/
















































































































































Автор: Буторин Н.В.
Теги: материальные основы жизни биохимия молекулярная биология биофизика биология экология водохранилища
Год: 1975




Текст
ЙЙ«ЖЛв4
АКАДЕМИЯ НАУК СССР
ИНСТИТУТ БИОЛОГИИ ВНУТРЕННИХ вод
Труды, вып. 27 (30)
ЭКОЛОГИЯ ОРГАНИЗМОВ
ВОДОХРАНИЛИЩ-ОХЛАДИТЕЛЕЙ
А
ж»* m
HbJW EKAI
ИЗДАТЕЛЬСТВО «НАУКА»
ЛЕНИНГРАДСКОЕ ОТДЕЛЕНИЕ
ЛЕНИНГРАД 1975
Academy of Sciences of the USSR
Institute for the biology of inland waters.
ECOLOGY OF THE-ORGANISMS
OF THE HEATED WATERS
IN RESERVOIRS
Transactions, volume 27(30)
Главный редактор - д-р географ, наук Н.В.Буторин
Ответственный редактор - д-р биолог, наук
проф. ф.Д. Мордухай-Болтовской
q 21009-648
а 055(02)-75®5в^74 © Издательство ,Наука", 1975
ИВАН ДМИТРИЕВИЧ ПАПАНИН -
ОРГАНИЗАТОР ИНСТИТУТА БИОЛОГЙИ
ВНУТРЕННИХ ВОД АН СССР
26 ноября 1974 г. исполнилось 80 лет со дня рождения
дважды Героя Советского Союза, доктора географических наук
Ивана Дмитриевича Папанина.
• Популярность имени И.Д.Папанина, его широкая известность
как в нашей стране, так и за рубежом связаны прежде всего
с освоением Северного'морского пути и исследованиями Аркти-
ки. Кульминационным этапом этих исследований было осущест-
вление первой в истории станции «Северный полюс' и героичес-
кий дрейф четырех мужественных советских исследователей -
И.Д.Папанина, Е.К.Федорова, П.П.Ширшова и Э.Т.Кренкеля во
льдах Арктики. Участие в этой.героической эпопее, руковод-
ство ею требовали не только мужества, .но и незаурядных орга-
низаторских способностей. Талант организатора, созидателя
нового И;Д.Папанин проявил не только в ходе работ по освое-
нию Арктики и Северного морского пути,- но и в годы Великой .
Отечественной войны, и при организации экспедиционных работ
И создании исследовательского флота в системе Академии наук
СССР. Эта сторона деятельности И.Д.Папанина широко извест-
на. Менее известна роль И.Д.Папанина в создании и развитии
одного из крупнейших биологических учреждений Академии наук
СССР - Института биологии внутренних вод.
\ В 1932 г. почетный академик, революционер-народиик
Н.А.Морозов передал Академии наук СССР часть своей усадь-
бы «Борок*, предоставленной ему Постановлением Совнаркома
(1923 г.)' в пожизненное пользование. В 1938 г, здесь была
основана Верхневолжская база АН СССР, преобразованная в
1943 г. в биологический стационар, а в 1947 г. - в биологичес-
кую станцию «Борок* им. Н.А.Морозова.
3
До 1062 г. задачей станции было изучение изменений фло-
ры и фауны, л также почвенных процессов под влиянием гидро-
логических и климатических условий, возникших в связи с об-
разованием Рыбинского водохранилища. В 19S2 г. была пере-
смотрена тематика работ станции и принято решение направить
ее дальнейшие исследования иа комплексное изучение Рыбинс-
кого водохранилища в свете задач его рыбохозяйственного ос-
воения. В январе 1952 г. И.Д.Папании назначается уполномо-
ченным Президиума Академии науи и директором биологической
станции «Борок* им. Н.А.Морозова.
В то время биологическая станция «Борок* представляла
собой весьма скромное научное учреждение. Научный коллектив
ее состоял из 12 человек при общем штате 30 единиц. Рабочие
помещения размешались в бывшем помещичьем доме и распола-
гали самым примитивным лабораторным оборудованием. Под
жилые помещения использовались служебные пристройки имения.
Для работ иа водоеме площадью 4550 км2 станция располагала
лишь одним фанерным катером.
Интенсивное гидротехническое строительство, развернув-
шееся в Советском Союзе в послевоенные годы, образование
крупных водохранилищ вызвало необходимость создания биологи-
ческого учреждения со специальной задачей изучения водохрани-
лищ. Биостанция «Борок*, расположенная на берегу одного из
них, была вполне подходящей базой для создания такого учреж-
дения. Однако в реализации этой идеи имелись серьезные за-
труднения. Борок находится в 18 им от ближайшей железнодо-
рожной станции. В течение всей весны, значительной части лета
и осени грунтовые дороги от станции до Борка были практичес-
ки непроходимы для автотранспорта. Зимой использованию его
мешали снежные заносы. Таким образом, связь с железной до-
рогой большую часть года осуществлялась на лошадях. Правда,
в навигационный период в Борок можно было доставлять грузы
по воде.
В таких условиях взять на себя руководство капитальным
строительство^ без которого немыслимо создание любого уч-
реждения, мог человек, обладающий хорошими организаторскими'
способностями, исключительной энергией и настойчивостью. Все
эти качества присуши И.Д.Папанину и характерны для него.
Приняв руководство станцией, Иван Дмитриевич приступил
к ее реорганизации. Уже к концу 1952 г. помещичий дом ста-
рой усадьбы был отремонтирован и переоборудован под разме-
щение лабораторий. На месте овсяного поля, примыкающего к
усадьбе, развернулось строительство жилых домов. Было уста-
новлено 10 немецких и 13 финских стандартных трехкомнатных
домиков. Старая конюшня перестроена под гараж и электростаи-
4
цию. Поп служебные помещения и пол жилье переоборудовались
усадебные постройки. Было начато сооружение водопровода и
канализации.
Понимая, что главной задачей нового учреждения являет-
ся биологическое изучение крупных водохранилищ, И.Д.Папании
с первых дней своей деятельности обратил особое внимание иа
оснащение его экспедиционными судами, способными работать
на етих водоемах. В Первый же год он приобрел два таких
судна, а в начале второго года - еще одно.
Не меиее важным делом было привлечение к работе в уч-
реждении квалифицированных специалистов. Из старого научного
персонала станции после ее реорганизации остался лишь один
кандидат наук и три младших научных сотрудника без ученой
степени. Таким образом, и в этом вопросе пришлось начинать
почти с нуля. И.Д.Папании в первый же год привлек к работе
трех докторов наук, двух кандидатов наук - старших научных
сотрудников и ряд способных молодых специалистов. Для руко-
водства работами по некоторым направлениям были приглашены
видные ученые из московских и ленинградских учреждений и
вузов.
С самых первых дней своей деятельности в Борке И.Д.Па-
панин принял все меры к оснащению лабораторий современным
научным оборудованием.
Вса это позволило уже в 1952 г* развернуть исследова-
тельские работы на Рыбинском, Угличском и Иваньковском во-
дохранилищах на совершенно ином уровне и в таких масштабах,
какие были немыслимы для станции год тому назад.
С' 1952 г. идет непрерывное научное, материально-техни-
ческое и хозяйственное укрепление учреждения. Пополняются и
качественно усиливаются научные кадрьь Организуются новые
лаборатории. Улучшается их оборудование и оснащение. Строят-
ся новые лабораторные, вспомогательные, хозяйственные в жи-
лые помещения, экспериментальная прудовая база.
В настоящее время в составе Института имеются лабора-
тории; гидрологии, продукционных процессов, микробиологии,
биологии низших организмов, биологии и систематики водных
организмов, экологии водных беспозвоночных, ихтиологии, фи-
зиологии и паразитологии пресноводных животных. При Инсти-
туте имеется Куйбышевская станция, находящаяся в г. Тольятти.
Лаборатории Института размешены в каменных корпусах,
оснащены современным оборудованием, контрольно-измеритель-
ными приборами и счетно-вычислительной техникой, включая
ЭВМ ,Минск- 22'. Флот Института насчитывает 10 экспедицион-
ных судов, имеющих от 8 до 20 мест дли персонала экспедиций.
Имеется ряд судов, обслуживающих хозяйственные нужды Инс-
титута.
8
При Институте имеется отличная библиотека, насчитываю-
щая свыше 100 тыс. библиотечных единиц. В специальном зда-
нии расположена мастерская для изготовления и ремонта раз-
личных приборов и инструментов. . .
Жилой поселок в настоящее время, кроме деревянных
домиков, сооруженных в 1952 г. и частично замененных на со-
временные коттеджи, состоит из 8 восьмиквартирных двухэтаж-
ных домов, 5 двадцатичетырехквартирных и 9 тридцатишести-
квартирных каменных домов. Институт имеет прекрасную гости-
ницу на 120 мест для приезжих ученых, аспирантов и студен-
тов, а также общежитие для строительных рабочих.
Отдельный комплекс составляет больничный городок с од-
ноэтажным каменным зданием, вмещающим больницу на 25 коек
и поликлинику, с аптекой и другими постройками. Начато строи-
тельство нового здания больницы на 75 коек.
Построены здания средней школы, детского комбината, от-
деления связи, пожарного депо, универсального магазина, цент-
ральной котельной-, складские -помещения.
Институт и поселок электрифицированы, имеют центральное
отопление, водопровод и канализациях Переведена на природ-
ный газ центральная котельная и идет газификация жилых домов.
Острая проблема бездорожья решена еще в 1958 г. постройкой -
дороги от Борка до ближайшей железнодорожной станции Шести-
хино, на которой выстроен прекрасный вокзал в современном
стиле.
Огромная работа, проделанная- под руководством И.Д.Па-
панина по строительству, материально-техническому оснащению,
подбору и воспитанию кадров, обеспечила создание и нормальное
функционирование большого исследовательского учреждения, ка-
ким является сегодня Институт биологии внутренних вод. Созда-
ние Института биологии внутренних вод АН СССР в необычных
условиях удаленнооти от крупных научных и культурных цент-
ров было бы немыслимо, если бы организация его в свое время
не была поручена Ивану Дмитриевичу Папанину.
И каждый иоследователь, который имел возможность рабо-
тать в лабораториях Института, остается навсегда благодарным
И.Д.Папанину за тот труд, который вложен им в создание уч-
реждения.
Ф. Д. Мордухай-Болтовской
ПРОБЛЕМА ВЛИЯНИЯ ТЕПЛОВЫХ
И АТОМНЫХ ЭЛЕКТРОСТАНЦИЙ
НА ГИДРОБИОЛОГИЧЕСКИЙ РЕЖИМ ВОДОЕМОВ
(ОБЗОР)'
Введение
Проблема влияния электростанций на экологические систе-
мы водоемов, использующихся ими для охлаждения, возникла
в связи с быстрым увеличением числа, и мощности тепловых и
атомных (ядерных) электростанций. Электростанции забирают
из водоемов большие массы воды, которые затем сбрасывают-
ся в водоемы в подогретом состоянии, внося большие количест-
ва дополнительного тепла. Это неизбежно ведет к изменению
экологических условий в водоемах.
Биологу, изучающему влияние подогретых, вод на биологию
водоемов, небеополезно ясно представлять, в чем заключается
работа энергетической станции и в частности откуда берется
добавочное тепло.
Генератором энергии в теплоэлектростанции (ТЭС) (рис.1)
служит турбина, приводимая в действие паром, который полу-
чается в результате подогревания специально подготовленной
(освобожденной от солей) воды. Отработанный, прошедший че-
рез турбину пар охлаждается и конденсируется в воду, которая
опять подогревается и снова превращается в пар, направляемый
в турбину. Этот процесс повторяется многократно, и для того,
чтобы турбина могла непрерывно работать, необходимо обес-
печить быстрое охлаждение пара. Охлаждение осуществляется
при помощи воды, накачиваемой из водоема насосом и пропускае-
мой через систему из нескольких сот или тысяч тонких латун-
ных трубок длиной в несколько метров, образующих так назы-
ваемый конденсатор. Окружающий конденсаторные трубки пар
охлаждается и превращается в воду, отдавая свое тепло воде,
проходящей по трубкам. Нагревшаяся в трубках вода выходит
из ТЭС в водосбросный канал, по которому сбрасывается в тот
же водоем, из которого она была взята, или в специальную
охладительную башню (градирню). Таким образом, пока ТЭС
работает, через нее непрерывно проходит поток воды, но вода
входит с естественной температурой, а выходит с температурой,
повышенной обычно на 8-11 °C (от 5 до 18°). Водоем же непре-
рывно получает с этой нагретой водой массы дополнительного
тепла.
Рис. 1. Схема энергетической электростанции
при открытом прямоточном (А) и закрытом (Б) спо-
собах охлаждения.
1 - водозабор, 2 - насосы, 3 - конденсатор, 4 -
котел, 5 - турбина, 6 - сбросный канал, 7 - охлади-
тельная башня, -р - топливо, t - тепло, е - электро-
энергия.
Существует несколько способов и систем водоснабжения
электростанций охлаждающей водой. При закрытом способе во-
да, выйдя из конденсаторов, поступает в охладительную башию-
традирню, где охлаждается испарением я возвращается обрат-
но в ТЭС, а потеря на испарение компенсируются добавлением
воды со стороны. Закрытый способ не использует для ох-
лаждения водоем и поэтому не представляет для нас интереса.
При открытом способе подогретая вода поступает в водоем,
где и охлаждается, но при этом есть две системы охлаждения.
При «оборотной' системе вода сбрасывается, в специально
сооружаемый для этой пели небольшой водоем-охладитель (во-
8
дохрани лише или пруд). По возможности место сброса отдаляет-
ся от места забора, и на электростанцию поступает уже охла-
дившаяся вода, но под влиянием постоянного притока теплых
вод в малых водоемах-охладителях может в большей или
меньшей мере нагреваться вся масса воды. При .прямоточной'
системе подогретая вода сбрасывается в большой водоем вдали
от места ее забора насосами. В водоеме образуется эона по-
стоянного подогрева около места сброса, постепенно переходя-
щая в область нормального термического режима. Для гидробио-
лога эта система наиболее важна, так как обычно именно
она применяется на мощных тепловых и атомных станциях,
расположенных на крупных водоемах многостороннего хозяйст-
венного использования.
Атомные (ядерные) электростанции (АЭС) построены по
такой же схеме, как и тепловые, но тепло для превращения во-
ды в пар в них получается не от сгорании ископаемого топли-
ва (угля, нефти, газа), а в результате реакции расщепления
атомов радиоактивных веществ. Объем воды, требуемой для
охлаждения тепловых и атомных электростанций, увеличивается
с увеличением мощности станций. Один .блок* тепловой элект-
ростанции, соответствующий мощности 300 МВт, требует для
охлаждения 10-11 м^/с воды. Поэтому крупные современные
ТЭС мощностью 2100-2400 МВт (2.1-2.4 млн кВт) забирают для
охлаждения 70-90 м3/с воды, создавая непрерывно поступающий
в водоем поток теплой воды примерно такого масштаба, как
реки Сена, Москва, Хопер, Южный Буг.
Атомные станции менее экономичны с точки зрения рас-
ходования воды, чем тепловые, так как при той же мощности
требуют в среднем в полтора раза (на 20-60%) больше охлаж-
дающей воды. Сбрасываемая атомными станциями вода подогре-
вается приблизительно так же, как и вода, сбрасываемая ТЭС.
В американских АЭС подогрев колеблется от 6 до 18°С, сос-
тавляя в среднем 10.8°С (Coutant, 1971).
' Так как температура представляет собой один из наибо-
лее мощных экологических факторов, поступление дополнительно-
го .тепла со сбрасываемой подогретой водой может приобрести
весьма важное гидробиологическое значение. Оно нарушает ес-
тественную экологическую обстановку в водоемах, поэтому по
аналогии. со сбросом бытовых и промышленных сточных вод,
изменяющих химизм водоемов, сброс теплых вод, изменяющих
их физические условия, в США часто называют .тепловым
загрязнением* (thermal pollution). Этим термином, однако,
лучше не пользоваться. Во-первых, следует иметь в виду прин-
ципиальное отличие сброса тепла, т.е. энергии, от сброса раз-
личных веществ в бытовых и промышленных отходах. Во-вторых,
8
поступление дополнительного тепла до известных пределов во-
обще не приносит прямого вреда, хотя и может вызвать неко-
торые изменения. Поэтому лучше говорить о сбросе подогретых
вод, или „калефакции' - нагревании (Merriman, 1971), а
может быть даже лучше о влиянии электростанций вообще, ко-
торое, как ниже будет показано, не ограничивается одним по-
догревом водоемов.
Рис. 3. Рост потребле-
ния воды для охлаждения
электростанций в США.
'Рис. 2. Перспективы
роста производства энергии
в США.
1 - станции на иско-
паемом горючем, 2 - ядер-
ные (атомные) станции,
3 - гидростанции (по:
Krenkel a. Parker,
1969).
1 - всего, 2 - от рек
и озер (по: Dysart а.
Krenkel, 1965). (амер,
галлон = 3.78 л).
Выяснение влияния сброса подогретых вод чрезвычайно
важно, так как этот сброс быстро растет в связи со строитель-
ством новых тепловых и атомных электростанций. Известно,
что в настоящее время более 80% электроэнергии в промышлен-
но развитых странах, в частности в СССР, вырабатывается
этими станциями. В СССР мощность всех ТЭС в 1970 г. сос-
тавляла более 120 млн кВт, и они сбрасьвали 225 млн м^ подо-
гретой воды в сутки. Очень быстрый рост производства элект-
роэнергии наблюдается в США, где каждые десять лет общая
мощность станций удваивается, причем увеличивается доля
ядерных (атомных) станций, сбрасывающих больше подогретых
вод. К 1980 г. атомные станции США будут сбрасывать до 40%
10
подогретых вод. К этому времени, по оценкам ряда авторов,
все электростанции США будут использовать для охлаждения
190-200 млрд галлонов воды (т.е. 730-760 млн м3) в день,
что составляет 1/6-1/5 всего годового стока рек этой страны.
А к 2000 г. уже 1/3 тодового стока (или весь меженный сток)
рек США будет проходить через конденсаторы электростанций
(Arnold, 1962; Clark, 1969; Levin е. а., 1972). В даль-
нейшем предусматривается еше больший рост производства
электроэнергии, и в основном за счет атомных станций, как
это хорошо видно из рис. 2 и 3.
В США, расположенных целиком южнее 49° с.ш., естест-
венная температура многих рек в летние месяцы достигает
30°, а иногда и более. Сброс подогретых вод повышает темпе-
; ратуру реки, а если на одной реке строится серия станций на
сравнительно небольшом расстоянии друг от друга, то вниз по
течению реки температура воды становится все выше и выше
(рис. 4). Поэтому в США возникло сильное беспокойство по.
поводу возможных тяжелых последствий сброса подогретых вод,
приведшее, как пишет Бошан (Beauchamp, 1969), даже к
враждебной позиции организаций по охране природы по отноше-
нию к компаниям по производству электроэнергии. 1 Многие аме-
риканские экологи считают, что интенсивное развитие энергети-
ки и особенно атомных электростанций представляет настолько
серьезную угрозу для жизни рек, что некоторые из них могут
стать практически необитаемыми.
В США уже давно начались работы по определению норм
подогрева, т.е, повышения температуры, которое можно допус-
тить, чтобы подогрев ие оказывал вредного влияния на жизнь .
водоема и в частности на рыб. В разных штатах были приняты
различные нормы; в последние годы Технический совещатель-
ный комитет порекомендовал установить допустимое повышение
средней температуры воды для рек не более чем на 2.8°С *
(5°Ф), для озер - на 1.7°С (3°Ф), для эстуарных и прибрежных
районов' летом - на 0.8°С (1.5°Ф), в остальное время - на
2.2°С (4°Ф).
Принятая в нашей стране одна санитарная норма допускает
подогрев не более чем на 3°С, а для зимнего периода - на
5 °C.
Естественно, что изучение проблемы влияния сброса подо-
гретых вод иа водоемы в США началось раньше и развернулось
Так, в связи с проектированием на оз. Кейюга второй
мошной ядерной электростанции в Итаке в 1968 г. был орга-
низован ,Гражданский комитет по спасению озера Кейюга**
(Thermal pollution of Cayuga Lake, 1968).
11
в больших масштабах, чем в других странах. Первые статьи
По отдельным вопросам этой проблемы стали появляться в кон-
це 50-х годов, но широкие исследования начались в 60-х годах.
В настоящее время они ведутся и в ряде других зарубежных
стран, особенно в Англии, Польше, Франции, Швеции.
В нашей стране первым научным учреждением, приступив-
шим к разработке рассматриваемой проблемы, был Институт
Рис. 4. Температура воды в августе в
реке Мононгахелла (США).
Пунктирные вертикали сд^ретствуют
местам поступления подог петых вод из про-
мышленных центров. По оси ординат - темпе-
ратура, по оси абсцисс - расстояние (по:
Clark, 1969).
гидробиологии АН УССР. В 1962 г. Институт начал, а с 1966 г.
развернул пр многосторонней программе исследования водое-
мов-охладителей теплоэлектростанций Украины - небольших
озер и водохранилищ площадью от 4 до 15 км^, имеющих спе-
циальную функцию охладителей.
Исследования влияния подогретых вод на больших водое-
мах многоцелевого назначения были начаты Институтом биоло-
гии внутренних вод АН СССР. В 1967 г. Институт приступил
к работам по многосторонней программе на крупном Иваньков-
ском водохранилище на Волге площадью S27 км2, в районе
мощной (достигшей к 1970 г. 2400 МВт) Конаковской ГРЭС.
В 1970 г. под руководством Института группой Горьковского
отделения .Теплоэлектропроекта* начались исследования на
еше более крупном (плош. 1300 км2) Горьковском водохрани-
лище в районе Костромской ГРЭС, которая к 1978 г. будет
самой мошной в стране (4800 МВт). Исследования на этих
двук водохранилищах, преимущественно по влиянию подогретых
вод на биологию рыб, ряд лет ведет также Институт озерного
И речного рыбного хозяйств^ (ГосНИОРХ).
12
Некоторые другие институты и вузы за последние годы
тоже начали работы по рассматриваемой проблеме (на Днепров-
ском водохранилище, Кучурганском лимане, на водоемах-охла-
дителях в районе Горького, в Литве, на Урале и в некоторых
других местах).
К настоящему времени по вопросу влияния подогретых
вод на экосистемы водоемов проведено уже много исследований,
породивших обширную литературу. Тем не менее эта проблема
в целом далеко еще не может считаться решенной. Многие воп-
росы ее до сих пор еше не ясны и для их разрешения необхо-
димы новые исследования.
По своему существу рассматриваемая проблема очень
сложна. Влияние сообщаемого водоему дополнительного тепла
зависит не только от количества последнего, но и от теплоза-
паса самого водоема, и от абсолютной величины температуры,
получающейся в результате подогрева воды, и от состава на-
селения и отношения различных населяющих водоем организмов
к этой температуре.
Деятельность каждой тепловой и атомной электростанции
неизбежно сопровождается не только подогреванием воды, но
и другими воздействиями на воду и содержащиеся в ней орга-
низмы. Наиболее важно влияние прохождения воды через агре-
гаты станции. Забираемая для охлаждения из водоема вода
вместе с планктоном и рыбами сначала проходит через загра-
дительные решетки, а затем подвергается действию значитель-
ного давления в насосах и высоких Скоростей в трубках кон-
денсатора и на водосбросе, а также в открытой части канала. Ско-
рости в трубках конденсатора составляют обычно t.5-2.0 м/с,
причем трубки относительно узкн (диаметр их обычно 15-30 мм)
и, несмотря на кратковременность пребывания воды в пределах
электростанции (всего 4-5 мин.), организмы, находящиеся в
забираемой воде, могут подвергаться механическому травмиро-
ванию и одновременно тепловому шоку от быстрого повышения
температуры. Травмирование может быть еше значительнее в
конце водосбросного канала, по которому вышедшая из конден-
саторов подогретая вода сбрасывается в водоем, В конце этого
канала обычно находится водослив с порогом, где, особенно
при понижении уровня водоема, образуется водопад. Вода рас-
пределяется по порогу тонким слоем и при этом обычно под-
вергается сильной аэрации.
Кроме того, вода перед поступлением в конденсатор во
многих станциях подвергается хлорированию, с целью предотвра-
щения бактериального и грибкового обрастания внутренней по-
верхности трубок.
13
Наконец, одновременно с подогретой водой в водоем посту-
пают в каком-то количестве сточные воды с промышленными,
в частности нефтяными, загрязнениями от электростанции, из
цеха химической водоочистки и сточные воды с бытовыми за-
грязнениями от примыкающего к ней поселка.
Для выяснения влияния главного фактора - Подогрева -
очень важно оценить влияние этих сопутствующих факторов.,
которые могут исказить или замаскировать влияние поступаю-
щего в водоем тепла.
Влияние энергетических станций
на гидролог о-г идрохимический
режим водоемов
Влияние сброса подогретых вод теплоэлектростанций на
гидрологический, особенно термический, режим водоемов под-
робно рассмотрено в работе Н.В.Буторина и Т.Н.Курдиной
(наст, сб.) на примере Конаковской ГРЭС и Иваньковского во-
дохранилища. Поэтому здесь я напомню только основные осо-
бенности гидрологии и гидрохимии принимающих эти сбросы
водоемов, важные для их биологии.
Так как естественные температуры водоемов в течение
года и в разных климатических условиях сильно колеблются,
абсолютная величина температуры сбрасываемой - воды тоже
сильно колеблется. В умеренном поясе, где естественная тем-
пература воды при подледном режиме составляет 0-4°С, в по-
догретых сбросах зимой температура обычно ,не выше 10-15°,
летом же, при естественной температуре 20° и более, в подо-
гретой воде она может превосходить 30°, достигая иногда при
сильном естественном подогреве 35-40°С и- даже более.
Поток подогретой воды уже начинает остывать, проходя
через открытый водосбросный отводящий канал, но температура
воды за это время (около .получаса) понижается не более, чем
на 0.5-1.0°, Основная теплоотдача происходит в водоеме, где
подогретая вода смешивается с неподогретыми. водами. В ма-
лых водоемах-охладителях вся масса воды в большей или мень-
шей степени подогревается. В крупных водоемах создается зона
подогрева, конфигурация, плопвдь и вертикальная структура
которой различны и зависят от ряда причин. В реках зона по-
догрева вытягивается в виде длинного языка или полосы, рас-
пространяющейся вниз по течению и способной захватить всю
14
ширину реки. В озерах она может иметь различные, часто не-
правильные очертания. В водохранилищах со слабой проточно-
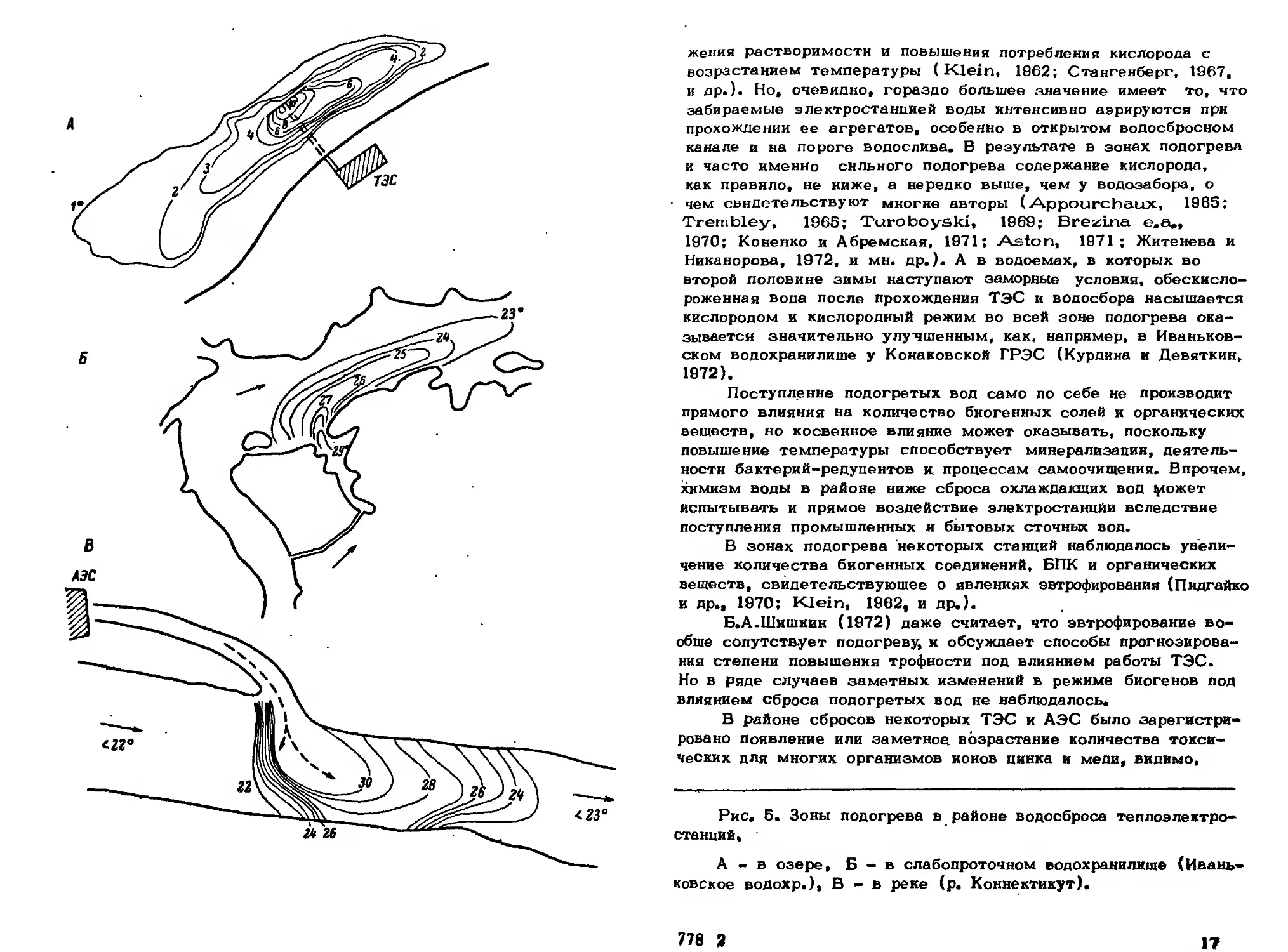
стью образуются промежуточные картины (рис. 5).
Под влиянием непрерывного поступления подогретых вод
в водоеме создается постоянное течение, скорость которого
в районе их сброса может быть очень значительной, но быстро
убывает по мере удаления от него.
Подогретые воды в летнее вреья обычно растекаются в
поверхностном слое, в результате чего создается вертикальная
термическая стратификация, но в районе их поступления захва-
тывают всю толщу воды. Осенью и весной с усилением тепло-
отдачи в более охлажденную атмосферу и ветрового перемеши-
вания стратификапия ослабевает й подогрев распространяется
глубже. В зимние месяцы в водоеме образуется более или ме-
нее обширная постоянная полынья, при обратной термической
стратификации: более высокие температуры располагаются не
у поверхности (где идет интенсивная теплоотдача), а на неко-
торой глубине, и наиболее высокие температуры (3-5°) могут
быть в придонных слоях, в связи с чем повышена и темпера-
тура грунта.
В водоемах или их частях, в которые сбрасываются по-
догретые воды, можно различать зоны разной степени подогре-
ва. За местом сброса начинается зона постоянного сильного
подогрева; при этом ее участок, расположенный непосредствен-
но у места поступления подогретых вод, отличается наиболее
сильной проточностью, перемешиванием всей толши воды и пол-
ным размьвом донных отложений. За сильно подогреваемой рас-
полагается зона более слабого или умеренного подогрева, за
.пределами которой начинается неподогреваемая область. Эти
зоны в течение всего года отличаются по температуре, как
хорошо видно, например, в р. Трент в районе ТЭС ХантерстоН
(рис. 6). При исследованиях водоемов-охладителей ТЭС Украи-
ны к зоне сильного подогрева относили воды, температура ко-
торых превышает естественную летом на 6°, к зоне умеренно-
го подогрева - воды, подогреваемые летом на 4-6°, а кроме
того выделяли эону минимального подогрева, переходную к
области естественного температурного режима, в которой тем-
пература повышена в пределах О.5-З.О°С (Пидгайко и др.,
1970). В крупных водоемах распределение подогретых вод
очень мобильно и изменчиво, находясь под влиянием силы и
направления ветра (Буторин и Курдина, наст. c6j.
Кислородный режим в зонах подогрева в большинстве
случаев оказывается благоприятным. Некоторые авторы при
прогнозировании условий эон подогрева предполагали ухудшение
в них кислородного режима, исходя из известных фактов поив—
15
же ни я растворимости и повышения потребления кислорода с
возрастанием температуры (Klein, 1962; Стангенберг, 1967,
и др.). Но, очевидно, гораздо большее значение имеет то, что
забираемые электростанцией воды интенсивно аэрируются прн
прохождении ее агрегатов, особенно в открытом водосбросном
канале и на пороге водослива. В результате в эонах подогрева
и часто именно сильного подогрева содержание кислорода,
как правило, не ниже, а нередко выше, чем у водозабора, о
чем свидетельствуют многие авторы (Appourchaux, 1965;
Trembley, 1965; Turoboyski, 1969; Brezina е.а»,
1970; Коненко и Абремская, 1971; Aston, 1971; Житенева и
Никанорова, 1972, и мн. др.). А в водоемах, в которых во
второй половине зимы наступают заморные условия, обескисло-
роженная вода после прохождения ТЭС и водосбора насыщается
кислородом и кислородный режим во всей зоне подогрева ока-
зывается значительно улучшенным, как, например, в Иваньков-
ском водохранилище у Конаковской ГРЭС (Курдина и Девяткин,
1972).
Поступление подогретых вод само по себе не производит
прямого влияния на количество биогенных солей и органических
веществ, но косвенное влияние может оказывать, поскольку
повышение температуры способствует минерализации, деятель-
ности бактерий-редуцентов и, процессам самоочищения. Впрочем,
химизм воды в районе ниже сброса охлаждакщих вод уожет
испытывать и прямое воздействие электростанции вследствие
поступления промышленных и бытовых сточньк вод.
В зонах подогрева некоторых станций наблюдалось увели-
чение количества биогенных соединений, БПК и органических
веществ, свидетельствующее о явлениях эвтрофирования (Пидгайко
и др., 1970; Klein, 1962, и др.).
Б.А.Шишкин (1972) даже считает, что эвтрофирование во-
обще сопутствует подогреву, и обсуждает способы прогнозирова-
ния степени повышения трофности под влиянием работы ТЭС.
Но в ряде случаев заметных изменений в режиме биогенов под
влиянием сброса подогретых вод не наблюдалось.
В районе сбросов некоторых ТЭС и АЭС было зарегистри-
ровано появление или заметное возрастание количества токси-
ческих для многих организмов ионов цинка и меди, видимо,
Рис. 5. Зоны подогрева в районе водосброса теплоэлектро-
станций,
А - в озере, Б - в слабопроточном водохранилище (Ивань-
ковское водохр.), В-в реке (р. Коннектикут).
778 2
1?
Рис. в. Головой ход температуры воды р.Трент
в районе ТЭС Хантерстон.
1 - в неподогреваемой зоне выше ТЭС, 2 - ниже
водосброса ТЭС (по: Langford, 1071).
поступавших из агрегатов станций ( Grimes, 1971; Davis,
1060; Rothwell, 1071; Thermal effects..., 1071).
Применяющееся иа некоторых станциях хлорирование (про
тив обрастания трубок) тоже может оказывать вредное влияние
на организмы.
Содержание нефтепродуктов повышается в районе некотор!
ГРЭС за счет их потерь в пуско-наладочный период, но и в
дальнейшем может оказаться довольно значительным. В районе
Конаковской и Костромской ГРЭС наблюдалась „замазученност:
донных отложений - накопление тяжелых фракций, распростра-
няющееся на некоторое расстояние вниз по течению. (Скальска
наст. сб.).
. В обшем изменении в химизме воды под влиянием сбро-
сов электростанций незначительны и при достаточно хорошей
очистке не оказывают существенного влияния на жизнь водоем
18
Влияние энергетических станций
на биологический режим
водоемов
Влияние собственно подогрева, повышения температуры
воды на органический мир водоема многообразно и сложно.
Это важнейший фактор, с которым по всесторонности и глуби-
не воздействия на организм не может сравниться никакой дру-
гой фактор среды.
По характеру своего воздействия тепло резко отличается
от промышленного загрязнения. В то время как с увеличением
концентрации фенола или другого ядовитого вещества усилива-
ется его отрицательное, вредное, влияние на организмы, увели-
чение количества тепла, выражаемое в повышении температуры,
в общем оказывает положительное воздействие, стимулируя
жизненные процессы. Достаточно напомнить известное правило
Ваит-Гоффа, выражаемое уравнением mQ«uti где ut “
скорость химических реакций при температуре t, a QTO обыч-
но составляет 2-3. Оно в общем хорошо применимо для биоло-
гических процессов, ускоряющихся в 2-3 раза при повышении
температуры на 10°.
Для процессов развития Белехрадеком
1935) было предложено другое уравнение: у
( Belehradek,
шГЙлЬ • где
в -
время (длительность) развития при температуре Ъ, а. - темпе-
ратура, при которой развитие прекращается, а - коэффициент,
зависящий от применяемых единиц. На основании этого уравне-
ния Мак Дарен (McLaren, 1963) предложил формулу для ско-
рости развития D=a(T+a)\ из которой следует, что скорость
развития организмов, вообще ’есть степенная функция темпера-
туры; показатель Ъ в большинстве случаев колеблется между
1.0 и 3.0. Если показатель равен 1, а развитие .прекращается
при 0°, что во многих случаях имеет место, формула упроща-
ется до I) = аТ. При графическом изображении зависимости ско-
рости процесса от температуры получается характерная вогну-
тая кривая, близкая к так называемой „нормальной кривой*
Крога (рис. 7, а).
Однако повышение температуры производит стимулирукщее
действие на пойкилотермные организмы до определенных гра-
ниц, по достижении которых начинает оказывать неблагоприят—
-ное влияние. Влияние подогрева меняет знак и вскоре, по до-
стижении верхнего порога, оказывается гибельным Для орга-
низма. Пороговые, или летальные, температуры неодинаковы
для разных видов и у теплолюбивых и эвритермных организмов
19
значительно выше, чем у холополюбивых. Реакция организма
иа изменение температуры может быть выражена в випе кри-
вой на графике, у которого по оси абсцисс откладываются тем-
пературы, а по оси ординат какие-нибудь биологические пока-
затели: распределение вида в градиентных условиях (в приро-
де это может быть встречаемостью), рост, интенсивность ды-
хания или питания и т.п. (рис. 7, б). Местоположение моды
кривой определяет степень хододо- или теплолюбивости вида.
Чем выше вершина и уже основание кривой, -тем более стено-
термичен вид. Степень эвритермичности можно оценить по отно-
шению площади между кривой и абсциссой к максимальной орди-
6
Рис. 7. Графики, иллюстрирующие отношение пойкило-
термного вида к температуре.
а - „нормальная кривая* Крога; б - схема реакции
ьойкилотермного вида на изменение температуры, v - ско-
рость биологического процесса, Ъ - биологические пока-
затели, • нижняя. летальная, tmax- верхняя леталь-
ная, - оптимальная температура (по: Ивлев и Лейэе-
рович, 1960).
нате (моде) (Ивлев'и Лейэерович, 1960). Интересно, что в
большинстве случаев эти кривые асимметричны: правая ветвь
кривой, приходящаяся на температуры выше оптимальных, явно
и иногда значительно короче левой, так что оптимальная тем-
пература приближена к максимальной пороговой.
В пресных водоемах умеренного пояса холодолюбивых ви-
дов организмов вообще мало. К иим относятся из рыб преиму-
щественно лососевые, из беспозвоночных - некоторые планктон-
ные формы и ледниковые реликты Глубоких северных озер,
реофильные формы (преимущественно личинки насекомых) холод-
ных горных ручьев и речек, также некоторые коловратки, копе-
поды и другие формы весенних временных водоемов. Преобладаю-
20
шее большинство видов имеет довольно высокий температур-
ный оптимум, причем он оказывается часто выше максималь-
ных летних температур в водоеме. Вследствие этого небольшое
повышение температуры воды должно способствовать жизнедея-
тельности большинства пресноводных организмов в условиях
умеренных широт. Однако в субтропических и тропических эонах
где большинство организмов живет при температуре, близкой
к оптимуму, уже незначительное повышение ее оказывается
опасным, приближаясь к пороговой.
Ранее высказывались предположения, что подогреваемые
водоемы и зоны, в которых температуры' в течение круглого
года не опускаются ниже 4-5° (или даже более высокого уров-
ня) и которые по лимнологической классификации должны счи-
таться субтропическими, должны были бы иметь и соответст-
вующий субтропический растительный и животный мир. Можно
было думать, что в подогреваемых эонах в средней и северной
полосе СССР разовьются виды, характерные для озер и прудов
стран Африки или Присредиэемноморья. Так, например, среди
кладоцер можно было бы ожидать появления „циркумтропических
Pseudosida bidentata, Diaphanosoma sarsi, Cerio-
daphnia cornuta и др. Однако скоро оказалось, что подо-
грев.аемые водоемы и зоны средних широт, даже сушествукшие
уже десятки лет, вообще не населяются термобионтными вида-
ми и представителями субтропической флоры и фауны. Известно
лишь несколько теплолюбивых видов, найденных в некоторых
эонах подогрева: олигохета Branchiura sowerbii (в Англии,
Швеции, США) (Mann, 1965; Wiederholm., 1971); высшие
растения Eichhornia (в США) и Vallisnieria spira-
lis (в Горьковской области) (Шахматова и др., 1971); некото-
рые виды диатомовых водорослей (в охладителях ТЭС Украины)
(Виноградская, 19716). В эстуариях Англии в эонах подогрева
встречаются теплолюбивые мшанка Bugula neritina, усо-
ногое Balanus amphitrite и краб Brachynotus sex-
dentatus, характерные для теплых и субтропических
морей (Naylor, 1965). Но основную массу флоры и
фауны в условиях даже сильного подогрева составляют
эвритермные или относительно более теплолюбивые (но не
термобионтные) виды из наличного набора видов данного водое-
ма или бассейна. Поэтому в зонах подогрева в зимнее время
наблюдается увеличенное число видов (приращение за счет
форм, которые в естественных водах исчезают при близких к
нулю температурах), но в летнее время, особенно при сильном
прогреве, число видов обычно уменьшается (за счет выпадения
менее эвритермных). Температуры между 25 и 35°С для очень
многих, даже большинства "видов Пресноводных организмов
21
оказываются близкими к пороговым. Так, Лангфорд ( Langford,
1971а), исследовавший реки Англии в районе ТЭС и АЭС, сре-
ди нескольких сотен видов донных беспозвоночных обнаружил
32 вида, встречавшихся при 30°С, выделив из них, как наиболее
выносливых к повышению температуры, только пять: Limno-
drilus’ hoffmeisteri, Asellus aquaticus, Ischnura ele-
gans, Limnaea pereger, Hydrobia jenkinsi. Среди
Рис. 8. Полигон
теплоустойчивости пой-
килотер,много вида ор-
ганизма.
А - эона терми-
ческой выносливости,
Бв- верхняя, Бн- ниж-
няя эона термического
сопротивления. 1 -
граница 10% смертнос-
ти, 2 - граница 50%
смертности, 3 - край-
няя начально-леталь-
ная температура, 4 -
температура „мгновен-
ной* смерти. По оси
ординат - начально-ле-
тальная температура,
по оси абсцисс - тем-
пература акклимации
(по: Coutant, 1970 ).
*С
Рис, 9. Полигон темпера-
турной устойчивости для хо-
лодолюбивого вида лососевых
(нерка) и эвритермичного ви-
да карповых рыб (карась).
А - полигон нерки: 1 -
зона размножения, 2 - эона
роста, 3 - зона переживания; •
Б - полигон карася, значения
осей те же, что и на рис. 8
(из Проссера и Брауна, 1967).
22
растений, особенно водорослей и тем более бактерий, выносли-
вых к высоким температурам видов больше, но и для них так
называемые «заколдованные тридцать' (Пайош, 1970) представ-
ляют собою в обшем порог, выше которого происходит уменьше-
ние видового разнообразия и смена доминирующих групп.
Рис. 10. Влияние температуры и времени ее
воздействия на выживаемость (50%) пойкилотерм-
ногб вида.
А - эона термической выносливости (неоп-
ределенно долгого выживания), Б - зона сопро-
тивления (выживания в течение некоторого време-
ни); 1 - крайняя начально-летальная температура,
2 - температура акклимапии (от 5 до 26.6°), По
оси ординат - температура шока, по оси абсписс'-
время выживания,по логарифмической шкале (по:
Coutant, 1071).
Подводя итоги исследованиям водоемов-охладителей юга
УССР, М.Л.Пидгайко с соавторами (1970) приходят к выводу,
что температуры 28-30° (в поверхностных слоях воды) представ-
ляют собою тот рубеж, по переходе которого начинаются отри-
цательные явления, т.е. ухудшение качества воды и понижение
продуктивности.
Очень большое значение для реакций организмов на повы-
шение температуры имеет их способность к температур-
23
ной акклимации. Это явление, довольно хорошо изученное у
многих рыб, может быть выражено в виде так называемых
„полигонов теплоустойчивости", примеры которых показаны
на рис. 8 и 9. Полигон строится на основании опытов по точкам
(температурам), при которых начинается гибель 50% животных
У эвритермичных организмов полигоны занимают большую пло-
щадь и более вытянуты: как видно из рис. 9, акклнмированный
(т.е. выдержанный сутки или более) при 5° лосось выносит
до 23.5°, а эвритермичный карась - до 28°; акклимированные
при 20° они выносят соответственно 25° и 35°; у лосося пре-
дел 25°, а карась при акклимании к более высоким температу-
рам может вынести повышение температуры до 42° (Brett,
1960; Leynaud, 1967; Проссер и Браун, 1967).
За пределами полигона гибель животных происходит тем
скорее, чем выше температура.
Рис. 11» Выносливость .1
к температуре на разных ста-
диях жизненного цикла пойкило-
термного вида.
1 - максимальная темпера-
тура, 2 - диапазон колебаний
температуры, 3 - скорость из-
менения температуры, 4 - час-
тота максимальной температуры
(по: Jensen, 1969).
Из рис. 10 видно, например,
что акклимированный при 5°
организм погибает через 10
минут при 25° и немедленно -
при 25.8°, т.е. еше до дости-
жения начально-летальной
температуры (26.6 ); аккли-
мированный же при 15° вооб-
ще не гибнет при 25°, поги-
бает через 3 часа при 26.6°
и немедленно - лишь при 29 о.
Большое значение имеет
также скорость изменения
температуры. Как было Пока-
зано опытами с различными
рыбами (Brett, 1960), они
хуже переносят более быстрое
изменение температуры, чем
медленное, причем особенно
ее понижение. Летальная для
рыбы Lepomis macrochi-
rus скорость понижения тем-
пературы в 20 раз меньше,
чем летальная скорость ее по-
вышения. „Холодовой шок*
Под акклимацией понимают приспособление к одному
фактору среды, акклиматизация же есть приспособление, „при-
живление" ко Всему комплексу условий среды. Впрочем, мно-
гие авторы пользуются в обоих случаях только вторым терми-
ном.
24
при быстром понижении температуры (например, при внезапной
остановке электростанции и прекращении сбросов) может быть
опаснее для рыб, чем „тепловой шок* (Speakman а, Кгеп-
kel, 1972; Coutant, 1972а, 1973).
Установление диапазона температур, при которых возмож-
но существование вида, затрудняется еще тем, что теплоустой-
чивость и способность к акклимапии неодинаковы на разных
стадиях развития. Во многих случаях они наименьшие в период
размножения. Йенсен (Jensen, 1969) попытался изобразить
это на схеме, которая приводится на рис. 11.
У водных беспозвоночных и растений явления акклимации
изучены гораздо хуже, чем у рыб, но также имеют место. Так,
оии были установлены у различных высших ракообразных (Куд-
рявцева и Горюнова, 1971; Sprague, 1963). Явления терми-
ческой акклимапии, конечно, очень осложняют понимание и про-
гнозирование пропессов, происходящих при подогреве воды.
Общей для всех биоценозов и групп организмов чертой
является сдвиг фенологических фаз на более раннее время и
удлинение вегетационного периода в эонах подогрева.- Естест-
венно, что температуры порядка 10-15°, при которых в уме-
ренном поясе происходит созревание, размножение, метаморфоз
многих организмов, наступают в подогретых водах значительно
раньше, вскоре после таяиия льдов, когда при естественном
температурном режиме прогрев только начинается и достигает
всего нескольких градусов выше нуля. Вследствие этого сезон-
ная динамика численности и биомассы изменяется и характерный
для многих организмов весенний и летний подъем (пик) насту-*
пает раньше. С другой стороны, осенью вегетация задерживает-
ся и прекращение размножении, метаморфоза и роста как след-
ствие похолодания наступает гораздо поэие. Схематизируя,
можно сказать, что к северу от Москвы в эонах подогрева
весна начинается вместо мая в апреле, а глубокая осень -
вместо октября в ноябре, приблизительно как на юге Украины
и иа Северном Кавказе, находящихся примерно на 10° южнее.
Существуют серьезные отличия в воздействиях электро-
станций на планктонное и донное население. Как указывалось
выше, планктон и рыбы, вовлекаемые вместе с водой в элект-
ростанцию, подвергаются при ее прохождении энергичному ме-
ханическому и термическому воздействию, которое не прихо-
дится испытывать бентосу. Прежде всего много рыб травми-
руется и погибает, ударяясь о сороудерживающие решетки или
под воздействием всасывающей силы насоса, прижимающей их
к решеткам. Молодь рыб и планктонные организмы, прошедшие
через решетки в конденсаторные трубки, подвергаются еще
большим механическим травмам в конце водосбросного канала.
25
если он заканчивается высоким порогом, на котором образует-
ся водопад (Ривьер, наст. сб.). Кроме того, попадая в конден-
саторные трубки, они испытывают тепловой шок от быстрого
нагревания в течение одной-двух минут. От ударов и теплового
шока может происходить гибель части организмов, но, как ука-
зывают Темплтон и Кутан (Templeton a. Coutant, 1971),
не меньшее, а иногда и большее значение имеют „сублетальные
эффекты* - потеря равновесия (дезориентировка движений,
иммобилизация), за которой следует повышение гибели от хищ-
ников. Повышение доступности молоди некоторых рыб хищникам
было доказано экспериментально. Результаты одного из опы-
тов с молодью радужной форели показаны на рис. 12, из кото-
рой видно, как доступность молоди хищникам возрастает с
увеличением длительности термического воздействия (Temple-
ton a. Coutant, 1971; Coutant, 1971, 1972с).
Рис. 12. Влияние повыше-
ния температуры и длительнос-
ти ее воздействия на гибель
от хищников, потерю равнове-
сия и смертность молоди ра-
дужной форепи ('акклимирован-
иой при 15°С).
1 - гибель от хищников,
2 - потеря равновесия, 3 -
смерть; по оси ординат -
температура опыта, по оси
абсцисс - время (по; Cou-
tant, 1971).
В изменении доступности
хищникам видно большое зна-
чение поведения животных в их
реакциях на температуру.
Способность к быстрым пове-
денческим реакциям в ответ
на термические воздействия
резко отличает животных от ’
растений, реагирующих медлен-
ными изменениями процессов
метаболизма, роста и т.п.
Рыбы могут приходить к ис-
точнику теплых вод и уходить
из эоны сильного подогрева,
избегать температурных барье-
ров во время миграции; планк-
тонные рачки - опускаться в
нижние, более холодные слои
воды. Таким образом, многие
подвижные животные могут
избегать неблагоприятных ус-
ловий, не акклимируясь к ним
(Мордухай-Болтовской, 1971;
Ривьер, 1972; Jensen, 1969;
Warren, 1971).
Все это показывает, что .
повышение температуры далеко
не всегда оказывает прямое
действие на организмы, но до
многих случаях действует кос-
23
веяно, через другие факторы. С другой стороны, одно-
временное воздействие других факторов может подавить
или замаскировать влияние температуры. Так, хорошо
известно, что токсическое действие многих загрязнителей
усиливается с' повышением температуры (Klein, 1962;
Hargis a. Warinner, 1971; Rothwell, 1971: Warren,
1971, и др.). В результате и незначительный подогрев мо-
жет вызвать летальный исход. Отсутствие некоторых пе-
лофильных форм непосредственно ниже водосброса объяс-
няется не сильным подогревом, а размывом или изме-
нением свойств отложений. Понижение фотосинтеза фито-
планктона в эоне сильного подогрева может быть след-
ствием и хлорирования, и механического травмирования,
и сильного течения. Иначе говоря, имеет место син-
ергизм, многие примеры которого будут приведены ниже.
Это явление во многих случаях чрезвычайно затрудняет
выяснение действия главного фактора - повышения тем-
пературы - и заставляет при сравнительных исследова-
ниях в водоеме и при проведении ' экспериментов тща-
тельно следить за однородностью остальных факторов или
по крайней мере учитывать степень их влияния.
Перехожу теперь к . краткому обзору накопившихся
к настоящему времени данных о влиянии работы терми-
ческих станций на отдельные группы организмов и био-
ценозы.
Планктон
Бактериальная флора в толще воды - бактериопланк-
тон, -видимо, не- страдая от механических воздействий
при прохождении электростанции, стимулируется подогревом.
Общая численность бактерий обычно заметно возрастает
с повышением температуры, что замечалось как в малых
охладителях (Ленчина, 1971а), так и в эонах подогрева
крупных водоемов (Вайнштейн, 1971; Кривенцова, 1972;
Спиглаэов, 1972; Рох a, Moyer, 1973). В районе Кост-
ромской ГРЭС, в Горысрвском водохранилище численность
бактерий максимальна при наиболее высоких температурах,
достигавших там 29-30°. В малых водоемах-охладителях
Украины гетеротрофные микроорганизмы находили темпера-
турный оптимум между 28 и 35° (Пидгайко и др.,
1970).
27
Известно, что бактерии вообще наиболее выносливы к вы-
соким температурам и некоторые группы их могут обитать в
горячих источниках при +73°С ( Kempner, 1963)Л
Иногда, однако, в эоне подогрева численность бактерио-
планктона не увеличивалась или даже понижалась; в таких
случаях, возможно, она ограничивалась недостатком органичес-
ких веществ .в воде (Романова, 1972; Спиглаэов, 1972; Flor-
czyk u, а,, 1972; Merriman, 1973). Бактерии играют очень'
важную роль в процессе самоочищения воды. Самоочищение во-
обще ускоряется с повышением температуры, но есть данные,
что его скорость максимальна при температуре около 30° и
при более высоких температурах,
возможно, наступает нарушение
биохимической активности неко-
торых групп микробов и замед-
ление минерализации, накопление
органики (Dysart a. Krenkel,
1965; Hawkes, 1971; Horvath
Рис. 13. Последователь-
ность развития разных групп
водорослей при повышении
температурь^
1 - диатомовые, 2 -
зеленые, 3 - синезеленые.
По оси ординат - число ви-
дов, по осн абсцисс - темпе-
ратура (по: Cairns, 1956).
а. Brent, 1972).
В общем микробиологичес-
кий режим эон подогрева изучен
недостаточно. Можно предполагать
усиление бактериальной деструк-
ции и размножение бактерий в
связи с гибелью части планкто-
на непосредственно ниже водо-
сброса.
Фитопланктон исследован
более подробно. В связи с удлине-
нием времени вегетации всех
групп водорослей число одновре-
менно или круглогодично вегети-
рующих видов увеличивается.
Видовой состав изменяется мало,
но с повышением температуры обычно уменьшается роль в фи-
топланктоне диатомовых и возрастает роль зеленых (протокок-
ковых) и пирофитовых, а также сйнезеленых. Последние оказы-
ваются наиболее выносливыми к высоким температурам. После-
довательность групп в общем соответствует схеме Кэрнса
(рис. 13), хотя едва ли число видов синезеленых максимально
По мнению этого автора, указанная температура - мак-
симальная, при которой возможна активная жизнедеятельность
организмов вообще и все указания на более высокие темпера-
туры относятся к переживанию без метаболизма.
28
при 40°. По данным некоторых авторов ( Patrick, 1969), в
культурах синезеленые заменяют диатомовых при 34-38°.
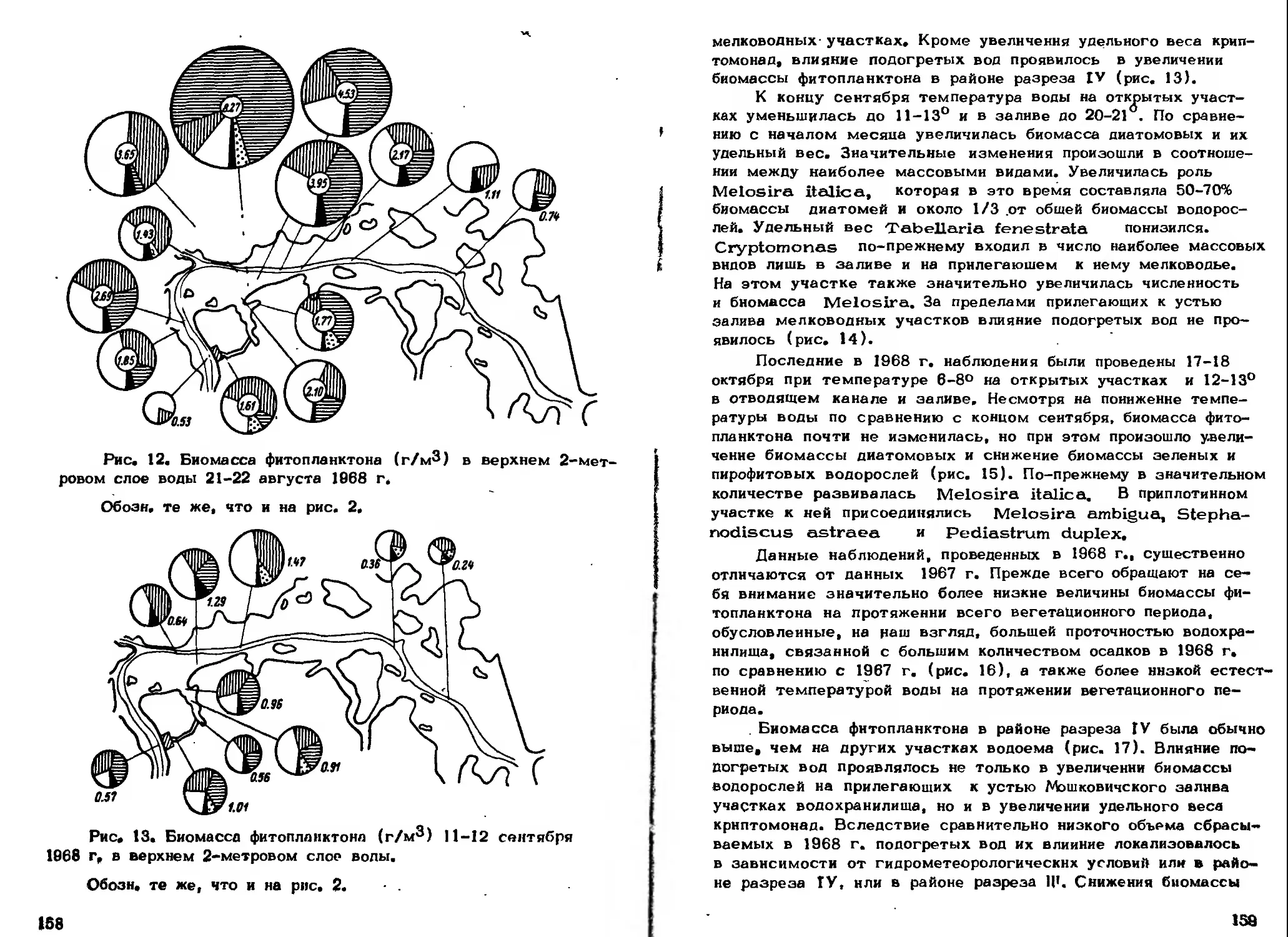
Прохождение фитопланктона через агрегаты ТЭС вообще
мало сказывается на нем. Разрушения клеток водорослей, по
всей видимости, не происходит. По некоторым наблюдениям
фотосинтез фитопланктона ослабевает при прохождении ТЭС в
летние месяцы (Пырина и Наумова, 1973; Рох a, Moyer,
1973). Случаи очень сильного (на 90%) подавления фотосинте-
за после прохождения ТЭС объясняются, видимо, отрицатель-
ным влиянием хлорирования воды ( Morgan a. Stross, 1969).
В других случаях понижение интенсивности фотосинтеза незна-
чительно, а иногда и вовсе не замечается.
В водоемах-охладителях в условиях сильного подогрева
нередко происходит массовое развитие водорослей, особенно
протококковых и сннеэеленых, при температурах выше 30°
(Гринь и Виноградская, 1968; Богатова, 1969). Однако все же
здесь не наблюдалось массового „цветения", как в крупных
днепровских водохранилищах. При подогреве же до 33-35° и бо-
лее начинается угнетение даже многих видов сннеэеленых и
биомасса фитопланктона понижается (Пидгайко и др., 1970; Ви-
ноградская, 1971). В водохранилищах Волги в районе Конаковс-
кой и Костромской ГРЭС тоже не происходило настоящего
„цветения* даже жарким летом 1972 г., когда температуры
приближались к 30°. В фитопланктоне здесь господствуют диа-
томовые. Видимо, для „цветения* сннеэеленых необходимы и
другие факторы (непроточность, определенный' уровень биогенов).
В эоне подогрева в летние месяцы при наиболее высоких тем-
пературах наблюдалось лишь усиленное развитие пирофитовьх
(Cryptomonas ovata). За счет пирофитовьх и диатомовых
биомасса фитопланктона увеличивается в среднем в 1.5-3.раза,
достигая максимума уже за пределами зоны сильного подогре-
ва, в районах,, где температура начинает уже понижаться. Еше
дальше от ТЭС,-в 10-15 км от водосброса, биомасса опять
уменьшается. Реакция фитопланктона на подогрев, таким обра-
зом, происходит с запозданием, что вполне естественно, так
как для размножения водорослей нужно время и, кроме того,
фитопланктон сносится вниз по течению (Девяткин, 1970а
наст, сб.) (рис. 14).
Влияние подогрева на интенсивность фотосинтеза водорос-
лей изучалось многими авторами. За немногими исключениями
( Brezina е. а. 1970) исследователи сходятся на том. что
первичная продукция повышается при подогреве воды со сравни-
тельно невысокими температурами, не выше 15-20°, ио тормо-
зится или подавляется, если исходная температура воды была
выше ( Warinner a. Brehmer, 1966; Morgan а.
29
Stress, 1969; Thorslund, 1971; Елизарова, 1973; Пырина
и Наумова, 1973; Шишкин и пр., 1972, и пр.). В обшем фото-
синтез фитопланктона при 30-35° ослабевает, а по некоторым
наблюдениям при 40-45° прекращается (Девяткин, 1973а). Это,
однако, не относится ко многим синезеленым и протококковым,
имеющим более высокие температурные оптимумы (Sorokin,
1971). Возможно, что в зонах подогрева наблюдается некоторая
Рис. 14. Изменение температуры, содержа-
ния кислорода и биомассы фитопланктона в райо-
не Конаковской ГРЭС в Иваньковском водохрани-
лище зимой.
1 - в поверхностном, 2 - в придонном слое
(по: Курдина и Девяткин, 1972).
акклимапии водорослей, приводящая к повышению термическо-
го оптимума фотосинтеза (Девяткин, 1973а). Вопрос о зависи-
мости фотосинтеза от температуры еще не окончательно выяс-
нен и его экспериментальное решение наталкивается иа значи-
тельные трудности.
Более сильное влияние на фитопланктон (как на видовое
разнообразие, так и на численность) подогрев оказывает в хо-
лодное время года. Здесь сказывается не только повышение
температуры, Но значительное улучшение светового режима за
счет отсутствия ледового покрова в зонах подогрева. Так, в
80
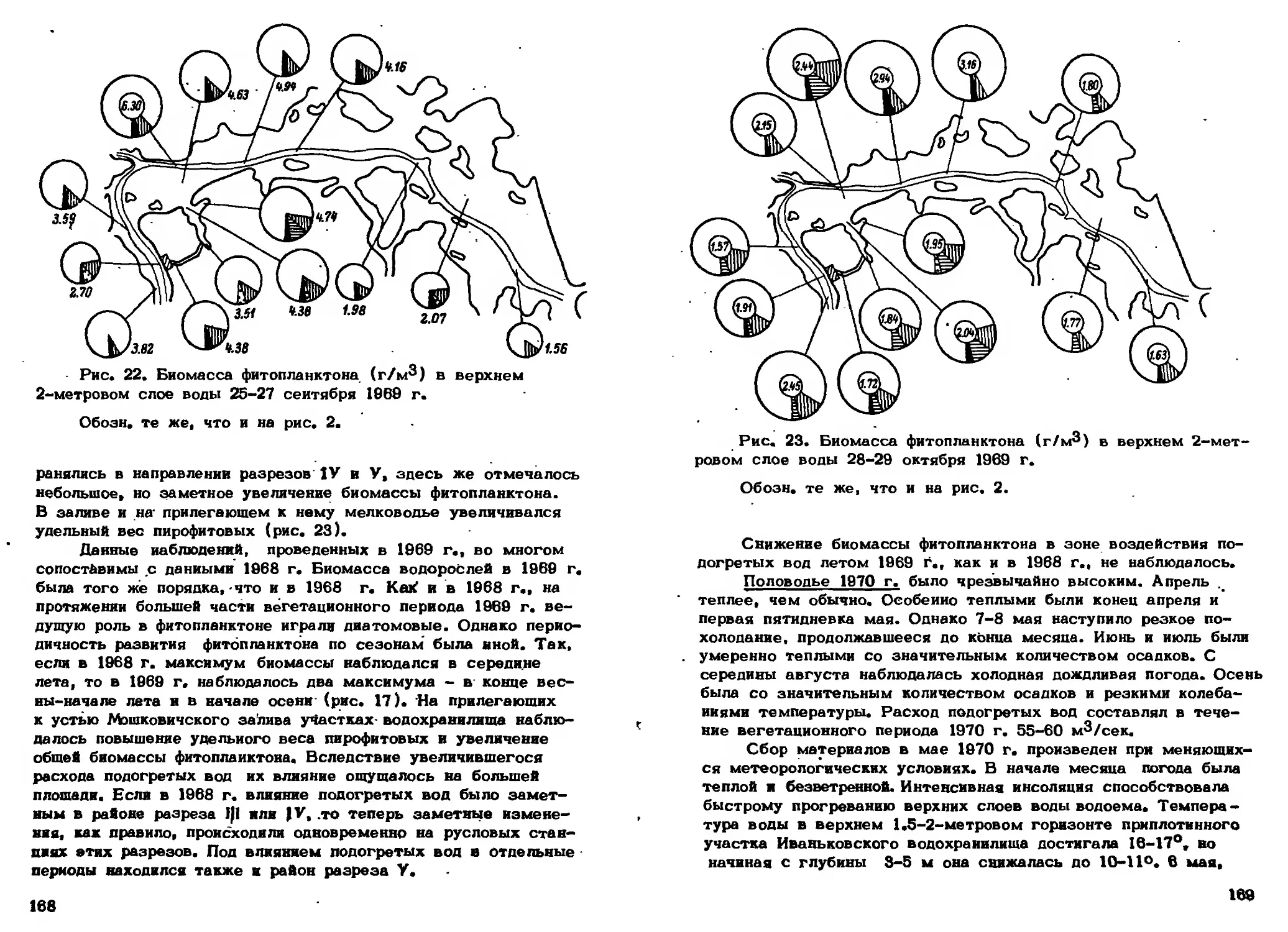
Иваньковском водохранилище биомасса фитопланктона возрастает в
5-10 раз и это возрастание распространяется на большое расстояние
вплоть до плотины ГЭС. Однако вследствие относительно низкой
численности фитопланктона зимой даже десятикратное повышение его
биомассы дает лишь небольшую абсолютную величину (Девяткин,
1971) (рис. 14).
Усиленное развитие водорослей и повышение фотосинтеза сопро-
вождаются увеличением количества кислорода, ио вместе с тем способ-
ствуют и вторичному загрязнению, как избыточная первичная продукция.
Зоопланктон.подвергается сильному воздействию в агрега-
тах ТЭС, однако их травмирующее влияние неодинаково в раз-
ных случаях и зависит как от Состава зоопланктона, так и от
абсолютной величины температуры. Вообше оно наблюдается,
видимо, во всех ТЭС независимо от характера водоема и кли-
мата местности и было зарегистрировано многими авторами в
Европе и Америке ( Heinle, 1969; Churchill a. Wojtalik,
1969; Bader a. Tabb, 1970; Поливанная и др., 1971;
Астраускас и др., 1971; Ривьер, 1971, наст, сб., и др.). Ниже
водосброса всегда имеется большее или меньшее количество
поврежденных или разрушенных планктонных животных. Обычно
травмируются ракообразные удлиненной формы тела или с высту-
пающими придатками и конечностями (особенно Leptodora, но
также Daphnia, многие из Calanoida, Cyciopoida), а
из коловраток крупные с тонкими покровами ( Asplanchna).
Округлые, с короткими конечностями, как Bosmina, Chy-
dorus, большинство коловраток повреждаются значительно
меньше. Из морской фауны, кроме ракообразных, легко поги-
бают медузы и гребневики, а личинки моллюсков и усоногих
выживают хорошо ( Cory a. Nauman, 1969; Barnett,
1972). При этом доля поврежденных и погибших организмов
выше при более высоких температурах.- Так, в Грин-Ривер при
подогреве с 27.5 до 36.3°С погибало более 90% зоопланктона
(Churchill a. Wojtalik, 1969); в заливе Бискайи во Флори-
де при 33° погибало около 10%, а при 40° - до 80% сетного
зоопланктона (Bader a, Tabb, 1970). В Иваньковском
водохранилище уже при 26° наблюдалась гибель 35%, а' при
32-33° - гибель 54% (а некоторых видов даже 70-86%) планк-
тонных рачков (Ривьер, 1971, иаст. сб.). Очевидно, здесь ска-
зывается „потеря равновесия", о которой говорит Кутан
(Coutant, 1971), но, возможно, что она приводит к повышенной
гибели и независимо от деятельности хищников. Кроме того,
не меньшее значение может иметь травмирование на водопаде
в конце водосбросного канала, о котором Кутан не упоминает.
В пробах планктона, взятых непосредственно у водосброса, мож-
но видеть массу рачков с поломанными антеннами и ногами,
очевидно, уже нежизнеспособных (Ривьер, наст. сб.).
31
Кутан (Coutant, 1071, 1972а) делает попытку расчета
вреда, приносимого планктону водоема электростанцией, по
соотношению стока водоема и объема вовлекаемой в станцию
воды Но прохождение агрегатов станции не всегда вызывает
сильную гибель зоопланктона: во-первых, при низких естест-
венных температурах она вообще понижена, .тепловой" шок
слабее; во-вторых, как указывалось, многие планктонные фор-
мы мало страдают от травмирования, и, вероятно, этим и
объясняется, что у ТЭС в эстуарии р. Патаксент (США) планк-
uSaptlpoc
Рис. 15. Изменение температуры и количества зоопланк-
тона в районе теплоэлектростанций.
а - в р. Грин-Ривер в районе Перэдайз ТЭС, в мае
1964 г., по оси Ординат слева - объем зоопланктона (по: Ch'ur-
chill' a. Wojtalik, 1969); б - в Иваньковском водохранили-
ще в районе Конаковской ГРЭС, в августе 1970 г„ по осн ор-
динат слева - биомасса зоопланктона (по: Ривьер и Куэичкин,
1972).
тонные личинки усоногих, моллюсков, мшанок и других форм
(большей частью округлые, в раковинках), видимо, не страда-
ли при прохождении станции и обеспечивали более высокую
продукцию обрастаний, чем у водозабора (Cary a. Nauman,
1969; Nauman a. Cory, 1969).
Вследствие гибели части зоопланктона его численность
и биомасса в районе сброса подогретых вод обычно, особенно
летом, понижены (иногда в несколько раз). Но планктонные
сборы в этом месте могут создать ложное впечатление повыше-
нии биомассы за счет того, что в уловы попадают поврежден-
ные нежизнеспособные или погибшие формы, так как в месте
сброса вод образуется водоворот. С удалением от места сбро-
са зоопланктон, уносимый потоком, постепенно восстанавлнвает-
82
ся и в районе, гае прекращается течение и температура уже
заметно понижается, постигает первоначального или более вы-
сокого уровня (Ривьер, наст. сб.). Это хорошо випио, напри-
мер, в районе Конаковской ГРЭС и у ТЭС Грин-Ривер (рис.15).
В зоне подогрева видовой состав зоопланктона, как упо-
миналось 'выше, изменяется мало, только при сильном подогре-
ве число видов уменьшается, отходят на задний план или исче-
зают менее эвритермные виды и господствующее положение
занимают самые эвритермные и относительно теплолюбивые.
В пресных водах умеренной полосы это копепода Mesocyc-
lops leuckarti (космополит, как известно, в массовых коли-
чествах встречающийся в Африке), из кладоцер Diaphano
-------------- 28 И
* * еР льное распределение температуры и зоо-
планктона в эоне сильного подогрева ниже Конаковской ГРЭС
летом 1970 г.
Столбики пропорциональны биомассе зоопланктона в поверх-
ностном и придонном слое, по оси ординат - глубина; сверху —
расстояние от водосброса (по: Литвинов и Ривьер, 1972).
soma braehyurum, Daphnia cuculiata, Bosmina longi-»
rostris; на юге к иим присоединяются Daphnia magna,
виды Moina (Сергеева, 1968; Богатова, 1969; Поливанная,
Сергеева, 19716; Ривьер, 1971, и др.).
При более слабом подогреве ие замечается и‘изменений
в соотношении форм. Так,в оз. Троусфинид (Англия) при тем-
778 3
33
пературе 26° (при подогреве на 5°) зоопланктон в подогревае-
мой и неподогреваемой эонах практически не различался, при-
чем в неподогреваемой численность теплолюбивой Diaphano-
soma была повышена, возможно вследствие более благоприят-
ных условий питания (Whitehouse, 1971). В оэ. Онтарио бы-
ло обнаружено значительное возрастание количества босмин
после начала действия АЭС, при подогреве до 23-24°
(McNaught a. Fenlon, • 1972); однако при обсуждении этого
сообщения было высказано сомнение в том, что это результат
подогрева, а не годовых флюктуаций по другим причинам.
Удлинение вегетационного периода и фенологические сдви-
ги в зоопланктоне обычно хорошо заметны. Так, в эоне подо-
грева Конаковской ГРЭС сильное весеннее размножение коло-
враток начинается уже в мае, отчасти в апреле, и сравнитель-
но богатый зоопланктон держится до ноября. При сильном повы-
шении температуры, достигающей 27-28° и более, количество
зоопланктона обычно понижается, что наблюдается в наиболее
жаркие месяцы в зонах не только сильного, ио и умеренного
подогрева (Сергеева, 1968; Ривьер, наст. сб.). В районах с
явственной термической стратификацией наблюдается характер-
ная быстрая поведенческая реакция планктонных рачков: они
уходят из сильно нагретых верхних слоев в более холодные
нижние, создавая значительное повышение биомассы в придонном
слое (рис. 16). Однако прогрев и придонных слоев до 26-28°
приводит к сильному уменьшению численности всех видов, а
при 28-30° исчезает большая часть зоопланктона вследствие
прекращения размножения и гибели значительной части ракооб-
разных (Ривьер, 1971 и наст, сб.; Поливанная и Сергеева,
1971; Елагина, наст, сб,). По-видимому, такой подогрев сильно
угнетает и инфузорий (Мамаева, наст. сб.).
Продукция зоопланктона, вообще говоря, может повыситься
с подогревом вследствие ускорения роста и увеличения числа
пометов и поколений животных с повышением температуры,
пока она не приблизилась к пороговым. Однако имеющиеся
наблюдения над продукцией противоречивы. В подогреваемых
озерах в Польше наблюдалось увеличение продукции зоопланк-
тона вдвое (при сравнении озера с температурами в У11-УП1
26-28°С с озером с температурами в это время 20-22°С)
( Patalas, 1970; Thorslund, 1971). В водохранилищах-ох-
ладителях Украины (Кураховское водохранилище) при слабом
подогреве продукция зоопланктона оказалась, напротив, вдвое
пониженной (Пидгайко и др., 1970). Очевидно,, при определении
продукции необходимо привлечь оценку условий питания, а так-
же степени постоянства температур.
Бентос
По микрофитобентрсу .есть лишь данные, относящиеся к ма-
лым прудам-охладителям Украины и к району р. Делавэр (США).
При подогреве наблюдается сильное разрастание нитчаток, осо-
бенно синезеленых Oscillatoria, наиболее устойчивых к Повы-
шению температур. По Тремблей (Trembley, 1965) они обиль-
ны даже при 45°, но по В.Г.Гринь (1971а, б) и М.Л.Пидгайко
и др- (1970), хотя при - умеренном подогреве количество микро-
фитобентоса возрастает в 6-8 раз, при сильном подогреве
(выше 25°) оно начинает понижаться. Сильное развитие сине-
зеленых и иных нитчаток и микрофлоры перифитона было обна-
ружено и в других случаях (Thermal effects... 1971; Flor-
czyk e. a,, 1972).
Макрофиты на подогрев реагируют по-разному, в зависи-
мости от того, к какой группе принадлежат. Морские водорос-
ли, судя по данным американских авторов, менее теплоустойчи-
вы, чем животные, и вымирают в районе тепловых сбросов, хо-
тя это несомненна лишь в тропических эстуариях, а в других
случаях возможно отрицательное влияние мутности или других
факторов (Reeves, 1970; Steidinger a. van Breedveld,
1971).
Высшие растения вообще интенсивно развиваются в эонах
подогрева. В районе электростанции Турецкий Мыс (Флорида),
т.е. в субтропической эоне, обширные ковры „черепаховой травы*-
Thalassia «разрастались при 28°С, но когда температура до-
стигла 40-41 , началось их отмирание (Bader’a. Tabb,
1970), Малые водоемы-охладители нередко, особенно на юге,
зарастают на 50% и более, что сильно ухудшает условия охлаж-
дения воды и приводит к засорению решеток и фильтров и к за-
грязнению водоема, особенно при массовом развитии погружен-
ных растений (Шиманский, 1963, 1968, 1970 и др., Шахматова
и др., 1971). В охладителях и других водоемах умеренной и
средней полосы в эонах сильного подогрева обычно- изменяется
соотношение видов,- разрастаются более теплолюбивые или юж-
ные формы (тростник, рдест курчавый, местами валлиснерия).
Хорошо замечается сдвиг фенологических фаз: более ран-
нее (на месяц и более) начало вегетации, цветения и плодоно-
шения. При этом во многих случаях общая длительность вегета-
ции не увеличивается и в зоне подогрева раньше начинается
и отмирание. Общая продукция макрофитов часто возрастает
(Trembley, 1965; Зубарева, 1971; Лисицына и Жукова, 1971;
Coutant, 1971; Шишкин и др., 1972). По некоторым данным
.(Coutant a, Goodyear, 1972), затормаживание развития
макрофитов наступает при 30-35°.
33
Но в водохранилищах часто не наблюдается возрастания’
продукции макрофитов и при меньшем подогреве, очевидно в
связи с тем, что для них большое значение имеют другие
факторы - освещенность (прозрачность воды), характер грун-
та и особенно колебание уровня. В районе Конаковской ГРЭС
полное обнажение и вымерзание мелководной эоны водохрани-
лища в течение поздней осени и зимы, очевидно, сдерживает
развитие макрофитов, н их продукция в подогреваемой зоне,
несмотря на более раннее появление и максимум биомассы,
почти такая же, как в иеподогреваемых районах. Имеет значе-
ние и то, что здесь, как и около других электростанций, естест-
венные донныё отложения во время строительства ГРЭС были
разрушены и растительный покров их до сих пор еше не пол-
ностью восстановился (Лисицына и Жукова, 1971; Экзернев
и Лисицына, наст. сб.).
Зообентос зон подогрева изучался чаще и более интенсив-
но, чем фнтобентос, однако вопрос о влиянии подогрева на зоо-
бентос с трудом поддается решению. Для бентоса, естествен-
но, не имеют значения условия прохождения вод через агрега-
ты станции (хотя некоторое количество донных (Организмов
вовлекается в них при водозаборе), поэтому вопрос может
ставиться лишь о влиянии сбрасываемых подогретых вод. Но
эти воды ие только могут нести с собой загрязнение, но, как
уже говорилось, воздействуют на грунты и размывают их не-
посредственно в месте сброса, что очень важно для бентоса. •
В некотором отдалении от водосброса, где создается термичес-
кая стратификация, придонные слои воды, и особенно грунт по-
догреваются Значительно меньше, чем толща воды и особенно
ее поверхностные слои. Поэтому бентос подвергается действию
высоких температур (в умеренных эонах 30° и выше) лишь
в ограниченных участках перед самым водосбросом и на силь-
но прогреваемых (и естественным путем, и сбросами электро-
станции) мелководьях. К этому следует добавить, что наибо-
лее распространенные в наших внутренних водоемах средних ши-
рот пелофильные беспозвоночные (некоторые виды хирономид,
тубифицид и моллюсков) - большей частью формы со сравнитель*
но' долгим (нередко год и более) жизненным циклом, выносли-
вые к органическому и химическому загрязнению; а такие
формы, как верно замечает Вурц (Wurtz, 1961), обычно вынос-
ливы и к изменениям физических факторов.
Все это очень затрудняет выяснение влияния подогрева
на зообентос, хотя казалось бы, что вследствие своей малопод-
вижности бентические беспозвоночные, особенно зарывающиеся
В грунт, могут быть наиболее удобными показателями степени
подогрева, и на них ₽ этом смысле возлагались надежды
(Wurtz, 1661; Jensen, 1969; Thermal effects.., 1971).
Зв
Неудивительно, что большинство исследователей не могли
обнаружить заметного влияния подогрева на бентос или обнару-
живали незначительные, статистически не всегда достоверные
изменения. Во многих случаях там, где. подогрев заметно влия-
ет на планктон, в бентосе изменений не замечается, как, на-
пример, в зонах подогрева Конаковской и Костромской ГРЭС,
где бентос по составу и биомассе приблизительно такой же,
как в иеподогреваемых частях водохранилищ. В районе Конаков-
ской ГРЭС на илах биомасса бентоса, по Т.Л.Поддубной
(1971а), примерно на треть повышена, а по В.И.Митропольско-
му (1.973), напротив, несколько ниже; Только в эоне сильного
(временами до 25-29°) подог|эева Костромской ГРЭС, по
И.А.Скальской (наст, сб.), бентос значительно беднее, но в
основном в связи с измененным в этом участке характером
грунта. В оз. Меларен (Швеция) незначительное повышение
количества олигохет может быть объяснено не только подогре-
вом, но и общим эвтрофированием озера (Wiederholm, 1971), а
в оз. Троусфинид (Англия) сильное обеднение бентоса при по-
догреве до 25°, по мнению автора (Rothwell, 1971), вызвано
скорее накоплением токсических ионов меди и цинка. Не было
замечено влияния подогрева на бентос и в море около некото-
рых ТЭС в Америке и Англии ( Markowski, 1960; Lyons
е. а., 1971).
Чаше всего замечаются сдвиги в фенологии, в моментах
наступления фаз жизненного цикла. Подогрев воды в течение
всего зимнего периода и особенно весной вызывает значительно
более раннее (почти иа месяц) созревание и размножение оли-
гохет (тубифицид), в результате чего становится возможным
появление к концу лета второго поколения (Поддубная, 1971а;
Aston, 1971). Раньше происходят и окукливание и вылет хи-
роиомид. Можно было ожидать нарушений в прохождении цикла
хироиомид (и других гетеротопов), имаго которых вылетают
слишком райо, в условиях еше холодной атмосферы, и могут
в массах погибать (Мордухай-Боптовской, 1971.; Brungs, 1970;
Rothwell, 1971). Однако закономерного понижения численнос-
ти личинок в зонах подогрева не наблюдалось, видимо, потому,
что сравнительно небольшая акватория последних легко заселя-
ется комарами из окружающих иеподогреваемых районов. Сдвиги
в сроках вылета веснянок и поденок в реках Англии при подо-
греве до 26-28° также не вызывали угнетения популяций личи-
нок (Langford, 1971b; Langford a, Aston, 1972). Для
вылета хироиомид, как было выяснено на двух видах Chiro-
nomus, температура играет ббльшую роль, чем фотопериод
(длительность освещения), но последний тоже имеет значение,
воздействуя на гистологию нервной системы и хромосомы
(Clarke a. Credland, 1971).
3?
В области распространения каспийских фаунистических
элементов, в общем сравнительно теплолюбивых, т.е. в бассей-
нах среднего и нижнего течения рек Понто-Каспийского бассей-
на, в подогреваемых эонах наблюдается усиленное развитие
некоторых каспийских видов. Так, в водоеме-охладителе Кура-
ховской ГРЭС (басе. р. Днепра) развился бокоплав Pontogam-
marus robustoides, который здесь (при повышении средне-
годовой температуры на 3-5°) размножается круглый год,
дает большее число генераций и пометов, более крупные разме-
ры и вдвое более высокую биомассу, чем в неподогреваемых
водоемах (Китицына, 1970, 1971а, 19716).
В подогреваемом участке Днепровского водохранилища,
в сбрасываемых водах Приднепровской ГРЭС, успешно развива-
ются несколько каспийских видов, в том числе искусственно
вселенная миэида Limnomysis benedeni, в этих условиях
перешедшая к круглогодичному размножению („ациклии")
(Журавель, 1971).
В водоеме-охладителе Северского Донца оз. Лиман силь-
но размножились акклиматизированные здесь каспийские мизи-
ды, у которых более чем на 1.5 месяца удлинился период
размножения, появились две новые генерации, а теплоустойчивость,
очевидно как следствие акклимации, повысилась (Дукина и
Кудрявцева , 1970; Кудрявцева и Горюнова, 1971). . .
Ясно выраженное вредное влияние подогрева проявляется
тогда, когда температура (у дна) приближается к 30-32°. Это
было показано еще на фауне порожистой части р. Делавэр,
где при подогреве до 37-42° наступало резкое обеднение бенто-
са (Coutant, 1962). Позже это же было обнаружено на р. Шуил-
килл, где температура ниже сброса ТЭС держалась на уровне
32-35°С, временами поднимаясь до 37-40°. Бентос в этом райо-
не сильно обеднел по числу видов и изменился по составу,
стали преобладать выносливые к сильному подогреву виды; та-
кие изменения авторы (Wurtz, 1961; Wurtz a. Dolan, 1961)
называют .биологической депрессией" и „биологическим иска-
жением' (distortion). Выражая эти показатели в процентах
(по отношению к числу видов при естественных условиях, и по
отношению числа выносливых видов к числу невыносливых),
авторы пользуются ими для характеристики не только теплово- .
го, но и другого загрязнения. Исследованная ими река подвер-
галась также сильному органическому загрязнению.
В тропических водах (в эстуариях Флориды) уже подогрев
на 3-5° вызывает сильное обеднение бентоса (G-looschenko,
1968, и др,).
По наблюдениям на водоемах-охладителях Украины незна-
чительный подогрев в общем ведет к увеличению числа видов
88
и некоторому увеличению биомассы, но только если в фауне
есть теплолюбивые виды. Сильньй же подогрев вызывает умень-
шение числа видов и значительное понижение численности и
биомассы, причем уяе летняя придонная температура около 25°
является тем пределом, превышение которого ведет к сильно-
му угнетению макрозообентоса (Пидгайко и др., 1970).
Продукция бентоса в этих же водоемах при незначитель-
ном подогреве в целом повышается приблизительно на 40%. Од-
нако у разных групп беспозвоночных продукция изменяется по-
разному: у олигохет она на 20% понижена, из хироиомид одни
дают повышение, а другие понижение продукции. Коэффициенты
_>же P/В (удельная продукция) оказываются в общем такими же
или более низкими в подогреваемой зоне, и лишь у упомянутого
выше каспийского бокоплава Pontogammarus robustoides
среднегодовая продукция и Р/В - коэффициент в подогреваемом
водоеме несколько повышены (Пидгайко, 1971а; Китицына,
1971а). В Иваньковском водохранилище в зоне умеренного по-
догрева продукция олигохет (тубифицид) оказалась, наоборот,
повышенной в 1.S-2 раза (Поддубная, 19716).
Этот разнобой в данных по продукции, очевидно, объясня-
ется, как и для зоопланктона, интерференцией различных факто-
ров; Для бентоса, очевидно, условия местообитания (субстрата)
и питания оказываются чаще более важными. Поэтому при
исследованиях продукции необходимо строго следить за одно-
родностью этих факторов в подогреваемом и неподогреваемом
участках.
Очень мало пока данных по обрастаниям и эпибиозам, хотя
эта весьма разнообразная фауна, не связанная с грунтами и
обитающая в условиях более сильного подогрева, заслуживает
подробного изучения. Имеющиеся данные говорят о том, что в
районе водосброса в эстуариях эпифауна развивается успешно
и даже в большем количестве, чем у водозабора (Markovski,
1960; Nauman a. Cory, 1969). В эстуарии р. Патаксент
(США) при подогреве в среднем на 6.8° продукция эпифауны
была втрое выше, чем у водозабора, несмотря на то что летом
температура достигала иногда 37-88°; правда, в период наиболь-
шего подогрева число видов и продукция понизились ( Cory
a. Nauman, 1969).. В р. Луаре при подогреве на 5-8° (до
28°) наблюдалось массовое развитие эпифитной микрофауны,
в частности сидячей коловратки Lacinularia ( Pourriot
е. а., 1972).
По' микрозообентосу, также заслуживающему большего
внимания, данных еше меньше. В Ку чу рга неком лимаие-охла-
дителе не было замечено влияния подогрева (в среднем на
4.2°) на популяцию гарпактицид (Набережный и Витковская,
39
1S72). Не удалось найти в литературе никаких сведений о влия-
нии подогрева на фитофильные биоценозы, представляющие наи-
больший интерес благодаря их богатству и связи с макрофитами.
Рыбы
Влияние энергетических станций и сбрасываемых ими вод
на рыб, естественно, было предметом особенно интенсивных
исследований. Не ставя перед собой цель дать полный обзор
обширной относящейся сюда литературы, ограничусь только не-
которыми общими замечаниями.
Многие рыбы, как упоминалось выше, сильно страдают
от травмирования на сороудерживаюших решетках у водозабора
станций, а молодь и личинки рыб - в трубках конденсатора и
на водосбросе, где подвергаются тепловым шокам и частью
гибнут, частью становятся добычей хищников. Воздействие
прохождения через станции на организмы изучено главным обра-
зом именно иа рыбах.
В зонах подогрева обычно наблюдается постоянное скоп-
ление рыб, привлекаемых повышенной температурой, а иногда
и обилием добычи. Здесь наблюдается преимущественная кон-
центрация более теплолюбивых форм, но для холодолюбивых
(главным образом лососевых) зоны подогрева могут быть
барьером, препятствующим, нерестовой миграции. В зоне подо--
грева хорошо выражено смешение фаз жизненного цикла, более
раннее созревание рыб и усиленный рост молоди. Общий период
роста- значительно удлиияетси и сеголетки рыб достигают к
осени значительно больших размеров, чем при естественном
температурном режиме.
Рыбы обладают хорошо разнитой способностью к терми-
ческой акклимацин, повышающей пороги у эвритермичных форм
иа 5-10°. Это позволяет многим видам обитать в зонах подо-
грева, но при етом, однако, нередко наблюдаются нарушения
в процессе воспроизводства вследствие несоответствия между
готовностью к нересту и состоянием нерестилищ и потребнос-
тью молоди в пище' и состоинием кормовой базы в зоне подо-
грева.
Температуры, приближающиеси к 30°, несмотря на аккли-
манионные возможности, оказывают отрицательное влияние
почти на всех рыб, вызывая уход их из зон подогрева или
депрессию размножения, роста и питания. В зонах подогрева, !
кроме того, наблюдается значительное повыщение интенсивности
заражения рыб различными паразитами (ремнецом и другими).
40
Влияние подогрева на рыб рассмотрено в ряде работ, среди
которых следует указать работы, имеющие также общее значе-
ние для рассматриваемой проблемы (Brett, 1960; Alabas-
ter, 1963; Leynaud, -1967; Coutant, 1971; Sylvester,
1972).
За счет выращивания теплолюбивых видов рыб из евро-
пейской и азиатской фауиы (а также некоторых промысловых
беспозвоночных) возможно хозяйственное использование по-
догретых зон и водоемов. Но вопрос об использовании подо-
гретых вод должен составлять предмет специальных статей.
Заключение
Подводя итоги вышеизложенному, можно сделать следую-
щее общее заключение.
Тепловые и атомные электростанции оказьвают влияние
на гидробиологический режим водоемов', воды которых они ис-
пользуют для охлаждения, во-первых, при пропускании забирае-
мой для охлаждения массы воды через свои агрегаты; во-вто-
рых, при выбрасывании в водоем той же массы воды в подо-
гретом (обычно на 8-11°) состоянии.
При прохождении через электростанцию (продолжающемся
всего несколько минут) организмы, находящиеся в забираемой
воде, подвергаются механическим травмам и тепловому шоку,
в результате которых часть рыб и планктонных животных поги-
бает немедленно или через некоторое время (частично от .
истребления хищниками).
Сброс охлаждающих вод приводит к образованию в водое-
ме зоны постоянного подогрева. Подогрев, в условиях умерен-
ного пояса, вызывает у водных организмов ускорение роста и
развития и сдвиг фенологических фаз на более раннее время,
удлинение вегетационного периода, изменение соотношения ви-
дов (увеличение роли эвритермных и теплолюбивых). Сильный
подогрев ведет к нарушениям размножения, развития, структу-
ры сообществ и к отмиранию организмов, по достижении их
летальной температуры.
Но для суждения о реакциях водных организмов на подо-
грев необходимо учитывать следукщие особенности.
1. Обычно отношение организма к температуре может быть
выражено в виде асимметричной кривой, вершина 'которой, соот-
ветствующая оптимуму, приближена к максимуму, т.е, верхне-
му летальному порогу.
41
2. У большей.части гидробионтов умеренного пояса верх-,
ний термический порог близок к 30° или отличается от него
на один-два градуса. •- .Немногие холодолюбивые виды имеют
пороги не выше 20°, а теплолюбивые (преимущественно из
бактерий и синезеленых водорослей) - между 30 и 40°С.
3. Большинство организмов способно к акклимации к бо-
лее высокой температуре, повышающей верхние и нижние поро-
ги на несколько градусов.
4. Многие водные животные могут реагировать на подогрев
не только физиологическими, но и поведенческими реакциями,
уходя от неблагоприятных температур в горизонтальном или
вертикальном направлении.
5. В природе почти всегда имеет место синэргизм, т.е.
одновременное действие температуры и других факторов. С по-
вышением температуры усиливается токсическое действие ядо-
витых веществ, повышается потребность в кислороде, пище.
Изменение субстрата, освещенности, течения и т.д. часто ока-
зывает более сильное влияние, подавляя действие незначитель-
ного изменения температуры.
Эти особенности сильно затрудняют выяснение влияния
подогрева на экосистемы водоемов. Но в общем имеющиеся г
к настоящему времени сведения показывают, что при подогре-
ве наблюдается накопление в водоеме органических веществ и
тенденция к эвтрофированию. Сильный подогрев, при котором .
температуры в летние месяцы приближаются к 30° или превос-
ходят их, оказывает вредное влияние на жизнь водоемов, сни-
жая видовое разнообразие, численность и продуктивность. Но
умеренный и слабый подогрев, хотя и вызывает еше недоста-
точно изученные изменения экосистем, в большинстве случаев,
видимо, не вызывает явственных отрицательных последствий
и часто способствует повышению продуктивности.
Степень влияния сброса подогретых вод энергетичеекими
станциями зависит от нескольких факторов:
1) мощности электростанций, их типа (атомные станции
при той же мощности расходуют больше воды) и взаимного
расположения (у близких станций зоны подогрева могут смы-
каться);
2) характера водоема, используемого для охлаждения: в
реках рассеивание тепла и перемешивание вод происходит быст-
рее;
3) естественной температуры водоема, определяющей воз-
можность достижения при подогреве температур около 30°С.
Эти температуры достигаются легче и держатся дольше в лет-
ние месяцы и на более южных широтах, особенно в субтропи-
ческом и тропическом поясах.
22
При современном состоянии энергетической промышлен-
ности в умеренном поясе вредное влияние электростанций на
жизнь водоемов пока невелико и локально, ограничиваясь не-
большими (площадью в несколько квадратных километров или
немногие десятки их) зонами сильного подогрева. Несомненно,
что прохождение охлаждающих вод через электростанции и их
сброс в подогретом виде в водоем приносят гораздо меньше
вреда жизни водоемов, чем сброс промышленных отходов. С
точки зрения экономики, деятельность ТЭС и АЭС гораздо
эффективнее, чем сооружение гидроэлектростанций с водохрани-
лищами, вызывающими затопление обширных площадей, но боль-
шей частью малопродуктивными.
Однако планируемое массовое строительство тепловых и
особенно мощных атомных станций, требующих все больших
количеств охлаждающей воды, может привести к расширению
зон сильного подогрева и областей с нарушенными экосистема-
ми. Нежелательные последствия будут усиливаться особенно в
более южных широтах. Поэтому нормы допустимого подогрева
должны зависеть прежде всего от климатических условий и мо-
гут быть неодинаковыми в разных областях. Принятая в СССР
стандартная санитарная норма подогрева 3° может быть увели-
чена в более холодном климате на 1-2° и настолько же умень-
шена в более теплом. В тропических же зонах подогрев вообще
недопустим. ,
Для уточнения воздействия электростанций на жизнь во-
доемов и обоснования норм подогрева необходимы дальнейшие
исследования, в основном в следующих направлениях:
1) изучение отношения к температуре наиболее массовых
форм водных растений и животных (особенно беспозвоночных)
и реакпий на подогрев неизученных с этой стороны биоценозов
(эпибионтных, фитофильных, микробеитических) ;
2) изучение, взаимовлияния различных факторов среды с
целью выявления действия одной температуры;
3) выявление «индикаторов теплового загрязнения" - бо- ‘
лее или менее стенотермичиых форм, по наличию и состоянию
популяции которых можно судить о степени подогрева в биоло-
гическом аспекте.
Исследования по всем трем направлениям должны прово-
диться при сочетании экспериментальных работ с наблюдениями
в7природе.
43
Библиография по влиянию
энергетических электростанций
иа гидробиологический
режим водоемов
Литература по вопросам влияния теплоэлектростанций и
сбрасываемых ими подогретых вод на водные организмы до
половины 60-х гг. была очень скудной. Это были небольшие
статьи преимушественно в технических журналах, содержавшие
или некоторые данные по специальным вопросам, или общие
соображения об экологических изменениях, вызываемых поступ-
лением дополнительного тепла в водоемы. В нашей стране появ-
лялись статьи о зарастании небольших водоемов-охладителей,
а также о возможностях выращивания в них. рыб. В двух анг-
лийских руководствах по загрязнению водоемов ( Hynes, I960;
Klein, 1962) ,термическому загрязнению" отводится всего
несколько страниц.
С половины и особенно с конца 60-х гг. положение меня-
ется. Литература по проблеме влияния тепловых, а затем и
атомных станций на биологию водоемов начинает быстро расти,
чему способствует особенно организация совещаний, посвящен-
ных этой проблеме.
В июне 1967 г. в нашей стране (в Киеве) было созвано
первое координационное совещание по биологическому и хими-
ческому изучению водоемов-охладителей. Некоторые вопросы
биологии этих водоемов рассматривались также во Львове в
1970 г. на* совещании по водохранилищам ТЭС.
В марте 1971 г. в 'Борке, в Институте биологии внутрен-
них вод АН СССР, был организован симпозиум по вопросам
влияния подогретых вод теплоэлектростанций. на гидрологию и
биологию водоемов.
Особенно активно проблема влияния подогретых сбросов
электростанций обсуждалась в США. В 1968 г. там были про-
ведены симпозиумы по гтепловому загрязнению" (по его инже-
нерным аспектам -.в Нэшвилле, по биологическим аспектам -
в Портленде) и.совещание ^Термической рабочей группы" МБП
в Чисепикской лаборатории; в. 1969 г. - конференция по прак-
тическому использованию подогретых сбросов, а в 1970 г. -
симпозиум по значению термальных сбросов при производстве
электроэнергии в Вашингтоне и симпозиум по экологическим
аспектам атомных электростанций в Нью-Йорке. Весной 1971 г.
был созван симпозиум по пресноводной биологии в связи с про-
изводством электроэнергии в Лондоне, а летом 1972 г. - сим-
позиум по влиянию подогретых вод ТЭС и АЭС на водоемы в
Вене.
44
Все перечисленные совещания выпускали к их открытию
или спустя некоторое время сборники (докладов или тезисов),
перечень которых приводится ниже.
Множество отдельных статей по рассматриваемой пробле-
ме публиковалось и продолжает публиковаться также в различ-
ных периодических изданиях, как технических, так и биологи-
ческих.
В настоящее время литература по влиянию тепловых и
атомных станций на водоемы насчитывает уже многие сотни
названий. Трудности в использовании этой разбросанной лите-
ратуры побудили к составлению библиографий и обзоров. Первая
попытка такого рода принадлежит Нейлору (Naylor, 1965),
опубликовавшему обзор по влиянию температуры и в частности
подогретых сбросов на морские и эстуарные организмы, с боль-
шим списком литературы; в том же году краткий обзор по
влиянию подогретых сбросов Hai биологию рек, тоже с большим
списком литературы, опубликовал Тремблей (Trembley, 1965).
Через два года в США появилась более обширная „Библиогра-
фия по тепловому загрязнению" (Bibliography..., 1967), сос-
тавленная Комитетом по тепловому загрязнению, и библиогра-
фия по влиянию температуры в водной среде В.Кеннеди и Ю.Ми-
хурского, позже ими же дополненная (Kennedy а.. Mihur-
sky, 1967, 1969). Обе охватывают и влияние подогретых сбро-
сов на гидрологию и гидрохимию водоемов.
Начиная с 1968 г., составление библиографий специально
по биологическим аспектам „теплового загрязнения', сопровож-
даемых обзорами, взял в свои руки Ч.Кутан (Ch. Coutant),
ежегодно публикующий их в журнале Journal WPCF ( Wa-
ter pollution control federation). Им опубликовано (в
последние годы - с соавторами) уже шесть таких литератур-
ных обзоров, объем которых, как и число упоминаемых работ,
непрерывно растет. В обзоре за 1972 г* приводится уже 427
названий, но в число их входят статьи по влиянию не только
подогретых сбросов, но и вообще температуры на организмы и
биоценозы.
В последнее время в этой работе деятельное участие при-
нимает Лаборатория Оук Ридж (Oak Ridge national labo-
ratory ) при Комиссии по атомной энергии США. В 'изданиях
этой лаборатории Кутаном *с соавторами были опубликованы
индексы для литературы с 1967 г. и аннотации к библиографии
за 1971-1972 гг.,а Ульриксоиом и Стокдэйлом ( Ulrikson
a. Stockdale, 1971) - перечень всех учреждений, исследо-
вательских программ и специалистов, занимающихся проблемой
изучения влияния температуры на водные организмы в США.
Широкий размах этих работ виден хотя бы из того, что в этом
перечне более 280 лиц.
45
На русском языке большие обзоры и библиографии не пуб-
ликовались, одна ко следует упомянуть очень сжатую, но содер-
жательную статью об итогах исследований малых водоемов-охла-
дителей юга СССР, составленную М.Л.Пидгайко с сотрудника-
ми (1970), и вышедшие в самое последнее время краткие об-
зоры по влиянию подогретых сбросов на режим и биологию
водоемов А.В.Быковой (1973) и Н.С.Золотаревой и др. (1973).
Последние содержат списки литературы (из S2-54 названий),
но оии охватывают меиее одной десятой всей литературы по
рассматриваемой проблеме.
Ниже приводится список всех работ по влиянию тепловых
и атомных электростанций и сбрасываемых ими подогретых вод
на гидробиологический режим, микро- и макрофлору и фауну
(за исключением рыб) водоемов, которые мне удалось иайти
в отечественной н заграничной литературе. Статьи по влиянию
температуры на организмы вообше как в экспериментах, так
и в природе, вне связи с электростанциями, не приводятся, за
исключением нескольких руководств и обобщающих работ, на
которые в тексте есть ссылки (таких работ не более десяти).
Таким образом, в этот список, состоящий более чем из
260 названий, не входят работы по влиянию ТЭС и АЭС на
гидрологию и гидрохимию водоемов и на их рыбное население.
Последних очень много, особенно в США, где вопросы влияния
энергетических станций на рыб интенсивно разрабатываются.
К настоящему времени накопилось и много работ по проблеме
хозяйственного использования подогретых вод, которая совсем
не затрагивалась в настоящем обзоре. Со включением литера-
туры по всем этим вопросам число работ по влиянию ТЭС* и
АЭС на экосистемы водоемов по ориентировочной оценке приб-
лижается к • тысяче.
Сборники и библиографии
О мерах борьбы с минеральными и органическими
отложениями в системах технического водоснабжения и с за-
растанием и .цветением* водохранилищ ТЭС. Тезисы докл.
совет., состоявшегося в 1970 г. при ОРГРЭС во Львове. М,
1970.
Симпозиум по влиянию подогретых вод теплоэлект-
ростанций на гидрологию и биологию водоемов. Тезисы докл.
Борок, 1971.
Гидрохимия и гидробиология водоемов-охладителей
тепловых электростанций СССР. Киев, 1971.
46
Hynes H.N. 1960. The biology of polluted wa-
ters. London.
Klein L. 1962. River pollution. London.
Proceedings of the 2 hd thermal workshop
of the US IBP. 1969. Chesapeake Science, 10, 3-4.
P.A.K r e n к e 1 a. F.L.P a г к e r, ed. 1969.
Biological aspects of thermal pollution. - Vanderbilt
Univ, Press.
Eisenbud M. a. G.G 1 e a s о n, ed, 1969,
Electric power and thermal discharges. - Gordon and
Breach publ,, N.Y.
S.M a t h u r a. R.S t e w a r t, ed. 1970. Pro-
ce edings of the conference on the beneficial uses of
thermal discharge. - N.Y. St, Dept, envir. conservation,
Albany.
Symposium on freshwater biology and electric
power generation. 1, 2. 1971. Centr, electr, generat.
board, research labor., London.
T.P.G r a h a m, ed. 1971, Ecological effects of
hot water discharged by an electric power generating
plant. — Univ, North Carolina, Asheville.
Environmental aspects of nuclear power
stations. Proceed, of Sympos,, 1971. Intern, atom. •
energy agency, Vienna.
C ol u m b i a river thermal effects study. 1971.
Envir. protect, agency, Washington.
Thermal effects and U.'S.Nuclear power sta-
tions. 1971. Divis. react, developm. technol. U.S.atom,
ener, commis.
Bibliography on thermal pollution. Journ..
Sanit. division, Amer, Soc, civ. eng,, June 1967.
Kennedy V.S. a. J.A.M i h u r s к y. 1967.
Bibliography on the effects of temperature in the
aquatic environment Univ. Maryland, Natur. Res. Inst.
Contr,, 326.
Kennedy V.S, a, J.A.M i h u r s к y. 1969.
Addendum to: Bibliography on the effects of temperatu-
re in the aquatic environment. Univ. Mar,, Natur. Res.
Inst, Contr. 326.
Raney E.C. a. Menzel B.W, 1969. Heated
effluents and effects on aquatic life with emphasis on
fish: A bibliography. Ichthyol. Assoc. Bull. N2, Cornel!
University water research center and Philadelphia
electric company.
47
Coutant C.C. Thermal pollution-biological
effects. Journal WCPF: 1968, 40, 6; 1969, 41, 6; 1970,
42, 6; 1971, 43, 6,
Coutant C.C. a, C,Ph.G о о d у e a r. 1972.
Thermal pollution-biological effects. Journal WPCF, 44,6
С о u t a n t C.C, a. H.A.P f u d e r e r. 1973. *
Thermal pollution-biological effects. Journal WPCF,
45, 6.
Coutant C.C, 1972. Index for annual reviews
1967 - 1970. „Thermal pollution-biological effects," Oak
Ridge nation, labor., Ecol, scien. divis.
Coutant C.C., E.E.H uber, H.A.P f u d e r e r.
1973. Thermal effects on aquatic organisms. Annotated
bibliography of 1971 literature. Oak Ridge nation.labor.,
Ecol. scien. divis.
Ulrikson G.U. a. W.G.S t о с к d a le. 1971.
Survey of thermal research programs, sponsored by
federal, state and private agencies (1970). — Oak
Ridge nation, laborat,, Ecol. ’scien. divis.
Coutant C.C. a, H.A.P f u d e r e r. 1973.
Thermal effects on aquatic organisms. Annotated bib-
liography of 1972 literature. Oak Ridge national labor,,
Environ, scien. divis.
Статьи
по отдельным вопросам
Аннотации докладов на Координационном совещании по
водохранилищам-охладителям тепловых электростанций.
1968. Гидробиол. журн., ТУ, 3.
Астраускас А.С. и Рачюиас Л.А. 1971. Эко-,
логические условия и состояние фауны водохранилища-
охладителя Литовской ГРЭС. Симпоз. по влияя, подо-
грет. вод ТЭС... Борок.
Богатова И.Б. 1969. О гидробиологическом режиме
водоемов-охладителей ГРЭС. - В кн.: Рыбоводство
в теплых водах. М.
Богатова И.Б. 1971. Планктон водоема-охладителя ГРЭС
им. Классона. Симпоз. по влиян. подогрет, вод ТЭС...
Борок.
Богатова И.Б. и Ле изо А.П. 1969. Культивиро-
вание Daphnia magna Str, в садках на\ теплых во-
д$х. М
48
БУ т OP И Н Н.В. и Курдина Т.Н. 1975. Особеннос-
ти температурного режима Иваньковского водохранили-
ща в условиях искусственного подогрева. Наст. сб.
Быкове А „В. 1973. Термальное загрязнение водоемов и
влияние повышенных температур на водные организмы.
Обзорная информ. Рыбохоз. использ. внутр, вод., З.ЦНИИГЭПРХ.
Вайнштейн М..Б. 1971. Действие подогретых вод ТЭС
на первичные продукционные процессы в Иваньковском
водохранилище. Симпоз. по влияи. подогрет, вод ТЭС...
Борок.
Васильчи.ко ва А.П. 1967. Зарастание прудов-охлади-
телей промышленного Урала и меры борьбы с ним.
Вопр. водн. хоз-ва Урала и мелиорация, 22. Свердловск.
В а сильчикова А.П. 1969. Влияние термического режи-
ма на развитие фитопланктона в водохранилищах-охла-
дителях среднего Урала. Второе Всесоюзн. совеш. по
вопр. кругов, вещ. и энергии в озерн. вод., ч. П.
Лиственничное на Байкале.
Васильчикова А.П. 1970. Фитопланктон водохранилищ-
охладителей Урала и методы борьбы с ним. - В кн.:
О мерах борьбы с минер, и орг. отложен, в сист. тех-
нич. водоснабж. и с зараст. и цвет, водохр. ТЭС. Те-
зисы докл. М.
Васильчикова А.П. 1971. Периодичность и динамика
биомассы фитопланктона водохранилищ-охладителей
Урала. Симпоз. по влиян. подогрет, вод ТЭС... Борок.
Веригин Б.В. 1962. Теплоэнергетика и рыбное хозяйст-
во. Рыбное хозяйство, 9.
Вииоградська’ Т.А. 1968, Сезонна динамхка фито-
планктону водоймищ-охолоджувач!в Кураховоькой та
Зухвськох ДРЭС. 3-я Наук, конфер. молод, учен. 1нст.
г£дроб. АН УРСР. Ки1в.
Виноградская Т.А. 1970. Влияние сброса подогретых
вод Луганской ГРЭС на развитие фитопланктона р. Се-
верский Донец. - В кн.: Вопр. рыбохоз. освоен, сан-
. биол. реж. водоемов Украины. 4, 1. Киёв.
Виноградская Т.А. 1971а. Некоторые итоги изучения
влияния подогрева иа развитие фитопланктона в водо-
хранилищах-охладителях ГРЭС юга Украины. Симпоз.
по влиян. подогрет, вод ТЭС...' Борок.
Виноградская Т.А. 19716. Влияние подогрева на
развитие фитопланктона водохранилища-охладителя Ку-
раховской ГРЭС. - В кн.: Гидрохимия и гидробиология
водоемов-охладителей ТЭС СССР. Киев.
Гасанов М.В. и Эйвазова Х.С. . 1971. Изучение
биологического обрастания теплообменной системы
778 4
40
НПЗ и сезонные изменения ее видового состава. Тр.
Бакин.филиала ВНИИ водоснабж., канализ. и гидротех.
сооруж., 6.
Г о р о б и й А.Н. 1973. Экологические условия развития
зоопланктона Иваньковского водохранилища в зоне
сбросных теплых вод Конаковской ГРЭС. - В кн.: Фор-
миров. и регулир. естеств. кормов, базы искусств,
водоемов. М.
Грииь В.Г. 1971а. Фитомикробентос и его продукция в
некоторых водоемах-охладителях ГРЭС юга Украины.
Симпоз. по влиянию подогрет, вод ТЭС... Борок.
Гринь В.Г. 19716. Донная альгофлора водоемов-охлади-
телей ГРЭС Украины. - В кн.: Гидрохим. и гидробиол.
водоемов-охладителей ТЭС СССР. Киев.
Гринь В.Г., Абре мекая С.И., Виног р адская
t Т.А., К о и е н к о А.Д., Пидгайко М.Л.,
Поливанная М.Ф., Сергеева О.А. 1970.
О влиянии сброса нагретых вод ГРЭС на процессы за-
грязнения и самоочищения в водоемах-охладителях. -
В кн.: Биолог, проц, в морских и континент, водоемах.
Кишинев.
Гринь В.Г. и Виноградская Т.А. 1968. Пере-
гретые воды тепловых электространций как фактор фор-
мирования фитопланктона в водоемах-охладителях. -
В кн.: Цветение воды, т. 1, Киев.
Гринь В.Г., Вино градская Т.А., Л е н ч и н а
Л.Г. 1970. Закономерности развития бактериопланкто-
на и водорослей в водоемах-охладителях ТЭС. - В кн.:
О мерах борьбы с минер, и орг. отложен, в сист.
технич. водоснабж. и с зараст. и цвет, водохр. ТЭС.
Тезисы докл. М.
Девяткин В.Г. 1970а. Влияние термальных вод на фито-
планктон приплотинного плеса Иваньковского водохрани-
лища. Гидробиол. журн., У1, 6.
Девяткин В.Г. 19706. Влияние термальных вод на фи-
топланктон Иваньковского водохранилища. - В кн.: Би-
олог. проц, в морских и континент, водоемах. Кишинев.
Девяткин В.Г. 1971. Динамика фитопланктона в зоне
влияния подогретых вод Конаковской ГРЭС. Симпоз.
по влиян. подогрет, вод ТЭС... Борок.'
Девяткин В.Г. 1973а. Влияние повышенной температуры
на фотосинтез фитопланктона. Информ, бюлл. Инет,
биол. внутр, вод АН СССР, 16.
Девяткин В.Г. 19736. О влиянии загрязнений на разви-
тие фитопланктона в зоне воздействия подогретых вод
60
Конаковской ГРЭС. Информ, бюлл. Иист. биол. внутр,
вод АН СССР. 17.
Девяткин В.Г. 1975. Влияние подогретых вод на фито-
планктон Иваньковского водохранилища. Наст. сб.
Д у к и н а В.В. и Кудрявцева Г.С. 1970. Неко-
торые биологические и эколого-физиологические изме-
нения мизид в процессе акклиматизации. Вести. Харь-
ковск. ун-та, 39, 2.
Д у к и н а В.В. и Кудрявцева Г.С. 1971. Опыт
акклиматизации ракообразных в водоеме-охладителе
Змиевской ГРЭС (оз. Лиман). Симпоз. по влиян. по-
догрет. вод ТЭС... Борок.
Д ы г а А.К. 1971. Изменение зоопланктона в Днепровском
водохраиилише под влиянием тепловой электростанции.
Симпоз. по влиян. подогрет, вод ТЭС... Борок.
Е л а г и и а Т.С. 1975. Зоопланктон Горьковского водохра-
, нилиша в районе Костромской ГРЭС. Наст. сб.
Елизарова В.А. 1971. Состояние фотосинтетических
пигментов фитопланктона в отработанных водах Кона-
ковской ГРЭС. Симпоз. по влиян. подогрет, вод ТЭС...
Борок.
Елизарова В.А. 1973. Влияние подогрева на состояние
фотосинтетических пигментов фитопланктона в сбросных
водах Конаковской ГРЭС. Информ, бюлл. Инет. биол.
внутр, вод АН СССР, 17.
Житенева Г.С. и Никанорова Е.А. 1972. Не-
которые итоги влияния подогретых вод Конаковской
ГРЭС на биологический режим Иваньковского водо-
хранилища. Рыбохоз. изуч. внутр, вод, 8.
Журавель П.А. 1971. Об изменениях в биологии и эко-
логии мизиды Limnomysis benedeni Cz, в теп-
лых водах Приднепровской ГРЭС в зоне Днепровского
водохранилища. Симпоз. по влиян. подогрет, вод ТЭС...
Борок.
Эв'Лота рева В.И. 1971. К вопросу о влиянии Приднеп-
ровской ГРЭС на биологию Pontogammarus cras-
sus (Grimm). Симпоз. по влиян. подогрет, вод
ТЭС... Борок. •
Золотарева Н.С., Мусатов А.П. и Санин
М.В. 1973. О характере влияния термальных сбросных
вод на биоценозы водоемов. Води, ресурсы, 1.
Зубдрвва Э.Л. 1971. Высшая водная растительность
и процессы зарастания в Егоршииском водохранилище-
охладителе. Симпоз. по влиян. подогрет, вод ТЭС...
Борок.
Зубарева Э./К и Васильчикова А.П. 1970.
Высшая водная растительность водохранилищ-охлади-
телей Урала и меры борьбы с ней. - В кн.: О мерах
борьбы с минер, и орг. отложен, в сист. технич.
водоснаб. и с зараст. и цвет, водохр. ТЭС. Тезисы
докл. М.
Ивлев В.С. и Лейзерович Х.А. 1960. Экологи-
ческий анализ распределения животных в градиентных
температурных условиях. Тр. Мурман, мор. биол. инет.,
5.
Кибальчич И.А., Стрижак Е.К., Артемова
Т.З. и др. 1969. Тепловые электростанции как ис-
точники загрязнения водохранилищ. Матер, научн. кон-
фер. по пробл. гигиены, водоснабж. и санит. охр. во-
доемов. Пермь.
Кхтхд и на Л.О. 1968. Темпи росту Pontogammarus
robustoides (G-r.) у водоймипа-охолоджувач!
Кураховськой ДРЭС. 3-я Наук, конфер. молод, учен.
Ihct. гйдроб. АН УРСР. Кихв.
Китицына Л.А. 1970. Размножение и плодовитость
Pontogammarus robustoides в водоеме-охлади-
теле Кураховской ГРЭС. Гидробиол. журн., VI, 2. .
Китицына Л.А. 1971а. Влияние температурного режима
водоема-охладителя ТЭС на рост, плодовитость и ин-
• уенсивность обмена бокоплава. - В кн.: Устойчивость
к экстрем, темпер, и температ. адаптации. Харьков.
Китицына Л.А. 19716. Жизненный цикл и продукция
бокоплава Pontogammarus robustoides (Gr.)
в водоеме-охладителе Кураховской ГРЭС при разных
температурных условиях. Симпоз. по влиян. подогрет,
вод. ТЭС... Борок.
Китицына Л.А. 1971в. Интенсивность обмена Ponto-
gammarus robustoides (Grimm) в водоеме-ох-
ладителе Кураховской ГРЭС. - В кн.: Гидрохим. и гид-
робиол. водоемов-охладителей ТЭС СССР. Киев.
Кондратюк В.Г. 1968. Влияние сброса подогретых вод
тепловых электростанций на реки. Матер. Х1У конфер.
по изуч. внутр, вод-Прибалтики, Рига.
Коне нк о А.Д. и Абремская С.И. 1971. Влияние
теплообменных вод на гидрохимический и санитарный
режим водоемов-охладителей ТЭС Украины. Симпоз.
по влиян. подогрет, .вод ТЭС... Борок.
Кри в ендова Т.Д. 1968. Бактериальная продукция
Кучурганского лимана-охладителя Молд. ГРЭС. Матер.
ХГУ Конфер, по изуч. внутр, вод Прибалтики, Рига.
52
Кривенцова Т.Д. 1872. Основная бактериофлора, раз-
рушающая органические вещества в Кучурганском ли-
мане-охладителе Молд. ГРЭС. - В кн.: Биолог, ресур-
сы водоемов Молдавии, 10. Кишинев.'
Кудрявцева Г.С. и Горюнова Т.И. 1971. К
вопросу о температурных адаптациях у мизид. - В кн.:
Устойчив, к экстрем, темпер, и температ. адаптации.
Харьков.
Кузьмич В.Н. и Семенюк Г.А. 1972.' Зоопланк-
тон озера Кенои. Зап. Забайкал. фил. Геогр. обш. СССР,
62.
К у р Д и и а Т.Н. и Девяткин В.Г. 1972. О влиянии
подогретых вод Конаковской ГРЭС иа содержание кис-
лорода и развитие фитопланктона в Иваньковском водо-
хранилище в зимний период. Гидробиол. журн., VII, 4.
Лавыгииа З.П. 1973. К вопросу о влиянии теплообмен-
ных вод электростанций на биологический режим водое-
мов. - В кн.: Предупредительная медицина. Кемерово.
Л а й о ш Ф. 1970. Загрязнение воды вследствие теплового
воздействия. Будапешт.
Л е и ч и н а Л.Г. 1971а. Численность бактерий группы кишеч-
’ ной палочки в водохранилище Кураховской ГРЭС.
Гидробиол. журн., VII, 1.
Л е н ч и н а Л.Г. 19716. Особенности развития бактериофло-
ры в водоемах-охладителях тепловых электростанций.
Симпоз. по влиян. подогрет, вод ТЭС... Борок.
Лисицына Л.И. и Жукова Г.А. 1971. О раститель-
ности Мошковичского залива Иваньковского водохрани-
лища. Симпоз. по влиян. подогрет, вод ТЭС... Борок.
Литвинов А.С. и Ривьер И.К. 1972. О верти-
кальном распределении зоопланктона в Мошковичском
заливе в связи с особенностями гидрологических усло-
вий. Информ, бюлл. Инет. биол. внутр, вод АН СССР,
13.
Ма ксимова Г.Д, 1973. Экологические условия развития
личинок хирономид (Chironomus f. 1. semireductus)
в отепленной и неотепленной зонах Иваньковского
водохранилиаЩ. - В кн.: Формиров. и регулир. естеств.
кормов, базы искусств, водоемов. М.
Максимова Г.Д., Ника норов Ю.И. и Ф л е й с
М.Л. 1971. Влияние сбросных теплых .вод Конаковской
ГРЭС на гидрохимический и биологический режим Ивань-
ковского водохранилища. Симпоз. по влиял, подогрет,
вод. ТЭС... Борок,
53
Ма ма ев a. Н.В. 1975. Влияние сбросных теплых вод
Конаковской ГРЭС на планктонных простейших Ивань-
ковского водохранилища. Наст. сб.
Мирошниченко А.З. 1971. Зоопланктон водоема-ох-
ладителя Змиевской ГРЭС. - В кн.: Гидрохим. и гид-
робиол. водоемов-охладителей ТЭС СССР. Киев.
Митропольский В.И. 1974. Зообентос Иваньковско-
го водохранилища в районе сброса подогретых вод Ко-
наковской ГРЭС. Информ, бюлл. Инет. бнол. внутр,
вод АН СССР, 24.
Мордухай-Болтовской Ф.Д. 1971. Состояние воп-
роса о влиянии подогретых вод теплоэлектростанций
на биологию водоемов. Симпоз. по влиян. подогрет,
вод ТЭС... Борок.
Набережный А.И. и Витковская Е.Д. 1972.
Состав, численность и некоторые стороны биологии
Harpacticoide в Кучурганском лимане-охладителе
Молд. ГРЭС. - В кн.: Биолог, ресурсы водоемов Мол-
давии. 10. Кишинев.
Пидгайко МЛ. 1967, Координационное совещание по
вопросам биологического, гидрохимического и санитар-
ного изучения водохранилищ-охладителей тепловых
электростанций. Гидробиол. журн., III, 6.
Пидгайко М.Л. 1971а. Влияние подогрева на развитие
и продуцирование зоомакробентоса в водоемах-охлади-
телях ГРЭС юга УССР. Симпоз. по влиян. подогрет,
вод ТЭС... Борок.
Пидгайко М.Л. 19716. Зоомакробентос Кураховского
водохранилища и влияние на него подогрева. - В кн.:
Гидрохим. и гидробиол. водоемов-охладителей ТЭС
СССР. Киев.
Пидгайко М.Л. 1971,в. Материалы к сравнительной физи-
ко-географической характеристике водоемов-охладите-
лей электростанций Украины. - В кн.: Гидрохим. и гид-
робиол. водоемов-охладителей ТЭС СССР. Киев.
Пидгай. ко М.Л., Г р и и ь В.Г., Поливанная М.Ф.
и др. 1967. Гидробиологический режим водоемов-охладителей
тепловых электростанций Украины. ГиДробиол. журн., Ш, 5.
Пидгайко М.Л., Гринь В.Г., Поливанная
М.Ф. и др. 1970. Итоги изучения гидробиологичес-
кого режима пресных водоемов-охладителей юга СССР.
Гидробиол. журн., VI, 2.
Пидгайко МьЛ., Поливанная М.Ф. и Сергее-
ва О.А. 1970. Значение зоопланктона и зообентоса
водоемов-охладителей в практике водопо'льзования
54
ТЭС. - В' кн.: О мерах борьбы с минер, и орг. отложен,
в сист. технич. водоснабж. и зараст. и цвет, водохр.
ТЭС. Тезисы докл. М.
Поддубная Т.Н. 1968. Донная фауна Иваньковского
водохранилиша в зоне сброса теплых вод Конаковской
ГРЭС. - В кн.: Волга-1. 1 конфер. по изуч. водоемов
басе. Волги. Тезисы докл. Тольятти.
ПоДДУбная Т.Л. 1971а. Донная фауна Иваньковского
водохранилища в районе сброса теплых вод Конаковской
ГРЭС. Тр. Инет. биол. внутр, вод АН СССР, 21 (24).
Поддубная Т.Л. 19716. Динамика донного населения
Иваньковского водохранилиша в районе сброса теплых
вод ГРЭС и за его пределами.’ Симпоз. по влиян.
подогрет, вод ТЭС... Борок.
Поливанная М.Ф. и Сергеева О.А. 1971а. К биологии
массовых видов Cladocera. водоема-охладителя Курахов-
ской ГРЭС. Гидробиол. журн., УН, 6.
Поливанная М.Ф. и Сергеева О.А. 19716.
Зоопланктон'водоемов-охладителей тепловых электро-
станций Украины. - В кн.: Гидрохим. и гидробиол. во-
доемоа-охладителей ТЭС СССР. Киев.
Поливанная М.Ф. и Сергеева О.А. 1971в.
Динамика зоопланктона и биология доминирующих видов
в водоемах-охладителях ГРЭС юга Украины. Симпоз.
по влиян. подогрет, вод ТЭС ... Борок.
Проссер Л. и Браун Ф. 1967. Сравнительная фи-
зиология животных. Гл. 9. Температура. М.
П ы р и н а И.Л. и Наумова Е.И. 1971. Фотосинте-
зирующая деятельность фитопланктона в отработанных
водах Конаковской ГРЭС. Симпоз. по влиян. подогрет,
вод ТЭС... Борок.
П ы р и н а И.Л. и Наумова Е.И. 1973. Интенсивность
фотосинтеза фитопланктона в сбросных водах Конаков-
ской ГРЭС. Информ, бюлл. Инет. биол. внутр, вод
АН СССР, 17.
Р а ч ю н а с Л.А. 1971а. Распределение беспозвоночных
в водохранилище-охладителе Литовской ГРЭС. Гидробиол.
журн., УП, 1.
Р а ч ю н а с Л.А. 19716. Распределение зообентоса в водо-
хранилище-охладителе Литовской ГРЭС. Информ, бюлл.
Инет. биол. внутр, вод АН СССР, 10.
Рачюнас Л.А. 1973. Состояние фауны беспозвоночных и
питание рыб водохранилища-охладителя Литовской ГРЭС
при новых экологических условиях. Автореф. дисс.
Вильнюс.
55
Ривьер И.К. 1968. Влияние сбросных теплых вод Кона-
ковской ГРЭС иа зоопланктон Иваньковского водохра-
нилища. - В кн.: Волга-1. 1 конфер. по изуч. водое-
мов басе. Волги. Тезисы док л. Тольятти.
Ривьер И.К. 1971. Зоопланктон Мэшковичского зали-
ва, подверженного влиянию подогретых сбросных вод
Конаковской ГРЭС. Симпоз. по влиян. подогрет, вод
ТЭС... Борок.
Ривьер И.К. 1972. Поведенческие реакции планктонных
рачков в зоне наибольшего влияния сбросных вод
Конаковской ГРЭС. - В кн.: Поведение водных беспозв.
Борок.
Ривьер И.К. 1975. Зоопланктон Иваньковского водохра-
нилища в зоне влияния подогретых вод Конаковской
ГРЭС. Наст. сб.
Ривьер И.К. и Гец Н.А. 1971. Зоопланктон Мошко-
вичского залива, подогреваемого сбросными водами
Конаковской ГРЭС. Информ, бюлл. Инет. биол. внутр,
вод АН СССР, 12.
Романова А.П. 1972. Бактериальная флора приплотинно-
го плеса Иваньковского водохранилища, испытывающе-
го влияние подогретых вод Конаковской ГРЭС. Рыбо-
хоз. изуч. внутр, вод., 8.
Сергеева О.А. 1968. Рхчна та сезониа д!намика зоо-
планктону водоймиша-охолоджувача Кураховськой
ДРЭС. 3-я Наук, конфер. молод. учен.‘1нст. г!дроб.
АН УРСР. Ки1в.
С ергеева О.А. 1971. Изменение биологических показа-
телей и продукция Ceriodaphnia reticulata (jur.)
и Diaphanosoma brachyurum (Liev.) в разных
температурных условиях водоема-охладителя К'урахов-
. ской ГРЭС. - В кн.: Устойчив, к экстрем, темпер, и
температ. адаптации. Харьков.
Скальская И.А. 1975. Состав и распределение зообен-
тоса Горьковского водохранилища в районе Костромской
ГРЭС. Наст. сб.
Смирнова Н.Ф. 1975. Рост и продукция унионид (Mol-
lusc а, Unionidae) в районе Костромской ГРЭС.
Наст. сб.
С пиглазов Л.П. 1972. Численность бактерий в озере
Кенон. Зап. Забайкал. фил. ГеОграф. общ. СССР, 62.
Стангенберг At 1967. Естественные следствия сбро-
са теплых вод в реки. - В кн.: Санитарн. и технич.
гидробиол. М
58
Сысолятина Т.Л. 1971. Первичная продукция и чис-
ленность бактерий в воде водоемов-охладителей ГРЭС’
№ 3. Симпоз. по влиян. подогрет, вод ТЭС... Борок.
'Тагирова Н.А. 1969. Опыт культивирования Cerio-
daphnia. reticulata (Jur.) на теплых водах ГРЭС. -
В кн.: Рыбоводство в теплых водах. М.
Топачевский А.В. и Пидгайко М.Л. 1971.
•Цели и задачи гидробиологического исследования водое-
мов-охладителей тепловых электростанций. - В кн.:
ГидроХим. и гидробиол. водоемов-охладителей ТЭС
СССР. Киев.
Федоров а Е.И. 1971. Некоторые черты гидрохимичес-
кого и биологического режима водоемов-охладителей.
Симпоз. по влиян. подогрет, вод ТЭС... Борок.
Федорова Е.И. 1972. О некоторых особенностях фито-
планктона водоемов-охладителей. Гидробиол. журн.,
УШ, 2.
Филимонов Л.А. и Соколова Г.В. 1971. О на-
хождении все ленца южной фауны Moina .weberi
Rich, в водоемах Костромской обл. Уч, зап. Костром,
пед. инет., 29.
Харин Н.Н. 1971. Развитие зоопланктона в каналах Ново-
черкасской ГРЭС. Симпоз. по влиян. подогрет, вод
ТЭС... Борок.
Чаплина А.М. и Махонина А.В. 1971. Влияние
подогретых вод Криворожской ГРЭС-2 на гидробиологи-
ческий режим водохранилища. Симпоз. по влиян. подо-
грет. вод ТЭС... Борок.
Ill а л а р ь В.М., Б ы з г у С., Усата М. 1968. Доев!
прхгничения синьозелених водоростей в Кучургансько-
му ли мани охолоджувач! Молдавськой ДРЭС. 3-я Наук,
конфер. молод, учен. 1нст. гхдроб. АН УРСР. Кихв.
111 а л а р ь В.М. и Яловицкая Н.И. 1971. Развитие
фитопланктона в Кучурганском лимане-охладителе
Молд. ГРЭС. - В кн.: Гидрохим. и гидробиол. водоемов- .
охладителей ТЭС СССР. Киев.
Шахматова Р.А., Тухсанова Н.Г. и Ю л о в а
Г.А. 1971. Биологическая характеристика термального
водоема ГОГРЭС им. А.В.Винтера. Симпоз. по влиян.
подогрет, вод ТЭС... Борок.
Шиманский Б.А. 1956. Борьба с биологическими поме-
хами, нарушающими нормальную эксплуатацию водохра-
нилищ,. Электрич. станции, 7.
57
Шиманский Б.А. 1962. Борьба с высшей водной растительностью
в водохранилищах-охладителях. Электрич. станции, 33, 1.
Шиманский Б.А. 1963. Активные Меры борьбы с зарастанием во-
дохранилищ-охладителей. Тр. Всесоюз. гидробиол. общ., 14.
Шиманский Б.А. 1965а. Развитие синезеленых водорослей в во-
дохранилищах-охладителях тепловых электростанций и меры
борьбы с ними. - В кн.: Экология и физиология синезеленых
водорослей. М-Л.
Шиманский Б.А. 19656. Технико-гидробиологическая характерис-
тика водохранилищ-охладителей тепловых электростанций.
Гидробиол. журн., '1, 2.
Шиманский Б.А. 1968. Биологические помехи в эксплуатации
систем технического водоснабжения тепловых электро-
станций и методы борьбы с ними. Гидробиол. журн.,1 У, 2.
Шиманский Б.А. 1970. Высшая и низшая водная раститель-
ность в водохранилищах-охладителях ТЭС, предотвраще-
ние ее развития и методы борьбы. - В кн.: О мерах
борьбы с минер, и орг. отложен, в сист. технич. водо-
снабж. и зараст, и цвет, водохр. ТЭС, Тезисы докл. М
Шишкин Б.А. 1972. Физико-географическая характеристика
озера Кенон (водоема-охладителя Читинской ГРЭС).
Зап. Забайкал. фил. Географ, общ. СССР, 62.
Шишкин Б.А., Вологдин МП., Гурова Л.И. и др. Ре-
жим водоема-охладителя Читинской ГРЭС (оз. Кенон).
Симпоз. по влиян. подогрет, вод ТЭС... Борок.
Шишкин Б.А., Спиглазова Г.Н. и Локоть Л.И.
1972. Первичная продукция озера Кенон. Зап. Забайкал.
фил. Географ, общ. СССР, 62.
Юревичюте В.С., Яскелявичене ' Б.П. и Вилене
В.В. 1971. Динамика фито- И зоопланктона теплых вод
Литовской ГРЭС. Симпоз. по влиян. подогрет, вод ТЭС...
Борок.
Ярошенко МФ., Шаларь В.М, Набережный А.И. и
Кубрик И.Ф. 1970. Биологические причины ухудшения
технических качеств воды в Кучурганском лимане-охлади-
теле Молд. ГРЭС й пути их устранения. - В кн.: Биолог,
ресурсы водоемов Молдавии, в. 6, Кишинев.
Экзерцев В.А. и Лисицына Л.И. 1975. Растительность'
Иваньковского водохранилища и влияние на нее подогре-
тых вод Конаковской ГРЭС. Наст. сб.
Adams J,R. 1969а. Ecological investigations
around some thermal power stations in Califor-
nia tidal waters, Chesapeake Science, 10.
Adams J.R. 1969b. Thermal power, aquatic life
and kilowatts on the /Pacific Coast. Nuclear
News, 75.
jabaster J.S. 1963. The effect of heated
effluents on fish. Int. Journ. air and water pollut.,
7.
Allen T.F. 1969. Research needs for thermal
pollution control. Biol. asp. therm, pollut.
дпй erso n R.R. 1969. Temperature and rooted
aquatic plants. Chesapeake Science, 10.
дпееН Ai), 1969. Thermal releases and shell-
fish culture: possibilities and limitations. Chesa-
peake Science, 10.
yxppourchaux M. 1965. Effects de la tempe-
rature de I’eau sur la faune et la flore aquati-
ques. L’eau, 5 2, 7.
/s/r n о 1 d G.E. 1962. Thermal pollution of surface
supplies. J.Americ, Wat. Work. Ass., v, 54.
As to n R.A. 1971. The effects of temperature
and dissolved oxygen concentration on repro-
duction in Limnodrilus hoffmeisteri (Clap.) and
Tubilex tubilex (Mui,). Sympos. freshwat. biol.
and electr. power gener,, II.
'Bader R.G., Roessler M.A., Thorhaug
A. 1970. Thermal effluents in a tropical mari-
ne estuary. Rosenst. School mar, atm. Sci.,
Univ. Miami,
Bader R.G. and Tabb D.C. 1970. An eco-
logical study of south Biscayne Bay in the
vicinity of Turkey Point, University of Miami.
Barnett P.R.O. 1971. Some changes in intertidal
sand communities due to thermal pollution. Pro-
ceed. Roy, Soc, London, B. 177, No 1048.
Barnett P.R.O. 1972. Effects of warm wafer
effluents from power stations on marine Life.
Proceed, Roy. Soc, London, B. 180, No 1061.
Barnett P.R.O, and Hardy B.L.S. 1969.
The. effects of temperature on the benthos near
the Hunterston generating station, Scotland.
Chesapeake Science, 10.
Beauchamp R.S.A, 1969. The central electri-
city generating board’s contribution to the stu-
dy of coarse fish. Proceed 4^ Brit, coarse x
fish conf,, Liverpool.
Beauchamp R.S.A., Ross F,F. and Whi-
te h о u s J,W, 1970. The thermal enrichment
of aquatic habitats. Proc. 5^ Intern, water poll,
res, conf,, 1-11.
58
50
Belehrddek J. 1935. Temperature and living
matter. Borntraeger, Berlin.
Brett J.R. 1960. Thermal requirements of fish.
Three decades of study. Biolog. prob], water
pollut., U.S.Public Health Service, Publ.
60-3.
Brezina E., Campbell R., Whitley J.
1970.- Thermal discharge and water quality in
a 1500-acre reservoir. Jour. W.P.C.F., 42, 1.
В r u n g s W.A. 1970. Effect of heated water from
nuclear plants on aquatic life. Nucl. Power and
public. Minneapol.
C air ns J., jr. 1956. The effects of increased tem-
perature upon aquatic organisms. Indust, was-
tes, 1(4).
Cairns J., jr. 1969. The response of fresh-water
protozoan communities to heated waste waters.
Chesapeake Science, 10.
Cairns J., jr. 1971. Thermal pollution — a cause
for concern. Jour. W.P.C.F., 43, 1.
Chang ТЛ. 1971. Thermal pollution and its effects
on aquatic life. Tech. Eng. News, 53, No 1.
Churchill M.A. and Wojtalik T.A. 1969,
Effects of heated discharges: the T.V.A. experi-
ence. Nucl. News, 12, 9.
Clark J.R. 1969. Thermal pollution and aquatic
life. Scient. Americ., 220.
Clark J.R. and Silverman ,M.J. 1969. . .
The thermal pollution controversy - ecological
comments. Proc ЗО^1 I nt, Wat. Confer., Pitt-
sbourgh.
Clarke K.U. and Credland P.F. 1971.
Some aspects of temperatire tolerance in Chiro-
nomus riparius M. and Ch. dorsalis M. Sympos.
freshwat. biol. and electr. power, gener., II.
Cory R.L. and Naumann J.W, 1969. Epi-
fauna and thermal additions in the upper Patu-
xent river estuary. Chesap. Science, 10.
Couta nt C.C, 1962. The effect of a heated water
effluent upon the macroinvertebrate riffle fauna
of the Delaware river. Proceed. Pennsylv. Acad.
Sc., XXXVI.
Couta nt C.C. 1969, Temperature, reproduction
and behavior. Chesapeake Science, 10, 3/4.
60
с ° u
tant ’C.C. 1970. Biological aspects of ther-
mal pollution. I. Entrainment and discharge canal
effects. CRC Crit, Rev, Environ. Contr., 1, 3.
r o u t a n t C.C. 1971. Effects on organisms of
G entrainement in cooling water: steps toward
predictability. Nuclear Safety, 12.
z- o u t a n t C.C. 1972a. Evaluating the ecological
impact of steam electric stations on aquatic
systems. Ann. meet. Amer, assoc. Advan. Sc.,
27-28 Dec. 1972, Wash.
Content C.C. 1972b. Biological aspects of ther-
mal pollution. II. Scientific basis for water tem-
perature standards at power plants. CRC Criti-
cal reviews in environm, control, 3, 1.
С о u t a n t C.C. 197 2c. Time-temperature relation-
ships for thermal resistances of aquatic orga-
nisms, principally fish. Oak Ridge Nation. La-
bqrat,, Tenn,, ORNL-EIS-27.
Coutant C.C. 1973. Thermal effects. Oak Ridge
nation, labor., Environ, sc. divis., Annual prog-
ress report;
С о u t a nt C.C. and Becker C.D. 1970.
Growth of the Columbia river limpet, Fischero la
nuttali (Hald.) in normal and reactor warmed wa-
ter. AEC research and development report.
Craven R.E. and Brown B.E. 1970, Power
plant heated discharge water and benthos in
Boomer lake. Jour. Kans. Entom, Soc„ 43.
D aj о z ,R. 1970. Les eaux polluees par la cha-
leur. Sci. progr. decouv,, No 3422.
Davis W. 1969. Conditions for coexisting of
aquatic communities with the expanding nuclear
power industry. - Nuclear Safety, 10, 4.
Drew H.R. and T i 11 о n J.R. 1970. Thermal
requirements to protect aquatic life in Texas
reservoirs. Jour. W.P.C.F., 42.
.Dysart B.C. and К r e n к e 1 P.A. 1965. The
effects of heat, on water quality. Proceed. 20^h
Industr. waste confer,, Purdue Univ.
Eicher GJ, 1967. Nuclear power generation and
aquatic life. Proc. 47^ Ann, Conf, west assoc,
state game and fish commis., Honolulu. S. 1.
Elster H.J. 1968. Einige limnologische und
fischereiliche Gesichtspunkte zur Aufheiiung
unserer Flieszgewasser durch Kiihlwasserein-
leitungen. Der Fischwirt, n, 12.
61
F enlon M.W., McNaught D.C. and S chro-
der GJD. 1971. Influence of thermal efflu-
ents upon aquatic production in lake Ontario. ~
Proc. 14-th confer> Great lakes res.
Florczyk H., Golowin S. and S о 1 s к i
A. 1972. Einige physische, chemische u. biolo-
gische Aspekte uber d. Einfluss erwSrmten
Wassers auf die Gewasser der Wisla. Fortbil-
dungs Kursus, Aktuelle Probl. Wassergutewirt.,
Dresden,
Fox J.L, and- Moyer M.S. 1973. Some effects
of a power plant on marine microbiota. Chesapea
ke Science, 14, No 1.
Gadkowski M. 1971. О dopuszczalnym podgrze-
waniu wody w rzece przez elektrownie ciepine
Gospod. wodna, 31, No 12.
Gammon K.M. 1968. Planning the use of water
for cooling purposes in power stations. Wat.
research, 2.
Gammon K.M, 1969. Planning cooling water for
power stations, Advan. wat, pollut., Proc. 4.
Int, Conf., Prague.
Gi no с c hi о . R. 1963. Les. besoins en eau pour
la production d’energie thermique et nucleaire.
L’eau, 50 е Annee., No 2.
Glooschenko W. 1968. Statement. Hearings of
the Committee on public works, United State
Senate XIX Congress. Thermal pollution.
Glooschenko W.A. and Glooschenko
K.A. 1969. Thermal pollution. Florida Natura-
list, 42, No 1,
G о u b e t A. 1965. Influence des centrales thermi-
ques sur les cours d'eau. Electricite de France,
No 12.
G r i m a s V. 1970a. Heat - a question of life and
death. Zool, Revy (Swed.), 32, 91.
Grim as V. 1970b. Warm water effluents in Sweden.
Marine poll. bull. (G.B.), 1, 151.
Grimes Ch.B. 1971. Thermal addition studies of
the Crystal River steam electric station. Profess.
Pap. Ser. Florida Dep. Natur. Resour., No 11.
Hamilton D.H., Flemer D.A., Keef C.W.,
Mihurski I,A. 1970. Power plants: effects
of chlorination on estuarine production. Science,
169, 197,
82
a rg i s Jr. and I.E.W a r i n n e r, 197,1.
Impacts of thermal alterations on estuarine and
coastal environments. Envir. aspects nucl. po-
wer stat., Proceed, Sympos,, Vienna.
H’awkes H.B. 1971. Water in electricity genera-
tion. Sympos. freshwat. biol. and electr, power
gener,, I.
Redden D.T. 1969. Utility company initiates
study of effects of „thermal pollution". Water and
Sewage works, v. 116, No 11.
H e i nl e D.R. 1969a. Temperature and zooplank-
ton. Chesapeake Science, 10, 3—4.
geinle D.R. 1969b. Patuxent thermal studies,
supplementary report. Thermal loading and the
zooplankton community. Natur. Resour. Instit,,
Ref. No 69-8. Univ, of Md.
pj © a к R.D. 1961. The thermal pollution problem.
Jour. W.P.C.F., 33, 12.
Horvath R.S. and Brent M.M. 1972. Ther-
mal pollution and the aquatic microbial commu-
nity: possible consequences. Environ.,pollut.,
3, No 2.
Hubbs C. 1972. Some thermal consequences
of environmental manipulations of water. Biol.
Conserv,, 4, No 3.
Hynes H.B.N., 1959. The biological effect of
water pollution. The effects of pollution on living
material, ed. by W.Yapp, London, 11.
Iles R. 1963. The subtropical waters of Britain.
Journ. Instit. electric, engineers, v, 6.
lohannes R.E. 1970. How to kill a coral reef.
Mar. PoUut. Bull., 1. No 12.
Jaske R.T., Templeton W.L. and Cou-
tant C.C. 1970, Methods for evaluating
effects of transient conditions in heavily loaded
and extensively regulated streams. Chemie,
engineer, progress, Sympos. ser., v. 67, No 107.
Jensen L.D. 1969. Spectrum of biological con-
cerns from power plant thermal discharges.
Environm, aspects nucl. power stat., Proceed.
Sympos., Vienna.
Jones M.L. 1964. Thermal pollution: the effects
of heated effluents. Fish and river pollution,
Butterworth, London.
Kempner E.S. 1963. Upper temperature limit
of life. Science, 142.
63
Kennedy V.S. and Mihursky J. A. 1971.
Upper temperature tolerance of some estuarine
bivalves. Chesapeake Science. v, 12, No 4.
Kinne O. 1970. Temperature. In: Marine ecology,
ed. by O,Kinne, Chapter 3.
Langford T.E. 1971a. The biological assessment
of thermal effects in sone british rivers. Symp,
freshwat. biol. and electr. pow. gener., 1.
Langford T.E, 1971b. The distribution, abundan-
ce and life-histories of stoneflies and mayflies
in a british river, warmed by cooling water from
a power station. Hydrobiologia, V» 3 8, is. 2,
Langford T.E. and Aston R.J. 197 2. The
ecology of some British rivers in relation to
warm water discharges from power stations.
Proceed, Roy. Soc, London, B180, No 1061.
Levin- A,, Birch T„ Hillman R„ Rai-
nes G. 1972. Thermal discharges: ecolo-
gical effects. Envir. Science abd Technology, 6,
No 3.
Leynaud G. 1967. Les pollutions thermiques.
Influence de la temperature sur la vie aquatique.
Bull, techn. inform., 24.
Lyons W,G., Cobb S.P., Camp D.K.,
Mountain J.A,, Savage Th., Lyons
L., Joyce E.A, 1971. Preliminary inventory
of marine invertebrates collected near the elect-
rical generating-plant Crystal River in 1969. -
Fla Dep. Natur. Resour. Mar. res. lab., prof.
pap. ser„ No 14.
Mann K.H, 1965. Heated effluents and their effects
on the invertebrate fauna of rivers. Proc, Soc.
Water treatment and examination, 14.
McErlean, Mihursky J.A. and Brink Ге у
H.T, 1969. Determination of upper temperature
tolerance triangle for aquatic organisms. Chesa-
peake Science, 10.
McLaren LA, 1963. Effects of temperature on
growth of zooplankton, and the adaptive value
of vertical migration, Journ. Fisheiy res. board
Canada, vol, 20, No 3.
McNaught D. and Fenlon N.W. 1972. The
effects of thermal effluents upon secondary
production Verh, Int. Verein. LimnoL» 18.
64
j^arlc°wski S. 1959. The cooling water of
power stations: a new factor in the environment
of marine and freshwater invertebrates. The
journ. anim. ecol., v. 28, No 2.
Markowski S. 1960. Observations on the res-
ponse of some benthonic organisms to power
station cooling water. The journ. anim. ecol.,
v. 29, No 2.
Merriman D. 1971. Does industrial calefaction
jeopardize the ecosystem of a long tidal river?
Environm, aspects nucl. power stat., Proceed.
Sympos,, Vienna.
Merriman D. 1972. Power stations and aquatic
ecology. Proceed. Amer, power confer., v. 34,
Chicago.
Merriman D. 1973. Calefaction of the Connecti-
cut River, U.S.A. Transact. New York Acad.Sci.,
35, No 1.
Mord ukh ai-B о Ito vs koi Ph. D. 1970. The
effect - of the heated water, discharged from the
cooling system of the Konakovskaja power
station, on the biology of Ivankovskoe reservoir.
Unesco, IBP Sympps, Productiv., II. Warszawa.
Morgan R.P. and Stross R.G, 1969. Des-
truption of phytoplankton in the cooling water
supply of steam electric station. Chesapeake
Science, 10, 3—4.
Mount D.D. 1971. Thermal standards in the Uni-
ted States of Amerika. Environm. aspects of
nuclear power stations, Proceed. Sympos.,
Vienna.
Nauman J.W. and Cory R.L. 1969. Thermal
additions and epifaunal organisms at Chalk
Point, Maryland. Chesapeake Science, 10,3-4.
Naylor E« 1965. Effects of heated effluents
upon marine and estuarine organisms. Advan.
marine bioL, v. 3.
N e b e ke r A.V. 1971. Effect of high winter water
temperature on adult emergence of aquatic in-
sects. Water Res. (G.B.), 5.
P a n n e 1 I.P.M., Johnson A.E. and R a y-
mo nt J.E. 1963. An investigation into the
effects of warmed water from Marchwood power
station into Southhampton water. Proc. Inst,
civil, engin., 23.
778 5
65
Pat alas К. 1970. Primary and secondary pro-
duction in a lake heated by thermal power plant.
Proc. Jnst. Environ. Sciences,
Patrick R. 1969. Some effects of temperature
on freshwater algaa Biologic. Aspects Therm.
Pollut», Proc. nat. sympos., ed. P.Krenkel and
F.Parker, Nashville, Tenn.
P e a r c e D.W. 1965. Plankton of an artifically
heated dock. Ann. Rep. Challenger soc. 3 (16).
Pearce J.B. 1969. Thermal addition and the ben-
thos, Cape Cod' canal, Chesap. Sci., 10, 3—4.
Pidgaiko M.L. et al. 1970. Biological producti-
vity of the Kurachov cooling reservoir. UNESCO,
IBP Sympos. on productiv,, V. II, Warszawa.
Poltoracka J. 1970, . Wplyw zrzutu wod podgrze-
wanych na sklad gatunkowy planktonu roslin-
nego w jeziorach okolic Konina. Gospod. rybna,
22, No 12.
Poltoracka J, 1968. Specific composition of
phyroplankton in a lake warmed up by wastewa-
ter from a thermal power station and in lakes
with normal temperatures. Acta Soc, Bot, Pol-
lut,, 37,
Pourriot R„ Rouyer G. et Peltier M.
1972. Proliferation de rotiferes epiphytes et
pollution thermique dans la Loire, - Bull, franc,
piscicult., 44, No 244.
Powell C.J. and Waugaman C.H. 1969.
Studies show need for thermal-effect data. -
Electr, world, Sept. 1969.
R a у m о n d M.P, and Raymond S.G, 1969.
Destruction of phytoplankton in the cooling water
supply of a steam electric station. Chesapeake
Science, No 10.
Reeve M.R, 1972. Seasonal changes in the zoo-
plankton of South Biscayne bay and some prob-
lems of assessing the effects on the zooplank-
ton of natural and artificial thermal and other
fluctuations. Bull mar. Scienc., -20, 4.
Reeves J.N, 1970. Effects of thermal discharge
from the San Onofre nuclear generating station.
Water and sewage works, v. 117, No 2.
Roessler M.A. 1971. Environmental changes
associated with a Florida power plant. - Marine
Poll, bullet., 2. 87.
ее
р о о s enburg W.H. 1969. Greening and cooper
accumulation in the american oyster, Crassostrea
virginica, in the vicinity of a steam electric ge-
nerating station. Chesap. Sci., 10, No 3-4.
Rose A.N., edit. 1967. Thermobiology. Acad.
Press, London.
Ross F.F. and Whitehouse J.W. 1973.
Cooling towers and water quality. Water Research
v. 7.
Rothwell D.R. 1971. The calefaction of lake
Trawsfynydd. Sympos. freshwat. biol. and electr.
power gener., It
Singer S.F. 1968. Waste heat menagement.
Science, v. 159, No 3820.
Snyder G.R. and Blahm Т.Н. 1971. Effects
of increased temperature on cold-water orga-
nisms. Journ. W.P.C.F., 43.
Sorokin C. 1971. Calefaction and phytoplankton.
Bioscience, v. 21, No 23.
Speakman J.N. and Krenkel P.A. 1972.
Quantification of the effects of rate of tempera-
ture change on aquatic biota. Water research,
v. 6.
Sprague J.B. 1963. Resistance of four freshwa-
ter crustaceans to lethal high temperature and
low oxygen. Journ, Fishery res. board Canada,
vol. 20. No 2,
Stangenberg M. and Pawloczyk M.
1969. The influence of a warm water influx from
a power staticn upon the formation of biocoeno—
tic communities in a river. Zesz. nauk. polit.
Wroczlaw, No 40.
Steidinger K. A. and Van Breedveld J.F.
1971. Benthic marine algae from waters adja-
cent to the Crystal River electric power plant
(1969 and 1970). - Fla Dep. natur resour. mar.
res. lab., prof. pap. ser„ No 16.
Sylvester J.R. 1972. Possible effects of therma
effluents on fish: a review. Environm, Pollut.,
3, No 3.
Tarzwell C1.M. 1970. Thermal requirements to
protect aquatic life. Journal W.P.C.F., 42, No 5.
Templeton W. and Coutant C.C. 1971.
Studies on the biological effects of thermal dis-
charges from nuclear reactors to the Columbia
67
river at Hanford. Environ, aspects, nucl, pow.
stat., Proceed. Sympos., Vienna.
Thermal effects and U.S. Nuclear power stations.
Jan. 1971. Divis. reactor develop, technol. U.S,
atom, energy commis.
Thermal pollution of Cayuga Lake by a proposed
power plant, 1968. Ithaca, N.Y.
Thorslund А.-Ч 1971. Potential uses of waste
and heated effluents. EIFAC Occasion, papers,
No 5, FAO, Rome.
Thorhaug A., Roessler M., S e g a r D.,
Bader R„ Gerchakov S. 1973.
The environmental impact of a power plant
on a subtropical bay. Transact. Amer. Nucl.
Soc,, 16, Suppl. No 1.
Trembley F.J. 1965. Effects . of cooling water
from steam-electric power plants on stream
biota. Biol, probl, in water pollut. U.S.Public
health service, Publ. No 999-WP-25.
Turoboyski L. 1969. Chemiczno-biologiczne
badania wod nad wplywem wod podgrzewanych
z elektrowni w Skawinie na Wisle i Skawinke.
Mater, bad., Instyt, Gob pod ar ki wodnej. Tom IV,
z. 2.
Vaillancourt G., С о u t u r e R., Dube J.
1972. Influence d*un' apport thermique sur une
biocenose fluviale. 6е Congr. int, tendances nouv.
radioprot., Bordeaux.
Warinner J.E, and Brehmer M.Z. 1966.
The effects of thermal effluents on marine orga-
nisms. Int. Jour. Air and water pollut., 10, No 4.
Warren Ch.E. 1971. Biology and water pollu-
tion control, W.Saunders Company.
Whitehouse J.W, 1971. Some aspects of the
biology of lake Trawsfynydd: a power station
cooling pond. Hydrobiologia (Neth.), 38, is. 2.
Wiederholm T. 1970. An exotic oligochaete,
Branchiura sowerbyi Bed,, in a warm water
effluent in lake Malaren. - Naturvardsverk. Lim-
nolog. undersokning., No 37.
Wiederholm T. 1971. Bottom fauna. and cooling
water discharges in a basin of lake Malaren.
Inst, freshwat. res. Drottningholm. Rep. No 51.
Wood L.J. and Z i e m a n J.C. 1969. The
68
effects of temperature on estuarine plant commu-
nity. Chesap. Sci., 10, 3/4.
Wright J.H., Champlin J.B.F., Davis O.H.
1971. The impact of environmental radiation
and discharge heat from nuclear power plants.
Environm. aspects nucl. power stat., Proceed.
Sympos., Vienna.
Wurtz C.B. 1967. Misunderstandings about heated
discharges. Ind. water Eng. No 9.
Wurtz C.B. 1969. The effects of heated dischar-
ges on freshwater benthos. Biol, aspects therm,
pollut., ed. Krenkel and Parker, 199.
Wurtz Ch.B. and Dolan Th. 1961. A. biolo-
gical method used in the evaluation of effects
of thermal discharge in the Schuylkill River. -
Proc. 15^ industr. waste confer. Purdue Univ.
Zhitenjowa T.S. and Nikanorow J.I.
197 2. Der Einfluss des vom Konakowschen
Warmekraftwerk abfliessenden Warmwasser auf
die biologischen Prozesse im Iwanjkowo—Stau-
see, Verh. Int. Verein. Limnol., 18, II.
Н.В.Б у т о р и н, Т.Н.К у р ди на
ОСОБЕННОСТИ ТЕМПЕРАТУРНОГО РЕЖИМА
ИВАНЬКОВСКОГО ВОДОХРАНИЛИЩА
В УСЛОВИЯХ ИСКУССТВЕННОГО ПОДОГРЕВА
Общая характеристика
водохранилища
»
Иваньковское водохранилище расположено в бассейне верх-
ней Волги. Климатические условия бассейна характеризуются
умеренно холодной зимой и умеренно теплым летом (Алисов,
1956). Средняя температура воздуха в январе у г. Калинина
около -10°, а в июле - 17.5°С. С ноября по март среднемесяч-
ная температура воздуха изменяется от —1.8 до -0.6°, а с ап-
реля по октябрь - от 4.2 до 18.2°. Максимальные дневные тем-
пературы достигают 34-36° (Справочник по климату СССР,
1964).
Основной рекой, питающей водохранилище, является Волга.
По расходам ее у г. Калинина видно, что колебания месячного
и годового стока реки весьма существенны. В годы средней
водности зимой приток волжских вод колеблется от 58 до 107
м3/сек., а летом - от ПО до 155 м3/сек. В маловодные сезо-
ны зимний расход снижается до 22-33, а летний - до 35-85
мЗ/сек. (Ресурсы поверхностных.вод СССР, 1967).
Иваньковское водохранилище принимает и регулирует сток
Волги от истока до Иваньковского гидроузла. Площадь водо-
сбора его по створу плотины 41 тыс. км2. Площадь зеркала
при нормальном подпорном уровне (уПУ) 327 км2, общий объем
1120 млн м3, полезный - 950 млн м . Геоморфологические осо-
бенности затопленной территории определили сложную конфигу-
70
к о S
Рис. 1. Схема Иваньковского водохранилища.
рацию водоема. А.В.Гавеман (1952) выделяет в нем три плеса _
Волжский, Шошинский и Иваньковский (рис. 1 ). Два последних
плеса характеризуются большой изрезанностью береговой линии,
большим количеством островов и мелководных заостровных
участков. В связи с этим при понижении уровня средняя глуби-
на данных плесов увеличивается, а площадь и объем заметно
уменьшаются. Так, при падении уровня на 3 м объем и плошадь
Иваньковского плеса уменьшаются более чем на '50%.
Гидрологический режим водоема в значительной степени
определяется его водным балансом. Среднегодовая сумма при-
ходной части баланса равняется 10.6 км^, основная его состав-
ляющая (речной сток) дает в год 10.2 км® (96.6%) с колебания-
ми по годам от 4.7 до 14.8 км^. Осадки дают 0.18 кмЗ (1.7%),
остальной приток идет за счет возврата воды гидросооружения-
ми и тающего на водоеме льда и снега.
Основная составляющая расходной части баланса - сброс
воды через гидроузел. Средняя величина ее 8.6 кмЗ (87.7%),
изменение по годам от 3.3 до Л 3.8 кмЗ. Забор воды в канал
им. Москвы в последнее десятилетие заметно увеличился и сос-
тавляет теперь 1.8-2.0 кмЗ, или 35-40% от объема сброса гид-
роузла. За счет испарения и на обсохший лед уходит около
0,23 кмЗ в год (Зайцева, 1966).
Небольшая полезная емкость водохранилища, равная 0.95
кмЗ, при среднем притоке, превышающем 10 км^, позволяет
ежегодно наполнять водохранилище до некоторой форсированной
ОТметки и. пропускать в половодье значительную часть вод
транзитом. Заполнение водохранилища начинается в апреле и
продолжается в среднем две недели. За это время уровень по-
вышается на 3.5 — 6.7 м. Максимальный весенний уровень,
как правило, выше НПУ на 0.3-0.7 м, обеспеченность НПУ в
мае 99.5% (Савина, 1966). Летом уровень водохранилища не-
сколько понижается, но не более чем на 1 м. Осенью уровень
Вновь повышается и обеспеченность НПУ в ноябре 71%. Зим-
няя сработка водоема начинается после ледостава и особенно
интенсивно проходит в феврале-марте. За зиму уровень понижа-
ется примерно на 5.5 м.
Особенности морфометрии водохранилища, искусственное ре-
гулирование его объема и связанные с ним изменения в соот-
ношении приходно-расходных элементов водного баланса опре-
деляют сложный режим стоковых течений. Наибольшие скорос-
ти течения наблюдаются в половодье, когда объем водохранили-
ща обновляется от 4 до 13 раз при годовых коэффициентах во-
дообмена 9.6-19.7. Наименьшие скорости течения отмечаются
летом и осенью. В эти сезоны коэффициент водообмена 1.6-3.0.
Зимой при сработке объема водохранилища коэффициент повыша-
ется в среднем до 3.2 (Зиминова, 1959).
72
По расчетам В.П.
Кудрина (1961) при объе-
ме половодья выше нор-
мы скорости течения в
Волжском плесе превы-
шают 1 м/сек., а в наи-
более узкой части Ивань-
ковского плеса достига-
ют 0.5 м/сек. В летне-
осенний период скорости
течения в Волжском пле-
се значительно ниже, чем
весной, и изменяются в
пределах 0.05-0.10 м/сек.,
лишь у г, Калинина они
повышаются до 0.15 м/сек.
В Иваньковском плесе
скорости течения меняют-
ся от 0.03 до 0.12 м/сек.,
при этом наибольшие зна-
чения скорости отмеча-
ются при пике нагрузки
на гидростанцию, а наи-
меньшие - при прекраще-
нии сброса воды.
Рис. 2. Распределение темпе-
ратуры в русловой части Иваньков-
ского водохранилища 15 июня
1961 г. (по: Рутковский, 1964).
Многосуточные измерения скорости и направления течения
самописцами БПВ-2р в июле 1963 г. в верхнем бьефе ГЭС и в
25 км выше нее на правом склоне русла Волги у дер. Городи-
ще показали, что в течение суток скорость течения на глуби-
не 2.5 м в верхнем бьефе изменялась от 0.03 до 0.16 м/сек.,
а в 25 км от ГЭС она составляла 0.03-0.07 м/сек. Направление
течения у плотины было более устойчивым, чем у дер. Городи-
ще (Буторин, 1969).
Зимой при сработке водохранилища скорости течения вновь
возрастают, особенно в верхней части Волжского плеса, кото-
рая постепенно выходит из подпора. У г. Конакова в .феврале -
марте скорости были порядка 0.06-0.08 м/сек. Направление течения
наименее устойчиво в Иваньковском плесе (Буторин, 1959, 1969).
В приплотинной части Иваньковского плеса наблюдаются
обратные течения. По нашим измерениям, выполненным в
летне-осенний период 1968, 1969 гг., величины их у бывшего
г. Корчева колебались от начальной скорости измерителя
течений ГР-42 до 0.06 .м/сек., а продолжительность дости-
гала 4-5 часов. Вероятно, что на характер перемещения
водной массы Иваньковского плеса оказывает влияние конструк-
73
пня водозаборных сооружений гидроузла и канала им, Москвы (
поскольку водоводы ГЭС берут воду из придонных слоев, а на-
сосные станции судоходного канала - из поверхностных (Рут-
ковский, 1864). Однако основной причиной возникновения обрат-
ных течений, как к в Рыбинском водохранилище, слуиат обрат-
ные уклоны водной поверхности, образующиеся при резком
уменьшении или полном прекращении сброса воды. через гидро-
узел (Буторин, Литвинов, 1963; Литвинов, 1970).
Различия в проточности и морфометрии отдельных участ-
ков водохранилиша определяют особенности распределения тем-
пературы воды по акватории и глубине водоема. Наибольшие
различия наблюдаются в температурном режиме Волжского и
Иваньковского плесов. Весной после прохождения пика поло-
водья прогрев Волжского и мелководного Шошинского плесов
идет гораздо быстрее, чем глубокого Иваньковского. Последний
имеет наибольший объем и заполняется более ранними и холод-
ными водами начала половодья, тогда как Волжский и Шошин-
ский плесы аккумулируют относительно теплые Вопы конца
половодья. В связи с этим обстоятельством температура воды
Волжского и Шошинского плесов весной на 2-3° выше, чем в
приплотинной части водохранилиша. Характер изменения темпе-
ратуры воды по длине водохранилиша в конце весны-начале ле-
та виден на рис. 2. В конце весны наиболее отчетливо прояв-
ляется и температурная неоднородность-водной толши: разность
температуры поверхность-дно на русловых участках Иваньковс-
кого плеса достигает 9°. Слой температурного скачка располо-
жен обычно между 2 и 6 м глубины.
Летом водохранилище прогрето относительно равномерно
и существенных различий в температуре между отдельными
участками водоема не наблюдается. Многолетняя средняя месяч-
ная температура воды с июня по август у водомерных постов
Калинин, Конаково, Дубна колеблется в пределах 18-20.4°, наи-
большая среднемесячная - 20-23.2°, наименьшая - 15-17.6°.
Средний многолетний максимум температуры воды колеблется
от 23 до 23.9°, а срочные максимальные температуры изменя-
лись от 19-21° в 1982 г, до 25.6-26.4° в 1960 г. (Ресурсы
поверхностных вод СССР, 1967). Максимум температуры воды
имеет наибольшую повторяемость в июле, наименьшую - в
июне (Тачалов, 1966).
Охлаждение водной толши начинается в августе при гомо-
термин и прежде всего отмечается в Шошинском и Волжском
плесах. По акватории водоема осенью температура воды неоди-
накова: в Иваньковском плесе она выше на 1-2° и ледовые ►
образования начинаются здесь несколько позднее, чем в других
плесах. Так, средняя дата ледостава у Конакова и Дубны со-
74
ответственно 21 и 24 ноября, ранняя - 7 ноября, поздняя - 0
января. Для Калинина средняя дата ледостава 19 ноября, ран-
няя - 3 ноября, поздняя - 15 декабря.
Некоторое превышение температуры воды приплотннного
плеса над температурой остальной части водоема заметно и
зимой. В декабре-январе в Иваньковском плесе придонные слои
воды имеют температуру до 2.0-2.50, тогда как на других
участках наибольшая ее величина около 1.5°. Основная же мас-
са воды охлаждена до 0.5-1.0°. Наименьшая температура в во-
доеме наблюдается в конце зимы, когда водохранилище запол-
няется зимними речными водами с температурой 0-0.5°, и в
начале половодья, когда первые снеговые воды идут с преиму-
щественной температурой менее 0.5°.
Различия в характере течений, перемешивания вод и их
температуры отражаются на мощности ледового покрова. Сред-
няя толщина льда в декабре в Волжском плесе 21-28, а в
Иваньковском 24-33 см. В феврале-марте толщина льда увеличи-
вается до 40-50 см. Очищение водоема от льда идет сверху
вниз. Средняя дата очищения водоема от льда у Калинина
15 апреля, у Конакова - 20, у Дубны - 26 апреля.
Таким образом, ледово-термический режим водохранили-
ща по сравнению с режимом реки характеризуется более позд-
ним вскрытием, заметными различиями в температуре воды
отдельных участков и отчетливо выраженной температурной
стратификацией водной толши в зоне подпора в весенне-летний
период.
Анализ температурных условий водохранилища показывает,
что сезонный и многолетний ход температуры воды в нем оп-
ределяется прежде всего метеорологическими условиями. Од-
нако в отличие от естественных водоемов на изменение темпе-
ратуры воды водохранилища существенное влияние оказывают
также антропогенные факторы. Для Иваньковского водохранили-
ща важнейшими из них являются искусственное регулирование
стока сооружениями гидроузла, забор воды насосными станция-
ми канала им. Москвы и сброс подогретых вод Конаковской
ГРЭС. Если роль первых двух факторов, обусловливающих свое-
образие водного баланса и лежащих в основе эксплуатации
водохранилища, освещена в литературе Достаточно полно, то
проблема влияния сброса подогретых вод на температурный
режим и экологические условия водоема разработана слабо.
Слабая изученность влияния сбросных вод тепловых и атомных
электростанций на физико-химические и биологические процессы,
протекающие в водоемах, затрудняет комплексное использование
водных ресурсов и осложняет решение некоторых вопросов,
связанных с водоснабжением.
75
Рис. 3. Расположение разрезов(римские цифры) и станций (араб-
ские цифры) в районе работ(нумерация, принятая в мае 1967 г.).
А - водозабор, Б - отводящий канал, В - Мошковичский
залив; пунктиром дано русло Волги.
Несомненно, что постоянный приток дополнительного теп-
ла в водоем с замедленным водообменом вызывает повышение
температуры воды, но чтобы определить, в каких масштабах
оно происходит, необходимы натурные исследования. Такие ис-
следования проведены нами в районе Конаковской ГРЭС, проект-
ная мощность которой 2.4 млн кВт. Для охлаждения конденсато-
ров пара и других теплообменников ГРЭС потребляет воды на
каждый из восьми блоков зимой по 8.4, летом 11,4, а в сред-
нем около 10 м°/сек. Забор воды производится по двум подво-
дящим каналам, глубина которых соответствует глубине русло-
вой части водоема. Погружение оси рабочих колес при НПУ
составляет 6.5 м. За годы исследований мощность станции
возрастала и водопотребление увеличивалось. В 1967 г. рабо-
тало в основном три блока и ГРЭС обслуживала одна насосная
станция с одним подводящим каналом. В 1968 г. работало уже
шесть блоков и были подключены вторая насосная и второй
подводящий канал. В конце 1969 г. введен в эксплуатацию
последний восьмой блок и водопотребление ГРЭС достигло оп-
тимальной величины 60-70 мЗ/сек.
76
Район и методика работ
С целью выявления влияния сброса подогретых вод
Конаковской ГРЭС на ледово-термический режим водохрани-
лища и биологические процессы, протекающие в нем, Инсти-
тутом биологии внутренних вод АН СССР с февраля 1967 г.
проводятся комплексные исследования водоема. Рекогносци-
ровочными обследованиями был определен район работ.
Первоначально он охватьвал участок водохранилища в райо-
не ГРЭС: 2 км выше водозабора и 5 км ниже водовыпуска.
На этом участке сбор материалов производился на шести
разрезах. На них было расположено 34 станции. Весной
1967 г. район работ расширен до плотины гидроузла, при
этом число постоянных разрезов увеличилось до 9, коли-
чество станций - до 55 и введена новая их нумерация
(рис. 3).
Программа работ включала измерения температуры и
электропроводности воды на каждой станции через метр
глубины, направление и скорость течения на стандартных
горизонтах, определение количества растворенного в воде
кислорода, прозрачности и цветности воды. С весны 1969 г.
измерялась температура грунта на глубине 5, 10, 20 см.
В безледный период регистрировались погодньв условия,
а зимой велись наблюдения за размерами полыньи и оп-
ределялась толщина ледяного покрова.
Измерение температуры воды и грунта производилось
электротермометрами с полупроводниковыми термисторами
МТ-4 и термокаппаметрами, которыми измерялась и элект-
ропроводность воды (Ершова, Эдельштейн, 1966). Элементы
течения измерялись вертушкой ВМ-М, измерителем течений
ГР-42 и самописцами течений БПВ-2р. Измерение про-
зрачности и цветности воды, а также определение количест-
ва растворенного в воде кислорода велись стандартными
методами. Ежегодно выполнялось 9-10 съемок выбранного
района водоема; кроме разовых измерений отдельных ха-
рактеристик проводились многосуточные наблюдения за их
изменчивостью. В ряде случаев осуществлялись повторные
съемки отдельных участков водоема. Объем полученной ин-
формации представлен в табл. 1.
Режим водохранилища за годы исследований в общих чер-
тах можно характеризовать величинами основных составляющих
Водного баланса, вычисленных за 1967 г. Рыбинской, а за
77
00
Таблица 1
Материалы съемок Иваньковского водохранилища
в районе Конаковской ГРЭС (1967-1970 гг. )*
Год Число рейсов. Число станций Число наблюдений над Число определе- ний кис- лорода
течением темпера- турой воды темпера- турой грунта электро- провод- ностью цвет- ностью
1967 2 8 70 439 1У 413 2562 - 178 1522 230 20 185
1968 3, 7 112 476 612 713 3260 - 615 2307 Тзо 70 88
1969 3. 6 158 454 865 1008. 3041 605 781 2287 - 116
1970 2. 7 _60 450 727 463 3170 132 360 364 2318 - 44
Числитель - измерения прн ледоставе, знаменатель - в безледный период
Таблица 2
Основные составляющие водного баланса
Иваньковского водохранилища за 1968-1970 гг. (млн м^)
Сезоны 1987-1968 гг. 1968-1969 гг. 1969-1970 гг,
Ул ¥4 АУ Ул У2 йУ Ул Уе
Зима (X1I-I11) 1191 1813 -622* 698 1350 -703 1474 2324 -702
Весна (ТУ, У) 5066' 4070 996* 2407 1497 841 6980 5780 727
Лето (УГ-УШ) 1431 1426 5* 1158 1437 -231 1332 1424 -138
Осень (IX-X!) 767 810 -54 1457 1211 242 980 976 58
Всего за год 8455 8119 325 5720 5495 149 10766 10504 -55
у,- сумма приходных компонентов, - сумма расходных компонентов, Ау - аккумуляция
воды; ж - вычислено без учета невязки баланса.
последующие годы Комсомольской гидрометеорологическими
обсерваториями (табл. 2).
Особенностью его в 1068 г. был пониженный осенний при-
ток (80% нормы), вследствие чего объем последующей зимней
сработки составил всего 65% нормы. Для 1969 г. характерна
маловодная весна, когда приток в апреле, равный 1.83 к№,
был близок к многолетнему минимуму. Лето этого года оказа-
лось засушливым, а осень - дождливой. В ноябре и декабре
приток в водохранилище составлял полторы месячных нормы,
благодаря чему зимой 1969/70 г. сброс воды из водохранили-
ша превысил норму на 40% и был выше объема сброса пред-
шествующих зим соответственно на 72 и 28%. Вследствие этого
объем воды в водохранилище зимой 1969/70 г. по нашим расче-
там менялся более трех раз, хотя предвесенняя сработка водое-
ма была неполной и уровень в марте превышал обычный для
этого времени на 1-1.5 м. Интенсивный водообмен оказал су-
щественное влияние на гидравлический режим водоема: сред-
няя месячная скорость течения, рассчитанная по объему сброса,
этой Эимой на IV разрезе (у Корчевы) менялась от 0.10 до
0.05 м/сек.., а в предыдущую зиму не превышала 0.05 м/сек.
Анализ материалов по температурному режиму водоема
в условиях его искусственного подогрева целесообразно начать
с того участка, куда первоначально поступают подогретые воды.
Этим участком является Мошковичский залив, который вместе
с отводящим каналом составляет трассу водовыпуска ГРЭС и
где наблюдаются первые существенные изменения характерис-
тик сбрасываемых вод.
Динамика вод
и температурные условия
в Мошковичском заливе
После прохождения через агрегаты ГРЭС вода, подогре-
тая на 6-10°, реже на 13°, сбрасывается в открытый отводя-
дий канал. Длина его 2.4 км, площадь поверхности 0.1 км^.
Канал заканчивается водосбросом (косой водослив длиной
100 м), через который вода переливается в Мошковичский залив
водохранилища. Длина залива 3 км, площадь 1.1 км2, глубина
у водослива до 8, на остальной части до 5 м. Зимой при сра-
ботке уровня водохранилища залив постепенно выходит из под-
пора и становится продолжением канала. Высота перепада воды
на водосбросе в это время превышает 2 м, тогда как летом
перепад около 0.5 м. Таким образом, зимой подогретые воды
80
Рис. 4. Ха-
рактер измене-
ния температу-
ры воды'Ь по оси
потока подогре-
тых вод в кон-
пе зимы 1967
(а), 1968 (б)
и 1969 (в) гг.
смешиваются с холодными водами водохранилиша вне залива,
летом — в заливе ниже водосброса.
Зимой при сработке водохранилиша уклон водной поверх-
ности в Мошковичском заливе весьма значителен и скорость
течения колеблется от 0.3 до 0.6 м/сек., а в марте-апреле на
отдельных участках повышается до 1.0-1.5 м/сек. При указан-
ных скоростях течения время добегания подогретых вод от на-
чала канала до водохранилиша ограничивается примерно 3 часа-
ми. Высокие скорости течения обусловливают резко выраженную
турбулентность потока и интенсивное перемешивание вод. Вер-
тикальные температурные градиенты отсутствуют. Понижение
температуры воды от начала открытого канала до устьевой
части Мошковичского залива составляет около 2.5° при наибо-
лее низких температурах воздуха и. 0.5-1.0 при значениях ее
около 0° и выше. Таким образом, даие зимой существенного
охлаждения потока в Мошковичском заливе не происходит, и
при работе 6-7 агрегатов ГРЭС температура его на выходе
из залива составляет 6-12°.
В предвесенний период залив заполнен исключительно
сбросными водами ГРЭС. Характер изменения температуры во-
ды по длине залива и распределения ее по глубине виден на
рис. 4. Смешение подогретых вод ГРЭС с водами водохранили-
ща происходит в это время вне залива. В начале весны воды
778 6
81
Рис. 5. Скорость течения (а) и температура воды “С (6)
в заливе и на прилегающей части водохранилища 20 апреля
1970 г. (по: Литвинов, 1973).
половодья пропускаются через Иваньковское водохранилище
транзитом. Скорость течения иа русле Волги в апреле дости-
гает 0.5-0.6 м/сек. При этом высокие скорости течения со-
храняются и в Мошковичском заливе (рис. 5, а). Теплые воды
при выходе из залива прижимаются транзитным потоком к пра-
вому берегу водохранилища. Характер распределения темпера-
туры воды в заливе сохраняется таким же, как показано, на
рис, 4. Вертикального расслоения водной толщи не наблюдает-
ся, но на выходе подогретых вод из залива в водохранилище
отмечаются существенные горизонтальные градиенты темпера-
туры (рис. 5, б).
В процессе дальнейшего наполнения водохранилища уклон
водной поверхности в заливе уменьшается и скорости течения
понижаются до 0.1-0.3 м/сек. Четко выраженное турбулентное
перемешивание воды' наблюдается лишь вблизи водосброса.
На большей части залива формируется двухслойное разнрнаправ-
лениое перемещение вод: поверхностный слой теплых вод
мощностью до 2 м перемещается ® сторону водохранилища,
а придонный - в противоположном направлении. Так, 9 и 10 мая
1939 г. при расходе ГРЭС порядка 40-45 м^/сек. и уровне на
8 см выие НПУ в Мошковичском заливе ниже водосброса наблю-
82
Рис. 6. Величина и направление переноса вод в Мэшко-
вичском заливе 9 и 10 мая 1969 г.
Рис. 7. Направление и
июня 1969 г. на глубине 1 м
скорость течения в заливе 7 и 8
(а) и в придонном слое (б).
далось четко выраженное стоковое течение. Скорость его на
поверхности равнялась 0.21 м/сек. и сравнительно быстро
уменьшалась с глубиной. На глубине 3 м она не превышала
0U3 м/сек., а на горизонте 4 м вертушкой не улавливалась.
По ходу воды на расстоянии около 1 км от рассмотренной вер-
тикали (ст. 4 ) поверхностный слой воды до глубины 2 м про-
должал перемещаться в сторону водохранилища. На поверхности
скорость течения была порядка 0.20-0,13 м/сек., а на глубине
3 м уменьшилась до 0.04 м/сек., и направление переноса вод
стало неустойчивым. В придонных горизонтах течение имело
направление, противоположное поверхностному, а скорость его
составляла 0.03-0.0.7 м/сек. Аналогичная картина в характере
перемещения вод по глубине наблюдалась по всей длине Мош-
ковичского залива (ст. 5, ст. 2 УШ разреза). Это хорошо видно
на эпюрах переноса вод (рис. 6), построенных по методу Хель-
стрёма (Hellstrom, 1963). Эпюры показывают, что по ходу
потока скорость переноса теплых вод уменьшается. Уменьшает-
ся она и с глубиной, особенно резко в слое от 2 до 3 м.
Подробная съемка течений в Мошковичском заливе и на
прилегающей к нему части водохранилища, выполненная 7-8
июня 1969 г., подтвердила описанные выше особенности переме-
щения вод в заливе (Бутории, Курдина, 19706). Как и в мае,
в начале июня верхний слой воды до 2.5-3.0 м, представляющий
собой подогретые воды ГРЭС, перемешался в сторону водохра-
нилища. Скорость течения на глубине 1 м равнялась 0.11-0.14
м/сек., а направление незначительно отклонялось от оси залива
(рис. 7, а). В придонных горизонтах по затопленному руслу
ручья четко прослеживалось противотечение. Скорость его ко-
лебалась около 0.02-0.03 м/сек. (рис. 7, б). В отличие от по-
верхностного направление течения в придонном слое менее
Рис. 8. Ход изотерм в заливе 9 июня 1969 г. на глубине
1 м (а) и в придонном слое (б).
84
устойчиво и в значительной степени определяется рельефом
дна. Противотечением охватывается слой воды примерно в два
раза меньший, чем поверхностным течением. В заливе он равен
1.0-1.5 м, а на выходе из него увеличивается до 2 м. В райо-
не выхода подогретых вод в водохранилище противотечение наи-
более устойчиво в русле ручья, а на мелководье оно возникает
и затухает в зависимости от характера течений в основном по-
токе по руслу Волги.
Отмеченные особенности структуры потока в Мошковичс-
ком заливе хорошо подтверждаются распределением температу-
ры воды (рис. 8). По ходу изотерм в верхнем метровом и при-
донном слоях видно, что теплые воды ГРЭС перемешаются по
более холодным водам водохранилища. Так, 9 июня 1969 г. на
глубине 0.5-1.0 м температура воды по акватории залива и при-
легающего участка водохранилища составляла 22-24° (рис.8,а).
Рис. 9. Расположение изотерм на продольной оси потока
подогретых вод в мае (а) и июле (б) 1967-1969 гг.
85
В придонном слое изотерма 22° оконтуривала лишь прибрежье,
а по руслу ручья в залив поступали воды водохранилиша с тем-
пературой 14-16° (рис. 8, б). Проникновение вод водохранили-
ща в Мошковичский залив четко прослеживается по расположе-
нию изотерм вдоль продольной оси потока (рис. 9). На рисунке
видно, что при такой структуре потока в заливе формируется
слой температурного скачка. Максимальные значения градиентов
температуры в нем достигают 7-8° на метр и наблюдаются в
мае-июие (рис. 9, а). В июле величина градиентов уменьшает-
ся, а эона выклинивания противопотока располагается ближе
к устью залива (рис. 9, б). Глубина залегания слоя скачка бли-
зка к глубине смены направлении течения. При расходах сброс-
ных вод 25-30 м3/сек. (1967-1968 гг.) температурное расслое-
ние вод летом прослеживалось до верховьев Мошковичского
залива. При увеличении расхода их до 50 м3/сек. и более
(1969-1970 гг.) стратификация вод по температуре в этом се-
зоне отмечалась преимущественно в нижней части залива. Не-
которые особенности формирования слоя скачка в заливе и его
устойчивость при различных условиях - рассмотрены А.С.Литвино-
вым (1973).
Несмотря на температурную стратификацию, в Мошковичс-
ком за ливе, происходит смешение подогретых вод с водами во-
дохранилиша и их охлаждение. Интенсивность этого процесса
обусловливается динамикой теплого и холодного потоков и за-
висит от соотношения объемов вод, участвующих в смешении.
Для условий Конаковской ГРЭС предполагалось , что расход
холодных вод водохранилища, участвующих в смешении с подо-
гретыми водами ГРЭС, составит.не менее половины расхода
последних и что в силу прыжкового сопряжения потоков смеше-
ние вод будет происходить непосредственно у водосброса. Расче-
ты, выполненные по уравнению баланса тепла в смешанном по-
токе, показали, что при расходе подогретых вод, равном 20-
35 мЗ/сек., расход вод водохранилища, участвующих в смешении
в заливе, составлял от 7 до 12 мЗ/сек., или 25-40% от расхо-
да ГРЭС. С увеличением расхода подогретых вод до 40-60
м^/сек. расход подмешиваемых холодных вод уменьшился до
7-25%. Наибольший расход вод водохранилиша, участвующих в
охлаждении подогретых вод ГРЭС, всегда был в мае-июне, а
наименьший - осенью.
Таким образом, в весенне-летний период в Мошковичском
заливе наблюдается четко выраженный двухслойный поток вод.
На границе раздела потоков температура смешанных вод резко
понижается и за счет этого средняя температура сбросных вод
уменьшается на 1-3°. В конце лета на выходе из Мошковичско-
№
1967г.
a
б
Рис. 10. Расположение изотерм на продольной оси потока
подогретых вод в августе (а) и сентябре (б) 1967-1969 гг.
го залива, (разрез УП1) отмечается уже незначительное расслое-
ние вод по температуре (рис. 10, а). С увеличением проточнос-
ти водоема и усилением ветрового перемешивания вод в осен-
ний период температура воды по глубине выравнивается и в
заливе наблюдается гомотермия (рис. 10, б).
Дальнейшее разбавление и охлаждение теплых вод ГРЭС
водами водохранилиша происходит на мелководье у выхода из
ЛЬшковичского залива. В этом районе также наблюдается дву-
слойное течение, но оно, как и температурное расслоение, ме-
нее устойчиво, чем в заливе. При изменении ветрового или
гидравлического режима водоема скорости течения к характер
переноса вод могут быстро изменяться (Буторин, Курдина,
1970а). Здесь в смешении участвует больший объем вод водо-
ет
хранилища, чем в заливе. Так, в 1969-1970 гт. на расстоянии
1.0-1.5 км от залива расход холодных вод водохранилища,
участвующих в смешении, составлял 50-80% от расхода подо-
гретых вод ГРЭС. В связи с этим охлаждение их в данном
районе происходит, интенсивнее, а температура смешанных вод
на 2-4° ниже, чем на выходе из залива.
Распространение подогретых вод
Конаковской ГРЭС
в водохранилище
Динамические процессы, протекающие в Мошковичском за-
ливе и на выходе из него, оказывают существенное влияние
ие только на охлаждение подогретых вод, но и на характер
распространения их в водоеме. При изучении влияния сброса
подогретых вод иа изменение экологических условий это имеет
большое значение. Наблюдении в районе Конаковской ГРЭС,
произведенные в феврале 1967 г., показали, что уже при расхо-
де подогретых вод около 30 м^/сек. они четко прослеживаются
в водоеме. При одинаковой температуре воды в подводящем ка-
нале и в прилегающей к нему русловой части водохранилища в
отводящий канал поступала вода с температурой 9.5°. За время
прохождения канала и Мошковичского залива температура ее по-
нижалась иа 2°, и в водохранилище подогретые воды поступали
с температурой 7.5° (Буторин и др., 1968).
После выхода из залива поток теплых вод придерживался
русла Волги, прижимаясь к приглубому правому берегу. Рас-
пространение их в водоеме прослеживалось по образовавшейся
полынье. Температура воды в ней (разрез IX) изменялась на
поверхности от 0.8° у левой кромки льда до 5.6 у правой. В
придонном слое она равнялась соответственно 3.2 и 5.5°. В
конце полыньи .температура воды была 0.5° на поверхности и
4.0° у дна. Подогретые воды ГРЭС отчетливо прослеживались
и в районе бывшего г. Корчева (1У разрез), расположенном
примерно в 4.5 км от залива, прежде всего по ледовому пок-
рову, толщина которого изменялась от 30 см у правого берега
до 50 см у левого. Кроме того, они прослеживались и по ха-
рактеру распределения температуры воды (рис. 11). Если на
русловой вертикали выше водозабора и в подводящем канале
средняя температура составляла около- 1°, а максимальная рав-
88
лилась 1.5°, то на данном разрезе их значения были 2.3 и 3°.
Это свидетельствует о том, что в феврале 1967 г. влияние по-
догретых вод на температурные условия водохранилища распро-
странялось и дальше Корчевы в сторону Иваньковской ГЭС.
В марте 1967 года
температура воды в русле
у водозабора понизилась
до 0.4°, а в подогретых
водах повысилась до 13.5°.
Несмотря на 2-метровый
перепад на водосливе и ин-
тенсивное перемешивание в
заливе, охлаждение подогре-
тых вод шло медленно, и в
водохранилище они поступа-
ли с температурой 11°. К
этому времени размеры по-
лыньи увеличились и кром-
ка льда оказалась в райо-
не Корчевы. Здесь на пра-
вом склоне русла от февра-
Рис. И. Распределение
температуры воды на IV (быв-
шем П) разрезе 10 февра jh
1967 г.
ля к марту температура воды повысилась на горизонте 1м-
иа 6°, на 4 м - на 3°. Присутствие подогретых вод в общем
потоке можно видеть по данным табл. 3, где представлено
распределение температуры на П разрезе, лежащем в 2 км ни-
же водозабора, и на дополнительном разрезе IV-A в 600 м
ниже. Корчевы (IV разрез). Первый участок зимой 1967 г. на-
ходился вне зоны влияния подогретых вод, в марте температу-
ра воды здесь только вблизи дна достигала 0.5°, В это время
ниже Корчевы с глубины 4 м температура воды была выше 1°,
а в придонных слоях русловой части водоема превышала -3°.
Как и в феврале, более теплые смешанные воды перемешались
преимущественно в нижней половине потока, придерживаясь
русла Волги (табл. 3).
В безледный период подогретые воды перемешаются в во-
дохранилище верхним, преимущественно 2-метровым слоем.
Район распространения их опредепяется в основном гидрометео-
рологическими условиями и прежде всего стоковыми и дрейфо-
выми течениями. Так, 5 мая 1967 г. при устойчивом южном
ветре теплые воды были отнесены к северному берегу водохра-
нилища, где температура воды повысилась на 1.5° по сравне-
нию с таковой вне зоны их влияния. При северо-восточном
ветре 21 мая поток подогретых вод прослеживался вверх от
АЪшковичского залива на 8-9 км и выклинивайся примерно
в 1.5 км выше водозабора ГРЭС. Температура воды здесь была
на 1.0-1,5° выше температуры воды, поступающей в водохрани-
88
Температура воды (ос) и толщина льда (см)
на русловых вертикалях разрезов 13,14 марта 1967
Тол- щи- на льда «о । Я to
Ч 1 QJ (TJ Ф к S а о. Я* К О) ф >> О Я h К Ь (О ОЗ О О'—
Горизонты наблюдений, м I о г-Ч 3.3
9-4 3.3
О (О СО О 1 СО
о ю оз о 1 о?
00 (О СО Г; О О «
г- о ос?
со м; со о о 6 04
(О со со f; ' 6 о
м* со и ® о d
со СЧ СО О 6 ©
сч СЧ СО ИЗ О О О
СЧ 03 Ч* о do
0.5 СЧ СЧ СО о do
Разре- зы II Водо- забор 1У-А
лише. Летом и осенью с увеличением забора воды из водо-
хранилища подогретые воды распространяются преимущественно
в сторону плотины, особенно при совпадении направления стоко-
вого и дрейфового течений. В 1967 г. наибольшее продвижение
их в сторону Иваньковской ГЭС было отмечено в конце июня
и составляло около 10 км.
Рис. 12. Распределение тем-
пературы воды на V разрезе 22
января 1968 г.
Осенью с усилением
теплоотдачи и ветрового
перемешивания различия
в температуре воды от-
дельных участков водо-
хранилища уменьшаются.
Так, если в сентябре
температура воды в зоне
влиянии подогретых вод
была на 2-3° выше, чем
в водохранилище, то в
октябре-ноябре эти раз-
личия не превышали 1°.
Максимальная разность
поверхностной температу-
ры подогретых вод на вы-
ходе из залива и воды
водохранилища в навигационный период 1967 г. составляла 8°.
В конце 1967-г. расход сбросных вод Конаковской ГРЭС
увеличился до 40-50 мЗ/сек. Естественно было ожидать усиле-
ния влияния их на температурные условия водоема. Наблюде-
ния, выполненные в январе 1968 г., подтвердили это. В полынье
температура верхнего метрового слоя не опускалась ниже 3°,
а в большей части потока -была около 5°. Район распростране-
ния подогретых вод по сравнению с зимой 1967 г. заметно
расширился, и они хорошо прослеживались ие только в районе
Корчевы, но и в 3 км нйже (разрез У). Из данных табл. 4
видно, что ниже Мошковичского залива температура большей
части потока превышает 3°, а средняя по вертикали значитель-
но выше, чем перед водозабором ГРЭС. Расположение изотерм
на рис, 12 подтверждает, что повышение температуры характер-
но для большей части потока.
Высокие температуры воды на разрезе У, расположенном
в 7.5 км ниже места поступления подогретых вод в водохрани-
лище, небольшое отличие их как на отдельных горизонтах,
так и по средним значениям от температуры на разрезе IV,
дают основание считать, что район влияния теплых вод в ян-
варе 1968 г. по сравнению с тем же периодом предшествующей
зимы значительно расширился. Принимая во внимание, что на
90
81
Температура воды (°C) и толщина льда (см)
на русловых вертикалях разрезов 20-22 января 1968
Т ол- щи- на льда 35 00 £ о in —1 со
Сред- няя тем- пе- ра- тура 1.1 N со 05 Г; Л СО со
Горизонты наблюдений, м ~ ~| 1 ‘ 1 — — со г- со СМ (О CD 1 1
12 6’1 э с М С 5.5 5.4
05 О О СО со S CM CM CD (D
10 О0 D 05 (D •-« М ID (D
05 00 05 оо со 05 4 J
00 СО 00 00 СО —• 4 -4 хГ тг
г- (D Г; 05 05 4 .4 -Г со СО
со тр СО СО О 05 S 4 4 со
СО У и ГО (D • Г- 00 4 СО со
’ф 0.5 ПК С 4 3.3 3.0
СО 0.1 0.2 0.2 3.0 2.7.
см С с 0.0 0.1 1.9 1.7
с с > с > о 0*0 CM CD ~ О
(Г с 1 1 с с 0.0 0.0 0.5 0.3
Раз- резы Водо- забор ►—< IV У
пути перемещения воды от IV к У разрезу понижение темпера-
туры в большей части потока не превышало 0.1° на километр,
можно утверждать, что влияние подогретых вод в это время
распространялось на большую часть Иваньковского плеса водо-
хранилища. Такое заключение подтверждается результатами
мартовских наблюдений. Данные табл. 5 показывают, что влия-
ние подогретых вод четко прослеживается на всей приплотинной
Рис. 13. Ход изотерм в русловой части водохра-
нилиша между Корчевой (IV разрез) и плотиной 15
марта 1968 г.
части водохранилиша. На расстоянии 2-3 км от плотины темпе-
ратура воды повысилась в среднем до 2.2°, а максимальная -
до 2.8°. Повышение температуры наблюдалось не только в рус-
ле, но и по всему поперечному сечению потока.
Таким образом, в 1968 г. с увеличением расхода подогре-
тых вод Конаковской ГРЭС акватория распространения их уве-
личилась. Зимой 1967 г. при расходе до 30 мЗ/сек. влияние
их ниже водовыпуска Прослеживалось на расстоянии 10-12 км.
Зимой 1968 г. с увеличением расхода до 40-50 мЗ/сек. оно
распространилось до плотины Иваньковского гидроузла, т.е.
на 25 км от водовыпуска. Если до сооружения ГРЭС в припло-
тиниой части водохранилища зимой средняя температура воды
редко достигала 1.5° (Зиминова, 1959; Тачалов, 1966), то в
1968 г. вся водная толща глубже 3 м имела температуру свы-
ше 2° (рис. 13).
Первый годовой цикл наблюдений позволил установить за-
метное влияние подогретых вод Конаковской ГРЭС на ледово-
термические условия района водохранилища, лежащего ниже
Мошковичского залива. Вблизи залива образовалась полынья
до 5 км длиной. Средняя температура воды в русловой части
водоема ниже водосброса (разрез IV) повысилась по сравнению
92
93
Температура воды (°C) и толщина льда (см)
на русловых вертикалях разрезов 14-21 марта 1968 г.
Тол- ШИ- на льда 50 30 со о 52 52 55 35
Сред- няя тем- пера- тура 0.6 0.8 9*0 3.6 3.5 3.2 2.3 2.2
1 1 1 1 1 1 3,0 I2-8
12 1 1 1 4.8 1 4.1 3.0 2.8
10 о 1.2 1 4.8 4.5 4.1 2.6 2.5
03 0.9 1.0 4.8 4.5 4.2 2.6 2.2
00 0.9 о о 4.7 4.5 4.2 2.6 2.2
X X О) ЕГ 2 с г- 8*0 о 8*0 4.5 4.1 СО сч 2.2 -
о <и X 2 со о a о о 4.6 4.5 4.0 2.6 2.2
Ь X о (9 X (О 9’0 8*0 0.5 4.5 4.4 4.0 2,5 СО СЧ
fp хг • 0.5 0.8 0.5 О Xf 3.8 О хг 2.3 2.3 |
СО 0.5 9*0 0.5 2.2 2.9 2.4 9*1 2.2 1
сч хг о (О о 0.3 (О 1.3 о о сч
0.3 0.5 0.3 0*1 О о 0.9 0.5 I 8*0
* 0.5 0.2 хг 6 о о 0.5 о 0.8 о СО о
1
£ £ к 3 е и иип ►И 1 О а о £0 а о W »-м X вм 1 •-ч > ь О
с температурой выше водозабора (разрез 1) при расходе ГРЭС
до 30 м°/сек. примерно на 1.5°, а при расходе порядка
40 мЗ/сек. - на 1.5-2.5°. Зимой влияние подогретых вод в во-
дохранилище прослеживалось до плотины Иваньковской ГЭС, а
летом присутствие -их отмечалось в 8-10 км выше и ниже Мош-
ковичского залива.
Зимой 1969 г. в связи с увеличением расхода подогретых
вод можно было ожидать дальнейшего повышения температуры
воды в водохранилище, осо-
бенно на участке ниже их
поступления в водоем.
Однако как в январе, так
и в марте 1969 г. темпе-
ратура воды здесь оказа-
лась заметно ниже, чем
в предшествующую зиму.
Как видно из данных
табл. 6, средняя величина
ее иа русловых вертикалях
IV и V разрезов составля-
ла 2-2.8°, а максимальная
не превышала 3.9°. Пони-
жение температуры воды
в районе Корчевы зимой
1969 г. вероятно связано
с изменением гидродина-
мических условий в иссле-
дованной части водоема.
С увеличением мощности
ГРЭЬ увеличился объем
вод, забираемых для ох-
лаждения ее агрегатов.
Это привело, с одной
стороны, к ослаблению
основного потока вод во-
Рис. 14. Ход изотерм в рус-
ловой части водохранилища зи-
мой 1969 г.
а - 15-19 января, б -
14-19 марта.
дохранилиша, а с другой -
создало предпосылки для
подсоса вод из района во-
довыпуска к водозабору. Доказательством этого является за-
метное повышение температуры воды выше водовыпуска. Так,
в январе 1969 г. большая часть врдной толщи в русле Волги
на разрезе 1П имела температуру свыше 2°, а максимальное
значение иа глубине 8 м равнялось 3.3°, тогда как на I и 11
разрезах средняя температура и значения ее по горизонтам
были ниже (табл. 6). В течение зимы температура воды на HI
95
разрезе продолжала повышаться и в марте в придонных слоях
русловой вертикали достигала 3.7 .
Несмотря на некоторое понижение температуры воды в
районе Корчевы по сравнению с предшествующей зимой, что
хорошо видно из сопоставления данных табл. 4, 5 и 6, допол-
нительный приток тепла в водоем продолжал увеличиваться.
На это указывает прежде всего дальнейшее повышение темпе-
ратуры воды в приплотинном участке водохранилища. Здесь
на отдельных вертикалях средняя температура повысилась до
2.7-3.30, а максимальная - до 3.9°. Зимой 1967 и 1968 гг. та-
ких температур в приплотинном участке не было. Характер рас-
пределения температуры в потоке на III разрезе 'зимой 1969 г.
виден на рис. 14. Здесь интересен тот факт, что, если в янва-
ре подогретые воды выше водовыпуска прослеживались в виде
отдельных очагов (рис. 14, а), то в марте, несмотря на про-
должавшееся предвесеннее опорожнение водохранилища, они за-
няли большую часть русла (рис. 14, б).
Водопотребление Конаковской ГРЭС зимой 1970 г. было
близко к проектной величине, поэтому распределение темпера-
туры воды в этом сезоне наиболее реально отражает термичес-
кие условия водоема при нормальной его эксплуатации. Несмот-
ря на повышенный водообмен и более низкую по сравнению с
предшествующими зимами естественную температуру воды в
водоеме, равную 0.2-0.70, четко прослеживается увеличение
влияния сброса подогретых вод ГРЭС на температурные усло-
вия водохранилища. На рис. 15 отчетливо наблюдается повыше-
ние температуры воды как в сторону плотины, так и к водо-
забору ГРЭС. Вниз по течению от Мошковичского залива повы-
шение температуры отмечалось по всей глубине потока. На рус-
ловых вертикалях в районе Корчевы средние ее значения сос-
тавляли 2.9-3.70, а максимальные - до 4.6° (табл. 7). В сто-
рону водозабора на разрезе III и против устья р. Созь на глу-
бинах свыше 5 м температура воды оказалась выше естествен-
ной на 1-2° (рис. 15, а). От февраля к марту тенденция повы-
шения температуры и особенности ее распределения на отме-
ченных участках сохранились (рис. 15, б).
Анализ материалов наблюдений четырех зим показал, что
с увеличением мощности Конаковской ГРЭС температура сбрасы-
ваемых вод повысилась от 8.5-10.0 зимой 1968 г. до 11.5-12°
в 1970 г. Соответственно изменялась температура воды на
трассе водовыпуска (табл. 8). В зависимости от величины рас-
хода ГРЭС и сработки водохранилища характер распространения
подогретых вод и их влияние на температурные условия водое--
ма изменялись (Буторин, Курдина, 1972а). Зимой 1967 и 1968гг.
при близких значениях расхода ГРЭС и сходном режиме регу-
96
Температура воды ( С) и толщина льда (ем)
на русловых вертикалях разрезов зимой 1969 г
Тол- щина льда 1 Ю СО (Л (О (О О СМ Ю СЧ (Л Г! «О ТГ о V со СО (Л
i 1 я « в к 2 а. а. а « g g b (_) Я Ь Е Ь ю. * О сч ~ « сч Л СО СЧ ~ СЧ СЧ и СО и
Горизонты наблюдений, м 14 о 1 IIIе01 СЧ 00 00 00 СО со СО со
12 со со и ° N, СЧ 1 СЧ СЧ W т СО 00 00 00 СО СО СО СО
10 СО Tf Я О ", СЧ 1 N N N я СО 00 00 00 СО СО СО СО
СТ) X С « —< М Г- О I СЧ СЧ СЧ 04. т « со СО 00 СО СО со СО
00 СМ со 05 00 СМ СМ СЧ СО °* О X со г- ГО СО СО СО
г- 1 ' ря , • О 05 05 сч < Г СЧ ~ СЧ СЧ D СО Ю СО О СО со со
со <0 oi-N"Nn'«nn <Я
со •—Ч Ю Г-СОГ-^ОТГ-ЮГ^С» ~ J J N - J о! Я oi И
чг ч1® cot°o5Wt~'rr~ ~ " oi " U w ci ci м
со с CO^OKJ^O^COIO J ci .гч OJ oi oj О)
см - a с —«сог-^г-Осо^сч «о^**-о^*^счсч
о ^•сЗоо^сэИ' 0 0 0 ^0° см со со J! Д
со о СО 6 Я N И О «5 С О О о — 6 с СР 00 О • • • о о •*
Разре- зы и стан- ции < а О о ч io < £8 s О > > УГ-А Ст. 7 Ст. 8
. 778 7
87
оо Таблица в (продолжение)
Разре- зы и стан- ция Горизонты наблюдений, м Сред- няя тем- пера- тура Тол- щина льда
O.S 1 2 3 4 5 6 7 8 9 10 12 14
I 0.0 0.0 0.0 0.0 14-1 0.0 ) мара 0.0 0.1 0.3 0.3 0.3 0.1 54
Водо- забор 0.0 0.0 0.0 0.0 0.1 0.2 0.3 — а. а» — — — 0.1 43
II 0.0 0.0 0.0 0.0 0.1 0.2 0.4 0.5 0.8 1.1 а. а. - 0.3 70
III 0.0 0.2 0.3 1.5 3.4 3.6 3.6 3.6 3.6 3.6 3.6 3.7 — 2.8 56
ТУ 0.6 0.6 2.7 2.7 3.2 3.2 3.2 3.3 3.5 3.5 3.5 а. — 2.8 0
У 0.6 2.2 2.6 2.6 2.9 3.0 3.1 3.1 3.1 3.1 3.2 — * 2.8 62
У-А 0.5 0.7 1.3 2.1 2.8 3.2 3.5 3.6 3.6 3.5 3.6 а. а» 2.7 65
У!-А — 1.0 1.9 2.5 3.1 3.8 3.8 3.8 3.8 3.8 3.9 3.9 - 3.3 68
Таблица 7
Температура воды (°C) и толщина льда (см)
на русловых вертикалях разрезов зимой 1970 г.
Раз- резы Горизонты наблюдений, м Сред- няя тем- пера- тура Тол- щина льда
0.5 1 2 3 4 5 6 7 8 9 10 12 14
1 0.1 0.1 0.1 0.1 0.1 12- ОЛ 15 фев 0.3 раля 0.3 0.3 0.3 0.3 0.3 0.2 32
Водо- забор 0.0 о’.о 0.0 0.1 0.1 0.2 0.2 0.2 0.2 0.2 0.2 0.2
II 0.1 0.1 0.1 0.1 0.2 0.4 0.4 0.5 0.5 0.5 0.5 1.4 2.0 0.5 51
Устье р.Созь 0.0 0.1 0.1 0.3 0.3 0.6 0.6 1.5 1.7 2.6 2.6 3.3 __ 1.4 41
III 0.1 0.2 0.3 0.4 0.9 1.2 1.6 2.2 2.5 2.6 2.6 2.7 2.8 1.8 30
ТУ 3.1 3.1 3.1 3.1 3.2 3.8 3.4 3.8 4.0 4.0 4.0 4.6 4.4 3.7 Нет
У 0.2 0.4 2.4 2.6 2.8 3.2 3.2 3.3 3.4 3.4 3.7 3.6 - 2.9 40
а 7 (продолжение) 1 ' ' 1 - I । 1 я о н шина льда со 1 г- to о £ о (О to СО ю
Сред-1 НЯЯ тем- пера- тура СО CD Г; О СМ Г; 00 q d о oi см см со
ч* гм CD О ( t t СО СО СО I
см со ш о | II СО СО СО 1
10 «-• Ю О) О Д см см со со см
О) 0) г- см см 05 о о Д Л СО со см тГ
>онты наблюдений, м 00 05 тг О0 О СМ 02 со d -I «4 со со см
г* СО Г- СО Г- 05 05 05 d d d см см см со
со ' 2 g СЧ СО СО СО U3 0) го «в О' О О -- СЧ СЧ СО 2 со со -и аз оо о . О ОО -- сч ч* _ сч
(О
S а о U о —« — 00 о d d d d «Л см
со О СЧ —< 00 02 d d d d d сч со
см о о СМ СО о о о о d сч со
г—« О О — СЧ — -И СО • • • • « • • О О О О О СЧ СЧ
0.5 о о — сч — —* С5 Йо О О СЧ
Я 1 R I Ю 1 Ь | назре- зы ’ а ® го 2 ° £ о 0 « О Ч - > «»CQroS>>Q.= <M>
Рис. 15. Ход изотерм в русловой части во-
дохранилища между 1 и У разрезами зимой
1970 г.
а - 12-15 февраля, б - 21-24 марта.
лирования объема водохранилища распространение подогретых
вод в нем имело сходный характер: поступая из Мошковичског
залива, они по руслу Волги перемещались в сторону плотины
Иваньковского гидроузла. Зимой 1967 г. при расходах ГРЭС
25-30 м^/сек. влияние их ограничивалось небольшим участком
водоема ниже водовыпуска протяженностью 10-12 км, а зимой
1968 г. при расходе ее 40-45 мЗ/сек. влияние подогретых вод
распространилось на всю глубоководную часть водоема от во-
довыпуска до плотины.
Зимой 1969 г. при расходе 50-55 мЗ/сек. наряду с даль-
нейшим повышением температуры воды в приплотиниой части
водохранилища отмечено и повышение ее на участке водоэабо
водовыпуск, поскольку часть подогретых вод стала перемещу
ся в сторону ГРЭС. Наблюдения, выполненные зимой 1970 г.,
при расходах ГРЭС порядка 55-60 мЗ/сек., подтвердили отме
ченные особенности распространения подогретых вод и покаэ-
100
Температура сбросных вод Конаковской ГРЭС
в отводяшем канале, Мошковичском заливе и на мелководье
водохранилиша (разрез IX) в зимний период
1970 г. 24 III о о о о О О О О 05 00 С5 00 г**
| 12 II О СО СО 05 С5 СО СО 00Г- Г-СЧ^СЧ сч Л Л Л ~ Л о о со Л 10 -т
1969 г. 16 III СО СО СО (Л СМ СО СМ — — 1 I I 1 ~ ~ о р о о
18 I СМ 05 оо о —< см ~ со см со со О 00 00 05 05 05 05 СО со ~ ~ —
1968 г. 17 III 00 а 05 СО 05 05 со 05 °, Х. °. а, О « 05 05 05 05 05 <35 005005
21 I «г см ч* о w R "2 м. Ч ® ® СО Л Л Л Л СО СО <0 О ТЧ Т Tf
Гори- зонт и и « ••••*• • •• л; 7? Ш О Д Q Ш о ДО Д О Д Q 0,2 о х о « о У ох о х о а ЕЧСПЕсЗ СЦ СПСЙ
Глубина, м ю. 7 Т Т 7 7 7 ХГ СО W СО со — со
Разрезы и станции = £ о со Ч" со сч ; о, см ч- п . а 0 . . ej Ь D ь 5 ь * s ь ь gesso о о о g к: о о 5 5 ' « ° * со й. ffl
ли, что при определенных условиях эти воды могут достигать
водозабора и повторно участвовать в охлаждении агрегатов
ГРЭС (рис. 15). Подобный характер распространения подогре-
тых вод в водохранилише зимой сохранится и в дальнейшем.
Присутствие их будет отмечаться по всей глубоководной части
Иваньковского'плеса. Температура основной массы воды здесь
составит 2-4°. Близка к этим значениям будет и температура
нижних слоев воды между водовыпуском и водозабором ГРЭС.
Только при продолжительных оттепелях с увеличением- Притока
и расхода гидроузла до значений, близких к половодным, ука-
занное распределение температуры может существенно изме-
ниться.
Весной при пропуске половодья водообмен в водохранили-
ше настолько высок, что его объем обновляется за сезон в
среднем 5.5, а в отдельные годы до 13.5 раза (Зиминова, 1959).
Многолетний среднемесячный расход воды через Иваньковский
гидроузел в апреле равен 945, а в мае 443 м^/сек. Эти расхо-
ды превышают максимальные летние расходы Конаковской ГРЭС
соответственно в 10 и 5 раз. Если же весной расход ГРЭС
будет равен 67 м^/сек. (максимальный зимний), а температура
сбросных вод в районе поступления их в водоем будет выше
естественной на 6-7°, то повышение температуры смешанного
потока на 1° произойдет лишь тогда, когда расход в водохрани-
лище будет менее 400 м^/сек. Расход, вдвое превышающий ука-
занный, имеет продолжительность до 30 дней. Следовательно,
в половодье около месяца соотношение расходов ГРЭС и гидро-
узла таково, что подогретые воды не могут оказать существен-
ного влияния на температуру воды водохранилища. Они отжи-
маются основным потоком на мелководий И перемешаются в
сторону Иваньковской ГЭС узкой полосой вдоль правого бере-
га водохранилища.
Это подтверждается данными наблюдений, выполненными с
16 по 20 апреля 1970 г. во время пропуска через гидроузел
пика половодья. В период наблюдений приток в водохранилише
составлял 1060-2100 м^/сек., а расход из него колебался от
350 до 1200 мЗ/сек. При расходе ГРЭС около 50 м^/сек. и
температуре подогретых вод 11-13° на мелководье у залива
температура их понижалась до 5-8°, а на периферии потока -
до 2°. В русловой части водоема разбавление подогретых вод
холодными водами водохранилища с температурой около 0.5°
было настолько значительным, что уже в районе Корчевы по-
вышение температуры до 1.0-1.50 отмечено только вблизи уре-
за правого берега. На спаде половодья влияние сброшенных вод
на температуру водной массы отмечается преимущественно на
небольшом участке, лежащем вблизи Мошковичского залива
103
(рис. 16). Ниже заметно лишь выравнивание температуры по
длине водоема.
В конце весны с установлением температурной стратифи-
кации и летом распространение подогретых вод в водохранили-
ще существенно меняется. Акватория и глубина проникновения
их в русловой части водоема определяются прежде всего взаи-
модействием различного рода течений: стоковых, дрейфовых и
Рис. 16, Расположение смешанных вод в русловой части
водоема 25 апреля 1968 г. (а) и 7 мая 1969 г. (б).
Цифры при кривых - температура воды, ®С.
компенсационных. При небольшом летнем притоке интенсивность
стоковых течений зависит от объема попусков воды в нижний
бьеф через турбины ищ!®зга также от величины забора воды
о канал им. Москвы. Поскольку гидростанция работает в часы
„пик* нагрузки в сети, шлюзование судов идет преимуществен-
но в дневные часы, а вода в канал забирается круглосуточно
с максимальным расходом в ночное время, режим стоковых
течений в приплотинном плесе неустойчив и характеризуется
наличием прямых и обратных течений, возникающих при резком
изменении расхода воды из водоема (Буторин, Литвинов, 1963).
Дрейфовые течения также неустойчивы, поскольку в теп-
лые месяцы из-за малых градиентов атмосферного давления
направление ветра в районе водохранилища быстро меняется.
По данным ближайшей к Иваньковскому плесу гидрометеороло-
гической станции г. Савелово с конца весны до начала осени
наибольшую повторяемость имеют ветры западных румбов.
Они составляют 15-25% от общего числа наблюдений. У восточ-
ных ветров наименьшая повторяемость. (4-9%) н лишь северо-
восточные отмечаются в 8-17% случаев. Средняя месячная ско-
рость ветра невелика: 2.6-3.0 м/сек. Вероятность ветра в
104
4-5 м/сек. около 20%, а 6-7 м/сек. - 8-12%. Наибольшие ско-
рости наблюдаются при западных и северо-западных ветрах
(Справочник по климату СССР, 1966). Следовательно, в летнее
время в дрейфовых течениях преобладает восточное, совпадаю-
щее по направлению со стоковым.
Анализ материалов наблюдений показывает также, что
распределение подогретых вод в рассматриваемом сезоне каж-
дый год имеет свои- особенности, обусловленные прежде всего
гидрометеорологическими условиями и режимом эксплуатации
водоема. Так, летом 1967 г. погода была более теплая и тихая,
чем летом 1968 и 1969 гг. В июле и августе днем температу-
ра воздуха достигала 25-27° при легком и слабом ветре. Сред-
несуточная температура воды с конца июня до середины авгус-
та у водомерного поста Конаково была выше 20°, а в отдель-
ные периоды достигала 22°. В этих условиях охлаждение подо-
гретых вод шло медленно и район распространения их выделял-
ся довольно четко. Лето 1968 г. было более ветреным и про-
хладным. Похолодание во второй декаде июля вызвало пониже-
ние температуры воды от 20-22° до 16-17°. Только в конце
августа температура воды вновь повысилась до ее значений в
начале июля. Охлаждение подогретых вод шло более интенсивно
и несмотря на увеличившийся расход теплых вод зона их посто-
янного влияния расширилась незначительно. Для лета 1969 г.
характерно малое количество осадков, благодаря чему приток
в водоем был невелик, а сброс близок к минимальному. Умень-
шение скорости стоковых течений при увеличении недопотреб-
ления ГРЭС вызвало продвижение теплых вод в сторону водоза-
бора, и район 1П разреза постепенно стал входить в зону по-
стоянного их влияния. Процесс отепления этого участка водо-
хранилища еще более четко прослеживался летом 1970 г., когда
сбросные воды на III разрезе находились постоянно й иеодно-
. кратно достигали II разреза.
Температурные съемки, проведенные в безледный период,
позволили проследить зависимость направления переноса подо-
гретых вод от конкретных гидрометеорологических условий. Из
сопоставления хода изотерм с преобладающим направлением
ветра, наблюдавшимся в течение двух-трех дней перед съемкой
(рис. 17), видно, что стоковым течением подогретые воды с
температурой на 2-5° превышающей естественную, от залива
относятся в сторону гидроузла (рис. 17, а, в). Их присутствие
четко прослеживается на расстоянии 10-15 км от залива. Пе-
ремешиваясь с водами водохранилища и теплыми водами, вы-
носимыми с заостровных мелководий, подогретые воды ГРЭС
постепенно охлаждаются и по температуре теряют свою инди-
видуальность. Иногда компенсационным течением, а также
105
б
Рис. 17. Акватория распространения подогретых вод при
ветрах разного направления в 1967 г.
а - 31 V, 1 VI, роза ветров за 27 V - 1 VI; 6-27
28 VI, роза ветров за 24-28 VI; в _ 23-24 VIII, рОЗа ветров
за 20-24 УШ; г - 27, 28 IX, роза ветров за 25-28 IX. Цифры
при кривых - температура воды,'С.
ветром северного и северо-восточного направлений подогретые
воды от залива относятся к водозабору ГРЭС и дане выше
его на расстояние 8-10 км от залива (рис. 17, б). В этом
случае граница распространения их выделяется более четко,
так как естественная температура воды в узком и глубоком
106
речном участке водохранилища летом ниже, чем в расширенной
части Иваньковского плеса. В ряде случаев ветровыми течения-
ми подогретые воды отжимались к одному из берегов водохра-
нилища, вдоль которого они перемещались узкой полосой, по-
степенно вовлекаясь в общий поток и охлаждаясь (рис. 17, г).
Сложный характер переноса подогретых вод в водохрани-
лище виден на таком примере: 22 августа 1968 г. в 20 час.
выше водозабора ГРЭС на русле Волги наблюдалось устойчивое
стоковое течение с максимальной скоростью 0.13 м/сек. на
глубине 8 м и направлением 12°. В 8 час. 23 августа при се-
веро-восточном ветре 2-5 м/сек. на этой же вертикали в верх-
нем 2-метровом слое зарегистрировано перемещение Вод в об-
ратном направлении со скоростью 0.06 м/сек. Аналогичный
характер перемещения вод отмечен в этот день и ниже водоза-
бора ГРЭС, где верхний 2-метровый слой воды перемещался
в направлении, противоположном переносу основной массы воды.
Двуслойный поток с незначительными изменениями скорости
течения наблюдался до 20 час. и вероятно сохранялся до смены
направления ветра. В противопотоке отмечалось’ повышение тем-
пературы воды на 1.5-2.0° по сравнению с измеренной до его
появления в 20 час. 22 августа.
Материалы наблюдений на многосуточных станциях показы-
вают, что даже в течение одних суток скорость и направление
переноса вод в Иваньковском водохранилище могут сильно ме-
няться, причем изменение охватывает иногда всю водную тол-
щу от поверхности до дна. В связи с этим и глубина проникно-
вения подогретых вод также непостоянна. Это хорошо просле-
живается на примере данных многосуточной станции, выпол-
ненной 7-10 июня 1970 г. в районе Корчевы (1У разрез). На-
блюдения велись при устойчивой погоде. Температура воздуха
имела четко выраженный суточный ход: ночью колебалась от
8 до 13°, а днем повышалась до 18-20°. Ветер с 21 часа 7
июня до 3 час. 9 июня был северо-восточным с преобладающей
скоростью 3-7 м/сек. К 6 час. 9 июня направление его смени-
лось на восточное, а затем на северное. Скорость ветра
уменьшалась до 2-4 м/сек., а с 20 до 24 час. наблюдался
штиль. Утром 10 июня зарегистрирован слабый ветер западно-
го направления. Большую часть времени наблюдений отмечалось
стоковое течение: от 5 м до дна оно было довольно устойчи-
вым, а выше 5 м устойчивость его нарушалась воздействием
ветра. Особенности переноса вод в отдельные сроки наблюдений
видны на рис. 18. Вечером 7 июня и в течение 8 июня на боль-
шей части глубины отмечено стоковое течение, направленное
к плотине Иваньковской ГЭС. В то же время верхний 3-мет-
ровый слой воды северо-восточным ветром перемешался в про—
107
8 VI О час. 6 час. 9 час. 15 час. 18 час. 21 час
009 0 0.10 -0.09 0 , 0.10 -0.04 0 0.10. -0.05 О 0.10 -0.05 0 0.10 0 20-005 п
Рис. 18. Направление и скорость переноса
вод на русловой станции IV разреза 7-10 июня
1970 г.
тивоположном направлении (рис. 18, а-е). К 18 час.
8 июня толщина слоя волы, захваченного противоречени-
ем, увеличилась до 6 м (рис. 18, д). Такая картина
переноса вод с некоторым уменьшением мощности слоя,
охваченного противотечением, и при ослаблении стоково-
го течения сохранялась до О час. 9 июня (рис. 18, ж).
С уменьшением скорости ветра и последующим затем
изменением его направления характер переноса вод изме-
нился. К 6 час. 9 июня по всей глубине наблюдалось
стоковое течение, при котором скорость- переноса вод
колебалась от 0.02 до 0.05 м/сек. (рис. 18, э). Такое
направление переноса с увеличением скорости в середи-
не потока до 0.07 м/сек. и постепенным ее уменьшени-
ем в поверхностном слое отмечалось до 15 час. (рис.
18, и, к). Около 18 час. на поверхности образовалось
слабое противотечение, которое сохранялось до 21 часа,
охватывая слой воды до 2 м (рис. 18, л, м). В ночь
на 10 июня с прекращением работы ГЭС и при штиле-
вой погоде- картина вновь резко изменяется: в 0 час.
в придонном горизонте зарегистрирован перенос вод в
направлении, обратном стоковому течению (рис. 18, н).
Мощность этого потока быстро возрастала и к 9 час.
10 июня он захватил практически всю водную толщу,
исключение составил лишь поверхностный слой до 1 м
глубины, который слабым западным ветром перемещался
в сторону гидроузла (рис. 18, о, п).
Все это свидетельствует о том, что в результате
взаимодействия течений различного происхождения в усло-
виях искусственного регулирования объема вод перемеще-
ние их в Иваньковском водохранилище .летом представля-
ет сложную картину. В общих чертах она сохраняется
и осенью. Характер распространения подогретых вод в на-
начале ‘ этого сезона мало отличается от летнего. Как
и . летом, преобладает перенос их в поверхностных слоях
водной толщи. Однако осенью с увеличением притока
все более существенную роль в динамике вод играет
стоковое течение. Это особенно четко проявляется при
сравнении характера течений в рабочие и нерабочие дни
недели, а также в течение рабочих суток. Результаты
наблюдений на многосуточной станции, выполненной в
районе Корчевы 27-30 сентября 1969 г., показали, что
в нерабочие дни недели (27 и 28 сентября) здесь про-
слеживалось слабое стоковое течение. Оно охватывало
всю водную толщу, имело скорость 0.02-0.04 м/сек. и
характеризовалось сменой направлений иа обратное (рис.
19, а, б). Такой режим скоростей сохранялся в течение
109
29IX 3 час. 9 час.
Рис. 19. Направление
станции 1У разреза 28, 29
и скорость переноса вод на русловой
сентября 1969 г.
всего дня 28 сентября (рис. 19, в, г, д, е). С на-
чалом рабочей недели картина резко меняется. Уже в 3
часа 29 сентября в поверхностном слое до 5 м заре-
гистрировано усиление стокового течения (рис. 19, ж).
Скорость его возросла до 0.06 м/сек. К 9 час. стоко-
вок течение распространилось на всю водную толщу
(рис. 19, з). Со временем в придонных слоях воды фор-
мируется компенсационное течение, которое к 18 час.
было выражено вполне отчетливо и постепенно усилива-
лось (рис. 19, и, к). К утру 30 сентября обратное тече-
ние охватило всю толщу воды.
При сложной динамике вод в безледный период трудно су-
дить о распространении подогретых вод в водохранилище по
характеру течений. С целью выявления более общих закономер-
ностей их распространения осуществлена математическая об-
работка данных температурных съемок за 1967-1970 гг. (Бу-
торин, Курдина, 19726). Анализ поля- температуры воды Ивань-
ковского водохранилища методом разложения -на естественные
ортогональные составляющие (Багров, 1958; Смирнов и Кондра-
нова, 1972) показал, что большая часть дисперсии поля темпе-
110
f
a
ратуры описывается двумя первыми векторами. На их долю при-
ходится от 75 до 90% суммарной дисперсии, в том числе на
долю первого вектора в поверхностном слое - от 69 до 83, а
в придонном - от 54 до 76%. Учитывая это, рассмотрим распре-
деление их характеристик по водоему. На рис. 20 представлено
поле первого вектора в поверхностном слое воды. Оно -весьма
неоднородно. На участке водохранилища, примыкающем к Мошко-
внчскому заливу, значения данного вектора оказались максималь-
ными по величине и противоположными по знаку значениям его
на большей части водоема. В отдельные годы характеристики
первого вектора в этом районе достигают +10.7, +10.8 (рнс.20,
а, б), тогда как у водозабора ГРЭС и в приплотинной части
их значения отрицательны и равны -1.0, -2.0. Очевидно', что
поле первого вектора и описывает ту особенность распределе-
ния температуры воды в водохранилище, которая связана с
поступлением и распространением подогретых вод Конаковской
ГРЭС. Следовательно, если проанализировать ход нулевой
изолинии первого вектора за годы наблюдений, то можно уста-
новить зону устойчивого влияния подогретых вод. Оказалось,
что несмотря на большие различия в расходе ГРЭС от года к
году, общая картина распространения теплых вод в водохрани-
лище меняется незначительно: лишь в 1970 г. при оптимальном
водопотреблении ГРЭС эта зона продвинулась к водозабору
(рис. 20, г). В придонном слое характеристики первого вектора
меняются с той же закономерностью, что и в поверхностном.
Что же касается зоны устойчивого влияния подогретых вод,
то в придонном слое она ограничивается частью мелководья,
прилегающего к Мошковичскому заливу.
Поле второго вектора имеет более однородный характер.
Это указывает на то, что причины, обусловливающие его изме-
нение, являются общими для всего водоема. На рис. 21 пока-
зано изменение характеристик второго вектора в 1970 г., ког-
да удельный-вес его был наиболее высоким и данный вектор
освещал в поверхностном слое 16, а в придонном - 25% диспер-
сии. Из рис. 21 видно, что наибольшие колебания значений ха-
рактеристик вектора отмечаются на выходе Мошковичского за-
лива - от 1.3 в поверхностном слое до 5.3 в придонном. На
прилегающем к заливу мелководье устойчиво располагается зо-
на минимальных значений вектора (до -2.4). В остальной час-
ти исследуемого района изменение характеристик идет более
монотонно от О до +1.5, причем вблизи водозабора ГРЭС значе-
ния их всегда отрицательны, а ниже залива преимущественно
положительны. Такое распределение характеристик вектора в
русловой части, очевидно, связано с теми особенностями тем-
пературного режима плеса, которые определяются его морфо-
метрией и метеорологическими условиями над водоемом. Так,
112
Рис. 21. Значение характеристик второго вектора в по-
верхностном (а) и придонном (б) слоях воды, 1970 г.
положительные значения вектора отмечаются в глубокой и ме-
нее проточной приплотинной части водохранилища, где аккумули-
руется больше тепла, чем на узком и проточном участке у
ГРЭС. Изменение характеристик второго вектора на выходе из
Мошковичского залива и прилегающем мелководье определяется
часто возникающим здесь расслоением и особенностями дина-
мики вод (Буторин, Курдина, 19706).
Таким образом, разложение поля температур иа естествен-
ные составляющие показалб, что зона устойчивого распростра-
нения подогретых вод в безледиый период ограничена, и на
большей части водоема ведущую роль в формировании темпера-
турных условий играет радиационньй фактор, С увеличением '
объема подогретых вод, сбрасываемых в водоем, влияние их
778 8
1 «4
на температуру воды в приплотинном плесе усиливается. Это
видно иа сопоставления значений характеристик первого векто-
ра у ГРЭС и вблизи плотины. У водозабора ГРЭС величина их
меняется от -2.0 до -2.7, а на остальной части плеса вне
района, прилегакшего к Мошковичскому заливу, от -0.3 до
-1.0. Если от характеристик вектора перейти к значениям тем-
пературы воды, поскольку эти величины функционально связаны^
то окажется, что за безледный период 1970 г. средняя темпе-
ратура воды вблизи плотины была примерно на 1.5° выше, чем
в створе ГРЭС.
Особенности верт
распределения те
в зоне распростр
подогретых вод
икального
мпературы воды
а н е н и я
Сложный характер динамики вод в Иваньковском водохра-
нилище оказывает большое влияние не только на распростране-
ние подогретых вод, но и иа вертикальное распределение тем-
пературы воды. Для характеристики особенностей температур-
ного поля и динамики температуры воды в зоне влияния теплых
вод ГРЭС использованы данные температурной съемки 8 и 9
мая н многосуточных стационарных наблюдений (производимых
четыре раза в сутки) с 9 по 14 мая 1968 г. Район работ и
схема расположения станций показаны на рис. 22. Станции 2
и 4 находятся вблизи потока подогретых вод, ст. 3 - в прото-
ке между берегом и грядой островов, а ст. 6 отделена от
оси потока теплых вод обширной отмелью и цепью островов.
Она считалась контрольной и в период наблюдений не подверга-
лась существенному воздействию теплых вод (Буторин, Курди-
на, 19706). Метеорологические" условия в районе исследований
были одинаковыми и изменялись одновременно.
В период наблюдений в район водозабора ГРЭС поступала
вода с температурой около 12.5° по всей глубине. По ходу
воды с уменьшением скорости течения и увеличением глубины
возникала стратификация водной толщи по температуре. Так,
в районе ст. 6 температура на поверхности оставалась около
12.5°, а у дна ие превышала 10.8°. Ниже Мошковичского зали-
ва под влиянием сбросных вод температура воды заметно по-
вышалась, а стратификация водной толщи усиливалась. Граница
Рис. 22. Направление потока подогретых вод 8, 9 мая
1968 г. В рамке - полигон многосуточных наблюдений и номера
станций.
Цифры при кривых - температура воды,°C.
влияния теплых вод четко прослеживалась на расстоянии 8 км
ниже аалива. У плотины температура воды оказалась такой же,
как на контрольной ст. 6. О том, что повышение температуры
воды в данном случае обусловлено влиянием подогретых вод
ГРЭС, свидетельствует тот факт, что все измерения температу-
ры были выполнены в один день и велись от плотины в направ-
лении ГРЭС.
Характер вертикального распределения температуры воды
в исследуемом районе и его изменение при разных погодных
условиях представлены на рис. 23. По ходу кривых отчетливо
видны различия в вертикальном распределении температуры на
отдельных станциях, особенно между контрольной ст. 6 и ст. 2
и 4. На последних двух четко выражено температурное расслое-
ние водной толщи, хорошо прослеживается слой температурного
скачка на глубине 1.0-1.5 м с разницей температур до 8°.
Ниже слоя скачка идет медленное понижение температуры на
1-2°.
Вне зоны влияния подогретых вод в подавляющем большин-
стве случаев наблюдается плавное понижение температуры воды
с глубиной или относительное постоянство ее в водной толще.
Только в двух случаях из 20 на ст. в отмечалась стратифика-
ция, причем слой скачка, как и на остальных станциях, расно-
115
Вертикальные градиенты температуры воды 9-14 мая 1968 г.
20,40 1.88 1 20.55 1.03 20.30 0.71 20.10 СО о • 2 q ф q а к <0 ко ф я о о ь ф к я 8 сч
11 мая О о со d О 1.07 g •м § Е м а d
сч tD •и tD «О d S 8.05 I 3
’ЧГ ГЧ
О1Ч1 а>|о 8|£ do 9,30 СЧ § 8 СО о сч о f
3.45 1.00 4.20 1.07 4.00 0.58 3.20 ег'о 154 1 OS’OS .
о 1.80 8. d 1.70 СО сч • 0.81 1;40 S чТ to co d
сч сч сч сч № о
§ >02 о 10 О со 2 8 8 00 сч 13 м * CD !•/
§ tO •-Ч сч to oi 2 ° ю d 8 3
о 9.40 1.28 9.55 0.80 СЧ|О g |<ю о |d V
did оэ to to co
.40 Ю о ю °. § 81 •4
* О о со о со ° a co
О со о о о • 18 _• <о 00 о о О о ф к я ф. • § о q si
К « 2 CD о ^*1 00 о о 2 мая 8.1 .02
а с «1
8.30 0.86 8.50 0.10 8.00 0.32 SCSI 10'0 • сэ • со я 3.55 0.70
№ стан- цин С ч 0 хг со S R КО 2 1 я в ь о 1 иип СЧ
. о 00 00*0 7.50 0.02 col 3 00’0 E- X Ф s ц «О Q.
ol СЧ 8 1
-ч-| о Ф
к
08 Q toi SI to СЧ ь CO
СЧ d о сч| d 81 d X Ф 2
CO
0
15.55 о d о co to ₽4 0.22 15.40 его Д s
X
oi о о о о to s
5 co co co co R
Ю
d о OD O uu <-> co
«—« X
a
о О CO Ю Ю 2
". * 4 ". ". a
О CO ° о Д 1
Ф
о [Se to ’Г о CO R
co co • Ф
сч о .0 СЧ о 8 о S R
О
о to |сч О to CQ
co 00 CO СЧ
t a
to 1 to о W1 о
Г-» Ф
и
to ICO о о ICO
to |4* cc CD
I
ОЭ 1° 00 |o |o
. Ф
2
to о 1° о 1° s
c c< co CO
|o
co Io Io c
лагалея на глубине около 1 м. О
степени стратификации вод на дан-
ном участке водоема можно судить
по температурным градиентам, рас-
считанным для всей толши воды
(табл, 9). Из данных табл. 9 вид-
но, что величина градиентов колеб-
лется в широких пределах, но наи-
меньшие значения их наблюдаются
на ст. 6. Поскольку район располо-
жения ст. 6 и ст. 4 характеризу-
ется примерно одинаковой проточно-
стью, а глубина последней даже
больше, чем первой, то повышение
значений градиента температуры
на ст. 4 по сравнению со ст. 6
обусловлено только влиянием подо-
гретых вод Конаковской ГРЭС
(Буторин,'1971). Влияние подогре-
тых вод ГРЭС на вертикальное
распределение температуры осо-
бенно хорошо прослеживается на
ст. 2, расположенной вблизи оси
потока; здесь на поверхности в от-
дельные сроки температура воды
достигала 21°, а у дна была около
13°. Естественно, что при глубине
3..5 м температурные градиенты
здесь особенно велики.
Сложная картина распределения
температуры воды по глубине в
зоне распространения подогретых
вод сохраняется и летом. Обычно
температурное расслоение вод в
равнинных водохранилищах явление
непродолжительное. При мелковод-
ности их оно нарушается ветром,
который способствует выравниванию
температуры й быстрому перерас-
пределению тепла по глубине.
Вследствие этого уже в начале лета
температуры поверхностных и при-
донных слоев близки между собой.
В Иваньковском водохранилише
вследствие поступления подогретых
117
Рис. 23. Вертикальное распределение температуры воды
мая 1968 г.
Цифры у кривых - порядковый номер наблюдения.
вод стратификация водной толщи по температуре усиливаетсяа
становится более устойчивой и продолжительной. Так в районе
Корчевы температурное расслоение вод наблюдается все лето,
118
причем характер и степень стра-
тификации меняются в зависимос-
ти от направления переноса
подогретых вод. Это видно из
сопоставления эпюр переноса
вод (рис. 18) с кривыми верти-
кального распределения темпера-
туры в те же сроки (рнс. 24).
По характеру кривых на рис.24,а
видно, что слой температурного
скачка чаше расположен на глу-
бине 4-5 м (рис. 24, 1, 4) или
ниже горизонта смены направле-
ния переноса вод. Иногда спой
скачка опускается до 9 м (рис.
24, 3), в отдельных случаях на-
блюдаются два слоя скачка
(рис. 24, 5), но и в этих слу-
чаях резкое изменение темпера-
туры чаше всего связано с ди-
намикой вод. Еще более резкое
расслоение водной толщи по тем-
пературе отмечено в этом районе
в период наиболее интенсивного
прогрева водоема (рис. 24, б).
В отдельные сроки наблюдений,
когда подогретые верхние слои
воды интенсивно нагреваются
солнцем, разность между темпе-
ратурой воды на поверхности и у
дна может превышать 8°.
Отмеченные особенности
вертикального распределения тем-
пературы воды в Иваньковском
водохранилище подтверждаются
обширными материалами наблю-
дений Московского отделения
института Теппоэпектропроект ,
которым изучалось влияние сброс-
ных вод Конаковской ГРЭС на
термический режид водоема в
наиболее теплый период года:
июль-первая половина августа 1970 г. На рис. 25 представлена
одна из многих серий кривых вертикального распределения тем-
пературы воды на отдельных участках водфхранилища за 14 июля.
119
Рис. 24. Вертикальное распределение температуры на рус-
ле IV разреза летом 1970 г.
а _ 7-Ю июня (1 - 21 час 7 VI; 2-9 час, 3-18 час
8 У1; 4 - 15 час 9 У1; 5-9 час 10 VI),' б - 3-6 июля.
Рис. 25. Вертикальное
распределение температуры
воды в характерных точках
района наблюдений 14 июля
1870 г.
1 - входной створ, 2 -
у водозабора, 3 - на выхо-
де из Мошковичского залива,
4 и 5 - мелководье у залива
(данные МОТЭП).
Характер кривых показывает,
что вне зоны влияния подо-,
гретых вод, в районе водо-
забора ГРЭС и выше, наблю-
дается незначительное пониже-
ние температуры воды с глу-
биной (рис. 25, 1, 2). Здесь
водная масса находится прак-
тически в состоянии гомотер-
мии. В зоне же распростране-
ния сбросных вод даже при
ветре до 5.4 м/сек. юго-за-
падного направления отчетли-
во прослеживается темпера-
турное расслоение водной тол-
ши (рис. 25, 3, 4, 5). О том,
что такое распределение тем-
пературы воды на рассматри-
ваемом участке водоема не
случайно, свидетельствует
многократная повторность
наблюдений у водозабора
(рис. 26, а) и залива (рис.26,
б). Таким образом, и летом
при одних и тех же метеоро-
логических условиях на от-
дельных участках Иванькове-
120
a
Рис. 26. Вертикальное распределение температуры у водо-
забора (а) и на выходе иа Мошковичского залива (б) 28-31
июля 1870 г. (данньБ МОТЭП).
а — в русловой части, 27-30 IX 1969 г., б - на правом скло-
не 2-4 XI 1970 г.
кого водохранилища наблюдаются существенные различия в рас-
пределении температуры воды по вертикали. Они обусловлены
влиянием сброса в водоем подогретых вод Конаковской ГРЭС.
Интересно отметить, что температурная стратификация
в зоне распространения подогретых вод наблюдается и в период
осеннего охлаждения водоема, несмотря на усиление ветровой
деятельности. В качестве примера могут служить данные мно-
госуточных станций в районе Корчевы, Здесь в конце сентября
в отдельные сроки прослеживается четко выраженный скачок
.при значительных колебаниях температуры воды (рис. 27,а).
121
Только в ноябре при ветре свыше 5 м/сек. в этом районе
наблюдается гомотермия или состояние близкое к ней, хотя
суточные колебания температуры превышают 4° (рис. 27, б).
Таким образом, в зоне распространения подогретых вод
вертикальное распределение температуры воды и ее динамика
имеют свои особенности. Прежде всего водная толша на этом
участке резко стратифицирована. Подогретые воды нарушают
естественный суточный ход температуры, значительно усложня-
ют ее динамику. Перенос подогретых вод ветровыми течениями
приводит к быстрому изменению температуры не только поверх-
ностных, но и придонных слоев. Несмотря на значительные
градиенты температуры, время полного перемешивания потока
теплых вод невелико - около 4-5 часов. Примерно такой же
срок требуется и для восстановления стратификации.
Однако решающая роль в формировании температурного
поля в районе распространения сбросных вод ГРЭС принадлежит
метеорологическим факторам, и прежде всего солнечной радиа-
ции и ветру. С метеорологическими условиями связана различ-
ная скорость прогрева водной толши на отдельных участках
водоема, а также абсолютная величина температуры воды.
Ветровое перемешивание перераспределяет тепло по акватории
и глубине водоема.
О влиянии сбросных вод
Конаковской ГРЭС
на температуру донных отложений
В тесной связи с температурой водной массы водоема на-
ходится температура донных отложений. Данные непосредствен-
ных измерений температуры донных отложений водохранилища
весьма ограничены и относятся преимущественно к верхнему
слою. Учитывая это, для общей характеристики температурных
условий дна использована температура придонного слоя воды.
Сопоставление температур придонного слоя воды и грунта пока-
зало, что в русловой части водоема, где донные отложения
состоят из серых и песчанистых илов, разность температур
вода-грунт редко превышает 1°. На пойменных участках и за-
островных мелководьях, где грунты представлены разновиднос-
тями почв или отложениями макрофитов (Курдин, 1961), эта
разность увеличивается до 2-3°. Различное соотношение темпе-
ратуры придонного слоя воды и Донных отложений объясняется
122
различием теплофизических свойств грунтов и особенностями
проточности (Бакастов, 1966). Учитывая эти обстоятельства,
для условий Иваньковского водохранилища можно принять, что
в глубоководных участках весной температура грунта верхнего
10-сантиметрового слоя в среднем на O.5-1.50 ниже придон-
ной температуры воды, летом эта разность уменьшается на 0-
0.5°. а осенью грунт теплее воды на 1-1.5°.
Поскольку приток тепла ко дну водоема и его отдача идут
через водную толщу, температура которой измеряется регуляр-
но на водомерных постах, предпринята попытка найти зависи-
мость между температурой воды и температурой грунта. С
этой целью осуществлена корреляция средней за предшествую-
щую декаду температуры воды по водомерному посту Конако-
во, наиболее полно отражающей температуру водной массы
водохранилища (Тачалов, 1966), с температурой грунта, изме-
ренной на глубине 10 см. На основе установленной зависимос-
ти нолучены кривые связи температуры грунта и средней тем-
пературы воды для ряда русловых станций (рис. 28). Оказалось,
что для отдельных лет кривые связи индивидуальны, так как
температуры грунта по календарным датам смежных лет су-
щественно различаются в зависимости от предшествующих ме-
теорологических условий. Так, в отдельные периоды весны и
пета 1970 г, температура дна Иваньковского плеса превышала
те же значения 1969 г. на 2.5-4°. Наряду с индивидуальностью
кривые имеют и обшие черты: раздельное расположение их вес-
ной при низких температурах грунта (рнс. 28, а), постепенное
сближение летом и тесное расположение точек на кривой в кон-
це лета и осенью (рис. 28, б). Это указывает на неравномер-
ный прогрев дна водоема весной, выравнивание температуры
донных отложений летом и относительно равномерное выхола-
живание дна осенью. .
Для оценки влияния подогретых вод Конаковской ГРЭС
иа температуру донных отложений водоема рассмотрим фоновый,
естественный годовой ход температуры грунта в Иваньковском
плесе водохранилища. Минимальные температуры донных отло-
жений наблюдаются в предвесенний период и в начале половодья,
когда идут воды с температурой около 0°. Весенний прогрев
начинается со второй и третьей декады апреля. К концу апре-
ля температура грунта может достигать 4-6°. В мае-июне
отмечается интенсивный прогрев дна водоема. Наблюдения в
середине мая 1969 г, показали, что к этому времени темпера-
тура дна в русле на входном створе вблизи г. Конаково дости-
гала 8-12°, а у плотины 6-8°. К началу июня на тех же стан-
циях температура повысилась до 12-15 и 11-12° соответственно
В период интенсивного прогрева водной массы наблюдаются
123
Рис. 28. Кривые связи температуры грунта, измеренной
на русловых станциях разрезов на глубине 10 см,и средней
декадной температуры воды у водомерного поста Конаково для
весенне-летнего (а) и осеннего (б) периодов 1969 г.
1 - развез I, 2 - разрезы HI, IV, V, 3 - разрез VI, 4 -
ст. 8. По оси ординат - температура грунта, по оси абсцисс -
температура воды.
и наиболее существенные различия в температуре донных отло-
жений. В верховьях водохранилища и в районе ГРЭС, где
водная масса интенсивно перемешивается стоковым течением,
дно водоема прогревается более равномерно. Но и в этих ус-
ловиях разность температур по поперечному сечению достигает
5°. В Иваньковском плесе при наличии обширных мелководий,
частично заостровных, разность температур на профиле дости-
гает 10°. Так, в июне 1970 г. при температуре дна на русле
от 11° у плотины до 14-15 у водозабора и Корчевы ла откры-
тых пойменных участках значения ее достигали 16-18° (рис.29,
а), а на заостровных - 16-21°. В июле-августе разность тем-
ператур донных отложений по акватории водоема уменьшается
124
Рис. 29. Ход изотерм в.грунте на глубине 10 см.
а — июнь, б — февраль 1970 г.
и редко превышает 2°. Максимальный прогрев дна наблюдается
во второй половине июля при температуре 18-21° на русловые
участках и 19-23° на мелководьях. Охлаждение дна водоема
начинается в августе. Первоначально идет равномерное пониже-
ние температуры грунта по'всему водоему, а поздней осенью
123
в русловой части водоема температура дна на 1-2° выше, чем
на мелководьях.
В естественных условиях водохранилища к январю темпе-
ратура дониых отложений на пойменных участках снижается
до 1-1.5°, а в русловых она примерно на 1° выше. При зимней
сработке объема водохранилища температура дна выравнивается
и в феврале колеблется около 1°, а в марте - от 0.5 до 1°.
С поступлением в водоем подогретых вод Конаковской
ГРЭС температурные условия дна в зоне их распространения
заметно изменились. Наиболее существенное влияние на темпе-
ратуру донных отложений подогретые воды оказывают зимой.
В этом сезоне величина температуры грунта в Иваньковском
плесе заметно повысилась: в Мошковичском заливе она дости-
гает 11°, на мелководье у выхода из залива колеблется от 5
до 9°, а по ходу воды в русловой части водоема - от 3 до 5°.
Более подогретым оказалось дно водохранилища к востоку от
Мошковичского залива разрезы ГУ и У. Менее прогрет учас-
ток от залива до водозабора ГРЭС - разрезы III и II (рис. 29,
б). Вблизи плотины большую часть зимы температура грунта
была около 3.5°, т.е. заметно выше той, что наблюдалась здесь
до поступления подогретых вод. Общее представление о темпе-
ратуре донных отложений на отдельных участках водоема в
зоне распространения подогретых вод позволяют получить дан-
ные табл. 10 и 11.
В безледный период в верховье Мошковичского залива тем-
пература дна превышает естественную на величину до 8°. Вы-
сокие температуры донных отложений залива наблюдаются вес-
ной и летом до границы распространения холодного придонного
течения. В зоне влияния последнего температура на 4-6° ниже,
чем у водосброса (табл. 10). При этом на некоторых участках
залива возникают значительные, иногда до 5°, перепады тем-
пературы как на границе вода-грунт, так и в толще донных от-
ложений. Осенью с выравниванием температуры водной толщи
температура донных отложений становится более однородной и
по своим значениям близкой к температуре воды.
Существенное влияние подогретые воды оказывают и на
температуру дна мелководий у Мошковичского залива. Ранней
весной при высокой проточности водоема в этом районе про-
исходит интенсивное смешение теплых вод с водами водохрани-
лища, и температура rpjHTa здесь выше, чем на мелководьях,
ие подверженных влиянию подогретых вод,до 10°. Так, в сере-
дине апреля 1969 г. при наполнении водохранилища температура
доииых отложений вие зоны влияния подогретых вод не превыша-
ла 1°, а на выходе из Мошковичского залива изменялась от 2°
128
Таблица 10 Температура грунта на глубине 10 см в зоне влияния подогретых вод и вне ее (Перетрусовский залив) в 1969 г. Перетрусовский залив защищенное мелководье ст. 4, 4.0 м to ео о оо - w’ 2
ст. 3, 2.0 м СХ О Ю Г- О —• Г* СО СМ СМ ~
открытое мелководье ст. 2, 2.0 м СО СО О (П ~ а —<
ст. 1, 3.0 м О О со ю СО •—« СО СО ~ см —
Водохранилище, мелководье ' Ш разрез ст. 4, 3,0 м Г- * — г- Г- со 05 to
ст. 3, 1.8 м •ч го —• сч О 00 05 СО СЧ — —
IX разрез 1 ст. 4, 6.8 м СО (О о —« ’Г о со СМ —'
ст, 2, 2.8 м со о со см со со —« CM W
Мошковичский залив, 1 | разрез УШ | левая пойма, э ч со со о о СО со Л см см см см
русло, 4.5 м 18.1 27.9 23.6 11.9
правая пойма, 2.5 м О Г- ’Г 00 со о см см см см
Дата измере- ния 6-7 VI 5-7 УШ 4-6 IX 1, 2 XI
Температура грунта на глубине 10 ем в Мошковичском заливе
и на отдельных разрезах зимой 1970 г.
ле- вый склон 5.0 м । 4.3 СО ’Г _
СО
& а рус- ло. 10. On ’Г СО X-
пра- вый склон в«0м 3.4 1 e’t-
(0 ле- вый склон, 5.0м ' 1
а (0 а рус- ло» 12.0м 4.1 4.5
> склон 6.0 м
пра- вый СО • со СО СО
ле- вый склон 6.0 м 3.8 3.3
ф О. (0 а рус- ло. 10.0м со со о
пра- вый склон. 5.0 м 3.3 3.7
О W 0. С9 2 ст. 4, 6.0 м 4.8 СО
IX I ст.2, 1.5 м со 00 1
Мош- КО- ВИ не- кий залив, ст.5, 2 о •<? 11.1 9.6
ле- вый склон 4.0 м 0.1 0.7
разрез рус- ло, । S о • со 1-4 2.4
пра- вый склон, 4.0 м 0.2 0.8
Дата изме- рения | SI-81 21-24 Е
128
да вновь залитой прнурезной полосе до 7-10° на участках, под-
верженных постоянному воздействию теплых вод.
С началом температурного расслоения водной толщи подо-
грев дна сброшенными водами прекращается и температура дон-
ных отложений имеет значения, близкие к температуре воды
водохранилища. Контрольные наблюдения, проведенные на мел-
ководье с естественным режимом прогрева дна, показали, что
летом значения температуры донных отложений в Перетру-
совском заливе и на мелководье у Мошковичского залива близки
между собой (табл. 10). Осенью при гомотермии выхолаживание
дна в зоне распространения подогретых вод идет медленнее,
чем вне этой зоны. При ледоставе (в естественных условиях)
температура грунта в русловой части водоема составляет пре-
имущественно 0.5-2.0; на выходе из залива значения ее меня-
ются от 4 до 7°, а в русле Волги равны 3.5-4.50 (табл. 11).
Таким образом, зимой, ранней весной и осенью в зоне
распространения подогретых вод Конаковской ГРЭС температу-
ра дна выше, чем на других участках водохранилища и в сосед-
них водоемах. Летом эта.разница минимальна. Различие в теп-
лосодержании Дна отдельных участков водоема хорошо просле-
живается по сумме градусо-дней, подсчитанной за один и тот
же промежуток времени. За зиму 1969/70 г. на станции, распо-
ложенной на склоне русла выше ГРЭС, она составила 70, а на
склоне русла у разреза V (в 7.5 км ниже Мошковичского зали-
ва, в зоне постоянного воздействия подогретых вод) - 410.
На русловых вертикалях указанных участков сумма градусо-
дней равнялась 180 и 480 соответственно. Следовательно, в зо-
не влияния теплых вод теплосодержание дна увеличилось по
сравнению с естественным в 2.5 раза на русле и в 6 раз на
склонах русла. На остальной части водоема температура донных
отложений близка к средней температуре водной массы и сход-
на с таковой соседних водохранилищ.
Изменение теплозапаса
водохранилища под влиянием
поступления сбросных вод
Конаковской ГРЭС
На основе температурных съемок водохранилища предпри-
няв попытка определить изменение теплосодержания вод, кото-
рое произошло на отдельных участках Иваньковского водохрани-
лища под воздействием сброшенного тепла. В 1967 г. при рас-
ходе 25-30 м^/сек. через конденсаторы ГРЭС было пропущено
778 9
129
1 а о л и n a12
Количество дополнительного тепла,
поступившего в Иваньковское водохранилище в 1968 г.
Месяц W, млн м® At Од, млн т/кал.
Январь .... 105.5 7.6 801.8
Февраль .... 74.3 8.7 646.4
Март 83.3 1О.З 858.0
Апрель .... 85.1 7.1 604.2
Май 116.6 5.7 664.6
Июнь 109.7 5.0 548.5
Июль 155.4 5.6 870.2
Август .... 182.5 4.8 876.0
Сентябрь . . . 150.7 6.3 949.4
Октябрь .... 102.8 7.9 812.1
Ноябрь .... 104.2 9.7 1011
Декабрь .... 122.5 9.3 1139
около 800 млн м^воды. В 1968 г. с увеличением мощности стан-
ции объем сброшенных вод составил 1400 млн м^. По проект-
ным данным, которые подтвердились натурными измерениями,
повышение температуры воды на выходе из ГРЭС по сравне-
нию с ее значениями на водозаборе колеблется от 6 до 10°,
иногда Достигает 13°. На трассе водовыпуска сбросные воды
частично охлаждаются, и на выходе в водохранилище температу-
ра их- превышает температуру водоема на 5-10, реже на 12°.
Количество дополнительного тепла, вносимого подогретыми во-
дами в водохранилище за месяц (Од.), определялось по месячному
объему сбросных вод (W) и среднему превышению температуры
их над естественной в нижнем створе Мошковичского залива
(At). Оказалось, что в зависимости от режима эксплуатации
ГРЭС и условий охлаждения подогретых вод количество допол-
нительного тепла, поступающего с ними в водоем, в течение
года существенно меняется (табл. 12).
Для оценки роли дополнительного тепла в термическом
режиме Иваньковского плеса водохранилища было подсчитано
месячное количество тепла, вносимого в плес проходящими че-
рез него водами с естественной температурой (Qe). Соотноше-
ние величин показало, что удельный вес дополнитель-
ного тепла возрастает по мере охлаждения вод водохранилища.
130
Рис. 30. Номера и границы участков водохранилища, для
которых подсчитывалось
теплосодержание вод.
Если в августе дополнительное тепло составляет менее 10%
отфтепла, приносимого водами с естественной температурой, то
В октябре на его долю приходилось 50%, а в декабре-марте со
сбросными водами ГРЭС в плес поступало тепла в 3-5 раз
больше, чем с водами водохранилища (Курдина, Буторин, 1971).
Естественно, что дополнительное тепло в водоеме распре-
деляется неравномерно. Для того чтобы выяснить этот вопрос,
район исследований был разбит на участки (рис. 30), а затем
для каждого из них подсчитано теплосодержание. Исследования
С.Н.Тачалова (1966), измерения температуры воды, выполнен-
ные до сооружения Конаковской ГРЭС, и наши измерения позво-
лили достаточно надежно определить естественное теплосодер-
жание каждого участка, а материалы температурных съемок
дали возможность установить его изменение под влиянием
подогретых вод (табл. 13).
Из данных табл. 13 видно, что наиболее существенно теп-
лосодержание повышается в районе выхода в водоем подогре-
тых вод. На участке 4 (между )П и IV разрезами) в январе
1969 г. количество тепла в воде увеличивалось на 60% от ес-
тественной величины, а в марте возрастало более чем в 5 раз.
Зимой 1970 г. в части водоема, лежащей от Мошковичского
залива по ходу воды, теплосодержание увеличилось в 3-3.5 ра-
за, а между водовыпуском и водозабором в 1.2-3.5 раза. Вес-
ной и осенью количество тепла в воде по отдельным участкам
131
Теплосодержание вод по отдельным участкам района
исследований в % к естественному
Весь район ж ж 1 СО СО — СЧ 5$ Г- CD ~ — 4“ со со сч СЧ О О о о ° СО СВ О о t- 05 о О О — — —> — — — СЧ — — СО СЧ — —, _
СО 1 О о О О й Ю О О О 1 1 со СМ СО о о О О ° СО ПО о О ООО „ О1 Г-1 —4
г- _ СМ О О О | | (О сч см 1Л О OI ~ С £ СО из о о ООО со О О ОО 5 —. см —« —« сч -*
со СО СО Г- СЧ СЧ ® тГ со сч сч ч- СО СО СЧ со СО О О О О ° со 05 О О СЧ — ООО Ч1 — — — — ” „Сч — — 05 СО — — —
(О 3 — о 3 3 ° о о со 8 о? еп со со — ч> — — — — 2! со со о — со со о О о — Ч< — со — — —
3 '8838« Й8ёё о«88о [Q
ь о <0 >> 2 со 1 СО О — СО СО Ч* СО — — СО г- сч ч* со оооо— ч* г- о о со г- о о о — — — — — —СО — — СО сч — — —
сч 1 2 2 2 2 2 со «о о о счч'счо — О о о О о о сч о О СО СЧ О О О F4 a-t «—« ч—с a-t а—> «-М а-t а-t a-t 1
'88888 8§88 8888S *“* г-» «М а-ч а—< а-^ а-Ч 1—«
Дата . 5 X -Е — . 1^5 — ?• Г* ?s „ СОГ- СО Ч* >. со — >• — о — — ю о — сч СО - > „СО 1 1 > Г- 1 1 * Г- сч а> со со ч> ч> оо со со ч> -оо сч— - i • — — СЧ оо СЧ СЧ „ — — — с- ч> — — сч со со —
Таблица 13 {продолжение)
132
Весь район Z? СО О О ° о с" —I •—«
со SsSS »—» •—* •"< /
г- N - О СО о о о о
со
ю SggS s _ ь о <0
участка >> о 00 « СО х о О о со h •м —• r-С о Ф 3
2 СО 103 104 105 122 ржания по
сч ф п о О о сч о о о о о о »- К с ф ь
ф а §§§§ i ~ ~ ~ S о
fl е а Е 3 1 4-6 VIII 3-5 IX 1, 2Х 1,2 XI * И
повышается на 20-40%, а летом -
примерно на 5%. По ходу воды
к плотине повышение ее тепло-
содержания за счет подогретых
вод постепенно уменьшается и
в приплотинном участке водо-
хранилища отмечается только
зимой. Так, на участке 8 в ян-
варе оно составляло 130%, а в
марте увеличивалось в 2.5 раза.
Повышение содержания тепла
на участках 3 и 2, лежащих по
ходу воды выше Мошковичского
залива, свидетельствует о том,
что зимой подогретые воды
подтягиваются в этот район
насосными станциями ГРЭС, а
летом приносятся ветровым те-
чением.
Таким образом, общее по-
вышение теплосодержания ис-
следуемого района водохранили-
ща за счет поступления подогре-
тых вод Конаковской ГРЭС наи-
более заметно зимой: в январе
оно составляет 30-40%, а в
феврале-марте увеличивается в
3-3.5 раза. В летний период из-
менение теплосодержания за
счет этого фактора незначитель-
но.
О некоторых
последствиях
поступления
подогретых вод
Конаковской ГРЭС
в Иваньковское
водохранилище
Как было показано выше, за
счет поступления подогретых
вод Конаковской ГРЭС заметно
133
Рис. ЗК Величина и контуры полыньи (незаштрихованная
часть) в январе-феврале и марте 1968 (а), 1969 (б), 1970(в)гг.
изменилось теплосодержание отдельных участков водоема. Это
в свою очередь вызвало изменения в ледовом режиме водохра-
нилища. Так, в Мошковичском заливе и прилегающем к нему
участке водоема лед. не образуется и на протяжении всей зимы
существует полынья. Размеры ее непостоянны и определяются
метеорологическими условиями сезона и особенностями динами-
ки вод. Наибольшие размеры полыньи в середине зимы отмече-
ны в январе 1969 г., когда длина ее достигала 4.5, а ширина
1.5 км (рис. 31). В начале и конце зимы длина полыньи может
достигать 9 км.
Уменьшилась толщина льда над большей частью акватории
Иваньковского плеса. На расстоянии 1-1.5 км от полыньи тол-
шина льда обычно не превышает 25 см. Если у водомерного
134
поста Конаково в середине зимы 1968 г. и зимой 1969/70 г.
толщина льда превышала норму, то в районе Дубны начиная с
1968 г. она стала ниже средних многолетних значений. Одно-
временно увеличилась повторяемость образования полыней и
промоин вблизи плотины Иваньковской ГЭС. Зимой 1970 и
1971 гг. в этом районе водохранилиша полыньи наблюдались на
протяжении всего сезона.
С поступлением подогретых вод связаны изменения в кис-
лородном режиме водохранилиша. Характерной особенностью
его до сооружения Конаковской ГРЭС являлась слабая насы-
щенность кислородом вод водохранилиша зимой, особенно в кон-
це ее. По данным Н.А.Трифоновой (1960), в феврале—марте ко-
личество кислорода выше и ниже г. Калинина в среднем около
7 мг/л, в Шошинском плесе - 4, в широком и расчлененном
приплотинном участке от 1.5 на мелководье до 6 мг/л в рус-
ловой части. Рядом авторов отмечалось, что в многоводные
годы, когда перед ледоставом водоем имеет полный объем и
воды его достаточно аэрированы, зимой дефицит кислорода
наблюдается преимущественно в Шошинском плесе. В маловод-
ные годы, когда перед ледоставом водоем уже частично срабо-
тан, а зимний приток невелик и представлен глубокими грунто-
. выми водами, бедные кислородом воды р. Шоши поступают в
водохранилише уже в феврале, и вместе с водами других прито-
ков вызывают резкое снижение содержания кислорода. В такие
годы при глубокой сработке водохранилиша недостаток кисло-
рода отмечается не только по всей акватории водоема, но и в
канале им. Москвы (Себенцов и др., 1940; Мейснер, 1968).
Особенности кислородного режима водохранилища в годы
исследований были в значительной мере связаны с интенсивно-
стью водообмена. Так, в 1967 г. к середине февраля сработка
водоема составила около 0.45 км®. При этом речные воды с
содержанием кислорода 1.13-0.72 мг/л до Ж разреза занимали
большую часть воднсй толши, а на IV разрезе прослеживались
только в придонном'слое. В янБ5р&19б8 и 1969 гг. при мень-
шей сработке водоема (порядка 0.3 км^) граница речных вод
с содержанием кислорода 2.1-2.7 мг/л располагалась выше -
в придонном слое они распространялись до створа ГРЭС и II
разреза. В поверхностных горизонтах содержание кислорода
здесь составляло 6.0-7.5 мг/л. К середине марта 1968 г. объем
водохранилиша уменьшился вдвое. *<В этом случае речные воды
в русловой части водоема продвинулись до плотины. Содержа-
ние кислорода в них колебалось в пределах 0.1-1.8 мг/л. При
менее интенсивном водообмене зимой 1968/69 г. до конца се-
зона в поверхностных горизонтах приплотиниого ' участка со-
хранилось кислорода 4.0-5.0 мг/л.
133
Таблица 14
Содержание кислорода в исходных водах
на водозаборе (А) и в подогретых водах
ниже водосброса (Б)
Дата Водозабор (А) Водосброс (Б) Б мг/л ' — Источник
А мг/л
мг/л 1% на- сыще- ния мг/л % на- сыще- ния «
1967 г. 9,10 II 2.60 18.1 9.64 82.1 3.6
3 III 2.13 14.0 8.44 75.5 4.0 М.Л.Флейс, 1070
14 Щ 1.55 10.6 8.79 81.6 5.7
1968 г. 2 I 4.74 32.5 9.92 85.8 2.1 Тот же
20,21 I 4.08 28.4 9.80 80.5 2.4
27 II 1.42 9.7 9.72 83.2 6.8
17,19 If! 1.46 10.1 8.58 74.5 5.9
21 Ш 2.98 20.3 9.46 84.2 3.2
1969 г. 16 i 5.14 36.1 9.98 85.0 1.9
28 II 1.22 8.7 10.59 97.3 8.7 Л.Пан, 1969
4 Ш 1.17 8.3 10.83 101.2 9.2 Тот же
6 1|1 0.96 6.8 10.75 97.8 11.2
1970 г. 12,13 II 1.91 13.0 9.0 81.5 4.7
24 Ц1 1.00 7.4 9.52 83.2 1 8.0
В водозаборном канале ГРЭС количество кислорода изме—
няется так же, как и в прилегаюшеЛ к нему русловой части во-
доема. Но благодаря забору воды с глубинных горизонтов (при-
мерно с 6-7 метров ) большую часть зимы ГРЭС забирает реч-
ные воды, бедные кислородом. Среднее насыщение их в январе
около 30-40%, а в марте - не более 10% (табл. 14), Аэрация
вод, забираемых електростанцией, очевидно, начинается в по-
лынье у водозабора и продолжается в отводящем канале. Одна-
ко обогащение кислородом сбросных вод в канале невелико и
составляет около 0.2 мг/л (Паи, 1060). При дальнейшем дви-
жении вод интенсивность аэрапии их определяется как содержа-
136
Рис. 32. Распределение
температуры ( С) и растворен-
ного кислорода (мг/л) на
У разрезе в марте 1968 г.
нием кислорода в исходной воде (скорость аэрации прямо про-
порциональна дефициту растворенного кислорода), так и высотой
перепада уровня на водосбросе. При этом известную роль игра-
ет к температура сбрасываемых вод. Поскольку перепад уровня
на водосбросе зимой увеличивается до двух метров и более, то
именно здесь наблюдается самая интенсивная аэрация подогре-
тых вод. Так, ниже водосброса, на ст. 3, по сравнению со ст.1,
количество кислорода увеличивается на 5-8, а в отдельных слу-
чаях на 10 мг/л. В Мошковичском заливе содержание кислоро-
да в подогретых водах меняется незначительно и на выходе
из залива количество его больше, чем у водозабора, в январе при-
мерно в 2 раза (до 10 мг/л), в феврале-марте это превышение ко-
леблется от 3 до 11 раз, а содержание составляет 8.5-10.5 мг/л
(табл. 14).
Расход подогретых вод составляет зимой от 20 до 50%
расхода воды из водохранилища, следовательно, аэрированные
сбросные воды оказывают существенное влияние на кислород-
ный режим Иваньковского плеса. Это особенно заметно во вто-
рой половине зимы. Так, в марте 1968 г. при закончившемся
предвесеннем опорожнении водохранилища присутствие подогре-
тых вод с повышенным содержанием кислорода прослеживается
по всей длине Иваньковского плеса. При этом подогрев и аэра-
ция вод были максимальными в иижней половине потока (рис.32).
Подсос подогретых вод насосными станциями ГРЭС обеспечива-
ет Повышение содержания кислорода в нижних слоях воды и на
участке водохранилища выше Мошковичского залива, что свиде-
тельствует о положительном влиянии подогретых вод иа кисло-
родный режим значительной части водоема.
Помимо зимнего увеличения количества растворенного кис-
лорода в зоне распространения подогретых вод, наблюдаются и
другие- изменения в химическом режиме водоема (Флейс. 1970).
137
.Яте'цШиши Mtipae
/ гои f rJL
Рис. 33. Мощность вто-
ричных отложений в русловой
части Волжского и Иваньков-
ского плесов в 1957 (1) и
1969-1970 (2) гг. (данные
В.П.Курдица).
По оси ординат - мощ-
ность отложений.
на 2-3 мг/л и незначительном
Хотя поступление подогретых
вод непосредственно не долж-
но оказывать влияния на со-
левой состав воды, содержа-
ние биогенных элементов н
органических веществ, однако
косвенное влияние повышенной
температуры на перечислен-
ные компоненты вполне воз-
можно. Общеизвестно, напри-
мер, что повышение темпера-
туры способствует ускорению
минерализации, активизирует
деятельность бактерий-реду-
центов и тем самым влияет
на процессы самоочищения.
На примере Конаковской
ГРЭС прослеживается прямое
воздействие на химизм воды
сточных вод с площадки ГРЭС
которое выражается в повы-
шении общей минерализации
еличении окисляемости и ко-
личества биогенных соединений азота и фосфора. В зоне пос-
тоянного подогрева это увеличение оценивается в 3-5% к нх
количеству выше водозабора.
Постоянный забор воды электростанцией и переброска ее по
трассе водовыпуска на расстояние около 10 км ниже водозабора
изменили гидравлику потока воды, проходящего по водохранилищу.
Выше водозабора скорости течения увеличиваются, ниже с уменьше-
нием проходящего расхода воды они несколько уменьшаются, а за-
тем вновь уменьшаются в районе водовыпуска, где существует обрат*
ный уклон водной поверхности и резко возрастает площадь водного
сечения. Изменение скоростей течения на указанных
участках в свою очередь вызвало перераспределение донных
отложений на всем протяжении водоема - от Шошинского плеса
до плотины гидроузла. Грунтовая съемка, произведенная В.П.
Курдиным в 1969-1970 гг., показала (рис. 33),* что после со-
здания Конаковской ГРЭС отложения серого н песчанистого се-
рого ила, достигавшие в Волжском плесе против устья Шоши
68 см, а против г. Конакова - 44 см, были в большей части
смыты и переотложены между Шошей и водозабором или выне-
сены в Иваньковский плес. Здесь значительная часть их седи-
ментировалась в районе водовыпуска и у Корчевы, захоронив
прежние донные отложения. Мощность вторичных отложений у
названных пунктов увеличилась соответственно на 90 и 30 см.
138
Всего из Волжского плеса в Иваньковский было вынесено грун-
та около 500 тыс. м3.
Влияние сброса подогретых вод на биологические процес-
сы сложно и многообразно. Общим для всех биоценозов в зоне
постоянного подогрева вод является удлинение вегетационного
периода и изменение сезонной динамики. Некоторые результаты
изучения влияния сброса подогретых вод Конаковской ГРЭС на
экосистему Иваньковского водохранилища освещены в специаль-
ных статьях настоящего сборника. Несомненно, что исследова-
ние этого вопроса неразрывно связано с дальнейшим изучением
климатических условий и гидрологического резшма водоема.
Заключение
Анализ материалов наблюдений, выполненных на Ивань-
ковском водохранилише за ряд лет при различных режимах
работы Конаковской ГРЭС, позволил оценить последствия сбро-
са ее подогретых вод на температурные условия водоема. Они
оказались отличными от бытовавших представлений. Исследова-
ниями установлено, что при всех наблюдавшихся расходах
ГРЭС в безледный период зона устойчивого распространения
подогретых вод ограничена и на большей части водоема веду-
щую роль в формировании температурных условий играет радиа-
ционный фактор. Наиболее существенное нзмененне температуры
воды отмечено лишь в Мошковичском заливе и на небольшом
участке примыкающей к нему акватория Иваньковского плеса.
Теплосодержание вод Иваньковского плеса водохранилища
за счет сбрасываемого ГРЭС тепла несколько повысилось.
Особенно это заметно поздней осенью и зимой, что, вероятно,
окажет благоприятное влияние на эксплуатацию водозаборных
сооружений'. Весной и летом увеличение теплозапаса за счет
искусственного подогрева наблюдается лишь в Мошковичском
заливе и прилегающих к нему мелководьях. В остальной части
плеса повышение температуры воды сказалось в пределах мно-
голетних колебаний среднемесячных значений ее до сооружения
Конаковской ГРЭС, которые определяются в 4-5.5°.
Близкий к естественному тепловой режим Иваньковского
плеса в условиях искусственного подогрева обеспечивается его
гидрологическим режимом. Транзитный пропуск половодья пол-
ностью освобождает водоем от подогретых зимних вод тепла,
аккумулированного грунтом. Летом при нормальной водности
объем плеса обновляется около 3 раз, и расход воды из водо-
139
хранилища превышает расход подогретых вод в 2-3 раза. Это
способствует быстрому распределению поступающего тепла в
толще воды и выносу его в нижний бьеф. При таком гидрологи-
ческом режиме водоема и существующих формах использования
его водных ресурсов не может произойти существенных измене-
ний температурных условий водохранилища.
Литература
Алисов Б.П. 1856. Климат СССР. М
Багров Н.А. 1858. Аналитическое представление После-
довательности метеорологических полей посредством
естественных ортогональных составляющих. Тр. ЦИП,
74. М.
Бакастов С.С. 1866. Теплофизические характеристики
затопленных грунтов. Тр. Инет, биол, внутр, вод АН
СССР, 13 (16).
Буторин Н.В. 1859. К вопросу о проточности Иваньков-
ского и Угличского водохранилищ. Бюлл. Инет, биол.
водохр. АН СССР, 3.
Буторин Н.В. 1868. Гидрологические процессы и динами-
ка водных масс водохранилищ Волжского каскада. Л.
Буторин Н.В. 1871. Особенности вертикального распре-
деления температуры воды в зоне влияния Конаковской
ГРЭС. Информ, бюлл. Инет. биол. внутр, вод АН СССР,
10.
Буторин Н.В., Бакастов С.С.,* Литвинов
А.С. 1868. Рекогноспировочное обследование Иваньковс-
кого водохранилища в районе Конаковской ГРЭС. Ин-
форм. бюлл. Инет, биол, внутр, вод АН СССР, 2.
Буторин Н.В., Курдина Т.Н. 1870а. О динамике
температуры воды Иваньковского водохранилища в зо-
не поступления подогретых вод Конаковской ГРЭС.
Информ, бюлл. Инет. биол. внутр, вод АН СССР, 7.
Буторин Н.В., Курдина Т.Н. 19706. Новые дан-
ные о характере переноса вод в Иваньковском водохра-
нилище. Информ, бюлл. Инет. биол. внутр, вод АН
СССР, 8.
Буторин Н.В., Курдина Т.Н,. 1972а. О распрост-
ранении сбросных вод Конаковской ГРЭС в зависимос-
ти от их расхода. Информ, бюлл. Инет. биол. внутр.
вод АН СССР, 14.
140
т о р и н Н.В., Курдина Т.Н. 19726. Статистичес-
*’ 7 кий анализ поля температуры в Иваньковском водохра-
нилище. Информ, бюлл. Инет. биол. внутр, вод АН
СССР, 15.
g у т о Р и н Н.В., Литвинов А.С. 1953. О течениях
в Рыбинском водохранилище. Тр. Инет. биол. внутр,
вод АН СССР, 6(9).
равеман А.В. 1952. Московское море. Калинин.
g р ш о в а М.Г. и Эдельштейн К.К. 1966. О ме-
тодике измерения электропроводности водных масс
волжских водохранилищ. Тр. Инет. биол. внутр, вод
АН СССР, 12(15).
Зайцева Е.А. 1966. Водный баланс Иваньковского водо-
хранилища. Сб. работ Рыбинской гидрометеорол. об-
серв., 3.
Зиминова Н.А. 1959. Элементы гидрологического режи-
ма и водный баланс Иваньковского водохранилища за
1951-1956 гг. Тр. Инет. биол. водохр. АН СССР, 2(5).
Курдин В.П. 1961. Грунты Иваньковского водохранили-
ща. Тр. Инет. биол. водохр. АН СССР, 4(7).
Сурдина Т.Н., Буторин Н.В. 1971. О влиянии
сбросных вод Конаковской ГРЭС на теплосодержание
Иваньковского водохранилища. Информ, бюлл. Инет,
биол. внутр, вод АН СССР, 11.
Литвинов А.С. 1970. Влияние Рыбинской ГЭС на режим
течений верхнего бьефа. Информ, бюлл. Инет. биол.
внутр, вод АН СССР, 5.
Литвинов А.С. 1973. Формирование и устойчивость слоя
скачка в Мошковнчском заливе Иваньковского водохра-
нилища. Информ, бюлл. Инет. биол. внутр, вод АН
СССР, 18.
Мейснер Е.В. 1958. Зимний кислородный режим Ивань-
ковского водохранилища и канала им. Москвы в связи
с годовыми различиями в их эксплуатации. - В кн.:
Волга-I. { конфер. по изуч. водоемов басе. Волги.
Тезисы докл. Тольятти.
Пан Л. 1969. Газовый режим охлаждающей воды Конаков-
ской ГРЭС и его влияние на Иваньковское водохрани-
лище. Матер, к совеш. по прогноз, содерж. биоген.
элемент, и органич. вещ-ва в водохр. Рыбинск.
Ресурсы поверхностных вод СССР. Основные гидрологи-
ческие характеристики, т. 10, 1967. Л.
Рутковский В.И. 1964. Опыт применения скоростной
съемки температуры и электропроводности для изуче-
ния температурного режима и перемещения водных
141
масс в водохранилищах. - В кн.: Методы изуч. и исполь-
зов. водн. ресурсов. М.
Савина В.Д. 1966. Уровенный режим Иваньковского во-
дохранилища. Сб. работ Рыбинской гидрометеорол.
обсерв., 3, Л.
Себенцов Б.М, Биек МД. и Мейснер Е.В.
1940. Режим и рыбы Иваньковского водохранилища
в первые два года его существования. Тр. Воронежск.
отд. Всеросс. инет. пруд.-рыбн. хоз., 3. 2.
Смирнов Н.П., Кондрацова О.Ф. 1972. Примене-
ние метода разложения по естественным ортогональ-
ным функциям к районированию стока Волги. Информ,
бюлл. Инет. биол. внутр, вод АН СССР, 13.
Справочник по климату СССР, выл. 8. 1964, 1966.
Тача лов С.Н. 1966. Особенности термического режима
Иваньковского и Угличского водохранилищ. Сб. работ
Рыбинской гидрометеорол. обсерв., 3. Л.
Трифонбва Н.А. 1960. О зимнем гидрохимическом ре-
жиме Иваньковского водохранилиша. Тр. Инет. биол.
водохр. АН СССР, 3(6).
Ф л е й с МЛ. 1970. Влияние сброса теплых вод Конаковской
ГРЭС на гидрохимический режим Иваньковского водо-
. хранилища. Гидробиол. журн. VI, 3.
Hellstrom В. 1963. Seiches and Wind currents
in lakes. Trans. Roy. Inst. Techn,, 211.
В.Г.Д е в я т к и н
ВЛИЯНИЕ ПОДОГРЕТЫХ ВОД НА ФИТОПЛАНКТОН
ИВАНЬКОВСКОГО ВОДОХРАНИЛИЩА
Крупные тепловые и атомные электростанции требуют для
охлаждения конденсаторов больших масс воды. Их водоснабже-
ние осуществляется нз различных источников - рек, озер, во-
дохранилищ, прибрежных участков морей. После прохождения
через охлаждающие системы электростанций сбрасываемая вода
имеет повышенную температуру.
Кроме этого в зоне воздействия подогретых вод тепло-
электростанций нередко наблюдается повышение минерализации
воды, смещение гндрокарбонатно-кальциевого равновесия, изме-
нение газового режима. Физические и химические явления, со-
путствующие сбросу подогретых вод в водоемы, приобретают
значение важного экологического фактора для развития гид-
робионтов.
Иваньковское водохранилище, первое в цепи крупных волж-
ских водохранилищ, является важнейшим источником водоснаб-
жения Москвы. Последствия сброса подогретых вод Конаковской
ГРЭС мощностью 2400 тыс. МВт на протекание гидробиологичес-
ких процессов привлекают внимание исследователей различных
специальностей.
Гидрологические особенности изучаемого участка Ивань-
ковского водохранилиша и его температурный режим в связи
со сбросом подогретых вод подробно рассматриваются в работе
Н.В.Буторина и Т.Н.Курднной (наст. сб.). Влйяние подогретых
вод на химические процессы в водохранилище изучено слабее.
Этому вопросу посвящены работы М.Л. Флейса ( 1870, 1972 ).
Влияние подогретых вод Конаковской ГРЭС на газовый режим
водохранилища рассмотрено в работе Л.Пана (1869), Т.Н.Курди-
ноЙ и В.Г.Девяткина (1872).
143
Таблица 1
Даты проведения рейсов (1) и число обработанных качественных
Годы Пока- зате- ян Месяцы
I II III IV У УТ
1 — — — 4-6;30 1;27-28
1967 2 — — — — 15 13
3 - - - — 40 18
1 19-21 - 16-20 24-26 - 1-2
1968 2 1 — 9 — 11
3 13 - 6 29 — 16
1 16-21 14-17 - 6-8 4-6
1969 2 — — — — . 3 17
3 28 - 29 - 49 37
1 — 21-24 6-8 5-6;30
1970 2 — — — — 10 —
3 - - 29 - 57 44
В предыдущем сообщения (Девяткин, 1970) нами кратко
рассмотрены некоторые результаты изучения влияния подогре-
тых вод Конаковской ГРЭС на развитие фитопланктона в 1967-
1968 гг. Предварительный анализ этих данных показал, что под
влиянием подогретых вод на участках водохранилища, прилегаю-
щих к устью Мошковичского залива, наблюдалось повышение
биомассы фитопланктона весной и 'осенью 1967 г., а также во
все сезоны вегетационного периода 1968 г. Летом 1967 г. в
зоне воздействия подогретых вод обилие водорослей было мень-
ше, чем за пределами этой зоны. Наряду с изменениями био-
массы в зоне воздействия подогретых вод наблюдалось увели-
чение удельного веса пирофитовых и зеленых водорослей. Наи-
более заметные изменения в составе и обилии фитопланктона
наблюдались за пределами Мошковичского залива в районах
распространения смешанных подогретых вод. В настоящем сооб-
J44
(2 ) и количественных ( 3) проб фитопланктона
Всего обрабо-
VII VIII IX X XI XII тано проб
качеств. количест
23-24 23-24 27 22-24 15-16 4-5
12 14 11 13 — 10 88 144
12 25 11 18 8 10
23-24 21-22 11-12; 18-21 — —
24-26
3 10 7 — — 40 180
8 31 48 29 - —
25-28 21-22 25-27 28-29 — —
й- 17 7 — — — 44 •262
68 18 15 18 - -
2 4-6 30 4;30 2
3 — 4 . 3 — 20 256
8 65 7 37 9 -
Всего обработа но проб в 1967 -1870 гг. . . . 192 842
шении более обстоятельно рассмотрены результаты наблюдений
за 1867-1970 гг.
Всего за вегетационные периоды 1867-1870 гг. на Ивань-
ковское водохранилище было проведено 28 рейсов, в которых
собирался фитопланктон (табл. 1).
• Пробы отбирались на разрезах и станциях, схема
расположения которых показана на рис; 1. Отбор проб про-
изводился батометром длиной 1 м конструкции А.В.Франце-
ва (Гусева, 1956) с борта экспедиционного судна с гори-
зонтов 0-2 м, 6-8 м и у дна. В Мошковичском заливе пробы
отбирались с лодки батометром Руттнера. В настоящем
сообщении в основном рассматриваются данные,' относящие-
ся к верхнему 2-метровому слою водоема, поскольку поток
Ц5
778 10
Пг/кпчп/смв
Рис. 1. Расположение разрезов и станций в Иваньковском
плесе водохранилища.
Римскими цифрами обозначены номера разрезов; арабски-
ми - станций.
подогретых вод в свободный ото льда период распростра-
няется в поверхностных горизонтах водохранилища.
Концентрация и обработка проб фитопланктона произ-
водились по принятой в. Институте биологии внутренних вод
АН СССР методике (Гусева, 1959; Кузьмин и Елизарова,
.1967). Пробы объемом 0.5 л концентрировали в сто раз
путем фильтрации через мембранный фильтр № 6. Подсчет
клеток водорослей проводили в камерах Нажотта объемом
0.01 и 0.05 мл. При подсчете биомассы фитопланктона' нс -
пользовали объемный метод, удельный вес водорослей при-
нят равным 1. При расчете объемов клеток водорослей тех
или иных видов их приравнивали к простейшим геометри-
ческим фигурам (шар, цилиндр, эллипсоид, параллелепипед,
конус). При обработке проб 1967 г. измеряли все встречен-
ные в счетной камере клетки. При обработке проб 1968-
1970 гг. для массовых видов и разновидностей водорослей
определяли их средние размеры и объем для каждого рейса.
Для определения средних размеров клеток того или иного
вида водорослей измеряли не менее 30 клеток. Размеры и
объем клеток редко встречающихся видов определяли не-
посредственно в камере.
146
Динамика биомассы фитопланктона
верхнего двухметрового слоя
в зоне воздействия
подогретых вод
К началу вегетационного периода 1967 г, объем сбрасы-
ваемых вод составлял около 30 м^/сек. В это время работали
3 блока ГРЭС из 8. Такой объем расхода подогретых вод со-
хранялся до сентября. В октябре и ноябре он возрос до 40,
а в декабре до 50 м^/сек. В связи с ограниченным расходом
подогретых вод сеть станций 1967 г, была реже, чем в после-
дующие годы наблюдений, а наблюдения на околоплотинном
участке проведены лишь в 3 рейсах из 8.
Первый рейс на Иваньковском водохранилище был проведен
4-6 мая 1967 г. Температура воды во время наблюдений сос-
тавляла 10-12° на русловых станциях водохранилища и 16-17°
в Мошковичском заливе. Отбор, проб производился при безвет-
ренной погоде , и совпал по времени с интенсивным естествен-
ным подогревом водоема. Подогретые воды распространялись
узкой струей в направлении разреза IV.
Наиболее массовыми формами фитопланктона были Melo-
sira italics, S urine 11 a ovata, Stephanodiscus hant-
zschii из диатомовых и С гур tom on as ovata из пирофи—
товых. Биомасса фитопланктона как в Мошковичском заливе,
так и в районе разреза 1У была выше, чем в районе водозабо-
ра. Особенно заметно возросла в районе воздействия подогре-
тых вод биомасса диатомовых, составлявших до 80% от обшей
биомассы водорослей. В связи с интенсивным прогревом водоема,
сравнительно большую роль в ранневесеннем фитопланктоне в
мае 1967 г. играли пирофитовые, в основном виды рода Сгур—
tomonas. заметном количестве развивались и зеленые во-
доросли, однако увеличения их биомассы в потоке теплых вод
не наблюдалось, в то время как биомасса пирофитовых в пото-
ке теплых вод увеличивалась (рис. 2).
В конце мая температура воды на русловых станциях сос-
тавляла 16-17°, а в заливе 24-25°. В связи с интенсивным
ветровым перемешиванием не наблюдалось повышения темпера-
туры под влиянием подогретых вод на открытых участках во-
дохранилища (русловые станции). Распределение фитопланктона
по акватории водоема не зависело от влияния подогретых вод.
Доминирующей группой фитопланктона по-прежнему были диато-
мовые водоросли с наиболее массовыми формами Melosira
italics и Fragilaria crotonensis. Присутствие в значитель-
ном количестве Fragilaria - летней формы фитопланктона
147
1 - синезеленые; 2 - золотистые; 3 - эвгленовые^ 4 -
пирофитовые; 5 - зеленые; 6 - диатомовые водоросли.
Иваньковского водохранилиша, - по-видимому, связано с жар-
кой погодой в мае. В пределах Мошковичского залива биомас-
са диатомовых заметно снизилась (рис. 3).
В течение июня стояла теплая погода. К концу этого ме-
сяца водохранилище прогревалось до 20-27° на русловых стан-
циях и до 29-30° в Мошковичском заливе. Температура потока
подогретых вод, распространяющихся в направлении плотины,
вследствие высокой температуры воздуха снижалась медленно.
Так, в районе разреза XV температура воды достигала 26-27°,
а в районе разреза VX - 23-24°. Общая биомасса фитопланкто-
на заметно понизилась в пределах отводящего канала и залива.
Однако в районе разрезов IV и VI биомасса водорослей была
заметно выше, чем в районе разреза I. По сравнению с пре-
дыдущими наблюдениями в планктоне возросла роль зеленых
Водорослей * криптомонад (ряс. 4). Наиболее массовыми вида-
ми в фитопланктоне были Pandorina morurii из зеленых,
148
Рис. 3. Биомасса фитопланктона (г/м^) в верхнем 2-мет-
ровом слое воды 30 мая-1 июня 1967 г.
Обози. те же, что и на рис. 2.
Cryptomonas ovata из пирофитовых и Fragilaria croto-
nensis из диатомовых.
Следующий рейс был проведен 23-24 июля при температу-
ре воды 21-23° на русловых станциях и 26-27° в заливе.
Предшествующая наблюдениям теплая погода и слабая проточ-
ность водохранилиша в это время обусловили массовое развитие
фитопланктона, биомасса которого на отдельных участках в верх-
нем 2-метровом горизонте превышала 60 г/м\ что является
максимальной величиной за годы наших наблюдений. Наиболее
массового развития достигли диатомеи Melosira italic а,
М. granulata, Fragilaria crotonensis, Tabdlaria fe-
nestrate, а также Cryptomonas ovata из пирофитовых.
Биомасса M. italic а и F, crotonensis составляла в это
время около 10 г/м3. Как в заливе, так и на русловых стан-
циях в районе распространения смешанных подогретых вод чис-
ленность и биомасса наиболее массовых форм фитопланктона
за исключением Tabellaria fenestrate были ниже, чем в
районе водозабора. Общая биомасса фитопланктона в районе
влияния подогретых вод также была значительно ниже, чем в
районе водозабора (рис. 5).
149
Рис. 4. Биомасса фитопланктона (г/м®)
ровом слое волы 27-28 июня 1967 г.
в верхнем 2-мет-
Обозн. те же, что и на рис. 2.
В августе температура воды в заливе составляла 27-28°,
а на открытых участках водохранилища - 20-22°. К этому вре-
мени, наряду со снижением обшей биомассы, в планктоне умень-
шился удельный вес пирофитовых и зеленых водорослей и воз-
росла роль диатомовых, а в приплотинном участке - синезеле-
ных. Наиболее массовыми видами в это время были Melosira
italic a, Tabellaria fenestrate и Fragilaria crotonen-
sis из диатомей, а также Crypt om on as ovata из .пирофи— .
товых. К устью Мошковичского залива по сравнению с его
началом наблюдалось заметное увеличение биомассы водорослей,
в основном за счет Melosira italica. На открытых участках
существенного изменения биомассы под влиянием подогретых
вод не происходило (рис. 6).
В сентябре наблюдения были проведены на Ъграниченном
числе станций (табл. 2). Более показательны данные октябрь-
ских наблюдений. Температура воды в заливе в это время была
около 15°, а на открытых участках - 8-9°. Как и в сентябре,
наиболее массовой формой в фитопланктоне была Melosira
150
Q
Рис. 5. Биомасса фитопланктона (г/м°) в верхнем 2-мет-
ровом слое воды 23-24 июля 1967 г.
Обоэн. те же, что и на рис. 2.
italic а. Несмотря на снижение биомассы фитопланктона в зали-
ве, на прилегающих к нему участках'водохранилища биомасса
водорослей увеличивалась по сравнению с районом водозабора
(рис. 7).
В ноябре и в декабре наблюдения были проведены на огра-
ниченном участке (табл. 2). Они свидетельствуют, что влияние
подогретых вод в это время года на развитие фитопланктона
проявлялось сравнительно слабо. Это связано с быстрым раз-
бавлением подогретых вод в результате ветрового перемешива-
ния, так как их объем в 1967 г. был сравнительно невелик
(Буторин и Курдина, наст. сб.). ъ
Таким образом, уже при сравнительно небольшом объеме
подогреваемых вод в 1967 г, наблюдались заметные различия
по величине биомассы фитопланктона в районе русла между
станциями, находящимися под влиянием подогретых вод, и сво-
бодными от него. Так, весной и осенью величина биомассы во-
дорослей в районе разреза IV была выше, а летом ниже, чем
в районе разреза I. В результате наблюдались различия в ди-
151
ев
ев
ф
в
х
X
ф
2
ю
I
2
ф
Т а б л и п а 2
Биомасса фитопланктона (г/м®) осенью 1967 г.
Разрезы и станции Сентябрь Нэябрь Декабрь
диато- зеле- пирофи L " всего диато- зеле- оинезе- всего днато- зеле- синезе- всего
мовые ные товые мовые ные ле ные мовые ные ле ные
I 2.175 1.233 1.128 4.950 0.388 0.073 0.087 0.548 0.099 0.002 0.022 0.123
Ст. 3 9.064 3.825 6.149 19.427 0.312 0.044 0.049 0.405 0.110 0.003 0.019 0.132
УП1 - - - - 0.379 0.071 0.035 0.485 0.135 0.006 0.050 0.191
JI1 13.414 1.143 1.758 16.541 6.375 0.114 0.007 0.499 - - — 1 -
IV 17.041 2.027 2.677 22.213 - - - — - - - -
Таблица 3
Среднемесячная температура воздуха
(данные метеостанции с.Тургиново)
isa
Год Месяцы Сред- него- довая
Г II Ш IV V VI VII VJU ТХ X XT XII
1967 -15.2 -11.3 -0.2 5.5 15.2 15.2 16.6 16.6 10.4 8.3 0.4 -11.4 4.2
1968 -17.5' -10.8 -2.6 5.0 11.2 16.8 14.7 16.5 9,7 2.3 -3.1 -5.4 3.1
1969 -18.2 -15.5 -9.7 4.2 10.1 13.9 16.9 14.9 9.0 3.8 1.1 -9.9 1.7
Рис. 8. Биомас-
са фитопланктона
(1, г/мЗ, логарифми-
ческая шкала) в райо-
не разреза I (сплош-
ная линия) и разреза
IV (прерывистая ли-
ния) и температура
воды (2, °C) в райо-
не разреза I (внизу)
и ТУ по данным
1967 г.
намике развития фитопланктона на этих участках (рис. 8). Уве-
личение биомассы фитопланктона в районе разреза IV весной
И осенью по сравнению с районом разреза Т, на наш взгляд;
связано с воздействием подогретых вод. Причины снижения
биомассы водорослей на этом участке летом нам не ясны. Ра-
нее (Девяткин, 1970) нами было высказано предположение, что
снижение биомассы фитопланктона в районе разреза TV связано
с отрицательным воздействием повышения температуры воды
на этом участке в результате сброса подогретых вод. Однако
последующие наблюдения, особенно в
1972 г., не подтвердили этого. Так,
в июле 1972 г. при температуре воды
в Мошковичском заливе до 33.4°, а в
районе разреза IV - до 29.5° не наблю-
далось существенного снижения биомас-
сы фитопланктона на этих участках
по сравнению с районом разреза I.
В период наблюдений 1967 г. темпера-
тура воды не превышала 30° в Мош ко-,
вичском заливе и 27° в районе разреза
IV, т.е. была заметно ниже, чем летом
1972 г.
Весна 1968 г. была ранней и теп-
лой с опережением многолетнего графи-
ка фенологических явлений на 9-10
дней. Однако в конце второй декады
мая и до конца месяца наблюдалось
сильное похолодание. Июнь 1968' г. был
очень теплым, но июль - холодным и
пасмурным. Погода в августе и начале
сентября была сравнительно теплой.
Резкбе похолодание наблюдалось с
конца первой декады сентября. Осень
1968 г. была холодной и со значитель-
ным количеством осадков. В целом лето
1968 г. было несколько холоднее нор-
мы (табл. 3).
Первый в 1968 г. рейс был прове-
ден в конце апреля. Температура воды
водохранилища составляла 7-10°, а в
Как и весной 1967 г., руководящей группой
иа открытых участках
заливе - 14-18°.
водорослей были диатомовые с-наиболее массовым видом Melo—
sira italic а. Биомасса фитопланктона снижалась в пределах
Мошковичского залива. Причиной заметного снижения биомассы
водорослей в заливе, по-видимому, явилось интенсивное подме-
154
Рис. 9. Биомасса фитопланктона .(г/м^) в верхнем 2-мет-
ровом слое воды - 24-26 апреля 1968 г.
Обозн. те же, что и на рис. 2.
шивание к теплому потоку холодных вод, более бедных фито-
планктоном. Об интенсивном разбавлении подогретых вод в за-
ливе в это время свидетельствует заметное снижение темпе-
ратуры воды в пределах этого участка. На открытых участках
водохранилища наблюдалось постепенное уменьшение биомассы
водорослей в направлении от разреза III к плотине (рис. 9).
Отсутствие влияния подогретых вод на развитие фитопланктона
на открытых участках, по-видимому, связано с их сильным
разбавлением, так как наблюдения проведены на спаде весенне-
го половодья.
К началу июня температура воды в водохранилище повы-
силась до 14-17° на открытых участках и до 19-21° в заливе.
В фитопланктоне, биомасса которого возросла до 2-3 г/м^,
по-прежнему руководящей группой были диатомовые, составляв-
шие свыше 90% обшей биомассы. Melosira italics доставляла
в это время около половины от обшей биомассы фитопланктона.
Численность и биомасса водорослей заметно возрастали в за-
ливе, На русловых станциях биомасса водорослей возрастала
по направлению от разреза I к плотине без заметного ее уве-
личения в районе разрезов III и ГУ (рис. 10).' Отсутствие за-
метного влияния подогретых вод на развитие фитопланктона
в июне связано, по-видимому, с тем, что наблюдения проводи-
лись в период интенсивного прогрева водохранилища, на фоне
которого влияние подогретых вод было мало ощутимым. Так,
153
Обозн. те же, что и на рис. 2.
I
в приплотинном участке, где отмечена наибольшая биомасса
фитопланктона, температура воды была выше, чем в районе
разрезов 111 и IV.
В конце июля температура воды на открытых участках
водохранилища была равна 16-18°, а в заливе - 23-24 , т.е.
гораздо ниже, чем в соответствующий период 1967 г. Несмотря
иа возрастание биомассы водорослей по сравнению с предыду -
щими наблюдениями, она была значительно ниже, чем в июле
1967 г. По сравнению с июнем в планктоне возрос удельный вес
пирофитовых и зеленых водорослей. Наиболее массовыми вида-
ми в это время были Melosira italica и Cryptomonas
ovata, В отличие от предыдущих наблюдений вегетационного
периода 1968 г, влияние подогретых вод на развитие фитопланк-
тона отчетливо проявилось как в заливе и на прилегающем к
нему мелководье (разрез IX), так и в районе разреза IV. В
потоке подогретых вод наряду с увеличением обшей биомассы
фитопланктона увеличилась относительная роль криптомонад
(рис. 11).
156
Рис.
11. Биомасса фитопланктона (г/м^)
в верхнем 2-мет-
ровом слое воды 23-24 июля 1968 г.
Обозн. те же, что и на рис. 2.
К концу августа температура воды повысилась до 19-21°
на открытых участках и до 24-25° в заливе. Набор наиболее
массовых видов не изменился. По-прежнему высокий удельный
вес приходился на долю пирофитовых и зеленых водорослей.
Влияние подогретых вод отчетливо проявилось на прилегающем
к устью залива мелководье, где увеличивалась биомасса фито-
планктона и возрастал удельный вес пирофитовых. Аналогичные
тенденции прослеживались и в районе разрезов III и IV (рис.12).
В сентябре материал был собран в двух рейсах. 11-12
сентября температура воды на открытых участках составляла
16-18°, а в заливе - 22-24°. Биомасса фитопланктона по срав-
нению с концом августа несколько уменьшилась. Снизился
удельный вес пирофитовых, а относительное значение зеленых
возросло. По-прежнему в число наиболее массовых видов вхо-
дила Melosira italica, к ней присоединилась Tabellaria
fenestrata, тогда как Cryptomonas составлял более 10%
от обшей биомассы лишь в заливе и иа прилегающих к нему
157
Рис. 12. Биомасса фитопланктона (г/мЗ) в верхнем 2-мет-
ровом слое воды 21-22 августа 1868 г.
Обозн. те же, что и на рис. 2.
Рис. 13. Биомасса фитопланктона (г/мЗ) 11-12 сентября
1868 г. в верхнем 2-метровом слое волы.
Обозн. те же, что и на рис. 2.
168
мелководных' участках. Кроме увеличения удельного веса крип-
томонад, влияние подогретых вод проявилось в увеличении
биомассы фитопланктона в районе разреза IV (рис. 13).
К концу сентября температура воды на открытых участ-
ках уменьшилась до 11-13° и в заливе до 20-21°. По сравне-
нию с началом месяца увеличилась биомасса диатомовых и их
удельный вес. Значительные изменения произошли в соотноше-
нии между наиболее массовыми видами. Увеличилась роль
Melosira italic а, которая в это время составляла 50-70%
биомассы диатомей и около 1/3 .от обшей биомассы водорос-
лей. Удельный вес Tabellaria fenestrata понизился.
Cryptomonas по-прежнему входил в число наиболее массовых
видов лишь в заливе и на прилегающем к нему мелководье.
На этом участке также значительно увеличилась численность
и биомасса Melosira. За пределами прилегающих к устью
залива мелководных участков влияние подогретых вод не про-
явилось (рис. 14).
Последние в 1968 г. наблюдения были проведены 17-18
октября при температуре 6-8° на открытых участках и 12-13°
в отводящем канале и заливе. Несмотря на понижение темпе-
ратуры воды по сравнению с концом сентября, биомасса фито-
планктона почти не изменилась, но при этом произошло увели-
чение биомассы диатомовых и снижение биомассы зеленых и
пирофитовых водорослей (рис. 15). По-прежнему в значительном
количестве развивалась Melosira italica. В приплотинном
участке к ней присоединялись Melosira ambigua, Stepha-
nodiscus astraea и Pediastrum duplex.
Данные наблюдений, проведенных в 1968 г., существенно
отличаются от данных 1967 г. Прежде всего обращают на се-
бя внимание значительно более низкие величины биомассы фи-
топланктона на протяжении всего вегетационного периода,
обусловленные, на наш взгляд, большей проточностью водохра-
нилища, связанной с большим количеством осадков в 1968 г.
по сравнению с 1967 г. (рис. 16), а также более низкой естест-
венной температурой воды на протяжении вегетационного пе-
риода.
. Биомасса фитопланктона в районе разреза IV была обычно
выше, чем на других участках водоема (рис. 17). Влияние по-
догретых вод проявлялось не только в увеличении биомассы
водорослей на прилегающих к устью Мошковичского залива
участках водохранилища, но и в увеличении удельного веса
крнптомонад. Вследствие сравнительно низкого объема сбрасы-
ваемых в 1968 г. подогретых вод их влииние локализовалось
в зависимости от гидрометеорологических условий или в райо-
не разреза ТУ, или в районе разреза Ш. Снижения биомассы
158
Рис. 14. Биомасса фитопланктона
ровом слое волы 24-26 сентября 1968
Q
(г/м°) в верхнем 2-мот-
Обо.ш. те же, что п па рис. 2.
ровом слое воды 18-21 октября 1968 г.
Обозн. те же, что и иа рис. 2.
100 \.................................................
/ Il III IУ У VI УЦ УШ IX X XI XII
Рис. 16. Суммарная при-
точность (млн м®/месяц, ло-
гарифмическая шкала) в Ивань-
ковском водохранилише в
1967-1970 гг.
Рис. 17. Биомасса фи-
топланктона (г/м^, лога-
рифмическая шкала) в райо-
не разрезов I (1) и IV (2)
в 1968-1970 гг.
1 - 1967 г„ 2 - 1968 г.»
3 - 1969 г.» 4 - 1970 г.
фитопланктона в зоне воздействия подогретых*вод летом 1968г.
не происходило.
В 1969 г. объем сбрасываемых с ГРЭС подогретых вод
колебался в пределах 40-50 м^/сек.
В мае 1969 г. температура воды в поверхностном слое
составляла 8-12° на открытых участках и 18-19° в заливе.
.Распределение фитопланктона было сравнительно равномерным:
его биомасса колебалась от 0.5 до 1.3 г/м®, причем иа долю
диатомовых ’ приходилось не менее 75% (рис. 18). Как и весной
1968 г., влияние подогретых вод на развитие фитопланктона
проявлялось слабо. В пределах залива при некотором снижении
обшей биомассы фитопланктона несколько возраста/ia биомасса
криптомонад. Некоторое повышение обшей биомассы наблюда-
лось в районе разрезов Ш и IV. Наиболее массовыми видами
в начале мая были Melosim italics и Diatoms elongatum.
В зоне распространения смешанных подогретых вод увеличива-
лась численность лишь - Melosira, а численность и биомасса
Diatoma оставались без существенных изменений.
В июне температура воды на открытых участках колеба-
лась от 13 до 16°, а в заливе составляла 23-24°. Биомасса
фитопланктона в июне варьировала в более широких пределах,
чем в мае. Максимальные величины ее отмечены в районе раз-
161
778 11
реза IV (4.7 г/м^), а минимальные - на пойменных, мелководьях
залива (рис. 19). Как и в июне 1968 г., по направлению от
разреза I к плотине наблюдалось постепенное увеличение био-
массы фитопланктона. Основную массу водорослей составляли
Рис. 18. Биомасса* фитопланктона (г/м®) в верхнем 2-мет-
ровом слое воды 6-8 мая 1969 г.
Обозн. те же, что и на рис. 2.
диатомеи: 85-99% на русловых станциях и несколько меньше в
прибрежье. К числу наиболее массовых видов присоединилась
Asterionella formosa. Удельный вес Melosira italica
сократился. Численность и биомасса наиболее массовых видов
возрастала в заливе и на прилегающем к нему мелководье.
За исключением Diatoma elongatum, увеличения биомассы
руководящих форм в районе распространения смешанных подо-
гретых вод не наблюдалось. В результате влияние подогретых
вод на открытых участках на величину биомассы водорослей-
проявлялось слабо. По-видимому, как и в июне предыдущего
года, влияние подогретых вод в это время маскировалось ес-
тественным прогревом водохранилища.
На пойменных расширениях залива биомасса фитопланкто-
на была ниже, чем иа русловых его участках» Здесь сравнитель-
но большую по сравнению с другими участками роль играли .
162
пирофитовые и зеленые водоросли, но состав наиболее массовых
видов диатомовых не изменился.
В июле состав фитопланктона по группам был менее мо-
нотонен, чем в весенние месяцы. Свыше половины обшей
Рис. 19. Биомасса фитопланктона (г/мЗ) в верхнем 2-мет-
ровом слое воды 5-6 июня 1969 г.
Обози. те же, что и на рис. 2.
биомассы водорослей составляли пирофитовые и зеленые. Из
числа наиболее массовых видов, отмеченных в июне, выпали
Asterionella formosa и Diatoma elongatum. Их мес-
то заняли Pandorina morum и Cryptomonas ovata.
Численность и биомасса наиболее массовых форм фитопланкто-
на, а также его общая биомасса в районе воздействия подогре-
тых вод были заметно выше, чем на других участках. Распро-
странение подогретых вод на открытых участках водохранили-
ща (русловые станции) зависело от метеорологических условий.
163
Рис. 20. Биомасса фитопланктона (г/мЗ) в верхнем 2-мет-
ровом слое воды 25-28 июля 1969 г.
Обозн. те же, что и на рис. 2.
В свою очередь состав и обилие фитопланктона на участках, на-
ходящихся под влиянием подогретых вод, зависели от направле-
ния распространения последних. Так, 26 июля, когда подогретые
воды распространялись в направлении разреза IV, биомасса
фитопланктона на этом участке "была около 1.8 г/мЗ. После
того как сильный северо-восточный ветер отклонил поток теп-
лых вод В направлении вышележащих участков и температура .
воды здесь к 28 июля снизилась на 3°, биомасса фитопланкто-
на понизилась до 0.9 г/м^ (рис. 20).
К вечеру 28 июля ветер стих и установилась штилевая
погода. Под воздействием стокового течения подогретые воды
вновь подошли к этому участку. В результате в поверхностном
слое воды резко повысилась температура. Это привело к суще-
ственным изменениям в соотношении между ведущими группами
водорослей и увеличению общей биомассы фитопланктона. Изме-
нения средневзвешенной биомассы в верхнем 10-метровом слое
воды были не столь значительны (табл. 4).
164
Рис. 21. Биомасса фитопланктона (г/м^) в верхнем
2-метровом слое воды 21-22 августа 1969 г.
Обозн. те же, что и на рис. 2.
В августе подогретые воды распространялись довольно
широким фронтом в направлении разрезов 1П и IV. Температу-
ра воды иа открытых участках составляла 17-19°, а в зали-
ве — 24-25°. Биомасса фитопланктона по сравнению с предыду-
щими наблюдениями несколько увеличилась. При этом возрос
удельный вес диатомей и сократилось относительное значение
криптомонад. Наиболее массовыми видами в августе были
Melosira italic a, Fragilaria crotonensis, Asterionella
formosa и Tabellaria fenestrate. Численность и биомасса
наиболее массовых видов, за исключением Asterionella for-
mosa возрастала на прилегающих к устью Мошковичского
залива мелководных участках и в районе разрезов 1J1 и IV,
здесь же наблюдалось увеличение общей численности и биомас-
сы водорослей, в то время как иа русловых участках залива
биомасса фитопланктона снижалась (рис. 21).
К концу сентября температура воды понизилась иа 6-7°
и на открытых участках была равна 11-13°, а в заливе -
16-18°. Однако при этом происходило дальнейшее нарастание
163
Таблица 4
Биомасса фитопланктона в верхнем 2-метровом (1)
(28-29 июля 1969 г.)
Показа- тели Слой 28 июля
19 ч. 21 ч. 23 ч. 01 ч. 05 ч.
Темпера- тура, град. 1 19.8 19.7 19.4 19.6 19.6
2 19.8 19.7 19.4 19.4 19.4
Диатомо- вые 1 0.373 0.317 0.208 0.366 0.442
2 0.100 0.456 0.178 0.394 0.294
Зеленые 1 0.102 0.097 0.115 0.252 0.168
2 0.065 0.398 0.060 0.146 0.279
Пирофито- вые 1 0.428 0.382 0.014 0.511 0.582
2 0.260 0.355 0.434 0.650 0.535
Прочие 1 0.022 0.015 0.000 0.014 0.000
2 0.000 0.000 0.097 0.000 0.048
Всего 1 0.911 0.811 0.337 1.143 1.192
2 0.425 1.209 0.769 1.190 1.156
биомассы фитопланктона, которая по сравнению с концом ав-
густа увеличилась в 1.5-2.0 раза. Особенно значительно возрос-
ла биомасса диатомей, которая составляла 75-93% от обшей.
Наиболее массовым видом была Melosira italica. Общая
биомасса фитопланктона в пределах залива изменялась незна-
чительно, однако под влиянием подогретых вод она несколько
увеличивалась в районе разрезов Щ и ТУ (рис. 22).
В конце октября температура воды составляла 3-6° на
открытых участках водоема и 11-12° в заливе. Несмотря на
заметное выхолаживание водоема, значительного уменьшения
биомассы водорослей по сравнению с предыдущими наблюдения-
ми не наблюдалось. По-прежнему в фитопланктоне доминирова-
166
и 10-метровом (2) слое воды в районе разреза IV
29 июля
07 ч. 09 ч. 11 ч. 13 ч. 15 ч. 17 ч. 19 ч.
19.6 19.4 21.3 19.5 23.5 19.9 24.6 20.6 24.6 20.8 24.8 20.8 25.9 21.1
0.253 0.400 0.451 0.411 0.572 0.651 0.549 0.417 0.536 0.423 0.988 0.917 1.248 0.462
0.128 0.053 0.332 0.236 0.296 0.730 0.207 0.221 0.559 0.202 0.312 0.486 0.333 0.188
0.955 0.685 0.460 0.590 1.745 1.390 1.750 0.650 2.290 0.474 2.600 1.060 2.830 0.775
0.000 0.094 0.005 0.043 0.024 0.097 0.000 0.000 0.000 0.021 0.000 0.049 0.000 0.000
1.336 1.232 1.248 1.180 2.637 2.868 2.506 1.288 3.385 1.120 3.900 1.412 4.411 1.425
ли диатомовые. Наиболее массовыми видами были Melosira
italica, Tabellaria fenestrate Diatom a elonga-
tum. Относительное значение Melosira italica по сравнению
с предыдущими наблюдениями уменьшилось. В пределах Мошко-
виче кого залива численность и биомасса Melosira оставались
без существенных изменений, биомасса Diatoma elongatum
уменьшалась, a Tabellaria fenestrate увеличивалась. На
прилегающем к заливу мелководье увеличивались численность
и биомасса Melosira Tabellaria, а биомасса Diatoma
оставалась без изменений. На открытых участках под влиянием
подогретых вод заметно увеличивались лишь численность в
биомасса Melosira и Tabellaria. Подогретые воды распрост-
167
Обозн. те же, что и на рис. 2.
ранились в направлении разрезов IV и У, здесь же отмечалось
небольшое, но заметное увеличение биомассы фитопланктона.
В заливе и на- прилегающем к нему мелководье увеличивался
удельный вес пирофитовых (рис. 23).
Данные наблюдений, проведенных в 1969 г., во многом
сопоставимы с данными 1968 г. Биомасса водорослей в 1969 г.
была того же порядка, -что и в 1968 г. КаК и в 1968 г., на
протяжении большей части вегетационного периода 1969 г. ве-
дущую роль в фитопланктоне играли диатомовые. Однако перио-
дичность развития фитопланктона по сезонам была иной. Так,
если в 1968 г. максимум биомассы наблюдался в середине
лета, то в 1969 г. наблюдалось два максимума - в конце вес-
ны-начале лета и в начале осени (рис. 17). На прилегающих
к устью Мэшковичского за'лива участках- водохранилища наблю-
далось повышение удельного веса пирофитовых и увеличение
обшей биомассы фитопланктона. Вследствие увеличившегося
расхода подогретых вод их влияние ощущалось на большей
плошади. Если в 1968 г. влияние подогретых вод было замет-
ным в районе разреза 1)1 или )У, .то теперь заметные измене-
ния, как правило, происходили одновременно на русловых стан-
циях этих разрезов. Под влиянием подогретых вод в отдельные
периоды находился также к район разреза У.
168
Рис. 23. Биомасса фитопланктона (г/м^) в верхнем 2-мет-
ровом слое волы 28-29 октября 1969 г.
Обозн. те же, что и на рис. 2.
Снижение биомассы фитопланктона в эоне воздействия по-
догретых вод летом 1969 г., как и в 1968 г., не наблюдалось.
Половодье 1970 г. было чрезвычайно высоким. Апрель
теплее, чем обычно. Особенно теплыми были конец апреля и
первая пятидневка мая. Однако 7-8 мая наступило резкое по-
холодание, продолжавшееся до конца месяца. Июнь и июль были
умеренно теплыми со значительным количеством осадков. С
середины августа наблюдалась холодная дождливая погода. Осень
была со значительным количеством осадков и резкими колеба-
ниями температуры. Расход подогретых вод составлял в тече-
ние вегетационного периода 1970 г. 55-60 м^/сек.
Сбор материалов в мае 1970 г. произведен при меняющих-
ся метеорологических условиях. В начале месяца погода была
теплой и безветренной. Интенсивная инсоляция способствовала
быстрому прогреванию верхних слоев воды водоема. Темпера-
тура воды в верхнем 1.5-2-метровом горизонте приплотинного
участка Иваньковского водохранилища достигала 16-17°, но
начиная с глубины 3-5 м она снижалась до 10-11°. 6 мая.
169
Рис. 24. Биомасса фитопланктона
ровом слое волы 6-8 мая 1970 г.
(г/м3) в верхнем 2-мет-
Обозн. те же, что и на рис. 2.
в .первый день проведения сбора материалов, при штилевой по-
годе были отобраны пробы на приплотинном участке водохрани-
лища. Однако уже на следующий день вследствие ветрового
- перемешивания произошло снижение температуры воды в поверх-
ностном слое до 13-14°, а в заливе температура воды была
около 17°. Таким образом, остальные материалы были собраны
при иной гидрометеорологической ситуации, и поэтому сравнен
ние данных приплотинного участка с другими должно произво-
диться с учетом этих изменений.
В начале проведения сборов в связи с устойчивой темпера-
турной стратификацией наблюдалось неравномерное вертикаль-
ное распределение планктонных водорослей, которое впоследст-
вии нарушилось. По мере продвижения от плотины к разрезу .
I биомасса водорослей в верхнем 2-метровом слое снижалась,
что может быть связано с постепенным перемешиванием толпш
* воды н нарушением установившейся в начале съемки стратифика-
ции водорослей по биомассе. Вследствие интенсивного ветрово-
170
го перемешивания подогретые воды быстро разбавлялись, поэто-
му их влияние на развитие фитопланктона открытых участков
водоема не ощущалось. В отводящем канале и заливе, а также
на прилегающих к заливу мелководных участках влияние повы-
шения температуры было более заметным. Здесь понизился
удельный вес диатомей и возросло относительное значение пи-
рофитовых, золотистых и эвгленовых водорослей. Биомасса
Рис. 25. Биомасса фитопланктона (г/м^) в верхнем 2-мет-
ровом слое воды 5-6 июня 1970 г.
Обозн. те же, что и на рис. 2.
водорослей в заливе и на прилегающем к нему мелководье
также была выше, чем в районе водозабора (рис. 24). Наибо-
лее массового развития в мае достигали Melosira italica,
Stephanodiscus hantzschii и Cryptomonas ovata.
К началу июня температура воды составляла на русловых
станциях 17-19°, т.е. была лишь ненамного выше, чем в нача-
ле мая. В заливе температура воды составляла 23-25°. Во
время проведения сборов преобладал ветер северо-восточного
направления, поэтому влияние потока теплых вод на развитие
фитопланктона в районе разрезов Ш и IV было ослаблено. В
связи с ветровым переносом подогретых вод в вышерасполо-
женные участки биомасса фитопланктона в районе разреза IV
уменьшилась 8 июля по сравнению с 6 июня более чем в два
раза (рис. 25). Ведущей группой в июньском фитопланктоне
Обозн. те же, что и на рис. 2.
были диатомовые. На мелководных участках наряду со сниже-
нием общей биомассы возрос удельный вес пирофитовых й
зеленых водорослей. Максимальная биомасса водорослей наблю-
'далась в районе разреза II, в направлении которого распрост- -
ранялись подогретые воды. Наиболее массовыми видами в фито-
планктоне были Melosira italic а' и Diatoma elongatum.
В районе разреза II, где влияние подогретых вод проявилось
б олее отчетливо,- к ним присоединялись Melosira varians
и М. granulate.
Во время наблюдений, проведенных в конце июня-начале
июля, поток подогретых вод, так же как и во время наблюдений,.
проведенных в начале июня, под влиянием северо-восточного
ветра уклонялся в направлении разрезов III и II, где темпера-
тура воды была около 21<>, тогда как в районе водозабора 18°.
В соответствии с таким распределением подогретых вод
распределялся и фитопланктон поверхностногогоризонта водо-
хранилища (рис. 26). Однако эта ситуация оказалась неустой-
чивой. После перемены направления ветра на западное поток
подогретых вод сместился в направлении разреза IV, где к
9 июля температура воды повысилась до 22-23°, а биомасса
водорослей увеличилась по сравнению с таковой 1 июля более
чем в 2 раза (рис. 26). Diatoma elongatum выпала к это-
172
Рис. 27. Биомасса фитопланктона (г/мЗ) в верхнем
2-метровОм слое волы 4-6 августа 1970 г.
Обозн. те же, что и на рис. 2.
му времени на числа наиболее массовых форм фитопланктона.
На ряде станций до 30% биомассы составляла Pandorina
morum. В число наиболее массовых форм входили также Me-
losira italic а и Cryptomonas ovata. Биомасса Melo-
sira, Cryptomonas и Pandorina на русловых станциях
в зоне воздействия теплых вод была выше, чем в районе раз-
реза I.
В начале августа температура воды составляла на откры-
тых участках 21-23.5°, а в Мошковичском заливе - 30-31°.
Подогретые воды распространялись в направлении разрезов Ш и
II, где отмечалось заметное увеличение биомассы водорослей.
Район разреза IV в начале проведения сбора материалов был
свободен от влияния подогретых вод, и биомасса фитопланкто-
на на этом участке была сравнительно низкой. Одиако после
173
перемены направления ветра и появления на этом участке по-
догретых вод биомасса водорослей в районе разреза ТУ воз-
росла примерно в 4 раза (рис. 27). На участках водохранили-
ща, расположенных вне зоны влияния подогретых вод, 50-70%
от общей биомассы составляли зеленые водоросли, а в районе
их воздействия - 25-40%. Зеленые водоросли были представле-
ны большим числом видов, в основном протококковых, однако
массового развития среди них достиг лишь Pediastrum dup-
lex. Кроме Pediastrum к числу наиболее массовых видов,
как и в предыдущем месяце, относились Melosira italica
и Cryptomonas ovata. В пределах Мошковичского залива
увеличилось лишь обилие Cryptomonas и Pediastrum, тог-
да как численность и биомасса Melosira изменялись незначи-
тельно. Заметное увеличение обилия всех наиболее массовых
видов отмечено на прилегающем к заливу мелководье. На рус-
ловых станциях под влиянием подогретых вод заметно возрос-
ли численность и биомасса Cryptomonas ovata и Pedi—
astrum duplex.
Биомасса пирофитовых водорослей в августе в районе
воздействия подогретых вод была значительно выше, чем в
районе разреза I и водозабора. Увеличение биомассы пирофи-
товых (преимущественно криптомонад) отмечено уже в конце
Мошковичского залива. За пределами Мошковичского залива
район воздействия подогретых вод легко отграничивался по
повышенному удельному весу крнптомонад. После появления
подогретых вод в районе разреза IV биомасса пирофитовых'
возросла на этом участке более чем в 10 раз. Общая биомас-
са фитопланктона в районе разрезов И, !П и IV была заметно
выше, чем на выше и ниже расположенных участках водохрани-
лища (рис. 27).
Во время наблюдений, приведенных в конце сентября-на-
чале октября, преобладала пасмурная погода с умеренным
ветром северо-восточного направления. Температура воды на
русловых станциях бЫдд равна 10-11°. Под влиянием подогре-
тых вод температура воды на русловых станциях повышалась
не более чем на 1°С, что связано с ветровым перемешиванием
толщи воды. От 40 до 80% биомассы фитопланктона состав-
ляли диатомовые с наиболее массовым видом Melosira itali-
ca. По направлению от разреза I к плотине возрастал удель-
ный вес зеленых. Пирофитовые с наиболее массовым видом
Cryptomonas ovata играли существенную роль в устьевых
участках Мошковичского залива и на прилегающем к нему мел-
ководье. Однако в районе водозабора также отмечено значитель-
ное количество криптомонад. На наш взгляд, в данном случае
не исключена возможность ветрового переноса воды из района
П4
Рис. 28. Биомасса фитопланктона (г/мЗ) в верхнем
2-метровом слое воды 30 сентября-4 октября 1970 г.
Обозн. те же, что и на рис. 2.
влияния подогретых вод в район водозабора, несмотря на то
что данные температурной съемки не подтверждают этой точки
зрения. В отдельных случаях состав фитопланктона может быть
более точным 'индикатором влияния подогретых вод, чем темпе-
ратура воды (Курдина, Девяткин, 1972). Влияние подогретых
вод на общую биомассу фитопланктона проявилось слабо (рис.28).
Последние наблюдения в 1970 г. были проведены в конце
октября-начале ноября, когда температура воды на открытых
участках составляла 3-6.5°. Основная масса подогретых вод
распространялась в направлении разрезов IV и V, где и отме*.
чена наибольшая биомасса фитопланктона. Как и месяц назад,
существенную роль в фитопланктоне играли зеленые водоросли,
с наиболее массовым видом Scenedesmus quadricaudа.
Среди диатомей, как и в конце сентября, наиболее массового
развития достигла Melosira italica. Пирофитовые с наибо-
лее массовым видом Cryptomonas ovata играли су-
щественную роль лишь в Мошковичском заливе и на прилегаю-
щем чк- нему мелководье, а также в районе водозабора. Под
влиянием подогретых вод отмечалось заметное увеличение био-
массы фитопланктона в районе разрезов IV и V (рис. 29).
175
Таким образом, в 1970 г., как и в предыдущие годы на-
блюдений, на прилегающем к Мошковичскому заливу мелководье
отмечено увеличение удельного веса пирофитовых водорослей
и увеличение общей биомассы фитопланктона. Увеличение био-
массы фитопланктона по сравнению с районом водозабора отме-
чено и на русловых станциях, находящихся под влиянием подо-
гретых вод. Заметное увеличение биомассы пирофитовых наблю-
далось здесь только летом.
Рис. 29. Биомасса фитопланктона (г/м^) в верхнем 2-мет-
ровом слое воды 30 октября-2 ноября 1970 г.
Обозн. те же, что и на рис. 2.
Во время наблюдений 1970 г. в 4 рейсах из 6 преоблада-
ли ветры северного и северо-восточного направлений. Частая
повторяемость ветров северных направлений вызвала заметные
различия в распределении фитопланктона по сравнению с 1969 г.
Наибольшая биомасса в среднем за вегетационный период отме-
чена в районе разреза И, что не вполне соответствует данным
температурных съемок (рис. 30)} На наш взгляд, главную роль
* При расчете средних величин биомассы и температуры
были исключены данные наблюдений, проведенных в начале мая, так
как они проведены при резко меняющихся гидрометеорологичес-
ких условиях (см. выше).
176
Рис. 30. Биомасса фи-
топланктона (г/мЗ) и темпе-
ратура воды (°C) в среднем
за вегетационные периоды
1969 и 1970 гг.
1 - биомасса; 2 - тем-
пература.
в повышении биомассы фитопланктона в районе разреза И ле-
том 1872 г. играл ветровой перенос воды с высокой биомассой
фитопланктона из районов, Прилегающих к устью Мошковичского
залива. Следует отметить, что район разреза II отделен от
устья залива группой островов с мелководными участками меж-
ду ними. При ветровом переносе подогретые воды могут ока-
заться в районе разреза II или в обход островов, или же про-
сачиваясь между ними. По-видимому, обычно в той или иной
пропорции имеют место оба
эти явления. Возможность сноса
воды с межостровных мелково-
дий подтверждается наблюдения-
ми, проведенными в июне 1970 г.
В это время в районе разреза
11 отмечена максимальная био-
масса (5.4 г/м^). В то же
время только в районе разреза
II отмечено значительное коли-
чество Melosira varians и
крупных клеток м. granu-
late. Как известно, М, va-
rians ~ форма, обитающая
преимущественно в обрастаниях
и фитобентосе.. М. granulate
также нередко в массе разви-
вается в обрастаниях (Скаби-
чевский, 1960). Появление этих
видов на русловой станции раз-
реза II и в то же время отсут-
ствие на других русловых стан-
циях свидетельствуют о возмож-
ности прохождения части подо-
гретых вод сквозь группу силь-
но зарастающих островов, расположенных к северу и северо-
западу от Мошковичского залива.
Отсутствие соответствия между средиесезонными величи-
нами температуры воды и биомассы фитопланктона в районе раз-
реза 11, наблюдавшегося иа других русловых станциях водо-
хранилища иа исследуемом участке, может быть объяснено
двумя причинами. Во-первых, подогретые воды при переносе
в район разреза II должны преодолеть довольно большое рас-
стояние, если они движутся в обход островов. В этом случае
их температура должна заметно понизиться. В случае прохож-
дения сквозь сильно зарастающее мелководье подогретые воды
разбиваются на отдельные струи и также сильно выхолаживают-
ся. Выхолаживание ускоряется ветровым перемешиванием, так
778 12
17?
как перенос подогретых вод в район разреза II при слабом, вет-
ре в свободный ото льда период до сих пор не наблюдался.
Во-вторых, не исключается возможность ветрового переноса
воды с зарастающих мелководий в район разреза II еще до
подхода подогретых вод. Однако в последнем случае, на наш
взгляд, не должно происходить столь заметного увеличения
биомассы фитопланктона, как это наблюдалось в 1870 г., так
как по нашим наблюдениям биомасса водорослей на зарастаю-
щих мелководьях обычно не выше или даже ниже, чем на откры-
тых участках водоема. Так, например, в июне 1970 г., когда,
по-видимому, наблюдалось ветровое перемещение подогретых
вод сквозь зарастающее мелководье, биомасса фитопланктона
в сильно зарастающих Кладбищенском (разрез X) и Перетрусов-
ском заливах (к северу от разреза VI) не превышала таковую
на русловых станциях (рис. 25).
Таким образом, необычно высокая биомасса фитопланкто-
на, отмеченная в районе разреза II в 1970 г., вероятно, в зна-
чительной степени связана с воздействием .подогретых вод. Од-
нако это предположение нуждается в более прочном фактичес-
ком обосновании с привлечением данных гидрологических на-
блюдений.
Если исключить разрез II, ход изменений биомассы в
среднем за вегетационный период 1970 г. на русловых станцнях
во многом сходен с таковым по данным 1869 г» и со средне-
сезонными значениями температуры в эти годы наблюдений
(рис. S0). Такое же соответствие между среднесезонными тем-
пературой я биомассой на русловых станциях наблюдалось и
по данным 1868 г. (Девяткин, 1870). Особенностью данных
1970 г. является очень низкая величина биомассы в предпло-
тинном участке (ст. 7). Величина биомассы в районе разреза
У! не превышала таковую в районе разреза I и водозабора.
Заметное влияние подогретых вод иа величину биомассы фито-
планктона ограничивалось, как и в 1868 г., районом разреза
У н не отмечалось в предплотиииом участке. В отличие от
1868 г. снижения биомассы фитопланктона в зоне воздействия
подогретых вод летом 1970 г., как и летом 1968 и 1869 гг.,
не наблюдалось.
178
Влияние прохождения воды
через агрегаты Конаковской ГРЭС
на биомассу фитопланктона
Как уже. отмечалось выше, работа тепловых электростан-
ций связана с изменением не только температуры, ио и ряда
других показателей, способных оказывать влияние на развитие
гидробионтов. Работа любой ГРЭС связана с возможностью
механического травмирования гидробионтов при прохождении
воды через охлаждающие /системы электростанции, насосные
станции и другие водотоки. Отдельные сведения о возможности
травмирования фитопланктона при прохождении через эти сис-
темы содержатся в работах ряда исследователей. Так, В.М.Ша-
ла рь и Н.И.Яловицкая (1871) сообщают о разрушении при про-
хождении конденсаторов Кучурганской ГРЭС в среднем 72%
фитопланктона, причем наиболее сильно при этом травмируются
синезелеиые водоросли, далее следуют протококковые и воль-
воксовые и почти ие разрушаются диатомовые. Наличие цирку-
ляции воды в водоемах-охладителях препятствует, по мнению
А.П.Васильчиковой (1871), массовому развитию сииезеленых в
водоемах-охладителях Урала. Разрушение при прохождении
через конденсаторы является одной из причин, сдерживающих
массовое развитие протококковых в Кураховском водохранили-
ще в летнее время (Виноградская, 1871). Прохождение через
конденсаторы в сочетании с хлорированием неблагоприятно от-
ражается иа физиологической активности фитопланктона, вызы-
вая снижение интенсивности фотосинтеза (Morgan a, Stross,
1868; Hamilton е. а,, 1870). С другой стороны, водоросли
обычно не страдают при прохождении через конденсаторы
(Patrick, 1868).
Прохождение фитопланктона через конденсаторы Конаков-
ской ГРЭС в сочетании с резким повышением температуры на
6-8° (хлорирование отсутствовало), по нашим наблюдениям,
вызывало временное снижение фотосинтетической активности
фитопланктона лишь в летнее время. Это явление, по-видимому,
не сопровождалось или почти ие сопровождалось разрушением
клеток водорослей, так как через несколько часов (ие более
чем через сутки) фотосинтетическая активность фитопланкто-
на полностью восстанавливалась (Вайнштейн и др., 187S).
Микроскопическое исследование нефиксированного фитопланкто-
на, состоящего преимущественно из диатомовых и протококко-
вых, до и после прохождения через водотоки Конаковской ГРЭС
проведенное нами в июле и августе 1868 г„ а также в июле
и в августе 1970 г., не обнаруживало видимых повреждений
179
клеток водорослей. Аналогичные данные были получены при об-
работке законсервированных модифицированным кислым фикса-
тором Утермеля (Кузьмин и Елизарова, 1967) материалов 1969
к 1970 гг. Фитопланктон в это время состоял преимущественно
из диатомовых, зеленых и пирофитовых водорослей. По-видн-
мому, степень травмирования фитопланктона при прохождении
его через конденсаторы и другие водотоки теплоэлектростанции
зависит от конструктивных особенностей последних, а также
от состава фитопланктона. Степень разрушения фитопланктона,
состоящего из диатомовых, зеленых и пирофитовых,* в конден-
саторах и других водотоках Конаковской ГРЭС в период прове-
дения наблюдений 1967-1970 гг., На наш взгляд, невелика.
В л и я н и е ГРЭС на периодичность
развития фитопланктона
в Мош ко вичском заливе
и прилегающем к нему
мелководье
Насосные станции ГРЭС оборудованы устройством для
глубиного забора воды, поэтому численность и биомасса во-
дорослей в, начале отводящего канала обычно несколько отли-
чаются в ту или иную сторону по сравнению с этими показате-
лями .в верхнем горизонте воды в районе водозабора. Ход се-
зонных изменений биомассы водорослей в начале отводящего
канала был несколько иным, чем в верхнем 2-метровом слое
района водозабора (рис. 31). Эти различия сохранялись н в
начале Мошковичского залива, где ход сезонных изменений
был почти таким же, как в начале отводящего канала. В пре-
делах канала биомасса фитопланктона и его состав, как прави-
ло, изменялись незначительно.
На .участке залива от его начала до ст. 5 изменения
биомассы фитопланктона и его состава также, как правило, не-
велики. Чаше всего здесь отмечалось небольшое повышение
биомассы водорослей. Ход сезонных изменений биомассы фито-
планктона в районе ст. 5 был почти таким же, как в начале
отводящего канала (рис. 32).
Отсутствие значительных изменений в составе и обилии
фитопланктона в канале и в начале залива связано с высокой
проточностью этого участка. В течение вегетационного перио-
да скорость течения в канале составляет около 0.5 м/сек. при
длине канала около 3 км, а в, начале Мошковичского залива -
около 0.2 м/сек. при длине участка около 1.5 км.
180
г)**
v vi vuvmix х xi v и vnmi'x x xi
1969г. 1970г.
Рис. 31. Биомасса фи-
топланктона (г/м3) в райо-
не водозабора (1) и в нача-
ле отводящего канала (2)
в 1969 и 1970 гг.
Рис. 32. Биомасса фи-
топланктона (г/м3) в по-
верхностном слое воды в
районе ст. 1 (1), ст. 3 (2)
и ст. 5 (3) в 1969 и 1970 гг.
ШШР ^3 ggg# Qf
Рис. 33. Биомасса фитопланктона (г/м3) в поверхностном
слое воды в районе ст.5, ст.5-а, разрезов УШ и IX в 1968 г.
" 1 - диатомовые, 2 - зеленые, 3 - пирофитовые, 4 - синезе—
леные, 5 — прочие.
Рис. 34. Биомасса фи-
топланктона (г/м^) в по-
верхностном слое волы в
начале отводящего канала
(а) и в конце Мошковичско-
го залива (б) в 1870 г.
Обозн. те же, что и
на рис. 33.
Ниже ст. 5 скорость тече-
ния заметно снижается вследст-
вие образования здесь обширных
пойменных расширений. Между
ними и русловыми участками за-
лива существует достаточно ин-
тенсивный водообмен, что обус-
ловливает, с одной стороны, по-
вышение температуры на поймен-
ных расширениях, а с другой -
вынос сформировавшегося на
пойменных расширениях планктона
в русловые участки залива. В
зависимости от интенсивности
водообмена между русловыми
участками и пойменными расши-
рениями залива фитопланктон
этих участков различается по
величине биомассы и соотноше-
нию между наиболее массовыми
видами водорослей. По сравнению
с началом залива на его поймен-
ных расширениях, как правило,
возрастает обилие криптомонад
и смешается сезонная периодич-
ность развития фитопланктона.
Удельный вес зеленых водорослей на пойменных расширениях
залива также обычно возрастает (рис. 33).
Вынос планктона с пойменных расширений вызывает замет-
ные различия в сезонной периодичности развития фитопланктона
и в соотношении между ведущими группами в конце Мошкович-
ского залива по сравнению с его началом и серединой (рис.33,
34). По сравнению с районом водозабора и разреза- I, т.е.
участком, ие подверженным влиянию подогретых вод, в течение
Вегетационного периода здесь возрастает удельный вес пиро-
фитовых водорослей (рис. 35), а в зимнее время - диатомовых.
Однако биомасса фитопланктона в пределах Мошковичского
залива В среднем за вегетационные периоды 1867-1870 гг.
изменялась незначительно. Она лишь несколько увеличивалась
по сравнению с таковой в начале залива (рис. 36).
После выхода из залива скорость перемещения потока
подогретых вод заметно снижается. По мере удаления от устья
залива снижается и температура воды. Однако в районе разре-
за IX она обычно на 2-6°, в зависимости от гидрометеороло-
гических условий, выше, чем в районе водозабора. Несмотря
182
Рис. 35. Соотношение наиболее массовых групп фитопланк-
тона на разных станциях Иваньковского водохранилища в 1969-
1870 гг.
1 - диатомовые, 2 - зеленые, 3 - пирофитовые, 4 - прочие.
f=j 2
ШШ)*
I I ♦
иа снижение температуры волы на этом участке, влияние подо-
гретых вод здесь сказывается более отчетливо, чем в заливе.
Биомасса фитопланктона на этом участке возрастает (рис.36).
По сравнению с устьем Мошковичского залива еще заметнее
возрастает удельный вес пирофитовых. Периодичность изменений
биомассы, как отдельных групп водорослей, так и всего фито-
планктона по сезонам отличается своеобразием, сохраняя при
этом ряд особенностей, присущих фитопланктону устьевых
участков Мошковичского залива (рис. 34, 37).
Связь между температурой
и биомассой фитопланктона
на открытых участках водохранилища
Тенденция к увеличению биомассы фитопланктона под
влиянием подогретых вод сохраняется и на более удаленных от
устья Мошковичского залива участках водохранилища. Направле-
ние влияния подогретых вод и размеры участка, на котором
это влияние регистрировалось, зависели прежде всего от метео-
183
Рис. 36. Температура
(°C) воды (1) и биомасса
(г/мЗ) фитопланктона (2) •
в среднем за вегетационные
периоды >1968, 1969 и 1970 гг.
на разных станциях Иваньковс-
кого водохранилиша.
Рис. 37. Биомасса фи-
топланктона (г/м3) в поверх-
ностном слое воды в райо-
не разрезов УШ (а) и IX
(б) в 1968 г.
Обозн. те же, что и
иа рис. 33.*
рологических условий, расхода воды в предплотинном плесе во-
дохранилища, объема сбрасываемой с ГРЭС воды и ее темпера-
туры (см. статью Н.В.Буторина и Т.Н.Курдиной, наст. сб.).
В тихую погоду наиболее заметное увеличение биомассы фито-
планктона под влиянием подогретых вод обычно наблюдалось в
районе разреза TV. С увеличением объема подогретых вод по-
вышение биомассы фитопланктона нередко одновременно реги-
стрировалось и в районе разреза Ш. Биомасса фитопланктона
на этих участках в таких случаях обычно была в 1.5-3.0 раза
выше, чем в районе водозабора и разреза I. С увеличением
расхода подогретых вод в сферу их воздействия, как правило,
вовлекался и район разреза V, Случаев достоверного увеличе-
ния биомассы водорослей в предплотинном участке под влияни-
ем подогретых вод в 1969 и 1970 гг. мы не наблюдали, хотя
ВТО, по-видимому, не исключено в условиях безветренной пого-
ды, при наличии стокового течения и максимальном расходе
подогретых вод.
1Я4
Степень воздействия
подогретых вод на разви-
тие фитопланктона в зна-
чительной степени зависе-
ла от условий их разбав-
ления. В меженный период
влияние подогретых вод
проявлялось наиболее от-
четливо. Весной при про-
хождении через водохра-
нилише вод весеннего
половодья влияние подо-
гретых вод было менее
заметным. Влияние подо-
гретых вод локализовалось
на более ограниченном
участке при сильном вет-
ровом перемешивании во-
доема. Осенью влияние
подогретых вод было- ме-
нее заметным вследствие'
отсутствия устойчивой
стратификации водоема
по температуре, частой
повторяемости ветреных
дней и более быстрого
выхолаживания подогре-
тых вбд' при низкой тем-
пературе воздуха. Наи-
более отчетливо влияние
подогретых вод на разви-
тие фитопланктона прояв-
лялось в летнее время
при солнечной безветрен-
ной погоде.
Наличие достаточно
отчетливой связи между
температурой воды и
биомассой фитопланктона
в верхнем 2-метровом
слое водохранилища на
открытых участках под-
тверждается при статис-
тической обработке данных
1968 г. (Девяткин, 1970).
Таблица 5 Корреляция между температурой воды и биомассой фитопланктона иа открытых участках водохранилища по данным 1969 г. 28-29 X 3-7 9 Значения Я
25-27 IX 11-14 10
21-22 -УШ 18-22 10
25-26 VII 19-23 И
4-6 VI 17-20 11
6-8 V 10-13 14
Дата 1 Температура воды, °C Число вариант
Диатомовые........... 0.81 -0.02 0.12 0.79 0.60 0.62
Зеленые........*...... 0.46 0.04 0.24 0.18 0.23 0.07
Пнрофитовые.....‘.... 0.62 -0.48 0.83 0.58 -0.52 -0.33
Общая биомасса........ 0.78 -0.07 0.79 0.78 0.60 0.59
185
Корреляция между температурой воды биомассой фитопланктона
иа открытых участках водохранилища по данным 1970 г.
в Z.-S- IX 1-Х 06 Ю Г- со Ч* О 00 ю «о co со -j о о о о । о
30 IX-2 X 16-18 в © оо из г- со о т 00 Ч* Ч* из — *— О © 6 © © <j> Cj>
4-7 УШ 21-27 11 со со со г- со со ч* o'oo'oo’^d
в kZ-61 ПА 6-1А ОБ t- С- СЧ Ч* Г- ч* г- из со СО — СО © © О © 1 ©
6-7 VI 17-20 8 Значения R со ч* со со сч — 4j сч ddo'dd 1 ©
5-8 V 13-16 10 СО © а— СО г- СО СО ID t*j СО — СО сч со © © © © ^ © ©
Дата Температура воды, °C Число вариант Диатомовые ... Зеленые ............ Эвгленовые .......... Пирофитовые .......... Синезеленые Золотистые Общая биомасса . .
Аналогичные данные были получены и для наблюдений 1969 и
1970 гг. (табл. 5.6). В ряде случаев, однако, корреляция меж-
ду биомассой водорослей и температурой воды, как, например,
в мае 1970 r.f была вызвана не влиянием подогретых вод, а
разной степенью естественного прогрева водоема на разных
его участках. Но независимо от причин повышения температу-
ры воды в водоеме связь между этим показателем и биомас-
сой водорослей часто была положительной. Статистически дос-
товерной отрицательной связи между температурой воды и
обшей биомассой фитопланктона на протяжении 1969-1970 гг.
мы не наблюдали. Ряды данных 1967 г. слишком коротки для
статистической обработки.
Между биомассой отдельных групп водорослей и темпера-
турой также нередко наблюдалась статистически достоверная
связь. Заслуживает внимания наличие в ряде случаев положи-
тельной связи между температурой воды и биомассой диатомо-
.вых, что не подтверждает распространенного представления
об их гХолодолюбивости*.
В периоды массового развития пирофитовых связь между
температурой и их биомассой также была положительной.
Положительная корреляция нередко наблюдалась между
температурой и биомассой зеленых. Корреляция между темпера-
турой и биомассой других групп водорослей была менее тес-
ной, что объясняется, на наш взгляд, менее достоверным опре-
делением биомассы водорослей этих групп, так как в количест-
венных пробах они были представлены небольшим числом клеток.
Заключение
Проведенные наблюдения показывают, что реакция фито-
планктона на повышение температуры воды в Иваньковском
водохранилище имеет ряд существенных отличий от таковой
в небольших водоемах-охладителях. Одно из них - запаздыва-
ние реакции фитопланктона по сравнению с ходом изменений
температуры, как уже отмечалось (Девяткин, 1970), определя-
ется высокой проточностью, создающейся в отводящем канале
и Мошковичском заливе. Другой особенностью является отсутст-
вие значительных сдвигов в сезонной периодичности развития
фитопланктона под влиянием подогретых вод, которые наблюда-
ются в небольших водоемах-охладителях с высокой тепловой
нагрузкой (Виноградская, 1971).
В отличие от небольших водоемов-охладителей влияние по**
Погретых вод на развитие планктонных организмов в крупных
186
187
водоемах в большей степени зависит от гидрометеорологичес-
ких условий. Поскольку в Иваньковском водохранилище весной
и осенью создаются более благоприятные условия для смешения
подогретых вод, влияние последних отчетливее проявлялось в
летнее время, что также нехарактерно для небольших водое-
мов-охладителей.
В связи с вопросом о влиянии подогрева на качество воды,
используемой для нужд водоснабжения Москвы, наибольший ин-
терес представляет предплотинный участок водохранилища. Про-
веденные нами наблюдения показывают, что на протяжении веге-
тационного периода подогретые воды, по-видимому, не оказыва-
ли заметного влияния на этом участке. Не отмечалось здесь
и заметных изменений в составе фитопланктона. Однако даль-
нейшее увеличение объема сбрасываемых в Иваньковское водо-,
хранилище подогретых вод может вызвать увеличение обилия
фитопланктона и в предплотинном участке.
Рассматривая изменения, наблюдающиеся в сообществе фи-
топланктона в зоне воздействия подогретых вод, нельзя не
рассмотреть вопрос о причинах этих изменений. Следует отме-
тить, что он почти не освещается в литературе, посвященной
влиянию подогретых вод ТЭС на водоемы (ф.Д.Мэрдухай-Бол-
товской, 1971, наст, сб.; Patrick, 1969).
В сипу „облигатности* температурного фактора для всех
гидробионтов и сбалансированности естественных биоценозов
влияние изменения температурных условий на какую-либо груп-
пу организмов, в частности на фитопланктон, нельзя рассматри-
вать изолированно. Достаточно очевидно, что при этом необхо-
димо учитывать изменения в составе и обилии, возникающие
под влиянием подогретых вод в других группах гидробионтов,
прежде всего планктонных. В силу того что изменения, проис-
ходящие в биоценозе, взаимосвязаны и обычно взаимозависимы,
влияние того .или иного фактора иа любую группу организмов
может проявляться не прямо, а опосредованно, через изменения,
протекающие в другой группе под воздействием данного факто-
ра. Это одно из условий, затрудняющих анализ причин измене-
ний, происходящих в естественных биоценозах под влиянием
подогретых вод и определяющих сравнительно небольшую цен-
ность громадной литературы, посвященной влиянию температур-
ного фактора на гидробионтов в условиях эксперимента, для
решения проблемы влияния подогретых вод на биологические
процессы в водоемах. Другим условием является комплексность
влияния теплоэлектростанций, так как оио, кроме собственно
повышения температуры, связано с изменениями химизма,
гидрологических условий и механическим воздействием на ор-
ганизмы.
188
Попытаемся рассмотреть, насколько это возможно в дан-
ный момент, некоторые побочные эффекты воздействия ТЭС
на фитопланктон, не связанные непосредственно с повышением
температуры. Наиболее важным из них нам представляется из-
менение гидрохимических условий в зоне воздействия подогре-
тых вод. Однако, к сожалению, этот вопрос до сих пор изучен
недостаточно. Данные наблюдений М.Л.Флейса (1970, 1972)
свидетельствуют, что в Мошковичском заливе по сравнению с
районом водозабора .в отдельных случаях увеличивалось содер-
жание минеральных форм азота, особенно нитритов. Однако
это увеличение было нерегулярным и особенно отчетливо прояв-
лялось зимой, когда уровень содержания биогенов не лимитиру-
ет развитие фитопланктона. За пределами Мошковичского зали-
ва в зоне распространения смешанных подогретых вод заметно-
го увеличения содержания биогенных элементов по сравнению
с районом водозабора не отмечено. Наблюдения, проведенные
на других водоемах-охладителях (Стангенберг, 1967; Карпова
и др., 1969; Кибальчич и др., 1969; Абремская, 1971; Вульф-
сон, 1971; Коненко и Абремская, 1971; Коненко и др., 1971;
Лаумянскас и Снукишкис,1971а; Moor, 1958; Krolewski,
1967; Edinger a. Geger, 1968; Pourriot e.a,, 1972).,
показывают, что влияние- ТЭС на гидрохимические условия
проявляется прежде всего в изменении кислородного режима.
Причем в водоемах с незначительным и умеренным загрязнени-
ем кислородный режим, как правило, улучшается, а в сильно
загрязненных водоемах ухудшается. В водоемах со щелочной
реакцией среды может происходить смещение карбонатио-каль- •
циевого равновесия и наблюдается пересыщение карбонатами.
Влияние сбрасываемых ТЭС вод иа содержание биогенных эле-
ментов в водоемах-охладителях незначительно. Увеличение со-
держания последних обычно не связано с температурным режи-
мом, а вызывается побочными загрязнениями. В отдельных
случаях в зоне повышения' температуры наблюдается снижение
концентрации биогенов вследствие более эффективного, их ис-
пользования фитопланктоном (Лаумянскас, Снукишкис, 19716).
Существенным моментом, могущим оказывать влияние на
содержание биогеиов в стратифицированных водоемах, может
оказаться оборудование большей части крупных ТЭС глубинны-
ми водозаборами. В этом случае вода с более богатых биоге—
нами глубинных горизонтов, будучи извлеченной на поверхность,
может стимулировать развитие фитопланктона и бактериальную
активность. Однако в таком - сравнительно слабо стратифициро-
ванном на протяжении большей части вегетационного периода
водоеме, как Иваньковское водохранилище, это явление, как
нам кажется, вряд ли имеет большое значение, хотя конкрет-
ных исследований этого вопроса не проводилось.
188
Важным обстоятельством для режима биогенов нам пред-
ставляется факт разрушения организмов зоопланктона в охлаж-
дающих системах Конаковской и других ТЭС (Астраускас и
Рачюнас, 1971: Поливанная и Сергеева, 1971; Ривьер, 1971,
и др.). Биомасса зоопланктона Иваньковского водохранилища
соизмерима с биомассой фитопланктона, причем в охлаждающих
системах Конаковской ГРЭС гибнет до 60% и более зоопланк-
тона (Ривьер, 1971). По некоторым данным, в результате ав-
толиза уже в течение первых 15-30 мин. после гибели в окру-
жающую среду выделяется примерно 30% содержащегося в теле
зоопланктеров азота (Zawach, 1971). Быть может, именно
это обстоятельство является причиной некоторого повышения
содержания азота, наблюдавшегося в отдельных случаях в на-
чале отводящего канала Конаковской ГРЭС (Флейс, 1970,1972).
В отводящий канал Конаковской ГРЭС сбрасываются сточ-
ные воды г. Конаково. В настоящее время соотношение их
объема и объема подогретых вод колеблется в пределах 1/600-
1/1200. Учитывая сравнительно высокое качество очистки этих
стоков, а также высокое фоновое содержание биогенов в воде
Иваньковского водохранилища, вряд ли следует предполагать,
что стоки г. Конаково оказывают в данное время существенное
влияние на развитие фитопланктона в зоне воздействия подогре-
тых вод. Однако в связи с ростом промышленности г. Конако-
во и его населения объем сточных вод, поступающих в отводя-
щий канал, будет возрастать.
Таким образом, вопрос о степени изменений гидрохими-
ческого режима в зоне воздействия подогретых вод Конаков-
ской ГРЭС остается открытым, хотя имеющиеся в настоящее
время данные свидетельствуют о том, что они не очень зна-
чительны. Проведенные нами наблюдения (Девяткин, 1973а) по-
казали, что химизм воды в отводящем канале и Мошковичском
заливе изменялся не столь значительно по сравнению с районом
водозабора, чтобы вызвать заметные различия в скорости на-
растания водорослей на этих участках при одинаковых темпера-
турных и световых условиях. Косвенно это подтверждается дан-
ными по интенсивности фотосинтеза водорослей, согласно кото-
рым не наблюдалось существенных изменений этого показате-
ля при сходных температурных условиях в пределах отводящего
канала и залива по сравнению с районом водозабора (Вайнштейн
и др.. 1973).
Другим побочным эффектом, связанным с работой ТЭС,
является травмирование гидробионтов в ее охлаждающих систе-
мах я других водотоках. Как уже отмечалось выше, данные
микроскопических наблюдений и данные по фотосинтетической
190
активности водорослей свидетельствуют, что фитопланктон
Иваньковского водохранилища, представленный на протяжении
большей части года преимущественно диатомеями и протококко-
выми, сравнительно мало травмируется при прохождении через
агрегаты Конаковской ГРЭС.
Собственно повышение температуры, как уже отмечалось,
может оказывать прямое н опосредованное влияние на разви-
тие фитопланктона. К числу опосредованных эффектов следует
отнести усиление бактериальной активности при повышении
температуры, которое согласно предположению Патрик (Patrick,
1869) может оказаться дополнительным источником биогенных
элементов для водорослей. Однако конкретные исследования
этого вопроса нам неизвестны. Можно сослаться лишь на дан-
ные Келлера и др. (нит. по: Coutant, 1970), согласно
которым рост Chlorella в безбактериальной культуре
в меньшей степени зависит от температурных условий, чем в
культурах с наличием бактерий. Наши наблюдения показали, что
повышение температуры на 10° вызывает заметное увеличение
интенсивности деструкции в опытных склянках с естественной
водой в первые сутки экспозиции, однако затем интенсивность
деструкции в „подогретых* склянках снижается, по сравнению
с контрольными склянками, что, по-видимому, связано с исчер-
пыванием запасов легкодоступной для бактерий органики (Девят-
кин, 19736). В Иваньковском водохранилише, по нашим данным,
интенсивность деструкции возрастает в начале отводящего ка-
нала, по сравнению с районом водозабора; однако к концу за-
лива она снижается До более низких, по сравнению с районом
водозабора, величин, что, по-видимому, связано с теми же,
что и в экспериментальных условиях, процессами. Если повы-
шение температуры ие увеличивает доступность для бактерий
связанных в прочные комплексы соединений, то увеличение
бактериальной активности при повышении температуры само по
себе не может оказаться источником поступления значительно-
го количества биогенов.
Непрямое влияние повышения температуры на развитие
фитопланктона может проявиться путем изменения численности
организмов, питающихся водорослями. Как уже отмечалось вы-
ше, в охлаждающих системах ГРЭС, а также иа водопаде в кон-
це отводящего канала гибнет значительная часть организмов
зоопланктона. Низкая численность зоопланктона сохраняется в
верхних горизонтах Мошковичского залива (Ривьер, 1971). Од-
нако, за исключением криптомонад и Surirella ovata, нэ
диатомовых наиболее массовые виды фитопланктона водохранили'
• ща являются колониальными организмами, обилие которых вряд
ли контролируется степенью выедания. Кроме того, заметное
181
повышение биомассы фитопланктона наблюдается за пределами
залива, где численность зоопланктона восстанавливается. По-
этому изменение численности организмов зоопланктона в зоне
воздействия подогретых вод едва ли может быть причиной уве-
личения биомассы водорослей.
Прямое воздействие повышения температуры на развитие
фитопланктона может проявляться в изменении скорости роста
и интеисивности фотосинтеза водорослей. Большая часть наи-
более массовых видов фитопланктона Иваньковского водохрани-
лища является эвритермными видами, поэтому повышение тем-
пературы на 1-5°, как это наблюдается на прилегающих к Мош-
ковичскому заливу участках, по-видимому, ускоряет скорость
их роста. Увеличение скорости роста энергетически обеспечива-
ется небольшим, но заметным увеличением интенсивности фото-
синтеза, которое отмечалось при повышении температуры до
20-26°, а при летнем составе фитопланктона - до 26-30° (Вайн-
штейн и др., 1973). При повышении температуры на 1-5° увели-
чение интенсивности фотосинтеза было, как правило, незначи-
тельным. Однако при достаточно длительном воздействии повы-
шения температуры дополнительно синтезируемая органика при
прочих равных условиях может накапливаться, обеспечивая тем
самым более интенсивный рост клеток.
Учитывая всю важность побочных и опосредованных эффек-
тов воздействия ТЭС и повышения температуры, роль прямо-
го воздействия повышения температуры при объяснении причин
увеличения биомассы фитопланктона нам представляется значи-
тельной. Косвенным подтверждением этого являются статисти-
ческие данные о наличии довольно высокой корреляции между
температурой воды и биомассой фитопланктона за пределами
Мошковичского залива. В случае опосредованного влияния повы-
шения температуры величины коэффициентов корреляции должны
были бы бЖть, на иаш взгляд, заметно ниже.
В целом вопрос о причинах повыденйя обилия фитопланк-
тона в зоне воздействия подогретых вод еще не может счи-
таться полностью выясненным. Очевидно, что для его оконча- '
тельного решения необходимы дальнейшие исследования с учас-
тием работников различных специальностей.
Таким образом, планктонные водоросли при прохождении
через охлаждающие системы Конаковской ГРЭС, и другие водо-
токи травмируются незначительно.
-Вследствие неравномерного вертикального распределе-
ния фитопланктона в районе водозабора и наличия уст-
ройства для глубинного забора охлаждающей воды се-
зонная динамика биомассы поверхностного слоя в отводящем
192
канапе Мошковичском заливе имеет ряд отличий по сравне-
нию с другими участками водохранилища. В пределах отводя-
щего канала и Мошковичского залива биомасса фитопланктона,
как правило, изменяется незначительно, однако в устьевых
участках залива в течение вегетационного периода возрастает
роль криптомонад. Сравнительно слабое проявление влияния
подогретых вод на развитие фитопланктона в канале и в зали-
ве связано с их. высокой проточностью и с запаздыванием
реакции водорослей на изменение температурных условий.
На мелководных участках, прилегающих к устью Мошко-
вичского залива, наблюдается повышение биомассы фитопланк-
тона по сравнению с заливом и увеличение относительной роли
криптомонад.
В районе распространения смешанных подогретых вод
(разрезы Н-У) наблюдается статистически достоверное увели-
чение биомассы фитопланктона по сравнению с районом водо-
забора и предплотинным участком водохранилища, а также уве-
личение удельного веса криптомоиад в летний период.
Степень воздействия подогретых вод на развитие фито^
планктона в районах распространения смешанных подогретых
вод зависит от метеорологических и гидрологических условий.
Наиболее отчетливо влияние подогретых вод проявляется в лет-
ний период при тихой погоде. Ветровое перемешивание ослабля-
ет влияние подогретых вод. Ветровые и стоковые течения ока-
зывают сильное влияние на распределение смешанных подогре-
тых вод и фитопланктона в водохранилище. При безветренной
погоде и ветрах западных направлений наиболее заметное уве-
личение биомассы фитопланктона наблюдается в районе разрезсв
Щ-У, а при достаточно длительном воздействии ветров восточ-
ных румбов - в районе разреза II. При изменении гидрометеоро-
логических* условий распределение фитопланктона может изме-
няться в течение суток.
Влияние подогретых вод на сезонную динамику биомассы
фитопланктона в зоне умеренного и слабого подогрева незна-
чительно. Снижение обилия фитопланктона в районе распростра-
нения смешанных подогретых вод происходило лишь летом
1867 г.( однако, по-видимому, это не было вызвано переходом
температуры за пределы оптимальной, так как при сходных
температурных условиях летом 1868-1871 гг. и при более вы-
сокой температуре летом 1872 г. снижения биомассы водорос-
лей в зоне умеренного и слабого повышения температуры не
происходило.
В результате воздействия подогретых вод не происходит
существенной перестройки сообщества фитопланктона в течение
вегетационного периода. Наблюдается лишь повышение роли
778 13
183
криптомонад, а соотношение между наиболее массовыми вида*,
ми фитопланктона в районе распространения подогретых вод
ие изменяется.
Влияние подогретых вод на предплотннный участок, отку-
да вода поступает в канал Москва-Волга, в течение вегета-
ционного периода незначительно.
Литература
Абремская С.И. 1871. Гидрохимический режим водое-
ма-охладителя Кураховской ГРЭС. Гидробиол. жури.,
VII, 6.
Астраускас А.С., Рачюнас Л.А. 1971. Экологи-
ческие условия и состояние фауны водохранилища-охла-
дителя Литовской ГРЭС. Симпоз. по влиянию подогре-
тых вод теплоэлектростанций на гидрологию и биоло-
гию водоемов. Тезисы докл. Борок.
Вайнштейн МБ., Девяткин В.Г., Митрополь-
ская И.В. 1973. Влияние подогретых вод Кона-
ковской ГРЭС иа фотосинтетическую активность фито-
планктона Иваньковского водохранилища. ГидробиоЬ.
журн., IX, 6.
Васильчикова А.П. 1971. Периодичность и динамика
биомассы фитопланктона водохранилиш-охладителей
Урала. Симпоз. по влиян. подогрет, вод ТЭС... Борок.
Виноградская Т.А. 1971. Влияние подогрева на раз-
витие фитопланктона водохранилища-охладителя Курахов-
ской ГРЭС. - В кн.: Гидрохим. и гидробиол. водоемов-
охладителей ТЭС СССР. Киев.
Вульфсои В.И. 1971. Физико-химическое воздействие
тепловых электростанций на охлаждающую воду и воп-
росы водоподготовки. Симпоз. по влиян. подогрет. .
вод ТЭС... Борок.
Гусева- К.А. 1956. Фитопланктон Рыбинского водохрани-
лища. Тр. биолог, станции .Борок", 2.
Гусева К.А. 1959. К методике учета фитопланктона. Тр.
Инет. биол. водохр. АН СССР, 2(5).
Девяткин В.Г. 1970. Влияние термальных вод на фито-
планктон приплотинного плеса Иваньковского водохра-
нилища. Гидробиол. журн., У1, 2.
Девяткин В.Г. 1973а. Влияние загрязнений на развитие
фитопланктона в эоне воздействия подогретых вод Ко-
164
ваковской ГРЭС. Информ, бюлл. Инет. биол. внутр, вод
АН СССР, 17.
Девяткин В.Г. 19736. Влияние кратковременного повы-
шения температуры на фотосинтез фитопланктона. Ин-
форм. бюлл. Инет. биол. внутр, вол АН СССР, 18.
Карпова З.Ф., Го ле ев а С.К., Шадиева К.Н.,
Заха рова Т.Н. 1969. Влияние сброса горячей
воды от охлаждения турбин Томь-Усинской ГРЭС на
р. Томь. Матер, республ. науч, конфер. по пробл.
гигиены, водоснабж. н санит. охраны вод. Тез. докл.
Пермь.
Кибальчич И.А., Стрижак Е.К., Артемова
Г.В., Ехина Г.В., Тача лов С.Н., Мель-
ниченко А.Г., Талин А.П. 1969. Тепловые
электростанции как источники загрязнения водохрани-
лищ. Матер, республ. науч, конфер. по пробл, гигиены,
водоснабж. н санит. охраны вод. Тез. докл. Пермь,
Коней ко А.Д., Абремская С.А. 1971. Влияние
теплообменных вод на гидрохимический и санитарный
режим некоторых водоемов-охладителей тепловых
электростанций. Симпоз. по влиянию подогретых вод.
ТЭС... Борок.
Коней ко А.Д., Абремская С.А., Кутовеико
В.М. 1971. Характеристика гидрохимического режима
водоемов-охладителей ГРЭС Украины. - В кн.: Гидро-
хим. и гидробиол. водоемов-охладителей ТЭС СССР.
Киев.
Кузьм и н Г. В., Елизарова В.А. 1967. Фитопланк-
тон Шекснннского плеса Рыбинского водохранилища
в 1963-1965 гг. Тр. Инет. биол. внутр, вод АН СССР,
19.
Курдина Т.Н., Девяткин В.Г. 1972. О некоторых
показателях распространения подогретых вод. Гидро-
биол. журн., УЩ, 4.
Лаумянскас Г.А., Снукишкис Ю.Ю. 1971а.
Изменение гидрохимического режима водоема-охладите-
ля под влиянием Литовской ГРЭС. Симпоз. по влияя,
подогрет, вод ТЭС... Борок.
Лаумянскас Г.А., Снукишкис Ю.Ю. 19716.
Режим биогенных элементов, органических веществ и
газового состава водохранилища-охладителя Литовской
ГРЭС. - В кн.: Гидрохим. и гидробиол. водоемов-охла-
дителей ТЭС СССР. Киев.
Мордухай-Болтовской Ф.Д. 1971. Состояние воп-
роса о влиянии подогретых вод теплоэлектростанций
193
на биологию водоемов. Симпоз. по влиян. подогрет,
вод ТЭС... Борок.
Пан Л. 1969. Газовый режим охлаждающей воды Конаковс-
кой ГРЭС и его влияние на Иваньковское водохранилище.
.Матер, к совет, по прогнозир.' биогенных и органичес-
ких веществ в водохр. Рыбинск.
Поливанная М.Ф., Сергеева О.А. 1971. Динами-
ка зоопланктона и биология некоторых доминирующих
видов в водоемах-охладителях ГРЭС юга Украины. Сим-
поз. по влиян. подогрет, вод ТЭС... Борок.
Ривьер И.К. 1971. Зоопланктон Мошковичского залива,
подверженного влиянию подогретых сбросных вод Ко-
наковской ГРЭС. Симпоз. по влиян. подогрет, вод
ТЭС... Борок.
Стангенберг М 1967. Естественные следствия сбро-
са теплых вод в реки. Материалы I съезда ВГБО.
Санит. и технич. гидробиол.
Ф л е й с МЛ. 1970. Влияние сброса теплых вод Коиаковс- .
кой ГРЭС на гидрохимический режим Иваньковского
водохранилиша. Гидробиол. журн., VJ , 3.
Ф л е й с МЛ. 1972. Изменение санитарного состояния водо-
хранилища в зоне сброса теплых вод. - В кн.: Теория
. и практика биологического самоочищения сточных вод.
М
Ш а л а р ь В.М, Яловицкая И. И. 1971. Развитие
фитопланктона в Кучурганском лимане-охладителе Мол-
давской ГРЭС. - В кн.: Гидрохим. и гидробиол. водое-
мов-охладителей ТЭС СССР. Киев.
Сои t ant Е.С. 1970. Thermal pollution-biological
effects. Jour. Water. Poll. Contr. Fed., 42, 6.
Edinger I.E. and Gr e g e r J.C. 1968, - Analy-
- sing steam electric power plant discharges. J.
Sanit. Engng, Div. Am, Soc. Civ. Engrs, No 94,
SA-4.
Hamilton: D.H., Flemer D.A., Keef Ca-
ro 1 i n W„ Mihurski I.A, 1970. Power
plants: effects of chlorinations on estuarine pro-
duction. Science, 169, 3941.
Krolewski H, 1967. Artificial heating of river
water and its effect, Wasserwirtschaft, Stuttg.,
57, '
Moor E.W, 1958. Thermal „pollution" of streams.
Industrial and Engng. Chern,, 50, 1-4, 87A.
196
Morgan R.P., Stress R.G, . 1969. Destru-
ction of phytoplankton in the cooling water
supply of steam electric station. Chesapeake
Science, 10, 3-4.
Patrick R. 1969. Some effects of temperature
on freshwater algae. Proc, of the Nat. Symp.
on Therm. Poll., ed. by P.Krenkel and F.Par-
ker. Vanderb. Univ. Press.
pourriot R., Royer G„ Peltier M. 1972.
Proliferation de rotiferes epiphytes et pollution
thermique • dans la Loire. Bull, franc, piscicult.,
44, 244.
Zawach W. 1971., Substancje organiezne rozpu-
szczone w wodzie, ich pochodzenie i udzial w
krazeniu materii. Wiad, ecoL* 17, 2.
В.А.Э к з е р ц е в и Л.И.Л и с и и в н а
РАСТИТЕЛЬНОСТЬ НИЖНЕГО ПЛЕСА
ИВАНЬКОВСКОГО ВОДОХРАНИЛИЩА
И ВЛИЯНИЕ НА НЕЕ ПОДОГРЕТЫХ ВОД
КОНАКОВСКОЙ ГРЭС
Иваньковское водохранилище создано первым в каскаде
Большой Волги. Естественно, что первый крупный искусствен-
ный водоем привлек внимание многих исследователей, в том
числе и ботаников. С момента его наполнения было начато изу-
чение процесса формирования гидрофильной растительности.
Большие работы в этом направлении выполнены. Ю.Д.Шмелевой
(1940, 1954), А.В.Калининой (1945а, б) и Н.Н.Третьяковым
(1947). В 50-х годах наиболее подробно изучил зарастание
мелководий искусственных водоемов А.А.Потапов (1954, 1959,
1960). Начиная с 1957 г. и до настоящего времени сотрудника-
ми Института биологии внутренних вод АН СССР проводились
Постоянные наблюдения за процессами формирования и смен рас-
тительности (Экзерцев, 1958, 1963, 1966, 1971; Экзерпев и
Трифонова, 1959; Экзерцев и Экзерцева, 1962; Экзерцев и
Солнцева, 1962; Экзерцев и Соколова, 1963).
Сброс теплых вод Конаковской ГРЭС в Мэшковичский
заЛив водохранилища вызвал необходимость изучения влияния
подогретых вод на естественный растительный покров мелково-
дий залива и иа сукцессионные процессы в нем. Эти исследо-
вания были начаты нами в 1968.г. Если*о развитии раститель-
ности в небольших водоемах-ойладителях теплостанпий имеются
некоторые сведения (Павлннова, 1939; Лукина, 1972), то о
влиянии теплых вод ГРЭС на крупные водохранилища почти
ничего не известно.
198
По данным гидрологических наблюдений заметное повыше-
ние температуры воды в Иваньковском водохранилище наблю-
дается в Мошковичском заливе и на близрасположеииых участ-
ках Иваньковского плеса. Только в этом районе через ряд
лет можно ожидать изменения естественного зарастания мелко-
водий, вызванного термическим фактором. Поэтому в настоя-
щем сообщении мы даем характеристику растительности мелко-
водий Иваньковского плеса водохранилища до строительства
ГРЭС (1966 г.) и кратко останавливаемся на тех естественных
ее изменениях, которые уже произошли в последние годы. .
Общая характеристика
растительности
Иваньковского плеса
Иваньковское водохранилище - довольно мелководный ис-
кусственный водоем. Почти половина его площади (46%) при-
ходится па участки с глубинами менее 2 м. Мелководен и его
Иваньковский (озерный) плес. Особенности морфометрии и уро-
венного режима водохранилища способствовали развитию гид-
рофильной растительности. За 35 лет его существования боль-
шая часть литорали плеса заросла. Растительный покров хоро-
шо развит. Фитоценозы имеют четкую ярусную структуру и рав-
номерное сложение. В прибрежье они расположены резко раз-
граниченными поясами, сменяющими друг друга в зависимости
от нарастания глубины и изменения грунтовых условий. Наи-
большее распространение на мелководьях плеса имеет воздушно-
водная растительность , представленная ассоциациями Glyce-
ria maxima purum, Glyceria maxima + Equisetum; flu-
viatile, Glyceria maxima + Rorippa amphibia, Glyceria
maxima — Spirodela polyrrhiza, Glyceria maxima ♦
Typha latifolia, Equisetum fluviatile purum, Equisetum
fluviatile + Carex acuta, Equisetum fluviatile + Slum
latifolium, Equisetum fluviatile + Galla palustris* Equi-
setum fluviatile - Spirodela polyrrhiza, Equisetum flu-
viatile — Nymphaea Candida, Phragmites communis
purum, Phragmites communis + Herbae mixtae aquatiles.
Phragmites communis - Spirodela polyrrhiza, Seirpus
lacustris purum, Scolochloa festucacea purum, Typha
angustifolia purum, Sagittaria sagittifolia purum, Carex
acuta purum. Фитоцеиозы воздуш ио-водных растений встре-
чаются от границы зоны' затопления до глубины 150 см. На не—
189
1 которых отшнурованных мелководьях воздушно-водные растения
образуют сплавины с господством Bquisetum fluviatile,
Glyceria maxima, Typha latifolia, Calls palustris, Me-
nyanthes trifoliata.
Значительные площади Иваньковского' плеса заняты погру-
женной растительностью, из которой наиболее широко распро-
странены ассоциации Potamogeton perfoliatus purum, Po-
tamogeton perfoliatus + Myriophyllum spicatum, Po-
tamogeton lucens purum, Potamogeton 'pectinatus
purum, Stratiotes aloides purum, Stratiotes aloides —
Spirodela polyrrhiza, Myriophyllum spicatum purum.
Фитопенозы погруженной растительности занимают участки с
. глубиной от 80 до 180 см.
В последнее десятилетие иа Иваньковском плесе произош-
ло увеличение площадей ассоциаций плавающей прикрепленной
растительности, особенно ассоциаций Nymphaea Candida
purum, Nymphaea Candida - Stratiotes aloides, Nuphar
lutea purum и Potamogeton natans purum. Заросли
плавающих растений встречаются преимушественио по защищен-
ным участкам, с илистыми грунтами на глубине от 80 до 200 см.
Типы зарастания мелководий
Зарастание отдельных участков литорали плеса различно
и зависит от гидрологических и грунтовых условий. Схематич-
но можно выделить следующие типы биотопа: 1) открытая лито-
раль, подверженная интенсивному прибою, с песчаными грунта-
ми; 2) открытая литораль, подверженная умеренному прибою,
с Песчаио-илистыми грунтами; 3) защищенная литораль с илис-
тыми грунтами; 4) защищенная литораль, изолированная от
основного плеса, с торфянистыми грунтами.
Зарастание первого типа биотопа незначительно. Постоян-"
иый прибой и перемещение грунта препятствуют поселению
макрофитов по границе воды. И только на глубине 30-40 см
появляются фитопенозы Potamogeton pectinatus. Этот вид
в сипу своих морфологических особенностей хорошо переносит
волвобой и способен в данных условиях сознавать устойчивые
сообщества. Ширина зарослей рдеста зависит от уклона дна,
но обычно не превышает 20 см. № глубине 60-80 см они
сменяются фитопеиозами Р. perfoliatus, гПубииное распро-
странение которых не превышает 120 см. Таким образом, в
•том типе зарастающего прибрежья представлен только один
200
Рис. 1. Профили зарастающего прибрежья Иваньковского
плеса.
а - открытая литораль, подверженная интенсивному при-
бою, с песчаными грунтами; б - открытая литораль, подвержен-
ная умеренному прибою, с песчано-илистыми грунтами; в -
защищенная литораль с илистыми грунтами; г - защищенная
литораль, изолированная от основного'плеса, с торфянистыми
грунтами; а — изолированные ^заболоченные мелководья без
заметного уклона дна.
пояс погруженных растений, состоящий из зарослей двух видов
рдеста (рис. I, а).
Открытая литораль, подверженная умеренному прибою
(второй тип), зарастает иначе (рис. 1, б). В этом случае при-
сутствует растительность двух поясов. Первый пояс, воздуШ”
201
но-водных растений, состоит из фитоценозов Carex acuta,
расположенных по урезу воды, и Phragmites communis,
занимающих участки с глубинами 20-100 см. Последние иногда
замешаются сообществами Glyceria maxima. Второй пояс,
погруженных растений, представлен участками ассоциаций ро_
tamogeton perfoliatus, проникающих на глубину до 180 см.
Третий тип биотопа (защищенные мелководья с илистыми
грунтами) наиболее благоприятен для развития всех групп вод-
ных растений. Благодаря этому экологический ряд ассоциаций
зарастающего прибрежья является в этом случае наиболее пол-
ным (рис. 1, в). Пояс воздушно-водных растений состоит из
односоставных фитоценозов Carex acuta, Glyceria maxima
и Sagittaria sagittifolia, последовательно сменяющих друг
друга с увеличением глубин от 0 до 80 см. При этом наиболь-
шие площади занимают ассоциации манника большого. Пояс
погруженных растений также хорошо развит и представлен
сплошными зарослями Potamogeton lucens, Р, perfolia-
tus или Myriophyllum splcatum, приуроченными к глуби-
нам от 90 до 150 см. Наконец, пояс растений с плавающими
листьями образован сплошным ковром зарослей Nymphaea
carfdida, занимающих участки с глубиной от 120 до 200 см.
Иная картина зарастания наблюдается на глухих изоли-
рованных участках с торфянистыми грунтами (четвертый тип).
Хотя в этом случае при пологом уклоне дна имеются тоже три
растительных пояса, но представлены они другими ассоциациями.
В первом поясе безраздельно господствуют фитоценозы Equi-
setum fluviatile, окаймленные со стороны берега узкой
полосой Carex acuta или Carex inflata. Довольно часто
в хвощах стало наблюдаться сплавинообразование с преоблада-
нием рогозо-цикуто-хвощовых ассоциаций. На глубинах 80 см
их сменяют сплошные заросли Stratiotes aloides, представ-
ляющие пояс погруженной растительности. Наконец, на глуби-
нах 150-200 см появляется пояс плавающих растений, который
состоит из пятнистых группировок Nuphar lute a, Nymphaea
Candida или Potamogeton natans (рис. 1, г).
Подобная зависимость размещения ассоциаций нарушается
на участках, где на больших площадях нет заметного уклона
диа, а глубина не превышает 1 м. Такие мелководья нацело за-
растают, растительный покров их пятнист, никакой поясности в
распределении фитоценозов не наблюдается (рис. 1, д). Преоб-
ладают группировки воздушно-водных растений, чередующиеся
со сплавинами, на которых доминируют Glyceria maxima,
Equisetuni fluviatile, Typha latifolia, Calla palustris.
Comarum palustre, Menyanthes trifoliate и Cicuta
viroaa.
202
До создания ГРЭС такие же закономерности распределе-
ния растительности можно было проследить в Мошковичском
заливе и на прилегающих к нему открытых участках плеса.
Верховья залива и верховья его отрогов зарастали по 4-му
типу - заболоченных биотопов. В них господствовали заросли
Equisetum fluviatile, Stratiotes aloides и Nuphar lu-
te a. В средних участках залива преобладали ассоциации Carex
acuta, Glyceria maxima, Potamogeton lucens и My-
riophyilum spicatum (3-й тип). На открытых устьевых
мелководьях доминировали Glyceria maxima и Potamoge-
ton perfoliatus (2-й тип). Литораль вокруг островов, рас-
положенных перед заливом, зарастала по типу открытых прибре-
жий,» подверженных постоянному прибою (1-й тип).
Фитоценозы всех растительных поясов имели четкую струк-
туру и равномерное сложение. Пятнистости в размещении груп-
пировок не наблюдалось. Кроме перечисленных господствующих
зарослей в заливе встречались несколько куртин Scirpus
lacustris, Phragmites communis и Typha angustifolia.
Влияние строительства ГРЭС
и сброса теплых вод
на растительность
Во время строительства электростанции в Мошковичском
заливе проводились земляные работы. Строился сбросной канал,
и земснаряд углублял русло залива, намывая грунты на левый
берег. Потоки мутной воды распространялись по заливу, что
ие могло не повлиять на растительность.' В результате за два
года многие ассоциации были нарушены или совсем замыты.
Наиболее пострадали заросли левого берега залива и участка,
прилегающего к сбросному каналу.
Общая картина состояния растительности в этот период
(1965-1966 гг,) была следующей. По левому берегу от сброса
до устья залива на намытых грунтах преобладали пятнистые
группировки Glyceria maxima, Polygonum amphibium,
Scirpus lacustris. Они не создавали сплошного покрова,
а располагались отдельными куртинами. Один из отрогов лево-
го берега был нацело замыт. Второй, предустьевой, был менее
подвержен действию мутных вод, л растительность в нем со-
хранила свой прежний облик, хотя сложение фитоцеиозов стало
меиее равномерным. В верховьях отрога преобладали группиров-
ки Equisetum fluviatile - Spirodela polyrrhiza, Nym-
203
SHI
phaea Candida — Potamogeton lucens и Potamo—
geton natans (purum). Там же имелось несколько куртин
чистых зарослей Potamogeton lucens и Scirpus lacu—
stris. Наконец, на мелководьях левого берега, расположенных
в устье залива, сохранился только пояс Carex acuta и по-
явились отдельные пятна Polygonum amphibium,
В верховьях Мошковичского залива выше сбросного кана-
ла по заболоченным участкам на глубинах более 60 см встре-
чались ассоциации Stratiotes aloides - Spirodela po-
lyrrhiza, а на глубинах до 90 см преобладали фитоценозы
Glyceria maxima - Spirodela polyrrhiza с пятнами
Typha latifolia и Rorippa amphibia. На открытых участ-
ках верховий большие площади мелководий занимали чистые
заросли Potamogeton lucens с единичными куртинами Nym-
phaea Candida.
В среднем участке залива у правого берега и в первом
отроге растительность тоже была частично нарушена. На не-
больших глубинах преобладали скопления Glyceria maxima,
Typha latifolia, Butomus umbellatus и Rorippa amphi-
bia. В самых верховьях отрога сохранился участок ассоциации
Equisetum fluviatile - Spirodela polyrrhiza,. На боль-
ших глубинах доминировали пятна Potamogeton lucens,
Nymphaea Candida, Polygonum amphibium и Sparga-
nium simplex. Наконец, растительность предустьевого отрога
правого бёрега залива сохраняла в основном свой прежний
облик. В поясе воздушно-водных растений господствовали Gly-
ceria maxima- и Equisetum fluviatile с вкраплениями
куртин Phragmites communis. В поясе погруженных и пла-
вающих растений большие площади занимали чистые заросли
Potamogeton lucens и ассоциация Nymphaea Candida -
Myriophyllum spicatum. Однако и здесь имела место не-
которая неравномерность в распределении растений и их груп-
пировок.
Рис. 2. Схематический план растительности Мошковичско-
го залива (1972 г.).
1 - крупноосочники; 2 - асе. с господством манника боль-
шого; 3 — асе. с господством тростника; 4 - асе. с господст-
вом рогоза широколистного; 5 — асе. с господством рогоза
узколистного; 6 - асе. с господством хвоща приречного; 7 -
куртины камыша озерного; 8 - комплексные заросли маниика
большого и рогоза широколистного; 9 - асе. с господством
рдеста блестящего; 10 - асе. с господством рдеста пронзениолист-
ного; 11 - асе. с господством кувшинки чистобелой; 12 -асе. с
господством телореза алоэвидного.
205
В залив с нарушенным растительным покровом, где раз-
личные участки его находились на разных этапах становления,
Начали сбрасывать подогретые воды. После этого на мелко-
водьях можно было наблюдать два процесса; восстановление
нарушенной растительности и изменения в развитии и размеще-
нии макрофитов, вызванные действием теплых вод.
В первые годы (1967-1968 гг.) действие теплых вод
на растительность проявилось только в сдвиге фаз развития
гидрофитов (Жукова и Лисицына, 1971). Особенно оно было'
ошутимо в весеннее время, когда разница температур воды за-
лива и водохранилища достигала значительных величин. Макро-
фиты в Мошковичском заливе начинали вегетировать раньше,
чем на других участках Иваньковского плеса. Но в залнве про-
грев вод был неравномерен. Более высокие температуры про-
слеживались на участках, расположенных ближе к руслу, а
мелководья с глубинами 10-90 см почти не нагревались. Раз-
личие температур по глубинам привело к тому, что первыми
начинали вегетировать растения, расположенные на больших
глубинах,, тогда как в естественных условиях весеннее разви-
тие растительности начиналось с хорошо прогреваемых мелко-
водий прибрежья. В середине лета разница в фенологическом
состоянии растительности залива и водохранилища сглаживалась.
Одиако отмирание погруженных растений в теплых водах
начиналось раньше и протекало быстрее. Только отдельные
экземпляры Potamogeton crispus, Potamogeton lucens
и Nymphaea Candida сохраняли небольшие вегетативные
побеги до конца сентября. В зимнее время уровень воды в во-
дохранилище и в заливе снижается на несколько метров, а за-
растающие мелководья обсыхают и таким образом выходят из-
под влияния теплых вод.
Интересно было выявить влияние подогрева и на продукцию
растительности (см. таблицу). Сравнив продуктивность, фито-
неиозов Мошковичского залива с продуктивностью их в водохранили
ше вне эоны подогрева, можно отметить, что основное дей-
ствие подогретых вод проявляется в смешении сроков достиже-
ния максимальной фитомассы. Обычно в водоемах лесной зо-
ны максимум фитомассы приходится на конец июля-начало ав-
густа. В Мошковичском заливе он у разных фитоценозов насту-
пает в разное время, что связано с неравномерным распределе-
нием температур по заливу и сдвигом фаз развития растений.
Если в первые три года изменений в сложении и соста-
ве растительности под влиянием теплых вод не наблюдалось,
то спустя пить лет они начали намечаться (рис. 2).
Коротко некоторые закономерности наблюдавшихся смен
сводятся к следующему. Прежде всего произошло сокращение
Таблица
Динамика фитомассы ассоциаций макрофитов
в заливах с подогретой водой (Мошковичский зал.)
и вне зоны-подогрева (залив по р. Бабня)
в 1960 г. (возд.-сух. вес, г/м^)
Ассоциация 25 V 26 У1 10 УШ 10 IX
i. Equisetum fluviati- le - Spirodela polyrrhiza . . . Мошкови» 117.3 1ский зали 982 В 688 433.8
Phragmites commu- nis (purum). . . 384 1338 1339 1234
Carex acuta (pu- rum) ...... 197.5 510 756.5 780
Stratiotes aloides - Spirpdela polyrr hiza ....... 70.6 177 461 387.4
Potamogeton lucens (purum) .... - 312 254.9 —
Equisetum fluviatile Spirodela poly- rrhiza ...... la лив no p Ab 40 Бабня 702 940 573.4
Phragmites commu- nis (purum) , . 315 1234 948.3
Carex acuta (pu- rum) ...... 187 423 649.5 517.5
Stratiotes aloides - Spirodela po- lyrrhiza .... 41.6 241 862 607
Potamogeton lucens (purum) ... 31 328.5 274.2
площадей зарослей Equisetum fluviatile и Stratiotes
aloides. С другой стороны, начала проявляться приспособлен-
ность некоторых видов к новому термическому режиму. Увели-
чились площади зарослей тростника, рогоза узколистного. Осо-
бенно увеличились площади зарослей тростника в устьевом
участке, где они сменили господствовавшие там фитоиенозы
Glyceria maxima. Кроме того, появились и начали рассе-
ляться в заливе три новых вида: Potamogeton crispus, Po-
tamogeton obtusifolius и Ricciocarpus natans,
Заключение
1. Растительность Иваньковского плеса к моменту строи-
тельства Конаковской ГРЭС вполне сформировалась и заняла
все доступные участки мелководий. В ее размещении в при-
брежье наблюдалась строгая поясность, обусловленная измене-
ниями глубинных и почвенно-грунтовых условий.
2. При строительстве сбросного канала в Мошковичском
заливе растительный покров был нарушен. В первые годы
поступления теплых вод пестрота растительности в заливе бы-
ла вызвана не их действием, а последствием земляных работ.
3. В настоящее время действие теплых вод на высшую
водную растительность водохранилища проявляется ли шь на
ограниченном участке Иваньковского плеса - в Мошковичском
заливе и прилегающих' к нему мелководьях.
4. За 5 лет сброса теплых вод не наблюдалось сколько-
нибудь заметного увеличения степени зарастания мелководий
водохранилиша и возрастания обшей годовой продукции макро-
фитов.
5. Непосредственное действие теплых вод. на раститель-
ность проявляется прежде всего в сдвиге фаз развития, в сдви-
ге достижения максимальной фитомассы и в наметившейся в
последние годы смене одних фитоцеиозов другими.
6. В дальнейшем можно ожидать увеличения площадей за-
рослей тростника, рогоза узколистного, роголистника, рдестов
курчавого и туполистного. Возможно, они займут господствую-
щее положение в Мошковичском заливе, вытеснив сообщества
манника большого, хвоша приречного и телореза алоэвидного.
7, В растительном покрове мелководий всего Иваньков-
ского водохранилиша и его продуктивности в настоящее вре-
мя ощутимого изменения не наблюдается и едва ли оно про-
явится в следующие годы при современном режиме работы
208
ГРЭС. Увеличение же мощности станции или строительство но-
вой может повлечь ал собой расширение сферы действия подо-
гретых вод на большую часть мелководий Иваньковского плеса,
что вызовет более интенсивные смены одних фитоцеиозов дру-
гими и увеличит степень эьтрофикаини водоема.
Литература
Жукова Г.А. и Л.И.Л и с и ц ы н а. 1971. Влияние
строительства Конаковской ГРЭС на растительность
Мошковичского залива. Информ, бюлл. Инет. биол.
•внутр, вод АН СССР, 12.
Калинина А.В. 1945а. Зарастание Иваньковского водо-
хранилиша. Реф. раб. учрежд. отд. биол. наук АН СССР
за 1941-1943 гг. М.-Л.
Калинина А.В. 19456. Первые стадии зарастания мел-
ководий Московского моря. Сов. ботан., XIII, 4.
Лукина Е.В. 1972. Водная и прибрежно-водная раститель-
ность водохранилища-охладителя ГоГРЭС им. А.В.Вин-
тера. Биология внутренних вод. Информ, бюлл. Инет,
бнол. внутр.вод^АИ СССР, 15.
Павликова Р.М. 1939. Кваттросуо зарастании водо-
хранилищ на примере водохранилища Горьковской
энергетической станции. Бюлл. МОИП, отд. биолог.,
X1.VIU, 4.
Потапов А. А. 1954. Распределение водных растений в
заливах Иваньковского и Истринского водохранилищ
в зависимости от состава воды и характера донных от-
ложений. - В кн.: Строительство водохранилищ и проб-
лема малярии. М.
Потапов А.А. 1959. Зарастание, водохранилищ при раз-
личном режиме, уровней. Бот. журн., 2.
Потапов А .А. 1960. О значении донных отложений при
прогнозах зарастания водохранилищ. Тр. Всесоюзн,
гидробиол. общ., 10.
Третьяков Н.Н. 1947. Материалы к формированию фау-
ны водоплавающих и болотных птиц Московского моря. '
В кн.: Очерки природы Подмосковья и Московской
области. М. ,
Шмелева Ю.Д. 1940. Зарастание Иваньковского водохранилища
канала Москва-Волга и заселение его личинками анофелеса
778 И 200
за три года его существования. - Медиц. паразитол. и пара-
зитарн. болезни, IX, 3.
Шмелева Ю.Д. 1954. Зарастание и анофелогенность
Иваньковского водохранилища. - В кн.: Строительство
водохранилищ и проблема малярии. М.
Экзерцев В.А ' 1958. Продукция прибрежно-водной рас-
тительности Иваньковского водохранилища. Бюлл. Инет,
биол. воцохр. АН СССР. 1.
Экзерц ев В.А. 1963. Зарастание литорали волжских
водохранилищ. Тр. Инет. Биол. внутр, вод АН СССР,
6(9).
Э кзерцев В.А. 1966. Флора Иваньковского водохрани-
лища. Тр. Инет. биол. внутр, вод АН СССР, 10(13).
Экзерцев В.А. 1971. Растительность Иваньковского
водохранилища. Тр. Инет. биол. внутр, вод АН СССР,
21(24).
Э к з е р.ц е в В.А. и Трифонова Н.А. 1959. О свя-
зи некоторых растительных груптировок Иваньковского
водохранилища с элементами окружающей среды. Бюлл.
Инет. биол. водохр. АН СССР, 4.
Экзерцев В.А. и Солнцева И.О. 1962. Эколо-
гические ряды ассоциаций Шошинского плеса Иваньков-
ского водохранилища. Бюлл. Инет. биол. водохр. АН
СССР, 13.
Экзерцев В.А. и Экзераева В. В. 1962. Про-
дукция макрофитов в заливах Иваньковского водохрани-
лища. Бюлц,Инст. биол. водохр. АН СССР, 12.
Экзерц ев В.А. и Соколова М.А. 1963. Сооб-
щества хвоща приречного на Иваньковском водохрани-
лище. Тр. Инет. биол. водохр. АН СССР, 5(8).
Н.В.М а м а е в
ВЛИЯНИЕ СБРОСНЫХ ТЕПЛЫХ ВОД
КОНАКОВСКОЙ ГРЭС
НА ПЛАНКТОННЫХ ПРОСТЕЙШИХ
ИВАНЬКОВСКОГО ВОДОХРАНИЛИЩА
•
Подогретые воды, сбрасываемые Конаковской ГРЭС, ока-
зывают существенное влияние на гидрологический режим, а так-
же на планктон и бентос Иваньковского водохранилища (Под-
дубная, 1968; Ривьер, 1968; Буторин и Курдина, 1970; Девят-
кин, 1970).
Влияние этих вод иа планктонных простейших до сих пор
не изучалось. А между тем простейшие - постоянный компо-
нент зоопланктона, составляющий существенное звено в трофи-
ческой цепи водоемов. Кроме того, в силу морфологических и
физиологических особенностей простейшие - чувствительные
индикаторы на изменение условий внешней среды.
Сбор материала производился с мая по октябрь
1972 г. на русловых стандартных станциях в зоне влияния
подогретых вод Иваньковского водохранилища (рис. 1).
Пробы отбирали метровым батометром системы А.В.Фран-
цева с каждого метра по вертикали до 10 м, глубже —
через 5 м. После этого все пробы перемешивали, из полу-
ченной смешанной пробы отбирали 0.5 л и концентрирова-
ли путем фильтрования через мембранный фильтр № 8 без
применения вакуума до 10 мл. Для учета мелких форм
0.5 мл полученного объема распределяли на стекле в виде
капель, в которых производили подсчет простейших при
увеличении микроскопах50-160. В оставшемся объеме учи-
тывали более крупные формы при меньшем увеличении. Од-
211
Рис. 1. Схема расположения станций в Иваньковском водо-
хранилище.
Римские цифры - разрезы, арабские - станции.
новременно производилось определение видовой принад-
лежности простейших.
Подогретые воды Конаковской ГРЭС сбрасываются в Мош-
ковичский ‘залив Иваньковского водохранилища (рис. 1). Зона
влияния этих вод охватывает участок водохранилища от разреза
III вверх по течению от Мошковичского залива до разреза V
вниз По течению. Станции в районе водозабора и II разреза
выше по течению, а также VI разреза и предплотинного пле-
са находились практически вне влияния или испытывали незна-
чительное влияние подогретых вод.
Численность простейших в планктоне, как показали наши
наблюдения, сильно колебалась в различные месяцы (рис. 2).
В мае в зоне влияния подогретых вод она была очень высокой,
до 4.8 тыс. экз./л, что заметно выше численности вне этой
зоны, где было отмечено не больше 1.5 тыс. экз./л. Только
в верховье Мошковичского залива, где перепад температуры
составлял 5° (от 12 до 17°С), численность резко сокращалась
до 800 экз./л.
В июне все выглядело совершенно иначе. Высокая числен-
ность простейших, наблюдавшаяся в районе водозабора, резко
уменьшилась в Мошковичском заливе и далее, после неболь-
шого подъема в районе IX разреза, она оставалась невысокой.
Таким образом, в мае в зоне влияния подогретых вод числен-
ность простейших значительно повышалась, а в июне она сни-
жалась.
212
В июле-августе- отмечалось
минимальное развитие простейших,
численность которых не превышала
500 экз./л. В Мошковичском зали-
ве таковые практически отсутст-
вовали. Температура здесь достига-
ла 32-33°, что, по-видимому, и
привело к гибели простейших. За
пределами залива численность по-
вышалась до прежнего уровня.
В сентябре влияние подогрева
сказывалось в меньшей степени,
чем в предыдущие месяцы. Перепад
температуры в 7° (16° вне зоны
подогрева и 23°С в верховье
Мошковичского залнва) вызывал
лишь небольшое сокращение числен-
ности простейших в зоне влияния
подогретых вод. Возможно, что ко-
лебание температуры в указанных
пределах не оказывает большого
влияния на простейших.
В октябре перепад температу-
ры в верховье МэшковичскогО;Зали—;
ва составлял 8° (от 7° вне зоны
влияния подогретых вод до 15РС
на водосбросе). При очень низкой
численности простейших вне зоны
влияния подогретых вод в районе
подогрева численность была в 10
раз больше, чем в районе водоза-
бора, и в 50 раз больше, чем в
предплотйнном плесе. Подобную
же картину мы наблюдали в мае.
В. p-С ,3 УШ, /У S 8
МешквВичыий
золиб
Рис. 2. Колебание
численности простейших в
мае-октябре 1072 г.
когда численность простейших
значительно повышалась в зоне
влияния подогретых вод. Однако
в октябре в Мошковичском за'ливе
численность простейших заметно
повышалась, а в мае она резко
снижалась в верховье залива, где
перепад температуры был наиболь-
шим.
T-IX - разрезы,
3-9 - станции, В - водо-
забор, р. С. - река Созь.
По оси ординат - колич.
простейших.
213
Средняя численность отдельных видов простейших
по месяцам в зоне влияния подогретых вод, экз./л
Виды Май Июнь 1 Июль, авгу ст Сентябрь Октябрь
Tintinnidium flu- viatile Stein, 1833 435 310 165 375 638
Codonella cra- ters Leidy, 1877 119 362 108 184 24
Strombidium viride Stein, 1859 181 100 60 108 0
Strobilidium ve- lox Faure-Fr„ 1924 7 36 4 18 4
Coleps hirtus Nitzsch, 1817 0 .1203 140 557 205
Tintinnopsis cylindrata Kof,-Cam., 1892 О 238 90 120 4
Holophrya sp. 201 205 80 0 0
Mesodinium pu- lex Clap, u. L„ 1858 75 0 98 60 0
Askenasia vol- vox Clap. u. L„ 1858 132 198 120 0 0
Didinium balbia- ni Fabre- Dom., 1888 21 101 60 0 0
Vorticella sp. 0 100 0 9 28'
Stokesia verna- lis (Wang, 1928) Wen- rich, 1929 12 0 0 0 8
Epistylis rotans Svec., 1897 0 116 60 0 0
Продолжение таблицы
Виды Май Июнь Июль, август Сентябрь Октябр!
Vorticella sphae-
rica d’Ude-
kem, 1664 .Amphileptus 0 94 0 120 0
trachelioid es Zach,, 1893 Condylostoma 6 0 0 0 7
vortic ella Ehrb., 1833 3 3 о
Vorticella ana- 0
baena Still., 1940 0 31 82 0 п
Tinti nnidium
fluviatile Stein f. cyli- ndrica Ga-
jew, 1933 Phascolodon 0 0 120 180 0
vorticella
Stein,, 1859 Dileptus a ns er 1237 0 О 0 0
O.F.Mull., 1786 0 0 0 0 10
Strobilidium sp. 264 0 0 0 0
Litonotus sp. Vorticella natans 79 0 0 0 о
Faure-Fr„ . 1924 80 0 0 0 0
P arur ole ptus
piscis Ko- ^zalewski, 1882 3 0 '0 0 0
Paramecium sp. Vorticella co- 2 0 0 0 0
no chili Sto- kes, 1888 0 43 0 0 0
Nassula aurea
Ehrb., 1833 0 7 0 0 0
Tintinnidium s p. 0 0 0 120 0
Продолжение таблицы
Виды Май Июнь Июль, август Сентябрь Октябрь
Paradileptus со- nicus Wen- rich, 1929 0 0 3 О 0
Urotricha pela- gica Kahl, 1935 0 0 0 60 0
Stentor globa- tor Stokes, 1885 0 0 0 3 0
Staurophrya ele- gans Zach. 0 0 0 0 2
Strombidium ' sulcatum Clap.u«L., 1859 0 0 0 0 20
Stentor roeseli Ehrb., 1835 0 0 0 о 2
Actinophrys sol. Ehrb., 1830 0 0 0 0 2
Простейшие в планктоне были представлены 35 видами
(см. таблицу). Этот список безусловно не исчерпывает всего
видового состава простейших, так как отбор проб производился
главным образом- в русловой части водохранилища. Таблица
дает представление о средней численности каждого вид»., про-
стейших в отдельные месяцы. Список составлен в порядке умень-
шения их встречаемости. Из 34 видов инфузорий 14 относятся
к Holotricha, 13 - к Spirotricha.B - к Peritricha, 1 вид “
к Sue tor ia. Кроме того, был обнаружен 1 вид солнечников
( Heliozoa). К числу массовых можно отнести И видов, 6 из
которых встречались практически постоинно в значительном
количестве: Tintinnidium fluviatile, Codonella cratera,
Coleps hirtus, Strombidium viride, Strobilidium velox,
Tintinnopsis cylindr ata.
Виды теплолюбивые, обитавшие только в эоне влияния
подогретых вод или получившие какие-то морфологические изме-
нения под действием повышенной температуры, нами не были .
216
обнаружены. По-видимому, это естественно в условиях большо-
го водообмена.
Состав видов по месяцам в зоне влияния подогретых вод
изменялся в общем так же, как и в водохранилище за ее пре-
делами. Май отличался от других месяцев большим видовым
.разнообразием при значительной численности основных видов.
На IX разрезе наибольшей численности (до 3200 экз./л) дости-
гал Phascolodon vorticella. На всех станциях в этом
месяце встречалась Stokesia vernalis - - крупный весенний
вид инфузорий'.
В июне видовой состав заметно изменился. Выпал ряд
форм, в том числе Phascolodon vorticella, Stokesia ver-
nalis, Mesodinium pulex, Amphileptus trachelioides.
На смену им пришли другие: Coleps hirtus, Epistylis ro-
tans, эпибионтные (поселяюциеся иа водорослях и планктонных
животных) инфузории, которые в этот период достигли большо-
го развития. В наибольшем количестве в июне развивался Co-
leps hirtus (до 3500 экз./л в районе водозабора).
В июле-августе видовой состав простейших при малой их
численности имел большое разнообразие. Преобладали: Tintin-
nidium fluviatile, Coleps hirtus, Askenasia volvox.
В сентябре видовое разнообразие несколько уменьшилось
при заметном увеличении численности. Coleps hirtus вновь
выходит в число преобладающих видов, достигая численности
1020 экз./л, но это вдвое меньше, чем в июне.
Октябрь отличался заметным своеобразием фауны. Появи-
лись Stokesia vernalis и Amphileptus trachelioides,
которые были отмечены только в мае. Tintinnidium fluvia-
tile достиг большей, чем в мае, численности причем инфузории
отличались крупными размерами. Появилась Strombidium
sulcatum,, ранее не отмеченная в Иваньковском водохранили-
ще. Описанная выше смена' форм по месяцам и сезонам года
происходит, как правило, в каждом водоеме. Заметного влияния
подогретых вод на смену видов простейших мы не обнаружили.
Однако влияние водогрева на отдельные виды было заметно.
На рис. 3. представлено колебание численности Tintinni-
dium fluviatile “ одного из самых многочисленных и постоян-
но встречаемых видов в зоне влияния подогретых вод в раз-
личные месяцы. В мае и июне в Мошковичском заливе, где
влияние подогрева сказывалось в наибольшей степени, числен-
ность этого вида резко сокращалась. В июле-августе этот
вид в зоне влияния подогретых вод практически отсутствовал.
В октябре картина была совершенно иной. Наибольшая числен-
ность Т. fluviatile наблюдалась именно в Мошковичском за-
ливе.
217
мз./л
В. р.С J УШ, IV 9 8
МошкоВцчский
залиВ
Рис. 4. Численность
Phascolodon vorticella
в Иваньковском водохрани-
лище.
По оси ординат - ко-
лич. простейших.
Рис. 3. Численность
Tintinnidium fluviatile
в. Иваньковском водохрани-
лище.
По оси ординат - ко-
лич. простейших.
Рис. 5. Численность
Coleps hirtus в Иваньков-
ском водохранилище.
По оси ординат — колич.
простейших.
Колебание численности Phascolodon vorticella, пре-
обладающего вида в мае, показано на рис. 4. У этого вида
можно отметить заметное сокращение численности вида в вер-
ховье Мошковичского залива и возрастание ее в близлежащей
к заливу зоне.
У Coleps hirtus, в массовом количестве развившего-
ся в июне, в Мошковичском заливе численность резко умень-
318
шилась, и, после небольшого подъема в устье залива, ниже по
течению была еще меньше (рис. 5).
Таким образом, подогретые воды, сбрасываемые Конаков-
ской ГРЭС, оказывают заметное влияние иа численность планк-
тонных простейших Иваньковского водохранилища. Весной и
осенью (в мае и октябре) численность простейших в зоне
влияния подогретых вод значительно повышалась, в июне -
снижалась, небольшое сокращение численности отмечено и в
сентябре. В июле-августе при самой высокой температуре в
Мошковичском заливе, куда.сбрасываются подогретые воды,
простейшие полностью отсутствовали.
Литература
Б у т о р и и Н.В., К у р д и н а Т.Н. 1970. О динамике
температуры воды Иваньковского водохранилища в зо-
не поступления подогретых вод Конаковской ГРЭС. Ин-
форм. бюлл. Инет. биол. внутр, вод АН СССР, 7.
Девяткин В.Г. 1970. Влияние термальных вод иа фито-
планктон приплотииного плеса Иваньковского водохра-
нилища. Гидробиол. жури., VI, 2.
Поддубная Т.Л. 1968. Донная фауна Иваньковского
водохранилища в зоне сброса теплых вод Конаковской
ГРЭС. - В кн.: Волга-1. | конфер. по изуч. водоемов
басе. Волги. Тезисы докл. Тольятти. *
Ривьер И.К. 1968. Влияние сбросных теплых вод Ко-
наковской ГРЭС на зоопланктон Иваньковского водохра-
нилища. - В кн.: Волга-}. I конфер. по изуч. водоемов
басе. Волги. . Тезисы докл. Тольятти.
И.К.Р и в ь е р
ЗООПЛАНКТОН ИВАНЬКОВСКОГО ВОДОХРАНИЛИША
В ЗОНЕ ВЛИЯНИЯ ПОДОГРЕТЫХ ВОД
КОНАКОВСКОЙ ГРЭС
Исследование воздействия теплых вод на планктонных жи-
вотных Иваньковского водохранилища началось Институтом био-
логии внутренних вод АН СССР в 1967 г. и продолжается до
настоящего времени. Основное внимание было уделено зоне
сильного подогрева вод - Мошковичскому заливу. В заливе изу-
чалось изменение состава зоопланктона, сезонная динамика ру-
ководящих форм и вертикальное распределение организмов при
различных гидрологических условиях и при прохождении живот-
ными охладительных агрегатов ГРЭС, водосбросного канала и
водопада, особенности биологии некоторых видов (Ривьер,
1968, 1971 ; Ривьер и Гец, 1971; Литвинов и Ривьер, 1972).
В настоящей работе более подробно изложены вопросы,
касающиеся трансформации зоопланктона при прохождении всей
охладительной системы ГРЭС, рассмотрены некоторые особен-
ности. вертикального распределения животных в зоне воздейст-
вия подогрева при наличии холодного придонного слоя, просле-
жены реакции избегания перегретых поверхностных слоев наи-
более крупными и Подвижными ракообразными. Отмечены более
ярко выразившиеся в последние годы изменения видового сос-
тава и соотношения отдельных видов, а также резкие вспышки
размножения отдельных видов в разные годы. Рассмотрена
динамика численности и биомассы зоопланктона в теплом пото-
ке вплоть до его полного охлаждения и смешивания с водами
водохранилища в разные сезоны. Прослежены изменения в
структуре популяций некоторых массовых видов в зоне сильно-
го воздействия теплых вод.
220
Материалами послужили 750 проб зоопланктона, со-
бранных автоматически захлопывающимся плаиктонобато-
метром объемом 10 л, батометром Руттиера, сетью Джуцая.
Собраны и обработаны также многочисленные пробы, в ко-
торых определяли лишь видовой состав животных.
При изучении степени травмирования животных в ох-
ладительной системе пробы просматривали непосредственно
после нх взятия без какой-либо фиксации. Такие исследо-
вания велись у самой ГРЭС в начале охладительного канала
и в районе водопада, в месте впадения - сбросного канала
в Мошковичский залив.
Вертикальное распределение зоопланктона в заливе
изучалось на 8 суточных станциях, для измерения измене-
ний температуры и течения применялись автоматические
приборы.
Обработку проб в лаборатории производили по обще-
принятой методике. Материалы, касающиеся изменений в
структуре популяций Daphnia cucullata и изменений
отдельных биологических показателей этого вида, обработа-
ны статистически.
- Трансформация зооплаиктоиа
в охладительной системе ГРЭС
Район водозабора ГРЭС представляет собой русловой учас-
ток Волги, углубленный после строительства электростанции.
Для охлаждения забираются глубинные, наиболее холодные слон
воды. Одиако, судя по составу планктона непосредственно
после прохождения охладительной системы ГРЭС, в водоворот
в водозаборном канале вовлекаются серединные и поверхностные
слои воды. Наиболее ярко это прослеживается весной при рез-
ком качественном различии населения поверхностных к придон-
ных слоев. Зоопланктон в районе сбросного канала в количест-
венном и качественном отношении более всего похож на планк-
тон слоя 0-8 м, а не придонных слоев. Летом при более рав-
номерном вертикальном распределении зоопланктона резкой
разницы в составе водного населения забираемой и сбрасывае-
мой воды не наблюдается.
Охладительную систему ГРЭС вода проходит за несколько
минут. Здесь организмы испытывают механическое воздействие
(давление в иасосах и высокие скорости течения - 2 м/сек. в
трубках), а также воздействие резкого возрастания температуры
221
волы иа 7-10°С. Однако воздействия эти настолько кратковре-
меины, что животные обычно не теряют подвижности сразу.
В пробах, взятых непосредственно после прохождения ГРЭС
на ст. 21 при температуре до 26-28°С и немедленно просмот-
ренных, погибших животных почти не обнаруживается. Но при
33°С (максимальная наблюдавшаяся за весь период исследова-
ния температура воды летом 1972 г.) полностью лишенной под-
вижности оказалась основная масса рачкового планктона. Не-
подвижны были почти все Leptodora kindtii, Daphnia lon-
gispina, D. cristata, среди D. cucullata - 75%. Рачков
с остановившимся сердцем особенно много среди молодых даф-
ний. Неподвижных Diaphanosoma - 83%. Меньше страдают
циклопы, неподвижных среди них - 24%; почти не страдают
мелкие круглые формы Chydorus и Bosmina. Среди коло-
враток сильно травмируется нежная крупная Asplanchna,
несколько менее - виды Synchaeta. Среди живых и не ли-
шенных подвижности форм многие оказываются сильно травми-
рованными и погибают через несколько часов. В трехлитровые
цилиндры были помешены пробы зоопланктона, взятые в водо-
заборном канале (ст. 1) и в сливе у ГРЭС (ст. 2). Через 6
часов в цилиндрах с планктоном от водозабора все животные
были живыми и в хорошем состоянии; в пробах, взятых в сли-
ве у ГРЭС, неподвижны почти все рачки (Daphnia cuculla-
ta, D. longispina, Diaphanosoma brachyurum, Leptodora
kindtii и Daphnia cristata); циклопов погибло лишь 39%.
Травмирование животных в водопаде (ст. 3) происходит
под воздействием механических причин и наблюдается при любой
температуре, значительно усиливаясь при максимальной (28-
33°С). Нами было произведено подробное обследование воздей-
ствия водопада на массовые формы рачков 20-21 июля 1971г.
при 26° и 20 июля 1972 г. при 32.5°С (табл. 1).
С увеличением температуры до 32.5° количество неподвиж-
ных животных увеличивается, однако оно достаточно велико и
при 26°. По-видимому, у водопада основная причина высокой
степени травмирования рачков - механическое воздействие па-
дающей воды. Неподвижными,- с уже остановившимися дыха-
тельными движениями конечностей оказываются рачки, потеряв-
шие способность плавать после охладителей ГРЭС. У значитель-
ной части рачков начинается разложение кишечника, растекает-
ся глазной пигмент. Кроме того, в пробах попадается масса
Схема Мошковичского .залива дана в статье Н.В.Бу-
торина и Т.Н.Курдиной .Особенности температурного режима
Ивеньковского водохранилища...' (иаст. сб., рис. 5).
222
Таблица 1
Травмирование планктонных рачков
после прохождения водопада (ст. 3). %
Виды 20-21 июля 1071 г., при 26° 20 июля 1972 г,. при 32.5°
Diaphanosoma brachyurum 23 86
Limnosida frontosa ..... БЗ
Daphnia cristata ...... 51 70
D. longispina 24 48
D. cucullata 35 61
Bosmina (в. coregoni, B. longirostris) ....... 14 43
Leptodora kindtii . 61 50
Copepoda (взрослые и ко- пеподиты) 15 17
Таблица 2
Биомасса травмированных рачков
при разной температуре, г/мЗ
Виды 20-21 июля 1871 г., при 26° 26 июля 1972 г.» при 32.5°
живые непод- вижные живые непод- вижные
Diaphanosoma brachyu- rum 0.16 0.14
Limnosida frontosa . . 0.21 0.11
Daphnia cristata . . . « 0,03 0.02 0.14 0.10
D. longispina ..... 0.03 0.01 0.14 0.07
D. cucullata ...... 2.73 0.95 0.92 0.56
Bosmina ........ 0.28 0.04 0.12 0.05
. Leptodora kindtii ... 0.35 0.21 1.56 0.78
Copepoda (взрослые и копеподиты) . . . . . . 0,78 0.12 1.00 0.02
Всего . . ' 4.41 1.46 4.04 1.72
223
Искалеченных рачков, с оторванными антеннами, фурками и
другими частями тела.
При обследовании такого рода соблюдение календарных
сроков особенно важно, так как степень травмирования планкто-
на зависит от его видового состава. Весной и осенью,, когда
основную биомассу зоопланктона образуют мелкие формы - ко-
ловратки (Keratella cochlearis и К. quadrate), босми-
ны и копеподы, - гибель зоопланктона незначительна. Летом
же пагубное воздействие падающей воды особенно проявляется
из-за преобладания в планктоне крупных, легко травмирующих-
ся, нежных ракообразных, образующих летом основную биомассу.
Нами была подсчитана биомасса травмированных в течение
суток рачков при температуре 26 и 32.5°С (табл. 2).
Таким образом, за сутки при 26° и при расходе воды
4500 тысЛяЗ в сутки травмируется 6.5 т рачкового планктона.
При современном расходе 7000 тыс./м^ и при температуре
32.5° за сутки травмируется 12 т рачков.
Распределение зоопланктона
в Мошковичском заливе
Мощковичский залив искусственно углублен и соединяется
с водосливом ГРЭС цементированным водосбросным каналом]
Верховье Мошковичского залива, куда изливаются теплые воды
из канала, представляет собой чашу глубиной 6-7 м. Направлен-
ное течение на атом участке существует только в поверхност-
ных слоях, глубже наблюдаются сложные циркуляционные токи,
затухающие у самого дна. Здесь происходит накопление поте-
рявших подвижность рачков. Вследствие этого биомасса рачков
здесь оказывается повышенной, особенно у дна.' Такое явление
наблюдается обычно летом, когда в планктоне преобладают круп-
ные наиболее легко травмируемые пелагические ракообразные
(рис. 1). При обработке фиксированных проб травмированные
животные часто иё отличаются от зафиксированных живых.
Биомасса в чаще у водопада (ст. 3) вследствие накопления
у дна неподвижных и разлагающихся рачков бывает часто зна-
чительно выше , чем у водозаборного канала.
Теплый поток, выходящий в залив, оказывается обеднен-
ным зоопланктоном, численность и биомасса животных в нем
значительно ниже. Оставшиеся в теплом потоке живые ракооб-
разные активно реагируют на неблагоприятные условия (тече-
ние и высокие температуры). Рачковое население придонных
224
5.0Ь
3.0
1.0 -
5.0
3.0
Водозабор Cm. 2
1969 УШ
_ I
1970И
Ст.З СтЛ Ст.5 Ст.2, РюрУШ
Рве. 1. Изменения биомассы зоопланктона от водозабора
до устья Мошковичского залива.
и поверхностных слоев значительно различается (табл. 3). Рач-
ки мигрируют в более холодные придонные слои воды. Здесь
происходит накопление их биомассы и складывается своеобраз-
ное планктонное сообщество, состоящее из холодолюбивого
778 15.
328
К 2
® а
а
226
«о
о со
я
ех 2
• °
0*0
к ю
сч
s«
сч
CI
о о
8
ю
о о
8
to
ю о
— со
ю
88
8°
ю
сч
т
«
si
к
с
ю
«
Ь
о о
о о
о
ф
о
о
и
о
я
*
со
S
л
с
S
9
£
ф
ей
со
а
go
С 00
s сч
со
2-«п
О 00
о сч
§
2
3
88
оз to
3 cd о
о
S
0)
я
S
к
3
ф
3
g
ф
я
а
Е
О
а
S
ф ф
о й
S
ё
§
8
0
О
_о
о to
ЪЧ —*
«сч
л о
о о
Е СЧ
2 «>
Я
88
со со
® 2
со
8 8
S*°
Cyclops vicinus и крупных ветвистоусых. У дна сосредото-
чиваются крупные половозрелые особи. Особенно неестественно
построены популяции дафний (Daphnia cucullata, D« cris-
tata, D, longispina), основную часть которых составляют
крупные половозрелые особи, а молоди почти нет, что в естест-
венных популяциях у этих видов обычно ие наблюдается. Ано-
мальную структуру популяций дафний у дна можно объяснить
сложными гидрологическими и гидробиологическими условиями
залива (табл. 3).
В 1972 г. при температуре воды 32°С нарушения в струк-
туре популяций у дна были особенно ясно выражены, так как
значительная часть молоди погибала уже в агрегатах ГРЭС.
Оставшиеся в живых рачки в Мошковичском заливе неодинаково
ведут себя в теплом потоке. Крупные, более сильные особи
активно мигрируют из теплого потока в поисках более благо-
приятных условий и скапливаются у дна. Молодь же, не способ-
ная преодолеть течение, остается в поверхностном теплом по-
токе и пассивно сносится к устью залива. Скопившиеся у дна
крупные особи дафний размножаются, однако молодь активно
поедается циклопами, численность которых у дна очень вели-
ка. Своеобразное водное сообщество, образующееся в придон-
ных слоях Мошковичского залива, обычно не Наблюдаемое в
естественных условиях, было подробно изучено петом 1971-
1972 гг. (Ривьер, 1972).
При падении уровня в Мошковичском заливе придонный
холодный слой исчезает (перемешается к устью залива) вмес-
те с присущим ему водным населением. При этом усиливается
водообмен между русловой, обладающей максимальной проточ-
ностью частью залива И его отрогами. Водные массы из отро-
гов вместе с их зоопланктоном поступают в русловую часть
залива и изливаются в водохранилише. Такая картина наблюда-
лась в июне 1972 г., когда прибрежные водные массы вместе
с Bosmina longirostris, размножившейся там в огромном
количестве, при резком падении уровня поступили в русло за-
лива и распространились вплоть до его устья. Рачки активно
мигрировали из быстро текущего поверхностного слоя и скопи-
лись у диа в устье залива. У дна образовались огромные кон-
центрации Bosmina longirostris (до 70 млн экэ./мЗ, при
этом биомасса составляла 400 г/м®! ).
При отсутствии холодного придонного слоя н гомотермии'
больших скоплений рачков у дна обычно ие образуется, но
миграции ко дну в некоторых случаях все же происходят. Это
явление, как и предыдущее, определяется уходом рачков •
менее освещенную зону при температуре выше оптимальной
(табл. 4) и избеганием зоны с большими скоростями течения.
22»
Численность Daphnia cucullata в Мошковичском заливе
при гомотермии и перегреве, экз./мЗ
о
1970 г. июнь ст. 5 со о СЧ 27.8 150С
в ст. 5 I 27.2 17300 26.8 12500
S а> ё S к | ст. 4 I 26.2 22500 26.2 33000
в о >> | ст. 5 26.2 ' 18300 00911 0*92
0 <Я 00 ® 8 °-8
в 6 Г- ю СЧ СЧ сч 8
со
£ ЧТ “Го ю.8
ё о со 8 СЧ ID со С сч оо
Я Я ст.. 3 28.5 2400 28.5 ' 5200
и август ст. 4 27.5 6600 27.7 3400
1967 июнь ст. 3 29.8 700 009 4'62
* •
о • ° ' о : °. •
Показатели Поверхность: S-e £ о £ о ag ® « S о ФЕ” ь о я «3 «• л о. в г? к £ о ® К £• ® Е с Z и Ф ж В sr
Кроме того, миграция в придонные слои как при наличии темпе-
ратурной стратификации, так и при гомотермии вызвана в ка-
кой-то степени неблагоприятными условиями в потоке вследст-
вие высоких скоростей течения. Пелагические рачки вынуждены
искать более спокойную зону. Этим, видимо, были вызваны
большие скопления В. longirdstris у дна, описанные выше л
наблюдавшиеся нами в июне 1972 г. Отрицательное влияние на
пелагических рачков циркуляции воды отмечается и в водоемах-
охладителях Украины (Поливанная и Сергеева, 1971).
Изменения в видовом составе
зоопланктона
Использование Иваньковского водохранилища в качестве
водоема-охладителя привело к изменениям в видовом составе
зоопланктона. В зоне сильного воздействия теплых вод исчезли
такие виды, как Cyclops vicinus и Bythotrephes longi-
manus, резко сократилась численность Diaptomus, Daph-
nia cristata, D. longispina; в разные по температурному
режиму годы отмечались вспышки численности отдельных видов
как в пелагиали, так и в прибрежной зоне: Peracantha trun—
cata, Bosmina longirostris.
Изменение в соотношении видов происходит вследствие
воздействия охладителей ГРЭС и водопада. Это сказывается
и в резких нарушениях структуры популяций отдельных видов,
когда разные возрастные группы страдают неодинаково. Многие
ракообразные, обладая значительной подвижностью, активно
избегают перегретой зоны.
Отсутствие Cyclops vicinus в зоне сильного влияния
теплых вод 'связано с гибелью взрослых особей в охладителях
ГРЭС и в самом заливе при температурах, близких к порого-
вым. Обнаружить этот вид в Мошковичском заливе можно лишь
у дна при наличии холодного придонного слоя, т.е. в зоне с
естественной температурой. Здесь же держится и молодь этого
вида. В перегретый поверхностный слой С.- vicinus ие миг-
рирует и локализуется у самого дна (рис. 2). При установлении
гомотермии он вообще исчезает из залива.
Положительную реакцию на изменение термики обнаружил
другой вид циклопов - Mesocyclops leuckarti. В зоне
влияния подогретых вод вплоть до IV разреза (район Корчевы)
это - самый массовый вид циклопид. Особенно многочисленным
был М. leuckarti в наиболее теплые за весь период исследо-
228
229
м 22.30 0.45 3.00 5.00 7.00 9.00 11.00 ' 15.00 tsoo
МААДАо^Лл
0 ZOOOOjkj./h3
Рис. 2. Вертикальное распределение Cyclops vicinus
в течение суток 22-23 УП 1871 г. в Мошковичском заливе
(ст. 2, разрез VIII) при температуре поверхностного слоя воды
26 и придонного 18°С.
Рис. 3. Распределение ко-
ловраток в теплом потоке.
1 - температура; 2 -
Synchaeta sp.; 3 -
Asplanchna.
на подогрев D. longispina и
температурного расслоения D. longispina и D. cristata
встречаются только в холодном придонном слое (Ривьер и Гец,
1871). Количество их в Мошковичском заливе резко падает при»
вания 1867 и 1872 годы, ког-
да температуры были близки
к пороговым (27-ЗО°С). Попу-
ляции этого вида'в Мошкович-
ском заливе содержат огром-
ное количество молоди, что
свидетельствует об интенсив-
ном размножении. В 1972 г.
при температуре в течение
всего лета около 30° числен-
ность М. leuckarti в Мош-
ковичском залнве составляла
в августе в среднем 720000
экз./мЗ.
Изменилось соотношение
видов и в группе ветвистоу-
сых. Среди дафний менее
страдает Daphnia cuculla-
ta; отрицательно реагируют
D. cristata. При наличии
температурах выШе 25°С. За пределами залива при нормализа-
ции температурных условий количество, этих двух видов обычно
восстанавливается (табл. 5).
В пелагическом планктоне в зоне влияния теплых вод
в более теплые годы (1967, 1972) в значительном количестве
размножается Diaphanosoma brachyurum; В прохладные
годы больше было Limnosida frontosa, a D. brachyu-
rum встречались единично.
Особенно, сказывается влияние теплых вод на крупные
весенние виды коловраток - Synchaeta sp. и виды As-
planchna. При массовом развитии их весной в районах с ес-
230
Таблица
5
Соотношение трех видов дафний
в различных районах Иваньковского водохранилища
(в % от обшей численности Daphnia )
Дата и район исследований Темпера- тура, °C Daphnia cucullata Daphnia longispi- na Daphnia cristata
Июль 1967 г.
Водозабор . . . . 21.2 39.5 3.5 57
Ст. 4 27.0 95 0 5
Ст. 5 27.2 95 0 5
VIII разрез . . . 26.7 87.5 .0 12.5
Июль 1 969 г.
Водозабор . . . . 20.1 60 38 2
Ст. 3 27.0 89.5 9 1.5
Ст. 4 26.2 90 10 0
VIII разрез . . . 26.0 79 16 5
IX разрез . . . . 19.7 51 37.5 2
Август 1969 г.
Водозабор . . . . 17.0 90.5 7 2.5
Ст. 3 24.5 90 6 3
Ст. 4 24.5 93 5 2
Ст. S 24.5 94.5 5.5 0
V111 разрез ; . . 24.1 78 6 16
IX разрез . . . . 22.8 81 5 4
Заборье 17.3 91 6 3
Июль 1970 г.
Водозабор . . . . 21.1 80.5 12.5 7
Ст. 3 27.8 77 14 9
Ст. 5 28.0 92 5 2
VIII разрез . . . 28.3 95 3 2
X разрез .... 23.6 82 12 7
Кладбищенский
залив . 24.8 73 7 20
тественными условиями в Мошковичском заливе наблюдается
резкое падение их численности — почти до полного исчезновения
(рис. 3). Это связано с травмированием особенно крупной и
нежной As planch па в охладителях и на водопаде, а также с
торможением развития в зоне сильного влияния теплых вод.
Только за предела'мн залива при снижении температур до естест-
венных и падении скоростей течения эти виды восстанавливают
свою численность. Исчезновение As planch па и Synchaeta
sp. резко сказывается на величине биомассы всего зоопланк-
тона, так как эти виды весной и в начале лета составляют ее
основу.
Мелкие с твердым панцирем формы Keratella quadrate
и К. cochlearis мало реагируют на неблагоприятные условия
в зоне сильного влияния теплых вод. За пределами залива (раз-
резы УШ, IX, X, IV)., где ослабляется течение, но темпера-
тура выше естественной в среднем за сезон на 3—4°С, в неко-
торые годы отмечалось массовое развитие К. quadrata.
Численность этого вида достигала 900000 особей в/мЗ, значи-
тельно превосходя количество этого вида в естественных усло-
виях (Ривьер и Гец, 1971).
Годы использования Иваньковского водохранилища как ох-
ладителя сбросных вод значительно отличались по температур-
ным условиям. Наиболее теплыми были 1967 в особенно 1972 гг.;
более прохладными 1968 и 1969 гг. За весь этот период были
отмечены вспышки численности отдельных видов. В 1967 и
1972 гг. создались условия для массового развития Bosmina
longirostris. Размножение этого вида в больших количествах
в зоне влияния теплых вод наблюдалось именно в наиболее
теплые за весь период исследования годы. Высокие биомассы
В. longirostris отмечаются ежегодно в южных водоемах-
охладителях на Украине (Мирошниченко, 1971; Поливанная, Сер-
геева, 1971). Это связано с чрезвычайной эвритермичностью
и теплолюбивостыо В. longirostris, развивающейся в массах
именно в южных водоемах, в частности, в нижнем Дону" в его
пойменных водоемах и близлежащих прудах (Мордухай-Болтовс-
кой, 1957). В 1970 г. в прибрежной зоне Мошковичского зали-
ва в массе размножались Peracantha truncate (Pleuro-
xus truncatus); численность этого вида составляла 887000
экз./м^, а биомасса - 17.7 г/м^.
Такие кратковременные вспышки численности отдельных
видов, когда количество их значительно превосходит обычно
наблюдаемое в естественных условиях (а затем эти виды поч-
ти исчезают из планктона на несколько лет), свидетельствуют
О нарушениях биотических и абиотических факторов среды, про-
исходящих под воздействием дополнительного тепла.
992
Динамика численности
и биомассы зоопланктона
ЛЮ
350
250
150
50
М>- tula йп.5 'tapix'/taplt
xfap Cm3 Ш Разр.т йвцХ
Рис. 4. Распределение
зоопланктона в зоне влияния
теплых вод в холодное время
года.
А - численность коловра-
ток, Б - биомасса всего зоо-
планктона: 1 - температура.
Воздействие повышенной температуры на зоопланктон по-
разному сказывается в разные сезоны года. В холодное время
года подогреваемые участки более богаты планктоном, чем
районы с естественной температурой воды. Здесь осенью и в
начале зимы продолжается жизнь зоопланктона в основном за
счет развития коловраток, количество которых в 5-10 раз вы-
ше по сравнению с неподогреваемыми участками (рис. 4). Тем-
пература воды у водозабора
в этот период колеблется от
1 до 9°С, в Мошковичском
заливе - от 6 до 14°С. В
феврале—марте при максималь-
ной сработке-уровня водохра-
нилища и в связи с этим
максимальными скоростями
течения в теплом потоке ко-
личество планктона в заливе
резко снижается по сравнению
с водозабором. В апреле начи-
нается подъем уровня, скорос-
ти течения падают и в зоне
воздействия теплых вод начи-
нается развитие Коловраток,
а затем ветвистоусых. В ап-
реле и начале мая при темпе-
ратуре воды в обогреваемых
участках 3-14°С здесь значи-
тельно больше планктона, чем
в водохранилище. Наибольшее
количество коловраток в это
время'наблюдается в зоне .
сильного влияния теплых вод,
к району Корчевы (разрез 1У)
количество их несколько сни-
жается (рис. 4, Б).
Обратная картина (резкое уменьшение численности в Мош-
ковичском заливе) наблюдается в самое жаркое время при тем-
пературах выше 25°C. В этот период количество зоопланктона
в заливе резко снижается по сравнению с водозабором и оста-
ется низким. на протяжении всего залива. В его устье оно
обычно ниже, чем у водозабора. Подъем численности, наблюдае-
233
о.ч
0.3
о.г
Вейо- СтЛ РазрУШ Разр.Х
забор Cm3 Ст. 5 Разр.К РазрСУ
Рис. 6. Распределение
биомассы зоопланктона в
теплом потоке весной и
осенью при оптимальных
температурах (16-23°С).
1 - температура, 2 -
биомасса.
Вейо- СтМ РшрМ Разр.Х
забор Cm3 Ст5 Разр.1Х
Рис. 5. Распределе-
ние зоопланктона в Мош-
ковичском заливе.
1 - температура, 2 -
биомасса.
мый иногда на ст. 3, связан, как указывалось выше, с накоп-
лением массы погибающих животных у дна. Чем ближе темпе-
ратура воды в заливе к 30°С, тем ниже биомассы животных
в теплом потоке (рис. 5).
Весной и осенью количество зоопланктона в заливе бывает
тоже несколько ниже, чем у водозабора. На выходе из залива
(разрез VIII) величины биомассы приближаются к наблюдаемым
у водозабора. Однако за пределами залива, в эоне слабого
влияния теплых вод, обычно наблюдается увеличение биомассы
планктонных животных (рис. в). Здесь (разрезы IX, X, IV)
количество зоопланктона в 5-10 раз выше, чем у водозабора.
Такая картина наблюдается в период оптимальной температуры
в потоке (от 16 до 23°С).
234
Некоторые стороны биологи в
Daphnia с u с u 11 a t а
в эоне влияния теплых вод
Daphnia cucullata - самый многочисленный вид круп-
ных пелагических .рачков в зоне влияния теплых вод летом.
Сезонная динамика численности этого вида, его вертикальное
распределение, некоторые особенности строения популяций в
Мошковичском заливе (малое количество новорожденных, более
ранние сроки наступления гамогенеза) были рассмотрены нами
ранее (Литвинов и Ривьер, 1972).
Размерный и половой состав популяций D. cucullata
был исследован в Мошковичском заливе, у села Заборье, у Го-
родищ, в районе водозаборного канала, а также (для сравнения
с более северными популяциями) в Рыбинском водохранилише.
В каждой пробе тщательно промеряли 100 особей, определяли
их пол и стадию зрелости. Вычисляли средний размер особей
в пробе, количество Новорожденных рачков, неполовозрелой
молоди и яйценосных самок. Кроме того, определяли минималь-
ный и максимальный размер яйценосных партеногенетических
и эфиппиальных самок, вычисляли их средний размер.
Наименьший средний размер (близкий к размеру новорож-
денных рачков) имеют дафнии в начале мая (табл. 6). К июлю-
августу средние размеры постепенно возрастают, а затем к
осени уменьшаются. Эта величина в естественных условиях в
течение лета не испытывает резких колебаний, что связайо с
устойчивостью строения популяций.
В зоне сильного влияния теплых вод (Мошковичский залив)
средний размер часто значительно выше, чем в естественных
условиях, и испытывает заметные колебания в течение лета.
Это связано с большими нарушениями в структуре популяций
Da cucullata в заливе, с малым количеством новорожден-
ных и мододых особей.
При подробном исследовании размерного состава полово-
зрелой части популяции оказалось, что минимальные размеры
половозрелых особей отмечены в июле в заливе, хотя макси-
мальные размеры везде близки (табл. 7). К осени минималь-
ные и средние размеры Партеногенетических самок везде ниже,
чем летом. Максимальные размеры эфиппиальных самок всегда
ниже, чем партеногенетических. Эта особенность присуща не-
которым другим видам дафнид (Ривьер, 19726) и определяется
тем, что самые крупные* самки в популяции размножаются пре-
имущественно партеногенетическим способом и в конце своей
жизни вообще не откладывают эфиппии. Судя по данным, прк-
235
Таблица в
Средние размеры Daphia cucullata,
в популяциях, мм
Сроки отбора проб Иваньковское водохранилище Рыбинское водо- хранилище
Район Д. Городищи Мошкович- ский за- лив 1969 г. 1870 г.
1868 г. 1868 г. 1868 г.
5-10 V - 0.54 - - *
1-5 Y1 0.75 - 0.64 - 0.72
6-15 VI 0.74 - 1.10. - 0.75
16-20 VI - - * * 0.88
21-30 VI - - * - 0.86
1-10 VII 0.97 - 0.84 0.91 -
11-20 VII — 0.86 1.10 0.90 -
21-30 УП 0.83 - * 1.05 -
1-15 У1П - - - - -
16-30 УШ 0.92 0.85 . 0.83 - -
1-10 IX - - - -
11-30 IX 0.80 0.80 1.17 0.88 -
веденным в табл. 7, существенных отклонений в размерах по-
ловозрелой части популяции в зоне воздействия теплых вод не
прослеживается, лишь в начале лета партеногенетические особи
несколько мельче, чем в естественных условиях. Видимо, в
начале лета в подогреваемых участках полово^ созревание про-
исходит при меньших размерах, чем в необогреваемых.
Имея данные по размерному и половому составу (коли-
чество половозрелых, яйценосных самок, количество молоди и
новорожденных рачков) И вычислив средний размер особей в
популяциях, мы попытались сравнить популяции D. cucullata,
обитающие в разных условиях. Если рост молоди сходен и
наступление половой зрелости происходит при одних и тех же
296
Т а б лица 7
Размеры партеногенетических и эфиппиальных самок
Daphnia cucullata, мм, в Иваньковском
водохранилище в 1868 г., мм
Месяц Мошковичский залив
В1 В В2 С1 С с2
Июль 0.85 1.15 1.5 0 0 ! о
Август 1.0 1.21 1.53 0 0 1 0
Сентябрь .... 0.8 1.03 1.4 0.85 1 1.05 1 1.27
Водоз а бор
Июль 11.03 | 1.17 1 1.5 1 0 1 0 1 0
Август 10.05 1 1.21 I 1.5 1 ° | 0 1 °
Сентябрь .... .10.85 1 .0.8 1 1,4 | 0.73 0.951 1.2
Район г. Городищи
Месяц В1 в в2 С1 С 1 С2 TV
Июль 1.0 1.18 1.4 0 э 0 1500
Август 1.05 1.17 1.45 0 0 0 2100
Сентябрь .... 0 0 0 1.0 1.05 1.2 1500
Примечание. В}, в « В2 - > лииимал ьный, средний и макси-
мальный размеры партеногенетических самок; cj, С -и С2 - со-
ответственные размеры эфиппиальных самок.
размерах, то популяции, имеющие одинаковую структуру, долж-
ны характеризоваться одинаковым средним размером особей.
Были произведены многочисленные (48) наблюдения за по-
пуляциями в природе. В каждом отдельном случае популяции
отличаются 6 какой-то степени структурой и имеют различные
средние размеры особей, так что сравнивать их очень трудно.
Средний размер особей данной популяции зависит, естественно^
от структуры. С увеличением количества половозрелых особей
он возрастает, а с увеличением количеотва новорожденных -
уменьшается.
Коэффициент корреляции (г ) для данных по Иваньковско-
- му водохранилищу, вычисленный по среднему размеру в коли-
честву половозрелых рачков (в %), равен 0.86, вычисленный по
среднему размеру и количеству новорожденных,- 0.93.
2ЭТ
Для Кураховского, Иваньковского и Рыбинского водохрани-
лищ были вычислены уравнения линий регрессии (рис. 7, 8).
При сравнении популяций D. cucullata из южного водое-
ма (Кураховское водохранилище, по данным М.Ф. Полива иной и
О.А.Сергеевой, 1971) с нашими по зависимости среднего раз-
мера особей от доли половозрелых самок в популяции оказа-
лось, что эти популяции существенно различны. Особенно отли-
4--1__I_I__I__I--1 ।_____
0.6 0.6 1.0 1.2Х.ИН
Рис. 7. Зависимость
среднего размера особей
от количества полово
зрелых самок в различ-
ных популяциях.
1 - Кураховский
водоем (у, “ -129.7+
+226x4); - Иваньковс-
кое водохранилище (yt я
• -56+102хг); 3 - Ры-
бинское водохранилище
(у4 • -38.6+82 Xj ). По
оси ординат - колич.
половозр. особей; по
оси абсцисс - ср. раз-
мер особей.
Рис. 8. Зависимость
среднего размера осо-
бей от количества ново-
рожденных в различных
популяциях.
1 - район водоза-
борного канала ( =
ж 8I-57X4); 2 - Мэшко-
вичский залив ( уа 106-
89 Ха ); 3 - район разр.
IX (у,- 130-116 ®s).
По оси ординат - колич.
новорожд. особей; по
оси абсцисс — ср, раз-
мер особей.
чается популяция D. cucullata из Кураховского водоема
(рис, 7), где по сравнению с Иваньковским и Рыбинским водо-
хранилищами при одном и том же количестве яйценосных
самок средний размер особей значительно ниже. Это, естест-
венно, свизано с более ранним наступлением половозрелости
при более высоких температурах.
238
Рис. 0. Поло-
возрелые особи и
молодь Daphnia
cucullata из од-
ной пробы, взятой
1 июня 1969 г.
близ Корчевы (Ивань-
ковское водохрани-
лище).
Мы пытались сравнить популяции D, cucullata, обитающие
в заливе, в эоне сильного воздействия теплых вод, в зоне слабого
воздействия их у разреза IX и при естественной температуре. В
данном случае уравнения были получены по зависимости среднего
размера особей от количества новорожденных рачков в популяциях.
Оказалось, что эти три популяции несколько различны, ио досто-
верно различаются они только до пересечения прямых (рис. 8). При
одном и том же количестве новорожденных рачков у популяций в
подогреваемых участках средние размеры особей выше (дафнии луч-
ше растут), чем в иеобогреваемых. Но подобная зависимость отме-
чается только до достижения в популяции средних размеров особей
0.76-0.8 мм, что наблюдается лишь в мае-начале июни (табл. 6).
В июле-августе картина становится обратной. Однако различия здесь
мало достоверны. Можно лишь предположить, что летний перегрев
тормозит рост особей в зоне воздействия теплых вод.
Постоянная жизнь популяции при повышенной- температуре
воды должна, казалось бы, отразиться иа морфологических при-
знаках пелагических форм. У дафний это скорее всего должно
сказаться на цикломорфозе. Были произведены многочисленные
промеры D. cucullata из зоны сильного воздействия теплых вод
239
Таблица 8 Относительная высота шлема у Daphnia cucullata, мм Рыбинское водохранилище яйце- носные со со со ** , | С? О О о о 900 экз,.
tfi а о о 2 g $ 3« 1 1 Q О ° о О
со сч ^2*0 о ’ ' * * 2 8 -
дата 9 июня 24 июня 14 июля И августа 22 сентября
Иваньковское водохранилище | яйце- носные 1- оо т со м сч оз ч; ч; o’ । । о о о о е Количество обследо- ванных особей 950 экэ.
молодь СО СО — Ч* со со t~ сч сч ч ч ч ч оз о о о о о о о
4* <0 03 СО СЗ из о 03 СО ХГ СО ч* ч* со >- —< •— сч сч •—
дата *« а сЛ& « К И tB S д « « ® 2 2 Д Ф 2 S 2 s и « о 2 2 » Г- СЧ Г- г~ Г- >- -X сч сч
Пробы из Мэшковичского залива
240
и из района с естественным температурным режимом. Для
сравнения были взяты D. cucullata из Волжского плеса Ры-
бинского водохранилища. Вычислялась относительная высота
шлема (рис. 9).
Высота головы до конца ростиума (К)
общая длина тела без спины (L )
Оказалось, что у молоди, обитающей при температуре в
естественных условиях 9°С и при температуре 19.5°С в Мош-
ковичском заливе, высота шлема одинакова (табл. 8). Макси-
мальная высота шлема, когда „голова' (Н) у рачка составляет
половину длины тела, отмечена в Рыбинском водохранилище
в июле при температуре всего 18-21 °C. Никаких отклонений в
проявлениях дикломорфоэа у дафний из подогреваемых районов
не обнаружено. Видимо, это служит лишним доказательством
того, что цикломорфные изменения не обусловливаются темпе-
ратурным фактором.
В процессе исследований выявилась особенность, прису-
щая Daphnia cucullata, а именно неравномерность роста
шлема в течение жизни, его преимущественный рост у молодо-
го рачка. У дафний как из Рыбинского, так и из Иваньковско-
го водохранилища (табл. 8, рис. 9) шлем у взрослых, полово-
зрелых особей относительно ниже, чем у молоди. Эта особен-
ность наиболее проявляется весной, в начале июня, когда
размножаются еше только самки, появившиеся из эфиппиев, а
молодь первого помета не достигла половой зрелости. На
рис. 9 представлены рачки из одной пробы. У молоди шлем в
полтора раза выше, чем у яйценосных самок. Увеличение вы-
соты шлема, по-видимому, происходит до наступления половой
зрелости. У взрослых самок он сохраняет свою высоту и форму,
хотя рачок продолжает линять при отрождении молоди и расти
уже при более высокой температуре воды.
Заключение
Приход дополнительного тепла оказывает большое влияние
на зоопланктонное сообщество близлежащих районов Пеньков-
ского водохранилиша. Изменяются сложившиеся взаимоотноше-
ния организмов со средой. Несомненно, некоторые более эври*
топные и эвритермичные виды, (например, Chydorus sphae-
ricus, Bosmina longirostris) лучше переносят изменившие-
ся условия и даже увеличивают свою численность, другие же,
более чувствительные к новым условиям, пелагические (на-
778 16
241
пример, Bythotrephes longimanus, Leptodora kindtii,
Daphnia longispina) ж стенотермичные (Cyclops vici-
nus, весенние коловратки Synchaeta и Asplanchna) ви-
ды отрицательно реагируют на дополнительное тепло.
Изменения в видовом составе сейчас проявились главным
образом в исчезновении отдельных видов лишь из зоны силь-
ного воздействия теплых вод и в нарушении естественного со-
отношения некоторых видов. В дальнейшем при длительном воз-
действии повышенной температуры и увеличившихся скоростей
течения эти изменения, постепенно накапливаясь, по-видимому,
приведут к еше большим нарушениям в сообшестве зоопланк-
тона.
Механическое воздействие охладительной системы ГРЭС
при одновременном воздействии температурного фактора наибо-
лее разрушительно сказьвается на крупных пелагических рачках,
самых ценных в кормовом отношении для рыб. При 26°C рач-
ковый планктон иа этом пути гибнет на 34.5%, при 33-32° -
на 53.3%.
Зона наибольшего воздействия сбросных вод (Мошковинс-
кий залив) мало благоприятна для развития пелагического
планктона, как следствие высокой температуры в потоке ле-
том, так и вследствие быстрого течения. Более благоприятен
для обитания придонный слой, сохраняющий естественную тем-
пературу, где скапливаются мигрирующие из теплого потока
рачки. В последние годы после запуска очередных агрегатов
ГРЭС и увеличения забора воды придонный холодный слой поч-
ти исчез, располагаясь только вблизи устья залива.
Сильнее всего воздействие повышенной температуры ска-
зывается летом. При Этом на температурный режим значитель-
но влияют метеорологические условия. В наиболее жаркие перио-
ды лета при установлении пороговой температуры и выше (29-
33°С) наблюдается массовое травмирование животных уже в
агрегатах ГРЭС. При этом теплый поток вплоть до выхода из
залива оказывается чрезвычайно бедным водным населением
(биомассы 0.05-0.1 г/м®).
При температуре в потоке до 25-26°С биомасса в заливе -
зоне наибольшего влияния теплых вод - падает не столь низко,
а в зоне слабого воздействия - разрезы IX, X и IV - наблю-
дается даже повышение биомассы зоопланктона.
Изучение самого массового вида ветвистоусых рачков
Daphnia cucullata показало, что значительных отклонений
в някломорфозе, в размерах партеногенетических и эфиппиаль-
ных самок не наблюдается. Лишь весной в зоне сильного воз-,
действия рачки лучше растут н быстрее достигают половой зре-
лости* Летом отмечаются значительные нарушения в структуре
242
популяции, происходящие вследствие исчезновения молоди, вы-
носа ее теплым потоком, а в придонном слое и вследствие
выедания циклопами.
Литература
Литвинов А.С. и Ривьер И.К. 1972. О вертикаль-
ном распределении зоопланктона в Мошковичском зали-
ве в связи о особенностями гидрологических условий.
Информ, бюлл. Инет. биол. внутр, вод АН СССР, 13.
Мирошниченко А.З. 1971. Зоопланктон водоема-охла-
дителя Змиевской ГРЭС. - В кн.: Гидрохим. и гидро-
биол. водоемов-охладителей ТЭС СССР. Киев.
Мордухай-Болтовской Ф»Д. 1957. Развитие фау-
ны беспозвоночных в нерестово-вырастных водоемах
на Дону в связи с выращиванием в них молоди рыб.
Тр. Совет, по рыбоводству. М.
Поливанная М>Ф. и Сергеева О.А. 1971а. К
биологии массовых видов Cladocera водоемат-охлади-
теля Кураховской ГРЭС. Гидробиол. журн., VII, 5.
Поливанная М.Ф. и Сергеева О.А. 19716.
Зоопланктон водоемов-охладителей электростанций
Украины, - В кн.: Гидрохимии гидробиол. водоемов-ох-
ладителей ТЭС СССР. Киев.
Ривьер И.К. 1968. Влияние сбросных теплых вод Кона-
ковской ТЭС на зоопланктон Иваньковского водохра-
нилища. - В кн.: Волга-Т. 1 конфер. по изуч. водоемов
басе. Волги. Тезисы докл, Тольятти.
Ривьер И.К. 1971. Зоопланктон Мошковичского залива,
подверженного влиянию подогретых сбросных вод Ко-
наковской ГРЭС. Симпоз. по влиян. подогрет, вод
ТЭС... Борок.
Ривьер И.К. 1972а. Поведенческие реакции планктонных
рачков в зоне наибольшего влияния сбросных вод Ко-
наковской ГРЭС. — В кн.: Поведение водных беспозв.
Борок.
Ривьер И.К. 19726. Особенности структуры популяций
Daphnia pulex во временных водоемах. Иифор.бюлл.
И нет.би о л. внутр, вод АН СССР, 13.
Ривьер И.К. и Ген Н.А. 1971. Зоопланктон Мошкович-
ского залива, подогреваемого сбросными водами Кона-
ковской ГРЭС. Информ.бюлл. Инст.биол.внутр.вод АН
СССР, 12.
243
Т.С.Е л а г и н а
Зоопланктон горьковского водохранилища
в районе костромской ГРЭС
Впервые гидробиологические съемки на Горьковском водо-
хранилище в районе Костромской ГРЭС были проведены отде-
лом изысканий ГОТЭПа в 1964 г., а затем в период пуска пер-
вых энергоблоков в июне-октябре 1969 г. Основное внимание
в этих работах уделялось вопросу обрастаний гидротехнических
сооружений старции. Специальное изучение зоопланктона данно-
го района Горьковского водохранилища было начато весной
1970 г. и проводилось на протяжении вегетационных сезонов
1970 и 1971 гг. Зимние и ранневесенние съемки были сделаны
лишь в 1971 г.
Сбор проб осуществлялся в 1970 г. через каждые де-
сять дней, а в 1971 г. - два раза в месяц с двух горизон-
тов (поверхностного и придонного) на Четырех створах
(разрезах), каждый с двумя или тремя станциями, - в во-
дохранилище и на 11 станциях в водозаборном канале
и заливе - в устье р. Шачи (рис. 1). Пробы отбирали
планктоночерпателем системы Вовка, а также видоизменен-
ным планктобатометром системы Дьяченко (оба - объе-
мом 10 л). Подсчет организмов производили в рэндом-каме-
ре, при этом делали промеры животных, отмечали наличие
науплиальных и копеподитных стадий у веслоногих и моло-
ди у ветвистоусых. Всего собрано и обработано в 1970 г.-
400, а в 1971 г. - 36 проб зоопланктона.
Собранный материал позволил составить подробную
характеристику зоопланктона исследуемого района водохра—
244
Рис. 1. Схема
расположения станций.
1 - станции отбо-
ра проб зоопланктона
в 1870 г., 2 - станции
отбора проб зоопланк-
тона в 1970 и 1971 гг.
нилища и проследить динамику его развития в различных
по термическому режиму зонах.
Общая характеристика
зоопланктона
По сообщению Е.Б.Каневской, в 1969 г. видовой состав
зоопланктона исследуемого района водохрацилища был представ-
лен 57 видами, относящимися к Cladocera, Copepoda и
Rotatoria. Наши сборы позволили расширить этот список до
65 видов: Cladocera - 26, Copepoda - 15, Rotatoria -
24 вида. Список всех видов приводится в приложении. Следует,
однако, иметь в виду, что это, собственно, список не планктон-
ных видов, а видов копепод, кладоцер и коловраток, найденных
в планктонных пробах, в которых, как правило, встречается
примесь и фитофипьных "и бентических форм (из копенод - виды
Macrocyclops, Paracyclops; из кладоцер - Ilyocryptus,
Leydigia, виды Pleuroxus, Alona, из коловраток - виды
Lecane к др.).
Доминировали на протяжении всего периода исследований
ракообразные, составлявшие в среднем 74.3% суммарной числен-
ности и 95.2% обшей биомассы. К числу руководящих видов от-
носились: Bosmina longirostris, В. coregoni, Daphnia
345
cucullata, D, longispina, Eudiaptomus gracilis, E, gra-
ciloides, Meso cyclops leuckarti, Cyclops vicinus.
Средн коловраток наиболее часто встречались: Asplanchna
priodonta, Keratella quadrata, Kellicottia longispina,
Polyarthra vulgaris и представители сем. Brachionidae.
Соотношение численности и биомассы массовых видов приведе-
но в табл. 1 и 2.
Зоопланктон водосбросного канала и приустьевого залива
р. Шачи представлен в основном теми же видами, что и в рус-
ловой части водохранилища. Вследствие недавно проведенных
дноуглубительных работ водная растительность в этом районе
в 1870-1971 гг. отсутствовала, что исключало массовое разви-
тие таких форм, как Polyphemus pediculus, Ceriodaph-
nia reticulata и представителей сем. Chydoridae, густо
населяющих прибрежную зону водохранилища.
Как видно из данных табл. 1 и 2, зоопланктон в различ-
ных участках исследуемого водоема характеризуется относитель-
но близкими средне сезонными показателями численности и био-
массы.
Сезонная динамика
зоопланктона
Наши наблюдения позволили проследить сезонные измене- .
иия количественных показателей зоопланктона в участках с
естественными температурными условиями и в подогреваемой
зоне водоема.
1. Русловая часть водохранилища: неподогреваемая зона
(створы 1, II и водозабор) и зона слабого подогрева (створ
Ш). В 1870 г. отбор контрольных проб в русловой части водо-
хранилища выше сброса подогретых вод осуществлялся на 3
станциях II створа, а в 1971 г. - на 2 станциях 1 створа.
В мае 1870 г. средняя температура воды в зоне естест-
ве иного температурного режима составляла 11 °C, а в зоне сла-
бого подогрева была лишь на 0.7°С выше. В этот период для
зоопланктона П створа'характерно преобладание коловраток,
составлявших 65% общёй численности. Среди них наиболее мно-
гочисленны были Asplanchna priodonta, Keratella qua-
drats, Brachionus calyciflorus. Общая биомасса зоопланк-
тона на атом участке была несколько ниже, чем в зоне слабо-
го подогрева, где биомасса на 81% создавалась веслоногими
рачками. Присутствие в планктоне III створа половозрелых осо-
348
Численность и биомасса зоопланктона и основных видов (% от общей)
h районе Костромской ГРЭС в 1970 г. (средние за вегетационный период)
3 -S Ф с 13 £ СМ СЧ -« — ~ ~ о d о do
ф з * а О СЧ О СО СЧ d ю со d о
£ С а . о СЧ со О со со со ~ сч со
AspL chna priod ta сч оо — ~ СО СО СЧ со
ipto- gra- E. loi- см О О Г; О IO ю Я о —' —' — OS
Eudie mus cilis, graci des *1 о со со со d со
Mesocyc- lops leu- ckarti см СО Г; О СО СО СО со со СО со
(0 сч св о СО ЧГ «5 * 03
Bosmina longirost- ris, B. coregoni сч СО О СО о о • • • • • сч со со ю о Ч* ’Г СО ч* сч
Q О <0 О О • • а ее Ч* 05 ЧГ О Ю СЧ $м сч со сч
si c <o сч •* О (0 -* 00 СО со со чг
Daph longj na ем О со п о со сч сч сч со ю
inia Llata сч чг о о сч с- а ю сч со ю
— 3 в 2 Q u *7 ‘Q ^5 со со
о ® о O Ci и со со ср — Г- СЧ 0В V со • • • • о — о
Весь г планкч , ГН <о «э е * * а -I а со со » ю 55 и «
Станции а«8 S 5 £ 2 а со о £ 9 2> * о ® г*» 6 -« сП г сл§ <5^ эг§ 5-23 и
Примечание. В таблицах 1 и 2 : 1 - численность (тыс. экз./м^), 2 - биомасса (г/м^
ф
н
ю
о
о
&
ф
к
г* м
3
* X
о
X
я
h
ф
L
Ф
Я
<0
я
ф
X
к
а
ф
о,
и
ffl
о
д
X
Л
X
Я
о
X
и
о
о
U
ф
СЧ
Я
л
к
ю
su
о ~
S2
! °
§О
„£
« U
и
о «
Q О
3 х
0 й
х 2
ю £
X ь
о
££
о •*•
|£
g«
h
у ®
о я I - - - - 8 N | d о о do
1 О со со СО - 1 СЧ О 00 ~ О
i о <2 « -D , 1 °. Ч ч 1 ** ** сч S СО
CL С Q <00.5 ’ - 1 О Ю О \р — 1 ** СО ci to
1 .2 о m < 1 о ~ °2 сч — ч I со со d <о со
з E 8 003 Cd ° uo Id ° " , 1 СО СО ОЗ СО тч 1 to сч со со
° J >» Й 0 <0 c о x 1 О СО со СО СО * 1 00 СО Г* J
It jot - 1 «0 оо X °0 « 1 оо оо со ~
c A 01 •g oo E fi) 1 *• *2 ® °Q 1 ОЗ £ о ~ ТГ
tn c 0 0 -s - (B 0 c оз сч со СЧ СО 1 СО ОЗ 00 со
M <0 W .£ 0 ci сч СЧ О о со со сч о ТГ сч I сч сч со сч сч
c .2 и -2 b I " Я 1 о С5 —< г- из 1 СЧ — сч — — .
<0 Lie N Г со со О со со 1 сч to сч U
& c-s 0 ° и I тг со г* оз со 1 *4 сч ~ *4
X 3 1 *7 °! Ч °i со -X СО чг
S-O ® 3 M — QoS 1 О 'Ч* со о Г; 1 о to
О Д О о сч <9 ь 1 Ю со О ОЗ СЧ I СО 00 г- со 1 о о о о о
Весь плат 1 1 I °° СМ V о 1 СО 00 СО со со СЧ СО СО СО
Станции а — о " g д । о «о °”a ® g 1 8 т 1*0 со ь s о о «зо Г М о с цо о ® g а г о из и в я а. • о а • вою Л ев —। вою (• в ю ь — —во О й я в о О — о О
бей относительно теплолюбивого Mesocyclops leuckarti
и значительного количества копеподитных стадий Cyclopoida,
видимо, объясняется выносом зоопланктона в этот участок
водохранилища из залива р. Шачи, где развитие копепод начи-
нается несколько раньше. И.К,Ривьер (1968) отмечала, что в
теплых водах Мошковичского залива Иваньковского водохрани-
лища в' районе Конаковской ГРЭС развитие весенних форм
планктонных организмов начинается значительно раньше, чем
в не подогревав мой зоне.
Майский зоопланктон 1971 г. на всем исследуемом участке
водохранилища характеризовался преобладанием веслоногих рач-
ков, но если выше водосброса (I створ) при температуре воды
9.3°С массовыми видами были Cyclops vicinus и Cyclops
strenuus (69% от общей численности), то в зоне влияния
подогретых вод при температуре 11.8°С - Mesocyclops leu-
ckarti и ювенильные стадии Copepoda (74% от общего чис-
ла зоопланктеров).
В первой половине июня 1970 г, начинается массовое
размножение ветвистоусых рачков, более интенсивное в районе
ниже водосброса (створы III и IV), где биомасса зоопланктона
была выше (рис. 2). Особенно многочисленны здесь были
Daphnia cucullata и Bosmina longirostris (65% от об-
щей биомассы). В зоопланктоне неподогреваемой зоны водохра-
нилища коловратки по-прежнему оставались руководящей труп-'
пой, хотя численность их несколько снизилась (до 45%) в ре-
зультате увеличения числа ветвистоусых; общие количествен-
ные показатели мало Изменились.
В июне 1971 г. среднемесячная температура воды была
на 1.5-2.0°С выше, чем в предыдущем году. В этот период био-
масса зоопланктона на 68-70% (I и III- створы соответственно)
создавалась ветвистоусыми рачками. В неподогреваемой зоне
водохранилища доминировали в основном Bosmina longirost-
ris и В, coregoni, а в зоне слабого подогрева ведущее
положение занимала Daphnia cucullata. В результате
массового развития этого рачкд количественные показатели
зоопланктона здесь достигают к концу июня наивысшего за се-
зон значения: биомасса - 1.93 г/мЗ и численность - 70.1 тыс.
экз./мЗ (рис. 3).
В июне 1970 г. максимум обилия зоопланктона наблюдал-
ся при среднемесячной температуре воды 20-’22°С; он опреде-
лялся главным образом Daphnia cucullata и Bosmina
longirostris (72.5% от обшей биомассы). Биомасса на III ство-
ре достигала 1.71 г/м3, а на II створе - 1.41 г/мЗ (рис. 2).
Июльский зоопланктон 1971 г. представлен теми же видами,
что и в предыдущем году, ио. в неподогреваемой и слабоподогре-
249
ваемой зонах количественно он был значительно беднее: сред-
няя биомасса в 4.5 раза, а численность в 3 раза ниже, чем в
1970 is По сравнению с июнем значительно снизилось количе-
ство Bosmina coregoni, В, longirostris, Daphnia Ion-
gispina и D. cucullata, но в неподогреваемой зоне водо-
хранилища последний вид остается столь же многочисленным.
г/чг
—1—1—।—।__। ।
v п п т а х
Рис. 2. Динамика
биомассы зоопланктона
в русловой части водо-
хранилиша в 1970 г.
1 - створ II (зо-
на естественного тем-
пературного режима),
2 - створ III (зона по-
догрева).
—।—।—<-----1-1__и
V V! VII пи и X
Рис. 3. Динамика
биомассы зоопланктона
в русловой части водо-
хранилиша в 1971 г.
1 - створ I (зона
естественного темпера-
турного режима), 2 -
створ BI (эона подогре-
ва).
В августе 1970 г. зоопланктон на различных по термичес-
кому режиму участках водохранилиша характеризовался одними
и теми же доминирующими формами: Daphnia cucullata
(40% от обшей численности) и Eudiaptomus gracilis и е.
graciloides (45% от обшей численности).
В августе 1971 г. зоопланктон был представлен теми же
видами, что и в предыдущем- году, численность их в зоне сла-
бого подогрева (Ш и IV створы) была значительно меньше,
чем в зоне естественного температурного режима (1 створ):
Daphnia cucuUata - в 1.5 раза, Eudiaptomus gracilis
к Е. graciloides - в 1,7 раза. Естественно, что и обшая
численность и биомасса зоопланктона' в районе подогрева была
меньше. Уменьшение численности этих рачков, возможно, явля-
ется результатом выедания „мирного' планктона рыбами и хищ-
ными ракообразными. Косвенно подтверждает это тот факт, что
численность популяции Mesocyclops leuckartt в августе
1970 г, повысилась в 2 раза, а в 1971 г. - в 3.8 раза.
250
Осенью (сентябрь и октябрь) динамика численности и
биомассы зоопланктона в оба года исследования одинакова,
В первой половине сентября с понижением температуры воды
до 13-14°С наблюдался подъем в развитии ветвистоусых рач-
ков Bosmina longirostris и В, coregoni (79% от обшей
биомассы), продолжавшийся в зоне подогрева до конца сентяб-
ря.
В октябре 1870 г. на различных по термическому режиму
участках водохранилища распределение зоопланктона было равно-
мерным, численность колебалась в пределах 16.6-19.6 тыс.
экз,/м3. В октябре 1971 г» как и в 1970 г.,'ведущее положе-
ние занимали ветвистоусые рачки Bosmina longirostris
(59% от общей биомассы), а более высокая численность зоо-
планктона была отмечена в слабоподогреваемой зоне водохра-
нилиша - 31.0 тыс. экз./м^.
2. Водосбросный канал и приустьевой залив р. Шачи (зо-
на значительного подогрева). Вода, забираемая насосами
ГРЭС из водохранилища, сбрасывается в залив р. Шачи, подо-
гретая в среднем на 5-8°С. При прохождении через о’хладитель-
ную систему станции и .водопад' (порог на водосбросе) проис-
ходит травмирование крупных, наиболее ценных в кормовом
отношении планктонных организмов: Leptodora kindtii,
взрослых Daphnia и Cyclopoida, что часто вызывает их -
гибель. Это явление отмечалось на Иваньковском водохранили-
ше в-районе Конаковской ГРЭС (Ривьер, 1968, 1971) и в во-
доемах-охладителях тепловых электростанций юга Украины
(Поливанная, Сергеева, 1971).
В январе 1971 г. температура воды в сбросном канале
превышала естественную на 8.3°С. В этот период наибольшее
количество зоопланктона обнаружено в незамерзающей части
залива (сТ. 8} - 3.22 тыс. экз./мЗ. В основном здесь присут-
ствовали копеподитные стадии веслоногих и коловратки, из
которых наиболее массовым видом была Keratella quadrate.
В марте при. температуре 8.1°С общая численность планк-
тонных организмов в подогреваемой зоне залива увеличилась
в 2 раза. Доминирующими видами были Cyclops vicirius,
Eudiaptomus gracilis и Keratella quadrate. Наиболее
высокая биомасса - 0.09 г/м^ - была на ст. 8 (на 67% за
счет веслоногих).
Майский зоопланктон сбросного канала и залива за два
года наблюдений состоял преимущественно из копепод и коло-
враток; доминирующее положение занимали: Mesocyclops
leuckarti, Cyclops vicinus и Asplanchna priodonta.
При этом количество зоопланктона на этом участке водохрани-
лища превышало его в районе водозабора в 2 раза.
251
Рис. 4. Динамика
численности Daphnia
cucullata в 1971 г. у
водозабора (ст. 3) и в
устьевом заливе р. Ша-
чи (ст. 4).
1 - ст. 3 (зона ес-
тественного температур-
ного режима), 2 - ст. 4
(зона подогрева).
Рис. 5. Динамика
биомассы зоопланктона
и ход температуры на
ст. 4 (ниже водосбросно-
го канала) в 1970 г.
1 - биомасса, 2 -
температура.
Г 'И VU VIII IX’ X'
Рис. 6. Динамика биомас-
сы зоопланктона и ход темпе-
ратуры на ст. 4 (ниже водо-
сбросного канала) в 1971 г.
1 - биомасса, 2 - темпера-
тура.
В конце мая-начале июня начинается интенсивное разви-
тие ветвистоусых рачков Bosmina longirostris, В. core-
goni Daphnia cucullata. Характерной особенностью лет-
него планктона подогреваемой зоны залива является более ран-
нее размножение Daphnia cucullata, численность которой
на ст. 4 в несколько раз выше, чем у водозабора (рис. 4). За
счет этого в июне на ст.' 4 наблюдалась максимальная биомас-
сы 8.85 г/м3 в 1970 г. и 1.58 г/м3 в 1971 г. ( рис. 5,8). ’
252
В июле 1970 г. температура сбрасываемой воды достигла
26°С, а в 1971 г. - 28.9°С._ По всей видимости, именно высо-.
кая температура воды привела к резкому уменьшению количест-
ва зоопланктона. В оба года его биомасса в июле сильно пони-
зилась (рнс. 5, 6). В июле 1971 г. на ст. 4 численность Bos-
mina longirostris снизилась в 16 раз, Daphnia cuculla-
ta - в 2 раза, совсем исчезли D, longispina и D, crista-
te, многие коловраткй, Основная масса зоопланктона была
сосредоточена в придонных слоях (табл. 3). В этот период
количество травмированных организмов в планктоне сбросного
канала было наиболее высоким и достигало 50%.
Таблица 3
Вертикальное распределение зоопланктона (тыс. экз./м^)
в районе Костромской ГРЭС 28 июля 1971 г.
‘Зона Горизонт лова t°C Clado- cera Cope- pod а Rota- toria Всего
1 i 1 1 1 и иида ч. подогрева (ст. 4) Поверх- ность Придонный слой 28.9 25.7 5.07 5.88 13.86 25.80 5.87 20.72 24.80 52.40
S с 0 9 < венного тем- пературного режима (ст. 3) Поверх- ность Придонный слой 20.1 19.4 6.37 2.15 15.49 9.31 7.29 1.41 30.16 12.87
Августовский зооплаиктои 1970 г. в зоне значительного
подогрева (при 24°С) был представлен преимущественно (66%
биомассы) веслоногими рачками Mesocyclops leuckarti,
Eudiaptomus gracilis и E. graciloides и их ювенильны-
ми стадиями. Численность и -биомасса зоопланктона понизились
еше больше. Наибольшее количество зоопланктона (62.96 тыс.)
наблюдалось на ст, 8, так как здесь наряду с вышеуказанны-
ми видами присутствовали Daphnia cucullata « Bosmina
longirostris (30% от обшей биомассы).' В 1971 г. в августе
биомасса зоопланктона повысилась в связи с возрастанием чис
ла ветвистоусых рачков, численность которых на ст, 4 по
сравнению с предыдущим годом увеличилась в 4.1 раза, а на
ст. в - в 2 раза.
253
В сентябре 1070 и 1971 гг, зоопланктон на различных
участках водоема характеризовался единством руководящих
форм и относительно близкими показателями численности и
биомассы. Но в зоне значительного подогрева наряду с рачка-
ми Bosmina longirostris и В. coregoni постоянным
компонентом зоопланктона была Daphnia cucullata.
В октябре 1871 г. температура сбрасываемой воды равня-
лась 13.8°С, что на 7.2°С превышало естественную. На всех
станциях преобладали ветвистоусые Bosmina coregoni и
В. longirostris (87.8% от обшей численности), но их числен-
ность в заливе была в 5-6 раз выше, чем у водозабора.
-Заключение
В составе зоопланктона исследованного участка Горьковс-
кого водохранилиша в районе Костромской ГРЭС - преобладают
виды эврибионтные, широко распространенные в водоемах евро-
пейской части СССР. Доминирующее положение занимают рако-
образные: Bosmina longirostris, В. coregoni. Daphnia
cucullata, Mesocyclops leuckarti, Cyclops vicinus,
C. strenuus, Eudiaptomus gracilis, E. graciloides.
Исследования динамики численности и биомассы зоопланктона
показали, что сброс подогретых вод Костромской ГРЭС оказы-
вает на него ощутимое влияние в устьевом заливе р. Шачи,
где температура воды превышает естественную в среднем иа
5-8°С.
Многие ракообразные при прохождении через охладитель-
ную систему станции и „водопад' травмируются или погибают.
Особенно страдают крупные формы: Leptodora kindtii, Da-
phnia и взрослые Cyclopoida.
В холодное время года повышение температуры воды в
заливе и сбросном канале способствует развитию зоопланкто-
на. Зимой 1971 г, в незамерзающей части залива численность
зоопланктона была в 2-2,5 pash выше, чем в водозаборном
канале. Весной в зоне подогрева массовое развитие планктон-
ных организмов начинается значительно раньше. Но повышение
температуры сбрасываемой воды до 26-28.9°С вызывает у них
отрицательную реакцию; общее количество организмов умень-
шается и основная масса зоопланктона сосредоточивается в
придонных, более холодных слоях. В осенние месяцы развитие
Зоопланктона в районе водосброса н залива ааканчнвается
позднее, чем в естественных условиях.
254
В русловой части водохранилища повышение температуры
воды ниже водосброса на 111 створе было небольшим (0.7-1.5°С)
и не оказывало значительного влияния на зоопланктон этого
участка. Однако здесь все же замечаются некоторые отличия:
появляются сравнительно более теплолюбивые копеподы в ве-
сеннее время, раньше достигают максимума количественные
показатели (1971 г.), удлиняется вегетационный период. Это,
видимо, объясняется сносом зоопланктона в этот район водо-
хранилища из залива р. Шачи.
Приложение
Список видов,
встреченных в планктонных пробах
в районе Костромской ГРЭС
Подкласс Copepoda
Отр. Calanoida
1. Eudiaptomus gracilis
Sars.
2. E. graciloides Lili.
3. Heterocope appendicu-
late Sars.
Отр. cyclopoida
1, Acanthocyclops bicus-
pidatus (Claus)
2. A, gigas (Claus)
3, A, viridis (Jurine)
4. Cyclops strenuus
Fisch.
5. C. vicinus Uljan.
6. C, lacustris Sars.
?. Mesocyclops leuckarti
Claus.
в. M. oithonoides Sars.
9. Macrocyclops albidus
(Jurine)
10. M. fuscus (Jurine)
11. Eucyclops serrulatus
(Fisch.)
12. Paracyclops fimbriatus
(Fisch.)
Подкласс Branchiopoda
Отр. Cladocera
1. Diaphanosoma brachyu-
rum (Lievin)
2. Limnosida frontosa
Sars.
3. Sida crystallina (O.F.
Muller)
4. Ceriodaphnia pulchella
Sars.
5. C. quadrangula (O.F.
Muller)
6. C. reticulata Jurine
?. C« affinis Lilljeborg
8. Daphnia longispina
O.F.Muller
9. D. cucullata Sars.
10. D. cristata Sars.
11. D. pulex (De Geer)
12. Simocephalus vetulus
(O.F.Muller)
13. Bosmina coregoni
(Baird)
14. B. longirostris (O.F.
Muller)
15. Ilyocryptus sp.
16. Alona quadrangularis
( O.F.Muller)
17. A, rectangula Bars.
18. A. affinis Leydig.
19. Chydorus sphaericus
(O.F,Muller)
20. Ch. globosus Baird.
21. Leydigia leydigii
(Leydig)
22. Pleuroxus aduncus
(jurine)
23. P, striatus Schoed-
ler
24. Polyphemus pediculus
(Linne)
25. Bythotrephes longi-
manus Leydig.
26. Leptodora kindtii
( Focke)
Kn; Rotatoria
1. Asplanchna priodonta
Gosse
2, Bractiionus angularis
Gosse
3, B. calyciflorus Pal-
las
4. B. diversicornis (Da-
day)
5. B. quad rid entatus Her-
mann
6. Kellicottia longispina
Kellicott.
7. Keratella cochlearis
Sosse
8. K. quad rata (o.F.
Muller)
9. Notholca cornuta Car-
lin.
10. Conochilus hippocre-
pis (Schrank)
11. Euchlanis dilatata
Ehrenberg
12. Filinia lengiseta
(Ehrenberg)
13. F. brachiata (jRousse-
let)
14. F. aseta Fadeev
15. Lecane luna (O.F..
Muller)
16. L. lunaris (Ehren-
berg)
17. La ludwigii. (Eckste-
in)
18. Polyarthra vulgaris
Carlin
19. P. minor Voigt.
20. P. major Burckhard^
21. Pleosoma sp.
22. Synchaeta sp.
23. Trichocerca capucina
(Wiez. et Zachar.)
24. Notommata sp.
Литература
пол«ваккая М,ф” Сергеев* О.А. 1071. Зоо-
планктон водоемов-охладителей ТЭС юга Украины. -
В кн.: Гидрохим. к гидробиол. водоемов-охладителей
ТЭС СССР. Киев.
ривьер И.К. 1868. Влияние сбросных теплых вод Кона-
ковской ГРЭС на зоопланктон Иваньковского водохра-
нилища, - В кн.: Волга-1. 1 конфер. изуч. водоемов
басе. Волги. Тезисы докл. Тольятти.
ривьер И.К. 1871. Зоопланктон Мошковичского залива,
подверженного влиянию подогретых сбросных вод Ко-
наковской ГРЭС. Симпоз. по влиян» подогрет, вод
ТЭС... Борок.
778 'А 17
И.А.С к а ль с к а я
СОСТАВ И РАСПРЕДЕЛЕНИЕ ЗООБЕНТОСА
ГОРЬКОВСКОГО ВОДОХРАНИЛИЩА
В РАЙОНЕ КОСТРОМСКОЙ ГРЭС
Изучение зообентода в районе Костромской ГРЭС, прово-
дившееся в 1970-1971 гг., является составной частью комплекс-
ного гидробиологического исследования влияния сброса теплых
вод этой электростанции на биологические процессы в Горьков-
ском водохранилише.
Пробы зообентоса отбирались с мая по октябрь через
каждые 10 дней в 1970 г. и два раза в месяц в 1971 г.
дночерпателем Петерсена (площадь сечения 1/40 м^). Про-
мывание грунта производилось через сито из шелкового
газа № 15-19. Организмы зообентоса, за исключением
моллюсков, фиксировали в 4% формалине. Моллюски храни-
лись в спирте. Определение видового состава и количест-
венную обработку проб проводили в Лаборатории экологии
водных беспозвоночных Института биологии внутренних вод
АН СССР. Считаю своим приятным долгом' выразить боль-
шую благодарность В.И.Митропольскому за помошь при об-
работке материалов.
Сборы бентоса производились в зоне с естественным
температурным режимом - на створах Г и II в русловой
части водохранилиша и в зоне подогрева - на ст. 4, 6, 8,
10 в приустьевом заливе р. Шачи и на Створе 111 в водо-
хранилише (рис. 1),
В 1970 Г. придонная температура в приустьевом заливе
р. Шачи превышала естественную в среднем за период нсследо-
258
ваний на 1.6-4.7°С. ^има
1970/71 г. была относи-
тельно теплой. В январе
отмечалась температура
воздуха выше нуля. В за-
ливе р. Шачи и в водохра-
нилише почти до III створа
оставалась полынья. В
1971 г. в связи с увеличе-
нием мощности ГРЭС подо-
грев усилился. В весенний
период на ст. 8 и 10 -
самых мелководных участ-
ках в приустьевом заливе
р. Шачи, где глубина сос-
тавляла 2.5-3.5 м - отме-
чен равномерный прогрев
толши воды. Во второй де-
каде мая 1971 г. придон-
ная температура здесь была
равна 16.2°С. (В 1970 г.
более чем на десять дней
позднее температура у дна
на этих станциях составляла
всего лишь 10.7-11.8°С). В районе ст. 4 и 6, расположённых
ближе к водосбросу, но более глубоководных (5-6 м), весной
прогрев у дна был меньше, чем на ст. 8 и 10. Теплая вода
в основном не охватывает придонные слои. Температурный ска- '
чок располагался на глубине 3-4 м, его величина составляла
6.0°С (рис. 2). Летом и осенью прогрев на ст. 4 более равно-
мерный (рис. 2), что,' вероятно, было связано с увеличением
объемов сбрасываемой воды, а следовательно, и интенсивности
перемешивания водных масс. В 1971 г. подогрев в среднем за
период наблюдений составлял на ст. 4 - 5.6, ст. 6 - 2.0, ст. 8 -
3.8, ст. 10 — 3.49С. Теплая вода по мере удаления от водосбро-
са движется верхним слоем и поэтому уже на створе Ш превы-
шение естественной придонной температуры составляло обычно
не более 1°С.
Грунты .в исследованном участке водохранилища представ-
лены заиленными мелко- и крупнозернистыми песками на стреж-
не, песчаными илами' у правого берега и кое-где серыми илами.
Район водосброса представляет собой небольшой залив в
устьевой части притока Волги - р. Шани, В этом районе наблю-
дается почти постоянно подпор со стороны водохранилища, сток
очень незначителен, практически скорость течения здесь близка
778 17
259
Рис. 2. Распределение температур по
глубинам в эоне смешения (продольный разрез)
в 1871 г.
А - 18 мая, Б - 17 августа.
к нулю. Вблизи водосброса ГРЭС расположена' пристань „Вол-
гореченск*. Летом 1870 г. проводились работы по углублению
участков, прилегающих к пристани. Землечерпалками были со-
вершенно нарушены естественные грунты и вся акватория это-
го района сильно взмучена, в результате чего естественные
биоценозы разрушены. Грунты здесь глинистые с осевшими
взвесями, вследствие размыва берегов прибрежная раститель-
ность почти не развивается.
Состав зообентоса
Видовой состав зообентоса исследуемого района за время
наблюдений был представлен 24 видами хирономид, 17 видами
олигохет. 18 видами моллюсков; кроме того, встречались в ви-
дов пиявок, по 1-2 вида личинок поденок, жуков, ракообразных.
Список всех найденных в сборах дночерпателя животных приво-
дится в приложении.
Наиболее распространены и чаше других встречаются оли-
гохеты: ими обильно заселены заиленные пески и песчаные
илы русла Волги, гораздо меньше их на серых илах и обна-
260
1970
л
Рис. 3. Соотношение доминирующих видов олигохет (в %
от обшей биомассы червей).
А - приустьевой залив р. Шачи, Б - створ II (1970 г.)
и створ 1 (1971 г.), В - створ III. 1 - Isochaetides ne-
waensis, 2 - Limnodrilus hoffmeisteri, 3 - Potamoth-
rix moldaviensis, 4 - Limnodrilus udekemianus, 5 ~
прочие олигохеты + молодь и неполовозрелые + фрагменты.
Площадь кругов пропорциональна общей биомассе олигохет.
же иных грунтах. Из олигохет Isochaetides newaensis в,
большом количестве встречается на песчаных илах и заиленных
песках водохранилища. Доля этой крупной олигохеты в биомассе
червей на разных участках водохранилища в 1970 и 1971 гг.
составляла от 32.4 до 74.6%, а в' приустьевом заливе р. Шачи -
всего лишь З.О-6.6% (рис. 3). Этот вид вообще, как известно
по Горьковскому и Рыбинскому водохранилищам, тяготеет к
илисто-песчаным грунтам и серым илам, к участкам, где имеет-
ся хотя бы временная проточность (Мордухай-Болтовской,
1963). На заиленных песках > песчаных илах водохранилища
встречались Peloscolex ferox, Arcteonais lomondi; *
приустьевом заливе р. Шачи эти олигохеты ие найдены. В от-
личие от указанных олигохет повсеместно были распростране-
ны Limnodrilus hoffmeisteri, Potamothrix moldavien- *
sis. По численности и биомассе эти черви относились к числу
281
доминирующих на всех обследованных участках. В приустьевом
заливе р. Шачи на долю L, hoffmeisteri приходилось до
42.7% биомассы олигохет, Р. moldaviensis - до 15.4%, а в
водохранилище соответственно - до 19.6 и 10.2%. Limnodrilus
udekemianus встречался в водохранилище редко, здесь на
его фолю приходилось не более 1.5%, в приустьевом заливе
р. Шачи значительно больше - 10.2-14.6% биомассы (рис. 3).
Хирономиды встречались повсеместно, но их численность
была невелика. В биомассе зообентоса приустьевого залива
р. Шачи они составляли 7.7%, а в водохранилище только 0.7%
(1971 Г.). Постоянно в пробах исследованного района находились
личинки Procladius sp., Cryptochironomus gr. defec-
tus, остальные хирономиды встречались очень редко и в неболь-
шом количестве.
Моллюски были найдены почти всюду. По числу представи-
телей богаче всего сем. Sphaeriidae, которых насчитывалось
И видов. Почти повсеместно встречаются Pisidiurn henslo—
wanum, Sphaerium scaldianum, Sph. solidum. Совер-
шенно отсутствовали сферииды на ст. 4 вблизи водосброса
ТЭС. Из других семейств встречались Dreissena poly-
morphs, Anodonta piscinalis, Uruo pictorum, U, tu-
midus, Viviparus viviparus, Valvata piscinalis. D.
polymorpha постоянно встречались в пробах только иа ст. 1
створа Ш. Осенью 1970 г. численность моллюсков здесь дости-
гала 8800 экз./м^ при биомассе 2.6 кг/м^. На следующий год
в ето же время их стало еще больше - 13 480 экз./м^ при
биомассе 3.2 кг/м2.
На ст. 1 створа III чаше, чем на других участках, обнару-
живались пиявки Herpobdella, Helobdella, Glossiphonia
и др. В октябре 1970 г. их насчитывалось 98 экз./м2, а в ок-
тябре 1971 г. — 400 экз. /м2.
Осенью 1971 г. в- районе створа III впервые была встрече-
на интродуцированная в Горьковское водохранилище в 1962-
1965 гг. байкальская гаммарида Gmelinoides fas ci atu s.
Ранее (1969 г.) этот вид был обнаружен не выше Плеса (Мор-
духай-Болтовской и Чиркова, 1971). Наши данные свидетельст-
вуют о дальнейшем расселении рачка вверх по течению Волги.
Обнаруженные в 1972 г. в пробе со ст. 4 (у водосброса ГРЭС)
G, fasciatus уже в конце апреля были яйценосными самками
и половозрелыми самцами. В открытой части Байкала этот вид
начинает размножаться в июне, а в лучше прогреваемом Посоль-
ском соре, где рачок образует скопления, массовая откладка
яиц наступает в мае (Бекман, 1962).
Средняя биомасса зообентоса в водохранилище в 1971 г. •
была равна 11.4 г/м2, в приустьевом заливе р. Шачи - 3.0г/м2,
262 *
Таким образом, водохранилище и приустьевой залив
р. Шачи существенно отличаются по составу и количеству бен-
тоса. В водохранилище в большом количестве обитает Isochae-
tides newaensis, а в приустьевом заливе р. Шачи он
встречается кое-где на бывшем русле н в небольшом количест-
ве. По этой причине средняя биомасса зообентоса в водохрани-
лище почти в 4 раза выше, чем в заливе. Только в водохрани-
лище обнаружены Peloscolex ferox, Arcteonais lomondi,
здесь же преимущественно живут и моллюски, в том числе
дрейссена.
В приустьевом заливе р. Шачи в соответствии с менее
благоприятными экологическими условиями для развития зообен-
тоса (глинистые грунты, слабый сток) обитают преимуществен-
но более эврибионтные виды, в том числе встречающиеся в по-
лисапробных водах - Limnodrilus hoffmeisteri, Potamo-
thrix moldaviensis. Преобладающее значение для зообентоса
имеет характер грунта, что замаскировывает действие темпера-
турного фактора. Следует отметить, что за время наблюдений
специфичных термофильных форм зообентоса в районе работ ие
обнаружено.
Динамика численности
и биомассы зообентоса
а) Неподогреваемая зона х естественным температурным
режимом - створы I и II в русловой части водохранилища.
Как уже отмечалось, основной группой зообентоса здесь
были олигохеты, из которых доминировали Isochaetides ne-
waensis, Limnodrilus hoffmeisteri, Potamothrix mol-
daviensis. Динамика численности этих тубифицид находится
в прямой/ связи с их жизненным- циклом. В общих чертах она
характеризовалась высокой* численностью олигохет перед раз-
множением (откладкой коконов), снижением количества червей
после размножения, очевидно, в результате гибели старых
особей, закончивших размножение, и увеличением численности
при появлении молоди.
В неподогреваемой зоне в 1970 и 1971 гг. олигохеты
размножались в июне. Для примера возьмем динамику числен-
ности олигохет иа ст. 2 створа I (1971 г.). Перед размноже-
нием основную часть червей составляли половозрелые особи.
В период размножения численность олигохет снизилась с
11088 экз./м2 до 910 экз,/м2 (рис. 4), первое появление иоко-
263
Рис; 4. Динамика чис-
ленности и биомассы оли-
гохет на ст. 2 створа I
в 1971 г.
1 —численность, 2 -
биомасса, 3 - коконы,
экз./м^ (1 мм - 5 экз./м^).
Рис. 7 Динамика чис-
ленности и биомассы оли-
гохет на ст. 10 (1971 г.).
Обозная. те же, что
ж на рис. 4.
•*«./** V**
1000
• П Щ ГШ ‘ IX ' X
364
нов отмечалось в первой декаде июня^ В конце июня появляет-
ся молодь, численность олигохет возрастает до 2140 экз./м2.
В этот период основную часть червей составляли неполовозре-
лые особи. В июле-августе численность олигохет колеблется
незначительно, черви растут и созревают. В сентябре часть
червей размножается, а в конце месяца появляется молодь,
к середине октября численность олигохет возрастает до 5740
экз./м^, а биомасса, наоборот, понижается до минимальных
величин 5.7 г/м2. Это объясняется тем, что основную часть
червей составляли чрезвычайно мелкие, ювенильные особи осен-
него рождения (второе поколение).
б) Зона со слабым подогревом - створ III.
В зоне слабого подогрева превышений естественной при-
донной температуры за два года наблюдений чаше всего сос-
тавляло не более 1°С. В 1970 г. на ст. 2 створа III первое
появление коконов в пробах было отмечено в конце мая. Молодь
олигохет появилась в конце июня-июле. В этот период здесь
находились мелкие черви, выходящие из разрушенных коконов,
численность олигохет достигала 7168 экз./м* (рис. 5). Осеннее
размножение происходило в сентябре, в конце месяца появля-
лась молодь, и к концу октября численность червей составляла
2520 экз./м2, основная масса - неполовозрелые. Аналогичный
ритм размножения олигохет наблюдался на этой станции и в
1971 г. с той разницей, что впервые коконы обнаружены в
июне. Осенью олигохеты 'размножаются менее активно, так как
размножается лишь часть особей. В осенних пробах коконы
нигде не встречались.
Таким образом, существенных различий в сроках размно-
жения олигохет в неотепленной и слабоотепленной зонах водо-
хранилиша не отмечено.
в) Зона значительного-недегрев^ - приустьевой залив
р,—Шачи.
В этой зоне непосредственно у водосброса на ст. 4 в 1
1970 г. доминировал Limnodrilus hoffmeisteri, ему сопут-
ствовали L, udek emi anus, Tubifex tubifex, L, ciapa-
redeanus, встречавшиеся непостоянно, их численность была
невелика. На следующий год в число ведущих организмов вклю-
чается Potamothrix moldaviensis. Численность олигохет
претерпевает здесь значительные колебания (рис. 6). В конце
июля 1970 г., в период самой высокой температуры (25.4°С),
наблюдалось полное исчезновение животньцс в пробах. М.Л.Пид-
гайко (1971) придонную температуру 25°С считает предельной
для нормального развития донных биоценозов. На протяжении
июня-августа половозрелые особи составляли основную часть
популяций, коконы в пробах не встречались, поэтому выявить
ритм размножения червей весьма трудно.
265
эк>./м*
Рис. 8. Суммарная численность хирономид (экз./м^) на
створе 1 и у водозабора (1) и в пруустьевом заливе р. Шачи
(II) в 1971 г.
1 - ст. 1, 2, 3 створа 1 + ст. 3 на водозаборе, II - ст.4,
6, 8, 10 в приустьевом заливе р. Шачи, а - личинки, б - пред-
ку колки, в - куколки.
Станции 6 и 8 находились в районе работ землечерпатель-
ных снарядов. Численность зообентоса здесь была очень низ-
кой, а средняя биомасса за сезон на ст. 6 составляла 1.1 г/м^,
на* ст. 8 - 1.6 г/м^.
В 1971 г. землечерпательные работы не проводились.
Грунты на этом участке стабилизировались, что, несомненно,
благоприятно отразилось на развитии зообентосе. Биомасса на
ст. 6 возросла до 3.4 г/м2, на ст. 8 - до 2.0 т7м2.
268
В 1971 г. на ст. 4 сильного загрязнения дна нефтепродук-
тами, как в прошлом году, не наблюдалось, Изменился характер
грунта; появились мелкозернистые включения, по-видимому по-
падающие в водоем со стоками промышленно-ливневой канали-
зации; увеличился подогрев. Средняя за сезон биомасса зоо-
бентоса была почти в 1.5 раза меньше, чем в 1970 г.
Отсутствие ледового покрова зимой 1970-71 г. в приустье-
вом заливе' р. Шачи. а также сильный весенний прогрев на мел-
ководных участках ускорили процесс созревания олигохет. Раз-
множение червей проходило в мае. Коконы червей обнаружива-
лись на ст. 10 с 18 мая; возможно, что они появились и рань-
ше, но, к сожалению, в более ранние сроки наблюдения не про-
водились. В конце июля отмечалась относительно высокая чис-
ленность олигохет - 1160 экз./м^ (рис. 7), большинство червей
были неполовозрелыми. В августе, очевидно, черви созревали и
размножались. Хотя в пробах коконы не обнаруживались, но от-
мечалось увеличение численности червей в .сентябре до 860
экз./м^ при одновременном снижении биомассы до 2.1 г/м2.
Таким образом, в зоне сильного подогрева, на мелковод-
ных хорошо прогреваемых участках размножение олигохет на-
чинается раньше, чем в неотепленной зоне. Возможно, второе
размножение червей наступает в августе*, а не в сентябре, как
это отмечено для зоны с естественным температурным режимом,
В Иваньковском водохранилище, где теплый поток воды
(район Конаковской ГРЭС) хорошо прогревает зимой придонный
слой, размножение доминирующего Limnodrilus hoffmeisteri
начинается в первых числах мая. В июне появляется молодь,
к концу августа она достигает половой зрелости. В сентябре
в размножении участвуют черви нового и старого поколений с
восстановленной половой системой, в октябре встречается мо-
лодь осеннего рождения (Поддубная, 1971). В приустьевом за-
ливе р. Шачи характер осеннего размножения червей недоста-
точно ясен и требует специального изучения.
Из хирономид в районе исследований повсеместно Встреча-
лись Procladius sp, и Cryptochirbnomus gr. defec-
tus; по численности преобладал Procladius sp. Вообще чис-
ленность хирономид в районе исследований невелика, поэтому
на рис, 8 даиа суммарная численность личинок, предкуколок и
куколок хирономид для ст. 1, 2, 3 створа 1 и ст, 3 на водоза-
боре (зона с естественным температурным режимом) и для
ст. 4, в, 8, 10 (зона сильного подогрева), В зоне сильного
подогрева развитие хирономид происходит раньше, чем в зоне
с естественным температурным режимом. Куколки хирономид
. в приустьевом заливе р. Шачи встречались в 1971 г, уже 18
мая; в этот период отмечена самая высокая суммарная числен-
28?
ность личинок - 1162 экэ./м^. Придонная температура в это
время была равна 16.0, а в неотепленной зоне только 9.9°С.
В зоне с естественным температурным режимом предкуколки
появляются в конце мая. Вылет, очевидно, начинается в июне.
В течение летних месяцев продолжается вылет имаго хироно-
мия в неподогреваемой и подогреваемой зонах, В сентябре-ок-
тябре в пробах присутствовали только личинки; предкуколки и
куколки отсутствовали в обеих зонах,
В 1972 г. первые сборы зообентоса были проведены зна-
чительно раньше, чем в предыдущие годы - 21 апреля. Резуль-
таты наблюдений показали, что в зоне естественного темпера-
турного режима на ст. 1 и 2 створа Т, а также на ст. 2 ство-
ра III при температуре у дна 2.2°С популяции червей на 62-85%
состояли из неполовозрелых особей. Численность и биомасса
зообентоса были высокими, доминировали те же виды, что и в
предыдущие годы,
В зоне подогрева картина была другая. В зоне слабого
подогрева на ст. 1 створа III придонная температура составля-
ла 5.3°С, в приустьевом заливе р. Шачи (зоне сильного подо-
грева) иа ст. 4, 8, 10 - от 8.1 до 11.1°С. Основная масса чер-
вей (50-69%) была половозрелыми особями.
Заключение ,
При изучении зообентоса Горьковского водохранилища в
районе Костромской ГРЭС основное внимание было уделено
характеристике его в зоне с естественным температурным ре-
жимом и в зоне наибольшего подогрева. Установлено, что зоо-
бентос этих участков существенно отличается по количеству
и по составу доминирующих видов. Средняя биомасса зообентоса
в водохранилише составляла 11.4 г/м2хав приустьевом зали-
ве р. Шачи - не более 3.0 г/м^. В водохранилище в большом
количестве встречался Isochaetides newaensis, а ® зали-
ве его было очень мало, он обнаруживался только на отдель-
ных участках старого русла р. Шачи. Причина этих различий
заключается в особенностях условий обитания организмов в
сравниваемых зонах. В водохранилише имеется постоянная
проточность, грунты представлены заиленными песками и ила-
ми, а в приустьевом заливе р. Шачи грунты глинистые вообще
бедны зообентосом, течение отсутствует. Основное зиачеиие
для развития зообентоса имеет характер грунта. Разный харак-
тер грунтов в сравниваемых зонах перекрывает действие тем-
268
пературного фактора и затрудняет выяснение его влияния. Спе-
цифических термофильных форм зообентоса в районе исследо-
ваний не обнаружено. Установлено, что в зоне наибольшего
подогрева созревание тубифицид и окукливание хироиомид про-
исходит раньше, чем в зоне с естественным прогревом. Вопрос
о состоянии популяций доминирующих аообентических организ-
мов в осенний период, в частности вопрос о второй генерации
олигохет, требует дальнейшего изучения.
Приложение-
Список зообентических организмов
Горьковского водохранилиша
в районе Костромской ГРЭС
К л. Oligochaeta
Сем. Tubificidae
1. Aulodrilus limnoblus
Bretscher
2, Isochaetides newaen-
sis Mich.
3. I. michaelseni Lasto-
ckln
4. Limnodrllus hoffmeiste-
ri Claparede
5. L. helvetlcus Piguet
6. L. clapared eanus Rat-
zel
7. L, udekemianus Cla-
‘ parede
B. Peloscolex ferox
(Bisen)
9. Potamothrix moldavien-
sis Vejdovsky
10. P, hammonlensis
Mich.
11, Ps ammo rye tides bar-
batus (Grube)
12, Tubifex tubifex Mull.
Сем. Naididae
1. Arcteonals lomondi
Martin
2. Ophidonals serpentina Mull.
3, Specaria Josinae Vej-
dovsky
4. Uncinais unclnata
О ersted »
Сем. Lumbriculidae
1. Stylodrilus heringianus
Claparede
2.
Kn. Hirudinea
1. Glos si pho nia compla-
nata (L,)
2. G, heterdciita (L.)
3. Batrachobdella sp.
4, Herpobdella sp.
5. HelobdeUa sp,
6. Piscicola* sp.
269
Кл, Gastropoda
Кл. Crustacea
Сем. Viviparidae
Отр. Iso pod а
1. Viviparus viviparus L.
Com, Vrdvatidae
1. Valvata piscinalis Mull.
2. Bithynia tentaculata
L.
Кл. Lamellibranchiata
Сем. Dreissenidae
1. Dreissena polymorpha
Pall.
Сем. Unionidae
1. Unio pictorum L.
2, U. tumidus Retz.
3. Anodonta piscinalis
Nilss.
Сем. Sphaeriidae
1. Sphaerium rivicola
Lam.
2. S. corneum L.
3, S. solid urn Norm.
4. S«, scaldianum Norm.
5. Pisldium amnicum
MU11.
6. P, casertanum Poli
7. P. moitesserianum Pa-
ladilhe
8. P. pond его sum Stelfox
9. P. henslowanum Shep.
10. P« subtruncatum
Malm,
11. P. tenuilineatum
Stelfox
1. Asellus aquaticus L.
Отр. Amphipoda
1, Gmelinoides fasciatus
Stebb.
Кл. Insecta
Отр. D i pt era
Сем. Chironomidae
1. Cryptochironomus gr.
conjugens Kieff.
2. C. gr. defectus Kieff.
3. C. gr. fuscimanus Kieff.
4. C. gr. pararostratus
Lenz.
5. Einfeldla gr. carbo na-
ria Mg.
6. Endochironomus gr. ten-
dens F.
7. Glyptotendipes gr. gri-
pekoveni Kieff.
6. Limnochironomus gr.
nervosus Staeg.
9. Pentapedilum gr. exsec-
tum Kieff.
10. Polypedilum gr. scalae»
num Schr.
11. P. gr. convictum
Walk.
12. P. gr. nubeculosum
Mg.
13, Chironomus f. 1. plu~
mosus L.
14, Ch. f. 1. semireductus
Lenz.
15. Ch. f. 1. reductus
Lipina
270
16. Tanytarsus gr. gre-
garlus Kieff.
17. T. gr. mane us v. d.
Wulp.
18, Cricotopus gr. alga-
rum Kieff.
19, Orthocladius gr. con-
sobrinus Holmgh
20. O. sp.
21, Psectrocladius gr.
psilopterus Kieff.
22. Ablabesmyia gr. mo-
nllis L.
23. Pelopia punctlpennis
Mg.
24. Procladius sp.
Сем. Culicidae
1, Chaoborus sp.
Сем. Heleldae
1» Culicoides sp,
Отр. Ephemeroptera
1. Ordella sp,
2, Caenls horaria L,
Отр. Coleoptera
1, Coleptera larvae sp.
Отр/Trfchoptera
1. Ryacophilia sp.
2, Tinodes sp.
Литература
Бекман М.Ю. 1962. Экология и продукция Mlcruropus
possolskil Sow. и Gmelinoides fasciatus Steb.
Тр. Лимнолог, инет. CO АН СССР, 2(22), 1.
Мордухай-Болтовской Ф.Д. 1863. Состояние
бентоса Горьковского водохранилища в 1860-1862 гг.
Тр. Инет, биол, внутр, вод АН СССР, 6(8).
М о р д у х а й-Болтовской Ф.Д., Чиркова З.Н.
1971. О распространении байкальского бокоплава Gme-
linoides fasciatus (Stebbing) в Горьковском
водохранилище. Информ, бюлл. Инет. биол. внутр, вод
АН СССР, 9.
Пидгайк'о М.Л. 1971. Влияние подогрева на развитие в
продуцирование зоомакробентоса в водоемах-охладите-
лях ГРЭС юга УССР. Симпоз, по влиян. подогретых
вод ТЭС... Борок.
Поддубная Т.Л. 1971. Донная фауна Иваньковского
водохранилища в районе сброса теплых вод Конаковской
ТЭС. Тр. Инет. биол. внутр, вод АН СССР, 21(24).
2?1
Н.Ф.С м и р и о в a
РОСТ И ПРОДУКЦИЯ УНИОНИД
(MOLLUSC A, UNIONIDAE)
В РАЙОНЕ КОСТРОМСКОЙ ГРЭС
Исследования проводились в районе Костромской ГРЭС, '
которая расположена на южном берегу Горьковского водохрани-
лища (правом берегу Волги) в устье р. Шачи, в 28 км ниже
г. Костромы. ГРЭС начала свою работу с июня 1969 г. и к де-
кабрю 1970 г, ее мощность составила 1 200 МВт. К 1975 г.
Костромская ГРЭС будет самой большой в стране, ее мощность
достигнет 4800 МВт. В результате сброса теплых конденсацион-
ных вод в водохранилище образуется участок с повышенной
температурой воды, границы которого находятся в определен-
ной зависимости от количества сбросных вод.
В задачу настоящей работы входило определение плотнос-
ти поселений, структуры популяции, закономерностей роста и
выяснение продукционных возможностей широко распространен-
ных в районе ГРЭС двустворчатых моллюсков Anodonta
piscinalis Nilss, Unlo pic to rum (L,)f Unio tumid us
Philips.
Материал и методика
Материал в районе Костромской ГРЭС собирался ежеме-
сячно с июня по ноябрь 1971 г. Первые пробы были взяты в
первых числах нюня, до появления молоди. В этот период мощ-
272
нос.ть станции не превы-
шала 1200 МВт. Исследо-
вания проводились в пяти
пунктах рассматриваемого
района: напротив водосбро-
са (ст. 5), в начале при-
устьевого залива р. Шачи
(ст. 10), выше по тече-
нию в р. Шаче (ст. 11),
в русловой части водохра-
нилища ниже устья р. Ша-
чи (ст. 2, разрез IV) и
выше района ГРЭС (ст. 4
разрез 1) (рис. 1). Ма- .
териал собирали на глуби-
не от одного до трех мет-
ров с помошью драги,
входное отверстие которой
составляло 60x30 см2.
Для количественного уче-
та моллюсков драгу про-
тягивали три раза на про-
Рис. 1. Схема расположения
станций.
тяжении 20 м между поставленными на этом расстоянии буями.
На каждой станции в момент взятия пробы определяли темпера-
туру и содержание кислорода. Характер грунта определяли ви-
зуально.
В начале работы, в июне, иа ст. 5 придонная температу-
ра воды составляла 19.8 , наиболее высокой она была в июле-
25.40, а к началу ноября она опустилась до 5.9°С. На ст. 10
в июне придонная температура была 17.5°, в июле - 23.5°, а в
конце октября - 3.5°С. На ст. 11 в июне температура в при-
донных слоях равиялась 20.2°, в августе 22.3°, в конце сен-
тября 2.3°С.
Температура воды в водохранилище была значительно нит
же: в июие на ст. 4 - 17.2°, иа ст. 2 - 19.8°С. Наиболее вы-
сокая температура наблюдалась в июле: 20.9 ° и 19.8°С, соот-
ветственно. К началу ноября иа обеих станциях температура
понизилась до 1.5°С.
Содержание кислорода на станциях определялось методом
Винклера. Из полученных результатов видно, что придовиые
сЛои воды во всех пунктах исследоваиия достаточно хорошо
аэрируются (рис. 2, б). Содержание кислорода за время наблю-
дений находилось в пределах от S.5 до 12.5 мг/л и в общих
чертах коррелировало с температурой. Лишь в июле, в период
наиболее высокой температуры, в придонных слоях всех рассмат-
273
Рис. 2. Температура (а) и со-
держание кислорода (б) в местах
сбора материала в период наблюдений.
♦ I—I—I___I__< I I
VI vn VIII ix X XI
риваемых станций на-
блюдается повышенное
содержание кислорода.
Грунт на ст. 10 и
11 до глубины 2-2.5 м
представлен заиленной
дерновиной (табл. 1).
Несколько глубже, до
3 м - жидким илом
с большим количеством
растительных остатков
и мертвой ракуши.
На ст. 4 грунт пред-
ставлен плотным серым
илом. На ст. 2 до глу-
бины 2м- глинистым
песком, а глубже -
серым плотным илом,
как и на ст. 4, но с
древесными остатками.
Для выяснения
структуры популяции
рассматриваемых видов в период исследования проводили изме-
рения всех собранных с мерных площадей особей и определе-
ние численности различных размерных групп. Вес определяли
прямым взвешиванием. Предварительно свежие моллюски с плот-
но закрытой раковиной подсушивали фильтровальной бумагой.
Возраст моллюсков устанавливали по годовым кольцам. Изме-
ряли длину каждого годового кольца, образовавшегося за счет
остановки роста в зимнее время. Получеииый таким образом
материал обрабатывался с помощью статистических методов.
Определяли среднюю длину каждой возрастной группы мол-
люсков, отклонение от среднего, величину дисперсии. Достовер-
ность различий в темпе роста моллюсков в разных участках
изучаемого района определяли по критерию Стыодента.
Для характеристики состояння популяции двустворчатых
моллюсков Anodonta piscinalis, Unio pictorum и U. tu—
nidus в районе Костромской ГРЭС изучали их продукционные
свойства. Для расчета продукции пользовались методом, пред-
ложенным А.Н.Голиковым (1970). Этот метод исходит из усло-
вий стационарного характера популяции. На основании анализа
размерно-весовой структуры популяции определяли ростовую
кривую особей популяции. Характеристика весового роста и
расчет изменений численности особей кайлой возрастной группы
послужили материалом для определения ростовой и подле ржнваю-
274
Таблица 1
Распределение грунтов в районе исследуемых станций
№ стан- ции Глубина, м Характер грунта
" 5 1.20-2.10 Заиленная дерновина
2.10-2.50 Глинистый, с примесью жидкого ила с большим количеством расти- тельных остатков и мертвой ра- куши
10 1.20-2.00 Заиленная дерновина
2.00-2.50 Твердый илистый, с большим коли- чеством мертвой ракуши
2.50-3.00 Жидкий илистый, с мертвой ракушей
1! 1.00-2.50 Заиленная дерновина
2.50-3.00 Жидкий ил
2, разр. IV 1.20-2.00 2.00-2.50 Глинистый песок с древесными и другими растительными остатка- ми Серый ил с древесными и раститель- ными остатками
4, разр.Х 1.00-2.50 Плотный серый ил
шей продукции. Под ростовой продукцией (Pg) понимается сумма
годовых приростов, умноженных на численность возрастной
группы:
TV t*4 t
гДе^+ Ni “ численность особей в данной возрастной группе;
(W^ — W4 ) - весовой прирост особи данного размера за
год (>W ). Среднегодовую ростовую продукцию определяли пу-
тем нахождения средней из рассчитанных ростовых продукций
за ‘'все 4 сезона я высчитывали по формуле
n WW
Pffcp’ 4
575
Такой способ расчета годовой продукции популяции позволяет
не принимать во внимание сезонные изменения скорости роста
и констатировать состояние популяции, к которому она прихо-
дит за год к моменту наблюдения.
Под поддерживающей продукцией (Pj) понимается сумма
биомассы всех особей популяции, достигших возраста одного
года (Bq), и произведения численностей всех остальных воз-
растных групп на приросты Их особей за прошедший год:
Среднегодовая поддерживающая продукция обозначалась как
сумма продукционного пропесса за год к моменту каждой
сезонной выборки, деленная на количество сезонов:
PSi*₽Se+pss+₽s4
4 •
Голиков и Мвншуткнн (1971) с помощью ЭВМ проверили
правомочность и точность такого метода определения продукции
путем моделирования продукционного процесса на .примере конк-
ретной популяции брюхоногого моллюска Epheria turrita (А.
Adams). Результат их расчетов показал, что поддерживаю-
щая продукция имеет иной смысл, чем чистая продукция ( Р )
в понимании других авторов. Поэтому использование величины
этой продукции в качестве оценки годовой продукции в устано-
вившемся понимании (Методы определения..., 1968) нецелесооб-
разно. Величина Pg дает представление о годовом приросте вы-
живших особей популяции, а ее коэффициент линейной за-
висимости связан с возрастом преобладающих в ней особей
(Голиков, 1970). Иными словами, поддерживающая продукция
отражает то количество органического вещества, которое об-
разуется за год и остается в пределах популяции, обеспечивая
ее существование на определенном, необходимом при данной
продолжительности жизни организмов уровне. Ростовая продук-
ция является несмещенной оценкой величины годовой чистой
продукции в общепринятом понимании авторов.
Описанный метод удобен, поскольку можно рассчитать
годовую продукцию вида по одному сбору. Однако необходимо
отметить, что при неблагоприятном стечении обстоятельств,
В случае одной выборки ошибка этого метода может достичь
величины около 80%. Но уже две пробы, взятые с интервалом
в полгода, позволяют определить годовую продукцию с макси-
мальной ошибкой 25%, а четыре выборки, равномерно распреде-
ленные в течение года, позволяют сократить ошибку в опреде— .
лении продукции менее чем до 5% (Голиков, Меншуткин, 1971 ).
27в
Рис. 3. Возрастной состав популяций A. piscinalis (а),
U, pictorum (б) и- U. tumidus (в). На оси ординат - ко-
личество моллюсков.
По оси абсцисс — возраст в годах.
Результа ты
и обсуждение
Наибольшая плотность поселений Anodonta piscinalis
наблюдается на станциях, находящихся на р. Шаче. Тах, на
ст. 5 эта величина составляет 1.15 экз./м2, на ст. 10 - 0.98
экз./м2, на ст. 11- 0.86 экз./м2. Причем наибольшим числом
экземпляров на этих станциях представлены ювенильные мол-
люски и особи в возрасте одного-двух лет: на ст. 5 они сос-
тавляют. 88%, на ст. 10 — 79%’ от общего числа моллюсков. На
ст. 11, помимо первых трех групп, хорошо представлены особи
трехлетнего возраста. Все эти группы вместе составляют 89%
от количества собранных моллюсков на этой станции (рис. 3, а).
На станциях водохранилища плотность поселений A, piscina-
lis значительно ниже. Так, на ст. 2 на 1 м2 приходится в
среднем 0.4 экз., а на ст. 4 - 0.45 экз. На этих станциях,
так же как и на предыдущих, лучше представлены молодые
моллюски (35 и 42% от общего числа собранных моллюсков, со-
ответственно).
778 18 отт
Рис. 4. Темп роста A. piscinalis (a), U. pic to rum
(б) и U« tumidus (в).
По оси ординат - длина моллюсков, по оси абсцисс -
возраст в годах..
Наибольшая плотность поселений Uni о pictorum наблю-
далась на ст. S (0.8 экз./м2) и ст. 10 (0.8 экз./м2), причем
на первой из этих станций преобладали молодь и особи первых
двух лет жизни (77%). На ст. 2 - особи первого года состав-
ляли 38%, остальные возрастные группы представлены равномер-
но (рис. 3, б). Достаточно высокая плотность U« pictorum
отмечалась и иа станциях в водохранилище." Их средняя числен-
ность на .ст. 2 равнялась 0.6 экз., а на ст, 4 — 0.7 экз./м^.
Все возрастные группы этого моллюска представлены в популя-
ции равномерно. Значитёльно меныие' плотность поселений U.
pictorum на ст. 11 (0.2 экз./м2). Количественное соотноше-
ние возрастных групп в этой популяции равномерное.
Наиболее массовым в водохранилище видом оказался ^
Unio tumidus (рис. 3, в). На ст. 2 его численность равна
0.8, на ст. 4 - 0.8 экз./м , причем все возрастные группы.в
указанных популяциях количественно распределяются равномер-
но. Несколько ниже плотность U. tumidus на ст. 5 - 0.6
экз./м^.'Эта популяция достаточно «молодая: 60% от числа веете
собранных особей приходится на мойодь и моллюсков первого "
года. На ст. 10 и ст. 11 на 1 м2 приходится 0.4 и 0.3 экз.
U. tumidus. соответственно.
' Изучение скорости роста моллюсков в различных популя-
циях позволяет отметить, что самые значительные различия в
278
росте имеются у Anodonta piscinalis (рис. 4, а). Наиболь-
ших размеров этот моллюск достигает в популяциях, располо-
женных в р. Шаче, причем с возрастом различия в темпе рос-
та увеличиваются.' Так, если ювенильные моллюски имеют дли-
ну раковины на ст. 5 — 22.2 мм, на ст. 10 - 24,5 мм, на ст.11 -
10.5 мм, на ст. 2 - 16.1 мм и на ст. 4 - 22.0 мм, то уже к
концу первого года жизни эти моллюски имеют размеры на
первых трех станциях в пределах от 40 до-44 мм, а на водо-
хранилишных - от 30 до 35 мм. На втором году жизни на стан-
циях, расположенных на р. Шаче, средняя длина A. piscinalis
равнялась 58-64 мм, а в водохранилище - 43-47 мм. На тре-f
тьем году — на первых трех станциях средняя длина моллюсков
составляла 71-76 мм, на ст. 2 и 4 - 53-55 мм; на четвертом
году - 77-85 и 63-64 мм, соответственно.
Статистическая обработка полученных результатов показа-
ла, что различия в скорости роста между популяциями A. pisci-
nalis, расположенными на ст. 5, 10 и 11, и популяциями ст. 2
и 4 почти для всех возрастных групп достоверны (табп. 2).
Темп роста Unio pictorum в исследуемых популяциях
почти повторяет закономерности роста Anodonta piscinalis
(рис. 4, б); На всех станциях размер ювенильных моллюсков
U. pictorum находится в пределах от 14 до 16 мм. Отличия
в средней длине моллюсков в значительной степени продляют-
ся с возрастом. Уже к концу первого года на ст. 5, 10 и Ц
средняя длина раковины моллюсков U. pictorum равна 27-30
мм, а на ст. 2 и 4 - 22 мм. На втором году жизни моллюски
вырастают до размеров 39-43 мм в популяциях р. Шачи, между
тем как в популяциях водохранилища лишь до 30-35 мм. На
третьем году жизни средняя длина моллюсков в первом случае
равна 47-55 мм, а во втором 40-45 мм. Проверив достоверность
полученных данных по критерию Стьюдента, можно с уверен-
ностью сказать, что темп роста U. pictorum на станциях
р. Шачи не различается, ио он выше и достоверно отличается
от темпа роста моллюсков, обитающих в водохраиилищнык по-
пуляциях (табц. 2). Заметим также, что наблюдаются достовер-
ные различия в темпе роста моллюсков старших возрастных
групп водохранилищных станций.
Исследование темпа роста U. tumidus показало, что
средняя длина раковины ювенильных моллюсков находится во
всех популяциях в пределах 12-14 мм (рис. 4, в), С возрастом
наиболее» значительно отличаются моллюски, собранные в райо-
не ‘ст. 2, поскольку они имеют наиболее ийзкий Темп роста
по сравнению с другими популяциями. Так, к концу первого
года средняя длина моллюсков здесь составляет 19 мм, второ-
го года - 28 мм, третьего - 35 мм, четвертого - 42 мм, пи-
279
Таблица 2
Опенка достоверности различий в темпе роста
исследуемых моллюсков в районе Костромской ГРЭС
№ станции Возраст, № станции
ГОДЫ 2 4 11 1 10
Anodonta piscinalix J —
5 <1 6,23 2.01 2.91 2.91
1 5.71 2.55 2.05 2.31
2 8,54 4.87 3,10 1.85
3 7,21 5.74 2.56 2.14
• /4 3,94 2.56 2.72 2.64
5 - 2.29 2.64 1.31
6 — — — -
10 <1 8.60 4.02 5.44
1 7.75 4.45 0.21
2 9.50 6.00 1.08
3 8.87 7.21 0.29
4 6.44 4,28 0.05
5 — 10.14 2.84
8 — — . — *
и <1 3,18 0.26
1 7.36 4.19
2 10.87 6.99
3 8.92 7.31 u
4 6.64 4,34
5 3.29
8 - -
4 <1 2,89
1 2.30
2 1.78
3 0.60
4 0.33
5
6 -
Таблица 2 (продолжение)
№ станции Возраст, годы № станции
2 4 1'1 10
X Jnlo plch >rum
5 <1 1 2 3 4 5 6 1.85 5.09 4,21 4.47 3.53 2.25 2.97 5.84 7.28 7.39 5.39 4.56 1.49 0.13 0.55 злз 1.33 1.61 0.22 0.73 0.12 0.47
10 <1 1 2 3 4 5 8 0.59 3.45 3,93 4.61 3,82 0.06 1.57 4.04 6,95 6,98 5.77 0.66 0.40 0.68 0.48
11 <1 1 2 3 4 5 6 0.10 2.21 0.85 2.21 0.83 2.30 1 36 7,62 •
4 <1 1 2 3 4 5 6 0.85 0.14 3,43 3,75 3.40 3.00 2.98
Unio tun ildus
5 <1 1 2 2,70 6,49 £23 1.11 4.47 2.52 2.28 1.35 0.63 0.51 1.78 0.10
Таблица 2 (продолжение)
№ станции Возраст, годы № станции
2 4 11 10
5 3 5.60 0.71 2.25 0.61
4 5.04 0.33 1.81 0.10
5 3.86 0.80 0.98 —
6 5,89 - - —
10 <1 3.08 1.55 2.65
1 4.38 2.45 0.47
2 4.88 2.31 0.71
3 4.29 1.13 1.87
4 2,85 . 0.09 1.70
5 — —
8 — - -
11 <1 0.06 1.23
1 5.12 3.07
2 4.39 2.55
3 3.87 2.49
4 3.59 1.96
5 2.13 1.24
6 - -
4 <1 1.47
1 2.41
2 3.55
4 4.94
о
4 5.69
5 3.74
6 *
Примечание. Подчеркнуты значения критерия Стыо-
дента выше уровня 99% обеспеченности.
того - 61 мм и шестого - 58 мм. Размеры этих же моллюсков
на других станциях значительно больше во всех возрастных
группах и достоверно между собой не различаются (табл. 2).
К концу первого года длина U. tumidus в популяпиях. рас-
положенных в районе ст. 5, 10, 11 и 4 составляет от 22 jjo
28 мм; второго - 32-39 мм; третьего - 42-54 мм; четверто- ,
282
Рис. S. Изменчивость длины раковины A. piscinalis
(a), U. pictorum (б) и U. tumidus (в).
По оси ординат - величина дисперсии, по оси абсцисс -
возраст в годах.
го - 51-61 мм; пятого - 58-67 мм; шестого - около 70 мм.
Необходимо отметить, что изменчивость в размерах мол-
люсков для всех рассмотренных видов с возрастом увеличива-
ется, о чем можно судить по величине дисперсии (рис. 5), Од-
нако резкие отклонения в ходе величин дисперсии для Апо—
donta piscinalis на ст. 4 (возр. группы четвертая и пятая,
рис. 5, а), для Unio pictorum на ст. 11 (все возрастные
группы, начиная с первой) и 10 (пятая возрастная группа),
(рис. 5, б) и для U. tumidus иа ст. 11 (начиная с третьей
возрастной группы, рис. 5,.в) связаны прежде всего с резким
уменьшением объема выборки, используемой дпя статистическо-
го анализа. Это в определенной степени сказалось и на оценке
достоверности различий в темпе роста, представленной в
табл.. 2. Так, например, сравнение размеров Anodonta pis-
cinalis на ст. 4 и ст. 5 (табл. 2) показало достоверные
различия только для второй и третьей возрастных групп, в то
время как для четвертой и пятой возрастные групп эти разли-
чия оказались несущественными. Но именно для четвертой и
пятой возрастных групп вследствие резкого сокращения коли-
чества исследуемых моллюсков возросла величина дисперсии,
что и привело к уменьшению критерия Стьюдента^ Однако
факт, что изменчивость в размерах моллюсков с возрастом
увеличивается, не есть только- результат статистической обра-
283
Станции.
Рис. 6. Биомасса (В), ростовая (Pg) й под-
держивакшая продукция (Р6 ) A. piscinalis (а),
U. pictorum (б) и U. tumidus (в). .
По оси ординат - биомасса, ростовая и
поддерживающая продукция, по оси абсцисс -
номера исследуемых станций.
ботки (уменьшение количества моллюсков старших возрастных
групп), поскольку такая закономерность наблюдается н в тех
случаях, когда количество собранных моллюсков равномерно
распределяется по всем возрастным группам. Можно заметить
также, что принятый критерий различия 99% обеспеченности
оказался в ряде случаев очень жестким. Тем не менее выбор
его оправдан, и во всех случаях, когда вычислительный крите-
рий Стьюдента оказывался выше табличного при 99% обеспе-
ченности, различия в темпе роста моллюсков действительно
достоверны.
Изучение продукционных возможностей ' Anodonta. pis-
cinalis, Unio pictorum и и. tumidus показало, что про-
дукция этих моллюсков, как и следовало ожидать, находится
в соответствии с численностью, темпом роста и составом по-
пуляции. Так, рассчитанная на'основании изложенного в начале
данной статьи принципа определения продукции, среднегодовая
ростовая и поддерживающая продукция A. piscinalis на ст.5,
10, 11 значительно выше, чем на ст. 4 и 2 (табл. 3, рис.6, а).
Для Unio pictorum, как и для Anodonta piscinalis,
наиболее благоприятны условия жизни на ст. 5 и 10, о чем так-
же свидетельствуют величины ростовой и поддерживакщей
продукции (табл. 3, рис. 6, б). На этих станциях, как отмеча-
лось выше, наблюдалась и наиболее значительная численность
284
рассматриваемых моллюсков, молодой состав популяции и высо-
кий темп роста. Значительно ниже величина среднегодовой про*
дукции U. pictorum на станциях в районе водохранилища.
Таблица 3
Ростовая ( Pg- ), поддерживающая ( Pg )_ продукции
(г/м2 в год) и соответствующие им ng- коэффициенты:
Вид № ‘ станции P9 $ P_S В
Anodonta piscinalis 5 10 11 13.45 20.25 18.2 7.13 15.0 12.12 1.1 0.6 0.6 0.6 0.5 0.4
4 1.37 0.75 0.9 0.5
2 1.55 0.8 1.5 0.6
Unio tu- 5 3.14 2.0 0.7 0.4
midus 10 3.4 2.58 0.4 0.3
11 2.05 1.29 0.9 0.5
4 • 3.3 2.19 0.6 0.4
2 3.46 2.54 0.6 0.4
Unio 5 3.51 2.12 0.6 0.4
pictorum 10 4.55 3.49 0.5 0.4
11 0.98 0.6 0.5 0.3
4 1.52 1.33 0.4 0.3
2 1.35 0.95 0.6 0.4
Такой низкий уровень продукции соответствует низкому
¥емпу роста этого моллюска в этом районе. Наиболее низкая
среднегодовая продукция U. pictorum наблюдалась в районе
ст. 11, где, несмотря на достаточно высокий темп роста, а
небольшая био-
также преобладание в популяции быстрорастущих молодых осо-
бей, ростовая продукция составляла 0.98 г/м^ в год, поддержи-
вающая - 0.6 г/м^ в год, а д- коэфф, равны соответственно
0.5 н 0.3. Причина такой низкой продукции - —*---------л
масса U,. pictorum в этой популяции.
Как уже отмечалось выше, численность
U. tunidus, его
темп роста, а также возрастной состав популяций - все эти
показатели значительно различаются между собой на шачинскнх
и водохранилншных станциях. Одиако значения продукции у»
285
tumidus на всех рассматриваемых станциях мало отличаются
(рис. 6, в). Исключение составляет лишь популяция этого мол-
люска, находящаяся на 11-й станции (табл. 3). Полученные ре-
зультаты показывают- такжэ, что продукция U. tumidus' на во-
дохранилищных станциях выше и по сравнению с продукцией
других двустворчатых моллюсков. Этот факт непосредственно
связан с относительно большей плотностью и, tumidus в по-
пуляциях водохранилища, чем A. piscinalis и U. pic to rum.
Таким образом, приустьевой залив р. Шачи, судя по чис-
ленности, темпу роста и продукции моллюсков, представляет
более благоприятные условия для A. piscinalis и и. picto-
rum (для U. tumidus этого не наблюдалось). Это можно бы-
ло бы объяснить более высокой температурой залива, чем во-
дохранилища. Однако увеличение темпа роста этих моллюсков
наблюдалось и в те годы (1966-1969), когда подогрева еше
ие было.* Очевидно, в мелководном заливе бы л всегда сильнее
естественный прогрев и лучшие условия питания.
По-видимому, полученные нами результаты следует при-
нять за контроль, с которым следует сравнить те изменения,
которые будут происходить в этом районе, когда мощность
ГРЭС будет значительно увеличена.
Лите ратура
Голиков А.Н. 1970. Метод определения продукционных
свойств популяции по размерной структуре и численнос-
ти. - ДАН СССР, 193, 3.
Голиков А.Н., Меншуткин В.В. 1971. Модельное
исследование ‘продукционного процесса популяции брю-
хоногого моллюска. - ДАН СССР, 197, 4.
Меишуткин В.В., Голиков А.Н. 1971. Моделирова-
ние популяции брюхоногого моллюска на электронно-
вычислительной машине. - Океанология, XI, 4.
Методы определения продукции водных животных. 1968
Под ред. Г.Г.Винберга. Минск.
286
РЕФЕРАТЫ
УДК 577.42
Проблема влияния тепловых и атомных электростанций на гидробиологи-
ческий режим водоемов (обзор). Мордухай-Болтовской
Ф.Д. В сб.: Экология организмов водохранилищ-охладителей. Тр. Инет,
биол. внутр, вод АН СССР, вып. 27. 1975. Изд-во' .Наука*, Леиингр.
отд., Л. 7-69.
Рассматривается современное состояние проблемы влияния деятельнос-
ти теплоэлектростанций, в особенности сброса ими подогретых вод, па
все стороны экосистемы водоемов, принимающих эти воды. Наиболее
подробно рассмотрены материалы, относящиеся к планктону и бентосу
водоемов. Высказываются предположения о направлении дальнейших
исследований. Дан обзор советской и иностранной литературы по вопро-
су о влиянии теплоэлектростанций на гидробиологию водоемов, в кото-
рые сбрасываются подогретые воды. В приводимом списке литературных
источников указываются также основные работы по влиянию температу-
ры на гидробионтов вообще. Илл. - 16, библ. - 295 назв.
УДК 627.81
Особенности температурного режима Иваньковского водохранилища в
условиях искусственного подогрева. Буторин Н.В. и К у р д и-
н а Т.Н. В сб.: Экология организмов водохранилищ-охладителей. Тр.
Инет. биол. внутр, вод АН СССР, вып. 22. 1975. Изд-во «Наука*, Ле-
нине р. отд., Л. 70-142.
Исследования температурного режима Иваньковского водохранилища В
условиях искусственного подогрева показали, что сброс подогретых вод
Конаковской ГРЭС оказал существенное влияние на температуру воды
лишь в Мошковичском заливе и на небольшом участке примыкающей
к нему акватории Иваньковского плеса. На остальной части плеса повы-
шение температуры воды не превышает многолетних колебаний средне-
месячных ее значений, наблюдавшихся до сооружения ГРЭС, Табл, - 4,
илл. - 33, библ. - 32 назв.
УДК 577.475 (28)
Влияние подогретых вод на фитопланктон 'Иваньковского водохранилища.
‘Девяткин В.Г. В сб.: Экология ергаиизмов водохранилищ-охла-
дителей. Тр. Инет. биоп. внутр, вод АН СССР, вып. 27. 1975. Изд-во
«Наука*, Леиингр. отд., Л. 143-187.
По данным 1967-1970 гг. рассматривается влияние подогретых вод Ко-
наковской ГРЭС на фитопланктон Иваньковского водохранилища. Отмеча-
ется, что в зоне распространения смешанных подогретых вод> наблюдает-
ся увеличение обилия фитопланктона в 1.5-3.0 раза по сравнению е вы-
ше- и ннжерасположениыми участками водоема. В отличие от вдвоем**-
287
охладителей с оборотной системой водоснабжения степень воздействия
подогретых вод Конаковской ГРЭС на фитопланктон Иваньковского водо-
хранилища в большей степени зависит от гидрометеорологических усло-
вий. Влияние подогретых вод на околоплотинный участок, откуда вода
поступает для водоснабжения г. Москвы, в течение вегетационного
периода было незначительным. Табл. - 6, илл. - 37, библ. - 36 назв.
УДК 58-1.526.3 .
Растительность Иваньковского водохранилиша и влияние на нее подогре-
тых вод Конаковской ГРЭС. Экзерцев В.А. и Лнсипнна
Л.И. В сб.: Экология организмов водохранилищ-охладителей. Тр. Инет,
биол. внутр, вод АН СССР, вып. 27. 1975. Изд-во „Наука*. Ленингр.
отд., Л. 198-210.
Характеристика растительности мелководий Иваньковского плеса и зако-
номерности зарастания мелководий. Влияние теплых вод, сбрасываемых
Конаковской ГРЭС, на развитие и распределение зарослей макрофитов
в Мошковичском заливе. Илл. - 2, библ. - 19 назв.
УДК 593.1:577.472(28)
Влияние сбросных теплых вод Конаковской ГРЭС на планктонных про-
стейших Иваньковского водохранилища. Ма м а е в а Н.В. В сб.:
Экология организмов водохранилиш-охладитепей. Тр. Инет. биол. внутр,
вод АН СССР, вып. 27, 1975. Изд-во „Наука", Ленингр. отд., Л. 211-219,
Изучение вопроса проводится впервые. Влияние подогретых вод Конаков-
ской ГРЭС на планктонных простейших Иваньковского водохранилища в
разные месяцы года проявляется различно. Весной и осенью количество
простейших увеличивается, летом - сокращается. В жаркое время года
в зоне влияния подогрева наблюдалась полная гибель простейших. Табл. -
1, илл, - 5, библ. - 4 назв.
УДК 577.475 (28)
Зоопланктон Иваньковского водохранилиша в зоне влияния подогретых
вод Конаковской ГРЭС. Ривьер И.К. В сб.: Экология организ-
мов водохранилиш-охладителей. Тр. Инет.' биол. внутр, вод АН СССР,
вып. 27, 1975. Изд-во „Наука", Ленингр. отд., Л. 220-243.
Рассмотрено влияние охладителей системы ГРЭС на различных планктон-
ных животных. Показано, что в наибольшем количестве гибнут крупные
пелагические ракообразные. Сильнее всего падает биомаоса животных
в самые теплые периоды лета. Оставшиеся в живых рачки активно миг-
рируют из теплого потока и скапливаются у дна. Весной, осенью и зи-
мой район, подверженный влиянию теплых вод, более богат зоопланк-
тоном. Весной и в начале лета Daphnia cucuHata в эоне влияния
теплых вод лучше растет и быстрее достигает половой зрелости. Летом
же развитие атого вида тормозится. Табл. - 8, илл. - 9, библ. - 10
назв.
288
УДК 577.475(28)
Зоопланктон Горьковского водохранилища в районе Костромской ГРЭС.
Елагина Т.С. В сб.: Экология организмов водохранилищ-охлади-
телей. Тр. Инет. биол. внутр, вод АН СССР, вып. 27, 1075. Изд-во
.Наука*, Ленингр. отд., Л. 244-257.
В зоне значительного подогрева (приустьевой залив р. Шачи) наблюда-
ется более раннее весеннее повышение .численности и биомассы зоо-
планктона и более позднее их осеннее понижение. В период наиболее
высоких температур (до 26-29°, июль-август) замечается угнетение
зоопланктона или уход его в нижние, более холодные слои воды. В это
время сильнее проявляется и гибель части планктонных ракообразных
вследствие травмирования на водосбросе. Табл. - 3, илл. - 6, библ. -
3 назв.
УДК 577.472(28)
Состав и распределение -зообентосаГорьковского водохранилиша в райо-
не Костромской ГРЭС. Скальская И.А. В сб.: Экология орга-
низмов водохранилищ-охладителей. Тр. Инет. биол. виутр. вод АН СССР,
вып. 27. 1975. Изд-во «Наука*, Ленингр. отд., Л. 258-271.
Средняя биомасса зообентоса в водохранилище (эоне с естественным
температурным режимом) составляла 11.4 г/м^, а В приустьевом зали-
ве р. Шачи (зона наибольшего подогрева) - 3.0 г/м^. Причина этих
отличий заключается, по всей видимости, в различиях не температурно-
го режима, а других условий обитания, особенно характера грунта. Спе-
цифических термофильных форм не обнаружено. В зоне наибольшего по-
догрева созревание тубифицид и окукливание- хироирмид происходит
раньше, чем в зоне с естественным температурным режимом. Илл. -
8, библ. — 5 назв.
УДК 594.1:577.472(28)
Рост и продукция уиионид (Mollusca, Unionidae) в районе Костром-
ской ГРЭС. Смирнова Н.Ф. В сб.: Экология организмов водохра-
нилищ-охладителей. Тр. Инет. бнол. внутр, вод АН СССР, вып. 27,
1975. Изд-во «Наука*, Ленингр. отд., Л. 272-^88,
Приустьевой залив р. Шачи, непосредственно прилегающий х ГРЭС, судя
по численности, темпу роста и продукции моллюсков Anodonta pis-
cinalis и Uni о pictorum (для U, tumidus этого ие наблюдалось)
имеет .наиболее благоприятные условия. Однако увеличение темпа роста
моллюсков в этом районе наблюдалось и до строительства ГРЭС. Поду-
ченные результаты следует принять за контроль для будущих исследо-
ваний, когда мощность ГРЭС будет значительно увеличена. Табл. - 3.
илл. - 6, библ. - 4 назв.
289
С одержание
Стр.
Иван Дмитриевич Папанин - организатор Института
биологии внутренних вод АН СССР ;............. 3
Ф.Д.М ордухай-Болтове к. ой. Проблема влияния
тепловых и атомных электростанций на гидробиоло-
гический режим водоемов (обзор).................... 7 •
Н.В.Буторин и Т.Н.Курдина. Особенности темпера-
турного режи ма Иваньковского водохранилища в
условиях искусственного подогрева ........... 70
В.Г.Девяткин. Влияние подогретых вод на фитопланк-
тон Иваньковского водохранилища ............. 143
В.А.Экзериев и Л.И.Л и си ни на. Растительность
Иваньковского водохранилища и влияние на нее по-
догретых вод Конаковской ГРЭС ........ •• . 198
Н.В.Мамаева. Влияние сбросных теплых вод Кона-
ковской ГРЭС на планктонных простейших Ивань-
ковского водохранилища..................... 211
И.К.Ривьер. Зоопланктон Иваньковского водохранили-
ща в зоне влияния подогретых вод Конаковской
ГРЭС........................ . .............. 220
Т.С.Елагина. Зоопланктон Горьковского водохранилища
в районе Костромской ГРЭС............. 244
И.А.Скальская. Состав и распределение зообентоса
Горьковского водохранилища в районе Костромской
ГРЭС......................................... 258
Н.Ф.С мир ио в а. Рост, и продукция унионид (Union!-
dae) в районе Костромской ГРЭС 272
Рефераты................................ . . . . 287
/
* «
Contents
Page
Ivan Dmitrievich Papanin - the organizer o£ the
Institut for the biology of inland waters
Academy of Sience of the USSR............. 3
Ph. D. Mo rdukhai-BoltovskoL The
problem of influence of power plants on
the • hydrobiological regime 'of water - bodies
(review)................................ 7
N.V.B u t о r i n and T.N.K u r d i n a. Peculiari-
ties of temperature regime of Ivankovskoie
Reservoir due to artificial heating.......'70
V.G.D evyatki n. Influence of heated water
on phytoplankton of Ivankovskoie Reservoir 143
V.A.E kzertzev and L.I.L isitzina.
Higher vegetation of Ivankovskoie Reser-
voir and influence on it of heated water
from Konakovskaya power plant............ 198
N.V.M a m a e v a. Influence of heated waters
from Konakovskaya power plant on plank-
tonic Protozoa of Ivankovskoie Reservoir 211
LK,Rivi er, Zooplankton of Ivankovskoie*
Reservoir in the zone influenced by heated
water from Konakovskaya power plant 220
T.S.E 1 agi na. Zooplankton of Gorkovskoie
Reservoir in the zone of Ko stroms к aya
power plant ......................... 224
LAS к alskay a. .Composition and distribution
of zoobenthos of Gorkovskoie Reservoir
in the zone of Kostromskaya power plant 258
N.F.S m i r n о v a. Growth and production of .
Onionidae (Molluspa, Bivalvia) in the zone .
of Kostromskaya power plant ........ 272
Abstracts............. ........ 287
ЭКОЛОГИЯ ОРГАНИЗМОВ
ВОДОХРАНИЛИЩ-ОХЛАДИТЕЛЕЙ
Труды Института биологии внутренних вод.
Том 27(30)
Утверждено х печати
Институтом биологии внутренних вод
Академии наук СССР
Редактор издательства И.П.Дубровская
Художник Д.С.Данилов
Технический редактор В.В.Шиханова
Корректоры Ж.Д.Андронова и Л.В.Субботина
Сдано в производство и подписано к печати 29/Х 1975 г.
Формат бумаги 60x90 1/16. Бумага № 1. Печ. л. 18 1 /4 „в
в 18.25 усл; печ, л. Уч.-изд. л. 16.10. Изд. №5521. Тип. зак. № 17
М-56195. Тираж 700. Цена 1 р. 85 к.
Ленинградское отделение издательства «Наука* 1
199164, Ленинград, В-164, Менделеевская линия, д. 1 I
1-я тип. издательства «Наука*, ' «
199034, Ленинград, В-34, 9 линия, д. 12
А.Н, <СС?
де 4MW4X «иу’уеви*
БИБЛИОТЕКА