.
612. Стрекалов А.А. Гемоглобин мышей некоторых инбредных линий. Характерис-
тика фракционного состава. Генетика, т. 3, №7, 39-47, 1967.
613. Stronkhorst J., Schipper С, Brils J. et al. Using marine bioassays to classify the toxicity
of Dutch harbor sediments. Envir. Toxicol, and Chemistry, 22, 1535-1547, 2003.
614. Судаков К.В. Биологические мотивации. М., Медицина, 1971.
615. Судаков К.В. Доминирующая мотивация в архитектуре функциональной систе-
мы. В кн.: Принципы системной организации функций. М., 68-75, 1973.
616. Судаков К.В. Приоритетные фундаментальные исследования интегративной де-
ятельности нервной системы. Вестник РАМН, 9, 3-6, 2003.
617. СудаковКВ. Устойчивость к психоэмоциональному стрессу как проблема био-
безопасности. Вестник РАМН, 11, 15, 2002.
618. Sun H, Pollock KG., BrewerJ. M. Analysis ofthe role ofvaccine adjuvants in modulating
dendritic cell activation and antigen presentation in vitro. Vaccine, 21, 849-855, 2003.
619. Суркова НИ., Малашенко A.M. Мутагенный эффект тиоТЭФ у лабораторных
мышей. Генетика, т. И, №1, 66-72, 1975.
620. Swenson M., Dukes J. Physiology of domestic animals. N.Y., Cornell University Press,
151, 1977.
621. Swindle M.M. Swine as replacement for dogs in the surgical teaching and research
laboratory. Lab. Anim. Sci., 34, №4, 383-385, 1984.
622. Swindle M.M., Horneffer P.J., Gardner T.J. Anatomic and anesthetic considerations in
experimental cardiopulmonary surgery in swine. Lab. Anim. Sci., 36, 357-361, 1986.
623. 51еШ-Суцг&1 A. Bioenergetics. Academic Press, N. Y, 1957.
624. Szent-Gywgyi A. Introduction to a Submolecular Biology. Academic Press, N. Y., 1960.
625. Сент-Дьёрдьи А. Биоэлектроника. Исследование в области клеточной регуля-
ции, защитных механизмов и рака. М., Мир, 1971.
626. Takeyoshi M., Sawaki M., Yamasaki К. et al. Assessment of statistic analysis in non-
radioisotopic local lymph node assay (non-RI-LLNA) with a-hexylcinnamic aldehyde
as an example. Toxicology, 191, 259-263, 2003.
627. Tarasov V.A.,AbilevS.K, Velibekov R.M. etal. Efficiency of batteries of tests for estimating
potential mutagenicity of chemicals. Russian Journal of Genetics, 39, 1191-1200, 2003.
Библиография
603
628. Tasker R.A., Bernard P.B., Doucette ТА. et al. Comparison of the in vitro and in vivo
neurotoxicity of three new sources of kainic acid. Amino Acids, 23, 45-54, 2002.
629. Tavelin S., Taipalensuu J., Hallbook F. et al. An improved cell culture model based on
2/4/A1 cell monolayers for studies of intestinal drug transport: characterization of
transport routes. Pharmaceutical Res., 20, 373-381, 2003.
630. Taylor B.A. Genetic relationships between inbred strains of mice. /. Hered, v. 63, 83-86,
1972.
631. Taylor B.A., Heinger H.J., Meier H. Genetic analysis of resistance to cadmium-induced
testicular damage in mice. Proc. Soc. Exp. Biol, and Med., v. 143, 629-633, 1973.
632. Tanaka K.,Goton F., Tomi S. et al. Inhibition of NO-synthesis induces a significant
reduction in local cerebral blood flow in the rat. Neurosci. Lett., v. 127, 129-132, 1991.
633. Tiozzo R., Boraldi F. Cell-matrix interactions of in vitro human. Tissue and Cell, 35, 37-
45, 2003.
634. Teesalu T, Orr S., Alexandre E. The establishment of a network of European research
tisse banks. Cell and Tissue Banking, 3, 133-137, 2002.
635. Ткаченко Б.И., Евлахов В.И., Поясов И.З. О роли сил vis afronte в формировании
венозного возврата крови к сердцу. Вестник РАМН, 12, 3-9, 2002.
636. Tong W., Welsh W.J., Shi L. et al. Structure-activity relationship approaches and
applications. Envir. Toxicol, and Chemistry, 22, 1680-1695, 2003.
637. Torok M., HuwylerJ., Gutmann H. et al. Modulation of transendothelial permeability
and expression of ATP-binding cassette transporters in cultured brain capillary
endothelial cells by astrocytic factors and cell-culture conditions. Exper. Brain Research,
153, 356-365, 2003.
638. Toth F.D., Vaczi L., Balogh M. Inheritance of susceptibility and resistance to Rauscher
leukaemia virus. Acta Microbiol. Acad. Sci. Hung., v. 20, 183-189, 1973/1974.
639. Труфакин В.А., Афтанас Л.И., Морозова Н.Б. Психонейроиммунологические
эффекты современных психонейротехнологий в преодолении постстрессовых
состояний. Руководство по реабилитации лиц, подвергшихся стрессорным нагруз-
кам. М., Медицина, 400, 121-133, 2004.
640. Цысина Э.Н., Ирлин И.С. Резистентность мышей линии AKR к онкогенному дей-
ствию вируса мышиной саркомы Молони. Бюл. эксп. биол. и мед., т. 72, №10, 85-
88, 1971.
641. Цысина Э.Н., Ирлин И.С. Спонтанная регрессия лейкемии Раушера у мышей
линии AKR. Бюл. эксп. биол. и мед., т. 72, №12, 68-71, 1971.
642. Tsung НС, Du Z. W., Rui R. et al. The culture and establishment of embryonic germ
(EG) cell lines from Chinese mini-swine. Cell Research, 13, 195-202, 2003.
643. Turton J., Shaw D., Bleby J. et al. Organ weights, plasma electrolyte values and blood
parametrs of three inbred strains of guinea-pig. Guinea-Pigs News Lett, 11, 10-31,
1977.
644. Тутельян В.А. Пища и биобезопасность. Вестник РАМН, 10, 14, 2002.
645. Utepbergenov D.L., Blasig I.E. NO and oxyradical metabolism in new lines of rat brain.
Microvascular Res., 62, 114-127, 2003.
646. Уёмов А.И. Логические основы метода моделирования. М., Мысль. 1971.
647. Vaiman M., Lenard Ch., La Fage P. et al. Evidence for a histocompatibility system in
swine (SI-A). Transplantation, 10, №2, 155-164, 1970.
604
Библиография
648. Vaz N. М., de Souza С. M., Hornbrook М. М. et al. Sensitivity to intravenous injections of
histamine and serotonin in inbred mouse strains. Int. Arch. Allergy andAppl. Immunol.,
v. 53, 545-554, 1977.
649. Ведерников А.А., Егоров И.К. Изучение мутаций Н-2 мышей. Генетика, т. 9, №2,
60-66, 1973.
650. Vega F. V, Olaisson Н. Distribution of carbonic anhydraseincells and membranes isolated
from pig gastric mucosa. Acta Physiol. Scand., 124, №4, 573-579, 1985.
651. Voipio N.M., Pekonen H., Nevalainen Т.О. A method for oral administration of tablets
to swine. Z. Versuchstierk, 37, №2, 115-116, 1985.
652. Воробьёв А.А. Современные проблемы микробиологической безопасности. Вест-
ник РАМН, 10, 9, 2002.
653. Walker J.D., Enache M., Dearden J.С. Quantitative cationic-activity relationships for
predicting toxicity of metals. Envir. Toxicol, and Chemistry, 22, 1916-1935, 2003.
654. Walker J.D., Jaworska J., Comber M.H.I, et al. Guidelines for developing and using
quantitative structure-activity relationships. Envir. Toxicol, and Chemistry, 22, 1653-
1665, 2003.
655. Wang Z., Gorski J.C., Hamman M.A. The effects of St. John's wort (Hypericum
perforatum) on human cytochrom P450 activity. Clin. Pharmacol. Ther., 70, 317-26,
2001.
656. Warren S.M., Nacamuli R.K., Song H.M. et al. Tissue-engineered bone using
mesenchymal stem cells and a biodegradable scaffold. J. of Craniofacial Surg., 15, 34-
37, 2004.
657. Wason S., Pohlmeyer-Esch G., Pallen С etal. 17a-methyltestosterone: 28-day oral toxicity
study in the rat based on the "Enhanced OECD Test Guideline 407" to detect endocrine
effects. Toxicology, 192, 119-137, 2003.
658. Wiebusch H, PoirierJ., Sevigny P. Further evidence for a synergistic association between
APOE epsilon-4 and BCHE. Hum. Genet., 104, 158-163, 1999.
659. Watson J., Riblet R. Genetic control of responses to bacterial lipopolysaccharides in
mice. I. Evidence of a single gene that influences mitogenic and immunogenic responses
to lipopolysaccharides. /. Exp. Med., v. 140, 1147-1161, 1974.
660. Weaver M.E., Sorenson F. W., Jump E.B. The miniature pig as an experimental animal
in dental research. Arch. Oral. Biol., 7, 17, 1962.
661. Вейн A.M., Судаков К.В., Левин Я.И. и др. Особенности структуры сна и личности
в условиях хронического эмоционального стресса и методы повышения адапта-
тивных возможностей человека. Вестник РАМН, 4, 13, 2003.
662. Weinstein Y., Givol D., Strausback P. Differences in the constitution of antibody
combining site in different mice strains. Europ. J. Immunol, v. 2, 186-188, 1972.
663. Weirich W.E., Will Y.A., Crumpton C.W. A technique for placing chronic indwelling
catheters in swine /. Appl. Physiol, 28, 117, 1970.
664. Whalsten D. Heritable aspects of anomalous myelinated fibre tracts in the forebrain of
the laboratory mouse. Brain Res., v. 68, 1-18, 1974.
665. White F.C., Roth D.M., Bloor CM. The pig as a model for myocardial ischemia and
exercise. Lab. Anim. Scl, 36, 351-356, 1986.
666. Wilkinson G.R., Shand D.G. Physiological approach to hepatic drug clearance. Clin.
Pharmac. Ther., 18, 377-390, 1975.
Библиография
605
667. Williams D. The engineering of an artery. Med. Device Technology, 14, 9-11, 2003.
668. Williamson I., Festing M. A note on some haematological parametrs in three strains of
guinea-pigs. Guinea-Pigs News Lett., 3, 7-11, 1971.
669. Wilson M.R. Immunological development of the neonatal pig. /. Anim. Sci., 38, 1018,
1974.
670. Wingate M.B., Wingate L. Experimental ileotubal anastomosis in the sow. Am. J. Obstet.
Gynecol., 105, 1176, 1969.
671. Wingfield W. E., Tumblesson M.E., Hicklin K. W. et al. An exteriorized cranial vena caval
catheter for serial blood sample collection from miniature swine. Lab. Anim. Sci., 24,
359, 1974.
672. Wise M.H., Gordon C., Johnson R. W.G. Intraportal autotransplantation ofcryopreserved
porcine islets of langerhans. Criobiol, 22, 359-366, 1985.
673. Witzel D.A., Litlledike E. Т., Cook H.M. Implanted catheters for blood sampling in swine.
Cornell. Vet., 63, 432, 1973.
61 A. Wolfe L., Griesmer R., Rohovsky M. Germfree cynomolgus monkeys. Lab. Animal Care.,
16, 364, 1966.
675. Wollensak G, Sporl E., Reber F. et al. Corneal endothelial cytotoxicity of riboflavin/
UVA treatment in vitro. Ophthalmic Res., 35, 324-328, 2003.
676. Wonigeit K., Gunter E. Differentiation antigens of T-lymphocytes. Rat. News Lett., 2,
12-13, 1977.
611. Wood J. D. Fat mobilization in large white pigs. Proc. Nutr. Soc, 33, 61, 1974.
678. Wood Т.К., Sullivan A.M., McDermott K.W. Viability of dopaminergic neurones is
influenced by serum and astroglial cells in vitro. J. of Neurocytology, 32, 97-103, 2003.
679. Woodle E.S., Hunter G.C., Eisele P. Ortotopic porcine liver transplantation. Operative
technique. J. Surg. Res., 39, 483-488, 1985.
680. Woodruff J. F., Woodruff J.J. Involvement of T-lymphocytes in the pathogenesis of
coxsackie virus B3 heart disease. /. Immunol, v. 113, 1726-1734, 1974.
681. Wright J.L., Farmer S.G., Churg A. A neutrophil elastase inhibitor reduces cigarette
smoke-induced remodelling of lung vessels. The European Respiratory Journ., 22, 77-
81, 2003.
682. Yamamoto A., Ishiguro H. et al. Ethanol induces fluid hypersecretion from guinea-pig
pancreatic duct cells. /. of Physiology, 551, 917-926, 2003.
683. Yang J.M., Arnush M., Chen Q. Y. et al. Cadmium-induced damage to primary cultures
of rat Leydig cells. Reproductive Toxicol, 17, 553-560, 2003.
684. Yang P., Hernandez M.R. Purification of astrocytes from adult human optic nerve heads
by immunop anning. Brain Res. Protocols, 12, 67- 76, 2003.
685. Ярыгин В.Н., Сухих Г. Т, Ситников В.Ф. и др. Ксенотрансплантация эмбриональ-
ных предшественников миогенеза человека для коррекции дистрофиопатии у
мышей с наследственной миодистрофией. Бюл. эксп. биол. и мед., 7, 100, 2003.
686. Yoon T.J., Lei ТС, Yamaguchi Y. et al. Reconstituted 3-dimensional human skin of
various ethnic origins as an in vitro model for studies of pigmentation. Analytical
Biochemistry, 318, 260-269, 2003.
687. Закусов В.В. Итоги и перспективы развития научных исследований в области
фармакологии. Вестник АМН СССР, 11, 47-53, 1977.
688. Zawia N.Н. Transcriptional involvement in neurotoxicity. Toxicol andAppl. Pharmacol,
606
Библиография
190, 177-188, 2003.
689. Зеленцов А.Г. Восприимчивость линейных мышей к гельминтам II. Развитие
Opistorchis felineus у мышей линий А/Не, CBA/Lac, CC57M/Mv, C57BL/6J,
DBA/2J, SWR/J. Мед. паразитол. и паразитарные болезни, т. 43, №1, 95-98,1974.
690. Zheng W., Aschner M., Ghersi-Egea J.F. Brain barrier systems: a new frontier in metal
neurotoxicological research. Toxicol. andAppl. Pharmacol, 192, 1-11, 2003.
691. Зилов В.Г., Миненко И.А., Смекалкин Л.М. Комплексная нелекарственная кор-
рекция посттравматических стрессовых расстройств у участников боевых дей-
ствий. Руководство по реабилитации лиц, подвергшихся стрессорным нагрузкам.
М., Медицина, 400, 194-206, 2004.
692. Зверев В.В., Семенов Б.Ф. Вакцинопрофилактика при биотерроризме и биоката-
строфах. Вестник РАМН, 10, 39, 2002.
693. Подопригора Г. И. Особенности асептического воспаления у безмикробных мор-
ских свинок. Бюл. эксп. биол. и мед., 11, 35-38, 1970.
694. Sutton К., Siok С, Stea A. et al. Inhibition of neuronal calcium channels by a novel
peptide spider toxin, DW13.3. Molec. Pharmacol., 54, 407-418, 1998.
695. Побопригора Г.И. Методологические аспекты гнотобиологии в медикобиологи-
ческих исследованиях. В кн.: Проблемы современной иммунологии. Новосибирск,
123-133, 1988.
696. Каркищенко Н.Н., Каркищенко В.Н., Степанов А.А. Способ получения раствора
нитроглицерина 0,1 % для иньекций. Патент на изобретение № 2229295, 2004.
697. Лазарев Н.В., Грех И.Ф. Пиримидины и терапия. Врачебное дело, №2,1959.
698. Teschemacher H, Koch G., Brantl V. Milk protein-derived opioid receptor ligands.
Biopolymers, v. 43(2), 99-117, 1997.
699. Williams A.J., Dave J.R., Phillips J.B. et al. Neuroprotective efficacy and therapeutic
window of the high-affinity N-methyl-D-aspartate antagonist conantokin-G: in vitro
(primary cerebellar neurons) and in vivo (rat model of transient focal brain ischemia)
studies. /. Pharmacol. Exp. Ther., 294(1), 378-86, 2000.
700. Yang S., Yunden J. et al. Rubiscolin, and selective opioid peptide derived from plant
Rubisko. FEBSLett., 509, 213-217, 2001.
701. Yatskin O.N., Philippova M.M., Blishchenko E.Yu. et al. LW- and W-hemorphins:
comparative levels in rat tissues. FEBS Lett., 428, №3, 286-90, 1998.
702. Zadina J.E., Martin-Schild S., GerallA.A. et al. Endomorphins: novel endogenous mu-
opiate receptor agonists in regions of high u.-opiate receptor density. Ann., N. Y, Acad.
ScL, v. 897, 136-144, 1999.
703. Zandstra P.W., Bauwens C, Yin T. et al. Scalable production of embryonic stem
cellderived cardiomyocytes. Tissue Engineering, 9, 767-778, 2003.
704. Zhou L., Li J., Goldsmith A.M. et al. Human bronchial smooth muscle cell lines show a
hypertrophic typical of severe asthma. Am. J. ofResp. and Critical Care Med., 169, 703-
711, 2004.
СПИСОК СОКРАЩЕНИЙ
АКАТОЛ — аденокарцинома толстой кишки
AT III — антитромбин III
БАВ — биологически активные вещества
ВМР-П — высокометастазирующий рак
ВПГ — вирус простого герпеса
ДМБА — индуцированная опухоль молочной железы
МФП+ — 1-метил-4-фенилпиридиний
МФТП — 1-метил-4-фенил-1,2,3,6-тетрагидропиридин
NMDA — N-метил-О-аспартат
НПВС — нестероидное противовоспалительное средство
НСТ — тиалактон гомоцистеин
ОА — окись азота
ПАФ — полный адьювант Фрейнда
ПС — паркинсонический синдром
РШМ — рак шейки матки
ТНХБ — 2,4,6-тринитрохлорбензол
УРАИ — условный рефлекс активного избегания
УРПИ — условный рефлекс пассивного избегания
ФС — фармакологическое средство
ХС — химическое средство
ХЯ — хвостатое ядро
Книгу можно приобрести по адресу: www.ozon.ru
Дополнительную информацию о книге можно
получить по адресу: www.labanimals.ru
Available at www.ozon.ru & www.labanimals.ru
Николай Николаевич Каркищенко
ОСНОВЫ БИОМОДЕЛИРОВАНИЯ
Научное редактирование: доц. Р.Г. Костогрызова
Подготовка рукописи и корректура: к.м.н. Е.В. Брайцева, Е.В. Иванова
Техническое редактирование
и компьютерная верстка: Б.М. Бороденков
Формат 70 х 100/16. Бумага офсетная. Гарнитура «Ньютон». Печать офсетная.
Усл.-печ. л. 49,2. Тираж 1500. Заказ тип. № 2098
Межакадемическое издательство «Вооружение. Политика. Конверсия»,
107564, г. Москва, 1-я Мясниковская ул., вл. 3, тел./факс (095) 169-51-97
ОАО «Чеховский полиграфический комбинат»
142300, г. Чехов Московской области тел: (272) 71-336, факс (272) 62-536