/











































































































































































































































































































































































































































































Текст
РОСТОВСКИЙ ОРДЕНА ТРУДОВОГО КРАСНОГО ЗНАМЕНИ
ГОСУДАРСТВЕННЫЙ УНИВЕРСИТЕТ
л. и. номоконов
ОБЩАЯ
БИОГЕОЦЕНОЛОГИЯ
Ответственный редактор
доктор биологических наук
И. X. Блюменталь
ИЗДАТЕЛЬСТВО РОСТОВСКОГО УНИВЕРСИТЕТА, 1989
Н81
Печатается по решению учебно-научного центра
биолого-почвенного факультета
Ростовского университета
Рецензенты:
кафедра геоботаники Днепропетровского ордена Трудового
Красного Знамени
государственного университета,
доктор биологических наук, академик АН КазССР
Б. Л. Быков
Номоконов Л. И.
Н81 Общая биогеоценология. Издательство Ростовского
университета, 1989. 456 с.
В монографии, в отличие от весьма распространенных взглядов, био-
геоценологня рассматривается не в качестве одного из разделов экологии,
а как самостоятельная синтетическая наука о сложных по составу и струк-
турно-функциональной организации бнокосных системах — биогеоценозах
и биогеоценотическом покрове Земля.
В книге описываются все стороны и основные свойства двуединого
объекта биогеоценологии: строение, функционирование, классификация,
взаимосвязь со средой, распределение на земной поверхности, изменение
во времени, развитие, в том числе и водных ценосистем — биогидроценозов.
Книга рассчитана на преподавателей, научных работников, учителей,
специалистов в области сельского, лесного и рыбного хозяйства. Она так-
же может служить учебным пособием для студентов и аспирантов вузов
соответствующей специальности.
Н
2001050000-051
Ml 75(03)—89
без объявления
ISBN 5-7507—0081-х
© Издательства Ростовского университета, 1989
ОГЛАВЛЕНИЕ
Пре ди с лов и е 7
Введение 9
Глава 1. ЭТАПЫ РАЗВИТИЯ БИОГЕОЦЕНОЛОГИЧЕСКНХ
ИДЕЙ 13
1. Возникновение биогеоцейологических идей в СССР и
за рубежом 13
2. Понятие экосистемы и биогеоценоза 15
3. Экология, бногеоценология и место их в системе наук 19
4. Развитие биогеоценологии в нашей стране и распро-
странение ее идей за рубежом 27
5. Научное и народнохозяйственное значение биогеоце-
нологни 30
Глава И. БНОГЕОЦЕНОЛОГИЯ И ЕЕ ОБЪЕКТ 32
1. Бногеоценология как наука 32
2. Непрерывность и дискретность биогеоценотического
покрова 32
3. Биогеоценоз и его состав по Сукачеву 35
*4. Биогеоценоз как система и взаимосвязь era составных
элементов 42
б. Развитие представлений о биогеоценозе и его составе 48
6. Границы биогеоценозов и ценокомплексы видовых це-
нопопуляций 52
7. Биосфера и ее планетарная роль по Вернадскому 55
8. Современное представление о биосфере и взаимоот-
ношении с ней человека 61
9. Бяогеосфера, типы ее строения и соотношение с био-
геоценозом 65
Глава III. ЖИВЫЕ КОМПОНЕНТЫ БИОГЕОЦЕНОЗА И ИХ
СОСТАВНЫЕ ЭЛЕМЕНТЫ 69
1. Виды и видовые популяции в составе живых компо-
нентов биогеоценоза 69
2. Жизненные формы (экобиоморфы) и системы эко-
биоморф растений и животных 71
3. Фитоценотипы, зооценотипы и биоценотипы 79
Глава IV. СОСТАВ И ФУНКЦИОНАЛЬНАЯ ДЕЯТЕЛЬНОСТЬ
ЖИВЫХ КОМПОНЕНТОВ БИОГЕОЦЕНОЗА 85
1. Состав и функциональная деятельность автотрофного
компонента биогеоценоза 85
3
2. Состав и функциональная деятельность биотрофного
компонента биогеоценоза 91
3. Состав и функциональная деятельность сапротроф-
иого компонента биогеоценоза 101
Глава V. КОСНЫЕ КОМПОНЕНТЫ БИОГЕОЦЕНОЗА, ИХ
СОСТАВ И ФУНКЦИОНАЛЬНАЯ РОЛЬ В БИОГЕОЦЕНОЗЕ 116
I, Аэротои и его материальная основа 117
2. Составные элементы и функциональная роль атмосфе-
ры в биогеоценозе 119
3. Преобразование атмосферы во внутреннюю воздуш-
ную среду биогеоценоза 132
4. Эдафотоп и его материальная основа 139
5. Почвообразование, его факторы и энергетика 142
6. Почва, ее состав, строение, мощность и характерные
черты 146
7. Генетические свойства и функциональная роль почвы
в биогеоценозе 150
8. Преобразование почвы во внутреннюю почвенную сре-
ду биогеоценоза 154
Глава VI. ВЗАИМОСВЯЗЬ БИОЛОГИЧЕСКИХ И БИОКОСНЫХ
СИСТЕМ СО СРЕДОЙ 158
1. Среда биологических и бнокосных систем и ее под-
разделение 158
2. Взаимосвязь биологических систем со средой 161
“3. Взаимосвязь бнокосных систем с природными факто-
рами среды 166
4. Взаимосвязь биокосных систем с социально-производ-
ственными факторами среды 178
Глава VII. СТРУКТУРНО-ФУНКЦИОНАЛЬНАЯ ОРГАНИЗА-
ЦИЯ БИОГЕОЦЕНОЗА 184
1. Структурно-функциональная организация биогеоце-
ноза, ее различные аспекты и назначение 184
2. Типы взаимоотношений между ценопопуляциями и
живыми компонентами биогеоценоза 186
‘3. Механизмы и условия сосуществования различных
видов в составе биогеоценоза 193
4. Пищевые цепи и сети, их состав, строение, формиро-
{ание и закономерности строения 197
. Понятие консорции, ее состав и строение в трактовке
разных авторов 206
4
*6. Многообразие консорций, их классификация и изме-
нение во времени 212
Консорция как основная функциональная структур-
ная единица биогеоценоза 215
8. Системообразующие единицы, их место и роль в ста-
новлении и функциональной организации биогеоценоза 219
Ярусность фитоценоза и пространственное размеще-
’Еие в нем животного и микробного населения 225
Морфологическое строение биогеоценоза, вертикаль-
ные и горизонтальные структурные части его 231
Глава VIII. МАТЕРИАЛЬНО-ЭНЕРГЕТИЧЕСКИЙ ОБМЕН И
ФУНКЦИОНИРОВАНИЕ БИОГЕОЦЕНОЗА 249
1. Материально-энергетический обмен в биогеоценозе и
круговорот веществ в природе 249
2. Биогеоценотический обмен веществ, циклы и типы
биологического круговорота 243
3. Биогеохимические циклы биофильных химических эле-
ментов в биогеосфере 249
4. Поступление солнечной энергии на Землю н ее ис-
пользование в биогеоценозе и биогеосфере 258
5. Трофическая структура, типы обмена веществ, носи-
тели и каналы переноса веществ и энергии в биогеоце-
нозе 263
6. Ход обмена веществ, поток энергии и роль живых
компонентов в обмене и функционировании биогеоце-
ноза 272
7. Биогеоценоз н биогеосфера как саморегулирующиеся
системы; механизмы и факторы саморегуляции 282:
Глава IX. КЛАССИФИКАЦИЯ БИОЦЕНОЗОВ И БИОГЕОЦЕ-
НОЗОВ И ТЕРРИТОРИАЛЬНОЕ ПОДРАЗДЕЛЕНИЕ БИОГЕО-
СФЕРЫ 290
1, Топологическая система классификации биоценозов
по А. Клюгу 290
2. Подход к классификации биогеоценозов н других
природных единств Сочавы, Долуханова и др. 291
3. Принципы классификации биогеоценозов по Сукачеву
и Дылису 293
4. Система классификации лесного типа биогеосферы 296
5. Межбиогеоценозные связи как основа единства, це-
лостности биогеосферы, их типы и механизмы 299
6, Территориальное объединение биогеоценозов в биогео-
с нс те мы и бногеомассивы по Бялловичу 301
7. Гео растительные системы Титова и топологические
геосистемы Сочавы 307
8, Представление Бялловича о пространственной струк-
туре биогеосферы н территориальном подразделении ее 309
9. Система пространственного подразделения биогеоце-
нотической оболочки Земли 310
10. Территориальное подразделение биогеосферы зару-
бежными экологами 316
Глава X. СТАНОВЛЕНИЕ И ДИНАМИКА БИОГЕОЦЕНОЗОВ
И ЭВОЛЮЦИЯ БИОГЕОЦЕНОЗОВ И БИОГЕОСФЕРЫ 318
1. Понятие о динамике биогеоценозов и формах ее
проявления 318
2. Циклические изменения биогеоценозов и понятие о
коренных изменениях и развитии их 319
3. Сингенез как процесс формирования биогеоценоза;
факторы формирования и стадии сингенеза 323
4. Эндогенеэ, эндогенные сукцессии биогеоценозов и
движущие силы развития биогеосферы 328
5. Экзогенные сукцессии и подразделения их по причи-
нам, скоротечности и продолжительности 334
6. Теория климакса, критика ее и современные взгляды
на устойчивость и динамику биогеоценозов 337
7. Понятие филоценогенеза и закономерности развития
растительности и биогеосферы 342
8. Современные представления о сопряженном характере
эволюции всех форм жизни на Земле 346
9. Становление и эволюция бнокосных систем разного
ранга 350
10. Факторы и механизмы органической эволюции и эво-
люционный прогресс биокосных систем 358
Глава XI. ВОДНАЯ СФЕРА ЖИЗНИ (БИОГИДРОСФЕРА) 363
1. Водная среда жизни и ее обитатели (гидробионты) 363
2. Континентальные водоемы и их классификация 367
3. Население континентальных водоемов и формирова-
ние живых компонентов биогидроценозов 371
4. Водные биокосные системы — бногидроценозы н их
особенности 376
5. Структурно-функциональная организация континен-
тальных биогидроцеиозов 378
6. Обмен веществ и энергии в биогидроценозах и типы
биотических круговоротов 384
7. О динамике континентальных биогидроцеиозов 388
6
8. Мировой океан, его гидрологическая структура и
стратиграфия водных масс и дна 389
9. Биотопы пелагяали и океанического дна 394
10. Население, зоны его концентрации в океане и осо-
бенности океанических биогидроценозов 397
11. Биогидроценозы планктонной пленки жизни, состав
н трофическая взаимосвязь их компонентов 400
12. Вертикальная структура бногидроценозов планктон-
ной пленки жизни океана 407
13. Биогидроценозы бентали, их компонентный состав и
население батиали, абиссали и коралловых рифов 409
14. Прибрежные и другие сгущения жизни и компонент-
ный состав их бногидроценозов 413
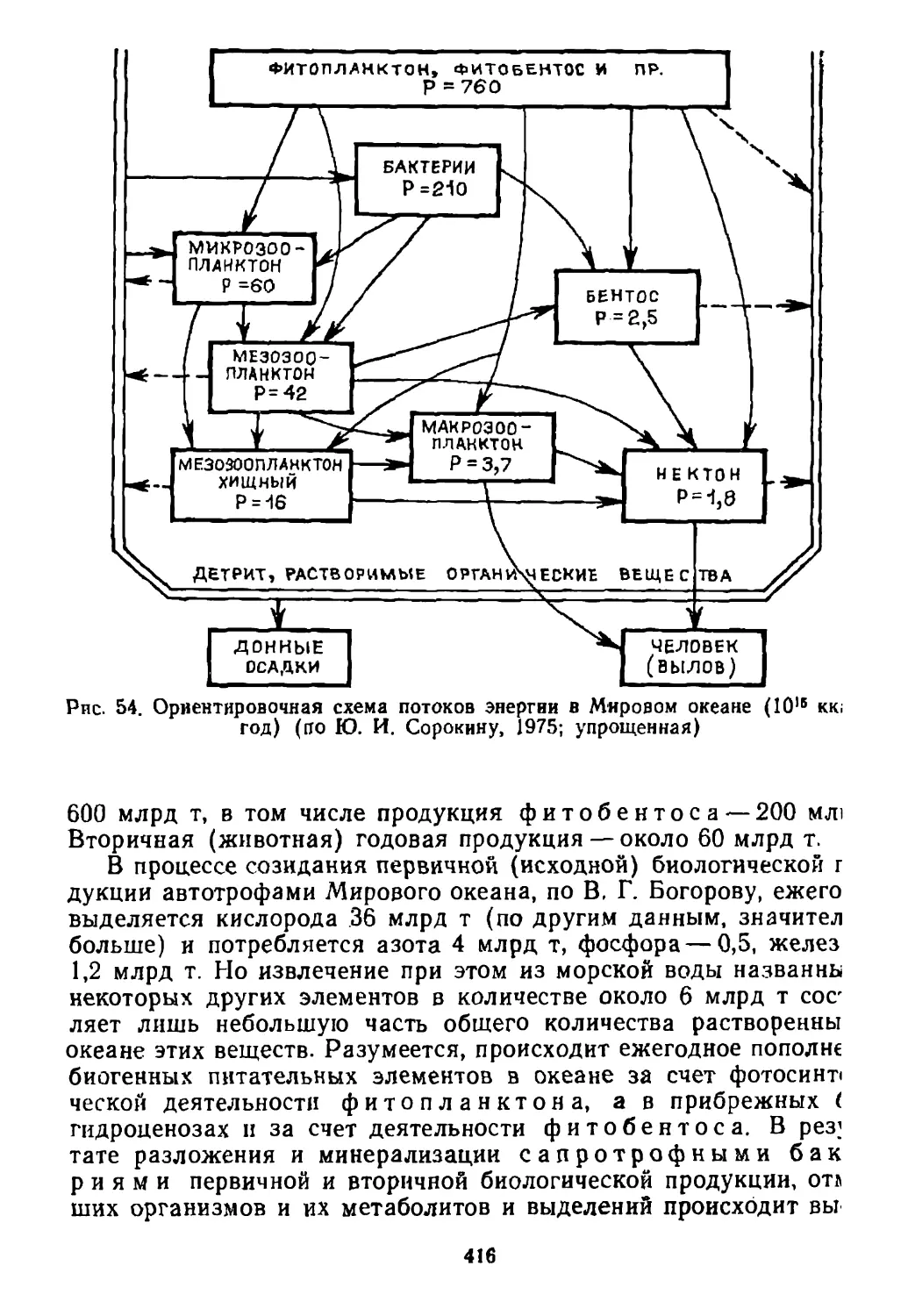
15. Обмен веществ и энергии в условиях Мирового
океана 415
16. Динамика бногидроценозов и этапы эволюции Ми-
рового океана 417
Заключение 420
Использованная литература 425
Предметный указатель 438
ПРЕДИСЛОВИЕ
Бногеоценология — наука молодая, возникшая в результате
развития и взаимодействия ряда биологических наук и наук о не-
живой (неорганической) природе. Обоснование биогеоценологии
как особой науки, науки о биокосных системах — биогеоценозах —
осуществил академик В. Н. Сукачев в начале 40-х годов текущего
столетия.
Идея о комплексном изучении природных явлений характерна
для русских и советских естествоиспытателей. В. В. Докучаев, осно-
ватель генетического почвоведения, рассматривал почвенный по-
кров как результат взаимодействия материнских пород раститель-
ности, животного населения и климата в их историческом развитии
с учетом хозяйственной деятельности человека. В. И. Вернадский
в начале 20-го века основал и развил учение о биосфере как особой
оболочке Земли, возникшей при активном взаимодействии живого
вещества с другими геосферами земного шара.
Автором книги систематизированы, обобщены и должным обра-
зом оценены современные достижения отечественных и зарубеж-
ных ученых в области биогеоценологии. Хорошо охарактеризованы
этапы развития биогерценологических идей у нас и за рубежом,
обсуждены вопросы о существе и объеме понятий «биогеоценоз» и
«бногеоценология» в сопоставлении с соответствующими понятиями
и терминами зарубежных ученых. Должным образом освещены
автором прогрессивные взгляды в этой области советских ученых —
В. И. Вернадского, В. Н. Сукачева, В. Р. Вильямса и др. В книге
детально рассмотрен состав биогеоценозов из живых и косных ком-
понентов, их взаимосвязь. Большое внимание уделено морфологи-
ческой структуре и функциональной организации биогеоценозов и
материально-энергетическому обмену в них. Особо рассмотрены
вопросы динамики биогеоценозов, их эволюции, принципов типо-
логии биогеоценозов и территориального подразделения биосферы.
Отдельная глава посвящена рассмотрению состава, структуры и
функциональной организации водного сектора биосферы (биогид-
росферы). Сложные вопросы биогеоценологии изложены автором
ясно и доступно. Книга будет прекрасным пособием для всех име-
ющих отношение к использованию и охране природных ресурсов.
Книга имеет и мировоззренческое значение, а потому она по-
лезна для всякого культурного человека.
И. X. Блюменталь
8
Посвящается
светлой памяти
Николая Владиславовича
ДЫЛИСА
ВВЕДЕНИЕ
В современной философской и биологической литературе при-
нято различать четыре основных уровня организации живой мате-
рии или, иначе, четыре формы жизни на Земле: организменную»
популяционно-видовую. биоценотическую и биостроматическую
(Лавренко, 1964, 1965; Завадский, 1966; Игнатов, 1966; Сетров,
1971). Некоторые биологи — Б. П. Ушаков (1963), Н. П. Наумов
(1964), Л. Берталанфи (1969) и др.— выделяют до десятка и более
уровней организации живой материи (молекулярный, клеточный,
органно-тканевый и т. п.). Но, как показал Завадский, в данном
случае смешиваются разные понятия: уровни организации живой
материи (формы жизни), структурные элементы каждой из этих
форм жизни и уровни исследования живой материи. Так, клетка
в многоклеточном организме — структурный элемент в его составе»
а не самостоятельная форма жизни; широко распространенные 9
природе одноклеточные живые существа функционируют по зако-
нам организменной формы жизни. Нечто подобное можно сказать
и о молекуле. Речь может идти, скажем, о молекулярном уровне
исследования организменной формы жизни, но не о молекуле как
самостоятельной специфической форме жизни. Названные выше
четыре формы жизни вслед за В. И. Вернадским (1928) большин-
ством ученых рассматриваются в настоящее время как основные
первичные носители жизни на Земле.
На протяжении многих столетий объектами исследования био-
логов всего мира были две формы жизни — организменная и по-
пуляционно-видовая. На первых этапах развития биологии это бы-
ло неизбежно и исторически оправданно. На этом пути биология
достигла больших успехов и еще далеко не исчерпала и, очевидно»
никогда не исчерпает всех возможностей, которые заложены в изу-
чении органического мира па этих уровнях организации живой
материи. Но с 1935 года за рубежом и с 1940 года в нашей стране
наряду с этим началось систематическое изучение биоценотической
формы жизни в виде образуемых ею таких биокосных систем, как
биогеоценоз и биогеоценотический покров земной
поверхности — по Сукачеву (1940) или экосистемы разного
размера —по Тенсли (Tansley, 1935). Несколько ранее, а именно
с 1926 года, началось изучение и биостроматической формы орга-
низации живой материи, ибо биострома, по Е. М. Лавренко (1964)
9
(фитострома, по Г Н. Вы-
соцкому, 1925), и представ-
ляет собою живое веще-
ство в трактовке В. И. Вер-
надского (1926),которое при
активном взаимодействии с
геофизическими оболочками
планеты и образует особую
оболочку земного шара —
биосферу. В связи с этим
напрашивается вывод: био-
логия в узком смысле мо-
жет рассматриваться как
наука о биологических си-
стемах (организмах и видо-
вых популяциях), биогеоце-
нология — как наука о био-
косных системах разного
ранга, а учение о биосфе-
ре— как особая самостоя-
тельная наука о глобальных
проявлениях живого веще-
ства на нашей планете в
широких вертикальных пре-
делах с учетом также дея-
тельности человеческого об-
щества.
Биогеоценология — мо-
лодая наука: со дня возник-
новения ее не прошло и пятидесяти лет. Она представляет собою
синтетическую науку о весьма сложных по составу и структурно-
функциональной организации биокосных системах — биогеоценозах
и образуемом ими биогеоценотическом покрове Земли. Основопо-
ложником биогеоценологии является выдающийся советский уче-
ный— академик В. Н. Сукачев.
Развитие биогеоценологии в нашей стране протекает довольно
успешно. За короткий срок она вышла на передовые позиции среди
наук о природе. Возникновение и относительно быстрый ход раз-
вития биогеоценологии подготовлен историей и логикой развития
классической биологии и диалектико-материалистической идеи о
взаимосвязи и взаимообусловленности всех явлений природы. Со-
циально-производственной базой такого хода ее развития являют-
ся научно-технический прогресс и порожденные им социальные,
эколого-ценотические и другие региональные и глобальные про-
блемы всего человечества.
ю
Разумеется, не по всем вопросам, определяющим содержание и
контуры науки биогеоценологии, имеются вполне ясные представ-
ления, не все основополагающие понятия данной науки, назначе-
ние которых — отражать реальные явления и протекающие в био-
геоценотических системах процессы, достаточно четко определены
и сформулированы. Многое еще является проблематичным, иногда
спорным и, чтобы стать общепризнанным, нуждается в дальнейшей
углубленной разработке. Все это вполне естественно и объясняет-
ся, с одной стороны, молодостью науки биогеоценологии, недоста-
точной разработанностью ряда стоящих перед ней проблем, с дру
гой — и это главное—чрезвычайной сложностью объекта исследо-
вания биогеоценологии, его многоранговостью и в связи с этим —
наличием множества разнообразных, сложно переплетающихся
взаимосвязей и взаимоотношений между его компонентами и сла-
гающими их элементами. Правда, некоторые из критических заме-
чаний и возражений биологов в адрес основоположника науки
биогеоценологии и его последователей, как будет видно, основань
на недоразумении, другие хотя и имеют некоторые основания, но
в свою очередь, не вполне верпы; высказываются некоторыми био
логами и такого рода возражения, которые методологически оши
бочны, а потому должны быть отвергнуты.
Из всего изложенного следует, что биогеоценологии, а через нес
и учению о биосфере в целом, должно принадлежать весьма важ
ное место в плане современной подготовки специалистов среднего
а через аспирантуру и высшего звена в области биогеоценологии
Она представляет собою фундаментальную научную основу под
готовки кадров в области охраны природы и природопользования
ибо основополагающие идеи и достижения биогеоценологии явля
ются теоретической базой многих мероприятий по охране и ради
овальному использованию природных ресурсов биосферы и реше
ния ряда народнохозяйственных проблем.
Возникает вопрос: каким должен быть подход к изложении
основного содержания книги? Очевидно, чтобы вызвать активно*
творческое отношение молодежи и зрелых специалистов к овладе
нию знаниями в области данной научной дисциплины и использо
ванию их в своей практической деятельности, необходимо сочетат
категоричное, утвердительное изложение материала с показом на
личных идей и теоретических построений представителей разны:
научных направлений в области биогеоценологии, различной трак
товки тех .или иных положений рассматриваемой проблемы.
Придерживаясь принципа преемственности биогеоценологиче
ских идей основоположника этой науки и его соратников и после
дователей, продолжающих дальнейшее развитие этих идей в со
временных условиях, мы стремимся при разработке, изложении ил1
трактовке тех или иных положений данной науки опираться в;
11
фундаментальные достижения современной биогеоценологии. В со-
ответствии со своими взглядами на содержание экологии и био-
геоденологии. их соотношение и место в системе наук о природе и
обществе (Номоконов, 1981) мы стремимся также, в меру наших
возможностей, использовать в данной монографии соответствую-
щую по содержанию позитивную информацию зарубежных ученых
о биокосных системах разного ранга и современные достижения их
в этой области.
Автор считает своим долгом выразить глубокую благодарность
академику Казахской АН Б. А. Быкову и профессорам Днепропет-
ровского университета А. П. Травлееву и А. Л. Бельгарду за це-
лый ряд замечаний, рекомендаций и пожеланий, сделанных при
рецензировании данной монографии в рукописи. Чувство особой
признательности и благодарности автор испытывает к профессору
Ленинградского университета И. X. Блюменталю за огромный труд
по научному редактированию при подготовке рукописи к изданию.
Все это вместе способствовало значительному улучшению содер-
жания предлагаемой читателю книги.
Вместе с тем, ввиду неясности и дискуссионного характера ряда
теоретических положений науки биогеоценологии, автор будет при-
знателен лицам за мотивированные высказывания и предложения,
способствующие более точному, отвечающему истине решению та-
кого рода вопросов.
Глава I
Этапы развития биогеоценологических идей
1. Возникновение биогеоценологических идей в СССР
и за рубежом
Всякая наука, как известно, характеризуется тремя чертами.
Во-первых, наличием присущего ей специфического объекта или
предмета исследования. Предмета исследования в том случае, ко-
гда объект исследования (растение в ботанике, животное в зооло-
гии и т. п.) изучается не в полном объеме, а лишь с одной какой-
либо стороны, например: внешнее или внутреннее строение объек-
та, взаимосвязь его со средой обитания, классификация объектов
или распределение их в пространстве и т. п. Во-вторых, всякая
наука обладает свойственными ей специфическими методами ис-
следования, обеспечивающими возможность реального и эффектив-
ного познания этого объекта или какой-либо из сторон его.
В-третьих, всякая наука имеет свою относительно самостоятельную
историю. Конечно, возникновение и развитие любой науки обуслов-
лено потребностями человеческого общества, уровнем развития
его производительных сил, состоянием и общим ходом его соци-
ально-экономического развития, но наука, как одна из форм обще-
ственного сознания, обладает относительной самостоятельностью
и логикой своего исторического развития.
История биогеоценологии весьма коротка. Ее возникновение в
нашей стране относится к началу сороковых годов текущего сто-
летия. Возникновение и довольно быстрое развитие биогеоценоло-
гии в послевоенный период было обусловлено, с одной стороны,
потребностями нашего общества в восстановлении разрушенного
войной народного хозяйства и дальнейшем развитии производи-
тельных сил страны, а также воздействием научно-технического
прогресса на окружающую среду, а с другой — созданием истори-
ческих и логических предпосылок развития биологической и смеж-
ных с ней наук о природе.
Лежащая в основе биогеоценологии идея о взаимосвязи и вза-
имообусловленности всех явлений природы — одна из фундамен-
тальных идей как в естественных, так и в гуманитарных науках, и
в частности" философии. Истоки ее уходят в историю науки неза-
памятных времен. В естественных пауках эта идея в отчетливой
форме проявилась во второй половине 19-го столетия в работах
Ч. Дарвина (1859), К- Мёбиуса (Mobius, 1877), В. В. Докучаева
(1899) и др. Ч. Дарвин, развивая свою теорию эволюции органи-
ческого мира» как известно, многократно подчеркивал решающую
13
роль различных форм взаимных отношений организмов в эволю-
ционном процессе, обращал внимание на сложное переплетение
взаимосвязей и взаимозависимостей между целым рядом вовлечен-
ных в этот круговорот живых существ. Широко известны также
представления К. Мёбиуса о существовании группировок тесно
взаимосвязанных организмов, образующих сообщества в опреде-
ленных условиях морских мелководий. Ему принадлежит и термин
для обозначения такого рода сообществ — биоценоз (biocoeno-
sis). В. В. Докучаев свои взгляды на наличие и значение всеобщих
взаимосвязей в природе выразил в следующих словах: «Изуча-
лись, главным образом, отдельные тела, — минералы, горные поро-
ды, растения и животные, — и явления, отдельные стихии, — огонь
(вулканизм), вода, земля, воздух... но не — их соотношения, не та
генетическая, вековечная, и всегда закономерная, связь, какая су-
ществует между силами, телами и явлениями, между мертвой и
живой природой, между растительными, животными и минераль-
ными царствами, с одной стороны, человеком, его бытом, и даже
духовным миром,—с другой. А между тем, именно эти соотноше-
ния, эти закономерные взаимодействия и составляют сущность по-
знания естества...—лучшую и высшую прелесть естествознания»
(1899, с. 5).
В начале 20-го столетия идея о взаимосвязи явлений природы
и образовании на этой основе разного рода внутренне взаимосвя-
занных биологических и природных комплексов, единств проникла
в лесоведение, географию и другие науки. Особенно плодотворны-
ми в этом плане оказались взгляды одного из учеников В. В. До-
кучаева— Г. Ф. Мопоэпвя (1912), нашедшие воплощение в ряде
его статей и в опубликованном в 1912 году труде «Учение о лесе»,
а также представления Р. И. Аболина (1914) об «эпигенеме».
Лес и как часть его — конкретное лесное насаждение — рас-
сматривались Г. Ф. Морозовым как сообщества, в состав ко-
торых входят не только древесные и другие растения, но и на-
селяющие их животные. Все они оказывают влияние друг на
друга, вследствие чего возникает ряд новых явлений, «не свой-
ственных одиноко растущим растениям» (1931, с. 55). В таком
сообществе, по его словам, обнаруживается также воздействие
древесных растений на занятую ими почву и атмосферу, что ведет
к изменению физико-географической среды и образованию внут-
ренней среды сообщества. Кроме того, вследствие воздействия
древесных растений друг на друга и на внешнюю среду происходит
также изменение формы и внутреннего строения самих растений.
Лес, как указывает Г. Ф. Морозов, нельзя понять вне зависимости,
существующей между деревьями, и вне физико-географической
обстановки, с которой они составляют «одно неотъемлемое целое»
(1931, с. 103). Отсюда лес как «географический индивидуум» рас-/
14
CMci'I’pHUaЛСИ ИМ hdft, MdLlD JlCCnULU UUnpUDd лалидлщи UFU/1
в закономерной связи со свойствами земной поверхности.
Таким образом, лес, лесное насаждение, по Морозову, — это не
просто сообщество древесных растений, а «общежитие более ши-
рокого порядка» (1931, с. 299), обладающее устойчивостью, дина-
мически равновесное, которое было названо им биоценодой.
«Итак, лес или часть его — лесное насаждение,— пишет он,— есть
некое социальное единство, есть сообщество, есть реальная сово-
купность, есть биоценода, есть, наконец, ландшафт» (1931, с. 300).
Не меньший интерес с биогеоценологической точки зрения пред-
ставляют взгляды на природу Р. И. Аболина, высказанные им в
работе по классификации болот, опубликованной в 1914 году. В
этой работе он изложил свои воззрения на растительность как на
один из компонентов поверхностной оболочки Земли, названной им
эпиген ем ой. В ее состав наряду с растительностью он включал
такие компоненты (эпигены), как рельеф, почва, подпочвенные
грунты, которые тесно связаны между собой и находятся в посто-
янном взаимодействии с растительностью. Эта сложная по составу
земная оболочка расчленялась Р. И. Аболиным па однородные по
компонентному составу участки, названные э л и морфами. Сход-
ные по ряду признаков, внешнему облику и генезису, эпиморфы
объединялись в эпи формацию, являющуюся основной таксоно-
мической единицей новой науки — э п и г е н о л о г и и.
Нетрудно заметить близость представлений Р. И. Аболина об
эпигенеме и ее конкретном представителе в природе — эпиморфе —
взглядам В. Н. Сукачева о биогеоценозе и биогеоценотическом
покрове Земли.
В географии идея о взаимообусловленности явлений природы
связана с учением о географических ландшафтах. Из географов
раннего периода, воспринявших эту идею, В. Н. Сукачев (1964)
упоминает Пассарга (Passarge, 1912, 1929)—на Западе и Л. С. Бер-
га (1913, 1931) —в России. Позднее в разработке ее в ландшаф-
товедении из отечественных географов принимали участие Б. Б. По-
лынов (1925, 1953), А. А. Григорьев (1938, 1956), С. В. Калеснив
(1940), Н. А. Солнцев (1948, 1961), А. Г Исаченко (1953),
Ф. Н. Мильков (1959) и др. Однако эти работы имели косвенное
отношение к учению о биогеоценозе, так как комплексные взгляды
на природу развивались в них, как отмечает Н. В. Дылис (1978),
несколько в ином плане, как бы параллельно с биогеоценологиче-
ским направлением.
2. Понятие экосистемы и биогеоценоза
Но все это была, так сказать, предыстория биогеоценологиче-
ской науки, характеризующаяся накоплением соответствующих
15
наблюдений, фактов и других материалов. С новой силой и в новом
качестве эта фундаментальная идея о взаимосвязи и взаимообу-
словленности всех явлений природы возродилась в естествознании
в 40-х годах 20-го столетия. Наибольшее развитие она получила в
США, Канаде и в Англии, а также в Советском Союзе. Как у нас,
так и за рубежом развитие ее шло через посредство геоботаники
(фитоценологии), сыгравшей роль промежуточного звена. У нас
□на привела к учению о биогеоценозе, а за рубежом — к учению
об экосистеме.
Учение об экосистеме
возникло несколько рань-
ше, чем учение о биогео-
ценозе. Основоположни-
ком его принято считать
английского геоботаника
А. Г. Тенсли (Tansley,
1935), опубликовавшего в
журнале «Ecology» ста-
тью, в которой и были
изложены основные поло-
жения этого учения и
дано понятие экосистемы.
В статье автором были
высказаны следующие
два положения: а) орга-
низмы нельзя отделить от
окружающей их среды,
ибо вместе с нею они со-
ставляют единую физиче-
скую систему; б) такого
рода системы являются
основными единицами
Ориг. рис. 1. Схема состава экосистемы
(в представлении западных экологов)
природы на земной поверхности. Эти основные единицы природы
Тенсли и предложил называть экосистемами. Сложное при-
родное образование — экосистема — в представлении Тенсли и дру-
гих зарубежных экологов состоит из двух компонентов: 1) ком-
плекса организмов, именуемого биосистемой (биота), и
2) комплекса факторов абиотической среды, так называемой фи-
яиосиетемы (рис. 1). Здесь же дается определение этого поня-
тия: экосистема — это «относительно устойчивая система динамиче-
ского равновесия, в которой организмы и неорганические факторы
являются полноправными компонентами» (цит. по В. Д. Александ-
ровой, 1971, с. 1225).
К сожалению, как заметил в 60-е годы В. Н. Сукачев, предло-
женное Тенсли понятие «экосистема» и сама идея о взаимосвязи
явлений природы долго не находили признания и не получали рас-
16
1ространения за рубежом. Однако за последние десятилетия эко-
югия, трактуемая как наука об экосистеме, стала интенсивно раз-
виваться. Опубликовано немало работ как по общим вопросам
1кологии, так и по конкретным исследованиям экосистем разного
Аипа и объема. Ряд работ, в которых их авторами излагаются
основы учения об экосистеме, переведены на русский язык: ГЕ Дю-
зиньо и М. Танг (1968), Р. Дажо (1975), Ю. Одум (1975), Р. Рик-
тефс (1979), Р. Уиттекер (1980) и др. В них также приводятся
шределения понятия экосистемы. Пожалуй, наиболее типично из
[их следующее: «Сообщество и его среда, рассматриваемые со-
вместно как функциональная система, образующая единое целое
1 характеризуемая присущим ей потоком энергии и круговоротом
вещества, называется экосистемой» (Уиттекер, 1980, с. 9). Следо-
вательно, в современных определениях экосистемы по сравнению
: первоначальным подчеркивается функциональный характер ее
сак системы и обращается внимание на обмен веществ и энергии
в нон как основной процесс ее функциональной деятельности.
Экосистемы в представлении зарубежных экологов бывают раз-
личных размеров, различного объема, протяженности, например:
озеро, лесной массив со всем населением живых существ, ас другой
стороны — аквариум с рыбками, водными растениями (элоде-
ей, валиснерией) или пень дерева в лесу с насекомыми, микроор-
ганизмами и пр. Следовательно, экосистема в трактовке зарубеж-
ных авторов — это неопределенное по объему понятие. Не вполне
однозначно оно и по содержанию, ибо в понятие экосистемы, на-
ряду с биологическими, входят также хозяйственные и прочие
объекты, например: фабрика, завод, мастерская, зернохранили-
ще и т. п.
Таким образом, под понятие экосистемы зарубежными эколо-
гами подводятся природные системы разного объема, ранга и раз-
ного содержания: от капли воды до. экосистемы, охватывающей
всю поверхность суши или водные массы Мирового океана; по
словам В. В. Мазинга (1966), с такой точки зрения как экосисте-
му можно рассматривать и отдельный муравейник, и всю биосферу
в целом.
Учитывая это, П. Дювиньо и М. Танг (1968) все экосистемы
мира по их объему, протяженности подразделили на три катего-
рии: а) микроэкосистемы (например, пень дерева, аквариум);
б) мезоэкосистемы (лесное сообщество, пруд); в) макроэкосисте-
мы (лесной покров Земли, Мировой океан). Объединение же всех
экосистем мира, по их выражению, составляет «гигантскую экосис-
тему земного шара» (с. 13). Нетрудно понять, что в основе такого
представления большинства зарубежных экологов об экосистеме,
независимо от ее пространственной протяженности, лежит идея
абсолютной непрерывности ее, идея так называемого кон тину-
ума, исключающего подразделения ее на иерархически соподчи-
ненные, объективно существующие в природе биокосные образо-
вания.
Правда, по утверждению В. Д. Федорова и Т. Г. Гильманова
(1980), Тенсли рассматривал экосистему как конкретное элемен-
тарное образование, хотя и не подчеркивал ее автономность и про-
странственную выраженность. Утвердившаяся в современной эко-
логии расшарительная трактовка понятия экосистемы, не имеющей
якобы горизонтальных пространственных границ, или с границами,
являющимися весьма условными, приписывается Ф. Эвансу
(Evans, 1956).
Возникновение и первые шаги развития биогеоценологии в на-
шей стране относятся к 1940—1947 годам. Рассматривая в своих
статьях, опубликованных в 1940 и 1942 годах, вопрос о развитии
растительности, В. Н. Сукачев пришел к выводу, что растительный
покров находится во взаимосвязи и взаимодействии с другими гео-
графическими явлениями, такими, как литосфера, атмосфера и гид-
росфера, вместе с которыми он и развивается. Взаимодействие
растительности, состоящей из фитоценозов, с другими явлениями
природы протекает в рамках так называемого «географического
комплекса», который В. И. Сукачевым (1940) был наименован
геоценозом. В этом внутренне противоречивом взаимодействии
всех явлений природы, по его словам, и лежит источник развития
географического комплекса как целого. В результате взаимодей-
ствия географических явлений на поверхности Земли, включая
растительный покров, составляющий вместе с животным населе-
нием биосферу, образуется еще один элемент географического ком-
плекса (геоценоза) — педосфера (почва), которая также вступает
во взаимосвязь и взаимодействие с остальными явлениями природы.
Взаимосвязь и соотношение явлений природы в геоценозе, на-
званных В. Н. Сукачевым его элементами, можно представить в
следующем виде:
Биосфера
Биоценоз:
фитоценоз
зооценоз
ФизикО'Географические элементы
Экотон:
эдафотоп — литосфера, гидросфера, педосфера
клнматоп — атмосфера
Геоценоз в тот период признавался В. Н. Сукачевым как объ-
ект изучения физической географии.
В дальнейшем, в процессе разработки идеи о природном един-
стве, овеществленном в геоценозе, В. Н. Сукачев убедился в том,
Что ведущая активная роль в процессах взаимодействия его ком-
понентов принадлежит живым существам, биоценозу. Исходя из
18
этого первона-
чальный термин
для обозначения
этого природного
единства — гео-
ценоз—в 1944—
1945 годах был
заменен им терми-
ном биогеоце-
ноз. Кроме того,
поскольку завер-
шение цикла об-
мена веществ в
биогеоценозе и
бесперебойное
функционирова-
ние его возможны
лишь при условии
разложения, ми-
нерализации от-
мерших расти-
тельных и живот-
ных остатков и
трупов, осуществ-
ляемых в основ-
ном микроорганизмами, В. Н. Сукачев в состав живых компонен-
тов биогеоценоза, наряду с растениями и животными, включил
также микроорганизмы сначала под названием микросинузии, а
затем—микробоценоза, В результате этого схема компонентного
состава биогеоценоза, соотношения и взаимосвязи живых и косных’
компонентов в нем приобрела вид, изображенный на рис. 2.
Биогеоценоз вследствие этого стал рассматриваться В. Н. Су-
качевым как объект изучения не физической географии, а особой
науки — биогеоценологии. Так произошло оформление био-
геоценологии как самостоятельной науки, занимающей, по мнению
В. Н. Сукачева, место на грани биологии и географии. Однако, как
считает Н. В. Дылис (1978), она все же ближе стоит к биологии.
3. Экология, биогеоценология и место их
в системе наук
Идея о биологических, а затем и о природных единствах заро-
дилась давно в различных странах, в разных отраслях знания и в
умах ученых разных специальностей. В связи с этим, естественно,
в науке появилось множество, самых разнообразных терминов для
19
•обозначения форм, в которых была воплощена эта идея. За рубе-
жом; экосистема (Tansley, 1935), холоден (Friederichs,
1930), биосистема (Thienemann, 1941), биохора (Palmann,
1948) и др.; в СССР: эп и м о р ф а (Аболин, 1914), биогеоценоз
(Сукачев, 1944, 1945), бйоэкос (Нестеров, 1954), б и о ц е н о то п
(Иоганзеп, 1964), цепоэкосистема (Быков, 1970а), геоэко-
биота (Герасимов, 1973). Однако самыми распространенными из
них в настоящее время являются два: экосистема — в США,
Канаде, а также в Англии и некоторых других странах Западной
Европы и биогеоценоз — в СССР и некоторых восточноевро-
пейских странах. Последние понятия и термины на IX Междуна-
родном ботаническом конгрессе в 1959 году в Канаде были при-
знаны синонимами. Первый из них стал рассматриваться основ-
ным объектом изучения науки экологии, а второй — биогео-
ценологии, получивших развитие в названных странах.
В действительности понятия и термины биогеоценоз и экосис-
тема хотя и близки друг другу, но не тождественны. В. Н. Сукачев,
как известно, различие между ними видел в том, что биогеоценоз—
это вполне определенное по объему понятие, его границы на суше
определяются границами фитоценоза как автотрофной базы био-
геоценоза в целом. Фитоценозы, как известно, сравнительно легко
опознаются в природе, ибо, как правило, физиономически и струк-
турно, морфологически довольно четко отграничены друг от друга,
даже при наличии иногда переходной полосы между ними. Экосис-
тема же в представлении зарубежных экологов, как было отмечено
выше,— это, так сказать, безразмерное, безранговое понятие, охва-
тывающее природные и другие объекты различного объема — от
мельчайших, микроскопических размеров до размеров глобального
масштаба. Биогеоценоз — вполне определенное понятие и по содер-
жанию: это природная бнокосная система (естественная или ис-
кусственная) сложного состава и структурно-функциональной орга-
низации. Экосистема же — неоднозначное по содержанию понятие,
ибо охватывает объекты не только природные, но и социальные,
хозяйственные и пр.
Различие понятий «экосистема» и «биогеоценоз» отмечается
рядом советских ученых. Ю. М. Свирежев и Е. Я. Елизаров (1972),
в частности, указывают на то обстоятельство, что па принципы
выделения экосистем не наложено никаких ограничений; так, при
выделении их природные границы считаются несущественными: в
качестве экосистем рассматриваются любые, даже неустойчивые
сообщества или не имеющие какой-либо целостности структуры.
Понятие же биогеоценоза опирается лишь «на реально существу-
ющие, пространственно раздельные природные сообщества» (с. 12).
К этому надо добавить, что вследствие безранговости понятия
экосистемы, без выделения элементарной экосистемы как исходной
20
ячейки, на которые естественно распадается любая вышестоящая
биокосная система, невозможна типизация их по таксономическим
градациям, ступеням иерархической лестницы, невозможно по-
стпоение неоаохиир'"™“ гйртрмм кпяггйгКйияпии ^клгигтр.м поллб-
нли тлтгтемр кляссигЬикяпии биогеоценозов^ составленной Н. В. Hvr?
лисом (1964). Экосистемы можно типизировать лишь по размерам,
протяженности: микро-, мезо- и макроэкосистемы, что и сделано
П. Дювиньо и М. Тангом (1968).
Но самое существенное различие этих понятий, на наш взгляд,
состоит в трактовке самой сущности понятия «экосистема» за ру-
бежом и понятия «биогеоценоз» в нашей стране. В основе сочета-
ния видовых популяций живых существ в экосистемы, в представ-
лении зарубежных экологов, лежат экологические связи, т. е. связи
их со средой обитания. Отсюда «эко» (экосистема, экология) —от
греческого слова oikos — дом, хозяйство, обиталище. В представ-
лении же советских биогеоценологов ведущую, решающую роль в
сочетании видовых популяций живых существ в биогеоценозы
играют ценотические связи и отношения между ними, как бы на-
кладывающиеся на экологические. Последние в сформировавшемся
биогеоценозе находятся в «снятом» виде, т. е. в соподчиненном по-
ложении по отношению к цепотическим. Отсюда «цено» (биогео-
ценоз, бногеоценология)—от греческого слова koinos (латинизи-
рованное coenos) —общий, община.ГПри этом имеются в виду це-
цотические связи двоякого рода: Межвидовые „взаимоотношения
организмов в форме «борьбы за существование» в широком дар-
виновском смысле и трофические (пищевые) связи и отношения
между видовыми популяциями и группами популяций, противопо-
ложными по типу питания, характеру используемых источников
энергии и роли в обмене веществ и энергии- (автотрофными и ге-
теротрофными). Трофические, точнее, трофоэнергетическне взаимо-
связи в силу этого становятся взаимно необходимыми, обеспечива-
ющими относительную устойчивость и длительность существования
биогеоценоза в целом (об этом более подробно в VII главе).
В связи с этим глубокий смысл приобретают слова А. П. Щен-
никова (1938), который, критикуя «механистически-экологическое»
направление в фитоценологии и ботанической географии, пишет:
«Вообще противоречит теории Дарвина широко распространенное
мнение о том, что проблема растительности целиком сводится к
проблеме отношения между растениями и внешней средой. Реше-
ние проблемы в духе дарвинизма... предполагает рассмотрение не
двухчленной (растение — среда), а трехчленной системы отношений
(растения — взаимные между ними отношения — среда)» (с. 14).
Деление организмов по их питанию на автотрофов, гетеротро-
фов и миксотрофов, по словам Вернадского, было предложено
в 1880-е годы немецким физиологом Пфеффером. Однако важное
21
значение этого деления организмов в процессах природы было по-
казано в 20-е годы текущего столетия самим В. И. Вернадским, а
разработка и раскрытие биологического механизма этой фунда-
ментальной, диалектической по существу закономерности, лежа-
щей в основе формирования и функционирования биокосной сис-
темы любого ранга, принадлежит В. Р. Вильямсу в его учении о
растительных формациях, опубликованном в «Почвоведении», вы-
шедшем в свет в первом издании в 1900 году, а в третьем — в 1936
(с. 87—100). В этом состоит глубоко принципиальное отличие на-
шей советской расшифровки существа биокосных систем от трак-
товки их существа большинством зарубежных экологов, что весьма
важно.
В связи с изложенным не только советские ученые соответству-
ющих специальностей, по и некоторые из зарубежных отдают пред-
почтение понятию и термину «биогеоценоз», поскольку, несмотря
на известную громоздкость этого термина, он наиболее точно и
полно отражает существо биокосной системы низшего ранга. Из
зарубежных ученых в пользу применения в науке понятия и тер-
мина «биогеоценоз» высказываются: канадский ученый В. Край-
ина (Krajina, 1960), немецкий (ГДР) —А. Скамони (Scamoni,
1960), югославский — В. Мишич (Misic, 1961); американский —
А. Кюхлер (КйсЫег, 1973) и др.
Что же касается наук экологии и биогеоценологии, то следует
отметить, что в последние десятилетия в ходе их развития произо-
шли значительные сдвиги в трактовке содержания и объема этих
наук, их объекта исследования, соотношения их друг с другом и т. п.
Вместе с тем в связи с беспредельно широким толкованием науки
экологии и известной утратой исторически сложившейся со времен
Э. Геккеля (1887) присущей ей специфики как науки о взаимоот-
ношении организмов и сообществ с окружающей средой появилась
тенденция к отрицанию объективного существования данной науки,
к сведению ее содержания до «экологического подхода» к изуче-
нию любого объекта природы и общества (И. П. Герасимов, 1978).
С другой стороны, в самое последнее время у нас в стране обна-
ружилась тенденция к сведению биогеоценологии в ранг науки,
находящейся в соподчиненном положении по отношению к эко-
логии, как одного из частных разделов ее. Все это свидетельствует
о произвольном, необоснованном подходе к трактовке существа
данных наук.
В действительности экология и биогеоценологии — разные науки
по своему существу. Экология, как видно на схеме (рис. 3), охва-
тывает в качестве предметов (а не объектов) исследования все
формы движения материи, включая социальную (общественную),
а также все формы жизни на Земле, включая биоценотическую и
22
Науки об объектах природы и общества
О МАКРОТЕЛАХ О МИКРОЧАСТИЦАХ
И ИХ СИСТЕМАХ И ИХ АГРЕГАТАХ
МОЛЕКУЛЯРНО-ФИЗИЧЕСКИЕ
(ОБ АГРЕГАТАХ МОЛЕКУЛ)
Физико-химические
(о молекулах)
Морфологические
ФУНКЦИОНАЛЬНЫЕ
Систематические
Экологические
Тополого-
географические
Историке-
ДИНАМИЧЕСКИЕ
Науки
об отдельных сторонах
И СВОЙСТВАХ объектов
Геологические
(о неживой природе)
Биогеоценологические
(о биокосных системах)
Биологические
(о живой природе)
БИОСОЦИО ЛОГИЧЕСКИЕ
( О ЧЕЛОВЕКЕ И ЕГО
ДЕЯТЕЛЬНОСТИ)
Социологические
(о человеческом обществе)
Химические
(06 атомах)
Химика-физические
( ОБ атомных ядрах)
субатомно-физические
(об элементарных частицах)
Предмет
исследования
Строение внутреннее
И ВНЕШНЕЕ
Функционирование
(обмен веществ
и энергии)
КЛАССИФИКАЦИЯ
ОБЪЕКТОВ
ВЗАИМОДЕЙСТВИЕ
С ОКРУЖАЮЩЕЙ СРЕДОЙ
Распределение
НА ЗЕМНОЙ ПОВЕРХНОСТИ
Изменение во времени,
РАЗВИТИЕ
Ориг. рис. 3. Схема соотношения экологии и биогео-
ценологии
биостроматическую, которые в сочетании со средой в процессе
активного преобразования ее формируют биокосные системы раз-
ного ранга. Но охватывает она объекты исследования наук о при-
роде и обществе не в полном объеме, а лишь с одной стороны, а
23
именно со стороны взаимоотношения этих объектов со средой. Это
как раз и находит отражение в самом наименовании данной науки.
Биогеоценологии же, как видно на той же схеме, изучает биогео-
ценозы и биогеосферу не с одной стороны, как экология, располо-
женная в горизонтальном ряду, а в полном объеме, как и другие
науки, расположенные в вертикальных рядах схемы, охватывая все
стороны и свойства названных биокосных систем (строение, функ-
ции, взаимосвязь со средой, распределение в пространстве и пр.).
Из всего изложенного с неизбежной необходимостью следует,
что экология по своему существу не может трактоваться как наука
об экосистеме, ибо экосистема, как и биогеоценоз, несмотря на
указанные выше их различия, рассматриваются как биокосные сис-
темы, представляющие собою, как мы видели на схеме, объект
исследования биогеоценологии, изучающей их в полном объеме, а
не с одной стороны, как экология. Из изложенного следует также,
что экология не может трактоваться как всеобъемлющая наука,
включающая биогеоценологию в качестве одного из разделов. На-
против, биогеоценология, как наука о биокосных системах разного
ранга, является подлинно синтетической наукой, включающей эко-
логию, точнее, экологию биокосных систем в качестве одного из
разделов, выражающего взаимосвязь этих систем с их внешней
средой. Из изложенного следует также и то, что весь фактический
материал, вся научная информация, накопленные за рубежом и в
нашей стране о компонентном составе, структуре, функциональной
организации и функционировании биокосных систем, представляет
собою содержание науки, изучающей эти объекты в полном объеме,
т. е. науки биогеоценологии. Содержанием же экологии должна
стать лишь научная информация, касающаяся проблемы взаимо-
связи разнообразных объектов природы и общества (в том числе
и биокосных систем) с окружающей средой. В этом состоит ее
весьма важное научное и народнохозяйственное значение.
Что же касается учения А. Тенсли об экосистеме, в основе ко-
торого лежит, по существу, все та же глубоко прогрессивная и
фундаментальная идея передовых естествоиспытателей конца 19-го
и начала 20-го столетия о взаимосвязи и взаимообусловленности
всех явлений природы, то оно должно было бы получить особое,
соответствующее этому новому содержанию название, например
экосистемология. Использовав для наименования своего уче-
ния термин «экология», предложенный Э. Геккелем для науки о
взаимоотношении организмов (а последующими учеными и других
объектов природы) со средой, А. Тенсли тем самым не только внес
путаницу в эту науку, но и принизил суть новой по своему объекту
и содержанию науки—науки о биокосных системах, одним из осно-
воположников которой он должен быть признан.
Возникает вопрос, чем объяснить этот достойный сожаления
24
факт? Что лежит в основе этого? Суть ошибочных представлений
о биогеоценологии как науке, находящейся якобы в соподчиненном
положении по отношению к экологии, или даже как одном из раз-
делов ее, на нашвзгляд, состоит в неправильном понимании соот-
ношения в природе такой парной категории, как внутреннее и
внешнее. Многие при анализе взаимосвязи внутреннего и внешнего,
-скажем, организма и среды или биоценоза и экотопа, примат от-
дают внешнему. Отсюда процессы становления, развития, эволю-
ции их и пр. они рассматривают именно с этих позиций. В дей-
ствительности же активная роль во взаимодействии их принадле-
жит организму или биоте в форме биоценоза («живому веществу»,
по В. И. Вернадскому), усвоение и ассимиляция материальных
ресурсов экотопа которыми осуществляются на основе своей вну-
тренней качественной специфики, изменяя и перестраивая их при
этом в соответствии со своей «природой», на свой лад. На той же
основе происходит становление и развитие биогеоценозов. Из всего
этого следует, что главным, определяющим целостность, сущность
биокосной системы (биогеоценоза) является не единство живого
и неживого само по себе, а структурно-функциональное единство
диаметрально противоположных по типу питания и обмена веществ
автотрофного и сапротрофного ценокомплексов, аккумуляторов
кинетической энергии Солнца в форме потенциальной энергии ор-
ганического вещества и деструкторов его, следствием чего является
биотический круговорот веществ, обеспечивающий существование
биогеоценозов и непрерывность жизни на Земле. Что же касается
материальных ресурсов местообитания, то они в том и другом слу-
чае выступают как средства, условия, обеспечивающие осущест-
вление данного впутриоргапизмеппого или впутриценозного про-
цесса. При этом изменение, развитие целого совершается на осно-
ве «природы», качественной специфики самого организма или био-
ты в форме биоценоза как совокупности необходимым образом
взаимосвязанных живых существ, безусловно, с включением в этот
процесс материальных ресурсов внешней среды.
Из всего изложенного с неизбежностью вытекает, что биокос-
ные системы, о которых идет речь, это не ЭКОсистемы, а ЦЕНО-
системы (ценозы, биогеоценозы), а наука, объектом которой они
являются, не экология, а цепология (биогеоценологии). Эко-
логия же, как показано на рис. 3, — это совершенно иная наука
со своим специфическим предметом исследования.
К сожалению, многие из современных естественников, как за-
рубежных, так и отечественных, не могут уяснить и принять тот
непреложный факт, что биогеоценология — это не одна из биоло-
гических наук (как ботаника, зоология, микробиология и т. п.) и
уж тем более не один из разделов экологии, а что это целый ком-
плекс наук, свидетелями «рождения» и первых этапов развития
25
которых мы являемся. Не могут или не в состоянии они понять
того, что речь идет о системе наук, объектом исследования которых
является особая форма организации живой природы — биоценоти-
веская, одна из переходных (по Ф. Энгельсу) форм движения ма-
терии — биокосная.
Полагаю, что здесь уместно вспомнить, что проблема соотно-
шения экологических и ценотических наук возникла в 20—30-х го-
дах текущего столетия. Правда, тогда она дискутировалась на
более низком уровне, а именно как соотношение экологии рас-
тений и фитоценологии—науки о растительных сообще-
ствах (фитоценозах).
В. В. Алехин в предисловии к переведенной на русский язык
книге Мак-Дуголла «Экология растений» (1935) писал: «Вообще
американцы не отделяют экологию от фитоценологии, что, конечно,
не может быть принято, так как фитоценология имеет особый
объект изучения — фитоценоз» (с. 3).
Более 40 лет спустя X. X. Трасс (1976) по этому же поводу
пишет: «На наш взгляд, положение американских геоботаников о
том, что наука о растительных сообществах должна входить в эко-
логию (plant ecology), неверно. Как известно, экология... это нау-
ка, изучающая взаимоотношения организмов и внешней среды, и
лишь одна ее часть — синэкология— исследует проблемы взаимо-
отношения растительных сообществ и условий внешней среды...
потому неправомерно называть науку о растительном покрове и
образующих его растительных сообществах экологией растений»
(с. 94—95).
В Северной Америке, констатирует X. X. Трасс, фитоценология
(геоботаника в узком смысле) до сих пор «втиснута в рамки эко-
логии» (там же, с. 94).
Таким образом, попытка американских биологов свести науку
о растительных сообществах — фитоценологию — в ранг одного из
разделов экологии растений была отвергнута передовыми биоло-
гами нашей страны. Отечественная наука фитоценология (геобо-
таника) весьма успешно развивалась и развивается как самосто-
ятельная наука, принося огромную пользу многим отраслям
сельскохозяйственного производства. Точно так же, по-видимому,
надлежит в настоящее время поступать советским биогеоценоло-
гам — прилагать максимум усилий к развитию биогеоценологии
как самостоятельной науки о биогеоценозах и биогеоценотическом
покрове (биогеосфере) Земли.
Между прочим, X. X. Трасс отмечает, что многие американские
исследователи уже поняли, что экология — слишком расплывча-
тая наука. Все больше появляется ученых, которые называют себя
фитосоциологами или фитоценологами. Не исключено, что недале-
ко то время, когда многие из современных зарубежных (да и на-
26
ших отечественных) экологов также поймут, чтонаука о биокосных
системах не укладывается в рамки «общей экологии», что это осо-
бая, самостоятельная наука и что в основе ее объекта ведущая,
решающая роль принадлежит не экологическим, а ценотическим
связям и взаимоотношениям. Это будет означать, что в историче-
ской борьбе дарвинизма и ламаркизма и на биогеоценотическом
уровне победа в конечном итоге окажется на стороне дарвинизма.
4. Развитие биогеоценологии
в нашей стране и распространение ее идей за рубежом
Центром развития биогеоценологических исследований в нашей
[стране длительное время был Институт леса АН СССР, созданный
fa возглавляемый В. Н. Сукачевым, а позднее созданные им же
лаборатория леса и лаборатория биогеоценологии Академии наук.
Полевые исследования велись на целом ряде стационаров Инсти-
тута леса и других научных учреждений и высших учебных заве-
дений, созданных в разное время и в разных ландшафтно-геогра-
фических зонах и провинциях страны. Исследования на первых
порах носили разрозненный характер и в большинстве случаев
были односторонними, посвященными изучению главным образом
растительного компонента лесных, степных, луговых и других зо-
нальных и азональных биогеоценозов; на некоторых из них пред-
почтение отдавалось изучению почвенного покрова либо животного
населения. Комплексные исследования с охватом всех компонен-
тов биогеоценозов велись лишь на немногих академических стаци-
онарах. При этом исследованию подвергались в основном видовой
состав, морфологическая структура биогеоценоза и т. п., а также
биологическая продуктивность и некоторые другие их стороны.
Мало внимания уделялось изучению функциональной организации
биогеоценозов и совершающихся в ших обменных процессов, что,
разумеется, снижало научную и практическую ценность этих ис-
следований. Это было обусловлено недостатком необходимого для
выполнения такого рода работ полевого и лабораторного научного
оборудования и слабой разработкой соответствующей методики
исследования.
Резкий перелом в консолидации биогеоценологических исследо-
ваний, а также в расширении и углублении их произошел в начале
70-х годов текущего столетия после Постановления президиума
АН СССР от 11.06.1970 г. по биогеоценологии и ряда всесоюзных
совещаний, .связанных с реализацией его решений. На этих сове-
щаниях рассматривались как научно-организационные вопросы,
так и ряд теоретических проблем биогеоценологии. Подъему био-
геоценологических исследований на более высокий уровень способ-
ствовал также выход в свет таких работ, как коллективная
27
монография «Основы лесной биогеоценологии» (под редакцией
В. Н. Сукачева и Н. В. Дылиса, 1964), «Программа_л методика
биогеоценологических исследований» (1966 и 1974), «Избранные
труды В. Н. Сукачева», т. 1, «Основы лесной типологии и биогео-
ценологии» ([972)3 «Проблемы биогеоценологии» (1973), учебные
руководства разных авторов по фитоценологии, общей экологии и
’биогеоценологии, в особенности «Основы биогеоценологии»
Н. В. Дылиса (1978а), переводные монографии зарубежных авто-
ров по общей экологии, монографии и сборники работ по биоло-
гической продуктивности биогеоценозов (по программе МБП), мо-
нографии и труды лаборатории лесоведения Академии наук, мно-
гочисленные региональные монографии и сборники статей по
биогеоценологическим исследованиям, периодические издания
«Экология», «Бюллетень МОИП. Отдел биологический» и др.
Важное значение для развития биогеоценологических исследо-
ваний имело расширение всесоюзной сети биогеоценологических
станций и стационаров по четырем меридианам, охватывающих все
ландшафтно-географические зоны, провинции и высотные пояса в
пределах страны, организация отделов и лабораторий биогеоцено-
логии в ряде научных учреждений и кафедр биогеоценологии и
охраны природы в высших учебных заведениях, увеличение выпус-
ка литературы по общим теоретическим и программно-методиче-
ским вопросам биогеоценологии, а также конкретным исследова-
ниям природных и культурных биогеоценозов и созданию высоко-
продуктивных и устойчивых агроценозов, издание Государственной
научно-технической библиотекой Сибирского отделения АН СССР
с 1973 года библиографического указателя литературы «Биогео-
цепология».
Общее состояние биогеоценологии характеризуется широким
размахом биогеоценологических исследований в нашей стране с
охватом во многих случаях всех живых и косных компонентов био-
геоценозов, с применением не только узкоспециальных, но и инте-
грированных, собственно биогеоценологических методов исследо-
вания, а также методов математического анализа и моделирования
биокосных систем разного ранга. В программы комплексных иссле-
дований биогеоценологических станций и стационаров, наряду с
вопросами морфологии биогеоценозов и их биологической продук-
тивности, стали включаться и вопросы, относящиеся к изучению
обменных процессов, в них происходящих, роли живых и косных
компонентов в этих процессах, баланса веществ в ходе биотиче-
ского круговорота, потока энергии при функционировании биогео-
ценозов и т. п. На биогеоценологических стационарах в индустри-
альных районах и в районах угольных, железорудных и других
месторождений с характерным техногенным ландшафтом ведется
экспериментальная разработка проблемы рекультивации нарушен-
28
пых земель и* озеленения промышленных отвалов, терриконов,
склонов карьеров и т. п. В районах же с развитым земледелием и
животноводством, в особенности в условиях орошения или осуше-
ния земель, ведутся полевые экспериментальные исследования по*
юзданию высокопродуктивных и устойчивых агроценозов различ-
ного хозяйственного назначения.
В постановлении секции химико-технологических и биологиче-
ских наук президиума АН СССР (1971) по вопросам биогеоцено-
логии констатировалось: «Имеются существенные достижения в
теоретической разработке вопросов структурно-функциональной
организации биогеоценотических систем, закономерностей про-
странственного перемещения и превращения в них масс вещества
н энергии, вертикальных .и горизонтальных границ биогеоценозов
и биогеосферы, вопросов целостности, гомеостаза и классификации
биогеоценозов, биогеохимической работы как отдельных слагае-
мых, так и биогеоценозов в целом, роли взаимных отношений
организмов в становлении, сложении, функциях и динамике био-
। еоценотнческих комплексов и др.»
Таким образом, состояние и развитие биогеоценологии как
одного из магистральных направлений современного естествозна-
ния и развитие биогеоценологических исследований в нашей стра-
не находятся на довольно высоком уровне. Вместе с тем в связи
с прогрессирующим воздействием вооруженного современной тех-
никой человека на природу биогеоценологические исследования
приобретают все большее значение. Поэтому в решениях секции
химико-технологических паук президиума АН СССР (1973) под-
черкивалась необходимость еще более широкого развертывания
биогеоценологических исследований в пашей стране, оснащения
биогеоценологических лабораторий, станций и стационаров совре-
менным оборудованием, всемерного совершенствования методики
полевых и лабораторных исследований, теоретического обобщения
их результатов и увязки с практическими задачами развития на-
родного хозяйства и защиты окружающей среды.
Успешное развитие биогеоценологии в нашей стране привело к.
признанию учения В, Н. Сукачева о биогеоценозе в странах соци-
алистического содружества (Болгария, Венгрия, Германская Де-
мократическая Республика, Польша, Чехословакия). Имеются
приверженцы его и в ряде капиталистических стран (Канада, США,
Финляндия, Индия й др.). Это нашло подтверждение в торжестве
биогеоценологических идей на соответствующих научных форумах
и в ряде научных публикаций зарубежных ученых, посвященных
вопросам биогеоценологии. Так, на IV Мировом лесном конгрессе
в 1954 году в Индии было принято решение о проведении иссле-
дований типов леса как типов лесных биогеоценозов по разрабо-
танной в СССР методике. На IX Международном ботаническом
29
конгрессе в 1959 году в Канаде было признано, что общей плат-
формой для различных направлений в исследовании лесов может
быть биогеоценологический подход к типам леса. Там же на спе-
циальном симпозиуме были обсуждены употребляемые в разных
странах термины для обозначения природных единств и выявлены
среди них наиболее распространенные, к которым отнесен и термин
«биогеоценоз» (Рысин, 1964). На I рабочем совещании стран —
участниц Совета Экономической Взаимопомощи в 1973 году в Мо-
скве были рассмотрены некоторые общие теоретические вопросы
биогеоценологии и ряд частных вопросов, касающихся методики
комплексного изучения биогеоценозов, подготовки специалистов по
охране окружающей среды и т. п.; были показаны также резуль-
таты ведущихся в странах — участницах СЭВ конкретных биогео-
ценологических исследований (Саблина, 1973).
К сожалению, проявившаяся с начала 80-х годов тенденция к
умалению научно-теоретических потенций биогеоценологии и ее
практического значения в решении народнохозяйственных и при-
родоохранных проблем отрицательно сказывается не только на ее
дальнейшем развитии в нашей стране, но и на распространении
биогеоценологических идей в зарубежных странах.
5. Научное и народнохозяйственное значение биогеоценологии
Научное и народнохозяйственное значение биогеоценологии и
биогеоценологических исследований чрезвычайно велико. В назван-
ном выше Постановлении президиума Академии наук указывается,
что бногеоценология (наряду с экологией) является теоретической
основой охраны природы от вредных выбросов промышленности и
применения стойких биоцидов, восстановления и расширения био-
логических ресурсов планеты, рационального использования био-
логических и других природных ресурсов и т. п. Обращается
внимание и на то, что «через биогеоценологию возможно наиболее
эффективное внедрение большинства достижений биологических
дисциплин в народное хозяйство, особенно в земледелие».
В качестве основной проблемы биогеоценологического изучения
природы выдвигается согласование дальнейшего научно-техниче-
ского прогресса с поддержанием в биогеосфере надлежащих усло-
вий для нормальной жизни и успешной производственной деятель-
ности людей.
Наряду с этим перед биогеоцепологией как наукой о биогеоце-
нозах и биогеосфере, по мнению Н. В. Дылиса (1978а) и других
отечественных биологов, встает целый ряд частных научных и на-
роднохозяйственных проблем: а) взаимоотношения организмов
между собой и с физико-химическими и почвенно-географическими
факторами в биогеоценозах различных ландшафтно-географиче-
30
Liuix зон; б) популяционно-видовой состав живых компонентов био-
геоценозов и механизмы регуляции численности, жизненности и
устойчивости видовых полуляций в составе различных типов био-
геоценозов; в) структурно-функциональная организация биогеоце-
нозов и обмен веществ и энергии в них на различных местополо-
жениях рельефа как в условиях нормального, так и нарушенного
метаболизма; г) биологическая продуктивность природных биогео-
ценозов,- в особенности в индустриальных районах и в районах
нового хозяйственного освоения земель — арктических, пустынных»
высокогорных; д) прогнозирование биопродуктивности сухопут-
ных и водных биогеоценозов, управление ею без подрыва произво-
дительности земельных угодий и промысловых водоемов; е) дина-
мика биогеоценозов и сопряженная эволюция биокосных систем
разного ранга; ж) классификация различных типов биогеоценозов,
сухопутного, водного и прибрежно-водного секторов биогеосферы
па основе типологии их метаболизма; з) межбиогеоценозные связи
п матсрипльпо-эпсргетический обмен между смежными и удален-
ными друг от други биогеоценозами и территориальное объедине-
ние их в соответствующие природные единицы на этой основе;
н) методы биогеоценологической съемки, энергетической оценки и
картирования биогеоцепотического покрова Земли с составлением
региональных и обзорных карт разного масштабами) интегрирован-
ные количественные и качественные методы исследования биогео-
ценозов как целостных биокосных систем; л) изменение структур-
но-функциональной организации различных типов биогеоценозов
под влиянием хозяйственной деятельности человека в индустри-
альных районах н в районах интенсивного земледелия и животно-
водства; м) научные основы охраны и преобразования биогеосфе-
ры Земли п рационального использования ее биологических ресур-
сов и др.
В связи с этим на основе научных достижений биогеоценологии
и смежных с нею наук представляется возможным решение ряда
важных практических народнохозяйственных задач: а) перестройка
технологических процессов в сторону резкого сокращения, вплоть
до полного устранения вредных отходов в окружающую среду и
переход на замкнутые циклы производства; б) совершенствование
биологической очистки сточных вод, стоячих водоемов и водных
коммуникаций от вредных промышленных и бытовых отходов;
в) защита почв от токсических веществ, водной и ветровой эрозии
и вторичного засоления; г) биологические способы борьбы с вред-
ными насекомыми и другими вредителями сельскохозяйственных
культур; д) рекультивация нарушенных хозяйственной деятельно-
стью биогеоценозов; е) создание высокопродуктивных и устойчи-
вых культурных биогеоценозов, а также замкнутых ценотических
систем типа космического корабля и т. п.
31
Глава II
Биогеоценология и ее объект
1. Биогеоценология как наука
Биогеоценологии, как было сказано ранее, — это наук;
'биокосных системах разного ранга — биогеоценозах и биогеоце,
тическом покрове Земли, материальной основой которых являю'
области концентрации живого вещества на земной поверхнос
пленки и сгущения жизни (по Вернадскому, 1926). Главной
целью биогеоценологии, как пишет Н. В. Дылис (1978а), являет
«расшифровка структурно-функциональной организации элем<
тарных ячеек биогеоценотического покрова, в смысле Сукачева
биогеоценозов, и анализ всего того разнообразия связей и B3anN
действий, которые существуют между составляющими их матер
альными компонентами и которые при ведущем значении жив!
компонентов создают сложную, но целостную и реальную... биокс
ную систему» (с. 65).
Учение В. И. Вернадского о биосфере — более широкая
объему наука, чем биогеоценология, так как охватывает своим i
следованием весь слой жизни па Земле, включая рассеянные в пр
странстве одиночные организмы, биосферу как особую специфи*
скую оболочку земного шара, находящуюся под непрерывным вс
действием социального человека со дня появления его на Зем;
Это, по-видимому, и послужило для А. П. Шенникова (196
основанием рассматривать биогеоценологию как «часть нового си
тетического учения о биосфере Земли» (с. 18).
2. Непрерывность и дискретность
биогеоценотического покрова
Объектом исследования биогеоценологии принято считать би
геоценозы как ячейки биогеоценотического шокрова Земли (по С
качеву, 1964а), биогеосферы (по Дылису, 19696). Но такое пре,
ставление об объекте науки биогеоценологии, по-видимому, буд
неполным. В действительности объект исследования биогеоценол
гии является как бы двуединым, включающим биогеоценоз и би
геосферу в целом, объединенные, как отмечено, единством матер,
алыюго субстрата. Биогеоценозы, образующие в своей совокупи
сти биогеосферу нашей планеты, обычно рассматриваются при это
как элементарные целостные, т. е. далее неделимые без нарушени
их функционального единства и жизнеспособности, иерархическ
соподчиненные единицы биогеосферы. Такого же мнения приде]
32
живается и Ю. А. Львов (1979), считающий, что объектами био-
геоценологии являются биогеосфера и ее индивидуальные едини-
цы — биогеоценозы.
Однако наряду с этим как в нашей стране, так и за рубежом
издавна бытуют и другого рода воззрения на растительный, а
позднее и биогеоцепотический покров Земли, суть которых состоит
в постулировании идеи непрерывности этих общепланетарных об-
разований, так называемого континуума, в отрицании дискретности
их, т. е. возможности выделения в их составе реально существую-
щих относительно обособленных единиц —фитоценозов и соответ-
ственно биогеоценозов. «Такой взгляд полностью отрицает самое
существование в биогеоценозах взаимоотношения между организ-
мами, закономерности их ассоциирования, авторегуляцию ценозов
и пр. Он, по существу, отрицает биоценозы и фитоценозы в совре-
менном их понимании и противоречит многочисленным фактам,
накопленным фитоценологией», — пишет Б. А. Быков (19706, с. 9).
Идея непрерывности растительного покрова была высказана
Л. Г. Раменским еще в 1910 году и развивалась им до конца дней
своих. В основе ее лежит выведенное им «правило» экологической
индивидуальности растений, в силу чего каждое растение имеет
свою кривую размещения в рельефе в соответствии с градиентом
факторов среды. Постепенное изменение условий увлажнения или
какого-либо другого экологического фактора по мере, скажем, по-
нижения положительного элемента рельефа, по мнению Раменского,
влечет за собой столь же постепенное изменение видового состава
растительности, что лишает возможности разделения ее на расти-
тельные группировки — фитоценозы. Однако представление о рас-
тительности как «зеркале местообитания», по мнению Дю-Рие (Dii-
Rietz, 1921) и В. Н. Сукачева (1931), является механистическим,
игнорирующим учение Ч. Дарвина о «борьбе за существование».
В действительности же вследствие конкуренции совместно произ-
растающих растительных видов происходит взаимное вытеснение
их с менее благоприятных для каждого из них позиций в более
благоприятные в пределах зоны контакта, что ведет ко все более
отчетливому разграничению их и пространственному обособлению
в более или менее экологически и ценотически однородные расти-
тельные группировки и, в конечном итоге, к формированию раз-
личных, соседствующих друг с другом растительных сообществ
(фптоценозов).
Концепция непрерывности (континуума) растительного покрова
независимо от Раменского возникла и издавна развивается в ряде
зарубежных стран — в США, Франции, Англии, Италии и др. Сто-
ронники этой концепции имеются и в нашей стране. Так, В. И. Ва-
силевич (1969) пишет о постепенном переливании одного фитоце-
ноза в другой и невозможности разделения растительности на
33
какие-либо единицы в силу экологической специфичности расти
тельных видов и якобы отсутствия в природе дискретных экотопое
что, кстати сказать, не соответствует действительности и, в частно
сти, опровергается указанием Ю. Одума (1975) на наличие «дис
кретности наземной среды» (с. 70). Б. М. Миркин и др. (1978)
своей краткой справке о развитии фитоценологии утверждают, чт<
концепция непрерывности растительности как ее коренного свой
ства является прогрессивной, единственно правильной, якобы зна
менующей собой рождение нового, современного этапа в развити!
этой науки, пришедшей на смену устаревших, по его словам, пред
ставлений фитоценологов о дискретности растительного покрова i
признании ими взаимоотношений между растительными видам!
в качестве главного фактора организации растительных сообществ
Из сказанного следует, в частности, что концепция континуум;
в растительном (да и в биогеоценотическом) покрове базируете*
на признании ее сторонниками примата внешних связей в сообще
ствах над внутренними, что не соответствует действительности
П. М. Рафес по этому поводу пишет: «...континуум можно видеть
либо в наличии экотонов (переходных полос. — Л. //.), либо е
градиентном расположении ассоциаций или популяций в соответ-
ствии с величиной появления какого-нибудь признака. Однакс
нельзя на этом основании отрицать реальность отграниченных е
природе ассоциаций» (1980, с. 83).
Противопоставление друг другу идеи непрерывности раститель-
ного покрова, с одной стороны, и представления о его дискретно-
сти, т. е. признания наличия в природе фитоценозов как элементар-
ных единиц его, — с другой, с нашей точки зрения является
методологически несостоятельным, а потому лишенным смысла.
Имеется достаточно оснований для признания того, что раститель-
ность, а равно и биогеоценотический покров обладают свойством
непрерывности если не в глобальном масштабе, то, по крайней
мере, в пределах того или иного региона, массива. Однако как
растительный покров, так и биогеосфера в целом — это не аморф-
ные (бесформенные) природные образования и не конгломерат из
хаотически разрозненных элементов, а определенным образом
структурно организованные живым веществом и космическими
воздействиями динамически устойчивые планетарные системы, сла-
гающиеся из иерархически соподчиненных элементов, каковыми
являются фитоценозы в первом случае и биогеоценозы — во вто-
ром, Следовательно, наряду и одновременно со свойством непре-
рывности названным планетарным системам присуще и свойство
дискретности, т. е. способности к образованию относительно само-
стоятельных, пространственно и качественно обособленных элемен-
тарных ценотических единиц разного ранга. «Сплошная живая
пленка Земли распадается на в значительной степени дискретные
34
1м и пн цы — биогеоценозы, — пишут Н. В. Тимофеев-Ресовский и
Д. II. Тюрюкаиов (1966). — Биогеоценозы — это структурные еди-
ницы («блоки»), из которых состоит биосфера и в которых проте-
ItHior вещественно-энергетические круговороты, вызванные жизне-
деятельностью организмов» (с. 120). Сходные мысли высказыва-
ются IO. М. Сннрсженым и Е. Я- Елизаровым (1972), по словам
Которых биогеоценоз как элементарная единица биосферы выделен
удпчно г применением определенных природных ограничений, «ко-
лнчегию и интенсивность связей внутри выделенной области го-
|)н |до иышс, чем между этой областью и окружающим ее простран-
ством» (с. 11). Л большая мощность внутренних связей по
гр и ипен ню с внешними, по мнению П. М. Рафеса (1970),— обяза-
тельное условие разграничения природных комплексов. Каждый
бноы’оценоэ. по слонам В. А. Абакумова (1975), обладает также
fin * нчитторнмой химической индивидуальностью, а Н. В. Дылис
(Г I укмзынаст на то, что «в каждом биогеоценозе создается
। и*. 1<чин||ичггь1н' поде излучений, отличное от генерализованно-
। с иод|1намнч<ч ion о поли всей биосферы» (с. 66).
Ihkiim обрнюм, методологически верной научной основой реше-
нии рассматриваемой проблемы является не противопоставление
друг другу непрерывности и дискретности растительного и биогео-
ценптического покровов Земли, а признание объективно существу-
ющего единства этих присущих им свойств.
Характерно, что сторонники концепции континуума все же не
могут обойтись без выделения тех или иных элементарных единиц
ра<11Г1елы1о< п| плп биогеосферы. Однако выделяемые ими с при-
менением различных статистических методов различные по объ-
ему । руппнровкп рассматриваются как собрания абсолютно авто-
номных. чисто случайных в данном местообитании организмов, а
потому объявляются условными, эмпирическими; условными объ-
являются и границы между ними.
Перейдем к более детальному рассмотрению первой составляю-
щей части объекта исследования науки биогеоценологии, т. е. био-
геоценоза.
3, Биогеоценоз и его состав по Сукачеву
Проблема биогеоценоза и его компонентного состава — цент-
ральная проблема биогеоценологии, и, возможно, поэтому она
является одной из самых сложных и спорных проблем этой науки.
От правильного решения коренных вопросов данной проблемы,
объективно отражающего реальное положение их существа, зави-
сит соответствующее современным задачам направление и успеш-
ное развитие науки биогеоценологии, действенная роль ее в реше-
нии местных народнохозяйственных и ряда глобальных проблем,
35
затрагивающих жизненно важные интересы всего человечества и
многое другое.
Биогеоценоз, как было сказано выше,.— это элементарная еди-
ница биогеосферы. Но вместе с тем биогеоценоз сам представляет
собою весьма сложную по составу, морфологической (физической)
структуре и функциональной организации биокосную систему. Как
всякая сложная система, биогеоценоз состоит из элементов разных
иерархических ступеней, объединенных в единое целое множеством
разного рода взаимных и односторонних связей и отношений меж-
ду ними. Биогеоценоз, как объективно существующая, обладающая
качественной определенностью единица биогеосферы, функциони-
рует и развивается в пространстве и во времени, иначе говоря, яв-
ляется не только хорологической, но и функциональной, относи-
тельно открытой, динамически устойчивой системой. Такого рода
характеристики биогеоценоза, насколько нам известно, не вызыва-
ют каких-либо сомнений и возражений.
Одпако при расшифровке этих общих положений, ввиду чрез-
вычайной сложности объекта исследования биогеоценологии, воз-
никает целый ряд вопросов, вызывающих различные, порой про-
тиворечащие друг другу толкования, а потому нуждаются в
тщательном рассмотрении и правильном научно обоснованном
решении каждого из них.
Основные положения учения о биогеоценозе и других пробле-
мах биогеоценологии изложены В. Н. Сукачевым в ряде работ,
опубликованных в различных периодических изданиях и коллек-
тивных монографиях, составивших содержание 1-го тома его «Из-
бранных трудов» (1972). Термин «биогеоценоз» происходит от
греческих слов: bios — жизнь, geo —земля, koinos — общин, общи-
на (отсюда латинизированное coenos). Следовательно, он может
быть истолкован как сообщество совместно живущих организмов
па определенном участке земли.
Биогеоценоз рассматривается В. Н. Сукачевым как весьма
сложная биокосная система, включающая в свой состав пять ком-
понентов (см. рис. 2). I. Растительный компонент, представленный
тем или иным растительным сообществом — фитоценозом:
древесным, кустарниковым, травяным, мохово-лишайниковым и т. п.
II. Животный, состоящий из позвоночных и беспочвоночпых форм,
обитающих как в наземной (воздушной), так и в почвенной или
водной среде в границах данного растительного сообщества, обра-
зующих зооценоз. III. Микробный — из бактерий, низших гри-
бов, актиномнцетов и других микроорганизмов, обитающих в почве,
в особенности в прикорневой зоне растений (ризосфере), и в
наземной части растительного сообщества или в водной среде, об-
разующих сухопутный (или водный) микробоценоз. Разуме-
ется, что разные биогеоценозы различаются видовым составом
Зв
пинит, составом видовых популяций) названных живых компо-
зитов; ипаче говоря, каждый биогеоценоз характеризуется вполне
Шределенпым набором видов растений, животных и микроорганиз-
<оп. В процессе совместного существования живых компонентов
определенного видового состава в определенных условиях обита-
ем между ними возникают разнообразные связи и отношения,
К’лсдствие чего и образуется биологическое единство более высо-
кий ранга — биоценоз.
Помимо перечисленных живых компонентов, в состав био-
гоцепоза, по крайней мере сухопутного, входят два косных
юмпопепта. IV Почва и подпочвенные слои горной поро-
ды, включая почвенно-грунтовые воды на ту глубину, на которую
грогтнрпется взаимодействие с ними других компонентов биогео-
р*поэ?|, в особенности растений и почвообитающих животных и
(пкроорганизмов, следствием чего является образование косного
биогенного происхождения) единства, названного э д а ф от о-
|ом V Атмосфера, включающая в свой состав биогенные газы
кнглороД| углекислый газ), атмосферную влагу, осадки в твердом
I жидком виде, движение воздушных масс (ветер, вертикальные
loiiBCKitiioHiiide токи) и пр., образующие в процессе взаимодей-
Tiiiin с другими компонентами биогеоценоза так называемый к л и-
1ИТОИ. Последние два компонента биогеоценоза, относящиеся к
(ежиной природе, также взаимодействуют друг с другом в зоне
[интактя, образуя косное единство более высокого ранга —
к отоп.
II постоянном взаимодействии друг с другом, как видно на схе-
|с, находятся по только все пять компонентов биогеоценоза, но и
нюцеиоз и целом как совокупность живых компонентов взаимо-
ойстиует с экотоном как совокупностью компонентов неживой
ip ироды. Так, в процессе взаимодействия живых компонентов био-
еоценоза между собою и активного воздействия их и биоценоза
целом па экотоп происходит формирование природного единства
ще более высокого ранга — биогеоценоза, представляющего
обой не биологическую, как уже говорилось выше, а биокосную
истему весьма сложного состава и строения.
Следовательно, биогеоценоз, по Сукачеву, — это многоранговая
•покосная система:
III ранг
I ранг
Фитоценоз
(ооценоз
Дикробоценоз
^дафотоп
(лиматоп
II ранг
Биоценоз
Экотоп
(биотоп)
Биогеоценоз
37
Рассматривая вопрос о направлении и характере взаимосвязей
и взаимодействий компонентов биогеоценоза, В. Н. Сукачев указы-
вает на их чрезвычайную сложность. Любой из компонентов био-
геоценоза находится в многообразных прямых и опосредованных
отношениях с остальными его компонентами. Так, характер и сте-
пень развития растительности в границах биогеоценоза находятся
в известной зависимости от почвенного покрова, гидрологического
режима почвы и материнских горных пород, от погодных условий
местного климата и воздействия животных и микроорганизмов.
В свою очередь, растительное сообщество оказывает разносторон-
нее и весьма существенное воздействие на все перечисленные ком-
поненты биогеоценоза, определяя многие их характерные черты и
свойства. То же самое можно сказать и о любом другом компонен-
те биогеоценоза, ибо каждый из них в своем существовании и
развитии находится в тесной взаимосвязи и зависимости от воздей-
ствия других компонентов биогеоценоза, оказывая, в свою очередь,
то или иное влияние на их судьбу.
В общем виде взаимодействие компонентов биогеоценоза выра-
жается главным образом в обмене веществом и энергией между
ними. Атмосфера, почва, материнская порода, вода, по словам
В. Н. Сукачева, служат первичным материалом для совершающих-
ся в биогеоценозе процессов. Растительный компонент выступает
как аккумулятор лучистой энергии Солнца, так как зеленые расте-
ния, содержащие в своих клетках хлорофилл, как известно, из
углекислоты, воды и минеральных солей синтезируют за счет этой
энергии первичную биологическую продукцию (углеводы, расти-
тельные жиры, белки и т. п.). За счет аккумулированной расти-
тельностью в виде первичной биологической продукции энергии
Солнца живет весь остальной органический мир, в том числе и че-
ловек. Поэтому-то растительное сообщество нередко и называют
«энергетической установкой» биогеоценоза.
Животные как компонент биогеоценоза, по крайней мере расти-
тельноядные, служат, по выражению В, Н, Сукачева, как бы транс-
форматором растительной массы в более высокомолекулярные
органические соединения — вторичную биологическую продукцию
(животные белки, жиры, углеводы и т. п.).
Основная же роль микроорганизмов, составляющих микробный
компонент биогеоценоза, заключается в том, что они, разлагая с
помощью ферментов сложные органические соединения раститель-
ного и животного происхождения, превращают их в простые мине-
ральные соединения, соли которых становятся доступными для
питания растений. Тем самым они завершают цикл биохимических
превращений веществ в биогеоценозе. За счет выделяемой при
этом заключенной в сложных веществах энергии совершаются раз-
нообразные жизненные процессы в живых компонентах биогеоце-
38
CO
<o
’ Грунтовые воды
,Л'^ЛАЛИЕ ОРГАНИЧЕСКИХ
И МИНЕРАЛЬНЫХ ВЕЩЕСТВ
П>С»ПГГЫ РАЗЛОЖЕН
КАПИЛЛЯРНОГО .ПСАЬЕМА
Почва и
ПОЧВЕННАЯ
► ВЛАГА
Зона аэрацк»
МДТВРИ НЕКАЯ
► ПОРОДА
Рис. 4. Взаимодействие компонентов биогеоценоза (составили А. А. Молчанов и Н. В. Дылис)
Атмосферные осадки
fMCJCOA
НА ТРАНСПИРАЦИЮ
ИСПАРЕНИЕ
С ВОДОЕМА
Сток
ПОВЕРХНОСТНЫХ
вод
Опад
Испарение
с покрова и почвы
ноза и функционирование его как целостной системы (рис. 4).
Таким образом, биогеоценоз, по Сукачеву, предстает перед на-
ми как сложная по составу, но целостная элементарная единица
биогеосферы, все компоненты которой тесно взаимно связаны в
единое целое как территориально (пространственно), так и функ-
ционально в ходе обмена веществ и энергии в ней. При этом все
компоненты биогеоценоза выступают как равнозначные, выполняю-
щие в «ем вполне определенную в ходе обменных процессов рабо-
ту. Тем не менее, как видно из сказанного, из всех пяти компонен-
тов особое место принадлежит растительному компоненту — фито-
ценозу, ибо им начинается биогеоценотический процесс, поскольку
лишь обладающие хлорофиллом зеленые растения способны акку-
мулировать кинетическую энергию Солнца; они при этом химиче-
ски связывают ее, превращая в потенциальную энергию органиче-
ских веществ, служащих источником существования и развития
биогеоценоза в целом.
В одной из своих последних работ В. Н. Сукачев дал такое
определение биогеоценоза: «Биогеоценоз— это совокупность на
известном протяжении земной поверхности однородных природных
явлений (атмосферы, горной породы, растительности, животного
мира и мира микроорганизмов, почвы и гидрологических условий),
имеющая свою особую специфику взаимодействий этих слагающих
ее компонентов и определенный тип обмена веществом и энергией
их между собой и с другими явлениями природы и представляю-
щая собой внутренне противоречивое диалектическое единство, на-
ходящееся в постоянном движении, развитии» (1964а, с. 23).
В этом емком по содержанию определении заключена вся суть
биогеоценоза. Прежде всего в определении подчеркивается слож-
ность состава биогеоценоза и разнообразие входящих в его состав
живых и косных компонентов (которые далее перечисляются); при
этом обращается внимание на относительную однородность самих
компонентов в пределах занимаемого биогеоценозом пространства.
Во-вторых, в определении указывается на наличие разнообразных
взаимодействий между компонентами и определенного типа обме-
на веществом и энергией как между самими компонентами внутри
биогеоценоза, так и с его окружением (другими биогеоценозами,
внешними источниками веществ и энергии и т. п.); обращается
внимание и на особый, специфичный для биогеоценоза характер
как взаимодействий, так и обменных процессов в нем. В-третьих,
подчеркивается единство, целостность биогеоценоза, несмотря на
большое разнообразие его компонентного состава и внутреннюю
противоречивость этого единства; однако именно противоречивость
сущности биогеоценоза и служит источником его постоянного дви-
жения, поступательного развития.
Таково в общих чертах представление В. Н. Сукачева о биогео-
40
mm। tv кик элементарной единице биогеоценотического покрова
. Ivm.'i и
l.iKHM образом, не вдаваясь в детали функциональной неодно-
родности названных В. Н. Сукачевым живых компонентов биогео-
цгнозп и приняв во внимание, что подавляющее большинство рас-
|гпн11 являются автотрофами, а животные — биотрофами, что в
инк’ еапротрофов главенствующая роль принадлежит микроор-
iiiiMiiM, можно сказать, что его представления о компонентном
'< биогеоценоза в общей форме не вступают в противоречие
И1нтслы1остью. Все три живых компонента—растительный,
,ьи пай н микробный — являются непременными членами био-
111" н биогеоценоза в целом. Они реально существуют в составе
би. цепозн, и не в виде хаотически разрозненных индивидов, а в
||н<1 ci |ivniiiiponaiiiiiiix в разного рода целостные образования,
lit" 1 i*im обрн.юм взаимосвязанные друг с другом в ходе его
i|n ipoit । к пн и обмена веществ и энергии в нем.
но компоненты биогеоценоза неодинаковы по своей
Н|н '"а '1ИЧйЮТгм но степени целостности и относительной
। в и, пн критерию, лсжищсму в основе их интегра-
iiiii । 1.111 и значению в составе биогеоценоза. В силу
IH.I. и! । ни целостности, относительной автономности и струк-
'1у|.и 1н|н>рмлеппости растительный компонент, в основе интегра-
ции которого лежат характеризующие его морфологические при-
ппикн и функциональные особенности, служит не только автотроф-
ной базой существования других живых компонентов, но и как бы
Маркисом для их вертикального и горизонтального размещения в
11||огтрп11стп<* и нормального функционирования во времени. Жи-
111О11ЫЙ н микробный компоненты биогеоценоза, в основе интегра-
ции которых лежат главным образом их функциональные особен-
ности, и отличие от растительного являются гетеротрофами, по-
требителями готовой органической продукции; большая часть
животных является потребителями живой биомассы, микроорга-
низмы мертвого органического вещества растительного и живот-
ного происхождения. Животный и микробный компоненты отлича-
ются также меньшей степенью целостности и относительной
самостоятельности в составе биогеоценоза по сравнению с расти-
тельным компонентом, значительной зависимостью от последнего
По многих отношениях. Отмечая качественное своеобразие расти-
тельного и животного мира и ведущую роль сообщества растений,
Г А. Новиков пишет: «Растительная группировка составляет фун-
дамент биологической макросистемы, тогда как животное население
представляет только, так сказать, надстройку, которая, впрочем,
тесно взаимодействует с. растительностью по принципу обратной
связи» (1979, с. 18).
Исходя из всего изложенного, можно согласиться с Г А. Нови-
41
ковым и другими биологами о замене терминов «зооценоз» и «мик
робоценоз» более нейтральными выражениями, например, такими
как животный или микробный компонент или ценокомплекс, вклю-
чающими в свой состав ряд соподчиненных им животных или
микробных биологических комплексов в разного рода других груп-
пировок и сочетаний. Что же касается понятия и термина «фитоце-
ноз», то особое положение растительного компонента в составе,
строении и функционировании биогеоценоза, а также успешное
развитие давно сложившейся науки фитоценологии, объектом ко-
торой он является, служат достаточным основанием для сохране-
ния этого термина в его установившемся значении.
Таким образом, представления В. Н. Сукачева о биогеоценозе
и его компонентном составе в своей основе являются прогрессив-
ными, методологически, по существу, верными. С. С. Шварц (1970),
анализируя взгляды В. Н. Сукачева на биогеоценоз и биоценоз,
указывает па специфичность и новизну их, заключающуюся в
геохимическом и энергетическом подходе его к биогеоценозу и его
исследованию, а также в подчеркивании им, в противоположность
В. Шелфорду и другим зарубежным биологам, активности биоце-
ноза в составе биогеоценоза по отношению к местообитанию.
П. М. Рафес (1970) при сопоставлении взглядов В. Н. Сукачева и
К. Мёбиуса на биоценоз также обращает внимание на то обстоя-
тельство, что растениям в составе биоценоза Мёбиусом отводилась
пассивная роль кормового ресурса для их потребителей, что нахо-
дится в явном противоречии с представлениями В. Н. Сукачева о
роли растительного компонента в составе биоценоза и биогеоцено-
за в целом.
4. Биогеоценоз как система и взаимосвязь
его составных элементов
Большинством современных биологов соответствующих специ-
альностей биогеоценоз рассматривается как система, которая по
сложности своего состава и строения полностью отвечает требова-
ниям, предъявляемым к «очень большим системам», по выражению
Эшби (цит. по В. Д. Александровой, 19646, с. 501), а потому допу-
скает применение системного подхода к его изучению. Необходи-
мость системного подхода к изучению тех или иных объектов или
явлений обусловлена, как известно, невозможностью охватить их
обычными методами со всей мыслимой полнотой в силу большой
сложности их или большой продолжительности существования во
времени. Основными предметами исследования при системном под-
ходе являются системы и их элементы и, очевидно, взаимосвязи
между элементами системы. Суть же системности, как пишет
К- А. Куркин (1981), заключается в том, что «динамика всех эле-
42
мгп।он скоординирована и направлена на поддержание существо*
IUHIIOI системы, ее относительной замкнутости, целостности, сфоку-
сирована на осуществление ее интегральных функций. Не менее
нлжно и то, что за счет внутрисистемных взаимодействий возни-
h/uoT особые интегральные,., системные качества, отсутствующие у
слагающих систему элементов» (с. 54). Им выделяются также про-
межуточные между системой и элементами структуры, более круп-
ные ил которых названы подсистемами, а мелкие — блоками.
Нс вызывает сомнения, что живая составная часть биогеоценоза
как бнокосной системы формируется из видовых популяций — рас-
тительных, животных, микробных, являющихся исходными элемен-
тами его, а косная — из взаимодействующих между собою различ-
ных факторов воздушной, почвенной или водной среды. Каким же
образом происходит объединение этих разнородных по своей при-г
роде живых и косных элементов в единства более высокого ран-
га биоценоза, с одной стороны, и экотопа (биотопа)—с другой, и
через какие соподчиненные промежуточные ступени этот процесс
ггекпет? Рассматривая этот вопрос, надо иметь в виду следую-
В; во-первых, всякая сложная система (или подсистема) отли-
лся от простого множества тем, что она представляет собою
юкупность совместно находящихся и между собою взаимодей-
|ующих элементов и их сочетаний, обладающая в силу этого
1Йством целостности, т. е. определенной устойчивостью, органи-
1анностью, упорядоченностью своего состава и строения.
Н. Садовский (1972) по этому поводу пишет, что хотя степень
целостности разных систем неодинакова, но нет систем, которые
вообще не обладали бы никакой целостностью и не имели бы ни-
каких общих свойств. Во-вторых, в природе известны разные типы
целостности и степень целостности различных систем. Наряду с
органическим целым (организм и т. п.), где части в составе целого
теряют свою самостоятельность, в природе существуют так назы-
ваемые системные единства, относительная самостоятельность со-
ставных элементов и структурных частей которых в их составе
сохраняется, хотя и в «снятом» (соподчиненном) виде. «Член того
или иного системного единства может быть сам системным един-
ством точно так же, как определенное системное единство может
быть членом охватывающего его системного единства», — пишет
Р. Рокгаузен (1959, с. 77).
Прежде всего рассмотрим с этих позиций объединение живых
существ в биоценоз. Многие из современных биологов соответству-
ющего профиля, вслед за Л. Г. Раменским и некоторыми амери-
канскими экологами, в противоположность В. Н. Сукачеву, счи-
тают, что объединение видовых популяций в биоценоз происходит
непосредственно, минуя какие-либо промежуточные структурно-
функциональные образования. Другие биологи (П. М. Рафес, 1970Г*
43
В. В. Меншуткин, 1971; Т. А. Работнов, 1978; и др,) признают
наличие промежуточных звеньев между видовыми популяциями и
биоценозом в виде трофических групп пищевых цепей и сетей, та-
ких, как автотрофы, и гетеротрофы (в том числе животные и мик-
роорганизмы), по Ю. Одуму (1975)—продуценты и консументы
(в том числе макроконсументы и микроконсументы). Взаимодей-
ствие же живых существ в биоценозе, по мнению многих современ-
ных биологов, осуществляется непосредственно между особями и
видовыми популяциями. Так, Т. А. Работнов, изложив взгляды
В. Н, Сукачева по этому вопросу, пишет: «Однако взаимодей-
ствие между организмами в биоценозе происходит не на уровне
сообществ растений, животных, микроорганизмов, а на уровне осо-
бей или „видовых популяций”» (1978, с. 6).
В действительности взаимодействие живых существ в биоценозе
происходит как на том, так и на другом уровне. Дело в том, что в
основе сочетания видовых популяций в биоценозы и функциониро-
вания биокосных систем, как отмечалось ранее, лежат взаимоотно-
шения двоякого рода. I. Дарвиновская межвидовая борьба за су-
ществование между видовыми популяциями: а) внутри, в рамках
автотрофного растительного компонента; б) в рамках трофических
биокомплексов (фитофагов, зоофагов I и II порядка) биотрофного
животного компонента; в) в рамках функционально обособленных
биокомплексов сапротрофного микробного компонента. Этот тип
взаимоотношений между живыми существами в биоценозе действи-
тельно осуществляется па организменном уровне видовых популя-
ций в острой межвидовой борьбе за позицию в рамках данного
компонента или трофического биокомплекса (точнее, ценокомплек-
са) за место в экологической нише в его составе. 2. Вильямсовская
трофоэнергетическая взаимосвязь между названными диаметраль-
но противоположными по типу питания и направлению функцио-
нальной деятельности автотрофными и гетеротрофными компонен-
тами, необходимым образом связанными друг с другом, обеспечи-
вающими в силу этого бесперебойное функционирование системы
в целом. Этот тип взаимоотношений живых существ в биоценозе
осуществляется в основном на уровне целостных структурно-функ-
циональных ценотических единств, а не на уровне особей или ви-
довых популяций.
Что же касается косной составной части биогеоценоза, то
объединение ее элементов в такие объективно существующие при-
родные единства, как климатоп, включающий весь комплекс атмо-
сферных явлений воздушной среды обитания живых компонентов,
эдафотоп, представленный на суше почвой и почвенно-грунтовыми
водами, и гидротоп в водной среде, не вызывает сомнений. Подвер-
гается критике со стороны некоторых отечественных биологов
разделение В, Н. Сукачевым биогеоценоза, как видно на схеме (см.
44
риг 2). на биоценоз как совокупность живых компонентов и эко-
ii>ri (биотоп) как совокупность косных компонентов и представле-
ние его о взаимодействии их на этом уровне. В этом они, во-пер-
иых, видят попытку рассматривать биоценоз и экотон (биотоп) в
отрыве друг от друга, подчеркивая при этом, что они немыслимы
друг без друга и в природе сливаются воедино. Так, Б. Г. Иоганзен
пишет: «...лес или луг и другие подобные биоценотопы представ-
ляют подлинные единства: лес одновременно является и биоцено-
зом и биотопом» (1964, с. 66). На этом основании они считают, что
в термине «биогеоценоз», отражающем взаимосвязь биоценоза и
экотона (биотопа), надобность отпадает, ибо термин «биоценоз»,
но нх мнению, полностью перекрывает, заменяет его, поскольку
последний мыслится лишь в конкретной среде обитания. Во-вто-
рых, как было отмечено ранее, взаимодействие живых существ с
косной средой, как и друг с другом в биогеоценозе, по их мнению,
осущестнлистся на уровне особей или видовых популяций с отдель-
ными же разрозненными факторами среды (светом, температурой
ноздухл и почны, нллгой и т. и.).
11ри анализе приведенных возражений против раздельного рас-
смо|рения биоценоза н экотопа (биотопа) в составе биогеоценоза
прежде всего следует иметь в виду, что наличие взаимосвязи и
в;п1нмообусловлен11ости тех или иных предметов или явлений при-
роды пе только не исключает, а, напротив, как было отмечено ра-
нее, предполагает известного рода обособленность и относительную
питоном пость взаимодействующих сторон. Не следует только забы-
11вт|», что они представляют собою части, стороны единого целого.
Во вторых, пл различие понятий и терминов «экотоп» и «биотоп»
до гпх пор ними не указывалось на том основании, что большин-
ством отечественных биологов (в том числе и В. Н. Сукачевым)
онп рассматриваются как синонимы; при этом первый термин упо-
требляется преимущественно в ботанической литературе, а вто-
рой в зоологической. Между тем, по мнению А. П. Шенникова
(1964), это разные понятия: экотоп — совокупность факторов
физической (неорганической) природы, лишенной жизни, не засе-
ленной живыми существами; биотоп же — экотоп, преобразован-
ный биоценозом в процессе взаимодействия их друг с другом во
внутреннюю его среду (биоценотическую среду, по терми-
нологии Б. А. Быкова, 1970а, с. 154).
Что же касается самого процесса взаимодействия косной со-
ставной части биогеоценоза с другими его компонентами, уровня,
на котором око происходит, то при решении этого вопроса необхо-
димо принять во внимание следующие обстоятельства: во-первых,
активной стороной в этом процессе выступают живые компоненты,
которые избирательно извлекают из входящих в его состав компо-
нентов косной среды необходимые для существования и развития
45
материальные ресурсы: во-вторых, экологические факторы среды
обитания воздействуют на живые существа и их сочетания комп-
лексно, совокупно, а не разрозненно, хотя это и не исключает вы-
деления средй них как ведущих, так и ограничивающих, лимити-
рующих факторов; в-третьих, эффективность воздействия совокуп-
ности почвенных или атмосферных факторов экотопа на организмы
и их биокомплексы и обратное воздействие живых компонентов на
экотоп, как правило, выше степени воздействия отдельных факто-
ров среды или отдельных организмов при такого рода взаимодей-
ствии их.
Из этого видно, что взаимодействие живой и косной составных
частей биогеоценоза, подобно взаимодействию живых компонентов
между собой, протекает не только па уровне особей и видовых
популяций с отдельными факторами среды, но и на уровне образу-
емых ими различного рода структурно-функциональных единств с
совокупностью почвенных или атмосферных факторов среды оби-
тания, в частности на уровне биоценоза и экотопа. В процессе та-
кого рода взаимодействия, как было отмечено выше, экотоп преоб-
разуется в биотоп, который становится в силу этого одной из
сторон биогеоценоза, другую сторону последнего составляет биоце-
ноз. Однако экотоп, как «физическое» окружение биоценоза в
границах биогеоценоза, не исчезает при этом бесследно, экотоп с
его факторами воздействия сохраняется в составе биогеоценоза,
хотя и в соподчиненном положении. Таким образом, все живое
«население» сформировавшегося биогеоценоза живет в условиях
преобразованной и постоянно преобразуемой под воздействием
живого «населения» биоценотической среды. Следовательно, био-
топ (биоценосреда) в составе биогеоценоза, как указывает
А. П. Шенннков (1964),— это не сумма факторов внешней (физи-
ческой) и внутренней (биогенной) среды, а весьма сложное, каче-
ственно своеобразное сочетание тех и других факторов в его гра-
ницах.
Итак, биогеоценоз выступает перед нами как единство живого
и косного вещества биогеосферы при активном воздействии живого
на разных уровнях его организации, преобразовании им косной
(абиотической) среды во внутреннюю (биоценосреду) в его соста-
ве и поддержании ее в относительно стабильном состоянии.
Таким образом, в биогеоценозе, как системе чрезвычайно слож-
ного состава и структуры, имеет место взаимодействие и отдельных
организмов внутри популяции, и самих видовых популяций, а так-
же образуемых ими разнообразных биологических и ценотических
групп как между собой, так и с абиотическими факторами среды.
Однако взаимодействие живых существ на этом уровне осуществ-
ляется, так сказать, под контролем тех ценотических комплексов
и единств более высокого ранга, в состав которых они входят. Мно-
46
пюбразные связи и отношения существуют также между расти-
тельным, животным и микробным компонентами биогеоценоза,
исторически обусловленные свойственными каждому из них свое-
образными функциями в его составе, а также между ними и
компонентами экотопа, как и между самими компонентами послед-
него. Взаимодействия разного рода обнаруживаются и между био-
ценозом как совокупностью живых компонентов биогеоценоза и
екотопом — совокупностью абиотических компонентов его.
Следовательно, биогеоценоз нельзя рассматривать только как
сочетание абсолютно автономных растений, животных и микроор-
ганизмов или соответствующих биологических видов с отдельными
же разрозненными абиотическими факторами среды. Сопряжен-
ность друг с другом различных растительных, животных и микроб-
ных видовых популяций сохраняется в составе соответствующих
биокомплсксов и образуемых ими живых компонентов. Биоценоз,
как и биогеоценоз в целом,— это не аморфные образования разоб-
щенных элементов. Качественное своеобразие, известного рода
целостность, организованность растительного, животного и микроб-
ного компонентов в составе биоценоза, обусловленные их биологи-
ческой и функциональной спецификой, как и целостность биоцено-
за и биотопа в составе биогеоценоза, также сохраняются, посколь-
ку они выступают как соподчиненные составные части, элементы
биологических и биокосных единств более высокого ранга. Этим
подчеркивается также качественная определенность биогеоценоза
как целостной системы. Кстати сказать, в общей форме, в методо-
логическом аспекте эта проблема была рассмотрена нами в статье,
опубликованной более двадцати лет тому назад (Номоконов, 1963).
Итак, применяя принципы системного анализа биогеоценоза как
Покосной системы сложного состава и строения, биоценоз и био-
|л можно рассматривать как составные части, как подсистемы
о; в то же время каждое из этих единств выступает как целое по
отношению к составляющим их растительному, животному и ми-
кробному компонентам в первом случае, или по отношению к эда-
фотопу и климатопу — во втором. Но последние биологические и
абиотические единства также могут рассматриваться как целост-
ные категории, как ценокомплексы («блоки», по К. А. Куркину,
1981) по отношению к составляющим их структурно-функциональ-
ным ценоэлементам и видовым популяциям или определенным со-
четаниям и отдельным факторам среды. Раздельное рассмотрение
всех этих слагаемых каждого биологического, абиотического или
биокосного единства вполне допустимо и целесообразно. Но под
каким бы углом зрения их ни рассматривали, мы всегда должны
видеть не только различия, но и тесную взаимосвязь, взаимообу-
словленность и специфическую роль каждого из них в составе и
функционировании того единства, членами которого они являются.
47
Вместе с тем биогеоценоз, как пишет Н. В. Дылис, хотя и со-
стоит из структурно и функционально очень разнородных компо-
нентов живой и косной природы, но «...он не механическая их
смесь, не сумма своих составляющих, а сложная интегрированная
биокосная система, действующая и развивающаяся по особым за-
кономерностям, отличным от законов, управляющих действием и
поведением его участников. Каждый компонент, входя в состав
биогеоценоза в качестве его части, подчинен биогеоценозу как це-
лому, скорректирован им в своих свойствах и работе с общей
структурно-функциональной организацией системы и потому отра-
жает в каждом конкретном биогеоценозе не только свою субстрат-
ную специфику, но и общие особенности биогеоценоза» (1978,
с. 16—17).
5. Развитие представлений о биогеоценозе
и его составе
По мере развития биогеоценологии и разностороннего изучения
биогеоценоза постепенно углубляется и уточняется представление
о нем как одной из составных частей двуединого объекта этой на-
уки. Биогеоценоз в настоящее время рассматривается в более ши-
роком плане: во-первых, как многокомпонентная и многомерная
биокосная система; во-вторых, как система весьма сложная по со-
ставу, морфологической структуре и функциональной организации,
состоящая из соподчиненных друг другу единств разного ранга и
обладающая многочисленными внутренними каналами связи в виде
передачи веществ, энергии и информации; в-третьих, как система не
только хорологическая, но и функциональная, термодинамически
открытая, имеющая «вход» и «выход» в его окружение, находясь
по внешним каналам связи в постоянном контакте с другими био-
геоценозами и внешними источниками веществ и энергии; в-четвер-
тых, природные коренные биогеоценозы рассматриваются как си-
стемы самоорганизующиеся, саморегулирующиеся и информацион-
ные, а потому относительно устойчивые, динамически равновесные.
Важнейшие коренные свойства биогеоценозов связаны главным
образом со множественностью и многообразием видового состава
живых существ, многообразием и сложным переплетением связей
и взаимоотношений их между собой и с внешним окружением, фор-
мированием множества разнообразных по составу, сложению и
объему биологических и ценотических комплексов и сочетаний ор-
ганизмов и, в конечном итоге, биокосных образований. Этим
обусловлены и многомерность, и информативность, и относительная
устойчивость, стабильность природных биогеоценозов, отрегулиро-
ванных на длительный срок существования, способность биогеоце-
нозов реагировать на внешние воздействия перестройкой своего
48
нндового состава, структуры, характера и сроков циклических
флюктуаций, а также восстанавливаться при нарушениях, наконец,
it силу сопряженного характера эволюции органического мира,
способность биогеоценозов к коренным преобразованиям и после-
довательным сменам в ходе поступательного развития биогеоцено-
тивеского покрова.
Из представления о биогеоценозе как открытой функциональной
системе и неоднородности с этой точки зрения выделенных
В. II. Сукачевым компонентов биогеоценоза вытекает необходи-
мость уточнения компонентного состава биогеоценоза и несколько
иной, чем у Сукачева, группировки его.
Некоторые отечественные биологи (Рафес, 1970; Дылис, 1978;
Работпов, 1978; и др.), вслед за В. Р. Вильямсом (1922) и
В. И. Вернадским (1926), выделяют в настоящее время в составе
биогеоценоза следующие живые компоненты, образующие в сово-
купности биоценоз: 1) автотрофы — фототрофные и хемо-
трофные растительные организмы (в том числе микроскопические
формы), синтезирующие органические вещества из неорганических
за счет лучистой энергии (Золнца или химической энергии, получае-
мой при окислении некоторых неорганических соединений (амми-
ака, сероводорода и т. п.); 2) гетеротрофы — животные и мик-
роорганизмы, потребляющие живую растительную или животную
биомассу и некоторые прижизненные выделения организмов или
мертвое органическое вещество, — биофаги и сапрофаги, а также
растения-паразиты, полупаразиты и сапрофиты. Правда, гетеро-
трофы П. М. Рафесом подразделяются на две самостоятельные ка-
тегории: биофаги — потребители живой биомассы и сапрофаги —
потребители биомассы отмершей.
Т. А. Работнов в качестве особой функциональной группы вы-
деляет свободпоживущие или симбиотически связанные с высшими
растениями микроорганизмы — азототрофы (азотфиксаторы),
обладающие способностью усваивать атмосферный азот. Они непо-
средственно или опосредованно обеспечивают азотом другие орга-
низмы, а потому играют важную роль в составе многих биогеоце-
нозов. П. М. Рафесом, наряду с сапротрофамп — потребителями
мертвого органического вещества, выделяются также микроорга-
низмы, вовлекающие в биотический круговорот минералы и неко-
юрые минеральные продукты обмена, которые названы им абио-
•р о ф а м и. Однако такие сочетания живых существ, как биофаги,
апрофаги п паразиты среди животных, паразиты, полупаразиты и
апрофиты среди растений, нитрофикаторы, сульфофикаторы, азо-
офнкаторы и подобные им микроорганизмы, представляют собой
(е ценотические, а биологические комплексы (биокомплексы), вхо-
;ящие в состав соответствующих цепокомплексов. Абиотрофы же
з микроорганизмов, использующие в качестве источника энергии
49
минералы, и в особенности минеральные продукты метаболизма,
может быть, следует отнести к автотрофам типа хемотрофов.
Основной, генеральной функцией биогеоценоза и биогеосферы,
обеспечивающей их существование и развитие, является матери-
ально-энергетический обмен, совершающийся при непосредственном
участии названных ценотических комплексов организмов, состоя-
щих из соответствующих биокомплексов. Отсюда подобного рода
подход к выделению компонентного состава биогеоценоза можно
считать вполне оправданным и научно обоснованным. Правда,
Ю. Одум (1975) и некоторые другие зарубежные биологи выра-
жают сомнения в том, что такого рода группы живых существ
представляют собою реальные и устойчивые объединения соответ-
ствующих биологических видов, считая их беспредметными аб-
страктными наименованиями функций в процессах метаболизма.
Второй непременной составной подсистемой биогеоценоза наря-
ду с биоценозом является экотоп, который, по Работнову
(1978), бывает представлен воздушной, почвенной или водной сре-
дами. В сухопутных биогеоценозах экотоп подразделяется на поч-
венную среду — эдафотоп и надземную (воздушную)—аэротоп
(вместо термина «климатоп»). Эдафотоп, по Работнову, — это
особая биогенная почвенная среда как результат преобразования
косной части почвы почвообитающими живыми компонентами, об-
ладающая определенными физико-химическими свойствами и ре-
жимами; аэротоп же — это биогенная воздушная среда как
результат преобразования надземного косного окружения назем-
ными живыми компонентами, в особенности растительным. Из
этого следует, что эдафотоп и аэротоп по существу являются ком-
понентами не косной среды — экотопа, а преобразованной биоцено-
зом внутренней среды — биотопа (по Шенникову). Среди вод-
ных биогеоценозов имеются такие, экотопы которых включают все
три среды: воздушную, водную и почвенную (грунт), но экотопы
некоторых водных биогеоценозов состоят лишь из одной водной
среды — гндротопа.
Признавая выделение компонентного состава биогеоценозов
названными отечественными биологами в основном правильным,
вместе с тем находим необходимым заметить следующее: посколь-
ку трофоэнергетическая взаимосвязь автотрофов и гетеротрофов в
биоценозе осуществляется в основном на уровне целостных струк-
турно-функциональных ценотических единств, то это обстоятель-
ство должно найти отражение в наименовании живых компонентов
биогеоценоза. Кроме того, в согласии с П. М. Рафесом (1970), счи-
таем необходимым группу гетеротрофов по приведенным им моти-
вам подразделить на два самостоятельных компонента — био-
трофы и сапротрофы. В результате перечень компонентного
состава биогеоценоза примет следующее выражение:
50
А. Живые компоненты биогеоценоза, образующие биоценоз
1. Ценокомплекс автотрофов — из ценопопуляций расте-
ний и других живых существ фото- и хемосинтетиков, созидателей
первичной биологической продукции из элементов косной среды:
1) Фототрофы — зеленые высшие и низшие растения и не-
которые виды жгутиковых;
2) хемотрофы—бесхлорофилльные хемотрофные бактерии,
|1. Ценокомплекс биотрофов—из ценопопуляций живот-
ных и отчасти растений и микроорганизмов — потребителей и
трансформаторов живой биомассы и прижизненных выделений и
Созидателей вторичной биологической продукции:
1) фитофаги — растительноядные животные, микроорга-
низмы и растения-паразиты и полупаразиты;
2) зоофаги — плотоядные (хищные) животные, микроор-
ганизмы, насекомоядные растения, животные и микроорга-
низмы-паразиты.
III. Ценокомплекс с а п р о т р о ф о в—из ценопопуляций мик-
роорглнмзмон, а также животных и растений, потребителей и де-
структоров отмерших остатков, отбросов и трупов растительного
и животного происхождения;
1) сапрофаги — микроорганизмы, беспозвоночные и неко-
торые виды позвоночных животных;
2) сапрофиты — грибы и некоторые виды высших сапро-
фитных растений.
Б. Косные компоненты биогеоценоза, составляющие экотоп,
преобразуемый биоценозом в биотоп
IV Аэротоп — преобразованная биоценозом воздушная среда.
V. Эда фотоп — преобразованная биоценозом почвенная среда.
Сопоставляя изложенные представления о компонентном соста-
ве биогеоценоза с представлением о нем В. Н. Сукачева, следует
заметить, что выделенные им компоненты биогеоценоза, по край-
ней мере живые, взяты в весьма обобщенном виде, без учета их
неоднородности. На первых порах это было неизбежно, да в общей
1>орме, как отмечалось, это и соответствует действительности.
Эднако при более тщательном рассмотрении их оказалось, что хотя
подавляющее большинство растений относится к категории авто-
грофов созидателей первичного органического вещества из неор-
ганического, среди них есть и гетеротрофы — потребители готового
органического вещества в виде живой биомассы и мертвой, функ-
ционально уподобляющиеся животным или микроорганизмам-де-
структорам. Животные в большинстве являются потребителями
•кивой биомассы — растительной или животной и трансформатора-
ми ее в более высокомолекулярные органические соединения, ио
:реди них есть и потребители отмершей биомассы, участвующие,
следовательно, в минерализации органики. Еще большей неодно-
61
родпостью отличается мир микроорганизмов, среди которых, наря-
ду с господствующей группой деструкторов — разрушителей орга-
нического вещества, имеются и азотфиксаторы атмосферного
азота, и синтетики различных биологически активных веществ,
окислители и восстановители минеральных соединений, и даже
потребители живой биомассы.
Учет всех этих частностей, деталей и нашел отражение в при-
лагаемой схеме компонентного состава биогеоценоза (рис. 5). Что
же касается наименования компонентов, в особенности живых, то,
как было отмечено выше, с функциональной точки зрения они точ-
нее определяют суть, роль и место каждого из них в обменных
процессах биогеоценотической системы, чем в схеме В. Н. Сукачева.
6. Границы биогеоценозов и ценокомплексы
видовых ценопопуляций
Биогеоценоз, как система функциональная и хорологическая
одновременно, занимает в том или ином ландшафте определенный
участок территории или акватории. Размеры биогеоценоза, по
62
данным Тимофеева-Ресовского и А. И. Тюрюканова (1966), в зави-
симости от степени физико-географической, почвенно-геохимиче-
ской и гидрологической однородности занимаемого им участка
варьируют от нескольких десятков и сотен квадратных метров до
нескольких квадратных километров. Конфигурация и границы био-
геоценоза определяются, по Сукачеву, границами свойственного
ему фитоценоза, как автотрофной базы его, физиономически более
отчетливо, чем другие компоненты, выражающие его в про-
странстве.
Горизонтальные границы между биогеоценозами, как и между
растительными сообществами, по утверждению Ж. Леме (1976),
бывают резкими, особенно в условиях вмешательства человека, но
они могут быть и расплывчатыми, как бы размазанными в случае
Взаимопроникновения компонентов соседствующих биогеоценозов,
образующих более или менее широкую переходную полосу, именуе-
мую экотоном (грсч. «экологическая связка»), при наличии ко-
lopoil бногеоценотичсскпй покров будет казаться непрерывным
(«континуум»). Однако экотоны пространственно всегда неизмери-
мо меньше, чем еооГяцества, которые они разграничивают, следо-
Нитмьпа, сообщества эти разные по своему существу и «...границы
между ними (пусть даже «размытые» экотонами) естественны и
потому реальны» (Рафес, 1980а, с. 83).
Б. А. Быков (1970а) различает следующие типы границ между
растительными сообществами и, следовательно, между биогеоце-
нозами: а) резкие границы наблюдаются при резком различии в
смежных ценозах экологических условий или при наличии доми-
нантой, обладающих мощными средообразующими свойствами;
б) мозаичные границы в отличие от резких характеризуются вклю-
чением в переходную полосу смежных ценозов их отдельных фраг-
ментов, образующих своего рода комплексность; в) каемчатые гра-
ницы— когда в полосе контакта смежных ценозов развивается
у ткан кайма ценоза, отличающегося от обоих из них; г) диффузные
границы между смежными ценозами характеризуются постепенной
пространственной сменой видового состава в зоне контакта при
переходе одного в другой.
Вертикальные границы биогеоценоза, как и горизонтальные,
определяются по размещению живой растительной биомассы фито-
ценоза в пространстве: верхняя граница определяется максималь-
ной высотой надземных органов растений — фототрофов — над
поверхностью почвы, нижняя— максимальной глубиной проникно-
вения корневой системы в почву. При этом в древесно-кустарнико-
вых биогеоценозах вертикальные границы, как пишет Т. А. Работ-
ное (1974а), не изменяются в течение вегетационного периода, в
травяных же биогеоценозах (луговых, степных и т. п.) они варьи-
руют по сезонам, так как происходит то нарастание травостоя, то
33
снижение его, то полное отчуждение на сенокосах и пастбищах,
лишь нижние границы их не подвергаются сезонным изменениям.
Работнов обращает внимание на наличие ряда серьезных затруд-
нений, возникающих при проведении горизонтальных и вертикаль-
ных границ в некоторых биогеоценозах, в особенности в водных и
в неполночленных биогеоценозах, образованных гетеротрофами
(животным населением и другими организмами), ввиду подвижно-
сти либо среды обитания, либо живых компонентов таких биогео-
ценозов.
Заканчивая раздел о биогеоценозе, уместно коснуться следую-
щего вопроса, имеющего отношение к рассматриваемой проблеме.
Некоторыми биологами выделяются в качестве самостоятельных
автономных систем такие биологические единства, как э нт омо-
цено з, п а р а з итоце н оз, микробоценоз и др. Однако из.
представления В. Н. Сукачева и современных представлений о су-
ществе биокосных систем типа биогеоценоза следует, что назван-
ные биологические единства не могут быть признаны самостоя-
тельно функционирующими системами, поскольку ввиду отсутствия
в их составе автотрофной энергетической базы возможность более
или менее длительного автономного существования их не обеспе-
чена. В действительности подобного рода сочетания насекомых,
микроорганизмов и других существ, как наземных, так и почвооби-
тающих,— это не самостоятельные ценозы, а взаимосвязанные
биокомплексы или ценокомплексы организмов, входящие в состав
биогеоценозов в качестве соответствующих компонентов или в фор-
ме трофических звеньев пищевой цепи.
Лишь в пустынях и некоторых других областях, по словам
В. Н. Сукачева, встречаются микробоценозы как автономные био-
косные системы, включающие в свой состав ценокомплекс хемо-
трофных бактерий, осуществляющих синтез органических веществ
за счет энергии, выделяющейся при окислении некоторых неорга-
нических соединений, и функционально взаимосвязанный с ними
ценокомплекс гетеротрофных микроорганизмов, производящих ми-
нерализацию органических веществ. Это вполне естественно, ибо
микробоценозы и микробиосфера — это колыбель жизни на Земле,
первый, исходный концентр биотического круговорота веществ как
основа непрерывности жизни на этом этапе эволюции органическо-
го мира. Их можно рассматривать как окостеневшие прообразы
древнейших биогеоценотических систем, сохранившихся в малоиз-
мененном состоянии в специфических условиях местообитания.
Что же касается эндо- и эктопаразитов, связанных в своем су-
ществовании с организмом-хозяином, то ввиду полной непосред-
ственной или опосредованной зависимости последнего от автотроф-
ной энергетической базы биоценоза (и биогеоценоза в целом) о
возможности длительного в геоисторическом смысле самостоятель-
54
иого существования такого рода сочетания живых существ не
может быть и речи.
Поскольку объект исследования биогеоценологии, как было
отмечено ранее, является двуединым, то после разностороннего
рассмотрения биогеоценоза необходимо столь же тщательно рас-
смотреть и вторую составную половину его: биогеоценотический
покров Земли — биогеосферу.
7. Биосфера и ее планетарная роль по Вернадскому
Биосфера—это верхняя, наружная оболочка земной коры,
•охваченная жизнью и качественно переработанная ею и космиче-
скими излучениями, прежде всего излучениями Солнца.
Термин «биосфера» введен в биологию в начале 19-го века
7К.-Б. Ламарком, а в геологию в конце того же века — Э. Зюссом.
<)д1Н1ко основоположником учения о биосфере по праву считается
гептильный советский ученый В. И. Вернадский (1863—1945).
В. И. Вернадский рассматривал эту оболочку нашей планеты
ъак «сферу жизни», как целостную планетарную структуру живого
вещества. под которым он понимал совокупность живых существ
ня Земле и в ее ближайшем окружении. Возникла она на опреде-
ленном этапе геологической истории Земли в тесном взаимодей-
ствии живой материи с косным веществом планеты под действием
лучистой энергии Солнца и других космических излучений. Живое
вещество, оказывая огромное влияние на косные оболочки Зем-
ли атмосферу, гидросферу и литосферу, определяет, по словам
Вернадского, состав, динамику и пути миграции химических эле-
ментов в биосфере, движение земных атомов на нашей планете
в течение многих миллионов лет, от архейской эры до наших дней.
Образующие живое вещество биосферы организмы рассмат-
риваются Вернадским как особые автономные разнокачественные
живые существа, обладающие собственным термодинамическим
полем, отличным от термодинамического поля биосферы.! Однако
свойственное им термодинамическое поле определяет лишь область,
пределы существования их, а не внутреннюю специфику. Основны-
ми характеристиками живых существ являются такие их свойства,
как вес, объем, химический состав, энергия, характер отвечающего
нм пространства, а также такие процессы, как питание, дыхание,
рост, размножение и пр. Находясь в окружении косной среды, жи-
вые организмы тесно взаимосвязаны с ней и в то же время резко
обособлены от нее, поддерживая свою температуру, внутреннее
давление и пр.
Под действием живого вещества совершаются самые разнооб-
разные процессы на Земле — геохимические, атмосферные, гидро-
логические, Почвенные и др. В результате жизнедеятельности жи-
55
вых существ во взаимодействии их со средой происходит накопле-
ние кислорода и углекислого газа, устанавливается определенное
соотношение их в атмосфере, происходит образование биогенных
осадочных пород, месторождений каменного угля, нефти, горючих
сланцев, торфа, сапропеля, а также образование почвенного покро-
ва Земли, накопление гумуса, минеральных (зольных) элементов
в верхних горизонтах почвы и многое другое.
Масштабы разносторонней деятельности живого вещества н ее
последствия в биосфере чрезвычайно велики. Так, кислород атмо-
сферы полностью обновляется благодаря деятельности зеленых
растений каждые две тысячи лет, а молекулы углекислоты, уча-
ствующие в процессе фотосинтеза, — через каждые 300 лет. По
данным А. А. Саукова (1966), в течение 13 лет организмы пропу-
скают через себя такое количество углерода, которое в 10 раз пре-
вышает все содержание его в земной коре. Вся вода, содержащая-
ся во всех водных вместилищах земного шара, разлагается и вос-
станавливается в биотическом круговороте примерно за два
миллиона лет. Таким образом, за период органической эволюции,
как указывает М. М. Камшилов (1970), «не только кислород и
углерод, но и вся вода прошли через живое вещество планеты не
одну сотню раз» (с. 126). Под действием живого вещества, по сло-
вам Вернадского, в корне изменяются химическое состояние и про-
цессы в морских и океанических водах, мощные геохимические
процессы совершаются также в земной коре. Словом, в течение
короткого времени через живые существа может пройти все веще-
ство биосферы
Эта огромная геохимическая работа живого вещества в биосфе-
ре определяется наличием особых присущих ему геохимических,
точнее, биогеохимических функций, выполняемых в течение дли-
тельного геологического периода: а) газовая функция—поступление
и выделение кислорода, углекислоты и пр.; б) окислительно-вос-
становительная функция — окисление аммиака и сероводорода в
нитраты и сульфаты и восстановление последних до свободного
азота и сероводорода; в) функция метаболизма, дыхания организ-
мов и прочие процессы их жизнедеятельности; г) функция разру-
шения, минерализации органических соединений (углеводов, бел-
ков и т. п.) до соединений, доступных растениям. Биосферные
функции живого вещества тесно связаны с указанными выше свой-
ствами организмов, а также с такими отправлениями и процесса-
ми, при которых химические элементы входят в них из окружаю-
щей среды, преобразуясь в соответствующие органические матери-
алы, и выходят из них по отмирании и разложении во внешнюю
среду.
Исходя из этого, В. И. Вернадский приходит к выводу, что
жизнь — не случайное явление на Земле, она связана со строением
56
‘Земной коры, входит в ее механизм, выполняя важнейшие функ-
ции. Прекращение жизни неизбежно привело бы к прекращению
химических изменений в биосфере, в ней установилось бы химиче-
ское равновесие, скоро исчез бы кислород и резко уменьшилось бы
количество углекислоты в атмосфере и пр.
Центральная идея учения Вернадского о биосфере, по словам
Ю. А. Жданова, состоит в том, что «высшая форма развития ма-
терин па Земле — жизнь — определяет, подчиняет себе другие пла-
нетарные процессы» (1973, с. 9). Вместе с тем, как указывает
В. И. Вернадский, биосфера должна рассматриваться как область
превращений космической энергии в земную, ибо непрерывно по-
ступающие из космоса, в особенности от Солнца, разнообразные
излучения пронизывают всю биосферу, регулируя через посредство
живых существ химическое проявление земной коры.
Носителем космической энергии является солнечный луч, а «ме-
ханизмами» превращения лучистой энергии Солнца в действенную
земную энергию (химическую, механическую, тепловую и т. п.)
являются живые организмы. При этом зеленые автотрофные рас-
тения улавливают лучистую энергию Солнца, а бесхлорофильные
гетеротрофные животные и микроорганизмы участвуют в удержа-
нии и преобразовании ее в другие формы эпергииЛВсе живое ве-
щество, таким образом, представляет, по словам Вернадского,
«единую, особую область накопления свободной химической энер-
гии в биосфере, превращения н нес световых излучений Солнца»
(1967, с. 243). В результате жизнедеятельности живых существ
происходит унелпненне дсПствеппой энергии земной коры. «Весь
поверхностный слой нлингты,—пишет В. И. Вернадский, — стано-
яитсн через посредство живого вещества полем проявления кинети-
ческой и химической энергии»((1967, с. 181).
Волыпиго внимания заслуживает замечание В. И. Вернадского
о том, что проникающий из космоса солнечный луч создает самую
форму «трансформаторов», т. е. живых организмов, придает им
разный вид на суше и в воде, меняет количественное соотношение
между автотрофными и гетеротрофными организмами; вызывая
«давление жизни», он ведет, по выражению Вернадского, к «расте-
канию» живого вещества, т. е. к расселению его по лику Земли.
Под действием солнечной радиации у растений и растительных
сообществ развиваются различные приспособления к улавливанию
солнечной энергии: огромная площадь листовой поверхности, в
400 раз превышающая площадь земной поверхности, «листовая мо-
заика» и типы ветвления стеблей, ярусное строение растительных
сообществ и т. п. Все это обеспечивает максимальную в данных
условиях степень фиксации растительностью поступающей на Зем-
лю солнечной энергии и наиболее полное использование ее в про-
весе созидания первичной органической биомассы. В результате,
57
по некоторым данным, на суше ежегодно образуется и разрушает-
ся до 55 млрд г растительного органического вещества, а общая
сумма суточной работы, совершаемая живыми организмами, дости-
гает 1875 млрд т-км.
В дальнейшем в ходе длительной геологической истории проис-
ходит захоронение громадных масс различных органических обра-
зований (торфа, углей, нефти и т. п.) в осадочных породах, акку-
мулирующих солнечную энергию в виде углерода и водорода,
неоднократное погружение осадочных пород на большие глубины
в геосинклиналях (прогибах) земной коры, обычно заполненных
морскими водами, и превращение их в метаморфизированные по-
роды и гранит. Затем при орогенезе (горообразовании) происходит
вынос этих пород на дневную поверхность, разрушение их и обра-
щение в новые осадочные породы. Одним словом, совершается так
называемый большой геологический круговорот веществ, связыва-
ющий эндогенные и экзогенные процессы земной коры и устанав-
ливающий взаимосвязь между осадкообразованием, тектонически-
ми явлениями, магматизмом и развитием жизни на Земле.
В состав биосферы В. И. Вернадский включал: а) тропосфе-
ру— воздушную оболочку Земли, обогащенную биогенными газа-
ми и пронизанную световыми и тепловыми излучениями, включая
и нижнюю часть стратосферы; б) гидросферу — Мировой
океан со всеми непосредственно связанными с ним морями; в) на-
ружную часть литосферы — кору выветривания с твердым со-
стоянием вещества осадочных и отчасти других пород (рис. 6).
Важнейшими чертами биосферы Вернадский считал: а) постоянный
материально-энергетический обмен ее с космосом; б) существова-
ние живого вещества в форме растений, животных и микроорганиз-
мов; в) тесную связь живого вещества с окружающей средой и
г) подвижное, динамическое равновесие, организованность, сла-
женность ее.
Большую ценность для развития биогеоценологии представляют
идеи В. И. Вернадского о распространении жизни, живого веще-
ства на нашей планете и распределении его в биосфере Земли.
Факторами, обусловливающими верхний предел возможного рас-
пространения жизни, он считал коротковолновые лучи солнечной
радиации, а нижний—высокую температуру (100°С). Помимо
этого он различал также область временного проникновения орга-
низмов в свое окружение и область длительного их существования,
неизбежно сопровождающегося размножением.
Одной из характерных особенностей жизни в биосфере являет-
ся, по выражению Вернадского, «всюдностъ» жизни, наличие ее во
всех геосферах (оболочках) земного шара: а) в тропосфере,
где она проявляется в виде временно поднимающихся в воздух
организмов суши (птицы, насекомые, бактерии), а также пыльцы,
58
земная корона
Экзосфера
Ионосфера (термосфера)
мезосфера
Стратосфера 40 -50 (юо) км
Рис, 6. Биосфера и другие геосферы планеты Земля (по В. И. Вернадскому.
1965; М- И. Голенкину, 1949; и др.)
спор и других мельчайших носителей жизни на высоту до 22 км и
более над земной поверхностью; б) в гидросфере, занимающей
около 71% поверхности Земли, где жизнь распространяется в слое
воды до 3,8—4 км, проникая местами, по косвенным показателям,
59
на глубину 5—5,5 км и даже до 10 км; в) в литосфере, на долю
которой приходится 29,2% земной поверхности, где охваченный
жизнью слой идет в глубь Земли на 10 м, редко более; лишь в
трещинах земной коры, в подземных водах и в сопровождающих
нефть пластовых водах жизнь обнаруживается на глубине 1,5—
1,7 км и в самом крайнем пределе — до 5 км. Таким образом, об-
щая мощность биосферы, по В. И. Вернадскому, составляет на
континентах около 25 км и в океанических областях 26—27,5 (до
32) км. Правда, по некоторым данным, мощность биосферы исчис-
ляется в 38—40 км, поскольку в ее состав включаются бывшие
биосферы геологического прошлого.
На протяжении длительного эволюционного развития жизни на
Земле выкристаллизовались две формы распределения живых су-
ществ в биосфере, которые, по утверждению Вернадского, не изме-
нялись в течение всего геологического времени (1,5—2 млрд лет) :
а) одиночные или немногочисленные нахождения организмов, рас-
сеянные среди основного субстрата биосферы, и б) многочислен-
ные, собранные в биоценозы скопления организмов, называемые
Вернадским то слоями или областями концентрации жизни, то про-
сто пленками и сгущениями жизни, неразрывно связанными с ок-
ружающей средой.
В. И. Вернадским (1926, 1967) выделены следующие пленки
и сгущения жизни, в которых сосредоточена основная масса
живых существ, обитающих па Земле.
А. В Мировом океане:
1) водная (планктонная) пленка — в поверхностных слоях океана;
2) донная (бентосовая) пленка — по дну океана;
3) прибрежные (береговые) сгущения — у берегов над мелями
морей и океанов;
4) саргассовые сгущения плавающих макроводорослей—на поверх-
ности Саргассового и других морей.
Б. На суше:
5) наземная пленка — из наземных биоценозов;
6) почвенные сгущения;
7) водные сгущения озер и других стоячих водоемов.
Мощность таких пленок и сгущений жизни измеряется несколь-
кими дециметрами, метрами или десятками метров, редко дости-
гая одной-полутора сотни метров. Несмотря на это, биогеохимиче-
ский эффект этих пленок и сгущепий, по выражению Вернадского,
колоссален. Особенно большое значение в энергетическом плане
он придает зеленому живому веществу. Он указывает, что по об-
щему запасу биомассы па суше преобладают зеленые автотрофные
растения, в особенности травянистая растительность, близка к пей
по массе древесная растительность, микроскопические же зеленые
организмы отходят на второй план. В океане, напротив, преобла-
60
дает животная жизнь, а среди зеленых организмов господствуют
микроскопические одноклеточные водоросли, протисты и пр.; травы
же и крупные многоклеточные водоросли сосредоточены лишь в
прибрежьях морских водоемов да в морских мелководьях.
Обобщая представления В. И. Вернадского о биосфере, можно
сказать, что в понятие биосферы он вкладывал в основном биогео-
химический смысл. Главным агентом биогеохимической деятельно-
сти в биосфере, выполняющих роль посредника между космосом и
нашей планетой, по Вернадскому, служит живое вещество как
совокупность живых организмов. Основной формой проявления
этой деятельности и распределения геохимической энергии по зем-
ной поверхности являются размножение и рост организмов, осу-
ществляющиеся в процессе газового и материально-энергетического
обмена их с окружающей средой. Скорость же размножения, рост
организмов и накопление биомассы при определенной плотности их
совместного обитания в определенных условиях среды рассматри-
вались Вернадским как показатель их геохимической энергии и
работы в биосфере, выражающейся в количестве созданного орга-
нического вещества.
8. Современное представление о биосфере
и взаимоотношении с ней человека
Во второй половине 20-го столетия продолжается дальнейшая
разработка учения Вернадского о биосфере как советскими биоло-
гами, географами, геологами и отчасти философами, так и зару-
бежными учеными разного профиля. За рубежом разработка
основных положений этого учения идет главным образом в общем
контексте учения об экосистеме. Биосфера выступает в таком слу-
чае как единая функциональная система Земли, как макро - или
мегаэкосистема, выстилающая всю поверхность земного ша-
ра, включающая в себя живые организмы и среду их обитания.
«Все организмы на Земле,— пишут П. Дювиньо и М. Танг (1968),—
можно рассматривать как мировое сообщество; вместе со своими
средами они образуют экосистему биосферы, которую можно пред-
ставить как объединение всех наземных и морских экосистем»
(с. 224). В другом месте об этом сказано еще яснее: «Взаимосвязь
(интеграция) всех экосистем мира приводит к представлению о
гигантской экосистеме мира «земной шар», то есть практически —
к представлению о биосфере» (с. 13). Близкие к этому взгляды на
биосферу высказывает и Ю. Одум (1975). Правда, наряду с
этим за рубежом имеют место и несколько иные трактовки этого
понятия, более отвечающие его существу.
В современной советской литературе выявилось несколько аспек-
тов в разработке учения о биосфере. Наряду с биогеохимическим
61
направлением, в основе которого лежат основополагающие идеи
В. И. Вернадского, идет разработка этого учения также с позиций
биогеоценологии, термодинамики, системологии и др. В связи с
этим большое внимание уделяется таким вопросам, как целост-
ность биосферы как планетарной системы, ее компонентный состав,
его взаимосвязь и соотношение с биосферой как целым, иерархич-
ность структуры и функциональной организации биосферы и меха-
низмы ее интеграции; вопросам энергетики биосферы, обменных
процессов внутри ее и со своим земным и ближайшим космиче-
ским окружением и пр. Для биосферы характерна «комплексность
происходящих в ней явлений, когда органически сочетаются осо-
бенности разных форм движения материи»,— пишет Г. Ф. Хильми
(1975, с. 94). Отсюда биосфера, по его словам, в равной степени
является геофизической, термодинамической, химической, биоло-
гической и кибернетической системой.
Биосфера, как указывает Э. В. Гирусов (1968), представляет
собою целостную общепланетарную систему, характеризующуюся
наличием у нее особых качеств и свойств, отсутствующих у состав-
ляющих ее компонентов как «частей» этого «целого»; при этом сами
компоненты являются продуктом саморазвития биосферы во вза-
имодействии ее с окружающей средой. Вместе с тем биосфера, как
всякое целостное образование, обладает определенной автономно-
стью, обособленностью по отношению к внешней среде. Повышение
уровня целостности и нарастание автономности бисферы по мере
ее развития рассматриваются как две стороны единого процесса.
Основная закономерность существования и развития биосферы, в
формулировке Э. В. Гирусова (1968), заключается в следующем:
«...живое вещество играет ведущую роль в развитии биосферы, а
главное противоречие биосферы — это взаимодействие живого ве-
щества с неорганическими условиями существования. Конкретным
выражением этого противоречия является основной для биосферы
процесс превращения неорганического вещества в органическое, с
одной стороны, и разложение его до минеральных элементов, с
другой. Этим главным противоречием определяются все остальные
процессы биосферы» (с. 252).
Компонентами, составными частями биосферы в настоящее вре-
мя принято считать: а) живое вещество планеты; б) наружный
слой литосферы; в) гидросферу и г) тропосферу. Э. В. Гирусов в
состав биосферы включает также д) почву и е) космические излу-
чения в зоне поверхности Земли. Им подчеркивается особое поло-
жение живого вещества в биосфере и биогенное происхождение
большинства ее неорганических «частей» (или «сред»). Возможно,
учитывая это обстоятельство, Г Ф. Хильми подразделяет «части»
биосферы на две категории: 1) живое вещество, выражаемое
его биомассой, и 2) биокосные среды, т. е. геофизические среды или
62
земные оболочки, преобразованные деятельностью живых существ
и космических излучений. К числу биокосных сред им отнесены:
а) современная атмосфера, содержащая свободный кислород;
б) почвы, т, е. измененные деятельностью абиотических и биотиче-
ских факторов поверхностные слои горных пород, и в) природные
воды, содержащие растворенный кислород и продукты минерали-
зации органических веществ. К числу биокосных сред Хильми отно-
сит также некоторые органогенные породы литосферы, так назы-
ваемые биолиты.
Г. Ф. Хильми обращает внимание на неустойчивость как живого
вещества, так и биокосных сред, которая выражается в том, что
предоставленные самим себе среды теряют свои биокосные свой-
ства, а для сохранения их они нуждаются в непрерывном поступ-
лении энергии извне, которую они получают в процессе взаимодей-
ствия с живым веществом биосферы, в особенности с раститель-
ностью. Таким образом, живое вещество не только создает себя из
ншчппей среды с использованием при этом солнечной энергии, но и
поддерживает се в преобразованном состоянии, как необходимое
условие своего существования и развития. По мере развития био-
сферы, заселения ее живыми существами, формирования биоген-
ной атмосферы и т. п., по утверждению Э. В. Гирусова, происходит
постепенное возрастание и усложнение взаимосвязи биосферы с
косной средой и космическим окружением, а по Н. В. Дылису
(1978)—возрастает также устойчивость и уравновешенность ее.
Биосфера, как отмечает В. А. Абакумов (1975), организована
по принципу иерархии: ее подсистемы и компоненты разного ранга
объединены в единое целое, находясь в соподчиненном по отноше-
нию к вышестоящим ступеням иерархической лестницы. Такого же
мнения по этому вопросу придерживается Ф. Я. Шипунов (1980),
который пишет: «В биосфере проявляется не просто совокупность
живых и косных тел как механическая смесь, а определенные
иерархически соподчиненные структуры (биокосные тела)» (с. 8).
В основе иерархической организации биосферы, по словам Абаку-
мова, лежат циклические колебательные процессы, представляю-
щие собою один, из важнейших механизмов ее организации.
Организованность биосферы, по Шипунову, заключается в том,
что все ее «части» — живые и косные — взаимопроникают друг в
друга и взаимодействуют между собой, образуя единое «целое»;
при этом ни одна из составных частей биосферы не может быть от
нее отделена без уничтожения «целого», т. е. биосферы.
Таким образом, организованность биосферы — это основное
свойство ее как системы весьма сложного состава и структуры, и,
вслед за Вернадским, она должна рассматриваться как подвижное,
динамическое, все время колеблющееся равновесие, непрерывна
стремящееся к устойчивости.
63
Подводя итоги всему изложенному о современном представле-
нии о биосфере, уместно привести определение биосферы, предло-
женное В. А. Ковдой (1973), которое гласит: «Все это дает основа-
ние понимать биосферу как сложную многокомпонентную общепла-
нетарную термодинамически открытую саморегулирующуюся систе-
му живого вещества и неживой материи, аккумулирующую и пере-
распределяющую огромные ресурсы энергии и определяющую со-
став и динамику земной коры, атмосферы и гидросферы» (с. 100).
Наряду с этим следует отметить, что многие факты свидетель-
ствуют об интенсивном в последние десятилетия нарушении при-
родной слаженности, организованности биосферы вследствие все
возрастающего воздействия человеческого общества на все ее
иерархически сопряженные подсистемы и компоненты, в особен-
ности в связи с бурным ростом населения и развитием научно-тех-
нического прогресса, в частности, в связи с включением в первич-
ную биосферу множества промышленных предприятий, мощных
технических установок и современных индустриальных центров и
городов. Все это послужило основанием для наименования новей-
шего этапа развития биосферы биотехносферой (Г Ф. Хиль-
ми, 1968).
Превращение современной биосферы в биотехносферу, по
словам Г. Ф. Хильми (1975), — процесс неизбежный и закономер-
ный. Однако при формировании биотехносферы человечеству не-
обходимо идти, как предлагает Ф. Я. Шипунов (1980), «не путем
замены биосферы совокупностью технических устройств, а путем
сознательного регулирования процессов в биосфере на основе со-
хранения ее организованности и «вписывания» техники в биогео-
химические циклы и энергетические потоки» (с. 10).
Еще ранее В. И. Вернадским (1944) для обозначения наступа-
ющего нового этапа в развитии и хозяйственном освоении биосферы
было предложено наименование ноосфера, т, е. сфера разума,
суть которой в подчинении дальнейшего развития нашей планеты
силе разума социального человека. Это наименование, на наш
взгляд, точнее выражает суть современного этапа развития био-
сферы и роль в этом процессе человека как социального существа.
«В ноосфере, — пишет И. А. Шилов (1981), — действует сложный
комплекс факторов, включающий хозяйственные, политические,
юридические, моральные и иные социальные аспекты и порожда-
ющий новые закономерности динамики природных комплексов»
(с. 11). Исходя из этого, он полагает, что настало время говорить
о рождении новой науки, цель которой — «...сознательное регули-
рование соотношения различных аспектов деятельности человече-
ства с поддержанием устойчивости природных систем, как источни-
ков его благосостояния» (с. И). Г. Ф. Хильми (1968) предложил
назвать современное учение о биосфере ноогеникой. Некото-
64
рыми другими авторами, например В. А. Абакумовым (1975), упо-
требляется и другое наименование этой науки, а именно биосфе-
р ол оги я. Однако ни то, ни другое название современного учения
о биосфере пока не получило всеобщего признания среди специ-
алистов соответствующей отрасли знания.
9. Биогеосфера, типы ее строения
и соотношение с биогеоценозом
Биогеоценологический подход к рассмотрению и анализу соста-
ва, структуры биосферы и 1Происходящих в ней явлений неизбежно
предопределяет сужение ее вертикальных границ, поскольку эти
явления наиболее отчетливо и ярко проявляются лишь в той при-
земной части биосферы, которая Вернадским называлась пленками
и сгущениями жизни. Это вполне естественно, ибо основные про-
цессы и внутренние качественные перестройки тех или иных струк-
турных образований в биосфере возможны лишь при определенной
концентрации, уплотненности живого вещества, обеспечивающей
взаимодействие ее составных частей. Речь, по существу, идет об
особом уровне организации живой материи — биоценотическом,
структурным выражением которого и являются такие биокосные
системы, как биогеоценозы и образованная ими биогеосфера (био-
геоценотический покров Земли), представляющие собою объект
изучения биогеоценологии. В противоположность этому объектом
изучения науки биосферологин (или ноогеники) является та-
кая интегрированная совокупность организмов, как живое вещество
(биострома) и образованная им во взаимодействии с неорганиче-
ской средой под действием солнечной радиации биосфера.
Таким образом, биогеоценологический аспект изучения биосфе-
ры заложен в самом учении В. И. Вернадского о ней, в его пред-
ставлении о наличии особых зон сосредоточения живого вещества
(сгущений и пленок жизни) в биосфере, а также в современном
представление о разных уровнях организации биосферы и одновре-
менном, в геологическом смысле, сопряженном возникновении био-
косных систем разного ранга, иерархически последовательно со-
подчиненных друг другу. С этой точки зрения учение В. Н. Сукаче-
ва и его соратников и последователей о биогеоценозе и биогеосфере
представляет собою дальнейшее развитие учения В. И. Вернадско-
го о биосфере. Это — особый биогеоценотический уровень изучения
биосферы, характеризующийся переходом от интегративных, обоб-
щенных представлений и трактовок В. И. Вернадского о биосфере,
ее строении и функционировании к более дробному, дифференци-
рованному изучению структур и процессов, происходящих в ее при-
земном, насыщенном жизнью слое—биогеосфере. С этим неизбежно
3. Зак. 161 в5
связана и дифференциация самих понятий, и соответствующая
терминология, выражающая эти понятия.
Биогеосфера в настоящее время рассматривается как осо-
бый, насыщенный жизнью слой биосферы, характеризующийся наи-
более плотной концентрацией организованных в биоценозы живых
существ, образующих во взаимодействии с окружающей средой
элементарные биокосные системы — биогеоценозы и биогидроце-
нозы. Последние в совокупности и составляют не вполне однород-
ную, местами прерывистую, сложную по составу и структуре био-
косную оболочку сухопутного и водного сектора земного шара.
Мощность биогеосферы варьирует в значительных пределах. На
суше верхняя граница биогеосферы определяется в основном высо-
той растительного покрова с охватом находящейся в постоянном
газообмене с живыми существами воздушной прослойки до 20—
30 м над его поверхностью, а нижняя — глубиной проникновения
в почву и подпочву корневых систем ценозообразующих растений
и почвообитающих животных и микроорганизмов, а также взаимо-
действующих с ними почвенно-грунтовых вод до глубины первого
водоупорного горизонта. В морях, океанах и континентальных во-
доемах вертикальные гранйцы биогеосферы определяются верхни-
ми пределами надводной воздушной прослойки той же мощности,
что и на суше, и глубиной проникновения в водную среду фотосин-
тетически активных солнечных лучей, а также растворенных и
взвешенных в воде минеральных и органических веществ и свобод-
ного кислорода. Отсюда общая мощность биогеосферы намного
меньше, чем мощность биосферы. На суше, без учета простираю-
щейся над растительным покровом воздушной прослойки, общая
мощность надземной и подземной частей биогеосферы измеряется
дециметрами на скалистых обнажениях и в тундрах, немногими
метрами в степях и пустынях и десятками метров, редко до 100—
150 м, в лесах различных частей света. В условиях водной среды
мощность биогеосферы местами достигает глубины в 380—400 м,
хотя наибольшее сосредоточение жизни наблюдается в поверхно-
стном слое воды до глубины в 20—30, реже— 100 м.
Биогеосфера — хотя и прерывистая, но все же общепланетар-
ная биокосная система, а потому она играет весьма важную роль
в выполнении глобальных функций биосферы в целом. В б ногео-
сфере преимущественно осуществляются: а) фиксация энергии
солнечной радиации и преобразование ее в различные формы зем-
ной потенциальной энергии; б) создание н накопление на поверх-
ности Земли и в ее недрах биогенного вещества в виде органиче-
ских и оргапоминеральных образований; в) создание биокосного
вещества в процессе взаимодействия живого и неживого на Земле;
г) преобразование структуры и химического состава верхних слоев
земной коры, а также химического состава подземных вод атмо-
66
сферного происхождения. В пределах биогеосферы протекает эво-
люция органического мира, а также развитие человека и челове-
ческого общества, которое в настоящее время становится главной
деятельной силой в биосфере. Следствием всего этого являются;
а) уменьшение энтропии, т. е. рассеивания солнечной энергии в
тепловой форме; б) возникновение в процессе эволюции многооб-
разия форм органической жизни и усложнение их организации;
в) биологизация биогеосферы, а в последние десятилетия и техни-
зация ее.
В результате, по словам Н. В. Дылиса, в биогеосфере, с одной
стороны, автоматически осуществляется контроль за состоянием и
перемещением энергетических ресурсов и ряда важнейших пока-
эптслсй физико-географической среды, что, как отмечалось, ведет
К возрастанию ее устойчивости по мере развития, а с другой, бла-
ro'i qoi нес увеличивающимся масштабам и интенсивности хозяй-
(I iioin ноздсйствпя вооруженного современной техникой чело-
мин нПщггrun на различные стороны биогеосферы, происхо-
ди мр у шеи up yi н1П1Н1И1ппегогя в пей динамического равновесия.
|l tan г whim перед чг.'кин'чргтпом возникает целый ряд весьма
hr ых и ПГН1ЛОЖПЫХ 1лоблл1.11ого характера теоретических науч-
h и при K i ичегких народнохозяйственных проблем и задач по
пению негативных форм воздействия на биогеосферу и раз,-
• • ।кс научных основ регулирования и управления ею в
интересах современного населения и грядущих людских поко-
лений.
Термины «биогеосфера» и «биогеоценотический покров Земли»,
как отмечалось рапсе, являются синонимами. Синонимами этим
терминам но своему содержанию и объему следует считать и пред-
ложенное Е. М. Лаврепко (1949) понятие и термин фитогео-
сфера, а также термин витасфера, предложенный В. Д. Алек-
сандровой и А. Н. Тюрюкановым (1969). Что же касается ряда
понятий и терминов, введенных в науку географами для обозначе-
ния поверхностных земных оболочек, иных по содержанию и по
объему, чем биогеосфера, то останавливаться на их характеристике
п данном случае нет необходимости.
После разностороннего и тщательного рассмотрения биогеоце-
ноза и бпогеосферы как составляющих двуединый объект исследо-
вания науки биогеоценологии следует подчеркнуть их органиче-
скую взаимосвязь, единство.
Биогеосфера, как отмечалось, представляет собою планетарную
систему взаимосвязанных биогеоценозов; биогеоценозы же рассма-
триваются как элементарные структурные единицы (ячейки) био-
геосферы. Как биогеоценоз, так и биогеосфера являются структур-
ным выражением единого — биоценотического — уровня организа-
ции живой материи в определенных условиях среды, материальный
67
субстрат которого представлен названными выше «пленками» и
«сгущениями» жизни нашей планеты.
Биогеоценоз и биогеосфера — это биокосные системы,
структурно и функционально взаимосвязанные как «часть» и «це-
лое», но находящиеся на разных ступенях иерархической лестницы.
При этом биогеосфера, состоящая из иерархически соподчиненных
ей структурных единиц—биогеоценозов, представляет собой каче-
ственно обособленную целостную биокосную систему, обладающую,
как мы видели, такими свойствами, которые отсутствуют у состав-
ляющих ее отдельно взятых биогеоценозов. В то же время биогео-
ценозы, находясь в пространственно-временном соподчинении по
отношению к биогеосфере, сохраняют свою относительную само-
стоятельность, т. е. некоторую степень автономности, как системы
весьма сложные по составу и структурно-функциональной органи-
зации, обладающие качественной определенностью.
Глава HI
Живые компоненты биогеоценоза
и их составные элементы
Очередные три главы будут посвящены всестороннему рассмо-
трению компонентов биогеоценоза как живых, образующих биоце-
ноз, так и косных, биогенного происхождения, составляющих эко-
гоп, который под воздействием живых компонентов и биоценоза в
целом, как было сказано ранее, преобразуется в биотоп — внутрен-
нюю среду биогеоценоза (биоценосреду).
При рассмотрении живых компонентов биогеоценоза основное
ннимапие должно быть обращено на такие стороны и свойства их,
как популяционно-видовой, экобиоморфный и ценотический состав
каждого из них; при этом должны быть насколько возможно тща-
к’лыю рассмотрены и определены сами эти понятия, характеризу-
ющие живые компоненты биогеоценоза. Следует также детально
рассмотреть основные функции каждого живого компонента в
составе биогеоценоза, направление их биоценотнческой функцио-
нальной деятельности и степень участия в биогеоценозе и пр. При
рассмотрении же косных компонентов сухопутного биогеоценоза
предметом анализа должны стать те элементы атмосферы и почвы,
которые в совокупности составляют материальную основу воздуш-
ной и почвенной биогенной среды живых компонентов, биогеоцено-
тлческая функциональная роль атмосферы и почвы и их составных
элементов, н также преобразования каждой из них при взаимодей-
ствии с другими компонентами в одну из сторон внутренней био-
генной среды биогеоценоза и пр.
I. Виды и видовые популяции в составе
живых компонентов биогеоценоза
В данной главе надлежит более или менее подробно рассмот-
реть живые компоненты биогеоценоза—автотрофный, биотрофный
и сапротрофный. Общим для живых компонентов является то, что
все они состоят из биологических видов, точнее, из видовых популя-
ций. Что такое биологический вид и видовая популяция?
С общебиологической точки зрения вид — это одна из форм
организации живой материи, одна из форм жизни на Земле. Ви-
ды— универсальное явление природы, ибо все живые существа,
организмы, как известно, представлены видами. Вид — это весьма
сложное и внутренне противоречивое явление природы. Внутренняя
противоречивость вида, как указывает К- М. Завадский (1961), на-
69
ходит отражение в противоречивости признаков вида, например:
вид — качественно обособленное образование, но в то же время это
сложная система, состоящая из ряда внутривидовых подразделе-
ний; любой вид представлен в природе бесчисленным множеством
особей, но одновременно с этим он имеет единую наследственную
основу и единый тип организации; вид характеризуется многообра-
зием форм (скажем, формы сосны обыкновенной—лесная, дюнная,
болотная, а на болотах — форма Литвинова, Вилькома, карлико-
вая, торфяная), по вместе с тем это целостное образование, обла-
дающее определенной структурой и внутривидовыми отношениями
особей, и т. д.
Итак, вид — это сложная целостная система, состоящая из
ряда внутривидовых единиц (подвид, экотип, биотип и др.), основ-
ной из которых является местная популяция. Местная видо-
вая популяция, по Завадскому, — это поселение (или группа
поселений) особей вида, основная единица существования, приспо-
собления и воспроизведения вида. Различные виды растений, жи-
вотных и микроорганизмов более или менее широко распростра-
нены по земной поверхности, занимают в пределах своего ареала
различные местообитания на суше, 'в морских и континентальных
водоемах. В различных местообитаниях виды живых существ обыч-
но представлены различными по своим свойствам группами особей,
образующими так называемые популяции. Следовательно, в раз-
личных местообитаниях каждый вид представлен разными популя-
циями.
По данным Т. А. Работнова (1975, 1978а), С. С. Шварца (1971,
1972), П. М. Рафеса (1972), Н. П. Наумова (1970) и других совет-
ских биологов, занимавшихся изучением популяционного состава
разных видов и ценозов, популяции обладают целым рядом при-
сущих нм характерных черт, свойств и признаков, имеющих не
только общебиологическое, но и весьма важное экологическое и
ценотическое значение.
Популяция прежде всего трактуется как форма существования
вида в определенных конкретных условиях обитания в составе того
или иного ценоза, как элементарная биохорологическая (простран-
ственно обособленная) единица вида, обладающая необходимыми
свойствами для поддержания жизни вида в постоянно изменяю-
щихся условиях среды. Во-вторых, популяция располагает способ-
ностью спаривания особей, дающих жизнеспособное потомство,
характеризующихся примерно одинаковыми возможностями при-
способления к среде обитания и относительно сходными морфоло-
гическими, физиологическими и другими свойствами и признаками.
С другой стороны, соседствующие популяции одного и того же
вида, развивающиеся в заметно отличающихся друг от друга усло-
виях, обнаруживают способность к дивергенции и образованию
70
чкоэлементов, изореагентов и других форм вяутрипопуляционной
дифференциации. В-третьих, обладая сложной внутренней струк>
турой, популяция получает возможность расселяться по различным
стациям экотопа, занимать различные экологические ниши в со-
обществе, образуя различной плотности поселения особен с соот-
ветствующей возрастной и половой структурой и общим запасок*
биомассы. Однако не отдельные поселения особей, а популяция в
целом может обеспечить поддержание оптимальной численности
особей и популяционной структуры, стабильности вида в изменя-
ющихся условиях среды. В-четвертых, популяции представляют
функциональное единство вида, которое обеспечивается сходством
их положения в функциональной организации ценотических систем
и выполнением их особями или группами особей определенных:
функций в их составе по трансформации пищевых ресурсов в соб-
ственную биомассу, по воспроизводству новых поколений особей
и пр. В-пятых, популяция рассматривается как элементарная еди-
Himtt эволюционного процесса органического мира на Земле, ибо
тпк нпзы1Н1Смаи «мнкрочнолюция» совершается на уровне по-
пуляции.
Вместе с тем видовые популяции представляют собой составные
элементы любого живого компонента биогеоценоза. Поэтому видо-
вые популяции в конкретных ценотических системах принято назы-
вать ценопопуляциями (А. А. Корчагин, 1964). Из этого следует»
что ценопопуляция может рассматриваться как основная единица
существования вида в соответствующем биогеоценозе.
Следовательно, видовые популяции живых существ в процессе
длительного исторического развития должны были выработать
приспособительные свойства, с одной стороны, к физической среде
витания, а с другой — к совместной жизни с другими видами в
□ставе соответствующего биогеоценоза. Однако следует иметь в ви-
Ь, что это не два разрозненных приспособительных процесса, а
випын, но двусторонний процесс приспособления видовых популя-
|ай разных видов к обитанию в составе биогеоценотической сис-
темы, сформировавшейся при их же активном разностороннем
взаимодействии друг с другом и с окружающей средой.
2. Жизненные формы (экобиоморфы) и системы экобиоморф
растений и животных
Любой регион биогеосферы, как целостной планетарной систе-
мы, состоит из большого количества взаимосвязанных в единое
целое элементарных структурных единиц—биогеоценозов, которые,
в свою очередь, как мы видели, состоят из множества видовых
ценопопуляций живых существ. Но видовые ценопопуляции орга-
низмов входят в состав биогеоценоза не хаотично, не разрозненно,
71
не в виде обособленных особей, а закономерно, в виде определен-
ных эволюционно выработанных взаимосвязанных биологических
групп и ценотических сочетаний. Пригнанность этих групп и соче-
таний видовых ценопопуляций к среде обитания и взаимосвязь их
друг с другом и придают биогеоценозу свойства целостности, сла-
женности, стабильности, а также качественной определенности и
количественной выраженности.
К таким эволюционно выработанным биологическим, точнее,
биоморфологическим группам ценопопуляций живых существ пре-
жде всего относятся так называемые жизненные формы (Е. Вар-
минг, 1902), или экобиоморфы (Б. А. Быков, 1962; Е. М. Лавренко
и В. М. Свешникова, 1968).
Представление о жизненных формах (экобиоморфах) возникло
в биологии в начале 19-го столетня, когда Гумбольдт в 1806 году
ввел понятие «основные физиономические формы» растений, а
позднее, в 1874 году, Гризебах при описании растительности зем-
ного шара установил и охарактеризовал 60 таких «основных фи-
зиономических форм». В конце 19-го столетия эти основные формы
растений Варминг предложил называть жизненными фор-
мами. Понятие «жизненные формы» оказалось весьма плодотвор-
ным. Разработка его в текущем столетии получила широкий раз-
мах как за рубежом, так и в нашей стране. Наряду с разработкой
этого понятия в экологии растений шла разработка его и в эколо-
гии животных, где оно отразило специфику животного объекта этой
науки.
На первых порах за рубежом разработка проблемы жизненных
форм растений производилась с чисто физиономических позиций.
Устанавливались и выделялись жизненные формы по внешним
морфологическим признакам растений, особенно большое внимание
уделялось их общему внешнему облику. Позднее Варминг в выде-
ление жизненных форм внес экологический принцип: характер мес-
тообитания, способ питания растений и пр. В дальнейшем разра-
ботка проблемы жизненных форм производилась на основе учета
не только внешних морфологических признаков растений, по также
и отношения их к важнейшим экологическим факторам среды, ха-
рактерных эколого-биологических особенностей их и т. п. Решаю-
щее значение за рубежом придавалось таким биологическим осо-
бенностям и приспособительным признакам растений к среде оби-
тания, как продолжительность их жизни или периода вегетации,
положение почек возобновления (в почве, на ее поверхности или
на некоторой высоте над ее поверхностью), способы и степень за-
щиты их в неблагоприятный период года от холода или жары
и т. п. Наиболее типичной системой растительных жизненных форм
такого рода и наиболее распространенной на Западе, в Америке,
да в недавнем прошлом и в нашей стране, является система К. Ра-
72
ункьера (Raunkiaer, 1905) с некоторыми дополнениями Браун-
Бланке (Braun-Blanquet, 1928).
В нашей стране разработка проблемы жизненных форм расте-
ний связана с именами таки* отечественных биологов, как Г. Н. Вы-
соцкий (1915), В. Р. Вильямс (1922), Л. И. Казакевич (1922),
Б. А. Келлер (1933), И. Г. Серебряков (1955, 1962, 1964), Г. М. Зо-
зулин (1961, 1968, 1976), В. Н. Голубев (1957, 1960), Б. А. Быков
(1970а), Т. И. Серебрякова (1971, 1972) и др, Советские биологи
в разработке этой проблемы, по словам А. П. Щенникова (1950),
в отличие от зарубежных биологов идут «не от анализа флоры во-
обще и не от климатического районирования или от физиономиче-
ских признаков... а от эколого-биологического анализа конкретных
форм растительного покрова» (с. 357).
Ввиду того, что подобное изложение различных систем жизнен-
ных форм (экобиоморф) растений имеется в курсах экологии рас-
тений, ограничимся общей характеристикой лишь некоторых оте-
чсчтнеппых систем, имеющих более четко выраженную ценотиче-
скую значимость.
Из ранних отечественных систем жизненных форм растений,
представляющих наибольший интерес, в согласии с А. П. Щенни-
ковым следует признать системы В. Р. Вильямса, Г. Н. Высоцкого
и Л. И. Казакевича, которые в известной мере логически связаны
между собой, взаимно дополняют друг друга. В. Р. Вильямс, как
известно, ценозообразующие растения европейской части Союза
сгруппировал в жизненные формы растений по способу питания и
морфологии вегетативного размножения; затем по типу питания и
взаимосвязи в биотическом круговороте веществ подразделил их
па две категории основных жизненных форм (точнее, групп или
секций жизненных форм). I. Зеленые автотрофные растения: 1) де-
ревянистые лесные, 2) травянистые степные и 3) травянистые лу-
говые. И. Бесхлорофилльные гетеротрофные растения: I) низшие
грибы, 2) бактерии аэробные и 3) бактерии анаэробные. Эти основ-
ные жизненные формы (группы жизненных форм) зеленых и бес-
хлорофилльных растений, по Вильямсу, будучи взаимосвязанными
в биотическом круговороте веществ, образуют разного типа расти-
тельные формации (точнее, типы растительности, а по существу
группы биогеоценозов).
Таким образом, система жизненных форм В. Р. Вильямса, как
видим, отражает исторически обусловленную взаимосвязь жизнен-
ных форм не только со средой обитания, но и друг с другом. Сле-
довательно, она является не только эколого-биологической, но и
биоценотической, причем динамической системой жизненных форм.
Системы травянистых форм Г. Н. Высоцкого и Л. И. Казаке-
вича, построенные на анализе степной флоры юга России, могут
рассматриваться как дополнение к системе Вильямса, поскольку
73
ь них наряду с жизненными формами злаков и осок довольно де-
тально рассматриваются и подразделяются жизненные формы
бобовых и представителей различных семейств двудольного и одно-
дольного разнотравья. В основе этих двух систем жизненных форм
лежат различия растений в характере подземных органов и в спо-
собности их к вегетативному размножению и расселению.
По мере более разностороннего изучения жизненных форм рас-
тений уточнялось и углублялось представление о их сущности,
морфологических, анатомо-физиологических и эколого-биологиче-
ских особенностях, о взаимосвязи со средой обитания и значении
их в изменении и преобразовании среды, о месте и роли в составе
ценотических систем разного ранга; углублялось также представ-
ление о направлении и путях эволюции жизненных форм растений,
и в соответствии с этим совершенствовались и методы выделения
и систематизация их и пр. Из отечественных систем жизненных
форм растений последнего периода наибольшего внимания, на наш
взгляд, заслуживают системы И. Г. Серебрякова, Б. А. Быкова,
Г М. Зозулина.
Система жизненных форм растений И. Г Серебрякова (1962)
построена на эколого-морфологической основе в эволюционном
аспекте. В качестве главного структурно-биологического признака
он выдвигает длительность большого жизненного цикла растения
(от прорастания семени до полного отмирания всего растительного
организма), которым, по его мнению, определяется как общий
облик (габитус) жизненной формы, так и направление ее эволю-
ции, выражающейся в сокращении длительности жизненного цикла
надземных скелетных осей растения от деревьев к травам.
С эколого-ценотической точки зрения жизненные формы, как
указывает И. Г Серебряков, исторически возникли как приспособ-
ления к наиболее полному использованию всего комплекса условий
местообитания, выражая способность определенных групп растений
к расселению и закреплению за собой данной территории и к уча-
стию их в формировании растительного покрова.
По-иному к трактовке содержания и роли жизненных форм
растений в природе, к выделению и систематизации их подходит
Б. А. Быков (1970а). Предпочитая именовать жизненные формы
растений экобиоморфами, он дает следующее определение:
«Экобиоморфа является совокупностью видов (а иногда и внутри-
видовых форм), имеющих сходные формы роста и биологические
ритмы, а также сходные приспособительные и средообразователь-
ные особенности» (с. 23). Возникновение экобиоморф растений, по
его утверждению, происходит путем естественного отбора видовых
популяций, наиболее соответствующих по своим природным каче-
ствам и свойствам условиям существования в сообществах, нахо-
дящихся в определенной среде обитания. Вследствие этого видовые
74
растительные популяции, относящиеся к одной и той же экобио
морфе, способны занимать относительно одинаковые экологически!
ниши в составе ценозов и играть в них однотипную роль в обмен!
веществ и энергии.
В системе жизненных форм (экобиоморф) растений Б. А. Бы
кова обращает на себя внимание то обстоятельство, что в выделя
емых им высших таксонах — отделах — подчеркивается тип пита
ния входящих в их состав экобиоморф, а именно автотрофность i
гетеротрофность. Это придает данной системе ясно выражении!
биоценотический характер, ибо функциональная взаимосвязь со
ответствующих автотрофных и гетеротрофных живых существ i
служит основой формирования биоценозов и биогеоценозов 1
целом.
Наиболее детально разработанной системой жизненных форл
растений, очевидно, следует признать систему Г. М. Зозулин;
(1961, 1968). И. Г. Серебряков (1959) считает ее наиболее выдер
жапиой в фитоцепотпческом плане.
Прежде всего Г М. Зозулиным подчеркивается конвергентное
происхождение жизненных форм растений в отличие от категори!
филогенетической систематики растений, возникающих путем ди
вергенции исходных форм. Отсюда ход эволюции жизненных форл
им рассматривается лишь как исторический процесс конвергентно!
выработки этих форм. Поэтому в одну жизненную форму им обь
едиияются, как и должно, различные по систематическому поло
жению виды растений, но сходные по исторически сложившими
приспособлениям к условиям существования. Исходным положе
пнем в становлении и развитии жизненных форм, по автору, явля
стся тенденция каждого организма путем размножения прочно за
крепиться в среде своего обитания и как можно шире распростра
питься в условиях окружения другими организмами. Поэтом]
основным принципом, положенным в основу установления и выде
ления жизненных форм растении, автором признается процесс раз
вития приспособлений, обеспечивающих удержание особями пло
щади обитания и распространения в ней.
Помимо этого, при выделении жизненных форм растений авто{
предлагает руководствоваться следующими положениями: 1. По
скольку становление и эволюция жизненных форм протекает i
условиях того или иного растительного сообщества, при выделени!
их необходимо учитывать фитоценотическое окружение раститель
ных организмов, ибо взаимоотношения растений в фитоценозе i
воздействие фитоценоза па отдельные растения играют решающук
роль в формировании и развитии жизненных форм, выражаясь i
различного рода приспособлениях к жизни в фитоценозе. 2. В свя
зи с тем, что у травянистых растений луговых, степных, пустынны:
и других подобного типа сообществ значительная часть года про
75
текает в подземной (почвенной) среде, где ограничивающим фак-
тором является главным образом влага, а иногда также и запас
элементов минерального питания, в основу выделения жизненных
форм должны быть положены приспособительные особенности под-
земных органов растений, значительно превышающих по массе их
периодически отмирающую надземную часть. У деревьев и кус-
тарников лесных и кустарниковых сообществ с развитыми, не отми-
рающими э онтогенезе надземными органами, где лимитирующим
фактором является главным образом свет, в основу выделения жиз-
ненных форм должны быть положены приспособительные особен-
ности скелетных наземных частей растений.
Позднее Г. М- Зозулин (1976) предпринял попытку подойти к
разработке проблемы жизненных форм растений с позиции учения
В. И. Вернадского о биосфере. Не вдаваясь в подробности такого
подхода автора к анализу жизненных форм, считаем возможным
обратить внимание лишь на некоторые принципиальные положения.
В аспекте учения о биосфере жизненные формы растений рассма-
триваются им как «продуценты, определяющие выход биомассы и
запасаемой энергии на единицу площади» и как особые этапы
(звенья) эволюции приспособлений у совместно обитающих расте-
ний «в направлении усиления биогенной миграции химических эле-
ментов в условиях той или иной среды» (с. 1510). Исходя из этого,
автор находит возможным связать различные подходы к разработ-
ке проблемы жизненных форм в единое целое и рассматривать
жизненные формы растений как «объединения принципиально
сходных по указанным приспособлениям организмов» (с. 1510).
В свете изложенного Г. М. Зозулин приводит следующее опре-
деление жизненных форм растений: «Под жизненными формами
следует понимать конвергентно возникшие объединения раститель-
ных организмов, обладающих принципиально сходными морфоло-
гическими и биологическими приспособлениями к удержанию осо-
бями площади их обитания и распространению по ней в условиях
сложной абиотической и биотической среды; вместе с тем жизнен-
ные формы являются качественно особыми этапами эволюции при-
способлений к максимальному проявлению биогенной миграции
химических элементов в биосфере» (с. 1510).
Проблема жизненных форм (экобиоморф) животных менее раз-
работана, чем жизненных форм растений как за рубежом, так н в
нашей стране, хотя разного рода приспособительные биолого-мор-
фологические признаки выражены в животном мире не менее, если
не более, ярко, чем в растительном.
Из отечественных экологов животных раннего периода наиболее
известен Д. Н. Кашкаров, который в монографиях «Среда и орга-
низм» (1933) и «Основы экологии животных» (1938, 1944) писал:
«...тип животного, находящегося в соответствии с окружающими
76
условиями, мы называем «жизненной формой». И далее: «В
жизненной форме, как в зеркале, отражаются главнейшие домини*
рующие черты местообитания видов и основные черты климата и...
почв и биотических отношений» (1944, с. 173). Руководствуясь
этими положениями, Д. Н. Кашкаров предложил следующую
систему форм животных.
I. Плавающие формы:
1. Чисто водные: а) нектон, б) планктон, в) бентос.
2. Полуводные: а) ныряющие, б) неныряющие, в) лишь
добывающие из воды пищу.
II. Роющие формы:
1. Абсолютные землерои (всю жизнь проводящие под
землей).
2. Относительные землерои (выходящие на поверхность).
Ш. Наземные формы:
1. Не делающие нор: а) бегающие, б) прыгающие, в) пол-
зающие.
2. Делающие поры: а) бегающие, б) прыгающие, в) пол-
зающие.
3. Животные скал.
IV. Древесные лазающие формы:
а) не сходящие с деревьев, б) лишь лазающие по де-
ревьям.
V. Воздушные формы:
а) добывающие пищу в воздухе, б) высматривающие пи-
щу с воздуха.
В соответствующих работах современных отечественных зооло-
гов и зооэкологов можно встретить ряд высказываний по поводу
жизненных форм животных или даже высказывания, касающиеся
жизненных форм вообще, включая животные и растительные орга-
низмы. Так, по М. П. Акимову (1954, 1959), жизненные формы жи-
вотных представляют собой тот или иной тип приспособления их к
определенной среде обитания и к определенному образу жизни, в
том числе и к способу питания. В. В. Яхонтов (1969) рассматривает
жизненные формы животных, в частности насекомых, как истори-
чески сложившиеся комплексы их, обладающие определенными
биологическими, физиологическими и морфологическими свойства-
ми, которые обусловливают соответствующую реакцию их на воз-
действия среды. По мнению Ю. И. Чернова (1975), в характери-
стику жизненных форм животных следует включать особенности
движения, способа добычи пищи, отношения к субстрату и формы
тела животного. Он считает также, что у насекомых и других жи-
вотных, развивающихся с метаморфозом, разные онтогенетические
стадии (личинки, куколки), наравне со взрослой особью, должны
относиться к разным жизненным формам, а для объединения фаз
77
жизненного цикла таких животных Г. А. Мазохин-ПоршнякоЕ
(1954) предложил понятие биологический тип, характери-
зующийся такими общевидовыми признаками, как способ расселе-
ния, плодовитость и пр.
Таким образом, при установлении, выделении и систематизации
жизненных форм (экобиоморф) животных, ведущих, в отличие 07
растений, в подавляющем большинстве подвижный образ жизни,
дополнительно принимается во внимание ряд таких биологических
особенностей и эколого-морфологических признаков и свойств, ко-
торые отражают специфику животного мира. Животные, в отличие
от растений, имеют возможность избегать неблагоприятных усло-
вий жизни, на определенный период года или на более длительный
срок перемещаться в более благоприятные места обитания или,
оставаясь на месте, произвольно или непроизвольно повышать
интенсивность своей жизнедеятельности, усиливать темпы размно-
жения и т. п. Поэтому с этой точки зрения весьма важное значе-
ние приобретает учет таких особенностей и приспособительных
черт и свойств животных, как способ передвижения и образ жизни
их, тип питания и способ добычи пищи, реакция на воздействие
различных факторов физической среды и проведение при различ-
ных ситуациях контакта с особями других видов и пр.
При сопоставлении изложенных взглядов и подходов соответ-
ствующих специалистов ботанической и зоологической наук к ре-
шению проблемы жизненных форм растительного и животного ми-
ра прежде всего следует обратить внимание на сходство состава
самых низших таксонов жизненных форм растений в системах по.-
следнего времени. Причем характерно, что, по крайней мере в груп-
пах травянистых жизненных форм, они являются в основном по-
вторением состава таковых в системах Высоцкого, Казакевича,
Вильямса с некоторыми дополнениями и уточнениями, что свиде-
тельствует о правильности по своей сущности и соответствии своей
природе выделенных в свое время названными авторами жизнен-
ных форм. Различия между рассмотренными выше системами жиз-
ненных форм последнего времени касаются главным образом со-
става таксонов, и связаны они с разными представлениями их
авторов о направлениях и путях эволюции жизненных форм и
положенными в основу их выделения и систематизации приспосо-
бительными признаками, свойствами и эколого-биологическими
особенностями организмов.
С биогеоценологической точки зрения в силу сопряженного ха-
рактера органической эволюции проблема жизненных форм — об-
щебиологическая проблема, несмотря на отмеченную выше специ-
фику их в мире растений и животных. Характерные качественные
приспособительные черты тех и других вырабатывались в филоце-
ногенетическом плане в условиях биогеоценоза как целостной сисх
78
темы, и обусловлены они главным образом характером выполняе-
мой каждым из живых компонентов функции в его составе. По-
скольку в основе сочетания живых существ в биогеоценозы, как
отмечалось, наряду с трофодинамическнми взаимоотношениями
функциональных живых компонентов лежат также межвидовые
взаимоотношения ценопопуляций в форме «борьбы за существова-
ние» в рамках каждого компонента за свои позиции в его составе,
то, следовательно, при анализе и систематизации жизненных форм
растительных и животных ценопопуляций необходимо учитывать
как общие для них эколого-морфологические и биологические чер-
ты, так и указанные выше специфические для тех и других приспо-
собительные свойства, обусловленные этим вторым типом взаимо-
связей.
Весьма важным исходным принципиальным положением при
выделении и систематизации жизненных форм растительных и жи-
вотных ценопопуляций, на наш взгляд, является также учет «стра-
тегии жизни» тех и других. Под стратегией жизни биологических
видов, формирующейся в ходе органической эволюции, Т. А. Ра-
ботнов (1978а) понимает «совокупность приспособлений, обеспечи-
вающих им возможность существовать совместно с другими вида-
ми в определенных условиях экотопа и занимать в биоценозах
соответствующее место» (с. 14). Помимо этого Р. С. Флейшман
(1978) различает также тактическую цель биологических систем,
которая в его представлении неодинакова на разных уровнях орга-
низации живой материи. Так, на уровне особей это хищничество,
паразитизм, мимикрия и т. п., на популяционно-видовом уровне —
свойство к увеличению численности особей и расширению своего
ареала, па биоценотическом уровне — достижение наибольшей
устойчивости, успешное прохождение своего цикла и сохранение
функции системы.
3. Фитоценотипы, зооценотипы и биоценотипы'
Возникает вопрос: что представляют собой выделяемые фито-
ценологами в нашей стране и за рубежом фитоценотипы и
некоторыми из них, в частности А. Л. Бельгардом (1950), цено-
морфы и в каком отношении они находятся к экобиоморфам жи-
вых существ?
Т. А. Работнов (1980) под фитоценотипами понимает
«группы видов растений со сходным изменением их цепотической
значимости в зависимости от условий произрастания или особен-
ностей их жизненного цикла» (с. 72). А. П. Шенников (1964) на-
звал такне группы видов фитоценотическими жизненными форма-
ми, которые характеризуют роль каждого вида «в использовании
и в создании среды фитоценоза, отчасти и во взаимоотношениях с
79
другими видами в нем» (с. 263). Следовательно, фитоценотипы, а
в более широком плане биоценотипы, можно рассматривать
как экобиоморфы растительных и животных ценопопуляций, сход-
ные по своей ценотнпической роли и степени участия в составе
и структуре соответствующего компонента и опосредованно в
структуре и функционировании биогеоценоза. При этом если цено-
зообразующие растительные и животные экобиоморфы характери-
зуют популяционно-видовой состав биогеоценоза с качественной
стороны, то биоценотипы выражают количественные соотношения
этих видовых ценопопуляций в его составе.
Что же касается ценоморф, то они, по А. Л. Бельгарду,
представляют собою группы растительных (добавим от себя: и жи-
вотных) ценопопуляций, отличающиеся своей ясно выраженной
приуроченностью, благодаря наличию соответствующих приспосо-
бительных признаков и свойств, к фитоценозам того или иного ти-
па растительности; например, сильванты— клееному, степанты —
к степному, пратанты — к луговому и т. д. Следовательно, цено-
морфы Бельгарда также можно отнести к категории фитоценоти-
пов, но выделенных не по роли их в строении фитоценозов, а по
принадлежности к тому или иному типу растительности.
В отечественной фитоценологии разработке проблемы фитоце-
нотипов издавна уделялось и уделяется большое внимание
(Г. Н. Высоцкий, 1915; И. К. Пачоский, 1917; А. Я. Гордягин, 1922;
Г. И. Поплавская, 1925; В. Н. Сукачев, 1926, 1928; Л. Г. Рамен-
ский, 1938; Е. М. Лавренко, 1940, 1947; С. Я. Соколов, 1947;
И. X. Блюменталь, 1950, 1951; Б. А. Быков, 1949, 1953, 1957;
Л. И. Номоконов, 1958; Т. А. Работнов, 1974а, 1978а, 1980; и др.).
Широко известной и получившей наибольшее распространение
в нашей стране является система феноценотипов» разработанная
Г. И. Поплавской и В. Н. Сукачевым в 1923 году (Г. И. Поплав-
ская, 1925), которая и приводится здесь.
I. Эдификаторы—растительные виды, строители, созидатели
фитоценоза и его среды (фитосреды), наиболее обильные, опреде-
ляющие внешний облик сообщества и в значительной степени его
видовой состав, а также количество и качество фитомассы и пр.
(например, ель в еловом лесу или сфагновые мхи на сфагновом
болоте).
1. Автохтонные, т. е. самобытные, природные эдификаторы (на-
пример, ковыль в степи).
2. Дигрессивные, появившиеся на месте автохтонных эдифика-
торов под влиянием хозяйственной деятельности человека (напри-
мер, береза на вырубках елового леса или типчак на интенсивно
выпасаемом участке Ковыльной степи).
II. Ассект а тор ы—-растительные виды, сопутствующие эдифи-
80
катору фитоценоза, участвующие в построении сообщества, но мал<
влияющие на создание его фитосреды.
1. Автохтонные, т. е. самобытные, природные ассектаторы, вхо
дящие в состав фитоценоза в его естественном состоянии:
а) эдификаторофилы— виды, приспособленные к жизни в уело
виях созданной эдификатором биогенной среды (фитосреды), н<
встречающиеся и без него;
б) эдификаторофобы— виды, избегающие густых зарослей эди
фикатора, обитающие в прогалинах, на разреженных участках, гд«
влияние его ослаблено.
2. Адвентивные ассектаторы — пришлые, не свойственные дл:
данного фитоценоза виды растений, случайно попавшие в него.
В последующие годы эта система фитоценотипов подверглас!
дальнейшей разработке и детализации. Так, С. Я. Соколов (1947)
наряду с ассектаторами и эдификаторами, ввел в систему понятие
субэдификаторы, а также среди этих трех основных катего
рий фитоцепотипов предложил различать устойчивые, прогрессив
ные и регрессивные фитоценотипы, занимающие то пли иное и:
названных положений в фитоценозах в зависимости от условий
среды обитания. И. X. Блюменталь по степени обилия и постоян
ства подразделил субэдификаторы на 4 ранга и ассектаторы — не
5 рангов. В результате рассматриваемая система фитоценотипоЕ
стала по содержанию более универсальной и динамичной, охваты
вающей разнообразные состояния жизненных форм, входящих е
состав фитоценоза видовых ценопопуляций.
Не менее известной, но не получившей в геоботанической прак-
тике сколько-нибудь широкого распространения является система
фитоценотипов Л. Г Раменского (1938), включающая три основ-
ных ценотипа растений, различающихся по особенностям их отно-
шений к другим компонентам и способности к совместному про-
израстанию в фитоценозах:
I. В иоленты (силовики)—«энергично развиваясь, они захва-
тывают территорию и удерживают ее за собой, подавляя, заглушая
соперников энергией жизнедеятельности и полнотой использования
ресурсов среды» (с. 279). К ним относятся основные ценозообра-
зующие растения, соответствующие эдификаторам.
II. П атиен ты (выносливцы) —«в борьбе за существование они
берут не энергией жизнедеятельности и роста, а своей выносливо-
стью к крайним, суровым условиям... к засолению, кислой реакции
почвы, резкой переменности увлажнения и т. д.» (с. 279). Напри-
мер, сосна.
III. Эксплеренты (выполняющие)—растения, которые «имею!
очень низкую конкурентную мощность, но зато они способны очень
быстро захватывать освободившиеся территории, восполняя про-
межутки между более сильными растениями; так же легко они и
81
вытесняются последними» (с. 280). К ним относятся возобновляю-
щиеся семенами одно-двулетники, стелющиеся, корнеотпрысковые
и другие многолетние растения.
Т. А. Работнов (1980) придает большое значение выделенным
Л. Г. Раменским фитоценотипам, подчеркивая ценологическую, а
не автол огическую их сущность. Он склонен рассматривать их как
три линии эволюционного развития стратегии жизни автотрофных
растении, имея в виду способность одних растений полно исполь-
зовать ресурсы среды и сильно воздействовать на своих конкурен-
тов (виолепты), способность других видов растений избегать кон-
куренции с виолентами, создавая ценозы в условиях, непригодных
для последних (патиенты), и способность некоторых растений из-
бегать конкуренции с растениями обеих названных групп, быстро
захватывая участки с нарушенным покровом (эксплерепты).
Выделенные Л. Г Раменским ценотипы растений по своему су-
ществу имеют много общего и с фитоценотипами, выделенными
В. Н. Сукачевым и Г И. Поплавской (1925).
Из отечественных систем фитоценотипов последних десятилетий,
на наш взгляд, наибольшего внимания заслуживает детально раз-
работанная система Б. А. Быкова (1957, 1970). Исходя из оценки
участия экобиоморф растительных ценопопуляций в сложении
главных и второстепенных слоев фитоценозов (включающих как
надземный, так и подземный ярусы) и из степени их влияния на
среду обитания, он предложил следующую систему основных кате-
горий фитоценотипов.
1. Кондоминанты, принадлежащие к одной экобиоморфе, дву-
и многовидовые ценопопуляций которых совместно образуют глав-
ные слои фитоценозов. II. Доминанты, ценопопуляция каждого
из которых образует главный слой фитоценоза и наибольшую фи-
томассу, оказывает наибольшее влияние на среду, определяя важ-
нейшие черты сообщества. III. Субдоминанты, ценопопуляций
которых образуют второстепенные слои фитоценозов. IV Эзодо-
м и и а нт ы, ценопопуляций которых являются наиболее значитель-
ными в консорциях доминантов и субдомипантов, а также в микро-
группировках сообщества. V. Ингредиенты (ассектаторы) —
остальные участники в составе главных и второстепенных слоев
фитоценоза.
Первые две и четвертая из основных категорий фитоценотипов
подразделяются автором по своим биоморфологическим особенно-
стям на ряд групп фитоценотипов (от 2 до 7). К сожалению, обыч-
но довольно многочисленная по видовому составу, хотя непосред-
ственно и маловлиятельная категория фитоцепотипов — ингреди-
енты или ассектаторы, — не получила у автора дальнейшей
детализации.
Из зарубежных синэкологов вопросам, связанным с определе-
82
нием роли и степени участия растительных вадов в сообществах»
в частности вопросам доминирования их, довольно значительно
внимание уделяли шведский ботаник Дю-Рие (Dii-Riets, 1921)
ботаники франко-швейцарской школы Браун-Бланке и Павийя;
(Braun-Blanquet, Pavillard, 1925), экологи англо-американско
школы Клементс, Уивер (Clements, 1936; Weaver, Clements, 1938
и др.
Подразделение животных по численности особей (обилию) i
значению их в структуре и в жизни сообщества принадлежит пер;
видного отечественного эколога Д. Н. Кашкарова (1933), которьп
различал среди них 3 группы:
1, Предо мин ан ты — виды многочисленные, пребывающие
сообществе в течение всего деятельного периода года, составляв
«ядро» животного населения, играющие наиболее важную роль i
его строении и функционировании.
2. Флюктуирующие предоминанты — виды, являющи
еся предоминантами в некоторые периоды вследствие значительно
го возрастания их численности и усиления активной деятельност]
(например, в периоды пролета, кочевок, выхода молодняка и т. п.)
3. Сезонные субдоминанты — виды более или мене
многочисленные, активные, влияющие на структуру и функцнони
рование сообщества в благоприятные для них сезоны года, обу
словленные их жизненным ритмом (рано весной, летом, под осень)
Выделенные Д. Н. Кашкаровым на указанных основаниях труп
пы животных фактически представляют собою основные категории
ценотипов в составе животного компонента биогеоценоза. Они
очевидно, могут быть подразделены на более мелкие градации це
нотипов вплоть до исходных форм, которые по аналогии с выделяе
мыми в составе растительного компонента фитоценотипами могу
быть названы зооценотипами.
Сопоставляя рассмотренные отечественные системы фитоцено
типов друг с другом и с системой зооценотипов Д. Н. Кашкарова
прежде всего приходится констатировать различную степень раз
работанности и детальности этих систем, а также неполную одно
родпость принципов, положенных в основу выделения ценотипоЕ
Следствием этого является неизбежный в таком случае разнобо!
как в трактовке содержания, так и в номенклатуре выделяемы
единиц.
Биоцеиотипы (включающие фито- и зооценотипы), как бы
ло отмечено ранее, выражают собою количественные соотнощени.
между растительными и животными ценопопуляциями, входящим!
в состав живых компонентов биогеоценоза. Сами же количествен
ные соотношения являются результатом различного рода взаимс
отношений, сложившихся между видовыми ценопопуляциями пр
становлении растительных и животных экобиоморф в процессе фи
83
логенеза биогеоценотических систем. Показателями количествен-
ных соотношений между видовыми ценопопуляциями в составе жи-
вых компонентов биогеоценоза, как известно, служат: а) соотно-
шение числа особей растений или животных на единице площади
(обилие); б) соотношение плотности стояния и степени покрытия
сомкнутости надземных органов растений или плотности нахожде-
ния животных в среде своего обитания (доминирование); в) весо-
вое (или объемное) соотношение биомассы растений или животных
(продуктивность, прирост или общий запас биомассы). К- А. Кур-
кин (1977) обращает внимание на то обстоятельство, что не чис-
ленность особей сама по себе определяет «позицию» ценопопуля-
ций в составе того или иного компонента биогеоценоза, а числен-
ность с учетом мощности особей; иначе говоря, интегральная био-
масса ценопопуляций, а также общее покрытие и, очевидно, плот-
ность, полнота ее.
Таким образом, количественные соотношения различных видо-
вых ценопопуляций в составе живых компонентов биогеоценоза с
учетом размеров их особей — одна из важнейших характеристик
как самих компонентов, так и биогеоценоза в целом. По ним мож-
но судить о многом: а) о характере взаимоотношений видовых це-
нопопуляций в составе живых компонентов биогеоценоза и взаимо-
связи их со средой обитания; б) об экобиоморфном и ценотическом
составе и структуре живых компонентов биогеоценоза и о «пози-
ции» видовых ценопопуляций в составе каждого компонента; в) о
физпономичности биогеоценозов и о сходстве или различии их как
целостных биокосных систем; г) о биологической продуктивности
и хозяйственной ценности того или иного биогеоценоза и народно-
хозяйственной значимости его.
Сопоставляя рассмотренные нами видовые ценопопуляций, эко-
биоморфы и биоценотипы, приходится констатировать, во-первых,
то, что биоценотипы — это не что иное, как экобиоморфы биогеоце-
нозов, в составе и под контролем которых они выработались; во-
вторых, биоценотипы как экобиоморфы биогеоценозов представ-
лены в их составе соответствующими видовыми ценопопуляциями
живых существ, образующими ценотические сочетания разного
ранга, облекаясь при этом в такие функциональные структуры био-
геоценозов, как трофические звенья пищевых цепей, трофические
концентры консорций и т. п.
Гл ав a IV
Состав и функциональная деятельность
живых компонентов биогеоценоза
1. Состав и функциональная деятельность
автотрофного компонента биогеоценоза
Перейдем к рассмотрению состава и функциональной деятель-
ности живых компонентов биогеоценоза, начиная с автотрофного,
поскольку с него начинается материально-энергетический обмен в
биогеоценозе.
Во второй главе было обосновано особое место и роль расти-
тельного сообщества в составе, структуре и обменных процессах
биогеоценоза. Оно выступает, с одной стороны, как автотрофная
база биогеоценоза, как источник питания и энергии гетеротрофных
его компонентов, а с другой — как материальная основа структур-
но-функциональной организации и пространственной протяженно-
сти и ограниченности биогеоценоза, а также как место укрытия,
убежище других компонентов от непогоды и от своих естественных
врагов. Поэтому автотрофный компонент биогеоценоза, в отличие
от других живых компонентов, будет рассмотрен нами в форме
фитоценоза как относительно самостоятельной целостной катего-
рии, в качестве реально существующей подсистемы, точнее — блока
биогеоценоза.
Большинством современных фитоценологов фитоценоз тракту-
ется как форма совместного существования растений в простран-
стве и во времени. Основу фитоценозов континентального типа
биогеосферы, состоящую в основном из наземной (сухопутной)
пленки жизни, составляют высшие автотрофные растения, а также
низшие гетеротрофные растительные и другие организмы. Соче-
таясь определенным образом друг с другом на основе присущих
данному системному уровню организации живой материи законо-
мерностей, эти растения образуют огромное количество разнооб-
разных фитоценозов зонального, азонального и провинциального
типов. В совокупности они образуют растительный покров нашей
планеты.
В различных провинциях лесной зоны умеренного и холодного
поясов наиболее важную ценозообразующую роль играют темно-
хвойные, светлохвойные, а также широколиственные и мелколист-
венные древесные породы; в арктической зоне — приземистые кус-
тарники, кустарнички, мхи и лишайники, особенно кустистые (так
называемые ягельные); в степной — многолетние травы, в особен-
85
ности ксерофильные дерновинные злаки; в пустынной — ксерофиль-
ные кустарники, кустарнички и различные травянистые галофиты
и геофиты. В азональных типах растительности ценозообразующая
роль принадлежит на лугах многолетним травам, в особенности
мезофильным злакам и осокам; на болотах — сфагновым мхам»
кустарничкам и оксилофильным осокам, местами—особым формам
древесных пород и кустарников либо гидрофильным осокам и зла-
кам. Значительно более многочислен и разнообразен популяцион-
но-видовой состав древесных, кустарниковых, травяных и других
форм ценозообразующих растений в различных областях жаркого
тропического пояса Земли.
В силу чрезвычайного разнообразия и широкого диапазона эко-
лого-географических условий в пределах континентального типа
биогеосферы современной геологической эпохи весьма разнообра-
зен не только популяционно-видовой, но и экобиоморфный состав
ценозообразующих растений. Разумеется, состав и общая числен-
ность растительных экобиоморф не столь велики по сравнению с
численностью популяционно-видового состава растений, в особен-
ности в пределах отдельных зон и провинций и тем более в преде-
лах отдельных типов местообитаний. Так, по данным Б. А. Быкова
(1970), в наземных (сухопутных) условиях на поверхности почвы
в приземном слое воздуха преобладают стеблекорневые и напоч-
венные формы растений—древесные, кустарниковые, травянистые»
мхи, лишайники (в том числе — паразиты, сапрофиты и т. п ); в
поверхностных горизонтах почвы господствуют почвенные гетеро-
трофные организмы, в частности симбиотрофные грибы, бактерии;
в нижних слоях атмосферы на деревьях и травах обитают эпифи-
ты, паразитные грибы и пр. В прибрежных мелководьях преобла-
дают укореняющиеся в грунте и пронизывающие водные слои стеб-
лекорневые земноводные гигрофиты и полностью погруженные в
воду гидрофиты. В поверхностных слоях континентальных водо-
емов распространены свободноплавающие гидрофильные формы,
одноклеточные планктонные водоросли и прочие организмы.
Что же касается цепотнческого состава фитоценозов, то мы
являемся сторонниками теории доминирования определенных рас-
тений в их составе, так как не считаем возможным закрывать глаза
на вполне очевидные факты, часто не нуждающиеся в каком-либо
подтверждении. Разумеется, при этом наряду с доминантами (де-
терминантами, эдификаторами) главного слоя фитоценоза должно
быть тщательно изучено и учтено участие в образовании фитоце-
нозов и других видов растений в качестве субэдификаторов, ассек-
таторов разного ранга и т. п. Не исключает все это и применения
соответствующих для данного случая математических методов
определения степени участия тех или иных видов в составе фито-
ценозов.
86
Основная функция ценокомплекса автотрофных растительных и
некоторых других фото- и хемосинтетиков в составе фитоценоза и
биогеоценоза в целом, по Т. А. Работнову (1980), состоит, во-пер-
вых, в усвоении солнечной энергии и создании в процессе фотосин-
теза из первичных неорганических материалов органических ве-
ществ, являющихся источником питания и энергии для ценопопу-
ляций биотрофов, сапротрофов, азотфиксирующих организмов и
микоризных грибов; во-вторых, в изменении в процессе своей жиз-
недеятельности косных компонентов биогеоценоза — почвенной и
воздушной среды, в приспособлении ее в первом приближении к
своим потребностям; однако среда со временем может перейти
в этом смысле в свою противоположность, т. е. стать неблагоприят
ной для их обитания.
Биоценотическая функциональная деятельность * ценокомплек-
са автотрофов (преимущественно фототрофных растительных цено-
лопуляций) в составе биогеоценоза проявляется, по Н. В. Дылису
с соавторами (1964), в целом ряде последовательно взаимосвязан-
ных процессов. 1. Поглощение функционально-биоценотическими
группами растительных видов фитоценоза необходимых для жизни
минеральных питательных веществ и воды из почвы, кислорода и
углекислого газа из приземного слоя воздуха, лучистой энергии
Солнца из космического пространства. 2. Синтез из поступивших
в растительное сообщество и поглощенных автотрофными расте-
ниями неорганических соединений, воды и углекислоты за счет
усвоенной посредством хлорофилла солнечной энергии разнооб-
разных органических веществ — углеводов, растительных жиров,
белков и т. п., составляющих так называемую первичную биологи-
ческую продукцию. В результате этого происходит накопление
живой растительной биомассы — фитомассы, которая служит
источником питания и энергии для животных, человека и некото-
рых микроорганизмов, а также строительным материалом для
гнезд и убежищем для многих позвоночных и беспозвоночных жи-
вотных. 3. Использование растениями фитоценоза аккумулирован-
ной в органическом веществе потенциальной энергии на процессы
своей жизнедеятельности (фотосинтез, дыхание, транспирация), а
также выделение продуктов метаболизма — воды, кислорода, угле-
кислого газа, различных химических соединений — в почву и атмо-
сферу. 4. Поступление на поверхность почвы отмерших органов
растений и их частей, а также полностью отмерших растений и
образование за счет этого на ее поверхности мертвого напочвенно-
го покрова, а в самой почве — гумуса и запасов минеральной пищи
растений. 5. Преобразование в процессе жизнедеятельности расте-
* Термины биоценотические функции и функционально-биоценотические группы,
выделяемые по типу питания, принадлежат Д. В. Панфилову (1966).
87
ний и функционирования растительного сообщества в целом
свойств и состояний других, в особенности косных, компонентов
биогеоценоза, в частности: изменение количества и качества сол-
нечной радиации и ослабление интенсивности освещения, снижение
прогревания почвы и воздуха и испарения влаги с поверхности
почвы, замедление поверхностного стока воды и понижение уровня
почвенно-грунтовых вод, перемещение химических элементов по
почвенному профилю и изменение структуры и физико-химических
свойств почвы и т. п.
Однако специфика биоценотическон функциональной деятель-
ности автотрофного ценокомплекса состоит в создании первичной
биологической продукции, ибо автотрофные растения — это основ-
ные продуценты, осуществляющие синтез органических веществ из
неорганических, благодаря чему они являются начальным звеном
всех так называемых пищевых (кормовых) цепей и большинства
консорций. Причем чем интенсивнее осуществляется фотосинтез,
тем энергичнее протекает обмен веществ и энергии в биогеоценозе,
так как при этом больше фиксируется лучистой энергии Солнца,
больше вовлекается в обмен углекислоты, воды и элементов мине-
ральной пищи растений, больше выделяется в атмосферу кислорода
и больше создается органических веществ. Поэтому-то раститель-
ное сообщество и называют то «энергетической установкой», то
«сердцем» биогеоценоза, то «солнечным комбинатом» по перера-
ботке энергии Солнца в органическое вещество.
Таким образом, функциональная деятельность растительного
сообщества в составе биогеоценоза весьма многогранна и специ-
фична. Опа играет важную, по выражению Н. В. Дылиса (1978),
системообразующую роль в биогеоценозе и ведет к разнообразным
и глубоким последствиям в нем и за его пределами. Так, по его
словам, она определяет размеры, конфигурацию и границы био-
геоценоза, его вертикальное и горизонтальное строение и внешний
облик, физиономичность, популяционно-видовой состав животного
и микробного населения и распределение тех и других по струк-
турным частям сообщества, в особенности в его наземной (воздуш-
ной) среде. Кроме того, она корректирует и преобразует воздей-
ствие на биогеоценоз внешних, в особенности атмосферных, фак-
торов среды, а также направляет процесс обмена веществ и
энергии в биогеоценозе как целостной биокосной системе.
Последствия разносторонней биоценотической функциональной
деятельности фитоценоза выражаются в том, что в результате фо-
тосинтеза, дыхания, транспирации, роста и прочих процессов жиз-
недеятельности ценозообразующих растений расширяется и услож-
няется весь ход обменных процессов в биогеоценозе и биогеосфере
в целом. В частности, ускоряется общая миграция химических
элементов, образуются новые цепи материально-энергетических
88
связей и биохимических превращений, формируются процессы об-
разования новых природных тел — почвы, торфа, биогенных газов
(кислорода, углекислого газа), происходит накопление в многолет-
них частях растений, особенно в древесине, значительного количе-
ства фитомассы и аккумулированной в ней солнечной энергии;
вследствие этого уменьшается энтропия, т. е. рассеивание энергии
в космическое пространство, и т. п.
Некоторая часть образовавшихся в процессе фотосинтеза орга-
нических веществ, как отмечалось, используется самими растения-
ми в количестве энергетического материала на процессы своей
жизнедеятельности; другая часть их превращается в растительную
биомассу — фитомассу, прирост которой за определенный проме-
жуток времени составляет продукцию, урожай растительного сооб-
щества. Продуктивность различных фитоценозов неодинакова: она
находится в зависимости от характера фитоценоза,возраста ценозо-
образующих растений, длительности периода их вегетации, погод-
ных условий вегетационного периода п пр. Наиболее продуктивны
лесные фитоценозы, особенно влажнотропические и субтропические
леса ввиду почти круглогодичной фотосинтетической деятельности
их. Минимальная продуктивность характерна для растительных
сообществ в условиях короткого периода вегетации (на севере, в
горах) либо в условиях недостаточного или избыточного увлажне-
ния, в особенности застойного (в сухих степях, пустынях, на неко-
торых типах болот).
Однако поскольку фотосинтез обычно протекает интенсивнее,
чем расход пластических веществ на процессы жизнедеятельности
растений, то довольно значительная часть образовавшейся фито-
массы идет в запас, достигая, например, в спелых древостоях от
200 до 500 т/га и более. Она как бы консервируется в многолетних
частях растений и на многие годы и даже десятилетня выходит из
биотического круговорота веществ. При этом, как указывает
Н. В. Дылис с соавторами (1964а,б), чем мощнее развивается фи-
томасса фитоценоза, тем сильнее проявляется ее преобразующая
роль в биогеоценозе. Этим фитоценоз резко отличается от биотроф-
ного животного компонента и сапротрофного микробного, биогео-
ценотическая роль которых определяется не столько биомассой,
сколько численностью особей, скоростью размножения и быстротой
смены поколений.
Вся эта огромная по масштабу и разнообразная по своему ха-
рактеру деятельность фитоценоза и продуцируемой им фитомассы,
во много раз превышающей по весу биомассу микроорганизмов, и
в особенности животных, определяет центральное место его среди
других живых компонентов биогеоценоза и неизмеримо более вы-
сокую долю участия его в трансформации веществ и энергии в
биогеосфере.
69
Здесь необходимо подчеркнуть значительную долю культурных
фитоценозов в общем балансе биотического круговорота веществ
в пределах того или иного региона биогеосферы, более высокую
продуктивность большинства из них по сравнению с природными
фнтоценозами.
Н. В. Дылис обращает внимание также на то обстоятельство,
что природное разнообразие функциональной деятельности фито-
ценозов, масштабы, ритм и биогеоценотические результаты ее
далеко не одинаковы в различных физико-географических усло-
виях земной поверхности. Это зависит, во-первых, от биохимиче-
ских и физических свойств продуцируемой фитомассы, которая
неодинакова в разных местах, поскольку различен популяционно-
видовой и экобиоморфный состав ценозообразующих растений фи-
тоценозов; во-вторых, от набора и состояния других живых компо-
нентов, факторов абиотической среды и притока солнечной радиа-
ции, которые также неодинаковы в разных пунктах биосферы.
В фитоценозах, как известно, наряду с формированием живых
растительных организмов и накоплением фитомассы, происходит
и естественный процесс периодического отмирания надземных и
подземных органов и отдельных частей растений (листьев, стеблей,
ветвей, коры, корней, корневищ и пр.), образующих так называе-
мый опад. Происходит отмирание и целых растений по истечении
присущего каждому виду срока жизни, именуемое отпадом. При
этом в опад и отпад в течение одного поколения цепозообразующих
растений некоторых лесных фитоценозов, по данным А. А. Молча-
нова (1961), уходит фитомассы в 3—4 раза больше, чем ее остает-
ся в живом состоянии. На поверхности почвы отмершие раститель-
ные материалы подвергаются под действием сапротрофов и абио-
тических агентов разложению, вовлекаясь в обменные процессы в
биогеоценозе. Следствием этого является пополнение запасов ми-
неральной пищи растений в почве, формирование почвенного гуму-
са и почвенной кислотности (pH), накопление в верхних горизон-
тах почвы ряда химических элементов, в частности, повышается
содержание углекислоты, подбор определенного состава почвенных
животных и микроорганизмов.
Неразложившиеся растительные остатки накапливаются на по-
верхности почвы в виде мертвого напочвенного покрова, образуя
подстилку в лесу или войлок («калдан») в травяных сообществах.
Биогеоценотическая роль мертвого напочвенного покрова не менее
значительна и многообразна, чем роль разложившихся раститель-
ных материалов. Лесная подстилка или степной войлок, в зависи-
мости от их мощности, плотности, влажности и прочих свойств, по
наблюдениям Н. В. Дылиса (1978) и ряда других авторов, оказы-
вают различное, весьма существенное влияние на целый ряд про-
цессов в биогеоценозах: поверхностный сток воды и проникновение
90
осадков в почву и физическое испарение влаги с ее поверхности,
тепловой режим почвы и приземного слоя атмосферы и газообмен
между ними, прорастание семян и развитие всходов растений,
взаимоотношение ценозообразующих растений путем положитель-
ного или отрицательного воздействия на их развитие, деятельность
почвообитающих животных и микроорганизмов и пр. При этом
важно отметить, что подобного рода влияние мертвого напочвен-
ного покрова на биогеоценозы в историческом прошлом сыграло
немаловажную роль в формировании ряда жизненных форм рас-
тений и животных, в частности лесных сапрофитов, подстилочных
форм беспозвоночных животных и пр.
Таким образом, биогеоценотическая роль фитоценозов выража-
ется не только в продуцировании живой растительной биомассы
ценокомплексом автотрофных ценопопуляций, но и в разнообраз-
ных формах воздействия как живой фитомассы, так и мертвого
напочвенного покрова на состояние и развитие других живых ком-
понентов и на состояние и деятельность косных компонентов —
почвы и приземного слоя атмосферы в его границах, существенным
образом перестраивая их во многих отношениях. Это обстоятель-
ство, надо полагать, и дало основание Т. А. Работнову (1980)
назвать фитоценозы коллективными продуцентами и коллективны-
ми средообразователями.
2. Состав и функциональная деятельность
биотрофного компонента биогеоценоза
Вторым из живых компонентов биогеоценоза надлежит рас-
смотреть ценокомплекс биотрофных ценопопуляций. являющихся
потребителями и трансформаторами растительной или живот-
ной и отчасти микробной биомассы и прижизненных выделений
организмов и созидателями так называемой вторичной био-
логической продукции. По сравнению с автотрофным ценокомп-
лексом живых существ он является более сложным по составу.
Основную массу его составляют животные — беспозвоночные, по-
звоночные. Кроме того, в состав ценокомплекса биотрофов входят
микроорганизмы и незначительное число растительных форм. Со-
став биотрофного ценокомплекса усложняется еще и в связи с тем,
что он охватывает два, а то и три трофических звена пищевых
(кормовых) цепей: а) звено фитофагов, включающее растительно-
ядных животных, микроорганизмы и растения-паразиты и полупа-
разиты, и б) звено зоофагов—плотоядных (хищных) животных и
микроорганизмы, насекомоядные растения и паразитарные формы
животных и микроорганизмов, обычно, в свою очередь, подразде-
ляемое на хищников 1-го и 2-го порядка.
Поскольку основная масса ценокомплекса биотрофов как в на-
91
земной (воздушной), так и в почвенной среде континентального
типа биогеосферы представлена позвоночными и беспозвоночными
животными, то прежде всего следует охарактеризовать популяци-
онно-видовой и экобиоморфный состав и функциональную деятель-
ность животного населения. Термин животное население,
предложенный В. А. Догелем еще в 20-х годах текущего столетия
и уточненный А. Н. Формозовым и А. Г. Вороновым в середине
его, в настоящее время получил широкое распространение в зооло-
гической и зооэкологической литературе. Он применяется, по сло-
вам Ю. И. Чернова (1971), в качестве аналога термина «расти-
тельность» в геоботанике и, в отличие от термина фауна, озна-
чает не состав видов и других систематических групп животных
той или иной территории, а степень разнообразия, пестроты топи-
ческих сочетаний животных или их группировок (лесных, степных»
луговых и т. д.).
Животные—самая разнообразная и динамичная группа живых
существ в составе биотрофного ценокомплекса. Видовое и экобио-
морфное разнообразие животных, составляющих наземные (сухо-
путные) биогеоценозы, во много раз превышает разнообразие
ценозообразующих высших растений, хотя по биомассе значитель-
но уступает им.
Популяционно-видовой состав животного населения биогеоце-
нозов, численность особей и устойчивость его определяются как
природой и биологическими особенностями видовых ценопопуля-
ций, так и физическими (экологическими) условиями среды обита-
ния, и прежде всего наличием и характером корма. Имеет значение
и характер исторически сложившихся взаимоотношений между
видами. Видовой состав и количественные соотношения видов по-
звоночных животных различных ландшафтно-географических зон
и типов биогеоценозов в европейской части Союза далеко не оди-
наков. По данным К. С, Ходашевой (1966) и других зоологов, в
травяных биогеоценозах открытых пространств (степных, луговых
и т. п.), ввиду высокой питательности травы и круглогодичной во
многих местах доступности ее, преобладают травоядные животные
(копытные, грызуны); многообразна и группа роющих позвоноч-
ных. В древесно-кустарниковых биогеоценозах лесной области,
вследствие богатства питательными веществами ветвей, корней,
коры деревьев и кустарников, широко представлена группа потре-
бителей вегетативных частей растений. Многообразна также и
группа позвоночных, питающихся семенами и плодами. В кустар-
ничково-моховых и мохово-лишайниковых биогеоценозах тундро-
вой зоны наиболее обширна группа позвоночных, употребляющая
в пищу почки, листья, плоды кустарничков, а также копытных жи-
вотных— потребителей ягельных лишайников. Всюду, разумеется,
обильно представлены соответствующие хищники и паразиты.
92
Важно отметить и то обстоятельство, что в степной и тундровой,
зонах позвоночные обитают на поверхности земли и в верхних го-
ризонтах почвы; в лесной же области они сосредоточены преиму-
щественно в наземной (воздушной) среде, размещаясь в различ-
ных ярусах и микрогруппировках (синузиях) древостоя.
Из беспозвоночных животных в составе биогеоценозов различ-
ных ландшафтно-географических зон в пределах Советского Союза
наиболее важное место, по данным Д. В. Панфилова (1966), зани-
мают дождевые черви, брюхоногие моллюски, паукообразные, кле-
щи, многоножки и особенно насекомые. Причем большая часть
беспозвоночных животных принадлежит к числу обитателей поч-
венной среды, а среди них преобладают потребители отмерших
органических остатков. Из почвообитающих беспозвоночных к це-
нокомплексу биотрофов относятся лишь виды, использующие в
пищу подземные органы растений (корни, корневища, клубни, лу-
ковицы) и мицелий грибов, а также трофически связанные с ними
хищные и паразитарные формы. В условиях наземной (воздушной)
среды в составе беспозвоночных животных всюду преобладают
растительноядные виды, в частности такие специализированные
формы, как листоеды, цветоеды, семя- и плодоеды, древесиноеды,
короеды и т. п. Трофически связанные с ними плотоядные беспо-
звоночные представлены в этих условиях хищными видами, исполь-
зующими в качестве корма живую животную биомассу, а также
эндо- и эктопаразитами, питающимися соками животных-хозяев, и
кровососущими видами беспозвоночных.
Степень верности, приуроченности, пригнанности различных
видов позвоночных и беспозвоночных животных к биогеоценозу, в
состав которого они входят, неодинакова. В каждом биогеоценозе,
как известно, формируется постоянное ядро из оседлых форм
животных, приспособленных к совместной жизни друг с другом и
с другими живыми компонентами в наличных условиях среды оби-
тания. Многие животные, в особенности беспозвоночные, обычно
функционально тесно связаны с биогеоценозом и не выходят за его
пределы. Наряду с этим другие, более подвижные виды животных,
в некоторые периоды охватывают в своей жизнедеятельности два
и более смежных биогеоценоза; при этом нередко они выполняют
в них разные жизненные функции: в одних они добывают пищу, в
других находят убежища, устраивают логовища, гнездовья и вы-
водят потомство (рис. 7). Такие видовые ценопопуляции животных,,
по мнению Г. А. Новикова (1979), следует относить не к одному, а
ко всем этим биогеоценозам. Существуют и такие особо подвиж-
ные виды животных, которые обитают в том или ином биогеоцено-
зе лишь в определенные сезоны года, или численность их по сезо-
нам сильно колеблется, что обусловлено главным образом
недостатком корма в некоторые сезоны и связанными с этим зако-
93
номерными или случайными, эпизодическими перемещениями их
(весенние и осенние перелеты птиц, миграции массово размножаю-
щихся насекомых, мелких грызунов и т. п.).
Все отмеченные различия разных групп животных в степени
приуроченности к биогеоценозу служат важным показателем био-
геоценотической значимости каждой из этих групп и в известной
мере предопределяют место их как в составе и структуре живот-
ного населения биотрофного компонента, так и биогеоценоза в
целом.
Биоценотическая функциональная деятельность позвоночных и
беспозвоночных животных, относящихся к ценокомплексу биотро-
фов, в составе биогеоценотических систем не менее многогранна,
чем деятельность растительного сообщества. По данным П. М. Ра-
феса с соавторами (1964), она включает в себя ряд следующих
процессов. 1. Получение от автотрофного растительного компонен-
та биогеоценоза разнообразных кормов как в виде живой расти-
тельной биомассы, так и в виде отмерших растений и их отдельных
частей вместе с заключенной в них потенциальной энергией, а из
почвы и атмосферы — кислорода, воды и прочих неорганических
веществ. 2. Синтез животными-фитофагами из усвоенной в процес-
се питания растительной биомассы, а животными-зоофагами жи-
вотной биомассы —вторичной биологической продукции (живот-
94
ных жиров, белков, мочевины, органических кислот и т. п.), а
также накопление в биогеоценозе в процессе роста и размножения
запасов животной биомассы[ 3. Внесение животными в атмосферу
и почву разнообразных продуктов синтеза и метаболизма: угле-
кислого газа, твердых и жидких отбросов, минеральных и мертвых
органических веществ; 4. Участие позвоночных и беспозвоночных
животных в формировании микро- и нанорельефа (сурчины, му-
равьиные кучи, выбросы кротов и других землероев), увеличиваю-
щее тем самым разнообразие экологических ниш биотопа^ 5. Уча-
стие некоторых животных в опылении растений и распространении
семян и плодов, а также в переносе растительной биомассы по
площади в границах биогеоценоза и за его пределы (в виде строи-
тельного материала и корма) и животной биомассы путем соб-
ственного перемещения.!
Однако специфика биоцеиотической функциональной деятель-
ности животных биотрофного ценокомплекса состоит в переработке
в процессе своей жизнедеятельности употребляемой ими биомассы
в более высокомолекулярную вторичную биологическую продук-
цию, так как животные—это основные консументы и трансформа-
торы первичной биологической продукции во вторичную, исполь-
зуемую частично и человеком для своих потребностей. |
Обладая известной подвижностью, животные в силу сказанного
значительно усложняют и расширяют обмен веществ как внутри
биогеоценоза, так и между смежными, а порой и весьма отдален-
ными друг от друга биогеоценозами:^ Помимо этого, животные воз-
действуют посредством своих выделений, отбросов, роющей дея-
тельности и т. п. на косные компоненты, а через них опосредован-
но и па состояние и развитие автотрофного растительного компо-
нента биогеоценоза, на его возобновительные процессы и пр. |
Наряду с этим необходимо хотя бы кратко остановиться на
некоторых специфических особенностях функциональной деятель-
ности позвоночных и беспозвоночных животных биотрофного цено-
комплекса.
В наземной (воздушной) среде наибольшее значение в жизни
и функционировании биогеоценозов имеет деятельность позвоноч-
ных фитофагов, а также видов со смешанным питанием (полнфа-
гов) и хищников^ Из растительноядных позвоночных особенно
велика роль мелких грызунов; крупные фитофаги (лоси, олени,
свиньи, сайгаки) сравнительно немногочисленны, а потому роль их
в биогеоценозах суши незначительна. Однако в последние десяти-
летия благодаря охранным мерам численность их быстро возра-
стает, и в некоторых местах лесной и степной зон они начинают
оказывать существенное воздействие как на устойчивость биогео-
ценозов путем нарушения их структуры и функциональной органи-
зации, так и на их кормовую базу. Известное значение имеет и
95
косвенное воздействие наземных позвоночных биотрофов на струк-
туру и функциональную организацию биогеоценозов посредством
изменения почвенно-грунтовых условий вследствие вытаптывания
травяного и мохового покрова, разбивания дернины злаков, уплот-
нения или рыхления почвы и удобрения ее за счет экскрементов и
прочих отбросов и т. д. |
В почвенной среде наибольшее значение в жизни и функциони-
ровании биогеоценозов имеет деятельность мелких роющих позво-
ночных (слепыши, сурки, кроты, полевки и пр.), которые роют
норы, ходы сообщения, склады корма, рыхлят при этом почву,
улучшая ее аэрацию, водопроницаемость, газообмен, а также вы-
носят на поверхность земли значительную массу грунта (слепы-
ши— до 13 м3 на 1 га, сурки — до 100 м3 и более); кроты в лесах
в поисках пищи проделывают ходы протяженностью до 15—18 км
на 1 га и выбрасывают на поверхность кучки земли до 0,5 т на
I га. Те и другие тем самым существенно разнообразят микро- и
нанорельеф степных и лесных местообитаний.
Важное биогеоценотическое значение имеет и функциональная
деятельность плотоядных позвоночных животных. В их составе во
всех географических зонах страны больше всего птиц и млекопи-
тающих. Выедая определенное количество позвоночных и отчасти
беспозвоночных фитофагов, или хищников первого порядка, они
оказывают не только прямое воздействие на структуру и состояние
животного населения биогеоценоза, ио и выполняют тем самым
важную роль регулятора численности соответствующих видов по-
звоночных животных, поддерживая таким образом сбалансирован-
ность видового состава в этих звеньях пищевых цепей. \
Биоценотическая функциональная деятельность беспозвоночных
биотрофов более разнообразна и эффективна по сравнению с дея-
тельностью позвоночных животных, что связано с их значительным
преобладанием над позвоночными как по числу видов и особей,
так и в особенности по биомассе. В наземных условиях особенно
большое значение в функционировании и динамике биогеоценозов
имеет деятельность беспозвоночных фитофагов.» Они поедают до
40% создаваемой зелеными растениями биомассы. Особенно про-
жорливы некоторые виды саранчовых, а также личинки и гусени-
цы таких массово размножающихся насекомых, как сибирский
шелкопряд, дубовая листовертка, луговой мотылек и пр., которые
нередко опустошают растительность на огромных площадях. По-
следствия деятельности надземных беспозвоночных фитофагов вы-
ражаются в осветлении растительной части биогеоценозов, особен-
но лесных, увеличении освещенности и прогреваемости поверхности
почвы и приземного слоя воздуха, ослаблении фотосинтеза и сни-
жении продуктивности биогеоценозов и пр. К специфическим фор-
мам биоценотической деятельности беспозвоночных относятся так-
96
же опыление насекомыми энтомофильных растений, распростране-
ние семян и плодов, перенос возбудителей разных болезней
растений и животных (вирусных, бактериальных и пр.).
Видовое разнообразие и биомасса почвообитающих беспозво-
ночных фитофагов сравнительно невелики, за исключением таких
массово размножающихся видов, как майский жук и др., а потому
и биоценотическая функциональная деятельность их не столь зна-
чительна, как деятельность наземных беспозвоночных фитофагов.
Что же касается плотоядных (хищных) беспозвоночных (пауки,
божьи коровки, жужелицы, муравьи и т. п.), то они в процессе
питания уничтожают большое количество растительноядных бес-
позвоночных животных, Так, население одного гнезда рыжего му-
равья за сезон истребляет от 3 до 5 млн насекомьц, что составляет
15—20 кг. Биогеоценотическое значение деятельности плотоядных,
а также паразитарных и кровососущих беспозвоночных выражает-
ся, таким образом, в регуляции численности беспозвоночных фито-
фагов и поддержании биологического равновесия в природе.. В
практике же эти особенности их биоценотической деятельности
используются в качестве биологического способа борьбы с вреди-
телями леса и сельскохозяйственных культур, значительно сни-
жающих прирост древесины в лесу и урожайность зерновых, кор-
мовых и других сельскохозяйственных растений на полях. Некото-
рые плотоядные беспозвоночные виды (муравьи, термиты и др.)
оказывают также косвенное воздействие на биогеоценозы, разрых-
ляя почву в процессе своей жизнедеятельности, удобряя и переме-
шивая ее с минеральной частью и т. п.
Таким образом, позвоночные и беспозвоночные животные явля-
ются основными потребителями живой растительной п животной
биомассы и основными трансформаторами ее в собственную био-
массу.; Помимо растительноядных и плотоядных животных в со-
став оиотрофного компонента биогеоценозов, как было сказано
выше, входят паразитирующие па растениях высшие растения-па-
разиты и полупаразиты и низшие растения-паразиты и симбионты,
а также насекомоядные растения, паразитирующие па животных
беспозвоночные эндо- и эктопаразиты и патогенные бактерии и
вирусы, вызывающие различные болезни растений и животных.
К высшим растениям-паразитам относятся лишенные зеленой
окраски бесхлорофилльные виды. Из них наиболее известными яв-
ляются повилики (Cuscuta)—около 100 видов из семейства повили-
ковых, распространенные на Евроазиатском континенте, паразити-
рующие па стеблях как диких, так и многих культурных растений;
заразихи (Orobanche)—около 130 видов из семейства заразиховых,
распространенные главным образом в Северном полушарии, труд-
понскорепимые паразиты таких культурных растений, как
подсолнечник, табак, томаты, конопля, клевер, люцерна и др.;
97
петров крест (Latraea) из этого же семейства, паразитирующий на
корнях многих древесных пород в лесу; на Суматре и некоторых
других островах Малайского архипелага встречается паразитирую-
щая на корнях тропических деревьев раффлезия (Rafflesia) с
огромными цветами (до метра в диаметре) и др.
Высшие растения-полупаразиты содержат в своих клетках хло-
рофилл, следовательно, обладают способностью осуществлять
фотосинтез; вместе с тем, прикрепляясь к корням других растений,
они используют и пластические вещества этих растений. Из них
наиболее распространены в Евразии мытники (Pedicularis) —око-
ло 250 видов из семейства норичниковых, встречающиеся на лугах
очанки (Euphrasia) и погремки (Rhinanthus) из того же семей-
ства, а также встречающиеся в лиственных лесах и кустарниках
мермекофильные растения иван-да-марья (Melampyrum пешого-
sum). На ветвях лиственных древесных пород, в особенности на
тополе и некоторых плодовых деревьях,распространен полупаразит
омела (Viscum album) из семейства омеловых и др.
Низшие растения-паразиты распространены главным образом
среди грибов *. Лишенные хлорофилла и возможности синтезиро-
вать органические вещества из неорганических, грибы в большин-
стве своем становятся сапрофитами, т. е. живут на мертвых расти-
тельных остатках, а другие виды — паразитами либо симбионтами,
т. е. органически соединяются с некоторыми видами растений в
особые самостоятельные организмы — микоризы и лишайники; при
этом паразиты, питаясь за счет живой ткани растений-хозяев, в
конечном итоге убивают ее, а у симбионтов ткань растения обык-
новенно остается живой.
Грибы чрезвычайно распространены в природе: в 1 г почвы
находится иногда до 100 тыс. и более грибных зачатков, а если
учесть большую энергию их роста и размножения, то станет ясным
весьма важное значение их деятельности в природе и в большин-
стве случаев отрицательная роль в хозяйстве и жизни человека.
Особенно большой вред природным биогеоценозам и народному
хозяйству наносят ржавчинные и головневые грибы-паразиты.
Ржавчинных грибов около 4 тыс. видов. Наиболее распространен-
ная из них на различных культурных и диких злаках хлебная
(линейная) ржавчина (Puccinia graminis) наносит огромный
ущерб хозяйству. Общие убытки мирового хозяйства от ржавчины
хлебных злаков достигают в некоторые годы 500 млн долларов.
Головневые грибы, в частности виды головни (Ustilago), парази-
тирующие на хлебных злаках, заражают проростки зерен в почве
и вызывают заболевание их, снижая тем самым урожай зерновых
(в дореволюционной России на 10% и более); правда, в настоящее
♦ Грибы в последнее время выделяются в особое «царство» органического мира.
98
время ущерб от головни у нас сократился до 1%. На злаках,
особенности на ржи, паразитирует гриб спорынья (Claviceps pui
purea), образующая в колосьях ржи на месте зерен рожки (скл<
роции), которая также значительно снижает тем самым урожа
зерновых. Из других паразитных грибов большой урон хозяйств
наносит картофельный гриб (Phytophthora infenstans), поражав
щий ботву и клубни картофеля, который уничтожает в сырые год
у пас до 30% урожая этой культуры. Наряду с этим имеются
полезные в хозяйстве грибы, к ним относятся различные видь
употребляемые в пищу съедобные грибы, а также в хлебопеч<
нии — дрожжевые грибы и др.
Микоризы, представляющие собою симбиотическое соединена
гиф грибов с корнями растений, как установлено в последнее вр<
мя, очень широко распространены в растительном мире.* Биолоп
ческой основой такого сожительства низших грибов и высших ра<
тений является, как известно, обоюдная зависимость их друг с
друга: микоризные грибы обеспечивают высшие растения извл<
каемыми из почвы водой, минеральными и некоторыми органич<
скими веществами (азотистыми, витаминами), получая, в сво
очередь, из клеток корней высших растений углеродистое питань
в виде углеводов. Лишайники же, соединяющие в своем таллом
большей частью сумчатые грибы с зелеными или синезеленым
водорослями с участием азотфиксирующих бактерий, оказываютс
способными осуществлять фотосинтез, обеспечивать себя водо
минеральными солями и азотом.
Биогеоценологическое значение микориз и лишайников состоь
в том, что первые обеспечивают существование и нормальное ра:
внтис высших растений в анаэробных условиях (па болотах, заб<
лоченных лугах, в лесах), а вторые, являясь пионерами растител]
ности на бесплодных субстратах (вулканических пеплах, лава
горных скалах), создают благоприятные условия для освоения г
высшей растительностью и формирования на начальных стадия
освоения примитивных по составу и строению биогеоценозов.
Относящиеся к ценокомплексу биотрофов насекомоядные ра
тения — весьма интересная и оригинальная группа живых сущест
Наиболее известными и широко распространенными из них явл:
ются пузырчатки (Utricularia)—около 200 видов из семейстг
пузырчатковых, произрастающие в тропиках, особенно в Браз]
лии, встречаются и в Евразии, растут на сырой земле, в воде; ж\
рянки (Pinguicula) из того же семейства; росянки (Drosera)
около 85 видов из семейства росянковых, распространенные в А
стралии, Новой Зеландии, обычны и в евроазиатской флоре, раст
на болотах, в воде; мухоловки (Dionaea) того же семейства, пр
израстающие на востоке Северной Америки. Особенно своеобра
ны и замысловато устроены виды рода Nepenthes семейства N
99
penthaceae, распространенные в тропиках Старого Света (о. Бор-
нео, о. Мадагаскар, Австралия).
Все названные растения являются плотоядными биотрофами,
питающимися насекомыми и другими мелкими беспозвоночными,
обладающими целым рядом оригинальных приспособлений для
привлечения, ловли, удержания насекомых, переваривания их тела
и усвоения растворенных веществ. Пластинки листьев насекомо-
ядных растений с одной стороны обычно снабжены железками,
выделяющими липкую жидкость, богатую кислотами и протеоли-
тическими ферментами, способными растворять биомассу насеко-
мых. Всасывание же растворенных веществ осуществляется
большей частью всей поверхностью листовой пластинки. Удержа-
ние прилипших к пластинкам листьев насекомых обеспечивается
у мухоловок складыванием обеих половинок пластинки около сред-
ней жилки при прикосновении жертвы к чувствительным волоскам,
расположенным по краям пластинок; у росянок — сгибанием чув-
ствительных головчатых щетинок и всей листовой пластинки
вокруг насекомого, а у непентесов — образованным сросшейся
листовой пластинкой ловчим кувшинообразным мешочком с кры-
шечкой, по краям которого выделяется нектар для привлечения
насекомых, а на дне его ферментативная жидкость для перевари-
вания их биомассы.
Входящие в состав биотрофного компонента животные-парази-
ты наиболее широко распространены среди насекомых, которые,
по данным В. В. Яхонтова (1969), паразитируют на многих жи-
вотных; на мокрицах, паукообразных, птицах, млекопитающих и
на человеке. В то же время сами насекомые являются жертвами
целого ряда паразитирующих на них живых существ; кроме гри-
бов и бактерий, на них паразитируют простейшие животные, со-
сальщики, круглые черви, клещи, наездники и пр. Большинство
паразитирующих на других животных насекомых являются экто-
паразитами: кровососущие комары, постельный клоп, блохи, личин-
ки некоторых перепончатокрылых и двукрылых. К эндопаразитам
относятся личинки желудочных оводов лошадей, некоторые дву-
крылые и перепончатокрылые насекомые. Важно отметить, что у
некоторых насекомых, например у комаров, паразитический образ
жизни ведут лишь самки; у других насекомых—наездников, неко-
торых видов мух —паразитируют лишь личинки, зато блохи и
комары — только во взрослом состоянии.
В. Н. Беклемишев (1970) различает три типа паразитарных си-
стем; двойные (паразит — хозяин), тройные (хозяин — паразит —
переносчик) и множественные (паразит — несколько видов хозяев,
играющих различную роль в жизненном цикле паразита), напри-
мер гельминты с главным и двумя промежуточными хозяевами,
Биогеоценологическое значение одних паразитарных систем, по его
too
мнению, состоит в регуляции численных взаимоотношений сочле-
нов биоценоза, в других — в защите биоценоза от вторжения чуж-
дых элементов. В выполнении первой функции принимают участие
высоко патогенные для своих хозяев паразиты, которые вызывают
у них остро протекающие заболевания с высоким процентом гибе-
ли их, что ведет к колебанию численности хозяина; вторая же
функция осуществляется паразитами, вызывающими хронические
паразитарные заболевания либо у позвоночпого-хозяина, либо у
членистоногого-передатчика. Из этого автор делает вывод: «Пара-
зиты являются нормальными сочленами природных биоценозов, и
вызываемые ими болезни и даже эпидемии (эпизоотии) большей
частью принадлежат к числу нормальных жизненных отправлений
биоценоза, поддерживающих качественное и количественное по-
стоянство его состава» (с. 255).
Что же касается микроорганазмов-биотрофов, то к ним следует
отнести распространенные в природных и культурных биогеоцено-
зах патогенные бактерии и вирусы. Являясь, по существу, парази-
тами, они вызывают разнообразные болезни у растений, животных
и человека. Вирусами, в частности, вызываются чума многих диких
животных и свиней, анемия лошадей, ящур рогатого скота и пр.,
а также более сотни заболеваний растений. Среди микроорганиз-
мов имеется множество форм, паразитирующих на водорослях
(морских, пресноводных, почвенных), нередко ведущих к гибели
пораженных особей. Встречаются среди микроорганизмов и хищ-
ные формы, заглатывающие одноклеточные водоросли и др.
3. Состав и функциональная деятельность сапротрофного
компонента биогеоценоза
Третьим из живых компонентов биогеоценоза является цено-
комплекс сапротрофов, состоящий главным образом из микроорга-
низмов и некоторых ценопопуляций животных и растений, являю-
щихся потребителями и деструкторами отмершего органического
вещества растительного и животного происхождения. В составе
ценокомплекса сапротрофов различаются: а) сапрофаги — микро-
организмы, беспозвоночные и немногие виды позвоночных живот-
ных и б) сапрофиты — грибы и некоторые виды высших бесхлоро-
филльных растений.
Основными деструкторами отмерших растительных остатков к
целых мертвых растений, отбросов, жидких выделений и трупов
животных, доводящими эту работу до конца, являются сапротроф-
ные микроорганизмы, последовательно взаимосвязанные в процес-
се осуществления этой основной своей функции в биогеоценозе в
соответствующие биокомплексы. Микроорганизмы — весьма
своеобразная и специфичная во многих отношениях группа живых
101
существ, обладающая целым рядом присущих ей особенностей по
сравнению с растительными и животными организмами. Это свое-
образие и специфика микроорганизмов, по словам Т. В. Аристов-
ской (1967), заключается в следующем: во-первых, исключитель-
ное разнообразие способов питания (от сапрофитизма до парази-
тизма и хищничества) и вещественно-энергетического обмена (от
автотрофии до гетеротрофного обмена); во-вторых, чрезвычайное
разнообразие биохимических функций микроорганизмов, связан-
ное как с обилием видов, обладающих различными физиологиче-
скими свойствами, так и с полифункциональностью многих из них,
т. е. способностью при необходимости переходить к иным способам
вещественно-энергетического обмена; в-третьих, микроорганизмы
способны осуществлять любые биохимические процессы, соверша-
емые высшими растениями и животными, а также такие химиче-
ские реакции и процессы, которые недоступны ни тем, ни другим
(например, фиксация свободного азота атмосферы и др.), и, нако-
нец, неимоверная скорость роста и размножения микроорганизмов,
многие виды которых потенциально способны менее чем за двое
суток покрыть сплошь всю поверхность земного шара.
Следствием указанных особенностей микроорганизмов и уни-
версальности их биохимических функций является, во-первых,
чрезвычайно высокая активность микроорганизмов в составе
биогеоценоза, они выступают как «необычайно мощный химический
реагент, способный с неимоверной скоростью преобразовывать,
аккумулировать и рассеивать химические элементы...» — пишет
Т. В. Аристовская (1967, с. 7); во-вторых, возможность самостоя-
тельного существования и устойчивость чисто микробных биогео-
ценозов как в геоисторическом прошлом, так и в настоящее время,
например на скалах, вулканических лавах, в пустынных областях
и -в так называемых пластовых водах нефтяных месторождений.
С другой стороны, ограниченность и однонаправленность биохими-
ческих функций многоклеточных растительных и животных орга-
низмов предопределяют невозможность автономного (без участия
микроорганизмов) существования растительных или животных
сообществ в силу необеспеченности минерализации органики и
фиксации азота атмосферы.
В состав биогеоценоза входят весьма разнообразные микроор-
ганизмы как по своему систематическому положению, так и по
характеру своей деятельности и общему направлению работы: бак-
терии, низшие грибы, актиномицеты, одноклеточные водоросли,
вирусы, а также некоторые простейшие животные — амебы, инфу-
зории и т. п. (рис. 8). Разнообразны и места обитания микроорга-
низмов, которые встречаются всюду: в почве, воздухе, воде; в
телах высших растений и животных и экскрементах последних; в
горных породах, на поверхности вечных льдов, в горячих источни-
102
1. Водоросли
г. Грибы
ЗАктиномицеты
несовершенные гги&ы
4. Бдктерии
Рис. 8, Почвенная микрофлора (Pochon, 1955; и др.) (по П. Дювиньсу
и М. Тангу, 1968; А. П. Шенннкову, 1964; и др.)
ках и месторождениях нефти. Однако больше всего микроорганиз-
мов в мертвом напочвенном покрове и в верхнем слое почвы./В
почве они размещаются в полостях между почвенными агрегата-
ми, а также в порах между микроагрегатами (рис. 9). Общая чис-
ленность микроорганизмов в биогеоценозе чрезвычайно велика. По
данным отечественных микробиологов, в 1 г лесной почвы насчи-
тывается водорослей около 100 тыс., плесневых грибов — до 1млн,
актиномицетов — до 15 млн, бактерий — несколько миллиардов;
правда, в зависимости от почвы, растительности и прочих обстоя-
тельств численность бактерий колеблется от нескольких десятков
тысяч до нескольких миллионов и миллиардов в 1 г верхнего гори-
зонта почвы. Численность микроорганизмов, как известно, изме-
ню
Рнс. 9. Распределение микроорганизмов в агрегированном комочке почвы (по
Н. А. Красильникову, 1958). 1 — агрегаты, состоящие из микроагрегатов 2:
а — клетки и б — колонии бактерий, грибов и актиномицетов (между агрегата-
ми); в — клетки бактерий (в порах между микроагрегатами)
няется по сезонам вегетационного периода в связи с изменением
экологических условий; изменяется она и в силу внутренних зако-
номерностей развития микробных ценопопуляций,» Последние изме-
нения носят характер пульсаций с суточной и даже внутрисуточной
периодичностью.
Довольно велика и биомасса микроорганизмов, несмотря на их
микроскопические размеры. Решающую роль в этом играет ско-
рость размножения микроорганизмов и соответствующая быстрота
смены поколений (от 6—8 до 14—18 за вегетационный период). По
данным Е. Н. Мишустина (1956), живая бактериальная биомасса
104
составляет в подзолистых и дерново-подзолистых почвах лесной
зоны 0,6—0,9 т/га, в черноземах степной полосы — 3,7 т/га и в се-
роземах пустыни — 2,5 т/га. На окультуренных подзолистых поч-
вах под клевером, по данным Н. А. Красильникова (1944, 1958),
живая биомасса бактерий достигает 1,5—4,0 т/га, а на окультурен-
ных сероземах под люцерной —5—8 т/га. Правда, по мнению
И. В. Тюрина (1946), приведенные данные о биомассе бактерий
завышены, ибо накопление ее в таких размерах не обеспечивается
наличием и приростом органической пищи. Однако Е. Н. Мишу-
стин находит, что Тюриным не учитываются некоторые дополни-
тельные источники углерода, используемые бактериями, в частно-
сти продукты распада гумуса, корневые выделения (около
2,5 и/га), образуемые некоторыми растениями гелеподобные сли-
зистые вещества (в количестве нескольких т/га), органические
вещества, продуцируемые автотрофными водорослями и бактерия-
ми. Биомасса других почвенных микроорганизмов менее значи-
тельна по сравнению с бактериальной биомассой. По данным
Ж. Пошона и Г. Де Баржака (I960), биомасса микроскопических
грибов составляет 1—1,5 т/га, актипомицетов — 0,7 т/га, простей-
ших животных — 0,1—0,3 т/га. Общая биомасса микроорганизмов
в несколько раз превышает вес живой биомассы животного насе-
ления биогеоценозов.
В силу колоссальной численности микроорганизмов и наличия
указанных выше таких свойств, как чрезвычайно высокая биоло-
гическая активность их биомассы и многообразие присущих им
биохимических функций, биоценотическая функциональная дея-
тельность микроорганизмов весьма значительна и многогранна.
Однако микроорганизмы хотя и обладают способностью при необ-
ходимости коренным образом перестраивать свой материально-
энергетический обмен, тем не менее вполне определенные специ-
фические функции их в составе биогеоценозов и биогеосферы в
целом с достаточной степенью эффективности могут выполняться
лишь соответствующими специализированными биологическими
группами, занимающими определенное место в составе биокосньи
систем. К таким специализированным биологическим группам
микроорганизмов относятся: а) автотрофные микроорганизмь
(фототрофные водоросли и хемотрофные бактерии)—созидателе
первичной биологической продукции, отнесенные ранее к авто
трофному компоненту биогеоценоза; б) паразиты и хищники, по
требляющие в пищу биомассу или соки живых существ, отнесен
ные к биотрофному компоненту биогеоценоза; в) сапрофаги и сап
рофиты, питающиеся мертвым органическим веществом, входят i
состав сапротрофного компонента биогеоценоза.
Помимо перечисленных биологических групп микроорганизмов
играющих основную роль в обменных процессах соответствующи:
105
биогеоценозов, выделяются также г) азотфиксаторы и микоризо-
образователи, являющиеся за малым исключением симбионтами с
высшими растениями, обеспечивающие живые существа нитратным
азотом и минеральным питанием в особо неблагоприятных усло-
виях; д) микроорганизмы — окислители и восстановители азоти-
стых, сернистых и других биогенных минеральных соединений, уча-
ствующих в биологическом круговороте веществ; е) микроорганиз-
мы— синтетики биологически активных веществ (ферментов,
антибиотиков, витаминов, ростовых веществ и т. п.), необходимых
для многих жизненно важных процессов высших растений и жи-
вотных. В последние десятилетия выявлены ж) группа микроорга-
низмов— синтетиков совершенно новых минералов в составе
почвообразующих материнских пород и з) группа деструкторов
минералов материнских пород, пополняющая запасы минеральных
веществ в почве, поддерживая тем самым биотический круговорот
на должном уровне и даже расширяя его, как правило, в сторону
поступательного развития биогеоценотического покрова данного
участка Земли.
Однако основная функция микроорганизмов в составе биогео-
ценоза и биокосных систем более высокого ранга, по свидетельству
С. А. Егорова с соавторами (1964), состоит в разложении находя-
щегося в почве, подстилке или в степном войлоке мертвого органи-
ческого вещества растительного и животного происхождения па
простые минеральные соединения, соли которых служат источни-
ком минерального питания автотрофных растений и частично ис-
пользуются на процессы жизнедеятельности самих микроорганиз-
мов. В минерализации органики и состоит специфика биоценотиче-
ской функциональной деятельности микроорганизмов в составе
сапротрофного компонента биогеоценоза, ибо микроорганизмы —
это основные биоредуценты, осуществляющие этот процесс до
исходных элементов. Дело в том, что в природных условиях боль-
шая часть питательных веществ растений находится в недоступных
для них органических и сложных минеральных соединениях. Мик-
роорганизмы же минерализуют органические вещества, а трудно-
растворимые минеральные соединения переводят в усвояемую
растениями форму. Следовательно, главная роль сапротрофных
групп микроорганизмов в составе биогеоценоза сводится к обеспе-
чению автотрофных растений элементами минерального питания.
Энергию для процессов своей жизнедеятельности и всех этих про-
цессов превращения веществ микроорганизмы получают из двух
источников: аэробные ценопопуляций при окислении различных
веществ, анаэробные — при процессах брожения некоторых из них.
В состав сапротрофного компонента биогеоценоза, как было
сказано, наряду с микроорганизмами входят животные-сап-
рофаги, источником питания которых являются огромные запасы
106
отмерших растительных остатков, во много раз превышающие по
массе Животную биомассу, особенно в почве и мертвом напочвен-
ном покрове. Верхние горизонты почвы и подстилка, по словам
Г. Ф. Курчевой (1971), являются средой обитания колоссального
М 8 3 О Ф A U НА
М И К РО ЮДИНА
Рис. 10. Почвенно-подстилочные беспозвоночные животные (по П. Дювиньо 1
М. Тангу, 1968; Д. А. Криволуцкому, 1969; и др.). Мезофауна: 1—дождево!
червь; 2 — мокрица; 3 — личинка пластинчатого жука; 4 — многоножки; 5 —
многоножка хищная; 6 — ложный скорпион; 7 — жук хищный. Микрофауна: I —
ногохвостки; 2 — тихоходка; 3 — нематоды; 4 — клещ хищный; 5 — клещи паи
цирные; б — амебы раковинные
107
количества разнообразных беспозвоночных животных: простейших,
коловраток, тихоходок, энхитреид, нематод, клещей, составляющих
так называемую микрофауну, а также дождевых червей, брюхо-
ногих моллюсков, мокриц, многоножек и других относительно
крупных членистоногих, которые относятся к мезофауне (рис. 10).
Г Ф. Курчевой приводятся данные разных авторов о численности
и биомассе почвенно-подстилочных беспозвоночных. По этим дан-
ным, на 1 м2 подстилки и почвы насчитывается до 900 тысяч осо-
бей мелких и более 2 тысяч крупных беспозвоночных, а живая
биомасса беспозвоночных составляет около 3,5 т/га (1,1 т/га сухо-
го вещества). По числу видов и особей среди них преобладает
микрофауна, а по биомассе—мезофауна. Основную массу почвен-
ных беспозвоночных составляют сапрофаги. Биомасса почвенных
беспозвоночных, в том числе и доля сапрофагов, возрастает от
тундры к зоне широколиственных лесов и затем падает к пустыне.
К животным сапрофагам, помимо собственно сапрофагов, по-
требителей растительного и животного опада, относятся также
беспозвоночные, питающиеся гниющей древесиной, — ксилофаги;
сильно разложившимися растительными преимущественно остат-
ками (детритом)—детритофаги; потребители экскрементов — коп-
рофаги и трупов животных — некрофаги.
По характеру питания одни из них являются первичными
сапрофагами, остальные — вторичными и последующими сапрофа-
гами, потребляющими в пищу мертвый растительный материал,
прошедший иногда не один раз через кишечный тракт беспозво-
ночных.
Беспозвоночные сапрофаги играют значительную роль в мине-
рализации растительных и животных остатков, в ускорении их
разложения. Достигается это прежде всего измельчением разла-
гаемого материала и, следовательно, увеличением его поверхности,
благодаря чему он становится более доступным для использования
основными биоредуцентами — микроорганизмами. К этому,
по мнению М. С. Гилярова и Б. Р. Стригановой (1978), следует
добавить перемещение почвенными беспозвоночными органических
остатков в более глубокие горизонты почвы и перемешивание их
с минеральными частицами, что ускоряет их минерализацию, а
также стимуляцию развития почвенной микрофлоры и распростра-
нение ее в почвенной толще. Наряду с этим, обладая разнообраз-
ными ферментами и выделяемыми кишечной микрофлорой энзи-
мами, почвенные беспозвоночные сапрофаги, по утверждению
Г. Ф. Курчевой, в состоянии подвергать растительный и животный
опад и отпад глубоким химическим изменениям, не только расщеп-
ляя его до исходных минеральных соединений, но и синтезируя из
них гумусовые вещества. Такого же мнения по этому вопросу при-
держивается и Л. С. Козловская (1959).
108
Входящие в состав сапротрофного компонента биогеоценоз
сапрофиты представлены главным образом грибами и немпс
гими видами высших растений, такими, как подъельник (из. сел
грушанковых), надбородник, гнездовка, ладъян (из семейства ор
хидных) и пр. Произрастающие преимущественно в хвойных
лиственных лесах, высшие сапрофитные растения лишены зелено
окраски и типичных листьев. Питаются они отмершими полураз
ложившимися растительными остатками подстилки и верхних гс
ризоптов почвы и отчасти минеральными веществами за счет микс
ризного гриба-симбионта. Грибььмакро- и микромицеты по способ
питания, как указывает Л. Г. Бурова (1976), обычно подраздели
ются на три группы: сапрофиты, паразиты и грибы-микоризообра
зователи, а по месту своего обитания—на внутрипочвенные, па
почвенные, стволовые, а также пневые и валежные группировку
Подобного рода группировки многими исследователями рассмат
риваются как синузии в составе сообщества. Местом обитани:
напочвенных грибов-сапрофитов в лесных биогеоценозах являете:
лесная подстилка, которая оказывает большое влияние на газо
вый, тепловой и водный режимы почвы. В пей осуществляютс:
процессы разложения растительного и животного опада и отпад,
и синтеза гумусовых веществ. Мицелий и плодовые тела стволовы.
макромицетов располагаются в слое древесины или пней и вале
жа, где происходит под их воздействием разложение таких трудно
растворимых веществ, как лигнин, целлюлоза и т. п.
Обитающие в почве микроорганизмы, беспозвоночные сапрофа
гн и сапрофитные грибы тесно взаимно связаны в составе биогео
цеиоза, оказывая друг па друга разнообразные воздействия. П(
наблюдениям Г. Ф. Курчевой (1971), микроорганизмы (бактерии
водоросли), мицелий грибов служат источником питания для мно
гих видов почвенных беспозвоночных; выделяемые микроорганнз
мами биологически активные вещества стимулируют развита
простейших животных, а антибиотики, напротив, отрицательш
влияют на почвенную микрофауну. С другой стороны, живые i
мертвые ткани беспозвоночных служат пищей патогенным и дру
гим микробам. Почвенные беспозвоночные стимулируют деятель
ность многих микроорганизмов путем улучшения аэрации почвы
заглатывания и обогащения микрофлоры в своем кишечнике, вы
деления некоторых веществ (например, муравьиной кислоты) либ<
оказывают угнетающее воздействие на развитие микроорганизме]
путем выделения веществ с бактерицидными свойствами и т. п.
Накопление опада и отпада растительного и животного проис
хождения в биогеоценозах, как отмечалось ранее, происходит ш
поверхности почвы в виде лесной подстилки или степного войлока
на болотах — в виде очеса, а отложение отмерших подземные
органов растений и мертвой биомассы почвенных животных и мик
109
рооргандзмов — в верхнем горизонте почвы в виде перегноя. По-
этому и процессы преобразования органического вещества совер-
шаются на поверхности почвы и в зоне ризосферы. Основную
массу мертвого органического материала как в надземной, так и в
почвенной среде составляют растительные остатки; животная и
микробная мертвая биомасса по сравнению с ними настолько ни-
чтожна по весу, что при рассмотрении подобного рода процессов
обычно не принимается в расчет.
Превращение, переработка опада и отпада в биогеоценозе, как
указывает Т. В. Аристовская (1980), — многоступенчатый биологи-
ческий процесс, который не сводится только к разложению, мине-
рализации растительных остатков и высвобождению ряда химиче-
ских элементов для дальнейших превращений их в биологическом
круговороте, но включает также синтез сложных органических
соединений, образование гумуса, определяющего многие физико-
химические свойства и плодородие почвы, и, наконец, консервацию
не полностью разложившихся остатков, ведущую к выпадению
значительного количества химических элементов из круговорота
веществ на длительный срок. При этом она обращает внимание на
то обстоятельство, что в случае преобладания минерализации над
другими названными процессами в почве растительный опад почти
полностью разлагается и образование гумуса сводится почти к
нулю (например, во влажных тропиках). Превалирование процес-
са гумификации (например, в степной полосе) сопровождается на-
коплением больших запасов перегноя и образованием черноземных
почв различной мощности. В условиях избыточного застойного
увлажнения и недостатка кислорода в почве на болотах лесной
полосы процесс минерализации растительных остатков настолько
замедляется, что происходит накопление огромных масс слабораз-
ложившихся растительных материалов и образование мощных за-
лежей торфа.
Разложение, минерализация природных органических веществ
в биогеоценозе, по словам В. Я. Частухина и М. А. Николаевской
(1969), — весьма сложный комплексный процесс, результативность
и скорость осуществления которого находятся в зависимости от
состава оргаиизмов-сапротрофов, участвующих в этом процессе,
от химического состава и других особенностей разлагаемого мате-
риала и от экологических условий, в которых этот процесс про-
текает.
В разложении органических остатков растительного и живот-
ного происхождения, как видно из всего изложенного, принимают
участие самые разнообразные организмы, относящиеся к цено-
комплексу сапротрофов: бактерии, грибы, актнномицеты, предста-
вители микро- и мезофауны и пр. Участие тех или иных организмов
в разложении различных растительных и других материалов опре-
110
деляется их физиологическими свойствами, в особенности хара
тером выделяемых ими ферментов. Необходимо отметить, что эт
весьма важный процесс, являющийся составной частью биотич
ского круговорота веществ в биогеоценозе, осуществляется п|
самом тесном взаимодействии перечисленных групп живых с
ществ, сменяющих друг друга в определенной последовательное!
Осуществляется процесс разложения органики обычно в к
сколько этапов: вначале разлагаются простые водорастворимь
более легко усвояемые вещества при активном участии быст
размножающихся неспороносных бактерий, плесневых и некотор!
других грибов; затем на полурналожившихся остатках поселяют
разрушители крахмала, клетчатки — спороносные бактерии и г
совершенные грибы и некоторые беспозвоночные с соответству
щей кишечной микрофлорой; на смену им приходят базидиальн
и другие грибы, актиномицеты, способные выделять более актг
ные ферменты и усваивать наиболее устойчивые формы органш
ских соединений (лигнин, целлюлозу и т. п.), разрушая их в пр
цессе усвоения; на этом этапе к микроорганизмам подключают
и некоторые беспозвоночные сапротрофы, участвующие в минер
лизации белков и их производных. Характерно, что при разлол
нии различных по химическому составу органических материал
комплектуются соответствующие специфичные биокрмплексы с<
ротрофных организмов, образующие сукцессионный ряд после;
вательно сменяющих друг друга биологических групп, способн
в совокупности довести процесс деструкции до исходных MHHepaj
ных элементов.
Рассмотрим разложение отмерших остатков (опада) травян
и лесной растительности, а также минералов материнских поч]
образующих пород. Ход этих процессов и участие в них различи
групп сапротрофных организмов освещаются в работах Т. В. А]
стовской (1967, 1972, 1980), В. Я- Частухина и М. А. Николаевск
(1969), Г. Ф. Курчевой (1971), Р. И. Злотина и К. С. Ходашев
(1973, 1974) и др.
В растительном опаде травянистых биогеоценозов (степш
луговых) отмечается высокое содержание минеральных хими
ских элементов, поэтому он довольно легко подвергается раз.
жению бактериями и другими микроорганизмами; участие бесп
воночных животных в этих процессах невелико. По наблюдени
Злотина и Ходашевой, значительная часть растительного опад?
степных условиях разрушается также абиогенным путем.
В составе опада лесных биогеоценозов почти 2/3 по весу П]
ходится на долю хвои или листьев, которые относительно ле!
поддаются разложению. В целом же лесная подстилка, по дани
Арйстовской (1980), разлагается медленно, в особенности подст
ка хвойных лесов, ибо в ее составе содержится больше лигнин:
111
других труднорастворимых соединений, чем в подстилке листвен-
ных лесов, больше в ней также бактерицидных веществ, угнетающе
действующих на почвенно-подстилочную микрофлору. В разложе-
нии растительного опада в хвойных лесах главную роль играют
микроскопические грибы и базидиомицеты, принимают участие и
беспозвоночные» главным образом клещи. В лиственных лесах в
минерализации органики значительная роль, наряду с бактериями
и микобактериями, принадлежит беспозвоночным животным, в
частности дождевым червям. Т. В. Арнстовская (1980) приводит
такую последовательность разложения растительного опада в
хвойном лесу: вначале бактерии и некоторые грибы (аскомицеты,
базидиомицеты) разлагают сахара, крахмал и клетчатку; затем на
базе продуктов распада их появляются и действуют фикомицеты,
гифомицеты и некоторые беспозвоночные сапротрофы; завершают
этот процесс мукоровые и другие грибы.
Основную роль в разложении древесины и коры в составе лес-
ной подстилки выполняют базидиальные грибы и беспозвоночные
животные, обладающие способностью выделять наиболее активные
ферменты, разрушающие содержащиеся в коре и древесине наи-
более устойчивые к разложению вещества. Разрушение отмираю-
щей древесины на живых деревьях, как пишут В. Я. Частухин и
М. А. Николаевская, начинается под действием паразитных грибов
(сумчатых, несовершенных), выделяющих ферменты, способные
разлагать как углеводы, так и лигнин, целлюлозу и пр. На базе
происшедших изменений древесины происходит развитие после-
дующих, сменяющих друг друга биологических групп микроорга-
низмов и беспозвоночных животных, способных довести разложе-
ние древесины до простых минеральных соединений и образования
соответствующих форм гумуса.
Минерализация корневого опада в травяных и лесных биогео-
ценозах протекает в почвенной толще, точнее, в зоне ризосферы,
где и происходит непрерывное частичное или полное отмирание
корневых систем. Разложение корневого опада происходит значи-
тельно медленнее, чем надземной отмершей массы, особенно в ана-
эробных условиях, где этот процесс длится годами. В минерализа-
ции его принимают участие бактерии, грибы, актиномицеты, про-
стейшие животные, дождевые черви и пр. По наблюдениям
В. Я. Частухина и М. А. Николаевской (1969), разложение корне-
вого опада осуществляется следующими сменяющими друг друга
биологическими группами сапротрофпых организмов: в начальном
этапе разложения корневых остатков принимают участие плесне-
вые грибы и неспороносные бактерии, на смену им приходят спо-
роносные бактерии, и на завершающем этапе минерализации их
включаются целлюлозоразлагающие бактерии и отчасти актино-
мицеты.
112
Наряду с минерализацией растительного и животного опада
мертвом напочвенном покрове и верхнем горизонте почвы биоге<
ценозов, как было сказано ранее, с различной степенью интенси!
ности идет процесс гумификации разлагающихся материалов, о(
разование гумусовых веществ (гуминов, гуминовых кислот, фул!
вокислот и т. п.). Различаются три формы гумуса: а) грубый гум1
(мор), который состоит из слаборазложившихся органически
остатков, кислой реакции, устойчив к воздействию микрооргани:
мов, он обычно достигает значительной мощности, распадаясь н
три слоя (L — свежий опад, F — полуразложившаяся масса, Н-
гумифицированный слой); б) средний гумус (модер), состоящи
из не полностью гумифицированных разложившихся органически
остатков растительного и животного происхождения; в) мягки
гумус (муль), представляющий собою гомогенную массу с мине
ральной частью почвы, слабокислой реакции, легко поддающийс
минерализации.
Образование грубого гумуса характерно для хвойных лесов
главная роль в этом процессе принадлежит различным форма!
грибной флоры. Средний гумус формируется обычно в условия
достаточно быстрой минерализации органических остатков по,
действием преимущественно беспозвоночных сапрофагов. Мягки
гумус образуется в лиственных лесах в результате интенсивно)
деятельности бактериальной микрофлоры при заметном участи
беспозвоночных животных, в особенности дождевых червей.
Из образовавшегося в ходе почвообразования различных форг
гумуса формируется особый горизонт в самой верхней части поч
венного профиля соответствующих типов почв, называемый гуму
совым горизонтом. Этот различной мощности почвенный горизош
перекрытый, как правило, войлоком и в некоторых типах поч1
дерниной, представляет собою по существу резервный фонд, своеп
рода кладовую непрерывно пополняемого потенциального почвен
ного плодородия. В нем сосредоточен огромный запас питательны,
веществ, реализуемый в процессе минерализации почвенного гуму
са. Минерализация гумусовых веществ, по словам Т. В. Аристов
ской (1980), является важной частью круговорота углерода,
результате чего высшие растения обеспечиваются элементами золь
ного минерального питания и азотом. Осуществляется этот процес
биокомплексом взаимосвязанных между собой разнообразны
групп микроорганизмов (бактерий, микроскопических грибов, акти
номицетов и пр.) при участии различных представителей микро-
мезофауны.
Из сопоставления функциональной биоценотической деятельнс
сти основных представителей всех трех живых компонентов бис
геоценоза видно, что если основная функция ценопопуляци
автотрофного ценокомплекса состоит в созидании первичной бис
нз
логической продукции из неорганических веществ, а функция цено-
популяций биотрофного ценокомплекса — в трансформации ее в
более высокомолекулярную вторичную биологическую продукцию,
представляющую их собственную биомассу, то главная функция
ценопопуляций сапротрофного ценокомплекса состоит в разложе-
нии той и другой на простые минеральные соединения (углекисло-
ту, воду и простые минеральные соли), вновь используемые авто-
трофными растениями на синтез органических веществ. Иначе
говоря, сапротрофный компонент выполняет в составе биогеоцено-
за функцию, как бы диаметрально противоположную функциям
первых двух компонентов, функцию разрушения синтезируемой
ими биомассы, замыкая тем самым цикл биохимических превра-
щений веществ в биогеоценозе.
Однако следует иметь в виду, что обменные процессы в биогео-
ценозе и бногеосфере в целом не сводятся только к чередующемуся
синтезу и разрушению органических веществ при непосредственном
участии живых организмов. Наряду с этим, как указывает
Т. В. Аристовская (1973), в биогеоценозах протекают процессы
глубокого преобразования и минеральной части материнских поч-
вообразующих пород, которая не остается вечно инертной, а во-
влекается в обменные процессы в биогеоценозах посредством раз-
рушения минералов породы и образования новых минералов. Ми-
неральная часть почвообразующих пород в действительности
является первоисточником зольной пищи растений, а через них и
всех других живых существ; освоение же ее осуществляется в
основном почвенной микрофлорой. Т. В. Аристовская придает чрез-
вычайно большое значение этому процессу, поскольку это основной
путь вовлечения минеральной части почвы в обменные процессы
биогеоценоза и, следовательно, создания условий для расширения
биотического круговорота в биогеоценозе и биогеосфере в целом.
По своему значению для биогеосферы она ставит этот процесс в
один ряд с такими процессами, как ассимиляция углекислоты авто-
трофами или фиксация азота атмосферы определенными группами
микроорганизмов.
Из состава почвенной микрофлоры в разрушении минералов
почвы и материнских почвообразующих пород, по данным
Т. В. Аристовской (1973, 1980), участвуют водоросли, бактерии,
микроскопические грибы, микобактерии, актиномицеты и пр. Про-
цесс разрушения минералов, содержащих серу, железо, марганец,
осуществляется тиобактериями, железобактериями и др. с по-
мощью выделяемых ими ферментов; при этом минералы из
закисной формы переходят в более подвижную окисную, становясь
доступными растениям. Пожалуй, чаще разрушение минералов
совершается почвенными бактериями и водорослями с помощью
образуемых ими внеклеточных полимеров в виде микробных сли-
114
зей; содержащиеся в слизи химические вещества вступают в реак*
цию с элементами минералов, вследствие чего происходит разру-
шение последних и переход химических элементов в почвенный
раствор. Наряду с этим широко распространен косвенный способ
воздействия микроорганизмов (тиобактерий, микроскопических
грибов и пр.) на минералы почвообразующих пород посредством
выделения в окружающую среду продуктов своего обмена — мета-
болитов, являющихся сильными химическими реагентами (органи-
ческие и минеральные кислоты, биогенные щелочи и т. п.).
Таким образом, микроорганизмы, разлагая минералы почвы и
почвообразующих пород, обеспечивают мобилизацию питательных
веществ из минеральной части почвы, а также вовлекают в обмен-
ные процессы биогеоценоза инертные материнские породы.
Что же касается образования микроорганизмами новых мине-
ралов, то значение этого рода их деятельности в жизни биогеоце-
ноза также достаточно велико, ибо вследствие этого значительно
изменяются условия обитания всего почвенного населения, в част-
ности, изменяются- механический состав и структура почвы, погло-
тительная способность почвы и химические свойства ее и т. п.
Г л ав a V
Косные компоненты биогеоценоза,
их состав и функциональная роль
в биогеоценозе
В качестве косных компонентов биогеоценоза В. Н. Сукачев,
как известно, называл эдафотоп, включающий в свой состав
почву и почвенно-грунтовые воды на определенную глубину, и
климатоп в составе таких атмосферных явлений, как световое
н тепловое излучения, атмосферные осадки, ветер и пр. Т. А. Ра-
ботновым (1978), как было сказано ранее, эдафотоп рассматрива-
ется как совокупность факторов биогенной почвенной среды в
границах биогеоценоза, климатоп же, переименованный в аэро-
топ, он представляет как совокупность факторов биогенной воз-
душной среды в пределах биогеоценоза. Иначе говоря, косные
компоненты биогеоценоза в трактовке Работнова выступают как
компоненты внутренней среды биогеоценоза, ограниченной его го-
ризонтальными и вертикальными пределами, которая представляет
собою результат преобразования живыми компонентами в процес-
се своей жизнедеятельности косной части почвы, с одной стороны,
и косного атмосферного окружения — с другой. Отсюда следует,
что сами компоненты становятся вследствие этого биогенными
образованиями, подобно каменному углю, нефти и др.
Правда, Т. А. Работнов (1974) считает почву биокосным ком-
понентом биогеоценоза, состоящим из косной среды и живых орга-
низмов, хотя последние, по его мнению, могут рассматриваться
составной частью как почвы, так и фитоценоза. К категории био-
косных систем, со ссылкой на В. И. Вернадского, относит почву и
А. И. Перельман (1974), наряду с илами, корой выветривания, во-
доносными горизонтами и ландшафтом. Однако В. А. Ковда
(1973), рассматривая компонентный состав биосферы, относит
почву, наряду с нефтью, торфом, сапропелем и др., к категории
биогенных тел, образовавшихся при непосредственном участии жи-
вого вещества. На наш взгляд, последняя трактовка почвы как
косного тела биогенного происхождения является более обоснован-
ной. Дело в том, что наличие в почвенной среде позвоночных и
беспозвоночных животных, микроорганизмов и подземных органов
растений не может служить основанием для рассмотрения их в
качестве составных элементов почвы. Почва является для них сре-
дой обитания, подобно тому как воздушная среда является средой
обитания для наземных (сухопутных) животных, микроорганиз-
мов и наземных органов растений. Ведь никто на основании по-
ив
следнего факта не считает возможным эти наземные живые суще-
ства и образуемые ими биокомплексы относить к составным эле-
ментам воздушной среды биогеоценозов, резонно рассматривая их
как самостоятельные природные образования. Да и В. В. Докучае-
вым, как известно, почвообитающие живые существа рассматри-
вались, наряду с климатом, горными породами, рельефом и возра-
стом страны, как факторы почвообразования, а не как составные
элементы почвы.
Что же представляет собою косная часть воздушной и почвен-
ной среды, преобразуемой живыми компонентами в биогенные ком-
поненты биогеоценоза, каков ее состав, в чем состоит и во что
выливается преобразование той и другой живыми компонентами
во внутреннюю среду биогеоценоза — биоценосреду?
1. Аэротоп и его материальная основа
Прежде всего об аэротопе, контактирующем с почвой, с назем-
ной частью автотрофного ценокомплекса живых существ и надзем-
ными сухопутными фитофагами и зоофагами биотрофного цено-
комплекса. Материальным субстратом, лежащим в основе форми-
рования этого биогенного по происхождению компонента биогео-
ценоза, является атмосфера (более узко — тропосфера), кото-
рая представляет собою надземную газовую (воздушную) оболоч-
ку земного шара, связанную с ним силой земного притяжения
(гравитацией). Верхняя граница атмосферы прослеживается до
1100 км над поверхностью Земли, но простирается она предполо-
жительно до 20 тыс. км, постепенно переходя в межпланетное про-
странство. Нижний слой атмосферы мощностью до 1Б—18 км у
экватора и до 6—8 км у полюсов называется тропосферой.
Это наиболее плотная часть атмосферы, в ней сосредоточено около
80% всей ее массы. Здесь происходит образование облаков и осад-
ков, движения воздушных масс и других связанных с погодой
атмосферных явлений.
Современная земная атмосфера, являющаяся физической
смесью газов, кислородно-азотная по своему составу: азота по
объему в ней содержится 78,08%, кислорода — 20,95; из других га-
зов в ее состав входит инертный газ аргон — 0,94, а также углекис-
лый газ, гелий и прочие — 0,03%. Такой состав атмосферы сфор-
мировался около 2 млрд лет назад с появлением свободного
кислорода, продуцируемого зелеными растениями, особенно мор-
ским фитопланктоном. Под действием кислорода коренным обра-
зом изменился состав первичной атмосферы Земли (протоатмо-
сферы), состоящей в основном из метана, аммиака и водорода с
парами воды. При этом произошло окисление метана и аммиака
до углекислого газа и свободного азота, водород же улетучился
117
в космическое пространство. Так сформировалась вторичная совре-
менная атмосфера, состав которой остается с тех пор относительно
постоянным на всей поверхности нашей планеты до высоты 400—
600 км.
Многими исследователями отмечаются следующие характерные
черты атмосферы, по крайней мере нижних слоев ее: а) преобла-
дание в ее составе азота, кислорода и наличие других газов; б) ко-
лебание температуры в пределах от 4-50 до —50° С; в) величина
атмосферного давления около 1 атм с незначительными отклоне-
ниями от этого среднего показателя в ту или другую сторону;
г) непрерывный поток световых, тепловых и других излучений из
космоса. К этому, очевидно, надо добавить д) наличие постоянной
циркуляции воздушных масс на земной поверхности, в особенно-
сти в системе суша — Мировой океан.
В биогеоценотическом отношении наиболее важное значение
имеет самый нижний слой атмосферы, точнее, тропосферы мощно-
стью в 30—50 (до 150) м. Этот приземной слой атмосферы, на-
званный А. И. Воейковым (1904) деятельным слоем, рассматри-
вался им как обусловленная радиационным режимом особая зона
теплооборота, находящаяся в зависимости от подстилающей по-
верхности, в частности от характера растительного покрова. Верх-
няя граница деятельного слоя определяется высотой растительного
покрова данного биогеоценоза, нижняя — либо глубиной распро-
странения в почве корневой системы доминирующих растений —
1—2 (до 10) м и более, либо глубиной затухания температурной
волны в почве. Деятельный слой атмосферы (тропосферы) обла-
дает рядом важных особенностей: а) наибольшей плотностью био-
генных и прочих газов (кислорода, углекислого газа и др.); б) вы-
сокой динамичностью всех атмосферных явлений (температуры,
осадков, ветра и т. п.); в) наиболее тесным контактом его с
другими компонентами биогеоценоза (растительностью, животным
населением, горными породами, почвой и пр.). В границах биогео-
ценоза деятельный слой подразделяется на два подслоя: а) расти-
тельный подслой (полог растительности) и б) почвенный подслой
(верхние охваченные обменными процессами горизонты почвы).
Составными элементами или факторами атмосферы, в частно-
сти ее деятельного слоя, являются: а) солнечная радиация —
источник лучистой энергии света и тепла; б) атмосферные осадки,
несущие живительную влагу на поверхность Земля; в) газовый
состав воздуха, обеспечивающий газовый обмен живых существ;
г) общая циркуляция атмосферы, формирующая климат и погод-
ные условия в отдельных пунктах земного шара.
118
2. Составные элементы и функциональная роль
атмосферы в биогеоценозе
Рассмотрим составные элементы (факторы) атмосферы и зна-
чение, роль каждого из них в жизни живых существ, в составе и
функционировании биогеоценозов и биогеосферы в целом.
Солнечная радиация представляет собою непрерывный нотой
лучистой энергии Солнца на Землю через атмосферу. Мощность
потока лучистой энергии называется интенсивностью радиации,
1 о rrz 1см3 ч
которая оценивается в калориях на 1 см2 в минуту (K-j—).
Интенсивность радиации составляет 1,98—2,0 кал/см2 мин. Коли-
чество поступающей на подстилающую поверхность Земли солнеч-
ной радиации находится в зависимости от целого ряда факторов:
от географической широты местности вследствие шарообразной
формы Земли, от толщины и плотности атмосферы, от облачности
и содержания водяных паров в атмосфере, от времени суток и
рельефа местности. Из поступающей на верхнюю границу атмо-
сферы солнечной радиации поверхности земного шара достигает
менее половины ее, а именно 44% в виде прямого излучения и
4%—рассеянного, ибо 14% поступающей радиации поглощается
атмосферой, до 33% ее отражается от облаков и 5% диффунди
рует в космическое пространство (рис. 11). В результате па по-
верхность Земли поступает солнечной энергии в среднем
5-1020 ккал/год, в том числе на континенты— 1,4-1020, на Мировой
океан—3,6-1020 ккал/год.
•Поступающая из космоса на земную поверхность солнечная ра-
диация имеет огромное значение в природе, ибо это основной
источник энергии на Земле, Другие источники энергии (гравита-
ционные приливы, обусловленные вращением Земли, энергия, по-
ступающая с магнитно-гидродинамическими волнами из ближай-
шего космоса и пр.) ничтожно малы. Солнечная радиация оказы-
вает то или иное воздействие на все процессы, протекающие на
земной поверхности и в ее воздушном окружении. Прежде всего,
она определяет световой и тепловой режимы местности, а путем
воздействия на движение воздушных масс, следовательно, влияет
и на выпадение осадков; она определяет режим увлажнения или,
иначе говоря, водный, гидрологический режим местности. Солнеч-
ная радиация прямо или косвенно воздействует на многие химиче-
ские реакции и физические явления, в частности, путем нагревания
поверхности суши и водных пространств влияет на физическое
испарение влаги и, следовательно, на круговорот воды на Земле.
Ею определяются и многие биологические процессы, и в первую
очередь такие жизненно важные, как фотосинтез и транспирация
растений и пр.
110
СОЛНЕЧНАЯ РАДИАЦИЯ, ПОСТУПАЮЩАЯ
падстилки
РАССЕЯННАЯ
РАДИАЦИЯ НЕБА
ТУРБУЛЕНГ
* ОБМЕН
/мж
Поверхность
ПОД СТИРАЮЩАЯ
Рис. 11. Баланс солнечной радиации (по А. А Молчанову, 1964)
л-
В процессе фотосинтеза зелеными автотрофными растениями
органических веществ из неорганических используется далеко не
вся поступающая на их листья солнечная энергия, а лишь только
физиологически активная часть ее, называемая фотосинтетически
активной радиацией (ФАР)^Она составляет в прямых солнечных
лучах около 35% от поступающей энергии, в рассеянных — 50—
60%. В пределах СССР количество ФАР за вегетационный период
(при температуре выше 5—10° С) колеблется по зонам от 1,5 до
4—5 млрд ккал/га коэффициент использования фототрофными
растениями лучистой энергии Солнца на процессы фотосинтеза
очень низок: в среднем 1% от полной радиации и около 3% от
120
ФАР. При этом важно иметь в виду, что поскольку различные
лучи солнечного спектра обладают разными свойствами, то, будучи
поглощенными растениями, они оказывают неодинаковое воздей-
ствие на них. Так, оранжево-красные лучи спектра наибольшую
активность проявляют в процессах фотосинтеза органических ве-
ществ зелеными растениями, сине-фиолетовые лучи участвуют в
синтезе белков и затормаживают переход растений в фазу цвете-
ния, ультрафиолетовые лучи предотвращают вытягивание стеблей
растений, а самые короткие из них гибельны для растений, длин-
новолновые инфракрасные лучи положительно влияют на тепловой
режим растений.
Световая энергия солнечной радиации необходима и для суще-
ствования животного населения биогеоценозов. Немногие живот-
ные, как известно, могут жить в отсутствие света —это обитатели
пещер, морских глубин, почвообитающие животные. У них разви-
ваются органы свечения или наблюдается усложнение глаз для
улавливания слабых количеств лучистой энергии, а то и просто
органы зрения за ненадобностью полностью атрофируются; у дру-
гих животных развиваются компенсационные органы (слух, осяза-
ние, обоняние). Интенсивность освещения некоторыми животными
воспринимается как сигнальный фактор, побуждающий к активной
деятельности утром дневных насекомых, птиц, вечером — ноч-
ных ц_Т- п.
Солнечная радиация, как было отмечено, является источником
не только света, но и тепла на земной поверхности, но не столько
путем непосредственного воздействия на тепловой режим нижних
слоев атмосферы, сколько в виде теплового излучения нагретой
солнечными лучами поверхности суши или воды. Дело в том, что
лишь незначительное количество поступающей на Землю солнеч-
ной радиации, превращаясь в тепло, расходуется на испарение
воды, большая же часть ее идет на нагрев земной поверхности, ко-
торая и излучает тепло в атмосферу. Некоторое значение в повы-
шении температуры воздуха имеет и так называемый турбулент-
ный (беспорядочный) теплообмен в самой атмосфере.
В нагревании воздуха атмосферы принимает участие как пря-
мая, так и рассеянная радиация Солнца. Однако степень притока
тепла на поверхность Земли и нагревания воздуха неодинакова в
различных пунктах земного шара. Это находится в зависимости
прежде всего от преобладания прямой или рассеянной радиации,
а также от высоты стояния Солнца над горизонтом и интенсивно-
сти солнечной радиации; имеет значение и ход циркуляции атмо-
сферы и ее прозрачность, а также господствующие формы рельефа
в данной местности и экспозиция склонов при изрезанном рельефе
и т. п. В любой местности, за исключением тропиков, тепловой ре-
жим неравномерен в течение года, он колеблется по временам года
121
с более или менее ярко выраженным чередованием теплого перио-
да года с холодным. Особенно четко периодичность в смене времен
года проявляется в странах умеренного пояса, где колебания тем-
пературы по временам года обычно бывдцэт довольно резкими, а
амплитуда колебаний весьма значительной^ Неравномерность теп-
лового режима в таких местах бывает достаточно хорошо выра-
жена и по годам, когда происходит чередование группы лет, харак-
теризующихся преобладанием теплого режима в течение большей
части года с группой лет, отличающихся преобладанием холодного
режима (с учетом, разумеется, в том и другом случае общей сум-
мьцюдовых температур).
Состояние теплового режима атмосферы, безусловно, имеет
большое значение не только в жизни живых существ, но и в функ-
ционировании биогеоценоза и биогеосферы в целом, так как нет в
природе процессов, которые не были бы связаны с тепловым ре-
жимом местности и не находились бы в той или иной зависимости
от него^ Тепловой режим оказывает существенное влияние на жиз-
недеятельность растений и растительных сообществ, в частности
на фотосинтез, дыхание, транспирацию и минеральное питание
растений, а следовательно, на их рост и развитие.^ Вследствие это-
го, как указывает А. А. Молчанов (1964), тепловой режим опреде-
ляет северные и верхние (в горах) пределы распространения
ценозообразующих растений и фитоцепозов, а также широтную
зональность и вертикальную поясность растительного покрова. От-
рицательно сказывается на состоянии, жизнедеятельности и про-
дуктивности растений и растительных сообществ резкая смена
заморозков оттепелями и обратно, и особенно весною. Это приво-
дит к образованию морозобойных трещин на стволах деревьев и
кустарников, обмерзанию молодых побегов и цветочных почек,
снижению продуктивности фитомассы и плодоношения, а иногда и
к гибели, по крайней мере, травянистых растений, в особенности
озимых зерновых, вследствие образования кристалликов льда в
клетках и последующего разрыва последних.
Тепловой режим сказывается определенным образом и па жиз-
недеятельности животного населения биогеоценозов. Он оказывает
влияние на состояние, размножение и развитие животных, а сле-
довательно, и на численность и распространение их в природе. Не-
посредственное воздействие температуры воздуха на животных
осуществляется путем поглощения тепла их телом и отдачей его
во внешнюю £реду Теплообмен у животных, как отмечает
А. А. Молчанов, регулируется с помощью рефлекторной деятельно-
сти нервной системы. Известны два типа теплообмена у животных:
а) холоднокровность, характеризующаяся неустойчивостью уровня
обмена и непостоянством температуры тела (земноводные, пресмы-
кающиеся и др.), которая мало отличается от температуры окру-
122
жающей среды, и б) теплокровность, отличающаяся высоким и
устойчивым уровнем обмена, регуляцией его и относительным по-
стоянством температуры тела животных. Имеется и промежуточ-
ная по характеру теплообмена группа животных, которые в силу
этого в неблагоприятные для жизни периоды (зимние морозы, лет-
няя засуха) впадают в длительную спячку либо в относительно
короткий, но глубокий сон. М. П. Акимов (1959) различает сле-
дующие градации состояния скрытой жизни животных в условиях
зимнего снижения температуры и ухудшения условий питания:
а) зимний покой некоторых млекопитающих (м'едведи, барсуки,
белки в норах, логовах), холоднокровных (пчелы в ульях); б) на-
стоящую зимнюю спячку млекопитающих, сопровождающуюся по-
нижением температуры тела и общим окоченением; в) зимнее
окоченение холоднокровных позвоночных и беспозвоночных живот-
ных с возможным переохлаждением тела и весьма слабым обме-
ном веществ. Наряду с этим известны и другие способы пережива-
ния неблагоприятного сезона года: а) в стадии яйца, куколки (на-
секомые и др.); б) сезонные миграции, кочевки в области с более
благоприятными условиями тепла, влаги, корма (некоторые мле-
копитащие, птицы); в) постройка убежищ (гнезд в дуплах деревь-
ев, мор и т. п.). Известны и летние спячки животных в жарких
засушливых областях земного шара (пустыни, саванны и т. п.).
Растения, кстати сказать, также, по наблюдениям В. В. Крюч-
кова (1960), обладают свойственным им внутренним тепловым
(и водным) режимом, своей энергетикой, которая пока еще слабо
изучена.
Таким образом, с наступлением неблагоприятного для жизни
периода года (зимнего холодного или жаркого сухого) понижается
жизнедеятельность растений, животных и даже микроорганизмов;
замедляются также многие физические процессы и химические ре-
акции в природе; нарушаются или затормаживаются обменные
процессы в биокосных системах разного ранга. Периодические ко-
лебания в тепловом режиме — суточные, сезонные, разногодние —
также сказываются на состоянии как живых, так и косных компо-
нентов биогеоценозов и самих биокосных систем.
Атмосферные осадки — второй весьма важный элемент воздуш-
ной оболочки Земли — атмосферы. Образуются атмосферные
осадки из водяных паров как за счет местного испарения влаги,
так и за счет влаги, принесенной с воздушными массами из других
областей земного шара, обычно из окружающих материки океани-
ческих водных пространств. Правда, из транспортируемой воздуш-
ными массами влаги, например, в европейской части Советского
Союза, по данным А. А. Молчанова (1964), выпадает лишь около
37% осадков от их общего количества. Но ввиду большой массы
транспортируемой влаги в конечном итоге основная масса атмо-
123
сферных осадков выпадает за счет ее; за счет местного испарения
осадков выпадает в 3 раза меньше, чем за счет влаги, принесенной
извне.
Атмосферные осадки выпадают в виде дождя, снега, града, ро-
сы, инея, изморози и т. п. Общее количество выпадающих в раз-
личных областях земного шара атмосферных осадков, по тем же
данным, колеблется в больших пределах: от 0 до 5000—10000 (и
даже 12000) мм в год.
В пределах Советского Союза годовое количество атмосферных
осадков колеблется от 50 до 150 мм в среднеазиатских пустынях
и сухих степях Заволжья, до 2000—2600 мм — на Черноморском
побережье Кавказа и Закавказья. В средней полосе европейской
части СССР осадков выпадает 500—600 мм в год, а в азиатской
части от 400—500 мм в Западной Сибири до 150—300 мм в Восточ-
ной Сибири. На Дальнем Востоке годовая сумма осадков дости-
гает_800—1000 мм (А. А. Борисов, 1948).
(^Выпадают атмосферные осадки, как правило, неравномерно по
сезонам вегетационного периода и года. |Гак, для тропических об-
ластей характерны два периода дожДёЙ, один из которых начина-
ется в разгар лета, и два сухих периода между ними. В областях
на широте близ 35-й параллели обоих полушарий типичны зимние
дожди и сухое лето. В местностях севернее 50-й параллели осадки
выпадают во все сезоны года, но наибольшее количество их внутри
материков выпадает летом, в океанах — зимой, а на побережьях,
преимущественно западных, — осенью. На большей части террито-
рии Советского Союза основная часть годового количества осадков
приходится на вторую половину лета и осень, весна и первая по-
ловина лета отличаются недостатком атмосферных осадков.
( Поступающая на Землю в виде жидких и твердых осадков вода
опадает весьма важными свойствами. Прежде всего, вода нахо-
дится в природе, как известно, в трех физических состояниях —
твердом, жидком и газообразном — и обладает способностью пере-
ходить из одного состояния в другое; она обладает большой по-
движностью, особенно в жидком и газообразном состояниях; вода
является универсальным растворителем разнообразных веществ,
что повышает их транспортабельность и химическую реактивность;
она способна переносить различные вещества в растворенном и
взмученном состояниях на большое расстояние.
Вода, как известно также, совершает в биосфере круговорот
(влагооборот), охватывающий нижнюю часть атмосферы (тропо-
сферу), литосферу (земную кору с почвенным и растительным по-
кровом) и гидросферу (Мировой океан). Он является основой так
называемого большого геологического круговорота веществ. Вла-
гооборот, по данным А. А. Молчанова (1964, 1974), слагается из
следующих звеньев: а) выпадение атмосферных осадков в твердом
124
в виде грунтовых и артезианских вод.
Рис. 12. Круговорот воды в биогеоценозах
(по* А. А. Молчанову, 1964)
и жидком состояниях на земную поверхность; б) задержка влаги
растениями всех ярусов фитоценоза, а также подстилкой, войло-
ком, испарение ее с поверхности древостоя (травостоя) и подстил-
ки, сток влаги по стволам (стеблям) растений; в) проникновение-
влаги до поверхности почвы и физическое испарение ее, а также
сток воды в низины, ложбины и в долины рек (поверхностный и
русловой сток); г) просачивание влаги в почву и использование
ее растениями, животными, микроорганизмами на процессы жиз-
недеятельности, а растениями в том числе и на транспирацию;
д) инфильтрация влаги из почвы в грунт до водоупорного гори-
зонта, внутрипочвенный и грунтовый сток ее и выход в подножии
склонов на дневную поверхность в виде ключей или накопление'
в подземных резервуарах
(рис. 12).
Количественные пока-
затели перечисленных пу-
тей передвижения (транс-
портировки) воды в био-
сфере сильно варьируют
в зависимости от количе-
ства и интенсивности ат-
мосферных осадков, тем-
пературы и движения воз-
духа (ветра), рельефа
местности и физических
свойств почвы, состава и
структуры растительного
покрова.
Таким образом, влага,
поступившая из атмосфе-
ры в биогеоценотический
покров Земли, вновь воз-
вращается в атмосферу
путем физического испа-
рения ее с поверхности
почвы, мертвого напоч-
венного покрова, откры-
тых водоемов путем
транспирации ее растениями и т, п. В этом и состоит круговорот
воды (влагооборот) в биосфере (рис. 13).
Наличие воды в том или ином физическом состоянии в биосфе-
ре имеет исключительно большое значение как в жизни организ-
мов, так и в состоянии и функционировании биогеоценозов и
биогеоценотического покрова Земли. Эта разносторонняя и важ-
нейшая роль воды в природе определяется теми свойствами, кото-
125
Рис. 13. Круговорот воды б биосфере (Clodius, Keller, 1951)
(по П. Дювиньо и М. Тангу, 1968). I — поверхностный
сток; И — подземный сток; III — просачивание. Количества
воды указаны в миллиметрах от среднегодовой нормы
осадков
рыми она обладает, а также глобальной циркуляцией ее в биосфе-
ре в виде влагооборота, состоящего из целого ряда взаимосвязан-
ных звеньев. Вода в состоянии водяных паров в атмосфере
определяет влажность воздуха — абсолютную, относительную и
так называемый дефицит влажности (разность между упругостью
насыщения и фактической упругостью водяного пара в воздухе
при данной температуре и давлении). Влажность воздуха оказы-
вает влияние на все компоненты биогеоценозов, но особенно тесно
связана с растительностью и почвой, оказывая сильное воздействие
на влагообмен их с атмосферой. Особенно большое значение в
природе имеет дефицит влажности, который оказывает прямое
влияние на физическое испарение воды, на транспирацию влаги
растениями, а при сочетании с высокой температурой может вы-
звать завядание и гибель растений. Правда, у растений, произрас-
тающих в сухих и жарких областях, выработался в процессе эво-
люции целый ряд приспособлений — анатомо-морфологических
(толстая кутикула, войлочное опушение, восковой налет на лнсть-
126
ях, свертывание и складывание листьев, развитие механической
ткани, суккулентность некоторых растений), а также физиологиче-
ских (высокое осмотическое давление клеточного сока, мощность
корневой системы и способность корней быстро и полно улавли-
вать воду, интенсивная транспирация влаги при известной обеспе-
ченности ею и пр.).
На животный мир недостаточная влажность воздушной среды
оказывает неблагоприятное воздействие. Но у животных, как и у
растений, также имеются разного рода приспособления, обеспечи-
вающие выживание их в условиях сухого и жаркогр климата: раз-
витие малопроницаемых покровов (ороговение эпителия, хитино-
вый панцирь, раковины, перья, чешуи и т. п.), а также развитие
внутренних органов дыхания (легкие, легочные мешки, трахеи)»
образование специальных органов выделения влаги и пр.
Вода, выпадающая на поверхность Земли в виде атмосферных
осадков, прежде всего оказывает непосредственное воздействие на
все живые компоненты биогеоценозов, обеспечивая их влагой, ко-
торая жизненно необходима им, тем более что тело их на 50—98%
состоит из воды; во-вторых, атмосферные осадки оказывают воз-
действие на почвенный покров, а следовательно, и на органический
мир путем растворения различных минеральных веществ в почве,
перемещения их в вертикальном и горизонтальном направлениях,
обогащения почвы органическими и минеральными веществами,
принесенными с водой из атмосферы или смытыми с поверхности
растений, путем изменения температурного режима почвы и ее
аэрации; в-третьих, количеством, интенсивностью и продолжитель-
ностью атмосферных осадков, а также физическим испарением
воды и транспирацией ее растениями определяется гидрологиче-
ский (водный) режим поверхностных и грунтовых вод местности
в пределах водосборного бассейна.
Рассмотрим третий составной элемент атмосферы — газовый
состав. Выше при общей характеристике атмосферы как воздушной:
оболочки нашей планеты были приведены данные о газовом соста-
ве современной атмосферы; при этом было подчеркнуто биогенное
происхождение входящих в ее .состав газов, а также постоянство
газового состава атмосферы. Vlo начала бурного развития всех
отраслей производства в связи с наступлением научно-технической
революции газовый состав атмосферы действительно был неизмен-
ным (азота — 78,08%, кислорода — 20,95, аргона — 0,94, углекис-
лоты и пр. — 0,03%). Постоянство его в указанном по объему со-
отношении всех составных компонентов обеспечивалось тем, что
поглощение организмами кислорода и углекислого газа при осу-
ществлении разнообразных жизненных процессов компенсирова-
лось выделением их в ходе других процессов (фотосинтеза, дыха-
ния и т. п.).
127
Однако за последние полвека, по утверждению Ф. Ф. Давитая
(1971), содержание в составе атмосферы биогенных газов и их
процентное соотношение в той или иной мере изменились. Это, по
его данным, обусловлено следующими обстоятельствами. Количе-
ство свободного кислорода в атмосфере в настоящее время состав-
ляет 1,5-1015 т. За все время существования человечества на сжига-
ние топлива израсходовано кислорода (по заниженным расчетам)
273 млрд т, из которых 246 млрд т—за последние 50 лет; за счет
частичного использования кислорода за последние полвека обра-
зовалось углекислого газа 288 млрд т, что увеличило содержание
его в воздухе на 12%. Следует принять во внимание и то, что в
начале 70-х годов расход кислорода на сжигание топлива во всем
мире ежегодно составлял 10 млрд т, а углекислого газа выделя-
лось в атмосферу 14 млрд т. В дальнейшем мировое потребление
кислорода будет возрастать в среднем на 10% ежегодно; будет
увеличиваться и образование углекислого газа, что, очевидно, при-
ведет к дальнейшему изменению газового состава атмосферы.
Различные по своим природным свойствам и процентному со-
отношению в составе атмосферы биогенные газы оказывают раз-
личное воздействие на живые компоненты и почвенный покров
биогеоценозов и играют неодинаковую роль в функционировании
биокосных систем. Роль газообразного азота, несмотря на высокий
процент содержания его в воздухе, в жизни организмов и проте-
кающих в биогеоценозе процессах невелика, так как непосред-
ственно он усваивается лишь азотфиксирующими бактериями. В
почву же азота, преимущественно в виде аммиака, поступает чрез-
вычайно мало — несколько кг/га в год.
Гораздо большее значение в жизни организмов и обменных
процессах биогеоценозов по сравнению с газообразным азотом, как
отмечает А. А. Молчанов (1964), имеют кислород и углекислый
газ. Кислород используется, как известно, всеми живыми суще-
ствами на дыхание во всех средах жизни — воздушной, почвенной,
водной; он вступает во множество химических реакций в живой
природе, участвует в минерализации органических веществ в био-
геоценозах и биогеосфере. Особенно сильно ощущается нужда в
кислороде для дыхания животными, ибо не многие из них могут
некоторое время пребывать в анаэробных условиях (черви в иле,
некоторые моллюски в грунте водоемов, личинки комаров в воде).
Кислород в теле животных обеспечивает окислительные процессы
с освобождением энергии, используемой ими на процессы своей
жизнедеятельности. Разные животные по-разному реагируют на
содержание кислорода в среде обитания: более стойки к недостат-
ку кислорода птицы, особенно перелетные, а также подземные и
водные млекопитающие. У некоторых животных развиваются спе-
циальные приспособления к обитанию в анаэробных условиях: у
128
китов высокое содержание гемоглобина в крови, альвеолы легких
запираются специальными мускулами для удержания воздуха в
них па глубине, у кашалота образуется запасной воздушный мешок
и г. п. К обитанию в замкнутых убежищах (в почве, ходах под
корой и древесине) приспособились и беспозвоночные животные.
Однако потребляется кислород организмами и расходуется в раз-
личных процессах неживой природы необратимо, лишь зеленые
растения способны не только поглощать его при дыхании, но и вы-
делять при фотосинтезе.
Из сказанного следует, что главным источником углекислого
газа & атмосфере является дыхание животных, растений и гниение
их органических остатков и отбросов, а также так называемое поч-
венное «дыхание», т. е. выделение углекислоты в процессе разло-
жения органических веществ в почве микроорганизмами Исполь-
зуется же углекислый газ как источник углерода только Зелеными
автотрофными растениями на процессы фотосинтеза органических
веществ. Количество углекислого газа в атмосфере, несмотря на
низкий процент содержания его, в общем достаточно для обеспе-
чения потребностей в нем. Но в связи с неравномерностью потреб-
ления углекислоты растениями в течение суток и по сезонам в
отдельные периоды имеет место недостаток ее, что ведет к сни-
жению интенсивности фотосинтеза растений.
Углекислый газ выполняет также весьма важную планетарную
функцию в биосфере, а именно регулирует температуру воздуха
на поверхности Земли, пропуская тепловые лучи из космоса на
Землю, но задерживая тепло на ее поверхности, препятствуя рас-
сеиванию его в космическом пространстве.^
Атмосферный воздух, окружающий сплошным мощным слоем
поверхность пашей планеты, далеко не везде и не всегда находит-
ся в первозданно чистом состоянии. Обычно он бывает в той или
иной мере загрязнен мельчайшими твердыми частицами различ-
ного происхождения (вулканического, транспортного, промышлен-
ного, бытового) либо газами, большей частью вредными, также
различного происхождения (окисью углерода, сернистым газом,
сернистым ангидридом, хлором и пр.).
Как твердые, так и газообразные примеси или включения в
состав естественной воздушной оболочки Земли оказывают, как
правило, неблагоприятные воздействия на живые компоненты био-
геоценозов, в особенности на растительный покров. По словам
А. А. Молчанова (1964), они отрицательно влияют на содержание
хлорофилла в вегетативных органах растений, вызывают увядание
растений или задерживают их рост, задерживают распускание по-
чек и развитие листьев у деревьев, уменьшают прирост древесных
пород, вызывают повреждения и даже гибель их. Особенно сильно
< 'Радают от различных вредных газов хвойные природные и ис-
129
кусственные лесные насаждения. Правда, растительность, в осо-
бенности лесная, содействует очищению воздуха от пыли и газов,
но за счет нарушения своего нормального состояния, снижения
жизненности и продуктивности.
Четвертый и последний составной элемент воздушной оболочки
Земли — это общая циркуляция атмосферы, представляющая собою
систему или совокупность крупномасштабных воздушных течений
над поверхностью земного шара. Она включает перемещение воз-
душных масс в горизонтальном направлении (ветер) и в верти-
кальном (конвекция). Основная причина циркуляции воздушных
масс по лику Земли и на ограниченных участках ее поверхности —
это неравномерность нагревания атмосферы за счет теплового
излучения солнечной радиации, а также разность атмосферного
давления на разных шпротах земного шара, с одной стороны, на
материках и в океанах — с другой. Обычно различаются следую-
щие типы воздушных течений; а) основные зональные переносы
воздуха, например западный перенос в тропосфере и стратосфере
умеренных широт и восточные ветры в нижней части тропосферы
тропических широт; б) возмущения зональных переносов, вызы-
ваемые влиянием суши и моря; в) крупные атмосферные вихри
(циклоны и антициклоны). Общая циркуляция атмосферы меняет-
ся в годовом цикле — по сезонам и в многолетнем — от года к году
(по годам).
Наряду с общей циркуляцией атмосферы известны также мест-
ные циркуляции ее, охватывающие сравнительно небольшие участ-
ки территории или акватории, например бризы, горно-долинные
ветры и т. л. Они обусловлены специфическими особенностями
самих этих участков (резким контрастом температуры, некоторыми
особенностями рельефа и т, п.).
На основе закономерностей общей циркуляции атмосферы в
Северном и Южном полушариях формируется система шпротно-
зональных типов воздушных масс; а) арктический воздух; б) по-
лярный (умеренный) воздух; в) тропический воздух; г) экватори-
альный воздух (общий для обоих полушарий). Названные типы
воздушных масс разделяются неширокими (в несколько десятков
километров) пограничными полосами, получившими название клн-
матические фронты.
Таким образом, общая циркуляция атмосферы — явление обще-
планетарного порядка. Отсюда и воздействие ее на природные
процессы также носит общепланетарный характер. Так, общая
циркуляция атмосферы, по словам А. А. Молчанова, а) играет ре-
шающую роль в формировании климатов земного шара; б) опре-
деляет погоду и ее суточные и сезонные изменения; в) смягчает
контрасты тепла и холода; г) выравнивает газовый состав воздуха
и т. п. Тем не менее местные циркуляции воздуха, обусловленные
130
местными же особенностями территории, оказывают влияние на
ход тех или иных процессов в пределах не только более или менее
крупных биогеоценотических массивов, но и отдельных биогеоце-
нозов Так, по свидетельству А. А. Молчанова (1964), ветер в лес-
ном "биогеоценозе усиливает физическое испарение и транспирацию
влаги растениями, способствует внутреннему и внешнему газооб-
мену; влияет на развитие корней и сбежистость стволов; иногда
обнажает корни растений, повреждает всходы и молодые посадки,
вызывает буреломы и ветровалы; зимой способствует накоплению
снега на лесных опушках, полянах, в понижениях рельефа; уча-
ствует в опылении некоторых растений и распространении семян
и плодов и пр.
После рассмотрения всех элементов воздушной оболочки Зем-
ли — атмосферы — и воздействия каждого из них на компоненты
и те или иные стороны биогеоценозов и биогеосферы следует поды-
тожить роль атмосферы в целом в жизни и функционировании
биокосных систем.
Н. В. Дылис (1978) так характеризует роль атмосферы в жиз-
ни и деятельности биогеоценозов и биогеосферы, которая проявля-
ется в трех направления^ Прежде всего, атмосфера участвует во
всех превращениях веществ и энергии на пашей планете, оказывая
влияние на все звенья биогеоценотнческого процесса на земной по-
верхности, на его масштабы, направление, ритм и конечный резуль-
тат erqf Во-вторых, атмосфера воздействует на все компоненты
биогеоценозов, в особенности на растительность, являющуюся
экраном, воспринимающим вещества и энергию из атмосферы и
ближайшего космоса. Так, она определяет физико-химическое вы-
ветривание горных пород, влияет на почвообразовательный процесс
и физико-химические свойства почв; она накладывает значитель-
ный отпечаток на экобиоморфный состав растений и животных,
характер и степень участия их в биогеоценотическом процессе,
режим и интенсивность биогеохимической работы растительности,
миграцию животных, активизацию и оцепенение их в определенные
сезоны года и т. п! В-третьих, атмосфера, по словам Н. В. Дылиса,
является условием существования всей биогеосферы Земли; по-
средством так называемого озонового экрана — слоя воздуха
с повышенной концентрацией молекулярного кислорода на высоте
22—25 км — защищает все живое на Земле от губительного дей-
ствия ультрафиолетовых и других излучений Солнца, а посред-
ством углекислого газа предохраняет земную поверхность от излу-
чений тепла в космическое пространство; она служит проводником
солнечной энергии в биогеосферу и является средоточием огром-
ных материальных ресурсов биогеосферы — кислорода, углекислого
газа, атмосферной влаги и пр.
131
3. Преобразование атмосферы
во внутреннюю воздушную среду биогеоценоза
Однако связь атмосферы с почвой и живыми компонентами
биогеоценоза является обоюдной, последние со своей стороны так-
же оказывают то или иное влияние на воздушную оболочку Земли.
Особенно существенное и разностороннее воздействие на деятель-
ный слой атмосферы оказывают растительность и почвенный по-
кров. В пределах биогеоценоза это воздействие фитоценоза и
почвы на приземной слой воздушной среды является настолько
сильным, чю коренным образом изменяет световом и тепловой ре-
жимы и режим влажности воздуха в надземной части биогеоцено-
за, тепловой и водный режимы и режим минерального питания
растений в подземной (почвенной) части его. В результате в пре-
делах биогеоценоза происходит постепенное преобразование мест-
ного климата в особый, специфичный для данного биогеоценоза
внутренний климат, называемый фитоклиматом.
Под климатом данной местности, по определению М. И. Щер-
баня (1968), следует понимать «многолетний режим погоды, фор-
мирующийся под воздействием солнечной радиации, земной по-
верхности и связанной с ними циркуляцией атмосферы...» (с. 9).
В зависимости от масштабов рассматриваемых частей поверхности
Земли различают еще мезо климат, или местный климат —
климат сравнительно небольших участков земной поверхности. В
качестве примеров приводится климат леса, морского побережья,
города. Наконец, микроклимат — климат малого участка вну-
три географического ландшафта, например климат склона холма,
опушки леса, городской площади и т. п. Особенности микроклима-
та проявляются обычно в приземных слоях воздуха, поскольку они
определяются мелкими специфическими особенностями данного
участка.
В чем же выражается воздействие живых компонентов и почвы
биогеоценоза на деятельный слой атмосферы? Каким образом про-
исходит видоизменение, преобразование составляющих местный
климат элементов и конкретных метеорологических факторов ат-
мосферы во внутреннюю среду биогеоценоза? Как преломляются
эти факторы в биогеоценозах и в каких свойствах проявляется
видоизменение местного климата в фитоклимат?
Прежде всего, об изменении солнечной радиации и ее интенсив-
ности в биогеоценозах. Ярче всего это проявляется в лесных био-
геоценозах. Так, суммарная солнечная радиация под пологом дре-
весных пород лесного биогеоценоза состоит пе только из прямых
солнечных лучей (в виде бликов, пятен) и рассеянных лучей, отра-
женных от неба и облаков, как на открытых местах, но также и
из лучей, отраженных вниз кронами деревьев и лучей, прошедших
132
сквозь листья. Интенсивность прямого света под пологом древо-
стоя, по данным А. А. Молчанова (1964), в 2—4 раза слабее, чем
на смежных открытых участках, ибо происходит поглощение сол-
нечных лучей, отражение части из них нижними ярусами и пр.
Рассеянная радиация в лесном биогеоценозе беднее активными
для фотосинтеза лучами, чем на открытом месте, да и общая ин-
тенсивность ее меньше во всех частях спектра. Интенсивность сол-
нечной радиации, по тем же данным, находится в зависимости от
ярусного строения лесного биогеоценоза. Так, суммарная радиация
в сосновом лесу, принятая над кронами древостоя за 100%, под
кронами составляет лишь 30%, над травяным покровом—10, в
травяном ярусе близ поверхности почвы—менее 5%. Изменяется
она и с возрастом ценозообразующих древесных пород. Так, в ду-
бовом лесу суммарная радиация под пологом древостоя в 20 лет
составляет 20% от полной, в 70 лет — 4, а в 230 лет — 6%.
В кронах деревьев отдельные листья, как указывает В. Лархер
(1978), неодинаково обеспечиваются световой энергией в зависи-
мости от формы кроны и густоты листвы, от положения листьев
относительно лучей света и их места внутри кроны, следствием
чего является «характерный градиент радиации от хорошо осве-
щенной периферии кроны через зоны снижающейся освещенности
до внутренней части кроны» (с. 20). Степень освещенности, изме-
ряемая в люксах, внутри древостоя, по данным А. А. Молчанова,
находится в зависимости от состава древесных пород и их возра-
ста, от сомкнутости крон и их высоты от поверхности почвы, от
экспозиции склона и пр. В хвойных лесах освещенность, по утвер-
ждению Р. Дажо (1975), сильно ослаблена, но качество мало
изменяется. В лиственных же лесах она подвергается избиратель-
ному поглощению, отсюда желто-зеленый оттенок их. Степень
освещенности под пологом древостоя отражается на состоянии»
развитии и продуктивности нижних ярусов. В густых биогеоцено-
зах, по наблюдениям В. Лархера, благодаря многократному отра-
жению и поглощению солнечных лучей до почвы доходит так мало
света, что так называемое световое довольствие (т. е. освещен-
ность, выраженная в процентах от общего количества поступаю-
щего извне света) составляет лишь несколько процентов по отно-
шению к освещенности открытой местности. Поэтому в составе
подлеска, например, при относительном световом довольствии
5—20% могут произрастать лишь теневыносливые растения.
Отличается солнечная радиация и в травяных биогеоценозах
(луговых, степных, травяно-болотных) по сравнению с таковой на
открытых местах, поскольку в них также происходит и поглощение
солнечных лучей ценозообразующими растениями, и пропускание
их через листья, и отражение части радиации напочвенным ярусом
ценоза и пр. Свет, проходя через травостой, претерпевает значи-
133
тельные изменения, ибо лучи видимой радиации, по данным
Р. Дажо, поглощаются гораздо сильнее, чем инфракрасные лучи,
а в пределах видимой части спектра сильнее поглощаются крас-
ные и синие и слабее — зеленые. Вследствие этого происходит зна-
чительное снижение лучистой энергии: так, в степи ее поток над
травостоем, по его данным, составляет 1,08 кал ——, а на поверх-
см2 МИН
ности почвы — лишь 0,19 кал ------. Это обстоятельство сказы-
мин
вается и па распространении насекомых, живущих под раститель-
ным покровом, и пр.
Солнечная радиация, как было сказано ранее, является источ-
ником не только энергии света, но и тепла. Около 20% световых
лучей при прохождении через атмосферу поглощается находящи-
мися в пей водяными парами, непосредственно участвуя в нагре-
вании воздуха, но большая часть световых лучей поступает на
земную поверхность, которая, нагреваясь сама, нагревает и воздух
посредством излучения тепла в атмосферу.
Тепловой режим биогеоценозов, в особенности лесных, подобно
световому, значительно отличается от такового смежных открытых
пространств, так как прогревание воздуха и поверхности почвы
под пологом древесного яруса заметно снижается ввиду излучения
тепла с поверхности крон деревьев. Средняя годовая температура
в лесу, по данным Р. Дажо (1975), примерно на 1,5°С ниже^ по
сравнению с соседними безлесными участками. Днем самыми теп-
лыми являются кроны деревьев, но в течение суток температурный
максимум в пределах крон перемещается: с верхней части крон на
восходе солнца в их среднюю часть в 13 ч и снова на верхушки
деревьев вечером, так как днем солнечные лучи глубже всего про-
никают в древесный полог; ночью же температура в лесу на всех
уровнях древостоя почти одинакова. Разумеется, преобразование
теплового режима под древесным пологом неодинаково в различ-
ных типах лесных биогеоценозов, так как находится в зависимости
от видового состава ценозообразующих древесных пород, их воз-
раста и от ярусного строения биогеоценозов. А. А. Молчанов обра-
щает внимание и на то обстоятельство, что преобразующее влия-
ние лесного биогеоценоза распространяется и на тепловой режим
смежных участков — лесных полян, сплошных вырубок, просек на
расстояние до 50—100 м от опушки леса.
Известное влияние на изменение теплового режима в своих
пределах оказывают и травяные биогеоценозы. Температура в тра-
востое луговых биогеоценозов, по данным Р. Дажо, находится в
зависимости от характера растительности, высоты и плотности
травостоя и ориентации листьев по отношению к поверхности поч-
вы. Так, летом на лугу с густым (плотным) травостоем темпера-
турные различия между его верхним пологом и поверхностью
почвы меньше, чем на лугу с разреженным осветленным травосто-
134
ем. Осенью температуры в них выравниваются. На лугу темпера-
тура понижается по вертикальному профилю травостоя свер-
ху вниз.
Что же касается видоизменения режима атмосферных осадков
и влажности воздуха в условиях биогеоценозов и биогеоценотиче-
ского покрова Земли, то следует иметь в виду, что выпадение
осадков, как было сказано ранее, определяется факторами более
общего планетарного и даже космического характера (солнечная
радиация и связанное с этим нагревание поверхности материков
н океанов, циркуляция атмосферы и транспортировка влаги по
земной поверхности и пр.). Однако известное значение в этом име-
ет и так называемая подстилающая поверхность мыгности, а имен-
но морфология и геологическое сложение данной местности, ха-
рактер растительного и почвенного покрова и пр., определяющие
в свою очередь поверхностный сток и инфильтрацию воды в грунт,
физическое испарение воды и транспирацию ее растениями, а сле-
довательно, и влажность воздуха и почвы.
В лесах годовое количество атмосферных осадков, по данным
Р. Дажо (1975), примерно на 17% выше, чем в безлесных местно-
стях, но в лесных биогеоценозах значительное количество осадков
задерживается кронами деревьев. Так, в хвойном лесу задержи-
вается от 20 до 60% выпадающих осадков, а в лиственном — еще
больше. Задерживаются атмосферные осадки и верхним пологом
травостоя на лугах. Физическое испарение и интенсивность транс-
пирации влаги в биогеоценозах обычно меньше, чем на открытых
пространствах, отчасти в связи с уменьшением скорости ветра. Эти
процессы находятся в. зависимости от типа биогеоценоза, его струк-
туры и густоты (плотности) стояния древостоя или стеблестоя, от
сомкнутости крои лесных или общего проективного покрытия тра-
вяных биогеоценозов. Так, в сосняках черничных расход влаги на
транспирацию, по наблюдениям А. А. Молчанова (1964), больше,
чем в сосняках лишайниковых или сосняках сфагновых, в сосняках
с двухъярусным древостоем транспирируется влаги в 2 раза боль-
ше, чем в сосняках простых (одноярусных). Больше расходуется
влаги на транспирацию и в густых, сложных по составу биогеоце-
нозах по сравнению с разреженными. В связи с этим значительно
выше оказывается и гидрологическое значение таких биогеоцено-
зов по сравнению с биогеоценозами разреженными, простыми по
составу и структуре и, как правило, низкопродуктивнымн.
Режим влажности воздуха внутри биогеоценозов, так же как и
световой и тепловой режимы, видоизменяется, преобразуется под
действием живых компонентов биогеоценоза, значительно отлича-
ясь от режима влажности открытых смежных территорий. Так,
абсолютная и относительная влажность воздуха под пологом дре-
востоя и подлеска лесных биогеоценозов обычно выше влажности
135
наружного воздуха вследствие поступления водяных паров с по-
верхности почвы и крон деревьев и кустарников. Характерно и то,
что с утра влажность воздуха одинакова во всех ярусах биогеоце-
ноза, а днем постепенно падает от верхних ярусов к нижним. В
травяных биогеоценозах влажность воздуха также выше наружно-
го, и чем гуще травостой, тем выше относительная влажность воз-
духа; варьирует она и по вертикальному профилю травостоя: в
верхнем ярусе, по данным А. А. Молчанова, она составляет 55—
60%, а в нижних достигает 94—98%. Влажность воздуха внутри
травяных биогеоценозов изменяется и в течение светового периода
суток: утром и вечером она обычно выше, чем днем.
Биогеоценозы и биогеоценотический покров Земли оказывали
большое влияние на газовый состав атмосферы в геологическом
прошлом, оказывают и в настоящее время. Основную массу воз-
душной оболочки пашей планеты, как известно, составляют газы
биогенного происхождения: азот, кислород, углекислый газ. Совер-
шающийся на протяжении многих тысячелетий газообмен в био-
геосфере — результат жизнедеятельности живых компонентов
биогеоценозов, поглощающих продуцируемый автотрофными орга-
низмами кислород и выдыхающих углекислый газ, вновь исполь-
зуемый автотрофами на созидание первичной органической био-
массы в процессе фотосинтеза. Однако содержание наиболее
интенсивно используемых живыми компонентами кислорода и
углекислого газа неодинаково в различных биогеоценозах. Кон-
центрация углекислого газа, как свидетельствует А. А. Молчанов
(1964), выше в воздушной среде лесных биогеоценозов, чем в тра-
вяных; выше концентрация его в воздухе дубовых типов леса, чем
сосновых. В тропических лесных биогеоценозах концентрация угле-
кислоты выше, чем в таежных. Содержание углекислого газа в
воздушной среде биогеоценоза колеблется в течение суток с мак-
симумом ночью и минимумом днем, поскольку в дневные часы он
интенсивно используется растениями на процессы фотосинтеза.
Содержание углекислоты в биогеоценозе, безусловно, важный фак-
тор, отражающийся на продуктивности фотосинтеза и, следова-
тельно, на выходе органической массы, поскольку она является
основным источником углерода.
Животные в составе биогеоценоза, как известно, поглощая в
процессе дыхания кислород, обогащают воздух внутри биогеоце-
ноза углекислотой, оказывая тем самым влияние на соотношение
биогенных газов в его составе. Известную роль в этом отношении
играет и микрофлора, в особенности почвенная. Установлено, что
под пологом биогеоценозов в результате редуцирующей деятель-
ности микроорганизмов увеличивается содержание углекислого
газа в воздухе, в особенности в приземном слое его, что также ска-
зывается на газовом составе воздушной среды биогеоценоза.
136
Не остается в стороне и такой элемент атмосферы, как цирку
ляция воздуха. Горизонтальное, параллельное земной поверхност!
движение воздуха — ветер—под пологом лесных биогеоценозе!
видоизменяется в своем проявлении. Вследствие трения о древес
ные стволы и ветви, траты энергии на раскачивание стволов и т. п
ветер снижает свою скорость в лесном биогеоценозе. При плотно&
стоянии стволов древостой не только гасит ветер, но и дробит его
и даже отклоняет его направление. Все это в известной мере отра
жается и на тепловом режиме, и на режиме влажности воздух*
внутри биогеоценоза. «Лесные биогеоценозы ослабляют силу ветр<
и на своей опушке, предохраняя древостой от буреломов и ветро
валов. На этих свойствах лесной растительности основано и устрой
ство государственных и местных лесных полос, защищающих по
севы сельскохозяйственных культур от пыльных бурь и сухих, по
рой горячих ветров из пустынных областей, а на севере — oi
холодных арктических ветров. Известное значение имеет и верти
кальное движение воздуха, способствующее, наряду с горизонталь-
ным, повышению обмена газов и более равномерному распредели
нию водяных паров внутри биогеоценоза.
Преобразующее влияние на ветер и вертикальные токи воздух*
травяных биогеоценозов по понятным причинам менее эффективне
по сравнению с лесными бйогеоценозами. Но и здесь раститель
ность снижает скорость ветра, причем вблизи поверхности почвь
скорость тем меньше, чем выше и гуще травостой.
Так, в общей форме, представляется изменение, преобразова
пне основных элементов и конкретных факторов местного климат*
в своеобразный физический режим метеорологических явлений 1
пределах данного биогеоценоза, названный фитоклиматом
Характерная особенность фитоклимата, по Ю. Л. Раунер?
(1972),— резкое отличие его от местного климата смежных откры
тых пространств, и главная черта его — резкая смена метеоэлемен
тов (света, тепла, влаги, ветра и пр.) внутри растительного под
слоя биогеоценоза по вертикали по причине взаимодействие
энергетических потоков с фитоэлементами. Фитоклимат складыва
ется, по его словам, под воздействием «общих метеорологически:
условий данной местности, а также особенностей геометрическое
структуры данного ценоза. Оба этих фактора обусловливают взаи
модействия энергетических потоков с элементами фитомассы,
результате чего формируются разные типы фитоклиматов» (1972
с. 25).
С. Я. Соколов (1956) так характеризует отличительные особен
ности фитоклимата: «Фитоклимат отличается от наружного клима
та следующими чертами: общая радиация сильно снижается; ия
тенсивность химических лучей падает, качественный состав свет
изменяется в сторону преобладания зеленых лучей спектра; тем
137
пературный режим становится более плавным, крайности сглажи-
ваются; дневная амплитуда температуры, особенно летом, уменьша-
ется; движение воздуха замедляется, благодаря этому выделяемая
растениями (и животными. — Л. Н.) углекислота задерживается
в биоценозе; количество осадков, достигающих почвы, значитель-
но падает по сравнению с открытым местом; снежный покров
рыхл, хорошо отепляет почву, стаивает поздно; влажность воздуха
возрастает» (с. 517—518).
Б. А. Быков (1970а) приводит чертеж, наглядно иллюстрирую-
щий своеобразие воздушного режима и режима влажности поч-
венного покрова в сосновом лесу Серебряноборского лесничества
Московской области, составленный по материалам Молчанова
(1950), Никитина и Гребенниковой (1961) (рис. 14).
Более 25'Л
№1111111111:
от Ш ло 20
от 5 ло ТО
от 4 м 5
от Зло4
менее 5%
Илл юность почвы
S */• ОТ СУХОГО ми
Июль Аогоет Сеитшь
Рис. 14. Фитоклимат в сосновом лесу (по материалам А. А. Молчанова и др.)
(по Б. А. Быкову, 1970). 1 —температура на поверхности почвы в лесу; 2— то
же на лугу; 3 — абсолютная влажность воздуха в лесу на h = l м (в мм); 4 —
атмосферные осадки над лесом
Однако из изложенного выше материала по вопросу изменения,
преобразования основных метеорологических элементов и факто-
ров местного климата (или микроклимата) видно, что в становле-
нии, формировании специфических особенностей фитоклимата по
сравнению с климатом смежных открытых участков, наряду с
138
фитоценозом, играющим в этом главную роль, принимают участи'
и обитающие в его границах позвоночные и беспозвоночные живот
ные, а также эпифитная и почвенно-подстилочная микрофлора
Отсюда следует, что коль речь идет не о фитоценозе, а о система
более высокого ранга — биогеоценозе, то сформировавшийся в еп
составе при участии всех живых компонентов особый своеобразны!
климат имеется достаточно оснований назвать биоценоклима
том. А из этого, в свою очередь, следует, что биоценоклимат —
это не что иное, как тот биогенный компонент данного биогеоцено
за, который получил наименование аэротопа, представляющей
собою одну из составных частей внутренней среды биогеоценоз;
(биотопа, по А. П. Щенникову, 1964), названной ранее биоцено
средой.
4. Эдафотоп и его материальная основа
Перейдем к рассмотрению второго биогенного компонента бис
геоценоза—эдафотопа, представляющего собою вторую составну!
часть биоцепосреды (биотопа). Эдафотоп, по Работнову (1978)
как видно, — это совокупность факторов биогенной почвенной ср(
ды в границах биогеоценоза, являющейся результатом преобразс
вания почвообитающими живыми существами в определенны
климатических и гидрологических условиях в течение определен
ного времени косной ее части, а именно минеральной основы по1
венного слоя и материнской горной породы в пределах верхни
горизонтов коры выветривания. Таким образом, материальны
субстратом, лежащим в основе формирования эдафотопа как кок
попента биогеоценоза, являются поверхностные горизонты кор
выветривания, представленные почвообразующими материнским
горными породами различного генезиса, и минеральная часть с;
мой почвы.
Кора выветривания — это особое геологическое образа
ваиие, возникшее в результате глубоких преобразований верхш
слоев литосферы под действием агентов атмосферы, гидросф
ры и отчасти биосферы. Кора выветривания, по утверждени
А. И. Перельмана (1974), родственна почве; для нее также сво
ственны и деятельность микрофлоры, и зональность окислительн
восстановительных процессов, и свой вертикальный профиль ра
личной мощности. Отличие коры выветривания от почвы, по е
словам, состоит лишь в отсутствии в ней биогенной аккумулящ
химических элементов под влиянием растительности. Мощное
коры выветривания варьирует в весьма больших пределах. Так, i
поверхности скал магматических горных пород опа представлен
тонкой пленкой выветривания в 1—2 мм, на тех же породах Сев
ро-Западной Европы, освободившейся от ледникового цокро
139
5—6 тыс. лет тому назад, мощность коры выветривания 10—20 см.
Мощность же красноцветной коры выветривания субтропической
области Западного Закавказья, по данным В. А. Ковды (1973),
достигает 7—10 м, а толща аллитной коры выветривания третич-
ного возраста во влажно-тропических областях Азии и Африки
достигает огромной мощности — до 150 м.
В зависимости от продолжительности и истории процесса вы-
ветривания поверхности земной коры различаются: а) современ-
ные коры выветривания, образовавшиеся в кайнозое, — в четвер-
тичный, третичный периоды и отчасти в верхнем мелу; б) древние
(ископаемые) коры выветривания, формирование которых отно-
сится к юрскому периоду мезозойской эры, к девону и кембрию
палеозойской и даже до палеозоя. Мощность последних колеблется
от 60 до 300 м. Древние коры выветривания в геологическом
прошлом были при опускании суши погребены осадочными поро-
дами последующих периодов, а позднее при поднятии были выве-
дены на поверхность и вскрыты эрозионными процессами, оказав-
шись таким образом субстратом для современного почвообразо-
вания.
Выветривание (г и п е р ге н е з, по А. Е. Ферсману) — это весьма
сложный процесс, сопровождающийся разрушением одних горных
пород и слагающих их минералов и образованием других пород и
минералов. Процессы выветривания различных по своему генезису
и минералогическому составу горных пород как в геоисторическом
плане, так и в современных условиях протекали и протекают в
определенной последовательности и при ведущем значении то
одних, то других факторов. По мнению В. А. Ковды (1973), при
выветривании магматических горных пород главная роль принад-
лежит физическим и физико-механическим факторам, а при вы-
ветривании более или менее раздробленных метаморфических и
осадочных пород — химическим и физико-химическим факторам.
С появлением же и дальнейшим развитием жизни на Земле боль-
шое значение в процессах выветривания горных пород приобре-
тают также биологические и биохимические факторы. Важно при
этом заметить, что определенным этапам или стадиям выветрива-
ния горных пород соответствует и образование определенного на-
бора минералов. Так, на первых стадиях физико-химического
выветривания, по словам В. А. Ковды, происходит окисление вос-
становленных соединений железа и марганца; на последующих
стадиях образуются под воздействием водорода калий, натрий,
кальций, магний и пр. Затем, в результате изменения электрова-
лентных связей, происходит образование растворов и целого ряда
аморфных минералов.
В современных условиях, как указывает В. А. Ковда, в зависи-
мости от геоморфологического строения местности и складываю-
140
щегося при выветривании горных пород в соответствующих фор-
мах рельефа баланса веществ, наблюдается формирование сле-
дующих типов коры выветривания. I. На равнинных
плато, древних плоских водоразделах и древних террасах, в усло-
виях промывного типа баланса веществ, вследствие преобладания
процессов выщелачивания и выноса подвижных продуктов вывет-
ривания формируются элювиальные (остаточные) коры вы-
ветривания, II. На пологих склонах, наклонных делювиальных
равнинах, сухих дельтах и конусах выноса в условиях переходного,
промежуточного типа баланса веществ, характеризующегося тран-
зитом водных растворов, выпадением части взвешенных и раство-
ренных в них веществ по пути в осадок, засолением и загипсовы-
вапием этих форм рельефа, формируются транзитно-аккуму-
лятивные коры выветривания. III. В малодренировапных и
бессточных низменностях, депрессиях, нижних надпойменных и
пойменных террасах, дельтовых равнинах и высыхающих озерах
в условиях накопительного типа баланса веществ, вследствие акку-
муляции образовавшихся на месте и поступивших из вышележа-
щих форм рельефа продуктов выветривания, формируются акку-
мулятивные коры выветривания. Все три названных типа коры
выветривания, разумеется, в пределах любого географического
ландшафта тесно связаны между собой единством генезиса и свое-
образием геохимических процессов.
К материальному субстрату, служащему основой эдафотопа,
наряду с поверхностными горизонтами коры выветривания, отно-
сится, как сказано ранее, минеральная часть самой почвы, вклю-
чая самый верхний слой материнской почвообразующей породы.
Опа представляет собой арену непосредственного контакта и не-
прерывного взаимодействия атмосферы, гидросферы и биосферы,
результатом чего и является возникновение почвенного покрова
(педосферы), представляющего собой, по выражению В. А. Ковды
(1973), «самостоятельную специфическую биогенную оболочку
земного шара, окутывающую сушу материков и мелководья морей
и озер» (с. И).
В связи с этим необходимо принять во внимание то обстоятель-
ство, что процессы выветривания поверхности земной коры (лито-
сферы) и процессы почвообразования сопряжены как в простран-
стве, так и во времени. Но ввиду того, что в процессах выветри-
вания горных пород ведущими являются факторы абиотические, а
в процессах почвообразования — биологические, связанные с жиз-
недеятельностью растений, животных и микроорганизмов, в на-
правленности тех и других и в существе полученных результатов,
как указывает В. А. Ковда, вскрываются весьма важные в соотно-
шении этих процессов закономерности. Так, в условиях формиро-
вания элювиального типа коры выветривания, как мы видели, гос-
141
подствует вынос продуктов выветривания; однако в верхних гори-
зонтах, охваченных почвообразовательными процессами, происхо-
дит биогенное накопление веществ, в особенности гумусовых. В
условиях же формирования аккумулятивного и транзитно-аккуму-
лятивного типов коры выветривания имеет место отложение меха-
нических, химических и прочих осадков; вместе с тем и процессы
почвообразования также сопровождаются аккумуляцией органиче-
ских и минеральных веществ, особенно в гумусовом горизонте.
Следовательно, как пишет В. А. Ковда (1973), «формирование
элювиальной коры выветривания и биогенный цикл почвообразо-
вательного процесса диаметрально противоположны в своей на-
правленности и в суммарном балансе веществ» (с. 116). Хотя
вынос веществ здесь все же преобладает. Во втором случае «на-
правленность почвообразования и геохимических процессов сум-
мируется и сочетается без противоречий, ведя к формированию
плодородных богатых питательными веществами осадочных пород
и почв» (с. 116).
5. Почвообразование, его факторы и энергетика
Процесс почвообразования, как и процесс выветривания по-
верхности земной коры, весьма сложный и многогранный процесс,
охватывающий длительные отрезки времени. В осуществлении его
принимает участие целый ряд факторов—абиотических и биоло-
гических. В. В. Докучаев (1889), как известно, к числу непремен-
ных почвообразующих факторов относил климат, горные породы,
рельеф, организмы и время. В. Р. Вильямс (1936) ведущим
фактором почвообразования считал живые организмы; позднее
большое значение он придавал также хозяйственной деятельности
населения. Всеми последователями В. В. Докучаева признавалось
и признается в настоящее время, что перечисленные факторы поч-
вообразования действуют не разобщенно, а совместно, совокупно,
постоянно взаимодействуя между собой; они осуществляют в силу
этого огромную биогеохимическую работу, имеющую общеплане-
тарное значение. Вместе с тем факторы почвообразования различ-
ны как по своей природе, так и по своей роли и значению в этом
процессе.
В. А. Ковда в своей фундаментальной монографии «Основы
учения о почвах» (1973) приводит детальную характеристику фак-
торов почвообразования, исходя из представлений В. В. Докучае-
ва, выразив их в виде формулы: П= (КОГР)Т, где П — почва,
К —климат, О — организмы, Г — горные породы, Р — рельеф и
Т — время. Горные породы (и минералы) В. А. Ковда рассматри-
вает как главные объекты почвообразовательного процесса, кото-
рые, подвергаясь выветриванию, разрушению и новому синтезу,
142
преобразуются под действием организмов в почвенный покров (
гумусовым горизонтом в его профиле. Общий климат, включая вод
но-воздушный, световой и тепловой режимы, автором рассматри
вается как фактор космического происхождения, зависящий ог
солнечной радиации и ее распределения по земной поверхности
Этому фактору почвообразования он придает исключительно боль
шое значение, поскольку климат определяет многие процессы ш
Земле: выпадение осадков, водную и ветровую эрозию, перемещени'
разнообразных веществ по поверхности Земли, условия существо
вания и жизнедеятельности живых организмов и пр. Изменена
климата в геологическом прошлом приводили к глубоким и разно
образным изменениям на нашей планете, охватывая раститель
ность, животный мир и почвенный покров.
Рельеф местности вполне резонно считается автором косвеннс
действующим фактором почвообразования, перераспределяющик
климатические и другие процессы в зависимости от местоположе
ния. Рельеф оказывает влияние на поверхностный и подземные
сток атмосферных осадков, глубину залегания почвенно-грунтовы?
вод, характер и степень минерализации поверхностных и подзем
ных вод, распределение тепла и влаги, скорость испарения г
транспирации влаги; рельеф определяет смыв подвижных продук
тов выветривания с положительных форм, отложение и накопление
их в отрицательных местоположениях, химизм почвообразующи?
пород и формирование различных типов почв, размещение живы;
существ, их состояние и развитие и многое другое.
Роли, значению и результатам деятельности живых организмов
как биологическому фактору почвообразования уделяется большое
внимание многими почвоведами и другими специалистами, в осо
бенности в связи с глубокими обобщениями и выводами В. И. Вер
надского, касающимися роли живого вещества в общепланетар
ных биогеохимических процессах в пределах биосферы Земли.
Прежде всего, большинство почвоведов и биологов широкой
профиля вслед за В. Р. Вильямсом подчеркивают ведущую рол]
живых организмов в осуществлении почвообразовательного про
цесса, в особенности автотрофных растений как аккумуляторе]
солнечной энергии, служащей источником существования всех ор
ганизмов на Земле и осуществления разного рода реакций, состав
ляющих основу почвообразовательного процесса (В. А. Ковда
1973; С. В. Зонн, 1956, 1960; Т. А. Работнов, 1974; Н. В. Дылис
1978; Ю. Одум, 1975; Р. Риклефс, 1979; и др.). Во-вторых, процес
почвообразования в настоящее время трактуется как непрерывны
циклический обмен веществом и энергией разных групп организ
мов между собой, а также с материнской почвообразующей поре
дой и минеральной частью почвы. Он выражается: а) в механич€
ской и физико-химической переработке минеральной массы поч
143
вообразующей породы и почвенного слоя; б) минерализацир
поступивших в почвенный слой органических остатков и продуктов
метаболизма с выделением свободной энергии; в) усвоении орга-
низмами минеральных соединений и утилизации их на процесс
синтеза гумусовых веществ, новых минералов, различных органо-
минеральных соединений специфических газов в почвенном возду-
хе и т. п.
Иначе говоря, процесс почвообразования осуществляется сово-
купной деятельностью живых организмов в форме биологического
круговорота веществ и потока энергии, ведущих к коренному изме-
нению почвообразующей породы и превращению ее в почву.
Фактор времени, как видно из приведенной выше формулы, за-
нимает особое место среди других факторов почвообразования. Он
выражает историю, продолжительность почвообразовательного
процесса, протекающего под действием названных выше факторов.
Фактор времени, называемый также возрастом страны, находит
отражение в эффективности почвообразования и его результатах, в
формировании на разных этапах в зависимости от сочетания усло-
вий различных типов и разновидностей почв, которые в силу этого
могут рассматриваться как представители стадий почвообразова-
тельного процесса.
Эффективность и характер действия остальных факторов поч-
вообразования в ходе исторического развития этого процесса от
этапа к этапу постепенно изменяется. Так, например, роль живого
вещества в процессах почвообразования, в геоисторическом аспек-
те, как утверждает В. А. Ковда, неизменно возрастает, так как с
течением времени возрастает видовое разнообразие и численность
организмов, а следовательно, и общая масса живого вещества;
расширяются границы биосферы, и большее количество солнечной
энергии вовлекается в различные процессы на Земле, поэтому все
большие массы земной коры подвергаются действию биологическо-
го круговорота, преобразуясь в почвенный покров.
Помимо перечисленных природных факторов почвообразова-
ния, в последнее время все больше внимания стало уделяться роли
хозяйственной деятельности местного населения в этом процессе.
Хозяйственная деятельность человека в настоящее время является
мощным фактором изменения и преобразования горной породы в
почву и изменения природных свойств самой почвы, вплоть до со-
здания так называемых «окультуренных» почв. Вследствие этого
почва, являясь природным телом, постепенно превратилась в объ-
ект труда, важнейшее средство производства. По мере обществен-
ного развития человечества мощность всестороннего хозяйственно-
го воздействия населения на горные породы и почвенный покров
все более высокими темпами возрастает, в особенности в связи с
изменением форм его хозяйственной деятельности, а также в связи
144
с быстрым развитием техники и колоссальным повышением энерго-
вооруженности.
Для получения более полного представления о почве и почвен-
ной среде необходимо хотя бы в самых общих чертах ознакомить-
ся с энергетикой почвообразовательного процесса. Основным ис-
точником энергии на Земле, как известно, является лучистая
энергия Солнца, которая в почву поступает в виде тепловой энер-
гии. Из других источников тепловой энергии известны: а) энергия,,
поступающая из глубинных слоев Земли (энергия тектонических
движений земной коры, вулканов, термальных источников, газовых
выделений при извержениях вулканов); б) энергия естественной
радиации радиоактивного распада некоторых элементов земной
коры. Однако значение их в тепловом балансе Земли ничтожно.
Поступление в почву энергии, по данным исследований В. Р. Во-
лобуева (1961), В. А. Ковды (1966, 1968), К. И. Лукашева (1958)
и др., происходит а) с водой в разных формах почвенной влаги;
б) с мертвым органическим веществом в виде химической энергии
органических соединений в почвенном гумусе за счет биологиче-
ского массообмена организмов и почвы; в) с почвообразующей
материнской породой при формировании почвы за счет энергии
глубинных процессов планеты и пр. В результате в почве, в особен-
ности в гумусовом слое, происходит накопление значительного ко-
личества потенциальной энергии, примерно равного запасу энер-
гии, заключенной в наземной биомассе сухопутных организмов,
главным образом растений. Причем в травяных типах раститель-
ности запас энергии в гумусовом слое в 20—30 раз выше, чем в
наземной биомассе; в лесных же, наоборот, в наземной биомассе
запаса энергии в 2—4 раза больше, чем в почвенном гумусе. Это
связано с тем, что в лесах потенциальная энергия сосредоточива-
ется преимущественно в древесине, а не в почве, как в травяни-
стых сообществах.
Основная часть поступившей на земную поверхность солнечной
энергии в виде тепловой энергии (95—99,5% всей тепловой энер-
гии почвообразования) расходуется, по данным В. Р. Волобуева
(1958, 1961, 1969), на процессы влагооборота и теплообмена, глав-
ным образом на физическое испарение и транспирацию влаги, при-
чем эта энергия в конечном итоге выбывает из почвы в виде
тепловой энергии вовне. От 0,5 до 5% от всей тепловой энергии
расходуется на циклические биологические процессы превращения
эрганических веществ в более высокомолекулярные органические
I затем тех и других —в простые минеральные соединения. Лишь
ютые и тысячные доли процента от общей тепловой энергии ухо-
1ят на разложение минералов в процессах физического и хнмиче-
:кого выветривания, и столь же ничтожное количество ее тратится
ia явления миграции веществ по вертикальному профилю почвы.
145
Таким образом, суммарные затраты энергии на процессы почво-
образования, по тем же данным, составляют в ккал/см2 в год: в
тундрах и пустынях 2—5, в лесах и степях умеренного пояса
10—40, во влажнотропических типах растительности 60—70, но в
тропических пустынях примерно в 7 раз меньше, чем во влажных
тропиках.
6. Почва, ее состав, строение,
мощность и характерные черты
С. В. Зонн (1964), рассматривая почву в качестве одного из
компонентов биогеоценоза, пишет, что с биогеоцепотической точки
зрения под почвой следует понимать «поверхностный слой лито-
сферы, участвующий в биологическом круговороте и обретший
свойства природного компонентного тела, с характерным для него
непрерывным обменом вещества и энергии, обуславливающим его
образование, плодородие, закономерности формирования и эволю-
цию» (с. 372).
По мнению В. А. Ковды (1973), почва, как самостоятельное
ограниченное в пространстве природное тело, может трактоваться
как определенная термодинамическая система, обладающая спо-
собность^ пропускать через себя потоки вещества и энергии, не
аккумулируя их в своих границах. Исходя из этого, он дает такое
определение почвы: «...с термодинамической точки зрения почва
представляет собой открытую гетерогенную многофазную систему,
характеризуемую необратимыми термодинамическими процессами»
(с. 129). Открытой системой почва названа потому, что, как ком-
понент биогеоценоза, она находится «в постоянном и массо- и
энергообмене с окружающей средой» (с. 127); гетерогенной систе-
мой ввиду того, что она состоит «из нескольких гомогенных систем,
имеющих разные свойства» (с. 127); почва — многофазная систе-
ма, очевидно, в связи с тем, что в ее состав входят вещества как
в твердом, так и в жидком и в газообразном состояниях.
Вместе с тем почва, по мнению В. А. Ковды, может рассматри-
ваться и как биохимическая система, поскольку в результате жиз-
недеятельности и обменных процессов в сообществе почвообитаю-
щих организмов с участием подземных органов растений в почве
создается, по его выражению, «своеобразная биохимическая обста-
новка». В почве непрерывно синтезируются, разрушаются и из
продуктов распада вновь синтезируются разнообразные вещества:
аминокислоты, спирты, органические кислоты, гумусовые вещества,
энзимы, активные биохимические продукты, антибиотики, витами-
ны групп В, РР, токсические вещества и пр. Продукты метаболиз-
ма и другие продуцируемые вещества оказывают существенное
воздействие па процессы почвообразования, плодородие почвы, на
146
течение различных биохимических реакций в почве (окисление,
восстановление, гидролиз, брожение и т. п.).
Из всего изложенного следует, что почва как природное тело
состоит из подвергшейся физико-механическому и биохимическому
выветриванию косной минеральной основы, биогенной органиче-
ской массы в виде гумусового слоя различной мощности, продук-
тов почвообразования и метаболизма в различных состояниях, а
также специфических почвенных, газов и воды с растворенными в
ней веществами. По Ю. Одуму (1975), в состав почвы входят под-
стилающий минеральный субстрат как исходный материал и орга-
нические «добавки», а также организмы и продукты жизнедеятель-
ности, перемешанные с мелкими частицами измененного исходного
материала, а между частицами — газы и вода. Почвообитающие
организмы Ю. Одум, как видно, в согласии с некоторыми другими
биологами и почвоведами считает составными элементами, а не
обитателями почвы, что с нашей точки зрения, как было отмечено
ранее, не имеет достаточных оснований.
Происходящие в почве процессы аккумуляции, превращения и
перемещения органических и минеральных веществ, как указывает
С. В. Зонн (1964), находят отражение в морфологическом строе-
нии почвенного профиля, в особенностях вертикального расчлене-
ния почвы на так называемые биогеогоризонты и слои
(Ю. П. Бяллович, 1960), которые обычно почти полностью совпа-
дают с выделяемыми почвоведами генетическими горизонтами
почвы. С. В. Зонн (1964, 1974) при вертикальном расчленении поч-
вы как компонента биогеоценоза выделяет следующие биогео-
горизонты и слои (подгоризопты):
АА — биогеогоризонт активной аккумуляции сырого энергетическо-
го материала в виде подстилки, войлока и т. п., соответству-
ющий генетическому горизонту почвы Ао. По степени разло-
жения он подразделяется на три слоя (подгоризонта):
AAi—свежий неразложившийся опад (и отпад);
АА2—полуразложившиеся органические остатки опада;
АА3 — полностью разложившаяся гомогенная перегнойная
масса.
А — биогеогоризонт аккумуляции гумусовых и зольпых веществ,
соответствующий гумусовому горизонту почвы А]. По убыва-
нию степени гумусированности, изменению структурности
и пр. он подразделяется на слои — Аь А2, А3.
ИТ — биогеогоризонт интенсивного (элювиального) транзита гуму-
совых и подвижных минеральных веществ (в молекулярных
и коллоидных формах) и обеднения ими, соответствующий
элювиальному горизонту почвы А2. В зависимости от форм
транзита веществ различаются два варианта:
ИТ]—когда происходит перемещение веществ, поступающих
147
из биогеогоризонта А, а также разрушение их и вы-
нос продуктов разрушения в нижележащий горизонт
(в подзолистых, дерново-подзолистых и подзолисто-
глеевых почвах);
ИТг — когда происходит вымывание из этого биогеогоризон-
та илистой фракции и образование конкреций железа
(в дерново-подзолистых почвах на богатых породах
и бурых лесных оподзоленных черноземах).
ВА — биогеогоризонт вторичкой (иллювиальной) аккумуляции ор-
ганоминеральных и минеральных веществ (с изменением и
без изменения их физико-химического состояния), соответ-
ствующий иллювиальному горизонту почвы В. Различаются
два варианта его:
BAi—когда в нем происходят новообразования и отложения
различных солей (хлоридов, сульфатов и т. п.);
В Аз — когда происходит оглинение горизонта за счет вмыто-
го или образовавшегося на месте ила.
ГТ — биогеогоризонт глубинного транзита органоминеральных и
минеральных веществ в подстилающие горные породы или
грунтовые воды или накопления выбывающих из биогеоце-
нотического обмена веществ (в виде карбонатов, гипса
и т. п.). Он соответствует горизонту С, представляющему
собой слабо измененные продуктами обмена материнские
породы (с. 216—217).
Распределение химических элементов по почвенному профилю,
как отмечает С. В. Зонн, определяется двумя взаимно противопо-
ложными процессами: а) биогенной аккумуляцией элементов в
почве вследствие перемещения корнями растений калия, кальция,
фосфора, серы и др. из нижних горизонтов в верхние, что наибо-
лее характерно для степных и отчасти пустынных биогеоценозов
(черноземы, сероземы), и б) выщелачиванием легкорастворимых
элементов из верхних горизонтов в нижние при промывании почв
атмосферными осадками, в особенности в лесных биогеоценозах
(подзолистые почвы).
Таким образом, вертикальный профиль почвы, как указывает
А. П. Шенников (1950), характеризуется существенными разли-
чиями почвенных горизонтов как по механическому составу, физи-
ческим и химическим свойствам, так и по функциональным режи-
мам— солевому, водно-воздушному и тепловому. Важность учета
расчленения почвы на биогеогоризонты подчеркивает и Н. В. Ды-
лис (1978), ибо взаимодействие других компонентов биогеоценоза
с почвой, в особенности живых, складывается дифференцированно
по биогеогоризонтам и даже слоям. Особенно большое значение
он придает подстилке и гумусовому горизонту почвы, наиболее
плотно заселенным живыми существами и заполненным корнями
148
растений, продуктами метаболизма и отмершими органическими
остатками. Т. А. Работное (1978) обращает внимание на важную
роль в травяных биогеоценозах верхнего дернового слоя почвы, так
называемой дернины.
Мощность, толща различных типов почв, границы их верти-
кального простирания неодинаковы. Верхней границей почвенного
профиля принято считать поверхность мертвого напочвенного по-
крова (лесной подстилки, войлока на лугах и в степях), а нижней
границей — предел распространения основной массы корневых
систем растений, так называемого корнеобитаемого слоя. Общая
мощность почвы обычно составляет 1,5—2 м. В этом слое расте-
ния, почвообитающие животные и микроорганизмы находят необ-
ходимые для своего существования, жизнедеятельности и развития
тепло, влагу, кислород воздуха и питательные вещества (мине-
ральный азот, зольные элементы и пр.). Мощность почвы, по
А. П. Шенникову, свидетельствует о степени близости или уда-
ленности от ее поверхности материнской почвообразующей поро-
ды. Наиболее маломощные почвы встречаются на скалистых маг-
матических и метаморфизированных горных породах либо в
суровых климатических условиях, ограничивающих распростране-
ние биологических процессов почвообразования в более глубокие
слои материнских пород. Маломощные почвы характерны также
для первых стадий почвообразования.
От мощности почвы, как указывает С. В. Зонн (1964), следует
отличать мощность подземного слоя биогеоценотического покрова
(по В. Н. Сукачеву), включающего как толщу самой почвы, так
и поверхностные слои коры выветривания ниже корнеобитаемого
слоя, в пределах которых протекают процессы почвообразования.
Однако изменения в этих горизонтах происходят не столько под
действием живых существ, сколько под влиянием физических и
физико-химических процессов, связанных с передвижением влаги
и веществ, не использованных организмами.
Почва как природное тело отличается от материнской горной
породы, на основе которой она сформировалась, рядом характер-
ных особенностей. Во-первых, почвы, как отмечают Ф. Я. Гаври-
люк с соавторами (1977), в отличие от почвообразующей породы
анизотропны, т. е. их состав п свойства неодинаковы по генетиче-
ским горизонтам почвенного профиля; во-вторых, почва обладает
плодородием, т. е. способностью обеспечить необходимые условия
для существования и развития растений и других организмов.
В-третьих, по В. И. Вернадскому (1967), почва характеризуется
скоплением, концентрацией бесчисленного количества организмов,
обладающих в совокупности огромной химической энергией, обес-
печивающей грандиозный геохимический эффект их деятельности
в биогеоценозе и в биосфере в целом. В-четвертых, по словам
149
С, В. Зоина (1964), для почвы характерна резкая смена физико-
химических свойств по биогеогоризонтам почвенного профиля (па-
пример, смена кислой реакции (pH) среды с глубины 20—50 см
на щелочную). В-пятых, наличие в почве окислительно-восстанови-
тельных процессов с преобладанием восстановительных в верхних
горизонтах, где происходит интенсивное разложение органики, и
окислительных в нижних — с образованием окисных соединений
железа, марганца или переход их в анаэробных условиях в закис-
ные соединения.
7. Генетические свойства
и функциональная роль почвы в биогеоценозе
Любая почва, как и всякое природное тело, обладает присущи-
ми ей свойствами, названными специалистами в этой области
генетическими свойствами. К числу их относятся: механический и
минералогический состав почвы, реакция (кислотность) почвы,
физические свойства (основные и функциональные) и физико-хи-
мические свойства органической и минеральной частей почвы. Не
останавливаясь на характеристике этих почвенных свойств, кото-
рая дается в подробном изложении в соответствующих руковод-
ствах и монографиях, обратим внимание главным образом на воз-
действие наиболее важных из названных свойств, которое они
оказывают на различные стороны составных частей других ком-
понентов биогеоценозов, на роль и значение этих свойств в составе
и функционировании последних.
Прежде всего, физические свойства почвы, а из них как эколо-
гически наиболее важные водный, воздушный и тепловой режимы
почвы. Почвенная вода» как указывает Н. В. Дылис (1978), исполь-
зуется корнями растений и почвообитающими животными и мик-
роорганизмами на удовлетворение своих потребностей в ней. Кро-
ме того, опа транспортирует растворенные в ней минеральные и
органические вещества по профилю почвы, а также по уклону
земной поверхности или водоупорного горизонта в подземной ча-
сти. При этом так называемая гравитационная вода, перемещаясь
по почвенному профилю сверху вниз, способствует выщелачиванию
веществ и выносу их из верхних горизонтов почвы в нижние, а
капиллярная вода, наоборот, вследствие испарения ее с поверхно-
сти перемещает вещества снизу вверх, обогащая верхние горизон-
ты почвы, наиболее плотно заполненные подземными органами
растений и населенные животными и микроорганизмами, содер-
жащие различные минеральные соли, органические и минеральные
кислоты, кислород и другие необходимые для них вещества.
В зависимости от гидрологического режима местности и глуби-
ны залегания грунтовых вод складываются различные типы ре-
150
жима влажности почвы. А. А. Молчановым (1964) различа-
ются: болотный и полуболотный режимы, характеризующиеся
избыточной влажностью почвы, таежный — с оптимальной влаж-
ностью таковой, степной и пустынный, отличающиеся недостаточ-
ной влажностью почвы, а последний даже чрезмерной сухостью
ее. В условиях названных типов режима влажности почвы, есте-
ственно, создаются и соответствующие типы обеспеченности био-
геоценозов влагой, что сказывается на их состоянии, развитии и
продуктивности.
Воздушный режим почвы, как известно, является антагонистом
водного режима. Чем больше скважины почвы заполнены водой,
пишет А. П. Шенников (1950), тем меньше в ней окажется возду-
ха, а следовательно, и кислорода; остается лишь воздух, раство-
ренный в воде. В тяжелых по механическому составу почвах
создаются в таких случаях анаэробные условия, что, помимо не-
обеспеченности живых существ кислородом, замедляет также про-
цесс разложения органических остатков.
Весьма важным условием нормальной жизни организмов и
бесперебойного функционирования биогеоценозов является тепло-
вой режим почвы, который находится в зависимости от целого ряда
факторов: климатических особенностей местности, теплопроводно-
сти почвы и почвообразующих пород, обеспеченности почвы вла-
гой, характера растительного покрова и т. п. Тепловой режим
почвы оказывает влияние на многие стороны функционирования
биогеоценозов. По словам Н. В. Дылиса, он определяет жизнедея-
тельность корней растений и почвообитающих животных и микро-
организмов, подвижность воды и почвенных растворов и доступ-
ность их для живых существ, скорость разнообразных химических
реакций, а в конечном итоге — продуктивность биогеоценотических
систем. Тяжелые по механическому составу глинистые и суглини-
стые почвы в таежной полосе, как отмечает А. П. Шенников, более
влажны и холодны, чем песчаные почвы. Но в сухих жарких пу-
стынях песчаные почвы влажнее глинистых, поэтому и раститель-
ный покров на них более развит.
Из физико-химических свойств почвы наибольшее значение для
жизни почвообитающих организмов и функционирования биогео-
ценозов, как указывает С. В. Зоин (1967), имеют: а) химический
состав минеральной части почвы, а именно валовое содержание
основных окислов макро- и микроэлементов и подвижных раство-
римых в воде и кислотах минеральных и органоминеральных со-
единений (азот, фосфор, калий, кальций, магний и др.); б) биохи-
мический состав органической части почвы в виде подстилки или
войлока и гумусового горизонта различного механического состава
и мощности. Поступление и запас химических элементов в почве
находится в зависимости от содержания их в составе почвообра-
151
зующих пород, от приноса их извне или выноса за пределы био-
геоценоза, а также от количества и качества химизма раститель-
ной, животной и микробной биомассы и от притока их под дей-
ствием корневых систем растений из нижних горизонтов почвы и
подпочвы.
Органическое вещество почвы, представленное в основном гу-
мусом в коллоидальной форме, состоит из малорастворимых гуми-
новых веществ и растворимых в воде креновой и апокреиовой
кислот. При разложении гумуса выделяются усваиваемые расте-
ниями минеральные соли, а также углекислота, которая, раство-
ряясь в воде, подкисляет почву и вместе с тем облегчает растворе-
ние минеральных частиц почвы и усвоение их растениями. Гумус„
таким образом, представляет собою весьма важный источник
минеральной пищи растений, а с другой стороны, как пишет
Н. В. Дылис, он увеличивает поглотительную способность почвы,
благодаря наличию в его составе гуминовых веществ и извести
способствует образованию прочной структуры почвы, вследствие
чего существенно меняются физические и химические свойства поч-
вы, улучшаются водно-воздушный, тепловой и солевой режимы, что
ведет к повышению плодородия почвы и продуктивности биогео-
ценозов в целом. Кроме того, гумусовый горизонт почвы, как было
отмечено ранее, является резерватором аккумулированной живым
веществом лучистой энергии Солнца. В гумусовом слое почвенного^
покрова Земли, по свидетельству Н. В. Дылиса (1978) со ссылкой
на данные В. А. Ковды, сосредоточено 1,2-1018 ккал энергии
(с. 103). Однако мощность и химический состав гумусового гори-
зонта почвы и запас потенциальной энергии в нем далеко не оди-
наковы в различных физико-географических условиях: так, черно-
земные почвы степной зоны весьма богаты гумусом и заключенной
в нем энергией; почвы же таежной полосы обычно выщелочены и
обеднены как гумусом, так и энергией.
Таким образом, роль почвы в происходящих в биогеоценозах
процессах чрезвычайно велика. Представляя собою целостное при*
родное образование, она находится в постоянном взаимодействии
со всеми другими компонентами и составными частями биогеоце-
ноза. На нижнем пределе своего распространения почва непосред-
ственно связана с почвообразующей породой, главным образом
посредством обмена продуктами разрушения материнских горных
пород и разнонаправленной миграции различных химических эле-
ментов. Сверху она контактирует с приземным слоем атмосферы,
а в границах биогеоценозов с их внутренней биогенной воздушной
средой преимущественно через посредство живых организмов и
продуктов их жизнедеятельности. Но особенно тесными узами поч-
ва связана с живыми компонентами биогеоценозов, а из них в
первую очередь с ценопопуляциями автотрофных растений и с
152
соответствующими
группами гетеро-
трофных микроорга-
низмов (рис. 15).
Почва, по словам
С. В. Зонна, Н. В.
Дылиса, Т. А. Работ-
нова, П. М. Рафеса,
В. А. Ковды, В. Р.
Вильямса и других
почвоведов и биоло-
гов, представляет со-
бою среду обитания
и условия существо-
вания сухопутной
растительности, жи-
вотного и микробно-
го населения биогео-
ценоза, является вме-
стилищем живого
вещества. Она — по-
ставщик и «кладо-
вая» необходимых
Рис. 15. Взаимодействие почв с другими компо-
нентами лесных биогеоценозов (по С. В. Зонну,
1964)
для живых существ материально-энергетических ресурсов: воды,
минеральных и органических веществ, газов и пр. Почва обеспе-
чивает растительные сообщества (через корневую систему) и
взаимосвязанные с ними ценокомплексы животных и микроорга-
низмов зольными элементами, минеральным азотом, водой в раз-
ных формах, углекислотой и некоторыми органическими вещества-
ми (ферментами, витаминами и т. п.).
Но это только одна сторона функциональной роли почвы в био-
геоценозе; вторая, с нею взаимосвязанная, заключается в следую-
щем: почва является приемником продуктов метаболизма, жизне-
деятельности живых существ и их посмертных органических остат-
ков, а также посредником между почвообитающими организмами
и земными (вода и элементы питания) факторами среды. Почва
участвует во всех важнейших обменных процессах, происходящих
в биогеоценозе, — водном, органоминеральном и энергетическом.
В почве сходятся и переплетаются все инти, тянущиеся от других
компонентов биогеоценоза, в ней, главным образом, протекает
биологический круговорот веществ, осуществляющийся между жи-
выми компонентами, но с непременным включением почвенных,
атмосферных и космических материально-энергетических ресурсов.
Почва, можно сказать, суммирует результаты функциональной
деятельности всех компонентов биогеоценоза. Являясь более кон-
153
сервативной составной частью биогеоценоза по сравнению с живы-
ми компонентами, она в наибольшей степени отражает в своих
свойствах и составе процессы и явления, совершающиеся в биогео-
ценозе, а потому нередко именуется «зеркалом» всего биогеоцено-
тического процесса. Таким образом, главная функция почвы в
составе биогеоценоза состоит в обеспечении благоприятных усло-
вий существования бесчисленному количеству разнообразных жи-
вых существ, как населяющих самоё почву, так и обитающих в
наземной, воздушной среде. Отсюда коренным природным свой-
ством ее является плодородие.
8. Преобразование почвы
во внутреннюю почвенную среду биогеоценоза
Связь почвы с другими компонентами биогеоценоза, как и
связь атмосферы,—это двусторонний процесс. Почва, воздействуя
на компоненты и составные части биогеоценоза, сама испытывает
с их стороны и биогеоценоза как целого разностороннее и суще-
ственное влияние.^Из материнской горной породы (при выветрива-
нии) и почвенно-грунтовых вод в почву поступает большое коли-
чество разнообразных минеральных веществ. Из атмосферы и
ближайшего космоса почва получает с атмосферными осадками и
аэрозолями воду, кислород, азот и ряд других веществ и химиче-
ских элементов; поступает также атмосферная пыль, энергия в
тепловой форме и т. п. Различные вещества приносятся в почву
ветром, смываются с крон и коры деревьев и кустарников, с лист-
вы и стеблей травянистых растений.
Особенно сильное и разнообразное влияние на почву оказы-
вают живые существа. Прежде всего, как видно из ранее сказан-
ного, они играют важную, ведущую роль в самом процессе почво-
образования. Но не менее мощное воздействие на почву и подпочву
продолжают оказывать и в качестве компонентов того или иного
сформировавшегося биогеоценоза. В почву непрерывно поступает
большое количество органических и минеральных веществ в виде
так называемого опада и отпада (т. е. отмерших отдельных частей
и целых растительных и животных организмов), а также в виде
прижизненных корневых выделений, отбросов и выделений живот-
ных (мочевина, энзимы) и пр. По данным С. В. Зонна (1964), в
почву таежных биогеоценозов ежегодно поступает опада 3—5 т/га,
содержащего значительные запасы различных веществ и энергии.
Образующийся на поверхности земли в виде подстилки в лесах
или войлока на лугах и в степях мертвый напочвенный покров под
действием микроорганизмов, беспозвоночных животных и отчасти
сапрофитных растений подвергается разложению, гумификации и
частичной минерализации. Все это ведет к обогащению почвы орга-
154
никой и целым рядом минеральных соединений, в том числе и
биогенного происхождения, к усилению и усложнению биологиче-
ского круговорота веществ и миграции веществ по горизонтам
почвенного профиля с аккумуляцией их в гумусовом и некоторых
других горизонтах, к существенному изменению основных физиче-
ских свойств (структура, плотность, скважность) и физико-хими-
ческих свойств почвы, в особенности обусловленных составом лег-
корастворимых подвижных соединений. В конечном итоге все это
приводит к преобразованию почвы, превращению ее во внутрен-
нюю почвенную среду биогеоценоза (эдафотоп), находящуюся в
данный отрезок времени в относительном соответствии с потребно-
стями живых компонентов биогеоценоза и присущей им избира-
тельной способностью в использовании материально-энергетических
ресурсов почвенной среды как компонента биогеоценоза.
Таким образом, становление почвы и ее функционирование про-
текают в составе биогеоценоза, в силу этого ее разнообразные
генетические свойства как бы преломляются в процессе взаимодей-
ствия с другими компонентами, и почва, качественно преобразуясь
при этом, превращается в биогенную среду данного биогеоценоза,'
становится его непременным компонентом. Взаимосвязь почвы с
другими компонентами биогеоценоза и с биогеоценозом как це-
лостной системой, в состав которого она входит, хорошо видна на
примерах, приведенных С. В. Зонном (1964), А. П. Щенниковым
(1964) и Н. В. Дылисом (1978). Так, в составе таежно-лесных био-
геоценозов почва, по их свидетельству, формируется и функциони-
рует в условиях преобразованного под их пологом климата; поч-
венные процессы находятся в тесной и наиболее четко выраженной
взаимосвязи с другими компонентами биогеоценоза. Для почвы в
этих условиях характерны достаточно хорошее увлажнение и аэра-
ция, интенсивный водно-минеральный обмен и образование назем-
ной биомассы, в особенности растительной, накопление большого
количества отмерших органических остатков в виде лесной под-
стилки, довольно полное разложение органики с участием в соста-
ве микрофлоры низших грибов и актиномицетов, выделение угле-
кислоты и тепла в приземной слой атмосферы и вынос раствори-
мых веществ в подпочву и грунтовые воды, выщелачивание
подвижных элементов из верхних горизонтов почвы и формирова-
ние относительно бедных органическим веществом подзолистых,
дерново-подзолистых и других подобного типа почв, как правило,
кислой реакции.
Степные травяные биогеоценозы ввиду сравнительно незначи-
тельной мощности биогеоценотического покрова мало влияют на
приземной слой атмосферы, но оказывают значительное воздей-
ствие на почву; почвенные процессы в силу тех же причин связаны
не столько с фитоклиматом, сколько с общими атмосферными
155
условиями местности; взаимодействие компонентов биогеоценоза
вследствие пониженного увлажнения и высокой температуры в ве-
гетационный период значительно ослаблено. Образование назем-
ной биомассы и мертвого напочвенного покрова в виде войлока в
этих условиях невелико, зато развивается огромная масса корней
и других подземных органов растений, ежегодно отмирающих в
большом количестве; при этих обстоятельствах происходит преоб-
ладание процессов гумификации органики над процессами минера-
лизации ее, все это ведет к накоплению огромной массы гумуса в
верхних горизонтах почвы и формированию богатых органическим
веществом, хорошо аэрируемых, структурных черноземных почв
различной мощности.
Отмеченные резкие различия подзолистых и черноземных почв,
формирующихся под соответствующими группами типов таежно-
лесных и травяных степных биогеоценозов, в известной степени
связаны с зональными физико-географическими особенностями
местности. Однако и в пределах одной и той же зоны, как извест-
но, под разными группами типов биогеоценозов, например ельни-
ков и сфагновых болот в таежно-лесной области, формирующиеся
почвы весьма сильно отличаются друг от друга по своим призна-
кам и свойствам. Так, для корнеобитаемого слоя сфагновых болот,
развивающихся на минеральном грунте, по данным С. В. Зонна и
А. П. Шенникова, в отличие от подзолистых почв характерно на-
личие торфяного или минерально-торфяного субстрата, насыщен-
ного застойной холодной водой, очень кислой реакции и крайне
бедной элементами минерального питания в доступном для выс-
ших растений состоянии, чрезвычайно плохая аэрация субстрата
и весьма слабая в анаэробных условиях деятельность микрофлоры,
накопление травяно-кустарничково-сфагновым покровом неразло-
жившейся органической массы и образование различной мощности
торфяного слоя, формирование в этих условиях торфянисто-гле-
евых и торфяных болотных почв.
Специфичность биогенной почвенной среды, связанной в своем
происхождении и последующем функционировании с определен*
ным биогеоценозом в качестве его компонента, может быть под*
тверждена и другими общеизвестными фактами, например, таким,
как постепенное преобразование разновидности почвы при есте-
ственной или антропогенной коренной смене одного биогеоценоза
другим; о том же свидетельствует и так называемый лугово-дер-
новый процесс в интерпретации В. Р. Вильямса (1949) и пр. Прав*
да, в связи с известной «консервативностью» почвы наблюдается
отставание ее от темпов развития живой части биогеоценоза, что
приводит к нарушению установившегося ранее относительного со*
ответствия их друг другу. Однако это несоответствие между ними
со временем преодолевается в ходе поступательного развития био-
геоценотического покрова, и исходная биогенная почвенная среда
под действием живой части биогеоценоза преобразуется в должном
направлении.
Из всего изложенного следует, что почва как тип, характерный
для данного типа биогеоценозов, не является, строго говоря, ком-
понентом какого-либо конкретного биогеоценоза. Подлинным ком-
понентом такого биогеоценоза может быть лишь почвенная разно-
видность, свойственная ему, находящаяся в относительном соот-
ветствии в данный отрезок времени с потребностями живых
компонентов биогеоценоза, в состав которого они входят, преобра-
зованная ими в соответствии с этими потребностями. Иначе гово-
ря, компонентом определенного биогеоценоза является сформиро-
вавшаяся в его составе в течение относительно длительного
отрезка времени биогенная почвенная разновидность, подогнанная
к выполнению вполне определенных функций в биологическом
круговороте веществ данной биогеоценотической системы. Все это
находится в полном соответствии с основным универсальным прин-
ципом Жизни: все живое само создает из материального окруже-
ния необходимые условия своего существования и поступательного»
развития.
Глава VI
Взаимосвязь биологических
и биокосных систем со средой
Прежде чем рассматривать вопрос о взаимосвязи биологиче-
ских и биокосных систем со средой, необходимо выяснить и уточ-
нить: что же представляют собою взаимодействующие стороны?
Живые компоненты биогеоценоза, как мы видели, состоят из био-
логических видов, точнее — видовых популяций (ценопопуляций),
каждая из которых включает бесчисленное множество относящихся
к ней организмов, особей. Сами живые компоненты, представляю-
щие собою ценокомплексы ценопопуляций автотрофов, биотрофов
и сапротрофов, в определенном сочетании образуют биоценозы.
Биоценозы и их последовательно соподчиненные составные эле-
менты представляют собою биологические системы различного
уровня организации и степени интеграции жизни. Во взаимосвязи
со свойственной им средой обитания они (биоценозы) образуют
биокосные системы — биогеоценозы, которые являются элементар-
ными единицами биогеоцепотического покрова Земли.
1. Среда биологических и биокосных систем
и ее подразделение
Что же представляет собою среда биологических и биокосных
систем? Б. Г. Иоганзен (1959) при обсуждении этого вопроса раз-
личает следующие понятия: среда (или среда жизни), среда оби-
тания и биотоп; каждому из них он дает соответствующее опреде-
ление. «Среда, среда жизни — наиболее общее понятие, опре-
деляющее всю совокупность внешних факторов, воздействующих
на живое в пределах биосферы. Среда есть физическое окружение
организма, в котором действуют различные абиотические, биотиче-
ские и антропические факторы» (с. 100) *. Понятие «среда жизни»
в известной мере соответствует распространенному в экологии жи-
вотных понятию арена жизни. Б. Г. Иоганзен различает четы-
ре среды жизни: воду, воздух, почву и организм (для эндопарази-
тов); при этом первые две из них он считает элементарными сре-
дами жизни, а третью и четвертую — сложными. Вода в то же
время, по его мнению, является первичной средой жизни, осталь-
ные же — вторичными. В каждой из названных сред жизни автором
выделяются (помимо качественно различной пищи) специфические
* Разрядка автора.
158
факторы: в воздухе, например, температура, свет, влажность; в.
воде — температура, кислород, соленость, давление; в почве — ре-
акция (pH) и др. Среды жизни, по его словам, отличаются не
только своей физической природой, но и условиями жизни.
«Среда обитания — подразделение среды жизни, характе-
ризующееся географической и экологической определенностью»
(с. 100) *. Например, вода — соленая и пресная, текучая и стоячая;
почвы в зависимости от географического положения — подзоли-
стые, черноземные и т. п. «Биотоп — участок среды обита-
ния с более или менее однородными условиями жизни, как абиоти-
ческими, так и биотическими» (с. 100) *.
В. Б, Сочава (1974), применяя системный подход при обсуж-
дении вопроса о среде природных геосистем разного ранга, счи-
тает, что в соответствии с этим и географическая среда должна
подразделяться па иерархически соподчиненные категории: среда
ландшафтно-географической области, зоны, провинции, округа
т. д. - до iuiii мсныпей единицы среды, соответствующей выделу
фации, ннзппнной элементарной геох о р о й. Понятие среды
в общей форме он рассматривает как внешние условия функциони-
рования геосистемы в конкретных границах; при этом он приводит
следующее определение понятия среды из переведенной на русский
язык работы Л. Д. Холла и Р. Е. Фейджина (1969): «Для данной
системы окружающая среда есть совокупность всех объектов, из-
менение. свойств которых влияет на систему, а также тех объектов^
чьи свойства меняются в результате поведения системы» (с. 258).
Близких взглядов ла среду биологических и биокосных систем
придерживается и Г Гохлернер (1971), который так же, как и
В. Б. Сочава, считает, что понятие «среда» на разных уровнях
организации жизни неодинаково. В планетарном масштабе среда
живого вещества (по В. И. Вернадскому) им трактуется как
«комплекс физико-химических условий (абиотические факторы), в
которых протекает жизнь» (с. 67). В понятие же среды биоцено-
зов, видов, популяций и особей, наряду с географическими (абио-
тическими) условиями обитания, он включает также и биотическое
окружение, т» е. другие биоценозы, виды, популяции и особи, ко-
торые по отношению к названным биосистемам выступают, по его*
мнению, как факторы среды. Автор полагает, что эволюция орга-
нического мира сопровождается эволюцией и внешней среды; в ней
появляются новые биосистемы и их сочетания, которые не могут
существовать в отрыве от других подобного рода биосистем, как
не могут они существовать и без воды, воздуха или температуры
в определенном диапазоне. Особого внимания заслуживает подчер-
кивание автором наличия у биосистем, в частности у организмов,.
♦ Разрядка автора.
159
внутренней среды и сложной системы регуляции, поддерживающей
ее в состоянии относительного постоянства. Правда, система регу-
ляции внутренней среды организма, по его словам, непрерывно со-
относится с внешней средой, оповещая, информируя о всех видах
энергии, с которыми организм вступает в контакт.
Б. А. Быков (1970) усматривает у биоценозов, а также у коло-
ний растений и животных, наличие внутренней среды, которая
создается организмами при их совместном существовании и, сле-
довательно, является биогенной. Что же касается внешней
среды биосистем, то, по мнению Б. А. Быкова, ее имеет вокруг себя
каждый организм, даже микроскопический. Она, как полагает ав-
тор, представляет собою сферу измененной организмом внешней
физической среды, названную им эд а сферой. Объем эдасферы,
разумеется, неодинаков у разных видов; наиболее велик он у выс-
ших растений, особенно у деревьев с их обширной кроной, мощной
корневой системой и значительной массой отмерших органических
остатков на поверхности почвы.
Среда, среда жизни или арена жизни как наиболее общие по-
нятия обычно подразделяются на более мелкие соподчиненные
топологические единицы. Немецким зооэкологом Р. Гессе (Hesse,
1924) арена жизни подразделяется па жизненные области —
биоциклы (например, суша, море, пресные воды); биоциклы
делятся на жизненные округа — биохоры (например, степь, пу-
стыня, тундра, морской берег); биохоры распадаются на биото-
п ы (например, пустыня песчаная, глинистая, каменистая, щеб-
нистая; морской берег илистый, песчаный, галечниковый, ра-
кушечный). Биотоп — это основная единица арены жизни. Среди
биотопов различаются фации, которые представляют собой мо-
дификации биотопа (например, пески бугристые, грядовые, бар-
ханные). Эта система арены жизни не получила широкого распро-
странения в экологии.
Советским зооэкологом В. В. Станчинским (1933) была разра-
ботана на основе экологического принципа система подразделения
арены жизни на следующие соподчиненные ступени: зоны
(арктическая, таежно-лесная, лиственно-лесная, степная, пустын-
ная); среды обитания (воды, болота, открытые пространства,
горные местности); урочища (воды пресные, соленые и пр.);
биотопы (болота моховые, тростниковые, осоковые и др.), пред-
ставляющие собою основную единицу арены жизни. Встречающиеся
в разных зонах разновидности тех или иных биотопов названы
стациями. Эта система арены жизни признается более конкрет-
ной и экологически обоснованной по сравнению с системой
Р. Гессе.
Сопоставляя между собой приведенные системы подразделения
арены или среды жизни, мы находим наиболее последовательной
160
в экологическом отношении систему, предложенную Б. Г. Иоганзе-
ном (1959). Однако биотоп как основную единицу среды жизни, на
наш взгляд, следует рассматривать не в качестве синонима экото-
на, как это принято в экологии, а как биогенную среду био-
ценоза, являющуюся результатом преобразования физической
внешней среды и процессе жизнедеятельности его компонентов.
2. Взаимосвязь биологических систем со средой
н с|к хы, нэиращсиием самого суще-
iimoi нч in. Суть ошибочного представ-
<и <>|>1.1пизма и среды заключалась в
.роблгму на основе такой парной фи-
фчрмн и содержание, они утверждали,
i .iKoM случае как форма, а среда — как
ПереЙдсм к самому главному — к вопросу о взаимосвязи био-
логических и биокосных систем со средой. В последнее время, как
мы видели, утвердилось представление, что на разных уровнях
органii.iaii.itii Ж1ПК1Й материи взаимосвязь разных форм жизни с
окружающей гр" |>)й протекает по-разному.
В пернут <•'
обитании. Пр"1
через всю in
острый лиг
в связи г
ваемого
стал, x.ij
лепии и
том, ми ।
лое<>''
что
соде
Иг>|
жаП'
аии
ней среды н этом процессе, к попытке существование и развитие
и, рассмотрим взаимосвязь организма со средой
hi «opriiiiu.iM it среда» красной нитью проходит
• энолюционных учений в биологии, но особенно
пый хпример она приняла в недавнем прошлом
।H.iiipiiiiniiiirM некоторыми биологами так назы-
npl инн
। н ч reriioli
<’ре u ni НМ'
l.llpllllllM и
h.l H'lopilll, I. Ih
Hill IM IIЫГ ry HIH III
nine, т. о. трнкHHi.iJiH их кик дне стороны единой сущности,
in тип», что no п ||Н1Модейстпии категории «форма и содер-
ш’лущпй стариной, кок известно, признается содержание,
U4 такого рода взглядов пришли к переоценке роли внеш-
> hi
организма представить как результат прямого и непосредственного
влипним нпеишей среды, определяющей существование организма
и и iMrticiHie его. Следовательно, ими полностью игнорировалась
антпнпин роль организма во взаимодействии с окружающей сре-
дой. 1Чо внутренняя наследственно обусловленная качественная
специфика, которая находит выражение в относительной целесо-
образности его организации, в приспособлении строения его орга-
нон и даже тканей к наиболее совершенному выполнению функ-
uin'l, необходимых для сохранения его как индивидуума и самовос-
Нро|13»СД1Ч1ИЯ.
Как же и действительности осуществляется взаимосвязь и взаи-
модействие организма с окружающей средой? Э. В. Волкова
(1974) обращает внимание на большую зависимость биологических
систем, в том числе и организмов, от внешней среды, ибо необхо-
димым условием существования организма является, как известно,
161
непрерывный обмен веществом и энергией с внешней средой, по-
стоянное обновление его элементов. В то же время она преду-
преждает от переоценки ее роли в существовании и развитии
организма. Развитие организма, по ее словам, автономно, незави-
симо в известных пределах от окружающих условий. Автономность
существования и развития организма выражается прежде всего в
наличии свойства избирательности в усвоении ресурсов среды па
основе своих внутренних потребностей, а также в активности орга-
низма в процессе взаимодействия со средой, выражающейся в
преобразовании ее, в приспособлении к своим потребностям. А жи-
вотные, как пишет М. П. Акимов (1959), благодаря присущей им
подвижности способны активно отыскивать необходимые им усло-
вия жизни и избегать неблагоприятных воздействий внешней сре-
ды. С другой стороны, организмы обладают свойством пластично-
сти, что дает им возможность выживать в условиях колебания
факторов среды в известных пределах.
Кроме того, у организмов в процессе длительной эволюции вы-
работался ряд специфических приспособлений к внешним услови-
ям— морфологических, анатомических, физиологических, а у жи-
вотных еще и этологических, т. е. определенного поведения в
поисках пищи, размножении, защиты потомства и т. п. Характер-
но, что организмы не только разных видов, но одного и того же
вида по-разному реагируют на воздействие света, тепла, влаги, по-
разному ассимилируют пищевые ресурсы в зависимости от своей
природы, своих внутренних качеств и свойств и даже в зависимо-
сти от стадии индивидуального развития (онтогенеза). В ходе
своего осуществления организмы как бы извлекают из внешнего
окружения необходимые для них условия существования и раз-
вития.
Таким образом, взаимодействия организма со средой обитания
протекают не в форме прямого влияния внешней среды, а в ходе
преобразования среды организмом, приспособления ее к собствен-
ным потребностям. При этом организм во взаимодействии с окру-
жающей средой выступает как основание развития, ибо
направление, характер и конечный результат развития определя-
ются его внутренней качественной спецификой. Среда же высту-
пает как источник свободной энергии и доступных для организма
материальных ресурсов, как необходимое условие осуще-
ствления процессов обмена веществ и энергии. Но действует она
на организм, так сказать, преломляясь через своеобразие его внут-
ренних свойств, и, будучи ассимилированной им, она также вносит
известный вклад в формирование его как биологической системы.
Следовательно, существование и индивидуальное развитие орга-
низма осуществляются, с одной стороны, на основе единства функ-
циональной деятельности и внутренней организации самого орга-
162
низма, а с другой — на основе единства организма как целого и
среды его обитания, точнее, извлекаемых и ассимилируемых орга-
низмом необходимых условий жизни.
На популяционно-видовом уровне организации жизни взаимо-
связь и взаимодействие такого рода биологических систем со сре-
дой складываются по-иному, чем на организменном уровне. Видо-
вая популяция, как было сказано ранее, является элементарной
единицей формообразования и эволюционного процесса; так назы-
ваемая микроэволюция совершается именно на этом уровне
организации живой материи. Поэтому результатом взаимодействия
биологического вида со средой должно стать выживание не только
организма, отдельных особей данного вида, но и конкретных посе-
лений вида, т. е. видовых популяций и вида в целом. Следователь-
но, в процессе взаимодействия видовых популяций со средой оби-
тания должно обеспечиваться не только существование и индиви-
дуальное развитие, онтогенез особей, но и эволюционное
преобразование, филогенез вида. Следует принять во внима-
ние и то обстоятельство, что в процессе эволюции органического
мира особое значение приобретает взаимосвязь видовых популяций
не только с абиотическими факторами внешней среды, но и с жи-
выми существами, как относящимися к тому же самому виду, так
и к другим видам, т. е. внутривидовые и межвидовые взаимоотно-
шения живых существ, протекающие, как известно, в форме «борь-
бы за существование» и взаимного благоприятствования, по
Дарвину.
Биологический вид как основная единица эволюционного про-
цесса представлен в природе множеством особей, но он не сумма
особей, а целостное образование, обладающее «племенной общно-
стью», однородностью химического состава биомассы, единством
воздействия на окружающую среду, наличием внутривидовых кор-
реляционных зависимостей в строении и функции органов — кон-
груэнций, по С. А. Северцову (1951), и объединенное внутривидо-
выми связями и особым строем внутривидовых отношений. Поэто-
му внутривидовые отношения и определенный уровень численности
особей, являющиеся важным фактором существования и выживае-
мости вида, входят в качественную характеристику его и должны
рассматриваться как видовое приспособление, возникающее пол
действием естественного отбора в ходе органической эволюции.
Внутривидовая конкуренция является следствием биологическо-
го неравенства, разнокачественности особей одного и того же вида
в морфологическом, физиологическом и других отношениях, что
находит выражение в различной степени сопротивляемости их не-
благоприятным факторам абиотической среды и в различном исхо-
де конкуренции с другими особями того же вида за свет, влагу,
пищу и т. п. Большая интенсивность конкуренции, по наблюдениям
163
В. Н. Сукачева (1966), имеет место между экологически близкими
особями, как относящимися к одному, так и к разным видам. Толь-
ко при условии глубокого различия в потребностях особей разных
видов борьба за существование ослабевает и осуществляется, по
выражению Ч. Дарвина, наибольшая сумма жизни при наиболь-
шем многообразии.
Межвидовая конкуренция живых существ, возникающая чаще
всего на основе питания и размножения, неизбежна. Отсюда выра-
ботка в процессе эволюции различного рода приспособлений к
борьбе с другими видами или к защите от врагов: у животных это
клыки, зубы, рога, копыта, жала, устройство нор, закрытых гнезд,
образование раковин, панцирей, игл и т. п.; у растений — шипы,
колючки, выделение ингибирующих и других химических веществ
и пр. Внутривидовая конкуренция для самого существования вида
не является непосредственно необходимой. Отсюда отсутствие спе-
циальных приспособлений для подавления особей того же вида.
Следовательно, хотя по форме внутри- и межвидовые взаимоотно-
шения организмов весьма сходны (ибо в том и другом случае
происходит борьба за «средства жизни»), однако по существу они
различны, так как ведут, каю подчеркивает В. Н. Сукачев, к био-
логически различным результатам, а именно — к сохранению и
процветанию вида, к расширению его ареала в одном случае, к
угнетению и уничтожению или вытеснению из данного ценоза —
в другом.
Указанные различия внутри- и межвидовых взаимоотношений
живых существ определяются тем, что в основе межвидовых взаи-
моотношений лежат антагонистические противоречия, а в основе
внутривидовых — хотя и острые порой, но не антагонистические,
ибо в пределах вида действуют не только внутривидовые различия
между особями, но и внутривидовая общность их.
Таким образом, на популяционно-видовом уровне организации
жизни наиболее существенное значение имеют те типы и формы
взаимоотношения организмов и видовых популяций между собой
и с окружающей внешней средой, которые обеспечивают, во-пер-
вых, воспроизводство вида, т. е. способствуют появлению и выжи-
ванию особей данного вида, увеличению их плодовитости и защите
потомства; во-вторых, само существование, развитие и преобразо-
вание вида, т. е. смену поколений, поддержание оптимального
уровня численности всех возрастных групп популяций и т. п. В
данном случае имеются в виду отношения между особями разных
полов (самцами и самками), отношения родителей к своему по-
томству, внутривидовые отношения особей на базе совместного
добывания пищи, защиты от врагов и неблагоприятных воздей-
ствий внешней среды и пр. Следовательно, взаимосвязь и взаимо-
действие живых существ на популяционно-видовом уровне со сре-
164
дой обитания и друг с другом становятся условием существованъ
и развития вида как целостной биологической системы и как одй<
го из элементов ценотического уровня организации живой матери]
Отличительной особенностью взаимосвязи и взаимодействи
живых систем (биосистем) с окружающей внешней средой и др}
с другом на биоценотическом уровне организации жизни являете
образование в результате этого биокосных систем, названных 6ut
геоценозами. В связи с этим особый смысл приобретает проблем
внутренней и внешней среды данной биокосной системы. Возникав
вопрос: есть ли «среда» у биогеоценоза? Ведь приземной ело
атмосферы, почва, атмосферные и почвенно-грунтовые воды «орг;
нически», так сказать, входят в состав биогеоценоза в качестг
таких биогенных компонентов, как аэротоп.и эдафотоп, составляй
щих в совокупности экотоп, по Т. А. Работнову.
Вопрос о соотношении понятий, отражающих реальность супц
гтионапин биоценоза как совокупности живых компонентов биогес
ценоза и экотона (или биотопа), как совокупности косных ег
компонентов, в общей форме был рассмотрен во II главе (раз
дел 4). При этом были сопоставлены взгляды В. Н. Сукачева и ег
оппонентов по этому вопросу и показано, что для отрицания р<
альпости существования такого рода ценотических сочетаний Ж1
вых компонентов, с одной стороны, и косных — с другой, нет оснс
вапий. Единство биоценоза и экотопа (биотопа) в составе биогес
ценоза, по словам П. М. Рафеса (1980), следует рассматривать в
как «некое материальное слияние живого и косного», а как фуш
циопальпое объединение, при котором «все биотические и абйотр
ческие компоненты системы остаются дискретными» (с. 85).
Там же было показано также, что в процессе формировани
биогеоценоза происходит преобразование физической биокосно
среды (экотопа) под воздействием биоценоза во внутреннюю бис
генную его среду-биотоп, называемую также биоценосредой. Веле/
ствие этого биотоп (биоценосреда) становится одной из сторо
биогеоценоза, вторую сторону которого представляет биоцено:
При этом А. П. Шенниковым подчеркнуто, что экотоп в смысл
физического субстрата биоценоза с его факторами воздействия н
исчезает бесследно, а сохраняется в границах биогеоценоза, хот
и в соподчиненном положении. Отсюда, как мы видели, он прихс
дит к выводу: биотоп — это не сумма факторов косной физическо
и нпутренней биогенной среды, а весьма сложное диалектическо
сочетание факторов той и другой. Следовательно, все живое «па
селение» сформировавшегося биогеоценоза существует в условия
именно такой синтетической биоценосреды, изменяясь и эволюцис
пируя вместе с нею.
Биоценотическая среда биоценоза, являющаяся в силу сказан
ного одновременно внутренней средой биогеоценоза в целом, вклк
165
чает в свой состав надземную (воздушную) и почвенно-грунтовую
части, которые Н. В. Дылисом (1978) трактуются как ее надзем-
ный и подземный (почвенный) блоки. Она, по словам Б. А. Быкова
(1970), характеризуется наличием специфического биогенного типа
почв, особого состава воздуха с определенным соотношением био-
генных газов (кислорода и углекислоты), особого радиационного
режима с уменьшенным количеством физиологически активной
радиации, особых теплового и водного режимов и т. п.
3. Взаимосвязь биокосных систем
с природными факторами среды
Однако биогеоценоз — это не замкнутая в своих границах, а
функционально относительно открытая биокосная система. Пред-
ставляя собою основную элементарную единицу биогеосферы, он
связан с другими биогеоценозами, как смежными, так и порой
весьма отдаленными. Кроме того, функционирование и развитие
биогеоценоза находятся в зависимости от непрерывного обмена
веществом и энергией с окружающей его абиотической внешней
средой, и, наконец, на него оказывает прямое или косвенное влия-
ние целый ряд природных явлений более крупного масштаба (пла-
нетарных, космических), а также хозяйственная деятельность мест-
ного населения.
Исходя из этого, В. Н. Сукачев (1964), как известно, помимо
компонентов биогеоценоза выделял условия, или факторы,
оказывающие то или иное воздействие на биогеоценоз, его компо-
нентный состав и функциональную деятельность. Среди них он
различал: а) космические факторы (лучистая энергия Солнца,
ультрафиолетовые и другие излучения ближайшего космоса);
б) планетарные факторы (земное притяжение — гравитация,
электромагнитные и другие физические явления); в) эндогенные
процессы земной коры (вулканические извержения, землетрясения,
поднятия и опускания земной коры и т. п.); г) экзогенные процес-
сы на земной поверхности (водная эрозия, дефляция, суховеи,
пыльные бури и пр.); д) рельеф и геологическое сложение местно-
сти, перераспределение прямодействующих факторов в связи с
этим; е) хозяйственная деятельность человека, позитивные и нега-
тивные последствия ее. Все эти факторы, за исключением послед-
него, являются природными, а последний можно назвать антропо-
генным или социально-производственным.
Взаимосвязь биогеоценоза с другими биокосными системами
подобного рода; а также с абиотическими факторами окружающей
среды в ходе обменных процессов с надлежащей подробностью
166
будет рассмотрена в последующих главах. Здесь же главное вни-
мание надлежит уделить перечисленным внешним факторам, опре-
деляющим в совокупности те условия, в которых происходит и
происходило в геологическом прошлом формирование, функциони-
рование и поступательное развитие биогеоценозов как целостных
структурных единиц биогеоценотического покрова Земли. Ввиду
широкомасштабности упомянутых факторов среды воздействие их
не ограничивается рамками биогеоценозов, а охватывает порой
значительные территории биогеоценотического покрова на земной
поверхности, а также значительные по длительности отрезки гео-
логической истории планеты, в особенности четвертичного периода
кайнозойской эры.
Прежде всего о рельефе местности и его геологическом сложе-
нии, которые оказывают преимущественно Косвенное воздействие
на биогеоценозы и па их составные ценотические элементы. Оно
цырижиетсн главным образом в перераспределении, функциональ-
ной дифференциации таких постоянно действующих экологических
факторов, как сиетоиой и тепловой режимы, режим влажности
почвы, режим минерального питания ценозообразующих видовых
ценопопуляций и пр.
Характер и формы рельефа в значительной степени определя-
ются составом и сложением горных пород, характеризующихся,
как выяснилось в последнее время, помимо внешних признаков и
химических особенностей, специфической микроструктурой. Наибо-
лее распространенными из них являются равнины, которые, по
свидетельству В. В. Добровольского (1972), резко преобладают
над горными областями, занимая около 90% суши земного шара.
Равнины довольно разнообразны. В пределах пашей страны встре-
чаются низинные заболоченные равнины (Западно-Сибирская),
всхолмленные равнины (Восточно-Европейская), возвышенные
плато, так называемые «поверхности выравнивания» (Хибины,
Крымская яйла, Устюрт и др.). Неоднородны равнины на земной
поверхности и по своему происхождению (генезису), по геологиче-
скому сложению и составу горных пород, по характеру четвертич-
ных покровных отложении и пр. Характерной особенностью об-
ширных равнин является отчетливо выраженная широтная ланд-
шафтно-географическая зональность. На территории европейской
части нашей страны обычно выделяются следующие ландшафтные
зоны: тундровая, лесная и степная с лесотундровой и лесостепной
подзонами между ними.
Однако поверхность равнин не является идеально ровной пло-
скостью. Они обычно бывают изрезаны речными долинами, овра-
гами и балками; среди равнин встречаются невысокие пологоскло-
новые останцы эродированных коренных пород, различные по
форме и простиранию, перекрытые различной мощности четвер-
167
-гичными и современными рыхлыми отложениями. На территории
былого распространения материкового оледенения встречаются
грядово-моренные слабые возвышенности и зандровые всхолмле-
ния, чередующиеся местами с гляциально-озерными заболоченными
понижениями и пр. В соответствии с этим и биогеоценотический
покров не является вполне однородным. В пределах хвойно-лесной
(таежной) полосы европейской части Союза на невысоких водо-
разделах прослеживается такой экологический ряд биогеоценозов;
плоские вершины и верхние части пологих склонов с бедными вы-
щелоченными почвами обычно бывают покрыты ельниками брус-
ничными, ниже по склону на более увлажненных и минерально
богатых почвах они сменяются ельниками черничными, а еще ниже
по склону по мере возрастания увлажнения и заболачивания —
ельниками долгомошными и затем ельниками сфагновыми, пере-
ходящими в замкнутых или полузамкнутых низинах в сфагновые
болота. Зандровые холмы и гряды с легкими песчаными и супес-
чаными почвами заняты вересковыми, лишайниковыми и другими
сосняками, а гляциально-озерные обводненные низины — болотами
с ельниками на едва возвышающихся над ними островках. В пре-
делах северной части степной зоны на черноземных почвах рас-
пространены злаково-разнотравные степные биогеоценозы, а на
юге — дерновинно-злаковые степные. В пограничной между ними
полосе в условиях слабоизрезанного рельефа первые из них обыч-
но оказываются расположенными на северных, более увлажнен-
ных склонах возвышений, а вторые — на более прогреваемых юж-
ных склонах.
Таким образом, даже в условиях слабоизрезанного рельефа
достаточно отчетливо выявляется его влияние на набор (состав)
типов биогеоценозов и размещение их по элементам рельефа, ибо
сочетание прямодействующих экологических режимов различных
типов местоположений неодинаково. Так, температурный режим
неодинаков на склонах северной и южной экспозиций, режим
влажности и минерального богатства почвы на вершине водораз-
дела, в различных частях склона или в подножии его также раз-
личен, не говоря уже о днище примыкающей низины или речной
поймы.
Однако наиболее отчетливо воздействие рельефа на биогеоце-
нотический покров сказывается в горных местностях. Основные
формы горного рельефа—это водораздельные гребни хребтов, реч-
ные долины различной ширины, узкие и глубокие ущелья, но наи-
большая площадь горных массивов приходится на долю горных
склонов различной крутизны и экспозиции. Среди них отвесные
скалы из твердых коренных пород, крутые склоны от 60 до 45—30°,
пологие склоны и террасы различной ширины и различного возра-
ста. Кроме того, встречаются такие производные формы рельефа,
168
как осыпи в подножии скал и обрывов, селевые конусы и оползни
в подножии крутых горных склонов и пр.
Названные формы горного рельефа характеризуются большим
разнообразием и специфическим сочетанием экологических режи-
мов и взаимосвязанными с ними особенностями растительного и
биогеоценотического покрова. Так, например, горные массивы
среднеазиатских горных систем с отвесными или ступенчатыми
скалами, по наблюдениям О. Е. Агаханянца (1970), с южной сто-
роны днем нагреваются порой до 70—80°, а ночью остывают ниже
нуля; северные же их склоны и склоны массивов, затененных
смежными хребтами, постоянно охлаждены. Неодинаков и режим
увлажнения тех и других местоположений рельефа. Поэтому ска-
листые обнажения коренных твердых пород либо вовсе лишены
растительности, либо находятся на разных этапах физического и
биологического разрушения и заселения их то одноклеточными
водорослями и бактериями, то накипными лишайниками и бессте-
бельными мхами, то узкоспециализированными или в известной
степени адаптированными видами высших растений. Последние
обладают мощными, прочными на разрыв корнями, выделяющими
различные кислоты, способствующие проникновению их в глубь
твердых пород и освоению их жизнью.
Для горных областей, как известно, характерна вертикальная
поясность в распределении растительных сообществ и биогеоцено-
тических систем в целом по высотным поясам, в соответствии с их
температурным градиентом и градиентом влажности воздуха.
Однако состав и высотное распределение основных единиц расти-
тельного и биогеоценотического покрова неодинаковы в разных
горных системах в зависимости от приуроченности последних к той
или иной зоне и провинции, а в пределах горной системы неодина-
ковы в зависимости от экспозиции и крутизны склона горных мас-
сивов. В общем, каждой ландшафтно-географической зоне и про-
винции соответствует свой специфический тип вертикальной пояс-
ности с характерным для него набором типов биогеоценозов и их
сочетаний и специфической последовательностью распределения
их по высотным поясам. В полном соответствии с отмеченными
особенностями в различных местоположениях рельефа природных
условий находится и размещение в горных странах сельскохозяй-
ственных культур, сенокосных, пастбищных и других угодий с уче-
том, разумеется, наличествующей местами инверсии темпе-
ратур.
С другой стороны, в природе имеет место и обратный процесс,
т. е. изменение рельефа и образование новых форм под действием
биогеоценозов, в особенности их живых компонентов. Так, на за-
болоченных лугах образуются кочки осокового происхождения, на
сфагновых болотах—торфяные бугры, гряды и мочажины между
169
ними, в лесах и степях—муравьиные и землеройные кучи, сурчи-
ны и другие формы микро- и нанорельефа.
Менее зримым, но столь же важным, преимущественно косвен-
но действующим фактором на биогеоценозы и биогеоценотический
покров Земли, являются эндогенные процессы земной ко-
ры, обусловленные действием внутриземных сил планеты. К ним
относятся: а) вулканические явления, связанные с движением в
недрах Земли расплавленной массы, так называемой магмы, под-
нятием и внедрением ее в земную кору по трещинам (разломам)
и последующим извержением на поверхность в виде лавы и сопут-
ствующих этому горячих газов, водяных паров и пр.; б) землетря-
сения, также связанные с движением расплавленной магмы в нед-
рах Земли, иногда они предваряют извержение вулканов; в) тек-
тонические движения земной коры, вызывающие медленные
поднятия и опускания ее, смятие ее верхних слоев в складки, обра-
зование разломов, надвигов и пр.
Прежде всего о средообразующей роли вулканов конца третич-
ного и современного (четвертичного) периода геологической исто-
рии Земли. Распространение их, по утверждению В. В. Белинского
(1969), связано с такими структурными элементами земной коры,
как зоны напряжения растяжения, характеризующимися трещино-
ватостью, разломами ее. Эти зоны тянутся па многие сотни и ты-
сячи километров как на континентах, так и в океанических
областях. В нашей стране зоны вулканической деятельности, как
известно, расположены на северо-востоке (Чукотка, Камчатка, Ку-
рильские острова) и на юге (Тянь-Шань, Кавказ и пр.). При из-
вержении вулканов вылившаяся на земную поверхность х магма
застывает, образуя так называемые эффузивные породы (базаль-
ты, андезиты, липариты), при застывании же магмы на некоторой
глубине от поверхности земли образуются интрузивные породы
(граниты, диориты, габбро и пр.), которые в соответствующих
условиях также могут оказаться на земной поверхности. Под дей-
ствием физико-химических агентов те и другие породы постепенно
превращаются в рухляк и становятся, таким образом, материаль-
ным субстратом для развития почвообразовательных процессов.
Выше было показано, что на застывших вулканических лавах с
течением времени формируются весьма своеобразные почвы, выде-
ленные С. В. Зонном в особый класс аэрально-вулканогенных почв,
входящих в состав соответствующих групп типов биогеоценозов.
Особого внимания среди эндогенных процессов заслуживают
тектонические движения земной коры, в особенности вертикальные
поднятия и опускания ее. С этим связано образование мощных
горных систем или обширных равнин и низменностей, развитие
эрозионных процессов и образование речных долин, оврагов и ба-
лок, повышение или понижение базиса эрозии рек (т. е. уровня
170
бассейна, в который они впадают) и скорость их течения, усиление
или ослабление глубинной эрозии русла рек или поперечной эро-
зии их берегов. Следствием этого является повышение или пони-
жение уровня почвенно-грунтовых вод, а следовательно, и степени
увлажнения, образование надпойменных террас, а в устьях рек —
лиманов (вытянутых мелководных заливов) либо, наоборот, дале-
ко углубляющихся в море (озеро) конусов выноса.
Многие районы нашей страны за несколько последних миллио-
нов лет, с конца третичного периода, по утверждению А. И. Пе-
рельмана (1970), пережили чрезвычайно сложную геолого-геохи-
мическую историю, обусловленную новейшими тектоническими
движениями земной коры и сопровождающуюся резкой сменой
климатов и природных комплексов — ландшафтов. За этот период
произошло образование большинства современных основных струк-
турных форм земной коры, как платформенных, равнинных, так и
складчатых горных систем.
Колебательные движения земной коры самого последнего —
послеледникового — времени четвертичного периода, по данным
В. Б. Сочавы (1950), нашли отражение в значительном изменении
климатической обстановки в пределах всей Северной Евразии и е
характере развития почвенно-растительного покрова и биогеоцено-
тического покрова в целом. Их влияние сказалось также и нг
взаимоотношении лесных биогеоценотических систем со степным?
в лесостепной полосе и с тундровыми — в лесотундровой. Развитие
этих процессов во второй половине четвертичного периода шло нг
базе современных форм рельефа и в разных районах нашей стра
ны складывалось по-разному. Так, на северо-востоке Русской рав-
нины в пределах тундровой зоны в периоды поднятия территори?
(сразу после отступания ледника и в самое последнее время) про
исходило, по словам В. Б. Сочавы, усиленное эрозионное расчле
нение рельефа и, в частности, углубление речных долин, улучшение
дренажа и теплового режима местности, а следовательно, прогре
вание поверхности земли и отступание мерзлоты к северу. След
ствием этого явилось значительное облесение тундры, сокращение
площади болот и тундровых биогеоценозов, продвижение север hoi
границы леса далеко в Арктику. В период же опускания террито
рии между этими двумя поднятиями ее происходило меандриро
вание речных русел и развитие долин в ширину, увеличение аллю
виальных отложений в поймах рек и озерных котловинах. След
ствием этого явилось заболачивание территории, расширены
площади тундры и смещение северной границы леса к югу. Прав
да, действие некоторых других факторов (летнее вторжение поляр
ных воздушных масс и пр.) несколько сдерживало развити<
указанных процессов.
Для таежной зоны Русской равнины, по данным В. Б. Сочавы
особенно характерно совпадение общего хода заболачивания тер
171
ритории с ритмом новейших тектонических движений земной коры.
В период поднятия данной местности после отступления ледника
образование и развитие болотных массивов происходило преиму-
щественно на месте озерных впадин путем зарастания озер или
нарастания па водной поверхности травяно-моховых сплавин. В
период же опускания суши в так называемое бореальное время
голоцена образование болот и расширение их площади происходи-
ло в основном путем заболачивания лесных массивов. В современ-
ный период в связи с незначительным тектоническим поднятием
данной территории процесс распространения болотных ценозов
сравнительно ограничен.
Более рельефно влияние новейших вертикальных тектонических
движений земной коры на характер биогеоценотического покрова,
как указывает В. Б. Сочава, проявляется в пределах лесостепной
полосы Русской платформы. Установлено, что эта территория в
последнее время находится в состоянии поднятия (Средне-Русская,
Волыно-Подольская, Приволжская и другие возвышенности). С
этим связано усиление эрозионных процессов на их поверхности,
некоторые микроклиматические особенности и процессы выщела-
чивания почвы. Таким образом, природные условия здесь благо-
приятствуют развитию и распространению лесных биогеоценозов
за счет степных. Однако вмешательство человека препятствует
развитию этого процесса.
Иное эколого-ценотическое выражение тектонические движения
земной коры приобретают в пределах горных систем, особенно в
зонах наиболее молодой альпийской складчатости. Так, например,
вследствие поднятия в четвертичный период Главного Кавказского
хребта, по свидетельству В. Б. Сочавы (1950), произошло а) общее
повышение влажности климата, в особенности в высокогорье, и
мезофитизация ксерофитной флоры в новых условиях; б) надвига-
ние лесных типов биогеоценозов на степные и вытеснение их либо
олуговение в условиях высокогорного климата (например, пестро-
овсяничники); в) расселение в нижнем поясе более влаголюбивых
лесных типов биогеоценозов — буковых за счет вытеснения ими
мезофитных дубовых биогеоценозов. В области более слабых вер-
тикальных поднятий в послеледникоцре время, например на Се-
верном Урале, произошло лишь снижение верхнего предела дре-
весной растительности из лиственничного редколесья и смена его
лишайниково-кустарничковыми сообществами горно-тундрового
типа.
Таким образом, эндогенные процессы земной коры, в особенно-
сти вертикальные тектонические движения ее, оказывают весьма
существенное влияние на формирование, развитие и смены природ-
ных биогеоценозов и биогеоценотического покрова в зонах актив-
ного проявления названных процессов. Осуществляется воздей-
172
ствие эндогенных процессов на биокосные системы опосредованно,
путем значительного преобразования макрорельефа и изменения
климатических условий данной территории. При этом, несмотря на
то что в основе тектонических движений земной коры и других
эндогенных явлений лежат глубинные процессы, происходящие в
недрах Земли, характер воздействия их на органический мир, как
отмечает В. Б. Сочава, песет на себе печать широтной зональности
и вертикальной поясности.
Значительное воздействие на биокосные системы оказывают и
экзогенные процессы, протекающие на поверхности нашей
планеты: водная эрозия поверхности суши и другие близкие к ней
процессы, ветры различного характера и интенсивности и пр. Воз-
действуют они на биогеоценозы и биогеоценотический покров как
непосредственно, так и косвенно, а их проявление и позитивные
или негативные последствия их деятельности во многих случаях
очевидны и не нуждаются в пояснениях.
Водная эрозия — это размыв водными потоками горных пород
на склонах возвышенностей и вынос продуктов их разрушения в
отрицательные формы рельефа и водоемы. Эрозия — это также
размыв талыми и дождевыми водами почвенного покрова и смыв
его частиц в русла рек и далее в моря и океаны. Близка к водной
эрозии по характеру и результатам деятельности дефляция, т. е.
обтачивание и шлифовка твердых горных пород минеральными
частицами под воздействием ветра и выдувание ветром мелкозема
почвы, а также ледниковое выпахивание движущейся массой лед-
ника в его ложе борозд и стирание твердых горных пород с обра-
зованием так называемых «бараньих лбов» и «курчавых скал».
Непосредственным результатом длительной механической и физи-
ко-химической деятельности водной эрозии и дефляции является
не только размыв и разрушение горных пород и смыв наиболее
плодородного слоя почвы, но и размывание горных склонов, обра-
зование оврагов и балок, катастрофическое расчленение равнин-
ного рельефа местности, запое и заиливание русел рек, озер и водо-
хранилищ, а на юге и засоление водоемов, образование мощных
толщ осадочных пород в морях и океанах и пр. Следствием этого
является исключение из сельскохозяйственного использования ко-
лоссального количества полезной площади, снижение продуктив-
ности сельскохозяйственных земель и рыбопродуктивности есте-
ственных водоемов и пр. С другой стороны, эоловое (ветровое)
отложение лёсса на огромных равнинных пространствах имеет
положительное значение, ибо способствует повышению плодородия
земельных угодий.
Воздействие ветров на биогеоценозы и биогеоценотический по-
кров определяется их характерными особенностями. На юго-восто-
ке европейской части Союза и в Казахстане формируются сухие и
173
горячие ветры — суховеи, дующие в теплое время года, темпера-
тура которых достигает 35—40°С, а относительная влажность сни-
жается до 15% и ниже. В пустынях Африки и Аравийского полу-
острова обычно в продолжение нескольких часов дует знойный и
сухой ветер — самум, который несет массу мелкого песка и пыли,
затемняющих Солнце, а в южных горах — теплый и сухой, нисхо-
дящий по склону ветер — фён, возникающий вследствие разницы
атмосферного давления по обе стороны хребта. Нетрудно предста-
вить себе, какой колоссальный вред приносят сухие и горячие вет-
ры всем живым компонентам степных и пустынных биогеоценозов
как непосредственно, так и путем иссушения воздуха и почвенного
покрова. Кроме того, на севере пашей страны нередко в зимний
период свирепствуют снежные бураны, достигающие огромной ско-
рости (до 40—50 м/сек и более), а на юге и юго-востоке ранней
весною — пыльные (черные) бури. Те и другие, обладая огромной
разрушительной силой и неся с собой массу сыпучего материала,
также наносят большой и разнообразный вред живым компонен-
там биогеоценозов, биогеоценотическому покрову и хозяйству
местного края, обнажая почву в одних местах и засыпая в других.
В целях предотвращения или смягчения губительных и вредных
воздействий названных экзогенных процессов на природные био-
косные системы и культурные сельскохозяйственные угодья соот-
ветствующими государственными органами и местным населением
принимаются надлежащие меры: а) создание государственных
защитных лесных полос от сухих горячих ветров на юге и юго-во-
стоке и от холодных ветров на севере страны; б) создание местных
полезащитных лесных полос на степных равнинах, противоэрози-
онных и водопоглощающих полос на склонах водораздельных ува-
лов; в) сооружение магистральных республиканских и сети мест-
ных оросительных каналов в сухих степях Прикаспия и Северного
Кавказа и в знойных пустынях Средней Азии.
Важными факторами воздействия на биокосные системы и их
составные компоненты и элементы являются такие планетар-
ные явления, как гравитационные, магнитные и электрические
поля, которые представляют собою особую форму материи, суще-
ствующую наравне с такой формой материи, как вещество, и кото-
рые в силу этого обладают определенными физическими свойства-
ми. Названные статические поля, как указывает В. Брагинский
(1976), жестко связаны с соответствующими объектами: гравита-
ционное поле с создавшей его массой тела, магнитное поле с по-
стоянным естественным или искусственным магнитом, электриче-
ское поле с электрическим зарядом. Статические поля при
определенных обстоятельствах могут быть преобразованы в пере-
менные поля, которые существуют уже самостоятельно в виде
электромагнитных или гравитационных волн.
174
Гравитация в общей форме — это всемирное тяготение
Гравитация свойственна, наряду с другими космическими телами
и нашей планете Земля, которая обладает также присущим ei
гравитационным полем. Сила тяжести (сила земного притяже
ния)—это равнодействующая двух сил: силы тяготения Земли i
центробежной силы, обусловленной вращением Земли вокруг свое<
оси. Сила тяжести наглядно проявляется при падении физически:
тел, предметов, веществ в направлении к центру Земли, например
при падении снежной лавины, потока воды и т. п. При этом осво
бождается энергия, затраченная в свое время при осуществлени!
соответствующих процессов на подъем этих физических тел н;
определенную высоту; иначе говоря, при падении тел совершается
переход потенциальной энергии в кинетическую. Перемещения тел
веществ под действием силы тяжести называются гравитационным
процессами.
Сила тяжести (земного притяжения), по словам Н. В. Бочуро
вой и Е, Г. Мининой (1968), представляет собою совершенно осо
бый по стихийности и значимости фактор внешней среды. Ош
играет важную роль в образовании различных форм рельефа, i
особенности в горных местностях. Велика ее роль и в жизнедея
тельности живых существ, а следовательно, и образуемых им>
цепокомплексов биогеоценозов. Известны определяемые силой тя
жести явления положительного и отрицательного геотропизма j
растений, различные типы ветвления древесных пород и травяни
стых растений, положение и угол наклона ветвей на стволах i
стеблях относительно земной оси (или поверхности земли), чт<
сказывается на закладывании цветочных почек и, следовательно
на плодоношении древесных пород и плодовых деревьев, а также
на интенсивности фотосинтетической продуктивности растений
Сила тяжести оказывает самое разнообразное влияние и на жизне
деятельность, а также состояние животного населения биогеоцено
зов, как прямое, непосредственное, так и косвенное.
Планета Земля обладает не только свойством гравитации i
гравитационным полем, но и свойством магнетизма и маг
нитным полем. Магнитное поле — это пространство, в которое
обнаруживается действие магнитных сил. Земля имеет Северный i
Южный полюсы, где сходятся силовые линии магнитного поля
Магнитные полюсы по своему расположению близки географиче
ским полюсам земного шара, но не совпадают с ними, а кроме
того, они дрейфуют. Причиной магнитного поля Земли, по мнении
большинства исследователей, служат конвективные движения i
жидкой, т. е. расплавленной массе в недрах Земли и ее вращение
хотя М. Штеенбек (1970) полагает, что основным источником маг
нитного поля Земли являются турбулентные вихревые движение
в жидкой зоне планеты. Дело в том, что оболочка и твердое ядре
175
Земли, как утверждает М. Штеенбек, не имеют жесткого соедине-
ния, поэтому они могут вращаться в жидкой массе с различной
угловой скоростью.
Магнитное поле Земли, как считают многие ученые, выполняет
защитную функцию всего живого от вредных космических лучей.
А. Б. Коган (1977) приводит данные о влиянии магнитного поля
Земли на состояние и поведение животных. Так, с повышением
напряжения магнитного поля снижается дыхание улитки, умень-
шается потребление кислорода термитами, изменяется также дви-
гательная активность мышей. Повышенная магнитная активность
планеты, обусловленная интенсивностью космических излучений,
по данным того же автора, оказывает влияние и на состояние че-
ловеческого организма. В магнитно-активные дни наблюдается
чаще обычного острое начало инфаркта миокарда, наиболее вы-
сокая смертность больных инфарктами, повышенная возбудимость
нервных и психически больных людей и пр.
Велико значение разного рода космических излучений
как факторов прямого или косвенного, опосредованного воздей-
ствия на биокосные системы разного ранга и их компоненты. Из
них, как отмечает А. Б. Коган (1977), только световые, ультрафио-
летовые и радиоволновые излучения проникают через ионосферу
и, достигая поверхности Земли, оказывают прямое воздействие на
живые существа. Но космические излучения, будучи поглощенны-
ми верхними слоями атмосферы, вызывают, по его словам, обра-
зование озона, нагревание и перемещение воздушных масс, выпа-
дение атмосферных осадков, оказывая косвенное воздействие на
живые компоненты биогеоценозов, в частности, влияют на рост
древесных пород и других ценозообразующих растений, на дина-
мику численности многих видов животных, на распространение
некоторых болезней животных и человека и пр.
Особо важное значение из названных видов космических излу-
чений имеет солнечная радиация, несущая лучистую энергию
Солнца на земную поверхность, за счет которой, как было отмече-
но ранее, осуществляется фотосинтез высшими зелеными растения-
ми и фитопланктоном огромного количества первичной биологиче-
ской продукции, измеряемой сотнями миллиардов тонн, и обогаще-
ние атмосферы сотнями миллиардов тонн кислорода ежегодно.
Влияние ультрафиолетовых лучей спектра на растительные и
животные компоненты биогеоценозов также достаточно велико. По
данным И. Знаменского (1935), ультрафиолетовые лучи с длиной
волны от 290 до 400 р оказывают стимулирующее действие на
жизнедеятельность растений; известно также, что они предотвра-
щают чрезмерное вытягивание стеблей растений, но более корот-
кие из них губительны для растений. Правда, эпидермис многих
растений обладает способностью поглощать ультрафиолетовые
176
лучи и тем самым предохраняет их от гибели. Ультрафиолетовые
лучи, по свидетельству И. Знаменского, обладают сильными бак*
терицидными свойствами, стерилизуя загрязненные воды до 30 см
в глубину. Однако наиболее короткие, вредные для жизнедеятель-
ности живых существ ультрафиолетовые лучи, как известно, погло-
щаются озоновым экраном атмосферы и почти не достигают по-
верхности Земли.
Что же касается взаимодействия с окружающей средой
биокосных систем на биостроматическом уровне организации жиз-
ни, то этого вопроса приходилось касаться в тех случаях, когда
речь шла о взаимосвязи со средой не только биогеоценозов, но и
биогеоценотического покрова того или иного участка территории.
В планетарном же масштабе о взаимосвязи и взаимодействии био-
геоценотического покрова (биогеосферы) Земли и биосферы в
целом (в трактовке В. И. Вернадского) с так называемыми «гео-
сферами» нашей планеты было сказано во II главе (раздел 8).
К числу геосфер, как было отмечено, относятся атмосфера, гид-
росфера, литосфера и педосфера (почвенный покров). Перечислен-
ные геофизические косные оболочки Земли, будучи в течение
длительной геологической истории преобразованными под актив-
ным воздействием живого вещества биосферы и космических излу-
чений, и составляют в совокупности его биокосную среду.
Г Ф. Хильми (1975) обращает внимание на то обстоятельство, что
входящие в состав биокоспой среды геосферы для сохранения и
поддержания своих биокосных свойств нуждаются в непрерывном
поступлении энергии, получаемой ими в процессе взаимодействия
с живым веществом, в особенности с зеленой автотрофной расти-
тельностью. Э. В. Гирусовым (1968) подчеркивается также, что по
мере развития биосферы, заселения ее живыми существами, обра-
зующими пленки и сгущения жизни, происходит постепенное воз-
растание и усложнение взаимосвязи биосферы (точнее, ее живого
вещества) с биокосной средой и ближайшим космическим окруже-
нием, что в конечном итоге ведет к возрастанию ее устойчивости
и уравновешенности.
Таково в общих чертах воздействие крупномасштабных факто-
ров на биогеоценозы, их компоненты и биогеоценотический покров
(биогеосферу) Земли в целом. Обратное же влияние биокосных
систем разного ранга на окружающую природную среду хотя и
ограничено, но также весьма существенно. Растительный, точнее*
биогеоценотический покров Земли в целом, в свою очередь, по
словам А. П. Шенникова (1964), оказывает влияние на горные по-
роды, преобразуя их в почвы, на атмосферу, на распределение
тепла и света по земной поверхности. Без биогеоценотического, в
особенности растительного, покрова не было бы ни земной атмо-
сферы, ни почвы и т. п.
177
4. Взаимосвязь биокосных систем
с социально-производственными факторами среды
Наряду с природными факторами, условиями среды, В. Н. Су-
качев (1964) придавал большое значение хозяйственной деятель-
ности людского населения, влияние которой сказывается не только
на функционировании природных биогеоценозов и развитии био-
геоценотического покрова, но и ведет к коренному изменению
природных ландшафтов, к возникновению совершенно новых —
культурных ландшафтов, отсутствовавших ранее на пашей
планете.
В. В. Кучерук (1976) следующим образом характеризует ре-
зультаты воздействия человека на окружающую природу: «Облик
земной поверхности существенно изменен людьми. В настоящее вре-
мя практически не сохранилось участков суши, не затронутых в той
или иной мере деятельностью человека. На месте природных ланд-
шафтов возникли пашни и плантации, населенные пункты, города
и индустриальные объекты. Это создает большое количество преж-
де не существовавших экологических ниш. Повсюду разрушаются
естественные биоценозы и формируются новые. Животные, прежде
немногочисленные, становятся массовыми. Другие же виды, на-
против, сокращаются в числе или исчезают совсем. Воздействие
человека на природу становится все более многообразным, и темпы
его стремительно возрастают» (с. 5). Автор приводит многочис-
ленные данные, подтверждающие изложенное. Так, человечеством
в настоящее время хозяйственно освоено 55% поверхности суши,
в том числе под пашнями занято 13% суши, под промышленными
объектами — 2%. а 3% земель уже превращены в пустоши. Под
сельскохозяйственные культуры используются черноземные почвы
на 37—54%, черноземовидные почвы прерий — на 44, черные тро-
пические почвы Индии — на 45, оподзоленные, выщелоченные и
типичные черноземы Советского Союза — на 80—85%.
Наряду с этим, по данным Р. Дажо (1975), в текущем столетии
человечеством потеряно 5 млн км2 культурных земель, лесные пло-
щади сократились более чем на 40%; широко распространенное
возделывание монокультур (сахарного тростника, кофейного дере-
ва и т. п.) привело к истощению почв на значительной площади.
В результате чрезмерно интенсивного рыбного промысла произо-
шло резкое сокращение численности многих морских видов рыб, а
в связи с хищническим истреблением человеком полностью исчезли
с лица Земли 162 вида птиц и более 100 видов охотничье-промыс-
ловых млекопитающих.
Вслед за Вернадским И. П. Герасимов (1973) пишет: «...произ-
водственная деятельность людей стала мощным фактором глобаль-
178
вого воздействия на природу, сопоставимым с деятельностью гео-
логических или космических сил» (с. 4).
В каких же формах выражается хозяйственное воздействие
современного людского населения на биокосные системы и природу
в целом? В. В. Кучерук называет следующие формы: а) лесные и
степные пожары; б) рубка леса; в) сенокошение; г) выпас скота;
д) сооружение искусственных водопоев; е) появление антропоген-
ного микрорельефа; ж) осушение болот; з) сооружение водохра-
нилищ и каналов; и) искусственное орошение; к) распашка, посе-
вы, плантации; л) возникновение человеческих сооружений и
населенных пунктов. К этому, очевидно, надо добавить: м) откры-
тые и шахтные горнорудные разработки; н) промышленные пред-
приятия; о) тепловые и гидроэлектростанции; п) атомные электро-
станции^) железнодорожные и портовые сооружения; с) железные
и шоссейные дорожные магистрали; т) устройство государственных
и местных лесных полос; у) интродукцию и акклиматизацию цен-
ных древесных пород, животных и пр.; ф) создание культурных
лесных, полевых и кормовых ценозов; х) организацию государ-
ственных заповедников, заказников и т. п.
Перечисленные формы производственной деятельности людско-
го населения можно подразделить на три категории. Большинство
из них относится к сельскохозяйственному производству и лесному
хозяйству. Другая группа — это предприятия горнодобывающей и
перерабатывающей промышленности, энергетические сооружения,
а также города и другие населенные пункты. Третью группу со-
ставляют различные устройства и мероприятия, предназначенные
для защиты, охраны и воспроизводства биологических ресурсов
планеты.
Не вдаваясь в детальное обсуждение существа, результатов и
последствий производственной деятельности населения, обратим
внимание па то, что она представляет собою исторически сложив-
шуюся форму общения человека с природой, активного освоения
и использования природных ресурсов, приспособления их посред-
ством применения орудий и средств труда к своим насущным по-
требностям; иначе говоря, она является необходимым условием
существования человека, условием его общественного развития.
Вместе с тем разнообразная производственная деятельность люд-
ского населения, неизбежно связанная с вмешательством в ход
природных процессов, сопровождается, как мы видели, целым ря-
дом не только запланированных позитивных результатов, но и не-
предвиденных негативных последствий.
По мере количественного роста и качественного преобразования
форм' общественного производства и его энерговооруженности,
обусловленных научно-техническим прогрессом, естественно, на-
растают также темпы и масштабы воздействия человека на биоло-
179
гические и биокосные системы и окружающую среду, а следова-
тельно, и негативные последствия этого процесса. Самыми
нежелательными и повсеместными последствиями производствен-
ной деятельности человека в настоящее время являются разного
рода загрязнения воздуха, воды, почвы и изменение вслед-
ствие этого их физических, химических или биологических свойств.
Последние оказывают неблагоприятное влияние на состояние и
продуктивность биологических и биокосных систем, на здоровье и
жизнь человека, а также на ход производственных процессов, на
орудия и средства производства, на материальные и культурные
ценности людей и т. п. Все это в конечном итоге ведет к истоще-
нию, сокращению и потере природных, в частности биологических
ресурсов, к ухудшению условий жизни и производственной дея-
тельности людского населения и к непроизводительным затратам
сил и средств на ликвидацию или нейтрализацию разного рода за-
грязнителей среды.
Наиболее распространенными и универсальными загрязнения-
ми и загрязнителями воздуха, воды, почвы являются: а) бытовые
отходы — твердые (металлические банки, склянки и т. п., посту-
пающие обычно на свалку), жидкие (канализационные сточные
воды), пищевые, которые могут быть использованы и во многих
городах используются в качестве дополнительного корма для до-
машних животных; б) промышленные и транспортные отходы и
отбросы — твердые (обломки, обрезки, стружка, шлаки, вырабо-
танные горные породы, образующие отвалы, терриконы, а также
загрязняющие воздух, — дым, копоть, угольная пыль, сажа и пр.),
жидкие (нефть, отработанные сточные воды, в том числе горячие,
содержащие соли ртути, фенольные соединения и пр., а потому
нередко вредные для здоровья человека и ядовитые), газообраз-
ные (сернистый газ, угарный и углекислый газы, загрязняющие
атмосферу) и радиоактивные вещества и отходы; в) сельскохозяй-
ственные отходы — пестициды (инсектициды, гербициды, фунгици-
ды и т. nJ, загрязняющие почву и сточные воды.
По свидетельству Ю. Одума (1975), в развивающихся странах
преобладают органические загрязнения, связанные с испражнения-
ми животных и человека, являющиеся причиной целого ряда
инфекционных заболеваний, а в развитых странах — агроиндустри-
алъные химические загрязнения. Все виды загрязнений с экологи-
ческой точки зрения, по мнению того же автора, целесообразно
подразделить на две категории: а) стойкие загрязнения, которые
в естественных условиях, по его словам, либо вовсе не разрушают-
ся, либо разрушаются очень медленно, так как не существует при-
родных процессов, которые могли бы их разлагать с такой же
скоростью, с какой они поступают в природные биогеоценозы, а
потому они имеют тенденцию накапливаться и даже биологически
180
усиливаться по мере прохождения в биохимических циклах и по
пищевым цепям; б) нестойкие загрязнения, разрушаемые биологи-
ческими процессами, так как в природе существуют естественные
механизмы переработки их, либо некоторые из них (например,
теплые водяные пары) рассеиваются в пространстве естественным
путем или вторично используются в производстве.
Как же должны быть построены взаимоотношения человека с
природой в современный период бурного роста населения и науч-
но-технической революции, ведущей к превращению биосферы в
биотехносферу? Нельзя, очевидно, в этой ситуации изме-
нить направление или замедлить темпы развития промышлен-
ного и сельскохозяйственного производства. Напротив, решениями
XXVII съезда КПСС поставлена задача всемерного ускорения про-
изводственной деятельности всех отраслей народного хозяйства па
основе дальнейшего развития научно-технического прогресса.
Однако при этом необходимо учитывать естественный ход происхо-
дящих в природе процессов, ибо игнорирование этого чревато по-
рой непоправимыми последствиями.
Ю. А. Жданов (1973) обращает внимание на социальную
обусловленность форм воздействия общества на природу. «Капи-
тализм, будучи формацией, основанной па антагонистических про-
тиворечиях,— пишет он,— с одной стороны, способствует развитию
новых очеловеченных (и обесчеловеченных) форм природы, а с
другой — приводит к их разрушению, к гибели» (с. 29). Более того,
он «создает такие природные объекты, которые направлены против
человека и грозят ему уничтожением» (с. 29). «Социалистическая
система хозяйства, — продолжает автор, — дает возможность под-
чинить контролю общего интеллекта все условия функционирова-
ния производства, в том числе и взаимоотношения с природой»
(с. 30). Задача заключается в том, как пишет Ю. А. Жданов, «что-
бы общественное производство не допускало стихийного искажения
или разрыва естественного природного цикла, а включалось в него
в качестве необходимого звена, подчиняя себе природу и вместе с
тем становясь условием ее дальнейшего развития» (с. 32).
Подобного рода идеи могут найти и уже частично находят кон-
кретное воплощение в следующих формах охраны природы и ра-
ционального использования природных ресурсов. Во-первых, рас-
четливое и бережное использование невозобновимых природных
ресурсов (каменный уголь, природный газ, нефть, подземные воды
и т. п.); во-вторых, рациональное использование возобновляемых
природных, в особенности биологических, ресурсов (растительных,
животных), сопровождаемое воспроизводством, а еще лучше —
расширенным их воспроизводством на качественно более высокой
сортовой и породной основе; в-третьих, устройство сооружений для
очистки загрязненных бытовых и промышленных сточных вод,
181
устройство дымоуловителей и пр., а также организация безотход-
ного производства и производства с замкнутым по воде циклом;
в-четвертых, рекультивация карьеров, отвалов, терриконов и дру-
гих нарушенных промышленностью земель, возвращение их в
народнохозяйственное использование путем террасирования и вы-
полаживания откосов, с последующим задернением, залужением
их многолетними травами, сидератами или насаждением на них
белой и желтой акации, серой ольхи и других древесно-кустарни-
ковых пород; в-пятых, устройство государственных и местных лес-
ных полос различного назначения, оросительных каналов, а также
создание высокопродуктивных и устойчивых полевых и кормовых
агроценозов; в-шестых, организация заповедников, национальных
парков и других особо охраняемых участков природы с целью со-
хранения уникальных (в том числе реликтовых) ландшафтов, ред-
ких и ценных представителей растительного и животного мира в
соответствующих условиях обитания. По данным А. М. Красниц-
кого (1983), в нашей стране в то время заповедников было 142, в
настоящее время их около 150. Кроме того, в различных регионах
страны создано заказников более 1500 и так называемых памятни-
ков природы — более 9 тыс.
Следует заметить, что в современных условиях вполне эффек-
тивных результатов по охране живой природы и окружающей сре-
ды, в особенности от разного рода загрязнений, возможно достиг-
нуть лишь при организации этого дела в мировом масштабе, что
при наличии большой пестроты общественно-политических систем
и уровня экономического развития стран потребует, очевидно, не-
мало усилий и времени.
Так в общей форме решается вопрос о взаимосвязи биогеоце-
нозов и биогеосферы с окружающими их природными и хозяй-
ственно-производственными факторами внешней среды. Физгеогра-
фы, как известно, рельеф местности, человека с его хозяйственной
деятельностью включают в состав компонентов элементарного
ландшафта. По В. Н. Сукачеву же к компонентам биогеоценоза
относятся лишь такие материальные тела, которые в процессе и в
результате взаимодействия друг с другом определяют сущность
биогеоценоза. Исходя из этого, рельеф местности, уровень грунто-
вых вод, сила земного притяжения и т. п. рассматриваются Сука-
чевым не как компоненты биогеоценозов, а как факторы, условия
их существования. Не считает он возможным относить к компонен-
там биогеоценоза и человека с его хозяйственной деятельностью,
несмотря на огромную силу воздействия его на природные биогео-
ценозы и биогеосферу в целом. Очевидно потому, что во взаимо-
действии с ними человек выступает не как биологическое существо;
а как существо социальное, подчиняющееся в своей деятельности
закономерностям иного порядка. Кстати сказать, включение гео-
182
графами в компонентный состав «элементарного ландшафта»
наряду с компонентами биогеоценоза также рельефа, человека со
всей его деятельностью и пр. В. Н. Сукачев считает основным кри-
терием, позволяющим различать биогеоценоз и элементар-
ный ландшафт друг от друга как объекты исследования био-
геоценологии, с одной стороны, и физической географии — с
другой.
Глава VII
Структурно-функциональная
организация биогеоценоза
Проблема структурно-функциональной организации биогеоце-
ноза— одна из самых трудных, если не самая трудная в биогео-
ценологии. Здесь еще много неясного, требующего дальнейшего
осмысливания и разработки. В особенности в отношении того, как,
в какой последовательности и какой логической взаимосвязи и
причинно-следственной взаимообусловленности происходит сочета-
ние известных элементов структурно-функциональной организации
биогеоцепотических систем, обеспечивающее непрерывность их су-
ществования в пространстве и во времени.
1. Структурно-функциональная организация биогеоценоза,
ее различные аспекты и назначение
Биогеоценоз — это не хаотическое сочетание его составных эле-
ментов и структурных частей, а соответствующим образом органи-
зованная и упорядоченная биокосная система. Эта упорядочен-
ность и организованность биогеоценозов, как указывает Н. В. Ды-
лис (1974), проявляется «в виде определенного порядка в
размещении и группировке взаимосвязанных своей работой масс
живых и косных тел» (с. 14). Суть упорядоченности и организован-
ности биогеоценотических систем состоит в обеспечении нормаль-
ного и бесперебойного функционирования как их составных эле-
ментов разного ранга, так и системы в целом. Иными словами,
обеспечить возможность биогеоценотической системе успешно
осуществлять, по выражению Дылиса, «свою генеральную функ-
цию — материально-энергетический обмен между составляющими
биогеоценоза и между биогеоценозом и окружающей его средой»
(с- 14).
Под структурой биологических и биокосных систем в биологи-
ческой литературе, по словам В. В. Мазинга (1973), понимается
«пространственно-временная организация целостных систем, выра-
жающая закономерные связи морфологических и функциональных
элементов...» (с. 148). Он различает три направления в трактовке
понятия структуры ценоза: а) структура как синоним состава це-
ноза, т. е. его популяционно-видовой, экобиоморфный и ценотиче-
ский состав; б) структура как синоним строения ценоза, т. е. эко-
лого-ценотическое пространственное расчленение его на вертикаль-
ные и горизонтальные по отношению к земной поверхности
184
структурные части; в) структура как совокупность связей между
ценопопуляциями и компонентами ценоза — трофические и другие,
обеспечивающие обмен веществ и энергии в нем.
Н. В. Дылис (1974, 1978) также различает три аспекта струк-
туры биогеоценотических систем: а) структурно-физический аспект,
характеризующий пространственную группировку и размещение
живых и косных компонентов в пределах биогеоценоза; б) функ-
циональный аспект, отражающий взаимоотношение и работу этих
живых и косных компонентов в процессе функционирования био-
геоценоза как целостной системы; в) временной аспект, под
которым имеется в виду динамика «работы» биогеоценоза и био-
геоиенотической деятельности его компонентов в течение суток,
вегетационного периода, года, ряда лет. Н. В. Дылис, как видно,
исключает из понятия структуры биогеоценоза данные, характери-
зующие его состав, но зато вводит понятие о временном аспекте,
отражающем изменение строения и работы биогеоценоза во
времени. В свою очередь В. В. Мазинг считает, что нет достаточных
оснований для выделения временного аспекта структуры биогео-
ценоза, поскольку все материальные структуры существуют в про-
странстве и во времени. Важно также отметить, что оба автора
подчеркивают тесную взаимную связь выделяемых ими аспектов в
строении и организации биогеоценозов, как разные стороны одного
и того же явления.
При рассмотрении изложенных представлений В. В. Мазипга
я Н. В. Дылиса о структурно-функциональной организации биогео-
ценозов, по нашему мнению, следует иметь в виду, что аспект
структуры, характеризующий популяционно-видовой, экобиоморф-
ный и ценотический состав биогеоценоза, нет оснований относить
к «структуре» ценоза. В данном случае речь идет именно о со-
ставе и составных элементах биогеоценоза, а не о структуре, не
о строении и не о структурных частях его. Составными элементами
биогеоценоза, как было сказано ранее, являются биологические
виды (точнее, видовые популяции), облекающиеся в процессе бы-
тия в соответствующие жизненные формы (экобиоморфы), и био-
ценотипы высших живых существ — растений и животных. К струк-
турным же частям морфологической (пространственной)
структуры биогеоценоза относятся так называемые биогеого-
рнзонты и парцеллы. От морфологической структуры биогеоценоза,
как резонно подчеркивают и Мазинг и Дылнс, следует отличать
его функциональную организацию, функциональными
структурными единицами которой являются пищевые (кормовые)
цепи и сети и так называемые консорции.
В то же время В. В. Мазинг (1973) обращает внимание на
взаимосвязь строения и функции биогеоценотических систем. «Кон^
фигурация и размещение структурных элементов,— пишет он,—
185
являются условием для их функционирования, а последнее, в свою
очередь, определяет в значительной мере морфологию ценозов»
(с. 150). Следовательно, структура и функции биогеоценоза долж-
ны рассматриваться в единстве. При этом в процессах взаимодей-
ствия структуры (как выразителя формы) и функции, надо пола-
гать, и на ценотическом уровне примат принадлежит последней»
хотя, как известно, возникшая под ее воздействием и во взаимо-
связи с ней форма не остается пассивной, а в соответствующих
условиях начинает (по принципу обратной связи) оказывать воз-
действие на функцию.
Исходя из сказанного, мы полагаем, что наиболее целесообраз-
но рассмотрение вопросов, связанных с проблемой функциональ-
ной организации биогеоценозов, предпослать рассмотрению их
морфологической (физической) структуры. «Функциональная
структура (организация. — Л. Я.), — пишет В. В. Мазинг (1980),—
это сеть взаимоотношений их элементов, раскрывающая сущность
и механизм «работы» этих систем» (с. 57). Следовательно, прежде
всего необходимо с возможной тщательностью и полнотой рас-
смотреть основные функциональные структурные единицы функ-
циональной организации биогеоценозов, т. е. пищевые (кормовые)
цепи и сети и консорции, а затем всесторонне проанализировать и
обсудить их место, роль, взаимосвязь и соотношение в составе и
функционировании биогеоцепотических систем.
2. Типы взаимоотношений между ценопопуляциями
и живыми компонентами биогеоценоза
Основным свойством живой материи, как известно, является
обмен веществ и энергии с неорганической природой и составных
элементов ее между собой. Первый этап обмена — питание, как
указывает М. П. Акимов (1959), является «узловым пунктом» мно-
гочисленных связей организмов друг с другом. Исключительно
важную роль пищи в жизни живых существ, в частности живот-
ных, подчеркивает также Р Дажо (1975). Он указывает, что ко-
личество и качество пищи способны оказывать влияние на плодо-
витость, продолжительность жизни, развитие и смерть животных.
Разнообразие пищевых рационов, по его словам, лежит также в
основе многочисленных морфологических, физиологических и эко-
логических приспособлений организмов. Автором приводятся и
такие фактические данные: количество съедаемой пищи хищными
видами животных влияет на размеры их особей, недостаточное
питание насекомых ведет к уменьшению числа откладываемых
ими яиц, при достаточном количестве пищи развитие животных
идет гораздо быстрее. Интересные сведения о значении количества
и качества потребляемой животными пищи в их жизни имеются
186
также в работе М. П. Акимова. Прежде всего он отмечает, что ко-
личество потребляемой ими пищи на единицу веса их тела зависит
от калорийности пищи, степени ее усвояемости и интенсивности
процесса обмена веществ. Он обращает внимание и на следующие
обстоятельства: малая питательность корма компенсируется пол-
нотой ее усвоения животными, обладающими удлиненным и ус лож -
ненным кишечным трактом, и интенсификацией пищеварительного
процесса; необходимость обеспечения себя достаточным количе-
ством корма в период его отсутствия или недостатка приводит к
развитию у некоторых животных способности отложения жира в
подкожной клетчатке или к развитию инстинкта запасания корма
впрок (муравьи, пчелы, жуки-навозники, а также многие мелкие
млекопитающие и даже некоторые птицы). Не менее важное зна-
чение полнота, интенсивность и регулярность обеспечения элемен-
тами углеродного и минерального питания имеют и в нормальном
росте и развитии высших растений.
В силу сказанного трофические (пищевые) связи и отношения
живых существ выступают как основные. Они лежат в основе
взаимосвязей и отношений живых компонентов биогеоценоза меж-
ду собою — автотрофного, биотрофного, сапротрофпого, а также в
основе целого ряда так называемых межвидовых взаимоотноше-
ний, возникающих на почве совместного обитания, питания и раз-
множения между видовыми ценопопуляциями внутри живого ком-
понента или трофического уровня некоторых из них.
Весьма важно различать эти два типа взаимосвязей и отноше-
ний между составляющими биоценоз живыми компонентами и их
составными элементами, так как по своему характеру и биоцено-
тичсской роли в составе, структурно-функциональной организации
и функционировании биогеоценоза в целом они далеко не одина-
ковы. Взаимные связи и отношения между видовыми ценопопуля-
циями в составе того или иного живого компонента или в составе
трофического уровня пищевой цепи, как было ранее отмечено, осу-
ществляются в форме «борьбы за существование» в широком дар-
виновском толковании, включая различные формы конкуренции и
«сотрудничества», «взаимопомощи» между ними. Этот тип взаимо-
отношений, протекающий на уровне видов и видовых популяций
живых существ, определяет в конечном итоге их место, позицию,
экологическую нишу в составе и строении соответствующе-
го живого компонента биогеоценоза, выполняющего вполне опре-
деленную роль в функционировании последнего. | Второй тип
взаимосвязей и отношений между живыми компонентами биогео-
ценоза, представляющими собою ценокомплексы автотрофных, био-
трофных и сапротрофных ценопопуляций и их ценотических соче-
таний (групп), основан на различиях в типе питания,используемых
источников энергии и образа жизни представителей названных
187
ценокомплексов и вследствие этого необходимым образом последо
вательно взаимосвязанных в единую подсистему — биоценоз.
Этот тип взаимоотношений, протекающий на уровне названных
ценокомплексов и соподчиненных им ценотических групп, опреде-
ляет функциональную организацию биоценоза и биогеоценоза а
целом в воздушной и почвенной средах, составляющих в совокуп-
ности биотоп, и тем самым обеспечивает нормальное функциони-
рование биогеоценоза и стабильность его как целостной биокосной
системы.
Межвидовые связи и взаимоотношения живых существ много-
образны и по своему существу и последствиям чрезвычайно разно-
родны. Возможно, поэтому и принципы объединения и классифи-
кации их также довольно разнообразны. В последнее время оте-
чественными и зарубежными биологами выделяются следующие
формы межвидовых взаимоотношений преимущественно среди жи-
вотного мира: 1) мутуализм — облигатная (обязательная)
взаимосвязь с обоюдной пользой для взаимодействующих видов;
2) протокооперация — взаимосвязь видов также с обоюдной
пользой для них, но она не является для них обязательной;
3) симбиоз — форма сожительства двух видов, обычно отожде-
ствляемая с мутуализмом, но И. Н. Пономаревой (1975) выделя-
ется в самостоятельную форму взаимосвязи, поскольку в отличие
от мутуализма, как она полагает, здесь имеют место некоторые
элементы паразитизма одного из взаимосвязанных видов; 4) ком-
менсализм— взаимосвязь, при которой один вид получает
пользу от этого, а для его партнера она является безразличной.
Эту форму взаимосвязи названный автор считает целесообразным
подразделить на три самостоятельные формы: а) сотрапезничество,
б) нахлеб качество и в) квартирантство; 5) сипойкия — форма
взаимосвязи, выделяемая немногими биологами, поскольку она
отличается от комменсализма лишь тем, что не обязательна для
второго вида и что между видами нет пищевых отношений;
6) хищничество — взаимосвязь хищника и жертвы, при кото-
рой один вид нападает на другой, питаясь им; 7) парази-
тизм— взаимосвязь, при которой один вид живет за счет пита-
тельных веществ другого (хозяина), поселяясь на нем или внутри
его, и тем самым угнетает его, доводя иногда до гибели; 8) полу-
паразитизм— форма взаимосвязи, встречающаяся только в
растительном мире, где некоторые виды зеленых растений при-
крепляются корнями к другим видам растений, частично питаясь
за счет их минеральных и пластических веществ; 9) аменса-
лизм— взаимосвязь, приносящая пользу одному виду, который
угнетающе действует па другой, не испытывая при этом отрица-
тельного влияния с его стороны; 10) конкуренция — взаимо-
188
связь в виде состязания, прямого взаимного подавления или в виде
борьбы за «средства жизни» (пищу, пространство и т. п.) при их
недостатке. Эта форма взаимосвязи Ю. Одумом (1975) подразде-
ляется на две самостоятельные формы: а) конкуренция как непо-
средственное взаимодействие видов живых существ и б) конкурен-
ция как взаимодействие их из-за ресурсов среды обитания;
11) антагонизм — форма взаимосвязи, встречающаяся среди
микроорганизмов, при которой присутствие в данном местообита-
нии одного вида исключает пребывание в нем другого; 12) ней-
трализм — когда ни один из совместно обитающих видов не
оказывает влияния на своего партнера.
Перечисленные формы межвидовых взаимоотношений живых
существ (преимущественно животных), входящих в состав биоцено-
зов, объединяются и классифицируются по-разному. Ю. Одум
(1975) находит возможным выделяемые им формы межвидовых
взаимодействий объединить по характеру и эффекту взаимодей-
ствия в три типа. I. Тип положительных взаимодействий — мутуа-
лизм, протокооперация и комменсализм. II. Тип отрицательных
взаимодействий — аменсализм и конкуренция (в двух формах).
III. Тип положительно-отрицательных взаимодействий — хищниче-
ство и паразитизм. Автор при этом замечает, что выделенные нм
формы взаимодействия видов можно обнаружить в любом сообще-
стве (биоценозе), даже в самом сложном; кроме того, форма взаи-
модействия той или иной пары видов в сообществе может изме-
няться в зависимости от условий или этапов онтогенеза (например,
паразитические взаимоотношения в другой момент могут смениться
комменсализмом, а затем — стать нейтральными). Ю. Одум обра-
щает также внимание на две важные закономерности: «...а) в ходе
эволюции и развития экосистем существует тенденция к уменьше-
нию роли отрицательных взаимодействий за счет положительных,
увеличивающих выживание взаимодействующих видов; б) в недав-
но сформировавшихся или в новых ассоциациях вероятность воз-
никновения сильных отрицательных взаимодействий больше, чем
в старых ассоциациях» (с. 274).
И. Н. Пономарева (1975) считает наиболее целесообразным
перечисленные формы межвидовых взаимоотношений в ценозах
объединить в основном на тех же принципах в четыре типа. I. Тип
взаимополезных отношений — мутуализм, протокооперация и сим-
биоз. II. Тип полезно-нейтральных отношений — комменсализм и
синойкия. III. Тип полезно-вредных отношений — хищничество, па-
разитизм, полупаразитизм и аменсализм. IV. Тип взаимовредных
отношений — конкуренция и антагонизм. Автор при этом подчер-
кивает, что один и тот же вид в одно и то же время может нахо-
диться в разных отношениях с окружающими видами в биоценозе:
с одними видами, скажем, в симбиотических отношениях, с други-
189
ми — в коименсалистских, а с третьими — в конкурентных.
Несколько по-иному и в иные формы сложились и складываются
межвидовые взаимоотношения в растительном мире, что нашло
отражение в соответствующей группировке и классификации их.
Отечественными биологами в последнее время выделяются следую-
щие формы межвидовых взаимоотношений в растительных сообще-
ствах: 1) механические взаимоотношения — трение стволов,
охлестывание ветвями крон других видов деревьев и кустарников,
смещение особей одного вида под влиянием разрастания особей
другого и пр.; 2) взаимоотношения между лианами н растения-
ми-опорами, а также эпифитами и их «хозяевами», являющие-
ся полезными для лиан и эпифитов и в основном безвредными для
их «хозяев»; 3) физиологические взаимоотношения сросших-
ся корнями или стволами особей разных видов растений, заклю-
чающиеся в обмене пластическими веществами между ними;
4) биохимические взаимоотношения (аллелопатия) — влия-
ние специфических прижизненных выделений (метаболитов) одного
вида растений на особи другого; 5) симбиотические взаимо-
отношения растительных видов (включая мутуализм) со взаимной
пользой для обоих симбионтов; 6) паразитические взаимоот-
ношения— это взаимоотношения паразита и «хозяина» с односто-
ронней пользой для паразитирующего вида растений и во вред для
растения-хозяина; 7) полупаразитические взаимоотношения
некоторых видов зеленых растений, которые частично используют
минеральные и другие питательные вещества растений-хозяев и
тем самым наносят им некоторый ущерб; 8) хищнические
взаимоотношения — это взаимоотношения хищника (насекомоядно-
го растения, растительноядного или плотоядного животного) и
жертвы, которая используется им в пищу в качестве источника
энергии; 9) конкурентные взаимоотношения растительных ви-
дов из-за «средств жизни» (питательные вещества, свет, влага
и т. п.) в условиях их недостатка; 10) средообразующие
взаимоотношения разных видов растений через изменение абиоти-
ческих факторов среды обитания, оказывающих влияние на про-
цессы их жизнедеятельности.
Перечисленные формы межвидовых взаимоотношений расти-
тельных видов, входящих в состав биоценозов, объединяются и
классифицируются на разных основаниях. По характеру воздей-
ствия одних видов на другие они объединяются в две группы:
1) прямые взаимоотношения, 2) косвенные взаимоотношения
(Е. М. Лавренко, 1971; А. А. Корчагин, 1956); по каналам взаим-
ных влияний они (вслед за В. Н. Сукачевым) объединяются в три
группы: 1) контактные взаимоотношения, 2) трансбиотические вза-
имоотношения, 3) трансабиотические взаимоотношения (Б. М. Мир-
кин, 1978; Т. А. Работнов, 1978); по трофоэнергетическому прин-
190
ципу Б. А. Быков (1970) объединяет их в две группы: 1) взаимо-
отношения с передачей веществ и энергии от одного вида к
другому; 2) взаимоотношения без передачи веществ и энергии.
В отечественной биологической литературе имеют место и
другие подходы к подразделению и классификации межвидовых
взаимоотношений живых существ. Так, В. Н. Беклемишев (1951)
межвидовые связи животных сгруппировал в следующие четыре
типа: 1) тип топических взаимосвязей (хорологических, территори-
альных, по Г А. Новикову, 1979), возникающих на почве совмест-
ного обитания и выражающихся в изменении животными условий
местообитания, в частности физико-химического состава почвы и
воздуха посредством дыхания, выделения твердых и жидких отбро-
сов и т. п.; 2) тип трофических взаимоотношений, возникающих по
линии питания между растениями и их потребителями — животны-
ми-фитофагами и сапрофагами, питающимися живой фитомассой
или отмершими растительными остатками; 3) тип форических
взаимосвязей — перенос пыльцы растений энтомофилов насекомы-
мп-опылителями, семян и плодов — млекопитающими и птицами,
мелких червей-нематод — жуками, клещами, дождевыми червями
и др.; 4) тип фабрических взаимосвязей, выражающихся в исполь-
зовании птицами, насекомыми разных частей растений (листьев,
ветвей и пр.) для постройки своих убежищ. Правда, по мнению
некоторых биологов, последний тип взаимосвязей целесообразнее
объединить с форическим, поскольку он также связан с переносом
в качестве строительного материала тех или иных частей растений.
Вслед за Беклемишевым Б. Г. Иоганзен (1979) также находит
возможным выделить таких два типа межвидовых взаимосвязей
организмов, как 1) тип топических взаимосвязей и 2) тип трофиче-
ских взаимоотношений. К первому типу он относит следующие
формы межвидовых взаимоотношений: а) охлестывание кроны де-
ревьев одного вида ветвями другого; б) использование некоторых
видов растений лианами, эпифитами в качестве субстрата; в) вы-
таптывание и механическое повреждение растений животными в
лесу, на пастбищах. Во второй тип межвидовых взаимоотношений
им включаются такие формы, как а) паразитизм, б) хищничество,
в) конкуренция и г) симбиоз.
Межвидовым взаимоотношениям представителей всех живых
компонентов как отечественными, так и зарубежными биологами
издавна придается большое значение в составе, строении и опосре-
дованно через соответствующие биологические группы и ценотиче-
ские сочетания и в функциональной организации биоценозов и
биокосных систем в целом. Б. Г. Иоганзен (1959) квалифицирует
их как закон существования биоценозов. Особенно важными в
этом смысле считаются хищничество, паразитизм и симбиоз, по-
скольку они наиболее распространены в живой природе и в той
191
или иной степени связаны с передачей веществ и энергии от одной
видовой популяции к другой. Некоторыми биологами (Б. А. Бы-
ков, 1970; С. Н. Абросов и Б. Г Ковров, 1977; Б. Г. Иоганзен и др.,
1979) они выделяются на этом основании в особую группу трофи-
ческих (пищевых) межвидовых взаимоотношений. Столь же
важной формой межвидовых взаимосвязей обычно признается и
конкуренция из-за «средств жизни», в особенности в борьбе за
пищу, которая обычно развертывается между видами, занимаю-
щими в биоценозе одну и ту же экологическую нишу. Биоценотиче-
ское значение других форм межвидовых взаимосвязей, по мнению
Абросова и Коврова, заключается в обеспечении выживания
видов, не выдерживающих пищевой конкуренции или натиска
хищников.
Межвидовые взаимоотношения живых существ по своей приро-
де являются большей частью антагонистическими, в особенности
хищничество и пищевая конкуренция. Взаимоотношение между
хищником и добычей (жертвой), по утверждению М. П. Акимова
(1959), представляет собой главную форму межвидовых взаимо-
связей в животном мире. Характерно, что как у хищников, так и у
их жертв развиваются общие (сходные) для представителей раз-
ных систематических групп приспособления, обеспечивающие наи-
более успешное выполнение соответствующих функций теми и
другими. У хищников для поиска, нападения на жертву, захвата и
утилизации добычи наблюдаются а) высокое развитие централь-
ной нервной системы и органов чувств (острота зрения и обоня-
ния); б) наличие клыков, соответствующего зубного аппарата и
кишечного тракта, а также ферментов, способных переваривать
животную биомассу; в) размножение хищников, молодь которых
питается молодью жертвы, предшествует размножению жертвы
и пр. У животных, служащих добычей для хищника, также разви-
ваются приспособления для избежания встречи с хищником, за-
щиты от него: а) развитие рогов, копыт, жала; б) образование
раковин, панцирей, игл, а также защитной подражательной и пре-
дупреждающей окраски и формы; в) создание убежищ (пор, за-
крытых гнезд); г) быстрота и маневренность бега; д) принятие
угрожающей позы, взъерошивание шерсти, оскал зубов; е) образо-
вание стад, стай и пр. У хищников на основе их поведения при
добывании пищи формируются даже своеобразные жизненные фор-
мы, так называемые этологические типьп 1) охотники — по-
движные хищники большей частью за крупной добычей; 2) огляд-
чики, осматривающие место охоты, ввиду мелкой по размеру
добычи, ведущей открытый образ жизни; 3) обишрщики, питающи-
еся наиболее мелкой, скрытой, часто неподвижной добычей; 4) ак-
тивные засадчики, высматривающие добычу из места засады и
быстро внезапно нападающие на нее; 5) пассивные засадчики,
192
схватывающие добычу в непосредственной близости или с незна-
чительного расстояния.
Последствием хищничества может стать исчезновение на дан-
ной территории используемого хищником вида. Однако чаще, как
указывает Ж. Леме (1976), оно вызывает уменьшение численности
особей вида-жертвы, так как они, по словам названного автора,
оказываются защищенными от полного истребления самим фактом
уменьшения их численности, или же в силу того, что оставшиеся
особи приобретают способность сопротивляться полному истребле-
нию благодаря иммунологическим процессам и, следовательно, да-
вать жизнеспособное потомство.
Что же касается межвидовой конкуренции животных из-за
«средств жизни*, в особенности пищи, то, как указывает М. П. Аки-
мов (1959), опа наиболее напряженна между видами, обитающими
в одних и тех же стациях, сходными по своему образу жизни, спо-
собам и объектам питания, подверженности нападения одних и тех
же врагов и т. п. В сложившихся биоценозах острота конкуренции
экологически близких видов заметно снижается; это он объясняет
тем, что вследствие длительной конкуренции происходит все более
узкая специализация и разобщение их потребностей. Разобщение
же пищевых и других потребностей экологически близких конку-
рирующих видов, как он считает, находит в конечном итоге выра-
жение а) в территориальном разобщении таких видов по стациям
или микростациям в пределах биогеоценоза; б) во временном
разобщении активного периода их жизнедеятельности в течение су-
ток или по сезонам года; в) в формировании у конкурирующих
видов морфологических, анатомических и физиологических разли-
чий, названное автором конституционным разобщением.
Подвижные животные со сходными потребностями, как утверж-
дает Ж- Леме (1976), обычно избегают конкуренции между собой
путем миграции в другие места или путем далежа занимаемой
территории на зоны охоты. Поэтому межвидовая конкуренция им
рассматривается как важный фактор, оказывающий влияние на
ареал видов, на распределение видов по местообитаниям и стаци-
ям внутри ареала и даже на состав и эволюцию сообществ (це-
нозов) .
3. Механизмы и условия сосуществования различных видов
в составе биогеоценоза
Разного рода межвидовые взаимоотношения и взаимодействия
живых существ, как было упомянуто ранее, лежат в основе под-
бора и формирования популяционно-видового состава биокосных
систем и их соподчиненных ценотических образований. Они ока-
зывают влияние на сосуществование различных видов в составе
193
живых компонентов или в составе отдельных трофических уровней
так называемых цепей и циклов (сетей) питания. Выявлению и
изучению механизмов и условий сосуществования различных видов
в составе биогеоценозов и видовой регуляции этого состава боль-
шое количество экспериментальных исследований и публикаций
было посвящено коллективом авторов — Н. С. Абросовым,
Б. Г Ковровым, О. А, Черепановым и др. (1982). Особое внимание
ими было обращено на влияние межвидовых форм взаимодействия
живых существ на эти процессы, в особенности на роль межвидо-
вой конкуренции и взаимоотношения хищника и жертвы в этом.
В результате многолетних исследований названными авторами
были выявлены некоторые закономерности сосуществования
различных видов в составе трофических уровней живых ком-
понентов биогеоценоза и механизмы видовой регуляции этого
состава.
Рассматривая вопрос о роли межвидовой конкуренции в этих
процессах, авторы прежде всего отмечают важное значение режи-
ма поступления элементов питания в формировании видового со-
става живых компонентов биогеоценоза. Ими установлено следую-
щее: а) при межвидовой конкуренции ценопопуляций автотрофно-
го компонента биогеоценоза за взаимопеэаменяемые ресурсы
(свет, влага, элементы минерального питания и пр.) и ценопопуля-
ций гетеротрофного компонента за взаимозаменяемые (источники
углерода и энергии) число видов не должно превышать числа эле-
ментов питания; б) качественный видовой состав компонентов био-
геоценоза или трофических уровней некоторых из них определя-
ется трофическими свойствами конкурирующих видовых популяций
и относительной скоростью поступления всех элементов питания;
в) изменение скорости поступления материальных ресурсов ведет
к перестройке видового состава ценоза; при этом устанавливается
такой состав, который обеспечивает наименьшую сумму потерь
всех этих ресурсов. Следовательно, основой межвидовой конкурен-
ции как одного из важнейших механизмов поддержания видового
разнообразия на трофическом уровне, по их словам, помимо нали-
чия определенных трофических свойств конкурентов, служит раз-
нообразие ресурсов питания в данном местообитании (биотопе).
Но наряду с этим в составе выделяемых в процессе жизнедеятель-
ности видовых популяций метаболитов могут оказаться дополни-
тельные ресурсы, которые способны обеспечить существование на
данном трофическом уровне дополнительных видов. «Таким обра-
зом,— пишут авторы, — выявились два вида экологических меха-
низмов сосуществования и видовой регуляции, в основе которых
лежат разнообразие ресурсов питания и межвидовые взаимодей-
ствия на трофическом уровне через продукты метаболизма. При
этом величина вклада последних определяется интенсивностью
194
воспроизводства ресурсов питания» (Н. С. Абросов и др., 1982,
с. 156).
Изучение тем же коллективом авторов влияния ценопопуляций
некоторых трофических уровней биотрофного ценокомплекса на
популяционно-видовой состав последующего трофического уровня
и обратного влияния потребителей на разнообразие и интенсив-
ность воспроизводства их пищевых ресурсов показало следующее:
а) хищники, как утверждают авторы, ограничивая численность
своих жертв, могут уменьшать конкуренцию между ними и тем
самым способствовать сосуществованию на данном трофическом
уровне нескольких конкурирующих видов; б) кроме того, хищни-
ки, по их словам, обычно увеличивают выедание наиболее обиль-
ной жертвы, что сопровождается уменьшением плотности основной
жертвы этого хищника вследствие усиления косвенной конкурен-
ции жертв. Отсюда авторами делается вывод: «...единственно ре-
альный путь увеличения численности популяции малочисленной
жертвы (в условиях конкуренции двух хищников и двух жертв) —
это повышение специализации к ней ее основного хищника»
(с. 210). Названными авторами установлено также, что конкурен-
ция жертв и хищников оказывает влияние на частоту колебаний
плотности популяций около стационарного состояния при наруше-
нии равновесия в ценозе. Причем, как они пишут, «...слабая конку-
ренция на уровне жертв приводит к уменьшению частот колебаний,
а увеличение конкуренции хищников за какие-либо неистребимые
ресурсы... вызывает соответственно увеличение частоты колебаний
плотности популяций» (с. 213). Таким образом, в возрастании ви-
дового разнообразия на трофическом уровне ценоза, наряду с раз-
нообразием ресурсов питания и взаимодействием популяций через
метаболиты, важнее значение имеет и межвидовая взаимосвязь
живых существ в форме хищничества.
Выявленные названным выше коллективом авторов в процессе
длительных исследований закономерности сосуществования конку-
рирующих видов и механизмы, регулирующие видовой состав от-
дельных трофических уровней ценоза, могут быть, по их мнению,
распространены и на биоценозы, состоящие из нескольких трофи-
ческих уровней. В биоценозе из нескольких трофических уровней,
как утверждают авторы, «...хищничество и метаболиты могут по-
зволить сосуществовать количеству конкурирующих популяций,
равному сумме количества выделяемых метаболитов, популяций
хищников и субстратов, причем только в том случае, если каждый
фактор (метаболит, популяция хищников, субстрат) ограничивает
стационарную численность только одной своей конкурирующей
популяции. Суммарное действие этих факторов определяется как
структурой сообщества, так и интенсивностью взаимодействий»
(Н. С. Абросов и др., 1982, с. 249). Авторами установлено также,
195
что в биоценозах из нескольких трофических уровней межвидовая
конкуренция играет относительно большую роль в регуляции и
поддержании видового разнообразия на верхних трофических уров-
нях, а хищничество и метаболистические взаимодействия — на
нижележащих трофических уровнях. Что же касается значения
пищевых ресурсов в видовой регуляции в подобного рода биогео-
ценозах, то, по предположению авторов, прирост первичной про-
дукции приводит к изменению видового разнообразия сначала на
нижнем трофическом уровне, затем на следующем и т. д., хотя па
самых верхних трофических уровнях это практически не приводит
к увеличению числа видов.
Названные авторы обращают внимание на следующие обстоя-
тельства, которые должны учитываться при рассмотрении вопро-
сов, связанных с формированием и регуляцией видового состава
биоценозов и, следовательно, биогеоценозов в целом: а) простран-
ственную неоднородность местообитания, вызывающую неодно-
родность распределения видовых популяций в биогеоценозе; б) пе-
риодические флюктуации среды обитания, ибо то и другое оказы-
вают определенное влияние на формирование и перестройку
видового состава биогеоценотнческих систем.
Таково существо межвидовых связей и отношений, определяю-
щих видовую специфику каждого трофического уровня живых су-
ществ в соответствии с выполняемой ими функцией в составе того
или иного живого компонента биогеоценоза. Но в основе функцио-
нальной организации биогеоценоза, как было сказано, лежат взаи-
мосвязи двоякого рода. Наряду с межвидовыми взаимоотношения-
ми ценопопуляций внутри каждого живого компонента или
трофических уровней фитофагов и зоофагов в составе биотрофпого
ценокомплекса, в биогеоценозе имеют место взаимосвязи и отно-
шения между самими живыми компонентами и соподчиненными
им биологическими и ценотическими группами. Взаимосвязи данно-
го типа в корне отличаются от межвидовых взаимоотношений жи-
вых существ, являющихся большей частью, как мы видели, анта-
гонистическими, направленными на завоевание в борьбе за
«средства жизни» наиболее прочной позиции, овладение и освоение
соответствующей своей природе экологической ниши в составе тро-
фического уровня или живого компонента бногеоценотической
системы.
В основе взаимосвязей между живыми компонентами биогео-
ценоза, представленными ценокомплексами автотрофных, био-
трофных и сапротрофных ценопопуляций, и соподчиненными каж-
дому из них ценотическими образованиями, как было сказано, ле-
жат глубокие различия в типе питания, используемых источников
энергии и образа жизни входящих в их состав живых существ. При
этом следует подчеркнуть, что такого рода взаимосвязи между
196
автотрофными и гетеротрофными живыми компонентами, в силу
их диаметральной противоположности по используемым пищевым
и энергетическим ресурсам и направлению функциональной дея-
тельности, не являются и не могут быть антагонистическими, а,
напротив, они становятся для каждого из них жизненно необ-
ходимыми, обеспечивающими в совокупности в ходе их осу-
ществления бесперебойное функционирование биогеоценоза в
целом. Из изложенного следует, что трофическим (пищевым) взаи-
мосвязям, осуществляющимся на уровне живых компонентов, пред-
ставленных ценокомплексами соответствующих ценопопуляций,
принадлежит основная, решающая роль в функциональной органи-
зации биогеоценозов.
4. Пищевые цепи и сети, их состав, строение,
формирование и закономерности строения
Функциональными структурными единицами функциональной
организации биогеоценоза, как известно, являются пищевые
цепи и сети и консорции. Представление о цепях и циклах
питания у зоологов и зооэкологов появилось давно. Это обусловле-
но, по всей вероятности, тем, что животное население, в отличие от
растительности и микробного населения, представлено в ценозах
не одним, а двумя-тремя трофическими звеньями, что и могло по-
служить основанием для возникновения среди них понятия о нали-
чии в природе последовательно взаимосвязанных по линии питания
различных групп животных как звеньев единой цепи питания,
названной впоследствии пищевой (кормовой) цепью. Эти
звенья пищевой цепи, составленные сходными по типу питания и
образу жизни группами организмов, получили название трофиче-
ских уровней. В пищевой цепи обычно выделяются следующие тро-
фические уровни и группы организмов, их составляющие.
I. Зеленые автотрофные растения.
II. Растительноядные животные-фитофаги.
III. Плотоядные животные-зоофаги (хищники 1-го порядка).
IV. Хищные животные 2-го порядка, обычно более крупные
хищники.
Позднее стали выделяться в особую группу (звено) позвоноч-
ные и беспозвоночные животные, питающиеся отбросами и трупа-
ми животных, — мусорщики.
Таково примерно было представление зоологов о цепях пита-
ния. Начальное звено каждой цепи питания, как видно, представ-
лено популяцией (или популяциями) автотрофного кормового
растения, а конечное — наиболее крупным хищником (или хищни-
ками) в данной зоне (белый медведь в Арктике, рысь и бурый
медведь в тайге, волк и шакал в аридной полосе, лев, тигр и лео-
197
пард в тропиках и субтропиках). Животные разных трофических
уровней так тесно связаны пищевыми отношениями в цепях пита-
ния, что стоит произвести какие-либо нарушения в одном звене
взаимосвязанных видов живых существ, как произойдет изменение
их пищевых взаимоотношений во всем биогеоценозе. Позднее вы-
явилось, что некоторые виды животных того или иного трофиче-
ского уровня одной цепи питания временно или постоянно исполь-
зуют также в качестве источника энергии биомассу живых су-
ществ, относящихся к соответствующему трофическому уровню
другой цепи питания данного биогеоценоза. Вследствие такого
переплетения пищевыми взаимоотношениями животных разных це-
пей питания в составе биогеоценоза образуются сложные сочета-
ния видовых популяций, названные зоологами циклами пита-
ния, получившими впоследствии название пищевых сетей.
По мере развития биогеоценотических идей происходило
изменение и усложнение представления о пищевых цепях. Цепи
питания в трактовке зоологов и зооэкологов представляли собой
то, что Дж. Вудвелл (1972) назвал цепью выедания, В такого рода
схемах пищевых цепей цикл обмена веществ оставался незавер-
шенным; отсутствовали в схеме и указания на источники матери-
альных и энергетических ресурсов. Поэтому в схеме пищевых
цепей Вудвелла параллельно цепи выедания показана цепь разло-
жения, представленная редуцентами, а снизу схемы — источники
поступления извне энергии, воды, минеральных элементов и био-
генных газов, а пунктирными стрелками — выход отработанной
энергии и отходов в сток.
С учетом последующих замечаний и предложений разных авто-
ров пищевая (кормовая) цепь представляется в еще более услож-
ненном виде (рис. 16). В ней, наряду с дополнениями Вудвелла,
показываются каналы сосредоточения и потока вещества: а) тро-
фический канал связи; б) канал отмершего органического вещества
и в) канал законсервированной живой биомассы. Стрелки разной
ширины приблизительно выражают различную мощность потока
веществ по трофическому каналу от одного трофического уровня
к другому. С большей полнотой расшифровывается и популяцион-
но-видовой состав живых компонентов каждого трофического уров-
ня пищевой цепи. В таком развернутом виде пищевая цепь пред-
ставляется весьма информативной и важной функциональной
структурной единицей функциональной организации биогеоценоза.
В биологической литературе встречаются следующие определе-
ния или краткие фрагментарные характеристики понятия пищевой
цепи и сети. По П. Дювиньо и М. Тангу (1968), ц?пи пита-
ния— это «...соподчиненные ряды организмов... Они создают по-
следовательность иерархических уровней созидания и потребления
продукции. Переплетаясь, они образуют сложную трофическую
198
УРОВНИ
|<|
fel
БИОМАССЫ
ЖИВОМ
+
5 с х
СВЯЗИ
S
С К И Е
НИЯ
Цепь
ВЫЕДА
Z
Канал
отмершего
ОРГАНИЧЕСКОГО *
ВЕЩЕСТВА 1
а
।
2
Цепь разложения
Микроорганизмы
ЖИВОТНЫЕ—САПРОФАГИ И РАСТЕНИЯ- САПРОФИТЫ
Ориг. рис. 16. Усложненная пищевая (кормовая) цепь (на основе пищевой цепи
Дж. Вудвелла, 1970; и др.)
(кормовую) сеть» (с. 10—11). Ю. Одум (1975) пишет: «Перенос
энергии пищи от ее источника — растений — через ряд организмов,
происходящий путем поедания одних организмов другими, назы-
вается пищевой цепью» (курсив наш—Л. Н.) (с. 85). Более
развернуто, с учетом более поздних представлений о пищевых це-
пях, формулирует это понятие Н. С. Абросов с соавторами (1982):
«Пути передачи вещества и энергии... через биологические состав-
ляющие (экосистему) за счет поглощения внешней, как правило,
световой энергии... получили название пищевых цепей; они
образуют в природных экосистемах сложную пищевую сеть, обес-
печивающую круговорот вещества» (с. 15).
Зарубежными экологами в зависимости от исходного источника
энергии различается несколько типов пищевых цепей. Ю. Одум
(1968, 1975) выделяет два типа. I. Пастбищная цепь в со-
ставе трофических уровней: 1) продуценты — зеленые растения;
2) первичные консументы — растительноядные животные; 3) вто-
ричные консументы — хищные животные; 4) третичные консумен-
ты — вторичные хищники. II. Детритная цепь: 1) мертвое
199
органическое вещество; 2) микроорганизмы, питающиеся мертвой
органикой; 3) детритофаги — потребители детрита; 4) хищники,
питающиеся детритофагами. П. Дювиньо и М. Танг (1968) разли-
чают три типа пищевых цепей. I. Цепи хищников в составе
четырех трофических уровней: 1) зеленые кормовые растения;
2) растительноядные животные; 3) мелкие хищники; 4) крупные
хищники. II. Цепи паразитов из двух трофических уровней:
1) живые растения или животные; 2) паразиты на них или в их
теле. III. Цепи сапрофагов также в составе двух трофических
уровней: 1) мертвое органическое вещество растительного и жи-
вотного происхождения; 2) некоторые беспозвоночные животные
и микроорганизмы — потребители органики. Р. Дажо (1975) нахо-
дит возможным цепь хищников и цепь паразитов слить в одну цепь
хищников и паразитов и, в отличие от предыдущих авторов, вводит
в ее состав еще один трофический уровень — 5) деструкторы или
биоредуценты, состоящий преимущественно из микроорганизмов
(бактерий, грибов), разлагающих органические вещества, замыкая
тем самым круговорот веществ в пищевой цепи. Выделяемая авто-
ром детритная цепь включает два трофических уровня:
1) детрит (мертвое органическое вещество), и 2) консументы — пи-
тающиеся детритом организмы (почвенные беспозвоночные живот-
ные и микроорганизмы), завершающие разложение органических
веществ.
Из отечественных биологов Н. С. Абросов с Б. Г Ковровым
(1977), как и Ю. Одум, различают два типа пищевых цепей.
I. Пастбищная цепь — цепь питания «с передачей энергии от
зеленых растений к последующим звеньям цепи путем поедания
живых организмов предыдущего звена организмами последующего
звена» (с. 10). II. Детритная цепь — цепь разложения, «ко-
торая начинается от мертвого органического вещества, проходит
через микроорганизмы, а затем может вновь соединяться с паст-
бищной цепью» (разрядка и курсив наши. — Л. Н.) (с. 10).
Сопоставляя между собой типы пищевых цепей разных авторов,
видим: во-первых, пищевая цепь хищников в интерпретации Дю-
виньо и Танга по составу трофических уровней и по существу
соответствует пастбищной пищевой цепи Ю. Одума; во-вторых, вы-
деляемая Р. Дажо детритная пищевая цепь во всех отношениях
соответствует цепи сапрофагов П. Дювиньо и М. Танга; в то же
время она отличается от детритной пищевой цепи Ю. Одума, ибо
в последней поток веществ и энергии не замыкается на уровне
микроорганизмов, живущих за счет мертвого органического веще-
ства, а продолжается и далее, заканчиваясь звеном хищников, пи-
тающихся детритофагами; в-третьих, все рассмотренные типы
пищевых цепей, за исключением пищевой цепи паразитов, входят
в качестве составных частей в усложненную схему пищевой цепи
200
Дж. Вудвелла (1972): пищевая цепь хищников (или пастбищная)
под названием «цепь выедания», а детритная (или цепь сапрофа-
гов)— под названием «цепь разложения». Тем не менее все они
могут рассматриваться и раздельно, поскольку в природе процес-
сы синтеза и деструкции органических веществ нередко бывают
разобщены в пространстве или на длительные периоды во времени.
Анализируя пищевые цепи и сети, зарубежные и отечественные
биологи высказывают ряд важных наблюдений и замечаний отно-
сительно набора и . состава трофических уровней пищевых цепей,
функционирования пищевых цепей разного типа, роли и значения
разных трофических уровней в функционировании пищевых цепей,
сетей и биогеоценозов в целом и пр.
Разнообразие пищевых цепей, по наблюдениям Р. Дажо (1975),
связано с разнообразием условий функционирования биокосных
систем, в состав которых они входят. Однако чаще, как он пола-
гает, представленные в экосистеме (биогеоценозе) типы пищевых
цепей функционируют одновременно, но одна из них при этом до-
минирует; лишь в особо специфических условиях, например в под-
земной и абиссальной средах, сохраняется только детритная пище-
вая цепь. В природных сообществах к одному трофическому
уровню, по словам Ю. Одума (1975), относятся виды организмов,
получающие свою пищу от растений «через одинаковое число
звеньев цепи» (с. 85). При этом при каждом переносе энергии с
одного трофического уровня пищевой цепи на другой большая
часть энергии (до 80-90%) теряется для системы, переходя в теп-
ло. Отсюда «чем короче пищевая цепь (или чем ближе организм
к ее началу), — пишет Ю. Одум, — тем больше количество доступ-
ной энергии» (с. 85). Вследствие этого ограничивается также и
число трофических уровней в пищевой цепи до четырех-пяти. В
цепях хищников (пастбищных пищевых цепях) по мере продви-
жения по цепи животные, по данным Р. Дажо, все более увеличи-
ваются в размерах и уменьшаются численно, а в цепях паразитов,
наоборот, животные уменьшаются в размерах, но численность их
возрастает.
Из отечественных биологов Н. С. Абросов и Б. Г Ковров (1977)
считают, что каждый трофический уровень пищевой цепи отражает
основные черты жизнедеятельности организмов, их место и роль
в выполнении определенной функции в переносе веществ и энергии
по цепям и в биологическом круговороте веществ всей системы.
Энергетическим материалом для функционирования трофических
уровней служат биомасса организмов предыдущего трофического
уровня или продукты их метаболизма и деструкции отмерших
остатков, отбросов и трупов. «Таким образом, — пишут Н. С. Абро-
сов с соавторами (1982), — каждый трофический уровень представ-
ляет собой открытую по веществу и энергии систему, внутренняя
201
структура которой определяется качественными и скоростными
характеристиками входа и выхода» (с. 15).
Вместе с тем теми же биологами приводятся факты, свидетель-
ствующие о некоторой условности деления пищевых цепей на тро-
фические уровни. Так, Р. Дажо (1975) отмечает, что всеядные
животные питаются одновременно растительной и животной пи-
щей, а хищные животные нередко используют в качестве источни-
ка энергии значительное количество разнообразных видов жертв.
Ю. Одум (1975) утверждает, что популяция одного вида животных
может занимать один и более трофических уровней в зависимости
от разнообразия пищевого рациона. Н. С. Абросов и Б. Г. Ковров
(1977) указывают на принадлежность некоторых видов животных
к нескольким трофическим уровням пищевой цепи. П. П. Второв
(1971) обращает внимание на то, что некоторые, в основном рас-
тительноядные, виды грызунов, копытных животных могут поедать
и животную пищу, а многие растительноядные птицы почти поло-
вину своих пищевых потребностей удовлетворяют за счет животной
биомассы. Такого рода факты дали повод для критики трофоэнер-
гетической концепции. Тем не менее, пишет он, «представление о
иерархии соподчиненных трофических уровней использования энер-
гии показывает разные стороны энергетической микроструктуры
биотической системы и допускает некоторые количественные сопо-
ставления как внутри одного сообщества, так и между разными
сообществами» (с. 43).
Известный интерес представляет вопрос о формировании пище-
вых цепей. Этот процесс весьма ярко представлен П. М. Рафесом
(1972), по мнению которого, он протекает как бы поэтапно. От-
дельные простые по составу пищевые (трофические) системы, со-
стоящие из двух-трех звеньев, можно наблюдать в природе. Наибо-
лее простую пищевую систему составляет взаимодействие в
процессе питания двух особей разных видов. Автором приводятся
следующие примеры:
а) листья березы гусеница березовой пяденицы;
б) гусеница березовой пяденицысиница.
Такая пищевая система названа биотрофом. При этом в про-
цессе питания, как вслед за Г. Ф. Хильми (1966) замечает автор,
происходит не простая передача вещества и энергии с пищей по-
требителю, а он (потребитель) перерабатывает ее в собственную
биомассу. Из этого следует, что возникшая в процессе такого рода
взаимодействия организмов трофическая система является откры-
той трофоэнергетической системой.
Развивая в этом направлении свои мысли далее, П. М. Рафес
пишет, что потребитель того и другого биотрофа представляет со-
бою объект нападения для других видов. В таком случае возни-
202
кают трофические системы из взаимодействующих особей трех
различных видов:
а) листья березы—>> гусеница березовой пяденицы -^синица;
б) гусеница пяденицы—^синица-> ястреб (хищник).
Такого рода трофическая система названа триотрофом.
Рассмотренные триотрофы, как показывает П. М. Рафес, пред-
ставляют собою отдельные отрезки цепей питания и, следователь-
но, могут быть объединены в единую полночленную пищевую цепь
в следующем составе трофических уровней: листья березы->гусе-
ницы березовой пяденицы —> синицы ястребы.
Однако далёко не всегда процесс формирования пищевых цепей
протекает гладко, без каких-либо коллизий. Прослеживая судьбу
возникших простых трофических образований — биотрофов и трио-
трофов, П. М. Рафес констатирует, что организмы взаимодействую-
щих видов находятся в зависимости от наличия корма и от своих
естественных врагов. Поэтому судьба потребителя в составе био-
трофа или триотрофа может сложиться по-разному. Потребитель
(гусеница насекомого или синица) при благополучном стечении
обстоятельств может, пройдя соответствующие стадии развития,
дать одно или несколько поколений потомства и дожить до есте-
ственной смерти, но может и стать объектом паразитизма и испы-
тывать угнетение либо подвергнуться нападению своего врага и
быть съеденным, не оставив потомства. Исходя из этого, автор
логично приходит к выводу: трофическая связь между двумя инди-
видуумами (особями) разных видов носит случайный характер,
так как успех реализации ее обусловлен множеством разного рода
обстоятельств, вследствие чего не всегда обеспечивается при этом
существование особи, а следовательно, и продление существования
вида. Отсюда и рассмотренные простые трофические системы, осно-
ванные на индивидуальных профических связях, также являются
случайными, а потому скоротечными.
Совершенно по-иному складывается судьба трофической систе-
мы, как указывает П. М. Рафес, когда в основе ее оказываются не
отдельные особи, а видовые популяции, так как при этом не все
входящие в их состав, скажем, гусеницы или синицы будут съеде-
ны их «врагами»; часть из них достигнет взрослого состояния и
доживет до естественной смерти, дав при жизни потомство. Следо-
вательно, трофические системы в таком случае становятся посто-
янными, долговечными. Этот вывод автора, на наш взгляд, имеет
весьма важное, глубоко принципиальное значение, ибо подтвер-
ждает, что только видовая популяция как совокупность особей,
преодолевая стохастичность случайности, приобретает форму необ-
ходимости, вследствие чего становится прочной и долговременной
основой состава, строения и функциональной организации биогео^
ценотической системы.
203
При анализе состава и строения цепей питания Ч. Элтоном
(1934) выявлен ряд закономерностей, названных им правилами.
1. Правило величины особей, суть которого заключается в том, что
цепи питания, как правило, идут от более мелких форм к более
крупным. Однако, поскольку размеры животных не могут увели-
чиваться беспредельно, число звеньев цепи не может превышать
пяти-шести. Правда, из этого правила имеются исключения, при-
веденные Р. Дажо (1975): например, нападение волков на более
крупных животных, чем сами, благодаря охоте сообща или умерщ-
вление змеями крупных животных посредством выделяемого яда
и пр. 2. Правило пирамиды чисел, сформулированное так: число
особей в каждом предыдущем звене бывает значительно больше,
чем в последующем. Только при этом условии смертность видов-
жертв от истребления хищниками будет компенсироваться и чис-
ленность их будет поддерживаться на определенном уровне. Пира-
мида чисел, как отмечает Р. Дажо, тем выше, чем большее число
уровней входит в состав данной пищевой цепи. Он приводит и
некоторые исключения, например пищевые цепи паразитов, а так-
же сапрофагов. 3. Правило биомассы, вытекающее из предыдущего
правила, заключается в том, что биомасса с каждым последующим
звеном цепи уменьшается в среднем в отношении 10:1, так как
далеко не вся масса какого-либо звена превращается в массу по-
следующего звена, часть ее расходуется на процессы жизнедея-
тельности организмов, па их воспроизводство и т. п. Исключение
из этого правила относится к водным биокосным системам, в кото-
рых, например, биомасса фитопланктона меньше биомассы питаю-
щегося ею зоопланктона; правда, она значительно быстрее возо-
бновляется (рис. 17).
Р. Дажо (1975) приводит еще одно правило — 4) правило пи-
рамиды энергии, где пирамида имеет вид треугольника с верши-
ной, обращенной вверх, в связи с потерей энергии при переходе от
одного трофического уровня к другому. Это правило наилучшим
образом выражает трофоэнергетическую суть биокосной системы,
но для этого в настоящее время часто не хватает необходимых
данных.
Итак, что же представляют собою пищевые цепи и трофические
уровни, их составляющие? ГО. Одум (1975) подчеркивает, что
«трофическая классификация делит на группы не сами виды, а их
типы жизнедеятельности» (с. 85), а Н. С. Абросов и Б. Г Ковров
(1977) утверждают, что «трофические уровни пищевых цепей —
это не что иное, как живые компоненты биогеоценоза» (с. 10).
На наш взгляд, из всего изложенного в этом разделе следует,
что взаимодействующими «субъектами», образующими трофиче-
ские уровни в пищевых целях, несомненно, являются видовые по-
пуляции (ценопопуляций) живых существ, вступающие при
204
совместном обита-
нии в составе ценоза
в сложные трофиче-
ские (пищевые) и
другие взаимоотно-
шения между собой.
Трофические уровни
(звенья) в составе
пастбищной цепи
(цепи выедания, по
Вудвеллу) формиру-
ются из ценопопуля-
ций автотрофного це-
нокомплекса (I уро-
вень) и из ценопопу-
ляций биотрофного
ценокомплекса (II
уровень фитофагов,
III и IV — уровни у
зоофагов 1-го и 2-го X.
порядка); ИСТОЧНИ-рис Упрощенная схема пирамиды числа особей Эл-
КОМ энергии тех и тона (по Г А. Новикову, 1979)
других служат во-
площенная в последовательно трансформируемой биомассе свето-
вая энергия солнечной радиации и материальные пищевые и другие
ресурсы местообитания. Трофический уровень детритной цепи (цепи
разложения, по Вудвеллу) состоит из ценопопуляций сапротроф-
ного ценокомплекса, обитающих как в наземной (воздушной), так
и в почвенной среде биотопа, и в качестве источника энергии ис-
пользуют отмершие остатки, отбросы и мертвые органические
вещества растительного и животного происхождения, осуществляя
одновременно и синтез гумусовых и других веществ.
Следовательно, живые компоненты биокосных систем, пред-
ставленные в природе ценокомплексами автотрофных ценопопуля-
ций, биотрофных ценопопуляций (фитофаги и зоофаги) и ценопо-
пуляций сапротрофов, нельзя отождествлять с трофическими
уровнями пищевых цепей, как нельзя и обезличивать сами трофи-
ческие уровни. Трофические уровни — это формы, в которые обле-
каются в ходе функционирования в составе пищевой цепи видовые
ценопопуляций соответствующих ценокомплексов биогеоценотиче-
ских систем. При этом не следует забывать, что в состав биогео-
ценоза входит не одна, а целый ряд пищевых цепей, базирующихся
на ценопопуляциях автотрофных кормовых ценозообразователей.
Пищевые же цепи (и сети) того или иного типа, следовательно,
должны рассматриваться как важнейшие функциональные струк-
205
турные единицы функциональной организации биоценоза и биогео-
ценоза в целой, весьма сложные по популяционно-видовому составу
живых существ и по выполняемой в биогеоценозе функции.
А
5. Понятие консорции, ее состав
и строение в трактовке разных авторов
В последнее времяТ^аряду с пищевыми цепями и сетями, совет-
скими биологами выявлена еще одна функциональная структурная
единица функциональной организации биогеоценоза — это так на-
зываемая консорция. Проблема консорции — многоплановая
проблема, свидетельством чему служит тот факт, что это понятие
и термин почти одновременно независимо друг от друга были пред-
ложены зоологом В. Н. Беклемишевым (1951) и ботаником
Л. Г. Раменским (1952). Л. Г Раменский дал такое, несколько
вольное, определение этого понятия: консорции—это «сочета-
ния разнородных организмов, тесно связанных друг с другом в их
жизнедеятельности известной общностью их судьбы» (с. 186). Он
приводит примеры, иллюстрирующие это понятие. Одна из консор-
ций елового сообщества, представленная популяцией ели и целым
рядом популяций других видов, тесно связанных с нею в своей
жизнедеятельности: а) микоризные грибы — иа ее корнях; б) ку-
стистый лишайник—на ветвях; в) корковые лишайники — на коре
стволов; г) жуки-короеды и усачи — в стволах; д) еловая огнев-
ка— в кроне деревьев; е) бурундуки и белки, питающиеся семена-
ми ели и пр. (рис. 18). Консорция дубового сообщества, состоящая
из популяции дуба и связанных с ним растительных и животных
паразитов, эпифитов, симбионтов, вредителей из насекомых и пр.
Таким образом, центром каждой консорции служит популяция
какого-либо высшего автотрофного растения, с которым тесно свя-
заны трофическими, симбиотическими, паразитическими и другими
отношениями разнообразные виды растений, преимущественно низ-
ших, позвоночных и беспозвоночных животных и микроорганизмов.
Эти биологические виды-партнеры названы консортами, а
связи и отношения консортов с популяцией автотрофного кормово-
го растения — консортивными связями.
В отличие от Л. Г. Раменского, В. Н. Беклемишев предложен-
ный им термин употребил в мужском роде — «консорций». По его
мнению, организмы входят в состав биогеоценоза не сами по себе,
а в составе какого-либо консорция (или иначе — консорции). Кон-
сорция состоит из особи-эдификатора и целого ряда поселяющихся
на теле или в теле эдификатора особей других видов. Автором
приводятся следующие примеры консорций: а) сосна с микориз-
ными грибами, мхами, насекомыми и пр.; б) полевка с эктопара-
зитами, гельминтами, простейшими, бактериями и т. п. Следова-
206
Ориг. рис. 18. Схема консорции популяции ели (по данным
Л. Г. Раменского, 1952, и В. В. Мазинга, 1966)
тельно, центром консорции, по Беклемишеву, является не популя-
ция вида как поселение особей, а одна особь какого-либо доми-
нирующего вида, причем это может быть как растение, так и
животное (полевка в данном случае).
В дальнейшем понятие консорции значительно расширилось.
В. В. Мазинг (1966) предложил в состав консорции включать как
виды организмов, непосредственно связанные с популяцией авто-
трофного кормового растения, образующие I круг (концентр) во-
круг нее, так и виды, связанные с нею опосредованно через орга-
низмы I концентра, составляющие II, III и IV концентры. В ре-
зультате консорция выглядит весьма сложной как по составу, так
и по своему строению единицей функциональной организации
биоценоза и биогеоценоза в целом (см. рис. 18). Консорция такой
усложненной структуры и такого объема полностью принимается
и Т. А. Работновым (1969, 1978) на том основании, что живые су-
щества II и III концентров оказывают значительное воздействие
207
на организмы I концентра, а через них и на центральное кормовое
растение консорции, являющееся основным ценозообразователем
сообщества.
В связи с различными представлениями о составе и структуре
консорции эти вопросы стали предметом неоднократного обсужде-
ния и дискуссий на специальных научных совещаниях (в 1972 году
в Москве и в 1975 году в Перми), а также в научной периодиче-
ской печати.
Любая консорция, как видно на рис. 18, состоит из централь-
ного организма или видовой популяции, называемых то ядром, то
эдификатором или детерминантом консорции, и группы организ-
мов-консортов, связанных в своей жизнедеятельности с централь-
ным видом. Прежде всего, что считать центром консорции?
Е. М. Лавренко (1959), вслед за Л. Г. Раменским, предлагает
центром консорции в сухопутных системах считать популяцию выс-
шего автотрофного зеленого растения, являющегося как бы «энер-
гетической установкой биоценоза» (с?^29). По Т. А. Работнову
(1969), центральным видом, детерминантом консорции могут быть
не только высшие, но всякие автотрофные растения, на суше, в
частности, мхи, водоросли и др. А. А. Корчагин (1977), в согласии
с В. Н. Беклемишевым, находит, что на положении центра, ядра
консорции может оказаться не только автотрофное растение, но и
гетеротрофное животное. И. А. Селиванов (1974) и П. М. Рафес
(1970) идут еще дальше, считая, что в качестве центра консорции
следует признать не только живые авто- и гетеротрофные суще-
ства, но и мертвые тела (пни, валежник, трупы животных). Точка
зрения последних авторов разделяется и Н. В. Дылисом (1978), но
консорции, развивающиеся на мертвых (отмерших) телах, по его
мнению, должны быть выделены в особую группу. Но Б. А. Быков
(1970) высказывается против включения в состав консорций орга-
низмов, использующих в качестве источника энергии разлагающи-
еся ткани мертвых растений и животных.
Еще более глубокие расхождения во взглядах биологов обна-
жились по вопросу о составе консортов и в связи с этим о струк-
туре и объеме самих консорций. К группе консортов принято
относить: а) потребителей живой биомассы — растительной, живот-
ной— это так называемые биотрофы (консументы) и б) потреби-
телей отмерших остатков и мертвого органического вещества
растительного и животного происхождения — эт» сапротрофы (био-
редуценты). Т. А. Работновым сюда же относятся: в) эккрисотро-
фы, использующие в качестве источника энергии прижизненные
выделения организмов, и г) эпифиты, для которых живые суще-
ства, образующие ядро консорции, служат лишь субстратом для
прикрепления. Иными словами, в состав консортов входят орга-
низмы или видовые популяции, связанные с детерминантом консор-
208
ции трофически или топически (эпифитно, по А. А. Корчагину,
1977).
В состав I концентра консорции усложненной структуры входят
консорты, непосредственно связанные с центральным видом кон-
сорции. Одни из них связаны с ним трофически, получая от
него вещества и энергию: а) животные-фитофаги, большей частью
беспозвоночные — симбионты, паразиты, вредители, в том числе
насекомые (жуки, чешуекрылые, перепончатокрылые); б) фитопа-
разиты (бактерии, грибы, актиномицеты, вирусы и др.); в) расте-
ния-полупаразиты, получающие от центрального вида консорции
только вещества; г) азотфиксирующие растения-симбионты, полу-
чающие от центрального вида энергию и вещества (за исключени-
ем азота). Другие консорты связаны с центральным видом тро-
фически и топически: а) растения-паразиты; б) животные,
поселяющиеся внутри его особей или вступающие с ними в
симбиотические отношения. У третьих консортов связь с детерми-
нантом консорции только топическая: а) растения-эпифиты (мхи,
лишайники, некоторые высшие растения), использующие его в
качестве субстрата для прикрепления; б) животные, которые на-
ходят здесь убежище для себя и для своего потомства.
II концентр консорции составляют видовые популяции, орга-
низмы которых в качестве источника энергии используют биомассу
организмов I концентра как в живом, так и в отмершем состоянии,
а также их экскременты. Сюда относятся: а) животные-зоофаги
(хищники 1-го порядка)—млекопитающие, птицы, некоторые на-
секомые (жужелицы, божьи коровки, пауки и пр.); б) зоопаразиты
животных-фитофагов (личинки насекомых, кровососущие насеко-
мые); в) беспозвоночные животные-сапрофаги, некрофаги, копро-
фаги, живущие за счет мертвого органического вещества и экскре-
ментов животных. В состав III и последующих концентров консор-
ций входят видовые популяции живых существ, использующих в
процессах своей жизнедеятельности энергию, заключенную в био-
массе организмов предшествующих концентров: а) животные-зоо-
фаги (хищники 2-го и 3-го порядка); б) зоопаразиты плотоядных
животных предшествующих концентров, в) животные-сапрофаги и
микроорганизмы, живущие за счет использования энергии мертво-
го органического вещества (насекомые, бактерии и пр.).
При определении места, позиции, занимаемой той или иной
ценопопуляцией в составе консорции, выявился ряд трудностей.
Так, мхи и лишайники обычно относятся к консортам, но на мохо-
вых болотах и, очевидно, в некоторых биогеоценозах тундры они
становятся основными продуцентами, а потому, как считает
А. А. Корчагин (1977), должны рассматриваться в таком случае
как центральные виды консорций. Автотрофные полупаразиты, ко-
торые обычно относятся к консортам-консументам, по мнению
209
Т. А. Работнова, следует считать консортами-эпифитами, посколь-
ку они получают от детерминанта лишь воду и элементы мине-
рального питания; последний же выступает в таком случае как
субстрат для их прикрепления. Насекомоядные хлорофиллоносные
растения являются продуцентами и одновременно зоофагами-кон-
сументами, получающими от своих жертв только азот и фосфор.
Поэтому Т. А. Работиов склонен относить их к числу центральных
растений соответствующих консорций. Лианы, особенно деревяни-
стые, нередко настолько угнетают растение-опору, что становятся
основными продуцентами консорции. Следовательно, вопрос о по-
зиции лианы в составе консорции, по мнению А. А. Корчагина,
должен решаться по-разному, в зависимости от степени угнетения
опоры-хозяина и от доли участия каждого из них в продуцирова-
нии органики.
Значение и доля участия консортов в потреблении и трансфор-
мации органических веществ в составе консорций неодинаковы. Из
растительных консортов наибольшее значение в этом, по словам
А. А. Корчагина (1977), имеют почвенные бактерии и грибы; из
наземных растений — мхи, лишайники и водоросли, а также пара-
зиты и полупаразиты — из высших растений. Из животных видов к
числу наиболее важных консортов относятся беспозвоночные, осо-
бенно насекомые, среди них фитофаги и фитопаразиты.
Для характеристики консорций в целом наибольшее значение,
как утверждает В. В. Мазинг (1966), имеет I концентр консорции,
состоящей из видов, непосредственно связанных с автотрофным
детерминантом. «Чем дальше он от центра, — пишет он, — тем
меньше специфичных для консорция и облигатных связей. Виды
высших звеньев пищевых цепей («хищники хищников») требуют
большего количества пищи, поступающего не только из других
консорций, но и из различных биоценозов, часто пространственно
разобщенных» (с. 121).
Обращает на себя внимание исключительное видовое богатство
консортов большинства автотрофных ценозообразователей, в осо-
бенности древесных и кустарниковых пород. Так, по данным
В. В. Мазинга (1966), численный состав консортов только I кон-
центра болотной формы сосны на верховых болотах Эстонии со-
ставляет не менее 100 видов, в том числе: грибов (аскомицетов,
базиомицетов и несовершенных)—30 видов, эпифитпых мхов и
лишайников — 32 вида, насекомых-фитофагов (жуков, чешуекры-
лых, перепончатокрылых и др.) —38 видов. Общее число консор-
тов I концентра пушистой и бородавчатой березы, по данным того
же автора (1976), достигает почти 1000 видов, в частности: грибов
(паразитов и симбионтов) — 147 видов, эпифитных мхов и лишай-
ников— 69, высших цветковых растений-паразитов—1, животных-
консументов (паукообразных, насекомых и других беспозвоноч-
210
них)—636, птиц (куриных, воробьиных, дятловых)—8, млекопи-
тающих (грызунов, парнокопытных) —12; всего 974 вида, не считая
водорослей и других низших организмов. Среди консортов беспоз-
воночных животных особенно много листогрызущих форм, а также
стволовых и подкоровых обитателей. Менее богаты видами-консор-
тами, по сравнению с деревьями и кустарниками, травянистые
цветковые детерминанты. К числу бедных по видовому составу
относятся консорции плаунов, хвощей и папоротникообразных цено-
зообразователей. Видовое богатство консорций, по словам В.В.Ма-
зинга (1976), определяется: а) филогенетическим возрастом детер-
минанта и динамикой его ареала; б) разнообразием поставляемого
детерминантом корма для консортов и экологических ниш для их
обитания.
В связи с изменившимся представлением о консорции по срав-
нению с первоначальным появился целый ряд новых определений
консорции отечественными биологами: Е. М. Лавренко (1959),
В. В. Мазинга (1966), Т. А. Работнова (1969), Б. А. Быкова
(1970), А. Г Воронова (1976), Н. В. Дылиса (1974 и 1978) и др.
В этих определениях консорции подчеркиваются следующие при-
сущие ей черты: а) центром, «ядром» консорции большинством
биологов признается видовая популяция, а не отдельно взятый
организм, причем популяция автотрофного вида (или, по Мазингу,
также группы экологически близких видов) растений, а из авто-
трофов— популяция самостоятельно существующего вида (т. е. не
эпифита и не полупаразита); б) объединение или сочетание цент-
ральной видовой популяции происходит с организмами всех входя-
щих в состав консорции видов, как растительных, так и животных,
как высших, так и низших; в) в основе такого рода сочетания
видовых популяций консорции лежат трофические или топические
связи и отношения или те и другие одновременно; г) сформировав-
шаяся таким образом консорция как совокупность разнообразных
видовых популяций большинством биологов рассматривается как
единица функциональной организации биогеоценоза, а не фитоце-
ноза, как считалось ранее.
Из всех современных определений консорции названных авто-
ров наиболее полным является определение Т. А. Работнова
(1969). «...Под консорцием (консорцией. — Л. Н.) следует пони-
мать сочетание популяции любого самостоятельно существующего
(т. е. не эпифита) в пределах определенного фитоценоза автотроф-
ного растения и совокупности организмов, связанных с ними тро-
фически (консументов, редуцентов), или использующих как суб-
страт для прикрепления (эпифиты), или как источник воды и
элементов минерального питания (автотрофные полупаразиты)»
(с. 12). Однако наиболее точно раскрывающим суть консорций как
функциональных структурных единиц биогеоценоза следует при-
211
.знать определение их Н. В. Дылисом (1978): «С биогеоценологи-
ческой точки зрения, — пишет он,— консорции есть функциональ-
ные структуры биоты, отражающие в своей совокупности разнооб-
разие путей перемещения и трансформации веществ и энергии от
первичных продуцентов биогеоценоза ко всем в нем участвующим
консументам и деструкторам» (с. 28).
6. Многообразие консорций, их классификация
и изменение во времени
В любом биогеоценозе консорций не одна, а много, ибо каждая
популяция самостоятельно существующего автотрофного вида рас-
тений, по крайне мере доминантного, может стать центром (ядром)
консорции со своим специфическим набором консортов. Формиро-
вание и обособление консорций, как указывает В. В. Мазинг
(1966),— длительный во времени процесс, связанный с экологиче-
ской и ценотической дифференциацией самого центрального вида
консорции. По имеющимся литературным данным, большинство
консорций и консортивных связей в составе биогеоценозов совре-
менного биогеоценотического покрова складывались в четвертич-
ный геологический период. Они филогенетически закреплены и для
каждого автотрофного ценозообразующего вида растений специ-
фичны. Следствием всего этого является большое разнообразие
консорций, что вызвало необходимость в подразделении их на раз-
ного рода группы, типы и т. п., ибо популяционно-видовой состав
каждой консорции в совокупности определяет специфику обмена
веществ и энергии в данном пункте (секторе) биогеоценотической
системы.
Б. А. Быков (1970) и В. В. Мазинг (1976) подразделяют кон-
сорции на группы по их объему и некоторым другим признакам.
Первый автор различает следующие три группы консорций: 1) ин-
дивидуальные, центром (ядром) которых является одна особь
автотрофного детерминанта; 2) популяционные, с центром, пред-
ставленным видовой популяцией детерминанта или видом в целом
в пределах всего ареала; 3) синузиальные, центральное ядро кото
рых охватывает виды, относящиеся к одной жизненной форм<
(экобиоморфе), например мезофильные темнохвойные древесные
породы (ель, пихта и др.). В. В. Мазинг, исходя из тех же прин-
ципов, различает шесть групп консорций. Наряду с индивидуаль-
ными и популяционными группами консорций он выделяет группу
видовых консорций, расчленив выделенную Быковым группу по-
пуляционных консорций на две самостоятельные группы. Помимо
этого, им выделена группа родовых консорций на том основании,
что в отдельных случаях центром консорций становится не один
вид растения, а род в целом, например сфагпумы на верховых бо-
212
лотах, «белые» березы в лесной полосе; кроме того, он счел воз-
можным выделить две группы консорций, занимающие промежу-
точное положение между названными группами консорций: кло-
нальные консорции, объединяющие консорты по клонам (по сово-
купности вегетативно разросшихся побегов одной материнской
особи), и группу региональных консорций, представляющую собою
совокупность популяционных консорций не всего ареала централь-
ного вида, а какой-либо части его.
По роли в ценозе консорции Е. М. Лавренко (1959) подразде-
ляются на следующие группы: а) эдификаторные консорции
(основные, по В. В. Мазингу, 1966), в которых центральная
видовая популяция является эдификатором сообщества; б) доми-
нантные консорции, центральная популяция которых — доминант
сообщества; в) консорции, центральная популяция которых нахо-
дится в подчиненном положении по отношению к эдификатору,
В. В. Мазингом названы производными. Помимо этого,
Т. А. Работиовым (1969) выделена группа консорций, образован-
ных несамостоятельно существующими автотрофными растениями
(эпифитами, полупаразитами), которые были названы зависи-
мыми.
По мнению Н. В. Дылиса (1978), с биогеоценологической точки
зрения весьма важно более дробное расчленение консорций на со-
ответствующие подразделения консортов, связанных с отдельными
органами центрального автотрофного вида растений, ибо значение
их в жизни детерминанта консорции и в функционировании био-
геоценоза неодинаково. В. В. Мазингом (1976) вся эта группа
консортов отдельных органов детерминанта и его частей названа
мероконсорцией. В составе этой группы названными автора-
ми выделяются следующие подгруппы консортов: а) листоеды,
воздействующие на ход газообмена; б) корнееды, оказывающие
влияние на миграцию минеральных веществ и воды по горизонтам
почвенного профиля; в) семяеды и плод о еды; г) опылители расте-
ний и д) разносчики плодов и семян. Консорты последних подгрупп
способствуют поддержанию в относительно стабильном состоянии
популяционно-видового состава автотрофного компонента биогео-
ценоза. ’ ’ г
Первую попытку классификации консорций как единиц функ-
циональной организации фитоценоза и биогеоценоза в целом пред-
принял Б. А. Быков (1970). Взяв за основу выделенные им
группы популяционных и синузиальных консорций, он объеди-
нил их в типы консорций, выделяемые по принадлежности их
детерминанта к тому или иному подклассу экобиоморф. Типы кон-
сорций объединяются им в классы консорций по типу питания
основных ценозообразователей. Всего автором выделено три клас-
са, подразделяемых на подклассы.
213
I. Класс консорций автотрофных видов, ядрами которых яв-
ляются автотрофные растения; накопленная ими энергия,
рассеивается гетеротрофными организмами.
1. Подкласс консорций высших растений, доминирующих
в растительном покрове суши.
2. Подкласс консорций макрофитных водорослей, домини-
рующих в растительном покрове морских шельфов.
II. Класс консорций гемиавтотрофных видов, их ядра пред-
ставлены зелеными насекомоядными растениями; при этом
происходит рассеивание накопленной в процессе фотосин-
теза энергии и одновременно аккумуляция рассеянной
энергии в гетеротрофных организмах. Значение этих кон-
сорций в преобразовании веществ и энергии сравнительна
небольшое.
III. Кла сс консорций гетеротрофных видов, их ядрами являют-
ся сравнительно крупные, доминирующие по биомассе, чис-
ленности и значению в ценозах животные; полученная от
гетеротрофных и автотрофных организмов энергия рассе-
ивается.
1. Подкласс консорций морских растительноядных (в осо-
бенности питающихся фитопланктоном) и плотоядных
животных.
2. Подкласс консорций наземных животных — фитофагов
и полифагов.
Несмотря на отмеченную выше устойчивость формирующихся
в течение длительного времени консорций, последняя все же явля-
ется относительной. В .действительности наблюдается изменение
консорций во времени, неоднородны, вариабельны они и в про-
странстве. Это обусловлено особенностями развития компонентов
консорции, в первую очередь центрального вида ее, динамикой
биогеоценоза, в состав которого она входит, и особенностями фи-
зико-географической среды, в условиях которой протекает функ-
ционирование биогеоценоза и консорции в его составе. Т. А. Работ-
новым (1969) различаются следующие формы изменения консор-
ции: а) сезонные изменения, связанные с годичным ритмом
жизни детерминанта консорций и других ее компонентов;
б) флюктуационные изменения в составе, численности и жиз-
ненном состоянии детерминанта и консортов, связанные с разно-
гбдичными различиями погодных условий и циклами развития
отдельных консортов, особенно массово размножающихся; в) сук-
цессионные изменения, связанные с коренными или обрати-
мыми сменами самих биогеоценозов; г) эволюционные изме-
нения, выражающиеся в коренной перестройке консорций в связи
с эволюционным преобразованием компонентов консорции и изме-
нениями копсортивпых связей между ними.
214
Что же касается неоднородности, вариабельности консорций в
пространстве, то она выражается в том, что один и тот же домини-
рующий автотрофный вид в различных ландшафтно-географиче-
ских зонах, по словам В. В. Мазинга и Т. А. Работнова, образует
неодинаковые по своему составу и структуре консорции. Вслед-
ствие этого формирующиеся в различных физико-географических
условиях консорции проявляют себя по-разному в составе соот-
ветствующих зональных биогеоценозов: от тропиков к умеренным
и арктическим условиям наблюдается обеднение видового состава
и упрощение структуры консорции.
Таковы основные положения, фактические данные и теоретиче-
ские суждения отечественных биологов по вопросу о консорции.
7. Консорция как основная функциональная структурная единица
биогеоценоза
Итак, что же представляет собою консорция при рассмотрении
ее во всех, так сказать, ипостасях? В каком соотношении она на-
ходится с пищевыми цепями и сетями? Каково место ее в струк-
турно-функциональной организации биогеоценоза и какова роль в
выполнении обменных процессов в этой основной материально-
энергетической ячейке биогеосферы?
Прежде всего следует рассмотреть некоторые спорные или не-
ясные вопросы в свете современных представлений о консорции, с
тем чтобы в процессе обсуждения попытаться уяснить их суть, и
если не найти правильного решения или подходов к нему, то хотя
бы приблизиться к этому. Во всех суждениях по этим вопросам
надо, очевидно, исходить из известных фактов. Первый вопрос: что
считать центром консорции? Только живые организмы или так-
же и остатки отмерших растений и трупы животных? Только авто-
трофные зеленые растения или также и гетеротрофные животные
организмы? Отдельные особи доминирующих видов или популяции
этих видов? Если исходить из признания консорции в качестве
одной из основных единиц функциональной организации биогеоце-
ноза, то центром, ядром консорции следует считать живые суще-
ства, а из них только самостоятельно существующие автотрофные
растения, и не отдельные особи, а видовые популяции (ценопопу-
лянии) растительных форм, ибо только такого рода детерминанты
способны обеспечить длительное устойчивое функционирование в
составе соответствующей биогеоценотической системы. В то же вре-
мя известны неопровержимые факты, свидетельствующие о том, что
наряду с названной категорией консорции бытуют, во-первых, та-
кие последовательно сменяющие друг друга виды и группы видов
сапротрофов, источником энергии которых служат отмершие расти-
тельные и животные организмы и их части; во-вторых, встречаются
215
такие преимущественно скрыто живущие животные, с которыми
тесно связаны в своем существовании экто- и эндопаразиты, ком-
менсалы, патогенные бактерии и прочие живые существа.
На наш взгляд, такого рода сочетания живых существ, тесно
связанных в своей жизнедеятельности с популяцией животных или
существующих за счет утилизации мертвых органических материа-
лов, должны быть отнесены к рангу консорций, ибо те и другие
естественно включаются в общий жизненный поток, выполняя в
соответствующих природных биогеоценозах вполне определенные
функции. Но, по аналогии с соответствующими категориями пище-
вых цепей — детритных или лишенных непосредственной автотроф-
ной базы,— они могут быть выделены в особую категорию не пол-
ной лен ных (детритных, сапротрофпых) консорций. В отличие
от них вышепоименованные консорции с автотрофным детерминан-
том в центре, базирующиеся на энергии солнечной радиации, могут
быть в таком случае названы полночленными (пастбищными»
биотрофными). Что же касается так называемых индивидуальных
консорций, ядро которых представлено отдельными особями авто-
трофного вида растений и популяционных с центром из популяции
самостоятельно существующего автотрофного детерминанта, то те
и другие наличествуют в природе. Но поскольку судьба первых,
подобно индивидуальным простым трофическим системам пищевых
цепей, непредсказуема и скоротечна, то их можно рассматривать
как элементарные консорции, исходные для формирования популя-
ционных консорций, длительно существующих, относительно ста-
бильных, а потому представляющих собою надежную ценотическую
основу функциональной организации биогеоценоза в целом.
Второй вопрос подобного рода: какой является в действитель-
ности структура консорции и в соответствии с этим ее популяцион-
но-видовой состав? То ли она должна представляться нами про-
стой, состоящей из центральной автотрофной растительной популя-
ции и группы непосредственно трофически и топически связанных
с нею разнообразных видов-консортов, т. е. в том виде, в каком это
понятие было предложено Л. Г. Раменским (1952), то ли она
должна войти в науку в виде усложненной формы, состоящей, в
представлении В. В. Мазинга (1966), из нескольких концентров.
Очевидно, никто из биологов не будет отрицать наличия непо-
средственных трофических и других консортивных связей, ежечасно
и ежеминутно естественным образом осуществляющихся в природе
между различными растительноядными формами живых существ»,
составляющих, по Мазингу, I концентр консорций, и множеством
насекомоядных, паразитных, мелких и крупных хищных животных
форм, образующих последующие концентры консорций. Последняя
категория живых существ оказывается, таким образом, опосредо-
ванно связанной и с центральной популяцией консорции, по мере
216
удаления от которой, в связи с принадлежностью консортов к раз-
ным концентрам, эти связи с нею, как отмечалось ранее, все более
ослабевают, но не исчезают полностью. Отсюда следует, что пред-
ложенная Мазингом схема сложного строения консорции графиче-
ски отображает реально существующие в природе связи и отноше-
ния между консортами разных концентров и последних — с цент-
ральной видовой популяцией. А это означает, что сколько бы мы
ни высказывали свои пожелания об ограничении представлений о
консорции в первоначальном объеме, на уровне первого концентра
от этого ничто не изменится, ибо это объективный природный про-
цесс, протекающий независимо от нас. Следовательно, нам ничего
не остается, как признать реальность сложного состава и строения
основных полночленных консорций, в полной мере отвечающих в
таком виде своему назначению. Несколько проще, очевидно, струк-
тура неполночленных консорций, консорты которых базируются на
использовании в качестве источника энергии мертвой биомассы или
жизненных ресурсов преимущественно животных организмов.
Достаточно беглого взгляда на схему структуры полночленной
консорции, чтобы заметить радиально расходящиеся от трофически
связанных с центральной растительной популяцией консортов через
все концентры «лучи» и установить, что это не что иное, как пи-
щевые цепи, а в случае переплетения трофических связей меж-
ду ними — и пищевые сети. В связи с этим возникает вопрос:
в каком же соотношении находятся консорции и входящие в их
состав пищевые цепи и сети?
Из изложенного следует, что консорция — более широкое по
объему понятие, чем пищевая цепь, ибо в ее состав входит несколь-
ко пищевых цепей, базирующихся на пищевых и энергетических
ресурсах одной и той же популяции автотрофного эдификатора или
того или иного ценозообразующего растительного доминанта. Из
этого следует также, что если пищевая цепь представляет собой
важнейшую структурную единицу функциональной организации
биогеоценоза, то консорция — это основная функциональная
структурная единица его. Консорция, таким образом, выступает
как весьма сложное по составу и структуре образование, как сво-
его рода система, точнее, форма системного единства соответствую-
щего ранга. Недаром в связи с этим среди некоторых биологов
возникло мнение, что консорция представляет собой абсолютно
автономную единицу; это будто бы микробиоценоз.
На самом же деле входящие в состав биогеоценоза консорции
находятся в постоянном взаимодействии друг с другом, и они да-
леко не одинаковы по своему назначению и выполняемой в нем
функции. При сильном эдификаторе автотрофного компонента био-
геоценоза, по словам В. В. Мазинга (1966), в его составе обычно
формируется основная (эдификаторная, по Е. М. Лавренко) кон-
217
сорция и целый ряд тесно связанных с нею так называемых произ-
водных консорций. При наличии же в составе автотрофного компо-
нента нескольких равносильных доминантных ценопопуляций
образуется несколько консорций, каждая с соответствующим ее
экологии набором консортов; в силу этого они менее тесно связаны
друг с другом, проявляя большую самостоятельность, что, кстати
сказать, характерно для ранних этапов становления биогеоценозов
или в условиях неоднородности их ценотической среды. Следова-
тельно, обособленность консорции (экологическая, ценотическая,
биогеохимическая) относительна, т. е. она не выходит за рамки
характерных, присущих биогеоценозу черт.
По составу консорций биоценозы и биогеоценозы в целом неоди-
наковы. Набор консорций и их соотношение характеризуют функ-
циональную специфику биогеоценоза и определяют его качествен-
ное своеобразие. С другой стороны, сам консортный состав биоце-
ноза определяется комплексом ценозообразующих растений как
автотрофной базой его. В полночленных консорциях биогеоценоза,
состоящих преимущественно из пастбищных пищевых цепей, обра-
зованных ценопопуляциями автотрофного компонента и фитофага-
ми и зоофагами биотрофного, господствующим процессом является
синтез и трансформация веществ и поток энергии по трофическим
уровнях пищевых цепей от одной консорции к другой. Среди них
основные эдификаторные консорции занимают ведущее положение
в составе и функционировании биогеоценоза, а так называемым
производным консорциям принадлежит второстепенная роль в
этом. В неполночленных консорциях, в частности состоящих из
детритных пищевых цепей (или, иначе, пищевых цепей сапрофа-
гов), образованных ценопопуляциями сапротрофного компонента,
осуществляется деструкция, разложение органики, следствием чего
является пополнение запаса минеральных элементов в почве (или
водной среде). Роль и значение в биогеоценозе других форм непол-
ночленных консорций, очевидно, невелики; лишь в случае после-
дующего вовлечения их на том или ином этапе функционирования в
общий круговорот веществ значение их заметно возрастает. Так
что вклад разных консорций в функциональную организацию био-
геоценоза и в те или иные обменные процессы в нем неодинаков.
Наличествующие в биогеоценозе консорции, как и входящие в
их состав пищевые цепи всех типов (пастбищные, детритные и
пр.), функционируют одновременно и постоянно. Этим обеспечива-
ется бесперебойное функционирование биогеоценозов, в состав ко-
торых они входят.
218
в. Системообразующие единицы, их место и роль в становлении
и функциональной организации биогеоценоза
Перейдем к самому главному: к рассмотрению вопроса о фор-
мировании биогеоценоза и его функциональной оргазации из соот-
ветствующих составных элементов и функциональных структурных
единиц.
«Элементарные функциональные единицы биогеоценоза — это
видовые популяции, интегрированное единство которых и состав-
ляет биогеоценоз», — пишет С. С. Шварц (1971, с. 6). Но этот про-
цесс интеграции ценопопуляций в биогеоценоз протекает, судя по
всему, через ряд ступеней, представляющих собой качественно
различающиеся биологические и ценотические систе-
м ы разного ранга и соответствующие им формы системного
единства, в которые они облекаются. Взяв за основу схему
двух параллельных рядов надорганизменной системы иерархии жи-
вого (автотрофпо-фитоценотического и консортивно-биоценотиче-
ского), предложенную К. А. Куркиным и А. Р. Матвеевым (1981),
мы нашли возможным построить схему, в некотором отношении
сходную с ней, но и отличающуюся от нее (табл. 1)
На схеме показаны два взаимосвязанных ряда биологических и
деистических образований разного ранга. Первый ряд состоит из
.последовательно усложняющихся биологических и ценотических
.систем: а) особь (организм); б) видовая ценопопуляция; в) цено*
комплексы авто-, био- и сапротрофов; г) биота как система живых
компонентов. Во втором ряду схемы в такой же последовательно-
сти расположены соответствующие каждой био- и ценосистеме
формы системного единства: а) индивидуальная консорция; б) по-
пуляционная консорция; в) биоценоз как совокупность популяци-
онных консорций; г) биогеоценоз. Нетрудно в каждой ступени это-
го двоякого рода биологических и ценотических образований
увидеть единство двух взаимосвязанных сторон такой парной кате-
гории, как содержание и форма,
Исходя из такой трактовки существа множества элементов раз-
ного типа и ранга, рассматриваемых многими в качестве функцио-
нальных структурных единиц функциональной организации биогео-
ценоза, представляется возможным, на наш взгляд, с большей
точностью определить действительное место и ранг каждого из них
в общей иерархии системообразующих единиц, а также соотноше-
ние существующих взаимосвязей их между собой. Так, доминирую-
щие особи автотрофов, являясь центрами индивидуальных кон-
сорций, входят в качестве элементов в виде экобиоморф (иначе —
фитоцепотипов) в состав видовых ценопопуляций автотрофов, уча-
ствуя при этом вместе со своей «свитой» разнообразных консортов
в формировании соответствующих популяционных консорций —
219
Таблица 1
220
Био-
и ирносистемы
Формы
системного
ЕДИНСТВА
Становление функциональной организации биогеоценоза
полночленных (пастбищных, биотрофных) и неполночленных (де-
тритных, сапротрофных). Ценозообразующие видовые цено-
популяции, являющиеся системами особей живых существ я
одновременно центрами или консортами популяционных консорций
входят в качестве элементов в состав автотрофных, биотрофных и
сапротрофных ценокомплексов, занимая в каждом из них опреде-
ленные позиции в трофических звеньях пастбищной или детритной
пищевой цепи. Ценокомплексы авто-, био- и сапротрофов,
являющихся системами соответствующих ценозообразующих видо-
вых ценопопуляций и одновременно живыми компонентами биоце-
ноза, входят в качестве элементов в состав биоты, выполняя при-
сущие им функции в обмене веществ и преобразовании энергии в
трофических звеньях пищевых цепей и сетей. Биота, являющаяся
целостной системой авто-, био- и сапротрофного ценокомплексов
и одновременно подсистемой биогеоценоза, входит в форме биоце-
ноза в его состав; второй подсистемой биогеоценоза как основной
структурно-функциональной единицы биогеосферы является биотоп.
Итак, все иерархически последовательно взаимосвязанные био-
логические и ценотические составные элементы функциональной
организации биогеоценоза и соответствующие им формы системно-
го единства оказываются задействованными. При этом каждый из
них занимает определенное место в общей системе биогеоценоза,
обеспечивающее выполнение свойственной ему функции.
Из изложенного следует, что формирование биогеоценоза и его
функциональной организации —многоступенчатый процесс, кото-
рый совершается при непосредственном участии различных по типу
питания и использованию источников энергии видовых ценопопу-
ляций, объединенных в соответствующие иенокомплексы, в сово-
купности образующие биоту. Популяционные же консорции, как и
пищевые цепи (и сети) в их составе, с одной стороны, биоценозы
и биогеоценозы — с другой, — это не что иное, как формы, выра-
жающие весьма сложные, многократно переплетающиеся связи и
отношения видовых ценопопуляций автотрофного, биотрофного и
сапротрофного ценокомплексов и биоты в целом. Занимая в их со-
ставе определенные позиции, они включаются, таким образом, в
форме соответствующих трофических уровней в состав пищевых
цепей той или иной популяционной консорции. Последняя же с по-
зиции системной трактовки организации биогеоценоза представ-
ляет собою функциональную структурную единицу более крупного
системного единства-биоценоза, в форме которого биота входит в
качестве подсистемы в состав биогеоценоза как материально-энер-
гетической ячейки биогеосферы.
Таким образом, несколько перефразируя отнесенные к фитоце-
нозу слова А. П. Шенникова (1964), можно сказать, что биоценоз
и биогеоценоз в целом есть оформление бесконечного многообра*
221
зия взаимных связей и отношений их составных элементов разного
ранга.
Так в общей форме представляется становление биогеоценоза
и формирование его функциональной организации из соподчинен-
ных биологических и ценотических образований разного ранга и
соответствующих форм системного единства. Более конкретно этот
процесс представлен П. М. Рафесом (1972) на материале исследо-
вания осиново-березовых рощиц в «блюдцах» лесостепной полосы
Западной Сибири, называемых «березовыми колками». Рассмат-
ривая биогеоценоз как систему консорций, автор обращает внима-
ние на то обстоятельство, что в биогеоценозе, как правило, дей-
ствуют несколько популяционных консорций, детерминированных
определенными ценопопуляциями автотрофных растительных доми-
нант, например березой, осиной, сосной и др. Вследствие количе-
ственного и качественного различия продуцируемой названными
детерминантами фитомассы подбирается и различный состав кон-
сортов I концентра в каждой из них, что определяет и состав кон-
сортов последующих концентров консорций. Из этого им делается
вывод: биогеоценоз нельзя изображать как систему, характеризую*
щуюся единым потоком веществ и энергии. В действительности он
складывается из нескольких вещественно-энергетических потоков,
входящих в состав биогеоценоза консорций. Более того, поскольку
консорции включают по несколько пищевых цепей, каждая из ко-
торых также характеризуется специфичным потоком веществ и
заключенной в них энергии, то количество и разнообразие веще-
ственно-энергетических потоков еще более возрастает. «В конечном
счете, — заключает автор, — функция биогеоценоза — это осуще-
ствление круговорота материи на занимаемой им территории»
(с. 45).
Изложенное представление автора о формировании биогеоце-
ноза и его функциональной организации из слагающих его популя-
ционных консорций весьма наглядно иллюстрируется схемой
Заслуживает внимания и оригинальная схема консорции
исусттд \рис. 20) f составленная автором не в виде концентрических
кругов вокруг детерминанта, а в виде развернутой в одном направ-
лении полосы. Это дало возможность более полно показать боль-
шое разнообразие консортивных связей между консортами и кон-
сортов с центром консорции, в том числе и связь потребителей
живой биомассы всех концентров и детерминанта консорции с
сапротрофами, т. е. показано завершение цикла обмена веществ в
рамках популяционной консорции.
Важное значение представляет включение П. М. Рафесом в
процесс функционирования биогеоценоза, наряду с биотическим
круговоротом веществ, особого цикла, названного им а биоса п-
ротрофным. Основанием для этого, как видно, послужили сле-
222
Детерминанты
I Кон
| Концентры: т
с о рты
J JT . I - М
Рис. 19. Схема функциональной организации биогеоценоза (по П. М. Рафесу,.
1972)
дующие факты. Детритные пищевые цели, как показано Ю. Оду-
мом (1975), не всегда заканчиваются звеном сапротрофов, а в
случае поедания последних их врагами в пищевую цепь включа-
ются новые трофические звенья. С другой стороны, как показала
223
Рис. 29, Схема консорции популяции березы (по П. М. Рафесу, 1972)
Т. В. Аристовская (1973), некоторые почвенные микроорганизмы,
названные П. М. Рафесом абиотрофами, способны разлагать
не только органические материалы, но и многие минералы, делая
продукты их разложения доступными для питания растений. Ис-
ходя из этого, автор приходит к выводу о наличии в природе осо-
бого цикла обменных процессов. Этот цикл начинается с потребле-
ния органических остатков и трансформации минеральных веществ
иочны, который вводит мертвую материю в особые (без фотосин-
тези) цепи питания, отходы которых возвращаются в почву, со-
здавая цикл.
224-
9. Ярусность фитоценоза и пространственное размещение в нем
животного и микробного населения
Для успешного функционирования биогеоценоза, нормального
и бесперебойного осуществления обменных процессов в нем, наря-
ду с надлежащей функциональной организацией, необходима так-
же оптимальная морфологическая (пространственная) структура
его. Под морфологической структурой биогеоценоза имеется в виду
определенная группировка ценозообразующих биологических и це-
нотических систем разного ранга, их размещение в воздушной и
почвенной средах, взаимосвязь и расположение их по отношению
друг к другу и к факторам внешней среды. При этом группировка
и пространственное размещение био- и ценосистем должны быть
такими (как в вертикальном,так и в горизонтальном направлении),
которые обеспечивали бы наиболее полное использование ими как
энергии солнечной радиации, так и материальных ресурсов био-
цепосрсды (углекислоты, кислорода, атмосферной и почвенной вла-
ги, элементов зольного и азотного минерального питания и пр.).
Строение и функции биогеоценоза тесно взаимосвязаны. Функ-
ции биогеоценоза, по словам В. В. Мазинга (1973), определяют
его морфологическое строение; с другой стороны, структура по-
следнего является условием его успешного функционирования. На
важное значение морфологической (пространственной) структуры
биогеоценоза в его функционировании обращает внимание и
Ю. И. Чернов (1971). В частности, он указывает на наличие опре-
деленной количественной зависимости между функциональными и
структурными характеристиками биогеоценоза; например, зависи-
мость продукции от его структуры. Вместе с тем тот и другой авто-
ры подчеркивают, что нельзя также смешивать и тем более ото-
ждествлять явления и понятия морфологической (пространствен-
ной) структуры биогеоценоза и его функциональной организации,
ибо, как отмечает Ю. И. Чернов, морфологические категории не
всегда адекватно отражают их функциональное значение в биогео-
ценозе; с другой стороны, функциональные ценотические образо-
вания в составе биогеоценоза, по словам В. В. Мазинга, не всегда
могут быть увязаны с соответствующими частями морфологической
структуры его. Несовпадение морфологических и функциональных
категорий в биогеоценозе, по мнению Ю. И. Чернова, объясняется
тем, что биоценотический уровень организации живого характери-
зуется невысокой степенью интегрированности, целостности всех
форм ценотических образований, но большей дифференциацией их
морфологических структурных частей по сравнению, например, с
организменной формой жизни.
Структурной основой, своего рода каркасом пространственного
размещения компонентов биогеоценоза в наземной (воздушной)
225
среде является ценокомплекс высшйх автотрофных растений (фи-
тоценоз). Это обусловлено не только тем.что растительное сообще-
ство— единственный компонент, способный синтезировать органи-
ческие вещества из неорганических, но и тем, что, ввиду относи-
тельно равномерного повсюду распределения в пространстве
материальных ресурсов для фотосинтеза, высшие растения, как
было сказано ранее, ведут прикрепленный образ жизни, а это опре-
делило хорошо выраженное структурное оформление растительных
сообществ, вертикальное и горизонтальное расчленение их на так
называемые ярусы (стратосинузии, поП. М. Рафесу, 1966)
и синузии-микрогруппировки.
Древнейшие сухопутные растительные сообщества, как полага-
ет Н. М. Григорьева с соавторами (1977), были «стратифицирован-
ными образованиями». Среди вышедших на сушу первичных выс-
ших растений, по их мнению, со временем появились виды
«вертикально устойчивые и всесторонне дихотомически разветвлен-
ные», сомкнувшиеся впоследствии в ярусы. Следовательно, ярус-
иость сообществ, как утверждают авторы, — «им по природе
присущее, с ними неразрывными узами связанное свойство» (с. 12).
Таким образом, дифференциация растительных сообществ по вы-
соте рассматривается названными авторами как всеобщий процесс
вертикальной анизотропности, вторым проявлением которой явля-
ется мозаичность. Последняя, как и ярусность, по их мнению,
представляет собой свойство изначальное, появившееся одновре-
менно с возникновением самих сообществ.
Структурной основой пространственного размещения живых
компонентов (подземных органов растений, почвообитающих поз-
воночных и беспозвоночных животных и микроорганизмов) в под-
земной (почвенной) среде является сама почва, а именно диффе-
ренциация почвенного профиля на так называемые генетические
горизонты.
Вертикальное расчленение фитоценоза на ярусы определяется
в надземной части размерами, долголетием, формой роста ценозо-
образующих растений, особенностями их ветвления и облиственно-
сти, видовым разнообразием растений, отношением их к свету,
атмосферным осадкам; в подземной части — механическими» физи-
ческими и химическими свойствами почвы, отношением растений
к влаге, минеральным веществам почвы и т. п. Горизонтальное
расчленение фитоценоза на микрогруппировкн обусловлено харак-
тером роста господствующих растений (латками, куртинами, дер-
нинами, одиночными побегами), густотой и размещением различ-
ных растительных экобиоморф по площади, характером поверхно-
сти почвы (микрорельефом), этапами формирования растительного
сообщества и т. п.
Не останавливаясь на изложении морфологической структуры
22В
растительных сообществ, поскольку подробная характеристика ее
дается в руководствах по геоботанике (фитоценологии), обратим
внимание лишь на два принципа выделения вертикальных и гори-
зонтальных структурных частей фитоценозов, предложенные
В. В. Мазингом (1973): а) принцип перекрытия структурных ча-
стей и б) принцип взаимоисключения структурных частей. На осно-
ве принципа перекрытия выделяются в вертикальном направлении
ярусы, в горизонтальном — микрогруппировки. На основе
же принципа взаимоисключения в вертикальном направлении мож-
но выделять биогеоценотические горизонты (в воздуш-
ной и почвенной средах), а в горизонтальном — парцеллы
(рис. 21).
Рис. 21. Вертикальное строение лесного фитоценоза (по В. В, Ма-
зингу, 1973; и др.). В — взаимоисключающие и П — перекрыва-
ющие элементы
При анализе морфологической структуры фитоценоза во всех
случаях, разумеется, необходимо особое внимание уделять про-
странственному размещению наиболее эффективно ассимилирую-
щих органов растений и адсорбирующих частей подземных
органов.
227
Растительный компонент биогеоценоза — фитоценоз — является
не только автотрофной базой для гетеротрофных живых компонен-
тов, но и местом гнездования и укрытия их от своих естественных
врагов. Высшие зеленые растения в сухопутных биогеоценозах
производят основную массу первичной органической продукции,
служащей средством питания для гетеротрофов, и свободный кис-
лород для дыхания. Многоярусная древесно-кустарниковая либо
травянистая и мохово-лишайниковая растительность, а также об-
разующаяся из опада и отпада подстилка служат местом обитания
и размножения разнообразных позвоночных и беспозвоночных жи-
вотных и микроорганизмов. Поэтому вертикальное и горизонталь-
ное размещение животного и микробного населения биогеоценоза
в наземной (воздушной) среде находится в значительной зависи-
мости от особенностей морфологической структуры фитоценоза, а
также от характера питания, образа жизни и связанной с этим
степени подвижности животных и микроорганизмов. Размещение
же почвообитающих животных и микроорганизмов по почвенному
профилю определяется главным образом глубиной залегания бога-
тых органикой почвенных горизонтов (почвенно-подстилочного, гу-
мусового и др.) и распространением корневой системы, служащих
местом обитания и источником энергии для названных живых су-
ществ.
О вертикальном размещении животного населения в биогеоце-
нозе н степени соответствия его с ярусностью растительного
покрова известны такого рода наблюдения и данные. Так. Ж- Леме
(1976) утверждает, что животные живут преимущественно или
исключительно в одном каком-либо ярусе растительного сообще-
ства; например, некоторые птицы предпочитают вить гнезда и пре-
бывать на вершинах деревьев, в их кроне, другие виды гнездятся
в подлеске. Из 240 видов беспозвоночных животных (насекомых,
паукообразных, многоножек), зарегистрированных нм в дубовом
лесу США, более 180 видов оказались обитателями одного какого-
либо яруса данного сообщества и только 3—5 видов были обнару-
жены во всех ярусах. Даже в травянистых сообществах, по наблю-
дениям данного автора, часть животных обитает преимущественно
в верхнем пологе, а часть — близ поверхности почвы. Правда, у
некоторых видов животных он отмечает перемещения из одного
яруса в другой в разное время суток или в разные сезоны года.
Столь же конкретные данные о вертикальном распределении
беспозвоночных животных в зависимости от особенностей морфо-
логической структуры растительного сообщества приводит
Г. А. Новиков (1979). Так, в Мариупольском лесничестве среди
насекомых оказалось видов, обитающих на деревьях и кустарни-
ках,— 41, в травостое — 76, па поверхности почвы и в подстилке —
67. В дубовом лесу заповедника «Лес на Ворскле» Белгородской
228
области из общего количества насекомых и пауков обнаружены: в
древесном ярусе — 28,9%; в подлеске и подросте—14,4; в травостое
и на поверхности почвы — 56,7%. По мнению Г. А. Новикова, раз-
мещение животных в биогеоценозе по ярусам свойственно не толь-
ко беспозвоночным, но также некоторым видам млекопитающих и
птиц. Это проявляется в устройстве ими убежищ (гнезд, нор) в
том или ином ярусе, в выборе места кормежки и т. п.
Однако А. Г. Воронов (1967) и ряд других биологов считают,
что у наземных позвоночных животных строгой приуроченности к
определенному ярусу фитоценоза не наблюдается. Большинство
наземных позвоночных ими относится к категории так называемых
ярусно-подвижных форм, которые строят свои убежища в
одном ярусе (например, на поверхности почвы), а в поисках корма
взбираются на деревья, или наоборот, т. е. охватывают своей жиз-
недеятельностью, таким образом, два яруса сообщества. Некото-
рые виды позвоночных в разные сезоны вегетационного периода
или года занимают в фитоценозе разные ярусы. У немногих видов
позвоночных—млекопитающих, пресмыкающихся, птиц можно кон-
статировать более или менее ясно выраженную приуроченность к
тому или иному ярусу растительного сообщества.
Зато у беспозвоночных животных, особенно у почвообптающих,
явление ярусности довольно хорошо выражено. Так, например, в
почве пустынных сообществ наблюдается такого рода размещение
насекомых по почвенному профилю: в приземном слое преобла-
дают прямокрылые и двукрылые; в поверхностных горизонтах поч-
вы господствуют перепончатокрылые, щетинохвостые, веснянки и
пр.; в глубинных слоях почвы встречаются лишь жуки, термиты,
ногохвостые и т. п. Некоторая дифференциация в распределении
по горизонтам почвенного профиля отмечается и у почвообитаю-
щих позвоночных, таких, как слепушонок, слепыш и пр., которые
строят свои гнездовья в норах на разной глубине от поверхности
земли; камеры же (кладовые) для хранения запасов корма строят
обычно еще глубже.
Не менее отчетливо ярусное распределение наблюдается и у
водных животных, где выделяется несколько ярусов: надводный»
водный, донный и внутригрунтовый.
Из микробного населения биогеоценоза наибольшее значение
имеют микроорганизмы, осуществляющие минерализацию органи-
ки (спороносные и неспороносные бактерии, микроскопические
грибы, актиномицеты). Вертикальное и горизонтальное размеще-
ние микроорганизмов воздушной среды в биогеоценозе определя-
ется составом и морфологической структурой автотрофного расти-
тельного компонента. Распределение же почвенной микрофлоры по
горизонтам почвенного профиля находится в зависимости главным
образом от характера распространения подземных органов ценозо-
229
образующих растений, а также от физико-химических свойств раз-
личных почвенных горизонтов и гидротермических условий в каж-
дом из них. Наибольшее скопление микроорганизмов-деструкторов
наблюдается в мертвом напочвенном покрове биогеоценоза (в
лесной подстилке, степном войлоке и т. п.), в дернине и верхнем
гумусовом горизонте почвы, в прикорневой части растений, в так
называемой ризосфере, что связано с выделением корнями
растений различных физиологически активных и питательных ве-
ществ (органических кислот, сахаров, фитонцидов и т. п.).
Такого же рода закономерности имеют место в пространствен-
ном размещении животного и микробного населения биогеоценоза
и в горизонтальном направлении. Материальной основой распреде-
ления биологических и ценотических групп биотрофов и сапротро-
фов по площади служат такие структурные части фитоценоза, как
сйнузии-микрогруппировки (рис. 22).
Рис. 22. Горизонтальное сложение березово-лиственничника (по П. Д. Ярошенко,
1961). Микрогруппировки: 1 — соссюрейно-березово-лиственничная (L. dahurica);
2 — бруснично-лиственничная (L. dahurica); 3 — вейниково-березовая (В. р!а-
typhylla)
Таким образом, на основе вертикального и горизонтального
расчленения растительного сообщества происходит пространствен-
ное размещение и других живых компонентов биогеоценоза и со-
здается основа для формирования морфологической структуры
биогеоценоза в целом. Но морфологическая (физическая) струк-
тура биогеоценоза — это не сумма структурных частей его компо-
230
нентов. Структурные части вертикального и горизонтального рас-
членения биогеоценоза — это комплексные структурные образова-
ния; каждая из них включает, как правило, все живые и косные
(биогенные) компоненты, находящиеся в постоянной функциональ-
ной связи и взаимодействии между собой. Через периферийные
структурные части биогеоценоз связан в горизонтальном направ-
лении со смежными биогеоценозами, а в вертикальном — с воздуш-
ными массами атмосферы либо с грунтовыми толщами земной
коры за его пределами.
10. Морфологическое строение биогеоценоза,
вертикальные и горизонтальные структурные части его
Анализируя строение сухопутных биогеоценозов в вертикальном
направлении, Ю. П. Бяллович (1960) выделил структурные части,
которые назвал бногеоценотическими горизонтами
(бпогеогоризонтами), и дал такое определение этого понятия:
«Биогеоценотический горизонт есть вертикально обособленная и
по вертикали далее нерасчленимая структурная часть биогеоцено-
за. Сверху донизу биогеоценотический горизонт однороден по со-
ставу биогеоценотических компонентов, по взаимосвязям, по проис-
ходящим в нем превращениям веществ и энергии...» (с. 45). Во
многих случаях биогеогоризонты пространственно совпадают со
структурными частями природных образований (с ярусами фито-
ценоза, генетическими горизонтами почвы, слоями тропосферы,
горизонтами почвенно-грунтовых вод). Но обычно биогеоценоти-
ческие горизонты — более дробные подразделения биогеоценоза
по вертикали, так как служащие автотрофной базой биогеоценоза
наземные ярусы растительного компонента, особенно верхние, не-
однородны в пределах своей мощности сами по себе, а также по
степени освещенности, результатам фотосинтетической деятельно-
сти и другим показателям. Поэтому в каждом древесном ярусе
лесного биогеоценоза автором выделяются верхний «деятельный»
горизонт фотосинтеза и нижний горизонт фотосинтеза; в качестве
самостоятельных биогеогоризонтов выделяются ярусы подлеска,
травяно-кустарничкового и мохово-лишайникового покровов; кро-
ме того, выделяются межпологовые и подпологовый биогеогори-
зонты. В результате в лесном биогеоценозе с трехъярусной расти-
тельной основой оказывается около десятка биогеоценотических
горизонтов (рис. 23).
Биогеогоризонты биогеоценотических систем хотя каче-
ственно и отличаются друг от друга, но, по свидетельству автора,
им свойственны и некоторые общие морфологические закономер-
ности: а) отсутствие абсолютно перпендикулярных поверхностей
биогеогоризонта на всем его протяжении по отношению к силе зем-
231
Ярусы
1, Верхний („деятельный")
БИОГЕОГОРИЗОНТ
ФОТОСИНТЕЗА
Биогеогоризонты
2. Нижний БИОГЕОГОРИЗОНТ
ФОТОСИНТЕЗА
з. Первый межпологовый
БИОГЕОГОРИЗОНТ
5. НИЖНИЙ БИОГЕОГОРИЗОНТ
ФОТОСИНТЕЗА
4. Верхний биогеогоризонт
ФОТОСИНТЕЗА
в. Второй межпологовый
БИОГЕОГОРИЗОНТ
Л
5Ч5ЯЯ
Рис. 23. Соотношение растительных ярусов и биогеогоризонтов в лесном биогео-
ценозе (по Ю. П. Бялловичу, 1960)
7 БИОГЕОГОРИЗОНТ
ФОТОСИНТЕЗА ПОДЛЕСКА
8. Подпологовый
БИОГЕОГОРИЗОНТ
ного притяжения; б) непостоянство границ биогеогоризонтов в
связи с изменением их в ту или иную сторону в процессе сукцес-
сий; в) вклинивание между некоторыми биогеогоризонтами на
сроки различной продолжительности временных биогеогоризонтов
(например, ярусы эфемеров в пустыне весной). Связанные с био-
геоценозами внешние по отношению к ним покрывающие и подсти-
лающие участки биосферы характеризуются, по словам автора, тем,
что в них протекают лишь геофизические и геохимические процес-
сы. Поэтому вычленяющиеся в их составе в вертикальном направ-
лении слои он назвал геогоризонтами. В отличие от биогео-
горнзонтов, они лишены постоянно обитающих специальных
жизненных форм. Здесь могут оказаться лишь случайно залетев-
232
шие птицы или занесенные воздушными течениями насекомые,
микроорганизмы, зачатки растений и т. п.
По мнению Н. В. Дылиса (1969, 1974, 1978), А. И. Уткина
(1969, 1970) и др., подразделение биогеоценозов на биогеогоризон-
гы по предложенному Ю. П. Бялловичем принципу является в
значительной степени условным. Проведенные названными авто-
рами с участием ряда других биогеоценологов всесторонние иссле-
дования по вертикально-фракционному распределению фитомассы
древесных, кустарниковых, травянистых и других растений в раз-
личных типах лесных биогеоценозов Подмосковья показали:
а) особо важное, превалирующее значение автотрофного расти-
тельного компонента в вертикальном расчленении биогеоценотиче-
ской толщи, поскольку с этим связано соответствующее расслоение
воздушной и почвенной среды обитания, животного и микробного
населения биогеоценоза; б) неоднородность в биогеоценотическом-
отношении фитомассы в пределах растительных ярусов, а именно:
наличие горизонтов с высокой концентрацией акцепторных (вос-
принимающих) частей надземных и подземных органов растений
(листьев и молодых побегов древесных и кустарниковых пород,
листьев трав, мхов, слоевищ лишайников, молодых окончаний кор-
ней и пр.), а также горизонтов наибольшей аккумуляции органи-
ческих веществ. !
Исходя из этого, Н. В. Дылис (1974) при выделении биогеого-
ризонтов в вертикальном профиле наземной части лесного биогео-
ценоза, в частности в пределах кроны, предлагает использовать
такие показатели, как распределение массы световых и теневых
листьев (с учетом их размеров и годичного прироста), генератив-
ных органов растений, отмерших побегов, сучьев и т. п. Показатели
эти должны быть увязаны с особенностями вертикального распре-
деления света, влажности воздуха, концентрации углекислоты,
размещения животных-фитофагов и пр. Все это, по мнению автора,
даст возможность более обоснованно подойти к морфологической
стратификации биогеоценоза.
Н. В. Дылис и др. (1969) на основе анализа вертикально-фрак-
ционного строения фитомассы ценозообразующих растений и свя-
занного с этим состояния живых и косных компонентов выделяют
в кроне 1-го яруса елового биогеоценоза три фотосинтетических
биогеогоризопта. I. Верхний биогеогоризонт (М — 3—4 м) со све-
товой хвоей, работающей в условиях полного освещения. II. Сред-
ний биогеогоризонт (М = 7—8 м) со световой -f- теневой хвоей в
условиях сниженной интенсивности освещения. III. Нижний б йо-
ге ого риз о нт (М = 4—5 м) с теневой хвоей, работающий в условиях
резко ослабленного освещения с отрицательным балансом. Кроме
того, в самой нижней части кроны выделяется слой, характеризую-
щийся преобладанием процессов отмирания частей растений над
233
их новообразованием. В кронах сосновых биогеоценозов тем же
автором выделяются два биогеогоризонта, в осинниках — один
ввиду однородности строения листвы и освещенности кроны, обла-
дающей незначительной мощностью.
А. И. Уткиным с соавторами (1969) в пределах всего древес-
ного яруса елового биогеоценоза на тех же основаниях выделяют-
ся четыре биогеогоризонта — два фотосинтезирующих и два акку-
мулятивных. I. Биогеогоризонт фотосинтеза с активным балансом
(М = 9 м) с густоохвоенными ветвями кроны, доля рабочей части
которой более 10%, хвоя световая. Л. Биогеогоризонт фотосинтеза
с пассивным балансом (М = 4 м) с охвоенностью ветвей кроны
около 1/3, доля рабочей части ее менее 10%, хвоя преимуществен-
но теневая. III. Биогеогоризонт кроново-стволовой аккумуляции
(М = 5 м) с резким преобладанием стволовой аккумуляции над
аккумуляцией скелета кроны. IV. Биогеогоризонт стволовой акку-
муляции (М = 7 м), охватывающий всю подкроновую зону де-
ревьев.
Тот и другой авторы в лесных биогеоценозах, в отличие от
Ю. П. Бялловича, на биогоризонты подразделяют лишь древостой.
Фотосинтетическая деятельность нижних ярусов автотрофного рас-
тительного компонента в подкроновой зоне определяется древесным
ярусом. Поэтому они объединяются в один комплексный биогео-
горизонт, подразделяемый па ряд слоев: а) слой фотосинтеза
подлеска (М = 4,5 м) и б) слой фотосинтеза травяного покрова
(М = 0,4 м), внутри которого обособляются слой фотосинтеза под-
роста и пятнистый слой фотосинтеза мохового покрова.
Проблема горизонтального сложения биогеоценозов разработа-
на Н. В. Дылисом (1964, 1969, 1974, 1978 и др.). В 1964 году со-
вместно с А. И. Уткиным и И. М. Успенской им были впервые
выделены горизонтальные структурные части биогеоценоза, на-
званные парцеллами, и дано определение этого понятия: «Пар-
целлы— это комплексные части столь же комплексного целого —
биогеоценоза, отличающиеся друг от друга как радиальным
сложением компонентов, так и спецификой их радиально направ-
ленного материально-энергетического метаболизма, и на этом
основании обособленные друг от друга в пространстве на всю ра-
диальную толщу биогеоценоза» (с. 65). В последующих работах
формулировка этого понятия автором была несколько упрощена:
«Биогеоценотическими парцеллами называются структурные части
горизонтального расчленения биогеоценозов, отличающиеся друг
от друга составом, структурой и свойствами компонентов, специ-
фикой их связей и материально-энергетического обмена. Обособ-
лены парцеллы друг от друга в пространстве на всю вертикальную
толщу биогеоценоза» (1969, с. 12).
В первом определении понятия парцеллы особо подчеркива-
234
ется, что парцеллы — это комплексные структурные части биогео-
ценоза, так как в их состав входят как все живые компоненты —
ценокомплексы автотрофных, биотрофных и сапротрофных цено-
популяций, так и биогенные—аэротоп и эдафотоп. Автотрофной
базой парцелл являются выявленные П. Д. Ярошенко (1953) в
составе фитоценоза микрогруппировки (синузии-микрогруп-
пировки, по А. П. Щенникову, 1964). В соответствии с этим по
границам растительных микрогруппировок устанавливаются и гра-
ницы парцелл, а по названию доминирующих в их составе растений
производится и наименование их. Каждая парцелла в границах
биогеоценоза бывает обычно представлена многими конкретными
участками, различными по площади и конфигурации. Поэтому пар-
целла рассматривается автором как понятие типологическое, объ-
единяющее сходные между собой участки по присущим им при-
знакам и свойствам (компонентному составу, структуре и внешнему
облику, составу и запасам живой и мертвой биомассы, обменным
процессам), а также по местоположению в микрорельефе, режи-
мам света, тепла, влаги и т. п. В качестве примера Н. В. Дылисом
приводится состав парцелл ельника зеленчуково-волосистоосоково-
го (Piceetum qaleobdolonoso-pilosae caricosum); а) елово-зеленчу-
ковая (27% от общей площади биогеоценоза); б) елово-волосисто-
осоковая (19%); в) елово-кисличная (8%); г) осиново-елово-меду-
ничная (7%); д) елово-костяничная (7%); е) елово-щитовниковая
(6%); ж) елово-липовая (6%); з) крупнопапоротниковая (в «ок-
нах» (5%) и др. Всего—14 (рис. 24).
Нами (Л. И. Номоконов и др., 1978) в процессе исследования
природных луговых биогеоценозов в пойме Нижнего Дона выявле-
ны следующие парцеллы пырейника чистого (Aqropyretum purum),
названные впоследствии эпипарцеллами: а) дербенниково-
пырейная (36,7%); б) бекманниево-дербенниково-пырейная
(26,6%); в) дербенниково-ясколково-пырейная (14,9%); г) осоко-
во-вербейниково-пырейная (4,4%); д) солодково-дербенниково-пы-
рейная (11,1%); е) солодково-мятликово-пырейная (2,5%) и
ж) бекманниево-жирухово-пырейная (3,8%) (рис. 25).
Хотя парцеллы — пространственно обособленные друг от друга
образования, вместе с тем они тесно связаны между собой как
структурные части единого целого — биогеоценоза. Связаны они
посредством множества каналов и с внешним окружением (поверх-
ностный сток атмосферных осадков и талых вод, поток солнечной
энергии, перемещение животных, рассеивание семяп и плодов рас-
тений и т. п.).
Являясь структурными частями горизонтального сложения раз-
личных биогеоценозов, парцеллы весьма разнообразны. Они раз-
личаются по происхождению, по своей роли в структуре и метабо-
лизме биогеоценоза, по тенденциям развития и пр. Н. В. Дылисом
235
Рис. 24. Парцеллярное сложение ельника (с липой) зеленчуково-волоснстоосоко-
вого (по Н. В. Дылису, 1978). Парцеллы: 1 — елово-волосистоосоковая; 2 — ело-
во-зеленчуковая; 3 — елово-кисличная; 4 — осиново-елово-медуничная; 5 — елово-
костяничная; 6 — елово-щитовниковая; 7 — елово-пальчатоосоковая; 8— елово-
липовая; 9 — крупнопапоротннковая (в «окнах» древостоя); 10 — осиново-сныте-
вая; 11—дубово-снытевая; 12 — елово-лещиновая; 13 — лещиново-волосистоосо-
ковая; 14 — групповой подрост ели
с соавторами (1964) они подразделяются на следующие категории.
По происхождению: а) на корен иные, по своим признакам и
свойствам находящиеся в относительном соответствии с наличны-
ми материально-энергетическими ресурсами экотопа; б) произ-
водные, возникающие под влиянием хозяйственной деятельности
местного населения или природных стихийных явлений (бури, на-
воднения, пожары и т. п.). По роли парцелл в горизонтальном
сложении и обменных процессах биогеоценоза: а) на основ-
ные— наиболее крупные по занимаемой площади в экотопе, со-
здающие основной фон биогеоценотического покрова и основное
направление обмена веществ в биогеоценозе; б) дополняющие,
занимающие в экотопе небольшие по площади участки в виде пя-
236
Рис. 25. Эпипарцеллы пырейника чистого (по Л. И. Номоконову и др., 1978):
] —дербенниково-пырейная; 2 — бекманниево-дербенниково-пырейная; 3 — дер-
»бенниково-ясколково-пырейная; 4 — осоково-вербейниково-пырейная; 5 — солодко-
во-дербенниково-пырейная; 6 — солодково-мятликово-пырейная; 7— бекманниево-
жерухово-пырейная
тен и играющие небольшую роль в обменных процессах биогеоце-
ноза. По тенденциям развития: а) на устойчивые, прогрессив-
ные с расширяющимися границами; б) отмирающие, реликто-
вые с сокращающимися границами.
Б. А. Быковым (1970) парцеллы подразделяются, кроме того, по
высоте доминирующих в составе их автотрофного растительного
компонента видовых ценопопуляций на следующие группы: а) нор-
мальные (ординарные) парцеллы, состоящие из доминирующих
237
растений обычного размера; б) превышающие парцеллы, представ-
ленные более высокорослыми растениями, возвышающимися над
главным слоем биогеоценоза, часто за счет особей иной ценопопу-
ляции; в) пониженные парцеллы, нарушающие главный слой рас-
тительного компонента благодаря господству более низкорослых
особей, обычно иной ценопопуляций; г) беспокровные парцеллы из
низших растений (водорослей, грибов, бактерий) при отсутствии
наземной растительности, встречающиеся в биогеоценозах пустын-
ной области.
Наряду с биогеогоризонтами и парцеллами, являющимися
основными, общими для всех биогеоценозов морфологическими
структурными частями, Н. В. Дылисом с соавторами и другими
биогеоценологами выявлен ряд пространственных структур, не
укладывающихся в рамки названных структурных образований;
они отнесены к категории микроструктур. В число таких
микроструктур обычно включают следующие мелкие морфологи-
ческие объекты растительного и животного происхождения:
а) мертвые стволы древесных пород на поверхности почвы; б) пни
от высоко срезанных деревьев; в) бугристые выбросы кротов и
других землеройных животных; г) муравьиные кучи; д) поры зве-
рей и гнезда птиц на земле. А. И. Уткин (1970) к числу микро-
структур относит даже плодовые тела шляпочных грибов, трупы
животных и т. п. Все перечисленные микроструктуры, как он счи-
тает, являются своего рода включениями в общем фоне парцеллы,
обладающими относительной автономностью в ее составе. Они, по
словам Н. В. Дылиса, разнообразят микросреду, способствуя обра-
зованию специфических группировок в составе травяно-мохового
покрова, сапрофитных грибов, насекомых и других сапротрофных
беспозвоночных животных и т. п.
3. Ф. Павлова (1977) на повышенных участках побережий со-
леных озер Юго-Западной Барабы выделила в качестве микро-
структур земляные муравьиные кочки, которые, по ее выражению,
«становятся центрами аккумуляции биоты» (с. 62). Они оказывают
существенное влияние на формирование и развитие растительного
покрова, почвообитающего животного населения, на процессы
почвообразования в условиях засоления и пр. А. А. Титлянова и
И. В. Стебаев (1978) в процессе исследований в условиях сухих
степей Юго-Восточного Забайкалья установили, что не только рас-
тительные группировки, но даже отдельные крупные растения
(чий, карагана) вносят существенные разносторонние изменения в
общий фон биогеоценоза. Такого рода образования ими названы
фитоге иными узлами и отнесены к категории микрострук-
тур биогеоценоза.
Причины горизонтальной неоднородности, парцеллярности био-
геоценозов весьма многообразны. Т. А. Работнов (1978), Б. А. Бы-
238
ков (1970) и др. видят их в следующем: а) неоднородности эда-
фотопа (неровность поверхности почвы, различная мощность
мелкозема на твердых грунтах); б) наличии конкурентных взаимо-
отношений видовых ценопопуляций главным образом посредством
изменения экотопа; в) особенностях вегетативного размножения
некоторых ценозообразующих растительных видов с образованием
клонов; г) воздействии внешних факторов, в особенности ветра,
воды; д) непосредственном или косвенном влиянии животных и
хозяйственной деятельности местного населения посредством ло-
кальных воздействий на биогеоценотический покров и, наконец,
е) некоторых явлениях случайного характера (в распределении
зачатков растений, нападении хищников на свои жертвы и т. п.).
Все формы пространственной структуры биогеоценоза, как ука-
зывает Н. В. Дылис, динамичны. В процессе становления и после-
дующего развития биогеоценоза протерпевает соответствующее
изменение и его морфологическая (физическая) структура. Неко-
торые структурные части вертикального и горизонтального строе-
ния биогеоценоза отмирают, исчезают, появляются новые структур-
ные образования, которые со временем также могут отмереть и
замениться новыми. Все эти структурные изменения, разумеется,
совершаются в рамках биогеоценоза, не нарушая его целостности.
Автором приводятся следующие примеры: а) под пологом верхнего
фотосинтетического биогеогоризонта через 10—15 лет формируется
нижний биогеогоризонт ослабленного фотосинтеза; б) с течением
времени может произойти отмирание части кроны древесного яру-
са и превращение ее в биогеогоризонт стволовой аккумуляции;
в) упавший ствол погибшего дерева под действием физико-хими-
ческих факторов и воздействием дереворазрушающих насекомых,
грибов и бактерий постепенно превращается в трухлявую массу.
Каково же биологическое и ценотическое значение вертикаль-
ной и горизонтальной дискретности морфологического строения
биогеоценоза? Суть этого, надо полагать, заключается в том, что
в процессе эволюции под действием биогеоценотического отбора
(в определенных пространственных рамках) выработалась наибо-
лее совершенная группировка и размещение в пространстве (и во
времени) биологических и ценотических составных элементов, ко-
торые обеспечивают наиболее полное и эффективное использова-
ние ценозами в процессе функционирования лучистой энергии
Солнца и материальных ресурсов экотопа. Этим самым подтверж-
дается и взаимозависимость морфологической структуры биогео-
ценоза и его функциональной организации. Это, кстати сказать,
открыло также пути к созданию высокопродуктивных и устойчивых
агроценозов различного хозяйственного назначения и создало
определенные предпосылки для хозяйственного освоения неисполь-
зуемых малоценных земель.
239
Глава Vlll
Материально-энергетический обмен
и функционирование биогеоценоза
1. Материально-энергетический обмен в биогеоценозе
и круговорот веществ в природе
Биогеоценоз представляет собой термодинамически относитель-
но открытую, пространственно обособленную функциональную био-
косную систему, имеющую вход и выход биогенных и других
веществ и энергии. Отсюда функционирование биогеоценоза — это
процесс непрерывного обмена веществом и энергией между его
компонентами (внутриценозный обмен) и биогеоценоза как целост-
ной системы с внешней физической средой и другими биогеоцено-
зами (внешний обмен). Материально-энергетический обмен, по
выражению Н. В. Дылиса, — это генеральная функция биогеоцено-
за, так как без обмена веществом и энергией внутри биогеоценоза
и без притока энергии извне существование его невозможно. Об-
менный процесс биогеоценоза носит циклический характер и про-
текает в форме круговорота веществ и одностороннего потока
энергии, непрерывно поступающей на земную поверхность из бли-
жайшего космоса в виде квантов солнечного света.
Материально-энергетический обмен биогеоценоза, как было
сказано ранее, протекает при участии всех его компонентов. Но
активную роль в этом процессе играют живые компоненты — цено-
комплексы автотрофных, биотрофных и сапротрофных ценопопуля-
ций живых существ. При этом следует иметь в виду, что выполне-
ние присущих каждому из них функций осуществляется не обособ-
ленно друг от друга, а в тесной взаимной связи между собой. В
тесной взаимосвязи живые компоненты находятся также и с био-
генными компонентами — аэротопом и эдафотопом, непрерывно
вовлекая их в обменные процессы биогеоценоза. Основное назна-
чение последних состоит в том, что они служат поставщиками
первичных материальных ресурсов неорганического преимуще-
ственно характера (биогенных газов, элементов минерального пи-
тания, влаги и т. п.), а также средой, обеспечивающей нормальный
ход этих процессов. В материально-энергетическом обмене биогео-
ценоза, как в фокусе, отражаются все взаимодействия между его
составляющими и с внешним окружением. Взаимодействие живого
и неживого проявляется, по Вернадскому, в биогенной миграции
атомов, осуществляющейся за счет энергии Солнца и выражаю-
щейся в питании, дыхании, размножении, смерти и разложении
240
организмов. Круговорот веществ, следовательно, можно трактовать
как основную форму проявления жизни. «Циклическая структура
жизни — одна из самых существенных особенностей ее организа-
ции»,— пишет М. М. Камшилов (1961, с. 30).
Жизнь осуществляется за счет постоянного притока извне
энергии, вещества и информации. Передача энергии по пищевой
цепи происходит, по словам Камшилова, с помощью различных
механизмов — от миграции электронов до пожирания хищниками
своей жертвы. Энерговооруженность жизни в процессе эволюции,
как он полагает (ссылаясь на мнение В. И. Вернадского и других
ученых), должна постепенно возрастать. В результате расширения
сферы жизни должно происходить при этом и увеличение массы
органического вещества на планете за счет включения в биологи-
ческий (биотический) круговорот все новых и новых минеральных
элементов.
Р. Н. Степанов (1966) полагает, что цикличность присуща не
только процессам жизни, по и всем природным процессам на Зем-
ле, обусловленным вращением Земли вокруг своей оси или вокруг
Солнца, колебанием солнечной активности или глобальной цирку-
ляции атмосферы и т. п. С этим связана не только смена дня и
ночи и смена времен года, но также зональная неоднородность
планеты, широтное и сезонное разнообразие поступления солнеч-
ной радиации, распределения ее по земной поверхности и, следо-
вательно, неоднородность освещения Земли. Автором приводятся
следующие закономерности, обнаруженные в цикличности развития
природных процессов: а) каждый цикл высшего порядка состоит
из циклов низшего порядка; б) качественные особенности каждого
цикла определяются количественными изменениями в круговороте
веществ и энергии, осуществляющимися в предшествующем цикле
(или ряде циклов), а также изменениями в поступлении космиче-
ской энергии; в) с переходом от одного цикла развития природы
к другому подвижное равновесие нарушается и характер связей и
взаимосвязей изменяется. Следовательно, заключает автор, «цик-
лическое развитие природных процессов соответствует спиралевид-
ному развитию материи» (с. 75). Циклы развития природных про-
цессов, выявленные закономерности в их реализации, безусловно
находят отражение и в развитии живой природы и, в частности,
сказываются на ходе обменных процессов биологических и био-
косных систем.
В. Р. Вильямс (1936), как известно, различал два типа круго-
воротов веществ на Земле, протекающих под воздействием энергии
солнечной радиации: а) геологический (абиотический) круго-
ворот и б) биологический (биотический, по Камшилову). Под
геологическим круговоротом веществ Вильямс понимает кругово-
рот, совершающийся между сушей и Мировым океаном с кругово-
241
ротом воды и циркуляцией атмосферы. Им выделяются следующие
этапы в ходе его осуществления. I. Горные породы суши под дей-
ствием тепловой энергии Солнца выветриваются до образования
рухляка, а под действием различных химических агентов разру-
шаются до растворимых минеральных солей. II. Образовавшиеся
минеральные соли растворяются пропитавшейся в землю водой
атмосферных осадков и сносятся вместе с нею в океан. III. Часть
унесенных в растворенном виде элементов пищи растений усваива-
ется планктоном моря, который поедается рыбами, а последние
вылавливаются птицами, животными и человеком и, таким обра-
зом, вновь возвращается на сушу. IV Гораздо большая часть
оставшихся в океане минеральных элементов в виде отмерших
организмов и их отбросов попадает в состав осадочных пород, ко-
торые долгие геологические периоды будут лежать на дне океана,
пока оно не станет сушей и снова подвергнется выветриванию в
рухляк.
Биологический круговорот зольной и азотной пищи растений
развертывается, по Вильямсу, на части траектории геологического
круговорота веществ в природе. (При этом часть минеральных пи-
тательных элементов вырывается из геологического круговорота и
включается в биологический круговорот. Последний состоит из
двух диаметрально противоположных, по взаимосвязанных процес-
сов: а) созидания органического вещества в основном автотрофны-
ми зелеными растениями из простых минеральных соединений,
углекислоты и воды под действием лучистой энергии Солнца и
б) разрушения, минерализации органического вещества соответ-
ствующими гетеротрофными бесхлорофилльными организмами
(бактериями, грибами, актиномицетами и другими обитателями
почвы) па простые минеральные соединения, вступающие затем в
новый цикл биологического круговорота.
Средний элементарный химический состав биомассы раститель-
ных и других организмов (в процентах от их живого веса), по
А. П. Виноградову (1946), представляется в следующем виде: на
долю кислорода приходится 65—70%, водорода — 10% и 20—25%
составляют большое количество (более 70) других элементов. Из
числа последних наиболее важное место в составе биомассы жи-
вых организмов и в составе зольной пищи растений занимают С,
N, Са (1—10%); S, Р, К, Si (0,1 — 1%); Fe, Na, Cl, Al, Mg, Mn, Zn,
Си и др. (0,1—0,01%). В биотический круговорот, по М. М. Камши-
лову (1970), вовлекается до 40 разнообразных минеральных
элементов, среди которых основная роль, по его словам, принад-
лежит углероду, кислороду, водороду, азоту, фосфору и сере.
«Жизнь зависит от химических свойств этих элементов» (с. 124).
В основе биологического круговорота веществ, как указывает
В. Р. Вильямс (1936), лежит количественная ограниченность всех
242
элементов на Земле, и в особенности тех, которые играют наиболее
существенную роль в жизни организмов. «Единственный способ
придать элементу конечному, количественно ограниченному, свой-
ство бесконечного, — пишет В. Р. Вильямс, — это придать ему цик-
лическое движение, заставить его вращаться в круговороте»
(с. 86). Вот природа, по его словам, и использует в данном случае
этот принцип цикличности, круговорота, который лежит в основе
трофических (пищевых) отношений автотрофных и гетеротрофных
живых существ.
Таким образом, происходящий в биогеоценозе и биогеосфере в
форме биологического круговорота обмен веществ между живым
и неживым осуществляется, по Вильямсу, противоположными по
типу питания и обмена веществ автотрофными и гетеротрофными
группами организмов, выступающих в этих процессах носителями
и трансформаторами вещества и заключенной в нем потенциаль-
ной энергии по пищевой цепи. Что же касается почвы и атмосфе-
ры, то они являются, по его словам, посредниками в этом процессе,
источниками веществ и энергии..'Сходные по этому вопросу взгля-
ды развивает и М. М. Камшилов (1961), который пишет: «В каче-
стве звеньев, с помощью которых осуществляется биотический
круговорот, выступают организмы различных систематических
групп, от микроорганизмов до высших представителей раститель-
ного и животного мира. Абиотические факторы являются источни-
ками вещества и энергии, условиями существования биотического
круговорота» (с. 60).
Г. Вальтер (1982) различает две разновидности биологического
круговорота — малый и большой. При малом биологическом
круговороте в процессе обмена веществ и энергии в биогеоценозе
участвуют две группы организмов: растения, осуществляющие син-
тез первичной биологической продукции, и микроорганизмы, про-
изводящие разложение, минерализацию ee.jB осуществлении же
большого биологического круговорота, помимо растений и микро-
организмов, принимают участие также животные — растительно-
ядные и плотоядные; среди последних различаются хищники 1-го
порядка и хищники 2-го порядка. На долю же микроорганизмов в
таком случае выпадает минерализация как отмерших растении и
их частей, так и отбросов и трупов животных.
2. Биогеоценотический обмен веществ,
циклы и типы биологического круговорота
Поскольку биогеоценоз представляет собою не замкнутую абсо-
лютно, а термодинамически открытую биокосную систему, то в
биологическом круговороте веществ на уровне биогеоценоза и био-
геосферы, по мнению Б. А. Быкова (1970), следует различать два
243
круга обменных процессов: а) внутриценозный круговорот веществ,
совершающийся между биотой как совокупностью живых существ
в составе биогеоценоза и биоценотической средой, биотопом, и
б) внешний круговорот, включающий обмен веществ между био-
ценозом (точнее, биогеоценозом в целом) и внешним окружением
его, т. е. с другими сопряженными с ним биогеоценозами и внеш-
ней по отношению к нему физической средой.
Еще ранее такого рода суждения были высказаны Н. В. Дыли-
сом (19696). Они вытекают из основополагающих представлений
В. Н. Сукачева о биогеоценозе и обменных процессах между его
компонентами и биогеоценоза как такового с «другими явлениями
природы». Круговорот веществ, осуществляющийся между компо-
нентами в рамках биогеоценоза, Н. В. Дылисом был назван био-
геоцепотическим (с. 8). Очевидно, этим автор намерен под-
черкнуть наряду со сходством и некоторые важные отличия
биогеоценотического обмена от обмена веществ биологического
(физиологического), протекающего на уровне организма.
Биологический (физиологический) обмен веществ, как известно,
является основой жизненного процесса. Следствием его, в отличие
от обмена веществ в неживой природе, является не разрушение
материи, а самообновление живого тела. В обмене веществ на
уровне организма В. А. Павлов (1960) также различает внутреннее
в внешнее звенья обмена. Внешнее звено в организменном обмене
включает в себя такие процессы, как восприятие и поглощение из
окружающй среды одних веществ и выделение в нее других. Внут-
реннее звено обмена—это совокупность биохимических превраще-
ний поглощенных растительным или животным организмом ве-
ществ в их телах, это основное звено обмена, собственно метабо-
лизм. Он включает процессы синтеза сложных соединений из более
простых, протекающие с затратой энергии на эти процессы и, сле-
довательно, увеличением ее запасов в биомассе организмов, и
процессы расщепления сложных соединений на более простые, со-
провождающиеся высвобождением и выделением энергии вовне
и, следовательно, уменьшением ее в организме. Внешним проявле-
нием обмена веществ на уровне организма, по словам В. А. Пав-
лова, является осуществление внутренних жизненных функций
организма, процессы размножения, роста, развития его, активность,
подвижность живых существ и пр.
Однако биологический (физиологический) обмен веществ в от-
личие от биогеоценотического не обладает циклической обратимо-
стью вещества внутри организма, а потому предполагает непрерыв-
ность поступления извне как энергии, так и соответствующих
неорганических веществ для автотрофных растений и живой или
мертвой биомассы для гетеротрофных живых существ — животных
и микроорганизмов. Это обусловлено тем, что организм как инди-
244
видуум в составе видовой ценопопуляций является обладателем
какого-либо одного типа обмена веществ, что исключает циклич-
ность обмена и оборачиваемость веществ в процессе его осуществ-
ления.
Все процессы на уровне биогеоценоза, протекающие как внутри
биогеоценоза, так и при взаимодействии его с внешним окруже-
нием, должны рассматриваться, по мнению К. А. Куркина (1980),
как «системная совокупность взаимосвязанных процессов, как
единый биогеоценотический процесс» (с. 40). Централь-
ным явлением, фокусом его, очевидно, следует считать биогеоцено-
тический обмен в форме биологического круговорота веществ и
потока энергии. Биогеоценотический процесс, как утверждает ав-
тор со ссылкой на В. Н. Сукачева, представляет собою особую
форму движенйя материи, характеризующуюся наличием своих
особых специфических закономерностей, которые еще надлежит
открыть. А все компоненты биогеоценоза, участвующие в этом про-
цессе, по приведенным Куркиным словам В. Н. Сукачева, высту-
пают «и материалом для него, и трансформатором, и обменным
аппаратом, и суммарным выразителем его» (с. 40). Нельзя не
отметить при этом, что особо важное значение в обменных процес-
сах биогеоценоза К. А. Куркиным придается все же внутриценоз-
ному круговороту, ибо в нем, по мнению автора, скрыты специфи-
ческие качества, сущность биогеоценоза, и на нем зиждется
целостность последнего как биокосной системы.
Биологический круговорот веществ, как форма биогеоценотиче-
ского обмена, в принципе должен быть замкнутым, ибо с этим свя-
зана оборачиваемость вовлекаемых в него химических элементов.
Однако многие отечественные ученые-естественники (В. А. Павлов,
1960; А. И. Перельман, 1966; И. И. Смольянинов и Е. В. Рябуха,
1971; В. А. Ковда, 1973; Ю. А. Жданов, 1973) приводят данные о
необратимости ряда веществ и химических элементов в ходе био-
логического круговорота, подчеркивая тем самым относительный
характер замкнутости его. И. И. Смольянинов, в частности, указы-
вает на то, что биологический круговорот имеет выход в геологи-
ческий круговорот. При этом обычно наблюдается некоторое пре-
вышение поступления веществ и энергии в биогеоценоз над
выходом их в окружающую среду. Кроме того, как отмечает
В. А. Павлов, в ходе впутриценозного обмена веществ происходят
те или иные изменения в количестве и качестве образующихся ве-
ществ, а именно часть веществ выключается из процесса, а остаю-
щаяся часть их приобретает новые качества и свойства.
По данным В. А. Ковды, такие химические элементы, как хлор,
натрий и отчасти кальций и магний, в значительном количестве
сносятся с поверхностным и подземным стоком воды в низины,
моря и океаны. Даже такие жизненно важные элементы, как угле-
245
род, кислород, водород, азот, фосфор, калий и сера, в процессе
минерализации не полностью утилизируются соответствующими
группами организмов в пищевых цепях консорций и, следователь-
но, не полностью вовлекаются в биологический круговорот. Необ-
ратимый характер носит, по его словам, накопление кислорода и
уменьшение (до последних десятилетий) содержания углекислоты
в атмосфере, образование целого ряда биогенных осадков (карбо-
натов, фосфоритов, кремнеземов, глин и т. п.), а также изменение
окислительно-восстановительных и щелочно-кислотных условий в
земной коре и пр. Вместе с тем с появлением жизни на Земле, как
указывает В. А. Ковда, происходит образование целого ряда новых
биогенных соединений (сероводорода, сернистых металлов, нитра-
тов, фосфатов), а также химически активных органических кислот
(гуминовых, фульвокислот и пр.).
Ж- Леме указывает на то, что наличие в экосистеме (биогео-
ценозе) внутренних циклов обмена веществ не превращает ее в
систему, полностью замкнутую. В ходе обмена веществ в ней про-
исходит ряд изменений, связанных с такими процессами, как по-
ступление в нее новых минеральных элементов, вынос некоторых
из них поверхностным стоком, образование ряда органических ве-
ществ и соединений, отчуждение их и заключенной в них энергии
человеком, обмен веществом и энергией с другими экосистемами.
Имеются данные, свидетельствующие о том, что в биогеоценозе
под действием микроорганизмов происходит не только разложение
органических и сложных минеральных соединений, но и синтез
целого ряда совершенно новых минералов (Т. В. Аристовская,
1973).
А. И. Перельман также утверждает, что биологический круго-
ворот веществ в природе не является абсолютно замкнутым. Часть
труднорастворимых веществ закрепляется в почве, в то время как
ряд других веществ вымывается из почвы и сносится в отрицатель-
ные формы рельефа и водные бассейны. Вследствие этого природ-
ные географические ландшафты после каждого цикла совершаю-
щихся в них обменных процессов приобретают некоторые новые
черты. Ю. А. Жданов пишет: эволюция биосферы свидетельствует
о том, что «при всей своей повторяемости циклические процессы
каждый раз выступают в несколько модифицированном виде. На
базе этих изменений формируется поступательное развитие био-
сферы, ее усложнение, расширение и прогресс» (с. 13). Биологиче-
ский круговорот веществ, в форме которого протекают обменные
процессы в биогеоценозе и биогеосфере, может рассматриваться,
таким образом, как основной, магистральный путь развития био-
геоценотического покрова Земли, развития его по спирали.
Биологический круговорот веществ в биогеоценозе и биогеосфе-
ре в целом — это сложный полициклический процесс, включающий»
246
по словам С. В. Зонна и Н. И. Базилевич (1966), циклы различной
продолжительности и глубины воздействия на среду. Обычно раз-
личаются сезонные, годовые, многолетние и вековые циклы биоло-
гического круговорота. Центральное место среди них занимают
годовые циклы, которые неодинаковы в разные годы по количеству
синтезируемого органического вещества в связи с различием по-
годных условии и возрастных особенностей ценозообразующих
растений. Годовые циклы круговорота характеризуются, по дан-
ным тех же авторов, следующими протекающими в течение года
взаимосвязанными в единое целое процессами: а) извлечением и
потреблением соответствующими группами организмов биофильных
элементов из почвы и атмосферы; б) удержанием части из них в
биомассе растений, животных и микроорганизмов; в) возвраще-
нием другой части в почву и атмосферу с опадом, отпадом, жидки-
ми и газообразными выделениями с последующим включением их
в новый цикл круговорота. В годовом цикле круговорота веществ
выделяются иногда сезонные циклы, например, на протяжении
одной из стадий развития ценоза.
Многолетние циклы биологического круговорота веществ охва-
тывают период жизни одного поколения основного ценозообразую-
щего растения-эдификатора; в бидоминантных и полидоминантных
биогеоценозах — период жизни ценозообразователей-соэдификато-
ров. В травяных биогеоценозах длительность многолетних циклов
составляет 15—20 лет, в лесных 100—200 и более лет. Вековыми
циклами биологического круговорота охватываются периоды вре-
мени, в течение которого происходит смена ценозообразующих
растений-эдификаторов, т. е. коренная смена одного биогеоценоза
другим. Этот процесс в лесных биогеоценозах может длиться от
нескольких столетий до нескольких тысячелетий.
Из изложенного следует, что надо различать такие понятия, как
биологический круговорот веществ в целом и цикл биологического
круговорота той или иной продолжительности. Цикл биологическо-
го круговорота веществ биогеоценоза или биогеосферы конечен, он
завершается в течение года, нескольких или даже многих лет и
столетий, а биологический круговорот» как форма обменных про-
цессов биокосной системы того или иного ранга, длится бесконеч-
но. Этим и обеспечивается непрерывность жизни па Земле.
Биологический круговорот веществ разных биокосных систем
неодинаков. Он различается по целому ряду показателей. Для ха-
рактеристики различных типов биологического круговорота ве-
ществ разными авторами выдвигается ряд показателей. А. И. Пе-
рельман (1966) к числу основных показателей биологического
круговорота относит: а) емкость биологического круговорота, имея
в виду весовое количество химических элементов в составе живого
вещества того или иного ландшафта (иными словами, биомассу,
247
ц/га), а также структуру биомассы, т. е. весовое соотношение зеле-
ной части, корней растений, биомассы животных и микроорганиз-
мов; б) скорость биологического круговорота веществ, т. е. коли-
чество живого вещества, образующегося и разлагающегося в
единицу времени, а именно: ежегодный абсолютный прирост био-
массы и абсолютный опад (ц/га) и их структуру, ежегодный относи-
тельный прирост биомассы и относительный опад в процентах от
общего веса биомассы и опада. И. П. Герасимов (1973) опери-
рует по существу теми же показателями «внутреннего кругооборо-
та» веществ в биогеоценозах, ландшафтах, что и предыдущий
автор, но несколько в ином толковании их: а) интенсивность или
скорость оборота веществ, подразумевая под этим отношение еже-
годной биологической продуктивности к общей биомассе; б) общая
структура оборота веществ, которая может быть охарактеризована,
по словам автора, степенью своей завершенности и сбалансирован-
ности веществ (т. е. использованием биотой ежегодного прироста
биомассы и соотношением первичной и вторичной биологической
продукции) и остаточной продуктивностью (соотношением масс
живого и мертвого органического вещества). Кроме того, для ха-
рактеристики некоторых биокосных систем автор использует и та-
кой показатель, как в) степень замкнутости оборота веществ, опре-
деляемая уровнем использования веществ биотой внутри системы.
Р. Дажо (1975) и Ж. Леме (1976) также одним из наиболее
важных показателей, характеризующих особенности круговорота
веществ экосистемы (биогеоценоза), считают а) скорость (интен-
сивность) оборота веществ. Но под скоростью оборота веществ в>
системе Р. Дажо понимает скорость разложения подстилки и вы-
свобождения химических элементов при этом, что может быть уста-
новлено по отношению сухого вещества подстилки к сухому веще-
ству годового опада зеленых частей растений, а Ж. Леме имеет
в виду отношение количества получаемых или потребляемых
веществ в единицу времени на разных трофических уровнях пище-
вой цепи. Последний автор, наряду со скоростью оборота, в каче-
стве показателей характерных особенностей круговорота веществ
выдвигает также б) время оборота, т. е. время, необходимое для
«возобновления количества вещества, равного живой биомассе эко-
системы» (с. 114), а также в) характер циркуляции элементов в
экосистеме, а именно: количество элементов, ежегодно поглощае-
мых живыми существами, удерживаемых в своем теле и возвра-
щаемых в почву. Н. И. Базилевич и А. А. Титлянова (1974), как и
Р. Дажо, к числу важнейших показателей биологического круго-
ворота веществ в биогеоценозе относят скорость высвобождения
химических элементов при процессах разложения отмерших расти-
тельных остатков. При этом учитывается различная продолжи-
тельность закрепления их у разных групп растений — у древесных
248
пород, травянистых многолетников, однолетних растений и эфеме-
ров, которая варьирует от одного года и даже нескольких месяцев
у эфемеров до нескольких десятков лет у деревьев и кустарников.
На основе тех или иных особенностей обменных процессов в
биокосных системах разного ранга биологами выделяется ряд ти-
пов биологического круговорота веществ. Так, И. И. Смольянинов
(1971) по охвату, масштабу процессов преобразования веществ
подразделяет биологический круговорот на следующие два типа:
1) глобальный тип биологического круговорота, когда циклы
преобразования, трансформации веществ рассматриваются во все-
объемлющем пространственном масштабе — в масштабе биосферы;
2) факультативный, частный (точнее, локальный) тип
круговорота в случае рассмотрения циклов трансформации ве-
ществ в узких рамках — в масштабе биогеоценоза или тех или
иных территориальных объединений биогеоценозов. Тем же авто-
ром по тенденциям развития биокосной системы различаются:
1) прогрессивный тип биологического круговорота, когда под
совокупным действием живых компонентов системы происходит
улучшение биоценотической среды, повышение ее производитель-
ных потенций; 2) консервативный тип круговорота веществ
в случае отсутствия позитивных изменений биотопа, которые сви-
детельствовали бы о повышении его продуктивности. Другими био-
логами, как будет сказано ниже, выделяются па соответствующих
основаниях и другие типы биологического круговорота веществ и
содержащихся в них биофильных химических элементов питания.
3. Биогеохимические циклы биофильных химических элементов
в биогеосфере
После ознакомления с целым рядом общих положений, связан-
ных с характеристикой биологического круговорота веществ как
формы обменных процессов в биогеоценозе и биогеосфере, следует
перейти к рассмотрению круговорота отдельных химических эле-
ментов, играющих наиболее важную роЛь в жизни живых компо-
нентов биогеоценоза и функционировании биокосных систем раз-
ного ранга. Такие химические элементы П. Дювиньо и М. Танг
(1968) назвали биогенными, Р. Дажо (1975) — биофильными, а
Ю. Одум— (1975) питательными. Циклы же круговорота этих хи-
мических элементов получили название биогеохимических.
К таким жизненно важным химическим элементам, как было
сказано ранее, относятся углерод, водород, азот, фосфор и сера.
Все эти биофильные химические элементы биосферы в ходе
круговорота в биогеоценотической системе многократно поступают
в ее живые компоненты («организуются», по выражению А. Й. Пе-
рельмана, 1966) и многократно выходят из них — минерализуются.
249
Во внешнем круговороте химических элементов на уровне биосфе-
ры (биогеосферы) с выходом его в геологический (абиотический)
круговорот Ю. Одум (1975) выделяет два фонда: а) резервный,
включающий огромную массу весьма медленно движущихся ве-
ществ, и б) обменный — меньший по массе вещества, но более
активный, характеризующийся быстрым обменом питательных эле-
ментов между живыми существами и их непосредственным окру-
жением.
Биогеохимические циклы круговорота биофильных элементов в
некоторых отношениях специфичны. Вместе с тем в циркуляции
этих элементов имеются и общие черты, что позволило биологам
и геохимикам сгруппировать их в два типа биогеоценотического об-
мена: а) газовый тип круговорота таких элементов, как углерод,
кислород, азот, водород, вода как химическое соединение Н и О,
главными резервуарами которых служат атмосфера и гидросфера,
и циркуляция их в крупных пространственных масштабах
происходит в газовой форме, а потому сравнительно быстро;
б) осадочный тип круговорота, элементы которого — фосфор и
сера — циркулируют в водном растворе; резервуарами этих эле-
ментов являются почва и земная кора (литосфера), где они нахо-
дятся в составе осадочных пород, вследствие чего маломобильны
и круговорот их в биосфере совершается крайне медленно.
В. Лархер (1978) считает целесообразным круговорот азота выде-
лить в самостоятельный тип круговорота на том основании, что он
занимает как бы промежуточное положение между газовым и оса-
дочным типами биогеохимического обмена элементов, поскольку
растениями используется в основном минеральный азот из почвы,
а основные его запасы находятся в газовой форме в атмосфере.
Однако вряд ли в этом есть необходимость. В таком случае встал
бы вопрос о выделении в самостоятельный тип круговорота и ряда
других химических элементов (водорода и пр.).
Не раскрывая полностью ход и совокупность процессов биогео-
химических циклов круговорота биофильных элементов, роль жи-
вых и косных компонентов биокосных систем в осуществлении
их, рассмотрим в общих чертах биогеохимические круговороты
наиболее важных из этих химических элементов. Прежде всего,
биогеохимический круговорот углерода, поскольку он лежит в
основе того материала, из которого построено все живое на Земле.
В. И. Вернадский (1926) в круговороте углерода различал два
цикла: а) первичный или биотический цикл углеродного обмена,
который совершается между тропосферой или гидросферой, с одной
стороны, и живым веществом — с другой, при непосредственном
активном участии последнего; процесс этот протекает быстро и
интенсивно; б) вторичный или геохимический цикл обмена угле-
рода, который охватывает, по словам В. А. Павлова (1960), всю
250
совокупность превращений углерода в биосфере и литосфере и осу-
ществляется в течение длительного (геологического) времени.
Общий запас соединений углерода на нашей планете, по дан-
ным В. Лархера (1978)» составляет 26-1015 т. Основная масса
химически связанного углерода содержится в неорганических ве-
ществах, и лишь около 0,05% —в органических. Как видно из при-
веденных автором данных (с. 137), запасы неорганического угле-
рода, представленные в миллиардах тонн, распределяются по
земному шару следующим образом: в атмосфере в виде углекисло-
ты сосредоточено 700, в гидросфере (преимущественно в морских
и океанических глубинах) —35250 и в литосфере в виде известня-
ков и других осадочных пород — 25007500. Запасы органического
углерода общим количеством 4130 млрд т находятся в биосфере и
верхних слоях литосферы. Наибольшая часть органического угле-
рода (64%) содержится в ископаемых отложениях (торф, уголь,
нефть и пр.), в отмерших органических остатках в почве и водо-
емах содержится 32%, и лишь около 4% (примерно 420 млрд т) —
в биомассе наземных и морских организмов.
Круговорот углерода совершается при непосредственном участии
живых существ на разных уровнях организации живой материи.
Так, в зеленых автотрофных растительных организмах из погло-
щенной ими из соответствующих источников углерода углекислоты
синтезируются углеводы, липиды, пептиды и другие органические
вещества, которые расходуются в ходе жизнедеятельности расте-
ний, окисляясь в процессе дыхания в более простые соединения;
другая часть органического материала в процессах роста и раз-
множения идет на построение собственной (растительной) биомас-
сы. В биогеоценозах круговорот углерода совершается в пищевых
цепях с участием всех живых компонентов путем трансформации на
соответствующих трофических уровнях растительной биомассы в
животную, той и другой — в простые минеральные соединения с
высвобождением углекислоты, которая служит исходным материа-
лом для последующих циклов круговорота.
Сходным образом совершается круговорот углерода и в преде-
лах всей биогеосферы, поскольку основными материально-энерге-
тическими ячейками ее являются биогеоценозы. Разница по суще-
ству лишь в масштабах этого процесса. По данным В. Лархера
(1978), автотрофными растениями земного шара ежегодно связы-
вается в виде органических веществ около 6—7% углекислоты, со-
держащейся в атмосфере и в растворенном виде в гидросфере, что,
по расчетам А. А. Ничипоровича (1955), составляет примерно
175 млрд т углерода в год. Около 30% заключенного в органике
углерода используется автотрофами на процессы своей жизнедея-
тельности, а остальная часть его (за исключением углерода, закон-
сервированного в разного рода отложениях) служит источником
251
питания и энергии гетеротрофам на последующих трофических
уровнях пищевых цепей и сетей в биогеоценозах. Продолжитель-
ность глобального круговорота углерода оценивается по-разному.
По данным Р. Дажо (1975), она колеблется в пределах 300—
400 лет, по другим данным, как он указывает, на это уходит 1000
и более лет (рис. 26).
Рис. 26. Круговорот углерода в биосфере (по П. Дювинью и М. Тангу, 1968)
С круговоротом углерода в природе, как известно, взаимосвя-
зан круговорот кислорода, содержащегося в углекислоте и высво-
бождающегося в процессе фотосинтеза автотрофных растений.
Кислород используется всеми живыми существами в процессе
дыхания, а выделяющийся при этом углекислый газ вновь исполь-
зуется на процессы фотосинтеза в последующем цикле кругооборо-
252
та. В атмосфере кислород содержится, по данным В. Лархера, в
количестве 1,2-1016 т. В процессе фотосинтеза на каждую тонну
связанного углерода, как указывает Б. А. Быков (1970а), выделя-
ется кислорода 2,7 т; по расчетам В. Лархера, за год выделяется
7> 109 т. В морских и океанических водах кислород сосредоточен
главным образом в самом верхнем слое, где концентрация его бы-
вает в 2—3 раза выше, чем в атмосфере. Но благодаря непрерыв-
но происходящему газообмену между атмосферой и гидросферой
содержание как кислорода, так и углекислого газа в атмосфере
поддерживается на относительно постоянном уровне. Содержащий-
ся в атмосфере кислород используется, как уже сказано, на дыха-
ние живых существ, а кроме того, при разложении отмерших орга-
низмов и их частей, при сжигании топлива человеком и т. п. На
эти процессы расходуется почти все количество кислорода, высво-
бождающегося в процессе фотосинтеза за год. В водах морей и
океанов расход кислорода на дыхание организмов и разложение
органики значительно меньше, чем в атмосфере, благодаря преоб-
ладанию здесь анаэробных условий, в которых эти процессы осу-
ществляются, и в силу некоторых других причин. На полное об-
новление всего состава кислорода атмосферы на нашей планете
уходит, по П. Клауду и др. (1972), 2 тыс. лет, а по расчетам
А. А. Ничипоровича— не менее 3 тыс. лет.
К числу важнейших питательных элементов наряду с углеро-
дом относится и азот, который входит в состав белка протоплазмы
и ядра клетки, являющегося не только основой жизни, но и энер-
гетически наиболее емким материалом. Главными источниками
азота в природе являются фотохимическая и биологическая фик-
сация азота микроорганизмами, грозовые электрические разряды,
сопровождающиеся синтезом окислов азота (NO3) в количестве, по
П. Дювиньо и М. Тангу (1968), 4—10 кг/га в год, выбросы газов,
в том числе и азота, при вулканических извержениях и пр. Основ-
ные запасы азота сосредоточены в атмосфере в молекулярной
форме, составляя 78% от общего объема воздуха. Кроме того, азот
находится в верхних слоях почвы в виде почвенного гумуса, отмер-
ших остатков растений и животных и продуктов их жизнедеятель-
ности в количестве 0,1—0,4% сухого вещества (по В. Лархеру); в
земной коре и подпочве — 0,03%; в морских и океанических водах;
в виде азотсодержащих соединений в биомассе растительных и
животных организмов и пр. В. Лархер (1978) приводит также дан-
ные о распределении по земному шару азота, непосредственно
участвующего в обменных процессах биогеосферы в целом. По
этим данным, в атмосфере находится 99,4% (3,8- 101в т), в гидро-
сфере— 0,5, в почве — 0,05 и в биомассе живых существ — 0,0005%
(с. 158).
Высшие растения неспособны усваивать молекулярный азот
253
атмосферы. Такой способностью обладают лишь некоторые микро-
организмы. К ним относятся клубеньковые бактерии рода Rhizo-
bium, находящиеся в симбиозе с бобовыми растениями, связываю-
щие атмосферный азот, по данным П. Дювиньо и М. Танга
(1968), в количестве от 150 до 400 кг/га в год; свободноживущие
микроорганизмы (Azotobacter — аэроб, Clostridium — анаэроб),
связывающие около 25 кг/га азота в год; синезеленые водоросли
рода Nostoc, Anabaena, обитающие в воде, во влажной почве, в
рисовых чеках; актиномицеты и некоторые бактерии, находящиеся
в симбиозе с серой ольхой. В последнее время, по свидетельству
Ю. Одума (1975), было обнаружено, что способностью фиксации
атмосферного азота обладают также пурпурные бактерии рода
Rhodospirillum, почвенные бактерии, близкие к роду Pseudomonas,
и многие другие. Было установлено также, что в симбиозе с азот-
фиксирующими микроорганизмами, помимо бобовых, находятся
также ольха, лох и др. Ю. Одум приводит также количественные
данные разных авторов по биологической фиксации азота в био-
сфере. По данным одних авторов, в течение года фиксируется
азота от 140—700 мг/м2 до 1 г/м2, по данным других — в благо-
приятных условиях даже до 20 г/м2.
В круговороте азота, как и в круговороте углерода, можно
усмотреть два цикла: первичный — биотический и вторичный —
геохимический. В ходе биотического цикла круговорота азота осу-
ществляется ряд взаимосвязанных процессов. 1. Усвоение расте-
ниями из почвы (или воды) минерального азота в виде азотнокис-
лых или аммониевых солей и образование белков, идущих на
построение растительной биомассы и в запас питательных мате-
риалов. 2. Построение растительноядными животными из расти-
тельных белков и плотоядными — из животных своей животной
биомассы. 3. Разложение (минерализация) отмершей растительной
и животной биомассы при участии аммонифицирующих бактерий
с выделением аммиака, который подвергается дальнейшему окис-
лению. Этот процесс, получивший название нитрификации аммиа-
ка, протекает в два этапа. На первом этапе аммиак окисляется
бактериями рода Nitromonas до азотистой кислоты; на втором
этапе азотистая кислота бактериями рода Nitrobacter окисляется
до азотной кислоты. Далее в процессе взаимодействия азотной
кислоты с углекислыми солями в почве образуются нитраты —
азотнокислые соли, которые оказываются пригодными для усвое-
ния растениями.
Геохимический цикл круговорота азота протекает в анаэробных
условиях, включая такие процессы, как преобразование денитри-
фицирующими бактериями азотной кислоты в азотистую, а послед-
ней— в свободный азот, который выделяется в атмосферу (в коли-
честве 50—60 кг/га в год) и частично выщелачивается из почвы и
254
сносится сточными водами в моря и океаны. Там он поглощается
фитопланктоном, переходит по пищевым цепям к рыбам и птицам,
отлагаясь затем на суше в залежах гуано, а в морских глубинах —
в осадочных породах, надолго выходя, таким образом, из жизнен-
ного цикла круговорота (рис. 27).
Рис. 27. Круговорот азота в биосфере (по К. Дельвичу, 1972)
Весьма важным и необходимым элементом, входящим в состав
протоплазмы и ядра, в частности ДНК и РНК живой клетки, явля-
ется фосфор. Основным резервуаром фосфора в природе служат
осадочные горные породы и другие отложения, образовавшиеся в
далекие геологические времена. Под действием механического вы-
ветривания и физико-химических факторов фосфоросодержащие
породы подвергаются разрушению. Высвободившиеся при этом
фосфаты (соли фосфорной кислоты) в наземных условиях по-
падают в сухопутные биогеоценозы, но, пожалуй, большая часть
их выщелачивается и уносится сточными водами в моря и океаны.
Таким образом, круговорот фосфора относится к осадочному типу
биогеохимических круговоротов. По своей структуре он проще по
сравнению с круговоротом азота.
Циркуляция фосфора в наземном биотическом цикле представ-
ляется в следующем виде. Поступившие в биогеоценоз фосфаты
потребляются растениями и используются в процессе синтеза на
построение своей биомассы. Растительная биомасса в последую-
щих звеньях пищевых цепей трансформируется в животную био-
массу. По отмирании та и другая биомасса подвергаются минера-
255
лизации при участии фосфороредуцирующих бактерий, плесневых
и других грибов в соли фосфорной кислоты, поглощаемые расте-
ниями в последующем цикле круговорота. Циркуляция фосфора,
поступившего в виде фосфатов в морские водоемы, протекает в
следующих пищевых цепях круговорота: а) планктон -►рыбы -►
птицы и образование гуано; б) планктон-►рыбы растительнояд-
ные-►хищные рыбы и отложение фосфоросодержащих осадков на
больших глубинах морей и океанов либо вылавливание раститель-
ноядных и хищных рыб человеком. В случае образования на по-
следнем этапе круговорота залежей гуано или мощных глубинных
осадков фосфор выводится из круговорота на многие десятки и
сотни тысяч лет. Вылов рыбы, и в особенности вынос фосфора на
Рис. 28. Круговорот фосфора в биосфере (по IL Дювиньо и
М. Тангу, 1968)
256
поверхность земли при горнорудных разработках, также ведет к
нарушению круговорота этого элемента вследствие смыва фосфа-
тов стоком воды (рис. 28).
Вторым биофильным элементом, круговорот которого соверша-
ется по осадочному типу, является сера. Основными источниками
серы в природе служат содержащиеся в осадочных горных породах
серный и медный колчеданы и входящие в состав растительной и
животной биомассы содержащие серу белковые органические со-
единения. Хотя содержание серы в составе живой биомассы не
столь велико по сравнению с содержанием азота и фосфора, био-
геохимический круговорот ее, по выражению Ю. Одума (1975),
является ключевым в общем процессе обмена веществ в биокос-
ных системах. По своей структуре он представляется более слож-
ным, чем круговорот фосфора, что, очевидно, связано с наличием
нескольких ступеней валентности п окисленности серы, подобно
азоту.
Круговорот серы в наземных условиях совершается в основном
в почвенной среде биокосных систем, охватывая подпочвенные слои
материнских осадочных пород. Биотический цикл биогеохимиче-
ского круговорота серы включает ряд процессов: 1) разложение
при участии микроорганизмов отмерших растений и животных и
их органических остатков и отбросов; 2) дальнейшее окисление
таких продуктов разложения белков, как сероводород и сера, в
процессе так называемой сульфофикации. Этот процесс, как и ни-
трификация азота, совершается в два этапа особой группой микро-
организмов— серобактериями (бесцветными и пурпурными) при
участии тиобактерий. На первом этапе сероводород окисляется до
минеральной серы, а на втором сера окисляется до серной кисло-
ты, соли которой — сульфаты — могут усваиваться растениями,
вступая в новый цикл круговорота как в сухопутных биогеоцено-
зах, так и, в случае выноса в мелководные водоемы, в водных био-
косных системах. В анаэробных условиях на дне водоемов проис-
ходит восстановление сульфатов при участии разнообразных бак-
терий, главным образом вибрионов, до сероводорода. Последний
накапливается в этих условиях, а впоследствии улетучивается в
атмосферу либо при наличии в придонных отложениях железа
образует во взаимодействии с ним пирит, выходя на длительное
время из круговорота.
Вывод серы и ее соединений из морских глубинных осадков и
наземных осадочных пород и включение их в биотический цикл
[круговорота происходят разными путями в ходе реализации гео-
химического цикла круговорота этого элемента, растягиваясь по-
рой на длительные геологические времена. На некоторых этапах
этого цикла круговорота серы вмешивается человек путем перера-
ботки извлекаемой из недр руды и сжигания в промышленных
257
предприятиях содержащего серу ископаемого топлива. Сера в
таком случае вместо включения в жизненные циклы сухопутных и
водных биокосных систем попадает в виде ядовитого сернистого
газа в атмосферу, нанося вред всему живому.
Для нормальной жизнедеятельности организмов и функциони-
рования биогеоценотических систем необходимы не только рас-
смотренные биофильные химические элементы, но и такие, как
калий, кальций, магний и микроэлементы: железо, марганец, цинк»
медь, кобальт, молибден и др. На суше эти макро- и микроэлемен-
ты находятся в почве и частично поступают в биогеоценозы с
атмосферными осадками. Круговорот этих химических элементов,
как указывают П. Дювиньо и М. Танг (1968), также совершается
в ходе общего биологического круговорота веществ через пищевые
цепи биокосных систем.
4. Поступление солнечной энергии на Землю и ее использование
в биогеоценозе и биогеосфере
Обменные процессы живых компонентов биогеоценоза проте-
кают с затратой энергии. Энергия рассматривается как способность
производить ту или иную работу. Это свойство ее определяется
двумя законами термодинамики. Первый закон в формулировке
Ю. Одума (1975) гласит: «...энергия может переходить из одной
формы в другую, но она не создается заново и не исчезает». Со-
гласно второму закону, «процессы, связанные с превращениями
энергии, могут происходить самопроизвольно только при условии,
что энергия переходит из концентрированной формы в рассеянную»
(с. 52). Следствием этих законов является то, что циклической
циркуляции, оборачиваемости в ходе внутриценозного круга био-
логического круговорота подвергаются лишь материальные ресур-
сы биотопа (органические и минеральные вещества и содержащие-
ся в них различные химические элементы). Что же касается
заключенной в органическом веществе потенциальной энергии, то
она расходуется в процессе функциональной деятельности живых
компонентов биогеоценоза необратимо для него, переходя в про-
цессе дыхания в тепловую форму и рассеиваясь в виде теплового
излучения в ближайшем космическом пространстве. Иначе говоря,
поток энергии в биогеоценозе ориентирован в одном направлении.
Израсходованная внутри биогеоценоза и в биогеосфере энергия не
восстанавливается, а потому для нормального функционирования
необходим непрерывный приток энергии извне. Все это, по словам
Ю. А. Жданова (1973), свидетельствует об ограниченной, конечной
природе биогеоценозов и биогеосферы. «Ни одно природное тело,
будучи вырвано из всеобщей связи, — пишет он, не обладает актив-
258
костью, самодвижением. Свойством бесконечного самодвижения
обладает лишь материя в целом» (с. 13). Однонаправленный по-
ток энергии и круговорот веществ в биокосных системах Е. П. Одум
(1968) объявляет «двумя великими принципами или законами об-
щей биологии, так как они одинаково применимы ко всем услови-
ям существования и ко всем организмам, включая и человека»
(курсив наш.—Л. И.) (с. 52).
Основным источником энергии на Земле, как известно, является
поступающая из ближайшего космоса лучистая энергия солнечной
радиации. Но, кроме того, как утверждает В. А. Ковда (1973) со
ссылкой на В. И. Вернадского, живым веществом биосферы ча-
стично используется космическая атомная энергия радиоактивного
распада и энергия рассеянных элементов, исходящих из галактики.
Вся эта поступившая из космоса энергия находится на нашей пла-
нете, по словам В. А. Ковды, в действенном состоянии, выполняя
целый ряд важнейших функций в биосфере, и в потенциальном
состоянии, которое под действием живого вещества постоянно
переходит в действенное. Однако способностью усваивать и акку-
мулировать солнечную энергию обладают только хлорофиллопос-
ные автотрофные растения. Поэтому источником энергии гетеро-
трофным организмам служит биомасса: для животных-фитофагов
и зоофагов — живая растительная или животная биомасса, а для
микроорганизмов и животных-сапрофагов — отмершая биомасса
живых существ и продукты их метаболизма и деструкции остатков
и отбросов.
Поступление солнечной энергии на земную поверхность и рас-
ходование ее на различного рода процессы в биокосных системах
измеряется в энергетических единицах — калориях. Основными
единицами энергии принято считать калорию (кал), которая,
как известно, представляет собою количество тепла, необходимого
для повышения температуры 1 г (1 мл) воды на ГС (при 15°С),
и килокалорию (ккал) — количество тепла, необходимого для
повышения температуры 1 кг (1 л) воды на ГС (при 15°С).
В специальной литературе имеются некоторые сведения об ин-
тенсивности солнечной радиации и общем количестве поступающей
на Землю лучистой энергии. Так, по данным П. Дювиньо и М. Тан-
га (1968), поступающая на поверхность земного шара солнечная
энергия составляет 5-1020 ккал/год (15,3-108 кал/1 м2 в год), в том
числе на континенты поступает в год 1,4* 1020 ккал и на Мировой
океан — 3,6• 1020 ккал. Большая часть этой энергии отражается
атмосферой и поглощается водяными парами. Количество энергии,
достигающей поверхности растительного покрова, зависит от гео-
графической широты местности: в средних широтах ее приходится
около 9 млрд кал/га в год. По данным Ю. Одума (1975), на верх-
нюю границу биосферы поступает солнечной энергии 2 кал на 1 см2
259
в 1 мин. Но после прохождения ее через слой атмосферы на зеле-
ный ковер автотрофных растений падает не более 1,34 кал/см2 в
минуту, или в среднем за сутки 300—400 кал/см2 (с варьированием
от 100 до 800 кал/см2). А. А. Ничипорович и И. А. Шульгин (1976)
приводят данные фотосинтетически активной радиации (ФАР) в
пределах Советского Союза. Количество ФАР, поступающей за
вегетационный период (при температуре выше 5—10° С), колеблет-
ся по физико-географическим (ландшафтным) зонам от 1—1,5 до
4—5 млрд ккал/га.
Достигающая земной поверхности лучистая энергия состоит
примерно на 45% из видимого света, на 45%—из инфракрасного
излучения и на 1О°/о—из ультрафиолетового излучения. Зеленая
автотрофная растительность, как указывает Ю. Одум, наиболее
интенсивно поглощает необходимые для фотосинтеза оранжево-
красные лучи, в особенности дальние; ближнее же красное излу-
чение, несущее основную часть тепловой энергии, по его выраже-
нию, как бы отбрасывается листвой, предохраняя тем самым рас-
тение от перегрева в жаркие летние дни.
Используется поступившая на земную поверхность световая
солнечная энергия разнообразными природными биогеоценозами и
человеком в сельскохозяйственном производстве и в лесном хозяй-
стве. Вносится эта лучистая энергия в природные биогеоценозы и
в культурные посевы и посадки через популяции автотрофных зе-
леных растений, обладающих способностью посредством хлоро-
филла фиксировать солнечную радиацию с длиной волны от 380
до 700 ммк и использовать ее на фотосинтез и другие процессы
жизнедеятельности. Но фиксируется автотрофами (фототрофами)
лишь небольшая часть поступающей на зеленый сухопутный или
голубой водный экран лучистой радиации Солнца. Наземной сухо-
путной растительностью, по данным Ж- Леме (1976), в среднем
улавливается 1% от падающей на ее поверхность солнечной радиа-
ции, в континентальных водоемах — от 0,1 до 1%, в морях и океа-
нах— лишь 0,2%. В частности, лесными сообществами умеренного
пояса, по данным Ю. Одума, фиксируется до I—1,5% от поступаю-
щей солнечной энергии, а влажно-тропическими лесами — до 5%.
Однако наибольшим количеством фиксируемой энергии, по тем же
данным, отличаются сообщества коралловых рифов, аллювиальных
равнин, эстуариев или лиманов (полузамкнутых прибрежных во-
доемов) и такие сельскохозяйственные культуры, как сахарный
тростник, кукуруза, просо и др. Некоторые из этих сообществ и
названных растений способны фиксировать солнечную энергию до
25 кал/1 м2 в год.
Коэффициент использования лучистой энергии Солнца природ-
ных ценозов и культурных растений, называемый коэффициен-
том полезного действия (КПД),—это. отношение акку-
260
мулированиой автотрофами в органическом веществе валовой
первичной продукции энергии к поступившей на их поверхность
лучистой энергии Солнца. По данным Н. А. Ефимовой (1977), КПД
фотосинтетически активной радиации (ФАР) естественной расти-
тельности составляет в пустынях менее 0,1%; в полупустынях уме-
ренного пояса, сухих саваннах и редколесьях — от 0,1 до 0,5%; в
зоне сухих степей умеренного пояса и тропических саваннах и ред-
колесьях— от 0,5 до 1%; в субполярных и полярных широтах
Евразии и Северной Америки — от 1 до 1,5—1,8% и в зоне субтро-
пических и тропических лесов — от 1,5 до 2%. КПД ФАР есте-
ственной растительности на различных материках земного шара
также далеко не одинаков; в Европе он равен 1,26%; в Южной
Америке—1,13; в Северной Америке — 0,94; в Азии — 0,88; в Аф-
рике он падает до 0,59, а в Австралии—до 0,44% (с. 197).
М. И. Будыко (1977) приводит данные об использовании ФАР
естественной растительностью на территории Советского Союза.
КПД ФАР естественной растительности в среднем по Союзу со-
ставляет 0,8% (по другим данным, 1,09%), варьируя по отдельным
провинциям и областям в зависимости от особенностей теплового
и водного режимов в значительных пределах. На Черноморском
побережье Кавказа КПД ФАР равен 1,8—2%; на территории евро-
пейском части СССР (за исключением засушливого юга и юго-во-
стока) — 1—1,2; в центральной части Западной Сибири, на юге
Хабаровского края и в Приморском крае— 1; в пустынях и полу-
пустынях Средней Азии он снижается до 0,2—0,1% (с. 179). КПД
ФАР сельскохозяйственных растений в пределах Советского Сою-
за, по А. А. Ничипоровичу, в среднем составляет 1 —1,5% с варьи-
рованием от 0,25 до 4—5% у таких культур, как кукуруза и
сахарная свекла, при хорошей обеспеченности их влагой и мине-
ральным питанием.
Вообще КПД световой энергии при фотосинтезе, как отмечает
В. Лархер, в действительности бывает значительно ниже теорети-
ческой величины, равной 33%. Но зато «растительная фабрика»,—
пишет Ж. Леме (1976),— покрывает огромные пространства и
функционирует многие дни в году или даже весь год» (с. 108).
«Фотосинтез в самой общей форме, — пишут А. М. и
Д. М. Гродзинские (1973), — определяется как процесс использо-
вания энергии квантов видимого света для восстановления двуоки-
си углерода (СО2) до уровня углеводов. Свободная энергия, исполь-
зуемая любой живой клеткой, в конечном счете связана с запаса-
нием солнечного света в химических связях соединений, возникаю-
щих при фотосинтезе» (с. 416). При этом на 1 грамм-атом
поглощенного углерода, по В. Лархеру (1978), фиксируется 114 кал
энергии (с. 39). Следовательно, как отмечает Б. А. Быков (1970),
«процесс фотосинтеза замечателен тем, что в противоположность
261
другим совершающимся на Земле энергетическим процессам он ве-
дет к аккумуляции энергии, к повышению энергетического уровня
биосферы» (с. 141).
По данным А. М. и Д. М. Гродзинских, на нашей планете зеле-
ными растениями ежегодно образуется около 100 млрд т органиче-
ских веществ, в которых заключается около 450-1016 ккал солнеч-
ной энергии, воплощенной в виде энергии химических связей. При
этом растениями ассимилируется около 170 млрд т углекислоты,
фотохимически разлагается около 130 млрд т воды, из которой
выделяется 115 млрд т свободного кислорода. Таким.образом, зе-
леный растительный покров Земли выступает производителем и
аккумулятором огромного количества органических веществ и за-
ключенной в них потенциальной энергии, за счет которой живет,
функционирует и развивается весь остальной органический мир, в
том числе и человек. Помимо того, он является поглотителем избы-
точной углекислоты атмосферы и поставщиком жизненно необхо-
димого для всех живых существ кислорода.
Разумеется, приведенные данные о количестве синтезируемых
растительными сообществами земного шара органических веществ,
составляющих так называемую первичную продукцию, весьма при-
близительны, ориентировочны. Об этом можно судить хотя бы по
тому, что количественные данные разных авторов о первичной про-
дуктивности Земли значительно расходятся. Так, если А. М. и
Д. М. Гродзинскими (1973) годовая первичная продуктивность
растительного покрова земного шара оценивается в 100 млрд т, то
по расчетам П. Дювиньо и М. Танга (1968) она составляет 83, а
no Р. Дажо (1975) —лишь 61 млрд т; с другой стороны, по дан-
ным американских исследователей, приведенным П. Дювиньо, го-
довая первичная продуктивность Земли достигает в среднем
140 млрд т (от 70 до 280 млрд т), а М. И. Будыко (1977) оцени-
вает ее даже в 200 млрд т.
Определенный интерес представляют данные П. Дювиньо и
М. Танга о распределении годовой первичной продуктивности суши
земного шара в количестве 52,9 млрд т по зональным экосистемам
(типам ландшафтов). Так, па долю леса приходится 28,4 млрд т;
степей и лугов—10,4; пустынь — 5,4; полярные земли бесплодны,
продукция обрабатываемых земель составляет 8,7 млрд т (с. ПО).
Формирование первичной продукции автотрофными растениями
в процессе фотосинтеза и отчасти хемосинтеза трактуется
Б. Д. Абатуровым (1979) как «первый и наиболее ответственный
этап биопродукционного процесса, с которым связано образование
всего объема биологической продукции в экосистеме» (с. 7). На
дальнейших этапах биопродукционного процесса, как он вполне
резонно отмечает, происходит лишь трансформация созданного ав-
тотрофами органического вещества. Сам же биопродукнионный
262
процесс он рассматривает как комплексный процесс, который осу-
ществляется в биокосных системах, в особенности в экосистемах
пастбищного типа, и который включает образование как первич-
ной, так и вторичной продукции, а также использование той и дру-
гой соответствующими ценопопуляциями живых существ на раз-
ных трофических уровнях пищевых цепей этих биокосных систем.
Такая трактовка биопродукционного процесса названным автором
представляется нам чрезвычайно важной с биогеоценотической
точки зрения, ибо она вскрывает самую суть, сердцевину матери-
ально-энергетических обменных процессов, совершающихся в био-
геоценозе, обеспечивающих его нормальное функционирование и
поступательное развитие.
5. Трофическая структура, типы обмена веществ, носители
и каналы переноса веществ и энергии в биогеоценозе
Перейдем к рассмотрению с возможной полнотой циклически
совершающихся обменных процессов в биогеоценозе, в частности
процессов созидания и трансформации биологической продукции,
запасания ее в различных формах живой и мертвой биомассы, от-
ложения в виде косной органики в почве и верхних слоях литосфе-
ры и выноса неусвоенных веществ в сток. Одновременно с этим
подлежит рассмотрению органически взаимосвязанный с этим
поток энергии в биогеоценозе, как поступившей извне, так и осво-
бождаемой при трансформации и разложении растительной, жи-
вотной и микробной биомассы, расходование части ее на процессы
жизнедеятельности живых компонентов биогеоценотических систем
и рассеивание отработанной энергии в тепловой форме в окружаю-
щем пространстве. При этом должно быть обращено внимание на
выявленные закономерности обмена веществ и потока энергии в
биогеоценозе. Разумеется, все это должно происходить с учетом
необычайной сложности структурно-функциональной организации
биогеоценотических систем, слабой изученности протекающих в них
вещественно-энергетических обменных процессов, недостаточности
конкретных данных, характеризующих эти процессы с качествен-
ной и в особености с количественной стороны. Ведь это как раз и
служит значительным препятствием, ограничивающим возмож-
ность на данном этапе до конца разобраться в этом сложном клуб-
ке взаимосвязанных природных явлений.
В одном из разделов предыдущей главы было показано, что
единственным активно действующим в природных биогеоценозах
аппаратом и носителем веществ и энергии являются видовые цено-
популяции автотрофных, биотрофных и сапротрофных живых орга-
низмов, облекающиеся в соответствующие формы совместного су-
ществования, а именно в трофические уровни (звенья) пищевых
263
цепей и сетей, являющихся соподчиненными элементами более
крупных функциональных структурных единиц — консорций. По-
следние, в свою очередь, представляют собою элементы биоцено-
зов, входящих в форме этой подсистемы в такую биокосную систе-
му, как биогеоценоз, наряду с другой подсистемой — биотопом
(биоценосредой).
Там же было показано, что пищевые цепи и консорции в услож-
ненном варианте представляют собою такие функциональные
структурные единицы биогеоценоза, в которых циклическая цирку-
ляция биофильных элементов в составе органических веществ че-
рез их трофические звенья находит полное завершение — от синте-
зированных автотрофами углеводов и других веществ до исходных
химических элементов, пригодных для использования фототрофами
в последующем цикле биологического круговорота. При этом было
подчеркнуто, что, несмотря на это, ни пищевые цепи, ни консорции
не являются самостоятельно функционирующими системами, а
представляют собою лишь функциональные структурные единицы,
элементы биоценоза и биогеоценоза в целом. Было обращено вни-
мание и на то, что в состав консорций входит не одна пищевая
цепь, а в состав биоценоза не одна консорция, а несколько, даже
много, а также на то, что эти пищевые цепи разные (пастбищные,
детритные), что консорции также неодинаковы (индивидуальные
и популяционные, полночленные и пеполпочлепные); неодинакова
и роль тех и других в структурно-функциональной организации и
функционировании биогеоценоза как основной материально-энерге-
тической ячейки биогеосферы.
Из этого следует, что от известной всем трофической структуры
пищевых цепей необходимо отличать трофическую структуру кон-
сорций и биоценозов и, следовательно, биогеоценозов и биогеосфе-
ры в целом. Различие трофических структур названных составных
единиц биогеоценоза заключается в следующем. Узловыми звень-
ями пищевой цепи, входящей в состав той или иной консор-
ции, являются, как известно, трофические уровни, каждый из
которых представлен сходными по типу питания и обмена веществ
организмами соответствующей видовой ценопопуляций. Узловыми
звеньями популяционной консорции, входящей в состав
того или иного биогеоценоза, по-видимому, будут трофические кон-
центры, состоящие из трофических уровней пищевых цепей, каж-
дый из которых представлен видовыми ценопопуляциями соответ-
ствующего ценокомплекса (автотрофного, биотрофного, сапротроф-
ного). Узловыми же звеньями биоценоза, образующего в сово-
купности с биотопом биогеоценоз, следует считать трофические
ступени (ярусы, по Г. Ф. Хильми, 1966), которые состоят из трофи-
ческих концентров консорций; каждая из этих ступеней представ-
лена соответствующими ценокомплексами видовых ценопопуляций.
264
Вместе с тем трофические структуры пищевых цепей, консорций и
биокосных систем, несмотря на указанные различия, сходны между
собой по числу и последовательности в функциональном смысле
расположения в поточных линиях трофических звеньев,близ-
ки они и по характеру протекающих в каждом звене обменных
процессов.
В основе трофической структуры биогеоценоза, по мнению
Р. Риклефса (1979) и Ю. Одума (1975), лежит аккумулированная
в первичной биологической продукции автотрофов солнечная энер-
гия, поток которой не только формирует трофическую структуру,
но и приводит в движение все механизмы биогеоценотической си-
стемы, в том числе и круговорот питательных веществ в ней. При
этом, как утверждает Ю. Одум, условия существования организ-
мов определяются общим потоком поступающей на Землю солнеч-
ной радиации, а для формирования первичной биологической про-
дукции и круговорота веществ наибольшее значение имеет сум-
марное количество солнечной энергии, получаемой зелеными
автотрофными растениями за педели, месяцы, за весь вегетацион-
ный период и весь год.
И. И. Смольянинов и Е. В. Рябуха (1971) различают три типа
биокосных систем (биогеоценозов) по некоторым особенностям
осуществления в них обменных процессов. 1. Замкнутые системы.
Разумеется, в природе абсолютно замкнутых биокосных систем не
бывает. Они будут (или уже есть) на космических кораблях, ба-
тискафах и т. п. 2. Относительно замкнутые системы. К ним отно-
сится подавляющее большинство природных биогеоценозов земного
шара (лесные, луговые, степные и др.). 3. Открытые (проточные)
системы. Это биогеоценозы, расположенные по берегам рек, ручьев
в непосредственной близости к ним. Сюда же, вероятно, можно
отнести искусственные гидропонные установки.
Функционирование биогеоценоза — это спонтанный непрерыв-
ный процесс осуществления целого ряда разнообразных биохими-
ческих превращений и преобразования веществ при непосредствен-
ном участии живых компонентов этой сложной биокосной системы
с использованием при этом солнечной энергии и материальных
ресурсов биотопа. Действующими «агентами», как было сказано»
участвующими в обменных процессах биогеоценоза путем усвоения
и трансформации веществ в процессе своей жизнедеятельности,
являются ценопопуляций автотрофных, биотрофных и сапротроф-
ных организмов и ценокомплексы этих ценопопуляций. Материа-
лом же, подвергающимся биохимическим превращениям на авто-
трофном трофическом уровне, служат первичные природные ре-
сурсы— углекислота, атмосферная и почвенная влага, элементы
минерального питания, на уровне биотрофов — живая биомасса
265
растений и животных и продукты их жизнедеятельности, на уровне
сапротрофов — отмершая биомасса организмов, их отбросы, выде-
ления и трупы, которые могут рассматриваться как вторичные ма-
териальные и одновременно энергетические ресурсы.
Общий обменный процесс в биогеоценозе как основной мате-
риально-энергетической ячейке биогеосферы — весьма сложный
процесс не только сам по себе, но и в силу того, что он базируется
на биологическом (физиологическом) обмене, осуществляющемся
на организменном уровне и приобретающем необходимую устойчи-
вость и ценотическую значимость на популяционно-видовом уровне
в составе соответствующих трофических звеньев консортивных
пищевых цепей биогеоценоза. Кроме того, круговорот веществ и
поток энергии в полночленном биогеоценозе — это многоканальный
процесс, включающий биотрофный канал, соответствующий
каналу выедания пищевых цепей, сапротрофный канал (это
канал разложения в пищевых цепях) и канал запасания орга-
нических веществ и потенциальной энергии в виде живой и мерт-
вой биомассы (в древесине, в телах позвоночных животных, в пнях
н лесном валежнике, в гумусовом горизонте почвы и т. п.), не го-
воря уж об отложениях органики и органоминеральных соединений
за пределами биогеоценоза — на дне водоемов, в глубоких слоях
литосферы, в осадочных горных породах и пр. Важно и то обстоя-
тельство, что вследствие сопряженности круговорота питательных
веществ и потока заключенной в них энергии ход обменных про-
цессов в биогеоценозе практически является не линейным, а круго-
вым, хотя и не замкнутым абсолютно в самом себе. Ведь суть
различия этих двух процессов, на наш взгляд, состоит не столько
в односторонности, однонаправленности одного из них и в циклич-
ности другого, сколько в одноразовости использования энергии и
многоразовост и утилизации биофильных элементов в составе ис-
пользуемой живыми существами пищи.
Б. А. Быков (19706) довольно подробно рассматривает каналы
внутренних и внешних коммуникаций, обеспечивающие как связь
живых компонентов между собой и с косными компонентами (био-
ценосредой), так и связь биогеоценоза с внешним окружением.
К внешним коммуникациям (путям связи) он относит: 1) энер-
гетические каналы (коммуникации), по которым осуществля-
ется а) ввод в биогеоценоз извне энергии солнечной радиации
посредством фотосинтеза фототрофов и поглощения тепловых лу-
чей подстилающей земной поверхностью и б) вывод энергии из
биогеоценоза во внешнюю среду в процессе дыхания организмов
и прямой теплоотдачи; 2) сырьевые каналы (коммуникации),
по которым происходит а) ввод в биоценотическую среду биогео-
ценоза материальных ресурсов (воды, минеральных веществ, азота
И углерода в виде углекислоты) и б) вывод из системы биогеоце-
266
ноза различных материалов вовне с воздушными потоками, а так-
же путем фильтрации, выщелачивания и т. п.
Наибольшее значение в осуществлении обменных процессов
функционирования биогеоценозов Б. А. Быков придает внутренним
коммуникациям, поскольку только они, по его мнению, способны
обеспечить регуляцию, относительную стабильность биогеоценозов
и, следовательно, их «самодвижение» и развитие. Среди них он
различает: 1) аллел агонические каналы (коммуникации),
по которым протекает поток энергии по пищевым цепям консорций
и происходит передача ее от автотрофов через фитофагов и зоофа-
гов к хищникам 2-го порядка; 2) аллелопатические каналы,
по которым осуществляется транспортировка органических и более
простых органогенных веществ, а также тепловой преимуществен-
но энергии от ценозообразующих видовых популяций в воздушную
или почвенную среду биогеоценоза; 3) аллелосполические
каналы, по которым транспортируются различные минеральные
вещества и биогенные газы, а также энергия из биоценотической
среды в ценопопуляций живых компонентов, в особенности гетеро-
трофных, Однако следует иметь в виду, что разного рода «каналы»
как внутренних, так и внешних коммуникаций — это лишь вообра-
жаемые пути, направления реального перемещения веществ и энер-
гии при участии соответствующих видовых ценопопуляций или
каких-либо абиотических агентов.
Обменные процессы на каждом трофическом уровне обладают
некоторой спецификой, что определяется целым рядом особенно-
стей образующих данное трофическое звено видовых ценопопуля-
ций. Эти особенности в строении, функционировании и образе
жизни ценозообразующих популяций и сопряженные с этим специ-
фические особенности трансформации веществ и энергии В. А. Пав-
лов (I960) видит в следующем. Для ценопопуляций автотроф-
ного компонента, представленных главным образом хлорофилло-
носными зелеными растениями, по его словам, характерны: а) фо-
тотроф ность, т. е. способность фиксации лучистой энергии Солнца
и аккумуляции ее в химических связях сложных органических
веществ; б) прикрепленный образ жизни подавляющего большин-
ства сухопутных автотрофных растений, обусловленный широким
распространением и относительно равномерным по земной поверх-
ности распределением необходимых для их жизни материальных
ресурсов; в) чрезвычайная расчлененность растений, в особенности
высших, на внешние органы, а самих органов — на бесчисленное
множество морфологически однотипных образований (листья, кор-
ни), обеспечивающая огромную площадь соприкосновения ассими-
лирующих и адсорбирующих частей этих органов с соответствую-
щим субстратом; г) преобладание ассимиляции в обмене веществ
фототрофных растений, осуществляющейся за счет энергии,, кота-
267
рая выделяется при окислении углеводов в процессе дыхания, и
отложение жиров и белков в репродуктивных органах растений,
используемых в качестве энергетического материала при прораста-
нии их.
Характерными особенностями биотрофного компонента на
последующих трофических уровнях, представленного в основном
ценопопуляциями растительноядных и плотоядных животных, явля-
ются: а) гетеротрофность питания и обусловленная этим потреб-
ность в непрерывном потреблении для поддержания их жизни
сложных высокомолекулярных органических веществ, заключенных
в биомассе растительного или животного происхождения; б) ак-
тивность и подвижный образ жизни большинства представителей
животного мира в поисках и добывании пищи (корма и воды),
обусловленных наличием более интенсивного и разнообразного у
них обмена веществ по сравнению с растениями, а также извест-
ной ограниченностью либо неравномерностью распределения пище-
вых ресурсов в биосфере; в) развитие у животных специальных
сложных систем органов для восприятия, поглощения и переработ-
ки разнообразного пищевого материала и кислорода и удаления
из организма продуктов обмена; г) преобладание диссимиляции
в процессах обмена веществ у животных, особенно у наиболее
активных форм, обусловленной более высокой потребностью их в
энергии, которая удовлетворяется главным образом за счет утили-
зации более энергоемких органических веществ — жиров и отчасти
белков.
Что же касается сапротрофного компонента, представлен-
ного главным образом микроорганизмами (спороносными и неспо-
роносными бактериями, низшими грибами, актиномицетами), то
для них, по словам названного автора, характерны: а) необычай-
ное многообразие типов обмена веществ, особенно у бактерий; мик-
роскопические размеры, простота и однообразие формы и чрезвы-
чайная изменчивость физиологических свойств; исключительная
быстрота размножения и приспособленность к самым разнообраз-
ным условиям существования; б) повсеместная распространенность
микроорганизмов, особенно бактерий, во всех средах жизни (в воз-
душной, почвенной, водной) в очень большом диапазоне темпера-
турного режима, обеспеченности кислородом, кислотности почвы
и при чрезвычайном разнообразии используемой пищи; в) наличие
наряду с этим узкой специализации некоторых форм микроорга-
низмов, в частности бактерий, как в отношении условий обитания,
так и в использовании пищевых ресурсов вследствие ограниченного
набора у них ферментов.
Нетрудно представить себе, что выработавшиеся в процессе
органической эволюции различные типы обмена веществ, с одной
стороны, определили позиции носителей этих типов обмена, а имен-
268
но автотрофных, биотрофных и сапротрофных ценопопуляций в
общей системе совместного бытия, а с другой — обусловили неиз-
бежную зависимость их друг от друга, неразрывную взаимосвязь,
взаимообусловленность друг другом. Это, в свою очередь, опреде-
ляло каждый раз целостность и относительную стабильность воз-
никающих и развивающихся в данных физико-географических и
геоисторических условиях в совокупности с преобразуемой ими
средой (биоценосредой) соответствующих биокосных систем —
биогеоценозов.
Живые компоненты сухопутных биогеоценозов в ходе эволюци-
онного развития заняли и функционально освоили, как известно,
две среды жизни — наземную (воздушную) и почвенную, вошед-
шие в силу этого в состав биогеоценоза в качестве самостоятель-
ных биогенных компонентов. Е. М. Лавренко и Н. В. Дылис (1968)
на основании этого сухопутный биогеоценоз подразделяют на два
блока — наземный и почвенный. При этом они отмечают, что при
осуществлении «первой биохимической функции» живого вещества
(по В. 11. Вернадскому), связанной с питанием, дыханием и раз-
множением организмов, охватываются оба блока биогеоценоза, а
«вторая биохимическая функция», связанная с разложением от-
мерших организмов, протекает в почвенном блоке. Близкие к этому
мысли высказывает и Ю. Одум (1975), который пишет, что авто-
трофный метаболизм наиболее интенсивно протекает в «зеленом
ярусе», т. е. в надземной части биогеоценоза, а гетеротрофный
метаболизм преобладает в «коричневом поясе», т. е. в почве. Осу-
ществление этих двоякого рода функций, по словам Ю. Одума,
часто бывает разделено и во времени, поскольку лишь небольшая
часть первичной продукции используется гетеротрофами сразу же
непосредственно на месте, большая часть ее попадает в подстилку
и в почву, где утилизация ее может надолго задержаться. След-
ствием этого явилось подразделение Ю. Одумом обменных процес-
сов в биогеоценозе на два потока — пастбищный и детритный
(биотрофный и сапротрофный каналы).
Р. Дажо (1975) находит, что и сами живые компоненты биогео-
ценоза в связи с этим также пространственно разобщены. Так, авто-
трофные растительные сообщества, растительноядные животные и
трофически связанные с ними хищники распространены на поверх-
ности земли, а детритоядные формы приурочены к почве и под-
стилке. Живые компоненты биогеоценоза, по свидетельству того
же автора, разобщены в отношении утилизации пищевых ресурсов
и во времени, а именно: животные фитофаги и фитопаразиты, ис-
точником энергии которых являются живые растения, используют
их непосредственно по мере нарастания фитомассы, а детритояд-
ные живые существа, живущие за счет мертвой растительной и
269
животной биомассы, потребляют ее не сразу, а лишь после отми-
рания, следовательно, значительно позже.
Возникает вопрос: что же является движущей силой совершаю-
щихся в живой природе, в биогеоценозе обменных процессов? Йз
представлений Ф. Энгельса (1930) о жизни как форме существо-
вания «белковых тел», как самопроизвольно совершающемся про-
цессе, присущем своему носителю — белку, вытекает, что такой
движущей силой предстает сама жизнь во всех ее проявлениях и
формах — организменной, популяционно-видовой, биоценотической
и биостроматической. Но это не внешняя по отношению к жизни
сила, а внутренне присущая ей как ее атрибут, и обмен веществ,
по словам Энгельса, является существенной функцией жизни. Но
обмен веществ имеет место и в неживой природе. Однако в живой
природе обмен веществ ведет не к разрушению ее объектов, как в
неживой, а является, по выражению Энгельса, необходимым усло-
вием их существования.
Такова суть и логика жизни, которые, надо полагать, опреде-
лили и соответствующую стратегию жизни, и формы, в которые
она облеклась, и средства, которые пускаются в ход в процессе
преодоления ею сопротивления неорганической природы. Как след-
ствие этого можно рассматривать и утвердившуюся в живой при-
роде в ходе органической эволюции систему отношений между
биологическими видами. Суть и своеобразие ее, по мнению
Г. Ф. Хильми (1966), состоит в том, что любой биологический вид
в биосфере (биогеосфере) выступает как источник питания и, сле-
довательно, доступной энергии для других видов либо сам стано-
вится потребителем биомассы и энергии, накопленной другими
видами. С этим он, кстати сказать, связывает и ярусное (ступенча-
тое) в функциональном смысле строение биокосных систем (био-
геоценоза, биогеосферы), которое выражается в том, что видовые
ценопопуляций живых компонентов, составляющие данную трофи-
ческую ступень биокосной системы, неизбежно являются потреби-
телями биомассы и энергии, накопленной ценопопуляциями ниже-
лежащих ступеней. Исключение составляет самая нижняя ступень
системы, представленная зелеными автотрофными растениями,
источником питания для которых служат элементы «физической
среды», а источником энергии — солнечная радиация.
Исходя из представления о ступенчатом строении биокосных
систем, в частности биогеосферы, Г. Ф. Хильми выявил некоторые
закономерности движения веществ и энергии в процессе их функ-
ционирования, поданные нами в незначительной перефразировке:
а) движение веществ и энергии по трофическим ступеням биогео-
ценоза или биогеосферы представляет собою не передачу их из
нижних ступеней в верхние, а превращение ассимилированной на
каждой нижележащей ступени биомассы и заключенной в ней
270
энергии в биомассу и соответствующие формы энергии вышерас-
положенной над ней ступени; б) поступление энергии в животное-
пожирателя вместе с поедаемой им биомассой жертвы всегда свя-
зано с затратой его собственной энергии на поиски пищи, охоту
и т. п.; в) чем меньше организмом той или иной трофической сту-
пени затрачивается энергии на добывание пищи (поиски, охоту
и пр.), тем выше так называемый коэффициент рентабельности, и
наоборот. Коэффициент рентабельности (КР)—это
отношение количества приобретенной организмом энергии к коли-
честву предварительно затраченной на ее приобретение. Он соот-
ветствует коэффициенту полезного действия (КПД) питания. Уста-
новлено, что КР у разных животных неодинаков: он находится в
зависимости от характера пищи, ее обилия, плотности и степени
подвижности объектов питания, а также от способа добывания
пищи потребителем и степени пищевой специализации его.
Формирование биологической продукции в любой биокосной
системе, по мнению Б. Д. Абатурова (1979), представляет собою
многоступенчатый процесс; имеется, очевидно, в виду ступенчатый
характер их трофической структуры, состоящей из ряда трофиче-
ских уровней в составе пищевых цепей. При этом он подчеркивает,
что на каждом этапе, трофическом уровне происходит взаимодей-
ствие контактирующих компонентов системы: автотрофного расти-
тельного компонента с абиотической средой и солнечной радиаци-
ей, животных-фитофагов с автотрофной растительностью, хищных
животных и паразитов с растительноядными животными и т. д. В
процессе эволюции биогеоценозов, по его словам, выработался
такой характер этих взаимоотношений, который «позволяет созда-
вать максимальное количество биологической продукции на всех
этапах биопродукционного процесса» (с. 115).
Далее автором приводится ряд выявившихся приспособитель-
ных особенностей ценопопуляций каждого трофического уровня,
которые, как он пишет, «обеспечивают, с одной стороны, макси-
мально возможное использование потребителями своих ресурсов
питания, а с другой, предупреждает катастрофическое снижение
продуктивности потребляемых организмов и деградацию всей си-
стемы» (с. 115). С этими приспособлениями живых существ автор
связывает выход продукции на каждом трофическом уровне и био-
геоценоза в целом, а также зависимость ее от материально-энер-
гетических ресурсов на первом трофическом уровне и от ресурсов
питания в виде живой или мертвой биомассы на последующих
трофических уровнях. Отсюда он приходит к выводу: «Продуктив-
ность всей экосистемы, в том числе и вторичная продуктивность^
задается количеством ресурсов среды» (с. 116).
271
6.. Ход обмена веществ, поток энергии и роль живых компонентов
в обмене и функционировании биогеоценоза
Перед тем как приступить к рассмотрению самого хода обмен-
ных процессов в биогеоценозе, полезно уточнить некоторые связан-
ные с этим понятия, в частности такие, как биомасса и продукция.
Под биомассой следует понимать количество живого вещества
в телах отдельных организмов, видовой популяции или ценокомп-
лекса популяций на момент учета, выражаемого обычно в весовых
или энергетических единицах (калориях или джоулях*) на едини-
це площади или объема. Различают живую и мертвую (отмершую)
биомассу, а также растительную (фитомассу), животную (зоомас-
су) и микробную биомассу. Продукция — это количество жи-
вого органического вещества, создаваемого организмом или цено-
популяцией трофического уровня за единицу времени (обычно за
год), это годичный (суточный, месячный) прирост, прибавка рас-
тительной или животной биомассы. Продукция живого вещества
подразделяется на первичную и вторичную продукцию, а среди
первичной продукции различают валовую и чистую. Первичная
продукция — количество живого органического вещества, созидае-
мого автотрофными зелеными растениями в процессе фотосинтеза
в единицу времени или, иначе, количество солнечной энергии, свя-
занной автотрофами в органическом веществе. Вторичная про-
дукция— количество живого органического вещества, созидаемого
биотрофами и сапротрофами в процессе трансформации потреб-
ляемой ими живой или мертвой (отмершей) биомассы и продуктов
метаболизма в единицу времени.
Различие валовой и чистой первичной продукции состоит в том,
что валовая первичная продукция — это общее количество син-
тезированного автотрофами органического вещества и связанной в
нем энергии за единицу времени; чистая же продукция — это
разница между валовой первичной продукцией и той частью ее, ко-
торая израсходована па процессы жизнедеятельности растений,
рассеяна в виде тепла в окружающем пространстве за данный
промежуток времени. Вторичная продуктивность на валовую и
чистую по некоторым соображениям обычно не подразделяется.
Количественный расчет всех перечисленных видов продукции в
биогеоценозе ведется на единицу площади (или объема) в весовых
или энергетических единицах. Предпочтительно вести расчет в
энергетических единицах, ибо калория — такая единица, которая
дает возможность количественно сопоставлять выход различной по
энергоемкости продукции на разных трофических уровнях биогео-
♦ Джоуль (Дж)—единица измерения энергии по Международной системе
(1 Дж = 0,2388 кал).
272
ценотических систем, а также скорость и интенсивность обменных
процессов в разных типах биогеоценозов и у разных организмов.
Между прочим, установлено, что интенсивность метаболизма
на единицу веса живого организма, как правило, тем больше, чем
меньше его размеры. Это объясняется тем, что с уменьшением
размеров организма возрастает относительная величина его по-
верхности и, следовательно, диффузия кислорода и углекислоты в
его ткани, да и характер обменных физиологических процессов
далеко не одинаков у разных представителей органического мира.
Наряду с размерами и величиной биомассы организмов в продук-
ционном процессе имеет значение и продолжительность их жизни,
и быстрота смены поколений. Так, организмы с коротким сроком
жизни (например, планктон) или многократно частично употреб-
ляемые в пищу (травы на пастбище), как утверждает Ж. Леме
(1976), могут оказаться более продуктивными, чем организмы,
обладающие большей биомассой. Здесь, пожалуй, уместно обра-
тить внимание па такое понятие, как коэффициент эколо-
гической эффективности — «отношение биомассы орга-
низмов к количеству потребленного ими органического вещества»
(М. И. Будыко, 1971, с. 189). Он, как правило, не превышает
10-20%.
Что же касается потока энергии через трофические уровни пи-
щевых цепей биогеоценотической системы и соотношения его с
биологической продукцией, то, по свидетельству зарубежных эко-
логов, он представляет собой общую, суммарную ассимиляцию на
каждом трофическом уровне, т. е. сумму продуцированной био-
массы и веществ, израсходованных в процессе продуцирования на
дыхание или, иначе, всю ассимилированную на этом уровне энер-
гию. О соотношении потока энергии и продуктивности можно по-
лучить представление также со следующих слов М. И. Будыко
(1971): «Количество энергии, передаваемой организмами по пище-
вой цепи, соответствует их продуктивности, т. е. массе создавае-
мого ими органического вещества» (с. 410).
Итак, обмен веществ—необходимое условие существования
всего живого на Земле, в том числе и биогеоценозов как матери-
ально-энергетических ячеек биогеосферы. В биогеоценозе он начи-
нается с автотрофного звена пастбищных пищевых цепей
в составе индивидуальных и популяционно-видовых консорций, ибо
в процессе взаимодействия ценопопуляций автотрофного ценокомп-
лекса с материально-энергетическими ресурсами биотопа и внеш-
ней среды происходит ввод в бногеоценотическую систему необхо-
димых зольных элементов минерального питания растений, угле-
рода и азота в соответствующих формах и лучистой энергии
Солнца. Завершается взаимодействие контактирующих компонен-
тов биогеоценоза на этом трофическом уровне образованием пер-
273.
винной биологической продукции в виде углеводов, растительных
жиров, белков и пр.
Формирование первичной продукции и ее величина определяют-
ся, как отмечает Б. Д. Абатуров, двумя противоположными про-
цессами— фотосинтезом и дыханием автотрофов. Роль хемосинте-
за, осуществляемого хемосинтезирующими бактериями (хемотро-
фами) за счет энергии окисления некоторых минеральных веществ
{аммиака, сероводорода и др.)» незначительна. Продуктивность
ценопопуляций зеленых автотрофных (фототрофных) растений
биогеоценоза, создающих органические вещества в процессе фото-
синтеза, зависит, по словам В. Лархера (1978), от целого ряда
свойств самих ценозообразующих растений: характера и густоты
облиствления растений, размеров и расположения по отношению
к потоку солнечного излучения ассимиляционных поверхностей
(листьев), интенсивности ассимиляции ими питательных веществ
и солнечной энергии, интенсивности газообмена растений и дли-
тельности периода с положительным балансом его. Большое зна-
чение, разумеется, имеет в этом и морфологическая структура
растительного слоя сообщества (вертикальная, горизонтальная), а
также степень обеспеченности растений элементами минерального
питания, влагой, надлежащим тепловым режимом и пр.
Наземные растения, по данным Р. Дажо, фиксируют в среднем
5—8% минеральных веществ от собственного веса (в расчете на
сухое вещество); правда, галофиты отличаются способностью по-
глощать значительно большее количество минеральных солей из
почвы. Что же касается усвоения фототрофными растениями сол-
нечной энергии, то, по данным Ю. Одума, они фиксируют в сред-
нем около 50% поступающей на их поверхность лучистой энергии,
но лишь незначительная часть ее, не более 5%, превращается в
процессе фотосинтеза в растительную биомассу. Большое значение
в усвоении фототрофами энергии солнечной радиации придается
гак называемому листовому индексу (индексу листовой по-
верхности), под которым понимается отношение освещаемой пло-
щади листьев к площади, занятой растениями. На приведенном
им графике (с. 66) видно, что наиболее высокий листовой индекс
(L), например, для злаков близок к 4, а для спелых древесных
пород достигает 8—10.
Усвоенная растениями солнечная энергия расходуется прежде
всего на поддержание их жизни и осуществление ими таких внут-
ренних жизненных функций, как питание, транспирация, фотосин-
тез и т. п. (основной обмен). В процессе фотосинтеза происходит
формирование валовой первичной продукции. Значительная часть
аккумулированной в ней потенциальной энергии расходуется на
осуществление таких жизненных явлений, как рост, размножение
растений, обновление отмирающих в процессе развития тканей»
274
образование в соответствующих органах углеводных запасов и пр.
Некоторая часть энергии теряется путем прямой теплоотдачи, а
также вместе с растительным опадом и отпадом, поступающим в
подстилку и почву, превращаясь в почвенный гумус. Небольшая
часть продуцируемой на этом трофическом уровне органики пре-
вращается в минеральные вещества при лесных и степных пожа-
рах, и столь же незначительная часть ее отлагается на болотах, на
дне водоемов и т. п. Протекающие в растительных организмах
внутренние жизненные процессы представляют собою чрезвычайно
сложные взаимосвязанные биохимические превращения веществ и
преобразование энергии. Наряду и одновременно с синтетическими
процессами в организмах совершаются и процессы разложения,
распада веществ, заключающиеся в окислении сложных органиче-
ских веществ в более простые соединения при дыхании растений,
сопровождающиеся высвобождением энергии и выделением ее
в виде тепла.
Чистая первичная продукция, остающаяся от валовой после
вычета из нее продукции и энергии, потерянных при дыхании рас-
тений, поступает на вторую трофическую ступень (уровень) био-
геоценотической системы. На этом трофическом уровне она служит
прямым источником питания (корма) и энергии для растительно-
ядных животных (фитофагов) и для фитопаразитов. Следователь-
но, взаимодействующими компонентами на втором трофическом
уровне пищевых цепей становятся продуцирующие растительную
биомассу ценопопуляций автотрофного ценокомплекса и использую-
щие ее в качестве источника энергии животные-фитофаги и фито-
паразиты. Завершается взаимодействие названных компонентов
биогеоценоза преобразованием первичной биологической продук-
ции в более высокомолекулярную вторичную биологическую про-
дукцию, т. е. в животные жиры, белки и т. п.
Трофические взаимоотношения животных-фитофагов с ценопо-
пуляцнями зеленых автотрофных растений Б. Д. Абатуров (1979)
относит «к самостоятельному разряду отношений типа „потреби-
тель— корм"» (с. 22), считая их аналогичными отношениям
«хищник — жертва» и «паразит — хозяин». Своеобразие взаимо-
действия контактирующих на данном трофическом уровне компо-
нентов биогеоценоза, по мнению Б. Д. Абатурова, состоит в том,
что количество потребляемого корма растительноядными животны-
ми определяется не столько их энергетическими потребностями
(как принято считать), сколько обилием и состоянием кормовых
ресурсов. «...Количество потребляемого корма, — пишет он, — в
этом случае в среднем соответствует потребностям животных, в
противном случае их существование здесь было бы невозможным»
(с. 91). Подчеркнем: «соответствует», но не «контролируется» и не
«определяется» потребностями животного в корме.
275
преобразование первичной продукции во вторичную, как ука-
зывает Б. Д. Абатуров, находится в зависимости от интенсивности
питания (т. е. количества потребляемого в единицу времени кор-
ма), и в особенности от эффективности переработки корма (т. е.
переваримости и усвояемости его животными). Интенсивность
потребления кормов, отнесенная к единице веса тела, как отмечает
автор, неодинакова у разных видов растительноядных животных:
«...чем мельче животное, тем выше относительный уровень пита-
ния» (с. 91). Переваримость и усвояемость корма у разных видов
животных-фитофагов также неодинакова, зависит она и от конси-
стенции и питательности корма. Так, по приведенным автором
данным, у крупных копытных животных, питающихся грубым рас-
тительным кормом, усвояемость его 60—70%; у грызунов, потреб-
ляющих в пищу более питательные корма, она достигает 75—80
(95) %, ио у грызунов, питающихся вегетативными частями расте-
ний, опа, наоборот, снижается до 43—55%. У беспозвоночных
фитофагов, в частности у гусениц насекомых, переваримость и
усвояемость потребляемой пищи всего лишь 26—39 (49)%- Зави-
симость формирования вторичной продукции от переваримости и
усвоения корма выражается, по Абатурову, в следующем: чем
меньше потери потребленной растительной массы с непереварен-
ными и неусвоенными остатками пищи и, следовательно, больше
усвоенной фитофагом в процессе питания пищи, тем выше эффек-
тивность создания вторичной продукции.
Неусвоенная часть потребленной фитофагами растительной
биомассы выводится из организмов наружу в виде экскрементов,
с мочой и т. п. Ассимилированная же растительная биомасса и
аккумулированная в ней энергия частично (.не менее 10%) исполь-
зуются ими па образование биомассы своего тела; большая часть
ее расходуется на внутренние процессы поддержания жизни расти-
тельноядных животных (основной обмен), на переваривание и
усвоение потребляемого корма, а главным образом на мышечную
работу—передвижение в пространстве, поиск воды, пищи, сбор
корма (дыхание); у диких животных на это тратится до 60% сверх
количества энергии, расходуемого па основной обмен. На величину
траты энергии дикими и домашними животными оказывают влия-
ние местные климатические условия, а также физиологическое
состояние животных (половая активность, спячка и т. п.).
Важнейшим показателем продуктивности животных-фитофагов
является эффективность переработки чистой первичной продукции
во вторичную. Этот показатель можно трактовать как КПД исполь-
зования энергии на образование фитофагами вторичной продук-
ции, который представляет собой отношение количества энергии,
заключенной в созданной фитофагами вторичной продукции, к
количеству энергии, аккумулированной в потребленной ими пер-
276
вмчной продукции. У диких теплокровных растительноядных жи-
вотных этот показатель (КПД), по данным Б. Д. Абатурова, со-
ставляет 1—2% от величины потребления; у холоднокровных, на-
пример насекомых и др., он достигает 10—16%, а у домашних
животных может быть доведен до 50% от усвоенной части
корма.
Значительная часть вторичной продукции в виде биомассы, со-
зданной растительноядными животными на втором трофическом
уровне, поступает на третью трофическую ступень (уро-
вень) биогеоценотической системы. На этом трофическом уровне
опа служит источником пищи и энергии для плотоядных животных
(зоофагов) и для зоопаразитов. Непосредственно взаимодействую-
щими функциональными группами на третьем трофическом уровне
пищевых цепей являются ценопопуляций биотрофного ценокомп-
лекса — растительноядные животные с ценопопуляциями пло-
тоядных животных (хищниками 1-го порядка). Трофические взаи-
модействия контактирующих на данном трофическом уровне
функциональных групп животных осуществляются по типу хищ-
ник— жертва и отчасти по типу паразит — хозяин. Завер-
шается взаимодействие этих функциональных групп животных
формированием биологической продукции, соответствующей по
своим качественным показателям биомассе хищных форм.
Биопродукционные процессы и процессы жизнедеятельности
плотоядных (хищных) животных на третьем трофическом уровне
пищевых цепей характеризуются некоторыми особенностями, обу-
словленными своеобразием их объектов питания и энергии. По-
требляемая в пищу зоофагами животная биомасса отличается от
растительной массы, служащей пищей фитофагам, во многих отно-
шениях, а главное — более высокой калорийностью, энергоемко-
стью и неравномерным распределением в пространстве. С другой
стороны, использование ее хищниками в пищу сопряжено со значи-
тельной затратой энергии на поиск поедаемых животных, охоту на
них и пр. Все это определило возникновение и развитие в процессе
эволюции целого ряда анатомо-морфологических, физиологических
и эколого-ценотических приспособлений как у хищников, так и у
их жертв, а также весьма важных характерных особенностей в
образе жизни и поведении тех и других.
Существование и развитие ценопопуляций плотоядных живот-
ных третьего трофического уровня пищевых цепей зиждется па
использовании энергии с поедаемой ими биомассой в виде некото-
рой части особей своих жертв. Разумеется, не вся потребленная в
пищу хищниками биомасса ассимилируется, усваивается в процес-
се питания. Некоторая часть ее используется паразитами или в ви-
де экскрементов и других отбросов выделяется наружу. Ассимили-
рованная же часть съеденной ими биомассы фитофагов и за-
277
ключенной в ней энергии расходуется, как и на предыдущем
трофическом уровне, прежде всего на внутренние физиологические
процессы, связанные с поддержанием их жизни и отправлением
свойственных им жизненных функций (основной обмен). Однако
большая часть ассимилированной энергии затрачивается на такие
процессы жизнедеятельности хищных животных, как ходьба, бег,
связанные с поиском, выслеживанием добычи, охотой и пожирани-
ем своих жертв (дыхание). Лишь незначительная часть энергии
(около 10%) используется на синтез новой протоплазмы, явления
роста и другие процессы, связанные с формированием биомассы
тела животных и элементов для их размножения, а также с накоп-
лением жировых запасов и пр.
Показателем продуктивности животных-зоофагов на третьем
трофическом уровне консортивных пищевых цепей биогеоценозов
является эффективность образования ими биологической продук-
ции в виде собственной биомассы. Это не что иное, как КПД ис-
пользования энергии на образование этой продукции, который
представляет собой отношение энергии, заключенной в созданной
животными данного трофического уровня продукции, к энергии,
содержащейся в потребленной ими биомассе растительноядных
животных. По данным С. С. Шварца (1972), КПД у животных-фи-
тофагов, питающихся растительной массой, составляет, как было
показано ранее, 5—10% от содержащейся в ней энергии; у плото-
ядных же животных, использующих в качестве источника энергии
более калорийную и энергоемкую животную биомассу, КПД до-
ходит до 30% от доступной им энергии. Но отнесенный к энергии,
содержащейся в растительной биомассе, КПД у животных-зоофа-
гов, по данным П. Дювиньо (1968), составляет 1%, а по отноше-
нию к солнечной энергии — лишь 0,001 %.
Созданная животными-зоофагами (хищниками 1-го порядка)
биологическая продукция определенного химического состава и
энергоемкости (за исключением израсходованной в процессе жиз-
недеятельности) становится источником питания и энергии для
ценопопуляций хищников 2-го порядка, составляющих в совокуп-
ности четвертое и, как правило, последнее трофиче-
ское звено пастбищных пищевых цепей. Здесь происходят
такого же характера обменные процессы, что и на предыдущем
трофическом уровне.
Отмершие растения, животные и их части, а также отбросы и
выделения на разных трофических уровнях пастбищных пищевых
целей поступают в подстилку и поверхностный слой почвы, где ста-
новятся объектами питания и энергии для сапротрофов. Поэтому
в некоторых графических моделях (схемах), иллюстрирующих ход
обменных процессов в биокосной системе в виде пастбищной пище-
вой цепи, выделяется особый резервуар, служащий местом стока
278
отмершей биомассы и отбросов, поступающих со всех трофических
звеньев, и вместилищем сапротрофов, которые осуществляют здесь
минерализацию их, Однако, по представлению других биологов,
минерализация органики осуществляется не в одном резервуаре, а
в детритной пищевой цепи различными по видовому набору и соче-
танию комплексами сапротрофов в зависимости от консистенции и
химического состава поступающей с каждого трофического звена
органики, отбросов и выделений.
Детритные пищевые цепи, по словам Г. А. Новикова
(1979), функционируют в составе биогеоценоза бок о бок с пище-
выми цепями пастбищного типа. В составе детритной пищевой
цепи, по его мнению, можно выделить две группы (два звена) сап-
ротрофов, L'Животные-сапрофаги— беспозвоночные и отчасти по-
звоночные (насекомые, грызуны и пр.), которые, потребляя в пищу
отмершую растительную или животную биомассу, механически
разрушают ее, способствуя тем самым более интенсивному и пол-
ному разложению на последующих этапах деструкции. II. Микроб-
ные сапрофиты — бактерии, низшие грибы, актиномицеты и пр.,
которые в процессе своей жизнедеятельности завершают минера-
лизацию сложных органических веществ до простых исходных
соединений, а также осуществляют биосинтез некоторых веществ
в почве (гумуса, биологически активных веществ и т. п.).
Таковы важнейшие процессы и перемещения минеральных и
органических веществ, атмосферной и почвенной влаги и биоген-
ных газов в биогеоценозе, совершающиеся при непосредственном
участии его живых компонентов (рис, 29).
Из изложенного поэтапного (раздельно по трофическим звень-
ям) рассмотрения материально-энергетических процессов в биогео-
ценозе, а также из изложенного представления об интегральном
характере трофической структуры его вытекает ряд важных поло-
жений. Во-первых, в основе материально-энергетических процессов
в биогеоценозе лежат обменные процессы, протекающие в трофи-
ческих звеньях (уровнях) пищевых цепей на организменном уров-
не организации жизни. Начавшись в организмах автотрофных
ценопопуляций, обмен веществ и энергии продолжается в организ-
мах биотрофных ценопопуляций, завершаясь в ценопопуляциях
сапротрофов. Это, как видно из предыдущего, обусловлено тем, что
продуцируемая на каждом трофическом уровне биомасса и про-
дукты метаболизма становятся пищей и источником энергии для
потребителей последующего трофического уровня. Потоком ве-
ществ и энергии осуществляется связь групп организмов каждого
трофического уровня пищевых цепей в единую функциональную
линию, вплетающуюся далее через трофические концентры консор-
ций в обменные процессы, протекающие на популяционно-видовом
уровне и через трофические ступени биоценоза в единую функцио-
279
Рис. 29. Важнейшие процессы и потоки веществ, воды и газов в био-
геоценозе (по Н. В. Дылису, 1975). 1 —солнечная радиация: 1а —
отраженная от растительности; 16 — поглощенная при фотосинтезе;
1в — прошедшая к почве. 2. Атмосферные осадки: 2а — задержанные
растениями; 26 — проникшие к почве; 2в — просочившиеся в почву;
2г — физическое испарение влаги; 2д — расход воды на транспира-
цию, 3. Поглощение углекислоты при фотосинтезе: За — выделение
углекислоты из почвы при дыхании организмов н разложении орга-
ники. 4. Выделение кислорода при фотосинтезе: 4а — поглощение
кислорода почвенными организмами; 46 — то же —наземными жи-
вотными и растениями. 5. Отчуждение фитомассы фитофагами: 5а —
то же — почвенными организмами; 56 — то же — зоофагами 1-го и
2-го порядка. 6. Фиксация атмосферного азота микроорганизмами.
7. Растительный опад и формирование подстилки: 7а — отмирание
подземных органов растений. 8. Разложение подстилки почвенными
животными и микроорганизмами и образование гумуса. 9. Прижиз-
ненные выделения корней (ко л ины). 10. Поглощение корнями рас-
тений из почвы воды и минеральных веществ
налъную систему биогеоценоза на биоценотическом уровне
(рис. 30).
Рис. 30. Упрощенная схема потока веществ и энергии в пищевой це-
пи (ми П. Дювиньо и М. Тангу, 1968). С — свет; Пв —продукция
валовая; Пч — продукция чистая; А — ассимилированный корм; Н —
неиспользованные вещества; Э — экскременты и выделения; К—корм
растительноядным животным; Д — потери энергии на дыхание
При этом, как видно на прилагаемой графической модели, про-
исходит резкое, каскадное снижение, обесценивание аккумулиро-
ванной в веществе энергии при переносе ее с одного трофического
уровня пищевой цепи на другой вследствие потери ее в процессе
дыхания в виде тепла. В соответствии с этим происходит также
уменьшение продуцируемой на каждом трофическом уровне био-
массы и численности особей ценозообразующих ценопопуляций
(рис. 31).
Во-вторых, на каждой трофической ступени биоценоза и биогео-
ценоза в целом, как единой функциональной системы, происходит
суммирование результатов метаболизма в виде первичной и вто-
ричной биологической продукции входящих в ее состав ценокомп-
лексов автотрофных, биотрофных и сапротрофпых ценопопуляций,
подобно тому как на каждом трофическом уровне пищевой цепи
суммируются результаты метаболизма всех организмов соответ-
ствующих видовых ценопопуляций.
В-третьих, внутриценозный материально-энергетический обмен
биогеоценоза, как весьма сложной интегрированной бнокосной си-
стемы, также представляет собой сложный интегративный
процесс. Он включает в себя обменные процессы, совершаю-
щиеся на нижележащих уровнях организации жизни, вбирает в
себя результаты метаболизма .всех трофических звеньев соподчи-
ненных ему функциональных структурных единиц (пищевых цепей,
консорций). Это находится в полном соответствии с отмеченным
ранее высказыванием П. М. Рафеса (1972) о том, что материаль-
281
Ккал/и2 в ДЕНЬ: LA-1000 PN-10 PS^-I PS2-0,1
Рис. 31. Поток веществ и энергии в пищевой цепи (по Р. Дажо, 1975, с допол-
нениями). LT — падающие на растения лучи солнечной радиации; LA — погло-
щенные растениями лучи солнечной радиации; NV1*2*3 —неиспользованная сол-
нечная энергия; СН — рассеянные, превратившиеся в теплоту солнечные лучи;
РВ — первичная валовая продукция продуцентов; PN — первичная чистая про-
дукция; PS1»2 — вторичная чистая продукция первичных и вторичных консумен-
тов; J1*2 — энергия, поглощенная консументами с пищей; А1*2—энергия, ассимили-
рованная консументами; NA2— энергия, выбрасываемая консументами в виде
экскрементов и выделений; Rl> *•3 — потери энергии на процессы жизнедеятель-
ности (на дыхание)
-у й /
но-энергетический обмен биогеоценоза представляет собой не еди-
ничный поток веществ и энергии, а множество разнообразных по
веществу и энергии потоков входящих в его состав консорций и
пищевых цепей на всей занятой биогеоценозом площади.
Но в биогеоценозе, как относительно открытой биокосной систе-
ме, наряду и одновременно с внутриценозным обменом имеет место
и внешний обмен веществ и энергии с другими биогеоценозами и
с физической внешней средой. Это означает, что биогеоце-
ноз— это не изолированная бнокосная система, а материаль-
но-энергетическая ячейка биогеосферы, структур-
но и функционально соподчиненная ей, как часть целому.
7. Биогеоценоз и биогеосфера как саморегулирующиеся системы;
механизмы и факторы саморегуляции
Со структурно-функциональной организацией, функционирова-
нием и функциональной деятельностью биогеоценоза и его компо-
нентов непосредственно связано осуществление авторегуля-
ц и и его, обеспечивающей его устойчивость, гомеостаз.
282
Г Ф. Хильми (1966), исходя из своего представления о ярус-
ном, ступенчатом в функциональном смысле строении биосферы
(точнее, биогеосферы) по типу цепей питания, приходит к заклю-
чению, что биогеосфера состоит из двух подсистем. Первую из
них он назвал фи то сферой, включающей в свой состав а) рас-
тительный покров (или фитоценоз) и б) физическую (абиотическую)
среду. В состав второй подсистемы входят а) растительноядные
животные (фитофаги) и б) плотоядные животные (зоофаги, хищ-
ники 1-го порядка). Эту подсистему по аналогии с первой можно
назвать зоосферой. Названные компоненты той и другой под-
системы биогеосферы и, будем считать, также биогеоценоза связа-
ны между собой «замкнутыми контурами воздействия» (по выра-
жению автора), которые, по его мнению, являются источниками,
основой саморегулирования и самоорганизации каждой из этих
подсистем и системы в целом (рис. 32).
БИО СФЕРА
Ориг. рис. 32. Ступенчатое строение биосферы и ее са
морегулярование (по данным Г. Ф. Хильми, 1966)
283
В процессе взаимодействия компонентов каждой из названные
подсистем биогеоценоза и биогеосферы осуществляется их саморе-
гулирование и самоорганизация, вследствие чего фитосфера и зоо-i
сфера, как полагает автор, приобретают свойство устойчивости,
превращаясь в подсистемы с надежным и устойчивым саморегули-
рованием. Суть саморегулирования первой подсистемы состоит в
том, что между фитоценозом или растительным покровом в целоц
и их средой обитания устанавливается прямая и обратная связью
т. е., находясь во взаимозависимости, они постоянно воздействуют
друг на друга. Физическая среда воздействует на растительность:
всей совокупностью физических и химических факторов (свет, теп-
ло, влажность почвы и воздуха, кислород, углекислота, кислот-,
ность почвы, элементы минерального питания и т. п.), создавая;
тем самым определенные условия существования для ценозообра-i
зующих растений. Растительность в ответ на это воздействие фи-
зической среды, с одной стороны, приспосабливается к существо-
ванию в наличных условиях, что находит выражение в различного
рода анатомо-морфологических изменениях и физиологических
реакциях растений, а также в таких явлениях совокупного харак-
тера, как листовая мозаика, ярусное строение и мозаичное сложе-
ние растительного покрова и пр. С другой стороны, путем актив-
ного воздействия па воздушную, почвенную или водную среду
растительность обогащает ее продуктами метаболизма и отмерши-
ми органическими остатками, изменяя ее в соответствии со своими
требованиями, преобразуя в специфическую внутреннюю среду —
фитосреду.
Следствием саморегулирования и самоорганизации первой под-
системы, по словам Г. Ф. Хильми, является формирование устой-
чивых, стабильных растительных сообществ, находящихся в отно-
сительном соответствии со средой обитания. Популяционно-видо-
вой состав растений и структура таких сообществ приспособлены
к усвоению солнечной энергии и материальных ресурсов в количе-
стве, достаточном для обеспечения их нормального функциониро-
вания и продуцирования необходимого количества фитомассы. Рас-
тительные сообщества обретают способность восстанавливать свой
состав и структуру при нарушениях. Иначе говоря, растительный
покров, как пишет автор, «оказывается достаточно устойчивым, а
его биомасса и запас превратимой энергии сохраняют относитель-
но постоянное значение или колеблются в допустимых пределах,
позволяющих видам сохранить свое место в природе» (с. 218).
Саморегулирование и самоорганизация второй подсистемы био-
геоценоза или биогеосферы, компонентами которых являются
растительноядные животные и плотоядные, хищные животные, на-
ходят выражение, по словам Г. Ф. Хильми, в следующем взаимо-
действии их между собой. Хищник, пожирая свои жертвы, будет
284
ограничивать их размножение, и поэтому численность особей по-
пуляции жертвы будет снижаться. Это приведет к ухудшению
условий питания популяции хищника и, следовательно, к вымира-
нию и снижению численности его особей. Уменьшение численности
особей популяции хищника вызовет интенсивное размножение их
жертвы, численность особей которой возрастет. Вследствие этого
условия питания хищника улучшатся, что приведет к увеличению
численности его особей. Таким образом, несмотря на потенциаль-
ную возможность животных к беспредельному размножению, этого
не происходит, ибо, как пишет Г. Ф. Хильми, «любой вид живот-
ных, полностью исчерпавший свои источники энергии, обречен па
гибель и не может сохраниться в природе» (с. 248). В силу этого
возрастание численности популяции хищника в действительности
прекращается задолго до полного исчерпания доступных ему
источников питания и энергии.
Вскрывая механизмы авторегуляции в подсистеме «зоосфера»,
Б. Д. Абатуров (1979) обращает внимание на то обстоятельство»
что у животных-фитофагов, являющихся жертвами хищных живот-
ных, в процессе совместной эволюции выработался ряд приспособ-
лений к максимальному изъятию их особей, на которое способны
потребляющие их хищники, например увеличение плодовитости»
снижение смертности и т. п. Это дает возможность фитофагам, пи-
шет он, «выносить без ущерба для своей продуктивности значи-
тельный пресс хищников» (с. 115). Более того, изъятие некоторого
количества особей жертвы (в пределах 5—10% от их общего чис-
ла) при высокой их плодовитости становится, по утверждению
Б. Д. Абатурова, необходимым условием их нормального суще-
ствования, так как позволяет избавиться от излишних особей и
тем самым предотвратить избыточное перенаселение их. С другой
стороны, хищники, по приведенным автором словам Н. В. Батени-
ной (1977), «не могут уничтожить кормовых животных полностью,
так как при приближении последних к минимуму численности за-
трата энергии на отлов не оправдывается. Хищники прекращают
его и переходят на замещающие корма, при общей депрессии
откочевывают или гибнут» (с. 109).
Таким образом, саморегуляция второй подсистемы биогеоцено-
за и биогеосферы в целом осуществляется в ходе взаимного воз-
действия друг на друга популяций хищника и их жертвы, т. е. жи-
вотных-фитофагов. Однако решающее значение в этом, как видно
из сказанного, имеет воздействие популяции жертвы на числен-
ность особей популяции хищных животных; это так называемая
обратная связь.
Следствием саморегулирования и самоорганизации подсистемы
«зоосфера», по словам Г Ф. Хильми, является поддержание чис-
ленности особей взаимодействующих популяций хищников и их
285
жертв — растительноядных животных — на определенном уровне с
колебанием ее в определенных пределах. Иначе говоря, в подси-
стеме благодаря этому постоянно поддерживается определенное
соотношение между численностью особей популяций хищника и
его жертвы, обеспечивая тем самым относительную устойчивость
подсистемы.
Отношения между растительноядными животными и раститель-
ным покровом (в частности, фитоценозом) отличаются от взаимо-
связи трофических ступеней, образующих названные подсистемы
биогеоценоза и биогеосферы. Животные-фитофаги, как указывает
Г. Ф. Хильми, либо вовсе не испытывают обратного воздействия со
стороны растительности в случае питания широко распространен-
ными растениями, не требующими затраты энергии на их поиск
(насекомые, грызуны и пр.), либо это воздействие слабо выражено
при специализированном питании (лось, белка и др.). Поэтому
растительный покров не в состоянии контролировать, сдерживать
размножение животных-фитофагов и предотвращать чрезмерно
высокий уровень численности их особей. Следовательно, между
компонентами этого звена биогеоценоза и бногеосферы в целом
«замкнутые контуры воздействия» отсутствуют, а потому оно явля-
ется слабым звеном, неспособным к саморегулированию. Между
тем роль этого звена весьма велика, так как посредством его под-
системы фитосфера и зоосфера соединяются в одну целостную био-
косную систему — биогеоценоз или бногеосферу.
Возникает вопрос: как же все-таки осуществляется регуляция
этих биокосных систем разного ранга? Рассуждения Г. Ф. Хильми
по этому вопросу примерно таковы: численность особей раститель-
ноядных животных не может достигать такого уровня, при котором
произошло бы полное, необратимое уничтожение растительности
на месте их обитания, так как это привело бы к полной гибели
данных видов фитофагов. Но этого не может произойти в силу
того, что отношения между ними и пожирающими их хищниками
сбалансированы, саморегулируются «замкнутыми контурами воз-
действия». Следовательно, отношения животных-фитофагов с рас-
тительным покровом, взаимосвязь их между собой и сбалансиро-
ванность их взаимоотношений регулируются косвенным путем
хищниками, предотвращающими тем самым полное уничтожение
растительности на данном участке.
Несколько по-иному к решению этого вопроса подходит
Б. Д. Абатуров (1979). По его мнению, в ходе совместной эволю-
ции животных-фитофагов и растительного покрова на том или ином
участке территории у последнего выработался целый ряд приспо-
соблений, которые позволяют ему смягчать неблагоприятное воз-
действие на него со стороны растительноядных животных. Воздей-
ствие животных-фитофагов на растительность выражается главным
286
образом в отчуждении фитомассы путем стравливания ее в про-
цессе питания. Помимо этого они оказывают и косвенное воздей-
ствие на растительность посредством уплотнения или рыхления
поверхности почвы при пастьбе или благодаря роющей деятельно-
сти некоторых видов грызунов, а также посредством обогащения
почвы элементами минерального питания за счет продуктов мета-
болизма, различных отбросов и выделений копытных, грызунов и
других растительноядных животных, повышающих тем самым ее
плодородие. Реакция популяций ценозообразующих растений и
растительного покрова на такого рода воздействия животных-фито-
фагов выражается, во-первых, в развитии защитных приспособле-
ний у многих видов (шипов, колючек, ядовитости, мимикрии под
ядовитые растения и пр.); во-вторых, в появлении ползучих, сте-
лющихся, приземистых и дерновинных форм, способных выдержи-
вать вытаптывание их пасущимися животными; в-третьих, в спо-
собности многих видов трав быстро, полно и многократно в течение
вегетационного периода отрастать после стравливания и, в-четвер-
тых, в способности растительного покрова выносить отчуждение
некоторой части растений фитофагами, не только не снижая свою
продуктивность, но даже повышая ее вследствие стимулирующего
действия стравливания травостоя на его отрастание и установле-
ния надлежащей плотности его при условии оптимальной нагрузки
участка животным населением.
Из изложенного следует, что, несмотря на значительный пресс
животных-фитофагов на растительный покров, у последнего выра-
боталась способность противостоять этому давлению фитофагов,
как бы вынуждая их к ограничению потребления корма путем
саморегулирования численности особей или «автоматически при
снижении количества потребляемого растительного корма до кри-
тического для животных уровня... обычно до 60—70% от урожая»
(с. 113—114).
Таким образом, соединение двух подсистем — фитосферы и зоо-
сферы— в единую целостную биокосную систему (биогеоценоз или
биогеосферу) осуществляется, по Г Ф. Хильми, не внутренними
регулирующими связями, а косвенным регулированием. В силу
этого, как отмечает автор, связующее звено этих подсистем не
обладает высокой надежностью, а потому в этом звене биокосных
систем чаще всего происходят разрывы, ведущие к нарушению их
целостности; свидетельством тому служат многочисленные случаи
нашествия мышевидных грызунов, налета саранчи, массовое раз-
множение вредных насекомых и т. п.
По представлениям же Б. Д. Абатурова (1979), как видно из
изложенного, взаимосвязь и сбалансированность отношений между
животными-фитофагами и растительным покровом, служащих свя-
зующим звеном двух подсистем в единую систему, регулируются
287
не только косвенным путем посредством саморегуляции взаимосвя-
зи хищников с фитофагами. Не менее важное значение в этом
имеет и прямая взаимосвязь их самих между собой, которая опре-
деляется, по его словам, «соотношением между силой трофическо-
го воздействия фитофагов и степенью устойчивости растений к
этому воздействию» (с. 113). Конкретным же выражением такого
сбалансированного соотношения, по-видимому, является историче-
ски сложившееся соответствие допустимого изъятия наземной фи-
томассы (без ущерба для ее продуктивности) тому наибольшему
количеству ее, которое может потребляться растительноядными
животными. «Именно этот механизм, — пишет автор, — регулирует
характер отношений фитофагов и растительности и определяет
устойчивость данной системы» (с. 114).
Нельзя в связи с этим не обратить внимания на высказывания
В. В. Алексеева (1973), который подчеркивает важную роль тро-
фического уровня растительноядных животных, в особенности узко-
специализированных видов, в определении устойчивости биогеоце-
нозов к смене внешних условий. Биогеоценозы без консументов, по
его словам, «вырождаются в конечном итоге в одновидовые систе-
мы и оказываются беззащитными к резким изменениям внешних
условий, происходящих вслед за периодами продолжительного их
постоянства» (с. 456—457).
Некоторыми отечественными и зарубежными биологами при-
дается большое значение в регулировании характера взаимоотно-
шений между компонентами трофических ступеней биокосных си-
стем и в определении их устойчивости в пространстве и во времени
влиянию внешних условий среды обитания. Но взаимосвязь живых
существ и их биологических и ценотических комплексов с внешней
средой, на наш взгляд, — проблема иного порядка, и суть ее, как
было изложено в одной из предыдущих глав, состоит в другом.
И. И. Шмальгаузен (1968), касаясь этого вопроса, подчеркивает;
«...все регуляторные процессы осуществляются всегда за счет сил,
действующих внутри данной системы» (с. 176). Иначе говоря, ре-
гуляция биологических и биокосных систем — это есть саморе-
гуляция. В процессах саморегуляции этих систем он решающее
значение придает так называемым отрицательным обратным свя-
зям. «Механизмы поддержания равновесия в биогеоценозе имеют
характер замкнутого цикла с отрицательной обратной связью»
(с. 175).
Итак, в основе саморегуляции и самоорганизации биогеоценоза
и биогеосферы, как следует из всего изложенного в последнем раз-
деле настоящей главы, лежат внутренние процессы, внутрисистем-
ные связи и отношения (трофические, информационные и другие),
в особенности обратные связи, представляющие собой ответные
действия одного из взаимодействующих компонентов на сильное
288
воздействие на него со стороны другого компонента. Решающее
значение из них, как утверждает В. Д. Александрова (19646),
имеют отрицательные обратные связи, которые, по ее
словам, регулируют состав, численность и продуктивность живых
компонентов биогеоценоза, поддерживая тем самым видовые по-
пуляции взаимодействующих на каждой трофической ступени на
определенном уровне численности, создавая предел беспредельно-
му возрастанию их численности и пр. Иначе говоря, благодаря
этому поддерживается динамическое равновесие, обеспечивается
гомеостаз, устойчивость биогеоценоза, а следовательно, и био-
геосферы, основной материально-энергетической единицей которой
биогеоценоз является.
Глава IX
Классификация биоценозов и биогеоценозов
и территориальное подразделение биогеосферы
Разработка проблемы классификации биогеоценозов в основном
принадлежит перу Н. В. Дылиса (1964, 1975, 1978) с участием
А. И. Уткина (1968) и др. Имеются в литературе и данные других
авторов по некоторым принципиальным вопросам этой проблемы;
к работам этих авторов мы будем обращаться в ходе рассмотрения
или обсуждения данной проблемы,
1. Топологическая система классификации
биоценозов по А. Клюгу
До появления в науке понятий «биогеоценоз» и «экосистема»
природные комплексы живых существ, как известно, с легкой ру-
ки К. Мёбиуса (1877) стали называться биоценозами. В со-
ответствии с этим одной из первых классификаций природных
единств была система классификации биоценозов, составленная
американским экологом Клюгом (Kliigh, 1923). Как и ранние систе-
мы классификации растительных сообществ, классификация биоце-
нозов А. Клюга была топологической. Это и понятно, так
как составление такой классификации гораздо проще, чем эколого-
ценологической классификации, поскольку не требует познания са-
мих растений, животных, их сочетаний в ценозы и др. А. Клюг уста-
новил такую иерархическую систему таксономических единиц
для классификации морских биоценозов: association (ассоциация),
systasis (группа ценозов), coenosis (ценозы).
В развернутом виде классификация морских биоценозов А. Клю-
га выглядела так:
J. Пелагическая ассоциация
1. Верхнепелагическая группа ценозов.
2. Среднепелагическая группа ценозов.
3. Нижнепелагнческая группа ценозов.
II. Бентическая ассоциация
1. Верхнебентическая группа ценозов:
а) ценоз скалистого дна; в) ценоз гравийного дна;
б) ценоз песчаного дна; г) ценоз илистого дна.
2. Среднебентическая группа ценозов:
ценозы такие же, что и в 1-й группе.
3. Абиссальная группа ценозов.
290
2. Подход к классификация биогеоценозов
и других природных единств Сочавы, Долуханова и др.
А. П. Шенников (1964), опираясь на мнение канадского бота-
ника В. Крайины (Krajina, I960),считавшего топологические клас-
сификации биологических и природных объектов первой ступенью
в разработке классификационной проблемы, пришел к заключению,
что второй ступенью является фитоценологическая классификация;
биогеоценологическая же классификация природных комплексов
явится третьей ступенью разработки этой проблемы. Специфич-
ность и сложность ее А. П. Шенников видел в том, что она должна
строиться на более совершенном изучении взаимосвязей между
типами ценозов и средой, на более глубоком познании различных
группировок живых существ как элементов биогеоценозов, являю-
щихся участками биосферы (фитогеосферы), как носителей разли-
чий в энергетическом балансе между «живой» и «мертвой» средой,
«обладающих различиями в процессе аккумуляции и трансформа-
ции энергии, в обмене энергии и веществ в биосфере» (с. 410).
Проблема классификации растительных сообществ (фитоцено-
зов) и отчасти некоторых природных систем наиболее тщательно-
му обсуждению в нашей стране подверглась в 1959 году на посвя-
щенном этой проблеме совещании в Институте биологии Ураль-
ского филиала АН СССР в Свердловске и в 1971 году на совеща-
нии по классификации растительности, созванном научным советом
по проблеме «Биологические основы рационального использова-
ния, преобразования и охраны растительного покрова АН СССР»
и Всесоюзным ботаническим обществом в Ленинграде.
Некоторые из опубликованных в периодических изданиях ра-
бот, затрагивающих вопросы классификации природных единств,
заслуживают пристального внимания. В частности, Н. В. Тимофе-
ев-Ресовский (1961) обращает внимание на то, что классификаци-
онная система в биогеоценологии должна основываться «на типах
геохимической работы биогеоценозов и типах сообществ» (с. 26),
а также на то, что такого рода классификационная система явля-
ется типологической, а не пространственной, т. е. не территориаль-
ным подразделением бногеоценотического покрова.
Своеобразный подход к классификации растительных сообществ
(фитоценозов) и ряда природных единств (комплексов) предло-
жил на первом из этих совещаний В. Б. Сочава (1961). Во-первых,
подчеркнуто рассматривая растительное сообщество как автотроф-
ную базу, с которой неразрывно связаны в своем существовании
многие беспозвоночные и позвоночные животные, В. Б. Сочава счи-
тает, что сформировавшиеся на этой основе биоценозы становятся
непременными компонентами таких природных единств, как физи-
ко-географическая фация и экосистема или биогеоценоз. Во-вто-
291
рых, между последними тремя понятиями он не склонен видеть
существенных различий. Экосистему Тенсли и биогеоценоз Сука-
чева В. Б. Сочава рассматривал как «физиографические комплек-
сы», одновременно с этим считая, что «территориально биогеоценоз
и элементарная экосистема — это та же физико-географическая
фация, но трактуемая в других аспектах» (с. 7). Правда, физико-
географическую фацию он считает более универсальным образова-
нием, что, по его словам, обязывает «к анализу взаимоотношений
не только между биотическими и абиотическими компонентами
комплекса, но и к изучению связей в пределах абиотической части
фации)» (с. 7), т. е. уделять внимание не только биоэнергетиче-
ским, но и локальным геофизическим процессам.
Исходя из этих теоретических посылок, В. Б. Сочава (1961)
нашел возможным построить сравнительную систему таксономи-
ческих единиц для параллельной классификации фитоценозов, био-
ценозов, физико-географических фаций и биогеоценозов (экоси-
стем), которая и приводится нами (табл. 2).
Таблица 2
Параллельная классификация природных единств
Фитоценоз Биоценоз Физико-географи- ческая фация Биогеоценоз (экосистема)
Растительная ассоциация Тип биоценоза Тип физико-геогра- фических фаций Тип биогеоценоза
Группа Группа типов Группа типов фи- Группа типов
ассоциаций биоценоза зико-географических фаций биогеоценоза
Формация Биом Геом —
Группа формаций — — —
С иных позиций, чем В. Б. Сочава, к вопросам классификации
биогеоценозов подошел А. Г. Долуханов (1961). При классифика-
ции биогеоценозов, по его словам, «одинаково важно отразить
степень их родства (сходства.—Л, Н.) как по растительности, так
и по условиям произрастания» (с. 40). Однако осуществить это
одновременно в одной шкале иерархически соподчиненных таксо-
номических единиц невозможно. Отсюда он пришел к выводу,
что при классификации биогеоценозов необходимо остановиться на
такой системе, в которой «объединение биогеоценозов в типологи-
ческие таксоны различного ранга идет одновременно по трем на-
правлениям: ...путем объединения биогеоценозов по степени родства
растительности... по условиям среды обитания... путем классифи-
кации сукцессионных рядов развития» (с. 40).
292
Таким образом, по предложению В. Б. Сочавы (1959, 1961) и,
как отмечает Н. В. Дылис (1964), ряда зарубежных ученых —
Хильса (Hills, 1952, 1961), Pay (Rowe, 1960), классификация био-
геоценозов должна строиться путем объединения важнейших ха-
рактеристик отдельных компонентов биогеоценоза, т. е. путем сум-
мирования показателей растительного покрова, животного населе-
ния и т. п. Особое внимание, по их мнению, должно быть уделено
растительным сообществам, поскольку различным фитоценозам
соответствуют и определенные биокомплексы животных, микро-
организмов, а также определенные типы почвенного покрова. Все
это и дает им возможность классифицировать как фитоценозы, так
и биогеоценозы и даже физико-географические фации на одних и
тех же основаниях. По мнению же А. Г. Долуханова (1959, 1961)
и др., классификация биогеоценозов, как мы видели, должна со-
стоять из ряда частных классификаций, т. е. должна быть расчле-
нена на самостоятельные части, раздельно отражающие черты
растительности, экотопов и т. п. Иначе говоря, классификация био-
геоценозов должна быть дробной, состоящей из нескольких клас-
сификационных систем, а не целостной, каковой ей полагается быть.
3. Принципы классификации биогеоценозов
по Сукачеву и Дылису
Н. В. Дылис (1964) отрицательно отнесся к изложенным пред-
ложениям В. Б. Сочавы и А. Г Долуханова по вопросам класси-
фикации биогеоценозов. Возражая В. Б. Сочаве и др., Н. В. Ды-
лис отмечает, что биогеоценозы и растительные сообщества (фито-
ценозы) нельзя классифицировать на одних и тех же основаниях;
принципы классификации тех и других должны быть разными, ибо
это разные объекты природы. Классификацию растительных со-
обществ, по его мнению, можно использовать лишь для построения
низших ступеней классификации биогеоценозов и других природ-
ных единств. И действительно, попытка В. Б. Сочавы построить
на этой основе классификацию биогеоценозов и физико-географи-
ческих фаций не вышла за рамки первых трех ступеней системы.
Возражая А. Г. Долуханову, Н. В. Дылис указывает, что клас-
сификация биогеоценозов не может быть раздробленной, состоя-
щей из отдельных обособленных частей, ибо «биогеоценоз — не
сумма биоценоза и среды, а целостное и качественно обособленное
явление природы, действующее и развивающееся по своим соб-
ственным закономерностям, основу которых составляет метаболизм
его компонентов, совершающийся под действием солнечной радиа-
ции» (с. 488). Поэтому классификация биогеоценозов, по Дылису,
может строиться только как классификация природных единств, а
не механических сумм отдельных их компонентов; причем строить-
ся она должна на основе признаков, присущих этим единствам, а
293
именно на основе обмена веществ и энергии, протекающего в био-
геоценозах. Помимо этого, классификация биогеоценозов должна
быть не только целостной, но и по принципу выдержанной во всех
звеньях, на всех ступенях иерархической лестницы.
Таким образом, основные положения классификации биогеоце-
нозов Н. В. Дылиса находятся в полном соответствии с принципи-
альными установками В. Н. Сукачева (1964) по вопросам класси-
фикации биогеоценозов, образующих биогеоценотический покров
Земли. Об этом он писал: «...биогеоценстическая классификация
типов леса должна строиться на другом принципе, а именно на
степени сходства процесса превращения вещества н энергии в лес-
ных биогеоценозах и обмена ими внутри биогеоценозов (между ком-
понентами его) и между биогеоценозами и другими явлениями при-
роды, отражением чего является сходство всех компонентов био-
геоценозов, их сложения и структуры, так как в этом лежит сущ-
ность биогеоценозов и от этого зависят в первую очередь и пути
практического использования их» (с. 40). Согласуются принципы
классификации биогеоценозов Н. В. Дылиса и с приведенными вы-
ше высказываниями Н. В. Тимофеева-Ресовского. (1961) и А. П. Щен-
никова (1964) на эту же тему. Из зарубежных биологов Н. В. Ды-
лис ссылается на Овингтона (Ovington, 1960), придерживающегося
сходных взглядов по данной проблеме, и некоторых других авторов.
Четверть века тому назад Н. В. Дылис (1964) высказывал со-
жаление по поводу того, что возможности для построения биогео-
ценотической классификации на основе типологии биогеоценотиче-
ского обмена еще слишком ограничены. В то время, как отмечает
он, в специальной литературе имелись данные, характеризующие
различные типы биологического круговорота элементов, типы фи-
зико-географической среды, отдельные типы территории, геохими-
ческие типы ландшафтов и пр. Появились также отечественные и
зарубежные работы по изучению материально-энергетического об-
мена между отдельными компонентами биогеосферы, в частности
между растительностью и почвой и др. Имеющиеся данные, как
он пишет, позволяют установить на этой основе различия таких
типов биогеосферы, как тундра, лес, степь, пустыня. Появляются
отдельные работы, позволяющие обнаруживать различия в обмене
веществ между более мелкими подразделениями биогеосферы, на-
пример между зеленомошными и сложными ельниками и т. п. Од-
нако в целом имевшихся в то время для фундаментального, по его
словам, решения проблемы классификации биогеоценозов на осно-
ве «непосредственных индексов биогеоценотического метаболизма»
еще явно недостаточно. Недостаточно такого рода данных для это-
го, по-видимому, и в настоящее время.
В связи с этим, по мнению Н. В. Дылиса, следует как можно
полнее использовать при построении системы классификации био-
294
геоценозов косвенные признаки материально-энергетического мета-
болизма, которые, будучи ясно и однозначно связанными с глав-
ным биогеоценотическим процессом обмена веществ, в то же время
внешне хорошо выражены, а потому легко учитываются. Таких
признаков много, но наиболее показательными и информативными
из них, по его мнению, являются:
а) радиальная мощность биогеоценотического слоя ценоза (мохо-
вого, травяного, древесно-кустарникового);
б) продолжительность и ритмика материально-энергетического об-
мена (вегетации растительности, активной деятельности живот-
ного населения, теплообмена и влагообмена);
в) годичная продуктивность (прирост) органического вещества, на-
копление его и общий запас биомассы;
г) скорость и специфика разложения отмершего органического ма-
териала растительного и животного происхождения;
д) форма и масштабы отчуждения части органического вещества
из текущего обращения и накопление его в древесине, торфе,
подстилке, гумусе и т. п,;
е) «дыхание почвы», соотношение и режимы нисходящих и восхо-
дящих токов в почве;
ж) направление, скорость и химизм выноса за пределы биогеоце-
ноза неорганических материалов и продуктов жизнедеятельно-
сти живых компонентов его с водой, ветром и пр.;
з) ритмика и масштабы притока в биогеоценоз извне различных
веществ (пыли, наилков, минеральных солей и т. п.).
Перечисленные косвенные признаки касаются разных по объему
сторон метаболизма — то крупных, то более мелких деталей этого
процесса. Это обстоятельство, по словам Н. В. Дылиса, дает воз-
можность построения системы классификации биогеоценозов на
одном принципе во всех звеньях.
Изложенные принципы биогеоценотической классификации ис-
пользованы Н. В. Дылисом при классификации лесных биогеоце-
нозов, которые являются предметом длительного всестороннего
стационарного изучения им с участием большого коллектива со-
трудников на биогеоценологической станции в Подмосковье. Раз-
работанная им система классификации лесных биогеоценозов была
изложена в 1964 году и в несколько измененном виде в 1978 г., но
в наиболее детальном и развернутом виде она представлена в со-
авторстве с А. И. Уткиным в 1968 году.
Классификация биогеоценозов, по словам Н. В. Дылиса, может
производиться как путем подразделения биосферы на более мел-
кие таксономические единицы, так и путем объединения биогеоце-
нозов в более крупные таксоны. Автор нашел целесообразным ис-
пользовать первый путь классификации. Биосферу он подразделил
по особенностям структуры и характеру обмена веществ и энергии
295
на три сектора: а) сухопутный (биогеосфера); б) водный (биогид-
росфера) и в) прибрежно-водный (переходный от сухопутного к
водному). Кстати сказать, В. Лархер (1978), назвавший биосферу
экосферой, также подразделил ее на сухопутную часть (био-
геосферу) и водную — моря и океаны (биогидросферу). Под эко-
сферой он понимает физическую среду, населенную организмами
•с момента появления жизни на Земле.
Из трех выделенных секторов биосферы Н. В. Дылис сосредо-
точил свое внимание на сухопутном секторе — биогеосфере,
которая в отличие от физико-географической оболочки, как он от-
мечает, не имеет глобального, повсеместного распространения, так
как биогеоценозы, ее составляющие, «не могут быть в годичном
цикле постоянно безжизненными» (с. 489). Поэтому в состав био-
геосферы он не считает возможным включать вечные льды и снега,
жаркие песчаные пустыни, сильносоленые водоемы, молодые лаво-
вые поля, сели, осыпи, прибрежные аллювии и т. п. Биогеосферу
он подразделяет по типу биогеоценотического метаболизма на
типы биогеосферы: а) лесн он; б) тундровый; в) степ-
ной; г) пустынный и др.
4. Система классификации лесного типа биогеосферы
Приведем предложенную Н. В. Дылисом систему классифика-
ции лесного типа биогеосферы (1964).
Лесной тип биогеосферы, по словам автора, характеризуется
специфическим (лесным) типом биогеоценотического метаболизма,
большой радиальной мощностью биогеоценотического слоя (от не»
скольких метров до десятков и даже до полутора сотен метров),
высокой горизонтальной плотностью слоя активного биогеоценоти-
ческого обмена. Далее им приводится перечень признаков, харак-
теризующих лесной тип биогеоценотического метаболизма, в част-
ности: а) максимальный контакт растительной части биогеоценоза
с атмосферой и почвой; б) более или менее сбалансированный теп-
ло- и влагообмен; в) максимальная аккумуляция органического
вещества в стволах, ветвях, коре, корнях, листве; г) преобладание
(в 3—5 раз) надземных запасов органического вещества над под-
земными запасами его; д) большой процент в составе первичной
продукции лигнина; е) господство в составе микробного компонен-
та грибов — разрушителей лигнина и целлюлозы и аммонифици-
рующих бактерий; ж) преобладание в составе животного населе-
ния видов, трофически и топически тесно связанных с древесно-кус-
тарниковой растительностью; з) наличие более или менее мощной
лесной подстилки; и) повышенная подвижность почвенных раство-
ров, обусловливающая вынос части легкорастворимых минераль-
ных соединений за пределы биогеоценоза и пр.
296
Лесной тип биогеосферы подразделяется по продолжительности
и ритмике активного биогеоценотического обмена в годичном цик-
ле на два подтипа, или порядка, лесных биогеоценозов.
1. Леса с равномерно-непрерывным в годичном цикле метаболиз-
мом вещества н энергии между всеми компонентами благодаря
более или менее непрерывному и равномерному поступлению
тепла и влаги. Это леса влажных тропиков.
2. Леса с отчетливо выраженными перерывами в обменных про-
цессах благодаря паузам в поступлении тепла и влаги. Это леса
умеренных широт и сухих тропиков.
Среди лесов первого подтипа (порядка) автором различаются:
а) влажно-тропические и экваториальные вечнозеленые леса, или
' гилеи, с самым интенсивным тепло- и влагообменом;
б) мангровые леса на ежесуточно затопляемых морской водой при-
брежных участках;
в) заболоченные тропические леса (или лесные тропические боло-
та) на кислых торфяных грунтах.
Леса второго подтипа (порядка) в зависимости от причины,
вызывающей спад или прекращение функционирования биогеоце-
нозов, подразделяются на две группы, или союза, лесных био-
геоценозов:
а) гидропериодические леса, в которых спад биогеоценотического
обмена обусловлен периодическим прекращением поступления
влаги в летние месяцы, — это саванновые и отчасти муссонные
тропические и субтропические леса;
б) термопериодические леса, в которых глубокая пауза в годич-
ном цикле биогеоценотического обмена вызвана сменой с на-
ступлением зимы теплого периода холодным,— это леса умерен-
ного и холодно-умеренного пояса ( в том числе все леса Совет-
ского Союза).
Термопериодические леса по продолжительности теплопаузы,
количеству тепла и влаги, биологической продуктивности и скоро-
сти разложения органики подразделяются на три класса.
1. Класс северных лесотундровых и северотаежных биогеоценозов
с наиболее коротким периодом метаболизма и низкой продук-
тивностью.
2. Класс таежных биогеоценозов с более длинным периодом актив-
ного метаболизма, большой его интенсивностью и значительно
более высокой продуктивностью и большими запасами органи-
ческого вещества в древесине.
3. Класс широколиственных биогеоценозов с наиболее длинным
периодом активного метаболизма, высшей в данной группе клас-
сов продуктивностью и запасом органического вещества.
Класс таежных биогеоценозов на основе различий в сезонной
ритмике функционирования биогеоценозов и в обмене веществ меж-
297
ду растительностью и почвой подразделяется на две группы
формаций.
1. Группа формаций вечнозеленой темнохвойной тайги с наличием
фазы «колодного фотосинтеза» в ранневесенний и позднеосеи-
ний периоды года с равномерным в течение года сложением
растительной массы древостоя и круглогодичным глубоким пре-
образованием почвы и фитоклимата биогеоценозов.
2* Группа формаций летне-зеленой тайги с отсутствием фазы «хо-
лодного фотосинтеза» с резко выраженной сменой в сложении
растительной массы древесного яруса и периодичностью про-
цессов трансформации почвы и фитоклимата биогеоценозов.
Группа формаций вечнозеленой темнохвойной тайги в зависи-
мости от светового, теплового, водно-воздушного и пищевого режи-
мов, обусловленных положением в рельефе, геологическим сложе-
нием и характером почвенного покрова, подразделяется на био-
г.еоценотические формации:
3) формация плакорной вечнозеленой тайги на дренированных во-
доразделах с суглинистыми почвами, с нормальным для зоны
ходом тепла и влаги, аэрации почвы, миграции почвенных ра-
створов, первичной продуктивности и скорости разложения от-
мершей органики;
б) формация вечнозеленой тайги на южных крутых склонах, более
освещенных и теплых, по менее увлажненных, с менее выщело-
ченными почвами и более богатой высокопродуктивной расти-
тельностью, с более быстрым разложением органического опа-
. да и менее мощной подстилкой;
в) формация вечнозеленой тайги на северных крутых склонах, ме-
нее освещенных и обогретых, с более коротким периодом био-
геоценотического метаболизма, с менее быстрым разложением
органического опада и довольно мощной подстилкой;
г) формация заболоченной вечнозеленой тайги в понижениях — на
плоских междуречьях и речных террасах, на анаэробных холод-
ных почвах с замедленной циркуляцией почвенной влаги, низ-
кой продуктивностью и неполным разложением органики, с на-
коплением на поверхности торфа, а в почве — фульвокислот, за-
кисного железа и водорода;
д) формация пойменной вечнозеленой тайги на плодородных поч-
вах, обогащенных аллювиальным и делювиальным притоком
минеральных и органических веществ и влаги, с богатой по со-
ставу и сложной по строению мощно развитой древесно-кустар-
никовой и травянистой растительностью.
Помимо названных формаций, местами в той или иной степени
распространены своеобразные формации вечнозеленой тайги» раз-
вивающиеся на карбонатных породах, на сыпучих кварцевых пес-
ках, на различных породах вулканического происхождения и т. п.
298
Биогеоценотические формации вечнозеленых таежных лесов на
основе систематической однородности эдификаторов, принадлежа-
щих к одному и тому же роду или даже виду, подразделяются на
группы типов биогеоценозов — еловых, пихтовых, кедро-
вых и пр., которые по-разному используют и по-разному трансфор-
мируют биоценосреду.
Группы типов биогеоценозов таежных лесов по целому комплек-
су разнообразных признаков подразделяются на типы биогео-
ценозов, являющихся основной, низшей таксономической еди-
ницей биогеосферы. В типы биогеоценозов, по Н. В. Дылису (1978),
объединяются «участки земной поверхности, однородные по составу
и структуре растительности, животному и микробному населению,
по комплексу условий внутренней среды, по материально-энергети-
ческому обмену между компонентами биогеоценоза и по направле-
нию динамики и восстановительным процессам при разного рода
разрушениях» (с. 137—138).
Таким образом, разработанная Н. В. Дылисом с сотрудниками
система классификации биогеоценозов лесного типа биогеосферы
является теоретически обоснованной, цельной, отвечающей основ-
ным принципам во всех звеньях. Она выгодно отличается от других
изложенных классификационных систем подобного рода. В основу
ее положены наиболее существенные свойства и признаки биогео-
ценозов, как прямые (обмен веществ и энергии), так и косвенные
(ритмика функционирования и развития, биологическая продуктив-
ность, тепловлагообмен и пр.). В силу этого положенные в ее осно-
ву принципы классификации лесных биогеоценозов с успехом мо-
гут быть использованы для разработки принципов и систем клас-
сификации других типов биогеосферы (степного, лугового, пустын-
ного, тундрового, болотного и пр.).
Вместе с тем необходимо как можно больше внимания уделить
накоплению прямых данных не только по составу, структуре и про-
дуктивности, но и по функциональной деятельности и обмену ве-
ществ и энергии в биогеоценозах разных типов биогеосферы. С этой
целью необходимо шире развернуть глубокие и разносторонние
биогеоценологические исследования в этом направлении на био-
геоценологических станциях и стационарах по всем физико-геогра-
фическим зонам, провинциям и высотным поясам на территории
нашей страны.
5. Межбиогеоценозные связи как основа единства,
целостности биогеосферы,
их типы и механизмы
Выше обращалось внимание на то, что, наряду с типологией,
классификацией биогеоценозов на основе сходства их по
290
наиболее существенным природным свойствам или тем или иным
искусственно выделенным признакам, биогеоценотический покров
Земли может быть подразделен также на территориальные едини-
цы разного ранга. Подразделение такого типа в геоботанике было
названо районированием растительного покрова. В основе
объединения биогеоценозов в территориальные единицы более вы-
сокого ранга или расчленения биогеосферы на более мелкие еди-
ницы лежат межбиогеоценозные связи и отношения,
в первую очередь между соседними биогеоценозами, а также то-
пографические и географические закономерно-
сти, связанные с геолого-геоморфологическим строением местно-
сти и изменением градиентов тепла от экватора к полюсам и влаги
от морских и океанических побережий в глубь континентов, а в
горных местностях, помимо того, в зависимости от высоты над
уровнем моря.
Исходя из представлений В. Н. Сукачева об обмене веществ и
энергии не только внутри биогеоценоза между его компонентами,
но и между биогеоценозами, Н. В. Дылис (1973—1978) довольно
основательно разработал проблему межбиогеоценозных связей и
отношений в биогеосфере. Изучению биогеоценбзных связей он
придает большое значение, так как считает, что это дает возмож-
ность глубже познать внутриценозный метаболизм, а главное, эти
связи, по его словам, «играют роль важнейшего механизма, обес-
печивающего как глобальную целостность биогеоценотической обо-
лочки Земли, так и связанность ее отдельных крупных частей»
.(с. 121).
Суть межбиогеоценозных связей Н. В. Дылис видит во взаим-
ном обмене как соседних, так и порой весьма удаленных друг
от друга биогеоценозов метаболитами, энергией, организмами или
их репродуктивными зачатками, а также различными материалами
органического и неорганического происхождения, растворенными в
воде веществами и т. п. В биогеоценозы, как относительно откры-
тые системы, как указывает Н. В. Дылис, постоянно поступают
различные вещества и энергия из окружения; в то же время из них
выносятся продукты метаболизма и некоторые отмершие органи-
ческие остатки за пределы биогеоценоза. Одним словом, пишет
Н. В. Дылис (19746), «нет биогеоценотических систем, ничего не
получающих от своего окружения, равно как и ничего не отдаю-
щих своим соседям. Различия касаются лишь масштаба, форм и
направления этого обмена» (с. 370).
Обмен веществом, энергией и различными органическими и не-
органическими материалами между биогеоценозами осуществляется
разными путями: при помощи воды, воздуха и диффузии биоген-
ных и других газов; путем расселения и перемещения растений,
Животных, микроорганизмов; благодаря наличию градиентов теп-
300
ла и различных концентраций веществ в воде, почве и пр. Масшта-
бы межбиогеоценозных обменных процессов довольно велики, не-
редко огромны как по массе перемещаемых веществ (тучи саранчи,
массовые нашествия грызунов), так и по расстоянию (на сотни и
тысячи километров). Известно, например, что вынос растворенных
в воде веществ с территории СССР местами составляет 40—
50 т/км2; вынос веществ в растворенном виде в Мировой океан со
всей суши достигает 3300 млн т в год, в том числе органических
веществ — около 720 млн т.
О вещественно-энергетическом обмене смежных биогеоценозов
в газообразной, жидкой и твердой фазах пишет и Н. В. Тимофеев-
Ресовский (1976); общим стоком веществ они объединяются в свое-
образные цепи по направлению стока. С биохимической точки зре-
ния миграция веществ в цепях биогеоценозов, по его словам, может
рассматриваться как «серия сопряженных процессов рассеивания
и концентрирования веществ в растениях, почвах, водах, грунтах
и атмосфере» (с. 129). Он отмечает также, что популяционно-видо-
вое разнообразие территориально связанных биогеоценозов (дубо-
вых, осиновых, сосновых и т. п.) отражается на вещественном и
химическом составе стока и что поступающие в почвенно-грунтовые
воды химические элементы проходят через серию сопряженных по
направлению стока биогеоценозов, при этом на их границах они,
как правило, претерпевают изменения в составе стока; это так на-
зываемый биогеоценотический барьер, представляющий
собой «основную преграду веществам, выходящим за пределы био—
геоценозов» (с. 129).
6. Территориальное объединение биогеоценозов в биогеосистемы
и биогеомассивы по Бялловичу
Перемещение (движение) энергии, веществ и различных мате-
риалов, непосредственно или косвенно связанных с метаболизмом
биогеоценоза, а также живых организмов, их зачатков, да и сами
перемещающиеся объекты, по Ю. П. Бялловичу (1960, 1969, 1973,
1980), составляют предмет изучения особой науки, названной им
кинетикой биогеоценозов. При этом имеется в виду перемеще-
ние названных объектов как внутри биогеоценоза между его мор-
фологическими структурами (биогеогоризонтами и парцеллами),
так и между биогеоценозами (смежными и удаленными) и между
биогеоценотическим покровом и природными системами, находя-
щимися за его пределами.
Пространство как форма существования биогеоценоза, указы-
вает Ю. П. Бяллович, обладает трехмерной анизотропией и в связи
с этим — тремя различными направлениями перемещения объектов:
радиальным и двумя латеральными — фронтальным, со-
30!
впадающим с пространственным направлением главных межбио-
геоценозных связей, и фланговыми, пересекающими фронтальный
под определенным углом. В каждом из названных трех направле-
ний перемещения объектов, по словам автора, «по-своему выраже-
на симметрия, по-своему происходит изменение свойств биогеоце-
ноза, по-своему совершается перенос веществ и энергии, по-своему
построены связи между структурными элементами биогеоценоза и
между биогеоценозами» (с. 51).
В отличие от морфологического расчленения биогеоценоза на
структурные части, при расчленении его в кинетическом отноше-
нии, т. е. на основе учета, циркуляции веществ, энергии и пр.,
Ю. П. Бяллович различает следующие структурные элементы:
а) радиали (R), включающие всё находящееся в движении в
радиальном направлении из одного биогеогоризонта в другой и за
пределы биогеоценоза; б) латерали (L), охватывающие всё пе-
ремещающееся в латеральных направлениях из одной парцеллы в
пределах биогеогоризонта в другую внутри биогеоценоза и из од-
ного биогеоценоза в другой; в) стационали (S)—всё покоя-
щееся относительно границ биогеогоризонтов, парцелл и биогеоце-
ноза. Но «неподвижность» стационали относительна, поскольку она
постоянно находится в движении вместе со всевозможными движе-
ниями биогеогоризонта и может также совершать колебательные
движения (например, качание деревьев в лесу).
Биогеогоризонты в составе биогеоценозов по соотношению ста-
ционалей и латералей Ю. П. Бялловичем (1969, 1973) подразделя-
ются на три категории: а) поточные биогеогоризонты, лишенные
стационалей, в них происходит движение всей их массы в ла-
теральном направлении, они как бы сплошь заполнены разно-
образными латералями и пронизаны взаимодействующими с ними
радиалями; б) непоточные биогеогоризонты, лишенные латералей,
движения масс в латеральном направлении в них нет; в) полу-
поточные биогеогоризонты, состоящие из относительно неподвиж-
ных стационалей и из движущихся сквозь стационали масс в лате-
ральном направлении. Множеством движущихся в поточных и
полупоточных биогеогоризонтах латералей, имеющих общий путь
и на нем взаимодействующих, образуется так называемый био-
геопоток (рис. 33).
Таким образом, стационали, по словам автора, определяют дис-
кретность биогеоценотического покрова и самих биогеоценозов, их
относительную статичность, а латерали и биогеопотоки обусловли-
вают непрерывность, целостность биогеоценозов и биогеоценотиче-
ского покрова. При этом смежные биогеоценозы межбиогеоценоз-
ными связями и отношениями, реализующимися в латералях и био-
геопотоках, объединяются, как пишет Ю. П. Бяллович (1969), «в
интенсивно функционирующие, целостные, объективно обособлен-
302
Ориг. рис. 33. Латералк и бногеопотоки в биогеоценозах и между ними (по дан-
ным Ю. П. Бялловича, 1973)
ные реальные системы» (с. 60). Такне относительно замкнутые,
пространственно ограниченные системы названы автором биогео-
системами.
Непосредственные связи между биогеоценозами, следовательно,
осуществляются только латералями и биогеопотоками. По своему
характеру и материальному содержанию латерали и биогеопотоки
весьма разнообразны: простые и сложные, вещественные и энерге-
тические, живые и косные и т. п. Но наряду с ними в природе
имеют место и такие связи и отношения, которые протекают вне
биогеосферы по внешним каналам связи и осуществляются лате-
ралями и так называемыми геопотоками. Возникающие при
этом в тропосфере и литосфере горизонтально простирающиеся
образования названы Ю. П. Бялловичем (1969) геогоризонта-
ми (рис. 34). Они могут достигать различной мощности и нахо-
диться на различной высоте за пределами внешних радиальных
границ биогеосферы (перелет птиц, перенос спор, микроорганиз-
мов) или на различной глубине (подземные воды различного типа
и т. п.). В совокупности ими образуются системы высшего ранга —
геосистемы, по терминологии автора. Следовательно, геосисте-
мы, по Ю. П. Бялловичу, в отличие от биогеосистем базируются на
связях между составляющими их биогеоценозами, протекающими
преимущественно по внешним каналам и осуществляющимися ла-
тералями и геопотоками.
Ю. П. Бяллович (1969—1980), разработавший в первом прибли-
жении схему пространственного строения биогеоценотического по-
крова сухопутного сектора Земли, различает следующие типы
биогеосистем: а) неразветвленные биогеосистемы, в которых
303
Геогориэонты
(в литосфере)
Геогоризонты
(в тропосфере)
Биогеогориэонты
БИОГЕОЦЕНОЗА
Рис. 34. Системы биогеогоризонтов и геогоризонтов (по данным
Ю. П. Бялловича, 1973)
срединный биогеоценоз непосредственно соединен латералями с
двумя соседними — начальным и финальным, эти же, в свою оче-
редь, соединены с соседними биогеоценозами других, смежных био-
геосистем; б) разветвленные биогеосистемы, в которых имеются
-«узловой» биогеоценоз н несколько ответвленных от него в разных
направлениях биогеоценозов, связанных с ним латералями и био-
геопотоками; в) контурные биогеосистемы, в которых связи между
биогеоценозами перекрещиваются по типу сети, некоторые из них
являются кольцевыми. В неразветвленных и разветвленных биогео-
системах тот или иной вид биогеопотока играет наиболее важную,
ведущую роль (рис. 35).
Весьма важное место в структуре и функционировании биогео-
сферы, по мнению Ю. П. Бялловича, занимают территориальные
совокупности водных ценозов — биогидросистемы, с которы-
ми взаимосвязаны соседние сухопутные биогеосистемы. Наиболее
существенные межбиогеоценозные связи в них, по Бялловичу, осу-
ществляются биогеопотоками склонового и ложбинного поверх-
ностного стока к верхнего горизонта подземного стока, а также
геопотоками более глубоких горизонтов подземного стока, речным
биогеопотоком и подрусловым геопотоком.
Граничащие друг с другом, обычно сходные биогеосистемы
образуют сплошной, относительно однородный массив биогеоси-
стем. Такого рода массивы Ю. П. Бяллович назвал биогеомас-
сивами; например, биогеомассив на водоразделе небольшой ре-
ки или ручья, образованный сходными биогеосистемами водосбор-
ного бассейна. Основанием для объединения смежных биогеосистем
304
Биогеосистемы
Комть/рные
Ориг. рис. 35. Типы биогеосистем (по данным Ю. П. Бял-
ловича, 1973). Биогеоценозы: Н — начальные; С — сред-
ние; Ф — финальные; Уз — узловые
в биогеомассы, по Ю. П. Бялловнчу (1973), служит то обстоятель-
ство, что рельеф местности, отражая геологическое строение и со-
став горных пород, перераспределяет геопотоки и радиали и тем
самым «упорядочивает... и жестко фиксирует латеральную диф-
ференциацию биогеосферы» (с. 11). На том же основании может
быть выделен биогеомассив вдоль речной или приручейной долины
и т. п. Биогеомассивы автором подразделяются на однородные
секторы, простирающиеся поперек водораздела или долины.
При этом в верховьях долины и водораздела (с прилегающими
склонами) в пределах того или другого биогеомассива выделяется
верховой сектор, образованный в том и другом случае соответству-
ющими биогеосистемами; в низовье соответственно выделяется ни-
зовой сектор из соответствующего набора и комбинации биогеоси-
стем и между ними — срединный сектор (рис. 36 и 37).
Биогеомассивы и секторы биогеомассивов по Ю. П. Бялловичу
могут быть различными по размерам и относиться к различным
рангам. Это определяется величиной речного потока и его водо-
сборного бассейна — от ручья и небольшой речки до огромной ре-
ки, пересекающей несколько ландшафтно-географических зон в ме-
ридиональном направлении или несколько провинций — в широт-
305
зов
Верховой сектор
Срединный сектор
Водораздельный
Прирученный
Ориг. рис. 36. Топологические биогеомасснвы ручья (продольный профиль) (по
данным Ю. П. Бялловича, 1973)
/ Низовой сектор
Ориг. рис. 37. Топологические биогеомассивы ручья (попереч-
ный профиль) (по данным Ю. П. Бялловича, 1973). Биогео-
массивы; А — водораздельные; Б — приручейные
ном. Из этого следует, что представления Ю. П. Бялловича о тер-
риториальных единицах биогеосферы во многих отношениях пере-
кликаются с представлениями И. А, Титова (1936, 1952) по данно-
му вопросу.
7. Георастительные системы Титова
и топологические геосистемы Сочавы
И. А. Титов, рассматривая проблему взаимодействия раститель-
ных сообществ и условий среды» выдвинул и обосновал термин
и понятие георастительная система как «совокупность
взаимодействующих растительных сообществ и условий среды во-
досборного геологического бассейна, развивающаяся как единое
целое» (1952, с. 22). Компонентами, слагающими геораститель-
ную систему, он считал а) водосборный бассейн с его стра-
тиграфической структурой и геохимическими ресурсами и б) ра-
стительные сообщества со всеми почвообразовательными
последствиями и накопленными продуктами их жизнедеятельности.
Георастительным системам И. А. Титов придавал очень боль-
шое значение, ибо сухопутный сектор биогеосферы, по его пред-
ставлениям, состоит в основном из занятых растительным, а теперь
можно сказать — биогеоценотическим покровом водосбор-
ных бассейнов больших и малых водотоков.
В самом деле, если взглянуть с большой высоты на тот или
иной отрезок земной поверхности, то можно увидеть, что он пред-
ставляет собой массив, изрезанный многочисленными различной
величины речными долинами с приручейными и безводными бал-
ками или обширными низинами, озерными котловинами и другими
понижениями, с одной стороны, и разграничивающими их возвы-
шенностями, пенипленизированными плоскогорьями или островер-
хими гребнями горных хребтов с крутыми обрывистыми или пологи-
ми террасированными склонами — с другой. Короче говоря, нашему
307
взору представится разграниченная различного типа водораздела-
ми более или менее густая, древовидно разветвленная гидрографи-
ческая сеть поверхностных водотоков с общей ориентацией их в
сторону прилегающих континентальных или морских водоемов.
В последние десятилетия в физической географии изучению
водосборных речных бассейнов уделяется большое внимание. Они
рассматриваются как самостоятельно функционирующие целост-
ные географические системы — геосистемы, Ссылаясь на работы
В. Б. Сочавы о геосистемах, Л. М. Корытный (1974) пишет, что в
этих работах «географическая оболочка Земли рассматривается в
виде иерархии геосистем, под которыми понимаются природно-гео-
графические единства всех категорий с присущими им своеобраз-
ными структурами, законами изменения и развития» (курсив
наш.— Л. Н.) (с. 33). Основной причиной единства геосистем, по
его словам, является «устойчивость процессов взаимодействия меж-
ду природными компонентами (биотой, почвой, водой и т. д.), ко-
торая выражается в различных природных режимах» (с. 33), Сум-
марный эффект циркуляции этих режимов является «движущей си-
лой саморазвития природной среды» (с. 33).
В. Б. Сочава (1970—1974), развивая учение о ландшафтах, ука-
зывает, что физико-географический ландшафт и его подразделения
в настоящее время рассматриваются как многокомпонентные от-
крытые структурно-динамические системы разного ранга, назван-
ные им геосистемами.
При этом, исходя из того, что присущие геосистемам закономер-
ности действуют в ограниченных пространственных пределах, он
подразделяет их на три порядка размерностей — плане-
тарный, региональный и топологический. К планетарному порядку
размерности автором относятся следующие подразделения физико-
географической земной оболочки: а) физико-географические пояса,
б) группы физико-географических областей, в) субконтиненты и
г) физико-географические области; к региональному порядку раз-
мерности: д) природные зоны и е) подзоны (подразделяемые, в
свою очередь, в широтном направлении на провинции); к тополо-
гическому порядку размерности: ж) округа, з) районы, и) местно-
сти и к) урочища. Территориальные единицы топологического ряда
размерности названы автором геохорами. Элементарной гео-
хорой он считает выдел фаций, которую рассматривает как
сочетание смежных биогеоценозов.
Из изложенного видно, что понятие «геосистема» в трактовке
Ю. П. Бялловича и В. Б. Сочавы как по содержанию и по форме,
так и по объему далеко не одинаково. Отличается от них и поня-
тие «георастительная система» И. А. Титова. Общим для этих
понятий является лишь то, что все они представляют собой вне-
ранговые территориальные единицы либо растительного покрова,
808
либо биогеоценотического, либо физико-географической (ландшафт-
ной) оболочки земного шара. Вытекающие из этой их общности
некоторые принципиальные положения и закономерности, по-види-
мому, могут быть положены в основу системы территориального
подразделения биогеоценотического покрова Земли.
К таким положениям принципиального характера можно от-
вести некоторые высказывания В. Б. Сочавы (1972), который фи-
зико-географическое районирование ландшафтной оболочки Земли
рассматривает как территориальное деление его. основанное «на
выявлении соподчиненных регионов» (с. 9). Исходной логической
основой районирования он считает общую теорию систем; при
этом зональный и азональный принципы районирования, по его
мнению, могут быть использованы как аргументы системного ана-
лиза. Правда, к районированию он относит подразделение терри-
тории лишь планетарного и регионального порядка размерности.
Подразделение же территории топологического порядка размер-
ности на геохоры он рассматривает как выявление при крупно-
масштабных полевых исследованиях «их пространственной мо-
заики» (с. 19).
8. Представления Бялловича
о пространственной структуре биогеосферы
и территориальном подразделении ее
Ю. П. Бяллович (1973) отличие территориальных единиц рай-
онирования биогеосферы от типологических таксонов видит в том,
что они устанавливаются не по сходству, а по контрастности, и
классификация их должна базироваться на особенностях не вну-
триценозного метаболизма, а межценозного обмена и других меж-
биогеоценозных связей.
В одной из последующих посвященных данной проблеме статей
Ю. П. Бяллович (1980) приводит схему пространственной струк-
туры биогеоценотического покрова Земли. В ней «совокупность всех
охваченных жизнью частей биосферы» (т. е. биогеосферу.— Л. Н.)
(с. 26) он назвал всеобителью. Правда, строгого соответствия все-
обители с биогеосферой, по его словам, нет. Для обеспечения функ-
ционального единства и устойчивости всеобители, по его словам,
необходима «всеобщая пространственно-временная упорядочен-
ность связей между биогеоценозами, т. е. структуированность по
типу иерархической многоуровневой системы» (с. 26). В качестве
наивысшей территориальной единицы биогеоценотического покрова
(всеобители) автор называет обитель, понимая под этим тер-
мином «один слой биогеоценозов, определенным образом располо-
женный относительно границ между геосферами» (в смысле
В. И. Вернадского. — Л. Z/.) (с. 28). При этом автор замечает, что
309
деление биогеосферы (всеобители) на обители за редким исклю-
чением не совпадает с другими подразделениями биосферы — био-
географическим, ландшафтоведческим, океанологическим и пр. Ни-
же он приводит перечень обителей в количестве 14 (7 основных и
7 промежуточных) с довольно подробной характеристикой каждой
из них.
К категории низших территориальных единиц сухопутного сек-
тора биогеосферы Ю. П. Бяллович (1973) относит биогеомас-
сивы речных (и озерных) водосборных бассейнов разной величи-
ны (разделяемые в поперечном по отношению к потоку направлении
на секторы) и биогеосистемы как разного рода комбинации
смежных биогеоценозов. Позднее Ю. П. Бяллович (1980) внес ко-
ренные изменения в предложенную им систему территориальных
единиц биогеоценотического покрова. Было введено множество но-
вых понятий и терминов, некоторые из ранее предложенных тер-
минов получили новую трактовку. В результате такого понятийного
и терминологического усложнения система пространственной струк-
туры биогеоценотического покрова Земли, на наш взгляд, стала
излишне громоздкой и, возможно, поэтому, насколько нам извест-
но, не получила широкого признания. По-видимому, эта проблема
нуждается в дальнейшем осмысливании и разработке.
9. Система пространственного подразделения биогеоценотической
оболочки Земли
Исходя из всего изложенного нам представляется, что в каче-
стве одного из предварительных вариантов территориального под-
разделения биогеоценотической оболочки Земли на иерархически
соподчиненные территориальные единицы может быть принята
следующая система такого рода единиц.
Биогеоценотическая оболочка Земли
А. Планетарный порядок размерности
На уровне планетарного порядка размерности основными при-
родными факторами, обусловливающими формирование высших
градаций пространственной структуры биогеоценотического покро-
ва Земли, являются глобальные планетарные и космические фак-
торы и явления (солнечная радиация, земная гравитация, эндоген-
ные процессы земной коры, в частности поднятия и опускания
земной коры, горообразовательные процессы, вулканическая дея-
тельность и пр.).
В качестве наивысшей территориальной единицы, соподчинен-
ной биогеоценотической оболочке Земли, мы считаем возможным
использовать термин мегатип, предложенный Ю. А. Львовым
310
•(1979) в качестве типологической единицы подразделения биогео-
сферы. При наличии всесторонне глубоко обоснованной классифи-
кации биогеоценозов Н. В. Дылиса (1964) необходимость в нем в
этом смысле, на наш взгляд, отпадает. В качестве второй единицы,
соподчиненной мегатипу, вместо малореспектабельного термина
«обитель» можно использовать термин биогеом (М. А. Глазов-
ская, 1972) или еще лучше термины — геобиом для крупных тер-
риториальных сочетаний сухопутных ценозов и гидробиом для
экваториальных сочетаний водных ценозов такого же масштаба.
Биогеоценотнческая оболочка Земли может быть подразделена
на следующие м е г а т и п ы.
I. Мегатип континентальных биогеоценозов и биогидроце-
нозов.
П. Мегатип морских и океанических биогидроценозов.
Ill. Мегатип подземных биогеоценозов.
IV. Мега тип культурных биогеоценозов и биогидроценозов
(лесных, полевых и т. п.).
Мегатипы подразделяются на геобиомы и гидробиомы.
К первому мегатипу относятся следующие геобиомы и гидробиомы,
соответствующие обителям Ю. П. Бялловича:
1) наземный геобиом у раздела атмосфера — литосфера;
2) земноводный гидробиом периодически затопляемых речных и
озерных пойменных и внепойменных участков, пересыхающих
рек и озер, обнажающихся мелей, временных русловых по-
токов;
3) налёдный геобиом у раздела атмосфера — ледник на покров-
ных и шельфовых ледниках, многолетнем припое снежного
льда и других незагрязненных участках ледников;
4) наземно-налёдный геобиом у раздела атмосфера — ледник в
местах поступления значительной массы минерального мате-
риала на горных ледниках;
5) озерно-речной гидробиом, свободный от полуволной, плаваю-
щей и мелкопогруженной растительности;
Ко второму мегатипу относятся гидробиомы:
6) пелагический гидробиом у раздела атмосфера — океаническая
гидросфера до глубины перевеса влияния поверхностной цир-
куляции над влиянием дна;
7) бентический гидробиом у раздела океаническая гидросфера —
литосфера;
6) неритический гидробиом у двух сближенных разделов атмосфе-
ра— океаническая гидросфера — литосфера (при глубине дна
200—500 м);
9) литоральный гидробиом;
1:0) неритово-подледный гидробиом у двух сближенных разделов
ледник на плаву — океаническая гидросфера — литосфера под
311
шельфовым ледником или под мощным антарктическим при-
паем;
11) пелаго-подледный гидробиом у раздела ледяная подвижная
масса — океаническая гидросфера (айсберги, ледяные острова
до уровня омывания волнами);
К третьему мегатипу относятся геобиомы:
12) подземный геогидробиом с одним или несколькими потоками
воды, нефти, газов до глубины 1—12 км;
13) пещерный геобиом;
14) подземно-подледный геобиом у раздела ледник — литосфера
под относительно теплыми горными ледниками.
В составе мегатипа культурных ценозов могут быть выделены
геобиомы: 15) наземно-лесной; 16) наземно-сельскохозяйствен-
ный; 17) садово-парково-тепличный, а также 18) прудово-рисово-
чековый мелководный гидробиом и др.
Б. Региональный порядок размерности
На уровне регионального порядка размерности формирование
средних градаций пространственной дифференциации биогеоцено-
тического покрова совершается под действием зональных и провин-
циальных! факторов, главным образом климатических, обусловлен-
ных вращением Земли вокруг Солнца и распределением океаниче-
ских водных масс по отношению к континентам (атмосферные
осадки, температура воздуха, циркуляция тепла и влаги по земной
поверхности и пр.).
Нетрудно представить себе, что распростертые в разных направ-
лениях по лику Земли геобиомы н гидробиом ы под дей-
ствием закона зональности и других факторов значительно
варьируют по многим существенным показателям. Так, например,
наземный геобиом естественно расчленяется на биогеоценоти-
ческие области, зоны и провинции.
На Евроазиатском континенте в пределах СССР наземный гео-
биом, вследствие повышения температуры воздуха на 1—1,5° С на
каждые 3 градуса географической широты в направлении с севера
на юг, может быть подразделен на следующие биогеоценоти-
ческие области.
I. Арктическая биогеоценотическая область в холодном гумид-
ном климате.
II. Лесная биогеоценотическая область в умеренно холодном и
умеренно теплом климате.
III. Степная биогеоценотическая область в умеренно теплом и се-
миаридном климате.
IV. Пустынная биогеоценотическая область в умеренно теплом к
теплом аридном климате.
312
Биогеоценотические области в том же направлении можно под-
разделить на биогеоценотические зоны, а в широтном
направлении (с запада на восток), вследствие изменения градиен-
тов влажности воздуха от океанических окраин в глубь континента,
на биогеоценотические провинции.
1. Арктическая биогеоценотическая область:
1. Тундровая биогеоценотическая зона кустарниковых и мохо-
во-лишайниковых тундр.
2. Лесотундровая биогеоценотическая зона субарктических дре-
весно-кустарниковых редколесий.
Провинции: а) европейско-западносибирская лесотунд-
ровая; б) центрально-сибирская лесотундровая; в) беренгий-
ская лесотундровая.
П. Лесная биогеоценотическая область:
3. Хвойно-лесная (таежная) биогеоценотическая зона вечнозе-
леной темнохвойной и листопадной светлохвойной тайги.
4. Лиственно-лесная биогеоценотическая эона хвойно-широко-
лиственных, широко- или мелколиственных летне-зеленых
лесов.
Провинции: а) европейско-западносибирская вечнозеле-
ная темнохвойная; б) средне- и восточносибирская летне-зеле-
ная светлохвойная; в) южноохотская темнохвойная; г) евро-
пейская широколиственная; д) западносибирская мелколист-
венная; е) дальневосточная хвойно-широколиственная.
III. Степная биогеоценотическая область:
5. Лесостепная биогеоценотическая зона остепненных лугов и
луговых степей с широко- или мелколиственными перелес-
ками.
6. Еустепная биогеоценотическая зона разнотравно-дерновин-
но-злаковых и дерновинно-злаковых степей.
Провинции: а) восточноевропейская лесостепная; б) за-
падносибирская лесостепная; в) восточноевропейская (понтий-
ская) степная; г) казахстанская степная; д) даурско-монголь-
ская степная.
IV. Пустынная биогеоценотическая область:
7. Полупустынная биогеоценотическая зона полынно-дерновин-
но-злаковых степей и злаково-полынных остепненных пус-
тынь.
8. Еупустынная биогеоценотическая зона полукустарниковых
полынных пустынь.
9. Эфемерово-пустынная биогеоценотическая зона эфемерово-
полынных (южных) пустынь.
Провинции: а) восточноевропейская полупустынная;
б) казахстанская полупустынная; в) мангышлакская пустын-
313
ная; г) арало-каспийская пустынная; д) туркестанская пустын-
ная; е) копет-дагская эфемерово-пустынная.
Что же касается вертикальной поясности горных местностей, то
по типологическому составу биогеоценозов и последовательности
размещения таковых в пространстве она, как известно, аналогична
(но не гомологична) широтной зональности и вместе с тем отра-
жает специфические особенности той зоны или провинции, а пре-
делах которой горная страна находится.
В. Топологический порядок размерности
На уровне топологического порядка размерности на первое ме-
сто выступают местные природные топоэкологические факторы,
связанные с особенностями мезо- и микрорельефа, а также с экзо-
генными процессами на поверхности земли (в частности, водно-
воздушный, тепловой режимы, режим минерального и азотного пи-
тания растений и пр.). Что же касается производственно-хозяй-
ственного воздействия людского населения на биогеоценотический
покров и его пространственную дифференциацию, то оно проявля-
ется на всех уровнях размерности, хотя на данном уровне наиболее
эффективно.
В пределах топологического порядка размерности под действием
названных природных факторов биогеоценотический покров обла-
сти, зоны (и провинции) подразделяется на следующие последова-
тельно соподчиненные территориальные единицы: б и о гео ре-
гион, биогеомассив и биогеосистема.
Биогеорегион (по своему характеру и объему соответству-
ющий геораститеЛьной системе И. А. Титова) представляет собой
совокупность биогеомассивов водосборного бассейна речной
системы любой величины. Биогеорегионы водосборных бассейнов
различаются по величине водотока (длине, водности, площади во-
досборного бассейна) и по общему направлению его (северное, юж-
ное, западное, восточное). По совокупности этих показателей био-
георегноны в пределах Советского Союза могут быть подразделе-
ны на градации.
I. Биогеорегионы, водосборных бассейнов великих рек северной
покати (Оби, Енисея, Лены), южной (Волги), восточной
(Амура).
II. Биогеорегионы водосборных бассейнов крупных рек северной
покати (Северная Двина, Печора, Индигирка, Колыма), юж-
ной (Днепр, Урал) и крупные притоки великих рек северной
покати (Иртыш, Ангара, Подкаменная и Нижняя Тунгуски,
Витим, Алдан и др.), южной покати (Кама) и восточной —
составляющие Амура (Аргунь и Шилка).
III. Биогеорегионы водосборных бассейнов средних рек северной
314
покати (Хатанга), южной (Дон) и внутриконтинентальных во-
доемов (Амударья, Сырдарья, Селенга) и средние притоки ве-
ликих и крупных рек разных покатей (Припять, Десна, Ока,
Сухона, Ишим, Тобол, Чулым, Кеть, Вилюй, Олёкма, Бурея
И др.).
IV. Биогеорегионы водосборных бассейнов малых рек разных по-
катей (типа Прут, Днестр, Кубань, Терек, Неман, Зап. Двина,
Южн. Буг, Мезень, Онега, Пур, Таз, Оленек, Яна и многие
другие), а также великое множество притоков рек разной ве-
личины.
V Биогеорегионы водосборных бассейнов бесчисленного множе-
ства речек, ручьев и прочих водотоков, служащих истоками
рек более крупного масштаба.
VI. Биогеорегионы водосборных бассейнов стоячих водоемов (озер,
прудов, водохранилищ и т. п.).
Биогеорегионы вдоль водотока водосборного бассейна расчле-
няются на биогеомассивы различной величины в зависимости
от величины водотока. Биогеомассив — это совокупность
смежных биогеосистем разного типа и различного размера по пло-
щади. Среди них, пользуясь терминологией Г Н. Высоцкого
(с. 192), можно различать: 1) плакориые водораздельные био-
геомассивы (невысокие пологосклоновые возвышенности, всхолм-
ленные плато, выравненные пенеплены); 2) надплакорные
биогеомассивы различной протяженности и высоты над уровнем
моря (горные цепи хребтов, обычно с отчетливо выраженной вер-
тикальной поясностью растительного покрова, крутые и обрывистые
склоны разной экспозиции); 3) плаккатные водосборные био-
геомассивы различного типа (обширные наклонные низменности,
слабодренированные впадины, речные поймы различной величины,
озерные котловины и т. п.). В поперечном относительно водотока
направлении биогеомассивы Ю. А. Бяллович разделяет на
секторы. При этом биогеомассивы водосборных бассейнов вели-
ких и крупных речных систем, пересекающих на своем протяжении
ряд областей, зон и провинций, очевидно, следует расчленять на
соответствующие зональные или провинциальные секторы, а био-
геомассивы более мелких водосборных бассейнов, умещающихся в
пределах одной какой-либо зоны или провинции, — на топологиче-
ские секторы (верховой, срединный, низовой). Разумеется, каждый
сектор любого биогеомассива специфичен по набору и комбинации
составляющих его биогеосистем.
Впрочем, биогеорегион в целом, по-видимому, также
может быть подразделен в поперечном относительно водотока на-
правлении на соответствующие секторы, охватывающие секто-
ры всех примыкающих друг к другу биогеомассивов данного био-
315
георегиона, поскольку поперечные границы их, как правило, со-
впадают.
Биогеомассивы, по Ю. П. Бялловичу (1973), разделяются на
биогеосистемы разного типа, которые представляют собой
совокупность смежных биогеоценозов как основных функциональ-
но-биохорологических единиц биогеоценотического покрова любой
местности. Он различает, как отмечено, несколько типов био-
геосистем: неразветвленные, разветвленные и контурные. Ха-
рактерные особенности каждого из названных типов биогеосистем,
приведены выше.
10. Территориальное подразделение биогеосферы
зарубежными экологами
Зарубежными экологами (Р. Дажо, Ю. Одум, Р. Уиттекер и др.)
в качестве крупных биогеоценотических территориальных единиц,
выделяемых по физиономическим признакам растительности и по
сходству среды обитания, называются биомы. Р. Дажо (1975)
приводит следующие примеры биомов: а) североамериканская пре-
рия со стадами бизонов; б) африканская саванна с зонтиковидны-
ми акациями, баобабами, крупными травоядными (жирафы, анти-
лопы, зебры) и хищными (львы) животными и др. Ю. Одум (1975)
образование и существование биомов связывает с процессами вза-
имодействия регионального климата с биотой и субстратом, отме-
чая при этом, что для каждого биома характерна определенная
жизненная форма климаксовой растительности. Р. Уиттекер (1980)
так определяет биом: «Биом — это группа наземных экосистем
данного континента, которые имеют сходную структуру или физио-
номию растительности и общий характер условий среды, что нахо-
дит отражение в этой структуре и характеристиках их животного
населения» (разрядка наша.—Л. Н.) (с. 148).
Сходные по физиономическим признакам и условиям среды
биомы разных континентов Р. Уиттекером объединяются в основ-
ные физиономические типы биомов, к числу которых
он относит: I) леса; II) злаков ник и; III) редколесья
(невысокие редкостойные древостои с развитым нижним ярусом
растительности); IV) кустарники (с общим покрытием не бо-
лее 50%); V) полупустынные кустарники; VI) пусты-
ни (с покрытием растений не более 10%) Однако ввиду значи-
тельной неоднородности приведенных основных типов биомов, в
особенности по условиям среды обитания, автор затем подразделил
их на более мелкие типы, доведя число наземных типов биомов
до 21. Кроме того, им выделено 11 типов водных биомов, а всего»,
следовательно, им выделено 32 типа биомов» перечень которых на-
ми приводится.
316
Наземные (сухопутные) типы биомов
I. Леса:
1. Тропические дождевые леса. 2. Тропические сезонные леса.
3. Дождевые леса умеренного пояса. 4. Листопадные леса уме-
ренного пояса. 5. Вечнозеленые леса умеренного пояса. 6. Суб-
аркто-субальпийские хвойные леса. 7. Субальпийские карли-
ковые леса. 8. Тропические широколиственные леса.
II. Редколесья:
9. Колючие редколесья. 10. Редколесья умеренного пояса.
III. Кустарники:
11. Кустарники умеренного пояса. 12. Альпийские кустарники.
IV. 3 л а к о в н и ки:
13. Саванны. 14. Злаковники умеренного пояса. 15. Альпийские
луга.
V. Полупустынные кустарники:
16. Полупустынные кустарники теплого климата. 17. Холодные
полупустыни. 18. Тундры. 19. Арктоальпийские полупустыни.
VI. Пустыни:
20. Настоящие пустыни (в субтропиках). 21. Арктоальпийские
пустыни (в крайне холодном климате).
Водные типы биомов
I. Континентальные (пресноводные);
22. Гигрофитные сообщества (болота и пр. — Л. Н.). 23. Сооб-
щества стоячих пресных вод (озера и пруды). 24. Сообщества
проточных пресных вод (потоков).
И. Морские и океанические:
25. Приморские скалистые побережья. 26. Приморские песча-
ные отмели. 27. Приморские илистые мелководья. 28. Корал-
ловые рифы. 29. Морская поверхностная пелагиаль. 30. Мор-
ские глубоководные пелагические сообщества. 31. Бентос
континентального шельфа (неритический бентос). 32. Глубоко-
водные океанические бентосные сообщества.
Сопоставляя приведенный перечень наземных (сухопутных) ти-
пов биомов Р. Уиттекера с изложенным выше пространственным
подразделением биогеосферы на территориальные единицы разного’
ранга, можно прийти к следующим выводам: во-первых, все града-
ции типов биомов Уиттекера укладываются в категории региональ-
ного порядка размерности; во-вторых, основные типы биомов близ-
ки или даже сходны как по объему, так и по содержанию с выде-
ленными нами биогеоценотическими областями регионального по-
рядка размерности; в-третьих, типы биомов более дробного расчле-
нения биогеосферы в большинстве случаев соответствуют выделен-
ным нами биогеоценотическим зонам.
317
Глава X
Становление и динамика биогеоценозов
и эволюция биогеоценозов и биогеосферы
1. Понятие о динамике биогеоценозов
и формах ее проявления
В предыдущей главе на базе структурно-функциональной орга-
низации, функционирования биогеоценоза и функциональной дея-
тельности его компонентов был рассмотрен вопрос об осуществле-
нии авторегуляции биогеоценоза, обеспечивающей его устойчивость,
гомеостаз. Но устойчивость и динамика биогеоценоза — это две
стороны единого явления, а именно процесса развития биогеоцено-
тического покрова нашей планеты; это два состояния, присущие
любому объекту природы. Динамика—это изменчивость био-
геоценоза, накопление количественных изменений в его составе,
структуре, функциональной организации; устойчивость же
постулирует качественную определенность биогеоценоза как ре-
зультат перехода количественных изменений его в качественные по
достижении ими определенной меры. Следовательно, динамика и
устойчивость биогеоценоза неразрывно связаны друг с другом,
как две противоположности, переходящие друг в друга в ходе его
поступательного развития.
Становление, динамика и эволюция биологических и биокосных
систем совершаются по законам материалистической диалектики.
Широко известно выражение В. И. Ленина, что и овес растет по
Гегелю. С тем большим основанием можно сказать о таких слож-
ных во многих отношениях, многокомпонентных и многогранных
объектах природы, как биогеоценоз и биогеосфера. Недаром
В. Н. Сукачев, рассматривая вопросы развития биогеоценотическо-
го покрова Земли, обратился к известным положениям В. И. Лени-
на по проблеме развития, изложенным в его фрагменте «К вопросу
о диалектике» (1932).
Н. В. Дылис вслед за В. Н. Сукачевым рассматривает биогео-
ценозы как весьма динамичные природные образования, которые
непрерывно изменяются и развиваются «на основе внутренних про-
тиворечивых тенденций его компонентов» (1969, т. 9). Все биогео-
ценозы, несмотря на относительную устойчивость, стабильность,
«испытывают со временем большую или меньшую перестройку
своего компонентного состава, структуры и метаболизма, — пишет
юн. — Причины этих изменений разные. Изменения идут с разными
скоростями, в различных направлениях, разной глубины и могут
318
начинаться с изменения любого составляющего, любой связи>
(1978, с. 113).
Изменения биогеоценозов В. Н. Сукачевым (19646), как извест-
но, подразделяются на две принципиально различающиеся, хотя и
взаимосвязанные категории: 1) циклические (периодические)
и 2) сукцессионные (коренные). Последние, в свою очередь,,
подразделяются им на а) автогенные, выражающие процесс раз-
вития биогеоценозов, и б) экзогенные, не вытекающие из их спон-
танного развития, в известной мере случайные. К категории авто-
генных сукцессий биогеоценозов В. Н. Сукачев относит и так назы-
ваемые филоценоеенетические сукцессии, хотя последние, пожалуй,
целесообразнее выделить в самостоятельную категорию, поскольку
они представляют собой чрезвычайно длительные этапы медленно
протекающего процесса эволюции биогеоценозов и биогеоценотиче-
ского покрова, сопряженные с эволюционным преобразованием ви-
дов, коренной перестройкой всей флоры и фауны порой колоссаль-
ных областей земной поверхности. В противоположность им дру-
гие типы автогенных сукцессий — не столь длительные смены био-
геоценозов, выражающие формирование и развитие биогеоценозов,
из видов с готовыми, ранее сложившимися и наследственно закреп-
ленными биологическими, экологическими и ценотическими свой-
ствами, а потому протекающие относительно быстро по сравнению
с эволюцией биогеоценозов и биогеосферы в целом.
2. Циклические изменения биогеоценозов
и понятие о коренных изменениях
и развитии их
.Для циклических изменений биогеоценозов характерно периоди-
ческое возвращение последних в состояние, практически однород-
ное с исходным, хотя и не тождественное. (Обратимый характер-
таких изменений-—одна из существенных особенностей, отличаю-
щих таковые от коренных смен биогеоценозов. Цикличность изме-
нений биогеоценозов обусловлена периодической сменой ряда гло-
бальных природных явлений с различной продолжительностью
периодов и их колебательных циклических изменений. В соответ-
ствии с этим В. Н. Сукачевым различаются следующие типы цик-
лических (флюктуационных) изменений биогеоценозов: 1) суточ-
ные, 2) сезонные, 3) погодичные (разногодние) и изменения био-
геоценозов, связанные с процессом возобновления биогеоценозов.
1. Суточные изменения биогеоценозов. Основным фактором,
определяющим суточные изменения биогеоценозов, являются смена
дня и ночи, обусловленная вращением Земли вокруг своей осн, и
связанные с этим изменения солнечной радиации, а именно про-
должительность светового периода в течение суток, интенсивность.
319
дневного освещения и пр. Это влечет за собой изменение свойств
воздушной и отчасти почвенной среды, в частности температурного
режима и пр. В соответствии с этим суточные изменения живых
компонентов биогеоценозов выражаются в периодичности процес-
сов фотосинтеза, транспирации, роста—у растений, в периодично-
сти кормления, покоя — у животных. В длительном геоисторическом
плане это находит выражение в формировании различных эколо-
гических групп, форм животных: дневных, к числу которых отно-
сится большинство животных, ночных (сова, летучая мышь икт. п.)
и сумеречных промежуточных форм.
2. Сезонные изменения биогеоценозов. Основным фактором,
определяющим сезонные изменения биогеоценозов в странах с уме-
ренным климатом, являются смена времен года, обусловленная
обращением Земли вокруг Солнца, и связанные с этим колебания
по временам года температурного режима, режима влажности воз-
духа и прочих метеорологических явлений.
Сезонные изменения биогеоценозов проявляются в так назы-
ваемой смене аспектов, т. е. в смене их внешнего облика и
некоторых сторон состава и строения. Смена аспектов биогеоценоза
определяется периодичностью (ритмикой) сезонного развития ра-
стений и животных, связанной с тем, что фенологические фазы
разных видов растений и соответствующие состояния и характер
деятельности разных видов животных (спаривание, выведение
птенцов или вынашивание и рождение детенышей, вылет птенцов
и собирание их в стаи или выход из нор и расселение молодняка
и т. п.) протекают в разные сроки в течение теплого времени года.
Для аспектов растительного компонента биогеоценоза, по словам
А. Г Воронова (1963), характерна определенная последователь-
ность смены их во времени и определенная продолжительность
фенологических фаз, тогда как аспекты животного населения био-
геоценозов менее определенны по продолжительности и срокам
проявления, например; появление в биогеоценозе на короткий срок
пролетных птиц, мигрирующих грызунов, насекомых или довольно
продолжительное пребывание в степных биогеоценозах копытных
млекопитающих в период подрастания детенышей и др.
Характер аспектов в разных биогеоценозах неодинаков. Наибо-
лее яркие, красочные аспекты наблюдаются в северных разнотрав-
но-злаковых луговых степях и бобово-разнотравных лугах. В тем-
нохвойных лесах, дерновинно-злаковых степях, злаково-осоковых
лугах и болотах аспекты монотонны и довольно постоянны. Лишь
древесный ярус лиственных лесов, в особенности широколиствен-
ных, имеет разную окраску кроны ранней весной, в летний период
и осенью. В широколиственных лесах, кроме того, ранней весной
до распускания листьев на деревьях под их пологом развивается
кратковременная синузия яркоцветущих эфемеров и эфемероидов.
320
3. Погодичные (разногодние) изменения биогеоценозов. Ос-
новным фактором, вызывающим такого рода изменения, является
чередование ряда лет, характеризующихся одним типом погодных
условий в вегетационный период, с последующим рядом лет с со-
вершенно иным типом погоды; например, годы с ранней теплой
весной и затяжной холодной весной или с сухим и жарким летом
и дождливым прохладным летом и т. п. Предполагается, что это
обусловлено периодическими колебаниями солнечной активности.
Погодичные изменения биогеоценозов наиболее резко проявля-
ются в растительной части. Так, в дождливые годы на влажных
осоковых лугах обильно развивается пушица узколистная, образуя
белый пятнистый аспект. Изменяется в той или иной мере популя-
ционно-видовой состав растительного компонента, его строение,
продуктивность. Заметные изменения по годам наблюдаются так-
же в жизнедеятельности и темпах развития животного населения
и микробного биокомплекса. Колебания по годам выхода фитомас-
сы и семенной продуктивности растительной части биогеоценоза
сказываются на численности животных-фитофагов, а также трофи-
чески связанных с ними хищников и паразитов. В неурожайные
годы семян хвойных пород и плодов рябины, калины наблюдаются
миграции, кочевки таких семеядных животных, как белка, соболь,
птица-кедровка, и питающихся плодами рябины свиристелей и др.
Все это может отразиться не только на изменении физионо-
мичности биогеоценозов по годам, но и вызвать варьирование ви-
дового состава, вертикального и горизонтального строения биогео-
ценоза и т. п. При сильных и резких изменениях погодных условий
по годам или длительности чередующихся периодов подобного рода
изменения в составе и структуре живых компонентов и биогеоце-
ноза в целом могут перейти в необратимые, т. е. привести к корен-
ной смене данного биогеоценоза другим.
К категории циклических (периодических) изменений биогеоце-
нозов В. Н. Сукачев относит также изменения, связанные с есте-
ственным процессом возобновления господствующей синузии авто-
трофного растительного компонента, например древостоя в лесном
биогеоценозе. Возобновительный процесс биогеоценоза протекает,
как правило, циклично и во всех случаях приводит к изменению
не только растительной части, но и других его компонентов. Одна-
ко другими отечественными биогеоценологами такого рода измене-
ния биогеоценозов не включаются в разряд циклических изменений.
Рассмотренные и кратко охарактеризованные нами внутрице-
нозные типы циклических изменений биогеоценологических систем
представляют собой изменения количественного порядка. Биогео-
ценоз при этом остается самим собой-, коренного изменения популя-
ционно-видового состава живых компонентов, качеств и свойств
биоценосреды, а также структурно-функциональной организации
321
биогеоценоза не происходит. Иначе говоря, эти изменения не вы-
ходят за рамки качественной специфики данного биогеоценоза, а
потому они обратимы.
В чем же состоит биологическое и ценотическое значение цик-
лических изменений биогеоценозов, в частности периодической
смены фенологических фаз и состояний ценозообразующих расте-
ний и животных? Во-первых, благодаря этому в одном и том же
биогеоценозе могут ужиться растительные и животные виды с раз-
личным ритмом развития и, следовательно, с различной периодич-
ностью использования материально-энергетических ресурсов био-
ценосреды. Во-вторых, на определенной площади вследствие этого
может разместиться больше особей и видов растений и животных,
чем при одновременном прохождении присущих им фаз и стадий
развития. В-третьих, благодаря этому обеспечивается физиологи-
ческое обособление систематически близких видов, поскольку их
органы размножения созревают в разное время, что ведет к фор-
мированию разных экологических рас, а затем и видов (например,
существование двух рас дуба в европейской части Союза).
Внутриценозные циклические изменения, как сказано выше,—
это количественные изменения биогеоценоза, не выходящие за рам-
ки присущих ему структурно-функциональных, метаболических и
других особенностей. Вместе с тем происходящие в биогеоценозе
количественные изменения (суточные, сезонные, погодичные), на-
капливаясь и с течением времени усиливаясь, могут при отмечен-
ных выше обстоятельствах перерасти в качественные и привести
к коренному изменению данного биогеоценоза. Следовательно, ко-
ренные изменения биогеоценоза •— это изменения качественного по-
рядка; биогеоценоз при этом перестает быть самим собой, стано-
вится иным, с иными качествами и свойствами. Иначе говоря, про-
исходит смена старого биогеоценоза новым биогеоценозом.
Такого рода смены биогеоценозов во времени получили название
сукцессии (от греч. succedo— быть или стать на место чего-то
другого). Ряд сменяющих друг друга биогеоценозов во времени
назван серией смен (сукцессий); например, серия смен биогеоце-
нозов елового леса при повышении уровня почвенно-грунтовых вод
на прилегающем к сфагновому болоту пологом склоне водораздела
в среднетаежной зоне европейской части Союза: ельник чернич-
ный-тельник долгомошный (с Polytrichum commune) тельник
сфагновый (со Sph. Girgensohnii и Sph. Warnstorfii).
Коренная смена одного биогеоценоза другим обычно сопровож-
дается сменой смежных биогеоценозов, а это в конечном итоге
ведет к изменению (смене) всего биогеоценотического покрова за-
нимаемого им участка территории. Иначе говоря, на данной терри-
тории происходит развитие биогеоценотического покрова в целом в
определенном направлении. Правда, по мнению В. Н. Сукачева, не
322
всякое изменение биогеоценоза можно подвести под понятие раз-
вития его и не всякую серию смен биогеоценозов можно рассмат-
ривать как развитие биогеоценотического покрова на данной тер-
ритории. Только те коренные изменения биогеоценозов и серии их
смен подойдут под понятие «развитие», которые являются, по его
словам, «следствием противоречивых процессов, протекающих
внутри самого этого явления» (с. 459).
Процесс формирования биогеоценозов, смены их во времени и
развития биогеоценотического покрова на той или иной территории
(или акватории) В. Н. Сукачев (19646) назвал биогеоценоге-
незом (или биогеоценотическим процессом), включающим два
взаимосвязанных этапа: а) сингенез и б) эндогенез.
3. Сингенез как процесс формирования биогеоценоза;
факторы формирования и стадии сингенеза
Под сингенезом В. Н. Сукачев понимал процесс формиро-
вания биогеоценозов на лишенных жизни участках Земли. Смены
же стадий в процессе формирования биогеоценоза были названы
им сингенетическими сукцессиями. Ранее при рассмо-
трении вопроса о формировании растительных сообществ (фитоце-
нозов) в процессе сингенеза В. Н. Сукачев (19646) различал три
стадии. Заимствуя терминологию у Ф. Клементса (Clements, 1936),
первую из этих стадий он назвал миграцией, вторую — эце-
зисом и третью — конкуренцией. В сингенетическом про-
цессе формирования биогеоценозов он выделяет лишь две стадии:
а) миграцию и 6) эцезис.
На первой стадии сингенеза, по В. Н. Сукачеву, происходит
заселение нового участка растениями и трофически связанными с
ними животными и микроорганизмами и приживание тех и других
к наличным условиям среды (экотопа), т. е. размножение и рассе-
ление их по субстрату. Подбор видов живых существ на этой стадии
происходит на основе экологических связей организмов со средой
обитания. Это ведет, по выражению В. Н. Сукачева, к экологиче-
скому выравниванию видового состава, т. е. к распределению их
в пространстве по соответствующим их природе элементам микро-
и мезорельефа. На второй стадии происходит сживание различных
видов растений, животных и микроорганизмов друг с другом как
внутри каждого компонента, так и в составе ценотнческих групп
разных компонентов формирующегося биогеоценоза, т. е. вступле-
ние их по мере увеличения численности и плотности «населения»
в прямые и опосредованные взаимные связи и отношения. Подбор
видов и подгонка их к совместному обитанию на данной стадии
сингенеза происходят на основе ценотнческих связей организ-
мов, выражающихся в межвидовой борьбе за средства жизни внут-
323
ри компонента и в трофических, симбиотических и других связях
между живыми компонентами. Это ведет к эколого-ценотической
дифференциации видового состава живых существ по синузиям,
биогеогоризонтам, парцеллам и к формированию пищевых цепей,
консорций и т. п. Протекающие одновременно с этим под воздей-
ствием биоты изменения компонентов косной природы (экотопа)
приводят с течением времени к образованию почвенного слоя
и соответствующего фитоклимата.
В классической форме сингенез протекает на так называемых
новых субстратах: а) на твердых, не подвергшихся еще механиче-
скому разрушению и физико-химическому «выветриванию» суб-
стратах, полностью лишенных жизни (остывших вулканических ла-
вах, скалистых обнажениях горных пород, обвалах и осыпях в
подножье отвесных скал); б) на рыхлых естественных практиче-
ски безжизненных субстратах (элювиальных отложениях на скло-
нах и делювиальных в подножье нижних надпойменных террас,
конусах выноса в устьях рек и ручьев, аллювиальных наносах
речных прирусловых отмелей); в) на лишенных жизни искусствен-
ных обнажениях (склонах железнодорожных и других насыпей и
выемок, терриконах и горнорудных отвалах, карьерах и т. п.).
Сингенетический процесс можно наблюдать также на участках,
освободившихся от бывшего биогеоценотического покрова, в ре-
зультате уничтожения его под действием природных факторов или
хозяйственной деятельности местного населения (сплошные выруб-
ки леса, лесные гари, заброшенные пахотные земли — так назы-
ваемые «залежи» и т. п.). Проявление некоторых элементов син-
генеза можно обнаружить и в сформировавшихся биогеоценозах
в виде перестройки их популяционно-видового состава и строения
вследствие внедрения новых видов илн исчезновения некоторых из
произрастающих и т. п.
Во всех этих случаях сингенез протекает несколько по-иному,
обладает некоторыми специфическими особенностями, но в основ-
ном он сходен как по существу, так и по последовательности про-
исходящих процессов и смены стадий» хотя сами стадии сингенеза
разными авторами именуются по-разному.
Сингенез на подвергшихся выветриванию выходах твердых гор-
ных пород, по И. В. Стебаеву (1978), проходит следующие ста-
дии: 1} стадия корковых и листоватых лишайников; 2) литофиль-
ных мхов; 3) зеленых гипновых мхов: 4) высших сосудистых расте-
ний. В каждой из названных стадий сингенеза автором выделяется
по две фазы, Для первой фазы характерно увеличение видового
разнообразия и мозаичный характер размещения ценопопуляций
автотрофного ценокомплекса. Во второй фазе происходит слияние
раздельных растительных пятен в характерный для каждой стадии
сплошной ковер растительности. Каждая стадия сингенеза харак-
324
теризуется соответствующим составом растительноядных и детри*
тоядных животных (ногохвосток и других низших насекомых на
первой стадии, панцирных клещей — на второй, дождевых червей,
высших насекомых и других крупных беспозвоночных — на после-
дующих стадиях), а также трофически связанных с ними хищников.
Должного развития на каждой стадии сингенеза достигает и со-
ответствующая по групповому составу гетеротрофная микрофлора.
В результате фотосинтеза, метаболизма и других процессов
жизнедеятельности живых существ происходит увеличение от ста-
дии к стадии количества продукции, общего запаса биомассы и
отмершего органического материала, усиливаются процессы мине-
рализации органики и образования гумусовых веществ. Вместе с
тем возрастает биологическое разрушение поверхности горной по-
роды и мощность рыхлого субстрата, происходит обогащение слоя
мелкозема гумусом и минеральными питательными веществами и
превращение его в сплошной слой почвенного покрова, именуемого
обычно на этой стадии маломощной неразвитой или малоразвитой
почвой. По мере возрастания мощности и плотности автотрофного
растительного компонента и животного населения усиливается
разностороннее взаимодействие живых компонентов и видовых по-
пуляций каждого из них, а также укрепляется взаимосвязь живых
существ со средой обитания. Следствием этого является усложне-
ние структурно-функциональной организации биогеоценотического
слоя, дифференциация его по элементам морфологической и тро-
фической структуры и в конечном итоге становление относительно
устойчивой биогеоценотической системы.
Сингенез, протекающий на рыхлых субстратах, большей частью
легкого механического состава (песчаных, супесчаных, иногда за-
иленных), отличается от сингенеза на твердых скальных породах.
Здесь отсутствуют стадии из примитивных лишайниковых и мохо-
вых сообществ, связанных с биологическим разложением скальных
пород и формированием примитивного почвенного слоя. В этих
условиях процесс сингенеза от начала до конца протекает на базе
высших сосудистых, преимущественно покрытосеменных растений,,
относящихся к различным экобиоморфам и фитоценотипам, и со-
ответствующих представителей животного и микробного населения.
Так, Б. А. Быков (19706) в сингенетическом процессе становле-
ния фитоценоза как основного компонента биогеоценотической си-
стемы различают следующие стадии: 1) проценоз-колония;
2) проценоз-группировка; 3) гиперпроценоз. Первая стадия синге-
неза представлена разбросанными по площади разобщенными по-
селениями одной, реже — немногих популяции так называемых
инициальных видов высших сосудистых растений, относящихся
обычно к одной экобиоморфе. Она характеризуется почти полным
отсутствием взаимодействия между поселениями особей и весьма
325
слабым воздействием их на среду обитания. На второй стадии фор-
мирующееся растительное сообщество представлено группировка-
ми нескольких взаимодействующих между собой видовых популя-
ций, относящихся к двум и более экобиоморфам. Среда обитания
под действием растений и связанных с ними гетеротрофных живых
существ подвергается заметному изменению. Третья стадия форми-
рования сообщества характеризуется не только значительным раз-
нообразием популяционно-видового состава растений и соответ-
ствующих гетеротрофов, но и дифференциацией их на ценозообра-
зующие фитоценотипы — доминанты, субдоминанты, сопутствую-
щие виды. Фитоценоз вследствие этого приобретает надлежащее
структурное оформление и внешний облик, хотя и не является еще
вполне сложившимся как в отношении видового состава, так и сре-
ды обитания, особенно почвенного покрова.
Завершается процесс сингенеза образованием относительно ста-
бильного сообщества, обладающего свойственным ему составом
живых и косных компонентов, структурно-функциональной органи-
зацией, сложной системой разносторонних связей и механизмов
саморегуляции.
Сходным образом сингенетический процесс формирования фито-
ценозов на не освоенных жизнью рыхлых субстратах трактует
А. П. Шенников (1964). Выделяемые им стадии сингенеза по су-
ществу близки тем, что выделяются Б. А. Быковым, но по-иному
названы: 1) экотопическая группировка растений раздельного сло-
жения; 2) открытый фитоценоз раздельно-зарослевого сложения:
3) сомкнутый фитоценоз относительно диффузного сложения. ?На
первых двух стадиях естественный процесс формирования фитбЦе-
ноза, по А. П. Шенникову, протекает под контролем экотоп и че-
ского отбора, так как на первой стадии взаимовлияние меж-
ду разобщенными куртинами растений отсутствует, а на второй
еще слабо выражено; поэтому подбор растительных видов происхо-
дит на основе способности каждого из них произрастать в данном
экотопе. На третьей стадии и отчасти на второй, в связи с возра-
станием сомкнутости растительного покрова и взаимовлияния
растений, экотопический отбор перерастает в фитоценотиче-
ский. Последний совершается на основе способности раститель-
ных видов произрастать совместно с другими видами, занимая в
сообществе экологические ниши в соответствии с присущими им
эколого-ценотическими свойствами; это ведет к увеличению видо-
вого разнообразия растений и сопутствующих. им гетеротрофов.
Следовательно, фитоценотический отбор, в отличие от экотопиче-
ского, способен, по словам А. П. Шенникова, не только ограничи-
вать набор видов в сообществе, но и увеличивать и даже заменять
его новым набором.
Следствием увеличения видового разнообразия и численности
326
особей видовых ценопопуляций в сообществе является а) возра-
стание взаимовлияния ценозообразующих растений и б) превра-
щение экотопа под их воздействием во вторичную среду — фито-
среду (биотоп). Следовательно, «формирование фитоценоза и
свойственного ему биотопа, — заключает А. П. Шенников (1964),—
происходит одновременно» (с. 32—33). Более того, поскольку син-
генез представляет собой процесс становления не только раститель-
ного сообщества, но и трофически связанного с ним гетеротрофно-
го населения, то «формирование фитоценоза, — продолжает
А. П. Шенников,— является одновременно и формированием био-
ценоза и обособлением биогеоценоза* (с. 35—36).
Большого внимания заслуживает и приведенный А. П. Щенни-
ковым перечень факторов формирования фитоценоза, а следова-
тельно, и биогеоценоза на свободных субстратах, действие кото-
рых, как он полагает, продолжается и в сформировавшемся ценозе:
а) геологическая и культурная история района; б) экологическая
специфика зарастающего участка (экотопа); в) видовой состав
исторически сложившейся и пришлой флоры района, особенно бли-
жайшей к зарастающему участку; г) способность растений к раз-
множению и расселению; д) биологические особенности и эколо-
гические потребности растительных видов; е) степень напряженно-
сти экотопического отбора; ж) степень и способы влияния растений
на экотоп; з) экологическая специфика возникающей вторичной
фитогенной или шире — биогенной среды (биотопа) ; и) напряжен-
ность фито- или биоценотического отбора; к) наличие новых
мигрантов, выживающих и размножающихся в новой фитосреде;
л) интенсивность вымирания видов из числа первых мигрантов,
оказавшихся нежизнеспособными в этой фитосреде; м) микрофло-
ра, микро-, мезо- и макрофауна беспозвоночных и позвоночные
животные, участвующие в ф°РмиРовании фитоценоза и биогеоце-
ноза в целом.
Что же касается формирования биогеоценозов на так называ-
емых вторичных экотопа х, освободившихся после уничтоже-
ния существовавших на них биогеоценозов, то этот процесс харак-
теризуется рядом особенностей по сравнению с сингенезом на
«новых субстратах» — первичных экотопах. Специфика его опреде-
ляется тем, что на вторичных экотопах после уничтожения биогео-
ценотического покрова остаются заметные следы его былого су-
ществования (обогащенный органикой и минеральными веществами
почвенный слой, семена и другие репродуктивные зачатки расте-
ний, почвенная микрофлора и почвообитающие беспозвоночные
животные и пр.). Поэтому формирование биогеоценоза в данном
случае сводится в основном к восстановлению прежнего биогеоце-
ноза, хотя и не тождественного с исходным. Причем в редких слу-
чаях прежний, коренной биогеоценоз восстанавливается сразу
327
в первом поколении. Чаще этот процесс протекает через целый
ряд сменяемых одного другим промежуточных биогеоценозов,
называемых производными. В силу этих особенностей этот
процесс, растягивающийся на многие десятилетия, а то и столетия,
связанный со сменой (сукцессией) разных биогеоценозов, выходит
за рамки сингенеза — процесса формирования одного определен-
ного биогеоценоза, а потому большинством биогеоценологов пере-
водится в категорию динамики биогеоценозов, называемую эндо-
генезом (или эндоэкогенезом) (рис. 38).
1-3 ГОДА
4 -9ЛЕТ
10-15 ЛЕТ
60-80 ЛЕТ
00 -120 ЛЕТ
Рис. 38. Возобновление елового леса (через смену березняка)
(по Г. А. Новикову, 1979)
4. Эндогенез, эндогенные сукцессии биогеоценозов
и движущие силы развития биогеосферы
Однако, по представлениям В. Н. Сукачева (19646), на этапе
сингенеза формирование биогеоценоза не заканчивается, ибо на
328
этом этапе, по его словам, изменение свойств косных компонентов
формирующегося биогеоценоза еще не столь велико, чтобы играть
существенную роль в изменении биогеоценотического процесса, и
в такой степени, чтобы можно было говорить о смене биогеоцено-
зов. Из этого замечания В. Н. Сукачева следует: процесс формиро-
вания биогеоценоза продолжается и на следующем этапе биогео-
ценотического процесса, получившего название эндогенеза. «С не-
которого времени, — пишет он, — эти изменения делаются столь
велики и начинают так сильно воздействовать на весь органический
мир, что и он изменяется, и движущей силой биогеоценотического
процесса становится совокупность всех тех изменений в биогеоце-
нозе, которые сопровождают взаимодействия всех его компонентов.
Этому длительному периоду в жизни биогеоценотического покрова
можно присвоить название эндогенеза» (с. 468).
Другими же отечественными ценологами, как мы видели, син-
генез рассматривается как процесс, конечным результатом которого
является становление растительного сообщества, а также связан-
ного с ним многообразными отношениями животного и микробного
«населения» и соответствующей внутренней среды — биотопа, иначе
говоря, — становление биогеоценоза со всеми присущими ему
атрибутами.
На наш взгляд, такая трактовка сингенеза правильна. На вто-
ром этапе биогеоценотического процесса, протекающего в форме
эндогенеза, по существу, происходит не продолжение формирова-
ния биогеоценоза, а дальнейшее развитие уже появившегося на
свет биогеоценоза, всестороннее совершенствование его.» В частно*
сти, происходит усложнение популяционно-видового состава, уста-
новление оптимального количественного соотношения ценозообра-
зующих видов, усложнение морфологической структуры и функцио-
нальной организации биогеоценоза и, следовательно, энерготрофи-
ческих, информационных и других взаимосвязей между живыми
компонентами системы, а также взаимосвязи их и биоты в целом
с биогенной средой (биоценосредой), установление циклической
динамики биогеоценоза, выражающейся в суточном и сезонном ре-
жимах биогеоценотического процесса, и, наконец, совершенству-
ются механизмы авторегуляции биогеоценоза, повышается устой-
чивость, гомеостаз его, и на этой основе возрастает созидательная
деятельность всей биокосной системы, степень вещественно-энерге-
тической сбалансированности, а также общий уровень и слажен-
ность функциональной деятельности ее. ^Перечисленные показатели,
совершенствования биогеоценоза, позитивного преобразования всех
его сторон составляют суть эндогенеза, внешним выражением ко-
торого является сукцессия, коренная смена биогеоценозов в опре-
деленной последовательности. |В процессе замещения одного био-
геоценоза другим и осуществляются отмеченные преобразования
329
биогеоценозов и прогрессивное развитие биогеоценотического по-
крова на данном отрезке земной поверхности.
Существенным отличием второго этапа биогеоценотического
процесса от первого заключается и в том, что в ходе сингенеза про-
исходит смена стадий становления, формирования биогеоценоза, а
при эндогенезе — смена сформировавшихся при сингенезе биогео-
ценозов как качественно обособленных в пространстве и во времени
биокосных систем» Биогеоценозы, образующие последовательный
ряд смены во времени, выступают, по словам В. Н. Сукачева, как
звенья поступательного развития (саморазвития) биогеоценотиче-
ского покрова. Происходящие в процессе эндогенеза смены биогео-
ценозов во времени названы им эндогенными сукцес-
сиями.
Биогеоценотический процесс, по В. Н. Сукачеву, определяется
четырьмя частными процессами: во-первых, внутренним процессом
взаимодействия компонентов биогеоценоза и составных элементов
каждого из них; во-вторых, заносом из окружения ветром, водой,
некоторыми организмами репродуктивных зачатков растений и все-
лением в ценоз новых видов гетеротрофов; в-третьих, приносом со
стороны минеральных и отчасти органических веществ с пылью,
водой и т. п.; в-четвертых, выносом из биогеоценоза минеральных
и органических веществ водой и другими агентами.
Основной причиной и движущей силой биогеоценотического
процесса, в частности эндогенеза, В. Н. Сукачев считал внутрен-
нюю противоречивость взаимных связей и взаимоотношений ком-
понентов биогеоценоза. Н. В. Дылис (1978) указывает, что в силу
относительной самостоятельности компонентов биогеоценоза в обра-
зе жизни, поведении, работе и динамике, изменения их не могут
совершаться абсолютно «синхронно и адекватно» (с. 116); нередки
случаи отставания изменений одних компонентов от соответствую-
щих изменений других (например, почвы от растительности); име-
ют место несоответствия и в глубине перестройки их и пр. «Отсут-
ствие в биогеоценотических системах полной гармонии в связях и
взаимооотношениях отдельных компонентов и, наоборот, наличие
известной несогласованности между ними во времени и масштабе
действий,— пишет он, — служит постоянным источником спонтан-
ного саморазвития биогеоценозов» (с. 117).
В качестве примера саморазвития биогеоценотического покрова,
протекающего в форме эндогенеза в силу внутренних, заложенных
в самом биогеоценозе причин, обычно приводится постепенное з а-
растание неглубокого пологосклонового континентального во-
доема (озера). Но, пожалуй, наиболее убедительным, хотя и менее
наглядным примером такого явления может служить так назы-
ваемый дерново-луговой процесс В. Р. Вильямса. Со-
гласно этой теории, возникший в лесолуговой (лесостепной) зоне
330
на месте леса луг проходит в своем развитии три периода
(стадии)—корневищный, рыхлокустовой и плотнокустовой, сме-
няясь затем болотом. Господствующее в каждый из этих периодов
луговое растительное сообщество в процессе своего существования
постепенно преобразует среду обитания (почву и пр.) в соответ-
ствии с потребностями доминирующих в нем растений. Но изме-
няющаяся биогенная среда, будучи вначале благоприятной, с те-
чением времени становится все менее и менее пригодной для них,
но зато благоприятной для луговых растений с другими эколого-
ценотическими свойствами, обилие которых в силу этого возра-
стает, и они с течением времени становятся господствующими в
данном сообществе. Иными словами, происходит коренная смена
одного лугового растительного сообщества другим. Следовательно,
луговое сообщество в процессе своего существования на определен-
ном участке территории само подготавливает гибель для себя и
создает благоприятные условия для существования и развития
другого, идущего на смену ему сообщества.
Эндогенный процесс смены лугового сообщества одного периода
сообществом другого, согласно теории В. Р. Вильямса, ярко и сжа-
то описан А. П. Шенниковым (1941). Приведем это описание пол-
ностью.
«В корневищной стадии господствуют автотрофные длиннокор-
невищные злаки (пырей ползучий,вейник наземный и т.п.—Л.Н.).
После отступления леса они находят условия, благоприятные для
их массового и пышного развития: рыхлую почву, хорошую аэра-
цию поверхностного слоя почвы, обилие минерального питания за
счет разлагающейся лесной подстилки. Отмирающие корни и дру-
гие мертвые остатки корневищных злаков в течение лета разлага-
ются при посредстве аэробных бактерий (аэробиозис). Осенью,
когда почва насыщена водой, аэробиозис сменяется анаэробиози-
сом, разложение замедляется, а с понижением температуры и
совсем прекращается. Весной, когда в просохшую почву снова про-
никнет воздух, кислород его будет перехвачен корнями нового по-
коления злаков, а прошлогодний остаток неразложившейся орга-
нической массы так и останется без дальнейшего разложения.
С годами масса органических веществ в почве увеличивается.
Минеральные соли частично переходят в органоминеральные
соединения, и почва делается беднее легкодоступными питатель-
ными веществами. Вдобавок новые корневища, развиваясь ежегод-
но выше прошлогодних, наконец оказываются слишком близки к
поверхности почвы и чаще страдают от чрезмерного высыхания
летом. Глубже для них не хватает кислорода. Это ведет к угнетению
и изреживанию корневищных злаков. Между ними поселяются
рыхлокустовые, как более приспособленные к создавшимся усло-
виям (овсяница луговая, мятлик луговой и пр.— Л. Н.). Их кор*
331
яевища, молодые корни, почки, низовые листья находятся у самой
поверхности почвы и не страдают от недостатка кислорода. Корни
достигают большей глубины, используют более глубокие слои поч-
вы. Между злаками поселяются бобовые, обогащающие почву азо-
том. Так формируется рыхлокустовой луг. Именно в эту стадию
луг имеет, по Вильямсу, типичную дерновую почву.
Однако накопление органических веществ, обладающих свой-
ствами коллоидов, продолжается. От этого влагоемкость почвы
увеличивается; весной и осенью, да и летом после дождей, в сырой
почве коллоиды разбухают и прекращают доступ воздуха в почву.
По мере увеличения массы гумуса высыхание его затрудняется и
периоды аэробиозиса делаются все короче. Дело доходит до того,
что почва почти все время остается в условиях анаэробиозиса.
В этих условиях рыхлокустовые злаки не могут более господство-
вать на лугу. Наступает новая стадия: преобладание растений,
снабженных системой межклетных ходов, по которым воздух, про-
никающий в растение через устьица, проникает к растущим кон-
чикам корней. Таковы плотнокустовые злаки (луговик дернистый,
белоус торчащий и др. — Л. Н.) ...Имея узлы кущения над поверх-
ностью анаэробной почвы, обладая глубокими корнями с проводя-
щими воздух межклетниками и микоризой, достаточно олиготроф-
ные плотнокустовые луговые злаки могут жить в условиях полного
анаэробиозиса почвы и без подтока минеральных соединений.
В плотнокустовой стадии органические вещества накопляются и
на поверхности почвы, что и приводит, наконец, к смене луга бо-
лотом» (курсив наш. — Л. Н.) (с. 262—263).
Из изложенного следует, что основным фактором саморазвития
луговой растительности в лесолуговой зоне, согласно теории
В. Р. Вильямса, являются накопление коллоидных органических
веществ при гумификации отмерших растительных остатков и свя-
занные с накоплением гумуса анаэробность и физиологическая бед-
ность почвы. Однако в разных физико-географических и эдафиче-
ских условиях этот эндодинамический процесс протекает по-разно-
му и сопровождается разными последствиями. В степной зоне свя-
занная с накоплением гумуса физиологическая сухость луговых
почв приводит, по А. П. Шенникову, к остепнению лугов и смене
их степной растительностью, а в северотаежной зоне, лесотундре
и тундре накопление сырого и холодного гумуса и связанное с этим
ухудшение теплового режима ведет к смене лугов психрофильной
растительностью (пустошью).
Нашими исследованиями пойменных лугов Енисея (Л. И. Но-
моконов, 1959), помимо названных трех периодов (стадий) дерно-
во-лугового процесса, выявлена еще корневищно-рыхлокустовая
стадия — промежуточная между первой и второй (из мятлика
узколистного, овсяницы красной, лисохвоста лугового и др.). Кроме
332
того, установлено, что в проявлении дерново-лугового процесса, в
зависимости от климатических особенностей и элементов рельефа
поймы, отчетливо выделяются шесть типов (рядов) задер-
ней и я их: 1) мезофитный тип (ряд) задернения на равнинных
местоположениях рельефа енисейской поймы, наиболее характер-
ный для лиственно-лесной зоны, средней тайги и северной лесо-
степи; 2) ксерофитный ряд на положительных формах рельефа,
осуществляется в пойме безлесно-степной зоны и южной лесостепи;
3) психрофитный ряд характерен для северотаежной зоны и
лесотундры, В отрицательных формах рельефа поймы в аэробно
заболоченных условиях развивается 4) гидромезофитный ряд за-
дернения почти на всем протяжении енисейской поймы; 5) гидро-
сксилофитный ряд встречается в тех же зональных пределах, но
в анаэробных условиях; 6) гидрогалофитный ряд задернения лугов
наиболее ярко проявляется в неглубоких карбонатно- и галоидно-
засоленных пойменных понижениях в безлесно-степной зоне и от-
части в южной лесостепи. Характерно, что в рядах задернения на
равнинных местоположениях и гривах наблюдается тенденция не-
посредственного перехода корневищных лугов в процессе эндоге-
неза в рыхлокустовые, тогда как в рядах задернения в понижениях
поймы этот переход (смена) всегда осуществляется через посред-
ство корневищно-рыхлокустовых луговых формаций. Очевидно,
природа корневищно-рыхлокустовой луговой экобиоморфы в своем
происхождении связана с режимом переменного увлажнения в ука-
занных условиях.
Таким образом, поступательное развитие биогеоценотического
покрова на том или ином участке земной поверхности, начавшись
с сингенеза, протекает далее в форме эндогенеза, т. е. последова-
тельной смены (сукцессии) биогеоценозов в определенном направ-
лении. Этот внутренне противоречивый процесс развития биогеоце-
нотического покрова В. Н. Сукачев рассматривал как непрерывный
процесс перестройки биогеоценозов, т. е. разрушения старых био-
геоценозов и возникновения на их месте новых биогеоценотических
систем.
Процесс качественной перестройки существующего биогеоценоза
и смена его новым, как мы видели, весьма сложный, строго детер-
минированный процесс. При этом перестройке подвергается не
только популяционно-видовой состав живых компонентов (в пер-
вую очередь автотрофного растительного), но и так называемых
косных (биогенных) компонентов, в особенности почвенно-грунто-
зого (эдафотопа). Смена состава живых компонентов биогеоценоза,
как было показано, протекает в форме конкуренции за средства
жизни видовых ценопопуляций каждого из них под углом наиболь-
шего соответствия наличной экологической нише в структурно-
функциональной системе формирующегося биогеоценоза. Иначе го-
333
воря, этот процесс протекает под контролем биоценотическо-
го отбора автотрофных, биотрофных и сапротрофных ценопо-
пуляций, наиболее отвечающих по своим качественным особенно-
стям и свойствам новой совокупности сложившихся под их воздей-
ствием условиям среды обитания. Таким образом, формирование
нового биогеоценоза в недрах старого — это, по существу, синге-
нетический процесс становления нового биогеоценоза на базе
наличной и отчасти пришлой флоры и фауны, в ходе которого мож-
но обнаружить и стадию приживания идущих на смену старым но-
вых видов к изменившимся условиям биоценосреды, и стадию ежи-
вания их друг с другом при совместном обитании, т. е. вступления
их в разнообразные связи и отношения, обеспечивающие слажен-
ность составных элементов и структурно-функциональных частей
биогеоценотической системы и нормальное функционирование ее.
5. Экзогенные сукцессии и подразделения их по причинам,
скоротечности и продолжительности
Но поступательный эндодинамический автогенный процесс раз-
вития биогеоценотического покрова время от времени нарушается
или даже прерывается изменениями его, происходящими под дей-
ствием природных или производственно-хозяйственных (антропо-
генных) факторов. Эти изменения биогеоценотического покрова,
вызванные внешними по отношению к нему причинами, В. Н. Су-
качев назвал экзогенными.'Они, как и эндогенные изменения,
сопровождаются сменой исходного биогеоценоза, правда, не только
постепенной, но и разовой (катастрофической). Экзогенные сукцес-
сии, как отмечает Н. В. Дылис (1978), необязательны для биогео-
ценоза, так как не вытекают из внутренних закономерностей его
поступательного развития. Они могут быть, по его словам, «обра-
тимыми и необратимыми — направленными, длительными и быстро-
течными, локальными, местного значения и могут охватывать об-
ширные пространства Земли» (с. 119). Разнообразны они и по
другим показателям (по форме, масштабу, скорости и т. п.).
Экзогенные сукцессии (смены) В. Н. Сукачевым подразделяют-
ся на две группы: 1) локальные (местные) сукцессии, происхо-
дящие вследствие природных процессов, явлений того же порядка
и антропогенных воздействий. Среди них имеются как необратимые
смены, так и обратимые, если факторы, вызвавшие их, исчезают
или действие их прекращается; 2) гологенетические
сукцессии, вызванные изменениями природных единств более об-
щего порядка, не контролируемых человеком. Они, подобно эндо-
генным сукцессиям, необратимы. Среди локальных сукцессий, в за-
висимости от вызвавших их причин, В. Н. Сукачевым и Н. В. Ды-
лисом различаются: а) зоогенные смены биогеоценозов, вызванные
334
выпасом диких и домашних животных, налетом саранчи или наше-
ствием мышевидных грызунов и других массово размножающихся
беспозвоночных и позвоночных животных; б) фитогенные смены,
вызванные внедрением в биогеоценозы некоторых быстро размно-
жающихся видов, растений (канадская элодея, водный гиацинт
и пр.); в) пирогенные смены, происходящие на участках, подверг-
шихся лесным пожарам или степным «палам»; г) ветровальные
смены лесных биогеоценозов на крутых склонах горных местностей
с малоразвитым почвенным покровом;; д) селевые и оползневые
смены по склонам древних речных террас; е) антропогенные смены,
связанные с рубками леса, распашкой или осушением и орошением
земель, внесением удобрений, различных пестицидов и пр. Б. А. Бы-
ков (19706) и Н. В. Дылис (1978) вполне обоснованно считают це-
лесообразным антропогенные смены, происходящие под влиянием
хозяйственной деятельности местного населения, выделить в осо-
бую, самостоятельную группу экзогенных сукцессий, поскольку по-
следние специфичны по своему характеру, а размах и интенсив-
ность воздействия их на биогеоценозы в настоящее время достига-
ют колоссальных размеров и сопровождаются глубокими измене-
ниями биогеоценотического покрова местности.
Гологенетические сукцессии менее разнообразны по сравнению
с локальными. К этой группе сукцессий названными выше автора-
ми относятся: а) климатогенные смены, обусловленные планетар-
ными или региональными изменениями климата, охватывающие,
как правило, значительные промежутки времени, а потому их обыч-
но относят к так называемым вековым сменам, например, смена
различных типов ельников березово-сосновыми биогеоценозами,
сменяющимися затем сосново-широколиственными, а южнее — ши-
роколиственными типами биогеоценозов в связи с отступанием по-
следнего оледенения в центральной части Русской равнины; б) гео-
морфогенные смены, являющиеся следствием тектонических и дру-
гих эндогенных процессов земной коры, а также суффузионных и
прочих процессов, изменяющих основные формы макрорельефа и
базис эрозии речных долин того или иного региона суши; напри-
мер, смена ивово-тополевого биогеоценоза в речной пойме лесостеп-
ной полосы ильмовым или ильмово-дубовым (из Ulmus scabra)
в связи с понижением уровня почвенно-грунтовых вод в результате
глубинной эрозии русла реки; такие смены также весьма долго-
срочны; в) к этой же группе сукцессий В. Н. Сукачев относит также
селектоценогенетические смены, связанные с расселением растений
и животных, которые, внедряясь при расселении в существующие
в данной местности биогеоценозы, нередко постепенно вытесняют
ценозообразующие аборигенные виды, формируя на их месте иные,
новые биогеоценозы.
Локальные экзогенные сукцессии являются большей частью вне-
335
запными, катастрофическими, так как связаны с уничтожением
(сведением) растительного компонента коренного биогеоценоза»
что, естественно, влечет за собой исчезновение свойственной ему
наземной фауны, а с течением времени и изменение состава
почвенной микрофлоры, микро- и мезофауны и самого почвенного
покрова, включая условия увлажнения, минерального питания
растений и т. п. Такие смены ценозов получили название ката-
строфических, П. Д. Ярошенко (1961) назвал их внезап-
ными. Лишь немногие смены этой группы сукцессий протекают
медленно, постепенно; например, в результате неумеренного выпаса
скота в степной зоне коренные крупнодерновинные ковыльные био-
геоценозы постепенно сменяются мелкодерновинными тонконогово-
типчаковымн, на смену которых приходят смешанные злаково-раз-
нотравные ценозы, сменяющиеся, в свою очередь, редкостойным
сорно-разнотравным «сбоем». Такого рода смены биогеоценозов
В. Д. Александровой (1964) названы сменами развития, а П. Д. Яро-
шенко— последовательными. Те и другие локальные экзо-
генные сукцессии являются, как правило, обратимыми. Процесс
разрушения или существенного нарушения состава и структуры
коренных биогеоценозов под действием природных или антропоген-
ных внешних для каждого из них факторов Г. Н. Высоцким (1915)
назван дигрессией, а процесс восстановления их до состояния,
близкого к исходному, — демутацией. Восстановительный про-
цесс биогеоценозов во всех случаях протекает относительно мед-
ленно, постепенно, через ряд промежуточных стадий, смен, назы-
ваемых демутацианными, которые по своему характеру являются
эндогенными сменами.
В. Н. Сукачев (19646), приведя схему классификации форм ди-
намики биогеоценозов (с. 482—483), обращает внимание на следу-
ющие обстоятельства: 1) процесс биогеоценогенеза — все-
объемлющий и непрерывный процесс, охватывающий как цикличе-
ские (периодические) изменения, так и коренные автогенные и
экзогенные сукцессии, смены их; 2) в каждом конкретном случае
изменения и смены биогеоценозов являются «результатом одновре-
менного действия нескольких причин, и различные формы смен
накладываются одна на другую» (с. 486). П. Д. Ярошенко (1961)
свою мысль о взаимосвязи всех форм динамики ценозов выразил
в следующих словах: «...собщество в каждый момент своей жизни:
1) проходит одну из фаз своего сезонного развития; 2) отражает
на себе влияние условий данного года; 3) находится на пути к
смене каким-либо другим сообществом; 4) находится под влиянием
общей смены растительного покрова всей местности; 5) проходит
одну из стадий своей эволюции» (с. 195).
Л. Г. Раменский (1938), Е. М. Лавренко (1959) и другие оте-
чественные биологи группируют сукцессии биогеоценозов также
336
по скорости» продолжительности процесса смены. Все типы сукцес-
сий биогеоценозов они подразделяют на этом основании на три
группы: 1) быстротечные смены, происходящие в течение не-
скольких десятилетий, например возрастные стадии древесных це-
нозов, обусловленные изменениями возраста доминирующих по-
род; 2) длительные смены продолжительностью в несколько со-
тен лет, например смены производных березовых ценозов еловыми
при восстановлении коренных типов ельников после их вырубки
или лесных пожарищ; 3) вековые смены биогеоценозов в про-
цессе эволюции биогеоценотического покрова, связанные с эволю-
цией флоры и фауны на огромных пространствах земного шара,,
длительность которых исчисляется тысячелетиями, а то и десятка-
ми и сотнями тысяч лет.
Наличие тех или иных вековых или общих (по П. Д. Ярошен-
ко) смен биогеоценозов при реконструкции биогеоценотического
покрова прошлых времен устанавливается различными способами:
по ископаемым растительным и животным остаткам (в виде ока-
менелостей, отпечатков и т. п.); по наличию в составе современных
биогеоценозов реликтов древнего растительного покрова и живот-
ного мира; по данным пыльцевого анализа и пр. Так, а) на основа-
нии находок в ископаемом состоянии дриады (Dryas octopetala) в
окрестностях Ленинграда ученые пришли к заключению о суще-
ствовании здесь в далеком прошлом тундровой растительности;
б) по распространению в современном биогеоценотическом покрове
советского Приморья, а также в Колхиде и Ленкорани Закавказья
реликтов третичной флоры и фауны сделано заключение о суще-
ствовании в прошлом в этих местах третичных ценозов.
6. Теория климакса, критика ее и современные взгляды
на устойчивость и динамику биогеоценозов
Стабильное сообщество, завершающее серию сукцессий (смен)»
американским экологом Ф. Клементсом (Clements, 1916) названо
климаксным, или просто климаксом, находящимся в равнове-
сии с физическим метоообитанием. Основные положения теории
климакса Ф. Клементса следующие. 1. Основной единицей расти-
тельности является формация, которая представляет собой «слож-
ный комплексный организм» («квазиорганизм», по А. Тенсли). По-
добно организму, формация возникает, растет, зреет и умирает,
проходя при этом стадии: детство, юность, зрелое состояние,
старость, дряхлость и смерть. 2. Развитие формации протекает в
процессе сукцессии от пионерной стадии через промежуточные к
конечной, заключительной — к климаксу. 3. Сукцессия с необхо-
димостью идет в направлении от ксерофитной и гигрофитной расти-
тельности к мезофитной. Климакс и представляет собой наиболее
337
мезофитный тип, который данный климат может нормально под-
держивать. 4. Климаксы изменяются вследствие существенного из-
менения климата той или иной физико-географической зоны. Такие
зональные климаксные сообщества названы климатическими
климаксами, а развиваемая Клементсом теория климакса —
теорией моноклимакса.
Таким образом, климакс в представлении Ф. Клементса и неко-
торых его современников за рубежом — это абсолютно устойчивая
конечная формация сукцессионной серии, с достижением которой
развитие растительности прекращается. Толчком к дальнейшему
развитию растительности может стать лишь глубокое изменение
климата.
Позднее было установлено, что в пределах той или иной физико-
географической зоны наряду с климатическим климаксом нали-
чествует целый ряд климаксных сообществ, связанных с различны-
ми формами рельефа, особенностями почвенного покрова и т. п.,
варьирующих в соответствии с градиентами условий среды. Такого
рода климаксные сообщества стали называться субклимакса-
ми. На основе этого сформировалось представление о мозаике
климаксных сообществ в пределах зоны или более или менее круп-
ного территориально обособленного географического ландшафта.
Это, в свою очередь, послужило основанием для появления теории
поликлимакса, нашедшей среди зарубежных биологов немало сто-
ронников.
По Ю. Одуму (1975), стратегия развития экосистемы (биогеоце-
ноза) определяется следующими тремя параметрами: 1) это есть
упорядоченный процесс развития сообщества (биоценоза), связан-
ный с изменениями во времени видового состава и протекающих в
сообществе процессов, он определенным образом направлен и пред-
сказуем; 2) развитие (или сукцессия) происходит в результате из-
менений физической среды под действием сообщества; иначе гово-
ря, сукцессия контролируется сообществом, т. е. совокупностью
живых компонентов, несмотря на то, что физическая среда опре-
деляет характер сукцессии, скорость изменения, а часто и пределы,
до которых может дойти развитие; 3) кульминацией развития яв-
ляется стабилизированная экосистема, в которой «на единицу име-
ющегося потока энергии приходится максимальная биомасса (или
высокое содержание информации) и максимальное количество сим-
биотических связей между организмами» (с. 324).
Заслуживает внимания и следующее замечание Ю. Одума: «За-
мещение видов в сукцессиях вызывается тем, что популяции, стре-
мясь модифицировать окружающую среду, создают условия, бла-
гоприятные для других популяций; это продолжается до тех пор,
пока не будет достигнуто равновесие (осносительное соответствие.—
Л. Н.) между биотическими и абиотическими компонентами»
338
(с. 324). «Одним словом, — заключает автор,— «стратегия» сукцес-
сии, как быстро протекающего процесса, в своей основе сходна со
«стратегией» длительного эволюционного развития биосферы: уси-
ление контроля над физической средой (или гомеостаз со средой)
в том смысле, что система достигает максимальной защищенности
от резких изменений среды» (с. 324).
Для климаксного сообщества, по Ю. Одуму, характерно то, что
годовая продукция и приток вещества извне уравновешиваются го-
довым потреблением, расходом и выносом веществ в окружающее
пространство; поэтому чистая годовая продукция в нем отсутствует.
Р. Дажо (1975) приводит ряд закономерностей изменения в хо-
де сукцессии некоторых свойств биоценозов и экосистем (биогеоце-
нозов) в целом в направлении к климаксному ценозу независимо
от типа сукцессии: а) первоначально линейные пищевые цепи с
преобладанием растительноядных животных превращаются в слож-
ные пищевые сети, где все большую роль начинают играть детрито-
ядные формы; б) экологические ниши все более сужаются и под-
вергаются специализации; размеры организмов растут, биологиче-
ские круговороты удлиняются и усложняются; в) общее количество
органических веществ, скопившихся в экосистеме, на первых порах
невелико, но неуклонно возрастает, одновременно увеличивается
видовое и биохимическое разнообразие; г) отношение «валовая
продуктивность — дыхание», обычно превышающее единицу в био-
ценозах-пионерах, стремится к единице в климаксных био-
ценозах; д) отношение «валовая продуктивность — биомасса», вна-
чале высокое, начинает уменьшаться, оно соответствует скорости
обновления биоценоза; е) отношение «биомасса — энергетический
поток», напротив, по мере приближения к климаксному биоценозу
увеличивается.
Таким образом, когда выяснилось, что биоценозы и образуемые
ими биокосные системы представляют собой открытые системы,
непрерывно изменяющиеся во времени, концепция климаксного со-
общества, как некоего неизменного «квазиорганизма», подверглась
критике и претерпела значительные изменения, а некоторые зару-
бежные экологи, по утверждению Р. Риклефса, вовсе отказались
от нее.
Еще более резкой критике концепция климакса Ф. Клементса
подверглась в недалеком прошлом со стороны советских фитоцено-
логов, в особенности представление о завершающем серию смен це-
нозе как заключительном, конечном, застывшем в неподвижности,
дальнейшее развитие которого возможно лишь под действием
резких климатических перемен или каких-либо других широко-
масштабных и кардинальных причин (например, внедрение в ценоз
новых, более конкурентомощных видов и т. п.). Критиковались и
такие стороны ее, как аналогизация развития ценоза с онтогенезом
339
отдельного организма н пр. В силу этого советские ценологи
В. Н. Сукачев (1942), П. Д. Ярошенко (1949), А. П. Шенников
(1964) и др. сочли целесообразным отказаться от термина «кли-
макс», заменив его такими терминами, как выработавшийся
или узловой ценоз. Этим самым подчеркивалось отрицатель-
ное отношение к заложенному в термине «климакс» представлению
об окончании развития растительного или биогеоценотического по-
крова по достижении этого этапа; с другой стороны, указывалось
на относительный характер стабильности, устойчивости ценозов на
этом этапе,
В представлении советских ценологов развитие растительного
или биогеоценотического покрова с завершением той или иной се-
рии смен фито- или биогеоценозов не прекращается, а замедляет-
ся, точнее, протекает как бы в скрытой форме. Растительное сооб-
щество или биогеоценоз в ходе фито- или биогеоценогенеза дости-
гает на стадии «узлового» («выработавшегося») ценоза своего
совершенства в компонентном составе, морфологической структуре,
функциональной организации, в деятельности механизмов авторе-
гуляции, обеспечивающей слаженное, устойчивое функционирова-
ние фитоценоза или биогеоценоза в течение неопределенно дли-
тельного времени, поскольку продолжительность их существования
не запрограммирована. На этом этапе в фито- или биогеоценозе
происходят разнообразные изменения количественного порядка:
а) в популяционно-видовом составе живых компонентов за счет
внедрения из окружения новых видов и исчезновения старых, ока-
завшихся менее приспособленными к изменившимся условиям био-
топа; б) в структуре и функциональной организации ценозов в свя-
зи с изменением количественного соотношения ценозообразующих
видов и характера взаимоотношений их между собой в составе
каждого ценокомплекса; в) во взаимосвязи биоценоза как совокуп-
ности живых компонентов с биоценосредой и всего биогеоценоза
с биотическим и абиотическим окружением и пр.
Все эти малозаметные сами по себе количественные изменения
в рамках существующего «узлового» растительного сообщества или
биогеоценоза, постепенно накапливаясь, все более и более наруша-
ют его относительное равновесие, и далее с течением времени, как
отмечает П. Д. Ярошенко, они становятся настолько значительны-
ми, что выработанность ценоза, слаженность всех его элементов
совершенно расшатывается. Все это в конечном итоге ведет к ка-
чественному изменению «узлового» фито- или биогеоценоза, т. е. к
коренной смене его иным, новым ценозом, начинаю-
щим собой новую серию смен растительных сообществ, или биогео-
ценозов. Следовательно, «узловой» фито- или биогеоце-
ноз — это не что иное, как достигшее наибольшего совершенства
и стабильности звено в поступательном процессе фито- или био-
340
Сингенез
(становление БГЦ)
Эндогенез
(усложнение и стабилизация БГЦ)
ЭКЗО ГЕ И ЕЭ
( Нарушение .
И ДИГРЕССИЯ БГЦ }
Ориг. рис. 39. Схема биогеоценотического процесса (биогеоценогенеза) (по данным В. Н. Сукачева и др.)
геоценогенеза, постулирующее прерывистость этого процесса
во времени и утверждающее тем самым качественную определен-
ность данного биогеоценоза.
Ход биогеоценотического процесса (биогеоценогенеза) можно
иллюстрировать следующей схемой (рис. 39). На схеме видно, что
биогеоценогенез, как общий и относительно медленный
и длительный процесс, слагается из более быстрых частных
процессов — неогенезов (по В. Б. Сочаве, 1944). Таким обра-
зом, биогеоценогенез можно рассматривать как содержа-
ние процесса формирования биогеоценоза и последующего его со-
вершенствования и стабилизации; сингенез, эндогенез и
экзогенез — как формы проявления этого процесса во времени,
а неогенезы — как скоротечные этапы, звенья его. В качестве
таких звеньев биогеоценогенеза, как показали наши исследования
природных луговых биогеоценозов в пойме Нижнего Дона (Л. И. Но?
моконов и др., 1978), могут выступать так называемые эпипар-
целлы, приуроченные к определенным элементам микрорельефа
со свойственными им режимами увлажнения, минерального пита-
ния и т. п. Их, следовательно,- можно считать временными катего-
риями, развернутыми в пространстве в виде горизонтальных струк-
тур биогеоценозов (рис. 40).
Рис. 40. Смены ассоциаций пырейника через ценогенетические ряды эпипарцелл
(по Л. И. Номоконову, 1973). Эпипарцеллы основные: 1—дербенниково-пырей-
ная; 2 — бекманниево-дербенниково-пырейная. Эпипарцеллы дополняющие: 3 —
дербенниково-ясколково-пырейная; 4 — осоково-вербейниково-пырейная; 5—солод-
ково-дербенниково-пырейная; 6 — солодково-мятликово-пырейная; 7 — бекманние-
во-жерухово-пырейная
7. Понятие филоценогенеза и закономерности развития
растительности и биогеосферы
Перейдем ко второму разделу главы — эволюции биогеоцено-
тического покрова (биогеосферы) и его составных единиц — био-
342
геоценозов. Этот геоисторический процесс становления биогеоце-
нозов, их типов и других таксономических единиц, связанный с
изменением видов и их приспособлений, В. Н. Сукачевым (1928)
назван филоценогенезом. Он включает также процесс исто-
рического преобразования, развития биогеоценотического покрова
какой-либо обширной территории, акватории или в масштабе всей
планеты. Этот процесс охватывает временные отрезки в десятки,
сотни тысяч и даже миллионы лет. Другими отечественными, а
также зарубежными биологами обращается внимание, во-первых,
на то, что филоценогенез тесно взаимно связан с параллельно иду-
щими процессами флорогенеза и фауногенеза, которыми
определяется качественный (видовой) состав биогеоценозов; во-вто-
рых, что в процессе филоценогенеза происходит эволюционная вы-
работка типов биогеоценозов и других таксонов разного ранга, ко-
торая выражается в подборе популяционно-видового состава био-
геоценозов из существующих и вновь образующихся видов, в
установлении определенных количественных соотношений между
видами, в выработке определенного вертикального строения и го-
ризонтального сложения биогеоценозов и в становлении их функ-
циональной организации и пр.
Развивая свое представление о филоценогенезе как длительном
геоисторическом процессе становления и развития растительного,
а также и биогеоценотического покрова в целом, В. Н. Сукачев
вместе с тем отрицательно относился к идее П. Д. Ярошенко об
едином процессе развития растительности на нашей планете, осно-
ванной на учении В. Р. Вильямса (1936) об едином почвообразо-
вательном процессе на поверхности Земли.
С другой стороны, по поводу реальности существования в при-
роде геоисторического процесса филоценогенеза, признаваемого
большинством отечественных фитоценологов, со стороны некоторых
ученых имело место высказывание сомнений по этому поводу и
даже критические высказывания. Основными доводами, служивши-
ми основанием для этого, были следующие: а) растительные сооб-
щества (фитоценозы) не обладают свойством наследственности,
как организмы, следовательно, между ними нет и не может быть
генетических родственных связей; б) растительные сообщества воз-
никают каждый раз заново из наличной флоры, следовательно, нет
оснований для признания генетической, родственной преемствен-
ности между вновь возникшими и ранее существовавшими фито-
ценозами. Доводы эти, как видим, не лишены основания. Да и сам
Сукачев высказывал сходные мысли, которые, возможно, и послу-
жили ему основанием для отрицательного отношения к идее
В. Р. Вильямса об едином почвообразовательном процессе и к
представлению П. Д. Ярошенко об едином процессе развития рас-
тительного покрова Земли.
343
Как же в действительности протекает процесс развития расти-
тельного и биогеоценотического покрова? На наш взгляд, при об-
суждении этого вопроса следует исходить из ряда следующих
положений и вытекающих из них выводов.
I. Всякий процесс ограничен в пространстве и во времени. Бес
конечна только материя. По отношению к развитию биогеоценоти
ческого покрова это означает следующее. А. Развитие флоры i
фауны, с одной стороны, и биогеоценотического покрова — с другой
(а эти два процесса, как было отмечено, взаимосвязаны), в раз*
ных условиях климата, т. е. в разных ландшафтно-географических
зонах и провинциях, шло и идет по-разному. И действительно, nq
данным А. Н. Криштофовича (1936, 1946), дифференциация расти-)
тельного и» следовательно, биогеоценотического покрова, скажем,)
в пределах Евразии на зоны существует издавна (леса, степи, пу-»
стыни). Обусловлено это явление, как известно, определенным
распределением тепла и влаги на поверхности Земли вследствие
ее шарообразной формы.
Б. Развитие флоры и фауны, с одной стороны, и биогеоценоти-
ческого покрова — с другой, в разные геологические периоды исто-
рии Земли шло и идет также по-разному. И действительно, с пе-
риодом расцвета папоротникообразных, рыб и земноводных в
каменноугольный период палеозойской эры связано развитие и
широкое распространение по лику Земли биогеоценозов из хвощей»
плаунов и папоротников, в том числе и заболоченных лесных (ле-
пидодендроновых, сигилляриевых и различных типов папоротнико-
вых до 40 м высотой) с соответствующим животным и микробным
населением. С периодом расцвета покрытосеменных растений и
млекопитающих в третичный период кайнозойской эры связано
развитие и распространение (включая современный холодный
пояс) пышных третичных лесных биогеоценозов субтропического
характера. В современный четвертичный период кайнозоя с чередо-
ванием ледниковых и межледниковых фаз и установлением сезон-
ной ритмики климата связано развитие и распространение летне-
зеленых листопадных лесных биогеоценозов, а также луговых и
степных с отмирающими на зиму побегами, населенных современ-
ными биокомплексами животных и микроорганизмов.
Таким образом, в каждый геологический период и в каждой
ландшафтно-географической зоне биогеоценозы и биогеоценотиче-
ский покров в целом развиваются на базе наличной флоры и фауны
и «осколков» растительности и животного населения предшеству-
ющего периода.
II. Развитие биогеоценотического покрова есть спонтанный, са-
мопроизвольный процесс, основанный на его самодвижении, т. е.
развитие в силу внутренних причин и закономерностей, заложен-
ных в самом биогеоценозе. Но этот процесс совершается в тесной
344
связи биогеоценоза с внешним биотическим и абиотическим окру-
жением. Иначе говоря, развитие биогеоценотического покрова со-
вершается по законам материалистической диалектики, путем раз-
двоения единого на противоречивые противоположности и взаимо-
действия их, а именно на основе взаимодействия автотрофных и
гетеротрофных компонентов биогеоценозов, осуществляющих сози-
дание и разрушение органических веществ в процессе биологиче-
ского круговорота с использованием энергии солнечной радиации,
а также путем перехода количественных сезонных, разногодних и
прочих изменений в качественные преобразования биогеоценозов
в процессе сукцессий (смен) и серий смен, причем сукцессий, про-
текающих не по замкнутому кругу, а по спирали, с прогрессив-
ным развитием биогеоценозов и биогеоценотического покрова в
целом.
III. Биогеоценотические системы и биогеоценотический покров
той или иной зоны и провинции — это часть биогеосферы всего
земного шара, в развитии которой важную роль играют не только
региональные природные и антропогенные (хозяйственно-производ-
ственные) факторы, но и факторы общепланетарного и даже гло-
бального порядка. Иначе говоря, всегда следует иметь в виду и
брать в расчет всеобщую связь и взаимообусловленность всех про-
цессов, явлений в природе, происходящих на разных уровнях орга-
низации биокосных систем и с разной степенью протяженности их
в пространстве и во времени.
Из всего изложенного следует: а) в пределах того или иного
геологического периода и той или иной ландшафтно-географиче-
ской зоны и провинции имеет место филоценогенез, т. е. гео-
исторический процесс формирования и развития биогеоценозов раз-
ного типа и систематической принадлежности и биогеоценотиче-
ского покрова в целом; б) филоценогенез, как неимоверно
длительный геоисторический процесс, расчленен во времени на
множество биогеоценогенезов как ценозообразующих процессов
становления стабилизированных «узловых» («выработавшихся»)
биогеоценотических систем, являющихся этапами, венчающими
последовательные серии смен на пути поступательного развития
биогеоценотического покрова Земли; в) филоценогенеза как
единого процесса развития биогеосферы от первичных протобиогео-
ценозов в архее или палеозое до современных биогеоценотических
систем не существует и не может существовать. В самом деле, хотя
историческое преобразование биогеоценозов и преобразование флор
и фаун протекают в единстве, но в основе развития флор и фаун
лежит генеалогия растительных и животных организмов, и потому
флоро- и фауногенез уходят своими корнями в глубь первых дней
появления живых существ на Земле. Генеалогия же биогеоценозов
ограничена рамками образовавшихся флор и фаун, и потому нет
345
оснований для того, чтобы выводить происхождение и развитие,
скажем, господствующих в мезозое различных типов хвойных лес-
ных биогеоценозов от тех или иных типов биогеоценозов, обра-
зуемых различными видами папоротникообразных в каменноуголь-
ном периоде палеозойской эры. Нет оснований и для заключения
о происхождении и развитии современных березовых и осиновых
мелколиственных лесных биогеоценозов Северного полушария от
господствующих здесь в третичном периоде кайнозоя магнолиевых,
каштановых, миртовых и других лесных биогеоценозов субтропи-
ческого типа.
Следовательно, развитие биогеоценотического покрова (биогео-
сферы) Земли — это непрерывный процесс по флорогенезу и
фауногенезу, но прерывистый по ценогенезу, так как он
расчленен на ограниченные в пространстве и во времени филоцено-
генетические отрезки.
В общем, весь процесс развития биогеоценотического покрова
(оболочки) Земли, очевидно, можно представить в таком виде.
Биогеоценогенез— процесс становления и стабилизации био-
геоценозов— складывается из сингенезов и эндогенезов, а послед-
ние— из неогенезов. Подобно этому, ограниченный рамками геоло-
гических периодов и ландшафтно-географических областей и зон,
филоценогенез складывается из биогеоценогенезов, а гео-
исторический процесс развития, эволюции биогео-
ценозов и биогеоценотического покрова Земли в целом — из таких
филоценогенезов.
8. Современные представления
о сопряженном характере эволюции
всех форм жизни на Земле
Современные представления о происхождении и эволюции орга-
нического мира зиждутся на известных высказываниях В. И. Вер-
надского по этой проблеме. Во-первых, исходя из своего представ-
ления о совместном существовании суши и океана начиная с отда-
леннейших геологических эпох, В. Й. Вернадский (1967) писал:
«...попытки объяснить происхождение наземных организмов из
морских несостоятельны и фантастичны. Воздушная жизнь в рам-
ках геологического времени так же стара, как и морская» (с. 348).
Во-вторых, исходя из выявленных им биохимических принципов
«биогенной миграции атомов биосферы», Вернадский (1928) счи-
тал, что происхождение жизни на нашей планете не было случай-
ным явлением. Из возникших видов, писал он, «мог бы выжить и
войти в сложнейший биоценоз биосферы... тот, который был бы в
нем устойчив, т. е. который увеличивал бы биогенную миграцию
атомов биосферы» (с. 46). Следовательно, происхождение и эво-
346
вюция биологических видов осуществлялись в составе биоценозов,
под контролем биоценозов и совместно с ними.
В настоящее время большинство биологов, по крайней мере
этечественных, вслед за Вернадским считают, что все формы орга-
низации жизни на Земле произошли (в геологическом смысле)
одновременно, т. е. одновременно и сопряженно формировались и
организмы, и биологические виды, и первичные биоценозы (прото-
биоценозы), и первичная биосфера (биогеосфера). Такого рода
представление о сопряженном характере возникновения и развития
всех форм организации живой материи В. И. Вернадским (1960)
мотивируется тем, что огромные по объему и весьма разнообраз-
ные по характеру геохимические функции, выпавшие на долю жи-
вых существ с первых дней их появления, не могли бы быть выпол-
нены отдельными возникшими организмами и видами организмов.
В действительности одновременно в геологическом смысле все эти
формы жизни возникли и, следовательно, одновременно они и эво-
люпровали в дальнейшем.
Таким образом, современное представление о сопряженном ха-
рактере органической эволюции, как справедливо утверждает
В. В. Мазинг (1970), расширяет прежнее представление о ней как
эволюции организмов, видовых популяций и видов. В настоящее
время речь идет об эволюции и таких форм организации живой
материи, как биоценозы и их биокосные образования — биогеоце-
нозы, а также биострома (живое вещество, по Вернадскому) и
образуемые ею пленки жизни (т. е. биогеосфера). Все это застав-
ляет по-иному в настоящее время смотреть на весь ход эволюци-
онного процесса, на взаимосвязь всех четырех форм организации
жизни в ходе эволюции, а также на те структуры и явления, в ко-
торых сопряженный характер эволюции находит отражение. Ста-
новится совершенно ясным, что эволюционное преобразование рас-
тительных и животных видов связано с изменением и преобразо-
ванием других биологических и ценотических систем. Бесспорным
становится и то, что возникавшие на базе флоры и фауны того
или иного геологического периода ценотические системы в ходе гео-
исторического развития преобразовались, по словам В. В. Мазинга,
из случайных и кратковременных совокупностей организмов в
сложные стабильные, все более целостные и самоорганизующиеся
системы.
И. И. Шмальгаузен (1968) также считает, что, поскольку био-
логические и биокосные системы образуют единый ряд взаимосвя-
занных и иерархически соподчиненных единиц, историко-эволюци-
онные изменения и преобразования их всегда взаимообусловлены.
Элементарной эволюирующей единицей, как известно, является
видовая популяция, в которой, по его словам, протекают наиболее
существенные первичные процессы эволюционных преобразований.
347
Эти процессы, как он указывает, определяются взаимодействием
с биоценозом, который также не замкнут в себе, а, будучи связав
с внешней средой, образует биокосную систему, называемую био-
геоценозом.
Свое представление о сопряженном характере органической эво-
люции И. И. Шмальгаузен (1968) выразил в следующих словах:
«Если эволюирует организация особей, то меняются и популяции!
и виды в целом, а также биоценозы, в состав которых входят дан-
ные виды. Поэтому можно говорить и об эволюции высших биоло-
гических систем — популяций, видов и биоценозов в целом. Более
того... мы рассматриваем именно биогеоценоз как арену первичных
эволюционных преобразований» (с. 166).
Общий ход эволюции органического мира И. И. Шмальгаузен
рассматривал, по существу, как иерархическую систему взаимо-
связанных циклов эволюционных преобразований
разного ранга: а) элементарный цикл эволюционных преобразова-
ний видовой популяции, осуществляющийся в процессе микро-
эволюции путем «индивидуального» естественного отбора фено-
типов (особей) на основе внутрипопуляционного соревнования их
(точнее, это отбор генотипов через посредство отбора феноти-
пов); б) цикл эволюционных преобразований вида, совершаю-
щийся в процессе макроэволюции путем группового от-
бора популяций на основе внутривидового соревнования их; в) цикл
эволюционных преобразований биоценоза (и биогеоценоза), про-
исходящий путем отбора видов на основе межвидового соревнова-
ния внутри авто-, био- и сапротрофного ценокомплексов биогеоце-
нотической системы; г) цикл эволюционных преобразований био-
стромы (живого вещества, по Вернадскому) и биогеосферы в
целом, протекающий путем отбора фито- п биоценозов (естествен-
ного отбора 2-го порядка, по В. В. Ревердатто, 1935; и др.) внутри
биостромы в процессе эволюции, которая может быть названа м е-
гаэволюцией.
Разумеется, перечисленные циклы эволюционных преобразова-
ний находятся в состоянии постоянного взаимодействия, чем и
обеспечивается сопряженность эволюционного процесса биологиче-
ских и биокосных систем. Этот единый процесс эволюции биостро-
мы, биоценозов и видов живых существ Б. А. Быков (1970) пред-
ложил назвать биотоценогенезом.
Из того обстоятельства, что элементарной единицей органиче-
ской эволюции является видовая популяция, И. И. Шмальгаузен
(1968) приходит к заключению: «...закономерности эволюции, дей-
ствующие на популяционном уровне, образуют как бы «ядро» за-
кономерностей эволюции... биоценозов и биосферы в целом» (с. 99).
Но наряду с этим в природе совершаются и специфические для
эволюции биоценозов процессы. К ним, в частности, относится
348
совместная эволюция целого ряда видов и групп видов живых су-
существ, тесно взаимосвязанных друг с другом экологическими,
структурно-морфологическими, функционально-ценотическими и
другими формами взаимозависимости, например, таких, как а) ра-
стения-энтомофилы и их опылители насекомые; б) микотрофные
высшие растения и микоризные грибы; в) организм-хозяин и его
паразиты; г) растительные или животные экобиоморфы, относя-
щиеся к одной и той же экологической нише в биоценозе (и био-
геоценозе); д) так называемые консорты — организмы, связанные
с автотрофным растением консорции пищевыми, симбиотическими
и другими отношениями; е) группы видов, входящие в состав одной
и той же структурной части биогеоценоза (биогеогоризонта, пар-
целлы); ж) функциональные группы живых существ, выполняющие
определенную функцию в обмене веществ и энергии в биогеоце-
нозе; з) виды и группы видов, трофически взаимосвязанные в пи-
щевых цепях (растения и животные-монофитофаги, хищник и жерт-
ва) и пр.
С. С. Шварц (1973), подчеркивая сопряженный характер орга-
нической эволюции, включающей наряду с филогенезом видов и
групп организмов также и эволюцию ценотических систем, обра-
щает внимание на то, что роль биогеоценоза в эволюционном про-
цессе нередко упрощается, поскольку он трактуется лишь как
среда, в которой протекает филогенез групп организмов и видов.
Между тем в действительности эволюционный процесс биологиче-
ских систем, по его мнению, в значительной степени детерминиру-
ется эволюцией биогеоценозов. Этот сопряженный процесс органи-
ческой эволюции он назвал синэволюцией (биотоценогенез,
по Быкову).
Синэволюция, по данным С. С. Шварца, проявляется, во-пер-
вых, в том, что виды — эдификаторы биогеоценоза— определяют
темпы и формы эволюции сопутствующих видов живых существ.
Например, к срокам фенологического развития эдификатора лесно-
го биогеоценоза вынуждены приспособиться насекомые и другие
беспозвоночные, а также растения нижних ярусов в прохождении
фаз своего развития. Во-вторых, с течением времени физиологиче-
ские реакции обитающих в данном биогеоценозе различных видов
растений и животных на изменение условий биоценосреды стано-
вились сходными; например, сходство скорости их развития и хода
смены фенологических фаз в зависимости от общего хода метео-
рологических явлений биотопа и пр. Этим обеспечивается слажен-
ность функционирования биогеоценоза как целостной системы, не-
смотря на колебание факторов внешней среды. Следствием же
слаженности функционирования, согласованности действия популя-
ционных и биогеоценотических механизмов регуляции, как утверж-
дает С. С. Шварц, является поддержание относительного равнове-
сия, стабильности биогеоценоза.
349
Таковы в общих чертах основные закономерности сопряженной
эволюции всех четырех форм организации живого (организменной,
популяционно-видовой, биоценотической и биостроматической), ко-
торые признаются, по выражению К. М. Завадского (1961), «пер-
вичными и основными носителями жизни» (с. 31). Однако когда
речь идет о сопряженном характере органической эволюции, то
следует иметь в виду, что возникновение и развитие ценотических
форм организации живой материи и образуемых ими биокосных
систем ограничено определенными временными и пространствен-
ными пределами. Это, как было сказано, обусловлено тем, что
качественная специфика их определяется наличным составом флор
и фаун, характерных для каждого периода геологической истории
Земли и для каждой ландшафтно-географической зоны и провин-
ции земной поверхности.
9. Становление и эволюция
биокосных систем разного ранга
Жизнь, по многочисленным и разносторонним научным данным,
существует на нашей планете более 3,5 млрд лет-. Такая дли-
тельность ее существования, как полагает М. М. Камшилов
(1969), обусловлена циклическим характером ее организации.
Вслед за В. И. Вернадским он считает, что жизнь возникла на
Земле не в виде отдельных простейших организмов, а в форме
«круговорота органического вещества, основанного на процессах
синтеза и деструкции» (с. 30). Как и В, И. Вернадский, М. М. Кам-
шнлов большое значение придает принципу неравномерности в
развитии жизни на Земле, обусловленной, по его мнению, местом
органических форм в циклической структуре жизни. Он, в частно-
сти, обращает внимание на то, что около миллиарда лет со времени
возникновения жизни она существовала в форме одноклеточ-
ных организмов. Следовательно, одноклеточными осуществлялись
биотический круговорот веществ и другие биогеохимические функ-
ции первичной биосферы (рис. 41). Возможность успешного выпол-
нения в геологической истории одноклеточными живыми существа-
ми весьма сложных и многообразных функций жизни в биосфере
связана, по мнению М. М. Камшилова, с тем, что они обладают
свойством приспособления к весьма различным условиям обитания
и способностью использовать в качестве источника углерода и энер-
гии самые разнообразные субстраты.
Между прочим, Т. В. Аристовская (1967) отмечает также, что
древние виды микроорганизмов обладали значительно более раз-
нообразными биохимическими свойствами по сравнению с совре-
менными видами, а потому относительно примитивный биологиче-
ский круговорот мог осуществляться биоценозом, состоящим даже
350
из однородных видов, а не обязательно из разнообразных видов
микроорганизмов.
Рис. 41. Циклический характер организации жизни (по
М. М. Камшилову, 1969). 1 —одноклеточные автотрофные
организмы; 2— одноклеточные сапротрофные организмы; 3 —
многоклеточные автотрофные растения; 4 — растительноядные
животные; 5 — плотоядные животные (хищники)
Как видно из прилагаемой схемы биотического круговорота ве-
ществ в биосфере М. М. Камшилова, одноклеточные организмы-
деструкторы продолжали выполнять функцию разложения и мине-
рализации органики и тогда, когда к началу кембрия в биосфере
появились многоклеточные организмы— автотрофные растения,
растительноядные и плотоядные (хищные) животные и пр. При
этом микроорганизмы-деструкторы стали осуществлять минерали-
зацию органики не только одноклеточных автотрофов, но и много-
образных по типу питания многоклеточных растительных и живот-
ных форм, включая их таким путем в биотический круговорот
биосферы. Таким образом, в основе жизни — исторически и в совре-
менности— лежит деятельность низших организмов — одноклеточ-
ных синтетиков и деструкторов. Они способны, по словам М. М. Кам-
шилова, «осуществлять биотический круговорот и одни — без уча-
стия многоклеточных. Высшие организмы... представляют собой
сравнительно поздно развившуюся надстройку на прочном фунда-
менте одноклеточных» (с. 37). Более того, само появление на жиз-
351
ненной арене многоклеточных, по мнению Т. В. Аристовской (1967),
в известной мере было подготовлено существованием одноклеточ-
ных организмов, которые, «вовлекая в биологический круговорот
все большее количество неживой материи, создали обстановку,
пригодную для возникновения более высоко организованных форм
жизни» (с. 48).
Подобно М. М. Камшилову, Т. В. Аристовская значительное ме-
сто в органической эволюции отводит биологическому круговороту
веществ. «Эволюция жизни — это прежде всего эволюция круго-
ворота веществ, обусловленного биохимической активностью живых
организмов» (с. 50). Исходя из этого, она солидаризируется с
Б. Л. Личковым, который, по ее словам, даже такие коллизии в
геологической истории жизни на Земле, как, например, массовая
гибель животных в некоторые геологические периоды, объясняет
нарушением сбалансированности между отдельными процессами в
круговоротах. Вследствие недостаточной в эти периоды обводнен-
ности материков происходило снижение выхода первичной биоло-
гической продукции, что подрывало кормовые ресурсы раститель-
ноядных животных, а это, в свою очередь, отрицательно сказыва-
лось и на численности плотоядных (хищных) животных. Важное
значение в органической эволюции, по мнению ряда ученых, имело
и колебание содержания кислорода и углекислоты в атмосфере в
разные геологические эпохи вследствие выпадения углерода из
обменных процессов и пр.
Анализируя циклический характер организации жизни,
М. М. Камшилов обращает внимание на то, что появление в ходе
органической эволюции в далеком геологическом прошлом более
высоко организованных многоклеточных организмов создало не-
преодолимый барьер на пути новых попыток одноклеточных к про-
грессивному развитию в многоклеточные формы жизни, подобно то-
му как возникновение первых живых существ воспрепятствовало
новым попыткам «самозарождения» жизни на нашей планете.
Ю. Одум (1975), обсуждая на основании разнообразных данных
проблему происхождения жизни на Земле и первых этапов ее раз-
вития, так же как и М. М. Камшилов, приходит к заключению, что
жизнь на протяжении многих миллионов лет существовала в виде
примитивных одноклеточных форм. Первые многоклеточные орга-
низмы, как полагает Ю. Одум, появились на Земле в самом начале
кембрия, т. е. около 600 млн лет тому назад, когда содержание
кислорода в атмосфере достигло примерно 3% его современного
уровня.
Как же протекал геоисторический процесс становления и эволю-
ции биокосных систем разного ранга — биогеоценоза, биогеосферы
и соподчиненных им ценотнческих структур?
По Ю. Одуму (1975), первичные сообщества живых существ
352
состояли из мельчайших анаэробных гетеротрофов, которые могли
существовать в то время за счет органического вещества, образу-
ющегося в процессах абиотического синтеза. Позднее, как сказано
выше, по его выражению, произошел «популяционный взрыв»
автотрофных водорослей, что с течением времени привело к пре-
образованию восстановительной воздушной среды в насыщенную
кислородом окислительную. Это способствовало возникновению в
последующем более сложных и разнообразных ценотических си-
стем, состоящих из все более крупных и высокоорганизованных
видов многоклеточных организмов.
По мнению Н. П. Наумова (1971), с возникновением в далеком
геологическом прошлом в морских мелководьях молекулярных
образований с функциями живых существ начали складываться
и различного рода взаимодействия между ними. В первую очередь
устанавливались трофические (пищевые) связи и отношения, вы-
ражавшиеся в использовании одних «архебионтов» или продуктов
их обмена другими в качестве источника энергии. Иначе говоря, в
процессе подобного рода взаимодействия первичных живых су-
ществ произошло становление биологического круговорота с диффе-
ренциацией их на функциональные группы — продуцентов и реду-
центов, а впоследствии и промежуточной группы — консументов.
В ходе дальнейшего геоисторического развития, как полагает
Н. П. Наумов, все более возрастало разнообразие жизненных форм
и увеличивалось их число, происходило повышение их структурно-
функциональной организации и усложнение возникших биогеоце-
нотических систем. Одним словом, биологический круговорот ве-
ществ биогеоценозов все более расширялся и ускорялся, совершен-
ствовались механизмы авторегуляции биогеоценотических систем,
что способствовало освоению ими новых мест обитания, расшире-
нию их арены жизни.
Б. А. Быков (1970), рассматривая первые этапы эволюции рас-
тительных сообществ (фитоценозов) как ведущего компонента
ценоэкосистем (биогеоценозов), рисует такую картину этого гео-
исторического процесса. Возникшие еще в кембрийскую эпоху
палеозойской эры первые примитивные наземные растительные
сообщества, по его мнению, состояли «из дихотомически ветвящихся
многолетних водорослей, стелющихся и приподнимающихся над
землей в полосе прибоя и в иных постоянно увлажняемых усло-
виях» (с. 169). Гетеротрофными компонентами, как он полагает,
были бактерии, простейшие и, возможно, грибы. Позднее, в силу-
рийскую эпоху, существовали уже двухъярусные сообщества, со-
стоящие из довольно высоких травянистых растений (Psylophyton,
Zosterophyllum, Rhynia и др.) в первом ярусе и многолетних на-
земных водорослей — во втором. В девонскую эпоху произошло
дальнейшее усложнение состава и морфологической структуры
353
растительных сообществ. Наряду с травянистыми растительными
сообществами появились древесные ценозы. Особенно широкое рас-
пространение последние получили в конце девона и в эпоху кар-
бона, когда псилофитовые сменились папоротникообразными типа
Eospermatopteris, Lepidodendron и др. Образованные ими лесные
ценозы до 30 и более метров высотой отличались большим разно-
образием жизненных форм и сложным вертикальным строением.
Под их пышно облиственными кронами развивались кустарниковый
подлесок, лианы н ярус корневищных видов трав. Состав гетеро-
трофного компонента лесных ценозов пополнился многочисленны-
ми видами насекомых и других членистоногих, а также земновод-
ными видами животных и пр.
В последующие геологические периоды и эпохи мезозойской и
кайнозойской эры, несмотря на весьма значительные изменения в
систематическом составе растительных и животных форм, сложная
структура растительных сообществ и ценотических систем в целом,
как пишет Б. А. Быков, «никогда более не утрачивалась» (с. 170).
В. В. Мазинг (1970, 1972 и др.) на основе использования дан-
ных палеонтологии и палеоэкологии предложил довольно строй-
ную, хотя в известной степени умозрительную и схематичную си-
стему поэтапного становления, развития и усложнения популяци-
онно-видового состава и структуры биогеоценотических систем с со-
ответствующей характеристикой каждого из установленных им
этапов эволюции (рис. 42). Воспроизведем в кратком изложении
эту систему эволюции ценотических систем.
I. Эоценотический этап, начавшийся с возникновения
жизни на Земле. Он характеризуется тем, что первичными обита-
телями мелководных прибрежных морских лагун на этом началь-
ном этапе были наипростейшие доклеточные существа с признака-
ми живого — эобионты. Первичный живой покров планеты того
времени представляет собой, таким образом, неоднородные, раз-
бросанные по обводненным местам земной поверхности скопления
весьма несовершенных сочетаний еще мало различающихся живых
существ.
II. Микроб и оценотический этап эволюции ценотиче-
ских систем был представлен одноклеточными биоморфами разного
типа питания. Среди них, наряду с примитивными гетеротрофами,
существующими за счет абиотически синтезированного органиче-
ского вещества, были и хищные формы типа амебы, встречались,
возможно, и паразитирующие вирусного типа формы. Позднее по-
явились и автотрофные фотосинтетики — дробянки и другие одно-
клеточные зеленые организмы. Однако на этом этапе, по мнению
автора, преобладали биохимические взаимодействия в условиях
водной среды. Ценотические взаимосвязи между живыми существа-
ми были относительно редкими и носили случайный характер.
354
МП -Микробоценозы
АГ - Агломерации
с - Синузии
ГЛАВНЫЕ ЭТАНЫ
ЭВОЛЮЦИИ БИОЦЕНОЗОВ
Комплексно-ярусно -
ДИФФЕРЕНЦИРОВАННЫЙ
Ярусно—
ДИФФЕРЕНЦИРОВАННЫЙ
Прикрепленно-
ЦЕНОТИЧЕСКИЙ
Подвижно-
ЦЕНОТИЧЕСКИЙ
МГ -Микрогруппировки, zs Микробо-
МйКРОЦЕНОЗЫ \£/ ЦЕНОТИЧЕСКИЙ
Лк ЭОЦЕНОТИЧЕСКИЙ
Рис, 42. Схема эволюции биоценозов (и биогеоценозов) (по
В. В. Мазннгу, 1972)
Представителями этого этапа эволюции в настоящее время
В. В, Мазинг считает морские микробиоценозы и почвенные био-
комплексы микроорганизмов в составе сухопутных биогеоценозов,
но находящихся на более высокой стадии их филогенетического
развития.
III. Подвижно-ценотический этап эволюции связан,
по мнению В. В. Мазинга, с возникновением у одноклеточных орга-
низмов средств активного перемещения в виде ворсинок, жгутиков
и т. п. Это давало им возможность попадать в более благоприят-
ные для существования участки, облегчало поиски пищи, а также
355
создавало условия для скопления в местах концентрации пищевых
ресурсов и образования на этой почве более или менее постоянных
группировок, в которых со временем могли вырабатываться про-
стые двучленные пищевые сочетания типа биотрофов. С возникно-
вением в ходе эволюции колониальных форм одноклеточных орга-
низмов, находящихся большей частью во взвешенном в воде со-
стоянии, происходило дальнейшее усложнение состава ценотических
группировок. Современными аналогами такого уровня ценотиче-
ской организации автор считает фито- и зоопланктон и большей
частью временные сочетания других растительных и животных
одноклеточных форм.
IV. Прикрепленно-ценотический этап связан с воз-
никновением еще в нижнем кембрии прикрепленных к субстрату
одноклеточных и многоклеточных форм растений (сифональных»
нитчатых, листоватых или талломных водорослей), а также живот-
ных (инфузорий, губок, кораллов, мшанок, брахиопод и др.). В свя-
зи с оседлым образом жизни организмов появились некоторые
новые черты состава, структуры и функциональной организации це-
нотических систем: усложнение межвидовых взаимоотношений»
расширение трофических связей (комменсализм, симбиоз, парази-
тизм), образование вокруг прикрепленных растений, кораллов, бо-
лее длительно ценотически взаимосвязанных группировок (консор-
ций), начинается горизонтальная (латеральная) дифференциация
ценотических систем, происходит также укрепление и усложнение
взаимосвязи организмов со средой, отложение органических остат-
ков и изменение среды обитания, образование внутренней среды
(биоценосреды). Следовательно, происходит становление биогео-
ценозов как «стационарных», по выражению автора, элементар-
ных ячеек, биогеосферы.
Дальнейшая эволюция биогеоценотических систем, по В. В. Ма-
зингу, шла разными путями в водной среде и на суше. В условиях
открытого океана, как он полагает, образовались своеобразные
морские биоценозы из скоплений малоподвижных одноклеточных
автотрофов в верхних освещенных слоях воды и активно движу-
щихся животных-гетеротрофов в толще воды. Между находящими-
ся в разных слоях водной среды автотрофами и гетеротрофами»
разумеется, происходил непрерывный вещественно-энергетический
обмен.
Такого рода ценотические образования морфологически (а не
исторически), как видно, больше тяготеют к подвижно-ценотиче-
скому эволюционному этапу.
V. Ярусно-дифференцированный ценотический
этап эволюции биогеоценотических систем В. В. Мазинга под-
разделяется на две ступени.
1. Ярусно-дифференцированная ступень характерна для цено-
356
зов, состоящих преимущественно из бурых водорослей или корал-
ловых участков, развивающихся в условиях литорали тропических
и субтропических областей. Те и другие ценотические системы, как
утверждает автор, отличаются прочными внутри- и межъярусными
взаимоотношениями живых компонентов.
2. Комплексно-ярусно-дифференцированная ступень эволюцион-
ного развития биогеоценозов возникла и оформилась на суше в
условиях воздушной и почвенной среды, коренным образом отли-
чающейся от водной, характеризуясь значительным колебанием
температуры и влажности, часто с резко выраженной мозаичностью
различных местообитаний.
Первые двухъярусные травяные ценозы на суше, как сказано
выше, по мнению Б. А. Быкова, появились еще в силуре на базе
псилофитовой флоры, а в девоне уже существовали и древесные
ценозы довольно сложного состава и строения. В каменноугольную
же эпоху палеозоя на базе флоры папоротникообразных в увлаж-
ненных понижениях развивались пышные многоярусные лесные
биогеоценозы, признаваемые некоторыми биологами предшествен-
никами современных влажно-тропических лесов («гилей»). В усло-
виях обильного водоснабжения и минерального богатства суб-
страта, как указывает В. В. Мазинг (1970), лимитирующим
фактором был свет, и в конкуренции за свет образуются высоко-
ствольные многоярусные лесные биогеоценозы (с лианами, эпифи-
тами и т. п.). Они заселились многочисленными потребителями
(насекомыми, лесными животными, грибами — разрушителями
клетчатки), что способствовало ускорению биологического круго-
ворота веществ.
В дальнейшем на базе господствующих в мезозое голосеменных
растений, отличающихся мощной разветвленной корневой системой
и высокой продуктивностью, произошло образование почвенного
покрова. Это было, по словам В. В. Мазинга, переломным момен-
том в становлении и развитии биогеоценозов и биогеоценотического
покрова суши. Однако особенно значительного совершенства в по-
пуляционно-видовом составе и структурном оформлении достигли
высокоствольные многокомпонентные и многоярусные лесные био-
геоценозы, образовавшиеся в кайнозое на базе широко распро-
странившихся покрытосеменных растений и млекопитающих жи-
вотных. В последующие геологические периоды кайнозойской эры,
в связи с выходом в изменившихся климатических условиях-цено-
зообразующих растений и животных из пониженных местоположе-
ний на плакорные, по мнению В. В. Мазинга, произошло формиро-
вание новых растительных и животных экобиоморф и новых типов
биогеоценотических систем разного ранга. Резкое же изменение
климатических условий и возникновение периодической смены вре-
мен года нашли отражение в установившейся сезонной динамике
биогеоценозов.
357
Таким образом, первые три этапа эволюции ценотических си-
стем, по В. В. Мазингу, протекают в водной среде на уровне про-
стейших доклеточных и одноклеточных форм жизни с весьма при-
митивными типами взаимоотношений и структурно-функциональ-
ных формирований. Только на четвертом этапе эволюции на базе
возникновения прикрепленных к подводному субстрату живых су-
ществ и появления многоклеточных форм жизни происходит замет-
ное усложнение видового состава и структурно-функциональной
организации ценотических систем и образование собственно био-
геоценозов. Но по существу лишь на второй ступени пятого
этапа эволюционного геоисторического развития, с выходом живых
существ на сушу, происходит становление и последующее поступа-
тельное развитие сложных по видовому н консортивному составу,
вертикальной и горизонтальной (латеральной) структуре и функ-
циональной организации относительно стабильных биогеоценозов-
и образуемого ими биогеоценотического покрова Земли.
10. Факторы и механизмы органической эволюции
и эволюционный прогресс биокосных систем
Рассматривая вопрос о факторах, механизмах эволюционного*
развития органического мира, и в частности эволюции ценотиче-
ских систем, Ю. Одум (1975) и другие биологи подразделяют их
на две группы: а) аллогенные (внешние) силы, а именно гео-
логические и климатические изменения, и б) автогенные (внут-
ренние) процессы, являющиеся результатом деятельности живых,
компонентов биогеоценозов. Однако, по мнению Б. А. Быкова
(1970), основная, ведущая роль в органической эволюции принад-
лежит внутренним силам, обеспечивающим саморазвитие эво-
люирующих биологических и ценотических систем; внешняя же
среда, по его выражению, лишь создает «режимные условия для
развития самоуправляющихся ценоэкосистем» (с. 170). Среди внут-
ренних сил автор главным фактором считает биоцбиотиче-
ский отбор, выражающийся в естественном отборе новых форм
и видов и в преобразовании и отборе сообществ, ценозов. Ю. Одум
также подчеркивает особо важную роль в эволюционном процессе
естественного отбора, совершающегося не только на популяционно-
видовом уровне, но и на уровне сообществ, который «ведет к сохра-
нению признаков, благоприятных для группы в целом, даже если
они неблагоприятны для конкретных носителей этих признаков»
(с. 350).
В. В. Мазинг (1970) основную, ведущую роль среди механизмов
сопряженной эволюции всех форм организации живой материи, и
в частности в эволюции биогеоценозов и биогеосферы, отводит
естественному отбору, протекающему на разных уровнях.
358
Он указывает, что в ходе органической эволюции происходил и
происходит отбор не только отдельных особей в видовых популя-
циях, но и популяций разных видов, и целых групп видов (био-
морф), а также фито- и биоценозов. Доказательством тому служит
наличие в природе сравнительно ограниченного количества повто-
ряющихся на земной поверхности относительно устойчивых, так
называемых «узловых» типов биоценозов и биогеоценозов.
Таким образом, представления разных авторов о механизмах,
или, иначе, движущих силах сопряженной эволюции разных форм
жизни, как видно из изложенного, почти идентичны. Всеми назван-
ными биологами среди механизмов эволюции главная роль при-
писывается естественному отбору, протекающему на разных уров-
нях, т. е. в несколько расширенной, точнее, в более расшифрован-
ной, чем у Ч. Дарвина, интерпретации. И это, на наш взгляд,
правильно. Только, пожалуй, надо принять во внимание, что
бшщенотический отбор (по Быкову), или отбор второго порядка (по
Ревердатто и др.), очевидно, обладает некоторыми особенностями
по сравнению с естественным отбором отдельных составных элемен-
тов биогеоценотических систем ввиду известной качественной спе-
цифики этих систем. «В биотическом круговороте, как в макроси-
стеме,— пишет М. М, Камшилов (19706), — естественный отбор
проявляется как неизбежный результат взаимодействия между
организмами. Это взаимодействие и выступает в качестве управля-
ющего фактора... Управление осуществляется с помощью механиз-
ма естественного отбора, благоприятствующего развитию одних
тенденций и, напротив, не дающего возможности реализоваться
другим» (с. 174).
Когда речь идет об эволюции органического мира, то, естествен-
но, возникает вопрос: в каком направлении идет это эволюционное
развитие, и в чем выражается его движение вперед, или, иначе, в
чем состоит его прогресс?
Критериями эволюционного прогресса ценотических систем
разного ранга, по мнению В. В. Мазинга (1972), могут быть:
а) сложность морфологической структуры и функциональной орга-
низации ценотических систем; б) эффективность использования ве-
ществ и энергии в процессах построения и расширения системы;
в) степень относительной независимости ценотических систем как
результат совершенства регуляционных механизмов. Н. В. Тимо-
феев-Ресовский (1966) считает, что одним из критериев биогеоце-
нотического прогресса является интенсивность работы биогеоценоза
в общей биогеохимической работе биосферы.
Общим, генеральным направлением эволюции ценотических си-
стем, как полагает Б. А. Быков (1970), является прогрессивное раз-
витие, в результате которого происходит следующее: а) усложнение
структуры сообщества и ценоэкосистемы в целом (в том числе и
359
консорций), увеличение общей интеграции и информативности;
б) увеличение количества живого вещества (общей биомассы) и
повышение производительности системы; в) максимальное исполь-
зование занимаемого пространства (площади) и материально-энер-
гетических ресурсов местообитания; г) усиление преобразования
живыми компонентами внешней среды во внутреннюю среду (био-
ценосреду); д) возрастание приспособлений ценозообразующих ор-
ганизмов к совместному существованию в условиях изменяющейся
среды обитания; е) усложнение и упорядочение ценотнческих свя-
зей живых компонентов и механизма авторегуляции ценоэкосисте-
мы (биогеоценоза) в целом.
По мнению С. С. Шварца (1973), в ходе органической эволюции
происходит совершенствование биогеоценозов, что выражается в.
повышении эффективности использования ими наличных ресурсов
среды и в освоении жизнью новых пространств. Важным следстви-
ем эволюции он считает также повышение морфофизиологической
организации животных и совершенствование их популяционной
структуры, что ведет к увеличению трансформации веществ и энер-
гии и укреплению организационного единства биогеоценоза, повы-
шению уровня его целостности. М. М. Камшилов (1973) важным
прогрессивным результатом органической эволюции считает утол-
щение (по его выражению) цикла жизни вследствие формирования
в процессе эволюции параллельных рядов трофических звеньев пи-
щевых цепей в разных таксонах (насекомых, позвоночных живот-
ных и растений), что ведет к повышению организованности жизни,
возрастанию запаса внутренней информации и ко все более полному
освоению жизненных материальных источников неорганической
среды. По В. Л. Комарову, как отмечает Н. В. Дылис (1975), в
связи с возникновением в ходе эволюции множества разнообраз-
ных форм жизни и усложнением их морфофизиологической орга-
низации происходит удлинение цикла превращения солнечной энер-
гии на земной поверхности и замедление ее энтропии.
Вместе с тем на фоне общего прогрессивного хода орга-
нической эволюции, как пишет Б. А. Быков (1970), возможны част-
ные проявления регрессивного порядка. В качестве примеров
он приводит: а) выпадение из растительного покрова каменноуголь-
ного периода хвощевых лесов; б) регрессивное развитие некоторых
типов ценозов в условиях ксерофитизации климата и т. п.
Р. Риклефс (1979) обращает внимание на то обстоятельство, что
на ранних стадиях дивергенции в процессе эволюции видовые по-
пуляции проявляют, по его словам, эволюционную гибкость и спо-
собность приспосабливаться к изменяющимся условиям среды, но'
с течением времени они утрачивают эти свойства, которые сменя-
ются эволюционным консерватизмом. В подтверждение
этих мыслей он приводит ряд примеров из геологического прошло-
360
го. Так, а) двоякодышащие рыбы за последние 250 млн лет мало
изменились по сравнению с формами, сложившимися в течение
первых 50 млн лет; б) встречающиеся в настоящее время в Юго-
Восточной Азии и Северной Америке изолированные друг от друга
участки реликтовых третичных субтропического типа лесов оста-
лись почти без изменения за последние 50 млн лет. Эволюционный
консерватизм на фоне неблагоприятного для существования орга-
низмов изменения окружающей среды, по утверждению Р. Риклеф-
са, может привести к вымиранию видов. Хотя главной причиной
вымирания видов признается конкурентная борьба между ними, а
также воздействие хищников и возбудителей болезней, в случае
относительно быстрого в геологическом смысле изменения условий
среды, пишет он, «механизмы реакции некоторых видовых популя-
ций не успевают сработать» (с. 322),
Из изложенного видно, что Б. А. Быков, по сравнению с выска-
зываниями других отечественных биологов, обращает внимание на
возрастание в ходе прогрессивного развития приспособлений орга-
низмов к совместному существованию в условиях изменяющейся
среды и усложнение и упорядочение ценотических связей компо-
нентов и механизмов авторегуляции ценоза. С. С. Шварц подчер-
кивает повышение в процессе эволюции морфофизиологической
организации живых существ, совершенствование их популяционной
структуры и укрепление на этой основе организационного единства
биогеоценоза в целом. В высказывании же В. Л. Комарова особого
внимания заслуживает мысль об удлинении в ходе органической
эволюции цикла превращения солнечной энергии в ценотических
системах и замедление ее энтропии.
Завершая раздел об эволюции биогеоценотических систем раз-
ного ранга, считаем необходимым обратить внимание на взгляды
В. И. Вернадского (1928) по этой проблеме, который подошел к ее
решению с биогеохимических позиций. Это дало ему возможность
раскрыть некоторые новые стороны эволюционного процесса жи-
вой природы, его общей направленности и результативности про-
грессивного развития.
Рассматривая биологический вид с биохимической стороны, т. е.
не только как форму, морфологическое образование, а как материю
и его энергию, как вещество, характеризуемое его биомассой,
В. И. Вернадский по-иному подошел к оценке и самого эволюцион-
ного процесса, и его конечных результатов. Исходя из непреложного
факта резкого изменения и преобразования живых существ и их
интегральных системных структур в ходе органической эволюции,
с одной стороны, и неизменности, устойчивости в биогеохимическом
отношении жизни как целого — с другой, он приходит к выводу,
что «в сложном механизме биосферы происходили в пределах жи-
361
ъого вещества только перегруппировки химических элементов, а не
коренные изменения их состава и количества» (с. 39).
Биогеохимический подход к проблеме сопряженной эволюции»
всех форм организации живой материи привел В. И. Вернадского
к тому, что в процессах общей миграции химических элементов в
биосфере он усмотрел биогенную миграцию атомов, совершающую-
ся в процессе жизнедеятельности живого вещества. Принимая
участие в круговоротах веществ, протекающих в «природных систе-
мах равновесия», пишет Вернадский, «биогенная миграция хими-
ческих элементов в биосфере стремится к максимальному своему
проявлению» (с. 42). Этот сформулированный Вернадским биогео-
химический принцип, по его мнению, регулирует биохимические
явления в биогеоценозе и биосфере (биогеосфере). Проявляется это
стремление биогенной миграции атомов к максимальному своему
выражению, по его словам, во «всюдности» жизни и в «давлении»
жизни, следствием чего является конкурентная межвидовая борьба
живых существ за свои позиции в составе трофических звеньев пи-
щевых цепей.
Завоевывая в ходе эволюции в течение геологического времени
новые области обитания, живое вещество, по словам Вернадско-
го, увеличивает таким путем биогенную миграцию атомов в био-
сфере. Этот второй биогеохимический принцип выражен В. И. Вер-
надским в следующих словах: «Эволюция видов, приводящая к
созданию форм жизни, устойчивых в биосфере, должна идти в на-
правлении, увеличивающем проявление биогенной миграции атомов
в биосфере» (с. 44). Правда, этот биогеохимический принцип, как
отмечает автор, не может объяснить эволюцию видов, но «он ло-
гически неизбежно указывает направление, в котором должен идти
эволюционный процесс» (с. 44). В подтверждение своих идей о ходе
сопряженной органической эволюции и сформулированных биогео-
химических принципов В. И. Вернадским приводится целый ряд
примеров из геологической истории жизни на Земле. Основные вехи
геоисторического эволюционного процесса живой природы в об-
щем виде были изложены выше.
Глава XI
Водная сфера жизни (биогидросфера)
Все изложенные в предыдущих главах основополагающие теоре-
тические положения биогеоценологии, касающиеся разных сторон
состава, строения, функциональной организации и обменных про-
цессов биогеоценозов, а также их изменения во времени, распреде-
ления в пространстве и т. п., основаны на фактическом материале
и закономерностях, выявленных при изучении наземных (сухопут-
ных) биокосных систем. Нетрудно понять без всякого объяснения,
чем это обусловлено. Но суша, по Дювиньо и Тангу (1968), зани-
мает около 148 млн км2 (29% поверхности земного шара), более
363 млн км2 (71%) приходится на долю водных пространств —
морских, океанических, а таже континентальных водоемов разного
типа, растительное, животное и микробное население которых резко
отличается во многих отношениях от наземного (сухопутного) на-
селения, обитающего в условиях воздушной среды. Поэтому мы
сочли необходимым хотя бы одну главу книги посвятить ознаком-
лению читателей с водными биокосными системами — биогидро-
ценозами, тем более что в силу известных всем обстоятельств
народнохозяйственное значение водных материально-энергетиче-
ских и биологических ресурсов и их практическое использование
с каждым годом все более возрастают.
1. Водная среда жизни
и ее обитатели (гидробионты)
По мнению гидробиологов, как существование и развитие био-
гидроценозов, так и качественная специфика каждого из них в
отличие от сухопутных биокосных систем определяются главным
образом совокупностью косных компонентов — биотопом. Отсюда
делается вывод, что в основе всестороннего анализа и обобщения
данных о биогидроценозах и биогидросфере должен лежать биото-
пический подход. Это означает, что в целях как можно более глу-
бокого познания водных биокосных систем разного ранга необхо-
димо в первую очередь выявить специфику водной среды жизни.
К общим свойствам водной среды жизни, отличающим ее от
наземной (воздушной) среды, М. П. Акимов (1959) относит: а) зна-
чительную плотность водной среды (так, удельный вес морской
воды достигает 1,025, мало уступая удельному весу протоплазмы).
Благодаря этому в водной среде значительно уменьшается тяжесть
тела ее обитателей — гидробионтов, что обусловило формиро-
вание в процессе эволюции тела с упрощенным скелетом пли даже
363
без такового, в этих условиях облегчается также плавание гидро-
бионтов и создается возможность для многих из них находиться
во взвешенном состоянии; б) расширение воды перед замерзанием
при температуре 4-4° С с образованием на поверхности водоемов
ледяного покрова с уменьшением при этом его удельного веса, что
обеспечивает водным животным благоприятные условия перезимов-
ки подо льдом в странах с умеренным и холодным климатом;
в) подвижность и высокую теплопроводность воды, способствующие
созданию в водоемах более или менее однородных термических
условии обитания во всем их объеме; г) постоянство водной сре-
ды, в особенности морской и океанической, по сравнению с воз-
душной, что обеспечивает относительную неизменность водных
живых существ на протяжении длительного геологического времен
ни, исчисляемого миллионами лет.
Б. Г. Иоганзеи (1959) к общим свойствам воды как среды жиз-
ни относит также а) большое разнообразие ее в отношении газово-
го и солевого режимов вследствие простирания от полюсов до
экватора и на большую глубину в морях и океанах; б) разнообра-
зие характера и степени минерализации воды, обусловленной спо-
собностью ее растворять многие другие вещества, оставаясь при
этом химически инертной.
Помимо общих характерных свойств водной среды, в особых
условиях некоторых типов водоемов проявляется воздействие на их
обитателей ряда специфических факторов: а) высокое давление
водной среды на больших морских и океанических глубинах, дости-
гающее нескольких сот (до 1000 и более) атмосфер, населенной
сравнительно немногими специфическими видами животных, не
встречающимися выше 6—7 тыс. м; б) прозрачность воды, завися-
щая от количества планктона и взвешенных в воде органических
и минеральных частиц, которые обусловливают проникновение све-
та в глубину. Из других отдельных факторов водной среды
М. П. Акимов приводит сапробность — степень загрязненности во-
ды разлагающимися органическими веществами, соленость воды в
аридных условиях на континентах и в морских и океанических об-
ластях, активная реакция (pH) водной среды (кислая, щелочная,
нейтральная), содержание в воде кислорода в растворенном виде
и количество минеральных и органических питательных веществ в
растворенном и взешенном состояниях, обепечивающих нормальное
существование и развитие обитателей водной среды, и пр.
Сопоставляя биологическую значимость основных факторов на-
земной (воздушной) среды для сухопутных организмов (аэробион-
тов) и водной среды для водных живых существ (гидробионтов),
Ф. Д. Мордухай-Болтовской (1975) констатирует, что гидробионты
постоянно испытывают воздействие множества таких абиотических
факторов, которые не имеют почти никакого значения для аэро-
364
бнонтов, и наоборот. Для аэробионтов жизненно необходимыми
факторами, как он считает, являются влажность и температура
почвы и воздуха, а также содержащиеся в почве элементы мине-
рального питания; для гидробионтов же к таковым относятся тем-
пература воды, наличие в ней кислорода и соответствующих мине-
ральных и органических веществ в растворенном и взвешенном
в воде состояниях. Лимитирующими же факторами для аэробион-
тов является нередко находящаяся в дефиците в наземных усло-
виях влажность почвы и воздуха, а для гидробионтов — низкое
содержание растворенного в воде кислорода.
Таким образом, водная среда жизни, обладающая целым рядом
перечисленных особенностей, по сравнению с наземной (воздушной)
средой представляет собой весьма благоприятную среду обитания
для многих современных живых существ (растений, животных,
микроорганизмов). Присущие же ей особые физико-химические
свойства способствовали также зарождению жизни на нашей пла-
нете в далеком геологическом прошлом.
Б. Г. Иоганзен (1959) указывает, что, по данным С. А. Зернова,
в водной среде в настоящее время обитает 68 классов и подклассов
животных и 26 классов и подклассов растений. Большая часть жи-
вых существ и возникла в водной среде, предположительно в при-
брежных неглубоких лагунах с перемешанной морской и пресной
водой, откуда они и проникли впоследствии на сушу. По М.П. Аки-
мову, из всех известных классов и подклассов животных не менее
82% возникли в водной среде и лишь 18% — на суше. Но в настоя-
щее время в водной среде представлены все типы животного мира,
тогда как на суше нашли пристанище немногие из них, и лишь
только членистоногие да позвоночные животные, как он указывает,
наиболее широко и прочно заселили наземную (воздушную) среду.
Зато большая часть животных, возникших на суше, обитает в на-
стоящее время в воде, исключение составляют лишь первичнотра-
хейные да многоножки. Жизнь в целом, по общему признанию
биологов, в настоящее время в водной среде, в особенности мор-
ской, значительно богаче и разнообразнее, чем в наземной; но в
Мировом океане преобладает животная жизнь, а на суше — расти-
тельная.
В составе растительного населения водной среды обитания гос-
подствуют низшие растения — одноклеточные и многоклеточные во-
доросли: зеленые, синезеленые, диатомовые и др., на больших глу-
бинах— бурые и красные водоросли, использующие на процессы
фотосинтеза сине-фиолетовый спектр лучей солнечной радиации.
Высшие (сосудистые) растения распространены в прибрежной зоне
морских и континентальных водоемов да в мелководьях, тогда как
в наземных местообитаниях высшие растения, как известно, значи-
тельно преобладают. В составе животного населения по происхож-
365
дению и образу жизни, по М. П. Акимову (1959), различаются две
группы: а) первичноводные животные, к числу которых относится
большинство водных обитателей, всегда со времени возникновения
живущих в ней; б) вторичноводные животные — это выходцы из
наземных местообитаний, проникшие некогда в водную среду и вто-
рично приспособившиеся к ее особенностям; к этой группе отно-
сятся брюхоногие моллюски, водные насекомые, из млекопитаю-
щих— сиреновые и китообразные. Но от вторичноводных, как ука-
зывает М. П. Акимов, следует отличать земноводных животных:
амфибий, некоторые виды рептилий, птиц, млекопитающих и раз-
вивающихся в воде некоторых наземных насекомых. В наземных
же условиях, как известно, преобладают насекомые, многочислен-
ны также паукообразные, нематоды, олигохеты и пр., из позвоноч-
ных животных — птицы и млекопитающие.
•Водные организмы—гидробионты, в отличие от наземных
организмов, обитающих в условиях воздушной среды — а э р о б и-
онтов, обладают целым рядом биологических и экологических
особенностей. Во-первых, в условиях недостатка кислорода или
значительного колебания содержания его в воде у многих гидро-
бионтов выработались такого рода приспособления, как сильно
рассеченные листья у растений, жабры у рыб, легочные мешки у
вторичноводных животных и т. if. Во-вторых, благодаря значитель-
но большей плотности воды по сравнению с воздухом многие вод-
ные беспозвоночные способны находиться в водоемах в свободно-
плавающем или парящем состоянии, поглощая взвешенные в воде
органические вещества (детрит) и микроорганизмы; активные же
пловцы в процессе эволюции приобрели обтекаемую форму тела,
нередко покрытого с поверхности слизью. В-третьих, в условиях
колебания солевого режима водных местообитаний у живых су-
ществ, по крайней мере растительных форм, выработались специ-
альные осморегуляторные механизмы. В-четвертых, на больших
глубинах водоемов и в подземных водах, лишенных света, где по-
этому отсутствуют зеленые растения, гетеротрофы могут существо-
вать лишь за счет поступления органических веществ извне. В связи
с этим в них ярко выражена вертикальная миграция живых су-
ществ. В-пятых, благодаря выделению гидробионтами при дыха-
нии, а растениями и при фотосинтезе, в воду биогенных газов (кис-
лорода и углекислоты), а также различных продуктов метаболизма
в водных ценозах из диффундирующих веществ создается как бы
сеть, которая служит средством связи гидробионтов друг с другом
без непосредственного контакта.
Водные организмы — гидробионты—в зависимости от занима-
емого ими положения в пространственной структуре водной среды
Б. Г. Иоганзеном (1959, 1979) и Ф. Д. Мордухай-Болтовским (1975)
подразделяются на следующие эколого-биологические группы.
368
I. Население толщи воды (пелагиали)—пела гос.
1. Нейстон — плавающие и бегающие обитатели поверхност-
ной пленки воды.
2. Планктон — взвешенные в воде «пассивные пловцы», пе-
ремещающиеся с течением воды.
3. Нектон — самостоятельно перемещающиеся активные
пловцы.
II. Население дна водоемов (бентали) — бентос.
4. Эпибентос—прикрепленные или покоящиеся обитатели
поверхности дна.
5. Ин бенто с — внутригрунтовые организмы, живущие в тол-
ще донных осадков.
Ф. Д. Мордухай-Болтовским, кроме того, выделяются промежу-
точная и две особые группы гидробионтов.
III. Промежуточная группа гидробионтов.
6. П л а н кт о б е н то с—планктон придонного слоя воды.
7. Нектобентос — нектон придонного слоя воды.
IV. Особые группы гидробионтов.
8. Эпибиоз (перифитон)—группа организмов «обрастаниям
плотных субстратов (скал, камней, потонувших и плавающих
кораблей и т. п.).
9. Фитофильные гидробионты — обитатели прибреж-
ных зарослей высших водных растений.
2. Континентальные водоемы и их классификация
Водная среда (гидросфера), занимающая почти три четверти
поверхности Земли, разумеется, неоднородна на всем своем протя-
жении. Она делится на две резко отличающиеся друг от друга об-
ласти: внутриконтинентальные пресноводные (за немно-
гим исключением) водоемы разного типа и Мировой океан
вместе со связанными с ним морями. В соответствии с этим и вся
образуемая водным населением живых существ биогидросфе-
ра также может быть подразделена на две области: континен-
тальную область биогидросферы и океаническую об-
ласть. Рассмотрим их с возможной полнотой в этой последова-
тельности.
Внутриконтинентальные или просто континентальные водоемы,
по данным Ф. Д. Мордухай-Болтовского (1975), отличаются от мо-
рей и океанов по целому ряду показателей. Во-первых, они гораздо
меньше по занимаемой площади и своей глубине, лишь в трех озе«-
рах мира глубина достигает 1000—1700 м, среди них находящееся
на территории нашей страны озеро Байкал. Во-вторых, вода в кон*-
тинентальных водоемах пресная, немногие озера, обычно в аридных
местностях, содержат соленую воду, но по составу солей она отлк-
367
чается от морской воды. В-третьих, континентальные водоемы тесно
связаны с окружающей их сушей, которая оказывает большое
влияние на количество водной массы и качественный состав ее.
В-четвертых, соотношение значений факторов водной среды в кон-
тинентальных водоемах иное, чем в морях и океанах, а именно:
плотность и давление воды в них не достигают столь высоких
величин, зона, лишенная света, значительно меньше или вовсе от-
сутствует, температура воды и содержание в ней кислорода, напро-
тив, колеблется сильнее, нередко угнетающе воздействуя на орга-
низмы. В-пятых, условия водной среды более изменчивы во времени
и в пространстве по сравнению с морской и океанической средой.
Приведем подразделение континентальных водоемов М. П. Аки-
мовым (1959), Ф. Д. Мордухай-Болтовским (1975) и Ю. Одумом
(1975) на типы с краткой характеристикой каждого из них.
Прежде всего, континентальные водоемы подразделяются на та-
кие две группы. I. Проточные (текучие) водоемы (реки,
ручьи и источники или родники). II. Стоячие водоемы (озера,
пруды,водохранилища и пр.).Общими характерными чертами рек,
ручьев являются; а) течение воды и подвижность донных отло-
жений, а также неоднородность продольного профиля водотоков,
выражающаяся в уменьшении угла падения (наклона) русла и,
следовательно, скорости течения воды от истоков к устью и в пле-
сах По сравнению с перекатами, порогами, водопадами; б) значи-
тельные сезонные колебания уровня воды, в частности весенние
полые воды и летние паводки, ведущие к затоплению речных пойм
и образованию временных водоемов в пойменных понижениях;
в) отсутствие или слабая выраженность вертикального расчленения
водной массы на слои вследствие более или менее равномерного
распределения температуры воды и растворенных в воде биоген-
ных газов (кислорода и др.); г) значительно большая по сравне-
нию со стоячими водоемами поверхность контакта воды с сушей
и интенсивность обмена между ними.
Источники (родники) несколько отличаются от других
водных потоков данной группы водоемов. Характеризуются они по-
стоянством химического состава, скорости течения и температуры
воды. Среди них различаются горячие источники в вулканических
местностях, жестководные — в районах известняковых отложений и
мягководные — в местах залегания глинистых сланцев и кристал-
лических пород. Подземные грунтовые воды разного типа, питаю-
щие проточные водоемы, характеризуются большими запасами; тем
не менее, по вполне понятным причинам, они нуждаются в чрезвы-
чайно бережливом и разумном использовании их в определенных
целях и ограниченных размерах.
Из группы стоячих континентальных водоемов наиболее круп-
ными по площади являются озера. Происхождение озер, по Ю. Оду-
368
му, связано с такими эндо- и экзогенными планетарными явления-
ми, как эпохи оледенения, тектоническая и вулканическая деятель-
ность и т. п. Поэтому они распределены по земной поверхности
неравномерно, обычно бывают сгруппированы в озерные регионы.
В озерах хорошо выражены как вертикальное, так и горизонталь-
ное (по площади) расчленение водоема. В горизонтальном и одно-
временно в вертикальном направлениях озера расчленяются на три
зоны: а) литоральная зона — это мелководная прибрежная зо-
на, в пределах которой свет проникает до дна; б) сублитораль-
ная зона (при вертикальном расчленении называемая лимни-
ческой), охватывающая толщу воды по склону водоема и далее
до глубины эффективного проникновения света, где фотосинтез еще
уравновешивает дыхание, и в) профундальная глубинная зо-
на, охватывающая толщу воды и дно водоема, куда не проникает
солнечный свет (рис. 43).
Рис. 43. Зоны континентальных стоячих водоемов (по данным Ю. Одума, 1975)
На основе первичной продуктивности и глубины озера под-
разделяются на 1) мелководные: а) эвтрофные — богатые био-
генными веществами; б) морфометрически эвтрофные — бедные ими
и 2) глубоководные: в) олиготрофные—бедные биогенными
веществами; г) морфометрически олиготрофные — с богатым запа-
сом таковых. Помимо того, выделяются специальные типы
озер: а) дистрофные озера — с высоким содержанием гуминовой
кислоты, некоторые с торфянистыми берегами, со временем пре-
вращающиеся в торфяные болота; б) глубокие древние озера с эн-
демичной фауной (например,оз. Байкал); в) соленые или щелочные
озера пустынь с высоким содержанием хлоридов, сульфатов или
карбонатов; г) вулканические озера — кислые или щелочные с во-
дой, поступающей из магмы, химически токсичной; д) озера, хими-
чески постоянно расчлененные на поверхностный и придонный слои
вследствие внедрения или вымывания солей, с отсутствием кисло-
369
рода в придонной воде; е) полярные озера с температурой воды
ниже 4° С.
Относящиеся к стоячим континентальным водоемам, пруды
отличаются от озер не только меньшими размерами занимаемой
площади и меньшей глубиной, но и хорошо развитой литоральной
зоной и, как правило, отсутствием других зон и водных слоев. Ха-
рактерно для прудов также распространение по всей площади
растений-макрофитов (тростник, камыш, рогоз и т. п.). Естествен-
ные пруды возникают в притеррасьях речных пойм («старицы»), в
карстовых опусканиях и провалах в районах залегания известняков
и в местах распространения бобров, строящих плотины и запруды.
Искусственные пруды сооружаются человеком — мельничные, ры-
боводные и т. п.
Водохранилища — это искусственные водоемы, сооружае-
мые человеком при строительстве гидроэлектростанций и других
народнохозяйственных объектов. Для водохранилищ, по Ф. Д. Мор-
духай-Болтовскому и Ю. Одуму, характерны высокая мутность во-
ды и наличие зоны колебания ее уровня. Обнажаемая при этом на
длительное время зона заселяется своеобразным сообществом жи-
вых существ. Тепловой режим и запас биогенных элементов водо-
хранилища определяются характером сброса воды, зависящим, в
свою очередь, от конструкции и назначения плотины: так, при сбро-
се богатой биогенами холодной придонной воды (например, на
гидроэлектростанциях) водохранилище, по словам Ю. Одума, ста-
новится аккумулятором тепла и экспортером биогенных веществ, а
ври сбросе теплой поверхностной воды — аккумулятором биогенных
веществ и экспортером тепла. Влияние плотины на водохранилище
и на нижележащий отрезок реки выражается также в следующем:
а) при выбросе из водохранилища бедной растворенным кислоро-
дом природной воды снижается способность реки производить со-
ответствующую переработку органических отходов; б) в водохра-
нилище усиливается физическое испарение воды, так как удаляется
холодная придонная вода, а остается более теплая поверхностная;
в) с потерей при сбросе с водой из водохранилища биогенных ве-
ществ происходит снижение его продуктивности с одновременным
повышением ее ниже по течению реки и пр.
Образующиеся весной временные водоемы в понижениях
речных пойм при затоплении полыми водами или в низинах водо-
разделов талыми водами, местами называемые лиманами, не-
долговечны, продолжительность существования их исчисляется
лишь несколькими месяцами. Для них, по словам Ф. Д. Мордухай-
Болтовского, характерно чрезвычайно быстрое развитие водной
фауны и краткость жизненных циклов организмов.
Встречающиеся в горных местностях пещерные водоемы
характеризуются отсутствием или очень слабым развитием расти-
370
цельности из-за недостатка света, а следовательно, и других живых
существ.
Таковы особенности среды обитания континентальных водо-
емов— текучих и стоячих, в условиях которых формируются рас-
тительное, животное и микробное население этих водоемов и
образуемые нми биокосные системы — биогидроценозы.
3. Население континентальных водоемов
и формирование живых компонентов биогидроценозов
Континентальные пресноводные местообитания, по данным
М. П. Акимова (1959), населены преимущественно вторичноводны-
ми животными, относящимися к более высокоорганизованным клас-
сам, но с менее пластичными видами живых существ, чем морские
обитатели. Наибольшую роль в их составе играют кольчатые черви,
моллюски, ракообразные и насекомые, из позвоночных животных —
пресноводные формы рыб. Континентальные водоемы характеризу-
ются значительно меньшим разнообразием жизненных форм по
сравнению с населением морских и океанических водных про-
странств.
В прибрежной зоне таких стоячих водоемов, как озера, пруды
и водохранилища, в особенности с пологими склонами, формиру-
ются несколько сменяющих друг друга в пространстве и во времени
поясов растений-макрофитов: а) заросли тростника, камышей
(озерного или Табернемонтана), рогозов (широко- или узколист-
ного) и др.; б) глубже — пояс плавающих и прикрепленных к
грунту растений с погруженными и плавающими на поверхности
воды листьями (стрелолист, кувшинка, кубышка, рдесты, роголист-
ник); в) еще глубже — пояс придонных водных растений (харовые
и другие водоросли, водные мохообразные и т. п.) (рис. 44). За
ними, в зависимости от крутизны склона, глубины водоема и рель-
ефа дна, обычно простираются открытые водные пространства, ли-
шенные зарослей макрофитов, исключение составляют лишь мелко-
водья водохранилищ да некоторых неглубоких прудов.
Заросли макрофитов наиболее богаты животным населением. Из
первичноводных животных, по М. П. Акимову, здесь распростране-
ны как свободноплавающие, так и ползающие и буравящие дно
бентические формы. Среди вторичноводных господствуют насеко-
мые, паукообразные, легочные моллюски; из позвоночных довольно
обычны амфибии, добывающие здесь пищу водоплавающие птицы,
а также цапли, кулики; встречаются некоторые рептилии и млеко-
питающие. Жизненные формы открытых пространств стоячих во-
доемов представлены планктонными организмами и нектоном, прав-
да, сильно обедненным по сравнению с морским. Более бедно здесь
и животное население профундали по сравнению с морским насе-
371
Рис. 44. Прибрежные растения пресноводных водоемов (по А. П. Шенникову,
1964; и др.). 1 — тростник, камыш и др. (на тростниково-камышовом торфе); 2—
стрелолист, кувшинка, рдесты и др. (на сапропелевом торфе); 3 — харовые во-
доросли и подводные мхк (на сапропелите)
лением бентали — бентосом, среди прикрепленных форм преобла-
дают вторичноводные животные. Соленые озера отличаются сла-
бым развитием зарослей растений макрофитов, а следовательно,
они бедны и животным населением.
Различного типа мелководные временные водоемы, пересыхаю-
щие в сухие сезоны или в засушливые годы, характеризуются
чрезвычайной скудостью редкостойного растительного покрова и
узкоспециализированного животного населения. Обитатели таких
водоемов, по данным М. П. Акимова, обладают целым рядом при-
способлений к переживанию сухих периодов: образование стойких
к высыханию яиц и покоящейся стадии (цисты), покрытой плотной
оболочкой, закапывание в такие периоды в землю, обладание спо-
собностью переносить значительное обезвоживание тела, не теряя
жизнеспособности, или впадать в анабиотическое состояние. При
наступлении же благоприятных для жизни условий они способны
быстро размножаться.
Реки, ручьи и другие типы текущих в одном направлении вод-
ных потоков неблагоприятны для существования и развития как
372
растительных, так и животных форм жизни. Особенно неблагопри-
ятны для жизни верховья рек, речные пороги и водопады, горные
потоки, несущие массу песка, гальки, иногда и крупные камни. Та-
кие участки текучих водоемов заселяются, по данным М. П. Аки-
мова и Ю. Одума, специально приспособленными формами живых
существ, обладающих присосками, крючкообразными придатками,
клейкой поверхностью нижней части тела или плоской обтекаемой
формой его; у многих из них в процессе эволюции выработалась
прирожденная ориентация против течения воды и прирожденная
способность цепляться за поверхность предметов и пр. В составе
животного населения здесь встречаются немногие виды рыб, раков,
моллюсков, а также личинки ручейников. Из позвоночных, кроме
рыб, к обитанию в таких условиях приспособилась лишь единствен-
ная птица — оляпка, добывающая пищу среди камней на быстри-
нах. Животное население в более спокойном среднем и нижнем
течении рек, в особенности крупных, в общих чертах сходно с оби-
тателями стоячих пресных водоемов.
Не менее своеобразно и микробное население различных типов
континентальных водоемов, как природных, так и искусственных..
Микробное население водной среды жизни представлено огромным:
количеством (по составу и биомассе) низших одноклеточных водо-
рослей (зеленых, диатомовых, десмидиевых, синезеленых и т. п.)г
образующих в поверхностном слое воды так называемый фито-
планктон, осуществляющий созидание Органического вещества
в процессе фотосинтеза. На базе его обильно развивается и зоо-
планктон из беспозвоночных животных, включающий значитель-
ное число и микроскопических форм — жгутиковых и других про-
стейших и т. п. Но особенно обильна и разнообразна в водной
среде, по данным А. Г. Родиной (1971), бактериальная флора, обра-
зующая на поверхности и в толще воды бактериопланктон.
Формы существования бактерий в воде, как она указывает, весьма
разнообразны. Некоторые из них существуют в виде отдельных
обособленных клеток — это палочкообразные неспороносные бак-
терии, вибрионы, спириллы, кокки; встречаются нитевидные формы
серобактерий, железобактерий и пр. Однако большая часть бакте-
рий существует в воде в виде микроколоний различной формы
(клубковидной, кольчато-ячеистой и др.) и различной величины
(от 8 до 48 клеток). Они обычно бывают покрыты общей слизистой
массой, но некоторые микроколонии, состоящие из бактериальных
клеток, заключенных в тонкие коллоидные футляры, образуют скоп-
ления неправильной формы.
Помимо естественных континентальных водоемов различного
типа, в последние десятилетия, как уже отмечалось, чрезвычайно
возросло количество искусственных водоемов различного хозяй-
ственного назначения: рыбоводных прудов, водохранилищ, связан-
373
ных с сооружением гидроэлектростанций, и мощных оросительных
систем и т. п. В связи с этим появилась масса научных работ как
в пашей стране, так и за рубежом, посвященных не только узко-
специальным вопросам лимнологии и гидрологии, но и вопросам
формирования водной флоры и фауны и возникающих на их основе
водных сообществ и ценотических биокосных систем. Особенно тща-
тельному и всестороннему исследованию подверглись крупные
волжские водохранилища (Горьковское, Куйбышевское, Волгоград-
ское), а также водохранилища: Днепропетровское, Цимлянское,
Новосибирское, Иркутское и множество менее крупных по площади
водохранилищ. При этом разными специалистами исследовались
разные стороны этих искусственных водоемов. Так, К- А. Гусева
и В. А. Экзерцев (1966) главное внимание уделили вопросам фор-
мирования фитопланктона и высшей прибрежно-водной раститель-
ности, Ф. Д. Мордухай-Болтовской и Н. А. Дзюбан (1966) исследо-
вали формирование фауны беспозвоночных животных, И. В. Шаро-
нов (1966) занимался изучением формирования ихтиофауны в усло-
виях водохранилищ.
В процессе подобного рода исследований были выявлены ход,
скорость течения и некоторые закономерности процесса формиро-
вания названных биолого-ценотических комплексов живых существ,
были установлены также стадии, этапы этого процесса и пр. Со-
поставление всех этих данных разных авторов позволяет составить
довольно обоснованное представление о том, что формирование
различных биоценокомплексов происходило не изолированно друг
от друга, а в тесной взаимосвязи и взаимообусловленности между
собой во многих отношениях: по линии питания, срокам и продол-
жительности прохождения стадий развития и т. п.
Так, формирование в водохранилищах продуцентов первичной
биологической продукции — водорослевого фитопланктона и выс-
шей водной растительности, по данным исследований К. А. Гусевой
и В. А. Экзерцева, протекает по-разному. Преобразование фито-
планктона, существовавшего в водоеме до зарегулирования его
стока, протекает довольно быстро, почти полностью заканчиваясь
за время заполнения водохранилища до проектного уровня. По-
ставщиками водорослей, приспособленных к обитанию в условиях
относительно спокойного течения воды в водохранилищах, являют-
ся мелководные плесы среди водохранилища, многочисленные зали-
вы в случае значительной изрезанности береговой линии, а также
притоки реки и впадающие в водохранилище речки, особенно бога-
тые разнообразными представителями фитопланктона. Полностью
сформировавшийся фитопланктон водохранилища довольно одно-
типный по видовому составу для большинства из них. Преимуще-
ственно диатомовый фитопланктон исходного водоема в водохра-
нилище обычно сменяется фитопланктоном, состоящим из зеленых
374
водорослей (евгленовых, пирофитовых), а также синезеленых, осо-
бенно обильных в летний период, нередко вызывая в это время
«цветение» воды. Формирование высшей водной растительности,
приуроченной к прибрежной полосе и мелководьям водохранилищ,
протекает довольно медленно, особенно в условиях значительного
колебания уровня воды при заполнении водохранилища, а также
на участках озерного и долинного типа. На небольших по площади
водохранилищах, по данным упомянутых авторов, становление выс-
шей водной растительности завершается примерно в течение 5 лет,
в водохранилищах же площадью в сотни квадратных километров
этот процесс растягивается на 10—20 лет. К особенностям высшей
водной растительности водохранилищ южных областей страны от-
носится обильное развитие в ее составе представителей сорной и
земноводной растительности.
Формирование трофически связанного с фитопланктоном зоо-
планктона водохранилищ, поданным Ф. Д. Мордухай-Болтовского
и Н. А. Дзюбана, также протекает довольно быстро. В первые го-
ды формирования зоопланктона на открытых участках водохрани-
лищ в составе его господствуют коловратки и различные виды
ракообразных, а в молодых прибрежных зарослях высших водных
растений преобладают так называемые фитофильные группировки
беспозвоночных. На последующих стадиях развития видовой состав
зоопланктона упрощается, снижается и численность особей его.
Так постепенно складывается довольно однообразный, характер-
ный для водохранилищ зоопланктон с преобладанием в его составе
дафний, циклопов и других планктонных форм. В отличие от этого
формирование бентосных сообществ водохранилищ, и в
частности их животного компонента, по данным тех же авторов,
протекает гораздо медленнее. По мере заполнения водохранилища
и снижения скорости течения воды происходит заиление дна, что
ведет к разрушению прежних донных сообществ и заселению дна
водохранилищ различными по экобиоморфологии видами беспоз-
воночных животных. Однако этот биоценокомплекс, сформировав-
шийся из фрагментов разных биоценозов, вскоре сменяется также
недолговечным животным компонентом, состоящим из личинок мо-
тыля и других видов тендипедид, личинок комаров и т. п., биомасса
которого достигает 10—15 г/м2. Через несколько лет после запол-
нения водохранилища до проектной отметки на месте временного
сообщества постепенно складывается типичный для бентоса водо-
хранилищ донный биогидроценоз с участием в его составе гамме-
рид, осликов, пиявок и других представителей придонной и внутри-
грунтовой фауны, отличающейся низкой продуктивностью и запа-
сом биомассы.
Для становления рыбного населения биогидроценозов водохра-
нилищ, по данным И. В. Шаронова, требуется еще более продол-
375
жительный период, чем для формирования населения беспозвоноч-
ных животных бентосных ценозов, а именно от 5—7 до 9 лет. Есте-
ственный процесс становления ихтиофауны протекает в тесной
взаимосвязи с ходом заполнения водохранилища, изменением его
уровневого и гидрологического режимов, развитием фито- и зоо-
планктона и прочих источников питания рыбного населения и пр.
В начальный период заполнения водохранилищ в прибрежной по-
лосе и мелководьях обычно создаются благоприятные условия для
нереста рыб и выживания молоди, а буйное развитие зоопланктона
и тендипедид способствует быстрому росту численности и упитан-
ности большинства представителей ихтиофауны. В последующие
годы ввиду резкого уменьшения площади нерестилищ по мере за-
полнения водохранилища и общего повышения уровня воды, а так-
же вследствие отмеченного выше резкого снижения биомассы зоо-
планктона и бентоса происходит замедление темпов роста фитофа-
гов и бентофагов и значительное снижение рыбного поголовья
водохранилища. С заполнением водохранилища до проектной от-
метки и установлением гидрологического режима, характеризую-
щегося значительным периодическим колебанием уровня воды, в
нем создаются условия, неблагоприятные для размножения, зимов-
ки и питания рыбного населения, в результате чего не только сни-
жается общая численность ихтиофауны, но и заметно возрастает
в ее составе доля малоценных и сорных видов рыб.
По влажным склонам водохранилищ и оросительных каналов,
в водах каналов, по наблюдениям Г. Н. Олейника (1971), обычно
интенсивно развиваются нитчатые зеленые водоросли с участием
некоторых одноклеточных форм. На поверхности и в зарослях нит-
чаток, являющихся созидателями первичной биологической продук-
ции, формируются так называемые бактериальные биогид-
роценозы. В составе гетеротрофного населения этих ценозов
господствует большое количество разнообразных видов бактерий,
дрожжевых и дрожжеподобных низших грибов, актиномицетов
и т. п.
4. Водные биокосные системы — биогидроценозы
и их особенности
Зарождение и многовековое существование и развитие жизни в
водной среде свидетельствуют о достаточно высокой степени ее
организованности и обеспеченности надежной системой механизмов
.авторегуляции. Жизнь, по М. М. Камшилову, как было сказано ра-
нее,— это круговорот веществ и информации, совершаемый живы-
ми организмами на основе синтеза органического вещества за счет
солнечной энергии и его деструкции. Из этого следует, что такого
рода фундаментальные жизненные процессы имеют место и в вод-
376
ной среде, а это означает» что в ней объективно существуют и функ-
ционируют соответствующие ценотические биокосные системы, осу-
ществляющие эти процессы.
Однако водные биокосные системы, называемые биогидро-
ценозами, сильно отличаются от наземных биогеоце-
нозов как по составу видовых популяций живых компонентов,
так и по характеру косных компонентов. Но, пожалуй, самое суще-
ственное различие между ними состоит в отмеченных выше свой-
ствах водной среды обитания, в особенности в свойстве подвижно-
сти водных масс — однонаправленной или беспорядочной внутри
континента и типа круговоротов разного масштаба в Мировом
океане. Дело в том, что в силу этого отсутствует четкая выражен-
ность дифференциации биогидросферы на объективно существую-
щие функционально обособленные структурные единицы и не обес-
печивается четкость и устойчивость границ между ними; последние
в большинстве случаев являются подвижными и расплывчатыми.
Да и структурно-морфологическая обособленность биогидроцено-
зов весьма слабо, а то и вовсе не выражена. Все это послужило
основанием для отрицания некоторыми отечественными биологами
наличия в биогидросфере биокосных ценотических систем типа био-
геоценозов.
Но все же большинство биологов вслед за Л. А. Зенкевичем
(1970), как отмечает Е. М. Лавренко (1971), «считают возможным
распространить биогеоценотическую концепцию Сукачева и на во-
доемы, включая и океаны» (с. 398). Основанием для такого пред-
ставления может служить ряд существенных обстоятельств. Во-пер-
вых, наземные и водные биокосные ценотические системы, разли-
чаясь по систематическому и экологическому составу видовых по-
пуляций и в известной степени по морфологической структуре,
обнаруживают удивительное сходство по количественному соотно-
шению ценозообразующих видов, а именно: в том и другом типе
ценосистем имеются один-два, реже несколько доминантных видов,
несколько субдоминантов и значительное число второстепенных ви-
дов. Во-вторых, функциональная организация биогеоценозов также
идентична, а именно: в тех и других ценотических системах наличе-
ствуют а) производители первичной биологической продукции —
продуценты или автотрофы (фото- и хемосинтетики); б) потре-
бители первичной продукции и создатели вторичной биологической
продукции — консументы или биотрофы (фито- и зоофаги);
в) биоредуценты или сапротрофы (сапрофаги), осуществляю-
щие разложение и минерализацию органического вещества расти-
тельного и животного происхождения, завершая тем самым очеред-
ной цикл биотического круговорота веществ и закладывая мате-
риальные основы для осуществления последующего цикла его.
В-третьих, как для биогеоценозов, так и для биогидроцеиозов ха-
377
рактерна относительная однородность по набору и сочетанию эко-
логических факторов занимаемого ими участка территории или
акватории.
К представлению об объективном существовании качественно
обособленных водных ценотических систем (сообществ) на основа-
нии анализа результатов многочисленных исследований гидробио-
логов приходит, в частности, Б. Я. Виленкин (1966). Обнаружение
многими гидробиологами «пространственной локализации» населе-
ния гидробионтов в континентальных и морских водоемах, достига-
ющей высокой плотности в местах скопления, а также установление
в такого рода скоплениях не только определенного видового со-
става и величины биомассы, но и степени доминирования тех пли
иных видов и сложности состава сообществ, по его мнению, свиде-
тельствует о том, что такая «естественная совокупность» живых
существ неизбежно должна проявлять свойства, отсутствующие у
входящих в ее состав видовых популяций. Иначе говоря, в данном
случае, как он указывает, речь идет об ином — биоценотическом
уровне организации живой природы, о «структуированной совокуп-
ности некоторого числа разных видов» (с. 70), обладающей особы-
ми присущими ей свойствами.
Итак, логика самой жизни и конкретные исследования гидро-
биологов дают достаточно оснований для признания того, что био-
косные ценотические системы объективно существуют не только в
воздушной среде — на суше, но и в водной — внутри континентов и
в Мировом океане.
5. Структурно-функциональная организация
континентальных биогидроценозов
Рассмотрим особенности функциональной организации и про-
странственной, морфологической структуры биогидроценозов, в
первую очередь пресноводных, по сравнению с сухопутными био-
косными системами того же ранга.
Стоячие континентальные водоемы (озера, пруды, водохрани-
лища), как было сказано ранее, подразделяются на три зоны (ли-
торальная, сублиторальная, или лимническая, и профундальная),
каждая из которых характеризуется целым рядом особенностей
среды обитания и водного населения. В соответствии с этим и фор-
мирующиеся в них биогидроценозы также не идентичны.
Прибрежная л ит о р а л ь н а я зона, называемая некоторыми био-
логами также фитальной, занимает довольно значительную пло-
щадь, в особенности в водоемах с пологими склонами. Авто-
трофный компонент биогидроценозов данной зоны представлен
высшими сосудистыми макрофитами и одноклеточными и
колониальными водорослями, образующими фитопланктон.
378
Правда, по мнению некоторых гидробиологов, роль макрофитов в
продуцировании первичной биологической продукции незначитель-
на, главным продуцентом они считают фитопланктон. Однако
Н. С. Гаевская (1966) на основании своих многолетних исследова-
ний и литературных данных пришла к иным представлениям по
этому вопросу. В процессе исследований автором было установлено,
что общее число высших водных и водно-болотных растений, по-
требляемых в пищу водными животными, составляет 314 видов
(в том числе 90% цветковых). Из них 68 видов автор относит
к группе целиком или почти целиком погруженных в воду
(рдесты, роголистник, уруть, пузырчатка, элодея, валиснерия
и др ); 44 вида — к группе погруженных в воду, укореняющихся в
грунте растений с плавающими на поверхности воды листьями (кув-
шинки, кубышки и т. п.); 123 вида — к группе полупогруженных,
у которых значительная часть стеблей и листьев располагается
над водой (рогозы, камыш, тростник, стрелолист, ежеголовка,
ситняг и пр.); 79 видов — к группе растений сырых, избыточно
увлажненных, эпизодически заливаемых водой местообитаний
(см. рис. 44).
Видовое разнообразие биотрофного компонента литоральных
бногидроценозов еще более велико. По данным Н. С. Гаевской, об-
щее число животных, питающихся прибрежными макрофитами, до-
стигает 600 видов,
подавляющее боль-
шинство из них—на-
секомые (452 вида).
К растительноядным
беспозвоночным жи-
вотным относятся
также брюхоногие
моллюски, ракооб-
разные, нематоды и
пр. Средн раститель-
ноядных животных
25 видов рыб, 18 ви-
дов птиц и 5 ви-
дов млекопитающих
(нутрия, ондатра и
др ). К плотоядным
(хищным) формам
принадлежат из бес-
2
РАСТИТЕЛЬНОЯДНЫЕ
хищники
Рис. 45. Беспозвоночные животные литоральной зоны
пресноводных водоемов (по Ю. Одуму, 1975). 1 —
брюхоногие моллюски; 2 — мелкие членистоногие; 3—
нимфы поденок; 4 — жук-плавунец и его личинка;
5 — хищные клопы; 6 — нимфы стрекоз
позвоночных жуки-
плавунцы, некоторые
виды клопов, нимфы
стрекоз, а из позво-
379
ночных — щука»
окунь и др. (рис. 45).
В составе зоопланк-
тона, развивающего-
ся на базе литораль-
ного фитопланктона,
по Ю. Одуму (1975),
преобладают ветви-
стоусые и веслоно-
гие рачки, коловрат-
ки, а в составе ней-
стона — жуки-вер-
тячки, водомерки и
многочисленные мик-
роскопические фор-
мы — простейшие и
пр. (рис. 46).
Наибольшая пло-
щадь в континен-
тальных водоемах
приходится на долю
открытых водных
пространств. Это —
сублитораль-
ная (или лимниче-
Рис. 46. Пресноводный зоопланктон, зоонейстон и
зообентос (по ГО. Одуму, 1975). Зоопланктон: 1—
коловратки; 2 —веслоногие рачки; 3 —ветвисто-
усые рачки. Зоонейстон: 4 — жук-вертячка, 5 —
водомерки. Зообентос: 6 — личинки комара и ко-
ретры; 7 — двустворчатый моллюск; 8 — красные
кольчатые черви
ская) зона, охваты-
вающая толщу воды до глубины проникновения света (около
350—380 м), но главным образом — поверхностный слой до 30
(максимум до 100 м), обеспечивающий достаточно эффективный
синтез органического вещества. Основными автотрофами субли-
торальных биогидроцеиозов, по Ю. Одуму, являются зеленые, диа-
томовые и синезеленые водоросли и жгутиковые (евгленовые, пан-
цирные и вольвоксовые). Первичная биологическая продукция
образуемого ими фитопланктона нередко достигает значи-
тельных размеров, особенно в ранневесенний период, вызывая так
называемое «цветение» воды (рис. 47).
Биотрофный компонент биогидроцеиозов данной зоны, пред-
ставленный зоопланктоном, как отмечает Ю. Одум, небогат
по видовому составу. Здесь численно изобилуют некоторые виды
рачков, коловраток и т. п., отличающиеся от обитателей литорали
преобладанием форм с прозрачным телом (медузы и пр.), с длин-
ными антеннами, рядами щетинок для отфильтровывания в процес-
се питания детрита, фито- и бактериопланктона; имеются среди
них и хищные формы. В водной толще сублиторали довольно обыч-
ны суточные вертикальные перемещения гидробионтов. Нектон суб-
380
Рис. 47. Пресноводный водорослевый фитопланктон (по Ю. Одуму,
1975). Водоросли: 1 — зеленые; 2 — диатомовые; 3 — синезеленые
литорали представлен рыбами, питающимися преимущественно
более или менее крупными формами живых существ.
Функциональная организация биогидроценозов профундаль-
ной зоны континентальных водоемов своеобразна. Ввиду почти
полного отсутствия света не развит здесь и фототрофный компо-
нент. Поэтому существование биотрофов (фито- и зоофагов) нахо-
дится в зависимости от количества и качества органической пищи,
поступающей из литоральной и лимнической зон. По Ю. Одуму,
они представлены в основном бентосными формами (личинки
хирономид, кольчатые черви, мелкие виды моллюсков и т. п.) и
одной планктонной формой (коретра), совершающей ежесуточные
вертикальные миграции со дна водоема до лимнической зоны и
обратно. Широко распространены здесь и детритоядные фор-
м ы. Для животного населения профундали характерна приспособ-
ленность к существованию в условиях пониженной концентрации
кислорода (см. рис. 46).
Среди сапротрофов, выполняющих в процессе своей жизне-
деятельности в стоячих континентальных водоемах функцию раз-
ложения и минерализации органических остатков, распространены
соответствующие группы преимущественно анаэробных бактерий и
низших грибов. Особенно широко они распространены в водоемах,
богатых взвешенными в воде или лежащими на дне частицами ор-
ганического детрита, и в загрязненных различными отходами во-
доемах.
К косным компонентам биогидроценозов сублиторальной
(лимнической) зоны континентальных водоемов относится вода,
381
которая, по П. М. Рафесу (19806), заменяет в них почву. В ней
живые компоненты находят в растворенном или во взвешенном
состоянии питательные вещества и необходимые для дыхания, а
фототрофам и для фотосинтеза биогенные газы (кислород, углекис-
лоту). В пределах литорали для одних организмов всеобъем-
лющим косным компонентом служит вода, для других — вода и
подводный грунт различного механического состава и сложения,
перекрытый более или менее мощным слоем полуразложившихся
органических и органоминеральных отложений в виде торфа, са-
пропелита или сапропеля. Для полупогруженных в воду макрофи-
тов и эпифитной мезофауны и микрофлоры, помимо воды и грунта,
к косным компонентам относится и частично или полностью окру-
жающая их воздушная среда. Косными компонентами биогидроце-
нозов профундали являются: материковый грунт (илистый,
песчаный, каменистый) и придонный слой воды, скудно насыщен-
ный биогенными элементами и газами.
Для проточных континентальных вод (рек, ручьев) характерны
два типа ценотнческих систем: 1) биогидроценозы речных
перекатов и других быстрин и 2) биогидроценозы речных
плесов. На речных перекатах в зависимости от характера дна рус-
ла реки, по Ю. Одуму (1975), формируются сообщества двоякого
рода, которые, надо полагать, тесно связаны в единую ценотиче-
скую систему: а) в составе сообщества на каменистом дне перека-
тов господствуют прикрепляющиеся к камням, цепляющиеся и при-
сасывающиеся к ним растительные и животные формы; б) на пес-
чаном или илистом дне заводей среди перекатов в сообществе пре-
обладают роющие беспозвоночные формы (двустворчатые моллюс-
ки, поденки, некоторые виды стрекоз), а в более глубоких местах —
нектон, планктон и некоторые речные виды рыб.
Биогидроценозы речных плесов с песчаным и илистым дном и
относительно спокойным течением воды по составу живых компо-
нентов и другим показателям мало отличаются от биогидроцено-
зов литоральной зоны стоячих водоемов.
Вместе с тем следует иметь в виду, что водное население круп-
ных речных потоков, по мнению ряда отечественных гидробиологов,
неоднородно на всем их протяжении. На разных отрезках про-
дольного профиля реки, в зависимости от геолого-геоморфологи-
ческого строения местности, характера прибрежного растительного
покрова, формы и интенсивности хозяйственной деятельности мест-
ного населения и т. п., систематический и эколого-биологический
состав водного населения значительно разнится. В соответствии с
этим неодинаковы и образуемые ими биогидроценозы на раз-
ных участках, по крайней мере среднего и нижнего течения реки.
Так, например, по наблюдениям А. П. Скабичевского (1974), в со-
ставе фитопланктона вдоль течения Иртыша имеют место следую-
382
щие изменения. В истоке Иртыша — озере Зайсан — относительно
бедный фитопланктон состоит из синезеленых и диатомовых водо-
рослей; несколько выше г. Семипалатинска он становится богаче,
и господствуют в нем зеленые водоросли (протококковые и др.);
ниже по течению реки численность зеленых водорослей резко па-
дает, но зато возрастает численность диатомовых; у Омска господ-
ство в составе фитопланктона переходит к синезеленым водорос-
лям. В нижнем течении Иртыша изменения в составе и численности
особей фитопланктона еще более значительны. В соответствии с
этим, разумеется, происходит изменение и состава других живых
компонентов водных ценозов, трофически связанных с фитопланк-
тоном каждого отрезка продольного профиля реки.
Морфологическая (пространственная) структура естественных
континентальных биогидроценозов отличается от структуры
наземных биогеоценозов как в вертикальном, так и в горизонталь-
ном направлении. По вертикали (сверху вниз) биогидроценозы
обычно расчленяются на следующие ярусы (биогидрогоризонты):
1) надводный (птицы, насекомые и т. п.); 2) нейстический (бега-
ющие и плавающие на поверхности воды организмы — вертячки,
водомерки, ряска и пр.); 3) планктический (фитопланктон и зоо-
планктон в поверхностных слоях водной толщи); 4) бентический
(рдесты, роголистник, харовые водоросли, донные виды беспозво-
ночных); 5) внутригрунтовый (ракообразные, некоторые виды рыб).
Но границы между биогидрогоризонтами, да и сами горизонты,
подвижны в связи с вертикальными токами воды и суточными ми-
грациями организмов. В связи с этим одни биологи (К- В. Бекле-
мишев, 1969; и др.) относят население всей толщи водоема к одно-
му биогидроценозу, расчлененному на ярусы, другие (американский
биолог А. Клюг, 1923) население гидробионтов каждого яруса при-
нимают за самостоятельный биогидроценоз. В основе правильного
решения данного вопроса, очевидно, должно лежать наличие, в
пределах признаваемой в качестве самостоятельного биогид-
роценоза пространственной структуры, совокупности живых и
косных компонентов, обеспечивающих во взаимосвязи биотиче-
ский круговорот веществ и поток энергии и непрерывность этого
процесса.
Лежащие в основе существования и развития биогидроценозов
энерготрофические взаимоотношения между живыми компонентами
и топоэкологические взаимосвязи биоценоза с биотопом существен-
но отличаются от системы взаимосвязей, сложившихся в наземных
биогеоценозах. Непосредственная связь между живыми компонен-
тами биогидроценозов, по Мордухай-Болтовскому (1966), выражена
слабо; чаще наблюдается косвенное воздействие водных организ-
мов друг на друга — через воду, посредством воды. Отчетливо
выражены лишь трофические отношения хищника и жертвы. Так,
383
основным условием для успешного осуществления пищевых вза-
имоотношений автотрофного компонента с гетеротроф-
ным в открытых водных пространствах, по словам Н. С. Гаевской
(1966), является необходимость совпадения их появления в месте
контакта. В зоне же прибрежных зарослей макрофитов для
этого необходимо, чтобы заросли высших водных растений служили
для животных-фитофагов не только источником питания, но и бла-
гоприятным во всех отношениях местом обитания, в частности,
обеспечивали бы надежное укрытие от врагов, сохранность потом-
ства, давали бы материал для постройки гнезд и другого типа убе-
жищ и пр.
Связь между водным биоценозом (как совокупностью жи-
вых компонентов) и биотопом, как отмечает Ф. Д. Мордухай-
Болтовской, является по существу односторонней. Биотоп оказывает'
мощное воздействие на состав, структуру и динамику водных це-
нозов, влияние же населения гидробионтов на среду обитания
почти незаметно. В водной среде, по мнению Е. А. Тимофеевой-
Ресовской (1963), ведущими являются другие механизмы, чем на
суше. В наземных биогеоценозах особую роль, по ее словам, игра-
ют почвообразование и взаимоотношения между почвой и стоком
воды; в водоемах же (озерах, прудах, водохранилищах) особую
роль играет концентрация химических элементов в донных отложе-
ниях, в особенности первичная биологическая продукция, соотно-
шение трофических уровней в ценозе, общий баланс веществ и
энергии и т, п.
Такова функциональная организация и популяционно-видовой
состав живых компонентов континентальных биогидроцеиозов, как
естественных (стоячих и проточных), так и искусственных (водо-
хранилищ и пр.). Основное свойство водных ценотических биокос-
ных систем, по словам Г Г. Винберга (1971),—это «большое фи-
зиологическое единство функций разнообразных форм жизни и ви-
дов» (с. 12).
6. Обмен веществ и энергии в биогидроценозах
и типы биотических круговоротов
В основе существования и развития биогидроцеиозов, осуще-
ствления присущих им биогеохимических функций в биогидросфере
лежит непрерывный процесс обмена веществ и энергии. Процесс
этот, как и в наземных биокосных системах, имеет циклический ха-
рактер, протекающий в форме биотического круговорота веществ
и энергии. Хотя непосредственными активными агентами, осуще-
ствляющими обменные процессы в биогидроценозе, являются жи-
384
вые компоненты (автотрофы, биотрофы и сапротрофы), в хо-
де его непременно включаются и косные компоненты, и прежде
всего природные воды, содержащие в том или ином виде
органические и минеральные вещества и биогенные газы (угле-
кислота, кислород), а в донных ценозах и подводные грунты
разного типа. Принципиальная схема биотического круговорота^
как отмечает Ф. Д. Мордухай-Болтовской, остается такой же, как
и в наземных биогеоценозах. Однако в биогидроценозах эта общая
схема биотического круговорота, по его словам, значительно услож-
няется и в некоторой степени трансформируется. При этом некото-
рые звенья круговорота выпадают или выносятся на прибрежную
зону или даже за пределы водоема — на сушу, превращая ценоз
в таком случае в наземно-водный.
Начальным звеном биотического круговорота, как и на суше,
является созидание автотрофами в процессе фото- и хемосин-
теза первичной биологической продукции в виде растительной био-
массы. Поставщиками ее в континентальных водах, как было ска-
зано, является главным образом фитопланктон, а в стоячих
водоемах, кроме того, прибрежная водная растительность из мак-
рофитов. Годичный прирост и общая фитомасса макрофитов на
территории СССР, по данным Н. С. Гаевской (1966), достигают
огромных размеров. Первое место среди укореняющихся прибреж-
но-водных растений занимают тростник, камыш, рогозы и другие
виды с возвышающимися над поверхностью воды побегами. Так,
например, общая площадь тростниковых зарослей на территории
СССР, по тем же данным, достигает 5 млн га, а годичный прирост
фитомассы (урожайность) в воздушно-сухом состоянии составляет
от 16—18 до 30—40 ц/га. На втором месте по площади в южных
водоемах и дельтах рек стоит рогоз узколистный с урожайностью
13—25 ц/га, а в северной половине — рогоз широколистный, уро-
жайность которого достигает 100 ц/га сухой фитомассы. В стоячих
водоемах лесной области имеются обширные заросли камыша озер-
ного с урожайностью от 6—8 до 18 ц/га сухой фитомассы. Велика
площадь и довольно высока урожайность и других прибрежно-вод-
ных цветковых и высших споровых видов растений. Многие тысячи
и десятки тысяч тонн фитомассы в стоячих водоемах приходятся
также на долю харовых водорослей, образующих в южных и гор-
ных районах страны настоящие подводные луга, иногда с участием
в их составе гидрофильных видов мха.
По данным исследований С. И. Кузнецова (1970) и А. Г Роди-
ной (19716), в стоячих континентальных водоемах важное место в
качестве пищевого ресурса зоопланктона занимает бактери-
альное население, образующее бактериопланктон, состоя-
щий из разнообразных палочек, кокков, спирилл, а также из кле-
ток дрожжей, серобактерий, нитчаток и пр. Через бактериальное
385
звено водными животными используется большое количество выс-
шей водной растительности, значительная часть отмирающего фи-
топланктона, а также органическое вещество, растворенное в воде,
и пр. Так, например, в Рыбинском водохранилище продукция фито-
планктона за летний период, по данным С. И. Кузнецова, соста-
вила 100—110 тыс. т, а продукция биомассы бактерий достигла
160 тыс. т. Не менее важное пищевое значение в континен-
тальных водоемах, по данным А. Г. Родиной, имеет и детрит,
на базе которого развивается огромное количество планктонных
и донных животных, а также и бактериального населения
(см. рис. 46).
Процессы образования органического вещества в процессе фото-
синтеза фототрофов, как указывает С. И. Кузнецов, совершаются
в верхних слоях континентальных водоемов, а процессы деструк-
ции.— в толще воды и в донных отложениях ила и других суб-
стратов. Процесс деструкции органических веществ в водоемах осу-
ществляется различными физиологическими группами водной
микрофлоры. Остатки высшей водной растительности и фито-
планктона разлагаются и минерализуются аэробными и анаэроб-
ными бактериями, отчасти низшими грибами и актиномицетами, а
органические вещества, находящиеся в растворенном или коллои-
дальном состоянии в воде, — неспороносными бактериями (палоч-
ками, кокками и пр.). Однако не все органическое вещество, обра-
зовавшееся в водоеме и поступившее извне, как утверждает
С. И. Кузнецов, подвергается деструкции и минерализации. Часть
его удаляется из водоема со стоком воды или выбывает из него в
виде вылета взрослых насекомых, вылова человеком или поедания
птицами рыб, выхода биогенных газов в атмосферу; часть органики
откладывается на дне водоема, входя в состав минеральных дон-
ных отложений, и пр. С другой стороны, согласно исследованиям
В. И. Романенко (1967), величина деструкции органического ве-
щества в стоячих водоемах в расчете на весь столб воды на еди-
ницу площади, как правило, превышает величину фотосинтеза фи-
топланктона. Из этого он делает вывод о поступлении орга-
нического вещества в том или ином виде в водоем извне —
с поверхностным стоком воды, с атмосферными осадками, в
результате выщелачивания его из зоны временного подтопле-
ния и т. п.
Исходя из всего изложенного, нетрудно понять и признать обос-
нованность представлений Ф. Д. Мордухай-Болтовского о ходе об-
менных процессов в водных ценозах. В континентальных водоемах
он различает два типа биогидроценозов и в соответствии с этим
два основных типа биотического круговорота веществ. Первый тип
биогидроценозов характеризуется следующим направлением тро-
фических связей между живыми компонентами в пищевой цели.
386
I. Зеленые растения (фитопланктон и заросли макрофитов).
II. Беспозвоночные животные (зоопланктон) и рыбы.
III. Микроорганизмы-деструкторы.
В биогидроценозах
такого типа обмен свет вылов
веществ и поток
энергии протекают
по классической схе-
ме круговорота с
замкнутым циклом
его (рис. 48). Прав-
да, такая схема кру-
говорота в чистом
виде, по словам Мор-
духай-Болтовского,
реализуется лишь в
морских условиях; в
континентальных же
биогидроценозах, на-
ряду с основным на-
правлением трофи-
ческих связей, имеет
место и второстепен-
ный поток веществ
и энергии, поступаю-
щих в ценоз с бере-
говых склонов, с во-
дами рек, ручьев
ДОННЫЕ v ОТЛОЖЕНИЯ
Рис. 48. Схема циклического круговорота веществ,
в континентальных биогидроценозах (по Ф. Д. Мор-
духай-Болтовскому, 1975)
и пр.
Второй тип биогидроценозов обычно формируется в таких есте-
ственных водоемах и искусственных прудах и водохранилищах, где
основным источником энергии являются органические вещества, по-
ступающие в ценоз из внешнего окружения, а продукция фито-
планктона, состоящего из синезеленых и крупных диатомовых
водорослей, вследствие низкого кормового достоинства ее имеет
второстепенное значение. Функциональная организация биогидро-
ценоза, по Мордухай-Болтовскому, приобретает в таком случае сле-
дующий вид.
I. Детрит (продукты неполного распада отмерших растительных
и животных тканей).
II. Бактерии, живущие за счет детрита.
III. Беспозвоночные животные, питающиеся бактериями.
Нетрудно убедиться в том, что в биогидроценозе такого типа, вви-
ду непрерывного потока веществ извне н отсутствия звена сапро-
387
трофов, обмен веществ и энергии перестает быть циклическим; он
назван упомянутым автором поточным,
В биогидроценозах проточных (текучих) континентальных во-
доемов (рек, ручьев и т. п.), где первичная биологическая про-
дукция— отмершие растительные остатки и органическое вещество
в растворенном или взвешенном состоянии — не используется на
месте, а сносится вниз по течению, обмен веществ и энергии
Ф. Д. Мордухай-Болтовской назвал транзитным.
7. О динамике континентальных бногидроценозов
Население трофически взаимосвязанных в единое целое живых
компонентов бногидроценозов, развивающихся в соответствующих
биотопах континентальных водоемов, подобно наземным биогеоце-
нозам обладает определенной степенью устойчивости, стабильно-
сти в составе, структуре, функциональной организации и пр. В то
же время под воздействием целого ряда внутренних процессов
своего бытия и влиянием внешних факторов они подвергаются раз-
ного рода изменениям количественного и качественного порядка.
Одни из них, как было сказано ранее, относятся к категории цик-
лических изменений — сезонных и погод ичных (разногодних),
которые обратимы, другие являются коренными, необратимы-
ми изменениями.
Сезонные и разногодние циклические изменения
водных ценозов — прибрежно-водных, планктонных — в общих чер-
тах сходны с циклическими изменениями наземных биогеоценозов,
поскольку в основе их лежат одни и те же причины; лишь измене-
ния бентосных ценозов бывают не столь ярко выраженными. Ко-
ренные изменения бногидроценозов, так называемые сукцес-
сии, также в принципе не отличаются от сукцессий биогеоценозов
зуши. Они происходят вследствие постепенного приспособления жи-
вых компонентов к изменяющимся условиям водной среды, а так-
же при коренном изменении уровневого и водного режимов есте-
ственного или искусственного водоема и пр. Но в отличие от
циклических изменений бногидроценозов коренные изменения явля-
ются качественными изменениями, которые сопровождаются изме-
нением популяционно-видового состава, структурно-функциональ-
ной организации и характера обмена веществ и энергии, что в ко-
нечном итоге приводит к смене данного биогидроценоза другим.
Смена же и других типов бногидроценозов водоема, следователь-
но, может привести к смене всего населения данного водоема.
388
8. Мировой океан, его гидрологическая структура
и стратиграфия водных масс и дна
Перейдем к рассмотрению характерных черт л особенностей
океанической области биогидросферы, а также морских и океан-
ских биогеоценозов (биогидроцеиозов), ее составляющих. Океани-
ческая область биогидросферы (Мировой океан) во многих отно-
шениях значительно отличается не только от наземного (сухопут-
ного) сектора биосферы, но и от континентальной области био-
гидросферы.
В разделе, посвященном континентальной области биогидро-
сферы, были рассмотрены особенности водной среды жизни в це-
лом, и в частности пресноводной среды обитания, по сравнению
с наземной (воздушной) средой, типы континентальных водоемов
и типы биотопов в их составе; были рассмотрены характерные чер-
ты и особенности обитателей водной среды — гидробионтов и
образуемых ими биокосных систем — пресноводных биогидро-
ценозов. Были обсуждены и некоторые принципиальные вопро-
сы, касающиеся самого существа биогидроцеиозов, их компонент-
ного состава, структурно-функциональной организации и внутри-
ценозных обменных процессов, а также циклические и коренные
изменения континентальных биогидроцеиозов во времени и пр.
Е. М. Лавренко (1965) и Б. А. Быков (1970) вслед за В. И. Вер-
надским (1926) считают, что наиболее благоприятными средами
жизни на Земле являются области соприкосновения, контакта раз-
ных геосфер: а) поверхность суши (соприкосновение атмосферы с
литосферой); б) поверхность водоемов разного типа и масштаба
(атмосферы с гидросферой); в) дно водных бассейнов (гидросферы
с литосферой). Эти участки планеты, как указывает Быков, явля-
ются самыми деятельными, подвижными, динамичными; здесь, по
его словам, в огромных масштабах происходит трансформация
солнечной и химической энергии и протекают процессы физическо-
го и химического выветривания. Из этого следует, что наиболее
благоприятными средами для существования и развития органиче-
ской жизни в Мировом океане являются: а) поверхностная часть
водной толщи, а в менее глубоких областях, помимо того, б) при-
донная часть и дно (ложе) морей и океанов.
Ю. Одум (1975), рассматривая Мировой океан как среду
жизни, прежде всего подчеркивает непрерывность его, поскольку
все океаны и моря (за исключением немногих, вследствие текто-
нических процессов обособившихся морей), связаны друг с другом
в единый водный бассейн с находящимися среди него разобщен-
ными между собой континентами суши. Основными барьерами,
ограничивающими распространение живых существ по морским и
океанским просторам, он считает их глубину, а также температуру
389
в соленость воды. В качестве важнейших характеристик Мирового
океана как среды жизни Ю. Одум приводит: а) постоянные, до-
вольно сильные ветры в одном и том же направлении — пассаты;
б) постоянную циркуляцию морской воды; в) определенные
поверхностные и глубинные течения водных масс; г) волны и
регулярные приливно-отливные движения воды; д) так на-
зываемые апвеллинги — подъемы богатых биогенными элемен-
тами глубинных холодных вод на поверхность; е) наличие раство-
ренных в морской воде, наряду с солями хлористого натрия (око-
ло 27%), таких биогенных питательных элементов, как азот, фос-
фор, калий, а также солей магния, кальция и др.
Приведем из работы В. Г Богорова (1969) некоторые количе-
ственные показатели, характеризующие водную оболочку
планеты и Мировой океан, а также баланс воды на Зем-
ле. Водная оболочка всей планеты составляет 412,5 млн км2
(80,9% от поверхности Земли), а водная поверхность Мирового
океана — 361 млн км2 (70,8%). Средняя глубина Мирового океа-
на— 3795 м, а максимальная глубина его (в Тихом океане) —
11022 м. Общий объем воды на планете составляет 1460 млн км3,
нз них в Мировом океане —1370 млн км3. Благодаря физическому
испарению воды с поверхности Мирового океана и циркуляции воз-
душных масс над поверхностью Земли происходит обмен между
водами океана и атмосферой. С поверхности океана, по данным
В. Г. Богорова, ежегодно испаряется 448900 км3 воды, из которых
411600 км3 выпадает в виде атмосферных осадков непосредственно
над океаном, а остальные 37300 км3 уносятся в сторону континен-
тов и выпадают в виде осадков над сушей, откуда большая часть
их с поверхностным речным стоком снова попадает в океан.
Горизонтальное и вертикальное расчленение рельефа дна и
водной оболочки океана, по Ю. Одуму и В. Г. Богорову, представ-
ляется в следующем виде. I. Материковый шельф — это от-
мель. являющаяся продолжением суши в глубь океана до глубины
200 м, которая (вместе с морями) занимает 7,6% Мирового океана.
II. Материковый склон — это цоколь фундамента континен-
та — в среднем до глубины 3000 м, площадь дна которого состав-
ляет 15,3%. III. Ложе океана—это дно океана, включающее
океанские равнины, впадины, хребты; глубина его от 3 до 6 тыс. м,
а площадь 75,9%. IV. Узкие желоба (каньоны) глубиной от 6
до 11 тыс. м (11,022 м), общей площадью 1,2% от площади Миро-
вого океана. Водная оболочка океана в горизонтальном направле-
нии подразделяется на две области. 1. Неритическая об-
ласть в пределах материкового шельфа, подразделяющаяся на
три вертикальные зоны; а) супралитораль, б) литораль и
в) сублитораль; донная же часть неритической области носит
название бенталь. 2. Пелагическая область океана,
390
омывающая ма'^'ериковый склон и ложе океана; в пределах ее вы-
деляются сЛедУ^ющие вертикальные зоны: а) поверхностная
(эвфотичесКаЯ) >ю Ю. Одуму) зона глубиною 100—200 м; б) пере-
ходная (w014 ическая, по Ю. Одуму) зона глубиною до 1000 и;
в) промежуточная зона глубиною до 4000 миг) глубоко-
водная зона _^о 6qqq—8000 м глубиной. Донная часть пелагиали,
т. е. рельеф дн^ открытого океана, подразделяется: а) на бати-
аль— дно ^а^рикового склона; б) абиссаль (верхняя и ниж-
няя)— это доя^е океана; в) у л ь тр а а б и сс а л ь — дно желобов
(каньонов) океена (рис. 49).
Одна изглавных особенностей воды как среды жизни, как бы-
ло сказано,^ э>о подвижность ее, а главными барьерами распро-
странения Живьмх существ в океане в горизонтальном направлении
являются гемп^ратура и соленость воды. Эти температурно-соле-
ностные бйРье^ы в океане названы фронтами. Фронты — это
естественнЖе п^»граничные зоны между реально существующими в
океане водРыми структурными отдельностями. Т. А. Айзатуллия
с соавторами (1984) рассматривает фронты как функционально
активные А0ГР^ничные слои, с одной стороны, разделяющие, а С
другой — объед.иняющие водные структуры океана в единое целост-
ное планетРРнае образование. Фронты сопряжены с главными мор-
скими и о^ан^кими течениями, вместе с которыми они представ-
ляют собой высокоупорядоченные системы в полосе контакта
встречных в0Д*1ых масс, различающихся температурным и соле-
ностным гради^нтами
Протяженн(^сть фронтов измеряется многими десятками и сот-
нями килоМет£ков. Они тянутся по периферии окаймляемых ими
водных стрУктУр довольно широкой полосой, обычно с отчетливо
выделяющейся центральной пенистой зоной. Зона эта бывает на-
столько бС’гатг^й планктонными организмами, что в воде кишат
питающиеся иМи различные рыбы, головоногие моллюски и пр., а
также кру0ны^ хищники (акулы и др ), а над водой парят, летают
стаи морсМих Ьтиц, вылавливающих из воды добычу. Особое зна-
чение в воАн0^ и биологической структуре океана имеют плане-
тарные ФР^нты, которые, по словам Т. А. Айзатуллина и др.,
могут рассматриваться как особые ленточные ландшафты «с ха-
рактерной для пограничных зон активностью всех природных про-
цессов и обогащением экологических систем (биогидроценозов.—
Л. Н.)» (С- 34) Некоторые фронты, как отмечают авторы, имеют
замкнутые контуры, т. е. принимают форму круговоротов
разного мдСШ*аба, с малоподвижными в центральной части и в
химико-би^логИческом отношении сравнительно слабоактивными и
обедненными Чсивым веществом водными массами.
Воды океаНов нашей планеты и примыкающих к ним морей в
391
Ряс, 49. Горизонтальное и вертикальное расчленение пелагической и бентической областей океана (по
В. Г Богороьу, 1969, и Ю. Одуму, 1975)
той или иной мере отличаются друг от друга по целому ряду гид-
рологических и других показателей (солевому составу, темпера-
турному режиму, плотности воды и т. п.). В пределах каждого из
них воды также неоднородны на всем их протяжении. Гидрологи
выделяют в морях и океанах достаточно изолированные и обособ-
ленные друг от друга естественные объемы воды, именуемые вод-
ны м и массами. Каждая водная масса, как отмечает К. В. Бек-
лемишев (1969), формируется в определенных физико-географиче-
ских условиях, приобретая при этом соответствующие климатиче-
ски обусловленные качественные черты. В силу этого и в особен-
ности благодаря различию в плотности воды они не смешиваются
полностью с соседними водными массами, устойчиво сохраняя свою
специфику в течение неопределенно длительного времени.
Водные массы подразделяются гидрологами на две категории—
первичные и вторичные. Первичные водные массы, как
указывает К. 'В. Беклемишев, занимают широтные климатические
зоны океана. Основная масса воды циркулирует в них в форме
круговоротов. Водные массы, образующиеся в местах частичного
смешения двух смежных океанских водных масс или одной из них
с проникающими в океан водами моря, называются вторнчны-
м и. Для них характерна однонаправленная, а не круговая цирку-
ляция воды и более быстрое в силу этого обновление ее по сравне-
нию с первичными водными массами.
Набор разного типа водных масс горизонтального простирания
и определенный порядок распределения их в пространстве назы-
вается гидрологической структурой вод. Но этот тер-
мин употребляется и в смысле вертикального расчленения столба
воды относительно стационарных первичных водных масс на гори-
зонтальные водные «пласты», последние в таком случае также не-
которые называют водными массами.
Первичные воды в морях и океанах, как было сказано, цирку-
лируют в форме круговоротов разного масштаба. Линии водного
тока соприкасающихся круговоротов, по К. В. Беклемишеву, обра-
зуют так называемое деформационное поле, а акватория
между круговоротами получила название нейтральной обла-
сти. В нейтральных областях происходит взаимное проникновение
первичных водных масс соприкасающихся круговоротов и преобра-
зование их во вторичные водные массы. Следовательно, деформа-
ционное поле состоит из ряда соприкасающихся горизонтальных
круговоротов крупного масштаба и нейтральных областей между
ними. В пределах последних близ берегов суши образуется боль-
шое количество достаточно устойчивых круговоротов мелкого мас-
штаба. В Тихом океане, например, в составе деформационного поля
К. В. Беклемишевым (1973) выделяются следующие крупно-
масштабные круговороты с нейтральными областями
393
между ними: ССП — северный субполярный (субарктический),
ЮСП — южный субполярный (субантарктический), ССТ — север-
ный субтропический, ЮСТ — южный субтропический, СТ — север-
ный тропический (экваториальный), ЮТ—южный тропический (эк-
ваториальный), НО —н ейтр а л ьные области (рис. 50).
90 -120 450 480 450 420 90 GO 30
Рнс. 50. Деформационное поле в Тихом океане (по
К. В, Беклемишеву, 1973). Крупномасштабные круговороты
первичных водных масс: 1 — северный субполярный (суб-
арктический); 2 — южный субполярный (антарктический);
3 — северный субтропический; 4 — южный субтропический;
5—северный тропический (северный экваториальный); 6—
южный тропический (южный экваториальный). НО — ней-
тральные области вторичных водных масс
9. Биотопы пелагиали и океанического дна
Морская вода в морях и океанах служит средой жизни для по-
давляющего большинства их обитателей (гидробионтов). В толще
морской воды они, по выражению К. В- Беклемишева, находят
394
«стол и дом». Из этого следует, что необходимыми условиями су-
ществования и развития морских и океанических обитателей и
образуемых ими сообществ (биоценозов) являются отно-
сительно замкнутая циркуляция в форме круговорота достаточно
обособленной модификации водной массы, отличающейся от со-
седних водных масс, относительная однородность водных масс по
своим физико-химическим свойствам и наличие в том или ином
виде в воде необходимых питательных биогенных элементов. Замк-
нутая циркуляция воды, по словам К. В. Беклемишева, опреде-
ляет возможность существования какого бы то ни было постоян-
ного населения, а различия в физических и химических усло-
виях участка акватории определяют состав населения живых
существ.
Выделяемые гидрологами в морях и океанах водные массы
определенного состава и структуры, циркулирующие в форме кру-
говоротов, служат средой обитания, точнее, биотопами свой-
ственного им населения морских гидробионтов; границы
же круговоротов являются границами этих водных масс и, следо-
вательно, биотопов. Наиболее устойчивыми и энергоемкими
К. В. Беклемишев (1973) считает крупномасштабные
круговороты, состоящие из постоянных течений, являющих-
ся сегментами такого рода круговоротов. Крупномасштабные
круговорты обычно простираются от одного берега океана
до другого, размеры их исчисляются сотнями и тысячами километ-
ров в диаметре, каждый из них, как правило, занимает целую ши-
ротную климатическую зону, характеризуясь определенными пока-
зателями температуры, солености, а следовательно, и плотности
воды. В глубину они прослеживаются примерно до 1500 (2000) м
от водной поверхности, глубже направление течений обычно
не совпадает с направлением поверхностных водных те-
чений.
Позднее, как указывает К- В. Беклемишев, выяснилось, что под
действием ветровых волн, так называемых инерционных колебаний
волн и таких факторов, как различия в степени нагрева поверхно-
сти океана, пассатные ветры и т. п., возрастает энергоемкость круп-
номасштабных круговоротов порядка 1000 км и мелкомасштаб-
ных—10 км и 10 м в диаметре. Исходя из этого, он приходит к
заключению, что в открытом океане круговороты диаметром 1000 км
являются самыми устойчивыми биотопами пелагических сообществ,
так как они, наряду с высокой энергоемкостью, отличаются по-
стоянством и относительной однородностью в своих частях, а по-
тому и население живых существ в их границах также относитель-
но однородно и постоянно. В местах соприкосновения течений
смежных нейтральных областей возникают, как было сказано, вто-
ричные структуры водных масс со своими круговоротами разного
395
масштаба и разной степени устойчивости. Наиболее устойчивые из
них в открытом океане оказываются биотопами так называемых
дальненеритических сообществ. В прибрежной полосе нейтральных
областей близ континентов и больших островов формируется мно-
жество круговоротов мелкого масштаба диаметром 10 км и 10 м,
которые под действием суши оказываются вполне обособленными
модификациями водных масс, обладающими высокой степенью
устойчивости и энергоемкости. В силу этого они становятся биото-
пами прибрежно-водных неритических сообществ.
Что же касается биотопов морского и океанического дна, то и
они в известной мере находятся под воздействием океанических
круговоротов соответствующего масштаба, так как общие контуры
их в пределах проникновения водных масс определенной гидроло-
гической структуры в океанические глубины определяются грани-
цами этих водных масс. Правда, расчленение среды обитания в
пределах участка дна, омываемого водной массой того или иного
круговорота, на более дробные биотопы определяется характером
дна (песчаного, илистого, каменистого и т. п.). В океанических
глубинах, находящихся вне досягаемости водных масс деформа-
ционного поля, среда обитания донного и придонного населения»
по-видимому, не дифференцируется на дробные биотопы, так как
глубинные воды и донные морские осадки оказываются относи-
тельно однородными на огромных пространствах, представляя, та-
ким образом, биотопы колоссального размера, разграничивающиеся
геолого-геоморфологическими особенностями морского дна.
Таким образом, роль круговоротов воды в качестве биотопов
весьма существенна. Во-первых, они, по словам К. В. Беклемишева
(1973), создают «прочную материальную основу биологической
структуры» морских и океанических биоценозов. Если бы не
было в океане круговоротов воды, отмечает он, то не было бы и
районов с обособленным населением живых существ, т. е. биоце-
нозов. Во-вторых, каждый круговорот переносит водную массу
определенного состава, обладающую особыми, отличными от дру-
гих водных масс свойствами, представляющими собой необходимые
условия существования определенных биоценозов в своих пределах.
«Биотоп есть часть среды обитания, занятая биоценозом
(сообществом)»,— пишет К. В. Беклемишев (1967, с. 7) Одно из
основных свойств биотопа — относительная однородность его в про-
странстве по важнейшим параметрам водной массы, являющейся
результатом турбулентного перемешивания ее. Но однородность
водного биотопа, как и наземного, следует представлять не в виде
аморфной массы воды на всем занимаемом им пространстве, а в
виде мозаики более мелких, иерархически соподчиненных водных
структур разного ранга в пределах как горизонтальной, так и вер-
тикальной его протяженности. Ко второму свойству биотопа следу-
396
ет отнести изменчивость его в пространстве и во времени, связан-
ную с непрерывной горизонтальной и вертикальной циркуляцией
воды, а также колебание в некоторых пределах границ самого
биотопа и входящих в его состав мелких структурных отдель-
ностей.
Таким образом, биотопы пелагических сообществ в открытом
океане окантовываются, по К. В. Беклемишеву, крупномасштаб-
ными круговоротами, преимущественно порядка 1000 км в диамет-
ре, а биотопы дальненеритических сообществ — круговоротами дру-
гих размеров, формирующимися в нейтральных областях. Биото-
пами прибрежно-водных неритических сообществ являются водные
массы в границах мелкомасштабных круговоротов диаметром 10 км
и 10 м. Биотопы морского и океанического дна в пределах мате-
рикового шельфа очерчиваются границами различных по механи-
ческому составу донных грунтов. На более значительных морских
и океанических глубинах как донная, так и придонная водная сре-
да обитания ввиду относительной однородности на значительном
протяжении выступает в виде биотопов крупного масштаба.
Таковы характерные черты Мирового океана как среды жизни
и среды обитания морских и океанических гидробионтов и особен-
ности основных биотопов толщи воды открытого океана, морских
и океанических глубин, а также прибрежных участков акватории и
подводных грунтов.
10. Население, зоны его концентрации в океане
и особенности океанических бногидроценозов
Исходя из особенностей Мирового океана как среды обитания
и биологических особенностей морских и океанических гидробион-
тов, способных существовать и развиваться в этих условиях,
В. Г Богоров (1969) дает такое определение его в качестве вод-
ного сектора биосферы: «...биосфера океана (биогидросфера.—
Л. И.)—это термодинамическая оболочка с температурой от +30
до —2° С и давлением от 1 до 1000, вернее, до 1100 атм» (с. 16).
Л. А. Зенкевич (1970) рассматривает Мировой океан как «це-
лостный природный комплекс», населенный богатой и разнообраз-
ной флорой и фауной. Общее количество обитающих в океане ра-
стений и животных, по его данным, — более 200 тыс. видов; среди
них, по данным В. Г. Богорова, более 150 тыс. видов животных.
В составе животного населения подавляющее большинство состав-
ляют беспозвоночные — около 120 тыс. видов, в том числе: губок —
5 тыс. видов, кинечнополостных — 9, свободноживущих червей — 6,
иглокожих — 6, но особенно много головоногих и других мол-
люсков— 65 и ракообразных — 25 тыс. видов. Среди позвоночных
животных преобладают рыбы—16 тыс. видов, млекопитающих всего
397
лишь 90 видов, в том числ 58 видов китообразных и 32 — ластоно-
гих. Великое множество в морской воде и простейших — около
10 тыс. видов. В составе растений господствуют водоросли —
15 тыс. видов, а среди них — одноклеточные, в том числе: зеленые
водоросли — более 5 тыс. видов, диатомовые — около 5 и синезеле-
ные— 1 тыс. видов. На долю многоклеточных водорослей прихо-
дится около 4 тыс. видов, в том числе бурых водорослей—1 и
красных — 2,5 тыс. видов. Количество высших сосудистых растений
в морях и океанах ничтожно мало. Среди них наиболее распро-
страненными являются взморник (Zostera), филлоспадикс, некото-
рые виды морских солянок и пр.
Богаты морские и океанические воды и микрофлорой, особенно
бактериальной, которая не только выполняет функции деструкции
и минерализации отмершей растительной и животной органики, но
и служит источником питания для различных животных-фильтра-
торов, а также детритоядных, поедающих вместе с детритом и бак-
териальную массу.
Основу водного сектора биосферы (биогидросферы) В. И. Вер-
надский (1926, 1967), как известно, видел в высокой плотности,
концентрации жизни. Такие богатые жизнью участки, как было
сказано ранее, он называл пленками и сгущениями жиз-
ни. В Мировом океане В. И. Вернадский выделял две пленки жиз-
ни: а) планктонную в поверхностной части океана мощностью
в 50—60 м и б) донную различной мощности на глубинах 4—
6 тыс. м. К сгущениям жизни он относил прибрежноводные
сгущения в местах контакта океана и суши и так называемое
саргассовое и ему подобные сгущения в поверхностных водах
океана. Пленки и сгущения жизни отличаются от остальной части
океана не только концентрацией жизни, но и наиболее высокой
степенью трансформации световой солнечной энергии в различные
формы органического вещества, выполняя тем самым важнейшие
биогеохимические функции планетарного масштаба.
Планктонная пленка жизни, а также прибрежные и другие сгу-
щения жизни в Мировом океане представляют собой участки аква-
тории, отличающиеся весьма высокой концентрацией живого ве-
щества, состоящего из организмов разнообразной систематической
принадлежности и эколого-ценотических приспособительных потен-
ций. Их можно рассматривать как постоянные, самостоятельно
функционирующие зоны жизни в океане, поскольку они обеспечены
собственной автотрофной базой. В противоположность им донная
пленка жизни и промежуточная между обеими пленками водная
толща с рассеянной формой живого вещества в материально-
энергетическом отношении находятся в большой зависимости от
поверхностной планктонной пленки жизни.
Рассмотрим, какими типами биогидроценозов представлены
398
названные пленки и сгущения жизни и каковы их особенности по
сравнению с наземными биогеоценозами, каков их компонентный
состав, структурно-функциональная организация и прочие их
атрибуты.
К. В. Беклемишев (1969) указывает, что океаническая планк-
тонная пленка жизни состоит из биогидроцеиозов полного состава
(полночленных, по Н. В. Дылису), включающих как автотроф-
н ы й, так и гетеротрофные ценокомплексы живых су-
ществ. Биогидроценозы же донной пленки жизни в наиболее глу-
боких местах океана представлены только гетеротрофами,
существующими за счет избытка продукции биогидроцеиозов
планктонной пленки жизни, продуктов жизнедеятельности планк-
тонных организмов и отмерших органических остатков и отбросов.
Наиболее существенными особенностями морских и океаниче-
ских биогидроцеиозов, отличающими их от наземных биогеоце-
нозов, по словам Л. А. Зенкевича (1970), являются: во-первых,
разобщенность в подавляющем большинстве случаев животного на-
селения и автотрофного растительного компонента за исключением
поверхностной эвфотической зоны океана; во-вторых, исключитель-
ная зависимость живых ценокомплексов биогидроцеиозов от мор-
ской воды как среды обитания, ибо в ней в растворенном состоянии
находится все необходимое для их существования и развития (пи-
тательные минеральные соли, различные аминокислоты, биологи-
чески активные вещества и микроэлементы, биогенные газы), толь-
ко необходимая растениям лучистая энергия Солнца поступает
извне; в-третьих, наличие активных и пассивных в вертикальном
и горизонтальном направлениях миграций животных, связанных с
питанием, размножением и т. п., иногда на огромные в горизон-
тальном направлении расстояния.
В связи с этим возникает вопрос о границах морских и океани-
ческих биогидроцеиозов, структурно-функциональной и про-
странственной обособленности их друг от друга. Границы океани-
ческих биогидроцеиозов, по П. М. Рафесу (1980), маркируются
постоянными течениями воды. Причем границы между биогидро*-
ценозами, как утверждает К. В. Беклемишев, даже в океане не
всегда бывают расплывчатыми. Одним словом, границы между
морскими и океаническими биогидроценозами, как и континенталь-
ными, существуют, и для отрицания наличия их нет оснований.
Они определяются границами биотопов, соответствующих каждому
сообществу. Речь, очевидно, может идти лишь о постоянстве и чет-
кости этих границ, которые действительно между биогидроценоза-
ми, по крайней мере, планктонной пленки жизни, ввиду подвиж-
ности водного субстрата и самого населения, также подвижны.
Кроме того, следует иметь в виду, что некоторые животные, в осо-
бенности крупные планктонёры и птицы, в процессе своей жизне-
399
деятельности охватывают не один, а несколько смежных, а иногда
и разобщенных бногидроценозов, являясь в таком случае членами
нескольких бногидроценозов.
11. Биогидроценозы планктонной пленки жизни,
состав и трофическая взаимосвязь
их компонентов
Прежде всего рассмотрим планктонную пленку жизни. По сло-
вам В. И. Вернадского, она покрывает тонким слоем всю поверх-
ность океана. Общая площадь пленки, по его данным, измеряется
сотнями миллионов квадратных километров, а биомасса выража-
ется числами порядка 1015—1016 т. Однако, по современным пред-
ставлениям, планктон распределяется в океане не вполне равно-
мерно, планктонная пленка фактически прерывиста» мощность ее
также неодинакова на ее протяжении. Во-вторых, к планктонной
пленке жизни, как видно, в действительности относится водное на-
селение не только поверхностной (эвфотической) зоны пелагиали
(глубиною в 100—200 м), но и население гидробионтов переходной
(афотической) и промежуточной зон (глубиною до 3800—4000 м).
Дело в том, что обитатели всех этих зон трофически тесно связаны
друг с другом. Бесперебойное существование гидробионтов этой
водной толщи обеспечивается путем утилизации животными, микро-
и макроорганизмами первичной биологической продукции фито-
планктона и путем использования последним на процессы фото-
синтеза минеральных элементов и биогенных газов, находящихся
в растворенном виде в морской воде.
Из изложенного следует, что в качестве структурно обособлен-
ных единиц (бногидроценозов) в пределах планктонной пленки
жизни следует считать трофически взаимосвязанное в единую це-
лостную функциональную систему население гидробионтов фрон-
тальных водных масс крупномасштабных океанических круговоро-
тов и население круговоротов в водах нейтральных областей. Иначе
говоря, биогидроценозы планктонной пленки жизни пред-
ставлены биоценозами основных типов биотопов открытого океана
на всю их глубину с охватом эвфотической и афотической зон
вплоть до нижней границы биотопов (до 3000—4000 м). В средин-
ной же части окаймляемых круговоротами океанических водных
масс ввиду значительной рассеянности организмов оформление био-
ценозов, очевидно, не происходит.
Биокосным компонентом в составе бногидроценозов
планктонной пленки жизни в открытом океане служит, по суще-
ству» вся водная толща пелагиали мощностью до 3800—4000 м.
Плодородие верхнего продуцирующего слоя океана обеспечивается,
400
по словам Л. А. Зенкевича (1970), выносом из глубинных слоен
биогенных элементов (фосфатов, нитратов, силикатов), где они
накапливаются в количестве, «в тысячи раз превышающем их го-
довой расход» (с. 16). В морской воде в растворенном виде содер-
жатся также азот, кислород, углекислый газ и другие необходи-
мые для существования гидробионтов биогенные газы. По данным
В. Г. Богорова (1971), фитопланктоном в процессе фотосин-
теза ежегодно выделяется 40 млрд т свободного кислорода. Кроме
того, в морской воде пелагиали в различном состоянии содержит-
ся огромное количество органического вещества, образующегося
в процессе жизнедеятельности как обитателей поверхностного про-
дуцирующего слоя планктонной пленки жизни, так и трофически
связанных с ними обитателей промежуточной между двумя плен-
ками жизни водной толщи пелагиали. Сюда относятся неразло-
жившиеся и полуразложившиеся остатки и трупы мезо- и макро-
планктона и других гидробионтов, различные продукты метаболиз-
ма и экскременты представителей животного планктона и нектона,
в особенности головоногих моллюсков и морских млекопитающих,
а также связанных с морем по линии питания различных видов
морских птиц.
Вся эта огромная масса содержащихся в морской воде мине-
ральных и органических веществ и биогенных газов в виде водных
растворов и физических смесей — эмульсий — в совокупности обра-
зует двуединый биокосный компонент биогидроценозов планктон-
ной пленки жизни. По своему ценотическому значению он в полной
мере соответствует почвенному и воздушному блокам наземных
биогеоценозов.
Автотрофный компонент планктонных биогидроценозов, по
словам В. И. Вернадского (1967), представлен в основном одно-
клеточным водорослевым фитопланктоном. Обращая внима-
ние на то, что солнечный луч придает различный вид трансформа-
торам космической энергии на суше и в океане, он подчеркивает,
что зеленые одноклеточные водоросли с коротким сроком жизни
могут создавать в определенный промежуток времени несравненно
больше живого вещества, чем высшие сосудистые и другие зеле-
ные растения суши. При этом следует отметить, что наиболее важ-
ную фотосинтетическую роль Ю. Одум отводит нанопланкто-
ну с участием в этом процессе жгутиковых (евгленовых, пан-
цирных, вольвоксовых) и других простейших, ибо они обладают
особенно высокой скоростью размножения и быстрой сменой поко-
лений. Благодаря этому достигаются чрезвычайно высокие темпы
созидания органического вещества. Так, хотя общая биомасса фи-
топланктона Мирового океана, по данным В. Г. Богорова (1971),
составляет лишь 1,5 млрд т, годовая продукция его достигает около
600 млрд т (рис. 51).
401
Л НАТО новые ВОДОЮСЛ*
Цлшрные кгитнкавые
Рис. 51. Морской фитопланктон и нанопланктон (по Ю. Одуму, 1975)
Первичными консументами, потребителями фитопланктоне
морских и океанических биогидроцеиозов, по Ю. Одуму, являютс*
находящиеся в непосредственном контакте с фитопланктоном раз
личные формы зоопланктона, как микроскопические существ?
(фораминиферы, радиолярии, инфузории), так и растительноядные
беспозвоночные животные, относящиеся к мезо- и макро-
планктону (мелкие рачки, черви, медузы, моллюски, гребневики
из кишечнополостных, оболочники и др.). В составе микроскопи-
ческого зоопланктона встречаются и бесцветные гетеротрофные
формы, широко распространенные в афотической зоне, питающиеся
растворенным в морской воде органическим веществом. Общая
годовая продукция зоопланктона, по В. Г. Богорову (1971), при
биомассе 22 млрд т составляет около 55 млрд т (рис. 52). Прямое
выедание фитопланктона животными, по данным Ю. И. Сорокина
(1975), составляет лишь около 25% от его продукции, в том числе
около 15% потребляется организмами мезо- и макропланктона, а
остальная часть ее—микроскопическим зоопланктоном. Отсюда
следует, что 75—80% продукции фитопланктона океанических био-
гидроценозов используется по детритному пути в виде мертвого
органического вещества.
402
эис. 52. Морской растительноядный и хищный зоопланктон и планктоноядный
зектон (по Ю. Одуму, 1975): 1 — мелкие рачки; 2 — мелкие медузы; 3 — крыло*
югие моллюски; 4 — гребневики; 5 — оболочники (сальпы); 6 — многощетин-
ковые черви; 7 — щетинкочелюстные; 8 — сельди, сардины и др.
Для организмов морского и океанического планктона характер-
но наличие целого ряда оригинальных приспособлений. У од-
ноклеточных водорослей наблюдается соединение их в нитевидные
цепочки, отложение в организме запасных веществ в виде жира,
а не крахмала, как у наземных растений. Облегчение пассивного
плавания представителей зоопланктона достигается посредством
образующихся на теле выростов, капелек жира, капсул воздушных
пузырьков, а также обладанием парашютообразной формой тела,
уменьшением размеров тела и т. п. Для активно плавающих форм
зоопланктона характерно образование жгутиков, ресничек, вёслиц
и т. п. Передвижение в воде различных форм зоопланктона осу-
ществляется путем использования названных специальных органов,
а также изгибанием тела, движением задней части его. Важным
приспособлением более крупных форм зоопланктона, способству-
ющим его выживанию, является прозрачность тела или голубая,
под цвет морской воды, окраска его и пр.
Со способностью активного передвижения животного планкто-
на— от зеленых и бесцветных жгутиковых форм нанопланктона
403
до более или менее крупных растительноядных и плотоядных форм
макропланктона — связано важнейшее эколого-биологическое свой-
ство его: суточные вертикальные миграции. Благодаря этому обес-
печивается тесная взаимосвязь гетеротрофных обитателей проме-
жуточной между двумя пленками жизни водной толщи пелагиали
с автотрофным фитопланктоном эвфотнческой зоны. Максимальное
скопление зоопланктона, в особенности микроскопического, есте-
ственно, находится в поверхностном продуцирующем слое эвфоти-
ческой зоны, наиболее богатой необходимым для его развития
органическим веществом. Установлено, что днем зоопланктон опу-
скается на десятки и сотни метров в глубь океана, в пределы почти
полностью лишенной света афотической зоны, где служит объектом
питания для многих обитающих здесь животных. Ночью зоопланк-
тон всплывает в поверхностные слои эвфотнческой зоны для пита-
ния накопившимся за день фитопланктоном. Вслед за ним мигри-
руют и многие рыбы и другие животные.
Из этого следует, что значительная часть обитателей афотиче-
ской зоны и водной толщи пелагиали на глубину основных
океанических биотопов (до 3800—4000 м) оказывается по линии
питания через зоопланктон неразрывно связанной с основным про-
дуцентом первичного органического вещества — водорослевым фи-
топланктоном. Эта промежуточная водная толща морских и
океанических биотопов населена весьма разнообразными биокомп-
лексамн беспозвоночных и позвоночных животных, в особенности
верхняя часть ее — афотическая зона глубиной до 1000 (2000) м.
Здесь обитают питающиеся микро- и мезозоопланктоном плотояд-
ные животные, в частности относящиеся к макропланктону
ракообразные и к мелкому нектону — сельди, сардины и
другие планктоноядные рыбы, обычно плотными стаями
регулярно мигрирующие в эвфотическую зону (см. рис. 52). По-
скольку годовая продукция одноклеточного зоопланктона значи-
тельно превышает его биомассу, то Ю. Одум считает, что он мо-
жет рассматриваться как одно из ключевых звеньев пищевой цепи
от фитопланктона к планктоноядным беспозвоночным и последую-
щим звеньям плотоядных позвоночных животных (рыб, морских
млекопитающих и птиц).
На базе представителей макропланктона и мелкого нектона
как источника питания и энергии в океанических биотопах проме-
жуточной водной толщи пелагиали развивается средний и
крупный нектон. Это главным образом крупные хищ-
ные формы, обладающие, по М. П. Акимову (1959), мощной
мускулатурой, обтекаемой формой тела двусторонней симметрии,
хорошо развитыми органами движения. Главным движущим аппа-
ратом у них является задняя часть тела с вертикальным, горизон-
тальным и боковыми парными плавниками» служащими рулями
404
глубины и стабилизаторами дорзовентрального положения тела.
Передвижение в воде осуществляется полукруговым вращением
или изгибом хвостового отдела, с помощью гребных плавников ве-
сельным способом и т. п. Сюда относятся прежде всего крупные
морские хищные рыбы, многие из которых обладают зубами, а
также некоторые морские млекопитающие. К этой же категории
пелагических обитателей М, П. Акимов относит ластоногих (мор-
жей, тюленей), некоторых головоногих моллюсков, кальмаров, мор-
ских черепах, обладающих укороченным телом, передвигающихся
с помощью различно устроенных конечностей, а также животных
без специальных органов передвижения — пелагических червей,
змей, плавающих путем волнообразного изгибания тела. Сюда же,
по мнению М. П. Акимова,’следует относить таких морских живот-
ных, как осьминоги, каракатицы, медузы, движущихся по принципу
ракеты, резко с силой выбрасывая воду через суженные отверстия
или жаберные щели. Из рыбного населения более 75% обитают в
пределах афотической зоны (рис. 53).
По-видимому, особое положение среди потребителей морского
зоопланктона и мелкого нектона занимают гигантского размера
млекопитающие — усатые киты, которые отфильтровывают до-
бычу из захваченного пастью объема воды через сеть длинных
узких пластинок, образующих так называемый китовый ус. Об-
щая биомасса среднего и крупного нектона Мирового океана, по
В. Г. Богорову (1971), составляет 1 млрд т, годовая же продукция
его не превышает 200 млн т. К этому же звену пищевой цепи био-
гидроценозов планктонной пленки жизни открытого океана, очевид-
но, относятся и питающиеся животным макропланктоном и мелким
нектоном морские птицы (пингвины, альбатросы, фрегаты, качурки,
тропические крачки и пр.). Большинство из них не связаны с су-
шей, за исключением периода размножения.
Следующее звено пищевой цепи биогидроцеиозов промежуточ-
ной водной толщи пелагиали представлено такими крупными
морскими хищниками, как акулы (акула обыкновенная,
морская собака и др.). Зубатые киты (кашалоты) и дельфины так-
же должны быть отнесены к этому же трофическому уровню. Все
они живут за счет пожирания обитающих в водах пелагиали раз-
нообразных представителей крупного и среднего по размерам нек-
тона и других морских животных (см. рис. 53).
Крупные морские животные, в особенности млекопитающие,
ежесуточно пожирают огромное количество морской рыбы, рако-
образных, моллюсков, беспозвоночных животных. Так, по данным
А. Г. Томилина (1970), морскими млекопитающими в Мировом
океане за сутки потребляется в пищу около 2 млн 717 тыс. т раз-
нообразных животных. В частности, усатые киты потребляют в
сутки около 300 тыс. т, зубатые киты (кашалоты) — 450 тыс. т,
405
Рис. 53. Плотоядный и хищный морской нектон различных вер-
тикальных зон пелагиали (без соблюдения масштаба) (по Ю. Оду-
му, 1975). Поверхностная и переходная зоны: 1 — усатый кит;
2 — тюлень; 3 — дельфин; 4 — тунец; 5 —пеламида; 6 — макрель;
7 — карифены; 8 — крупные кальмары; 9 — осьминоги. Промежу-
точная зона: 10 — гигантская акула; 11—белая акула; 12 — меч-
рыба; 13 — кашалот (зубастый кит); 14 — рыбы-топорики; 15 —
светящиеся рыбы. Глубинная зона: 16 — удильщики; 17 — черный
пожиратель; 18 — большероты
дельфины— I млн 700 тыс. т, ластоногие — 267 тыс. т. Следователь
но, морские млекопитающие представляют собой весьма существен
ный элемент бногидроценозов открытого океана. При этом следуе'
иметь в виду, что в их пищевой рацион попадает немало и промыс
ловых морских животных, в том числе и промысловых видов рыб
Выше было сказано, что примерно три четверти биологическо!
продукции фитопланктона эвфотической зоны биогидроценозо!
океана утилизируется по детритному пути. Наибольшая массг
406
детрита скапливается в промежуточной водной толще пелагиали,
в особенности в афотической зоне. Формирование детрита проис-
ходит, во-первых, за счет отмерших органических остатков, трупов
и экскрементов обитающих в этой водной толще позвоночных и
беспозвоночных животных и парящих над ней морских птиц; во-
вторых, за счет органических веществ, находящихся в морской воде
в растворенном состоянии («пенистый детрит»); в-третьих, за счет
органики и продуктов метаболизма, опускающихся из продуциру-
ющего слоя эвфотнческой зоны. Вся эта масса сформировавшегося
детрита служит источником энергии для развивающегося в вод-
ной толще пелагиали сапротрофного бактериального
планктона, общая биомасса которого, по данным В. Г. Богоро-
ва (1971), составляет в океане около 10 млн т, а годовая продук-
ция достигает 100 млрд т.
Таким образом, функциональная организация биогидроценозов
планктонной пленки жизни открытого океана схематично может
быть представлена в следующем виде.
1. Автотрофный компонент.
1. Фотосинтезирующий микро- и нанофитопланктон.
II. Биотрофный компонент.
2. Растительноядный микро- и мезозоопланктон.
3. Хищники 1-го порядка — макрозоопланктон и мелкий нектон.
4. Хищники 2-го порядка — средний и крупный нектон.
5. Хищники 3-го порядка — наиболее крупный нектон.
III. Сапротрофный компонент.
6. Бактериальный планктон.
12. Вертикальная структура биогидроценозов
планктонной пленки жизни океана
Какова же вертикальная биологическая структура биогидроце-
нозов планктонной пленки жизни и водной толщи пелагиали Ми-
рового океана вообще?
Материальной основой, своего рода физическим каркасом вер-
тикальной биологической структуры биогидроценозов открытого
океана служит приведенная выше вертикальная гидрологическая
структура океана (см. рис. 49). На основе данных отечественных
и зарубежных гидробиологов о гидрологической вертикальной
структуре и компонентном составе населения морских и океаниче-
ских биогидроценозов пелагиали можно составить следующее пред-
ставление о вертикальной биологической структуре всей охвачен-
ной жизнью водной толщи его. Так, от поверхности пелагических
биогидроценозов ко дну океана выделяется: 1) поверхностная
(эвфотическая, по Ю. Одуму) зона — единственная продуцирующая
первичное органическое вещество зона мощностью 100—200 м,
объем вод ее, по В. Г. Богорову, составляет лишь 2,5% от всей тол-
407
щи океанических вод, а биомасса фито- и зоопланктона достигает
31% от всей биомассы планктона. Ниже в пределах промежуточ-
ной водной толщи пелагиали выделяются: 2) переходная (афо-
тическая, по Ю. Одуму) зона мощностью 800—900 м (от 100—200
до 1000 м), объем вод 22,5%, биомасса зоопланктона 32%; 3) про-
межуточная (батиопелагиальная, по Богорову) зона мощ-
ностью 3000 м (от 1000 до 4000 м) и 4) глубоководная (абис-
сопелагиальная) зона мощностью 2000—4000 м (от 4000 до 8000 м);
объем вод 3-й и 4-н зон—75% от всей толщи океанических вод,
а биомасса зоопланктона составляет лишь 37% от общей биомас-
сы всего планктона. Следовательно, биомасса планктона, по словам
В. Г Богорова, уменьшается на единицу объема от поверхности до
абиссальных глубин океана «в тысячи раз». Значительно уменье
шается с глубиной и рыбное население. По данным того же авто-
ра, на глубине от поверхности до 500 м рыбное население по весу
составляет более 50% от всего населения рыб, на глубине последу-
ющих 500 м (от 500 до 1000 м) оно составляет лишь 25%, а в
огромном объеме глубинных океанических вод — от 1000 до 6000 м
и более, т. е. на глубине последующих 5 тыс. м биомасса рыбного
населения не превышает 25% от всей его биомассы.
Поименованные зоны вертикального строения биогидроценозов
планктонной пленки жизни, характеризующиеся составом опреде-
ленных по типу питания и образу жизни биокомплексов гидро-
бионтов, могут рассматриваться как ярусы (биогидрогори-
зонты) этих водных биокосных систем. Характерная особенность
их по сравнению с наземными биогеоценозами состоит в том, что
отмеченная ранее разобщенность автотрофного и гетеротрофного
компонентов единой пищевой цепи находит выражение именно в
размещении последних по разным биогидрогоризонтам данного
биогидроценоза.
Что же касается населения глубоководных океанических про-
странств, превышающих 4—6 тыс. м, то оно, во-первых, чрезвычай-
но малочисленно как по видовому составу, так и по численности
особей каждого вида, во-вторых, весьма своеобразно. Это объяс-
няется тем, что глубоководная океаническая среда отличается це-
лым рядом малоблагоприятных для существования гидробионтов
экологических факторов. Известно, что с глубиной температура во-
ды понижается, давление же воды, напротив, значительно воз-
растает, в связи с этим возрастает и плотность воды; соленость же
морской воды изменяется с глубиной по-разному. Кроме того, на
больших глубинах постоянно царит полная тьма, содержание кис-
лорода в воде низкое, а главное — это чрезвычайная скудость пи-
щевых ресурсов, поскольку основная масса их потребляется некто-
ном и детритофагами нижних биогидрогоризонтов планктонной
пленки жизни. Естественно, что в таких аномальных условиях мо-
408
гут жить немногие виды живых существ, обладающие специфиче-
скими приспособлениями.
Поскольку в условиях темных океанических глубин отсутствуют
производители первичной биологической продукции— автотрофы,
население их, по К. В. Беклемишеву (1969), представлено лишь
гетеротрофными группировками (биокомплексами)
преимущественно беспозвоночных животных и бактерий (бактерио-
планктоном), существующих за счет органических веществ, посту-
пающих в том или ином виде из поверхностного слоя планктонной
пленки жизни. На глубине 6—7 тыс. м, по свидетельству В. Г. Бо-
горова (1969), встречаются фораминиферы, губки, кишечнополост-
ные, черви, моллюски, иглокожие, погонофоры и асцидии. На
глубине 10 тыс. м обнаружены лишь фораминиферы, актинии, не-
матоды, некоторые ракообразные, мелкие моллюски, эхиуриды и
голотурии (из иглокожих). Из состава населения темных океани-
ческих глубин, главным образом из-за недостатка пищи и кисло-
рода, полностью исчезли рыбы, крупные ракообразные, головоногие
моллюски и многие другие плотоядные животные. У оставшихся
плотоядных животных развились приспособления, способствующие
снижению энергетических затрат на процессы жизнедеятельности:
уменьшение размеров тела, переход от активного преследования
добычи к пассивному выжиданию ее. В связи с этим у них проис-
ходит редукция мускулатуры, скелета, дыхательной и выделитель-
ной систем, а с другой стороны — увеличение размеров пасти для
схватывания и заглатывания добычи и пр. У многих обитателей
больших глубин развиваются приспособления к отысканию друг
друга (органы свечения, длинные усикообразные придатки); у не-
которых видов (рыб-удильщиков, эхиуровых червей) — самцовый
паразитизм и пр. (см. рис. 53)
Таким образом, обширные глубоководные пространства морей
и океанов в силу указанных обстоятельств по существу представ-
ляют собой океанические «пустыни». Вследствие рассеянности и
разобщенности глубоководных обитателей, неимоверной разрежен-
ности всего их населения при отсутствии собственной автотрофной
энергетической базы и скудости пищевых ресурсов не обеспечива-
ются необходимые условия для формирования здесь трофически
взаимосвязанных целостных биокосных систем (биогидроцеиозов).
13. Биогидроценозы бентали, их компонентный состав
и население батиали, абиссали и коралловых рифов
Перейдем к рассмотрению донной пленки жизни, охватывающей
пограничную полосу соприкасающихся окраин литосферы и гидро-
сферы. «Донная живая пленка,— пишет В. И. Вернадский (1967),—
наблюдается в морской грязи и донном слое воды, ее пронизыва-
409
ющем и к ней прилегающем» (с. 327). По размеру и объему она,
по его расчетам, примерно равна планктонной пленке, по весу же
значительно превышает ее; значительно превышает она планктон-
ную пленку и по своей поверхности ввиду неровностей рельефа дна.
Мощность насыщенной жизнью донной пленки он считает равной
в среднем 10—60 м, но не превышающей 100 м. Донную пленку
жизни Вернадский подразделяет на две части: а) верхнюю, содер-
жащую кислород придонную часть пелагиали, названную пела-
геном, в составе населения которой довольно велика доля много-
клеточных животных; б) нижнюю, собственно донную
часть, представляющую, по его выражению, «слой грязи», насе-
ленный одноклеточными организмами, преимущественно анаэроб-
ными бактериями и отчасти проникающими из пелагена роющими
многоклеточными существами.
Морское и океаническое дно, по данным М. П. Акимова (1959),
В. Г. Богорова (1969), Л. А. Зенкевича (1970), Ю. Одума (1975)
и др., неоднородно по своей поверхности (рельефу), характеру
осадочных пород и грунтов, составу минеральных, скелетных и ор-
ганических отложений (кремневых, известковых и т. п.). В соответ-
ствии с этим М. П. Акимов в пределах бентали выделяет следу-
ющие группы типов и типы донных биотопов: а) прибрежные
галечниковые, каменистые и ракушечые — на мелководье; б) илис-
тые и песчаные — на средних глубинах; в) подводные и береговые
скалы и коралловые рифы; г) заросли морских трав и многокле-
точных водорослей как участки среды обитания фитофильных ор-
ганизмов.
Биотопы донных грунтов батиали и абиссали харак-
теризуются большим количеством кремневых, известковых и прочих
скелетных остатков организмов и различных минеральных веществ,
которые, как отмечает Л. А. Зенкевич (1970), резко преобладают
над органическими. Тем не менее выпадающее в осадок органиче-
ское вещество все же содержит в себе достаточное количество по-
тенциальной энергии, которая служит основой для существования
и развития донных беспозвоночных животных и анаэробных бак-
терий. Поступающие из поверхностного слоя планктонной пленки,
а также из переходной и промежуточной зон органические остатки
и экскременты сосредоточиваются в придонном слое воды, на по-
верхности и в толще грунта. Названные условно детритом, они
также используются беспозвоночными животными и бактериями в
качестве источника питания и энергии на процессы своей жизне-
деятельности. В наиболее значительных глубинах дно океана, по
свидетельству гидробиологов, состоит из обнажений красной гли-
ны, которая, очевидно, малопригодна для обитания живых существ.
Таким образом, биотопы донной пленки жизни в пределах бен-
тали, омываемой водами неритической области океана, и в преде-
410
лах батиали и абиссали, по крайней мере до глубины 3—4 тыс. м,.
обладают запасом пищевых ресурсов, достаточным, чтобы обеспе-
чить существование и развитие населяющих их гидробионтов.
Население донной пленки жизни довольно разнообразно по
видовому составу, по крайней мере в верхней окисленной зоне, в
особенности в пределах бентали, наиболее богатой питательны-
ми веществами. Основными обитателями этой зоны считают много-
щетинковых червей, двустворчатых моллюсков и множество мелких
животных (плоские черви, турбеллярии, тихоходки, коловратки,
нематоды, инфузории и пр,); в местах, освещаемых лучами солнеч-
ного света, развиваются и многоклеточные морские водоросли.
В восстановленном слое грунта донной пленки встречаются лишь
хемосинтезирующие бактерии, анаэробные инфузории да некото-
рые нематоды. Все эти мелкие бентосные формы автором рассмат-
риваются как объекты питания более крупных плотоядных жи-
вотных.
Выделяемые М. П. Акимовым и другими биологами типы и
группы типов биотопов донной плёнки жизни населены соответству-
ющим набором жизненных форм животных. Так, к обитателям
группы илистых типов биотопов, да отчасти и песчаных
и гравийных, автор относит: а) роющие и сверлящие формы
животных, живущие в грунте, иле, в отверстиях (ходах) горных
пород — многие черви, пластинчатожаберные, немногие губки, ра-
кообразные, некоторые голотурии и морские ежи из иглокожих,
ланцетник и немногие рыбы из хордовых; б) ползающие и бегаю-
щие по дну формы — черви, моллюски, ракообразные, иглокожие,
бентические рыбы (камбалы, скаты), некоторые высшие раки;
в) прикрепленные формы животных — губки, полипы, некоторые
пластинчатожаберные, кишечнополостные, моллюски, морские ли-
лии, устрицы, усоногие раки, простейшие.
Подводные биотопы каменистых, галечниковых и
скалистых берегов, а также ракушечников, богатые пи-
тательными веществами за счет фитобентоса и притока их
из открытого океана и суши, населены, по данным М. П. Акимова
(1959) и В. Г. Богорова (1969), такими жизненными форма-
ми животных, как а) прикрепляющиеся и прирастающие формы
из разных систематических категорий — простейшие, губки, кишеч-
нополостные, пластинчатожаберные, мелкие ракообразные и хор-
довые; б) буравящие или сверлящие животные формы, живущие
в отверстиях (ходах) горных пород, — черви, некоторые моллюски,
иглокожие, ракообразные и др.; в) подвижные и свободнолежа-
щие, укрывающиеся среди камней и в расщелинах скал живот-
ные, — гастроподы, цефалоподы с присосками, некоторые иглоко-
жие, пластинчатожаберные и крабы с цепкими конечностями,
многие ракообразные, черви, хищные рыбы и пр.
411
Наиболее характерные для донной пленки жизни виды живых
существ обладают целым рядом приспособительных особенностей:
наличие различных органов прикрепления к субстрату при обита-
нии в условиях прибойных волн, развитие чехлов, трубок, панци-
рей, обеспечивающих защиту их от нападения хищников, или вы-
сокой способности регенерации, способствующих выживанию их в
случае потери при этом некоторых частей тела; развитие способ-
ности к неполовому способу размножения, к образованию колоний
и пр. Особенно глубокие приспособительные изменения в строении
наблюдаются у глубинных бентосных форм животных, обитающих
в пределах абиссали: например, радиальное (лучевое) строение
тела у животных, ведущих сидячий образ жизни, билатеральная
(двубоковая) симметрия тела у ползающих животных и даже на-
рушение симметрии у лежащих форм, обладающих к тому же по-
кровительственной окраской (камбалы и др.).
Жизнь морского и океанического дна, по свидетельству В. Г. Бо-
горова, начинается с берега. Однако, исходя из ранее изложенных
соображений, следует иметь в виду, что донное и в особенности
придонное население бентали в границах неритической об-
ласти находится под определенным воздействием биотических и
-абиотических факторов планктонной пленки жизни. Проникновение
световых лучей, деятельность фито- и зоопланктона накладывают
известный отпечаток на состав, структуру и функциональную орга-
низацию донных биогидроценозов.
Автотрофными продуцентами в составе б и о г и д-
роценозов бентали являются синезеленые и диатомовые во-
доросли и другие одноклеточные организмы, составляющие так
•называемый фитобентос. За счет созидаемой ими первичной
биологической продукции живут животные фитофаги (мор-
ские звезды, ежи, некоторые ракообразные, брюхоногие моллюски
и пр.), но преобладают фильтраторы и детритофаги. Все
они, в свою очередь, становятся источниками питания и энергии
хищных бентосных и придонных животных, в частности хищных
рыб во время морских приливов. Видовой состав животных, отно-
сящихся к зообентосу, по свидетельству Ю. Одума, далеко не
одинаков на разных типах подводных грунтов. Это служит одним
из оснований для выделения различных типов бногидроценозов в
пределах бентали. Сапротрофный компонент донных биогид-
роценозов представлен преимущественно анаэробными бактерия-
ли, численность которых, по данным Ю. Одума, достигает 108 осо-
бей на 1 г грунта. Роль бактерий в минерализации отмершей био-
массы в донных осадках, по его мнению, не менее велика, чем в
наземных биогеоценозах.
Далее в глубь океана распространяется население донной плен-
ки жизни (бентос) материкового склона — батиали, а еще глуб-
412
«же — бентос ложа океана — абиссали. По мере увеличения глу-
бины плотность донного населения, как указывает Ю. Одум (1975),
резко снижается — от 6000 особей на 1 м2 в пределах бентали до
25—100 особей на абиссальном ложе. В соответствии с этим резко
‘Снижается и биомасса бентоса с глубиной. Так, поданным Л.А.Зен-
кевича (1970), биомасса его на глубине 0—200 м составляет
200 г/м2, на глубине 200—3000 м —лишь 20 г/м2, а на глубине
3000—6000 м — менее 0,2 г/м2. Из приведенных данных о числен-
ности и биомассе беспозвоночных обитателей донных отложений
-океана следует, что ложе абиссали» не говоря уж об ультраабисса-
ли, представляет собой зону рассеянной жизни.
Весьма своеобразный тип биогидроценозов донной пленки жиз-
ни небольших морских и океанических глубин представляют собой
коралловые рифы. Они состоят из сцементированных одно-
клеточными водорослями кораллов и раковин моллюсков. Такого
рода донные биотопы, по В. Г Богорову (1969), населены огром-
ным количеством разнообразных по типу питания гидробионтов:
различных губок, гидроидных полипов, кораллов, червей, мшанок,
моллюсков, иглокожих, рыб, в том числе и хищных, и пр. Они тро-
фически взаимосвязаны в соответствующие водные ценотиче-
ские системы.
14. Прибрежные и другие сгущения жизни
и компонентный состав их биогидроценозов
Помимо планктонной и донной пленок жизни, к участкам высо-
кой концентрации живого вещества в биогидросфере В. И. Вер-
надский (1967) относил так называемые сгущения жизни —
прибрежные сгущения и сгущения над мелями и на поверхности
более или менее глубоких морских и океанических вод.
Прибрежные сгущения жизни распространены, как было отме-
чено, в нейтральных областях близ континентов и больших
островов, где формируются достаточно устойчивые и энергоемкие
мелкомасштабные круговороты водных масс диаметром 10 км и
10 м. Мощность их, по К- В. Беклемишеву (1969), в среднем около
500 м. Местами они охватывают всю толщу воды в пределах не-
ритической области океана, соединяясь в таком случае в
единое целое с планктонной и донной пленками жизни. Общая
площадь их довольно значительна: почти 8% Мирового океана.
Биотопы прибрежных мелкомасштабных круговоротов, прони-
зываемые на значительную глубину солнечным светом, характери-
зуются достаточным количеством лучистой энергии Солнца, несу-
щей не только свет, но и тепло. Богаты они также минеральными
и органическими питательными веществами, поступающими с при-
легающих берегов суши. В этих благоприятных для жизни усло-
413
виях обычно развиваются мощные заросли многоклеточных,
морских водорослей и высших сосудистых трав, как прикрепленных
к субстрату, так и плавающих в поверхностных слоях морской
воды. Среди них господствуют взморник, зеленые ламинарии, а
дальше от берега в менее освещенных глубоких местах — бурые
и красные водоросли, использующие на процессы фотосинтеза си-
не-фиолетовую часть светового спектра. Названные многоклеточные
водные растения, наряду с одноклеточными водорослями, и явля-
ются основными созидателями первичной биологической продукции
прибрежных бногидроценозов, годичный прирост которой достигает
веса их биомассы.
На материально-энергетической базе прибрежных зарослей мор-
ских водорослей и трав и изобилующих в этих условиях однокле-
точных водорослей и других микроскопических организмов наблю-
дается большое скопление беспозвоночных животных (моллюсков,
коралловых полипов, мшанок и т. п.), а также многочисленных
мельчайших обитателей на самих многоклеточных растениях. Во
многих случаях немаловажную роль в прибрежных биогидроцено-
зах играют позвоночные животные, преимущественно птицы, гнез-
дящиеся на суше, но использующие в качестве источника пищи и
энергии мелкие виды рыб и других обитателей поверхностных слоев
морской воды.
Расположенное у юго-восточного атлантического побережья
Северной Америки саргассовое сгущение и другие подоб-
ного рода сгущения жизни приурочены к морским мелководьям и
к поверхностно обособленным водным массам относительно глубо-
ких частей океана. От прибрежных сгущений жизни они отлича-
ются, по словам В. И. Вернадского, независимостью от притока
минеральных и органических материалов и живых существ конти-
нентов, а также отсутствием связи с донной пленкой жизни. Для
них, как и для прибрежных сгущений, характерно мощное скоп-
ление плавающих в морской воде крупных многоклеточных во-
дорослей и некоторых видов высших растений, представляющих
собой источник энергии и среду обитания, плотно заселенную раз-
личными формами животного мира и водной микрофлоры. Проис-
хождение растительного и животного населения бногидроценозов
саргассового типа сгущений обычно связывается с бентосом нери-
тической области и с населением прибрежных водных сгущений
жизни, представители которого в процессе эволюции в новых усло-
виях приобрели некоторые новые приспособительные черты.
Таково разнообразие морских и океанических биогидроце-
н о з о в, составляющих живой покров наиболее плотно насыщен-
ных жизнью областей и вертикальных зон Мирового океана.
414
15. Обмен веществ и энергии
в условиях Мирового океана
Таким образом, Мировой океан в ценотическом .плане представ-
лен на земном шаре двумя пленками жизни и разного рода сгуще-
ниями жизни. Те и другие характеризуются определенным по-
пуляционно-видовым составом водного населения, особенностями
структурно-функциональной организации образуемых им биогидро-
ценозов и т. п. Однако в условиях постоянной циркуляции во все-
возможных направлениях колоссального средоточия на нашей
планете водных масс жизненно необходимые процессы обмена ве-
ществ и энергии в Мировом океане значительно отличаются от
таковых в сухопутных и водных условиях континента. «В Мировом
океане,— пишет В. Г. Богоров (1969),— взаимодействие космиче-
ских, земных небиологических и биологических процессов столь
тесное, что физические, химические, биологические и геологические
процессы в «чистом» виде почти не протекают. Все они взаимообус-
ловлены. Этому содействует и то, что само «тело» океана — вода —
всегда находится в движении и происходит перемешивание всей
массы воды со всем в ней находящимся» (с. 6). Из этого следует,
что межбиогидроценозные связи в океане накладывают столь зна-
чительный отпечаток на трофическое взаимодействие живых компо-
нентов биогидроцеиозов в ходе обмена веществ и энергии между
ними и со средой обитания, что сильно маскируют их, затрудняя
тем самым их дробное изучение. Поэтому отечественные гидробио-
логи обычно ограничиваются приведением данных по обмену ве-
ществ и энергии для океана в целом (рис. 54).
Утилизация солнечной энергии фитопланктоном в сред-
нем для Мирового океана, по данным В. Г Богорова (1969), равна
0,04% от энергии, поступающей на его поверхность, достигая 0,33%
лишь в высокопродуктивных районах океана. Однако если принять
во внимание фактор времени, в течение которого в процессе фото-
синтеза происходит освоение океаном энергии солнечной радиации,
то нетрудно представить себе, как велико общее количество ее,
ушедшее на формирование природы океана за истекшие миллиар-
ды лет. Годовая первичная продуктивность Мирового океана в ве-
совых единицах углерода, по данным Г. Г Винберга и О. И. Коб-
ленца-Мишке (1966), составляет 22-10эт, или 61 г углерода на
1 м2. Чистая годовая продукция фитопланктона (исключая фито-
массу, затраченную на процессы его жизнедеятельности) равна
13-109 т углерода. В наиболее продуктивных водах Мирового
океана ассимилируется углерода 237 г/м2, в менее продуктивных —
91 г/м2, а в центральных областях океана с рассеянной формой
жизни — лишь 28 г/м2. Годичный прирост фитомассы Мирового
океана в целом, по данным В. Г. Богорова (1971), составляет
415
Рис. 54. Ориентировочная схема потоков энергии в Мировом океане (10,в кк;
год) (по Ю. И. Сорокину, 1975; упрощенная)
600 млрд т, в том числе продукция фитобентоса — 200 МЛ1
Вторичная (животная) годовая продукция — около 60 млрд т.
В процессе созидания первичной (исходной) биологической г
дукции автотрофами Мирового океана, по В, Г. Богорову, ежего
выделяется кислорода 36 млрд т (по другим данным, значител
больше) и потребляется азота 4 млрд т, фосфора — 0,5, желез
1,2 млрд т. Но извлечение при этом из морской воды названнь
некоторых других элементов в количестве около 6 млрд т сосг
ляет лишь небольшую часть общего количества растворенны
океане этих веществ. Разумеется, происходит ежегодное пополни
биогенных питательных элементов в океане за счет фотосинт<
ческой деятельности фитопланктона, а в прибрежных (
гидроценозах и за счет деятельности фитобентоса. В рез;
тате разложения и минерализации сапротрофными бак
р и я м и первичной и вторичной биологической продукции, ота
ших организмов и их метаболитов и выделений происходит вы
416
бождение биогенных элементов, которые вновь и вновь вступают
в очередные циклы биотического круговорота. Обогащение поверх-
ностных вод океана биогенными элементами происходит также за
счет огромных запасов их в донных осадках при поднятии на по-
верхность холодных глубинных вод.
Таким образом, вследствие цикличности биотического кругово-
рота веществ и однонаправленности потока энергии при активном
участии трофически взаимосвязанных живых компонентов биогид-
роценозов биогенные питательные элементы довольно быстро воз-
вращаются в их водную среду обитания. Гораздо медленнее, как
указывает В. Г. Богоров, обмен веществ и энергии протекает в слу-
чае поступления биогенных элементов из глубинных слоев воды, а
некоторая часть из них превращается в осадочные породы к в та-
ком случае вообще на долгие геологические периоды выключаются
из биотического круговорота.
Следует также иметь в виду, что ход обменных процессов в био-
гидроценоза1Х прибрежных сгущений находится под большим влия-
нием притока различных веществ, поступающих с поверхностны-
ми водами с материка. Установлено, что реки ежегодно вносят в
океан более 12 млрд т твердых минеральных частиц и около 3 млрд т
растворимых веществ (азота, фосфора, кальция, кремния и раз-
личных органических соединений). Большинство из них вовлека-
ются живыми компонентами бногидроценозов в биотический кру-
говорот, благодаря чему поддерживается постоянство солевого
состава океанических вод. Углекислые соединения кальция откла-
дываются в раковинах и скелетах различных организмов (корал-
лов, моллюсков, корненожек и др.), а соединения кремния — в ске-
летах радиолярий и диатомей.
16. Динамика бногидроценозов
и этапы эволюции Мирового океана
В состоянии и развитии морских и океанических биогидроцено-
зов разного типа происходят различного рода периодические изме-
нения во времени, называемые временными аспектами, которые
рассматриваются как формы существования водных сообществ и
бногидроценозов в целом. К. В. Беклемишев (1969) различает су-
точные, приливно-отливные, сезонные и годичные аспекты. Времен-
ные аспекты биогидроценозов, по его словам, характеризуются чис-
ленностью, активностью и распределением гидробионтов во време-
ни, а иногда и частичным периодическим изменением, колебанием
видового состава данного биогидроценоза. Периодическая смена
аспектов, т. е. переход сообщества и биогидроценоза как та-
кового, из одного состояния в другое происходит в результате дей-
ствия целого ряда внутренних и внешних факторов и явлений:
417
популяционных процессов гидробионтов, периодического изменения
их состава и численности особей, периодичности в развитии послед-
них, вертикальных миграций и пространственных перемещений
животных, изменения внешних условий вследствие смены дня и
ночи или времен года и пр. В пелагиали, как отмечает К. В. Бекле-
мишев, смена аспектов нередко происходит в результате частичной
смены или модификации океанических вод. В арктических областях
она может быть обусловлена циклонами, а в тропиках — времен-
ными перестройками самих биотопов.
Происходящие в умеренных и высоких широтах сезонные
изменения в количестве и массе фито- и зоопланктона В. Г. Бо-
горов называет биологическими сезонами. В открытом
океане он различает четыре биологических сезона. Наиболее ярко
внешне выраженными из них он считает а) биологическую весну,
характеризующуюся бурным развитием фитопланктона, вызываю-
щим «цветение» воды, и б) биологическую осень, для которой
характерно обильное развитие зоопланктона; другие сезоны —
в) биологическое лето и г) биологическая зима — не столь ярки
и красочны, как первые два. Установлено также, что биологические
сезоны по-разному проявляются в разных широтах. Так, в аркти-
ческих морских водах вегетация фитопланктона длится 1—2 меся-
ца, в тропиках же — почти круглый год, захватывая н «биологиче-
скую зиму». Фито- и зоопланктон в Арктике развивается лишь в
период «биологического лета».
Сезонные изменения морских и океанических биогидроцеиозов
разного типа внешне выражаются в закономерном чередовании
определенных аспектов в течение года. Однако характер сезонной
периодической смены аспектов биогидроцеиозов в годичном цикле
при определенных условиях может быть настолько существенным,
что в конечном итоге может привести к коренной смене био-
гидроценозов данного типа другими биогидроценозами того же
типа и, следовательно, к смене всего трофически взаимосвязанного
населения на занимаемом ими водном пространстве.
Что же касается развития биогидросферы, то оно рас-
сматривается В. Г Богоровым на фоне эволюции Мирового океана
в целом. Автор выделяет такие стадии существования океана,
как а) абиотическая, характеризующаяся химическим этапом
возникновения и эволюции жизни на Земле, и б) биотическая,
для которой характерно все усиливающееся воздействие на при-
роду океана развивающейся живой материи. По масштабу событий
в эволюции Мирового океана В. Г Богоров различает два этапа.
Первый этап характеризуется восстановительными условиями вод-
ной среды и господством анаэробных гетеротрофных организмов
в составе его обитателей. Второй этап отличается преобразованием
водной среды океана из восстановительной в окислительную в ре-
418
зультате образования хлорофилла и воздействия хлорофиллонос-
ных организмов (типа синезеленых водорослей); разнится он также
развитием в связи с этим аэробных гетеротрофов (типа простей-
ших животных форм).
Следовательно, на втором этапе эволюции Мирового океана
наряду с гетеротрофным получает развитие автотрофный тип пита-
ния живых существ. Иначе говоря, развиваются простейшие формы
растительного и животного мира и примитивные формы водных
ценотических систем — протоценозы. Все это привело к обога-
щению водной и надводной воздушной среды океана кислородом
и азотом и к образованию в верхних слоях атмосферы озоново-
го э кр а на, что создало необходимые условия для развития жиз-
ни и на континентах. Имеющиеся данные о ходе эволюции биогео-
ценозов и биогеоценотического покрова на суше изложены в
VII главе.
ЗАКЛЮЧЕНИЕ
Для полноты курса биогеоценологии следовало бы поместить
по крайней мере еще две главы: главу о культурных биогеоцено-
зах, в частности об агроценозах, а также главу о коренных вопро-
сах методики биогеоценологических исследований. Но запланиро-
ванный объем книги не позволяет сделать это.
Правда, в отечественной литературе имеется немало работ,
посвященных вопросам агроценологии. Имеются даже учебные
пособия по агрофитоценологии (М. В. Марков, 1972; А. А. Часовен-
ная, 1975). Имеются и переводные работы по так называемой сель-
скохозяйственной экологии {Г Ацци, 1932; В. Тишлер, 1971). Од-
нако, признавая бесспорные высокие достоинства названных моно-
графий, нельзя не учитывать того обстоятельства, что в качестве
объекта агрофитоценологии или сельскохозяйственной экологии
принимается сельскохозяйственное растительное сообщество, агро-
фитоценоз. Между тем, как показали стационарные полевые иссле-
дования в нашей стране, в частности исследования лаборатории
биогеоценологии Ростовского университета на Нижне-Донском био-
геоценологическом стационаре (1973—1983), агро ценозы целесооб-
разно рассматривать не только как растительные объекты, но и как
сложные биокосные системы, как агробиогеоценозы (сокращенно —
агроценозы). Проблемы культурных биокосных систем, в частно-
сти сельскохозяйственных, в этом плане нуждаются в дальнейшей
тщательной экспериментальной и теоретической разработке, ибо
это один из самых эффективных путей утилизации даровой сол-
нечной энергии.
Сходные в некотором отношении мысли возникают и по вопро-
сам методики биогеоценологических исследований. Методика лабо-
раторных и полевых исследований растительных, животных и
микробных объектов и их сообществ, а также среды их обитания —
экотопов — разработана достаточно хорошо. Более того, еще в
1966 году под руководством основоположника биогеоценологии
В. Н, Сукачева была опубликована «Программа и методика био-
геоценологических исследований». Это ценнейшее программно-ме-
тодическое руководство во втором издании в 1974 году было пере-
работано и значительно расширено и дополнено такими важней-
шими разделами, как структурно-функциональная организация
биогеоценозов, обменные процессы в них, а также межбиогеоце-
нозные связи и пр. Тем не менее поскольку биогеоценоз — это не
сумма слагающих его живых и косных компонентов, а особое спе-
цифическое природное образование, функционирующее на основе
особых свойственных ему закономерностей, то, следовательно, и.
420
методы изучения их должны быть особыми, интегральными. Это
означает, что объектом исследования должен быть биогеоценоз как
целостная биокосная система, определенным образом устроенная
и определенным образом функционирующая. Компоненты же био-
геоценоза должны рассматриваться и изучаться не сами по себе»
а как части этого целостного системного единства.
Одним словом, следует иметь в виду, что биогеоценологические
исследования по своей природе — комплексные исследования и в
выполнении их должны принимать участие специалисты разного
профиля (фитоценологи, зооэкологи, микробиологи, почвоведы, фи-
токлиматологи и др.). Однако не всякие комплексные исследова-
ния по своему содержанию являются подлинно биогеоценологиче-
скими. Они могут стать таковыми лишь в том случае, если испол-
нители их будут обладать биогеоценологическим мышлением;,
иначе говоря, если они будут биогеоценологами с тем или иным
уклоном, совместно обеспечивающими изучение всех компонентов
биогеоценозов и основной роли последних в осуществлении гло-
бальных геохимических функций биогеосферы Земли, подобно тому
как среди физгеографов имеются фитогеографы, зоогеографы и т. п._
Подготовка биогеоценологов такого типа, как средней, так и выс-
шей квалификации, с должным успехом может осуществляться на
специальных эколого-биогеоценологических кафедрах крупных уни-
верситетов, в том числе и через аспирантуру.
Правда, высказываются такого рода суждения, что биогеоцено-
логия якобы не фундаментальная, а прикладная наука и что место
ее не в учреждениях академического или университетского типа,
а только в отраслевых институтах — сельскохозяйственном, лесном
и т. и. Очевидно, это просто недоразумение. Тот факт, что биогеоце-
нология наряду с разработкой множества широкомасштабных тео-
ретических проблем решает на основе этого целый ряд весьма
важных практических народнохозяйственных задач, не служит
основанием относить ее в разряд прикладных наук.
Биогеоценология президиумом Академии наук СССР в упомя-
нутом ранее постановлении по ее проблемам трактуется как «выс-
шее обобщение всех классических направлений биологических
наук». Наряду с молекулярной биологией она рассматривается как
одна из фланговых наук, которые в настоящее время в значитель-
ной мере определяют состояние и развитие других промежуточных
разделов биологии. Но биогеоценология, в отличие от молекуляр-
ной биологии, «изучает жизнь в ее макропроявлении». Исходя из
этого, президиум Академии наук постановляет одним из главных
направлений современной биологии считать разработку теоретиче-
ских основ учения о биогеоценозах и биогеосфере. Ставится также
задача разработки в рамках биогеоценологии научных основ охра-
ны и преобразования биогеосферы Земли и рационального исполь-
421
зования ее биологических ресурсов, разумного управления эволю-
цией и биопродуктивностью биогеоценозов без подрыва произво-
дительности земельных угодий и промысловых водоёмов.
Приведем высказывания В. Н. Сукачева о биогеоценологии, ее
проблемах и значении для современного бытия и дальнейших су-
деб человечества. В 1964 году он писал: «Бногеоценология — новая
научная дисциплина, возникшая... на стыке нескольких наук. Она
находится еще в первоначальной стадии своего развития. В даль-
нейшем она должна и будет применять новые исследовательские
методы... будет вскрывать новые еще не используемые биологиче-
ские ресурсы... а также содействовать общей перестройке биосферы
в интересах человечества» (1964, с. 517). По мере развития раз-
личных отраслей естествознания бногеоценология, по его словам,
«будет все более полно объединять достижения многих других
наук, перерабатывать их под своим углом зрения и создавать но-
вые обобщения большого теоретического и практического значе-
ния» (там же, с. 512).
Все это свидетельствует о том, что бногеоценология — одна из
основных, ведущих наук современного естествознания, разрабаты-
вающая фундаментальные проблемы, стоящие перед человечеством.
Попытка умалить ее роль и значение в решении важнейших гло-
бальных и региональных проблем охраны окружающей среды, вы-
явлении и рациональном использовании биологических ресурсов
планеты чревата многими непредвиденными и, возможно, непопра-
вимыми последствиями. Глубоко ошибочно полагать, что все по-
добного рода проблемы лежат в рамках науки экологии и разре-
шимы ее методами. Можно прямо сказать, что все проблемы и
практические мероприятия, связанные с нарушением и более или
менее глубокой перестройкой тех или иных объектов биогеосферы,
являются предметом соответствующих разделов биогеоценологии.
Это обусловлено тем, что в основе функционирования биокосных
систем разного ранга, как известно, лежат многообразные взаимо-
связи как между компонентами, так и между самими биокосными
системами, составляющими биогеоценотическую оболочку (биогео-
сферу) Земли.
Возьмем в качестве примера такую животрепещущую в настоя-
щее время проблему всего человечества, как охрана окружающей
среды. Это по существу не что иное, как охрана от нарушения
человеком в процессе хозяйственной деятельности существующего
в природе относительно устойчивого динамического равновесия, на-
зываемого по недоразумению экологическим. В действительности
же природное равновесие — это выработавшаяся в процессе дли-
тельного геоисторического развития сбалансированность многооб-
разных и разносторонних взаимосвязей и взаимоотношений живых
компонентов биогеоценозов и биогеосферы в целом в условиях
422
непрерывно изменяющейся под их воздействием внешней среды.
Экологическим, следовательно, является не природное равновесие,
а сам процесс бездумного нарушения его человеком.
Одной нз важнейших глобальных проблем нашего времени яв-
ляется, в частности, также борьба с различного рода загрязнения-
ми природной среды промышленными, бытовыми и другими отхо-
дами в твердом, жидком и газообразном состояниях. Практика
устройства специальных очистных сооружений показывает, что лишь
включение в эту систему наряду с механическими и химическими
биологических способов (в виде создания ценотических систем в со-
ставе трофически сопряженных компонентов живых существ) спо-
собно обеспечить полную очистку загрязненных отходами сточ-
ных вод.
Можно взять и такую проблему прикладного характера, как
борьба с сорняками или насекомыми-вредителями сельскохозяй-
ственных культур. Применение разнообразных пестицидов, в част-
ности инсектицидов, в борьбе с вредными насекомыми, как извест-
но, не приводит к полному избавлению от них. После некоторого
снижения их численности через непродолжительное время она
снова возрастает нередко в гораздо больших размерах, чем преж-
де, ибо вместе с ними погибают их естественные враги и паразиты.
Широко известны многочисленные факты неимоверно быстрого
распространения по тем же причинам завезенных из других стран
некоторых растений, позвоночных и беспозвоночных животных, на-
пример элодеи канадской, в последние десятилетия — жука коло-
радского по странам Западной Европы, ранее — кроликов в Австра-
лии и пр. Жук колорадский, опаснейший вредитель картофеля и
других культур из семейства пасленовых, семимильными шагами
почти беспрепятственно продвигается на восток и по нашей стране.
И, очевидно, лишь выявление использующего его в качестве источ-
ника энергии плотоядного потребителя способно радикально сокра-
тить его вредоносную деятельность *.
Не менее важна проблема повышения продуктивности сельско-
хозяйственных культур, которая веками осуществляется человече-
ством путем создания наиболее благоприятных условий для их
выращивания (осушение или орошение, внесение удобрений, раз-
личные способы обработки почвы и т. п.). Такие меры, которые
можно назвать собственно экологическими, давали и дают возмож-
ность повысить урожайность зерновых, кормовых и других культур
на несколько центнеров или даже десятков центнеров с гектара.
Между тем, как показали наши полевые исследования в совхозах
Ростовской области, применение ценотических методов, выразив-
* Недавно в периодической печати появилось сообщение, что такими плотоядны-
ми потребителями колорадского жука являются фазаны.
423
шееся в создании двух-трехъярусных многолетних и однолетних
кормовых агроценозов из биохимически совместимых видов расте-
ний, позволяет повысить их продуктивность в два раза и более по
сравнению с традиционными хозяйственными посевами тех же
культур. Это обусловлено тем, что сложные по вертикальному
строению и горизонтальному сложению агроценозы способны наи-
более полно использовать лучистую энергию Солнца и материаль-
ные ресурсы поля (углекислоту, воду, элементы минерального пи-
тания и пр.). Этим самым создается возможность внести немалый
вклад в решение продовольственной программы нашей страны.
Даже в основе решения такой проблемы, как мирное освоение
космоса, лежат ценотические принципы, ибо достаточно эффектив-
ной, долговременно и бесперебойно действующей системой жизне-
обеспечения космонавтов на космических кораблях и орбитальных
или в будущем межпланетных станциях может быть лишь биоцено-
тическая регенерация пищи, воды, воздуха. Она должна представ-
лять собой циклически функционирующую ценосистему, осно-
ванную на использовании в качестве компонентов нескольких био-
химических совместимых видов пригодных в пищу автотрофных
растений и взаимосвязанных с ними салротрофных микроорганиз-
мов с включением в нее в качестве биотрофов космонавтов. Такая
ценосистема, разумеется, должна быть по всем параметрам сбалан-
сированной и саморегулирующейся.
Одним словом, какую бы проблему прикладного характера мы
ни взяли из области охраны окружающей среды, рационального
использования биологических ресурсов планеты, создания искус-
ственных сооружений разного назначения и т. п., эффективное ре-
шение каждой из них базируется, как мы видим, на закономерно-
стях биогеоценологии. В основе связанных с этим практических мер
ведущее значение имеют не экологические связи (т. е. связи живых
существ со средой их обитания), а ценотические (т. е. разнообраз-
ные взаимоотношения между видовыми ценопопуляциями и энерго-
трофические связи между авто-, био- и сапротрофными компонен-
тами). Экологические же связи, как было сказано ранее, не исче-
зают при этом, а находятся, по выражению философов, в «снятом»
виде, т. е. в соподчиненном положении.
Таким образом, бногеоценология, являясь фундаментальной
наукой о биокосных системах разного ранга, характеризуется тем
что ей присущи и прикладные, практические аспекты локальногс
регионального и планетарного масштаба. Важно и то, что по мер»
общественного развития человечества по пути к социализму и ком
мунизму и повышения уровня и ускорения темпов хозяйственно^
деятельности человека роль и значение биогеоценологии в ниц
ально-экономическом и культурном развитии общества, по слон
В. Н. Сукачева, будет неуклонно возрастать.
424
ИСПОЛЬЗОВАННАЯ ЛИТЕРАТУРА
Абакумов В. А. Иерархичность организации биосферы//Методолог. аспекты
нсследов. биосферы. М., 1975.
Абатуров Б. Д. Биопродукционный процесс в наземных экосистемах. М.» 1979.
Аболин Р. И. Опыт эпигенологической классификации болот // Болотоведе-
ние. 1914. № 3. '
Абросов Н. С., Ковров Б. Г. Анализ видовой структуры трофического уровня
одноклеточных. Новосибирск, 1977.
Абросов Н. Ковров Б. Г. Черепанов О. А. Экологические механизмы со-
существования и видовой регуляции. Новосибирск, 1982.
Аганянц О. Е. Жизнь на склоне//Природа. 1970. № 11.
Айзатуллин Т. А., Лебедев В. Л.г Хайлов К М. Океан. Фронты, дисперсии,
жизнь. Л., 1984.
Акимов М. П. Понятие жизненной формы и его использование в экологиче-
ских исследования//Тез. докл. 3-й эколог, конф. Т. 1. Киев, 1954.
Акимов М. П. Экология животных. Киев, 1959.
Александрова В. Д. Изучение смены растительного покрова // Полев. геобот.
Т. 3. М.; Л., 1964а.
Александрова В. Д. О возможности применения идей я методов кибернетики
в лесной биогеоценологии//Основы леей, биогеоценол. М., 19646.
Александрова В. Д. Об объектах биогеоценологии//Ботан. журя. 1971.
Т. 56. № 9.
Алексеев В. В. О роли консументов в формировании разнообразия видов в
биогеоценозах//Журн. общ. биол. 1973. Т. 34. № 3.
Аристовская Т. В. Теоретические аспекты проблемы численности, биомассы
и продуктивности почвенных микроорганизмов // Вопр. числен., биом, и продукт,
почв, мнкроорг. Л., 1972.
Аристовская Т. В. О некоторых аспектах геохимической деятельности поч-
венных микроорганизмов как составной части биогеоценозов // Пробл. биогео-
ценол. М., 1973.
Аристовская Т. В. Микробиология процессов почвообразования. Л., 1980.
Арнольди К. В., Арнольда Л. В, О биоценозе как одном из основных поня-
тий экологии, его структуре и объеме//Зоолог, журн. 1963. Т. 42. № 2.
Ацци Г. Сельскохозяйственная экология. М.; Л., 1932.
Базилевич Н. И., Титлянова А. А. Изучение обменных процессов в биогео-
ценозах // Прогр. и методика биогеоценол. исследов. М., 1974.
Беклемишев В. Н. Основные понятия биоценологии в приложения к живот-
ным компонентам наземных сообществ//Тр. по защите растений. 1931. Т. 1.
Вып. 2.
Беклемишев В. И. О классификации биоценологических (симфизиологиче-
ских) связей//Бюл. МОИП. Отд. бнол. 1951. Т. 56. Вып. 5.
Беклемишев В. Н. Биоценологяческие основы сравнительной паразитологии.
М.. 1970.
Беклемишев К. В. Экология и биогеография пелагиали. М., 1969.
Беклемишев К. В. Биотопы морских биоценозов // Пробл. биогеоценол.
М., 1973.
Белоусова Л. С., Борисов В. А. Винокуров А. А. Заповедники и националь-
ные парки мира: Краткий справочник. М„ 1969.
Бельеард А, Л. Лесная растительность юго-востока УССР. Киев, 1950.
Бельгард А. Л. Биогеоценологические принципы степного лесоведения
(К истории степного лесоведения)//Струит.-функц. организ. биогеоценол,
М„ 1980.
Берг Л. С. Опыт разделения Сибири и Туркестана на ландшафтные и мор-
фологические зоны//Сб. в честь 75-летия Д. Н. Аиучина. М., 1913.
425
Берг Л. С. Ландшафтно-географические зоны СССР. Ч. 1. М., 1931.
Берталанфи Л. Общая теория систем (Обзор проблемы и регуляторов) //
Системные исследов. М.» 1969.
Биогеоценология. Текущий указатель литературы. Новосибирск (с 1973).
Биогеоценозы в пойме Нижнего Дона. Ростов н/Д, 1978.
Блюменталь И. X., Попова Т. А. Геоботанический очерк формации овсяницы
джамильской на Северо-Западном Кавказе//Тр. Ленингр. об-ва естественен ыт.
1950. Т. 70. Вып. 3.
Блюменталь И. Х„ Петровичева О. Л, Геоботанический очерк высокогорных
мелкоосоковых пастбищ Северо-Западного Кавказа // Учен. зап. Леиннгр. ун-та.
1951. № 143.
Богоров В, Г. Жизнь океана. М., 1969.
Богоров В. Г. Океаническая часть биосферы//Сб. тез. докл. по биогеоценоз
М., 1971.
Богоров В. Г., Зенкевич Л. А. Биологическая структура океана // Экология
водн. организмов. М., 1966.
Борисов А. Д. Климаты СССР. М., 1948.
Бочурова Н. В.» Минина Е. Г. О значении гравитации в жизнедеятельности
лесных пород//Лесоведение. 1968. Ns 6.
Брагинский В. В поисках гравитационных волн//Наука и жизнь. 1976. № 10.
Будыко М. И. Климат и жизнь. Л., 1971.
Будыко М. И. Глобальная экология. М., 1977.
Бурова Л. Г. Грибы в лесных биогеоценозах//Бюл. МОИП. Отд. биол. 1976.
Т. 81. Вып. 1.
Быков Б. А. Эдификаторы растительных формаций Советского Союза//
Вестник АН КазССР. 1949. № 3.
Быков Б. А. Геоботаника. Алма-Ата, 1953, 1957.
Быков Б. А. Доминанты растительного покрова СССР. Алма-Ата, 1962. Т. 2.
Быков Б. А. Введение в фитоценологию. Алма-Ата, 1970а.
Быков Б. А. Биоценоз как ценоэкосистема//Экология. 19706. Ns 3.
Бяллович Ю. П. Биогеоценотические горизонты // Сб. по геоботан., ботан.
геогр., систем, растений и палеогеогр. М., 1960.
Бяллович Ю. П. О кинетике биогеоценоза//Жури. общ. биол. 1969.
Т. 30. Ns 2.
Бяллович Ю. П. Системы биогеоценозов // Пробл. биогеоцеиол. М., 1973.
Бяллович Ю. П. О биогеоценотической структуре центрального слоя био-
сферы//Бюл. МОИП. 1980. Т. 85. Вып. 3.
Вальтер Г. Общая геоботаника. М., 1982.
Варминг Е. Распределение растений в зависимости от внешних условий (Эко-
логическая география растений). СПб., 1902.
Василевич В. И. Статистические методы в геоботанике. Л., 1969.
Василевич В. И. Фитоценотические объекты как системы//Пробл. эколог.»
геоботан., ботан. геогр. и флористики. Л., 1977.
Белинский В. В. Как и где образуются магмы//Природа. 1969. № 2.
Вернадский В. И. Биосфера. Л., 1926.
Вернадский В. И. Эволюция видов и живое вещество//Природа. 1928. № 3.
Вернадский В. И. Об условиях появления жизни на Земле//Изв. АН СССР.
Сер. мат. и естеств. наук. 1931. Ns 5.
Вернадский В. И. Несколько слов о ноосфере//Успехи соврем, биол. 1944.
Т. 18. Вып. 2.
Вернадский В. И. Химическое строение биосферы Земли и ее окружения.
М., 1965.
Вернадский В. И. Биосфера (Избранные труды по биогеохимии). 3-е изд.
М._ 1967
426
Виленкин Б, Я. Кибернетический и энергетический подход к проблемам био-
ценологии//Экология води, организмов. М., 1966.
Вильямс В. Р. Общее земледелие. Естественноисторические основы луговод-
ства или луговедение. М., 1922.
Вильямс В. Р. Почвоведение (Общее земледелие с основами почвоведения).
М., 1936.
Вильямс В. Р. Естественнонаучные основы луговодства или луговедение //
Собр. соч. М., 1949. Т. 4.
Винберг Г. Г. Бногеоценология водной среды//Природа. 1971. № 6.
Винберг Г. Г., Кобленц-Мишке О. И. Проблемы первичной продукции водо-
емов//Экология води, организмов. М., 1966.
Виноградов А. П. Биогеохимические провинции//Тр. юбил. сессии В. В. До-
кучаева. М., 1946.
Виноградов А. П. Биогеохимические провинции и их роль в органической
эволюции И Геохимия. 1963. № 3.
Воейков А. Я. Кругооборот тепла в оболочке земного шара//Сб. статей по
физике. СПб., 1904.
Волкова Э. В. Биологические системы и среда//Вопр. философии. 1974. № 12.
Волобуев В. Р. Некоторые вопросы энергетики почвообразования // Почвове-
дение. 1958. № 7.
Волобуев В. Р. Компонентная вода географической оболочки // Почвоведе-
ние. 1961. №11.
Волобуев В. Р. Биогеоэнергетика и ее задач и//Вестник АН СССР. 1969. №4.
Волобуев В. Р. Некоторые энергетические критерии в экологии гумусообра-
зования // Жури. общ. биол. 1974. Т. 35. № 2.
Воронов А. Г. Биогеография. М., 1963.
Воронов А. Г. Формирование некоторых зональных особенностей консор-
ций//Знач. консорт, связей в организ. биогеоценозов. Пермь, 1976.
Второв П. П. Проблемы изучения наземных экосистем и нх животных ком-
понентов. Фрунзе, 1971.
Вудвелл Дж. Круговорот энергии в биосфере//Биосфера. М., 1972.
Высоцкий Г. Я. Ергеня. Культур-фитологический очерк//Тр. Бюро прикл.
ботаники. 1915. Т. 8. № 10—11.
Высоцкий Г Н. Покрововедение // Зап. Белорусок, ин-та сельск. хоз-ва.
1925. № 4.
Гаврилюк Ф. Я., Вальков В. Ф., Садименко П. А. Почвы // Человек и био-
сфера. Ростов н/Д, 1977.
Гаевская Н. С. Роль высших водных растений в питании животных пресных
водоемов. М., 1966.
Герасимов И. П. Учение о природных экосистемах (геоэкобиотах) как син-
тез ландшафтоведения и биогеоценологии в советской географической и биологи-
ческой науках//Жури. общ. биол. 1973. Т. 34. № 5.
Герасимов И. п. Методологические проблемы экологизации современной
науки //Вопр. философии. 1978. X? 11.
Гиляров М. С., Стриганова Б. Р. Роль почвенных беспозвоночных в разло-
жении растительных остатков и круговороте веществ//Зоология беспозвоноч-
ных. М., 1978.
Гирусов Э. В. Биосфера как целое // Пробл. целостности в соврем, биол.
М., 1968.
Глазовская М. А. Технобиогеомы— исходные физико-географические объек-
ты ландшафтно-геохимического прогноза // Вестник Моск, ун-та. Сер. геогр.
1972. № 6.
Голубев В. Н, Материалы к эколого-морфологической и генетической харак-
теристике жизненных форм травянистых растений//Ботан. журн. 1957. Т. 42. №7.
Голубев В. Н. К вопросу о классификации жизненных форм // Тр. Центр.-
Черноз. госуд. запов. Курск, 1960. Вып. 7.
427
Г орд яг ин А. Я. Растительность Татарской республики//Геогр. описание Та-
тарской респ. Казань, 1922.
Гохлернер Г. Проблема <внутреннего> и «внешнего» в эволюции органиче-
ского мира//Наука и жизнь. 1971. № 10.
Григорьев А. А. Опыт характеристики основных типов физико-географической
среды//Пробл. физ. геогр. Ч. 1. 1938.
Григорьев А. А. О взаимосвязи и взаимообусловленности компонентов гео-
графической среды и о роли в них обмена веществ и энергии // Изв. АН СССР.
Сер. геогр. 1956. № 4.
Григорьева Н. Заугольнова Л. Б., Смирнова О. В. Особенности простран-
ственной структуры ценопопуляций некоторых видов растений // Ценопопуляцик
растений. М„ 1977.
Гризебах А. Растительность земного шара. СПб., 1874. Т. 1.
Гродзияский А. М.» Гродзинский Д. М. Краткий справочник по физиологии
растений. Киев, 1973.
Гумбольдт А. Идеи физиономичности растений//Геогр. растений. М.; Л., 1936.
Гусева К. А., Экзерцев В. А. Формирование планктона и высшей водной
растительности в равнинных водохранилищах//Экология води, организмов.
М., 1966.
Довитая Ф. Ф. Динамика газового состава атмосферы//Наука и жизнь.
1971. Хе 10.
Дажо Р. Основы экологии. М., 1975.
Дарвин Ч. Происхождение видов. М.; Л., 1935.
Добровольский В. В. Равнина под микроскопом//Природа. 1972. № 6.
Докучаев В. В. К учению о зонах природы. Горизонтальные и вертикальные
почвенные зоны. СПб., 1899.
Долуханов А. Г. Принципы классификации растительности//Тр. Ин-та биол.
Уральск, фил. АН СССР. 1961. Вып. 27. •
Дохман Г. И. Обсуждение книги И. А. Титова «Взаимодействие раститель-
ных сообществ и условий среды»//Организм и прир. среда. М., 1966.
Дылис Н. В. Принципы построения классификации лесных биогеоценозов //
Основы лесн. биогеоценол. М.. 1964а.
Дылис Н. В. Учение об экосистемах за рубежом // Лесоведение. 1967. 3.
Дылис Н. В. Структура лесного биогеоценоза//Комаровские чтения, 2L
М.» 1969а.
Дылис Н. В. Биогеосфера, ее свойства и особенности // Изв. АН СССР. Сер.
биол. 19696. № 4.
Дылис Н. В. Структура и функция биогеоценозов//Природа. 1971. № 6.
Дылис Н. В. О структуре консорции//Журн. общ. биол. Г973а. № 4.
Дылис Н. В. Межбиогеоценозные связи, их механизмы и изучение // Пробл.
биогеоценол. М., 19736.
Дылис Н. В. Структурно-функциональная организация биогеоценотических
систем и ее изучение // Прогр. и методика биогеоценол. исследов. М., 1974а.
Дылис Н. В. Межбиогеоценозные связи, их механизмы и изучение // Прогр.
и методика биогеоценол. нсследов. М., 19746.
Дылис Н. В. Учение о биогеоценозе и его проблемы. М., 1975.
Дылис Н. В. Основы биогеоценологии. М„ 1978а.
Дылис Н. В. Учение В. И. Вернадского о биосфере и его значение для раз-
вития биогеоценологии//Бюл. МОИП. Отд. биол. 19786. Вып. 1.
Дылис Н. B.t Цельникер Ю. Л„ Карпов В. Г. Фитоценоз как компонент лес-
ного биогеоценоза//Основы лесн. биогеоценол. М., 19646.
Дылис Н. В., Уткин А. И., Успенская И. М. О горизонтальной структуре
лесных биогеоценозов//Бюл. МОИП. Отд. биол. 1964в. Т. 69. Вып. 4.
Дылис Н. В,, Карпов В, Г.» Цельникер Ю, Л. Изучение высшей растительно-
сти как компонента биогеоценоза // Прогр. и методика биогеоценол. исследов.
М., 1974.
428
Дювиньо П., Танг М. Биосфера и место в ней человека (Экологические си-
стемы и биосфера). М., 1968.
Егорова С. А., Е никее в а М. Г., Большакова В. С. Микроорганизмы как ком-
понент лесного биогеоценоза // Основы лесн. биогеоцеиол. М., 1964.
Еленкин А. А. Закон подвижного равновесия в сожительствах и сообществах
растений//Изв. Главн. ботан. сада. 1921. Т. 20. Вып. 2.
Ефимова Н. А. Радиационные факторы продуктивности растительного покро-
ва. Л., 1977.
Жданов Ю. А. Учение о биосфере в свете диалектической концепции разви-
тия//Человек и биосфера. Ростов н/Д, 1973.
Завадский К. М. Учение о виде. Л., 1961.
Завадский К. М. Основные формы организации живого и их подразделе-
ние//Филос. пробл. соврем, бнол. М.; Л., 1966.
Зенкевич Л. А. Общая характеристика биогеоценозов океана и сравнение их
с биогеоценозами суши//Прогр. и методика изучения биогеоцеиол. водн. среды.
М., 1970.
Зенкевич Л. 4., Барсанова Н. Г., Беляев Г. М. Количественное распределение
донной фауны в абиссали Мирового океана//Докл. АН СССР. 1960. Т. 130. №1,
Злотин Р. И., Ходашева К. С. Влияние животных на автотрофный цикл био-
логического круговорота // Пробл. биогеоцеиол. М., 1973.
Злотин Р. И., Ходашева К. С. Роль животных в биологическом круговороте
лесостепных экосистем. М., 1974.
Знаменский И. Влияние ультрафиолетовых лучей на высшие растения //
Ботан. журн. 1935. Т. 20. № 4.
Зозулин Г М. Система жизненных форм высших растений // Ботан. журн.
1961. Т. 46. № 1.
Зозулин Г. М. Схема основных направлений и путей эволюции жизненных
форм семенных растений//Ботан. журн. 1968. Т. 53. № 2.
Зозулин Г. М. Жизненные формы растений в аспекте учения о биосфере//
Ботан. журн. 1976. Т. 61. № 11.
Зонн С. В. Биогеоценология и почвоведение//С б. акад. В. Н. Сукачеву к
75-летию со дия рожд. М.; Л., 1956.
Зонн С. В. О взаимодействии лесной растительности с почвами в свете но-
вых биогеоценологических исследований//Тр. МОИП. Отд. биол. 1960. Т. 3.
Зонн С. В. Почва как компонент лесного биогеоценоза//Основы лесн. био-
геоценол. М., 1964.
Зонн С. В. Задачи и направления изучения почвы как компонента лесного
биогеоценоза//Лесоведение. 1967. № 3.
Зонн С. В. Изучение почвы как компонента биогеоценоза // Прогр. и мето-
дика биогеоцеиол. нсследов. М., 1974.
Зонн С. В., Базилевич И. И. Изучение почвы как компонента биогеоценоза //
Прогр. и методика биогеоцеиол. исследов. М., 1966.
Игнатов А. И. Жизнь как система форм движения материн. М., 1966.
Иоганзен Б. Г. Основы экологии. Томск, 1959.
Иоганзен Б. Г. Закон единства биогеоценоза и его биотопа//Докл. зоолог,
совещ. Томск, 1964.
Иоганзен Б. Г. Экология. Ч. 1 // Экология, биогеоцеиол. и охрана природы.
Томск, 1979.
Исаченко А. Г. Основные вопросы физической географии. Л., 1953.
Казакевич Л. И. Материалы к биологии растений юго-востока России // Изв.
Саратовск. сельскохоэ. оп. станции. 1922. Т. 3. Вып. 2—3.
Калесник С. В. Задачи географии и полевые географические исследования//
Учен. зап. Ленингр. ун-та. Сер. геогр, наук. 1940. Вып. 2.
Камшилов М. М. Значение взаимных отношений между организмами в эво-
люции. М.; Л., 1961.
Камшилов М. М. Принципы организации живой природы // Природа.
1969. № 2.
429
Камшилов At М. Организованность н эволюция // Журн. общ. биол. 1970а.
Т. 31. № 2.
Камшилов М. М. Биологический круговорот. М., 19706.
Кашка ров Д. Н. Среда и сообщество. М.. 1933.
Кашкаров Д. Н. Основы экологии животных. М., 1938.
Кашкаров Д. Н. Основы экологии животных. Л., 1944.
Келлер Б. А. Об изучении жизненных форм при геоботанических исследова-
ниях//Сов. ботаника. 1933. № 2.
Ковда В. А. Проблема биологической и хозяйственной продуктивности су-
ши//Сел ьскохоз. биология. 1966. Т. 1. № 2.
Ковда В. А. Основы учения о почвах. Общая теория почвообразовательного
яроцесса. Кн. I. М., 1973.
Ковда В. А., Самойлова Е. М. О возможности определения возраста гидро-
морфных почв по содержанию СаСО3//Докл. АН СССР. Отд. геол. 1968.
Т. 182. № 5.
Коган А. Б. Биосфера и космос//Человек и биосфера. Ростов н/Д, 1977.
Козловская Л. С. Роль почвенной фауны в разложении органических остат-
ков заболоченных лесных почв//Тр. Ин-та леса АН СССР. 1959. Т. 49.
Коржинский С. И. Северная граница черноземно-степной области восточной
полосы Европейской России в ботанико-географическом отношении//Тр. об-ва
естествоиспыт. при Казанск. ун-те, 1891. Т. 22. Вып. 6.
Корчагин А. А. К вопросу о характере взаимоотношений растений в сообще-
стве//Сб. акад. В. Н. Сукачеву к 75-летию со дня рожд. М.; Л., 1956.
Корчагин А. А. Внутривидовой (популяционный) состав растительных сооб-
ществ и методы его изучения//Полев. геоботан. М.; Л., 1964.
Корчагин А. А. Понятие о консорции // Пробл. экол., геоботан., ботан., геогр.
и флористики. Л., 1977.
Корытный Л. М. Речной бассейн как геосистема//Докл. Ин-та геогр. Сиби-
ри и Дальнего Востока, 1974.
Красильников Н. А. Бактериальная масса почв//Микробиология. 1944. Т. 13.
Вып. 4.
Красильников Н. А. Микроорганизмы почвы н высшие растения. М., 1958.
Красницкий А. М. Проблемы заповедного дела. М., 1983.
Краснов А. Н. Опыт истории развития флоры южной части восточного Тянь-
Шаня. 1888. Вып. 19.
Краснов А. В. География растений. Законы распределения растений и опи-
сание растительности земного шара. Харьков, 1899.
Криштофович А. Н. Развитие ботанико-географических провинций Северного
полушария с конца мелового периода//Сов. ботаника. 1936. № 3.
Криштофович А. Н, Эволюция растительного покрова в геологическом про-
шлом и ее основные факторы//Матер, по истории флоры и растит. СССР. М.;
Л., 1946. Т. 2.
Крючков В. В. О микроклимате растений//Ботан. журя. 1960. Т. 45. № 3.
Кузнецов С. И. Микрофлора и ее геохимическая деятельность. Л., 1970.
Куркин К. А. Вклад А. А. Уранова в учение о жизненном состоянии видов
в фитоценозах и системный подход в фитоценологии // Бюл. МОИП. Отд. биол.
1977. Т. 82. Вып. 3.
Куркин К. А. Параметры биогеоценозов и системный подход к их определе-
нию И Бюл. МОИП. Отд. биол. 1980. Т. 85. Вып. 3.
Куркин К. А.. Матвеев А. Р. Ценопопуляций как системы особей и как эле-
менты фитоценозов (Системно-иерархический подход) //Бюл. МОИП. Отд. биол.
1981. Т. 86. Вып. 4.
Курчева Г Ф. Роль почвенных животных в разложении и гумификации рас-
тительных остатков. М., 1971.
Куче рук В. В. Антропогенная трансформация окружающей среды и грызу-
ны//Бюл. МОИП. Отд. биол. 1976. Т. 81. Выл. 2.
430
Лавренко Е. М. Об изучении эдификаторов растительного покрова // Сов.
ботаника. 1947. № 1.
Лавренко Е. М. О фитогеосфере//Вопр. географии. М., 1949. Вып. 15.
Лавренко Е. М. Основные закономерности растительных сообществ н пути их
изучения//Полев. геобот. М.; Л., 1959. Т. 1.
Лавренко Е. М. Об уровнях изучения органического мира // Изв. АН СССР.
Сер. биол. наук. 1964. № 1.
Левренко Е. М. Об уровнях изучения органического мира в связи с позна-
нием растительного покрова//Пробл. соврем, ботаники. М.; Л., 1965. Т. 2.
Лавренко Е. М. Основные проблемы биогеоценология и задачи биогеоцеио-
логических исследований в СССР//Журн. общ. бнол. 1971. Т. 32. № 4.
Лавренко Е. М., Дылис Н. В. Успехи и очередные задачи в изучении био-
геоценозов суши в СССР//Ботан. журн. 1968. Т. 53. № 2.
Лавренко Е. М., Свешникова В. М. Об основных направлениях изучения эко-
биоморф в растительном покрове//Осн. пробл. соврем, геоботан. Л., 1968.
Лархер В. Экология растений. М., 1978.
Леме Ж. Основы биогеографии. М., 1976.
Линков Б. Л. К основам современной теории Земли. Л., 1965.
Лукашев К. И. Основы литологии н геохимии коры выветривания.
Минск, 1958.
Львов Ю. А. Биогеоценология. Ч. 2//Экология, биогеоцеиол. я охрана при-
роды. Томск, 1979.
Мазинг В. В. Консорции как элементы функциональной структуры биогеоце-
нозов//Естеств. корм, угодья СССР. М.г 1966.
Мазинг В. В. Теоретические и методические проблемы изучения структуры
растительности: Автореф. дис. д-ра биол. наук. Тарту, 1969.
Мазинг В. В. К вопросу эволюции бноценотических систем//Теор. пробл.
фитоценол. и биогеоцеиол. М., 1970.
Мазинг В. В. Системы биоцен отнческого уровня и их усложнение в эволю-
ции//Развитие концепции структ. уровней в биол. М., 1972.
Мазинг В. В. Что такое структура биогеоценоза // Пробл. биогеоцеиол;
М„ 1973.
Мазинг В. В. Проблемы изучения консорций//Знач. консорт, связей в ор-
ганнз. биогеоценозов. Пермь; 1976.
Мазинг В. В. О функциональной структуре растительности (На примере вер-
ховых болот) //Бюл. МОИП. Отд. бнол. 1980. Т. 85. Вып. 3.
Мазохин-Поршняков Г. А. Основные приспособительные типы чешуекрылых
(Lepidoptera) //Зоолог, журн. 1954. Т. 33. Вып. 4.
Макфедьен Э. Экология животных. Цели и методы. М., 1965.
Меншуткин В. В. Математическое моделирование популяций и сообществ
водных животных. Л., 1971.
Методика изучения биогеоценозов внутренних водоемов/Отв. ред. Ф. Д. Мор-
ду хай-Болтовской. М., 1975.
Ми ль ко в Ф. Н. Основные проблемы физической географии (Избр. лекции),
Воронеж, 1959.
Миркин Б. М., Розенберг Г. С. Фитоценология. Принципы и методы. М., 19781
Мишустин Е. Н. Микроорганизмы и плодородие почвы. М„ 1956.
Молчанов А. А. Круговорот органического вещества в процессе роста сосня-
ка черничника/Сообщ. лаб. лесоведения АН СССР. 1961. Вып. 5.
Молчанов А. А. Атмосфера как компонент лесного биогеоценоза // Основы
лесн. биогеоцеиол. М., 1964.
Молчанов А. А. Изучение воды как компонента биогеоценоза//Прогр. и ме-
тодика биогеоцеиол. исследов. М., 1974.
Молчанов А. А., Грубова В. А. Взаимосвязи в лесном биогеоценозе. М„ 1980,
Морд у хай-Болтовской Ф. Д. Изучение пресноводных биогеоценозов // Прогр*
и методика биогеоцеиол. исследов. М., 1974.
431
Мордухай-Болтовской Ф, Д. Биологический круговорот в водоемах//Мето-
дика изучения биогеоценозов внутр, водоемов. М., 1975.
Мордухай-Болтовской Ф. Д„ Дзюбан Н. А. Формирование фауны беспозво-
ночных крупных водохранилищ//Экология водн. организмов. М., 1966.
Морозов Г, Ф. Учение о лесе. Введение в биологию леса. СПб., 1912. Вып. 1.
Морозов Г. Ф. Учение о лесе. М.; Л., 1931.
Наумов Н. Л. Экология животных. М„ 1955.
Наумов Н. П. О методологических проблемах биологии // Филос. науки.
1964. № 1.
Наумов Н. Л. Развитие представления о популяциях в экологии животных //
Очерки по истории экологии. М., 1970.
Наумов Н. Л. Уровни организации живой материи и популяционная биоло-
гия//Журн. общ. биол. 1971. Т. 32. № 6.
Нестеров В. Г. Общее лесоводство. М., 1954.
Ничипорович А. А. Световое и углеродное питание растений (фотосинтез).
Й4., 1955.
Ничипорович А. А. Фотосинтетическая деятельность и первичная продуктив-
ность фитоценозов на современном этапе эволюции биосферы // Пробл. биогео-
ценол. М., 1973.
Ничипорович А. А., Шульгин И. А. Фотосинтез и использование энергии сол-
нечной радиации //Ресурсы биосферы. Л., 1976.
Новиков Г А. Основы общей экологии и охраны природы. Л., 1979.
Номоконов Л. И. Пойменные луга Енисея. М., 1958.
Номоконов Л. И. Некоторые методологические вопросы биоценологии // Изв.
Сиб. отд. АН СССР. Сер. биол.-мед. наук. 1963. Вып. 2.
Номоконов Л. И. Опыт комплексного стационарного изучения природных
биогеоценозов в пойме Нижнего Дона//Ботан. журн. 1973. Т. 58. № 4.
Номоконов Л. И. Экология, биогеоценология и место их в системе наук //
Журн. общ. биол. 1981. Т. 42. № 6.
Номоконов Л. И., Бананова В. А., Лысенко С. В. Горизонтальное сложение
пырейника чистого//Биогеоценозы в пойме Нижнего Дона. Ростов н/Д, 1978.
Обсуждение доклада А. И. Толмачева «К критике учения о биогеоценоозе»
в комиссии флоры н растительности ВБО// Ботан. журн. 1953. Т. 38. № 6.
Одум Е. П. Экология. М., 1968.
Одум Ю. Основы экологии. М., 1975.
Олейник Г Н. Бактериальные ценозы и количественный состав эпифитной
микрофлоры зеленых нитчатых водорослей в канале Северский Донец —Дон-
басс//Пробл. микробиол. внутр, вод. М., 1971.
Основы лесной биогеоценологии/Под. ред. В. Н. Сукачева и Н. В. Дылиса
М, 1964.
Павлинин В. Н. Обсуждение в президиуме Академии наук СССР проблем
развития биогеоценологии//Экология. 1971. № 3.
Павлов В. А. Обмен веществ и биологический круговорот. М., 1960.
Павлова 3. Ф. Земляные кочки, заселенные муравьями, как узловые микро-
структуры биогеоценозов побережий озер//Экология. 1977. № 5.
Панфилов Д. В. Географическое распространение функционально-биоцеиоти-
ческих групп насекомых на территории СССР//Зональн. особен, населения на-
эеми. животных. М., 1966.
Паттен Б. Концепция информации в экологии. Некоторые аспекты поведения
планктонных сообществ//Концепция информ, и биолог, системы. М.» 1966.
Пачоский И. К. Стадии развития флоры//Вестник естествозн. 1891. № 8.
Пачоский И. К. Описание растительности Херсонской губернии. Степи.
Херсон, 1917.
Перельман А. И. Геохимия ландшафта. М., 1966»
Перельман А. И. Геохимическое прошлое ландшафтов СССР//Природа.
1970. Хз 7.
432
Перельман А. И. Геохимия биосферы//Природа. 1971. № 9.
Перельман А. И, В. В. Докучаев и учение о биокосных системах // Природа.
1974. № 5.
Полынов Б. Б. Ландшафт и почва//Природа. 1925. № 1, 2.
Полынов Б. Б. Учение о ландшафтах//Вопр. географии. 1953. Вып. 33.
Пономарева И. Н. Общая экология: Учебное пособие. Л., 1975.
Поплавская Г. И. Опыт фитосоциологического анализа растительности це*
лянной заповедной степи Аскания-Нова // Журн. Рус. ботан. об-ва. 1925. № 9^
Пошон Ж, Де Баржак Г Почвенная микробиология. М., 1960.
Проблемы биогеоценологии // Отв. ред. Е. М, Лавреико и Т. А. Работков.
М., 1973.
Программа и методика биогеоценологических исследований / Под ред.
В. Н. Сукачева и Н. В. Дылиса. М., 1966.
Программа и методика биогеоценологических исследований / Отв. ред.
Н. В. Дылис. М„ 1974.
Программа и методика изучения биогеоценозов водной среды. Биогеоцено-
зы морей и океанов/Отв. ред. Л. А. Зенкевич. М., 1970.
> Работное Т. А, О консорциях//Бюл. МОИП. Отд. биол. 1969. Т. 7. Вып. 4.
Работное Т. А. О значении консортивных связей в определении взаимных
отношений растений в фитоценозах//Бюл. МОИП. Отд. биол. 1970. Т. 74. Вып. 2.
Работное Т. А. Некоторые вопросы изучения консорций // Жури. общ. биол.
1973. Т. 34. № 3.
Работное Т. А. Некоторые вопросы изучения почвы как компонента биогео-
ценоза//Вестник Моск, ун-та. Биология и почвовед. 1974а. № 3.
Работное Т. А. Консорция как структурная единица биогеоценоза // Приро-
да. 19746. Хе 2.
Работное Т А. Изучение ценотических популяций в целях выяснения «стра-
тегии жизни» видов растений//Бюл. МОИП. Отд. биол. 1975. Т. 80. Вып. 2.
Работное Т. А. Фитоценология. М., 1978а.
Работное Т А. Еще раз о консорциях//Бюл. МОИП. Отд. биол. 19786,
Т. 83. Вып. 2.
Работное Т. А. Некоторые вопросы изучения автотрофных растений как ком-
понентов наземных биогеоценозов//Бюл. МОИП. Отд. биол. 1980. Т. 85. Вып. 3.
Раменский Л. Г. О сравнительном методе экологического изучения расти-
тельных сообществ//Дневник 12-го съезда русск. естествоиспыт. и врачей. 1910.
Вып. 9.
Раменский Л. Г. Основные закономерности растительного покрова и их изу-
чение//Вестник опыт. дела. Воронеж. 1924.
Раменский Л. Г. Введение в комплексное почвенно-геоботаническое исследо-
вание земель. М., 1938.
Раменский Л. Г О некоторых принципиальных положениях современной гео-
ботаники И Ботан. журн. 1952. Т. 37 № 2.
Раунер Ю. Л. Тепловой баланс растительного покрова. Л.» 1972.
Рафес П. М. О роли и значении растительноядных насекомых в лесном био-
геоценозе // Влияние животных на продукт, лесн. биогеоцен. М., 1966.
Рафес П. М. Развитие учения о биогеоценозах//Очерки истории экологии.
М., 1970.
Рафес П. М. Популяция и биогеоценоз//Природа. 1972. № 5.
Рафес П. М. Спорные вопросы и пути развития биогеоценологии//Бюл.
МОИП. Отд. биол. 1980а. Т. 85. Вып. 3.
Рафес П. М. Биологические исследования растительноядных лесных насеко-
мых. М., 19806.
Рафес П. М., Динесман Л. Г., Перель Т. С. Животный мир к;ак компонент
лесного биогеоценоза // Основы леей, биогеоценол. М., 1964.
Ревердатто В. В. Введение в фитоценологию. Томск, 1935.
Риклефс Р. Е. Основы общей экологии. М., 1979.
433
Родина А. Г О формах существования бактерий в водной толще водое-
мов//Пробл. микробиол. внутр, вод. М., 1971а.
Родина А. Г Продукционная микробиология пресных вод // Пробл. микро-
биол. внутр, вод. М., 19716.
Рокгаузен Р. Проблема целостности в биологии // Вопр. философии.
1959. № 3.
Романенко В. И. Соотношение между фотосинтезом фитопланктона и де-
струкцией органического вещества в водохранилищах//Микрофлора, фитопланк-
тон и высшая растит, внутр, водоемов. Л., 1967.
Рысин Л. П. Идеи биогеоценологии за рубежом. Краткий литературный об-
зор//Изв. Сиб. отд. АН СССР. Сер. бнол.-мед. наук. 1964. 12. Вып. 3.
Саблина Т. Б. Первое рабочее совещание стран — участниц СЭВ по теме
«Разработка общей теории биогеоценологии» // Журн. общ. биол. 1973.
Т. 34. № 5.
Садовский В. Н. Парадоксы системного^ мышления // Системы исследов. (еже-
годник). 1972.
Сауков А. А. Геохимия. М., 1966.
Свирежев Ю. M.t Елизаров Е. Я. Математическое моделирование биологиче-
ских систем//Пробл. космич. бнол. М., 1972. Т. 20.
Северцев С. А. Проблемы экологии животных//Неопу б ликов, работы. М.г
Селиванов И. А. Некоторые вопросы учения о консорциях//Учен. зап.
Пермск. пед. ин-та. 1974. Т. 133.
Серебряков И. Г. Основные направления эволюции жизненных форм у по-
крытосеменных растений // Бюл. МОИП. Отд. биол. 1955. Т. 60. Вып. 3.
Серебряков И. Г. Экологическая морфология растений. М., 1962.
Серебряков И. Г. Жизненные формы высших растений и их изучение//По-
лев. геоботан. М., 1964. Т. 3.
Серебряков К. К. История растительного мира СССР. Л., 1936.
Серебрякова Т. И. Морфогенез побегов и эволюция жизненных форм зла-
ков. М., 1971.
Серебрякова Т. И. Учение о жизненных формах на современном этапе // Бо-
таника. М., 1972. Т. 1.
Сетров М. И. Организация биосистем. Методологический очерк принципов
организации живых систем. Л., 1971.
Скабичевский А. П. Условия формирования речного фитопланктона // Водн.
и наземи. сообщ. низш. растений Сибири. Новосибирск, 1974.
Смольянинов И. И., Рябуха Е. В. Круговорот веществ в природе. Киев, 1971.
Соколов С. Я. Фитоценотические типы//Докл. АН СССР. 1947. Т. 55. № 2.
Соколов С. Я. Типы борьбы за существование средн растений //В. Н. Сука-
чеву к 75-летию со дня рожд. М.; Л., 1956. _
Солнцев Н. А. Природный географический ландшафт и некоторые его общие
закономерности//Тр. 2-го Всесоюз. геогр. съезда. 1948.
Солнцев Н. А. Некоторые дополнения и уточнения в вопросе о морфологии
ландшафта//Вестник Моск, ун-та. Сер. 1. География. 1961. № 3.
Сочава В. Б. Опыт филоценогенетической систематики растительных ассоциа-
ций // Сов. ботаника. 1944. № 1.
Сочава В. Б. Новейшие вертикальные движения земной коры н растительный
покров//Землеведение. МОИП. 1950. Т. 48.
Сочава В. Б. Классификация растительности и топология физико-географиче-
ских фаций //Матер, по классиф. растит. Урала. Свердловск, 1959.
Сочава В. Б. Вопросы классификации растительности, типология физико-гео-
графических фаций//Тр. Ин-та биол. Уральск, фил. АН СССР. 1961. Вып. 27,
Сочава В. Б. Теоретические положения топологии степных геосистем. Л., 1970.
Сочава В. Б, К теории классификации геосистем с наземной жизнью // Докл.
Ин-та геогр. Сибири и Дальнего Востока. 1972. Вып. 4.
434
Сочава В. Б. Топология как раздел учения о геосистемах//Тополог, аспек-
ты учения о геосист. Новосибирск, 1974.
Станчинский В. В. Среда жизни и ее подразделение//Тр. Науч.-исследов.
зоол.-биол. ин-та Харьк. ун-та. 1933. Т. 1.
Стебаев И. В. Об иерархическом строении систем биогеоценозов суши//
Мат. моделир. в экологии. М., 1978.
Степанов В. Н. Круговорот энергии и веществ и изменение природы Земли//
Вопр. философии. 1966. № 6.
Сукачев В. Н, Растительные сообщества (Введение в фитоценологию). М.;
Л., 1928.
Сукачев В. Н. Руководство к исследованию типов лесов. М.; Л., 1931.
Сукачев В. Н. Развитие растительности как элемента географической среды
в соотношении с развитием общества//О геогр. среде в лесн. произв. Л., 1940.
Сукачев В. Н. Идея развития растительности в фитоценологии // Сов. бота-
ника. 1942. № 1, 2.
Сукачев В. Н. О принципах генетической классификации в биогеоценологии//
Журн. общ. биол. 1944. Т. 5. № 4.
Сукачев В. Н. Биогеоценология и фитоценология//Докл. АН СССР. 1945.
Т. 47. № 6.
Сукачев В. Н. О лесной биогеоценологии и ее основных задачах // Ботан.
журн. 1955. Т. 40, № 3.
Сукачев В. Н. Основные понятия лесной биогеоценологии // Основы лесн.
биогеоцеиол. М., 1964а.
Сукачев В. Н. Динамика лесных биогеоценозов//Основы лесн. биогеоцеиол.
М., 19646.
Сукачев В. Н. Теоретическое и практическое значение лесной биогеоценоли-
гии//Основы лесн. биогеоцеиол. М„ 1964в.
Сукачев В. Н. Основные понятия о биогеоценозах и общее направление их
изучения//Прогр. и методика биогеоцеиол. исследов. М., 1966.
Сукачев В. Н. Основы лесной типологии и биогеоценологии // Избр. тр. Л.г
1972. Т. 1.
Тараненко Л. И. Роль позвоночных животных в биогеоценозах//Биогеоце-
нозы в пойме Нижнего Дона. Ростов н/Д, 1978.
Тимофее в-Ресовский Н. В. О некоторых принципах классификации биохоро-
логических единиц; Вопр. классиф. растит. // Тр. Ии-та биол. Уральск, фил.
АН СССР. 1961. Вып. 27.
Тимофеев-Ресовский Н. В. Структурные уровни биологических систем//Си-
стемные исследов. М., 1970.
Тимофеев-Ресовский Н. В. Популяции, биогеоценозы и биосфера Земли //
Мат. моделирование в биологии. М.. 1975.
Тимофеев-Ресовский Н. В., Тюрюканов А. Н. Об элементарных хорологиче-
ских подразделениях биосферы//Бюл. МОИП. Отд. биол. 1966. Т. 71. Вып. 1.
Тимофеев-Ресовский Н. В., Тюрюканов А. И. Биогеоценология и почвоведе-
ние//Бюл. МОИП. Отд. биол. 1967. Т. 72. Вып. 2.
Тимофеева-Ресовская Е. А. Распределение радиоизотопов по основным ком-
понентам пресноводных водоемов//Тр. Ин-та биол. Уральск, фил. АН СССР.
1963. Выл. 3.
М 19523 Взаимодействие растительных сообществ и условий среды.
Томилин А. Г. Морские млекопитающие и их биогеоценологические связи с
другими группами морской фауны // Прогр. и методика изуч. биогеоцеиол. водн„
среды. М., 1970.
Травлеев А. П. Роль личинок садовой мушки в разложении лесиой подстил-
ки в степи//Науч. докл. высш, школы. Биол. науки. 1961. № 1.
Травлеев А. 77. Взаимоотношение растительности с почвами в лесных био-
геоценозах степной зоны Украины//Лесоведение. 1976. Xs 6.
435
Тюрин И. В. О количественном участии живого вещества в составе органи-
ческой части почв//Почвоведение. 1946. № 1.
Тюрюканов А. Н.. Александрова В. Д. Витасфера Земли//Бюл. МОИП. Отд.
биол. 1969. Т. 74. Вып. 4.
Уиттекер Р. Сообщества и экосистемы. М., 1980.
Уткин А. И. Актуальные вопросы лесной биогеоценологии//Теор. лробл.
леей, биогеоценол. М.» 1970.
Уткин А. И, О показателях лесных биогеоценозов//Бюл. МОИП. Отд. биол.
1975. Т. 80. Вып. 2.
Уткин А. И. Об энергетике лесных биогеоценозов // Структ.-функц. организ.
биогеоценозов. М., 1980.
Уткин А. И., Дылис Н. В. Принципы построения биогеоценотической клас-
сификации лесов//Лесоведение. 1968. № 3.
Уткин А. И., Блэров Л. Г„ Дылис Н. В., Солнцева О. Н. Вертикально'фр ак-
ционное распределение фитомассы и принципы выделения биогеогоризонтов в
лесных биогеоценозах//Бюл. МОИП. Отд. биол. 1969. Т. 74. Вып. 1.
Ушаков Б. П. О классификации приспособлений животных и растений и о
роли цитоэкологии в разработке проблем адаптации//Пробл. цятоэкол. живот-
ных. М., 1963.
Федоров В. Д., Гильманов Т. Г Экология, М., 1980.
Флейшман В. С. Системные методы в экологии//Статист, методы анализа
почв, растит, и их связи. Уфа, 1978.
Хайлов К. М. Экологический метаболизм//Экология. 1970. № 2.
Хильми Г. Ф. Основы физики биосферы. Л., 196.
Хильми Г. Ф. Уроки биосферы // Методол. аспекты исследов. биосферы.
М., 1975.
Ходашева К. С. О географических особенностях структуры населения назем-
ных позвоночных животных // Зоиальи. особен, насел, наземн. животных.
М., 1966.
Холл А Д., Фейджин Р. Е. Определение понятия системы // Исследов. по
общей теории систем. М., 1969.
Часовенная А. А, Основы агрофитоценологии. Л., 1975.
Частухин В. Я., Николаевская М. А. Биологический распад и ресинтез орга-
нических веществ в природе. Л., 1969.
Чернов Ю. И. Понятие «животное население» и принципы геозоологических
исследований//Журн. общ. биол. 1971. Т. 32. № 4.
Чернов Ю. И. Природная зональность и животный мир суши. М., 1975.
Шаронов И. В, Формирование ихтиофауны водохранилищ//Экология води,
организмов, М., 1966.
Шварц С. С. К истории основных понятий современной экологии // Очерки
по истории экологии. М., 1970.
Шварц С. С. Популяционная структура биогеоценозов//Природа. 1971. № 6.
Шварц С. С. Характер функциональных связей и элементов биологических
систем на популяционном и организменном уровнях//Экология. 1972. № 4.
Шварц С. С. Эволюция и биосфера.//Пробл. биогеоценол. М.. 1973.
Шварц С. С„ Гурвич. Э. Д., Ищенко В. Г., Сосин В. Ф. Функциональное
единство популяции//Журн. общ. биол. 1972. Т. 33. № 1.
Шенников А. П. Дарвинизм и фитоценология//Сов. ботаника. 1938. № 3.
Шенников А. П. Луговедение. Л.» 1941.
Шенников А. П. Экология растений. М., 1950.
Шенников .4, П. Введение в геоботанику. Л., 1964.
Шилов И. А. Биосфера, уровни организации жизни и проблемы экологии //
Экология. 1981. № 1.
Шипунов Ф. Я- Организованность биосферы. М., 1980.
Шмальеауэен И. И. Кибернетические вопросы биологии. Новосибирск, 1968.
Штеенбек М. Почему Земля обладает магнитным полем//Природа. 1970. Хе 7.
Щербань М. И. Микроклимате логи я. Киев, 1968.
436
Элтон Ч, Экология животных. М„ 1934.
Энгельс Ф. Диалектика природы. М.; Л., 1930.
Яковлев В. П. Учение В. И. Вернадского о биосфере//Человек и биосфера.
Ростов н/Д, 1973.
Ярошенко П. Д. Основы учения о растительном покрове. М., 1950.
Ярошенко П. Д. Основы учения о растительном покрове. 2-е изд. М., 1953.
Ярошенко П. Д. Геоботаника. Основные понятия, направления и методы. М.;
Л., 1961.
Яхонтов В. В. Экология насекомых. М., 1969.
Braun-Blanquet /. Pflancensociologle. Berlin, 1928.
Braun-Blanquet /., Pavillard I. Vocabulaire der Sociologie vegetale. Montpel-
lier, 1925.
Clemenis F. Plant succession: an analisis of the developments of vegetation //
Carnegie Inst. PubL, 1916.
Clements F. Nature and structure of the dim ax//Jour, of Ecology. 1936. 24, 1.
Du-Rietz G. Zur metodologlschen Grudlage der modernen Pflanzensociologie//
Akademische Abhandlung. Upsala. 1921.
Ellenberg H. Ziele und Stand der Okosystemforschung // Okosystemforschung.
Berlin; New York, 1973.
Evans F. Ecosystem as the basis unit in ecology Science. 1956. 123, N 3208.
Friederichs K. Die Grundfragen und Gesetzmassigkelten der landund forst-
wirtschaftlichen Zoologie//Okologischer Teil. Berlin, 1930. Bd. 1.
Gleason H. The individualistic concept of the plant association//Bull. Torrey.
Bst Club. 1926. Vol. 53. N 1.
Haeckel E. Generell Morphologic der organismen // Allgemeine Entwickelungs-
.geschichte der Organismen. Berlin, 1866. Bd. 2.
Hesse R. Tier geographic auf okologischer Grundlage. Jena, 1924.
Hills G. The classification and evaluation of site for forestry//Ontario Dept
Lands and Forests. Res. Rept. 1952. N 24.
Krajina V. Can we find common platform for the different schools of the
forest type classification? //Silva fennica, 1960. N 105.
Kuchler A. Problems in classifying and mapping vegetation for ecological
regionaliration//Ecology. 1973. Vol. 54. N 3.
Lindeman R. The trophic-dinamic aspekt of ecology//Ecology. 1942. 23. 4.
Misic V. Poreclo sukcesija i degradaeija sumske vegetaeije Srbije//Zbornik
radova. Beograb. 1961. Kn. 5. N 3.
Mobius K. Die Austen und die Austernwirtschaft. Berlin, 1877,
Ovington J. The ecosystem concepts as and aid to forest classification//Silva
fennica. 1960. N 105.
Passarge S. Physiologlsche Morphologic//Mitt geogr. Ges. 1912. N 26.
Passarge S. Lander, reale Landschaften, ideale Landschaftstypen //Naturwis-
senschaften. 1929. 17. 36.
Patten B. In Introduction to the cybernetics of the ecosystem the trophiedina-
mic aspect//Ecology. 1959. Vol. 40. N 2.
Raunkiaer C. Types biologique pour la geographic botanique//Bull. Acad. R.
Sc. Danemark, 1905.
Rowe J. Forest regions of Canada. Ottawa. 1959.
Shelf ord V. Principes and Problems of ecology as illustrated by animals //
Ecology. 1915. 3. N 1.
Scamoni A. Konnen wir eine gemeinsame Grundlage ffir die verschiedenen
Schule der Waldtypeenklassification finden. //Silva fennica. 1960, N 105.
Tansley A. The use and abuse of vegetational concepts and terms//Ecology.
1935. 16,
Thienemann A. Grundzuge einen allgemeinen Okologie//Okologie Arch. Hyd-
robiologie. 1939. Vol. 35.
Weaver L, Clements F, Plant ecology. N. Y.; L„ 1938.
437
ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ
Абиосапротрофный круговорот веществ 222
Абиотическая (косная) среда 16, 76, 90, 163, 271
Абиотическое единство 47
Абиотрофы 49, 224
Абиссаль 391, 409—411, 413
Автогенные (внутренние) процессы 358
Автогенные сукцессии 319, 336
Авторегуляция 282, 285, 318
Автотрофная энергетическая база 41, 54, 288, 231, 409
Автотрофные водоросли 353
Автотрофные микроорганизмы 105
Автотрофные популяции 21
Автотрофные продуценты 412
Автотрофные растения 57, 73, 85, 87, 88, 106, 114, 120» 129, 197, 208, 210, 215,
244, 251, 259, 260, 267, 269, 272, 349, 424
Автотрофные фотосинтетики 354
Автотрофный компонент биогеоценоза 44, 69, 85; 95, 187, 217, 233, 237, 325, 345
Автотрофный тип питания 419
Автотрофный трофический уровень 265
Автотрофный фитопланктон 404
Автотрофный ценокомплекс 25, 88, 91, 221, 348, 399
Автотрофы 21, 41, 44, 49—51, 57, 75, 114, 136, 243, 261, 265, 274, 356,385; 409, 416
Агробиогеоценоз 420
Агрофитоценоз 420
Агрофитоценология 420
Агроценозы 28, 29, 239, 420, 424
Агроценология 420
Азототрофы 49
Азотофикаторы 49
Азотофиксаторы 49, 52, 87, 99, 106, 128, 209
Аккумулятивный тип коры выветривания 141, 142
Аллелагонические каналы (коммуникации) 267
Аллелогенные (внешние) движущие силы 358
Аллелопатические каналы (коммуникации) 267
Аллелопатия 190
Аллелослолические каналы (коммуникации) 267
Амменсализм 189
Анаэробиозис 332
Анаэробные гетеротрофы 418
Анаэробные ценопонуляции 106
Антагонизм 189
Апвеллинги 390
Арена жизни 158, 160, 353
Архебионты 353
Ассектаторы 80, 82, 86
Атмосфера 18, 37, 38, 55—57, 63, 69, 88, 91, 102, 117—Н9, 121, 123, 124, 126—
128, 130—132, 136, 137, 139, 141, 154, 15Я 165, 176, 177, 247, 251, 253,
254, 258, 259, 262, 352, 386, 389, 419
Афотическая зона пелагиали 404, 405
Аэробиозис 331, 332
Аэробионты 365, 366
Аэробные гетеротрофы 419
Аэробные ценопопуляций 106
Аэротоп 50, 51, 117, 139, 165, 235. 240
438
Бактериальное население 386
Бактериальная масса 398
Бактериальные бногидроценозы 376
Бактериоплаиктон 373, 380, 385, 407, 409
Батиаль 391, 409—412
Бенталь 372, 390, 409—412
Бентическая ассоциация 290
Бентические (бентосные) формы 371, 381, 411
Бентос 367, 372, 412—414
Бентосные сообщества (ценозы) 375, 376
Бентосные ценозы 388
Бентофаги 376
Бидоминантные биогеоценозы 247
Биогенная миграция атомов 346, 362
Биогенная миграция химических элементов 76, 240, 362
Биогенная почвенная среда 116, 156, 157, 327
Биогенная среда (биоценоза) 50, 155, 160, 161, 331
Биогенное вещество 66, 370
Биогенные газы 37, 58, 89, 118, 128, 166, 240, 300, 366, 382, 385, 386, 399—401
Биогенные осадочные породы 56
Биогенные циклы почвообразования 142
Биогенные элементы веществ 370, 382, 390, 395, 401, 417
Биогеогоризонты почвы 147, 148, 150, 231
Биогеомассивы 131, 301, 304, 305, 310, 314—316
Биогеопоток 302—304
Биогеорегион 314—316
Биогеосистемы 301, 303, 310, 315, 316
Биогеосфера 24, 26, 30, 31, 33—36, 46, 50, 55, 65—69, 71, 85, 86, 89, 92, 105,
114, 119, 122, 128, 131, 136, 166, 177, 182, 215, 221, 246, 247, 249—251,
258, 264, 266, 270, 273, 282—289, 294, 296, 297, 299, 300, 303—305, 307,
309, 310, 316—319, 345—347, 352, 356, 358. 362, 421
Биогеохимические циклы биофильных элементов 64, 249, 250
Биогеохимический круговорот серы 257
Биогеохимический круговорот углерода 250
Биогеоценогенез (биогеоценотический процесс) 323, 336, 340, 342, 346
Биогеоценоз 8—10, 15, 16, 19—22, 24—38, 40—42, 44-55, 65—69, 71, 72, 75,
78—80, 83—85, 87—96, 106, 109, 111, 115—119, 121, 122, 125, 128, 131 —
137, 139, 147, 149—152, 154—160, 165—167, 172—174, 176—178, 182—185,
187, 188, 193, 194. 196—198, 201, 207, 209, 211, 212, 214, 215, 218, 219, 221,
222, 225, 229—231, 233—235, 239, 240, 243—245, 248, 249, 251, 252, 258,
263—266, 269—274, 279, 281, 282, 284—287, 289—292, 293—304, 309, 310,
316, 318—323, 327, 329, 330, 334, 335—340, 342, 344—346, 348, 349, 352,
353, 355—363, 377, 383, 388, 389, 399, 408, 412, 419—422
Бногеоценология 8, 10—13, 18—22, 24—30, 32, 33, 35, 48, 55, 58, 62, 65, 67, 183,.
184, 291, 363, 420—422, 424
Биогеоценологическая классификация 291, 294
Биогеоценотическая зона 313, 317
Биогеоценотическая оболочка Земли 300, 309—311, 422
Биогеоценотическая провинция 313
Биогеоценотическая среда (биоценосреда) 267, 299, 321, 340, 360
Биогеоценотическая (функциональная) роль компонентов 69, 90
Биогеоценотические горизонты (биогеогоризонты) 185, 227, 231—234
Биогеоценотические области 312, 317
Биогеоценотические системы 11, 52, 54, 94, 151, 171, 184—186, 196, 203, 212,
215, 249, 258, 265, 273, 275, 277, 300, 303, 314, 325, 330, 333, 334, 353. 354,
356—358, 361.
439
Биогеоценотический барьер 301
Биогеоценотический внутренний обмен (веществ и энергии) 244
Биогеоценотический круговорот 244
Биогеоценотический отбор 239, 327, 358
Биогеоценотический прогресс 359
Биогеоценотический уровень изучения биосферы 65
Биогидрогоризонты 383, 408
Биогндросистемы 304
Биогидросфера 8, 296, 363, 377, 397, 398, 413, 418
Биогидроценоз 66, 363, 371, 375, 377, 378. 382, 383, 387—389, 391, 398—400, 402,
405—409, 413, 414, 416, 418
Биокосная система 10, 12, 20—25, 27, 28, 31, 32, 36, 37, 44, 47, 48, 54, 65, 66,
68, 84, 88, 106, 123, 131, 158—184, 193, 240, 241, 243, 245, 247—250, 257,
259, 263—265, 269—271, 278, 281, 282, 286—288, 318, 329, 330, 339, 345,
348, 350, 352, 363, 371, 374, 377, 378, 384, 389, 408, 409, 420—422, 424
Биокосная среда 62, 63, 177
Биокосное вещество 66
Биолиты 63
Биологическая система (биосистема) 16, 20, 79, 158, 159, 161, 165, 180, 219г
225, 241, 318, 347—349, 358
Биологическая структура океанических биогидроценозов 396, 407
Биологические комплексы (биокомплексы) 44, 48—50, 117, 288, 374
Биологические сезоны 418
Биологический вид 69, 158, 163, 185, 270, 347, 361
Биологический круговорот веществ 144, 146, 153, 155, 201, 241, 242, 245—249,
258, 264, 294, 339, 344, 345, 350, 352, 353, 357
Биологический (физиологический) обмен 244, 266
Биологический тип 78
Биом 292, 316
Биомасса 71, 76, 84, 89, 94, 96, 97, 104—106, 108, 109, 114, 136, 145, 152, 156,
198, 201, 202, 204, 217, 222, 235), 242, 244, 247, 248, 251, 254, 255, 259,
263, 265, 270—282, 360, 361, 376, 378, 401, 404, 407, 408, 413, 414
Биоморфы 354, 359
Биопродукционный процесс 262, 263, 271, 272, 277
Биоредуценты 106, 108, 200, 208, 377
Биострома 9, 65, 347
Биостроматическая форма жизни 9, 22, 270, 350
Биостроматический уровень организации жизни 177
Биосфера 8, 10, 11, 17, 18, 32, 35, 55—64, 67, 76. 90, 116, 124, 129, 139, 141,
143, 144, 158, 177, 181, 232, 246, 249—251, 259, 270, 291, 295, 296, 309,
310, 339, 346, 350, 351, 359, 361, 362, 422
Биосферология 65
Биота 16, 212, 219, 221» 238, 244, 248, 308» 324, 329
Биотехносфера 64, 181
Биотический круговорот веществ 49, 56, 73, 89, 90, 222, 243, 351, 359, 377, 383—
385, 417
Биотоп 37, 43, 45, 46, 50, 51, 69, 139, 159—161, 165, 188, 221, 244, 264, 265, 273,
327, 329, 349, 363, 384, 388, 396, 400, 404, 410, 411, 413
Биотопы дальнеиеритических сообществ 396, 397
Биотопы донной пленки жизни 410, 411
Биотопы морского и океанического дна 396-, 397
Биотопы морских гидробионтов 395, 418
Биотопы населения морских гидробионтов 395
Биотопы пелагических сообществ 395, 396
Биотопы прибрежных неритических сообществ 396, 397
Биотоценогенеэ 348, 349
440
Бнотрофные (полночленные) консорции 216, 221
Биотрофные ценопопуляций 279, 334
Биотрофиый канал перемещения веществ 266, 269
Бкотрофный (животный) компонент биогеоценоза 44, 69, 89, 94, 100
Биотрофный компонент биогидроценоза 379, 407
Биотрофы 41, 50, 96, 202, 208, 230, 272, 356, 377, 381, 385, 424
Биофаги 49
Биофильные химические элементы 249, 257, 258
Биохора 20, 160
Биоценода 15
Биоценоз 15—19, 25, 37, 41—45, 47, 49—^51, 60, 66, 69, 79, 101, 138, 158, 165„
187—189, 191—193, 195, 196, 207, 219, 221, 264, 279, 280, 290, 292, 327,
338, 339, 346—349, 356, 359, 384, 395, 396, 400
Биоценоклимат 139
Биоценотип 79, 80, 83, 84, 185
Биоценотическая роль компонентов биогеоценоза 187
Биоценотическая регенерация пиши 424
Биоценотическая среда (биоценосреда) 45, 46, 69, 117, 139, 165, 225, 244, 249,
264, 334, 349, 356
Биоценотическая форма жизни 9, 22, 25, 270, 350
Биоценотическая (функциональная) деятельность 69, 87, 88, 94—97, 105, ИЗ, 185
Биоценотическнй отбор 334, 359
Биоценотический уровень организации живой материи 65, 67, 79, 165, 225, 378
Биоценотоп 20, 45
Биоциды 30
Биоциклы 160
Биоэкос 20
Блоки 35, 43, 47, 166, 401
Большой биологический круговорот 58, 124, 243
Валовая первичная биологическая продукция 272, 274
Вертикальные зоны неритической области 390
Вертикальные зоны пелагической области 391
Вертикальные миграции живых существ 366, 368
Вещественно-энергетический поток 222
Видовые популяции 10, 21, 37, 43—46, 69—71, 93, 158, 163, 164, 185, 203, 204»
208, 215, 267, 272, 289, 325, 326, 347, 348, 360, 361, 377, 378
Видовые ценопопуляций 71, 83, 84, 187, 219, 221, 239, 245, 281, 327, 333
Витасфера 67
Внешние коммуникации (связи) 267
Внешний круговорот веществ 244, 250
Ruprrfwu& ОДА ОЯО
Внешняя среда 160—162, 165, 182, 225, 240, 244, 273, 282, 360, 423
Внутренние коммуникации (связи) 267
Внутренняя (биогенная) среда 46, 116, 160, 165
Внутривидовые единицы 70
Внутрисистемные связи и отношения 288
Внутриценозный биогеоценотический обмен 240, 245, 282
Внутриценозный круговорот веществ 244
Внутриценозный материально-энергетический обмен 281
Внутриценозный метаболизм (обмен) 300, 309
Внутриценозный обменный процесс 389
Водная биокосная система 204, 376, 384
Водно-воздушный режим почвы 148, 150, 152
Водная микрофлора 386, 414
Водная оболочка планеты 390
441
Водная среда жизни (гидросфера) 50, 66, 268, 269, 358, 363, 364. 367, 368 373
378, 384, 389, 391 ’
Водная среда обитания 377, 395, 417
Водная ценотическая система 378, 413, 419
Водная эрозия 173
Водные массы в океане 389, 391, 393, 395, 400, 413
Воздушная среда жизни 44, 50, 69, 117, 132, 188, 225, zo/—26У, 320, 357, 378, 382
Воздушный режим 138, 151
Войлок (<калдан>) в травянистых фитоценозах 90, 106, 149, 151, 154, 156, 230
Всеобитель 309, 310
Вторичная биологическая продукция (продуктивность) 38, 51, 91, 94, 95, 114,
263, 271, 272, 275—277, 281, 377, 416
Вторичноводные животные 366, 371
Вторичные водные массы (горизонтальные н вертикальные) 393
Вторичные консументы 199
Вторичный (геохимический) цикл в круговороте азота 254
Вторичный (геохимический) цикл углеродного обмена 250
Газовый тип круговорота химических элементов 250
Гемиавтотрофные виды 214
Генетические горизонты (почвы) 149, 226, 231
Геобиом 311, 312
Геогоризонты 232, 303
Географический комплекс 18
Геологический (абиотический) круговорот веществ 58, 124, 241, 242, 245, 250
Геом 292
Геопотоки 303, 305
Георастнтельная система 307, 308
Геосистема 159, 303, 308
Геосфера 58, 59, 177, 309, 389
Геохоры 308, 309
Геоценоз 18, 19
Геоэкобиота 20
Гетеротрофная микрофлора 57, 153
Гетеротрофное население 327, 376
Гетеротрофные животные 57, 208
Гетеротрофные микроорганизмы 57, 153
Гетеротрофные популяции 21
Гетеротрофные растения 51, 73, 85
Гетеротрофный компонент биогидроценоза 384, 408
Гетеротрофный компонент биоценоза 44, 197, 228, 345, 353, 354
Гетеротрофный тнп метаболизма 368
Гетеротрофы 21, 41, 44, 49—51, 54, 57, 75, 86, 214, 242—244, 251, 252, 269, 326,
330, 353, 356, 399, 404, 409
Гидробиом 311, 312
Гидробионты 363—366, 378, 380, 389, 394, 400, 401, 408, 411, 417, 418
Гидрологическая структура вод 389, 393, 407
Гидрологический режим 119, 150
Гидросфера 18, 55, 58, 59, 62, 64, 124, 139, 141, 177, 251, 253, 311, 389
Гидротоп 44, 50
Гипергенез (выветривание пород) 140
Гиперпроценоз 325
Глубоководная (абиссопелагиальная) зона 391, 408
Гологенетические сукцессии (смены) 334, 335
Гомеостаз (гомеостазис) биогеоценоза 29, 282, 289, 318, 329, 339
Гравитационная вода в почве 150
442
Гравитационное поле 174t 175
Гравитационные процессы 174, 175
Гравитация 166, 175
Демутационные смены 336
Демутацня 336
Деструкторы 52, 200, 212, 351, 387
Деструкция 201, 259, 279, 376, 386
Детерминант 86, 208, 210—212, 215, 216
Детрит 200, 366, 380, 381, 387, 407, 410
Детритная пищевая цепь 199—201, 218, 221, 223, 279, 402, 406
Детритные (неполночлеиные) консорции 216, 221
Детритный поток веществ и энергии 269, 402, 406
Детритофаги 108, 200, 269, 339, 381, 408, 412
Дефляция 166, 173
Деформационное поле (в океане) 393
Деятельный слой атмосферы 118, 132
Дистрофные озера 369
Доминантные консорции 213
Доминантные ценопопуляций 218
Доминанты 82, 86, 212, 377, 217, 326, 377
Донная пленка жизни 60, 399, 409, 411—413
Единицы функциональной организации биогеоценоза 211
Емкость биологического круговорота веществ 247
Естественный отбор на разных уровнях организации жизни 348, 358, 359
Живое вещество 8, 10, 32, 34, 55, 56, 58, 60-65, 143, 152, 153, 159, 177, 259,
347, 360, 362
Животное население 41, 83, 88, 92, 96, 121, 153, 175; 230, 238, 287, 296, 316, 325;
329, 363, 365, 371—373, 397
Жизненные формы 71, 72, 74, 75, 77, 91, 185, 212, 353, 354, 371, 411
Жизненные формы животных 77, 78
Жизненные формы растений 75, 76, 78
Законы термодинамики 258
Замкнутые контуры воздействия 283, 286
Запас биомассы 295, 325, 375
Звенья пищевой цепи 197, 202, 204, 210
Земная корона 59
Зимнее окоченение холоднокровных животных 123
Зимний покой 123
Зимняя спячка 123
Зообентос 412
Зоомасса 272
Зоопаразиты 209, 277
Зоопланктон 204, 356, 375, 376, 380, 385, 402—404, 408, 412, 418
Зоосфера 283, 285—287
Зоофаги 44, 51, 91, 94, 117, 196, 197, 205, 209, 210, 218, 259, 267, 277, 278, 283,
377, 381
Зооценоз 18, 36, 37, 42
Зооценотип 79, 83
Изореагент 71
Ин бентос 367
Ингредиенты 82
443
Индивидуальные консорции 212. 216. 219. 264
Интенсивность метаболизма 273
Интенсивность (скорость) оборота веществ 248
Интенсивность солнечной радиации 119. 133, 259
Интрузивные породы 170
Инфильтрация влаги 125
Информационные взаимосвязи 329
Инфракрасные лучи (излучения) 121. 134, 260
Ионосфера (термосфера) 59, 76
Канал выедания 266
Канал запасания органических веществ 266
Канал разложения органических веществ 266
Каналы коммуникации (перемещения веществ) 266
Квартнрантство 188
Кинетика биогеоценозов 301
Климакс (клнмаксовое сообщество) 337, 339
Климатические фронты 130
Климатический климакс 338
Климатоп 18, 37, 44, 47, 50, 116, 117
Клональные консорции 213
Комменсализм 188, 189, 356
Комменсалы 216
Компонентный состав биогеоценоза 340
Компоненты биогидроценоза 382, 389, 413
Компоненты биосферы 62, 63
Конвекция 130
Конгруэнция 163
Коидоминанты 82
Конкуренция 188, 189, 191—195
Ковсортивные связи 206, 212, 214, 216, 222
Консортивный состав биогеоценоза 218, 358
Консорты 206, 208—213, 216—222
Консорции 88, 185, 186, 197, 206—218, 264, 265, 267, 323, 349, 356, 360
Конституционное разобщение видов 193
Консументы 44, 200, 208—212, 353, 377, 402
Контактные взаимоотношения 190
Континентальная область бногидросферы 367, 389
Континуум 17, 33—35, 53
Концентры 207, 209, 216, 217, 222, 264
Копрофаги 108, 209
Кора выветривания 139, 140
Коралловые рифы 413
Коренная смена биогеоценозов 321, 322, 331, 340
Коренной биогеоценоз 48, 327, 336
Косвенное регулирование 287
Космические излучения 176
Косная среда 46, 55, 63
Косные компоненты биогеоценоза 88, 91, 95, 116, 123, 329, 377
Коэффициент полезного действия (КПД) питания 260, 271
Коэффициент рентабельности (КР) 271
Коэффициент фотосинтетически активной радиации (ФАР) 260
Коэффициент экологической эффективности 273
Круговорот азота 254
Круговорот биофильных химических элементов 249
Круговорот веществ 240, 241, 243, 244, 246, 265, 266, 350, 352, 362
444
Круговорот воды (влагооборот) 119, 124, 125, 241
Круговорот воды в океане 391, 393, 395
Круговорот серы 257
Круговорот углерода 251, 252
Круговорот фосфора 255
Крупномасштабные круговороты (в океане) 393, 395, 397, 400
Латерали 302—304
Латеральное направление перемещения объектов 301
Ленточные ландшафты в океане 391
Лиманы 171, 260, 370
Лимническая зона (озер н т. п.) 369, 378, 380, 381
Листовая мозаика 57, 284
Листовой индекс 274
Литораль (иеритической области) 390
Литоральная зона 369, 378, 382
Литосфера 18, 55, 58-60, 62, 63, 124, 139, 146, 177, 251, 303, 311, 389
Ложе океана 390, 391
Локальные сукцессии 334, 335
Лучистая энергия Солнца 121, 145, 152, 166, 176, 242, 259, 399, 424
Магнетизм 175
Магнитное поле 174—176
Макрозоопланктон 407
Макроконсументы 44
Макропланктон 401, 402, 404, 405
Макросистема 41, 359
Макрофауна 32
Макрофиты (высшие сосудистые растения) 371, 372, 379, 384, 385, 387
Макроэволюцня 348
Макроэкосистема 17, 21, 61
Материковый склон 390, 391
Материковый шельф 390
Материально-энергетическая база 414
Материально-энергетические ресурсы 360
Материально-энергетический обмен 240, 263, 279, 282, 294
Мегатип 310, 311
Мегаэволюция 348
Мега экосистема 61
Межбиогеоценозные связи и отношения 31, 299, 300, 302, 304, 309, 415, 420
Межбиогеоценозный обменный процесс 301
Межгидроценозные связи в океане 415
Мезозоопланктон 407
Мезоклимат (местный климат) 132, 138
Мезопланктон 401, 402, 404
Мезофауна 107, 108, ПО, 327, 336
Мезоэкосистема 17, 21
Мелкомасштабные (прибрежные) круговороты 395, 413
Мероконсорция 213
Мертвый напочвенный покров 90, 91, 103, 107, 113, 125, 154, 156, 230
Местная видовая популяция 70
Метаболизм 87, 95, 144, 147, 149, 153, 194, 201, 244, 259, 269, 272, 279, 281, 287,
294—297, 301, 318, 325, 407
Метаболиты 190, 194, 195, 300, 416
Механизмы авторегуляции 326, 329, 340, 349, 353, 359, 360, 361
Механизмы видовой регуляции 194, 195
445
Миграция организмов 123, 399
Микориза 98, 99, 332
Микоризообразователн 106, 109
Микоризные грибы 87, 99, 109, 206, 349
Микробиосфера 54
Мнкробиотическнй этап эволюции ценотнческих систем 354
Микробиоцеиоз 217, 354, 355
Микробное население 88, 153, 197, 230, 325, 329, 363, 371, 373
Микробные сапрофиты 279
Микробный компонент 38, 41, 42
Микробоценоз 19, 36, 37, 42, 54
Микрогруппировки 227, 235
Микрозоопланктон 4Q7
Микроклимат 132, 138
Микроконсументы 44
Микропланктон 402, 404
Микрорельеф 96, 170
Микросинузия 19
Микросреда 238
Микростации 193
Микроструктура биогеоценоза 238
Микрофауна 107, 108, НО, 327, 336
Микрофитопланктон 407
Микрофлора 111, 112, 114, 136, 139, 155
Микрофлора водная 382
Микроэволюция 71, 163, 348
Микроэкосистема 17, 21
Миксотрофы 21
Мировой океан 118, 119, 124
Мозаичное сложение (растительного, биогеоценотического) покрова 284
Мозаичность сообществ 226
Монофитофагн 349
Морфологическая (пространственная) структура биогеоценоза 225
Морфологическая (физическая) структура биогеоценозов 185, 186, 230, 239, 325,
329, 340, 353, 359
Морфологическая структура биогидроцеиоза 378, 383
Морфометрически олиготрофные озера 369
Морфометрически эвтрофные озера 369
Морфоструктура фитоцеиозов 274
Мусорщики 197
Мутуализм 188, 189
Наземная (воздушная) среда 88, 93, 95, 228, 363—365, 389
Наземная пленка жизни 60, 85
Наземный (воздушный) блок среды биогеоценоза 269
Надплакорные биогеомассивы 315
Нанопланктон 401, 403
Нанорельеф 95, 96, 170
Нанофитопланктон 407
Население гидробионтов 400
Нахлебничество 188
Нейстон 367, 380
Нейтрализм 189
Нейтральные области (в океане) 393, 394, 413
Некрофаги 108, 209
Нектобентос 367
446
Нектон 367, 371, 380, 404, 405, 407, 408
Неогенез 342, 346
Неполночленные консорции (детритные, сапротрофные) 216—218, 264
Неритическая область океана 390, 410, 412—414
Нитраты 56, 246, 401
Нитрофикаторы 49
Новые субстраты 324
Ноогеника 64, 65
Ноосфера 64
Области концентрации жизни 60
Обмен веществ и энергии 294, 299, 384, 387, 415, 417
Обменные процессы в биогеоценозе 52, 240, 241, 244—246, 263, 266, 268, 270,
273, 420
Обменный фонд в круговороте химических элементов 250
Обратная связь 284, 285
Обший запас биомассы 84, 325
Озоновый экран 131, 177, 419
Океаническая область биогидросферы 367, 389
Олиготрофные озера 369
Онтогенез 162, 163, 189
Опад 90, 108, 109, 111, 113, 154, 228, 247, 248, 275
Оранжево-красные лучи 121
Организменная форма жизни 9, 270, 350
Организменный уровень организации живой материи 9, 266, 279
Органическое целое 43
Орогенез 58
Осадочные породы 58
Осадочный тип круговорота химических элементов 250, 255
Основная (эдификаторная) консорция 218
Огпад 90, 108, 109, 112, 154, 223, 247, 275
Отраженная радиация 132, 133
Отрицательная обратная связь 288, 289
Паразитизм 79. 102, 188—191, 356
Паразитоценоз 54
Паразиты 98—101, 206, 210, 271, 321
Парцеллы 185, 227, 234, 235, 237, 238, 301, 302, 324, 349
Парцеллярность 238
Пассаты 390
Пастбищные (полночленные) консорции 216, 221
Пастбищная пищевая цепь 199, 201, 205, 218, 221, 278, 279
Пастбищный поток веществ и энергии 269
Педосфера (почвенный покров Земли) 18, 141, 177
Пелаген 410
Пелагиаль 400, 401, 404, 405, 407, 408
Пелагическая ассоциация 290
Пелагическая область океана 390
Пелагос 367
Пенистый детрит 407
Первичная биологическая продукция 38, 51, 88, 95. 114, 176, 243, 262, 263, 265,
272, 274, 276, 277, 281, 352, 377, 385, 388, 400. 409. 414—416
Первнчнсводные животные 366, 371
Первичные водные массы 393
Первичные консументы 199
Первичный (жизненный) цикл в круговороте азота 254
447
Первичный (жизненный) цикл углеродного обмена 250
Переходная (афотическая) зона 391, 400, 407, 408, 410
Периодичность (ритмика) сезонного развития 320
Пестициды (инсектициды, гербициды н др.) 180, 423
Пищевые сети 44, 185, 186, 197—199, 217
Пищевые (кормовые) цепи 44, 88, 185, 186, 197—206, 217, 223, 243—273, 277
278, 323, 349, 386
Пищевые цепи н сети 205, 264
Плаккатные водосборные биогеомассивы 315
Плакорные водораздельные биогеомассивы 315
Планетарные фронты в океане 391
Планетарный порядок размерности 308, 310
Планктобентос 367
Планктон континентальный и морской 242, 256, 273, 367, 371, 387, 400, 403, 408
Планктонная пленка жизни 60, 398—401, 405, 407, 409, 410, 412, 413
Планктоноядные животные 404
Поверхностная (эвфотическая) зона 391, 400, 406, 407
Погодичные (разногодные) изменения биогеоценозов 319, 321, 322
Подвижно-ценотнческий этап эволюции биогеоценозов 355, 356
Подземная (почвенная) среда 117, 188, 225, 267—269, 320, 357
Подсистема биосферы 63, 64
Подсистемы 43, 47, 63, 85, 188, 221, 283, 284, 286, 287
Подстилающая поверхность 119, 135
Подстилка (в лесу) 90, 106, 111, 112, 148, 149, 151, 154, 228» 230, 275, 278, 296,
331
Полидоминантные биогеоценозы 247
Полифаги 95, 214
Полициклический процесс 246
Полночленные биогеоценозы 266
Полночленные биогидроценозы 398
Полночленные консорции 217, 218, 221, 264
Полупаразитизм 189, 190, 209
Популяционная консорция 212, 216, 219, 221, 222, 264
Популяционно-видовая форма жизни 9, 270, 350
Популяционно-видовой состав биогеоценозов 69, 80, 184, 185, 193, 324, 329, 333,
340, 343, 354, 357, 358
Популяциоиио-видовой состав биогидроценозов 384, 388, 408, 415
Популяционно-видовой состав консорции 212
Популяционно-видовой состав растительного компонента 9, 284, 326
Популяционно-видовой уровень организации живой материи 163, 164, 266, 279,
358
Популяционный состав видов 70, 71
Популяционный состав ценозов 70, 71
Популяция автотрофного растения 206, 211, 260
Популяция вида 70, 71, 206, 207, 216, 217, 285—287, 348
Порядки размерностей (планетарный, региональный, топологический) 308
Поток веществ и энергии 240, 263, 266, 267, 272, 273, 282, 417
Поточные линии 265
Почвенные сгущения жизни 60
Почвенный подслой 118
Правило биомассы 204
Правило величины особей 204
Правило пирамиды чисел 204
Правило пирамиды энергии 204
Предоминанты 83
448
Прибрежные сгущения жизни 60, 398, 413, 414, 417
Прикреплеино-ценотический этап эволюции биогеоценозов 356
Принцип обратной связи 41, 186
Природные воды 63, 385
Протобноценоз 347
Продукция живого вещества 272, 325, 401
Продуценты 44, 76, 88, 199. 209, 210, 212, 353, 377, 379, 404
Производный биогеоценоз 328
Производные консорции 213, 218
Промежуточная (батиопелагиальная) зона 391, 400, 401, 404, 405, 408, 410
Протоатмосфера 117
Протокооперация 188, 189
Протоценоз 345, 419
Профундальная зона, профундаль 369, 371, 378, 381, 382
Проценоз-группировка 325
Прямая радиация 121, 132
Прямая связь 284, 288
Ради а ли 302, 305
Радиальное направление перемещения (объектов) 301
Радиоволновые излучения 176
Рассеянная радиация 121, 132, 133
Растительное сообщество 33, 34, 53, 57, 85—91, 94, 122, 153, 226, 227, 262, 284,
290, 291, 293, 307, 326, 327, 331, 353, 354
Растительный подслой 118
Растительный покров планеты 85, 125, 135, 151, 169, 177, 259. 262. 284, 286, 287,
291, 293, 340, 343, 360
Региональный порядок размерности 308, 312, 317
Регресс, регрессивное развитие 360
Редуценты 211, 353
Режим минерального питания растений 132, 167
Режим увлажнения (влажности) 132, 135, 137, 138, 167, 168, 320
Резервный фонд в круговороте химических элементов 250
Рекультивация нарушенных биогеоценозов 31
Рекультивация нарушенных земель 182
Ризосфера 112, 230
Родовые консорции 212
Самоорганизация 283—285, 288
Саморегуляция, саморегулирование 283—288.
Самум 174
Сапропелит 382
Сапропель 56, 116, 382
Сапротрофные беспозвоночные животные 238
Сапротрофные группы микроорганизмов 113, 268
Сапротрофный бактериальный планктон 407, 416
Сапротрофные неполночленные консорции 216, 221
Сапротрофные ценопопуляций 269, 334
Сапротрофный канал перемещения веществ 266, 269
Сапротрофный компонент биогеоценоза 69, 108, 114, 187, 268, 424
Сапротрофный компонент биогидроценоза 407—412
Сапротрофный микробный компонент 44, 89
Сапротрофный ценокомплекс биогеоценоза 25, 114, 221, 348
Сапротрофы 41, 50, 111, 112, 208, 215. 223, 230, 272, 278, 279, 377, 381, 385, 387,
424
Сапрофаги 49. 51, 101, 105, 106, 108, 191, 209, 218, 222, 259, 279, 377
449
Сапрофитные грибы 238, 323
Сапрофиты 49, 51, 91, 98, 101, 105, 109, 154
Саргассовое сгущение макроводорослей 60, 398, 414
Световая солнечная энергия 260, 398, 415
Световое довольствие 133
Световое излучение 176
Световой режим 132, 135, 167
Сгущения жизни, 60, 68, 398, 399
Сезонные изменения биогеоценозов 320
Секторы биогеомассивов 305, 315
Селектогснетические сукцессий 335
Серия смен биогеоценозов 322, 336
Серия сукцессий (смен) 338
Силикаты 401
Симбиоз 188, 189, 191, 254, 356
Симбионты 98, 109, 206, 209, 210
Симбиотические связи и отношения 189, 190, 206, 324, 338, 349
Симбиотрофные грибы 86
Сингенез 323—325, 328—330, 342, 346
Сингенетические сукцессии 323
Сингенетический процесс 324, 325, 334
Сииойкия 188, 189
Синузиальные консорции 212
Синузни-микрогруппнровки 93, 226, 230
Синузия 109, 320, 321, 324
Синэволюцня 349
Синэкология 26
Система 42, 43, 48, 159, 161, 165, 201, 221, 283, 284, 288, 303
Системные единства 43
Системные качества 43
Системология 62
Системообразующие единицы 219
Смена аспектов 320, 417
Солнечная радиация 119, 121, 132—135, 143, 176, 205, 225, 260, 270, 271, 274,
310 345 415
Сообщество 15—19, 35—48, 74, 75, 80, 83, 145, 172, 189, 195, 202, 206, 213, 291,
326, 327, 338, 352, 358, 368, 374, 378, 394, 396, 417, 420
Сотрапезничество 188
Среда, среда жизни 158, 160, 364, 391, 389
Среда обитания 153, 158—161, 233, 284, 288, 326» 331, 334, 356, 360, 365, 389,
395, 414, 415, 417, 420
Средообразующие взаимоотношения 190
Стадия сингенеза биогеоценозов 323
Стадии сингенеза растительных сообществ 323
Стадии эволюции Мирового океана 418
Стадия миграции биогеоценоза 323
Стадия эцезиса биогеоценоза 323
Стадионали 302
Стация экотопа (биотопа) 71, 193
Стратосинузии 226
Стратосфера 58
Структурно-функциональная организация биогеоценоза (и биогеосферы) 85, 185,
321, 263, 264, 325, 326, 353, 358, 420
Структурно-функциональная организация биогидроцеиозов 378, 388, 389, 399,415
Структурные единицы биогеоценозов 377
Ступенчатое (ярусное) строение биосферы (биогеосферы) 270, 283
450
Субдоминанты 82, 377
Субклимаксы 338
Сублитораль (неритической области) 390
Сублиторальная зона (озер) 369, 378, 380, 381
Субэдификаторы 81, 86
Сукцессии (смены) биогеоценозов 319, 322, 328, 329, 333, 338, 345
Сукцессионные (коренные) изменения биогидроценозов 388, 418
Сульфаты 56, 257
Сульфофикаторы 49
Сульфофикация 257
Суммарная (солнечная) радиация 132, 133
Супралитораль (неритической области) 390
Суточная вертикальная миграция животного планктона 383, 404
Сырьевые каналы коммуникации 266
Тектонические движения земной коры 58, 170, 172, 173
Температурный градиент 169
Теория климакса 337
Теория моноклимакса 338
Теория поликлимакса 338
Тепловая энергия 242, 260
Тепловой режим воды 370
Тепловой режим воздуха 122, 132, 134, 135
Тепловой режим почвы 148, 150—152, 167
Тепловой режим среды 166, 274
Теплокровность 123
Термодинамическое поле 55
Типы биогеоценозов 273, 343, 346
Типы биогидроценозов 386
Типы биокосных систем 265
Типы биологического (биотического) круговорота веществ 249
Типы биотопов бентали 410
Типы (ряды) задернения луговых фитоценозов 333
Типы континентальных водоемов 389
Типы коры выветривания 141
Типы обмена веществ и энергии биогидроценозов 387
Типы озер (по первичной продуктивности) 369
Типы пищевых цепей 199, 200
Типы структуры круговорота 249
Топические связи 191, 209, 211, 216
Топологическая классификация биоценозов 290, 291
Топологический порядок размерности 308, 314
Транзитно-аккумулятивный тип коры выветрявання 141, 142
Трансабиотические взаимоотношения 190
Трансбиотические взаимоотношения 190
Третичные консументы 199
Триотрофы 203
Тропосфера 58, 62, 117, 118, 303
Трофическая структура биогеоценозов 264, 325
Трофическая структура консорции 264
Трофическая структура пищевых цепей 264»
Трофические биокомплексы 44
Трофические группы 44
Трофические звенья пищевых цепей 54, 84, 197, 205, 221, 223, 266, 279, 281, 360
Трофические концентры консорции 84, 264
451
Трофические (пищевые) связи и отношения 21, 187, 191, 192, 209, 211, 216, 217,.
324, 353, 356, 383, 415, 417
Трофические ступени биосферы 264, 271, 286, 288, 289
Трофические уровни (звенья) пищевых цепей (и сетей) 187, 194—198, 201, 202^
204, 205, 218, 221, 252, 263, 264, 271—273, 275, 277—279, 281, 349, 362, 384
Трофоэнергетическая связь 21, 44, 50
Турбулентный обмен 121
Узкие желоба (каньоны) 390
Узловой (выработавшийся) ценоз 340, 345
Ультраабиссаль 391, 413
Ультрафиолетовые лучи (излучение) 121, 131, 166, 176, 177
Уровень биотрофов 265
Уровень сапротрофов 266
Уровни организации живой материи 9
Урочище 160, 308
Фабрические связи 191
Фауна 92
Фауногенез 343, 345, 346
Физико-географические фации 292, 293
Физиономические типы биомов 316
Физиосистема 16
Филогенез 84, 163, 349, 355
Филоценогенез 342, 343, 345, 346
Филоценогенетнческне сукцессии 319
Фильтраторы 398, 412
Фнтальная зона (озер и т. п.) 378
Фитобентос 411, 412, 416
Фитогенные узлы 238
Фитогеосфера 67, 291
Фитоклимат 132, 137, 138, 155, 298, 324
Фнтомасса 80, 87, 89—91, 122, 137, 233, 272, 287, 288, 385, 415
Фитопаразиты 209, 210, 269, 275
Фитопланктон 117, 176, 204, 214, 255, 356, 373—380, 382, 383, 385—387, 400—
402, 404, 406, 408, 412, 415, 416, 418
Фитосреда 80, 284, 327
Фитострома 10
Фитосфера 283. 286, 287
Фитофаги 44, 51, 91, 94—97, 117, 191, 196, 197, 205, 209, 210, 214, 218, 233, 259,
267, 269. 271, 275—278, 283, 285—288, 321, 377, 381, 384, 412
Фитофильные гидробионты 367, 410
Фитофильиые группы беспозвоночных 375
Фитоценоз 18, 20, 26, 33, 34, 36, 37, 40, 53, 75, 80—82, 85, 87, 89—91, 122, 125,
132, 139, 226—229, 283, 284, 286, 292, 325—327, 340, 343, 359
Фитоценология 21, 26, 28, 33, 34
Фитоценотнпы 79, 80, 82, 83, 219, 325, 326
Фитоценотическая классификация 291
Фитоценотнческие жизненные формы 79
Фитоценотический отбор 326, 327
Флорогенез 343, 345, 346
Форические связи 191
Форма системного единства 219, 221, 222
Фосфаты 246, 255—257
Фотосинтетнки 354
Фотосинтетически активная радиация (ФАР) 120, 260, 261
452
Фотосинтетические бногеогоризонты 233; 239
Фототрофность 267
Фототрофы 49, 51, 53, 120, 260, 267, 274, 386
Фототрофные водоросли 105
Фототрофные растительные организмы 49, 274
Фототрофные ценопопуляций 87
Фототрофный компонент 381
Фронтальные водные массы 400
«Фронтыэ в океане 391
Функциональная деятельность живых компонентов биогеоценоза 85, 282, 31К
Функциональная деятельность фитоценоза 90
Функциональная организация биогеоценоза 48, 71, 186, 188, 196—198, 203, 206г
216, 218, 219, 221, 225, 239, 329, 340, 356, 359
Функциональная организация биогидроценоза 377, 378, 381, 384, 387, 407, 412
Функциональная роль атмосферы 119
Функциональная роль почвы 150
Функционально-биоцеиотические группы 87
Функциональные структурные единицы 185, 186, 198, 205, 206, 215, 217, 219, 221г
264, 281
Хемосинтетики 87
Хемотрофные бактерии 105
Хемотрофные растительные организмы 49
Хемотрофы 50, 51, 274
Хищничество 79, 102, 188—191, 193, 195, 196
Холоднокровность 122
Холоцен 20
Хорологическая система 36, 48
Ценогенез (фнлоценогенез) 346
Ценоз 25, 53, 70, 75, 179, 184, 185, 193, 195, 197, 213, 290, 291, 327, 330, 336,
339, 340, 354, 357, 358, 361, 384, 385
Ценокомплекс автотрофных ценопопуляций 87, 91, 187, 196, 235, 240, 264, 265,
269, 275, 281
Ценокомплекс автотрофов 51, 87, 158, 219, 226
Ценокомплекс биотрофных ценопопуляций 91, 187, 196, 235, 240, 264, 265, 269»
281
Ценокомплекс биотрофов 51, 91, 93, 94, 99, 117, 158, 195, 196, 219, 221, 348
Ценокомплекс гетеротрофных микроорганизмов 54
Ценокомплекс сапротрофвых ценопопуляций 196, 235, 240, 264, 265, 269, 281
Ценокомплекс сапротрофов 51, 101, ПО, 158, 187, 219
Ценокомплекс хемотрофных бактерий 54
Ценокомплексы 42, 44, 47, 48, 50, 54, 175, 197, 288, 340, 374, 399
Ценология 25
Цсноморфы 79, 80
Ценопопуляций 71, 72, 79, 81—84, 106, 158, 185, 186, 196, 197, 204, 209, 218, 219и
263, 267, 271
Ценопопуляций автотрофного компонента 194, 218, 267
Ценопопуляций автотрофного ценокомплекса 113, 205, 221, 273
Ценопопуляций автотрофных растений 152, 222, 263, 270, 274
Ценопопуляций биотрофного компонента 218
Ценопопуляций биотрофного ценокомплекса 114, 205, 221, 277
Ценопопуляций биотрофов 87, 263
Ценопопуляций гетеротрофного компонента 194
Ценопопуляций сапротрофного компонента 218
Ценопопуляций еапрот^офиого ценокомидекса. 114, 20б> 221
453
Ценопопуляций сапротрофов 87, 263, 279
Ценотические б ио косные системы 377
Ценотические группировки (живых существ) 356
Ценотические единства 51
Ценотические связи организмов (или компонентов) 21, 323, 349, 354» 356. 361,
424
Ценотические системы (ценоскстемы) 25, 74, 219, 225, 347, 354, 356—359. 361,
382, 424
Ценотические элементы (ценоэлементы) 47
Ценотический состав биогеоценоза 184, 185
Ценотический состав живых компонентов биогеоценоза 69, 84
Ценотический уровень организации жизни 165, 186
Ценоэкосистема 20, 353, 358, 360
Центр (ядро) консорции 211, 212, 215, 219, 221
Цели паразитов 200, 201, 204
Цепи питания 198, 200, 224, 283
Цепи сапрофагов 200, 201
Цепи хищников 200, 201
Цели и «циклы* питания 194, 197
Цель выедания 198, 201, 205
Цепь разложения 198, 200, 201
Циклическая циркуляция биофильных элементов 264
Циклические (периодические) изменения биогеоценозов 319, 321, 329, 366, 388,
389
Циклический характер организации жизни 350, 352
Цикличная структура жизни 241, 350
Цикличность биотического круговорота веществ 417
Цикличность обмена веществ и энергии 384
Циклы биологического круговорота веществ 247
Циклы питания 198
Циклы эволюционных преобразований жизни 348
Циркуляция атмосферы 130, 135, 137, 242
Чистая первичная биологическая продукция 272, 275, 276
Эволюция ценотических систем 349, 358, 361
Эволюционный консерватизм 360, 361
Эволюционный прогресс ценотических систем 358, 359
Эвтрофные озера 369
Эвфотическая зона пелагиали 391, 400, 404, 406, 407
Эдасфера 160
Эдафотоп 18. 37, 47, 50, 51, 116, 139, 141, 165, 235, 240
Эдификаторы 80, 81, 86, 206, 208, 213, 217, 239, 247, 299, 333, 349
Эдификаторная консорция 213, 218
Экзогенез 342
Экзогенные процессы на земной поверхности 58, 166, 173, 174
Экзогенные сукцессии 319, 334, 335, о36
Экзосфера 59
Эккрисотрофы 208
Экобиоморфный состав (живых компонентов) 69, 84, 92, 131, 184, 185
Экобиом орфный состав растений 90
Экобиоморфы 71—76, 78, 80, 82—84, 86, 185, 219, 226, 325, 326, 349, 357
Экология 12. 17, 19, 20, 22, 24, 25, 28, 30, 160, 161, 420, 422
Экология животных 76
Экологическая ниша 44, 71, 75, 95, 178, 187, 192, 196, 211, 326, 333, 339, 349.
Экологические связи 21, 323, 348, 424
454
Экологический ряд 168
Экологическое выравнивание видового состава 323
Эколого-ценотическая дифференциация видового состава 323
Экосистема 9, 15—18, 20. 21, 24, 61, 189, 199, 201, 246’, 248, 262, 263 271 290—
292, 316, 338, 391 ’
Экосистемология 24
Экосфера 296
Экотон 34 53
Экотоп 181 25, 34, 37, 43, 45—47, 50, 51, 79, 165, 239, 293, 323, 324, 327, 420
Экотопический отбор 326
Экскременты 209, 276, 410
Эктопаразиты 54, 93. 97, 100, 216
Электрическое поле 174
Элементарная геохора 159
Элементарная консорция 216
Элементарная функциональная единица биогеоценоза 219
Элювиальный тип коры выветривания 141, 142
Эндогенез (эндоэкогенез) 323, 328—330, 333, 342, 346
Эндогенные планетарные явления 369
Эндогенные сукцессии 328, 330, 334, 336
Эндодинамнческий (автогенный) процесс 332, 334
Эндопаразиты 54, 93, 97, 100, 216
Энергетический канал коммуникации 266
Энергетические потоки 64
Энерготрофические связи и взаимоотношения 329, 424
Энтомоценоз 54
Энтропия 67, 89, 360, 361
Эобионты 354
Эоцен оти чески й этап эволюции цеяотических систем 354
Эпибентос 367
Эпибиос (перифитон) 367
Эпигенема 14, 15
Эпигенология 15
Эпигены 15
Эпиморфа 15, 20
Эпипарцеллы 235, 342
Эпифиты 86, 190, 191, 206, 208—211
Эпифитная мезофауна 382
Эпифитная микрофлора 139, 382
Эпиформация 15
Этапы эволюции Мирового океана 418
Этологические типы хищников 192
Эфемеры 249
Эффузивные породы 170
Ярусы 226, 227, 229, 231
Ярусно-дифференцированный ценотический этап эволюции биогеоценозов 356
Ярусное строение растительного покрова 57, 284
Ярусно-подвижные формы 229
Ярусность фитоценоза 225, 226, 228
Леоятв* Иванович НОМОКОНОВ
ОБЩАЯ
БИОГЕОЦЕНОЛОГИЯ
Технический редактор Н. П. Соловьева
Корректор Н. С. Атланова
Обложка М. А. Белозора
ИБ № 1312
Изд. Ns 51/1950. Сдано в набор 31.10.88. Подписано к печа-
ти 29.12.88. ПК 04176. Формат 60x84/16. Бумага тип. № 2. Гар-
нитура литературная. Печать высокая. Уел. п. л. 26.50. Уч.-
изд. л. 30,0. Тираж 1000. экз. Заказ 151. Цена 4 р. 50 к.
Издательство Ростовского университета. 344006, Ростов-на-Дону,
Пушкинская, 160.
Типография нм. М. И. Калинина Ростовского областного управ-
ления по делам издательств, полиграфии и книжной торговли,
344081, Ростовка-Дону, Советская, 57.