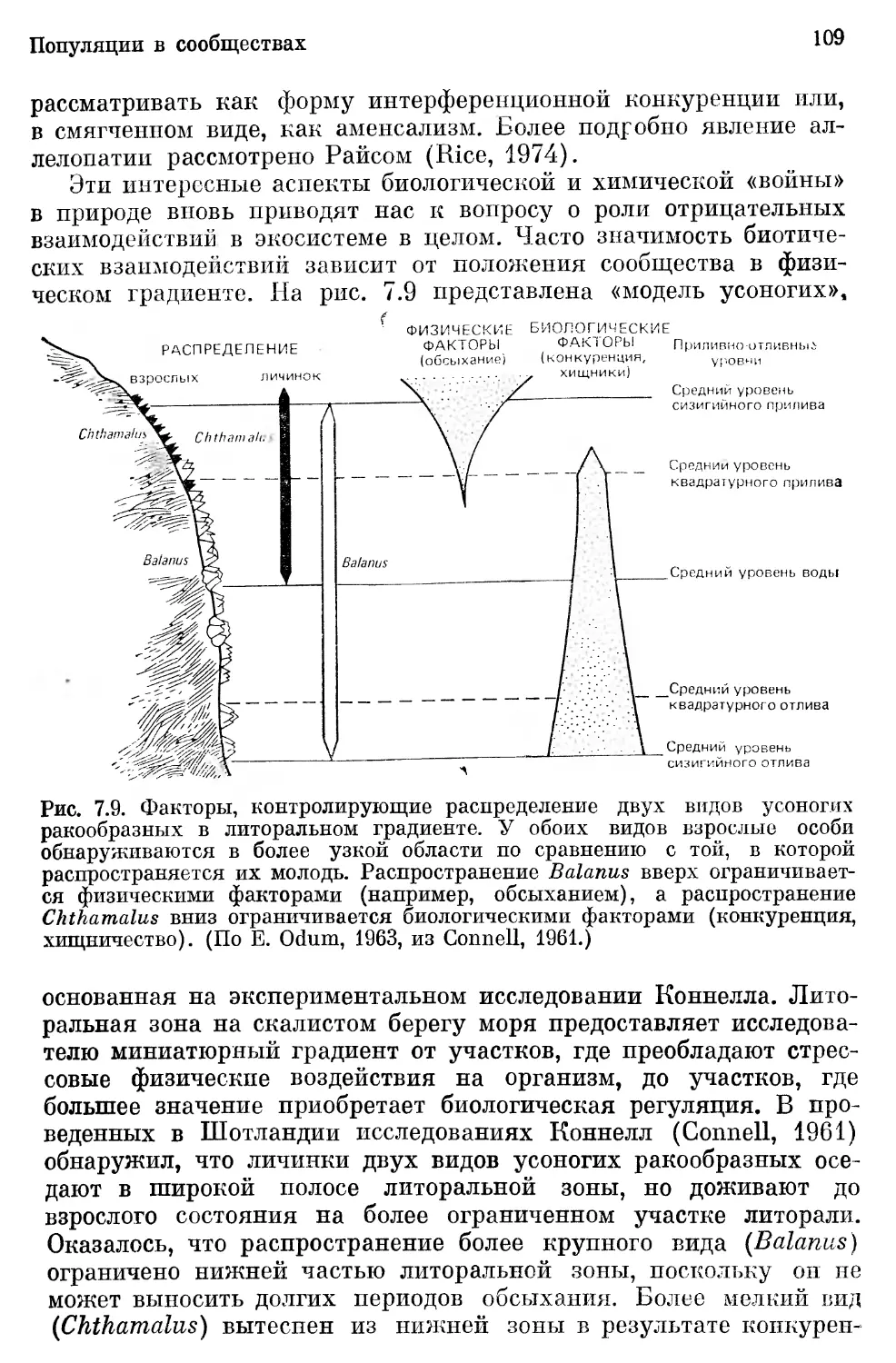
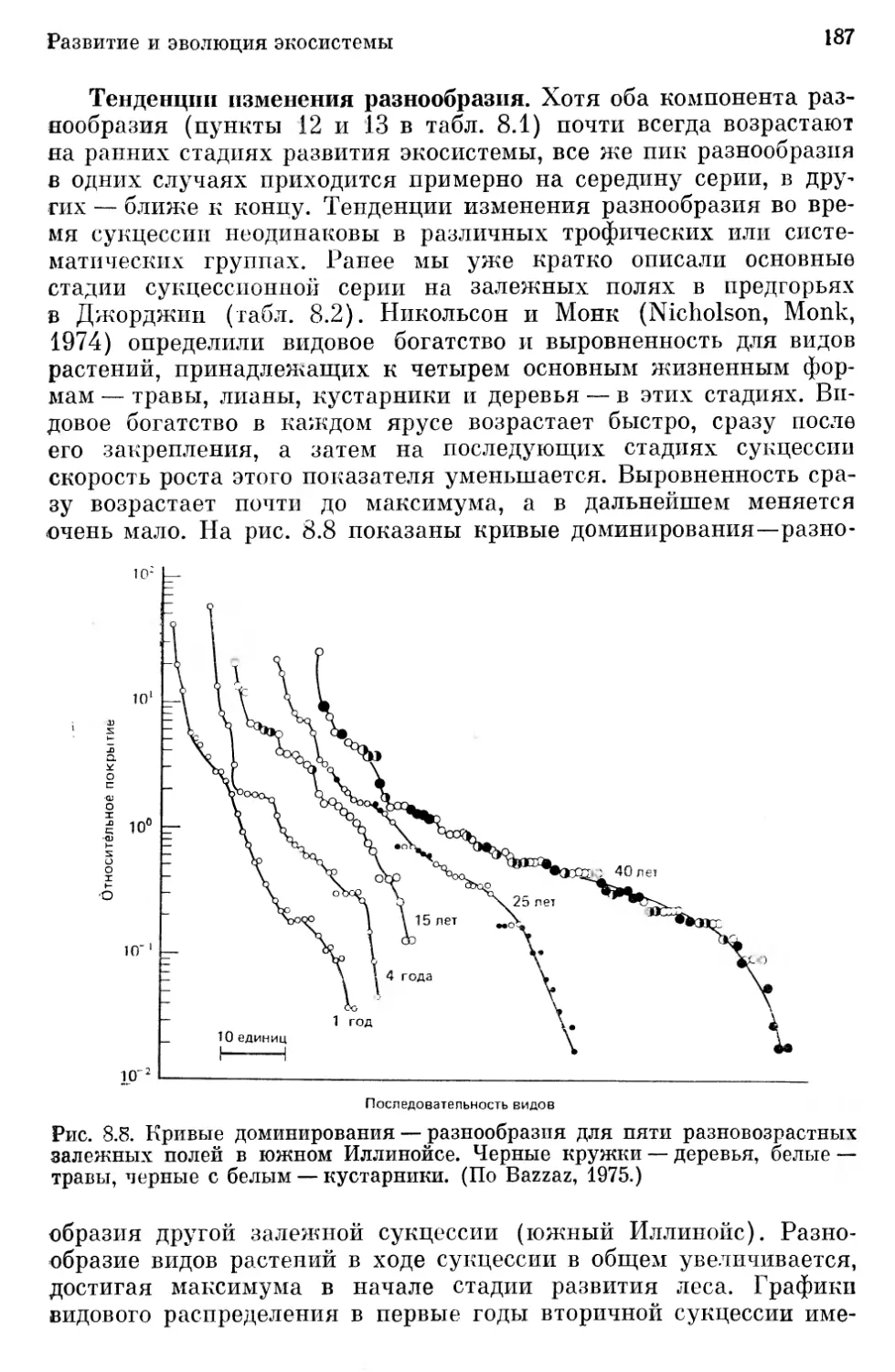
/


























































































































































































































































































































































































Текст
ю.одум ЭКО/1ОГИ7
экология
B/SIC EOLOGY
Eugene P. Odum
Calloway Professor of Ecology
and Director of the Institute of Ecology
University of Georgia
Saunders College Publishing
Philadelphia New York Chicago
San Francisco Montreal Toronto
London Sydney Tokyo Mexico City
Rio de Janeiro Madrid
Ю. Одум
ЭКОЛОГИЯ 2
Б двух томах
Перевод с английского
канд. биол. наук Б. Я. Виленкина
под редакцией
академика В. Е. Соколова
Москва «Мир» 1986
ББК 28.081
044
УДК 577.4
Одум 10.
044 Экология: В 2-х т. Т. 2. Пер. с англ. — М.: Мир, 1986.—
376 с., ил.
Книга представляет собой переработанное и сокращенное автором изда-
ние ранее опубликованных «Основ экологии» (М.: Мир, 1975).
Второй том содержит главы, в которых рассмотрены вопросы динамики
популяций; взаимоотношения между популяциями, сообществами и экоси-
стемами; динамики экосистем и эволюционной экологии; а также вопросы,
связанные с перспективами на будущее всего человечества. В конце книги
дана краткая сводка основных типов экосистем биосферы.
Предназначена для биологов всех специальностей, студентов и препо-
давателей биологических вузов, а также для лиц, интересующихся пробле-
мами использования природных ресурсов и охраны окружающей среды.
Ж1050000-Ж ч1
041(01)—86
ББК 28.081
Редакция литературы по биологии
© 1983 by CBS College Publishing. All rights
reserved. Printed in the United States of
America.
© перевод иа русский язык, «Мир», 1986.
. 6
Динамика популяций
Предмет этой и последующих глав составляют более биологпч-
ные аспекты экологии. Речь пойдет о взаимодействии организмов
с организмами при их функционировании в экосистеме. В гл. 2—5
обсуждались долгодействующие мощные физические и химиче-
ские факторы, выступающие как главные внешние ведущие си-
лы. Однако организмы не подстраиваются к этим силам путем
пассивной адаптации, а активно модифицируют и изменяют окру-
жающую среду, воздействуя на нее в пределах законов природы,
которые определяют преобразование энергии и круговороты ве-
ществ. Иными словами, человечество не единственная популя-
ция, которая изменяет окружающую среду и пытается взять ее
под контроль. В соответствии со схемой уровней организации
(рис. 1.1) в этой и следующей главах внимание будет сосредо-
точено на биотических уровнях популяций и сообществ. На этих
уровнях взаимодействие между генетическими системами и окру-
жающей средой определяет ход естественного отбора и тем самым
не только то, как индивидуальный организм оптимизирует свое
существование, но и то, как на протяжении эволюционных отрез-
ков времени должны были меняться и менялись целостные эко-
системы.
1. Свойства популяционной группы
Определения
Популяцию можно определить как любую группу организмов од-
ного вида (или иную группу, внутри которой особи могут обме-
ниваться генетической информацией), занимающую определенное
6
Глава 6
пространство и функционирующую как часть биотического со-
общества. Биотическое сообщество в свою очередь определяется
как совокупность популяций, которая в результате коэволюции
метаболических превращений функционирует как целостная еди-
ница в отведенном ему пространстве физической среды обитания.
Популяция характеризуется рядом признаков; хотя эти признаки
лучше всего выражаются статистическими функциями, единст-
венным их носителем является группа, но не особи в этой груп-
пе. Плотность, рождаемость, смертность, возрастная структура,
биотический потенциал, распределение в пространстве (диспер-
сия) и кривая роста — вот некоторые из таких свойств популя-
ции. Популяции обладают также генетическими характеристика-
ми, непосредственно связанными с их экологией. Это адаптив-
ность, репродуктивная (дарвиновская) приспособленность и не-
прерывность (т. е. вероятность оставления потомков на протяже-
нии длительного периода времени).
Объяснения
Популяция, как это хорошо выразил один из пионеров современ-
ной популяционной экологии Томас Парк (см. Allee et al., 1949),
обладает «биологическими свойствами», присущими как популя-
ции, так и составляющим ее организмам, и «групповыми свойст-
вами», присущими только группе в целом. Биологические свойст-
ва характеризуют жизненный цикл популяции: популяция, так
же как и отдельный организм, растет, дифференцируется и под-
держивает сама себя. Популяция имеет определенную организа-
цию и структуру, которые можно описать. В отличие от этого
групповые свойства, такие, как рождаемость, смертность, возраст-
ная структура и генетическая приспособленность, могут характе-
ризовать только популяцию в целом. Таким образом, особь рож-
дается, стареет, умирает, но применительно к особи нельзя гово-
рить о рождаемости, смертности, возрастной структуре — харак-
теристиках, имеющих смысл только на групповом уровне.
Ниже даются определения и краткие описания основных
свойств популяции.
Плотность. Плотность популяции — это величина популяции,
отнесенная к некоторой единице пространства. Ее обычно изме-
ряют и выражают числом особей или биомассой популяции на
единицу площади пли объема, например 500 деревьев на 1 га,
5 млн. диатомей на 1 м3 воды или 200 кг рыбы на 1 га поверх-
ности водоема. Иногда бывает важно различать среднюю плотность,
т. е. численность (или биомассу) на единицу всего пространства,
и удельную, или экологическую, плотность, т. е. численность (или
биомассу) па единицу обитаемого пространства (доступной пло-
щади или объема, которые фактически могут быть заняты попу-
Динамика популяций
7
ляцией). Часто важнее знать не конкретную величину популяции
в тот или иной момент времени, а ее динамику, т. е. возрастает
она или уменьшается. В таких случаях полезны показатели от-
носительного обилия. Они могут быть отнесены к определенным
промежуткам времени, например число птиц, отмеченных в те-
чение часа. Их можно выразить также в процентах, например,
частота встречаемости — это процент проб, в которых обнаружен
данный вид. В описательных работах по геоботанике плотность
и частоту встречаемости часто объединяют, получая показатель
значимости для каждого вида.
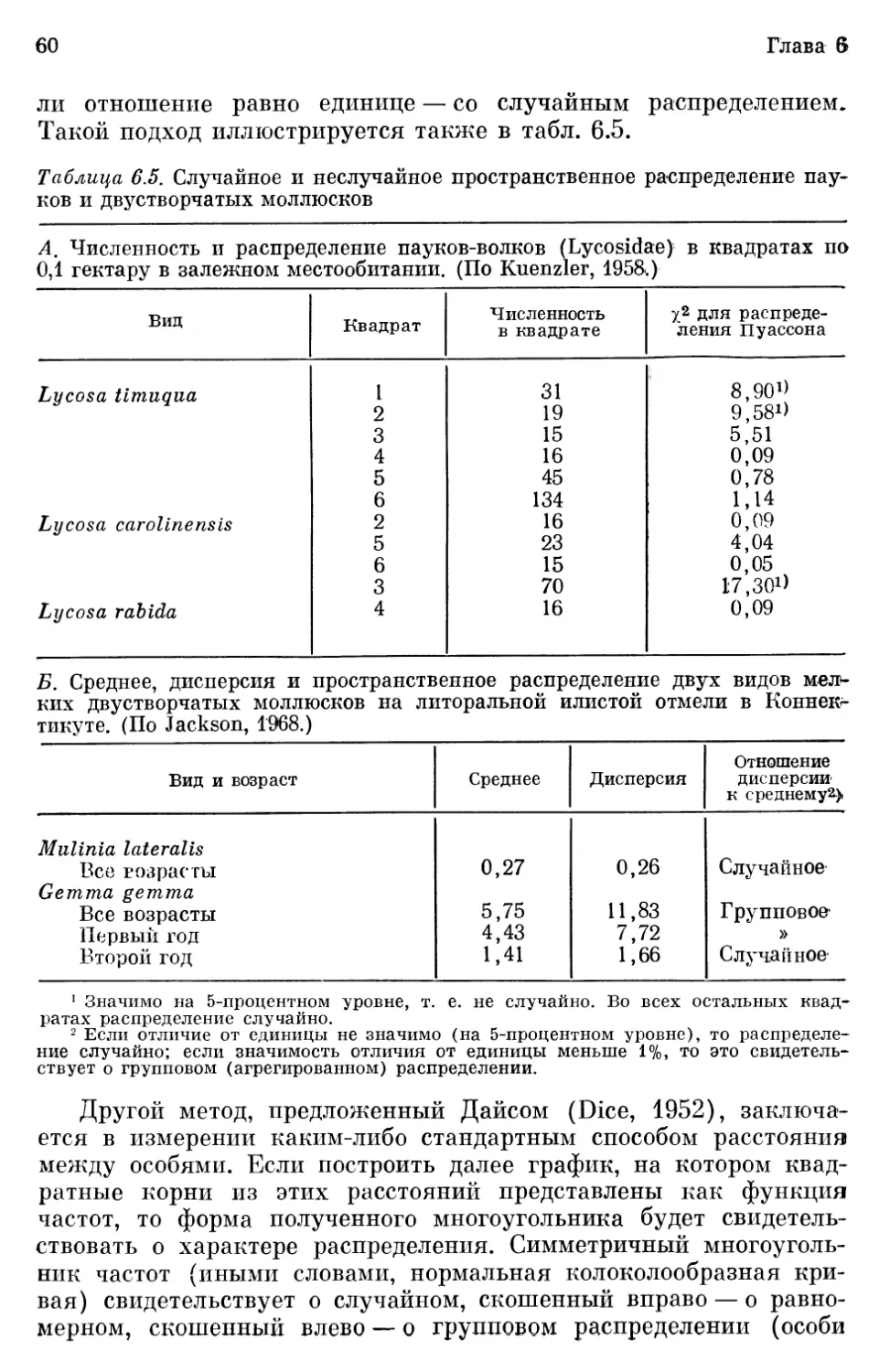
На рис. 6.1 п в табл. 6.1 показано соотношение между плот-
Г орностай
ПЛОТОЯДНЫЕ
Рысь ।
•'.•I Пума ।
Скунс
, Г-Лисица ВСЕЯДНЫЕ
I 1 • •..u-.i Черный медведь
0,001
I Хомячок (Peromyscus) t..'_з I
ПИТАЮЩИЕСЯ СЕМЕНАМИ И ( Бурундук г-;„- zzzzj I
ПЛОДАМИ । 1 !
I । Бел кап-- •и.. I
----------’----------7й—-— ! .
I П ол ев к и (Мicro rm /..- „• •.
I . I................. | ,,
ПИТАЮЩИЕСЯ ЛИСТЬЯМИ ' Лесной сурок
I । 1
I I Олень к’.-.-агг-у
inn
0,1 1,0
Биомасса.кг/га
Рис. 6.1. Диапазон изменений плотностей популяций (выраженных в био-
массе на гектар) у разных видов млекопитающих. Результаты получены
в предпочитаемых данным видом местообитаниях, где антропогенное влия-
ние выражено незначительно. Виды расположены в соответствии с их тро-
фическим уровнем, а в пределах каждого уровня — в соответствии с раз-
мерами животного, что иллюстрирует влияние на ожидаемую биомассу раз-
меров организма и его положения в пищевой цепи. (Схема составлена по
данным Mohr, 1940, и дополнена результатами позднейших исследований.)
ностями популяций млекопитающих, их трофическими (энерге-
тическими) уровнями и величиной особей. Хотя плотности попу-
ляций разных представителей млекопитающих как класса охва-
тывают диапазон почти в пять порядков величины, диапазон
колебаний плотности популяции для каждого вида или трофиче-
ской группы намного меньше (рис. 6.1). Чем ниже трофический
8
Глава 6
уровень, тем выше плотность, а чем крупнее животные внутри
данного уровня, тем больше их биомасса. Так как у крупных
животных интенсивность метаболизма на единицу массы меньше,
чем у мелких, на данной энергетической базе может поддержи-
ваться большая биомасса крупных животных.
Если размеры особей в популяции и интенсивности их мета-
болизма примерно одинаковы, то плотность удобно выражать чис-
лом особей, однако часто встречается ситуация, показанная в
табл. 6.1, Б. Относительные достоинства таких показателей, как
численность, биомасса и поток энергии, обсуждались в гл. 3 (см.,
в частности, табл. 3.6). Вспомним следующее утверждение из
этой главы: «данные по численности приводят к переоценке зна-
чения мелких организмов, а данные по биомассе — к переоценке
роли крупных организмов». Поток энергии «служит более под-
ходящим показателем для сравнения любого компонента с другим
и всех компонентов экосистемы между собой».
Многие специальные количественные показатели и термины
широко применяются только при работе с определенными попу-
ляциями или группами популяций. Лесные экологи, например,
часто пользуются термином «площадь оснований» (суммарная
площадь поперечных срезов стволов) как мерой плотности де-
ревьев. Лесники же определяют число бордсовых, или досковых,
футов на 1 акр как меру полезной с коммерческой точки зрения
части всего древостоя. Эти и другие меры могут служить оценка-
ми плотности, поскольку они в известной степени отражают раз-
меры «урожая на корню» и, следовательно, соответствуют данно-
му ранее определению плотности в широком смысле.
Мы уже указывали, что если необходимо знать характер из-
менения популяции или если в данных условиях нельзя опреде-
лить абсолютную плотность, то часто бывают полезными оценки
относительного обилия. Термины «обильный», «обычный», «ред-
кий» п т. п. применимы в тех случаях, когда они соответствуют
измерениям или оценкам, полученным таким способом, что срав-
нение имеет смысл.
Легко догадаться, что «индексы» относительного обилия ши-
роко используются для популяций крупных животных и назем-
ных растений, когда необходимо проводить оценку численности
на больших площадях без чрезмерных затрат времени и денег.
Например, административные органы, на которые возложена обя-
занность ежегодно регулировать охоту па перелетную водоплаваю-
щую дичь, чтобы соблюсти интересы и охотников,
и птиц, должны знать, в каких местах популяции стали меньше,
больше или остались такими же, как в предыдущем году. Это
можно сделать путем оценки относительного обилия, получен-
ной в результате обследования угодий егерями, опроса и учета
гнездовий. Данные суммируют и представляют как число птиц,
Динамика популяций
Таблица 6.1. Влияние трофического уровня на биомассу рыб (4) и взаимо-
связь между численностью и биомассой в популяции с быстро меняющейся
возрастной и размерной структурой (Б)
А. Смешанные популяции, биомасса на единицу площади
Рыба в искусственных прудах шт. Иллинойс. (Данные из Thompson, Ben-
nett, 1939.) Расположение групп рыб приблизительно соответствует их по-
ложению в пищевой цепи; рыбы, питающиеся «грубыми» кормами, зани-
мают нижний трофический уровень, объекты спортивной ловли — верхний
Количество рыбы, кг на 1 га
пруд №1 пруд №2 пруд №3
Объекты спортивного и хозяйственного рыбо-
ловства (окунь, ушастый окунь и др.) 260 50 10
Сомики (сомик-кошка, канальный сомик) Промысловые рыбы (нотрописы, сельдевые 0 45 70
и др.) Потребители грубых кормов (чукучаны, кар- 0 265 3,5
пы и др.) 0 97 1300
Всего 260 457 1383,5
Б. Сравнение численностей и биомасс в случае, когда размеры организма
сильно меняются с возрастом
Молодь нерки в одном из озер Британской Колумбии. Мальки вылупляются
в реках и в апреле мигрируют в озеро, где остаются до полового созрева-
ния. Обратите внимание, что с мая по октябрь особи быстро увеличиваются
в размерах, поэтому биомасса возрастает втрое, даже несмотря на значи-
тельное уменьшение численности рыб. С октября по апрель следующего
года рост очень замедлен, и общая биомасса снижается из-за продолжаю-
щейся гибели рыб. (Данные из Ricker, Forester, 1948.)
Май Октябрь Апрель
Число особей в озере, тыс. Биомасса, т 4000 1,0 500 3,3 150 2,0
увиденных или убитых за единицу учетного усилия. Показатели,
выраженные в процентах, широко используются при изучении
растительности; общеупотребительны такие специальные показа-
тели, как частота — процент пробных площадок, на которых встре-
чается данный вид; обилие — процент особей в пробе; покрытие -
процент поверхности почвы, оказавшейся закрытой при проекции
на нее надземных частей растений. Следует быть осторожным и
не путать эти индексы с истинной плотностью, которая всегда
выражается по отношению к определенному пространству.
10
Глава 6
Различие между средней плотностью и экологической плот-
ностью можно иллюстрировать данными Коля (Kahl, 1964) по
лесному аисту в Национальном парке Эверглейдс (Флорида). На
рис. 6.2 показано, что в этой местности плотность мелких рыб с
Рис. 6.2. Экологическая и средняя плотность рыб, служащих добычей аисту
в период его размножения. По мере падения уровня воды в течение сухого
сезона в водоемах на юге Флориды средняя плотность мелких рыб умень-
шается (т. е. уменьшается число рыб на 1 км3 поверхности), так как со-
кращаются размеры и число водоемов, но экологическая плотность (т. е. ко-
личество рыбы на 1 м2 сохранившейся водной поверхности) возрастает,
поскольку рыба скапливается в водоемах с меньшей площадью. Гнездова-
ние аиста приурочено к тому времени, когда максимальная доступность
пищи совпадает с наибольшими пищевыми потребностями подрастающих
птенцов. (По Kahl, 1964.)
падением уровня воды в течение сухого зимнего сезона в целом
снижается, но экологическая плотность при этом возрастает, так
как по мере сокращения зеркала воды увеличивается число рыб,
приходящихся на единицу водной поверхности. Аисты отклады-
вают яйца в такое время, что вылупление птенцов приходится
на период пика экологической плотности рыб. Это облегчает ро-
дителям ловлю рыбы, которая составляет основную пищу птенцов.
В мире лесного аиста в расчет принимается экологическая, а не
средняя плотность пищевых объектов. В мире людей наблюдается
во многом сходная картина; в расчет принимаются главным об-
разом высококачественная энергия и пищевая продукция, скон-
центрированные на 15 или 20 процентах земли, где возможен
высокий урожай, по не рассеянные и трудно доступные для до-
бывания ресурсы.
Для измерения плотности популяций испытано много разных
методов, и сама по себе методология составляет важную область
Динамика популяций
11
исследований. Было бы нецелесообразно подробно рассматривать
здесь эти методы; их лучше постигать, пользуясь методическими
и полевыми руководствами, а еще лучше советоваться с опытным
исследователем, который вначале изучил оригинальную литерату-
ру, а потом модифицировал и усовершенствовал существующие
методы, приспособив их к конкретной ситуации. Ничто не может
заменить опыт, приходящий при проведении полевых исследо-
ваний.
Практикуемые методы можно подразделить на несколько об-
ширных категорий:
1. Тотальный учет, возможный иногда для крупных, хорошо
заметных организмов (например, бизоны на открытых равнинах)
или для организмов, собирающихся на период размножения в
большие группы (например, морские птицы и тюлени).
2. Метод пробных площадок. Этот метод состоит в подсчете
и взвешивании организмов на пробных площадках или на тран-
септах. Размеры и число пробных площадок должны быть доста-
точными для получения оценки численности в обследуемой тер-
ритории.
3. Методы мечения с повторным отловом (для подвижных
животных). Из популяции отлавливают часть животных, метят их
и выпускают. Доля меченых животных в последующей выборке
используется для определения общей численности (см. Zippin,
1958).
4. Метод изъятия, при котором число организмов, собранных
с некоторой площади при последовательных выборках, отклады-
вается по оси ординат, а число из собранных ранее — по осн
абсцисс. Если вероятность поимки относительно постоянна,
то точки ложатся вдоль прямой линии. Эту линию можно про-
длить до нулевой точки (на оси абсцисс), теоретически соответ-
ствующей 100%-ному изъятию с данной площади (см. Menhinick,
1963).
5. Методы без взятия проб (применимые к неподвижным ор-
ганизмам, таким, как деревья). Примером может служить метод
случайных точек, при котором измеряют расстояние от ряда про-
извольно выбранных точек до ближайших особей по всем четы-
рем направлениям. Плотность на единицу площади оценивают по
среднему расстоянию (см. Phillips, 1959).
Рождаемость. Рождаемость — это способность популяции к
увеличению численности. Термин рождаемость используется так-
же и при анализе популяций человека (в демографии) 1. Фактп-
1 В оригинале сказано, что для обозначения рождаемости экологи и
демографы используют разные термины (соответственно «natality» и «birth
rate»). В русской литературе в обоих случаях используется один термин.—
Прим, пер ев.
12
Глава 6
чески в популяционной экологии это просто более широкий тер-
мин; он характеризует появление на свет новых особей любого
организма независимо от того, рождаются ли они, вылупляются
из яиц, прорастают из семян или появляются в результате деле-
ния. Максимальная рождаемость (иногда ее называют абсолют-
ной или физиологической) — это теоретический максимум скоро-
сти образования новых особей в идеальных условиях (когда от-
сутствуют лимитирующие экологические факторы, размножение
сдерживается только физиологическими). Максимальная рождае-
мость постоянна для данной популяции. Экологическая, или реа-
лизованная, рождаемость (или просто «рождаемость» без какого-
либо определения) обозначает увеличение численности популяции
при фактических или специфических условиях среды. Эта вели-
чина не постоянна и варьирует в завимости от размерного и воз-
растного состава популяции и физических условий среды. Обычно
рождаемость выражают либо как скорость, определяемую путем
деления общего числа вновь появившихся особей на время (абсо-
лютная или общая рождаемость), либо как число вновь появив-
шихся особей в единицу времени на 1 особь в популяции (удель-
ная рождаемость).
Различие между абсолютной и удельной рождаемостью легко
проиллюстрировать на таком примере: предположим, что попу-
ляция из 50 простейших в некотором объеме воды увеличивается
путем деления. Через час численность ее возросла до 150 особей.
Абсолютная рождаемость при этом равна 100 особям в час,
а удельная рождаемость (средняя скорость изменения численно-
сти на особь в популяции) составляет 2 особи в час при 50 исход-
ных. Или же предположим, что в городе с населением 10 000 по-
явилось 400 новорожденных. Абсолютная рождаемость составит
400 в год, а удельная — 0,04 (4 на 100, или 4%). В демографии
рождаемость принято рассчитывать на одну женщину репродук-
тивного возраста, а не на все население.
Различие между максимальной и реализованной рождаемостью
можно проиллюстрировать данными, полученными Лески (Laskey,
1939) при изучении природной популяции певчей птицы Sialis
stalls на искусственных гнездовьях городского парка в Нашвилле,
Теннесси, и данными, полученными Парком (Park, 1934) на ла-
бораторной популяции мучного хрущака. Птицы отложили
510 яиц (около 15 яиц на самку в трех последовательных клад-
ках) ; это число соответствует максимальной рождаемости. Опе-
рилось всего только 265 птенцов, поэтому экологическая или реа-
лизованная рождаемость составляет 52% максимальной (около
8 птенцов па самку или 4 па гнездящуюся пару). В опытах на
мучном хрущаке были получены иные данные; так, в одном из
контейнеров жуками было отложено 12 000 яиц (см. Park, Gins-
burg, Horwitz, 1945), из которых вылупились только 773 (или
Динамика популяций
13
6%) личинки. В общем для видов, которые не охраняют яйца и
не заботятся о потомстве, характерна высокая потенциальная и
низкая реализованная рождаемость. В последующих разделах бу-
дут обсуждаться другие обстоятельства, влияющие на рождае-
мость.
Смертность. Смертность отражает гибель особей в популяции.
В определенной степени это понятие является антитезой рождае-
мости. Смертность эквивалентна уровню смертности в демографии.
Подобно рождаемости, смертность можно выразить числом осо-
бей, погибших за данный период (число смертей в единицу вре-
мени), или же в виде удельной смертности для всей популяции
или любой ее части. Экологическая, или реализованная, смерт-
ность — гибель особей в данных условиях среды. Эта величина,
как и экологическая рождаемость, не постоянна и изменяется в
зависимости от условий среды и состояния самой популяции.
Теоретическая минимальная смертность — величина, постоянная
для популяции; она представляет собой гибель особей в идеаль-
ных условиях, при которых популяция не подвергается лимити-
рующим воздействиям. Даже в самых лучших условиях особи
будут умирать от старости. Этот возраст определяется физиоло-
гической продолжительностью жизни, которая, конечно же, часто
намного превышает среднюю экологическую продолжительность
жизни. Нередко значительно больший интерес представляет не
смертность, а выживаемость. Если число погибших особей выра-
зить через М, то выживаемость будет равна 1—М.
Как и рождаемость, смертность, особенно у высших организ-
мов, широко варьирует с возрастом. В связи с этим большое
значение имеет определение удельной смертности для возможно
большего числа возрастных групп или стадий развития, посколь-
ку это позволяет экологам выяснить механизмы, определяющие
общую смертность в популяции. Полная картина смертности в
популяции последовательно описывается статистическими табли-
цами выживания, специально разработанными в демографических
исследованиях. Раймонд Пирл впервые использовал таблицы вы-
живания для решения некоторых проблем общей биологии, при-
менив их к данным, полученным в лабораторных исследованиях
плодовой мушки Drosophila (Pearl, Parker, 1921). Диви (Deevy,
1947, 1950) обобщил в виде таких таблиц данные по многим при-
родным популяциям от коловраток до горного барана. Со време-
ни появления обзоров Диви таблицы выживания были составлены
для самых разных природных и экспериментальных популяций.
В табл. 6.2 и 6.3 суммированы данные, полученные для аляскин-
ской популяции дикого горного барана, популяции рисового дол-
гоносика и одной гипотетической популяции. В табл. 6.3 включе-
на также удельная рождаемость для разных возрастов. Возраст
баранов определяли по рогам (чем старше баран, тем больше
14
Глава 6
костяных колец). Когда барана убивает волк или он гибнет от
других причин, его рога сохраняются еще долгое время. Адольф
Мьюри в течение нескольких лет изучал взаимоотношения волка
и дикого барана в национальном парке Мак-Кинли на Аляске.
Он собрал много рогов, получив, таким образом, интересные дан-
ные о возрасте, в котором гибнут бараны в природных условиях,
когда они подвержены опасностям, в том числе и нападению хищ-
ников— волков (человек их не трогает постольку, поскольку охо-
та на барана в Национальном парке Мак-Кинли запрещена).
Таблица выживания состоит из нескольких столбцов, обозна-
ченных следующим образом: 1Х— число особей из 1000 (или лю-
бого другого удобного числа) представителей данной популяции,
которые выживают к концу определенного интервала времени
(день, месяц, год и т. д.), указываемого в столбце х; dx— число
Таблица 6.2. Таблица выживания для популяции горного барана Далла
(Ovis d. dalli) (Данные Murie, 1944, из Deevey, 1947), построенная на основе
данных о возрасте 608 особей к моменту смерти. Учитывались бараны, по-
гибшие до 1937 г. Данные для обоих полов объединены. Средняя продолжи-
тельность жизни — 7,09 лет. Небольшое число черепов было лишено рогов,
но, по остеологическим данным, они принадлежали особям 9 лет и старше
и поэтому были распределены пропорционально среди старших возрастных
групп
О X • - к „ Р ’S >> 5 ® h и К Р О 10 н
р га р 3 s й в £ 3 О га О сД
3 о га £ Q, <*. га ОС , И « Р о р о Снй S Е s :обей, поги м возрасты» :е на 1000 г обей, пыжи каждого в нтервала н денных :ть в нача/ возрастног на 1000 жн: 1ая продол: ь жизни, и время пред тзни, у ОСО1 dx соответс озраста
га Ко О к Р р н и ” Н ло ОС ачале ого и ) рож о о « £ о Р SK х. е< га о идаел ьност днее ей Ж1 ТИГШ1 его в
о CQ О tr И S S и о 2 + с,с *= га о о О « н о О h О к 2
X л' dx 1х 1000^х ех
0—0,5 —100 54 1000 54,0 7,06
0,5—1 —93,0 145 946 153,0 —
1—2 —85,9 12 801 15,0 7,7
2—3 —71,8 13 789 16,5 6,8
3—4 —57,7 12 776 15,5 5,9
4—5 —43,5 30 764 39,3 5,0
5—6 —29,5 46 734 62,6 4,2
6—7 —15,4 48 688 69,9 3,4
7—8 — 1,1 69 640 108,0 2,6
8—9 + 13,0 132 571 231,0 1,9
9—10 + 27,0 187 439 426,0 1,3
10—11 + 41,0 156 252 619,0 0,9
11—12 + 55,0 90 96 937,0 0,6
12—13 +69,0 3 6 500,0 1,2
13—14 +84,0 3 3 1000 0,7
Динамика популяций
15
Таблица 6.3. Таблицы выживания с удельными величинами выживания и
рождаемости
А. Таблица выживания гипотетической популяции с простым жизненным
циклом
Возраст, годы Удельное выживание Удельная смертность Удельная рождаемость (число потомков на самку в возрасте х)
X 1х т х 1 г
X
0 1,00 0,20 0 0,00
1 0,80 0,20 0 0,00
2 0,60 0,20 1 0,60
3 0,40 0,20 2 0,80
4 0,20 0,10 2 0,40
5 0,10 0,05 1 0,10
6 0,05 0,05 0 0,00
7 0,00
2 /хтпх=7?0 (чистая скорость роста) = 1,90
Б. Таблица выживания лабораторной популяции рисового долгоносика
(Calandra oryzae) при оптимальных условиях (29 °C и влажности риса 14%).
Соотношение полов равно единице
j, Середина возраст- ного интервала, нед Удельное выживание Удельная рождаемость, число потомков на 1 самку возраста к
к 1х т х 1хтх
4,5 0,87 20,0 17,400
5,5 0,83 23,0 19,090
6,5 0,81 15,0 12,150
7,5 0,80 12,5 10,000
8,5 0,79 12,5 9,875
9,5 0,77 14,0 10,780
10,5 0,74 12,5 9,250
11,5 0,66 14,5 9,570
12,5 0,59 11,0 6,490
13,5 0,52 9,5 4,940
14,5 0,45 2,5 1,125
15,5 0,36 2,5 0,900
16,5 0,29 2,5 0,800
17,5 0,25 4,0 1,000
18,5 0,19 1,0 0,190
Ro = 113,560
особей, погибших в последовательные интервалы времени; qx —
смертность в течение последовательных интервалов по отношению
к величине исходной популяции и ех — ожидаемая продолжитель-
ность жизни в конце каждого интервала. Как показано в табл. 6.2,
средний возраст горных баранов превышает 7 лет, и если баран
16
Глава 6
смог выжить в течение первого года, то его шансы на выживание
до относительно позднего возраста, несмотря на обилие волков и
другие превратности судьбы, остаются довольно высокими.
Кривые, построенные на основе таблиц выживания, могут быть
весьма информативными. Если данные столбца 1Х отложены на
графике, где горизонтальная координата соответствует интервалу
времени, а вертикальная — числу выживших, то полученная кри-
вая называется кривой выживания. Такие кривые, построенные
в полулогарифмическом масштабе, причем интервалы времени по
оси абсцисс отложены как процент средней продолжительности
жизни (см. столбец х' в табл. 6.2) или как процент общей про-
должительности жизни, позволяют сравнивать виды с очень раз-
ной продолжительностью жизни. Кроме того, прямая линия, по-
строенная в полулогарифмическом масштабе, свидетельствует о
постоянстве удельного выживания.
Кривые выживания подразделяют на три общих типа, пока-
занных на рис. 6.3. Сильно выпуклая кривая (Z на рис. 6.3) ха-
рактерна для таких видов, как, например, баран Далла, в попу-
ляциях которых смертность почти до конца жизненного цикла
остается низкой. (Мы предлагаем нанести данные из столбца
табл. 6.3 на полулогарифмическую миллиметровую бумагу и срав-
нить форму полученной кривой с кривыми на рис. 6.3.) Этот тип
кривой выживания характерен для многих видов крупных жи-
вотных и, конечно, для человека. Другой крайний вариант, сильно
вогнутая кривая (V на рис. 6.3), получается, если смертность вы-
сока на ранних стадиях. Хорошей иллюстрацией этого служат
устрицы и другие двустворчатые моллюски, а также дубы: смерт-
ность очень велика у свободноплавающих личинок и прорастаю-
щих желудей, но, как только особь хорошо приживется на под-
ходящем субстрате, ожидаемая продолжительность жизни сильно
увеличивается. К промежуточному типу относятся кривые вы-
живания для тех видов, у которых удельная выживаемость для
каждой возрастной группы более или менее одинакова, так что
в полулогарифмическом масштабе кривые приближаются к диа-
гоналям (Z/Z, /У на рис. 6.3). Ступенчатый тип кривой выжива-
ния характерен для видов, у которых выживаемость сильно варь-
ирует на последовательных стадиях жизненного цикла, как это
часто бывает у насекомых с полным превращением (голометабо-
лических), например у бабочек. На изображенной кривой {II на
рис. 6.3) крутые участки соответствуют стадиям яйца, окуклива-
ния и короткоживущему имаго: пологие участки соответствуют
стадиям личинки и куколки, у которых смертность ниже (смв
Ito, 1959). Вероятно, в природе не существует популяций, у ко-
торых выживаемость постоянна на протяжении всего жизненного
цикла (кривая III на рис. 6.3); однако слегка вогнутые или сиг-
моидные кривые (IV на рис. 6.3) всегда характерны для многих
Динамика популяций
17
Рис. 6.3. Разные типы кривых выживания. По оси ординат отложено число
выживших на тысячу особей (логарифмическая шкала), а по оси абсцисс —
возраст в процентах от продолжительности жизни. / — выпуклая кривая
(наибольшая смертность приходится на конец жизни). II — ступенчатая
кривая (выживание резко меняется при переходе от одной стадии жиз-
ненного цикла к другой). III — теоретическая кривая (прямая; выживание
для разных возрастов остается постоянным). IV — сигмоидная кривая,
приближающаяся к III. V — вогнутая кривая (смертность очень высока на
ранних стадиях).
птиц, мышей и кроликов. В этих случаях смертность высока у
молодых особей, а у взрослых (в возрасте 1 года и старше) она
ниже и более или менее постоянна.
Форма кривой выживания связана со степенью заботы о по-
томстве и другими способами защиты молоди. Так, кривые вы-
живания пчел и дроздов (которые заботятся о потомстве) зна-
чительно менее вогнуты, чем у кузнечиков и сардин (которые
не заботятся о потомстве). У последних видов это обстоятельство,
без сомнения, компенсируется значительно большим числом от-
кладываемых яиц (отношение максимальной к реализованной
рождаемости велико, как отмечалось в предыдущем разделе).
Форма кривой выживания очень часто варьирует также и в
зависимости от плотности популяции. На рис. 6.4 показаны кри-
вые выживания двух популяций оленя, обитающих в зарослях
чапараля в Калифорнии. Кривая выживания более плотной по-
пуляции заметно вогнута. Иными словами, у оленей, живущих
на освоенной территории, где количество пищи больше в резуль-
тате планомерного выжигания старых зарослей, ожидаемая про-
должительность жизни меньше, чем у оленей, живущих на неосво -
2 Заказ № 1483
18
Глава 6
Рис. 6.4. Кривые выживания для
двух стабильных популяций черно-
хвостого оленя, обитающих в зарос-
лях чапараля в Калифорнии. Попу-
ляция с высокой плотностью (при-
мерно 40 оленей на 1 км2) обитает
на территории, где покров открытых
кустарников п травянистых расте-
ний путем регулярных палов посто-
янно поддерживается в таком со-
стоянии, что имеется большое коли-
чество новых побегов, которые
олени обгрызают. Популяция с низ-
кой плотностью (примерно 17 оле-
ней на 1 км2) обитает на террито-
рии, где не ведется хозяйства; ста-
рые заросли кустарников не выжи-
гались уже 10 лет. Свежие гари
могут прокормить до 57 оленей на
1 км2, но эта популяция нестабиль-
на, и поэтому кривые выживания
нельзя строить по данным о возраст-
ном распределении. (По Taber и
Dasmann, 1957.)
енной территории; это, по-видимому, связано с более интенсивной
охотой, внутривидовой конкуренцией и т. д. С точки зрения охот-
ника, освоенная территория более благоприятна, но с точки зре-
ния оленя, на менее населенной территории больше шансов вы-
жить. То же самое можно сказать и относительно человеческих
популяций, где высокая плотность не всегда бывает благоприятна
для отдельного индивидуума. Многие экологи считают, что быст-
рый рост и высокая плотность в популяциях человека опасны
не столько для выживания, сколько для качества жизни инди-
видуума. И это несмотря на то, что «экологическая» продолжи-
тельность жизни человека значительно увеличилась благодаря до-
стижениям современной медицины, улучшению питания и т. д.
Кривая выживания для человека приближается к остроугольному
типу с минимумом смертности. Однако максимальная, или «фи-
зиологическая», продолжительность жизни человека, по-видимому,
не увеличилась, поскольку в наше время до 100 лет доживает
не больше людей, чем в минувшие столетия. Эти парадоксы еще
будут обсуждаться в гл. 8.
Чтобы подготовить основу для создания математических моде-
лей роста популяций, которые будут рассматриваться в следую-
щих разделах, целесообразно дополнить таблицы выживания дан-
ными по удельной рождаемости (числом потомков, производимых
самкой в единицу времени) в каждом возрастном классе.
В табл. 6.3 это столбец тх, так что теперь это не просто таблица
«смертей».
Динамика популяций
19
Если перемножить величины 1Х и тх и результаты для разных
возрастных классов сложить, то получится чистая скорость раз-
множения (7?о).
Таким образом,
(чистая скорость размножения) ~^1х-тх
(в данном случае 1Х относится только к самкам). Величина чис-
той скорости размножения 1,9 в табл. 6.3 означает, что число
потомков на каждую самку составляет 1,9. Если число самок рав-
но числу самцов, то получается, что примерно за одно поколение
популяция полностью обновляется. Лабораторная популяция ри-
сового долгоносика при самых оптимальных условиях увеличива-
ется через каждое поколение в 113,6 раз (т. е. 7?о= 113,6;
табл. 6.3,Б). В природе в стабильных условиях величина 7?о для
популяции в целом должна составлять около 1. Перис и Пителка
(Paris, Pitelka, 1962), использовав таблицы выживания и данные
по 1Х и тх по годовым классам, высчитали величину Ro для по-
пуляции мокриц в злаковнике. Она оказалась равной 1,02, что
свидетельствует о приблизительном равновесии между числом
рождений и числом смертей.
Особенности размножения сильно влияют на рост популяции
и другие ее характеристики. Естественный отбор может воздейст-
вовать на разные типы изменений жизненного цикла, что приво-
дит к появлению адаптивных свойств. Так, давление отбора мо-
жет изменить сроки начала размножения, не затрагивая общее
число производимых потомков, или же отбор может влиять па
продукцию, или «величину выводка», не меняя сроков размноже-
ния. Эти и многие другие аспекты динамики популяции можно
выявить при анализе таблиц выживания.
Возрастной состав популяции. Возрастная структура популя-
ции является ее важной характеристикой, которая, как было по-
казано на примерах, обсуждавшихся в предыдущем разделе, влия-
ет как на рождаемость, так и на смертность. Соотношение разных
возрастных групп в популяции определяет ее способность к раз-
множению в данный момент и показывает, чего можно ожидать
в будущем. Обычно в быстро растущих популяциях значительную
долю составляют молодые особи, в популяции, находящейся в
стационарном состоянии, возрастное распределение более равно-
мерно, а в популяции, численность которой снижается, будет
содержаться большая доля старых особей (рис. 6.5 и 6.6). Воз-
растная структура популяции может изменяться и без изменения
ее численности. По имеющимся данным, для каждой популяции
характерно «нормальное», или стабильное, возрастное распреде-
ление, к которому направлены изменения реальных распределе-
ний; теоретически это впервые предсказал Лотка (Lolka, 1925).
Коль скоро достигается стабильное возрастное распределение, вре-
2*
20
Глава 6
20
18
ё 16
S 14
£ 12
S. ю
3 8
со 6
4
2
ПОПУЛЯЦИИ ПОЛЕВКИ
Растущая Стабильная о
□
Относительное количество особей в разных
возрастных классах, %
ОНДАТРЫ
Крайние значения из 14 учетов
| ~~| 48%
| | 52%
Наименьшее
ФАЗАНЫ
Взрослые | | 48%
Ювенильные | j 52%
1945
Взрослые I _~125%
Ювениль-1 175%
ные 1947
| | 15%
) | 85%
Наибольшее
Рис. 6.5. Возрастные пирамиды. Вверху. Три типа возрастных пирамид,
характеризующих популяции с высокой, умеренной и малой численностью
(в %) молодых особей. В середине. Возрастные пирамиды лабораторных
популяций полевки Microtus agrestis. Слева. Экспоненциальный рост попу-
ляции в нелимитирующей среде. Справа. Рождаемость и смертность равны.
(Leslie, Ranson, 1940). Внизу. Крайние случаи соотношений численности
ювенильных и взрослых особей у фазана в Северной Дакоте (Kimball,
1948) и ондатры на востоке США (Petrides, 1950).
менные отклонения от него могут быть вызваны лишь необычны-
ми увеличениями рождаемости или смертности, после чего проис-
ходит спонтанный возврат к стабильной ситуации. По мере раз-
вития человеческого общества от состояния освоения новых
территорий и быстрого роста численности к зрелому состоянию
стабильной популяции процент населения в младших возрастных
классах уменьшается (рис. 6.6). Это изменение возрастной струк-
туры и увеличение процента населения старших возрастных клас-
сов оказывает глубокое воздействие на жизненный уклад общества
и его экономику.
Упрощенно в популяции можно выделить три экологические
возрастные группы: пререпродуктиеную^ репродуктивную и пост-
Динамика популяций
21
Рис. 6.6. Возрастная структура популяции человека, рассчитанная отдельно
для мужчин и женщин в Швеции (1965 г.) и Коста-Рике (1963 г.). Насе-
ление Швеции увеличивалось медленно, поэтому основание пирамиды уз-
кое. Велик процент старших возрастных классов (40—60 лет). Расширение
возрастного класса 15—23 года явилось следствием резкого увеличения
рождаемости после второй мировой войны. Быстрый рост населения Коста-
Рики сформировал пирамиду с широким основанием и высоким процентом
населения моложе 25 лет. (Ricklefs, 1979, по данным Keyfitz, Fieger, 1968.)
репродуктивную. Длительность этих возрастов по отношению к
общей продолжительности жизни сильно варьирует у разных ор-
ганизмов. У современного человека три этих «возраста» прибли-
зительно одинаковы, на каждый из них приходится около трети
жизни. У первобытных людей пострепродуктивный период был
намного короче. Для многих животных и растений характерен
очень длительный пререпродуктивный период. У некоторых жи-
вотных, в особенности у насекомых, пререпродуктивный период
чрезвычайно длителен, репродуктивный период очень короткий,
а пострепродуктивный период отсутствует совсем. Классические
примеры — некоторые виды поденок (Ephemeridae) и 17-летняя
цикада. У поденок личиночное развитие в воде занимает от года
до нескольких лет, а во взрослом состоянии они живут всего
несколько дней. У цикад необычайно продолжителен период раз-
вития (хотя и не обязательно 17 лет), по взрослые насекомые
живут менее одного сезона.
У промысловых птиц и пушных зверей отношение сеголетков
к старшим возрастным группам служит показателем тенденции
изменения численности популяции. Это отношение определяют во
время охотничьего сезона (осенью или зимой) по выборке из
популяции, полученной охотниками или промысловиками. В об-
22
Глава 6
щем высокая численность молоди по сравнению с численностью
взрослых, как это видно из нижних схем на рис. 6.5, указывает
на то, что сезон размножения сложился успешно; и если смерт-
ность молоди не будет чрезмерной, то можно ожидать увеличения
популяции в следующем году. Наибольший процент молоди
(85%) отмечался в популяции ондатры (рис. 6.5 внизу справа),
которая несколько предшествующих лет подвергалась интенсив-
ной промысловой нагрузке. Вызванное таким воздействием общее
снижение численности популяции привело, по-видимому, к уве-
личению рождаемости у выживших особей, что указывает на
гибкость, свойственную популяциям ондатры, — они быстро вос-
станавливают свою численность после периодов высокой смерт-
ности.
Явление, известное под названием «доминирование возрастного
класса», часто наблюдается в популяциях рыб, для которых ха-
рактерна высокая потенциальная рождаемость. Когда в резуль-
тате чрезвычайно высокой выживаемости икры и личинок зна-
чительную часть популяции составляют особи определенного воз-
растного класса, в последующие несколько лет размножение по-
давляется. Классический пример этого — данные Йорта по сельди
в Северном море (рис. 6.7). Рыбы возрастного класса 1904 года
доминируют в уловах с 1910 года (когда особи этой группы до-
стигли 6-летнего возраста; к этому времени они уже достаточно
крупны и эффективно вылавливаются промысловыми орудиями)
до 1918 года (когда в 14-летнем возрасте они еще превосходили
по численности рыб младших возрастных групп). Такая ситуация
порождает иногда периодичность общих уловов, которые были
велики в 1910 г. и снижались в последующие годы по мере сни-
жения численности доминирующего возрастного класса до тех
пор, пока его не заменили другие классы. Мы наблюдаем здесь
нечто вроде компенсаторного механизма, когда высокая выживае-
мость с большой вероятностью сопровождается годами низкой
выживаемости. Ихтиологи еще не пришли к единому мнению
относительно того, какие внешние факторы обусловливают на-
блюдающийся время от времени необычный характер выживания.
2. Основные представления о скоростях
Популяции изменчивы по своей сути. Даже если сообщество и
экосистема кажутся неизменными, плотность, рождаемость, вы-
живание, возрастная структура, скорость роста и другие харак-
теристики популяции обычно изменяются по мере того, как вид
постоянно подстраивается к сезонам, физическим факторам и
другим видам. Экологов соответственно в большей мере интере-
сует не величине! и структура той или иной популяции в некото-
рый момент времени, а то, как часто и с какой скоростью в по-
Возраст, годы
Рис. 6.7. Возрастное распределение сельди Северного моря в промысловых
уловах за период с 1907 по 1919 г. иллюстрирует феномен доминирующего
возрастного класса. Особи 1904 г. очень многочисленны и доминируют
в популяции на протяжении многих лет. Рыбы моложе 5 лет не попадают
в сети, поэтому класс 1904 г. не выявляется до 1909 г. Возраст рыб опре-
деляли по кольцам роста на чешуе, которые образуются ежегодно подобно
годичным кольцам на деревьях. (По Hjort, 1926.)
24
Глава 6
пуляциях происходят изменения. Поэтому при изучении экологии
популяции все чаще используют дифференциальное исчисление —
раздел математики, оперирующий не самими величинами, а ско-
ростью их изменения. Краткий обзор представлений о скоростях
может помочь тем, кто еще не знаком с этой областью.
Скорость можно определить, разделив изменение некоторой
величины на интервал времени, в течение которого произошло
это изменение. Скорость, выраженная таким образом, будет ха-
рактеризовать быстроту изменения чего-то во времени. Так, чис-
ло километров, которое проехал автомобиль за час, — это скорость
его движения, а число особей, родившихся за год,— это скорость
рождаемости. Предлоги «в» или «за» означают «деленное на».
Вспомним (гл. 3, разд. 3), что продуктивность — это скорость,
а не неподвижное состояние.
«Изменение» иногда принято сокращенно обозначать симво-
лом Д (дельта) перед буквой, обозначающей изменяющийся па-
раметр. Так, если N представляет собой число организмов, a t —
время, то
ДАТ = изменение числа организмов,
— или AN/At = средняя скорость изменения числа ор-
д* ганизмов за (деленное на) определен-
ное время. (Это и есть скорость роста.)
или ДА7(2УД£) — средняя скорость изменения числа ор-
ганизмов в расчете на 1 особь (ско-
рость роста, деленная на исходное чис-
ло организмов или же на среднее их
число за определенный период време-
ни). (Этот параметр часто называют
удельной скоростью роста п использу-
ют при сравнении популяций разной
величины. Умножая удельную скорость
на 100, т. е. AN/(NAt) Х100, мы полу-
чаем скорость роста популяции в про-
центах.)
Часто нам необходимо знать не только среднюю скорость,
но п теоретическую мгновенную скорость в тот или иной момент
времени; иными словами, такую скорость, когда Д/ стремится к
нулю. В математическом анализе в тех случаях, когда рассмат-
ривают мгновенную скорость, Д заменяют буквой d (символ для
производной). В этом случае предыдущие выражения приобрета-
ют следующий вид:
dN/dt — скорость изменения числа организмов за единицу
времени в некоторый момент,
Динамика популяций
25
dN/(N dt) = скорость изменения числа организмов в расчете
на 1 особь в единицу времени в некоторый мо-
мент.
Рис. 6.8. Кривые роста популяции (вверху) и кривые скорости роста (вни-
зу) двух колоний пчел на одной и той же пасеке. I — итальянские пчелы.
II — кипрские пчелы. (Из Bodenheimer, 1937.)
На рис. 6.8 показано различие между кривой роста и кривой
скорости роста. Как будет показано в разд. 4, S-образные кривые
роста и одновершинные кривые скорости роста часто характери-
зуют ранние стадии роста популяций.
В любой точке кривой роста скорость роста равна наклону
этой кривой (тангенсу касательной). Так, в случае популяции А
на рис. 6.8 скорость роста максимальна между 4-й и 11-й неде-
лями и равна нулю после 16-й недели. В популяции Б макси-
мальная скорость наблюдается между 9- и 16-й неделями и нуле-
вая — после 18-й недели. С величиной AN/kt удобнее оперировать
при построении модели на основе исходных данных, но при ма-
тематическом манипулировании над моделями ее следует заме-
нить величиной dNldt.
Мгновенную скорость dNldt нельзя измерить непосредственно;
точно так же нельзя непосредственно вычислить величину
dN/(Ndt), исходя из данных по учету популяций. Для вычисле-
26
Глава 6
нпя мгновенной скорости роста с использованием уравнений,
приведенных в разд. 4, необходимо знать тип кривой роста попу-
ляции. Мгновенную скорость роста популяции нельзя измерить
так, как измеряют скорость движения автомобиля. Конечно, мож-
но получить приближенную оценку скорости, производя измере-
ния через очень малые интервалы. Мы можем отложить на гра-
фике любые полученные результаты, соединить точки линиями и
тогда определить, какой тип уравнения в большей степени отве-
чает реальной кривой роста или, точнее, какое уравнение лучше
соответствует факторам, контролирующим рост конкретной по-
пуляции (таким образом мы избегаем процедуры «подгонки кри-
вой» в явном виде).
3. Внутренняя скорость естественного роста
Определения
Если среда не налагает никаких ограничений (пространство, пи-
ща и другие организмы не оказывают лимитирующего действия),
удельная скорость роста (скорость роста популяции в расчете на
1 особь) для данных микроклиматических условий становится
постоянной и максимальной. Значение скорости роста в этих
благоприятных для популяции условиях характеризует соответ-
ствующую возрастную структуру популяции и представляет со-
бой единственный показатель наследственно обусловленной спо-
собности популяции к росту. Этот показатель обозначают симво-
лом г, являющимся экспонентой в дифференциальном уравнении
роста популяции в нелимитирующей среде при определенных фи-
зических условиях:
dNjdt=rN\ r=dN!(Ndt). (1)
Это выражение аналогично тому, которое было использовано в
разд. 2. Параметр г можно рассматривать как коэффициент мгно-
венного роста популяции. Путем элементарного интегрирования
получаем экспоненциальную зависимость
Nt=Noe'\ (2)
где TVo — численность в начальный момент времени, Nt — числен-
ность в момент времени t и е — основание натуральных логариф-
мов. Логарифмируя обе части равенства, получаем уравнение в
форме, удобной для проведения расчетов:
1пМ— 1пМ)
in^z=in^+^; г = —5—• (3)
Пользуясь последним уравнением, можно вычислить показатель г
по двум измерениям численности популяции [7V0 и Nt или в лю-
Динамика популяций
27
бые два момента времени в фазе нелимитируемого роста; в этом
случае в приведенном уравнении величины и Nt2 подставляют
вместо No и Nt, а вместо t — разность (£2—£i)].
Показатель г — это фактически разность между удельной мгно-
венной скоростью рождаемости Ъ (т. е. скоростью рождаемости в
единицу времени на 1 особь) и мгновенной скоростью смертно-
сти d и может быть выражен так:
r—b—d. (4)
Общая скорость роста популяции в нелимитирующей среде (г)
зависит от возрастного состава и удельной скорости роста, обус-
ловленных вкладом в размножение различных составляющих по-
пуляцию возрастных групп. Таким образом, показатели скорости
роста для одного вида могут быть различными в зависимости от
структуры популяций. Когда устанавливается стационарное и ста-
бильное возрастное распределение, удельную скорость роста на-
зывают внутренней скоростью естественного роста или гтах. Мак-
симальное значение г часто называют биотическим или репродук-
тивным потенциалом. Название это менее специально, но более
употребительно. Разницу между максимальным г, или биотиче-
ским потенциалом, и скоростью роста, наблюдаемой в реальных
полевых или лабораторных условиях, часто используют как меру
сопротивления среды, которая характеризует сумму всех лимити-
рующих факторов среды, препятствующих реализации биотиче-
ского потенциала.
Объяснения
Такие показатели, как рождаемость, смертность и возрастное рас-
пределение, весьма важны, но каждый из них сам по себе мало
что дает; в частности, по каждому из них нельзя судить о росте
популяции как целого, о характере изменений, которые произой-
дут при изменении условий среды, о максимальных возможностях
популяции и отличии их от ее реальных возможностей. Для обо-
значения максимального репродуктивного усилия Чепмен (Chap-
man, 1928) предложил термин биотический потенциал. Он опре-
делил его как «врожденное свойство организма к размножению
и выживанию, т. е. к увеличению численности. Это своего рода
алгебраическая сумма числа молодых особей, производимых за
каждый цикл размножения, числа циклов размножения за дан-
ный период времени, соотношения полов и общей способности ор-
ганизма выживать при данных физических условиях». Однако
весьма общий характер определения биотического, или репродук-
тивного, потенциала, данного выше, привел к тому, что разные
исследователи вкладывают в него разный смысл. Для одних он
означает некую туманную способность к размножению, скрытую
28
Глава 6
Рис. 6.9. Кривая роста одной и той же популяции, построенная двумя раз-
ными способами. Число особей (7V) отложено в арифметическом (слева) и
логарифмическом масштабе (справа). Рассматривается гипотетическая по-
пуляция микроорганизмов, растущая экспоненциально в течение 6 суток;
в этот период каждые два дня численность возрастала в 10 раз. Объяснения
уравнений см. в тексте.
в популяциях и, к счастью, никогда не проявляющуюся в полной
мере благодаря воздействию окружающей среды («в неконтроли-
руемых условиях потомство одной пары мух через несколько лет
весило бы больше, чем земной шар»). Другие представляют его
просто и более конкретно, как то максимальное число яиц, семян,
спор и т. д., которое может продуцировать наиболее плодовитая
особь, хотя обычно такое представление имеет мало значения
применительно к популяции, поскольку в большинстве популяций
все особи не способны постоянно поддерживать максимальную
продуктивность.
Лотка (Lotka, 1925), Даблин и Лотка (Dublin, Lotka, 1925),
Лесли и Рэнсон (Leslie, Ranson, 1940), Бирч (Birch, 1940) и дру-
гие исследователи выразили это довольно широкое представление
о биотическом потенциале в математических терминах, понятных
на любом языке (однако подчас лишь с помощью хороших ма-
тематиков). Хорошо сказал по этому поводу Бирч: «Если попы-
таться количественно выразить «биотический потенциал» Чепме-
на с помощью всего лишь одного показателя, то, по-видимому,
лучше всего для этого использовать параметр г, поскольку он
характеризует внутренне присущую животным способность к уве-
личению численности в отсутствие лимитирующих факторов сре-
ды». Как будет отмечено далее, показатель г часто используется
также для количественного выражения «репродуктивной приспо-
собленности» в генетическом смысле.
Динамика популяций
29
Если вспомнить о кривых роста, обсуждавшихся в разд. 2,
то г представляет собой удельную скорость роста (&N/NAt) толь-
ко для случая экспоненциального роста популяции. Уравнение
(3), приведенное в разд. «Определения», есть уравнение прямой.
Следовательно, величину г можно определить графически. Если
кривую роста вычерчивать в логарифмическом или полулогариф-
мическом масштабе, то при экспоненциальном росте график за-
висимости логарифма численности популяции от времени будет
иметь вид прямой линии, а г будет наклоном этой прямой. Чем
круче наклон, тем выше внутренняя скорость роста. На рис. 6.9
одна и та же кривая роста построена двумя способами: слева
численности (N) отложены на арифметической, а справа — на ло-
гарифмической шкале (правый график на полулогарифмической
шкале). В этом примере гипотетическая популяция микроорга-
низмов шесть дней находилась в состоянии экспоненциального
роста, причем каждые двое суток происходило десяшкратное уве-
личение численности. Наклон прямой на полулогарифмическом
графике равен 1,15; это и есть величина г. В этом нетрудно убе-
диться, подставляя в уравнение (3) две любые величины числен-
ности популяции, например, на 2- и 4-е сутки:
1пЛГ/2 — 1п№
г=-----7^1—’
1 пЮО — liilO
Г =----2 •
Воспользовавшись таблицей натуральных логарифмов, получаем
Чрезвычайно широкий диапазон значений биотического потен-
циала выявляется особенно отчетливо, если выражать его с по-
мощью величины, показывающей, во сколько раз увеличится чис-
ленность при экспоненциальном росте или какое время требуется
для удвоения популяции. Эти два параметра математически вы-
водятся из внутренней скорости роста:
Конечная скорость прироста: X=er; logeX=r; %=antiloger.
Время удвоения: ^=loge2/r==0,6931/r
(эту величину мы получаем из уравнения (2), принимая
Nt/N0=2).
При максимальной внутренней скорости роста время удвоения
для популяции мучного хрущака в оптимальных лабораторных
условиях составляет меньше недели (см. Leslie, Park, 1949). Для
человека время удвоения в 1968 г. составляло 35 лет (Ehrlich,
Ehrlich, 1970), однако в некоторых странах население удваива-
30
Глава 6
ется быстрее. Смит (Smith, 1954) полагает, что г и соответствен-
но периоды удвоения в живой природе различаются более чем на
шесть порядков.
Коэффициент роста популяции г не следует путать с чистой
скоростью размножения /?о, рассмотренной в разд. 1 (см.
табл. 6.3). 7?о связано со временем генерации и не подходит для
сравнения разных популяций, характеризующихся различным
временем генерации. Однако среднее время генерации (Т) свя-
зано с Ro и г следующим образом:
г. „т
R0 = erJ, отсюда Т= ~ •
Зависимость между 7, Ro и г для популяций различных животных
графически представлена на стр. 52 книги Слободкина (Slobodkin,
1962). В природных популяциях в непродолжительные периоды
времени, когда пища имеется в изобилии, отсутствуют скучен-
ность, враги и т. д., нередко проявляется экспоненциальный ха-
рактер роста. В таких условиях популяция в целом растет с
огромной скоростью, хотя при этом каждый организм размножа-
ется с той же скоростью, что и раньше, а удельная скорость роста
постоянна. «Цветение» планктона, упомянутое в предыдущих
главах, вспышки численности вредителей, рост бактерий на све-
жей культуральной среде — служат примерами ситуаций, в ко-
торых рост может быть логарифмическим. Подобный характер
изменений отмечается и для многих других явлений, например
поглощения света, мономолекулярных химических реакций, роста
сложных процентов. Очевидно, что экспоненциальный рост не мо-
жет продолжаться очень долго, а часто и вовсе не реализуется.
Взаимодействия с другими популяциями, а также сопротивление
внешней среды быстро снижают скорость роста п различным об-
разом влияют на характер роста популяции.
4. Кривая роста популяции
Определения
Характер увеличения численности популяции может быть раз-
личным, и в связи с этим выделяют различные типы роста попу-
ляции. По форме кривых роста, построенных в арифметическом
масштабе, можно выделить два основных типа: рост, описываемый
J-образной и S-образной, пли сигмоидной, кривой. Эти противо-
положные типы в соответствии с особенностями разных организ-
мов и условий среды могут усложняться, модифицироваться или
сочетаться различным образом. При J-образной кривой плотность
быстро возрастает по экспоненте (как показано на рис. 6.9),
Динамика популяций
31
но затем, когда начинают действовать сопротивление среды пли
другой лимитирующий фактор (обычно более или менее резко),
рост быстро прекращается. Этот тип роста может быть описан
простым экспоненциальным уравнением, рассмотренным в преды-
дущем разделе:
dN
dt —rN при заданном ограничении на величину N.
При описываемом сигмоидной кривой типе роста популяция вна-
чале увеличивается медленно (фаза положительного ускорения
или становления), затем быстрее (возможно, приближаясь к ло-
гарифмической фазе), но вскоре под влиянием сопротивления
среды (возрастающем в процентном отношении) рост постепенно
замедляется (фаза отрицательного ускорения); в конце концов
будет достигнуто равновесие, которое и сохраняется. Такой тип
роста можно описать простым логистическим уравнением:
dN „ (K-N)
dt ~~rN к
Верхний предел увеличения численности популяции, соответст-
вующий константе К, называется верхней асимптотой сигмоидной
кривой. Ее называют также пределом роста. При J-образном типе
роста равновесие не достигается, напротив, плотность может ко-
лебаться в соответствии с ограничениями ресурсов, физическими
факторами и взаимодействием с другими популяциями (напри-
мер, хищниками). Два описанных типа роста и некоторые их ва-
рианты схематически показаны на рис. 6.10.
Объяснения
Когда тех или иных животных интродуцируют в незанятые об-
ласти или они сами распространяются туда (как это бывает, на-
пример, в начале сезона), часто можно наблюдать характерный
тип роста популяции. На графике, построенном в арифметическом
масштабе, участок кривой, соответствующей росту популяции,
часто имеет S- пли J-образную форму (рис. 6.10, Л, Б). Интерес-
но отметить, что два основных типа роста, описываемые этими
кривыми, сходны с двумя типами метаболизма или роста, кото-
рые были описаны для отдельных особей (Bertalanffy, 1957)
Однако неясно, существует ли причинная связь между ростом
популяций и ростом особей. При нынешнем уровне наших зна-
ний можно только отметить сам факт некоторого сходства в
форме кривых. Популяции и сообщества не являются «сверхор-
ганизмами», но могут обладать аналогичными организмам свой-
ствами (гл. 2, разд. 6).
32
Глава 6
Рис. 6.10. Некоторые особенности формы кривых роста популяции при изо-
бражении в линейном масштабе. Показаны J-образная кривая (экспонен-
циальный рост) (А), S-образная кривая (сигмоидный рост) (Б) и некото-
рые их варианты. Л-1 и А-2— колебания, выявляемые при экспоненциаль-
ном росте. Б-1, Б-2 и Б-3 — некоторые (но далеко не все) возможные вари-
анты роста, когда имеется разрыв во времени между увеличением плот-
ности организмов, вызванным появлением новых особей, и проявлением
тормозящего влияния этого увеличения плотности на рост популяции (такой
рост характерен для высших растений и животных). Если питательные ве-
щества и другие факторы накоплены до начала роста популяции, то может
произойти «овершут», как показывают кривые А-2 и Б-2 (этим объясняет-
ся, почему новые пруды и озера часто богаче рыбой, чем старые!). (Из
Nicholson, 1954.)
Уравнение, приведенное выше для описания J-образного роста,
представляет собой то же самое экспоненциальное уравнение, ко-
торое было рассмотрено в разд. 3, с той лишь разницей, что на
величину N налагается ограничение. Относительно свободный
рост внезапно прекращается, как только популяция исчерпывает
какой-либо ресурс (пищу или пространство, когда наступают вне-
запные заморозки или вмешивается другой сезонный фактор или
когда внезапно заканчивается сезон размножения (возможно, на-
пример, из-за наступления диапаузы, как описано в гл. 5.
Динамика популяций 33
стр. 267). По достижении верхнего предела для N плотность
может некоторое время оставаться на этом уровне либо сразу же
резко падает, что приводит к релаксационно-колебательному ха-
рактеру изменений плотности, показанному на рис. 6.10, АЛ и
Л-2. Такое изменение численности, названное Никольсоном (Ni-
cholson, 1954) «триггерным», свойственно, по-видимому, многим
природным популяциям, в частности одноклеточным планктон-
ным водорослям во время «цветения», однолетним растениям,
некоторым насекомым и, быть может, леммингам в тундре.
Другой, также нередко наблюдаемый тип роста может быть
выражен в арифметическом масштабе по отношению к времени
S-образной, или сигмоидной, кривой. Характерная форма кривой
обусловлена постепенным усилением — по мере нарастания плот-
ности популяции — действия неблагоприятных факторов (сопро-
тивления среды). В этом отношении такой тип роста является
прямой противоположностью ранее рассмотренному, при котором
популяция начинает испытывать сопротивление среды почти в
конце своего роста. По этой причине Никольсон (Nicholson, 1954),
говоря о сигмоидном росте, назвал его «обусловленным плот-
ностью». В простейшем случае усиление действия сдерживающих
факторов прямо пропорционально плотности. Такой тип роста на-
зывают логистическим \ и он соответствует логистическому урав-
нению, которое характеризует сигмоидный рост.
Логистическое уравнение можно записать несколькими спосо-
бами; три из них вместе с интегральной формой таковы:
dN
~ТГ — Г1\ , или
at А
Г
— rN—~2У2,ИЛИ
А
/ N \
где dNjdt — скорость роста популяции (изменение численности во
времени); г — удельная, или внутренняя, скорость роста (обсуж-
далась в разд. 3); N — величина популяции (численность);
К — максимально возможная величина популяции, или верхняя
"асимптота; е — основание натуральных логарифмов и а — посто-
янная интегрирования, определяющая положение кривой относи-
1 Логистическое уравнение было впервые предложено Ферхюльстом
(Р. F. Verhulst) в 1838 г.; им широко пользовался Лотка, а затем его «пере-
открыли» Пирл и Рид (Pearl, Reed, 1930).
3 Заказ № 1483
34
Глава б
тельно начала координат. Она численно равна (K—N)/N при
£=0.
Это уравнение отличается от экспоненциального уравнения,
приведенного в предыдущем разделе, лишь тем, что оно содержит
величину (К—N)/K, (r/K)N2 или (1—N/K). Эти три величины
соответствуют трем способам обозначения сопротивления среды,
которое порождается вследствие роста самой популяции и по
мере того, как величина популяции приближается к емкости сре-
ды, вызывает все большее уменьшение потенциальной скорости
размножения. В словесной форме эти уравнения означают сле-
дующее:
Скорость
роста по-
пуляции
равна
Максимальной
возможной скорос-
ти прироста (не-
ограниченной
удельной скорости
роста), умножен-
ной па числен-
ность популяции
умножен-
ной на
или
минус
Степень реали-
зации макси-
мальной скоро-
сти
Нереализован-
ный прирост
В общем, эта простая модель выводится из трех компонент: кон-
станты скорости (г), меры величины популяции (7V) и меры того,
какая часть доступных лимитирующих факторов не используется
популяцией (1—N/K).
Логистическое уравнение можно записать также в форме, ха-
рактеризующей скорость прироста на одно поколение, R:
dN
dt
=NlnR^
K — N
К
Хотя рост многих популяции микроорганизмов, растений и
животных как в лабораторных, так и природных условиях может
быть описан сигмоидной кривой, это вовсе не означает, что такие
популяции растут согласно логистическому уравнению. Сущест-
вует много математических уравнений, решение которых можно
представить графически в форме сигмоидных кривых. Почти лю-
бое уравнение, в котором увеличение отрицательного воздействия
находится в какой-то зависимости от плотности, дает сигмоидную
кривую. Как отметил Вигерт (Wiegert, 1974), логистическая кри-
вая представляет собой своего рода минимальный сигмоидный
рост, поскольку лимитирующие эффекты и пространства, и ресур-
сов сказываются с самого начала роста (т. е. максимальная удель-
ная скорость роста достигается только при нулевой плотности).
В большинстве случаев менее ограниченный рост ожидается в
начальной фазе, а затем по мере увеличения плотности скорость
роста медленно понижается. Рис. 6.11 иллюстрирует представле-
Динамика популяций
35
ние о том, что логистическая
кривая отражает наиболее
медленный, а экспоненциаль-
ная кривая — наиболее быст-
рый тип роста. Следует ожи-
дать, что рост большинства
популяций занимает проме-
жуточное положение.
В популяциях высших
растений и животных, имею-
щих сложные жизненные
циклы и длительный период
индивидуального развития,
наблюдается несовпадение во
времени увеличения плотно-
сти и воздействия лимити-
рующих факторов. Говоря
словами Никольсона (Nichol-
son, 1954), этот тип роста
можно назвать «обусловлен-
ным плотностью с запазды-
ванием». В таких случаях
получается более вогнутая
кривая роста (необходимо
больше времени, чтобы про-
явился эффект рождаемости),
Рис. 6.11. Кривые, показывающие тео-
ретически самое быстрое (экспоненци-
альное) и самое медленное (логистиче-
ское) увеличение численности популя-
ции. Максимальные скорости роста и
минимальные плотности одинаковы.
Между кривыми находится область
(заштрихована), соответствующая рос-
ту большинства популяций. (По Wie-
gert, 1974.)
популяция почти всегда «перепры-
гивает» верхнюю асимптоту и испытывает колебания численно-
сти, прежде чем стабилизироваться на уровне емкости среды (см.
рис. 6.10, кривая Б-2). Для того чтобы учесть два типа времен-
ных задержек: 1) время до начала прироста при благоприятных
^условиях и 2) время, нужное для того, чтобы в неблагоприятных
условиях перенаселения начали изменяться рождаемость и смерт-
ность, было предложено большое число модифицированных логи-
стических уравнений. Обозначив эти задержки соответственно как
\t~-1\ и t—t2, получаем
dN(t)
dt
При исследовании этого варианта уравнения на аналоговом уст-
ройстве или мини-ЭВМ получается, что плотность возрастает с
ювершутом («перепрыгивает» асимптоту) и колеблется с умень-
шающейся во времени амплитудой, как показано на рис. 6.10,
^кривая Б-2.
3*
36
Глава 6
Обозначения dt (мгновенное время) и (дискретный ин-
тервал времени), использовавшиеся в разделах 2, 3 и 4,
отражают два распространенных математических подхода
при моделировании популяций и экосистем: один из них
основан на применении систем дифференциальных уравне-
ний, а другой — систем разностных уравнений. Дифферен-
циальные уравнения более удобны в чисто математическом
плане, вероятно, поэтому им уделяется столько внимания в
учебных курсах вычислительной математики. В то же вре-
мя разностные уравнения лучше соответствуют способу по-
лучения исходных данных (в дискретные интервалы време-
ни) и структурам кривых роста популяций у видов с
дискретными поколениями. Для сравнения повторим диффе-
ренциальные уравнения J- и S-образных кривых роста по-
пуляций и запишем для каждого случая соответствующее
разностное уравнение. Дифференциальное уравнение непре-
рывного экспоненциального роста имеет вид
dNjdt^rN, (1)
а разностное уравнение дискретного экспоненциального роста
Nt+i=\Nt(r), или (2)
Nt^=e'Nt, (3)
где М+1 — изменение за данный временной интервал;
Z — мультипликативный коэффициент роста на поколение
или на какой-то другой выделенный период (это есть ко-
нечная скорость роста, введенная в разделе 3, с. 29, она
аналогична г, внутренней скорости роста). r=lnZ, так что
Z=er, что и отражает замена Z в уравнении (3). При росте
численности популяции X должна быть больше 1, а г боль-
ше 0, в противном случае численность популяции экспонен-
циально уменьшается и в конце концов она вымирает.
Дифференциальное уравнение для логистического (зави-
симого от плотности или сигмоидного роста) имеет вид
dN )dt — rN(\ — N (4)
а разностное уравнение для зависимого от плотности роста —
NM=Nte^~N^ (5)
В качестве аналогов уравнений логистического и других ти-
пов зависимого от плотности роста было предложено нес-
колько типов разностных уравнений (см. Мау, 1976; Мау
и Oster, 1976). Примечательно, что разностные уравнения
можно использовать в том виде, в каком они есть, не ин-
тегрируя их как дифференциальные уравнения. Сравните
Динамика популяций
37
уравнение (5) с интегральной формой логистического урав-
нения на с. 33.
Выбор подхода зависит от того, что интересует в данной
задаче: характер изменения некоторой величины или ско-
рость ее изменения. В последнем случае лучше использо-
вать дифференциальные уравнения.
Почти все математические модели роста популяций сильно
уязвимы вследствие того, что они оперируют в закрытых систе-
мах, не имеющих входа и выхода. Моделируется только само пе-
ренаселение и другие внутренние факторы. Как не раз обсуж-
далось в гл. 2 и 3, реальный мир состоит из открытых систем,
в которых большую роль играют среда на входе и среда на вы-
ходе (см. рис. 2.1). Эта ошибка особенно заметна при моделиро-
вании или предсказаниях характера роста популяций человека.
Эдвард Диви (Deevey, 1958) много лет назад отметил, что при
отсутствии «внешнего» управления, такого, как систематическое
планирование семьи, рост народонаселения должен был бы, ско-
рее всего, следовать некоей кривой с овершутом и колебаниями,
поскольку механизмы ограничения перенаселения срабатывают с
временными задержками. По-видимому, быстро растущие города,
зависящие от мощных внешних источников энергии, поступления
пищи, воды и общих ресурсов жизнеобеспечения, особенно под-
вержены в разной степени чередованию взлетов и кризисов: это
определяется факторами на входе и тем, насколько отдельные
граждане и правительственные органы могут предвидеть гряду-
щие условия и осуществлять соответствующее планирование. Так,
на ранних стадиях роста города, когда экологические условия
благоприятны (пространство и ресурсы доступны и недороги)
и когда потребность в обслуживании (вода, переработка отходов,
улицы, школы и т. д.) мала, численность населения растет быстро
(при этом часто главным образом за счет иммиграции), что отве-
чает J-образной кривой роста. Но спустя некоторое время (до сра-
батывания временной задержки) школы и жилой фонд оказыва-
ются перегруженными, увеличиваются потребности в обслужива-
нии, наблюдается рост цен и становятся ощутимыми общие
нарушения в масштабах всего хозяйства. В отсутствие отрица-
тельной обратной связи, такой, какая встроена в простую логи-
стическую зависимость или какую можно включить в рациональ-
ное планирование, города будут расти слишком быстро, что небла-
гоприятно скажется на их состоянии и приведет к упадку.
Рис. 6.12. Рост дрожжей в культуре. Простейший случай сигмоидного рос-
та, при котором сопротивление среды (в этом случае вредные факторы
обусловлены жизнедеятельностью самих организмов) прямо пропорцио-
нально плотности. Светлые кружки — наблюдаемые величины, сплошные
линии — кривые, построенные по уравнениям. На верхнем графике рост
дрожжей представлен в линейном масштабе и точки ложатся на логисти-
ческую кривую. На нижнем графике те же данные представлены в логариф-
мическом (L) масштабе; приведена также экспоненциальная кривая (Е),
показывающая, каким был бы рост при отсутствии самоограничивающих
влияний. (Нижний график — по данным Pearl, 1927, верхний — из Allee
et al., 1949.)
Динамика популяций
39
Примеры
На рис. 6.12 показана простейшая сигмоидная кривая, а на
рис. 6.13 — J-образные кривые. Рис. 6.13 показывает, что чис-
Рис. 6.13.
на розах.
1948.)
Сезонные изменения численности популяции взрослых трипсов
(График построен по данным из работы Davidson, Andrewartha,
ленность трипсов (мелкие насекомые) в благоприятные годы
быстро увеличивается, вплоть до конца сезона, после чего их
плотность так же быстро снижается. В менее благоприятные годы
кривая роста больше похожа на сигмоидную. В общем, J-образ-
ную кривую можно рассматривать как неполную сигмоидную
кривую, просто лимитирующие факторы среды в этом случае на-
чинают ограничивать рождаемость еще до того, как существенную
роль начнут играть внутренние факторы ограничения численно-
сти.
На рис. 6.12 представлены графики роста дрожжей в ариф-
метическом (вверху) и в полулогарифмическом (внизу) масшта-
бах, соответствующие логистическому уравнению. Обратите вни-
мание на то, что на полулогарифмическом графике кривая боль-
ше похожа на перевернутую букву J, чем на букву S. На нижнем
графике проведена также экспоненциальная кривая (Е), показы-
вающая, каким мог бы быть рост, не ограниченный размерами
культивационного сосуда и плотностью популяции. Эти уравне-
40
Глава 6
ния представляют собой уже описанные интегральные формы
логистического и экспоненциального (с J-образной кривой) урав-
нений, в которые подставлены реальные значения величин К, а
и г. Фактический (наблюдаемый в опыте) рост дрожжей хорошо
соответствует логистическому росту, что свидетельствует о ран-
нем проявлении эффекта скученности и его линейной связи с
плотностью. Площадь между двумя кривыми на нижнем графике
можно принять за количественную меру сопротивления среды.
Преимущество полулогарифмического построения заключается в
том, что по любому отклонению от прямой линии можно судить
об изменении скорости роста популяции (dNIdt). Чем больше
изгиб кривой, тем значительнее изменение.
Хотя простой логистический рост, вероятно, характерен лишь
для мелких организмов или для организмов с простыми жизнен-
ными циклами, S-образные кривые роста наблюдаются и у круп-
ных организмов, когда они интродуцируются на незанятые преж-
де острова. Пример этому — рост поголовья овец на о. Тасмания
(Davidson, 1938) или рост популяции фазанов, интродуцирован-
ных на один из островов в Пьюджет-Саунд, шт. Вашингтон
(Einarsen, 1945).
Вернемся вновь к не один раз упомянутой в этой книге теме:
открытым системам. Расселение — перемещение организмов или
их подвижных стадий (семян, спор, личинок и т. п.) на терри-
торию, уже занятую популяцией, или с этой территории — наряду
с рождаемостью и смертностью определяет характер роста попу-
ляции. Эмиграция — выселение с занимаемой территории — ана-
логична смертности; иммиграция — аналогична рождаемости.
Миграция — периодические уход и возвращение на данную тер-
риторию — в разные сезоны влияет на характер кривой роста то
как рождаемость, то как смертность. Расселение в большой сте-
пени зависит от различных преград и врожденной способности
к перемещениям, или подвижности, взрослых особей или стадий
их расселения. И, конечно, расселение — это средство колониза-
ции новых или освободившихся пространств. Расселение мелких
организмов и пассивных стадий обычно подчиняется экспоненци-
альной зависимости: в местах, удаленных от места расселения
на кратные расстояния, плотность снижается на постоянную ве-
личину. Расселение крупных активных животных носит иной ха-
рактер; оно может закончиться на некотором заданном расстоя-
нии, может иметь форму нормального распределения или прини-
мать другие формы. В исследовании Стюарта (Stewart, 1952),
выполненном на окольцованных сипухах, дан хороший пример
сочетания случайного расселения с тенденцией миграции к югу.
В пределах 100 миль расселение было ненаправленным (т. е. ве-
роятность встретить окольцованную птицу была примерно одина-
ковой в любой точке и в любом направлении), но за пределами
Динамика популяций
41
этого расстояния птицы имели явную тенденцию расселяться к
югу. Общие обзоры о характере и закономерностях расселения
см. в работах Wolfenbarger, 1946, и MacArthur, Wilson, 1967.
5. Флуктуации численности популяций
и «циклические» колебания
Определения
Когда популяция завершает рост и AN/At в среднем за длитель-
ное время становится равным нулю, плотность ее проявляет тен-
денцию к флуктуациям относительно стационарного уровня, даже
если в популяции работают разного рода механизмы обратной
связи. Часто флуктуации бывают обусловлены сезонными или
годовыми изменениями доступности ресурсов или же могут быть
стохастическими (случайными). Однако у некоторых популяций
флуктуации столь регулярны, что их можно рассматривать как
«циклические».
Объяснения
Для природных популяций важно различать: 1) сезонные изме-
нения численности, которые регулируются преимущественно
адаптациями жизненного цикла, сопряженными с сезонными из-
менениями факторов среды, и 2) годичные флуктуации. В ана-
литических целях удобно рассматривать две группы годичных
флуктуаций: а) флуктуации, контролируемые в первую очередь
годичными различиями внешних факторов (факторы, такие, как
температура и осадки, лежащие вне сферы популяционных взаи-
модействий), и б) флуктуации, связанные прежде всего с самой
динамикой популяции (биотическими факторами, такими, как
доступная пища или энергия, болезни и т. п.). Во многих случаях
представляется, что изменения численности из года в год хорошо
коррелируют с вариациями одного или нескольких внешних ли-
митирующих факторов; однако у некоторых видов изменения чис-
ленности столь регулярны и, по-видимому, не зависят от очевид-
ных внешних причин, что к этим изменениям вполне применим
термин «циклы». (Виды с такими регулярными изменениями ве-
личины популяции часто называют «циклическими».) Теории, ко-
торые были предложены для объяснения этих циклов, будут рас-
смотрены после соответствующих примеров.
В предыдущих главах не раз подчеркивалось, что популяции
модифицируют окружающую их среду и компенсируют наруше-
ния, вызванные внешними физическими факторами. Таким обра-
зом, чем более высокоорганизованно и зрело сообщество и чем
42
Глава 6
стабильнее физическая среда, или чем лучше совпадают оба эти
условия, тем меньше должна быть амплитуда флуктуаций плот-
ности популяции во времени.
Йримеры
Всем нам знакомы сезонные измененпя величины популяции.
Тучи москитов и комаров, леса, полные птиц, поля, заросшие
амброзией, — все это бывает в свое время, в другие сезоны попу-
ляции этих организмов могут сходить практически на нет. Хотя
в природе трудно найти популяции животных, микроорганизмов
и травянистых растений, численность которых не изменялась бы
по сезонам, наиболее выражены эти флуктуации у организмов с
ограниченным периодом размножения, особенно у организмов с
коротким жизненным циклом или со значительными сезонными
различиями расселения в пространстве (например, у мигрирую-
щих животных). На рис. 6.13, как уже упоминалось, показана
не только J-образная кривая роста, но и сезонные и годичные
флуктуации, выявленные в результате многолетних систематиче-
ских исследований. Изменения такого рода, по-видимому, типич-
ны для большинства насекомых, большинства однолетних расте-
ний и «однолетних» животных. Сезонные циклы планктонных
популяций интенсивно изучались и моделировались как в прес-
ных водах, так и в океане (один пример был описан в гл. 5,
рис. 5.2).
Пример довольно нерегулярного изменения величины попу-
ляции, коррелирующего, видимо, с погодными условиями, пока-
зан на рис. 6.14. На протяжении многих лет популяция цапель
в двух местностях в Великобритании оставалась относительно
постоянной. Очевидно, местные условиях обеспечивали достаточ-
но устойчивую несущую емкость среды для цапель. Однако в
годы, следовавшие после лет с суровыми зимами (указаны на
рис. 6.14 вверху), происходило резкое снижение плотности с по-
следующим ее восстановлением. Синхронность изменении плот-
ности в обеих местностях свидетельствует о том, что их причи-
на __ увеличение смертности в зимнее время. Всегда проводить
исследование, не ограничиваясь только одним районом, — вот хо-
рошее правило для полевого эколога! Популяции птиц оказались
одними из наиболее хорошо изученных, поэтому именно резуль-
таты этих исследований внесли большой вклад в популяционную
теорию. Обзор работ по исследованиям популяций птиц принад-
лежит Лэку (Lack, 1966).
К наиболее известным примерам «циклических» колебаний
можно отнести колебания численности у некоторых видов север-
ных млекопитающих и птиц, у которых наблюдаются либо 9—10-,
либо 3—4-летние циклы. Классическим примером 9—10-летних
Динамика популяций
43
Рис. 6.14. Изменения численности цапли Ardea cinerea в двух местностях
в Великобритании в период с 1933 по 1963 гг. Показана зависимость между
холодными зимами и снижением численности. (Из Lack, 1966.)
Рис. 6.15. Изменения численности рыси и зайца-беляка, регистрируемые по
числу шкурок, заготовленных Компанией Гудзонова залива. Это классиче-
ский пример циклических колебаний плотности популяций. (Рисунок из
работы MacLulich, 1937.)
колебаний служит изменение численности зайца-беляка и рыси
(рис. 6.15). Канадская Компания Гудзонова залива располагает
начиная примерно с 1800 года ежегодными отчетами о заготовке
пушнины. Если представить эти данные графически, то оказыва-
ется, что, например, у рыси пики численности популяции за весь
этот длительный период повторяются каждые 9—10 лет, в сред-
нем через 9,6 года. Пики численности часто сменяются резкими
спадами, когда рысь в течение нескольких лет встречается крайне
44
Глава 6
Рис. 6.16. Заяц-беляк, знаменитый в истории экологии в связи с резко вы-
раженными колебаниями численности (см. рис. 6.15). Показано животное
в белом зимнем наряде. Было выявлено, что смена коричневого летнего
наряда на белый зимний контролируется длительностью фотопериода.
(Фотография службы охраны почв США.)
редко. У зайца-беляка наблюдаются такие же циклы, но пики его
численности, как правило, на год или более предшествуют пикам
рыси. Поскольку рысь в большой степени зависит от зайца, слу-
жащего ей пищей, очевидно, что цикл хищника связан с циклом
жертвы. Но эти два цикла связаны не строгими причинно-след-
ственными взаимодействиями хищник—жертва, так как циклы
численности зайца отмечаются и в тех местах, где рысь не водит-
ся. Более короткие 3—4-летние циклы характерны для многих
Динамика популяций
45
северных мышевидных грызунов (леммингов, мышей, полевок)
и их хищников (особенно полярной совы и песцов). Цикл лем-
минга в тундре и циклы песца и полярной совы впервые на
фактическом материале показал Элтон (Elton, 1942). Каждые три
или четыре года на огромных пространствах тундры двух кон-
тинентов численность леммингов (два вида рода Lemmus
в Евразии и один в Северной Америке и еще один вид рода
Dicrostonyx в Северной Америке) катастрофически возрастает,
вслед за чем, часто за один сезон, резко падает. Численность
песцов и сов, увеличивавшаяся по мере увеличения количества
пищи, также очень скоро снижается. Совы при этом в поисках
пищи могут мигрировать на юг в США (иногда достигая
шт. Джорджия). Эта взрывная миграция избытка птиц происхо-
дит, по-видимому, в одном направлении; если обратный перелет
и совершается, то лишь единичными особями. Таким образом,
популяция сов резко уменьшается в результате миграции. Эти
колебания так регулярны, что орнитологи в США могут точно
предсказать нашествие полярных сов каждые три или четыре
года. Поскольку эти птицы хорошо заметны и появляются по-
всюду вокруг городов, они привлекают к себе большое внимание:
их фотографии появляются в газетах, чучела — в местных мага-
зинах. В периоды между нашествиями ни в США, ни на юге
Канады полярная сова не встречается или очень редка. Гросс
(Gross, 1947) и Шелфорд (Shelford, 1943) проанализировали
данные о нашествии сов и показали, что они коррелируют с пе-
риодическими уменьшениями числа леммингов — основной пи-
щи сов.
В Европе (но, по-видимому, не в Северной Америке) лемминги
порой достигают такой высокой плотности, что начинают мигри-
ровать из своих перенаселенных местообитаний. Элтон (Elton,
1942) живо описал знаменитую миграцию леммингов в Норвегии.
Животные проходили через деревни в таком количестве, что
кошки и собаки, которые вначале нападали на них, просто пере-
ставали обращать внимание на орды грызунов. Достигнув моря,
многие лемминги тонули. Движение леммингов, подобно движе-
нию сов, при взрывном возрастании их численности происходит в
одном направлении. Такие впечатляющие миграции происходят
не при каждом четырехлетием пике плотности, а только в годы
исключительно высоких плотностей. Часто сокращение числен-
ности популяции происходит без переселения животных из тунд-
ры или с гор.
Кребс и Мейерс (Krebs, Meyers, 1974) подготовили обзор по-
пуляционных циклов у мелких млекопитающих, а Файнерти (Fi-
nerty, 1980) применил некоторые новые статистические подходы,
пытаясь ответить на вопрос: «Существуют ли циклы?» Ответ гла-
сит «да» для одних видов и ситуаций и «нет» для других.
46
Глава 6
На рис. 6.17 приведены два примера многолетней регистрации
значительных колебаний численности листогрызущих насекомых
в европейских лесах. Такие хорошо выраженные циклы отмеча-
лись преимущественно в северных лесах, особенно в чистых хвой-
ных древостоях. Плотность может меняться больше чем на пять
Рис. 6.17. Колебания численности двух видов бабочек, личинки которых
питаются хвоей. Вверху — сосновая пяденица Bupalus в сосновом бору
близ Литцлингена, Германия. В этом лесу поддерживается чистое сосновое
насаждение. (Varley, 1949). Внизу — лиственничная листовертка Zeiraphera
griseana в лиственничном лесу долины Энгадин, Швейцария.
порядков (логарифмы циклов), от менее чем одной до более чем
10 000 особей на 1000 м2 (верхний график на рис. 6.17). Легко
представить, что 10 000 потенциальных бабочек сосновой пяде-
ницы на каждые 1000 м2, способных дать несколько поколений
за сезон, более чем достаточно для появления такого числа гусе-
ниц, чтобы уничтожить всю хвою и даже погубить деревья, как
это часто и случается. Циклы у этих насекомых не так регуляр-
ны, как у зайца-беляка, и повторяются примерно через 4—10 лет
(в двух приведенных примерах пики численности повторялись
через 7,8 и 8,8 года). У разных видов циклы не синхронны.
Хорошо известные примеры сходной периодичности — цикли-
ческие вспышки численности еловой листовертки-почкоеда и ко-
Динамика популяций
47
конопряда в северных районах Северной Америки. Исследования
по коконопряду провел Веллингтон (Wellington, 1957, 1960), а по
листовертке-почкоеду — Мак-Дональд (MacDonald, 1975) и Хол-
динг с сотрудниками (Ludwig et al., 1978; Clark et al., 1979;
Holling, 1980). Считается, что чередование «сильных» и «слабых»
физиологических рас у коконопряда (Malacosoma) имеет генети-
ческую основу. На подъеме цикла гусеницы строят удлиненную
паутину, особи активно питаются и перемещаются в ветвях на
большие расстояния. В пике численности гусеницы становятся
неактивными, строят более компактную паутину, едят меньше и
оказываются более восприимчивыми к болезням. Таким образом,
в популяции развился ограничительный контролирующий меха-
низм, благодаря которому предотвращается повторяющаяся дефо-
лиация деревьев и достигается длительное сосуществование насе-
комых и их растений-хозяев.
Можно с уверенностью сказать, что циклы листовертки-поч-
коеда представляют собой явления на экосистемном уровне, по-
скольку эволюция насекомых-листоедов, их паразитов и хищни-
ков, а также хвойных деревьев (ели и бальзамической пихты,
которые часто растут в чистых насаждениях) строго сопряжена
или протекает совместно. По мере возрастания биомассы деревьев
в лесу наиболее крупные и старые деревья становятся восприим-
чивыми к гусеницам листовертки-почкоеда, и многие из этих
деревьев погибают от повторяющейся дефолиации. Отмирание и
разложение древесины, фекалий и остатков насекомых возвраща-
ет в лесную почву питательные вещества. Менее чувствительные
к нападению насекомых молодые деревья избавляются от затене-
ния и быстро растут, за несколько лет заполняя полог. Тем вре-
менем паразиты и птицы, питающиеся насекомыми, совместными
усилиями снижают экологическую плотность листовертки-почко-
еда. Если рассматривать существование хвойных лесов на боль-
ших промежутках времени, то очевидно, что листовертка-почкоед,
периодически омолаживая экосистему хвойного леса, представля-
ет co6oii неотъемлемую часть этой целостной системы, а не яв-
ляет собой катастрофу, как могло бы показаться всякому, кто
видит только мертвые и умирающие деревья в пике цикла. Дей-
ствительно, Питермен (Peterman, 1978), изучив роль короедов в
полегании жердняка в горных лесах, пришел к заключению, что
благодаря деятельности жуков, которые изреживают загущенный
древостой, малопригодный для лесозаготовок, обитания в нем жи-
вотных и отдыха, создаются более ценные для человека леса. По-
жалуй, Питермен заставляет нас рассматривать этих жуков как
одно из средств ведения хозяйства, а не как вредителей леса.
По его мнению, в изолированных лесах следовало бы допускать
периодические вспышки численности жуков. Такой взгляд, естест-
венно, противоречит традиционным представлениям о мерах борь-
48
Глава 6
бы с вредителями, поскольку эти меры предусматривают контроль
численности насекомых только в периоды, когда их становится
слишком много и они способны полностью уничтожить деревья.
Другая стратегия может заключаться просто в том, чтобы ис-
пользовать в хозяйстве старые рослые деревья, прежде чем их
истребят жуки или листоеды. Сейчас такая стратегия входит в
практику, поскольку созданы модели, на основе которых можно
предсказать время вероятной вспышки численности насекомых.
Периодические бури выполняют функцию, подобную той, что вы-
полняют насекомые: вываливая старые деревья и разрежая загу-
щенные древостои горного леса, бури создают пятнистость в раз-
мещении молодых и более старых лесов, которые постоянно сме-
щаются по склону (см. гл. 8, рис. 8.3).
Во всех описанных до сих пор случаях взаимодействующие
популяции животных и растений длительное время эволюциони-
ровали совместно и, таким образом, выработали стратегии не толь-
ко для выживания, но и для взаимной выгоды.
Пожалуй, наиболее известные примеры колебаний численно-
сти у насекомых — ее циклы у саранчовых. Сведения о нашест*
виях странствующей саранчи (Locusta migratoria} в Евразии вос-
ходят к далекой древности (Carpenter, 1940а). Саранча живет в
пустынях и полуаридных зонах и на протяжении многих лет
не совершает миграций, не пожирает посевов и не привлекает к
себе внимания. Однако время от времени плотность популяции
саранчи достигает чудовищных размеров. Под влиянием скучен-
ности насекомые претерпевают морфологические изменения 1 (на-
пример, у них развиваются более длинные крылья) и начинают
мигрировать в земледельческие районы, съедая все на своем пу-
ти. Уваров (Uvarov, 1957) отметил, что такая практика возделы-
вания земель, как севообороты и перевыпас скота, не уменьшают,
а увеличивают вероятность вспышки, потому что мозаичное че-
редование растительности и обнаженной земли (куда саранча от-
кладывает яйца) благоприятствует экспоненциальному росту чис-
ленности популяции. По-видимому, мы имеем здесь дело с попу-
ляционным взрывом, обусловленным и нестабильностью
окружающей среды, и простотой условий. Вероятно, как и у лем-
мингов, не каждый популяционный максимум сопровождается
эмиграцией, поэтому частота повторения этих бедствий не обяза-
тельно отражает точную периодичность колебаний плотности
И даже при этом за период между 1695 и 1895 гг. вспышки чис-
ленности регистрировались по крайней мере каждые 40 лет. Бо-
лее современный исторический обзор, посвященный подъемам и
1 Одиночные и мигрирующие формы обнаружены у нескольких видов
саранчи; их часто описывали как разные виды, пока не были установлены
их истинные взаимоотношения.
Динамика популяций
49
спадам численности пустынной саранчи, принадлежит Валову
(Waloff, 1966).
Известен интересный пример колебаний численности типа
хищник — жертва с участием и животных, и растений. Продук-
ция семян у хвойных часто циклична, и соответствующие колеба-
ния часто наблюдаются у зерноядных птиц и других животных..
Регулярные циклы численности, такие, как у рыси и зайца
или у леммингов, представляются парадоксальными и загадочны-
ми: ведь в природе изменения часто далеко не столь регулярны.
Эти циклы имеют две поразительные особенности: 1) сильнее
всего они выражены в менее сложных экосистемах северных
районов, как явствует из приведенных примеров; и 2) хотя пики
численности могут одновременно отмечаться на обширных прост-
ранствах, у одного вида, обитающего в разных районах, ее пики
отнюдь не всегда совпадают по времени. Теории, выдвинутые для
объяснения этих регулярных циклов, относятся ко всем уровням
иерархической организации, и их можно подразделить на: 1) ме-
теорологические теории; 2) теории случайных флуктуаций;
3) теории взаимодействия популяций и 4) теории взаимодействия
трофических уровней. Ниже будут даны краткие характеристики
каждой из этих теорий.
Все попытки связать регулярные колебания численности с
климатическими факторами остаются пока безуспешными, несмот-
ря на то, что синхронность и четкая выраженность этих колеба-
ний в северных районах, казалось бы, свидетельствуют о перио-
дических событиях за пределами локальной экосистемы. Одно
время полагали, что циклы численности рыси и другие 10-летние
циклы объясняются цикличностью появления солнечных пятен,
с которым связаны основные изменения погоды. Однако, Мак-Ла-
лич (MacLulich, 1937) и другие показали, что на самом деле*
корреляция здесь отсутствует. До сих пор не выявлено широко-
распространенной климатической периодичности с 3—4-летними
интервалами.
Палмгрен (Palmgren, 1949) и Коул (Cole, 1951, 1954) выска-
зали предположение, что колебания, кажущиеся нам регулярны-
ми, возможно, являются следствием случайных изменений комп-
лекса биотических и абиотических условий среды, в которых
обитает популяция. Если эта гипотеза верна, то ни один из фак-
торов нельзя считать важнее остальных. Однако Кейт (Keith,
1963), проведя детальный статистический анализ циклов северных
млекопитающих и птиц, пришел к выводу, что десятилетние цик-
лы неслучайны, хотя доказать, что более короткие циклы не яв-
ляются следствием случайных флуктуаций, все же трудно.
Если будет доказано, что случайные и иные климатические
факторы не являются основной причиной резких колебаний чис-
ленности, то вполне естественно искать причины этих колебаний;
4 Заказ № 1483
Глава 6
в самих популяциях («внутренние факторы»). Имеются некоторые
данные, свидетельствующие о том, что, по-видимому, существуют
какие-то механизмы, которые могут действовать сопряженно с
изменениями погоды или других физических факторов.
Основываясь на медицинской теории стресса (общего адапта-
ционного синдрома), созданной Гансом Селье (Hans Selye), Крис-
тиан и его сотрудники (см. Christian, 1950, 1961, 1963; Christian,
Davis, 1964) собрали многочисленные данные как для природ-
ных, так и для лабораторных популяций, показывающие, что у
высших позвоночных в условиях перенаселенности происходит
увеличение надпочечников. Это один из симптомов сдвига нейро-
эндокринного равновесия, который в свою очередь сказывается
на поведении животных, их репродуктивном потенциале, устой-
чивости к болезпям и другим видам стресса. Комплекс таких
изменений часто бывает причиной резкого снижения плотности
популяции. Например, зайцы-беляки в периоды пика численности
часто внезапно погибают от «шоковой болезни», которая, как
было показано, связана с увеличением надпочечников и другими
признаками нарушения гормонального равновесия.
Чптти (Chitty, 1960, 1967) высказал предположение, что раз-
личия в агрессивном поведении и выживаемости, наблюдающиеся
в различные фазы цикла у полевок, обусловлены генетическими
сдвигами — ситуация, сходная с той, которая отмечена у коконо-
пряда. Таким образом, совершенно очевидно, что адаптационный
синдром сглаживает колебания.
Теории четвертой группы основываются на представлении,
согласно которому регуляция циклов численности осуществляется
на экосистемном, а не на популяционном уровне. Разумеется,
изменения, которые охватывают несколько порядков величин,
должны распространяться не только на взаимодействия высших
трофических уровней, например на взаимодействие хищник—
жертва, но и на взаимодействия, характерные для первичного
звена пищевой цепи, такого, как растения—травоядные. Приме-
ром служит описанная ранее гипотеза Холдинга, основанная на
исследованиях листовертки-почкоеда. Другим примером служит
предложенная для объяснения циклов численности полевок в
тундре гипотеза «возврата биогенных элементов» (Schultz, 1964,
I960; Pitelka, 1964, 1973). Согласно этой гипотезе, подтвержден-
ной данными изучения круговоротов биогенных элементов, в ре-
зультате сильного выедания растительности леммингами в годы
пика их численности в последующие годы происходит связывание
и уменьшение доступности биогенных элементов (особенно фос-
фора), в результате чего питательная ценность пищи леммингов
сильно снижается. В связи с этим рост и выживание молодых
особей ограничиваются. На протяжении третьего и четвертого года
биогенные элементы возвращаются в круговорот, растительный
Динамйка популяций
51
покров восстанавливается, и экосистема опять может поддержи-
вать высокую плотность леммингов.
Мы уделили столь большое внимание случаям резкого измене-
ния численности не потому, что они так уж часто встречаются в
природе, а потому, что при их исследовании выявляются функции
и взаимодействия, вероятно, имеющие более общий характер,
но не всегда с очевидностью проявляющиеся в тех популяциях,
где плотность меняется менее резко. Проблема циклических ко-
лебаний в каждом конкретном случае может быть сведена к уста-
новлению того, определяются ли они в основном одним или
несколькими факторами или же их причины столь многочислен-
ны, что разобраться в них не представляется возможным, хотя и
ясно, что общая структура взаимосвязей может быть понята на
основе того, что Коул (Cole, 1957) назвал «вторичной простотой»»
имея в виду, что закономерностей здесь «не больше, чем встре-
чается в последовательности случайных чисел». Первая ситуация,
конечно, возможна в простых экосистемах, как эксперименталь-
ных, так и природных; последняя более вероятна для сложных
экосистем.
Рассмотрев несколько интересных конкретных случаев, пе-
рейдем к анализу более общей проблемы регуляции численности
популяций.
6. Независимая и зависимая от плотности
регуляция численности популяций
Определения
В экосистемах с низким уровнем разнообразия, подверженных
воздействию физических стрессов или находящихся под влиянием
других нерегулярных или непредсказуемых внешних факторов,
размеры популяции обычно зависят от погоды, течений, лимити-
рующих химических факторов, загрязнения и т. п. В экосистемах
с высоким уровнем разнообразия, функционирующих в благопри-
ятной среде (вероятность периодического физического стресса,
такого, как буря или пожар, низка), регуляция осуществляется,
как правило, за счет биологических факторов; по крайней мере
в известных пределах их плотность подвержена саморегуляции.
Любой фактор — неважно лимитирующий или благоприятный
(отрицательный пли положительный) — может быть; 1) незави-
симым от плотности, если его влияние не зависит от величины
популяции, или 2) зависимым от плотности, если его влияние на
популяцию есть функция плотности. Влияние факторов второй
группы может быть прямым, т. е. оно обычно усиливается по
мере приближения к верхнему пределу плотности, но оно может
4*
52 Глава 6
подчиняться и обратной зависимости: с увеличением плотности
влияние уменьшается. Факторы, влияние которых прямо зависит
от плотности, действуют подобно регулятору числа оборотов дви-
гателя (следовательно, их можно назвать «управляющими плот-
ностью»); по этой причине их рассматривают как один из глав-
ных механизмов, предотвращающих перенаселение. Действие
климатических факторов часто (но отнюдь не всегда) не зависит
от плотности, тогда как действие биотических факторов (конку-
ренция, паразиты, патогенные организмы и т. д.) часто (но опять-
таки не всегда) зависит от плотности.
Объяснения
Общая теория регуляции численности популяций логически вы-
текает из представления о биотическом потенциале, типах роста
и колебаниях относительно уровня несущей емкости среды. Так,
рост, соответствующий J-образной кривой, наблюдается тогда,
когда независимые от плотности или внешние факторы вызывают
замедление или остановку роста. Вместе с тем сигмоидный рост
зависит от плотности, так как в данном случае рост популяции
регулируется переуплотнением и другими внутренними факто-
рами.
Поведение любой популяции, произвольно выбранной для
изучения, зависит от типа экосистемы, частью которой эта по-
пуляция является. Противопоставление (как мы это делали) «фи-
зически контролируемых» и «биологически контролируемых»
экосистем произвольно, и хотя в данном случае мы пользуемся
чересчур упрощенной моделью, такой подход вполне оправдан,
особенно в связи с тем, что человек на протяжении большей
части промышленной революции замещает самоподдерживаю-
щиеся экосистемы монокультурами и подвергает их все более
сильным стрессовым воздействиям; это требует от него значи-
тельных усилий для сохранения таких экосистем. По мере того
как росла стоимость (в энергетическом и денежном выражении)
физического и химического контроля, повышалась устойчивость
возбудителей к пестицидам, а концентрации побочных продуктов
токсичных химических веществ в пище, воде и воздухе станови-
лись все более опасными для здоровья, мы были вынуждены под-
вергнуть серьезному пересмотру наше отношение к биологиче-
скому контролю. Свидетельством этому служит все возрастающий
интерес к новому подходу, названному «интегральным методом
борьбы с вредителями», при котором основные усилия направле-
ны на восстановление природного, зависимого от плотности конт-
роля на экосистемном уровне в сельскохозяйственных и лесных
экосистемах, как это было подробно описано в конце гл. 5.
В предыдущем разделе мы описали, как физиологические и
генетические сдвиги или смены экотипов во времени могут смяг-
Динамика популяций
53
чать колебания и ускорить возврат плотности к более низкому
уровню, после скачка за пределы насыщения емкости экосистемы.
Остается, однако, вопрос о том, происходит ли в действительности
©волюция саморегуляции на популяционном уровне, если естест-
венный отбор действует на уровне особей. Винн-Эдвардс (Wynne-
Edwards, 1962, 1965), чьи обзорные статьи и книги пользовались
широкой популярностью, предложил два механизма стабилизации
плотности в пределах ниже уровня насыщения: 1) территориаль-
ное поведение, представляющее собой форму внутривидовой кон-
куренции, которая ограничивает рост популяции посредством ре-
гулирования «землепользования» (более подробно это будет об-
суждаться в разд. 9), и 2) групповое поведение, выражающееся,
например, в «очередности клевания», «половом доминировании»
я других формах поведения, повышающих приспособленность по-
томства, но уменьшающих его численность. Эти механизмы на-
правлены на повышение качества среды обитания особи и умень-
шение вероятности вымирания, которое могло бы произойти при
чрезмерном увеличении численности и, как следствие этого, ис-
тощении доступного количества ресурсов. Значение этих соци-
альных и поведенческих особенностей трудно проверить экспе-
риментально; в настоящее время вопрос этот привлекает к себе
усиленное внимание, о чем свидетельствует очень большая обзор-
ная книга Коэна и др. (Cohen et al., 1980). Мы же обсудим это
далее, в гл. 8, разд. 5 и 6.
Итак, независимые от плотности факторы среды вызывают
подчас драматические изменения плотности популяции и смеще-
ние уровня верхней асимптоты или уровня несущей емкости.
В то же время зависимые от плотности рождаемость и смертность
удерживают популяцию в стационарном состоянии или ускоряют
возврат в это состояние. Независимые от плотности факторы сре-
ды играют большую роль в регуляции экосистем, подвергающихся
сильному внешнему стрессовому воздействию, тогда как зависи-
мые от плотности факторы приобретают большее значение, когда
влияние этих стрессовых воздействий уменьшается. Как во вся-
кой отрегулированной кибернетической системе, дополнительная
регуляция по принципу обратной связи обеспечивается взаимо-
действием (как фенотипическим, так и генотипическим) между
популяциями разных видов, которые связаны друг с другом в
пищевые цепи, или другими существенными экологическими взаи-
моотношениями.
Примеры
Сильные бури, внезапные падения температуры и другие резкие
изменения физических факторов — наиболее характерные приме-
ры независимых от плотности воздействий. Изучая в течение трех
54
Глава 6
Нередко наблюдается явная корреляция между теорией
и той средой, которую изучает теоретик. Так, экологи, ра-
ботающие в местностях с резко неблагоприятными условия-
ми (например, в аридных областях) или имеющие дело с
мелкими организмами (такими, как насекомые или планк-
тон, для которых характерны короткие жизненные циклы,
высокий биотический потенциал, высокая интенсивность ме-
таболизма на 1 г массы, а следовательно, и небольшая био-
масса на единицу пространства в единицу времени), прежде
всего обращают внимание на: 1) значение периода времени,
когда скорость прироста (г) положительна; 2) значение
независимых от плотности факторов, таких, как погода,
в определении продолжительности благоприятных периодов;
3) второстепенную роль внутрипопуляционных факторов
самоограничения; 4) общее отсутствие стабильности в ди-
намике плотности любых видов, даже в стабильных экосис-
темах. Эти моменты подчеркнуты в хорошо известной книге
Андреварты и Бирча (Andrewartha, Birch, 1954).
Экологи, работающие в мягких, благоприятных условиях
(таких, как английские сады, коралловые рифы или тропи-
ческие леса) или с более крупными организмами (птицы,
млекопитающие или лесные деревья, у которых жизненные
циклы более продолжительны, а численность и биомасса ‘
более четко соответствуют потоку энергии), обращают вни-
мание на: 1) значение зависимых от плотности факторов,
и прежде всего — лимитирующее значение внутривидовой
конкуренции (как в уравнении S-образного роста) и различ-
ных межвидовых регулирующих влияний (межвидовая кон-
куренция, паразиты и т. д.); 2) стабильность или по мень-
шей мере постоянство характера изменений плотности;
3) значение биологических механизмов регуляции. Харак-
терно, что исследователи, работающие с отдельными замкну-
тыми популяциями мелких организмов, такими, как куль-
туры бактерий или лабораторные популяции мучных хру-
щаков, также обращают особое внимание на эти последнпе
аспекты, что, в общем, неудивительно, поскольку биомасса
па единицу объема в культурах больше, чем это обычно на-
блюдается в природе. В равной мере не следует удивляться
и тому, что на исследователей, работающих с монокульту-
рами зерновых и древесных растений, не производит впе-
чатления эффективность биологического контроля.
Анализ теорий зависимой от плотности регуляции содер-
жится в основополагающих работах Никольсона (Nicholson, |
1954, 1957, 1958), Лэка (Lack, 1954); см. также приложение ।
в работе Lack, 1966. :
Динамика популяций
55
лет моллюска Астаеа. живущего на камнях в литоральной зоне,
Фрэнк (Frank, 1965) обнаружил, что большая часть изменений,
происходящих в популяции, зависит от плотности; исключение
составляют случаи резкого увеличения смертности после суровых
морозных зим, когда поверхность камней разрушается и удаля-
ется вместе с моллюсками независимо от их численности. Хоро-
ший случай «совершенной» зависимости от плотности уже был
нами рассмотрен на примере роста дрожжей (рис. 6.12). Как
отмечалось в обсуждении, линейная зависимость от плотности
маловероятна в природных открытых системах. Проведя тщатель-
ное исследование васильковой орехотворки, Варли (Varley, 1947)
показал, что влияние ее основного паразита, насекомого Eurytoma
eurta, зависит от плотности, поскольку в условиях, когда плот-
ность популяции хозяина велика, паразит уничтожает большее
(как в процентном, так и в абсолютном выражении) число особей
хозяина (табл. 6.4). Однако в другом исследовании Варли и Эд-
вардс (Varley, Edwards, 1957) сообщили, что когда «площадь
поиска» мала, как в случае паразитической осы Mormoniella,
влияние паразита на своего двукрылого хозяина не обязательно
зависит от плотности. Таким образом, важную роль могут играть
различия в поведении. Холлинг (Holling, 1965, 1966) предложил
серию математических моделей, в которые ввел поведенческие
признаки. Эти модели позволяют прогнозировать эффективность
регулирующего влияния данного насекомого-паразита на насеко-
мое-хозяина при различных уровнях плотности.
Таблица 6.4. Зависимое от плотности влияние паразитического перепон-
чатокрылого на хозяина — личинку галловой мухи. (По Varley, 1947.)
Уровень численности популяции Популяция личи- нок в начале сезо- на, численность на 1 Число уничтожен- ных паразитом личинок на 1 м2 Смертность от паразита, %
Год с низкой численно- стью 43 6 14
Год с высокой числен- ностью 148 66 45
Все аспекты проблемы регулирования популяций можно пред-
ставить в виде графической модели, приведенной на рис. 6.18.
В основу графика положены данные о динамике популяции кон-
кретного вида (австралийской листоблошки сем. Psyllidae, жи-
вущей на эвкалиптах), однако он иллюстрирует многие принци-
пы, обсуждавшиеся выше. В норме наблюдается стабилизация
популяций на уровне, соответствующем низкой численности (уро-
вень 2, время I на рис. 6.18); этот уровень намного ниже уров-
ня 2, которого достигает популяция при использовании всех пи-
66
Глава 6
щевых и пространственных ресурсов. Регуляция на уровне ниже
насыщающего осуществляется метеорологическими условиями и
независимыми от плотности паразитизмом и хищничеством на
нимфах этого насекомого, а также зависимым от плотности хищ-
ничеством птиц на взрослых особях. Иногда стабилизирующие
процессы ослабевают; обычно это происходит под влиянием
необычайно низких температур, вызывающих снижение относи-
тельной численности паразитирующих организмов, в результате
чего плотность увеличивается выше контролируемого уровня
(т. е. «ускользает» от контроля; время II на рис. 6.18). Тогда
происходит стремительный рост (время III) по J-образной кривой
вследствие того, что 1) паразитизм на нимфах становится неэф-
фективным из-за быстрого увеличения сверхпаразитов (парази-
тов, которые паразитируют на паразитах листоблошки) и 2) чис-
ленность насекомоядных птиц растет медленнее, чем численность
насекомых (вспомните значительные различия рождаемости у
насекомых и птиц). Неограниченный какое-то время рост пре-
кращается на уровне 2, когда нимфы выедают всю пищу, а взрос-
лые особи не находят более мест, пригодных для откладки яиц.
Затем, когда деревья остаются без листьев, а популяции хищни-
ков (птиц и муравьев) и паразитов возрастают, происходит рез-
кий спад численности листоблошек (время IV). Если их числен-
ность опускается ниже уровня 7, то популяция опять попадает
в зону действия механизмов регуляции, и ее численность в тече-
ние ряда лет, вероятно, будет небольшой (рис. 6.18, а). Если же?
Рис. 6.18. Динамика численности насекомого Cardiaspina albitextura
(Psyllidae), которое кормится на деревьях ^Eucalyptus blakelyi. В норме
плотность популяции при совместном воздействии зависимых и независи-
мых от плотности факторов (погоды, паразитов и хищников) стабилизиру-
ется на низком уровне. Иногда популяция ускользает от естественного
контроля и вырывается на очень высокий уровень плотности; следствием
этого является дефолиация деревьев на обширных участках. При плотности
ниже уровня 1 эффективно действует механизм природного контроля. При
плотности на уровне 2 строго лимитирующими становятся пища и места
для откладки яиц. (Из Clark, 1964, см. также Clark et al., 1967, с. 158.)
Динамика популяций
57
плотность не снизится ниже контролируемого уровня, то может
возникнуть новая внезапная вспышка численности (рис. 6.18,6).
Эта модель иллюстрирует также некоторые трудности, кото-
рые возникают в практике контроля вспышек численности насе-
комых-вредителей. Частота вспышек увеличивается в результате
хозяйственной деятельности человека, который, подобно «необыч-
ным» погодным условиям, нарушает работу природных регули-
рующих механизмов. Нередко численность популяции возрастает
так быстро, что это обнаруживается лишь в фазе экспоненциаль-
ного роста, когда проводить обработку уже слишком поздно. Оче-
видно, контролирующие мероприятия могут предотвратить вторую
вспышку, если обработка была направлена на уничтожение имен-
но данного насекомого-вредителя. Однако применение инсектици-
дов с широким спектром действия в период IV может принести
больше вреда, чем пользы, поскольку при этом будут уничтожены
также паразиты и хищники, что скорее увеличит, а не уменьшит
вероятность новой вспышки. Во многих случаях, если нам не из-
вестны фаза популяционного цикла или состояние других попу-
ляций, участвующих в функционировании механизма естествен-
ного контроля, лучше воздержаться от каких бы то ни было
истребительных мероприятий.
7. Структура популяции: характер распределения
организмов в пространстве
Определения
Распределение особей в популяции может быть 1) случайным,
2) равномерным (более регулярным, чем при случайном распре-
делении) и 3) групповым (нерегулярным и неслучайным). Слу-
чайное распределение наблюдается тогда, когда среда очень од-
нородна, а организмы не стремятся объединиться в группы. Рав-
номерное распределение встречается там, где между особями
очень сильна конкуренция или существует антагонизм, способст-
вующий равномерному распределению в пространстве. Чаще всего
наблюдается образование различного рода скоплений. Однако ес-
ли особи в популяции обладают тенденцией образовывать группы
определенной величины (например, пары у животных или веге-
тативные клоны у растений), то распределение самих групп мо-
жет оказаться более близким к случайному или даже равномер-
ному. Выяснение типа распределения важно при выборе метода
взятия проб.
Объяснения п примеры
Три типа распределения особей внутри популяции упрощенно
показаны на рис. 6.19. В каждом прямоугольнике находится при-
мерно одинаковое число особей. При групповом распределении
58
Глава G
Рис. 6.19. Три основных типа
распределения особей, пар или
групп особей в популяции. А.
Равномерное распределение.
Б. Случайное распределение^
Б. Групповое распределение
(распределение групп нерегу-
лярно).
(В) группы могут быть
одинакового или разного-
размера (как на рисунке)
и распределяться случай-
но (как показано) или в
свою очередь объединять-
ся в скопления, между
которыми остаются об-
ширные незанятые прост-
ранства. Иными словами,
существует пять типов
распределения: 1) равно-
мерное; 2) случайное;
3) случайное групповое;
4) равномерное групповое
и 5) групповое, с образо-
ванием скоплений групп.
Все эти типы, несомненно’,
встречаются в природе.
Как явствует из рис. 6.19, малые выборки, взятые из трех пред-
ставленных популяций, дадут совершенно различные результаты.
При малой выборке из популяции с групповым распределением
будут получены либо слишком низкие, либо слишком высокие
величины плотности в пересчете на всю популяцию. Таким обра-
зом, прп изучении популяций с групповым распределением необ-
ходимо более тщательное планирование эксперимента, а число
выборок должно быть большим, чем в случае, если распределение
не групповое.
Случайному распределению соответствует нормальная, или
колоколообразная, кривая, на которой основаны стандартные ста-
тистические методы. Распределение такого типа можно ожидать
в природе в тех случаях, когда на популяцию одновременно воз-
действуют многочисленные, но слабые факторы. (Вспомните
разд. 5, в котором обсуждалась возможная случайная основа цик-
лических колебаний.) Если же, как это обычно бывает, преобла-
дает лишь несколько основных факторов (вспомните принцип ли-
митирующих факторов) или если у растений и животных наблю-
дается сильная тенденция к образованию скоплений для (или
Динамика популяций
59
вследствие) размножения и других целей, то вряд ли следует
ожидать полностью случайного распределения. Однако встречае-
мые иногда в природе неслучайные или «контактные» распреде-
ления можно рассматривать как результат случайного распреде-
ления групп, содержащих разное число особей (как на
рис. 6.19, В), или же группы могут быть распределены равно-
мерно (или по крайней мере не совсем случайно). Иными словами
(мы берем крайний случай), лучше определить каким-либо вы-
борочным методом число муравейников (используя муравейник
как популяционную единицу), а затем определить число муравьев
на 1 муравейник, нежели пытаться определить количество му-
равьев непосредственно, используя метод случайных выборок.
Предложено несколько методов, позволяющих определить тип
распределения особей в пространстве и степень скученности
особей в популяции (когда это не самоочевидно), однако здесь
предстоит решить еще много задач. В качестве примеров можно
упомянуть два метода. Один из них состоит в сравнении факти-
ческих частот встречаемости групп животных с разной числен-
ностью, выявленной в серии выборок, с членами ряда Пуассона,
задающего частоты, с которыми должны встречаться группы из
О, 1, 2, 3, 4, ...п особей, при случайном распределении. Так, если
частота малых групп (включая пустые выборки) и обильных
групп выше, а частота средних по величине групп ниже, чем
ожидаемая, то мы имеем дело с групповым распределением. Про-
тивоположная ситуация характеризует равномерное распределе-
ние. При определении значимости наблюдаемого отклонения от
кривой Пуассона проводят статистический анализ, однако этот
общий метод неудобен тем, что на результат может влиять вели-
чина выборки. Один из примеров использования метода Пуассона
для проверки случайного распределения пауков показан в
табл. 6.5. На восьми из одиннадцати площадок пауки распреде-
лялись случайно. Неслучайное распределение имело место на
площадках с наименее однородной растительностью.
Для случайных распределений характерно одно общее свой-
ство, состоящее в том, что дисперсия (V) равняется среднему
(т). Если дисперсия больше среднего, то распределение группо-
вое, если же дисперсия меньше среднего, то распределение рав-
номерное (регулярное). Таким образом, при случайном распре-
делении
У/т—1; стандартная ошибка = 1 •
Если при использовании стандартных критериев значимости ока-
зывается, что отношение дисперсия/среднее больше единицы,
то распределение групповое, если это отношение значимо меньше
единицы, то мы имеем дело с равномерным распределением, и ес-
60
Глава 6
ли отношение равно единице — со случайным распределением.
Такой подход иллюстрируется также в табл. 6.5.
Таблица 6.5. Случайное и неслучайное пространственное распределение пау-
ков и двустворчатых моллюсков
А. Численность и распределение пауков-волков (Lycosidae) в квадратах по
0,1 гектару в залежном местообитании. (По Kuenzler, 1958.)
Вид Квадрат Численность в квадрате X2 для распреде- ления Пуассона
Lycosa tlmuqua 1 31 8,904
2 19 9,584
3 15 5,51
4 16 0,09
5 45 0,78
6 134 1,14
Lycosa carolinensis 2 16 0,09
5 23 4,04
6 15 0,05
3 70 17,304
Lycosa rabida 4 16 0,09
Б. Среднее, дисперсия и пространственное распределение двух видов мел-
ких двустворчатых моллюсков на литоральной илистой отмели в Коннек-
тикуте. (По Jackson, 1968.)
Вид и возраст Среднее Дисперсия Отношение дисперсии к среднему^-),
Mulinia lateralis Все возрасты Gemma gemma Все возрасты Первый год Второй год 0,27 5,75 4,43 1,41 0,26 11,83 7,72 1,66 Случайное Групповое » Случайное
1 Значимо на 5-процентном уровне, т. е. не случайно. Во всех остальных квад-
ратах распределение случайно.
2 Если отличие от единицы не значимо (на 5-процентном уровне), то распределе-
ние случайно; если значимость отличия от единицы меньше 1%, то это свидетель-
ствует о групповом (агрегированном) распределении.
Другой метод, предложенный Дайсом (Dice, 1952), заключа-
ется в измерении каким-либо стандартным способом расстояния!
между особями. Если построить далее график, на котором квад-
ратные корни из этих расстояний представлены как функция
частот, то форма полученного многоугольника будет свидетель-
ствовать о характере распределения. Симметричный многоуголь-
ник частот (иными словами, нормальная колоколообразная кри-
вая) свидетельствует о случайном, скошенный вправо — о равно-
мерном, скошенный влево — о групповом распределении (особи
Динамика популяций 6i'
находятся ближе друг к другу, чем ожидалось). Численную меру
степени «скошенности» можно оценить количественно. Этот ме-
тод, разумеется, лучше применять для растений или неподвиж-
ных животных, но его можно использовать и для оценки распре-
деления в пространстве колоний или жилищ животных (нор лис
и грызунов, гнезд птиц и т. д.).
Личинки мучных хрущаков обычно случайно распределены в
своей очень однородной среде: их наблюдаемое распределение со-
ответствует распределению Пуассона (Park, 1934). Одиночные
паразиты или хищники, такие, как пауки, рассматриваемые в
табл. 6.5, характеризуются иногда случайным распределением
(и они часто проводят время, занимаясь случайным поиском хо-
зяина или жертвы). Джексон (Jackson, 1968) сообщил о случай-
ном характере распределения особей двустворчатого моллюска
Mulinia lateralis на илистой литорали. Так же были распределены
двухлетние особи Gemma gemma, однако для однолетних особей
и всей популяции Gemma закономерность оказалась иной: в связи
с яйцеживорождением (т. е. личинки развиваются в теле самки)
распределение этих животных было групповым. Результаты этого
исследования приведены в табл. 6.5, Б. Обратите внимание на то,
что в популяции со случайным распределением дисперсия близка
к среднему, а в популяции с групповым распределением — значи-
мо больше среднего. Илистое дно — очень гомогенная среда оби-
тания, и межвидовая конкуренция здесь выражена слабо. Оба
этих обстоятельства благоприятствуют случайному распределению.
У деревьев в лесу, достаточно высоких, чтобы их кроны обра-
зовали часть общего полога, распределение может быть равно-
мерным, поскольку конкуренция за свет у деревьев столь велика,
что у них наблюдается тенденция отстоять друг от друга на рас-
стояния, которые «скорее регулярны, нежели случайны». Поля
зерновых культур, фруктовые сады и посадки сосны могли бы
служить еще лучшими примерами. Кустарники в пустыне часто
распределяются очень равномерно, почти так, словно их специ-
ально сажали рядами. По-видимому, дело здесь в интенсивной’
конкуренции (быть может, связанной, в частности, с выделением:
антибиотиков) в среде с недостаточной влажностью. Тенденция к
более равномерному распределению у некоторых животных мо-
жет быть обусловлена территориальностью (разд. 9).
За примерами группового распределения и пятнистой среды
обитания читателю далеко ходить не придется. Из многих изу-
ченных Коулом (Cole, 1946, 1946а) беспозвоночных, обитающих
в лесной подстилке, только для пауков было характерно случай-
ное распределение. В другой работе Коул отметил, что случайное1
распределение удалось установить только для 4 из 44 видов расте-
ний. Все остальные виды были в той или иной степени объединены
в группы.
62
Глава 6
Более подробно статистика пространственного распределения
элементов популяций рассмотрена в работах Скеллама (Skellam,
1952), Гудолла (Goodall, 1970) и Пиелу (Pielou, 1975).
8. Структура популяции: агрегация,
принцип Олли и безопасные поселения
Определения
Как было отмечено в предыдущем разделе, для внутренней струк-
туры большинства популяций в разное время характерно обра-
зование групп разных размеров. Такие группы представляют со-
бой результат агрегирования особей, происходящего по разным
причинам: 1) вследствие локальных различий в местообитаниях;
2) под влиянием суточных и сезонных изменений погодных усло-
вий; 3) в связи с процессами размножения или 4) в результате
социального привлечения (у высших животных). Агрегация мо-
жет усиливать конкуренцию между особями за компоненты ми-
нерального питания, пищу или пространство, но это часто более
чем уравновешивается повышением жизнеспособности группы, по-
скольку группа обладает большими возможностями для своей за-
щиты, обнаружения ресурсов или изменения микроклимата или
микроместообитания. Степень агрегации, так же как общая плот-
ность, при которой наблюдаются оптимальный рост и выживание
популяции, варьирует у разных видов и в разных условиях; по-
этому как «недонаселенность» (или отсутствие агрегации), так и
перенаселенность могут оказывать лимитирующее влияние. Это
и есть принцип Олли.
Особый тип агрегации был назван «образованием безопасных
поселений». В этом случае обширные, социально организованные
группы животных обосновываются в благоприятном центрально
расположенном участке, откуда они регулярно расходятся для
удовлетворения потребностей в пищевой или других видах энер-
гии и куда возвращаются. Некоторые из наиболее успешно адап-
тирующихся животных на земле, в том числе скворцы и человек,
используют эту стратегию.
Объяснения и примеры
У растений агрегация может возникать под влиянием первых трех
пз перечисленных выше факторов. У высших животных сильно
выраженная агрегация может быть результатом действия всех
четырех факторов, но особенно социального поведения; хорошими
примерами служат разные виды оленей, в частности карибу, кото-
рые в очень монотонной арктической среде образуют огромные
Динамика популяций
63
стада, птицы, образующие большие мигрирующие стаи, или ан-
тилопы в саваннах восточной Африки, кочующие табунами с од-
ного пастбища на другое и предупреждающие таким образом ис-
тощение любого участка своей территории.
В целом для растений и, вероятно, для некоторых низших
животных применимо экологическое правило, согласно которому
тенденция к агрегации находится в обратной зависимости от по-
движности стадий расселения (семян, спор и т. д.). Впервые это
высказали Уивер и Клементс (Weaver, Clements, 1929) в своем
руководстве по экологии растений. В заброшенных насаждениях
кедры, хурма и другие растения, у которых семена не имеют
приспособлений, способствующих их переносу, всегда сгруппи-
рованы около родительских растений или вдоль заборов и в дру-
гих местах, где птицы и другие животные запасают семена. Вмес-
те с тем амброзия, злаки и даже сосны, легкие семена которых
далеко разносятся ветром, растут на залежах сравнительно более
равномерно.
Высокая выживаемость в группе — важный признак, который
может быть результатом агрегации. Группа растений способна
лучше противостоять ветру или эффективнее уменьшать потери
воды, чем отдельные особи. Однако у зеленых растений отрица-
тельные последствия конкуренции за свет и минеральное питании
в общем скоро перевешивают преимущества от объединения в-
группы. Лучше всего положительное влияние объединения в груп-
пу выражено у животных. Олли (Allee, 1931, 1938, 1951) провел
множество соответствующих экспериментов и подытожил полу-
ченные им данные в печатных работах. Он обнаружил, например^
что рыбы в группе могут лучше переносить введенную в воду
определенную дозу яда, чем изолированные особи. В то же время
отдельные особи оказываются более устойчивыми к яду, если их
посадить в воду, где раньше находилась группа рыб, чем если
их держать в предварительно не «кондиционированной» воде.
В первом случае слизь и другие выделения способствуют проти-
водействию ядам, выявляя в данной ситуации некоторые меха-
низмы эффекта группы. Другой пример положительного влияния
объединения в группу на выживание — пчелы. Пчелы в улье или
просто в скоплении выделяют и сохраняют достаточно тепла для
выживания всех oco6eii при температуре, при которой гибнут
изолированные особи. Колониальные птицы часто не могут ус-
пешно размножаться, если колония становится маленькой (Dar-
ling, 1938). Олли отметил, что такая примитивная кооперация
(протокооперация), встречающаяся даже у представителей при-
митивных типов организмов, есть начало социальной организации,
в разной степени развитой в животном царстве и достигающей
кульминации у человека. Принцип Олли схематически иллюстри-
руется на рис. 6.20.
64
Глава 6
Плотность--------
Рис. 6.20. Иллюстрация принципа Олли. В одних популяциях скорость рос-
та и выживание максимальны, когда величина популяции мала (И), тогда
как в других внутривидовая протокооперация приводит к тому, что наибо-
лее благополучны популяции промежуточных размеров (Б). В последнем
случае «недонаселение» так же вредно, как и «перенаселение». (По Allee
et al., 1949.)
Настоящая социальная агрегация в том виде, в каком она
встречается у общественных насекомых и позвоночных (в про-
тивоположность пассивной агрегации в ответ на некоторый об-
щий внешний фактор), характеризуется определенной организа-
цией, в частности социальной иерархией и специализацией
особей.
При социальной иерархии, которая может выражаться,
например, просто в «очередности клевания» (названа так потому,
что это явление было впервые описано у кур), особи часто свя-
заны между собой четкими связями подчинения—доминирования
(подобно армейской цепочке от генерала до рядового); она может
принимать и более сложную форму, при которой особи связаны
отношениями лидерства, доминирования и кооперации, как, на-
пример, в группах некоторых птиц и насекомых, ведущих себя
почти как единое целое. В таких группах может быть признан-
ный лидер, однако столь же часто ни одна особь не занимает
главенствующего положения, и члены группы следуют за дейст-
виями особи, которая ведет себя решительно, так, словно она
«знает, что делать». Подобные типы социальной организации бла-
гоприятны для популяции в целом, потому что они предотвраща-
ют перенаселение.
Очень успешная стратегия агрегаций у высших животных
была названа У. Дж. Гамильтоном (Hamilton, Watt, 1970) «об-
разованием безопасных поселений». В этом случае большое число
особей располагается в благоприятном центральном участке, или
ядре, как это наблюдается, например, в колониях скворцов или
гнездовых колониях морских птиц. Практически ежедневно особи
Динамика популяций
65
покидают центральный участок, добывая себе пищу на обширном
пространстве. Агрегация в центральном участке, если хороших
участков недостаточно, дает преимущества в том, что для каждой
особи в этих условиях гарантировано получение необходимого
количества чистой энергии. К неблагоприятным факторам следует
отнести стресс, обусловленный загрязнением участка экскремен-
тами и интенсивным вытаптыванием растительности или грунта.
Очевидно, что «городская агрегация» человека организована та-
ким же образом. Разработка горючих ископаемых фактически
уничтожила границы зон жизнеобеспечения; и поскольку города
и другие центральные поселения не имеют больших запасов энер-
гии, величина популяции в них ограничивается доступными ис-
точниками топлива. Однако по мере роста плотности населения
все большую роль, как лимитирующих факторов, начинают играть
загрязнение окружающей среды и уплотнение почвы.
Удивительные поселения общественных насекомых уникальны
в том отношении, что составляющие их особи крайне специали-
зированы. К числу наиболее высокоразвитых общественных на-
секомых следует отнести термитов (Isoptera), а также муравьев
и пчел (Hynienoptera). У наиболее специализированных видов
разделение труда обеспечивается существованием трех каст: про-
изводителей (например, царицы у пчел), рабочих и солдат. Каж-
дая из каст морфологически специализирована для выполнения
соответствующей функции: размножения, заготовки пищи или
защиты. В следующей главе будет показано, что такая адаптация
ведет к групповому отбору не только внутри вида, но и в группе
тесно связанных видов.
Принцип Олли вполне приложим к человеку. Вероятно, «го-
родская агрегация» (стратегия «безопасных поселений», обсуж-
давшаяся только что) благоприятна для человека, но лишь до
определенного предела (см. гл. 2, разд. 7, и гл. 3, разд. 8). Если
построить график зависимости выгоды (ось у) от величины го-
рода (ось х), то полученная кривая будет иметь такой же горб,
как кривая Б на рис. 6.20 и кривая «субсидия — стресс» на
рис. 3.5. Таким образом, города, как и колонии пчел или терми-
тов, могут оказаться слишком большими в ущерб собственному
благополучию. Оптимальная величина агрегации общественных
насекомых определяется пробами и ошибками естественного от-
бора.
Поскольку оптимальная величина города пока еще не под-
дается объективному определению, население городов следует
уменьшать, как только стоимость жизни в городе начинает пре-
вышать выгоду. С точки зрения экологических принципов оши-
бочно поддерживать или субсидировать город, который стал слиш-
ком большим для своего жизнеобеспечения. Кто должен сформу-
лировать это, исходя из интересов общественного благополучия?
5 Заказ Кз 1483
66
Глава 6
9. Структура популяции: изоляция
и территориальность
Определения
Факторы, способствующие изоляции или разделению в простран-
стве особей, пар или мелких групп, составляющих популяцию,
вероятно, распространены не так широко, как факторы, способ-
ствующие агрегации. Тем не менее эти факторы очень важны для
улучшения приспособленности и, возможно, для регуляции раз-
меров популяций. Обычно изоляция возникает как следствие:
1) конкуренции между особями за дефицитные ресурсы или
2) прямого антагонизма, включающего у высших животных по-
веденческие реакции, а у растений, микроорганизмов и низших
животных — химические изолирующие механизмы (антибиотики
и аллелопатические вещества). В обоих случаях это может при-
вести, как было показано в разд. 7, к случайному пли равномер-
ному распределению, поскольку ближайшие соседи уничтожаются
или изгоняются. Активность особей, пар или семейных групп
позвоночных и высших беспозвоночных обычно ограничена опре-
деленным пространством, которое называется индивидуальным,
или семейным участком. Если этот участок активно охраняется,
так что он не перекрывается с соседним участком антагонистиче-
ской особи, пары и т. д., то его называют территорией. По-види-
мому, территориальность наиболее выражена у позвоночных и
некоторых членистоногих, характеризующихся сложным репро-
дуктивным поведением, проявляющимся, в частности, в строи-
тельстве гнезд, откладке яиц, заботе о потомстве и его защите.
Объяснения и примеры
Точно так же, как агрегация, которая может усилить конкурен-
цию, но дает другие преимущества, разобщение особей в популя-
ции может уменьшить конкуренцию за жизненно необходимые
ресурсы или обеспечить «приватность», необходимую при слож-
ных репродуктивных циклах (как у птиц), но достигается это,
вероятно, за счет преимуществ, даваемых кооперированным по-
ведением группы. Полагают, что сохранение в процессе эволю-
ции какой-либо структуры зависит от того, дает ли эта структура
долговременные преимущества. Во всяком случае, в природе часто
встречается организация обоих типов; фактически в популяциях
некоторых видов одна форма поведения сменяется другой. Дроз-
ды, например, занимают изолированные территории в период раз-
множения и собираются в стаи к зиме, получая, таким образом,
преимущества от обоих типов распределения. Кроме того, особи
разного возраста и пола могут вести себя по-разному в один и
тот же период (например, взрослые живут изолированно, а мо-
лодые образуют скопления).
Динамика популяций
67
Роль внутривидовой конкуренции и «химической войны» за
пространство у деревьев в лесу и кустарников в пустыне мы уже
обсуждали в разд. 6. Такого рода изолирующие механизмы ши-
роко распространены у микроорганизмов и высших растений. Те-
перь необходимо подробно остановиться на механизмах разобще-
ния, основанных на активном антагонизме, наблюдаемом у выс-
ших животных. Многие животные ограничивают свою основную
деятельность определенной областью или индивидуальным участ-
ком, площадь которого может варьировать от нескольких квад-
ратных сантиметров до многих квадратных километров (таковы
размеры участка пумы). Поскольку индивидуальные участки час-
то перекрываются, достигается только частичная изоляция; край-
нее выражение разобщенности — территориальность (сравните че-
репах и птиц на рис. 6.21).
Рис. 6.21. Индивидуальные участки и территории (т. е. охраняемые инди-
видуальные участки) у различных животных. А, Б. Индивидуальные участ-
ки 15 черепах (Terrapene Carolina) на площади в 2 га (4 — самцы, Б — сам-
ки). Этот вид не охраняет свои участки, и они перекрываются. (Из Stickel,
1950.) В. Г. Территории певчего дрозда (Turdus philomelos) в 1955 и 1956 гг.
в Оксфордском ботаническом саду, Англия. Цифрами обозначены окольцо-
ванные самцы из территориальных пар. Обратите внимание на то. что у
особей 1, 6 и 7 одни и те же участки сохраняются из года в год, две другие
птицы на следующий год не вернулись, и на их территории на следующий
год поселились три новые особи. Обычно преуспевающие особи занимают
из года в год одну и ту же территорию на протяжении всей жизни. (Из
Lack, 1966.)
Термин «территория» в том смысле, как он определен в этом
разделе, был впервые использован Эллиотом Говардом (Elliot
Howard) в книге «Территория в жизни птиц», опубликованной
в 1920 г. Появившиеся с тех пор работы по этому вопросу посвя-
5*
Глава 6
6€
щены главным образом птицам. Однако «изоляция посредством
антагонистического поведения» широко распространена среди
других позвоночных и некоторых членистоногих, особенно у ви-
дов, у которых один или оба родителя охраняют гнезда и молодь.
Найс (Nice, 1941) подразделил территориальное поведение птиц
на несколько основных типов: 1) охраняется весь участок, где
происходит кормление и спаривание, выведение птенцов; 2) ох-
раняется участок, где происходит спаривание и гнездование, но
не кормление; 3) охраняется только тот участок, на котором
происходит спаривание; 4) охраняется только гнездо и 5) охра-
няются участки, не связанные с размножением. В первом случае
охраняемый участок может быть достаточно обширным, большим,
чем нужно для обеспечения пищей родителей и потомства. На-
пример, крошечный комаролов (Polioptila) весом всего около
7 граммов охраняет территорию, достигающую 1,8 га, но добывает
пищу со значительно меньшей площади вокруг гнезда (Root,
1969). В большей части случаев территориального поведения на-
стоящие столкновения на границах сведены к минимуму. Вла-
дельцы заявляют о своих правах на участок или о месте своего
нахождения с помощью песни или демонстративного поведения,
и потенциальный захватчик в общем избегает вторжения в заня-
тое владение. Многие птицы, а также рыбы и рептилии обладают
заметной, бросающейся в глаза раскраской головы, тела или при-
датков, которая демонстрируется для отпугивания незваного
гостя. У большинства перелетных певчих птиц самцы прилетают
в места гнездования раньше самок и проводят время, утвержда-
ясь на своей территории и заявляя об этом громким пением. Лю-
ди же, как правило, разочаровываются, когда узнают, что первый
весенний дрозд поет вовсе не для того, чтобы доставить удоволь-
ствие своей подруге, а для того, чтобы заявить о своих правах
на землю.
Поскольку в начале периода гнездования участок, охраняемый
птицами, часто больше, чем в конце, когда потребность в пище
максимальна, и поскольку многие виды птиц, рыб и рептилий,
проявляющие территориальное поведение, вообще не защищают
участков, на которых они кормятся, репродуктивная изоляция и
территориальный контроль, по-видимому, имеют большее значе-
ние для выживания, чем изоляция, связанная только с добыва-
нием пищи.
Территориальность определенно влияет на генетическую при-
способленность (вероятность оставить потомков), поскольку у
видов, характеризующихся территориальным поведением, особи,
которые не могут обеспечить своей территории, не размножаются.
Вопрос о том, предотвращает ли территориальность перенаселение
л эволюционирует ли она именно по этой причине, как это энер-
гично доказывает Винн-Эдвардс (Wynne-Edwards, 1962), до сих
Динамика популяций
69
пор не решен. Джеррем Браун (Jerram Brown, 1969) обобщил
аргументы, противоречащие гипотезе об ограничении популяции,
в том числе и такую идею, что большая, чем это необходимо,
энергетическая стоимость защиты участка не могла бы дать ка-
кие-либо селективные преимущества. Вернер (Verner, 1977) же
утверждает, что обладание территорией, большей чем та, которая
определяется сиюминутными потребностями, может иметь адап-
тивный смысл, поскольку, даже если наступит засуха или вслед-
ствие иных суровых условий ограничится запас доступной пи-
щи, ресурсы, необходимые для размножения, будут все-таки
гарантированы. Эта точка зрения подтверждается эксперимен-
тальным исследованием Рихерта (Riechert, 1981), выполненным
на одном из территориальных видов пустынных пауков (Agelenop-
sis operta). Рихерт обнаружил, что размер территории у этих
пауков фиксирован (на опытной площадке обычно находится
определенное число пауков) и, кроме того, величина территории
достаточна для того, чтобы паук сумел прокормиться даже при
очень сильном снижении численности жертв. В соответствии с
этим плотность пауков не может превзойти предел, определяемый
наличием соответствующих территорий, независимо от количества
пищи в наиболее благоприятные периоды. Особи, неспособные
занять территорию, теряют в весе и в конце концов погибают,
как показано на рис. 6.22. Особи, занимающие более выгодные
территории, успешнее размножаются, что особенно заметно в
суровых условиях (при неблагоприятной погоде и недостатке пи-
Рис. 6.22. Приспособленность, оцениваемая в суточном приросте или потере
веса тела, у территориальных пауков (7) и у особей, которые не могут
занять и поддерживать территорию (бродяг, 77). В неблагоприятные сезоны
бродяги оказываются в крайне тяжелом положении, и лишь немногие из
них доживают до возраста, в котором возможно размножение. (Из Riechert,
1981.)
70
Глава 6
щи). В этом случае реализуются возможности такого признака,
как территориальность, в ограничении численности популяции и
отборе наиболее приспособленных особей. Проблема, связанная
с гипотезой о регуляции, частично включает вопрос о групповом
отборе, который мы рассмотрим в гл. 8.
Полагают, что территориальное поведение позволяет избегать
давления хищников и предотвращает распространение болезней
вследствие пространственного разобщения особей; кроме того, оно
способствует разделению п сохранению ресурсов и облегчает
встречу особей при размножении.
Вопросы о том, насколько человеку свойственно врожденное
территориальное поведение, и о том, в каких пределах он может
научиться планировать землепользование, так чтобы избежать
перенаселения, чрезвычайно интересны. В поведении человека
обнаруживаются определенные черты территориальности, такие,
как императив личной собственности и законы и обычаи, пред-
писывающие ему защищать свой дом, — если нужно, то и с ору-
жием в руках. В популярной книге «Территориальный импера-
тив» Роберт Ардри (R. Ardrey) оптимистично утверждает, что
человеку от рождения присущи черты территориального поведе-
ния, и поэтому он неизбежно будет противостоять чрезмерной
скученности, избегнув тем самым гибели от перенаселения. К со-
жалению, человек слишком хорошо адаптируется к чрезмерной
скученности, как это можно видеть на примере Гонконга, одной
из самых густонаселенных точек Земли.
10. Распределение энергии и оптимизация: г- и /С-отбор
Определения
Концепция чистой энергии экосистемы как целого (обсуждалась
в гл. 3) основывается на распределении энергии между продук-
цией (Р) и дыханием, или поддержанием (7?). Сходным образом
отдельный организм и популяция могут расти или размножаться
только в том случае, если они получают энергии больше, чем ее
нужно для поддержания. Энергия для поддержания представля-
ет собой сумму энергии, образующейся в результате основного
обмена (т. е. обмена, который характеризует особь в состоянии
покоя; этот обмен можно измерить в лаборатории, и он меняется
в зависимости от типа организма п температуры) и кратной ему
величины энергии, обеспечивающей минимальную активность,
необходимую для выживания в полевых условиях. Такая энергия
для поддержания определяется при длительных наблюдениях в
поле, поскольку ее величина широко варьирует в зависимости от
того, является ли данный организм подвижным или сидячим.
Для размножения требуется дополнительная, пли чистая, энергия,
Динамика популяций 71
обеспечивающая выживание будущих поколений. Эта энергия
расходуется на развитие репродуктивных структур, спаривание,
продуцирование семян, яиц, молоди, заботу о потомстве и т. д.
В процессе естественного отбора организмы достигают наиболее
благоприятного соотношения между выгодами и затратами, или
соотношения чистой энергии и времени. У автотрофов эта эф-
фективность складывается из полезной части световой энергии
(преобразуемой в пищу) минус энергия, необходимая для под-
держания энергоулавливающих структур (например, листьев).
Эта величина зависит от времени, в течение которого доступна
световая энергия. Для животных критическим является отноше-
ние разницы между полезной энергией пищи и энергетической
стоимостью поиска и поедания пищевых объектов ко времени,
требующемуся для поиска и поедания. Оптимизация может до-
стигаться двумя основными путями: 1) минимизацией времени
(например, при эффективном поиске и преобразовании энергии
пищи) или 2) максимизацией чистой энергии (например, путем
выбора крупных пищевых объектов или легко перевариваемой
пищи) или одновременным использованием обеих возможностей.
Большинство оптимизационных моделей предсказывает, что, чем
меньше абсолютное обилие пищи (или другого источника энер-
гии), тем обширней должна быть область ее заготовки и тем шире
должен быть диапазон объектов питания, чтобы соотношение до-
ход/стоимость было оптимальным. Эта тенденция, однако, может
меняться под действием внешних факторов, таких, как конку-
ренция или кооперация с другими видами.
Отношение энергии размножения к энергии поддержания
варьирует не только в соответствии с размером организма и ха-
рактером его жизненного цикла, но также и в соответствии с
плотностью популяции и емкостью среды. В непересыщенной
среде давление отбора благоприятствует видам с высоким репро-
дуктивным потенциалом (высоким отношением усилий на раз-
множение к усилиям по поддержанию). Напротив, условия за-
полненной среды благоприятны организмам, характеризующимся
низким потенциалом роста, но более высокой способностью кон-
курировать за скудные ресурсы и использовать их (больше энер-
гии вкладывается в поддержание и выживание особи). Соответст-
венно эти два способа известны под названиями r-отбора и Й>от-
бора, по г- и /^-константам в уравнении роста (разд. 4). Точно
так же виды, подверженные одному или другому типу отбора,
называются соответственно г- или К-стратегами.
Объяснения
Согласно Коуди (Cody, 1966), распределение или подразделение
(Allocation) энергии между основными видами активности ор-
ганизма отражает балансы между преимуществами и стоимостью
72
Глава 6
каждого вида активности, которые обеспечивают изменения гтах—
внутренней (генетически детерминированной) скорости роста, по-
вышающие в будущем выживание или приспособленность. Преж-
де всего оплачивается, конечно, выживание и поддержание особи
(компонент дыхания) с необходимым добавлением в виде чистой
энергии, ассигнованной на рост и размножение (компонент про-
дукции). Крупные организмы (как и большие города) должны
направлять большую часть метаболизированной энергии на вход
блока поддержания; у мелких организмов эта доля меньше, так
как они не обладают большими структурами, требующими под-
держания. Естественный отбор, этот бескомпромиссный мастер
навязывать функции, требует, чтобы все организмы нашли опти-
мальное равновесие между расходами энергии на выживание в
будущем и расходами на выживание в настоящем.
На рис. 6.23 показаны четыре гипотетических способа распре-
Рис. 6.23. Гипотетическое распределение энергии между тремя основными
видами жизнедеятельности, которое необходимо для выживания в четырех
резко различающихся ситуациях, когда относительное значение каждого
вида жизнедеятельности меняется. (Из Cody, 1966, с изменениями.)
деления энергии между тремя основными статьями расхода:
1) энергия для оплаты конкуренции с другими видами за общие
ресурсы; 2) энергия, оплачивающая возможность быть не съеден-
ным хищником (или травоядным, если речь идет о растительном
организме); и 3) энергия для производства потомства. Если
ущерб от конкуренции и выедания невелик, то большая часть
потока энергии может быть направлена на размножение и про-
дуцирование молоди, как показано в случае А. Напротив, избе-
гание конкуренции или выедания может потребовать больше
Динамика популяций
73
энергии, как показано в случаях Б и В. В случае Г энергия при-
мерно поровну распределяется между этими тремя типами ак-
тивности. Л, Б, В и Г могут обозначать четыре разных вида,
у которых давление отбора приводит к такому характеру под-
разделения энергии, или четыре разных сообщества, в которых
такое подразделение присуще большинству видов, или же, как
это будет показано в гл. 8, А — обычная ситуация на стадиях
колонизации или начальных стадиях сукцессии, когда преоблада-
ет r-отбор, тогда как В и Г — ситуация на более зрелых стадиях,
когда преобладает Я-отбор.
Шенер (Schoener, 1971), Коуди (Cody, 1974), Пайк и др.
(Pyke et al., 1977), обобщив методы анализа путей распределе-
ния и оптимизации энергии, высказали предположение, что в
данном случае можно пользоваться методами, аналогичными при-
меняемым при анализе прибыли и затрат в экономике, причем
прибыль в экологии — это повышение приспособленности, а за-
траты — энергия и время, необходимые для обеспечения размно-
жения. Хищник, например, под давлением отбора вынужден уве-
личивать соотношение между чистой энергией (ассимилированная
энергия минус затраты на поимку жертвы) и временем, необхо-
димым для поиска, преследования и поедания добычи. Теорети-
чески увеличить количество энергии, используемой для размно-
жения, можно, либо 1) выбирая более крупную или более пита-
тельную жертву или жертву, которую легче поймать, либо
2) уменьшая время и усилия, направленные на поиск и пресле-
дование.
Для того чтобы определить оптимальное распределение энер-
гии для данного вида, популяции или данной ситуации в место-
обитании, было предложено несколько подходов. Один из подхо-
дов состоит в том, чтобы выявить, каким образом экологически
эквивалентные виды используют морфологические, метаболиче-
ские и поведенческие адаптации для сохранения приспособлен-
ности в сходных местообитаниях, но в различных географических
областях. Если параметры приспособленности очень сходны, т. е.
наблюдается конвергенция, то можно заключить, что отбор привел
к оптимальному решению, несмотря на различия в истории, мас-
штабе времени и генетическом происхождении. Примером служит
проведенное Коуди (Cody, 1974) сравнение степных птиц Канзаса
и Чили, результаты которого представлены в табл. 7.4.
Другой, более специальный подход состоит в графическом
«анализе стратегий» (Levins, 1968), как это показано на рис. 6.24.
На рисунке приведены две модели стратегий добывания пищи
гипотетическим видом, перед которым стоит задача выбора либо
числа пищевых объектов из шести возможных (Л), либо опре-
деленного числа изолированных участков («пятен») для добыва-
ния пищи (5). Если особи этого вида питаются каким-то одним
74
Глава 6
Рис. 6.24. Модели оптимизации стоимости и доходов. Объяснения см. в
тексте.
пищевым объектом из многих возможных, то им потребуется за-
трачивать больше усилий на отыскание каждой жертвы, чем если
бы они питались любым из шести объектов (кривая AS на
рис. 6.24,Л). Но при увеличении числа объектов питания растут
затраты на преследование (одни объекты трудно поймать, другие
слишком малы) (кривая АР). Оптимальное соотношение дохода
и затрат достигается в точке пересечения восходящей и нисходя-
щей кривых, в нашем гипотетическом случае — при питании на
четырех объектах (рис. 6.24,Л). Взаимодействия с другими ви-
дами или другими факторами среды могут сдвинуть оптимум в
любом направлении. Конкуренция с другими видами может при-
вести наше гипотетическое животное к «специализации» на пи-
тании только одним объектом, если это дает ему преимущество
в конкуренции. Или же такой отбор может давать преимущество,
когда пищи много. Тогда условия будут диктовать, что лучшая
стратегия — это стратегия «генералиста». Пример смещения пи-
щевой избирательности будет приведен ниже.
Из рис. 6.24, Б видно, что охотничье усилие АН возрастает по
Мере увеличения числа используемых участков для поиска пищи,
но это уравновешивается уменьшением времени перехода АТ для
поимки одного объекта. И вновь оптимум представляет собой
компромисс между противоположными тенденциями; в описывае-
мой ситуации это три изолированных охотничьих участка.
Динамика популяций
75
Более общая модель приведена на рис. 6.24, В. Здесь по мере
увеличения времени, затрачиваемого на добывание энергии (ни-
щи), репродуктивный выход единицы энергии снижается, а нере-
продуктивный — растет (в обоих случаях монотонно). Область
максимального репродуктивного выхода на рисунке покрыта точ-
ками, а оптимальное время питания, обеспечивающее благопри-
ятный баланс между двумя необходимыми путями расхода энер-
гии, вновь соответствует пересечению кривых.
Как отмечалось в разд. «Определения», виды с высоким био-
тическим потенциалом (г) отбираются обычно в ненасыщенной
или неопределенной среде, подверженной периодическим стрес-
сам (таким, как штормы, засухи и т. д.). Виды, у которых энер-
гия распределяется в пользу поддержания и повышения конку-
рентной способности, чувствуют себя лучше при плотности, рав-
ной К (насыщение), или при стабильных физических условиях
(мала вероятность сильных неблагоприятных возмущений). По
другому пути идут виды, рост популяций которых описывается
J-образной кривой. Это хорошие пионерные виды, которые могут
быстро использовать нетронутые или недавно накопленные ре-
сурсы, они упруго реагируют на различного рода нарушения.,
Медленно растущие виды и популяции лучше адаптированы к
зрелым сообществам и более устойчивы, но менее упруго реаги-
руют на нарушения (вспомните обсуждение резистентной и упру-
гой устойчивости в гл. 2, разд. 6).
Общая модель г- и A-отбора, предложенная Мак-Артуром
(MacArthur, 1972), показана на рис. 6.25. На рисунке Х\ и Х2
обозначают два конкурирующих генетических аллеля, однако они
могут также обозначать виды. В области Л (слева от точки С),
где плотность низка, а пищи (или света и биогенных элементов
в случае растений) много, побеждает быстрее размножающийся
вид или аллель Х2 (dX/Xdt — удельная скорость роста, как описа-
но в разд. 2), т. е. мы имеем дело с r-отбором. В области В (справа
от точки С) вид Xi растет быстрее, чем X2i и поэтому побеждает,
т. е. имеет место A-отбор. Мак-Артур отмечает, что A-отбор пре-
обладает в тропиках с относительно слабо выраженной сезон-
ностью, тогда как r-отбор преобладает в умеренных зонах, где
сезонность хорошо выражена; в таких условиях численность по-
пуляции растет экспоненциально, а затем следует ее катастрофи
ческий спад.
На величину выводка у птиц (число яиц или молоди за ре-
продуктивный период) влияют не только смертность и выжива-
ние, она зависит также от г- и К-отбора (см. Lack, 1966, 1968;
Cody, 1966; Hutchinson, 1975, гл. 3). У оппортунистических видов
выводки больше, чем у сбалансированных, что видно при сравне-
нии птиц умеренной зоны и тропиков.
Обозначение вида как г- или A-стратега может быть ошибоч-
76
Глава 6
Высокая -е— ПЛОТНОСТЬ ПИЩИ (РЕСУРСА) ---Низкая
Рпс. 6.25. Модель г- и A-отбора Мак-Артура (MacArthur, 1972). Скорости
размножения двух аллелей (или видов) изображены в виде функций плот-
ности популяции и ресурса. Когда плотность превышает С, а ресурсов ма-
ло, Х\ растет быстрее, чем Х2, но Х2 растет быстрее и «победит» в естест-
венном отборе, если плотность ниже С. В соответствии с этим А представ-
ляет собой область r-отбора, а В — область /^-отбора.
ным из-за чрезмерного упрощения классификации, поскольку
многие популяции изменчивы в этом смысле или занимают про-
межуточное положение. Однако Пианка (Pianka, 1970) считает,
что в природе наблюдается бимодальность относительна г- и К-
отбора, связанная с размерами тела и временем генерации. Он
утверждает, что любая из стратегий (либо г, либо К) обладает
преимуществом по сравнению с любым компромиссом. Шеффер
(Schaeffer, 1976) попытался уточнить эту концепцию, оценивая
преимущества и недостатки размножения на разных стадиях и в
разное время жизненного цикла.
Левине (Levins, 1968) в книге «Эволюция в меняющейся сре-
де» пришел к заключению, что специализация в эволюции видов
ограничивается неопределенностью среды обитания. Для отбора
в нестабильных условиях лучше быть «генералистом» и обладать
высокой Гщах. Сообщества в таких условиях могут быть только
очень слабо организованными. Специализация и организация мо-
гут подняться до высокого уровня лишь тогда, когда непредска-
зуемость среды мала (согласно Левинсу). До каких пределов
могут совместно действующие группы популяций и сообществ
Динамика популяций
17
снизить вариабельность среды и открыть тем самым путь для
формирования организации более высокого уровня, как это иног-
да (но не всегда) происходит в человеческом обществе? Этот
вопрос остается пока без ответа.
Примеры
Аспекты распределения энергии в связи с трофическими уровня-
ми, размером организма, наличием или отсутствием терморегу-
ляции обсуждались в гл. 3, разд. 4 (см., в особенности, модель
универсального потока энергии на рис. 3.12). Более частный
случай представлен в табл. 6.6, где сравниваются пути распреде-
ления ассимилированной энергии (А на рис. 3.12) между про-
дукцией Р (рост и размножение) и дыханием R (поддержание)
у четырех видов, представляющих хищников и растительнояд-
ных, а также позвоночных и членистоногих. В общем хищники
(болотные крапивники и пауки-охотники) тратят больше ассими-
лированной энергии на поддержание (добывание пищи, защита
территории и т. д.), чем растительноядные (хлопковые крысы и
гусеницы). Сходным образом крупные эндотермные животные
(теплокровные позвоночные) в отличие от мелких экзотермных
(членистоногих) направляют большую часть ассимилированной
энергии на дыхание. Мак-Нейл и Лоутон (McNeil, Lawton, 1970)
отмечают, что у водных видов стоимость дыхания выше, чем у
наземных видов, однако на вопрос о том, является ли это разли-
чие основным и почему так должно быть, ответа до сих пор не по-
лучено.
Таблица 6.6. Распределение ассимилированной энергии между продукцией
(рост и размножение) и дыханием (поддержание)
Количество ассимилиро- ванной энергии (%), идущей на Источник данных
дыхание продукцию
Эндотермные позвоночные Болотный крапивник (на- Kale, 1965
секомоядный) Хлопковая крыса (раститель- 99 1
ноядная) Экзотермные членистоногие 87 13 Randolph et al., 1977
Пауки-волки (хищники) Гороховая тля (раститель- 75 25 Hymphreys, 1978
ноядная) 42 58 Randolph et al., 1977
78
Глава 6
Интересно сравнить распределение энергии у пауков-охотни-
ков и тенетников. Поскольку паутина содержит много белка,
на синтез шелка затрачивается много энергии, но многие пауки
используют шелк повторно, съедая старую паутину при постройке
новой. Таким образом, цена паутины снижается. Пиколл и Витт
(Peacall, Witt, 1976) установили, что на образование шелка у
одного из пауков, который реутилизирует свою паутину, тратится
примерно четвертая часть того количества калорий, которые из-
расходованы на работу по строительству паутины и ее регуляр-
ному ремонту. Общая энергетическая стоимость паутины состав-
ляет около половины основного потребления энергии, и это мень-
ше, чем затраты на охоту у некоторых не строящих паутины
видов. Возможно, это урок для человека: вид, который конструи-
рует дорогостоящие и сложные механизмы, может удешевить их
путем повторного использования материалов.
Теория, согласно которой хищник оптимизирует энергетику
дохода и затрат, выбирая наиболее благоприятный размер жертвы
в зависимости от общего обилия пищи, была экспериментально
проверена и подтверждена Вернером и Холлом (Werner, Hall,
1974). Эти исследователи предлагали ушастым окуням в разных
комбинациях ветвистоусых рачков разной величины и регистри-
ровали размеры избираемой окунями жертвы. Когда абсолютное
количество пищи было невелико, съедались подряд жертвы всех
размеров. При возрастании количества пищи рыбы выедали наи-
более крупных рачков, не трогая представителей мелких размер-
ных классов. Таким образом, при увеличении количества пищи
(и наоборот, когда пищи становится мало) рыбы способны ме-
нять стратегию, становясь из «генералистов» «специалистами».
Для иллюстрации г- и К-отбора приведем два примера: амбро-
зию, которая растет по залежам и другим недавно нарушенным
местообитаниям, и травянистое растение Dentaria laclniata, оби-
тающее в относительно стабильном нижнем ярусе леса. Эти рас-
тения сравниваются по продукции семян и максимальному ре-
продуктивному усилию в табл. 6.7. Амброзия продуцирует при-
мерно в 50 раз больше семян, чем Dentaria, и тратит значительно*
большую долю чистой энергии на размножение.
Золотарник дает пример целого ряда репродуктивных страте-
гий, промежуточных между крайними стратегиями г- и ЙГ-отбора.
На рис. 6.26 приведены данные, характеризующие репродуктив-
ное усилие и накопление биомассы для шести популяций четы-
рех видов золотарника рода Solidago. У популяции вида 1, кото-
рый растет на сухих открытых полях и нарушенных местообита-
ниях, поддерживающая биомасса листьев мала и около 45 %
чистой продукции направлено на образование репродуктивных
тканей. Популяция 6, встречающаяся во влажных лиственных
лесах, напротив, вкладывает большую часть энергии в листья и
Динамика популяций
79
Рис. 6.26. Репродуктивное усилие (отношение сухой массы репродуктивных
тканей к общей сухой массе надземных тканей) как функция отношения
массы листьев к общей массе в шести популяциях четырех видов золотар-
ника (Solldago). Популяция 1 принадлежит виду, который растет на сухих
открытых полях или нарушенных почвах; популяция 6 встречается во
влажных лиственных лесах, остальные популяции занимают промежуточ-
ные по влажности и стабильности местообитания. (По Abrahamson, Gadgil,
1973.)
только 5% затрачивает на размножение. Другие популяции встре-
чаются в промежуточных по влажности и стабильности местооби-
таниях и в соответствии с этим распределяют энергию в проме-
жуточных пропорциях.
Таблица 6.7. Противоположные стратегии размножения двух травянистых
растений, а также полевого и лесного сообществ, в которых они растут:
пример г- и A-отбора (Newell, Тгатег, 1978)
Сообщество Среднее число семян в расчете . на 1 особь Наибольшее репро- дуктивное усилие (сухая масса реп- родуктивных струк- тур по отношению к общей сухой массе>
Однолетнее поле 1190 0,30
Ambrosia arte mis if olia
Среднее для доминантов — 0,21
Остальное сообщество — 0,24
Лес
Dentaria laciniata 24 0,01
Среднее для доминантов — 0,05
Остальное сообщество — 0.05
80
Глава 6
Солбриг (Solbrig, 1971) описал случай, когда и г-, и 7^-стра-
теги встречаются у одного и того же вида. У обычного одуван-
чика (Тaraxacum officinale) имеется несколько линий, или варие-
тетов, различающихся по генотипам, контролирующим распреде-
ление энергии. Растения одной линии первыми занимают
нарушенные местообитания и продуцируют много мелких рано
созревающих семян. Растения другой линии встречаются в менее
нарушенных местообитаниях, больше энергии направляют в
листья и стебли и продуцируют относительно меньше поздно со-
зревающих семян. Если обе линии произрастают на хорошей
почве, то более плодовитая линия оказывается затененной менее
плодовитой. Таким образом, первая линия лучше приспособлена
для заселения новых местообитаний и считается г-стратегом;
вторая линия характеризуется лучшей конкурентоспособностью,
и ее следует считать К-стратегом.
Хотя вариабельность и нарушение местообитаний должны бла-
гоприятствовать r-стратегам, это не значит, что Я-стратеги исче-
зают.
В адаптированных к пожарам сообществах, например в со-
обществах калифорнийского чапараля (гл. 5, разд. 5), растения
запасающие большое количество энергии в подземных частях
и размножающиеся побегами, приспособлены к переживанию пе-
риодических пожаров так же хорошо, как растения, вкладываю-
щие свое будущее в семена (см. Carpenter, Recher, 1979).
Другие примеры распределения энергии в растительном цар-
стве приведены в работах Харпера (Harper, 1977) и Гэджила и
Солбрига (Gadgil, Solbrig, 1972).
11. Интеграция: особенности и тактика
жизненного цикла
Определения
Давление отбора, обусловленное влиянием физических факторов
окружающей среды и биотическими взаимодействиями, форми-
рует определенный тип жизненного цикла так, что каждый вид
приобретает адаптивную комбинацию популяционных особенно-
стей, о которых мы уже говорили в предыдущих разделах этой
главы. Каждый вид характеризуется уникальным, присущим
только ему жизненным циклом, тем не менее мы можем выде-
лить несколько основных типов жизненных циклов и предсказать
с определенной точностью, какой комбинацией специфических
особенностей будут обладать организмы, обитающие в определен-
ных условиях.
Динамика популяций
8Б
Объяснения и примеры
Стирнс (Stearns, 1967) приводит четыре характерные особенности
жизненного цикла, которые играют ключевую роль в тактике вы-
живания: 1) величина выводка (число семян, яиц, молоди или
других потомков); 2) размеры молодой особи (при рождении^
вылуплении или прорастании); 3) возрастное распределение ре-
продуктивного усилия и 4) соотношение репродуктивного усилия
и смертности взрослых (особенно соотношение смертности молоди
и взрослых). Теории, позволяющие предсказывать ту или иную»
тактику выживания, были предложены Гэджилом и Боссертом:
(Gadgil, Bossert, 1970), Стирнсом (Stearns, 1976) и другими:
1. В тех случаях, когда смертность взрослых выше, чем смерт-
ность молоди, вид может размножаться только один раз на про-
тяжении жизненного цикла, и соответственно в тех случаях, когда
смертность выше у молоди, организм должен размножаться не-
сколько раз.
2. Величина выводка должна максимизировать число потомков;,
доживших до половой зрелости за среднее время жизни родите-
ля. Так, птицам, гнездящимся на земле, необходима кладка ви
20 яиц, чтобы обеспечить непрерывность поколений, тогда как.
птицам, гнездящимся в пещерах и других защищенных местах,,
достаточно намного меньшей кладки.
3. В увеличивающейся популяции (на восходящей ветви кривой^
роста) отбор должен минимизировать возраст полового созрева-
ния (организмы должны размножаться в более раннем возрасте).,
В стабильных популяциях (на уровне несущей емкости среды
или К) процесс полового созревания должен быть более длитель-
ным. Этот принцип соблюдается в популяциях человека; в стра-
нах с быстро растущим населением семья обзаводится детьми в?
более раннем возрасте, тогда как в странах со стабильной чис-
ленностью населения обзаведение детьми откладывается на более-t
поздний возраст.
4. Там, где существует риск быть съеденным хищником или
имеется очень мало ресурсов, или действуют одновременно два
этих фактора, новорожденные должны быть крупными; если же-
ресурсы легкодоступны, давление хищников или конкурентов,
незначительно, то размеры молоди уменьшаются.
5. В растущей популяции, в общем, не только снижается воз-
раст начала размножения, которое приурочено к ранним перио-
дам жизненного цикла, но также увеличивается число потомков^
в помете и большая часть потока энергии направляется на раз-
множение. Это сочетание биологических особенностей рассматри-
вается как тактика /’-отбора, описанная в разд. 10. В стабильной
популяции следует ожидать противоположной комбинации при-
знаков, иными словами, в такой популяции действует 7^-отбор.,
6 Заказ № 1483
«2
Глава 6
6. Если ресурсы жестко не лимитированы, то размножение
начинается в раннем возрасте.
7. Сложные жизненные циклы с личиночными стадиями по-
зволяют видам использовать больше чем одно местообитание или
один пищевой ресурс.
Сравнение двух крайних типов флор — пустынной флоры и
*флоры влажных тропических лесов — дает нам представление о
преобладании основных черт жизненных циклов в различных ти-
пах экосистем. В пустынях преобладают однолетние растения,
поскольку выживание многолетников очень затруднено из-за дли-
тельного засушливого периода. Напротив, в дождевых тропиче-
ских лесах условия благоприятствуют жизненным циклам много-
летников, так как интенсивная конкуренция и выедание семян
сильно снижают выживание сеянцев. Этот случай можно рассмат-
ривать как пример первой из перечисленных выше прогностиче-
ских теорий.
Связь между размерами новорожденных особей и давлением
хищников иллюстрируется сравнением антилоп и медведей: мо-
лодые особи у антилоп крупнее (по отношению к размерам тела
^взрослого животного), чем у медведей. Молодые антилопы могут
следовать за табуном уже через несколько часов после рождения,
тогда как медвежата должны долгое время находиться под защи-
той матери.
Интерес экологов к стратегиям жизненных циклов восходит к
»одной из первых в этой области работ Ла Монт Коула (LaMont
‘Cole, 1954), озаглавленной «Популяционные следствия феномена
жизненного цикла». Эта работа рекомендуется для чтения наряду
с работой Чарнова и Шеффера (Charnov, Schaffer, 1973), в кото-
рой повторно рассматривается теория Коула.
Популяции в сообществах
1. Типы взаимодействия между двумя видами
Определения
Взаимодействия популяций двух видов теоретически можно вы-
разить в виде следующих комбинаций символов: 00,----, 4~ + -,
+0, —0 и 4—. Три из них (4—Н и 4—) в свою очередь
обычно подразделяются, и в результате мы получаем девять ос-
новных типов взаимодействий (см. схему классификации HaskeL
1949, в адаптации Barkholder, 1952): 1) нейтрализм, при котором
ассоциация двух популяций не сказывается ни на одной пз них;
2) взаимное конкурентное подавление, при котором обе популя-
ции активно подавляют друг друга; 3) конкуренция за общий
ресурс, при которой каждая популяция косвенно отрицательно’
воздействует на другую в борьбе за дефицитный ресурс; 4) амен-
сализм, при котором одна популяция подавляет другую, но сама
не испытывает отрицательного влияния; 5) паразитизм и 6) хищ-
ничество, при которых одна популяция неблагоприятно воздейст-
вует на другую, нападая непосредственно на нее, но тем не менее-
сама зависит от объекта своего нападения; 7) комменсализм, при'
котором одна популяция извлекает пользу из объединения, а для
другой это объединение безразлично; 8) протокооперация, при*
которой обе популяции получают от ассоциации выгоду, но эти
отношения не обязательны, и 9) мутуализм, при котором связь
популяций благоприятна для роста и выживания обеих, причем в>
естественных условиях ни одна из них не может существовать-
без другой. Все эти типы взаимодействий приведены в табл. 7.1..
Следует подчеркнуть три принципа, основанные на этих кате-
гориях:
1. Негативные взаимодействия проявляются на начальных ста-
диях развития сообщества или в нарушенных природных усло-
виях, где высокая смертность нейтрализуется г-отбором.
6*
84
Глава 7
Таблица 7.1. Анализ взаимодействий популяций двух видов1
Тип взаимодействий2 Виды Общий характер взаимодействия
1 1 2
4. Нейтрализм 0 0 Ни одна популяция не влияет на другую
2. Конкуренция, непо- средственное взаи- модействие Прямое взаимное подавление обоих видов
3. Конкуренция, взаи- модействия из-за ре- сурсов Непрямое подавление при дефиците общего ресурса
4. Аменсализм 0 Популяция 2 подавляет популяцию 1, но сама не испытывает отрица- тельного воздействия
5. Паразитизм + — Популяция паразита (1) состоит из меньших по величине особей, чем популяция хозяина (2)
<5. Хищничество (и по- едание растений) + — Особи хищников (1) обычно круп- нее, чем особи жертвы (2) Популяция 1, комменсал, получает пользу от объединения; популяции 2 это объединение безразлично
7. Комменсализм + 0
8. Протокооперация + + Взаимодействие благоприятно для обоих видов, но не обязательно
*9. Мутуализм + + Взаимодействие благоприятно для обоих видов и обязательно
1 0 означает отсутствие значимых взаимодействий; + означает улучшение роста,
«выживания и другие выгоды для популяции (к уравнению роста прибавляется по-
ложительный член); — означает замедление роста и ухудшение других характери-
стик (к уравнению роста добавляется отрицательный член).
2 Типы 2—4 можно считать «отрицательными взаимоотношениями», типы 7—9—
«положительными взаимоотношениями», а типы 5 и 6 можно относить к обеим этим
«группам.
2. В процессе эволюции и развития экосистем обнаруживает-
ся тенденция к уменьшению роли отрицательных взаимодействий
за счет положительных, повышающих выживание взаимодейству-
ющих видов.
3. В недавно сформировавшихся или новых ассоциациях веро-
ятность возникновения сильных отрицательных взаимодействий
’больше, чем в старых ассоциациях.
Объяснения
Одна популяция часто влияет на скорость роста или смертность
другой. Так, члены одной популяции могут поедать членов дру-
гой популяции, конкурировать с ними за пищу, выделять вред-
ные вещества или взаимодействовать с ними другим способом.
Точно так же популяции могут быть полезными друг для друга,
Популяции в сообществах 85
причем польза в одних случаях оказывается обоюдной, а в дру-
гих — односторонней. Как было отмечено в предыдущем разделе
и показано в табл. 7.1, взаимодействия такого рода подразделя-
ются на несколько категорий.
Все эти популяционные взаимодействия можно изучать на
примере некоего среднего сообщества. Тип взаимодействий кон-
кретной пары видов может изменяться в различных условиях или
в зависимости от стадии жизненных циклов. Так, в одно время у
двух видов может быть выявлен паразитизм, в другое время —
комменсализм, а в какой-то третий период жизни они могут быть
полностью нейтральны друг к другу. Упрощенные сообщества и
лабораторные эксперименты позволяют экологам получать всего
один из многих типов взаимодействий и изучать его количест-
венно. Дедуктивные математические модели, построенные на осно-
ве таких исследований, дают возможность анализировать факто-
ры, которые обычно трудно отдифференцировать один от другого.
Мы уже говорили в предыдущей главе о том, что для выяс-
нения действий различных факторов в сложных природных си-
туациях, а также для более точного определения понятий и боль-
шей четкости рассуждений полезно пользоваться «моделями»
в форме уравнений. Если рост одной популяции можно описать с
помощью уравнения, то влияние другой популяции можно вы-
разить членом, который изменяет рост первой популяции. В за-
висимости от типа взаимодействия в уравнение можно подстав-
лять разные члены. Например, при наличии конкуренции ско-
рость роста каждой популяции равна скорости нелимитируемого
роста минус влияние собственной численности (которое растет с
ростом численности популяции) и минус величина, характери-
зующая отрицательное влияние другого вида, N2 (которая также
растет по мере роста численностей обоих видов и N2), или
dVj лг г
- CN^
Скорость — Пелимитируе- — Влияние собст- — Отрицательное
роста мый рост венной числен- влияние друго-
нос ти го вида
Нетрудно видеть, что это по существу то же логистическое
уравнение, которое было приведено на с. 34, с той лишь разни-
цей, что в нем имеется дополнительный член «минус отрицатель-
ное влияние другого вида». Взаимодействие такого рода может
привести к разным последствиям. Если величина С мала для
обоих видов, так что ингибирующее межвидовое влияние меньше
соответствующих внутривидовых влияний, то скорость роста и,
вероятно, окончательная плотность обоих видов будут всего лишь
несколько снижены, но оба вида, по-видимому, будут сосущест-
вовать; иными словами, межвидовые взаимодействия будут иметь
«6
Глава 7
меньшее значение, чем внутривидовая конкуренция. Точно так
же если для видов характерен экспоненциальный рост (в урав-
нении отсутствует член, соответствующий самоограничению чис-
ленности), то сглаживание кривой роста может произойти вслед-
ствие межвидовой конкуренции; популяция, следовательно,
ускользает от закономерности собственного роста. Однако если С
велико, то вид, оказывающий наибольшее влияние, либо уничто-
жит своего конкурента, либо вынудит его занять другое место-
обитание. Таким образом, теоретически виды, обладающие сход-
ными потребностями, не могут жить совместно, так как велика
вероятность возникновения сильной конкуренции и элиминации,
одного из них. В следующем разделе мы рассмотрим, как реали-
зуются те возможности, на существование которых указывают
наши модели.
Когда виды двух взаимодействующих популяций оказывают
друг на друга благоприятное, а не повреждающее влияние, в урав-
нение вводят положительный член. В таких случаях обе попу-
ляции растут и процветают, достигая равновесных уровней, что
благоприятно для обоих видов. Если для роста и выживания
каждой из популяций необходимо их взаимное влияние друг на
друга, то такие взаимоотношения называются мутуализмом. Если
же, с другой стороны, эти благоприятные влияния лишь вызы-
вают увеличение размеров популяции или скорости ее роста,
но не являются необходимыми для ее роста и выживания, то та-
кое взаимодействие соответствует кооперации или протокоопера-
ции. (Поскольку такая кооперация не есть следствие сознатель-
ной или «разумной» деятельности, предпочтительнее пользоваться
последним термином.) И мутуализм, и протокооперация приводят
к сходному результату: рост популяции в отсутствие другой либо
замедлен, либо равен нулю. По достижении равновесия обе по-
пуляции продолжают сосуществовать, обычно сохраняя опреде-
ленное соотношение.
Рассмотрение популяционных взаимодействий так, как это
сделано в табл. 7.1 или на основе уравнений роста, позволяет
избежать противоречий, нередко возникающих при обсуждении
терминов или определений как таковых. Например, термин «сим-
биоз» иногда употребляют в том же смысле, что и мутуализм,
а иногда его понимают более широко, включая в него также
комменсализм и паразитизм. Поскольку слово симбиоз буквально
означает «жить вместе», мы используем его в этой книге в ши-
роком смысле, безотносительно к природе взаимосвязи. Термин
«паразит» многие используют применительно к любому мелкому
организму, живущему на поверхности или внутри другого орга-
низма, независимо от того, положительно, отрицательно или без-
различно его влияние. В таком же смысле трактуется и предмет
науки паразитологии. Для одного и того же типа взаимодействий
Популяции в ^сообществах
87
предлагались разные термины, что только увеличивало путаницу.
Однако, когда отношения представлены в виде схемы, обычно
не остается сомнений, к какому типу относить рассматриваемое
взаимодействие; слово или «ярлык» оказывается в таком случае
вторичным относительно механизма и его результата.
Обратите внимание на то, что слово «вредоносный» не ис-
пользовалось в описании отрицательных взаимодействий. Конку-
ренция и хищничество снижают скорость роста популяций, ис-
пытывающих эти влияния, но этот эффект не обязательно вреден,
<если рассматривать его с точки зрения выживания популяции на
протяжении длительного времени или с точки зрения ее эволю-
ции. Отрицательные взаимодействия фактически могут ускорять
естественный отбор, приводя к возникновению новых адаптаций.
Хищники и паразиты часто полезны для популяций, не имеющих
механизмов саморегуляции для предотвращения перенаселения,
следствием которого могло бы быть самоуничтожение таких по-
пуляций.
2. Конкуренция и сосуществование видов
Определения
В самом широком смысле конкуренция — это взаимодействие двух
организмов, стремящихся получить один и тот же ресурс. Меж-
видовая конкуренция — это любое взаимодействие между популя-
циями двух или более видов, которое неблагоприятно сказывается
на их росте и выживании. Как показано в табл. 7.1, она может
проявляться в двух формах. Тенденция к экологическому разде-
лению, наблюдаемая при конкуренции близкородственных или
сходных в иных отношениях видов, известна как принцип кон-
курентного исключения. Вместе с тем конкуренция способствует
возникновению в процессе отбора многих адаптаций, что приво-
дит к увеличению разнообразия видов, сосуществующих в данном
пространстве или сообществе.
Объяснения
О межвидовой конкуренции написано много и экологами, и гене-
тиками, и эволюционистами. В большинстве случаев слово «кон-
куренция» применяется в ситуациях, в которых отрицательные
взаимодействия являются следствием уменьшения ресурсов, ис-
пользуемых обоими видами. Более непосредственные взаимодей-
ствия, такие, как взаимное хищничество или выделение вредных
веществ, следует отнести к другой категории (табл. 7.1), хотя
для ее обозначения и нет общепринятого термина. Однако термин
88
Глава 7
аллелопатия (введенный в разд. 4, гл. 2) теперь широко приме-
няется для обозначения такого типа взаимодействия, когда один
из конкурирующих видов выделяет химические посредники (Уит-
такер (Whittaker, 1970) назвал их аллелопатическими вещества-
ми), которые обеспечивают конкурентное преимущество одного
вида перед другим.
Конкурентное взаимодействие может касаться пространства,
пищи или биогенных элементов, света, неиспользованных ве-
ществ, зависимости от хищников, подверженности болезням и т. д.,.
и многих других типов взаимодействий. Результаты конкуренции
представляют огромный интерес; они были многократно исследо-
ваны как один из механизмов естественного отбора. Межвидовая
конкуренция, независимо от того, что лежит в ее основе, может
привести либо к установлению равновесия между двумя видами,
либо, при более жестокой конкуренции, к замене популяции од-
ного вида популяцией другого, либо к тому, что один вид вы-
теснит другой в иное место или же заставит его перейти на ис-
пользование иной пищи. Неоднократно отмечалось, что близко-
родственные организмы, ведущие сходный образ жизни и
обладающие сходной морфологией, не обитают в одних и тех же
местах. Если же они живут в одном месте, то часто используют
разные ресурсы или активны в разное время. Объяснение эколо-
гического разделения близкородственных (или сходных в иных
отношениях) видов получило известность как принцип Гаузе*
по имени русского биолога, который в 1932 г. впервые подтвер-
дил его существование экспериментально (см. рис. 7.2), или как
принцип конкурентного исключения (Harden, 1940).
К числу наиболее широко обсуждавшихся в литературе тео-
ретических проблем, относящихся к явлению конкуренции, от-
носятся закономерности, получившие название уравнений Лот-
ки—Вольтерра (эти модельные уравнения были предложены Лот-
кой и Вольтерра независимо друг от друга в 1925 и 1926 гг.)~
Эти уравнения представляют пару дифференциальных уравнений*,
сходных с уравнением, приведенным в предыдущем разделе. Та-
кие уравнения можно использовать для моделирования систем!
хищник—жертва, паразит—хозяин, конкуренции и других типов
взаимодействия между двумя видами. В случае конкуренции в=
ограниченном пространстве, где для каждой популяции имеется
определенный уровень равновесия, характеризуемый величиной
Л, уравнения одновременного роста двух популяций, основанные
на логистическом уравнении, можно записать так:
dM Ki —м —
—,
dNa „ Kg-Vg-₽^
—>
Популяции в сообществах
89
где TVi и N2 — соответственно численности видов 1 и 2. а — коэф-
фициент конкуренции, характеризующий ингибирующее воздей-
ствие вида 2 на вид 1, р соответствует коэффициенту конкурен-
ции, означающему ингибирование вида 2 видом 1. В более общем
виде уравнения конкуренции были предложены Вигертом (Wie-
gert, 1974). В отсутствие убежищ (рефугий) или в других усло-
виях вид, который сильнее ингибирует своего партнера, вытес-
нит его независимо от того, что коэффициенты конкуренции очень
малы по сравнению с соотношениями насыщающих плотностей
(КХ1К2 или К2/Лд).
Чтобы понять причины конкуренции, необходимо рассматри-
вать не только особенности популяций и условия, определяющие
конкурентное исключение, но также ситуации, в которых сосу-
ществуют сходные виды, поскольку в открытых природных систе-
мах большое число видов фактически пользуется общими ресур-
сами. В табл. 7.2 и на рис. 7.1 показана ситуация, которую мож-
но было бы назвать моделью Tribolium—Trijolium-. эта модель
наглядно демонстрирует конкурентное исключение в паре видов
Рис. 7.1. Сосуществование видов клевера (Trifolium). На графике показан
рост популяций двух видов в чистых (т. е. представленных только одним-
видом) и смешанных травостоях. Обратите внимание на то, что кривые
роста двух видов в чистых травостоях различны и в разное время дости-
гают зрелости. Это и другие различия позволяют видам сосуществовать
в смешанных травостоях, но при этом их плотность понижена, так как
они в одинаковой степени мешают друг другу. Листовой индекс, который
используется для оценки плотности биомассы, представляет собой отноше-
ние площади поверхности листьев к поверхности почвы (см2/см2). I — Т.
Jragiferum, чистый травостой; II — Т. repens, чистый травостой; III — Т.
repens, смешанный травостой; IV — Т. fragiferum, смешанный травостой.
90
Глава 7
жуков (Tribolium) и сосуществование двух видов клевера (ТгЦо-
lium).
Одно из наиболее тщательных и длительных эксперименталь-
ных исследований межвидовой конкуренции было проведено в
лаборатории покойного доктора Томаса Парка в Чикагском уни-
верситете. Парк, его студенты и сотрудники работали с мучным
хрущаком, в частности с видами рода Tribolium. Эти мелкие жуки
могут завершить весь свой жизненный цикл в очень простой и
однородной среде — в банке с мукой или пшеничными отрубями.
В этом случае среда служит одновременно и пищей, и местооби-
танием личинок и имаго. Если регулярно добавлять свежую сре-
ду, то популяция жуков может поддерживаться долгое время.
С точки зрения представлений о потоке энергии такую экспери-
ментальную систему можно описать как стабилизированную ге-
теротрофную экосистему, в которой импорт энергии пищи урав-
новешивается затратами на дыхание.
Таблица 7.2. Случай конкурентного исключения в популяциях мучного хру-
щака (Tribolium). (По Park, 1954).
«Климат» Темпера- тура, С Относитель- ная влаж- ность, % Исход межвидовой конкуренции!, %
Побеждает Т. castaneum Побеждает Т. confи sun
Жаркий влажный 34 70 100 0
Жаркий сухой 34 30 10 90
Теплый влажный 29 70 86 14
Теплый сухой 29 30 13 87
Холодный влажный 24 70 31 69
Холодный сухой 24 30 0 100
1 Каждый из 6 вариантов опытов проведен в 20—30 повторностях. В чистой1
культуре каждый вид выживает при любых сочетаниях условий, однако при сов-
местном культивировании двух видов выживает только один. В процентах выраже-
но относительное число повторностей, в которых сохранялся только один вид а
другой элиминировался.
Оказалось, что если в этот однородный мирок поместить два
разных вида Tribolium, то раньше или позже один из видов неиз-
бежно будет элиминирован, тогда как другой будет успешно раз-
множаться. Один вид всегда «побеждает», или, иными словами,,
два вида Tribolium не могут выжить в этом специфическом «од-
номестном» микрокосме. Относительное число особей каждого ви-
да, первоначально внесенных в культуру (т. е. исходное соотно-
шение особей) не влияет на окончательный результат, однако*
«климат» экосистемы существенно сказывается на исходе борь-
бы. Как показано в табл. 7.2, один из видов (7. castaneum) всег-
да побеждает в условиях высокой температуры и влажности,.
Популяции в сообществах
91
тогда как другой (Т. confusum) всегда побеждает в прохладном
и сухом «климате», хотя и тот и другой при любом из испытан-
ных сочетаний условий может неопределенно долго существовать
в чистой культуре. В промежуточных условиях для каждого вида
имеется определенная вероятность оказаться победителем (напри-
мер, для Т. castaneum эта вероятность в теплой и влажной среде
составляет 0,86). Понять некоторые результаты конкурентного
взаимодействия помогает анализ свойств популяции каждого вида
в чистой культуре. Например, было обнаружено, что победителем
обычно оказывается вид, обладающий в изучаемых условиях на-
много большей скоростью роста (/'), чем другой. Если же величи-
ны г для обоих видов различаются не слишком сильно, то вид с
более высоким показателем не всегда окажется победителем. Ви-
русное заболевание в одной из популяций легко может решить
исход дела. Финер (Feener, 1981) описал случай, когда конку-
рентное равновесие между двумя видами муравьев было наруше-
но паразитическим двукрылым. Кроме того, конкурентоспособ-
ность может в значительной степени различаться у различных
генетических линий.
Пользуясь данными, полученными в модельных эксперимен-
тах на Tribolium, легко создать условия, в которых виды нр ис-
ключали бы друг друга, а сосуществовали. Если культуры по-
переменно помещать то в жаркие и влажные, то в сухие и хо-
лодные условия (для имитации сезонных изменений погоды),
преимущество одного вида перед другим будет недостаточно дли-
тельным для уничтожения другого. Если бы система культивиро-
вания была «открытой» и особей доминирующего вида вынуждали
эмигрировать (или их бы удаляли, как это делает хищник) до-
статочно быстро, то конкуренция оказалась бы настолько слабой,
что оба вида могли бы сосуществовать. Существованию могли бы
благоприятствовать и многие другие условия.
Интересные эксперименты по конкуренции у растений провел
Дж. Л. Харпер и его коллеги из Университетского колледжа Се-
верного Уэльса (см. Нагрет, 1961; Нагрет и Clatworthy, 1963;
Clatworthy, Нагрет, 1962). Результаты одного из этих исследова-
ний представлены на рис. 7.1. Вследствие различий в характере
роста два вида клевера могут сосуществовать в одной и той же
среде (при одинаковых условиях освещенности, температуры,
на одной и той же почве и т. д.). Из этих двух видов Trifolium
repens растет быстрее и скорее достигает максимальной облист-
венное™. Однако Т, fragiferum имеет более длинные черешки и
выше посаженные листья, поэтому он может перейти в верхний
ярус раньше, чем быстро растущий вид (особенно после того,
как скорость роста Т. repens пошла на убыль), и таким образом
избежать затенения. В смешанных травостоях из-за этих особен-
ностей каждый вид ингибирует развитие другого, но оба они
92
Глава 7
оказываются в состоянии завершить жизненный цикл и дать се-
мена, хотя плотность каждого вида понижена (впрочем, общая
плотность в смешанных травостоях двух видов была примерно
равна плотности в чистых травостоях). В этом случае оба вида»,
несмотря на сильную конкуренцию за свет, могут сосущество-
вать, и это сосуществование обусловлено особенностями морфо-
логии и различиями во времени максимального роста. Харпер,
(Нагрет, 1961) пришел к заключению, что два вида растений
могут длительно сосуществовать, если их популяции независимо
регулируются посредством одного или нескольких из следующих
механизмов: 1) различия потребностей в питании (например, бо-
бовые и небобовые); 2) различия причин смертности (например»
разная чувствительность к выеданию скотом); 3) чувствительно-
сти к различным токсинам и 4) чувствительности к одному и то-
му же регулирующему фактору (свет, вода и т. д.) в разное
время (как в случае только что описанного примера с клеве-
ром).
Парк (Park, 1954), Брайен (Brian, 1956) и Кромби (Crombie,
1947) (все работали с мучными хрущаками) впервые стали про-
водить различие между непрямой конкуренцией, или эксплуата-
ционной конкуренцией, и прямой конкуренцией, или интерфе-
ренционной конкуренцией. На основании данных, полученных
при изучении головастиков, Стейнуошер (Steinwasher, 1978) при-
шел к заключению, что эксплуатационная конкуренция сменяется
интерференционной конкуренцией при уменьшении количества
пищи. Хатчинсон (Hutchinson, 1978) полагает, что интерферен-
ционная конкуренция встречается все чаще по мере продвижения
вверх по филогенетическому «древу» от примитивных фильтрую-
щих простейших и ветвистоусых, которые обычно конкурируют за
захват пищевых частиц, к позвоночным с их развитыми поведен-
ческими механизмами агрессии, территориальности и т. д. Он за-
ключает (как это сделал Слободкин (Slobodkin, 1964) на основе
экспериментов по конкуренции у Hydra), что оба указанных типа
конкуренции перекрываются, но что по теоретическим соображе-
ниям полезно различать эти два процесса, так как уравнениям
эксплуатационной конкуренции соответствуют графики с прямо-
угольными изоклинами, тогда как для интерференционной кон-
куренции характерны криволинейные изоклины. Таким образом,
сосуществование более вероятно во втором случае. Более подроб-
ные сведения, касающиеся конкуренции, можно найти в книге
Хатчинсона (Hutchinson, 1978, гл. 4). Хороший обзор экспери-
ментальных и генетических исследований, выполненных на по-
пуляциях Drosophila, имеется у Айалы (Ayala, 1972) и Ричмон-
да и др. (Richmond et al., 1975).
При изучении литературы, посвященной конкуренции, скла-
дывается общее впечатление, что в системах, где отсутствуют или
Популяции в сообществах
93;
понижены иммиграция и эмиграция, происходит более жестокая
конкуренция и более вероятно конкурентное исключение. К чис-
лу таких систем следует отнести лабораторные культуры, острова-
или другие природные ситуации с трудно преодолимыми прегра-
дами для входа и выхода. В обычных природных открытых сис-
темах вероятность сосуществования выше.
Примеры
Результаты одного из оригинальных экспериментов Гаузе пред-
ставлены на рис. 7.2. Перед нами классический пример кон-
Рис. 7.2. Конкуренция между двумя близкородственными видами простей-
ших, занимающих сходные ниши. В изолированных культурах при посто-
янном обеспечении пищей рост Paramecium caudatum (I) и Paramecium,
aurelia (III) происходит в соответствии с обычной S-образной зависимостью^
но при совместном культивировании (II и IV) Р, caudatum (II) элимини-
руется. (По Гаузе из Allee et al., 1949.)
курентного исключения. Два близкородственных вида инфузорий»
Paramecium caudatum и Paramecium aurelia в раздельной куль-
туре с постоянной средой и плотностью организмов, служащих пи-
щей (инфузории питаются бактериями, которые в этих средах не?
размножаются, и поэтому бактерии вносили в среду через неболь-
шие промежутки времени, поддерживая их плотность на постоян-
94
Глава 7
ном уровне), росли в соответствии с типичной S-образной зави-
симостью, причем численность парамеций оставалась постоянной.
Однако когда оба вида простейших выращивали в одной культу-
ре, через 16 сут в ней обнаруживалась только Р. aurelia. При
атом ни один пз организмов не нападал на другой и не выделял
вредных веществ, просто популяции Р. aurelia отличаются более
высокой скоростью роста (более высокой внутренней скоростью
естественного роста), п поэтому при заданных условиях ограни-
ченного количества пищи побеждают Р. caudatum (явная эксплуа-
тационная конкуренция). Однако Paramecium caudatum и Para-
mecium bursaria могут выжить и достичь устойчивого равновесия
при их совместном культивировании в одной и той же среде.
Происходит это потому, что, хотя оба вида и питаются одной
и той же пищей, Р. bursaria занимала часть культурального со-
суда, где она могла питаться, не конкурируя с Р. caudatum. Та-
ким образом, было показано, что местообитания у этих двух видов
существенно различались, что позволяло им сосуществовать,
несмотря на одинаковую пищу.
Пример конкуренции с прямым подавлением описан Кромби
(1947). Он обнаружил, что совместное культивирование в муке
Tribolium и Oryzaephilus (другой род мучных хрущаков) закан-
чивается уничтожением Oryzaephilus. поскольку Tribolium актив-
нее истребляет неполовозрелые стадии Oryzaephilus. Однако если
в муку положить стеклянные трубочки, в которых могут прятать-
ся неполовозрелые особи меньшего по размерам Oryzdephilus,
то выживут обе популяции. Таким образом, если в среде имеются
убежища, позволяющие скрываться от прямого воздействия
(в данном случае хищничества), то конкуренция снижается на-
столько, что сохраняются оба вида.
Но довольно лабораторных примеров. Совершенно очевидно,
что скученность в лабораторных экспериментах может быть весь-
ма значительной, что приводит к чрезмерной конкуренции. В по-
левых исследованиях межвидовая конкуренция была подробно
изучена у растений; результаты этих исследований позволили
прийти к выводу (в настоящее время этот вывод является обще-
принятым) , что конкуренция — важный фактор, вызывающий
смену впдов^(как будет показано в гл. 8). Кивер (Keever. 19b5}
описал случай, когда залежь первого года была почти целиком
занята чистым травостоем высокого сорняка, в дальнейшем его
постепенно вытеснил отсюда другой вид, ранее не известный в
этих местах. Эти два вида, хотя и принадлежат к разным родам,
имеют очень сходные жизненные циклы (время цветения и со-
зревания семян) и жизненные формы, попадали в условия ин-
тенсивной конкуренции. Тщательные последующие исследования
этих залежных полей показали, что новый пришелец не вытеснял
растущий здесь ранее вид; оказалось, что оба вида сосуществуют,
Популяции в сообществах
95-
но их численное соотношение зависит от почвы, времени и сте-
пени нарушений.
Мы уже отмечали, что конкуренция между особями одного
вида — один из наиболее важных природных факторов, завися-
щих от плотности. То же самое можно сказать о межвидовой кон-
куренции. Конкуренция, по-видимому, оказывает огромное влия-
ние на распределение близких видов, хотя зачастую об этом сви-
детельствуют лишь косвенные данные. Правило Гаузе трудно
проверить в природе, используя стандартные методики, упрощаю-
щие реальную ситуацию, поскольку на исход конкуренция — со-
существование или исключение — могут влиять многие другие пе-
ременные. Тем не менее близкородственные виды или виды с
очень сходными потребностями обитают обычно в разных геогра-
фических областях или разных местообитаниях в одной и той
же области либо избегают конкуренции каким-либо иным спосо-
бом, например благодаря различиям в суточной или сезонной
активности или различиям в пище. Вероятно, действие естествен-
ного отбора на протяжении длительного времени направлено на
то, чтобы исключить или предотвратить продолжительную кон-
фронтацию видов (гл. 8, разд. 4) со сходным образом жизни.
Эволюционный процесс, названный «смещением признаков»,
может быть причиной морфологических различий, усиливающих
экологическое разобщение. Например, в Центральной Европе со-
существует шесть видов синиц (мелкие птицы рода Parus); их
изоляция друг от друга обусловлена отчасти различиями в место-
обитаниях, отчасти же различиями в местах кормежки и разме-
рах добычи, что находит отражение в небольших изменениях
длины и толщины клюва. В Северной Америке встретить в одной
местности более двух видов можно редко, хотя в целом на кон-
тиненте обитает семь видов. Лэк (Lack, 1969) высказал предпо-
ложение, что «американские виды синиц находятся на более ран-
них стадиях эволюции, чем европейские, и имеющиеся у них
различия в строении клюва, размерах тела и пищевом поведе-
нии — это адаптации к соответствующим местообитаниям, а не*
адаптации, которые позволили бы им сосуществовать в одном
местообитании ».
Роль конкуренции в разделении местообитаний схематически
иллюстрируется на рис. 7.3. Кривыми показаны границы место-
обитаний, внутри которых вид может существовать; отмечены так-
же оптимальные и краевые зоны. При наличии конкуреншш с
близкородственными или экологически сходными видами диапа-
зон местообитаний, возможных для данного вида, сокращается
до оптимальных границ (т. е. вид распространяется в наиболее
благоприятных для него местообитаниях, в которых он обладает
какими-либо преимуществами по сравнению со своими конкурен-
тами). Если же межвидовая конкуренция выражена слабо, то под
Глава 7
Краевые Опги Краевые
Межвидовая конкуренция
Внутривидовая конкуренция
’Рпс. 7.3. Влияние конкуренции на распространение птиц в местообитаниях.
Щели преобладает внутривидовая конкуренция, то вид широко распростра-
няется, занимая и менее благоприятные (краевые) области. Если же ин-
тенсивна межвидовая конкуренция, то распространение ограничивается
•узкими пределами, где имеются оптимальные условия. (Из Svardson, 1949,
с изменениями.)
влиянием внутривидовой конкуренции популяции данного вида
расширяют границы своего местообитания.
На островах, которые не заселены потенциальными конкурен-
тами, особенно хорошо прослеживается тенденция к расширению
местообитаний. Например, на островах, где отсутствуют лесные
•полевки (Clethrionomys), луговые серые полевки (Microtus) часто
занимают лесные местообитания (см. Cameron, 1964). Кроуэлл
(Crowell, 1962) обнаружил, что на Бермудских островах карди-
налы более многочисленны и занимают более краевые по усло-
жним местообитания, так как здесь нет многих из его материко-
вых конкурентов.
Хорошим примером экологического разделения сфер питания
могут служить две родственные рыбоядные птицы Великобрита-
нии, большой баклан (Phalacrocorax carbo) и длинноносый, или
хохлатый, баклан (Р. aristotelis), исследованные Лэком (Lack,
1945). Эти два вида обычно кормятся в одних и тех же водах
ги гнездятся на одних и тех же обрывах, однако более глубокий
анализ показывает, что на самом деле места их гнездовий раз-
.личаются, а, кроме того, существенные различия выявляются и в
составе их пищи (рпс. 7.4): длинноносый баклан ловит рыбу,
плавающую в верхних слоях воды, тогда как большой баклан
добывает пищу в основном у дна, где он ловит камбал и донных
’беспозвоночных (креветок и др.).
Тот факт, что близкородственные виды в природе четкэ раз-
граничены, вовсе не означает, что для поддержания этой разоб-
щенности между ними непрерывно должна поддерживаться кон-
куренция. В процессе эволюции у этих видов могут выработаться
различные потребности и предпочтения, и тогда конкурентные
отношения между ними прекращаются.
Популяции в сообществах
97
Процентное содержание в пище
Рис. 7.4 Состав пищи двух близкородственных видов морских птиц, боль-
шого баклана (Phalacrocorax carbo) и длинноносого, или хохлатого, бакла-
на (Р. aristotelis), живущих в период размножения в одних и тех же мес-
тах. Данные исследований показывают, что, несмотря на сходство место-
обитаний, пища этих бакланов различна. Таким образом, два вида зани-
мают разные ниши и фактически не конкурируют за пищевые ресурсы.
(Из Lack, 1945.)
В Европе, например, один из видов рододендрона Rhododen-
dron hirsutum растет на известковых почвах, а другой — R. farru-
gineum — на кислых; ни один из них не может жить на почве
противоположного типа, поэтому конкуренция между ними прак-
тически отсутствует (Braun-Blanquet, 1932). Тил (Teal, 1958) экс-
периментально изучил выбор местообитания у манящих крабов
(С7са), которые встречаются на засоленных маршах обычно по-
рознь. Один вид U. pugilator был обнаружен на открытых песча-
ных отмелях, другой, U. pugnax, на илистом грунте, поросшем
болотной травой. Тил выявил, что ни один из видов не стремится
вторгнуться в местообитание другого, даже если оно и не заселе-
но, потому что каждый из них может рыть норы только в пред-
почитаемом им субстрате. Отсутствие активной конкуренции, ко-
нечно, не означает, что ее следует исключить и в прошлом, когда
она, по-видимому, служила фактором, который привел к выра-
ботке поведения, поддерживающего сейчас видовую изоляцию.
Три пробные модели, предложенные Филипом (Philip, 1955),
могут служить основой для будущих наблюдений, анализа и экс-
периментальных работ: 1) несовершенная конкуренция, при ко-
торой межвидовая конкуренция служит лимитирующим фактором,
но не приводит к полной элиминации одного из конкурентов из
арены взаимодействия; 2) совершенная конкуренция, описывае-
мая моделями Гаузе и Лотки—Вольтерра, при которой один вид
постепенно элиминируется в процессе конкуренции за общий
ресурс, и 3) сверхсовершенная конкуренция, при которой эффект
7 Заказ № 1483
98
Глава 7
подавления очень силен и проявляется немедленно, как, напри-
мер, при выделении антибиотиков. Наглядный пример прямого
подавления или «сверхсильной» конкуренции у растений приве-
ден в следующем разделе.
3. Хищничество, растительноядность, паразитизм
и аллелопатия (антибиоз)
Определения
Хищничество и паразитизм — это хорошо известные примеры
таких взаимодействий между двумя популяциями, результаты ко-
торых отрицательно сказываются на росте и выживании одной по-
пуляции и положительно или благоприятно — на другой. Если
хищник — первичный консумент (обычно животное), а жертва
или «хозяин» — первичный продуцент (растение), то взаимодей-
ствие называют растителъноядностъю. Если же одна популяция
продуцирует вещества, вредные для конкурирующей популяции,
то для такого взаимодействия обычно применяют термин аллело-
патия, или антибиоз. Следовательно, взаимодействия типа Ч--
разнообразны.
В тех случаях, когда взаимодействующие популяции эволю-
ционируют синхронно в относительно стабильной экосистеме, от-
рицательные эффекты проявляют тенденцию становиться слабее.
Иными словами, естественный отбор стремится уменьшить от-
рицательные влияния или устранить взаимодействие популяций,
поскольку продолжительное и сильное подавление популяции
жертвы или хозяина популяцией хищника или паразита может
привести к вымиранию одной или обеих популяций. Следователь-
но, наиболее тяжелые разрушительные взаимодействия чаще все-
го наблюдаются там, где контакт между популяциями возник
недавно (две популяции встретились впервые) или когда в эко-
системе произошли крупномасштабные или внезапные изменения
(может быть, временные), нередко в результате деятельности
человека.
Объяснения и примеры
Объективный подход к вопросу о паразитизме и хищничестве
(как и к вопросу о других отрицательных взаимодействиях) найти
нелегко. Все мы испытываем естественное отвращение к парази-
там, будь то бактерии или ленточные глисты. Люди настроены
и против всех хищников, хотя человек — самый типичный хищ-
ник и основной виновник природных эпидемий. К тому же обви-
нительный приговор хищнику часто выносится наобум, без вы-
Популяции в сообществах
99
яснения того, действительно ли он вредит интересам человека.
«Волк хорош только мертвым» — эта широко распространенная
точка зрения, как мы увидим далее, оказывается весьма далекой
от истины.
Популяционный подход наиболее объективен при анализе хищ-
ничества, паразитизма, питания растениями и аллелопатии. Хищ-
ники, паразиты и растительноядные, конечно же, убивают или
наносят вред тем особям, которыми они питаются или против
которых выделяют токсичные химические вещества, они замедля-
ют до некоторой степени скорость роста популяции или умень-
шают ее численность. Но означает ли это, что без хищников или
химического ингибирования популяции всегда были бы в лучшем
состоянии? И только ли хищники получают выгоду от связи с
организмами, за счет которых они существуют, если взглянуть
на эту связь с точки зрения отдаленных результатов? Как уже
отмечалось при обсуждении регуляции численности популяции
(гл. 6, разд. 5; см., в частности, рис. 6.20), хищники и паразиты
помогают поддерживать плотность популяций растительноядных
насекомых на низком уровне, так что эти последние не уничто-
жают собственные источники пищи и не разрушают собственные
местообитания (хотя хищники могут оказаться неэффективными
и в тех случаях, когда происходит вспышка численности хозяина
и он ускользает от влияния зависимых от плотности регуляторных
механизмов). В гл. 3, разд. 4 мы говорили о том, что эволюция
растительноядных животных и растений привела к тому, что
между этими организмами возникли почти мутуалистические
взаимоотношения (+ + ). Валерио (Valerio, 1975) описал случай
обоюдного «хищничества» при взаимоотношениях паразит—хо-
зяин у пауков: молодые паучки поедают паразитов, как только
те выходят из яиц пауков, это позволяет паукам избежать зара-
жения паразитами. Более полно положительные стороны парази-
тизма описаны Смитом (Smith, 1968).
В качестве примера популяции, которая дает вспышку чис-
ленности в ответ на понижение давления хищников, обычно при-
водят ситуацию с оленями на плато Кайбаб, численность которых,
как впервые отметил Леополд (Leopold, 1943) на основе подсче-
тов Расмуссена (Rasmussen, 1941), увеличилась якобы с 4000 го-
лов (на площади 280 000 га в северной части Большого Каньона,
Аризона) в 1907 г. до 100 000 в 1924 г., что совпало с органи-
зованной правительством кампанией по борьбе с хищниками.
Коли (Caughley, 1970) повторно проанализировал имеющиеся
материалы и пришел к выводу, что, хотя численность оленей дей-
ствительно сильно увеличилась, популяция истощила свою кор-
мовую базу и затем уменьшилась, вызывают сомнения степень
перенаселенности, а также утверждение, что единственной его
причиной явилось уничтожение хищников. Определенную роль
7*
100
Глава 7
здесь сыграли также выпас крупного рогатого скота и пожары.
Коли полагает, что вспышки численности парнокопытных есть
следствие таких изменений среды и условий питания, которые
позволяют популяции ускользнуть от влияния обычного механиз-
ма регуляции численности.
Ясно одно: наибольшие вспышки численности отмечаются в
тех случаях, когда вид интродуцируется на новую территорию,
где имеются неиспользованные ресурсы и отсутствуют отрица-
тельные взаимодействия. «Взрыв» популяции кроликов, ввезен-
ных в Австралию, — самый известный случай такого рода среди
тысяч случаев резких колебаний численности, происходящих при
вселении вида с высоким биотическим потенциалом в новые места
обитания. Интересное следствие попытки контроля размножения
кроликов путем интродукции болезнетворных организмов предо-
ставило нам данные, свидетельствующие о наличии группового
отбора в системе паразит—хозяин (гл. 8, разд. 6).
Наиболее важное обобщение состоит в том, что отрицательные
взаимодействия со временем ослабевают, если стабильность и
пространственное разнообразие экосистемы достаточны для уста-
новления взаимных адаптаций. В популяциях паразитов и хозяев
или хищников и жертв, интродуцируемых в экспериментальные
микроэкосистемы, обычно наблюдаются резкие колебания чис-
ленности с определенной вероятностью вымирания. В модельных
системах хищник—жертва, описываемых уравнениями Лотки—
Вольтерра, происходят постоянные незатухающие колебания, если
в уравнения не введены дополнительные члены второго порядка,
характеризующие действие факторов самоограничения численно-
сти, которые могут вызвать затухание колебаний. Пиментел и
Стоун (Pimentel, Stone, 1968) экспериментально показали, что
такие члены второго порядка могут отражать взаимные адапта-
ции.
Как видно на рис. 7.5, сильные колебания численности про-
исходят в том случае, когда хозяина (комнатную муху) и пара-
зита (осу) впервые вместе помещают в замкнутую культиваци-
онную систему. Если же новые культуры создавали из особей,
ранее на протяжении двух лет совместно существовавших в куль-
туре, где их численность подвергалась значительным колебаниям,
то оказалось, что у них посредством отбора выработался эколо-
гический гомеостаз, при котором каждая из популяций была «по-
давлена» другой в такой степени, что оказалось возможным их
сосуществование при значительно более стабильном равновесии.
Однако в реальном мире обстоятельства могут складываться
таким образом, что взаимная адаптация в новых ассоциациях
не возникает. Всегда есть опасность, что отрицательная реакция
может стать необратимой и хозяин будет элиминирован. История
с заболеванием каштана в Америке служит примером такого взаи-
Популяции в сообществах
101
Рис. 7.5. Развитие гомеостаза в системе паразит — хозяин (хозяин — ком-
натная муха Musca domestica (I), паразит — оса Nasonia vitropennis (II)
содержались в многокамерном садке, состоявшем из 30 пластиковых камер,
соединенных друг с другом трубочками, которые замедляли распростране-
ние паразита. А. В популяциях, объединенных недавно (дикие особи
впервые посажены вместе), наблюдаются сильные колебания плотности,
причем эти колебания в популяции хозяина (мухи) происходят с некото-
рым опережением. Б. Популяции, которые получены из колоний, в которых
оба вида сосуществовали в течение двух лет; отмечается более устойчивое
равновесие, резкие спады численности отсутствуют. О возникновении у хо-
зяина адаптивной устойчивости свидетельствуют сильное снижение рож-
даемости у паразита (46 потомков на самку вместо 133 в начале опыта) и
уменьшение плотности его популяции. Этот эксперимент показывает регу-
ляторную и стабилизирующую функции генетического механизма обратной
связи в системе популяций. (А — из Pimentel, Stone, 1968; Б — по данным
для двух экспериментальных популяций, приведенным на рис. 2 и 3 их
работы; плотность выражена числом особей на 1 камеру в 30-камерном
садке.)
содействия популяции, которое с равной вероятностью может за-
кончиться и адаптацией, и вымиранием.
Американский каштан раньше был важным компонентом лесов
в районе Аппалачских гор на востоке Северной Америки, состав-
ляя часто до 40% биомассы леса. У него были свои паразиты,
хищники и болезни. Родственный ему вид каштана, произраста-
ющий в Китае, также имел своих паразитов, в том числе пара-
зитический гриб Endothia parasitica, поражающий кору на стволе.
В 1904 г. этот гриб был случайно завезен в США. Американский
102
Глава 7
Рис. 7.6. Последствие заболевания каштанов в Южных Аппалачах (Джорд-
жия). Пример того, сколь роковым может оказаться воздействие паразити-
ческого организма (гриба), завезенного из Старого Света, на нового, ин-
фицированного им хозяина (американский каштан).
каштан оказался неустойчивым к этому новому паразиту, так что
к 1952 г. все крупные деревья погибли, их мрачные серые стволы
стали характерной чертой аппалачских лесов (рис. 7.6). Каштан
продолжает давать корневые побеги, и эти побеги, прежде чем их
снова убьет гриб, часто плодоносят, но никто не может сказать,
каким будет окончательный исход этой борьбы — вымирание или
адаптация. Таким образом, популяция каштана, по крайней мере
на время, утратила свою былую доминирующую роль в лесной
экосистеме. Есть, однако, некоторая надежда на то, что недавно
обнаруженное вирусное заболевание гриба можно будет исполь-
зовать для борьбы с болезнью каштанов (см. Anagnostakis, 1982).
(Реакция леса на изъятие главного доминанта показана в
табл. 7.7.)
Приведенные выше примеры не подбирались специально, что-
бы подтвердить определенную точку зрения. Тот, кто интересу-
ется этим вопросом, легко может обнаружить множество подобных
примеров, проведя совсем немного времени в библиотеке. Ему
станет ясно, что: 1) при длительном контакте паразитов и хищ-
Популяции в сообществах
103
ников с их хозяевами или жертвой влияние на последних стано-
вится умеренным, нейтральным или даже благоприятным (ко
нечно, при рассмотрении длительных промежутков времени) и
2) наиболее сильное повреждающее действие оказывают новые
паразиты и хищники. Действительно, если составить список бо-
лезней, паразитов и насекомых-вредителей, причиняющих наи-
больший ущерб сельскому и лесному хозяйству, то окажется, что
многие из них недавно занесены в новые для них районы, как,
например, гриб Endotia parasitica, паразитирующий на каштане,
или недавно приобрели новых хозяев или жертв. Европейская
хлопковая совка, непарный шелкопряд, японский жук, средизем-
номорская плодовая мушка — это лишь немногие из интродуци-
рованных насекомых-вредителей, принадлежащих к этой катего-
рии. Тот же самый принцип применим во многом и к болезням
человека; наиболее опасны новые, завезенные впервые возбуди-
тели заболеваний.
В общем то, что мы называем «принципом внезапного усиле-
ния патогенности», можно сформулировать следующим образом:
эпидемии, эпизоотии или эпифитотии часто вызываются 1) внезап-
ным или быстрым вселением организма, обладающего потенци-
ально высокой скоростью собственного роста, в экосистему, в ко-
торой механизмы регуляции численности для этого нового вида
или отсутствуют или малоэффективны; 2) резкими или очень
сильными изменениями окружающей среды, приводящими к
уменьшению энергии, необходимой для регуляции по принципу
обратной связи, или каким-либо иным образом нарушающими
способность системы к саморегуляции. Урок из всего этого таков:
не следует допускать интродукцию новых потенциальных вреди-
телей и избегать по возможности стресса в экосистемах, возник-
новению которого способствуют ядохимикаты, уничтожающие
как вредные, так и полезные организмы.
Хотя с экологической точки зрения хищничество и паразитизм
очень сходны, между их крайними формами — крупным хищни-
ком и мелким внутренним паразитом — имеются весьма сущест-
венные различия, касающиеся не только размеров. Паразитиче-
ские или патогенные организмы обычно обладают более высоким
биотическим потенциалом, чем хищники. Их строение, обмен, вы-
бор хозяина и жизненные циклы часто более специализированы,
что связано со своеобразием их среды обитания и проблемой рас-
пространения от одного хозяина к другому.
Особенно интересны организмы, занимающие промежуточное
положение между хищниками и паразитами, например парази-
тические насекомые, которые нередко, подобно хищникам, пожи-
рают свою добычу целиком, но вместе с тем напоминают парази-
тов своим высоким биотическим потенциалом н высокой специ-
фичностью к хозяину. Энтомологи научились искусственно
104
Глава 7
разводить некоторых паразитических насекомых и использовать
их для борьбы с вредными насекомыми. Попытки использовать с
той же целью крупных неспециализированных хищников в общем
оказались неудачными. Например, мангусты, которых завезли на
острова Карибского моря для уничтожения крыс на полях сахар-
ного тростника, значительно сильнее понизили численность гнез-
дящихся на земле птиц, чем численность крыс. Такой способ
борьбы с вредителями может быть успешным при условии, что
хищник мал, специализирован в выборе пищи и обладает высоким
биотическим потенциалом. Хороший пример такого рода приводят
в своей работе Хаффейкер и Кеннетт (Huffaker, Kennett, 1956).
Эти авторы описали мелкого хищного клеща, который эффективно
контролирует численность растительноядного клеща на посадках
земляники в Калифорнии. Растительноядные насекомые иногда
успешно используются в борьбе с сорняками (Huffaker, 1957,
1959).
Человек, к сожалению, еще очень медленно учится быть,
по выражению Слободкина (Slobodkin, 1962), «расчетливым хищ-
ником», т. е. не истреблять свою жертву полностью при эксплуа-
тации ее как ресурса. Проблема оптимального изъятия из экс-
плуатируемой популяции обсуждалась Бивертоном и Холтом (Ве-
verton, Holt, 1957), Риккером (Ricker, 1958), Меншуткиным
(1962), Слободкином (Slobodkin, 1962), Силлименом (Silliman,
1969), Вагнером (Wagner, 1969), Мак-Калохом (McCullought,
1979) и многими другими. Теоретически, если кривая сигмоид-
ного роста симметрична, как в логистической модели, то ско-
рость роста dNIdt будет максимальной при плотности, равной
К/2 (при половине величины плотности насыщения). Эта точка
перегиба сигмоидной кривой была обозначена как уровень под-
держивающей емкости I на рис. 3.19. Однако вершина «горба-
той» пли параболической кривой скорости роста вследствие ско-
шенности вправо (из-за временных задержек) часто может и не
находиться точно посередине между 0 и К, поэтому плотность,
при которой обеспечивается максимальный выход живой продук-
ции, часто составляет несколько меньше половины равновесной
плотности неэксплуатируемой популяции.
К анализу проблемы можно подойти экспериментально, созда-
вая опытные популяции в мпкроэкосистемах. Одна из таких экс-
периментальных моделей представлена на рис. 7.7. Аквариумную
рыбку гуппи (Lebistes reticulatus) использовали для имитации
популяции промысловой рыбы, облавливаемой человеком. Можно
видеть, что максимальный урожай получали в том случае, когда
в каждый репродуктивный цикл изымалась одна треть популя-
ции, при этом равновесная плотность оказывалась несколько мень-
шем, чем половина плотности необлавливавшейся популяции.
В пределах данного эксперимента это соотношение оставалось
Популяции в сообществах
105
Рис. 7.7. Биомасса и выход продукции в экспериментальных популяциях
гуппи, которые содержались при трех режимах питания и разных уров-
нях эксплуатации (в процентах изъятия за репродуктивный период; уро-
вень доступной пищи в опыте II принят за единицу; в опыте I он в 1,5 раза
больше, а в опыте III — в два раза меньше). Наибольший выход продук-
ции получен в том случае, когда в каждый репродуктивный период изыма-
ли около трети популяции, а средняя биомасса снижалась менее чем на
половину по сравнению с биомассой в неэксплуатируемой популяции
(кривые выхода продукции скошены влево). (По Silliman, 1969.)
независимым от несущей емкости системы, которая поддержива-
лась на трех разных уровнях путем изменения количества корма.
Одновидовые модели часто слишком упрощают реальные от-
ношения, поскольку в них не учитывается роль конкурирующих
видов; между тем плотность последних может увеличиваться из-за
уменьшения плотности изымаемого вида, и, кроме того, они ис-
пользуют пищу и другие ресурсы, необходимые для поддержания
эксплуатируемого вида. Хищнику высшего порядка, например
человеку (или наиболее эффективному пастбищному животному,
например корове), очень легко нарушить конкурентное равнове-
сие, что может привести к замещению эксплуатируемого вида
на другой, непригодный для данного хищника или пастбищного
животного. Случаи таких «сдвигов» все множатся, по мере того,
как активизируется рыболовная, охотничья и т. п. деятельность
человека. Это дает основание высказать следующее утверждение,
которое следует рассматривать и как вызов и как предупрежде-
ние: одновидовые системы, как и монокультуры в сельском хо-
зяйстве, неустойчивы по своей природе, поскольку в условиях
стресса они сильно уязвимы для конкурентов, возбудителей бо-
106
Глава 7
лезней, паразитов, хищников и других факторов, оказывающих
отрицательное влияние. Несколько хороших примеров этого об-
щего принципа можно найти в промышленном рыболовстве.
Мерфи (Murphy, 1966, 1967) отметил уменьшение численно-
сти интенсивно облавливаемой популяции тихоокеанской сардины
(Sardinops caerulea) и последующее возрастание численности
близкого по экологии анчоуса (Engraulis mordox); в данном слу-
чае произошло замещение чрезмерно эксплуатируемого вида его
конкурентом, не подвергавшимся эксплуатации.
В другом случае Смит (Smith, 1966) описал, как последова-
тельная эксплуатация разных видов в оз. Мичиган в сочетании
с интродукцией новых видов п эвтрофикацией привела к чередо-
ванию вспышек и падений численности промысловых рыб. Пер-
воначально на протяжении полустолетия неизменным объектом
рыболовства оставалась озерная форель, однако в результате
чрезмерного облова, интродукции паразитической миноги п эв-
трофикации форель была фактически уничтожена. Вслед за этим,
быстро сменяя друг друга, здесь появлялись ряпушка, сиг, го-
лавль и один экзотический вид сельдевых. Их численность в свою
очередь сначала возрастала, а затем под влиянием усиленного
отлова, конкуренции, хищников и паразитов резко снижалась.
В 1960 г. в озеро был интродуцирован кижуч, который, к радости
рыболовов-спортсменов, сильно размножился, питаясь экзотиче-
ским видом сельди Pomolobus pseudoharengus. Однако эта неожи-
данная удача может быть сведена на нет, если не будет налажен
лучший контроль за количеством отлавливаемой рыбы и сбросом
сточных вод.
Давление хищников или промысел часто влияют на размеры
особей эксплуатируемой популяции. Так, отлов рыбы на уровне
максимального урожая обычно приводит к уменьшению среднего
размера рыб, а вырубка леса, при которой достигается макси-
мальный выход пиломатериалов на единицу объема древесины,
вызывает уменьшение величины деревьев, ухудшение качества
древесины. Как мы неоднократно повторяли, нельзя одновременно
добиться и максимального количества, и наилучшего качества.
Брукс и Додсон (Brooks, Dodson, 1965) описали случай, когда в
озерах крупные виды зоопланктеров заменялись более мелкими
после вселения питающихся зоопланктоном рыб, которых ранее
в этих озерах не было. В подобных случаях, когда экосистема от-
носительно мала, один или несколько видов хищников способны
контролировать как размерный, так и видовой состав всего тро-
фического уровня.
Хорошие примеры аллелопатии можно найти в работе Мулле-
ра и сотрудников, которые изучали ингибиторы, продуцируемые
ароматическими кустарниками в Калифорнийском чапарале. Эти
исследователи пе только выявили химическую природу и физио-
Популяции в сообществах
107
логическое действие ингибирующих веществ, но показали также
их важность в регуляции состава и динамики сообщества (см.
Muller, 1966, 1969; Muller et al., 1964, 1968). На рис. 7.8 показа-
но, как летучие терпены, выделяемые двумя видами ароматиче-
ских кустарников, ингибируют рост травянистых растений. Лету-
чие токсины (особенно эвкалиптол и камфора) продуцируются в
листьях и во время сухого сезона накапливаются в почве в таком
количестве, что с наступлением сезона дождей они подавляют
прорастание семян или последующий рост проростков в обширной
воне вокруг каждой группы кустов. Другие виды кустарников
образуют водорастворимые антибиотические вещества различной
химической природы (например, фенолы или алкалоиды), которые
также дают им возможность занять доминирующее положение в
сообществе. Однако периодические пожары, являющиеся неотъ-
емлемой частью экосистемы чапараля, эффективно уничтожают
источники токсинов, обезвреживают токсины, накопившиеся в
почве, и тем самым дают возможность появиться здесь растениям,
семена которых устойчивы к пожарам. В результате в дождливый
сезон, следующий после пожара, наблюдается пышное цветение
однолетников, которое происходит каждую весну до тех пор, по-
ка кустарники опять не разрастутся и их токсическое действие
не достигнет достаточной эффективности. В зрелом сообществе
чапараля выживает очень немного травянистых растений. Таким
образом, за счет попеременного действия пожаров и антибиотиков
происходят циклические изменения видового состава раститель-
ности, что представляет собой адаптивную черту экосистемы этого
типа.
Уиттэкер (Whittaker, 1970) в своем обзоре растительных ин-
гибиторов приходит к следующему заключению:
«Высшие растения синтезируют значительное число веществ,
являющихся репеллентами или ингибиторами для других орга-
низмов. Аллелопатические воздействия оказывают существенное
влияние на скорость растительных сукцессий и на видовой состав
стабильных сообществ. Химические взаимодействия двояко ска-
зываются на видовом разнообразии природных сообществ; сильное
доминирование и интенсивные аллелопатические эффекты поддер-
живают низкое видовое разнообразие в одних сообществах, тогда
как в других сообществах благодаря различным химическим адап-
тациям (вследствие того, что эти адаптации лежат в основе диф-
ференциации ниш) поддерживается высокое видовое разнообра-
зие».
Антибиоз, разумеется, не является достоянием только высших
растений, известны многочисленные примеры его среди микроор-
ганизмов. Иллюстрацией этого служит гриб Penicillium, образу-
ющий бактериальный ингибитор пенициллин, в настоящее время
широко используемый в медицине. Химический антибиоз можно
Рис. 7.8. Вверху. Аэрофотоснимок, на котором отчетливо видно вторжение
ароматических кустарников Salvia leucophylla и Artemisia californica в за-
росли однолетних степных растений в долине Санта-Инес, Калифорния.
Виден эффект биохимического ингибирования. Внизу. Куст Salvia крупным
планом (слева от /), выделяющий летучие токсины и подавляющий вокруг
себя рост других растений. Между 1 и 11 находится зона шириной в 2 м,
лишенная всякой травянистой растительности, за исключением нескольких
маленьких, чахлых проростков. Таким образом, корневая система кустар-
ника, простирающаяся под этой зоной, не испытывает конкуренции со сто-
роны других видов. В зоне между 11 и Ш находятся низкорослые растения,
и видов здесь меньше, чем па участках справа от III, па которые не рас-
пространяется ингибирующее влияние. (Фотографии с разрешения
д-ра С. II. Muller из Калифорнийского университета, Санта-Барбара.)
Популяции в сообществах
109
рассматривать как форму интерференционной конкуренции или,
в смягченном виде, как аменсализм. Более подробно явление ал-
лелопатии рассмотрено Райсом (Rice, 1974).
Эти интересные аспекты биологической и химической «войны»
в природе вновь приводят нас к вопросу о роли отрицательных
взаимодействий в экосистеме в целом. Часто значимость биотиче-
ских взаимодействий зависит от положения сообщества в физи-
ческом градиенте. Иа рис. 7.9 представлена «модель усоногих»,
Рис. 7.9. Факторы, контролирующие распределение двух видов усоногих
ракообразных в литоральном градиенте. У обоих видов взрослые особи
обнаруживаются в более узкой области по сравнению с той, в которой
распространяется их молодь. Распространение Balanus вверх ограничивает-
ся физическими факторами (например, обсыханием), а распространение
Chthamalus вниз ограничивается биологическими факторами (конкуренция,
хищничество). (По Е. Odum, 1963, из Connell, 1961.)
основанная на экспериментальном исследовании Коннелла. Лито-
ральная зона на скалистом берегу моря предоставляет исследова-
телю миниатюрный градиент от участков, где преобладают стрес-
совые физические воздействия на организм, до участков, где
большее значение приобретает биологическая регуляция. В про-
веденных в Шотландии исследованиях Коннелл (Connell, 1961)
обнаружил, что личинки двух видов усоногих ракообразных осе-
дают в широкой полосе литоральной зоны, но доживают до
взрослого состояния на более ограниченном участке литорали.
Оказалось, что распространение более крупного вида {Balanus)
ограничено нижней частью литоральной зоны, поскольку он не
может выносить долгих периодов обсыхания. Более мелкий вид
(Chthamalus) вытеснен из нижней зоны в результате копкурен-
по
Глава 7
ции с крупным видом и более активными в нижней литорали
хищниками. Таким образом, физическое стрессовое воздействие —
обсыхание — оказывается основным контролирующим фактором
в верхней части градиента, тогда как межвидовая конкуренция
и хищничество служат главными контролирующими факторами в
нижних зонах. Эту модель можно применить и к значительно
более широким градиентам, таким, как Арктика—тропики или
градиент от высокогорных областей до уровня моря; необходимо
только иметь в виду, что все модели, хотя и в разной степени,
чрезвычайно упрощают реальные отношения.
4. Положительные взаимодействия: комменсализм,
кооперация и мутуализм
Определения
Ассоциация двух видовых популяций, сказывающаяся поло-
жительно на обеих, чрезвычайно широко распространена в при-
роде и, вероятно, имеет не менее важное значение для определе-
ния облика популяций и сообществ, чем конкуренция, паразитизм
и т. д. Положительные взаимодействия удобно рассматривать в
той последовательности, в какой они формировались в ходе эво-
люции: комменсализм (преимущество имеет одна популяция),
протокооперация (пользу получают обе популяции) и мутуализм
(пользу получают обе популяции, причем они полностью зависят
друг от друга).
Объяснения
Широкое восприятие дарвиновской идеи о «выживании наибо-
лее приспособленных» как важном механизме естественного от-
бора послужило причиной того, что внимание исследователей со-
средоточивалось па различных проявлениях конкуренции в жи-
вой природе. В результате значение кооперации между видами
оказалось недооцененным. Во всяком случае, до настоящего вре-
мени! положительные взаимодействия изучены количественно да-
леко не столь подробно, как отрицательные взаимодействия.
С полным основанием можно предположить, что в конечном итоге
отрицательные и положительные взаимодействия между видами
в природе уравновешивают друг друга и что оба типа взаимодей-
ствий в равной степени важны для эволюции видов и стабилиза-
ции экосистемы.
Наиболее простой тип положительных взаимодействий, кото-
рый, по-видимому, представляет собой первый шаг на пути их
развития, — комменсализм. Оп характерен, в частности, для взаи-
моотношений между неподвижными растениями и животными,
Популяции в сообществах
111
с одной стороны, и подвижными организмами — с другой. Особен-
но часто можно наблюдать комменсализм в океане. Практически
в каждой норке червя, в каждой раковине двустворчатого мол-
люска, в теле каждой губки находятся «незваные гости», которые
получают здесь укрытие, но в свою очередь не приносят ему ни
пользы, ни вреда. В мантийной полости устриц, например, иногда
обитает небольшой изящный краб. Эти крабы — обычные ком-
менсалы; впрочем иногда они нарушают свой статус «гостя» и
закусывают хозяином (Christensen, McDermott, 1958). Дейле (Da-
les, 1957) в своем обзоре, посвященном комменсализму у морских
животных, перечисляет 13 видов, живущих как «гости» в норках
крупных морских червей (Erechis) и роющих креветок (Callia-
nassa и Upogebta). К их числу относятся рыбы, двустворчатые
моллюски, многощетинковые черви и крабы; они питаются ос-
татками пищи хозяина или продуктами его жизнедеятельности.
Многие комменсалы используют в качестве хозяина один опре-
деленный вид, другие же — разные виды.
Лишь один шаг отделяет комменсализм от такой ситуации,
при которой оба организма получают преимущества от объедине-
ния или какого-либо иного взаимодействия друг с другом; в этом
случае мы имеем дело с протокооперацией. Много работал в этой
области и писал по этому вопросу покойный Олли (Alee, 1938,
1951). Он полагал, что начальные стадии кооперации между ви-
дами встречаются в природе повсюду. Он выявил многие из этих
случаев и экспериментально доказал ее пользу для обоих видов.
За примером вновь обратимся к морю, где такая взаимная польза
часто возникает при объединении крабов и кишечнополостных.
Кишечнополостные прикрепляются к спине крабов (иногда крабы
«сажают» их сами), маскируя и защищая их (у кишечнополост-
ных имеются стрекательные клетки), в свою очередь получая от
крабов кусочки пищи, остающиеся от их трапезы, и используя их
как транспортное средство.
В этом примере ни крабы, ни кишечнополостные не находятся
в полной зависимости друг от друга. Следующий этап — развитие
зависимости популяций друг от друга. Такие случаи получили
название мутуализма, или облигатного симбиоза. Часто в ассо-
циацию входят совсем разные животные. Фактически случаи му-
туализма чаще встречаются у организмов с очень разными по-
требностями. (Организмы со сходными потребностями чаще
вступают в конкуренцию.) Наиболее важные мутуалистические
системы возникают между автотрофами и гетеротрофами, что
неудивительно, поскольку эти два компонента экосистемы долж-
ны в конце концов достичь некоторого сбалансированного сим-
биоза. Однако общая взаимозависимость, часто наблюдаемая в
сообществах, не относится к категории явлений, которые мы
обозначили как мутуализм; последний имеет место лишь тогда,
112
Глава 7
когда какой-либо определенный гетеротроф становится полностью
зависимым от некоторого автотрофа в отношении пищи, а суще-
ствование этого автотрофа зависит от защиты, минерального об-
мена и других жизненно важных функций гетеротрофа. Приме-
ром может служить сотрудничество между азотфиксирующими
бактериями и высшими растениями (подробнее об этом см. гл. 4).
Мутуализм обычен также между микроорганизмами, способными
переваривать целлюлозу и другие стойкие растительные остатки,
и животными, у которых нет необходимых для этого ферментных
систем.
Как отмечалось ранее, мутуализм, по-видимому, приходит на
смену паразитизму по ходу эволюции экосистем в сторону боль-
шей зрелости; и он, вероятно, особенно важен в тех случаях,
когда лимитированы некоторые ресурсы среды (например, на не-
плодородной почве); в такой ситуации взаимная кооперация мо-
жет обеспечить весьма существенное селективное преимущество.
Вопрос о том, каким образом возникают и закрепляются генети-
чески такие взаимовыгодные отношения в мире дарвиновской
борьбы за существование, до сих пор вызывает оживленные дис-
куссии среди популяционных генетиков; речь об этом пойдет в
гл. 8.
Примеры
Хорошо изучен облигатный симбиоз между жвачными (ко-
ровы) и бактериями, обитающими в их рубце (см. Hungate, 1963,
1966, 1975). Анаэробные условия в рубце неблагоприятны для
роста бактерий (только 10 процентов энергии травы или сена,
съеденных коровой, ассимилируются бактериями), но именно бла-
годаря этим неблагоприятным условиям жвачные вообще могут
существовать на таком субстрате, как клетчатка. Основная часть
энергии, полученной в результате деятельности микроорганизмов,
запасается в жирных кислотах, которые образуются из клетчатки,
но не разлагаются дальше. Жвачные могут непосредственно ас-
симилировать эти конечные продукты. Таким образом, партнер-
ство очень полезно для жвачных, которые получают большую
часть энергии в виде клетчатки; эту энергию они не смогли бы
использовать без помощи симбионтов. Бактерии в свою очередь
обеспечиваются «культуральной средой» с постоянными темпера-
турными условиями.
Достаточно хорошо изучены мутуалистические отношения
между переваривающими клетчатку микроорганизмами и чле-
нистоногими; часто такие мутуалистические отношения являются
ведущими в детритных пищевых цепях (гл. 3, разд. 4). Наиболее
известно сотрудничество между термитами и обитающими в их
кишечнике жгутиковыми; впервые этот случай мутуализма был
Популяции в сообществах
113
исследован Кливлендом (Cleveland, 1924, 1926). Многие виды
термитов не могут переваривать древесину, которой они питают-
ся, без этих специализированных жгутиковых (группа видов от-
ряда Hypermastigina). Это было доказано экспериментально: тер-
миты погибали от голода, если у них искусственно удаляли жгу-
тиковых из кишечника. Жизненный цикл симбионтов точно
скоординирован с жизнедеятельностью хозяина; в частности, при
выделении термитами гормонов линьки жгутиковые образуют цис-
ты. Это очень важно для передачи, а также для повторного за-
ражения термитов, теряющих при линьке выстилку кишечника
и затем снова заглатывающих ее. Симбионты термитов живут
внутри тела хозяина. Однако еще более тесная взаимосвязь может
возникать между хозяевами и микроорганизмами, живущими вне
животного-хозяина; подобные ассоциации фактически представ-
ляют собой следующую ступень в эволюции мутуализма (здесь
меньше вероятность того, что такие взаимоотношения могут пре-
вратиться в паразитизм!). Интересны в этом отношении некото-
рые тропические муравьи, которые разводят целые грибные сады
на листьях, принесенных в муравейник. Муравьи удобряют «по-
севы» грибов, заботятся о них и собирают урожай точно так же,
как это делал бы рачительный фермер. Мартин (Martin, 1970)
показал, что в системе муравьи—грибы детритная цепь укора-
чивается, что ускоряет естественное разложение листьев. Для
разложения листьев в норме требуется определенная последова-
тельная смена микроорганизмов, причем грибы базидиомицеты
появляются на последних стадиях разложения. Однако если листья
в грибных садах «удобряются» выделениями муравьев, то грибы,
подобно быстрорастущей монокультуре, оказываются способными
быстро размножаться на свежих листьях, обеспечивая муравьев
пищей. Для поддержания этой монокультуры так же, как при
интенсивном возделывании зерновых человеком, требуются боль-
шие затраты энергии, в данном случае «муравьиной энергии».
Мартин следующим образом резюмирует описанные факты:
«Культивируя как источник пищи организм, разрушающий клет-
чатку, муравьи получают доступ к огромным запасам клетчатки
в тропическом дождевом лесу, используя ее опосредованно в
качестве пищи. То, что у термитов осуществляется на основе эн-
досимбиотической связи с микроорганизмами, разрушающими
клетчатку, у муравьев достигается посредством более сложной
эктосимбиотической ассоциации с разрушающими клетчатку гри-
бами. Если говорить о биохимическом аспекте этой связи, то гриб
можно рассматривать как поставщика ферментного аппарата,
служащего для разложения клетчатки. В фекалиях муравьев со-
держатся протеолитические ферменты, которых нет у грибов,
и муравьи, таким образом, являются поставщиками ферментного
аппарата, служащего для разрушения белков. Этот симбиоз, еле-
8 Заказ № 1483
Рис. 7.10. Типы микоризы. А. Трехлетние сеянцы белой сосны (Pinus stro-
bus), лишенные микоризы (7) и с сильно развитой эктотрофной микоризой
(2). Б. Эндотрофная микориза; мицелий гриба проникает в клетки корня.
В. Перитрофная (внематричная) микориза образует нечто наподобие чехла
или оболочки вокруг корня сеянца ели (Picea pungens). Г, Перитрофная
микориза, образующая плотную обертку или плотную мантию вокруг кор-
ней, погруженных в богатую органикой подстилку тропического леса.
(Фотографии с разрешения проф. S. A. Wilde из Висконсинского универ-
ситета.)
116
Глава 7
довательно, можно рассматривать как метаболический альянс,
при котором происходит интеграция углеродного и азотного об-
менов двух организмов».
Копрофагию, или поедание экскрементов, которое, по-видимо-
му, является характерной чертой детритофагов, можно, вероятно,
рассматривать как значительно менее сложный, но гораздо более
распространенный случай мутуализма, объединяющий углерод-
ный п азотный обмен микроорганизмов и животных, своего рода
«наружный рубец».
Эколог Джензен (Jansen, 1965, 1966) описал замечательный
пример мутуалистической связи между определенными видами
муравьев и одним из видов акации в тропиках. Акация предо-
ставляет муравьям пищу и жилье (муравьи гнездятся в основании
раздутых колючек). Муравьи же в свою очередь защищают дерево
от возможных посягательств растительноядных насекомых. Если
в опыте убрать муравьев (это иногда встречается в природе при
обработке деревьев инсектицидами), то на дерево сразу же на-
падают листогрызущие насекомые, оно лишается листвы и часто
погибает.
Благодаря симбиозу между микроорганизмами и растениями
ускоряются круговороты минеральных веществ и продукция пи-
щи. Наиболее яркий пример этого — микориза (т. е. грибы—кор-
ни), которая представляет собой мицелий гриба, находящийся в
отношениях мутуализма с живыми корнями растений (не путать
с паразитическими грибами, которые убивают корни). Как и в
случае азотфиксирующих бактерий и бобовых, грибы при взаи-
модействии с тканью корня образуют своего рода сложные «ор-
ганы», повышающие способность растения извлекать из почвы
минеральные вещества. В свою очередь грибы получают некото-
рые продукты фотосинтетической деятельности растения. Поток
энергии, идущий через микоризу, настолько важен, что его мож-
но считать главным направлением пищевой цепи, как было по-
казано на рис. 3.11, А.
Микориза может иметь несколько форм (рис. 7.10):
1. Эктотрофная микориза, чаще всего образуемая базидиомп-
цетами; при этом формируются корневидные структуры, выходя-
щие наружу через кору корня (рис. 7.10, А).
2. Эндотрофная микориза, чаще всего образуемая фикомице-
тами, которые проникают в клетки корня (рис. 7.10,Б). Микори-
за этого типа широко распространена на корнях деревьев, но куль-
тивировать и исследовать ее весьма сложно.
3. Перитрофная (или внематричная) микориза, образующая
вокруг корней обертку или скопления (рис. 7.10,В, Г), но мице-
лий при этом не проникает через эпидермис корня. Полагают,
что микориза этого типа играет важную роль в создании благо-
приятной химической «ризосферной среды», способствующей пре-
Популяции в сообществах
117
вращению нерастворимых или недоступных минеральных веществ
в форму, которая может использоваться корнями.
Многие деревья не могут расти без микоризы. Лесные деревья,
пересаженные на почву прерий или интродуцированные в другие
районы, часто не способны расти до тех пор, пока не произойдет
заражения грибными симбионтами. Сосны со здоровой микоризой
прекрасно растут на такой бедной почве, которая по принятым
сельскохозяйственным стандартам не пригодна для посевов зер-
новых культур. Грибы могут метаболизировать «недоступный»
фосфор п другие минеральные элементы путем образования хе-
латных соединений (объяснение этого процесса см. т. 1, с. 54)
или иными, пока еще неясными путями. Если в почву добавить
меченые минеральные соединения (например, радиоактивный
фосфор), то до 90% этих веществ быстро поглощается микоризой;
затем происходит медленное перемещение этих веществ в рас-
тение.
Весьма кстати, что такая мутуалистическая система, как сос-
на—микориза, способна выживать на миллионах гектаров юга
США, где верхний слой почвы был уничтожен в результате дли-
тельного возделывания монокультуры и сложившейся там прак-
тики ведения сельского хозяйства, при которой владельцы не жи-
вут на собственных землях; не будь сосны, все эти земли пре-
вратились бы в пустыню. Замечательные свойства системы
сосна—микориза были использованы также и в Коппер-Хилле
(гл. 2, разд. 4).
Роль микоризы в возвращении минеральных веществ в кру-
говорот, ее значение в тропиках и необходимость для культур,
зависящих от таких механизмов возврата, рассматриваются в
гл. 4, разд. 7 и 8. Дополнительные сведения о мутуализме между
микроорганизмами и растениями см. Harley, 1959; Rovira, 1965;
Wilde, 1968; Marks, Kozlowski, 1973; Sanders et al., 1975 и Marx,
Ruehle, 1979.
Лишайники — это симбиотическая система определенных гри-
бов и водорослей; функциональная и морфологическая связь этих
организмов настолько тесна, что их можно рассматривать как
своего рода особый организм, не похожий ни на один из слага-
ющих его компонентов. Лишайники обычно классифицируют как
отдельные «виды», несмотря на то что они состоят из двух нерод-
ственных видов. Компоненты лишайников нередко можно изоли-
рованно культивировать в лаборатории, однако сами лишайники
трудно вырастить в культуре, хотя в природе они могут сущест-
вовать в очень суровых условиях. Лишайники интересны тем, что
на их примере можно наблюдать постепенную эволюцию от па-
разитизма к мутуализму. У некоторых более примитивных ли-
шайников гриб фактически проникает в клетки водоросли, как
показано на рис. 7.11, А, и по существу является паразитом.
118
Глава 7
проникают в
.летки водоросли
Б
Гифы гриба
переплетаются с
нитями водоросли
Гифы f риба тесно
примыкают к клеткам
водорослей, но не
проникают в них
Рис. 7.11. Направление эволюции от паразитизма к мутуализму у лишай-
ников. А. У некоторых примитивных лишайников грибы проникают в клет-
ки водорослей. Б и В. У эволюционно более развитых видов водоросль и
гриб живут в гармоничных отношениях, принося друг другу взаимную
пользу. (Е. Odum, 1963.)
У эволюционно более развитых видов мицелий гриба (его гифы)
не проникает в клетки водоросли, но оба организма живут в пол-
ной гармонии (рис. 7.11,Б и В).
Общие сведения о симбиотических ассоциациях см. в трудах
симпозиума под редакцией Натмена и Месси (Natman, Masse,
1963), а также в работах Henry, 1966 и Cheng, 1971. Взаимоот-
ношения между человеком, культурными растениями и домашни-
ми животными, которые можно рассматривать как особую форму
мутуализма, обсуждаются в гл. 8. В гл. 2 была описана ассоциация
между кораллами и водорослями, способствующая увеличению
скорости возврата биогенных элементов и повышению продуктив-
ности всей экосистемы.
Модель лишайника, представленная на рис. 7.11, симво-
лична для человека. До сих пор человек существовал в своей
автотрофной среде как паразит, потребляя то, что ему нуж-
но и не заботясь о благосостоянии своего хозяина, т. е. пла-
неты. Непомерно разрастающиеся города становятся парази-
тами сельской местности, которая так или иначе должна
снабжать их нищей, водой и воздухом и перерабатывать
огромные количества отходов. Ясно, что человек должен
установить мутуалистический союз с природой. Если он
не научится этому, то, подобно «неразумному» или «неадап-
тированному» паразиту, может довести эксплуатацию своего
хозяина до такой степени, что погубит и себя.
Популяции в сообществах
119
5. Концепции местообитания, экологической
ниши и гильдии
Определения
Местообитание организма — это место, где он живет, или мес-
то, где его обычно можно найти. Экологическая ниша — понятие
более емкое, включающее в себя не только физическое простран-
ство, занимаемое организмом, но и функциональную роль орга-
низма в сообществе (например, его трофическое положение) и его
положение относительно градиентов внешних факторов — темпе-
ратуры, влажности, pH, почвы и других условий существования.
Эти три аспекта экологической ниши удобно обозначить как про-
странственную нишу, или нишу места, трофическую нишу и мно-
гомерную нишу, или нишу как гиперобъем. Следовательно, эко-
логическая ниша организма зависит не только от того, где он
живет, но включает также общую сумму его требований к окру-
жающей среде. Концепция ниши оказывается наиболее полезной
для количественных оценок различий между видами (или внутри
вида в разных местах и в разное время) по одному или немногим
основным (т. е. операционально значимым) признакам. Для ха-
рактеристики ниши чаще всего используются два измерения —
ширина ниши и перекрывание ниши с соседними. Группы видов
в сообществе, обладающих сходными функциями и нишами оди-
накового размера, называются гильдиями. Виды, занимающие
одинаковые ниши в разных географических областях, называют-
ся экологическими эквивалентами.
Объяснения и примеры
Термин «местообитание» широко используется не только в эко-
логии, но и в других областях науки. Так, местообитание водных
насекомых: клопа-гладыша (Notonecta) и плавта (Corixa), пока-
занных на рис. 7.12, — это мелководные, покрытые раститель-
ностью пространства (литоральная зона) прудов и озер, куда
можно пойти, чтобы собрать этих замечательных водных клопов.
Эти виды занимают, однако, очень разные трофические ниши.
Гладыш — активный хищник, а плавт питается преимущественно
разлагающейся растительностью. Другой пример разделения ниш,
основанного на способах питания, показан на рис. 7.4. В эколо-
гической литературе можно найти множество подобных примеров
сосуществования видов, использующих разные источники энергии.
Местообитание может также обозначать место, занимаемое це-
лым сообществом. Например, местообитание шалфейного сообще-
ства песчаной степи — это гряды холмов с песчаной почвой, тя-
нущиеся вдоль северных берегов рек на юге Великих равнин в
120
Глава 7
Рис. 7.12. Два водяных клопа (Hemiptera) — Notonecta (Л) и Corixa (Б),
которые живут в одном и том же местообитании, но из-за различий в пи-
тании занимают разные трофические ниши.
США. В данном случае местообитание слагается преимуществен-
но из физических, или абиотических, комплексов, тогда как мес-
тообитание упомянутых выше водных клопов включает живые и
неживые объекты. Таким образом, местообитание одного организ-
ма или группы организмов (популяции) включает помимо абио-
тической среды также и другие организмы. Для описания место-
обитания сообщества достаточно только последнего. Во избежание
недоразумений важно различать эти два возможных применения
термина «местообитание».
Экологическая ниша — понятие более новое и оно не имеет
столь широкого применения вне экологии. Расплывчатые, хотя и
полезные термины, такие, как ниша, трудно определить п выра-
зить количественно; в данном случае лучше всего рассматривать
историю возникновения понятия. Джозеф Гриннелл (Joseph
Grinnell, 1917, 1928) пользовался словом ниша «для обозначения
основной единицы распределения, в пределах которой данный вид
удерживают его структурные и инстинктивные ограничения...;
на одной и той же территории не может быть двух видов, кото-
рые долго занимали бы совершенно идентичную экологическую
нишу». (По случайному совпадению, последнее утверждение пред-
шествовало экспериментам Гаузе, продемонстрировавшим прин-
цип конкурентного исключения; см. рис. 7.2.) Таким образом,
Гриннелл трактовал пишу преимущественно как микроместооби-
тание или то, что мы назвали бы теперь пространственной нишей.
Чарлз Элтон (Ch. Elton, 1927, и последующие публикации) в Анг-
лии был одним из первых, кто придал термину «ниша» смысл
«функционального статуса организма в сообществе». Поскольку
Элтон имеет очень большое влияние на экологическое мышление,
представление о том, что ниша не есть синоним местообитания,
получило широкое распространение, а поскольку он уделял ос-
новное внимание энергетическим отношениям, его вариант этого
понятия можно обозначить как трофическую нишу.
Популяции в сообществах
121
В 1957 г. Хатчинсон показал, что нишу можно рассматривать
как многомерное пространство или гиперобъем, в пределах кото-
рого условия среды позволяют существовать особи или виду
неопределенно долго. Нишу Хатчинсона, которую мы будем на-
зывать многомерной, или гиперпространственной, нишей, можно
описать с помощью количественных характеристик и оперировать
с ней математически. Например, двумерные климаграммы
(рис. 5.15), изображающие климатическую нишу одного вида
птиц и одного вида плодовой мухи, можно расширить путем вве-
дения ряда новых координат, учитывающих другие параметры
окружающей среды. Хатчинсон (Hutchinson, 1965) различал так-
же фундаментальную нишу — наибольший «абстрактно заселен-
ный гиперобъем», когда вид не ограничен конкуренцией с другими
видами, и реализованную нишу — меньший гиперобъем, занятый
в условиях биотических ограничений. Эти два понятия двумерных
ниш иллюстрируются на рис. 7.13, Л и В.
Вандермеер (Vandermeer, 1972) несколько расширил концеп-
цию реализованной ниши Хатчинсона, включив в нее допущение
о том, что если в данном местообитании имеется 0, 1, 2, 3,... п
взаимодействующих видов, то их реализованные ниши будут раз-
личными. В отсутствие потенциальных конкурентов вид занимает
оптимальную (или фундаментальную, по Хатчинсону) нишу,
но при наличии других сходных видов следует говорить не об
одной реализованной нише, а о целом ряде частных ниш, число
которых зависит от числа потенциальных конкурентов. Итак,
Вандермеер рекомендует пользоваться термином частная ниша
вместо термина «реализованная ниша». Простая аналогия из по-
вседневной жизни, вероятно, может помочь нам разобраться в
этих перекрывающихся и иногда противоречивых вариантах при-
менения в экологии термина «ниша». Если в нашем человеческом
обществе мы хотим с кем-то познакомиться, то для начала нам
необходимо узнать адрес этого человека, т. е. место, где его мож-
но найти. «Адрес» здесь соответствует местообитанию организма.
Однако, чтобы узнать человека как следует, желательно знать о
роде его занятий, его интересы, знакомых и вообще роль, которую
он играет в жизни общества. В данном случае профессия будет
аналогом ниши.
Сходная ситуация наблюдается при изучении организмов:
знание местообитания — это только начало. Для определения ста-
туса организма в природном сообществе мы должны располагать
некоторыми сведениями о его активности, в частности о питании,
источниках энергии и путях ее распределения; нужно, кроме
того, знать соответствующие популяционные характеристики, та-
кие, как внутренняя скорость роста, приспособленность и т. д.,
и, наконец, нужно знать о влиянии данного организма на другие
организмы, с которыми он взаимодействует, и о той степени, в ка-
122
Глава 7
Рис. 7.13. Схематическое представление концепций ниши. А и В — ниша
как гиперобъем. На верхней диаграмме В два вида занимают не перекры-
вающиеся ниши, на нижней диаграмме ниши перекрываются настолько
сильно, что возникает жестокая конкуренция, приводящая к дивергенции
(показано стрелками). Б. Распределение активности двух видов вдоль оси,
имеющей размерность ресурса, иллюстрирует концепцию ширины ниши и
перекрывания ниш.
кой он воздействует или может воздействовать на важные собы-
тия в экосистеме.
Мак-Артур (MacArthur, 1968) отметил, что экологический
термин «ниша» и генетический термин «фенотип» — понятия ана-
логичные; оба они связаны с неопределенным числом признаков,
имеют некоторые или даже все общие переменные и оба весьма
полезны при установлении различий между особями и видами.
Таким образом, ниши сходных видов, объединенных одним общим
местообитанием, можно точно сравнить, если проводить сравнение
Популяции в сообществах
123
лишь по нескольким операционально значимым переменным. Мак-
Артур сравнивает ниши четырех видов американских лесных пе-
вунов (Parulidae). Все они размножаются в одном макроместо-
обитанин — еловом лесу, но собирают корм и гнездятся па разных
частях елей. Для этой ситуации Мак-Артур построил математи-
ческую модель, представляющую собой систему уравнений конку-
ренции в матричной форме. Коэффициенты конкуренции в модели
вычислялись для взаимодействий каждого вида с каждым из трех
других. В одной из пар видов наблюдалась особенно жестокая
конкуренция, поэтому весьма вероятно, что в отсутствие любого
из этих двух видов другой займет вакантное пространство ниши.
Общая тенденция уменьшения ниши при конкуренции между ви-
дами иллюстрируется на рис. 7.3.
Термин гильдия в современной экологической литературе час-
то используется в применении к группам видов, таким, как изу-
ченные Мак-Артуром лесные певуны, роль которых в сообществе
одинакова или сравнима (см. Root, 1967, который первым указал
на возможность такого использования термина). Можно привести
и другие примеры гильдий — это осы, паразитирующие на попу-
ляциях питающихся нектаром насекомых, улитки, живущие в лис-
товой подстилке леса, и лианы, проникающие в полог тропическо-
го леса. Гильдия — удобная единица для изучения взаимодействий
между видами, однако при анализе сообщества ее можно рассмат-
ривать как функциональную единицу, в результате чего отпадает
необходимость рассматривать все до единого виды по отдельности.
Разделение микроместообитаний или пространственных ниш
внутри гильдий широко распространено, являясь почти правилом.
Два примера показаны на рис. 7.14. У шести из семи видов лес-
ных слизней более четко разделяются ниши в областях питания,
тогда как у пяти видов бокоплавов наблюдается значительное пе-
рекрывание ниш вдоль градиента солености.
В качестве показателей при сравнении ниш крупных растений
и животных часто пользуются результатами промеров морфоло-
гических признаков. Например, Ван Вален (Van Valen, 1965)
нашел, что показателем «ширины ниши» могут служить длина п
толщина клюва у птиц (клюв, конечно, отражает вид потребляе-
мой пищи). Оказалось, что коэффициенты вариации толщины
клювов у островных популяций шести видов птиц больше, чем у
материковых; это соответствует большей ширине ниш (большему
разнообразию занимаемых местообитаний и типов пищи) на ост-
ровах, где меньше конкурирующих видов. Конкуренция внутри
вида часто сильно уменьшается, если разные стадии жизненного
цикла занимают разные ниши. Головастики, например, — расти-
тельноядные, а взрослые лягушки, обитающие в том же самом
пруду, — хищники. Разделение ниш можно наблюдать даже меж-
ду разными полами. У дятлов рода Picoides (старое название —
124
Глава 7
Рпс. 7.14. Л. Разделение ниш по высоте обитания у лесных слизней. (По
Junnings, Barkham, 1979.) Б. Расположение местообитаний бокоплавов
вдоль градиента солености. (По Fenchel, Raiding, 1979.)
Dendrocopus) самки и самцы различаются по величине клюва и
пищевому поведению (Ligon, 1968). У ястребов, как и у многих
насекомых, особи разного пола заметно различаются по размерам
тела и, следовательно, по характеристикам пищевых ниш.
Можно ожидать, что введение в природные экосистемы как
биогенных элементов, так и токсичных химических веществ в
большей степени изменяет соотношение ниш тех видов, которые
наиболее подвержены этому воздействию. Так, Бакелар и Одум
(Bakelaar, Odum, 1978) при изучении влияния производимого в
промышленных масштабах удобрения, содержащего N, Р и К,
на растительность залежи обнаружили, что в результате приме-
нения этого удобрения ширина ниши и перекрывание ниш не из-
менились у большинства видов, но у главного доминанта — золо-
Популяции в сообществах
125
тарника (один из видов Soltdago) ширина ниши существенно
увеличилась, численность его возросла за счет растения-кодоми-
нанта (одного из видов Aster).
В следующем разделе мы рассмотрим, в какой степени рас-
пределение видов по обилию внутри трофических уровней, так-
сономических групп (птицы, насекомые и т. д.) и сообществ в
целом позволяет нам лучше понять природу взаимодействия ниш.
Виды, занимающие одну и ту же или похожие ниши в разных
географических областях, называются экологически эквивалент-
ными, причем в соприкасающихся областях эти виды обычно
близкородственны таксономически, тогда как в несоприкасато-
щихся областях они часто не имеют родственных связей. В раз-
ных флористических и фаунистических областях сообщества силь-
но различаются по видовому составу, но во всех тех местах, где
независимо от географического положения физические условия
одинаковы, развиваются сходные экосистемы. Эквивалентные
функциональные ниши оказываются занятыми теми биологиче-
скими группами, которые имеются в фауне и флоре данной об-
ласти. Так, экосистемы типа злаковников развиваются повсюду,
где имеются соответствующие климатические условия, но виды
злаков и травоядных могут быть совершенно различными, особен-
но если эти области разделены мощными барьерами. Крупные
кенгуру Австралии экологически эквивалентны бизонам и анти-
лопам Северной Америки (на обоих континентах этих животных
теперь в значительной степени замещают коровы и овцы).
Табл. 7.3 и 7.4 иллюстрируют два примера экологической эк-
вивалентности в местообитаниях морского берега и степи. Брюхо-
ногие моллюски, сменяющие друг друга по высоте берега на по-
бережье Северной и Южной Америки, все принадлежат к одному
Таблица 7.3. Экологические эквиваленты в трех главных нишах четырех зон
побережья Северной и Центральной Америки
Ниша Тропики Северо-западное побережье Мексиканский залив Северо-вос- точное побе- режье
Питание растени- ями на литораль- ных камнях Littorina ziczac L. danaxis L. scutelata L. irrorata | L. littorea
Хищничество на Лангуст Королевский Каменный краб Омар
Дне (Panuliriis) краб (Paralithodes) (Menippe) (Но marus)
Планктоноядная рыба Анчоус Тихооке анская сельдь, сардина Менхеден, пальцепер Атлантичес- кая сельдь, помоло- бус
126
Глава 7
роду, но птицы в степях Канзаса и Чили относятся к разным
семействам.
Таблица 7.4. Экологические эквиваленты среди видов степных птиц в Кан-
засе и Чили1
Экологические эквиваленты (пары видов) Размеры тела, мм Длина клю- ва, мм Отношение толщины клюва к его длине
Восточный трупиал (Sturnella magna) Канзас 236 32,1 0,36
Крас ног рудный трупиал (Pezites inilttaris) Чили 264 33,3 0,40
Саванный воробей (Ammatramus savannarum) Канзас 118 6,5 0,60
Желтый степной вьюрок (Sicalis luteula) Чили 125 7,1 0,73
Рогатый жаворонок (Eromorphila alpestris) Канзас 157 И,2 0,50
Чилийский конек (Anthus correnderas) Чили 153 13,0 0,42
1 Три вида из каждого региона различаются своими нишами питания, на что
указывают различия величины тела и размеров клюва, но каждая пара эквивален-
тов очень сходна морфологически, что указывает на большое сходство ниш. Тру-
пиалы таксономически близко родственны, вторая пара родственна только на уров-
не семейства, виды третьей пары принадлежат разным семействам (По Cody, 1974.)
6. Видовое, структурное и генетическое
разнообразие в сообществах
Определения
Из общего числа видов какого-либо трофического уровня или со-
общества в целом обычно лишь немногие бывают доминирующи-
ми, т. е. имеют значительную численность (большую биомассу,
продуктивность или другие показатели «значимости»); большая
же часть относится к редким видам (т. е. имеет низкие показате-
ли «значимости»). Правда, иногда доминантные виды отсутству-
ют, а многие виды характеризуются промежуточным обилием.
Видовое разнообразие слагается из двух компонентов: 1) видового
богатства, или плотности видов, которое характеризуется общим
числом имеющихся видов, и 2) выровненности, основанной
на относительном обилии или другом показателе значимости вида
Популяции в сообществах
127
и положении его в структуре доминирования. Видовое разнообра-
зие увеличивается при увеличении размеров площади и при про-
движении от высоких широт к экватору. В сообществах же. под-
вергающихся стрессовым воздействиям, видовое разнообразие не-
велико; но, кроме того, оно может снижаться в результате
конкуренции в старых сообществах, существующих в стабильной
физической среде. Большую роль играют два других типа разно-
образия: 1) структурное разнообразие, которое является следстви-
ем зональности, стратифицпрованности, периодичности, пятнисто-
сти, наличия пищевых сетей и других способов ранжирования
компонентов популяций микроместообитаний, и 2) генетическое
разнообразие, т. е. поддержание генотипических гетерозпготности,
полиморфизма и другой генотипической изменчивости, которая
вызвана адаптационной необходимостью в природных популяци-
ях. Многие экологи уже знают, что уменьшение видового и гено-
типического разнообразия, происходящее вследствие деятельности
человека, ставит на грань риска возможность будущих адаптаций
как в природных экосистемах, так и в агроэкосистемах.
Объяснения
Характерной чертой структуры сообществ в северных широтах,
а также сообществ тропических областей с чередованием дожд-
ливых и сухих сезонов является наличие немногих обычных,
или доминантных, видов, представленных большим числом
особей, и многих редких видов, представленных малым числом
особей. Во влажных тропиках, где нет смены сезонов, обычно
насчитывается множество видов с низким относительным обили-
ем. Общую тенденцию увеличения числа видов с севера на юг
иллюстрирует рис. 7.15. Другая общая тенденция, или закон, при-
роды состоит в том, что число видов возрастает с увеличением
площади и, вероятно, также с увеличением эволюционного време-
ни, в течение которого происходили колонизация, специализация
ниши видообразование (см. Preston, 1960; Sanders, 1968).
Для анализа видового разнообразия в разных ситуациях приме-
няют два широких подхода: 1) сравнения, основанные па формах
кривых относительного обилия или доминирования—разнообразия,
и 2) сравнения, основанные на индексах разнообразия, представ-
ляющих собой отношения или другие математические выражения
зависимости между числом видов и их значимостью. Позднее мы
более подробно рассмотрим эти подходы, но прежде всего важно
уметь различать два основных компонента видового разнообразия,
которые могут по-разному реагировать на географические, физи-
ческие и эволюционные факторы. Один из главных компонентов —
видовое богатство (многообразие), или плотность, видов; это прос-
то общее число видов, которое в сравнительных целях обычно
128
Глава 7
Рис. 7.15. Широтный градиент числа видов гнездящихся наземных птиц (Л)
и числа видов муравьев (Б). (Fischer, 1960.)
выражается как отношение числа видов к площади или числа
видов к числу особей. Часто, но далеко не всегда, существует
логарифмическая зависимость между числом видов и числом осо-
бей (прямая линия в полулогарифмических координатах), и чис-
ло видов представляет собой степенную функцию от площади
(прямая линия в двойных логарифмических координатах), как
показано на рис. 7.16,Л, Б, Из рис. 7.16,5 видно, что на увели-
чение числа видов с увеличением площади оказывают влияние
как разнообразие местообитаний, так и площадь per se. Соотно-
шения виды/площадь обычно используются для определения оп-
тимальной величины пробы и для предсказания числа видов на
большей площади, чем та, где брались пробы. По данным Конно-
ра и Мак-Коя (Connor, McCoy, 1979), хорошее приближение
кривых виды/площадь получается при использовании нескольких
различных уравнений (в том числе степенной функции в двойной
логарифмической шкале), но ни одно из них не имеет биологиче-
ского смысла.
200 ----1-------।-------1----------------'-------'---
1 10 10? 103 104 10s
Площадь, квадратные мили (логарифм)
Рис. 7.16. А. Соотношение между числом видов и числом особей на засо-
ленных маршах и залежных землях в Джорджии. При логарифмической
зависимости в качестве индекса разнообразия можно использовать число
видов на 1000 особей. Б. Зависимость между площадью и числом видов
растений (Великобритания). Линии регрессии проходят с наклоном от 0,1
до 0,35. Низкие величины характерны для континентов, высокие — для
островов. (См. Connor, McCoy, 1979.)
9 Заказ Ks 1483
130
Глава 7
Второй важный аспект разнообразия — выровненностъ относи-
тельного распределения особей среди видов. Например, две систе-
мы, каждая из которых состоит из десяти видов и 100 особей,
могут иметь одинаковые индексы разнообразия *$7А, но совер-
шенно различные индексы выровненности в зависимости от рас-
пределения 100 особей между десятью видами. Так, в одном край-
нем случае распределение может иметь вид: 91—1—1—1—1—
1 — 1 — 1 1 । (минимальная выровненностъ и максимальное до-
минирование), а в другом крайнем случае — по десяти особей на
каждый вид (максимальная выровненностъ и отсутствие домини-
рования). Выровненностъ, как правило, высока и постоянна среди
популяций птиц (вероятно, это обусловлено их территориальным
поведением), а различия этого показателя в разных сообществах
и географических зонах определяются главным образом видовым
богатством (Tramer, 1969). Напротив, у растений и фитопланкто-
на выровненностъ в среднем низка, и оба компонента подвержены
значительным вариациям (Austin, Tomoff, 1978).
Наилучший способ представить оба компонента разнообразия—
построить график, где в логарифмическом масштабе отложено
число особей (биомасса, продуктивность или другие подходящие
меры значимости) каждого вида, а на оси х — ранжированная
последовательность видов от наиболее обильного до наименее
обильного. Линия, соединяющая точки или проходящая близко
от них, названа Уиттэкером (Whittaker, 1965, 1972) кривой до-
минирования—разнообразия. Еще одно удачное название—«кри-
вая значимости видов» — было предложено Пианкой (Pianka,
1978). На рис. 7.17 показаны три основные формы, которые мо-
жет иметь такая кривая, построенная для гипотетической выбор-
ки из 1000 особей и 20 видов. Если наиболее обильный вид в два
раза многочисленнее следующего за ним по обилию, а этот в
свою очередь вдвое многочисленнее третьего и т. д., то на графике
мы получаем прямую линию (рис. 7.17,7). Отсюда можно предпо-
ложить, что первый вид занимает половину доступного простран-
ства ниш, второй — половину оставшегося пространства (25% ис-
ходного) и т. д. Иными словами, каждый вид занимает прежде
всего свободную нишу, не перекрывающуюся с другими. Если,
однако, пространство ниш поделено на случайные, соприкасаю-
щиеся и неперекрывающиеся участки, то получится совершенно
другая кривая (рис. 7.17,77). Эта кривая изображает модель «раз-
ломанного стержня», предложенную Мак-Артуром (MacArthur,
1957). Эти две возможности представляют крайние случаи, обыч-
но же распределение в природе имеет вид промежуточной S-об-
разной кривой (рис. 7.17,777), что указывает на более сложный
характер дифференциации и перекрывания ниш. Об этом свиде-
тельствует логнормальное распределение видов по значимости
(рпс. (См. Preston, 1948; Whittaker, 1972, и
Популяции в сообществах
131
Рис. 7.17. Кривые доминирования—разнообразия для гипотетической вы-
борки, состоящей из 1000 особей, относящихся к 20 видам. По оси ординат
отложено число особей каждого вида, а по оси абсцисс — порядковый номер
вида в последовательности от более обильного к менее обильному. I — гео-
метрический ряд с приоритетами в захвате ниш; II — ниши не перекры-
ваются, их величина случайна; III — промежуточная система, соответст-
вующая 5-образной кривой; ниши многомерны и перекрываются, что обус-
ловливает логнормальное распределение видов по значимости. (Whittaker,
1965.) ..
рис. 7.21,5.) У групп с интенсивной межвидовой конкуренцией
и территориальным поведением, например у лесных птиц, харак-
тер распределения соответствует представлению о неперекрываю-
щихся случайных нишах, как на кривой II; простые геометриче-
ские последовательности (кривая /) характерны для некоторых
растительных сообществ в суровых условиях окружающей среды.
Как было подчеркнуто в разд. 2, большинство видов в природных
открытых системах сосуществует в условиях соревнования за ре-
сурсы, а не в условиях прямой конкуренции; множество адапта-
ций дает возможность делить ниши без конкурентного исключе-
ния из местообитания. Таким образом, наиболее вероятную мо-
дель изображает кривая III, особенно для ненарушенных сооб-
ществ.
В общем расположение видов по степени значимости, как на
рис. 7.47 и 7.18, не только точно отражает богатство и относи-
тельное обилие видов как компонентов видового разнообразия,
но и объясняет, каким образом подразделяется пространство ниш.
9*
132
Глава 7
Ранг вида
Рис. 7.18. Сравнение кривых доминирования—разнообразия для двух тро-
пических лесов и двух лесов умеренной зоны. Оценки значимости видов в
лесах умеренной зоны основаны на чистой годовой продукции, в сухом лесу
Коста-Рики — на площади оснований (площадь сечений всех стволов дан-
ного вида), в лесу Амазонки — на биомассе надземных частей. (Hubbel,
1979.) I — влажный тропический лес (Манаус, Бразилия); II — сухой тро-
пический лес (Гуанакасте, Коста-Рика); III — сырой лес умеренной зоны
(Смоки-Маунтинс, Теннесси); IV — горный (субальпийский) лес умеренной
зоны (Смоки-Маунтинс, Теннесси).
Чем выше кривая и чем более она уплощена, тем больше при дан-
ном числе видов общее разнообразие; так, на рис. 7.17 разнообразие
11>разнообразие 777 >разнообразие I. Чем круче идет кривая,
тем меньше общее разнообразие и сильнее доминирование одного
или нескольких видов. В стрессовых ситуациях независимо от
того, вызваны ли они естественными причинами (например, суро-
выми погодными условиями) или антропогенным воздействием
(например, загрязнения), кривая становится более крутой. Таким
образом, кривую доминирования—разнообразия можно исполь-
зовать для оценки влияния нарушений на видовую структуру.
При другом подходе используются индексы разнообразия.
В табл. 7.5 приведены четыре наиболее употребительных индекса
и некоторые их варианты. А — индекс видового богатства; Г — ин-
декс выровненности; Б и В содержат комбинацию обоих компо-
нентов и, таким образом, представляют собой обобщенные индек-
сы разнообразия. Индекс Симпсона показывает «концентрацию»
доминирования, так как его величина тем больше, чем сильнее
Популяции в сообществах
133
доминирование одного или немногих видов. Единица минус ин-
декс Симпсона (или просто обратная ему величина) становится
индексом разнообразия, сравнимым с другими.
Таблица 7.5. Некоторые полезные показатели разнообразия
А. Индекс видового богатства (d)
g______________________I
---- (также S/N и 5 на 100 особей),
IgN
где S — число видов, N — число особей.
Б. Индекс Симпсона 1 2 (с)
с = 2(пг7Я)2, или c=2n< J индекс доминирования
и
1
1—Щпг/N)2 и ./дт)2~ ’ инДекс разнообразия.
где Пг — оценка значимости каждого вида (численность, биомасса и т. д.),
N — сумма оценок значимости.
В. Индекс Шеннона3 (Я)
H=—^niIN\ng(niIN), или Я=-—SPJogPf,
где гц — оценка значимости каждого вида, Л7 — сумма оценок значимости,
Pi — вероятность вклада каждого вида.
Г. Индекс выровненности Пиелу4 * * (е)
Я
е~ logs’
где Н — индекс Шеннона, S — число видов.
1 Simpson, 1949.
2 См. Margalef 1958, — логарифмический индекс; Menhinick, 1964, — индекс
квадратного корня; Н. Т. Odum, Cantion, Kornicker, 1960, — индекс 8/1000 особей.
3 Shannon, Weawer. 1949; Margalef, 1968.
4 См. Pielou, 1966; другие типы показателя выровненности — см. Lloyd, Ghelardi,
1964. __
Примечание. При расчете величин d, Н_п е обычно пользуются натуральными ло-
гарифмами (loge), но для вычисления Н иногда применяют логарифм с основани-
ем 2 (log2), чтобы сразу получить эту величину в битах на особь.
Из двух обобщенных индексов индекс Симпсона придает обыч-
ным видам больший вес (поскольку при возведении в квадрат
малых отношений Пг/N получаются очень малые величины). Ин-
декс Шеннона придает больший вес редким видам. Так как ин-
декс Шеннона заимствован из теории информации и представляет
собой формализацию, которая широко используется при оценке
сложности и содержания информации в любых типах систем, он
лучше всего подходит для целей сравнения в тех случаях, когда
нас не интересуют компоненты разнообразия по отдельности.
134
Глава 7
И коль скоро Н вычислено, мы можем, поделив эту величину на
логарифм числа видов, легко получить индекс выровненности
(Г в табл. 7.5). Индекс Шеннона к тому же почти не зависит от
величины пробы и вследствие того, что N — всегда целое число,
характеризуется нормальным распределением (Bowman et al.,
1970; Hutcheson, 1970); это обстоятельство позволяет использо-
вать обычные статистические методы для проверки значимости
различий между средними. Лайонс (Lyons, 1981) показал, что
если известно число особей, то в таких статистических тестах
могут быть использованы показатели биомассы или продуктивно-
сти (нецелые величины), которые с экологической точки зрения
часто оказываются более подходящими.
Поскольку стабильные экосистемы, такие, как дождевой лес
или коралловые рифы, обладают высоким видовым разнообразием,
возникло искушение сделать вывод, что разнообразие повышает
стабильность. Маргалеф (Margalef, 1968) выразил это так: «Эко-
лог видит в разнообразии, измеренном любым способом, проявле-
ние возможности построить системы с обратной связью». Однако
результаты более поздних исследований и критические обзоры,
начиная с симпозиума 1969 г. «Разнообразие и устойчивость эко-
логических систем», материалы которого вышли под редакцией
Вудвелла и Смита (Woodwell, Smith, 1969), и кончая статьями в
Трудах I международного экологического конгресса под редак-
цией Ван Доблена и Лоу-Мак-Коннелла (Van Doblen, Lowe-Mc-
Connell, 1975), показали, что связь между видовым разнообразием
п устойчивостью сложнее, что положительная корреляция может
быть вторичной п что, напротив, стабильность экосистемы может
обусловливать высокое разнообразие. Хастон (Huston, 1979) при-
шел к выводу, что экосистемы, названные им «неравновесными»,
т. е. экосистемы, претерпевающие периодические возмущения, как
правило, характеризуются большим разнообразием, чем «равно-
весные» экосистемы, в которых сильнее выражены доминирова-
ние и конкурентное исключение. Вместе с тем Мак-Нотон
(McNaughton, 1978) на основе изучения залежей и восточноаф-
риканских злаковнпков заключил, что на уровне первичных про-
дуцентов (растительности) видовое разнообразие служит средст-
вом функциональной стабильности сообщества. Однако, как гово-
рится, хорошенького понемножку; многочисленные виды, вступая
в конкурентную борьбу друг с другом, могут дестабилизировать
экосистему (см. Мау, 1973). В природе нигде и никогда не до-
стигается максимальное теоретическое разнообразие, т. е. не бы-
вает так, что одновременно одинаково значимы многие виды; как
правило, одни виды всегда более редки, чем другие. В тех случа-
ях, когда разнообразие высоко, в среднем, по-видимому, достига-
ется 80% максимальной выровненности (Odum, 1975). В ходе
экологической сукцессии видовое разнообразие обычно увеличи-
Популяции в сообществах
135
вается. Однако это увеличение не всегда затрагивает старые пли
зрелые стадии, о чем будет сказано в следующей главе (разд. 1
и 2).
Основная трудность при изучении видового разнообразия со-
стоит в том, что исследователи имеют дело только с частями
сообществ, обычно с таксономическими группами (например, пти-
цами или насекомыми) или, в лучшем случае, с одним трофиче-
ским уровнем. Для оценки разнообразия всего сообщества требу-
ется, чтобы роли разных ниш и организмов разных размеров
были «взвешены» каким-то образом на основе общего знаменате-
ля, такого, например, как энергия. Некоторые возможные способы
оценки см. Odum, 1982.
Представляется вероятным, что стабильность более тесно свя-
зана с функциональным, а не со структурным («урожай на кор-
ню») разнообразием (см. гл. 6, разд. 2).
Для того чтобы разнообразие внутри одного местообитания или
типа сообщества не путать с разнообразием ландшафта или ре-
гиона, который содержит смесь местообитаний, Уиттэкер (Whitta-
ker, 1960) предложил следующие термины: 1) альфа-разнообра-
зие для разнообразия внутри местообитания пли внутри сообще-
ства; 2) бета-разнообразие для разнообразия между местообита-
ниями и 3) гамма-разнообразие для разнообразия в обширных
регионах биома, континента, острова и т. д.
Рассматривая трофические уровни, отдельные хорошо изучен-
ные зоны, такие, как донное население водоемов, или другие
части сообщества, мы обнаруживаем, что большое влияние на
видовое разнообразие оказывают функциональные связи между
этими уровнями. Например, количество травоядных пли хищников
сильно влияет на разнообразие травостоя или жертв. Умеренное
«хищничество» часто снижает плотность доминантов, предостав-
ляя, таким образом, менее конкурентоспособным видам большие
возможности для использования пространства и ресурсов. Разно-
образие травянистой растительности на меловых возвышенностях
Англии снизилось после того, как были огорожены пастбища для
кроликов (Harper, 1969). Вместе с тем чрезмерный выпас ока-
зывает стрессовое воздействие, уменьшая число видов до немно-
гих несъедобных. Пейн (Paine, 1969) обнаружил, что как в тропи-
ческих, так и в умеренных областях прикрепленные организмы
литорали (где пространство в общем более ограничено, чем пища)
характеризуются большим видовым разнообразием в тех местах,
где активно действуют хищники и первого, и второго порядка.
В таких ситуациях искусственное удаление хищников снижает
видовое разнообразие всех прикрепленных организмов независимо
от того, питались ими хищники или нет. Пейн приходит к вы-
воду, что «локальное видовое разнообразие непосредственно за-
висит от эффективности, с которой хищники предотвращают мо-
136
Глава 7
нополизацию одним видом основных необходимых ресурсов сре-
ды». Этот вывод не всегда обязателен для местообитаний, где
конкуренция за пространство менее напряженная. Деятельность
человека, как правило, направлена в сторону уменьшения разно-
образия и поощрения монокультур, однако часто человек увели-
чивает разнообразие местообитаний в ландшафте в целом, созда-
вая открытые пространства в лесу, сажая деревья в прерии, ин-
тродуцируя новые виды и т. д. Разнообразие растений и мелких
певчих птиц намного больше в старых, давно заселенных районах,
чем во многих природных местообитаниях (гл. 2, разд. 7).
Виды не всегда служат лучшей экологической единицей для
оценки разнообразия, так как различные стадии жизненного цик-
ла или различные жизненные формы одного и того же вида часто
занимают разные местообитания и экологические ниши и вносят,
таким образом, свой вклад в разнообразие экосистемы. Гусеница
и бабочка или лягушка и головастик играют значительно более
разнообразные роли в сообществе, чем два вида гусениц или
взрослых лягушек. Харпер (Harper, 1977) заметил, что «различ-
ные стадии жизненного цикла растений в такой же степени обус-
ловливают разнообразие экосистемы, как и многочисленные ви-
ды». Следовательно, для оценки разнообразия не нужно быть
опытным систематиком (или нуждаться в его помощи). Напри-
мер, в коллекции насекомых, собранных сачком, нужно только
разобрать особей, у которых есть четкие морфологические и раз-
мерные отличия, при этом нет необходимости знать видовые на-
звания или выяснять, принадлежат ли разные «сорта» к одному
виду или нет. Иными словами, щ в формулах разнообразия могут
быть представлены разными морфологическими типами, которые
будут скорее всего представлять разные экологические типы (по-
скольку, как уже отмечалось, различия в размерах и морфологии
указывают на различия ниш).
Рассмотрим генотипическое разнообразие, которое остается
скрытым, если характеризовать сообщество только на уровне ви-
дов. Как поясняет Вайда (Vida, 1978), классическая теория осно-
вывалась на том, что особи в популяции должны быть гомозигот-
ными по аллелям, дающим наибольшую приспособленность. Это
видно на приведенной ниже схеме, где некая гипотетическая
особь гомозиготна по всем, за исключением одной из шести пар
аллелей:
aibic1d1e1f1
Альтернативная теория утверждает, что особи гетерозиготны по
большинству локусов, и высокий полиморфизм поддерживается
разными формами сбалансированного отбора, как показано на
Популяции в сообществах
137
следующей схеме, где наблюдается гетерозиготность по всем ал-
лелям, за исключением одного. Разнообразие аллелей здесь намно-
го больше:
Теория «сбалансированного полиморфизма» представляется более
приемлемой, что подтверждается современными биохимическими
методами, которые могут выявить скрытую генетическую измен-
чивость; вероятно также, что полиморфизм поддерживается ес-
тественным отбором. В отсутствие такой генотипической изменчи-
вости виды оказались бы неспособными адаптироваться к новым
ситуациям и, следовательно, должны были бы вымереть в изме-
няющейся среде.
Разнообразие видов, стадий жизненных циклов и генетических
типов не исчерпывает собой набор элементов, составляющих раз-
нообразие сообщества. Структуры, формирование которых обус-
ловливается распределением организмов в среде или их взаимо-
действием со средой, рассмотрены Хатчинсоном (Hutchinson,
1953). Различные способы аранжировки одновременно представ-
ленных в сообществе организмов характеризуют его структурное
разнообразие L Например:
1. Стратификационные принципы (вертикальная слоистость,
ярусность растительного покрова и структура почвенных профи-
лей).
2. Зональность (горизонтальная разобщенность, вертикальная
поясность в горах или в литоральной зоне).
3. Характер активности (периодичность).
4. Структура пищевой сети (сетевая организация пищевых це-
пей).
5. Репродуктивные системы (ассоциации родителей и потомст-
ва, клоны растений и т. д.).
6. Социальные структуры (стада и табуны).
7. Системы взаимодействия (возникают в результате конку-
ренции, антибиоза, мутуализма и т. д.).
8. Стохастические структуры (возникают в результате дейст-
вия случайных сил).
Увеличение разнообразия обусловлено также «краевыми эф-
фектами» — контактами между пятнами контрастирующих типов
растительности или физическими местообитаниями. Этот вопрос
будет обсуждаться в следующем разделе.
1 Пиелу (Pielou, 1966а) использует термин «структурное разнообразие»
в более узком смысле для обозначения степени разобщения особей одной
популяции от особей другой.
138
Глава 7
Примеры
На рпс. 7.18 приведены кривые доминирования—разнообразия,
построенные для деревьев в лесах, простирающихся от влажных
тропиков до субальпийских горных районов. Очень хорошо виден
градиент видового разнообразия от Арктики до тропиков. Различ-
ные типы кривых (геометрическая, логнормальная) также хорошо
показаны на обобщенной модели рис. 7.17.
Использование кривых и индексов разнообразия для оценки
влияния антропогенного стресса (сброс сточных вод) на речной
бентос (организмы, живущие на дне) показано на рис. 7.19, А,
где приведены кривые доминирования—разнообразия и индексы
Шеннона для донной фауны трех небольших параллельных ре-
чек, протекающих через пригородную зону Атланты. Одна река
протекает через густо заселенный пригородный район, и в нее
сбрасывается большое количество сточных вод; другая расположе-
на дальше от города, и в нее попадает умеренное количество
стоков; третья пересекает сельскую местность. И кривые, и ин-
дексы показывают, как снижается разнообразие при увеличении
количества бытовых стоков. На рис. 7.19,5 показано влияние
бытовых и промышленных сточных вод на разнообразие бентос-
ных организмов в одной из рек штата Оклахома. Бентос был
обеднен на протяжении примерно 100 км вниз по течению от
места загрязнения (Wilhm, 1967). Результаты этих и многих дру-
гих исследований свидетельствуют о том, что определение индек-
сов разнообразия бентоса — хороший способ для оценки загряз-
нений воды.
Прежде чем использовать индексы разнообразия для сравне-
ния разных ситуаций, следует определить влияние, оказываемое
на них размером проб. Один из способов определения показан на
рис. 7.20,А. Индексы вычислены здесь для накопленных проб
членистоногих, отловленных с нескольких участков (размером по
0,1 м2) на довольно однородном поле зерновых. Индекс Шеннона
выходит на постоянный уровень при 4—5 пробах, несмотря на
то что при этом в пробы попала лишь небольшая часть имеющих-
ся видов, тогда как индекс разнообразия (S—1/logN) изменялся
вплоть до суммы 9—10 проб.
На рис. 7.20,5 показано влияние острого инсектицидного
стресса на разнообразие членистоногих просяного поля. Хотя
индекс разнообразия значительно снизился в результате обработ-
ки, показатель выровненности увеличился и оставался на доволь-
но высоком уровне на протяжении большей части вегетационного
периода. В этом примере разнообразие членистоногих до обработ-
ки было низким, причем на каждом трофическом уровне домини-
ровало всего по нескольку видов. Гибель от инсектицида многих
видов-доминантов привела к большей выровненности распреде-
Расстояние по течению от места сброса, км
Рис. 7.19. А. Профили доминирования—разнообразия для трех речек в
одном и том же водосборном бассейне. Речки в разной степени загрязнены
городскими бытовыми стоками. Индексы разнообразия Шеннона составля-
ют: 3,31 для незагрязненной речки (1);2,80 для умеренно загрязненной (//);
2,45 для загрязненной (Z//) (Е. Р. Odum, Cooley, 1980). Б. Изменения индекса
разнообразия Шеннона (Я) для бентоса (организмы, живущие на дне) иллю-
стрируют эффект хронического загрязнения речки недостаточно очищен-
ными стоками. Индексы рассчитаны для разных участков вниз по течению
от места сброса бытовых и промышленных сточных вод маленького городка
(Wilhm, 1967).
140
Глава 7
Рпс. 7.20. А. Влияние увеличения размера проб на два индекса разнообра-
зия. Графики построены в полулогарифмическом масштабе; изображены
кривые числа видов членистоногих и индексов разнообразия в последова-
тельных суммирующихся пробах, которые брали с площадок размером
0,1 м2 в посеве проса (Panicum) в Джорджии. Б. Влияние однократной об-
работки инсектицидом севином (фосфорорганическое соединение, сохра-
няющее токсичность всего около 10 дней) на население членистоногих,
обитающих на участке просяного поля площадью 0,4 га. Пробы обработан-
ной и контрольной площадок брались через одну-две недели с начала июля
до сентября. Две составляющие разнообразия (d и е) и общий индекс раз-
нообразия (Н) вычислялись по десяти пробам, взятым с площадок размером
0,1 м2. Применение полулогарифмического масштаба облегчает непосред-
ственное сравнение относительных отклонений, обусловленных острым ин-
сектицидным стрессом. (Barrett, 1969.)
ленпя по численности выживших популяций членистоногих. Ин-
декс Шеннона выражает взаимодействие этих двух компонентов
разнообразия и поэтому показывает некую промежуточную реак-
цию. Хотя использованный в этих опытах инсектицид сохранял
токсичность лишь в течение десяти дней, а резко выраженная
депрессия длилась всего около двух недель, овершуты и колеба-
ния коэффициентов разнообразия отмечались еще достаточно дол-
го. Эта работа освещает несколько интересных моментов: 1) же-
лательно рассматривать порознь видовое богатство и относитель-
ное обилие; 2) не строго направленное или умеренное нарушение
при наличии сильного доминирования вызывает увеличение, а не
понижение разнообразия; 3) восстановление происходит быстро,
если стресс затрагивает небольшую область, поскольку с соседних
участков быстро поступает пополнение. Применение инсектици-
Популяции в сообществах
141
дов на больших пространствах влечет за собой совершенно иные
последствия.
На рис. 7.21 показаны два других типа графического представ-
ления видового разнообразия. На верхнем графике с помощью
метода «разрежения» сравнивается разнообразие основных видов
беспозвоночных, обитающих в донных осадках («инфауна») раз-
личных морских местообитаний. В этом случае строится кривая
зависимости между суммарным числом видов и суммарным чис-
лом особей, и по форме этой кривой оцениваются различия в
разнообразии (Sanders, 1968). Неожиданно выяснилось, что глу-
боководные морские осадки дают приют большему числу видов
животных, чем прилежащие участки континентального шельфа,
хотя плотность населения (число особей на 1 м2) в осадках на-
много ниже. Для объяснения этого факта Сандерс предложил
гипотезу «стабильности во времени»: конкурентные взаимодейст-
вия в физически стабильной среде на протяжении длительного
эволюционного времени привели к сосуществованию многих ви-
дов, характеризующихся узкими нишами. Однако Абеле и Уол-
терс (Abele, Walters, 1979) считают, что высокое разнообразие в
морских глубинах также хорошо можно объяснить огромной пло-
щадью этого местообитания (гипотеза виды—площадь).
Если построить график зависимости числа видов от числа осо-
бей, взятых с интервалами, образующими геометрическую про-
грессию (1—2, 2—4, 4—8, 8—16 и т. д.), то часто получается,
как на нижнем графике рис. 7.21, усеченная нормальная кривая.
Это такое же логнормальное распределение, о котором упомина-
лось в этом разделе раньше. Здесь опять, как и в других случаях,
под влиянием загрязнения или иных стрессовых воздействий кри-
вая становится более плоской и снижается (т. е. в данном случае
уменьшается модальная высота).
В табл. 7.6 сравниваются плотность и разнообразие популяций
членистоногих на просяном поле и сменившем его в следующем
году природном сообществе разнотравья. Приведенные значения
представляют собой средние для 10 проб, взятых на протяжении
вегетационного периода. Уже через год «хозяйничанья природы»
произошли следующие изменения:
1. Значительно уменьшилась численность растительноядных
насекомых (фитофагов), а также общая плотность членистоно-
гих.
2. Существенно увеличились компонент видового разнообразия
и индексы общего разнообразия для каждой гильдии и для всего
сообщества членистоногих.
3. Увеличился компонент выровненности.
4. Значительно возросли численность, разнообразие и соотно-
шение хищников и паразитов; хищники и паразиты на просяном
поле составляли всего 17% плотности всего населения, тогда как
Рпс. 7.21. Использование кривых видового обилия для сравнения видового
разнообразия в различных местообитаниях. А. Метод разрежения, основан-
ный на построении кривых процентного соотношения видов в отдельных
пробах донных осадков достаточной величины (от 500 до 3000 особей дву-
створчатых моллюсков и полихет). Метод используется для определения
гипотетического числа видов в последовательно уменьшающихся пробах.
В соответствии со снижением видового разнообразия исследованные место-
обитания распределяются следующим образом: I — тропическое мелководье^
II — глубоководные участки моря, III — континентальный шельф, IV —
бореальное мелководье, V — эстуарий бореальной зоны. (Sanders, 1968.)
Б. Видовая структура диатомовых водорослей в двух эстуарных сообщест-
вах в шт. Техас. Структура представлена усеченными нормальными кри-
выми, которые получаются, если откладывать на графике число видов в
классах обилия, образующих геометрическую прогрессию. Например, 1—
2 особи — первый интервал, 2—4 — второй, 4—8 — третий, 8—16 — четвер-
тый п т. д. В загрязненном канале (судоходный канал Хьюстона) число
видов во всех классах обилия резко снижено. (Patrick, 1967.)
Популяции в сообществах
143
на лугу —47%, т. е. здесь они практически превосходили по
численности фитофагов.
Это сравнение до некоторой степени позволяет понять, почему
в искусственных сообществах часто приходится применять хими-
ческие или другие средства борьбы с растительноядными насеко-
мыми, тогда как в природных сообществах в этом нет необходи-
мости, если только дать природе возможность развивать собствен-
ные средства защиты.
Таблица 7.6. Сравнение плотности и разнообразия населения членистоногих
на несжатом поле проса и сменившем его через год природном сукцессион-
ном сообществе 1}.
1 [Показатели Популяции Посев проса Природное сукцессион- ное сообще- ство
Плотность, число особей на 1 м2 Индекс разнообра- зия (d) Выровненность Общий индекс_раз- нообразия (Н)2) Растительноядные Хищники Паразиты Все членистоногие Растительноядные Хищники Паразиты Все членистоногие Растительноядные Хищники Паразиты Все членистоногие Растительноядные Хищники Паразиты Все членистоногие 482 82 24 624 7,2 3,9 6,3 15,6 0,65 0,77 0,89 0,68 2,58 2,37 2,91 3,26 1563) 117 513) 3553) 10,63) 11,43) 12,4 30,93) 0,793) 0,80 0,90 0,843) 3,283) 3,323) 3,693) 4,493)
1) Поле проса (Рапгсит) представляет собой участок в 0,4 га; данные по этому
участку послужили основой для построения графика рис. 7.20. Удобрения во время
посева вносились как обычно, но инсектицидов и другой химической обработки не
применяли, урожай собран не был. Все цифры представляют собой средние еже-
недельных проб в сезон роста, с июля по сентябрь.
2) Формулы индексов разнообразия приведены в табл. 7.5.
3) Различия между сообществами значимы на 99%-ном уровне.
Табл. 7.7 иллюстрирует изменение в разнообразии видов де-
ревьев после того, как каштановый гриб уничтожил главный до-
минирующий вид в лесах юга Аппалачей. Каштаны, составляв-
шие от 30 до 40% биомассы исходного насаждения, оказались
замененными не одним, а несколькими видами дуба, и в резуль-
тате открытия полога появилось несколько видов субдоминантов,
или пионерных, деревьев (таких, как тюльпанное дерево). Соче-
144
Глава 7
тание этих изменений привело к снижению доминирования и
увеличению общего разнообразия. В 1970 г., спустя 25 лет после
исчезновения каштанов, суммарная площадь оснований раститель-
ности вернулась к уровню, бывшему до эпифитотии, разнообразие
же так и осталось на низком уровне.
Таблица 7.7. Разнообразие лесных деревьев до распространения заболевания
каштанов и после выпадения главного доминанта в лесах Южных Аппала-
чей. (По Nelson, 1955; Day, Monk, 1974.)
Площадь оснований Индекс Симпсона
доминиро- вания разнообра- зия
1934, до болезни 25 0,25 0,75
1953, после болезни 22 0,13 0,87
1970, после болезни 26 0,17 0,83
Стратификацию и ее вклад в разнообразие удобно изучать в
лесах и озерах. Интересным примером стратификации популяций
служит вертикальное распределение трех видов рыб — объектов
спортивного лова в искусственных водоемах в долине Теннесси
в середине лета (рис. 7.22). Как и во многих глубоких озерах
умеренного пояса, летом здесь можно четко различить два слоя:
верхний теплый, богатый кислородом слой, где происходит цир-
куляция воды, и лежащий под ним более глубокий и холодный
слой, где циркуляция воды отсутствует и содержание кислорода
часто бывает очень низким. Как указано на рисунке, большеро-
тый окунь лучше других видов переносит высокую температуру
(это подтверждается также тем, что он встречается в природе
значительно южнее двух других видов) и держится ближе к
поверхности. Два других вида держатся в более глубоких слоях
воды, причем канадский судак — в самых глубоких (и потому
особенно холодных), содержащих достаточное количество кисло-
рода.
Определив вертикальное распределение кислорода и темпе-
ратуры, можно предсказать, где рыба должна быть особенно
многочисленна. В местных газетах даже публикуют диаграммы,
подобные изображенной на рис. 7.22, чтобы помочь рыболовам
решить, на какой глубине нужно ловить данный вид рыбы. Од-
нако, как хорошо известно каждому рыболову, одного лишь зна-
ния того, где находится рыба, еще недостаточно для успеха, хотя
это и может помочь.
В океане косяки рыб часто настолько четко приурочены к
разным глубинам, что создают хорошо ограниченные звукорассеи-
вающие слои или ложное дно на эхограмме судового эхолота.
Рис. 7.22. Распределение по глубине трех непромысловых видов рыб в се-
редине лета в трех водохранилищах долины Теннесси. На графиках справа
показаны вертикальное содержание кислорода и температура, определяю-
щие глубину, к которой приурочены особи того или иного вида. (Dendy,
1945.)
Ю Заказ Ks 1483
146
Глава 7
Озабоченность исчезновением видов
и утратой генетического разнообразия
На протяжении истории уменьшение видового и генетического
разнообразия приносило кратковременную выгоду человеку при
ведении лесного п сельского хозяйства. Об этом свидетельствует
распространение на больших площадях зерновых и лесных на-
саждений специализированных, высокоурожайных сортов. Слиш-
ком сильная зависимость от этих сортов может, однако, в буду-
щем привести к катастрофам. Эти катастрофы могут быть вызва-
ны изменениями климата, уменьшением химических и энергети-
ческих субсидий, необходимых для поддержания этих сортов, или
новыми болезнями и вредителями, к которым эти сорта воспри-
имчивы. О глубокой озабоченности, вызванной утратой разнообра-
зия сельскохозяйственных культур, свидетельствуют предприни-
маемые в настоящее время попытки создания питомников и
банков посевного материала, которые призваны служить храни-
лищами как можно большего числа сортов сельскохозяйственных
культур. Для привлечения внимания к глобальной угрозе потери
разнообразия некоторые общественные деятели создают програм-
мы сохранения генетических ресурсов. Приводим выдержку из
отчета одной из таких программ \
«Биологическое разнообразие животных, растений и микро-
организмов представляет собой фактор фундаментальной важно-
сти для выживания человечества. Термин «генетические ресурсы»
можно определить как генетическое разнообразие, играющее ре-
шающую роль во все времена и для удовлетворения всех нужд
общества. Это разнообразие выражается как в различиях между
видами, так и в различиях между особями, составляющими тот
или иной вид. Генетические ресурсы слагаются из генофондов
диких и одомашненных видов, включая многие виды, не имеющие
непосредственной потребительской ценности, но существенно важ-
ные для выживания экономически ценных. Каждый год эти ре-
сурсы используются для производства новых и уже знакомых нам
продуктов стоимостью в миллиарды долларов: пищи, одежды,
жилищ, лекарств, энергии п сотен промышленных товаров. Ши-
рокий набор видов и их продуктов требуется для медицинских
и других исследований. Сельское и лесное хозяйство и связанные
с ними отрасли промышленности становятся зависимыми, как
только в этом возникает нужда, от необходимого разнообразия
(например, от растений, устойчивых к болезням). Именно это
разнообразие определяет пределы, в которых дикие и домашние
виды могут адаптироваться к различным изменчивым факторам,
1 Программа сохранения генетических ресурсов, 1583А Арч Стрит,
Беркли, Калифорния 94708, Дэвид Кофтой, руководитель.
Популяции в сообществах
147
в том числе к 1) погодным условиям, насекомым и болезнямv
2) технологическим приемам, 3) потребностям и 4) предпочте-
ниям людей. Большая часть биологического разнообразия сосре-
доточена пока в природных экосистемах, существование которых,
зависит в значительной степени от их внутреннего разнообразия.
В США и во всем мире растет серьезное беспокойство по по-
воду быстрого истощения генетических ресурсов, о чем свидетель-
ствуют многочисленные быстро накапливающиеся данные. Основ-
ной резервуар генетических ресурсов — природные экосистемы —
оказался значительно измененным или разрушенным. Большое
разнообразие сортов растений и пород животных, используемых я
«примитивном» сельском хозяйстве менее развитых стран, без
всякой необходимости теряется, замещаясь сравнительно малым
числом высокоурожайных сортов и пород. К тому же существую-
щие коллекции и хранилища недостаточны и отсутствует необ-
ходимая поддержка для их сохранения. Несмотря на то что утра-
та генетических ресурсов в основном необратима, до сих пор
не ведется соответствующей национальной политики, не разра-
батываются системы или программы, которые гарантировалп бы
сохранение важных для всех нас генетических ресурсов в безопас-
ности. Долговременная цель программы сохранения генетических
ресурсов — помочь в получении всемирной поддержки, необходи-
мой для гарантии безопасности разнообразия, требующегося для
нужд общества во все времена, и установить национальную по-
литику и программу для достижения этой цели».
То, что американский народ действительно озабочен сохране-
нием генетических ресурсов, хотя, быть может, еще и не прояв-
ляет серьезной озабоченности, видно по тому, какую реакцию
вызывают в нем попытки спасения угрожаемых видов от выми-
рания. О силе общественного мнения свидетельствует законода-
тельный акт конгресса об охране редких видов, особенно если они
интересны п эффектны, как американский журавль; однако этот
акт не затрагивает обычных видов или их местообитаний, если
меры по сохранению этих видов приходят в конфликт с планами
развития, сулящими немедленную экономическую выгоду (на па-
мять приходит случай с одним из окуней и строительством боль-
шой плотины в долине Теннесси). Поскольку такой подход к
охране, при котором перебирается вид за видом, неизбежно по-
терпит крах при любом подобном конфликте, сейчас самое время
перейти к охране разнообразия экосистемы в целом. В пользу
этого можно привести более убедительные доводы, кроме того,
это позволит сохранить большее разнообразие видов и генофондов.
Поль и Энн Эрлихи (Paul Ehrlich, Ann Ehrlich, 1981) написали
по этому поводу доступную, но основанную на строгих докумен-
тах книгу, которую мы рекомендуем для чтения студентам и всем
тем, кто интересуется проблемами охраны природы.
10*
<48
Глава 7
Хорошее пространственное и региональное планирование мо-
жет до некоторой степени компенсировать снижение локального
(или альфа-) разнообразия, которое обычно сопровождает интен-
сивное развитие сельского хозяйства, лесной промышленности и
городов. Если сельскохозяйственные и лесные монокультуры,
а также дома вдоль дорог (многочисленные ряды одинаковых до-
мов на небольших покрытых травой лужайках) чередуются с бо-
лее разнообразными естественными и полуестественными экосис-
темами (парки, природные центры и т. д.) и если остаются неза-
строенными речные долины, крутые склоны, озера, болота и
овраги, то мы сохраняем не только прекрасные уголки природы,
предоставляющие нам большие возможности для отдыха, но и
гарантируем высокий уровень бета-разнообразия. Примером мо-
жет служить Национальный парк исследований в области окру-
жающей среды при министерстве энергетики, расположенный
примерно па 80 000 га при электростанции в Саванна-Ривер. При-
мерно треть этой площади, раньше занятой фермами, теперь пре-
вращена в плантацию сосен, так что альфа-разнообразие здесь
приближается к нулю (в целом на плантации только один вид).
Однако поскольку речные долины и другие прибрежные зоны
вдоль рек, возвышенности, смешанные лиственные леса, полосы
живых изгородей и прочая первоначальная растительность оста-
лись на двух третях территории более или менее нетронутыми,
разнообразие местообитаний, или бета-разнообразие, на террито-
рии в целом достаточно велико и сравнимо с разнообразием при-
родных типов почв (см. Odum, Kroodsma, 1975). Служба леса
США, ведущая хозяйство на этой территории, надеется расширить
лесные плантации за счет других типов растительности, что по-
зволит увеличить выход древесной массы для бумажной промыш-
ленности и древесины. Такое изменение было бы, однако, доро-
гостоящим экономически (сомнительное соотношение затраты-
выгоды) и резко понизило бы бета-разнообразие, подвергнув рис-
ку постоянство и стабильность в будущем.
На рис. 7.23 показано, как можно планировать ландшафты с
сохранением разнообразия и в согласии с городским и промыш-
ленным развитием. Новый город Колумбия в шт. Мэриленд яв-
ляется хорошим примером успешного планирования, осуществ-
ленного в пределах частного сектора со свободным рынком с
благословения федерального правительства и правительства шта-
та, но с минимальной финансовой помощью, оказываемой с их
стороны. Этот пример будет более подробно рассмотрен в гл. 8,
разд. 7.
Многие экологические концепции разнообразия противоречи-
вы и требуют дальнейших исследований, но все согласны с тем,
что разнообразие необходимо для выживания всего человечества
и сохранения природы.
Рис. 7.23. Планирование ландшафта, сохраняющее его разнообразие (на
бета- и гамма-уровнях) п качество среды. А. План городского поселения
(11000л жителей, 18 000 га), в котором сохраняются качество среды и есте-
ственная красота окружающей природы и предусматривается обширное
пространство для отдыха и уничтожения загрязнений. Обширные свобод-
ные пространства (на схеме показаны белым) сохраняются 1) благодаря
концентрации жилых массивов (показаны две степени плотности застрой-
ки) вокруг городских и сельских центров (черные кружки); жилые зоны
разделены широкими зелеными поясами или парками и 2) благодаря тому,
что речные долины, озера и местности с красивыми пейзажами остаются
незанятыми. Промышленные предприятия расположены в обширной зоне
для переработки отходов (правый край схемы). Здесь есть место для
предприятий по переработке отходов, прудов-окислителей, земляных насы-
пей и других устройств для разрушения, восстановления, хранения и по-
вторного включения в цикл отходов и воды. (Wallace, McHarg, Roberts,
Todd, «Plan for Volleys», c. 59). Б. Схема зоны переработки отходов при
электростанции (а) или, с необходимыми изменениями, при химическом
или другом крупном промышленном предприятии. Токсичные отходы в
твердом состоянии захороняются в специальных участках (г), легко раз-
рушающиеся отходы и теплую воду (в) можно утилизировать в ряде пру-
дов-отстойников, а также в природных или сельскохозяйственных зонах.
Состав речных и грунтовых вод и газов из дымовых труб непрерывно ре-
гистрируется у плотин (д), в специальных скважинах (е) и при помощи
специальных установок в трубах. Эта система имеет следующие основные
входы и выходы (см. номера у стрелок на схеме): 1) поступление солнеч-
ного света и атмосферных осадков; 2) вывоз радиоактивных отходов в
места захоронения; 3) электрическая энергия для городов; 4) ввод ядерно-
150
Глава 7
го и прочего горючего; 5) выход — пищевые продукты, волокно, чистый
воздух; 6) поступление чистой воды для сельского хозяйства, промышлен-
ности и городов; 7) использование территории для отдыха, а также для
обучения и научных исследований в области окружающей среды.
В гл. 2, использовав убедительные аргументы, мы показали,
что природные ландшафты необходимо сохранить, поскольку они
играют особую роль в жизнеобеспечении. Теперь можно добавить
еще одну причину, вынуждающую нас делать это, — сохранение
разнообразия, обеспечивающее адаптацию и выживание в буду-
щем.
7. Популяции и сообщества в географических
градиентах; экотоны и понятие краевого эффекта
Определения
При описании расположения популяций и сообществ в пределах
данной географической области или в данном участке ландшаф-
та пользуются двумя различными подходами: 1) зональным под-
ходом, при котором идентифицируют и классифицируют отдель-
ные сообщества, располагая их в виде списка типов сообществ,
и 2) градиентным подходом, при котором популяции располагают
вдоль одно- или многомерного градиента внешней среды или со-
Популяции в сообществах
151
ответствующих осей. В этом случае идентификация сообществ
основана на распределениях частот, коэффициентах сходства или
других статистических методах сравнения. Для обозначения упо-
рядоченности в расположении видовых популяций и сообществ
по градиенту часто применяется термин ординация. Термином
континуум обозначают весь градиент расположенных по порядку
популяций или сообществ. В общем, чем резче изменяется гради-
ент внешней среды, тем более отчетливы и дискретны сообщест-
ва. Это объясняется не только большей вероятностью резких
изменений физических условий среды, но и тем, что границы
сообществ становятся более четкими в результате процессов кон-
куренции и коэволюции взаимодействующих и взаимозависимых
видо®.
Экотон представляет собой резкий переход между двумя или
более различными сообществами, например между лесом и степью
или между твердым и мягким грунтами на дне моря. Эта зона
контакта, или зона «напряжения», может иметь значительную
линейную протяженность, но всегда бывает уже территории са-
мих соседних сообществ. Сообщество экотона содержит обычно
многие организмы из контактирующих сообществ и, кроме того,
организмы, характерные только для экотона и нередко ограничен-
ные им в своем распространении. Часто число видов и плотность
популяций некоторых из них в экотоне выше, чем в лежащих по
обе стороны от него сообществах. Тенденция к увеличению раз-
нообразия и плотности организмов на границе сообществ известна
под названием краевого эффекта.
Объяснения и примеры
Определение границ экосистем — не слишком сложная проблема
при их анализе (гл. 2, разд. 2). До тех пор пока среда на входе и
среда на выходе будут рассматриваться как части системы,
не имеет значения, как ее ограничить: при помощи естественных
ориентиров или произвольно — так, как это удобно. Сложность
возникает, однако, при попытке ограничить биотическое сообще-
ство, используя для этого слагающие его видовые популяции.
На протяжении последних 50 лет экологи растений вели ожив-
ленную дискуссию по поводу того, следует ли рассматривать на-
земные растительные сообщества как дискретные единицы с
определенными границами (Clements, 1905, 1916; Braun-Blanquet,
1932, 1951; Daubenmire, 1966) или же как континуум, поскольку
отдельные популяции реагируют на градиенты окружающей среды
настолько независимо, что сообщества постепенно переходят друг
в друга, и всякое выделение дискретных единиц произвольно
(Gleason, 1926; Curtis, McIntosh, 1951; Whittaker, 1951; Goodall,
1963 и др.). Уиттэкер (Whittaker, 1967) иллюстрирует эти раз-
152
Глава 7
личные точки зрения следующим примером. Если бы в разгар
осеннего разноцветья в Национальном парке Грейт-Смоки-Маун-
тинс нам предложили выбрать удобную точку, с которой можно
видеть весь градиент высот — от долины до гребней хребтов,
то мы бы заметили пять по-разному окрашенных зон: 1) много-
цветный лес на площадках среди скал; 2) темно-зеленый лес из
тсуги; 3) темно-красный дубовый лес; 4) красновато-коричневые
заросли из дуба и вереска и 5) светло-зеленый сосновый бор на
гребнях. Эти пять зон можно рассматривать как дискретные типы
сообществ, а можно считать их частями одного континуума, ко-
торые подлежат какой-либо форме градиентного анализа; при этом
выявятся распределение популяций отдельных видов и их реак-
ция на изменение условий внешней среды в соответствии с гра-
диентом. Эту ситуацию иллюстрирует рис. 7.24, на котором по-
Рис. 7.24. Распределение популяций доминирующих видов деревьев вдоль
гипотетического градиента (от 0 до 10), иллюстрирующее размещение
популяций в сообществе типа континуум. Кривые распределения каждого
вида имеют колоколообразную форму, причем пики относительного обилия
(в процентах от общего числа деревьев) приходятся на разные точки вдоль
градиента. Некоторые виды устойчивы к более широкому диапазону внеш-
них условий (степень доминирования обычно меньше) по сравнению с
другими. Внутри этого большого сообщества можно, используя сочетания
двух или более доминирующих видов (виды-индикаторы) и другие при-
знаки, выделить подсообщества (Л—Г в верхней части графика). Такое
подразделение до некоторой степени условно, но полезно для целей опи-
сания и сравнения. Кривые построены по данным ряда исследований рас-
пределения видов деревьев по градиенту высоты. (Whittaker, 1954.)
казано частотное распределение (в виде гипотетических колоко-
лообразных кривых) 15 видов доминирующих деревьев (от а
до п), перекрывающихся вдоль градиента, а также 5 до некоторой
степени условных типов сообществ (Л, Б, В\, В2 и Г), выделе-
ние которых основано на пиках одного или нескольких доминан-
тов. Можно привести много доводов в пользу того, чтобы рассмат-
Популяции в сообществах
153
ривать склон в целом как одно большое сообщество, поскольку
все эти леса связаны друг с другом обменом биогенными элемен-
тами, энергией и животными в одну экосистему водосборного
бассейна. Как подчеркивалось в гл. 2, такая экосистема представ-
ляет собой минимальную единицу экосистемы, поддающуюся
функциональному исследованию и регулированию со стороны че-
ловека. Однако лесникам или землеустроителям удобнее рассмат-
ривать каждую зону как отдельное сообщество, поскольку каждая
из них отличается по скорости прироста древесины, качеству
древесины, рекреационной ценности, восприимчивости к пожарам
и болезням и по многим другим показателям.
Как это часто бывает, концепции и подходы зависят от гео-
графии; так, экологи, работающие в областях с постепенно из-
меняющимися условиями среды и однообразными почвами или
субстратами, предпочитают концепцию континуума и разные ме-
тоды ординации (т. е. статистические методы распределения по-
пуляций и сообществ по градиентам). Вместе с тем экологи, ра-
ботающие в областях с резкими изменениями условий среды и
неоднородным рельефом, отдают предпочтение зональной концеп-
ции. Методы ординации часто требуют от экологов сравнения
сходства (или несходства) последовательных проб, взятых вдоль
градиента внешней среды. При этом используется в общей форме
такой индекс:
2С
Индекс сходства (5) = д
где А — число видов в пробе А; В — число видов в пробе В и
С — число общих видов в обеих пробах. В работе Уиттэкера
(Whittaker, 1967) и Мак-Интоша (McIntosh, 1967) рассматрива-
ются методы ординации и другие приемы градиентного анализа.
Билз (Beals, 1969) непосредственно сравнил изменение расти-
тельности на крутых и пологих склонах гор в Эфиопии; резуль-
таты этой работы приведены на рис. 7.25. На крутом склоне на-
блюдается большая разрывность, о чем свидетельствуют несколь-
ко острых пиков на кривой индекса несходства (1—5, как было
показано выше), рассчитанной для каждой пары соседних проб
(рис. 7.25, вверху}. Кроме того, на крутом склоне в большей
степени, чем на пологих, выражена тенденция к внезапному по-
явлению и исчезновению видов. Как показывает рис. 7.25,
внизу, распределения доминирующих видов по частоте при по-
степенном изменении высоты имеют колоколообразную форму
нормального распределения (напоминая гипотетические кривые
рис. 7.24). При резком же изменении высоты эти кривые харак-
теризуются резким подъемом и спадом и усеченной вершиной.
Билз пришел к выводу, что «вдоль крутых склонов сама расти-
тельность может образовывать разрывы, несмотря на непрерыв-
154
Глава 7
Крутой склон
Рис. 7.25. Градиентный анализ изменения растительности на крутом
(сплошные липни) и пологом (штриховые линии) склонах одинаковой
высоты (Эфиопия). Вверху. График показателей различия между сосед-
ними участками вдоль склона. Внизу. Частотное распределение двух видов,
Acacia Senegal (I) и Carissa edulis (II). На крутом склоне виды и группы
сообществ разграничены более четко. (Beals, 1969.)
ность градиента среды, тогда как на пологих склонах этого может
и не быть». Обособлению одного сообщества от другого могут
способствовать различные типы взаимодействия между популя-
циями, например: 1) конкурентное исключение; 2) симбиоз меж-
ду группами зависящих друг от друга видов и 3) коэволюция
групп видов (это будет подробно рассмотрено в следующей гла-
ве). Такие факторы, как пожары и антибиотики, также могут
создавать четкие границы. Бьюэлл (Buell, 1956) описал ситуацию
в парке Итаска (шт. Миннесота), где в пределах основного мас-
сива кленово-липового леса имеются довольно хорошо отграни-
ченные островки елово-пихтового леса, не связанные с изменения-
ми рельефа местности. В морских донных сообществах на крутых
обрывах также возникает четко выраженная зональность, подоб-
ная зональности на горных склонах.
Популяции в сообществах 155
В тех местах, где происходят резкие изменения по градиенту
или где соприкасаются границы двух хорошо отличающихся мес-
тообитаний или сообществ, образуется экотон или переходная
зона. Часто в этой переходной зоне развивается сообщество, от-
личающееся от прилежащих сообществ; объясняется это тем, что
многим видам нужны в качестве частей местообитаний или мест,
где проходят разные стадии жизненного цикла, такие условия,
какие имеются в двух или более соприкасающихся и часто силь-
но различающихся по структуре сообществах. Например, амери-
канскому дрозду нужны деревья, на которых он гнездится, и от-
крытые травянистые пространства, где он питается. В хорошо
развитые сообщества экотонов входят виды, характерные для
каждого из перекрывающихся сообществ, и, кроме того, виды, ха-
рактерные только для экотона, поэтому число видов и плотность
популяций в экотоне выше. Тенденция к увеличению разнообра-
зия и плотности живых организмов на границах сообществ из-
вестна под названием краевого эффекта.
В одной из первых работ, где был показан краевой эффект,
Бичер (Beecher, 1942) обнаружил, что плотность населения птиц
увеличивается с увеличением отношения длины опушки к пло-
щади, занимаемой сообществом. Хорошо известно, что плотность
певчих птиц выше на территории разного рода усадеб, в окрест-
ностях колледжей и тому подобных местах, которые состоят из
«смешанных местообитаний и, следовательно, характеризуются
большей протяженностью «границ» по сравнению с большими од-
нородными участками леса или полей.
Экотоны могут иметь характерные для них виды, не встречаю-
щиеся в соседних сообществах, участвующих в образовании этих
экотонов. Например, при изучении населения птиц вдоль гради-
ента развития сообщества (см. табл. 8.2) пробные площадки были
выбраны с расчетом, чтобы свести к минимуму влияние контак-
тов с другими сообществами. В некоторых из этих стадий разви-
тия сообщества плотность 30 видов птиц составляла не менее
пяти пар на 40 га. Однако известно еще около 20 видов птиц,
которые обычно гнездятся в сообществах внутренних областей
этого региона в целом. Семь из них были встречены в неболь-
шом числе, а остальные тринадцать видов вообще не были заре-
гистрированы на обследованной однородной территории, в том
числе такие обычные виды, как дрозд, синяя птица, многоголосый
пересмешник, овсянка индиговая, воробьиная овсянка и садовая
иволга. Многим из них деревья необходимы как места для устрой-
ства гнезд или в качестве наблюдательных постов, питаются же
они преимущественно на земле, в траве или на других открытых
участках. Поэтому их требования к местообитаниям лучше всего
удовлетворяются в экотонах между лесными и луговыми или
кустарниковыми сообществами, а не в местах, занятых лишь од-
156
Глава 7
ним из этих сообществ. Таким образом, в рассмотренном случае
около 40% обычных видов птиц (20 из 50), размножающихся в
этом районе, можно считать в основном илп полностью экотон-
ными.
Дэвид Пэттон (David Patton, 1975) предложил один из вари-
антов индекса разнообразия для сравнения отношения протяжен-
ности границ к площади в разных ситуациях. Поскольку наимень-
шее отношение к площади характерно для окружности, именно
это отношение принимается за единицу. Исходя из этого, краевой
индекс (EI) можно вычислить следующим образом:
где ТР — общий периметр площади плюс длина всех линейных
границ внутри этой площади, А—площадь и л=3,14. Квадрат-
ная площадка с одним типом растительности характеризуется
индексом 1,13. Если на этой площадке четыре разных типа рас-
тительности, занимающие одинаковые площади, то дополнитель-
ные внутренние границы увеличат значение индекса до 1,69. Если
два из четырех типов растительности разделить еще дополнитель-
ной границей, то индекс возрастет до 1,97. Два других варианта
краевого индекса рассматриваются в работе Тейлора (Taylor,
1977).
Одним из обычных типов экотонов, наиболее важных для че-
ловека, является опушка леса. Опушку можно определить как
экотон между лесным и травянистым или кустарниковым сооб-
ществами. Где бы ни поселился человек, он стремится сохранить
поблизости от своих жилищ сообщества опушек. Так, если он
селится в лесу, он сокращает лес до отдельных небольших участ-
ков, перемежающихся лугами, сельскохозяйственными угодьями
и другими, более открытыми местообитаниями. В работе Хокинса
(Hawkins, 1940) приводятся карты, на которых зарегистрированы
изменения, происшедшие в шт. Висконсин за столетие после по-
явления здесь в 1838 г. первых поселенцев. Если человек селится
на открытом месте, то он сажает деревья, создавая подобную же
мозаичную структуру ландшафта. Можно сказать, что предпочи-
таемое местообитание Homo sapiens — это опушка леса, поскольку
он не прочь понежиться в тени деревьев и кустарников, но «добы-
вает» себе пищу на лугах и полях. Некоторые виды, обычные
для лесов и степей, способны жить на созданных человеком опуш-
ках. Другие же, хорошо адаптировавшиеся к жизни на опушках,
особенно многие виды трав, насекомых, птиц и млекопитающих,
в таких условиях часто увеличиваются в числе и расширяют свои
границы обитания по мере создания человеком новых местооби-
таний типа опушек.
Популяции в сообществах 15?
Вообще говоря, охотничье-промысловые виды животных, та-
кие, как олени, кролики, куропатки, фазаны и т. д., предпочитают
держаться на опушках. Поэтому человек искусственно создает
опушки, сажая съедобные для этих видов растения или растения,
дающие им приют и укрытие. Пятнистые местообитания создают-
ся также путем расчисток, выжигания (см. рис. 5.25) и других
мероприятий.
Олдо Леополд, которому приписывают честь введения понятия
краевого эффекта, писал в своей новаторской работе по охот-
ничьему хозяйству (Leopold, 1933), что «дикая природа — это
феномен опушек». Не так давно Хэнссон (Hansson, 1979) рас-
смотрел значение гетерогенности ландшафта в жизни северных
теплокровных животных, которые активны круглый год. Сельско-
хозяйственные и другие нарушенные пространства предоставляют
этим животным зимой больше пищи, чем зрелые ненарушенные
леса. Весной и летом, однако, эти последние предоставляют боль-
ше пищи.
Не следует думать, что увеличение плотности в экотонах —
явление универсальное. Многие организмы ведут себя противо-
положным образом. Так, не вызывает сомнений, что плотность
деревьев в экотоне лесной опушки ниже, чем в лесу. Вырубка
на обширных пространствах дождевого тропического леса почти
наверняка уменьшает видовое разнообразие и вызывает вымира-
ние многих видов, адаптированных к большим площадям одно-
родного местообитания. Возникает предположение, что наиболь-
шее значение экотоны приобретают там, где люди на протяжении
многих столетий сильно модифицировали природные сообщества
и осваивали ландшафты, предоставив развитию адаптаций эволю-
ционное время. Например, в Европе, где от лесов остались лишь
узкие полоски, дрозды и другие лесные птицы охотнее живут в
городах и пригородах, чем родственные им виды в Северной Аме-
рике. Но, конечно же, многие другие европейские виды не адап-
тировались и стали редкими или вымерли.
Как и в большинстве случаев положительных реакций или
выгоды, горбатая кривая субсидия—стресс (см. рис. 3.5) приме-
нима и к соотношению граница—разнообразие. Чрезмерное уве-
личение границ (много мелких блоков местообитания) вызывает
перегиб кривой разнообразия в сторону снижения. Хотя увеличе-
ние границ часто повышает разнообразие, уменьшение площади
местообитания снижает разнообразие (тенденция разнообразие-
площадь). Теоретический максимум видового бета-разнообразия
должен находиться там, где велики или достаточно велики блоки
местообитания и велика также общая протяженность границ в
регионе (Thomas et al., 1979). Эти противоположные тенденции
нужно учитывать при ведении лесного и охотничьего хозяйства
и общем планировании ландшафтов. Островная теория биогеогра-
158
Глава 7
фии (будет обсуждаться в следующей главе) может до некоторой
степени помочь в определении величины блоков, необходимой для
сохранения как отдельных видов, так и разнообразия в целом.
8. Палеоэкология: структура сообществ
в прошлые века
Определения
Ископаемые остатки и другие материалы свидетельствуют о том,
что в прошлом организмы были иными и достигли современного
состояния в процессе эволюции. Отсюда естественно предполо-
жить, что структура сообществ и характер окружающей среды
также были иными. Знания о сообществах и климате прошлого
могут внести значительный вклад в наше понимание современных
сообществ. Это предмет исследования палеоэкологии — науки, по-
граничной между экологией и палеонтологией, которую С. Кейн
(S. Cain, 1944) определил как «изучение биот прошлого с исполь-
зованием в той мере, в какой это возможно, экологических кон-
цепций и методов» пли более широкого — как изучение «взаимо-
действия земли, атмосферы п биосферы в прошлом». Палеоэколо-
гия исходит из следующих основных допущений: 1) в различные
геологические периоды действовали по существу одинаковые эко-
логические принципы; 2) об экологии ископаемых организмов
можно судить на основе того, что известно об эквивалентных
или родственных современных видах.
Объяснения
С тех пор как Чарлз Дарвин сформулировал теорию эволюции,
многие ученые увлеклись реконструкцией жизни прошлого на
основе изучения обнаруженных ископаемых остатков. Теперь ста-
ло возможным представить полную картину эволюции многих
видов, родов и более высоких таксономических групп. Например,
во всех элементарных учебниках биологии описана история эво-
люции лошади от четырехпалого животного размером с лисицу
до современного состояния. Но как была связана с окружающей
средой лошадь на разных стадиях ее эволюционного развития?
Чем она питалась, в каких условиях обитала, какова была плот-
ность ее популяций? Какие хищники ей угрожали и с кем она
конкурировала? Какой в то время был климат? Как эти эколо-
гические факторы содействовали естественному отбору, играюще-
му формирующую роль в структурной эволюции? Конечно, неко-
торые из этих вопросов так и останутся без ответа. Однако, свя-
зав количественные данные по ископаемым остаткам с
Популяции в сообществах
159’
определенным местом и временем, ученые могут получить неко-
торое представление о природе сообществ прошлого и формах,
которые в них доминировали. А сопоставление этих данных с
геологическими данными может помочь в реконструкции клима-
тических и других физических условий, существовавших в то
время. Благодаря развитию методов радиоактивной датировки и
использованию других современных геологических методов мы
получили возможность установить время, когда жила та или иная
группа ископаемых организмов.
До недавнего времени вопросам, перечисленным в предыдущем
абзаце, уделялось мало внимания. Палеонтологи были заняты опи-
санием своих находок, определением их таксономического поло-
жения и интерпретацией этих данных в свете эволюционной тео-
рии. Однако по мере накопления такой информации, которая
становилась все более обширной, возрастал интерес к эволюции
групп. В связи с этим и возникла новая ветвь науки — палеоэко-
логия.
Короче говоря, палеоэкологи по ископаемым остаткам пыта-
ются определить, как организмы прошлого взаимодействовали
друг с другом, как они реагировали на существовавшие тогда
физические условия среды и как происходило изменение сооб-
ществ во времени. В основе палеонтологии лежит допущение, что
«законы природы» в прошлом были такими же, как и сейчас,
и что организмы прошлого и настоящего, имеющие сходный тип
организации, должны быть сходными также по характеру пове-
дения и по экологическим признакам. Так, если ископаемые на-
ходки свидетельствуют о том, что 10 000 лет назад на месте со-
временного леса из дуба и гикори рос еловый лес, то у нас есть
все основания считать, что 10 000 лет назад климат был холоднее,
поскольку современные виды ели адаптированы к более холодно-
му климату, чем виды дуба и гикори.
Примеры
Ископаемая пыльца растений дает превосходный материал для
реконструкции наземных сообществ, существовавших со времен
плейстоцена. Диаграмма на рис. 7.26 показывает, как путем опре-
деления доминирующих видов деревьев можно реконструировать
характер послеледниковых сообществ и климат тех времен. От-
ступая, ледник часто оставлял после себя котлованы, которые
затем превратились в озера. Пыльца растущих вокруг озера рас-
тений опускалась на дно и фоссилизировалась в донном иле. Озе-
ро постепенно заполнялось осадками и превращалось в болото.
Проба в виде колонки, полученная на болоте пли со дна озера,
дает нам хронологическую запись, по которой можно определить
процентное соотношение разных типов пыльцы. Так, на рис. 7.26
160
Глава 7
А. СОДЕРЖАНИЕ ПЫЛЬЦЫ В ПРОБЕ, %
О 60%
Li ,I .l U.-J
10 000
5 000
5 000
ТИП РАСТИТЕЛЬНОСТИ
15 000
10 000
Б.СКОРОСТЬ НАКОПЛЕНИЯ ПЫЛЬЦЫ
Сельскохозяйственные И
лесные участки
Лиственный лес
Сосновый и лиственный леса
Разреженные еловые леса
паркового типа
Тундра
0 , W х 103 зерен пыльцы/ смх В ГОД
Рис. 7.26. Профили ископаемой пыльцы из датированных горизонтов в про-
бах донных осадков из озера на юге Новой Англии. А. Число пыльцевых
зерен каждой группы видов, выраженное в процентах от общего числа
зерен в пробе. Б. Оценки скорости отложения пыльцы для каждой группы
растений. Диаграмма, основанная на скорости отложения, дает лучшее
представление о количественном облике постплейстоценовой растительно-
сти, чем диаграмма, основанная на процентных соотношениях. (Davis, 1969.)
в пробах «старейшей» пыльцы представлена главным образом
пыльца ели, пихты, лиственницы, березы и сосны, что свидетель-
ствует о холодном климате. Смена пыльцы этих пород пыльцой
дуба, тсуги и бука указывает на то, что несколько тысяч лет
спустя климат потеплел и стал более влажным; появление позд-
нее пыльцы дуба и гикори свидетельствует о теплом сухом пе-
риоде, сменившемся еще позднее несколько более холодным и
влажным периодом (современная часть профиля). Наконец,
в пыльцевом «календаре» ясно отражены результаты недавней
вырубки лесов человеком — в пробах повысилось содержание
Популяции в сообществах
161
пыльцы травянистых растений. По данным Девиса (Davis, 1969),
пыльцевые диаграммы Европы зарегистрировали даже последст-
вия эпидемии чумы («черной смерти»), приведшей к упадку
сельского хозяйства. Это выразилось в уменьшении доли пыльцы
травянистых растений в отложениях, датированных тем же вре-
менем, к которому относится массовая гибель людей.
Рис. 7.26 иллюстрирует также, каким образом совершенство-
вание количественных методов может повлиять на интерпретацию
ископаемых данных. Если изобразить графически относительное
содержание пыльцы разных видов в общем содержании пыльцы
в пробе (обычный метод, применявшийся, во всяком случае, до
недавнего времени), то создается впечатление, что 10—12 тыс. лет
назад Новая Англия была покрыта густыми еловыми лесами. Од-
нако после того, как радиоуглеродный метод дал возможность
определить скорость отложения пыльцы в датированных слоях
п были построены графики этих скоростей (рис. 7.26,Б), стало
очевидным, что 10 000 лет назад деревьев в Новой Англии вооб-
ще было мало п ее растительность представляла собой открытый
парковый ельник, вероятно схожий с тем, который тянется в на-
стоящее время вдоль южной границы тундры. Это служит хоро-
шим примером того, от чего постоянно предостерегают статисти-
ки: не увлекайтесь процентами — они могут ввести в заблужде-
ние!
В океане наплучшие сведения часто дают раковины и скелеты
животных. Особенно удобны для оценки видового разнообразия в
прошлом ракушечники. Валентайн (Valentine, 1968) подчерки-
вает, как важно отличать разнообразие «в сообществе» от «гео-
графического» разнообразия или разнообразия «в градиенте». Так,
в прошлые геологические периоды, когда Северный полюс не был
покрыт льдами, придонная фауна северных морей была гораздо
богаче видами, чем теперь. Однако если рассматривать весь гра-
диент от полюса до экватора в целом, то сейчас, когда полюса
покрыты льдом, на дне моря обитает вдвое больше видов моллюс-
ков. Причина этого, видимо, в том, что на более крутом градиенте
увеличивается разнообразие местообитаний и ниш.
Изучение колонок донных отложений в озере — путь к про-
чтению недавней истории антропогенных нарушений на площади
водосбора. Брагем (Brugem, 1978), изучая фоссилизированных
диатомей, личинок комаров-звонцов, зоопланктон и химический
состав датированных слоев колонки из Линдсли-Понд, небольшого
озера в шт. Коннектикут, выявил три стадии антропогенных на-
рушений: 1) раннее фермерское хозяйство в конце XVIII — нача-
ле XIX вв. мало отразилось на озере; 2) в результате интенси-
фикации сельского хозяйства примерно после 1915 г. возрос сток
в озеро химикалий, что послужило причиной увеличения доли
эвтрофных видов диатомей и звонцов; 3) приближение к озеру
И Заказ № 1483
162
Глава 7
пригородной зоны с 1960-х годов до настоящего времени привело
к еще большему обогащению его биогенными элементами (гипер-
эвтрофикации), а эрозия почвы обусловила приток в осадки
больших количеств минералов и металлов (Fe, Си). Эти измене-
ния вызвали значительные изменения в составе биоты, особенно
в составе зоопланктона.
9. От популяций к сообществам и экосистемам
Существует два подхода к изучению и пониманию экосистем,
а когда это необходимо, и к управлению ими (гл. 2, разд. 3) —
холистический и мерологический. При первом подходе интересу-
ющую нас область или систему ограничивают каким-либо удоб-
ным способом, получая тем самым систему типа «черного ящи-
ка». После этого исследуют энергетические и прочие входы п
выходы (см. рис. 2.2) и оценивают основные функциональные
процессы внутри системы (см. рис. 2.1). Следуя принципу береж-
ливости (экономии усилий), изучают затем операционально зна-
чимые популяции и факторы, которые определены посредством
наблюдений, моделирования и возмущений самой экосистемы.
При таком подходе детали популяционных компонентов внутри
«ящика» анализируют только постольку, поскольку это необхо-
димо для понимания или управления экосистемой как целым.
Нам предстоит еще выяснить столько захватывающе интерес-
ных вопросов, касающихся отдельных популяций и взаимодей-
ствий между ними, что само по себе предложение перейти от
изучения этих уровней организации к изучению организации на
экосистемном уровне может восприниматься как вызов. Совер-
шенно ясно, что подробно изучать каждую популяцию непрак-
тично. К тому же, как об этом неоднократно говорилось в на-
стоящей главе, популяции ведут себя очень по-разному, если они
функционируют в сообществе и если они изолированы в лабора-
тории пли огорожены в поле. Однако коль скоро компоненты
изучены, то как вновь объединить их в сообщества и экосистемы
и как рассматривать новые холистические свойства, которые мо-
гут проявиться, если компоненты функционируют совместно в
интактной экосистеме? Коуди (Cody, 1974) предлагает два под-
хода: 1) исследование экспериментально вызванных изменений
окружающей среды для определения операциональной значимо-
сти популяционных структур, наблюдавшихся в ненарушенной
системе, и 2) изучение изменений, происходящих в естественных
условиях, для выявления значимых структур. Фойн и Джейн
(Foin, Jain, 1977) предложили нечто вроде «дорожной карты» для
объединения популяционной биологии с наукой, занимающейся
изучением экосистемы: 1) провести поверхностный описательный
анализ тех свойств на уровне сообщества, которые имеют отно-
Популяции б сообществах
163
шение к цели исследования (разнообразие, видовой состав, доми-
нирование, биомасса, продуктивность); 2) предложить некую ги-
потезу, объясняющую структуру и функцию сообщества, на осно-
ве тех свойств, которые считаются наиболее важными; 3) выбрать
для подробного изучения те популяции, которые включены в вы-
явленные на этапах 1 и 2 процессы в сообществе: 4) на основе
полученных для популяции данных построить математическую
модель и проверить, насколько хорошо она имитирует изучаемую
экосистему. Независимо от выбранной процедуры в ходе исследо-
вания могут возникать различные конструктивные идеи (напри-
мер, решение о том, какие популяции важны), и эти идеи долж-
ны быть основаны на данных, полученных непосредственно в
полевых исследованиях.
Взаимоотношение частей и целого может в значительной сте-
пени зависеть от уровня сложности системы. Одну крайность
представляют экосистемы, находящиеся в суровых условиях сре-
ды (как, например, экосистемы арктической тундры или горячих
источников), где биотических компонентов мало. Такие «системы,
характеризующиеся малой размерностью», можно изучить и по-
нять на основе изучения составляющих их частей, поскольку
весьма возможно, что целое здесь просто представляет собой сум-
му этих частей и обладает либо очень небольшим числом эмер-
джентных свойств, либо вообще не обладает ими. Напротив, «сис-
темы, характеризующиеся большой размерностью» (как биосфе-
ра), состоят из огромного множества компонентов, которые, дей-
ствуя синергически, обусловливают появление эмерджентных
свойств; в данном случае целое, безусловно, не равно сумме
его частей. Об изучении всех частей по отдельности не может
быть и речи, поэтому приходится сосредоточить внимание на
свойствах целого. Большая часть реально существующих экосис-
тем (например, озеро или лес) представляет собой «системы с
промежуточной размерностью»; эти системы лучше изучать по-
средством многоуровневого подхода или как «черный ящик»,
о чем говорилось в первом абзаце этого раздела. Более подробные
сведения относительно теории уровней сложности содержатся в
работе Аллена и Старра (Allen, Starr, 1982).
Как соотнести части и целое? Этот вопрос долгое время ста-
вил в тупик философов и сбивал с толку общество. Во все време-
на между учеными всех специальностей существует раскол на
почве атомизма и холизма. Насколько трудно иметь дело одно-
временно п с частью, и с целым, отражает лучше всего, по-ви-
димому, конфликт между «благом» для индивидуума и «благом»
для общества. Было предложено или испытано множество эконо-
мических и политических подходов, призванных решить этот
конфликт, но пока все они имели мало успеха. В США политика
выборных правительств из года в год колеблется между препму-
11*
164
Глава 7
шественным вниманием к индивидууму (консервативная пози-
ция) и заботой об общественном благосостоянии (либеральная
позиция), так что внимание уделяется и частям (индивидуумам),
и целому (обществу), но не одновременно. В решении этой проб-
лемы может помочь, видимо, познание законов развития природ-
ных экосистем, о чем пойдет речь в гл. 8.
В семидесятые годы были опубликованы труды многочислен-
ных симпозиумов по экологии популяций и сообществ, в том чис-
ле мемориальный том, посвященный Роберту Мак-Артуру под
редакцией Коуди и Даймонда (Cody, Diamond, 1975), и том,
посвященный Дэвиду Лэку под редакцией Стоунхауса и Перрин-
са (Stonehouse, Perrins, 1977). Другие ссылки — в библиографи-
ях изданий Goulden, 1977; Bernardi, 1979; Soule, Wilcox, 1980,
и Taunsend, Calow, 1981. Поскольку на симпозиумы авторы обыч-
но представляют всеобъемлющие и подробные обзоры в своей
узкой области, эти книги и рекомендуются в первую очередь для
углубленного изучения этого предмета. Для более общих справок
рекомендуются работы Хатчинсона (Hutchinson, 1978) и Бигона
и Мортимера (Begon, Mortimer, 1981).
ШЕИ 8
Развитие и эволюция экосистемы
1. Стратегия развития экосистемы
Определения
Развитие экосистемы, более известное под названием экологиче-
ской сукцессии, состоит в изменении во времени видовой струк-
туры и биоценотических процессов. В отсутствие внешних нару-
шающих процессов сукцессия представляет собой направленный
и, следовательно, предсказуемый процесс. Сукцессия происходит
в результате изменений сообществом физической среды и взаи-
модействий конкуренция—сосуществование на популяционном
уровне. Таким образом, сукцессия контролируется сообществом,
несмотря на то что физическая среда определяет характер и
скорость изменения, а часто и ограничивает пределы развития.
Если сукцессионные изменения определяются преимущественно
внутренними взаимодействиями, то говорят об аутогенной (т. е.
самопорож дающейся) сукцессии. Если же на изменения регуляр-
но воздействуют или контролируют их внешние силы среды на
входе (например, штормы и пожары), то такую сукцессию назы-
вают аллогенной (порожденной извне). Аутогенная сукцессия при
заселении новых территорий (возникших в результате изверже-
ния вулкана, строительства новых водохранилищ) обычно начи-
нается с несбалансированного сообщества. Метаболизм в таком
сообществе характеризуется тем, что валовая продукция (Р) ли-
бо больше, либо меньше дыхания (/?), и процесс направлен в
сторону более сбалансированного состояния, когда P=R. Отно-
шение биомассы к продукции (В/P) возрастает в течение сукцес-
сии до тех пор, пока не произойдет стабилизации экосистемы,
в которой на единицу доступного потока энергии будет прихо-
диться максимум биомассы (или высокое содержание информа-
ции) и симбиотических связей между организмами.
Последовательность сообществ, сменяющих друг друга в дан-
ном пространстве, называется серией’, относительно недолговеч-
166
Глава 8
ные переходные сообщества называются по-разному: сериальными
стадиями, или стадиями развития, или пионерными стадиями.
Терминальная стабилизированная система известна под названи-
ем климакса, который теоретически должен быть постоянным во
времени и существовать до тех пор, пока его не нарушат силь-
ные внешние возмущения. Сукцессия, начинающаяся с состояния
P>R,— это автотрофная сукцессия-, напротив, если сукцессия
начинается с состояния P<ZR, то это гетеротрофная сукцессия.
Сукцессия, начинающаяся на участке, который прежде не был
занят (на потоке застывшей лавы, например), называется пер-
вичной сукцессией, тогда как сукцессия, начинающаяся на пло-
щади, с которой удалено прежнее сообщество (например, на лес-
ной вырубке или заброшенном поле), называется вторичной сук-
цессией.
Мы уже не раз подчеркивали, что экосистема — это не «сверх-
организм», тем не менее между развитием экосистемы, развитием
организмов, а также развитием человеческого общества сущест-
вует множество параллелей.
Объяснения и примеры
Описательные исследования сукцессий на песчаных дюнах, в сте-
пях, лесах, на морских берегах или в других местах наряду с
более новыми функциональными исследованиями позволили по-
строить ряд теорий, частично объясняющих процесс развития и
его причины. Г. Т. Одум и Пинкертон (Н. Т. Odum, Pinkerton,
1955), опираясь на сформулированный Лоткой (Lotka, 1925) «за-
кон максимума энергии в биологических системах», впервые ука-
зали на то, что сукцессия связана с фундаментальным сдвигом
потока энергии в сторону увеличения количества энергии, на-
правленной на поддержание (дыхание) системы, по мере того,
как накапливаются биомасса и органическое вещество. Позднее
Маргалеф (Margalef, 1963, 1968) подкрепил эту концепцию о
биоэнергетических основах сукцессии фактическими данными и
расширил ее. В последние 15 лет интенсивно обсуждается роль,
которую играют межпопуляционные взаимодействия в формиро-
вании последовательности смены видов, характерном признаке
экологической сукцессии. (См. соответствующие обзоры Connell,
Slayter, 1977; McIntosh, 1980.)
Изменения основных структурных п функциональных харак-
теристик аутогенной сукцессии перечислены в табл. 8.1. Для удоб-
ства обсуждения двадцать признаков экологических систем раз-
биты на пять групп. Начальные и конечные стадии развития
характеризуются различными тенденциями. Абсолютная величина
изменений, скорость изменений и время, необходимое для дости-
жения стационарного состояния, могут варьировать не только
Развитие и эволюция экосистемы
167
различных климатических и физико-географических ситуациях,
но и для разных признаков экосистемы в одной и той же физи-
ческой среде. При наличии хороших исходных данных кривые
скорости изменений имеют обычно выпуклую форму (причем в
начале изменения происходят быстрее). Известны также двувер-
шинные или периодические кривые.
Таблица 8.1. Тенденции изменения основных характеристик экосистемы, ко-
торых следует ожидать в ходе аутогенной сукцессии
Энергетика экосистемы
1. Возрастают биомасса (В) и количество органического детрита
2. Возрастает валовая продукция (Р) за счет первичной; вторичная про-
дукция изменяется мало
3. Уменьшается чистая продукция
4. Увеличивается дыхание (R)
5. Соотношение PIR приближается к единице (равновесию)
6. Соотношение В)Р возрастает (P/В соответственно уменьшается)
Круговороты биогенных элементов
7. Круговороты элементов становятся все более замкнутыми
8. Увеличиваются время оборота и запас важных элементов
9. Возрастает коэффициент цикличности (возобновление/вход)
10. Удерживается и сохраняется больше биогенных элементов1
Виды и структура сообщества
11. Меняется видовой состав сообщества (флористические и фаунистиче-
ские эстафеты)
12. Возрастает богатство как компонент разнообразия
13. Возрастает выровненность как компонент разнообразия
14. r-Стратеги в широких масштабах замещаются /^-стратегами
15. Усложняются и удлиняются жизненные циклы
16. Увеличивается величина организмов и (или) их стадий расселения (се-
мян, молоди и т. д.)
17. В значительной степени развивается взаимовыгодный симбиоз1
Стабильность
18. Возрастает резистентная устойчивость 1
19. Снижается упругая устойчивость 1
Общая стратегия
20. Возрастает эффективность использования энергии и биогенных элемен-
тов1
1 Тенденции обоснованы теоретически и еще не подтверждены в полевых усло-
виях.
Перечисленные в табл. 8.1 тенденции наблюдаются в том
случае, если в сообществе преобладают внутренние, аутогенные,
процессы. Влияние внешних, аллогенных, возмущений может по-
вернуть вспять пли иным образом изменить эти тенденции раз-
вития. Эти вопросы мы обсудим позднее.
Биоэнергетика развития экосистемы. Биоэнергетику экосисте-
мы отражают первые шесть пунктов из табл. 8.1. На ранних
стадиях автотрофной сукцессии в среде, лишенной органического
вещества, скорость образования первичной продукции, или общий
168
Глава 8
(валовой) фотосинтез (Р), превышает скорость дыхания сообще-
ства (Р), так что соотношение Р/Р, как правило, больше 1.
В особых условиях, когда сукцессия протекает в богатой органи-
кой среде (например, в очистном отстойнике), соотношение Р/Р
бывает меньше 1. Такую среду первыми заселяют бактерии и дру-
гие гетеротрофные организмы, поэтому сукцессия называется ге-
теротрофной. Однако в обоих случаях, согласно теории, величина
Р/Р приближается к единице по мере развития сукцессии. Иными
словами, в зрелой, или климаксной, экосистеме наблюдается тен-
денция к равновесию между связанной энергией и энергией, за-
трачиваемой на поддержание биомассы (т. е. суммарным дыха-
нием сообщества). Таким образом, отношение Р/Р является функ-
циональным показателем относительной зрелости экосистемы. От-
ношения Р/Р для ряда хорошо изученных экосистем приведены
на рис. 8.1, А, где указано также направление автотрофной и
гетеротрофной сукцессий.
Пока Р больше Р, в системе будут накапливаться органиче-
ское вещество и биомасса (Р), в результате чего отношения Р/Р,
Р/Р или В/Е (где Е=Р+Р) будут увеличиваться или соответ-
ственно отношение Р/Р будет уменьшаться. Напомним, что эти
отношения уже обсуждались в связи с концепцией термодинами-
ческой упорядоченности (т. 1, с. 109). Теоретически, следователь-
но, урожай на корню, поддерживаемый доступным потоком энер-
гии (Р), достигает максимума на зрелых, или климаксных, ста-
диях. Как следствие этого, чистая продукция сообщества (или
урожай за годовой цикл) велика на ранних стадиях и мала или
равна нулю в зрелой системе.
Сравнение сукцессии в лабораторном микрокосме и в лесу.
Биоэнергетические изменения легко наблюдать, вызвав сукцес-
сию в экспериментальных лабораторных микроэкосистемах, про-
исходящих от природных систем; такие микроэкосистемы были опи-
саны в гл. 2 (см. рис. 2.17, Р). На рис. 8.2 общее течение 100-днев-
ной автотрофной сукцессии в микрокосме (данные Cooke, 1967)
сравнивается с гипотетической моделью 100-летней сукцес-
сии леса (данные Kira, Shidei, 1967). В течение первых 40—
60 дней типичного эксперимента с микрокосмом дневная чистая
продукция (Р) превосходит дыхание ночью (Р), поэтому в систе-
ме накапливается биомасса (Р). После начального периода «цве-
тения» примерно к 30-му дню скорости обоих процессов начинают
снижаться и на 60—80-й день становятся приблизительно равны-
ми. По мере приближения к стационарному состоянию отношение
Р/Р (в граммах суточной продукции углерода, поддерживающих
1 г углерода биомассы) возрастает от менее 20 до более 100.
В климаксном сообществе не только сбалансирован автотрофный
и гетеротрофный метаболизм, но крупные органические структу-
ры поддерживаются невысокой суточной продукцией и дыханием.
A
P/R > 1
Исходная культура водорослей
(оптимум биогенных элементов)
Коралловые рифы
Продуктивные лиманы
Богатые леса
5
5
10
Сельское хозяйство
с применением удобрений
Вторичная
Первичная
Океаны
Автотрофная ___
сукцессия
Богатые озера
Степи
Загрязненные
реки (зона
пониженного
содержания О2)
Болотные
воды
Пруды
Г етеротрофная
сукцессия
Бедные озера
Пустыни
Неочищенные
сточные воды
10; 100
Дыхание сообщества, г]*м“2-сут‘
Рис. 8.1. А. Положение разных сообществ в системе классификации, осно-
ванной на метаболизме сообщества. Слева от диагональной линии валовая
продукция (Р) превышает дыхание (R); Р/R больше единицы = автотроф-
ность. Справа от диагонали — обратная ситуация: Р/R меньше едини-
цы — гетеротрофность. В последнем случае сообщества живут за счет орга-
нического вещества, получаемого извне, или за счет его предварительного
запасания или накопления. Стрелками показано направление автотрофной
и гетеротрофной сукцессий. Сообщества, расположенные вдоль диагонали,
в среднем за год потребляют примерно столько, сколько создают, и их
можно считать метаболическими климаксами. (Из Н. Т. Odum, 1956.)
Б, Общесистемная модель развития экосистемы.
170
Глава 8
Рис. 8.2. Сравнение энергетики развития экосистем леса (из Kira, Shidei,
1967) и лабораторного микрокосма (из Cooke, 1967). Pg — валовая продук-
ция; PN — чистая продукция; R — суммарное дыхание сообщества; В —
суммарная биомасса.
Меняется и относительное обилие видов, так что в конце 100-
дневной сукцессии доминирующими становятся другие бактерии,
водоросли, простейшие и мелкие ракообразные, а не те, которые
доминировали вначале (Gorden et al., 1969).
В проточном лабораторном микрокосме, где культуральная
среда медленно протекает через контейнер с организмами, имити-
руя вход и выход среды для воды и питательных веществ, сук-
цессия замедленна; для накопления биомассы и приближения
PJB к 1 требуется в 2—3 с лишним раза больше времени, чем в
закрытом микрокосме (Hendrix et al., 1981).
Непосредственная экстраполяция данных, полученных для ла-
бораторного микрокосма, на природные системы может быть и
не вполне правомерна, поскольку в первом случае в системе су-
ществуют только мелкие организмы с простыми жизненными
циклами и неизбежно снижено видовое и химическое разнообра-
зие. Тем не менее в микрокосме можно наблюдать такие же
основные тенденции, которые характерны для сукцессии на су-
ше (рис. 8.2, верхний график) и в крупных водоемах. Сезонные
сукцессии часто происходят по той же самой схеме: вслед за
«цветением» в начале сезона, которое характеризуется быстрым
ростом нескольких доминирующих видов, к концу сезона уста-
Развитие и эволюция экосистемы
171
навливается высокое отношение BIP, увеличиваются разнообразие
и относительная, хотя и временная, устойчивость, как это уста-
новлено в терминах Р и R (Margalef, 1963). В открытых систе-
мах на зрелых стадиях понижения общей, или валовой, продук-
ции, наблюдаемой в пространственно ограниченном микрокосме,
может и не происходить, но общая схема биоэнергетических из-
менений в последнем, судя по всему, достаточно хорошо имитиру-
ет природу.
Интересно также отметить, что пик чистой первичной продук-
ции (PN) соответствующий максимально возможному урожаю,
приходится на 30-й день в микрокосме и 30-й год в лесу. Коротко-
цикличное лесное хозяйство ориентировано на пик Ду, который
во многих случаях повторяется через 20—40 лет.
Аллогенные и аутогенные воздействия. До сих пор мы рассмат-
ривали изменения, происходящие в результате процессов, проте-
кающих в самой экосистеме. Привнесенные вещества или энер-
гия, геологические силы, шторма, антропогенные нарушения
и т. п. могут изменить, остановить или повернуть вспять тенден-
ции, указанные в табл. 8.1 (так обычно и бывает на самом деле).
Эвтрофикация озера, например (неважно — естественная или ан-
тропогенная), происходит в результате поступления в озеро био-
генных элементов и почвы извне, т. е. с водосбора. Это равнознач-
но добавлению биогенов в лабораторную микроэкосистему или
удобрению поля; в сукцессионном отношении такая система от-
брасывается назад, к более молодому состоянию. Аллогенная сук-
цессия такого типа во многих аспектах обратна аутогенной сук-
цессии. Если влияние аллогенных процессов намного сильнее
влияния аутогенных процессов, как во многих прудах и малых
озерах, то экосистема не только не в состоянии стабилизировать-
ся, но и «угасает», т. е. заполняется органическим веществом и
осадками, превращаясь в болото или наземное сообщество. Такова
неизбежная судьба искусственных озер, берега которых подвер-
жены ускоряющейся искусственной эрозии.
Исследования озерных осадков (Mackereth, 1965; Cowgill, Hut-
chinson, 1964; Harrison, 1962) наряду с теоретическими сообра-
жениями указывают, что озера могут развиваться и действитель-
но развиваются в сторону более олиготрофного (менее обогащен-
ного) состояния, если поток биогенных элементов с водосбора
замедляется пли прекращается. Таким образом, есть надежда, что
вызывающую беспокойство антропогенную эвтрофикацию, кото-
рая ухудшает качество воды и укорачивает жизнь водоема, мож-
но повернуть вспять, если удастся сильно уменьшить приток
биогенов с водосбора. Примером служит описанное Эдмондсоном
(Edmondson, 1968, 1979) «выздоровление» озера Вашингтон в
Сиэтле. В течение 20 лет очищенные, богатые биогенамп сточные
воды сливались в озеро, которое становилось все более мутным,
172
Глава 8
и в нем все чаще наблюдались обильные «цветения» водорослей.
В результате призывов общественности сточные воды отвели от
озера, и оно быстро вернулось к более олиготрофному состоянию
(вода стала чище, и прекратились «цветения» водорослей).
Взаимодействие внешних и внутренних сил можно выразить
при помощи обобщенной системной модели (рис. 8.1,Б). Впервые
модель такого типа была приведена на рис. 1.4. Аутогенные силы
изображены как внутренний вход или обратная связь, которая
теоретически способствует сдвигу системы к своего рода равно-
весному состоянию. Аллогенные силы изображены как периоди-
ческое внешнее возмущение на входе, которое возвращает назад
или иным способом меняет траекторию развития системы.
В тех системах, которые развиваются длительное время, как
это бывает, например, при зарастании лесом голой почвы, сук-
цессионный процесс подвержен влиянию периодических возмуще-
ний, особенно в изменчивой среде умеренной зоны. Оливер и
Стефенс (Oliver, Stefens, 1977) описали изменения растительного
покрова на небольшой площади Гарвардского леса (Массачусетс).
Растительность этой территории 14 раз подвергалась воздействи-
ям природных и антропогенных стрессов разной силы с нерегу-
лярными промежутками за период с 1803 по 1952 г. Имеются
также данные о том, что до 1803 г. здесь пронеслось два урагана
и возник один пожар. Слабые воздействия не вызывают появления
новых видов деревьев, но часто позволяют видам, уже представ-
ленным в нижнем ярусе (черная береза, красный клен, тсуга),
достигать полога. Крупномасштабные возмущения, такие, как
ураган пли сильный пожар, создают открытые пространства,
прогалы, в которые вторгаются виды, свойственные ранним ста-
диям сукцессии (например, белая береза или пенсильванская че-
ремуха). В таких прогалах развивается также молодая поросль
из находившихся в почве семян и проростков. Северный дуб часто
заполняет таким образом прогалы и через несколько десятилетий
начинает доминировать в пологе. Смена видов и сукцессия в лес-
ных прогалах была названа Брэем (Bray, 1956) «сукцессией фазы
заполнения пробела». Оливер и Стефенс на основе полученных
ими данных пришли к выводу, что существующий видовой состав
леса представляет собой в большей степени результат аллогенных
процессов, нежели аутогенного развития. В более поздней обзор-
ной работе Оливер (Oliver, 1981) высказал предположение, что
основным фактором, определяющим структуру леса и видовой
состав деревьев во многих районах Северной Америки, являются
сила и частота внешних возмущений.
Если в силу периодических возмущений внешней среды или
вследствие характера развития самого сообщества нарушения
происходят через более пли менее регулярные промежутки вре-
мени, то экосистема претерпевает то, что логично было бы на-
Развитие и эволюция экосистемы
173
Рис. 8.3. Циклическая («волнообразная») сукцессия в лесу бальзамической
пихты.
звать циклической сукцессией (Уатт, 1947). Хороший пример са-
могенерируемой циклической сукцессии дает описанный ранее
(гл. 5, разд. 4) цикл пожаров в растительности чапараля, где сла-
боразложившаяся подстилка образует запас горючего материала
для периодических пожаров в сухой сезон.
Другой пример «волнообразной» сукцессии — высокогорные
леса из бальзамической пихты на северо-востоке США (Sprugel,
Bormann, 1981). Деревья здесь растут на тонком слое почвы,
и по мере достижения максимальной высоты они теряют устойчи-
вость к штормовым ветрам, которые вырывают с корнем и таким
образом губят старые экземпляры. Таким образом, возникает
возможность вторичной сукцессии. Как показано на рис. 8.3,
склон горы покрыт рядами полос молодых, зрелых п мертвых
деревьев (последние образуют на фотографии светлые полосы).
Вследствие непрерывной циклической сукцессии эти полосы дви-
жутся, подобно волнам, через весь ландшафт в направлении до-
минирующих ветров. В любой момент времени здесь представле-
ны все стадии сукцессии, предоставляющие множество местооби-
таний животным и более мелким растениям. Склон горы в целом
представляет собой «циклический климакс», своего рода стацио-
нарное состояние, находящееся в равновесии со своим окруже-
нием;.
174
Глава 3
Природная структура, состоящая из чередующихся полос
молодого и зрелого древостоя, наводит на мысль о том, что вы-
рубки в виде лент или пятен могут быть хорошим способом ве-
дения лесного хозяйства на обширных пространствах, поскольку
в таком случае легче происходило бы естественное возобновление*
(и не нужно было бы проводить дорогостоящих лесопосадок).
Почвы и популяции животных также оказывались бы менее на-
рушенными, чем при сплошных рубках в больших лесных мас-
сивах. Кроме того, наличие разных стадий сукцессии создает
множество пограничных зон, что способствует увеличению видо-
вого разнообразия диких животных.
Другой пример циклической сукцессии — цикл листовертки-
почкоеда и ели, описанный в гл. 6, разд. 5. В этом случае перио-
дические нарушения вызываются не физическими силами, а рас-
тительноядными животными, которые лишают хвои и уничтожают
старые деревья, что дает возможность расти молодым деревьям.
Термин «зависимый от возмущений» часто используется для
обозначения таких экосистем, которые специально адаптированы
к регулярно повторяющимся возмущениям благодаря тому, что
они располагают видами и процессами, способными к быстрому
восстановлению (см. обзор Vogl, 1980). Для того чтобы предска-
зать восстановление экосистемы и управлять им после таких, на-
пример, нарушении, как открытые разработки ископаемых, необ-
ходимо в деталях знать структуру сукцессионного процесса и
восстановительный потенциал конкретной экосистемы, иначе уси-
лия по ее восстановлению могут мешать, а не способствовать ес-
тественному восстановлению (см. McIntosh, 1980).
Теоретические траектории экологической сукцессии в благо-
приятных и неопределенных физических условиях среды показа-
ны на рис. 8.4. По мере развития биотического сообщества слож-
ность организации быстро увеличивается. На увеличение содер-
жания информации указывает возрастание биомассы и отношения
В/P, Это же говорит об увеличении разнообразия и эффективно-
сти функциональных процессов, связанных с потоком энергии и
циркуляцией биогенных элементов. Если физические условия сре-
ды благоприятны и вероятность катастрофических нарушений
мала, то достигается высокий уровень организации и длительное
время сохраняется стационарное состояние или климакс
(рис. 8.4, 7). Траектория, ведущая к более низкому уровню орга-
низации, характерна, напротив, для неблагоприятной среды, под-
верженной периодическим возмущениям, которые прерывают и
возвращают вспять процесс сукцессии (рис. 8.4, II). На кривой II
показаны две возвращающие сукцессию вспять петли, которые
следуют после двух возмущений, обозначенных стрелками. По-ви-
димому, этот более низкий уровень лучше адаптирован к возму-
щениям и более устойчив. Можно высказать рабочую гипотезу,
Развитие и эволюция экосистемы
175
Рис. 8.4. Траектория экологической сукцессии в благоприятной среде с низ-
кой вероятностью катастрофических возмущений (/) ив стрессовой среде
с периодическими возмущениями, нарушающими и отбрасывающими на-
зад процесс развития (II). На кривой II показаны две гипотетические петли
траектории возврата сукцессии назад. Эти события происходят вслед за
нарушениями, показанными стрелками; биотическое сообщество адапти-
руется к нарушениям, и в нем поддерживается уровень организации ниже
той, которой оно бы достигло в отсутствие стресса на входе. (По Odum,
1981.)
что в общем более зрелые стадии сукцессии ведут себя стабиль-
нее при кратковременном стрессе или стрессе, вызванном опреде-
ленным фактором (как, например, засушливый год), тогда как
ранние стадии устойчивее (восстанавливаются быстрее) при ка-
тастрофических стрессах, таких, как сильная буря или пожар
(см. пункты 18 и 19 в табл. 8.1).
Круговороты биогенных элементов. Важной тенденцией в раз-
витии сукцессии являются увеличение времени оборота и повы-
шение способности к сохранению и регенерации веществ. Все это
приводит к замыканию или «уплотнению» биогеохимических цик-
лов основных биогенных элементов, таких, как азот, фосфор и
кальций (табл. 8.1, пункты 7—10). Исследователи не пришли к
единому мнению в оценке тех пределов, в которых сохранение
биогенов представляет собой главное направление или стратегию
развития экосистемы. Отчасти это противоречие обусловлено тем,
что способы выявления соответствующих процессов различны.
Эту проблему хорошо иллюстрирует рис. 8.5. Витусек и Рейнере
(Vitousek, Reiners, 1975) высказали предположение, что в системе,
в которой биомасса накапливается на ранних стадиях сукцессии,
с большей вероятностью будет идти процесс запасания бпогенов.
Согласно теории этих авторов, при переходе биогенов в накоп-
ленную биомассу отношение выхода к входу падает ниже еди-
ницы. В зрелой климаксной системе, где рост больше не проис-
176
Глава 8
Сукцессия------------>-
Рис. 8.5. Гипотетические тенденции изменений отношения выход,'вход, ин-
декс цикличности (СГ) и отношение запас/выход (S/О) для биогенных
элементов в процессе сукцессии. I — вход; О — выход; S — запас: С — во-
зобновление; BG — обнаженная почва; ES — ранние стадии; MS — проме-
жуточные стадии; SS — стационарное состояние. (Основано на схеме
J. Т. Finn.)
ходит, вход и выход уравновешиваются, и это отношение вновь
повышается до единицы (рис. 8.5). Вудманси (Woodmansee, 1978)
отметил, однако, что даже после того, как растения перестали
давать прирост живой биомассы, накопление биогенов в почве
может продолжаться. К тому же Гендерсон (Henderson, 1975),
Уэстмен (Westman, 1975) и Финн (Finn, неопубликованная ру-
копись) считают, что определение отношения выход/вход не
единственный и, видимо, не лучший способ оценки поведения
биогенов. Как показано на рис. 8.5, индекс возобновления
(С1= отношению регенерированной части на входе к выходу, см.
гл. 4, разд. 7) устойчиво возрастает по мере созревания системы.
Биогены, следовательно, удерживаются более долгое время и ис-
пользуются повторно, при этом даже в случае сбалансированных
входа и выхода необходимость притока уменьшается. Точно так
же отношение запаса (S) к потерям (О), по-видимому, мало на
ранних стадиях и возрастает на поздних стадиях. В общем име-
ются экспериментальные данные и можно также теоретически
обосновать то, что в процессе развития экосистемы возрастают
запас и степень возобновляемости биогенных элементов, в резуль-
тате необходимый уровень поступления этих элементов на вход
для поддержания единицы биомассы снижается. У нас нет осно-
ваний считать, что так же сохраняются второстепенные и токсич-
ные вещества.
Райс и Пэнколи (Rice, Pancholy, 1971, 1972, 1974) предста-
вили в серии работ косвенные доказательства того, что на зрелых,
Развитие и эволюция экосистемы
177
или климаксных, стадиях происходит аллелохимическое ингиби-
рование нитрификации. Таким образом, в ходе сукцессии проис-
ходит смена источника азота с нитрата на аммоний. Пионерные
растения используют в первую очередь нитрат, тогда как на
поздних стадиях, в частности на стадиях леса, в качестве источ-
ника азота они используют аммоний. Питер Мак-Рой (личное
сообщение) наблюдал, что в зрелых стадиях сукцессии морских
трав азот регенерируется в форме аммиака благодаря деятельно-
сти анаэробных микробов. Факт этот очень интересен, так как
при переходе с нитрата на аммоний уменьшается количество
энергии, расходуемой на регенерацию азота (см. обсуждение кру-
говорота азота в гл. 4), и возрастает, следовательно, эффектив-
ность использования энергии. Вопрос, однако, остается открытым,
потому что Робертсон и Витусек (Robertson, Vitousek. 1981)
не получили в экспериментальных данных подтверждающий сдвиг
от потребления нитрата к потреблению аммония.
Не решен также вопрос о том, увеличивается пли снижается
в течение сукцессии интенсивность азотфиксации, микоризного
симбиоза и других форм мутуализма, которые повышают эффек-
тивность регенерации биогенов (пункт 17 в табл. 8.1). Эти формы
мутуализма проявляются и широко распространены па всех ста-
диях сукцессии, но до сих пор было предпринято мало попыток
количественно оценить их деятельность на разных сукцессионных
стадиях. По-видимому, мутуалистические отношения, способству-
ющие сохранению биогенов, в большей степени подвержены влия-
нию «текущих запросов» (бедность биогенами), чем процессов
развития экосистемы на таковых.
Замещение видов. Более или менее непрерывная смена видов
во времени — характерная черта большинства сукцессионных се-
рий. Смена видового состава растительности была названа Эгле-
ром (Egler, 1954) «флористической эстафетой». Несомненно, су-
ществует и «фаунистическая эстафета», поскольку виды живот-
ных также сменяют друг друга в сериях.
Если развитие начинается на участке, который до этого не быль
занят каким-либо сообществом (на обнажившихся скалах, на пес-
ке или на потоке лавы), то первичная сукцессия на ранних ста-
диях протекает медленно и для достижения стационарного со-
стояния, характеризующего зрелость системы, потребуется много
времени. Классический пример первичной сукцессии — дюны в
штате Индиана у южной оконечности озера Мичиган, которое
некогда было гораздо больше, чем сейчас. Отступая к своим со-
временным границам, озеро оставляло все более и более молодые
песчаные дюны. Поскольку субстрат песчаный, сукцессия проте-
кает медленно, и можно наблюдать серию сообществ разного воз-
раста: пионерные стадии у берегов и все более старые стадии по
мере удаления от берега. Именно в этой «природной лаборатории
12 Заказ № 1483
478
Глава 8
сукцессии» Каулес (Cowles, 1899) провел свои основополагающие
исследования растительной, а Шелфорд (Shelford, 1913) —живот-
ной сукцессии. Оба автора показали, что виды как растений, так
и животных сменяют друг друга по мере увеличения возраста
дюн; существующие вначале виды в старых сообществах заме-
няются другими, совершенно отличными от них. Олсон (Olson,
1958) повторно исследовал развитие экосистемы на этих дюнах и
представил современные данные о происходящих при этом процес-
сах и их скоростях. Из-за вторжения в эти области тяжелой про-
мышленности специалисты по охране природы, которые пытаются
сохранить «дюны Индианы», оказались в затруднительном положе-
нии. Однако, к счастью, некоторая часть этих дюн входит теперь
в состав национальных парков и парков штата. Общественность
должна поддержать их усилия по защите дюн, поскольку этот
район не только бесценен своей природной красотой, которой мо-
гут наслаждаться жители городов, но и представляет собой ес-
тественную учебную лабораторию с поразительно наглядной эко-
логической сукцессией.
Первые поселенцы на дюнах — это прибрежные злаки (Ammo-
phila, Agropyron, Calamovilfa), ива, песчаная вишня, хлопковое
дерево и животные, такие, как бегающие по песку длинноногие
жуки-скакуны, живущие в норках пауки, кузнечики. Вслед за
сообществом первых поселенцев следуют открытые сухие леса
из сосны Банкса, затем черного дуба и, наконец, на самых старых
дюнах — влажные леса из дуба и гикори или бука и клена.
Несмотря на то что развитие сообщества начинается в очень су-
хом и бесплодном местообитании, оно в конечном счете приводит
к влажному и холодному сомкнутому лесу, резко отличающемуся
от голых дюн. Мощная богатая гумусом почва, населенная дожде-
выми червями и улитками, контрастирует с сухим песком, на ко-
тором она образовалась. Так сравнительно негостеприимная вна-
чале масса песка в конце концов совершенно преобразуется под
влиянием сукцессии сообщества.
На ранних стадиях сукцессия на дюнах часто приостанавли-
вается, если ветер засыпает песком растения, и дюны приходят в
движение, погребая всю растительность на своем пути. Это при-
мер аллогенных возмущений, которые задерживают или обраща-
ют сукцессию вспять, о чем говорилось в начале данного раздела.
Однако по мере продвижения дюны от берега она стабилизирует-
ся, и ее первые поселенцы, злаки и деревья, вновь восстанавли-
ваются. Использовав современные методы радиоуглеродного да-
тирования, Олсон (Olson, 1958) установил, что для достижения
климаксного лесного сообщества на дюнах озера Мичиган требу-
ется около 1000 лет. Это примерно в пять раз больше, чем нужно
для развития зрелого леса в более гостеприимном месте, как будет
видно из следующего примера.
Развитие и эволюция экосистемы
179*
Пример вторичной сукцессии дан в табл. 8.2, где показаны
растительные сообщества и популяции птиц, сменяющие друг
друга на месте заброшенных сельскохозяйственных угодий, нахо-
дящихся в возвышенном предгорье на Юго-Востоке США. Пер-
вые поселенцы — это r-стратеги, однолетние растения, такие, как
Digitaria, Erigeron и Ambrosia, затрачивающие большую часть
своей энергии на расселение и размножение (см. табл. 6.7). Через
два-три года их место занимают многолетнее разнотравье, пред-
ставленное астрами и золотарником, злаки, в особенности бородач
(Andropogon), и такие кустарники, как ежевика (Rubus). Если
поблизости есть обильный источник семенного материала, то по-
являются сосны, которые вскоре образуют сомкнутый полог и
затеняют ранних поселенцев. Часто вместе с соснами появляется
несколько видов быстро растущих листопадных деревьев, как, на-
пример амброво и тюльпанное деревья. Поскольку все эти виды
живут подолгу, стадия сосны (с редкими широколиственными
деревьями) существует длительное время, но в нижнем ярусе по-
степенно развиваются теневыносливые дубы и гикори. Молодые
сосны не могут развиваться в тени взрослых деревьев, поэтому,
по мере того как старые сосны погибают от болезней, старости
и ураганов, в пологе начинают доминировать дубы и гикори.
В табл. 8.2 показано, что население птиц сменяется на каждой
крупной сукцессионной стадии; эти изменения проявляются силь-
нее всего при смене доминирующих жизненных форм растений
(травы—кустарники—сосны—лиственные породы). Птицы выби-
рают местообитания, ориентируясь больше на жизненные формы,
а не на виды растений. Ни один вид растений или птиц не может
процветать на протяжении всей сукцессии. Максимумы разных
видов приходятся на разные точки временной шкалы.
Животные принимают не только пассивное участие в измене-
нии сообщества. Птицы разносят семена, необходимые для уста-
новления стадий кустарников и широколиственных деревьев, рас-
тительноядные организмы, паразиты и хищники часто контроли-
руют последовательность видов. И, конечно же, в мелководных
морских местообитаниях именно крупные животные, а не расте-
ния образуют структурную матрицу сообщества. Глемарек (Gle-
marec, 1979) описал вторичную сукцессию донных животных вда-
ли от побережья Бретани (Франция). После штормов, вызываю-
щих перераспределение осадков и нарушающих донную фауну,
наступает период относительного затишья. В это время при от-
сутствии внешних воздействий происходит более пли менее пред-
сказуемая смена доминирующего населения. Первыми поселяются
двустворчатые моллюски, питающиеся взвесью, затем такие же
моллюски, но добывающие пищу в донных отложениях, и, нако-
нец, в бентосе начинают преобладать питающиеся детритом мно-
гощетинковые черви. Таким образом, подтверждается теория, что*
12=
180
Глава 8
Таблица 8.2. Вторичная сукцессия в предгорной области на Юго-Востоке
США1
|С,
' Возраст, годы
7 ип сообщества
1 1 1 2 | 3-20 25 - 100 | 150+
1 Голое попе Степь 1 Злаки и j кустарники) Сосновый лес (Климаксный лес из дуба и | гикори
Гикори
Сорняки
гг <??? т
Бородач Кустарники 0СНЭ Подлесок из Дуб
лиственных пород
ЯКП Yttt
Мелко- Астра
лечестник
Дом пнирующая растительность Разнотравье Злаки Злаки и кустар- ники Сосновый лес Климаксный лес из дуба и гикори
Возраст стадии в месте изучения, годы 1—2 2—3 15 20 25 35 60 100 150— 200
Виды птиц (имеющие на данной стадии плотность популяции не ниже 5 особей)
Саванный воробей {Ammodramus savannarum) Луговой трупиал (Sturnella neg- lecta) Овсянка Spizella pusilia Американская славка (Geothlipus trichas) Желтогрудая славка (Icteria virens) Кардинал Richmondena cardinalis Тауи (Pipilio erythrophthalmus) Юнко (Aimophila aestivalis) Древесница Dendroica discolor Виреон белоглазый (Virco griseus) Древесница Dendroica pinus Танагра (Piranga rubra) Крапивник каролинский (Troglodytes lydovicianus) Синица каролинская (Parus carolinensis) Комаролов (Polioptila caerulea) 10 5 30 10 25 15 35 15 5 5 5 2 48 18 16 4 8 8 6 8 25 9 13 6 6 16 6 8 10 10 4 4 34 13 4 2 2 3 14 15 5 43 13 5 5 13 20 15 55 15 20 5 23 10 10 5 13
Развитие и эволюция экосистемы
181
Продолжение
Доминирующая растительность Разнотравье Злаки Злаки и кустар- ники Сосновый лес Климаксный лее из дуба и гикори
Поползень Sitta pus ilia 2 5
Тиранн С onto pus virens 10 1 3
Колибри 9 10 10
Синила Parus bicolor 6 10 15
Виреон желтолобый
(Vireo jlavifrons) 3 5 7
Древесница Wilsonia citrina 3 30 И
Виреон красноглазый
(Vireo olivaceus) 3 10 43
Дятел Dendrocopus villosus 1 3 5
Дятел Dendrocopus pubescens 1 2 5
Тиранн Myiarchus crinitis 1 10 6
Дрозд Hylocichla mustelina 1 5 23
Американская кукушка
(Coccysus americanus) 1 9
Древесница (Mniotilta varia) 8
Древесница (Oporornis formosus) 5
Тиранн зеленый
(Empiodonax virescens) 5
Всего (вместе с не приведенными
в таблице редкими видами) 15 40 но 136 87 93 158 239 228
а По Johnston, Е. Р. Odum, 1956. Цифры показывают число занятых участков
или оценку числа пар на 40 га. Главные растительные доминанты в серии, следую-
щей за пустошами на месте полей, нарисованы над таблицей. Сукцессия была под-
робно описана Устингом (Oosting, 1942). Кивер (Keever, 1950) изучил некоторые
взаимодействия между растениями. В таблице показана сукцессия обычно гнездя-
щихся птиц.
при непрерывной сукцессии в среде увеличивается количество
органического вещества.
Вторичная растительная сукцессия в степных районах так же
поразительна, как и лесная, несмотря на то что в ней участвуют
только травянистые растения. В 1917 году Шанц (Sliantz) описал
сукцессию на старых заброшенных фургонных дорогах, по кото-
рым первые поселенцы пересекали степи центральных и западных
штатов; впоследствии такую же в сущности последовательность
видов описывали не один раз. Хотя в разных географических
областях встречаются разные виды, общий характер сукцессии
всюду одинаков. Сукцессия слагается из четырех последователь-
ных стадий: 1) стадия однолетних сорняков (2—5 лет); 2) ста-
дия короткоживущих злаков (3—10 лет); 3) ранняя стадия мно-
182
Глава 8
голетних злаков (10—20 лет); 4) климаксная стадия многолетних
злаков (достигается за 20—40 лет). Таким образом, природе нуж-
но от 20 до 40 лет, чтобы, начав на голом или перепаханном
грунте, «построить» степной климакс; точные сроки зависят от
лимитирующих воздействий таких факторов, как влажность, вы-
пас и т. п. Несколько засушливых лет подряд или чрезмерный
выпас вызывают возврат сукцессии к стадии однолетних сорня-
ков. Насколько далеко может зайти такой возврат, зависит от си-
лы воздействия.
В водных местообитаниях, как и в наземных, можно наблю-
дать сукцессию. Но, как уже подчеркивалось, процесс развития
сообщества в мелководных экосистемах (прудах и небольших
озерах) обычно осложняется сильным притоком веществ и энер-
гии, который может ускорить, остановить или повернуть вспять
нормальное течение развития сообщества, т. е. такое развитие,
какое наблюдалось бы без этого сильного аллогенного воздейст-
вия. Сложное взаимодействие аутогенной и аллогенной сукцессии
выступает более наглядно в искусственных прудах и водохрани-
лищах. Когда при создании водохранилища затопляют тучные
почвы или участок с большим количеством органического вещест-
ва (например, при затоплении лесистой местности), первой ста-
дией развития бывает высокопродуктивная стадия «цветения»,
для которой характерны интенсивное разложение, высокая ак-
тивность микроорганизмов, обилие биогенных элементов, низкое
содержание кислорода в придонном слое воды. При этом чаете
наблюдается быстрый и интенсивный рост рыбы. На этой стадии
рыболовы очень счастливы. Однако после того, как запасенные
биогенные элементы рассеиваются, а запасы пищи оказываются
израсходованными, водоем стабилизируется на некотором более
низком уровне продуктивности, более высоком содержании кис-
лорода в придонном слое и пониженном урожае рыбы. Эта стадия
приносит рыболовам огорчение (см. рис. 8.6). Такая стабилизи-
рованная стадия, своего рода климакс, может существовать неко-
торое время, если водосбор хорошо защищен зрелой раститель-
ностью или если почвы на площади водосбора неплодородны.
Однако эрозия и различные усиливаемые человеком источники
эвтрофикации обычно порождают длительную серию «переходных
состояний», продолжающуюся до тех пор, пока водоем не окажет-
ся заполненным осадками. Водохранилища на истощенных водо-
сборах пли на первично стерильных участках дают, конечно,
обратную картину — низкую продуктивность вначале. Непонима-
ние основных принципов экологической сукцессии и зависимости
между водосбором и водоемом привело к многочисленным неуда-
чам и разочарованиям при попытках человека поддерживать та-
кие искусственные экосистемы. Мелководные экосистемы могут
быть «импульсно стабилизированы» на высоком уровне продук-
Развитие и эволюция экосистемы
183
Рис. 8.6. Изменение обилия рыбы (средние по двум методам сбора) в новом
русловом водохранилище в верховьях Миссури со второго по пятнадцатый
годы после постройки плотины и заполнения водой образовавшегося озера
Френсис-Кейз, Южная Дакота. (Gasaway, 1970.)
тивности в результате поступления большого количества энергии
за счет колебаний уровня воды (это будет описано в разд. 3 на-
стоящей главы).
Так как океаны, вообще говоря, находятся в стационарном
состоянии и на протяжении столетий химически и биологически
стабилизированы, океанографов не интересовали вопросы эко-
логической сукцессии. Однако по мере того, как в результате
загрязнений равновесие в океанах начинает нарушаться, взаимо-
действие аутогенных и аллогенных процессов будет, несомненно,
привлекать большее внимание морских биологов. Более очевидны
сукцессионные изменения в прибрежных водах, как это уже от-
мечалось на примере развития донных сообществ после сильных
штормов, разрушающих поверхность морского дна. Маргалеф
(Margalef, 1967) следующим образом подытожил свои наблюдения
над сукцессионными изменениями в толще прибрежных вод:
1. Возрастают средний размер клеток п относительное обилие
подвижных форм в фитопланктоне.
2. Продуктивность или скорость размножения снижаются.
184
Глава 8
3. Меняется химический состав фитопланктона, о чем можно
судить, в частности, ио растительным пигментам.
4. Меняется видовой состав зоопланктона: вместо пассивных
фильтраторов развиваются более активные и питающиеся выбо-
рочно охотники. Это происходит вследствие сдвига от многочис-
ленных мелких взвешенных пищевых частиц к более рассеянной
пище, сконцентрированной в более крупные частицы и распре-
деленной в более организованной (стратифицированной) среде.
5. На поздних стадиях сукцессии общий перенос энергии мо-
жет быть снижен, но его эффективность, вероятно, повышается.
Джессби и Голдмен (Gassby, Goldman, 1974) использовали
скорость смены видов планктона для построения диаграммы се-
зонной сукцессии в озерах. Виды быстро сменяют друг друга
весной п намного медленнее летом. Если в июне добавить в озеро
биогенные элементы, то скорость смены видов достигнет весеннего
уровня п останется такой примерно в теченпе месяца.
Сукцессия организмов на искусственных субстратах в водной
среде привлекает больше внимания, что объясняется практиче-
ской важностью обрастания днищ судов и причалов усоногими
ракообразными и другими сессильными морскими организмами.
К тому же небольшие, используемые в нескольких повторностях
образцы субстратов, такие, как стеклянные пластинки или квад-
раты из пластика, дерева или других материалов, широко приме-
няются для оценки влияния загрязнений на биоту пресных и
соленых вод (см. Patrick, 1954; Cairns, Dickson, 1971). Такпе
субстраты представляют собой своего рода микрокосм, на кото-
ром можно ожидать развития экологической сукцессии. Но это
только ограниченная и упрощенная модель, поэтому экстраполи-
ровать полученные с ее помощью данные на более обширные,
менее ограниченные в пространстве открытые системы со многи-
ми типами субстратов следует с осторожностью. В общем, первы-
ми заселяют субстрат виды, у которых в том месте и в то время,
где и когда появилась доступная поверхность, в воде имеются
многочисленные стадии расселения. Иногда эти первые поселенцы
так меняют химическую и физическую природу субстрата, что
облегчается вторжение других видов, но чаще они сопротивля-
ются вторжению других видов и существуют до тех пор, пока их
не сменят более удачливые конкуренты. Как уже отмечалось при
обсуждении литоральных сообществ на скалистых берегах, в про-
цессе распределения видов в местообитаниях с определенными
ограничениями или в условиях нехватки свободного пространства
отрицательные взаимодействия (конкуренция, хищничество) иг-
рают большую роль, чем положительные (сосуществование, мутуа-
лизм) .
Гетеротрофная сукцессия. В качестве примера гетеротрофной
сукцессии можно привести данные ранних экспериментов Вуд-
Развитие и эволюция экосистемы
185
Рис. 8.7. Сукцессия в сенном отваре с последовательным доминированием
разных видов. Это лабораторный пример гетеротрофной сукцессии. (По
Woodruff, 1912, с добавлением гипотетической кривой для гетеротрофных
бактерии.)
раффа (Woodruff, 1912) с сенным отваром. Если дать постоять
культуральной среде, приготовленной из отвара сена, то разви-
вается пышная культура бактерий. Если затем добавить к ней
немного прудовой воды, содержащей «посевной материал» раз-
нообразных простейших, то произойдет вполне определенная сук-
цессия населения простейших со сменой доминантных форм
(рис. 8.7). В этой ситуации количество энергии достигает макси-
мума вначале, а затем уменьшается. Если не добавлять свежую
культуральную среду или не переводить систему в автотрофный
режим, то она в конечном счете прекращает свое существование
и все организмы погибают или образуют покоящиеся стадии (спо-
ры, цисты и т. п.). Такая система радикально отличается от авто-
трофной сукцессии, в которой поток энергии поддерживается
неопределенно долго. Микрокосм в сенном отваре представляет
собой хорошую модель сукцессии, происходящей в разлагающей-
ся древесине, трупах животных, фекальных пеллетках и на вто-
ричных стадиях очистки сточных вод. Эту сукцессию можно рас-
сматривать также как модель сукцессии, «скатывающейся по на-
клонной плоскости», которую следует ассоциировать с обществом,
зависящим от ископаемого горючего. Во всех этих примерах мы
имеем дело с рядом переходных состояний в понижающемся энер-
186
Глава 8
гетическом градиенте, причем стационарного состояния достичь
невозможно.
В лабораторной модели микроэкосистемы можно скомбиниро-
вать гетеротрофную и автотрофную сукцессии. Для этого пробы
из каждой системы нужно добавить в среду, обогащенную орга-
ническим веществом. Горден и др. (Gorden et al., 1969) описали
сукцессию в такой системе и измерили в ней поток энергии. Вна-
чале, когда «цветут» гетеротрофные бактерии, система становится
мутной; затем, когда в результате активности бактерий в среде
накапливаются биогены и ростовые вещества (в частности, вита-
мин тиамин), нужные водорослям, система становится ярко-зеле-
ной. Эта сукцессия служит, конечно же, хорошей моделью куль-
турной эвтрофикации, вызываемой органическим загрязнением^
таким, как неполностью очищенные городские сточные воды.
Давление отбора: количество против качества. Стадии заселе-
ния островов, описанные Мак-Артуром и Уилсоном (McArthur^
Wilson, 1967), прямо соответствуют стадиям экологической сук-
цессии на континентах. На ранней стадии колонизации острова^
когда плотность населения мала, преобладает, как и на ранних
стадиях сукцессии, r-отбор, т. е. успешнее вселяются виды с вы-
сокой скоростью размножения и роста. Напротив, при равновес-
ной плотности, характерной для поздних стадий заселения острова
п экологической сукцессии, давление отбора благоприятствует
A-стратегам, т. е. видам с низким потенциалом роста, но более
высокой способностью к выживанию в условиях конкуренции
(пункт 14 в табл. 8.1).
Можно предположить, что происходящий при сукцессии пе-
реход от количества продукции к качеству сопровождается гене-
тическими изменениями, охватывающими всю биоту, на что ука-
зывает тенденция к увеличению размеров организмов (пункт 16
в табл. 8.1). Изменение размеров растений, по-видимому, связано
с возникновением адаптации к переходу на другой источник пи-
щи, от неорганических биогенных элементов к органическим ве-
ществам. Мелкие организмы обладают селективным преимуще-
ством в среде, богатой биогенами и компонентами минерального
питания. Это справедливо, в частности, для автотрофов, поскольку
у них велико отношение поверхности к объему. Однако по мере
развития экосистемы неорганические биогенные элементы оказы-
ваются все более и более связанными в биомассе (т. е. становятся
интрабиотическими), так что селективное преимущество перехо-
дит к более крупным организмам (либо к более крупным особям
того же вида, либо к более крупным видам, либо и к тем и к
другим), способность к запасанию веществ у которых выше и
жизненные циклы сложнее. Благодаря этим свойствам они адап-
тированы к использованию сезонных или периодических поступ-
лений биогенов пли других ресурсов.
Развитие и эволюция экосистемы
187
Тенденции изменения разнообразия. Хотя оба компонента раз-
нообразия (пункты 12 и 13 в табл. 8.1) почти всегда возрастают
на ранних стадиях развития экосистемы, все же пик разнообразия
в одних случаях приходится примерно на середину серии, в дру-
гих — ближе к концу. Тенденции изменения разнообразия во вре-
мя сукцессии неодинаковы в различных трофических или систе-
матических группах. Ранее мы уже кратко описали основные
стадии сукцессионной серии на залежных полях в предгорьях
в Джорджии (табл. 8.2). Никольсон и Монк (Nicholson, Monk,
1974) определили видовое богатство и выровненность для видов
растений, принадлежащих к четырем основным жизненным фор-
мам — травы, лианы, кустарники и деревья — в этих стадиях. Ви-
довое богатство в каждом ярусе возрастает быстро, сразу после
его закрепления, а затем на последующих стадиях сукцессии
скорость роста этого показателя уменьшается. Выровненность сра-
зу возрастает почти до максимума, а в дальнейшем меняется
очень мало. На рис. 8.8 показаны кривые доминирования—разно-
Последовательность видов
Рис. 8.8. Кривые доминирования — разнообразия для пяти разновозрастных
залежных полей в южном Иллинойсе. Черные кружки — деревья, белые —
травы, черные с белым — кустарники. (По Bazzaz, 1975.)
образия другой залежной сукцессии (южный Иллинойс). Разно-
образие видов растений в ходе сукцессии в общем увеличивается,
достигая максимума в начале стадии развития леса. Графики
видового распределения в первые годы вторичной сукцессии име-
188
Глава 8
ют характер геометрических кривых (приближаются к прямым в
полулогарифмических координатах), затем, по мере увеличения
числа новых видов, постепенно становятся логнормальными. Ре-
зультатом этого процесса оказывается высокий уровень выровнен-
ности (см. гл. 7, разд. 6). Последовательность кривых на рис. 8.8
очень напоминает последовательность от Арктики до тропиков,
показанную на рис. 7.18.
Оклер и Гофф (Auclair, Goff, 1971) провели одно из наиболее
полных исследований тенденций изменения разнообразия в ходе
продолжительной сукцессии. Они измеряли видовое разнообразие
деревьев в лесах разного возраста на западе района Великих
озер. В сырых местах большее разнообразие наблюдалось в лесах
среднего возраста и меньшее — в самых старых лесах. В сухих
местах, напротив, разнообразие непрерывно повышалось по мере
увеличения возраста леса.
Характер изменения видового разнообразия во время сукцес-
сии — непрерывный рост или прохождение пика в промежуточ-
ной стадии — зависит от соотношения двух процессов, а именно
от возрастания числа потенциальных ниш в результате увеличе-
ния биомассы, стратифицированное™ и других последствий био-
логической организации, и от противоположных эффектов увели-
чения размеров организмов (пункт 16 в табл. 8.1) и конкурент-
ного исключения среди хорошо адаптированных долго живущих
доминантов, которые снижают видовое разнообразие. Никому еще
не удалось составить полный перечень всех видов на площади
сколько-нибудь значительных размеров, не говоря уже о том,
чтобы проследить за всем разнообразием видов в сукцессионной
серии. Мы располагаем лишь данными для отдельных частей со-
общества (деревья, птицы и т. д.). Можно ожидать, что характер
изменения видового состава в значительной степени зависит от
изучаемой группы и географического положения, что определяет
набор видов, способных к вселению.
Как указывалось ранее (гл. 2, разд. 6), экологи единодушны
в своем мнении относительно того, что изменение видового раз-
нообразия представляет собой в большей степени косвенное след-
ствие увеличения количества органического вещества и сложно-
сти, а не прямое следствие причинных факторов сукцессии. До-
стигнутый уровень разнообразия может сильно зависеть от
энергетики, поскольку поддержание высокого разнообразия тре-
бует энергетических затрат и этот уровень может быть дестаби-
лизирован (опять синдром «хорошенького понемножку»).
Другие аспекты биохимического разнообразия в целом изуче-
ны далеко не столь систематически, как видовое разнообразие,
тем не менее было бы логично выявить их тенденции в ходе ауто-
генного развития экосистемы. Например, Джеффрис (Jeffries,
1979) обнаружил, что по мере созревания и усложнения морского
Развитие и эволюция экосистемы
189-
сообщества становится разнообразнее состав жирных кислот
планктона и бентоса. Это может служить примером увеличения
биохимического разнообразия.
Некоторые теоретические соображения. В начале этого разде-
ла было установлено, что развитие экосистемы происходит в ре-
зультате 1) изменений физической среды под воздействием сооб-
щества как целого и 2) конкурентных взаимодействий и сосуще-
ствования между слагающими сообщество популяциями. Логично
было бы предположить, что в многоплановое сукцессионное раз-
витие, описанное в этом разделе, вносят вклад процессы, проте-
кающие и на экосистемном, и на популяционном уровнях, однако
некоторые экологи считают, что такое развитие обусловлено ка-
кой-либо одной из этих причин, но не обеими. Коннелл и Слейтер
(Connell, Slayter, 1977) сравнили две теории: 1) «модель облег-
чения», в соответствии с которой виды ранних сукцессионных
стадий меняют условия существования и подготавливают, таким
образом, почву для последующих вселенцев, и 2) «модель инги-
бирования», согласно которой первые виды противостоят вторже-
ниям и сохраняются до тех пор, пока не окажутся вытесненными
в результате конкуренции, выедания и внешних возмущений. Кон-
нелл и Слейтер явно предпочитают вторую модель, по крайней
мере для вторичной сукцессии. Сущность аргументов сторонников
популяционных теорий причин сукцессии сводится к тому, что
если наблюдаемые сукцессионные тенденции можно объяснить
взаимодействиями на видовом уровне, то нет необходимости при-
влекать процессы на высших уровнях. Другие теоретики, напро-
тив, считают, что видовая сукцессия — это только часть процесса,
что самоорганизованное развитие — это свойство целых экосистем5
и что, следовательно, нет необходимости объяснять фундаменталь-
ные тенденции взаимодействиями составляющих популяций.
Идея о том, что экологическую сукцессию следует рассматри-
вать с холистической точки зрения, восходит к Фридерику Е. Кле-
ментсу и его монографии 1916 г. (F. Е. Clements, 1916), озаглав-
ленной «Растительная сукцессия» (переизданной в 1928 г. поц
названием «Растительная сукцессия и индикаторы»). Клементс
считал, что сообщество в своем развитии повторяет последова-
тельные стадии развития организма и что все сообщества в одной
климатической области развиваются в направлении одного и тоге
же климакса (идея «моноклимакса», см. следующий раздел); од-
нако в настоящее время эти идеи потеряли свою привлекатель-
ность или оказались сильно модифицированными. Тем не менее
его основной тезис — экологическая сукцессия — это процесс раз-
вития, а не только смена функционирующих изолированно ви-
дов — остается в основе одной из самых важных объединяющих
теорий экологии. Маргалеф (Margalef, 1963, 1963а, 1968)
и Ю. П. Одум (Е. Р. Odum, 1969) переработали и расширили?
190
Глава 8
фундаментальную теорию Клементса, дополнив ее такими функ-
циональными характеристиками, как метаболизм сообщества.
Главным для холистического подхода является вопрос о самоор-
ганизации как свойстве неравновесных термодинамических систем
(т. 1, с. 109). Ниже дается подборка современных утверждений,
принадлежащих авторам, которые нашли по крайней мере теоре-
тические основы для того, чтобы рассматривать сукцессию как
стратегию самоорганизации: «Полагают, что процесс в целом [сук-
цессия] протекает таким образом, что система накапливает раз-
нообразие и специализацию до тех пор, пока уровень неопреде-
ленности окружающей среды не обратит дальнейшее усложнение
организации во вред системе». (Valiela, 1971.)
«Твердо установлено, что в стабильных условиях окружающей
среды экологические системы с большим разнообразием успешно
конкурируют с более простыми и замещают их. Если особый
поток энергии не направлен на предотвращение сукцессии,
то сложная специализированная система характеризуется боль-
шей работой выхода, направленной в русло общих усилий, и вы-
тесняет те системы, энергия которых запасается, а не использу-
ется для полезной работы, направленной на выживание в конку-
ренции». (Н. Т. Odum, 1971.)
«Смена направлений приложения энергии при ее превращени-
ях объясняет относительные эффективности видов, которые сме-
няют друг друга во вторичной сукцессионной серии наземных
растений. Применение принципа смены способностей видов в се-
рии позволяет предсказать самые общие свойства роста и раз-
множения в ходе сукцессии». (Christopher Smith, 1976.)
«Ход и особенности вторичной сукцессии выводятся из теоре-
тической динамической модели взаимообусловленных обратных
связей. Эта модель [соответствует] табличной модели экологиче-
ской сукцессии Одума». (Gutierrez, Fey, 1975).
«Популяции сменяются в ходе сукцессии, но продуктивность
системы можно измерять на протяжении всего процесса. Сукцес-
сия— это не просто последовательность разных систем, а единая
система, которая меняет во времени переходящие виды и попу-
ляции». (O'Neil, Reichle, 1977).
«Предполагается, что самоорганизующиеся диссипативные сис-
темы развиваются во времени так, чтобы оптимизировать свою
независимость (сложность биомассы и потоков*в сетях). Это пред-
положение подтверждается тенденциями развития экосистем».
(Ulanowicz, 1980).
Противоположная концепция состоит в том, что экологическая
сукцессия не имеет оптимизирующей стратегии и происходит в
результате борьбы особей и видов за пространство. Эту концепцию
высказывает в своих работах Глисон (Н. A. Gleason), в частности
в статье 1926 г., озаглавленной «Индивидуалистическая концеп-
Развитие и эволюция экосистемы
191.
цпя растительной сукцессии». Работы Глисона, рассмотренные
недавно Мак-Интошем (McIntosh, 1975), служат отправной точ-
кой построения новых популяционных теорий сукцессии. Эти
теории характеризуются новыми подходами к эволюционной био-
логии и оценками тех влияний, которые оказывают друг на друга
консументы и продуценты, и реакций на такие воздействия. Тео-
рии сукцессии, основывающиеся на свойствах организмов, а не на
эмерджентных свойствах экосистем, разработаны в обзорах Друри
и Нисбета (Drury, Nisbet, 1973) и Хорна (Horn, 1974, 1975).
Главная предпосылка, согласно Пикетту (Pickett, 1976), состоит
в том, что меняющееся в зависимости от внешних возмущений и
физических градиентов положение вида в сукцессионной после-
довательности определяется его эволюционной стратегией (дар-
виновский отбор, конкурентное исключение и т. д.) п характери-
стиками жизненного цикла. С тех пор как холистическая теория
Клементса была признана эволюционной теорией сообщества, по-
пуляции и экосистемы, экологи не смогли далее оставаться в
стороне от этой проблемы, о чем свидетельствуют их работы. Эту
позицию занимал в общем покойный Роберт Уиттэкер (Whittaker,
1957, 1975; Whittaker, Woodwell, 1971). Глессер (Glasser, 1982)
в недавнем обзоре также отметил, что, хотя в фазе ранней ко-
лонизации закрепляются случайные организмы, которым повезло,
позднейшие стадии намного более организованны и упорядоченны.
Раньше или позже любая теория проверяется в практическом
мире прикладной науки, например в лесном хозяйстве. Лесоводы,
вообще говоря, считают, что лесная сукцессия направлена и пред-
сказуема. Для того чтобы оценить будущие запасы древесины,
они часто строят модели, в которых сочетаются тенденции при-
родной сукцессии с нарушениями и хозяйственной деятельностью,
меняющими природный ход процесса. В предгорных районах
Джорджии, например, естественная лесная сукцессия направлена-
от сосен к широколиственным деревьям. Поскольку сосны в на-
стоящее время представляют большую промышленную ценность,
чем широколиственные породы, были предприняты попытки оста-
новить сукцессию на стадии сосен, чтобы эта стадия сохранялась
и восстанавливалась, особенно в районах с развитыми заготовками
древесины. Джонсон и Шарп (Johnson, Sharpe, 1976) пишут, что,
несмотря на усилия по освоению лесов и сохранению сосны,
с 1961 по 1972 г. увеличилась площадь произрастания широко-
лиственных деревьев (как амбрового и тюльпанного деревьев на
ранних стадиях, так и дуба и гикори на поздних). Основываясь
на модели, построенной на 30-летнюю перспективу, эти авторы
считают, что площадь, занятая широколиственными породами,
будет увеличиваться и дальше, хотя скорость такого увеличения
будет меньше, чем в ходе естественной сукцессии. Важное значе-
ние в этой прогностической модели придается таким факторам.
192
Глава 8
как урбанизация и борьба с пожарами, которые дают широко-
лиственным деревьям преимущество перед соснами. Джонсон и
Шарп пришли к выводу, что хотя состав предгорного леса и
находится под сильным влиянием хозяйственной деятельности
человека, будущие изменения будут следовать тенденциям при-
родной сукцессии.
2. Концепция климакса
Определения
Терминальным, или стабильным, сообществом развивающейся се-
рии является климаксное сообщество. Теоретически климаксное
сообщество поддерживает само себя неопределенно долго, все
внутренние его компоненты уравновешены друг с другом, и оно
находится в равновесии с физической средой. В климаксном со-
обществе в отличие от развивающихся стадий и других переход-
ных сообществ годовая продукция и «импорт» уравновешиваются
годовым потреблением и «экспортом» (см. рис. 8.1 и 8.2). Удобно,
хотя и несколько произвольно, различать для данной области:
1) единственный региональный, или климатический, климакс, на-
ходящийся в равновесии с общими климатическими условиями,
п 2) различное число локальных, или эдафических, климаксов,
которые представляют собой модификации стационарных состоя-
ний, соответствующие особым местным условиям субстрата. В тех
местах, где рельеф местности, почва, водный режим и регуляр-
ные возмущения, например пожары, препятствуют развитию эко-
системы до теоретического конечного состояния, сукцессия закан-
чивается эдафпческпм климаксом.
Объяснения и примеры
Представление о том, что аутогенное развитие неизбежно приво-
дит к стабильному сообществу, широко принято, поскольку оно
основано на здравой теории и глубоких наблюдениях. Однако по
интерпретации этого представления экологи делятся на две шко-
лы. Согласно концепции моноклимакса (которая, как отмечалось
в предыдущем разделе, восходит к Клементсу), в любой области
теоретически возможен лишь один климакс, в направлении ко-
торого, хотя и медленно, развиваются все сообщества. Согласно
более реалистичной концепции поликлимакса, не следует ожи-
дать, что все сообщества в данной климатической области придут
к одному и тому же, несмотря на все разнообразие физических
условий. Нельзя также рассчитывать, что под влиянием данного
сообщества все местообитания достигнут какого-то единого уровня
Развитие и эволюция экосистемы 193
за приемлемые отрезки времени, сопоставимые с продолжитель-
ностью жизни человека (или в несколько раз большими!). Удоб-
ный компромисс между этими двумя точками зрения — призна-
ние единственного теоретического климатического климакса и
разного числа эдафических климаксов, зависящих от изменчиво-
сти субстрата.
Концепцию поликлимакса лучше всего проиллюстрировать на
частном примере. На рис. 8.9 показаны особенности рельефа юж-
ной части провинции Онтарио и стабильные сообщества, связан-
ные с различными физическими условиями. В равнинных или
слабохолмистых местностях с хорошо дренированной, но влажной
почвой терминальная стадия сукцессии представлена кленово-бу-
ковым сообществом (доминирующие растения — сахарный клен и
бук). Так как этот тип сообщества в данном регионе можно
встретить повсюду, где рельеф и дренирование выражены умерен-
но, сообщество клена и бука можно произвольно принять за кли-
матический климакс региона. Там, где, несмотря на деятельность
сообщества, почва более влажная или более сухая, чем в норме,
мы встречаем несколько отличные типы терминальных сообществ
(см. рис. 8.9). Еще большие отклонения от климатического кли-
макса наблюдаются на крутых южных склонах, где микроклимат
теплее, или на северных склонах и в глубоких лощинах, где
микроклимат холоднее. Эти климаксы часто похожи на климати-
ческие климаксы, находящиеся соответственно южнее или север-
нее. Так что если вы живете на востоке США или в гористой
местности на западе и заинтересуетесь, каким был бы климакс-
ный лес севернее, найдите ненарушенный северный склон или
лощину. Южный склон соответственно покажет вам, каким был
бы тип леса южнее.
Теоретически если предоставить лесному сообществу на сухой
почве развиваться неопределенно долго, то почва постепенно обо-
гатилась бы органическим веществом и соответственно возросла
бы ее влагоемкость; со временем это создало бы условия для
более влаголюбивого леса, например кленово-букового. Мы не зна-
ем, произойдет ли это на самом деле, так как у нас очень мало
данных о такого рода изменениях и так как наблюдения в нетро-
нутых областях не проводились в течение достаточно длительного
времени, что, по-видимому, было бы необходимо. В любом случае
вопрос этот чисто академический, поскольку задолго до того, как
могли бы произойти любые аутогенные изменения, скажется вме-
шательство климатических, геологических или антропогенных воз-
действий. Альтернативой рассмотрению ряда климаксов или серий,
связанных в мозаичном ландшафте, показанном на рис. 8.9, с част-
ными физико-географическими условиями, была бы некоторая
разновидность градиентного анализа (см. гл. 7, разд. 7). Экологи-
ческая сукцессия — это, в сущности, градиент во времени, кото-
13 Заказ № 1483
Местообитание: микроклимат и почва
Климаксное
биотическое
сообщество
Нормальный микроклимат
над сырой почвой
Клен— бук
} КЛИМАТИЧЕСКИЕ
КЛИМАКС
Нормальный микроклимат
над влажной почвой
Нормальный микроклимат
над сухой почвой
Дуб—ясень
Дуб—гикори
IIIH
W1
Теплый микроклимат
над сырой почвой
Тюльпанное дерево-
орех
Теплый микроклимат
над влажной почвой
Явор —тюльпанное
дерево
I f f Теплый микроклимат
над сухой почвой
Дуб—каштан
I ЭДАФИЧЕСКИЕ
[ КЛИМАКСЫ
41 111
МВ
Холодный микроклимат
над сырой почвой
Вяз—ясень—дуб
IIIII
Холодный микроклимат
над влажной почвой
Белая ель—
бальзамическая пихта
11111
ЕЕШ
Холодный микроклимат
над сухой почвой
Тсуга—желтая береза
Рис. 8.9. Климатический и эдафические
тарио. (Из Hills, 1952, с упрощениями.)
климаксы на юге провинции Он-
Развитие и эволюция экосистемы 195
рый взаимодействует с градиентом рельефа местности и климати-
ческим градиентом в пространстве.
Аутогенная экологическая сукцессия происходит в результате
изменений окружающей среды, вызванных самими организмами.
Поэтому, чем более экстремальны физические условия среды, тем
труднее становится модификация окружающей среды и тем боль-
ше вероятность того, что развитие сообщества прекратится, не до-
стигнув теоретического регионального климакса. Процент площа-
дей, на которых может развиться климатический климакс, сильно
варьирует в разных областях. Когда первые поселенцы достигли
Великих равнин, большая часть этого региона с его мощными
почвами была покрыта климаксной степью. Напротив, на песча-
ных, геологически более молодых прибрежных равнинах Юго-Вос-
тока климатический климакс (вечнозеленый широколиственный
лес) и в то время был такой же редкостью, как и теперь. Боль-
шая часть прибрежной равнины занята эдафическими климакса-
ми сосны или влаголюбивых сообществ, или их последовательны-
ми стадиями. Напротив, можно считать, что океаны, занимающие
геологически древние бассейны, представляют собой климаксы в
той мере, в какой это касается развития сообщества. Однако, как
уже упоминалось, здесь наблюдаются сезонные сукцессии и сук-
цессии, происходящие вслед за возмущениями и особенно выра-
женные в прибрежных водах.
На рис. 8.10 показан наглядный пример различия между ре-
гиональным и эдафическим климаксом. На одном прибрежном
участке в северной части Калифорнии леса из гигантской вечно-
зеленой секвойи соседствуют бок о бок с карликовыми лесами из
крошечных, чахлых деревьев. Как видно на рисунке, оба типа
лесов подстилает один и тот же песчаник, но карликовые леса
растут там, где водонепроницаемый горизонт залегает близко к
поверхности, сильно ограничивая развитие корней и препятствуя
перемещению воды и биогенов. Растительность, которая в этих
условиях достигает равновесия или климакса, почти полностью
отличается по своей структуре и видовому составу от раститель-
ности, покрывающей соседние участки, где отсутствует водонепро-
ницаемый горизонт.
Там, где преобладают описанные в предыдущем разделе цик-
лические сукцессии, климакс отсутствует в том смысле, что мы
не наблюдаем самоподдерживающегося стационарного состояния.
Существует, однако, тенденция рассматривать конец каждого цик-
ла как короткоживущий климакс или, по некоторым утверждени-
ям, «катастрофический климакс», разрушение которого «запро-
граммировано».
Обычно критерием того, является ли данное сообщество кли-
максным или нет, служит видовой состав. Однако одного этого
критерия часто оказывается недостаточно, поскольку видовой со-
13*
196
Глава 8
Ркг. 8.10. Эдафические климаксы на побережье северной Калифорнии
(район Мендосино). На примыкающих друг к другу береговых террасах
с одинаковой материнской породой (горизонт С) — береговые отложения и
песчаники — по соседству растут леса гигантской секвойи и карликовый
хвойный лес. Хвойный лес чахлый и низкорослый, потому что сцементиро-
ванный окислами железа горизонт В залегает примерно на глубине 45 см
от поверхности. Почва над непроницаемым горизонтом очень кисла»
(pH 2,8—3,9) и бедна Са, Mg, К, Р и другими минеральными компонентами
питания. По крайней мере одна из доминирующих карликовых сосен пред-
ставлена экотипом, специально адаптированным к этим крайним почвенным
условиям. (По Jenny, Arkley, Schultz, 1969.)
став заметно меняется в ответ на сезонные п кратковременные
колебания погоды, даже если экосистема в целом остается ста-
бильной. Как уже указывалось, хорошим показателем может слу-
жить отношение P/R или другие функциональные критерии. По-
лезны также различные статистические оценки, например время
оборота (в климаксе дольше, чем в серии), коэффициенты вариа-
ции, индексы сходства и другие показатели устойчивости.
Человек, конечно, сильно влияет на ход сукцессии и дости-
жение климакса. Когда стабильное сообщество, не представляю-
щее собой климатический или эдафический климакс для данной
местности, поддерживается человеком или домашними животны-
ми, его удобно было бы обозначить как дисклимакс (т. е. нару-
шенный климакс) или антропогенный субклимакс (т. е. порож-
денный человеком). Например, чрезмерный выпас может привести
к образованию пустынного сообщества с зарослями креозотового
кустарника, мескита и кактусов там, где по условиям локального
климата могла бы сохраниться степь. Пустынное сообщество в
данном случае — дисклимакс, а степь — климатический климакс.
Пустынное сообщество в таких условиях — показатель бесхозяй-
ственности человека, тогда как в регионе с настоящим пустынным
климатом его следует считать естественным. Интересное сочетание
эдафического климакса и дисклимакса наблюдается на обширных
Развитие и эволюция экосистемы
197
пространствах степных районов Калифорнии, где интродуцирован-
ные виды однолетников почти полностью вытеснили местные зла-
ки прерий (более подробно об этом дисклимаксе см. McNaughton,
1968).
Сельскохозяйственные экосистемы, остающиеся долгое время
стабильными, определенно можно рассматривать как климаксы
(или дисклимаксы), поскольку в среднем за год «импорт» и про-
дукция уравновешивают дыхание и «экспорт» (урожай), и ланд-
шафт при этом из года в год не меняется. Примерами длительно-
го антропогенного стационарного состояния являются сельское
хозяйство в Голландии и Бельгии и древняя культура риса на
Востоке. К сожалению, многие системы ведения сельского хозяй-
ства, особенно современные системы в тропиках и орошаемых
пустынях, никоим образом нельзя считать стабильными, так как
эти системы подвержены эрозии, выщелачиванию, засолению и
нашествиям вредителей. Поддержание высокой продуктивности
таких систем требует все возрастающих энергетических субсидий,
а слишком крупные субсидии могут оказаться стрессовыми для
системы (гл. 3, разд. 3, рис. 3.5).
3. Эволюция биосферы
Определения
Как и при описанном в разд. 1 этой главы развитии в течение
коротких промежутков времени, долговременная эволюция био-
сферы формируется под влиянием 1) аллогенных (внешних) сил,
таких, как геологические и климатические изменения, и 2) ауто-
генных (внутренних) процессов, обусловленных активностью жи-
вых компонентов экосистемы. Первые экосистемы, существовав-
шие 3 млрд, лет тому назад, были населены крошечными анаэроб-
ными гетеротрофными организмами, существовавшими за счет
органического вещества, синтезировавшегося в абиотических про-
цессах. Затем последовали возникновение и популяционный взрыв
автотрофных водорослей, которые, как полагают, сыграли одну
из главных ролей в превращении восстановительной атмосферы
в кислородную. С этого момента на протяжении длительного гео-
логического времени эволюция организмов шла по пути создания
все более сложных и разнообразных систем, которые 1) контро-
лировали состав атмосферы и 2) содержали в себе все более круп-
ные и высокоорганизованные виды многоклеточных. Принято
считать, что эволюционные изменения происходят в принципе
путем естественного отбора на видовом или более низком уровнях.
Однако возможно, что естественный отбор на более высоких уров-
нях также играет важную роль, особенно 1) коэволюция, т. е.
198
Глава 8
взаимный отбор зависящих друг от друга автотрофов и гетеро-
трофов, и 2) групповой отбор, или отбор на уровне сообществу
который ведет к сохранению признаков, благоприятных для груп-
пы в целом, даже если они неблагоприятны для конкретных но-
сителей этих признаков внутри группы.
Объяснения
На рис. 8.11 изображена грубая схема эволюции организмов и
кислородной атмосферы. Эти два фактора сделали биосферу уни-
кальной среди планет нашей Солнечной системы. Сейчас считает-
ся общепринятым, что, когда свыше 3 млрд, лет назад на Земле
зародилась жизнь, атмосфера содержала азот, аммиак, водород,
окись углерода, метан и водяной пар, но свободный кислород в
ней отсутствовал (см. Berkner, Marshall, 1964, 1965; Drake, 1968;
Tappen, 1968; Calvin, 1969). В атмосфере содержались также
хлор, сероводород и другие газы, ядовитые для большинства со-
временных организмов. Состав атмосферы в то время в значи-
тельной степени определялся вулканическими газами; вулканы
тогда были намного более активными, чем сейчас. Из-за отсут-
ствия кислорода не существовало и озонового слоя (под действи-
ем коротковолнового излучения Ог превращается в Оз, в свою
очередь поглощающего ультрафиолетовое излучение), экраниру-
ющего губительное ультрафиолетовое излучение Солнца, которое,
таким образом, достигало поверхности суши и воды. Это излуче-
ние убило бы любые незащищенные от него организмы, но, как
это ни странно, считается, что именно оно породило химическую
эволюцию, приведшую к возникновению сложных органических
молекул, таких, как аминокислоты, которые послужили блоками
для построения примитивных живых систем. Очень малое коли-
чество кислорода, образуемого за счет абиотических процессов,
например при диссоциации водяного пара под действием ультра-
фиолета, могло обеспечить достаточное количество озона, чтобы
создать некоторую защиту от самого ультрафиолетового излуче-
ния. Все же, пока атмосферного кислорода и озона было мало,
жизнь могла развиваться только под защитой слоя воды. Первы-
ми живыми организмами были, следовательно, дрожжеподобные
анаэробы, которые получали необходимую для дыхания энергию
путем брожения. Так как брожение гораздо менее эффективно,
чем кислородное дыхание (см. т. 1, с. 50), примитивная жизнь не
могла эволюционировать дальше одноклеточной стадии прокариот
(организмов, не имеющих оформленного ядра). Снабжение при-
митивных организмов пищей также было очень ограничено; их
питание, по-видимому, зависело от медленно опускавшихся на дно
органических веществ, синтезировавшихся под действием радиа-
ции в верхних слоях воды, куда не рисковали подниматься эти
Возникн- еение
цветковых растении
Очень низка — Быстро Стационарна на ч Стационарна на Колеблющееся* *- Гмищ-ДРТСИ WKMIkTlM ^.ЙПЧ.ПЛЛ.ЛРТ-^СТЯПИЛНЯПНПГ ПЕРВИЧНАЯ
возоастает высоком уровне уровне состоя ние продукция
Преимущественно анаэробы Зеленые и сине-зеленые водоросли с целлюлозной оболочкой Преи му щественно известковые олитофориды и др. Известко- вые и кремне- вые диа- томовые и др. ОКЕАНИЧЕСКИЙ ПЛАНКТОН
Только одноклеточные организмы Только Возникновениеi анаэробы фотосинтези Восстановительная атмосфера 1 Возникновение и быстрая ^^воноТных'и назем- эволюция многоклеточных н^^^ительности® Эволюция высших животных и растений Популя - ЦИОННЫЙ взрыв человека ОБЩАЯ ЭВОЛЮЦИЯ БИОСФЕРЫ
Увеличение числа л популяционный взрыв растительноядных Популяционный взрыв наземных теплокровных на суше
эующих автотрофов живо отло - -Окислительная атмосфера5^£|?! (тных и биогенное жение кальция ние запасов
ого горючего
ДОКЕМБРИЙ — Кембрий Ордовик . Силур Девон 1 со с g С Пермь Триас Юра 5 < Третичным ГЕОЛОГИЧЕСКИЕ ПЕРИОДЫ
ПАЛЕОЗОЙ МЕЗОЗОЙ КАЙНО- ЗОЙ
1000 500 400 300
200 100
Миллионы лет назад
Рис. 8.11. Эволюция биосферы и ее кислородной атмосферы. Л. Датировка
событий биологической эволюции на фоне предполагаемого изменения
уровня кислорода. (Cloud, 1978.) Б. Геологические периоды и возникновение
основных биотических компонентов по данным палеонтологической ле-
тописи.
200
Глава 8
голодные микробы! Так на протяжении миллионов лет жизнь
вынуждена была существовать в очень неподходящих условиях,
подвергаясь множеству опасностей. Беркнер и Маршалл (Berk-
ner, Marshall, 1969) обрисовали эту ситуацию: «Для этой мо-
дели примитивной экологии необходим водоем, достаточно глу-
бокий, чтобы поглощался губительный ультрафиолет, но не столь
глубокий, чтобы не ослабить чрезмерно видимый свет. Жизнь
могла зародиться на дне небольших водоемов или мелководных
закрытых морей, питавшихся, по-видимому, горячими источника-
ми, богатыми пригодными в пищу химическими веществами».
Возникновение фотосинтеза покрыто тайной. По-видимому,
определенную роль сыграло давление отбора, обусловленное ску-
достью органической пищи. Постепенное увеличение в воде ко-
личества кислорода за счет жизнедеятельности организмов и его
диффузия в атмосферу около 2 млрд, лет назад (рис. 8.11, Б)
вызвали громадные изменения в химии Земли и сделали возмож-
ным быстрое распространение жизни и развитие эукариотических
(обладающих оформленным ядром) клеток, что привело в свою
очередь к эволюции более крупных и более сложных живых сис-
тем. Многие минералы, такие, как железо, выпали из воды в
осадок и образовали характерные геологические формации.
По мере увеличения содержания кислорода в атмосфере слой
озона в ее верхней части становился все более мощным и мог
экранировать разрушающее ДНК ультрафиолетовое излучение.
Жизнь теперь могла свободно распространиться к поверхности
моря. Затем последовало то, что Клауд (Cloud, 1978) назвал «по-
зеленением суши». Аэробное дыхание сделало возможным разви-
тие сложных многоклеточных организмов. Считается, что первые
ядерные клетки появились после того, как содержание кислорода
в атмосфере достигло 3—4% (или около 0,6% атмосферы) его
современного уровня (20%). Сейчас полагают, что это произошло
примерно 1 млрд, лет назад. Маргулис (Margulis, 1981, 1982)
приводит достаточно убедительные доводы в пользу теории, пред-
полагающей, что эукариотические клетки возникли как мутуа-
листическое объединение независимых прежде микробов анало-
гично современной эволюции лишайников, показанной на
рис. 7.11.
Когда содержание кислорода около 700 млн. лет назад достиг-
ло примерно 8%, появились первые многоклеточные организмы
(Metazoa) (рис. 8.11, Б). Термин «докембрий» используется для
обозначения того огромного периода времени, когда существовали
только мелкие, прокариотические одноклеточные формы жизни.
В кембрии произошел эволюционный взрыв новых форм жизни,
таких, как губки, кораллы, черви, моллюски, морские водоросли
и предки семенных растений и позвоночных. Так, благодаря спо-
собности мельчайших зеленых растений моря продуцировать та-
Развитие и эволюция экосистемы
201
кое количество кислорода, которое превышало потребности в нем
всех организмов, оказалось возможным заселение живыми суще-
ствами за сравнительно короткое время всей Земли. В течение
последующих периодов палеозойской эры жизнь не только за-
полнила все моря, но и вышла на сушу. Развитие зеленой назем-
ной растительности обеспечило большие количества кислорода и
пищи, которые были необходимы для последующей эволюции
таких крупных животных, как динозавры, млекопитающие и. на-
конец, человек. В это же самое время к океаническому планкто-
ну, ранее состоявшему из форм с клеточными оболочками из
органических веществ, прибавились формы с известковыми,
а позднее и с кремневыми оболочками (рис. 8.11, Л).
Когда примерно в середине палеозоя, около 400 млн. лет на-
зад, продукция кислорода сравнялась с его потреблением, содер-
жание кислорода в атмосфере достигло современного уровня,
т. е. 20 % • Следовательно, с экологической точки зрения эволю-
цию биосферы можно сравнить с гетеротрофной сукцессией, на
смену которой пришел автотрофный режим, подобной той сукцес-
сии, которую можно воспроизвести в лабораторном микрокосме,
начав с культуральной среды, обогащенной органическим вещест-
вом. В конце палеозоя произошло снижение содержания О2 и
повышение содержания СО2, сопровождавшееся изменениями кли-
мата и, по-видимому, послужившее толчком к обширному «авто-
трофному цветению», создавшему запасы ископаемого топлива,
на которых основана современная промышленная цивилизация.
Затем последовало постепенное возвращение к атмосфере с высо-
ким уровнем О2 и низким уровнем СО2, после чего отношение
О2/СО2 остается в состоянии, которое можно назвать осциллирую-
щим стационарным состоянием. Антропогенное загрязнение ат-
мосферы пылью и СО2 может сделать это ненадежное равновесие
еще более «нестационарным» (см. гл. 2 и гл. 4).
Между прочим подобную краткую историю развития атмосфе-
ры следовало бы довести до сведения каждого гражданина и
каждого школьника, так как она ярко показывает абсолютную
зависимость человека от других организмов, населяющих среду,
в которой обитает он сам. Согласно гипотезе Геи (гл. 2, разд. 4),
регуляция состояния среды организмами, особенно микроорганиз-
мами, установилась в очень ранний период истории биосферы.
Противоположная гипотеза утверждает, что жизнь на ранних ста-
диях просто была в состоянии адаптироваться к условиям вы-
званных радиацией физико-химических изменений. Иными сло-
вами, вопрос ставится так: была ли ранняя эволюция в большей
степени аутогенной или аллогенной? Вызывает оживленные дис-
куссии также вопрос о том, эволюционировала ли жизнь посте-
пенно или в процессе эволюции наблюдались четко выраженные
импульсы (короткие периоды быстрых изменений сменялись дли-
202
Глава 8
тельными периодами, когда изменения были невелики или их
вовсе не было), как об этом свидетельствует изучение ископаемых
остатков. Этот вопрос будет рассмотрен в следующем разделе.
На протяжении всей органической эволюции на развитие форм
жизни значительное влияние оказывал «дрейф континентов». Этот
процесс подробно обсуждается Уилсоном (Wilson, 1972).
Очень хорошо читается книга об эволюции биосферы, напи-
санная Клаудом (Cloud, 1978). Прекрасное изложение эволюции
жизни можно найти в книге Маргулис, озаглавленной «Ранняя
жизнь» (Margulis, 1982).
4. Естественный отбор: аллопатрическое
и симпатрическое видообразование;
микроэволюция и макроэволюция
Определения
Вид — это естественная биологическая единица, всех членов ко-
торой связывает воедино участие в общем генофонде. Эволюция
состоит в изменении частот генов, которое происходит вследствие
1) давления отбора, вызванного окружающей средой и взаимо-
действующими видами; 2) повторяющихся мутаций и 3) генети-
ческого дрейфа (стохастических или «случайных» изменений ге-
нетической структуры). Видообразование, или формирование но-
вого вида и развитие видового разнообразия, происходит тогда,
когда поток генов в пределах общего генофонда прерывается
каким-либо изолирующим механизмом. Если изоляция возникает
вследствие географического разделения популяций, происходящих
от общего предка, то происходит аллопатрическое (разные районы
происхождения) видообразование. Если изоляция осуществляется
экологическими или генетическими средствами в пределах данной
области, то можно говорить о симпатрическом (одинаковые райо-
ны происхождения) видообразовании. В настоящее время неясно,
в какой степени видообразование представляет собой медленный,
градуальный процесс (микроэволюция), или же это процесс пе-
риодических быстрых изменений (макроэволюция).
Объяснения
Биологическая эволюция, включающая дарвиновский естествен-
ный отбор и генные мутации на видовом уровне, широко призна-
ется учеными как факт, а не только как теория. Однако до сих
пор нет единого мнения относительно ее механизмов. Особенно
неясно, каковы относительные роли трех перечисленных механиз-
мов: отбора, мутации и случайности — и какова роль отбора на
Развитие и эволюция экосистемы
203
высших уровнях биологической организации (коэволюция и груп-
повой отбор). Эти последние аспекты будут рассмотрены в разд 6
и 7.
Со времени Дарвина биологи в общем придерживались теории,
что эволюционное изменение — это медленный, постепенный про-
цесс, включающий множество мелких мутаций и непрерывный ес-
тественный отбор, который обеспечивает преимущество в конку-
ренции на уровне особи. Однако пробелы в палеонтологической
летописи и неудачи попыток найти промежуточные формы («не-
достающие звенья») заставили многих палеонтологов допустить
существование «прерывистого равновесия», как это назвали Гоулд
и Элдридж (Gould, Eldredge, 1972, 1977). Согласно этой теории,
на протяжении долгих периодов времени виды остаются неизмен-
ными в состоянии своего рода эволюционного равновесия. Время
от времени это равновесие «прерывается», тогда небольшая попу
ляция отделяется от родительского вида и быстро развивается в
новый вид, при этом в палеонтологической летописи не остается
промежуточных форм. Новый вид может настолько отличаться от
родительского, что будет сосуществовать с ним, не вытесняя его,
либо оба вида вымрут. Теория прерывистой эволюции не припи-
сывает исключительной роли конкуренции на индивидуальном
уровне как движущей силе процесса; однако до сих пор неясно,
что может быть причиной внезапного разделения популяции,
формирующего новую, генетически изолированную единицу.
Дальнейшие сведения о макроэволюции как альтернативе микро-
эволюции содержатся в работах Гоулда (Gould, 1977), Ренсбер-
гера (Rensberger, 1981, 1982) и Стенли (Stanley, 1979, 1981).
Виды, обитающие в разных географических областях или раз-
деленные пространственными барьерами, называются аллопатри-
ческими. Если виды обитают в одной и той же местности, то их
называют симпатрическими. Аллопатрическое видообразование
принято считать главным механизмом возникновения новых ви-
дов. Согласно этой традиционной точке зрения, две части какой-
либо свободно скрещивающейся популяции становятся разделен-
ными пространственно (как на острове или по обе стороны гор-
ного хребта). Со временем такие изолиты накапливают достаточ-
но генетических различий, так что если они вновь придут в
контакт, то уже не смогут обмениваться генами (скрещиваться)
и будут сосуществовать как отдельные виды в разных нишах.
Иногда эти различия могут усилиться за счет смещения призна-
ков. Если ареалы двух близкородственных видов перекрываются,
у них наблюдаются тенденция к дивергенции по одному или
нескольким морфологическим, физиологическим или поведенче-
ским признакам в зоне перекрывания и тенденция к конверген-
ции (остаться такими же или стать похожими друг на друга)
в частях ареала, где каждый вид встречается поодиночке. Браун
204
Глава 8
и Уилсон (Brown, Wilson, 1956) приводят подробные объяснения
и примеры явления смещения признаков.
Накапливается все больше данных о том, что для видообра-
зования не обязательно необходима строгая географическая изо-
ляция и что симпатрическое видообразование распространено и
шире и играет более важную роль, чем полагали прежде. Внутри
одной и топ же географической области популяции могут стать
генетически изолированными вследствие особенностей поведения и
размножения, таких, как колонизация, ограниченное расселение
стадий размножения, бесполое размножение, отбор, выедание хищ-
никами и т. и. В локальных частях популяции со временем на-
капливается достаточно генетических различий, чтобы воспрепят-
ствовать скрещиванию.
Примеры
Галапагосские вьюрки представляют собой классический пример
аллопатрического видообразования (вследствие географической
изоляции) с последующим смещением признаков. Этот случай
был впервые описан Дарвином, который посетил Галапагосские
острова во время своего знаменитого плавания на «Бигле». В ус-
ловиях изоляции на разных островах от общего предка возникла
целая группа видов; они претерпели столь сильную адаптивную
радиацию, что при повторных заселениях различных островов за-
полняли самые разнообразные потенциальные ниши. Среди совре-
менных видов есть тонкоклювые насекомоядные, толстоклювые
зерноядные, питающиеся на земле и на деревьях, крупные и
мелкие вьюрки и даже один вьюрок, похожий на дятла, хотя он
вряд ли выдержал бы конкуренцию с настоящим дятлом и может
выжить лишь в отсутствие этих птиц (см. Lack, 1947). На
рис. 8.12 показаны вариации размера клюва одного из галапагос-
ских вьюрков в зависимости от того, обитает ли он на острове
один или же вместе с двумя близкородственными видами на
другом, более крупном острове. В этой ситуации (Б на рис. 8.12)
происходит «смещение» размера клюва в направлении увеличения
его толщины, так что он не перекрывается по размерам с клю-
вами двух конкурентов. В результате конкуренция за пищу умень-
шается, так как каждый из трех видов адаптирован к питанию
семенами разной величины.
Засоленные марши Британских островов дают нам пример
симпатрического видообразования в результате гибридизации и
полиплоидии. Когда Spartina alternvflora, злак, произрастающий
на засоленных маршах Америки, попал на Британские острова,
он стал скрещиваться с местным видом Spartina maritima. Воз-
ник новый полиплоидный вид Spartina townsendii, который засе-
Развитие и эволюция экосистемы
205
Рис. 8.12. Величина клюва у Дарвинова выорка Geospiza fortis, живущего
на одном из Галапагосских островов в отсутствие конкурентов (Л) и на
других островах вместе с конкурентами — другими видами Geospiza
(В). Увеличение размеров клюва в присутствии конкурентов — пример
смещения признаков. (Lack, 1947.)
лил теперь голые, прежде илистые отмели, где не росли мест-
ные виды.
Другой пример быстрого естественного отбора, вызванного
вмешательством человека, — это так называемый «индустриаль-
ный меланизм»: возникновение темной окраски у бабочек в про-
мышленных районах Англии, где стволы деревьев заметно потем-
нели от копоти, которая погубила лишайники, придававшие
стволам светлый оттенок. Кеттлуэлл (Kettlewell, 1956) экспери-
ментально показал, что темные бабочки лучше выживают на тем-
ных (закопченных) стволах, а светлые — на обычных; вероятно,
это вызвано тем, что птицы выборочно истребляют особей, не име-
ющих покровительственной окраски. Этот пример, естественно,
приводит нас к проблеме прямого, или целенаправленного, отбора,
осуществляемого человеком.
Искусственный отбор: доместикация. Отбор, который произ-
водит человек с целью приспособить растения и животных к
своим нуждам, называют искусственным отбором. Доместикация 1
растений и животных — это не только изменение генетики вида,
так как при этом необходима взаимная адаптация одомашнивае-
мого вида и доместикатора. Доместикация, следовательно, ведет
к особой форме мутуализма. Будучи эгоцентричным, человек впа-
1 Для искусственного отбора растений многие предпочитают термин
«окультуривание». Здесь мы используем термин «доместикация» (одомаш-
нивание) в общем смысле для растений и для животных. — Прим, автора.
206
Глава 8
дает в заблуждение и считает, что, одомашнивая другой организм
путем искусственного отбора, он просто «покоряет» природу, при-
способляя ее для наших целей. На самом же деле доместикация
вызывает у человека такие же изменения (если не генетические,
то, во всяком случае, экологические и социальные), как п у одо-
машненных видов. Поэтому человек в той же мере зависит от
сельскохозяйственных растений, как и сельскохозяйственные рас-
тения зависят от нас. Общество, хозяйство которого зависит от
кукурузы, развивается в культурном отношении совершенно по-
иному, нежели общество, занятое пастбищным скотоводством.
Еще вопрос: кто у кого в рабстве? Этот же вопрос можно отнести
к взаимоотношениям человека и его машин. Взаимоотношения
человека с трактором, по-видимому, не слишком отличаются от
взаимоотношений человека с лошадью; разница только в том, что
трактор требует гораздо больше высококачественной энергии (го-
рючего) и выделяет ядовитые отходы!
Хорошим примером взаимозависимости между одомашнивае-
мым видом и доместикатором-человеком служит искусственный
отбор зерновых — основа «зеленой революции». Повышение уро-
жайности получено путем отбора на большее отношение зерна
(или других съедобных частей) к поддерживающим тканям
(листья, корни, стебли и т. д.), иными словами, повышению уро-
жайности выше определенного уровня приносится в жертву адап-
тивная способность растения к самоподдержанию. Таким образом,
высокоинбредные линии требуют мощных вкладов энергии, удоб-
рений и пестицидов, что вызывает глубокие изменения в социаль-
ной, экономической и политической структуре общества. Для
многих отсталых стран эти требования к социально-экономиче-
скому укладу и ресурсам являются наибольшим препятствием
для использования высокоурожайных сортов, необходимых для
повышения производства продуктов питания.
Некоторые из наиболее серьезных проблем, стоящих перед
человеком, связаны с одомашненными растениями и животными,
которые «вырвались из плена» (т. е. одичали) и, вернувшись в
природу, превратились в настоящий бич. Одичавший организм
отличается от своего дикого предка тем, что он перенес период
искусственного отбора, приобретя некоторые новые свойства и
утратив некоторые «дикие». Вернувшись к жизни в природе,
одичавший организм вновь подпадает под действие естественного
отбора, который благоприятствует свойствам, необходимым для
самостоятельного выживания. Например, отбор работает против
пятнистой или светлой окраски и крупных размеров домашних
сзиней, если они попадают в дикую природу. Поэтому одичавшие
свиньи становятся более мелкими и темными. Сочетание искус-
ственного и естественного отбора приводит к тому, что такие
растения и животные процветают в частично измененных или
Развитие и эволюция экосистемы
207
нарушенных местообитаниях. Например, одичавшие свиньи и ко-
зы могут произвести опустошительные разрушения растительно-
сти островов или районов, из которых изъяты крупные хищники.
Островная биогеография. Острова представляют собой природ-
ные лаборатории для изучения эволюции, зачаровывая биологов и
экологов со времени посещения Дарвином Галапагосских остро-
вов. Сложная взаимосвязь изоляции, естественного отбора и ви-
Рис. 8.13. Теория островной биогеографии. Число видов на острове опреде-
ляется равновесием между скоростями иммиграции и вымирания. Показаны
четыре точки равновесия, соответствующие разным комбинациям размеров
острова и удаления его от материка. (MacArthur, Wilson, 1963.)
дообразования привлекает внимание и сейчас, особенно после
того, как Мак-Артур и Уилсон (MacArthur, Wilson, 1963, 1967)
опубликовали свою теорию островной биогеографии. В простей-
шей формулировке эта теория состоит в том, что число видов на
острове определяется равновесием между иммиграцией новых ви-
дов и вымиранием уже существующих. Поскольку число видов
увеличивается и уменьшается примерно логарифмически и по-
скольку скорости иммиграции и вымирания зависят от величины
острова и расстояния до материка, служащего источником видов,
общая модель равновесия может быть изображена графически
так, как это сделано на рис. 8.13. Показаны четыре точки рав-
новесия, соответствующие 1) небольшому удаленному острову,
на котором предсказывается существование немногих видов;
2) большому, близкому к материку острову, который должен быть
населен большим числом видов; 3) большому удаленному острову
и 4) небольшому близкому острову, которые занимают промежу-
точное положение в отношении видового богатства. Теория рав-
новесия была проверена Симберлоффом и Уилсоном (Simberloff,
Wilson, 1969), которые уничтожили всех членистоногих (путем
обработки инсектицидом) на одном из небольших мангровых
208
Глава 8
островов из группы Флорида-Кис и наблюдали реколонизацию.
В обзоре Симберлоффа (Simberloif, 1974) содержатся более по-
дробные сведения об островной биогеографии.
Уилсон и Уиллис (Wilson, Willis, 1975), Даймонд п Мей
(Diamond, Мау, 1981) и другие исследователи предположили, что
теория островной биогеографии дает основу для планирования
резерватов, предназначенных для сохранения природного разно-
образия и (или) находящихся под угрозой исчезновения видов.
В соответствии с этой теорией один большой резерват предпочти-
тельнее группы мелких равной площади. Если приходится разби-
вать небольшие парки, то они должны быть расположены близко
друг от друга или должны быть соединены коридорами, облегча-
ющими расселение животных. Резерват в форме круга, когда
максимизируется отношение площади к периметру, предпочти-
тельнее вытянутого резервата. Резерваты или парки, находящиеся
внутри континентальных областей, редко бывают изолированы в
такой степени, как океанические или прибрежные острова, по-
этому неясно, применима ли к ним модель Мак-Артура—Уилсона.
Некоторые другие проблемы, возникающие при применении ост-
ровной теории к природным заповедникам, рассмотрены в работах
Симберлоффа и Эбиля (Simberloff, Abele, 1976) и Кашлена (Kush-
Ian, 1979).
5. Коэволюция
Определения
Коэволюция — это тип эволюции сообщества (т. е. эволюционных
взаимодействий между организмами, при которых обмен генети-
ческой информацией между группами минимален или отсутству-
ет), заключающийся во взаимных селективных взаимодействиях
друг на друга двух больших групп организмов, находящихся в
тесной экологической взаимозависимости, таких, как растения и
травоядные, крупные организмы и их микроскопические симби-
онты, паразиты и их хозяева.
Объяснения
Теория коэволюции в том виде, в каком она принимается сейчас
исследователями-эволюционистами, была сформулирована в об-
щих чертах Эрлихом и Равеном (Ehrlich, Raven, 1965), которые
использовали в качестве основы для этой теории свои исследова-
ния, проведенные на бабочках и растениях. Их гипотеза вкратце
сводится к следующему: в результате случайных мутаций и ре-
комбинаций растения начинают синтезировать химические веще-
Развитие и эволюция экосистемы 209"
ства, которые не связаны непосредственно с основными метабо-
лическими путями (или образуются в этих путях как побочные*
продукты метаболизма). Эти вещества не мешают нормальному
росту и развитию, но некоторые из них случайно уменьшают
съедобность растений для травоядных. Такие защищенные от на-
секомых-фитофагов растения попадают в известном смысле в
новую адаптивную зону. Затем может последовать эволюционная
радиация растений и то, что возникло как случайная мутация
или рекомбинация, может при известном стечении обстоятельств
стать признаком целого семейства или группы родственных се-
мейств. Однако насекомые-фитофаги также способны эволюиро-
вать в ответ на возникшие физиологические препятствия, о чем
свидетельствует широкое распространение устойчивых линий..
В самом деле, создается впечатление, что реакция на побочные
вещества растений и развитие устойчивости к инсектицидам тесно
связаны друг с другом (Gordon, 1961). Если в популяции насе-
комых появляется такой мутант или рекомбинант, способный пи-
таться на растениях, которые прежде были устойчивы к данному
насекомому, то отбор вынесет эту линию в новую адаптивную
зону, что даст ей возможность дивергировать в отсутствие других,
растительных конкурентов. Таким образом, не только разнообра-
зие растений способствует увеличению разнообразия животных-
фитофагов, но может оказаться верным и обратное. Иными сло-
вами, растения и фитофаги эволюируют совместно в том смысле,,
что эволюция каждого из них зависит от эволюции другого.
Для обозначения этого типа эволюции Пиментел (Pimentel,
1968) использовал выражение «генетическая обратная связь»; эта
обратная связь приводит к гомеостазу популяции и сообщества-
внутри экосистемы.
Примеры
Многочисленные примеры коэволюции обсуждаются и анализиру-
ются в трудах симпозиума, опубликованных под редакцией Гилы
берта и Равена (Gilbert, Raven, eds, 1980). Один пример из этой'
книги наглядно показывает такую совместную эволюцию растений*
и животных.
Широко распространенные виды шмелей рода Bombus извест-
ны как важные опылители и диких растений, и таких культур-
ных форм, как люцерна, клевер, бобы и черника. Хейнрих (Hein-
rich, 1979, 1980) исследовал энергетику взаимодействия цветков*
и шмелей. Он измерил продукцию нектара (количество доступ-
ного сахара в одном цветке), подсчитал число посещений цветков-
шмелями и скорость потребления нектара в зависимости от вре-
мени дня и температуры. Шмели в отличие от бабочек должны-
посещать цветки часто, чтобы получить энергетическую выгоду,,
14 Заказ № 1483
210
Глава 8
так как у них высок уровень метаболизма. Для привлечения
этих необходимых опылителей и обеспечения выживания многие
виды цветковых выработали определенную «стратегию»: они либо
цветут одновременно, либо растут группами.
Как показал Колуэлл (Colwell, 1973), реципрокный естествен-
ный отбор не ограничивается двухвидовыми взаимодействиями.
Он описал интересный случай, когда в тропическом субсообщест-
ве совместно эволюировали десять разных видов — четыре цветко-
вых растения, три колибри, еще одна птица и два клеща. Коэво-
люция может захватывать больше одной связи в пищевой цепи.
Например, Брауэр и др. (Brower et al., 1968) изучали бабочку-
монарха (Danaus plextppus), которую, как хорошо известно,
не едят позвоночные хищники. Они обнаружили, что эта бабочка
может ассимилировать очень ядовитые гликозиды, содержащиеся
в млечном соке растений, которыми она питается, обеспечивая
себя таким образом очень эффективной защитой от птиц (защи-
щенными оказываются не только гусеницы, но и взрослые бабоч-
ки). Таким образом, бабочка не только приобрела в процессе
эволюции способность питаться растениями, не поедаемыми дру-
гими насекомыми, но и способность «использовать» яд растения
для собственной защиты от хищников.
6. Эволюция кооперации и сложности;
групповой отбор
Определения
Для того чтобы описать удивительное разнообразие и сложность
биосферы, ученые вынуждены были постулировать, что естествен-
ный отбор действует вне видового уровня и вне коэволюции. В со-
ответствии с этим групповой отбор — это естественный отбор в
труппах организмов, которые не обязательно тесно связаны в му-
туалистических ассоциациях. Теоретически групповой отбор ведет
к сохранению благоприятных для популяции и сообщества при-
знаков, которые могут быть селективно невыгодны для их отдель-
ных генетических носителей внутри популяций. Справедливо и
обратное: групповой отбор элиминирует или поддерживает на
уровне низких частот признаки, неблагоприятные для выживания
вида, но селективно ценные внутри популяций. Групповой отбор
включает выгоды, которые получает организм, совершенствуя ор-
ганизацию сообщества, необходимую для длительного существо-
вания самого организма.
Развитие и эволюция экосистемы 21Ь
Объяснения и примеры
«Борьба за существование» и «выживание наиболее приспособ-
ленных» (по Дарвину) вовсе не значит, что «человек человеку
волк». Выживание во многих случаях основывается на коопера-
ции, а не на конкуренции. С помощью эволюционной теории труд-
но объяснить, как возникают и генетически закрепляются коопе-
рация и развитые мутуалистические отношения. Трудность со-
стоит в том, что исходно взаимодействия почти всегда дают
преимущество особям, действующим в собственных интересах,,
а не в интересах кооперации. Аксельрод и Гамильтон (Axelrod,
Hamilton, 1981) проанализировали эволюцию кооперации и пред-
ложили модель, основанную на «игре в дилемму заключенного»
(см. Rapoport, Chammah, 1970). Другая предпосылка модели —
теория взаимности как расширение общепринятой, основанной на>
конкуренции генетической теории выживания наиболее приспо-
собленных. В игре в дилемму заключенного два игрока решают,
вступить им в кооперацию или нет в интересах ближайшей выго-
ды. При первой попытке решение играть в свою пользу п не ко-
оперироваться обеспечивает наибольший выигрыш каждому игро-
ку независимо от того, что делает партнер. Однако если оба ре-
шают играть каждый за себя, то каждый из них теряет больше,,
чем в случае, если бы они объединились. Если взаимодействие
особей продолжается (т. е. продолжается «игра»), то при повтор-
ных попытках и оценке их преимуществ может возникнуть отбор
на кооперацию. Выводы, сделанные на основе этой модели, и ре-
зультаты компьютерного турнира показывают, как может возник-
нуть основанная на такой взаимности кооперация в асоциальной
среде, как она развивается дальше и продолжает существовать,,
будучи полностью закрепленной. Способность высших животных
распознавать особей повышает шансы установления взаимовыгод-
ных отношений, а постоянный тесный контакт между многочис-
ленными особями микроорганизмов или между микроорганизмами
и растениями повышает вероятность обоюдовыгодных взаимодей-
ствий вроде тех, что развились в процессе эволюции азотфикси-
рующих бактерий и бобовых.
Аксельрод и Гамильтон показали также, что исходной вехой
в эволюции кооперации даже неродственных особей может быть
альтруизм — принесение в жертву приспособленности одной особи
ради выгоды другой, — который встречается среди родственных
особей (родители и потомство, например). Гены, определяющие
этот признак, будучи однажды закрепленными отбором родичей
(kin selection), могут обеспечить распространение кооперации
среди все более дальних родственников.
Д. С. Уилсон (Wilson, 1975, 1977, 1980) следующим образом
сформулировал представление о групповом отборе (см. с. 97 его*
14*
212
Глава 8
книги 1980 г.). «Обычно популяции эволюируют таким образом,
чтобы стимулировать или придерживать другие популяции, от ко-
торых зависит их собственная приспособленность. В таком случае
приспособленность организма на эволюционных промежутках вре-
мени в значительной степени отражает его собственное влияние
на сообщество и реакцию сообщества на присутствие этого орга-
низма. Если эта реакция достаточно сильна, то длительно суще-
ствовать будут только те организмы, которые положительно влия-
ют на сообщество». Уилсон приводит аргументы, что групповой
отбор облегчается отбором между «структурными демами» (тесно
связанными генетическими подразделениями популяции). Он при-
водит также аналогию между парадоксом противоречия приспо-
собленности особи и сообщества в биологическом сообществе и
противоречием между частной выгодой и общественной пользой
в обществе людей.
Так как напряженность отрицательных взаимоотношений хищ-
ник — жертва и паразит — хозяин со временем уменьшается
(гл. 7, разд. 3), Джилпин (Gilpin, 1975) предположил, что в
процессе группового отбора развиваются черты «предусмотритель-
ности», удерживающие хищников и паразитов от чрезмерной экс-
плуатации своих жертв или хозяев. Левин и Пиментел (Levin,
Pimentel, 1981) в качестве примера отбора на понижение виру-
лентности приводят историю интродукции возбудителя миксома-
тоза в Австралию для борьбы с ввезенными туда из Европы
кроликами. Сразу после интродукции вирус убивал кролика в те-
чение нескольких дней. Затем вирулентный штамм был вытеснен
менее вирулентным, которому нужно было в два-три раза больше
времени, чтобы убить хозяина, при этом комары, которые пере-
носят возбудителя, получили возможность дольше питаться на
инфицированном кролике. Так как авирулентные штаммы разру-
шали свой пищевой ресурс не так быстро, как вирулентные, то их
становилось все больше и больше, и они распространялись шире.
Таким образом, отбор между демами дал преимущество авиру-
лентному штамму перед вирулентным, в противном случае и па-
разит, и хозяин должны были неизбежно вымереть.
Хотя само существование группового отбора вряд ли вызывает
сомнения, степень его влияния на эволюцию остается весьма
неясной. В числе других Саундерс и Хо (Saunders, Но, 1976)
утверждают, что сложность окружающего нас мира невозможно
•объяснить отбором только на видовом и организменном уровнях,
поэтому главную роль в эволюции должен играть отбор на высших
уровнях. Напротив, Уильямс (Williams, 1966) и Левин и Майр
(Levin, Мауг, 1981), как и некоторые другие исследователи, счи-
тают, что все можно объяснить в рамках традиционного дарвинов-
ского естественного отбора; отсюда вытекает, что вряд ли группо-
вой отбор служит одним из основных механизмов эволюции.
Развитие и эволюция экосистемы
213
Более подробные сведения о групповом отборе приведены в
работах Уилсона (Е. О. Wilson, 1973), Мэйнарда Смита (Maynard
Smith, 1976) и Александера и Борджиа (Alexander, Borgia, 1978).
7. Применимость теории развития
экосистемы и эволюции биосферы
к экологии человека
Определения
Принципы развития экосистемы имеют важное отношение к взаи-
модействиям между человеком и природой, потому что тенденции
развития природных систем, заключающиеся в повышении уров-
ня структурированности и сложности на единицу потока энергии
(стратегия максимальной сохранности), часто вступают в проти-
воречие со стремлением человека получить максимум продукции
(иными словами, получить наибольший возможный урожай). Осо-
знание экологической основы этого конфликта между человеком и
природой — первый шаг в установлении рациональной системы
землепользования.
Объяснения
Основной конфликт между стратегиями человека и природы изо-
бражен на рис. 8.14 (и на рис. 8.2). Способ подразделения потока
энергии на ранних стадиях развития экосистемы (30-дневный
Молодые (развивающиеся) экосистемы
Поток
энергии
Поток
энергии
Рис. 8.14. Противоположные пути распределения энергии в развивающейся
и зрелой системах.
214
Глава 8
микрокосм или 30-летний лес) иллюстрирует современные пред-
ставления политических и экономических лидеров о том, как сле-
дует управлять природой. Например, цель современного сельского
или интенсивного лесного хозяйства состоит в получении высоких
урожаев легко убираемых продуктов, с тем чтобы доля урожая
на корню, которая остается и накапливается в ландшафте, была
как можно меньше; иными словами, хозяйство направлено на
достижение высокого отношения PIB. Человек всегда старается
получить как можно больше продукции с ландшафта, развивая
и поддерживая экосистемы ранних стадий сукцессии, обычно мо-
нокультуры. Но ведь человеку нужны не только пища и одежда;
ему необходимы также сбалансированная по СОг и О2 атмосфера,,
мягкий климат, который обеспечивают океаны и зеленые массивы,
чистая (т. е. непродуктивная) вода для бытовых, культурных и
промышленных нужд. Многие необходимые для жизни ресурсы,
не говоря уже о тех, которые необходимы для удовлетворения
рекреационных и эстетических потребностей, лучше обеспечивают
менее продуктивные ландшафты. Иными словами, ландшафт —
это не только склад продовольствия и товаров, но еще и «ойкос» —
дом, в котором мы должны жить. До последнего времени чело-
вечество принимало более или менее как должное, что природа
обеспечивает ему газообмен, очистку воды, круговороты биогенных
элементов и другие защитные функции самоподдерживающихся
экосистем. Так было до тех пор, пока численность населения зем-
ного шара и вмешательство человека в окружающую среду не воз-
росли до такой степени, что начали влиять на региональное и
глобальное равновесие. Самый благоприятный для жизни ланд-
шафт — это такой, в котором имеются разнообразные сельскохо-
зяйственные угодья, леса, озера, реки, обочины дорог, болота,,
морские берега и места для переработки отходов, — иными слова-
ми, смесь сообществ разных экологических возрастов. Каждый из
нас как индивидуум более или менее инстинктивно старается
окружить свой дом защитным несъедобным покровом (деревья,
кусты, травы), и в то же время мы стараемся выжать лишний
бушель из хлебного поля. Пашня — это, конечно, хорошая вещь,
но большинство из нас не согласилось бы жить на ней, а занять
под пашни всю обширную поверхность биосферы было бы проста
самоубийством. Мы лишились бы «несъедобного» буфера жизне-
обеспечения, который жизненно необходим для стабильности био-
сферы, и открыли бы путь различным бедствиям от эпидемиче-
ских заболеваний.
Поскольку одну и ту же систему невозможно оптимизировать
по двум несовместимым критериям, вырисовываются два прием-
лемых решения дилеммы: либо мы должны все время искать
какой-то компромисс между количеством урожая и качеством
жизненного пространства, либо должны продумать такой план
Развитие и эволюция экосистемы
215
расчленения ландшафта, при котором можно было бы на разных
участках поддерживать высокопродуктивный и преимущественно
протективный типы в качестве отдельных единить к которым при-
меняется совершенно различная стратегия ведения хозяйства
(стратегия при этом варьирует от, например, интенсивного сель-
ского хозяйства с одной стороны, до сохранения совершенно
нетронутой природы — с другой). Если теория развития экосистем
состоятельна и применима при планировании, то стратегия мно-
гоцелевого использования, о которой мы столько слышим, осуще-
ствима только в рамках одного или обоих этих подходов, посколь-
ку в большинстве случаев множество заложенных в проект целей
противоречат друг другу. Например, плотины на больших реках
часто расхваливают как источник многих благ, —- это и производ-
ство электроэнергии, и контроль над наводнениями, и водопотреб-
ление, и рыболовство, и отдых и т. д. Но все эти цели на самом
деле противоречат одна другой; так, если осуществлять контроль
над наводнениями, то нужно спускать воду еще до сезона разли-
ва, а это понижает выработку электроэнергии и служит помехой
для рекреационного использования водохранилища. Можно, таким
образом, максимизировать систему для одной цели (или, возмож-
но, для немногих тесно сопряженных целей), поступившись ос-
тальными, или можно принять решение в пользу нескольких це-
лей, т. е. пойти на компромисс. Теперь уместно проанализиро-
вать несколько примеров стратегий компромисса и расчленения.
Импульсная стабильность. Более или менее регулярные, но рез-
кие физические возмущения, поступающие извне, могут поддер-
живать экосистему на некоторой промежуточной стадии развития,
так сказать, в состоянии компромисса между молодостью и зре-
лостью. Хорошим примером этого служат так называемые «эко-
системы с колеблющимся уровнем воды». Лиманы и литораль в
общем поддерживаются на ранних относительно плодородных
стадиях благодаря приливам, поставляющим энергию для быстро-
го круговорота биогенных элементов. Сходным образом пресновод-
ные болота, например, в Национальном парке Эверглейдс во Фло-
риде поддерживаются на ранней стадии сукцессии сезонными ко-
лебаниями уровня воды. Падение уровня воды в сухой сезон
ускоряет аэробное разложение скопившегося органического веще-
ства, в результате чего высвобождаются биогенные элементы, ко-
торые при паводковом затоплении поддерживают расцвет продук-
тивности. Жизненные циклы многих организмов тесно связаны с
этой периодичностью, как, например, у лесного аиста (см.
рис. 6.2). Стабилизация уровня воды в болотах парка Эверглейдс
при помощи плотин, шлюзов и водохранилищ не сохраняет эти
водоемы в их настоящем виде, а способствует их разрушению,
как и при максимальном дренировании. Без периодического спус-
ка воды и пожаров мелководные бассейны заполнились бы орга-
216
Глава 8
ническим материалом и произошла бы сукцессия от современного
прудово-прерийного ландшафта к кустарникам или заболоченному
лесу.
Вызывает удивление, что человеку так трудно оценить всю
важность периодических изменений уровня воды в природной
ситуации, такой, как в парке Эверглейдс, когда подобные измене-
ния лежат в основе одного из наиболее древних способов произ-
водства пищи человеком. Периодическое заполнение и осушение
прудов на протяжении многих веков было одним из обычных
приемов в рыбоводстве как в Европе, так и на Востоке. Другой
пример — затопление, осушение и аэрация почвы при выращива-
нии риса. В этом смысле рисовое поле — культурный аналог при-
родного болота или литоральной экосистемы.
Другой физический фактор, периодические воздействия ко-
торого имели жизненно важное значение на протяжении столе-
тий, — это пожары. Как было описано в гл. 4, целые биоты, та-
кие, как африканские степи и калифорнийский чапараль, адапти-
ровались к периодическим пожарам, сформировав то, что экологи
часто называют «пирогенным климаксом». На протяжении веков
человек сознательно использовал пожары для поддержания таких
климаксов или для возвращения сукцессии на какую-либо из
желаемых стадий. Контролируемый пожарами лес дает меньший
урожай древесины, чем интенсивные лесные хозяйства (где рас-
тут молодые деревья приблизительно одного возраста, располо-
женные рядами с коротким циклом воспроизведения), но он
лучше защищает ландшафт, дает древесину более высокого ка-
чества и служит убежищем для птиц — объектов спортивной охо-
ты (перепелов, диких индеек и т. п.), которые не выживают в
условиях интенсивного лесного хозяйства. Таким образом, пиро-
генный климакс представляет собой пример компромисса между
продуктивностью и простотой с одной стороны и защитой и раз-
нообразием — с другой.
Импульсная стабильность срабатывает только в том случае,,
если сообщество в целом (т. е. не только растения, но и животные-
и микроорганизмы) адаптировано к некоторой определенной час-
тоте и интенсивности возмущений. Адаптация (возникающая под
действием отбора) требует времени, измеряемого в эволюционном
масштабе. Большинство антропогенных физических стрессов
слишком внезапны, слишком разрушительны или слишком арит-
мичны, чтобы к ним можно было адаптироваться; поэтому они
приводят к сильным колебаниям, а не к стабильности. По крайней
мере во многих случаях попытки приспособить естественно адап-
тированные экосистемы к хозяйственным нуждам, вероятно, дадут
лучшие результаты, чем их полная переделка.
Перспективы ведения сельского хозяйства на основе детрита^
Гетеротрофное использование первичной продукции в зрелых
Развитие и эволюция экосистемы
217
экосистемах в значительной степени связано с потреблением раз-
латаемого детрита. Нет причин, по которым человек не мог бы
(интенсивнее использовать детритную пищевую цепь для получе-
ния пищи и других продуктов, сохраняя в то же время защитные
-функции экосистем. Такое решение вновь означало бы компро-
мисс, так как «сиюминутный» урожай не может быть таким же
большим, как при непосредственной эксплуатации пастбищной
пищевой цепи. Однако у детритного сельского хозяйства имелись
бы некоторые компенсирующие недобор урожая преимущества.
Современное сельское хозяйство ориентировано на селекцию рас-
тений, быстрый рост и высокую пищевую ценность, что, конечно,
делает их восприимчивыми к насекомым и болезням. Следователь-
но, чем интенсивнее мы ведем отбор на сочность и скорость роста,
тем больше сил нам приходится затрачивать на химические сред-
ства борьбы с вредителями. Это в свою очередь увеличивает ве-
роятность отравления полезных животных, не говоря уже о самом
человеке.
Почему бы нам не практиковать также и обратную страте-
гию — отбор малосъедобных растений или растений, вырабаты-
вающих в процессе роста собственные системные инсектициды с
последующей переработкой чистой продукции в продукты пита-
ния путем микробиологического или химического обогащения на
пищевых фабриках? Тогда мы могли бы направить биохимические
исследования на изучение процессов обогащения, вместо того что-
бы отравлять наше жизненное пространство химическими ядами.
Производство силоса из малоценного корма путем ферментации
служит примером такого рода процесса, уже широко используе-
мого человеком. Другой пример — разведение детритоядных рыб
на Востоке. Производство органики и сельское хозяйство с огра-
ниченной вспашкой (гл. 2, разд. 7) — также шаги в этом направ-
лении.
Блоковые модели землепользования. Анализируя вопрос о том,
каким образом принципы развития экосистемы связаны с ланд-
шафтом в целом, рассмотрим блоковые модели, изображенные на
рис. 8.15. На верхней диаграмме изображены три типа окружаю-
щей среды, служащие системами жизнеобеспечения для четверто-
го блока, представляющего собой гетеротрофную систему промыш-
ленно-городского типа. Продуктивная «окружающая среда» че-
ловека представлена экосистемами на ранних стадиях сукцессии,
такими, как посевы, пастбища, древесные плантации и интенсив-
но эксплуатируемые леса. Все эти экосистемы обеспечивают че-
ловека продуктами питания и волокном. Зрелые экосистемы, та-
кие, как старые леса, климаксные степи и океаны, выполняют в
большей степени защитную, чем продуктивную функцию. Они
стабилизируют субстраты, служат буферами в круговоротах воз-
духа и воды, смягчают колебания температуры и другие физиче-
218
Глава 8
Рис. 8.15. Блоковые модели для планирования использования окружающей
среды. А. Разделение в соответствии с теорией экосистемы. Б. Разделение
с точки зрения ландшафтного архитектора и дизайнера. Объяснения см.
в тексте.
ские факторы и при этом часто дают полезные продукты. Третья
категория естественных или полуестественных экосистем прини-
мает на себя главную нагрузку по ассимиляции большого коли-
чества отходов, производимых городскими промышленными и
сельскохозяйственными системами. Эти экосистемы представлены
водными путями (континентальными и береговыми), болотами и
другими местами, подвергающимися сильной нагрузке. Экосисте-
мы этой произвольной категории находятся преимущественно на
промежуточных, эвтрофицированных или задержанных стадиях
Развитие и эволюция экосистемы
219
развития сукцессии. Все они непрерывно взаимодействуют, будучи
объединены друг с другом входами и выходами (на рисунке по-
казаны стрелками).
Подразделение ландшафта на три компонента окружающей
среды—«природный», «освоенный» и «искусственный», как это
традиционно делается в ландшафтной архитектуре (рис. 8.15,Б),—
открывает другой удобный подход к изучению потребностей и
взаимосвязей нашего хозяйства.
Хотя урбанизированная, или искусственная, среда «парази-
тирует» на среде жизнеобеспечения (природной и «одомашнен-
ной»), получая биологическую продукцию для своих нужд, она
создает и экспортирует другие, преимущественно небиологические
ресурсы. Это удобрения, капитал, произведенная энергия и това-
ры, которые могут быть как полезными, так и опасными для
биологической среды. Можно было бы сделать значительно боль-
ше для увеличения полезных субсидий и снижения стрессовых
воздействий на выходе с этой заряженной энергией и плотно на-
селенной «горячей точки», и отныне это должно стать главной
целью человечества. Неизвестно, может ли соответствующая тех-
нология заменить в глобальном масштабе блага, предоставляемые
биотическим комплексом жизнеобеспечения, и услуги, выполняе-
мые природными экосистемами (вспомните обсуждение автоном-
ного космического корабля в гл. 2, разд. 7).
Прежде чем рассматривать перспективный план землепользо-
вания, или «систематизированного развития», вместо существую-
щего, которое ведется от случая к случаю, преимущественно ради
«сиюминутных» политических и экономических выгод, определя-
ющих пользование землей и водой (см. Watt, 1977, гл. 2, для
сравнения случайного и систематизированного развития), остано-
вимся вкратце на подразделении, или расчленении, окружающей
среды в США в 1980 г. Около 24% площади суши (рис. 8.16,А)
составляют агроэкоспстемы и 6 % — городские промышленные сис-
темы (включая транспортную сеть). Оставшиеся примерно 70% —
это природные экосистемы, которые в разной степени подвержены
выпасу, заготовкам древесины, использованию в качестве рекреа-
ционной зоны и загрязнению. 4% входит в состав национальных
парков или парков штатов, убежищ дикой природы и диких жи-
вотных. Эти 4% лучше всего защищены от антропогенных нару-
шений, хотя некоторые из них контролируются, с тем чтобы со-
хранить определенную растительность, виды диких животных или
их научную ценность. Общественные леса и открытые простран-
ства (степи и пустыни) составляют около 30% поверхности
48 штатов (кроме Аляски и Гавайев). Предполагается, что здесь
хозяйство ведется так, чтобы сохранялось устойчивое состояние
растительных структур и чтобы собираемый урожай и выпас
уравновешивались компенсирующим приростом. Однако перед ли-
Рис. 8.16. Землепользование в США в 1980 г. А. Земельная плошадь кон-
тинентальной части США. Б. Площадь с включением вод над континен-
тальным шельфом.
Развитие и эволюция экосистемы
221’
цом растущей численности и запросов населения становится все-
труднее следовать этой национальной политике.
Воды океана над континентальным шельфом функционируют
как большие системы ассимиляции отходов и смягчения климата.
Они служат также источником пищи, поэтому представляют собой
существенную часть системы жизнеобеспечения прилежащих кон-
тинентальных районов. Если воды над шельфом включить в об-
щую схему (рис. 8.16,Б), то получим, что 75% поверхности США
находится в природном или полуприродном состоянии (в разных
случаях под воздействиями разной силы) и 25% составляют го-
родские и сельскохозяйственные системы.
Прежде чем делать вывод, что соотношение от 3:1 до 5:1,.
характеризующее доли природной и окультуренной среды, указы-
вает на достаточное (по-видимому, более чем достаточное) раз-
витие среды жизнеобеспечения, подумаем о следующих трех огра-
ничениях:
1. Поскольку уровень мощности городских промышленных сис-
тем примерно в 100 раз выше уровня в любой природной экосис-
теме (см. табл. 3.18), для рассеяния неупорядоченности на выхо-
де городской системы малой площади нужна очень большая пло-
щадь природной системы, которая, как уже отмечалось, должна
обладать определенной диссипативной емкостью, чтобы могли
развиваться и поддерживаться высокоорганизованные структуры.
2. Жизнеобеспечивающая способность природной среды может
меняться на несколько порядков в зависимости от ее продуктив-
ности (рабочего потенциала) и степени уже действующего стрес-
са. Так, 1 га пустыни никогда не может быть столь же эффек-
тивным, как 1 га плодородного прибрежного болота, а уже сильна
загрязненное озеро имеет очень немного дополнительной возмож-
ности для жизнеобеспечения.
3. Страны с высокой плотностью населения и развитой про-
мышленностью, такие, как Япония и многие европейские страны,,
зависят в обеспечении необходимого притока энергии, материалов,,
продуктов питания и общих благ и услуг от обширных про-
странств вне собственной территории. Эти потребности в окру-
жающей среде на входе Боргстром (Borgstrom, 1969) назвал «ак-
рами-приведениями». Он подсчитал, что в Японии только для
обеспечения населения страны одной пищей на каждый акр сель-
скохозяйственных угодий в пределах государства требуется 5 ак-
ров моря и суши вне страны. Примерно так же высоко отношение
акров-привидений к акрам внутри страны в Бельгии, Голландии
и повсюду в Европе. США, напротив, не только сами кормят себя,,
но и экспортируют продукты питания, обеспечивая тем самым
акрами-привидениями другие страны, которые не в состоянии
прокормиться сами. Следовательно, нельзя сравнивать схемы зем-
лепользования и соответствующие им коэффициенты в малых.,
Й22
Глава 8
плотно населенных странах с такими же показателями в больших
странах с низкой плотностью населения, если не учитывать внеш-
ние площади жизнеобеспечения, в которых нуждаются страны
первой категории. Если включить в рассмотрение эти площади,
то отношение площади жизнеобеспечения к площади городов и
промышленных районов окажется во всех случаях высоким.
Все это делает крайне трудным объективное определение того,
как много природной среды нужно сохранить в пределах каждой
данной политической единицы (штата или страны), чтобы обес-
печить заданный уровень развития человеческого общества. Один
подход, предложенный Одумом и Одумом (Odum, Odum, 1972),
состоит в построении такой модели землепользования, в которой
можно было бы менять соотношение природных и окультуренных
земель, как показано на рис. 8.17. Если все входы и выходы
компонентов модели количественно оценить в единицах энергии,
то разные варианты планирования землепользования можно про-
игрывать на цифровой или аналоговой машине. «Природные зем-
ли» определяются в такой модели как часть ландшафта, которая
сама себя обеспечивает и для поддержания себя нуждается лишь
в минимальном вмешательстве человека. В функциональном от-
ношении «природная среда» — это такая часть системы жизне-
обеспечения человека, которая работает без включения на вход
человеческой энергетики или экономики. На рис. 8.17 показаны
кривые, характеризующие (в величинах чистой энергии) одно из
решений такой модели. Если площадь обрабатываемой и застроен-
ной земли превышает 40%, то, как только возможности природ-
ного жизнеобеспечения оказываются не в состоянии удовлетво-
рить потребности интенсивного городского и сельскохозяйственно-
го развития, стоимость всего ландшафта резко падает. В этой
ситуации пропорционально уменьшился оборот (гл. 3, разд. 8,
см., в частности, рис. 3.20). По словам Одума и Одума, «даже
простейшие модели ясно показывают, что высокоэнергетические
•системы, такие, как города, нуждаются в обильном природном
жизнеобеспечении. Если для обеспечения необходимых входов от
природы не сохраняются большие пространства природной среды,
то качество жизни в городе снижается, и этот город не может
больше экономически конкурировать с другими городами, где
имеются мощные входы системы жизнеобеспечения. Часто именно
это, а не энергия сама по себе становится лимитирующим факто-
ром. При этом некоторые основные природные ресурсы нуждают-
ся в высоком уровне потока энергии для своего поддержания.
Непрерывный городской, промышленный и (или) сельскохозяйст-
венный рост во многих частях США (и мира в целом) будет
зависеть от создания новых водных ресурсов (опреснение морской
воды, выкачивание подземных вод или использование удаленных
источников). Все это требует больших энергетических затрат. Ес-
Рис. 8.17. А. Модель земельного хозяйства, допускающая изменение соот-
ношения природных и освоенных земель для определения оптимального’
с точки зрения среды в целом их баланса. Б. Характеристические кривые,
основанные на данных, полученных с помощью модели для гипотетического’
региона с экстенсивным развитием городов. (Odum, Odum, 1972.) I — общая
стоимость; II — стоимость освоенных земель; Ill — стоимость природы;
IV — стоимость взаимодействия.
224
Глава $
ли создавать такие источники, то энергетическая цена города
шозрастет настолько, что он не сможет более конкурировать с
теми городами, которые не оплачивают столь большие энергети-
ческие затраты. Складывается грустная ситуация, когда город
перерос (свои средства и не может больше оплачивать свое суще-
ствование. Эти города занимают деньги или требуют федеральные
дотации, чтобы расти еще больше и соответственно предъявлять
еще большие требования к системам жизнеобеспечения, тогда как
им следовало бы направлять все больше энергии на сохранение
качества и эффективности уже освоенной окружающей среды и
на уменьшение нагрузки на жизненно важную поддерживающую
1часть окружающей среды».
Определение оптимального соотношения разных типов ланд-
шафта — это, конечно же, самый первый и простой шаг в разум-
ном планировании. В такой же мере для достижения нужного
взаимодействия важна пространственная структура, т. е. размеще-
ние энергетических станций, промышленных районов, сельскохо-
зяйственных угодий и участков природы. Иными словами, если
электростанции, аэропорты и промышленные предприятия окру-
жены зелеными поясами естественных природных участков, то бу-
ферная емкость этих участков оказывается более эффективной,
чем если эти же самые площади природных зон расположены в
удалении. Огромное значение приобретают социальные проблемы,
такие, как неравенство, экономическое положение и социальная
•справедливость, особенно если плотность населения велика и про-
мышленность интенсивно развита. Последствия выбранного ва-
рианта планирования нужно определять не только в целом (на
душу населения), но и по отношению к разным социальным, ра-
совым и экономическим группам, воздействие на каждую из ко-
торых данного варианта плана может быть очень различным.
'Так, скоростная дорога, проектируемая на благо местному хозяй-
ству, в долговременном аспекте может оказаться чистым убытком,
если она будет отрицательно воздействовать на соседние жилые
районы. Наряду с планируемой экономической выгодой необхо-
димо учитывать и стоимость разрушения общественной струк-
туры.
Моих учеников, моих коллег и меня самого несколько раз про-
сили построить холистические модели для принятия решения от-
носительно строительства сверхскоростных дорог. В одном случае
нужно было выбрать между альтернативными трассами, в дру-
гом — отказаться от строительства пригородной дороги через одно
из населенных предместий Атланты, в третьем — изменить на-
правление скоростной дороги с интенсивным движением, чтобы
снизить ее отрицательное воздействие на природные участки,
.обладавшие рекреационной и эстетической ценностью и важные
для жизнеобеспечения. В каждом случае все доступные велпчи-
Развитие и эволюция экосистемы
225
ны — экономические, экологические, социальные и т. п. — были
даны в числовом масштабе общих единиц измерения и взвеши-
вались в соответствии с лучшими из имевшихся экспертных оце-
нок (см. Е. Р. Odum et al., 1976). В каждом случае попыткам
оценить тотальный вред противопоставлялись деловые и прочие
частные интересы, но когда «все карты были раскрыты», оказа-
лось довольно просто согласовать эти интересы в соответствии с
решением, выданным холистической моделью. Редакционная
статья одной из газет выразила это так: «Это единственный путь
к согласию на благо целого, которое может быть достигнуто в
условиях демократии».
Если вернуться к структуре землепользования в США, которая
показана на рис. 8.16, то в свете уже обсуждавшихся теоретиче-
ских соображений представляется вполне приемлемым отношение
7з освоенной территории : 2/з природной среды. Для страны в це-
лом можно сказать, что американцам повезло в обладании таким
обилием природных благ и услуг. Однако большая часть США
аридна и не обладает такой поддерживающей способностью, как
хорошо увлажненные районы. Об этом свидетельствуют острый
недостаток воды и увеличение ее цены в большинстве Западных
штатов. Таким образом, в США никоим образом нет излишков
площади жизнеобеспечения, и американцам следует заботиться
о постоянной защите существующей природной среды, о ее ка-
честве п емкости. Кроме того, увеличивающееся распространение
кислых дождей, смога и других загрязнений говорит о том, что
уже исчерпана емкость некоторых регионов, таких, как Южная
Флорида, Южная Калифорния, промышленное Северо-Восточное
побережье и индустриальный район Великих озер.
Всеобъемлющее планирование с учетом перспективы экологи-
ческого и эволюционного развития. Некоторые новаторские под-
ходы к конструированию ландшафта, базирующиеся на холисти-
ческих экологических принципах, будут проиллюстрированы тремя
примерами.
Прекрасно иллюстрированная книга Мак-Харга (McHarg,
1969) «Конструируем с природой» стала классической и вдохно-
вила много попыток проектирования, согласованного с природ-
ными особенностями ландшафта. Впервые в одной книге в до-
ступной форме приведены факты в пользу целостного планиро
вания землепользования как альтернативы неконтролируемого
развития. Перефразируя Мак-Харга, можно сказать, что неконтро-
лируемое развитие, распространяясь широко, уничтожает ланд-
шафт вследствие перенаселения и загрязнения; оно необратимо
разрушает все красивое и памятное независимо от того, насколь-
ко хорошо спланированы отдельные дома или участки территорий.
В табл. 8.3 сравниваются последствия беспланового землепользо-
вания с плановым развитием городского района, предложенным
15 Заказ № 1483
226
Глава 8
Мак-Харгом. В этом районе предусмотрен рост населения от
20 000 до 100 000 жителей. При разумном планировании развития
жилых и прочих массивов треть всей площади, включающая хол-
мы и все речные долины, остается в виде свободного пространст-
ва. Желательно также строить промышленные предприятия по-
среди обширных зон, предназначенных для переработки отходов.,
где расположены пруды и другие полуестественные системы тре-
тичной обработки, которая была бы эффективной и недорогой
(рис. 7.23,Б). Очень удачный «новый город», в котором сочета-
ется все лучшее из промышленного развития с типом устройства
открытых пространств, показанным в табл. 8.3, — это Колумбия
в шт. Мэриленд. Этот город — итог размышлений мастера плани-
рования Джеймса Роуза. Роуз начал с проектирования и строи-
тельства закрытых торговых рядов, но скоро стал с энтузиазмом
заниматься новаторскими проектами более широкого масштаба?
включая реконструкцию внутренних городских районов, и новыми
городами. В ходе этой работы Роуз и его сотрудники стали очень
пылкими, но практичными консервационистамиКолумбия в
Мэриленде занимает площадь в 22 квадратные мили (примерно
величиной с Манхэттен), здесь живет 60 000 человек, из них
30 000 работает в городе. В городе имеется пять автономных
центров (вместо одного), каждый из которых включает в себя
три-четыре жилых массива со смешанной застройкой, где есть
высоко- и малодоходные дома. Треть города — постоянное, не под-
лежащее застройке «открытое пространство», состоящее из реч-
ных долин, старых лесных массивов и других живописных мест.
Этот город — целиком частное предприятие, была выдана всего
лишь одна небольшая правительственная субсидия для строитель-
ства дешевых домов. Колумбия Ассошиэйшн, ведущая коммуналь-
ное хозяйство, содержит в своем штате эколога, наблюдающего за
растительностью, реками, озерами, дорогами и дикими животными
и птицами, в том числе за сипухами, которые живут в старых
постройках, превращенных в склады или подсобные помещения
в местах отдыха. Сохранение открытых пространств стоит меньше
10 000 долларов в год, поскольку сенокос и другие дорогостоящие
мероприятия по поддержанию среды проводятся только в местах,
где расположены игровые площадки1 2.
Третий пример новаторских размышлений об использовании
окружающей среды дает небольшая книга под названием «План
завтрашней Калифорнии», изданная Альфредом Хеллером (Heller,
1972). В этом отчете, составленном группой авторов по данным
1 Очерк о Джеймсе Роузе напечатан в журнале «Таймс» от 24 августа
1981 г. (т. 118, № 8, с. 42—53); на обложке журнала помещен его портрет.
2 Письмо от 24 июля 1981 г. от эколога Колумбия Ассошиэйшн Чарлза
Роудхеймела.
Развитие и эволюция экосистемы
227
Таблица 8.3. Сравнение беспланового (бесконтрольного) и планового (опти-
мального землепользования) развития быстро растущего городского района
с пригородами 1>
1970 г., 20СОО жителей В 2000 г. планируется 110 000 жителей
Бесплановое развитие Оптималь- ное исполь- зование зе- мель
Застроенный район Освоенная площадь 5 200 га 15 200 га 12 000 га
Жилые дома 3 000 10 400 8 520
Торговые предприятия 200 280 252
Промышленные предприятия 28 120 28
Учреждения 1 000 2 200 1 200
Дороги «Свободное пространство 1 000 2 200 2 000
(«неосвоенное пространство») 12 800 2 8002) 6 000
Зоны переработки отходов 0 0 4003>
Парки для отдыха 200 8 00 2 000
•Сельское и лесное хозяйство 4 600 0 8004)
Нетронутая природа 8 000 2 000 2 8005)
€’бщая площадь 18 000 18 000 18 000
•Свободное пространство, % 71 16 33
1 Данные взяты из плана городского развития Мэриленда, подготовленного фир-
мой «Уоллес—Мак-Харг», 2121 Уолнут-Стрит, Филадельфия; дополнено новыми кон-
цепциями Ю. П. Одума о свободном пространстве.
2 При бесплановом развитии и при населении в 150 000 человек свободное про-
странство свелось бы к нулю!
8 Земли, примыкающие к промышленным и городским предприятиям по перера-
ботке отходов или обезвреживанию их. На этих предприятиях очистные пруды и дру-
гие устройства по снижению загрязнений могут быть использованы для эффективной и
дешевой третичной обработки всех отходов.
4 Сюда можно включить не только овощные хозяйства, но и показательные фер-
мы и леса в качестве учебных лабораторий для школ и колледжей.
5 Включая все крутые склоны, овраги, речные долины, болота и озера (по состо-
янию на 1963 г.), а также участки спелого леса и сельскохозяйственные угодья,
восстановленные до первоначального естественного состояния.
разных групп специалистов, предлагается установить законода-
тельно комиссию штата для разработки плана землепользования,
который должен быть представлен для общественного обсуждения
с внесением необходимых поправок. Цель работы — выявление н
охрана природной среды жизнеобеспечения, для того чтобы со-
хранить качество жизни в городах и регулировать, насколько это
возможно, способы и интенсивность эксплуатации среды, не пре-
вышая ее несущую емкость. Уникальная идея проекта состоит в
том, чтобы поселения устраивались в стороне от больших резер-
ватов нетронутых земель. Решения об использовании этих зе-
мель должны быть отложены на будущее, что обеспечивает гиб-
кость в случае возникновения в будущем непредвиденных об-
стоятельств. В отчете Хеллера отмечается, что ни один подобный
15*
228
Глава 8
план не может быть реализован до тех пор, пока на смену су-
ществующему фрагментарному бюрократическому контролю со*
стороны дюжин комиссий городов и округов не придет координи-
рованное региональное управление землепользованием. И если
общий контроль над использованием окружающей среды перейдет
на региональный уровень или к штату, то контроль над жилищ-
ным строительством и другими подобными структурами должен
оставаться в руках местных властей, что обеспечит личную сво-
боду принятия решений о локальном использовании частной соб-
ственности, работе школ, жилищном строительстве, деятельности
гражданских обществ и т. д.
Некоторые соображения об интеграции экологических и эко-
номических предприятий. При наличии сильного общественного
мнения можно, вероятно, преодолеть политические трудности?
но экономические соображения остаются главным препятствием
для любого рода разумного планирования с целью долговремен-
ного использования окружающей среды. Эта проблема возникает
из-за резкого несовпадения рыночных и нерыночных ценностей.
Независимо от политической системы в разных странах промыш-
ленные товары и услуги, такие, как автомобили или электроэнер-
гия, оцениваются очень высоко, тогда как не менее важные для
жизни блага и услуги природного происхождения вроде очистки
воды и воздуха и их возобновления остаются обычно вне эконо-
мической системы и обладают очень низкой денежной стоимостью
или не обладают ею вовсе (следовательно, соответствуют «неры-
ночным» ценностям). Причины этого различия иллюстрируются
оценкой общей стоимости одного из эстуариев, как показано на
рис. 3.23,Л. Лестер Браун так комментирует это в обзоре о «гло-
бальных экономических перспективах» (Brown, 1978): «Эконо-
мисты не приучены думать о роли биологических систем в эко-
номике, еще меньше они думают о состоянии этих систем. Стол
экономиста может быть завален ссылками на последние данные
о состоянии здоровья экономики, но экономист на самом деле
редко бывает озабочен состоянием здоровья главных биологиче-
ских систем Земли. Отсутствие экологической осведомленности
вносит свой вклад в недостатки экономического анализа и фор-
мирование политики».
Браун выбрал в качестве фундамента глобальной экономики
четыре биологические системы — водоемы, леса, степи и сель-
скохозяйственные угодья. Продолжая свой комментарий, он пи-
шет: «Здоровье экономики не может быть отделено от здоровья
биологических систем. При развитии глобальной экономики воз-
растает давление на биологические системы Земли. На обширных
пространствах всего мира требовании человека к этим системам
достигли уже невыносимого уровня, той точки, где их продук-
тивность нарушается. Когда это происходит, то рыболовство тер-
Развитие и эволюция экосистемы
229
пит крах, леса исчезают, степи превращаются в голые пустыни,
и сельское хозяйство разрушается, падает качество воздуха, воды
и других ресурсов жизнеобеспечения».
Проект Брауна для «Построения устойчивого общества»
(Brown, 1981) призывает к систематическому, опирающемуся на
поддержку правительства глобальному сохранению ресурсов, ко-
торое состоит в том, что ресурсы следует расходовать экономнее,
с большей эффективностью, всячески расширяя их повторное ис-
пользование. Резко контрастирует этому сетование Саймона (Si-
mon, 1981), что люди вроде Брауна продлевают «миф» о нараста-
ющем оскудении ресурсов. Саймон принижает роль биологиче-
ских факторов и утверждает, что «запасы природных ресурсов
бесконечны в любом экономическом смысле» и что «нет оснований
полагать, что когда-либо иссякнет человеческая изобретательность
и предприимчивость, поэтому мы всегда сможем решить проблемы,
связанные с грядущими дефицитами ресурсов, используя различ-
ные открытия, и после некоторого периода адаптации станем еще
сильнее, чем до возникновения этих проблем». Этой крайней точ-
ки зрения придерживаются очень немногие исследователи. Кен-
нет Боулдинг (Boulding, 1982) написал резкую статью по поводу
книг Брауна и Саймона, которую не лишне прочесть.
Большинство экономистов придерживаются мнения, что рынок
начинает давать сбои, когда он сталкивается с распределением
многих природных ресурсов. В экономической литературе от-
мечено, что недостатки в работе рынка происходят тогда, когда
общество считает некоторые ценности более или менее желатель-
ными, чем это обозначается рыночными ценами. Бейтор (Bator,
1958), исходя из «теории распределения», определяет несостоя-
тельность рынка как «неспособность более или менее идеализи-
рованной системы институтов цены — продажи поддерживать «же-
лательную» активность и приостанавливать «нежелательную» ак-
тивность» (это относится, конечно, к обществу в целом). По тра-
диционному мнению экономистов, вторжение политики в рыноч-
ную сферу требуется для защиты человеческих ценностей и для
распределения недостаточных ресурсов или ресурсов, которым
нет замены (например, земли и воды). Показательны три при-
мера из прошлого: 1) законодательное запрещение потогонной
системы производства, которая была предпочтительнее с промыш-
ленной точки зрения, но нежелательна с общественной; 2) уста-
новление зон землепользования, которое ограничило землеполь-
зование в общественных интересах; 3) установление системы
национальных парков, при которой земля либо изымалась из
сферы действия рынка, либо никогда не предназначалась для
этого. Экономисты подчеркивают, кроме того, что люди не долж-
ны только потреблять или использовать ресурсы, но должны так-
же создавать новые ресурсы, чтобы не быть полностью ограни-
230
Глава 8
ченными имеющимися ресурсами природы (Straffa, 1973). Это
утверждение, несомненно, верно относительно некоторых природ-
ных ресурсов и услуг, однако основные ресурсы жизнеобеспече-
ния мы не можем создать искусственно.
Проведенный через конгресс США Национальный акт об охра-
не окружающей среды во многих отношениях стал первой по-
пыткой подвести в национальном масштабе правовую основу под
распространение системы ценностей на природную среду. Акт
требует, чтобы при каждом планируемом антропогенном наруше-
нии составлялся «официальный отчет об ущербе». Медленно,
иногда мучительно, этот временный подход должен привести к
улучшению процедуры установления общей оценки, включающей
оценки затрат и прибылей для природных и общественных собы-
тий наряду с принятыми экономическими.
В табл. 8.4 показана в общих чертах классификация ценно-
стей применительно к проблеме развала рынка. Нерыночные цен-
ности поделены на две категории: характеризуемые и нехаракте-
ризуемые. По мнению большинства экономистов, характеризуемым
нерыночным ценностям можно приписать денежную стоимость на
общепринятом языке рыночной экономики. Например, стоимость
изъятия природной среды можно было бы определить исходя из
того, что стоило бы обеспечение искусственной замены бесплат-
ных благ и услуг (например, переработки отходов), предлагае-
мых природной экосистемой (обзор попыток найти пути к тому,
чтобы «включить» природные услуги в антропогенную экономику,
дан Уэстменом (Westman, 1977). Напротив, неосязаемые или
нехарактеризуемые ценности не могут быть включены в обычный
в экономике расчет стоимости или подвергнуты анализу затраты—
прибыли.
Одним из первых экономистов, бросивших вызов свободному
рынку как средству эффективного распределения ресурсов, был
Пигоу (А. С. Pigou, 1920). Он очень точно заострил внимание на
недостатках рынка, которые проявляют себя, если бизнес пресле-
дует только свои интересы, не заботясь об общественных. Пигоу
был впереди своего времени, высказывая опасения по поводу
разрушения городов; он писал, что только государство может
«установить обязательные правила и использовать их для защиты
воздуха и воды от опасности загрязнения, что вряд ли можно
компенсировать». Он рассматривал налоги и субсидии как сред-
ство уравнения частных и общественных издержек. (Обсуждение
проведенного Пигоу анализа общественных благ см. Britt, 1971.)
Так как действия «государства» становятся все более затруд-
нительными или, хуже того, слишком запоздалыми перед лицом
возрастающего давления рынка на землепользование и так как
экономисты, экологи и законоведы стремятся иметь дело только
с частью проблемы (и стремятся обвинить в неудачах друг дру-
Развитие и эволюция экосистемы 231
Таблица 8.4. Сравнение рыночных и нерыночных ценностей. (Farnwortli et
al., 1981.)
Рыночные ценности
Сюда относятся главным образом производимые товары и услуги — продук-
ция фабрик и ферм и коммерческие услуги. На рынке свободного предпри-
нимательства они распределяются по законам спроса и предложения по-
средством неограниченной конкуренции. В теории рыночная стоимость от-
ражает общественную опенку товара и услуг, что приводит к эффективно-
му распределению ресурсов. На практике это не всегда так, поэтому допус-
кается необходимость некоторой регуляции со стороны правительства. Ос-
новы «рыночной модели» заложены в книге Адама Смита «Исследования о
природе и причинах богатства народов», изданной в 1776 г.
Нерыночные ценности
Это главным образом природные товары и услуги, их иногда называют
«свободными» или «общими» или «общественными» товарами и услугами.
Обычно эти «бесплатные» ценности существуют вне рыночной экономики.
Характеризуемые и приписываемые нерыночные ценности
Ценность реки для ассимиляции отходов. Здесь можно применять кон-
цепции и язык рыночной экономики, денежная стоимость может быть оп-
ределена.
Неуловимые, неприписываемые или нехарактеризуемые ценности
Ценность для жизнеобеспечения природных экосистем. Леса, степи, реки,
озера и океаны осуществляют, смягчают и стабилизируют атмосферные и
гидрологические циклы и круговороты минеральных элементов. К этой же
категории относится присущая видам ценность, ценность туземной куль-
туры, красоты природы и множество эстетических ценностей, которые со
временем получают признание людей. Нехарактеризуемые категории яв-
ляются личными и общественными ценностями, а не частными рыночны-
ми, с которыми они очень часто приходят в конфликт.
га), то, по-видимому, есть некий смысл в идее разработать новую
дисциплину, которая имела бы дело с проблемой рынок—не ры-
нок в целом. Для обозначения дисциплины, которая должна рас-
сматривать роль биологических и равным образом небиологиче-
ских, созданных людьми систем, в поддержание всеобщего хозяй-
ства был предложен термин биоэкономика (Georgescu-Roegen,
1977; Clark, 1981). Более подходящее название — голоэкономика,
которое свыше полувека назад предложила группа экономистов
(см. Grunchy, 1947, и эпилог).
Многообещающей основой для такой новой дисциплины явля-
ется использование в качестве общей единицы измерения «затра-
ченной» энергии (см. гл. 3, разд. 10). Другой обещающий под-
ход — это превращение современной линейной экономики в кру-
говую экономику, как показано на рис. 8.18. В таком случае
экономическая система стала бы соответствовать общей системной
модели с внутренней обратной связью, как схематически показано
на рис. 1.4. Антрополог Бернштейн отмечает, что многие изоли-
рованные культуры, которые обречены существовать на местных
232
Глава 8
ЛИНЕЙНАЯ ЭКОНОМИКА
Изъятие ресурсов
(ведет *: снижению незозоб-
ноЕЛЯемых ресурсов:
Максимизация производства
\ (максимизируется
' поток вещества*
Максимизация
потребления
Загрязнение'
Выбрасывание
остатков:
загрязнение
КРУГОВАЯ ЭКОНОМИКА
Акцент на производство
Рис. 8.18. Использование ресурсов можно в значительной степени улучшить,
если современную линейную экономику заменить круговой экономикой,
(Т. Е. Jones, 1977.)
ресурсах, умеют отличать действия, которые в будущем могут
оказаться вредными для окружающей среды, и избегают таких
действий. Такой способ локальной обратной связи при принятии
решения был утрачен, когда изолированные культуры вошли в
состав больших и сложных индустриальных обществ. Бернштейн
заметил, что «экономисты должны развить согласованную теорию
принятия решений, применимую на всех уровнях групповой орга-
низации. При этом возникнет необходимость определения собст-
венных интересов в терминах выживания, а не потребления».
Такое смещение подвергло бы экономику своего рода «естествен-
ному отбору», который работает достаточно хорошо, чтобы обес-
печивать продолжение жизни на Земле на протяжении многих
эон.
Для дополнительного чтения мы рекомендуем следующие кни-
ги и статьи:
Об экономическом росте: Galbraith, 1958; Mishan, 1967, 1970; Kru-
tilla, 1967; Wagner, 1970; Barkley, Seckier, 1972; Ridder, 1972.
О нерыночных ценностях и неудачах рынка: Coase, 1960; Barnett,
Morse, 1963; Costle, 1965; Pearse, 1968; Alfred Kahn, 1966; Ran-
del, 1974; Lugo, Brinson, 1978; Sinden, Worrell, 1979; Farnworth
et al., 1981.
Об экономике ресурсов: Kneese, Ayres, d’Arge, 1970; Schurr (ed.),
1972; Fisher, Krutilla, Cicchetti, 1972; Kneese, 1973, 1979; Page,
1977; Ayres, 1978; Smith, Krutilla, 1979.
Развитие и эволюция экосистемы
233
Об экономических индексах (GNP, NEW): Samuelson, 1954; Nord-
haus, Tobin, 1972; Nordhaus, 1979.
Об экономике стационарного состояния: Ayres, Kneese, 1971; Daly,
1973.
Об энергетическом анализе экономики: H. Т. Odum, 1973; Hannon,
1973; Gilliland, 1975, 1978; Huetner, 1976; Slesser, 1974; Costanga,
1980; Odell, 1980.
Работы Кеннета Боулдинга и Николаса Джорджеску-Регена,
являющихся основоположниками синтеза экономики и экологии,
приведены в библиографии.
Эпилог. Трудности, стоящие перед
человечеством: взгляд в будущее
Увлекательная игра — предсказание будущего; она становится
особенно популярной во времена кризисов. На самом деле никто
не может реально предсказать, что случится в следующем году,
еще меньше можно сказать о том, что будет через 25 лет или
позже. Слишком многого мы не знаем и не можем предвидеть
многочисленные новые события, технологические новшества и
другие факторы. Поучительно, тем не менее, рассмотреть ряд
альтернативных вариантов того, что могло бы произойти. Тогда
мы были бы в состоянии оценить вероятность будущих событий,
исходя из современных условий нашего понимания и знаний.
Еще важнее то, что мы смогли бы предпринять кое-какие меры
сейчас, чтобы уменьшить вероятность нежелательных последствий
в будущем.
Единственно, что мы знаем определенно, приближаясь к
2000 г., это то, что численность населения будет продолжать рас-
ти, и что индустриально развитым странам придется по мере
уменьшения количества, снижения качества ископаемого топлива
п роста цен на него совершить значительный и очень болезненный
сдвиг в использовании энергии. Этот сдвиг уже начался, и мы
можем по крайней мере строить какие-то гипотезы относительно
«волновых эффектов», которые будут сопровождать радикальные
изменения в энергетической среде на входе. Большинство футу-
рологов считает, что мы должны уменьшить современное чудо-
вищное количество отходов и стать более экономными и береж-
ливыми, чтобы обойтись меньшим количеством высококачествен-
ной энергии. Мнения большинства исследователей сходятся также
в том, что рост потребления энергии на душу населения сверх
современного уровня в промышленно развитых странах не улуч-
шит качества жизни, а фактически может вызвать обратный эф-
234
Трудности, стоящие перед человечеством
235
фект (см. Nader, Beckerman, 1978). Сходным образом большин-
ство предсказателей видит необходимость получать в мировом
масштабе больше продуктов питания с меньшими энергетически-
ми затратами на ограниченной площади с благоприятными для
сельского хозяйства почвами, качество которых уже сейчас ухуд-
шается из-за непомерно тяжелой нагрузки (эрозия, засоление).
Нет недостатка в исследованиях, отчетах и популярных кни-
гах, содержащих разбор тех затруднений, с которыми сталкива-
ется современное человечество. В большинстве этих изданий на-
рисована мрачная картина существующих глобальных проблем,
но многие полны оптимизма относительно будущего. Мы можем
быть оптимистами, но только в том случае, если (и это очень
большое если) нам есть что противопоставить современным труд-
ностям и есть, что предпринять. Среди исчерпывающих отчетов
следует отметить отчеты правительственных агентств США. и в
частности два, подготовленных Советом по качеству среды (СКС)
в 1980 и 1981 гг. и отчет Отдела сельского хозяйства 1979 г.
Очень интересен отчет СКС 1980 г., озаглавленный «Мир в
2000 г., отчет для Президента; переход в XXI век». В трех томах
детально рассмотрены вопросы, связанные с ростом численности
населения, доходами, использованием ресурсов и нагрузками на
окружающую среду. Включена также глава, где эти данные срав-
ниваются с другими глобальными оценками, и «Правительствен-
ная модель», где исследуются перспективы на будущее.
Позиции, с которых рассматривается будущее, и которые раз-
деляют как ученые, так и общество в целом, различаются от пол-
ного доверия к новой технологии до убежденности в том, что
общество должно полностью перестроиться, умерить свою власть
над природой, создать новые международные всеохватывающие
политические и экономические механизмы, чтобы работать в мире
ограниченных ресурсов. Хорошо известный сторонник первой точ-
ки зрения — Герман Кан (см. Kahn, Wiener, 1967; Kahn et al.,
1976). Вторая точка зрения предполагает необходимость принять
«стратегию кораллового рифа или тропического леса на выщело-
ченной почве», чтобы преуспеть в условиях ограниченных ресур-
сов (т. 1, с. 16, 17, 240—241). В гл. 8, разд. 7, разбирая книги спе-
циалиста по ресурсам Лестера Брауна и экономиста Джулиана
Саймона, вышедшие в свет в 1981 г., мы противопоставляли эти
два возможных варианта. Уатт (Watt et al., 1977) посвятил целую
главу (гл. 2) сравнению того, что он назвал «верой в технокуль-
туру» и «противоположной позицией». Противоположная позиция
по сути дела представляет собой экологическую парадигму, со-
гласно которой человечество как часть глобальной экосистемы
подвержено действию законов природы в условиях ограничения
ресурсов. В своей широко известной книге «Красота малого» Шу-
мейкер (Schumacher, 1973) приводит хорошие доводы в пользу
236
Эпилог
увеличения вкладов в маломасштабную технологию, гармонирую-
щую с человеческими и природными ценностями (в этой связи в
обиход вошел термин «подходящая технология» — appropriate
technology). Маргалеф (Margalef, 1979) заметил, однако, что,
хотя малое часто бывает прекрасным, большое — могущественно!
Впрочем, чем нам руководствоваться в решении вопроса о том,
что должно быть большим, и что должно быть малым? Когда и
где, например, следует строить мощнейшую централизованную
электростанцию или же, напротив, устанавливать на крышах фо-
тоэлектрические батареи, строить мини-гидростанции, ветроэнер-
гогенераторы и прочие децентрализованные устройства? Анализ
затрат и доходов столь сложен, что прежде чем найти лучшее
сочетание для долговременной выгоды, мы должны эксперименти-
ровать с многими вариантами. Здравый смысл подсказывает нам,
что мы слишком много черпали из нефтяных скважин, и теперь
следует, фигурально выражаясь, черпать из многих других «сква-
жин».
Другой подход к той сложной ситуации, в которой находится
человечество, — анализ различий, которые необходимо преодолеть,
чтобы люди и окружающая среда, так же как промышленно раз-
витые и не развитые страны, находились в гармоничном равно-
весии. В этой книге уже говорилось об этих различиях:
1. Различия в доходах: в одной и той же стране есть богатые
и бедные; различия существуют между 30% промышленно раз-
витых стран и 70% неиндустриализованных стран (т. 1, с. 135).
2. Различия в питании (т. 1, с. 136).
3. Различия в ценностях: рыночные и нерыночные товары и
услуги (с. 228).
4. Чудовищные различия в уровне образования: грамотные и
неграмотные, охваченные и неохваченные школьным обучением
(а также квалифицированные и неквалифицированные рабочие);
ранее об этом не упоминалось, но мы обсудим данный вопрос
ниже.
Ни одно из этих различий не может быть ликвидировано в
ближайшем будущем. Ни усилия Организации Объединенных
Наций (которая затрачивает их в основном на попытки остано-
вить военные столкновения), ни выделение процветающими стра-
нами больших средств на помощь слаборазвитым странам не име-
ли большого эффекта. Фактически различия в доходах и различия
в ценностях стали за несколько последних десятилетий еще боль-
ше. Морхаус и Сигурдсон (Moorehouse, Sigurdson, 1977) отмечают,
что передача промышленной технологии бедным странам часто
приносит пользу только небольшому современному сектору, но не
массам городской бедноты. Благополучие не может упасть с неба
при наличии глубоких различий в культуре, образовании и ресур-
сах. Нельзя перенести энергоемкую технологию в бедную страну
Трудности, стоящие перед человечеством
237
без того, чтобы не снабдить ее источником высококачественной
энергии для поддержания этой технологии. Фарвар и Милтон
(Farvar, Milton, 1972) приводят многочисленные случаи неудач
при передаче технологии.
Рене Дюбо, ученый, гуманист и убежденный оптимист, крас-
норечиво пишет о том, что путь к ликвидации противоречий меж-
ду человеком и окружающей средой — «одомашнивание» всей био-
сферы, с тем чтобы сохранить не только пригодность ферм, полей,
лесов, водоемов, парков и т. д., но и использовать их с толком,
соблюдая гармонию между творениями человеческих рук и при-
родой. Идея Дюбо — это утопическая идея глобальной экосистемы,
которая питалась бы от возобновляемых ресурсов и была бы орга-
низована наподобие европейских деревень, так глубоко запавших
в его память со времен детства во Франции (Dubos, 1976, 1980).
Если цивилизация — это система, а не организм, тогда в от-
личие от того, что утверждает Тойнби в «Изучении истории»
(Toynbee, 1961), цивилизация не должна подобно организму рас-
ти, становиться зрелой, стареть и умирать, хотя такие процессы
и случались в прошлом (например, подъем и падение Римской
Империи). Согласно Батцеру (Butzer, 1980), цивилизация стано-
вится нестабильной, когда сильно повышается стоимость ее соб-
ственного поддержания, причиной чего является бюрократия,
предъявляющая чрезмерные требования к сектору производства.
Следующий за этим крах является результатом «случайного пе-
реплетения взаимно усиливающих процессов, а не результатом
старения и упадка». Такая точка зрения совпадает с экологиче-
ской теорией потока энергии и сложности (гл. 3).
Доклады Римского клуба
В апреле 1968 г. группа в 30 человек из десяти стран собра-
лась в Риме по инициативе итальянского специалиста в области
управления промышленностью, экономиста и просто дальновид-
ного человека доктора Аурелио Печчеи. В эту группу входили
ученые-естественники, математики, экономисты, социологи, пре-
подаватели, промышленники, гражданские служащие националь-
ного и международного значения. Они собрались, чтобы в гло-
бальном масштабе обсудить настоящие и будущие трудности че-
ловечества; эта группа получила название «Римский клуб».
С этого времени группа заказала серию «Докладов Римскому клу-
бу» под общим названием «Затруднения человечества». Первый из
этих докладов — «Пределы роста» (1972) — был подготовлен груп-
пой ученых из Массачусетского технологического института, воз-
главляемой Деннисом и Донелл ой Медоузами. Группа строила
глобальные модели по новаторской методике системного анализа,
разработанной Джеем Форрестером (P^orrester, 1968, 1971). Эти
238
Эпилог
модели должны были предсказать, что ожидает нас в будущем
при сохранении современных экономических и политических ме-
тодов. Результаты моделирования показали, что во многих слу-
чаях или в связи со многими жизненно важными ресурсами бу-
дут наблюдаться циклы взлетов и падений. Промышленный рост
и потребление ресурсов будут продолжать увеличиваться ускоря-
ющимися темпами наряду с ростом численности населения и
увеличением потребления энергии и т. д. до тех пор, пока не бу-
дет достигнут некий предел; затем произойдет катастрофа, или
«банкротство». Этот отчет осуждает общественную манию роста?
при которой цель на каждом уровне — индивидуальном, семейном?
классовом, национальном — стать богаче и могущественнее без
учета окончательной платы за экспоненциальный рост.
Хотя «Пределы роста» претендовали только на то, чтобы по-
казать, что может случиться, если мы не изменим наши методы
и цели, многие, включая большинство политических лидеров и
большие группы общественности, восприняли отчет так, как если
бы он предсказывал день Страшного суда для цивилизации. Ро-
дилась, естественно, буря критики. Многие обозреватели отмечали?
что в моделях не были приняты во внимание новая технология?
открытие новых ресурсов, замена истощенных ресурсов новыми
и т. д. Казалось, большинство людей осознало, что для человече-
ства было бы слишком болезненно претерпеть любой цикл взлета
и падения, поэтому следует прекратить промышленный рост или
изменить весь стиль нашей жизни еще до достижения этого мо-
мента.
Несмотря на то, что книга вызвала серьезные критические
замечания, а большинство политических деятелей не признало
возможность чего-то похожего на взлет и падение, книга произве-
ла ошеломляющее впечатление. Она предупреждала нас о том, что
мы должны с большим вниманием отнестись к тому, куда направ-
лено развитие человечества.
«Пределы роста» можно найти во многих книжных магазинах
наряду с более старыми классическими трудами, служащими пре-
дупреждением человечеству: Vogt, 1948; Osborn, 1948; Leopold.,
1949; Carson, 1962.
За первым докладом последовал ряд дополнительных докладов,
в которых были сделаны попытки не только более подробно опи-
сать существующую ситуацию и рассмотреть возможные вариан-
ты, но и обосновать действия, которые необходимо предпринять,
чтобы избежать Страшного суда. С этой целью Медоуз опублико-
вал вторую книгу (Meadows, 1973).
Второй доклад Римскому клубу называется «Человечество на
перепутье». Доклад подготовлен Михайло Месаровичем, директо-
ром и профессором Центра системных исследований при Универ-
ситете Кейс Уэстерн Резерв, и Эдуардом Пестелем, профессором
Трудности, стоящие перед человечеством
239
Института механики из Ганновера (ФРГ). Пытаясь устранить
недостатки «Пределов роста», авторы этого доклада делят Землю
на десять взаимозависящих регионов. Логическим обоснованием
такого деления послужило то, что решать глобальные проблемы
на региональной основе более практично и более эффективно,
поскольку в разных частях Земли проблемы, стоящие перед че-
ловечеством, существенно различаются. В этом докладе вновь
анализируются природа глобального кризиса и опасность промед-
ления. Авторы доклада пришли к заключению, что разные регио-
ны мира в целом должны быстро перейти от «недифференцирован-
ного (стихийного) роста» к тому, что они назвали «органическим
ростом». Иными словами, мир больше не может развиваться сти-
хийно, ибо пассивное следование такому стихийному развитию
ведет просто-напросто к гибели.
Во втором докладе также особое внимание обращается на две
постоянно расширяющиеся пропасти, которые, видимо, находятся
в самой сердцевине современного кризиса. Это пропасти между
человеком и природой и между богатыми и бедными. Между
прочим, именно на эти различия в течение многих лет указы-
вали экологи. Если мы хотим избежать глобальной катастрофы,
то эти пропасти должны быть уничтожены, однако достигнуть
этого можно, только обеспечив некое единство всего мира, при
котором вновь была бы подтверждена взаимозависимость человека
п природы, и народы всех стран четко представили бы себе, что
Земля не бесконечна.
Идею органического длительного роста мировой системы
не следует истолковывать как упрощенную идею развития «мо-
нолитной» системы, устроенной по единому образцу. В докладе
подчеркивается, что концепция единого мира, много раз предла-
гавшаяся утопистами, как раз и не будет работать в сложной
для человечества ситуации. Напротив, концепция одного гомоген-
ного мира по существу не совместима с истинно глобальным
подходом к облегчению затруднений человечества. На самом деле
рекомендуемый подход должен начинаться с определения на-
правлений, которых следует придерживаться, и с сохранения ре-
гионального разнообразия в мире. Необходимо, чтобы развитие
каждого региона шло своим специфическим путем, но при этом
нужно исходить из глобальных, а не узких национальных инте-
ресов. Таким образом, модель мира выглядит как гибкий, бази-
рующийся на вычислительной технике инструмент планирования,
содержащий в себе многоуровневые региональные модели мировой
системы. Эта модель имеет фундаментальные отличия от любой
другой предлагавшейся ранее в «Пределах роста» модели, потому
что она распознает существующее в мире разнообразие, имеющее
глубокие корни в прошлых культурных традициях, которые, не-
сомненно, будут преобладать и в будущем. И вновь мир рассмат-
240
Эпилог
ривается как система, т. е. собрание взаимодействующих и взап-
мозависящих частей. Различные географические регионы с прису-
щими им культурными традициями являются блоками этой!
предложенной модели, которая может обеспечить лиц, ответствен-
ных за принятие решений в разных частях мира, всесторонним
глобальным средством планирования.
Региональные подходы, предложенные Месаровичем и Песте-
лем, очень похожи на социологическую теорию регионализма, раз-
витую несколькими десятилетиями ранее покойным Говардом
У. Одумом (Odum, 1935, 1951; Odum, Moore, 1938). Регионализм
как подход к изучению общества основан на признании четких
различий в культурных и природных характеристиках разных об-
ластей, которые тем не менее зависят друг от друга. Региональное
социальное исследование побуждается отчасти надеждой поднять
выше «отстающие» регионы (как Юг США в 30-е годы), чтобы
они смогли вносить свой вклад в экономическое благосостояние
нации, а не только черпать из него. Концепция биолога о тесной
связи всей природы и о взаимозависимости между человеком и
остальной биотой насчитывает уже более 100 лет. Об этом писал,
например, Т. Гексли в своем эссе «Доказательство места человека
в природе» (Huxley, 1863, переиздание в 1959 г.). Эта тема много
раз поднималась экологами животных, писавшими об «экологии
человека», например Чарлзом С. Адамсом (Adams, 1935),
Ф. Фрейзером Дарлингом (Darling, 1951), Марстеном Бейтсом
(Bates, 1952) и У. К. Олли (Allee, 1931, 1951). Связь между по-
ведением человека и других животных послужила также основой
для новой и противоречивой теории социобиологии, развиваемой
Эдвардом О. Уилсоном (Wilson, 1975, 1980) и другими исследова-
телями.
Следовательно, и тема второго доклада Римскому клубу не но-
ва, но она ставится в расширенном контексте: все наши кризи-
сы, которые один за другим привлекают внимание общественно-
сти, на самом деле являются компонентами одного синдрома и
сигнализируют о необходимости найти способ постепенного отхо-
да от недифференцированного роста и приблизиться к многоуров-
невому органическому росту с обратными связями, достаточными
для действий, предотвращающих циклы взлетов и падений. До-
клад позволяет предположить, что мы используем все эти кризи-
сы как детекторы ошибок и начинаем действовать, как действует
хорошая органическая система — кибернетически, с сильной отри-
цательной обратной связью, которая срабатывает, когда рост при-
ближается к состоянию с уменьшающейся отдачей (см. гл. 3,
разд. 3). Хирш (Hirsch, 1978) оптимистично полагает, что соци-
альные ограничения п стоимость поддержания обеспечат обратную
связь, которая ограничит рост до того, как возникнет нехватка
материальных благ.
Трудности, стоящие перед человечеством
24Э
Третий доклад Римскому клубу называется «РИО — перестрой-
ка международного порядка» (1977). В этом докладе, координа-
цией которого занимался Нобелевский лауреат, голландский эко-
номист Ян Тинберген, особое внимание уделяется слаборазвитым
странам, руководители которых отвергают в общем любую идею
об ограничениях роста. Тинберген и соавторы доклада пришли
к выводу, что для того чтобы провести в жизнь идею органиче-
ского мирового порядка, предложенную во втором докладе, все
страны и все регионы должны согласовывать свои локальные пе-
ли с определенными глобальными целями, которые будут работать
на благо человечества в целом. Этот доклад должен был показать,
что сочетание локальных и глобальных целей возможно. Напри-
мер, какой-то стране необходимо достичь определенной степени
индустриализации для того, чтобы обеспечить разумно высокий
уровень жизни своих граждан — это достойная локальная цель.
Если это достигается путем по возможности полного повторного
использования скудных ресурсов и по возможности полной пере-
работки отходов, то «общее», глобальное, т. е. воздух, океаны
и т. д., будет повреждено в меньшей степени. Конечно, чрезвы-
чайно трудно убедить граждан и политических лидеров в том, что
соответствие их собственных национальных целей целям челове-
чества как целого отвечает их же интересам. Жизненно важное1
значение имеет ослабление гонки вооружений, так как трата энер-
гии в этом направлении затрудняет возможность всех стран на-
править свою волю, энергию и деньги на сотрудничество друг с
другом ради общей цели. Когда люди заняты военными конфлик-
тами, они мало думают о сотрудничестве для взаимной выгоды..
На память приходит эволюционная тенденция в природных систе-
мах—переход от конкуренции к мутуализму (гл. 7, разд. 4 и
гл. 8, разд. 6).
Четвертый доклад под названием «Цели для глобального обще-
ства» (1977) составлен Эврином Ласло, профессором философии,
который известен своими книгами «Системный взгляд на мир»
и «Стратегия для будущего» (цитировались в гл. 2 этой книги).
Этот доклад посвящен двум фундаментальным вопросам, которые
лежали в основе предыдущих докладов: 1) в чем на самом деле
заключаются цели человечества? и 2) согласны ли мы предпо-
честь материальному росту развитие человеческих качеств? Ины-
ми словами, будет ли человечество на протяжении длительного
времени использовать свою чудовищную научно-техническую
мощь на благо всех людей, не причиняя существенного вреда
планете? Для этого нам придется умерить некоторые свои жела-
ния в ближайшее время ради стабильности и приемлемых условий
жизни будущих поколений.
«Цели для глобального общества» базируются на переписи на-
циональных и региональных целей, предложенных рабочими
16 Заказ № 1483
242
Эпилог
группами из разных стран и регионов. Так, в этом докладе име-
ются главы о целях США и Канады, Западной Европы, Восточной
Европы, Латинской Америки, Африки и Среднего Востока. За эти-
ми главами следуют главы о целях в понимании международных
групп, таких, как Организация Объединенных Наций, межнацио-
нальные корпорации и Всемирный Совет Церквей. Затем рассмат-
риваются цели, направленные на сохранение глобальной безопас-
ности, цели, связанные с использованием энергии, пищи и ресур-
сов. В гл. 16 доклада обсуждается «текущее глубокое расхождение
целей»; эта глава содержит ряд диаграмм в виде строк и столб-
цов, в которых цели оцениваются от 0 (усилия сосредоточены
исключительно на ближайших национальных интересах) до 10
(цели включают долговременные глобальные проблемы). В США
в отношении долговременных проблем охраны природы, экономи-
ки, энергетики и сельскохозяйственной политики деятельность де-
ловых кругов оценена низко, а деятельность правительства —
сравнительно высоко. Политика в отношении «стиля жизни» оце-
нена, как и можно было ожидать, низко. Во всех случаях выше
оцениваются устремления интеллектуальных и религиозных или
духовных групп (больше глобальных интересов). Средние нацио-
нальные оценки по шкале от 0 до 10 для ведущих стран таковы:
США — 4,0; Канада — 4,7; Бельгия — 3,7; Нидерланды — 4,7; Шве-
ция — 4,6; Япония — 5,5; Китай — 7,0; Алжир — 6,4; Саудовская
Аравия — 4,2; Бразилия — 4,0; Египет — 4,6; Гана — 6,5; Паки-
стан — 4,7 и Индия — 4,3. Интересно, что народы слаборазвитых
стран оптимистичнее смотрят в будущее, чем народы развитых
стран, но долговременные глобальные интересы оценены высоко
только для Китая. В последних главах книги авторы приходят к
заключению, что мы можем достичь глобальных целей и солидар-
ности всех народов мира, и начало этому движению уже положе-
но. Обсуждаются и сравниваются проекты, проведенные совмест-
но религиозными и научными группами, деловыми и научными
группами в правительственной группой. В докладе отмечено, од-
нако, что мы не имеем определенного ответа на вопрос, который
<о временем обязательно возникнет: каким образом прийти к ми-
ровой солидарности?
Последующие доклады Римскому клубу, опубликованные меж-
ду 1978 и 1980 гг., посвящены важным составным частям гло-
бальных проблем, например проблеме переработки отходов (Ga-
bor, 1987), использования энергии (de Montbrial, 1979), организа-
ции общества (Hawrylyshyn, 1980) и достижению изобилия и
благосостояния (Giarini, 1980). Ключевой доклад посвящен проб-
лемам образования и называется «Нет пределов обучению: наве-
дение мостов через разделяющую людей пропасть» (Botkin et al.,
1980). В этом отчете противопоставляется то, что авторы назвали
микро- п макрообучепием. Они отмечают, что в большинстве слу-
Трудности, стоящие перед человечеством
243
чаев все усилия направлены на индивидуальное обучение, цел1
которого — сделать индивидуума знающим и разумным. Это —
уровень микрообучения. Напротив, мало внимания уделяется кол-
лективному или общественному обучению, т. е. макрообучению.
Иными словами, как сделать, чтобы осведомленность о региональ-
ных и глобальных трудностях и понимание этих трудностей стали
доступны людям, которые мало или вовсе не учились в школе?
Как научить общество в целом избегать циклы взлетов и падении,
энтропийную ловушку и другие опасности, понятные сейчас толь-
ко немногим высокообразованным его представителям? И самое
важное, как сделать, чтобы общество было осведомленным об об-
разе действий, соответствующем органическому росту, так красно-
речиво обоснованному во втором и третьем докладах, и чтобы оно
захотело воспринять этот образ действий? Те, кто надеялся, что
телевидение могло бы стать средством макрообучения, до сих пор
были разочарованы, но, по-видимому, в данном случае исчерпаны
еще далеко не все возможности. Разработка методов социального
обучения, несомненно, заслуживает того, чтобы ей уделили самое*
пристальное внимание.
Ласло (Laszlo, 1977а) так оценивает общее действие докладов
Римскому клубу: «Главным образом благодаря усилиям Римского
клуба быстро возросла международная осведомленность о мировой
проблематике. Если продолжить аналогию с медициной, то можно
сказать, что Клуб первым перешел от постановки диагноза (Ме-
доуз, Месарович, Пестель) к предписанию определенных средств
(Тинберген, Ласло и другие доклады). Но не в укор героическим
усилиям группы Аурелио Печчеи, в области терапии было достиг-
нуто сравнительно мало. Используя другую метафору можно ска-
зать, что Клуб помог наметить путь, но мало сделал, чтобы воз-
никло желание следовать по этому пути. Если верна пословипа.
что где хотение, там и умение, то телега оказалась впереди ло-
шади. Конечно, для группы обеспокоенных граждан мира легче
наметить путь, чем вызвать волевой подъем, необходимый, чтобы
следовать этим путем. Тем не менее есть реальная опасность, что
единственный результат проделанной до сих пор работы будет в
том, что от длительного неупотребления она покроется тонкой
изящной патиной. Было бы лучше — и здесь пищущий эти стро-
ки полагает, что его мнение совпадает с мнением других членов
Римского клуба — если бы отчеты попадали в горнило конструк-
тивной дискуссии, а пе пылились в качестве респектабельных до-
кументов эпохи, которым никто не следует».
Глобальные модели и варианты будущего мира
Между 1971 и 1981 г. было создано около десятка крупномас-
штабных глобальных моделей. В этих моделях обсчитывались ма-
тематические имитации физических и социально-экономические
16*
244
Эпилог
систем мира. Результаты проецировались в будущее, которое пред-
ставлялось логическим следствием заложенных в модель данных
и предположений, и следует подчеркнуть, что в этом отношении
все модели различались. Обзор и сравнение группы этих моделей
даны в докладе, изданном Службой Конгресса по технологическим
оценкам (СКТО) (1982), в книге Медоуза (Meadows, 1983) и
статье Донеллы Медоуз (Meadows, 1982). Особенно рекомендует-
ся последняя работа, поскольку в ней содержится обзор методики
моделирования, а также имеется иллюстрированный раздел под
названием «Инструкция для детей для овладения системным
взглядом».
В этом Эпилоге уже упоминались четыре из глобальных моде-
лей — это модели Форрестера, Медоуза с соавторами, Месарови-
ча—Пестеля и Глобал-2000. Кроме того, предложены и такие
модели, в основе которых лежат преимущественно социоэкономи-
ческие исследования, а не данные по ресурсам и населению как
таковым. Это в особенности Латиноамериканская модель мира
(ЛАММ), Британская SARUM-модель, японская модель FUGI и
мировая модель ООН (UNWM). Построение этих моделей в дей-
ствительности вызвано критикой ранних перспектив предельного
роста, и эти модели основывались на допущениях, что в наших
несчастьях так же повинны политическое, социальное и экономи-
ческое неравенство, как и ограниченность ресурсов и пресс наро-
донаселения.
Несмотря на различие допущений и акцентов, вся последова-
тельность моделей приводит к единодушному согласию по неко-
торым моментам. Вот те точки согласия, которые собраны в отче-
те СКТО и обзоре Д. Медоуза (Meadows, 1982):
1. Технологический прогресс желателен и жизненно важен,
но необходимо, чтобы при этом происходили также социальные,
экономические и политические изменения.
2. Народонаселение и ресурсы не могут расти беспрестанно на
конечной планете.
3. Мы не имеем надежной и полной информации относительно
того, до какой степени физическая среда Земли и система жизне-
обеспечения могут удовлетворить нужды и потребности будуще-
го роста населения (т. е. нам неизвестна несущая емкость среды),
однако резкое снижение скорости роста сильно уменьшит вероят-
ность превышения допустимого уровня или больших катастроф
(экобедствпй).
4. Продолжение практики «обычного бизнеса» не приведет к
желаемому будущему; результатом этого будет скорее углубление
нежелательных противоречий (например, между богатством и бед-
ностью).
5. Долговременное сотрудничество было бы выгоднее для всех
партий, чем политика сиюминутной конкуренции.
Трудности, стоящие перед человечеством
245
6. Поскольку народы, страны и окружающая среда находятся
в более тесной зависимости друг от друга, чем это обычно пред-
ставляют, все решения должны приниматься в контексте холи-
стического подхода. Действия, направленные на достижение узко
ограниченных целей, скорее всего контрпродуктивны.
7. Природа будущего глобального устройства, хуже оно или
лучше настоящего, но предопределена. Многое зависит от того,
как скоро изменятся существующие нежелательные тенденции.
Действия, предпринятые быстро (за ближайшие два десятилетия),
окажутся, по-видимому, более эффективными и менее дорогостоя-
щими, чем те же действия, предпринятые с запозданием. Это тре-
бует сильного руководства и более широкой макрообразованности
(о чем говорилось в связи с докладом Римскому клубу «Нет пре-
делов обучению»), поскольку к тому времени, когда проблема
станет очевидной каждому, предпринимать какие бы то ни было
действия будет уже слишком поздно.
Итак, для тех, кто «молод душой и телом», есть чему будить
воображение и есть цели, достичь которые стоит немалого.
Экологическая оценка
Философия многих составителей докладов Римского клуба
также как и данные, полученные с помощью глобальных моделей,
хорошо соответствуют основам теории экосистемы, особенно двум
ее парадигмам: 1) при работе со сложными системами необходимо
использовать холистический подход и 2) при приближении систе-
мы к пределам в отношении ресурсов и проч, сотрудничество име-
ет большую ценность для выживания, чем конкуренция. Все упо-
мянутые ранее трактаты подтверждают также древнюю мудрость
здравого смысла, выраженную в пословицах, таких как «семь раз
примерь, а один отрежь», «не ставь все на карту», «поспешишь —
людей насмешишь», «легче предупредить болезнь, чем ее выле-
чить», «сила развращает» и многих других. Что касается здравого
смысла и экологии, см. Е. Р. Odum, 1977.
Главный источник неопределенности, влияющий на модельные
предсказания — будущий рост народонаселения. На рис. 3.21 гра-
фически изображены три тенденции, основанные на предположе-
нии, что рождаемость и смертность в будущем столетии уравнове-
сятся. Большинство футурологов считают, что все наши проблемы
было бы легче решить, если бы снизилась скорость роста населе-
ния, хотя чтобы достичь этого в глобальном масштабе необходимо
изменить глубоко укоренившиеся религиозные и культурные убеж-
дения относительно контроля над рождаемостью, абортов и других
противоречивых материй. В 1971 г. Национальная академия наук
издала отчет, знаменующий собой веху в истории и озаглавленный
«Быстрый рост народонаселения; последствия и смысл политики».
246
Эпилог
В отчете сделан вывод, что быстрый рост не приносит экономи-
ческой и какой-либо другой выгоды, а, напротив, порождает со-
циальные проблемы, связанные с окружающей средой, причем эти
проблемы возникают гораздо быстрее, чем решаются. Быстрый
рост может обладать инерцией, которая ведет к перескоку через
допустимый уровень, и избежать этого трудно. Многие демографы
верят в демографический переход — в теорию, согласно которой
рост населения замедляется, когда люди становятся богаче в
меньше зависят от детского труда; однако все это весьма проти-
воречиво (см. Teitelbaum, 1975).
Экологи считают, что одно из препятствий в достижении ка-
кого-либо порядка в мире — это слишком узкая экономическая
теория и представления, преобладающие в мировой политике (см.
гл. 3, разд. 7 и гл. 8, разд. 7). Примерно в середине нашего сто-
летия группа ученых, которые называли себя «холистическими
экономистами», сформировала активную «школу», критически на-
строенную по отношению к экономическим моделям того времени.
Гранчи (Grunchy, 1947) посвятил истории этой школы, в состав
которой входили Торстейн Веблен, Дж. Р. Коммонс, Уэсли Мит-
челл, Джон М. Кларк, Рексфорд Тагуэлл и Гардинер Минс, кни-
гу. Перечисленные имена мало знакомы тем, кто изучает эконо-
мику сегодня. Холисты отрицательно относились к тому, что эко-
номисты-классики уделяют мало внимания экономике как
динамично функционирующему целому, рассматривая лишь ее
аспекты по отдельности. В результате этого модели носят жесткий
механистический характер и плохо предсказывают реальный мир.
Дальнейшее развитие холистической экономики было «затоп-
лено» потоком нефти, породившим быстрый рост денежного и ма-
териального изобилия. Классическая теория роста служит доста-
точно хорошо до тех пор, пока снабжение нефтью существенна
превосходит потребности в ней. Теперь, когда нефтяной бум спал,
самое время взяться вновь за развитие этой теории, но так, чтобы
она наряду с денежными включила в себя культурные ценности и
ценности окружающей среды. Историк Джон Хааг (Haag. 1981)
в обзоре, посвященном развитию экономических идей после
1945 г., заметил появление нового поколения экономистов, кото-
рое не «заражено» «материалистической этикой роста любой це-
ной». Лауреат Нобелевской премии экономист Василий Леонтьев
(Leontief, 1982) сетует на то, что экономисты не способны уви-
деть, что экономика — это «саморегулируемая система, образуемая
очень большим числом разных, но взаимосвязанных п тем самым
взаимозависящих действий». Он задает вопрос, «как долго иссле-
дователи в таких соприкасающихся областях, как демография,,
социология и политика, с одной стороны, и экология, биология,,
медицина, техника и другие прикладные физические науки — с:
другой, будут воздерживаться от выражения озабоченности той
Трудности, стоящие перед человечеством 247
блестящей изоляцией, в которой находится сейчас академическая
экономика?» Критика Леонтьева во многом совпадает с той, ко-
торую высказывали представители упомянутой выше школы се-
редины века. Это же недовольство слишком узкой точкой зрения
высказывают и многие академические экологи. Однако экологи
начинают вносить все больший вклад в экономику. В результате
экологические концепции проникают в современные экономиче-
ские труды, и наоборот. Знаменательно, что исследователи в
разных областях начинают применять одни и те же аналитиче-
ские методы. Например, использованный Леонтьевым подход с
построением открытых моделей применяется, среди прочих, и эко-
логами Хенноном и Финном, а системный экологический подход
Г. Т. Одума привлекает внимание экономистов.
Многие футурологи полагают, что необходимо расширить и
укрепить этику, включив в нее представление о жизнеобеспече-
нии и других нерыночных ценностях, для того чтобы преодолеть
пропасть между «знаю как» науки и желанием политических ли-
деров принимать широкомасштабные решения на благо целому.
В гл. 1 подчеркивалось важное значение «Этики земли» Леополь-
да. Однако никто не знает, как можно личный интерес распро-
странить на озабоченность всех глобальными проблемами, но мно-
гие об этом пишут.
Когда «наука о доме» (Экология) и наука о «ведении домаш-
него хозяйства» (Экономика) сольются, и когда предмет Этики
расширит свои границы, и включит в себя наряду с ценностями,
произведенными человеком, ценности, создаваемые окружающей
средой, тогда мы на самом деле сможем стать оптимистами отно-
сительно будущего человечества.
Приложение. Краткое описание основных
типов природных экосистем биосферы
1. Наземные биомы
Ключевая характеристика, позволяющая разграничивать и узна-
вать наземные биомы, — жизненная форма (травы, кустарники,
листопадные деревья, хвойные деревья и т. д.) растительности
климатического климакса. Так, климаксная растительность степ-
ного биома (злаковника) — злаки, хотя в разных частях биома
и на разных континентах могут произрастать разные виды зла-
ков. Итак, основа классификации — растительность климатическо-
го климакса, однако биом включает в себя и эдафические климак-
сы, и стадии развития, в которых во многих случаях доминируют
иные жизненные формы. Например, степные сообщества пред-
ставляют собой стадии развития лесного биома, а прибрежные
леса — составная часть степного биома. Подвижные животные,
для которых Шелфорд предложил термин «пермеанты», связыва-
ют между собой различные ярусы и стадии развития раститель-
ности. Птицы, млекопитающие, рептилии и многие насекомые
свободно передвигаются между подсистемами и между развиваю-
щимися и зрелыми стадиями растительности, а перелетные птицы
в определенные сезоны — между биомами разных континентов.
Во многих случаях жизненные циклы и поведение в разные сезо-
Рис. П.1. Схематическая карта основных биомов земного шара. Обратите
внимание на то, что только тундра и северные хвойные леса тянутся более
или менее непрерывно вокруг всего земного шара. Другие сравнимые
биомы (например, степи умеренной зоны или тропический дождевой лес)
занимают изолированные пространства, следовательно, можно ожидать,
что в них обитают экологически эквивалентные, но неродственные виды.
Распределение главных биомов сходно с распределением основных типов
почв, показанным на рис. 5.22, но не совпадает с ним. (Схема основана на
карте первичной растительности из книги: Finch V. S., Trewartha G. Т.,
Physical Elements of Geography. New York, McGraw-Hill, 1949.)
250
Приложение
ны организованы так, что данный вид будет занимать несколько
часто совершенно различных типов растительности. Для наземных
биомов характерны крупные растительноядные млекопитающие —
олени, антилопы, бизоны, домашний скот. Многие из них отно-
сятся к жвачным, обладающим замечательной микроэкосистемой—
рубцом, где анаэробные микроорганизмы разрушают и обогащают
лигноцеллюлозу, составляющую большую часть растительной био-
массы. Сходным образом, одним из основных путей потока энер-
гии служат детритная пищевая цепь, характеризующаяся нали-
чием грибов и почвенных животных — редуцентов, а также му-
туалистические системы между корнями растений и микоризой,
азотфиксаторами и другими микроорганизмами.
Биомы мира схематически показаны на рис. П.1; сравнение
климаграмм шести основных биомов приводится на рис. П.2.
Рис. П.2. Климаграммы шести основных биомов по среднегодовым темпе-
ратурам и среднегодовому количеству осадков. (С разрешения National
Science Foundation.)
Некоторые из основных биомов мира (степи, листопадные
леса умеренной зоны, северные хвойные леса, тундра и пустыня)
были в 70—80-е годы объектом междисциплинарного изучения.
Эти исследования представляют собой часть вклада США в Меж-
дународную биологическую программу (МБП). Результаты этих
длительных интенсивных работ опубликованы в многочисленных
статьях и сериях книг. Широкий обзор исследований МБП дан
Блейром (Blair, 1977) и Луксом (Loucks, 1983). По составу фло-
ры и фауны биогеографы разделяют мир на пять или шесть глав-
Краткое описание основных типов природных экосистем
251
ных областей, которые приблизительно соответствуют основным
континентам. Самые изолированные континенты — Австралия и
Южная Америка; можно соответственно ожидать, что экологиче-
ски эквивалентные виды в биомах этих континентов будут таксо-
номически совершенно различными (см. гл. 7, разд. 5 и табл. 7.3).
Тундры — арктическая и альпийская (рис. П. 3)
Между лесами на юге и шапкой полярных льдов на севере цир-
кумполярно расположена полоса безлесной местности площадью
примерно 2Х104 км2 (рис. П.З). Небольшие, но экологически
сходные районы расположены в высоких горах выше границы
леса. Такие районы, встречающиеся даже в тропиках, называются
альпийскими тундрами. В Северной Америке и Евразии граница
между тундрой и лесотундрой сдвинута к северу на западе, где
теплые западные ветры смягчают климат.
Основными лимитирующими факторами служат низкие тем-
пературы и короткий сезон вегетации; осадков тоже может быть
мало, но они не относятся к лимитирующим факторам, посколь-
ку скорость испарения здесь невелика. Грунт, за исключением
верхнего слоя толщиной в несколько сантиметров, на протяжении
всего лета остается промерзшим. Эти постоянно промерзшие глу-
бокие слои грунта называются вечной мерзлотой. Тундра — это в
сущности влажная арктическая степь с растительностью, состоя-
щей из злаков, осок, карликовых древесных растений и на более
сухих - местах — лишайников («олений мох»). Для «низменной
тундры» (как на прибрежных равнинах Аляски) характерны
толстая рыхлая подушка из живых и очень слабо разложившихся
растений, часто пропитанная водой и испещренная небольшими
озерцами. «Высокая тундра», особенно если рельеф хорошо вы-
ражен, покрыта скудным покровом лишайников и злаков. Хотя
вегетационный сезон краток, длинные летние фотопериоды спо-
собствуют образованию в благоприятных местах, например на
береговой равнине мыса Барроу (Аляска), довольно большого ко-
личества первичной продукции (до 5 г сухого вещества в день).
Продуктивные водные (в том числе плодородный арктический
океан) н наземные экосистемы поддерживают не только большое
число перелетных птиц и насекомых, активных только в бесснеж-
ный сезон, но и постоянных обитателей, которые остаются ак-
тивными на протяжении всего года. Сюда относятся: овцебык,
европейский северный олень, карибу, белый медведь, волки, пес-
цы, хищные птицы, а также лемминги и другие мелкие млеко-
питающие, строящие ходы в толще растительности. Для крупных
животных характерны обширные миграции; «циклы» обилия мно-
гих более мелких животных обсуждались в гл. 6, разд. 5. Особую
уязвимость тундры следует иметь в виду при все возрастающей
Рис. П.З. Вид тундры в июле (прибрежная равнина у арктической исследо-
вательской лаборатории, мыс Барроу, Аляска). А. Видна широкая болоти-
стая низменность в верховьях речной системы, тянущаяся примерно в 3 км
от берега. Доминирующие растения — арктический злак Dupontia fischeri
и осока Carex aquatalis, укоренившиеся на пласте торфа в полузаболочен-
ной почве. Эту местность с полным основанием можно считать типичной
климаксной низинной прибрежной тундрой. На фотографии показаны проб-
ные квадраты и «выгородка», защищающая участок от леммингов.
Б. Участок примерно в 16 км от берега. Видна характерная полигональная
структура грунта. Образованию полигональных структур способствуют
выклинивания льда под грунтом. Белые плоды, заметные на переднем
плане, — пушица Eriophorum scheuzchert. (Фотографии R. Е. Shanks,
Е. Е. Clebsch, Koranda.)
Краткое описание основных типов природных экосистем
253
эксплуатации минеральных ресурсов и другой деятельности че-
ловека, вызывающей нарушения окружающей среды. Тонкий жи-
вой покров легко разрушается и медленно восстанавливается.
Строительство трубопровода на Аляске преподало нам много на-
глядных уроков.
Биомы северных хвойных лесов (рис. П. 4)
Широким поясом вытянулись через всю Северную Америку п
Евразию области северных вечнозеленых лесов. Горные районы,
занятые таким лесом, встречаются даже в тропиках. Доминирую-
щая жизненная форма представлена здесь хвойными вечнозеле-
ными деревьями, особенно елью, пихтой и сосной (Picea. Abies,
Pinus). На протяжении всего года в таких лесах царит густая
тень, поэтому кустарниковый и травяной ярусы обычно развиты
слабо. Однако сплошной зеленый покров, содержащий хлорофилл,,
сохраняется круглый год, и поэтому, несмотря на низкую темпе-
ратуру в течение половины года, для этого биома характерен до-
статочно высокий уровень годовой продукции. Хвойные леса —
самые крупные в мире поставщики лесоматериалов. Хвоя разла-
гается очень медленно, поэтому почва имеет характерный подзо-
листый профиль (см. рис. 5.18). В почве обитает многочисленное
население мелких животных, но крупных животных мало (по
сравнению с почвами листопадных лесов или степей). Многие из
крупных травоядных позвоночных, таких, как лось, заяц-беляк и
тетерев, зависят, по крайней мере частично, от развивающихся
широколиственных сообществ. Семена хвойных служат важным,
источником существования многих животных, таких, как белки,
чижи, клесты.
Как и в тундре, здесь сильно выражены сезонная периодич-
ность и колебания численности многих популяций. Классический
пример — циклы численности зайца-беляка и рыси (см. рис. 6.15).
В хвойных лесах наблюдаются вспышки численности жуков-ко-
роедов и насекомых-дефолиантов (пилильщиков, почкоедов
и т. п.), особенно там, где в древостое доминируют всего один
или два вида. Такие вспышки, однако, как описано в гл. 6, разд. 5,
являются частью непрерывного цикла развития, к которому адап-
тирована экосистема хвойного леса.
Другой тип хвойного леса встречается вдоль западного побе-
режья Северной Америки, от центральной Калифорнии до Аляски,
где температуры выше, сезонные ее колебания относительно ма-
лы, а влажность очень высока. Эти леса, хотя в них и доминируют
хвойные, флористически совершенно отличаются от обычной се-
верной тайги. Следовательно, эти «дождевые леса умеренной зо-
вы», как их часто называют, могут с достаточным основанием'
рассматриваться как отдельный тип биома.
Гис. П.4. Три типа хвойных лесов. А. Высокогорный лес из ели Энгельман-
на и пихты субальпийской в Колорадо. Обратите внимание на мощный
слой подстилки, который накапливается вследствие низких температур и
длительного снежного периода. Б. Ельник в шт. Айдахо с одной из основ-
ных стадий развития — осинником. Осенью листья осин становятся золо-
сгпсто-желтыми (светлые деревья слева и в центре фотографии).
Краткое описание основных типов природных экосистем
25S
В. Пример влажного хвойного леса, который часто называют дождевым
лесом умеренной зоны (заповедник Олимпик, шт. Вашингтон). Обратите
внимание на величину деревьев, пышный ковер папоротников и других
травянистых растений и эпифитные мхи, обрамляющие сучья деревьев
(С разрешения службы леса США.)
Этот лес достигает наибольшего развития в районе Пьюджет-
Саунда, где доминируют в основном четыре вида — западная тсу-
га {Tsuga heterophylld), западная туя {Thuja plicata), пихта ве-
ликая {Abies grandis) и лжетсуга {Pseudotsuga douglasii}. По-
следний вид преобладает в более сухих местообитаниях или
субклимаксах влажных местообитаний. Южнее произрастают ве-
личественные секвойи Sequoia, а севернее — ситхинская ель Pices
sitchensis. В отличие от более сухих и более северных хвойных
лесов в этом лесу всюду, куда проникает хоть немного света,
хорошо развита растительность нижних ярусов, обильны мхи п
другие влаголюбивые низшие растения. Эпифитные мхи пред-
ставляют здесь экологический эквивалент эпифитных бромелис-
вых влажных тропических лесов. Биомасса продуцентов в этом
биоме огромна, и, как легко себе представить, продукция строевого
леса с единицы площади потенциально может быть очень велика,
если поддерживать возобновление растений и круговороты био-
генных элементов. Как и во всех экосистемах, где большой про-
цент биогенных элементов связан в биомассе, существует серьез
Рис. П.5. Климатический, эдафический и пирогенный климаксы на востоке
США. А. Девственный листопадный лес на западе Северной Каролины —
климатический климакс. Б. Эдафический климакс — болотистый лес камед-
ного дерева (Nyssa) с бахромой эпифитного «испанского мха» Tillandsta
в затопляемой долине реки Саванна.
Краткое описание основных типов природных экосистем
257
В. Остатки пирогенного климаксного леса длиннохвойной сосны близ То-
масвилла, шт. Джорджия. Использование частых контролируемых пожаров
сохраняет открытый, парковый облик и предотвращает вторжение лист-
венных деревьев, не выносящих пожара.
ная опасность истощения этих ресурсов в будущем вследствие
чрезмерной эксплуатации.
Другие подразделения биома северных хвойных лесов, которые
можно рассматривать как отдельные биомы, — это леса карлико-
вой сосны и можжевельника, или «карликовых хвойных», и леса
желтой сосны, растущие на промежуточных высотах между сте-
пями и зоной елей в Колорадо, Юте, Нью-Мексико и Аризоне
(см. рис. П.11). Это в общем редколесья, в которых количество
травянистых растений велико, особенно там, где пожары пред-
ставляют собой естественный фактор окружающей среды.
Листопадные леса умеренной зоны (рис. П. 5)
Сообщества листопадных лесов занимают области с большим
количеством равномерно распределенных осадков (750—1500 мм)
и умеренной температурой, для которой характерны четкие се-
зонные колебания. Первоначально умеренные листопадные леса
J7 Заказ № 1483
258
Приложение
покрывали весь восток Северной Америки, всю Европу, часть
Японии, Австралии и южную оконечность Южной Америки. Та-
ким образом, биомы листопадных лесов в большей степени изо-
лированы друг от друга, чем тундра и северные хвойные леса.
Их видовой состав, несомненно, должен отражать эту степень изо-
ляции. Поскольку в таких лесах деревья и кустарники в течение
части года лишены листьев, контраст между зимой и летом очень
велик. Травянистый и кустарниковый ярусы, а также почвенная
биота развиты хорошо. Многие растения дают сочные мясистые
плоды и орехи, например желуди или буковые орешки. Животное
население первоначальных лесов Северной Америки представлено
виргинским оленем, медведем, серой и черной белкой, серой ли-
сицей, рыжей рысью и дикой индейкой. Характерные мелкие
птицы зрелых стадий — красноглазый виреон, большой американ-
ский дрозд, американская хохлатая синица, желтоголовая амери-
канская славка и несколько видов дятлов. Развитие листопадных
лесов умеренной зоны представляет собой длительную сукцессию,
которая рассматривалась в гл. 8, разд. 1 (см. табл. 8.2). Для
ранних сукцессионных стадий характерна растительность пере-
ходных степей или залежей, на промежуточных стадиях часто
выделяются сосны.
Листопадные леса умеренной зоны представляют собой наи-
более важные биотические области мира, так как именно в этих
районах цивилизация достигла наибольшего развития. В резуль-
тате этот биом сильно изменился под влиянием человеческой
деятельности, его большая часть замещена культурными сообще-
ствами или сообществами лесных опушек.
Листопадные леса Северной Америки подразделяются на мно-
го важных типов, являющихся разными климакснымп типами ле-
сов. Вот некоторые из них:
буково-кленовый лес на севере центральных районов США:
кленово-липовый лес в шт. Висконсин и Миннесота;
дубово-гикориевый лес в западных и южных штатах;
дубово-каштановый лес Аппалачских гор (теперь это главным
образом дубовый лес, так как каштан уничтожен паразитическим
грибом);
различные смешанные мезофитные леса Аппалачского плато;
эдафический сосновый лес па юго-востоке прибрежной равнины,
который сохраняется благодаря пожарам и бедной песчаной поч-
ве (на бедных почвах вечнозеленые леса обладают конкурентным
преимуществом перед листопадными лесами).
Каждый из этих типов леса имеет свои отличительные черты,
но многие организмы, особенно крупные животные, распростра-
нены в пределах двух или более типов («связующие» виды). Лис-
топадные леса Западной Европы относительно бедны видами (ис-
ходное разнообразие снизилось в результате оледенения). На-
Краткое описание основных типов природных экосистем 259
против, смешанный мезофитный лес на юге Аппалачей (здесь
не было оледенения) и сравнимые с ним леса Восточной Азии —
самые богатые видами среди лесов умеренной зоны мира.
Степи умеренной зоны (рис. П. 6)
Степи умеренной зоны расположены там, где выпадает проме-
жуточное между пустынями и лесами количество осадков (250—
750 мм); существование степей в этих условиях зависит от тем-
пературы, сезонного распределения осадков и влагоемкости поч-
вы. Упоминавшиеся уже исследования в рамках МБП показали,
что ключевым фактором здесь является влажность почвы, в част-
ности потому, что опа лимитирует микробное разложение и воз-
врат в круговорот биогенных элементов. Обширные степные про-
странства занимают внутренние части Евразийского и Североаме-
риканского континентов, юг Южной Америки (аргентинские пам-
пасы) и Австралию.
В Северной Америке биом степей подразделяется (с востока
на запад) на следующие зоны: высокотравные, смешанные, низ-
котравные и кустисто-злаковые прерии. Эта зональность опреде-
ляется градиентом количества осадков, который одновременно
является и градиентом уменьшения первичной продуктивности.
Ниже приводится перечень некоторых наиболее важных видов
многолетников, сгруппированных по высоте надземных частей.
1. Высокие злаки (от 150 до 240 см) — бородач (Andropogon
gerardi), просо (Panicum virgatum), индейская трава (Sorghastrum
nutans) и в низинах Spartina pectinata.
2. Средние злаки (от 60 до 120 см) — малый бородач (Andro-
pogon scoparius), ковыль (Stipa spartea), спороболус (Sporobolus
heterolepis), пырей (Agropyron smithii), тонконог (Koeleria crista-
ta), индейский рис (Oryzopsis) и многие другие.
3. Низкие злаки (от 15 до 45 см) — бизонова трава (Buchloe
dactyloides), голубая грамова трава (Bouteloua gracilis), другие
виды грамовой травы (Bouteloua. sp.), интродуцированные мятлик
(Роа) и костер (Bromus sp.).
Корни большинства видов глубоко (до 2 м) проникают в поч-
ву, п масса корней климаксных многолетников в несколько раз
превышает массу их надземных частей. Большое значение имеет
форма роста корней. Некоторые из упомянутых видов, например
бородач, бизонова трава и пырей, образуют подземные корневища
и формируют таким образом дернину. Другие виды, такие, как
малый бородач, тонконог, ковыль, кустятся и растут куртинами.
Эти две жизненные формы можно найти во всех зонах, но кусто-
вые злаки преобладают в более засушливых областях, где степь
переходит в пустыню.
17'
Рис. П.6. Естественный злаковник умеренной зоны в центральной части
Северной Америки с двумя видами травоядных млекопитающих. А. Зла-
ковник с умеренным выпасом в Национальном резервате Ред-Род-Лейк,
Монтана; видно небольшое стадо вилорогих антилоп. Б. Низкотравшлй
злаковник в Национальном парке Уэйнрайт, пров. Альберта, Канада, со
стадом бизонов. Животное в центре «принимает пылевую ванну». Старые
«бизоньи ванны» можно обнаружить в прерии через много лет после ис-
требления бизонов.
Краткое описание основных типов природных экосистем
261
Разнотравье (сложноцветные, бобовые и т. и.) составляют
обычно лишь малую часть биомассы продуцентов климаксной сте-
пи, но встречаются неизменно. Некоторые виды особенно интерес-
ны как индикаторы стресса. Усиление выпаса, засуха или обе эти
причины увеличивают процент разнотравья, что выражено также
на ранних стадиях сукцессии. Вторичная сукцессия в степном
биоме и ритмические изменения растительности на протяжении
влажного и сухого сезонов были описаны ранее в гл. 8, разд. 1.
Те, кто путешествует на автомобиле по континентальным районам
США, должны помнить, что живописные однолетние травы на
обочинах дорог, такие, как солянка (Salsola) и подсолнечник
(Helianthus), обязаны своим «процветанием» постоянному раз-
рушению почвы, чему способствуют ремонтные работы на ско-
ростных дорогах.
Обширные пространства степей, особенно высокотравных пре-
рий, в настоящее время заняты посевами зерновых, культурными
пастбищами или древесной растительностью. Сейчас трудно найти
первоначальную, или девственную, высокотравную прерию, и там,
где она сохраняется для научных и учебных целей (как в арборе-
туме Висконсинского университета), ее нужно время от времени
выжигать, чтобы сохранить специфику этого биома.
В хорошо развитом степном сообществе есть виды с различ-
ными температурными адаптациями; одна группа растет в холод-
ные сезоны (весной и осенью), другая — в теплый (летом). Степь
в целом «скомпенсирована» по температуре, что удлиняет период
первичного продуцирования. Роль в этой адаптации Сз- и Сатинов
фотосинтеза обсуждалась в гл. 2, разд. 5 (см. табл. 2.2).
В степном сообществе развивается почва, полностью отличная
от лесной, даже если оба сообщества стартовали на одной и той
же материнской породе. Так как злаки по сравнению с деревьями
живут недолго, почву обогащает большое количество органическо-
го вещества. Первая фаза разложения протекает быстро, в резуль-
тате образуется мало подстилки, но много гумуса, иными словами,
скорость гумификации велика, а минерализации мала. В итоге
степная почва может содержать в 5—10 раз больше гумуса, чем
лесная, как видно при сравнении профилей на рис. 5.18. Темная
степная почва лучше других подходит для произрастания куку-
рузы, пшеницы и других зерновых, представляющих собой виды
культурных злаков.
Пожары помогают степной растительности конкурировать с
лесной в теплых или влажных районах. Характерная особенность
степей — наличие крупных травоядных. «Экологические эквива-
ленты» бизонов, антилоп и кенгуру в степях разных биогеогра-
фических областей отмечались на с. 125. Крупные травоядные
представлены двумя «жизненными формами»: бегающей, к кото-
рой относятся только что упомянутые животные, и роющей, к ко-
262
Приложение
торой относятся суслики и луговые собачки. Когда природные
степи превращаются в пастбища, то присущие им виды травояд-
ных животных заменяются домашними: коровами, лошадьми, ов-
цами и козами. Поскольку степи адаптированы к мощному пото-
ку энергии, проходящему через пастбищную пищевую цепь, такое
переключение экологически безвредно. Однако люди постоянно
страдают от «трагедии общин», допуская чрезмерный выпас и
выпахивание. Таким образом, многие степи превратились в на-
стоящее время в созданные руками человека пустыни, которые
трудно вернуть вновь к состоянию степей. Например, Морелло
(Morello, 1970) сообщает, что в аргентинских пампасах в резуль-
тате интенсивного выпаса коров количество способных гореть ве-
ществ сократилось настолько, что не осталось пищи пожарам,
необходимым для поддержания покрова злаков. Это привело к
преобладанию колючих кустарников, рост которых прежде сдер-
живался периодическими пожарами. Единственный путь восста-
новления продуктивности пастбищ — затрата энергии топлива для
механического удаления и выжигания древесной растительности.
Описанная ситуация — пример такого антропогенного изменения
растительности, которое можно сделать обратимым только доро-
гой ценой.
Тропические степи и саванны (рис. П. 7)
Тропические саванны (степи с редкими деревьями или груп-
пами деревьев) расположены в теплых областях, где в год выпа-
дает большое количество осадков (1000—1500 мм), но имеется
один пли два продолжительных сухих сезона, когда возникают
пожары, представляющие собой важный фактор окружающей
среды. Самая обширная область этого типа находится в Централь-
ной и Восточной Африке, но довольно значительные области тро-
пических саванн (grassland) встречаются также в Южной Аме-
рике п Австралии. Поскольку и деревья, и злаки должны быть
устойчивыми к засухе и пожару, растительность состоит из не-
большого числа видов, что составляет разительный контраст с
расположенными рядом экваториальными лесами. В покрове до-
минируют знаки, принадлежащие к таким родам, как Panicum,
Pennisetum, Andropogon, Imperata.
Вместе с тем редкие разбросанные деревья принадлежат к
видам, отличным от тех, что растут в дождевом лесу. Ландшафт
африканской саванны усеян колючими и живописными акациями
и другими деревьями и кустарниками, относящимися к семейству
бобовых, баобабами (Adansonia), древовидными молочаями (эко-
логические эквиваленты кактусов) и пальмами. Часто на боль-
ших пространствах доминирует один вид деревьев и злаков.
Краткое описание основных типов природных экосистем
263
Рис. П.7. Тропическая саванна в Восточной Африке (Найроби). Характер-
ные черты этого биома — травы, редкие деревья причудливой формы,
пожары в сухой сезон и обилие видов крупных травоядных млекопитаю-
щих (показана газель Томсона). (Фотографии Donald I. Ker; Ker, Downey
Safaris, Ltd.)
Африканская саванна не имеет себе равных по численности
и разнообразию популяции копытных. Здесь пасутся или объеда-
ют деревья многочисленные виды антилоп (включая гну), зебры
и жирафы, а в тех местах, где эту «крупную дичь» не вытеснил
человек со своим домашним скотом, на нее охотятся львы п дру-
гие хищники. Мутуализм между травоядными и травами довольно
подробно обсуждался в гл. 3 на с. 163. Насекомые наиболее
обильны во время влажного сезона, когда гнездится большинство
птиц; рептилии же активнее во время сухого сезона. Следова-
тельно, сезонность определяется дождями, а не температурой,
как в степях умеренной зоны.
Самые древние ископаемые остатки человека найдены в Вос-
точной Африке, однако мы не знаем, была ли эта область на
заре развития человечества засушливее или более влажной, чем
сейчас.
Чапараль и жестколистные леса (рис. П. 8)
В областях с мягким умеренным климатом, где обильны зимние
дожди, но лето сухое, растительность состоит из деревьев или
кустарников (или тех и других) с жесткими толстыми вечнозеле-
ными листьями (рис. П.8). В данный раздел мы включили рас-
Рис. П.8. А. Общий вид поросших чапаралем холмов в Калифорнии. Б. Борь-
ба с небольшим пожаром в растительности пирогенного типа.
Краткое описание основных типов природных экосистем
265
тительность от прибрежного чапараля, в котором преобладают
кустарники, до «широколиственно-склерофильных» лесов, в кото-
рых преобладают вечнозеленые деревья небольшой или средней
величины. «Картина» климата одной из областей с зимними дож-
дями дана на рис. 5.14. Сообщества чапараля широко распростра-
нены в Калифорнии и Мексике, по берегам Средиземного моря,
в Чили и вдоль южного берега Австралии. В зависимости от
области и местных условий доминировать могут многие виды
растений. Все виды обладают микоризой, и у многих имеются
клубеньки с азотфиксирующими актиномицетами. Оба варианта
мутуализма способствуют выживанию в жестких условиях среды.
Пожары — важный фактор, способствующий доминированию кус-
тарника, а не деревьев. Чапараль как пример «пирогенной эко-
системы» и циклического климакса рассматривался в гл. 5, с. 314,
гл. 7, с. 107, и гл. 8, с. 172.
В Калифорнии чапараль покрывает склоны и каньоны на
площади 2—3 млн. га. Обычными кустарниками являются чамисо
(Adenostoma) и толокнянка (Arctostaphylos'), которые образуют
густые заросли; характерны также разные виды вечнозеленых
дубов и других кустарников и деревьев. Дождливый сезон, когда
происходит рост, продолжается обычно с ноября по май. В чапа-
рале в этот период живут чернохвостый олень и многие птицы;
с наступлением жаркого сухого лета они откочевывают к северу
или в горные районы. Постоянно живущие в чапарале позвоноч-
ные невелики и имеют тусклую окраску, гармонирующую с низ-
корослым лесом. Характерны мелкий кролик Бахмана (Sylvilagus
bachmani), древесные крысы, бурундуки, ящерицы, мелкие во-
робьиные и тауи. К концу вегетационного сезона плотность по-
пуляции гнездящихся птиц и размножающихся насекомых бывает
довольно высокой; в конце сезона роста, когда высыхает расти-
тельность, численность популяций уменьшается. Именно в этот
сезон часто возникают пожары, которые с невероятной быстротой
очищают склоны (и пригородные районы в южной Калифорнии).
После пожара с первыми дождями в чапарале начинают усилен-
но расти кустарники, достигая за 15—20 лет максимальной вели-
чины.
Жестколистные леса в средиземноморских областях с зимними
дождями носят местное название «маки»; в Австралии подобная
растительность с доминирующими деревьями и кустарниками из
рода Eucalyptus называется «мали-скрэб». Не удивительно, что
австралийские эвкалипты хорошо себя чувствуют в Калифорнии,
куда их активно интродуцировали и где они в значительной сте-
пени заменили в городах местные виды древесной растительности.
Рис. П.9. Три типа пустынь на западе Северной Америки. А. Низинная
«жаркая» пустыня на юге Аризоны с преобладанием креозотового кустар-
ника (Larrea). Обратите внимание на характерную форму роста пустынных
кустарников (многочисленные ветви ответвляются на уровне земли) и на
довольно равномерное распределение кустов. (Фотография R. R. Humphrey.)
Б. Пустыня в Аризоне, расположенная на несколько большей высоте с не-
сколькими формами кактусов, разнообразными пустынными кустарниками
и небольшими деревьями. Это одно из мест, выбранных для междисципли-
нарного изучения по МБП. (Фотография R. Н. Chew.)
Краткое описание основных типов природных экосистем
267
В. «Холодная» пустыня на востоке шт. Вашингтон с преобладанием полы-
ни {Artemisia). Фотография сделана весной во время максимума первичной
продукции, когда почва между кустами и вокруг них покрыта ковром одно-
летних злаков и трав. Показан момент эксперимента с введением радиоак-
тивной метки, проводимого для получения сравнительных оценок потреб-
ления элементов питания кустарниками и однолетниками. (Фотография
Hanford Atomic Products Operation.)
Пустыни (рис. П. 9)
Пустыни встречаются в общем в тех областях, где в год выпадает
меньше 250 мм осадков. Иногда к пустыням относят области и с
большим количеством осадков, но эти осадки распределены очень
неравномерно (рис. П.9). Скудость осадков может быть вызвана:
1) высоким субтропическим давлением (Сахара и австралийские
пустыни); 2) географическим положением в области «дождевой
тени» (пустыни на западе Северной Америки; см. рис. 5.11);
3) большой высотой местности (пустыни Тибета и Боливии, Го-
би). В большинстве пустынь в течение года выпадает какое-то
количество дождей, и имеется хотя бы редкая растительность,
если эдафические условия субстрата не оказываются очень небла-
гоприятными (например, подвижные песчаные дюны, барханы).
По-видимому, единственные места, где дождей выпадает очень
мало или они не выпадают совсем, — это центральная Сахара и
пустыня на севере Чили. Характеристики климата пустыни
Ныо-Мексико приведены на рис. 5.14 (см. также рис. П.2).
268
Приложение
Вальтер (Walter, 1954) измерил чистую продукцию ряда пус-
тынных и полупустынных сообществ, расположенных вдоль гра-
диента количества осадков в юго-западной Африке. Оказалось,
что годовая продукция сухого вещества представляет собой ли-
нейную функцию количества осадков. Это иллюстрирует «жест-
кость» действия влажности как главного (ведущего) лимитирую-
щего фактора. Чистая годовая первичная продукция составляет
в пустынях менее 1000 кг сухого вещества с гектара, или около
200 ккал/м2 (см. рис. 3.7 и табл. 3.7).
В условиях орошения пустынь, когда вода перестает быть
лимитирующим фактором, первостепенное значение приобретает
тип почвы. Если текстура почвы и содержание элементов пита-
ния в ней благоприятны, то вследствие обилия солнечного света
оводненные пустыни могут быть крайне продуктивными. Однако
из-за высокой стоимости строительства и эксплуатации иррига-
ционных систем удельная стоимость выращенной продукции мо-
жет быть очень высокой. Через такую экосистему нужно пропус-
кать очень большие количества воды, в противном случае в почье
будут накапливаться соли (вследствие высокой скорости испаре-
ния), которые станут лимитировать продуктивность. По мере
«старения» орошаемой экосистемы потребности в воде растут. Это
вызывает «спираль инфляции»; возникает необходимость в строи-
тельстве новых водоводов, что увеличивает стоимость продукции
и ведет к более интенсивной эксплуатации подземных и горных
водных ресурсов. В пустынях Старого Света легко найти разва-
лины древних оросительных систем. Во многих случаях никто
не знает, почему они пришли в упадок и почему «райские сады»
опять превратились в пустыню. Эти руины должны служить нам
по меньшей мере напоминанием того, что орошенная пустыня
не будет процветать бесконечно, если игнорировать основные за-
коны развития экосистемы.
Существуют три жизненные формы растений, адаптированные
к пустыням: 1) однолетники, избегающие засухи тем, что они
растут только при достаточной влажности (см. т. 1, с. 267);
2) суккуленты, такие, как кактусы, характеризующиеся сохраня-
ющим влагу CAM-фотосинтезом (т. 1, с. 46) и накапливающие
воду; 3) пустынные кустарники, у которых многочисленные от-
ходящие от короткого основного ствола ветви несут мелкие тол-
стые листья. Во время длительного засушливого сезона эти листья
опадают. Пустынные кустарники всего мира весьма сходны по
внешнему виду, хотя могут принадлежать к различным таксоно-
мическим группам (еще один четкий пример экологической экви-
валентности).
Для пустынных кустарников характерно очень своеобразное
распределение: редкие отдельные растения разбросаны далеко
друг от друга с обширными пустынными" Пространствами между
Краткое описание основных типов природных экосистем
269
ними. В некоторых случаях определенную роль в поддержании
дистанции между растениями играют антибиотики. Однако всегда
разреженность снижает конкуренцию за скудные ресурсы, иначе
интенсивная конкуренция за воду могла бы привести к гибели
или задержке роста всех растений.
На основе температурного режима удобно различать два тина
пустынь: жаркие и холодные, хотя такая классификация несколь-
ко произвольна. Доминирующий вид в жаркой пустыне на юго-
западе Северной Америки представлен креозотовым кустарником
Larrea, а в расположенных севернее холодных пустынях Большого
Бассейна — полынью (Artemisia), В южных областях широко рас-
пространен также колючий шалфей (Franseria), тогда как на
высокогорьях, где влаги немного больше, характерными компо-
нентами являются гигантский кактус Sahuaro и пало верде. Вос-
точнее к пустынным кустарникам примешивается большое коли-
чество злаков и пустыня переходит в полупустыню. В холодных
пустынях, особенно на щелочных почвах внутренних водосборных
бассейнов, обширные площади заняты солянками сем. Chenopodia-
сеа, в частности видами родов Atriplex, Grayia, Eurotia, Sarco-
batus.
Солянки не менее широко распространены в аридных районах
и других частей света. В мексиканской пустыне наибольшего
развития достигает жизненная форма суккулентов, включая как-
тусы, древовидные юкки и агавы. Некоторые их виды проникают
в кустарниковые пустыни Аризоны и Калифорнии, но в холодных
пустынях такая жизненная форма не имеет смысла. Однолетние
травы и злаки создают замечательное зрелище во всех пустынях
во время короткого влажного периода. Обширные участки «голо-
го грунта» пустынь не обязательно лишены растений. Здесь могут
существовать мхи, водоросли и лишайники; на песках и других
типах тонкодисперсной почвы они образуют стабилизирующую
корку. Сине-зеленые водоросли (часто в составе лишайников)
имеют большое значение как фиксаторы азота.
Животные пустыни, как и растения, по-разному адаптированы
к недостатку воды. Пресмыкающиеся и некоторые насекомые
«преадаптированы», так как они обладают непроницаемыми по-
кровами и выделяют сухие экскреты (мочевую кислоту и гуанин).
Пустынные насекомые защищены от испарения особыми водоне-
проницаемыми при высокой температуре веществами. Полностью
устранить испарение с поверхности дыхательных путей невоз-
можно, однако у пустынных насекомых оно сведено к минимуму
благодаря особой системе внутренних дыхалец. Следует заметить,
что образование метаболической воды (в результате расщепления
углеводов), которое часто бывает единственным ее источником,
(само по себе не является адаптацией. Адаптация состоит в спо-
собности сохранять эту воду или, как это наблюдается у жуков-
Рис. П.10. Тропические леса. А. Вид сверху на листопадный низинный лес
в Панаме. Выделяются высокие деревья с белыми стволами, теряющие
листву в сухой сезон; они возвышаются над пологом широколиственных
вечнозеленых деревьев и пальм. (Фотография G. Child, Институт экологии,
университет шт. Джорджия.) Б. Горный дождевой лес в Пуэрто-Рико.
Видно обилие эпифитов, что характерно для дождевых горных лесов во
влажных тропиках. (Фотография Е. Р. Odum.) В. Крупным планом пока-
зано основание ствола дерева в том же горном лесу в Пуэрто-Рико; видна
масса мелких надземных корней, которые проникают в подстилку (а не
в почву, как в лесах умеренной зоны). (Фотография Е. Р. Odum.)
Краткое описание основных типов природных экосистем 271
чернотелок (характерных обитателей пустыни), в способности к
усиленному образованию метаболической воды при низкой влаж-
ности. Млекопитающих как группу нельзя считать хорошо адап-
тированными к условиям пустыни, так как они выделяют моче-
вину, что связано с потерей большого количества воды. Все же
некоторые виды приобрели удивительные вторичные адаптации.
К таким млекопитающим в Новом Свете принадлежат грызуны
из сем. Heteromyidae, особенно кенгуровая крыса (Dipodomys)
и карманчиковая мышь (Perognathus), а в пустынях Старого
Света — тушканчики (Dtpus сем. Dipodidae). Эти животные могут
неопределенно долго существовать на сухих семенах и вовсе
не нуждаться в воде для питья. Днем они остаются в норах и
сохраняют воду, выделяя очень концентрированную мочу и не
расходуя воду на терморегуляцию. Следовательно, адаптация этих
животных к жизни в пустыне носит не только физиологический,
но и поведенческий характер. Другие пустынные грызуны, на-
пример древесные крысы (Neotoma), не могут существовать ис-
ключительно на сухой пище, но выживают в некоторых частях
пустыни, поедая суккулентные кактусы или другие запасающие
воду растения. Должен пить даже верблюд, но верблюды могут
длительное время обходиться без воды потому, что их ткани
устойчивы к повышению температуры тела и к такой степени
дегидратации, которая была бы смертельна для большинства жи-
вотных. (Кстати, верблюды не запасают воду в своих горбах,
как это принято считать.)
Полувечнозеленые сезонные тропические
леса (рис. П. 10, Л)
Сезонные тропические леса, в том числе муссонные леса тропиче-
ской Азии, произрастают в областях с влажным тропическим кли-
матом, где выражен сухой сезон, во время которого некоторые или
все деревья теряют листву (в зависимости от продолжительности
и резкости сухого сезона). Ключевым фактором здесь являются
строгие сезонные колебания в выпадении довольно обильных в
течение года осадков. Там, где продолжительность сухого и влаж-
ного периодов примерно одинакова, сезонность проявляется так
же, как в листопадном лесу умеренной зоны, причем «зима» здесь
соответствует сухому сезону. В сезонном лесу Панамы, показан-
ном на рис. П.10, А, крупные, возвышающиеся над пологом де-
ревья теряют листву во время сухого сезона, а пальмы и другие
деревья из нижних ярусов сохраняют листья (отсюда термин
«полувечнозеленый»). По своему видовому богатству сезонные
тропические леса занимают второе место после дождевых
лесов.
272
Приложение
Тропические дождевые леса (рис. П. 10, Б и В)
Разнообразие жизни достигает, по-видимому, максимума в широ-
колиственных вечнозеленых дождевых тропических лесах, ко-
торые расположены на малых высотах полосой вдоль экватора.
Дождей выпадает от 2000 до 2250 мм в год; в течение года от-
мечается один или несколько относительно «сухих» сезонов
(120—130 мм в месяц или меньше). Дождевые леса встречаются
в трех главных областях: 1) бассейны Амазонки и Ориноко в
Южной Америке (крупнейший сплошной лесной массив) и Цент-
ральноамериканский перешеек; 2) бассейны Конго, Нигера и
Замбези в Центральной и Западной Африке и Мадагаскар; 3) об-
ласти Индо-Малайская и Борнео—Новая Гвинея. Эти области
отличаются друг от друга по видовому составу (так как находят-
ся в разных биогеографических областях), но сходны по структу-
ре и экологии лесов. Различия в температуре между зимой и
летом выражены не столь сильно, как между ночью и днем. Се-
зонная периодичность размножения и других жизненных функций
растений и животных в значительной степени связана с колеба-
ниями количества осадков или же регулируется внутренними
ритмами. Например, некоторые деревья сем. Winteraceae растут
непрерывно, тогда как для других видов этого же семейства ха-
рактерен периодический рост с образованием годичных колец.
Птицы дождевого леса также, по-видимому, нуждаются в периоде
«покоя», поскольку размножение у них часто происходит с пе-
риодичностью, которая не связана с сезонами. Дождевой лес
характеризуется очень сильно выраженной ярусностью. Деревья
обычно формируют три яруса: 1) редкие очень высокие деревья,
которые возвышаются над общим уровнем полога; 2) полог, об-
разующий сплошной вечнозеленый покров на высоте 25—35 м;
3) нижний ярус, который становится густым только там, где
имеются просветы в пологе. Корни у высоких деревьев неглубо-
кие; утолщенная нижняя часть ствола выполняет функцию
«контрфорса». Изобилие лазящих растений, в частности древес-
ных лиан и эпифитов, таково, что они часто скрывают контурьз
деревьев. Следует особо упомянуть «фикус-душитель» и другие
деревянистые лианы. Число видов растений очень велико; часто
на нескольких гектарах можно встретить больше видов деревьев,
чем во флоре всей Европы.
В отличие от лесов умеренной зоны, где жизнь приурочена к
надпочвенному уровню, в дождевом лесу значительно большая
доля животных обитает в верхних ярусах растительности. Напри-
мер, в Британской Гвиане на деревьях живет больше 50% мле-
копитающих. Кроме древесных млекопитающих здесь в изобилии
встречаются хамелеоны, игуаны, гекконы, древесные змеи, лягуш-
ки и птицы. Важную экологическую роль играют муравьи и
Краткое описание основных типов природных экосистем
273
прямокрылые, а также дневные и ночные бабочки. В таких лесах
широко распространен симбиоз между животными и эпифитами.
По отношению к фауне также справедливо утверждение, что она,
как и флора, необычайно богата видами. Например, на 15 км2'
в Барро-Колорадо, хорошо изученном участке дождевого леса в
зоне Панамского канала, насчитывается 20 000 видов насекомых^
тогда как на территории всей Франции их всего несколько сотен.
Разнообразные архаичные типы животных и растений выжили в
многочисленных экологических нишах неизменяющейся среды.
Многие ученые полагают, что для тропического леса характерна
высокая скорость эволюционных изменений и видообразования,
поэтому такие леса стали местом возникновения многих видов,
вошедших позднее в состав более северных сообществ. В настоя-
щее время ученые всего мира озабочены необходимостью сохра-
нения больших пространств тропических лесов в качестве «ре-
сурса генов» (см. с. 146—150).
Основной источник пищи животных в тропических лесах —
плоды и термиты. Обилие птиц в этих лесах объясняется тем, что
многие из них растительноядные; это питающиеся плодами по-
пугаи, птица-носорог, туканы, котинги, трогоны и райские птицы.
Поскольку «чердаки» джунглей перенаселены, многие птицы вьют
себе висячие гнезда, а насекомые строят висячие коконы, что
спасает их от армии муравьев и других хищников. Хотя известно
несколько видов ярких птиц и насекомых, обитающих в более
открытых местах, основная масса животных дождевого леса не-
приметна, и многие из них ведут ночной образ жизни.
В горных районах тропиков расположен горный дождевой лес^
который представляет собой разновидность низинного дождевого
леса с некоторыми характерными особенностями. По мере про-
движения в горы лес становится все более низкорослым, и эпи-
фиты составляют все большую долю автотрофной биомассы; эта
тенденция достигает кульминации в карликовом лесу горной ги-
леи. Функциональная классификация дождевых лесов может быть
основана на дефиците насыщения, так как он определяет транс-
пирацию, от которой в свою очередь зависит биомасса корней и
высоты деревьев- Еще одна разновидность дождевого леса встре-
чается вдоль берегов затапливаемых речных долин; это так на-
зываемый галерейный лес, или прибрежный лес.
Замечательным свойством дождевых лесов является эффек-
тивный прямой возврат в круговороты биогенных элементов, осу-
ществляемый мутуалистическими микроорганизмами. Именно
благодаря этому на скудных почвах леса растут так же пышно,
как и на более плодородных. Эта особенность более подробно об-
суждалась в гл. 4, разд. 7.
На местах, где дождевой лес уничтожен, часто развивается
вторичный лес, в состав которого входят хвойные деревья, такиеь
18 Заказ № 1483
Приложение
*274
как Musangcb (Африка), Cecropia (Америка) и Macoranga (Ма-
лайзия). Вторичный лес выглядит пышным, но совершенно от-
личается от девственного леса как экологически, так и флористи-
чески. «Климакс» возвращается обычно очень медленно, особенно
на песчаных и других бедных минеральным питанием почвах, так
как все элементы питания первоначального леса были утрачены
.в результате изъятия биомассы и нарушения микробиологической
сети их возврата в круговорот.
Вопрос о том, как осваивать дождевые леса, продолжает по-
рождать противоречия и крушения планов как тех, кто видит в
этих лесах последнюю границу на пути колонизации, так и тех,
кто рассматривает их как потенциальный источник богатства.
^Огромная величина деревьев ввела в заблуждение ранних евро-
пейских исследователей Амазонки, предположивших, что это сви-
детельствует о плодородии почвы. Поэтому последовали много-
численные безуспешные попытки создать на месте дождевого
.леса сельскохозяйственную или лесную индустрию. Несмотря на
неудачи, предприниматели и правительства продолжают попытки
приспособить сельскохозяйственную и лесную технологию уме-
ренной зоны к районам дождевого тропического леса, хотя эта
технология здесь определенно не годится. В конце 60-х годов
^миллиардер Д. К. Людвиг заполучил в Бразильской Амазонии
(Жара) участок площадью примерно с Коннектикут. Он сплавил
;зрелый лес на целлюлозно-бумажные фабрики и превратил его в
плантации экзотических видов. Джордан (Jordan, 1982) так ком-
ментирует это дорогостоящее предприятие: «Теперь, спустя менее
45 лет после начала осуществления проекта, он потерпел провал.
'Еще есть возможность отказаться от мнения, что проблема связа-
ла с почвами. В неудаче повинно скверное (экономическое)
управление». По мнению Джордана, лучший способ использова-
ния лесных ресурсов — организация вырубок в виде полос, так,
чтобы не оказался сильно нарушенным запас биогенов в корне-
вых подушках и чтобы обсеменение от ненарушенных прилежа-
щих участков позволило быстро восстановиться лесу на местах
.вырубок. Сходным образом можно организовать и садоводство.
Ясно одно, что человек должен строить свои проекты с исполь-
зованием адаптаций региональных экосистем, а не в противоре-
чие им.
Тропический скрэб, или колючее редколесье
Там, где условия влажности промежуточны между пустыней и
саванной, с одной стороны, и сезонным или дождевым лесом —
с другой, можно встретить тропический скрэб, или колючее ред-
колесье. Такая растительность занимает большие площади в
Краткое описание основных типов природных экосистем
275
центре Южной Африки, в юго-западной Африке и в части юго-
западной Азии. Ключевой климатический фактор здесь — небла-
гоприятное распределение вполне достаточных в целом осадков.
Колючие леса, которые в Африке или Австралии часто называют
«бушами», а в Бразилии—«каатингами», состоят из некрупных
лиственных деревьев, которые нередко бывают колючими и при-
чудливо изогнутыми: эти деревья покрыты мелкими листьями,,
опадающими в сухие сезоны. Деревья могут образовывать как
густые насаждения, так и быть редко рассеянными или образовы-
вать группу. В некоторых случаях не всегда можно точно опре-
делить, является ли колючее редколесье в данной области при-
родным или оно возникло благодаря многолетним выпасам скота.
Зональность в горах (рис. П. 11)
Распределение биотических сообществ в горных областях носит
своеобразный характер, как этого и следовало ожидать, учитывая
разнообразие физических условий. Основные сообщества распо-
ложены обычно в виде неправильных поясов, часто с очень уз-
кими экотонами. Мелкомасштабная карта, подобная приведен-
ной на рис. П.1, не может передать эти особенности. На какой-
3900 - БИОМ ЗОНА
СКЛОН
СКЛОН
Т ундра
Зона осок и злаков
2. Зона ели и пихты
3. Зона туи и тсуги
4. Зона лжетсуги тиссолистной
Б. Зона желтой сосны
2 2400
8 2100
Степь
7. Зона овсяницы и пырею
8. Зона пырея и мятлика
9. Зона ковылей и пастбищных трав
НЮ. Зона полыни
11. Зона креозотового кустарника:
Рис. П.11. Вертикальная зональность в горах на западе Северной Америки.
Схема не относится к какой-либо определенной горе, а показывает общую
картину, характерную для всего центрального (так называемого «меж-
горного») района Скалистых гор в шт. Юта. К северу, югу, востоку или
западу этого района зональность может быть несколько иной. Так, к северу
и востоку отсутствуют зона креозотового кустарника и зона карликовой
сосны и можжевельника. Между степью и северным хвойным лесом на
западе располагаются различные зоны биома чапараля и на востоке — зона
дуба и Cercocarpus montanus.
18*
£76
Приложение
либо конкретной горе могут быть представлены четыре или пять
главных биомов с многими зональными подразделениями
(рис. П.И). Следовательно, здесь имеется более тесный контакт
между биомами и более активный обмен между биотами, чем в
равнинных областях. Вместе с тем сходные сообщества в горах
изолированы друг от друга в большей степени, так как горы
редко образуют непрерывную цепь.
В общем многие виды, характерные для широких равнинных
вариантов биомов, столь же характерны и для вытянутых напо-
добие поясов горных «фаз». Вследствие изоляции и особенностей
рельефа многие другие виды и вариететы присущи только горным
сообществам. Как показано на рис. П.11, высотные границы зон
расположены выше на более теплом южном склоне и ниже —
на северном.
2. Пресноводные экосистемы
Пресноводные местообитания удобно подразделять на три
группы:
1. Стоячие воды или лентические (лат. lenis — спокойный)
экосистемы — озера и пруды.
2. Текучие воды или логические (лат. lotus — омывающий)
экосистемы — родники, ручьи и реки.
3. Заболоченные участки с колеблющимся уровнем воды часто
по сезонам, ио возможно и от года к году — марши и болота.
Типы этих местообитаний приведены на рис. П.12. Грунтовые
воды, хотя они и образуют очень большой пресноводный резер-
вуар и представляют собой очень важный для людей ресурс
(гл. 5, разд. 4), в общем не считаются экосистемой, так как они
безжизненны или населены очень слабо (иногда бактериями).
Грунтовые воды связаны с тремя главными поверхностными эко-
системами (см. рис. 4.10) и, таким образом, представляют собой
среду на входе и среду на выходе для этих экосистем.
По сравнению с морскими и наземными местообитаниями
пресноводные местообитания занимают сравнительно малую часть
поверхности Земли, ио их значение для человека поистине огром-
но. Это объясняется рядом причин.
1. Пресные воды — самый удобный и самый дешевый источник
воды для бытовых и промышленных нужд. (Мы можем и, по-ви-
димому, будем получать большую часть пресной воды из моря,
но стоимость такой воды чрезвычайно высока, если учесть расход
энергии для опреснения и возрастающее при этом засоление
среды.)
2. Пресные воды — узкое место планетарного гидрологическо-
го цикла (см. рис. 4.9,Б).
Краткое описание основных типов природных экосистем 277
3. Пресноводные экосистемы наряду с лиманами и эстуариями
представляют собой самую удобную и дешевую систему третичной
переработки отходов. Почти все без исключения крупнейшие го-
рода мира расположены на больших реках, озерах или лиманах,
которые служат бесплатными коллекторами. Человек настолько
злоупотреблял использованием этого природного ресурса, что те-
перь уже стала очевидной необходимость прилагать значительные
усилия для немедленного уменьшения возникшего стресса. В про-
тивном случае вода станет основным лимитирующим фактором
для человека как биологического вида (т. 1, с. 289).
Вода обладает рядом уникальных термодинамических свойств,
способствующих уменьшению температурных колебаний; таким
образом, диапазон различных изменений в воде меньше и проис-
ходят они медленнее, чем в воздухе. Самые важные из этих
термодинамических свойств воды следующие:
1. Высокая удельная теплоемкость. Это значит, что для изме-
нения температуры воды требуется относительно большое коли-
чество тепла. Для повышения температуры 1 мл воды (1 г) на
1 °C (от 15 до 16 °C) необходимо затратить 1 кал тепла. Только
аммиак и немногие другие вещества обладают удельной тепло-
емкостью больше 1.
2. Большая скрытая теплота плавления. Для превращения 1 г
льда в воду необходимо затратить 80 кал (столько же тепла вы-
деляется при замерзании воды без изменения температуры).
3. Самая высокая из известных скрытая теплота парообразо-
вания. При испарении (а оно происходит более или менее по-
стоянно с поверхности растений, воды и льда) 1 г воды поглоща-
ется 536 кал. Значительная часть поступающей солнечной энергии
в экосистемах всего мира расходуется на испарение воды (см.
гл. 3, разд. 1). Этот поток энергии смягчает климат и делает воз-
можным существование жизни во всем ее фантастическом разно-
образии.
4. Наибольшей плотностью вода обладает при 4 °C; при тем-
пературах выше или ниже 4 °C вода расширяется и, следователь-
но, становится легче. Это уникальное свойство предохраняет озера
от промерзания до дна.
Лентическпе экосистемы (озе^а и пруды)
Большинство пресных водоемов, которые заполнены сейчас стоя-
чей водой, в геологическом смысле молоды. Жизнь прудов про-
должается от нескольких недель или месяцев (маленькие времен-
ные пруды) до нескольких сотен лет (большие водоемы). Очень
немногие озера, например озеро Байкал в России, являются древ-
ними, большинство же крупных озер образовалось не раньше
ледникового периода. Можно ожидать, что пресноводные экосис-
278
Приложение
темы изменяются во времени со скоростью, более или менее обрат-
но пропорциональной их размеру и глубине. Географическая
разобщенность пресноводных водоемов способствует видообразова-
нию, однако отсутствие изоляции во времени препятствует этому
процессу. Вообще говоря, видовое разнообразие в пресноводных
сообществах невелико, и многие таксоны (виды, роды, семейства)
широко распространены в пределах континентов и даже на смеж-
ных континентах. Более подробно пруд как экосистема, удобная
вследствие малых размеров при изучении вводного курса эколо-
гии, рассматривался в гл. 2, разд. 7. Характерной чертой озер
и крупных прудов является четкая зональность и стратификация.
В типичном случае мы можем различить литоральную зону, где
встречаются прибрежные укореняющиеся растения, лимническую
зону, или зону открытой воды, где доминирует планктон, и глу-
боководную профундальную зону, где живут только гетеротрофы.
В областях с умеренным климатом озера часто становятся стра-
тифицированными летом и зимой, что вызывается нагреванием и
охлаждением воды. Более теплые верхние слои в озере, или эпи-
лимнион (греч. limnion — озеро), временно изолируются от охлаж-
денных глубинных вод, или гиполимниона, зоной термоклина, ко-
торая служит преградой для обмена различными веществами.
Вследствие этого запасы кислорода в гиполимнионе и биогенных
элементов в эпилимнионе истощаются. Весной и осенью, когда
температура всей водной массы выравнивается, происходит пе-
ремешивание. «Цветение» фитопланктона часто происходит имен-
но после этих перемешиваний, поскольку в фотической (освещен-
ной) зоне становятся доступными придонные биогены. В странах
с теплым климатом перемешивание происходит только один раз
в году (зимой), в тропиках процесс перемешивания происходит
постепенно или нерегулярно.
Первичная продукция экосистем со стоячей водой зависит от
химической природы бассейна и от природы поступающих с при-
токами и с суши веществ; обычно она связана с глубиной обрат-
ной зависимостью. Следовательно, урожай рыбы с единицы вод-
ной поверхности выше в мелких, чем в глубоких озерах (см.
табл. 3.10), однако в глубоких рыба бывает крупнее. На основе
продуктивности озера часто подразделяются на олиготрофные
(малокормные) и эвтрофные (кормные). Парадокс состоит в том„
что с точки зрения хозяйственного и рекреационного использова-
ния биологически бедные озера предпочтительнее высокопродук-
тивных. В одних частях биосферы люди увеличивают плодородие
озер для нужд собственного питания, тогда как в других частях
они сдерживают плодородие (удаляя биогены, отравляя растения
и т. д.) для сохранения красоты окружающей природы. Плодород-
ные зеленые пруды, которые могут давать много рыбы, не счи-
таются подходящими для отдыха и купания.
‘Краткое описание основных типов природных экосистем
279
При строительстве искусственных прудов и озер (водохрани-
лищ) человек изменяет ландшафт в тех регионах, где нет при-
родных водоемов. В США почти при каждой ферме сейчас имеет-
ся пруд, и почти на каждой реке построено водохранилище.
Многие такие искусственные водоемы приносят доход, но идея
строительства водохранилищ может завести слишком далеко; за-
топление плодородных земель, которые дают намного больше про-
дуктов питания, чем водохранилища, не всегда лучший способ
длительного землепользования. В стоячих водах загрязнения окис-
ляются в общем менее эффективно, чем в проточных. Если водо-
сборный бассейн не покрыт хорошей растительностью, осадки,
образующиеся в результате эрозии, могут заполнить весь водоем
за время жизни одного поколения. Поэтому, прежде чем строить
новые водохранилища, следует проводить очень тщательный ана-
лиз всех затрат и выгод.
Тепловой бюджет водохранилищ может сильно отличаться от
«бюджета природных озер, что зависит от конструкции плотины.
Если водосброс, как на плотинах гидроэлектростанций, придон-
ный, то вниз по течению экспортируется холодная вода, богатая
«биогенными элементами и бедная кислородом, тогда как теплая
вода задерживается в водохранилище. Водохранилище в этом
«случае работает как ловушка тепла и экспортер биогенных эле-
ментов в противоположность природным озерам с поверхностным
водосливом, которые, таким образом, функционируют как ловуш-
ка биогенов и экспортер тепла. Характер водосброса, следователь-
но, сильно влияет на условия вниз по течению.
.Логические экосистемы (ручьи и реки)
В общем различия между текучими и стоячими водами скон-
центрированы вокруг триады условий: 1) течения, являющегося
значительно более важным контролирующим и лимитирующим
фактором; 2) обмена между сушей и водой, относительно более
интенсивно выраженного в реках, в результате чего в реке со-
здаются более «открытая» экосистема и «гетеротрофный» тип
метаболизма сообщества, особенно если река невелика; 3) кон-
центрации кислорода, которая в реке обычно выше, его более
равномерного распределения и более слабо выраженной (пли
отсутствующей вовсе) термической стратификации; исключения
составляют большие медленно текущие реки.
Вдоль «речного континуума» происходят изменения метаболиз-
ма сообщества, биотического разнообразия и размера частиц от
истоков до устья, как показано на рис. 4.11. На протяжении
конкретного речного потока выявляются обычно две зоны:
1. Перекаты: участки с достаточно быстрым течением. Дно
свободно от ила и обломочных материалов, вследствие чего по-
280
Приложение
верхность его твердая. В этой зоне обитают специализированные
организмы, которые прочно прикрепляются или прилипают к
субстрату (как личинки веснянок и ручейников), или хорошие
пловцы, способные плыть против течения (форель, дартер и т. д.).
2. Плёсы: глубоководные участки с медленным течением, в ре-
зультате чего ил и песок оседают на дно, образуя мягкий суб-
страт. Такие условия благоприятны для жизни роющих и пла-
вающих животных, укореняющихся растений и на больших плё-
сах — развития планктона. Сообщества плёсов крупных рек
напоминают прудовые.
В верховьях реки обычно вымывают породу, врезаясь в суб-
страт, в результате чего дно чаще бывает твердым. В нижнем
течении реки взвесь обычно оседает, формируются заливные до-
лины и дельта, которые часто бывают крайне плодородными.
По химическому составу воды Ливингстон (Livingstone, 1963)
подразделяет реки мира на два типа: 1) жестководные, или кар-
бонатные, реки, содержащие 1ОО°/оо или более растворенных неор-
ганических веществ, и 2) мягководные, или хлоридные, реки, где
растворенных веществ менее 25%о.
Химизм воды карбонатных рек определяется преимущественно
выветриванием скальных пород, а доминирующий фактор, опре-
деляющий химизм хлоридных рек, — атмосферные осадки. Еще
один тип рек, характерный для теплых низменностей, — гумусо-
вые, или «черные», реки с высокой концентрацией растворенных
органических веществ.
Ряд превосходных исследований, проведенных преимуществен-
но на рыбах, по энергетике пищевых цепей в реках был выпол-
нен, например, Алленом (Allen, 1951), Хортоном (Horton, 1961),
Камминсом (Cummins, 1974) и на р. Темзе Манном (Mann,
1965).
Несмотря на малочисленность родников и источников и не-
большое пространство, занимаемое ими, изучение их чрезвычайно
важно. Источники послужили объектом изучения целостных эко-
систем; к числу лучших исследований следует отнести исследова-
ния крупных известковых источников во Флориде (Н. Т. Odum
и сотрудники), маленьких холодных родников в Новой Англии
(Teal, Tilly) и горячих источниках в Йеллоустоне (Wiegert, Brock
и сотрудники).
Заболоченные пресноводные участки
(марши и болота) (рис. П. 12, А)
Заболоченными участками называются любые пространства, ко-
торые хотя бы часть года покрыты пресной водой. Почвы здесь
насыщенны водой постоянно или на протяжении части года. Клю-
чевой фактор, определяющий продуктивность и видовой состав
Рис. П.12. Три пресноводных экосистемы. А. Болото; пресноводный марш
в национальном резервате Сакраменто, шт. Калифорния, где в зарослях
водной и полуводной растительности находят убежища и обильный корм
стаи диких гусей. Б. Лентическая экосистема; природный пруд в зоне сте-
пей в западной Канаде. В. Лотические экосистемы; слияние двух рек на
севере Нью-Джерси. Водосбор реки, находящейся внизу в центре фотогра-
фии, защищен травянистой и древесной растительностью; река, текущая
слева, загрязнена илом в результате плохого ведения сельского хозяйства
(А и Б — Department of Interior, Fich and Wildlife Service; B — U.S. Soil
Conservation Service.)
282
Приложение'
этих сообществ, — гидропериод, т. е. периодичность колебаний
уровня воды. Роль потока воды как вклада в энергетику обсужда-
лась в гл. 3, разд. 3, и в гл. 5, разд. 4. Связь между продуктив-
ностью и гидропериодом в заболоченных лесах была схематически
показана на рис. 5.16. В соответствии с этим заболоченные место-
обитания можно отнести к «экосистемам, импульсная стабиль-
ность которых обеспечивается колебаниями уровня воды» (как и
морские литоральные и эстуарные экосистемы, гл. 8, разд. 7).
Болота представляют собой очень открытые системы; их удоб-
но классифицировать по степени связи либо с более глубокими
водоемами, либо с наземными экосистемами, либо с теми и други-
ми вместе.
1. Речные болота, расположенные в низких депрессиях и за-
ливных долинах, связанных с реками. Одни из самых продуктив-
ных природных экосистем — низинные лиственные леса в долинах
больших рек и пресноводные литоральные марши вдоль низовьев
больших равнинных рек у берега моря.
2. Озерные болота, которые соединены с озерами, прудами или
каналами запруженных рек. Они периодически заполняются во-
дой, когда ее уровень повышается в этих глубоководных водоемах.
3. Собственно болота — это верховые и низинные болота, мар-
ши, топи, сырые луга и временные пруды, встречающиеся в де-
прессиях, не связанных непосредственно с озерами и реками (хо-
тя они и могут располагаться в речных старицах и занесенных
ложах прудов или озер). Такие болота широко разбросаны в
областях бывшего оледенения. Обычно они покрыты погруженны-
ми водными макрофитами, воздушными болотными растениями,
кустарниками. Часто болота служат наиболее подходящими мес-
тообитаниями для размножения водоплавающих птиц и других
водных или полуводных позвоночных.
Хотя болота занимают лишь около 2 % поверхности Земли,
по существующим оценкам здесь содержится от 10 до 14% угле-
рода (Armentano, 1980). Некоторые болотные почвы содержат до
20% углерода по массе, а торфяники — еще больше. Дренирование
таких почв и их использование для нужд сельского хозяйства
способствуют высвобождению в атмосферу больших количеств
СО2, что усугубляет «СОг-проблему» (гл. 4, разд. 4). Несмотря на
малую площадь болот, аэробно-анаэробная стратификация в бо-
лотных осадках (включая солоноводные марши) очень важна, что
объясняется ролью, которую играют эти осадки в глобальных
круговоротах серы, азота, фосфора и углерода (см. гл. 2, разд. 5).
Важнейшие аспекты микробного разложения и возврата биоген-
ных элементов в круговорот в болотных и мелководных морских
осадках показаны на рис. ПЛЗ.
Отношение человека к болотам коренным образом изменилось
в 70-е годы, когда исследования экологов и экономистов выявили
Краткое описание основных типов природных экосистем
283
Рис. П.13. Микробное разложение и возврат биогенных элементов в болот--
ных осадках. Четыре основных типа бактерий, осуществляющих процессы
разложения в анаэробных условиях, возвращают в атмосферу в газообраз-
ной форме углерод, азот и серу. Фосфаты также превращаются из нерас-
творимой формы (сульфиды) в растворимую и вновь становятся доступ-
ными организмам. (По Е. Р. Odum, 1979.)
их скрытую раньше ценность. Болота теперь уже не рассматри-
ваются как безусловно бросовые земли, которые нужно уничто-
жать или переделать. В деле сохранения болот, особенно при-
брежных, уже достигнуты некоторые успехи, тем не менее в этой
области предстоит еще огромная работа с привлечением законода-
тельных и политических органов. Многочисленные сведения, ка-
сающиеся значения болот, собраны и опубликованы в Трудах
симпозиума, изданных под редакцией Грисона, Кларка и Кларка
(Greeson, Clark, Clark, 1979).
3. Морские экосистемы (рис. П. 14)
Наиболее интересны с экологической точки зрения следующие ха-
рактеристики морской среды:
1. Море занимает 70% поверхности Земли.
2. Глубина моря огромна, и жизнь обнаруживается на всей
этой глубине. В океане, по-видимому, не существует безжизнен-
ных зон, однако вблизи материков и островов воды заселены зна-
чительно гуще.
3. Море непрерывно и не подразделяется на изолированные
области, подобно суше и пресным водам. Все океаны соединены
друг с другом. Основными барьерами для свободного передвиже-
Рис. П. 14. Зональность в мировом океане, выявленная по оценкам первичной продукции фитопланктона. Прибрежные
воды континентального шельфа и мелководья вокруг островов во много раз продуктивнее, чем воды открытого океана.
Более темным на карте указаны, области с большей плотностью жизни в море и с большей рыбопродуктивностью Неко-
ТЖ^Л3 этих прибрежных зон процветают благодаря апвеллингу (см. рис. П.17). (Из Атласа живых ресурсов морей,
FAO Fisheries Circular 126, Rev. 1, 1972. Rome, FAO.) F p ’
Краткое описание основных типов природных экосистем
28Б
ния морских организмов служат температура, соленость и глу-
бина.
4. В море происходит постоянная циркуляция. Разница в тем-
пературе воздуха на полюсах и у экватора порождает сильные
ветры, дующие на протяжении всего года в одном направлении
(пассаты). В результате совместного действия этих ветров и вра-
щения Земли образуются определенные течения. Заслуживают
внимания экваториальные течения, направленные к востоку и
западу, и прибрежные течения, направленные к северу и югу.
Хорошо известны Гольфстрим и Североатлантическое течение,
которые несут теплую воду и смягчают климат высоких широт в
Европе. Калифорнийское течение, напротив, несет на юг массы
холодной воды, отчею образуется характерный для побережья
Калифорнии пояс туманов (что важно для секвойи и других ги-
гантских хвойных). Основные течения действуют подобно гигант-
ским вертушкам (или волчкам), которые вращаются по часовой
стрелке в северном полушарии и против часовой стрелки —
в южном.
Кроме поверхностных ветровых течений имеются глубинные
течения, наличие которых обусловлено различиями в плотности
воды, что в свою очередь обусловлено различиями в температуре-
и солености. Взаимодействие ветрового давления, силы Кориоли-
са, термохалинных течений и физической конфигурации бассейна
очень сложно и является центральной проблемой широких иссле-
дований в области физической океанографии. Перемешивание в
море настолько эффективно, что часто встречающийся в пресно-
водных водоемах дефицит кислорода, или «стагнация», сравни-
тельно редок в океанских глубинах.
5. На море постоянно господствуют разного рода волны, про-
исходят приливы и отливы, вызванные притяжением Луны п
Солнца. Роль приливов особенно велика в прибрежной зоне, где-
жизнь часто особенно богата и разнообразна. Именно приливы
обусловливают заметную периодичность в жизни сообществ, «за-
водя» лунные, а не солнечные биологические «часы» литоральных
животных. Эти «часы» согласовывают активность животных с
периодичностью приливов. Поскольку приливы повторяются при-
мерно через 12,5 ч, в большинстве мест они происходят дважды
в сутки, сдвигаясь в каждые последующие сутки примерно на
50 мин. Каждые две недели, когда силы солнечного и лунного
притяжений складываются, амплитуда приливных колебаний до-
стигает наибольшей величины, при этом приливы бывают особен-
но высоки, а отливы — особенно низки. Это так называемые си-
зигийные приливы. В середине этого двухнедельного периода раз-
ница между высокой и низкой водой бывает наименьшей — это
так называемые квадратурные приливы, когда силы притяжения
Солнца и Луны направлены в противоположные стороны. Ампли-
286
Приложение
туда приливов варьирует от менее чем 30,5 см в открытом море
до 3,5 м в некоторых узких или полузамкнутых заливах. На при-
ливы влияют многие факторы, поэтому картина может различать-
ся в разных частях планеты. Самое первое, что должен сделать
морской эколог, работающий в прибрежной зоне, — это получить
графики приливов и отливов для данной местности.
6. Одна из характеристик морской среды — ее соленость.
Средняя соленость, или содержание солей, составляет 35 частей
соли на 1000 частей воды по массе, или 3,5%. Обычно это запи-
сывается так: 35%о ( = частей на тысячу; вспомните, что соле-
ность пресной воды меньше О,5°/оо). Около 27°/оо приходится на
долю хлористого натрия, а остальное главным образом на соли
магния, кальция и калия. Поскольку соли диссоциированы на
ионы, химический состав морской воды указывают обычно в ионах
(табл. П.1). Соотношение основных компонентов в воде остается
фактически постоянным, поэтому общую соленость можно рас-
считать по содержанию хлорида (это легче, чем определить об-
щую соленость). Таким образом, 19%о содержания хлора соответ-
ствует 35°/оо солености.
Таблица П.1. Химический состав морской воды (величины даны в частях на
тысячу=граммы на 1 кг).
Положительные ионы (катионы) Отрицательные ионы (анионы)
Натрий 10,7 Хлорид 19,3
Магний 1,3 Сульфат 2,7
Кальций 0,4 Бикарбонат 0,1
Калий 0,4 Карбонат 0,007
Бромид 0,07
7. Морская вода имеет щелочную реакцию, сильно забуфере-
на, но часто бедна жизненно важными элементами питания. Си-
лы электролитической диссоциации катионов больше этих сил
анионов (примерно на 2,4 миллиэквивалента), что и определяет
щелочную природу морской воды (в норме рН=8,2). Морская
вода устойчива к изменению pH. Помимо перечисленных выше
ионов морская вода содержит множество других элементов (тео-
ретически все известные). К сожалению, в морской воде возраста-
ет содержание токсичных химических веществ промышленного
происхождения. Концентрация биогенных ионов, однако, часто
бывает такой низкой, что лимитирует первичную продукцию. Все
эти прочие ионы составляют менее 1% солености океана. Как и
следовало ожидать, время пребывания для солей намного больше
Краткое описание основных типов природных экосистем
287
'Литораль
Рис. П.15. Горизонтальная и вертикальная зональность в море. На попе-
речном разрезе в районе Западной Атлантики показаны некоторые геоло-
гические особенности строения морского дна: желоба (которые могут ухо
дить в глубину более чем на 6000 м), каньоны, хребты, абиссальная рав-
нина и срединно-океанические хребты, которые вздымаются подобно ог-
ромным горным вершинам. (По Heezen, Tarp, Ewing, 1959.)
времени пребывания самой воды; по оценке Вейля (Weyl, 1970),.
эти величины имеют порядок 107 и 104 лет соответственно.
На рис. П.15 показаны зональность в море и подчас сложная
природа морского дна. В общем континентальный шельф прости-
рается на некотором расстоянии от берега, на границе шельфа
дно круто обрывается вниз, образуя континентальный склон: за-
тем несколько выравнивается — это континентальное подножие,
и опускается к более глубокому и ровному ложу — абиссальной
равнине. Мелководная зона континентального шельфа называется
неритической (прибрежной) зоной, а зона берега между отмет-
ками высокой и низкой воды — зоной прилива (или литоральной
зоной). Область открытого океана за пределами континентального
шельфа называется океанической областью, область континен-
тального склона и подножия — батиальной зоной', эта последняя
зона может быть геологически активной, пересеченной желобами
и каньонами, подверженными действию подводной эрозии и об-
валов. Область океанических глубин, или абиссальная область,
может находиться в разных местах на глубине от 2000 до 5000 м.
Абиссальную область перерезают желоба, достигающие в глубину
6000 м (эти очень глубоководные области иногда называют ха-
бальной зоной). Абиссаль можно рассматривать как крупнейшую
в мире экологическую единицу. Это, конечно, гетеротрофная сис-
тема, поскольку первичный источник энергии расположен намного
выше (за исключением районов геотермальных рифтов, описанных
288
Приложение
в т. 1, с. 47). Считается, что срединно-океанические хребты оста-
лись после того, как разошлись континенты, прежде плотно сом-
кнутые.
По количеству проникающего света море подразделяют на две
горизонтальные зоны: верхнюю, или эефотическую зону первич-
ного продуцирования, и нижнюю, простирающуюся до больших
глубин афотическую зону, где света для фотосинтеза недостаточ-
но. Эвфотическая зона достигает больших глубин (до 100—200 м)
в прозрачных водах океанической области. В более мутных (и бо-
гатых) прибрежных водах глубина эффективного проникновения
света редко превосходит 30 м.
Океанические области
Из-за низкой концентрации биогенов в фотической зоне
(рис. 3.7) большая часть открытого океана представляет собой
«пустыню» по сравнению с прибрежными водами и лиманами.
Арктические и антарктические моря продуктивнее морей средних
широт, на это указывает большое количество рыбы и китов, оби-
тающих в полярных областях.
До последнего времени считалось, что первичный источник
энергии в морских пищевых цепях образуют диатомеи, динофла-
геллаты и мелкие ракообразные, составляющие макропланктон
(называемый также сетным планктоном, потому что эти организ-
мы хорошо ловятся планктонными сетями, сделанными из шелко-
вой ткани или нейлона). Однако позднее было обнаружено, что
основу пищевых сетей в море образует микропланктон, или нан-
нопланктон (панно — карликовый, крошечный, слишком мелкий,
чтобы задержаться в планктонной сети), который может быть
автотрофным и гетеротрофным. В его состав входят мельчайшие
зеленые и бесцветные флагеллаты и многочисленные типы бак-
терий (см. Pomeroy, 1974). Связь между автотрофами и гетеро-
трофами, как и в большинстве других типов экосистем, обеспечи-
вается детритом — взвешенным органическим веществом (ВОВ)
и растворенным органическим веществом (РОВ). Однако в море
не только ВОВ может превращаться в РОВ, но и РОВ под дей-
ствием волн может превращаться в ВОВ. Это происходит также
при образовании фекальных пеллеток и посредством других не-
полностью выясненных механизмов. Образующиеся агрегаты и
населяющие их бактерии обеспечивают, видимо, важный источ-
ник питания для многочисленных и разнообразных фильтраторов.
Об удивительно высоком разнообразии глубоководной фауны
мы уже говорили. Две теории, объясняющие это явление, обсуж-
дались в гл. 7, разд. 6. Глубоководные рыбы во многом курьезны,
некоторые из них светятся (рыбы-лампы), у других светятся кон-
чики подвижных лучей, которые служат приманкой для привле-
чения добычи (рыба-ангел), у многих огромные рты, и они могут
Краткое описание основных типов природных экосистем 289
заглатывать добычу, превосходящую по размерам их самих (ры-
ба-гадюка, хаулиод). Нищи мало, и она редка в темных глубинах,
но рыбы паилучшпм образом адаптировались к использованию
счастлпвых возможностей.
Область континентального шельфа
Жизнь в море сконцентрирована около берега, где благоприятны
условия питания. Пн в каком другом месте, даже в дождевых
тропических лесах, нет такого разнообразия жизни. Значительную
часть морского прибрежного зоопланктона составляет меропланк-
тон (временный пли сезонный планктон), представленный пела-
гическими личинками донных организмов (крабов, морских чер-
вей, моллюсков и т. д.); эта особенность резко отличает его от
планктона пресных вод и открытого океана, где большую часть
взвешенных организмов составляет голопланктон (т. е. организмы,
которые на протяжении всего жизненного цикла остаются в
планктоне). Выло показано, что пелагические личинки обладают
замечательпой способностью определять тип субстрата, пригодный
для жизни прикрепленных взрослых организмов. Когда прибли-
жается время метаморфоза, личинки не оседают где попало,
а только при наличии конкретных химических свойств субстрата.
Бентос состоит из двух вертикальных компонентов: 1) эпифау-
ны— организмов, которые живут на поверхности, прикрепившись
к пей или свободно передвигаясь по субстрату, и 2) инфауны,
представители которой закапываются в субстрат пли строят труб-
ки и поры. Агрегации бентоса распространены всюду в виде так
называемых «параллельных донных сообществ» (Thorson, 1955),
в которых доминируют экологически эквивалентные виды, часто
относящиеся к одному роду.
Все крупное промысловое рыболовство мира почти полностью
сосредоточено на континентальном шельфе или вблизи него, осо-
бенно в районах поднятия глубинных холодных вод, или в райо-
нах апвеллинга (см. следующий раздел и рис. П.17). Основу
уловов составляют сравнительно немного видов промысловых рыб,
куда входят анчоусы, сельдь, треска, макрель, сайда, сардина,
камбалообразные (камбалы, палтус), лососи и тунец. Большин-
ство специалистов по промысловой ихтиологии считают, что пик
мировых уловов уже прошел и в настоящее время во многих
районах наблюдается перелов. Рыболовство, особенно с использо-
ванием траулеров и сейнеров, работающих вдали от баз, требует
больших затрат энергии. Рост добычи пищевых продуктов из моря
будет в значительной степени зависеть от марикультуры (рыбные
хозяйства в замкнутых заливах и лиманах).
Доступность и богатство жизни на морских берегах привели
к тому, что эти области стали наиболее изученной частью конти-
19 Заказ № 1483
Средний уровень отлива _________________
Камбалы(Pleuronectes). Fundulus (рыбы)
голубые крабы раки-отшельники
(Callinectes), (Pagurus)^^
Menidia^ из дт ерник-
Морской еж
{Echinarachnius ).
морские перья
(Ren dial
Средний уровень прилива
Прибрежные
птицы
Краб-крот
( Emerita),
моллюск
Donax
Морской еж
( Met Uta ),
закапывающаяся
креветка (Ogyris),
голотурия (Thyone),
моллюск-
сердцевидка
(Cardium),
моллюск-олива
(Oliva) ,
песчаные
полуводные
веслоногие
Рак-привидение
(Callianassa).
закапывающийся
равноногий рачок
( Chiridotea),
закапывающийся
бокоплав
(Haustorius),
многощетин ковые
черви
Краб-привидение
* (О сур ode),
| морские блохи
| {Ta/orchestia,
Orchestia),
I жуки-скакуны и
| другие наземные
। насекомые
I Бактерии - Диатомовые Одноклеточные водоросли
I КРУТОЙ БЕРЕГ ПОЛОГИЙ БЕРЕГ !
I
СУПРАЛИТОРАЛЬНАЯ
кораллы( Leptogorgia,
О си Нпа)
СУБЛИТОРАЛЬНАЯ ЗОНА
(зона водорослей)
ЛИТОРАЛЬНАЯ ЗОНА
(зона усоногих рачков)
IСУПРАЛИТОРАЛЬНАЯ
I ЗОНА
(зона литторин.или
"черная зона")
Рис. П.16. Схематические разрезы песчаного пляжа (вверху) и каменис-
того берега (внизу) у Бофорта, Сев. Каролина. Показаны зоны и характер-
ные доминирующие формы. (Верхняя схема по данным Pearse, Hunn,
Wharton, 1942, нижняя — по данным Stephenson, Stephenson, 1952.)
Краткое описание основных типов природных экосистем
291
пентильного шельфа. Нет такого биолога, не говоря уже о на-
туралистах-любителях, который считал бы свое образование за-
конченным, если он не прошел хорошей «школы» на берегу моря.
Сообщества в литоральной зоне, как и в горах, характеризуются
четкой зональностью. На рис. 11.16 показаны некоторые аспекты
этой зональности па берегах двух разных типов — на песчаном
пляже и скалистом берегу. На экологов, изучающих организацию
сообществ в плотно заселенной зоне литорали, сильное впечатле-
ние производят конкуренция и хищничество (см. гл. 5, разд. 1).
Энергия волн, прибоя и приливов — это главный фактор па
входе, к которому должны адаптироваться организмы. Низкоэнер-
гетический берег, подверженный слабому воздействию волн, будет
заселен больше и другими видами, нежели высокоэпергетический
берег, па который воздействуют сильные волны.
Области апвеллинга
Большую роль в жизни океана играет процесс, названный апвел-
лингом. Он происходит там, где ветры постоянно отгоняют по-
верхностную воду от крутого берегового склона. В этих местах па
поверхность поднимается холодная глубинная вода, богатая на-
копленными биогенными элементами. Наиболее продуктивные
области оксанов сосредоточены в районах апвеллинга, о чем сви-
детельствует развитый здесь рыболовный промысел. Расположены
апвеллинги, как показано па рис. П.17, вдоль западных берегов.
Апвеллинги поддерживают, кроме того, большие популяции мор-
ских птиц, откладывающих па берегах и островах бесчисленные
тонны богатого нитратом и фосфатом гуано.
Приведем некоторые характерные черты биома апвеллингов:
1. Высокая концентрация биогенных элементов и организмов;
преобладают пелагические, а не демерсальные (донные) рыбы.
2. Колоссальные популяции рыб (и птиц) характеризуют
не только высокую продуктивность, по и короткие пищевые цепи.
Некоторые виды ракообразных и рыб, которые в океанических
областях хищничают, в областях апвеллинга становятся расти-
тельноядными. В короткой пищевой цепи доминируют диатомеи
и сельдевые рыбы.
3. Отложения осадков на морском дне содержат много орга-
нических веществ и фосфата.
4. В противоположность богатству моря прилегающие районы
суши часто представляют собой пустыню, так как формирующие
апвеллинг ветры дуют большей частью с суши, унося прочь вла-
гу. Однако частые туманы поддерживают развитие некоторой
растительности.
5. Продуктивное «цветение» подвержено периодическим спа-
дам, когда меняется направление ветра, развиваются бескисло-
19*
292
Приложение
Рис. П.17. Пять наиболее продуктивных зон прибрежных апвеллингов (об-
ласти, покрытые точками) и системы атмосферного давления на уровне мо-
ря (антициклоны), влияющие на апвеллинги. Стрелками показано пример-
ное положение основных течений. Потенциальный урожай рыбы (млн. т
в год): Перуанско-Чилийский район —12, Юго-Западная Африка —5,
Орегон—Калифорния — 5, Северо-Западная Африка — 4, Аравийское мо-
ре — 4. Различия между районами в значительной степени определяются
числом месяцев в году, когда эффективно работает апвеллинг. Например,
12 мес в Перуанском районе и только б мес в Аравийском море. (Карта
воспроизведена с разрешения Richardi Т. Barber.)
родные условия или когда происходит «цветение» ядовитых ди-
нофлагеллят (красные приливы). При этом наблюдается массовая
гибель рыбы, отрицательно сказывающаяся на промысловом ры-
боловстве. Такая катастрофа происходила несколько раз в зоне
перуанского апвеллинга, который обычно оценивается как самый
продуктивный район мирового рыболовства. Человек и природа
нс в состоянии постоянно поддерживать очень высокий уровень
продукции, циклическое поведение может быть основным свойст-
вом эвтрофицированпых систем (включая индустриализованное
сельское хозяйство).
Лиманы (рис. П. 18)
Лиман — это полузамкнутый прибрежный водоем, свободно со-
единяющийся с открытым морем; таким образом, лиман сильно
подвержен воздействию прилива, а морская вода в нем смешана
(и обычно заметно опреснена) с пресной водой материкового сто-
Краткое описание основных типов природных экосистем 293
ка. Примерами лиманов служат устья рек, заливы, литоральные
марши и водоемы между берегом и барьерными отмелями. Эстуа-
рии и лиманы можно было бы рассматривать как переходные
зоны или экотоны между пресноводными и морскими местооби-
таниями, но многие из их важнейших физических и биологиче-
ских признаков являются не промежуточными, а уникальными.
Более того, человек настолько злоупотреблял использованием
этой зоны, что при сложившемся сейчас критическом положении
крайне важно, чтобы уникальные признаки и свойства лиманов
стали понятными всем.
С точки зрения геоморфологии удобно рассматривать пять ти-
пов лиманов:
1. Затопленные речные долины наиболее распространены вдоль
береговой линии относительно низких и широких прибрежных
равнин. Хороший пример — Чесапикский залив на среднеатлан-
тическом побережье США.
2. Лиманы типа фьордов — глубокие, U-образные выемки бе-
рега, пропаханные ледниками, обычно с мелководным порогом в
устьевой части, образованным ледниковыми отложениями. При-
мерами могут служить знаменитые фьорды Норвегии и сходные
образования вдоль побережья Британской Колумбии и Аляски.
3. Лиманы, ограниченные отмелями или барьерными острова-
ми, представляют собой мелководные бассейны, нередко частично
обсыхающие в отлив; они отгорожены от моря цепочкой отмелей
пли барьерных островов, разделенных проливами, которые и обес-
печивают «свободную связь с морем». В одних случаях песчаные
отмели образованы отнесенными от берега отложениями, в дру-
гих — барьеры — это бывшие береговые дюны, которые стали изо-
лированными вследствие постепенного повышения уровня моря.
Хорошо изученными примерами лиманов этого типа служат «про-
ливы» позади «внешней отмели» в Северной Каролине (Нацио-
нальный парк на берегу мыса Хаттерас), засоленные марши в
Южной Каролине, Джорджии и «морские острова» вдоль побе-
режья Мексиканского залива.
4. Лиманы, образованные тектоническими процессами, — это
береговые выемки, сформировавшиеся в результате геологических
сдвигов или локальных опусканий, часто с большим притоком
пресной воды. Залив Сан-Франциско представляет собой хороший
пример такого типа лимана.
5. Лиманы речных дельт часто образуются в устьях больших
рек, таких, как Миссисипи или Нил. В таких ситуациях при пе-
ремещении илистых осадков часто формируются полузакрытые
заливы, протоки и солоновато-водные марши. Если речной сток
преобладает над приливами, то устанавливается характерная сис-
тема циркуляции в стратифицированной воде с соленым клином.
Пресная вода разливается по поверхности более тяжелой соленой
20 Заказ № 1483
Краткое описание основных типов природных экосистем 295
воды, которая образует клин, простирающийся над дном на зна-
чительное расстояние вверх по течению. Если пресноводный по-
ток и приливные течения более или менее уравновешивают друг
друга, то перемешивание осуществляется главным образом за счет
турбулентности. В этом случае возникает слабосоленый, частично
перемешанный и слабо стратифицированный лиман. Наконец, ес-
ли преобладают сильные приливы, то вода хорошо перемешивается
от дна до поверхности и соленость приближается к солености от-
крытого моря.
Лиманы характеризуются некоторыми важными свойствами,
которые к тому же служат основанием для их сохранения:
1. Лиманы обычно продуктивны вследствие поступления боль-
ших энергетических субсидий в виде приливов и обилия биоген-
ных элементов. Вертикальное перемешивание п турбулентность
образуют своего рода ловушку для этих элементов, так что ли-
манные растения получают больше фосфора и других биогенов,
чем растения из прилежащих пресноводных пли морских место-
обитаний.
2. На протяжении круглого года гетеротрофы обеспечиваются
пищей за счет активности автотрофов трех типов: макрофитов
(болотные травы, морские травы, водоросли), донных водорослей,
развивающихся на поверхности осадка и в его толще (в том числе
диатомей, которые живут в трубочках из ила и передвигаются в
них вверх и вниз в соответствии с режимом освещенности и при-
ливов), и фитопланктона.
3. Лиманы служат местом откорма для молоди многих морских
промысловых видов. Устрицы и промысловые крабы остаются в
лиманах на протяжении всего жизненного цикла, а различные
креветки и многие рыбы — объекты промыслового п спортивного
Рис. П.18. Ограниченный островами лиман в Джорджии (Уоссо-Айленд)
А. Вид с моря на берег барьерного острова, ряд дюн (самые старые искры
ты спелым лесом) и широкую ленту засоленных маршей, расположенных
между внешним барьером и материком. (Фотография Floyd Jillson, Atlanta
Journal и Constitution Magazine.) Б. Вид с близкого расстояния зарослей
Spartina на засоленном марше. Видны высокая трава, многочисленные
бухты, протоки на литорали и участки богатого водорослями ила (один
такой небольшой участок показан на переднем плане в центре). Обратите
также внимание на ранние стадии образования детрита из Spartina (спра-
ва на переднем плане); этот детрит, возможно, будет кормить многие
квадратные километры воды. На верхней фотографии отчетливо видна
взаимосвязь моря, берега, острова и лимана. Засоленный марш улавливает
осадки и биогенные элементы, это происходит постоянно благодаря тече-
ниям и избытку осадков; без этого процесса красивый белый песчаный
пляж на острове был бы размыт и занесен илом. Барьерные острова по-
стоянно изменяются, размываются в одних местах (обратите внимание на
северную оконечность острова справа) и достраиваются в других (на пе-
реднем плане слева). Если бы растительность не защищала дюны, скорость
эрозии легко превысила бы скорость образования нового берега.
20’
296
Приложение
рыболовства, — лов которых ведется вдали от берега, проводят
ранние стадии жизненного цикла в лиманах, где они находят
изобилие пищи и защиту от хищников, благодаря чему лучше
выживают и быстрее растут на критических стадиях жизненного
цикла.
Мутуализм между манящими крабами и растениями, состав-
ляющими их источник пищи, отмечался в т. 1, с. 163. Лиман слу-
жит хорошим примером краха рынка, если подходить к нему с
экономическими оценками, как обсуждалось в т. 1, с. 197—199.
При планировании строительства и изменений лиманов нуж-
но иметь в виду следующее. Во-первых, необходимо стараться
не нарушать нормального водообмена. Например, дороги должны
проходить по эстакадам и мостам, а не по земляным насыпям,
преграждающим путь воде. Во-вторых, нельзя нарушать мелко-
водные продуктивные зоны, такие, как рифы, заросли морских
трав и марши, которые служат местом откорма ценных морских
промысловых видов. Слишком часто в прошлом изменения были
недальновидными и без необходимости вредоносными (см.
W. Е. Odum, 1970).
Литература1
Биогеография
Daubenmire R., 1978. Plant Geography, with Special Reference to North Ame-
rica, New York, Academic Press.
Good R. D., 1964. The Geography of Flowering Plants (3rd. ed.), London,
Longmans, 518 pp.
Hunt Ch. B., 1973. Natural Regions of the United States and Canada, San
Francisco, W. H. Freeman.
Neill W. T., 1969. The Geography of Life, New York, Columbia University
Press, 480 pp.
Pielou E. C., 1979. Biogeography, New York, Wiley-Interscience, 351 pp.
Polunin N., 1960. Introduction to Plant Geography, New York, McGraw-Hill,
640 pp.
Udvardy M. F. D., 1969. Dynamic Zoogeography, with Special Reference to
Land Animals, New York, Van Nostrand Reinhold, 446 pp.
Биосфера и Международная Биологическая Программа (МБП)
* Blair W. F., 1977. Hig Biology; The US/IBP, Stroudsburg, Pa., Dowden,
Hutchinson and Ross.
Botkin D. B., 1977. Bits, bytes, and IBP. BioScience, 27, 385—386.
Flessa K. W., 1980. Biological effects of plate tectonics and continental drift,
BioScience 30, 518—523.
Hallam A., 1972. Continental drift and the fossil record. Sci. Am. 226 (4),
42-52.
Hutchinson G. E. (cd.), 1970. The Biosphere, Special issue of Scientific Ame-
rican (Vol. 223, No 3) (Also published in book form by W. H. Freeman
and Co., San Francisco.)
Lieth H., R. II. Whittaker (eds.), 1975. Primary Productivity of the Biosphere,
Berlin and New York, Springer-Verlag.
* Loucks O. L., 1983. The U. S. IBP, in perspective after 10 years. In: Eco-
system Theory and Application, C.-A. Knox, ed. New York, John Wiley
and Sons.
Наземные биомы (работы общего характера)
Carpenter J. R., 1939. The biome, Am. Midi. Nat., 21, 75—91
Holdridge L. R., 1947. Determination of world plant formations from simple
climatic data, Science, 105, 267—368.
Jordan C. F., 1971. A world pattern of plant energetics, Am. Sci., 59, 425—
433.
Merriam С. H., 1894. Laws of temperature control of the geographical distri-
bution of terrestrial animals and plants, National Geographic, 6, 229—238.
(См. также Life Zones and Crop Zones, U. S. D. A. Bull. No 10, 1899).
Odum E. P., 1945. The concept of the biome as applied to the distribution of
North American birds, Wilson Bull, 57, 191—201.
Pitelka F. A., 1941. Distribution of birds in relation to major biotic commu-
nities, Am. Midi. Nat., 25, 113—137.
Riley D., Young A., 1968. World Vegetation, Cambridge, England, Cambridge
University Press.
1 Непосредственно в приложении цитировались только работы, отме-
ченные звездочкой.
298 Литература
Shelf ord V. Е., 1963. The Ecology of North Ameruca, Urbana, University of
Illinois Press.
Walter H., Box E., 1976. Global classification of natural terrestrial ecosystems,
Vegetatio, 32, 75—81.
Тундра
Bliss L. C., 1971. Arctic and alpine plant life cycles, Ann. Rev. Ecol. Syst., 2,
405—438.
Brown J., Miller P. C., Tiezen L. L., Bunnell F. L. (eds.), 1981. An Arctic
Ecosystem, The Coastal Tundra at Barrow, Alaska, New York, Academic
Press.
Johnson P. L., 1969. Arctic plants, ecosystems and strategies, Arctic, 22, 341—
355.
Северный хвойный лес
Denison W. C., 1973. Life in tall tress, Sci. Am., 228(6), 74—90.
Edmonds B. L. (ed.), 1981. Analysis of Coniferous Forests Ecosystems in
the Western United States, New York, Academic Press.
Shelf ord V. K., Olson S., 1935. Sere, climax and influent animals with special
reference to the transcontinental coniferous forest of North America,
Ecology, 16, 375—402.
Warning В. H., Franklin J. F., 1979. The evergreen coniferous forests of the
Pacific Northwest, Science, 204, 1380—1386.
Листопадный лес умеренной зоны
Braun Е. L., 1950. Deciduous Forests of Eastern North America, Philadelphia,
Blakiston Co.
McCormick J., 1966. The Life of the Forest (Our Living World of Nature
Series), New York, McGraw-Hill.
Reichle D. E. fed.), 1970. Analysis of Temperature Forest Ecosystem, Berlin
and New York, Springer-Verlag.
Степи умеренной зоны
Allen D. L., 1967. The Life of Prairies and Plains (Our Living World of Na-
ture Series), New York, McGraw-Hill.
Carpenter J. R., 1940. The grassland biome, Ecol. Monogr., 10, 617—684.
Clements F. E., Shelf ord V. E., 1939. The North American grassland. In: Bio-
Ecology, F. E. Clements and V. E. Shelf ord, New York, John Wiley and
Sons.
Hanson H. D., 1950. Ecology of the grassland. II. Bot. Rev., 16, 283—360.
Love R. M., 1970. The rangelands of the western United Stales, Sci. Am., 272
(2), 89—96.
* Morello J., 1970. Modelo de relaciones entra pastizales у lenosas colonzodo-
ras en el Chaco — Argentine. (A model of relationship between grassland
and wood colonizer plank in the Argentine Chaca.), Idia, 276, 31—51
(Dec. 1970).
Weaver J. E., Albertson. F. W., 1956. Grasslands of the Great Plains: Their
Nature and Use, Lincoln, Neb., Johnsen Publ. Co.
Тропические степи и саванна
Bell R. H V., 1971. A grazing ecosystem in the Serengeti, Sci. Am., 225(1),
86—93.
Bourliere F., Hadley M., 1970. The ecology of tropical savannas, Ann. Rev.
Ecol. Syst., 7, 125—152.
Sinclair A. R. E., Norton-Griffiths M. (eds.), 1979. Serengeti: Dynamics of an
Ecosystem, Chicago, University of Chicago Press.
Литература
299
Чапарраль
Castri F. di, II. A. Mooney H. A. (eds.), 1973. Mediterranean Type Ecosystems,
Ecol. Studies, Vol. 7, Berlin and New York, Springer-Verlag.
Cogswell H. L., 1947. Chaparral country, Audubon, 49, 75—81.
Cooper W. S., 1922. The broad-sclerophyll vegetation of California: An eco-
logical study of the chaparral and its related communities. Washington,
D. C., Carnegie Inst., 319, 1—124.
Пустыни
Gore R., 1979. The desert; an age-old challenge grows, Nat. Georg., 156, 594—
639.
McGinnis Ж G., Goldman B. J., Paylore P. (eds.), 1969. Deserts of the World,
Tucson, University of Arizona Press.
Niering W. A., Whittaker R. H., Lowe С. H., 1963. The saguaro: a population
in relation to environment, Science, 142, 15—23.
Noy-Meir I., 1973. Desert ecosystems: Environment and producers, Ann. Rev.
Ecol. Syst., 4, 25—51.
Noy-Meir I., 1974. Desert ecosystems: Higher trophic levels, Ann. Rev. Ecol.
Syst., 5, 195—214.
Shantz H. L., 1942. The desert vegetation of North America, Bot. Rev.. 8, 195—
246.
Sutton A., Sutton M., 1966. The Life of the Desert (Our Living World of Na-
ture Series), New York, McGraw-Hill.
* Walter H., 1954. Le facteur eau dans les regions arides et sa signification
pour 1’organisation de la vegetation dans les contrees sub-tropicales. In:
Les Divisions Ecologiques du Monde, Paris, Centre Nationale de le Re-
cherche Scientifique, pp. 27—39.
Тропические леса
Golley F. B., McGinnis J. T.. Clements R. G., Child G. I., Duever M. I.,
1969. The structure of tropical forests in Panama and Columbia. BioSci-
ence, 19, 693—696; 696—700.
Golley F. B., Medina E. (eds.), 1975. Tropical Ecological Systems, Ecol. Stu-
dies, № 11, Berlin and New York, Springer-Verlag.
Gomez-Pompa A., Vasquez-Yanes C., Guaria S., 1972. The tropical rain forest,
a non-renewable resource, Science, 177, 762—765.
* Jordan C. F., 1982. Amazon rain forest, Am. Sci., 79, 394—401.
National Academy of Science, 1980. Conversion of Tropical Forests, Committee
Report, Norman Meyers, Chairman. Washington, D. C., National Academy
Press.
Richards P. W,, 1952. The Tropical Rain Forest, New York, Cambridge Uni-
versity Press.
Richards P. W., 1973. The tropical rain forest, Sci. Am., 229 (6), 58—67.
Водные экосистемы — общие сведения
Barnes R. S. К., Mann К. H. (eds), 1980. Fundamentals of Aquatic Ecosystems,
Oxford, Blackwell, 229 pp.
Mann К. H., 1969. The dynamics of aquatic ecosystems. In: Advances in
Ecological Research, J. B. Cragg, ed., Vol. 6, New York, Academic Press,
pp. 1—81.
Russel-Hunter W. D., 1970. Aquatic Productivity: An Introduction to Some
Basic Aspects of Biological Oceanography and Limnology, New York and
Toronto, Macmillan.
300
Литература
Пресноводные экосистемы (работы общего характера)
Coker R. Е., 1954. Streams, Lakes, Ponds, Chapel Hill, University of North
Carolina Press.
Gerking S. C. (ed.), 1967. The Biological Basis of Freshwater Fish Production,
Oxford, Blackwell, 495 pp.
Gibbs R. J., 1970. Mechanisms controlling water chemistry, Science, 170,
1088—1090.
Haynes H. B. N., 1969. The Biology of Polluted Waters, Liverpool, England,
Liverpool University Press.
* Livinstone D. A., 1963. Chemical composition of rivers and lakes, U. S.
Geological Survey Prof., Paper 440-G.
Porter K., 1977. The plant-animal interface in freshAvater ecosystems, Am,
Sci, 65, 159—170.
Лентические экосистемы
Beeton A. M., 1969. Changes in the environment and biota of the Great Lakes.
In: Eutrophication: Causes, Consequences, and Corrections, Washington,
D. C, National Academy of Science, pp. 150—187.
Bennett G. W., 1962. Management of Artificial Lakes and Ponds, New York,
Reinhold.
Brinkhurst R. O., 1974. The Benthos of Lakes, London and New York, Mac-
millan.
Brylinsky M., Mann К. H., 1973. An analysis of factors governing productivity
in lakes and reservoirs, Limnol. Oceanogr, 18, 1—14.
Deevy E. S., Jr., 1951. Life in the depths of a pond, Sci. Am, 185, 68—72.
Lowe-McConnell R. H. (ed.), 1966. Man-Made Lakes, New York, Academic
Press.
Ragotzkie R. A., 1974. The Great Lakes rediscovered, Am. Sci, 62, 454—464.
Richey J. E., et al., 1978. Carbon flow in four lake ecosystems: A structural
approach, Science, 202, 1183—1186.
Логические системы
* Allen К. R., 1951. The Horokiwi stream: A study of a trout population. New
Zealand Mar. Dept. Fish Bull, № 10.
Cummins K. W., 1974. Structure and function of stream ecosystems, BioSci-
ence, 24, 631—641.
Fisher S. G., Likens G. E., 1972. Stream ecosystem: Organic energy budget,
BioScience, 22, 33—35.
* Horton P. A., 1961. The bionomics of brown trout in a Detmoor stream,
J. Anim. Ecol, 30, 331—338.
Hynes H. B. N., 1970. The Ecology of Running Waters, Toronto, University
of Toronto Press.
* Mann К. H., 1964. The pattern of energy flow in the fish and invertebrate
fauna of the River Thames, Vert. I nt. Ver. Limnol, 15, 485—495. (Cm,.
также J. Am. Ecol, 34, 253-275, 1965.)
Patrie R. 1970. Benthic stream communities, Am. Sci, 58, 546—549.
Vannote R. L. et al, 1980. The rever continuum concept, Can. J. Fish. AquaL
Sci, 37, 130—137.
Whitten B. A. (ed.), 1975. River Ecology, Two vols. Berkeley, University of
California Press.
Болота
* Armentano Th. V., 1980. Drainage of organic soils as a factor in the world
carbon cycle, BioScience, 30, 825—830.
Brinson M. M. et al, 1981. Primary productivity, decomposition and consumer
activity in freshwater Avetlands, Ann. Rev. Ecol. Syst, 12, 123—162.
Литература
301
Cowardin L. M., Carter V., Golet F. C., 1979. Classification of wetlands and
deepwater habitats of the United States, FWS/OBS—79/31, Washington,
D. C., Fish and Wildlife Service, U. S. Dept, of Interior.
Good R. E., et ah, 1978. Freshwater Wetlands. Ecological Processes and Mana-
gement Potential, New York, Academic Press.
* Greeson P. E., Clark J. R., Clark J. E. (eds.), 1979. Wetland Functions and
Values: The State of Our Understanding, Minneapolis, American Water
Resources Assoc.
* Odum E. P., 1979. The value of wetlands; a hierarchical approach. In: Wet-
land Functions and Values, Greeson, Clark and Clark eds. Minneapolis,
American Water Resources Assoc.
Niering W. A., 1966. The Life of the Marsh (Our Living World of Nature
Series), New York, McGraw-Hill.
Wharton С. H. et al., 1982. The ecology of bottomland hardwood awamps of
the Southeast: A community profile, FWS/OBS-81/37, Washington, D. C.,
U. S. Fish and Wildlife Service, Biol. Serv. Program.
Океаны (работы общего характера)
Berrill N. J., 1966. The Ocean (Our Living World of Nature Series), New York..
McGraw-Hill.
Coker R. E., 1947. This Great and Wide Sea, Chapel Hill, University of North
Carolina Press.
Cushing D. H., Walsh J. J. (eds.), 1976. The Ecology of the Seas, Phila-
delphia, W. B. Saunders.
Falkowski P. G. (ed.), 1980. Primary Productivity in the Sea, New York,.
Plenum Press. (См. Review in Science, 212, 794, 1981).
Goldreich P., 1972. Tides and the earth-moon system, Sci., Am., 226(4). 42—52.
Heezen R. C., Tarp M., Ewing M., 1959. The floors of the ocean, I. North
Atlantic, Geol. Soc. Amer. Spec. Paper. 65, 122 pp.
MacIntyre F., 1970. Why the sea is salt, Sci. Am., 223(5), 104—115.
Ravelie R. (ed.), 1969. The Oceans, Special issue of Scientific American (Vol
221, № 3).
Steele J. H., 1974. The Structure of Marine Ecosystems, Cambridge Mass..
Harvard University Press, 128 pp.
Sverdrup H. U., Johnson M. W., Fleming R., 1942. The Oceans: Their Physics,
Chemistry, and General Biology, Englewood Cliffs, N. J., Prentice-HalL
Walsh J. J., 1972. Implications of a system approach to oceanography, Science
176, 969—975.
Океаническая область
Hardy A. C., 1957. The Open Sea: The World of Plankton, New York, Hough-
ton-Mifflin.
Menzies R. J.. George R. Y., Rowe G. T., 1973. Abyssal Environment and Eco-
logy of the World Oceans, New York, John Wiley and Sons, 488 pp.
* Pomeroy L. R., 1974. The ocean’s food web, a changing paradigm. BioSci-
ence, 24, 299—504.
Rex M. A., 1981. Community structure in the deep-sea benthos, Ann. Rev. Ecol
Syst., 12, 331—354.
Континентальный шельф, прибрежная зона и побережье
Amos W. Н., 1966. The Life of the Seashore (Our Living World of Nature
Series), New York, McGraw-Hill.
Bahr L. M., Lanier W. P., 1981. The ecology of intertidal oyster reefs of
the South Atlantic coast: A community Profile, PWS/OBS-81/15, Washing-
ton, D. C., U. S, Fish and Wildlife Service, Office of Biological Services.
Barnes R. S. K. (ed.), 1977. The Coastline, New York, Wiley-Interscience„356 pp’
Carson R., 1955. The Edge of the Sea, New York, Houghton-Mifflin.
302
Литература
Dolan R.. Hayden В., Lins H., 1980. Barrier islands, Am. Sci., G5, 16—26.
Jackson T. C., Reische D., 1981. Coast Alert: Scientists Speak Out, San Fran-
cisco, Friends of the Earth.
Mann К. H., 1982. Ecology of Coastal Waters: A Systems Approach, Berkeley^
University of California Press.
Mann К. H., 1973. Seaweeds: Their productivity and strategy for growth,
Science, 182, 975—981.
Odum H. T., Copeland B. J., McMahan E. A., 1974. Coastal Ecological Systems
of the United States, Washington, D. C., The Conservation Foundation.
Odum W. E., Mcluor С. C., Smith T. Ill, 1982. The ecology of mangroves
of South Florida: A community profile, Washington, D. C., U. S. Fish and
Wildlife Services, Office of Biological Services.
Paulson A. C.. 1980. The corall atoll; An oasis in the desert, San Frontiers,
Jan.-Feb., pp. 37—43.
Pearse A. S., Humm H. J.. Wharton G. W., 1942. Ecology of sand beaches at
Beaufort, N. C. Ecol. Monogr., 12, 136—190.
Peterson С. H., Peterson N. M., 1979. The ecology of intertidal flats of North
Carolina: A community profile, FWS/OBS-79/39. Washington, D. C., U. S.
Fish and Wildlife Service, Office of Biological Services.
Stephenson T. A., Stephenson A., 1949. The universal features of zonation
between tidemarks on rocky coasts, J. Ecol., 37, 289—305.
Stephenson T. A., Stephenson A., 1973. Life Between Tidemarks on Rocky
Shores, San Francisco, W. H. Freeman.
* Thorson G., 1956. Marine level-bottom communities of recent seas, their
temperature adaptation, and their ’’balance11 between predators and food
animals, Trans. N. Y. Acad. Sci., 18, 693—700.
Области апвеллинга
Barber R. T., Smith R. L., 1980. Coastal upwelling ecosystems. In: Analysis of
Marine Ecosystems, A. R. Longhurst, ed., New York, Academic Press.
Boje R., M. Tomczak (ed.), 1978. Upwelling Ecosystem, New York, Springer-
V erlag.
Лиманы
Bronin L. E. (ed.), 1975. Estuarine Research, Two volumes, New York, Aca-
demic Press.
Kennedy V. S. (ed.), 1980. Estuarine Perspectives, New York, Academic Press.
Laufj G. A. (ed.), 1967. Estuaries, Publ. No 83, Washington, D. C., American
Association for the Advancement of Science, 757 pp.
McHugh J. L., 1966. Management of estuarine fisheries. In: A Symposium on
Estuarine Fisheries, Washington, D. C., Am. Fish. Soc.
McLusky D., 1981. The Estuarine Ecosystem, New York, Halsted Press, John
Wiley and Sons.
Odum E. P., 1961. The role of tidal marshes in estuarine production, The Con-
servationists, June-July, pp. 12—15, Albany, N. Y. Dept, of Conservation.
Odum E. P., 1980. The status of three ecosystem-level hypotheses regarding
salt marsh estuaries. In: Estuarine Perspectives, V. S. Kennedy, ed., New
York, Academic Press.
* Odum W. E., 1970. Insidious alteration of the estuarine environment, Trans.
Am. Fish. Soc., 99, 838—847.
Ragotzkie R. A., 1960. Marine marsh. In: McGraw-Hill, Encyclopedia of Sci-
ence and Technology, New York, McGraw-Hill, pp. 217—218.
Turner R. E., 1977. Intertidal vegetation and commercial yields of penaeid
shrimp, Trans. Am. Fish. Soc., 106, 411—416.
Warner W. W., 1976. Beautiful Swimmers; Watermen, Crabs and Chesapeake
Bay., New York, Penguin Books.
Литература
В списке литературы приведены не только источники, непосредственно ци-
тированные в этой книге, но также ряд других книг, обзорных статей и
трудов смпозиумов. Литература, которая рекомендуется для использования
при изучении основных типов экосистем, приведена в приложении.
Aaronson S., 1970. Experimental Microbial Ecology, New York, Academic
Press, 598 pp.
Abele L. G., Walters K., 1979. The stability-time hypothesis: Reevaluation of
the data, Am. Nat., 114, 559—568.
Abrahamson W. G., Gadgil M., 1973. Growth form and reproductive effort in
goldenrods (Solidago, Compositae), Am. Nat., 107, 651—661.
Adams С. C., 1935. The relation of general ecology to human ecology, Ecology,
16, 316—335.
Adkisson P. L., Niles G. A., Walker J. K., Bird L. S., Scott H. B., 1982.
Controlling cotton’s insect pests: A new system, Science, 216, 19—22.
Agren G. I., Axelsson B., 1980. Population respiration: A theoretical approach,
Ecol. Modelling, 11, 39—54.
Ahlgren I. F., Ahlgren С. E., 1960. Ecological effects of forest fires, Bot. Rev.,
26, 483—533.
Albertson F. W., Tomanek G. W., Riegel A., 1957. Ecology of drought cycles
and grazing intensity on grasslands of Central Great Plains, Ecol. Monogr.,
27, 27—44.
Alexander M., 1964. Biochemical ecology of soil micro-organisms, Am. Rev.
Microbiol., 18, 217—252.
Alexander M., 1971. Microbial Ecology, New York, John Wiley and Sons,
511 pp.
Alexander R, D., Borgia G., 1978. Group selection, altruism, and the levels of
organization of life, zAnn. Rev. Ecol. Syst., 9, 449—474.
Allee W. C., 1931. Animal Aggregations: A Study in General Sociology. Chi-
cago, University of Chicago Press.
Allee W. C., 1951. Cooperation Among Animals with Human Implications,
New York, Schuman. (Revised edition of Social Life of Animals, New
York, W. W. Norton, 1938.)
Allee W, C., Emerson A. E., Park O., Park Th., Schmidt K. P.t 1949. Prin-
ciples of Animal Ecology, Philadelphia, W. B. Saunders Co., 837 pp.
Allen K. R., 1951. The Horokiwi Stream: A study of a trout population. New
Zealand Mar. Dept. Fish. Bull., No. 10, 238 pp.
American Public Health Association, 1975. Standard Methods for Examination
of Water and Wastewater Including Bottom Sediments and Sludges
(14th ed.), New York, American Public Health Association.
Anagnostakis S. L., 1982. Biological control of chestnut blight, Science, 215,
466-471.
304
Литература
Anderson F. R. et al., 1977. Environmental Improvement Through Economic
Incentives, A Resources for the Future Book, Baltimore, Johns Hopkins
Press.
Anderson G. R., 1955. Nitrogen fixation by pseudomonas-like soil bacteria,
J. Bacterio!., 70, 129—133.
Anderson R. M., Turner B. D., Taylor L. R. (eds.), 1979. Population Dyna-
mics, Oxford, Blackwell.
Andrewartha H. G., 1961. Introduction to the Study of Animal Populations,
Chicago, University of Chicago Press, 281 pp.
Andrewartha H. G., Birch L. C., 1954. The Distribution and Abundance of
Animals, Chicago, University of Chicago Press.
Ardrey R., 1967. The Territorial Imperative, New York, Atheneum, 390 pp.
Aruga Y., Monsi M., 1963. Chlorophyll amount as an indicator of matter
production in biocommuniti.es, Plant Cell Physiol., 4, 29—39.
Ashby W. R., 1963. An Introduction to Cybernetics, New York, John Wiley
and Sons, 295 pp.
Auclair A. N., 1976. Ecological factors in the development of intensive ma-
nagement ecosystems in the midwestern United States, Ecology, 57, 431—
444.
Auclair A. N., Goff F. G., 1971. Diversity relations of upland forests in the
western Great Lakes area. Am. Nat., 105, 499—528.
Auerbach S. I., Crossley D. A., Engelman M. D., 1957. Effects of gamma ra-
diation on collembola population growth, Science, 126, 614.
Auerbach S. J., Olson J. S., Waller H. D., 1964. Landscape investigations using
cesium 137, Nature, 201, 761—764.
Austin G. T., Tomoff C. S., 1978. Relative abundance in bird populations, Am.
Nat., 112, 695—699.
Axelrod R.. Hamilton W. D., 1981. The evolution of cooperation, Science, 211,
1390—1396.
Ayala F. J.. 1968. Genotype, environment and population numbers, Science,
162, 1453—1459.
Ayala F. J., 1969. Experimental invalidation of the principle of competitive
exclusion, Nature. 224, 1076—1079.
Ayala F. J., 1972. Compel ition between species, Am. Sci., 60, 348—357.
Ayres R. V., 1978. Resources, Environment and Economics. Applications of
the Materia Is/Energy Balance Principle, New York, Wiley-Inter science,
207 pp.
Ayres R. V., Kneese A. V., 1971. Economic and ecological effects of a sta-
tionary economy, Ann. Rev. Ecol. Syst., 2, 1—22.
Azevedo J., Morgan D. T... 1974. Fog precipitation in coastal California forests,
Ecology, 55, 1135—1141.
Baes C. F., Jr., Goeller H. E.. Olson J. S., Rotty R. M., 1977. Carbon dioxide
and climate: The uncontrolled experiment, Am. Sci., 65, 310—320.
Bakelaar R. Gary, Odum E. P., 1978. Community and population level res-
ponses io fertilization in an oil-field ecosystem, Ecology, 59, 660—665.
Baker H. G., 1961. The adaptation of flowering plants to nocturnal and cre-
puscular pollinators, Quart, Rev. Biol., 36, 64—73.
Baker H. G., 1963. Evolutionary mechanisms in pollination biology, Science
139, 877—883.
Bakuzis E. V., 1969. Forestry viewed in an ecosystem perspective. In: The Eco-
system Concept in Natural Resource Management, G. Van Dyne, ed. New
York, Academic Press, pp. 189—257.
Ballard R. D., Grassle J. F., 1979. Return to oases of the deep; strange world
without sun, Nat, Geogr., 156, 680—703.
Baltensweiler W., 1964. Zeiraphera griseana Hubner (Lepidoptera: Tortricidae)
in the European Alps. A contribution to the problem of cycles, Can..
Entomol., 96, 792—800.
Bardach J. E., 1968. Harvest of the Sea, New York, Harper and Row.
Литература
305
Bardach J. E., 1969. Aquaculture, Science, 161, 1098—1106.
Barick F. B., 19a0. The edge effect of the lesser vegetation of certain Adiron-
dack forest types with particular reference to deer and grouse. Roosevelt
Wildl. Bull, 1—146.
Barkley P. W., Seckier D. W., 1972. Economic Growth and Environmental
Decay, New lork, Harcourt Brace Jovanovich.
Barney G. O., 1980. The Global 2000 Report, See under Council on Environ-
mental Quality.
Barr T. N., 1У81. the world food situation and global grain prospects, Science,
214, 1087—1095.
Barrett G. W., 1969. The effects of an acute insecticide stress on a semienclo-
sed grassland ecosystem, Ecology, 49, 1019—1035.
Barrett G. W., Rosenberg R. (eds.), 1981. Stress Effects on Natural Ecosys-
tems, New York, John Wiley and Sons.
Bates M., 1964. Man in Nature (2nd ed.), Englewood Cliffs, N. J., Prentice-
Hall.
Batie S. S., Healy R. G. (eds.), 1980. The Future of American Agriculture as
a Strategic Resource, Washington, D. G., The Conservation Foundation,
291 pp.
Bator F. M., 1958. The anatomy of market failure, Quart. J. Econ., 72, 351—
379.
Batra S. W. T1982. Biological control in agroecosystems, Science, 215, 134—
139.
Baylor E. R., Sutcliffe W. IL, Jr., 1963. Dissolved organic matter in seawater
as a source of particulate food, Limnol. Oceanogr., 8, 369—371.
Bazzaz F. A., 1975. Plant species diversity in old-field successional ecosystems
in southern Illinois, Ecology, 56, 485—488.
Beals E. W., 1969. Vegetational change along altitudinal gradients, Science,
165, 981—985.
Beck S. D., 1960. Insects and the length of the day, Sci Am., 202(2), 108—
118.
Beecher W. J., 1942. Nesting birds and the vegetation substrate, Chicago,
Chicago Ornithological Society.
Beery J. A., 1975. Adaptation of photosynthetic processes to strees, Science,
188, 644—650.
Beeton A. M., 1961. Environmental changes in Lake Erie, Trans. Am. Fish.
Soc, 90, 153—159.
Begon M., Mortimer M., 1981. Population Ecology: A Unified Study of Ani-
mals and Plants, Sunderland, Mass, Sinauer Associates.
Belt С. B., Jr., 1975. The 1973 flood and man’s constriction of the Mississippi
River, Science, 189, 681—684.
Bennett G. W., 1962. Management of Artificial Lakes and Ponds, New York,
Reinhold.
Bennett I. L. (Panel Chairman), 1967. The World Food Problem. Report of
the Panel on World Food Supply to President’s Science Advisory Com-
mittee, Three vols, Washington, D. C, Supt. of Documents.
Benton G. S., Blackburn R. T., Snead V. O., 1950. The role of the atmosphere
in the hydrological cycle, Trans. Am. Geophys, Union, 31, 61—73.
Bergmann G., 1944. Holism, historicism and emergence, Philos. Sci, 11, 209—
221.
Berkner L. V., Marshall L. C., 1964. The history of growth of oxygen in the
earth’s atmosphere. In: The Origin and Evolution of Atmospheres and
Oceans, D. J. Brancazio and A. G. W. Cameron, eds. New York, John
Wiley and Sons, pp. 102—126.
Berkner L. V., Marshall L. C., 1965. History of major atmospheric components,
Proc. Natl. Acad. Sci. USA, 53, 1215—1226.
Berkner L. V., Marshall L. C., 1966. The role of oxygen. Saturday Review,
May 7, 1966, pp. 30—33.
306
Литература*
Berlinski D., 1976. On Systems Analysis, Cambridge, Mass., MIT Press.
Bernardi R. (ed.), 1979. Biological and Mathematical Aspects in Population
Dynamics, Memorie dell ’’Institute di Idrobiologia,“ Vol. 37, Suppl. Pallan-
za, Italy.
Bernstein В. B., 1981. Ecology and economics: Complex systems in changing
environments, Ann. Rev. Ecol. Syst., 12, 309—330.
Bertalanjjy L. von, 1950. An outline of general systems theory, Brit. J. Philos.
Sci., 1, 139—164.
Bertalanjjy L. von, 1957. Quantitative laws in metabolism and growth, Quart.
Rev. Biol., 32, 217—231.
Bertalanjjy L. von, 1968. General Systems Theory: Foundations, Development,
Application, New York, George Braziller, 295 pp. (Revised edition, 1975.)
Best R., 1962. Production factors in the tropics. In: ’’Fundamentals of Dry-
Matter Production and Distribution,“ Special issue of Neth. J. Agr. Sci.t
10(5), 347—353.
В everton R. J. H., Holt S. J., 1957. On the dynamics of exploited fish populati-
ons, Great Brit. Min. Agr. Fish, Food, Fish. Invest. Ser. 2—19, 1—533.
Beyers R. J., 1962. Relationship between temperature and the metabolism of
experimental ecosystems, Science, 136, 980—982.
Beyers R. J., 1963. The metabolism of twelve aquatic laboratory microecosys-
tems, Ecol. Monogr., 33, 281—306.
Beyers R. J., 1964. The microcosm approach to ecosystem biology, Am. Biol.
Teacher, 26. 491—498.
Billings W. D., 1952. The environment complex in relation to plant growth and
distribution, Quart. Rev. Biol., 27, 251—265.
Billings W. D., 1957. Physiological ecology, Ann. Rev. Plant Physiol., 8, 375—
392.
Billings W. D., 1968. Plants, Man and the Ecosystem, Belmont, Calif., Wads-
worth, 154 pp.
Birch L. C. 1948. The intrinsic rate of natural increase of an insect population?
J. Anim. Ecol., 17, 15—26.
Birge E. A., 1915. The heat budgets of American and European lakes, Trans.
Wisconsin Acad. Sci. Arts Letters, 18. 166—213.
Bjorkman J., 1966. The effect of oxygen concentration on photosynthesis in
higher plants, Physiol. Plantarium, 19, 618—633.
Bjorkman O., 1973. Comparative studies on photosynthesis in higher plants.
In: Photophysiology, A. G. Giese, ed. Vol. 8, New York, Academic Press.
Bjorkman O., Beery J., 1973. High efficiency photosynthesis, Sci. Am., 229
(4), 80—93.
Black C. A., 1968. Soil—Plant Relationships (2nd ed.), New York, John Wiley
and Sons, 792 pp.
Black С. C., 1971. Ecological implications of dividing plants into groups with
distinct photosynthetic capacities, Adv. Ecol. Res., 7, 87—114.
Black J. N., 1971. Energy relations in crop production: A preliminary survey?
Ann. Appl. Biol., 67, 272—278.
Bliss L. C., 1966. Plant productivity in Alpine microenvironments, Ecol. Mo-
nogr., 36, 125—155.
Blum M. S,, 1969. Alarm pheromones, Ann. Rev. Entomol., 14, 57—80.
Bodenheimer F. S., 1937. Population problems of social insects, Biol. Rev., 12?
393—430.
Bolin B., 1977. Changes of land biota and their importance for the carbon
cycle, Science, 196, 613—615.
Borgstrom G., 1965. The Hungry Planet, New York, Macmillan, 487 pp.
Borgstrom G., 1969. Too Many: An Ecological Overview of Earth’s Limitations,
New York, Collier Books, 400 pp.
Borgstrom G., 1979. Ecological constraints on global food production. In:
Growth without Ecodisasters, Nicholus Polumin, ed. London, Macmillan
Press, pp. 293—323.
Литература 307
Bormann F. H., Likens G. E., 1967. Nutrient cycling, Science, 155, 424—429.
Bormann F. H., Likens G. E., 1979. Pattern and Process in a Forested Eco-
system, New York. Springer-Verlag, 253 pp.
Bormann F. H., Likens G. E., Melillo J. M., 1977. Nitrogen budget for an
aggrading northern hardwood ecosystem, Science, 196, 981—983.
Botkin D. B., Woodwell G. M., Tempel Neal, 1970. Forest productivity estima-
ted from carbon dioxide uptake, Ecology, 51, 1057—1060.
Botkin J. W., Elmandjra M., Malitza M., 1980. No Limits to Learning. Brid-
ging the Human Gap, A Report to the Club of Rome, New York, Perga-
mon Press, 159 pp.
Boulding. К. E., 1962. A Reconstruction of Economics, New York. Science
Editions.
Boulding К. E., 1964. The Meaning of the 20th Century: The Great Transition,
New York, Harper and Row.
Boulding К. E., 1966. The economics of the coming spaceship earth. In: Envi-
ronmental Quality in a Growing Economy, A Resources for the Future
Book, Baltimore, Johns Hopkins Press, 314 pp.
Boulding К. E., 1966a. Economics and ecology. In: Future Environments of
North America, F. F. Darling and J. P. Milton, eds. Garden City, N. Y.,
The Natural History Press, pp. 225—234.
Boulding К. E., 1978. Ecodynamics: A New Theory of Societal Evolution.
Beverly Hills, Calif., Sage.
Boulding. К. E., 1982. Knowledge, resources and the future, A Review of Si-
mon's The Ultimate Resource and Brown’s Building a Sustainable Society,
BioScience, 32, 343—344.
Bouvonder B., 1979. Impact of green revolution in India, Tech. Forecasting
and Social Change, 13, 297—313.
Bouwer H., 1968. Returning wastes to the land, a new role for agriculture,
J. Soil Water Cons, 23, 164—168.
Bowen G. D., 1973. Mineral nutrition of ectomycorrhizae. In: Ectomycorrhizae:
Their Ecology and Physiology, G. C. Marks and T. T. Kozlowski, eds..
New York, Academic Press.
Bowman К. O., Hutcheson K., Odum E. P., Shenton L. B., 1970. Comments
on the distribution of indices of diversity. In: International Symposium
on Statistical Ecology, Vol. 3. University Park, The Pennsylvania State
University Press.
Box E., 1978. Geographical dimensions of terrestrial net and gross producti-
vity, Radiat. Environ, Biophys., 15, 305—322.
Braun-Blanquet J., 1932. Plant Sociology: The Study of Plant Communities.
G. D. Fuller and H. C. Conard, trans, and eds.. New York, McGraw-Hill
Braun-Blanquet J., 1951. Pflanzensoziologie, Vienna, Springer-Verlag.
Bray J. B., 1956. Gap phase replacement in a maple-basswood forest, Ecology
37, 598—600.
Bray J. B., 1958. Notes towards an ecologic theory, Ecology, 30, 770—776.
Bray J. H., Gorham E., 1964. Litter production in forests of the world. Adv
Ecol. Res., 2, 101—157.
Breit W., 1982. The Academic Scribblers (2nd ed.), Hinsdale, Ill.. Dryden
Press.
Brian M. W., 1956. Exploitation and interference in interspecies competition.
J. Anim. Ecol., 25, 335—347.
Brill W. J., 1977. Biological nitrogen fixation, Sci. Am., 236(3). 68—81.
Brill W. J., 1979. Nitrogen fixation: Basic to applied, Am. Sci., 67, 458—
466.
Brillouin L., 1949. Life, thermodynamics and cybernetics, Am. Sci., 37, 354—
368.
Brink B. A., Densmore J. W., Hill G. A., 1977. Soil deterioration and the
growing world demand for food, Science, 197, 625—630.
308
Литература
Brock Т. D., 1967. Relationship between primary productivity and standing
crop along a hot spring thermal gradient, Ecology, 48, 566—571.
Brock T. D., Brock M. L., 1966. Temperature options for algal development
in Yellowstone and Iceland hot springs, Nature, 209, 733—734.
Brody S., 1945. Bioenergetics and Growth, New York, Reinhold.
Broecker W. S., Takahashi T., Simpson H. J., Peng T. H., 1979. Fate of fossil
fuel carbon dioxide and the global carbon budget, Science, 206, 409—
418.
Brooks J. L., Dodson S. I., 1965. Predation, body size and composition of
plankton, Science, 150, 28—35.
Brower L. P., Ryerson IF. N., Coppinger L. L., Glazier S. C., 1968. Ecological
chemistry and the palatability spectrum, Science, 161, 1349—1350. (See
also Zoologica 49, 137; 1964.)
Brown F. A., Hastings J. W., Palmer J. D., 1970. The Biological Clock—Two
Views, New York, Academic Press, 94 pp.
Brown H. S., 1977. World Food and Nutrition Study, See under National Aca-
demy of Science.
Brown H. S., 1978. The Human Future Revisited, New York, Norton.
Brown J. L., 1969. Territorial behavior and population regulation in birds,
a review and reevaluation, Wilson Bull., 81, 293—329.
Brown J. L., Arians G. H., 1970. Spacing patterns in mobile animals, Ann.
Rev. Ecol. Syst., 1, 239—262.
Brown L. L., Wilson E. O., 1956. Character displacement, Syst. ZooL, 5, 49—64.
Brown L. R., 1975. The world food prospect, Science, 190, 1053—1059.
Brown L. R., 1978. The global economic prospect: New sources of economic
stress, Worldwatch Paper No. 20, Washington, D. C., Worldwatch Institute,
56 pp.
Brown L. R., 1978a. The worldwide loss of cropland, Worldwatch Paper No. 24,
Washington, D. C., Worldwatch Institute, 48 pp.
Brown L. R., 1980. Food or fuel: New competition for the world croplands,
Worldwatch Paper No. 35, Washington, D. C., Worldwatch Institute.
Brown L. R., 1981. Building a Sustainable Society, New York, Norton, 433 pp.
Brown L. R., 1981a. World population growth, soil erosion and food security,
Science, 214, 995—1002.
Brown S., 1981. A comparison of the structure, primary productivity and
transpiration of cypress ecosystems in Florida, Ecol. Monogr., 51(4), 403—
427.
Brown IF. L., 1961. Mass insect control programs: Four case histories, Psyche,
68, 75—109.
Brugam R. B., 1978. Human disturbance and the historical development of
Linsley Pond, Ecology, 59, 19—36.
Bryson R. A., 1974. A perspective on climatic change, Science, 184, 753—
760.
Buckman H. O., Brody N. C., 1974. The Nature and Properties of Soil (8th
ed.), New York, Macmillan.
Budyko M. I., 1955. Atlas of the Heat Balance, Leningrad, See: The heat
balance of the earth’s surface, Translated by N. A. Stepannova.
Buell M. F., 1956. Spruce-fir and maple-basswood competition in Itasca Park
Minn. Ecology, 37, 606.
Bullock T. H., 1955. Compensation for temperature in the metabolism and
activity of poikotherms, Biol. Rev., 30, 311—342.
Burkholder P. R., 1952. Cooperation and conflict among primitive organisms
Am. Sci., 40, 601—631. ’
Burris R. H. (ed.), 1978. Future of biological N2 fixation, BioScience, 28.
563—592.
Butzer K. W., 1964. Environment and Archaelogy, An Introduction to Pleisto-
cene Geography, Chicago, Aldine Press, 524 pp.
Литература
309
Bulzer К. IV, 1980. Civilizations: Organisms or systems?, Am. Sci., 68, 517—
523.
Calin R., 1978. Footprints on the Planet; A Search tor an Environmental
Ethic., New York, Universe Books.
Cain S. A., 1944. Foundations of Plant Geography, New York, Harper and
Bros., 556 pp.
Cain S. A., 1950. Lifeforms and phytoclimate. Bot. Rev., 16, 1—32.
Cairns J. (ed). 1970. The structure and function of freshwater microbial com-
munities, Research Division Monograph 3. Blacksburg, Va, Va. Polytech
nic Institute and Slate University, 300 pp.
Cairns J. (cd.), 1980. The Recovery Process in Damaged Ecosystems, Ann Ar
bor, Mich., Ann Arbor Science, 167 pp.
Cairns J. (ed.), 1982. Artificial Substrates, Ann Arbor, Mich., Ann Arbor
Science, 300 pp.
Cairns J., Jr., Dickson K. L., 1971. A simple method for the biological as-
sessment of the effects of water discharges on aquatic bottom dwelling or-
ganisms, J. Water Pollution Control Fed., 43, 755—772.
Cairns J., Dickson K. L., Herrick E. E. (eds.), 1977. Recovery and Restoration
of Damaged Ecosystems, Charlottesville, University of Virginia Press,
531 pp.
Calhoun J. B., 1962. Population density and social pathology, Sci. Am., 206(2),
1399—1408.
Callahan D., 1972. Ethics and population limitations, Science, 175, 487—494.
Callicott J. B., 1979. Elements of an environmental ethic: Moral considerability
and the biotic community, Environ. Ethics, 1, 71—81.
Calvin M., 1969. Chemical Evolution; Molecular Evolution Towards the Origin
of Living Systems on the Earth and Elsewhere, New York, Oxford Uni-
versity Press, 278 pp.
Cameron A. W., 1964. Competitive exclusion between the rodent genera Mic-
rotus and Clethrionomys, Evolution. 18, 630—634.
Cannon W. B., 1939. The Wisdom of the Body, New York. Norton.
Carpenter E. J.. 1969. A simple, inexpensive algal chemostat, Limnol. Ocea
nogr., 14, 720—721.
Carpenter F. L., Rec.her II. F., 1979. Pollination, reproduction and fire, Am.
Nat, 113, 871.
Carpenter J. R., 1940. Insect outbreaks in Europe, J. Anim. Ecol, 9, 108—
147.
Carson II. L., 1958. Response to selection under different conditions of recoin
bination of Drosophila, Cold Spring Harbor Symp. Quant. Biol, 23, 291—
306.
Carson R., 1962. Silent Spring, Boston, Houghton Mifflin.
Castle E. M.. 1964. The market mechanism, externalities and land economics,
J. Farm Economics, 47(3), 546.
Caswell IE, Reed F., Stephenson S. N., Werner P. J, 1973. Photosynthetic
pathways and selected herbivory: A hypothesis, Am. Nat., 107, 46)5—480.
Caughley G., 1970. Eruption of ungulate populations with emphasis on Hi-
malayan th or in New Zealand, Ecology, 51, 53—72.
Chambers K. L. (ed.), 1970. Biochemical Coevolution, Twenty-ninth Biology
Colloquium. Corvallis, Oregon State University Press.
Chambers R. S., Herendeen R. A., Joyce J. J., Penner P. S.. 1979. Gasohol:
Does it or doesn’t it produce positive net energy?, Science, 206, 789—
795.
Chapman R. N„ 1928. The quantitative analysis of environmental factors,
Ecology, 9, 111 — 122.
Chapman R. N., 1931. Animal Ecology, with Special Reference to Insects, New
York, McGraw-Hill.
Ch arnov E. L., Schaffer W. M., 1973. Life-history resequences of natural se-
lection: Cole’s result re-visited, Am. Nat, 107, 791,
ЗЮ Литература
Cheng Т. С. (ed.), 1971. Aspects of the Biology of Symbiosis, Baltimore, Uni-
versity Park Press
Chew R. M., 1974. Consumers as regulators of ecosystems, Ohio J. Sci., 74,
359—370.
Chilly I)., 1900. Population processes in the vole and their relevance to ge-
neral theory. Can. J. Zool., 38, 99—113.
Chilly D., 1907. The natural selection of self regulatory behavior in animal
populations, Proc. Ecol. Soc. Austr., 2, 51—78.
Christensen J. M., McDermott, 1958. Life history and biology of the oyster
crab, Pinnotheres ostreum. Say. Biol. Bull., 144, 140—179.
Christian J. J., 1950. The adreno-pituitary system and population cycles in
mammals, J. Mammal., 31, 247—259.
Christian J. J., 1903. Endocrine adaptive mechanisms and the physiologic
regulation of population growth. In: Physiological Mammalogy, W. V. May-
er and R. G. van Gelder, eds. New York, Academic Press, pp. 189—353.
Christian J. J., 1970. Social subordination, population density, and mammalian
evolution, Science, 168, 84—90.
Christian J. J., Davis D. E., 1964. Endocrines, behavior and populations, Sci-
ence, 146, 1550—1560.
Clark C. W.. 1981. Bioeconomics. In: Theoretical Ecology (2nd ed.), R. M. May,
ed. Sunderland, Mass., Sinauer Associates, pp. 387—418.
Clark L. R., 1964. The population dynamics of Cardiaspina albitextura (Psylli-
dae), Austr. J. ZooL, 12, 362—380.
Clark L. R., Geir P. W., Hughes R. D., Morris R. F., 1967. The Ecology of
Insect Populations in Theory and Practice, London, Methuen, 232 pp.
Clark W. C., Jones D. D., Rolling C. S., 1979. Lessons for ecological policy
design: A case study of ecosystem management, Ecol. Model., 7, 153.
Clarke F. W., 1924. The data of geochemistry. U. S. Geol. Surv. Bull., No. 228.
Clarke G. L., 1954. Elements of Ecology, New York, John Wiley and Sons.
(Revised printing, 1965).
Clalworlhy J. N., Harper J. L., 1962. The comparative biology of closely re-
lated species living in the same area, J. Exp. Bot., 13. 307—324.
Clements F. E., 1905. Research Methods in Ecology, Lincoln, Neb., Univ. Publ.
Co., 199 pp.
Clements F. E., 1916. Plant succession: Analysis of the development of ve-
getation, Washington, D. C., Publ. Carnegie Inst., 242, 1512. (See also
reprinted edition, 1928, entitled Plant Succession and Indicators. New
York. Wilson.)
Clements F. E., Sh el ford E. E., 1939. Bioecology, New York, John Wiley and
Sons.
Cleveland L. R., 1924. The physiological and symbiotic relationships between
the intestinal protozoa of termites and their host, with special reference
Io Reticulilermes fluipes Kollsx, Biol. Bull., 46, 177—225.
Cleveland L. R.. 1926. Sumbiosis among animals with special reference to ter-
mites and their intestinal flagellates, Quart. Rev. Biol., 1, 51—60.
Cloud P. E., Jr. (ed.), 1969. Resources and Man, San Francisco, W. II. Freeman,
259 pp.
Cloud P. E., Jr. (ed.), 1978. Cosmos, Earth, and Man: A Short History of the
Universe, New Haven, Yale University Press.
Coase R. IE, 1960. The problem of social cost, J. Law Econ., 3, 1—44.
Cobh J. C., Dyer T., 1979. Environmental protection or economic prosperity:
Georgia. Tennessee and the Tennessee Copper Companies, 1903—1975,
Presented at Annual Meeting, Organization of American Historians, New
Orleans, 26 pp. г notes (manuscript).
Cody M. L., 1966. A general theory of clutch size, Evolution, 20, 174—184.
Cody M. L., 1974. Optimization in ecology, Science, 183, 1156—1164.
Cody M. L., Diamond M. (eds.), 1975. Ecology and Evolution of Communities,
Cambridge, Mass., Belknap Press, Harvard University.
Литература
311
Coffin С, С., Hayes F, R., Jodrey L. H., Whileway S. C., 1949. Exchanges of
materials in a lake as studied by the addition of radioactive phosphorus.
Can. J. Res., Section C, 27, 207—222.
Cohen M. N., Malpass R. S., Klein II. G., 1980. Bisocial Mechanisms of Po-
pulation Regulation, New Haven, Yale University Press.
Cole LaMont C., 1946. A theory for analyzing contagiously distributed popu-
lations, Ecology, 27, 329—341.
Cole LaMont C., 1946a. A study of the cryptozoa of an Illinois woodland,
Ecol. Monogr., 16, 49—86.
Cole LaMont C., 1951. Population cycles and random oscillations, J. Wildl.
Mgmt., 15, 233—251.
Cole LaMont C., 1954. Some features of random cycles, J. Wildl. Mgmt. 18,
107—109.
Cole LaMont C., 1954a. The population consequences of life history pheno-
mena, Quart. Rev. Biol., 29, 103—137.
Cole LaMont C., 1957. Sketches of general and comparative demography, Cold
Spring Harbor Symp. Quant. Biol., 22, 1—15.
Cole LaMont C., 1966. Protect the friendly microbes. In: ’’The Fragile Breath
of Life,“ 10th Anniversary Issue, Science and Humanity Supplement, Sa-
turday Review, May 7, 1966, pp. 46—47.
Coleman D. C., 1976. Energy flow and partitioning in selected man-managed
and natural ecosystems, Agro-Ecosystems, 3, 45—54.
Coleman D. C., Cole С. V., Runt H. W., Klein D. A., 1978. Trophic interactions
in soil as they affect energy and nutrient dynamics, Microb. Ecol., 4,
345—349, 373—380.
Coleman D. C., Reid С. P. P., Cole С. V., 1983. Biological strategies of nutrient
cycling in soil systems, Adv. Ecol. Res., 13 (in press).
Colwell R. K., 1973. Competition and coexistence in a simple tropical commu-
nity, Am. Nat., 107, 737—760.
Colwell R. N. et al., 1963. Basic matter and energy relationships involved in
remote reconnaissance, Photogr. Engr., 29, 761—799.
Comar C. L., 1955. Radioisotopes in Biology and Agriculture: Principles and
Practice, New York, McGraw-Hill.
Comar C. L., 1965. The movement of fallout radionuclides through the bio-
sphere and man, Ann. Rev. Nucl. Sci., 15, 175—206.
Commoner B., 1971. The Closing Circle: Nature, Man and Technology, New
York, Alfred A. Knopf.
Connell J. H., 1961. The influence of interspecific competition and other fac-
tors on the distribution of the barnacle, Chthamalus stellatus, Ecology,
42, 133—146.
Connell J. H.> 1972. Community interactions on marine rocky intertidal shores,
Ann. Rev. Ecol. Syst., 3, 169—192.
Connell J. H., 1978. Diversity in tropical rain forests and coral reefs, Science,
199, 1302—1310.
Connell J. H., Orias E., 1964. The ecological regulation of species diversity,
Am. Nat., 98, 399—414.
Connell J. H., Slayter R. O., 1977. Mechanism of succession in natural commu-
nities and their role in community stability and organization, Am. Nat.,
Ill, 1119—1144.
Conner W. H., Day J. W., 1976. Productivity and composition of a bald cypress
water tupelo site and a bottomland hardwood site in a Louisiana swamp,
Am. J. Bot, 63, 1354-1364.
Connor E. F., McCoy E. D., 1979. The statistics and biology of the species-area
relationship, Am. Nat., 113, 791—833.
Cooke G. D., 1967. The pattern of autotrophic succession in laboratory mic-
roecosystems, BioScience, 17, 717, 721,
312
Литература
Cooke G. D., Beyers R. J., Odum E. P., 1968. The case for the multi-species
ecological system, with special reference to succession and stability. In:
Bioregenerative Systems, NASA Special Publication, 165, 129—139.
Cooper Ch,. R., 1961. The ecology of fire, Sci. Am., 204(4), 150—160.
Copeland B. J., Dorris T. C., 1962. Photosynthetic productivity in oil refinery
effluent holding ponds, J. Water Poll. Control Fed., 34, 1104—1111.
Costanza R., 1980. Embodied energy and economic valuation, Science, 210,
1219—1224.
Council on Environmental Quality (CEQ), 1972. Integrated Pest Management,
Washington, D. C., U. S. Governing Printing Office.
Council on Environmental Quality, 1978. Ninth Annual Report; 1979, Tenth
Annual Report, 1980, Eleventh Annual Report; 1981, Twelfth Annual Re-
port, Washington, D. C., U. S. Government Printing Office.
Council on Environmental Quality, 1980. The Global 2000 Report to the Pre-
sident. Entering the Twenty-first Century. Vols. 1, 2, and 3, Washington,
I). С., U. S. Government Printing Office. (Textbook edition published by
Pergamon Press, Elmsford, N. Y.)
Council on Environmental Quality, 1981. Global Future — Time to Act,
G. Speth, ed. Washington, D. C., U. S. Government Printing Office.
Council on Environmental Quality, 1981. Global Energy Futures and the
Carbon Dioxide Problem, Washington, 1). C., U. S. Government Printing
Office, 92 pp.
Council on Environmental Quality, 1981. Contamination of Ground Water by
Toxic Organic Chemicals, Washington, D. C., U. S. Government Printing
Office.
Cowgill U. 71/., Hutchinson G. E., 1964. Cultural eutrophication in Lago Mon-
terosi during Roman antiquity, Proc. Ink Assoc. Thooret. Appl. Limnol.,
15(12), 6Vi—645.
Cowles В. C., 1899. The ecological relations of the vegetation of the sand
dunes of Lake Michigan, Bot. Gaz., 27, 95—117, 167—202, 281—308, 361 —
391.
Cox G. IF., Atkins M. D., 1979. Agricultural Ecology, San Francisco, W. IL Fre-
eman, 721 pp.
Craighead J. Craighead F. C., Jr., 1956. Hawks, Owls and Wildlife, Harris-
burg Pa., The Stackpole Co.
Crisp D. (ed.), 1964. Grazing in terrestrial and marine environments, Oxford,
Blackwell, 322 pp.
Crocker R. L., 1952. Soil genesis and the pedogenic factors, Quart. Rev. Biol.,
27, 139—168.
Crombie Л. C., 1947. Interspecific competition, J. Anim. Ecol., 16, 44—73.
Crossley D. A., 1963. Consumption of vegetation by insects. In: Radioecology,
V. Schultz and A. W. Klement, eds. New York, Reinhold, pp. 427—430.
Crossley D. A., 1964. Biological elimination of radionuclides, Nuclear Safety,
5, 265—268.
Crossley D. A., Bohnsack К. K., 1960. The oribated mite fauna in pine litter,
Ecology, 41, 785—790.
Crossley D. A., Coulson R. N., Gist C. S., 1973. Trophic level effects on spe-
cies diversity in arthropod communities of forest canopies, Environ. Ent.,
2, 1097—1100.
Crossley D. A., Hoglund M. P., 1962. A litterbag method for the study of
microarthropods inhabiting leaf litter, Ecology, 43, 571—573.
Crossley D. A., Witkamp M., 1964. Forest soil mites and mineral cycling, Aca-
rolgia 137—145.
Crowe B. L., 1969. The tragedy of the commons revisited. Science 166, 1103.
Crowell K. L., 1962. Reduced interspecific competition among the birds of
Bermuda, Ecology, 43, 75—88.
Currie R. I., 1958. Some observations on organic production in the northeast
Atlantic, Rapp. Proc. Verb. Cons. Int. Explor. Mer., 144, 96—102.
Литература 313
Curtis J. T,, 1955. A prairie continuum in Wisconsin, Ecology, 36, 558—566.
Curtis J. T., McIntosh R. P., 1951. An upland forest continuum in the prairie
forest border region of Wisconsin, Ecology, 32, 476—496.
Dale M. B., 1970. Systems analysis and ecology, Ecology, 51, 2—16.
Dales R. P., 1957. Commensalism. In: Treatise on Marine Ecology and Paleo-
ecology, J. W. Hedgpeth. ed. Vol. 1. Boulder, Col., Geological Society of
America, pp. 391—412.
Daly H. E., 1973. Toward a Steady-State Economy, San Francisco, W. H. Free-
man.
Darling F. F., 1938. Bird Flocks and the Breeding Cycle, Cambridge, England,
Cambridge University Press.
Darling F. F., 1951. The ecological approach to the social sciences, Am. Set,
39, 244—254.
Darling F. F., Milton J. P. (eds.), 1966. Future Environments of North Ame-
rica, Garden City, N. Y, Natural History Press, 265 pp.
Darmstadler J., Dunkerley J., Alter man J., 1977. How Industrial Societies
Use Energy: A Comparative Analysis, A Resources for the Future Book,
Baltimore, Johns Hopkins Press.
Dasmann R. F., 1968. A Different Kind of Country, New York, Macmillan.
Daubenmire R., 1966. Vegetation: Identification of typal communities, Science,
151, 291—298.
Daubenmire R., 1968. Ecology of fire in grasslands. In: Advances in Ecologi-
cal Research, J. B. Cragg, ed. Vol. V. New York, Academic Press,
pp. 209—266.
Daubenmire R., 1968a. Plant Communities, New York, Harper and Row.
Daubenmire R., 1974. Plants and Environment (3rd ed.), New York, John
Wiley and Sons.
Daubenmire R., 1978. Plant. Geography; with Special Reference to North
America, New York, Academic Press.
Davidson J., 1938. On the growth of the sheep population in Tasmania, Trans.
R. Soc. S. Austr, 62, 342—346.
Davidson J., Andrewartha H. G., 1948. Annual trends in a natural population
of Thrips imaginis (Thysanoptera), J. Anim. Ecol., 17, 193—199, 200—
222.
Davis J. J., Foster R. F., 1958. Bioaccumulation of radioisotopes through aquatic
food chains, Ecology, 39, 530—535.
Davis K. (ed.), 1965. Cities, Sci. Am. (Special Issue), 213(3), 1—280.
Davis M. B., 1969. Palynology and environmental history during the Quater-
nary Period, Am. Sci., 57, 317—332.
Day F. P., Monk C. D., 1974. Vegetation patterns on a southern Appalachian
watershed, Ecology, 55, 1064—1074.
Dayton R. K., 1971. Competition, disturbance and community organization:
The provision and subsequent utilization of space in a rocky intertidal
community, Ecol. Monogr, 41, 351—389.
Dayton R. K., 1975. Experimental evaluation of ecological dominance in
a rocky intertidal algal community, Ecol. Monogr., 45, 137—389.
DeBach P., 1964. Biological Control of Insect Pests and Weeds, New York,
Reinhold.
DeBenedictis P. A., 1974. Interspecific competition between tadpoles of R. pi-
piens and R. sylvatica: An experimental field study, Ecol. Monogr., 44,
129—151.
Deevey E. S., Jr., 1947. Life tables for natural populations of animals, Quart.
Rev. Biol, 22, 283—314.
Deevey E. S., Jr., 1950. The probability of death, Sci. Am, 182, 58—60.
Deevey E. S., Jr., 1958. The equilibrium population. In: The Population
Ahead, R. G. Francis, ed. Minneapolis, University of Minnesota Press,
pp, 64—86.
21 Заказ № 1483
314 Литература
de la Cruz A. A., 1975. Proximate nutritive value changes during decomposi-
tion of salt marsh plants, Hydrobiologia, 47, 475—480.
de la Cruz A. A., 1979. Production and transport of detritus in wetlands. In:
Wetland Functions and Values: The State of Our Understanding, Minnea-
polis, Am. Water Resources Assoc., pp. 162—173.
Delwiche C. C., 1970. The nitrogen cycle, Sci. Am., 223(5), 137—146.
Dendy J. S., 1945. Predicting depth distribution in three TVA storage type
reservoirs, Trans. Am. Fish. Soc., 75, 65—71.
Diamond J. M., May R. M., 1981. Island biogeography and the design of na-
tural reserves. In: Theoretical Ecology (2nd ed.), R. M. May, ed. Sun-
derland, Mass., Sinauer Associates.
Dice L. R., 1952. Measure of spacing between individuals within a population,
Contr. Lab. Vert. Biol., U. of Michigan, 55, 1—23.
Dice L. R., 1952a. Natural Communities, Ann Arbor, University of Michigan
Press.
Dickinson R. E., 1970. Regional Ecology: The Study of Man’s Environment,
New York, John Wiley and Sons.
Dobzhansky T., 1968. Adaptedness and fitness. In: Population Biology and
Evolution, R. C. Lewontin, ed. Syracuse, N. Y., Syracuse University Press,
pp. 109—121.
Dokuchaev V. V., 1889. The zones of nature, (In Russian.), Akad. Nauk
Moscow, Vol. 6.
Dommergues Y. R., Krupa S. V., 1978. Interactions Between Non-pathogenic
Soil Microorganisms and Plants, New York, Elsevier.
Dougherty E. C., 1959. Introduction to axenic culture of invertebrate metazoa:
A goal, Ann. N. Y. Acad. Sci., 77, 27—54.
Drake E. T. (ed.), 1968. Evolution and Environment, New Haven, Yale Uni-
versity Press, 478 pp.
Drury W. H., Nisbet I. С. T., 1973. Succession. J. Arnold Arboretum, 54, 331—
368.
Dublin L. L, Lolka A. J., 1925. On the true rate of natural increase as exem-
plified by the population of the United States, 1920, J. Am. Statist.
Assoc., 20, 305—339.
Dubos R., 1976. Symbiosis between the earth and humankind, Science, 193,
459—462.
Dubos R., 1980. The Wooing of Earth: New Perspective on Man’s Use of
Nature, New York. Charles Scribner’s Sons, 183 pp.
Duckham A. N.. Jones J. G. W., Robert E. H. (eds.), 1976. Food Production
and Consumption. The Efficiency of Human Food Chains and Nutrient
Cycles, Amsterdam, North-Holland; New York, Elsevier; 542 pp.
Dugger W. M., Jr., Koukol J., Palmer R. L., 1966. Physiological and bioche-
mical effects of atmospheric oxidants on plants, J. Air Poll. Control
Assoc., 16, 467—471.
Dugsdale R. C., 1967. Nutrient limitation in the sea: Dynamics, identification,
and significance, Liranol. Oceanogr., 12, 685—695.
Duncan O. D., 1964. Social organizations and the ecosystem. In: Handbook
of Modern Sociology, R. E. L. Faris, ed. Chicago, Rand McNally.
Duvlgneaud P., 1971. La Synthese Ecologique. Populations, Communantes,
Ecosystemes, Biosphere, Noosphere, Paris, Boin.
Dyer M. 1., Bokhari U. G.. 1976. Plant-animal interactions: Studies of effects
of grasshopper grazing on blue grama grass Ecology, 57, 762—772.
Dyksterhuis E. J., 1958. Ecological principles in range evaluation, Bot. Rev.,
24, 253—272.
Echholm E., 1975. The other energy crisis: Firewood, Worldwatch Paper No. 1,
Washington, D. C., Worldwatch Institute, 22 pp.
Edmondson W. T.. 1968. Water-quality management and lake eutrophication:
The Lake Washington case. In: Water Resources Management and Public
Литература
315
Policy, Т. Н. Campbell and R. О. Sylvester, eds. Seattle, University of
Washington Press, pp. 139—178.
Edmondson W. T., 1970. Phosphorus, nitrogen and algae in Lake Washington
after diversion of sewage, Science, 169, 690—691.
Edmondson W. T., 1979. Lake Washington and the predictability of limno-
logical events, Arch. Hydrobiol. Bech. Ergobn. Limnol., 13, 234—241.
Edson M. M., Foin T. C, Knapp С. M., 1981. Emergent properties and ecolo-
gical research, Am. ft at., 118, 593—596.
Edwards C. A., 1969. Soil pollutants and soil animals, Sci. Am., 220(4), 88—92,
97—99.
Edwards C. A., Reichle D. E., Crossley D. A., 1970. The role of soil inver-
tebrates in turnover of organic matter and nutrients. In: Analysis of
Temperate Forest Ecosystems, D. E. Reichle, ed. Berlin, Springer-Verlag,
pp. 147—172.
Egerton F. N., 1968. Leeuwenhoek as a founder of animal demography, J.
Hist. Biol., 1, 1—22.
Egerton F. N., 1969. Richard Bradley’s understanding of biological productivi-
ty; a study of eighteenth-century ecological ideas, J Hist. Biol., 2(2),
391—410.
Egler F. E., 1954. Vegetation science concepts. 1. Initial floristic composition—
a factor, in old-field vegetation development, Vegetatio, 4, 412—417.
Ehrenfeld D., 1978. The Arrogance of Humanism, New York, Oxford Univer-
sity Press.
Ehrlich P. R., 1968. The Population Bomb, New York, Ballantine Books.
Ehrlich P. R., Ehrlich A., 1981. Extinction: The Causes and Consequences of
Disappearance of Species, New York, Random House, 305 pp.
Ehrlich P. R., Ehrlich A. H., Holdren J. P.., 1977. Ecoscience: Population,
Resources, Environment. San Francisco, W. H. Freeman.
Ehrlich P. R., Raven P. H., 1965. Butterflies and plants: A study of coevolu-
tion, Evolution, 18, 586—608.
Einarsen A. S., 1945. Some factors affecting ring-necked pheasant population
density, Murrelet, 26, 39—44.
Eldredge N., Gould S. J., 1972. Punctuated equilibria: An alternative to phy-
letic gradualism. In: Models in Paleobiology, T. J. M. Schopf, ed. San
Francisco, Freeman, Cooper, pp. 82—115.
Eliassen R., 1952. Stream pallution, Sci. Am., 186, 17—21.
Ellul J., 1967. The Technological Society, New York, Random House.
Elton Ch., 1927. Animal Ecology, New York, Mcmillan. (2nd ed., 1935; 3rd ed.,
1947).
Elton Ch., 1933. The Ecology of Animals, London, Methuen.
Elton Ch., 1942. Voles, Mice and Lemmings: Problems in Population Dynamics,
London, Oxford University Press.
Elton Ch., 1958. The Ecology of Invasions by Animals and Plants, London,
Methuen, 181 pp.
Elton Ch., 1966. The Pattern of Animal Communities, London, Methuen,
432 pp.
Elwood J. W., Nelson D. J., 1972. Periphyton production and grazing rates in
a stream measured with a 32P material balance method, Oikos, 23, 295—
303.
Emanuel W. R., Mulholland R. J., 1976. Linear periodic control with applica-
tions to environmental systems, Int. J. Control, 24(6), 807—820.
Emerson A. E., 1954. Dynamic homeostasis: A unifying principle in organic,
social and ethical evolution, Sci. Monthly, 78(2), 67—85.
Emerson F. C. (ed.), 1973. The economics of environmental problems, Mich.
Business Papers No. 58, Ann Arbor, University of Michigan, 98 pp.
Emery К. O., Iselin С. O. D., 1967. Human food from ocean and land, Science,
157, 1279—1281.
21*
316
Литература
Emlen J. M., 1966. The role of time and energy in food preference, Am. Nat.,
100, 611—617.
Emlen J. M., 1970. Age specificity and ecological theory, Ecology, 51. 588—
601.
Emlen J. T., Dejong M. J., 1981. Intrinsic factors in the selection of foraging
substrates by pine warblers; a test of a hypothesis, Auk, 98, 294—298.
Engellberg J., Boyarsky L, L., 1979. The noncybernetic nature of ecosystems,
Am. Nat., 114, 317—324.
Engelmann M. D., 1968. The role of soil arthropods in community energetics,
Am. Zool., 8, 61—69.
Enright J. T., 1970. Ecological aspects of endogenous rhythmicity, Ann. Rev.
Ecol. Sys I., 1, 221—238.
Epstein S. S., 1974. Environmental determinants of human cancer. Cancer
Res., 34, 2425—2435.
Erringlon P. L., 1946. Predation and vertebrate populations, Quart. Rev. Biol.,
21, 144—177, 221—245.
Esch G. W., Hazen T. C., 1978. Thermal ecology and stress: A case history
for red-sore disease in large-mouth bass. In: Energy and Environmental
Stress in Aquatic Systems, Thorp and Gibbons, eds. U. S. Dept, of Energy,
Symp. No. 48, Springfield, Va., National Technical Info. Center, pp. 331—
363.
Esch G. W., McFarlane R. W. (eds.), 1975. Thermal Ecology II. Energy Re-
search and Development Administration, Springfield, Va., National Tech-
nical Info. Center, 404 pp.
Estep M. F., Dabrowski H., 1980. Tracing food webs with stable hydrogen
isotopes, Science, 109, 1537—1538.
Etkins R., Epstein E. S., 1982. The rise of global mean sea level as an indi-
cation of climate change, Science, 215, 287—289.
Evans F. C., 1956. Ecosvstem as the basic unit in ecology, Science, 123, 1227—
1228.
Evans F. C., Cain S. A., 1952. Preliminary studies on the vegetation of an
old field community in southeastern Michigan, Contr. Lab. Vert. Biol.,
U. of Michigan, 51, 1—17.
Evans L. T. (ed.), 1963. Environmental Control fo Plant Growth, New York,
Academic Press, 449 pp.
Evans L. T. (ed.), 1980. The natural history of crop field, Am. Sci., 68. 388—
397.
Ewel K., Odum H. T., 1978. Cypress swamps for nutrient removal and waste-
water recycling. In: Advances in Water and Wastewater Treatment;
Biological Nutrient Removal, Ann Arbor, Mich., Ann Arbor Science.
Fager E. W., 1972. Diversity: A sampling study, Am. Nat., 106, 293—310.
Falk J. H., 1976. Energetics of a suburban lawn ecosystem, Ecology, 57, 141—
150.
Falk J. H., 1977. The frenetic life forms that flourish in suburban lawns,
Smithsonian, 8, 90—96.
Famer D. S., 1964. The photoperiodic control of reproductive cycles in birds,
Am. Sci., 52, 137—156.
Famer D S., 1964a. Time measurement in vertebrate photoperiodism, Am.
Nat, 98, 375-386.
Farnworth E. G., Tidrick T. H., Jordan C. F., S mothers W. M., 1981. The va-
lue of natural ecosystems. An economic and ecological framework,
Environ. Conserv., 8, 275—282.
Farvar M. T., Milton J. P. (eds.), 1972. The Careless Technology; Ecology
and International Development, Garden City, N. Y., Natural History Press,
1030 pp.
Faulkner E. H., 1943. Plowman’s Folly, Norman, University of Oklahoma
Press, 161 pp.
Литература
317
Feener D. H., 1981. Competition between and species: Outcome controlled by
parasitic flies, Science, 214, 815—817.
Fenchel T. M., 1969. The ecology of marine microbenthos, Part IV, Ophelia,
6, 1—182.
Fenchel T. M., 1978. The ecology of micro- and meiobenthos, Ann. Rev. Ecol.
Syst., 9, 99—122.
Fenchel T. M., Raiding S., 1979. Habitat selection and distribution patteerns
of 5 species of the amphropod genus Gammarus, Oikos, 33. 316—322.
Fernandez L., 1980. Managing chemilcal wastes. In: Nat. Jour. Special Report
’’Chemical Issues in the News," Washington, D. C.
Fiebleman J. K., 1954. Theory of integrative levels, Brit. J. Philos. Sci., 5,
59-66.
Filzer P., 1956. Pflanzengemeinschaft und Umwelt, Ergebnisse und Probleme
der Botanischen Standortforschung, Stuttgart, Enke.
Flnerty J. P., 1980. The Population Ecology of Cycles in Small Mammals,
New Haven, Yale University Press.
Finn J. T., 1976. Measures of ecosystem structure and function derived from
the analysis of flows, J. Theoret. Biol., 56, 363—380.
Finn J. T., 1978. Cycling index: A general definition for cycling in compart-
ment models. In: Environmental Chemistry and Cycling Processes, D. C.
Adriano and I. L. Brisbin, eds., U. S. Dept, of Energy Symp., No. 45,
Springfield, Va., National Technical Info. Center, pp. 138—164.
Fischer A. G., 1960. Latitudinal variations in organic diversity, Evolution, 14,
64-81.
Fisher A. C., Krutilla J. V., Cicchetti C. J., 1972. The economies of environ-
mental preservation: A theoretical and empirical analysis, Am. Econ. Rev.,
62, 605-619.
Flader S. L., 1979. Thinking Like a Mountain: Aldo Leopold and the Evolu-
tion of an Ecological Attitude Towards Deer, Wolves and Forest, Lincoln
University of Nebraska Press.
Fleischer W. E., 1935. The relation between chlorophyll content and rate of
photosynthesis, J. Gen. Physiol., 18, 573—597.
Flint M. L., Bosch R. van den, 1981. Introduction to Integrated Pest Ma-
nagement, New York, Plenum, 256 pp.
Fogg G. E., 1965. Algal Cultures and Phytoplankton Ecology, Madison, Uni-
versity of Wisconsin Press, 126 pp.
Foin T. C., Jain S. K., 1977. Ecosystem analysis and population biology: Les-
sons for the development of community ecology, BioScience, 27, 532—539.
Forbes S. A., 1887. The lake as a microcosm, Bull. Sc. A. Peoria, Reprinted
in Ill. Nat. Hist. Surv. Bull., 15, 537—550, 1925.
Forrester J. W., 1968. Principles of Systems, Cambridge, Mass., Wright-Allen
Press.
Forrester J. W., 1971. World Dynamics, Cambridge, Mass., Wright-Allen Press,
142 pp.
Fortescue J. A. C., 1980. Environmental Geochemistry: A Holistic Approach,
Ecol. Series 35, New York, Springer-Verlag, 346 pp.
Foster R. F., Davis J. J., 1956. The accumulation of radioactive substances in
aquatic forms, Proc. Int. Conf. Peaceful Uses Atomic Energy, Geneva,
13, 364—367.
Frank P. W., 1957. Coactions in laboratory populations of two species of
Daphnia, Ecology, 38, 510—518.
Frank P. W., 1965. The biodemography of an intertidal snail population,
Ecology, 46, 831—844.
Frank P. W., 1968. Life histories and community stability, Ecology, 49, 355—
357.
Frankenberg D., Smith K. L., Jr., 1967. Coprophagy in marine animals, Limnol.
Oceanogr., 12, 443—450.
318
Литература
Freeze R. A., Cherry J. A., 1979. Groundwater, Englewood Cliffs, N. J., Pren-
tice-Hall.
French N. R., 1965. Radiation and animal population: Problems, progress and
projections, Health Physics, 11, 1557—1568.
Fretwell S. D., 1977. The regulation of plant communities by the food chains
exploiting them, Perspect. Biol. Med., 20, 169—185.
Fried M., Broeshart П. 1967. The Soil-Plant System, New York, Academic
Press.
Frieden E., 1972. The chemical elements of life, Sci. Am., 227(1), 52—60.
Fredericks K., 1930. Die Grundfragen und Gesetzmassigkeiten der landund
fortswirtschaf (lichen Zoologie, Two vols, Berlin, Paul Parey.
Fritsch A. J., 1980. Environmental Ethics: Choices for Concerned Citizens,
New York, Anchor Books.
Fry F. E. E, 1958. Temperature compensation, Ann. Rev. Physiol., 20, 207—
227.
Gabor D., Columbo V., King A., Galli R., 1978. Beyond the Age of Waste,
A Report to the Club of Rome, New York, Pergamon Press.
Gadgil EL, Bossert W, H., 1970. Life historical consequences of natural selec-
tion, Am. Nat., 104, 1—24.
Gadgil AL, Solbrig О. T.. 1972. The concept of r and К selection: Evidence
from wild flowers and some theoretical considerations, Am. Nat., 106,
14—31.
Galbraith J. K., 1958. The Affluent Society, New York, The New American
Library.
Galbraith J. K., 1967. The New Industrial State, Boston, Houghton Mifflin,
427 pp.
Gardner M. R., Ashby W. R., 1970. Connectance of large dynamic cybernetic
systems: Critical values for stability, Nature, 288, 784.
Garrets R. M., Mackenzie F. T., Hunt C., 1975. Chemical cycles and the glo-
bal environments; assessing human influences, Los Altos, Calif., William
Kaufmann, 206 pp.
Garren К. H., 1943. Effects of fire on the vegetation of the southeastern
United States, Bot. Rev., 9, 617—654.
Gasaway Ch. R., 1970. Changes in the fish population of Lake Francis Case
in South Dakota in the first 16 years of impoundment, Tech. Paper
No. 56, Washington, D. C., Bureau of Sport Fisheries and Wildlife.
Gates D. M., 1962. Energy Exchange in the Biosphere, New York, Harper and
Row, 151 pp.
Gates D. M., 1963. The energy environment in which we live, Am. Sci., 51,
327—348.
Gates D. M., 1965. Radiant energy, its receipt and disposal, Metero, Monogr.,
6. 1—26.
Gates D. M., 1965a. Energy, plants and ecology, Ecology, 46, 1—13.
Gates D. M., 1971. The flow of energy in the biosphere, Sci. Am., 224(3),
88—99.
Gates D. M., 1980. Biophysical Ecology, New York, Springer-Verlag, 640 pp.
Gates J. E., Gysel L. W., 1978. Avian nest dispersion and fledgling success in
a field-forest ecotone, Ecology, 59, 871—883.
Gause G. F., 1932. Ecology of populations, Quart. Rev. Biol., 7, 27—46. (See
also Quart. Rev. Biol., 11, 320—336, 1936.)
Gause G. F., 1934. The Struggle for Existence, Baltimore, Williams and
Wilkins, 163 pp.
Georgescu-Roegen N., 1971. The Entropy Law and the Economic Process,
Cambridge, Mass., Harvard University Press.
Georgescu-Roegen N., 1977. Bioeconomics. In: The Political Economy of Food
and Energy, L. Junker, ed. Michigan Business Papers No. 62, Ann Arbor,
University of Michigan, pp. 105—134.
Литература
319
Georgescu-Roegen N., 1977a. The steady-state and ecological salvation: A ther-
modynamic analysis, BioScience, 27, 266—271.
Georgescu-Roegen N., 1977b. Energy and Economic Myths, Elmsford, N. Y.,
Pergamon Press.
Gessner F., 1949. Der chlorophyllgehalt in see and seine photosynthetische
Valenz als geophysikalishes problem, Schweizer Z. Hydrologie, 11. 378—
410.
Giarini 0., 1980. Dialogue on Wealth and Welfare: An Alternate View of
World Capital Formation, A Report to the Club of Rome, New York,
Pergamon Press.
Gibbons J. W., Sharitz R. R., 1974. Thermal alteration of aquatic ecosystems,
Am. Sci., 62, 660—670.
Gibbons J. W., Sharitz R. R. (eds.), 1981. Thermal Ecology, U. S. Atomic
Energy Commission, Springfield, Va., National Technical Info. Center,
670 pp.
Gibson D. T., 1968. Microbial degradation of aromatic compounds, Science,
161, 1093—1097.
Giesy J. P. (ed.), 1980. Microcosms in Ecological Research, U. S. Dept, of
Energy Symp. No. 52, Springfield, Va., National Technical Info. Center,
1110 pp.
Gifford С. E., Odum E. P., 1961. Chlorophyll — a content of intertidal zones
on a rocky seashore, Limnol. Oceanogr., 6, 83—85.
Gilbert L. E., Raven P. H. (eds.), 1975. Coevolution of Animals and Plants,
Austin, University of Texas Press. (Revised edition, 1980).
Gilliland M. W., 1975. Energy analysis and public policy, Science, 180, 1051—
1056.
Gilliland M. W. (ed.), 1978. Energy Analysis: A New Public Policy Tool,
Selected Symposium 9, American Association for the Advancement of
Science, Boulder, Col., Westview Press.
Gilpin M. E., 1975. Group Selection in Predator-Prey Communities, Princeton,
N. J., Princeton University Press.
Glasser J. W., 1979. The role of predation in shaping and maintaining the
structure of communities, Am. Nat., 113, 631—641.
Glasser J. W., 1982. On the causes of temporal change in communities: Modi-
fication of the biotic environment, Am. Nat., 119, 375—390.
Glasstone S. (ed.), 1957. The Effects of Nuclear Weapons, Washington, D. C.,
U. S. Atomic Energy Commission.
Gleason H. A., 1917. The structure and development of the plant association.
Bull. Torrey Bot. Club, 44, 463—481.
Gleason H. A., 1926. The individualistic concept of the plant association, Bull.
Torrey Bot. Club, 53, 7—26.
Glemarec M., 1979. Problemes d’ecologie dynamique et de succession en Baie
de Concerneau, Vie Milou, Vol. 28—29, Fasc. 1, Ser. AB, pp. 1—20.
Goldman C. R., 1960. Molybdenum as a factor limiting primary productivity
in Castle Lake, California, Science, 132, 1016—1017.
Goldman C. R. (ed.), 1965. Primary productivity in aquatic environments,
Berkeley, University of California Press, 464 pp.
Goldschmidt V. M., 1954. Geochemistry, Oxford, Clarendon Press, 730 pp.
Golley F. B., 1960. Energy dynamics of a food chain of a old-field communi-
ty, Ecol. Monogr., 30, 187—206.
Golley F. B.', 1968. Secondary productivity in terrestrial communities, Am.
ZooL, 8, 53—59.
Golley F. B., 1983. The historical origins of the ecosystem concept in biology,
In press.
Golley F. B., McGinnis J. T., Clements R. G., Childs G. I., Duever M. J., 1975.
Mineral cycling in a tropical moist forest ecosystem, Athens, University
of Georgia Press.
'320 Литература
Goodall D. W., 1963. The continuum and the individualistic association, Ve-
getatio, 11, 297—316.
Gorden R. W., 1969. A proposed energy budget of a soybean field, Bull. Ga.
Acad. Sci., 27, 4152.
Gorden R. W., Beyers R. J.. Odum E. P., Eagon E. G., 1969. Studies of a sim-
ple laboratory microecosystem: Bacterial activities in a heterothrophic
succession, Ecology, 50, 86—100.
Gordon H. T., 1961. Nutritional factors in insect resistance to chemicals, Ann.
Rev. EntomoL, 6, 27—54.
Gorham E., VHouse к P. M., Reiners W. A., 1979. The regulation of chemical
budgets over the course of terrestrial succession, Ann. Rev. Ecol. Syst.,
10, 53—84.
Gornitz V., Lebedeff S., Hansen J., 1982. Global sea level trend in the past
century, Science, 125, 1611—1614.
Gosselink J. G., Odum E. P., Pope R. M., 1974. The Value of the Tidal Marsh,
LSU-SG-74-03, Center Wetland Resources, Louisiana State University,
Baton Rouge, 33 pp.
Gould S. J., 1977. The return of hopeful monsters, Nat. Hist., 86, 22—30.
Gould S. 1982. Darwinism and the expansion of evolutionary theory, Sci-
ence, 216, 380—387.
Gould S. J., Eldredge N., 1977. Punctuated equilibria: The tempo and mode
of evolution reconsidered, Paleobiology, 3, 115—151.
Goulden C. (ed.), 1977. Changing Scenes in Natural Sciences, Special Publi-
cation 12, Philadelphia. Academy of Natural Sciences.
Graham S. A.. 1951. Developing forests resistant to insect injury, Sci. Monthly,
73, 235—244.
Grinnell J., 1917. Field test of theories concerning distributional control, Am.
Nat., 51, 115—128.
Grinnell J., 1928. Presence and absence of animals, Univ. Calif. Chron., 30,
429—450.
Gross A. O., 1947. Cyclic invasion of the snowy owl and the migration of
1945—46, Auk, 64, 584—601.
Grunchy A. G., 1947. Modern Economic Thought: The American Contribution,
Englewood Cliffs, N. J., Prentice-Hall.
Grzenda A., Caver G. J., Nicholson H. P., 1964. Water pollution by insecticides
in an agricultural river basin. II. The zooplankton, bottom fauna, and
fish, Limnol. Oceanogr., 9, 318—323.
Gutierrez L. T., Fey W. R., 1975. Feedback dynamics analysis of secondary
successional transients in ecosystems, Proc. Natl. Acad. Sci. USA, 72,
2733—2737.
Gutschick V. P., 1978. Energy and nitrogen fixation, BioScience, 28, 571—
575.
Haag J., 1981. A yearning for synthesis: Organic thoughts since 1945, Int. J.
Social Econ., 8, 87—111.
Haagen-Smith A. J., 1963. The control of air pollution., Sci. Am., 209(1),
24-29.
Haagen-Smith A. J., Darley E. F., Zaitlin E. F., Hull M., Noble W., 1952.
Investigation of injury to plants from air pollution in the Los Angeles
area, Plant Physiol., 27, 18—34.
Haines E. B., Hanson R. B., 1979. Experimental degradation of detritus made
from salt marsh plants Spartina alterniflora, Salicornus Virginia and
J uncus roemerianus, J. Exp. Marine Biol. Ecol., 40, 27—40.
Haines E. B., Montague C. L., 1979. Food sources of estuarine invertebrates
analyzed using 13C/12C rations, Ecology, 60, 48—56.
Hairston N. G., 1959. Species abundance and community organization, Ecolo-
gy, 40, 404—416.
Hairston N. G., Smith F. K., Slobodkin L. B., 1960. Community structure,
population control and competition, Am. Nat., 94, 421—425.
Литература
321
Half on E. (ed.), 1979. Theoretical Systems Ecology: Advances and Case Stu-*,
dies, New York, Academic Press, 516 pp.
Hall Ch. A. S., Day J. W., 1977. Systems and models: Terms and basic prin-
ciples. In: Ecosystem Modeling in Theory and Practice, C. Hall and
J. Day, eds., New York, John Wiley and Sons, pp. 5—36.
Hamilton W. J. Ill, Watt К. E. F., 1970. Refuging, Ann. Rev. Ecol. Syst., 1,
263—286.
Hannon B., 1973. The structure of ecosystems, J. Theoret. Biol., 41, 535—546.
Hannon B., 1973a. An energy standard of value, Ann. Am. Acad. Political
Social Sci., 410, 139—153.
Hannon B., 1974. Energy, manpower and the highway trust fund, Science,
185, 669—675.
Bannon B., 1979. Total energy costs in ecosystems, J. Theoret. Biol., 80. 271—
293.
Hansson L., 1979. On the importance of landscape heterogeneity in northern
regions for breeding population density of homeotherms: A general hy-
pothesis, Oikos, 33, 182—189.
Hardin G., 1960. The competitive exclusion principle, Science, 131, 1292—
1297.
Hardin G., 1963. The cybernetics of competition. A biologist’s view of society,
Perspect. Biol. Med., 7, 58—84.
Hardin G., 1968. The tragedy of the commons, Science, 162, 1243—1248.
Hardin G., 1977. The Limits of Altruism: An Ecologist’s View of Survival,
Bloomington, Indiana University Press.
Hardin G., Boden J., 1977. Managing the Commons, San Francisco, W. H. Free-
man.
Harlan J. R., 1974. Our vanishing genetic resources, Science, 188, 618—621.
Harley J. L., 1952. Associations between microorganisms and higher plants
(mycorrhiza), Ann. Rev. Microbiol., 6, 367—386.
Harley J. L., 1959. The Biology of Mycorrhiza, Plant Science Monographs.
London, Leonard Hill; New York, Interscience, 233 pp.
Harper J. L., 1961. Approaches to the study of plant competition. In: ’’Mecha-
nisms in Biological Competition.", Symp. Soc. Exp. Biol., Number XV.
pp. 1—268.
Harper J. L., 1969. The role of predation in vegetational diversify. In: Diver-
sity and Stability in Ecological Systems, G. M. Woodwell and H. H. Smith,
eds., Brookhaven Symp. Biol., Number 22. Upton, N. Y., Brookhaven Na-
tional Laboratory, pp. 48—62.
Harper J. L., 1974. Agricultural ecosystems, Agro-E co systems, 1, 1—6.
Harper J. L., 1977. Population Biology of Plants, New York and London, Aca-
demic Press, 892 pp. (Paperback edition, 1981.)
Harper J. L., 1977a. The contributions of terrestrial plant studies to the de-
velopment of the theory of ecology. In: Changing Scenes in Natural
Science, С. E. Goulden, ed. Special Publication 12, Academy of Natural
Sciences, Philadelphia, Lancaster, Pa., Fulton Press, pp. 139—157.
Harper J. L., Clatworthy J. N., 1963. The comparative biology of closely re-
lated species. VI, Analysis of the growth of Trifolium repens and T, fra-
gifesum in pure and mixed populations, J. Exp. Bot., 14, 172—190.
Harper J. L., White J., 1974. The demography of plants, Ann. Rev. Ecol. Syst..
5, 419—463.
Harris E., 1959. The nitrogen cycle in Long Island Sound, Bull. Bingham
Oceanogr. Coll., 17, 31—65.
Harris P., 1974. A possible explanation of plant yield increases following
insect damage, YgroEcosyslems, 1, 219—225.
Harrison A. D., 1962. Hydrobiological studies of all saline and acid still
waters in Western Cape Province, Trans. R. Soc. S. Afr., 36, 213.
Hart J. S.. 1952. Lethal temperatures of fish from different latitudes, Publ.
Ontario Fish. Res. Lab., 72, 1—79.
322 Литература
Hart М. L. (ed.), 1962. Fundamentals of dry-matter production and distributi-
on, Neth. J. Agr. Sci., 10, 309—444 (special issue).
Hartenstein R., 1981. Sludge decomposition and stabilization, Science, 212,
743—749.
Harvey H. W., 1950. On the production of living matter in the sea off Ply-
mouth, J. Marine Biol. Assoc. U. K. n. s., 29, 97—137.
Harwell M. A., Cropper W. P., Ragsdale H. L., 1977. Nutrient cycling and
stability: A reevaluation, Ecology, 58, 660—666.
Haskell E. F., 1940. Mathematical systemization of ’’environment," ’’organism"
and ’’habitat.", Ecology, 21, 1—16.
Haskell E. F.3 1942. The religious force of a unified science, Sci Monthly, 54,
545-551.
Haskell E. F., 1949. A clarification of social science, Main Currents in Modern
Thought, 7, 45—51.
Haskell E. F., 1972. Full Circle: The Moral Force of Unified Science, New
York, Gordon and Breach.
Hasler Л. D., 1947. Eutrophication of lakes by domestic drainage, Ecology, 28,
383—395.
Hasler A. D., 1969. Cultural eutrophication is reversible, BioScience, 19, 425—
431.
Hasler A. D. (ed.), 1975. Coupling of Land and Water Systems, New York,
Springer-Verlag, 309 pp.
Hawkes H. A., 1963. The Ecology of Waste Water Treatment, New York,
Pergamon Press, 203 pp.
Hawkins A. S,, 1940. A wildlife history of Faville Grove, Wisconsin. Trans.
Wise. Acad. Sci. Arts Letters, 32, 39—65.
Ha wryly shy n B., 1980. Road Maps to the Future: Toward More Effective
Societies, A Report to the Club of Rome, New York, Pergamon Press.
Hayes F. R., Coffin С. C., 1951. Radioactive phosphorus and exchange of lake
nutrients, Endeavour, 10, 78—81.
Hazard T. P.3 Eddy R. E., 1950. Modification of the sexual cycle in the brook
trout (Salvelinus fontinalis) by control of light, Trans. Am. Fish. Soc., 80,
158—162.
Heald E. J., Odum W. E., 1970. The contribution of mangrove swamps to
Florida fisheries, Proc. Gulf Carib. Fish. Inst., 22, 130—135.
Heath R. T., 1979. Holistic study of an aquatic microcosm: Theoretical and
practical implications, Int. J. Environ. Studies, 13(2), 87—93.
Heatwold IL, Levine K., 1972. Trophic structure, stability and faunal change
during recolonization, Ecology, 53, 531—534.
Hedgpeth J. W. (ed.), 1957. Treatise on Marine Ecology and Paleoecology,
Vol. 1, Ecology, New York, The Geological Society of America, 1296 pp.
Hegner R., 1938. Big Fleas Have Little Fleas, or Who’s Who Among the Pro-
tozoa, Baltimore, Williams and Wilkins.
Heinrich B., 1979. Bumblebee Economics, Cambridge, Mass., Harvard Univer-
sity Press, 224 pp.
Heinrich B., 1980. The role of energetics in bumblebee-flower interrelation-
ships. In: Coevolution of Animals and Plants, L. E. Gilbert a nd P. H. Ra-
ven, eds. Austin, University of Texas Press, pp. 141—158.
Heller A. (ed.), 1972. The California Tomorrow Plan, Los Altos, Calif., William
Kaufmann.
Henderson G. S., 1975. Letter to editor, BioScience, 25, 770.
Henderson IL, 1978. Creating Alternate Futures: The End of Economics, New
York, Berkley, 403 pp.
Henderson J. V., 1974. Optimum city size: The external diseconomy questions,
J. Polit. Econ., Chicago, (82) n. r. 2, p. 373.
Henle P., 1942. The status of emergence, J. Philos., 39, 483—493.
Henry S. M. (ed.), 1966. Symbiosis, Two volumes, New York, Academic Press.,
Vol. 1, 478 pp.; Vol. 2, 400 pp.
Литература
323
Herrera R., Jordan C. F., Klinge H., Medina E., 1978. Amazon ecosystems.
Their structure and functioning with particular emphasis on nutrients,
Intersciencia, 3, 223—232.
Hibbert A. R., 1967. Forest treatment effects on water yield. In: International
Symposium on Forest Hydrology, W. E. Sopper and H. W. Lull, eds.,
New York, Pergamon Press, pp. 527—543.
Hickey J. J., Anderson D. W., 1968. Chlorinated hydrocarbons and egg shell chan-
ges in raptorial and fish-eating birds, Science, 162, 271—272.
Rickling C. F., 1948. Fish farming in the Middle and Far East, Nature, 161,
743—751.
Hill J., Durham S. L., 1978. Input, signals and control in ecosystems. In: Proc.
1978 Conference on Acoustics, Speech and Signal Processing. Tulsa, Okla,
New York, Inst. Electrical and Electronics Engineers, pp. 391—397.
Hill J., Wiegert R. G., 1980. Microcosms in ecological modelling. In: Micro-
cosms in Ecological Research, J. P. Giesy, ed. U. S. Dept, of Energy
Symp. 52, Springfield, Va., National Technical Information Service,
pp. 138—163.
Hills G. A., 1952. The classification and evaluation of site for forestry, Res.
Rep. No. 24, Ontario Dept, of Lands and Forests.
Hirsch F., 1978. Social Limits to Growth, Cambridge, Mass., Harvard Univer-
sity Press, 208 pp.
Hjort J., 1926. Fluctuations in the year classes of important food fishes,
J. du Conseil Permanent Internationale pour 1’Exploration de la Мег, 1,
1—38.
Ho M. W., Saunders P. T., 1979. Beyond neo-Darwinism, an epigenetic appro-
ach to evolution, J. Theoret. Biol., 78, 573—591.
Hoch I., 1976. City size: Effects, trends and policies, Science, 193, 856—863.
Holdgate M. W., Woodman M. J. (eds.), 1978. The Breakdown and Restoration
of Ecosystems, New York, Plenum, 496 pp.
Holdren J. P., Ehrlich P. R., 1974. Human population and the global environ-
ment, Am. Sci., 62, 282—292.
Holeman J. N„ 1968. The sediment yield of major rivers of the world. Water
Res., 4, 737—747.
Rolling C. S., 1961. Principles of insect predation, Ann. Rev. Entomol., 6
163—182.
Rolling C. S., 1965. The functional response of predators to prey density and
its role in mimicry and population regulation, Mem. Entomol. Soc. Ca-
nada, No. 45, 60 pp.
Rolling C. S., 1966. The functional response of invertebrate predators to prey
density, Mem. Entomol. Soc. Canada, No. 48, 86 pp.
Rolling C. S., 1973. Resilience and stability of ecological systems, Ann. Rev.
Ecol. Syst., 4, 1—23.
Rolling C. S. (ed.), 1978. Adaptive Environmental Assessment and Manage-
ment, New York, Wiley-Interscience.
Rolling C. S., 1980. Forest insects, forest fires, and resilience. In: Fire Regi-
mes and Ecosystem Properties, H. Mooney, J. M. Bonnicksen, N. L. Chris-
tensen, J. E. Lotan, and W. E. Reiners, eds. USDA Forest Service Gen.
Tech. Rep., WO-26.
Holt S. J., 1969. The food resources of the ocean, Sci. Am., 221(3), 178—194.
Hopkinson C. S., Day J. W., 1980. Net energy analysis of alcohol production
from sugarcance, Science, 207, 302—304.
Horn H. S., 1974. The ecology of secondary succession, Ann. Rev. Ecol. Syst.,
5 25_______37
HornH. S., 1975. Forest succession, Sci. Am., 232, 90—98.
Horn H. S., 1981. Succession. In: Theoretical Ecology (2nd ed.), R. M. May,
ed. Sunderland, Mass., Sinauer Associates.
Howarth R. W., Teal J. M., 1979. Sulfate reduction in a New England salt
marsh, Limnol. Oceanogr., 24, 999—1113.
324
Литература
Howarth R. W., Teal J. M., 1980. Energy flow in a salt marsh ecosystem:
The role of reduced inorganic sulfur compounds, Am. Nat., 116, 862—
872.
Hubbell S. P., 1979. Tree dispersion, abundance and diversity in a tropical
dry forest, Science, 203, 1299—1309.
Huber D. M., Warren H. L., Nelson D. W., Tsai C. Y., 1977. Nitrification in-
hibitors — new tools for food production, BioScience, 27, 523—529.
Huettner D. A., 1976. Net energy analysis: An economic assessment, Science,
192, 101—104.
Huff a her С. B., 1957. Fundamentals of biological control of weeds, Hilgardia,
27, 101—167.
Huffaker С. B., 1959. Biological control of weeds with insects, Ann. Rev.
Entomol., 4, 251—276.
Huffaker С. B., Kennett C. E., 1956. Experimental studies on predation: Pre-
dation and cyclamen-mite populations on strawberries in California,
Hilgardia, 26, 191—222.
Hulbert M. K., 1971. The energy resources of the earth, Sci. Am., 224(3).
Humphreys W. F., 1978. Ecological energetics of Geolycosa godeffroyi (Aru-
neae: Lycosidae) with an appraisal of production efficiency of ectother-
mic animals, J. Anim. Ecol., 47, 627—652.
Hungate F. P., 1963. Symbiotic associations: The rumen bacteria. In: Symbiotic
Associations, 13th Symp. Soc. Gen. Microbiol., New York, Cambridge
University Press, pp. 266—297.
Hungate F, P., 1966. The Rumen and Ils Microbes. New York. Academic
Press, 533 pp.
Hungate F. P., 1975. The rumen microbial ecosystem, Ann. Rev. Ecol. Syst.,
6, 39—66.
Hunter W. S., Vernberg IE. B., 1955. Studies on oxygen consumption of dige-
netic trematodes. 11. Effects of two extremes in oxygen tension, Exp.
Parasit., 4, 427—434.
Huston M., 1979. A general hypothesis of species diversity, Am. Nat., 113,
81—101
Hutcheson K., 1970. A test for comparing diversities based on the Shannon
formula, .1. Theoret. Biol., 29, 151—154.
Hutchinson G. E.. 1944. Nitrogen and biogeochemistry of the atmosphere, Am.
Sci., 32, 178—195.
Hutchinson G. E., 1948. On living in the biosphere, Sci, Monthly, 67, 393—
398.
Hutchinson G. E., 1948a. Circular causal systems in ecology, Ann. N. Y. Acad.
Sci., 50, 221—246.
Hutch inson G. E., 1950. Survey of contemporary knowledge of biogeochemistry.
HI The biogeochemistry of vertebrate excretion, Bull. Am. Mus. Nat.
Hist., 95, 554.
Hutchinson G. E., 1953. The concept of pattern in ecology, Proc. Acad. Nat.
Sci. (Phila.). 105, 1—12.
Hutchinson G. E., 1957. A Treatise on Limnology. Vol. I, Geography, Physics
and Chemistry, New York, John Wiley and Sons, 1015 pp.
Hutchinson G. E.. 1957a. Concluding remarks, Cold Spring Harbor Symp.
Quant. Biol., 22, 415—427.
Hutchinson G. E., 1959. Homage to Santa Rosalina. or why are there so many
kinds of animals?. Am. Nat., 93. 145—159.
Hutchinson G. E., 1961. The paradox of the plankton, Am. Nat., 107, 406—
425.
Hutchinson G. E.. 1964. The lacustrine microcosm reconsidered, Am. Sci., 52
331—341.
Hutchinson G. E.. 1965. The niche: An abstractly inhabited hyper-volume.
In: The Ecological Theatre and the Evolutionary Play, New Haven, Yale
University Press, pp. 26—78.
Литература
325
Hutchinson G. E., 1967. A Treatise on Limnology, Vol. II, Introduction to Lake
Biology and the Limnoplankton, New York, John Wiley and Sons, 1115 pp.
Hutchinson G. E., 1967a. Ecological biology in relation to the maintenance
and improvement ot* the human environment. In: Applied Science and
Technical Progress, Proc. Natl. Acad. Sci. USA, pp. 171—184.
Hutchinson G. E., 1978. An Introduction to Population Biology, New Haven,
Yale University Press, 260 pp.
Hutchinson G. E., Bowen V. T., 1948. A direct demonstration of phosphorus
cycle in a small lake, Proc. Natl. Acad. Sci. USA, 33, 148—153.
Hutner S. H., Provasoli L., 1964. Nutrition of algae, Am. Rev. Plant Physiol.,
15, 37—56.
Huxley J., 1935. Chemical regulation and the hormone concept, Biol. Rev., 10,
427.
Huxley T. H., 1863. Evidence as to Man’s Place in Nature. (Reprinted with
an introduction by Ashley Montagu, Ann Arbor Paperbacks, University
of Michigan Press, 1959, 184 pp.)
Hynes H. B. N., 1960. The Biology of Polluted Waters, Liverpool, Liverpool
University Press, 202 pp.
Ito У., 1959. A comparative study on survivorship curves for natural insect
populations, Jpn. J. Ecol., 9, 107—115.
Ito У., 1971. Some notes on the competitive exclusion principle, Res. Pop.
Ecol., XIII, 46-54.
Ivlev V. S., 1945. The biological productivity of waters. In. Uspekhi Soure-
minnoi Biologii (Advances in Modern Biology), 19, 98—120. (In Russian;
English translation by W. E. Ricker.) J. Fish. Res. Bd. Can., 23, 1727—
1759,. 1966.
Jackson J. В. C., 1968. Bivalves; spatial and size-frequency distributions of
two intertidal species, Science, 161, 479—480.
Jackson W., Bender M., 1981. New roots for American agriculture, J. Soil
Water Cons., Nov. — Dec., pp. 320—324.
Jahoda M., 1982. Wholes and parts; meaning and mechanisms, Nature, 296,
8—14.
Jannasch H. W., 1969. Estimations of bacterial growth rates in natural waters,
J. Bacterio!., 99, 156—160.
Jannasch H. W., Jones G. E., 1959. Bacterial populations in seawater as deter-
mined by different methods of enumeration, Limnol. Oceanogr., 4, 128—
138.
Jansen D. H., 1966. Coevolution of mutualism between ants and acacias in
Central America, Evolution, 20, 249—275.
Jansen D. H., 1967. Interaction of the bull’s horn acacia (Acacia cornigera L.)
with an ant inhabitant (Pseudomyrmex fermginea F. Smith) in eastern
Mexico, Univ. Kansas Sci. Bull., 57, 315—558. (See also Ecology, 48, 26—
35, 1967.)
Jantsch E., 1980. The Self-Organizing Universe, Oxford, Pergamon Press,
343 pp.
lass by A. D., Goldman Ch. R., 1974. A quantitative measure of succession role
and its application to the phytoplankton of lakes, Am. Nat., 108, 688—
693.
Jeffries H. P., 1979. Biochemical correlates of a seasonal change in marine
communities Am. Nat., 113, 643—658.
Jenkins J. IL, Fendley T. T., 1968, The extent of contamination, detention and
health significance of high accumulation of radioactivity in southeastern
game populations, Proc. 22nd Annual Conf. Southeast Assoc. Game and
Fish Commissioners, 22. 89—95.
Jenny H.. 1980. Alcohol or humus?, Science, 209, 444.
Jenny H.. Arkley R. J., Schultz A. M.. 1969. The pygmy forest-podsol ecosys-
tem ami ils dune as^>< noes Г an Mendocino coast, Madrono, 20, 60—75.
326
Литература
Johannes R. E., 1964. Phosphorus excretion in marine animals; microzooplank-
ton and nutrient regeneration, Science, 146,, 923—924.
Johannes R. E., 1965. The influence of marine protozoa on nutrient regenera-
tion, Limnol. Oceanogr., 10, 434—442.
Johannes R. E., 1968. Nutrient regeneration in lakes and oceans. In: Advan-
ces in Microbiology of the Sea, M. Droop and E. J. Ferguson Wood,
eds. Vol. I, New York, Academic Press, pp. 203—213.
Johnson L. E., Albee L. R., Smith R. 0., Moxon A. L., 1951. Cows, calves and
grass, South Dakota Agr. Expt. Sta.. Bull., No. 412.
Johnson M. P., Raven P. H., 1970. Natural regulation of plant species diver-
sity, Evol. Biol., 4, 127—162.
Johnson P. L. (ed.), 1969. Remote Sensing in Ecology, Athens, University of
Georgia Press, 244 pp.
Johnson P. L., 1970. Dynamics of carbon dioxide and productivity in an arctic
biosphere, Ecology, 51, 73—80.
Johnson W. C., Sharpe D. M., 1976. An analysis of forest dynamics in the
northern Georgia piedmont, Forest. Sci., 22, 307—322.
Johnston D. W., Odum E. P., 1956. Breeding bird populations in relation to-
plant succession on the Piedmont in Georgia, Ecology, 37, 50—62.
Jones Jt R. E., 1949. A further ecological study of a calcareous stream in the
’’Black Mountain" district of south Wales, J. Anim. Ecol., 18, 142—159.
Jones T. E., 1977. Current prospects of sustainable economic growth. In:
Goals in a Global Community. (The original background papers for
Goals of Mankind, A Report to the Club of Rome.) E. Laszlo and J. Bier-
man, eds. New York, Oxford, Frankfurt, Pergamon Press, pp. 117—179.
Jordan C. F., 1971. A world pattern in plant energetics, Am. Sci., 59(4), 425—
433.
Jordan C. F., Herrera R., 1981. Tropical rain forests: Are nutrients really
critical?, Am. Nat., 117, 167—180.
Jordan C. F., Kline J. R., 1972. Mineral cycling: Some basic concepts and
their application in a tropical rain forest, Ann. Rev. Ecol. Syst, 3, 33—50.
Jordan C. F., Kline J. R., Sasscer D. E., 1972. Relative stability of mineral
cycles in forest ecosystems. Am. Nat, 106, 237—253.
Jorgenson В. B., Fenchel T. M.s 1974. The sulfur cycle of a marine sediment
model system, Marine Biol., 24, 189—201.
Juday C., 1940. The annual energy budget of an inland lake, Ecology, 21,
438—450.
Juday C., 1942. The Summer standing crop of plants and animals in four
Wisconsin lakes, Trans. Wise. Acad. Sci., 34, 103—135.
Junnings T. J., Barkham J. P., 1979. Niche separation in woodland slugs,
Oikos, 33, 127-131.
Kahl M. P.. 1964. The food ecology of the wood stork, Ecol. Monogr., 34,
97—117.'
Kahn A, E., 1966. The tyranny of small decisions: Market failures, imperfec-
tions and the limits of economies, Kyklos, 19, 23—47.
Kahn H., Wiener A., 1967. The Year 2000, New York, William Morrow.
Kahn H., Broun W., Leon M., 1976. The Next 200 years, New York, William
Morrow.
Kale H. W., 1965. Ecology and bioenergetics of the long-billed marsh wren
Telmatodytes palustris griseus (Brewster) in Georgia salt marshes, Publ.
No. 5, Cambridge, Mass.. Nuttall Ornithol. Club, 142 pp.
Kamen M. D., 1953. Discoveries in nitrogen fixation, Sci. Am., 188, 38—42.
Kamen M. D., Gest H., 1949. Evidence for a nitrogenase system in the pho-
tosynthetic bacterium Rh о do spirillum rubrum, Science, 109, 560.
Kardos L. T., 1967. Waste water renovation by the land: A living filter. In:
Agriculture and Quality of Our Environment, Brady, ed. Publ. No. 85,.
Washington, D. C., American Association for the Advancement of Science*
pp. 241—250.
Литература 327
Karl D. M., Wirsen С. О., Jamnasch Н. W., 1980. Deep-sea primary produc-
tion at the Galapagos hydrothermal vents, Science, 207, 1345—1347.
Kaushik N. K., Hynes H. B. N., 1968. Experimental study on the role of au-
tumn—shed leaves in aquatic environments. J. Ecol., 56, 229—243.
Keever C., 1950. Causes of succession on old bields on the Piedmont
North Carolina, Ecol. Monogr., 20, 229—250.
Keever C., 1955. Heterotheca latifolia, a new and aggressive exotic dominant
in Piedmont old-field succession, Ecology, 36, 732—739.
Keith L. B., 1963. Wildlife’s Ten-year Cycle, Madison, University of Wisconsin
Press, 201 pp.
Kellogg Ch. E., Agricultural Development, Soil, Food, People, Work.
Madison, Wis., Soil Sci. Soc. of America, Inc.
Kellogg W. W., Cadle R. D., Allen E. R., Lazrus A. L., Martell E. A, 1972.
The sulfur cycle, Science, 175, 587—596.
Kendeigh S. Ch., 1934. The role of environment in the life of birds, Ecol.
Monogr., 4, 299—417.
Kendeigh S. Ch., 1949. Effect of temperatire and season on energy resources
of the English Sparrow, Auk, 66, 113—127.
Kendeigh S. Ch., 1974. Ecology Englewood Cliffs, N. J., Prentice-Hall, 468 pp.
Kennedy J. S., 1968. The motivation of integrated control, J. Appl. Ecol., 4,
492—499.
Kercher J. R., Shugart H. H., 1975. Trophic structure, effective trophic posi-
tion, and connectivity in food webs, Am. Nat., 109, 191—206.
Kettle well H. B. D., 1956. Further selection experiments on industrial mela-
nism in the Lepidoptera, Heredity, 10, 287—301.
Keyfitz N., Flieger W., 1968. World Population: An Analysis of Vital Data,
Chicago, University of Chicago Press.
Kimball J. W., 1948. Pheasant population characteristics and trends in the
Dakotas, Trans. 13th N. A. Wildl. Conf., 13, 291—314.
Kinne O., 1956. Uber den Einfluss des Salzgehaltes und der Temperatur auf
Wachstum, Form und Vermehrung bei dem Hydroidpolypen Cordylopho-
ral caspia (Pallas), Thecata, Clavidae. Zool. Jahrb., 66, 565—638.
Kira T.. Shidei T., 1967. Primary production and turnover of organic matter
in different forest ecosystems of the western Pacific, Jpn. J. Ecol., 17,
70—87.
Kit ch ell J. F., O’Neill R. V., Webb D., Gallepp G. W., Bartell S. M., Ke-
once J. F., Ausmus B. S., 1979. Consumer regulation of nutrient cycling,
BioScience, 29, 28—34.
Klages К. H. W., 1942. Ecological Crop Geography, New York, Macmillan.
Kleiber M., 1961. The Fire of Life, New York, John Wiley and Sons.
Kleiber M., Dougherty J. E., 1934. The influence of environmental temperature
on the utilization of food energy in baby chicks, J. Gen. Physiol., 17,
701—726.
Klopjer P. H., Hailman J. P., 1967. An Introduction to Animal Behavior;
Ethology’s First Century, Englewood Cliffs, N. J., Prentice-Hall, 297 pp.
Kneese A. V.. 1977. Economics and Environment, New York, Penguin Books.
Koestler A., 1969. Beyond atomism and holism — the concept of holon. In:
Beyond Reductionism, The Alpbach Symposium 1968, London. Hutchinson,
pp 192—232.
Komarek E. F., 1967. Fire and the ecology of man. Proc. 6th Annual Tall
Timbers Fire Ecol. Conf., Tallahassee, Fla., pp. 143—170.
Komarek E. V. (ed.), 1969. Ecological Animal Control by Habitat Management,
Tallahassee. Fla., Tall Timbers Research Station, 244 pp.
Kononova M. M., 1961. Soil Organic Matter: Its Nature, Its Role in Soil
Formation and in Soil Fertility. (Translated from Russian by T. Z. Now-
ankowski and G. A. Greenwood.) New York, Pergamon Press, 450 pp.
Kozlowski D. G., 1968. A critical evaluation of the trophic level concept,
(1) Ecological efficiencies, Ecology, 49, 48—116.
328
Литература
Kozlowski Т. T.f 1964. Water Metabolism in Plants, New York, Harper and
Row.
Kozlowski T. T. (ed.), 1968. Water Deficits and Plant Growth, Two vols,
New York, Academic Press.
Kozlowski T. T., 1980. Impacts of air pollution on forest ecosystems, Bio-
Science, 30, 88—93.
Kozlowski T. T., Ahlgren С. E. (eds.), 1974. Fire and Ecosystems, New York,
Academic Press, 538 pp.
Krebs C. J., 1978. A review of the Chitty hypothesis of population regulation,
Can. J. Zook, 56, 2463-2480.
Krebs J. R., Davies N. B. (eds.), 1981. An Introduction to Behavioral Ecology,
Sunderland, Mass., Sinauer Press.
Krebs C. J., Meyers J. IL, 1974. Population cycles in small mammals, Adv.
Ecol. Res., 8, 267—349.
Krenkel R. A., Parker F. L. (eds.), 1969. Biological Aspects of Thermal Pollu-
tion, Nashville, Tenn., Vanderbilt University Press.
Kuentzel L. E., 1969. Bacteria, carbon dioxide and algal blooms, J. Water
Poll. Control Fed., 41, 1737—1747.
Kuenzler E. J., 1958. Niche relations of three species of Lycosid spiders,
Ecology, 39, 494—500.
Kuenzler E. J., 1961. Phosphorus budget of a mussel population, Limnok
Oceanogr., 6, 400—415.
Kuenzler E. J., 1961a. Structure and energy flow of a mussel population,
Limnok Oceanogr., 6, 191—204.
Kushlan J. A., 1979. Design and management of continental wildlife reserves:
Lesions from the Everglades, Biol. Conserv., 15, 281—290.
Kyke G. H., Pulliam H. P., Charnov E. L., 1977. Optimal foraging: A selective
review of theory and test, Quart. Rev. Biol., 52, 137—154.
Lack D. L., 1945. Ecology of closely related species with special reference to
cormorant (Phalacrocorax carbo) and shag (P. aristotelis), J. Anim. Ecol.,
14, 12—16.
Lack D. L., 1947. Darwin’s Finches, New York, Cambridge University Press.
Lack D. L., 1947. The significance of clutch size. Ibis, 89, 302—352 (continued,
90, 25—45, 1948).
Lack D. L., 1954. The Natural Regulation of Animal Numbers, New York,
Oxford University Press.
Lack D. L., 1966. Population Studies of Birds, Oxford, Clarendon Press,
341 pp.
Lack D. L., 1969. Tit niches in two worlds; or homage to Evelyn Hutchinson,
Am. Nat., 103, 43—49.
Landsberg H. (ed.), 1979. Energy: The Next 20 Years, Report Ford Foundation
and Resources for Future, Cambridge, Mass., Ballinger.
Lange O. L., Kock W., Schulze E. D., 1969. CO2-gas exchange and water rela-
tionships of plants in the Negev Desert at the end of the dry period, Ber.
Dtsch. Bot. Ges., 82, 39—61.
Lange W., 1967. Effect of carbohydrates on the symbiotic growth of plank-
tonic blue-green algae with bacteria, Nature, 215, 2177.
LaPorte L. F. (ed.), 1972. The Earth and Human Affairs, Rept. Comm. Geok
Sci. Nat. Acad. Sci., Nat. Res. Council, San Francisco, Canfield Press.
Larkin P. A., 1963. Interspecific competition and exploitation, J. Fish. Res. Bd.
Canada, 20, 647—678.
Laskey A. R., 1939. A study of nesting eastern bluebirds, Birdbanding, 10,
23—32.
Laszlo E., 1972. The Systems View of the World, New York, George Braziller,
131 pp.
Laszlo E. (ed.), 1977. Goals for Mankind: A Report to the Club of Rome on
the New Horizons of Global Community, New York, Dutton, 374 pp.
Литература
329”
Laszlo Е., 1977а. The Club of Rome of the future vs. the future of the Club
of Rome. In: Goals in a Global Community, E. Laszlo and J. Bierman,
eds., New York, Pergamon Press, pp. 281—285.
Laszlo E., Margenau H., 1972. The emergence of integrating concepts in con-
temporary science, Philos. Sci., 39, 252—259.
Lave L. B., Seskin E. P., 1970. Air pollution and human health, Science, 169.
723—733.
Lawton J. H., 1981. Moose, wolves, daphnia and hydra: On the ecological
efficiency of endotherms and ectothcrms, Am. Nat., 117, 782—783.
Leffler J. W., 1978. Ecosystem responses to stress in aquatic microcosms. In:
Energy and Environmental Stress in Aquatic Systems, J. H. Thorp and
J. W. Gibbons, eds. U. S. Dept, of Energy, Springfield, Va., National
Technical Info. Center.
Leffler J. W., 1980. Microcosmology: Theoretical applications of biological
models. In: Microcosms in Ecological Research, John Giesy, ed. U. S.
Dept, of Energy, Springfield, Va., National Technical Info. Center, pp. 14—
29.
Lemon E. R., 1960. Photosynthesis under field conditions. II. An aerodynamic
method for determining the turbulent carbon dioxide exchange between
the atmosphere and a corn field, Agron. J., 52, 697—703.
Lent С. M, (ed.), 1969. Adaptations of intertidal organisms, Am. Zook, 9.
Leontief W., 1966. Input-Output Economics, New York, Oxford University
Press
Leontief W., 1980. The world economy of the year 2000, Sci. Am., 243(3),
206—231.
Leontief W., 1982. Academic economics, Letter to the editor, Science, 217.
104, 107.
Leopold A., 1933. Game Management, New York, Charles Scribner’s Sons.
Leopold A., 1933a. The conservation ethic, J. Forestry, 31, 634—643.
Leopold A., 1943. Deer irruptions. Wise. Cons. Bull., August 1943, Reprinted
in Wise. Cons. Dept. Publ., 321, 3—11.
Leopold A., 1949. The land ethic. In: A Sand County Almanac, New York,
Oxford University Press.
Leopold L. B., 1974. Water: A Primer, San Francisco, W. H. Freeman.
Leopold L. B., Langbein W. B., 1962. The concept of entropy in landscape
evolution, U. S. Geological Survey Paper 500 A.
Leopold L. B., Maddock Th-, 1954. The Flood Control Controversy: Big Dams,.
Little Dams, and Land Management, New York, The Ronald Press.
Lephowski W., 1979. The social thermodynamics of Ilya Prigogine, Chem.
Engr. News, 57(16), 30—33.
Leslie P. H., 1945. On the use of matrices in certain population mathematics.
Biometrika, 33, 183—212.
Leslie P. H., Park Th., 1949. The intrinsic rate of natural increase of Triboli-
um castaneum Herbst, Ecology, 30, 469—477.
Leslie P. H., Ranson R. M., 1940. The mortality, fertility, and rate of natural
increase of the vole (Microtus agretis) as observed in the laboratory, J.
Anim. Ecol., 9, 27—52.
Levin D. A.. 1976. Alkaloid-bearing plants: An ecogeographic perspective, Am.
Nat, 110, 261—284.
Levin S., Pimentel D., 1981. Selection of intermediate rates of increase in
parasite-host systems, Am. Nat, 117, 308—315.
Levins R., 1966. The strategy of model building in population biology, Am.
Sci., 54, 421—431.
Levins R., 1968. Evolution in Changing Environments, Princeton, N. J., Prin-
ceton University Press.
Lewin J. C., 1963. Heterotrophy in marine diatoms. In: Marine Microbiology,
С. H. Oppenheimer, ed. Springfield, Ill., Charles C. Thomas, pp. 229—
235.
22 Заказ № 1483
330 Литература
Le wont in R. C. (ed.), 1968. Population Biology and Evolution, Syracuse,
N. Y., Syracuse University Press, 205 pp.
Liebig J.f 1840. Chemistry in Its Application to Agriculture and Physiology,
London, Taylor and Walton. (4th ed., 1847.)
Lieth H. (ed.), 1962. Die Stoffproduktion der Pflanzendecke, Stuttgart, Gustav
Fischer Verlag, 156 pp.
Leith H., Whittaker R. H. (eds.), 1975. Primary Productivity of the Biosphere,
New York, Springer-Verlag, 340 pp.
Ligon J. D., 1968. Sexual differences in foraging behavior in two species of
Dendrocopus woodpeckers, Auk, 85, 203—215.
Likens G. E., Bormann F. H., 1974. Linkages between terrestrial and aquatic
ecosystems, BioScience, 24, 447—456.
Likens G. E., Bormann F. H., 1974a. Acid rain: A serious regional environ-
mental problem, Science, 184, 1176—1179.
Likens G. E., Bormann F. IL, Pierce R. S., Eaton J. S., Johnson N. M., 1977.
Biogeochemistry of a Forested Ecosystem, New York, Springer-Verlag,
146 pp.
Likens G. E., Wright R. F., Galloway J. N., Butler Th. J., 1979. Acid rain, Sci.
Am. 241(4), 43—51.
Lloyd M., Ghelardi R. J., 1964. A table for calculating the equitability com-
ponent of species diversity, J. Anim. Ecol., 33, 421—425.
Lorenz K. Z., 1966. Evolution and Modification of Behavior, London, Methuen.
Lotka A, J., 1925. Elements of Physical Biology, Baltimore, Williams and
Wilkins, 460 pp. (Reprinted as Elements of Mathematical Biology, New
York, Dover, 1956.)
Lovelock J. E., 1979. Gaia: A New Look at Life on Earth, New York, Oxford
LTni versify Press, 157 pp.
Lovelock J. E., Epton S. R., 1975. The quest for Gaia, New Scientist, 65,
304—306.
Lovelock J. E., Margulis L., 1973. Atmospheric homeostasis by and for the
biosphere: The Gaia hypothesis, Tellus, 26, 1—10.
Lovins A. B., 1977. Soft Energy Paths, Cambridge, Mass., Ballinger.
Lowe C. IL, Heath W. G., 1969. Behavioral and physiological responses to
temperature in the desert pupfish С у pr inodon maculariusi, Physiol. Zook,
42, 53—59.
Lowry A., 1974. A note on emergence, Mind, 83, 276—277.
Lowry W. P., 1967. The climate of cities, Sci. Am., 217(2), 15—23.
Lucas С. E., 1947. The ecological effects of external metabolites, Biol. Rev.
Cambridge Philos. Soc., 22, 270—295.
Ludwig D., Jones D. D., Hulling C. S., 1978. Qualitative analysis of insect
outbreak systems: The spruce budworm and forest, J. Anim. Ecol., 47,
315—332.
Lugo A. E., Brinson M. H., 1978. Calculations of the value of salt water
wetlands. In: Wetland Functions and Value: The State of Our Understan-
ding, P. E. Greeson, J. R. Clark, and J. E. Clark, eds., Minneapolis, Am.
Water Resources Assoc., pp. 120—130.
Lugo A. E., Farnworth E. G., Pool D., Jerez P. Kaufman G., 1973. The impact
of the leaf cutter ant Attica Colombia on the energy flow of a tropical
wet forest, Ecology, 54, 1292—1301.
Lynch K., 1965. The city as environment, Sci. Am.. 213(3), 209.
Lyons N. L, 1981. Comparing diversity indices based on counts weighted by
biomass or other importance values, Am. Nag, 118, 438—442.
MacArthur R. IL. 1955. Fluctuations of animal populations and a measure
of community stability. Ecology. 36, 533—536
MacArthur R. IL, MMLl. On ' i e relative abundance of bird species. Proc. Natl.
Acad. Sci. USA, 45. 293—295
MacArthur R. IL, 1958. po. nla i«m or< h« у m some warblers of northeastern
con Fcrous fores’. E- J -gy. 39. 5'. 9—619.
Литература 33 Ь
MacArthur R. Н., 1960. On the relative abundance of species, Am. Nat. 94,
25—36.
MacArthur R. H., 1965. Patterns of species diversity, Biol. Rev., 40, 410—
533.
MacArthur R. H., 1968. The theory of the niche. In: Population Biology and
Evolution, R. C. Lewontin, ed. Syracuse, N. Y., Syracuse University Press,
pp. 159—176.
MacArthur R. H., 1972.. Geographical Ecology: Patterns in the Distribution of
Species, New York, Harper and Row, 269 pp.
MacArthur R. H., Connell J., 1966. The Biology of Populations, New York,.
John Wiley and Sons, 200 pp.
MacArthur R. H., Pianka E. R., 1966. On optimal use of a patchy environ-
ment, Am. Nat., 100, 603.
MacArthur R. H., Wilson E. O., 1963. An equilibrium theory of insular zoogeo-
graphy, Evolution, 17, 373—387.
MacArthur R. H., Wilson E. O., 1967. The Theory of Island Biogeography,
Princeton, N. J., Princeton University Press, 208 pp.
MacDonald D. R., 1975. Biological interactions associated with spruce budworm
infestations. In: Ecological Effects of Nuclear War, G. Woodwell, ed.
Brookhaven National Laboratory Publ. No. 917, pp. 61—68.
MacElroy R. D., Averner M. M., 1978. Space ecosynthesis: An approach to
the design of closed ecosystems for use in space, NASA Tech. Memo.
78491, Moffet Field, Calif., National Aeronautics and Space Administration,
Ames Research Center, 31 pp.
MacFadyen A., 1949. The meaning of productivity in biological systems, J.
Anim. Ecol., 17, 75—80.
MacFadyen A., 1963. Animal Ecology: Aims and Methods (2nd ed.), London.
Pitman, 344 pp.
Mackereth F. J. H., 1965. Chemical investigations of lake sediments and their
interpretation, Proc. Soc. London, Series B, 161, 295—309.
MacLulich D. A., 1937. Fluctuations in the numbers of the varying hare
(Lepus americanus), U. of Toronto Studies, Biol. Ser., No. 43.
Major J., 1969. Historical development of the ecosystem concept. In: The Eco-
system Concept in Natural Resource Management, G. M. Van Dyne, ed.
New York, Academic Press, pp. 9—22.
Malthus T. R., 1798. An Essay on the Principle of Population, London, John-
son. (Reprinted in Everyman’s Library, 1914).
Margalej R., 1958. Temporal succession and spatial heterogeneity in phyto-
plankton. In: Perspectives in Marine Biology, Buzzati-Traverso, ed. Ber-
keley, University of California Press, pp. 323—347.
Margalej R., 1958a. Information theory in ecology, Gen. Syst., 3, 36—71.
Margalej R., 1961. Communication of structure in plankton populations, Lim-
nol. Oceanogr., 6(2), 124—128.
Margalej R., 1963. Successions of populations, Adv. Frontiers Plant Sci. (In-
stitute Adv. Sci. and Culture, New Delhi, India), 2, 137—188.
Margalej R., 1963a. On certain unifying principles in ecology, Am. Nat., 97,
357—374.
Margalej R., 1967. Concepts relative to the organization of plankton, Oceanogr.
Marine Biol. Ann. Rev., 5, 257—289.
Margalej R., 1968. Perspectives in Ecological Theory, Chicago, University of
Chicago Press, 112 pp.
Margalej R., 1974. Ecologia, Barcelona, Ediciones Omega.
Margalej R., 1979. The organization of space, Oikos, 33, 152—159.
Margulis L., 1981. Symbiosis in Cell Evolution; Life and Its Environment on
the Early Earth, San Francisco, W. H. Freeman, 419 pp.
Margulis L., 1982. Early Life, Boston, Science Books International, 160 pp.
Margulis L., Lovelock J. E., 1974. Biological modulation of the earth’s atmo-
sphere, Icarus, 21, 471—489.
221
332
Литература
Margulis L., Lovelock J. E., 1975. The atmosphere as circulatory system of
the^ biosphere — The Gaia hypothesis, Goevolulion Quarterly, Summer
Margulis L., Schwartz K., 1982. Five Kingdoms: An Illustrated Guide to the
Phyla of Life on Earth, San Francisco, W. H. Freeman.
Marks G. C., Kozlowski T. T. (eds.), 1973. Eclomycorrhizae, Their Ecology
and Physiology, New York, Academic Press.
Marks P. L., 1974. The role of the pin cherry (Prunus Pennsylvania) in the
maintenance of stability in northern hardwood ecosystems, Ecol. Monogr.,
44, 73-8S.
Marsh G. P., 1864. Man and Nature; or Physical Geography as Modified by
Human Action, Reprinted by Harvard University Press, Cambridge,
Mass., 1965 (D. Lowen thal, cd.). For an evaluation of Marsh’s classic,
see Franklin Russell, Horizon, 10, 17—23, 1968.
Martin M. M., 1970. The biochemical basis of the fungus-attine ant symbiosis,
Science, 1969, 16—20.
Maruyama ML, 1963. The second cybernetics. Deviation-amplifying mutual
causal processes, Am. Sci., 51. 164—179.
Marx D. IL, Ruel de J. L., 1979. Fiber, food, fuel and fungal symbionts, Science,
106, 419—422.
Mason W. IL, Odum E. P., 1969. The effect of coprophagy on retention and
bioelimination of radionuclides of detritus-feeding animals, Proc. 2nd
Nat. Symposium, on Radioecology, D. J. Nelson and F. C. Evans, eds.
U. S. Dept, of Commerce., Springfield, Va., Clearinghouse Fed. Sci. Tech.
Info., pp. 721—724.
Mattson W. J. (ed.), 1977. The Role of Arthropods in Forest Ecosystems,
New York, Springer-Verlag.
Mattson IV. J., Addy A., 1975. Phytophagous insects as regulators of forest
primary production, Science, 190, 515—521.
Mauldin W. P., 1980. Population trends and prospects. In: Science Centennial
Issue, P. H. Abelson and R. Kulstad, eds., Science, 209, 148—157.
May R. M., 1973. Stability and Complexity in Model Ecosystems Monographs
in Population Biology, Princeton, N. J., Princeton University Press, 235 pp.
May R. M., 1979. Production and respiration in animal communities, Nature,
282, 443—444.
May R. M. (ed.), 1981. Theoretical Ecology: Principles and Applications (2nd
ed.), Sunderland, Mass., Sinauer Associates, 489 pp.
Maynard Smith L, 1976. Group selection — a commentary, Quart. Rev. Biol.,
51, 277—283.
McCarthy J. J., Goldman J. C., 1979. Nitrogenous nutrition of marine phyto-
plankton in nulrien [-depleted waters, Science, 203, 670—672.
McCormick F. J., 1969. Effects of ionizing radiation on a pine forest. In:
Proceedings of 2nd National Symposium on Radioecology, D. Nelson and
F. Evans, eds. U. S. Dept, of Commerce, Springfield, Va., Clearinghouse
Fed. Sci. Tech. Info., pp. 78—87.
McCormick F. L, Golley F. B., 1966. Irradiation of natural vegetation — an ex-
perimental facility, procedures and dosimetry, Health Physics. 12, 1467—
1474.
McCullough D. IL. 1979. The George Reserve Deer Herd. Population Ecology
of a K-soIemed Species, Ann Arbor, I Diversity of Michigan Press,
271 pp.
McIIarg I. L., 1969. Design with Nature, Garden City, N. Y., Natural History
Press, 197 pp.
McIntosh R. P. 1967. The coBlinimm concept of vegetation, Bot. Rev., 33,
130—187.
McIntosh R. P., 1975. H. A. Gleason —’’individualistic ecologist,“ 1882—1975,
Bull. Torrey Bot. Club. 102, 253—273.
Литература
333
McIntosh R. P., 1980. The relationship between succession and the recovery
process in ecosystems. In: The Recovery Process in Damaged Ecosystems,
John Cairns, ed. Ann Arbor, Mich., Ann Arbor Sciences, pp. 11—62.
McIntosh R. P., 1980a. The background and some current problems of theore-
tical ecology, Synthese, 43, 195—255.
McLeese D. W1956. Effects of temperature, salinity and oxygen on the sur-
vival of the American lobster, J. Fish. Res. Board Canada, 13, 247—272.
McMillan C., 1956. Nature of the plant community. 1. Uniform garden and
light period studies of five grass taxa in Nebraska, Ecology, 37, 330—
340.
McMillan C., 1969. Ecotypes and ecosystem function, BioScience, 19, 131—
134.
McNab В. K., 1963. Bioenergetics and the determinations of home range size,
Am. Nat., 97, 135—140.
McNaughton S. L, 1966. Ecotype function in the Tijpha community-type,
Ecol. Monogr., 36, 297—325.
McNaughton S. J., 1968. Structure and function in California grasslands. Eco-
logy, 49, 962—972.
McNaughton S. J., 1975. r- and К-selection in Typha, Am. Nat., 129, 251—
261.
McNaugthon S. J., 1976. Serengeti migratory wildebeest: Facilitation of energy
flowT by grazing, Science, 191, 92—91.
McNaughton S. L, 1978. Stability and diversity in grassland communities,
Nature, 279, 351—352.
McNaughton S. L, 1979. Grazing as an optimization process: Grass—ungulate
relationships in the Serengeti, Am. Nat., 113, 691—703.
McNeill S., Lawton J. II., 1970. Annual production and respiration in animal
populations, Nature, 225, 472.
Meadows D. L., 1974. Dynamics of Growth in a Finite World, Cambridge,
Mass. MIT Press.
Meadows D. L., Meadows D. Il. (eds.), 1973. Toward Global Equilibrium:
Collected Papers, Cambridge, Mass., Wright-Allen Press, 358 pp.
Meadows D. H., 1982. Whole earth models and systems, CoEvolution Quarterly,
Summer 1982, pp. 98—108.
Meadows D. H., Meadows D. L., Randers J., Behrens IT. W., 1972. The Limits
to Growth: A Report for the Club of Rome’s Project on the Predicament
of Mankind, New York, Universe Books, 205 pp.
Meadows D. IL, Richardson J., Bruckmann G. (eds.), 1983. Groping in the
Dark; The First Decade of Global Modeling, New York, John Wiley and
Sons.
Meentemeyer V., 1978. Macroclimate and lignin control of litter decomposition
rates, Ecology, 59, 465—472.
Meentemeyer V., Box E. O., Thompson R., 1982. World patterns and amounts
of terrestrial plant litter production, BioScience, 32, 125—128.
Menhinick E. F., 1963. Estimation of insect population density in herbaceous
vegetation with emphasis on removal sweeping, Ecology, 44, 617—622.
Menhinick E. F., 1964. A comparison of some species diversity indices applied
to samples of field insects, Ecology, 45, 858—861.
Menshutkin Г. Г.. 1962. The realization of elementary models for fish popu-
lations on electronic computers (in Russian), Vopr. Ilshtiol., 4, 625—631.
Menzel D. W., 1980. Applying results derived from experimental microcosms
to study of natural pelagic ecosystems. In: Microcosms in Ecological
Research, J. P. Giesy, ed. U. S. Dept, of Energy Symposium No. 52.
Springfield, Va., National Technical Info. Center, pp. 742—752.
Menzel D. W., Hulbert E. M., Ryther J. IL, 1963 The effects of enriching
Sargasso Sea water on the production and species composition of the
phytoplankton, Deep-Sea Res., 10, 209—219.
334
Литература
Menzel D. W., Ryther J. IL. 1961. Nutrients limiting the production of phyto-
plankton in the Sargasso Sea, with special reference to iron, Deep-Sea
Res., 7, 276—281.
Mertz W., 1981. The essential trace elements, Science, 213, 1332—1338.
Mesarovic M., Pestel Ed. (eds.), 1974. Mankind at the Turning Point: The Se-
cond Report to the Club of Rome, New York, Signet, 210 pp.
Mesarovic M. D.f Takahara Y., 1975. General Systems Theory: Mathematical
Foundations, New York, Academic Press.
Metcalf R. L., Luckman W. (eds.), 1975. Introduction to Pest Management,
New York, Wiley-Interscience.
Middleton J. T., 1961. Photochemical air pollution damage to plants, Ann.
Rev. Plant Physiol., 12, 431—448.
Middleton J. T., 1964. Trends in air pollution damage, Arch. Environ. Health,.
8, 19—24.
Miles J., 1979. Vegetation Dynamics, New York, John Wiley and Sons; Lon-
don, Chapman and Hall; 80 pp.
Miller M. W., Berg G. G. (eds.), 1969. Chemical Fallout, Springfield, Ill.,,
Charles C Thomas.
Miller R. S., 1967. Pattern and process in competition, Adv. Ecol. Res., 4,
1—74.
Milthrope F. L., 1956. The Growth of Leaves, London, Butterworth.
Mishan E. J., 1967. The Cost of Economic Growth, London, Stapes Press;
New York, Praeger.
Mishan E. J., 1970. Technology and Growth: The Price We Pay, New York,.
Praeger.
Misra R., Singh J. S., Singh К. P., 1968. A new hypothesis to account for
the opposite trophic-biomass structure on land and in water, Current Sci.
(India), 37, 382—383.
Mitchell R., 1979. The Analysis of Indian Agroecosystems, New Delhi, Inter-
print
Mohr C. 0., 1940. Comparative populations of game, fur and other mammals^
Am. Midi. Nat., 24, 581—584.
Monk C. D., 1966. Ecological importance of root/shoot ratios, Bull. Torrey Bot.
Club, 93, 402—406.
Monk C. D., 1966a. An ecological significance of ever-greenness, Ecology, 47,
504—505.
Monk C. D., 1966b. Effects of short-term gamma irradiation on an old field,
Rad. Bot, 6, 329—335.
Montague C. L., 1980. A natural history of temperate western Atlantic fiddler
crabs (genus Uca) with reference to their impact of the salt marsh, Contr.
Mar. Sci., 23, 25—55.
deMontbrial T1979. Energy: The Countdown, A Report to the Club of Rome,
New York, Pergamon Press.
Monteith J. L., 1962. Measurement and interpretation of carbon dioxide fluxes
in the field, Netherlands J. Agr. Sci., 10 (special issue), 334—346.
Mooney H. A., Ehleringer O. J., Berry L, 1976. Photosynthetic capacity of
in situ Death Valley plants, Carnegie Inst. Year Book, 75, 410—413.
Moore A. W., 1969. Azola: Biology and agronomic significance, Bot. Rev., 35,
17—34.
Moore N. W. (ed.), 1966. Pesticides in the environment and their effect on
wildlife. (A supplement to the Journal of Applied Ecology, Vol. 3.)
Oxford, Blackwell, 312 pp.
Morehouse W„ Sigurdson L, 1977. Science, technology and poverty, Bull. Ato-
mic Sci., 33, 21—28.
Mudd J. B., Kozlowski T. T. (eds.), 1975. Responses of Plants to Air Pollution,
Newr York, Academic Press.
Muller С. H., 1966. The role of chemical inhibition (allelopathy) in vegeta-
tional composition, Bull. Torrey Bot. Club, 93, 332—351.
Литература
335
.Muller С. Н., 1969. Allelopathy as a factor in ecological process, Vegetatio,
18, 348-357.
Muller С. H., Hanawalt R. B., McPherson J. K., 1968. Allelopathic control of
herb growth in the fire cycle of California chaparral, Bull. Torrey Bot.
Club, 95, 225—231.
Muller С. H., Muller W. H., Haines B. L., 1964. Volatile growth inhibitors
produced by aromatic shrubs, Science, 143, 471—473.
Mumford L., 1967. Quality in the control of quantity. In: Natural Resources,
Quality and Quantity, Ciriacy-Wantrup and Parsons, eds. Berkeley, Uni-
versity of California Press, pp. 7—18.
Munson R. D., Doll J. P., 1959. The economics of fertilizer use in crop pro-
duction. In: Advances in Agriculture XI, A. G. Norman, ed.
Murie A., 1944. Dall sheep. In: Wolves of Mount McKinley, Washington,
D. C., Natl. Parks Service Fauna No. 5.
Murphy G. I., 1967. Vital statistics of the Pacific sardine (Sardinops caeru-
lea) and the population consequences, Ecology, 48, 731—736.
Murphy G. I., Isaacs J. D., 1964. Species replacement in marine ecosystems
with reference to the California Current, Calif. Coop. Oceanic Fish Invest.
Marine Res. Comm. Mtg., San Pedro, Doc. VII, 1—6.
Murray B. G., Jr., 1979. Population Dynamics — Alternative Models, New York,
Academic Press, 224 pp.
Muscatine L. C., 1961. Symbiosis in marine and freshwater coelenterates.
In: The Biology of Hydra and Some Other Coelenterates, H. M. Lenhoff
and W. F. Loomis, eds. Miami, University of Miami Press, pp. 255—268.
Muscatine L. C., Porter J., 1977. Reef corals: Mutualistic symbioses adapted
to nutrient-poor environments, BioScience, 27, 454—456.
Myers N., 1979. The Sinking Ark: A New Look at the Problem of Disappearing
Species, Oxford and New York, Pergamon Press.
Nader L., Beckerman S., 1978. Energy as it relates to the quality and style
of life, Ann. Rev. Energy, 3, 28. Palo Alto, Calif. Annual Reviews, Inc.
National Academy of Science, 1966. Waste Management and Control, Publ.
No. 1400, A. Spilhaus, ed., 1966.
National Academy of Science, 1969. Resources and Man, P. E. Cloud, Jr., ed.
San Francisco, W. H. Freeman.
National Academy of Science, 1969. Eutrophication: Causes, Consequences and
Correctives, Int. Symp. Eutrophication, Washington, D. C., 661 pp.
National Academy of Science, 1971. Rapid Population Growth, R. Revelle,
ed. Baltimore, Johns Hopkins Press.
National Academy of Science, 1972. The Earth and Human Affairs, L. F. La-
Porte, ed. San Francisco, Canfield Press.
National Academy of Science, 1973. Geographical perspective and urban prob-
lems, Committee on Geography, Division Earth Sciences, Nat. Res. Coun-
cil, Washington, D. C.
National Academy of Science, 1977. World Food and Nutrition Study; The Po-
tential Contributions of Research, Harrison S. Brown, Chairman of Study
Committee, Washington, D. C., National Research Council, 192 pp. (Also
5 volumes of supporting papers.)
National Academy of Science, 1978. Nitrates: An Environmental Assessment,
Environ. Studies Board. Panel on Nitrates, P. L. Brezonik, Chairman.
Washington, D. C., National Research Council.
National Academy of Science, 1979. Carbon Dioxide and Climate, A Scientifie
Assessment, Washington, D. C., National Research Council.
National Academy of Science, 1981. Testing for the effects of chemical on
ecosy>tems, Committee to Review Methods for Ecotoxicology, John Cairns,
Chairman. Washington, D. C., Commission on Natural Resources, National
Research Council, Nat. Acad. Press.
Neess J. C., 1946. Development and status of pond fertilization in central
Eiir.pe. Trans. Am. Fish. Soc., 76, 335—358.
336
Литература
Nelson D. J., 1967. Microchemical constituents in contemporary and рге-Сот
lumbian clam shells. In: Quaternary Paleoecology, Proc. VII Congress
Int. Assoc. Quaternary Research, Vol. 7, pp. 185—204.
Nelson D. J., Scott D. C., 1962. Role of detritus in the productivity of a rock-
outcrop community in a Piedmont stream, Limnol. Oceanogr., 7, 396—
413.
Nelson T. C., 1955. Chestnut replacement in the southern highlands, Ecology.
36, 353—354.
Newell R., 1965. The role of detritus in the nutrition of two marine deposit
feeders, the prosobranch Hydrobia ulvae and the bivalve Macoma balthica,
Proc. Zool. Soc. London, 144, 25—45.
Newell S. J., Tram,er E. J., 1978. Reproductive strategies in herbaceous plant
communities in succession, Ecology, 59, 228—234.
Newland K., 1980. City limits; emerging constraints on urban growth, World-
watch Paper 38. Washington, D. C., Worldwatch Institute, 31 pp.
Nice M. M., 1941. The role of territory m bird life, Am. Midi. Nat., 26, 441—
487.
Nichols J. D., Conley W., Batt B., Tipton A. R., 1976. Temporally dynamic
reproductive strategies and the concept of r- and А-selection, Am. Nat.,
110, 990—1005.
Nicholson A. J., 1954. An outline of the dynamics of animal populations, Austr.
J. Zool., 2, 9—65.
Nicholson A. J., 1957. The self-adjustment of populations to change, Cold
Spring Harbor Symp. Quant. Biol., 22, 153—173.
Nicholson A. J., 1958. Dynamics of insect populations, Ann. Rev. Entomol., 3,
107—136.
Nicholson S. A., Monk C. D., 1974. Plant species diversity in old-field successi-
on on the Georgia piedmont, Ecology, 55, 1075—1085.
Nicol is G., Prigogine L, 1977. Sell-organization in Non-equilibrium Systems,
New York, John Wiley and Sons.
Nixon S. W., 1969. A synthetic microcosm, Limnol. Oceanogr., 14, 142—145.
Nordhaus W., 1979. The Efficient Use of Energy Resources, New Haven, Yale
LTniversity Press.
Nordhaus W., Tobin J., 1972. Economic Growth: Is Growth Obsolete?, Collo-
quium V, National Bureau of Economic Research, New York, Columbia
University Press, 92 pp.
Norman A. G., 1957. Soil-plant relationships and plant nutrition, Am. J. Bot.„
44, 67—73.
Novikojj A. B., 1945. The concept of integrative levels in biology, Science,
101, 209—215.
Nutman P. S, (ed.), 1976. Symbiotic Nitrogen Fixation, Cambridge, England,
Cambridge University Press, 211 pp.
Nutman P. S., Masse B. (eds.), 1963. Symbiotic Associations, 13th Symposium
for Gen. Microbiology, New York, Cambridge University Press, 356 pp.
Oberlander G. T., 1956. Summer fog precipitation on the San Francisco Pe-
ninsula, Ecology, 37, 851—852.
Odell R., 1980. Environmental Awakening, Washington, D. C., The Conserva-
tion Foundation.
Odum E. P., 1957. The ecosystem approach in the teaching of ecology illustra-
ted with sample class data, Ecology, 38, 531—535.
Odum E. P., 1961. The role of tidal marshes in estuarine production, The Con-
servationist (New York State Conservation Dept., Albany), 15(6), 12—15.
Odum E. P., 1963. Primary and secondary energy flow in relation to ecosys-
tem structure, Proc. XVI Int. Congr. Zool., Washington, D. C., pp. 336—
338.
Odum E. P., 1964. The new ecology, BioScience, 14, 14—16.
Odum E. P., 1968. Energy flow in ecosystems: A historical review, Am. Zool.,
8, 11—18.
Литература
337
Odum Е. Р., 1968а. A research challenge: Evaluating the productivity of coas-
tal and estuarine water, Proc. 2nd Sea Grant, Conf., Grad. School Oceano-
graphy, University of Rhode Island, Newport, pp. 63—64.
Odum E. P., 1969. The strategy of ecosystem development, Science, 164, 262—
270.
Odum E. P., 1969a. Air-land-water = an ecological whole, J. Soil Water Cons.,
24, 4—7.
Odum E. P., 1970. Optimum population and environment: A Georgian micro-
cosm, Curr. Hist., 58, 355—359, 365.
Odum E. P., 1971. Ecological principles and the urban forest. In: Proc. Symp.
on Role of Trees in the South’s Urban Environment. Athens, Ga., Ga.
Center for Continuing Education, pp. 78—81.
Odum E. P., 1972. Ecosystem theory in relation to man. In: Ecosystems:
Structure and Function, 31st Biol. Coll. J. Wiens, ed. Corvallis, Oregon
State University Press.
Odum E. P., 1975. Ecology: The Link Between the Natural and the Social
Sciences (2nd ed.). New York, Holt, Rinehart and Winston, 244 pp.
Odum E. P., 1975a. Harmony between man and nature: An ecological view.
In: Essays on Alternate Futures, Yale University School of Forestry,
Bull. 88, pp. 43—55.
Odum E. P., 1976. Diversity as a function of energy flow. In: Unifying Con-
cepts in Ecology (Proc. 1st International Congress of Ecology), The Ha-
gue, W. Junk Publ.
Odum E. P., 1976a. Energy, ecosystem development and environmental risk,
Risk Insurance, 43(1), 1—16.
Odum E. P., 1977. The emergence of ecology as a new integrative discipline,
Science, 195, 1289—1293.
Odum E. P., 1977a. Ecology—the common sense approach, The Ecologist,
7(7), 250—253.
Odum E. P., 1977b. The life support value of forests. In: Forest for People,
Washington, D. C., Society of American Foresters, pp. 101—105.
Odum E. P., 1978. Ecological importance of the riparian zone. In: Strategies
for Protection of Floodplain Wetlands and Other Riparian Ecosystems,
B. R. Johnson and J. F. McCormick, eds. Gen. Tech. Rept. WO-12,
Washington, D. C., U. S. Forest Service, pp. 2—4.
Odum E. P., 1978a. The value of wetlands: A hierarchical approach. In: Wet-
land Functions and Values: The State of Our Understanding, P. E. Gree-
son, J. R. Clark, and J. E. Clark, eds. Minneapolis, American Water Resources
Association, pp. 16—25.
Odum. E. P., 1979. Rebuttal of “Economic value of natural coastal wetlands:
A critique/4, Coastal Zone Mgmt. Jr., 5, 231—237.
Odum E. P., 1980. The status of three ecosystem-level hypotheses regarding
salt marsh estuaries: Tidal subsidy, outwelling and detritus-based food
chains. In: Estuarine Perspectives, Kimbel, ed. New York, Academic
Press, pp. 485—495.
Odum E. P., 1980a. Radiation ecology at Oak Ridge. In: Environmental Scien-
ces Laboratory Dedication, Oak Ridge, Tenn., Oak Ridge National Labora-
tory, pp. 53—57.
Odum E. P., 1981. The effects of stress on the trajectory of ecological successi-
on. In: Stress Effects on Natural Ecosystems, G. W. Barrett and R. Ro-
senberg, eds. London, John Wiley and Sons, pp. 43—47.
Odum E. P., 1982. Species diversity of whole communities with importance
values weighted according to energy quality, (In press.)
Odum E. P., Bramlett G. A., Ike A., Champlin J. R., Zieman J. C., Shu-
gart H. H., 1976. Totality indices for evaluating environmental impacts
of highway alternates, Transportation Record No. 561, Washington, D. C.,
National Academy of Science, pp. 57—67.
338 Литература
Odum Е. Р., Burleigh Th. D., 1946. Southward invasion in Georgia, Auk, 63
388—401.
Odum E. P., Connell С. E., Davenport L. B., 1962. Population energy flow of
three ^primary consumer components of old-field ecosystems, Ecology, 43,
Odum E. P., Cooley J. L., 1980. Ecosystem profile analysis and performance
curves as tools for assessing environmental impacts. In: Biological Eva-
luation of Environmental Impacts, Washington, D. C., Council on Envi-
ronmental Quality and Fish and Wildlife Service, pp. 94—102.
Odum E. P., de la Cruz A. A., 1963. Detritus as a major component of ecosys-
tems, AIBS Bulletin (now BioScience), 13, 39—40.
Odum E. P., de la Cruz A. A., 1967. Particulate organic detritus in a Georgia
salt marsh-estuarine ecosystem. In: Estuarines, G. Lauff, ed. Am. Assoc.
Adv. Sci. Publ., 83, 383—388.
Odum E. P., Fanning M. E., 1973. Comparison of the productivity of Spartina
alterniflora and S. cynosuroides in Georgia coastal marshes, Bull. Ga.
Acad. Sci., 31, 1—12.
Odum E. P., Finn J. T., Franz E. H., 1979. Perturbation theory and the sub-
sidy-stress gradient, BioScience, 29, 349—352.
Odum E. P., Franz E. H., 1977. Whither the life-support system. In: Growth
Without Ecodisaslers, N. Polunin, ed. London, MacMillan Press, pp. 264—
274.
Odum E. P., Golley F. B., 1963. Radioactive tracers as an aid to the measure-
ment of energy flow at the population level in nature. In: Radioecology,.
V. Schultz and A. W. Element, eds., New York, Reinhold, pp. 403—410.
Odum E. P., Johnston D. W., 1951. The house wren breeding in Georgia:
An analysis of a range extension. Auk, 68, 357—366.
Odum E. P., Kroodsma R. L., 1977. The power park concept: Ameliorating
man’s disorder with nature’s order. In: Thermal Ecology II, G. W. Esch
and R. W. McFarlane, eds. Springfield, Va., National Technical Info.
Service, pp. 1—9.
Odum E. P., Kuenzler E. J.., 1963. Experimental isolation of food chains in
an old-field ecosystem with use of phosphorus-32. In: Radioecology,
V. Schultz and A. W. Element, eds. New York, Reinhold, pp. 113—120.
Odum E. P., Kuenzler E. J., Blunt M. X., 1958. Uptake of P32 and primary
productivity in marine benthic algae, Limnol. Oceanogr., 3, 340—345.
Odum E. P., Odum H. T., 1972. Natural areas as necessary components of
man’s total environment, Proc. 37th N. A. Wildl. and Nat. Res. Conf.
Washington, D. C., Wildlife Mgmt. Institute, pp. 178—189.
Odum E. P., Pomeroy S. E., Dickinson J. C., Hutcheson K., 1973. The effects
of late winter burn on the composition, productivity and diversity of
a 4-year-old fallow field in Georgia. In: Proc. Annual Tall Timbers Fire
Ecol. Conf., E. V. Eomarek, ed. Tallahassee, Fla., Tall Timbers Research
Station.
Odum E. P., Smalley A. E., 1957. Comparison of population energy flow of
a herbivorous and a deposit-feeding invertebrate in a salt marsh ecosys-
tem, Proc. Natl. Acad. Sci., 45, 617—622.
Odum H. T., 1956. Primary production in flowing waters, Limnol. Oceanogr.,
1, 102—117.
Odum H. T1956a. Efficiencies, size of organisms, and community structure,
Ecology, 37, 592—597.
Odum H. T., 1957. Trophic structure and productivity of Silver Springs,
Florida. Ecol. Monogr., 27, 55—112.
Odum H. T., 1960. Ecological potential and analogue circuits for the ecosys-
tem, Am. Sci., 48, 1—8.
Odum H. T„ 1962. Ecological tools and their use — Man and the ecosystem.
In: Proceedings of the Lock wood Conference on the Suburban Forest
Литература 339
and Ecology, Paul E. Waggoner and J. D. Ovington, eds. The Connecticut
Agricultural Experiment Station Bull., 652, pp. 57—75.
Odum H. T., 1963. Limits of remote ecosystems containing man, Am. Biol.
Teacher, 25, 429—443..
Odum H. T., 1967. Biological circuits and the marine systems of Texas. In:
Pollution and Marine Ecology, T. A. Olson and F. J. Burgess, eds.
New York, Wiley Interscience, pp. 99—157.
Odum II. T., 1967a. Energetics of world food production. In: The World Food
Problem. A Report of the President’s Science Advisory Committee, Panel
on World Food Supply (I. L. Bennett, Chairman), Vol. 3, Washington,
D. C., The White House, pp. 55—94.
Odum II. T., 1968. Work circuits and systems stress. In: Symposium on Pri-
mary Productivity and Mineral Cycling in Natural Ecosystems,
H. E. Young, ed. Orono, University of Maine Press, pp. 81—138.
Odum II. T., 1971. Environment, Power and Society, New York, John Wiley
and Sons, 331 pp.
Odum H. T., 1973. Energy, ecology and economics, Ambio, 2(6), 220—227.
Odum H. T., 1975. Energy quality and carrying capacity of the Earth, Trop.
Ecol, 16(1), 1—8.
Odum H. T., 1978. Energy analysis, energy quality, and environment. In:
Energy Analysis: A New Public Policy Tool, Martha Gilliland, ed. AAAS
Selected Symp, 9, Washington, D. C, American Association for the Ad-
vancement of Science, pp. 55—87.
Odum H. T., 1982. Systems Ecology, New York, John Wiley and Sons.
Odum II. T., Cantion J. E., Kornicker L. S., 1960. An organizational hierarchy
postulate for the interpretation of species-individuals distribution, species
entropy and ecosystem evolution and the meaning of a species-variety
index, Ecology, 41, 395—399.
Odum H. T., Hoskin Ch. M., 1957. Metabolism of a laboratory stream micro-
cosm, University of Texas Publ. Inst. Marine Sci, 4, 115—133.
Odum H. T., McConnell W. M., Abbott W., 1958. The chlorophyll ”A“ of
communities, University of Texas Publ. Inst. Marine Sci, 5, 65—97.
Odum II. T., Odum E. C., 1981. Energy Basis for Man and Nature (2nd ed.),
New York, McGraw-Hill, 337 pp.
Odum H. T., Odum E. P., 1955. Tropic structure and productivity of a wind-
ward coral reef community on Eniwetok Atoll, Ecol. Monogr, 25, 291—
320.
Odum H. T., Pigeon B. F. (eds.), 1970. A tropical Rain Forest. A study of
irradiation and ecology at El Verge, Puerto Rico, Springfield, Va, Natio-
nal Technical Info. Service, 1678 pp.
Odum H. T., Pinkerton R. C., 1955. Times speed regulator, the optimum effi-
ciency for maximum output in physical and biological systems, Am. Sci,
43, 331—343.
Odum H. W., 1936. Southern Regions of the United States. Chapel Hill, Uni-
versity of North Carolina Press, 664 pp.
Odum H. W., 1951. The promise of regionalism. In: Regionalism in America,
Merrill Jensen, ed. Madison, University of Wisconsin Press.
Odum H. W., Moore H. E., 1938. American Regionalism, New York, Henry
Holt and Co.
Odum W. E., 1968. Pesticide pollution in estuaries, Sea Frontiers (Internatio-
nal Oceanographic Foundation, Miami, Fla.), 14, 234—245.
Odum W. E., 1970. Insidious alteration of the estuarine environment, Trans.
Am. Fish. Soc, 99, 836—847.
Odum W. E., Heald E J., 1972. Tropic analysis of an estuarine mangrove
community, Bull. Marine Sci, 22(3), 671—738.
Odum W. E., Heald E. J., 1975. The detritus-based food web of an estuarine
mangrove community. In: Estuarine Research, G. E. Cronin, ed. Vol. 1,
New York, Academic Press, pp. 265—286.
340 Литература
Odum W. E., Woodwell G. M., Wurster C. F., 1969. DDT residues absorbed
from organic detritus by fiddler crabs, Science, 164, 576—577.
Office of Technology Assessment (OTA), U. S. Congress, 1982. Global Models,
World Futures and Public Policy, A Critique, Washington, D. C., Office
of Technology Assessment.
Ogburn W. F., 1922. Social change with respect to culture and original na-
ture, New York, Henbsch.
Oliver C. D., 1981. Forest development in North America following major
disturbances, Forest Ecol. Mgmt., 3, 153—168.
Oliver C. D., Stephens E. P., 1977. Reconstruction of a mixed-species forest in
central New England, Ecology, 58, 562—572.
Olson J. S., 1958. Rates of succession and soil changes on southern Lake
Michigan sand dunes, Bot. Gaz., 119, 125—170.
Olson J. S., 1963. Energy storage and the balance of producers and decom-
posers in ecological systems, Ecology, 44, 322—332.
Olson J. S., 1964. Gross and net production of terrestrial vegetation, J. Ecol.,
62, 99—118.
Omernik J. M., 1977. Non-point source stream nutrient level relationship:
A nationwide study, EPA Ecol. Res. Ser. 600/3-77-105, Corvallis, Oregon.
O'Neill G. K., 1977. The High Frontier; Human Colonies in Space, New York,
William Morrow, 288 pp.
O'Neill R. V., 1967. Niche segregation in seven species of diplopods, Ecology,
48, 983.
O'Neill R. V., 1976. Ecosystem persistence and heterotrophic regulation, Eco-
logy, 57, 1244—1253.
Oosting H. J., 1942. An ecological analysis of the plant communities of Pied-
mont, North Carolina, Am. Midi. Nat., 28, 126.
Oosting H. L, 1956. The Study of the Plant Communities (2nd ed.), San Fran-
cisco, W. H. Freeman.
Oparin A. I., 1938. The Origin of Life, New York, Macmillan.
Ophel I. L., 1963. The fate of radio strontium in a freshwater community.
In: Radioecology, V. Schultz and W. Klement, eds., New York, Reinhold,
pp. 213—216.
Osborn F., 1948. Our Plundered Planet, Boston, Little, Brown.
Overgaard-Neilsen C., 1949. Feeliving nematodes and soil microbiology, Proc.
4 th I nt. Congr. Microbiology, Copenhagen, 1947, pp. 283—284.
Ovington J. D., 1957. Dry matter production by Pinus sylvestris, Ann. Bot
n. s., 21, 287—314.
Ovington J. D., 1961. Some aspects of energy flow in plantations of Pinus
sylvestris, Ann. Bot. n. s., 25, 12—20.
Ovington J. D., 1962. Quantitative ecology and the woodland ecosystem con-
cept. In: Advances in Ecological Research, J. B. Cragg, ed. Vol. 1, New York,
Academic Press, pp. 103—192.
Ovington J. D., 1965. Organic production, turnover and mineral cycling in
woodlands, Biol. Rev., Cambridge Phil. Soc., 40, 295—336.
Owen D. F., Wiegert R. G., 1976. Do consumers maximize plant fitness?,
Oikos, 27, 489—492.
Page T., 1977. Conservation and Economic Efficiency, A Resources for the Fu-
ture Book, Baltimore, Johns Hopkins Press.
Paine R. T., 1966. Food web diversity and species diversity, Am. Nat., 100.
Palmgren P., 1949. Some remarks on the short-term fluctuations in the num-
bers of northern birds and mammals, Oikos, 1, 114—121.
Paris О. H., Pitelka F. A., 1962. Population characteristics of the terrestrial
isopod Armedillidium vulgare in California grassland, Ecology, 43, 229—
248.
Park Th., 1934. Studies in population physiology: Effect of conditioned flour
upon the productivity and population decline of Tribolium confusum*
J. Exp. Zook, 68, 167-182.
Литература
34Г
Park Th., 1954. Experimental studies of interspecific competition. (I, Tempe-
rature, humidity and competition in two species of Tribolium, PhvsioL
Zool, 27, 177—238.
Parker J. R., 1930. Some effects of temperature and moisture upon M elan о plus
mexicanus and Cannula pellucida Scudder (Orthoptera), Bull. Univ. Mon-
tana Agr. Exp. Sta., 223, 1—132.
Patrick R,, 1954. Diatoms as an indication of river change, Proc. 9th Industrial
Waste Conference, Purdue University Engineering Extension Series. 87,
325—330.
Patrick R., 1967. Diatom communities in estuaries. In: Estuaries, G. H. Lauff,
ed. Publication No. 83, Washington, D. C.. American Association for
the Advancement of Science, pp. 311—315.
Patten R. C., 1963. Plankton: Optimum diversity structure of a summer com-
munity, Science, 140, 894—898.
Patten В. C., 1963a. Information processing behavior of a natural plankton
community, Am. Biol. Teacher, 25, 489—501.
Patten В. C., 1966. Systems ecology; a course sequence in mathematical ecolo-
gy, BioScience, 16, 593—598.
Patten В. C. (ed.), 1971. Systems Analysis and Simulation in Ecologv. Vol. I,
New York, Academic Press. (See also Vol. II, 1972; Vol. Ill, 1974; Vol. IV,
1976.)
Patten В. C., 1978. Systems approach to the concept of environment. Ohio J.
Sci., 78, 206—222.
Patten В. C., 1981. Environs: The superniches of ecosystems, Am. Zool., 21,
845—852. (See also Am. Nat., 119, 179—219, 1982).
Patten В. C., Anuble G T1981. The systems theory of the ecological niche,
Am. Nat, 118, 345—369.
Patten В. C., Odum E. P., 1981. The cybernetic nature of ecosystems, Am.
Nat, 118, 886—895.
Patten В. C., Witkamp M., 1967. Systems analysis of 134cesium kinetics in
terrestrial microcosms, Ecology, 48, 813—824.
Patton D. R., 1975. A diversity index for quantifying habitat ”edge.“, WildL
Soc. Bull, 3, 171—173.
Paul E. A., Kucey R. M. N., 1981. Carbon flow in plant microbial associations,
Science, 213, 473—474.
Payne W. J., 1970. Energy yields and growth of heterotrophs, Ann. Rev.
Microbiol, 24, 17—51.
Payne W. J., 1981. Denitrification, New York, John Wiley and Sons.
Peakail D. B., 1967. Pesticide-induced enzyme breakdown of steroids in birds,
Nature, 216, 505—506.
Peakail D. B., Witt P. N., 1976. The energy budget of an orb web-building
spider, Comp. Biochem. Physiol.. 54A, 187—190.
Pearl R., 1927. The growth of populations, Quart. Rev. Biol, 2. 532—548.
Pearl R., Parker S. L., 1921. Epberimental studies on the duration of life:
Introductory discussion of the duration of life in Drosophila, Am. Nat,
55, 481—509.
Pearl R., Reed L. J., 1930. On the rate of growth of the population of the Uni-
ted States since 1790 and its mathematical representation, Proc. Natl.
Acad. Sci, (Wash.) 6, 275-288.
Pearse A. S., 1939. Animal Ecology (2nd ed.), New York, McGraw-Hill.
Pearse P. H., 1968. A new approach to the evaluation of non-priced recrea-
tional resources. Land Economics, 44(1), 87—99.
Peet R. K., 1974. The measurement of species diversity, Ann. Rev. Ecol. Syst,
5, 285—308.
Peet R. K., 1978. Ecosystem convergence, Am. Nat, 112, 441—444.
Penfound W. T., 1956. Primary production of vascular aquatic plants, Limnol.
Oceanogr, 1, 92—101.
542 Литература
Penman Н. L., 1956. Weather and water in the growth of grass. In: The
Growth of Leaves, F. L. Milthrope, ed. London, Butterworth.
Peterman R. M., 1978. The ecological role of mountain pine beetle in lod-
gepole pine forests, and the insect as a management tool. In: Mountain
Pine Beetle Management in Lodgepole Pine Forests, Berryman, Stark,
and Amman, eds. Moscow, University of Idaho Press.
Peters G. A., 1978. Blue-green algae and algal associations, BioScience, 28,
580—585.
Peterson B. J. et al., 1980. Salt marsh detritus: An alternative interpretation
of stable carbon isotope ratios and the fate of Spartina alterniflora, Oikos,
34, 173—177.
Petrides G. A., 1950. The determination of sex and age ratios in fur animals,
Am. Midi. Nat., 43, 355—382.
.Petrides G. A., 1956. Big game densities and range carrying capacity in East
Africa, N. A. Wildl. Conf. Trans., 21, 525—537.
Petrusewicz K. (ed.), 1967. Secondary productivity of terrestrial ecosystems,
Inst. Ecol. Polish Acad. Sci. Int. Biol. Program (Warsaw). Vol. I, 367 pp.;
Vol. II, 879 pp.
.Pfennig N., 1967. Photosynthetic bacteria, Ann. Rev. Microbiol., 21, 285’—
324.
Philip ]. P., 1955. Note on the mathematical theory of animal population
dynamics and a recent fallacy, Austr. J. Zool., 3, 287—294.
Phillips E. A., 1959. Methods of Vegetation Study, New York, Holt, Rinehart
and Winston.
Pianka E. R., 1967. Lizard species diversity, Ecology, 48, 333—351.
Pianka E. R., 1970. On r- and А-selection, Am. Nat., 104, 592—597.
Pianka E. R., 1978. Evolutionary Ecology (2nd ed.), New York, Harper and
Row.
Pickett S. T. A., 1976. Succession: An evolutionary interpretation., Am. Nat.,
110, 107—119.
Picozzi N., 1968. Grouse bags in relation to the management and geology of
heather moors, J. Appl. Ecol., 5, 483—488.
Pielou E. C., 1966. The measurement of diversity in different types of biolo-
gical collections, L Theoret. Biol., 13, 131—144.
Pielou E. C., 1966a. Species-diversity and pattern-diversity in the study of
ecological succession. J. Theoret. Biol., 10, 370—383.
Pielou E. C., 1975. Ecological Diversity, New York, Wiley-Interscience, 165 pp.
.Pigou A. C., 1920. The Economics of Welfare (4th ed., 1932), London, Mac-
millan, 976 pp.
.Pilson M. E. Q., Nixon S. W., 1980. Marine microcosms in ecological research.
In: Microcosms in Ecological Research, J. P. Giesy, ed. U. S. Dept, of
Energy Symp. No. 52, Springfield, Va., National Technical Info. Center
pp. 724—741.
Pimentel D., 1961. Animal population regulation by the genetic feedback
mechanism, Am. Nat., 95, 65—79.
Pimentel D., 1961a. Species diversity and insect population outbreaks, Ann.
Entomol. Soc. Am., 54, 76—86.
Pimentel D., 1968. Population regulation and genetic feedback, Science, 159,
1432—1437.
.Pimentel D., Hurd L. E., Bellotti A. C., et al., 1973. Food Production and
the energy crisis, Science, 182, 443—449.
Pimentel D., Moran M. A., Fast S., et al., 1981. Biomass energy from crop and
forest residues, Science, 212, 1110—1115.
Pimentel D., Pritschilo W., Krummel J., Kutzman J., 1975. Energy and land
constraints in food protein production, Science, 190, 754—760.
Pimental D., Stone F. A., 1968. Evolution and poulation ecology of parasite-
host systems, Can. Entomol., 100, 655—662.
Литература 343
Pimentel D., Terhume E. C., Dyson-Hudson R., et al., 1976. Land degradation:
Effects on food and energy resources, Science, 194, 149—155.
Pippenger N., 1978. Complexity theory, Sci. Am., 238(6), 114—124.
Pirt S. J., 1957. The oxygen requirement of growing cultures of an Aerobacter
species determined by means of the continuous culture technique, J. Gen.
Microbiol., 16, 59—75.
Pitelka F. A., 1964. The nutrient recovery hypothesis for arctic microtine
cycles. In: Grazing in Terrestrial and Marine Environments, A. J. Crisp,
ed. Oxford, Blackwell.
Pitelka F. A., 1973. Cyclic patterns in lemming populations near Barrow,
Alaska. In: Alaska Arctic Tundra, M. E. Britton, ed. Tech. Paper 25,
Washington, D. C., Arctic Inst. N. Am., pp. 119—215.
Plass G. N., 1955. The carbon dioxide theory of climate change, Tellus, 8,
140—154.
Plass G. N„ 1959. Carbon dioxide and climate, Sci. Am., 20(1), 41—47.
Platt R. B., 1965. Ionizing radiation and homeostasis of ecosystems. In: Eco-
logical Effects of Nuclear War, G. M. Woodwell, ed. Upton, N. Y., Brook-
haven National Laboratory Publ. No. 917, pp. 39—60.
Pollard H. P., Gorenstein S., 1980. Agrarian potential, population, and the Ta-
rascan state, Science, 209, 274—277.
Polumin N. (ed.), 1980. Growth Without Ecodisasters, Proc. 2nd Int. Conf,
on Environ, Future, Iceland, 1977, London, Macmillan Press, 675 pp.
Polynov В. B., 1937. The Cycle of Weathering, Trans, from Russian by Mu-
ir A. Murby, London, 230 pp.
Pomeroy L. R., 1959. Algal productivity in Georgia salt marshes, Limnol.
Oceanogr., 4, 386—397.
Pomeroy L. R., 1960. Residence time of dissolved phosphate in natural waters,
Science, 131, 1731—1732.
Pomeroy L. R., 1970. The strategy of mineral cycling, Ann. Rev. Ecol. Sy st.,
1, 171—190.
Pomeroy L. R. (ed.), 1974. Cycles of Essential Elements, Benchmark Papers
in Ecology, Stroudsburg, Pa., Dowden, Hutchinson and Ross; New York,
Academic Press, 373 pp.
Pomeroy L. R., 1979. Secondary production mechanisms of continental shelf
communities. In: Ecological Processes in Coastal and Marine Systems,
R. J. Livingston, ed. New York, Plenum, pp. 163—186.
Pomeroy L. R., 1980. Detritus and its role as a food source. In: Fundamentals
of Aquatic Ecosystems, Barnes and Mann, eds. Oxford, Blackwell, pp. 84—
192.
Pomeroy L. R., Johannes R. E., Odum E. P., Roffman B., 1969. The phospho-
rus and zinc cycles and productivity of a salt marsh. In: Proc. 2nd Symp.
Radioecology, D. J. Nelson and F. C. Evans, eds. Springfield, Va., Clea-
ringhouse Fed. Sci. Tech. Info., pp. 412—419.
Pomeroy L. R., Mathews H. M., Min H. S., 1963. Excretion of phosphate and
soluble organic phoshporus compounds by zooplankton, Limnol. Oceanogr.,
8, 50—55.
Pomeroy L. R., Odum E. P., Johannes R. E., Roff man B., 1966. Flux of 32P
and 65Zn through a salt marsh ecosystem, Proceedings of the Symposium
on the Dispersal of Radioactive Wastes into Seas, Oceans and Surface
Waters, Vienna, pp. 177—188.
President’s Science Advisory Committee, 1967. The World Food Problem, Three
vols., Washington, D. C., U. S. Government Printing Office.
Preston F. W., 1948. The commonness and rarity of species, Ecology. 29,
254—283.
Preston F. W., 1960. Time and space and the variation of species, Ecology. 41,
611—627.
Prigogine I., 1962. Introduction to Nonequilibrium Thermodynamics, New York,.
John Wiley and Sons.
:344
Литература
Prigogine I., 1978. Time structure and fluctuations, Science, 201, 777—785.
Prigogine I., Nicolis G., Babloyantz A., 1972. Thermodynamics and evolution,
Physics Today, 25(11), 23—38; 25(12), 138—144.
Prigogine N. G., Prigogine I., 1977. Self Organization in Nonequilibrium
Systems, New York, John Wiley and Sons.
Primm S. L., Lawton J. H., 1977. Number of trophic levels in ecological
communities, Nature. 268, 329—331.
Pritchard D. W1967. What is an estuary: Physical viewpoint. In: Estuaries,
G. H. Lauff, ed. Publ. No. 83, Washington, D. C., American Association
for the Advancement of Science, pp. 3—5..
Prosser C. L. (ed.), 1967. Molecular Mechanisms of Temperature Adaptation,
Washington, D. G., American Association for the Advancement of Science.
Provasoli L., 1958. Nutrition and ecology of protozoa and algae, Ann. Rev.
Microbiol., 12, 279—308.
ProvasoU L., 1963. Organic regulation of phytoplankton fertility. In: The Sea,
M. N. Hill, ed. Vol. 2, New York, Wiley Interscience, pp. 165—219.
Pyke G. H., Pulliam H. R., Charnov E. L., 1977. Optimal foraging: A selective
review of theory and tests, Quart. Rev. Biol., 52, 137—154.
Randolph P. A., Randolph J. C., Barlow C. A., 1975. Age-specific energetics
of the pea aphid, Ecology, 56, 359—369.
Randolph P. A., Randolph J. C., Mattingly K., Foster M. M., 1977. Energy
costs of reproduction in the cotton rat, Sigmodon hispidus, Ecology, 58,
31—45.
Rapoport A., Chammah A. M., 1970. Prisoner’s Dilemma, Ann Arbor. Univer-
sity of Michigan Press.
Rappaporl R. A., 1967. Pigs for the Ancestors. New Haven, Yale University
Press.
Rappaport R. A., 1971. The flow of energy in an agricultural society, Sci.
Am., 224(3), 116—132.
.Rasmussen D. I., 1941. Biotic communities of Kaibab Plateau, Arizona. Ecol.
Monogr., 11, 229—275.
Raunkaier C., 1934. The Life Form of Plants and Statistical Plant Geography,
Oxford, Clarendon Press.
Ravera O., 1969. Seasonal variation of the biomass and biocoenotic structure
of plankton of the Bay of Ispra (Lago Maggiore), Verh. Int. Ver. Limnol.,
17, 237—254.
. Rawson D. S., 1952. Mean depth and the fish production of large lakes, Eco-
logy, 33, 513—521.
Reardon P. O., Leinweber C. L., Merrill L. B., 1972. The effect of bovine saliva
on grasses, J. Anim. Sci., 34, 897—898.
Reardon P. O., Leinweber C. L., Merrill L. B., 1974. Response of sideoats gra-
ma to animal saliva and thiamine, J. Range Mgmt., 27, 400—401.
Redfield A. C., 1958. The biological control of chemical factors in the envi-
ronment, Am. Sci., 46, 205—221.
Reichle D. (ed.), 1970. Analyses of Temperate Forest Ecosystems, Heidelberg,
Berlin, Springer-Verlag. 304 pp.
Reichle D. (ed.), 1975. Advances in ecosystem analysis, BioScience, 25, 257—
264.
Reichle D. E., Franklin J. F., Goodall D. W. (eds.), 1975. Productivity of
World Ecosystems, Washington, D. G., National Academy of Science.
Reif A. E., 1981. The causes of cancer, Am. Sci., 69. 437—447.
. Reijsynder W. E., Lull H. W., 1965. Radiant energy in relation to forests,
Tech. Bull. No. 1344, Washington, D. C., U S. Dept, of Agriculture, Forest,
Service, 111 pp.
Rensberger B., 1981. Evolution of evolution, Mosaic (NSF Journal) 12(5),
14—22.
Rensberger B., 1982. Evolution since Darwin, Science-82, 3(3), 41—45.
Литература
345
Revelle R. (ed.), 1966. Population and food supplies: The edge of the knife.
In: Prospects of the World Food Supply: A Symposium, Washington,
D. C., National Academy of Science, pp. 24—47.
Revelle R. (ed ), 1971. Rapid Population Growth, National Academy of Science
Study, Baltimore, Johns Hopkins Press.
Rice E. L., 1974. Allelopathy, New York, Academic Press, 368 pp.
Rice E. L., Pancholy S. K., 1972. Inhibition of nitrification by climax vege-
tation, Am. J. Bot.. 59, 1033—1040; 60, 691—702 (1973); 61, 1094—1103
(1974).
Rich P. H., 1978. Reviewing bioenergetics, BioScience, 28, 80.
Rich P. H., Wetzel R. G., 1978. Detritus in the lake ecosystem, Am. Nat., 112,
57—71.
Richards B. N., 1974. Introduction to the Soil Ecosystem, New York, Longman,
266 pp.
Richmond R. C., Gilpin M. E., Salas S. P., Hyala F. J., 1975. A search for
emergent competitive phenomena: The dynamics of multispecies Drosophi-
la systems, Ecology. 56, 709—714.
Ricker W. E., 1946. Production and utilization of fish populations, Ecol.
Monogr., 16, 373—391.
Ricker W. E., 1958. Handbook of computations for biological statistics of fish
populations, Fish. Res. Bd. Canada Bull., 119, 1—300.
Ricker W. E., Forester R. E., 1948. Computation of fish production, Bull.
Bingham Oceanogr. Coll., 11, 173—211.
Ricklejs R. E., 1970. Clutch size in birds; outcome of opposing predator and
prey adaptations, Science, 168, 599—600.
Rickiefs R. E., 1979. Ecology (2nd ed.), New York, Chiron Press, 966 pp.
Ridker R. G., 1973. To grow or not to grow: That’s not the relevant question,
Science, 182, 1315—1318.
Richert S. E., 1981. The consequences of being territorial: Spiders, a case
study, Am. Nat.. 117, 871—892.
Rifkin J., 1980. Entropy, Entropy, Entropy; A New World Review, New York
The Viking Press, 305 pp.
Rigler F. H., 1961. The uptake and release of inorganic phosphorus by
Daphnia magna Straus, Limnol. Oceanogr., 6, 165—174.
Riley G, A., 1944. The carbon metabolism and photosynthetic efficiency of
the earth, Am. Sci., 32, 132—134.
Riley G. A., 1952. Phytoplankton of Block Island Sound, 1949, Bull. Bingham
Oceanogr. Coll.. 13, 40—64.
Riley G. A., 1956. Oceanography of Long Island Sound, 1952—54. IX. Produc-
tion and utilization of organic matter, Bull. Bingham Oceanogr. Coll.,
15, 324—344.
Riley G. A., 1963. Organic aggregates in sea water and the dynamics of their
formation and utilization, Limnol. Oceanogr., 8. 372—381.
Ripper W. E., 1956. Effect of pesticides on the balance of arthropod popula-
tions, Ann. Rev. Entomol., 1, 403—438.
Robertson G. P., Vitousek P. M., 1981. Nitrification potential in primary and
secondary succession, Ecology, 62, 376—386.
Rodhe W., 1955. Can plankton production proceed during winter darkness in
subarctic lakes?, Proc. Int. Assoc. Theoret. Appl. Limnol., 12, 117—122.
Root R. B., 1967. The niche exploitation pattern of the blue-gray gnatcatcher,
Ecol. Monogr., 37, 317—350.
Root R. B., 1969. The behavior and reproductive success of the blue-gray
gnatcatcher, Condor, 71, 16—31.
Rosenzweig M. L., 1968. Net primary production of terrestrial communities;
prediction from climatological data, Am. Nat., 102, 67—74.
Roughgarden J., 1971. Density-dependent natural selection, Ecology, 52, 453—
468.
23 Заказ № 1483
646 Литература
Rounsefell G. A., 1946. Fish production in lakes as a guide for estimating
production in proposed reservoirs, Copeia, 1946, 29—40.
Rovira A. D., 1965. Interaction between plant roots and soil microorganisms,
Ann. Rev. Microbiol., 19, 241—266.
Rudd R. L, 1964. Pesticides and the Living Landscape, Madison, University
of Wisconsin Press, 320 pp.
Ruehle J. L.f Marx D. H., 1979. Fiber, food, fuel and fungal symbionts,
Science, 206, 419—422.
Russell E. W., 1961. Soil Conditions and Plant Growth (9th ed.). London,
Longmans.
Russell F., 1968. The Vermont Prophet: George Perkins Marsh., Horizon, 10,
17—23.
Ruttner F., 1963. Fundamentals of Limnology (3rd ed.), Frey and Fry, trans.,
Toronto, University of Toronto Press, 295 pp.
Ryther J. H., 1954. The ecology of phytoplankton blooms in Moriches Bay
and Great South Bay, Long Island, New York., Biol. Bull., 106, 198—209.
Ryther J. H., 1956. Photosynthesis in the ocean as a function of light inten-
sity, Limnol. Oceanogr., 1, 61—70.
Ryther J. H., 1969. Photosynthesis and fish production in the sea, Science,
166, 72—76.
Ryther J. H., Yentsch. C. S., 1957. The estimation of photoplankton produc-
tion in the ocean from chlorophyll and light data, Limnol. Oceanogr., 2,
281—286.
Saarinen E., 1943. The City: Its Growth, Its Decay, Its Future, Cambridge,
Mass., MIT Press, 380 pp. •
Sale K., 1978. The polis perplexity: An inquiry into the size of cities. Wor-
king Papers, Jan.-Feb. 1978.
Salt G. W., 1979. A comment on the use of the term emergent properties,
Am. Nat., 113, 145—148.
Sanders F. E., Mosse B., Tinker P. B. (eds.), 1975. Endomycorrhizas, New York
and London, Academic Press.
Sanders H. L., 1968. Marine benthic diversity: A comparative study, Am. Nat.,
102, 243—282.
San Pietro A., Greer F. A., Army T. J. (eds.), 1967. Harvesting the Sun,
New York, Academic Press, 342 pp.
Santos P. F., Phillips J., Witford W. G., 1981. The role of mites and nema-
todes in early stages of buried litter decomposition in a desert, Ecology,
62, 664—669.
Santos P. F., Whitford W. G., 1981. The effects of microarthropods on litter
decomposition in a Chihuahuan desert ecosystem, Ecology, 62, 654—663.
Sauer С. O., 1950. Grassland, climax, fire and man, J. Range Mgmt., 3, 16—21.
Sauer С. O., 1963. Fire and early man. In: Land and Life: A Selection from
the Writings of Carl Ortwin Sauer, J. Leighly, ed. Berkeley, University
of California Press, pp. 228—299.
Saunders D. S., 1976. The biological clock of insects, Sci. Am., 234(2), 114—
121.
Saunders G. W., 1957. Interrelationships of dissolved organic matter and phy-
toplankton, Bot. Rev., 23, 389—409.
Saunders J. F. (ed.), 1968. Bioregenerative Systems, NASA. SP165. Washington,
D. C., Superintendent of Documents, U. S. Govt. Printing Office, 153 pp.
Saunders P. T., 1978. Population dynamics and length of food chains, Nature,
272, 189—190.
Saunders P. T., Ho M. W., 1976. On the increase in complexity in evolution,
J. Theoret Biol., 63, 375—384.
Schaffer W. M., 1974. Selection for optimal life histories: The effects of age
structure, Ecology, 55, 291—303.
Литература
347
Schelske С. L., 1977. Trophic status and nutrient leaching for Lake Michigan.
In: North American Project — A Study of U. S. Water Bodies, EPA Re-
port 600/3-77-086, Corvallis, Oregon.
Schelske C. L., Odum E. P., 1961. Mechanisms maintaining high productivity
in Georgia estuaries, Proc. Gulf Carib. Fish. Inst., 14. 75—80.
Schindler D. W.. 1977. Evolution of phosphorus limitation in lakes, Science,
195, 260—262.
Schmidt-Nielsen B., Schmidt-Nielsen K., 1952. Water metabolism of desert
mammals, Physiol. Rev., 32, 135—166.
Schmidt-Nielsen K., 1964. Desert Animals: Physiological Problems of Heat
and Water, London, Oxford University Press, 270 pp.
Schoener T. W., 1969. Optimal size and specialization in constant and fluc-
tuating environments: An energy-time approach. In: Diversity and Stabi-
lity in Ecological Systems, G. M. Woodwell and H. H. Smith, eds. Brook-
haven Symp. Biol. No. 22. Upton, N. Y., Brookhaven National Laboratory,
pp. 103—114.
Schoener T. W., 1971. Theory of feeding strategies, Ann. Rev. Ecol. Syst., 2,
369—404.
Schoener T. W., 1974. Resource partitioning in ecological communities, Science,
185, 27—39.
Schoener T. W., 1974a. Competition and the form of habitat shift, Theor. Pop.
Biol., 6, 265—307.
Schroder G. D., Rosenzweig M. L., 1975. Perturbation analysis of competitions
and overlap in habitat utilization between Dipodomys ordi and Dipodo-
mys mernami, Oecologia, 19, 9—28.
Schrodinger E., 1945. What Is Life? The Physical Aspects of the Living Cell,
Cambridge, England, Cambridge University Press, 91 pp.
Schultz A. M., 1964. The nutrient recovery hypothesis for arctic microtine
cycles. II. Ecosystem variables in relation to arctic micro tine cycles. In:
Grazing in Terrestrial and Marine Environments, D. J. Crisp, ed. Oxford,
Blackwell, pp. 57—68.
Schultz A. M., 1969. A study of an ecosystem: The arctic tundra. In: The Eco-
system Concept in Natural Resource Management, G. Van Dyne, ed.
New York, Academic Press, pp. 77—93.
Schumacher G. F., 1973. Small Is Beautiful, New York, Harper and Row.
Schurr S. H. (ed.), 1972. Energy, Economic Growth and the Environment,
A Resources for the Future Book, Baltimore, Johns Hopkins Press.
Schurr S. H. (Project Director), 1979. Energy in America’s Future: The Choi-
ces Before Us., A Resources for the Future Book., Baltimore, Johns
Hopkins Press.
Scott J. O. (ed.), 1956. Allee memorial number, Series of papers on social
organization in animals, Ecology, 37, 211—273.
Sears P. B., 1935. Deserts on the March, Normal, University of Oklahoma
Press.
Sears P. B., 1957. The Ecology of Man, Condon Lectures, Corvallis, University
of Oregon Press, 61 pp.
Selye H., 1955. Stress and disease, Science, 122, 625—631.
Shabman L., Batie S. S., 1978. Economic value of natural coastal wetlands:
A critique, Coastal Zone Mgmt. J., 4, 231—247.
Shannon C., 1950. Memory requirements in a telephone exchange, Bell Tech.
J., 29, 343—347.
Shannon С. E., Weaver W., 1949. The Mathematical Theory of Communication,
Urbana, University of Illinois Press, 117 pp.
Shantz H. L., 1917. Plant succession on abandoned roads in eastern Colorado,
J. Ecol., 5, 19-42.
Shelford V. E., 1911. Physiological animal geography, J. Morphol., 22, 551—
618.
23*
348
Литература
Shelford V. Е., 1911a. Ecological succession: Stream fishes and the method of
physiographic analysis, Biol. Bull., 21, 9—34.
Shelf ord V. E., 1911b. Ecological succession: Pond fishes, Biol. Bull., 21, 127—
151.
Shelford V. E., 1913. Animal Communities in Temperate America, Chicago,
University of Chicago Press.
Shelf ord V. E., 1943. The abundance of the collared lemming in the Churchill
area, 1929—1940, Ecology, 24, 472—484.
Sheridan D., 1981. The underwatered west: Overdrawn at the well, Environ-
ment, 23, 6—12, 30—33.
Shugart H. H., O’Neill R. V. (eds.), 1979. Systems Ecology, Benchmark Papers
in Ecology No. 9, Stroudsburg, Pa., Dowden, Hutchinson and Ross.
S illi man R. P., 1969. Population models and test populations as research tools,
BioScience, 19, 524—528.
Silverstein R. M., 1981. Pheromones: Background and potential for use in
insect control, Science, 213, 1326—1332.
Simberloff D. S., 1974. Equilibrium theory of island biogeography and ecology,
Ann. Rev. Ecol. Syst., 5, 161—182.
Simberloff D. S., 1976. Trophic structure determination and equilibrium in
an arthropod community, Ecology, 57, 395—398.
Simberloff D. S., Abele L. G., 1976. Island biogeography theory and conserva-
tion practice, Science, 191, 285—286, 193—1032.
Simberloff D. S., Wilson E. 0., 1969. Experimental zoogeography of islands:
The colonization of empty islands, Ecology, 50, 278—296.
Simon H. A., 1962. The architecture of complexity, Proc. Am. Phil. Soc., 106,
467—482. Reprinted in Sciences of the Artificial (2nd ed.), 1981, Cam-
bridge, Mass., MIT Press.
Simon H. A., 1973. The organization of complex systems. In: Hierarchy
Theory: The Challenge of Complex Systems, H. H. Pattee, ed. New York,
George Braziller, pp. 3—27.
Simon J. L., 1981. Ultimate Resource, Princeton, N. J., Princeton University
Press, 415 pp.
Simpson E. H., 1949. Measurement of diversity, Nature, 163, 688.
Simpson G. G., 1969. The first three billion years of community evolution.
In: Diversity and Stability in Ecological Systems, G. M. Woodwell and
H. H. Smith, eds. Brookhaven Symposium in Biology, No. 22, Upton,
N. Y., Brookhaven National Laboratory, pp. 162—177.
Sinclair A. R. E., Norton-Griffiths M. (eds.), 1979. Serengeti: Dynamics of
an Ecosystem, Chicago, University of Chicago Press, 389 pp.
Sindcn J. A., Worrell A. C., 1979. Unpriced Values: Decisions Without Market
Prices, New York, Wiley-Interscience, 511 pp.
Skellum J. G., 1952. Studies in statistical ecology, I-spatial pattern, Biometrica,
39, 346—362.
Slesser M., 1978. Energy in the Economy, New York, Macmillan.
Slobodkin L. B., 1954. Population dynamics in Daphnia obtusa, Kurz. Ecol.
Monogr., 24, 69—88.
Slobodkin L. B., 1960. Ecological energy relationships at the population level,
Am. Nat., 95, 213—236.
Slobodkin L. B., 1962. Growth and Regulation of Animal Populations, New
York, Holt, Rinehart and Winston, 184 pp.
Slobodkin L. B., 1964. Experimental populations of hydrida. In: Brit. Ecol.
Soc. Jubilee Symp. Suppl. to J. Ecol. 52 and J. Anim. Ecol. 33, 1—244,
Oxford, Blackwell, pp. 131—148.
Slobodkin L. B., 1968. How to be a predator, Am. Zool., 8, 43—51.
Small M., 1976. Marsh/pond sewage treatment plants. In: Proceedings of
Symposium on Freshwater Wetlands and Sewage Effuent Disposal,
D. L. Tilton, R. H. Kodlec, and C. J. Richardson, eds., Ann Arbor, Uni-
versity of Michigan Press, pp. 197—213.
Литература 349
Smith A. D., 1940. A discussion of applications of climatological diagrams,
the hythergraph, to distribution of natural vegetation types, Ecology, 21,
Smith C., 1976. When and how much to reproduce: The trade-off between
power and efficiency, Am. ZooL, 16, 763—774.
Smith F. E., 1954. Quantitative aspects of population growth. In: Dynamics
of Growth Processes, E. J. Boell, ed. Princeton, N. J., Princeton University
Press.
Smith F. E., 1969. Today the environment, tomorrow the world, BioScience, 19,
317—320.
Smith F. E., 1969a. Effects of enrichment in mathematical models. In: Eutro-
phication, Washington, D. C., National, Academy of Science, pp. 631—
645.
Smith F. E., 1970. Ecological demand and environmental response, J. Forestry
68, 752—755.
Smith&N. G., 1968. The advantage of being parasitized, Nature, 219, 690^
Smith R. F., van den Bosch R., 1967. Integrated control. In: Pest Control:
Biological, Physical and Selected Chemical Methods, Kilgore and Doutt,
eds., New York, Academic Press, pp. 295—340.
Smith R. F. (ed.), 1966. A symposium on estuarine fisheries, Trans. Am.
Fish. Soc. (Suppl.), 95(4), 1—154.
Smith S. H., 1966. Species succession and fishery exploitations in the Great
Lakes, Symposium on Overexploited Animal Populations, Washington,
D. C., American Association for the Advancement of Science, 28 pp.
(mimeographed).
Smith V. K., Krutilla J. V., 1979. Resource and environmental constraints to
growth, Am. J. Agr. Econ., 61, 395—408.
Smith W. H., 1981. Air Pollution and Forests: Interaction Between Air Con-
taminants and Forest Ecosystems, New York, Springer-Verlag.
Snow С. P., 1959. The Two Cultures and the Scientific Revolution, New York,
Cambridge University Press.
Solbrig О. T., 1971. The population biology of dandelions, Am Sci., 59. 686—
694.
Somero G. N., 1969. Enzymic mechanisms of temperature compensation, Am.
Nat., 103, 517—530.
Sopper W. E., 1968. Waste water renovations for reuse: Key to optimum use
of water resources. In: Water Research, Vol. 2. New York, Pergamon
Press, pp. 471—480.
Sorokin J. T., 1964. On the trophic role of chemosynthesis in water bodies,
Int. Rev. Ges. Hydrobiol., 49, 307—324.
Soule 1/. E., Wilcox B. A. (eds.), 1980. Conservation Biology: An Evolutiona-
ry-Ecological Perspectivef Sunderland, Mass., Sinauer Associates, 396 pp.
Sparrow Л. H., 1962. The role of the cell nucleus in determining radiosensiti-
vity, Brookhaven Lecture Series No. 17, Brookhaven National Laboratory
Publication No. 766.
Sparrow A. H.. Schairer L. A.. Sparrow R. C., 1963. Relationship between
nuclear volumes, chromosome numbers, and relative radiosensitivities,
Science, 141, 163—166.
Sparrow A. H., Woodwell G. M., 1962. Prediction of the sensitivity of plants
to chronic gamma irradiation, Rad. Bot., 2, 9—26.
Spilhaus A. (ed.), 1966. Waste Management and Control, Publication No. 1400,
Washington, D. C., National Academy of Science, 257 pp.
Spring D., Spring E. (eds.), 1974. Ecology and Religion in History, New York,
Harper Torchbooks, Harper and Row, 154 pp.
Sprugel D. G., Bormann F. H., 1981. Natural disturbance and the steady state
in high altitude balsam fir forests, Science, 211, 390—393.
350
Литература
Spurlock J. M., Modell M., 1978. Technology requirements and planning cri-
teria for closed life support systems for manned space missions, Washing-
ton, D. C., Office of Life Sciences, NASA.
Stanford Research Institute, 1975. City Size and the Quality of Life, Report
of Subcommittee on Rural Development of Committee on Agriculture and
Forestry, U. S. Senate. (Published as a Congressional Hearing in 1978.)
Stanley S. M., 1979. Macroevolution: Pattern and Process, San Francisco,
W. H. Freeman, 332 pp.
Stanley S. M., 1981. The New Evolutionary Timetable, New York, Basic Books,
222 pp.
Starr C. (ed.), 1971. Energy and Power, Sci. Am. Special Issue, Vol. 224,
No. 3.
Stearns S. C., 1976. Life-history tactics; a review of ideas, Quart. Rev. Biol.,
51, 3—47.
Stearns S. C., 1977. The evolution of life history traits, Ann. Rev. Ecol. Syst.,
8, 145—172.
Steele J. H. (ed.), 1970. Marine Food Chains, London, Oliver and Boyd, 552 pp.
Steeman-Nielsen E., 1952. The use of radioactive carbon (C!4) for measuring
organic production in the sea, J. Cons. Int. Explor. Mer., 18, 117—140.
Steemann-Nielsen E., 1954. On organic production in the ocean, J. Cons. Int.
Explor. Mer., 49, 309—328.
Steinhart J. S., Steinhart С. E., 1974. Energy use in the U. S. food system,
Science, 184, 307—316.
Steinwascher K., 1978. Interference and exploitation competition among tad-
poles of Rana utricularia, Ecology, 59, 1039—1046.
Stern A. C. (ed.), 1968. Air Pollution (2nd ed.), Three vols, New York, Aca-
demic Press.
Stewart P. A., 1952. Dispersal, breeding, behavior, and longevity of banded
barn owls in North America, Auk, 69, 277—285.
Stickel L. F., 1950. Populations and home range relationships of the box
turtle, Terrapene c. Carolina (Linnaeus), Ecol. Monogr., 20, 351—378.
Stobaugh R., Yergin D. (eds.), 1979. Energy Future: Report of the Energy
Project at the Harvard Business School, New York, Random House.
Stoddard D. R., 1965. Geography and the ecological approach. The ecosystem
as a geographical principle and method, Geography, 50, 242—251.
Stoddard H. L., 1936. Relation of burning to timber and wildlife, Proc. 1st.
N. A. Wildl. Conf., 1, 1—4.
Stokes A. W. (ed.), 1974. Territory, Benchmark Papers in Animal Behavior,
Stroudsburg, Pa., Dowden. Hutchinson and Ross, 416 pp.
Stonehouse B., Perrins C. (eds.), 1977. Evolutionary Ecology, London and
New York, Macmillan Press.
Straffa P., 1973. The Production of Commodities by Means of Commodities.
Prelude to a Critique of Economic Theory, Cambridge, England, Camb-
ridge University Press.
Stugren, Von Dr. Bogdan, 1965. Ecologie generala, Bucharest, Editura De-
dochea Sipedogogua.
Stugren, Von Dr. Bogdan, 1972. Grundlagen der allgemernen Okologie, Jena,
Veb Gustav Fischer Verlag.
Sukachev V. N., 1944. On principles of genetic classification in biocenology,
(In Russian.) Zur Obshchei Biol., 5, 213—227, Translated by F. Raney
and R. Daubenmire, Ecology, 39, 364—376.
Sukachev V. N., 1959. The correlation between the concepts ’’forest ecosystem'*
and ’’forest biogeocoenose“ and their importance for the classification of
forests, Proc. IX Int. Bot. Congr., Vol. II, page 387 (abstract). (See also
Silva Fennica, 105, 94, 1960).
Sukachev V. N., Dylis N. V. (eds.), 1964. Fundamentals of Forest Biogeocoeno-
logy, Moscow, Bot. Inst. Lab. For. Sci., 474 pp. (In Russian; see review in
Science, 148, 868, 1965).
Литература
351
Sutherland J. P., 1974. Multiple stable points in natural communities, Am.
Nat, 108, 859-873.
Svardson G., 1949. Competition and habitat selection in birds. Oikos, 1, 157—
174.
Svenson В. H., Soderlund R. (eds.), 1976. Nitrogen, phosphorus and sulfur —
global cycles, Ecol. Bull./NFR 22, Stockholm, Royal Swedish Academy
of Science.
Swank W. T.. Miner N. H., 1968. Conversion of hardwood-covered watershed
to white pine reduces wafer yield, Water Resources Res., 4, 947—954.
Sweeney J. R., 1956. Responses of vegetation to fire, Univ. Calif. Publ. Bot.,
28, 143—250.
Swift M. J., Heal 0. W., Anderson J. M., 1979. Decomposition in Terrestrial
Ecosystems. Studies in Ecology, Vol. 5, Berkeley, University of California
Press, 372 pp.
Swingle H. S.. Smith E. V., 1947. Management of farm fish ponds (rev. ed.),
Alabama Polytechnic Inst. Agr. Exp. Sta. Bull. No. 254.
Taber R. D.. Dasmann R., 1957. The dynamics of three natural populations
of the deer, Odocoileus hemionus columbianus, Ecology, 38, 233—246.
Takahashi M., Ichimura S.. 1968. Vertical distribution and organic matter
production of photosynthetic sulfur bacteria in Japanese lakes, Limnol.
Oceanogr., 13, 644—655.
Talbot L. M.. Talbon M. H., 1963. The high biomass of wild ungulates on East
African savanna, Trans. N. A. Wildl. Conf., 28, 465—476.
Tansley A. G.. 1935. The use and abuse of vegetational concepts and terms,
Ecology, 16, 284—307.
Tappen H., 1968. Primary production, isotopes and the atmosphere In: Pa-
laeogeography, Palaeoclimatology, Palaeoecology, Vol. 4, Amsterdam, El-
sevier, pp. 187—210.
Taub F. B., 1969. A biological model of a freshwater community in a gno-
tobiotic ecosystem, Limnol. Oceanogr.. 14, 136—142.
Taub F. B., 1974. Closed ecological systems, Ann. Rev. Ecol. Syst., 5. 139—
160.
Taylor M. W., 1977. A comparison of three edge indexes, Wildl. Soc. Bull.. 5,
192—193.
Taylor О. C., Dugger W. M.. Jr., Cardiff E. A., Darley E. F., 1961. Interaction
of light and atmospheric photochemical (smog) within plants. Nature, 192,
814-816.
Taylor W. R., 1964. Light and photosynthesis in intertidal benthic diatoms,
Helgd. Wiss. Meeresunters, 10. 29—37.
Teal J. M., 1957. Community metabolism in a temperate cold spring, Ecol.
Monogr., 27, 283—302.
Teal J. M., 1958. Distribution of fiddler crabs in Georgia salt marshes. Ecolo-
gy, 39, 185—193.
Teal J. M.. 1962. Energy flow in the salt marsh ecosystem of Georgia, Ecology,
43, 614—624.
Teitelbaum M. S., 1975. Relevance of demographic transition theory for de-
veloping coutries, Science, 188, 420—425.
Theobald R. (ed.). 1979. Challenge in Renewable Natural Resources. A Guide
to Alternate Futures, Washington, D. C., U. S. Department of Agriculture,
123 pp.
Thienemann A.. 1929. Der Nahrungskreislauf im Wasser, Verh. deutsch. Zool.
Ges., 31, 29—79.
Thienemann A., 1939. Grundzuge einer allgemeinen Oekologie, Arch. Hydro-
biol., 35, 267—285.
Thomas J. W., Maser C., Rodiek E., 1979. Edges. In: Wildlife Habitat in Ma-
naged Forests, J. W. Thomas, ed. Agricultural Handbook No. 53. USDA
Forest Service, Washington, D. C., Supt. of Documents, U. S. Govt. Prin-
ting Office, pp. 48—59.
352
Литература
Thomas М. D., Hill G. R., 1949. Photosynthesis under field conditions. In:
Photosynthesis in Plants, James Franck and Walter E. Loomis, eds.
Ames, Iowa State College Press, pp. 19—52.
Thomas JU. A., 1969. Accumulation and cycling of calcium by dogwood trees,
Ecol. Monogr., 39, 101—120.
Thomas W. L. (ed.), 1956. Man’s Role in Changing the Face of the Earth,
Chicago, University of Chicago Press.
Thompson D. II., Bennett G. W., 1930. The fishes of Champaign Country:
A study of the distribution and abundance of fishes in small streams.
Ill. Nat. Hist. Surv. Bull., 19, 1—101.
Thornthwaite C. W., 1931. The climates of North America according to a new
classification, Geogr. Rev., 21, 633—655.
Thornthwaite C. W., 1948. An approach to a rational classification of climate,
Geogr. Rev., 38, 55—94.
Thornthwaite C. W., Mather J. R., 1957. Instructions and tables for computing
potential evapotranspiration and water balance, Drexel Inst. Technol. Lab.
Climatol. Publ. Glimatol., 17, 231—615.
Thorson G., 1955. Modern aspects of marine level-bottom animal communi-
ties, J. Marine Res., 14, 387—397.
Tilly L. J., 1968. The structure and dynamics of Cone Spring, Ecol. Monogr.,
38, 169—197.
Tilton D. L., Kedlec R. H., Richardson C. J. (eds.), 1976. Proceedings of Sym-
posium on Freshwater Wetlands and Sewage Effluent Disposal, Ann.
Arbor, University of Michigan, 343 pp.
Tinbergen J. (ed.), 1977. The Rio: Reshaping the International Order, A Report
to the Club of Rome Coordinator, New York, Signet, 432 pp.
Tinbergen N., 1968. On war and peace in animals and man, Science, 160,
1411-1418.
Tjephema J. D., Winship L. J., 1980. Energy requirement for nitrogen fixation
in actinorhizal and legume root nodules, Science, 209, 279—281.
Torrey J. G., 1978. Nitrogen fixation by actinomycetenodulated angiosperms,
BioScience, 28, 586—592.
Tounsend C. R., Calow P. (eds.), 1981. Physiological Ecology: An Evolutionary
Approach to Resource Use, Sunderland, Mass., Sinauer Associates, 401 pp.
Toynbee A., 1961. A Study of History, New York, Oxford University Press.
Tramer E. J., 1969. Bird species diversity; components of Shannon’s formula,
Ecology, 50, 927—929.
Transeau Ё. N.. 1926. The accumulation of energy by plants, Ohio J. Sci., 26,
1—10.
Trautman M. B., 1942. Fish distribution and abundance correlated with stream
gradients as a consideration in stocking programs, Trans. 7th N. A. Wildl.
Conf., 7, 221—223.
Tribe H. T., 1957. Ecology of microorganisms in soil as observed during their
development upon buried cellulose film. In: Microbial Ecology, Williams
and Spicer, eds. Cambridge, Cambridge University Press.
Tribe H. T., 1963 The microbial component of humus. In: Soil Organisms,
J. Packser and J. van der Drift, eds. Amsterdam, North-Holland Publi-
shing Co.
Triplett G. B., Jr., VanDoren D. M., Jr., 1977. Agriculture without tillage, Sci.
Am., 636(1), 28-33.
Turner F. B. (ed.), 1968. Energy flow in ecosystems (Refresher Course, Am.
Soc. ZooL). Am. Zool., 8, 10—69.
Udall S. L., 1965. The Quiet Crisis, New York, Holt, Rinehart and Winston,
209 pp.
Udvardy M. F. D., 1959. Notes on the ecological concepts of habitat biotope
and niche, Ecology, 40, 725—728.
Ulanowicz R. E., 1980. A hypothesis on the development of natural commu-
nities, J. Theoret. Biol., 85, 223—245.
Литература
353
United States Department of Agriculture, 1979. Challenge in Renewable Na-
tural Resources. N Guide to Alternate Futures, R. Theobald, ed. Washing
ton, D. C., U. S. Dept, of Agriculture, 123 pp.
United States Water Resources Council, 1978. The Nation’s Water Resources —
1975—2000, Two vols, Washington, D. C., U. S. Govt. Printing Office.
Uvarov В. P., 1957. The aridity factor in the ecology of locust and grasshop-
pers of the old world. In: Human and Animal Ecology, Arid Zone Res.,
VIII. Paris, UNESCO.
Valentine J. W., 1968. Climatic regulation of species diversification and ex-
tinction, Bull. Geol. Soc. Am., 79, 273—276.
Valerio С. E., 1975. A unique case of mutualism, Am. Nat., 109, 235—238.
Valiela I., 1971. Food specificity and community succession: Preliminary or-
nithological evidence for a general framework, Gen. Syst., 16, 77—84.
Vallentyne /. R., 1960. Geochemistry of the biosphere. In: McGraw-Hill En-
cyclopedia of Science and Technology, Vol. 2, New York, McGraw-Hill,
pp. 239—245.
Vallentyne J. R., 1962. Solubility and the decomposition of organic matter in
nature, Arch. Hydrobiol., 58, 423—434.
Van den Bosch R., 1971. Biological control of insects, Ann. Rev. Ecol. Syst., 2,
45-66.
Vandermeer J. IL, 1981. Elementary Mathematical Ecology, New York, Wiley-
Interscience.
Van Doblcn W. EL, Lowe-McConnell R. H. (eds.), 1975. Unifying Concepts in
Ecology, Report of First International Congress of Ecology, The Hague,
W. Junk В. V. Publishers.
Van Dyne G. M. (ed.), 1969. The Ecosystem Concept in Natural Resource
Management, New York, Academic Press, 383 pp.
Vannote R. L., M Inshall G. W., Cummins K. W., Sedell J. R., Cushing С. E.,
1980. The river continuum concept. Can. J. Fish. Aquat. Sci., 37, 130—
137.
Van Valen L., 1965. Morphological variation and width of ecological niche,
Am. Nat., 99, 377—390.
Van Voris P., O’Neill R. V., Emanuel W. R., Shugart H. EL, 1980. Functional
complexity and ecosystem stability, Ecology, 61, 1352—1360.
Varley G. C., 1947. The natural control of population balance in the knap-
weed gall-fly {Urophora faceana), J. Anim. Ecol., 16, 139—187.
Varley G. C., 1949. Population changes in German forest pests, J. Anim. Ecol.,
18, 117—122.
Varley G. C., 1970. The concept of energy flow applied to a woodland commu-
nity. In: Quality and Quantity of Food, Symp. Brit. Ecol. Soc. Oxford,
Blackwell, pp. 389—405.
Varley G. C., Edwards R. L, 1957. The bearing of parasite behaviour on the
dynamics of insect host and parasite populations, J. Anim Ecol 26
471—477. ' ’
Verhulst P. F., 1838. Notice sur la loi que la population suit dans son accro-
issement, Corresp. Math. Phys., 10, 113—121.
Vernadsky V. L, 1945. The biosphere and the noosphere, Am. Sci., 33, 1—12.
Vernberg F. J., 1978. Multiple-factor and synergistic stresses in aquatic sys-
tems. In: Energy and Environmental Stress in Aquatic Systems,
T. H. Thorp and J. W. Gibbons, eds. U. S. Dept, of Energy, Symp. No. 48,
Springfield, Va., Technical Info, Center, pp. 726—747.
Vernberg F. J., Vernberg W. B., 1970. The Animal and the Environment,
New York, Holt, Rinehart and Winston, 416 pp.
Vernberg W. B., De Coursey P. J., Hara J. O., 1974. Multiple environment
factor effects on physiology and behavior of the fiddler crab, Uca pugi-
Литература
lator. In: Pollution and Physiology of Marine Organisms, New York,
Academic Press, pp. 381—425.
7.^1977. On the adaptive significance of territoriality, Am. Nat., Ill,
Viskery P. J., 1972. Grazing and net primary production of a temperate gras-
sland, J. Appl. Ecol., 9, 307—314.
Vida G., 1978. Genetic diversity and environmental future, Environ Cons., 5.
127—132.
V io sea P., Jr.s 1936. Statistics on the productivity of inland waters, the master
key to better fish culture, Trans. Am. Fish. Soc., 65, 350—358.
I'House к P. AL, Reiners W. A., 1975. Ecosystem succession and nutrient re-
tention: A hypothesis, BioScience, 25, 376—381.
Vogl R. J., 1980. The ecological factors that produce perturbation dependent
ecosystems. In: The Recovery Process in Damaged Ecosystems, J. Cairns,
Jr., ed. Ann Arbor, Mich., Ann Arbor Science, pp. 63—69.
Vogt W., 1948. Road to Survival, New York, Sloane.
Volterra V., 1926. Variations and fluctuations of the number of individuals
in animal species living together. In: Animal Ecology, R. N. Chapman,
ed. New York, McGraw-Hill, pp. 409—448.
Volterra V., 1928. Variations and fluctuations of the number of individuals
in animal species living together, J. Cons. Inst. Explor. Mer., 3, 3—51.
Wager J. A., 1970. Growth versus the quality of life, Science, 168, 1170—
1184.
Wagner F. H., 1969. Ecosystem concepts in fish and game management. In:
The Ecosystem Concept in Natural Resource Management, G. M. Van
Dyne, ed. New York, Academic Press, pp. 259—307.
Waksman S. A., 1952. Soil Microbiology, New York, John Wiley and Sons.
Walker J. C. G., 1977. Evolution of the Ahmosphere, London, Collier and
MacMillan, 318 pp.
Wallace B., Srb A. M., 1961. Adaptation, Englewood Cliffs, N. J., Prentice-Hall,
113 pp.
Waloff Z., 1966. The upsurges and recessions of the desert locust; an histori-
cal survey, Antilocust Mem. No. 8, London, 111 pp.
W angersky P. J., Cunningham W. J., 1956. On time lags in equations of
growth, Proc. Nat. Acad. Sci. (Wash.), 42, 699—702.
Ward B., 1966. Spaceship Earth, New York, Columbia University Press.
Ward B., 1976. The Home of Man, New York, W. W. Norton.
Warrington R., 1851. Notice of observation on the adjustment between animal
and vegetable kingdoms, Quart. J. Chem. Soc. London, 3, 5254.
Warrington R., 1857. On the aquarium, Notices Proc. Royal Inst., 2, 403—
408.
Watson V,, Loucks O. L., 1979. An analysis of turnover times in a lake eco-
system and some implications for system properties. In: Theoretical
Systems Ecology: Advances and Case Studies, E. Halfon, ed. New York,
Academic Press, pp. 355—383.
Watt A. S., 1947. Pattern and process in the plant community, J. Ecol., 35,
1—22.
Watt К. E. F., 1963. How closely does the model mimic reality?, Can. Ento-
mol. Mem., 31, 109—111.
Watt К. E. F., 1963a. Dynamic programming, ’’look ahead programming,44
and the strategy of insect pest control, Can. EntomoL, 95, 525—536.
Watt К, E F., 1966. Systems Analysis in Ecology, New York, Academic Press,
276 pp.
Watt K. E I' , 1968. Ecology and Resource Management: A Quantity live
Approach, New York. McGraw-Hill, 450 pp.
HkzK К. E. F., Molloy L. F., Varshney С. K., "Weeks D., Wirosardjono S.,
1977. The Unsteady Stale; Environmental Problems, Growth, and Culture,
Honolulu, East-West Center, The University Press of Hawaii, 287 pp.
Литература
355
Weaver J. E., Clements F. E., 1929. Plant Ecology (2nd ed., 1938), New York,
McGraw-Hill.
Wrebster J. R., Patten В. C., 1979. Effect of watershed perturbation on stream
potassium and calcium dynamics, Ecol. Monogr., 49, 51—72.
Weinberg A. M., Hammond R. P.f 1970. Limits to the use of energy, Am. Sci.,
58, 412-418.
Weisz P. B., Marshall I. F., 1979. High-grade fuels from biomass farming:
Potentials and constraints, Science, 206, 24—29.
Welch IL, 1967. Energy flow through the major macroscopic components of
an aquatic ecosystem, Ph. D. Dissertation, University of Georgia, Athens.
Welch IL, 1968. Relationships between assimilation efficiencies and growth
efficiencies for aquatic consumers, Ecology, 49, 755—759.
Wellington W. G., 1957. Individual differences as a factor in population dy-
namics: The development of a problem, Can. J. Zool., 35, 293—323.
Wellington W. G., 1960. Qualitative changes in natural populations during
changes in abundance, Can. J. ZooL, 38, 289—314.
Wells B. W., 1928. Plant communities of the coastal plain of North Carolina
and their successional relations, Ecology, 9, 230—242.
Went F. W., 1957. The Experimental Control of Plant Growth, Waltham
Mass., Chronica Botanica Co.
Went F. W., Stark N., 1968. Mycorrhiza, BioScience, 18, 1035—1039.
Werner E. E., Hall D. J., 1974. Optimal foraging and size selection of prey
by bluegill sunfish, Ecology, 55, 1042—1052.
Wesley J. P., 1974. Ecophysics: the Application of Physics to Ecology, Spring-
field, Ill., Charles C Thomas, 340 pp.
West D. C., Shugart II. IL, Botkin D. (eds.), 1981. Forest Succession: Concepts
and Applications, Ecol. Studies Vol. 41, New York, Springer-Verlag.
Westman W. E., 1975. Leiter to editor, BioScience, 25, 770.
West man W. E., 1977. How much are nature’s services worth?, Science, 1977
960—964.
Westman W. E., 1978. Measuring the inertia and resilience of ecosystems,
BioScience, 28, 705—710.
White G. F., 1980. Environment. In: Science Centennial Issue, P. H. Abelson
and R. Kulstad, eds. Science, 209, 183—190.
White L., 1967. The historical roots of our ecological crisis, Science, 155P
1203—1207.
White L., 1980. The ecology of our science, Science-80, 1, 72—76.
Whitehead F. IL, 1957. Productivity in alpine vegetation (abstract), J. Anim..
Ecol., 26, 241.
Whittaker R. II., 1951. A criticism of the plant association and climatic climax
concepts, Northwest Sci., 25, 17—31.
Whittaker R. IL, 1954. Plant populations and the basis of plant indication.
In: Angewandte Pflanzensoziologie, Veroffentlichungen des Karntner
Landesinstituts fur angewandte Pflanzensoziologie in Klagenfurt, Fest-
schrift Aichinger, Vol. 1.
Whittaker R. IL, 1957. Becent evolution of ecological concepts in relation to
eastern forests of North America. Am. J. BoL, 44, 197—206.
Whittaker R. IL, 1960. Vegetation of the Siskiyou Mountains, Oregon and
California, Ecol. Monogr., 30, 279—338,
Whittaker R. IL, 1965. Dominance and diversity in land plant communities,
Science, 147, 250—260.
Whittaker R. IL, 1967. Gradient analysis of vegetation, Biol. Rev., 42, 207—
264.
Whittaker R. H., 1969. New concepts of kingdoms of organisms. Science, 163,
150—160.
Whittaker R. IL, 1970. The biochemical ecology of higher plants. Tn: Chemical
Ecology, E. Sundheimer and J. B. Simeone, eds. New York, Academic
Press, pp. 43—70,
°00 Литература
Whittaker R. IL, 1972. Evolution and measurement of species diversity, Tar-
pon, 21, 213—251.
Whittaker R. II., 1975. Communities and Ecosystems (2nd ed.), New York,
Macmillan, 385 pp.
Whittaker R, H., Feeny P, P.f 1971. Allelechemics; chemical interaction
between species, Science, 171, 757—770.
Whittaker R. H., Levin S. A., Root R. B., 1973. Niche, habitat and ecotope,
Am. Nat., 107, 321—338.
Whittaker R. H., Likens G. E. (eds.), 1973. The primary production of the
biosphere, Human Ecol., 1, 301—369.
Whittaker R. H., Woodwell G. M., 1969. Structure, production and diversity of
the oak-pine forest at Brookhaven, New York, J. Ecol., 57, 155—174.
Whittaker R. H., Woodwell G. M., 1972. Evolution of natural communities.
In: Ecosystem Structure and Function, J. A. Wiens, ed. Corvallis, Oregon
State University Press, pp. 137—156.
Wiebes J. T., 1979. Coevolution of figs and their insect pollinators, Ann. Rev.
Ecol. Syst., 10, 1—12.
Wiegert R. G., 1965. Energy dynamics of the grasshopper populations in old-
field and alfalfa field ecosystems, Oikos, 16, 161—176.
Wiegert R. G., 1968. Thermodynamic considerations in animal nutrition, Am.
Zool., 8, 71—81.
Wiegert R. G., 1974. Competition: A theory based on realistic, general equa-
tions of population growth, Science, 185, 539—542.
Wiegert R. G., 1975. Simulation models of ecosystems, Ann. Rev. Ecol. Syst.,
6, 311—338.
Wiegert R. G., Coleman D. C., Odum E. P., 1970. Energetics of the litter-soil
subsystem. In: Methods of Study in Soil Ecology, Proc. Paris Symp. Int.
Biol. Prog. Paris, UNESCO, pp. 93—98.
Wiegert R. G.. Evans F. C.. 1967. Investigations of secondary productivity in
grasslands. In: Secondary Productivity of Terrestrial Ecosystems, K. Petru-
sewicz, ed. Vol. II, Warsaw. Polish Acad. Sci., pp. 499—518.
Wiegert R. G., Odum E. P., 1969. Radionuclide tracer measurement of food
web diversity in nature. In: Proceedings of 2nd National Symposium on
Radioecology, D. J. Nelson and Evans F. C., eds. Springfield, Va., Clea-
ringhouse Fed. Sci. Tech. Info., pp. 709—710.
Wiegert R. G., Odum E. P., Schnell J. FL, 1967. Forb-anthropod food chains
in a one-year experimental field, Ecology, 48, 75—83.
Wiegert R. G., Owen D. F., 1971. Trophic structure, available resources and
population density in terrestrial vs. aquatic ecosystems, J. Theoret. Biol.,
30, 69—81.
Wiener N., 1948. Cybernetics (2nd ed., 1961) Cambridge, Mass., MIT Press.
Vie ns J. A., 1976. Population responses to patchy environments. Ann. Rev.
Ecol. Syst., 7, 81—120.
Wiens J. A., 1977. On competition and variable environments, Am. Sci., 65,
590—591.
Wilbur H. M., Tinkle D. W., Collins J. P., 1974. Environmental and resource
availability in life history evolution, Am. Nat.. 108, 805—817.
Wilde S. A., 1968. Mycorrhizae and tree nutrition, BioScience, 18, 482—484.
JU/77г т/г L L., 1967. Comparison of some diversity indices applied to populations
of benthic macroinvertebrates in a stream receiving organic wastes, J.
Water Poll. Control Fed., 39, 1673—1683.
Wilhm J. L., Dorris T. C., 1968. Biological parameters for water quality crite-
ria, BioScience, 18, 477—481.
Williams С. B., 1964. Patterns in the Balance of Nature; and Related Problems
in Quantitative Ecology, New York, Academic Press, 324 pp.
Williams С. M., 1967. Third-generation pesticides, Sci. Am., 217(1), 13—17.
Williams G. C., 1966. Adaptation and Natural Selection, Princeton, N. J.,
Princeton University Press.
Литература
357
Wilson С. L., 1979. Nuclear energy: What went wrong?, Bull. Atomic Set,
35(6), 13—17.
Wilson D. S.., 1975. Evolution on the level of communities, Science, 192,
1358—1360.
Wilson D. S~, 1977. Structured denes and the evolution of group-advantageous
traits, Am. Nat., Ill, 157—185.
Wilson D. S,, 1980. The Natural Selection of Populations and Communities,
Menlo Park, Calif., and Reading, Mass., Benjamin/Cummings.
Wilson E. 0., 1965. Chemical communications in the social insects, Science,
149, 1064—1071.
Wilson E. O., 1969. The species equilibrium. In: Diversity and Stability in
Ecological Systems, G. M. Woodwell and H. H. Smith, eds. Brookhaven
Symposia in Biology, No. 22, Upton, N. Y., Brookhaven National Labora-
tory, pp. 38—47.
Wilson E. O., 1973. Group selection and its significance for ecology, BioSci-
ence, 23, 631—638.
Wilson E. O., 1975. Sociobiology: The New Synthesis. Cambridge, Mass.,
The Belknap Press of Harvard University Press. (Abridged edition, 1980).
Wilson E. O., Bossert W. H., 1971. A Primer of Population Biology, Stamford,
Conn., Sinauer Associates.
Wilson E. O., Willis E. O., 1975. Applied biogeography. In: Ecology and Evo-
lution of Communities M. L. Cody and J. M. Diamond, eds., Cambridge,
Mass., Harvard University Press, pp. 522—534.
Wilson J. T. (ed.), 1972. Continents Adrift. Readings from Scientific Ameri-
can, San Francisco, W. H. Freeman.
Winogradsky S., 1949. Microbiologie du Sol: Problemes et Methodes, Paris,
Masson et Cie., 861 pp.
Witherspoon J. P., 1965. Radiation damage to forest surrounding an unshielded
fast reactor, Health Physics, 11, 1637—1642.
Witherspoon /. P., 1969. Radiosensitivity of forest tree species to acute fast
neutron radiation. In: Proceedings of 2nd National Symposium on Ra-
dioecology, D. J. Nelson and F. C. Evans, eds. Springfield, Va., Clearing-
house Fed. Sci. Tech. Info, pp. 120—126.
Witkamp M., 1963. Microbial populations of leaf litter in relation to environ-
mental conditions and decomposition, Ecology, 44, 370—377.
Witkamp M., 1966. Decomposition of leaf litter in relation to environmental
conditions, microflora and microbial respiration, Ecology, 47, 194—201.
Witkamp M., 1971. Soils as components of ecosystems, Ann. Rev. Ecol. Syst.,
2„ 85—110.
Witkamp M., Olson J. S., 1963. Breakdown of confined and nonconfined oak
litter, Oikos, 14, 138—147.
Witt J. M., Gilett S. W. (eds.), 1978. Terrestrial Microcosms and Environmen-
tal Chemistry, Corvallis, Oregon State University Press.
Wohlschlag D. E., 1960. Metabolism of an antarctic fish and the phenomenon
of cold adaptation, Ecology, 41, 287—292.
Wolcott G. N., 1937. An animal census of two pastures and a meadow in
northern New York, Ecol. Monogr., 7, 1—90.
Wolfanger L. A., 1930. The Major Soil Divisions of the United States,
New York, John Wiley and Sons.
Wolfenbarger D. O., 1946. Dispersion of small organisms, Am. Midi. Nat., 35,
1—152.
Wollast R., Billen G., Mackenzie F. T., 1975. Behavior of mercury m natural
systems and its global cycle. In: Ecological Toxicity Research, A. D. McIn-
tyre and C. F. Mills, eds. New York, Plenum Press.
Wolman A., 1965. The metabolism of cities, Sci. Am., 213, 179—190.
Wood E. J. F. Microbiology of Oceans and Estuaries, Amsterdam, Elsevier,
319( pp.
358
Литература
Wood J. M., 1974. Biological cycles for toxic elements in the environments
Science, 183, 1049—1058.
W oodmansee R. G., 1978. Additions and losses of nitrogen in grassland eco-
systems, BioScience, 28, 448—453.
Woodruff L. L., 1912. Observations on the origin and sequence of the proto-
zoan fauna of hay infusions, J. Exp. Zool., 12, 205—264.
Woodwell G. M., 1962. Effects of ionizing radiation on terrestrial ecosvstems,.
Science, 138, 572—577.
Woodwell G. M., 1963. The ecological effects of radiation, Sci. Am., 208(6).
1—11.
Woodwell G. M.f (ed.), 1965. Ecological Effects of Nuclear War, Upton, N. Y.,
Brookhaven National Laboratory Publ. No. 917, 72 pp.
Woodwell G. M., 1967. Toxic substances and ecological cycles, Sci. Am.. 216(3).
24—31.
Woodwell G. M., 1977. Recycling sewage through plant communities, Am. Sci.,.
65, 556—562.
Woodwell G. M., Dyke man W. D., 1966. Respiration of a forest measured by
CO2 accumulation during temperature inversions, Science, 154, 1031—
1034.
Woodwell G. M., Pecan E. E. (eds.), 1973. Carbon and the Biosphere, U. So.
Atomic Energy Commission, Springfield, Va., National Technical Info,
Center.
Woodwell G. M., Smith II. H. (eds.), 1969. Diversity and Stability in Ecolo-
gical Systems, Upton, N. Y., Brookhaven National Laboratory Publ.
No. 22, 264 pp.
Woodwell G. M., Whittaker R. II., 1968. Primary production in terrestrial
communities, Am. Zool., 8. 19—30.
Wo<-dwell G. M., Whittaker R. II., Reiners W. A., Likens G. E., Delwiche C. C.^
Rotkin D. R., 1978. The biota and the world carbon budget, Science, 199.
141—146.
Woodwell G. M., Wurster С. F., Isaacson P. A., 1965. DDT residues in an east
coast estuary: A case of biological concentration of a persistent insecti-
cide, Science, 156, 821—824.
Wright S., 1945. Tempo and mode in evolution; a critical review, Ecology, 26,.
415—419.
Wurster Ch. F., 1969. Chlorinated hydrocarbon insecticides and the world
ecosystem, Biol. Cons., 1, 123—129.
Wynne-Edwards V. C., 1959. The control of population through social beha-
vior: A hypothesis, Ibis, 101, 436—441.
Wynne-Edwards V. C., 1962. Animal Dispersion in Relation to Social Behavior,.
New York, Hafner.
Wynne-Edwards V. C., 1964. Population control in animals, Sci. Am., 211(2),
68—72.
Wynne-Edwards V. C., 1965. Self-regulating systems in populations of animals,
Science, 147, 1543—1548.
Wynne-Edwards V. C., 1965a. Social organization as a population regulator,
Symp. Zool. Soc. London, 14, 173—178.
Zelitch I.. 1975. Improving the efficiency of photosynthesis, Science, 188,.
626—633.
Zeuthen E.. 1953. Oxygen uptake and body size in organisms, Quart. Rev.
Biol., 28, 1—12.
Zippin C., 1958. The removal method of population estimation, J. Wild!.
Mgmt., 22, 82—90.
Zunuska J. A., 1971. Cooperation or chaos: The challenge to natural resource
biologists, BioScience, 21, 726—728.
Предметный указатель
Термины, помещенные на страницах, выделенных курсивом, относятся
к рисункам. Римскими цифрами обозначены тома.
Абиотическая среда I: 24, 28, 30, 68
---- биологический контроль I:
36—41. См. также Лимитирую-
щие факторы; Элементы пита-
ния
Абиссальная область (абиссаль) II:
287
Автодороги, планирование II: 224
Автолиз I: 244
Автомобили и загрязнение воздуха I:
213, 229
Автотрофная сукцессия см. Сукцес-
сия экологическая
Автотрофы I: 26, 28, 29—30, 42—48,
70, 231—232
Агрегация II: 62—65
Агроэкосистемы I: 97—102
— доля, занимаемая на суше II:
219—220
— типы I: 98
Аденозинтрифосфат (АТФ) I: 31
Азот в болотных осадках II: 282, 283
— в лесах II: 239 (табл.)
— в ручьях I: 221, 223, 224
— и бактерии 1: 204—207
— и фосфор 1: 223, 295—296
— круговорот I: 203—209
— о кислы и загрязнение воздуха I:
213—214, 321
— удобрения 1: 208—209
— фиксация I: 205—209
Азот/фосфор, соотношение I: 223, 296
Азотфиксирующие актиноризные
растений I: 206—207 II: 265
Аквариум, равновесие I: 82
Аксенические культуры I: 80
Актиномицеты I: 207
Аллелопатия I: 58; II: 98, 106—108
Аллелопатические вещества I: 88.
См. также Аллелопатия
Аллопатрическое видообразование
II: 202—204
Альфа-излучение см. Ионизирующее
излучение
Аменсализм II: 83, 84 (табл.)
Аммиак и круговорот азота I: 204,
209
Анаэробная зона в водных местооби-
таниях I: 49—50; II: 283
Анаэробное дыхание I: 48—51
Анаэробы гетеротрофные I: 48—51
— фотосинтезирующие I: 43
Антибиоз II: 98, 107. См. также Ал-
лелопатия
Антропогенный стресс и токсичные
отходы I: 316—321. См. также
Стресс
Апвеллинг I: 103, 131 (табл.): И:
291, 202
Ассиммиляционное число I: 134
Ассимиляция I: 143, 148, 150, 151, 152
— эффективность I: 161 (табл.)
Атмосфера Земли, буферная способ-
ность I: 87—88
----сравнение с другими планета-
ми I: 37 (табл.)
----газы I: 293—294
----СО2 I: 60, 227—228, 293—295
----эволюция II: 197—202
Атмосферные газы как лимитирую-
щие факторы I: 293—295
Атомизм I: 18; II: 163
360
Предметный указатель
Атомная энергия как источник энер-
гии для человека I: 194, 195. См.
также Ионизирующее излучение
АТФ (Лдепозинтрифосфат) I: 31
Африка I: 163; II: 262—263, 272, 274,
275
Аэробное дыхание 1: 48; II: 200. См.
также Разложение; Дыхание
Бабочки, колебания численности II:
46, 47
Бактерии см. Разложение; Микроор-
ганизмы; Сапротрофы
— и круговорот азота I: 204, 205—
209;"II: 283
-------серы I: 46-47, 211—213; II:
283
— как деструкторы I: 29, 30 (рис.),
33, 51
— метаболизм I: 178 (табл.)
— метановые I: 48—49
— хемолитотрофные (хемосинтези-
рующие) I: 46—47, 147, 206
Батиальная зона (батиаль) II: 287
Безопасные поселения, образование
II: 64—65
Бентос (бентосные организмы) I: 70,
153; II: 289
Беспахотное земледелие см. Сельское
хозяйство беспахотное
Бета-излучение см. Ионизирующее
излучение
Биогенные элементы I: 295—296
Биогеография островная II: 207—208
Биогеохимические циклы I: 200—232
----количественное излучение I:
214—215
Биогеоценоз I: 14, 25
Биологический контроль геохимиче-
ской среды I: 36—38
Биологическое накопление в пище-
вых сетях I: 165—167, 279—280
Биологические часы I: 265—267; II:
285
Биомы наземные I: 14, 24, 103; II:
248—276
Биомасса I: 69, 91, 119, 129, 149, 164,
171—178
— и дыхание (соотношение R/В) I:
109
— и плотность популяции II: 6—7
— и поток энергии I: 175—177
— как топливо I: 138—140
— калорийность I: 110 (табл.)
— накопление в ходе сукцессии II:
167—168
— пирамиды I: 174, 175, 176
Биосистема I: 14—15, 17. См. также
Системы, Экосистема
Биосфера I: 15, 24, 67, 86, 88, 105
— как кибернетическая система I:
37
— первичная продукция I: 130, 131
(табл.)
— эволюция II: 197—202
— энергетические характеристики I:
112—117
Биофаги I: 29
Биохимическое потребление кисло-
рода (ВПК) I: 73
Биохимическое разнообразие II:
188—189
Биоценоз I: 14, 25
Биоэнергетика см. Энергетика, Поток
энергии
Блоковые модели см. Модели
Бобовые, фиксация азота I: 206—208,
267
Болезни и взаимодействие популя-
ций II: 103. См. также Парази-
тизм
Болота II: 282—283
Борьба с вредителями I: 324—325
----интегрированный метод I: 325
Брожение I: 48, 49; II: 198
Будущее мира, экологические оцен-
ки II: 245—247
Бумага, вторичное использование I:
246—247
Бури и ураганы как физические фак-
торы среды I: 299
Валовая продукция см. Первичная
продукция валовая
Валовый национальный продукт I:
135
Ветер как лимитирующий фактор I:
298—299
Вечная мерзлота II: 251
Взаимоотношения между видами II:
83—87
----паразит — хозяин II: 100
-------эволюция II: 100—103, 212
Взвешенное органическое вещество
(BOB) I: 31
Видовое разнообразие II: 126—136.
См. также Разнообразие
Видообразование II: 202—205, 278
Виды/площадь соотношение Иг
128—129
— показатель значимости II: 7, 126
— распределение по значимости II:
130—131
— угрожаемые II: 147
Винклера метод определения кисло-
рода I: 71
Влажность I: 283—284. См. также
Вода
Предметный указатель
— и температура I: 290—293
Внутренняя скорость естественного
роста II: 26, 27, 29
Вода, глобальный круговорот I: 225,
226
— как лимитирующий фактор I:
281—286
— охлаждающая способность I:
322—323
— тепловое загрязнение I: 322—323
— термодинамические свойства II:
277
— течения I: 297—298
Водные экосистемы, сравнение с на-
земными I: 74—77
----типы II: 276—296
Водоносные горизонты (слои) I:
230-231; 286—288
Водоросли I: 42, 47—48, 69—70, 74—
75, 81, 167—168
— сине-зеленые (цианобактерии) I:
207
Водосборный бассейн, антропогенные
нарушения II: 161
----биогеохимия I: 220—225
----как экосистема I: 77, 78, 79
----урбанизация I: 223, 286
Водохранилища II: 279
Возврат веществ в круговорот I: 203,
219, 242—246
----------коэффициент циркуля-
ции I: 247
Воздух, загрязнение I: 213—214,
320-321
Возрастные пирамиды II: 20, 21
Время пребывания I: 216
— удвоение II: 29
Вспомогательный поток энергии I:
123. См. также Энергетическая
субсидия
Вулканы и круговорот элементов
питания I: 209, 210
Выжигание контролируемое (плано-
вое) I: 312, 313—315.
Выровненностъ II: 130, 133 (табл.),
134
Газохол I: 140
Гамма-излучение см. Ионизирующее
излучение
Гаузе правило см. Конкурентного
исключения принцип
Геи гипотеза I: 35—38; II: 201
Генетика естественного отбора II:
202—212
Генетический дрейф II: 202
Генетическое разнообразие II: 146—
150
Геологические периоды II: 199
361
Геотермальная глубоководная экоси
стема I: 47
Гетеротрофы I: 26, 28, 231—232
Гидрологический цикл I: 230
Гидропериод II: 282
Гильдия II: 119, 123
Глобальные модели II: 237, 243—245
Гнотобиотическпе микрокосмы I: 80.
81
Голоэкономика II: 231
Гомеостаз I: 62, 63
Гомотаксис конгенерический I: 64
Горные экосистемы II: 251, 273, 275>
Город, его размеры и качество жиз-
ни I: 183—185
— зависимость от крупных водое-
мов II: 277
— как гетеротрофная система 1г.
89-97
— климат I: 94—95, 116
— метаболизм I: 96
— потребление энергии I: 94, 192—
193
— сравнение с устричной банкой 1г
90
— среда на входе и на выходе 1г
89, 183
Градиенты в реках I: 231—232
— высотные II: 152
— географические II: 150—156
— анализ II: 151, 153—155
— на литорали II: 109—110
— от полюса до экватора II: 161
— растительности II: 152—154
Грибы I: 32, 33, 58, 70. См. также?
Микориза, Лишайники
Групповой отбор II: 210—213
Грызуны II: 45, 77, 96, 262, 271
Гумификация I: 55, 301
Гумус (гумусовые вещества) I: 31,,
53—55, 227, 301
Давление барометрическое как ли-
митирующий фактор I: 299; Пг
267
Дарвинизм (теория эволюции) II:
158, 202—205, 210—211
Дарвиновы вьюрки II: 204—205
Двуокись углерода, влияние на кли-
мат I: 60, 228; II: 36, 42, 60, 225—
229, 293-294
----круговорот см. Углерод, гло-
бальный круговорот
----содержание в атмосфере I: 36.,
42, 60, 225—229, 293—294
-------------и фотосинтез I: 41—
47
------- в воде I: 294—295
94 Заказ № 1483
362
Предметный указатель
Двустворчатые моллюски I: 227; II:
60
ДДТ в пищевых цепях I: 165—167
Демографический переход II: 246
Денитрификация I: 204, 205, 209
Деньги и энергия I: 194—199
Деструкторы (разрушители) I: 29.
33, 48
Детрит органический I: 31
— в сельском хозяйстве II: 217
— и круговорот элементов питания
I: 218, 219, 243—244
Детритные пищевые цепи I: 142, 144,
145, 153, 158—160
Детритофаги II: 116
Дефицит давления пара I: 283
Диапауза I: 265, 267; II: 32
Дисклимакс II: 196—197
Диспергированное органическое ве-
щество (ДОВ) I: 147, 148 См.
также Детрит органический
Диссипативные структуры в экоси-
стемах I: 108; II: 218
Дифференциальные уравнения II:
31, 33, 36, 85, 88
Дпэлдрин I: 166
Длина светового дня см. Фотоперио-
дизм
ДИК (дезоксирибонуклеиновая кис-
лота) I: 31
Дождевая тень I: 281, 282
Дождевые осадки и эвапотранспира-
цпя I: 290—291
— распределение I: 281—282
Докембрийские формы жизни I: 50
Домашние животные см. Доместика-
ция
----и запасы пищи I: 141
----пастьба I: 145—146
----эквивалент популяции I: 138,
141
Доместикация II: 205—206
Доминирование II: 127, 130—134
— возрастного класса II: 22, 23
— индекс II: 133, 144 (табл.)
Древесина для топлива, глобальная
нехватка I: 138
Дрожжи I: 48
— рост в лабораторных культурах
II: 38, 39, 55
Дрейф континентов II: 202
Дыхание I: 48—50
— в пищевых цепях I: 143, 150, 151
— и биомасса, соотношение I: 109
— и продукция II: 77 (табл.), 168,
169. 170, 213
— сообщества I: 71, 109, 126, 128
(табл.); II: 169
Естественный отбор II: 202—205
Жвачные II: 112, 250
Жгутиковые I: 70; II: ИЗ. См. также
Простейшие
Железо как микроэлемент I: 259, 297
Животные. См. также Консументы;
названия отдельных групп
— домашние см. Домашние живот-
ные
— роль в биомах II: 248
— роль в пищевых цепях I: 162—165
— царство I: 32—33
Жизненная форма II: 248, 261
Жизненный цикл, особенности и
тактика II: 81—82
Жуки II: 47—48, 90, 94
Загрязнение
— воды I: 289, 317—318
— воздуха I: 320—321
— и видовое разнообразие II: 138,
139, 142
— тепловое I: 322—323
— токсичными отходами I: 316—318,
319
Засоление орошаемых земель I: 309
Заяц-беляк, изменения численности
II: 43, 44
«Зеленая революция» I: 124—125; II:
206
Земледелие см. также Сельское хо-
зяйство
— в тропиках I: 238—240
Землепользование
— блоковые модели II: 217—223
— в США II: 219, 220
Значимость вида II: 7, 126, 130, 131
Зональность II: 127, 137, 152—153.
См. также Стратифицирован-
пость
— в горах II: 542—543
— в море II: 284, 287—288
— в озерах и прудах II: 278
Зоопланктон I: 30, 70, 75 (табл.),
145, 254
— и круговорот элементов питания
I: 243-244
— морской II: 289
Иерархия I: 13—15
Избавление от отходов I: 318, 319
Избыточность и устойчивость (ста-
бильность) I: 60, 62, 64
Излучение
— ионизирующее см. Ионизирующее
излучение
— световое I: 270—272
— солнечное см. Солнечное излуче-
ние
— тепловое I: 114
Предметный указатель
— электромагнитное, спектр I: 270,
271
Импульсная стабильность, концеп-
ция II: 215—216
Индекс возобновления II: 176
— выровненное ти Пиелу II: 133
(табл.), 134
— Симпсона (разнообразия) II: 133
(табл.), 144 (табл.)
— сходства II: 153
— цикличности II: 176
Индивидуальный (семейный) учас-
ток II: 66—67
Инсектициды I: 122, 323—325. См.
также Пестициды
Интегрированный контроль вредите-
лей I: 325
Инфауна II: 288
Инфракрасное излучение I: ИЗ—114
(рис. 3.2), 270 1
Ионизирующее излучение I: 272—
281. См. также Радионуклиды
— — влияние на сообщество в це
лом I: 278—279
----единицы измерения I: 275—276
----сравнительная чувствитель-
ность организмов I: 276—277
— типы I: 273—275
— фоновое I: 275
Ископаемое топливо I: 12, 31; II: 234
----в будущем II: 234
----и атмосферная СО2 I: 59—60,
226, 228; II: 282
----и солнечная энергия I: 170—
171
----потребление городами I: 193
----эра горючих ископаемых I:
195
Искусственное обогащение, экспери-
менты I: 258—259
Искусственный отбор II: 205—206
Источники (родники) II: 280
Кадмий I: 260
Калий I: 296
Калория I: 110 (табл.)
Кальций I: 236, 279; II: 286
— как лимитирующий фактор I:
296
— круговорот на облесенном водо-
сборном бассейне I: 220—222
Карбонатные реки II: 280
Качество жизни II: 234. См. также
Энергия, качество
----и размер города I: 183—185
— ресурса I: 148, 170
Кислород I: 200, 202, 208, 251
— в атмосфере I: 42, 55, 227—228;
II: 200—201
363
— и деструкция (разложение) I:
48—49
— и фотосинтез I: 42—44, 73
— и хлорофилл I: 134
— содержания в воде, изменение I:
71
Кислый дождь I: 40, 96, 213
Клевер, сосуществование видов II:
89, 91_92
Климаграммы I: 291, 292, 293, II: 25G’
Климакс II: 192—197
Климат, влияние атмосферной СО2 I:
60, 228
— влияние водных масс I: 290
— городов I: 89—96
— классификация I: 290
Климатические камеры I: 293
Ковита, экспериментальный водо-
сборный бассейн (Северная Ка-
ролина) I: 220, 223
Комменсализм II: 83, 84 (табл.)г>
110—111
Компенсационный уровень I: 73
Компьютерное моделирование I: 19..
219
Конкурентного исключения принцип;
II: 88—90, 93
Конкуренция II: 83—84, 87—98
— интерференционная II: 92
— коэффициент II: 89
— и сосуществование видов I: 87,.
90—94
— межвидовая и внутривидовая II:
94—96
— роль в эволюции II: 203, 211
— эксплуатационная II: 92, 94
Конструирование ландшафта II: 225?
Конструкционизм I: 18
Консументы I: 29, 70, 75 (табл.)
Континентальный шельф II: 287
---как часть системы жизнеобес-
печения II: 221
Континуум в лесу II: 152—153
— в реках I: 231—232
— концепция II: 151, 153
Контроль численности насекомых-
вредителей II: 57
Кооперация II: 110—112
— эволюция II: 210—211
Копперхилл, бассейн (шт. Теннесси)
I: 38, 39, 40
Копрофагия I: 56, 158, 159
Коралловые рифы I: 16, 264: II: 235
Космическая колония I: 86—88
Космические полеты I: 86, 256
Космическое излучение I: 275
Коэволюция II: 197, 208—209
Коэффициент накопления I: 279—280
24*
364
Предметный указатель
Коэффициент рециркуляции (возвра-
та) I: 245, 246, 247
Краевой эффект II: 151, 155
— индекс II: 156
Кривые выживания II: 16, 17—18
— доминирования — разнообразия
II: 127, 130—132, 138, 139, 187
— роста популяции II: 25, 28, 32, 35,
38, 39
-------уравнения II: 26, 29, 30, 31
Круговорот воды I: 225, 226, 229—
” 232, 286
— — энергетика I: 229—230
— второстепенных элементов I:
235—238
— ртути I: 237—238
— серы I: 211—214
— элементов питания I: 200—247.
См. также Биогеохимические
циклы, Элементы питания
-------в тропиках I: 238—241
—------и экологическая сукцессия
II: 173—177
Крупный рогатый скот (жвачные)
II: 100, 112, 262, 272. См. также
Домашние животные
-------модель интенсивности вы-
паса I: 146
Культура риса, значение Azolla I:
207
Культуры сельскохозяйственные II:
135—141
— — отношение к энергетическим
субсидиям и удобрению I: 122—
123
Ландшафтная архитектура II: 219
Лемминги I: 152; II: 45, 251
Лентические экосистемы I: 103; II:
276—277
Леса, годовой опад листьев I: 302
— городские I: 89, 91 (табл.)
— заболоченные, продуктивность I:
298
— продуктивность I: 128 (табл.), 129,
130, 131 (табл.)
— северные хвойные II: 253—255,
257
— тропические дождевые II: 272—
274
----круговорот элементов питания
I: 238—240
----сезонные полувечнозеленые
II: 271
— хвойные, циклические колебания
численности насекомых в них
II: 46—47
.Лесные певуны (Parulidae), ниши II:
123
Либиха «закон» минимума I: 248—
252
Лигнин I: 51, 53
Лимитирующие факторы, концепция
I: 258—260
---биотические I: 256, 259
---физические I: 267—315
Лимническая зона II: 278
Листовой индекс II: 89
— опад I: 301, 302
Литоральная зона I: 259; II: 109, 278,
287, 290
Лишайники (мутуализм водоросли и
гриба) II: 117—418, 251
— как пример для человека II: 118
Логистическое уравнение II: 31, 33,
35, 36
Лотки — Вольтерра уравнения кон-
куренции II: 88, 100
Луг как экосистема I: 68, 70, 74—77
Лучистая энергия I: 117
Лэнгли I: 117
Люцерна I: 126; II: 209
Магний I: 296
Макроконсументы I: 29, 70
Макроэлементы I: 296
Мангры, детритная пищевая цепь I:
158, 159
Манящие крабы I: 166; II: 97
Марс, атмосфера I: 37
Математические модели см. Модели
Международная Биологическая Про-
грамма (МБП) II: 250, 259
Мезокосмы I: 82—84
Мейофауна I: 158
Меропланктон II: 289
Местообитание II: 119—120
Метаболизм и размеры особей I:
171-173
— GAM-типа (кислотный метабо-
лизм толстянковых) I: 46, 285
Метан и стабильность озонового слоя
I: 229
— как горючее I: 139
Микориза (микоризные грибы) I:
241; II: 114, 115, 116
— в пищевых цепях I: 148—149
Микрокосмы I: 79—83
— и сукцессия в них II: 168, 170
Микроконсументы I: 29
Микроорганизмы I: 33, 46, 50, 55,
56. См. также Бактерии, Грибы,
Сапротрофы
— автохтонные I: 55
— зимогенные I: 55
— и гипотеза Геи I: 35—36
— размягчающие целлюлозу II: 113
Микроэкосистема см. Микрокосмы
Предметный указатель
365
Микроэлементы I: 227
Минерализация органического ве-
щества I: 53, 58
Модели (моделирование) I: 8—12,
219; II: 163
— блоковые I: 20—21, 26, 115, 159,
218; II: 218
— глобальные II: 243—245
— графические I: 19, 23
— землепользования II: 217, 218
— круговорота фосфора I: 218—219
— общие системные I: 22
— пищевых цепей I: 143—147
— продукция фитопланктона I: 254
— рыбного промысла II: 104—105
— экологической сукцессии II: 167
(табл.), 169
Молибден I: 297
Монокультуры (одновидовые систе-
мы) II: 105
— в сельском хозяйстве I: 101, 141
Море II: 283—292. См. также Океан;
Морские экосистемы
Морские экосистемы II: 283—296
Мощность I: 189, 192
— единицы I: 110
Муравьи
— колонии, поток энергии в них I:
186
— мутуализм с грибами и деревья-
ми II: ИЗ, 116
Мутуализм I: 208, 241; II: 84 (табл.),
110—118
— и экологическая сукцессия II:
177
Мучной хрущак (Tribolium), экспе-
римент по межвидовой конку-
ренции II: 90—91
Наземные экосистемы (биомы) II:
248—276
---сравнение с водными I: 74—
77
Наннопланктон II: 288
Нарушения в экосистеме I: 35, 126,
179; II: 161
Насекомые I: 267, 277; II: 16, 46—
47, 248, 265, 269, 273
Нейтрализм 83, 84
Нейтроны I: 274, 275
Неритическая зона II: 287
Нерыночные товары и услуги II:
228, 230, 231 (табл.), 236
Неупорядоченность см. Энтропия
Нефть см. Ископаемое топливо
— влияние на морские экосистемы
I: 84
Ниша и конкуренция II: 95—96
— многомерная (гиперпространст-
венная) II: 121
— морфологические показатели II:
123
— разделение микроместообитаний
II: 123—124
— трофическая II: 119, 120
— экологическая II: 119—126
Ноосфера Г. 67
Обогащение См. также Эвтрофика-
ция
— культуральной среды I: 256
— эксперименты 1: 259
Оборот, концепция I: 74, 109, 216
— и соотношение биомасса/продук-
ция I: 129, 130
Обратная связь I: 21—22, 26, 60—
63, 64, 209
---в экономических системах II:
231—232
---отрицательная I: 62
---положительная I: 61, 162—163
---системы управления I: 61
Общественное поведение см. Поведе-
ние
Объем хромосом и чувствительность
к ионизирующему излучению I:
278
Огаллала, водоносный горизонт
(США) I: 231, 288
Одичавшие животные II: 206
Озера II: 276—279. См. также Водо-
хранилища, Пруды
— влияние развития сельского хо-
зяйства и городов I: 161—162
— круговорот фосфора I: 217 (табл.)
— стратификация II: 144—145, 278
Озон I: ИЗ, 229
— и загрязнение воздуха I: 321
— и эволюция биосферы II: 198—
200
Океан, буферная способность I: 86
— зональность II: 287—288
— и глобальные круговороты I:
225—226, 226, 229, 230
— продуктивность I: 129, 130, 131
(табл.), 132; II: 284
Океанические области II: 288
Окислы азота I: 213—214, 321
— серы I: 213
Окись углерода I: 228—229
Олень северный II: 152
— карибу II: 152
Олиготрофность I: 164; II: 171—172
Оптимизация потока энергии I: 70—
74
— урожая в рыболовстве II: 104—
105
366
Предметный указатель
Орлппапия II: 151, 153
Орошение I: 99, 286, 309; II: 268
— п подземные воды, истощение за-
пасов I: 287
Осадки болотные, разрез II: 283
— радиоактивные I: 234, 273, 281
(табл.)
— (отложения), снос I: 234—235
Осадочный цикл I: 233—235
Осмотрофы I: 29
Отбор групповой II: 210—212
— естественный см. Естественный
отбор
— искусственный II: 205—206
— К~ II: 71, 73, 75—80
— г- II: 71, 73, 75—76, 78—79
Относительная влажность I: 283—
284, 292
Палеоэкология II: 158—160
Паразитизм II: 83—84, 98—106
Паразитические насекомые I: 62; II:
103—104, 141
Паразиты II: 86—87. См. также Па-
разитизм
Пастбищная пищевая цепь I: 142—
145
Пауки II: 61, 77—78
— территориальное поведение II: 69
Первичная продукция (продуктив-
ность) I: 117—121
-----биосферы I: 131 (табл.), 132
-----валовая и частая I: 118, 124,
126, 128 (табл.)
-----вертикальное распределение I:
129—130
-----заболоченных лесов I: 298
-----зависимость от Солнца I: 120
(табл.)
-----и дыхание (соотношение про-
дукция/дыхапие) I: 151, 231—232
— и эвапотранспирация I: 136
— — использование человеком I:
135—141
-----моря II: 284
-----сельскохозяйственных куль-
тур I: 121 (табл.), 136
-----топлива и волокна I: 138
— и хлорофилл I: 133—135
Перевыпас I: 145—146. См. также
Пастбищная пищевая цепь
Переменные состояния I: 20
Перепончатокрылые паразитические
I: 62
Пермеапты II: 248
Пероксиацетилпитрат (ПАН) I: 321
Пестициды I: 323—325. См. также
Инсектициды
— и урожай сельскохозяйственных
культур I: 122
Пирамиды возрастные II: 20—21
— экологические I: 174—179
Пищевая сеть I: 26, 27, 142, 153
— радионуклиды в ней I: 280
— роль консументов I: 162—163
— цепь I: 142—148, 152—159
-----восстановленной серы I: 212
— — графические модели I: 143,144*
147
-----длина 1: 164
-----изотопные метки для ее изу-
чения I: 167—168
-----и размер организмов I: 157—
158
---- качество ресурса в ней I: 148
----- концентрирование токсичных
веществ I: 165—167
Планирование ландшафта II: 148.
149, 222—225
-----и города II: 225—227
Планктон I: 70; II: 288
Плейстоцен, растительность II:
159—160
Плотность видов II: 126—127
— популяций II: 6—11
-----и выживание II: 17—18
-----измерение II: 10—11
-----показатели II: 7—8
-----средняя и экологическая II: 6.
10
-----человека I: 184—186; II: 18
— потребления энергии I: 193
(табл.)
— энергии I: 189
Площадь оснований II: 8, 144 (табл.)
Поведение и регуляция температу-
ры I: 262
— территориальное II: 66—70
Поддерживающая емкость среды I:
180—181; II: 35
— -----для популяций животных
I: 185—186
-------для человека I: 187, II:
244
Пожар (ы) взаимодействие с анти-
биотиками II: 107
— в зарослях чапараля I: 314; II:
107
— как экологический фактор I:
310—315
— контролируемый или плановый I:
313—314
— и управление ресурсами I: 314—
315
— типы I: 311—313
Предметный указатель
367
Подземные (грунтовые) воды I: 224,
286—289
— — загрязнение I: 289
----истощение запасов I: 287—289
----пополнение I: 231
Полевки II: 45, 96
Полиморфизм сбалансированный II:
137
Популяция, возрастной состав II:
19—22
— вспышки численности II: 56—57,
99
— градиентный подход II: 150
— групповые свойства II: 6—22
— межвидовые взаимодействия II:
83—88
— плотность II: 6—И
— — методы измерения II: 11
----энергии II: 70—73, 77—80
— регуляция численности II: 50—
56
— — — зависимая от плотности II:
51-55
-------независимая от плотности
II: 51—53, 56
— рождаемость II: 11—13
— рост II: 26—40
— смертность II: 13
— типы распределения особей II:
57—59
— циклические колебания II: 41—
43, 51
Потенциал биотический (репродук-
тивный) II: 27
Поток энергии I: 25, 26, 111—112, 177
— — диаграммы I: 143, 144, 150
— — и круговорот элементов пита-
ния I: 202
----и размеры организмов I: 177
----и трофические уровни I: 150
----распределение в экосистемах
II: 213
Почва I: 299—305
— засоление I: 309
— и рельеф местности I: 303,304,305
— профили I: 300, 301, 302, 304
— серпентинитовая I: 255
— сцементированный слой I: 303
— типы I: 303, 305 (табл.), 306
— уплотнение I: 299—305
— эрозия I: 235
Пределы толерантности I: 249—253
----размножающихся особей и
стадий развития I: 250—251
Пресноводные экосистемы I: 103; II:
276-277
----сравнение с наземными I:
68-75
Прибрежная зона II: 289—296
--- экологические эквиваленты в
ней II: 125 (табл.)
Приливы II: 285, 292
— как энергетическая субсидия I:
118, 123, 125
Принцип Олли II: 62, 63, 64
Природные ландшафты, роль в под-
держании жизни II: 214
---сохранение разнообразия II:
148, 149, 150
— товары и услуги I: 12, 199; II:
288, 230, 231 (табл.)
Продолжительность жизни II: 13, 18
Продуктивность I: 117—141
— баланс продукции и разложения
I: 59
— вторичная I: 118, 157 (табл.)
— глобальная I: 41
— первичная см. Первичная продук-
ция
Продукция/дыхание, соотношение I:
151, 231—232
— — изменение в ходе сукцессии
II: 166—170
Продуценты I: 29
Прокариоты I: 50, 208; II: 198
Простейшие I: 48, 56, 57, 81, 178. См.
также Жгутиковые
— конкуренция в лабораторных
культурах II: 93—94
— сукцессия в лабораторных куль-
турах II: 185
Протокооперация II: 84 (табл.),
110—111
Профили ископаемой пыльцы II: 160
Профундальная зона II: 278
Пруды как пример экосистемы I:
68—77
— пищевые цепи в них I: 152—156
Птицы II: 11, 12, 21, 42, 45
— в экологической сукцессии II:
179—181
— и краевой эффект II: 155
— и круговорот фосфора I: 210
— разделение ниш II: 96, 97
— расширение ареалов I: 257
— смещение признаков II: 204, 205
— территориальность II: 67—68
— циклические колебания численно-
сти II: 41, 45
— экологические эквиваленты II:
II: 126 (табл.)
Пуассона распределение II: 50—60
Пустыни II: 269, 271
— антропогенные II: 262
Пчелы, кривая роста колонии II: 25
Пшеница I: 123, 137 (табл.)
368
Предметный указатель
Радиационная экология I: 217
Радиация чистая I: 116
Радиоактивные изотопы см. Радио-
нуклиды
Радиоактивные индикаторы
— — и биогеохимические циклы I:
214—218
---и пищевые цепи I: 167
— продукты деления ядер I: 279
Радионуклиды I: 273
— как изотопные метки в пищевых
цепях I: 165, 280
— как индикаторы в биогеохимиче-
ских круговоротах I: 214—218
— накопление в пищевых цепях I:
165, 280
Радиочувствительность сравнитель-
ная I: 276—278
Развитие городов, планирование II:
149, 227 (табл.)
— плановое и неконтролируемое II:
225—227 (табл.)
— экосистемы II: 165. См. также
Сукцессия экологическая
Разложение I: 48—59
— лесного опада I: 53
— общий баланс с продукцией I: 59
— роль микрофауны I: 56, 57
Размножение и возраст II: 21—22
— скорость внутренняя II: 26—27,
29
— скорость чистая II: 19
Разнообразие II: 126—148
— биохимическое II: 188—189
— видовое II: 126—137
— генетическое II: 127, 136, 146—
148
— в глубоководных районах II:
141—142
— и географическая широта II: 127
— и жизненный цикл II: 136—137
— и загрязнение II: 138, 189, 140—
142
— и инсектицидный стресс II: 138,
140
— и сукцессия II: 187—188
— п устойчивость II: 134
— индексы II: 132—133
— ландшафта II: 148—150
— кривые см. Кривые доминирова-
ния — разнообразия
— структурное I: 127, 137
— а, |3 и у II: 135, 148
Разнотравье II: 261
Распределение II: 57—62
— биомов II: 248
— и численность организмов I: 257
— особей в популяции II: 57—62
----------групповое II: 57, 58, 611
----------равномерное II: 57—61
----------случайное II: 57—61
— типов почв I: 306
Расселение II: 40
Растения С3- и С4- I: 44—46, 167
Растительность I: 78, 102, 103, 222,
II: 248.
Растительноядные I: 80, 70; II: 143
— в пищевых цепях I: 143, 145, 156
164—165; II: 248, 250
Расходы, связанные с масштабами I:.
180, 183—184
Редукционизм I: 18
Регионализм II: 240
Рентгеновские лучи I: 275
Ресурсы жизнеобеспечения см. Си-
стема жизнеобеспечения
Резистентная устойчивость см
Устойчивость
Рис, культура I: 99, 207, II: 216
Ритмы циркадные I: 265
Рождаемость I: 187; II: 12—13,
Рост городов I: 93—94
— народонаселения I: 188; II: 37.,
234, 245
— недифференцированный и орга-
нический II: 239
— популяции логистический II г
33—35
----сигмоидный (S-образная кри-
вая) I: 181, 182; II: 30—36
----экспоненциальный (J-образная
кривая) II: 28—29, 30—32, 36, 39
— скорость II: 24—31
— форма II: 31—40
Ртуть, круговорот I: 237—238
Рубец и бактерии-симбионты жвач-
ных II: 112
Рыба, возрастное распределение II:
22—23
— в пищевых цепях I: 153—156, 159
Рыболовство в водохранилищах II:
182, 183
— как пищевой ресурс для челове-
ка I: 154—157
— морское II: 289
— оптимальный размер вылова II:
104—105
Рыночные товары и услуги I: 197:
П: 228—231
Рысь, циклы изменения численности
II: 43
Саванна I: 282; II: 262—263
Сальпы (пелагические оболочники)
I: 56
Саранча, вспышки численности II: 48
Сапротрофы I: 29, 31, 33, 58, 70
Предметный указатель
369
Сапрофаги I: 29, 49, 75 (табл.)
Свет I: 270—272. См. также Солнеч-
ное излучение; Фотопериодизм
— как источник энергии I: 105—106
— как лимитирующий фактор I:
270—272
— спектральное распределение 1:
112—113, 271
Светлых и темных сосудов метод I:
71—73
Световое насыщение I: 272
Свойства экосистем I: 20, 21
----коллективные I: 16, 17
----на уровне сообщества П:
162—163
----несводимые I: 16
----эмерджентные I: 16—17, 34,
250; II: 163
Сельское хозяйство см. также Земле-
делие, Культуры сельскохозяй-
ственные
----беспахотное I: 101 (табл.), 125
----детритное II: 216—217
----и круговорот элементов пита-
ния I: 222—224, 227
----и эрозия почвы I: 305—309
----тенденции развития I: 101
(табл.)
----эффективность I: 99
Сера и загрязнение воздуха I: 213
См. также Кислый дождь
— как лимитирующий фактор I: 296
— круговорот I: 211—214; II: 283
— окислы загрязняющие I: 213
Сервомеханизмы I: 62
Сериальные стадии II: 166, 187
Серия II: 165, 177, 187, 193
Серные бактерии см. Бактерии
Сигмоидная (J-образная) кривая
роста численности популяции I:
182; II: 30—36, 38, 39—40
Симбиоз II: 86, 117
— облигатный II: 111. См. также Му-
туализм
Сине-зеленые водоросли (цианобак-
терии) I: 207, 268, 296
Синергизм I: 321
Система жизнеобеспечения (процес
сы и ресурсы среды) I: 12, 38,
88, 103, 190
----как нерыночная ценность II:
228—232, 247. См. также Природ-
ные товары и услуги
Системы I: 14, 20—22, 35. См. также
Модели
— обобщенная модель I: 22; II: 169
— уровень сложности II: 163
— с промежуточной размерностью
II: 163
Скорость, основные представления
II: 22, 24—27
Следовые элементы I: 227, 296—297
Сложность и соотношение между ча-
стями и целыми II: 163
— теория I: 179
— эволюция II: 210—212
Смена видов в сукцессии II: 166,
177—184
Смертность II: 13—18
Смещение признаков II: 95, 203—205
Смог фотохимический I: 21, 321
Соевые бобы I: 121, 136, 137 (табл.)
Соленость океана II: 286
— и температура I: 260—261
Солнечная постоянная I: 112
Солнечное излучение I: 107—108,
112—117, 234
— — рассеяние I: 116 (табл.)
---- спектральное распределение I:
113, 271
Сообщества биотические I: 14, 24, 26,
35, 272. См. также Биомы, Экоси-
стема
----и популяции II: 162—164
----и сукцессия см. Сукцессия эко-
логическая
----классификация на основе ме-
таболизма II: 169
— — содержание хлорофилла I:
133—135
— — структура в прошлом II: 158—
161
— — структурное разнообразие II:
137
Сосны II: 179—180, 191, II: 253, 257
— и микориза I: 40; II: 117
Спартина (морская трава) I: 51, 52,
218, 219; II: 294-295
Спектр электромагнитного излуче-
ния I: 270, 271
Среда см. также Экосистема
— биологический контроль I: 36—41
— и энергия I: 112—117
— на входе и на выходе I: 24, 27
— энергетические характеристики
I: 112—117
Стабильность (устойчивость) экоси-
стем I: 60, 63—66
— и разнообразие II: 135
— и экологическая сукцессия II:
166, 167 (табл.)
— импульсная II: 215—216
— резистентная и упругая I: 66—67
Стандартный урбанизированный рай-
он (СУР) I: 89. См. также Город
370
Предметный указатель
Стеногалинность I: 251
Стеногидрия I: 251
Стеноойкность I: 251
Стенотермия I: 251, 252—253
Стенофагия I: 251
Стоки бытовые, разложение I: 56—57
-------влияние на разнообразие в
ручье II: 138, 139
Стратифицированность в сообщест-
вах II: 127. См. также Зональ-
ность
Стресс антропогенный I: 316—318
— и разнообразие II: 127, 138—139
— и субсидия I: 125—127
— острый I: 316
— тепловой I: 322, 323
— хронический I: 316
Стронций радиоактивный I: 237
---накопление в пищевых цепях
I: 279—280
Структурное разнообразие в сообще-
ствах II: 126—127, 137
Субклимакс антропогенный II: 196
Субсидия и стресс I: 125—127 \ II:
157
Сукцессия экологическая II: 165—192
— автотрофная II: 166—169, 186
— аллогенная II: 165, 171, 182
— аутогенная II: 165—167, 171, 182
— в водных местообитаниях II:
182—184
— влияние возмущений II: 174, 175
— вторичная II: 166, 179—181
— гетеротрофная II: 166, 168, 169,
184—186.
— донных животных II: 179
— и давление отбора II: 186
— и тенденции изменения разнооб-
разия II: 187—188
— на искусственных субстратах II:
184
— на песчаных дюнах II: 177—178
— птиц II: 179
— первичная II: 166, 177
— - теоретические соображения II:
189—191
— циклическая II: 173—174
Сульфат в круговороте серы I: 212
Таблицы выживания II: 14, 15
Температура и влажность I: 290—
293
— и соленость I: 260
— как лимитирующий фактор I:
268—269
Температурная компенсация I: 262,
263
Тепловое загрязнение I: 322—323
Термодинамика, законы I: 104—111
— неравновесная I: 105, 108
— пищевых цепей I: 144
Термоклин II: 278
Территориальность II: 53, 66—70
Течения I: 297—299
Токсичные (ядовитые) вещества I:
55, 316-321
----в грунтовых водах I: 317—318
----избавление I: 319
----круговорот I: 236—238
----накопление в пищевых цепях
I: 165—167
----синтезируемые растениями II:
106
Толерантность, пределы I: 249—252
Топливо из биомассы I: 138, 140
— ископаемое см. Ископаемое топ-
ливо
Транспирация I: 77, 284
Трипсы, характер роста численно-
сти популяции II: 39
Трофическая структура I: 28
----и экологические пирамиды I:
174—178
----реакция на стрессовое взаимо-
действие I: 179
----уровни I: 142—144
Туман и прибрежная растительность
I: 286; II: 291
Тундра II: 251—253, 275
— пищевые цепи I: 152
Тяжелые металлы I: 236—238
Углерод, глобальный круговорот I:
225—229
Уголь I: 55
— и загрязнение воздуха I: 213
Удобрения и урожай сельскохозяйст-
венных культур I: 122, 127
Ультрафиолетовое излучение I: 113г
229, 270
•---и эволюция биосферы II: 198—
200
Управление осадками I: 286
Ураганы I: 299
Уран, продукты деления ядер I::
279—280"
Урбанизация I: 93. См. также Город
— и налоги I: 184, 185
— и элементы питания в ручьях I:
223, 224
Уровни организации I: 13—14; II:.
163
— смертности II: 13
— сложности II: 163
Урожай см. также Первичная про-
дукция
— максимальный II: 104, 106
— на корню I: 119
Предметный указатель
371
— пищи для людей I: 135—137
— устойчивый I: 182
Усиливающий эффект I: 21
Условия существования как регули-
рующие факторы I: 264—267
Усоногие ракообразные, распределе-
ние II: 109
Устойчивость резистентная I: 66
— упругая I: 66
Фаготрофы I: 29
Факторы, взаимодействие I: 249,
259—260
— компенсация I: 261—164, 272
— лимитирующие см. Лимитирую
щие факторы
— регулирующие I: 264—267
— функционально важные I: 252
Фенолы (фенольные соединения) I:
54
Феромоны I: 325
Физические факторы (обзор) I: 267—
315
Фиксация азота, метод восстановле-
ния ацетилена I: 207
Фитопланктон I: 30, 70, 129, 272
— лимитирующие факторы I: 254
— морской II: 288
— эволюция II: 199
Фотопериодизм I: 265—267
Фотосинтез, происхождение II: 200—
201
— типы I: 42—47. См. также Первич-
ная продукция
Фосфат в круговороте фосфора I:
210—211
Фосфор, взаимодействие с азотом I:
223, 296
— как лимитирующий фактор I: 211,
296
— круговорот I: 210—211, 219
— радиоактивный (32Р) I: 215—216
Функции взаимодействий: 20
Хаббард-Брук (шт. Ныо-Гэмпшир),
экспериментальный водосборный
бассейн I: 220
Хелатирование I: 54—55
Хемолитотрофы I: 46, 147
Хемосинтезирующие бактерии I: 46
Хемостат I: 81
Хирономиды (мотыль) I: 155
Хищники I: 30, 70. См. также Хищ-
ничество
— в нишевых цепях I: 143—144, 156
Хищник — жертва, взаимоотноше-
ния I: 186; II: 84 (табл.), 99, 103,
212
Хищничество II: 83, 98—106
— как лимитирующий фактор I:
257, 259; II: 106
— размеры жертвы II: 106
Хлорелла I: 256
Хлорофилл I: 32, 114
— и первичная продукция I: 133—
135, 296
Холизм и экономика и политика I:
18, 34; II: 163, 246
Хрущаки мучные
----межвидовая конкуренция II:
89—91
Царства таксономические I: 32—33
«Цветение» (период высокой продук-
тивности) I: 126; II: 30, 292
Цезий радиоактивный (137Cs) I: 237
----в оленях I: 280—281
Целлюлоза I: 51, 71, 74. См. также
Лигнин
Ценности рыночные п нерыночные
II: 228—233, 236, 247
Цианобактерии. См. также Сине-зе-
леные водоросли
Цивилизация деньги и энергия I:
194—195
— зависимость от окружающей сре-
ды I: 12. См. также Город, Систе-
ме! жизнеобеспечения
— и энергия I: 194—196
— как система II: 237
Цинк I: 248, 297
Циркадный ритм I: 265
Чапараль I: 67, 314, II: 80, 107, 108,
173, 263—265
«Черный ящик», концепция, исполь-
зуемая в моделировании I: 34;
II: 162, 163
Численность относительная II: 6—8.
См. также Плотность популяции
— населения земного шара, прогноз
на будущее I: 188
Число видов/число особей, соотно-
шение II: 128, 129
Чистая первичная продукция см.
Первичная продукция
Чистая энергия, концепция I: 196
Членистоногие см. также Насекомые
п др. группы
— и мутуалистические микроорга-
низмы II: 113
— плотность и разнообразие I: 75
(табл.); II: 141, 143 (табл.)
— роль в деструкции I: 57
Шмели п цветки, коэволюция II: 209
Шредингера соотношение I: 109
Шеннона индекс II: 133—134
Эвапотринспирация I: 133, 290—291
372
Предметный указатель
— атмосферные осадки и осадки I:
291
— и первичная продукция I: 284
— и разложение подстилки I: 53
Эверглейдс (шт. Флорида), нацио-
нальный парк I: 78; II: 216
Эволюция II: 197—213
— атмосферы I: 59, 227—229
— биосферы II: 197—202
— и экология человека II: 213—228
— микро- и макро- II: 202
— прерывистая II: 203
— ранней жизни I: 50; II: 197—201
— сложности II: 210—212
Эвригалинность I: 251
Эвригидричность I: 251
Эвриойкпость I: 261
Эвритермность 1: 251, 253
Эврифагия I: 251
Эвтрофикация антропогенная I: 78,
258—259; II: 106, 171
— и пищевые цепи I: 164
— как лимитирующий фактор I:
255, 258—259
— экспериментальная I: 258—259
Эвфотическая зона I: 73; II: 288
Эдафический климакс II: 192—194,
195, 196, 248
Эквивалент популяции домашнего
скота I: 100, 138, 141
Экологическая пиша см. Ниши эко-
логические
Экологические возрастные группы
II: 20—21
— пирамиды I: 174—178
— эквиваленты II: 119, 125—126
(табл.)
Экологическое доминирование см.
Домипи рование
Экология, изменение интересов ака-
демической экологии I: 12—13
— история I: 12—13
— определение I: И
— популяционная см. Популяция
— предмет I: 11—23
— радиационная I: 217, 272—281
— «стрессовая» I: 35, 249
— человек см. Человек
— экосистем I: 25
Экономика, определение I: 11
— п разрушение окружающей среды
I: 40*
— и экологические соображения I:
199; II: 228—232, 245—246
— и энергия I: 194—199
— рыночная и нерыночная II: 231
(табл.)
Экосистема (ы) I: 14, 24—28
— водосборного бассейна I: 77, 78
— движимые топливом 1: 89—90,
190, 192
— зависимые от возмущений II: 174
— залежи I: 68, 80; II: 91, 178—179
— избыточность компонентов I: 63,
64
— изучение, мерологический подход
I: 34-35; 82; II: 162
— и ископаемое топливо I: 170
— кибернетическая природа I: 60—
63
— классис^икация I: 188—193
— компоненты 1: 20, 68—70
— логические I: 103; II: 276—280
— приводимые в действие Солнцем
I: 188—191
— примеры I: 68—102; Приложение
— развитие II: 165—192. См. также
Сукцессия экологическая
— сельскохозяйственные I: 97—102
— структура I: 28—33
— устойчивость I: 65—67
— холистический подход I: 34—35;
II: 162
Экосистемный уровень организации
I: 24
Экотипы I: 261
Экотоны II: 150, 155—157
Экосфера I: 15
Экспоненциальный рост II: 28, 299
30, 34
Эктокрины I: 58
Электромагнитный спектр I: 270, 271
Элементы питания I: 296
---время оборота I: 216, 217
(табл.)
-------пребывания I: 216
--- скорость оборота I: 216
Эмерджентные свойства I: 16—173>
34; II: 163, 250
Энергетика. См. также Поток энер-
гии
— круговорота воды (гидрологиче-
ского цикла) I: 229, 230
— экологической сукцессии II: 167
(табл.), 168—171
Энергетическая субсидия I: 121—127
— — и классификация экосистем I:
188, 190 (табл.)
Энергия в экосистемах I: 104—199
— единицы измерения I: 110—111
(табл.)
— затраченная 1: 107, 170, 199
— и деньги I: 194—199
— и качество жизни II: 234
— качество I: 108, 168—171
— количество и качество I: 170, 194
Предметный указатель
373
— потребление I: 192—193
— распределение II: 70—80, 213
— фундаментальные концепции (за-
коны) I: 104—111
— чистая I: 196
Энтропия I: 104—105, 108—109
Эпилимнион II: 278
Эпифауна II: 289
Эпифиты тропического леса II: 272
Эрозия см. Почва
Эстуарии I: 125, 133; II: 293
Эукариоты I: 32; II: 200
Эффективность экологическая 1г.
160-162
Оглавление
’Глава 6. Динамика популяций.................................... 5
1. Свойства популяционной группы................................ 5
Определения.......................................... 5
Объяснения........................................... 6
2. Основные представления о скоростях ..........................22
3. Внутренняя скорость естественного роста......................26
Определения..........................................26
Объяснения...........................................27
4. Кривая роста популяции.......................................30
Определения......................................... 30
Объяснения...........................................31
Примеры..............................................30
5. Флуктуации численности популяций и «циклические» колебания 41
Определения..........................................41
Объяснения...........................................41
Примеры . . . ‘.................................42
6. Независимая и зависимая от плотности регуляция численности
популяций.......................................................31
Определения..........................................31
Объяснения...........................................52
Примеры..............................................53
7. Структура популяции: характер распределения организмов в про-
странстве ......................................................57
Определения..........................................57
Объяснения и примеры.................................57
8. Структура популяции: агрегация, принцип Олли и безопасные
поселения.......................................................52
Определения..........................................52
Объяснения и примеры.................................52
9. Структура популяции: изоляция и территориальность .... 66
Определения..........................................56
Объяснения и примеры................................ 56
10. Распределение энергии и оптимизация: г- и К-отбор .... 70
Определения..........................................70
Объяснения...........................................71
Примеры..............................................77
Оглавление
375
11. Интеграция: особенности и тактика жизненного цикла .... 80
Определения..........................................80
Объяснения и примеры.................................81
Глава 7. Популяции в сообществах................................83
1. Типы взаимодействия между двумя видами.......................83
Определения..........................................83
Объяснения...........................................84
2. Конкуренция и сосуществование видов..........................87
Определения......................................... 87
Объяснения...........................................87
Примеры..............................................93
3. Хищничество, растительноядность, паразитизм и аллелопатия (ан-
тибиоз) .....................................................98
Определения..........................................98
Объяснения и примеры ................................98
4. Положительные взаимодействия: комменсализм, кооперация и му-
туализм ................................................... 110
Определения..........................................ПО
Объяснения....................................... . ПО
Примеры.............................................112
5. Концепции местообитания, экологической пиши и гильдии . . 119
Определения.........................................119
Объяснения и примеры................................119
6. Видовое, структурное и генетическое разнообразие в сообществах. 126
Определения.........................................126
Объяснения......................................... 127
Примеры.............................................138
Озабоченность исчезновением видов и утратой генетиче-
ского разнообразия..................................146
7. Популяции и сообщества в географических градиентах; экотоны и
понятие краевого эффекта........................................150
Определения.........................................150
Объяснения и примеры................................151
8. Палеоэкология: структура сообществ в прошлые века . . . . 158
Определения.........................................158
Объяснения..........................................158
Примеры.............................................159
9. От популяций к сообществам и экосистемам....................162
Глава 8. Развитие и эволюция экосистемы .... . . 165
1. Стратегия развития экосистемы...............................166
Определения.........................................166
Объяснения и примеры................................166
2. Концепция климакса..........................................192
Определения.........................................192
Объяснения и примеры................................192
3. Эволюция биосферы...........................................197
Определения.........................................197
Объяснения..........................................198
4. Естественный отбор: аллопатрическое и симпатрическсе видообразо-
вание; микроэволюция и макроэволюция.................... . 202
Определения................................... . . 202
Объяснения..........................................202
Примеры.............................................204
5. Коэволюция..................................................208
Определения.........................................208
376
Оглавление
Объяснения........................................208
Примеры...........................................209
6. Эволюция кооперации и сложности; групповой отбор . . . . 210
Определения.......................................210
Объяснения и примеры........................... . . 211
7. Применимость теории развития экосистемы и эволюции биосферы
к экологии человека.............................................213
Определения..........................................213
Объяснения...........................................213
Эпилог. Трудности, стоящие перед человечеством: взгляд в будущее 234
Отчеты Римского клуба................................237
Глобальные модели и варианты будущего мира . . . 243
Экологическая оценка.................................245
Приложение. Краткое описан не основных типов природных экосис-
тем биосферы.........................................248
1. Наземные биомы...............................................248
Тундры — арктическая и альпийская (рис. П. 3) . . . 251
Биомы северных хвойных лесов (рис. П. 4) .... 253
Листопадные леса умеренной зоны (рпс. П.5) .... 257
Степи умеренной зоны (рис. П. 6)....................259
Тропические степи и саванны (рис. П. 7)..............262
Чапараль и жестколистные леса (рис. П. 8) . . . . 263
Пустыни (рис. П. 9)..................................267
Полувечнозеленые сезонные тропические леса
(рис. П. 10,А):......................................271
Тропические дождевые леса (рис. П. 10, Б и В) . . . 272
Тропический скрэб, или колючее редколесье . . . . 274
Зональность в горах (рис. П. И)......................275
2. Пресноводные экосистемы.....................................276
Леитические экосистемы (озера и пруды)...............277
Лотические экосистемы (ручьи и реки).................279
Заболоченные пресноводные участки (марши и болота)
(рис. П. 12, А).....................................230
3. Морские экосистемы (рис. П. 14)..............................283
Область континентального шельфа......................289
Области апвеллинга...................................291
Лиманы (рис. П. 18)..................................292
Литература..........................................297
Литература......................................................303
Предметный указатель............................................359
Юджин п. Одум
ЭКОЛОГИЯ
Научный редактор М. Б. Николаева. Мл. научи, редактор О. В. Шагинян. Художник
В. С. Стуликов. Художественный редактор Л. М. Кузнецова. Технический редактор
А. Л. Гулина. Корректор В. И. Киселева.
ИБ № 5491
Сдано в набор 26.11.85. Подписано к печати 8.04.86. Формат бОХЭО’Ле. Бумага
типографская № 1. Печать высокая. Гарнитура обыкновенная. Объем 11,75 бум. л.
Усл. печ. л. 23,50. Усл. кр.-отт. 23,50. Уч.-изд. л. 26,80. Изд. № 4/4136. Тираж
13 500 экз. Зак. 1483. Цена 2 р. 30 к.
Издательство «МИР». 129820, ГСП, Москва, И-110, 1-й Рижский пер., 2.
Московская типография № 11 Союзполиграфпрома при Государственном комитете
СССР по делам издательств, полиграфии и книжной торговли. Москва, 113105,
Нагатинская, 1.