/







































































































































































































































































































































Автор: Лемеза Н.А. Лисов Н.Д. Камлюк Л.В.
Теги: общая и теоретическая биология общая биология биология учебное пособие 10 класс
ISBN: 985-03-0814-1
Год: 1998




Текст
1 » : щШк
Ш- Щ л 'ч
' % f !‘Ж
Ж
1
Ш г! т Ш Ш Ш
§ 1
S i
1 1*1
1Н51Р«Ш Г
$ «■Н
*\л^Т 1аР
у 1РИ • уД
Iyi
rF IT ^ t
1 1
' - /-■-
.-
« « КАЛ T K r « « ? * S s ааЛгЭБ;
*л
^ .ч л я т ш
шт
*#ГЙ$ *
Ж
• ,<%
| ife.Зи
^ ~ S
Ш
Ё Ы
Ш
'Ш
Ш
Ш
Ш
Ш
Ш
Ж
Н. А. Лемеза
Н. Д. Лисов
Л. В. Камлюк
ОБЩАЯ
Н \ М J [•
Учебное пособие для 10 класса
с углубленным изучением биологии
общеобразовательных школ
с русским языком обучения
Под редакцией Н. Д. Л и с о в а
Допущено Министерством образования
Республики Беларусь
Минск «Народная асвета» 1998
УДК 573(075.3)
ББК 28.0я721
Л 44
Р е ц е н з е н т ы :/'. А. Бавтуто, проф. БГПУ им. М. Танка, доктор биол. наук; А. Г. Песнякевич, доц. Б ГУ,
канд. биол. наук; 3. И. Шелег, учитель биологии сред. шк. № 34 г. Минска, канд. биол. наук
Л 44
Лемеза Н. А. и др.
Общая биология: Учеб, пособие для 10-го кл. с углубл.
изуч. биологии общеобразоват. шк. с рус. языком обуче
ния / Н. А. Лемеза, Н. Д. Лисов, Л. В. Камлюк; Под ред.
Н. Д. Лисова.— Мн.: Нар. асвета, 1998.— 366 с.: ил.
ISBN 985-03-0814-1
УДК 573(075.3)
ББК 28.0я721
ISBN 985-03-0814-1
© Н. А. Лемеза, Н. Д. Лисов,
Л. В. Камлюк. 1998
© Б. Г. Клюйко, оформление, 1998
© ИГП «Народная асвета», 1998
Введение
История Земли и многообразие усло
вий на нашей планете определили уни
кальное свойство — жизнь, создали ог
ромное по размаху биологическое разно
образие жизни. Что же такое жизнь? До
настоящего времени нет достаточно чет
кого и однозначного ее определения как
реально существующего явления. При
описании живых систем обычно выделя
ют отдельные черты или свойства, при
знаки или критерии, на основе которых
можно отличить живое от неживого.
Более или менее строгое определение
жизни появилось во второй половине
XIX в. Жизнь определялась как способ
существования белковых тел, состоящий,
по существу, в постоянном самообновле
нии химических составных частей этих
тел. Исходя из этого, жизнь — это спо
соб существования материальных белко
вых тел. Жизнь — это и непрерывное
движение, заключающееся в постоянном
самообновлении белковых тел, которое
осуществляется за счет обмена веществ
биологических систем с окружающей
средой. Именно обмен веществ, по су
ществу, и есть особая форма движения
материи — биологическая. Обмен ве
ществ составляет основу всех других
свойств жизни: раздражимости, способ
ности к росту и размножению, наслед
ственности и изменчивости, внутреннего
движения в процессах ассимиляции пита
тельных веществ и т. д. Вне движения нет
и не может быть жизни.
Изучение свойств жизни показало,
что жизнь связана со сложным коллоид
ным состоянием протопласта (внутрен
него содержимого клетки), для которого
характерны обмен веществ и энергии,
обусловленные особым способом реали
зации наследственной информации, за
ключенной в нуклеиновых кислотах. Ж и
вые системы от клетки до биосферы в це
лом представляют собой системы, асси
милирующие энергию из внешней среды
таким образом, что могут активно проти
востоять разрушению сложившейся ор
ганизованности, т. е. противостоять про
цессу, характерному для всех тел неорга
нической природы.
Таким образом, жизнь можно опреде
лить как способ существования откры
тых коллоидных систем, обладающих
свойствами саморегуляции, воспроиз
ведения и развития на основе биохими
ческого взаимодействия белков, нукле
иновых кислот и других соединений
вследствие преобразования веществ и
энергии из внешней среды.
Проявления жизни на нашей планете
чрезвычайно многообразны. В связи с
этим выделяют различные уровни или
ступени организации жизни: молекуляр
ный, клеточный, организменный, популя
ционно-видовой, биогеоценотический и
биосферный.
М о лекуляр н ы й — самый низкий
уровень организации жизни. Элементар
ными единицами этого уровня являются
химические вещества: нуклеиновые кис
лоты, белки, углеводы, липиды и др. На
этом уровне в основном проявляются
такие процессы жизнедеятельности, как
реализация и передача наследственной
информации, биосинтез, физико-хими
ческие реакции и др. Выяснение сути
процессов, происходящих на молекуляр
ном уровне, помогает понять некоторые
явления, происходящие на последующих
уровнях организации. Основная страте
гия жизни на молекулярном уровне —
способность создавать живое вещество и
кодировать информацию, приобретенную
в меняющихся условиях среды.
На клет очном уровне организации
структурными элементами выступают
различные органеллы. Способность к
воспроизведению себе подобных, вклю
чение различных химических элементов
Земли в содержимое клетки, регуляция
химических реакций, запасание и расхо
дование энергии — главные процессы
данного уровня. Основная стратегия
3
жизни на клеточном уровне — вовлече
ние химических элементов Земли и энер
гии солнечного излучения в живые био
системы.
Организменный уровень организа
ции присущ многоклеточным и однокле
точным биосистемам. Здесь жизнь пред
ставлена растениями, грибами, протеста
ми и разнообразными микроорганизма
ми, животными, в том числе человеком.
Все они являются структурными едини
цами этого уровня. У представителей организменного уровня проявляются такие
свойства, как питание, дыхание, раздра
жимость, подвижность, выделение, раз
множение, рост и развитие, поведение,
продолжительность жизни, плодови
тость, образ жизни, взаимоотношения с
окружающей средой. Все вместе пере
численные процессы характеризуют ор
ганизм как целостную саморегулирующу
юся биосистему. Основная стратегия
жизни на этом уровне — ориентация ор
ганизма (особи) на выживание в постоян
но меняющихся условиях среды, что за
висит главным образом от его состояния
здоровья, а в связи с этим обеспечение
длительного существования популя
ции вида.
П опуляц и о н но -ви д о во й уровень
организации характеризуется объедине
нием родственных особей в популяции, а
популяций — в виды, что приводит к воз
никновению новых свойств системы, от
личных от предыдущих уровней. Основ
ные свойства этого уровня: рождаемость,
смертность, выживаемость, структура
(половая, возрастная, экологическая),
плотность, численность, функционирова
ние в природе. Основная стратегия попу
ляционно-видового уровня проявляется в
более полном использовании возможнос
тей среды обитания, в стремлении к воз
можно более длительному существова
нию, в сохранении свойств вида и самос
тоятельном развитии.
На биогеоценот ическом уровне
организации основными структурными
4
элементами являются популяции и ви
ды — целостные природные образова
ния. Однако они как структурные едини
цы включаются в биосистемы более вы
сокого ранга — биогеоценозы. Данный
уровень характеризуется множеством
надорганизменных свойств. К ним отно
сятся: структура экосистемы, видовой и
количественный состав ее населения, ти
пы биотических связей, пищевые цепи и
сети, трофические уровни, продуктив
ность, энергетика, устойчивость и многое
другое. Организующие свойства прояв
ляются в круговороте веществ и потоке
энергии, саморегулировании и устойчи
вости, автономности, открытости систе
мы, сезонных изменениях. Основными
функциональными единицами здесь вы
ступают популяции, основная страте
гия — активное использование всего
многообразия возможностей окружаю
щей среды и создание благоприятных ус
ловий развития и процветания жизни во
всем ее многообразии.
Основными структурными единицами
наиболее высокого биосферного уров
ня организации являются биогеоценозы и
окружающая их среда, т. е. географичес
кая оболочка Земли (атмосфера, гидро
сфера, почва, солнечная радиация идр.),
и антропогенное воздействие. Для этого
уровня организации характерны: актив
ное взаимодействие живого и неживого
вещества планеты; биологический круго
ворот веществ и потоки энергии с входя
щими в него геохимическими циклами;
хозяйственная и этнокультурная деятель
ность человека. Основная стратегия жиз
ни биосферного уровня — стремление
обеспечить динамическую устойчивость
биосферы как самой большой экосисте
мы нашей планеты.
Жизнь во всем ее многообразии
изучается биологией. Современная
биология представляет собой комплекс
биологических наук. Общая биология
исследует наиболее общие свойства и за
кономерности живой материи, проявляе
мые на разных уровнях организации, и
объединяет ряд частных биологических
наук, которые изучают отдельные сторо
ны существования и развития объектов
живой природы.
Клетку как структурную и функцио
нальную единицу живых организмов из
учает цитология, ткани — гистология,
общие закономерности индивидуального
развития организмов исследуют эмбрио
логия и биология развития.
Химический состав клеток, структуру,
функции, распределение и превращение
химических веществ в клетке изучают
биохимия и молекулярная биология-,
физические и химико-физические явле
ния в клетках и организмах исследует
биофизика, из которой в самостоятель
ные разделы выделились радиобиоло
гия, биомеханика, биоакустика, био
метеорология и др.
Закономерности наследственности и
изменчивости живых систем изучает ге
нетика-, историческое развитие живой
природы и многообразие органического
мира — эволюционное учение.
Важнейшим разделом общей биоло
гии является биометрия, которая разра
батывает методы обработки результатов
биологических экспериментов и оценки
их достоверности.
Изучением коллективной жизни со
обществ живых организмов занимаются
этология — наука о поведении живот
ных и экология — наука о взаимоотно
шениях различных организмов в образуе
мых ими сообществах между собой и с
окружающей средой. Отдельными разде
лами экологии являются популяционная
биология — отрасль знаний, изучающая
структуру и свойства популяций, и био
ценология — наука о сообществах жи
вых организмов.
Биогеография изучает общие за
кономерности географического рас
пространения живых организмов на
Земле.
В зависимости от того, в какой облас
ти практической деятельности человека
используются биологические знания, вы
деляют такие дисциплины, как биотех
нологию — совокупность промышлен
ных методов, позволяющих с высокой
эффективностью использовать живые
организмы и отдельные их части для про
изводства ценных продуктов (антибиоти
ков, аминокислот, белков, витаминов,
гормонов, ферментов и др.), для защиты
растений от вредителей и болезней, для
борьбы с загрязнением окружающей сре
ды, в очистных сооружениях и т. п.; агро
биологию — комплекс знаний о возде
лывании сельскохозяйственных культур;
селекцию — науку о методах создания
сортов и гибридов растений, пород жи
вотных, штаммов микроорганизмов с
нужными человеку признаками. В этот
же ряд следует поставить также живот
новодство, ветеринарию, медицин
скую биологию, ф ит опат ологию,
биологию охраны природы.
В данном учебном пособии изложены
основные сведения о свойствах и законо
мерностях, проявляющихся на клеточном
и организменном уровнях организации
жизни.
Р А З Д Е /1 ^
7П П Г М 5
КЛЕТКА —
СТРУКТУРНАЯ И ФУНКЦИОНАЛЬНАЯ
ЕДИНИЦА ЖИЗНИ
Клетка в системе живой природы.
Клеточная теория
Клетка в системе живой природы.
Все живые организмы построены из кле
ток. Одноклеточные организмы (бакте
рии, некоторые цианобактерии и протес
ты) состоят из одной клетки, многокле
точные (грибы, растения, животные) —
из множества клеток.
Клетка обладает всеми свойствами
живой системы: она осуществляет обмен
веществ и энергии, размножается и пере
дает по наследству свои признаки, реаги
рует на действие внешних раздражите
лей. Она является наименьшей структур
ной и функциональной единицей всего
живого.
Многоклеточные организмы состоят
из огромного количества клеток, которые
выполняют разнообразные функции и на
этой основе объединяются в ткани. Ком
6
плексы тканей в свою очередь формиру
ют органы. Будучи связанными функцио
нально, органы образуют живой орга
низм. В большинстве случаев в результа
те такой «узкой специализации» клетки
не могут существовать отдельно, вне ор
ганизма.
История создания клеточной тео
рии. Открытие клетки связано с именем
английского естествоиспытателя Р. Гу
ка, который в 1665 г. впервые рассмот
рел тонкий срез пробки в усовершенство
ванном им микроскопе. На срезе четко
просматривалась структура, похожая на
пчелиные соты, построенные из ячеек.
Элементы тонкого слоя пробки Гук на
звал латинским словом cellula, что оз
начает «ячейка», или «клетка». Вслед
за Гуком клеточное строение расте
ний подтвердили ботаники: итальянец
М . М а л ь п и г и (1675) и англичанин
Н. Гр ю (1682). Их внимание привлекли
форма клеток и строение их оболочек.
В результате было дано представление о
клетках как о «мешочках» или «пузырь
ках», наполненных вязким содержимым.
Значительный вклад в изучение клет
ки внес голландский ученый А . Л е в е н
гук, открывший в 1674 г. одноклеточные
организмы, в том числе и бактерии. Он
также впервые наблюдал под микроско
пом животные клетки — эритроциты
крови и сперматозоиды.
Дальнейшее усовершенствование
микроскопа привело к открытию фран
цузским ученым Ш . Ф. Б р и с с о М и р б е (1802, 1808) того факта, что все
растительные организмы образованы
тканями, которые в свою очередь состоят
из клеток. Еще дальше в обобщениях по
шел французский ученый Ж . Б. Л а
м а р к (1809), который распространил
идею Бриссо-Мирбе о клеточном строе
нии и на животные организмы.
В начале XIX в. предпринимаются по
пытки изучения внутреннего содержимо
го клетки. В 1825 г. чешский ученый
Я. П у р к и н е открыл ядро в яйцеклетке
птиц, а в 1831 г. английский ботаник
Р. Б р о у н впервые описал ядро в расти
тельных клетках. Таким образом, к 30-м
годам XX в. были накоплены некоторые
конкретные данные о структурной орга
низации клетки.
В 1839 г. немецкий биолог
Т. Ш в а н н опубликовал книгу «Мик
роскопические исследования о соответ
ствии в структуре и росте животных и
растений», в которой он обобщил имев
шиеся знания о клетке, в том числе ре
зультаты исследований немецкого бота
ника М . Ш л е й д е н а о роли ядра в
клетках растений. Главная идея книги
(потрясающая по своей простоте) своди
лась к тому, что жизнь сосредоточена в
клетках. Это вызвало революцию в био
логии. Иными словами, Т. Шванн сфор
мулировал клеточную теорию.
Основными положениями клеточной
теории были следующие:
1. Как растительные, так и живот
ные организмы состоят из клеток.
2. Клетки растительных и животных
организмов развиваются аналогично
и близки друг к другу по строению и
функциональному назначению.
3. Каждая клетка способна к самос
тоятельной жизнедеятельности.
Клеточная теория — одно из выдаю
щихся обобщений биологии XIX в., став
шее основой для понимания жизни и рас
крытия эволюционных связей между ор
ганизмами.
Дальнейшее развитие клеточная тео
рия получила в трудах немецкого ученого
Р. Вирхова, который в 1858 г. внес сущес
твенное дополнение в теорию: клетка мо
жет возникнуть только из предшествую
щей клетки в результате ее деления.
В 30-х годах XIX в. русский ученый
К ■ Б э р открыл яйцеклетку млекопитаю
щих и установил, что все многоклеточные
организмы начинают свое развитие из од
ной клетки. Это открытие показало, что
клетка является единицей не только
строения, но и развития всех живых орга
низмов.
Идея о том, что все организмы по
строены из клеток, стала одним из наибо
лее важных теоретических достижений в
истории биологии, поскольку создала
единую основу для изучения всех живых
существ. На клеточном уровне даже наи
более отдаленные виды живых организ
мов оказались весьма схожи по строению
и биохимическим свойствам, что указы
вало на общность их происхождения и ис
торического развития.
Дальнейшие успехи науки о клетке —
ц и т о л о г и и — связаны с усовершенст
вованием приборов и развитием физиче
ских и химических методов исследования.
Методы изучения клетки. Развитие
знаний о клеточном строении живых ор
ганизмов основывалось на данных све
тового микроскопирования.
Человеческий глаз обладает разре
шающей способностью около 100 мкм
(1 мкм = 0,001 мм). Это означает, что ес
ли вы смотрите на две точки, которые на
ходятся на расстоянии менее чем
100 мкм, то они кажутся одной расплыв
чатой точкой. Чтобы различить более
мелкие структуры, применяют оптичес
кие приборы, например микроскопы.
Один из лучших световых микроскопов
имеет разрешающую способность около
0,2 мкм, т. е. позволяет увеличивать ис7
слелуемый объект примерно в 1500—
2000 раз. Создать световой микроскоп с
большим разрешением невозможно, по
тому что разрешающая способность свя
зана с длиной волн световых лучей, а не
только с качеством увеличительных сте
кол и другими техническими проблемами.
Разрешающая способность элек
тронного микроскопа, изобретенного в
30-е годы XX в., почти в 400 раз больше,
чем светового. В принципе электронный
микроскоп устроен так же, как и свето
вой, только роль светового пучка выпол
няет в нем пучок электронов, и фокусиру
ется он не линзами, а электромагнитами.
В трансмиссионном электронном мик
роскопе электроны проходят сквозь
объект подобно тому, как в световом мик
роскопе сквозь объект проходит свет.
Современные электронные микроскопы
позволяют увеличивать тонкие структуры
клетки в 200 000 раз и больше.
Сканирующий электронный м ик
роскоп стали применять сравнительно
недавно — в 50-е годы. В нем очень точ
но сфокусированный пучок электронов
движется взад и вперед по поверхности
образца. Отраженные от поверхности
электроны формируют изображение на
подобие того, которое возникает на экра
не телевизора. Преимущество этого ме
тода заключается в том, что благодаря
глубине резкости создается эффект трех
мерности. Сканирующий электронный
микроскоп позволяет получать превос
ходные фотографии, воспроизводящие в
мельчайших деталях строение поверхнос
ти некоторых органелл клетки.
Современные
микрохимические
( цитохимические) методы анализа
позволяют установить химический состав
клетки и отдельных ее компонентов —
ядра, митохондрий, хлоропластов, рибо
сом и т. д.
Комплексное использование физи
ческих и химических методов исследова
ния, таких, как дифференциальное цен
трифугирование, метод меченых
8
атомов, автография, генная и кле
точная инженерия и др., позволило
установить тончайшую организацию
клетки, а также доказать неразрывную
связь между ее структурой и функцией.
Благодаря этому были развиты и углуб
лены основные положения клеточной
теории:
1. Клетка как элементарная живая
система, способная к самообновле
нию, саморегуляции и самовоспроиз
ведению, является основой строения
и развития всех живых организмов.
2. Клетки всех организмов построе
ны по единому принципу,сходны (го
мологичны) по химическому составу,
основным проявлениям жизнедея
тельности и обмену веществ.
3. Размножение клеток происходит
путем их деления, и каждая новая
клетка образуется в результате деле
ния исходной (материнской) клетки.
4. В многоклеточных организмах
клетки специализированы по выпол
няемым функциям и образуют ткани.
Из тканей состоят органы и системы
органов.
Цитология стремительно развивается
и в наши дни. Сегодня наука имеет доста
точно точные представления о химичес
ком составе, строении и функциях всех
структур клетки.
О
1. В чем сущность клеточной теории и кто ее
авторы? 2. С помощью каких приборов изучалась
клетка в XIX и XX вв.? 3. Каково значение клеточ
ной теории? 4. Каковы основные положения совре
менной клеточной теории?
ГЛАВА I
Молекулярные основы
жизни
Структурная организация и функцио
нальные особенности живой клетки оп
ределяются постоянно протекающими в
ней многочисленными химическими ре
акциями, в которых участвуют десятки
элементов и сотни простых и сложных со
единений.
1. Элементы и атомы
По химическому составу клетки раз
личных организмов могут существенно
отличаться друг от друга, но химические
элементы, входящие в их состав, одина
ковы.
Химическим элементом называется
совокупность атомов с одинаковым заря
дом ядра и одинаковым распределением
электронов по оболочкам и подоболоч
кам. В клетках обнаружено более 20 эле
ментов периодической таблицы Менде
леева, причем живая клетка состоит из
тех же элементов, что и неживые объек
ты. Это указывает на тесную связь и
единство живой и неживой природы.
Макро- и микроэлементы. Наиболь
ший удельный вес в элементном составе
живых организмов приходится на кисло
род, углерод, водород и азот. На долю
этих элементов приходится почти 98 %
массы клеток, причем их относительное
содержание в живом веществе гораздо
выше, чем в земной коре. Жизненно важ
ными являются также такие химические
элементы, как кальций, фосфор, сера,
калий, хлор, натрий, магний, йод, железо.
На их долю приходится немногим более
1 % массы клеток. Перечисленные хими
ческие элементы относятся к группе
макроэлементов.
Другие химические элементы, входя
щие в состав клетки, — медь, марганец,
молибден, кобальт, бор, цинк, фтор, се
лен, хром — содержатся в исключитель
но малых количествах (менее 0,01 %
массы клетки) и относятся к группе м и к
р о э л е м е н т о в . Содержание и значение
для жизнедеятельности клеток и организ
ма в целом различных химических эле
ментов приведено в табл. 1.
Т а б л и ц а 1 . Важнейшие химические элементы клетки
Элемент
Символ
Примерное
содержание,
% сухой мас
сы
Кислород
Углерод
Водород
Азот
О
62
с
20
н
10
N
3
Кальций
Са
2,5
Фосфор
Р
1
Сера
S
0,25
Калий
К
0,25
Хлор
С1
0,2
Натрий
Na
0,10
Значение для клетки и организма
Входит в состав воды и органических веществ
Входит в состав всех органических веществ
Компонент воды и органических веществ
Входит в состав аминокислот, белков, нуклеиновых кислот,
АТФ, хлорофилла, витаминов
Входит в состав клеточной стенки растений; костей и зубов;
повышает свертываемость крови и сократимость мышечных
волокон
Входит в состав костной ткани и зубной эмали, нуклеино
вых кислот, АТФ, некоторых ферментов
Входит в состав важнейших аминокислот (цистеина, цисти
на, метионина), некоторых витаминов, участвует в образова
нии дисульфидных связей в третичной структуре белка
Содержится в клетке только в виде ионов; повышает актив
ность ферментов белкового синтеза, обусловливает нормаль
ный ритм сердечной деятельности, участвует в процессах фо
тосинтеза, в генерации биоэлектрических потенциалов
Содержится преимущественно в виде отрицательного иона
в организме животных; компонент соляной кислоты в желу
дочном соке
Содержится в клетке только в виде ионов; обусловливает
нормальный ритм сердечной деятельности, влияет на синтез
9
Продолжение
Элемент
Символ
Примерное
содержание,
% сухой мас
сы
Магний
Mg
0,07
Йод
Железо
J
Fe
0,01
0,01
Медь
Си
следы
Марганец
Мп
Молибден
Мо
Кобальт
Со
Бор
В
Цинк
Zn
Фтор
F
—
Значение для клетки и организма
гормонов, участвует в поддержании и регулировании кислот
но-щелочного равновесия организма
Входит в состав молекул хлорофилла, а также костей и зу
бов; активизирует энергетический обмен и синтез ДНК
Входит в состав гормонов щитовидной железы
Входит в состав многих ферментов, гемоглобина и миоглобина, участвует в биосинтезе хлорофилла, в транспорте элек
тронов в процессах дыхания и фотосинтеза
Входит в состав гемоцианинов у беспозвоночных и некото
рых ферментов; участвует в процессах кроветворения, фото
синтеза, синтеза гемоглобина
Входит в состав или повышает активность некоторых фер
ментов; участвует в развитии костей, ассимиляции азота и
процессах фотосинтеза
Входит в состав некоторых ферментов (нитратредуктазы),
участвует в процессах связывания атмосферного азота клу
беньковыми бактериями
Входит в состав витамина В12, участвует в фиксации атмос
ферного азота клубеньковыми бактериями и образовании
эритроцитов
Влияет на ростовые процессы растений, активирует вос
становительные ферменты дыхания
Входит в состав некоторых ферментов, расщепляющих по
липептиды; участвует в синтезе растительных гормонов (аук
синов) и гликолизе
Входит в состав костей и эмали зубов
Строение атома. Все химические
элементы состоят из атомов. Атом —
это мельчайшая частица элемента, спо
собная принимать участие в химических
реакциях. В состав атомов входят три ви
да элементарных частиц: протоны
(электрический заряд + 1), нейтроны
(электрический заряд 0) и электроны
(электрический заряд —1). Массы прото
нов и нейтронов почти равны: в сум
ме они составляют массу ядра, рас
положенного в центре атома. Осталь
ное пространство атома принадле
жит электронам, каждый из которых
движется вокруг ядра по определенной
орбите.
Атом электрически нейтрален, потому
что число протонов в его ядре равно чис
лу электронов, движущихся вокруг ядра.
10
Такая система весьма устойчива, по
скольку положительно заряженные ядра
удерживают отрицательно заряженные
электроны, благодаря чему атом не рас
падается.
Электроны движутся вокруг ядра, за
полняя несколько электронных оболочек.
Каждая оболочка может содержать стро
го определенное число электронов: пер
вая, ближайшая к ядру оболочка, —
2 электрона, вторая — 8, третья — 18,
четвертая — 32 и т. д. Легко подсчитать,
что количество электронов в каждой обо
лочке равно 2«2, где п — номер оболоч
ки. На рисунке 1 представлено размеще
ние электронов в электронных оболочках
первых восьми элементов периодической
таблицы Менделеева. Атом водорода со
держит 1 электрон, атом гелия — 2, атом
Рис. 1. Размещение электронов в электронных оболочках атомов: 1 — водород: а — электрон, б —
ядро; 2 — гелий, 3 — литий, 4 — бериллий, 5 — бор, 6 — углерод, 7 — азот, 8 — кислород.
лития — 3, атом бериллия — 4, атом
кислорода — 8 электронов.
Связи между атомами. Любой хими
ческий элемент, у которого внешняя
электронная оболочка полностью запол
нена, практически не способен вступать в
реакции с другими элементами. Именно
этим объясняется инертность гелия, ко
торый крайне редко встречается в соста
ве химических соединений.
Все прочие элементы стремятся за
полнить свои внешние электронные обо
лочки, вступая в химические реакции с
другими элементами. Реакции сопровож
даются перегруппировкой электронов, в
результате которой внешняя электронная
оболочка каждого атома оказывается
полностью заполненной, и атом стано
вится более стабильным. При этом меж
ду атомами возникают связи одного из
следующих типов;
1)
и о н н ы е связи образуются в том
случае, когда один или несколько элек
тронов переходят от одного атома к дру
гому. В качестве примера рассмотрим ре
акцию между натрием и хлором (рис. 2 ).
Атом натрия, теряя единственный элек
трон своей внешней оболочки, получает
положительный заряд +1. Атом хлора, на
внешней оболочке которого 7 электро-
Рис. 2
11
нов, приобретая еще 1 электрон, получа
ет отрицательный заряд —1. У обоих ато
мов внешние электронные оболочки ока
зываются заполненными и, следователь
но, стабильными. Такие заряженные час
тицы называются ионами. Например, ион
натрия обозначают Na+, а ион хлора —
С1_. Ионы натрия и хлора, несущие про
тивоположные заряды, притягиваются
друг к другу и образуют соединение, на
зываемое хлоридом натрия (NaCl).
Другим типичным примером ионного
соединения является хлорид кальция
(СаС12). В этом случае атом кальция те
ряет два электрона, а каждый атом хлора,
наоборот, по одному приобретает. Ион
кальция обозначают Са++ или Са2+;
2)
к о в а л е н т н ы е связи возникают
в том случае, если атомы, участвующие в
их образовании, не отдают и не принима
ют электронов; вместо этого электроны
оказываются общими для двух атомов.
Например, два атома водорода, имеющие
по одному электрону, могут «обобщес
твить» их, и тогда каждый из атомов бу
дет иметь заполненную внешнюю обо
лочку с двумя электронами (рис. 3). В ре
зультате образуется молекулярный водород(Н2).
Между двумя атомами могут быть по
делены две пары электронов; в таком
случае образуется двойная ковалентная
связь, как, например, между двумя ато
мами кислорода (рис. 4). В некоторых со
единениях, например в ацетилене (С2Н2),
имеется тройная связь, указывающая
12
на три пары обобществленных элек
тронов.
В том случае, если ковалентная связь
образуется между двумя атомами одного
и того же элемента, эти атомы с равной
силой оттягивают на себя пару электро
нов, в результате чего общие электроны
находятся как бы посредине между ато
мами (см. рис. 4). Если же два атома при
надлежат разным элементам, то один из
них обычно притягивает электроны силь
нее, чем другой, и обобществленная пара
электронов смещается, вызывая смеще
ние и электрического заряда. Такая связь
называется полярной. На рисунке 5 изоб
ражена полярная молекула хлорида во
дорода (НС1).
Ковалентные связи встречаются в
биологических системах гораздо чаще,
чем ионные;
3)
в о д о р о д н ы е связи образуются
при участии атомов водорода, соединен
ных с сильно электроотрицательным ато
мом (обычно кислородом или азотом).
При этом атом водорода приобретает та
кой сильный положительный заряд, что
рода одной молекулы воды притягивается
к положительно заряженному атому во
дорода другой молекулы с образованием
водородной связи (сплошной линией
обозначены ковалентные связи, пунктир
ной — водородные):
н
0 -— н1
может образовать вторую связь с другим
электроотрицательным атомом (О, N).
Подобным образом связываются друг с
другом молекулы воды, у которых один
конец несет частично положительный за
ряд, а другой — частично отрицательный.
Такую молекулу называют диполем. Бо
лее электроотрицательный атом кисло
н
0——н----0——Н--- 0
н
н
4)
г и д р о ф о б н ы е взаимодейст
вия (рис. 6) — это наиболее слабые из
Рис. 6. Схема гидрофобного взаимодействия между двумя неполярными группами аланина и лейци
на — синий цвет; молекулы воды — голубой цвет.
13
нековалентных (дополнительных) связей,
встречающихся в органических соедине
ниях. Они представляют собой сближе
ние и взаимодействие неполярных моле
кул и частей молекул (например, поли
пептидных цепочек), которые сопровож
даются ослаблением их взаимодействия с
окружающей водой.
На рисунке 6 видно, что гидрофобное
взаимодействие возникает благодаря
сближению двух неполярных групп до тех
пор, пока они не соприкоснутся. Подоб
ное сближение сопровождается умень
шением числа окружающих их молекул
воды, т. е. молекулы воды как бы вытал
киваются из той среды, в которой возни
кает гидрофобное взаимодействие. Спо
собностью к таким взаимодействиям
обладают, например, остатки валина,
лейцина, изолейцина, фенилаланина,
триптофана, цистина и других амино
кислот.
В водном растворе нековалентные
связи (ионные, водородные и гидрофоб
ные) примерно в 100 раз слабее кова
лентных. В совокупности эти связи
определяют пространственную струк
туру
макромолекул
органических
соединений и, как следствие, их
функции.
?«
1. Что такое элемент? 2. Какие химические
элементы входят в состав клетки? 3. Каково строе
ние атомов? 4. В чем заключается различие между
атомом и ионом натрия? 5. Какая связь образуется
в результате обобществления электронов, принад
лежащих разным атомам? 6. Каков механизм об
разования водородных, ионных и гидрофобных
связей?
2. Химические соединения
и молекулы
Соединением называют вещество,
состоящее из химически связанных ато
мов одного или нескольких различных хи
14
мических элементов. Соединения харак
теризуются не только определенным сос
тавом, но и иными свойствами, чем ис
ходные вещества. В химическом соеди
нении уже нельзя различить свой
ства отдельных компонентов. Напри
мер, нельзя обнаружить свойства
железа или хлора в хлориде желе
за FeCl3.
Химические соединения образуются
либо при взаимодействии простых ве
ществ, либо в результате превращения
других соединений. Например, при взаи
модействии железа и хлора получается
хлорид железа:
2Fe + ЗС12 = 2FeCl3.
Это же соединение может образо
ваться при взаимодействии гидроксида
трехвалентного железа с соляной кис
лотой:
2Fe(OH)3 + 6 НС1 = 2 FeCl3 + 6 Н20 .
Молекула — это мельчайшая части
ца какого-нибудь соединения (вещес
тва), сохраняющая все его химические
свойства.
Свойства соединений определяются
не только составом, но и взаимным рас
положением атомов, а также типом хими
ческой связи. Поэтому для изображения
химического состава соединений исполь
зуют как общие, так и структурные фор
мулы. Последние более громоздки, но
они дают представление не только о рас
положении атомов и связей, но и об их
природе и числе. Более того, если два ка
ких-либо соединения содержат одинако
вое число одних и тех же атомов, то раз
личить их можно лишь по структур
ным формулам. Например, ниже при
ведены структурные формулы двух соеди
нений, у которых одинаковы общие фор
мулы:
н
I
с=о
I
н - с - он
I
н о -с -н
I
н - с - он
I
н - с - он
I
н -с -о н
I
н
н
н-с -о н
I
С= О
I
но - с - н
I
н - с -о н
I
н -с -о н
I
н -с -о н
I
н
глюкоза
фруктоза
Все имеющиеся в клетке соединения
можно разделить на две группы — неор
ганические и органические. К неоргани
ческим соединениям относятся вода и ми
неральные соли, а к органическим — ве
щества, в состав которых входят атомы
углерода (углеводы, липиды, белки, нук
леиновые кислоты и др.). Органические
соединения характерны для живых орга
низмов, в то время как неорганические
существуют и в неживой природе. Поэто
му не случайно на долю органических со
единений приходится большая часть су
хого вещества клетки (табл. 2).
Т а б л и ц а 2. Примерный химический состав ти
пичных клеток бактерий и млекопитающих
Дол Я от о б щей
массы
клетки %
Компонент
Б ак
тер ии
Вода
Неорганические ионы
Разнообразные низкомолекуляр
ные органические соединения
Белки
Нуклеиновые кислоты
Липиды
Углеводы
Млекопитаю
щие
70
1
70
1
3
15
3
18
7
2
2
1
5
2
Макромолекулы собираются из низко
молекулярных субъединиц, которые, при
соединяясь друг кдругу, образуют длинную
полимерную цепь с молекулярной массой
до нескольких сотен тысяч. Так, аминокис
лоты, связываясь с другими аминокисло
тами, образуют белки; нуклеотиды, связы
ваясь, образуют нуклеиновые кислоты, а
сахара — полисахариды.
9
. 1. Что такое химическое соединение и как оно
образуется? 2. В чем различия между неорганичес
ким и органическим соединениями? 3. В чем преи
мущество структурных формул соединений по срав
нению с общими формулами? 4. Каков примерный
химический состав типичной клетки?
3. Вода и другие неорганические
вещества. Их роль в клетке
В количественном отношении первое
место среди химических соединений клет
ки занимает вода. Ее содержание колеб
лется в зависимости от вида организма, ус
ловий его обитания, типа клеток и их функ
ционального состояния. Так, например, в
костной ткани содержится не более 20 %
воды, в жировой ткани — около 40, в мы
шечных клетках — 76, а в клетках разви
вающегося зародыша — более 90 %.
С возрастом количество воды в клет
ках любого организма заметно снижает
ся. Отсюда слелует, что, чем выше фун
кциональная активность клеток и орга
низма в целом, тем выше содержание в
них воды, и наоборот.
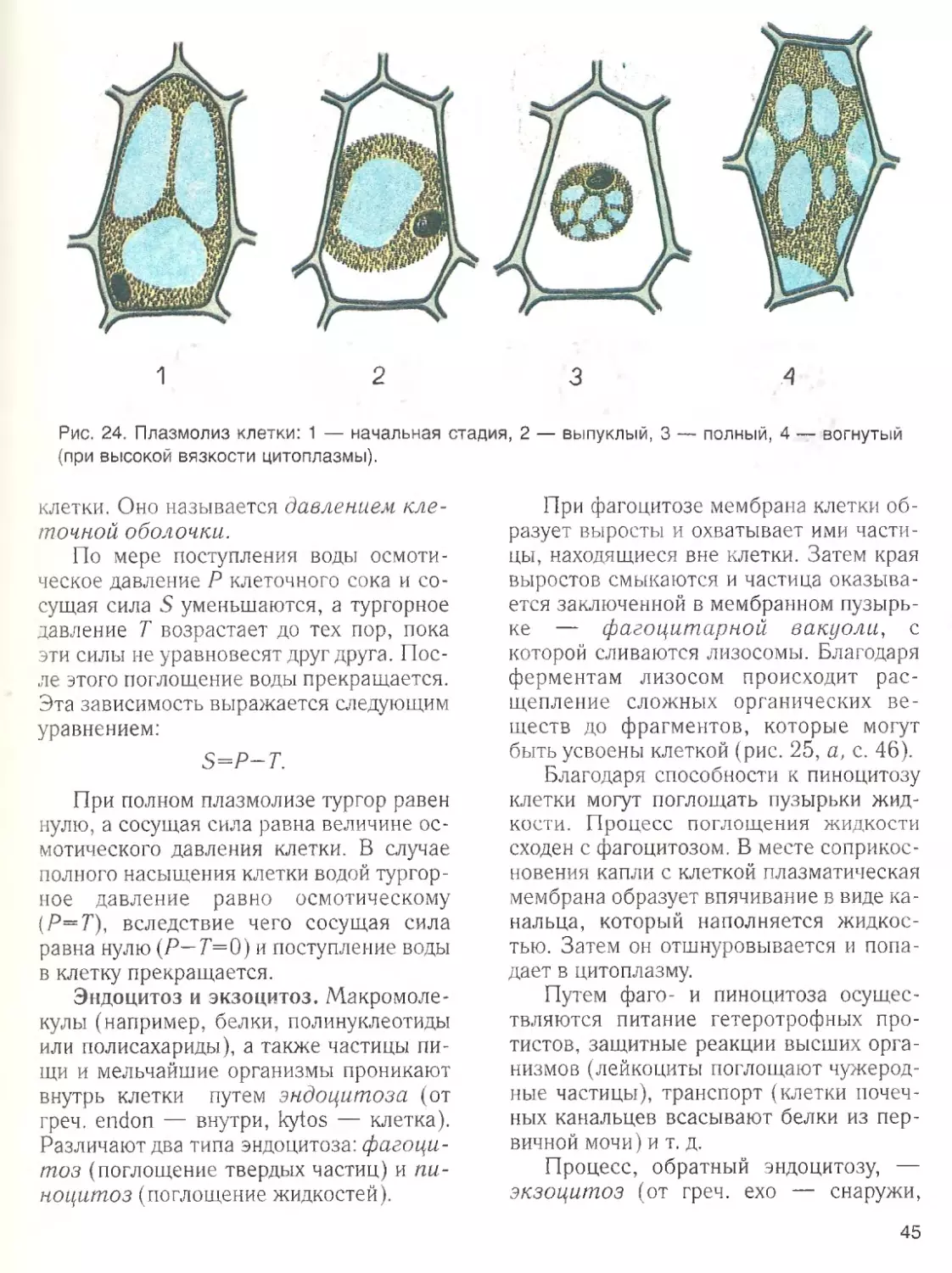
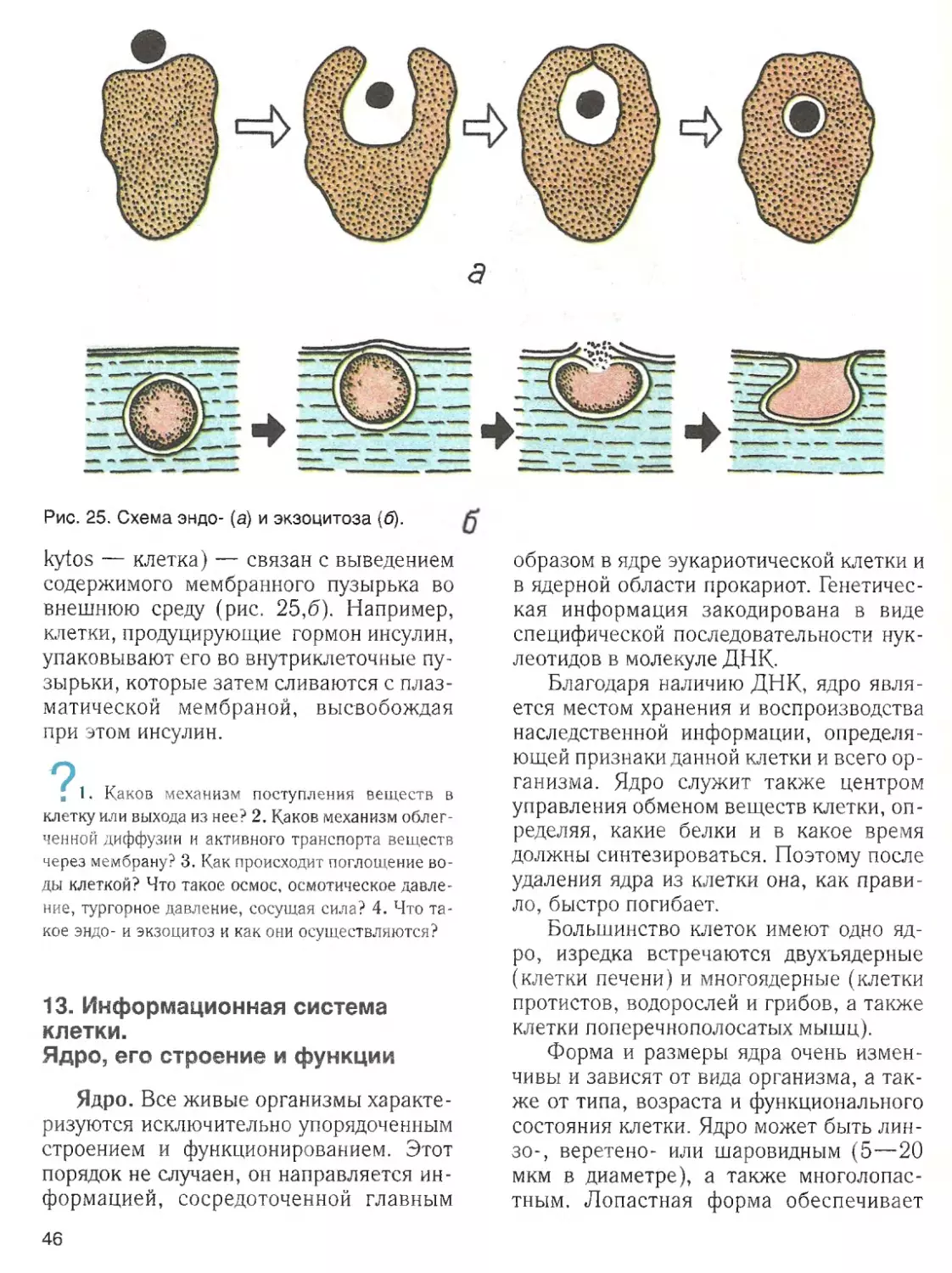
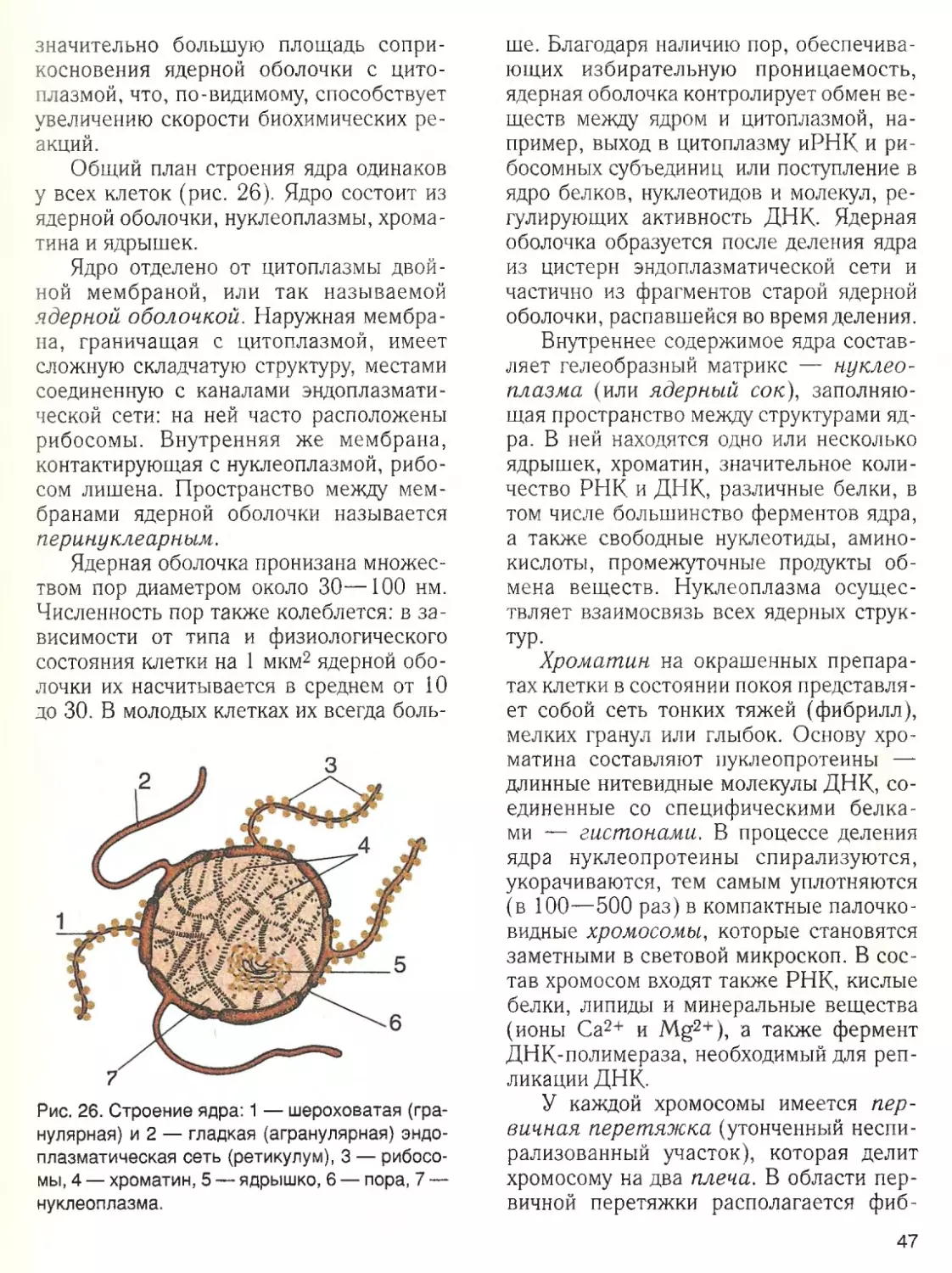
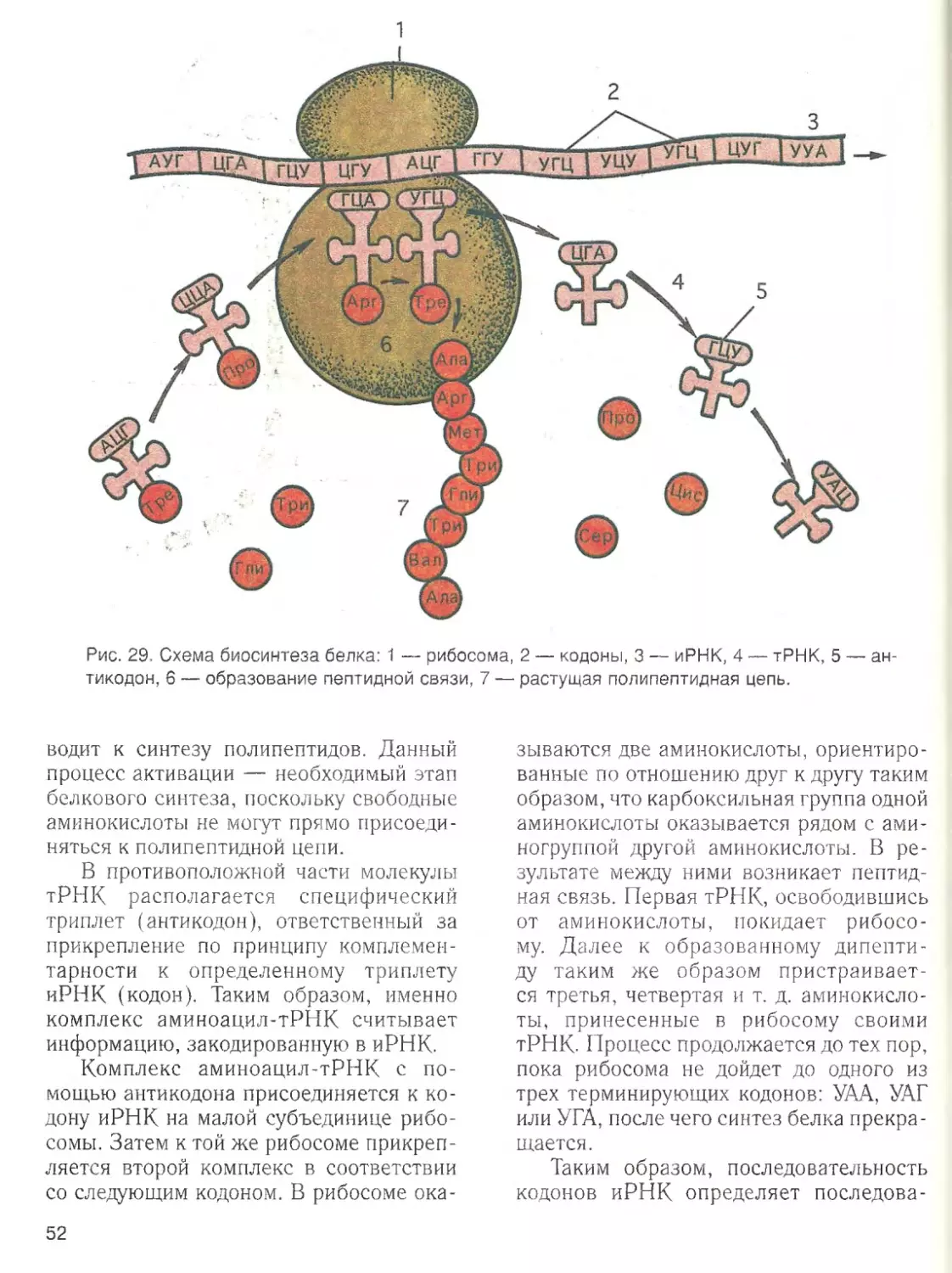
Наличие воды ■
— обязательное усло
вие жизненной активности клетки. Вода
составляет основную часть внутреннего
содержимого клетки — цитоплазмы,
поддерживает ее структуру и устойчи
вость входящих в ее состав коллоидов.
Роль воды в клетке определяется ее
химическими и структурными свойства
ми. Эти свойства связаны с малыми раз
мерами молекул воды, их полярностью и
способностью соединяться друг с другом
водородными связями.
15
Благодаря этим свойствам вода явля
ется хорошим растворителем для поляр
ных веществ, называемых гидрофильны
ми. К ним относятся ионные соединения,
у которых заряженные частицы (ионы)
диссоциируют (отделяются друг от друга)
в воде, когда вещество растворяется
(например, соли). Такой способностью
обладают и некоторые неионные соеди
нения, в молекуле которых присутствуют
заряженные (полярные) группы. У саха
ров, аминокислот, простых спиртов это
ОН-группы.
Когда вещество переходит в раствор,
его молекулы или ионы получают воз
можность двигаться свободно, и реакци
онная способность вещества возрастает.
По этой причине вода является основной
средой, где протекает большинство хими
ческих реакций. Все реакции гидролиза и
многочисленные окислительно-восстано
вительные реакции идут при непосред
ственном участии воды.
Неполярные вещества, например ли
пиды, практически нерастворимы в воде,
т. е. они гидрофобны.
Вода обладает наивысшей удельной
теплоемкостью из всех известных жидкос
тей. Это означает, что существенное уве
личение тепловой энергии вызывает срав
нительно небольшое повышение темпе
ратуры воды. Такое явление обусловлено
тем, что значительная часть данной энер
гии расходуется на разрыв водородных
связей, ограничивающих подвижность
молекул воды. Большая теплоемкость во
ды защищает ткани растений и животных
от быстрого и сильного повышения тем
пературы, тогда как высокая теплота па
рообразования надежно обеспечивает
стабильную температуру тела организма.
Необходимость значительных количеств
энергии для испарения (транспирации)
воды обусловлена наличием водородных
связей между ее молекулами. Эта энергия
черпается из окружающей среды, поэтому
испарение сопровождается охлаждением.
Указанное явление наблюдается у живот
16
ных при потоотделении, при одышке у со
бак, or о играет важную физиологическую
роль и в охлаждении испаряющих влагу
органов растений, особенно в условиях
пустынь и сухих степей.
Вода обладает также высокой тепло
проводностью, что обеспечивает равно
мерное распределение тепла по всему ор
ганизму. В результате устраняется риск
возникновения локальных «горячих то
чек», которые могли бы послужить при
чиной повреждения клетки. Следова
тельно, высокая удельная теплоемкость и
высокая теплопроводность делают воду
идеальной средой для поддержания опти
мального теплового режима организма.
Для воды характерно исключительно
высокое поверхностное натяжение. Это
свойство имеет очень важное значение
для адсорбционных процессов, передви
жения растворов по тканям (кровообра
щение, восходящий и нисходящий токи по
растению и т. д.).
Вода используется в качестве источ
ника кислорода и ионов водорода, выде
ляемых в световую фазу фотосинтеза.
К числу важных в физиологическом
отношении свойств воды относится ее
способность растворять газы ( 0 2, С 0 2
и др.). Кроме того, вода как растворитель
принимает участие в явлениях осмоса,
играющего важную роль в жизнедеятель
ности клетки и организма.
Соли. Неорганические соединения
клетки чаще всего находятся в виде со
лей, диссоциированных в растворе на ио
ны, реже — в твердом состоянии (карбо
нат и фосфат кальция костной ткани, из
вестковые или кремниевые панцири гу
бок, раковин моллюсков, диатомовых
протистов и красных водорослей). Наи
более важны для клетки катионы К+,
Na+, Са2+, Mg2+ и анионы Cl- , НСО3,
Н2РС>4, Н Р О |_, H SO 4 и др. (см.
табл. 1).
Концентрации ионов в клетке и в сре
де ее обитания, как правило, резко раз
личаются. Так, на внутренней поверхнос
ти клеточной мембраны довольно высо
кая концентрация ионов калия, а на
внешней — высокая ионов натрия и низ
кая — калия. По этой причине создается
разность потенциалов между внутренней
и внешней поверхностями клеточной
мембраны, что обусловливает передачу
возбуждения по нерву или мышце.
От концентрации ионов внутри клет
ки зависят также ее буферные свойства,
т. е. способность клетки сохранять реак
цию своего содержимого на постоянном
уровне, несмотря на то, что в процессе
жизнедеятельности непрерывно образу
ются ионы водорода, кислоты и щелочи.
Роль буфера играют анионы слабых кис
лот (НСО3, Н РО |~) и слабые кислоты
(Н2С 0 3), которые связывают и отдают
ионы водорода, благодаря чему реакция
внутри клетки практически не меняется и
остается слабо щелочной, почти ней
тральной.
Ряд важных функций в живых орга
низмах выполняют неорганические кис
лоты и их соли. В частности, клетки же
лудка образуют значительное количество
соляной кислоты, которая в составе ж е
лудочного сока участвует в процессе пе
реваривания пищи.
О
; 1. Чем отличается химический состав тел жи
вой и неживой природы? 2. Какова биологическая
роль воды в клетке? 3. Каково значение для клетки
и организма в целом важнейших химических эле
ментов? 4. Какую роль играют содержащиеся в
клетке катионы и анионы? 5. Какие соли входят в
состав клеток?
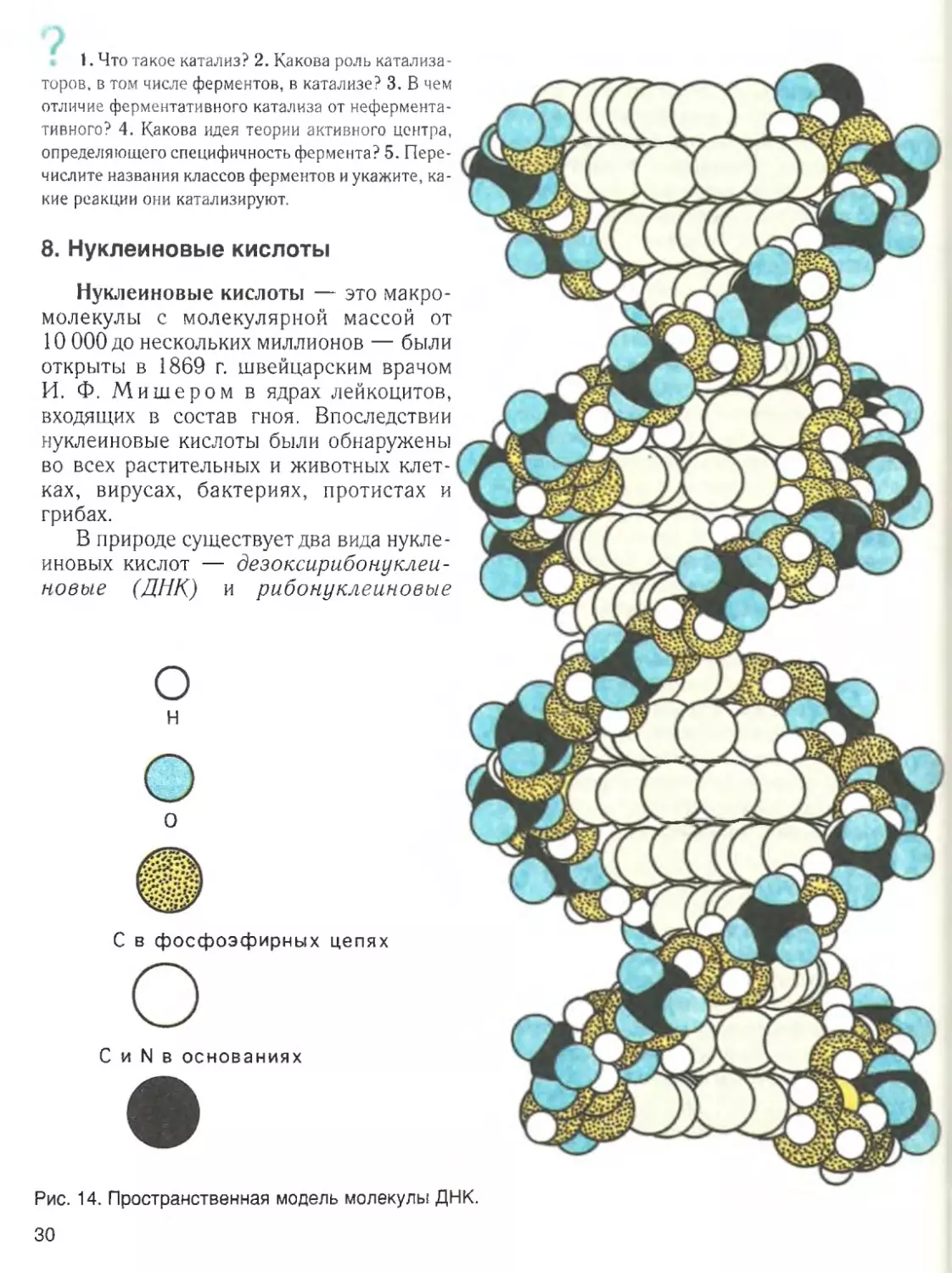
4. Органические соединения
Углеродсодержащие соединения —
химическая основа жизни. Не считая во
ды, почти все молекулы клетки — это со
единения углерода, или органические ве
щества. Углерод имеет ряд уникальных
химических свойств, благодаря которым
возможна жизнь. Малый размер атома
углерода и наличие на его внешней обо
лочке четырех электронов дает ему воз
можность образовывать четыре прочные
ковалентные связи с другими атомами.
Наиболее важное значение имеет
способность атомов углерода соединять
ся друг с другом, образуя цепи и кольца и
создавая в результате большие и слож
ные молекулы. Именно этой особеннос
тью углерода и объясняется чрезвычай
ное разнообразие органических соедине
ний. Другими словами, С — С-связи
можно рассматривать как скелет органи
ческих молекул.
Простые правила образования кова
лентной связи между углеродом и други
ми элементами (Н, N, О, Р и S) обеспе
чивают существование астрономического
числа разнообразных органических со
единений.
Разнообразие проявляется в разме
рах молекул, их химических свойствах и
зависит от степени насыщенности угле
родного скелета и различной формы мо
лекул, определяемой углами связей.
Биополимеры. Так называемые ма
лые органические молекулы клетки пред
ставляют собой соединения углерода с
молекулярной массой от 100 до 1000.
Они содержат немногим более 20 атомов
углерода. Такие молекулы обычно нахо
дятся в свободном состоянии в цитоплаз
ме, являясь промежуточным продуктом в
химических реакциях энергетического
обмена, дающих начало макромолекулам,
или полимерам.
Биологические полимеры — это ор
ганические соединения, молекулы кото
рых состоят из большого числа повторя
ющихся звеньев — мономеров. Макро
молекулы полимеров образуются в ре
зультате реакций полимеризации или
конденсации мономеров — низкомолеку
лярных соединений, содержание различ
ных видов которых в клетке около 1000.
Процесс этот обратим: полимеры могут
быть разрушены до мономеров, из кото
рых они состоят. Именно это и происхо
дит в любой живой клетке, например в
желудочно-кишечном тракте животных и
17
человека: макромолекулы пищи разру
шаются здесь до малых молекул, которые
затем всасываются в кровь и использу
ются организмом для образования новых
макромолекул, принадлежащих уже са
мому организму,
По форме биополимеры — это ли
нейные либо ветвящиеся цепи. Благода
ря такой структуре они характеризуются
рядом замечательных свойств.
Во-первых, их взаимодействие отли
чается кооперативностью, т. е. тесной
взаимосвязанностью всех функциональ
ных групп. Это означает, что взаимодей
ствие одних групп полимера изменяет ха
рактер взаимодействия других его групп.
Пример такого кооперативного взаимо
действия — связывание молекулы кис
лорода белком эритроцитов крови — ге
моглобином.
Во-вторых, биополимеры способны
образовывать так называемые интерпо
лимерные комплексы, которые могут
возникать как между отдельными частя
ми молекулы, так и между разными моле
кулами. Благодаря образованию ком
плексов и другим свойствам биополиме
ров осуществляются биосинтез белков,
нуклеиновых кислот, регуляция обмена
веществ, реакция иммунитета и другие
важнейшие биологические процессы.
Все живые организмы содержат четы
ре главных класса органических соедине
ний: углеводы, липиды, белки и нуклеи
новые кислоты. Соединения каждого
класса построены из особых мономеров,
объединяющихся в полимеры.
Углеводы (сахариды) представляют
собой первичные продукты фотосинтеза
и исходные продукты биосинтеза других
веществ (органических кислот, амино
кислот) в растениях. Углеводы содержат
ся в клетках всех живых организмов. В
животной клетке содержание углеводов
колеблется в пределах 1—2 %, в расти
тельной в некоторых случаях оно может
достигать 85— 90 % массы сухого ве
щества.
18
Углеводы состоят из углерода, водо
рода и кислорода, причем у большинства
углеводов водород и кислород содержатся
в таком же соотношении, как и в воде (от
сюда их название — углеводы). Таковы,
например, глюкоза С6Н 120 6 или сахароза
С 12Н22 0 ц.
Углеводы делятся на простые (моно
сахариды) и сложные (полисахариды).
Среди моносахаридов по числу угле
родных атомов различают триозы (ЗС),
тетрозы(4С), пентозы(БС), гексозы(бС)
и гептозы (7С). Моносахариды с пятью и
более атомами углерода, растворяясь в
воде, могут приобретать кольцевую цик
лическую структуру. В циклических фор
мах атомы углерода не обозначаются —
их присутствие подразумевается в каж
дом углу углеродного кольца (рис. 7.).
В природе чаще встречаются пентозы (напр., рибоза, дезоксирибоза, рибулоза) и гексозы (напр., глюкоза, фрукто
за, галактоза). Рибоза и дезоксирибоза
входят в состав важнейших соединений
клетки — нуклеиновых кислот и АТФ.
Глюкоза служит универсальным источни
ком энергии в клетке. С превращениями
моносахаридов связано и обезврежива
ние и выведение из организма ядовитых
веществ, проникающих извне или обра
зующихся в процессе обмена веществ,
например при распаде белков.
Ди- и полисахариды образуются путем
соединения двух и более моносахаридов,
таких, как глюкоза, галактоза, манноза,
арабиноза или' ксилоза. Так, соединяясь
между собой с выделением молекулы во
ды, две молекулы моносахаридов образу
ют молекулу дисахарида (рис. 8). Типич
ными представителями этой группы ве
ществ являются сахароза (тростниковый
сахар), мальтоза (солодовый сахар),
лакт оза (молочный сахар). Дисахариды
по своим свойствам близки к моносахари
дам. Например, и те и другие хорошо рас
творимы в воде и имеют сладкий вкус.
С увеличением количества моно
меров растворимость полисахаридов
с=о
Линейная
форма
I
н -с -о н
I
н -с -о н
н
Глицеральдегвд
н
I
г=о
I
н -с -о н
I
но—с—н
I
н -с -о н
I
н -с -о н
н -с -о н
н
I
с=о
I
н -с—он
I
н -с—он
I
н -с -о н
I
н -с -о н
I
с=о
I
н о -с -н
I
н -с -о н
I
н -с —он
I
н -с —он
А
с 3н6о 3
Проекции
циклических
форм
он н
Глюкоза
СйН ,,О й
Рвбоза
Фруктоза
C3HinO.
с«н12о 6
Рис. 7 Примеры биологически важных моносахаридов.
Глюкоза -— -----,
#---------- - Фруктоза
Рис. 8
уменьшается, исчезает сладкий вкус.
К числу полисахаридов принадлежат
крахмал, гликоген, целлюлоза, инулин
и др.
Основная функция углеводов —
энергетическая. При их ферментативном
расщеплении и окислении выделяется
энергия, которая используется клеткой.
Полисахариды главным образом играют
роль запасных продуктов и легко доступ
ных источников энергии (например, крах
мал и гликоген), а также используются в
качестве строительного материала (цел
люлоза, хитин).
Полисахариды являются прекрасны
ми запасными веществами по ряду при
чин: во-первых, будучи нерастворимы в
воде, они не оказывают на клетку ни ос
мотического, ни химического влияния,
что весьма важно при длительном хране
нии их в живой клетке. Во-вторых, твер
дое, обезвоженное состояние полисаха
ридов увеличивает полезную массу про
дуктов запаса за счет экономии их объ
ема. В-третьих, в твердом виде запасные
вещества не доступны для болезнетвор
ных бактерий, грибов, которые, как из
вестно, не могут заглатывать пищу, а вса
сывают ее всей поверхностью тела. И на
конец, при необходимости путем гидро
лиза они легко могут быть превращены в
сахара.
Целлюлоза является одним из важ
нейших структурных компонентов кле
19
точных стенок некоторых протистов, гри
бов и растений: в среднем 20— 40 %
материала клеточной стенки состав
ляет именно этот сахарид, а хло
пок состоит почти полностью из целлю
лозы.
Целлюлоза имеет большое народно
хозяйственное значение. Ее используют
для производства бумаги, вискозного
шелка, различных тканей. Целлюлоза
служит также пищей многим животным,
бактериям и грибам. Однако большин
ство животных, в том числе и человек, не
могут использовать целлюлозу, посколь
ку в их желудочно-кишечном тракте от
сутствует фермент целлюлаза, расщеп
ляющий целлюлозу до глюкозы. Целлю
лозные волокна играют важную роль в
нашем рационе по другой причине. Они
придают пище объемность и грубую кон
систенцию, что стимулирует перисталь
тику кишечника. У жвачных животных в
пищеварительном тракте целлюлозу рас
щепляют бактерии.
Хитин входит в состав клеточных сте
нок некоторых протистов и грибов, вы
полняя опорную функцию, а также встре
чается у отдельных групп животных
(чаще у членистоногих) в качестве
основного компонента их наружного ске
лета.
Большинство полисахаридов живот
ных соединено с белками или липидами.
При этом образуются гликопротеины и
гликолипиды, выполняющие многочис
ленные важные функции. Исключитель
но велико биологическое значение гли
копротеина гепарина. Он препятствует
свертыванию крови в кровеносных сосу
дах, а также участвует в регуляции обме
на липидов и иммунных реакциях орга
низма.
Гликолипиды принимают участие в
построении клеточных мембран. Особен
но богата гликолипидами нервная ткань
млекопитающих. При нарушении обмена
гликолипидов могут возникать заболева
ния нервной системы.
20
f 1. В чем выражаются уникальные свойства уг
лерода и какова его биологическая роль? 2. Какие
углеводы содержатся в клетках растений и живот
ных? 3. Какова роль углеводов в клетках? 4. Оха
рактеризуйте структуру и функции полисахаридов.
5. Липиды
Это обширная группа органических
соединений, которые содержатся во всех
живых клетках. Они нерастворимы в во
де, но хорошо растворяются в непо
лярных органических растворителях
(эфир, бензин, бензол, хлороформ
и др.).
Липиды отличаются исключительно
большим химическим разнообразием, од
нако можно все же сказать, что на
стоящие липиды — это сложные эфи
ры жирных кислот и какого-либо
спирта.
Жирные кислоты — это молекулы с
длинной цепью, состоящей из атомов уг
лерода (чаще всего 16 или 18), водорода
и карбоксильной группы ( — СООН). Уг
леводородные хвосты гидрофобны, а кар
боксильная группа крайне гидрофильна и
легко образует эфиры.
Иногда в жирных кислотах имеются
одна или несколько двойных связей
(С = С). В этом случае жирные кислоты,
а также содержащие их липиды называ
ются ненасыщенными. Жирные кислоты
и липиды, в молекулах которых нет двой
ных связей, называются насыщенными.
Последние образуются путем присоеди
нения дополнительной пары атомов водо
рода по двойной связи ненасыщенной
кислоты:
Н
Н
I I
—С= С—
---- >
Н
Н
I I
— С— С—
I
I
н н
ненасыщенная
насыщенная
Ненасыщенные жирные кислоты пла
вятся при более низких температурах,
чем насыщенные. Например, олеиновая
кислота (Тпл = 13,4 °С) при комнатнай
температуре бывает жидкой, тогда как
пальмитиновая (Тпл = 63,1 °С) и стеари
новая (Тпл = 69,6 °С) кислоты в этих ус
ловиях остаются твердыми.
Большинство липидов являются
сложными эфирами трехатомного спирта
глицерола и трех остатков жирных кис
лот. Такие соединения называются триацилглицеролами, или триглицерида
ми (рис. 9).
Среди соединений этой группы разли
чают жиры и масла в зависимости от то
го, в каком состоянии — жидком (масло)
или твердом (жиры) — они находятся
при комнатной температуре. Известно,
что температура плавления липидов тем
ниже, чем выше в них доля ненасыщен
ных жирных кислот. В маслах, как прави
ло, присутствует больше ненасыщенных
жирных кислот, чем в жирах. В организме
животных, живущих в холодном климате,
например у р ы б . арктических морей,
обычно содержится больше ненасыщен
ных триацилглицеролов, чем у обитате
лей южных широт. По этой причине тело
их остается гибким и при низких темпера
турах воды.
Важнейшими группами липидов явля
ются также с т е р о и д ы (желчные кис
лоты, холестерол, половые гормоны,
витамин D и др.), т е р п е н ы (ростовые
вещества растений — гиббереллины,
каротиноиды, витамин К), в о с к а ,
ф о с ф о - и г л и к о л и п и д ы, л и п о п р о
теины.
Липиды играют важную роль как ис
точники энергии. При окислении они да
ют более чем в два раза больше энергии,
чем углеводы и белки, и, таким образом,
более экономичны как форма хранения
запасов питательных веществ. Это связа
но с тем, что в липидах по сравнению с уг
леводами и белками больше атомов водо
рода и совсем мало атомов кислорода.
н
н
II
н -
с
IJ
с
II
-
I
=
сн2
I
сн 2
сн 2
II
сн 2
II
сн 2
II
сн 2
II
сн 2
II
сн 2
II
сн 2
II
сн2
j
сн3
Рис. 9
—
о
I
О
С
н
О
С
=
сн 2
I
сн2
О
с —
I
I
о
I
с =
сн 2
I
сн 2
сн2
I
сн 2
I
сн 2
I
сн2
СН 2
II
сн2
II
сн2
II
сн2
I
сн 2
II
сн 2
II
сн2
I
сн
II
сн
II
сн
II
сн
сн3
2
сн2
I
2
2
2
сн3
Нерастворимость в воде делает липи
ды важнейшими структурными компо
нентами клеточных мембран, состоящих
главным образом из фосфолипидов, а
также гликолипидов и липопротеинов.
Животные, впадающие в спячку, на
капливают жиры и расходуют их в про
21
цессе жизнедеятельности. Высокое со
держание липидов в семенах растений
обеспечивает энергией процесс развития
зародыша и проростка. Семена кокосо
вой пальмы, клещевины, сои, подсолнеч
ника служат сырьем для промышленного
получения растительного масла.
Благодаря низкой теплопроводности,
липиды выполняют защитные функции,
т. е. служат для теплоизоляции организ
мов. Например, у многих позвоночных
животных хорошо выражен подкожный
жировой слой, что позволяет им жить в
условиях холодного климата, переносить
зиму. У китов он играет еще и другую
роль — способствует плавучести.
Важно отметить также значение ли
пидов как источников воды. При окисле
нии 100 г жира образуется примерно
105 г воды. Эта метаболическая вода
очень важна для обитателей пустыни, в
частности для верблюдов, способных об
ходиться без воды 10— 12 дней. Жир, за
пасаемый в горбе, используется именно
для этой цели. Необходимую для жизне
деятельности воду находящиеся в спячке
медведи, сурки и другие животные тоже
получают в результате окисления жира.
?■ , !.ч Чем отличаются липиды от других веществ
клетки? 2. Как можно объяснить, что в клетках пойкилотермных животных содержание ненасыщенных
жирных кислот обычно выше, чем в клетках гомойотермных животных? 3, Какова биологическая роль
липидов?
6. Белки (протеины)
В жизни всех организмов белки име
ют первостепенное значение. На их долю
приходится 50-—80 % сухой массы клет
ки. По своей химической природе бел
ки — это сложные органические соеди
нения (полимеры), состоящие из атомов
углерода, водорода, кислорода и азота.
В некоторых белках содержится еще и
сера. Мономерами белков являются ами
нокислоты.
22
Аминокислоты — низкомолекуляр
ные органические соединения, в состав
которых входят одна или две аминогруп
пы (— NH2) и одна или две карбоксиль
ные группы ( —СООН), обладающие ще
лочными и кислотными свойствами соот
ветственно. Этим объясняются амфотер
ные свойства аминокислот, благодаря че
му в клетке они играют роль буферных
соединений. Общая формула аминокис
лот имеет вид:
н
\
R
О
//
N— С—с
/
\
1
Н
н
он
1
Все известные аминокислоты разли
чаются между собой только радикалами.
В природных белках содержится 20 раз
личных аминокислот. Кроме них, извес
тно еще более 150 аминокислот, которые
встречаются в некоторых пептидах, гор
монах, антибиотиках или находятся в сво
бодном состоянии.
У растений все необходимые амино
кислоты синтезируются из простых со
единений — С 0 2, Н20 и NH3. Однако
человек и многие животные в процессе
эволюции утратили способность синтези
ровать ряд белковых аминокислот, став
ших незаменимыми в питании: они обяза
тельно должны поступать с пищей,
то есть в готовом виде. К ним относят
ся лизин, валин, лейцин, изолейцин,
треонин, фенилаланин, триптофан, мети
онин.
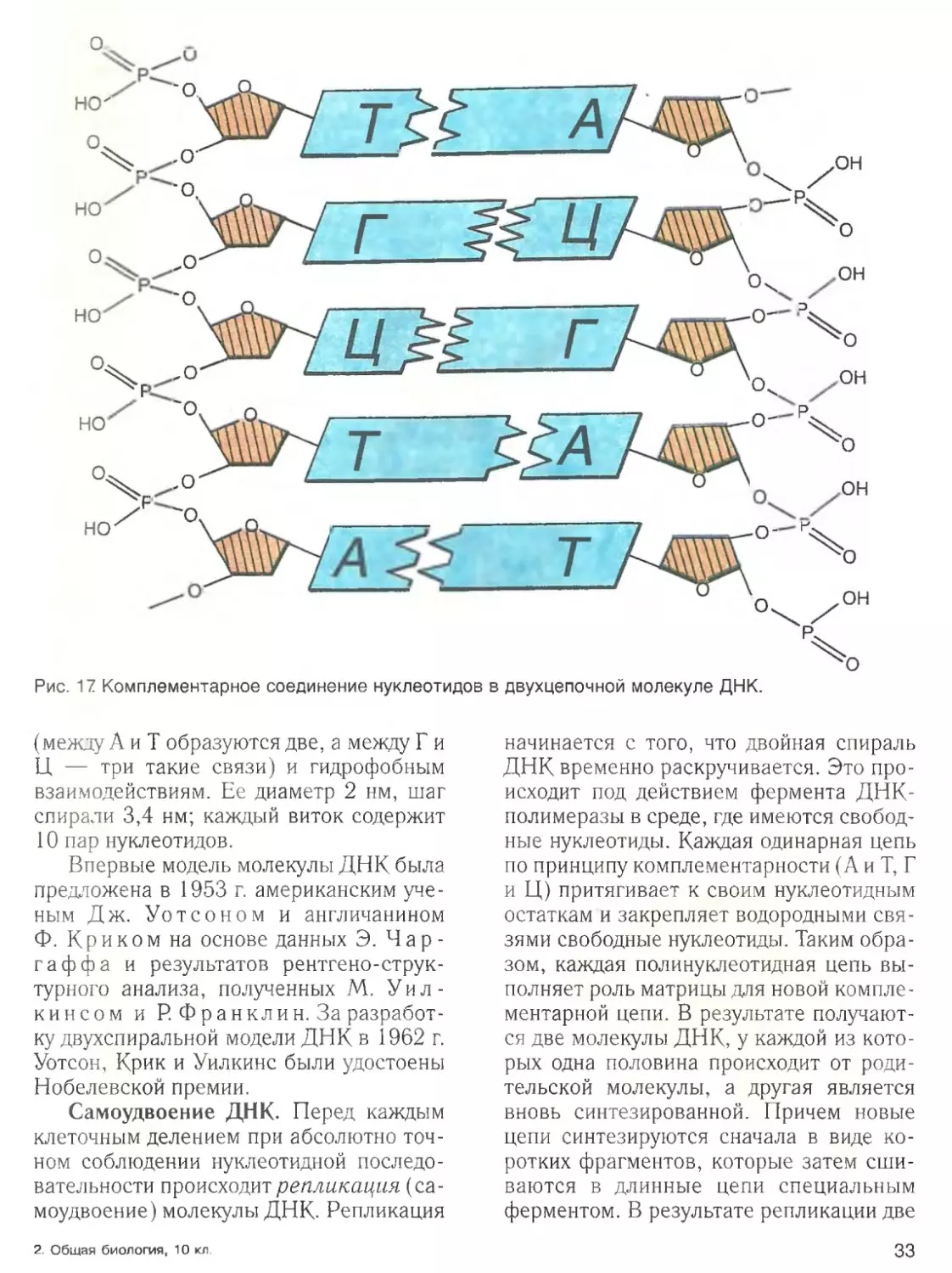
Строение белков. Аминогруппа од
ной аминокислоты способна вступать в
реакцию с карбоксильной группой другой
аминокислоты. В результате выделяется
молекула воды, а две аминокислоты ока
зываются соединенными прочной кова
лентной азот-углеродной связью, назы
ваемой пептидной ( — СО — NH — ).
Соединение, образующееся в результате
конденсации двух аминокислот, пред
ставляет собой дипептид:
Н
Ri О н r2 о
\ 1 //
\
1 /
—с —с—>
N—С—с + N■
1 \
/
1
/
он
н
н £>Н HJ н
н
R, О н r2 0
\
1 I 1 1 //
-- >Н90 + N--с-- с - -N—С--с\
/ [
1 \
н он
н н
Дальнейшее соединение аминокислот
с помощью пептидных связей приводит к
образованию полипептидной цепоч
ки — первичной структуры белка
(рис. 10). Таким образом, молекула белка
является полипептидом, в состав кото
рого входит от 50 до нескольких тысяч
аминокислот с молекулярной массой
свыше 10 000 и диаметром молекулы от 5
до 10 нм (1 нм — 0,001 мкм).
Все химические, морфологические и
функциональные свойства клеток зави
сят от содержащихся в них специфичес-
23
ких белков. Специфичность белков опре
деляется набором, количеством и после
довательностью расположения амино
кислот в полипептидной цепочке. Замена
одной-единственной аминокислоты в сос
таве молекулы белка или изменение по
следовательности их расположения, как
правило, приводит к изменению его фун
кций. Этим достигается огромное разно
образие первичной структуры белковых
молекул. Поэтому не удивительно, что
живой организм может использовать для
выполнения каждой своей функции осо
бый вид белка и что его возможности в
этом отношении неограниченны. Напри
мер, в организме человека встречается
более 5 млн различных белков, а в клетке
бактерии около 3 тыс. Следовательно,
огромное многообразие живых существ в
значительной степени определяется раз
личиями в строении их белков.
Свойства белка определяются также
пространственным расположением поли
пептидных цепочек. В живой клетке по
липептидные цепочки сложены или со
гнуты, приобретая вторичную и третич
ную структуры (см. рис. 10).
Вторичная структура представля
ет собой спирально закрученную белко
вую цепочку. Витки спирали удержива
ются водородными связями (обозначены
пунктиром), которые образуются между
СО- и NH-группами, расположенными
на соседних витках.
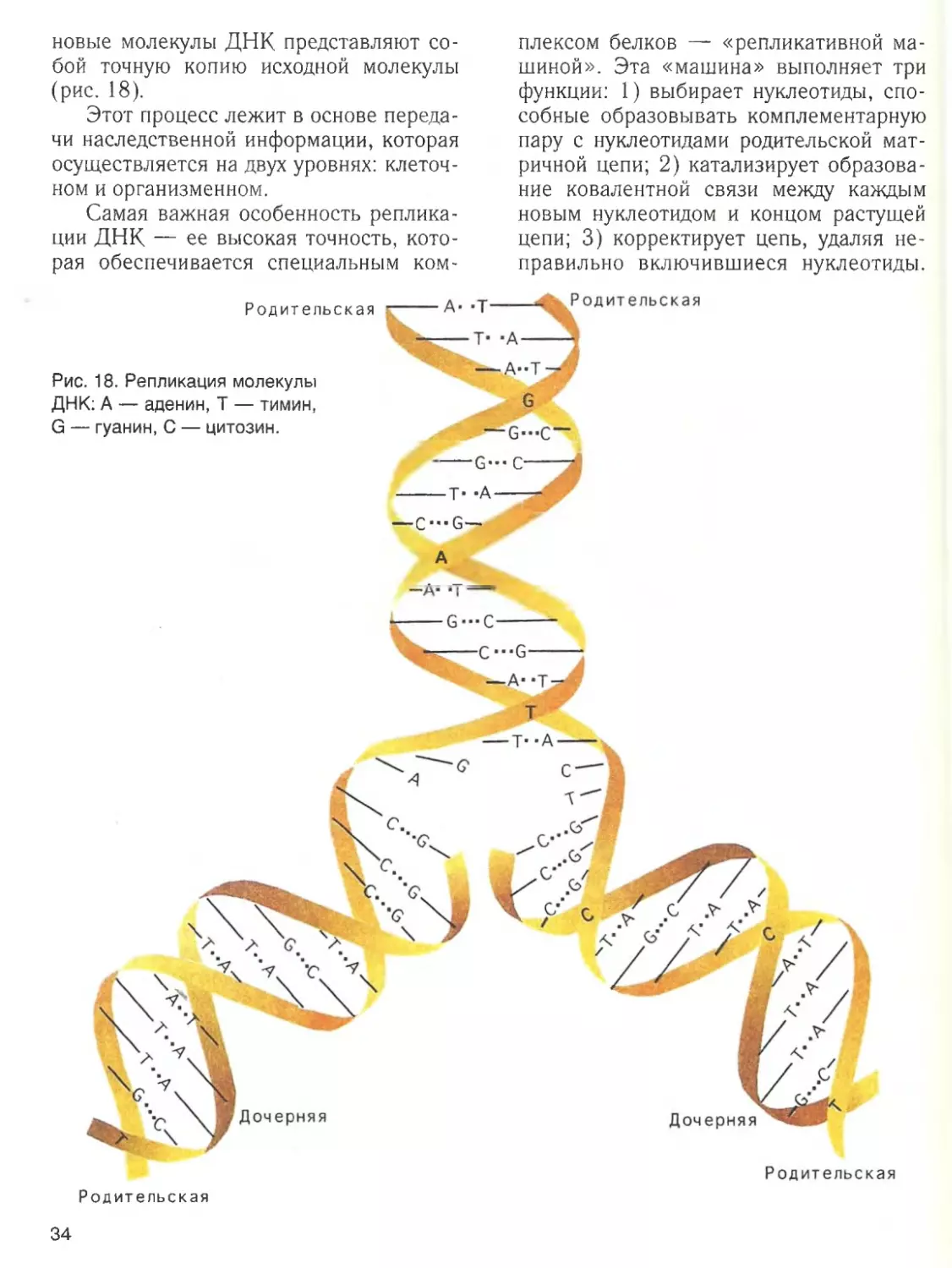
В результате дальнейшей укладки
спирали возникает специфичная для каж
дого белка конфигурация, называемая
третичной структурой. Она стабили
зируется за счет связей ме>кду входящими
в состав данного полипептида аминокис
лотными остатками — ковалентных ди
сульфидных связей ( — S — S — ) между
остатками цистеина, а также водородных,
ионных и гидрофобных взаимодействий.
Дисульфидные связи могут соединить
между собой не только различные учас
тки одной и той же пептидной цепи (рис.
11), но и отдельные пептидные цепи в
24
единую молекулу белка. Например, в мо
лекуле инсулина (гормона белковой при
роды) две полипептидные цепи, состоя
щие из 21 и 30 остатков аминокислот,
связаны между собой двумя дисульфид
ными связями. На рисунке 12 показана
структура молекулы инсулина крупного
рогатого скота, причем аминокислотные
остатки обозначены: Гли — глицин,
Изол — изолейцин, Вал — валин,
Глю — глютаминовая кислота, ГлюNH2 — глютамин, Цис — цистеин, Ал —
аланин, Сер — серин, Лей — лейцин,
Тир — тирозин, Acn-NH2 — аспарагин,
Лиз — лизин, Пр — пролин, Тр — трео
нин, Apr — аргинин, Гис — гисти
дин, Фен — фенилаланин. Черным цве
том на рисунке выделены дисульфидные
связи.
В количественном отношении наибо
лее важны гидрофобные взаимодействия,
обусловленные стремлением неполярных
боковых цепей аминокислот объединять
ся друг с другом, а не смешиваться с
окружающей водной средой. При этом
молекула принимает в растворе такую
форму, чтобы ее гидрофобные боковые
цепи были скрыты внутри молекулы, то
есть защищены от соприкосновения с во
дой, а гидрофильные боковые цепи, на
оборот, направлены наружу. Следует
помнить, что количество молекул цистеи
на и аминокислот с гидрофобными ради
калами, а также порядок их расположе
ния в полипептидной цепочке характерны
для каждого белка.
Сохранение спиральной формы моле
кулы обеспечивает также взаимное рас
положение групп атомов, которое необ
ходимо для проявления каталитической
активности белка, его гормональной фун
кции и др. Поэтому устойчивость глобул
(молекул) — не случайное свойство, а
одно из важнейших средств стабилизации
организма.
Биологическую активность белок
проявляет только в состоянии третичной
структуры, поэтому замена даже одной
Рис. 11. Схема первичной структуры молекулы лизоцима белка куриного яйца (цифры обозначают но
мера аминокислотных остатков).
аминокислоты в полипептидной цепочке
может привести к изменению конфигура
ции белка и к снижению или утрате в це
лом его биологической активности.
Иногда две, три, четыре и более бел
ковые молекулы с третичной организаци
ей объединяются в комплекс. Подобные
агрегаты представляют собой четвер
тичную структуру белка (см. рис. 10).
Примером такого сложного белка может
служить гемоглобин, состоящий из четы
рех белковых субъединиц и небелковой
части — гема. Только в такой форме ге
моглобин способен выполнять свои фун
кции. В четвертичной структуре белко
вые субъединицы, как правило, не связа
ны друг с другом ковалентно, однако вся
структура достаточно прочна за счет дей
ствия слабых межмолекулярных сил.
По строению белки делятся на п р о с ты е и с л о ж н ы е . В состав простых
белков входят только аминокислоты, а
сложные, кроме аминокислот, могут
включать нуклеиновые кислоты (нуклеопротеины), липиды (липопротеины), уг
леводы (гликопротеины) и др.
Под влиянием различных химических
и физических факторов (обработка спир
25
том, ацетоном, кислотами, щелочами,
высокой температурой, облучением, вы
соким давлением и т. д.) происходит изме
нение третичной и четвертичной структур
белка вследствие разрыва водородных и
ионных связей. Процесс нарушения на
тивной (естественной) структуры белка
называется денатурацией. При этом
наблюдается уменьшение растворимости
белка, изменение формы и размеров мо
лекул, потеря ферментативной активнос
ти и т. д.
Процесс денатурации иногда обра
тим, т. е. возвращение нормальных усло
вий среды может сопровождаться само
произвольным восстановлением естес
твенной структуры белка. Такой процесс
называется ренатурацией. Отсюда сле
дует, что все особенности строения и
функционирования макромолекулы бел
ка определяются его первичной струк
турой.
Функции белков. Белки в клетке вы
полняют чрезвычайно важные и многооб
разные функции, обусловленные слож
ностью и разнообразием структуры и сос
тава самих белков.
Одной из важнейших функций белко
вых молекул является ст руктурная
(строительная). Белки входят в состав
всех мембран и органелл клетки. Стенки
кровеносных сосудов, хрящи, сухожилия
у высших позвоночных животных состоят
преимущественно из белка.
Двигательная функция обеспечива
ется особыми сократительными белками,
которые обусловливают движение ресни
чек и жгутиков, сокращение мускулату
ры, перемещение хромосом при делении
клетки, движение органов растений и т. д.
Велика роль белков в транспорте
веществ в организме. Например, гемо
глобин переносит с током крови кислород
из легких к клеткам других тканей и орга
нов. В мышцах эту функцию берет на се
бя белок миоглобин. В этом случае четко
проявляется кооперативное взаимодей
ствие всех функциональных групп био
26
полимера.
Молекула
гемоглобина
( C 738H 1166O 208N 203S 2F e ) 4 СОСТОИТ ИЗ Д в у х
пар полипептидных цепочек, каждая из
которых связана с гемом ■
— железосо
держащим соединением, придающим
цвет гемоглобину. Каждый гем способен
присоединять одну молекулу кислорода, а
весь гемоглобин — до четырех молекул
0 2. Присоединение каждой молекулы об
легчает присоединение следующей, а по
теря одной из них — потерю следующей.
Благодаря этому гемоглобин легко насы
щается кислородом, когда кровь течет по
легочным капиллярам, и легко отдает его
в тканях, где концентрация кислорода
ниже.
Белки плазмы крови способствуют
переносу липидов и жирных кислот раз
личных биологически активных веществ.
Молекулы белков, входящие в состав
плазматической мембраны, принимают
участие в транспорте веществ в клетку.
Белки выполняют и защитную фун
кцию. В клетке в ответ на проникновение
в нее чужеродных веществ ( а н т и г е н о в )
вырабатываются а н т и т е л а — особые
белки — иммуноглобулины, которые
нейтрализуют чужеродные вещества и
обеспечивают иммунитет.
Такие белки-антигены могут входить в
состав вирусов, бактерий, спор грибов и
растений или чужеродных клеток (напри
мер, в случае переливания крови или пе
ресадки органов). Иммунная система ор
ганизма узнает антиген по характерным
для него участкам молекулы — антиген
ным детерминантам. Такое специфичес
кое связывание сходно со специфичнос
тью ферментов по отношению к своим
субстратам.
Антитела у человека синтезируются
лимфоцитами, причем каждый из них, как
правило, синтезирует только один вид ан
тител. В ответ на введение антигена в ор
ганизме резко возрастает число лимфо
цитов, синтезирующих соответствующие
антитела.
Белкам присуща также сигнальная
функция. В поверхностную мембрану
клетки встроены молекулы белков, спо
собных изменять свою третичную струк
туру в ответ на действие факторов внеш
ней среды. Так происходит прием сигна
лов из внешней среды и передача команд
в клетку.
Регуляторная функция принадлежит
белкам-гормонам, которые оказывают
влияние на обмен веществ. Гормоны под
держивают постоянные концентрации
веществ в крови и в клетках, участвуют в
росте, размножении и других жизненно
важных процессах. Одним из наиболее
известных белков-гормонов является
инсулин, который снижает содержание
сахара в крови путем повышения прони
цаемости клеточных мембран для глюко
зы, то есть способствует ее переходу из
крови в клетку. При недостатке инсулина
содержание сахара в крови увеличивает
ся, и он начинает выделяться с мочой —
развивается сахарный диабет.
Для некоторых белков характерна
токсическая функция. Токсические бел
ки обнаружены в яде змей, насекомых, в
грибах, растениях, у микроорганизмов.
Белки являются также энергетичес
ким материалом. При расщеплении 1 г
белка до конечных продуктов выделяется
17,6 кДж энергии, необходимой для мно
гих жизненно важных процессов клетки.
?I
1. Каковы химическая структура и свойства
аминокислот? 2. Какие аминокислоты называются
незаменимыми? 3. Каково строение белков? 4. Чем
определяются специфичность и свойства белков?
5. Охарактеризуйте структурные особенности пер
вичной, вторичной, третичной и четвертичной
структур белка. 6. Каковы функции белков?
7. Ферменты
Химические реакции в живой клетке
протекают при умеренной температуре,
нормальном давлении и в нейтральной
среде. В таких условиях реакции органи
ческого синтеза или распада идут очень
медленно, если они не подвергаются воз
действию биологических катализаторов
белковой природы — ферментов, или
энзимов.
Все процессы в живом организме осу
ществляются с помощью ферментов.
Например, под действием различных
ферментов составные компоненты жизни
(белки, углеводы, липиды и др.) расщеп
ляются до более простых соединений, за
тем в организме синтезируются новые
макромолекулы, свойственные данному
организму. Поэтому нарушение деятель
ности ферментов ведет к возникновению
тяжелых патологий.
Катализом называется явление
ускорения реакции без изменения ее об
щего результата, а катализатором —
вещество, которое способно ускорять эту
реакцию. Роль катализаторов, в том чис
ле и ферментов, заключается в снижении
энергии активации, т. е. в присутствии
катализатора требуется меньше энергии
для придания реакционной способности
молекулам, вступающим в реакцию.
Ферментативный катализ подчиняет
ся тем же законам, что и неферментатив
ный катализ в химической промышлен
ности. Однако биологический катализ от
личается необычайно высокой степенью
специфичности (фермент катализирует
только одну реакцию или действует на
один тип связи), что позволяет осущес
твлять тонкую регуляцию всех процессов
(дыхание, пищеварение, фотосинтез и
др.), протекающих в клетке и организме.
Например, фермент уреаза катализирует
расщепление лишь одного вещества —
мочевины (H2N—C 0 - N H 2+ H 20 ---- >
---- >2NH3+ С 0 2), не оказывая катали
тического действия на сходно построен
ные соединения.
Чтобы понять механизм действия
ферментов, обладающих высокой специ
фичностью, следует рассмотреть теорию
активного центра. Согласно этой тео
рии, в молекуле фермента имеется один
или более участков, в которых происхо
27
дит катализ за счет тесного (во многих
точках) контакта между молекулами фер
мента и специфического вещества (суб
страта). Активным центром выступает
или функциональная группа (например,
ОН-группа серина), или отдельная ами
нокислота. Обычно же для каталитичес
кого действия необходимо несколько
аминокислотных остатков (в среднем от 3
до 12), расположенных в определенном
порядке. Активный центр также может
формироваться металлами, витаминами и
другими соединениями небелковой при
роды — коферментами, связанными с
ферментом. Причем форма и химическое
строение активного центра таковы, что с
ним могут связываться только опреде
ленные субстраты в силу их идеального
соответствия друг другу (рис. 13).
б
____
_
_____ _________
Рис. 13. Активный центр фермента: а — суб
страт, б —■поверхность фермента.
28
Роль остальных аминокислотных ос
татков в крупной молекуле фермента сос
тоит в том, чтобы обеспечить его молеку
ле соответствующую глобулярную фор
му, которая нужна для эффективной ра
боты активного центра. Кроме того, вок
руг крупной молекулы фермента возни
кает сильное электрическое поле. В та
ком поле становится возможной ориента
ция молекул субстрата и приобретение
ими ассиметричной формы. Это приводит
к ослаблению химических связей, и ката
лизируемая реакция происходит с мень
шей начальной затратой энергии, а сле
довательно, с намного большей скорос
тью. Например, одна молекула фермента
каталазы за 1 мин может расщепить бо
лее 5 млн молекул пероксида водорода
(Н20 2), который возникает при окисле
нии в организме различных соединений.
У некоторых ферментов в присут
ствии субстрата конфигурация активного
центра претерпевает изменения, т. е.
фермент ориентирует свои функциональ
ные группы таким образом, чтобы обес
печить наибольшую каталитическую ак
тивность. Молекулы субстрата, присо
единяясь к ферменту, тоже в определен
ных пределах изменяют свою конфигура
цию для увеличения реакционности фун
кциональных групп центра. Эти пред
ставления о ферментно-субстратном вза
имодействии отличаются от принятой ра
нее теории, согласно которой считалось,
что молекула субстрата точно соответ
ствует активному центру фермента и под
ходит к нему как ключ к замку.
Фермент-субстратный комплекс пос
ле химической реакции распадается с об
разованием конечных продуктов и сво
бодного фермента. Освободившийся при
этом активный центр может принимать
новые молекулы субстрата.
Скорость ферментативных реакций
зависит от многих факторов: природы и
концентрации фермента и субстрата,
температуры, давления, кислотности сре
ды, наличия ингибиторов и т. д. Напри
мер, при температурах, близких к 0° С,
скорость биохимических реакций замед
ляется до минимума. Это свойство широ
ко используется в различных отраслях
народного хозяйства, особенно в сельс
ком хозяйстве и медицине. В частности,
охлаждением сохраняют различные орга
ны (почки, сердце, селезенку, печень) пе
ред их пересадкой больному, чтобы сни
зить интенсивность биохимических реак
ций и тем самым продлить время жизни
органов.
Международный биохимический съезд
(Москва, 1961) утвердил классификацию
ферментов, в основу которой был поло
жен тип реакции, катализируемой данным
ферментом. В названии фермента обяза
тельно присутствует название субстрата,
на который воздействует данный фер
мент, и окончание -аза. Например, целлюлаза катализирует гидролиз целлюло
зы до моносахаридов, а протеаза — гид
ролиз белков до аминокислот.
По этому принципу все ферменты бы
ли разделены на шесть классов.
1. Оксидоредуктазы катализируют
окислительно-восстановительные реак
ции, осуществляя перенос атомов Н и О
или электронов от одного вещества к дру
гому, окисляя при этом первый и восста
навливая второй. Эта группа ферментов
участвует во всех процессах биологичес
кого окисления, например в дыхании:
АН + В
А + ВН (окисленный) или
А + О s=s АО (восстановленный).
2. Трансферазы катализируют пере
нос группы атомов (метальной, ациль
ной, фосфатной или аминогруппы) от од
ного вещества к другому. Например, под
действием фосфотрансфераз происходит
перенос остатков фосфорной кислоты от
АТФ на глюкозу или фруктозу:
АТФ + глюкоза *=* глюкоза-6-фосфат +
+АДФ.
3. Гидролизы ускоряют реакции рас
щепления сложных органических соеди
нений до простых путем присоединения
молекул воды в месте разрыва химичес
кой связи. Подобное расщепление назы
вается гидролизом. К гидролазам отно
сятся амилаза (гидролизует крахмал), ли
паза (расщепляет жиры) и др.:
АВ + Н20 ^ АОН + ВН.
4. Лиазы катализируют негидролити
ческое присоединение к субстрату или от
щепление от него группы атомов. При
этом могут разрываться связи С — С,
С —N, С— О, С— S. Примером может
служить отщепление карбоксильной
группы декарбоксилазой:
О
II
СН3— С— С
II
I
о он
О
//
со2+сн3—с
\
н.
5. Изомеразы осуществляют внутри
молекулярные перестройки, т. е. катали
зируют превращение одного изомера в
другой:
глюкоза-6-фосфат
глюкоза- 1-фосфат.
6.
Лигазы (синтетазы) катализиру
ют реакции соединения двух молекул с об
разованием новых связей С— О, С— S,
С— N, С—С, используя энергию АТФ.
К лигазам, например, относится группа
ферментов, катализирующих присоедине
ние остатков аминокислот к транспортной
РНК (тРНК). Эти синтетазы играют роль
в процессе синтеза белка. Примером
такой синтетазы служит фермент лейцинтРНК-синтетаза, под действием ко
торого образуется комплекс лейцинтРНК:
АТФ + лейцин + тРНК = АДФ +
+ Н3Р 0 4 + лейцин-тРНК.
Большая часть ферментов связана с
определенными клеточными структурами
(ядро, митохондрии, пластиды, лизосомы
и др.), где и осуществляется их функция.
29
1. Что такое катализ? 2. Какова роль катализа
торов, в том числе ферментов, в катализе? 3. В чем
отличие ферментативного катализа от нефермента
тивного? 4. Какова идея теории активного центра,
определяющего специфичность фермента? 5. Пере
числите названия классов ферментов и укажите, ка
кие реакции они катализируют.
8. Нуклеиновые кислоты
Нуклеиновые кислоты — это макро
молекулы с молекулярной массой от
10 000 до нескольких миллионов — были
открыты в 1869 г. швейцарским врачом
И. Ф. М и ш е р о м в ядрах лейкоцитов,
входящих в состав гноя. Впоследствии
нуклеиновые кислоты были обнаружены
во всех растительных и животных клет
ках, вирусах, бактериях, протистах и
грибах.
В природе существует два вида нукле
иновых кислот — дезоксирибонуклеи
новые (ДНК) и рибонуклеиновы е
Н
О
С в ф о сф о эф и р н ы х цепях
С и N в основаниях
Рис. 14. Пространственная модель молекулы ДНК.
30
(РНК). Различие в названиях объясня
ется тем. что молекула ДНК содержит са
хар дезоксирибозу, а молекула РНК —
рибозу. В настоящее время известно
бол! лое число разновидностей ДНК и
РНК. отличающихся друг от друга по
строению и значению в метаболизме.
Приче? каждый вид организма содержит
свой, характерный только для него одно
го набор этих кислот. Нуклеиновые кис
лоты являются исключительно важными
веществами клетки, обеспечивающими
хранение и передачу генетической ин
формации в живых организмах. ДНК на
ходится преимущественно в хромосомах
клеточного ядра (99 % всей ДНК клет
ки ). а также в митохондриях и хлороплас
тах. РНК входит в состав ядрышек, ри
босом. митохондрий, пластид и цитоп
лазмы.
Молекула ДНК состоит из двух поли
нуклеотидных цепочек, спирально за
крученных относительно друг друга
(рис. 14).
Структурными компонентами (моно
мерами) каждой такой цепочки служат
нуклеотиды. Количество нуклеотидов в
молекулах нуклеиновых кислот бывает
разным — от 80 в молекуле тРНКдо не
скольких миллионов в ДНК- В состав лю
бого нуклеотида ДНК входит одно из че
тырех азотистых оснований (аденин, гу
анин, тимин или цитозин), пентозный
сахар дезоксирибоза (С5Н 10О4) и остаток
фосфорной кислоты (рис. 15).
Нуклеотиды отличаются только азо
тистыми основаниями, между которыми
имеется близкая родственная связь. Ци
тозин, урацил и тимин называются пири
мидиновыми, а аденин и гуанин — п у
риновыми основаниями.
В полинуклеотидной цепочке сосед
ние нуклеотиды связаны между собой ко
валентными связями, которые образуют
ся между дезоксирибозой одного и остат
ком фосфорной кислоты другого нуклео
тида (рис. 16, с. 32).
И хотя в молекуле ДНК всего четыре
типа нуклеотидов, различная последова
тельность их расположения в цепочке да
ет огромное разнообразие молекул ДНК.
Объединяются две цепи в единую мо
лекулу ДНК при помощи водородных
связей, возникающих между азотистыми
основаниями нуклеотидов разных цепей
(рис. 17, с. 33). При этом аденин (А) со
единяется только с тимином (Т), а гуанин
(Г) — с цитозином (Ц). В результате у
Рис. 15. Общая формула нуклеоти
да — мономера ДНК.
Азотистое
основание
NH2
А
N
НО
ОН
Фосфатная
группа
ОН
Пентозный сахар
31
НО — Р =
о
НО — Р = 0
О
Рис. 16
каждого организма число адениловых
нуклеотидов равно числу тимидиловых, а
число гуаниловых — числу цитидиловых.
Эта закономерность получила название
«правило Чаргаффа». Благодаря этому
свойству последовательность нуклеоти
дов в одной цепочке определяет их после
довательность в другой, т. е. цепи ДНК
32
являются как бы зеркальным отражени
ем одна другой. Такая способность к из
бирательному соединению нуклеотидов
называется комплементарностью. Этс
свойство лежит в основе образования но
вых молекул ДНК на базе исходной.
Двойная спираль стабильна благода
ря многочисленным водородным связям
о
но
о
■ ч /-°
.о н
но
о
о
о \
г
но
о.
оо
'О.
т
но
о.
но
он
о
он
о.
А^
он
Т
/.4
о
о
о
он
о
Рис. 17 Комплементарное соединение нуклеотидов в двухцепочной молекуле ДНК.
(между А и Т образуются две, а между Г и
Ц — три такие связи) и гидрофобным
взаимодействиям. Ее диаметр 2 нм, шаг
спирали 3,4 нм; каждый виток содержит
10 пар нуклеотидов.
Впервые модель молекулы ДНК была
предложена в 1953 г. американским уче
ным Дж. У о т с о н о м и англичанином
Ф. К р и к о м на основе данных Э. Ч а р г а ф ф а и результатов рентгено-струк
турного анализа, полученных М. У и л
к и н с о м и Р. Фр а н к л и н . За разработ
ку двухспиральной модели ДНК в 1962 г.
Уотсон, Крик и Уилкинс были удостоены
Нобелевской премии.
Самоудвоение ДНК. Перед каждым
клеточным делением при абсолютно точ
ном соблюдении нуклеотидной последо
вательности происходит репликация (са
моудвоение) молекулы ДНК. Репликация
2. Общая биология, 10 кл.
начинается с того, что двойная спираль
ДНК временно раскручивается. Это про
исходит под действием фермента ДНКполимеразы в среде, где имеются свобод
ные нуклеотиды. Каждая одинарная цепь
по принципу комплементарности (А и Т, Г
и Ц) притягивает к своим нуклеотидным
остаткам и закрепляет водородными свя
зями свободные нуклеотиды. Таким обра
зом, каждая полинуклеотидная цепь вы
полняет роль матрицы для новой компле
ментарной цепи. В результате получают
ся две молекулы ДНК, у каждой из кото
рых одна половина происходит от роди
тельской молекулы, а другая является
вновь синтезированной. Причем новые
цепи синтезируются сначала в виде ко
ротких фрагментов, которые затем сши
ваются в длинные цепи специальным
ферментом. В результате репликации две
33
новые молекулы ДНК представляют со
бой точную копию исходной молекулы
(рис. 18).
Этот процесс лежит в основе переда
чи наследственной информации, которая
осуществляется на двух уровнях: клеточ
ном и организменном.
Самая важная особенность реплика
ции ДНК — ее высокая точность, кото
рая обеспечивается специальным ком
плексом белков — «репликативной ма
шиной». Эта «машина» выполняет три
функции: 1) выбирает нуклеотиды, спо
собные образовывать комплементарную
пару с нуклеотидами родительской мат
ричной цепи; 2) катализирует образова
ние ковалентной связи между каждым
новым нуклеотидом и концом растущей
цепи; 3) корректирует цепь, удаляя не
правильно включившиеся нуклеотиды.
Р о д и тел ьск ая
Рис. 18. Репликация молекулы
ДНК: А — аденин, Т — тимин,
G — гуанин, С — цитозин.
G - - С-
С ■■■G
А
Р о д и тел ьск ая
34
Число ошибок «репликативной машины»
составляет ничтожно малую величину:
менее 1 на 109 нуклеотидов.
Однако бывают случаи, когда «репли
кативная машина» пропускает или встав
ляет несколько лишних оснований, вклю
чает Ц вместо Т или А вместо Г. Каждое
такое изменение последовательности
нуклеотидов в молекуле ДНК — генети
ческая ошибка, называемая мутацией.
Естественно, такие ошибки будут вос
производиться во всех последующих по
колениях клеток, что может привести к
заметным негативным последствиям.
Рибонуклеиновые кислоты (РНК).
Молекулы РНК состоят из одной поли
нуклеотидной цепочки, которая синтези
руется на молекуле ДНК и является ком
плементарной копией участка одной из
цепочек ДНК, только вместо тимина в
РНК входит похожее по своим свойствам
азотистое основание урацил (У). Кроме
того, во всех нуклеотидах молекулы РНК
находится не дезоксирибоза, а рибоза
• С5Н 10О5).
Существуют различные типы РНК,
различающиеся по величине молекулы,
структуре, местонахождению в клетке и
функциям. Низкомолекулярные тРНК
составляют примерно 10 % от всей кле
точной РНК. При реализации генетичес
кой информации каждая тРНК присоеди
няет и переносит определенную амино
кислоту к рибосомам — месту синтеза
белка. Следовательно, существует боль
ше двадцати различных тРНК, которые
различаются по своей первичной струк
туре (последовательности нуклеотидов).
Рибосомные РНК (рРНК) составля
ют до 85 % всей РНК клетки. Они входят
в состав рибосом, выполняя структурную
функцию. Кроме того, рРНК участвуют в
формировании активного центра рибосо
мы, где происходит образование пептид
ных связей между молекулами аминокис
лот в процессе биосинтеза белка.
Информационные, или матричные,
РНК(иРНК, мРНК) программируют син
тез белков клетки. Несмотря на относи
тельно низкое процентное содержание
(около 5 %) в общей массе РНК клетки,
иРНК по значению стоят на первом мес
те. Информационная РНК осуществляет
непосредственную передачу кода ДНК
для синтеза белков. При этом каждый бе
лок клетки кодируется специфической
иРНК. Это обусловлено тем, что иРНК
во время своего синтеза получают ин
формацию о структуре белка от ДНК в
форме транскрибированной последова
тельности нуклеотидов и переносят ее на
рибосомы, где эта информация реализу
ется.
Таким образом, значение всех типов
РНК определяется тем, что они представ
ляют собой функционально объединен
ную систему, направленную на осущес
твление синтеза в клетке специфических
для нее белков.
1. Какие виды нуклеиновых кислот обнаруже
ны в клетке? 2. Где в клетке находятся ДНК и РНК?
3. Какова химическая структура нуклеотидов?
4. Как связаны между собой нуклеотиды? 5. В чем
сущность принципа комплементарности? 6. Как
осуществляется самоудвоение (репликация) ДНК?
7. Чем отличаются по строению РНК и ДНК? 8. Ка
кие виды РНК имеются в клетке и какие функции
они выполняют?
9. АТФ и ее роль в клетке
Аденозинтрифосфорная
кислота
(АТФ) содержится в каждой клетке в
растворимой фракции цитоплазмы (гиа
лоплазме), митохондриях, хлоропластах и
ядрах. Она снабжает энергией большин
ство реакций, происходящих в клетке.
Молекула АТФ состоит из азотистого
основания аденина, пятиуглеродного са
хара рибозы и трех остатков фосфорной
кислоты. Фосфатные группы соединены
между собой высокоэнергетическими
(макроэргическими) связями (в формуле
обозначены со , рис. 19, с. 36).
35
Рис. 19
NHj
Аденин
С ^
N
\
N
I
С- н
Н - С
Г'
он
ОН
I
НО — Р и О —
II
О
он
I
I
Р «Ло - Р - о
II
О
II
Рибоза
О
я I_ йI *
он он
Фосфатные группы
В результате гидролитического от
щепления концевой фосфатной группы
АТФ превращается в аденозиндифосфор
ную кислоту (АЦФ) и высвобождается
энергия. При отщеплении второй фос
фатной группы получается аденозинмо
нофосфорная кислота (АМФ) и вновь вы
деляется энергия:
АТФ + Н20 = АДФ + Н3Р 0 4 + 40 кДж,
АДФ + Н20 = АМФ + Н3Р 0 4 + 40 кДж.
АТФ образуется из АДФ и неоргани
ческого фосфата за счет энергии, осво
бождающейся при окислении органичес
ких веществ, а также в процессе фото
синтеза. При этом должно быть затраче
но не менее 40 кДж энергии на 1 моль
АТФ, которая аккумулируется в ее макроэргических связях:
АДФ + Н3РО4 + 40 кДж —АТФ + Н20 .
АТФ чрезвычайно быстро обновляет
ся. У человека, например, каждая моле
кула АТФ расщепляется и вновь восста
навливается 2400 раз в сутки, так что
средняя продолжительность ее жизни ме
нее 1 мин. Синтез АТФ осуществляется,
главным образом, в митохондриях и хло
ропластах.
^
АТФ играет главную роль в энергети
ческом обмене клетки. Это означает, что
36
любые виды клеточной активности (дви
жение, биосинтез, деление и др.) совер
шаются за счет энергии, высвобождаю
щейся при гидролизе АТФ. Остальная
энергия (около 50 %), которая выделяет
ся при расщеплении молекул углеводов,
жиров, белков и других органических со
единений, рассеивается в виде тепла и не
имеет существенного значения для жиз
недеятельности клетки.
О
J 1. Какова химическая структура АТФ? 2. Како
во значение АТФ для жизнедеятельности клетки?
3. Где и каким образом синтезируется АТФ в клет
ке? 4. Почему именно АТФ, а не макромолекулы
является единым и универсальным источником
энергии в клетке?
ГЛАВА II
Строение и системы
жизнеобеспечения клетки
Клетка представляет собой самовос
производящуюся систему. Для того чтобы
поддерживать необходимую концентра
цию химических веществ, эта система
должна быть отделена от окружающей
среды и вместе с тем должна обладать
способностью к обмену с этой средой.
Другими словами, она должна быть спо-
собна поглощать те вещества, которые
-ребуются ей в качестве «сырья», и вы
водить наружу накапливающиеся «отхо
ды». Таким путем эта система может со
хранить стабильность (гомеостаз). Роль
5арьера между клеткой и ее окружением
играет плазматическая мембрана.
В каждой клетке имеются цитоплазма
а генетический материал в форме ДНК.
ДНК регулирует жизнедеятельность
-тетки и воспроизводит самое себя, бла
годаря чему образуются новые клетки.
Каждая органелла клетки выполняет
свою особую функцию, а в совокупности
все они определяют жизнедеятельность
клетки в целом, ее приспособленность к
_.словиям окружающей среды.
10. Строение клетки
Клетки различных организмов отли
чаются друг от друга размерами, формой,
происхождением и функциями. Напри
мер, бактериальные клетки имеют диа
метр около 0,2 мкм. Клетки других орга
низмов достигают 10— 100 мкм, не
сколько реже — 1— 10 мм, очень ред
ко — 10— 20 см (яйцеклетки страусов,
пингвинов, гусей) и даже 1 м и более
(длина отростков нервных клеток живот
ных и млечных сосудов растений).
По форме различают округлые,
овальные, многогранные, звездчатые,
дисковидные и другие клетки (рис. 20).
Форма клетки часто зависит от выполня
емой функции.
Несмотря на многообразие форм, ор
ганизация клеток всех живых организмов
подчинена единым структурным прин
ципам.
Живое содержимое клетки — про
топласт — отделено от окружающей
среды плазматической мембраной, или
плазмалеммой. Внутри клетка запол-
Ркс. 20. Различные формы клеток одноклеточных и многоклеточных организмов: протисты: 1 — инсЬузория-туфелька; 2 — эвглена зеленая; бактерии: 3 — бактерии со жгутиками; 4 — кокки, дипло
кокки, стрептококки; животные: 5 — клетки эпителия трахеи, 6 — нервная клетка, 7 — эритроциты;
застения: 8 — эпидермис клеток чешуи лука.
37
йена цит оплазмой, представляющей
собой полужидкую студенистую массу
сложного физико-химического состава, в
которой расположены ядро, многочис
ленные органеллы и включения.
Прокариоты и эукариоты. Все извес
тные одно- и многоклеточные организмы
делятся на две группы: прокариоты и эу
кариоты. К прокариотам относят бакте
рий и цианобактерий, а к эукарио
там — протистов, грибы, растения и
животных.
Клетки прокариот в отличие от клеток
эукариот устроены относительно просто
(рис. 21). В таблице 3 дана сравнитель
ная характеристика структурной органи
зации прокариотической и эукариотичес
кой клеток.
ТаблицаЗ. Структурные элементы клетки
Эукариотическая клетка
Протопласт
Плазмалемма
Ядро
Цитоплазма
Гиалоплазма, матрикс
Пластиды' | имеют
Прокариотическая клетка
Протопласт
Плазмалемма и ее про
изводные: впячивания
мембраны, мезосомы,
тилакоиды
Эквивалент ядра (нук
леоид)
Цитоплазма
Матрикс
Рибосомы
Митохондрии )Рибосомы
Эндоплазматический
ретикулум
Комплекс Гольджи
Вакуоли
Микрофиламенты
Микротрубочки
Центриоли
Реснички, жгутики2
Рибосомы
Производные протопласта:
клеточная оболочка2' 3
включения (гранулы,
кристаллы)
Нет
Нет
Нет
Нет
Нет
Нет
Жгутики
Рибосомы
Производные
пласта:
прото-
клеточная оболочка,
включения (гранулы)
1 Характерны для клеток растений и зеленых
протистов.
2 Одноименные структуры у про- и эукариоти
ческих клеток не гомологичны.
3 Отсутствует у клеток животных и некоторых
протистов.
Рис. 21. Строение прокариотической клетки:
1 — клеточная стенка, 2 — плазматическая
мембрана, 3 — стопка мембран, 4 — мезосома,
5 — кольцевая молекула ДНК, 6 — вакуоли,
7 — рибосомы, 8 — впячивания цитоплазмати
ческой мембраны.
38
В прокариотической клетке нет ис
тинного ядра, ядрышек и хромосом.
Вместо ядра имеется его эквивалент —
нуклеоид, лишенный оболочки и состоя
щий из одной-единственной кольцевой
молекулы ДНК. Именно этот признак яв
ляется решающим при делении клеток на
лочкой, состоящей из двух мембран. В
ядре находится генетический материал
клетки: ДНК и связанные с ней вещес
тва. Пластиды служат главным образом
для улавливания энергии солнечного
света и превращения ее в процессе фото
синтеза в химическую энергию углево
дов, а митохондрии — для выработки
энергии путем расщепления углеводов,
жиров и белков.
Мембранные системы цитоплаз
мы — эндоплазмат ическая сеть и
комплекс Гольджи — участвуют в син
тезе и упаковке макромолекул, необхо
димых для осуществления жизнедеятель
ности клетки. Вакуоли, лизосомы и пероксисомы выполняют специфические
для каждой из этих органелл функции и
также имеют мембранное строение.
И только у рибосом, хромосом, м ик
рот рубочек и микрофиламентов
мембраны отсутствуют.
Типы и разнообразие эукариотичес
ких клеток. Несмотря на сходство в стро
ении и химическом составе между клет
ками организмов различных царств жи
вой природы имеются существенные раз
личия. Клетки одноклеточных протистов
и многоклеточных грибов, растений и жи
вотных часто отличаются не только
структурной, но и физиолого-биохими
ческими особенностями (см. табл. 4).
прокариотические (доядерные) и эукари
отические (ядерные).
Прокариотические клетки не имеют
внутренних мембран, кроме впячиваний
илазмалеммы. Это значит, что у них нет
органелл, имеющих мембранное строе
ние, — митохондрий, эндоплазматичес
кой сети, хлоропластов, лизосом и ком
плекса Гольджи, характерных для эукари
отической клетки. Из органелл прокари
отическая клетка содержит лишь более
мелкие, чем в клетках эукариот, рибо
сомы.
Клетки прокариот покрыты жесткой
клеточной оболочкой и часто — слизис
той капсулой. Плазматическая мембрана
обычно образует впячивания внутрь ци
топлазмы — мезосомы. На мембранах
мезосом располагаются окислительнозисстановительные ферменты, а у фото
синтезирующих прокариот — пигменты:
'Пктериохлорофилл у бактерий, хлороэилл и фикоцианин — у цианобактерий.
Благодаря этому мембраны прокариот
способны выполнять функции митохон
дрий, хлоропластов и других органелл.
Функции в эукариотической клетке
распределены между ядром и различны
ми органеллами, такими, как митохон
дрии, рибосомы, комплекс Гольджи и др.
Клеточное ядро, митохондрии и плас_йды четко отделены от цитоплазмы обо
Т аб л и ц а 4. Сравнительная характеристика различных типов эукариотических клеток
Клетки
Признаки
протистов
грибов
растений
Ж ИВОТНЫ Х
Состоит в основном Состоит из целлю Нет
лозы
из хитина
Нет
Бывает редко
Есть
Есть
Крупная вакуоль
Есть
Нет
Бывают часто
Нет
Хлоропласты
Гетеротрофный
Автотрофный
Авто- и гетеротроф Гетеротрофный
Способ питания
ный
Только у некоторых Есть
Бывают редко
Бывают часто
Центриоли
мхов и папоротни
ков
Крахмал
Гликоген
Резервный
пита Крахмал, гликоген, Гликоген
парамилон, хризотельный углевод
ламинарин
Клеточная стенка
У многих есть
39
Цитоплазма. Как уже отмечалось,
цитоплазма состоит из полужидкого со
держимого и органелл. Кроме того, в ней
могут присутствовать различные включе
ния — временно выведенные из обмена
вещества (запасные питательные и стро
ительные) или нерастворимые отходы
метаболических процессов. Правда, в
большинстве случаев запасные вещества
и конечные продукты обмена расположе
ны не в основном веществе цитоплазмы,
а в определенных органеллах — пласти
дах, вакуолях и др.
Основное полужидкое вещество ци
топлазмы называют гиалоплазмой (от
греч. hyalos — стекло), или матриксом.
Гиалоплазма представляет собой очень
важную часть клетки, ее внутреннюю
среду. Гиалоплазма имеет вид гомогенно
го или тонкозернистого вещества. Она
представляет собой сложную коллоидную
систему, включающую различные биопо
лимеры: белки, нуклеиновые кислоты,
полисахариды и др. Эта система способна
переходить из золеобразного (жидкого)
состояния в гелеобразное (более плот
ное), и наоборот. Так, например, при вы
соком гидростатическом давлении цито
плазма не уплотняется, а обратимо раз
жижается. Это явление объясняется на
рушением связей между молекулами или
мицеллами (коллоидными частицами) в
составе гиалоплазмы. Переходы золя в
гель и обратно могут быть вызваны изме
нением кислотности, концентрации опре
деленных ионов, различными химически
ми реакциями, изменением температуры
и т. п. Таким образом, цитоплазма может
менять свое агрегатное состояние, а
вместе с ним вязкость, текучесть в зави
симости от изменения физических и хи
мических факторов.
Несмотря на то что в электронном
микроскопе гиалоплазма выглядит гомо
генным веществом, она не является од
нородной. В гиалоплазме обнаружена
микро трабекулярная система, т. е.
система тонких белковых нитей (около
40
2 нм толщиной), пересекающих цито
плазму в различных направлениях. Нити
микротрабекулярной системы состоят из
разных белков, молекулы которых обра
зуют друг с другом сложные комплексы.
Микротрабекулярная система связы
вает все внутриклеточные структуры
клетки: мембранные органеллы, различ
ные фибриллярные и трубчатые структу
ры и плазматическую мембрану. В местах
пересечения или соединения концов тра
бекул располагаются группы рибосом.
Вместе с фибриллярными (микрофиламенты) и трубчатыми (микрот ру
бочки) органеллами микротрабекуляр
ная система образует внутриклеточный
цитоплазматический скелет (цито
скелет) (рис. 22), который упорядочивает
размещение всех структурных компонен
тов клетки. Кроме того, микротрубочки
обеспечивают определенную форму кле
ток и отвечают за направленное движе
ние некоторых клеточных структур.
Микротрабекулярная система разде
ляет гиалоплазму как бы на две фазы: по-
Рис. 22. Цитоскелет: 1 — трабекулярные нити,
2 — микротрубочка, 3 — полисома, 4 — клеточ
ная мембрана, 5 — эндоплазматический рети
кулум, 6 — митохондрия, 7 — микрофиламенты.
лимерную, богатую белками (микротра
бекулы), и жидкую, заполняющую про
межутки между трабекулами.
Функциональная роль этой системы
■ ключается в создании и поддержании
цитоплазматического скелета, а также в
способности гиалоплазмы изменять агре
гатное состояние отдельных участков ци
топлазмы, ее вязкость и текучесть. Микротрабекулярная система очень динамич
на. Она может распадаться на отдельные
молекулы белка, которые переходят в
раствор и изменяют физические свойства
гиалоплазмы. Это происходит обычно
при изменении внешних и внутренних ус
ловий.
С распадом и сборкой микротрабекул
связывают также движение цитоплазмы,
которое имеет очень важное значение в
перемещении веществ и структурных
элементов клетки. В движении цитоплаз
мы принимают участие также микрофиламенты (актиновые и миозиновые во
локна).
Гиалоплазма выполняет различные
функции в клетке. Являясь основным ве
ществом цитоплазмы, ее внутренней сре
дой, она объединяет все клеточные
структуры и обеспечивает химическое
взаимодействие между ними.
Через гиалоплазму осуществляется
большая часть внутриклеточных тран
спортных процессов: перенос аминокис
лот, жирных кислот, нуклеотидов, саха
ров и т. д. Через гиалоплазму идет посто
янный поток ионов, растворенных в воде
газов от плазматической мембраны и об
ратно, к митохондриям, ядру, вакуолям.
Гиалоплазма — основное вместилище и
зона перемещения молекул АТФ. Она яв
ляется также резервуаром воды для рас
творения веществ и химических реакций.
?
, 1. Какие свойства характеризуют клетку как
живую систему? 2. В чем принципиальное отличие
чежду клетками прокариот и эукариот? 3. В чем
сходство и различие между основными типами эука
риотических клеток? 4. Что такое цитоплазма?
5. Что представляет собой гиалоплазма? 6. Какие
функции выполняет гиалоплазма в клетке? 7. Что
такое цитоскелет, какими структурами он образо
ван и каковы его функции в клетке?
11. Биологические мембраны
эукариотической клетки
Одной из основных особенностей всех
эукариотических клеток является разно
образие и сложность строения внутрен
них мембран. Мембраны окружают не
только цитоплазму, но и ядро, митохон
дрии и пластиды. Они образуют лабиринт
эндоплазматического ретикулума и стоп
ки уплощенных пузырьков, составляю
щих комплекс Гольджи. Мембраны огра
ничивают лизосомы, пероксисомы и
крупные вакуоли. Некоторые химические
реакции, в частности световые реакции
фотосинтеза в хлоропластах или окисли
тельное фосфорилирование при дыхании
в митохондриях, протекают на самих
мембранах. Здесь же, на мембранах, рас
положены и рецепторные участки для
распознавания веществ (гормонов и др.),
поступающих из окружающей среды или
из других частей самого организма.
Именно плазматическая мембрана, окру
жающая каждую клетку, обеспечивает
сохранение существенных различий меж
ду клеточным содержимым и окружаю
щей средой. Следовательно, без мембран
существование клетки было бы невоз
можным.
Плазматическая мембрана, или
плазмалемма, — это основная, универ
сальная для всех клеток система. Она
представляет собой тончайшее (6—
10 нм) образование, покрывающее всю
клетку и состоящее из упорядоченно рас
положенных молекул фосфолипидов и
белков. Эти молекулы удерживаются
вместе с помощью нековалентных взаи
модействий.
Внутренний слой мембраны состоит
из двух рядов липидов, молекулы которых
расположены таким образом, что их не41
полярные гидрофобные концы находятся
в глубине мембраны, а полярные гидро
фильные обращены к внутренней и внеш
ней водной среде. Липидный слой не яв
ляется сплошным. В отдельных местах
мембрана пронизывается белковыми мо
лекулами, образуя гидрофильные поры,
через которые проходят водораствори
мые вещества. Другие белковые молеку
лы находятся на внешней и внутренней
стороне мембраны (рис. 23).
На поверхности мембран всех эукари
отических клеток имеются углеводы.
Чаще они ковалентно присоединены к
мембранным белкам, образуя гликопро
теины, и в меньшей степени к липидам —
с образованием гликолипидов. В целом
содержание углеводов в плазматических
мембранах варьирует от 2 до 10 % по
массе. Полисахаридный слой толщиной
от 10 до 20 нм, покрывающий сверху
плазмалемму животных клеток, получил
название гликокаликс.
Все клеточные мембраны представ
ляют собой подвижные, текучие структу
ры: большая часть составляющих их моБ
Рис. 23. Строение цитоплазматической мем
браны: А — трехмерная модель, Б — плоскост
ное изображение: 1 — белки, примыкающие к
липидному слою (а), погруженные в него (б),
пронизывающие его насквозь (в); 2 — слои мо
лекул липидов, 3 — гликопротеины, 4 — глико
липиды, 5 — гидрофильный канал (пора).
42
лекул белков, липидов, полисахаридов,
воды, ионов калия, натрия, кальция и др.
способна достаточно быстро переме
щаться в плоскости мембраны, меняя в
ней свое расположение. При этом мигра
ция указанных веществ осуществляется
как диффузно, так и активным путем с
потреблением энергии. Кроме того, мем
браны быстро восстанавливаются после
повреждений, а также растягиваются и
сжимаются при клеточных движениях.
Все это свидетельствует об их динамичес
кой природе.
Поверхностные рецепторы. Мем
браны разных типов клеток существенно
различаются как по химическому соста
ву белков, гликопротеинов и липидов,
так и по их относительному содержа
нию. Следовательно, каждый тип кле
ток индивидуален, причем определяет
ся это преимущественно гликопротеи
нами.
Разветвленные цепи гликопротеинов,
выступающие из клеточной мембраны,
участвуют в распознавании определен
ных факторов внешней среды и в реакции
клеток на эти факторы. Например, яй
цеклетка и сперматозоид узнают друг
друга по гликопротеинам клеточной по
верхности, которые подходят друг другу
как отдельные элементы цельной струк
туры. Такое взаимное узнавание — необ
ходимый этап, предшествующий опло
дотворению.
Подобное явление наблюдается и в
процессе дифференцировки тканей.
В этом случае сходные по строению клет
ки с помощью распознающих участков
плазмалеммы правильно ориентируются
по отношению друг к другу, обеспечивая
тем самым сцепление и образование
тканей.
С распознаванием связана и регуля
ция транспорта молекул и ионов через
мембрану, а также иммунный ответ, в ко
тором гликопротеины играют роль анти
генов. Сахара, таким образом, могут фун
кционировать как информационные мо
лекулы (подобно белкам и нуклеиновым
кислотам).
В мембранах содержатся специфи
ческие рецепторы, переносчики электро
нов, преобразователи энергии, фермен
тные белки. Белки участвуют в обеспече
нии транспорта молекул внутрь клетки
или из нее, осуществляют структурную
связь цитоскелета с клеточными мембра
нами или же служат рецепторами для по
лучения и преобразования химических
сигналов из окружающей среды.
Столь четкая структурная организа
ция и упорядоченность плазмалеммы
обусловливает такую жизненно важную
функцию, как избирательная проница
емость, т. е. способность избирательно
го пропускания в клетку и выхода из нее
различных молекул и ионов. Благодаря
этому в клетке создается и поддержива
ется определенная концентрация ионов и
осуществляются осмотические явления.
Мембраны как основные структурные
элементы клетки ограничивают боль
шинство ее органелл. Они служат не
просто физическими границами, а пред
ставляют собой динамичные функцио
нальные поверхности. На мембранах осу
ществляются многочисленные биохими
ческие процессы, такие, как активное
поглощение неорганических и органичес
ких веществ, синтез АТФ, преобразова
ние энергии при фотосинтезе и др.
Клеточные (внешние) оболочки неко
торых клеток животных образованы це
ментоподобными соединениями. Это ха
рактерно для яиц морских ежей, пресмы
кающихся. Их кожистые оболочки состо
ят из гликопротеинового вещества —
муцина. Кроме того, плазматическая
мембрана часто покрывается дополни
тельными слоями слизи, хитина и др.
, 1. Каковы химический состав, структура и
свойства плазматической мембраны? 2. Что такое
поверхностные рецепторы клеточной мембраны?
3. На чем основано утверждение, что сахара могут
функционировать как информационные молекулы?
12. Молекулярный транспорт
через биологическую мембрану
Главной функцией плазматических
мембран является регулирование обмена
веществ между клеткой и внешней сре
дой. Существуют четыре основных меха
низма поступления и выведения веществ
из клетки: диффузия, осмос, активный
транспорт и экзо- или эндоцитоз.
Диффузия. Известно, что в растворах
происходит перемещение растворенных
веществ из области с высокой концен
трацией в область с более низкой. Этот
поток веществ (транспорт по градиенту
концентрации) существует до тех пор, по
ка концентрации веществ во всем объеме
раствора не выравняются. Перемещение
вещества, движущей силой которого яв
ляется градиент концентрации, называ
ется диффузионным, а процесс — диф
фузией.
Скорость дйффузии вещества через
мембрану зависит, главным образом, от
размера молекул и их относительной рас
творимости в жирах. Это означает, что,
чем меньше молекула и чем легче она
растворяется в липидах, тем быстрее она
будет диффундировать через мембрану.
Например, малые неполярные молекулы
легко растворяются в липидном слое
мембраны, проходят через нее и оказыва
ются по другую сторону. Незаряженные
молекулы небольших размеров (С 0 2,
этанол, мочевина) проходят через мем
брану быстро, а глюкоза, аминокислоты,
жирные кислоты, глицерол — медленно.
Интересно отметить, что вода легко диф
фундирует через липидный слой, несмот
ря на то что молекулы воды относительно
нерастворимы в жирах. Большая ско
рость диффузии воды отчасти объясняет
ся тем, что ее молекулы малы и не заря
жены. Напротив, для всех заряженных
молекул (ионов) независимо от их разме
ров липидные слои плазматической мем
браны служат серьезным препятствием
для проникновения в клетку.
43
За перенос через клеточные мембра
ны различных полярных молекул, таких,
как ионы, сахара, аминокислоты, нуклео
тиды и многие другие метаболиты, ответ
ственны специфические белки, называе
мые мембранными транспортными
белками. Каждый конкретный белок
предназначен для транспорта строго оп
ределенного соединения. Такие белкипереносчики способны соединяться с мо
лекулой или ионом и без затрат энергии,
т. е. пассивно транспортировать их через
мембрану по градиенту концентрации.
Этот процесс, получивший название об
легченной диффузии, является главным
механизмом избирательной проницае
мости мембран.
Активный транспорт. В отличие от
облегченной диффузии активный тран
спорт веществ осуществляется против их
градиентов концентрации, т. е. вещества
переходят из области низкой концентра
ции в область более высокой. В этом слу
чае для переноса протонов или неоргани
ческих ионов через мембрану требуется
энергия, источником которой является
АТФ. Например, необходимый растениям
катион К+ благодаря затратам энергии
может проникнуть в клетки корня даже в
том случае, если его концентрация в поч
венном растворе в 100 раз ниже, чем в
клеточном соке. Наоборот, не столь нуж
ный растениям катион Na+ удаляется
клеткой в окружающую среду даже при
более высокой концентрации в них этого
элемента.
Известно, что механизмы активного
транспорта действуют только для некото
рых ионов, например К+, Са2+. Следова
тельно, клетка обладает избирательной
способностью к различным ионам. Ос
тальные ионы проникают в клетку в зави
симости от градиента их электрохимичес
кого потенциала и проницаемости мем
браны.
Осмос. Вода поглощается клеткой
преимущественно путем осмоса. Ос
мос — это диффузия воды через полу
44
проницаемую мембрану, вызванная раз
ностью концентраций. Если клетку по
местить в воду, то создается градиент вод
ного потенциала: вода поступает в клетку
по градиенту-концентрации и при этом
мембрана избирательно пропускает
только молекулы воды.
В гипертоническом растворе вода под
действием осмотических сил выходит из
клетки. Например, эритроциты в таком
растворе сморщиваются. В растительной
клетке наблюдается явление плазмоли
за — уменьшения вакуоли и отделения
цитоплазмы от клеточной стенки (рис.
24), что приводит к увяданию растений.
Движение воды через мембрану мож
но приостановить, приложив к раствору
определенное внешнее давление, полу
чившее название осмотического давле
ния. Оно вызывается стремлением моле
кул воды пройти сквозь полупроницае
мую мембрану и выровнять концентра
цию по обе ее стороны. Следовательно,
чем выше концентрация раствора, тем
большую силу надо приложить для того,
чтобы воспрепятствовать проникнове
нию воды в раствор через полупроницае
мую мембрану. Поэтому более концен
трированный раствор обладает большим
осмотическим давлением по сравнению с
разбавленным и сильнее притягивает во
ду из окружающего раствора. Другими
словами, осмотическое давление опреде
ляется числом частиц растворенного
вещества в единице объема раство
рителя.
Поскольку концентрация ионов и мо
лекул различных соединений в расти
тельной клетке выше, чем в окружающей
среде (например, в почве), в ней развива
ется сосущая сила, которая и приводит к
поглощению воды. В результате клетка
набухает и создает внутреннее гидроста
тическое давление, направленное на кле
точную стенку. Этому давлению, называ
емому тургорным, противостоит равное
ему по величине механическое давление
клеточной стенки, направленное внутрь
1
2
3
А
Рис. 24. Плазмолиз клетки: 1 — начальная стадия, 2 — выпуклый, 3 — полный, 4 — вогнутый
(при высокой вязкости цитоплазмы).
клетки. Оно называется давлением кле
точной оболочки.
По мере поступления воды осмоти
ческое давление Р клеточного сока и со
сущая сила S уменьшаются, а тургорное
давление Т возрастает до тех пор, пока
эти силы не уравновесят друг друга. Пос
ле этого поглощение воды прекращается.
Эта зависимость выражается следующим
уравнением:
S= P —T.
При полном плазмолизе тургор равен
нулю, а сосущая сила равна величине ос
мотического.давления клетки. В случае
полного насыщения клетки водой тургор
ное давление равно осмотическому
(Р=Т), вследствие чего сосущая сила
равна нулю (Р—7=0) и поступление воды
в клетку прекращается.
Эндоцитози экзоцитоз. Макромоле
кулы (например, белки, полинуклеотиды
или полисахариды), а также частицы пи
щи и мельчайшие организмы проникают
внутрь клетки путем эндоцитоза (от
греч. endon — внутри, kytos — клетка).
Различают два типа эндоцитоза: фагоци
тоз (поглощение твердых частиц) я пи ноцитоз (поглощение жидкостей).
При фагоцитозе мембрана клетки об
разует выросты и охватывает ими части
цы, находящиеся вне клетки. Затем края
выростов смыкаются и частица оказыва
ется заключенной в мембранном пузырь
ке — фагоцитарной вакуоли, с
которой сливаются лизосомы. Благодаря
ферментам лизосом происходит рас
щепление сложных органических ве
ществ до фрагментов, которые могут
быть усвоены клеткой (рис. 25, а, с. 46).
Благодаря способности к пиноцитозу
клетки могут поглощать пузырьки жид
кости. Процесс поглощения жидкости
сходен с фагоцитозом. В месте соприкос
новения капли с клеткой плазматическая
мембрана образует впячивание в виде ка
нальца, который наполняется жидкос
тью. Затем он отшнуровывается и попа
дает в цитоплазму.
Путем фаго- и пиноцитоза осущес
твляются питание гетеротрофных протистов, защитные реакции высших орга
низмов (лейкоциты поглощают чужерод
ные частицы), транспорт (клетки почеч
ных канальцев всасывают белки из пер
вичной мочи) и т. д.
Процесс, обратный эндоцитозу, —
экзоцитоз (от греч. ехо — снаружи,
45
Рис. 25. Схема эндо- (а) и экзоцитоза (б).
kytos — клетка) — связан с выведением
содержимого мембранного пузырька во
внешнюю среду (рис. 25,6). Например,
клетки, продуцирующие гормон инсулин,
упаковывают его во внутриклеточные пу
зырьки, которые затем сливаются с плаз
матической мембраной, высвобождая
при этом инсулин.
9
I I . Каков механизм поступления веществ в
клетку или выхода из нее? 2. Каков механизм облег
ченной диффузии и активного транспорта веществ
через мембрану? 3. Как происходит поглощение во
ды клеткой? Что такое осмос, осмотическое давле
ние, тургорное давление, сосущая сила? 4. Что та
кое эндо- и экзоцитоз и как они осуществляются?
13.
И нф орм ационная систем а
кл е тки .
Я д р о , е го с тр о е н и е и ф у н кц и и
Ядро. Все живые организмы характе
ризуются исключительно упорядоченным
строением и функционированием. Этот
порядок не случаен, он направляется ин
формацией, сосредоточенной главным
46
образом в ядре эукариотической клетки и
в ядерной области прокариот. Генетичес
кая информация закодирована в виде
специфической последовательности нук
леотидов в молекуле ДНК.
Благодаря наличию ДНК, ядро явля
ется местом хранения и воспроизводства
наследственной информации, определя
ющей признаки данной клетки и всего ор
ганизма. Ядро служит также центром
управления обменом веществ клетки, оп
ределяя, какие белки и в какое время
должны синтезироваться. Поэтому после
удаления ядра из клетки она, как прави
ло, быстро погибает.
Большинство клеток имеют одно яд
ро, изредка встречаются двухъядерные
(клетки печени) и многоядерные (клетки
протистов, водорослей и грибов, а также
клетки поперечнополосатых мышц).
Форма и размеры ядра очень измен
чивы и зависят от вида организма, а так
же от типа, возраста и функционального
состояния клетки. Ядро может быть линзо-, веретено- или шаровидным (5—20
мкм в диаметре), а также многолопаетным. Лопастная форма обеспечивает
значительно большую площадь сопри
косновения ядерной оболочки с цито
плазмой, что, по-видимому, способствует
увеличению скорости биохимических ре
акций.
Общий план строения ядра одинаков
у всех клеток (рис. 26). Ядро состоит из
ядерной оболочки, нуклеоплазмы, хрома
тина и ядрышек.
Ядро отделено от цитоплазмы двой
ной мембраной, или так называемой
ядерной оболочкой. Наружная мембра
на, граничащая с цитоплазмой, имеет
сложную складчатую структуру, местами
соединенную с каналами эндоплазмати
ческой сети: на ней часто расположены
рибосомы. Внутренняя же мембрана,
контактирующая с нуклеоплазмой, рибо
сом лишена. Пространство между мем
бранами ядерной оболочки называется
перину клеарным.
Ядерная оболочка пронизана множес
твом пор диаметром около 30— 100 нм.
Численность пор также колеблется: в за
висимости от типа и физиологического
состояния клетки на 1 мкм2 ядерной обо
лочки их насчитывается в среднем от 10
до 30. В молодых клетках их всегда боль-
Рис. 26. Строение ядра: 1 — шероховатая (гра
нулярная) и 2 — гладкая (агранулярная) эндо
плазматическая сеть (ретикулум), 3 — рибосо
мы, 4 — хроматин, 5 — ядрышко, 6 — пора, 7 —
нуклеоплазма.
ше. Благодаря наличию пор, обеспечива
ющих избирательную проницаемость,
ядерная оболочка контролирует обмен ве
ществ между ядром и цитоплазмой, на
пример, выход в цитоплазму иРНК и рибосомных субъединиц или поступление в
ядро белков, нуклеотидов и молекул, ре
гулирующих активность ДНК. Ядерная
оболочка образуется после деления ядра
из цистерн эндоплазматической сети и
частично из фрагментов старой ядерной
оболочки, распавшейся во время деления.
Внутреннее содержимое ядра состав
ляет гелеобразный матрикс — нуклео
плазма (или ядерный сок), заполняю
щая пространство между структурами яд
ра. В ней находятся одно или несколько
ядрышек, хроматин, значительное коли
чество РНК и ДНК, различные белки, в
том числе большинство ферментов ядра,
а также свободные нуклеотиды, амино
кислоты, промежуточные продукты об
мена веществ. Нуклеоплазма осущес
твляет взаимосвязь всех ядерных струк
тур.
Хроматин на окрашенных препара
тах клетки в состоянии покоя представля
ет собой сеть тонких тяжей (фибрилл),
мелких гранул или глыбок. Основу хро
матина составляют нуклеопротеины —
длинные нитевидные молекулы ДНК, со
единенные со специфическими белка
ми — гистонами. В процессе деления
ядра нуклеопротеины спирализуются,
укорачиваются, тем самым уплотняются
(в 100— 500 раз) в компактные палочко
видные хромосомы, которые становятся
заметными в световой микроскоп. В сос
тав хромосом входят также РНК, кислые
белки, липиды и минеральные вещества
(ионы Са2+ и Mg2+), а также фермент
ДНК-полимераза, необходимый для реп
ликации ДНК.
У каждой хромосомы имеется пер
вичная перетяжка (утонченный неспирализованный участок), которая делит
хромосому на два плеча. В области пер
вичной перетяжки располагается фиб
47
риллярное тельце — центромера (кинет охор), которая регулирует движение
хромосом при клеточном делении: к ней
прикрепляются нити веретена деления,
разводящие хромосомы к полюсам.
В зависимости от расположения пе
ретяжки определяют три основных вида
хромосом: 1) равноплечие — с плечами
равной длины; 2) неравноплечие — с
плечами неравной длины; 3) палоч
ковидные с одним длинным и другим
очень коротким, едва заметным плечом
(рис. 27).
Некоторые хромосомы имеют вт о
ричную перетяжку, не связанную с
прикреплением нити веретена деления.
В области вторичной перетяжки обычно
формируется ядрышко. Поэтому этот
•
«г
1
Ц
2
3
Рис. 27 Типы хромосом: 1 — равноплечая, 2 —
неравноплечая, 3 — палочковидная, 4 — цен
тромера, 5 — спирально закрученная нить
ДНК.
48
участок хромосомы называется ядрыш
ковым организатором.
Каждой клетке того или иного вида
живых организмов свойственно опреде
ленное число, размеры и форма хромо
сом. Совокупность хромосом соматичес
кой клетки (клетки тела многоклеточного
организма), типичную для данной систе
матической группы протистов, животных
или растений, называют хромосомным
набором, или кариотипом.
Число хромосом в зрелых половых
клетках называют гаплоидным набором
и обозначают п. Соматические клетки со
держат двойное число хромосом (дипло
идный набор), обозначаемое 2п. Клетки,
имеющие более двух наборов хромосом,
называют полиплоидными (4п, 8п и
т. д.). Парные хромосомы, т. е. одинако
вые по форме, структуре и размерам, но
имеющие разное происхождение (одна
материнская, другая отцовская), называ
ются гомологичными.
Количество хромосом в кариотипе не
связано с уровнем организации живых
организмов: примитивные формы могут
иметь большее число хромосом, чем вы
сокоорганизованные, и наоборот. Напри
мер, клетки радиолярий (морских про
тистов) содержат 1000— 1600 хромосом,
а клетки шимпанзе — всего 48. Однако
следует помнить, что все организмы од
ного вида имеют одинаковое количество
хромосом, т. е. для них характерна видо
вая специфичность кариотипа. В клетках
человека диплоидный набор составляет
46 хромосом, пшеницы мягкой — 42,
картофеля — 18, мухи домашней — 12,
плодовой мушки дрозофилы — 8. Прав
да, клетки разных тканей даже одного ор
ганизма в зависимости от выполняемой
функции могут иногда содержать разное
число хромосом. Так, например, в клетках
печени животных бывает разное число
наборов хромосом (4л, 8л). По этой при
чине понятия «кариотип» и «хромосом
ный набор» не совсем идентичны.
Ядрышки — это округлые, сильно
уплотненные участки клеточного ядра ди
аметром 1— 5 мкм и больше. В состав
ядрышек входит около 80 % белка, 10—
15 % РНК, некоторое количество ДНК и
другие химические компоненты.
Во время деления ядра ядрышки раз
рушаются, а затем в конце деления фор
мируются вновь. Под контролем генов
ядрышковых организаторов осуществля
ется синтез рибосомной РНК. В ядрышке
происходит объединение РНК с белком, в
результате чего образуются рибонуклеопротеидные субъединицы рибосом. По
следние через поры ядерной оболочки пе
реходят в цитоплазму, где заканчивается
их формирование.
Таким образом, ядрышко является
местом синтеза рибосомной РНК и са
мосборки субъединиц рибосом.
ется кодоном. Нетрудно подсчитать, что
число возможных комбинаций из четырех
нуклеотидов по три составит 64, что бо
лее чем достаточно для кодирования
20 аминокислот, входящих в состав
белка.
Выяснилось, что код является м но
жественным, или «вырожденным», т. е.
одна и та же аминокислота может коди
роваться несколькими триплетами (от 2
до 6), в то время как каждый триплет ко
дирует только одну аминокислоту, напри
мер:
фенилаланин — УУУ, УУЦ;
изолейцин — АУУ, АУЦ, АУА;
пролин — ЦЦУ, ЦЦЦ, ЦЦА, ЦЦГ;
серин — УЦУ, УЦЦ, УЦА, УЦГ, АГУ,
АГЦ.
?■ 1. Из каких компонентов состоит система ядра?
Помимо этого, код является неперекрывающимся, т. е. один и тот же нукле
отид не может входить одновременно в
состав двух соседних триплетов. И нако
нец, это код без знаков препинания.
Это значит, что если произойдет выпаде
ние одного нуклеотида, то при считыва
нии его место займет ближайший нуклео
тид из соседнего кодона, из-за чего изме
нится весь порядок считывания. Поэтому
правильное считывание кода с иРНК
обеспечивается только в том случае, если
он считывается со строго определенного
пункта. Стартовыми кодонами в мо
лекуле иРНК являются триплеты АУГ
иГУГ.
Нуклеотидный код универсален для
всех живых организмов и вирусов: одина
ковые триплеты кодируют одинаковые
аминокислоты. Это открытие представ
ляет собой серьезный шаг на пути к более
глубокому познанию сущности живой ма
терии, ибо универсальность генетичес
кого кода свидетельствует о единстве
происхождения всех живых орга
низмов.
К настоящему времени расшифрова
ны триплеты для всех 20 аминокислот,
входящих в состав природных белков.
2. Каковы основные функции ядра? 3. Как устроена
ядерная оболочка? 4. Какие структуры ядра содер
жат молекулы ДНК? б. Как устроена хромосома?
6. Что такое кариотип? 7. Каковы строение и фун
кции ядрышка?
14. Хранение наследственной ин
формации.
Генетический код
Последовательность нуклеотидов в
полинуклеотидной цепи специфична для
каждой клетки и представляет собой ге
нетический код, посредством которого
записана информация о синтезе опреде
ленных белков. Это значит, что в молеку
ле ДНК каждое сообщение закодировано
специфической последовательностью из
четырех знаков — А, Г, Т, Ц, подобно то
му, как письменные сообщения кодиру
ются знаками (буквами) алфавита или аз
буки Морзе. Код является триплетным,
т. е. каждая аминокислота кодируется из
вестным сочетанием из трех расположен
ных рядом нуклеотидов, которое называ
49
Например, аминокислота лизин кодиру
ется триплетом ТТТ, валин ЦАА, трипто
фан — АЦЦ и т. д. Таким образом, зная
порядок расположения триплетов в мо
лекуле ДНК (генетический код), можно
установить порядок расположения ами
нокислот в белке. В одной молекуле ДНК
может быть закодирована последова
тельность аминокислот для многих бел
ков. Функциональный отрезок молеку
лы ДНК, несущий в себе информацию о
структуре одного белка, называется ге
ном. Различают структурные гены, в ко
торых закодирована информация для
синтеза структурных и ферментных бел
ков, и гены с информацией для синтеза
тРНК, рРНК и др.
Итак, план построения белка закоди
рован в ДНК, которая непосредственного
участия в синтезе белковых молекул не
принимает. Она служит лишь матрицей
для синтеза иРНК.
Процесс биосинтеза белка осущес
твляется на рибосомах, расположенных
преимущественно в цитоплазме. Следо
вательно, для передачи генетической ин
формации с ДНК к месту синтеза белка
требуется посредник. Таким посредником
является иРНК, которая на основе прин
ципа комплементарности синтезируется
на одной из цепей молекулы ДНК. Этот
процесс получил название транскрип
ции или переписывания-.
ц
. . .Г
1
А.
1
1
Т
1
1
.А
1
. .
1
Т . .
1
1 1
Г ...ц
1
1. . . А11
т г
1. . . г1
ц
1
1
ДНК
50
ц
1
1
А
1
1
У
1
Г1
1
У
1
ц
11
иРНК
Транскрипция происходит не на всей
молекуле ДНК одновременно, а лишь на
небольшом ее участке, отвечающем оп
ределенному гену. При этом часть двой
ной спирали ДНК раскручивается, обна
жая короткий участок одной из цепей, ко
торый теперь будет служить матрицей
для синтеза иРНК. Затем вдоль этой цепи
движется фермент РНК-полимераза, со
единяя между собой нуклеотиды в расту
щую цепь иРНК, последовательность
нуклеотидов которой является точной ко
пией последовательности нуклеотидов
матрицы —- одного или группы рядом
расположенных генов. Так, если в моле
куле ДНК имеется азотистое основание
цитозин, то в РНК — гуанин, и наоборот.
В ДНК комплементарной парой являют
ся аденин и тимин. Однако в состав иРНК
вместо тимина входитурацил. Транскрип
ция может происходить одновременно на
нескольких генах одной хромосомы и на
генах, расположенных на разных хромо
сомах.
На специальных генах синтезируются
и два других типа РНК — тРНК и рРНКНачало и конец синтеза всех типов РНК
на матрице ДНК строго фиксирован спе
циальными триплетами, которые контро
лируют запуск (инициирующие) и оста
новку синтеза (терминирующие) РНК.
Триплеты выполняют функцию знаков
препинания между генами.
1. Что такое генетический ход и каковы его
свойства? 2. Что такое транскрипция?
15. Реализация наследственной
информации.
Синтез белка на рибосомах
Синтезированная в ядре иРНК отде
ляется от ДНК и через поры ядерной обо
лочки поступает в цитоплазму, где при
крепляется к рибосоме.
Рибосомы —■это сферические грану
лы диаметром 15— 35 нм, являющиеся
местом синтеза белка из аминокислот.
Они обнаружены в клетках всех организ
мов, в том числе прокариотических. Каж
дая рибосома состоит из двух нуклеопротеидных субъединиц разной величины,
формы и химического строения, удержи
вающихся вместе благодаря присутствию
в них ионов магния. Рибосомы состоят из
примерно равных количеств белка и
рРНК.
В цитоплазме десятки тысяч рибосом
расположены свободно (поодиночке или
группами) или прикреплены к наружной
поверхности мембраны ядра и эндоплаз
матической сети. Они обнаружены также
в митохондриях и хлоропластах.
В процессе синтеза белка рибосома
защищает иРНК и синтезируемый белок
от разрушающего действия клеточных
ферментов типа РНКаз и протеаз. Меха
низм защитного действия заключается в
том, что нить иРНК проходит между
большой и малой субъединицами рибосо
мы, а начальная часть вновь синтезируе
мого белка находится в каналоподоб
ной структуре большой субъединицы
(рис. 28).
На одной молекуле иРНК может раз
мещаться несколько рибосом. Их число
определяется длиной иРНК. Комплекс из
иРНК и рибосом (от 5— 6 до нескольких
десятков) называется полисомой. Обра
зование полисом повышает эффектив
ность функционирования иРНК за счет
того, что одновременно протекает синтез
нескольких идентичных полипсптидных
цепей. Именно па полисомах происхо
дит синтез белка, или трансляция
(рис. 29, с. 52).
Трансляция начинается со стартового
кодона АУГ. Отсюда молекула иРНК пре
рывисто, триплет за триплетом продвига
ется через рибосомы, что сопровождает
ся ростом полипептидной цепочки. Число
аминокислот в таком белке равно числу
триплетов иРНК.
Выстраивание аминокислот в соот
ветствии с кодонами иРНК осуществля
ется на рибосомах при помощи тРНК —
Рис. 28. Схема строения рибосомы: 1 — малая
субъединица, 2 — иРНК, 3 — тРНК, 4 — амино
кислота, 5 — большая субъединица, 6 — мем
брана эндоплазматического ретикулума, 7 —
синтезируемая полипептидная цепь.
главных агентов синтеза белка. Благода
ря определенному расположению ком
плементарных нуклеотидов цепочка
тРНК имеет форму, напоминающую лист
клевера (рис. 30, с. 53). При этом тРНК
имеет акцепторный конец, к которому
присоединяется активированная амино
кислота. Активацию аминокислот осу
ществляют специфичные ферменты аминоацил-тРНК-синтетазы, т. е. для каж
дой аминокислоты существует свой фер
мент. Механизм активации заключается в
том, что фермент одновременно взаимо
действует с соответствующей аминокис
лотой и АТФ, которая теряет при этом пи
рофосфат. Тройной комплекс из фермен
та, аминокислоты и АТФ называется ак
тивированной (богатой энергией) ами
нокислотой, которая способна спонтан
но образовать пептидную связь, что при51
1
Рис. 29. Схема биосинтеза белка: 1 — рибосома, 2 — кодоны, 3 — иРНК, 4 — тРНК, 5 — ан
тикодон, в — образование пептидной связи, 7 — растущая полипептидная цепь.
водит к синтезу полипептидов. Данный
процесс активации — необходимый этап
белкового синтеза, поскольку свободные
аминокислоты не могут прямо присоеди
няться к полипептидной цепи.
В противоположной части молекулы
тРНК располагается специфический
триплет (антикодон), ответственный за
прикрепление по принципу комплемен
тарное™ к определенному триплету
иРНК (кодон). Таким образом, именно
комплекс аминоацил-тРНК считывает
информацию, закодированную в иРНК.
Комплекс аминоацил-тРНК с по
мощью антикодона присоединяется к ко
дону иРНК на малой субъединице рибо
сомы. Затем к той же рибосоме прикреп
ляется второй комплекс в соответствии
со следующим кодоном. В рибосоме ока
52
зываются две аминокислоты, ориентиро
ванные по отношению друг к другу таким
образом, что карбоксильная группа одной
аминокислоты оказывается рядом с ами
ногруппой другой аминокислоты. В ре
зультате мещцу ними возникает пептид
ная связь. Первая тРНК, освободившись
от аминокислоты, покидает рибосо
му. Далее к образованному дипепти
ду таким же образом пристраивает
ся третья, четвертая и т. д. аминокисло
ты, принесенные в рибосому своими
тРНК. Процесс продолжается до тех пор,
пока рибосома не дойдет до одного из
трех терминирующих кодонов: УАА, УАГ
или УГА, после чего синтез белка прекра
щается.
Таким образом, последовательность
кодонов иРНК определяет последова-
Е
80 дней. За всю жизнь человек обновляет
свои белки около 200 раз.
Каждый этап биосинтеза катализиру
ется соответствующими ферментами и
снабжается энергией за счет расщепле
ния АТФ. Синтезированные белки посту
пают в каналы эндоплазматической сети,
по которым транспортируются к опреде
ленным участкам клетки.
?
■ 1. Каковы особенности строения рибосом?
2. Как осуществляется биосинтез белка в клетке и
какова роль нуклеиновых кислот в этом процессе?
16.
Рис. 30. Структура тРНК: А, Б, В, Г — участки
комплементарного соединения, Д — участок
соединения с аминокислотой, Е — антикодон.
тельность включения аминокислот в цепь
белка.
Большое значение в данном процессе
имеет участие цепочки рибосом (поли
сом), что дает возможность молекуле
иРНК последовательно присоединиться к
ним и служить матрицей для синтеза не
скольких одинаковых молекул белка.
После завершения синтеза белка иРНК
под действием ферментов распадается на
отдельные нуклеотиды.
Следовательно, роль нуклеиновых
кислот в биосинтезе белка заключается в
преобразовании генетической информа
ции, представленной в виде последова
тельности нуклеотидов ДНК, в структуру
молекулы иРНК- Затем происходит син
тез белков из аминокислот.
Синтез белковых молекул происходит
непрерывно и идет с большой скоростью:
за 1 мин образуется от 50 до 60 тыс. пеп
тидных связей. Синтез одной молекулы
белка длится всего 3—4 с. В результате
половина белков нашего тела (в среднем
в нем около 17 кг белка) обновляется за
Э н е р ге ти ч е с ка я с и сте м а кл е тки
Любая живая клетка представляет
собой активную, динамичную систему.
В ней происходит преобразование энер
гии, синтез, распад и перенос веществ.
Весь этот комплекс реакций живой клет
ки носит название метаболизма или об
мена веществ и энергии. Процессы
синтеза, которые приводят к образова
нию составных частей клетки, называют
ся анаболизмом, а процессы распада,
обычно связанные с высвобождением
энергии, катаболизмом. Анаболизм
часто называют ассимиляцией или плас
тическим обменом, а катаболизм —
диссимиляцией или энергетическим
обменом.
Для осуществления любых проявле
ний жизнедеятельности необходима
энергия. Энергия требуется для процес
сов химического синтеза, для всех видов
движения, в том числе и мышечного; для
передачи нервных импульсов. Энергия
тратится и на процесс активного перено
са веществ через плазматическую мем
брану (в клетку и из клетки), причем на
этот процесс расходуется весьма значи
тельная часть энергетических ресурсов
клетки. Энергия требуется также для об
разования теплоты и поддержания посто
янной температуры тела у птиц и млеко
питающих и т. д.
Откуда же берется энергия, необхо
димая для осуществления жизнедеятель
53
ности клетки? Она поступает из окружа
ющей среды. Первичным источником
энергии для всего живого служит та часть
солнечной радиации, которая называется
видимым светом. Эта энергия улавлива
ется зелеными растениями и в процессе
фотосинтеза превращается сначала в
электрохимическую, а затем в химичес
кую энергию, которая запасается в орга
нических веществах, образующихся при
фотосинтезе. Животные организмы, гри
бы, большинство бактерий и протистов,
не способных к фотосинтезу, зависят (в
смысле снабжения энергией) от веществ,
синтезируемых растениями. Эта зависи
мость может быть прямой, как у травояд
ных, 11ли непрямой, как у плотоядных, ко
торые питаются другими животными, в
том числе травоядными.
Далее запасенная энергия переводит
ся в форму, в которой она может исполь
зоваться растительными и животными
клетками, клетками других организмов
для выполнения какой-либо работы, на
пример для синтеза определенных моле
кул, необходимых клетке, или же для ме
ханической, электрической и осмотичес
кой работы.
Авто- и гетеротрофные клетки.
Жизнь на нашей планете зависит от
энергии, излучаемой Солнцем. При фо
тосинтезе происходит улавливание и за
пасание солнечной энергии в молекулах
органических веществ. Исходными со
единениями для фотосинтеза служат
простые, бедные энергией неорганичес
кие вещества — диоксид углерода (С 0 2)
и вода (Н20 ). Их этих соединений строят
ся более сложные органические вещес
тва. Клетки, способные к самостоятель
ному синтезу необходимых для них орга
нических соединений за счет энергии све
та (фотосинтез) или энергии, выделив
шейся при окислении неорганических со
единений (хемосинтез), называются а в
тотрофными или авт от роф ами (от
греч. autos — сам и trophe — пища, пи
тание). К автотрофным принадлежат
54
клетки зеленых растений, зеленых и пур
пурных бактерий, хемоавтотрофные бак
терии, цианобактерии и некоторые протисты.
Гетеротрофные клетки не могут синте
зировать органические вещества из неор
ганических, поэтому для процессов асси
миляции они нуждаются в поступлении ор
ганических веществ извне в виде пищи, по
лучая с ней готовые углеводы, жиры, бел
ки. Гетеротрофными являются все клетки
животных, большинство бактерий, клетки
грибов и высших растений — сапротрофов и паразитов (например, подъельник,
заразиха, Петров крест), а также клетки
растений, не содержащие хлорофилл (на
пример, подземные органы).
Можно выделить также авт огет еротрофные клетки, которые в зависи
мости от условий могут осуществлять фо
тосинтез либо потреблять готовые орга
нические вещества из окружающей сре
ды; например, такие протесты, как хла
мидомонада, эвглена и др.
Синтез первичного органического
вещества автотрофами. Различают два
типа автотрофных организмов — фоm o
rn хемотрофные. Большую часть авто
трофных организмов составляют фототрофы, роль которых в биосфере трудно
переоценить. Достаточно сказать, что
процесс фотосинтеза — это основной'
путь, по которому практически вся энер
гия входит в нашу биосферу. При этом
каждый год в процессе фотосинтеза на
земном
шаре
образуется
более
150 млрдт органических веществ, необ
ходимых для всех гетеротрофных орга
низмов. Вклад же хемотрофов в синтез
питательных веществ на Земле ничто
жен, т. к. имеющиеся в их распоряжении
ресурсы химической энергии крайне ма
лы по сравнению с потоком солнечной
энергии.
9
1. Что называется метаболизмом? 2. Что такое
анаболизм, катаболизм? 3. На какие процессы за
трачивается энергия в клетке? 4. Откуда клетки бе
рут энергию для обеспечения процессов жизнедея
тельности? 5. Чем отличаются автотрофные клетки
от гетеротрофных?
17. Фотосинтез
Фотосинтез — это процесс преобра
зования энергии света в энергию хими
ческих связей органических веществ.
Процесс фотосинтеза обычно описывают
уравнением:
свет
6 С 0 2+ 6 Н 20 -----> С6Н 120 6+ 6 0 2.
хлорофилл
Такое преобразование происходит в
зеленых пластидах — хлоропластах.
Пластиды — это характерные органеллы клеток автотрофных протистов и
растений. Их окраска, форма и размеры
весьма разнообразны. Классифицируют
пластиды главным образом по окраске:
хлоропласты, хромопласты и лейко
пласты.
Хлоропласты имеют зеленый цвет,
обусловленный присутствием основного
пигмента — хлорофилла и вспомогатель
ных — желтых пигментов — каротинои
дов.
Хлоропласты — это овальные тельца
размером 5—'10 х 2—4 мкм. В одной
клетке листа может находиться 15— 20 и
более хлоропластов, а у некоторых водо
рослей — лишь 1—2 гигантских хлоро
пласта (хроматофора) различной формы.
Хлоропласты покрыты оболочкой,
состоящей из наружной и внутренней
мембраны (рис. 31).
Оболочка ограничивает внутреннюю
гомогенную среду хлоропласта — с т р о
му (матрикс). В строме содержатся бел
ки, липиды, ДНК (кольцевая молекула),
РНК, рибосомы и запасные вещества
(липиды, крахмальные и белковые зер-
Рис. 31. Строение хлоропласта: 1 — липидная капля, 2 — зерна крахмала, 3 — наружная
мембрана, 4 — грана, 5 — ДНК, 6 — рибосома, 7 — строма, 8 — тилакоид граны, 9 — тилакоиды стромы, 10 — внутренняя мембрана.
55
на), а также ферменты, участвующие в
фиксации диоксида углерода.
Внутренняя мембрана образует утол
щенные замкнутые впячивания — т и л а ко иды, которые имеют форму дис
ков. Несколько таких тилакоидов, лежа
щих друг над другом, образуют г ра ну, и
в этом случае они называются т и л а к о ид а ми г р а н .
Другие тилакоиды, связывающие
между собой граны или не контактирую
щие с ними, называются ти л а ко идам и
с т р о м ы . Именно в мембранах тилакои
дов локализованы светочувствительные
зеленые (хлорофиллы) и желтые (каро
тиноиды) пигменты, а также переносчики
электронов и протонов, которые участву
ют в поглощении и преобразовании энер
гии света. Биохимические системы синте
за и превращения углеводов функциони
руют в строме хлоропластов. В ней же от
кладывается запасной крахмал.
В зависимости от степени освещен
ности хлоропласта перемещаются в тол
ще цитоплазмы таким образом, чтобы
слабый свет воздействовал на возможно
большую фотосинтезирующую повер
хность (усиление фотосинтеза), а силь
ный — на минимальную (защита от раз
рушительного действия прямых солнеч
ных лучей). В последнем случае хлоро
пласта располагаются вдоль клеточных
стенок, параллельно световому потоку.
Кроме хлоропластов, в растительных
клетках имеются бесцветные пласти
ды — пропластиды, лейкопласт ы,
этиопласты и окрашенные — хромо
пласты (рис. 32). В меристематических
тканях присутствуют пропластиды, у ко
торых внутренняя мембрана имеет лишь
небольшие впячивания. Если структура
пропластид сохраняется в органеллах
зрелых клеток, их называют лейкоплас
тами (рис. 33). В лейкопластах отклады
ваются запасные вещества, и названия
они получают в зависимости от этих со
единений: если запасается крахмал —
амилопласты, если жиры — элайо56
Рис. 32. Хромопласты
Рис. 33. Лейкопласты в клетке: 1 — на фоне
ядра, 2 — в цитоплазме.
пласты, белки — протеинопласты.
Этиопласты формируются при выращи
вании зеленых растений в темноте. При
освещении они превращаются в хлоро
пласта.
Хромопласты отличаются от других
пластид своеобразной формой (дисковид
ной, зубчатой, серповидной,треугольной,
ромбической и др.) и окраской. В пузырь
ках стромы они содержат кристалличес
кие каротиноиды, которые и придают им
желтую, оранжевую и красную окраску.
Их присутствием объясняется окраска
плодов и корнеплодов некоторых расте
ний — томатов, рябины, ландыша, ши
повника, моркови, свеклы и др. С хро
мопластами связан также синтез некото
рых витаминов.
Все типы пластид генетически род
ственны друг другу, и одни их виды могут
превращаться в другие, как это показано
на рисунке 34.
Таким образом, весь процесс взаи
мопревращений пластид можно предста
вить в виде ряда изменений, идущих в од
ном направлении — от пропластид до
хромопластов. Последнее рассматрива
ется как дегенерирующая (конечная)
форма.
Фотосинтезирующие
пигменты.
Важнейшую роль в фотосинтезе играют
фотосинтезирующие пигменты: хлоро
филлы, каротиноиды ( карот ины,
ксантофиллы), а у цианобактерий и
красных водорослей еще фикобилины
(фикоцианин, фикоэритрин).
Известно около 10 хлорофиллов (а, Ь,
с, d, е и др.), которые отличаются друг от
друга по химическому строению, окраске,
распространению. У всех высших расте
ний содержатся хлорофиллы а и Ь. Хло
рофиллы e n d обнаружены у ряда водо
рослей и протистов, бактериохлорофиллы а и Ь —-в клетках фотосинте
зирующих зеленых и пурпурных бак
терий.
Основным пигментом, участвующим
в фотохимических реакциях, является
хлорофилл а (у бактерий — бактериохлорофилл а). По химическому строению
хлорофилл — это сложный эфир двухос
новной хлорофиллиновой кислоты и двух
остатков спиртов — фитола (С2оН39ОН)
и метанола (СН3ОН) (см. с. 58):
Рис. 34
57
COOCH,
Mg
n 4o h 30c 32
'COOConH
20п 39
Центральное место в молекуле хлоро
филла занимает атом магния. Свет по
глощают все пигменты, но из них только
фотосинтетически активные (основные)
принимают непосредственное участие в
фотохимических реакциях. Вспомога
тельные пигменты (хлорофилл d, кароти
ноиды, фикобилины) передают погло
щенную энергию квантов света активным
пигментам (хлорофилл а), расширяя тем
самым спектр действия фотосинтеза. Та
ким образом, молекула хлорофилла а по
лучает энергию от окружающих ее много
численных молекул других пигментов,
выполняя чрезвычайно важную роль ак
тивного (реакционного) центра.
Хлорофиллы поглощают свет в синей
и красной областях спектра, каротинои
ды — в синей и сине-зеленой. В зеленой
и желтой областях свет высшими расте
ниями почти не поглощается и фотосин
тез не происходит.
Световая фаза фотосинтеза. Про
цесс фотосинтеза представляет собой
цепь окислительно-восстановительных
реакций, где происходит восстановление
углекислого газа до уровня углеводов.
Всю совокупность фотосинтетических
реакций принято подразделять на две фа
зы — световую и темповую.
Для световой фазы фотосинтеза ха
рактерно то, что энергия солнечной ради
ации, поглощенная хлоропластами, пре
образуется в электрохимическую. Это
достигается путем переноса электронов и
ионов водорода с помощью специальных
переносчиков через мембрану тилакоидов. Такой процесс переноса ионов Н+ и
электронов получил название хемиосмоса. Выяснен он еще не до конца, но в
общих чертах этот процесс можно пред
ставить следующим образом (рис. 35).
58
При попадании кванта света на моле
кулу хлорофилла один из его электронов
поглощает эту энергию и переходит на
более высокий энергетический уровень,
т. е. оказывается в возбужденном состоя
нии. Возбужденный электрон может пе
рейти в основное состояние, и в этом слу
чае его избыточная энергия может выде
ляться в виде флюоресценции (красное
свечение) либо теплоты, или передавать
ся в качестве энергии возбуждения дру
гим молекулам. Кроме того, электрон мо
жет также отрываться от молекулы хло
рофилла. В последнем случае электрон,
использовав запас энергии, передается
от одного переносчика к другому по электронтранспортной цепи. Переносчиками
электрона в этом случае являются слож
ные органические соединения, встроен
ные в мембраны хлоропласта.
Таким образом, при действии света на
хлоропласт начинается электронный по
ток по системе переносчиков, встроен
ных в мембраны тилакоидов. С перено
сом электронов по электронтранспортной цепи сопряжено поступление прото
нов из стромы внутрь тилакоида. В ре
зультате активного переноса протонов из
стромы во внутритилакоидное простран
ство на мембране создается электрохи
мический потенциал водорода (Д цН +),
имеющий две составляющие: концентра
ционную ДрН, возникающую в результа
те неравномерного распределения ионов
Н+ по разные стороны мембраны, и элек
трохимическую, обусловленную противо
положными зарядами разных сторон
мембраны (благодаря накоплению прото
нов с внутренней стороны мембраны на
ее внешней стороне скапливаются отри
цательно заряженные частицы). Раз
ность потенциалов на мембранах дости
гает 50 мВ. Соответственно и энергия,
накапливаемая на мембране в результате
концентрации протонов на одной сторо
не, имеет две составляющие — концен
трационную и электрическую. Чтобы
использовать эту энергию, необходимо
ФОТОЭЛЕКТРОННЫЙ
- ПЕРЕНОС
Рис. 35. Схема хемиосмоса.
59
обеспечить обратный поток протонов.
Выделяющаяся при этом энергия и ис
пользуется для образования АТФ. Когда
ионы водорода Н+ проходят через каналы
мембраны, на ее наружной поверхности е
участием фермента АТФ-синтетазы из
АДФ и фосфорной кислоты синтезирует
ся АТФ. Синтезированная АТФ переходит
в строму хлоропласта.
Чем же заканчивается передача элек
тронов? Достигнув внешней поверхности
мембраны тилакоида, электрон соединя
ется с ионом водорода. Еще один элек
трон из электронтранспортной цепи вос
станавливает молекулу переносчика ато
мов водорода НАДФ+ (никотинамидадениндинуклеотидфосфат), которые в дос
таточном количестве присутствуют в
строме хлоропласта. При этом атом водо
рода соединяется с молекулой НАДФ:
НАДФ++2Н ++ 2 е - ---- => НАДФ • Н +Н +.
Таким образом, активированные све
товой энергией электроны используются
на присоединение атома водорода к его
переносчику, т. е. на восстановление
НАДФ+ в НАДФ • Н + Н +.
Вернемся теперь к молекулам хлоро
филла, утратившим свои электроны. Об
разовавшаяся в их молекулах электрон
ная «дырка» действует как сильный окис
литель и отрывает электроны от молекул
воды внутри тилакоида.; Через ряд пере
носчиков при участии ионов марганца и
хлора эти электроны передаются на мо
лекулы хлорофилла••и заполняют «дыр ку». Происходит фотоокисление (фо
т олиз) воды, в результате которого вы
деляется свободный кислород, а внутри
тилакоида накапливаются ионы водорода
Н +:
2 Н , 0 ---- > 4Н+ + 4е~ + 0 2.
Ионы водорода остаются внутри тилакоидов и увеличивают их концентрацию.
В результате фотоокисления воды обра
зуется свободный кислород, а также по
полняется запас положительно заряжен
60
ных ионов водорода на внутренней сторо
не тилакоида.
Таким образом, в результате переноса
электронов и протонов через мембрану
тилакоида происходит превращение све
товой энергии в электрохимическую и да
лее в химическую энергию макроэргических связей молекулы АТФ. При этом про
исходит восстановление НАДФ и выделе
ние свободного кислорода.
Кислород, образующийся при разло
жении воды, является побочным продук
том фотосинтеза. В дальнейшем он мо
жет использоваться растительными клет
ками для дыхания или диффундировать в
окружающую среду.
Темновая фаза фотосинтеза осущес
твляется в строме хлоропластов без не
посредственного поглощения света. Цепь
реакций, приводящих к восстановлению
С 0 2 до уровня органических веществ,
идет за счет использования энергии АТФ
и НАДФ-Н + Н+, синтезированных в све
товую фазу (рис. 36).
Восстановление молекул С 0 2 начи
нается с их фиксации молекулами акцеп
тора — пятиуглеродного сахара рибулозодифосфата. При взаимодействии рибулозодифосфата и С 0 2 сначала образует
ся нестойкое шестиуглеродное соедине
ние, которое затем ферментативным пу
тем распадается на две трехуглеродные
молекулы фосфоглицериновой кислоты
(ФГК). Дальнейшее превращение ФГК
требует участия продуктов световой фазы
фотосинтеза — АТФ и НАДФ ■Н + Н+. В
конечном итоге через ряд промежуточных
соединений образуются прежде всего
шеетиуглеродные углеводы, а также дру
гие органические вещества (амино- и ор
ганические кислоты, спирты, в том числе
глицерол и др.).
С 0 2 + С5 --------5я- С6 ------- з»Рибулозодифосфат
Промежуточное
вещество
2С3+АТФ+НАЦФ ' Н+Н+-^
+АДФ + НАДФ+ + Н3Р 0 4.
2С3.
ФГК
С6Н120 6+
ф
- акцептор электрона
ф
- акцептор С 0 2
Следует учесть, что в этих реакциях
одновременно участвуют многие одно
типные молекулы. На определенном эта
пе судьба трехуглеродных молекул ФГК
может оказаться различной. Одни из них
соединяются друг с другом и образуют
шестиуглеродные сахара, которые в свою
очередь могут полимеризоваться в крах-
Рис. 36. Общая схема фотосинтеза.
мал, целлюлозу и другие вещества, или
перемещаться в другие клетки (в виде са
харозы) и полимеризоваться там, или ис
пользоваться на энергетические и другие
нужды.
Другие молекулы ФГК используются
для синтеза аминокислот путем присо
единения аминогрупп или синтеза карбо
61
новых кислот, спиртов, в том числе глицерола и др.
Наконец, третья часть молекул ФГК
вовлекается в длинный ряд реакций, ко
торые приводят к превращению трехуг
леродных молекул в молекулы исходного
пятиуглеродного сахара, которые снова
могут акцептировать углекислый газ.
Поскольку часть трехуглеродных ко
нечных продуктов превращается в новые
молекулы исходного пятиуглеродного са
хара, процесс фиксации углерода по су
ществу представляет собой цикл (рис.
37), называемый С3-циклом (по его С3продуктам) или циклом Кальвина (в
честь американского ученого М . К а л ь
вина, открывшего этот процесс).
В темновой фазе, фотосинтеза энер
гия макроэргических связей АТФ преоб
разуется в химическую энергию органи
ческих веществ, т. е., по сути дела, энер
гия как бы консервируется в химических
связях между атомами органических ве
ществ.
Если реакции световой и темновой
фазы объединить, исключив все проме
жуточные стадии и вещества, то можно
получить суммарное уравнение процесса
фотосинтеза, которое было приведено в
начале параграфа.
Таким образом, фотосинтез у расте
ний — это процесс, при котором проис
ходит поглощение электромагнитной
энергии солнца хлорофиллом и вспомо
гательными пигментами, поглощение уг
лекислого газа из атмосферы, восстанов
ление его в органические соединения и
выделение кислорода в атмосферу.
Факторы, влияющие на фотосинтез.
На скорость фотосинтеза оказывают вли
яние различные факторы окружающей
среды: количество падающего света, на
личие влаги, минеральных веществ, тем
пература, концентрация С 0 2 и др.
Для растений умеренного пояса мак
симальная интенсивность фотосинтеза
наблюдается при 20— 25 °С. Дальнейшее
повышение температуры до 40 °С приво
дит к резкому снижению его интенсив
ности. В то же время некоторые растения
пустынь способны осуществлять фото
синтез при температуре 58 °С.
-►ЦЕЛЛЮЛОЗА
■-КРАХМАЛ
•■АМИНОКИСЛОТЫ
КАРБОНОВЫЕ
•-кислоты
— СПИРТ и др.
ШЕСТИУГЛЕРОДНЫЕ
САХАРА
Рис. 37 Цикл Кальвина.
62
(запасенная энергия)
Минимальная температура, при кото
рой еще может осуществляться фотосин
тез, -15 °С (при такой температуре фото
синтезируют некоторые морозостойкие
растения — в частности, ель и сосна, но
интенсивность фотосинтеза у них при та
кой температуре очень низкая.
Вода непосредственно участвует в фо
тосинтезе как источник атомов водорода.
Кроме того, величина оводненности лис
тьев определяет степень открывания ус
тьиц и, следовательно, поступления С 0 2
в лист.
Существуют так называемые С4-растения, у которых фотосинтез может про
текать при низких концентрациях С 0 2.
В клетках мезофилла этих растений име
ется особый фермент, который способ
ствует присоединению С 0 2 к трехугле
родной молекуле фосфоенолпировиноградной кислоты с образованием четырех
углеродного соединения — щавелевоук
сусной кислоты (ЩУК), в составе кото
рой временно удерживается С 0 2. Затем
ЩУК переходит в другие клетки листа
(например, в клетки обкладки проводя
щих пучков), где С 0 2 отщепляется и на
капливается в количествах, необходимых
для нормального протекания фотосинте
за при высоких температурах. Такой про
цесс получил название С4-фотосинтеза и выработался в процессе эволюции у
растений жарких, засушливых мест. К та
ким растениям относятся, например, ку
куруза, сахарный тростник, сорго.
Существует еще один путь фиксации
С 0 2 — так называемый кислот ны й
м ет аболизм т олст янковы х (сокра
щенно САМ, от англ. Crassulaceae acid
metabolism). К CAM-растениям относят
ся кактусы, молочаи и другие суккуленты,
которые живут в условиях пустынь. Эти
растения запасают С 0 2 в виде органичес
ких кислот ночью, когда открыты устьи
ца, а днем осуществляют фотосинтез, от
щепляя С 0 2 от этих кислот.
Уникальность и общебиологическое
значение фотосинтеза определяются тем,
что ему обязано своим существованием
все живое на нашей планете. Этот про
цесс является основным источником об
разования органических веществ, а так
же единственным источником свободного
кислорода на Земле. Из кислорода обра
зовался и поддерживается озоновый
слой, защищающий живые организмы от
коротковолновой ультрафиолетовой ра
диации. Кроме того, благодаря фотосин
тезу регулируется содержание С 0 2 в ат
мосфере.
В изучение процесса фотосинтеза,
раскрытие его механизма большой вклад
внесли русский ученый К. А. Т и м и р я
зев, американский М . К а л ь в и н , авс
тралийские М. Д. Хет ч и К. Р. Слэк,
а также наши белорусские ученые
Т. Н . Годнев и А. А. Шлык.
Т. Н. Годнев — белорусский физиолог
растений. Его основные научные работы
посвящены биохимии фотосинтезирую
щих пигментов, в частности, хлорофиллу
и его предшественникам, условиям их об
разования в растении. Годневу принадле
жит теория образования хлорофилла.
Им была создана белорусская шко
ла исследователей биосинтеза хлоро
филла.
Т. Н. Годнев (1893— 1982)
63
• 1. Что такое фотосинтез и в каких органеллах
осуществляется этот процесс? 2. Каковы особен
ности строения хлоропластов, хромопластов и лей
копластов? 3. В каких структурах хлоропластов про
ходит световая фаза фотосинтеза? Каков механизм
световой фазы? 4. Где осуществляется темновая фа
за фотосинтеза и что происходит в этой фазе? 5. Ка
кая связь между световой и темновой фазами фото
синтеза? 6. Какое вещество является источником
кислорода и в какой фазе фотосинтеза происходит
его образование? 7. Каково значение фотосинтеза в
природе? 8. Расскажите о С4-фотосинтезе. 9. Како
вы особенности фиксации С 02 суккулентами?
18. Бактериальный фотосинтез
Фотосинтезирующие бактерии (зеле
ные серные, пурпурные серные и пурпур
ные несерные) обитают в пресной и мор
ской воде, во влажной и илистой почве, в
прудах и озерах со стоячей водой, в сер
ных источниках и т. д. Для них характер
ны примитивные, древнейшие формы фо
тосинтеза. Бактерии в отличие от расте
ний не способны использовать воду в ка
честве субстрата для синтеза, поэтому
бактериальный фотосинтез протекает без
выделения кислорода.
В клетках фотосинтезирующих бакте
рий нет пластид, они заменены тилакоидами, образующимися в результате выпя
чивания цитоплазматической мембраны.
В тилакоидах находятся фотосинтезирую
щие пигменты и осуществляется свето
вая фаза фотосинтеза, а темновая фаза
проходит в цитоплазме.
Пигментные системы фотосинтезиру
ющих бактерий несколько отличаются от
таковых у растений. Их называют бактериохлорофиллами. По своей структу
ре эти пигменты подобны хлорофиллам а
и Ь, отличаясь от них лишь природой бо
ковых цепей при некоторых атомах угле
рода. В настоящее время известно пять
типов бактериохлорофиллов: a, b, с, d, е.
Кроме того, в реакционных центрах всех
бактерий обнаружен бактериофеофитин,
который отличается от бактериохлоро64
филла заменой центрального атома маг
ния на два атома водорода. Основные
каротиноидные пигменты бактерий так
же несколько отличаются от каротинои
дов растений.
Энергия света поглощается молеку
лами бактериохлорофилла и каротинои
дов, а затем путем миграции электронно
го возбуждения передается реакционно
му центру, содержащему 2 или 4 особым
образом упакованные молекулы бакте
риохлорофилла. Разделенные заряды пе
реносятся через мембрану их молекул,
запуская
электронный
транспорт,
обусловливающий образование АТФ,
НАД • Н + Н* или восстановленного
ферредоксина. Следовательно, фотосин
тезирующие бактерии способны синтези
ровать как АТФ, таки НАД • Н + Н+, т. е.
запасать энергию и «восстановительную
силу» для фиксации С 0 2. Почти у всех
видов фотосинтезирующих бактерий най
дены ферменты цикла Кальвина, откуда
следует, что данные организмы способны
фиксировать С 0 2 в реакциях этого
цикла.
Зеленые бактерии используют в ка
честве доноров электронов сероводород,
серу или в некоторых случаях тиосуль
фат, а пурпурные бактерии —• карбоно
вые и дикарбоновые кислоты, спирты и
др. Наиболее распространенным доно
ром электронов у фотосинтезирую
щих бактерий является сероводород
(H2S).
свет
6 С 0 2+ 12H9S ---- > С6Н 120 6+ 6Н 20 +
+ 12S.
При недостатке H2S сера, которая
часто накапливается в клетке в виде ка
пель, может утилизироваться как донор
электрона. Суммарное уравнение этого
процесса имеет следующий вид:
свет
6С 02 + 4S + 16Н20 ---- > С6Н ,90 6 +
+ 6Н20 + 4H2S 0 4.
В этой реакции используются прото
ны воды, однако происходит не фотоокис
ление (Н20 ---- э- 2Н+ + 2е- + */20 2), а
лишь не нуждающаяся в энергии диссо
циация (Н20 ---- > Н+ + ОН- ).
Подобным образом происходит реак
ция, в которой донором электронов слу
жит тиосульфат (H2S20 3):
1
свет
6C 02+3H2S20 3+ 5Н20 —
+6Н 20 + H2S 0 4.
^ с6н12о6+
Углеводы не являются единственным
и даже не всегда главным продуктом этих
форм бактериального фотосинтеза. Со
единения, образующиеся в клетках зеле
ных и пурпурных бактерий, могут быть в
дальнейшем использованы в качестве
субстратов хемосинтезирующими анаэ
робами, которые в свою очередь проду
цируют вещества, играющие роль пита
тельных веществ у фототрофных бакте
рий. Следовательно, в анаэробных усло
3. Общая биология, 10 кл.
виях бактерии этих типов могут сосущес
твовать.
Зеленые и пурпурные фотосинтезиру
ющие бактерии можно считать предста
вителями очень древних генеалогических
линий организмов, появившихся около 3
млрд лет назад. В глубокой древности, в
условиях бескислородной атмосферы,
эти бактерии были единственными орга
низмами, способными использовать для
своего роста и развития солнечную энер
гию.
Результаты современных исследова
ний дают'основание думать, что в эволю
ционном ряду фотосинтезирующие бак
терии занимают промежуточное положе
ние между древними анаэробными бакте
риями, осуществлявшими брожение, и
появившимися позднее водорослями и
растениями (см. схему):
?
а 1. Каковы особенности бактериального фото
синтеза? 2. Как можно представить эволюцию фо
тосинтеза?
65
19. Хемосинтез
Кроме фотосинтеза, существует еще
одна форма автотрофной ассимиляции —
хемосинтез, свойственный некоторым
бактериям. В отличие от фотосинтеза ис
точником энергии в данном случае явля
ется не свет, а процесс окисления неко
торых неорганических соединений —
например сероводорода, серы, аммиака,
водорода, азотистой кислоты, оксид
ных соединений железа и марганца
и др.
Открытие бактериального хемосин
теза принадлежит русскому ученомумикробиологу С. Н . В и н о г р а д
скому.
Важнейшей группой хемосинтезиру
ющих организмов являются нитрифици
рующие бактерии, способные окислять
образующийся при гниении органических
остатков аммиак до нитрита, а затем и до
нитрата:
2NH3+ 3 0 2=2H N 02+2H 20 + 6 3 2 кД ж ,
2H N 02 + 0 2 = 2H N 03 + 152 кДж.
Азотная кислота, реагируя с мине
ральными соединениями почвы, превра
щается в соли азотной кислоты, которые
хорошо усваиваются растениями.
Бесцветные серобактерии окисляют
сероводород и накапливают в своих клет
ках серу:
2H2S + 0 2 = Н20 + 2S + 272 кДж.
При недостатке сероводорода бакте
рии производят дальнейшее окисление
накопившейся в них серы до серной кис
лоты.
2S + 3 0 2 + 2Н20 = 2H2S 0 4 + 636 кДж.
Железобактерии переводят железо
Fe2+ в железо Fe3+:
4FeC03 + 0 2 + 6Н20 = 4Fe(OH)3 +
+ 4 С 0 2 + 324 кДж.
Водородные бактерии используют в
качестве источника энергии реакции
66
окисления молекулярного водорода, а в
качестве единственного источника угле
рода — углекислый газ. Реакция окисле
ния происходит по схеме:
2Н2 + 0 2 ---- > 2Н20 + 235 кДж.
Энергия, выделяемая при окислении
указанных выше соединений, использует
ся бактериями-хемосинтетиками для вос
становления С 0 2 до органических ве
ществ.
Экологическая роль хемосинтеза.
Хемотрофные нитрифицирующие бакте
рии широко распространены в природе.
Они встречаются и в почве, и в разных
водоемах. Масштаб осуществляемых ими
процессов достаточно велик, поэтому хе
мосинтетики играют существенную роль
в круговороте азота в биосфере.
Серобактерии способствуют посте
пенному разрушению и выветриванию
горных пород вследствие образования
ими серной кислоты, являются причиной
порчи каменных и металлических соору
жений, выщелачивания руд и серных
месторождений. Многие виды серобакте
рий, окисляя до сульфатов различные со
единения серы, играют большую роль в
процессах очищения промышленных
сточных вод.
В результате деятельности некоторых
железобактерий
вырабатывается
Fe(OFI)3, скопления которого образуют
болотную железную руду.
Водородные бактерии используются
для получения дешевого пищевого и кор
мового белка, а также для регенерации
(восстановления) атмосферы в замкну
тых системах жизнеобеспечения. Напри
мер, система «Оазис-2» была испытана
на космическом корабле «Союз» в
1973 г. Кроме того, в природных услови
ях водородные бактерии участвуют в
окислении водорода, который может на
капливаться при действии некоторых
микроорганизмов, размельчающих орга
нические вещества почвы, донные отло
жения водоемов и т. д.
i 1. Что такое хемосинтез? 2. Чем отличается хе
мосинтез от фотосинтеза? 3. В чем сходство процес
сов хемосинтеза и фотосинтеза? 4. Какова экологи
ческая роль хемосинтеза?
20. Клеточное дыхание
Высвобождение
потенциальной
энергии химических связей. Образую
щиеся в процессе фотосинтеза органи
ческие вещества и заключенная в них хи
мическая энергия служат источником ве
ществ и энергии для осуществления жиз
недеятельности всех организмов. Однако
использование животными, грибами,
многими протистами и бактериями созда
ваемых зелеными растениями органичес
ких веществ, синтез на их основе специ
фических для каждого вида соединений
возможны лишь после предварительных
преобразований, которые заключаются в
расщеплении этих сложных веществ до
простых: углеводов — до моносахаров,
жиров — до высших карбоновых кислот
и глицерина, белков — до аминокислот,
нуклеиновых кислот — до нуклеотидов.
То же касается и содержащейся в ор
ганических веществах энергии. Будучи
заключенной в химических связях, она
недоступна для непосредственного ис
пользования клеткой, в том числе и клет
ками растений, которые преобразовали
эту энергию из световой в химическую.
Для этого потенциальная энергия ор
ганических молекул должна быть высво
бождена и переведена в пригодную для
использования форму.
Образование и накопление энергии,
доступной клетке, происходит в процессе
клеточного дыхания. Для осуществле
ния клеточного дыхания большинству ор
ганизмов необходим кислород — в этом
случае говорят об аэробном дыхании,
или аэробном высвобождении энергии.
Однако некоторые формы живых орга
низмов могут получать энергию из пищи
без использования свободного атмосфер
ного кислорода, т. е. в процессе так назы
ваемого анаэробного дыхания (анаэ
робного высвобождения энергии).
Таким образом, исходными вещества
ми для дыхания служат богатые энергией
органические молекулы, на образование
которых в свое время была затрачена
энергия.
Основным веществом, используемым
клетками для получения энергии, являет
ся глюкоза. Процесс расщепления глю
козы выражается уравнением:
С6Н 120 6 + 6 0 2 ---- > 6С 02 + 6Н20 +
+ энергия.
Этим уравнением можно также опи
сать сжигание глюкозы вне клетки. Ко
личество теплоты, выделяющееся при
сжигании, очень велико. Если бы приды
хании происходил такой же мгновенный
распад глюкозы и выделялась вся запа
сенная в ней энергия, то такой процесс
привел бы к разрушению клетки. Тем не
менее этого не происходит, так как вы
свобождение энергии в клетке осущест
вляется постепенно, ступенчато, в не
сколько этапов. Все превращения осу
ществляются в ходе ферментативных ре
акций.
Аэробное (кислородное) дыхание.
Процесс аэробного дыхания можно ус
ловно разделить на два последовательных
этапа.
П е р в ы й э т а п — бескислород
ный, или неполный. Он протекает в ци
топлазме клеток в анаэробных (бескис
лородных) условиях.
На данном этапе дыхательный суб
страт подвергается ферментативному
расщеплению. Примером такого процес
са является гли к о ли з — многоступен
чатое бескислородное расщепление глю
козы.
В реакциях гликолиза шестиуглерод
ная молекула гексозы (С6) расщепляется
на две молекулы пировиноградной кисло
ты (С3). При этом образуются две моле
кулы АТФ и атомы водорода. Водородные
67
2
3
Рис. 38. Строение митохондрии: а — трехм ерная с т р у к т у р а , б — продольный разрез, в — часть
кр и сты с вы ростам и: 1 — внутренняя мембрана, 2 — матрикс, 3 — межмембранное пространство,
4 — наружная мембрана, 5 — АТФсомы, 6 — гранулы, 7 — ДНК, 8 — рибосома.
атомы присоединяются к переносчику
НДД+ (никотинамидадениндинуклеотиду), который переходит в свою восста
новленную форму НАД- Н+ Н+ (НАД
очень сходен с НАДФ, т. е. с переносчи
ком атомов водорода при фотосинтезе).
Суммарная реакция гликолиза имеет вид:
С6Н 120 6 + 2АДФ + 2Н3Р 0 4 +
+ 2НАД+ ---- > 2С3Н40 3 + 2АТФ +
т 211АД - Н т Н ' .
Таким образом, продуктами гликоли
за являются пировиноградная кислота,
2 молекулы АТФ и восстановленный
НАД ■Н + Н+.
В т о р о й э т а п аэробного дыха
ния — кислородный. Он протекает в ми
тохондриях и требует присутствия кисло
рода.
Митохондрии являются неотъемле
мыми компонентами всех эукариотичес
ких клеток и представляют собой грану
лярные или нитеподобные структуры
толщиной 0,5 мкм и длиной до 7—
10 мкм. Митохондрии очень динамичны:
они могут изменяться, ветвиться, делить
ся. Непостоянно и число митохондрий в
клетках — оно колеблется в зависимости
от типа клетки, а также фазы развития и
68
функционального состояния от несколь
ких единиц до нескольких сотен, иногда
тысяч штук.
Митохондрии имеют оболочку, кото
рая состоит из двух мембран — наружной
и внутренней (рис. 38).
Между внешней и внутренней мем
бранами имеется так называемое перимитохондриальное пространство, ко
торое является местом скопления ионов
водорода Н+. Внутренняя мембрана об
разует множество впячиваний внутрь ми
тохондрии — так называемых крист. На
мембране крист или внутри нее распола
гаются ферменты, в том числе перенос
чики электронов и ионов водорода Н+,
которые участвуют в кислородном дыха
нии. Наружная мембрана отличается
высокой проницаемостью, и многие со
единения легко проходят через нее. Внут
ренняя мембрана менее проницаема.
Ограниченный ею внутренний компонент
заполнен близким по составу к цитоплаз
ме веществом — матриксом, который
содержит ДНК (кольцевые молекулы),
все типы РНК, аминокислоты, рибосомы,
ряд витаминов. ДНК обеспечивает неко
торую генетическую автономность мито
хондрий, хотя в целом их работа коорди
нируется ДНК ядра. Кроме того, в мат
риксе содержатся почти все дыхательные
ферменты.
?
, 1. В каком виде энергия используется в клетке?
2. Какие процессы в клетке обеспечивают ее энер
гией? 3. Что представляет собой гликолиз? 4. Где
осуществляются реакции гликолиза? 5. В чем сущ
ность аэробного дыхания? 6. Что представляют со
бой митохондрии?
21. Дальнейшее превращение
пировиноградной кислоты
Продукт гликолиза — пиро ви н о
градная кислота — заключает в себе
значительную часть энергии, и дальней
шее ее высвобождение осуществляется в
митохондриях. Здесь происходит полное
окисление пировиноградной кислоты до
С 0 2 и Н20 . Этот процесс проходит три
основные стадии:
1) окислительное декарбоксилирование пировиноградной кислоты;
2) цикл трикарбоновых кислот
(цикл Кребса, названный так в честь
открывшего его английского учено
го Г. Кре б с а ) ;
3) заключительная стадия окисле
ния — элект ронт ранспорт ная
цепь.
На первой стадии пировиноградная
кислота взаимодействует с веществом,
которое называют коферментом А
(КоА), в результате чего образуется ацетилкофермент А с высокоэнергетичес
кой связью. При этом от молекулы пиро
виноградной кислоты отщепляется моле
кула С 0 2 и атомы водорода, которые за
пасаются в форме НДЦ ■Н + Н+.
В цикл Кребса вступает ацетил-КоА,
образованный на предыдущей стадии.
Ацетил-КоА взаимодействует со щавеле
воуксусной кислотой (рис. 39, с. 70). Д а
лее превращение идет через ряд органи
ческих кислот, в результате чего щавеле
воуксусная кислота регенерирует в
прежнем виде. В процессе цикла участву
ют три молекулы воды, выделяются две
молекулы 0 2 и четыре пары атомов водо
рода, которые восстанавливают соответ
ствующие коферменты: флавинадениндинуклеотид (ФАД) и НАД. Суммарная ре
акция цикла Кребса может быть выраже
на следующим уравнением:
ацетил-КоА + ЗН20 + ЗНАД + ФАД +
+ АДФ + Н3Р 0 4' — > КоА + 2 С 0 2 +
+ ЗНАД ■Н + Н+ + ФАД ■Н + Н+ + АТФ.
Таким образом, в результате распада
одной молекулы пировиноградной кисло
ты в аэробной фазе (декарбоксилирование ПВК и цикл Кребса) выделяется
ЗС 0 2, 4НАД • Н+ Н+, ФАД • Н + Н+.
Суммарную реакцию гликолиза,
окислительного декарбоксилирования и
цикла Кребса можно записать в следую
щем виде:
C6H i20 6 + 6Н20 -----> 6 С 0 2+ 4АТФ +
+ ЗНАД • Н + Н+ + ФАД • Н + Н+.
На третьей стадии пары водородных
атомов, отщепляемые от промежуточных
продуктов в реакциях дегидрирования при
гликолизе и в цикле Кребса, в конце кон
цов окисляются молекулярным кислоро
дом до Н20 с одновременным фосфорили
рованием АДФ в АТФ. Происходит это,
когда водород, отделившийся от НАД • Н +
+ Н+ и ФАД • Н + Н+, передается по цепи
переносчиков, встроенных во внутреннюю
мембрану митохондрий. Пары водорода
можно рассматривать как 2Н+ + 2е_ .
Именно в таком виде они и передаются по
цепи переносчиков. Путь переноса водоро
да и электронов от одной молекулы пере
носчика к другой представляет собой окис
лительно-восстановительный процесс.
При этом молекула, отдающая электрон
или водород, окисляется, а молекула, вос
принимающая электрон или ион водорода,
восстанавливается. Движущей силой
транспорта водорода в дыхательной цепи
является разность потенциалов.
С помощью переносчиков ионы водо
рода Н+ переносятся с внутренней сторо
ны внутренней мембраны на ее внешнюю
69
COOH
сторону, иначе говоря, из матрикса мито
хондрии в межмембранное пространство
(рис. 40).
Переносчики в мембране митохон
дрий расположены неоднородно. Одни из
них переносят целые атомы водорода, а
другие лишь электроны. Молекула пере
носчика, несущая целый атом водорода,
взаимодействует с переносчиком, вос
принимающим только электроны, и про
тоны высвобождаются в межмембранное
пространство. При переносе пары элек
тронов с НАД на кислород они пересека
ют мембрану 3 раза и этот перенос со
провождается выделением на внешнюю
сторону мембраны 6 протонов. На заклю
чительном этапе электроны переносятся
на внутреннюю сторону мембраны и ак
цептируются кислородом:
1/2 0 2 + 2е“ ---- > О2-.
70
Перестройка
Кислород, заряжаясь, воспринимает
2Н+ из внутренней среды и образуется
Н20:
2Н+ + 2е~ + 1 /2 0 2 ---- > Н20 .
В результате такого переноса ионов
водорода на внешнюю сторону мембраны
митохондрий в перимитохондриальном
пространстве создается их повышенная
концентрация, т. е. возникает электро
химический
градиент
протонов
(Др.Н+).
Протонный градиент представляет
собой как бы резервуар свободной энер
гии. Эта энергия используется при обрат
ном потоке протонов через мембрану для
синтеза АТФ. В ряде случаев может на
блюдаться непосредственное использо
вание энергии протонного градиента
(Д(дН+). Он может обеспечивать осмоти-
Э л е ктр и ч е с ки й потенциал
НАДФ -Н+Н +
Рис. 40. Окислительное фосфорилирование в митохондриях.
ческую работу и транспорт веществ через
мембрану против градиента их концен
трации, механическую работу и др. Таким
образом, клетка располагает двумя фор
мами используемой энергии — АТФ и
Др.Н+. Первая форма — химическая.
АТФ растворяется в воде и легко исполь
зуется в водной фазе. Вторая (Д цН +) —
электрохимическая — неразрывно свя
зана с мембранами. Обе формы энергии
71
могут переходить одна в другую. При об
разовании АТФ участвует энергия Д|дН+,
при распаде АТФ энергия может аккуму
лироваться в виде Д цН +.
Ионы водорода из Н+-резервуара
движутся по специальным каналам в
мембране, и их запас энергии использу
ется для синтеза АТФ.
Процесс образования АТФ в резуль
тате переноса ионов Н+ через мембрану
митохондрии получил название окисли
тельного фосфорилирования. Он осу
ществляется при участии фермента АТФсинтетазы.
В результате расщепления двух моле
кул пировиноградной кислоты и переноса
ионов водорода через мембрану по спе
циальным каналам синтезируется в це
лом 36 молекул АТФ (2 молекулы в цикле
Кребса и 34 молекулы в результате пере
носа ионов Н+ через мембрану).
Можно заметить, что ферментные
системы ориентированы в митохондриях
противоположно тому, как мы наблюдали
это в хлоропластах: там Н+-резервуар
находится с внутренней стороны внутрен
ней мембраны, а в митохондриях — с на
ружной. Кроме того, при фотосинтезе
электроны движутся в основном от воды
к переносчикам атомов водорода, а при
дыхании переносчики водорода, передаю
щие электроны на электронтранспортную цепь, — с внутренней стороны мем
браны. Электроны же в конечном счете
включаются в образующиеся молекулы
воды.
Кислородный этап, таким образом,
дает энергии в 18 раз больше, чем ее за
пасается в результате гликолиза.
Суммарное уравнение аэробного ды
хания можно выразить следующим обра
зом:
С6Н 120 6 + 6 0 2 + 6Н20 + 38АДФ +
+38Н3Р 0 4 — ^ 6 С 0 2 + 12Н20 +
+38АТФ.
Совершенно очевидно, что аэробное
дыхание в отсутствие кислорода прекра
72
тится, поскольку именно кислород слу
жит конечным акцептором водорода. Ес
ли клетки не получают достаточного ко
личества кислорода, все переносчики во
дорода вскоре полностью насытятся и не
смогут передавать его дальше. В резуль
тате основной источник энергии для об
разования АТФ окажется блокирован
ным.
« 1. В каких органеллах осуществляется кисло
родный этап дыхания? 2. Из каких стадий состоит
кислородный этап дыхания? 3. Что происходит на
каждой стадии? 4. Какова функция кислорода в
процессе дыхания?
22. Анаэробное дыхание.
Брожение.
При отсутствии или недостатке кис
лорода, играющего роль конечного ак
цептора электронов, цепь передачи элек
тронов через мембрану не осуществляет
ся, а значит, нет и. Н+-резервуара, спо
собного обеспечить энергией синтез
АТФ. В этих условиях многие клетки син
тезируют АТФ, расщепляя питательные
вещества в процессе анаэробного дыха
ния.
Для анаэробного дыхания характерен
гликолиз, в процессе которого глюкоза
расщепляется дО пировиноградной кис
лоты и высвобождаются атомы водорода.
В условиях, когда отсутствует кислород, а
значит, ему не могут быть переданы ато
мы водорода, все молекулы НАД+ вскоре
оказываются нагруженными атомами во
дорода. Для того чтобы процесс гликоли
за осуществлялся дальше, нужен другой
акцептор. Таким акцептором становится
пировиноградная кислота. Кислота при
соединяет атомы водорода и высвобожда
ет НАД+, который может снова акцепти
ровать атомы водорода, отщепляемые в
результате гликолиза, и, значит, гликолиз
может продолжаться, т. е. может синте
зироваться хотя бы некоторое количество
АТФ.
У разных организмов дальнейшая
судьба пировиноградной кислоты в зави
симости от метаболических путей может
быть различной. Например, дрожжи в от
сутствие кислорода образуют этиловый
спирт и С 0 2. Вначале от каждой молеку
лы пировиноградной кислоты отщепляет
ся молекула С 0 2 и образуется молекула
двууглеродного соединения — уксусного
альдегида, который затем восстанавли
вается до этилового спирта:
С6Н 120 6+2ДЦФ + 2Н3Р 0 4+2НАД---- >
---- > 2СН3СОСООН + 2АТФ +
+ 2 НАД • Н + Н+
СН3С О С О О Н ---- > СН3СОН +
СН3СОН + НАД ■Н + Н+ — ->
---- >С Н 3СН2ОН + НАД+
С6Н 120 6 — -э* 2СН3СН2ОН + 2АТФ
со2
Такой процесс называется спирто
вым брожением. Спиртовое брожение
наблюдается также в растительных клет
ках в отсутствие кислорода.
У некоторых бактерий, а также в жи
вотных клетках, испытывающих времен
ный недостаток кислорода, происходит
молочнокислое брожение, приводящее к
образованию молочной кислоты.
О
О
II //
СН3— С— С + НАД • Н + Н+ ---- >
\
н
он о
I
//
---- > СН3— СН— С+НАДА
ОН
Ни тот ни другой процесс не дают до
полнительного количества АТФ, так что
выход на одну молекулу глюкозы, рас
щепленную путем анаэробного дыхания,
соответствует двум молекулам АТФ. Зна
чительная часть энергии, заключенная в
молекуле глюкозы, при этом не извлека
ется и остается в конечных продуктах
брожения — этиловом спирте и молоч
ной кислоте. Поэтому анаэробное дыха
ние по сравнению с аэробным является
процессом малоэффективным.
Энергия, заключенная в этиловом
спирте, для дрожжей всегда недоступна.
Из молочной же кислоты в животных ор
ганизмах в присутствии кислорода может
быть извлечено довольно большое коли
чество энергии. В присутствии кислорода
молочная кислота в клетках печени пре
вращается в пировиноградную. Послед
няя вступает затем в цикл Кребса и окис
ляется до С 0 2 и Н20 , в результате чего
образуется АТФ. Пировиноградная кис
лота может превращаться также в глюко
зу, которая хранится в клетках печени или
мышц в форме гликогена; правда, эти
превращения требуют дополнительных
затрат энергии в виде АТФ.
Наиболее распространенным дыха
тельным субстратом является глюкоза.
Однако следует помнить, что любое орга
ническое вещество при соответствующих
условиях может выступать источником
энергии для синтеза АТФ.
При недостатке в клетке глюкозы в
дыхание могут вовлекаться жиры и
белки.
Жиры могут превращаться в углево
ды, а в клетках, богатых жирами (клетки
печени, эндосперма растений), они могут
использоваться в качестве дыхательного
субстрата без предварительного превра
щения в углеводы. При этом жиры гидро
лизуются до глицерина и высших карбо
новых кислот (рис. 41, с. 74).
Дальнейшее превращение жиров (де
гидрирование и декарбоксилирование)
сопровождается выделением более 160
молекул АТФ: 19 молекул АТФ при окис
лении глицерина и 147 — при окислении
стеариновой кислоты. Как видим, карбо
новые кислоты — важный источник
энергии. Считается, что около половины
энергетических затрат сердечной и ске
летных мышц (в покое), печени и почек
восполняется за счет окисления высших
карбоновых кислот.
73
Рис. 41. Упрощенная схема превращения белков, углеводов и жиров в организме: ФГА фосфоглицериновый альдегид, Щ УК — щавелевоуксусная кислота.
Белки же как дыхательный субстрат
используются крайне редко. Белки клет
ки начинают расщепляться только в слу
чае полного отсутствия запасов углево
дов и жиров.
Если с пищей поступает избыток ами
нокислот, то организм может использо
вать их как источник энергии.
Аминокислоты подвергаются окисли
тельному дезаминированию — отщеп
лению NH2-группы и атомов водоро
да, при котором выделяется аммиак и
74
образуются органические кислоты,
которые участвуют затем в цикле
Кребса. Аммиак выводится из организ
ма в виде мочевой кислоты или мочевины.
Иногда в клетках происходит трансаминирование — при участии фермен
тов трансаминаз осуществляется перенос
аминогруппы с аминокислоты на органи
ческую кислоту. При этом образуется
другая аминокислота и пировиноградная
кислота, которая дальше вовлекается в
цикл Кребса.
• 1. Чем отличается брожение от дыхания?
2. Что происходит в мышечных клетках при кисло
родном голодании? 3. Почему фотосинтезирующие
органы нуждаются в клеточном дыхании? Все ли
клетки растений дышат? 4. Чем отличаются про
цессы горения и биологического окисления — ды
хания? 5. В чем сходство и различие процессов фо
тосинтеза и дыхания?
23. Транспортная система клетки
В клетке осуществляется синтез ог
ромного количества различных веществ.
Часть из них потребляется на собствен
ные нужды — синтез АТФ, построение
органелл, накопление питательных запа
сов; часть выводится из клетки — по
строение оболочки (клетки растений,
грибов, некоторых протистов), гликокаликса (животные клетки), клеточные сек
реты (ферменты, гормоны, коллаген, ке
ратин и т. д.). 1де же осуществляется син
тез этих веществ и каким образом они пе
ремещаются из одной части клетки в дру
гую либо выводятся за ее пределы? Это
происходит в системе замкнутых цито
плазматических мембран — эндоплазма
тической сети, или эндоплазматичес
ком ретикулуме, и комплексе Голь
джи, составляющих экспортную систему
клеток.
Эндоплазматический ретикулум был
открыт с помощью электронного микрос
копа в 1945 г. Он представляет собой
систему разветвленных каналов, цистерн
(вакуолей), пузырьков, создающих подо
бие рыхлой сети в цитоплазме (рис. 42).
Стенки каналов и полостей образованы
элементарными мембранами.
В клетке существует два типа эндо
плазматического ретикулума: грануляр
ный (шероховатый) и агранулярный
(гладкий). Гранулярный эндоплазмати
ческий ретикулум густо усеян рибосома
ми, а гладкий, состоящий в основном из
длинных узких трубчатых каналов, не
связан с ними. Наличие рибосом на мем
бранах гранулярного эндоплазматическо
го ретикулума свидетельствует о том, что
этот тип характерен для биосинтеза бел
ка. Как правило, на рибосомах грануляр
ного эндоплазматического ретикулума
синтезируются белки, не используе
мые самой клеткой, — они выводятся
за ее пределы. Белки, используе
мые на внутренние потребности клет
ки, синтезируются на рибосомах цито
плазмы.
Гранулярный эндоплазматический ре
тикулум особенно хорошо развит в клет
ках, которые вырабатывают большое ко
личество белковых секретов: некоторые
клетки соединительной ткани, вырабаты-
нспорт
Конденсация
Концентрация
Рис. 42. Схема строения гранулярного эндоплазматического ретикулума.
75
вающей антитела, клетки слюнных же
лез, поджелудочной железы и т. д. Синте
зируемые белки проходят через мембрану
в каналы и полости эндоплазматического
ретикулума, изолируются от цитоплазмы,
накапливаются здесь и перемещаются по
каналам от места синтеза в другие части
клетки либо выводятся за ее пределы.
Таким образом, в мембранах грану
лярного эндоплазматического ретикулу
ма накапливаются и изолируются белки,
которые после их синтеза могли оказать
ся вредными для клетки. Например, син
тез гидролитических ферментов и их сво
бодный выход в цитоплазму привел бы к
самоперевариванию клетки и ее гибели.
Однако этого не происходит, потому что
подобные белки надежно изолированы в
полостях эндоплазматического ретику
лума.
Необходимо подчеркнуть, что на ри
босомах гранулярного эндоплазматичес
кого ретикулума синтезируются также
интегральные и периферические белки
мембран клетки и некоторая часть белков
цитоплазмы.
Цистерны шероховатого эндоплазма
тического ретикулума связаны с ядерной
оболочкой, причем некоторые из них яв
ляются прямым продолжением послед
ней. Считается, что после деления клетки
оболочки новых ядер образуются из мем
бран цистерн эндоплазматического рети
кулума.
В гладком эндоплазматическом рети
кулуме протекают процессы синтеза ли
пидов и некоторых углеводов (например,
гликогена).
Комплекс Гольджи. Выведение ве
ществ, синтезированных в эндоплазмати
ческом ретикулуме, из клетки осущес
твляется с участием другой мембранной
структуры — комплекса Гольджи
(рис. 43). Комплекс Гольджи был открыт
в 1898 г. итальянским ученым К . Го л ь дж и и назван его именем.
Комплекс Гольджи представляет со
бой систему плоских дискообразных цис-
Рис. 43. Схема строения комплекса Гольджи: а — цистерны в виде дисков (диктиосома),
б — перфорированные цистерны.
76
терн, ограниченных мембраной. Цистер
ны располагаются стопками одна над дру
гой и образуют диктиосому. По краям
от цистерн отшнуровываются крупные и
мелкие пузырьки. Число диктиосом в
клетках варьирует от одной до десятков и
сотен в зависимости от типа клеток и фа
зы их развития.
К комплексу Гольджи доставляются
вещества, синтезируемые в эндоплазма
тическом ретикулуме. От цистерн эндо
плазматического ретикулума отшнуровы
ваются пузырьки, которые соединяются с
цистернами комплекса Гольджи. В ком
плексе Гольджи эти вещества дорабаты
ваются и созревают. Одновременно с
поступлением веществ из эндоплазмати
ческого ретикулума происходит пере
стройка мембран: тонкая мембрана пу
зырька эндоплазматического ретикулума,
вошедшая в состав комплекса Гольджи,
превращается в более плотную мембрану
с иным белково-липидным составом,
сходную с цитоплазматической мембра
ной. Липиды поступают из гладкого эн
доплазматического ретикулума, а бел
ки — частично из гранулярного эндо
плазматического ретикулума и частично
от свободных рибосом цитоплазмы.
Пузырьки комплекса Гольджи учас
твуют в формировании цитоплазматичес
кой мембраны и стенок клеток растений
после деления, а также в образовании ва
куолей и первичных лизосом.
Зрелые цистерны диктиосомы отшнуровывают пузырьки, или вакуоли Голь
джи, заполненные секретом. Содержи
мое таких пузырьков либо используется
самой клеткой, либо выводится за ее пре
делы. В последнем случае пузырьки Голь
джи подходят к плазматической мембра
не, сливаются с ней и изливают свое со
держимое наружу (рис. 44), а их мембра
на включается в плазматическую мем
брану и таким образом происходит ее об
новление.
Цистерны комплекса Гольджи актив
но извлекают моносахариды из цитоплаз-
Рис. 44. Схема связи эндоплазматического ре
тикулума и комплекса Гольджи с образованием
и выведением веществ из клетки: 1 — грану
лярный эндоплазматический ретикулум, 2 —
комплекс Гольджи, 3 — отделившиеся от ком
плекса Гольджи пузырьки, 4 — выход (экзоцитоз) содержимого пузырьков Гольджи за пре
делы клетки.
мы и синтезируют из них более сложные
олиго- и полисахариды. У растений в ре
зультате этого образуются пектиновые
вещества, гемицеллюлоза и целлюлоза,
используемые для построения клеточной
стенки, слизь корневого чехлика. У жи
вотных подобным образом синтезируют
ся гликопротеины и гликолипиды гликокаликса, вырабатываются секрет подже
лудочной железы, амилаза слюны, пеп
тидные гормоны гипофиза, коллаген.
Комплекс Гольджи участвует в обра
зовании белков молока в молочных желе
зах, желчи в печени, веществ хрусталика,
зубной эмали и т. п.
Комплекс Гольджи и эндоплазмати
ческий ретикулум тесно связаны между
77
собой: их совместная деятельность обес
печивает синтез и преобразование ве
ществ в клетке, их изоляцию, накопление
и транспорт.
?
». 1. Что представляет собой эндоплазматический
ретикулум? 2. Чем отличаются шероховатый и глад
кий эндоплазматические ретикулумы? 3. Какова
роль эндоплазматического ретикулума в синтезе
белков? 4. Какие функции выполняет комплекс
Гольджи? 5. Каким образом осуществляется выве
дение веществ из клетки? 6. Какие вещества тран
спортируются за пределы клетки и какова их даль
нейшая судьба?
24. Внутриклеточное
переваривание
Некоторые клетки поглощают пищу
путем эндоцитоза. Возникающие при
этом внутри клетки мембранные пузырь
ки, содержащие частички или капельки
веществ, называются соответственно
фагосомами или пиносомами.
Эндоцитоз — довольно распростра
ненный способ поступления в клетку ве
ществ, находящихся в виде крупных агре
гатов — частичек или капелек.
С помощью эндоцитоза осуществля
ется, например, питание яйцеклеток
желточными белками. Пищеварительные
вакуоли некоторых протистов представ
ляют собой не что иное, как фагосомы.
С помощью эндоцитоза лейкоциты
крови поглощают чужеродные частицы
и иммуноглобулины, а почечные ка
нальцы всасывают белки из первичной
мочи.
Вещества, попадающие в клетку пу
тем эндоцитоза, расщепляются (напри
мер, содержимое пищеварительных ваку
олей), накапливаются (например, жел
точные белки) или снова выводятся из
клетки путем экзоцитоза.
Переваривание веществ, оказавших
ся внутри эндоцитозных пузырьков, осу
ществляется с помощью лизосом. Лизо
78
сомы — это пузырьки величиной до
2 мкм, окруженные мембраной. Внутри
лизосом содержатся гидролитические
ферменты, способные переваривать бел
ки, липиды, углевод,ы, нуклеиновые кис
лоты. Лизосомы образуются из пузырь
ков, отделяющихся от комплекса Гольджи
или гладкого эндоплазматического рети
кулума, причем предварительно на шеро
ховатом эндоплазматическом ретикулуме
синтезируются гидролитические фер
менты.
Сливаясь с эндоцитозными пузырька
ми, лизосомы образуют пищеваритель
ную вакуоль (вторичная лизосома),
где происходит расщепление органичес
ких веществ до слагающих их мономеров.
Последние через мембрану вторичной
лизосомы поступают в цитоплазму клет
ки. Именно так происходит, например,
обезвреживание бактерий в клетках кро
ви — нейтрофилах.
Вторичные лизосомы, закончившие
процесс переваривания, практически не
содержат ферментов. В них находятся
лишь непереваренные остатки, т. е. не
гидролизуемый материал, который либо
выводится за пределы клетки, либо на
капливается в цитоплазме.
Расщепление лизосомами чужерод
ного, поступившего путем эндоцитоза
материала называется гетерофагией.
Лизосомы участвуют также в разрушении
собственных материалов клетки, напри
мер запасных питательных веществ, а
также макромолекул и целых органелл,
утративших функциональную активность
(аутофагия). При патологических изме
нениях в клетке или ее старении мембра
ны лизосом могут разрушаться: фермен
ты выходят в цитоплазму и осуществля
ется самопереваривание клетки — а в
толиз. Иногда с помощью лизосом унич
тожаются целые комплексы клеток и ор
ганы. Например, когда головастик пре
вращается в лягушку, лизосомы, находя
щиеся в клетках хвоста, переваривают
его: хвост исчезает, а образовавшиеся во
время этого процесса вещества всасыва
ются и используются другими клетками
тела.
■ 1. Каким образом происходит поглощение час
тичек клетками? 2. Какова судьба веществ, посту
пивших в клетку путем эндоцитоза? 3. Что пред
ставляют собой лизосомы, где они образуются и ка
ковы их функции? 4. Чем отличаются вторичные ва
куоли от первичных? 5. Что такое гетеро- и аутофагия, автолиз? 6. В каких случаях происходит пере
варивание органелл и самопереваривание клеток?
25. Вакуоли
В клетках растений, грибов и многих
протистов содержатся вакуоли — круп
ные пузырьки или полости в цитоплазме,
заполненные преимущественно водным
содержимым. Вакуоли образуются из пу
зыревидных расширений эндоплазмати
ческого ретикулума или из пузырьков
комплекса Гольджи.
Вакуоли растительных клеток. В меристематических клетках растений воз
никает много небольших вакуолей из пу
зыревидных расширений эндоплазмати
ческого ретикулума. Увеличиваясь, они
сливаются в центральную вакуоль, ко
торая занимает до 70— 90 % объема
клетки и может быть пронизана тяжами
цитоплазмы (рис. 45). Окружающая ее
мембрана — тонопласт — имеет тол
щину мембраны эндоплазматического
ретикулума (4— 6 нм) в отличие от более
толстой, плотной и менее проницаемой
плазмалеммы.
Содержимое вакуолей — клеточный
сок. Он представляет собой водный рас
твор различных неорганических и органи
ческих веществ. Большинство из них от
носится к группе продуктов метаболизма
протопласта, которые могут появляться и
исчезать в различные периоды жизни
клетки. Химический состав и концентра
ция клеточного сока очень изменчивы и
зависят от вида растений, органа, ткани и
состояния клетки. В клеточном соке со
держатся соли, сахара (прежде всего са
хароза, глюкоза, фруктоза), органичес
кие кислоты (яблочная, лимонная, щаве
левая, уксусная и др.), аминокислоты,
белки. Эти вещества являются промежу
точными продуктами метаболизма, вре
менно выведенными из обмена веществ
клетки и изолированными тонопластом.
Р ис. 45
79
Рис. 46
Они являются запасными веществами
клетки.
Помимо запасных веществ, которые
могут вторично использоваться в обмене
веществ, клеточный сок содержит фено
лы, танины (дубильные вещества), алка
лоиды, антоцианы, которые выводятся из
обмена в вакуоль и таким путем изолиру
ются от цитоплазмы.
Танины особенно часто встречаются в
клеточном соке (а также в цитоплазме и
оболочках) клеток листьев, коры, древе
сины, незрелых плодов и семенных обо
лочек. Алкалоиды присутствуют, напри
мер, в семенах кофе (кофеин), плодах ма
ка (морфин) и белены (атропин), стеблях
и листьях люпина (люпинин) и др. Счита
ется, что танины с их вяжущим вкусом,
алкалоиды и токсичные полифенолы вы
полняют защитную функцию: их ядови
тый (чаще горький) вкус и неприятный
запах отталкивают травоядных живот
ных, что предотвращает поедание этих
растений.
В вакуолях также часто накапливают
ся конечные продукты жизнедеятельнос
ти клеток {отходы). Таким веществом
для клеток растений является щавелево
кислый кальций, который откладывается
80
в вакуолях в виде кристаллов различной
формы (рис. 46).
В клеточном соке многих растений со
держатся пигменты. Самыми распрос
траненными из них являются ант оциа
ны, которые придают клеточному соку
пурпурный, красный, синий или фиолето
вый цвета. Близкие к антоцианам флавоны и флавонолы окрашивают клеточный
сок в желтый или кремовый цвета. Имен
но эти пигменты и определяют окраску
венчиков цветков, плодов, почек и листь
ев, а также корнеплодов некоторых рас
тений (например, свеклы).
Цвет антоцианов может изменяться в
зависимости от кислотности среды: в кис
лой среде он может быть красным, в ней
тральной — фиолетовым, в щелочной —
синим. Кроме того, могут наблюдаться
все переходные оттенки. Реакция клеточ
ного сока в процессе жизнедеятельности
растений может меняться от сильнокис
лой до слабокислой или даже слабоще
лочной, что вызывает соответствующие
изменения цвета пигментов. Поэтому
цветки некоторых растений (например,
медуницы неясной) во время цветения
могут изменять свою окраску от розовой
до синей.
Клеточный сок некоторых растений
содержит физиологически активные ве
щества — фитогормоны (регуляторы
роста), фитонциды, ферменты. В пос
леднем случае вакуоли действуют как лизосомы. После гибели клетки тонопласт,
как и другие мембраны, теряет свою из
бирательную проницаемость, и фермен
ты, высвобождаясь из вакуолей, вызыва
ют автолиз клетки.
Вакуоли играют главную роль в по
глощении воды растительными клетками.
Вода путем осмоса через тонопласт по
ступает в вакуоль, клеточный сок которой
является более концентрированным, чем
цитоплазма, и оказывает давление на ци
топлазму, а тем самым и на оболочку
клетки. В результате в клетке развивает
ся т у р г о р н о е давление, которое под
держивает относительную жесткость
растительных клеток, а также обусловли
вает растяжение клеток во время их,
роста.
В запасающих тканях растений вмес
то одной центральной часто бывает не
сколько вакуолей, в которых скаплива
ются запасные питательные вещества.
Это, например, жировые (содержащие
растительные масла) или белковые
(алейроновые зерна) вакуоли.
Сократительные (пульсирующие)
вакуоли служат для осмотической регу
ляции, прежде всего, у пресноводных
протистов, так как в их клетки путем
осмоса непрерывно поступает вода из ок
ружающего гипотонического раствора
(концентрация веществ в речной или
озерной воде значительно ниже, чем кон
центрация веществ в клетках протистов).
Эту воду и воду, поглощенную путем пиноцитоза, сократительные вакуоли вса
сывают осмотическим путем и затем вы
водят наружу с помощью сокращений.
Сокращения осуществляются благодаря
наличию в мембране вакуолей пучков
эластичных волокон.
Таким образом, вакуоли выполняют в
клетках важные функции, связанные с
регуляцией водного баланса и под
держанием тургора клетки, а так
же с изоляцией запасных питатель
ных веществ и конечных продуктов
обмена.
Взаимосвязь между мембранами
клетки. Рассмотрев мембранные систе
мы клетки — эндоплазматический рети
кулум, комплекс Гольджи, лизосомы,
вакуоли, — вы убедились, что все
они представляют собой ограни
ченные элементарными мембранами
пространства клетки, специализиро
ванные для выполнения определенных
функций.
Между мембранами различных струк
тур клетки существует вполне определен
ная взаимосвязь, которая выражается в
способности мембран одних органелл пе
реходить (трансформироваться) в мем
браны других (рис. 47, с. 82).
Из мембран эндоплазматического ре
тикулума формируются вакуоли; они же в
виде пузырьковых образований могут
входить в состав цистерн комплекса Голь
джи, а последние, при участии пузырьков
эндоплазматического ретикулума, обра
зуют лизосомы. Кроме того, экзоцитозные пузырьки комплекса Гольджи, выно
ся свое содержимое за пределы цито
плазматической мембраны, сами включа
ются в ее состав. Таким образом, мож
но считать, что в клетке существует пе
реход мембранных систем от эндоплаз
матического ретикулума в сторону ци
топлазматической мембраны, причем
каждая мембранная система модифи
цируется на выполнение определенных
функций.
■ 1. Что представляет собой вакуоль? 2. Какие
функции выполняют вакуоли в растительных клет
ках? 3. Какие вещества содержатся в клеточнрм со
ке? 4. Как возникают вакуоли в клетке? 5. У каких
организмов присутствуют пульсирующие вакуоли,
какова их функция и работа? 6. Чем отличаются пи
щеварительные вакуоли от других вакуолей клетки?
81
Рис. 47 Взаимосвязь между мембранными системами клеток: 1 — ядерная оболочка, 2 — шерохова
тый эндоплазматический ретикулум, 3 — гладкий эндоплазматический ретикулум, 4 — лизосомы,
5 — комплекс Гольджи, 6 — пузырьки комплекса Гольджи.
26. Опорно-двигательная система
клетки
Для клетки характерны разнообраз
ные типы как внутренних, так и внешних
движений. Внутренние — это движение
цитоплазмы, перемещение хлоропластов
и других органелл, внешние — переме
щение клеток некоторых протестов и бак
терий с помощью ресничек и жгутиков. В
организмах животных и человека имеются
специализированные клетки, мышечная
работа которых позволяет осуществлять
различные движения органов, отдельных
частей тела или всего организма.
Выявлено, что наличие любых двига
тельных аппаратов структурно связано с
существованием опорных (каркасных,
или скелетных) внутриклеточных образо
82
ваний. Поэтому можно говорить о нали
чии в клетке опорно-двигательной сис
темы.
Микротрубочки. К собственно двига
тельным компонентам клеток относятся
микротрубочки и микрофиламенты, кото
рые были выявлены в цитоплазме при
помощи электронного микроскопа.
Микротрубочки содержатся во всех
эукариотических клетках и представляют
собой полые неразветвленные цилиндры
(рис. 48). Это очень тонкие структуры.
Их внешний диаметр не превышает
30 нм, а толщина стенки микротрубоч
ки — около 5 нм. В длину они могут дос
тигать нескольких микрометров. Цито
плазматические микротрубочки легко
распадаются (разбираются) и собирают
ся вновь.
Сборка микротрубочек из молекул гло
булярного белка тубулина (одну субъеди
ницу образуют две молекулы белка) осу
ществляется в присутствии ионов Mgf+,
молекул АТФ и в кислой среде. Разборка
ускоряется повышением концентрации ио
нов Са2+ и понижением температуры.
Сборка микротрубочек, вероятно, мо
жет начаться при наличии матрицы. Счи
тают, что роль матрицы (организатора
микротрубочек) могут играть центриоли,
базальные тельца ресничек и жгутиков
(см. далее), а также особые структуры
хромосом в области первичной перетяж
ки — кинетохоры (центромеры).
Микротрубочки выполняют опорную
функцию в клетке, придавая ей опреде
ленную форму. Известно, что при обра
ботке клеток колхицином происходит
разрушение микротрубочек и, например,
животные клетки, лишенные такой внут
ренней опоры, приобретают шаровидную
форму. Микротрубочки образуют верете
но деления и обеспечивают расхождение
хромосом к полюсам клетки. Перемеще
ние хромосом (хроматид) во время деле
ния осуществляется благодаря способ
ности микротрубочек скользить одна по
другой. Это скольжение обеспечивается
благодаря энергии АТФ. Одни микротру
бочки (хромосомные) прикрепляются к
хромосомам и скользят по другим мик
д
ротрубочкам (полюсным), в результате
чего хромосомы растаскиваются к полю
сам клетки.
Микротрубочки отвечают за переме
щение клеточных органелл, подобно то
му, как поезд следует в определенном
направлении по рельсам.
Центриоли и базальные тельца. У
большинства животных клеток, а также у
некоторых грибов, водорослей, мхов и
папоротников, имеются центриоли. Это
мелкие полые цилиндры длиной не более
0,5 мкм. Они располагаются парами пер
пендикулярно одна к другой. Накануне
деления клетки в ней содержится две па
ры центриолей (по одной паре у каждого
полюса).
Каждая центриоль построена из 9
триплетов микротрубочек (рис. 49, с. 84).
Основная функция центриолей — ини
циация образования микротрубочек ве
ретена деления клетки.
Центриолям по структуре идентичны
базальные тельца, которые всегда об
наруживаются в основании жгутиков и
ресничек. По всей вероятности, базаль
ные тельца образуются путем удвоения
центриолей.
Базальные тельца, как и центриоли,
являются центрами организации микро
трубочек, входящих в состав жгутиков и
ресничек.
б
Рис. 48. Схема строения микротрубочки: а — спиральное расположение тубулиновых единиц (1), б — поперечное сечение.
83
Рис. 49. Схема строения центриолей: 1 — мате
ринская центриоль, 2 — триплеты микротрубо
чек, 3 — дочерняя центриоль, 4 — микротру
бочки.
Жгутики и реснички представляют
собой подвижные цитоплазматические
отростки, служащие либо для передви
жения всего организма (протесты, рес
ничные черви) или репродуктивных кле
ток, либо для транспорта частиц и жид
костей (например, реснички мерцатель
ных клеток слизистой оболочки носовых
полостей и трахеи, яйцеводов и т. д.).
Ж гутики эукариотических клеток по
всей длине содержат 20 микротрубочек: 9
периферических дублетов и 2 централь
ные одиночные. У основания жгутика в
цитоплазме располагается базальное
тельце (рис. 50).
Жгутики эукариотических клеток
имеют длину около 100 мкм и больше.
Короткие (10—20 мкм) жгутики, кото
рых бывает много на одной клетке, назы
ваются ресничками.
Скольжение микротрубочек, входя
щих в состав жгутиков или ресничек, вы
зывает их б ие н и е , что обеспечивает пе
ремещение клетки либо продвижение
частиц.
Микрофиламенты представляют со
бой тонкие нити, встречающиеся во всей
цитоплазме клеток. Особенно много их в
поверхностном слое цитоплазмы, в лож
84
ноножках подвижных клеток, где они об
разуют густую сеть тонких нитей, кото
рые пересекаются в разных направлени
ях. Пучки микрофиламентов обнаружи
ваются в микроворсинках эпителия ки
шечника.
Микрофиламенты образованы бел
ком актином, молекулы которого полимеризуются в длинную фибриллу, состоя
щую из двух закрученных относительно
друг друга спиралей. Содержание актина
составляет 10— 15 % от общего коли
чества клеточных белков. Обнаружива
ются в клетках и нити другого важного
сократительного белка — миозина, хотя
количество его значительно меньше, чем
Рис. 50. Схема строения жгутика: 1 — микро
трубочка, 2 — плазматическая мембрана, 3 —
базальное тельце.
актина. Взаимодействие актина и миози
на лежит в основе сокращения мышц.
Микрофиламенты актина взаимодей
ствуют с микротрубочками поверхнос
тного слоя цитоплазмы и с плазмалеммой
и обеспечивают двигательную активность
цитоплазмы. Считается также, что мик
рофиламенты участвуют в эндоцитозе,
образовании перетяжки при делении кле
ток животных, а также обеспечивают
амебоидные движения.
Строение мышечных фибрилл и мы
шечные сокращения. Специализирован
ные сократительные клетки животных
организмов имеют в цитоплазме сократи
тельные фибриллы — миофибриллы.
Особенно много миофибрилл в скелет
ных мышечных волокнах (слиянии мы
шечных клеток •—■миобластов), в клет
ках сердечной мышцы и гладкой мускула
туры.
В световой микроскоп можно видеть,
что пучки миофибрилл окрашены нерав
номерно: через равные промежутки чере
дуются темные и светлые участки
(рис. 51, с. 86).
Темные участки были названы ани
зотропными дисками (A-диски), свет
лые — изотропными (1-диски). Свет
лый 1-диск пересекается темным Z-диском (Z -полоска). Таким образом, миофибрилла представляет собой нить тол
щиной около 0,5 мкм с чередующимися
участками:
А + 1/2 I + Z + 1/2 I + А + 1/2 I + Z... .
Единицей строения и функционирова
ния миофибриллы является саркомер —
участок между двумя Z-дисками. Величи
на саркомера в расслабленном состоянии
всегда одинаковая: 1,8—2,8 мкм в зави
симости от вида животного.
Под электронным микроскопом было
выявлено, что миофибрилла состоит из
ряда более тонких нитей — протофиб
рилл. Их диаметр в разных частях сарко
мера разный. В 1-дисках встречаются
тонкие нити (7 нм), а в A-дисках, кроме
тонких, присутствуют и толстые нити
(16 нм). Все протофибриллы расположе
ны параллельно и одна в другую не пере
ходят (см. рис. 51).
Тонкие нити представляют собой актиновые нити, а толстые — миозиновые.
В состав последних входит около 300 мо
лекул белка миозина.
Нити актина и миозина могут сколь
зить одна относительно другой, в резуль
тате чего происходит суммарное сокра
щение саркомеров, приводящее к сокра
щению мышечного волокна.
Скольжение нитей актина и миозина,
приводящее к сокращению или расслаб
лению саркомеров, осуществляется с
затратой большего количества АТФ и в
присутствии ионов Са2+. Периодичное
скольжение нитей в одну и другую сторо
ну приводит к сокращению и расслаб
лению миофибрилл и мышц в целом,
обеспечивая таким образом работу
мышц.
? lf
■. 1. С помощью каких структур осуществляются
внутриклеточные движения? 2. Как организованы
микротрубочки и при каких условиях осуществля
ются их сборка и разрушение? 3. Какую функцию
выполняют микротрубочки в клетке? 4. Как органи
зованы реснички и жгутики и как они способствуют
перемещению клеток? 5. Что представляют со
бой центриоли? Какие функции они выполняют в
клетке? 6. Каков механизм сокращения мышечных
волокон?
27. Клеточная стенка
Клеточная стенка (оболочка) явля
ется неотъемлемым компонентом клеток
растений и грибов и представляет собой
продукт их жизнедеятельности. Она при
дает клеткам механическую прочность,
защищает их содержимое от поврежде
ний и избыточной потери воды, поддер
живает форму клеток и их размер, а так
же препятствует разрыву клеток в гипо
тонических условиях. Клеточная стенка
участвует в поглощении и обмене различ85
Рис. 51. Строение саркомера: 1 — мышца, 2 — сухожилие, 3 — кость, 4 — мышечные
волокна, 5 — мышечное волокно, 6 — саркомер, 7 — Н-полоска, 8 — 4-диск,
9 — /-диск, 10 — актиновые нити, 11 — миозиновые нити.
86
ных ионов, т. е. является ионообменником. Через клеточную оболочку осущес
твляется транспорт веществ.
Клеточная стенка, формирующаяся
во время деления клеток и их роста путем
растяжения, называется первичной.
После прекращения роста клетки на пер
вичную клеточную стенку изнутри откла
дываются новые слои и образуется проч
ная вторичная клеточная оболочка.
В состав клеточной стенки входят
структурные компоненты (целлюлоза у
растений и хитин у грибов), компоненты
матрикса (гемицеллюлоза, пектин, белки),
инкрустирующие компоненты (лигнин, су
берин) и вещества, откладывающиеся на
поверхности оболочки (кутин и воск).
Целлюлоза представляет собой длин
ные неразветвленные цепочки, состоя
щие из 3— 10 тыс. остатков a -D -глюкозы. Молекулы целлюлозы взаимодей
ствуют между собой с образованием во
дородных связей и собраны в микрофиб
риллы (рис. 52). Переплетенные микро
фибриллы составляют каркас клеточной
оболочки.
У большинства грибов микрофибрил
лы клеточной стенки состоят из хит и
на — полимера, построенного из мо
лекул P-D-глюкозы, у которой при
втором атоме углерода гидроксильная
( — ОН) группа замещена на группу
— NH — СО — СН3.
Микрофибриллы погружены в мат
рикс клеточной стенки. Матрикс состоит
из смеси различных химических веществ,
среди которых преобладают полисахари
ды (гемицеллюлозы и пектиновые вещес
тва).
Гемицеллюлозы — это группа поли
сахаридов (полимеры пентоз и гексоз —
ксилозы, галактозы, маннозы, глюкозы и
др.). Молекулы гемицеллюлоз, как и цел
люлозы, имеют форму цепи, но в отличие
от последней их цепи короче, менее упо
рядочены и сильно разветвлены. Они
Рис. 52. Строение клеточной оболочки: 1 — микрофибрилла, 2 — агрегаты из цепочек целлюлозы,
3 _ цепочки целлюлозы, 4 — поры с плазмодесмами, 5 — первичная оболочка, 6 — вторичная обо
лочка, 7 — срединная пластинка.
87
легче растворяются и разрушаются фер
ментами.
Пектиновые вещества — это поли
меры, построенные из моносахаридов
(арабиноза и галактоза), галактуроновой
кислоты (сахарная кислота) и метилового
спирта. Длинные молекулы пектиновых
веществ могут быть линейными или раз
ветвленными. Молекулы пектиновых ве
ществ содержат большое количество
карбоксильных групп и поэтому способ
ны соединяться с ионами Mg2+ и Са2+.
При этом образуются клейкие, студнеоб
разные пектаты магния и кальция, из ко
торых затем складываются срединные
пластинки, скрепляющие стенки двух со
седних клеток.
Ионы двухвалентных металлов могут
обмениваться на другие катионы (К+, Н+
и т. д.). Это обусловливает катионооб
менную способность клеточных обо
лочек.
Пектиновыми веществами и пектатами богаты оболочки клеток многих пло
дов. Так как при их извлечении из оболо
чек и добавлении сахара образуются ге
ли, пектины используют как желеобразующие вещества для изготовления мар
мелада и др.
Помимо углеводных компонентов, в
состав матрикса клеточной стенки входит
также структурный белок экстенеин —
гликопротеин, который по своему составу
близок к межклеточным белкам живот
ных — коллагенам.
На долю матрикса приходится до
60 % сухого вещества клеточной оболоч
ки. Матрикс оболочки не просто запол
няет промежутки между микрофибрилла
ми, а образует прочные химические (во
дородные и ковалентные) связи между
макромолекулами и микрофибриллами,
что обеспечивает прочность клеточной
стенки, ее эластичность и пластичность.
Основным инкрустирующим вещес
твом оболочки клеток растений является
лигнин — полимер с неразветвленной
молекулой, состоящей из ароматических
88
спиртов (кумарового, кониферилового,
синапового).
Интенсивная лигнификация (пропит
ка слоев целлюлозы лигнином) клеточ
ных оболочек начинается после прекра
щения роста клетки.
Лигнин может откладываться отдель
ными участками — в виде колец, спира
лей или сетки, как это наблюдается в
оболочках клеток проводящей ткани —
ксилемы (рис. 53) или сплошным слоем,
за исключением тех мест, где осущес
твляются контакты между соседними
клетками в виде плазмодесм.
Лигнин скрепляет целлюлозные во
локна. Он действует как очень твердый и
жесткий матрикс, усиливающий проч
ность клеточных стенок на растяжение и
сжатие. Он же обеспечивает клеткам до
полнительную защиту от физических и
химических воздействий, снижает водо
проницаемость. Содержание лигнина в
Рис. 53. Вторичная оболочка сосудов ксилемы.
оболочке достигает 30 %. Инкрустация
им клеточных оболочек приводит к их од
ревеснению. Одревеснение часто влечет
за собой отмирание живого содержимого
клетки.
Лигнин в сочетании с целлюлозой
придает особые свойства древесине, ко
торые делают ее незаменимым строи
тельным материалом.
На клеточную оболочку могут откла
дываться также жироподобные вещес
тва — суберин, кутин и воск.
Суберин откладывается на оболочку
изнутри и делает ее практически непро
ницаемой для воды и растворов. В ре
зультате протопласт клетки отмирает и
клетка заполняется воздухом. Такой про
цесс называется о п р о б к о в е н и е м .
Опробковение оболочки клеток на
блюдается в покровных тканях много
летних древесных растений — пе
ридерме, корке, а также в эндодерме
корня.
Поверхность эпидермальных клеток
растений защищена гидрофобными ве
ществами — кутином и восками. Пред
шественники этих соединений секретируются из цитоплазмы на поверхность, где
и происходит их полимеризация. Слой ку
тана обычно пронизан полисахаридными
компонентами (целлюлозой и пектином)
и образует кутикулу. Воск часто откла
дывается в кристаллической форме на
поверхности частей растений (листья,
плоды), образуя восковой налет.
Кутикула и восковой налет защищают
клетки от повреждений и проникновения
инфекции, уменьшают испарение воды с
поверхности органов.
В оболочках эпидермальных клеток
некоторых растений (злаки, осоки и др.)
накапливается большое количество ми
неральных веществ (минерализация), в
первую очередь карбоната кальция и
кремнезема. При минерализации листья
и стебли растений становятся жесткими,
твердыми и в меньшей степени поедают
ся животными.
Таким образом, клеточная стенка иг
рает важную роль в жизни клеток расте
ний и грибов и выполняет ряд специфи
ческих функций.
Существенно отличается по строению
клеточная стенка прокариот. По строе
нию клеточной стенки бактерий делят на
две группы —- г р а м п о л о ж и т е л ь н ы е
(окрашиваются по Граму) и г р а м о т р и ц а т е л ь н ы е (обесцвечиваются при от
мывке красителя). В клеточной стенке
тех и других есть особая жесткая решет
ка, состоящая из пептидогликана. Пептидогликан — это полисахарид, со
стоящий из чередующихся остатков двух
аминосахаров (N-ацетилглюкозамина и
близкого к нему азотсодержащего поли
сахарида — N-ацетилмурамовой кисло
ты). Молекула пептидогликана представ
ляет собой правильную сеть из парал
лельно расположенных полисахаридных
цепей, сшитых друг с другом короткими
пептидными цепочками (рис. 54, с. 90).
Таким образом, каждая бактериальная
клетка окружена сетевидным мешком,
образованным, по сути дела, одной моле
кулой.
У грамположительных бактерий в
пептидогликановую сетку встроены дру
гие вещества, главным образом белки и
полисахариды. Так вокруг пептидогликановой сетки создается сравнительно тол
стая и жесткая упаковка.
У грамотрицательных бактерий кле
точная стенка тоньше, чем у грамполо
жительных, но устроена она сложнее
(рис. 55, с. 90). У этих бактерий снаружи
от пептидогликанового слоя расположен
дополнительный слой клеточной стен
ки — наружная мембрана. Она состоит
из фосфолипидов, типичных для элемен
тарных мембран, белков, липопротеина и
липополисахарида. Наличие наружной
мембраны у грамотрицательных бакте
рий значительно расширяет круг функций
клеточной стенки. Наружная мембрана
осуществляет регуляцию транспорта ве
ществ и ионов, необходимых клетке. Она
89
/
/
,
/
/
/ г
/ г
x
/
M
/
\
Mr
. '4 f /
/
/
*
/
/
/
/
/
IVLw~~M
/ 5' 5 //
/
7 7 77
/ /
M
/
/ г
M
/
/
/
‘
Рис. 54. Структура пептидогликана: кружочками обозначены аминокислоты пептидных цепочек; ли
ниями — полимерные цепочки, образованные аминосахарами (Г — N-ацетилглюкозамин, М — N-ацетилмурамовая кислота), соединенные между собой гликозидными связями.
препятствует проникновению в клетку
токсичных веществ, что делает более
устойчивыми грамотрицательные бакте
рии по сравнению с грамположительными к действию некоторых ядов, химиче
ских веществ, ферментов, антибиотиков.
Между наружной мембраной и пептидогликановой сеткой образуется обособ
ленная полость, где локализуются раз
личные вещества, в том числе транспор
тные белки, обеспечивающие транспорт
веществ от наружной мембраны к цито
плазматической, и гидролитические фер
менты.
Снаружи клеточная стенка прокариот
часто бывает окружена слизистым чех
лом, или капсулой. Это результат
биосинтеза прокариотами органиче
ских полимеров и отложения их вокруг
клетки.
2
3
4
Рис. 55. Клеточная стенка: а — грамположительных и б — грам отрицательны х бактерий: 1 — ци
топлазматическая мембрана, 2 — пептидогликан, 3 — периплазматическое пространство, 4 — на
ружная мембрана, 5 — цитоплазма, в центре которой расположена ДНК.
90
1. Клетки каких организмов имеют оболочку?
2. Какие функции выполняет оболочка клетки?
3. Из каких компонентов состоит клеточная оболоч
ка? 4. Что является структурным компонентом обо
лочки клетки? 5. Из каких веществ складывается
матрикс оболочки? 6. Благодаря чему достигается
высокая прочность, эластичность и пластичность
клеточных оболочек? 7. В клетках каких органов
наблюдаются одревеснение, опробковение, кутинизация клеточных оболочек? К чему это приводит?
8. Чем отличается по строению клеточная оболочка
прокариот от оболочки эукариот? 9. Каковы основ
ные различия в строении оболочек у грамположительных и грамотрицательных бактерий?
28. Система связи
и взаимодействия между клетками
в многоклеточном организме
Для того чтобы многоклеточный орга
низм мог нормально функционировать,
его активность каким-то образом должна
координироваться. Иными словами,
между клетками должна существовать
система связи и координации.
Существует несколько способов вза
имосвязи между клетками. Растительные
<3
клетки связываются между собой плазмодесмами — тонкими трубчатыми ка
налами. Как вы помните, растительные
клетки покрыты целлюлозными оболоч
ками. В некоторых местах клеточные
стенки растений пронизаны отверстия
ми — порами диаметром до 1 мкм. Через
поры проходят плазмодесмы (рис. 56),
каждая из которых представляет собой
канал, выстланный плазмалеммой, не
прерывно переходящей из клетки в клет
ку. Центральную часть плазмодесмы за
нимает десмотрубочка, состоящая из
спирально расположенных белковых
субъединиц. Десмотрубочка сообщается
с мембранами эндоплазматического ре
тикулума соседних клеток. Вокруг дес
мотрубочки имеется слой цитоплазмы,
которая соединяется с цитоплазмой со
седних клеток. Таким образом, посред
ством плазмодесм клетки связываются
друг с другом через эндоплазматический
ретикулум и цитоплазму. Единая система
цитоплазмы клеток тканей и органов на
зывается симпластом.
Аналогичные связи в животных клет
ках называются щелевыми контактами.
В области щелевых контактов мембраны
б
Рис. 56. Схема строения плазмодесмы: а — продольный разрез, 6 — поперечный
разрез: 1 — эндоплазматический ретикулум, 2 — десмотрубочка, 3 — цитоплаз
ма, 4 — плазмалемма, 5 — клеточная стенка, 6 — срединная пластинка.
91
2
Рис. 57 Щелевой контакт в животных клетках: 1 — цитоплазма; 2 — субъединицы, образующие ще
левой контакт; 3 — цитоплазматическая поверхность мембраны; 4 — межклеточное пространство;
5 — липидный бислой; 6 — наружная поверхность мембраны; 7 — гидрофильные каналы (коннексоны); 8 — интегральный мембранный белок; 9 — белки, нуклеиновые кислоты; 10 — ионы, аминокис
лоты, сахара, нуклеотиды.
соседних клеток находятся на расстоянии
2 — 3 нм друг от друга. Эти мембраны
пронизаны тонкими каналами диамет
ром 44 нм — коннексонами, кото
рые образованы белком коннектином
(рис. 57).
Одному коннексону на плазматичес
кой мембране клетки точно противостоит
коннексон на плазматической мембране
соседней клетки, так что каналы двух кле
ток образуют единое целое. Коннексоны
играют роль прямых межклеточных кана
лов, по которым ионы и низкомолекуляр
ные вещества могут диффундировать из
клетки в клетку. Коннексоны могут со
кращаться, изменяя диаметр внутренне
го канала и тем самым участвовать в
регуляции транспорта молекул между
клетками.
В нервной системе особый интерес
представляют контакты между клетками,
осуществляющиеся при помощи особых
структур, называемых синапсами (рис.
58). Синапс представляет собой тип кон
такта между'двумя нейронами (нервны
ми клетками) либо между нейроном и ка
ким-либо другим элементом — рецеп
92
тором или эффектором. Число синап
сов, как правило, очень велико, что обес
печивает большую площадь для передачи
информации.
Существует два типа синапсов — х и
мические и электрические — в зависи-
Рис. 58. Схема синаптического контакта: 1 —
митохондрия, 2 — синаптический пузырек, 3 —пресинаптическая мембрана, 4 — синаптичес
кая щель, 5 — постсинаптическая мембрана.
мости от природы проходящих через них
сигналов.
Синапс — это структура, образующа
яся между участками двух клеток — дендритов и аксонов. Межнейронные си
напсы имеют обычно вид грушевидных
(луковицевидных) расширений на конце
отростка нервной клетки. Такие расшире
ния называются синаптическими бляш
ками.
Цитоплазма синаптической бляшки
содержит митохондрии, гладкий эндо
плазматический ретикулум и многочис
ленные синаптические пузырьки. Каж
дый пузырек содержит медиатор — ве
щество, с помощью которого нервный
сигнал передается через синапс. Основ
ными медиаторами нервной системы поз
воночных являются ацетилхолин и норад
реналин. Мембраны контактирующих
клеток разделены межклеточным прос
транством — синаптической щелью.
Мембрана в области синаптического
контакта одной клетки называется пресинаптической, а в другой, воспринима
ющей импульс,— постсинаптической.
Синаптические пузырьки в момент про
хождения нервного импульса выбрасыва
ют свое содержимое в синаптическую
щель. Молекулы медиатора, диффунди
руя через синаптическую щель, связыва
ются с находящимися на постсинаптичес
кой мембране рецепторами, способными
узнавать структуру медиатора. Это в
свою очередь изменяет проницаемость
постсинаптической мембраны, что и спо
собствует передаче нервного импульса.
У многих животных передача импуль
сов через некоторые синапсы осущес
твляется путем прохождения электричес
кого тока между пре- и постсинапти
ческими нейронами. Ширина щели меж
ду этими нейронами составляет около
2 нм, кроме того, суммарное сопротивле
ние тока со стороны мембран и межкле
точной жидкости очень мало, что и спо
собствует передаче электрического им
пульса.
I 1. Каковы основные способы связи между
клетками? 2. Что представляют собой плазмодесмы
и как взаимодействуют клетки с помощью плазмодесм? 3. Каким образом осуществляются контакты
между клетками в организмах животных? 4. Расска
жите о клеточных контактах в нервной системе жи
вотных.
Глава III
ВОСПРОИЗВЕДЕНИЕ
КЛЕТКИ
29. Деление клетки.
Клеточный цикл
Все новые клетки в организмах воз
никают в результате деления надвое уже
существующих. Если делится однокле
точный организм, то из старого организ
ма образуется два новых. Многоклеточ
ный организм также начинает свое раз
витие чаще всего с одной-единственной
клетки. Путем многократных делений об
разуется огромное количество клеток,
которые и составляют организм. В мно
гоклеточных организмах не все клетки
имеют способность к делению по причине
их высокой специализации.
Животные клетки в связи с высоким
уровнем специализации имеют разную
продолжительность жизни. Например,
нервные и мышечные клетки после за
вершения эмбрионального периода раз
вития организма перестают делиться и
функционируют на протяжении всей по
следующей жизни организма. Многие
специализированные клетки не размно
жаются в обычных условиях, но при по
вреждении органов и тканей, в состав ко
торых они входят, их способность к деле
нию восстанавливается. К таким клеткам
относятся клетки печени, эндокринных
желез и других органов. Другие клетки, в
частности костного мозга, эпителия тон
кого кишечника, эпидермиса, в процессе
выполнения своей специфической фун
кции погибают, поэтому на смену им при
93
ходят новые, образованные путем де
ления.
Постоянное деление клеток верху
шечной и боковых меристем растений
(камбий, пробковый камбий) обеспечи
вает длительный рост побегов и корней в
длину и толщину.
Вновь образованные клетки приобре
тают способность к делению после неко
торого периода роста. Кроме того, деле
нию предшествует удвоение клеточных
структур: хлоропластов, митохондрий,
центриолей и др.
Перед началом клеточного деления
должна реплицироваться ДНК, посколь
ку она несет в себе информацию, необхо
димую клетке для синтеза белков. Если
бы дочерние клетки не получали точных
копий ДНК материнской клетки, они пе
рестали бы быть похожими друг на друга.
Это в конечном итоге привело бы к не
возможности существования определен
ных видов организмов. Чтобы этого не
случилось, ДНК должна идеально репли
цироваться и каждая дочерняя клетка при
клеточном делении должна получать ее
копию.
Совокупность последовательных и
взаимосвязанных процессов в период
подготовки клетки к делению и в период
деления называется митотическим цик
лом (от названия основного типа деле
ния — митоза).
Жизнь клетки от одного деления до
следующего деления или до смерти назы
вается клеточным, или жизненным цик
лом клетки. У одноклеточных организмов
(протистов) клеточный цикл совпадает с
жизнью особи. В непрерывно размножа
ющихся тканевых клетках клеточный
цикл совпадает с митотическим циклом и
состоит из четырех периодов со строгой
последовательностью смен друг друга:
постмитотического (G[ — от англ,
grow — расти, увеличиваться), синте
тического (S — от англ, synthesis —
синтез), премитотического (G2) и м и
тоза (М) (рис. 59). Первые три перио94
Gi
Рис. 59. Жизненный цикл клетки многоклеточ
ного организма: 2л •— диплоидный набор хро
мосом, 2с, 4 с — количество сестринских хроматид в наборе.
да — это и н т е р ф а з а . По продолжи
тельности она составляет большую часть
митотического цикла клетки.
В период Gj клетка растет, в ней уси
ленно образуются РНК и белки, в первую
очередь ферменты, катализирующие об
разование предшественников ДНК, фер
менты метаболизма РНК и строительных
белков. В этот период синтезируется зна
чительное количество гистоновых бел
ков. После завершения периода G[ клет
ка приступает к синтезу ДНК, или ее
репликации — удвоению. Две спирали
материнской молекулы ДНК расходятся,
становясь матрицей для воспроизводства
новых цепей ДНК- Каждая из двух дочер
них молекул обязательно включает две
цепочки: старую и новую. Удвоение ДНК
происходит с поразительной точностью,
что обеспечивает в дальнейшем развитие
присущих только данному организму
признаков. Однако под воздействием хи
мического и физических факторов (иони
зирующее и ультрафиолетовое излуче
ние, повышенная температура, химичес
кие вещества и др.) может нарушаться
правильность вновь синтезированных це
почек ДНК- Эти нарушения в большин-
стве случаев ликвидируются с помощью
специальных репарирующих ферментов,
которые «узнают» участок молекулы
ДНК, не сходный с матрицей, и «выре
зают» его, после чего недостающий
участок достраивается. Таким обра
зом, точная передача наследственной
информации обеспечивается матричным
синтезом ДНК и системой репарирова
ния поврежденных участков моле
кул ДНК.
Продолжительность S-периода в раз
ных клетках неодинакова: от нескольких
минут до нескольких часов (табл. 5).
Т а б л и ц а 5. Продолжительность митотического цикла у разных видов организмов
Вид организма и клеток
Вика посевная (корень)
Горох (корень)
Кукуруза (корень)
Мышь (эпителий тонкой кишки)
Мышь (эпителий кожи)
Митоти
ческий
цикл,ч
Митоз,
ч
G„
ч
15,0
19,3
22,5
18,8
585,6
1,2
2,3
2,2
1,0
3,8
0,8
6,7
6,3
9,5
528,0
В результате удвоения молекул ДНК в
каждой хромосоме оказывается вдвое
больше ДНК, чем было до S-периода,
т. е. количество ДНК в диплоидных клет
ках соответствует тетраплоидному набо
ру. Однако количество хромосом считает
ся диплоидным, потому что образовавши
еся дочерние хромосомы (хромонемы)
тесно переплетены между собой и сос
тавляют единую хромосому с четырьмя
сестринскими цепочками ДНК13 S-период, кроме репликации ДНК,
продолжается синтез РНК и белков, в
том числе и гистоновых, осуществляется
синтез рРНК, которые используются в
следующем периоде для синтеза белков,
обеспечивающих митоз, осуществляется
удвоение центриолей.
Время от окончания синтеза ДНК и до
начала митоза называется 0 2-периодом.
В этот период завершается подготовка
клетки к митозу, активно синтезируются
иРНК, различные белки, в том числе и
тубулиновые, из которых образуются ни
ти веретена деления, а также иРНК и
белки для осуществления G j-периода
после митоза. Далее наступает деление
клетки.
Клеточный цикл и его периоды сос
тавляют в размножающихся клетках
10—50 ч и зависят от типа клетки, воз
S,
ч
7,8
8,0
6,7
7,5
30,0
g 2.
Ч
5,2
2,3
7,3
0,8
4,6
раста, количества ДНК в ядре, темпера
туры, времени суток и других факторов.
Наиболее вариабельны Gr и (32-периоды; они могут значительно удлиняться,
особенно у так называемых покоящихся
клеток. В этом случае выделяют Go-пе
риод (от англ, gap — промежуток, интер
вал), или период покоя. С учетом перио
да покоя клеточный цикл может длиться
недели и даже месяцы (например, у кле
ток печени), а у нейронов клеточный цикл
равен продолжительности жизни орга
низма.
?
■ 1. Что такое клеточный цикл? 2. Что называет
ся интерфазой? 3. Из каких периодов состоит ин
терфаза? 4. Какие основные события осуществля
ются в G!-, S-, С2-периодах? 5. Все ли клетки спо
собны к делению? 6. Каков набор хромосом клеток,
находящихся в S -периоде?
30. Митоз
После того как в клетке завершаются
биохимические процессы подготовки к
делению, начинается таинственный и до
конца не изученный процесс. В настоя
щее время описано несколько способов
деления клетки: митоз, амитоз и мейоз.
Митоз (от греч. mitos — нить), т. е.
непрямое деление — основной способ
95
деления эукариотических клеток. М и
тоз — это деление ядра, приводящее к
образованию двух дочерних ядер, в каж
дом из которых имеется точно такой же
(аналогичный) набор хромосом, как и в
родительском ядре. Вслед за делением
ядра обычно следует деление самой клет
ки, поэтому часто термин «митоз» озна
чает деление клетки в целом.
Впервые митоз в спорах плаунов
наблюдал русский ученый И . Д . Ч и с
т я к о в в 1874 г. Детальные исследова
ния поведения хромосом в ходе митоза
были выполнены немецким ботаником
Э . С т р а с б у р г е р о м в 1876 —
1879 гг. на растениях и немецким гисто
логом В. Ф л е м м и н г о м в 1882 г. на
животных.
Митоз — процесс непрерывный, но
для удобства изучения биологи делят его
на четыре стадии в зависимости от того,
как выглядят в это время хромосомы в
световом микроскопе. В митозе выделя
ют профазу, метафазу, анафазу и телофазу (рис. 60, 61).
В п р о ф а з е происходит укорочение
Pgsppgj
1
2
3
7
Рис. 60. Схема митоза в растительной
клетке: 1 — интерфаза, 2— 5 — профаза,
6, 7 — метафаза, 8, 9 — анафаза, 10—
12 — телофаза.
96
и утолщение хромосом вследствие их
спирализации. В это время хромосомы
двойные — состоят из двух хроматид,
связанных между собой в области пер
вичной перетяжки особой структурой —
центромерой. Удвоение хромосом, как
вы помните, произошло в S-периоде ин
терфазы. Одновременно с утолщением
хромосом исчезает ядрышко и фрагмен
4. Общая биология, 10 кл.
тируется (распадается на отдельные цис
терны) ядерная оболочка. После этого
хромосомы лежат в цитоплазме свободно
и беспорядочно.
В профазе центриоли (в тех клетках,
где они есть) расходятся к полюсам клет
ки. В конце профазы начинает образо
вываться верет ено деления, кото
рое формируется из микротрубочек пу97
тем полимеризации белковых субъ
единиц.
Микротрубочки начинают формиро
ваться со стороны центриолей либо со
стороны хромосом (в клетках растений).
В м е т а ф а з е завершается образо
вание веретена деления, которое состоит
из микротрубочек двух типов: хромосом
ных, которые связываются с центроме
рами хромосом, и центросомных (по
люсных), которые тянутся от полюса к
полюсу клетки. Каждая двойная хромосо
ма прикрепляется к микротрубочкам ве
ретена деления. Хромосомы как бы вы
талкиваются микротрубочками в область
экватора клетки, т. е. располагаются на
равном расстоянии от полюсов. Они ле
жат в одной плоскости и образуют так на
зываемую экваториальную, или метафазную, пластинку. В метафазе отчет
ливо видно двойное строение хромосом,
соединенных только в области центроме
ры. В этот период легко подсчитывать
число хромосом, изучать их морфологи
ческие особенности.
В а н а ф а з е центромеры делятся и
из каждой удвоенной хромосомы образу
ются две отдельные, совершенно иден
тичные дочерние хромосомы. Разделив
шись, эти хромосомы с помощью микро
трубочек веретена деления движутся к
полюсам клетки. Во время движения до
черние хромосомы несколько изгибаются
наподобие шпильки, концы которой по
вернуты в сторону экватора клетки. Та
ким образом, в анафазе хроматиды удво
енных в интерфазе хромосом точно рас
ходятся к полюсам клетки. В этот момент
в клетке находятся два диплоидных набо
ра хромосом.
В т е л о ф а з е происходят процессы,
обратные тем, которые наблюдаются в
профазе: начинается деспирализация
(раскручивание)хромосом, они набухают
и становятся плохо различимы в микрос
коп. Вокруг хромосом у каждого полюса
из мембранных структур цитоплазмы
формируется ядерная оболочка, в ядрах
98
возникают ядрышки. Разрушается вере
тено деления.
На стадии телофазы происходит раз
деление цитоплазмы (цитотомия) с об
разованием двух клеток. В клетках жи
вотных плазматическая мембрана начи
нает впячиваться внутрь, в области, где
располагался экватор веретена деления.
По-видимому, это происходит благодаря
сокращению микрофиламентов, которые
здесь находятся. В результате впячивания
образуется непрерывная борозда, опоя
сывающая клетку по экватору. В конце
концов клеточные мембраны в области
борозды смыкаются, полностью разделяя
две клетки.
В клетках растений в области эквато
ра из остатков нитей веретена деления
возникает бочковидное образование —
фрагмопласт. В эту область со стороны
полюсов клетки устремляются многочис
ленные пузырьки комплекса Гольджи, ко
торые сливаются друг с другом. Содержи
мое пузырьков образует срединную плас
тинку, которая делит клетку на две дочер
ние, а мембрана пузырьков комплекса
Гольджи образует недостающие цито
плазматические мембраны этих клеток.
Впоследствии на срединную пластинку со
стороны каждой из дочерних клеток
откладываются элементы клеточных обо
лочек.
В результате митоза из одной клетки
возникают две дочерние с тем же набо
ром хромосом.
Таким образом, биологическое значе
ние митоза состоит в строго одинаковом
распределении между дочерними клетка
ми материальных носителей наслед
ственности — молекул ДНК, входящих в
состав хромосом материнской клетки.
Благодаря равномерному разделению
реплицированных хромосом между до
черними клетками обеспечивается обра
зование генетически равноценных клеток
и сохраняется преемственность в ряду
клеточных поколений. Это обеспечивает
такие важные моменты жизнедеятель
ности, как эмбриональное развитие и
рост организмов, восстановление орга
нов и тканей после повреждения. Мито
тическое деление клеток является цито
логической основой бесполого размно
жения организмов.
9
1. В чем заключается сущность митоза? 2. Ка
кие клетки делятся митозом? 3. Чем характеризу
ются фазы митоза? 4. Начало какой фазы митоза
можно непосредственно наблюдать в микроскоп?
5. В чем заключается биологическое значение ми
тоза? 6. Чем определяется упорядоченное распре
деление хромосом в митозе между дочерними клет
ками? 7. Какие функции жизни обеспечивает деле
ние клеток?
31. Амитоз.
Клеточное деление прокариот
Амитоз, или прямое деление, — это
деление интерфазного ядра путем пере
тяжки. При амитозе веретено деления не
образуется и хромосомы в световом мик
роскопе неразличимы. Такое деление
встречается у одноклеточных организмов
(например, амитозом делятся большие
полиплоидные ядра инфузорий), а также
в некоторых высокоспециализированных
с ослабленной физиологической актив
ностью, дегенерирующих, обреченных на
гибель клетках растений и животных, ли
бо при различных патологических про
цессах, таких, как злокачественный рост,
воспаление и т. п.
Амитоз можно наблюдать в тканях
растущего клубня картофеля, эндоспер
ме семян, стенках завязи пестика и па
ренхиме черешков листьев. У животных и
человека такой тип деления характерен
для клеток печени, хрящей, роговицы
глаза.
При амитозе часто наблюдается толь
ко деление ядра: в этом случае могут воз
никнуть двух- и многоядерные клетки.
Если же за делением ядра следует деле
ние цитоплазмы, то распределение кле
точных компонентов, как и ДНК, осущес
твляется произвольно.
Амитоз в отличие от митоза является
самым экономичным способом деления,
так как энергетические затраты при этом
весьма незначительны.
Клеточное деление прокариот. Бак
териальная клетка содержит только одну,
чаще всего кольцевую молекулу ДНК,
прикрепленную к клеточной мембране.
Перед делением клетки ДНК реплициру
ется и образуются две идентичные моле
кулы ДНК, каждая из которых также
прикреплена к клеточной мембране.
При делении клетки клеточная мем
брана врастает между двумя молекулами
ДНК так, что в конечном итоге в каждой
дочерней клетке оказывается по одной
идентичной молекуле ДНК. Такой про
цесс получил название прямого бинар
ного деления.
9
1. Чем отличается амитоз от митоза? 2. В чем
принципиальное различие между делением клеток
прокариот и амитозом эукариот? Что общего между
этими типами деления? 3. Какой биологический
смысл заключается в том, что амитоз характерен
для клеток с ослабленной физиологической актив
ностью?
32. Мейоз и его биологическое
значение
Мейоз (от греч. meiosis — уменьше
ние) — это особый способ деления кле
ток, в результате которого происходит ре
дукция (уменьшение) числа хромосом
вдвое и переход клеток из диплоидного
состояния 2п в гаплоидное п (рис. 62). С
помощью мейоза образуются споры и по
ловые клетки — гаметы. В результате
редукции хромосомного набора в каждую
гаплоидную спору и гамету попадает по
одной хромосоме из каждой пары хромо
сом, имеющихся в данной клетке. В ходе
дальнейшего процесса оплодотворения
(слияния гамет) организм нового поко
ления получит также диплоидный набор
99
Рис. 62. Схема мейоза (I и II деления): 1 — интерфаза, 2— 6 — профаза I, 7 — метафаза I, 8 — ана
фаза I, 9 — телофаза I, 10 — метафаза II, 11 — профаза II, 12 — анафаза II, 13 — телофаза II.
100
II
101
хромосом, т. е. кариотип организмов
данного вида в ряду поколений остается
постоянным. Важнейшее значение мейоза заключается в обеспечении постоян
ства кариотипа в ряду поколений орга
низмов данного вида.
Мейоз
впервые
был
описан
В. Ф л е м м и н г о м в 1882 г. у животных
и Э. С т р а с б у р г е р о м в 1888 г. у рас
тений.
Мейоз включает два быстро следую
щих одно за другим деления. Перед нача
лом мейоза каждая хромосома реплици
руется (удваивается). В течение некото
рого времени две ее образовавшиеся ко
пии остаются связанными друг с другом
центромерой. Следовательно, в каждом
ядре, в котором начинается мейоз, содер
жится эквивалент четырех наборов гомо
логичных хромосом 4с. Поэтому, для того
чтобы образовались ядра гамет, содержа
щие одинарный (гаплоидный) набор хро
мосом, необходимы два ядерных деления.
Эти деления так и называются: первое
деление мейоза и второе деление мей
оза. Второе деление мейоза следует прак
тически сразу же за первым, и синтеза
ДНК в промежутке между ними не проис
ходит (т. е., по сути дела, между первым и
вторым делением отсутствует интерфаза).
Первое мейотическое (редукцион
ное) деление приводит к образованию из
диплоидных клеток (2л) гаплоидных кле
ток (л). Оно начинается с п р о ф а з ы 1,
в которой, как и в митозе, осуществляет
ся упаковка наследственного материала
(спирализация хромосом). Одновре
менно происходит сближение гомологич
ных (парных) хромосом своими одинако
выми участками — конъюгация (в мито
зе не наблюдается). В результате конъю
гации образуются хромосомные пары —
биваленты (см. рис. 62). Каждая хромо
сома, вступая в мейоз, как уже отмеча
лось, имеет удвоенное количество нас
ледственного материала и состоит из двух
хроматид, поэтому бивалент включает
уже 4 нити.
102
Когда хромосомы находятся в конъю
гированном состоянии, продолжается их
дальнейшая спирализация. При этом от
дельные хроматиды гомологичных хромо
сом переплетаются между собой. В по
следующем гомологичные хромосомы от
талкиваются и несколько отходят одна от
другой. В результате этого в местах пе
реплетения хроматид может происходить
их разрыв, и, как следствие, в процессе
воссоединения этих разрывов гомологич
ные хромосомы обмениваются соответ
ствующими участками. В результате хро
мосома, перешедшая к данному организ
му от отца, включает участок материн
ской хромосомы, и наоборот. Перекрест
гомологичных хромосом, сопровождаю
щийся обменом соответствующими учас
тками между иххроматидами, называется
кроссинговером. После кроссинговера
расходятся уже измененные хромосомы,
т. е. с другим сочетанием генов. Являясь
процессом закономерным, кроссинговер
приводит каждый раз к обмену разными
по величине участками и обеспечивает
эффективную рекомбинацию материала
хромосом в гаметах.
К концу профазы хромосомы в бива
лентах, сильно спирализуясь, укорачива
ются. Как и в митозе, в конце профазы I
разрушается ядерная оболочка и начина
ет формироваться веретено деления.
В м е т а ф а з е I завершается форми
рование веретена деления. Его нити
прикрепляются к центромерам хромосом,
объединенных в биваленты таким обра
зом, что от каждой центромеры идет
лишь одна нить к одному из полюсов
клетки. В результате нити, связанные с
центромерами гомологичных хромосом,
устанавливают биваленты в плоскости
экватора веретена деления.
В а н а ф а з е I гомологичные хромо
сомы разделяются и расходятся к полю
сам клетки. При этом к каждому полюсу
отходит гаплоидный набор хромосом,
состоящих из двух хроматид.
В т е л о ф а з е I у полюсов веретена
деления собирается одиночный, гаплоид
ный набор хромосом, в котором каждый
вид хромосом представлен уже не парой,
а одной хромосомой, состоящей из двух
хроматид. В короткой по продолжитель
ности телофазе I восстанавливается
ядерная оболочка, после чего материн
ская клетка делится на две дочерние.
Таким образом, образование бива
лентов при конъюгации гомологичных
хромосом в профазе I мейоза создает ус
ловия для последующей редукции числа
хромосом. Формирование гаплоидного
набора в гаметах обеспечивается рас
хождением в анафазе I не хроматид, как в
митозе, а гомологичных хромосом, кото
рые ранее были объединены в бива
ленты.
Второе мейотическое деление следу
ет сразу за первым и сходно с обычным
митозом (поэтому его часто называют
митозом мейоза), но в отличие от митоза
клетки, вступающие в него, имеют гапло
идный набор хромосом. П р о ф а з а II
непродолжительная. В м е т а ф а з е II
снова образуется веретено деления, хро
мосомы выстраиваются в экваториаль
ной плоскости и центромерами прикреп
ляются к микротрубочкам веретена деле
ния. В а н а ф а з е II осуществляется
разделение их центромер, и каждая хроматида становится самостоятельной хро
мосомой. Отделившиеся друг от друга до
черние хромосомы направляются к полю
сам веретена деления. В т е л о ф а з е II
завершается расхождение сестринских
хромосом к полюсам и наступает деление
клеток: из двух гаплоидных клеток обра
зуются четыре гаплоидные дочерние
клетки. Таким образом, в результате мей
оза из одной диплоидной клетки образу
ются четыре клетки с гаплоидным набо
ром хромосом.
Редукционное деление является как
бы регулятором, препятствующим непре
рывному увеличению числа хромосом при
слиянии гамет. Не будь такого механиз
ма, при половом размножении число хро
мосом удваивалось бы в каждом новом
поколении. Иными словами, благодаря
мейозу поддерживается определенное и
постоянное число хромосом во всех поко
лениях каждого вида растений, живот
ных, протистов и грибов.
Другое важное значение мейоза за
ключается в обеспечении чрезвычайного
разнообразия генетического состава га
мет в результате как кроссинговера, так и
различного сочетания отцовских и мате
ринских хромосом при их расхождении в
анафазе I. Это обеспечивает появление
разнообразного и разнокачественного
потомства при половом размножении ор
ганизмов.
1. Что такое мейоз? 2. Каковы отличия мейоза
от митоза? 3. Какие события в мейозе обеспечива
ют уменьшение числа хромосом в гаметах вдвое по
сравнению с соматическими клетками? 4. Что такое
конъюгация? В чем биологическое значение этого
процесса? 5. Какие механизмы обусловливают ге
нетическое разнообразие гамет, образуемых орга
низмом? 6. Что такое кроссинговер, когда он проис
ходит и каково его биологическое значение?
7. В чем заключается биологическое значение мей
оза как особой формы клеточного деления?
33. Клетка — структурная
и функциональная единица жизни
Все системы клетки взаимосвязаны и
функционируют как единое целое
(рис. 63, с. 104). Гетеротрофные клетки
получают углеводы извне, а автотрофные
создают их в хлоропластах путем фото
синтеза. Большая часть углеводов рас
щепляется с целью высвобождения энер
гии — аэробно или реже анаэробно, т. е.
путем окисления (дыхания) сначала по
пути гликолиза, а затем в цикле Кребса и
другими путями. Получаемая энергия
связывается в форме АТФ, причем боль
шая часть — в цепи переноса электронов
(цепи дыхания), которая, как и цикл
Кребса, локализована в митохондриях.
Энергию АТФ клетка использует на раз103
личные жизненные процессы — синтез,
разрушение, движение и т. д.
Глюкоза и другие углеводы использу
ются также для биосинтеза полисахари
дов, которые в форме гликолипидов и
гликопротеинов включаются в гликокаликс (у животных), в форме гемицеллю
лозы и пектиновых веществ — в клеточ
ную стенку растений, в форме хитина —
в клеточную стенку грибов. Целлюлоза
оболочек растительных клеток синтези
руется в плазмалемме или в самой кле
точной стенке. Автотрофные клетки пе
редают большую часть синтезируемых
ими углеводов гетеротрофным клеткам, в
основном в виде сахарозы.
Растительные клетки сами синтезиру
ют все аминокислоты, входящие в состав
белков: происходит это в хлоропластах.
Синтез некоторых аминокислот может
осуществляться в митохондриях и цито
плазме, в том числе и в животных клет
ках. Однако часть аминокислот (незаме
нимых) животным клеткам приходится
получать из окружающей среды; они по
глощают и белки, в основном путем эндоцитоза, и расщепляют их затем в лизосомах до аминокислот.
Белки, в том числе и ферменты, син
тезируются на рибосомах с участием
иРНК и тРНК. Этот синтез идет главным
образом в цитоплазме, а также в хлоро
пластах и митохондриях. Из цитоплазмы
белки переходят в клеточное ядро (гисто
новые и негистоновые белки хромосом,
белки субъединиц рибосом и др.), в мито
хондрии и хлоропласты.
На рибосомах, связанных с эндоплаз
матическим ретикулумом, синтезируются
резервные и экспортные белки, которые
через комплекс Гольджи путем экзоцитоза покидают клетку, а также через глад
кий ретикулум или систему Гольджи пос
тавляют ферменты для лизосом. Клеточ
ные и резервные белки в конце концов в
результате автофагии расщепляются до
аминокислот.
Все эти и другие процессы осущес
твляются путем реализации генетической
информации, которая сосредоточена в
ядре клетки в виде молекул ДНК, а также
в пластидах и митохондриях. Там проис
ходит репликация ДНК — необходимая
предпосылка идентичного деления ука
занных органелл и клетки в целом, а так
же транскрипция, обеспечивающая по
явление различных видов РНК. На рибо
сомах при участии всех типов РНК осу
ществляется трансляция — конечная
реализация генетической информации —
биосинтез белков, посредством которых
регулируются процессы синтеза и рас
щепления веществ в клетке, синтез АТФ,
клеточный рост, подготовка и осущес
твление деления клетки и другие про
цессы.
Таким образом, клетка является наи
меньшей единицей жизни — единицей
строения, функционирования и размно
жения организмов.
Рис. 63. Взаимосвязь систем клетки: 1 — ядро, 2 — ядрышко, 3 — митохондрии, 4 — хлоропласт, 5 —
полисома, 6 — гранулярный эндоплазматический ретикулум, 7 — гладкий эндоплазматический рети
кулум, 8 — система Гольджи, 9 — пузырьки Гольджи, 10 — первичные лизосомы, 11 — фагосома,
12 — аутофагосома, 13 — цитоплазматическая мембрана, 14 — экзоцитоз, 15 — эндоцитоз, 16 —
клеточная стенка; Ф — фермент, Угл — углеводы, Лип — липиды, ЦЛК — цикл Кребса.
раздел!
Я
ОРГАНИЗМ —
БИОЛОГИЧЕСКАЯ СИСТЕМА
1
Основные свойства живых
организмов
Всякое живое тело, живое существо,
которое является реальным носителем
жизни, называется организмом (от позднелат. organizo — устраиваю, сообщаю
стройный вид). Живые организмы проис
ходят от одного зачатка: семени, споры,
зиготы и т. д. Кроме того, все живые ор
ганизмы характеризуются рядом общих
свойств и признаков.
Так, живым организмам присуще вы
сокоупорядоченное строение: их струк
турной и функциональной единицей явля
ется клетка.
Все организмы представляют собой
открытые системы, являющиеся устойчи
выми лишь при условии непрерывного
доступа к ним веществ и энергии извне.
Живые организмы извлекают, преобра
зуют и используют вещества и энергию
106
окружающей среды и возвращают в нее
продукты распада и преобразованную
энергию, например, в виде тепла. Таким
образом, для организмов характерен об
мен веществ с окружающей средой и
энергозависимость.
Живые организмы обладают способ
ностью поддерживать постоянство своего
химического состава и интенсивность об
менных процессов. Недостаток поступле
ния каких-либо питательных веществ мо
билизует внутренние ресурсы организма,
а избыток — вызывает прекращение
синтеза этих веществ. Это свойство орга
низма называется саморегуляцией.
В течение жизни организмы претер
певают ряд количественных (возрастает
число клеток, масса) и качественных
(идет дифференцировка клеток, образо
вание тканей и органов, старение) изме
нений, т. е. происходит их рост и разви
тие.
Живые организмы размножаются.
При размножении организмы воспроиз
водят себе подобных.
Воспроизведение себе подобных (са
мовоспроизведение) теснейшим образом
связано с наследственностью — способ
ностью организмов передавать свои
признаки и свойства в неизменном виде
потомкам. Это осуществляется с по
мощью носителей генетической инфор
мации — ДНК и РНК. Генетический ма
териал детерминирует возможные преде
лы развития организма, его структур,
функций и реакций на окружающую сре
ду. В то же время потомки обычно быва
ют похожи на своих родителей, а не иден
тичными им. Способность организмов
приобретать новые свойства и признаки
называется изменчивостью.
Живые организмы адаптированы к
среде обитания. Особенности строения,
функций и поведения данного организма,
соответствующие его образу жизни, на
зывают адаптациями.
Живые организмы характеризуются
раздражимостью — способностью отве
чать на определенные внешние воздей
ствия специфическими реакциями. Лю
бое изменение в окружающей среде яв
ляется раздражителем, а реакция орга
низма — проявлением раздражимости.
Сочетания раздражитель — реакция мо
гут накапливаться в виде опыта и исполь
зоваться в дальнейшем.
ГЛАВА IV
Обмен веществ и энергии
в живых организмах
34. Структурная организация
живых организмов
Для современных организмов, насе
ляющих нашу планету, характерны раз
личные формы структурной организации.
Среди них встречаются мелкие высо
кодифференцированные одноклеточ
ные организмы, тело которых представ
лено одной-единственной клеткой. Тако
вы бактерии, протесты и др. (рис. 64).
Другие организмы имеют более круп
ное, многоядерное, со сложной морфоло
гической дифференцировкой тело, не
разделенное на клетки: например водо
росль каулерпа (рис. 65, с. 108). Она
представляет собой гигантскую много ядерную клетку, расчлененную на листо
видную и корневидную части. Такая
структурная организация называется си-
1
2
Рис. 64. Одноклеточные организмы: 1 — хло
релла, 2 — хламидомонада.
фоновой. Она характерна для сифоновых
водорослей и некоторых грибов. У других
водорослей со сходной организацией
(кладофора) тело разделено поперечны
ми перегородками на многоядерные учас
тки. В этом случае говорят о сифонокладной организации.
Существуют колониальные формы,
представляющие собой объединения кле
ток, например вольвокс (рис. 66, с. 108).
Колонии возникают путем клеточного де
ления. Клетки в них образуют морфоло
гическое и функциональное единство. Их
цитоплазмы соединены между собой
плазмодесмами, вследствие чего клетки
могут координировать свои реакции.
Например, благодаря согласованному
синхронному биению жгутиков, колония
вольвокса волчкообразно перемещается.
В таких высокоорганизованных коло
ниях имеет место разделение функций
между клетками: есть вегетативные
клет ки, обеспечивающие движение и
питание, и генеративные, служащие
для размножения. Такие колониальные
организмы можно считать простейшими
многоклеточными организмами.
107
Рис. 65. Организмы с сифоновой организацией: 1 — каулерпа, 2 — кладофора.
жит тканей. Оно называется талломом
(рис. 67).
Большинство организмов являются
многоклеточными. Тело многоклеточных
организмов состоит из огромного мно
жества клеток, которые дифференциро
ваны по строению и выполняемым фун
кциям. Многоклеточные организмы, в
свою очередь, подразделяются на орга
низмы б е з и с т и н н ы х т к а н е й и ор
ганизмы с и с т и н н ы м и т к а н я м и .
Тело многоклеточных организмов без
истинных тканей не дифференцирова
но на вегетативные органы и не содер108
Талломную структурную организацию имеют
грибы, большинство водорослей и некоторые мохо
видные, например печеночные мхи. У грибов т а л ло м н ы е н и т и , или гиф ы , могут объединяться в
ложные ткани: переплетаясь, они образуют плете
ную ткань, а срастаясь,— п с ев д о п а р е н х и м у , или
п л е к т е н х и м у (рис. 68). Из такой ложной ткани у
шляпочных грибов образуются плодовые тела.
В нитчатых талломах клетки, размножаясь попе
речным делением, образуют один ряд (спирогира,
улотрикс). Иногда возникают плоские талломы,
когда клетки делятся в продольном и поперечном
направлениях (ульва). У некоторых форм появляет
ся полярная дифференцировка: у них есть растущий
конец таллома и прикрепляющийся к субстрату ба
зальный р и з о и д (улотрикс). У более высокооргани
зованных форм наблюдается ветвление таллома.
Например, таллом грибов — м и ц е л и й — чаще все
го образован ветвящимися гифами.
Тело многоклеточных организмов с
истинными тканями дифференцировано
на вегетативные органы, которые состоят
из различных тканей. Такую структурную
организацию имеют высшие растения и
большинство животных.
У высших растений, за исключением
моховидных, выделяют корень и побег
Рис. 67 Талломные организмы: 1 — ульва, 2 — макроцистис, 3 — маршанция, 4 — улотрикс.
Рис. 68. Плектенхима в ножке белого гриба.
(стебель с листьями и почками). У мо
ховидных функцию корня выполняют р и
зоиды — нитевидные вытянутые клетки
нижней части стебля.
Почти все высшие растения автотрофы, поэтому им необходима большая по
верхность, поглощающая свет. Это дос
тигается благодаря ветвлению стеблей и
значительному количеству листьев, кото
рые обычно располагаются в простран
стве таким образом, чтобы не затенять
друг друга. Развитие большой листовой
поверхности у наземных растений приво
дит к огромным потерям воды в результа
те транспирации. Эти потери постоянно
восполняются из почвы. Кроме того, с
помощью корней растения поглощают из
почвы минеральные вещества, необходи
мые для нормальной жизнедеятельности.
Чтобы обеспечить всасывание воды и ми
неральных веществ из почвы, корневая
система растений часто сильно развет
влена. Стебель выполняет опорную и
проводящую функции. На стеблях разви
ваются почки (зачатки новых побегов и
листьев)и генеративные органы (цветки
и плоды). Растения ведут прикрепленный
образ жизни, поэтому для них характерна
радиальная симметрия.
Животные ведут подвижный, редко
прикрепленный (только водные формы),
или паразитический образ жизни. Для
свободы перемещения им в отличие от
растений необходима минимальная по
верхность и компактная форма тела.
Большинству животных свойственна
109
двусторонняя (билатеральная) сим
метрия и вытянутость тела в направлении
движения.
Для прикрепленных форм характерна
радиальная симметрия.
Клетки и ткани у животных и челове
ка образуют многочисленные органы,
объединенные в системы органов, согла
сованная работа которых обеспечивает
слаженное функционирование всего ор
ганизма. Для координации и управления
функциями у высших животных есть две
взаимосвязанные системы — нервная и
эндокринная.
Прочность тела животного обеспечи
вается скелетными образованиями,
которые находятся либо на поверхности
тела (у моллюсков, членистоногих и др.),
либо внутри него (хордовые животные).
Передвижению животных в поисках пи
щи, партнера для размножения, спасения
от хищников способствует хорошо разви,тая мышечная система, которая вместе
со скелетными образованиями формиру
ет опорно-двигательную систему.
Все животные — гетеротрофы. В за
висимости от способа питания у них
сформировались разнообразные формы
ротовых аппаратов и сложная пищевари
тельная система.
Кроме того, у большинства животных
есть кровеносная, дыхательная, выде
лительная и половая системы.
У животных имеются различные ор
ганы чувств, с помощью которых они
воспринимают различные раздражения
из окружающей среды. Это органы зре
ния, слуха, обоняния, вкуса, осязания
и др.
9
I. Какие формы структурной организации жи
вых организмов существуют и в чем их принципи
альное различие? 2. Почему высокоорганизован
ные колониальные организмы (типа вольвокса) счи
таются многоклеточными организмами? 3. Какие
органы выделяют у растений, каковы их функции?
4. Перечислите органы животных, которые образу
ют кровеносную, дыхательную, пищеварительную
110
системы. 5. Почему для растений и животных, веду
щих прикрепленный образ жизни, характерна ради
альная симметрия? 6. С чем связано возникновение
билатеральной симметрии?
35. Растительные ткани
Тканью называется группа клеток,
имеющих общее происхождение, сход
ное строение и выполняющих одинако
вые функции.
Понятие «ткань» в биологии ввел в XVII в.
английский натуралист Н. Грю, который изучал
внутреннее строение растений и обнаружил, что они
состоят из переплетений сходных элементов — во
локон. По аналогии с кружевами и тканями, выра
батываемыми человеком, переплетения раститель
ных волокон Н. Грю также назвал тканями.
Микроструктуру листьев, стеблей и корней
изучал в XVII в. итальянский ученый М. Мальпиги.
Он обнаружил и достаточно подробно описал стро
ение некоторых тканей растений (например, прово
дящих тканей и их элементов).
Ткани возникли в процессе исторического раз
вития растений и животных. У растений это проис
ходило при переходе от водного образа жизни к на
земному.
Находясь в воде, растения находятся под ее за
щитой. Окружающая вода снабжает их минераль
ными веществами, предохраняет клетки от высыха
ния, поддерживает тело растений на своей повер
хности или в толще (что устраняет необходимость в
опорных системах), служит подходящей средой для
встречи гамет при половом размножении и сред
ством для распространения спор — при бесполом.
Покинув гостеприимную водную среду, растения
должны были приспособиться к жизни на суше. Для
этого им пришлось образовать ряд новых структур,
которые бы взяли на себя выполнение многочис
ленных функций, осуществлявшихся прежде водой.
Завоевание суши было, по-видимому, длительным и
трудным процессом. В процессе становления на
земных растений тело их постепенно дифференци
ровалось на органы. Корни обеспечивали мине
ральное питание и водоснабжение, а листья — воз
душное питание. У водорослей, обитающих в водной
среде, не было необходимости в специализирован
ных органах для снабжения водой и минеральными
солями, поскольку и то и другое находилось вокруг
в достаточном количестве.
Для нормального функционирования двух прос
транственно разделенных органов — корня и лис
та — у наземных растений необходимо было быст
рое передвижение воды и минеральных веществ к
листьям, а органических веществ — из листьев в
другие части растения. Это обусловило развитие
проводящих тканей.
В наземной среде у растений развилась также
сложная система покровных тканей, защищающая
их от чрезмерной потери воды, от резких колебаний
температуры и одновременно — сложный устьичный аппарат, регулирующий газообмен и испаре
ние воды.
В процессе эволюции у высших растений воз
никали механические ткани. Водоросли же, живя в
воде, не нуждаются в опоре, поскольку вода поддер
живает их тело в расправленном состоянии даже в
том случае, когда тело достигает 20 м и более в
длину.
У современных высших растений в за
висимости от уровня сложности выделя
ют до 80 (у покрытосеменных) видов тка
ней. Однако все их многообразие можно
свести к следующим основным типам:
образовательные, покровные, прово
дящие, механические и основные
ткани.
Ткани могут быть простыми или
сложными. Простые ткани состоят толь
ко из одного вида клеток и характерны
для листа и молодого корня, сложные —
из различных по строению клеток, вы
полняющих, кроме основных, и дополни
тельные функции. Например, клетки
сложной проводящей ткани ксилемы
(древесины) наряду с проводящей выпол
няют также опорную функцию.
Ткани растений делят на временные и
постоянные. Временные, или меристемы
(от греч. meristos — делящийся), состоят
из активно делящихся клеток. Часть кле
ток меристемы сохраняет способность
делиться, а часть постепенно дифферен
цируется, превращаясь в клетки различ
ных постоянных тканей.
Образовательные ткани (меристе
мы) являются эмбриональными тканями.
Рис. 69. Образовательная ткань в конусе на
растания стебля элодеи канадской.
Благодаря постоянному делению образо
вательные ткани принимают участие в
строительстве всех постоянных тканей
растения и фактически формируют его
тело.
Клетки образовательной ткани тон
костенные, с густой цитоплазмой и очень
мелкими вакуолями, с крупным, располо
женным в центре ядром (рис. 69). Они
многогранные, плотно прилегают друг к
другу и могут делиться в разных направ
лениях. Меристемы находятся на кончике
корня и стебля (верхушечные меристе
мы), у основания цветоносных побегов и
междоузлий некоторых растений, напри
мер злаков (вставочные меристемы),
внутри стеблей и корней (боковые ме
ристемы — прокамбий, камбий, проб
ковый камбий, перицикл).
Тело растения одето покровной
тканью (эпидермис, пробка, корка).
Эпидермисом покрыты однолетние стеб
ли и листья. Клетки эпидермиса сомкну
ты, их наружные стенки утолщены
(рис. 70, с. 112). Снаружи клетки эпидер
миса покрыты особой пленкой — кут и
кулой, состоящей из жироподобных ве111
Рис. 70. Эпидермальная ткань
листа ириса: 1 — клетки эпидер
миса, 2, 3 — устьице.
ществ, вырабатываемых клетками эпи
дермиса (кут ин). На эпидермисе неред
ко развиваются многочисленные волос
ки, образуется восковой налет. Все эти
приспособления надежно защищают рас
тения от излишних потерь воды.
Для сообщения растительных орга
низмов с внешней средой в эпидермисе
имеются специальные образования —
устьица. Через устьица происходит га
зообмен и выделяется вода в парообраз
ном состоянии. Устьице представляет со
бой щелевидное отверстие, которое с
обеих сторон ограничено двумя клетками
бобовидной формы. Эти клетки называ
ются замыкающими. Они могут изме
нять свою форму и регулировать ширину
устьичной щели.
Пробка (перидерма) покрывает стеб
ли и корни многолетних двудольных рас
тений и голосеменных. Ее образование
связано с деятельностью боковой образо
вательной ткани — пробкового камбия.
Оболочки клеток пробки пропитаны осо
бым жироподобным веществом — субе
рином — и не пропускают ни воды, ни
воздуха, поэтому их цитоплазма и ядро
быстро отмирают, а клетки заполняются
воздухом. У березы в клетках пробки
112
присутствует белое зернистое вещес
тво — бетулин, которое и придает ство
лам белую окраску. Эти мертвые клетки
предохраняют растения от неблагоприят
ных воздействий среды. Уже осенью пер
вого года жизни у древесных и кустарни
ковых растений все их органы, которым
предстоит зимовать, бывают покрыты
пробкой. Окраска этих частей растения
вместо зеленой становится бурой или се
рой. Это свидетельствует о том, что уже
произошла смена эпидермиса на пробку.
Весной для обеспечения газообмена и
испарения в пробке образуются разрывы,
заполненные рыхло расположенными
клетками — чечевички (рис. 71). На по
верхности стеблей чечевички выглядят
небольшими округлыми или овальными
бугорками.
Многочисленные слои пробки, форми
рующиеся в течение многих лет на стеблях
древесных растений, образуют корку.
Проводящие ткани обеспечивают пе
редвижение воды и питательных веществ
по растению. От корней к другим органам
растения передвигается вода с раство
ренными в ней минеральными солями.
Она движется по сосудам (рис. 72) —
сплошным полым трубкам, образован-
Рис. 71. Чечевичка на ветке бузи
ны: 1 — эпидермис, 2 — пробка,
3 — феллоген, 4 — феллодерма,
5 — выполняющая ткань.
— 4
Рис. 72. Сосуды и трахеиды: 1, 2 — кольчатые сосуды, 3 — 5 — спиральные, 6 —
сетчатый. 7 — лестничный, 8 — сосуд с окаймленными порами, 9 — 11 — тра
хеиды.
ным из лишенных живого содержимого
клеток, между которыми разрушены по
перечные перегородки. Такая ткань назы
вается ксилемой (древесиной). Клетки
ксилемы имеют утолщенные вторичные
оболочки, пропитанные лигнином, поэто
му сосуды, кроме проводящей функции,
выполняют также и механическую. У го
лосеменных, папоротников, хвощей и
плаунов проводящие элементы ксилемы
представлены не сосудами, а отдельными
вытянутыми одревесневшими клетками
со скошенными концами — трахеидами. Проникновение растворов из одной
113
трахеиды в другую происходит путем
фильтрации через поры. В состав ксиле
мы, кроме собственно проводящих эле
ментов, входят паренхимные клетки (дре
весинная паренхима) и клетки механи
ческой ткани.
От листьев органические вещест
ва — продукты фотосинтеза — переме
щаются по ситовидным трубкам (рис.
73). Эти трубки представляют собой ряд
вытянутых живых клеток, в которых нет
ядра. Цитоплазма расположена в цен
тральной части, а их поперечные перего
родки имеют сквозные округлые отвер
стия (наподобие сита), через которые в
соседние клетки проходят тонкие тяжи
цитоплазмы.
Благодаря этому соседние клетки со-
А
Б
Рис. 73. Ситовидные трубки и клетки-спутни
цы: А — продольный разрез, Б — поперечный
разрез через ситовидную перегородку: 1 —
клетки-спутницы, 2 — поперечная перегородка
ситовидной трубки, 3 — поры.
114
общаются друг с другом. Ситовидные
трубки проходят по всей длине растения,
что обеспечивает передвижение органи
ческих веществ. Рядом с ситовидными
трубками залегают клетки-спутницы.
Они имеют типичное для растительных
клеток строение и, очевидно, играют
вспомогательную роль, помогая передви
жению органических веществ. Ситовид
ные трубки вместе с клетками-спутницами (лубяной паренхимой и лубяными
волокнами) образуют сложную проводя
щую ткань флоэму (лу б ).
Ксилема и флоэма находятся в тесном
взаимодействии друг с другом и обычно
образуют в органах растения особые тка
невые комплексы — проводящие пучки.
Прочность растению придают меха
нические ткани. Растению приходится
оказывать сопротивление ветру и дождю,
выдерживать тяжесть ветвей и листьев.
Механические ткани составляют как бы
каркас растения, который заполнен мяг
кими и тонкостенными клетками осталь
ных тканей. Для клеток механических
тканей,, обеспечивающих их прочность и
упругость, характерно мощное утолще
ние и одревеснение оболочек, тесное
смыкание между клетками.
В зависимости от формы клеток, их
строения, физиологического состояния и
способа утолщения клеточных оболочек
различают два вида механической ткани:
колленхиму и склеренхиму.
Колленхима представлена живыми
паренхимными клетками с неравномерно
утолщенными оболочками, что особенно
важно для укрепления молодых растущих
органов. Будучи первичными, клетки кол
ленхимы легко растягиваются и практи
чески не мешают удлинению той части
растения, в которой находятся. Обычно
колленхима располагается отдельными
тяжами или непрерывным цилиндром под
эпидермисом молодого стебля и череш
ков листьев, а также окаймляет жилки в
листьях двудольных.
Склеренхима состоит из вытянутых
клеток с равномерно утолщенными, часто
одревесневшими оболочками, содержи
мое которых отмирает на ранних стадиях.
Оболочки склеренхимных клеток облада
ют высокой прочностью, близкой к проч
ности стали. Эта ткань широко пред
ставлена в вегетативных органах назем
ных растений и составляет их осевую
опору.
Различают два типа склеренхимных
клеток: в о л о к н а и с к л е р е и д ы . Во
локна — это длинные тонкие клетки,
обычно собранные в тяжи или пучки
(например, лубяные или древесинные во
локна) (рис. 74).
Склереиды — это округлые мертвые
клетки с очень толстыми одревесневши
ми оболочками. Ими образованы семен
ная кожура, скорлупа орехов, косточки
вишни, сливы, абрикоса; иногда они при
дают мякоти груш крупитчатый характер.
Основная ткань, или паренхима, —
Рис. 74. Волокна: а — древесинные, б — лубя
ные.
составляет основную часть тела расте
ния. Клетки паренхимы изодиаметричны
(длина клетки равна ширине) или имеют
таблитчатую форму (длина не более чем в
два раза превышает ширину). Паренхи
ма — одна из немногих тканей, которая
в зависимости от расположения в рас
тении может выполнять различные
функции: хлорофиллоносную (хлоренхима), запасающую, воздухоносную (аэрен
хима).
Хлорофиллоносная паренхима, или
ассимиляционная ткань, образуется в
зеленых листьях и стеблях растений и вы
полняет функцию фотосинтеза. В клетках
этой ткани в большом количестве содер
жатся хлоропласты.
В клетках запасающей паренхимы
откладываются в твердом или растворен
ном виде запасные питательные вещес
тва (крахмал, сахара, белки), которые
впоследствии используются растением в
процессе жизнедеятельности. У расте
ний, периодически испытывающих недос
таток воды, в запасающих тканях стебля
(кактусы, молочаи) или листьев (алоэ,
очитки) накапливается вода. Большое ко
личество запасающей паренхимы имеет
ся в стеблях древесных растений, корне
вищах, клубнях, луковицах, корнепло
дах, плодах. Иногда в запасающих
тканях накапливаются продукты обме
на веществ: смолы, органические
кислоты, кристаллы щавелевокислого
кальция.
У высших растений, обитающих в во
де (кувшинки, кубышки, рдесты и др.),
развивается особый тип основной тка
ни — воздухоносная паренхима, или
аэренхима. Ее основная функция —
обеспечение нормального газообмена в
теле растения в условиях пониженной
аэрации. Клетки воздухоносной паренхи
мы могут иметь разнообразную форму
(округлую, звездчатую) и располагаться
рыхло, так что между ними образуются
крупные межклетники, по которым цир
кулируют газы.
115
I
1. Чем было обусловлено возникновение тка
ней у высших растений? 2. Какую функцию выпол
няет эпидермис и как это отражено в строении клет
ки? 3. Почему, на ваш взгляд, происходит замена
эпидермиса перидермой? 4. Почему стволы берез
белые? 5. Каковы принципиальные различия в
строении ксилемы и флоэмы? 6. В каких условиях у
растений формируется аэренхима?
36. Животные ткани
У животных различают четыре основ
ных типа тканей: эпителиальную (по
кровную), соединительную, мышечную и
нервную. В состав животных тканей, кро
ме клеток, часто входит межклеточное
вещество, продуцируемое клетками.
Эпителиальная ткань, или эпите
лий, — является пограничной тканью и
покрывает организм снаружи, а также
выстилает полости внутренних органов.
Через слой эпителия покровов многих
животных происходит газообмен между
организмом и окружающей средой. Кро
ме того, он защищает животное от про
никновения извне вредных веществ и
микроорганизмов и предохраняет его от
потери веществ, необходимых для жизне
деятельности (например, воды). В неко
торых органах клетки эпителия выраба
тывают тот или иной секрет. Эпителий,
содержащий такие секреторные клетки,
называется железистым.
Клеточные элементы эпителия обыч
но располагаются в виде пласта, состоя
щего из клеток различной формы. Иногда
клетки плотно прилегают друг к другу, в
других случаях между ними имеются тон
кие ходы, по которым циркулирует ткане
вая жидкость. Межклеточное вещество,
как правило, неразвито. Клетки эпителия
почти всегда имеют одно ядро. В зависи
мости от числа слоев клеток в пласте
эпителий бывает однослойным и много
слойным. Однослойный эпителий быва
ет плоский, кубический, цилиндрический
(столбчатый) и др. (рис. 75).
116
Рис. 75. Типы эпителиальной ткани: а — плос
кий эпителий, б — кубический, в — стол б ча
ты й , г — ресничный (мерцательный), g — сен
сорный (кл е тки из вы стилки носовой полос
т и ), е — ж елезисты й: 1 — реснички, 2 — чув
ствительные волоски, 3 — бокаловидные
клетки.
П л о с к и й эпителий состоит из кле
ток, имеющих многоугольную форму, и
образует поверхностный слой кожи и
выстилку ротовой полости, пищевода,
мочевого пузыря. Почти у всех беспозво
ночных животных встречается только од
нослойный эпителий. У человека и выс
ших позвоночных животных плоский эпи
телий обычно состоит из нескольких сло
ев клеток, накладывающихся друг на дру
га. Такая ткань называется многослой
ным эпителием. В многослойном эпите
лии клетки основного слоя имеют обычно
кубическую или цилиндрическую форму,
вышележащие клетки несколько упло
щены, а поверхностные становятся плос
кими. Нередко наружные клетки ороговевают и отмирают. Из ороговевших кле
ток эпителия у позвоночных животных
образуется поверхностный слой кожи,
рога, копыта, ногти и т. д. У большинства
беспозвоночных животных эпителий по
кровов выделяет на своей поверхности
плотную, а иногда твердую оболочку —
кутикулу.
К у б и ч е с к и й эпителий состоит из
кубовидных клеток. Он выстилает почеч
ные канальцы у человека.
Клетки
цилиндрического
( с т о л б ч а т о г о ) эпителия имеют про
долговатую форму и напоминают столби
ки или колонны. Цилиндрическим эпите
лием выстланы желудок и кишечник
млекопитающих.
Столбчатые клетки на своей повер
хности могут иметь мельчайшие цито
плазматические выросты — реснички,
ритмическое сокращение которых про
двигает какие-либо, находящиеся у по
верхности клеток, частички в одном на
правлении. Такая ткань называется рес
ничным эпителием. Ресничный эпителий
выстилает большую часть дыхательных
путей. Ресничным эпителием покрыта
вся поверхность тела плоских червей, по
верхность жабер у моллюска беззубки
и т. д.
Ч у в с т в и т е л ь н ы й ( с е н с о р н ый )
эпителий состоит из клеток, которые вос
принимают внешние раздражения. На
пример, в носовой полости имеются
клетки, которые воспринимают запахи.
Клетки ж е л е з и с т о г о эпителия вы
рабатывают различные вещества — сек
реты, например молоко, ушную серу, пот,
пищеварительные ферменты.
Таким образом, эпителиальная ткань
характерна для большинства животных
и человека, играя в их жизни важную
роль, выполняя функции защиты, вса
сывания, секреции и восприятия раздра
жений.
Соединительная ткань. Это рыхлые
объединения клеток, которые участвуют
в образовании связок и прослоек между
органами, а также внутреннего скелета
многих животных: некоторые виды этой
ткани (кровь, лимфа) осуществляют пе
ренос веществ по всему телу.
Строение различных видов соедини
тельной ткани разнообразно, но сходство
в том, что их клетки выделяют межкле
точное ( основное) вещество, которое
в некоторых видах соединительной ткани
мягкое и может содержать коллагено
вые, или эластические, волокна, рас
положенные беспорядочно, параллельно
друг другу (в сухожилиях) или крест на
крест (в фасциях). В состав коллагеновых
волокон входит фибриллярный белок —
коллаген, который отличается высокой
прочностью и после гидратации превра
щается в желатин. При вываривании
коллагеновые волокна дают клей.
Эластические волокна более тонкие.
Они содержат белок эластин.
Для других видов соединительной тка
ни характерно прочное и плотное меж
клеточное вещество.
Различают следующие основные виды
соединительной ткани: рыхлая волокнис
тая, плотная волокнистая, хрящевая и
костная.
Рыхлая волокнистая ткань слага
ется из редко расположенных звездчатых
клеток, переплетающихся волокон и тка
невой жидкости, заполняющей проме
жутки между клетками и волокнами (рис.
76,а, с. 118). Такая ткань находится
обычно в прослойках между органами.
Разновидностью рыхлой соединитель
ной ткани является жировая ткань, ко
торая состоит из накапливающих капель
ки жира клеток. Жировая ткань входит в
состав подкожной клетчатки.
Плотная волокнистая ткань сос
тоит в основном из пучков коллагеновых
волокон. Аморфного межклеточного ве
щества мало, немногочисленные клетки
расположены между пучками волокон.
Такое строение данной ткани придает
прочность структурам, в которые она
входит, и позволяет им выдерживать
большие нагрузки. Плотная волокнистая
ткань образует связки, сухожилия, глубо
кие слои кожи (дерму) позвоночных жи
вотных и человека.
117
Рис. 76. Типы соединительной ткани: а — волокнистая, б — хрящ, в — кость, г — микроскопическая
структура кости.
Кроме того, плотной соединительной
тканью являются: твердая оболочка го
ловного и спинного мозга, выстилающая
изнутри череп и позвоночный канал у че
ловека; надкостница, покрывающая кос
ти; плотные слои фасций, отделяющих
отдельные мышцы друг от друга; фиброз
ный перикард и склера(одна из оболочек
глазного яблока).
Хрящевая ткань состоит из округ
лых или овальных клеток, лежащих в
капсулах в количестве от одной до
четырех штук среди мощно развитого
плотного и твердого межклеточного ве
щества, содержащего тонкие волокна
(рис. 76, б). Межклеточное вещество
этой ткани эластичное, гибкое, его мож
но резать. В нем нет кровеносных сосу
дов, и питание хряща осуществляет
ся диффузно из кровеносных сосудов
надхрящницы, покрывающей хрящ сна
ружи.
Хрящевая ткань входит главным об
разом в состав суставов, развивающихся
костей скелета многих животных и чело
века.
Хрящ образует хрящевую часть ре
бер, носа, ушной раковины, гортани, тра
хеи и бронхов.
Разновидность хрящевой ткани —
эластический хрящ (в его состав входят
118
многочисленные эластичные волокна)
встречается в ушной раковине, надгор
таннике и хрящевой части слуховой (ев
стахиевой) трубы. При сжатии или изги
бе эти структуры легко изменяют свою
форму, быстро возвращаясь в исходное
положение.
Костная ткань отличается тем, что
в ее межклеточном веществе откладыва
ются соли кальция, придающие ей твер
дость. Кроме того, она содержит гаверсо
вы каналы с кровеносными сосудами и
нервами. Костные клетки (остеоциты)
располагаются концентрическими круга
ми вокруг гаверсовых каналов и связаны
между собой плазматическими отростка
ми (рис. 76, в, г). Костная ткань свой
ственна позвоночным животным и чело
веку, у которых она образует кости.
Мышечная ткань является основным
элементом мышц животных и человека.
Ее клетки способны к обратному сокра
щению под действием различных раздра
жителей, что обусловливает способность
к движению. Мышечная ткань слагается
из отдельных мышечных волокон, в
которых расположены тончайшие со
кратительные волоконца — миофибриллы.
Различают три типа мышечной ткани:
поперечнополосат ую (скелет ную ),
сердечную и гладкую. Как следует из
названий, скелетные мышцы прикрепля
ются к костям скелета; сердечная мышца
образует основную массу ткани сердца, а
гладкие мышцы — мышечные слои внут
ренних органов (пищеварительного трак
та, кровеносных сосудов, матки, мочево
го пузыря и др.). У низших многоклеточ
ных животных гладкая ткань образует
всю массу их мышц.
Скелетные мышцы характеризуются
поперечной исчерченностью и состоят из
пучков, образуемых множеством много
ядерных волокон диаметром от 0,01 до
0,1 мм и длиной от 1 до 40 мм. Эти волок
на в свою очередь состоят из более тон
ких мышечных фибрилл (миофибрилл)
(рис. 77, а).
Сердечная мышца состоит также из
поперечно исчерченных мышечных кле
ток, но обладает иными свойствами, что
связано с ее структурой — волокна рас
положены не параллельным пучком, а
ветвятся (рис. 77, б). Кроме того, сосед
ние волокна соединены между собой ко
нец в конец. Благодаря этому все волок
на сердечной мышцы образуют единую
сеть, хотя каждое волокно заключено в
отдельную мембрану. Между волокнами,
соединенными своими концами, образу
ется множество щелевых контактов, ко
торые позволяют нервным импульсам
поступать от одного волокна к другому.
Поэтому сокращение сердечной мышцы в
одном участке быстро распространяется
по всей мышечной массе, обеспечивая
одновременное сокращение, а затем рас
слабление.
Гладкие мышцы лишены поперечной
исчерченности, так как у них отсутствует
упорядоченное расположение нитей ак
тина и миозина (рис. 77, в). Клетки глад
ких мышц веретенообразные, длиной
около 0,1 мм, с одним ядром в центре.
Источником энергии для мышечного сокраще
ния служат АТФ, креатинфосфат, аргининфосфат, а
также — при интенсивной мышечной работе — за-
j : j к; •1'; р"'
. И :•и ;; \ : •. •-1;*.'i f : к п
fj?* '
::
’ ■
б
Рис. 77 Типы мышечной ткани: а — поперечно
полосатая, б — сердечная, в — гладкая.
пасы углеводов в форме гликогена и жирные кис
лоты.
Аргининфосфат и креатинфосфат выполняют
роль своеобразных аккумуляторов химической
энергии, которые используются для быстрого фос
форилирования АДФ при ресинтезе АТФ во время
энергичного мышечного сокращения. В мышцах
позвоночных и в организме некоторых беспозвоноч
ных осуществляется следующая реакция трансфос
форилирования:
фермент
трансфосфорилаза
креатинфосфат + АДФ ----- ^-креатин + АТФ.
У позвоночных обнаружен только креатинфос
фат, а аргининфосфат широко распространен в
мышцах и тканях беспозвоночных.
Сокращение скелетных мышц осу
ществляется произвольно посредством
соматических нервов в отличие от сер
119
дечной и гладких мышц, управляемых ве
гетативной нервной системой.
Следует отметить, что скелетные
мышцы позвоночных состоят из волокон
по меньшей мере двух типов — «быс
трых» и «медленных». «Быстрые» во
локна содержат меньше миоглобина
(транспортный белок, который переносит
кислород в мышцах), поэтому называют
ся б е л ыми , а «медленные», с большим
количеством миоглобина,— к р а с н ы
ми. Мышца может состоять только из
«быстрых», только из «медленных» во
локон или из тех и других одновременно.
Нервная ткань выполняет функции
восприятия, переработки, хранения и пе
редачи информации, поступающей как из
окружающей среды, так и изнутри орга
низма. Деятельность нервной системы
обеспечивает реакцию организма на раз
личные раздражения и координацию ра
боты всех его органов.
Нервные клет ки — нейроны —
обычно имеют звездчатую или веретено
видную форму (рис. 78). Тело нейрона
снабжено одним или несколькими ветвя
щимися отростками. В этом случае, как
правило, короткие, толстые, сильно раз
ветвленные отростки (дендриты) прово
дят возбуждение к телу нейрона, а один
очень длинный отросток (нервное во
локно, или аксон) — от тела нейрона. В
большинстве случаев тела нейронов рас
полагаются в нервных центрах серого ве
щества головного и спинного мозга, и на
периферии в ганглиях. Аксоны могут
объединяться в тонкие пучки, а те в свою
очередь — в более толстые пучки, обра
зуя нерв. Афферентные (чувствитель
ные) нервы проводят возбуждение от пе
риферии к центру, а эфферентные (эффекторные) — в обратном направле
нии. Смешанные нервы содержат как
афферентные, так и эфферентные волок
на. На разветвленных концах аксонов на
ходятся концевые пуговки. Это места
контактов (синапсы) для передачи воз
буждения на следующий нейрон или иную
120
Рис. 78. Схема строения нервной клетки: 1 —
тело клетки, 2 — клеточное ядро, 3 — дендри
ты, 4 — голый участок аксона, 5 — миелиновая
оболочка, 6 — неврилемма, 7 — коллатераль,
8 — ядро неврилеммы, 9 — часть аксона выре
зана, 10 — перехват Ранвье, 11 — аксон, 12 —
концевые разветвления.
клетку (например, железистую или мы
шечную). Нервные клетки, связанные
синапсами, образуют нервную систему,
в которую входят также глиальные
клетки (опорные и защитные элемен
ты) и соединит ельнот канные обо
лочки.
1.
Какие функции выполняет эпителий и како
вы в связи с этим особенности строения его клеток?
2. Каково происхождение рогов, копыт, ногтей?
3. Что принципиально отличает соединительную
ткань от других тканей животных? 4. Чем различа
ются разные типы мышечной ткани? 5. Вспомните,
каков механизм передачи нервного импульса от ре
цептора на эффектор.
37. Питание как способ получения
вещества и энергии
из окружающей среды
Обязательным условием существова
ния любого организма является постоян
ный приток питательных веществ и пос
тоянное выделение конечных продуктов
обмена. Питательные вещества исполь
зуются организмами в качестве источни
ка энергии и атомов химических элемен
тов (прежде всего атомов углерода), из
которых строятся либо обновляются все
структуры организма. Для создания орга
нических веществ (построения присущих
данному виду клеточных структур), под
держания высокоупорядоченной органи
зации и на другие цели организмам тре
буется энергия. Все живые организмы в
связи с этим можно рассматривать как
активно работающие машины, которые
не могут жить (работать) без постоянно
го притока энергии, необходимой для
синтеза веществ и восстановления кле
ток и тканей; активного транспорта ве
ществ внутри клетки, из одних клеток в
другие, из одной части организма в дру
гую; передачи нервных импульсов; дви
жения организмов; поддержания посто
янной температуры тела (у птиц и млеко
питающих); биолюминесценции (у свет
ляков, некоторых морских животных);
электрических разрядов (у скатов, элек
трического угря) и т. д.
Энергия, необходимая для осущес
твления процессов жизнедеятельности
организмов, выделяется в процессе кле
точного дыхания. При этом недоступная
для выполнения работы энергия хими
ческих связей органических веществ
превращается в доступные для непосред
ственного использования формы —
энергию макроэргических связей АТФ и
энергию мембранного потенциала.
Процесс приобретения организмами
вещества и энергии называется питани
ем. Главным элементом, из которого жи
вые организмы строят органические ве
щества, является углерод. В зависимости
от того, какой источник атомов углерода
потребляют организмы из окружающей
среды, их можно разделить по типу пита
ния на несколько групп.
Как уже говорилось, одни организмы
в качестве источников атомов углерода
для синтеза органических веществ по
требляют диоксид углерода (С 0 2). Такой
тип питания называется автотрофным
(от греч. autos— сам, trophe — питание),
а организмы автотрофными, или автотрофами. Другие организмы исполь
зуют в качестве источников атомов угле
рода и других элементов готовые органи
ческие вещества. Такой тип питания на
зывается гетеротрофным (от греч. heteros — иной, разный и trophe — пита
ние), а организмы, питающиеся готовой
органической пищей, — гетеротроф
ными, или гетеротрофами.
Для осуществления процессов синте
за органических веществ также необхо
дима энергия. Те организмы, которые
осуществляют первичный синтез органи
ческих веществ, используя энергию сол
нечного света, называются фототрофами (от греч. photos — свет и trophe —
питание), а те, которым для этого нужна
химическая энергия, — хемотрофами.
Для фототрофов характерно наличие в их
121
клетках пигментов (в том числе какойлибо формы хлорофилла). Пигменты
поглощают солнечную энергию и способ
ствуют превращению ее в химическую
энергию; за счет второй и осуществляет
ся синтез органических веществ. Таким
образом, по типу питания — источнику
атомов углерода и источнику энергии,
используемой в создании органических
веществ, все живые организмы делятся
на четыре группы: фотоавтотрофы, фотогетеротрофы, хемоавтотрофы и хемогетеротрофы.
К ф от оавт от роф ам относятся все
зеленые растения, некоторые протесты,
цианобактерии, зеленые и пурпурные
бактерии. Эти организмы осуществляют
фотосинтез — синтезируют первичные
органические вещества из неорганичес
ких с использованием солнечной энергии.
Хемоавт отрофные организмы осу
ществляют синтез органических веществ,
используя диоксид углерода и химичес
кую энергию. В отличие от фотоавтотрофов, данная группа организмов использу
ет энергию окисления некоторых неорга
нических соединений — сероводорода,
серы, аммиака, водорода и др. К этой
группе относятся бактерии, осуществля
ющие хемосинтез, — нитрифицирующие,
бесцветные серобактерии, железобакте
рии, водородные и некоторые другие.
Некоторые бактерии, например пур
пурные несерные, содержат бактериюхлорофилл и способны фотосинтезиро
вать, но углерод при этом берут из орга
нических источников. Такие организмы
называются фотогетеротрофными.
Хемогет ерот роф ы используют в
пищу готовые органические вещества,
которые дают гетеротрофам энергию, не
обходимую для их жизнедеятельности, а
также служат источником специфических
атомов и молекул, идущих на поддержа
ние и возобновление клеточных структур
и образование протопласта в процессе их
роста. Вместе с пищей гетеротрофы по
лучают также коферменты и витамины,
122
которые не синтезируются в их организ
ме. К хемогетеротрофам относятся все
животные, грибы, часть протестов, боль
шинство бактерий, некоторые паразити
ческие растения.
Самые важные группы организмов —
фотоавтотрофы и хемогетеротрофы.
Именно фотоавтотрофы, улавливая сол
нечную энергию, создают запасы органи
ческого вещества на Земле. Хемогете
ротрофы потребляют органические ве
щества, создаваемые фотоавтотрофами,
и всецело зависят от них. В то же время,
расщепляя органические вещества в про
цессе своей жизнедеятельности, они
превращают их в простые неорганичес
кие вещества и способствуют таким об
разом круговороту веществ в приро
де, что является одним из обязательных
условий существования жизни на Земле.
Поскольку фотоавтотрофы и хемоге
теротрофы составляют подавляющее
большинство среди всех организмов, к
ним часто применяют определение «ав
тотрофные» и «гетеротрофные».
Способы добывания и поглощения
пищи у гетеротрофных организмов весь
ма разнообразны, но путь превращения
питательных веществ у большинства из
них сходен. По существу, это превраще
ние состоит из двух процессов: расщеп
ления больших и сложных молекул
(полимеров) на более простые (мономе
ры) — переваривания, всасывания прос
тых молекул и их транспорта ко всем
клеткам и тканям организма.
Известны четыре способа гетеро
трофного питания: голозойный, сапротрофный, симбиотический и паразити
ческий.
Голозойный тип питания (от греч.
holos — целый, полный, zoo — живот
ное) характерен для животных и насеко
моядных растений. Свободноживущие
голозойные организмы обладают специ
альным пищеварительным трактом,
Голозойный тип питания состоит из
следующих процессов: поглощение пи
щи, ее переваривание (ферментативное
расщепление), всасывание, транспорт
простых органических веществ к клеткам
и тканям, ассимиляция (использование
всосавшихся молекул для получения
энергии и синтеза собственных органи
ческих веществ) и экскреция (выделение
из организма в окружающую среду непе
реваренных остатков пищи).
Сапротрофный тип питания (от греч.
sapros — гнилой) характерен для грибов
и большинства бактерий. Сапротрофы
питаются мертвым или разлагающимся
органическим материалом, выделяя фер
менты непосредственно на него. Раство
римые конечные продукты такого внеорганизменного переваривания всасывают
ся и ассимилируются автотрофами.
Симбиотрофный тип питания харак
терен для симбиотических организмов.
Симбиоз (от греч. symbiosis — сожи
тельство) — одна из форм тесного со
вместного существования двух или более
видов, которое, как правило, стало необ
ходимым и полезным для обоих партне
ров (симбионтов). Существует две раз
новидности симбиоза — мутуализм и
комменсализм, которые подробнее будут
рассмотрены дальше.
При паразитическом типе питания
организмы получают органические ве
щества от организма хозяина, причиняя
последнему существенный вред. Парази
тический способ питания характерен для
некоторых бактерий (дифтерийная и
столбнячная палочки, стафилококк, хо
лерный вибрион и др.), протистов (маля
рийный плазмодий, дизентерийная аме
ба, лейшмании, трихомонады, лямблии),
животных (сосальщики, ленточные чер
ви, аскариды и др.), высших растений
(повилика европейская, заразиха, Пет
ров крест и др.).
Существует группа организмов, кото
рых нельзя всецело отнести по типу пита
ния ни к автотрофам, ни к гетеротрофам.
В зависимости от условий обитания они
могут себя вести по-разному. На свету
это типичные автотрофы, но, если имеет
ся источник органического углерода, они
ведут себя как гетеротрофы. Эту группу
составляют автогетеротрофные протисты (в первую очередь эвгленовые).
Таким образом, по типу питания по
давляющее большинство растений (за
исключением растений-паразитов и насе
комоядных) являются автотрофами, а все
животные и грибы — гетеротрофами.
Среди протистов есть автотрофы, гете
ротрофы и автогетеротрофы, среди бак
терий — гетеротрофы и автотрофы.
Следует отметить, что все живые ор
ганизмы нуждаются не только в органи
ческих веществах. Для их нормального
существования необходим целый ряд не
органических веществ: вода, кислород,
минеральные соли, содержащие натрий,
калий, кальций, магний, азот, фосфор,
серу, железо, а также бор, марганец,
цинк, молибден, медь, кобальт и др.
?;
1. На какие цели организмы расходуют энер
гию? 2. Каковы источники энергии, потребляемой
организмами? 3. Какова форма энергии, доступной
для непосредственного использования клетками?
4. На какие цели живые организмы расходуют по
требляемые из окружающей среды вещества?
5. Охарактеризуйте фотоавтотрофное и хемогетеротрофное питание. 6. Чем различаются способы
хемогетеротрофного питания?
38. Поступление и транспорт
питательных веществ
в живых организмах
Бактерии, протисты и грибы погло
щают все питательные вещества, необхо
димые им для поддержания жизни, из
окружающей среды путем всасывания,
диффузии, облегченной диффузии или
активного переноса. Причем грибы и
гетеротрофные бактерии выделяют в
окружающую среду гидролитические фер
менты, которые расщепляют сложные ор
ганические вещества на более простые.
Последние всасываются клетками бакте123
рий и грибов. В клетки этих организмов
диффузно поступают также молекулы во
ды, кислорода и других веществ.
Некоторые гетеротрофные протесты
поглощают питательные вещества с по
мощью фаго- и пиноцитоза. Например,
амебы, которые питаются мелкими про
тестами, бактериями, мельчайшими час
тичками мертвого органического вещес
тва, обтекают пищевые частички или од
ноклеточные организмы псевдоподиями и
включают их в цитоплазму с небольшим
количеством воды. Образуется пищева
рительная вакуоль. Эта вакуоль окру
жается множеством мельчайших лизосом, которые в конечном итоге сливают
ся с ее мембраной и изливают свое содер
жимое в вакуоль. Образуется вторичная
лизосома, в которой и осуществляется
расщепление сложных органических ве
ществ на более простые. Среди вливаю
щихся в вакуоль ферментов содержатся
различные гидролазы. Эти ферменты
поступают в вакуоль в разное время, по
этому их расщепляющее действие разде
лено во времени, а не в пространстве, как
это наблюдается у животных. Во время
расщепления органических веществ из
меняется pH содержимого вакуоли от
кислой до слабощелочной (рис. 79). Об
разовавшиеся в результате расщепления
более простые органические вещества с
помощью диффузии или облегченной
диффузии переносятся в цитоплазму, где
и используются.
Автотрофные протесты поглощают
все вещества из окружающей среды так
же диффузным путем.
В тело растений вода и минеральные
вещества поступают из почвы посред
ством всасывания их клетками корня.
Всасывание воды и растворенных в ней
минеральных веществ происходит глав
ным образом в области кончика корня,
где имеется множество мелких корневых
волосков.
Независимо от продолжительности жизни рас
тения кончик корня все время растет, хотя по мере
старения корни становятся толще и снаружи покры-
Рис. 79. Схема пищеварения у инфузории: 1 — порошица,
2 — pH 6, 3 — -рот, 4 — глотка, 5 — pH 1, 6 — сократитель
ная вакуоль, 7 — пищеварительная вакуоль (pH 4), 8 —
ротовая впадина (перистом). Белой стрелкой показан т о к
воды.
124
Рис. 80. Схема строения кончика корня
пшеницы: 1 — корневой чехлик, 2 — зона
деления, 3 — зона роста, 4 — зона вса
сывания, 5 — зона проведения, 6 — кор
невые волоски.
ваются пробковым слоем. В кончике корня можно
выделить несколько зон (рис. 80). На верхушке кор
ня размещается зона деления, где сосредоточена
верхушечная меристема. Благодаря делению кле
ток верхушечной меристемы корень постоянно рас
тет в длину. Кончик корня покрыт корневым чехликом, который защищает нежные клетки апикальной
(верхушечной) меристемы от механических по
вреждений и способствует продвижению корня в
почве. Клетки чехлика не прочно связаны друг с
другом и легко отделяются — слущиваются, благо
даря чему облегчается продвижение корня между
комочками почвы. Этому же способствует обильная
слизь, продуцируемая клетками чехлика. Чехлик
125
непрерывно обновляется. По мере того как с его
поверхности слущиваются клетки, меристема обра
зует новые клетки.
За зоной деления располагается зона роста и
дифференциации, где клетки достигают своих по
стоянных размеров и приобретают вполне опреде
ленное строение в связи с теми функциями, которые
они будут выполнять. Некоторые клетки начинают
удлиняться и дифференцироваться еще в зоне деле
ния, в то время как другие достигают зрелости и
специализации в зоне роста. За зоной роста распо
лагается зона всасывания (зона корневых волос
ков), где различают ризодерму (систему покровной
ткани), первичную кору (систему основной ткани)
и центральный цилиндр (систему проводящих тка
ней) (рис. 81). Клетки покровной ткани (эпиблема,
ризодерма) образуют корневые волоски — вырос
ты длиной 0,1 —8 мм.
Корневые волоски тесно соприкаса
ются с отдельными частицами почвы.
Благодаря этому осуществляется их кон
такт с водой и растворимыми минераль
ными солями.
Поглощение воды корнями происхо
дит, по-видимому, главным образом пу
тем осмоса, хотя есть данные, указываю
щие на активный перенос. Как правило,
концентрация растворенных веществ в
цитоплазме выше, чем в окружающей
корни воде. По мере проникновения воды
в клетки концентрация солей в них пони
жается, поэтому вода из этих клеток пе
реходит в соседние, пока не достигнет
центрального цилиндра. Центральный
цилиндр содержит проводящую ткань —
ксилему, по которой вода поднимается
вверх к листьям.
Проникновение солей в корневые во
лоски и далее в клетки коры и централь
ного цилиндра происходит путем диффу
зии и активного переноса. Эксперимен
тально установлено, что всасывание ми
неральных солей не прекращается и тогРис. 81. Поперечный срез кор
ня в зоне всасывания: 1 — ко
жица, 2 — корневые волоски,
3 — эндодерма, 4 — сосуды
(стрелками показан путь воды
от корневых волосков к сосу
дам).
126
да, когда концентрация их-в клетке стано
вится выше, чем в почве.
Передвижение воды из корневых во
лосков по клеткам корня к центральному
цилиндру может осуществляться разны
ми путями: по цитоплазме клеток через
плазмодесмы (симпластный путь) либо
через вакуоли (вакуолярный путь). Мо
лекулы воды могут также диффундиро
вать по оболочкам между клетками
(апопластный путь). Во всех случаях
вода может передвигаться от клетки к
клетке либо по клеточным оболочкам до
тех пор, пока не достигнет эндодермы —
ткани на границе первичной коры и цен
трального цилиндра (рис. 82). Клетки эн
додермы выполняют регулирующую фун
кцию — они препятствуют свободному
проникновению воды в центральный ци
линдр. Это достигается благодаря водо
непроницаемому веществу суберину, ко
торое откладывается в клеточных стенках
в виде полосок, называемых поясками
Каспари. Эти пояски не проницаемы для
воды, и поэтому вода и растворенные в
ней соли должны проходить через плаз
матическую мембрану под контролем
протопласта эндодермальной клетки.
Следует отметить, что клеток эндодермы
с поясками Каспари (пропускные клет
ки) не много: все другие клетки этой тка
ни имеют суберинизированные с трех
сторон оболочки. Именно поэтому вода
не может пройти свободно по апоплас-
Рис. 82. Схема передвижения во
ды и минеральных веществ в кор
не: а — передвижение воды и ио
нов, б — к л е тки эндодермы на
поперечном срезе, в — трехм ер
ное изображение соседних кле
т о к эндодермы: 1 — частичка
почвы; 2 — пленка воды; 3 — кор
невой волосок; 4 — вода и воз
дух; 5 — клеточная стенка; 6 —
вакуоль; 7 — цитоплазма; 8 —
слой клеток, образующих корне
вые волоски; 9 — эндодерма;
10 — перицикл; 11 — сосуды кси
лемы; 12 — поясок Каспари; 13 —
пропускная клетка; 14 — субе
рин. Черная стрелка — апоплас
тн ы й п у т ь (прерывается в эндо
дерме), красная стрелка — сим
пл астн ы й п у т ь , белая стрел
ка — вакуолярный п у ть .
127
тному пути или через любую клетку эндо
дермы. Вода может проходить только че
рез немногочисленные пропускные клет
ки с поясками Каспари. Как полагают,
благодаря этому живые клетки эндодер
мы могут регулировать передвижение во
ды и минеральных солей из почвы в кси
лему. Возможно, этот механизм служит
также для защиты от проникновения в
растения ядов, патогенных бактерий и
грибов и т. д.
Дальнейший транспорт воды и мине
ральных веществ, а также органических
веществ, синтезируемых в листьях расте
ний, осуществляется попроводящим тка
ням. Из корня через стебель к листьям
идут многочисленные проводящие пучки.
Часть пучка, обычно внутренняя (ксиле
м а, или древесина), содержит сосуды для
транспорта воды и минеральных веществ.
Растения не имеют никакого «насос
ного» механизма для передвижения ве
ществ, что резко отличает их от живот
ных с развитой сердечно-сосудистой сис
темой. Поэтому проблема переноса ве
ществ стоит у растений достаточно остро.
Некоторые растения достигают 120—
150 м в высоту, и общий диаметр сосудов
ксилемы у них больше, чем диаметр ма
гистральной трубы городского водопро
вода. Девичий виноград может подняться
по стене дома до пятого этажа и даже вы
ше, а вся вода, необходимая для поддер
жания его роста, поднимется по сосудам
ксилемы из почвы. Каким же образом у
растений осуществляется процесс пере
носа веществ?
Вода перемещается из участка с бо
лее высоким водным потенциалом к учас
тку с более низким. Условно принято счи
тать, что при обычном атмосферном дав
лении водный потенциал чистой воды ра
вен нулю. В этих условиях все растворы
будут иметь более низкий водный потен
циал, а, следовательно, величина его бу
дет отрицательной. Значит, можно опре
делить понятие «осмос» по-другому: это
перемещение молекул воды через полу
128
проницаемую мембрану, направленное из
области с более высоким водным потен
циалом в область с более низким. Если
контактируют две системы с разными
водными потенциалами, например клетка
и окружающая ее чистая вода, то вода
поступает в клетку, т. е. в систему с более
низким водным потенциалом.
Сухому воздуху свойствен очень низ
кий водный потенциал, а почве — высо
кий. Растение же, обладая средним вод
ным потенциалом, занимает среднее по
ложение в градиенте водных потенциа
лов, и вода проходит от почвы к воздуху,
т. е. в растении столб воды движется
вверх. Разрыву водяного столба в корне,
стебле и листьях препятствует сила сцеп
ления между молекулами воды. Вода из
почвы проходит через кору корня в кси
лему, поднимается по ней к листьям и ис
паряется с поверхности клеток мезофил
ла, а затем диффундирует наружу через
устьица. Этот процесс называется тран
спирацией. С наружных клеток эпидер
миса листьев и стеблей вода испаряется
через кутикулу — восковой налет, по
крывающий листья и зеленые стебли.
У листопадных растений после сбрасыва
ния листьев испарение может происхо
дить через чечевички.
Потери воды растением при транспирации мо
гут быть очень велики. Подсолнечник, например,
ежедневно теряет 1— 2 л воды, а старый дуб — бо
лее 60 л. Одна яблоня за сутки испаряет до 100 л
воды; за сезон эта величина достигает 18 000 л,
причем в это количество не входит вода, используе
мая в фотосинтезе (на создание 1 г сухого вещества
растения тратят от 200 до 1000 г воды, что состав
ляет 0,5— 1 % всей воды, проходящей через расте
ние).
Транспирация увеличивается при уменьшении
влажности воздуха, повышении температуры и вет
реной погоде. Транспирация может регулироваться
с помощью замыкающих клеток устьиц. В усло
виях дефицита влаги устьица могут закрываться и
транспирация снижается. Кроме того, для борьбы с
излишними потерями воды многие растения имеют
специальные приспособления: сворачивание или
складывание листьев, увеличение толщины кутику
лы, уменьшение площади листовой пластинки, гус
тое опушение и т. д.
Для перехода воды из жидкого состояния в па
рообразное нужна дополнительная энергия. Ее дает
Солнце. Именно солнечная энергия- обеспечивает
общий поток воды через растение.
При транспирации происходит существенное
понижение температуры на поверхности листьев,
что предотвращает перегрев растений. Таким обра
зом транспирация способствует осуществлению
двух важнейших процессов, происходящих в расте
ниях: подъему воды из корней в листья и борьбе с
перегревом.
Вместе с током воды перемещаются
растворенные в ней вещества: соли, не
которые органические соединения азота
и гормоны. Главным потребителем мине
ральных элементов, т. е. местом их ис
пользования, являются растущие части
растения — верхушечные и пазушные
меристемы, молодые листья, развиваю
щиеся цветки, плоды и запасающие орга
ны. Растворенные вещества «выделяют
ся» из ксилемы на концах тончайших жи
лок; при этом они поступают в клетки пу
тем диффузии и активного поглощения.
Другая часть проводящего пучка,
обычно наружная — флоэма, или луб.
По флоэме растворы органических ве
ществ, прежде всего продукты фотосин
теза, передвигаются ко всем органам рас
тения. Необходимо отметить, что органи
ческие вещества по флоэме транспорти
руются и вниз, и вверх, т. е. в двух нап
равлениях. Это отличает флоэму от кси
лемы, по которой вещества передвигают
ся только вверх.
Кроме органических веществ, по фло
эме в различной форме переносятся азот
и сера (в форме аминокислот), фосфор (в
виде фосфорилированных сахаров и ио
нов неорганического фосфата), калий (в
виде ионов). С током веществ по флоэме
в небольшом количестве перемещаются
витамины, ростовые вещества (ауксины,
гиббереллины и др.), вирусы, поступив
шие через листья из воздуха химические
5. Общая биология, 10 кл.
вещества и т. д. Передвижение веществ
по флоэме осуществляется активно, с
затратами энергии АТФ, которая, по всей
вероятности, вырабатывается в клеткахспутницах.
Диоксид углерода поступает из возду
ха через устьица и далее по межклетни
кам доходит до клеток, где осуществляет
ся процесс фотосинтеза. К фотосинтези
рующим клеткам стебля диоксид углеро
да может также проникать через чечевич
ки. В каждую клетку растения, где проис
ходит фотосинтез, диоксид углерода по
ступает с помощью диффузии.
9■
1. Каковы механизмы поглощения питатель
ных веществ одноклеточными организмами? 2. Ка
кова судьба веществ, поступивших в одноклеточные
организмы путем эндоцитоза? 3. Назовите особен
ности поглощения питательных веществ клетками
грибов и гетеротрофных бактерий. 4. Как передви
гаются вещества в организме растений? 5. Какие
особенности ксилемы обеспечивают транспорт во
ды и растворенных в ней веществ на большие рас
стояния? 6. Почему транспирация осуществляется
главным образом через листья? 7. Перечислите ос
новные структурные и функциональные отличия
ксилемы и флоэмы.
39. Поступление питательных в е
ществ в животный организм (спо
собы питания)
В отличие от растений, автотрофных
протестов и бактерий животные сами не
создают питательных веществ, а получа
ют их из окружающей среды и превраща
ют в те соединения, которые организм
способен усваивать. В организм живот
ного питательные вещества поступают
вместе с пищей, которую животные за
глатывают. В связи с этим все животные
имеют рот и специальный мускульный
орган — глотку.
Разные животные способны потреб
лять пищевые частицы различной вели
чины. Для животных, потребляющих
мелкие пищевые частицы, характерен
129
фильтрационный способ питания.
Фильтрация осуществляется с помощью
щетинок, ресничек, роговых пластинок и
т. п. Фильтрационный способ питания
наблюдается у мелких ракообразных
(дафний и др.), двустворчатых моллюс
ков, сельдевых рыб, усатых китов, неко
торых птиц (фламинго, буревестники и
ДР-)У дафний (рис. 83), например, имеет
ся ряд листовидных грудных конечностей,
покрытых жесткими щетинками. Движу
щиеся одновременно вперед конечности
захватывают воду со взвешенными пище
выми частицами. Щетинки отфильтровы
вают эти частицы, а когда конечности
движутся в обратном направлении, пи-
Рис. 83. Дафния: 1 — сложный глаз, 2 — пер
вая антенна, 3 — рот, 4 — первый туловищный
отросток, 5 — щиток, 6 — анальное отверстие,
7 — вторая антенна, 8 — мозг, 9 — челюсть,
10 — сердце, 11 — яичник, 12 — эмбрионы в
выводковой камере.
130
Рис. 84. Садовая улитка: 1 — терка (радула),
2 — голова, 3 — челюсть, 4 — рот, 5 — хрящ,
6— нога, 7— мешочек радулы, 8— втягивающий
мускул глотки, 9 — пищеварительная трубка.
щевые частицы проталкиваются по
брюшному желобку к ротовому отвер
стию под действием других щетинок, рас
положенных у основания каждой ноги.
У ротового отверстия пищевые частицы
обволакиваются липким слизистым ве
ществом и заглатываются.
Для животных, питающихся крупны
ми частицами, способы питания более
разнообразны. Так, животные, перераба
тывающие детрит (свежий или разлагаю
щийся органический материал — отмер
шие части растений или животных), за
хватывают кусочки пищи, находящиеся в
почве или на ее поверхности, и проталки
вают в глотку. Т. е. для них характерно
заглат ы вание пассивных пищевых
масс. Таковы, например, дождевые чер
ви, мокрицы, ногохвостки, нематоды идр.
Другие животные питаются путем
соскабливания, перетирания, жева
ния и сверления. Таковы морские ежи,
улитки, некоторые насекомые и позво
ночные.
Например, садовая улитка (рис. 84)
питается с помощью специального скре
бущего органа — радулы, или терки, и
роговой челюсти. Радула представляет
собой пластинку, на которой расположе
но до 150 поперечных рядов зубчиков,
направленных вершинами назад. В каж
дом ряду содержится около 100 зубчиков.
Радула лежит на особом выступе на дне
ротовой полости; внутри выступа нахо
дится хрящ, дающий опору всему аппара
ту терки. Благодаря особой мускулатуре
терка может перемещаться вперед или
назад внутри ротовой полости, а кроме
того, может немного выдвигаться через
ротовое отверстие. Когда улитка захва
тывает листья, радула движется взад и
вперед по листьям, прижимая их к челюс
тной пластинке, и ее зубчики перетирают
пищу. В конечном счете образуются
мельчайшие фрагменты растения, кото
рые постепенно проталкиваются в глотку.
У некоторых насекомых (например, у
жуков-листоедов) имеется специальный
грызущий ротовой аппарат, с помощью
которого животные отрывают кусочки
листьев или других частей растений, раз
грызают и перетирают их.
У многих хордовых функцию захвата и
перетирания пищи выполняют зубы, при
чем у жвачных животных вместо резцов и
клыков имеется роговая пластинка.
Для таких животных, как кишечнопо
лостные, рыбы, змеи, летучие мыши, ха
рактерны ловля и заглатывание жер
твы, когда пища не подвергается пред
варительному измельчению, а заглатыва
ется целиком. Существует ряд животных,
которые в процессе их исторического
развития приспособились к питанию
жидкой пищей или мягкими тканями. Для
этого у них имеются специальные при
способления. У насекомых это лижущий
(мухи), сосущий (бабочки) (рис. 85, а, б)
или лакающий (пчелы) ротовые аппа
раты, с помощью которых насекомые
собирают нектар и сок растений. У ко
либри функцию всасывания нектара вы
полняет свернутый в трубочку тонкий
язык.
Животные-кровососы
питаются
кровью других животных и человека. Для
них характерно наличие специальных ко
лющих частей ротового аппарата, или
зубчиков, и мускульных приспособлений
для всасывания крови: колюще-сосущий
аппарат у комаров (рис. 85, в), мускулис
тый хоботок у пиявок, сосательный же
лобчатый конус у клещей и т. п.
Некоторые паразитические живот
ные, которые живут внутри других жи
вотных и человека, поглощают питатель
ные вещества всей поверхностью тела.
Таковы, например, ленточные черви.
Животные различаются не только
Рис. 85. Ротовые аппараты насекомых: а — ком натная муха: 1 — глаз, 2 — антенна, 3 — максилярный щупик, 4 — нижняя губа; б — бабочка капустница: 1 — простой глазок, 2 — антенна, 3 —
сложный глаз, 4 — щупик нижней губы, 5 — хоботок; в — комар анофелес: 1 — антенна, 2 — верх
няя губа, 3 — верхняя челюсть, 4 — подбородок, 5 — нижняя челюсть, 6 — нижняя губа, 7 — щупик,
8 — глаз.
131
способами питания, но и пищевой специ
ализацией, т. е. тем, какой пищей они пи
таются.
В зависимости от склонности живот
ных к тому или иному пищевому рациону
их можно разделить на три группы: фито
фаги, зоофаги и сапрофаги. Ф итофаги
(от греч. phytos — растение, phagos —
пожиратель) — растительноядные жи
вотные (парнокопытные, непарнокопыт
ные, многие насекомые и т. д.). Зооф а
ги — плотоядные организмы, питаю
щиеся животными других (или своих)
видов.
. Среди зоофагов выделяют хищников,
поедающих других животных (жертв), и
паразитов, питающихся за счет пита
тельных веществ и живых тканей хозяи
на. Сапрофаги — животные, питающи
еся отмершим органическим веществом.
Среди них различают некрофагов, копрофагов и детритофагов.
Иекрофаги (от греч. nekros — мер
твый), трупоеды — питаются трупами
животных. К этой группе сапрофагов от
носятся некоторые птицы (грифы, мара
бу), млекопитающие (гиены, шакалы),
насекомые (жуки-могильщики, мерт
воеды).
Крпрофаги (от греч. kopros — помет,
кал) — животные, питающиеся экскре
ментами главным образом млекопитаю
щих животных. Это жуки-навозники, жу
ки рода стафилинид, личинки многих
двукрылых.
Детритофаги (от лат. detritus — ис
тертый) — организмы, питающиеся дет
ритом — мертвым или частично разло
жившимся органическим веществом. К
этой группе животных относятся ры
бы — лини, бычки, подусты; личинки
многих насекомых; малощетинковые и
многощетинковые черви и др.
Пищевая специализация может до
стигать крайних пределов, когда опреде
ленный вид животного может питаться
только строго определенной пищей. В за
висимости от степени требовательности к
132
определенному виду пищи животных де
лят на несколько групп.
Монофаги (от греч. monos — один),
одноядные — животные, которые пита
ются представителями одного вида (на
пример, одного вида растений или близ
кими видами одного рода). Примером та
кой крайней пищевой специализации яв
ляется тля филлоксера, живущая только
на виноградных лозах, или гороховая зер
новка, обитающая в основном в семенах
гороха.
Олигофаги (от греч. oligos — немно
гий) — животные, которые питаются
родственными видами, относящимися к
одному или немногим близким родам.
Например, колорадский жук питается
растениями семейства пасленовых, бе
лянки — крестоцветными.
Полифаги (от греч. polys — мно
гий) — питаются многочисленными ви
дами. Например, хищные млекопитаю
щие поедают разнообразных животных,
травоядные копытные — всевозможные
виды трав. Так, мигрирующая саранча
уничтожает более 70 видов растений, гу
сеницы лугового мотылька — около 200,
а муфлон на острове Корсика питается
более чем 100 видами растений. По срав
нению с олиго- и монофагами полифаги
легко добывают пищу, хотя иногда на по
иски тратят много энергии.
Пантофаги (от греч. pan (pantos) —•
все) — всеядные организмы, питающие
ся организмами разных царств — живот
ными, растениями,грибами.
9в.
1. Как осуществляется поступление питатель
ных веществ в организм животных? 2. Какие спосо
бы питания характерны для животных, какова прин
ципиальная разница между ними? 3. Как вы думае
те, с чем связано возникновение разных способов
питания у животных? 4. Каким образом осущес
твляется фильтрация пищевых частицу разных жи
вотных? 5. Какими приспособлениями обладают
кровососы? 6. Что такое пищевая специализация и
какие группы животных выделяют по отношению к
пище?
40. Пищеварение
Пищеварение — это совокупность
процессов, обеспечивающих механичес
кое измельчение пищи, химическое рас
щепление макромолекул питательных ве
ществ на компоненты, пригодные для
всасывания и участия в обмене веществ.
У большинства животных есть специали
зированный орган, называемый пищева
рительным, или ж елудочно-киш еч
ным, трактом. Он предназначен для то
го, чтобы извлекать пищу из окружаю
щей среды, временно запасать и перера
батывать ее прежде, чем она всосется че
рез клеточные мембраны. Поступающая
в организм пища перерабатывается под
действием гидролитических ферментов.
Пищеварительные ферменты рас
щепляют питательные вещества либо в
клетках (внут риклет очное пищ ева
рение), либо вне клеток, в полостях пи
щеварительного тракта (внеклет очное
пищ еварение).
У многих низших животных, напри
мер у губок (рис. 86), преобладает внут
риклеточное пищеварение. Однако в
процессе исторического развития все бо
лее важную роль приобретает внеклеточ
ное пищеварение, свойственное боль
шинству животных, а также грибам и
бактериям. Оно характеризуется тем, что
синтезирующиеся в клетках ферменты
выделяются во внешнюю среду и осу
ществляют расщепление макромолекул
за пределами клетки, иногда на значи
тельном расстоянии.
У кишечнополостных осуществляется
частично внеклеточное, а частично внут
риклеточное пищеварение. Гидры, меду
зы и актинии (рис. 87) могут захватывать
поразительно крупную добычу, а затем
долго ее переваривать, прежде чем захва
тить новую порцию. У этих организмов
пища поступает через рот в кишечную
полость. Здесь под действием пищевари
тельных ферментов, выделившихся из
железистых клеток энтодермы, начинает-
Рис. 86. Схема строения губки: 1 — устье, 2 —
эпидермальная клетка, 3 — воротничковая
клетка, 4 — спикула, 5 — вода, 6 — породит,
7 — амебоидная клетка, 8 — поры.
Рис. 87 Схема строения актинии: 1 — щупаль
ца с стрекательными клетками, 2 — рот, 3 —
остии, 4 — глотка, 5 — перегородка, 6 — стен
ка тела, 7 — пищеварительная полость, 8 — го
нады, 9 — волокна септ, 10 — стрекательные
нити, 11 — мышцы, 12 — подошва.
133
ся процесс внеклеточного переварива
ния. Как только комочек пищи распада
ется на мелкие полупереваренные час
тички, пищеварительные клетки энтодер
мы с помощью псевдоподий захватывают
их и окончательно переваривают. Раство
римые продукты переваривания далее
диффундируют через мезоглею в клетки
эктодермы.
У коралловых полипов глотка образуется пу
тем заворота эктодермы, вдающейся в виде трубки
внутрь тела, так что кишечник примыкает к ее внут
реннему концу. Наружным ртом становится наруж
ное отверстие глотки. Начиная с коралловых поли
пов, у всех животных пищеварительный аппарат пе
рестает быть чисто эктодермальным производным.
Его энтодермальная часть с этого момента получает
название средней кишки. Вновь образованная эк
тодермальная часть, глотка, представляет собой
переднюю кишку. Эти два отдела кишечника ха
рактерны и для плоских червей.
Смешанное пищеварение, свойствен
ное кишечнополостным, также наблюда
ется у плоских червей. Свободноживущие плоские черви (планарии) засасыва
ют пищу с помощью мускулистой глот
ки (рис. 88). У многих из них кишечник
сильно разветвлен, так как, помимо пи
щеварительной, кишечник выполняет
также функцию распределения. Ответ
вления кишечника практически пронизы
вают все участки тела, что облегчает дос
тавку и обмен веществ между клетками
тела и содержимым кишечника. Клетки
кишечника выделяют ферменты для
внеклеточного пищеварения; далее ма
ленькие пищевые частички поглощаются
фагоцитирующими клетками, и процесс
пищеварения заканчивается внутриклеточно. И у кишечнополостных, и у плос
ких червей непереваренные остатки вы
водятся через то же отверстие, через ко
торое поступила пища, т. е. через рот.
Внеклеточное пищеварение наблюда
ется у круглых и кольчатых червей, рако
образных, насекомых и хордовых, а также
у бактерий и грибов. Например, некото134
Рис. 88. Пищеварительная система планарии:
2 — глотка, 3 — рот, 4 — пищеварительный
тракт. 1 — глаз.
рые насекомые и паукообразные вводят
пищеварительные ферменты в пойман
ную добычу, а бактерии и грибы выделя
ют их в окружающую среду.
У высших многоклеточных животных
внеклеточное пищеварение происходит в
специализированных полостях желудоч
но-кишечного тракта (полостное пище
варение). Полостное пищеварение свой
ственно круглым червям, большинству
кольчатых червей, членистоногих, мол
люскам и хордовым. У сосальщиков, не
которых кольчатых червей после предва
рительного внеклеточного переварива
ния осуществляется более полное внут
риклеточное.
У круглых червей пищеварительный
тракт становится сквозным, так как по
является задняя эктодермальная киш
ка, образованная впячиванием экто
дермы зародыша и срастанием его с
нижним концом средней кишки. Начи
ная с круглых червей, непереваренная
пища у всех животных удаляется через
анальное отверстие.
У высокоразвитых животных пищева
рительный тракт — это, в сущности,
трубка, на одном конце которой располо
жен рот, куда поступает пища, а на дру
гом — анальное отверстие, через кото
рое выделяются непереваренные остат
ки. Наличие сквозного кишечника позво
ляет осуществлять «конвейерный» про
цесс пищеварения. Пища может переме
щаться лишь в одном направлении, так
что животное продолжает поглощать пи
щу, в то время как пища, съеденная ра
нее, еще переваривается. При этом раз
ные участки пищеварительного тракта
специализируются и выполняют различ
ные функции.
У большинства животных на перед
нем конце пищеварительного тракта рас
положены структуры, выполняющие
функцию измельчения пищи. У млекопи
тающих — это зубы, у черепах и птиц —клюв, у большинства моллюсков (кроме
двустворчатых) — терка, у дождевых
червей, насекомых, большинства птиц и
крокодилов — мускульный желудок со
специальными выростами или набитый
камешками. У дождевых червей и у неко
торых членистоногих, например, имеется
не только глотка, втягивающая пищу, но
и зоб позади глотки, который представ
ляет собой расширенную часть пищево
да, где накапливается пища (рис. 89). За
ним расположен жевательный желу
док — мускулистая тонкостенная каме
ра, выстилка которой снабжена тверды
ми выступами, напоминающими зубы;
при сокращении мышечных стенок они
дробят пищу на мелкие частицы. Пере
тертая или раздробленная пища подвер
гается затем действию пищеварительных
ферментов. Механическое измельчение
пищи увеличивает поверхность пищевых
частиц, повышая эффективность воздей
ствия пищеварительных ферментов.
4
11
10
5
э
6
7
8
Рис. 89. Внутреннее строение дождевого чер
вя: 1 — рот, 2 — мозговой ганглий, 3 — сердца,
4 — пищевод, 5 — зоб, 6 — желудок, 7 — неф
ридий, 8 — кишечник, 9 — брюшная нервная
цепочка, 10 — глотка, 11 — ротовая полость.
Пищеварительные ферменты у жи
вотных с полостным пищеварением обра
зуются в кишечном эпителии и крупных
пищеварительных железах (слюнных и
поджелудочной — у позвоночных, в ж е
лезах средней кишки — у беспозвоноч
ных). У многих моллюсков и членистоно
гих имеются крупные пищеварительные
железы, в которых происходит внутри
клеточное пищеварение. Эпителий ки
шечника, кроме пищеварительных фер
ментов, выделяет в кровь гормоны, кото
рые регулируют процесс переваривания,
а также слизистый секрет муцин, предот
вращающий самопереваривание стенок
пищеварительного тракта.
Пищеварительные тракты многих жи
вотных снабжены небольшими слепыми
выростами, в которых пища задержива
ется для более полного усвоения.
У ряда организмов наблюдается так
же мембранное, или пристенное, пи
щеварение. Мембранное пищеварение
осуществляется ферментами, локализо
ванными на структурах клеточной мем
браны. У большинства высокоорганизо
ванных животных такое переваривание
происходит на поверхности эпителиаль
ных клеток кишечника. Оно обеспечива
ет промежуточные и конечные этапы пе
реваривания и начальные этапы всасыва135
ния. Мембранное переваривание свой
ственно многим беспозвоночным (черви,
ракообразные, насекомые, моллюски) и
всем позвоночным. У некоторых организ
мов, например у кишечных паразитов —
аскарид, развито только мембранное пи
щеварение, в то время как полостное ре
дуцировано. У многих организмов сочета
ются все три типа пищеварения, что по
вышает эффективность и экономичность
работы пищеварительной системы. Про
дукты переваривания по мере образова
ния всасываются стенками кишечника.
В верхних отделах кишечника выделяют
ся ферменты, а следующие его отделы
специализированы для всасывания пита
тельных веществ. Увеличение всасываю
щей поверхности достигается развитием
мелкой складчатости слизистой или обра
зованием мелких выростов —- ворсинок.
Особенно многочисленны и хорошо раз
виты ворсинки в тонком кишечнике птиц
и млекопитающих.
?>
1. Какое значение имеет пищеварение в жизни
живых организмов? 2. Присуще ли пищеварение
растениям и грибам? 3. Чем различаются между со
бой разные типы пищеварения? 4. Почему у аска
рид осуществляется только мембранное пищеваре
ние? 5. Где вырабатываются пищеварительные
ферменты у разных организмов и почему они не пе
реваривают собственные клетки? 6. Каков смысл
механического измельчения пищи? Каким образом
измельчается пища у ракообразных, насекомых,
пресмыкающихся и птиц?
41. Пищеварение у человека
Рис. 90. Пищеварительный тракт человека:
1 — ротовая полость, 2 — глотка, 3 — пище
вод, 4 — желудок; то н ки й кишечник-. 5 — две
надцатиперстная, 6 — тощая, 7 — подвздош
ная
кишка;
толстый
кишечник: 8 —
червеобразный отросток слепой кишки, 9 —
поперечная, 10 — нисходящая, 11 —
восходящая ободочные, 12 — сигмовидная,
13 — слепая и 14 — прямая кишки, 15 — зад
ний проход.
Человек — существо всеядное. Он
питается как растительной, так и живот
ной пищей. Именно поэтому его пищева
рительный тракт избежал крайней специ
ализации, характерной для многих позво
ночных животных. Так, например, все зу
бы у человека примерно одной величины.
Передние зубы, или резцы, по форме на
поминающие долото или стамеску, отку
сывают кусочки пищи. Заостренные клы-
ки помогают удерживать пищу. Располо
женные в глубине рта уплощенные ко
ренные зубы перетирают пищу, превра
щая ее в кашицу. Во рту размельченная
пища смешивается со слюной, ферменты
которой начинают переваривание. Кроме
того, слюна смачивает пищу и тем самым
облегчает ее проглатывание. Переже
ванная влажная пища проходит через
глотку (горло) в пищевод (рис. 90), со
136
скальзывая по надгортаннику, который
закрывает вход в дыхательные пути и не
позволяет попадать туда пище.
В слюне содержатся ферменты птиа
лин и мальтаза (табл. 6), а также бел
ки муцин и лизоцим. Лизоцим обла
дает антибактериальными свойствами —
это вещество убивает многие виды
бактерий, которые могут оказаться в
пище.
Т а б л и ц а 6. Пищеварительные соки человека и их действие
П ищ еварительн ы й сок
Слюна
Птиалин (амилаза)
Мальтаза
Желудочный сок
Д ей ствие ф ерм ента
Ф ерм ент
П епсиноген(пепсин)
Реннин
Липаза
Панкреатический сок Липаза
(сок поджелудочной
Трипсиноген (трип
железы)
син)
Химотрипсиноген
(химотрипсин)
Д руги е вещ еств а, со д ер ж ащ и е
ся в п и щ еварительном соке, и
их ф у н кци и
Превращает крахмал и глико Слизь (муцин) обволакива
ет пишу
ген в мальтозу
Превращает мальтозу в глюкозу Лизоцим (антибактериаль
ное вещество)
Расщепляет белки до олигопеп Соляная кислота способ
ствует превращению пепситидов
ногена в активный пепсин
Свертывает молоко
Расщепляет жир до высших Трипсин превращает химо
карбоновых кислот и глицерола трипсиноген в химотрипсин
Частично расщепляет белки (до
пептонов и полипептидов)
Расщепляет белки (до пептонов
и полипептидов)
Амилопсин (амилаза) Превращает крахмал в мальто
зу
Карбоксипептидаза Отщепляет аминокислоты от
полипептидов
Рибонуклеаза
Расщепляет РНК
Дезоксирибонуклеаза Расщепляет ДНК
Желчь
«
Кишечный сок
Эмульгирует
жиры;
NaHC03 нейтрализует кис
лую реакцию желудочного
сока
Эрепсин (аминопеп- Расщепляет пептоны до амино
кислот
тидазы)
Расщепляет жир до высших
Липаза
карбоновых кислот и глицерола
Мальтаза
Сахараза
Лактаза
Энтерокиназа превраща
ет трипсиноген в трипсин
Кишечный сок смачивает
пищу и обволакивает ее
слизью
Расщепляет мальтозу с образо Секретин стимулирует вы
ванием глюкозы
деление сока поджелудоч
ной железой
Расщепляет сахарозу с образо
ванием глюкозы и фруктозы
Расщепляет лактозу с образо
ванием галактозы и глюкозы
Из пищевода пища поступает в желу
док, выполняющий несколько функций.
Продвижение пищи осуществляется бла
годаря непроизвольным перистальтичес
ким движениям пищевода. Перистальти
ческие движения происходят на всем про
тяжении пищеварительного тракта. Они
представляют собой последовательное,
волнообразное сокращение мышц пище
варительного тракта, постепенно протал
137
кивающих пищу к нижнему концу тракта.
Верхнее отверстие желудка открывается
в момент приближения перистальтичес
кой волны — пища входит в желудок, на
капливается и на некоторое время задер
живается. В результате сокращения мы
шечных стенок желудка пища перетира
ется, пока не приобретает консистенцию
жидкого супа. Железы, расположенные в
стенках желудка, секретируют желудоч
ный сок, который содержит соляную кис
лоту и поэтому имеет сильно кислую ре
акцию. Соляная кислота действует как
антисептик и дезинфицирующее вещес
тво, обезвреживающее большинство
микроорганизмов, поступающих с пищей,
а также создает pH среды, при которой
возможно переваривание белков. Ж еле
зы желудка секретируют пепсиноген, ко
торый в присутствии соляной кислоты
превращается в пепсин, расщепляющий
белки. Пепсин расщепляет коллаген, со
держащийся в соединительнотканных
компонентах мяса. Только позвоночные
способны синтезировать пепсин, благо
даря чему они могут переваривать боль
шую часть потребляемой ими мясной пи
щи. Вместе с тем пепсин и соляная кис
лота обычно не повреждают стенки само
го желудка, т. к. их защищает слой слизи.
В состав слизи входит муцин, выделяе
мый клетками стенок желудка. В состав
пищеварительного сока желудка входит
также липаза, однако она оказывает на
жиры слабый гидролизирующий эффект.
Она расщепляет лишь жиры молока,
поскольку они эмульгированы и легко
доступны действию липазы.
Жизнь клетки слизистой оболочки желудка,
несмотря на муциновую защиту, очень коротка. Эта
оболочка теряет около полумиллиона клеток в ми
нуту, а каждые трое суток происходит ее полное об
новление. Если клетки желудка выделяют недоста
точно слизи, чтобы защитить себя от действия соля
ной кислоты и пепсина, то возникает язвенная бо
лезнь. Язвы могут образовываться также на пище
воде и двенадцатиперстной кишке, если их участки
138
подвергаются чрезмерному воздействию соляной
кислоты. Риск возникновения этого заболевания
возрастает в стрессовых ситуациях. Ученые предпо
лагают, что страх, нервозность, беспокойство угне
тают секрецию желудочного сока, в том числе и вы
деление муцина.
Через некоторое время после попада
ния пищи в желудок она перистальтичес
кими движениями стенок проталкивается
небольшими порциями (примерно по
14 г) через периодически открывающий
ся пилорический сфинктер в двенад
цатиперстную кишку (начальный от
дел тонкого кишечника). Здесь пища сме
шивается с желчью, выделяемой пе
ченью, соком поджелудочной железы и
кишечным соком, секретируемым клет
ками слизистой оболочки тонких кишок.
В двенадцатиперстной кишке выделяе
мый печенью бикарбонат натрия нейтра
лизует кислую реакцию содержимого же
лудка. Пока пища нейтрализуется, сфин
ктер привратника желудка закрывается.
После нейтрализации сфинктер вновь
расслабляется, и в двенадцатиперстную
кишку поступает новая порция содержи
мого желудка.
Поступающая в двенадцатиперстную
кишку желчь эмульгирует жиры,
увеличивая тем самым поверхность, на
которой осуществляется их гидролиз
липазой. Желчь не содержит пище
варительных ферментов, но, несмотря на
это, она облегчает процесс пищеварения,
активизируя панкреатические (секрета руемые поджелудочной железой) и
кишечные ферменты. Кроме того, в ней
растворяются продукты гидролиза жи
ров, что способствует их последующему
всасыванию. Желчь активизирует мотор
ную и секреторную деятельность тонкого
кишечника. Соли желчных кислот спо
собствуют всасыванию витамина К, не
обходимого для свертывания крови.
Печень является
многофункциональным
органом. Кроме желчеобразующей функции, она
выполняет и ряд других важнейших функций.
В клетках печени из глюкозы, поступающей по
воротной вене от кишечника, синтезируется за
пасной углевод — гликоген. У взрослого человека
его содержание в печени может достигать 200 г.
При необходимости под действием адреналина он
превращается в глюкозу и выводится в кровь.
Печень выполняет также барьерную функцию,
которая заключается в обеззараживании ядовитых
веществ, как поступивших из внешней среды, так и
образовавшихся в ходе обмена веществ и пи
щеварения. В клетках печени, например, осу
ществляется дезаминирование аминокислот, в
результате чего образуется ядовитый аммиак, из
которого в клетках печени синтезируется менее
токсичная мочевина. В печени разрушается
этиловый спирт путем его ферментативного
окисления.
При нарушении деятельности кишечника и
активации гнилостных процессов, осуществляемых
бактериями, в толстой кишке из аминокислот
образуются токсичные фенол, индол, скатол. Они
всасываются в кровь и по воротной вене попадают в
печень, где из них образуются нетоксичные
соединения.
В клетках печени синтезируются белки
протромбин и фибриноген, участвующие в сверты
вании крови.
Под действием ферментов пища по
мере продвижения по кишечнику почти
полностью переваривается.
Выделение пищеварительных ферментов под
чиняется нервной и гуморальной регуляции. Напри
мер, выделение слюны и желудочного сока начина
ется во время приема пищи под действием ее запа
хов и вида. Эта фаза называется «психической».
Вкус пищи стимулирует «нервную» фазу образова
ния желудочного сока. Попадание пищи в желудок
вызывает химическую стимуляцию -— выделение
стенкой желудка гормона, который называется гастрином. Гастрин стимулирует выделение желудоч
ного сока.
При попадании пищи из желудка в двенадцати
перстную кишку соляная кислота стимулирует вы
деление клетками кишечника гормона секретина.
Секретин поступает в кровь и разносится по всему
телу. При этом часть его попадает в поджелудочную
железу и стимулирует выделение панкреатичес
кого сока.
Но какая-то часть пищи обычно не
переваривается. Так, клетчатка, содер
жащаяся в оболочках растительных кле
ток, и жесткие волокна мяса проходят че
рез желудочно-кишечный тракт почти не
изменяясь. Кроме того, в ряде случаев
пища не переваривается в тонком кишеч
нике из-за недостаточно полного контак
та с пищеварительными ферментами.
Остатки пищи проходят в начальный от
дел толстого кишечника — слепую киш
ку (см. рис. 90). Это своего рода мешок,
который заканчивается маленьким, так
называемым червеобразным, отрос
тком., или аппендиксом.
Иногда остатки пищи попадают в аппендикс, и
довольно часто по тем или иным причинам в этом
отростке начинается воспалительный процесс. Тог
да аппендикс приходится удалять хирургическим пу
тем. Считается, что червеобразный отросток не не
сет никаких определенных функций в пищеварении
и представляет собой рудиментарный остаток отде
ла толстого кишечника, функционировавшего у лю
дей в те далекие времена, когда рацион их был со
вершенно иным.
Пищевые массы, попадающие в тол
стый кишечник, имеют жидкую консис
тенцию. В слепой кишке и в следующем
отделе — так называемой ободоч
ной кишке — происходит интенсивное
всасывание воды и минеральных ве
ществ.
В процессе образования и выделения пищева
рительных соков экзокринными тканями пищевари
тельного тракта и его вспомогательными органами
расходуется большое количество воды с растворен
ными в ней солями. У человека это обычно около 8 л
в сутки. Ясно, что, если бы такое количество воды и
солей ушло из организма вместе с фекалиями, про
изошла бы настоящая осмотическая катастрофа.
Но этого не происходит, потому что почти вся вода и
соли в кишечнике всасываются обратно — главным
139
образом в нижних отделах тонкого кишечника
(рис. 91). Здесь же наблюдается наиболее активное
поглощение солей.
В толстом кишечнике обитает мно
жество бактерий, использующих остатки
пищи и, очевидно, играющих важную
роль в их переработке. Бактерии синте
зируют и выделяют в кишечник вита
мин К, который необходим для образова
ния протромбина, участвующего в свер
тывании крови.
Суточное потребление
1 000 г твердой пищи
и 1 500 мл ж идкости С Л Ю Н Н Ы в
Остатки пищи проталкиваются в пря
мую кишку. И наконец при сокращении
стенок прямой кишки и расслаблении
анального сфинктера — кольцевой
мышцы, окружающей дистальный конец
прямой кишки, — в виде фекалий выде
ляются наружу. Фекалии состоят на три
четверти из воды и на одну четверть из
твердого материала: 30 % его составля
ют обитающие в кишечнике бактерии,
15 % — неорганические вещества,
3 % — белки, 20 % — жиры, осталь
ное — непереваренные остатки пищи.
9
, г
.
1. Где начинается процесс переваривания пи
щи у человека? 2. Какие функции выполняет желу
док? 3. Каким образом осуществляется разделение
кислой среды желудка и слабощелочной среды две
надцатиперстной кишки? 4. Каковы причины появ
ления язв желудка и двенадцатиперстной кишки?
5. Каковы функции печени в пищеварении? 6. Ка
ковы функции толстого кишечника?
42. Всасывание питательных
веществ
Рис. 91. Всасывание воды в кишечнике чело
века.
140
По мере продвижения пищи по ки
шечнику она не только переваривается,
но и всасывается. Ежедневно из кишеч
ника в кровь и лимфу поступает около
10 л жидкости, содержащей воду и пита
тельные вещества. Всасывание происхо
дит в основном в тонком кишечнике.
Стенки желудка всасывают только неко
торые вещества, такие, например, как
алкоголь и некоторые лекарственные
препараты. Стенки кишечника очень хо
рошо приспособлены для всасывания. На
поверхности слизистой оболочки кишеч
ника расположены многочисленные ки
шечные ворсинки, которые в свою оче
редь покрыты микроворсинками — паль
цевидными выростами плазматической
мембраны. Последние вместе с гликокаликсом образуют щеточную кайму, что
создает огромную всасывающую повер
хность.
Рис. 92. Строение ворсинки: 1 — эпителий, 2 — капиллярная сеть, 3 — центральный
млечный капилляр, 4 — венула, 5 — лимфатическое сплетение, 6 — артериола, 7 —
мышечная пластинка слизистой оболочки.
Клетки, выстилающие кишечник, содержат
большое количество пищеварительных ферментов,
которые, по-видимому, обеспечивают пристеночное
пищеварение и расщепляют молекулы питательных
веществ при прохождении их через эти клетки.
Здесь короткие молекулы сахаров и пептидов гид
ролитически расщепляются на мономеры и перено
сятся через клеточную мембрану. Транспорт неко
торых сахаров может происходить путем облегчен
ной диффузии или активного переноса. Важную
роль в поглощении мелких полипептидов играет эндоцитоз.
Ворсинки состоят из соединительной
ткани с небольшим количеством гладко
мышечных клеток и покрыты эпителием
(рис. 92). В их состав входят также лим
фатические и кровеносные капилляры. В
центре ворсинки расположен лимфати
ческий капилляр — центральный млеч
ный синус, который слепо начинается и
окружен сетью кровеносных капилляров.
В лимфатические капилляры через клет
ки эпителия ворсинок из кашицеобразно
го содержимого тонкой кишки — химу
са — происходит всасывание продуктов
расщепления жиров (рис. 93, с. 142), ко
торые проходят по лимфатическим сосу
дам и через общий проток поступают в
ток крови.
Легче других жиров всасываются
продукты расщепления рливочного мас
ла, свиного жира. Глицерин растворим в
воде и легко всасывается через эпителий
слизистой оболочки кишечника. Высшие
карбоновые кислоты, нерастворимые в
воде, при всасывании соединяются с
желчными кислотами и солями, образуя
комплексы (растворимые мыла), которые
проходят через кишечную стенку. Пройдя
через эпителиальные клетки кишечника,
комплексы разрушаются.
Из продуктов переваривания жиров — моно
глицеридов, жирных кислот и глицерола — внутри
всасывающих клеток кишечника синтезируются но
вые триглицериды. Это происходит в гладком эн
доплазматическом ретикулуме, Затем триглицери
ды переходят в шероховатый эндоплазматический
ретикулум, где часть их превращается в фосфоли
пиды и холестерол. Триглицериды, фосфолипиды и
холестерол образуют мельчайшие капли, покрытые
слоем белка. Эти капли называются хиломикронами. Их диаметр около 1500 мкм. Хиломикроны на
капливаются в аппарате Гольджи. Из всасывающих
клеток они постепенно удаляются посредством экзоцитоза в тканевую жидкость, а из нее поступают в
лимфатическую систему и кровь. Около 80 % хиломикронов попадает в кровь через лимфатическую
систему.
Всосавшиеся жиры минуют печень и поступают
в венозную кровь через лимфатический проток.
141
Рис. 93. Всасывание липидов из просвета ки
шечника в лимфатический сосуд: 1 — мицелла,
2 — просвет кишечника, 3 — капля жира, 4 —
межклеточное пространство, 5 — центральный
лимфатический сосуд, 6 — аппарат Гольджи,
7 — шероховатый и 8 — гладкий эндоплазма
тический ретикулумы, 9 — микроворсинка. Ус
ловные обозначения: а — панкреатическая ли
паза, б — триглицерид, в — диглицерид, г —
моноглицерид, д — жирная кислота, е — соли
жирных кислот, ж — глицерол, з — хиломикрон.
выми частицами. Поэтому лимфа, оттекающая от
кишечника, по цвету напоминает молоко и часто на
зывается млечным соком. Отсюда и название сосу
да — центральный млечный синус.
Жиры являются основным энергетическим депо ор
ганизма. В нормальных условиях, когда организм
получает достаточно глюкозы для образования
энергии, жиры не требуются. В этой ситуации они
откладываются в жировых депо: подкожной жиро
вой клетчатке, жировой ткани, окружающей сер
дце, почки, и в брыжейке. Часть жиров встраивает
ся в плазматические и ядерные мембраны клеток.
Ворсинка кишечника, сокращаясь, выдавливает
лимфу с содержащимися в ней мельчайшими жиро142
Все другие питательные вещества по
падают непосредственно в кровеносные
капилляры ворсинок и через воротную
вену поступают в печень, где подвергают
ся дальнейшим биохимическим превра
щениям. Большая часть глюкозы откла
дывается в печени в виде гликогена, но
какая-то часть покидает печень через пе
ченочную вену и разносится по всем
клеткам организма, где подвергается
окислению в процессе клеточного дыха
ния. Часть глюкозы может откладывать
ся про запас в мышцах в виде гликогена.
Если между приемами пищи организму
требуется значительное количество энер
гии, гликоген превращается в глюкозу, и
последняя транспортируется с кровью к
тем тканям, которые в ней нуждаются.
Аминокислоты используются для син
теза новых белков цитоплазмы клеток,
восстановления поврежденных участков
тела, образования ферментов и гормо
нов, а также на другие нужды. Лишние
аминокислоты в запас не откладываются
и подвергаются в печени дезаминирова
нию. При этом от них отщепляются ами
ногруппы, которые превращаются в мо
чевину. Мочевина доставляется с кровью
в почки и выделяется с мочой. Остатки
молекул аминокислот превращаются в
гликоген и в таком виде депонируются.
Кроме того, в печени происходит дето
ксикация — расщепление ядовитых ве
ществ, всосавшихся в кишечнике, либо
перевод их в нерастворимое состояние с
последующим выделением в составе жел
чи в пищеварительный тракт.
?
!. Каково строение и функции кишечных вор
синок? 2. Каков механизм, обеспечивающий по
ступление питательных веществ из тонкого кишеч
ника в кровяное русло? 3. Объясните нецелесооб
разность дальнейшего расщепления аминокислот,
гексоз и жирных кислот в пищеварительном тракте,
хотя это могло бы облегчить диффузию веществ че
рез эпителий кишечника. 4. Какова судьба посту
пивших в кровь веществ? 5. Накапливаются ли в
организме человека запасные белки и что
происходит с избыточными аминокислотами? 6. Как
организм освобождается от ядовитых веществ,
попавших в пищеварительный тракт, а затем в
кровь?
43. Витамины
Кроме углеводов, жиров, белков и
минеральных веществ, в пище должны
содержаться витамины. Они являются
незаменимым компонентом пищевого ра
циона, так как обеспечивают нормальное
течение обменных процессов в организ
ме, поддерживая его жизнедеятельность
и рост. Витамины служат исходными ве
ществами для синтеза ферментных сис
тем клетки, поэтому организм очень
чувствителен к недостатку хотя бы
одного из них. При длительном от
сутствии витаминов в пище разви
ваются заболевания — авит ам ино
зы, при их недостатке — гиповитаминозы.
В витаминах нуждаются все живые
клетки, но лишь некоторые организмы
способны сами их синтезировать. Так, ряд
бактерий и дрожжей способны синтези
ровать все витамины из простых хими
ческих соединений. Такой же способнос
тью обладает и большинство высших рас
тений.
В настоящее время известно несколь
ко десятков витаминов. Они подразделя
ются на две группы: жирорастворимые
и водорастворимые витамины (табл. 7).
Всасывание их происходит главным об
разом в тонком кишечнике.
Т а б л и ц а 7. Витамины
В итам ин
П родукты ,
содерж ащ и е витам ин
Р о л ь витам ина
в орган и зм е
П р о я в л ен и я гипои ави там и н о за
Жирораствор! мые витамины
А
Ретинол
Каротин (предшественник витамина
А)
Печень трески, растительное и животное масла, молоко, кисломолочные про
дукты
Петрушка, укроп, лук, морковь, томаты и др.
d2
Эргокальциферол
Рыбий жир, яйца, сливочное масло, жирная рыба.
Синтезируется в коже под
действием ультрафиолета
Е
Токоферол
Растительное масло, цельное пшеничное зерно, яичный желток, молоко, овощи, зелень
Стимулятор роста, проти- Замедление роста молодого
вовоспалительное действие организма, повреждение
роговицы глаз (ксерофтальмия), нарушение зре
В организме превращается ния — «куриная слепота»,
эпителия,
в витамин А (в стенке ки- повреждение
предрасположенность к иншечника и в печени)
фекциям
Необходим для роста кос- Рахит у детей, остеомалятей и зубов, т. к. усиливает ция (размягчение костей) у
всасывание кальция и фос- взрослых
фора — антирахитный ви
тамин
Регулирует половую потен- Дистрофия
скелетных
цию у животных. Функция у мышц, ослабление половой
человека неизвестна (пред- функции (бесплодие), анеполагается антиоксидан- мия, нарушение зрения у
детей
тное действие)
143
Продолжение
Витамин
к
Филлохинон
Продукты,
содержащие витамин
Родь витамина
в организме
Проявления гипои авитаминоза
Люцерна, шпинат, соевые Способствует образованию Нарушение свертываемос
бобы, свиная печень. Син протромбина
ти крови; увеличение вре
тезируется бактериями, зе
мени свертываемости, же
леными частями растений
лудочно-кишечные крово
течения, подкожные крово
излияния
Водораствори мые витамины
В,
Аневрин, тиамин
В2
Рибофлавин
В3(РР)
Никотиновая
кислота, ниацин,
РР-фактор
в5
Пантотеновая
кислота
в6
Пиридоксин
М или Вс
Фолиевая кислота,
фолацин
В,2
Цианокобаламин
Н
Биотин
С
Аскорбиновая
кислота
Р
Гесперидин
144
Цельное зерно пшеницы, Участвует в регуляции об Болезнь бери-бери (пери
риса, ржи, бобовых; свиная мена углеводов
ферические полиневриты),
печень, яйца, дрожжи
сердечная недостаточность
Цельное зерно, молоко, пе Участвует в транспорте во Задержка роста молодого
чень, соя, бобы, горох, че дорода в процессе тканево организма, поражение глаз
чевица
го движения, в обмене ами (катаракта), дерматиты и
нокислот
трещины вокруг губ и носа
Мясо, печень, пшеничное Незаменимый компонент Пеллагра (поражение ко
зерно, хлеб грубого помола, коферментов НАД и НДЦФ жи), фотодермиты, сыпь,
зеленые овощи и др.
диарея
Во всех пищевых продуктах Входит в состав кофермен- Замедление роста, наруше
та А
ние нервномышечной коор
динации, утомляемость, су
дороги
Яйца, печень, почки, мясо, Кофермент в обмене ами Депрессия и повышенная
рыба, мука грубого помола, нокислот и жирных кислот, раздражительность, сонли
свежие овощи, дрожжи
имеет отношение к образо вость, диарея, анемия, дер
ванию эритроцитов
матиты
Печень, белая рыба, зеле Участвует в образовании Анемия
ные овощи
эритроцитов и синтезе нуклеопротеинов
Мясо, молоко, яйца, рыба, Участвует в синтезе РНК
сыр
Злокачественная анемия
Дрожжи, печень, почки,
цветная капуста, яичный
белок, молоко, орехи; син
тезируется бактериями ки
шечника
Играет роль кофермента в Дерматиты, конъюнктиви
реакциях карбоксилиро- ты, мышечные боли
вания; участвует в синте
зе белка и трансаминировании
Цитрусовые, черная сморо
дина, плоды шиповника,
картофель, томаты, зеле
ные овощи, шпинат
Важен для нормального
развития всех структурных
производных соединитель
ной ткани, здоровой кожи.
Необходим для синтеза
коллагеновых волокон; по
вышает сопротивляемость
организма, способствует
заживлению ран
Цинга: десны становятся
слабыми и кровоточат; не
заживают раны; не образу
ются волокна соединитель
ной ткани. Анемия; сердеч
ная недостаточность; быс
трая утомляемость; ослаб
ление иммунитета
Цитрусовые, черная сморо Способствует укреплению Подкожные кровоизлияния
дина, плоды шиповника, зе стенок кровеносных капил
леные овощи
ляров
Витамины должны поступать в орга
низм постоянно и в достаточном количес
тве. Однако их содержание в пищевых
продуктах колеблется (в зависимости от
сроков хранения и технологии приготов
ления пищи) и не всегда обеспечивает
потребность организма. При длительном
хранении овощей и фруктов содержание
витаминов в них снижается. Разрушают
ся витамины и при действии на продукты
высоких температур. Например, при вар
ке мяса теряется от 15 до 60 % витами
нов группы В, при варке овощей — до
20 % витаминов группы В и до 50 % ви
тамина С. Кроме того, витамин С разру
шается при соприкосновении с воздухом.
Поэтому овощи следует очищать и наре
зать только перед самой варкой, опускать
в кипящую воду и недолго варить в кас
трюле с закрытой крышкой. При кипении
воды растворенный в ней воздух удаляет
ся, что способствует сохранению витами
нов в пище. Овощи необходимо готовить
в эмалированной посуде, так как при со
прикосновении с металлом витамин С
разрушается.
Для предупреждения авитаминозов,
повышения устойчивости организма к ин
фекционным заболеваниям необходимо в
зимне-весенний период принимать спе
циальные витаминные препараты.
?
« 1. Какова роль витаминов в организме? 2. Ка
ким образом поступают витамины в организм?
3. Что является основным источником витаминов?
4. Какие авитаминозы, вам известны? 5. Как
избежать разрушения витаминов в пищевых про
дуктах?
44. Особенности пищеварения
у растительноядных животных
Растительная пища в сыром виде
обычно содержит много воды и грубых,
плохо перевариваемых волокон. Ее пи
щевая ценность сравнительно низкая.
Главной особенностью растительной пи
щи является ее насыщенность целлю ло
зой — основным компонентом клеточ
ных стенок растений. Связи, которыми
соединены глюкозные единицы в молеку
ле целлюлозы, отличаются от аналогич
ных связей в молекулах крахмала и гли
когена. Относительно немногие организ
мы синтезируют ферменты, способные
расщеплять эти связи, что необходимо
для усвоения целлюлозы. Поэтому расти
тельноядные животные, для того чтобы
получить достаточное количество пита
тельных веществ, должны тратить на еду
очень много сил и времени. Неудивитель
но, что некоторые из них употребляют в
пищу лишь те части растений, которые
наиболее богаты питательными вещес
твами — плоды, семена, клубни, лукови
цы, корни. Животные же, извлекающие
питательные вещества в основном из
листьев и стеблей, должны питаться поч
ти непрерывно. Усвоение целлюлозы
представляет для них важнейшую проб
лему, которая у разных животных реша
ется по-разному.
Большинство растительноядных на
секомых не могут переваривать целлюло
зу. Зато их ротовой аппарат приспособ
лен к тому, чтобы прокалывать или раз
рушать клеточные стенки, с тем чтобы
добраться до содержимого клеток. Одной
из причин прожорливости саранчи и дру
гих прямокрылых является малая усваиваемость поедаемой ими пищи, в связи с
чем для получения необходимого коли
чества питательных веществ они должны
съедать очень много. Причем почти поло
вина съедаемой ими растительной массы,
прежде всего клеточные стенки и крах
мальные зерна, выделяется с фекалиями,
не претерпев практически йикаких изме
нений.
Некоторые группы животных способ
ны вырабатывать фермент целлюлазу,
которая расщепляет целлюлозу. Такой
способностью обладают сверлящие дере
во двустворчатые моллюски (корабель
ный червь), некоторые ракообразные,
кольчатые черви и иглокожие. Расти
145
тельноядные брюхоногие моллюски сами
вырабатывают целлюлазу или получают
ее от симбиотических бактерий, обитаю
щих в их кишечнике.
Среди насекомых и других беспозво
ночных существуют животные, которые
приспособились к использованию целлю
лозы за счет расщепления ее организмами-симбионтами, обитающими в их пи
щеварительном тракте. Таковы, напри
мер, термиты, питающиеся древесиной.
Функцию переваривания целлюлозы у
них выполняют одноклеточные жгутико
вые и целлюлозолитические бактерии,
обитающие в задней кишке.
У большинства растительноядных
млекопитающих в специальных отделах
кишечника обитает, как и у некоторых
беспозвоночных, множество микроорга
низмов. Среди них есть бактерии, спо
собные расщеплять целлюлозу на более
простые сахара. Затем бактерии расщеп
ляют эти сахара в процессе брожения до
малых органических молекул, потребляе
мых как бактериями, так и самими жи
вотными.
У некоторых растительноядных жи
вотных целлюлоза переваривается в сле
пой кишке, отходящей от пищеваритель
ного тракта на границе между тонким и
толстым кишечником, или в прямой киш
ке, которые могут быть сильно увеличе
ны. Такой тип переваривания целлюлозы
встречается у животных с простым же
лудком — непарнокопытных, зайцеоб
разных, хоботных.
Самой высокой сложности процесс
пищеварения с помощью симбиотиче
ских микроорганизмов достигает у жвач
ных млекопитающих, таких, как крупный
и мелкий рогатый скот, олени, лоси и др.
Жвачные — это животные, у которых
желудок имеет сложное, многокамерное
строение. Он состоит из четырех отделов:
рубца, сетки, книжки и сычуга (рис.
94). Первые три отдела образуют преджелудок. Его стенки лишены пищевари
тельных желез. Слегка пережеванная пи-
Рис. 94. Желудок жвачного животного: 1 — привратник, 2 — сычуг, 3 — книжка,
4 — пищевод, 5 — рубец, 6 — сетка.
146
ща поступает сначала в рубец и сетку. Ру
бец — это самая объемная часть пище
варительного тракта. Стенки его выстла
ны прочной рогоподобной кутикулой.
Вместе с пищей в рубец изливается боль
шое количество слюны, обладающей сла
бощелочной реакцией. Поэтому содер
жимое рубца тоже слабощелочное.
В рубце пища подвергается предвари
тельной обработке в результате действия
ферментов слюны, а также деятельности
огромного количества микроорганизмов
(анаэробных бактерий и инфузорий). Их
ферменты расщепляют клетчатку, сос
тавляющую значительную часть пищи
жвачных, а также пектиновые вещества и
лигнин, а получающиеся продукты сбра
живают. Продукты брожения — в основ
ном органические кислоты — нейтрали
зуются обильно выделяемой слюной (у
коровы, например, в сутки ее выделяется
десятки литров), что создает подходящую
ферментативную среду для жизнедея
тельности микроорганизмов.
Микроорганизмы рубца синтезируют
также белок, используя в качестве источ
ника азота соли аммония.
Переваривание растительных воло
кон облегчается и измельчением в про
цессе дополнительного жевания. Живот
ное отрыгивает содержимое рубца в ро
товую полость и вновь начинает переже
вывать («жевать жвачку»).
Обработанная таким образом пища
вторично проглатывается и на этот раз
попадает сначала в книжку, а потом в сы
чуг, который представляет собой настоя
щий желудок; реакция в нем, как и у всех
млекопитающих, кислая. Вместе с пищей
(продукты брожения растительной пищи,
белки, синтезированные бактериями и
др.) в сычуг попадают и сами микроорга
низмы. В сычуге происходит дальнейшее
переваривание пищи под действием вы
деляемых его железами соляной кислоты
и пепсина. Таким образом, в желудке под
действием ферментов происходит пере
варивание не только пищи, но и тел мик
роорганизмов, что дает некоторое допол
нительное количество белковых веществ.
Корова, например, получает с телами
микроорганизмов более 100 г белка в
сутки. Благодаря использованию белка
симбиотических организмов, жвачные
практически не нуждаются в белковой
пище. Кроме того, жвачные получают от
микроорганизмов и некоторые витамины
группы В. Из сычуга через его пилори
ческую часть пища поступает в кишеч
ник, где происходит окончательное ее пе
реваривание и всасывание.
Таким образом, благодаря наличию
сложного желудка и обитанию в нем сим
биотических микроорганизмов пищева
рение жвачных является наиболее эф
фективным среди растительноядных жи
вотных. Полагают, что именно это пре
имущество предоставило жвачным воз
можность широкого распространения за
счет вытеснения нежвачных животных.
?
> 1. Каковы особенности растительной пищи?
2. Какие трудности испытывают животные при пе
реваривании растительной пищи? 3. Каким образом
решаются проблемы переваривания растительной
пищи у разных организмов? 4. Каковы принципи
альные отличия в пищеварении жвачных и
нежвачных животных? 5. Почему жвачные живот
ные не испытывают нужды в высокобелковой
пище?
45. Транспорт веществ
в организмах животных
и человека
Почти у всех животных имеется спе
циальная система для транспорта и рас
пределения веществ в организме. Ис
ключение составляют кишечнополос
тные, плоские и круглые черви. Внутри
тела этих организмов вещества проходят
настолько незначительные расстояния,
что вполне могут перемещаться путем
диффузии. У планарии, например, ки
шечник сильно разветвлен, и его ветви
отходят ко всем участкам тела (см.
147
рис. 88). Это сокращает путь питатель
ных веществ ко всем клеткам животного.
С увеличением размеров тела и слож
ности строения организма возрастает и
количество веществ,
поступающих
внутрь. Расстояния, которые приходится
проходить веществам в организме, тоже
увеличиваются, и для их распределения
уже недостаточно простой диффузии.
При значительном удалении клеток тела
от места всасывания питательных ве
ществ (кишечника) пассивная диффузия
их слишком медленна и не обеспечивает
своевременного поступления веществ.
В связи с этим в процессе исторического
развития животных сформировалась си
стема кровообращения. Транспорт ве
ществ осуществляется здесь главным об
разом путем конвекции в системе трубок
(кровеносная система), по которой
циркулирует жидкость (кровь, гемолим
фа). Кровеносная система служит для
транспорта газов, участвующих в процес
се дыхания, питательных веществ, про
дуктов распада, гормонов, антител и со
лей. Кроме того, у теплокровных живот
ных с током крови переносится и равно
мерно распределяется в организме тепло,
что способствует сохранению постоянной
температуры тела и предупреждает пе
регрев глубоко расположенных органов.
У животных различных групп наблю
даются разные типы кровеносных систем.
Однако, несмотря на разнообразие этих
типов, в любой кровеносной системе
можно выделить основные элементы, вы
полняющие у всех животных сходные
функции.
1. Главный сократительный орган, слу
жащий для проталкивания крови по
организму. В большинстве случаев та
ким органом является сердце.
2. Артериальная система, отвечающая
за распределение крови и играющая
роль напорного резервуара.
3. Капилляры, благодаря которым про
исходит перенос веществ между
148
кровью и клетками других тканей ор
ганизма.
4. Венозная система, представляющая
собой резервуар для крови и обеспе
чивающая ее возврат к сердцу.
Кровь движется по организму под
действием сил, создаваемых ритмичными
сокращениями сердца, сдавливанием со
судов при движении тела и (или) перис
тальтическими сокращениями гладких
мышц в стенках сосудов. Вклад каждого
из этих факторов в создание тока крови
различен у разных животных. Так, у поз
воночных главную роль в кровообраще
нии играет сердце, расположенное с
брюшной стороны, у членистоногих столь
же важны движения конечностей и со
кращения сердца, расположенного у них
на спинной стороне. У кольчатых червей
продвижение крови из задней части тела
в переднюю и наполнение боковых сер
дец обеспечивается главным образом за
счет перистальтических сокращений
спинного кровеносного сосуда. У всех
животных однонаправленный ток крови
обеспечивается клапанами или перего
родками, а просвет сосудов регулируется
окружающими эти сосуды гладкими
мышцами, что дает возможность управ
лять количеством крови, протекающей
по тому или иному сосудистому руслу и
тем самым перераспределять кровоток в
организме.
S.
Что обусловило возникновение кровенос
системы? 2. Каковы основные функции кровенос
ной системы? 3. Охарактеризуйте роль каждого из
элементов системы кровообращения. 4. Благодаря
чему осуществляется движение крови у разных
групп животных? 5. Почему кровь по сосудам дви
жется в одном направлении?
46. Типы кровеносных систем
У животных встречается два типа кро
веносной системы — замкнутый и от
крытый (незамкнутый).
Впервые кровеносная система возникает у
кольчатых червей, относящихся к вторичнополос
тным животным. Благодаря наличию вторичной по
лости тела (целома) стенки тела отделены у них от
внутренних органов. Однако с появлением вторич
ной полости тела появляется необходимость в при
способлении, которое связывало бы разделенные
ею области и обеспечивало обмен питательными
веществами, газами и продуктами метаболизма
между кишкой и окружающей средой. Именно поэ
тому на уровне вторичнополостных появляется кро
веносная система, осуществляющая связь между
пищеварительной трубкой и стенками тела.
При замкнутой кровеносной системе
кровь циркулирует в замкнутой полости,
в пределах которой она разносится от
сердца (сердец) по проводящим каналам
к органам и тканям и затем, не выходя из
этой полости, возвращается к сердцу.
У кольчатых червей кровеносная сис
тема состоит из спинного и брюшного со
судов, соединенных между собой на пе
реднем конце тела пятью парами так на
зываемых «сердец» (рис. 95). Эти сердца
представляют собой сокращающиеся
участки кольцевых кровеносных сосудов,
снабженные клапанами и действующие
как насосы. Каждое сердце имеет по че
тыре клапана и пропускает кровь только
в сторону брюшного сосуда. Кровотоку
способствуют также перистальтические
движения самих сосудов, в первую оче
редь спинного. Мелкие разветвления со
судов — капилляры ■
— пронизывают все
ткани организма червя. Хотя у дождевых
1
2
з
Рис. 95. Замкнутая система кровообращения
дождевого червя'. 1, 2 — сердца, 3 — спинной
сосуд, 4 — брюшной сосуд.
2
Рис. 96. Кровеносная система рыб: 1 — жабер
ные сосуды, 2 — спинная аорта, 3 — сердце,
4 — брюшная аорта.
червей кровь красная (что придает соот
ветствующую окраску телу червя), одна
ко у них в отличие от позвоночных нет
эритроцитов: гемоглобин находится в
крови в растворенном состоянии. Погло
щение питательных веществ осуществля
ется капиллярами, густо оплетающими
кишечник, а кислорода — капиллярами,
пронизывающими кожу.
Кроме кольчатых червей, замкнутая
кровеносная система характерна для го
ловоногих моллюсков, иглокожих и хор
довых животных (у последних она устрое
на наиболее сложно). Главным напорным
органом является сердце, которое выбра
сывает кровь в артерии и поддерживает в
них высокое кровяное давление. Артери
альная система в свою очередь играет
роль напорного резервуара, подающего
кровь в капилляры.
У низших позвоночных, например рыб, кровь
по артериям проталкивается к жабрам. Здесь кровь
обогащается кислородом, а затем попадает в другой
сосуд (спинную аорту), который, разветвляясь, раз
носит кровь к остальным органам и тканям. К сер
дцу кровь возвращается по венам. Таким образом, у
этих животных имеется один круг кровообраще
ния (рис. 96). Эта система обладает тем преиму
ществом, что вся кровь, приходящая к клеткам те
ла, обогащена кислородом. Недостатком ее являет
ся то, что кровь, проходящая через капиллярное
русло в жабрах, находится под низким давлением, а
149
в остальной части системы давление еще ниже. Это
в целом снижает скорость поступления кислорода и
питательных веществ в клетки и ограничивает ско
рость клеточного метаболизма, т. е. скорость всех
биохимических процессов, поскольку клеточное ды
хание поставляет энергию, необходимую для всех
остальных метаболических процессов.
У животных, дышащих легкими, силы кровяно
го давления недостаточно для проталкивания крови
по организму после прохождения ее по узким сосу
дам легких. Поэтому проблема низкого кровяного
давления в капиллярах устраняется благодаря нали
чию двух кругов кровообращения: из легких кровь
направляется снова к сердцу, оттуда уже разносится
по всему организму. Естественно, что у таких жи
вотных сердце должно состоять из большего числа
камер, чем у рыб. У земноводных и пресмыкающих
ся сердце трехкамерное, с двумя предсердиями и од
ним желудочком. В левое предсердие поступает
кровь из легких, а в правое — от всего тела. У жи
вотных с двумя кругами кровообращения сердце,
таким образом, разделено на две половины: в одну
поступает бедная кислородом кровь от организма, а
в другую — богатая кислородом кровь из легких
(рис. 97). Без такого разделения кровь, поступаю
щая из легких и остальных органов в сердце, сме
шивалась бы, и в результате часть обогащенной
кислородом крови возвращалась бы снова к легким,
тогда как бедная кислородом кровь разносилась бы
к органам и тканям. У земноводных и пресмыкаю
щихся в сердце один желудочек, однако благодаря
особенностям строения желудочка и кровеносных
сосудов, отходящих от сердца, кровь полностью не
смешивается. У птиц, млекопитающих и человека
появляется второй желудочек, происходит полное
разделение сердца на правую и левую половины
(рис. 98) и соответственно разделение кровотоков
на венозный и артериальный.
Наличие двух кругов кровообращения дает два
преимущества. Во-первых, кровь, проходя по ле
гочным капиллярам, находится под очень низким
давлением, так как мышечные стенки правого ж е
лудочка примерно втрое тоньше стенок левого же
лудочка. У человека кровь, поступающая в легоч
ную артерию, находится под давлением около
16 мм рт. ст. Если бы давление в легочных капилля
рах было высоким, из крови в легкие поступало бы
очень много жидкости, наполняя легкие водой. Во150
вторых, кровь от легких снова проходит через сер
дце, где ее давление значительно повышается, что и
обеспечивает продвижение крови ко всем клеткам
тела. Высокое давление необходимо также для того,
чтобы проталкивать плазму с растворенными в ней
веществами из капилляров во внеклеточную жид
кость, а также нагнетать в почки, где кровь очища
ется.
Суммарная скорость циркуляции кро
ви определяется главным образом силой
и частотой сердечных сокращений. Через
сердце человека прокачивается около 5 л
крови в минуту во время сна и в пять раз
больше при тяжелой физической нагруз
ке. У большинства людей сердце сокра
щается от 60 до 80 раз в минуту (частота
пульса). Однако у спортсменов в резуль
тате тренировоксобъем крови, проходя
щей через сердце при каждом сокраще
нии, возрастает так, что у бегуна частота
пульса может достигать в состоянии по
коя только 40 ударов в минуту, и при этом
через сердце будет проходить, как обыч
но, 5 л крови.
Кровяное давление в момент сокра
щения желудочков сердца называется
систолическим давлением, а минималь
ное давление в промежутке между сокра
щениями диастолическим. У взрослого
человека в нормальном состоянии систо
лическое давление составляет около
120 мм рт. ст., а диастолическое — около
80. Самое высокое давление — в артери
ях, отходящих от сердца; затем по мере
замедления движения крови вследствие
трения о стенки сосудов в процессе кро
вообращения давление постепенно пони
жается.
Стенки капилляров тонкие, поэтому в
них может происходить быстрый обмен
веществами между кровью и тканями.
Плотность капилляров в тканях велика, и
кащцая клетка отделена от ближайшего
капилляра не более чем двумя-тремя
клетками. Стенки капилляров состоят из
одного слоя клеток, причем между сосед
ними клетками есть узкие щели, благода-
Рис. 97 Общая схема кровеносной систе
мы млекопитающих: ПП — правое пред
сердие, ЛП — левое предсердие, ПЖ —
правый желудочек, ЛЖ — левый желу
дочек.
13
Рис. 98. Внутреннее строение сердца чело
века: 1 — аорта, 2 — клапан аорты, 3 — ле
вое предсердие, 4 — передняя (обрезана) и
5 — задняя створки двустворчатого клапа
на, 6 — левый желудочек, 7 — межжелудоч
ковая перегородка, 8 — сосочковая мышца,
9 — правый желудочек, 10 — сухожильные
нити, 11 — нижняя полая вена, 12 — задние
створки трехстворчатого клапана, 13 •—
правое предсердие, 14 — клапан легочного
ствола, 15 — верхняя полая вена.
151
ря чему вода с растворенными в ней соля
ми, питательными и другими веществами
поступает в межклеточные пространства
и далее в клетки.
Капиллярные сети соединены между
собой параллельно, и благодаря этому
возможна точная регуляция перераспре
деления крови между различными тка
нями.
По венозной системе кровь оттекает
от капилляров к сердцу. Давление в венах
обычно низкое, а стенки вен весьма элас
тичны, и поэтому большие изменения
объема крови в этих сосудах лишь незна
чительно сказываются на давлении в них.
Таким образом, венозная система играет
роль вместительного резервуара. Именно
из этого резервуара берут кровь у доно
ров при переливании или вводят в него
лекарства во время лечения,а поскольку
при этом давление в венах лишь незначи
тельно изменяется, объем крови и ско
рость ее протекания в других местах сис
темы кровообращения также не претер
певает существенных изменений. Стенки
вен содержат сравнительно мало мышеч
ных волокон и легко растягиваются. По
а
этому сами вены не принимают активно
го участия в возврате крови в сердце.
Кровь движется по венам отчасти под
действием кровяного давления, отчасти
под присасывающим действием предсер
дий, но в основном в результате сокраще
ния прилегающих мышц. При сокраще
нии мышц (например, конечностей) вя
лые стенют вен сжимаются и кровь про
талкивается по направлению к сердцу.
Когда же мышцы расслабляются, имею
щиеся в венах полулунные клапаны
препятствуют движению крови в обрат
ном направлении (рис. 99).
У многих беспозвоночных кровенос
ная система открытого типа (незамкну
тая). В такого рода системах кровь, или
гемолимфа, выбрасывается сердцем че
рез артерию в гемоцель, который пред
ставляет собой сеть заполненных кровью
незамкнутых полостей, называемых си
нусами, в которых лежат внутренние ор
ганы, Гемолимфа, попавшая в гемоцель,
не поступает в капилляры, а непосред
ственно омывает ткани. Вены, по кото
рым кровь поступала -бы в сердце, отсут
ствуют. Кровь из гемоцеля возвращается
б
в
Рис. 99. Схема работы клапанов в венозных сосудах: а — мышцы расслаблены, клапаны закрыты;
б — мышцы сокращаются, верхний клапан открывается, и кровь выталкивается вверх, нижний кла
пан закрыт; в — мышцы расслабляются, верхний клапан закрывается, а нижний открывается вслед
ствие сокращения мышц в нижележащих участках, и кровь проходит: 1 — верхний клапан, 2 — ниж
ний клапан, 3 — мышца, 4 — вена.
152
даря непосредственному контакту крови с
клетками эффективность выполнения
функций кровообращения достаточно ве
лика, что обеспечивает своевременный
приток питательных веществ ко всем
клеткам тела.
?9
Рис. 100. Незамкнутая система кровообраще
ния рака: 1 — артерии, 2 — сердце, 3 —
клапан.
к сердцу и поступает в него через клапа
ны, открывающиеся в момент расслабле
ния сердца. Такая система кровообраще
ния развилась у большинства членистоногих(рис. 100), моллюсков (кроме голо
воногих) и оболочников.
Важнейшими особенностями незам
кнутой системы кровообращения явля
ются обычно высокое давление крови,
трудности регулирования ее распределе
ния и медленное возвращение крови к
сердцу.
Давление крови в открытых системах не превы
шает 4,5— 13,5 мм рт. ст. Более высокое давление в
таких системах встречается лишь в исключительных
случаях, например, в определенных частях тела у
наземных улиток, некоторых водных моллюсков и
др. У этих животных высокое давление создается
благодаря сокращениям сердца либо мышц, его
окружающих. Физиологический смысл высокого
давления в некоторых отделах кровеносной системы
открытого типа состоит в том, что благодаря такому
давлению некоторым органам и отдельным частям
тела придается тургор, т. е. кровь выполняет фун
кцию гидростатического скелета. Так, например,
движение беззубки было бы невозможным без пе
риодического нагнетания в промежутки тканей ноги
гемолимфы. У пауков только давление гемолимфы
обеспечивает разгибание члеников конечностей,
так как мышцы-разгибатели отсутствуют.
Замкнутая система кровообращения
более динамична. Однако это не означа
ет, что открытая система кровообраще
ния не имеет своих преимуществ. Благо
1. Что такое кровяное давление (систоличес
кое, диастолическое), как оно возникает? 2. Где и
каким образом осуществляется обмен веществ
между кровью и другими тканями организма? 3. По
чему изменение количества крови в венах не вызы
вает существенного изменения давления в них?
4. Благодаря чему кровь передвигается по венам?
47. Кровь, тканевая жидкость
и лимфа
Кровь — это жидкая ткань организ
ма, которая состоит из жидкой среды —
плазмы и взвешенных в ней клеток —
форменных элементов крови. Состав
крови у разных организмов различен.
У человека кровь составляет около
8 % от общей массы тела. Так, у челове
ка массой 80 кг содержится примерно
6,5 л крови.
Клетки (форменные элементы) сос
тавляют примерно 45 % от общего объе
ма крови у мужчин и 40 % — у женщин;
остальной объем приходится на плазму.
Плазма — это бледно-желтая жид
кость, 90 % которой составляет вода, а
10 % — растворенные и взвешенные в
ней вещества (9 % — белки и 0,87 % —
минеральные соли). Именно плазма осу
ществляет перенос различных веществ и
клеток (pH плазмы составляет 7,3).
К белкам плазмы относятся фибриноген
и протромбин — два компонента, необ
ходимые для нормального свертывания
крови. Кроме них, в плазме имеются гло
булины. а-глобулин связывает тироксин
и билирубин, ф-глобулин связывает же
лезо, холестерол и витамины A, D и К.
у-глобулин связывает антигены и играет
важную роль в иммунологических реак
153
циях организма, у-глобулины обычно на
зывают антителами.
Наконец, в плазме содержится еще
один белок — альбумин, придающий ей
вязкость. Кроме того, альбумин связыва
ет присутствующий в крови кальций.
Клетки крови (рис. 101). Среди них
больше всего красных кровяных кле
ток — эритроцитов. У человека это
мелкие клетки, лишенные ядра и имею
щие форму двояковогнутых дисков. Диа
метр эритроцитов составляет в среднем
около 7— 8 мкм и приблизительно равен
диаметру самых узких капилляров. Спе
цифическая форма эритроцитов обуслов
ливает более высокое отношение повер
хности к объему, что увеличивает воз
можность газообмена. Благодаря элас
тичности мембран эритроциты могут про
ходить через капилляры, просвет кото
рых даже меньше диаметра эритроцитов.
В кубическом миллиметре крови муж
чины содержится примерно 5 млн эрит
роцитов, а в крови женщины — 4,5 млн,
но эти величины могут варьировать в за
висимости от возраста и состояния здо
ровья человека. Эритроциты вырабаты
ваются в красном костном мозге плоских
костей грудины, черепа, в ребрах, поз
вонках, ключицах и лопатках, а также в
головках длинных трубчатых костей. У
эмбриона на той стадии развития, когда
кости еще не сформировались, эритроци
ты образуются в печени и селезенке.
Важнейшей особенностью эритроци
тов является присутствие в них гемогло
Рис. 101. Форменные элементы крови.
154
бина, который связывается с кислородом
(превращаясь в оксигемоглобин) в учас
тках с высокой концентрацией 0 2 (лег
ких) и отдает его в участках с низкой кон
центрацией 0 2 (ткани). В эритроцитах
содержится также фермент карбоангидраза, участвующий в транспорте углекис
лого газа.
Продолжительность жизни эритроци
тов у взрослых людей составляет около
трех месяцев, после чего они разрушают
ся в печени или селезенке. Белковые
компоненты эритроцитов расщепляются
на аминокислоты, а железо, входящее в
состав гема, удерживается печенью и
хранится в ней в составе белка ферритина. Железо может в дальнейшем исполь
зоваться при образовании новых эритро
цитов и при синтезе цитохромов. Осталь
ная часть гемоглобина расщепляется с
образованием пигментов билирубина и
биливердина, которые вместе с желчью
выводятся в кишечник и придают окраску
каловым массам.
В среднем в организме человека со
держится около 20 млрд эритроцитов; их
образование и разрушение происходит с
постоянной скоростью: примерно 115
млн клеток в минуту. Правда, в условиях
низкого содержания кислорода в красном
костном мозге может образовываться
большее количество эритроцитов, неже
ли разрушается. Этот механизм лежит в
основе акклиматизации млекопитающих
к пониженному содержанию кислорода в
высокогорье.
Уменьшение числа эритроцитов или
снижение содержания в них гемоглобина
на 10 % ведет к развитию анемии: возни
кает кислородная недостаточность и, как
следствие, снижается интенсивность
синтеза АТФ.
Лейкоциты — белые клетки кро
ви — по размеру больше эритроцитов, но
в крови их содержится значительно мень
ше. Обычно в 1 мм3 крови насчитывается
в среднем около 8000 лейкоцитов. Они
играют важную роль в защите организма
от болезней. Все лейкоциты имеют ядро и
способны к делению, однако продолжи
тельность их жизни, как правило, неве
лика и составляет лишь несколько дней.
В то же время некоторые виды лейко
цитов (например, лимфоциты) мо
гут жить на протяжении десяти и бо
лее лет.
Лейкоциты способны к амебоидному
движению, что позволяет им проходить
сквозь небольшие отверстия в стенках
капилляров. Поэтому лейкоциты обнару
живаются не только в крови, но и в меж
клеточном пространстве тканей. Сущес
твует два основных типа лейкоцитов: зер
нистые (гранулоциты) и незернистые (агранулоциты).
Зернистые лейкоциты, или грану
лоциты, содержат в цитоплазме харак
терные гранулы (зерна) и ядра, разделен
ные на лопасти (доли). Лопасти в числе
двух или трех связаны между собой тон
кими перемычками. Кроме того, в гранулоцитах у женщин от одной из долей ядра
отходит отросток, имеющий форму бара
банной палочки; по этому признаку мож
но отличить кровь женщины от крови
мужчины. Гранулоциты играют особенно
важную роль в борьбе с проникающими в
организм микроорганизмами: обычно
гной, образующийся в очагах воспале
ния, содержит множество гранулоцитов.
Гранулоциты, как и эритроциты, образу
ются в красном костном мозге.
Гранулоциты можно разделить на три
вида в зависимости от окрашивания гра
нул цитоплазмы определенными красите
лями.
Н е й т р о ф и л ы составляют около
70 % всех лейкоцитов. Содержащиеся в
них мелкие гранулы окрашиваются обыч
но основными (синими) и кислыми (крас
ными) красителями в фиолетовый цвет.
Функция нейтрофилов — захват и пере
варивание бактерий.
Э о з и н о ф и л ы содержат в цито
плазме крупные гранулы, которые окра
шиваются при обработке кислым краси
телем эозином в красный цвет. Они сос
тавляют 1,5 % от общего числа лейкоци
тов, но при аллергических состояниях
(например, при астме или сенной лихо
радке) их количество резко возрастает.
Функция эозинофилов —• поглощение
комплексов антиген — антитело Б, при
чем делают они это эффективнее других
клеток.
Б а з о ф и л ы содержат крупные гра
нулы, которые окрашиваются в синий
цвет основными красителями типа мети
ленового синего. Базофилы составляют
около 0,5 % всех лейкоцитов. Они выра
батывают гепарин (сульфатированный
мукополисахарид, природный ингибитор
системы свертывания крови) и гистамин
(гормон — важнейшее звено в механиз
мах регуляции тонуса гладких мышц и
выделения желудочного сока). Нейтро
филы, эозинофилы и базофилы еще на
зывают микрофагами.
Незернистые лейкоциты, или агранулоциты, содержат крупное округ
лое или овальное ядро, которое может за
нимать почти всю клетку, и незернистую
цитоплазму. Существует два типа агранулоцитов: моноциты и лимфоциты.
М о н о ц и т ы ( м а к р о ф а г и ) — са
мые крупные из лейкоцитов; подобно
другим лейкоцитам, они вырабатываются
в красном костном мозге. Количество их
обычно невелико — около 4— 5 % от об
щего числа лейкоцитов, но при некото
рых заболеваниях их количество увели
чивается. Моноциты активно фагоцити
155
руют бактерии и другие крупные частицы.
Они способны мигрировать через стенки
капилляров в очаги воспаления, где дей
ствуют так же, как нейтрофилы.
Л и м ф о ц и т ы — самые мелкие из
лейкоцитов: по размеру они немногим
больше эритроцитов. Предшественники
лимфоцитов образуются в красном кос
тном мозге. Лимфоциты имеют округлую
форму и содержат очень мало цитоплаз
мы. Функция лимфоцитов состоит в том,
что они вырабатывают антитела в ответ
на попадание в организм чужеродного
белка. Лимфоциты составляют около
24 % от общего числа лейкоцитов.
В редких случаях в красном костном
мозге начинается неконтролируемое об
разование патологически измененных
лейкоцитов. Содержание их в 1 мм3 кро
ви может достигать 500 000 и более. Это
заболевание называется лейкозом.
Тромбоциты (кровяные пластинки).
Если приготовить мазок крови и исследо
вать его под микроскопом, то обычно в
нем видны лишь два типа клеток — эрит
роциты и лейкоциты. Существует, одна
ко, еще один тип форменных элемен
тов крови — тромбоциты; в мазке
они не видны потому, что при сопри
косновении с поверхностью стекла разру
шаются.
Тромбоциты представляют собой
фрагменты клеток, имеющие неправиль
ную форму, окруженные мембраной и
обычно лишенные ядра. Образуются они
в красном костном мозге из крупных кле
ток, называемых мегакариоцитами.
В 1 мм3 крови находится примерно
250 000 тромбоцитов. В тромбоцитах со
держатся вещества, которые играют
важную роль в свертывании крови.
Тканевая жидкость и лимфа. Ткане
вая жидкость окружает все клетки орга
низма. По составу эта жидкость сходна с
плазмой, однако в ней намного ниже со
держание белка. Образуется тканевая
жидкость из плазмы крови. Когда кровь
из мелких артерий попадает в капилляры,
156
в них сохраняется достаточно высокое
давление (около 32 мм рт. ст.), в резуль
тате чего вода и растворенные в ней ве
щества выходят из капилляров в мель
чайшие промежутки между клетками.
Стенки капилляров проницаемы для всех
компонентов крови, за исключением
эритроцитов и белков плазмы.
Роль тканевой жидкости состоит в
обеспечении клеток питательными ве
ществами, кислородом, гормонами и др.,
а также в удалении конечных продуктов
клеточного обмена. Капиллярная сеть
подходит не ко всем клеткам, но каждая
клетка омывается тканевой жидкостью.
Благодаря такому непосредственному
контакту обмен между кровью и клетками
тела происходит значительно более эф
фективно, чем только через стенки ка
пилляров.
Кровь не может постоянно терять
часть плазмы в связи с образованием
тканевой жидкости, и значительная часть
этой жидкости возвращается обратно в
кровяное русло. Это происходит двумя
путями. У венозного конца капиллярной
сети кровяное давление становится зна
чительно ниже, чем со стороны мелких
артерий (там оно падает до 12 мм рт. ст.),
что способствует обратному поступлению
воды и растворенных в ней веществ из
тканевой жидкости путем диффузии.
Большая же часть тканевой жидкости
диффундирует в замкнутые с одного кон
ца лимфатические капилляры и образует
лимфу. Лимфатические капилляры, со
единяясь между собой, образуют более
крупные лимфатические сосуды, которые
в конечном итоге открываются в кровя
ное русло. Совокупность лимфатических
сосудов образует лимфатическую систе
му. Лимфатические сосуды, идущие от
ног, объединяясь с лимфатическими со
судами пищеварительного тракта, обра
зуют грудной проток. Последний от
крывается в кровяное русло в области
шеи через левую подключичную вену.
Правый лимфатический проток изли
вает лимфу в кровь через правую под
ключичную вену (рис. 102).
По ходу лимфатических сосудов в не
которых местах расположены лимфати
ческие железы, или лимфоузлы. Лейко
циты, содержащиеся в лимфоузлах, по
глощают микроорганизмы и различные
чужеродные вещества, попавшие в орга
низм. При проникновении в организм
большого количества бактерий лимфа
тические узлы увеличиваются и уплотня
ются.
Лимфатическая система служит не
только для возврата жидкости, но и бел
ков, которые отфильтровались в ткани
через стенки капилляров. Через эту сис
тему в кровь поступают вещества с круп
ными молекулами, в частности жиры,
ной проток, 3 — лимфатические узлы.
всосавшиеся в кишечнике, а также высо
комолекулярные гормоны. Кроме того,
лимфатическая система участвует в за
щите организма от инфекций.
Движение лимфы осуществляется за
счет сокращения мышц, окружающих
лимфатические сосуды. Вот почему физи
ческая активность совершенно необходи
ма для нормальной циркуляции тканевой
жидкости и лимфы. Застой этих жидкос
тей нарушает правильный обмен ве
ществ. Обратному току лимфы в сосудах
препятствуют клапаны, которые имеются
в крупных сосудах и действуют подобно
клапанам вен.
Величина лимфотока может быть различной.
Обычно она не превышает 2 мл/мин. Однако, хотя
лимфоток и невелик, он очень важен для освобож
дения тканей от избыточной жидкости. Если лимфы
образуется больше, чем оттекает, то жидкость за
держивается в тканях и возникает отек.
Особенности крови животных.
У кольчатых червей, как мы уже отме
чали, гемоглобин находится в крови в
растворенном виде. Кроме того, в крови
циркулируют бесцветные амебоидные
клетки, выполняющие защитную фун
кцию.
У членистоногих кровь (гемолимфа)
бесцветная; она не содержит гемоглоби
на и служит только для транспорта пита
тельных веществ и продуктов метаболиз
ма, подлежащих экскреции. В крови чле
нистоногих циркулируют бесцветные
амебоидные лейкоциты. Крабы и омары
имеют сине-зеленый пигмент крови, ге
моцианин, который, в отличие от гемо
глобина, вместо железа содержит медь.
Гемоцианин присутствует также в крови
моллюсков.
У рыб, амфибий, рептилий и птиц в
крови имеются эритроциты, которые со
держат гемоглобин. Их отличительной
особенностью является наличие ядра (в
отличие от эритроцитов человека). Кроме
того, у данных организмов, как и у млеко
157
питающих, имеется лимфатическая сис
тема. Причем для многих рыб, земновод
ных и пресмыкающихся характерны лим
фатические сердца, способствующие
продвижению лимфы. У млекопитающих
и большинства птиц таких сердец нет, и
лимфа продвигается благодаря сокраще
нию прилегающих мышц.
7
„
1. Что представляет собой кровь и каков ее хи
мический состав? 2. Каковы особенности строения
и функции эритроцитов? 3. Каковы особенности
строения и функции разных видов лейкоцитов? 4.
Проследите, каким образом осуществляется тран
спорт питательных веществ из кишечника к клеткам
тела. 5. Что представляет собой тканевая жидкость
и каковы ее функции? 6. Каково назначение лимфа
тической системы в организме человека?
48. Защитные функции крови
Одной из важнейших функций крови
является защитная, в которой участвуют
три механизма:
а) свертывание крови, предотвра
щение кровопотери при травмах кро
веносных сосудов и проникновении в
организм болезнетворных агентов;
б) фагоцитоз, осуществляемый
клетками крови, захватывающими и
обезвреживающими попавших в кро
вяное русло микроорганизмов;
в) иммунная защита, осуществляе
мая антителами и лимфоцитами.
Свертывание крови. При поврежде
нии кровеносных сосудов из них вытекает
кровь, которая вскоре свертывается, об
разуя кровяной сгусток. Последний
препятствует дальнейшей потере крови и
проникновению микроорганизмов в кро
вяное русло. Очевидно, что этот процесс
имеет большое значение для выживания
животного. Не менее важно и то, что в
неповрежденных сосудах кровь не свер
тывается. Механизм свертывания крови
очень сложен. Его многостадийность слу
158
жит, вероятно, и для того, чтобы этот
процесс не происходил тогда, когда в нем
нет необходимости. Для его осуществле
ния требуется тринадцать различных
факторов крови, действующих согласо
ванно. Эти факторы обозначаются рим
скими цифрами от I до XIII в порядке их
хронологического открытия. Мы рас
смотрим только важнейшие из этих фак
торов.
Сущность ферментативного процесса
свертывания крови заключается в пере
ходе растворимого белка плазмы крови
фибриногена в нерастворимый белок
фибрин, образующий основу кровяного
сгустка — тромба.
При травмировании тканей происхо
дит разрыв стенок сосудов. Кровь, выте
кающая из раны, соприкасается с возду
хом и смешивается с веществами, выде
ляющимися из поврежденных клеток и
разрушенных при этом тромбоцитов.
Высвобождающимся из поврежденных
клеток веществом является липопротеин
тромбопластин, который совместно с
определенными плазменными факторами
(V, VII, X) и ионами Са2+ способствует об
разованию фермента протромбиназы
( фа з а I). Протромбиназа при участии
факторов V, X и Са2+ превращает неак
тивный фермент протромбин, образуе
мый клетками печени при участии вита
мина К, в активный фермент тромбин
( фа з а II). Тромбин — протеолитичес
кий фермент; он расщепляет крупные мо
лекулы растворимого глобулярного бел
ка плазмы фибриногена на более мелкие
единицы, которые затем полимеризуются
и образуют сеть, состоящую из длинных
перепутанных нитей фибрина — нерас
творимого фибриллярного белка ( фа з а
I I I ) (рис. 103). В сети, образованной во
локнами фибрина, задерживаются фор
менные элементы крови и в результате
образуется кровяной сгусток. Позднее он
сжимается, подсыхает и образует
ся струп, который препятствует поте
ре крови и создает механический
г
Ранение
-<
Разрушение
тромбоцитов
Соприкоснове
ние с воздухом
Повреждение /
ткани
Тромбопластин
Факторы
плазмы VII,V,X!
\L z .
Протромбиназа
Ш
ш
•S '
Протромбин
плазмы (образуется
в печени)
Тромбин
Факторы
плазмы V,X
2+
оа
Фибриноген
(образуется в печени
Фибрин
(сгусток)
Рис. 103. Основные этапы процесса свертывания крови.
барьер для проникновения микроорга
низмов.
В неповрежденных кровеносных со
судах кровь не свертывается, поскольку
изнутри поверхность сосудов очень глад
кая и не происходит разрушения формен
ных элементов крови. Кроме того, в кро
ви имеется и особая антисвертывающая
система, представленная активными ве
ществами, предотвращающими сверты
вание. Одно из них — гепарин — сульфатированный мукополисахарид, образу
емый базофилами и тучными клетками
соединительной ткани, расположен
ными главным образом вдоль стенок
кровеносных сосудов. Гепарин препят
ствует превращению фибриногена в
фибрин.
Если сгусток образуется внутри кровеносного
сосуда, он называется тромбом, и его образование
ведет к острому нарушению кровообращения —
тромбозу. Тромбоз может возникать как след
ствие повреждения внутренней стенки сосудов,
так как шероховатость поврежденного участка
способствует разрушению тромбоцитов, что ини
циирует процесс свертывания крови. Особенно
опасен и может привести к внезапной смерти
тромбоз венечной артерии сердца (инфаркт мио
карда), мозговых сосудов (инсульт), легочных ар
терий.
159
Фагоцитоз. Как уже отмечалось, фа
гоцитоз осуществляется фагоцитами,
главным образом нейтрофилами. Защит
ную функцию клеток, способных погло
щать и переваривать микробы, впервые
показалИ. И. М еч ни ко в . Среди фа
гоцитов он различал микрофаги (нейтро
филы, эозинофилы и базофилы) и макро
фаги (моноциты). Фагоциты представля
ют собой амебоидные клетки, способные
активно мигрировать в места поврежде
ния клеток и тканей организма, где обна
руживаются микроорганизмы. Стимулом
для их передвижения являются, вероят
но, какие-то вещества, выделяющиеся
при разрушении клеток. Фагоцит при
ближается к микробу и прилипает к нему.
Далее цитоплазматическая мембрана фа
гоцита впячивается и образуется фагосома, содержащая микроб. К фагосоме
присоединяются лизосомы, и под дей
ствием гидролитических ферментов мик
роб переваривается. Нейтрофилы погло
щают и переваривают в основном бакте
рий. Макрофаги фагоцитируют спирохет,
актиномицеты, грибы, протистов, виру
сы, атрофирующиеся, омертвевшие или
злокачественно перерожденные клетки.
Воспаление. При травмах (например,
сильном ушибе или разрыве тканей) ка
кого-либо участка тела возникает мес
тная реакция окружающих тканей, кото
рая проявляется в опухании, болезнен
ности и часто даже в повышении темпе
ратуры. Такое состояние называется вос
палением. Оно связано с выделением из
поврежденных тканей определенных ве
ществ, таких, как гистамин и серотонин.
Эти вещества вызывают местное расши
рение кровеносных сосудов, что ведет к
усилению притока крови к поврежденно
му участку. И, как следствие, повышению
температуры. Возрастает также прони
цаемость капилляров, в результате чего
плазма выходит в окружающие ткани и
вызывает их набухание — образуется
отек. В плазме содержатся бактерицид
ные факторы, антитела и нейтрофилы,
160
которые совместно противодействуют
распространению инфекции. В плазме
также присутствует фибриноген, кото
рый в случае повреждения кровеносных
сосудов участвует в свертывании крови.
Довольно часто во время ушиба травмируются
кровеносные сосуды и наблюдается подкожное кро
воизлияние. При этом разрушаются эритроциты и
образуются пигменты — продукты распада гема —
биливердин (зеленый) и билирубин (желтый).
Возникает синяк. Последовательное изменение
цвета синяка объясняется изменением соотноше
ния концентраций этих пигментов в месте распада
эритроцитов и их выведением.
Заживление ран. На завершающей
стадии воспалительного процесса в месте
ранения появляются клетки соединитель
ной ткани, называемые фибробластами,
которые секретируют коллаген. Колла
ген — это фибриллярный белок. Он со
единяется с полисахаридами и образует
сеть беспорядочно переплетающихся во
локон рубцовой ткани. Для образова
ния коллагена необходим витамин С.
Примерно через 2— 3 недели после ране
ния волокна рубцовой ткани реорганизу
ются и собираются в пучки, располагаю
щиеся вдоль линий натяжения в области
раны. Здесь начинают прорастать много
численные мелкие кровеносные сосуды,
которые снабжают кислородом и пита
тельными веществами клетки, участвую
щие в заживлении раны. Кроме того, в
заживление включаются эпидермальные
клетки — они мигрируют в рану. Встре
чаясь друг с другом, эпидермальные клет
ки соединяются, образуя сплошной слой
под струпом. Когда формирование этого
слоя заканчивается, струп отпадает.
7
> 1. В чем выражается защитная функция крови?
2. Какими механизмами обеспечивается защитная
функция крови? 3. Какова роль тромбоцитов в
свертывании крови? 4. Что представляет собой
отек, возникающий при ушибе? 5. Опишите после
довательность процесса заживления раны. Какова
роль крови в этом процессе?
49. Иммунная система
Иммунитет. Защитные функции кро
ви реализуются также через иммунную
систему. Иммунитет (от лат. immunitas — невосприимчивость) — это спо
собность распознавать появившийся в
организме чужеродный материал и моби
лизовать специальные клетки и образуе
мые ими вещества на быстрое и эффек
тивное удаление этого материала.
Чужеродным материалом могут быть
молекулы белков или полисахаридов, ко
торые находятся на поверхности клеток
микроорганизмов либо в свободном виде
(в этом случае эти вещества являются,
как правило, продуктами жизнедеятель
ности микроорганизмов). Кроме того, чу
жеродным материалом являются посто
ронние клетки, ткани, хирургически пе
ресаживаемые органы или изменившиеся
клетки собственного организма (напри
мер, раковые).
Иммунная система представлена в
организме тимусом, или вилочковой
железой, и красным костным мозгом
(центральные органы), лимфоузлами, се
лезенкой и лимфатическими скопления
ми в слизистых оболочках и внутренних
органах (периферические органы). Обес
печивают иммунитет находящиеся в этой
системе так называемые иммунокомпетентные клетки. Центральными назы
ваются те органы, в которых происходит
формирование и созревание иммунокомпетентных клеток, а периферичес
кими — органы, где эти клетки функцио
нируют.
Все позвоночные реагируют на чуже
родные тела, попадающие в организм
(помимо пищеварительного тракта), дву
мя основными способами (рис. 104, с.
162). Первый из них — гум оральны й
им м унит ет — это синтез и секреция
лимфоцитами и их потомками специфи
ческих белков — антител, или имму
ноглобулинов, которые перемещаются
по кровеносному руслу и действуют про
6 . Общая биология, 10 кл.
тив чужеродных субстанций и молекул —
антигенов, или иммуногенов.
Пространственная форма молекул
антител и распределение электрического
заряда по ее поверхности делают молеку
лу способной связывать антиген, компле
ментарный ей по форме и распределению
заряда. Такое специфическое связывание
сходно со специфичностью ферментов по
отношению к своим субстратам.
Второй способ —■ клет очный им
м унит ет , который осуществляется
прямым контактом лимфоцитов с чуже
родными антигенами, разрушающими их.
Такое разделение функций иммунной сис
темы связано с существованием двух ти
пов лимфоцитов — В-клеток(бурсазависимых) и Т-клеток (тимусзависимых).
Первые вырабатывают антитела, кото
рые служат эффекторами гуморального
иммунитета. Вторые антитела не выраба
тывают, а с помощью рецепторов, нахо
дящихся на их поверхности, распознают
антигены и либо уничтожают их с по
мощью специальных веществ, либо пере
дают сигнал и включают В- и Т-клетки в
антителообразование или другие эффекторные процессы.
Оба вида лимфоцитов образуются в
красном костном мозге из стволовых кле
ток-предшественниц. Окончательная
дифференциация в Т-лимфоциты проис
ходит в вилочковой железе (тимусе).
Тимус расположен в грудной клетке под груди
ной, у человека около передней стороны сердца. Он
начинает закладываться на шестой неделе эмбрио
нального развития и к моменту рождения ребенка
полностью созревает, что само по себе говорит о его
особом назначении.
Будущие Т-лимфоциты приобретают способ
ность выполнять свои защитные функции лишь пос
ле того, как пройдут через ткань тимуса.
В процессе созревания Т-лимфоциты
через лимфу и кровь перемещаются в
лимфатические узлы и селезенку, где в
случае встречи со специфическим аген
том узнают его и начинают делиться; так
161
А н ти ге н
.К л е т к и
пам яти
А ктивированны е
Т - лим ф оциты
(и м м у н о ц и т ы )
П л а зм ати чески е
кл е тки
(В -и м м у н о ц и т ы )
Р еакции,
осущ ествляем ы е
кл е ткам и
А нтитела
ГУМО РАЛЬНЫ Й
ИММУНИТЕТ
КЛЕТОЧНЫ Й
ИММУНИТЕТ
Рис. 104. Схема клеточного и гуморального иммунитетов.
образуются клоны идентичных клеток,
способных узнавать этот антиген и реаги
ровать с ним. Т-клетки регулярно перехо
дят из лимфоидных тканей в циркулирую
щую кровь и тканевую жидкость, благо
даря чему увеличивается вероятность их
встречи с антигеном и, следовательно,
его уничтожения.
По функциональным свойствам Тлимфоциты неоднородны. В зависимости
от реакций, которые они вызывают, раз
личают Т-киллеры (клетки-убийцы), Тхелперы (помощники) и Т-супрессоры
(регуляторы).
Т-киллеры способны разрушать ин
162
фицированные или злокачественно пере
рожденные клетки, поэтому их называют
еще цитотоксическими лимфоцитами.
Т-хелперы выполняют посредничес
кую сигнальную функцию, передавая ин
формацию об антигенах иммунокомпетентным клеткам. В гуморальном иммун
ном ответе Т-хелперы взаимодействуют с
антигеном и индуцируют превращение Влимфоцитов в плазмоциты (основные
продуценты антител). При этом хелперы
выделяют цитокины — вещества белко
вой природы, которые активируют деле
ние и трансформацию В-лимфоцитов.
В присутствии хелперов синтез антител
увеличивается на один-два порядка. Тхелперы индуцируют также образование
Т-киллеров. При СПИДе поражаются Тхелперы, что вызывает подавление имму
нитета.
Т-супрессоры являются регулятора
ми антителообразования и других иммун
ных процессов.
Стволовые клетки, которым предсто
ит стать В-лимфоцитами, должны подвер
гнуться дальнейшей дифференцировке за
пределами костного мозга. У птиц это
осуществляется в сумке (бурсе) Фабриция, которая располагается на задней по
верхности клоаки (отсюда и название
В-лимфоцитов — бурсозависимые). Уда
ление у цыплят сумки Фабриция приводит
к избирательному выключению способ
ности птиц синтезировать антитела, т. е.
блокирует В-систему иммунитета (при
сохранении клеточного иммунитета).
У млекопитающих В-лимфоциты прохо
дят дифференциацию, вероятно, в лимфо
идной ткани кишечника, аппендикса, мин
далинах, небно-глоточного кольца.
Основная функция В-лимфоцитов —
создание гуморального иммунитета путем
выработки антител. Когда поверхностные
рецепторы (иммуноглобулины) В-лимфоцитов узнают комплементарные им ан
тигены, В-лимфоциты начинают делиться
и дифференцироваться, образуя клоны
плазматических клеток (плазмоциты) и
«клетки памяти». Плазмоциты, иден
тичные друг другу, синтезируют большое
количество антител одного вида. И хотя
живут эти клетки всего лишь несколько
дней, но за это время они могут синтези
ровать около 2000 идентичных молекул
антител в секунду.
Лимфоциты позвоночных вырабаты
вают огромное количество различных ан
тител — число их типов оценивается в
сотни тысяч и даже миллионы.
Клетки памяти играют важную роль в
активации иммунного ответа организма
при повторном внедрении антигена, вы
звавшего формирование этих клеток. Об
этих клетках известно немного: живут
они в отличие от других лейкоцитов дол
го — 20 и более лет (некоторые на про
тяжении всей жизни организма) и позво
ляют организму очень быстро и энергич
но реагировать на повторное внедрение
любого антигена. Эта реакция носит наз
вание вторичного иммунного ответа
и приводит к массовому образованию ан
тител, которые быстро нейтрализуют
действие антигена.
Таким образом, в процессе иммунного
ответа, помимо эффекторных клеток,
формируется группа клеток иммунной
памяти, которые долго сохраняются и
распространяют информацию об антиге
не по всей иммунной системе.
Следует отметить, что существует
еще один, третий, класс клеток иммунной
системы — макрофагальный, который
также играет важную роль в иммунном
ответе.
Этот класс представлен моноцитами
крови, макрофагами тканей, лимфоузлов
и селезенки. Эта группа называется Аклетками (от англ, accessory — вспомо
гательный). А-клетки представляют со
бой крупные долгоживущие клетки с вы
соким содержанием лизосом. На их по
верхности имеются специфические ре
цепторы к В- и Т-лимфоцитам. Различа
ют подвижные (моноциты крови) и фик
сированные макрофаги. Последние лока
лизуются в дыхательных путях, печени,
брюшине, селезенке, лимфатических
узлах.
Значение макрофагов состоит в том,
что они накапливают и подвергают пере
работке проникающие в организм анти
гены и представляют их для распознава
ния Т-клеткам, вслед за чем стимулиру
ется деление и дифференциация В-лимфоцитов в антителообразующие плазмо
циты. При определенных условиях мак
рофаги проявляют цитотоксическое дей
ствие на опухолевые клетки. Макрофаги
также выделяют интерферон, лизоцим,
вещества, способствующие дифференци
163
ации стволовых клеток в гранулоциты,
которые стимулируют размножение и
созревание Т-лимфоцитов.
Необходимо отметить, что обычно в
иммунном ответе в какой-то степени
представлены оба типа иммунитета —
клеточный и гуморальный. Кроме того,
как было показано выше, осуществляет
ся тесное взаимодействие всех типов иммунокомпетентных клеток.
Типы и формы иммунитета. По про
исхождению выделяют наследственный и
приобретенный антимикробный иммуни
теты.
Наследственный иммунитет пред
ставляет собой невосприимчивость к то
му или иному заболеванию, полученному
ребенком по наследству. По направле
нию действия на чужеродные молеку
лы (клетки) наследственный иммуни
тет подразделяют на противоопухоле
вый, антимикробный и трансплантаци
онный.
Приобретенный антимикробный
иммунитет вырабатывается в процессе
жизни организма в природных условиях
(естественный иммунитет) или формиру
ется искусственным путем (искусствен
ный иммунитет).
Одной из форм естественного имму
нитета является врожденный. Он фор
мируется благодаря переданным от мате
ри через плаценту антителам, количество
которых ежедневно пополняется с моло
ком матери при грудном вскармливании.
(Вот почему так важно грудное вскар
мливание детей!) Продолжительность
врожденного иммунитета определяется
длительностью кормления
ребенка
грудью.
Другая форма приобретенного в
естественных условиях иммунитета —
постинфекционный — возникает у че
ловека в результате заболевания. Этот
вид иммунитета предохраняет организм
от повторного заболевания в результате
гибели патогенных микроорганизмов.
Появляется он вскоре после заболевания
164
и сохраняется в течение нескольких ме
сяцев, лет, а иногда и всю жизнь.
Приобретенный искусственный
иммунитет имеет две формы. При од
ной из них в организм вводят ослаблен
ных или убитых возбудителей той или
иной болезни — вакцину (антигены).
В этом случае организм переносит бо
лезнь в легкой форме благодаря активной
выработке антител против данного воз
будителя. Эта форма искусственного им
мунитета называется активной и сохра
няется в зависимости от качества вакци
ны и индивидуальных особенностей орга
низма от нескольких месяцев (грипп, ди
зентерия) до 5 и более лет (оспа, туляре
мия). Инъекция антигена стимулирует
иммунологическую память на выработку
большего количества антител и лейкоци
тов, что гарантирует надежную защиту на
случай проникновения в организм бакте
рий и вирусов — возбудителей болезней.
Выработка такой формы иммунитета яв
ляется ведущей в профилактике многих
тяжелейших инфекционных заболеваний,
таких, как столбняк, коклюш, паротит,
дифтерия и др., и осуществляется в ходе
проведения профилактических прививок,
о важности которых постоянно говорят
медики.
В медицинской практике широко ис
пользуется пассивный искусственный
иммунитет, когда заболевшему челове
ку вводят лечебные сыворотки с уже со
держащимися в них готовыми антитела
ми против определенного заболевания
или у-глобулины. Такой иммунитет буд^т
сохраняться до тех пор, пока не погибнут
все антигены (1 —2 месяца). К пассивной
иммунизации прибегают в лечебных
целях.
Система интерферона. Интерфе
рон — групповое название, которым
обозначают ряд белков со сходными
свойствами. У человека содержится три
группы интерферонов — а, р и у — в за
висимости от типа клеток, в которых они
образуются.
Наиболее изученным свойством ин
терферона является его способность пре
пятствовать размножению вирусов. Он
образуется в клетках млекопитающих и
птиц в ответ на вирусную инфекцию.
Считается, что интерферон — это актив
ный противовирусный агент, характер
ный для большинства типов клеток и дей
ствующий в большей или меньшей степе
ни против всех вирусов.
При заражении клетки вирусом собы
тия развиваются следующим образом.
Вирус начинает размножаться, и одно
временно клетка-хозяин начинает проду
цировать интерферон. Интерферон выхо
дит из клетки, вступает в контакт с сосед
ними клетками и делает их невосприим
чивыми к вирусу. Интерферон действует,
запуская цепь событий, приводящих к по
давлению синтеза вирусных белков и в
некоторых случаях — „сборки и выхода
вирусных частиц. Таким образом, интер
ферон не обладает прямым противови
русным действием, но вызывает такие из
менения в клетках, которые препятству
ют размножению вирусных частиц.
? , 1. ч
Чем
представлена в организме иммунная
система? 2. Чем отличаются клеточный и гумораль
ный иммунитеты? 3. Как происходит узнавание чу
жеродных тел в организме? 4. Как осуществляется
иммунный ответ организма? 5. Как вы думаете, по
чему не вырабатывается стойкий иммунитет против
гриппа? 6. Какова роль интерферона в организме
человека?
50. Газообмен
Для аэробного расщепления пита
тельных веществ с целью высвобождения
энергии всем живым клеткам организмов
необходим кислород, который служит ко
нечным акцептором водорода. Организ
мы получают кислород либо непосред
ственно из воздуха, либо из воды, в кото
рой он растворен. Одним из конечных
продуктов окисления питательных ве
ществ является диоксид углерода, кото
рый должен быть выведен из организма.
Процесс поступления кислорода в орга
низм и выведение диоксида углерода на
зывается дыханием (его нельзя путать с
клеточным дыханием, в процессе которо
го идет расщепление органических ве
ществ и высвобождение энергии). Дыха
ние на организменном уровне более пра
вильно называют газообменом.
Кислород и углекислый1газ перено
сятся через дыхательные поверхности
(т. е. через кожу или специальный дыха
тельный эпителий) путем пассивной диф
фузии. Основной движущей силой этого
процесса является разность концентра
ций газов. Для того чтобы при данной
разнице концентраций процесс переноса
шел быстрее, площадь поверхности ды
хательного эпителия должна быть как
можно больше, а диффузные расстоя
ния — как можно меньше.
Потребность в кислороде и скорость образова
ния углекислого газа прямо пропорциональны мас
се организма, а скорость переноса газов через по
верхность тела в значительной степени зависит от
величины этой поверхности. Если два тела геомет
рически подобны, то у большего из них площадь по
верхности, приходящаяся на единицу объема, мень
ше, чем у меньшего. Иными словами, отношение
поверхности к объему уменьшается с увеличением
размеров при сохранении формы. У очень мелких
организмов диффузные расстояния малы, а отноше
ние поверхности к объему тела достаточно велико.
Поэтому у организмов, диаметр тела которых не
превышает 0,5 мм, перенос газов вполне может
обеспечиваться одной лишь диффузией.
Одноклеточные организмы и очень
мелкие животные (например, плоские
черви, коловратки, клещи) удовлетворя
ют свои потребности в кислороде тем ко
личеством, которое поступает через по
верхность их тела (кожное дыхание).
Всей поверхностью тела дышат также
грибы и многие водоросли, у которых
кислород поступает диффузно непосред
ственно в кахддую клетку. Увеличение же
размеров тела животных приводит к воз
165
растанию диффузных расстояний и сни
жению отношения величины поверхности
к объему тела. Для того чтобы это отно
шение было достаточно большим, у круп
ных животных возникли специальные га
зообменные структуры (рис. 105). У од
них видов в переносе газов участвует вся
поверхность тела, однако у более круп
ных и ведущих активный образ жизни жи
вотных имеются особые участки, приспо
собленные специально для обмена газов.
У рыб и млекопитающих ■
— это специа
лизированные органы — жабры и лег
кие; у насекомых, пауков и многоно
жек — трахеи. Эти участки выстланы
тонким (0,5— 15 мкм) однослойным ды
хательным эпителием, на который
приходится большая часть поверхности
тела. Так, у человека, например, площадь
дыхательной поверхности легких состав
ляет 50— 100 м2, тогда как площадь по
верхности тела меньше 2 м2.
Для поддержания высокого градиента
диффузии кислорода на обменной мем
бране необходимо движение среды (вен
тиляция). Постоянное обновление среды
у поверхности дыхательного эпителия
обеспечивается у большинства животных
движением воды или воздуха в процессе
дыхания. У некоторых животных этому
способствуют дыхательные ритмические
движения всего тела (малощетинковый
червь трубочник, пиявки) либо работа
ресничного эпителия (моллюски, ланцет
ник). Кроме того, у крупных животных
возникла система кровообращения,
благодаря которой обеспечивается пере
нос 0 2 и С 0 2 между тканями и дыхатель
ным эпителием. В органах дыхания кровь
протекает по чрезвычайно разветвленной
Червь
Рис. 105. Основные типы газообменных структур у животных.
166
Рис. 106. Общая схема транспорта газов у млекопитающих.
капиллярной сети, образуя как бы тон
кую пленку, непосредственно гранича
щую с дыхательным эпителием; тем са
мым диффузионные расстояния для газов
сводятся к минимуму. Перенос газов от
дыхательной поверхности к тканям (и об
ратно) осуществляется с током крови, а
обмен газов между кровью и тканями (че
рез стенки капилляров) обеспечивается
диффузией. При этом в тканях, как и в
дыхательных органах, перенос газов об
легчается тем, что площадь диффузион
ной поверхности достаточно велика, а
диффузионное расстояние между любой
клеткой и ближайшим к ней капилляром
чрезвычайно мало.
Таким образом, у большинства жи
вотных газообмен и перенос газов проте
кает в несколько стадий (рис. 106):12
1. Дыхательные движения, благодаря
которым обеспечивается постоянный
приток свежей среды (воздуха или во
ды) к дыхательным поверхностям,
например жабрам или легким.
2. Диффузия 0 2 и С 0 2 через дыха
тельный эпителий.
3. Перенос газов кровью.
4. Обмен 0 2 и С 0 2 между кровью и
клеткой, осуществляемый путем диф
фузии через стенки капилляров.
Как уже отмечалось, специализиро
ванными дыхательными органами в вод
ной среде являются ж а б р ы , а в назем
но-воздушной — л е г к и е и т р а х е и.
Правда, ряд животных не имеет специа
лизированных органов дыхания. У них га
зообмен осуществляется через влажные
кожные покровы, снабженные обильной
сетью кровеносных сосудов (доледевой
червь). Кожное дыхание в качестве до
полнительного присутствует и у тех жи
вотных, которые обладают специализи
рованными органами дыхания. Напри
мер, у угрей, имеющих жабры, потреб
ность в кислороде на 60 % обеспечивает
ся за счет кожного дыхания; лягушки,
имеющие легкие, на 50 % дышат через
кожу.
Жабры представляют собой тонко
стенные, нитевидные или перистые вы
росты поверхности тела, пронизанные
густой сетью капилляров. Вторично жаб
167
ры могут быть заключены в полость, как,
например, у рыб, но это не меняет того
факта, что жабры в своей основе — вы
пячивания. Жаберное дыхание свой
ственно многощетинковым кольчатым
червям, большинству моллюсков, рако
образным, рыбам, личинкам земновод
ных. Наиболее эффективно происходит
извлечение кислорода из воды жабрами
рыб, причем в основе его лежит явление
противотока: кровь в капиллярах жа
берных лепестков течет в направлении,
противоположном току воды, омываю
щей жабры.
У наземных животных органы воз
душного дыхания (легкие) представляют
собой впячивания — тонкостенные по
лости, в которые свободно проникает
воздух. У беспозвоночных это эктодер
мальные впячивания поверхности тела, у
позвоночных — энтодермальные выпя
чивания кишки. Различают два типа лег
ких: диффузионные и вентиляционные.
В легких первого типа газообмен осущес
твляется только путем диффузии. Такие
легкие имеют относительно небольшие
животные: легочные моллюски, пауки,
скорпионы. Вентиляционные легкие име
ют только наземные позвоночные.
Для наземных позвоночных характерны доста
точно большие объемы тела и, кроме того, высокий
уровень интенсивности обменных процессов, что
требует большого количества кислорода. В связи с
этим у данных организмов происходит постепенное
усложнение строения легких от земноводных к мле
копитающим. Это усложнение связано с возраста
нием площади дыхательного эпителия. Так, у земно
водных 1 см3 легочной ткани имеет общую газооб
менную поверхность 20 см2. Аналогичный показа
тель для дыхательного эпителия легких человека,
представленных многочисленными мешковидными
альвеолами, окруженными снаружи кровеносными
капиллярами, равен 300 см2. Одновременно с уве
личением дыхательной поверхности происходит со
вершенствование механизма вентиляции легких.
Лягушки и тритоны просто заглатывают воздух:
когда ротовое дно опускается, воздух входит в рото
168
вую полость, когда оно поднимается при закрытых
ноздрях, воздух проталкивается в легкие. Сокраще
ние брюшных мышц выталкивает воздух назад в ро
товую полость и при открытых ноздрях — наружу.
У большинства рептилий воздух нагнетается в лег
кие путем расширения грудной клетки. Легкие при
этом также, растягиваются, и воздух засасывается
через ноздри. Исключение составляют черепахи,
обладающие твердым панцирем. Они утратили спо
собность к такому дыханию; заполнение легких про
исходит у нцх так же, как у амфибий, путем заглаты
вания воздуха.
Дыхание путем расширения грудной клетки
наблюдается и у птиц, когда они не летают; в поле
те, когда стенки грудной клетки фиксированы, ис
пользуются воздушные мешки, которые при взма
хах крыльев сжимаются и расправляются. Воздуш
ные мешки играют роль мехов, продувающих воздух
через легкие. Такая система вентиляции легких
очень эффективна. Известно, что птицы могут нахо
диться в полете очень долго, не проявляя признаков
усталости.
У млекопитающих вентиляция легких осущес
твляется двумя способами — за счет расширения
грудной клетки и за счет сокращения диафрагмы.
Трахеи представляют собой тонко
стенные заполненные воздухом и ветвя
щиеся трубчатые кутикулярные впячива
ния внутрь тела. Внутри трахеи находятся
кольца, благодаря которым она не сжи
мается. Трахеи сообщаются с наружной
средой через отверстия в кутикуле —
дыхальца. У насекомых их чаще всего
12 пар: 3 пары на груди и 9 пар на брюш
ке. Они могут закрываться клапанами в
зависимости от потребности в кислороде.
У насекомых трахейная система опле
тает все внутренние органы, так что воз
дух проникает непосредственно к каждой
клетке. Подобная система весьма эффек
тивна, поскольку она не связана с необ
ходимостью растворения кислорода в
крови для снабжения им клеток. В этом
заключается принципиальное отличие
трахейного дыхания от легочного и жа
берного, при которых 0 2 и С 0 2 тран
спортируются исключительно кровью.
В этом смысле трахейнодышащие живот
ные имеют определенные преимущества
перед другими, поддерживая достаточно
высокий уровень клеточного дыхания в
течение длительного периода и обеспечи
вая высокую физиологическую актив
ность. Правда этот тип дыхания эффекти
вен только при небольших размерах жи
вотных (насекомые). Работающая пчела,
например, совершает более сотни взма
хов крыльями в секунду на протяжении
дня без передышки. Однако система ды
хания у этих животных имеет и свои не
достатки. Любой яд из воздуха легко про
никает ко всем клеткам. Человек пользу
ется этим свойством, когда пытается из
бавиться от вредных насекомых, распы
ляя различные инсектициды.
Водные личинки некоторых насеко
мых дышат при помощи трахейных
жабр. У них трахейная система замкнута,
не имеет дыхалец и соединена с «жабра
ми» — придатками с большой повер
хностью. «Жабры» покрыты тонкой ку
тикулой, что позволяет осуществлять га
зообмен между водой и воздухом трахей
ной системы. Такие трахейные жабры
есть, например, у личинок поденок. У ли
чинок ряда стрекоз трахейные жабры
расположены в полости прямой кишки, и
насекомое вентилирует их, набирая воду
внутрь кишки и выталкивая ее обратно.
Газообмен у высших растений.
У высших растений нет специализиро
ванной дыхательной системы. У водных
растений развивается специальная
ткань — аэренхима, в которой накапли
вается и хранится кислород, используе
мый для дыхания. Кислород поступает в
тело наземных растений через устьица в
эпидермисе листьев и зеленых стеблей (у
травянистых растений), диффузно через
ризодерму корня, а также через чечевич
ки и трещины в коре на одревесневших
стеблях и корнях и перемещается по
межклетникам. Далее кислород раство
ряется в воде, содержащейся в клеточных
оболочках, оттуда он диффундирует
внутрь клеток. В растворенном виде кис
лород может перемещаться вместе с во
дой по оболочкам клеток (апопластный
путь) либо из клетки в клетку по плазмодесмам (симпластный путь). В клет
ках, содержащих хлорофилл, выделяе
мый при фотосинтезе кислород может
сразу же потребляться митохондриями.
7
» 1. Что представляет собой газообмен у живых
организмов? 2. Как дышат грибы, водоросли, протисты? 3. Что общего между жабрами и легкими?
4. Каковы преимущества и недостатки трахейной
дыхательной системы? 5. Как вы считаете, какими
должны быть дыхательные поверхности — влаж
ными или сухими? Почему? 6. Как осуществляется
снабжение кислородом корня и стебля, цветков,
плодов и семян растений?
51. Транспорт газов
в организме человека.
Перенос кислорода
и углекислого газа кровью
Из легочных альвеол кислород прони
кает в плазму крови. Однако потребности
организма в кислороде настолько велики,
что плазма, если бы она была единствен
ным переносчиком кислорода, никогда не
справилась бы с такой нагрузкой. Поэто
му основная часть кислорода — пример
но в 16 раз больше, чем растворено в
плазме, — непрочно связывается содер
жащимся в эритроцитах гемоглобином.
Гемоглобин представляет собой тетрамерный
белок: в его состав входят четыре молекулы гема,
которые присоединены к четырем полипептидным
цепям глобина и придают крови красный цвет. Каж
дый гем содержит один атом двухвалентного желе
за, непрочно связывающего одну молекулу кисло
рода. Когда молекула кислорода связывается с ато
мом железа одного из гемов, конфигурация этого
гема слегка изменяется. Это изменение отражается
на конфигурации всей молекулы гемоглобина, кото
рая определенным образом меняется. По мере при
соединения кислорода ко второй и третьей группам
169
гема происходит дальнейшее изменение конфигура
ции гемоглобина, в результате чего каждая последу
ющая молекула 0 2 присоединяется легче, чем пре
дыдущая, так что последний гем связывает кислород
в несколько сот раз быстрее, чем первый.
Присоединение кислорода к гемогло
бину с образованием оксигемоглобина
происходит при высоком парциальном
давлении кислорода — таком, как в ка
пиллярах легочных альвеол. При низком
давлении (как в капиллярах метаболи
чески активных тканей) связь между кис
лородом и гемоглобином становится не
прочной, кислород высвобождается и
диффундирует в окружающие клетки.
Способность крови переносить кис
лород снижается под воздействием двух
широко распространенных загрязнителей
воздуха: оксида углерода (II) — угарного
газа и азотистых соединений, молекулы
которых связываются с атомами железа и
блокируют их. Угарный газ и оксиды азо
та содержатся в выхлопных газах автомо
билей и попадают в кровь вместе с возду
хом. Нитраты могут попасть в организм
вместе с пищей и водой через пищевари
тельный тракт; обитающие в кишечнике
бактерии превращают азотные соедине
ния в ядовитые азотистые, которые и вса
сываются в кровь.
В отличие от кислорода большая
часть С 0 2 транспортируется кровью в
растворенном состоянии (в виде С 0 2 и
НСО3), и лишь небольшая его доля свя
зывается с гемоглобином и другими бел
ками крови. В этом случае С 0 2 присоеди
няется к аминогруппам с образованием, в
частности, карбаминогемоглобина:
Н
/
Белок — N
+ С 0 2?±
\
Н
Н
/
^ Н + + Белок — N
\з о с г .
170
Количество С 0 2, способное связать
ся с гемоглобином, зависит от количества
уже связанного гемоглобином кислорода.
Чем меньше кислорода связано с гемо
глобином, тем больше С 0 2 может быть
перенесено этим способом. Обычно, од
нако, таким способом транспортируется
от 10 до 20 % С 0 2.
Основная масса С 0 2 переносится
кровью в виде бикарбоната. Образую
щийся в тканях С 0 2 поступает в кро
вяное русло и далее в эритроциты,
где взаимодействует с водой, обра
зуя угольную кислоту, которая диссоци
ирует:
С 0 2 + Н20 = Н2С 0 3 = Н+ +НСО3.
Ионы бикарбоната выходят затем в
плазму и переносятся (в виде NaH0O3) в
легкие.
Этот процесс катализируется содер
жащимся в эритроцитах ферментом —
карбоангидразой (угольной ангидразой).
Реакция обратима; в легких, где концен
трация углекислого газа низкая, реакция
сдвигается влево и освободившийся С 0 2
переходит в альвеолы.
Приведенное описание переноса С 0 2 неполно;
необходимо остановиться еще на двух вопросах. Как
видно из реакции, образование иона Н С 03 сопро
вождается появлением одного иона водорода. По
скольку ионы водорода снижают значение pH, мож
но ожидать, что после поступления в кровь С 0 2 из
тканей плазма приобретает кислую реакцию. Из
вестно, однако, что этого не происходит; величина
pH крови остается постоянной (около 7,3); сниже
ние pH крови до 7,0 несовместимо с жизнью. Под
держание pH крови на постоянном уровне происхо
дит следующим образом.
Когда эритроциты с кровью покидают легкие,
содержащийся в них оксигемоглобин Н Ь02 имеет
слабокислую реакцию и связан с ионами калия.
В тканях оксигемоглобин легко отдает кислород,
приобретая при этом сильнощелочную реакцию.
В этом состоянии он теряет ионы калия и присоеди
няет ионы водорода (от угольной кислоты), превра
щаясь в ННЬ. В свою очередь ионы калия связыва-
Плазма
N aHC0 3
Плазма
;.>NaCI
М ито хон дрия
Рис. 107 Транспорт углекислого газа плазмой крови и эритроцитами.
ются с ионами бикарбоната, образуя бикарбонат
калия (рис. 107):
КНЮ 2 = КНЬ + 0 2
Н+ + НСОу + КНЬ = ННЬ + КНСОТ
Присоединяя ион водорода, гемоглобин дей
ствует как буферная система: поэтому большое ко
личество угольной кислоты может переноситься к
легким без значительного изменения pH крови. Ос
новная масса иона НСОу из Эритроцитов выходит в
плазму путем диффузии поТрадиенту концентрации
и, соединясь здесь с иОнами натрия, образует би
карбонат натрия. Однако в этом случае эритроцит
приобретает положительный заряд (за счет остав
шегося там связанного с гемоглобином иона Н+).
Этого, однако") не происходит, поскольку выход ио
нов бикарбоната компенсируется поступлением ту
да ионов хлора из плазмы. Таким образом поддер
живается электронейтральность. Это явление но
сит название хлоридного сдвига.
? 1.tRВ каком виде осуществляется транспорт кис
лорода в организмах, имеющих кровь? Почему?
2. Опишите, каким образом содержащаяся в крови
углекислота удаляется из легких в газообразной
форме. 3. Каким образом поддерживается на строго
определенном уровне кислотность крови?
52. Выделительная система
у животных и растений
В процессе обмена веществ в клетках
живых организмов образуются конечные
продукты. Среди них могут быть и ядови
тые для клеток вещества. При распаде
аминокислот, нуклеиновых кислот и дру
гих азотистых соединений образуются
токсические вещества — аммиак, моче
вина и мочевая кислота, которые по
мере их образования подлежат выведению из организма.
Н
I
I
N
О
/ \
II
h 2n — С — NH;
Н
Н
аммиак
мочевина
н
I
о
II
с
N
/ '\ / \
\
Н—N С
I
О= С
II
С
\ V
N
С= О
/
\
/
N
I
I
Н
Н
мочевая кислота
Токсический эффект конечных продуктов азо
тистого обмена снижается в ряду: аммиак — моче
вина — мочевая кислота. В зависимости от того, в
какой из этих трех форм преимущественно выделя
ется азот, животных подразделяют на три группы:
171
аммониотелические (выделяющие свободный ам
миак), уреотелические (выделяющие мочевину) и
урикотелические (выделяющие мочевую кислоту).
Форма выделения продуктов азотистого обмена
тесно связана с условиями жизни животного и обес
печенностью водой. Аммиак весьма токсичен даже в
малых концентрациях. Благодаря хорошей раство
римости и небольшой молекулярной массе он легко
диффундирует через любую поверхность, соприка
сающуюся с водой. Вот почему аммиак (обычно в
виде иона NHJ является конечным продуктом азо
тистого обмена у водных беспозвоночных, костис
тых рыб, личинок и постоянно живущих в воде зем
новодных. Наземные животные, для того чтобы ам
миак не накапливался в клетках и жидкостях тела,
должны обезвреживать его, преобразуя в менее
ядовитые продукты. Так, наземные ресничные чер
ви, взрослые земноводные и млекопитающие выде
ляют мочевину. Низкая растворимость мочевой кис
лоты (которая является к тому же практически не
ядовитой), выпадение ее в осадок делают это ве
щество осмотически неактивным. Для ее выведения
из организма вода практически не нужна. Поэтому
урикотелия в основном характерна для животных,
которые осваивают наземную, в том числе засушли
вую среду (наземные насекомые, пауки, выделяю
щие гуанин, чешуйчатые пресмыкающиеся, птицы).
Из организма выводится также избы
ток воды, минеральных солей, углекис
лый газ, а также яды, которые поступили
вместе с воздухом, пищей и водой, избы
ток витаминов, гормонов, лекарственные
препараты и т. п.
У протистов легкорастворимые экс
креты (аммиак, мочевина, углекислый газ
и др.) выводятся путем диффузии. У низ
ших водных многоклеточных (например,
у кишечнополостных) и малоактивных
морских животных (иглокожие) продукты
обмена также удаляются с помощью диф
фузии через поверхность тела.
По мере усложнения организации
животных у них развиваются специаль
ные выделительные органы, осуществля
ющие выведение отходов и избытков ве
ществ из организма в окружающую среду
через протоки и поры.
172
Такими органами выделения являют
ся протонефридии (у плоских червей,
некоторых групп круглых червей, у личи
нок моллюсков и кольчатых червей), ме
танефридии (у кольчатых червей, чле
нистоногих, моллюсков), антеннальные
и максиллярные железы (у членистоно
гих), мальпигиевы сосуды (у насекомых
и паукообразных). Открывающиеся на
ружу вышеперечисленные выделитель
ные органы характерны, таким образом,
для беспозвоночных животных. У позво
ночных органами выделения являются
почки с большим количеством плотных
сплетений кровеносных сосудов с каналь
цами выделительной системы.
При всем многообразии органов вы
деления в основе их функционирования
лежат два основных процесса: ультра
фильтрация и активный транспорт.
При ультрафильтрации жидкость поддавлением проходит через полупроницаемую
мембрану, которая задерживает белки и
другие крупные молекулы, но пропускает
воду и низкомолекулярные растворенные
вещества. Активный транспорт, как вам
уже известно, представляет собой движе
ние растворенных веществ против элек
трохимического градиента, что связано с
расходами энергии. Если эти вещества
переносятся из внутренней среды живот
ного в полость экскреторного органа, то
этот процесс называют активной секре
цией. Если же перенос осуществляется в
обратном направлении, то он называется
активной реабсорбцией. Как правило,
в выделительных органах фильтрацион
ная система дополняется процессами ак
тивного транспорта.
Протонефридии встречаются у жи
вотных, не имеющих полости тела. Они
представляют собой систему ветвящихся
по всему телу канальцев, открывающихся
наружу отверстием (одним или несколь
кими). Внутренние многочисленные кон
цы канальцев слепо замыкаются конеч
ной, расширенной в виде луковицы звезд
чатой клеткой с отростками (рис. 108, а).
Рис. 108. Органы выделения: а — планарии и б — кузнечика: 1 — реснички, 2 — звездчатая клетка,
3 — выделительные протоки, 4 — выделительные поры, 5 — мальпигиевы сосуды.
От клетки в полость канала выступает
одна либо пучок ресничек. Если ресничка
одна, конечную клетку называют соленоцитом; если их несколько (пучок) —
пламенной клеткой, так как биение
пучка ресничек напоминает мерцающее
пламя свечи.
Метанефридии имеются у животных,
обладающих вторичной полостью тела
(целомом). Для них характерно отсут
ствие ветвления канальца. На внутрен
нем конце канал открывается воронкой,
обращенной в полость целома. Как про
тонефридии, так и метанефридии фун
кционируют как фильтрационно-реабсорбционная почка, в которой жидкость
сначала образуется путем фильтрации, а
затем ее состав видоизменяется при про
хождении по мочевому канальцу. Разли
чие заключается в деталях. Так, вещества
для выведения протонефридиями посту
пают в канал из окружающих пламенную
клетку тканей и органов за счет градиен
та концентрации, создаваемого работой
ресничек. Вещества для выведения метанефридиями поступают из целомической
жидкости.
Парные компактные почки моллюс
ков и зеленые железы ракообразных
сходны по строению с метанефридиями и
удаляют продукты обмена из полостной
жидкости. Помимо фильтрационно-аб
сорбционных процессов, в мочевые ка
нальцы осуществляется активная секре
ция некоторых веществ. У ряда морских и
пресноводных моллюсков фильтрация
мочи идет через стенку сердца и около
сердечную полость. Далее фильтрат по
ступает по специальному каналу в «поч
ку», где реабсорбируются глюкоза, ами
нокислоты и важнейшие ионы.
Мальпигиевы сосуды насекомых
(рис. 108, б, 109, с. 174) и паукообраз
ных состоят из длинных трубочек, кото
рые впадают в пищеварительный канал в
месте соединения средней и задней киш
ки. Слепые концы мальпигиевых сосудов
находятся в гемоцеле. В механизме
работы мальпигиевых сосудов отсутству
ет начальная ультрафильтрация.
С развитием у насекомых трахейного дыхания
значение кровеносной системы уменьшается. В ре
зультате мальпигиевы сосуды не получают артери173
Рис. 109. Упрощенная схема выделительной
системы насекомого: 1 — средняя кишка, 2 —
задняя кишка, 3 — прямая кишка, 4 — мальпи
гиев сосуд.
альную кровь непрерывно под давлением, как, на
пример, нефроны млекопитающих. Зато они окру
жены кровью, давление которой, по существу, не
превышает давления в сосудах. Поскольку у насеко
мых значительной разницы давлений по обе сторо
ны стенки мальпигиевых сосудов нет, фильтрация
не может играть у них заметной роли в образовании
мочи. Поэтому данный процесс у насекомых и пау
кообразных должен идти полностью за счет секре
ции веществ, с последующим обратным всасывани
ем некоторых компонентов секретируемой жидкос
ти. Внутренняя поверхность мальпигиева сосуда
сплошь покрыта микроворсинками, и клетки, обра
зующие эти сосуды, богаты митохондриями (рис.
110). Такие особенности структуры органа связаны
с высокой активностью секреторного эпителия.
В просвет сосуда активно выделяется
калий и в меньшей степени натрий вмес
те с конечными продуктами азотистого
обмена (мочевой кислотой, аллантоином). Активный перенос К+ в просвет со
суда служит главной движущей силой
процесса образования мочи. Мочевая
кислота попадает в сосуд в виде раство
ренного в воде урата калия. Жидкость,
богатая калием, переходит в заднюю
кишку. Там нужные организму растворен
ные вещества, в том числе и К+, а также
вода, реабсорбируются, а мочевая кисло
174
Рис. 110. Секреторная клетка в стенке мальпи
гиева сосуда таракана: 1 — поверхность, обра
щенная к моче; 2 — поверхность, обращенная к
гемолимфе; 3 — микроворсинки; 4 — митохон
дрии.
та выпадает в осадок. Этим объясняется
дальнейшее извлечение воды, так как
осажденная мочевая кислота не участву
ет в осмотической активности содержи
мого кишки. Мочевая кислота затем уда
ляется из кишечника в смеси с остатками
непереваренной пищи.
Почки позвоночных функционируют
по принципу фильтрации-реабсорбции, к
которому добавляется секреция в ка
нальцах.
Почки водных позвоночных представ
ляют собой длинные органы, располо
женные вдоль полости тела. Они имеют
трубочки, открывающиеся в полость тела
и извлекающие конечные продукты обме
на веществ из целомической жидкости.
Почки наземных позвоночных — реп
тилий,
птиц
и
млекопитающих
(рис. 111) — представляют собой пар
ные органы бобовидной формы, располо
женные в полости тела, ближе к спинной
стороне. От каждой почки к мочевому пу
зырю направляется мочеточник. В моче
вом пузыре моча накапливается и затем
выводится наружу через мочеиспуска
тельный канал, иначе называемый
уретрой.
Функциональная единица почки —
нув коркового слоя, извивается рядом с капсулой
своего же нефрона и образует извитой каналец II
порядка, который переходит в собирательную
трубочку. Собирательные трубочки от нескольких
рядом расположенных нефронов объединяются в
более крупные собирательные протоки и открыва
ются в воронкообразную сплющенную полость с
тонкими стенками — почечную лоханку. Почеч
ные лоханки открываются в мочеточник.
Рис. 111. Внутреннее строение почки млекопи
тающих: 1 — нефрон, 2 — корковое вещество,
3 — междольковая артерия и клубочки, 4 — пи
рамида, 5 — мозговое вещество, 6 — сосочек,
7 -— капсула, 8 — мочеточник, 9 — лоханка,
10 — почечная вена, 11 — почечная артерия.
нефрон — сложная эпителиальная тру
бочка, один конец которой слепой, а вто
рой открывается в почечную лоханку че
рез собирательный проток (рис. 112,
с. 176).
В области слепого конца нефрон расширен и
напоминает шарик, вдавленный с одной стороны, в
результате чего образуется чашевидная структу
ра — капсула Боумена — Шумлянского. Полость
между стенками капсулы соединена с узким просве
том извитого почечного канальца. Внутри капсу
лы, тесно прилегая к ее стенкам, расположен пучок
кровеносных капилляров, образующих клубочек.
В мозговом слое почки каналец выпрямляется и в
середине изгибается на 180”, образуя петлю Генле.
В петле две части: нисходящее и восходящее коле
на. Конец восходящего колена петли Генле, достиг
В основе мочеобразования лежат че
тыре процесса: клубочковая фильтрация,
канальцевая реабсорбция, канальцевый
синтез и канальцевая секреция (рис. 113,
с. 177).
Клубочковая фильтрация представ
ляет собой процесс перехода плазмы кро
ви со всеми растворенными в ней вещес
твами, за исключением белков с молеку
лярной массой выше 7000 в полость кап
сулы Боумена — Шумлянского и далее в
извитой каналец.
К капсуле Боумена — Шумлянского
подходит тонкая приносящая артериола, распадающаяся на сеть мельчайших
капилляров — почечный югубочек. За
тем капилляры вновь соединяются и об
разуют выносящую артериолу. По
скольку диаметр выносящей артериолы
примерно в 2 раза меньше, чем принося
щей, в капиллярах поддерживается по
вышенное давление, под влиянием кото
рого вода и растворенные в ней вещества
выходят из крови в просвет капсулы. Так
образуется первичная моча, которая
поступает в почечный каналец. В сутки у
человека образуется 150— 180 л первич
ной мочи. Такой объем первичной мочи
образуется за счет прохождения через
почки больших объемов крови: 1700—■
1800 л в сутки; это примерно четвертая
часть крови, проходящей через сердце.
Если учесть, что у человека содержится
около 6 л крови, то можно рассчитать,
что каждая ее капля проходит через поч
ки почти 500 раз в сутки, причем всякий
раз ее состав контролируется и изменяет
ся. Ежесуточно из крови в нефроны
175
2
3
4
1
16
17
Рис. 112. Нефрон почки и его кровоснабжение:
I — приносящая артериола, 2 — капсула Боу
мена — Шумлянского, 3 — клубочковые капил
ляры, 4 — извитой каналец I порядка, 5 — из
витой каналец II порядка, 6 — корковое вещес
тво, 7 — собирательная трубочка, 8 — наруж
ное мозговое вещество, 9 — внутреннее мозго
вое вещество, 10 — выход в почечную лоханку,
I I — петля Генле, 12 — нисходящее колено,
13 — капилляры, 14 — восходящее колено пет
ли Г енле, 15 — выносящая артериола, 16 — ар
терия, 17 — вена.
фильтруется около 180 л жидкости. Ко
нечно, организм человека не производит
180 л мочи в сутки; большая часть воды,
фильтруемой в почках, вновь поступает в
организм через нефроны, а мочи образу
ется лишь 1— 1,5 л.
Канальцевая реабсорбция. В почеч
ном канальце происходит обратное вса
сывание (реабсорбция) воды, аминокис
лот, глюкозы, витаминов, нужных орга
низму солей. Это достигается благодаря
тому, что артериола, выходящая из капсу
лы, снова образует сплетение капилля
ров вокруг извитого канальца, отходягце176
10
го от капсулы. Именно там происходит
избирательное обратное всасывание. В
процессе реабсорбции значительную
роль играет активный перенос. Напри
мер, концентрация глюкозы в первичном
фильтрате и крови одинакова; тем не ме
нее вся глюкоза переходит из фильтрата в
кровь, так что в моче глюкозы практичес
ки не остается. Активно поглощаются
также соли, например NaCl. Мочевина
и другие продукты обмена реабсорбируются очень плохо, поэтому их кон
центрация увеличивается в просвете
нефрона.
Рис. 113. Четыре процесса, из которых склады
вается процесс образования мочи в нефроне:
1 — клубочковая фильтрация, 2 — канальце
вая реабсорбция, 3 — канальцевый синтез,
4 — канальцевая секреция, 5 — почечный ка
налец, 6 — капилляр.
Канальцевый синтез имеет место как
в клетках канальцевого эпителия, так и в
самом просвете почечного канальца.
К примеру, в почечном эпителии некото
рые аминокислоты дезаминируются (те
ряют NH3). Полученный аммиак диффун
дирует в просвет почечного канальца,
где, соединяясь с Н+, образует NHJ, ко
торый в свою очередь удерживается мо
чой и в виде азотсодержащих экскретов
выводится из организма.
Канальцевая секреция избирательна
и служит ддя регуляции содержания К+,
Н+ и бикарбонатов в крови, а также для
удаления из организма инородных ве
ществ, например лекарственных препа
ратов, ядов и т. п. Канальцевая секреция
почти всегда происходит за счет активно
го транспорта.
В результате всех рассмотренных вы
ше процессов образуется вторичная мо
ча, которая поступает в лоханку и далее
по мочевыводящим путям покидает орга
низм.
Выделительная система животных,
таким образом, выполняет множество
функций, таких, как поддержание опреде
ленной концентрации воды в клетках и в
теле, поддержание на определенном
уровне давления крови в кровеносной
системе и др. Все эти функции связаны с
обеспечением сохранения постоянства
внутренней среды организма. Можно вы
пить много воды или совсем не пить, а ко
личество воды в крови не изменится. И з
бирательное поглощение воды в нефронах протекает всегда с такой скоростью,
которая обеспечивает постоянство со
держания воды в крови. Почки, кроме то
го, поддерживают на постоянном уровне
ионный состав крови, что необходимо для
нормальной жизнедеятельности организ
ма. Если в рационе содержится большое
количество поваренной соли, то выделе
ние ионов натрия и хлора с мочой увели
чивается. Для выведения этих ионов тре
буется, однако, большое количество во
ды, и именно поэтому можно умереть от
жажды, выпив много морской воды.
Регуляция деятельности почек осуществляется
нервно-рефлекторным и гуморальным механизма
ми. Так, возбуждение симпатических нервов почки
приводит к сужению почечных сосудов. Если проис
ходит сужение приносящих артериол, то фильтра
ция плазмы замедляется; если сужаются вынося
щие артериолы, то фильтрация плазмы усили
вается.
Гормон задней доли гипофиза — вазопрессин
(антидиуретический гормон) уменьшает мочеотде
ление путем увеличения обратного всасывания во
ды при поступлении в- организм избыточного коли
чества солей. Гормон щитовидной железы — т и
роксин — усиливает мочеотделение. Противопо
177
ложное действие оказывает гормон мозгового ве
щества надпочечников — адреналин.
Кроме почек, у высших позвоночных
животных органами выделения являются
также кожа, легкие и печень. Вода, моче
вина и соли активно выводятся из кожных
кровеносных капилляров в протоки пото
вых желез. При выходе пота на повер
хность кожи вода испаряется, что приво
дит к потере тепла и способствует термо
регуляции. Через печень осуществляется
выведение желчных пигментов, образую
щихся при расщеплении гемоглобина
старых эритроцитов. В составе желчи эти
пигменты поступают в двенадцатипер
стную кишку и далее выходят из организ
ма по кишечному тракту с каловыми мас
сами, придавая им характерный цвет.
Кроме того, в печени происходит деза
минирование аминокислот — фермен
тативное отщепление аминогруппы
(—NH2) о т аминокислот, которые не ис
пользуются в данный момент клетками
для синтеза белка. Результатом этого
процесса является образование аммиака,
который идет либо на синтез других ами
нокислот или азотистых оснований, либо
превращается в мочевину, которая затем
с кровью доставляется к почкам и выво
дится из организма.
Диоксид углерода — один из основ
ных конечных продуктов дыхания — вы
деляется диффузно в окружающую среду
(у одноклеточных и мелких многоклеточ
ных — кишечнополостных, некоторых
червей ) или в кровь и далее через органы
дыхания — в окружающую среду.
У растений процессы выделения су
щественно отличаются от таковых у жи
вотных, т. к. растения синтезируют в
нужном количестве все необходимые им
органические вещества. Например, бел
ка в растениях образуется столько,
сколько его необходимо в данный мо
мент, и поэтому растения выделяют мало
азотистых отходов, образующихся при
расщеплении белков. Диоксид углерода,
178
кислород и вода — конечные продукты
обмена веществ — вновь используются
растениями как исходные вещества для
других реакций. Это в особенности отно
сится к диоксиду углерода и воде. У рас
тений нет выделительной системы, как у
животных. Избыток диоксида углерода из
клеток диффузно поступает в межклеточ
ное пространство и через устьица, чече
вички или трещины в корке выводится в
окружающую среду. Через устьица и че
чевички осуществляется также испаре
ние воды и выделенных в межклетники
спиртов, альдегидов, терпенов.
Многие органические отходы метаболиза откладываются у растений в отмер
ших тканях (например, в древесине), а
также в листьях и коре, которые периоди
чески сбрасываются. Многолетние рас
тения состоят в основном из омертвев
ших тканей. Экскреты скапливаются в
этих тканях и уже не могут оказывать
вредное воздействие на активность жи
вых клеток. Аналогичным образом могут
накапливаться многие минеральные со
ли, поглощаемые растениями в виде ио
нов. Накопление их связано с разным ис
пользованием катионов и анионов. Орга
нические кислоты, способные нанести
вред растению, часто связываются с из
быточными катионами и выпадают в виде
нерастворимых кристаллов, которые мо
гут сохраняться в клетках растения, не
причиняя им вреда. Например, ионы
кальция и сульфата одновременно погло
щаются растением, но сульфат сразу же
используется в синтезе аминокислот, а
ионы кальция остаются в избытке и свя
зываются со щавелевой или пектиновой
кислотами, образуя нерастворимые окса
лат и пектат кальция. Особенно много
этих кристаллов накапливается в вакуо
лях листьев. Ионы железа, марганца, не
которые органические кислоты (напри
мер, никотиновая и танниновая) также
поступают в листья, где они накаплива
ются и сбрасываются вместе с листвой во
время листопада. Подлежащие удалению
вещества удаляются не только с листвой,
но и с лепестками, плодами и семенами. У
водных растений основная масса отходов
метаболизма выводится путем диффузии
прямо в окружающую среду.
?
1. Каково назначение выделительной системы у
животных? 2. Как осуществляется функция выделе
ния у разных животных организмов и с чем это свя
зано? 3. Как у насекомых образуется моча и экскре
менты? 4. Назовите и опишите три основных про
цесса, происходящие в почке позвоночных и опре
деляющие конечный состав мочи. 5. Какие факторы
определяют скорость ультрафильтрации в почечном
клубочке? 6. Какую роль играют почки в регуляции
кровяного давления? 7. В процессе эволюции на
земные животные перешли на экскрецию в основ
ном мочевой кислоты и мочевины, а не аммиака.
В чем причина подобных изменений? 8. Како
вы особенности выделительной системы у рас
тений?
53. Гомеостаз — поддержание
постоянства внутренней среды
Любой живой организм подвергается
воздействию самых разных колеблю
щихся факторов окружающей среды. В то
же время для протекания всех процессов
в его клетках необходимы строго опреде
ленные условия. Вследствие этого у жи
вых организмов выработались различные
саморегулирующиеся системы, позволя
ющие поддерживать благоприятную
внутреннюю среду, несмотря на измене
ние внешних условий. Чтобы в этом убе
диться, достаточно вспомнить все те
приспособительные реакции, какими об
ладает организм человека. Когда мы вхо
дим с улицы в темное помещение, наши
глаза благодаря автоматической внутрен
ней регуляции быстро приспосабливают
ся к резкому уменьшению освещения.
Работаете ли вы зимой на севере или за
гораете летом на горячем песке у моря на
юге — во всех случаях температура ва
шего тела остается фактически постоян
ной, изменяясь не более чем на несколь
ко десятых долей градуса. Или другой
пример. Кровяное давление в головном
мозге должно оставаться всегда постоян
ным. Если оно падает, то человек теряет
сознание. При резком повышении давле
ния из-за разрывов мелких капилляров
может произойти кровоизлияние в мозг
(инсульт), которое остро нарушает фун
кцию головного мозга. При различных
изменениях положения тела (вертикаль
ное, горизонтальное, вниз головой) под
действием силы тяжести изменяется при
ток крови к голове; однако, несмотря на
это, комплекс приспособительных реак
ций поддерживает кровяное давление
мозга на строго определенном уровне.
Эти примеры иллюстрируют способность
организма поддерживать постоянство
внутренней среды с помощью специаль
ных механизмов регуляции. Поддержание
постоянства внутренней среды называет
ся гомеостазом.
Гомеостаз свойствен всем формам ор
ганизмов — от одноклеточных до круп
ных многоклеточных животных и расте
ний, масса которых может достигать со
тен килограммов.
Сократительная вакуоль, характерная
для протистов, поддерживает на постоян
ном уровне содержание воды в клетке. С
помощью специальных механизмов у
многоклеточных животных сохраняется
постоянный уровень воды в клетках, кон
центрация различных веществ, постоян
ное количество кислорода и углекислого
газа, кислотность, кровяное давление,
концентрация солей в крови, температу
ра тела и т. п.
Реакция организма на физическую
нагрузку. При интенсивной физической
нагрузке нервная система посылает сиг
налы в мозговое вещество надпочечни
ков, которые в ответ выделяют адрена
лин. Под действием адреналина из селе
зенки в сосуды поступает некоторое ко
личество депонированной в ней крови.
В результате этого объем крови в сосудах
увеличивается. Адреналин также вызы
179
вает расширение капилляров кожи,
мышц и сердца, увеличивая их крово
снабжение. Кроме того, под действием
адреналина кровеносные сосуды брюш
ной полости и почек сужаются, уменьша
ется их кровенаполнение. Такое перерас
пределение крови позволяет поддержи
вать кровяное давление на нормальном
уровне (при расширенном русле крови
для этого оказывается недостаточно).
Адреналин повышает также частоту
дыхания и сокращений сердца. В резуль
тате поступление в кровь кислорода и вы
ведение из нее диоксида углерода проис
ходит быстрее, кровь движется по сосу
дам с более высокой скоростью, достав
ляя больше кислорода работающим
мышцам и ускоряя удаление конечных
продуктов обмена.
При физической нагрузке мышцы вы
деляют больше углекислого газа, чем
обычно, и это само по себе обладает ре
гуляторным воздействием. Углекислый
газ повышает кислотность крови, что
влечет за собой усиление снабжения
мышц кислородом и расширение крове
носных сосудов мышц, а также через нер
вную систему стимулирует выделение ад
реналина, что в свою очередь увеличива
ет частоту дыхания и пульса (рис. 114).
На первый взгляд все приспособле
ния к физической нагрузке должны изме-
К ора го л о в н о го м о зга
(п р о и зв о л ь н а я р е гул я ц и я )
Учащ ение
ды ха ния
4
С и м п а ти ч е с к а я не р вн а я
“ ^ с и с т е м а (с п и н н о й м о з г )
(н е п р о и зв о л ь н а я р е гу л я ц и я )
i
А
-t> i.
У в е л иче ние
У сил е ние га зо о б м е н а
__________________________
а с т о т ы — £> 2 . У с к о р е н и е кро воо б р ащ е ния
с о кр а щ е н и й
у-. • ■ ~ f. се р д ц а
К ора н а д п о ч е ч н и ко в
А дреналин
В ы свобож дение
д е п о н и р о - __ ~
в ан но й крови- ^
Л ока льн ое
р асш ир ен ие — £> 4 .
сосудов
Усил ение к р о в о с н а б ж е н и я
И зм е н е н и е к р о в о с н а б ж е н и я р азл ичны х
о р га н о в
А
К р о в о с н а б ж е н и е , м л /м и н
В норм е
М инутны й объем
се рд ц а
П р и то к кр о в и
-£>М ыш ечная а к т и в н о с т ь -
Повыш ение
^ с о д е р ж а н и я СО|С> 5 .
17500
М озг
750
750
Брю ш ная п о л о с ть
1400
600
П о чки
1100
600
Мышцы
850
12500
Кожа
450
1900
С ердце
250
750
К р о в ь о т д а е т больше 0 2
и МОЛОЧНОЙ
ки сл о ты в кр о в и
Рис. 114. Реакция организма человека на физическую нагрузку.
180
сипл
При большой
н а гр у зке
нять состояние организма, однако в дей
ствительности они обеспечивают сохра
нение того же состава в неклеточной
жидкости, омывающей клетки организма
(что обеспечивает их бесперебойную ра
боту), и в особенности мозг, каким он был
бы в состоянии покоя. Если бы не было
этих приспособлений, физическая на
грузка приводила бы к повышению тем
пературы внеклеточной жидкости, к
уменьшению концентрации в ней кисло
рода и к повышению ее кислотности. При
крайне тяжелой интенсивной работе в те
чение длительного времени все это и про
исходит; в мышцах накапливается молоч
ная кислота, вызывающая судороги, при
которых, разумеется, дальнейшая физи
ческая работа становится невозможной,
что помогает организму вернуться в нор
мальное состояние.
Терморегуляция. Гомеостаз — это не
только постоянство состава жидкостей
организма, но и постоянство их темпера
туры. Большинство ферментов работает
достаточно эффективно лишь при опре
деленной, оптимальной для них или близ
кой к ней, температуре.
Все организмы вырабатывают тепло в резуль
тате происходящих в их клетках химических реак
ций. У млекопитающих и птиц метаболические ре
акции протекают с высокой скоростью, и это позво
ляет им поддерживать высокую температуру тела.
Высокая скорость метаболизма требует для клеточ
ного дыхания больших количеств кислорода. Для
обеспечения быстрой доставки кислорода к клеткам
у птиц и млекопитающих имеются разного рода
приспособления, такие, как обширная дыхательная
поверхность, где происходит поглощение кислоро
да, огромное число эритроцитов, переносящих кис
лород, высокое кровяное давление, нагнетающее
богатую кислородом кровь в капилляры.
Как же осуществляется поддержание
постоянной температуры тела в особен
ности при колебании температуры окру
жающей среды? Главный терморегулиру
ющий механизм расположен в головном
мозге, в области гипоталамуса. Когда в
морозный день человек выходит на улицу,
холодный воздух вызывает охлаждение
кожи рук, щек, носа, ушей. Происходит
небольшое уменьшение температуры
крови в этих органах. Охлажденная кровь
разносится по организму и уже через не
сколько секунд попадает в гипоталамус.
Этот мозговой центр, чувствительный к
малейшему понижению температуры, не
медленно посылает импульсы, мобилизу
ющие механизмы накопления и выработ
ки тепла (рис. 115, с. 182). Происходит
сокращение стенок капилляров и суже
ние их просвета; в результате уменьшает
ся ток крови через охлажденную повер
хность кожи. Благодаря этому тепло, вы
рабатываемое в клетках, сохраняется,
накапливается и температура тела не
сколько повышается. Если сужения ка
пилляров оказалось недостаточно для по
вышения температуры тела, то включа
ется второй механизм: человек начинает
дрожать. Под влиянием поступающих им
пульсов основные мышцы тела непроиз
вольно сокращаются. В процессе сокра
щения вырабатывается дополнительное
тепло, которое способствует повышению
температуры тела.
Возможна и еще одна реакция — так
называемая «гусиная кожа». Каждый во
лосок на теле имеет крошечную мышцу;
под влиянием импульсов, идущих от моз
га при понижении температуры тела, эти
мышцы сокращаются и волоски поднима
ются; выпрямившиеся волосяные сумки
вызывают появление на коже пупыры
шек. У кошки со вздыбленной шерстью
образуется воздушная прокладка, изоли
рующая кожу и предохраняющая ее от
холода. У человека волоски на теле на
столько малы, что они не могут создать
подобной воздушной изоляции, но тем не
менее эта унаследованная реакция на хо
лод сохранилась.
В организме животных и человека
имеется также механизм охлаждения,
благодаря которому они могут жить при
181
новном двумя способами. Во-первых,
возрастает приток крови к коже, и в ре
зультате увеличивается теплоотдача пу
тем излучения тепла. Во-вторых, что бо
лее важно, большинство птиц и млекопи
тающих используют для охлаждения ор
ганизма испарение, поскольку вода, пе
реходя из жидкого состояния в газооб
разное, поглощает большое количество
тепла. У человека при перегреве влага
выделяется через кожу в виде пота. За
тем содержащаяся в поте вода испаряет
ся, охлаждая тело. Млекопитающие до
стигают той же цели посредством т еп
ловой одышки — учащенного неглубо
кого дыхания, увеличивающего испаре
ние со слизистой оболочки верхних дыха
тельных путей. Всем, очевидно, приходи
лось наблюдать тепловую одышку собак:
частое дыхание, сильно высунутый язык.
Кроме того, птицы и звери могут изме
нять положение перьев и шерсти, что
усиливает приток воздуха к поверхности
кожи и ускоряет теплоотдачу.
Рис. 115. Гомеостатические реакции организ
ма человека при действии низких температур:
1 — мозг, 2 — охлажденная кровь, 3 — капил
ляр, 4 — холодный воздух, 5 — сужение про
света капилляров, 6 — мышца, 7 — повышение
интенсивности обмена веществ, 8 — «гусиная
кожа».
температурах, превосходящих нормаль
ную температуру тела (рис. 116). Если
температура тела выше нормальной,
включаются механизмы, увеличивающие
теплоотдачу. Это осуществляется в ос
182
Описанные механизмы терморегуляции спо
собны выравнивать отклонения температуры в пре
делах долей градуса. Однако в отдельных случаях в
организме происходят более резкие изменения тем
пературы. При некоторых заболеваниях механизмы
терморегуляции поддерживают температуру тела
на более высоком, чем в норме, уровне. Это проис
ходит вследствие повышения интенсивности обмена
веществ, которое сопровождается прекращением
секреторной активности потовых желез и сокраще
нием просветов капилляров кожи. В результате
происходит повышение температуры тела на нес
колько градусов. Причины подобного лихорадочно
го состояния заключаются в том, что в организме
начинают действовать иные гомеостатические меха
низмы. Болезнетворные микроорганизмы и выделя
емые ими токсины нарушают постоянство внутрен
ней среды организма, и, как следствие, повышается
температура. По выздоровлении термостатирующие механизмы устанавливаются вновь на нор
мальную температуру, свойственную здоровому ор
ганизму.
Не все животные в одинаковой степени способ-
Рис. 116. Гомеостатические реакции организ
ма человека при действии высоких темпера
тур: 1 — мозг, 2 — нагретая кровь, 3 — капил
ляр, 4 — нагретый воздух, 5 — расширение ка
пилляров, 6 — мышцы, 7 — понижение интен
сивности обмена веществ, 8 — потовые желе
зы, 9 — пот, 10 — охлаждение за счет испа
рения.
ны осуществлять терморегуляцию. Среди животных
различают гомойотермных (от греч. homoios —
одинаковый, therme — тепло) — способных под
держивать постоянную температуру тела независи
мо от температуры окружающей среды, и пойкилотермных (от греч. poikilos — изменяющийся,
therme — тепло), температура тела которых изме
няется в зависимости от температуры окружающей
среды. Часто считают, что все животные, за исклю
чением птиц и млекопитающих, являются пойкилотермными, поскольку они не способны к столь вы
сокому уровню теплопродукции, как птицы и звери.
Это не совсем верно. Основную часть пойкилотермных животных составляют водные беспозвоноч
ные. Большинство из них, начиная от медуз, обита
ющих в тропических морях с температурой около
20 °С, и кончая раками, населяющими водоемы се-,
верных широт, температура которых меняется от
4 °С зимой до 20 °С летом, хорошо переносят коле
бания температуры, характерные для их среды. По
мере того как при переходе от зимы к лету темпера
тура воды повышается, некоторые из этих живот
ных синтезируют новый набор ферментов, которые
активны при более высоких температурах. Боль
шинство рыб, земноводных и некоторые пресмыка
ющиеся также пойкилотермны, однако у большин
ства пресмыкающихся и у некоторых крупных рыб
температура тела заметно выше температуры окру
жающей среды. Так, вмерзшая в лед рыба не поги
бает, ибо ее кровь не замерзает. Ящерицы тоже
способны регулировать температуру тела, находясь
в своей природной среде, хотя возможности для та
кой регуляции неизбежно ограничены доступнос
тью для животных тепла из окружающей среды.
С другой стороны, было выявлено, что температура
тела у многих птиц и млекопитающих колеблется в
широких пределах относительно разных участков
тела либо во времени. Из этого ясно, что провести
четкую границу между пойкилотермными и гомойотермными животными нелегко.
Эти расхождения способствовали поиску более
точных терминов. В связи с этим животные, кото
рые сами обеспечивают нагрев своего тела, были
названы зндотермными, а животные, почти пол
ностью зависящие от внешних источников тепла, —
эктотермными. Необходимо подчеркнуть, что по
нятия «гомойотермные» и «пойкилотермиые», а
также «эндо- и «эктотермные» представляют собой
183
идеализированные крайности, лишь изредка пол
ностью совпадающие с тем, что на самом деле име
ет место в природе. Большинство организмов зани
мает промежуточное положение между приведен
ными крайностями.
К эндотермным относят животных, температу
ра тела которых обычно превышает окружающую и
обусловлена генерацией собственного тепла. Они
продуцируют метаболическое тепло с разной ско
ростью. У многих тело обладает относительно низ
кой теплопроводностью (т. е. хорошей теплоизоля
цией), позволяющей им сохранять тепло в организ
ме, несмотря на высокий градиент температуры по
отношению к окружающей среде. К эндотермам от
носятся птицы и млекопитающие, а также ряд круп
ных рыб и некоторые летающие насекомые. По
скольку эндотермы в холодном климате поддержи
вают температуру тела намного выше температуры
окружающей среды, им доступны более холодные
места обитания, чем большинству эктотермов.
И хотя у большинства эндотермных животных обес
печена хорошая теплоизоляция тела благодаря на
личию перьев или шерсти, сохранение тепла дости
гается за счет солидных метаболических затрат. Так,
интенсивность энергетического обмена у эндотермного животного, находящегося в состоянии по
коя, по меньшей мере в 5 раз выше, чем у эктотермного с такими же размерами и температурой
тела.
У эктотермных животных метаболическая теп
лопродукция относительно невелика, поэтому тем
пература тела у них определяется в основном тепло
обменом с окружающей средой, т. е. для повышения
температуры тела они должны поглощать тепло из
окружающей среды. К эктотермам относятся боль
шинство беспозвоночных, земноводных, пресмыка
ющихся.
Выделяют также гетеротермных животных,
которые могут- изменять интенсивность собствен
ной теплопродукции, но, как правило, не способны
поддерживать температуру тела в узких рамках.
Примерами гетеротермов могут служить летающие
насекомые, питоны и некоторые рыбы. Температура
тела этих животных может намного превышать тем
пературу окружающей среды, что сопряжено с вы
делением тепла при-интенсивной мышечной работе.
Так, некоторые насекомые в течение какого-то вре
мени перед полетом «разминают» свои летатель
184
ные мышцы, чтобы к моменту подъема в воздух ра
зогреть их.
На выработку метаболического тепла
в количестве, достаточном для поддержа
ния высокой температуры тела (напри
мер, около 40 °С), требуется много энер
гии. В определенные периоды жизни жи
вотного, когда нет потенциальных брач
ных партнеров, добычи и др., бессмыс
ленно поддерживать температуру тела на
уровне, необходимом для сохранения ак
тивного образа жизни. В этом случае
многие животные экономят энергию, до
пуская сильное понижение температуры
тела. Так, например, многие наземные
беспозвоночные, земноводные, пресмы
кающиеся, некоторые птицы и мелкие
млекопитающие ежедневно на некоторое
время впадают в оцепенение: животное
перестает двигаться, и температура его
тела изменяется вместе с температурой
окружающего воздуха. Так поступают ма
ленькие колибри, которые днем для под
держания высокой интенсивности обмен
ных процессов должны почти постоянно
летать в поисках корма. Чтобы избежать
истощения запасов энергии в ночной пе
риод, когда питаться нет возможности,
они впадают в похожее на сон состояние
(оцепенение), во время которого темпе
ратура тела падает, приближаясь к тем
пературе окружающей среды. Некоторые
крупные эндотермные животные впадают
в состояние, похожее на оцепенение на
длительные периоды. Такие периоды, в
течение которых температура тела жи
вотного понижена, называются спячкой.
Спячка бывает зимняя и летняя. Мед
ведь, например, впадает в зимнюю спяч
ку. За лето он нагуливает толстый слой
подкожного жира; во время спячки этот
жир используется организмом как источ
ник питательных веществ, а также пре
дохраняет животное от переохлаждения.
Температура тела медведя падает, благо
даря чему снижается расходование пита
тельных веществ. В летнюю спячку впа
дают некоторые степные животные, на
пример грызуны, что помогает им пере
жить сухой и жаркий сезон года.
? 1. Объясните, почему организмы должны под
держивать постоянство внутренней среды. 2. При
ведите примеры гомеостаза. 3. Как осуществляется
поддержание pH крови на определенном уровне?
4. Каким образом осуществляется регуляция кровя
ного давления? 5. Как осуществляется терморегу
ляция у разных животных? 6. Почему некоторые
животные впадают в оцепенение и спячку? 7. Пти
цы и млекопитающие обладают четырехкамерным
сердцем и гомойотермны. Какова связь между эти
ми признаками?
54. Передвижение организмов.
Опорные системы и движение
растений
Движение относится к основным
свойствам живого и играет важную роль
в жизни живых организмов, которые пе
редвигаются в поисках пищи и воды; спа
саются от хищников, расселяются и осва
ивают новые благоприятные местообита
ния, ишут партнеров для размножения.
Движение может происходить на
уровне клетки (например, движение ци
топлазмы, перемещение хлоропластов и
т. п.), на уровне органа (сокращение сер
дца, движение конечностей, складывание
листочков сложного листа и т. д.) или на
уровне целого организма. Передвижение
всего организма с одного места на другое
называется локомоцией. Растениям
свойственно движение на клеточном и
органном уровнях, локомоторная же ак
тивность у них, как правило, отсутствует,
поскольку растения ведут прикреплен
ный образ жизни. Для грибов характерны
клеточные движения. Многим бактери
ям, протистам и большинству животных
свойственны различные типы локомоции.
Многие бактерии передвигаются с
помощью жгутиков. Жгутики у бактерий
устроены гораздо проще, чем у эукариот.
Жгутики бактерий состоят из одинаковых сфе
рических субъединиц белка флагеллина (похожего
на мышечный актин), которые расположены по
спирали и образуют полый цилиндр диаметром око
ло 10— 20 нм. Т. е. по строению жгутик напоминает
одну из микротрубочек жгутика эукариот. Жгутики
одним концом встроены в цитоплазматическую
мембрану клетки. В движение они приводятся по
средством уникального механизма. Основание жгу
тика, встроенное в цитоплазматическую мембрану,
вращается так, что жгутик как бы ввинчивается в
среду, не совершая беспорядочных биений, и таким
образом продвигает клетку вперед. Это, видимо,
единственная известная в природе структура, где
используется принцип колеса.
Бактерии передвигаются часто в от
вет на определенные раздражители, т. е.
они способны к таксису. Так, например,
аэробные бактерии обладают положи
тельным аэротаксисом — движутся туда,
где среда богаче кислородом. Подвижные
фотосинтезирующие бактерии обладают
положительным фототаксисом — пере
мещаются к свету.
Некоторые протисты также способны
к передвижению. Амебы, например, пе
редвигаются с помощью псевдоподий —
временных выступов. Образование псев
доподий связано с изменением агрегатно
го состояния цитоплазмы (золь — гель),
а также, возможно, с участием актиновых
и миозиновых микрофиламентов. Амебо
идное движение, как вам известно, харак
терно также для лейкоцитов позво
ночных.
Эвглена перемещается с помощью
жгутика, расположенного на переднем
конце тела. Жгутик тянет животное впе
ред. Волны движения регенерируются в
самом жгутике, и по мере прохождения
волн от основания жгутика к его концу их
скорость и амплитуда увеличиваются
(рис. 117,а, с. 186). Работа жгутика за
ставляет тело эвглены вращаться вокруг
своей оси со скоростью около одного обо
рота в секунду и в то же время обеспечи
вает продвижение в воде вперед по спи185
Рис. 117 Последовательные стадии движения:
а — ж гутика и б — реснички. Рабочее положе
ние реснички и жгутика начинается из положе
ния 1.
рали. С помощью жгутиков передвигают
ся также хламидомонада, трипаносомы,
мужские половые клетки — сперматозо
иды и т. д.
Инфузории передвигаются с по
мощью ресничек, которые на теле орга
низма размещаются рядами (рис. 117, б).
Опорные системы у растений. Ж и
вое содержимое клеток организмов пред
ставляет собой мягкое образование. По
этому, для того чтобы клетки, ткани и це
лые организмы имели определенную
форму и сохраняли ее, необходимы спе
циальные приспособления. Такие опор
ные приспособления хорошо развиты у
растений и животных, причем они состав
ляют значительную часть массы живого
организма.
Важнейшими опорными приспособ
лениями у растений являются твердые
стенки отдельных клеток. Даже у боль
ших деревьев стенки каждой отдельной
клетки играют существенную роль в под
держании формы. Живые клетки в стволе
дерева образуют сплошной цилиндр во
круг древесины. Внутри этого цилиндра
постоянно образуются новые клетки кси
лемы — ткани, осуществляющей, как вы
знаете, транспорт воды и минеральных
186
веществ. Старые клетки ксилемы с тол
стыми стенками из целлюлозы и лигнина
постепенно перестают функционировать
и заполняются вязким секретом. Эти
клетки составляют основную часть дре
весины. Они сохраняются в течение всей
жизни дерева и служат ему прочной опо
рой. Кроме того, в состав древесины вхо
дят также вытянутые мертвые клетки
собственно механической ткани — скле
ренхимы — с толстыми одревесневшими
оболочками.
Клетки травянистых растений не об
ладают такими жесткими стенками. Кро
ме того, в стеблях травянистых растений
содержится намного меньше клеток с од
ревесневшими стенками, поскольку эти
растения долго не живут. Поэтому в под
держании их формы основная роль при
надлежит тургорному давлению. Если,
например, растение томата или огурца не
получает достаточно воды, то по мере
снижения в клетках тургорного давления
растение вянет. Если же его обильно по
лить, то благодаря осмосу клетки расте
ния начинают всасывать воду и набуха
ют; тургор повышается, и растение вновь
выпрямляется. Этот процесс можно на
блюдать на срезах клубня картофеля и
корнеплода свеклы. Если срезы помес
тить в чистую воду, то клетки останутся
«жесткими» — они сохраняют тургор, в
концентрированном солевом растворе
вода по законам осмоса выходит из кле
ток и из-за мягкости клеточных стенок
срез становится вялым (его клетки при
этом плазмолизируются). Стенки клеток
травянистого растения не такие прочные,
как у древесного, и поэтому они не могут
создать достаточной опоры для растения
при снижении тургорного давления.
Необходимо отметить, что у травя
нистых растений также имеются специа
лизированные опорные системы (механи
ческие ткани), к которым относятся, на
пример, лубяные волокна. Эти волок
на располагаются вокруг проводящих
пучков.
Движение растений. Мы обычно
представляем себе растения как относи
тельно неподвижные организмы. Однако
для них характерны некоторые типы дви
жений, в первую очередь ростовые и тургорные.
Ростовые движения довольно мед
ленны; они обусловлены неравномерным
ростом противоположных сторон различ
ных органов растения. Так, колебатель
ные движения верхушки побега обуслов
лены неодинаковой скоростью роста его
разных сторон. Этим же объясняется
развертывание листьев и распускание
цветков. Многие ростовые движения из
вестны под названием тропизмов. При
мером может служить фототропизм —
направленная реакция искривления, вы
зываемая односторонним освещением;
побеги растений искривляются в сторону
света.
Одностороннее освещение вызывает приток
ростового гормона ауксина на менее освещенную
сторону и обеднение ауксином освещенной сторо
ны, что ведет к торможению роста клеток на осве
щенной стороне. Обогащение же ауксином затем
ненной стороны, напротив, стимулирует рост кле
ток, что и ведет к искривлению.
Тургорные движения — это обрати
мые искривления, возникающие у расте
ний из-за неравномерного изменения
тургорного давления на противополож
ных сторонах органа.
У многих растений, особенно у бобо
вых (клевер, акация), в листьях и листоч
ках имеются структуры, называемые лис
товыми подушечками — особые взду
тия у основания черешка или листочка, в
которых находятся крупные паренхимные
клетки. Быстрое изменение тургорного
давления в таких клетках приводит к то
му, что листовая подушечка начинает ра
ботать как шарнир, с помощью которого
и происходит движение. Так, в ответ на
механическое раздражение складывают
ся листочки мимозы стыдливой (рис.
118).
Известно также, что изменение тур
горного давления в клетках листьев насе
комоядных растений под влиянием при
косновения к чувствительным волоскам
вызывает их складывание. Прикоснове
ние насекомого вызывает потерю тургор
ного давления клеток на верхней стороне
листа, в то же время в клетках нижней
стороны тургорное давление сохраняет
ся. Благодаря этому лист быстро склады
вается, и насекомое оказывается пойман
ным. Далее клетки листа секретируют
ферменты, которые переваривают мяг
кие ткани насекомого.
?,к
1. Какое значение имеет передвижение в жизни
живых организмов? 2. Как поставить опыт, чтобы
определить по фототаксису, какую интенсивность
света предпочитает эвглена или хламидомонада?
3. Чем представлены опорные системы у растений и
каково их значение? 4. Какова основная причина
того, что животные могут передвигаться, а расте
ния нет?
55. Опорные системы и движение
животных
У огромного большинства животных
имеются разного рода опорные структу
ры, начиная от плотных иглообразных
образований у губок до сложноустроен
ного скелета у членистоногих и позвоноч
187
ных. Опорные системы, как правило,
обеспечивают характерную форму тела, а
она, в свою очередь, обусловлена опреде
ленными потребностями организма, поэ
тому у разных животных опорные систе
мы имеют разное строение.
Скелет выполняет несколько фун
кций в организме. Прежде всего — это
опорная структура. Скелет любого типа
является жестким, устойчивым к сжатию
каркасом тела. У наземных организмов
скелет обеспечивает опору для всей мас
сы тела, противодействуя силе тяжести, и
во многих случаях приподнимает тело над
землей. Это облегчает передвижение жи
вотных на суше.
У некоторых животных наружный
скелет защищает нежные внутренние
органы (членистоногие, моллюски). У
других эту функцию выполняют части
внутреннего скелета. У человека, напри
мер, черепная коробка обеспечивает за
щиту головного мозга и некоторых орга
нов чувств; позвоночник защищает спин
ной мозг, а ребра и грудина — сердце,
легкие, крупные кровеносные сосуды.
Кроме того, скелет служит местом при
крепления мышц. При сокращении мышц
части скелета работают как рычаги, что
вызывает движение различных органов.
У мягкотелых животных при движении
опорой для сокращения мышц служит по
лостная жидкость.
Различают три основных типа скеле
та: гидростатический, внешний (экзоске
лет) и внутренний (эндоскелет).
Гидростатический скелет характерен
для мягкотелых животных (круглые и
кольчатые черви). У них имеется полос
тная жидкость, заключенная внутри мы
шечных стенок тела. Эта жидкость ока
зывает давление на мышцы, и те в свою
очередь способны сокращаться, преодо
левая давление. Мышцы не прикреплены
к каким-либо структурам, и поэтому при
сокращении они тянут лишь друг друга.
Животное сохраняет определенные раз
меры и форму тела благодаря давлению
188
полостной жидкости, с одной стороны, и
сокращающимся мышцам — с другой.
Обычно мышечные волокна образуют
два слоя — продольную и кольцевую
мускулатуру. Движение происходит бла
годаря тому, что эти два слоя работают
как антагонисты. У несегментированных
животных (круглые черви) давление на
жидкость передается во все части тела.
У сегментированных (дождевой червь,
нереида) этот эффект локализован, и
лишь определенные сегменты приводятся
в движение или изменяют форму.
Наружный скелет (экзоскелет) ха
рактерен для членистоногих. Экзоскелет
(кутикула) секретируется эпидермисом.
Он состоит в основном из хитина. Это
многослойный твердый наружный покров
тела, который построен из сочлененных
между собой пластинок или трубчатых
образований. Хитин — прочный и легкий
материал, однако он может приобретать
твердость при встраивании в него «задубленных» белков или при пропитке из
вестью.
В участках скелета, которые должны
сохранять подвижность, хитин остается
неизменным. Такая конструкция из плас
тинок или трубочек, соединенных гибки
ми пленками, обеспечивает одновремен
но и защиту, и подвижность.
Хитин проницаем для воды, и поэтому
тело наземных членистоногих могло бы
терять ее слишком много. Этого, однако,
не происходит, поскольку поверх хитино
вого слоя откладывается эпикутикула,
состоящая из воскоподобных веществ,
секретируемых клетками эпидермиса.
Поэтому экзоскелет служит не только
опорой и защитой для внутренних орга
нов, но и предохраняет тело от обезвожи
вания.
Членистоногие — это животные,
имеющие членистые конечности, кото
рые состоят из рычагов, соединенных по
добием шарниров. Эти рычаги приводят
ся в движение мышцами-сгибателями и
разгибателями, прикрепленными к внут
ренним выступам экзоскелета. Мышцы
этих животных имеют форму полосок или
лент, расположенных внутри тела. Со
кращая отдельные мышцы, членистоно
гое животное может двигать любой ко
нечностью в различных направлениях.
Как средство защиты наружный ске
лет обладает рядом преимуществ перед
внутренним, но он ограничивает рост.
Рак, например, заключенный в свой пан
цирь, не может расти под ним. Поэтому у
этих животных происходит линька. Сбро
сив старый панцирь, рак впитывает воду
и быстро растет, пока вокруг его тела не
образуется новый, постепенно затверде
вающий панцирь.
До того момента, пока экзоскелет не
станет жестким, животное уязвимо для
хищников и практически не может пере
двигаться. Именно поэтому перед линь
кой такие животные прячутся в укрытиях.
У большинства моллюсков тело одето
очень твердой раковиной, которая слу
жит им прекрасной защитой, но сильно
ограничивает подвижность. Улитке, на
пример, приходится выдвигать из ракови
ны свою мускулистую ногу, с помощью
которой она передвигается. Однако для
защиты нежного тела моллюска прочный
наружный скелет совершенно необходим.
Внутренний скелет (эндоскелет)
имеется у некоторых протистов (радиоля
рий) и губок — в виде кремниевых или
известковых игл (спикул), у головоногих
моллюсков в виде внутренней раковины,
но особенно характерен для позвоноч
ных, у которых он построен из хрящевой
или костной ткани и расположен в теле
под слоем мышц. Эндоскелет хотя и не
является столь надежной защитой от
повреждений, но зато не препятствует
росту животного. Именно поэтому позво
ночные оказались самыми крупными жи
вотными на Земле. У более примитивных
позвоночных нет костного скелета. Так, у
акул, например, скелет состоит из хряща.
Хрящ — это вещество, из которого по
строена, например, ушная раковина и
кончик носа у человека. Он обладает дос
таточной твердостью, но в то же время
эластичен, благодаря чему способен лег
ко изгибаться И быстро принимать преж
нюю форму. У эмбрионов всех позвоноч
ных скелет первоначально закладывается
в виде хряща или плотных мембран, и
только позже у животных, которым свой
ственен костный скелет, хрящ замещает
ся костной тканью, хотя отдельные части
скелета так и остаются хрящевыми.
Почти все части скелета человека закладыва
ются на седьмой неделе эмбрионального развития.
Некоторые части костей лица и черепной коробки
состоят на этой стадии из мембран, весь же осталь
ной скелет образован хрящом. В течение восьмой
недели появляются центры окостенения, где обра
зуются особые клетки — остеобласты. В резуль
тате деятельности остеобластов происходит заме
щение хрящевой ткани на костную путем отложения
карбоната и фосфата кальция. К моменту рождения
процесс окостенения охватывает многие участки
скелета, но отнюдь не все. На темени, например,
еще остается хорошо заметное мягкое место —
родничок. Неполное окостенение черепа ребенка
имеет очень большое значение при родах. Дело в
том, что диаметр таза женщины недостаточно велик
для того, чтобы через него могла свободно пройти
голова младенца. Поэтому при родах происходит не
которое сжатие не полностью окостеневшего чере
па новорожденного. Эта деформация черепа исче
зает через несколько дней.
Отдельные части скелета ребенка остаются не
окостеневшими в течение всего периода детства, и
благодаря этому кости растут одновременно с рос
том всего организма. На эпифизах (суставных кон
цах) длинных трубчатых костей рук и ног сохраня
ются хрящевые диски — в этих местах происходит
рост костей (рис. 119, с. 190). С наступлением зре
лости хрящевые диски окостеневают, и рост в высо
ту прекращается. Если происходит нарушение фун
кции передней доли гипофиза до исчезновения эпи
физарных хрящей в трубчатых костях, это приводит
к гигантизму, если же патология развивается пос
ле завершения полного отвердения костей, то это
приводит к акромегалии (увеличение кистей и
стоп, а также выступающих частей лица).
189
Рис. 119. Эпифизарные диски в длинных труб
чатых костях ребенка: 1 — эпифиз, 2 —
суставная поверхность, 3 — суставная по
лость, 4 — зоны роста.
Некоторые части скелета вообще не подверга
ются окостенению и остаются хрящевыми до ста
рости: это, например, передние концы ребер, благо
даря чему грудная клетка может расширяться при
дыхании. Не окостеневают и межпозвоночные дис
ки. Хрящ сохраняется также в местах соприкосно
вения трущихся поверхностей костей. С возрастом
некоторые из этих хрящей подвергаются окостене
нию, что нередко бывает причиной сильных болей.
Такое заболевание называется артрозом.
Скелет обычно состоит из множества
костей. Например, в состав скелета чело
190
века их входит более 200 (рис. 120). Кос
ти по-разному могут соединяться друг с
другом. Некоторые из них сочленены не
подвижно с помощью швов (кости чере
па). Многие кости скелета соединены
друг с другом с помощью подвижных со
единений, или суставов, что обеспечи
вает подвижность тела и отдельных его
частей. Кости удерживаются в суставе с
помощью прочных связок, которые идут
от одной кости к другой. Суставы окруже
ны синовиальной сумкой, содержащей
вязкую синовиальную жидкость, которая
действует как смазка и уменьшает трение
суставных поверхностей костей.
Суставы различаются по типу устрой
ства и характеру движения.
Цилиндрические суставы (локтевой)
допускают движение в основном в одной
плоскости. Шаровидные суставы (плече
вой и тазобедренный) имеют три оси дви
жения. Плоские суставы (запястье и
предплечье) допускают скользящие дви
жения костей.
Кости могут сочленяться также при
помощи хрящей (хрящевые соедине
ния). Такие соединения называются по
лу подвижными. При этом поверхности
костей разделяются небольшим количес
твом промежуточного вещества, физи
ческие свойства которого делают воз
можным только незначительные движе
ния. Примером хрящевого соединения
могут служить межпозвоночные диски —
участки волокнистого хряща между тела
ми соседних позвонков.
У некоторых позвоночных животных,
помимо внутреннего скелета, развивает
ся также и наружный.
Так, у черепах имеется панцирь, обра
зовавшийся из отдельных пластинок, ко
торые сильно разрослись и затем слились
между собой. Панцирь ограничивает под
вижность этих животных, но зато служит
надежным средством защиты.
У броненосцев кожа покрыта чешуями, образующими на спине животного
подобие брони.
20 — копчик, 21 —■запястье, 22 — пясть, 23 —
фаланги, 24 — бедренная кость, 25 — колен
ная чашечка, 26 — большая берцовая кость,
27 — малая берцовая кость, 28 — предплюсна,
29 — плюсна, 30 — фаланги.
Рис. 120. Скелет человека: 1 — теменная
кость, 2 — височная кость, 3 — скуловая кость,
4 — верхняя челюсть, 5 — нижняя челюсть,
6— 7-й шейный позвонок, 7 — 1-й грудной поз
вонок, 8 — I ребро, 9 — ключица, 10 — лопат
ка, 11 — грудина, 12 — плечевая кость, 13 —
лучевая кость, 14 — XII ребро, 15 — локтевая
кость, 16 — подвздошная кость, 17 — лобковая
кость, 18 — седалищная кость, 19 — крестец,
Движение позвоночных. Движение
позвоночных животных и человека обес
печивается работой мышц. У человека,
например, мышцы составляют около
40 % массы тела. Каждая клетка этой
громадной массы мышечной ткани вы
полняет функцию сокращения при воз
буждении. Как уже отмечалось, у позво
ночных имеется три вида мышечной тка
ни: поперечнополосатая, гладкая и
сердечная. В передвижении позвоноч
ных главная роль принадлежит скелет
ным поперечнополосатым мышцам. Ти
пичным примером такой мышцы может
служить двуглавая (бицепс), контур ко
торой хорошо виден на плече, если со
гнуть руку в локте. От обоих концов ске
летной мышцы обычно отходят плотные
сухожилия, которые прикрепляются, как
правило, к двум разным костям. При со
кращении мышцы происходит движение
одной из этих костей. Тот конец мышцы,
который при ее сокращении остается от
носительно неподвижным, называется
началом мышцы, а тот, который переме
щается, — местом прикрепления. На
пример, начало бицепса находится на ло
патке, а место прикрепления — на лу
чевой кости с внутренней стороны пред
плечья. Сокращение бицепса вызывает
сгиб руки в локтевом суставе. Таким об
разом, эта мышца относится к категории
сгибателей (рис. 121, с. 192). Противо
положная по действию мышца (антаго
нист), которая выпрямляет руку (трех
главая мышца — трицепс), принадлежит
к группе разгибателей. Мышцы, с по
мощью которых верхние конечности дви
жутся от тела, называются отводящими
(например, дельтовидная мышца, отводя
щая руку в сторону). Антагонистами от
водящих мышц являются мышцы, при191
а также в поддержании равновесия тела.
Действие каждой мышцы достигает
максимального эффекта лишь при одно
временном расслаблении мышцы-антагониста. Такая согласованность называ
ется мышечной координацией.
?,
1. Какие функции выполняет скелет в организ
ме? 2. Какие типы скелета выделяют и в чем заклю
чается принципиальная разница между ними?
3. Каково происхождение экзоскелета у членисто
ногих? 4. Благодаря чему осуществляется движение
позвоночных? 5. Мышцы прикрепляются к костям
посредством сухожилий. Как выдумаете, какие пре
имущества имеет этот способ по сравнению с не
посредственным прикреплением мышц к кости?
6. Каков механизм сокращения мышц? 7. Вспомни
те, как отличаются по строению и- функциям попе
речно-полосатые, гладкие и сердечная мышцы.
Рис. 121. Действие мышцы-сгибателя: а — рас
слабление, б — сокращение.
жимающие руку к телу, — приводящие.
Существуют мышцы, вращающие ту или
иную часть тела, например голову, плечо
или бедро.
Мышцы обычно образуют пары анта
гонистов — одна тянет кость в одну сто
рону, другая — в противоположную. В
зависимости от характера использования
этих мышц они обладают различной си
лой. Так, мышцы, опускающие крыло
птицы, значительно сильнее мышц, под
нимающих крыло. Именно поэтому на ки
ле у птиц значительно большая мышеч
ная масса, чем на спине. Мышцы, смыка
ющие челюсти, во много раз сильнее
мышц, размыкающих их и т. д.
Таким образом, движение позвоноч
ных животных и человека обеспечивает
ся благодаря тому, что кости, соединен
ные подвижными суставами, приводятся
в движение мышцами и работают как ры
чаги. Необходимо также отметить, что
поперечнополосатые мышцы участвуют
не только в передвижении организмов, но
и одновременно в фиксации суставов,
предотвращая ненужные движения,
192
ГЛАВА V
Размножение
й индивидуальное развитие
организмов
56. Типы размножения организмов.
Бесполое размножение
Способность размножаться, то есть
производить новое поколение особей то
го же вида, — одно и з. обязательных и
важнейших свойств всех без исключения
живых организмов. Это обусловлено тем,
что каждую секунду на Земле гибнут от
старости, болезней, хищников и природ
ных катаклизмов (вулканические извер
жения, пожары, наводнения и пр.) мил
лиарды живых существ. И только благо
даря размножению жизнь на Земле не
прекращается. При размножении проис
ходит передача генетического материала
(хромосом) от родительских форм дочер
ним, что обеспечивает воспроизведение
признаков не только данного вида, но и
конкретных родительских особей. Следо
вательно, размножение поддерживает
длительное существование вида, сохра
няя при этом преемственность между ро
дителями и их потомством в ряду многих
поколений, приводит к увеличению чис
ленности особей вида и способствует его
расселению. У растений, которые ведут
прикрепленный образ жизни, расселение
в процессе размножения — единствен
ный способ занять большую территорию
обитания.
Различают два типа размножения ор
ганизмов: бесполое и половое. Бесполое
размножение происходит без образова
ния гамет, и в нем участвует лишь одна
особь, которая делится, почкуется или
формирует споры. В случае полового
размножения особи нового поколения
появляются, как правило, при участии
двух организмов — материнского и от
цовского.
Бесполое размножение. Существует
несколько способов бесполого размно
жения, среди которых можно выделить
вегетативное и собственно бесполое
(спорообразование, или споруляция).
Вегетативное размножение осно
вано на способности организмов восста
навливать (регенерировать) недостаю
щие части. Этот способ. размножения
широко распространен в природе, но на
ибольшего разнообразия он достигает у
растений, особенно у цветковых.
У многих одноклеточных организмов
(бактерии, протесты) бесполое размно
жение сводится к простому бинарному
делению на две более мелкие идентичные
клетки. Делению клетки предшествует
репликация ДНК, а у эукариот — также
деление ядра. У многоклеточных циано
бактерий, водорослей, грибов, лишайни
ков размножение осуществляется фраг
ментами нитей, гиф и обломками слое
вищ соответственно.'При благоприятных
условиях это приводит к быстрому росту
популяций.
Одной из форм бесполого размноже
ния является почкование. В этом случае
новая особь образуется в виде выроста
(почки) на теле родительской особи, а за
тем отделяется от нее, превращаясь в са
7. Общая биология, 10 кл.
мостоятельный организм, совершенно
идентичный родительскому. Почкование
характерно для некоторых кишечнопо
лостных, например гидры (рис. 122, с.
194) и дрожжевых грибов. Если при этом
дочерние особи не отделяются от мате
ринской, могут возникать колонии.
У цветковых растений новые особи
часто формируются из вегетативных ор
ганов: стебля (кактус, элодея, ряска, ро
голистник), листа (фиалка, бегония, ли
лия, гиацинт), корня (малина, крыжов
ник, осот, одуванчик), видоизмененных
побегов: клубня (картофель, топинам
бур), луковицы (лук, чеснок, тюльпан,
нарцисс), клубнелуковицы (гладиолус,
шафран), корневища (пырей, мята, иванчай), усов (земляника, лютик ползучий).
Вегетативное размножение растений ши
роко распространено в природе (рис. 123,
с. 194) и часто используется в сельскохо
зяйственной практике при разведении
плодово-ягодных, овощных и цветочных
растений.
У животных в силу высокой специа
лизации клеток организма вегетативное
размножение встречается значительно
реже. Кроме кишечнополостных, оно
наблюдается у губок, плоских и некото
рых кольчатых червей. Например, тело
ряда примитивных морских червей и мор
ских звезд, разделенное на части, может
дать в результате регенерации каждой из
частей новую особь.
У отдельных видов млекопитающих
(южноамериканский броненосец) встре
чается вегетативное размножение заро
дышей, когда в ранний период эмбрио
нального развития делящийся зародыше
вый диск дает начало нескольким особям
(от 4 до 8). Подобное явление можно на
блюдать и у человека — однояйцевые
близнецы, которые поразительно похожи
друг на друга.
У многих организмов для воспроиз
водства потомства образуются специали
зированные клетки — споры, каяедая из
которых в благоприятных условиях мо193
Рис. 122. Почкующаяся гидра: 1 — взрос
лая особь, 2 — молодые гидры, 3 — щу
пальца, 4 — подошва.
Рис. 123. Вегетативное размножение растений: 1 — корневищем, 2
— усами, 3 — черенками, 4 — отводками, 5 — прививкой, 6 — лис
тьями.
194
жет прорасти и дать начало новому орга
низму. Спорообразование встречается у
протистов, грибов, водорослей и у выс
ших растений.
Споры образуются путем митоза или
мейоза в обычных вегетативных клетках
материнского организма или специаль
ных органах — спорангиях’, микроспоры
и мегаспоры у семенных растений обра
зуются в особых спорангиях, называемых
пыльцевым мешком и семязачатком
соответственно.
Споры представляют собой микро
скопические одноклеточные образова
ния, состоящие из небольшого количес
тва цитоплазмы и ядра с минимальным
запасом питательных веществ. Главное
достоинство таких спор — возможность
быстрого размножения и расселения ви
дов, в особенности грибов.
Преимущество бесполого размноже
ния в его простоте и эффективности —
не нужно искать партнера, потомство мо
жет оставить любая особь в любом мес
те, удачные сочетания генов не теряются.
При переходе к половому размножению
эти преимущества утрачиваются.
При любой форме бесполого размно
жения — частями тела или спорами —
наблюдается увеличение численности
особей данного вида без повышения их
генетического разнообразия: все особи
являются точной копией материнского
организма. Эта особенность использует
ся человеком для получения однородного,
с хорошими признаками потомства у пло
дово-ягодных, декоративных и других
групп растений. Новые признаки у таких
организмов появляются только в резуль
тате мутаций.
Вид, размножающийся только беспо
лым путем, может процветать достаточно
длительное время, но в конечном итоге он
обречен на вымирание, что для него го
раздо более вероятно, чем для вида с по
ловым размножением. Если в определен
ном местообитании возникает измене
ние, вызывающее гибель отдельных осо
бей того или иного вида живых организ
мов, то погибнут все особи, потому что
они генетически сходны.
?
■ I. Каково биологическое значение размноже
ния организмов? 2. Как осуществляется бесполое
размножение у бактерий, протистов, грибов, расте
ний и животных? 3. Каковы особенности и преиму
щества бесполого размножения?
57. Половое размножение
Половое размножение существенно
отличается от бесполого тем, что в дан
ном случае наследственные признаки по
томков возникают в результате перекомбинации генов, принадлежащих обоим
родителям. Это повышает возможности
организмов в приспособлении к меняю
щимся условиям среды. В результате по
томство может быть более жизнеспособ
ным, чем каждая из родительских особей,
что имеет первостепенное значение в
эволюции организмов.
Половое размножение характеризу
ется наличием полового процесса, одним
из важнейших этапов которого является
оплодотворение — слияние половых
клеток, или гамет. Гаметы различают
ся по строению и физиологическим свой
ствам и делятся на мужские (подвиж
ные — сперматозоиды, неподвиж
ные — спермии) и женские (яй ц е
клет ки).
В отличие от спор, одна гамета, как
правило, не может дать начало новой
особи. Этому предшествует оплодотворе
ние, в результате которого образуется зи
гота. В дальнейшем из зиготы развивает
ся зародыш нового организма.
Процесс образования половых кле
ток (гаметогенез) у водорослей, многих
грибов и высших споровых растений про
исходит путем мейоза или митоза в спе
циальных органах полового размно
жения: яйцеклеток — в гаметангиях,
оогониях или архегониях, сперматозои
дов и спермиев — в антеридиях, а у
195
протестов и некоторых водорослей — в
вегетативной клетке,
У животных и человека развитие яй
цеклеток (оогенез) и сперматозоидов
(сперматогенез) осуществляется в гона
дах, или половых железах — яичниках
и семенниках соответственно. В процес
се формирования половых клеток выде
ляют три периода — размножение, рост
и созревание (рис. 124).
Сперматогенез. Первичные мужские
половые клетки (сперматогонии) мно
гократно делятся путем митоза (период
размножения), в результате чего их коли
чество постоянно возрастает. В период
роста деление клеток прекращается, они
начинают усиленно расти и становятся
сперматоцитами первого порядка.
Последние в период созревания в резуль
тате первого деления мейоза образуют
гаплоидные сперматоциты второго
порядка, а после второго мейотического
деления превращаются в сперматиды.
Таким образом, из одной исходной клетки
в результате мейоза образуются четыре
сперматиды, из которых затем формиру
ются сперматозоиды (стадия формиро
вания). При этом ядро уменьшается в
размерах за счет плотной упаковки хро
мосом, а также сокращается объем ци
топлазмы, и формируются цитоплазмати
ческие структуры, характерные только
для сперматозоидов. Процесс образова
ния сперматозоида у человека занимает
примерно 70 дней; на 1 г массы яичка об
разуется 107 сперматозоидов в сутки.
Следует отметить, что образование
сперматозоидов продолжается в мужском
организме все время, независимо от того,
существует ли вероятность оплодотворе
ния яйцеклетки или не существует.
Типичный сперматозоид млекопитаю
щих состоит из головки, шейки и хвости
ка (рис. 125, с. 198). Головка содержит
ядро и небольшое количество цитоплаз
мы. На кончике головки располагается
аппарат Гольджи, преобразованный в
кольцевое тельце — акросому. В ней об
196
разуются ферменты, растворяющие мем
брану яйцеклетки при оплодотворении;
следовательно, функционально акросому
можно рассматривать как увеличенную
лизосому. В цитоплазме шейки сосредо
точены 4— 10 митохондрий, генерирую
щих энергию для движения жгутиков и
центриоли.
Сперматозоиды хорошо сохраняют
свою целостность вне организма и доста
точно устойчивы к воздействию небла
гоприятных условий внешней среды. Это
свойство широко применяется в сельс
ком хозяйстве, в частности, при разведе
нии крупного рогатого скота методом ис
кусственного осеменения. Сперму элит
ных пород животных собирают, замора
живают и хранят при очень низких темпе
ратурах (обычно в жидком азоте). В та
ком состоянии сперму можно сохранять
продолжительное время, а после размо
раживания использовать для получения
высокопродуктивного потомства.
Оогенез. В отличие от образования
сперматозоидов, которое начинается у
млекопитающих, в том числе и у челове
ка, только при половом созревании, об
разование яйцеклеток у женщин начина
ется еще до их рождения и завершается
только после оплодотворения.
Во время развития плода первичные
половые клетки многократно делятся пу
тем митоза (период размножения), обра
зуя множество клеток, называемых оогониями. Оогонии дифференцируются в
ооциты первого порядка, которые со
храняются без изменения долгие годы.
С наступлением половой зрелости от
дельные ооциты периодически вступают в
стадию роста, которая может продол
жаться несколько месяцев. За это время
ооцит достигает гигантских размеров за
счет синтеза РНК и белков в нем самом,
а также благодаря активному транспорту
веществ из окружающих ооцит клеток, из
крови или гемолимфы. В этом транспор
те активно участвуют окружающие ооцит
фолликулярные клетки. Плод женского
Рис. 124. Схема сперматогенеза (а) и оогенеза (б) у человека.
197
Рис. 125. Строение сперматозоида млекопита
ющих: а — головка, б — шейка, в — средняя
часть, г — хвостик, д — нить: 1 — акросома,
2 — головной чехлик, 3 — ядро, 4— 5 — центриоли, 6 — митохондрия, 7 — нить.
пола непосредственно перед ро>вдением
содержит около 2x 106 фолликулов, но
лишь около 450 из них достигают стадии
ооцитов второго порядка и выходят из
яичника (овуляция).
Перед овуляцией ооцит первого по
рядка делится путем мейоза (первое де
ление), образуя крупную гаплоидную
клетку (ооцит второго порядка),
состоящую почти целиком из цито
плазмы, и мелкую клетку, состоящую по
существу из ядра и минимального коли
чества цитоплазмы. Эту клетку называют
направительным (полярным, или р е
дукционным) тельцем. Второе деление
мейоза у человека доходит до стадии ме
тафазы и на этом останавливается до тех
пор, пока ооцит не сольется со спермато
198
зоидом. При оплодотворении ооцит вто
рого порядка совершает второе мейотическое деление, образуя крупную яйце
клетку и второе направительное тельце.
Первое направительное тельце, как пра
вило, тоже делится.
Таким образом, в отличие от сперма
тогенеза, где в ходе мейоза образуются
четыре гаплоидные, равноценные друг
другу клетки, при оогенезе развиваются
одна крупная яйцеклетка и три маленькие
клетки (направительные тельца), кото
рые вскоре погибают. Биологический
смысл формирования направительных
телец заключается в сохранении в яйце
клетке максимального количества желт
ка, необходимого для развития будущего
зародыша. Достигается это за счет гибе
ли трех полноценных (с генетической
точки зрения) направительных телец.
Яйцеклетки организмов в зависимос
ти от количества желтка имеют разную
величину (у морского ежа — 0,085 мм, у
человека — 0,2 мм, у страуса — 80 мм, у
сельдевой акулы — 22 мм). Типичное яд
ро яйцеклетки содержит гаплоидный на
бор хромосом. В цитоплазме функциони
руют митохондрии, рибосомы, аппарат
Гольджи и слабо развитая эндоплазмати
ческая сеть; накапливается значительное
количество нуклеотидов, аминокислот,
белков и других веществ, необходимых
для ранних стадий развития зародыша.
Яйцеклетка имеет чаще всего округ
лую форму и всегда окружена одной или
несколькими оболочками, имеющими
сложное строение (рис. 126). Их функция
заключается в защите яйцеклетки и раз
вивающегося зародыша от неблагопри
ятных условий среды.
Гермафродитизм. Виды, у которых
одна и та же особь способна производить
и мужские и женские гаметы, называют
двуполыми, или гермафродитами. К их
числу относятся многие протесты, ки
шечнополостные, плоские и кольчатые
черви, ракообразные, моллюски (улит
ка), некоторые рыбы и пресмыкающиеся
3
6
Рис. 126. Строение яйца курицы: 1 — желток,
2 — желточная оболочка, 3 — зародышевый
диск, 4 — скорлупа, 5 — белок, 6 — канатики,
удерживающие яйцеклетку в центре яйца.
(ящерица), а также большинство рас
тений.
Гермафродитизм развился как при
способление к сидячему, малоподвижно
му или паразитическому образу жизни.
Одним из его преимуществ является воз
можность самооплодотворения при нали
чии только одной особи. Кроме того, гер
мафродитизм увеличивает продолжи
тельность периода размножения и веро
ятность осеменения особей, особенно у
мелких животных с относительно круп
ными яйцами и низкой плодовитостью.
Необходимо отметить, что у большин
ства гермафродитных видов в оплодотво
рении участвуют гаметы, происходящие
от разных особей. У них имеются много
численные генетические, морфологичес
кие и физиологические адаптации, кото
рые препятствуют самооплодотворению
и способствуют перекрестному оплодо
творению. Например, у многих протистов
самооплодотворение предотвращается
генетической несовместимостью, у мор
ских ракообразных — неодновременным
развитием мужских и женских половых
желез, у ресничных и дождевых червей,
моллюсков — строением полового аппа
рата, не допускающего поступления се
мени в женские органы той же особи. По
добные приспособления имеются у мно
гих цветковых растений (например, неод
новременное созревание тычинок и пес
тиков и др.).
Аномальный гермафродитизм наблю
дается у всех крупных животных и у чело
века и обусловлен генетически. Он мо
жет быть истинным, когда у одной особи
имеются одновременно мужские и жен
ские половые железы, или ложным, ког
да у особи имеются половые железы од
ного пола, а наружные половые органы и
вторичные половые признаки соответ
ствуют признакам другого пола. Приме
ром подобного явления служит мужеподобие (мускулинизация) самок и жено
подобие (феминизация) самцов.
Партеногенез (от греч. parthenos —
девственница и genesis — происхожде
ние) — одна из модификаций полового
размножения, при которой развитие но
вой особи происходит из неоплодотворенной яйцеклетки. Партеногенетическое размножение встречается как у жи
вотных, так и у растений; преимущество
его состоит в том, что в некоторых случа
ях оно повышает скорость размножения.
В зависимости от числа хромосом в
яйцеклетке различают два вида партено
генеза — гаплоидный и диплоидный.
При гаплоидном партеногенезе у многих
насекомых (пчел, муравьев и ос) из неоплодотворенных яйцеклеток развива
ются гаплоидные самцы, а из оплодо
творенной — диплоидные самки, что
приводит к возникновению в данном со
обществе различных каст организмов.
Например, у медоносной пчелы матка
(царица) откладывает оплодотворенные
яйца (2п=32), из которых развиваются
самки (матки или рабочие пчелы), и неоплодотворенные яйца (п=16), которые
дают самцов (трутней), производящих
199
спермин путем митоза, а не мейоза. Такой
механизм размножения у общественных
насекомых позволяет регулировать чис
ленность потомков разного пола.
У тлей, дафний, коловраток, некото
рых ящериц и змей происходит диплоид
ный партеногенез, при котором ооциты
самки претерпевают особую форму мей
оза без расхождения хромосом. При этом
все хромосомы переходят в яйцеклетку, а
направительные тельца не получают их
вовсе. Яйцеклетки развиваются в мате
ринском организме, так что молодые сам
ки рождаются вполне сформировавши
мися, а не вылупляются из яиц. Такой
процесс называется живорождением.
Он может продолжаться в течение не
скольких поколений, особенно летом
(например, тли летом обычно представ
лены только бескрылыми самками, рож
дающими живых детенышей), до тех пор,
пока в одной из клеток не произойдет
почти полное нерасхождение хромосом.
Из этой клетки осенью партеногенетически развивается самец. Осенние самцы
и партеногенетические самки формируют
гаплоидные гаметы, участвующие в поло
вом размножении. Оплодотворенные
самки откладывают диплоидные яйца, из
которых весной вылупляются самки, раз
множающиеся партеногенетически и
рождающие живых потомков. Главное
преимущество, которое дает диплоидный
партеногенез, — быстрый рост числен
ности популяции, так как при этом все
половозрелые особи способны к откладке
яиц.
У растений партеногенез представлен
различными формами. Одна из них —
апомиксис — имитация полового раз
множения. Апомиксис наблюдается у не
которых цветковых растений (семейства
крестоцветные, розоцветные, сложно
цветные, злаки и др.), у которых диплоид
ная клетка семязачатка развивается в
функциональный зародыш без участия
спермиев. Из остального семязачатка об
разуется семя, а из завязи развивается
200
плод. В других случаях требуется присут
ствие пыльцевого зерна, которое стиму
лирует партеногенез, хотя и не прораста
ет; пыльцевое зерно индуцирует гормо
нальные изменения, необходимые для
развития зародыша. На практике такие
случаи трудно отличить от настоящего
полового размножения.
?
1. Каковы отличия и преимущества полового
размножения в сравнении с бесполым? 2. Где и как
осуществляется образование женских и мужских
гамет? 3. Каков биологический смысл формирова
ния при оогенезе направительных телец? 4. Каковы
особенности строения яйцеклеток и сперматозои
дов? 5. Что такое гермафродитизм и партеногенез и
каково их биологическое значение?
58. Оплодотворение
Процесс оплодотворения состоит в
слиянии женской и мужской гамет с об
разованием зиготы.
Оплодотворение у животных. Извес
тна закономерность: чем меньше вероят
ность встречи яйцеклетки и сперматозои
да, тем большее число половых клеток
образуется в организме. Например, у
большинства водных животных мужские
и женские гаметы выделяются в воду, где
соединяются в значительной мере слу
чайно, поэтому рыбы выметывают сотни
тысяч и даже миллионы икринок (яйце
клеток). Этот примитивный и довольно
ненадежный способ соединения гамет на
зывается наружным оплодот воре
нием.
Другие животные, преимущественно
наземные, имеют наружные половые ор
ганы для переноса спермальной жидкос
ти из тела самца в тело самки, где и про
исходит оплодотворение. В данном случае
речь идет о внутреннем оплодотворе
нии, при котором вероятность слияния
яйцеклетки и сперматозоида очень высо
ка и, как следствие, количество половых
клеток резко уменьшается.
У животных в спермальной жидкости
находятся миллионы сперматозоидов,
каждый из которых активно движется по
направлению к яйцеклетке. Как только
первый сперматозоид проникает сквозь
мембрану яйцеклетки, тут же образуется
оболочка оплодотворения, которая не до
пускает проникновения в яйцеклетку дру
гих сперматозоидов. Затем ядро сперма
тозоида начинает движение к ядру яйце
клетки и сливается с ним. Так образуется
зигота с диплоидным набором хромосом.
В ядре зиготы все хромосомы вновь
становятся парными: в каждой паре го
мологичных хромосом одна из них — от
цовская, другая — материнская. Следо
вательно, диплоидный набор хромосом,
характерный для соматических клеток
каждого вида организмов, восстанавли
вается именно при оплодотворении.
Развитие женских и мужских гамет и
оплодотворение у растений происходит
так же, как и у животных, — в женских и
мужских органах полового размножения.
Оплодотворение у растений. Рас
смотрим процесс оплодотворения у по
крытосеменных (цветковых) растений.
(Вспомните строение и функции цветка
из курса ботаники.)
Оплодотворению у цветковых расте
ний предшествует формирование жен
ского и мужского гаметофитов — поло
вого поколения в жизненном цикле выс
ших растений (рис. 127, с. 202). Муж
ской гаметофит образуется таким обра
зом. В пыльцевых мешках (спорангиях)
пыльников тычинок имеются особые дип
лоидные клетки — микроспороциты,
или материнские клетки микроспор.
Каждый микроспороцит претерпевает
мейоз и образует четыре микроспоры.
Затем гаплоидное ядро каждой микро
споры делится митотически,и образуется
пыльцевое зерно (пыльца) — мужской
гаметофит цветковых растений. Он
состоит из двух клеток: большой
(вегетативной) и малой (генеративной),
покрытых двойной оболочкой. У одно
дольных растений генеративная клетка
пыльцевого зерна делится митотически с
образованием двух мужских половых
клеток, лишенных жгутиков, — спермиев. У двудольных растений образование
половых клеток происходит позже, когда
пыльца попадает на рыльце пестика.
Таким образом, зрелая пыльца состо
ит из двух (вегетативной и генеративной)
или из трех (вегетативной и двух спермиев) клеток, покрытых двойной оболочкой.
Образование женского гаметофита
происходит в семяпочке, находящейся
внутри завязи пестика. Семяпочка, или
семязачаток, — это видоизмененный
мегаспорангий (нуцеллус), защищен
ный покровами. Покровы одевают семя
зачаток со всех сторон, но на верхушке не
срастаются и образуют узкий канат —
пыльцевход. В семяпочке, вблизи пыль
цевхода, начинает развиваться одна дип
лоидная клетка, которая становится
единственным макро-, или мегаспороцитом. Он делится мейотически, давая
четыре гаплоидные макро-, или мега
споры, обычно расположенные линейно,
т. е. друг за другом. Три мегаспоры вско
ре разрушаются, а четвертая, наиболее
удаленная от пыльцевхода, развивается в
зародышевый мешок.
Зародышевый мешок растет, его ядро
трижды делится митотически, в результа
те чего образуется восемь дочерних ядер.
Они располагаются по четыре двумя
группами — вблизи пыльцевхода гамето
фита и на противоположном конце. Два
полярных ядра перемещаются к центру
зародышевого мешка и сливаются, пре
вращаясь в одно диплоидное ядро (в не
которых случаях их слияние происходит
позднее, при оплодотворении).
Остальные шесть ядер, по три на
каждом полюсе, разделяются тонкими
клеточными стенками. При этом на по
люсе у пыльцевхода образуется яйцевой
аппарат, состоящий из яйцеклетки и двух
клеток-синергид. На противоположном
полюсе возникают так называемые
клетки-антиподы. Такая восьмиядерt
201
Т ы ч и н ка
Т е тр а д а
м и кр о с п о р
Три м е га спор ы
д е ге н е р и р у ю т
М итоз
в м икро
спо р а х
Я й ц е кл е тка
Полярные
ядра
А н ти п о д ы
З ароды ш евы й меш ок
б
Рис. 127 Образование у цветковых растений: а — пыльцевых зерен, б — зародышевых мешков.
202
ная семиклеточная структура — заро
дышевый мешок — и является зрелым
женским гаметофитом, готовым к опло
дотворению, которому предшествует
опыление.
Опыление — это процесс переноса
пыльцы из пыльников тычинок на рыльце
пестика. Пыльца может переноситься на
секомыми, птицами, ветром, водой, но в
основном — насекомыми и ветром.
Попав на рыльце пестика, пыльцевое
зерно начинает прорастать (рис. 128). Из
вегетативной клетки развивается длин
ная пыльцевая трубка, которая по тканям
столбика дорастает до завязи и далее до
семяпочки. Из генеративной клетки к
Рис. 128. Схема двойного оплодотворения у цветковых растений: а — продольный раз
рез пестика, б — прорастание пыльцевого зерна, в — проникновение пыльцевой труб
ки в зародышевый мешок, г — излияние содержимого пыльцевой трубки (двух спермиев) в зародышевый мешок, g — зародышевый мешок после оплодотворения: 1 — про
растающее пыльцевое зерно, 2 — пыльцевая трубка, 3 — завязь, 4 — зрелый зароды
шевый мешок, 5 — спермин, 6 — вегетативное ядро, 7 — яйцеклетка, 8 — полярные яд
ра, 9 — антиподы, 10 — синергиды, 11 — зигота, 12 — триплоидное ядро эндосперма.
203
этому моменту образуются два спермин,
которые спускаются в пыльцевую трубку.
Пыльцевая трубка входит в семяпочку
через пыльцевход, ее ядро дегенерирует,
а кончик трубки разрывается, освобож
дая спермин, которые проникают в заро
дышевый мешок в синергиду или в щель
между яйцеклеткой и центральным яд
ром. Вскоре после вхождения пыльцевой
трубки в зародышевый мешок синергиды
погибают.
После этого один из спермиев опло
дотворяет яйцеклетку. В результате обра
зуется диплоидная зигота, из которой
развивается зародыш нового раститель
ного организма. Второй спермий слива
ется с двумя полярными ядрами (или с
центральным диплоидным ядром), обра
зуя триплоидную клетку. Впоследствии из
этой клетки возникает питательная
ткань — эндосперм. В его клетках содер
жится запас питательных веществ, необ
ходимых для развития зародыша.
Слияние одного спермия с яйцеклет
кой, а другого — с полярными ядрами —
двойное оплодотворение — представля
ет собой уникальную особенность покры
тосеменных растений. Такой способ
оплодотворения был открыт в 1898 г.
русским
цитологом и эмбриологом
С. Г. Навашиным.
После оплодотворения семяпочка
разрастается и превращается в семя; в
результате же разрастания завязи пести
ка формируется плод. Стенки завязи ста
новятся стенками плода — околоплод
ником, внутри которого находятся се
мена.
Благодаря двойному оплодотворению
происходит очень быстрое образование
питательной ткани, что возможно только
после оплодотворения. Двойное оплодо
творение ускоряет весь процесс форми
рования семяпочки и семени.
Биологическое значение оплодотво
рения. При слиянии мужских и женских
половых клеток образуется новый орга
204
низм, несущий признаки отца и матери.
Образование половых клеток — гаметогенез — сопровождается формирова
нием гамет с разным сочетанием хромо
сом, поэтому после оплодотворения до
черние организмы могут сочетать в себе
признаки обоих родителей в самых раз
личных (в каждом случае уникальных)
комбинациях. В результате этого проис
ходит колоссальное увеличение наслед
ственного многообразия организмов, ко
торое служит материалом для естествен
ного отбора и эволюции вида.
9« 1. Проведите сравнительный анализ процесса
формирования половых клеток у животных и расте
ний. В чем выражается основное сходство и разли
чие этих процессов? 2. Какую роль выполняют про
цессы митоза и мейоза в спорогенезе и гаметогенезе у растений? 3. В чем преимущество внутреннего
оплодотворения по сравнению с наружным? 4. Что
такое мужской и женский гаметофиты у растений и
каков механизм их образования? 5. Что такое двой
ное оплодотворение и каково его биологическое
значение?
59. Индивидуальное развитие
организма. Онтогенез животных
Из оплодотворенной яйцеклетки на
чинается индивидуальное развитие орга
низмов — онтогенез, которое заключа
ется в постепенной реализации наслед
ственной информации, полученной от ро
дителей. Онтогенез включает весь пери
од жизни особи от зиготы до смерти орга
низма (жизненный цикл). За это время
организмы проходят определенный путь
развития, в котором различают два пери
ода — эмбриональный и постэмбрио
нальный.
Развитие зародыша. Эмбриональ
ное, или зародышевое, развитие охва
тывает промежуток времени от первого
деления зиготы до выхода из яйца или
рождения молодой особи у животных и до
прорастания семян — у растений.
Эмбриональное развитие большин
ства животных проходит по единому пла
ну и включает три основных этапа: дроб
ление, гаструляцию и органогенез.
В результате семи-восьми последова
тельных митотических делений оплодот
воренной яйцеклетки образуются много
численные (128 и более) бластомеры.
При делении дочерние клетки не расхо
дятся и не увеличиваются в размерах.
С каждым последующим делением они
становятся все меньше, поэтому процесс
деления в этом случае носит название
дробления.
У бедных желтком яиц происходит
полное дробление, т. е. делится вся масса
яйца, например у ланцетника (рис. 129).
В случае высокого содержания желтка
наблюдается частичное дробление —
дробится только диск цитоплазмы с яд
ром, а сам желток остается без измене
ний (головоногие моллюски, насекомые,
костистые рыбы, пресмыкающиеся, пти-
Рис. 129. Эмбриональное развитие лан
цетника: а — е — начальная фаза разви
тия оплодотворенного яйца, ж — образо
вание бластулы, з — бластула в разрезе,
и — начало образования гаструлы, к —
гаструла, л — ранняя нейрула, м — нейрула: 1 — сперматозоид, 2 — полость
бластулы (бластоцель), 3 — эктодерма,
4 — энтодерма, 5 — полость первичной
кишки, 6 — мезодерма, 7 — нервная
пластинка, 8 — хорда.
205
цы). В результате дробления образуется
однослойный многоклеточный заро
дыш — бластула. В типичном случае
она напоминает полый шарик с эпителие
подобной стенкой (бластодермой) из
одного ряда мелких бластомеров и цен
тральной полостью (бластоцелем, или
первичной полостью тела), заполнен
ной жидкостью.
После дробления идет процесс гаструляции, который характеризуется пе
ремещением части клеточного материала
с поверхности бластулы вовнутрь на мес
та будущих органов. В результате этих
перемещений образуется гаструла —
чашевидный зародыш, состоящий из двух
слоев, или зародышевых листков: наруж
ного — ■эктодермы и внутреннего —
энтодермы. У ланцетника гаструла воз
никает путем впячивания бластодермы в
бластоцель. Внутренняя полость, назы
ваемая первичной кишкой, связана с
внешней средой через отверстие, которое
становится первичным ртом. Существу
ют и другие способы образования гаструлы.
Губки и кишечнополостные заканчи
вают свое развитие на стадии двух заро
дышевых листков. У всех остальных жи
вотных в процессе гаструляции или после
ее завершения (у ланцетника) образуется
еще и третий зародышевый листок —
мезодерма. У иглокожих и хордовых жи
вотных она формируется в виде парных
карманоподобных выпячиваний эпите
лия первичного кишечника в бластоцель.
В итоге от первичной кишки отшнуровываются замкнутые пузырьки, или мешоч
ки, с полостью внутри. Они и представля
ют собой зачатки мезодермы, а полость
внутри пузырьков — вторичную по
лость тела, или целом.
Во время гаструляции клетки диффе
ренцируются, т. е. становятся различны
ми как по биохимическому составу, так и
по структуре. Биохимическая специали
зация клеток обеспечивается дифферен
цированной активностью их генов. Гене
206
тическая информация реализуется в ко
нечном итоге через специфические бел
ки, присутствие которых в клетке опреде
ляет, какие реакции будут в ней проте
кать, будет ли эта клетка подвижной или
нет, и др.
Дальнейшая дифференцировка кле
ток каждого зародышевого листка приво
дит к образованию одних и тех же тканей
и органов у подавляющего большинства
организмов животного мира, т. е. к орга
ногенезу.
Из эктодермы у позвоночных живот
ных образуется нервная система, органы
чувств, покровный эпителий с его желе
зами и производными структурами (воло
сы, перья, когти, копыта и т. п.).
Из энтодермы формируется эпителий
средней кишки с его придаточными желе
зами (печень и поджелудочная железа,
жабры и их производные — легкие, пла
вательный пузырь и др., а также щито
видная железа), из мезодермы — мы
шечная, все виды соединительной ткани,
кровеносная, выделительная и половая
системы.
Одновременно с мезодермой из энто
дермы образуется хорда — гибкий ске
летный тяж, расположенный у эмбрио
нов всех хордовых на спинной стороне.
Впоследствии хорда у всех позвоночных
замещается позвоночником, и только у
некоторых низших позвоночных ее остат
ки сохраняются между позвонками даже
во взрослом состоянии.
Из эктодермы, расположенной над
самой хордой, образуется нервная плас
тинка (см. рис. 129). В дальнейшем бо
ковые края пластинки приподнимаются, а
центральная часть опускается, образуя
нервный желобок. Постепенно верхние
края этих складок смыкаются, а желобок
превращается в лежащую под эктодер
мой нервную трубку — зачаток цен
тральной нервной системы. Из энтодер
мы возникают хорда, мезодерма, кишеч
ник и связанные с ним ткани и органы.
Нервная трубка, хорда и кишечник созда
ют осевой комплекс органов зародыша,
который определяет двустороннюю сим
метрию тела.
Зародыш животных развивается как
единый организм, в котором все клетки,
ткани и органы находятся в тесном взаи
модействии. При этом один зачаток ока
зывает влияние на развитие другого, в
значительной степени определяя его
дальнейший путь. Кроме того, на темпы
роста и развития зародыша воздействуют
внешние условия.
Особенности эмбриогенеза позво
ночных животных. Эмбриональное раз
витие организмов с разным типом онто
генеза протекает в разных условиях. При
этом необходимая связь зародыша со
средой обеспечивается специальными,
временно функционирующими внезародышевыми органами, называемыми про
визорными. Для всех позвоночных с не
личиночным типом развития, яйца кото
рых богаты желтком (рыбы, рептилии,
птицы), характерен такой провизорный
орган, как желточный мешок. У рыб он
образуется из клеточного материала всех
трех зародышевых листков, нарастающих
на желток. У пресмыкающихся, птиц и
млекопитающих желточный мешок имеет
энто- и мезодермальное происхождение.
В стенки желточного мешка врастают
кровеносные сосуды, образующие густую
капиллярную сеть. Клетки стенки жел
точного мешка выделяют ферменты, ко
торые расщепляют питательные вещес
тва желтка (белки, углеводы, жиры и др.)
до более простых соединений. Последние
поступают в кровеносные капилляры и
далее в организм зародыша. Желточный
мешок — это также первый кроветвор
ный орган зародыша.
У млекопитающих редуцированный
желточный мешок входит в состав пла
центы.
Важной особенностью эмбриональ
ного развития амниот — настоящих на
земных позвоночных (пресмыкающихся,
птиц, млекопитающих и человека) — яв
ляется развитие трех зародышевых обо
лочек: амниона, хориона и аллантоиса
(рис. 130, с. 208).
Амнион — внутренняя зародышевая
оболочка — представляет собой напол
ненный жидкостью мешок, окружающий
зародыш и защищающий его от механи
ческих повреждений и высыхания. Амни
отическая жидкость представляет собой
водный раствор белков, сахаров, мине
ральных солей, содержащий также гор
моны и мочевину. В акушерской практике
амниотическую жидкость, отходящую при
родах, называют водами.
Функцию наружной зародышевой
оболочки выполняет хорион, или вор
синчатая оболочка, названная так изза наличия на ее поверхности множества
пальцеобразных выростов — ворсинок.
Ворсинки хориона врастают в слизистую
оболочку матки — мешковидного органа
материнского организма, присущего
только млекопитающим.
Аллантоис представляет собой ме
шок, связанный с задней кишкой зароды
ша. У пресмыкающихся и птиц в нем на
капливаются продукты жизнедеятельнос
ти зародыша до вылупления из яйца.
У млекопитающих и человека аллантоис
разрастается и соединяется с хорионом,
образуя богатую кровеносными сосудами
структуру — хориоаллантоис. Послед
ний участвует в формировании детского
места, или плаценты, — органа, более
эффективно осуществляющего обмен
между зародышем и материнским орга
низмом.
Постэмбриональное
развитие.
В постэмбриональном, или послезародышевом, периоде осуществляются фор
мообразовательные процессы,определя
емые прежде всего генотипом организма,
а также факторами внешней среды.
Постэмбриональный период начина
ется с момента выхода организма из яй
цевых оболочек или с момента рождения.
Различают два способа постэмбриональ
ного развития: прямое, когда рождаю207
Рис. 130. Ранние (а) и поздняя (б) стадии развития внезародышевых оболочек у человека: 1 — тело
зародыша, 2 — амниотическая полость, 3 — желточный мешок, 4 — амниотическая ножка, 5 — вы
рост аллантоиса, 6 — хорион, 7 — ворсинки хориона, 8 — пупочный канатик, 9 — амниотическая обо
лочка.
щийся организм имеет все основные ор
ганы, свойственные взрослому животно
му (пресмыкающиеся, птицы, млекопи
тающие), и непрямое, когда эмбриональ
ное развитие приводит к образованию
личинки, которая по внешним и внутрен
ним признакам значительно отличается
208
от взрослого организма (плоские и коль
чатые черви, моллюски, ракообразные,
насекомые, земноводные). Например, из
яиц бабочек развиваются гусеницы, а из
яиц лягушки — головастики, которые
резко отличаются по строению, образу
жизни и среде обитания от взрослых жи
вотных. Так, у головастика имеются жа
берные щели, орган боковой линии,
хвост, двухкамерное сердце, один, как и у
рыб, круг кровообращения. Когда личин
ка достигает определенного уровня раз
вития, происходит ее метаморфоз, в
процессе которого вырабатываются
признаки взрослого организма и головас
тик превращается в лягушку.
Наличие личиночной стадии в разви
тии земноводных обеспечивает им воз
можность жить в разных средах и исполь
зовать разные источники пищи: головас
тик живет в воде и питается растительной
пищей, а лягушка ведет водно-наземный
образ жизни и питается животной пищей.
Такое явление наблюдается и у многих
насекомых. Смена среды обитания и, как
следствие, смена образа жизни животно
го при переходе его от личиночной стадии
к взрослому организму снижает интен
сивность борьбы за существование внут
ри вида. Кроме того, у некоторых сидя
чих, малоподвижных или паразитических
животных свободноживущие личинки
способствуют расселению вида, расши
рению ареала.
точный зародыш превращается в плот
ный многоклеточный шар (без обособ
ленной полости), называемый морулой.
Наружный слой клеток морулы называ
ется трофобластом, который участвует
в образовании плаценты. Из централь
ных бластомеров, или эмбриобласта,
развиваются органы будущего организма
и защитные оболочки. В моруле накапли
вается жидкость, за счет которой форми
руется центральная полость. Постепен
но полость увеличивается и превраща
ется в плодный пузырь, или бласто
цисту.
На 6— 7-е сутки после оплодотворе
ния бластоциста начинает внедряться в
слизистую оболочку матки. К концу вто
рой недели на трофобласте образуются
ворсинки, которые способствуют интен
сивному разрушению слизистой оболочки
матки и погружению в нее зародыша.
Этот процесс называется имплантаци
ей зародыша (рис. 131).
На 11 — 12-е сутки бластоциста пол
ностью погружается в стенку матки. Хори-
1. Что такое индивидуальное развитие? Назо
вите его периоды. 2. Охарактеризуйте стадии эм
брионального развития. 3. Назовите производные
трех зародышевых листков. 4. Каковы особенности
эмбриогенеза позвоночных животных? 5. Чем отли
чается прямое развитие от непрямого? Приведите
примеры.
60. Онтогенез человека
Эмбриогенез. Ранние стадии эмбрио
генеза человека и последовательность
их прохождения сходны с развитием
всех позвоночных животных, что от
ражает общность их эволюционной исто
рии.
Развитие зародыша человека начина
ется сразу же после оплодотворения с
дробления зиготы на бластомеры. В ре
зультате нескольких дроблений однокле
Рис. 131. Имплантация зародыша млекопитаю
щего в стенку матки: 1 — полость матки, 2 —
зародыш, 3 — стенка матки, 4 — развивающа
яся плацента.
209
он зародыша и ткани стенки матки форми
руют важный орган — детское место,
или плаценту, через которую осущес
твляется связь зародыша с организмом
матери. Внедряясь в слизистую оболочку
матки, ворсинки хориона разрушают ее
кровеносные сосуды. В местах таких раз
рушений образуются лакуны — неболь
шие полости, заполненные материнской
кровью, в которые погружены ворсинки.
Через плаценту зародыш (с девятой неде
ли развития он называется плодом) полу
чает необходимые питательные вещества
и кислород из крови матери и отдает угле
кислый газ и другие продукты жизнедея
тельности, подлежащие удалению.
Связь зародыша с плацентой осущес
твляется через кровеносные капилляры
пупочного канатика, или пуповины.
Кровь матери никогда и нигде не смеши
вается с кровью плода, поскольку она не
проникает в капилляры плода, а только
омывает ворсинки хориона.
Плацента, являясь барьером для мик
робов и ряда веществ из кровотока мате
ри, пропускает, однако, многие лекар
ственные и наркотические вещества, про
изводственные и пищевые яды, чужерод
ные белки и антитела. Особенно чувстви
телен зародыш человека к концу третьей
недели развития, когда у него формируют
ся основные системы органов — нервная,
пищеварительная, кровеносная. Потреб
ление в этот период лекарственных пре
паратов, алкоголя, никотина, а также пе
ренесение инфекционных заболеваний
матерью создают угрозу для нормального
развития эмбриона. Так, например, по
требление алкоголя в этот период чаще
всего приводит к тяжелым нарушениям
развития нервной системы плода. Заболе
вание матери краснухой может вызвать у
будущего ребенка серьезные пороки в
строении сердца, органов слуха и зрения,
развитие «волчьей пасти».
Причиной ряда уродств могут быть
также лекарственные препараты. Оказа
лось, что антибиотик актиномицин D, не
210
причиняя вреда материнскому организму,
у зародышей нарушает формирование
глаз и мозга. Используемое ранее в меди
цинской практике снотворное талидомид
приводило к недоразвитию у новорож
денных конечностей и другим нарушени
ям скелета.
Период внутриутробного развития (бе
ременность) у человека составляет около
280 суток. Беременность протекает под
контролем гормонов, выделяемых плацен
той, которые препятствуют наступлению
следующего после оплодотворения мен
струального цикла. Поэтому после того,
как началось развитие одного зародыша,
новый зародыш появиться уже не может.
Момент родов также определяется
гормонами, которые выделяет развиваю
щийся плод. Надпочечники зародыша,
достигшего соответствующей стадии раз
вития, выделяют гормоны, и это служит
сигналом к началу родов.
К моменту рождения масса плода до
стигает 3—4 кг, а длина тела составляет
около 50 см.
Рождение ребенка начинается с со
кращений мышц матки (схватки) и
брюшной стенки, благодаря которым
плод выталкивается наружу. В этот же
период происходит разрыв амниотичес
кого мешка и отхождение околоплодных
вод. Новорожденный после перевязыва
ния и перерезания пуповины начинает
самостоятельно дышать, и с этого момен
та начинается постэмбриональное разви
тие человека.
Постэмбриональное развитие. Пос
ле рождения в развитии человека выде
ляют следующие периоды:
новорожденный
грудной возраст
детский возраст
— 1— 10 дней
— 11 дней — 1 год
— 1 год — 11 лет
(девочки)
1 год — 12 лет
(мальчики)
подростковый возраст— 12— 15 лет
(девочки)
юношеский возраст
—
зрелый возраст
—
пожилой возраст
—
старческий возраст
—
долгожители
—
13— 16 лет
(мальчики)
16 — 20 лет
(девушки)
17 — 21 год
(юноши)
21 — 55 лет
(женщины)
22 — 60 лет
(мужчины)
56 — 74 года
(женщины)
61 — 74 года
(мужчины)
75— 90 лет
(женщины и
мужчины)
90 лет
и старше.
Приведенная возрастная периодиза
ция достаточно условна, поскольку раз
витие человека определяется как наслед
ственными факторами, так и условиями
жизни.
Особенностью постэмбрионального
развития человека является относитель
но более продолжительный, чем у других
видов млекопитающих, период детства.
Это имеет большое значение, так как в
этот период происходит не только физи
ческое развитие организма, но и станов
ление личности, т. е. развитие мышления,
речи, двигательной активности, связан
ной с трудовой деятельностью, и т. д.
На индивидуальное развитие любого
организма оказывают влияние факторы
внешней среды (свет, температура, соле
вой и газовый состав среды обитания, пи
щевые ресурсы и др.); Среди них разли
чают факторы, необходимые для нор
мального развития и нежелательные или
даже вредные. К числу вредных внешних
факторов следует в первую очередь от
нести алкогольные напитки, наркотичес
кие вещества и курение.
Употребление спиртных напитков и
курение, а также бесконтрольное приме
нение лекарственных препаратов прино
сят огромный вред на любом этапе онто
генеза человека и особенно опасны в под
ростковом возрасте. Алкоголь губитель
но действует на все системы органов че
ловека, поражая прежде всего централь
ную нервную систему, сердце и кровенос
ные сосуды, легкие, почки, печень. Алко
голь тормозит функциональное созрева
ние нервных клеток, а содержащийся в
табачном дыме никотин отравляет нер
вные клетки и служит причиной возник
новения различного рода функциональ
ных расстройств. У лиц, склонных к алко
голизму, гораздо чаще, чем у людей, веду
щих трезвый образ жизни, рождаются де
ти с врожденными уродствами.
?J 1. Что такое плацента? Как она образуется и
какова ее роль в организме матери? 2. Как осущес
твляется гормональная регуляция внутриутробного
развития человека? 3. Какие периоды выделяют в
развитии человека после рождения? 4. В чем выра
жается вредное воздействие алкоголя, наркотиков,
курения и других вредных факторов на эмбриональ
ное и постэмбриональное развитие человека?
61. Индивидуальное развитие
растений
Особенности онтогенеза растений.
Процессы развития животных и растений
радикально отличаются. Развитие живот
ных идет по определенному плану в точ
ной последовательности. При этом на
каждой стадии осуществляется строгий
генетический контроль. У животных ус
ловия внешней среды, как правило, не
сильно влияют на ход развития, которое у
них заканчивается по достижению взрос
лого состояния.
У растений, напротив, процесс разви
тия непрерывен: растения развиваются
на протяжении всей своей жизни. На их
развитие непосредственно влияют как
внешние факторы (свет, тепло, влага и
пр.), так и внутренние. Специфическая
для разных видов растений дифференци211
ровка на ткани управляется гормонами,
синтез которых чувствителен к изменени
ям условий окружающей среды. Способ
ность растений реагировать на изменения
внешних факторов способствует адапта
ции их к конкретным местообитаниям.
У растений почти всякая дифференцировка обратима при условии, что клет
ка жива. С этим связано отсутствие у рас
тений жесткого плана развития. У живот
ных такая обратимость встречается очень
редко.
Обратимость развития растений
обусловлена тем, что некоторые их клет
ки содержат всю генетическую информа
цию для развития целого растения. Под
тверждением этого является возмож
ность выращивания в пробирке целого
растения из отдельных клеток или кусоч
ков ткани (рис. 132).
Эмбриогенез. Эмбриональный период
у растений, как и у животных, начинается
с деления оплодотворенной яйцеклетки,
что приводит к постепенному формирова
нию зародыша нового организма. Ранние
стадии развития зародыша у двудольных и
однодольных очень сходны. Наиболее де
тально этот процесс изучен у покрытосе
менных (рис. 133, с. 214).
Первое поперечное деление зиготы в
зародышевом мешке семязачатка завер
шается образованием двух клеток: вер
хней (апикальной) и нижней (базаль
ной).. У большинства растений из базаль
ной клетки развивается своеобразная
ножка — одноклеточный или многокле
точный подвесок, или суспензор, кото
рый продвигает развивающийся зародыш
в глубь зародышевого мешка, где идет
образование особой питательной тка
ни — эндосперма или перисперма.
Кроме того, подвески цветковых достав
ляют зародышу питательные вещества из
эндосперма, а в некоторых случаях бел
ковые вещества, синтезированные в суспензоре, используются собственно заро
дышем в период его быстрого роста.
Из апикальной клетки после ряда
212
продольных, поперечных и тангентальных митотических делений развивается
шаровидный предзародыш. Когда предЗародыш достигает определенных разме
ров, начинается дифференцировка орга
нов, что приводит к превращению его в
зародыш.
Будущая эпидерма формируется в хо
де делений наружных клеток собственно
зародыша. В дальнейшем различия в сте
пени вакуолизации и плотности клеток
зародыша приводят к образованию про
камбия и основной меристемы. По
следняя, сильнее вакуолизированная и
менее плотная, дает начало основной
ткани, окружающей более плотный про
камбий, предшественник проводящих
тканей — ксилемы и флоэмы. Из верху
шечных частей эмбриона формируются
зародышевые листья (семядоли) и по
чечка, состоящая из зародышевого
стебелька и конуса нарастания с рас
положенными на нем зачаточными листь
ями, а из базальных частей — зароды
шевый корешок с конусом нарастания
и корневым чехликом. Таким образом, в
зародыше возникают все органы будуще
го растения.
В течение всего периода формирова
ния зародыша питательные вещества
непрерывно поступают от родительского
организма к тканям семязачатка, в ре
зультате чего значительный их запас на
капливается в эндосперме, перисперме
или в семядолях развивающегося семени.
Параллельно с развитием зародыша идет
преобразование всех частей семяпочки,
которая отделяется от семяножки, связы
вающей ее со стенкой завязи, и превра
щается в семя, а завязь пестика разрас
тается и превращается в плод.
В типичном случае семя состоит из
зародыша, запасной пит ат ельной
ткани (эндосперм, перисперм или семя
доли) и семенной кожуры. Последняя
предохраняет зародыш от механических
повреждений и неблагоприятных условий
в различные периоды жизни растения, в
Рис. 132. Развитие растения моркови из нескольких клеток зрелого корня: 1 — растение моркови;
2 — клетки корня, выращенные для получения каллуса (ткани ); 3 — отдельные клетки каллуса, вы
ращиваемые в жидкой среде; 4 — деление клеток в культуре тканей с образованием зародыша; 5 —
зародыш на ранней стадии развития; 6 — более поздние стадии развития зародыша; 7 — образова
ние зародышем корней и превращение в маленькое растеньице.
213
Рис. 133. Зародыш двудольного растения: а — развитие'. 1— 16 — последователь
ные фазы; б — строение семени куколя: 1 — зародыш, 2 — перисперм.
том числе в ходе расселения на значи
тельные расстояния.
Распространение плодов и семян осу
ществляется при помощи ветра, воды,
животных. Чем дальше разносятся семе
на, тем менее выражена конкуренция со
стороны родственного растения. Это дает
больше шансов занять новую террито
рию, что со временем приводит к увели
чению размеров популяции данного вида
в целом.
214
Условия прорастания семян. При по
падании в благоприятные условия семенаначинают прорастать. Однако некоторые
семена не прорастают даже в подходящих
для этого условиях. Их называют покоя
щимися. Причинами покоя семян явля
ются физиологическая незрелость заро
дыша и непроницаемость семенной кожу
ры для воды и кислорода, а также нали
чие в семенах веществ, тормозящих про
растание. Чтобы прорасти, такие семена
должны дозреть. Необходимость в дозре
вании помогает предотвратить прораста
ние в холодное время, когда выживание
проростка практически невозможно.
Период покоя очень важен для вы
живания растений. Он является механиз
мом, обеспечивающим прорастание се
мян только в условиях, благоприятных
для дальнейшего развития. Некоторые
семена должны пройти через пищевари
тельный тракт животных. У ряда расте
ний семя прорастает только после по
вреждения семенной кожуры или после
вымывания дождем Ингибиторов.
Прорастание семян — это переход
семян от состояния покоя к вегетативному
росту зародыша и формирующегося из не
го проростка. Оно начинается при опти
мальном для каждого вида и сорта расте
ний сочетании внутренних и внешних (эко
логических) факторов — влажности, теплаи свободного доступа кислорода. Кроме
того, для прорастания мелких семян, на
пример салата, мятликов и многих сорня
ков, обычно требуется воздействие света.
Зрелые семена в большинстве случа
ев крайне сухие (их влажность обычно
5— 20 %), вследствие чего прорастание
невозможно до тех пор, пока они не впи
тают определенное количество воды, не
обходимой для метаболической активнос
ти. Потребность в воде для набухания за
висит от состава семян. Семена с высо
ким содержанием жиров поглощают
30— 40 % воды по отношению к своей
массе, богатые крахмалом — 50—70 %,
а с большим количеством белка — около
90 % и более.
При набухании семян в клеточных органеллах активизируются окислительновосстановительные ферменты, усилива
ющие дыхание и гидролиз белков, угле
водов, жиров и других органических со
единений запасающих тканей зародыша.
В результате образуются простые водо
растворимые соединения, доступные для
поглощения клетками развивающегося
зародыша. Кроме того, набухание семян
сопровождается активированием фитогормонов, регулирующих их прорастание,
а также освобождением энергии, которая
используется в биохимических процессах
зародыша и проростка (синтез органи
ческих веществ, формирование клеточ
ных органелл, деление клетки и т. д.).
На ранних стадиях прорастание мо
жет быть полностью анаэробным, но, как
только семенная кожура лопается, оно
становится аэробным и требует кислоро
да. Свободный доступ кислорода усили
вает интенсивность дыхания прорастаю
щих семян в сотни раз. Если почва пе
ренасыщена водой, доступное семени ко
личество кислорода может оказаться не
достаточным для такого дыхания и про
растание станет невозможным. Только у
немногих растений (рис, тимофеевка) се
мена могут прорастать при пониженной
аэрации.
Интенсивность дыхания увеличивает
ся также при повышении температуры
окружающей среды. Согласно закону
Я. В а н т - Г о ф ф а , скорость большин
ства химических реакций возрастает в
2— 4 раза при повышении температуры
на каждые 10 °С. Однако у живых орга
низмов подобное повышение имеет свои
пределы. Например, при 45— 48 °С дыха
ние семян практически прекращается и
они теряют способность к прорастанию.
Оптимальная температура для прораста
ния семян многих растений 25—30 °С, а
минимальная, при которой семена еще
могут прорастать, колеблется в широких
пределах — от 0 °С до 15— 18 °С. Обыч
но семена растений южного происхожде
ния прорастают при более высокой тем
пературе, чем семена растений северных
и умеренных широт.
Развитие проростка. При прораста
нии семени первым появляется корешок,
или зародышевый корень. После того
как он укрепится в субстрате и начнет
поставлять зародышу воду, трогается в
рост зародышевая почка и формируется
надземная часть растения — система по
215
бегов. В результате ростовых и формооб
разовательных процессов зародыш семе
ни превращается в проросток.
В ходе прорастания и последующего
развития питательные вещества семени
переводятся в растворимую форму и
транспортируются в растущие ткани про
ростка. У двудольных семядоли при этом
постепенно уменьшаются, вянут и в кон
це концов погибают. К тому времени про
росток укореняется и его питание пере
стает зависеть от запасных веществ се
мени. Он становится фототрофным орга
низмом, т. е. с помощью вегетативных
органов (корня, стебля и листа) растение
полностью обеспечивает себя питатель
ными веществами.
Период роста и развития растения
сопровождается сложными физиолого
биохимическими изменениями в конусе
нарастания стебля, в результате чего
происходит переход от вегетативной ста
дии развития к репродуктивной — на
растении начинают формироваться орга
ны размножения.
У большинства растений онтогенез
состоит из периодически повторяющихся
жизненных циклов, в которых происходит
чередование поколений — гаметофита и
спорофита. Продолжительность онтоге
неза различна: у однолетних семенных
растений развитие завершается в течение
одного вегетационного периода, у двулет
них — в течение двух лет, а у многолет
них оно длится на протяжении десятков и
даже сотен лет.
Познание тонких механизмов зависи
мости роста и развития растений от раз
личных факторов внешней среды (темпе
ратура, свет, влажность, питание и т. д.)
позволяет целенаправленно влиять на
ход их онтогенеза, добиваться значитель
ного повышения урожайности. Напри
мер, предпосевное облучение клубней
картофеля различными источниками све
та приводит к увеличению количества,
длины и массы стеблей, площади листь
ев, накоплению сухих веществ во всех ор
216
ганах растений. Прибавка урожая клуб
ней составляет до 30 % и более.
?« 1. Каковы особенности эмбриогенеза у расте
ний? 2. Что такое покой семян и каково его биоло
гическое значение? 3. Почему прорастание семян
возможно только при оптимальных для каждого ви
да и сорта растений условиях влажности, тепла и
доступа кислорода? 4. Каковы особенности форми
рования и развития проростка? Как можно регули
ровать рост и развитие проростков?
62. Старение и смерть организмов
Старение — общебиологическая за
кономерность, свойственная всем живым
организмам. Старость представляет со
бой заключительный этап онтогенеза, ко
торый характеризуется существенными
структурными, функциональными и био
химическими изменениями в организме,
ограничивающими его адаптационные
возможности. Признаки старения прояв
ляются на всех уровнях организации жи
вого организма — от молекулярно-гене
тического до организменного.
Признаки старения. На молекуляр
но-генетическом уровне у животных
и человека ведущими механизмами
старения являются необратимые на
рушения ДНК, неравномерные измене
ния в синтезе РНК и белков разных
классов.
В цитоплазме накапливаются свобод
ные радикалы, и, как следствие, ассими
ляция уже не может полностью восстано
вить потерь, связанных с диссимиляци
ей. Снижается интенсивность синтеза ря
да гормонов.
На клеточном уровне наблюдается
уменьшение содержания воды в цито
плазме, изменение активного транспорта
ионов, что сказывается на важнейших
физиологических свойствах клетки, сни
жении ее электрического потенциала.
Ведущие механизмы на клеточном уров
не: деградация и гибель части клеток,
снижение их митотической активности,
уменьшение количества митохондрий,
разрушение лизосом и изменение струк
туры эндоплазматического ретикулума,
который нередко распадается на фраг
менты. Кроме того, происходит падение
лабильности клеток и их реакции на воз
действие физиологически активных ве
ществ.
На организменном уровне изменения
при старении выражаются прежде всего
во внешних признаках: у человека, на
пример, изменяется осанка, форма тела,
появляется седина, кожа теряет эластич
ность, что приводит к образованию мор
щин. Наблюдается ослабление зрения и
слуха, ухудшение памяти.
У пожилых людей уменьшается жиз
ненная емкость легких, увеличивается
артериальное давление, отмечаются из
менения в стенках сосудов, развивается
атеросклероз. Ослабляются функции ос
новных физиологических систем организ
ма (нервной, эндокринной, сердечно-со
судистой, пищеварительной и др.), сни
жается нервный контроль над их деятель
ностью. Наблюдается падение активнос
ти щитовидной железы, происходит инво
люция половых желез и снижение про
дукции половых гормонов.
Складывается впечатление, что ста
рение сопровождается ослаблением
всех функций организма. Однако это не
совсем так. Согласно современным
представлениям, возрастные изменения
могут быть разнонаправленными. Одни
функции прогрессивно снижаются с
возрастом, другие — заметно не изме
няются, а некоторые показатели даже
возрастают. Например, сократительная
способность сердца, гормональная ак
тивность многих эндокринных желез,
острота зрения и слуха, падение актив
ности многих ферментов с возрастом
прогрессивно снижаются. В то же время
уровень сахара в крови, количество
эритроцитов, лейкоцитов, гемоглобина
практически не изменяется, а вот синтез
гормонов гипофиза, чувствительность
клеток к гуморальным факторам повы
шается.
Возрастные изменения характеризу
ются также гетерохронностью, т. е.
протекают с различной скоростью в раз
ных клетках, тканях и органах. Напри
мер, атрофия тимуса у человека начина
ется в возрасте 13— 15 лет, угасание
функции яичников в 48— 52 года, а от
дельные функции гипофиза могут сохра
няться на высоком уровне и в старческом
возрасте. Отмечаются неодинаковые
проявления старения в различных угол
ках мозга.
У растений, как и у животных, наступ
ление старения связано со снижением
интенсивности основных функций в клет
ках и тканях, повышением активности
гидролитических ферментов. В результа
те накапливаются конечные продукты
метаболизма и наступают структурные
разрушения органелл клеток (хлоропластов, митохондрий, комплекса Гольджи
и др.).
Одной из удивительных особенностей
растений является то, что старение кле
ток, тканей и целых органов не ведет к
старению всего организма сразу, по
скольку в течение всей жизни одно
временно с отмиранием органов у рас
тения идет их новообразование (лис
топад, опадение веток, зрелых плодов и
семян компенсируется ростом новых по
бегов).
Смерть. Процесс старения животных
и растений приводит к смерти, т. е. к
прекращению жизнедеятельности орга
низма. В зависимости от причин, вызвав
ших наступление смерти, у животных и
человека различают смерть естественную
(физиологическую) и преждевременную.
Естественная смерть наступает в резуль
тате длительного постепенного угасания
основных жизненных отправлений орга
низма в процессе старения. Причинами
преждевременной смерти организма мо
гут быть болезни, поражения его жизнен
217
но важных органов, прекращение дыха
ния и кровообращения, гибель.
Каковы же основные причины
старения? К сожалению, единой точки
зрения на этот счет на сегодняшний день
нет. Изучением закономерностей старе
ния, начиная от молекулярного и клеточ
ного уровня до целого организма, зани
мается геронтология (от греч. geron —
старик) — наука о старости.
Проблемы геронтологии. Геронтоло
гия располагает сейчас огромным коли
чеством фактов об изменении различных
структур и функций организма в процессе
старения. Выдвинуто свыше 300 гипотез
о причинах старения, однако, согласно
большинству из них, первичные механиз
мы старения связаны с изменением сос
тояния генетического аппарата клетки.
По мнению одних авторов, старение и
смерть генетически запрограммированы.
Для каждого вида характерна определен
ная продолжительность жизни, которая
может изменяться под действием отбора.
Дубы и секвойи живут до 2000 лет и бо
лее, слон умирает от старости в возрасте
около 150 лет, человек — между 70 и 100
годами, многие насекомые живут не
сколько дней или недель, а некоторые од
ноклеточные организмы — всего не
сколько часов. Таким образом, процесс
старения приводит к смерти особей каж
дого вида в определенном возрасте.
Другие ученые считают, что старение
является результатом повреждения гене
тического аппарата в ходе онтогенеза,
нарушения его регуляции, появления и
накопления мутаций во всех клетках те
ла, что ведет к вышеописанным необра
тимым изменениям в организме.
В то же время, согласно адаптацион
но-регуляторной гипотезе, старение не
рассматривается как процесс равномер
ного угасания структурно-функциональ
ной организации живого, а как более
сложное явление. В ходе старения проис
ходит мобилизация важных приспособи
тельных механизмов, которые тормозят
218
процессы старения и тем самым способ
ствуют увеличению продолжительности
жизни.
Практически в каждой отрасли меди
цины и биологии появились геронтологи
ческие проблемы. На современном этапе
задача геронтологии состоит не только в
том, чтобы продлить жизнь человека, но
и дать возможность людям старших воз
растных групп принимать активное учас
тие в общественной и трудовой деятель
ности. Ученые полагают, что у человека
высокий уровень социально-трудовой ак
тивности и постоянные тренировки спо
собствуют сохранению умственной и фи
зической работоспособности до глубокой
старости.
?,ч
1. Что такое старение и каковы его признаки на
молекулярно-генетическом, клеточном и организменном уровнях? 2. Какие процессы в организме
человека с возрастом замедляются, не изменяются
и какие даже ускоряются? 3. Каковы особенности
старения у растений? 4. Какие задачи решает герон
тология?
ГЛАВА VI
Наследственность
и изменчивость
организмов
63. Гибридологический метод
изучения наследственности
Н а сл ед с т в е н н о ст ь — это свой
ство живых организмов сохранять и пе
редавать в ряду поколений характерные
для вида или популяции особенности
строения, функционирования и разви
тия. Наследственность обеспечивает
постоянство и многообразие форм жизни
и лежит в основе передачи наследствен
ных факторов, ответственных за форми
рование признаков и свойств организма.
Именно благодаря наследственности не
которые виды организмов оставались
почти неизменными в течение сотен мил
лионов лет, воспроизведя за это время
огромное количество поколений. Напри
мер, современная кистеперая рыба латимерия мало чем отличается от своих де
вонских предков, живших около 400 млн
лет тому назад.
Как известно, организмы объединяют
в определенные систематические катего
рии: виды, роды, семейства, отряды (по
рядки) и т. д. Эта системность возможна
лишь при наличии наследственности, ко
торая сохраняет не только черты сходства
внутри каждой группы организмов, но и
различия между ними по морфологичес
ким, физиологическим, биохимическим и
другим признакам и свойствам.
На протяжении нескольких тысячеле
тий человека волновал вопрос о причинах
сходства потомков и родителей, а также о
способности организмов передавать свои
признаки от одного поколения к другому.
Уже в XIX в. ученые начали понимать,
что передачу признаков по наследству
осуществляют какие-то имеющиеся в
клетке частицы — наследственные за
датки того огромного множества призна
ков, из которых слагается каждый орга
низм.
Первый серьезный научный шаг в
познании закономерностей наследствен
ности был сделан выдающимся чешским
ученым Г. Ме н д е л е м . На основании
многочисленных опытов по гибридизации
(скрещиванию) растений гороха с точным
количественным учетом всех типов полу
ченных гибридов он впервые открыл,
обосновал и сформулировал в книге
«Опыты над растительными гибридами»
(1865) основные законы наследственнос
ти. Мендель установил, что наследствен
ные задатки не смешиваются, а переда
ются от родителей потомкам в виде дис
кретных
(обособленных)
единиц.
В 1909 г. датский ботаник В. И о г а н с е н
назвал эти единицы генами, а в 1912 г.
219
американский генетик Т. М о р г а н пока
зал, что они находятся в хромосомах.
Выявленные Менделем закономер
ности наследования были по достоинству
оценены только через 35 лет, когда в
1900 г. Д е Ф р и з в Голландии, К. К о р
ре и с в Германии и Э. Ч е р м а к в Ав
стрии независимо друг от друга подтвер
дили законы Менделя и признали его
приоритет. С этого момента всем стало
очевидно, насколько велико значение ра
бот Менделя; именно им был открыт путь
к разгадке тайны наследственности. Переоткрытие законов Менделя вызвало
стремительное развитие генетики.
Гибридологический метод. Среди
множества современных методов генети
ческого анализа центральное место при
надлежит гибридологическому методу.
Суть его заключается в гибридизации ор
ганизмов, отличающихся друг от друга по
одному или нескольким признакам, и в
последующем анализе потомства.
Поскольку в дальнейшем речь пойдет
о закономерностях наследования призна
ков, следует определить понятие «при
знак». Под признаком принято пони
мать любую особенность организма,
т. е. любое отдельное его качество или
свойство, по которым можно различить
две особи. У растений это форма венчика
(например, симметричный — ассиметричный) или окраска цветка (пурпур
ный — белый), скорость созревания
растений (скороспелость — поздне
спелость). Кроме того, это способность
или неспособность организма к синте
зу какого-либо вещества, например
крахмала, гликогена, масла и др.;
устойчивость или восприимчивость к
заболеванию и т. д.
К числу признаков, наследование которых мож
но изучать, относится длина рестрикционных фраг
ментов ДНК. Их получают с помощью фермента
рестриктазы, которая узнает определенную после
довательность нуклеотидов в молекулах ДНК и раз
рывает ее именно в этих участках. Последние явля
220
ются уникальными для каждого индивидуума. На
званный признак имеет большое практическое зна
чение: он используется для диагностики наслед
ственных заболеваний человека как в эмбри
ональный период, так и после рождения. К числу та
ких заболеваний относятся серповидноклеточная
анемия, гемофилия, дефицит гормона роста и др.
Этот метод используется также в криминалистике.
Совокупность всех признаков орга
низма, начиная с внешних и кончая осо
бенностями строения и функциониро
вания клеток и органов, называется фе
нот ипом. Этот термин может употреб
ляться и по отношению к одному из аль
тернативных
(взаимоисключающих)
признаков.
До Менделя многие ученые пытались
понять, как наследуются биологические
признаки. Они скрещивали различные
виды растений, животных и наблюдали
сходство между родителями и потомками.
Результаты были обескураживающими:
одни признаки оказывались общими у по
томка с одним родителем, другие — с
другим, третьи — отличались от обоих
родителей. Эти наблюдения не позволили
ученым сделать хотя бы предположение о
закономерности наследования призна
ков. Причиной подобных неудач было от
сутствие четкой гипотезы и методологии
ее решения.
Главное достижение Менделя заклю
чается в том, что он сформулировал и
применил принципы гибридологического
анализа для проверки конкретной гипо
т е з ы— г и п о т е з ы о н а с л е д с т в е н
ной п е р е д а ч е д и с к р е т н ы х ф а к
т о р о в . Решению этой проблемы спо
собствовали принципиально новые мето
дические подходы Менделя в изучении
закономерностей наследования призна
ков, которым и по сей день следуют всё
генетики.
1. Скрещиваемые организмы должны
принадлежать к одному виду.
2. Скрещиваемые организмы должны
четко различаться по одной, двум и
более парам контрастных, альтер
нативных признаков.
3. Изучаемые признаки должны быть
константны, т. е. воспроизводиться
из поколения в поколение при скре
щивании в пределах родственной
формы.
4. Должен применяться индивидуаль
ный анализ потомства от каждого
гибридного организма.
5. Необходимо использовать количес
твенный учет гибридных организмов,
различающихся по отдельным парам
альтернативных признаков, в ряду
последовательных поколений.
Перечисленные приемы исследова
ния составили принципиально новый
гибридологический метод, открывший
целую эпоху в изучении наследственнос
ти и изменчивости.
Кроме разработки замечательной ме
тодологии, научная гениальность Менде
ля проявилась в способности сформули
ровать теорию, объясняющую данные эк
спериментов, и поставить эксперименты,
подтверждающие эту теорию. Время убе
дительно доказало фундаментальную
полноту и правильность всех предполо
жений и выводов Менделя.
Со времен Менделя генетический
анализ обогатился целым рядом методов.
В частности, м е т о д о т д а л е н н о й
г и б р и д и з а ц и и (скрещивание особей
различных видов или родов) позволяет
выяснить степень эволюционного род
ства между видами и родами. В последние
годы широкое распространение получили
методы г и б р и д и з а ц и и с о м а т и
ческих клеток животных и р а с
тений.
I . Что такое наследственность и каково ее био
логическое значение? 2. Что мешало предшествен
никам Менделя подойти к анализу наследования
признаков? 3. Какие непреложные правила поло
жены Менделем в основу метода гибридологическо
го анализа?
64. Моногибридное скрещивание.
Первый и второй законы Менделя
Моногибридным называется такое
скрещивание, при котором родительские
организмы отличаются друг от друга по
одной паре альтернативных признаков.
Мендель проводил опыты с горо
хом — самоопыляющимся растением. До
начала скрещивания он отобрал из
34 сортов гороха только 7 пар растений,
которые различались по семи отдельным,
хорошо заметным признакам: пурпурные
или белые цветки, их пазушное или вер
хушечное расположение, высокие (до 2
м) или низкие (не более 60 см) стебли,
желтая или зеленая окраска незрелых
плодов (бобов), выпуклые или с
перехватами бобы, гладкая или морщи
нистая поверхность семян, желтая или
зеленая их окраска (рис. 134, с. 222).
В результате длительного самоопыле
ния внутри каждого из отобранных сор
тов Мендель создал то, что позднее
В. Иогансен назвал «чистыми линия
ми» — группу особей, у которых иссле
дуемый признак постоянно воспроизво
дился в потомстве из поколения в поко
ления.
Доминирование. Закон единообра
зия гибридов первого поколения. В од
ном из опытов Мендель изучал наследо
вание окраски цветка, скрещивая расте
ния с пурпурными и белыми цветками.
В результате у всех гибридных растений
первого поколения (/у ) цветки оказались
пурпурными независимо от того, мате
ринским или отцовским было растение с
такими цветками. При этом белая окрас
ка цветков не проявлялась.
Мендель обнаружил также, что все
гибриды F/ оказались единообразными
(однородными) по каждому из семи
перечисленных выше исследуемых при
знаков (рис. 135, с. 223).
Следовательно, у гибридов первого
поколения из пары родительских альтер
нативных признаков проявляется только
221
один, а признак другого родителя как бы
исчезает. Явление преобладания у гибри
дов первого поколения признаков одного
из родителей Мендель назвал доминиро
ванием, а соответствующий признак —
доминантным. Признаки, не проявляю
щиеся у гибридов//, он назвал рецессив
ными. Это явление оказалось универ
222
сальным для всех живых организмов и
потому было возведено в ранг правила
доминирования.
Поскольку все гибриды первого поко
ления единообразны, это явление было
названо К. Корренсом первым законом
Менделя, или законом единообразия
гибридов первого поколения.
Закон расщепления. Из гибридных
семян гороха Мендель вырастил расте
ния, которые подверг самоопылению, и
образовавшиеся семена вновь высеял.
В результате было получено второе поко
ление гибридов, или F2- Среди гибридов
F2 обнаружилось расщепление по каждой
паре признаков в соотношении 3:1, т. е.
три четверти растений имели доминан
тные признаки (пурпурные цветки, желРодители
тые семена, высокие стебли и т. д.) и одна
четверть — рецессивные (белые цветки,
зеленые семена, низкие стебли и т. д.).
Следовательно, рецессивный признак у
гибрида С, не исчез, а только был подав
лен и вновь проявился во втором поколе
нии. Это обобщение позднее было назва
но вторым законом Менделя, или зако
ном расщепления.
Гомозиготные и гетерозиготные осо-
Р
Гибрид первого
поколения F1
Второе
поколение F2
1 1 I
1
Третье
поколение F3
Рис. 135. Ход моногибридного
скрещивания.
223
би. Чтобы выяснить, как будет осущест
вляться наследование признаков в треть
ем, четвертом и последующих поколени
ях, Мендель путем самоопыления вырас
тил гибриды этих поколений и проанали
зировал полученное потомство. Он выяс
нил, что растения, обладающие рецес
сивными признаками, например белые
цветки в следующих поколениях (F3, F4 и
т. д.), воспроизводят потомство только с
белыми цветками.
Иначе вели себя гибриды второго по
коления, обладающие доминантными
признаками (например, пурпурные цвет
ки). Среди них при анализе потомства
Мендель обнаружил две группы расте
ний, внешне совершенно неразличимых
по каждому конкретному признаку.
Первая группа, составляющая 1/ г от
общего числа растений с доминантным
признаком, далее не расщеплялась, т. е.
во всех последующих поколениях у них
обнаруживалась только пурпурная окрас
ка цветков. Оставшиеся 2/ 3 растений вто
рого поколения с доминантным призна
ком в F3 снова давали расщепление такое
же, как в F2, т. е. на 3 растения с пурпур
ными цветками появлялось 1 с белыми.
Особи, которые не дают в потомстве
расщепления и сохраняют свои признаки
в «чистом» виде, называют гомозигот
ными (от греч. gomo — равный, одинако
вый, zygoios — соединенный вместе) по
данному признаку. Особи, у которых в по
томстве обнаруживается явление рас
щепления, называют гетерозиготными
(от греч. getero — иной, другой). Поня
тие «гомозиготность» и «гетерозиготность» ввел в генетику У. Б э т с о н в
1902 г.
Таким образом, Менделем впервые
было установлено, что растения, сходные
по внешним признакам (по фенотипу),
могут обладать различными наследствен
ными свойствами.
Статистический анализ расщепле
ния. Для установления причины расщеп
ления, причем в строго определенных
224
численных отношениях доминантных и
рецессивных признаков, следует вспом
нить, что связь между поколениями при
половом размножении осуществляется
через половые клетки (гаметы). Очевид
но, гаметы несут материальные наслед
ственные задатки, или факторы, опреде
ляющие развитие того или иного призна
ка. Эти факторы позже и были названы
генами.
Под термином «ген» принято пони
мать функционально неделимую едини
цу наследственного материала, опреде
ляющую проявление отдельного эле
ментарного признака или свойства ор
ганизма, а совокупность всех генов ор
ганизма называют ген о т и п о м .
В соматических клетках почти каждо
го организма содержится пара одинако
вых генов: один получен от отцовского
организма, а другой — от материнского.
Мендель предложил обозначать доми
нантные наследственные задатки боль
шой буквой (например, Л), а соответству
ющие им рецессивные задатки — малой
буквой (а).
А теперь результаты опыта Менделя
по моногибридному скрещиванию пред
ставим в виде схемы (рис. 136). Символы
Р, Fu F2 и т . д . обозначают родительское,
1-е и 2-е поколения соответственно, знак
умножения указывает скрещивание, сим
вол </ обозначает мужской пол, a J —
женский пол. Из схемы видно, что в роди
тельском поколении (Р) материнская и
отцовская формы гомозиготны по иссле
дуемому признаку, поэтому производят
гаметы только с геном А или только с а.
При оплодотворении эти гаметы об
разуют зиготу, которая имеет оба гена,
как доминантный, так и рецессивный —
Аа. В результате все гибриды F{ единооб
разны по конкретному признаку, по
скольку доминантный ген А подавляет
действие рецессивного гена а. Во время
образования половых клеток гены А м а
попадают в них по одному. Следователь
но, гибридные организмы способны про-
изводить гаметы двух типов, несущие ге
ны А на. Организмы, имеющие одновре
менно доминантный и рецессивный гены
и формирующие гаметы двух типов (толь
ко с Л или только с а), называют гетеро
зиготными.
Для облегчения расчета сочетаний
разных типов гамет английский генетик
Р. П ен н е т предложил производить за
пись в виде решетки, которая и вошла в
литературу под названием решетки
Пеннета (см. рис. 136). Слева по верти
кали располагаются женские гаметы,
сверху по горизонтали — мужские.
р
ГАМ ЕТЫ
Рис. 136. Наследование пурпурной и белой
окраски цветков гороха.
8. Общая биология,
10 кл.
В квадраты решетки вписывают образу
ющиеся сочетания гамет, которые соот
ветствуют генотипам зигот.
При самоопылении в F2 получается
расщепление по генотипу в отношении
\AA\2Aa: \аа, т. е. одна гомозигота по до
минантным генам, две гетерозиготы и од
на гомозигота по рецессивным генам. Так
как генотипам АА и Аа соответствует
один и тот же фенотип — пурпурная
окраска цветка, расщепление по феноти
пу будет 3 пурпурных : 1 белый. Следова
тельно, расщепление по фенотипу не
совпадает с расщеплением по генотипу.
Теперь легко объяснить, почему гомо
зиготные белоцветковые растения второ
го поколения с рецессивными генами аа
при самоопылении в F3 дают только себе
подобных. Такие растения производят га
меты одного типа, и, как следствие, рас
щепления не наблюдается. Ясно также,
что среди пурпурноцветковых */3 доми
нантных гомозигот (АА) также не будет
давать расщепления, а 2/ 3 гетерозигот
ных растений (Аа) будут давать в F3 рас
щепление 3:1, как и у гибридов F2.
На основании анализа результатов
моногибридного скрещивания были
сформулированы не только первый и вто
рой законы Менделя и правило домини
рования, но и правило чистоты гамет.
Правило чистоты гамет. При моногибридном скрещивании в случае полно
го доминирования у гетерозиготных гиб
ридов (Аа) первого поколения проявля
ется только доминантный ген (Л); рецес
сивный же (а) не теряется и не смешива
ется с доминантным. В F2 как рецессив
ный, так и доминантный гены могут про
являться в своем «чистом» виде, т. е. в
гомозиготном состоянии. При этом гены
не только не смешиваются, но и не пре
терпевают изменений после совместного
пребывания в гибридном организме.
В результате гаметы, образуемые такой
гетерозиготой (Аа), являются «чистыми»
в том смысле, что гамета А «чиста» и не
содержит ничего от гена а, а гамета а
225
«чиста» от Л. Это явление несмешивания
генов пары альтернативных признаков в
гаметах гибрида получило название пра
вила чистоты гамет. Данное правило,
установленное Менделем, указывает на
дискретность гена, несмешиваемость ге
нов друг с другом.
Многочисленными наблюдениями
многих исследователей в последующие
годы был установлен универсальный ха
рактер законов Менделя. Им подчиняют
ся все живые организмы, в том числе че
ловек, у которого изучено и описано мно
го пар альтернативных признаков.
Установление общих законов насле
дования признаков позволило Менделю
сделать вывод о дискретности признаков
и их связи с определенными наследствен
ными структурами — генами.
9
1. Какие правила и законы установил Мендель
на основе моногибридного скрещивания? 2. Что та
кое доминантный и рецессивный признаки, гомози
гота и гетерозигота, генотип и фенотип? 3. Из жел
того семени гороха получено растение, которое дало 218 семян, из которых 159 желтых и 59 зеленых.
Каковы генотипы всех форм? Докажите соответ
ствие теории и экспериментальных данных.
65. Аллельные гены
и их взаимодействие.
Анализирующее скрещивание
Аллельные гены. На примере насле
дования у гороха видно, что гены, опреде
ляющие развитие взаимоисключающих
признаков, составляют пары. Пару генов,
определяющих альтернативные призна
ки, называют аллеломорфной парой, а
само явление парности — аллеломорфизмом, или аллелизмом. Каждый ген
имеет два состояния — А и а, поэтому
они составляют одну пару, а каждого из
членов пары называют аллелем. Таким
образом, гены, расположенные в одних и
тех же локусах (участках) гомологичных
226
хромосом и определяющие альтернатив
ное развитие одного и того же признака,
называются аллельными.
В самом простом случае ген представ
лен двумя аллелями. Например, пурпур
ная и белая окраска цветка гороха явля
ется доминантным и рецессивным при
знаками соответственно двум аллелям
одного гена. Пример трехаллельного ге
на — ген, определяющий у человека сис
тему групп крови АВО. Аллелей бывает и
больше: для гена, контролирующего син
тез гемоглобина человека, их известно
много десятков. Однако, сколькими алле
лями ни был бы представлен ген, в поло
вой клетке- имеется только один аллель
(правило чистоты гамет), а в диплоидной
клетке организма не больше двух — от
каждого из родителей.
Взаимодействие аллельных генов.
Установлено, что существует огромное
количество признаков и свойств живых
организмов, которые определяются дву
мя, тремя и даже многими парами генов,
и, наоборот, один ген часто влияет на
многие признаки. Кроме того, действие
гена может быть изменено соседством
других генов и условиями внешней сре
ды. Таким образом, в онтогенезе действу
ют скорее не отдельные гены, а весь ге
нотип как целостная система со сложны
ми связями и взаимодействиями между
генами.
Явление, когда за один признак отве
чает несколько генов (аллелей), называ
ется взаимодействием генов. Причем ес
ли это аллели одного и того же гена, то
такие взаимодействия называются ал
лельными, а в случае разных генов — неаллельными.
Выделяют следующие основные типы
аллельных взаимодействий: доминирова
ние, неполное доминирование, сверхдо
минирование и кодоминирование.
Доминирование — это такой тип
взаимодействия двух аллелей одного ге
на, когда один из них полностью исключа
ет проявление действия другого. В ре
зультате гетерозиготные организмы по
фенотипу точно соответствуют родителю,
гомозиготному по доминантным аллелям.
Такое явление возможно при условии,
что: 1) доминантный аллель в гетерози
готном состоянии обеспечивает синтез
продуктов, достаточный для проявления
признака такого же качества и интенсив
ности, как и в состоянии доминантной го
мозиготы у родительской формы, 2) ре
цессивный аллель совсем неактивен либо
продукты его активности не взаимодей
ствуют с продуктами активности доми
нантного аллеля. Примерами полного до
минирования могут служить доминирова
ние пурпурной окраски цветков у гороха
над белой, гладкой формы семян над мор
щинистой, темных волос над светлыми и
карих глаз над голубыми у человека и т. д.
Неполное доминирование, или про
межуточный характер наследования,
наблюдается в том случае, когда фенотип
гибрида (гетерозиготы) отличается от
фенотипа обеих родительских гомозигот,
т. е. выражение признака оказывается
промежуточным, с большим или мень
шим уклонением в сторону одного или
другого родителя. Механизм этого явле
ния состоит в том, что рецессивный ал
лель неактивен, а степень активности до
минантного аллеля недостаточна для то
го, чтобы обеспечить нужный уровень
проявления признака доминантной роди
тельской гомозиготы.
Примером неполного доминирования
является наследование окраски цветков у
растений ночной красавицы (рис. 137).
Как видно из схемы, гомозиготные расте
ния имеют либо красные (АА), либо бе
лые (аа) цветки, а гетерозиготные
(Аа) — розовые. При скрещивании рас
тения с красными цветками и растения с
белыми цветками в Fl у всех растений
цветки розовые, т. е. наблюдается проме
жуточный характер наследования. При
скрещивании гибридов с розовой окрас
кой цветков в F2 имеет место совпадение
расщепления по фенотипу и генотипу, так
как доминантная гомозигота (АА) отлича
ется от гетерозиготы (Аа). Так, в разби
раемом нами примере с растениями ноч
ной красавицы расщепление в Р2 по
окраске цветков обычно следующее —
1красная (АА) : 2розовые (Аа) \ 1белая(аа).
Неполное доминирование оказалось
широко распространенным явлением.
Оно наблюдается в наследовании курча
вости волос у человека, масти крупного
рогатого скота, окраски оперения у кур,
многих других морфологических и физио227
логических признаков у растений, живот
ных и человека.
Сверхдоминирование — это более
сильное проявление признака у гетерози
готной особи (Ла), чем у любой из гомо
зигот (АА и аа). Г
КодоминироЗание — участие обоих
аллелей (отцовского и материнского) в
определении признака у гетерозиготной
особи. Ярким и хорошо изученным при
мером кодоминирования может служить
наследование антигенных групп крови
человека: А, В, АВ и 0, детерминируемых
геном /.
Известны три типа аллелей этого ге
на: 1А, 1В, /°. При гомозиготности 1А1А
эритроциты имеют только антиген
А (группа крови А, или II). При гомози
готности 1В1В эритроциты несут только
антиген В (группа В, или III). В случае го
мозиготности /°/° эритроциты лишены
антигенов типа А я В (группа 0, или I).
При гетерозиготности 1АР или /в/° группа
крови определяется соответственно А (II)
или В (III). Эритроциты имеют антигены
только А или только В соответственно.
Это уже известный случай полного доми
нирования.
Если же человек гетерозиготен 1А1В,
его эритроциты несут оба антигена: А я В
(группа крови АВ, или IV). Это и есть
случай кодоминирования. Аллели 1А и 1В
работают в гетерозиготе как бы незави
симо друг от друга.
Пример наследования групп крови
иллюстрирует и проявление множест
венного аллелизма: ген / может быть
представлен тремя разными аллелями, а
есть гены, имеющие десятки аллелей.
Все аллели одного гена получили назва
ние серии множественных аллелей, из
которых каждый диплоидный организм
может иметь любые, но только два алле
ля. Между этими аллелями возможны все
перечисленные варианты аллельных вза
имодействий.
Явление множественного аллелизма
широко распростанено в природе. Извес
228
тны обширные серии множественных ал
лелей, определяющих тип совместимости
при оплодотворении у грибов, опылении
у семенных растений, детерминирующих
окраску шерсти животных и т. д.
Таким образом, разные аллели одного
гена могут проявлять свое действие неза
висимо друг от друга. Проявление алле
лей при этом может быть разным (полное
или неполное доминирование).
Реципрокные скрещивания. В рас
смотренных нами выше примерах доми
нантный признак имело материнское рас
тение, а рецессивный — отцовское. Воз
никает вопрос: изменится ли характер
наследования, если материнский орга
низм будет нести рецессивный признак, а
отцовский —- доминантный? Для ответа
на этот вопрос необходимо провести реципрокное скрещивание. Реципрокными
называют такую пару скрещиваний, при
которых организмы с доминантными и
рецессивными признаками используются
и как материнские, и как отцовские:
1 АА
х
с / аа и J
аа
х o ' АА.
Оказалось, что в подавляющем боль
шинстве случаев реципрокные скрещи
вания дают одинаковые результаты.
Реципрокные скрещивания использу
ют в генетическом анализе для выявле
ния наследственных факторов, локализо
ванных в половых хромосомах. В этом
случае в одном из реципрокных скрещи
ваний наблюдают «крисс-кросс» (крестнакрест) наследование, когда материн
ский признак передается только сыновь
ям, а отцовский — только дочерям.
Возвратные скрещивания. Скрещи
вание гибрида первого поколения с одной
из родительских форм, несущих данную
пару аллелей, в гомозиготном состоянии
называют возвратным скрещиванием,
или беккроссом, а потомство обозначают
FB. Эти два скрещивания имеют неодина
ковую ценность для генетического ана
лиза.
При возвратном скрещивании гибри
да F{(Aa) с одной из родительских форм,
например АА, все гаметы родительского
организма будут нести доминантный ал
лель Л, а у гибрида образуются гаметы
двух типов — Л и а. Поэтому в результа
те случайного сочетания этих гамет при
оплодотворении в потомстве имеет место
расщепление по генотипу в отношении
1:1 (2Ла:2ЛЛ), в то время как расщепле
ния по фенотипу не наблюдается (рис.
138). Беккросс используется для выявле
ния генотипа особи (анализирующее
скрещивание), а также для преодоления
стерильности отдаленных гибридов пер
вого поколения.
Анализирующее
скрещивание.
В случае полного доминирования по
внешнему проявлению признака (по фе
нотипу) судить о генотипе организма не
возможно, так так и доминантная гомози
гота (ЛЛ), и гетерозигота (Аа) обладают
1Аа
1аа
Рис. 138. Возвратное скрещивание гибрида F,
с рецессивной родительской формой (анализи
рую щ ее скрещивание).
фенотипически доминантным признаком.
Для того чтобы отличить доминантную
гомозиготу от гетерозиготы, применяют
анализирующее скрещивание. Скрещи
вание организма неизвестного генотипа с
организмом, гомозиготным по рецессив
ным аллелям, называется анализирую
щим. В этом случае рецессивная форма
образует только один сорт гамет с алле
лем а, что позволяет проявиться любому
из двух аллелей исследуемого организма
уже в первом поколении.
Например, у плодовой мушки дрозо
филы длинные крылья доминируют над
зачаточными. Особь с длинными крылья
ми может быть гомозиготной (LL) или ге
терозиготной (LI). Для установления ее
генотипа надо провести анализирующее
скрещивание между этой мухой и мухой,
гомозиготной по рецессивному аллелю.
Если у всех потомков от этого скрещива
ния будут длинные крылья, то особь с не
известным генотипом гомозиготна по до
минантному аллелю (LL). Численное со
отношение потомков с длинными и ко
роткими крыльями указывает на гетерозиготность особи с неизвестным гено
типом.
Таким образом, по характеру расщеп
ления можно проанализировать генотип
гибрида, типы гамет, которые он образу
ет, и их соотношение. Поэтому анализи
рующее скрещивание является очень
важным приемом генетического анализа
и широко используется в генетике и се
лекции.
?
* 1. Какие гены называются аллельными? 2. Что
такое генотип? 3. Какие взаимодействия генов на
зываются аллельными? 4. Объясните механизмы
таких аллельных взаимодействий, как доминирова
ние, неполное доминирование, кодоминирование. 5.
Что такое множественный аллелизм? 6. Что такое
реципрокные и возвратные скрещивания и какова
их роль в генетическом анализе и селекции? 7. Мо
жет ли у матери с группой крови А и отца с группой
крови 0 родиться ребенок с группой крови В?
Объясните ответ.
229
66. Дигибридное и полигибридное
скрещивания.
Третий закон Менделя
Существование дискретных наслед
ственных факторов, или генов, следует из
анализа моно гибридного скрещивания,
рассмотренного нами ранее. Однако оче
видно, что в большинстве случаев орга
низмы различаются по многим генам.
Чтобы одновременно проанализировать
наследование нескольких признаков, не
обходимо изучить наследование каждой
пары признаков в отдельности, не обра
щая внимания на другие пары, а затем со
поставить и объединить все эти наблюде
ния. Именно так поступил Мендель.
Скрещивание, при котором роди
тельские формы отличаются по двум
парам альтернативных признаков (по
двум парам аллелей), называется дигибридным. Гибриды, гетерозиготные по
двум генам, называются дигетерозиготными, а в случае отличия их по трем
и многим генам — три- и полигетерозиготными соответственно.
Поскольку каждый организм характе
ризуется огромным разнообразием при
знаков, а число хромосом ограничено
(например, у человека всего 46), то каж
дая из них должна нести большое число
генов. Результаты дигибридного и полигибридного скрещивания зависят от того,
располагаются ли гены, определяющие
рассмотренные признаки, в одной хромо
соме или в разных.
Независимое наследование. Для ди
гибридного скрещивания Мендель ис
пользовал гомозиготные растения горо
ха, различающиеся одновременно по
двум парам признаков. Одно из скрещи
ваемых растений имело гладкие желтые
семена, другое — морщинистые зеленые
(рис. 139).
Все гибриды первого поколения этого
скрещивания имели гладкие желтые се
мена. Следовательно, гладкая форма се
мян доминирует над морщинистой и жел
230
тая окраска над зеленой. Обозначим ал
лели гладкой формы В, морщинистой —
Ь, а аллели желтой окраски Л, зеленой —
а. Гены, определяющие развитие разных
пар признаков, называются неаллельны
ми. В данном случае гены формы семени
В и b неаллельны генам окраски Л и а.
Неаллельные гены обозначают разными
буквами алфавита. Родительские расте
ния будут иметь генотипы ААВВ и aabb.
Генотип гибридов F\ будет АаВЬ, то есть
характеризуется дигетерозиготностью.
Во втором поколении после самоопы
ления гибридов F\ в соответствии с зако
ном расщепления вновь появились мор
щинистые и зеленые семена. При этом
наблюдались все возможные сочетания
численных признаков: 315 гладких жел
тых, 101 морщинистое желтое, 108 глад
ких зеленых и 32 морщинистых зеленых
семени. Это соотношение очень близко к
9:3:3:1.
Чтобы выяснить, как ведет себя каж
дая пара аллелей в потомстве дигетеро
зиготы, целесообразно провести раздель
ный учет каждой пары признаков — по
форме и окраске семян. Из 556 семян
Менделем получено 423 гладких и 133
морщинистых, а также 416 желтых и 140
зеленых. Таким образом, и в этом случае
соотношение доминантных и рецессив
ных форм по каждой паре признаков сви
детельствует о моногибридном расщеп
лении по фенотипу 3:1. Отсюда следует,
что дигибридное расщепление представ
ляет собой по существу два независимо
идущих моногибридных, которые как бы
накладываются друг на друга.
Проведенные наблюдения убедитель
но показывают, что отдельные пары
признаков ведут себя в наследовании не
зависимо. Этот феномен отражает сущ
ность третьего закона Менделя — зако
на независимого наследования призна
ков, или независимого комбинирования
генов.
Он формулируется так: каждая пара
аллельных генов (и альтернативных
р
Зеленые
морщинистые
aabb
Желтые
гладкие
ААВВ
Желтые гладкие АаВЬ
Гаметы
с/©
V
©
©
00 >
00 >
Желтые
гладкие
Желтые
гладкие
®
»
Желтые
гладкие
®
Желтые
морщинистые
i:
Желтые
гладкие
®
f2
О
O' >
СГ >
Желтые
гладкие
©
*
Желтые
гладкие
"
Желтые
гладкие
®
Зеленые
гладкие
:
:
:
Зеленые
гладкие
©
:
Зеленые
гладкие
ф
__
О
Желтые
морщинистые
эЭ
-
Желтые
морщинистые
-
‘»
----v.
®
®
О
®
Желтые
гладкие
Желтые
гладкие
вь
Зеленые
морщинистые
аа
вь
ЬЬ
Рис. 139. Дигибридное скрещивание Горохов, различающихся по форме и окраске семян: А — жел
тые, а — зеленые, В — гладкие, в — морщинистые.
231
признаков, контролируемых ими) на
следуется независимо друг от друга.
Закон независимого комбинирования
генов составляет основу комбинативной изменчивости, наблюдаемой при
скрещивании у всех живых организмов.
Отметим также, что этот закон действи
телен только для генов, локализованных
в разных парах гомологичных хромосом.
Это обусловлено тем, что негомологич
ные хромосомы комбинируются в клетке
независимо друг от друга, что было дока
зано не только при изучении характера
наследования признаков, но и прямым
цитологическим методом. Поведение
хромосом при дигибридном скрещивании
показано на рисунке 140.
Цитологические основы дигибридного скрещивания. Как известно, в про
фазе I мейоза гомологичные хромосомы
конъюгируют, а в анафазе одна из гомо
логичных хромосом отходит к одному по
люсу, а другая — к другому. При расхож
дении к разным полюсам негомологичные
хромосомы комбинируются свободно и
независимо друг от друга. При оплодо
творении в зиготе восстанавливается
диплоидный набор хромосом и гомоло
гичные хромосомы, оказавшиеся в про
цессе мейоза в разных половых клетках
родителей, соединяются вновь.
Предположим, что каждая хромосома
содержит только один ген. Палочковид
ные хромосомы несут аллель А или а,
сферические — В или Ь, то есть эти две
пары аллелей находятся в негомологич
ных хромосомах.
Гомозиготные родители (ААВВ и
aabb) формируют только один тип гамет с
доминантными (АВ) или с рецессивными
(ab) аллелями. При слиянии гамет обра
зуется единообразное первое поколение
гибридов. Этот гибрид дигетерозиготен
(АаВЬ), но так как у него присутствуют
гены Л и Б, то по фенотипу он сходен с од
ним из родителей.
В тех случаях, когда необходимо ука
зать, что те или иные гены находятся в го
232
мологичных хромосомах, в генетических
формулах зигот хромосомы принято
изображать в виде двух или одной черточ
ки с указанием обоих аллелей гена. Фор
мула дигетерозиготы может быть записа
на так: = = . Поскольку гаметы несут
а Ь
только по одной из гомологичных хромо
сом и соответственно по одному аллелю
каждого гена, то формулы гамет могут
быть записаны так: А В, а Ь_и т. д.
При мейозе у гибридных организмов
происходит расхождение к полюсам го
мологичных хромосом каждой пары. При
этом по причине случайности расхожде
ния отцовских и материнских хромосом в
процессе мейоза ген А может попасть в
одну гамету с геном В или с геном Ь. Точ
но так же ген а может оказаться в одной
гамете с геном В или с геном Ь. Поэтому
гибриды образуют четыре типа гамет: две
родительские (отцовскую и материнскую)
и две рекомбинантные, содержащие один
отцовский ген и один материнский (АЬ и
аВ). Это соответствует двум равноверо
ятным вариантам расположения хромо
сом в мейозе и их расхождения к полю
сам: = = и = = . Поэтому образование
всех четырех типов гамет равновероятно,
то есть все они образуются в равных ко
личествах. Свободное сочетание таких
гамет в процессах оплодотворения закан
чивается образованием 16 типов зигот, а
значит, и потомков (см. рис. 140).
Они распадаются на четыре феноти
пических класса: доминантные по обоим
признакам — 9 частей, доминантные по
первому и рецессивные по второму при
знаку — 3, рецессивные по первому и до
минантные по второму — 3, рецессивные
по обоим признакам — 1 часть. Геноти
пических классов 9: 1ААВВ, 2ААВЬ,
lAAbb, 2Aabb, 4АаВЬ, 2АаВВ, laaBB,
2aaBb, laabb.
<
CD
Гаметы F1
? Г
А 00а
А 0 0А
1 В о о в B o o b В о оВ B o o b
А [Ш а А П Па
А0 0 А А 0 0 А
1 b о о В b @© b b ©ОВ b o o b
ч а П П А а П ПА
а П Па а П П а
| В О 0 в B o o b В ОО В B o o b
\
аП ПА
1 Ьоов
П П А а 0 Па
а
boob
а 00а
Ь о о в b ©@ь
Рис. 140. Цитологические основы расщепления признаков при дигибридном скрещивании.
233
Полигибридное скрещивание. Рас
суждая аналогично, можно представить
расщепление при тригибридном и полигибридном скрещиваниях, то есть когда
родители различаются по аллелям трех и
более генов, а в F\ образуются тригетерозиготы и полигетерозиготы. Соотно
шения генотипических и фенотипических
классов в F2 три- и полигибридных скре
щиваний, а также число типов гамет (и
число фенотипов) у гибридов /д опре
деляются простыми формулами: при
моногибридном скрещивании число
типов гамет равно 2, при дигибридном —
4 (22), а при полигибридном — 2"; число
генотипов равно соответственно 3,
9(32) и 3".
Опираясь на независимость наследо
вания разных пар аллелей, можно также
любые сложные расщепления предста
вить как произведение от соответствую
щего числа независимых моногибридных скрещиваний. Общая формула
определения фенотипических классов
при полигибридном скрещивании имеет
вид (3:1)", где п равно числу пар призна
ков, по которым идет расщепление. Для
моногибрида эта формула имеет вид
(3:1), для дигибрида — 9:3:3:1 или
(3:1)2, тригибрида — (3:1)3 и т. д. Рас
щепление по генотипу имеет вид ( 1:2:1)",
где п — число расщепляющихся пар ал
лелей.
Известно, что каждый организм гете
розиготен по многим генам. Если предпо
ложить, что человек, у которого отдель
ные пары хромосом содержат не одну, а
сотни пар аллелей, гетерозиготен хотя бы
по 20 генам, то число типов гамет у
такой полигетерозиготы составит 220 =
= 1 048 576. Эта цифра дает определен
ное представление о потенциальных воз
можностях комбинативной изменчивос
ти. Поэтому каждый человек обладает
неповторимой индивидуальностью. На
Земле нет двух людей, тождественных по
их наследственности, за исключением од
нояйцевых близнецов.
234
■ 1. В чем заключается смысл закона независи
мого наследования признаков? 2. Какова связь
между первым, вторым и третьим законами Менде
ля? 3. Каковы цитологические основы дигибридного скрещивания? 4. Напишите общие формулы для
числа классов фенотипов, генотипов, типов гамет,
формулы расщепления по генотипу и фенотипу в за
висимости от числа пар расщепляющихся аллелей.
5. Что такое комбинативная изменчивость и какие
факторы вызывают ее появление у растений и жи
вотных? Приведите примеры получения новых ком
бинаций при дигибридном скрещивании. 6. Выпи
шите все типы гамет, образуемые особью
AAbbCcDdEe. 7. Какое расщепление по генотипу и
фенотипу будет в потомстве от скрещивания расте
ний генотипа: DdEEAa х ddEeaa, если D — высо
кое растение, d — низкое, Е — гладкий корнеплод,
е — морщинистый, А — простое соцветие, а —
сложное?
67. Неаллельные взаимодействия
генов
Закон независимого наследования
признаков еще раз демонстрирует дис
кретный характер генетического матери
ала. Это проявляется в независимом ком
бинировании аллелей разных генов
(неаллельных генов) и в их независи
мом действии — фенотипическом выра
жении.
Ген как единица наследственности,
детерминирующая признаки организма,
характеризуется определенными функци
ональными свойствами, т. е. в своем дей
ствии ген дискретен. Дискретность гена
определяется тем, что он контролирует
присутствие или отсутствие отдельной
биохимической реакции, от которой зави
сит развитие или подавление определен
ного признака организма. Очевидно, если
несколько генов определяют какое-либо
одно свойство или один признак (форма
гребня у кур, окраска глаз у дрозофилы,
длина колоса у пшеницы и т. д.), они дол
жны взаимодействовать между собой.
Отсюда следует, что понятие «наследова
ние признаков» употребляется скорее
всего как образное выражение, посколь
ку в действительности наследуются не са
ми признаки, а гены. Признаки формиру
ются в ходе индивидуального развития
организма, обусловливаются генотипом и
влиянием многочисленных факторов
внешней среды.
Взаимодействие неаллельных генов
описано у многих растений и животных.
Оно приводит к появлению в потомстве
дигетерозиготы необычного расщепле
ния по фенотипу: 9:3:4; 9:7; 9:6:1; 13:3;
12:3:1; 15:1, т. е. модификации общей
менделевской формулы 9:3:3:1. Извес
тны случаи взаимодействия двух, трех и
большего числа неаллельных генов. Сре
ди них можно выделить следующие ос
новные типы: комплементарность, эпис
таз и полимерию.
Комплементарное взаимодействие
генов. Комплементарным, или допол
нительным, называется такое взаимо
действие неаллельных доминантных ге
нов, в результате которого появляется
новый признак, не определявшийся ни
одним, ни другим геном. Например, при
скрещивании двух сортов душистого го
рошка с белыми цветками появляется по
томство с пурпурными цветками. Если
обозначить генотип одной белой расы
ААаа, а другой — ааВВ, то
Р: ААвв х ааВВ
Гаметы: Ав
аВ
Fp
АаВв
Гибрид первого поколения с двумя до
минантными генами (А и В) получил био
химическую основу для выработки пур
пурного пигмента антоциана, в то время
как поодиночке ни ген А, ни ген В не
обеспечивали синтеза этого пигмента.
Синтез антоциана представляет собой
сложную цепь последовательных биохи
мических реакций, контролируемых не
сколькими неаллельными генами, и толь
ко при наличии как минимум двух генов
(А-В-) развивается пурпурная окраска.
В остальных случаях (ааВ- и А -b'b) цвет
ки у растения белые (знак «-» в формуле
генотипа означает, что это место может
занять как доминантный, так и рецессив
ный аллель).
При самоопылении растений душис
того горошка из /д в F2 наблюдалось рас
щепление на пурпурно- и белоцветковые
формы в соотношении, близком к 9:7.
Пурпурные цветки были обнаружены
9
7
16
16
у — растений, белые — у — . Решетка
Пеннета наглядно показывает причину
этого (рис. 141, с. 236).
Эпистаз. Сущность эпистаза сводит
ся к тому, что доминантный аллель одно
го гена подавляет действие аллельной
пары другого гена. Гены, подавляющие
действие других генов, называются эпистатическими генами, ингибиторами,
или супрессорами. Подавляемый ген но
сит название гипостатического.
По изменению числа и соотношения
фенотипических классов при дигибридном расщеплении в F2 рассматривают
несколько типов эпистатических взаимо
действий: доминантный эпистаз (А>В или
В>А) с расщеплением 12:3:1; рецессив
ный эпистаз (а>В или Ь>А), который вы
ражается в расщеплении 9:3:4, и т. д.
Рассмотрим проявления доминантно
го эпистаза на примере наследования ок
раски у кур (рис. 142, с. 237). Ген С кон
тролирует окраску оперения (сс — белое
оперение), а ген i — ген-ингибитор гена
окраски (тоже белое оперение). При
скрещивании белых кур породы леггорн
(ССП) с белыми курами породы виандот
(ccii) в Fl все потомство белое, а в F2 —
13 частей белых и 3 части окрашенных.
Окрашенное оперение только у тех, кото
рые имеют доминантный ген окраски С,
не блокированный наличием гена-инги
битора i. Белое оперение наблюдается у
кур, у которых отсутствует ген окраски С
(сс), а также у кур, имеющих доминан
тный аллель гена-ингибитора.
Эпистаз можно сравнить с полным
235
ААЬЬ
белый
X
ааВВ
ЬкША
белый
АаВЬ
F1 :
пурпурный
ГАМЕТЫ
АВ
АЬ
ААВВ
ААВЬ
AAEfo
ААЬЬ
аВ
ab
3 *
АВ
АаВЬ
АЬ
ф
АаЬЬ
F2 '
аВ
Аг^ВВ
ааВВ
ааВЬ
ааВЬ
aabb
ab
Рис. 141
а2
АаЬЬ
доминированием. И в том и в другом слу
чае наблюдается подавление одного ал
леля другим. Однако при эпистазе это ал
лели разных генов, а при доминирова
нии — аллели одного гена.
Полимерия. Когда один признак фор
мируется под влиянием сразу нескольких
генов с одинаковым фенотипическим вы
ражением, говорят о явлении полиме
рии, а сами гены называются полимер
236
ными. В этом случае принят принцип од
нозначного действия генов на развитие
признака. Например, при скрещивании
растений пастушьей сумки с треугольны
ми и с овальными плодами (стручочками)
в Fy образуются растения с плодами тре
угольной формы. При их самоопылении в
F2 наблюдается расщепление на растения
с треугольными и овальными стручочка
ми в соотношении 15:1. Это объясняется
CCii
CCII
Белая
P:
Белые
F,:
C -l-
c cl-
ccii
4---------v --------------------- '
С- i i
Окрашенные
Белые
3 /|6
щих однозначно. В этих случаях их обо
значают одинаково (Л[ и А2). Тогда все ге
нотипы (А г А2 А г а2а2, а {а ;А2-) будут
иметь одинаковый фенотип — треуголь
ные стручочки, и только растения
а^а{а2а2 будут отличаться — образовы
вать овальные стручочки. Это случай не
кумулятивной полимерии.
Полимерные гены могут действовать
и по типу кумулятивной полимерии.
Чем больше подобных генов в генотипе
организма, тем сильнее проявление дан
ного признака, т. е. с увеличением дозы
гена (Л[, А2, А3 и т. д.) его действие сум
мируется, или кумулируется. Например,
интенсивность окраски эндосперма зерен
пшеницы пропорциональна числу доми
237
нантных аллелей разных генов в тригибридном скрещивании. Наиболее окра
шенными были зерна А 1А 1А 2А2А3А3, а
зерна а 1а1а2а2а3а3 не имели пигмента.
По типу кумулятивной полимерии
наследуются такие количественные при
знаки, как молочность, яйценоскость,
масса и другие признаки сельскохозяй
ственных животных; многие важные па
раметры физической силы, здоровья и
умственных способностей человека; дли
на колоса у злаков, содержание сахара в
корнеплодах сахарной свеклы или липи
дов в семенах подсолнечника и т. д.
Таким образом, многочисленные на
блюдения свидетельствуют о том, что
проявление большей части признаков
представляет собой результат влияния
комплекса взаимодействующих генов и
условий внешней среды на формирование
каждого конкретного признака.
?
> 1. Какова биохимическая основа комплемен
тарное™? 2. Чем отличается явление доминирова
ния от эпистаза? 3. Как можно объяснить, что изу
чение явления полимерии особенно важно для по
нимания наследования количественных признаков?
4. При скрещивании между собой черных мышей
всегда получается черное потомство. В случае скре
щивания между собой желтых мышей одна треть
потомства оказывается черной, а две трети — жел
той. Как это можно объяснить? Как проверить пра
вильность предположения?
68. Хромосомная теория
наследственности.
Сцепленное наследование
В 1902— 1903 гг. американский цито
лог У. С е т т о н и немецкий цитолог и эм
бриолог Т. Б о в е р и независимо друг от
друга отметили параллелизм в поведении
генов и хромосом в ходе формирования
гамет и оплодотворения. Эти наблюдения
послужили основой для предположения,
что гены расположены в хромосомах. Од
нако экспериментальное доказательство
238
локализации конкретных генов в кон
кретных хромосомах было получено толь
ко в 1910 г. американским генетиком
Т. М о р г а н о м , который в последующие
годы (1911 — 1926/ обосновал хромо
сомную теорию наследственности. Со
гласно этой теории передача наслед
ственной информации связана с хро
мосомами, в которых линейно, в опре
деленной последовательности ло к а
лизованы гены. Таким образом, именно
хромосомы представляют собой матери
альную основу наследственности.
Разработка хромосомной теории и, в
частности, открытие эффекта положения
гена (т. е. зависимости проявления гена
от места расположения его в хромосоме)
позволили сформулировать один из важ
нейших принципов генетики о единстве
дискретности и непрерывности генети
ческого материала.
Группы сцепления. Независимое
комбинирование признаков (третий за
кон Менделя) осуществляется при усло
вии, что гены, определяющие эти призна
ки, находятся в разных парах гомологич
ных хромосом. Следовательно, у каждого
организма число генов, способных неза
висимо комбинироваться в мейозе, огра
ничено числом хромосом. Однако в орга
низме число генов, как правило, зна
чительно превышает количество хро
мосом.
Например, у кукурузы изучено более
500, у мухи дрозофилы — более 1000, а у
человека — около 2000 генов, тогда как
число хромосом у них 10, 4 и 23 пары со
ответственно. Это дает основание пред
положить, что в каждой хромосоме лока
лизовано множество генов. Гены, ло
кализованные в одной хромосоме, обра
зуют группу сцепления и наследуются
вместе.
Совместное наследование генов
Т. Морган предложил назвать сцеплен
ным наследованием. Число групп сцеп
ления соответствует гаплоидному набору
хромосом.
Сцепленное наследование и кроссинговер. Способ наследования сцеп
ленных генов отличается от насле
дования генов, локализованных в
разных парах гомологичных хро
мосом. Так,
если
при
незави
симом комбинировании дигибрид = =
образует четыре типа гамет (.А В, АЬ,аВ и
ab) в равных количествах, то такой же дигибрид
А
В
образует только два типа
гамет: АВ и аЬ тоже в равных количес
твах. Последние повторяют комбинацию
генов в хромосоме родителя.
Было установлено, однако, что, кроме
обычных гамет, возникают и другие —
АЬ и аВ — с новыми комбинациями ге
нов, отличающимися от родительской га
меты. Причиной возникновения новых
гамет является обмен участками гомо
логичных хромосом, или кроссинговер.
Кроссинговер происходит в профазе I
мейоза во время конъюгации гомологич
ных хромосом (рис. 143). В это время
части двух хромосом могут перекрещи
ваться и обмениваться своими участками.
В результате возникают качественно но
вые хромосомы, содержащие участки (ге
ны) как материнских, так и отцовских
хромосом. Особи, которые получаются из
таких гамет с новым сочетанием аллелей,
получили название кроссинговерных,
или рекомбинантных.
Рассмотрим один из первых экспери
ментов Т. Моргана по изучению сцеплен
ного наследования. При скрещивании
дрозофил, различающихся по двум парам
признаков — серых с зачаточными
I b* vg \
крыльями ~'7 7 " I и черных с нормальны/ b vg+\
ми крыльями!-
, — были получены
особи дигетерозиготные по этим генам.
Все они были нормальными по обоим
признакам в соответствии с правилом до
минирования и законом единообразия
гибридов первого поколения. Далее были
проведены два типа анализирующих
скрещиваний. В первом из них брали
самцов и скрещивали с гомозиготными
по рецессивным аллелям самками, а во
втором — дигетерозиготную самку скре
щивали с самцами, рецессивными по
обоим признакам (черное тело и зачаточ
ные крылья). Результаты этих скрещива
ний оказались разными (рис. 144, с. 240).
В п е р в о м случае были получены все
потомки только родительского типа, т. е.
серые с зачаточными крыльями, в отно
шении 1:1. Следовательно, данная диге
терозигота образует только два типа га-
Рис. 143. Схема перекреста гомологичных хромосом.
Рекомбинанты
239
?
240
мет (b+ vg и b vg+) вместо четырех, при
чем сочетание генов в гаметах самца бы
ло такое же, как у его родителей. Исходя
из указанного расщепления, следует
предположить, что у самца наблюдается
полное сцепление генов и не происходит
перемешивания (рекомбинации) призна
ков по причине отсутствия у него обмена
участками гомологичных хромосом.
Во в т о р о м случае в F2 наблюдалось
иное расщепление. Кроме родительских
комбинаций признаков, появились но
вые — мухи с черным телом и зачаточны
ми крыльями, а также с серым телом и
нормальными крыльями. Правда, коли
чество рекомбинантных потомков неве
лико и составляет 17 %, а родительс
ких — 83 %. Причиной появления не
большого количества мух с новыми соче
таниями признаков является кроссинговер, который приводит к новому реком
бинантному сочетанию аллелей генов b и
vg в гомологичных хромосомах. Эти об
мены происходят с вероятностью 17 % и
в итоге дают два класса рекомбинантов с
равной вероятностью — по 8,5 %.
Частота (процент) перекреста между
двумя генами, расположенными в одной
хромосоме, пропорциональна расстоя
нию между ними. Кроссинговер между
двумя генами происходит тем реже, чем
ближе друг к другу они расположены. По
мере увеличения расстояния между гена
ми все больше возрастает вероятность
того, что кроссинговер разведет их по
двум разным гомологичным хромосомам.
Расстояние между генами характери
зует силу сцепления и выражается в морганидах (в честь Т. Моргана) или в про
центах рекомбинации (кроссинговера).
Генетическое расстояние, на. котором
кроссинговер происходит с вероятностью
1 %, представляет собой сантиморган
^
(сМ). Имеются гены с высоким процен
том сцепления и такие, где сцепление
почти не обнаруживается. Однако при
сцепленном наследовании максимальная
величина кроссинговера не превышает
50 %. Если же она выше, то наблюдается
свободное комбинирование между пара
ми аллелей, неотличимое от независимо
го наследования.
Биологическое значение кроссинго
вера чрезвычайно велико, поскольку ге
нетическая рекомбинация позволяет соз
давать новые, ранее не существовавшие
комбинации генов и тем самым обеспе
чить повышение выживаемости организ
мов в процессе эволюции. Человек спе
циально проводит гибридизацию с целью
получения в результате комбинативной
изменчивости необходимых вариантов
комбинаций для использования в селек
ционной работе.
9а 1. Что такое группа сцепления и сколько групп
сцепления может быть у каждого из видов организ
мов? 2. Какие факты, полученные при изучении
сцепления и кроссинговера между генами, подтвер
ждают хромосомную теорию наследственности?
3. Объясните, почему на рисунке 144 вместо обыч
ного менделевского расщепления 1:1:1:1 появляет
ся отношение 41,5:41,5:8,5:8,5 %. 4. Какого потом
ства следует ожидать от скрещивания серых мух с
нормальными крыльями из Р2 (рис. 144) с черными
бескрылыми самцами из Fj?
69. Генетические карты
Сцепление генов, локализованных в
одной хромосоме, не бывает абсолют
ным. Кроссинговер, происходящий в про
цессе мейоза между гомологичными хро
мосомами, приводит к рекомбинации (пе
рераспределению) генов. Т. Морган и его
Рис. 144. Наследование сцепленных признаков у дрозофилы: а — полное сцепление (дигетерозиготен самец, у которого отсутствует кроссинговер); б — сцепление с кроссинговером (дигетерозиготна самка, у которой кроссинговер не подавлен);
b — серая и черная окраска тела; vg+, vg — длин
ные и зачаточные крылья соответственно.
241
сотрудники К - Б р и д ж е с , А. С т е р т е в а н г и Г. М е л л е р экспериментально
показали, что знание явлений сцепления
и кроссинговера позволяет не только
установить группу сцепления генов, но и
построить генетические карты хромосом,
на которых указаны порядок расположе
ния генов в хромосоме и относительные
расстояния между ними.
Генетической картой хромосом на
зывают схему взаимного расположения
генов, находящихся в одной группе
сцепления. Генетические карты состав
ляются для каждой пары гомологичных
хромосом.
Возможность такого картирования
основана на постоянстве процента крос
синговера между определенными генами.
Если известно взаимное расположение
генов на хромосоме (их порядок и рассто
яние между ними), то его можно изобра
зить в виде схемы (рис. 145).
Построение генетических карт. Пос
ле определения группы сцепления, в сос
тав которой входит ген, приступают к
следующему этапу анализа и устанавли
вают конкретное место гена в данной
группе сцепления. Локализация гена
устанавливается путем учета результатов
кроссинговера, т. е. учета данных о часто
тах рекомбинаций. При этом хромосом
ные карты строятся путем прямого пере
вода частоты рекомбинаций между гена
ми в предполагаемые расстояния на хро
мосоме. Относительное расстояние меж
ду двумя генами, лежащими в одной паре
хромосом, можно определить, проведя
большое число скрещиваний, сосчитав
число потомков с каждым фенотипом и
вычислив процент потомков с новыми
кроссинговерными комбинациями генов.
Если частота рекомбинации между гена
ми Л и В равна 3 %, то это означает, что
они расположены в одной и той же хро
мосоме на расстоянии трех морганиддруг
от друга; если частота рекомбинаций
между генами В и С равна 7 %, то они
разделены расстоянием в семь морганид.
242
Но по этим показателям еще нельзя ска
зать, где находится ген С, слева или спра
ва от гена Л, нельзя судить и о положении
гена Л по отношению к гену В:
С?
А
В
С
?
Решение этого вопроса возможно пу
тем определения частот рекомбинаций по
меньшей мере для трех генов одновре
менно. Именно такой метод позволяет
определить не только расстояние между
генами, но и их порядок расположения в
хромосоме. Если в указанном нами при
мере процент перекреста между третьей
парой генов Л и С составляет 10 %, то
ген В должен находиться обязательно
между генами Л и С:
А
в
с
✓ 0 Ч
е------- ----- 1 0 ------------------ >
На схеме цифры указывают процент
кроссинговера между генами.
Генетические и цитологические кар
ты. Для установления локализации гена
иногда привлекают и цитологические ме
тоды, которые позволяют получить дока
зательство расположения отдельных ге
нов в определенных участках хромосомы.
На рисунке 146 (с. 244) показан участок
политенной хромосомы личинки дрозо
филы, в которой в отдельных дисках, или
полосах, локализованы гены.
Политенные, или гигантские, хро
мосомы содержат во много раз больше
ДНК, чем обычные. Они не изменяют
своей формы на протяжении цикла
деления, достигая 0,5 мм в длину и
0,025 мм в толщину. Они встречаются,
например, в клетках слюнных желез
личинок насекомых (мух, комаров), в
ядрах инфузорий и в тканях завязи бобов.
Сравнение генетических карт, полу
ченных при изучении кроссинговера меж
ду генами, и цитологических карт, пред
ставляющих хромосому как физическое
тело, показывает соответствие положе-
4 0.0,----- 1spa
2.01-----1ey
ru
ve
jv
se
h
D
th
st
cu
Sb
ss
sr
Di
H
e
cd
ro
ca
Рис. 145. Генетические карты четырех хромо
сом дрозофилы с указанием ряда генов. 1 хро
мосома'. у — желтое тело, sc — бесщетинковые, w — белые глаза, fa — фасеточные глаза,
ес — увеличенные глаза, rb — рубиновые гла
за, cw — отсутствие поперечной жилки крыла,
ct — обрезанный край крыла, sn — опаленные
щетинки, 1— желтоватое тело и антенны, !z —
ромбовидные глаза, v — киноварный цвет
глаз, m — миниатюрные крылья, s — темное
тело, g — гранатовые глаза, г — рудиментар
ные крылья, f — вильчатые щетинки, В — по
лосковидные глаза, fu — слившиеся жилки,
саг — алые глаза, bb — подстриженные волос
ки; 2 хромосома: al — без усов, S — звездча
тые глаза, dp — короткие крылья, b — черное
тело, rd — короткие щетинки, рг — пурпурные
глаза, сп — киноварные глаза, vg — зачаточ
ные крылья, L — лопастные глаза, с — тонкие
растопыренные крылья, рх — добавочные пе
реплетенные жилки, bw — коричневые глаза,
sp — пятнистое тело; 3 хромосома: ru — гру
бые глаза, ve — укороченные продольные жил
ки, se — красно-коричневые глаза, h — волоса
тое тело, D — двущетинковые, th — нитевид
ные аристы, st — багряно-красные глаза, си —
загнутые крылья, Sb — остриженные щетинки,
ss — бесщетинковые, sr — полосатое тело,
DI — дельтовидные жилки, Н — бесщетинко
вые, е — черное тело, cd — ярко-красные гла
за, го — грубоглазые, са — рубиновые глаза;
4 хромосома: spa — грубые глаза, еу — безгла
зые.
243
mi
pd
mr Ins bs M33
Рис. 146. Генетическая (а) и цитологическая (б) карты гигантской (политенной) третьей хромосомы
личинки дрозофилы: рх — сплетение жилок, b w — коричневые глаза.
ния генов. Гены, удаленные друг от друга
на генетической карте, действительно
располагаются на хромосоме на значи
тельном расстоянии. Та же закономер
ность наблюдается и в отношении близко
расположенных генов. Таким образом,
генетические карты точно отражают по
рядок расположения генов в хромосоме.
Установленные связи определенных
генов с определенными физическими ло
кусами в хромосоме имеют исключитель
но важное значение. В этом случае при
помощи цитологического анализа хромо
сомная теория получила свое полное ис
черпывающее доказательство.
Генетические карты хромосом состав
лены для многих организмов: насекомых
(дрозофила, комар, таракан и др.), грибов
(дрожжи, аспергилл), для многих протистов, бактерий и вирусов.
Использование генетических карт.
Наличие генетической карты свидетель
ствует о высокой степени изученности то
го или иного вида организма и представля
ет большой научный интерес. Такой орга
низм является прекрасным объектом для
244
проведения дальнейших эксперименталь
ных работ, имеющих не только научное, но
и практическое значение. В частности,
знание генетических карт позволяет пла
нировать работы по получению организ
мов с определенными сочетаниями приз
наков, что теперь широко используется в
селекционной практике. Так, создание
штаммов микроорганизмов, способных
синтезировать необходимые для фармако
логии и сельского хозяйства белки, гормо
ны и другие сложные органические вещес
тва, возможно только на основе методов
генной инженерии, которые, в свою оче
редь, базируются на знании генетических
карт соответствующих микроорганизмов.
Генетические карты человека также
могут оказаться полезными в развитии
здравоохранения и медицины. Знания о
локализации гена на определенной хро
мосоме уже теперь используются при ди
агностике ряда тяжелых наследственных
заболеваний человека. Уже теперь по
явилась возможность для генной тера
пии, т. е. для исправления структуры или
функции генов.
Сравнение генетических карт хромо
сом разных видов живых организмов спо
собствует пониманию эволюционного
процесса.
Основные положения хромосомной
теории наследственности. Анализ явле
ний сцепленного наследования, кроссинговера, сравнение генетической и цито
логической карт позволяют сформулиро
вать основные положения хромосомной
теории наследственности:
1) гены локализованы в хромосомах.
При этом различные хромосомы
содержат неодинаковое число генов.
Кроме того, набор генов каждой из
негомологичных хромосом уникален;
2) аллельные гены занимают одина
ковые локусы в гомологичных хромо
сомах;
3) гены расположены в хромосоме в
определенной линейной последова
тельности;
4) гены одной хромосомы образуют
группу сцепления, благодаря чему
имеет место сцепленное наследова
ние некоторых признаков. При этом
сила сцепления находится в обратной
зависимости от расстояния между ге
нами;
5) каждый биологический вид харак
теризуется определенным набором
хромосом — кариотипом.
9m
1. Что такое генетическая карта и какова по
следовательность работы по ее составлению?
2. В чем сходство и отличие генетической и цитоло
гической карт? 3. Какие положения хромосомной
теории наследственности помогли обнаружить и
подтвердить генетические и цитологические карты?
4. Где и как используются генетические карты?
70. Генетика пола
У многих видов животных самым за
метным фенотипическим различием меж
ду особями является пол. Более того, это
одно из глубоких различий, поскольку
пол влияет на развитие и функциониро
вание многих органов, непосредственно с
половым размножением не связанных, а
половые гормоны накладывают отпеча
ток на экспрессию (проявление) многих
генов.
Пол, как и любой другой признак ор
ганизма, наследственно детерминирован.
Важнейшая роль в генетической детер
минации пола и в поддержании законо
мерного соотношения полов принадле
жит хромосомному аппарату.
Хромосомное определение пола.
Очень давно было замечено, что у раз
дельнополых организмов (животных и
двудомных растений) соотношение полов
обычно составляет 1:1, т. е. мужские и
женские особи встречаются одинаково
часто. Это соотношение совпадает с рас
щеплением в анализирующем скрещива
нии, когда одна из скрещиваемых форм
является гетерозиготной (Аа), а дру
гая — гомозиготной по рецессивным ал
лелям (аа). В потомстве в этом случае
наблюдается расщепление в отношении
\Аа\ 1аа. Если пол наследуется по такому
же принципу, то впол'не логично было бы
предположить, что один пол должен быть
гомозиготным, а другой — гетерозигот
ным. Тогда расщепление по полу должно
быть в каждом поколении равным 1:1,
что и наблюдается в действительности.
При изучении хромосом самцов и са
мок ряда животных между ними были об
наружены некоторые различия. Как у
мужских, так и у женских особей во всех
клетках имеются пары одинаковых (гомо
логичных) хромосом, но по одной паре
хромосом они различаются. Так, у самки
дрозофилы имеются две палочковидные
хромосомы, а у самца — одна такая же
палочковидная, а вторая, парная первой,
изогнутая. Такие хромосомы, по которым
самцы и самки отличаются друг от друга,
называют половыми хромосомами. Те из
них, которые являются парными у одного
из полов, называют Х-хромосомами. Не-'
парная половая хромосома, имеющаяся у
особей только одного пола, была названа
245
У-хромосомой. Хромосомы, в отношении
которых между самцами и самками нет
различий, называют аутосомами. Сле
довательно, у дрозофилы особи обоих по
лов имеют по 6 одинаковых аутосом плюс
половые хромосомы XX у самок и ХУ у
самцов.
Пол, содержащий различные половые
хромосомы (X и У), образует гаметы двух
типов (половина с Х-хромосомой и поло
вина с У-хромосомой) и называется гетерогаметным, а пол, содержащий в
канедой клетке одинаковые половые хро
мосомы (Х-хромосомы), гомогаметным.
От чего же зависит рождение муж
ских и женских особей? Рассмотрим это
на примере определения пола у дрозофи
лы (рис. 147). В ходе мейоза у самок об-
Рис. 147 Определение пола у дрозофилы: хро
мосомные наборы соматических клеток (1),
сперматозоидов (2), яйцеклеток (3), потом
ства (4).
246
разуется один тип гамет, содержащий
гаплоидный набор аутосом и однуХ-хромосому. Самцы образуют два типа гамет,
половина из которых содержит три ауто
сомы и одну Х-хромосому (ЗЛ+Х), а по
ловина — три аутосомы и одну У-хромосому (3A+Y). При оплодотворении яй
цеклеток (ЗЛ+Х) сперматозоидами с
X-хромосомами будут формироваться
самки (6Л+ХХ), а от слияния яйцеклеток
со сперматозоидами, несущими У-хромосому, будут развиваться самцы (6Л+ХУ).
Поскольку число мужских гамет с X- и Ухромосомами одинаково, то и количество
самцов и самок тоже одинаково. В дан
ном случае пол организма определяется в
момент оплодотворения и зависит от хро
мосомного набора зиготы.
Сходный способ определения пола
(ХУ-тип) присущ всем млекопитающим, в
том числе и человеку, клетки которого со
держат 44 аутосомы и две Х-хромосомы у
женщин либо ХУ-хромосомы у мужчин.
Соответственно у мужчин образуются
сперматозоиды двух типов — с X- и Ухромосомами. Правда, в отношении ме
ханизмов определения пола человек от
личается от дрозофилы тем, что для фор
мирования нормального мужского фено
типа необходимо присутствие У-хромосомы.
Установлено, что у человека У-хромосома контролирует дифференцировку се
менников, которая в дальнейшем влияет
на развитие половых органов и мужских
признаков. Однако у большинства орга
низмов У-хромосома не содержит генов,
имеющих отношение к полу. Ее даже на
зывают генетически инертной или гене
тически пустой, поскольку в ней очень
мало генов. Предполагают, что гены, оп
ределяющие мужские признаки дрозофи
лы, локализованы в аутосомах и их фено
типические эффекты маскируются нали
чием пары Х-хромосом. Об этом свиде
тельствует тот факт, что у дрозофилы пол
определяется отношением числа Х-хро
мосом к числу аутосом: при Х/А = 1 , 0
развиваются самки; если Х/А = 0,5, раз
виваются самцы. Другими словами, в
присутствии только одной X-хромосомы
мужские признаки у мух проявляются.
Это пример наследования, ограниченного
полом (в отличие от наследования, сцеп
ленного с полом), при котором, напри
мер, у женщин подавляются гены, детер
минирующие рост бороды.
Таким образом, ХУ-тип определения
пола, или тип дрозофилы и человека, —
самый распространенный способ опреде
ления пола, характерный для большин
ства позвоночных и некоторых беспозво
ночных (рис. 148, с. 248). Х0-тип встре
чается у большинства прямокрылых, кло
пов, жуков, пауков, у которых У-хромосо
мы нет вовсе, так что самец имеет гено
тип Х0, а самки XX.
У птиц, бабочек и пресмыкающихся
самцы являются гомогаметным полом, а
самки — гетерогаметным (типа XY или
ХО). Половые хромосомы у этих видов
иногда обозначают буквами Z и W, выде
ляя таким образом данный способ опре
деления пола, при этом самцы обозна
чаются символом ZZ, а самки — ZW,
или Z0.
Другие варианты определения пола.
Совершенно другой механизм определе
ния пола известен у пчел и муравьев. У
этих организмов нет половых хромосом:
самки — это диплоидные особи, а самцы
(трутни) — гаплоидные. Самки развива
ются из оплодотворенных яиц, а трут
ни — из неоплодотворенных. В процессе
сперматогенеза у трутней не происходит
редукции числа хромосом.
Из оплодотворенной яйцеклетки мо
жет развиваться либо «матка» — круп
ная, способная к размножению самка,
либо стерильная рабочая пчела. Это за
висит от условий выкармливания личин
ки рабочими особями.
Наконец, у некоторых животных оп
ределение пола зависит от внешних усло
вий. У морского червя Bonellia самцы и
самки имеют одинаковый генотип. Осо
би, которые в личиночной стадии остают
ся свободноплавающими, становятся
самками, а личинки, прикрепившиеся к
телу взрослой самки, развиваются в мик
роскопических самцов. Самцы мигриру
ют в половые пути самки, где они жи
вут в качестве паразитов, выполняя
главную функцию — оплодотворение
яйцеклеток.
■ 1. Почему рождается примерно одинаковое ко
личество особей мужского и женского пола? 2. Ка
кие существуют типы определения пола? 3. Как ге
нетически определяется пол у человека? 4. Объяс
ните цитологический механизм расщепления по по
лу. 5. Какой пол будет у дрозофил, имеющих следу
ющие наборы хромосом: ЗА+ЗХ, 2А+ЗХ, ЗА+2Х,
2А+2Х, 2A+XXY, 2А+Х0? Выводы объясните.
71. Наследование признаков,
сцепленных с полом
Признаки, сцепленные с полом.
В том случае, когда гены, контролирую
щие формирование того или иного приз
нака, локализованы в аутосомах, насле
дование осуществляется независимо от
того, кто из родителей (мать или отец) яв
ляется носителем изучаемого признака.
Если же гены находятся в половых хро
мосомах, характер наследования призна
ков резко изменяется. Например, у дро
зофилы У-хромосома, в отличие отХ-хромосомы, не содержит генов, т. е. генети
чески пуста. Поэтому гены, локализован
ные в Х-хромосоме, как правило,
не имеют аллелей в У-хромосоме.
По этой причине рецессивные гены в
Х-хромосоме гетерозиготного пола мо
гут проявляться будучи в единственном
числе.
Признаки, гены которых локализо
ваны в половых хромосомах, называ
ются признаками, сцепленными с по
лом. Это явление было открыто Т. Мор
ганом у дрозофилы.
Наследование, сцепленное с полом.
247
Рис. 148. Основные типы определения пола: 1 — человек (ХУ), 2 — кузнечик (ХО), 3 — курица (ZW),
4 — пчела (самец гаплоид).
248
Т. Морган и его сотрудники заметили, что
наследование окраски глаз у дрозофилы
зависит от пола родительских особей, не
сущих альтернативные аллели.
При скрещивании красноглазых са
мок и белоглазых самцов в F{ все мухи
имели красные глаза (рис. 149, а), что
соответствует предположению о доми
нантности этого признака. В f 2 происхо
дит расщепление в отношении 3:1, т. е.
три четверти потомства были красногла
зыми, а одна четверть — белоглазыми.
Но в F2 все самки были красноглазыми,
среди самцов половина имели красные
глаза, а половина — белые. Это не сов
падало с менделевскими законами нас
ледственности.
При скрещивании белоглазых самок с
красноглазыми самцами результаты бы
ли другими (рис. 149, б). Уже в F\ наблю
далось расщепление по окраске глаз 1:1.
При этом белоглазыми оказались только
самцы, а все самки — красноглазыми,
т. е. дочери наследуют красную окраску
глаз от отцов, а сыновья — белый цвет
глаз от матерей. Такой тип передачи
признаков от матерей сыновьям, а от от
цов дочерям получил название наследо
вания крест -накрест , или крисскросс. В F2 этого скрещивания потомство
состояло наполовину из белоглазых мух и
наполовину из красноглазых, причем в
равном отношении у обоих полов.
Морган показал, что все эти резуль
таты можно объяснить, предположив,
что только Х-хромосома несет ген окрас
ки глаз, а в К-хромосоме соответствую
щего локуса вообще нет. Следовательно,
белый цвет глаз у дрозофилы — это
признак сцепления с полом. Обнаружен
ное в этих опытах соответствие между по
ведением конкретного гена и конкретной
хромосомы послужило веским доказа
тельством справедливости хромосомной
теории наследственности.
Признаки, сцепленные с полом, из-
Рис. 149. Наследование, сцепленное с полом, у дрозофилы: а — скрещивание красноглазых самок с
белоглазыми самцами, б — белоглазых самок с красноглазыми самцами.
249
вестны и у человека. Например, развитие
первичных{половые железы и наружные
половые органы) и вторичных половых
признаков мужского пола определяют ге
ны, локализованные в У-хромосоме. При
наличии в клетках только одной половой
хромосомы (44+Х) развивается особь,
которая называется интерсекс (от лат.
inter — между, sexus — пол) — сущес
тво с промежуточным полом (синдром
Шерешевского — Тернера). Человек,
имеющий две и более Х-хромосомы, при
наличии У-хромосомы (44+XXY) будет
человеком мужского пола (синдром
Клайнфелътера).
X- и У-хромосомы имеют общие гомо
логичные участки (рис. 150). В этих учас
тках локализованы гены, определяющие
признаки, которые наследуются одинако
во как у мужчин, так и у женщин (по
добно признакам, сцепленным с аутосомами).
Помимо гомологичных участков, X- и
У-хромосомы имеют негомологичные
участки. Негомологичный участок У-хро
мосомы, кроме генов, определяющих
мужской пол, содержит гены перепонок
между пальцами ног и волосатых ушей у
Рис. 150. Гомологичные участки в X- и У-хромосомах человека: а — Х-хромосома, б — У-хромосома: 1 — негомологичные участки, 2 — цен
тромера, 3 — гомологичные участки.
250
человека. Патологические признаки,
сцепленные с негомологичным участком
У-хромосомы, передаются всем сыновь
ям, поскольку они получают от отца
У-хромосому.
Негомологичный участок Х-хромосомы содержит в своем составе ряд важных
для жизнедеятельности организмов
генов. Поскольку у гетерогаметного
пола (ХУ) Х-хромосома представлена в
единственном числе, то признаки,
определяемые генами негомологичного
участка Х-хромосомы, будут проявляться
даже в том случае, если они рецессивны.
Такое состояние генов называется
гемизиготным. Примером такого рода
наследования у человека являются гемо
филия, дистрофия Дюшена, атрофия зри
тельного нерва, несахарный диабет, даль
тонизм, облысение.
Гемофилия — это наследственная бо
лезнь, при которой кровь теряет способ
ность свертываться. Ранение, даже цара
пина или ушиб, могут вызвать обильные
наружные или внутренние кровотечения,
которые нередко заканчиваются смер
тью. Поэтому больных гемофилией сле
дует тщательно оберегать от всякого рода
травм. В некоторых странах для таких де
тей созданы специальные школы. Это за
болевание встречается, за редким исклю
чением, только у мужчин. Было установ
лено, что гемофилия обусловлена рецес
сивным геном, локализованным в Х-хромосоме, поэтому гетерозиготные по дан
ному гену женщины обладают нормаль
ной свертываемостью крови. Рассмот
рим, какое потомство может появиться у
женщины, вступающей в брак с нормаль
ным по этому признаку мужчиной. Ген,
определяющий нормальную свертывае
мость крови,обозначим Я, а ген,который
не определяет ее свертываемость, — h.
Учитывая, что в генотипе женщин име
ются две Х-хромосомы, а у мужчин — од
на Х-хромосома и одна У-хромосома,
запишем схему наследования гемо
филии:
Х"ХЬ
Р:
X
носительница
гена
гемофилии
Гаметы:
© ©
@>©
ХНХ» X"Xh
Д:
XHY
здоровый
мужчина
XPYXW
Как видно из схемы, потомки данного
брака проявляют расщепление признака:
половина дочерей (XHXh) являются носи
тельницами гена гемофилии, а половина
сыновей (XhY) — гемофиликами; вторая
половина — дочери (ХНХН) и сыновья
(.XHY) — окажутся здоровыми.
Таким образом, гемофилия, передава
емая через женщин, проявляется у поло
вины их сыновей. Известным примером
такого наследования является генеалогия
нескольких королевских семей в Европе,
которые восходят к английской королеве
Виктории. Предполагают, что ген гемо
филии возник в результате мутации у са
мой королевы Виктории или у одного из
ее родителей.
Фенотипическое проявление гемофи
лии у девочек наблюдается в том случае,
если мать девочки является носительни
цей гена гемофилии, а отец — гемофили
ком:
Р:
Xй Xh
Гаметы:
@
Fp.
х
X^Y
®
XHXh XhXh
XHY XhY
носительница гемофилик здоровый гемофилик
гена
девочка
мальчик мальчик
гемофилии
Ген, вызывающий дальтонизм, так
же сцеплен сХ-хромосомой. Что же каса
ется гена облысения, то он локализован в
аутосоме, но его экспрессия зависит от
мужских половых гормонов. У мужчин
этот ген доминирует из-за присутствия
мужских половых гормонов; у женщин
же он ведет себя как рецессивный
ген, так что женщина лысеет только в
том случае, если у нее имеются две дозы
этого гена.
« 1. Какие признаки считаются сцепленными с
полом? 2. Каковы особенности наследования приз
наков, сцепленных с полом? 3. Какой пол будет у
дрозофилы и человека с набором половых хромосом
XXY? 4. Дочь дальтоника вышла замуж за сына дру
гого дальтоника, у которого цветное зрение нор
мальное. Каковы генотипы мужа и жены? Какой
фенотип будет у их детей? 5. Мужчина-дальтоник
женился на женщине — носителе цветной слепоты.
Можно ли ожидать в этом браке здорового сына?
Дочь с цветной слепотой? Какова вероятность од
ного и другого события? 6. Могут ли нормальные по
гемофилии мужчина и женщина иметь гемофиликасына?
72, Цитоплазматическая наслед
ственность
Ранее приведенные данные свиде
тельствуют о том, что ведущая роль в ге
нетических процессах принадлежит ядру
и хромосоме. Вместе с тем носителями
наследственной информации могут быть
и некоторые органеллы цитоплазмы, в
которых содержится собственная ДНК.
В эукариотических клетках внехромосомная ДНК содержится в хлоропластах и
митохондриях, а также представлена ви
русоподобными частицами. Молекулы
ДНК этих органелл несут информацию о
собственных белках, а также об иРНК и
тРНК, участвующих в их синтезе. Такая
информация передается с цитоплазмой,
поэтому она получила название цит оп
лазматической (внеядерной, внехромосомной) наследственности. Причем
эта информация передается только через
материнский организм, в связи с чем на
зывается материнской наследствен
ностью. Это обусловлено тем, что и у
растений, и у животных яйцеклетка со
держит много цитоплазмы, а сперматозо
251
ид (спермий) ее почти лишен (у некото
рых организмов содержит только ядро).
Плазмиды. Факторы наследствен
ности, расположенные в клетке вне
хромосом, получили название плаз
мид. Они представляют собой коль
цевые двухцепочные молекулы ДНК
с молекулярной массой 106— 108 дальтон.
Плазмиды способны существовать
и размножаться в клетке автономно
от хромосомной ДНК. Однако есть и
такие плазмиды, которые включают
ся в определенные участки геномной
ДНК и реплицируются вместе с ней.
Такие плазмиды называются эписомами.
В клетках прокариот обнаружены
плазмиды, наследственный материал ко
торых определяет способность бактерий
к конъюгации (половой фактор F), а так
же их устойчивость к некоторым лекар
ственным веществам (фактор R). Кроме
того, плазмиды-колициногены (Со1-плазмиды) контролируют синтез особых бел
ков колицинов, уничтожающих бактерий
того же вида, но не обладающих этими
плазмидами.
Пластидное (хлоропластное) насле
дование. Пластиды содержат ДНК, вос
производятся делением и распределяют
ся в дочерние клетки. Совокупность
пластид клетки как структур, передаю
щих наследственную информацию, бы
ла названа пластидомом, а сово
купность генов, расположенных в коль
цевых молекулах ДНК пластид, — пластомом.
Пластидное наследование обнаруже
но у ряда растений: львиного зева, ночной
красавицы, кипрея и др. У них наряду с
формами, имеющими зеленые листья, су
ществуют пестролистные формы, у кото
рых в точках роста были обнаружены
разные группы клеток: с нормальными
пластидами и с пластидами, неспособны
ми к синтезу хлорофилла. Вследствие
этого пестролистные формы образуют
252
иногда чисто зеленые или совершенно
белые побеги.
При опылении цветков бесхлорофилльного побега пыльцой зеленого расте
ния в Fj появляются только бесхлорофилльные растения, которые вскоре по
гибают, так как не способны к фотосин
тезу. При реципрокном скрещивании в F{
все растения оказываются зелены
ми. При опылении цветков пестро
листного побега пыльцой зеленой фор
мы в F{ образуются белые, пестролис
тные и зеленые растения. При реципрок
ном скрещивании — вновь только зе
леные.
На основании этих наблюдений был
сделан вывод, что наследование пестро
листное™ связано с передачей при кле
точных делениях двух типов пластид —
зеленых (хлоропластов) и бесцветных
(лейкопластов). Пластиды воспроизво
дятся в клетке автономно и случайно рас
пределяются между дочерними клетками.
Клетки, содержащие смесь зеленых и
бесцветных пластид, при делении дают
клетки всех трех типов, что приводит к
образованию зеленых, пестрых и белых
участков растения. Поскольку единствен
ный способ проникновения пластид
в зиготу — через яйцеклетку, то насле
дование осуществляется по материн
ской линии — материнское наследо
вание.
Митохондриальное наследование. У некото
рых грибов (дрожжи, нейроспора) была обнаруже
на дыхательная недостаточность, которая обуслов
лена необратимыми наследственными изменениями
функций митохондрий — органелл, обеспечиваю
щих дыхание клеток подавляющего большинства
организмов.
У дрожжей известны мутантные формы, обра
зующие на глюкозе мелкие колонии с дыхательной
недостаточностью. Поскольку колонии возникают
при вегетативном размножении гаплоидных дрож
жей, они получили название «вегетативные карли
ковые». Другая форма, сходная с первой по феноти
пу — росту и дыхательной недостаточности, — бы
ла названа генеративным карликовым штаммом,
поскольку он давал расщепление по признаку кар
ликовости.
Генетический анализ вегетативного и генера
тивного карликовых штаммов показал, что фенотип
генеративной карликовости определяется ядерным
геном, поскольку при скрещивании наблюдается
расщепление спор дрожжей в отношении 1:1. При
скрещивании вегетативных карликов и нормальных
дрожжей диплоидная зигота, в которой есть мито
хондрии от нормальной формы, не дает расщепле
ния — из спор мелкие колонии не формируются.
Следовательно, у этих форм генотипы одинаковы,
различалась лишь цитоплазма. Расщепления по ти
пу цитоплазмы в мейозе не происходит. Значит, по
лученные данные свидетельствуют о цитоплазмати
ческом наследовании через митохондрии.
Влияние ядерных генов через ци
топлазму яйцеклетки. Самым наглядным
примером предопределения генами мате
ри свойств цитоплазмы яйцеклетки явля
ется наследование направления завитка
раковины у пресноводной улитки пру
довика. Существуют прудовики с пра
возакрученной (D ) и левозакрученной
(d) раковинами. Реципрокные скрещива
ния:
J D D x с/ dd и % ddx o ' DD
дают в F{ различный результат: гибриды
наследуют признак матери (рис. 151,
с. 254). При первом скрещивании они все
правозакрученные, а во втором — лево
закрученные.
Благодаря тому что прудовики гер
мафродиты, у них возможно самоопло
дотворение. От самооплодотворения ге
терозиготных особей из С/ все потомки F%
от обоих скрещиваний имеют правоза
крученную раковину. И только в F3, полу
ченном также путем самооплодотворе
ния, наблюдается менделевское расщеп
ление — 3 правозакрученных : 1 лево
закрученная. Благодаря материнскому
эффекту проявление признаков запазды
вает на одно поколение, и расщепление
3:1 наступает не во втором, а в третьем
поколении.
На рисунке 151 видно, что в F2 име
ются особи с генотипом dd, однако они
правозакрученные, и лишь в следующем
поколении они дают чисто левозакручен
ное потомство. На данном примере мож
но видеть, что в определенных случаях
фенотип потомков не соответствует их
генотипу, а определяется материнским
генотипом через цитоплазму яйцеклетки,
от которой и зависит направление закру
чивания раковины.
Этот признак формируется в раннем
эмбриогенезе направлением веретена
второго дробления оплодотворенного яй
ца. Положение веретена деления опреде
ляется материнским организмом перед
созреванием яйца. Такое явление получи
ло название предетерминации цитоп
лазмы генотипом матери, или мате
ринского эффекта.
Непосредственное влияние материн
ского организма на развитие зародыша
часто приводит к большему сходству по
томства с матерью, поскольку условия
эмбрионального развития организма
полностью зависят от матери. Влияние
самки особенно четко сказывается в реципрокных скрещиваниях при получении
межвидовых гибридов. Так, например,
мул — гибрид кобылы и осла — сильно
отличается от лошака — гибрида ослицы
и жеребца. Мул крупнее и больше напо
минает лошадь, а лошак — небольшой и
внешне больше похож на ослицу.
Цитоплазматическая мужская сте
рильность. Примером наличия цитоплаз
матической наследственности служит
также явление цитоплазматической
мужской стерильности (ЦМС). Оно
было обнаружено в 30-е годы одновре
менно в СССР М . И. Х а д ж и н о в ы м
и в США М. Р о д с о м у кукурузы, свек
лы, льна и др.
Кукуруза — однодомное растение,
мужские цветки которого собраны на
верхушке стебля в метелку, а женские —
в початок. У некоторых сортов кукурузы
были обнаружены растения, имевшие в
метелках недоразвитые пыльники со сте253
р
F,
F,
F3
Рис. 151. Наследование направления закручивания раковины у прудовика: D — фенотип правозавитковости, d — фенотип левозавитковости.
рильной пыльцой. Опыление растений с
ЦМС нормальной пыльцой, взятой от
других растений, давало в потомстве фор
мы также со стерильной пыльцой. Зна
чит, признак мужской стерильности пе
редавался по материнской линии.
254
Даже в том случае, когда все 10 пар
хромосом растения со стерильной пыль
цой замещали хромосомами от растения с
нормальной пыльцой, мужская стериль
ность сохранялась (рис. 152). Это послу
жило убедительным доказательством то-
73. Современные представления
о гене
Ген, или наследственный фактор, —
это участок молекулы ДНК (у некоторых
вирусов РНК), кодирующий первичную
структуру полипептида, молекулы тран
спортной или рибосомной РНК или
взаимодействующий с регуляторным
белком.
Вз
I
В4
I
I
I
П о сте п е н н о е зам ещ ение
геном а сте р и л ь н о й линии
геном ом линии с ф ер тил ьно й
пыльцой
Вп
Рис. 152. Наследование мужской
стерильности у кукурузы по мате
ринской линии: а, б — растения с
мужской стерильностью; в, г —
растения с фертильной пыльцой:
1 — цитоплазма, 2 — ядро, 3 —
образуются редко.
го, что наследование данного признака
осуществляется через цитоплазму и что к
этому процессу хромосомы не имеют ни
какого отношения.
ш. 1. Что такое плазмиды? 2. Каковы основные
отличия цитоплазматической наследственности от
хромосомной? 3. Чем обусловлена преимуществен
ная роль женских организмов при цитоплазмати
ческой наследственности? 4. В чем суть явления ци
топлазматической мужской стерильности у расте
ний?
Краткая история открытия гена. Предположе
ние о существовании дискретных наследственных
факторов в половых клетках впервые было выска
зано Г. Менделем в 1865 г., а в 1909 г. В. Иогансен
назвал их генами. Дальнейшие представления о ге
не связаны с развитием хромосомной теории на
следственности.
Первые эксперименты, доказавшие сложное
строение гена у дрозофилы, были выполнены в
20— 30-х гг. XX в. советскими учеными А . С. С е р е б р о в с к и м , Н. П. Д у б и н и н ы м и др.
В 1953 г. Д ж . Уо т с о н о м и Ф . К р и к о м была
раскрыта структура ДНК- Это открытие позволи
ло говорить о том, каким образом особенности
данной структуры определяют функции ДНК в ка
честве материального носителя наследственной ин
формации.
В 60-х гг. американский ученый С. Б е н з е р
доказал, что ген одного из бактериофагов кишечной
палочки состоит из линейно расположенных, неза
висимо мутирующих (изменяющихся) элементов.
Чуть позже он показал, что наименьшими мутирую
щими элементами гена являются отдельные пары
нуклеотидов ДНК.
К концу 60-х гг. благодаря работам Ф. Крика,
С. О ч о а , М. Н и р е н б е р г а и др. утвердилось
представление об универсальности основных черт
строения и функции гена как сложной линейной
структуры участка ДНК, который в результате тран
скрипции и последующей трансляции определяет
первичную структуру белков.
Дальнейшее развитие представлений б гене
связано с выявлением отличий в организации гене
тического материала у организмов разных си
стематических групп (начиная от вирусов и
прокариот и кончая высшими растениями и жи
вотными).
255
Организация генома. Молекула
ДНК представляет собой чрезвычайно
длинный неразветвленный биополимер, в
состав которого входят миллионы и даже
миллиарды нуклеотидов, расположенных
в определенной последовательности. Эта
последовательность нуклеотидов в цепи
ДНК является кодовой формой записи
наследственной информации, или гене
тическим кодом.
Каждая молекула ДНК, состоящая из
множества генов, входит в состав отдель
ной хромосомы. Совокупность генов, ха
рактерных для гаплоидного набора хро
мосом данного вида организмов, назы
вается геномом. В отличие от генотипа
геном представляет собой характеристи
ку вида, а не определенной особи.
Различные виды организмов резко от
личаются по количеству и составу нукле
отидов ДНК, составляющей их геномы. У
вирусов размер генома колеблется от
нескольких сотен до нескольких тысяч
пар нуклеотидов. Гены в таких геномах
расположены один за другим и занимают
практически всю длину ДНК или РНК. У
бактерий геном содержит несколько мил
лионов нуклеотидных пар, составляющих
единственную молекулу ДНК (одна хро
мосома). При этом более половины пар
нуклеотидов представлены структурными
генами. Остальную часть бактериальной
хромосомы составляют неспособные
транскрибироваться нуклеотидные пос
ледовательности.
Геном высших эукариот заметно пре
вышает по размерам геном прокариот.
Например, геном человека представлен
примерно 3,6x109 парами нуклеотидов,
распределенных в 46 хромосомах. Другие
млекопитающие имеют геном примерно
такого же размера. В каждой отдельной
хромосоме человека содержится от 50 до
250 млн нуклеотидных пар. Однако сле
дует отметить, что количество структур
ных генов, контролирующих синтез жиз
ненно важных белков, у млекопитающих
возрастает не очень сильно в сравнении с
256
прокариотами. Количество ДНК в геноме
человека достаточно для кодирования
почти 3 млн видов белков средних разме
ров. Реально имеющееся число таких ге
нов оценивается как 50—-60 тыс.
Главной функцией генома является
образование молекул РНК- Определен
ные участки нуклеотидных последова
тельностей ДНК копируются с образова
нием соответствующих молекул РНК.
Одни из них кодируют белки (иРНК),
другие являются структурными (тРНК и
рРНК).
Структура гена. Общее число генов в
клетке высших организмов по разным
оценкам составляет 50—200 тыс. Каж
дому структурному гену соответствует
белок. Гены, кодирующие синтез белка, у
эукариот состоят из нескольких постоян
ных элементов (рис. 153, а).
Прежде всего это регуляторная об
ласть, которая оказывает заметное влия
ние на активность гена в разные периоды
жизни организма. Регуляторная область
включает промотор — последователь
ность ДНК длиной до 80— 100 пар нукле
отидов, которую узнает молекула фер
мента РНК-полимеразы и соединяется с
ней, чтобы начать транскрипцию данного
гена. При этом промотор определяет, ка
кая из цепей двойной спирали ДНК при
соединит к себе РНК-полимеразу и тем
самым будет служить матрицей для син
теза иРНК.
За промотором расположена струк
турная часть гена, которая содержит в се
бе информацию о первичной структуре
соответствующего белка.
У прокариот за промотором располо
жен оператор — участок ДНК, «узна
ваемый» специфическими белками-реп
рессорами и регулирующий транскрип
цию отдельных генов. Оператор и управ
ляемые им структурные гены в совокуп
ности образуют функциональную едини
цу, называемую опероном (рис. 153, б).
Кроме того, в хромосомах были най
дены участки — усилители транскрип-
Р егул яторн а я
обл асть
_А_
днк
С т р у к т у р н ы й ген
________ А ________
ЛГ
П ро м отор
Э кзо н
Э кзо н
И нтрон
И нтрон
Э кзо н
__________
Р Н К -п о л и м е р а за
Р егул яторн а я
обл асть
_________/V _______
ДНК
П ро м отор
а
Терм и
н ато р
С т р у к т у р н ы е гены
_________ А _________
О ператор
Р Н К -п о л и м е р а за
Терм и
н а то р
Рис. 153. Схема строения гена и его регуляторных элементов у эукариот (а) и оперона у прокари
от (б).
ции, расположенные на большом рассто
янии от кодирующей последовательности
гена и его промотора. Для нормальной
работы гена очень важны также участки,
расположенные в конце кодирующей
последовательности и служащие сигна
лом остановки транскрипции. Их называ
ют терминаторами. В некоторых генах
терминатор можно обнаружить и внутри
кодирующей последовательности. Такой
ген может полностью транскрибировать
ся только в присутствии так называемого
белка-антит ерминат ора, позволяю
щего РНК-полимеразе «проскочить»
внутренний терминатор.
Важная особенность генов эукарио
тических организмов — их прерывис
тость. Это значит, что если у прокариот в
молекуле иРНК представлена вся нукле
отидная последовательность структурно
го гена, то у эукариот кодирующие учас
тки гена — экзоны — чередуются с не
кодирующими вставками — нитронами
(см. рис. 153).
Какова же причина такого явления?
Дело в том, что гены, входящие в состав
9. Общая биология, 10 кл.
хромосом эукариот, содержат десятки и
даже сотни тысяч нуклеотидных пар.
Вместе с тем для кодирования белка
среднего размера (содержащего от 300 до
400 аминокислотных остатков) требуется
всего лишь 900— 1200 нуклеотидных
пар, которые в совокупности и представ
ляют собой экзон. По размерам интроны
варьируют от нескольких единиц до нес
кольких тысяч нуклеотидных пар; при
этом их длина может быть больше длины
экзонов, а число интронов в составе гена
может достигать двух-трех десятков.
Молекула мРНК, синтезированная с
такого гена (содержащего не только эк
зоны, но и интроны), называется первич
ным транскриптом. Для того чтобы
первичный транскрипт превратился в
зрелую мРНК, требуется его предвари
тельное созревание — процессинг (рис.
154, с. 258). Для этого необходимо уда
лить (вырезать) некодирующие последо
вательности, или интроны, а кодирующие
участки (экзоны) сшить (срастить) в еди
ную молекулу. Этот процесс получил на
звание сплайсинга РНК■
257
Структурный ген
Ядро
ДНК
Э кзон
И н тр о н
ция ДНК
Э к з о н Интро$£-Э{<зон{
J
РНК- полимераза
Интрон
Первичный
Интрон
~~А А л—
транскрипт РНК
4 Сплайсинг РНК \ л / ч А
м РНК
Интроны
4 + Транспортные
белки
Мембраны
ядра
Цитоплазма
мРНК
Я д е р н а я пора
Рис. 154. Схема процессинга мРНК в ядре.
Экзоны ковалентно соединяются в
молекулу транслируемой иРНК в том же
порядке, в котором они располагаются в
ДНК.
Структура гена вирусов отражает ге
нетическую организацию клетки хозяина.
Так, гены бактериофагов собраны в опероны и не имеют нитронов, а вирусы
эукариот их имеют. В то же время в гене
тическом материале вирусов прокариот и
эукариот обнаружена общая характерная
черта — перекрывание генов. Напри
мер, в геноме фага ФХ174 ген В распо
ложен внутри гена А, а ген Е — внутри
гена D.
Свойства генов. Характер проявле
ния действия гена может изменяться в
различных ситуациях и под влиянием раз
личных факторов. В этом можно легко
убедиться, если рассмотреть свойства ге
нов и особенности их проявления в при
знаках:
258
1) ген дискретен в своем действии,
т, е. обособлен в своей активности от
других генов;
2) ген специфичен в своем проявле
нии, т. е. отвечает за строго определен
ный признак или свойство организма;
3) ген может действовать градуально,
т. е. усиливать степень проявления
признака при увеличении числа доми
нантных аллелей (дозы гена);
4) один ген может влиять на развитие
разных признаков — это множес
твенное, или плейотронное, действие
гена;
5) разные гены могут оказывать оди
наковое действие на развитие одного
и того же признака (часто количес
твенных признаков) — это множес
твенные гены, или полигены;
6) ген может взаимодействовать с
другими генами, что приводит к появ
лению новых признаков. Такое взаи-
модействие осуществляется опосре
дованно — через синтезированные
под их контролем продукты своих ре
акций;
7) действие гена может быть модифи
цировано изменением его местона
хождения в хромосоме (эффект поло
жения) или взаимодействием различ
ных факторов.
?
■ 1. Дайте определения понятиям: г е н , а л л е л ь ,
2. Что такое геном и каковы особенности его
организации у прокариот и эукариот? 3. Каково
строение гена? 4. В чем сходство и отличие струк
турной организации гена у вирусов, прокариот(бак
терий) н эукариот? 5. Каковы свойства генов?
локус.
74. Изменчивость организмов.
Норма р е а к ц и и
Изменчивость
это одно из важ
нейших свойств всех живых организмов,
связанное с их способностью приобре
тать новые признаки и свойства и терять
старые. Все многообразие живого и его
постоянное совершенство были бы невоз
можны без изменчивости. Это связано с
тем, что генотип последовательно реали
зуется в фенотип в ходе индивидуального
развития и в определенных условиях внут
ренней и внешней среды. Именно этим оп
ределяется тот факт, что организмы, име
ющие одинаковые генотипы, могут очень
резко отличаться по фенотипу.
Роль генотипа и среды в формиро
вании фенотипа. Возможность проявле
ния и развития признака зависит не толь
ко от генотипической среды (вспомните
взаимодействие генов), но и от влияния
регуляторных систем организма, в пер
вую очередь эндокринной. Такие призна
ки у петухов, как яркое оперение, боль
шой гребень, характер пения и тембр го
лоса, определяются действием мужских
половых гормонов (рис. 155, с. 260).
Женские половые гормоны, введенные
петухам, стимулируют работу генов, кон
тролирующих синтез белков в печени,
входящих в состав яйцеклетки. В норме
эти гены функционируют только у кур.
Следовательно, внутренняя среда орга
низма оказывает сильное влияние на
проявление наследственной информации
в форме соответствующих признаков.
Каждый организм развивается в оп
ределенных условиях, испытывая на себе
действие таких важнейших факторов
внешней среды, как колебания темпера
туры, освещенности, влажности, условий
питания, взаимоотношений с другими ор
ганизмами. Все эти факторы могут изме
нять морфологические и физиологичес
кие признаки и свойства организмов, т. е.
их фенотип. Приведем несколько приме
ров.
Если растения примулы, которые в
обычных условиях имели красные цветки
(Р-), перенести в оранжерею и содержать
их там при температуре 30.— 35 °С и вы
сокой влажности, то через некоторое
время все цветки у них оказываются бе
лыми. Если эти же растения вновь вер
нуть в обычные комнатные условия, то
распустившиеся цветки будут красными.
Напомним, что у гомозиготы по рецес
сивному аллелю (р р ) цветки будут белы
ми при любой температуре.
Другим классическим примером, ил
люстрирующим влияние условий внеш
ней среды на проявление качественных
признаков, служит изменение окраски
шерсти у гималайского кролика. Обычно
при 20 °С у этого, кролика шерсть белая,
за исключением черных ушей, лап, хвос
та и мордочки. При 30 °С такие кролики
вырастают полностью белыми. Если же у
гималайского кролика сбрить шерсть на
боку или спине и содержать при темпера
туре воздуха ниже +2 °С, то вместо белой
шерсти вырастет черная. Но если сбрить
шерсть на ухе, то в обычных условиях там
снова вырастет черная шерсть.
Для каждой области тела у кролика
есть свой порог температуры, выше кото
рого вырастает белая шерсть, а ниже —
черная (рис. 156, с. 261).
259
Рис. 155. Изменение вторичных половых признаков у кур после кастрации и пе
ресадки половых желез: 1 — нормальный петух, 2 — нормальная курица, 3 —
кастрированный петух, 4 — кастрированная курица, 5 — кастрированный петух
с пересаженным яичником, 6 — кастрированная курица с пересаженным се
менником.
260
25-27°
ментации шерсти у гималайского кролика
(цифры — пороговая тем п ера тура, выше ко
торой шерсть на данном участке тела белая, а
ниже — черная).
Эти наблюдения объясняют, почему
гималайские кролики рождаются белы
ми, без участков черной шерсти: их эм
бриональное развитие происходит в усло
виях высокой температуры.
Норма реакции. Все признаки и
свойства организма наследственно детер
минированы, однако организмы наследу
ют не сами признаки и свойства, а лишь
возможность их развития. Для проявле
ния и развития признака необходимы со
ответствующие условия внешней среды.
Если семена растений, напрцмер,
злаков (ржи, пшеницы, ячменя и Др./) или
клубни картофеля проращивать на свету,
то образующиеся проростки имеют зеле
ный цвет из-за присутствия в их клетках
хлоропластов. Такие же проростки, но
выращенные в темноте, становятся эт и
олированными (бесцветными или с жел
тым оттенком). На свету этиолированные
проростки вновь приобретают зеленый
цвет. Таким образом, возможность обра
зования хлорофилла и, как следствие,
хлоропластов в клетках была в обоих слу
чаях, но для реализации ее был необхо
дим свет. Другими словами, проявление
гена, контролирующего биосинтез хлоро
филла в клетках растений, осуществляет
ся только на свету.
Но даже в том случае, когда появле
ние и развитие того или иного признака
происходит, степень его выраженности
может быть разной в зависимости от ус
ловий внешней среды: при одних она мо
жет быть усилена, при других — ослаб
лена. Пределы этих изменений определе
ны возможностями, заложенными в гено
типе.
Степень варьирования признака,
ограниченная действием генотипа, на
зывается нормой реакции.
Для разных признаков и свойств орга
низмов границы, определяемые нормой
реакции, неодинаковы. Наибольшей
пластичностью и изменчивостью харак
теризуются количественные признаки:
семенная продуктивность злаков, вели
чины удоя у крупного рогатого скота,
масса животных, число и размеры листь
ев и колосков у растений и т. д. В то же
время качественные признаки (масть жи
вотных, окраска семян, цветков и плодов,
остистость и опушенность колоса у рас
тений) мало варьируют при изменении
условий среды.
Так, у крупного рогатого скота удой во
многом зависит от качества кормления и
ухода, т. е. от условий содержания. При
хорошем подборе кормов нужного качес
тва и количества ценная порода скота мо
жет дать 5000— 6000 кг молока в год, а в
случае ухудшения условий содержания их
продуктивность снижается до 2500 кг и
даже ниже. Что касается масти живот
ных, то при самых разных условиях она
почти не изменяется.
Подобную закономерность можно
наблюдать и у растений. Например, рас
тения элитного сорта пшеницы, выра
щенные в условиях высокой культуры аг
ротехники, дают урожай зерна 50— 60
ц/га. Ухудшение условий (тяжелые гли
261
нистые почвы, низкая культура земледе
лия и ухода за растениями) может при
вести к тому, что урожайность этого
прекрасного сорта пшеницы снизится до
10— 15 ц/га и ниже. Правда, окраска се
мян и остистость колоса при этом изме
няются в очень малой степени.
Рассмотренные примеры свидетельс
твуют о том, что количественные призна
ки обладают очень широкой нормой реак
ции, а качественные (окраска цветков у
примулы и шерсти у гималайского кроли
ка) гораздо более узкой.
Таким образом, все признаки и свой
ства каждого сорта растений и породы
животных генетически детерминированы
и могут изменяться только в пределах
нормы реакции. Поэтому попытки до
биться высоких урожаев, резко повысить
продуктивность сельскохозяйственных
животных только за счет повышения
уровня агротехники, улучшения условий
кормления и содержания животных не
состоятельны. В первую очередь необхо
димо получить новые сорта растений и
породы животных улучшенного генотипа,
отзывчивых на хорошие условия возде
лывания и выращивания.
?
» 1. Почему разнообразие качественных призна
ков в малой степени зависит от влияния условий
среды? 2. Чем определяется широта нормы реак
ции? 3. Можно ли, повысив уровень культуры агро
техники, превратить мягкую пшеницу в твердую? 4.
Какое практическое значение в сельском хозяйстве
имеет знание нормы реакции животных и растений?
75. Типы изменчивости.
Модификационная изменчивость
Типы изменчивости. По механизмам
возникновения и характеру изменений
признаков различают два типа изменчи
вости: наследственную и ненаследствен
ную.
Под наследственной изменчивостью
понимают способность к изменениям са
мого генетического материала, а под не
262
наследственной — способность орга
низмов реагировать на условия окружаю
щей среды, изменяться в пределах нормы
реакции, определяемой генотипом.
Наследственную изменчивость в свою
очередь подразделяют на мутационную и
комбинативную. Комбинативная измен
чивость возникает при скрещивании орга
низмов и представляет собой результат
перекомбинации генов или хромосом, не
сущих различные аллели. Эти перекомби
нации выражаются в появлении у потом
ков новых признаков, обусловленных дру
гим сочетанием генов, уже существовав
ших у родительских форм. В то же время в
результате мутационной изменчивости
появляются качественно новые признаки,
которых ранее не существовало.
Изменчивость может реализоваться у
отдельных организмов или клеток в ходе
их индивидуального развития, или онто
генеза; такая изменчивость называется
онтогенетической. Примеры онтоге
нетической изменчивости можно привес
ти из личного опыта, вспомнив, как зако
номерно и постепенно происходит физи
ческое и умственное развитие человека.
При этом типе изменчивости генотип
остается неизменным, хотя в данном слу
чае онтогенетические изменения детер
минированы и предопределены генети
ческими факторами. Причиной онтогене
тической изменчивости является функци
онирование различных наборов генов на
разных этапах онтогенеза организма в
пределах одного генома, причем порядок
«включения» или «выключения» опреде
ленных генов наследуется при половом
размножении организмов. По этому кри
терию она относится к ненаследственной
изменчивости.
Однако имеется ряд фактов, указыва
ющих и на изменения самого генотипа в
ходе онтогенеза, что приближает онтоге
нетическую изменчивость к наследствен
ной. Поэтому онтогенетическая изменчи
вость перекрывается наследственной и
ненаследственной изменчивостью:
Изменчивость
Наследственная
Комбинативная
Мутационная
Модификационная изменчивость.
Модификационной (ненаследственной,
или фенотипической) называют изменчи
вость, которая возникает в организме в
процессе его роста и развития в разных
условиях среды. Размах модификацион
ной изменчивости определяется нормой
реакции. Возникшее конкретное модифи
цированное изменение признака не на
следуется, но диапазон модификационной
изменчивости, или норма реакции, гене
тически обусловлен и наследуется. Мо
дификации не влекут за собой изменений
генотипа и сохраняются лишь на протя
жении жизни данного организма.
Норма реакции, лежащая в основе
модификационной изменчивости, скла
дывалась исторически в результате ес
тественного отбора. Способность орга
низмов реагировать на факторы среды,
как правило, имеет приспособительный
характер, и это составляет важнейшую
сторону всего учения о норме реакции.
Модификационной
изменчивости
подвержены как количественные, так и
качественные признаки. Возникновение
модификаций связано с тем, что такие
важнейшие факторы среды, как свет,
тепло, влага, воздействуют на активность
ферментов и в известной мере изменяют
течение биохимических реакций, проте
кающих в развивающемся организме.
Этим, в частности, объясняется появле
ние различной окраски цветков у приму
лы и шерсти у гималайских кроликов, о
чем говорилось выше.
Примерами модификационной измен
чивости у человека могут служить усиле
ние пигментации кожи — загар, под вли
Ненаследственная
Онтогенетическая
янием ультрафиолетовых лучей, развитие
костно-мышечной системы в результате
физических нагрузок и т. д. К модифика
ционной изменчивости следует отнести
также и явление физиологического гоме
остаза — способность организмов про
тивостоять колеблющимся условиям сре
ды путем приспособительного реагиро
вания.
У человека при жизни на разных вы
сотах над уровнем моря вырабатывается
разное количество красных кровяных те
лец — эритроцитов. Количество эритро
цитов в 1 мм3 крови у людей, живущих на
уровне моря, в два раза меньше, чем у
людей, живущих высоко в горах (табли
ца 8).
Та бл и ца 8.
Высота над уровнем
моря, м
Количество эритроцитов,
млн на 1 мм3крови
0
1300
2400
3100
3700
4100
4880
5600
4,2
5,2
6,0
6,6
6,8
7,5
7,8
8,3
Число эритроцитов растет пропорци
онально подъему над уровнем моря. Это
явление можно легко объяснить, если
вспомнить, что главная функция эритро
цитов — перенос кислорода от легких к
тканям и С 0 2 от тканей к легким. Увели
чение высоты над уровнем моря сопро
вождается снижением концентрации кис
лорода в атмосфере, что приводит к не
достатку 0 2 в тканях. Поэтому насущная
263
потребность в кислороде заставляет че
ловека и животных адаптивно реагиро
вать путем изменения числа эритроцитов
на разных высотах.
Эта реакция обратима: переезд в мес
та, расположенные на уровне моря, при
водит к снижению числа эритроцитов в
крови.
В некоторых случаях роль среды тако
ва, что особи, обладающие гомозиготным
по рецессивным аллелям генотипом, не
проявляют никаких признаков, отличаю
щих их от нормы. Например, в семье, где
отец и мать имеют голубой цвет глаз,
рождаются только голубоглазые дети.
При этом не имеет значения, в каких ус
ловиях живет данная семья. В некоторых
странах мира это учитывают при паспор
тизации граждан: в паспорте, помимо ос
новных сведений о гражданине, указыва
ется цвет его глаз.
Статистические закономерности модификационной изменчивости. Для из
учения закономерностей модификационной изменчивости, а также выявления ее
вклада в фенотип наряду с вкладами ге
нотипа необходимо придерживаться од
ного непременного условия: исследова
ния должны проводиться с генотипически
однородным материалом, развитие кото
рого проходит в имеющихся условиях
внешней среды. Только в этом случае ис
следователь может быть уверен в том,
что любые изменения признаков, возник
шие в ответ на действие различных фак
торов, являются проявлением именно
модификационной изменчивости, а не
обусловлены разным генотипом особей.
Такие условия среды, как влажность,
температура, освещенность, физические
свойства почвы и ее плодородие, глубина
заделки семян, взаимодействие и конку
ренция растений с другими сообитателями, никогда не бывают тождественными
даже на одном поле. Поэтому длина ко
лосьев пшеницы на одном поле может ко
лебаться от 6 до 14 см, а размеры листь
ев одного дерева иногда варьируют в еще
264
более широких пределах, хотя генотип их
одинаков. Если листья или колосья рас
положить в порядке нарастания или убы
вания их длины, то получается в а р и а
ц и о н н ы й ря д и з м е н ч и в о с т и дан
ного признака, слагающийся из отдель
ных вариант.
Как показывают подсчеты, частота
встречаемости отдельных вариант в вари
ационном ряду неодинакова. Чаще всего
встречается среднее значение признака,
а к обоим концам вариационного ряда
частота встречаемости закономерно сни
жается. Рассмотрим это на примере из
менчивости числа колосков в колосе
пшеницы. Возьмем произвольно (не вы
бирая) 100 колосьев одного сорта и под
считаем в каждом из них число колосков.
Полученные цифры (варианты) располо
жим в порядке нарастания признака и
подсчитаем, сколько раз каждая варианта
v встречается в данном ряду р, затем
сгруппируем их, т. е. составим вариаци
онный ряд:
Число колосков в колосе (о): 14, 15,
16, 17, 18, 19, 20.
Количество колосьев (р ): 2, 7, 22, 32,
24, 8, 5.
Эти цифры отражают степень измен
чивости признака. Ее среднее значение М
рассчитываем по формуле:
Е — сумма, п — общее число вариант
данного вариационного ряда. В нашем
примере среднее значение признака ва
риационного ряда составляет 20, 19.
Распределение вариант в этом ряду
можно выразить наглядно на графике.
Для этого на оси абсцисс откладывают
значения вариант v в порядке их увеличе
ния, на оси ординат — частоту встречае
мости р каждой варианты.
Графическое выражение изменчивос
ти признака, отражающее как размах ва
риаций, так и частоты встречаемости от
дельных вариант, называют вариацион-
Рис. 157 Вариационная кривая числа колосков
в колосе пшеницы.
ной кривой (рис. 157). Установлено, что
модификационная изменчивость самых
разнообразных признаков у растений,
животных и человека имеет общие черты,
что хорошо видно на графике.
Кривая на графике бывает, как пра
вило, симметричной, особенно когда
изучается большое число особей. Это
значит, что вариации, как большие, так
и меньшие, но отличающиеся от средне
го арифметического на одну и ту же ве
личину, встречаются одинаково часто.
Отсюда следует, что минимальные и
максимальные величины должны встре
чаться очень редко, но с одинаковой
частотой.
Значение модификаций. Модифика
ционная изменчивость в естественных ус
ловиях носит приспособительный харак
тер и в этом смысле имеет важное значе
ние в эволюции. Обусловленные нормой
реакции адаптивные модификации дают
возможность организму выжить и оста
вить потомство.
Знание закономерностей модификационной изменчивости имеет очень боль
шое практическое значение, так как поз
воляет предвидеть и заранее планировать
многие показатели. В частности, созда
ние оптимальных условий для реализации
генотипа (высокая культура агротехники,
грамотный уход за животными) позволя
ет получать высокие урожаи и высокую
продуктивность сельскохозяйственных
животных.
Такой подход в равной мере относится
и к человеку. Каждый ребенок обладает
большими способностями, иногда даже в
нескольких областях. Задача психологии
и педагогики состоит в том, чтобы как
можно раньше найти эту область и обес
печить максимальное развитие ребенка в
этом направлении наряду с общим обра
зованием, т. е. в пределах нормы реакции
достичь максимальной величины реали
зации его генотипа. Утверждение о воз
можности обучить всех всему в одинако
вой степени лишено всяких оснований.
?
I 1. Какова современная классификация типов
изменчивости? 2. Что такое онтогенетическая из
менчивость и какова ее причина? Приведите приме
ры онтогенетической изменчивости у растений,жи
вотных и человека. 3. Что такое модификационная
изменчивость? Приведите примеры. 4. Дайте опре
деление физиологического гомеостаза. Какова
главная особенность в реагировании организмов на
изменения условий среды? 5. Каковы статистичес
кие закономерности, отражающие модификационную изменчивость? 6. Каково практическое значе
ние модификаций?
76. Комбинативная изменчивость.
Мутации
Как уже отмечалось, наследственную,
или генотипическую, изменчивость при
нято делить на комбинативную и мутаци
онную.
Комбинативная изменчивость. В ос
нове комбинативной изменчивости лежит
половое размножение организмов,
вследствие которого возникает огромное
разнообразие генотипов. Практически
неограниченными источниками генети
ческой изменчивости служат три про
цесса:
1) независимое расхождение гомоло
гичных хромосом в первом мейотическом делении — важнейший источ
ник комбинативной изменчивости.
265
Именно независимое комбинирова
ние хромосом при мейозе является
основой третьего закона Менделя.
Появление зеленых гладких и желтых
морщинистых семян гороха во втором
поколении от скрещивания растений
с желтыми гладкими и зелеными мор
щинистыми семенами — пример
комбинативной изменчивости;
2) реципрокный обмен генами между
хроматидами гомологичных хромо
сом, или кроссинговер. Он создает
новые группы сцепления, т. е. служит
важным источником генетической ре
комбинации аллелей. Рекомбинан
тные хромосомы, оказавшись в зиго
те, способствуют появлению призна
ков, не типичных для родителей;
3) случайное сочетание гамет при
оплодотворении.
Эти три источника комбинативной из
менчивости действуют независимо и од
новременно, обеспечивая при этом по
стоянную «перетасовку» генов, что при
водит к появлению организмов с другими
генотипом и фенотипом (сами гены при
этом не изменяются). Однако новые ком
бинации генов довольно легко распада
ются при передаче из поколения в поко
ление. Поэтому для закрепления важных
признаков селекционеры используют
близкородственные скрещивания, при
которых возрастает вероятность встречи
одинаковых гамет. В результате могут
возникнуть потомки с комбинацией ге
нов, близкой к родительской комбина
ции.
Крмбинативная изменчивость являет
ся важнейшим источником всего колос
сального наследственного разнообразия,
характерного для живых организмов. Од
нако перечисленные выше источники из
менчивости не порождают крупных изме
нений в генотипе, которые необходимы,
согласно эволюционной теории, для воз
никновения новых видов. Такие измене
ния возникают в результате мутаций.
266
Мутационная изменчивость. М ут а
ционной называется изменчивость само
го генотипа. Мутации — это внезапные
скачкообразные и ненаправленные изме
нения исходной формы с появлением у
живых организмов качественно новых
наследственных признаков и свойств, ко
торых ранее в природе не существовало.
Основные положения мутационной
теории разработаны Г. Д е Ф р и з о м
в 1901 — 1903 гг. и сводятся к следую
щему:
1) мутации возникают внезапно,
скачкообразно как дискретные изме
нения признаков;
2 ) новые формы устойчивы;
3) в отличие от ненаследственных из
менений мутации не образуют непре
рывных рядов. Они представляют со
бой качественные изменения;
4) мутации проявляются по-разному
и могут быть как полезными, так и
вредными, как доминантными, так и
рецессивными;
5) вероятность обнаружения мутаций
зависит от числа исследованных осо
бей;
6) сходные мутации могут возникать
повторно;
7) мутации ненаправленны (спонтан
ны), т. е. мутировать может любой
участок хромосомы, вызывая измене
ния как незначительных, так и жиз
ненно важных признаков.
Классификация мутаций. Существу
ет несколько способов классификации
мутаций.
I. По характеру изменения генома.
1. Геномные мутации — изменение
числа хромосом.
2. Хромосомные мутации — изме
нение структуры хромосом.
3. Генные мутации — изменение ге
нов.
II. По проявлению в гетерозиготе.
1. Доминантные мутации.
2. Рецессивные мутации.
III. По уклонению от нормы.
1. Прямые мутации.
2. Реверсии — восстановление у
мутантного организма нормаль
ного фенотипа в результате пов
торной мутации.
IV. По причинам, вызывающим му
тации.
1. Спонтанные, возникающие без
видимой причины.
2. Индуцированные мутации.
V. По локализации в клетке.
1. Ядерные мутации.
2. Цитоплазматические, или внеядерные, мутации.
VI. По отношению к возможности
наследования.
1. Генеративные мутации, происхо
дящие в половых клетках.
2. Соматические мутации, происхо
дящие в соматических клетках.
Кроме того, очень часто мутации
классифицируют по их фенотипическому
проявлению. В этом случае рассматрива
ют летальные, морфологические, биохи
мические, поведенческие мутации, а
также мутации устойчивости или чув
ствительности к повреждающим агентам
и т. д.
Соматические и генеративные мута
ции. Мутации могут возникать в любых
. клетках многоклеточного организма и
Йа разных стадиях его развития. Му
тации, возникающие в половых клет
ках, называют генеративными, а в
клетках других тканей — соматиче
скими.
Соматические мутации по своей при
роде ничем не отличаются от генератив
ных. Такое разделение вызвано тем, что
биологическое значение их неравноценно
и связано с типом размножения организ
мов. У организмов с бесполым размно
жением соматические мутации могут
иметь огромное значение и в эволюции, и
в селекции.
При делении соматической клетки,
несущей мутантный ген, новые свойства
передаются ее потомкам. Если у растения
мутирует клетка, из которой образуется
почка, а затем побег, то последний при
обретает новые свойства. Так, у черной
смородины может появиться ветка с бе
лыми ягодами. При вегетативном
размножении (например, при помо
щи отводков и прививок) новый приз
нак, возникший в результате сома
тической мутации, сохраняется у по
томков.
При половом размножении признаки,
появившиеся в результате соматических
мутаций, потомкам не передаются и в
процессе эволюции никакой роли не иг
рают. Однако в индивидуальном развитии
они могут влиять на формирование приз
нака, причем чем раньше в онтогенезе
возникает соматическая мутация, тем
большим оказывается участок ткани, не
сущий данную мутацию, и чем позднее —
тем меньшим. Такие особи называются
мозаиками, или химерами. Например,
мозаиками являются люди, у которых
цвет одного глаза отличается от другого;
появляются темные родинки, седая прядь
волос и др.
Генеративные мутации возникают в
половых клетках и передаются по наслед
ству следующим поколениям организмов
(потомкам).
?
> 1. Какие биологические явления лежат в осно
ве комбинативной изменчивости? 2. Что такое му
тация и каковы ее свойства? 3. Какие существуют
способы классификации мутаций? 4. Чем отличает
ся наследование соматических мутаций от наследо
вания генеративных мутаций?
77. Мутационная изменчивость.
Геномные мутации
Почти любое изменение в структуре
или количестве хромосом, при котором
клетка сохраняет способность к самовос
произведению, обусловливает наслед
ственное изменение признаков организ
ма. По характеру изменения генотипа
267
различают геномные, хромосомные и
генные мутации.
Геномные мутации — это изменение
числа хромосом в геноме клеток организ
ма, сопровождающееся изменением его
признаков и свойств.
Организмы с отклонениями от нор
мального числа хромосом называются
хромосомными мутантами. Измене
ние числа хромосом происходит в двух
направлениях: в сторону увеличения или
уменьшения числа целых гаплоидных
наборов (полиплоидия) и в сторону по
тери или включения отдельных хромосом
(анеуплоидия) (рис. 158).
Полиплоидия — это кратное увели
чение гаплоидного набора хромосом.
Клетки с разным числом гаплоидных на
боров хромосом называются триплоидными (3га), тетраплоидными (4я), гексаплоидными (6п), октаплоидными (8п)
и т. д.
Чаще всего полиплоиды образуются
при нарушении расхождения хромосом к
полюсам клетки при мейозе или митозе.
Это может быть вызвано действием фи
зических (высокая и низкая температура,
радиоактивное излучение) и химических
(колхицин, винбластин, аценафтен, хло
роформ, эфир, хлоргидрид) факторов.
Химические вещества типа колхицина
подавляют образование митотического
веретена в клетках, приступивших к де
лению, в результате чего удвоенные хро
мосомы не расходятся, и клетка оказыва
ется тетраплоидной.
Для многих растений известны так на
зываемые полиплоидные ряды. Они
включают формы от 2 до Юге и более.
Например, полиплоидный ряд из наборов
в 12, 24, 36, 48, 60, 72, 96, 108 и 144 хро
мосомы составляют представители рода
паслен
(Solanum).
Род пшеница
(Triticum) представляет ряд, члены кото
рого имеют 14, 28 и 42 хромосомы.
Различают две формы полиплои
дии — автополиплоидию и аллополипло
идию.
268
Автополиплоидия возникает на ос
нове кратного увеличения гаплоидного
набора хромосом индивидуума опреде
ленного вида. Если обозначить основное
число хромосом (геном) буквой А, то А
будет соответствовать гаплоиду, АА —
автодиплоиду, ААА — автотриплоиду
ит. д.
Аллополиплоидией, или амфиплоидией, называется явление, возникающее
на основе умножения геномов разных ви
дов. Так, если у межвидового гибрида
совмещаются геномы Л и В, то получен
ный от него аллотетраплоид будет ААВВ.
Известно, что при отдаленной гибри
дизации наблюдается бесплодие особей,
поскольку их генеративные клетки не
имеют гомологичных хромосом для
конъюгации и образуются нежизнеспо
собные гаметы, которые гибнут при пер
вом же делении. Для преодоления бес
плодия гибридов в этом случае использу
ют полиплоидию. При кратном увеличе
нии числа хромосом каждая из них имеет
гомолога. Гаметы такого полиплоидного
гибрида образуют тетраплоидные зиготы
(2ге хромосом от одного и 2п хромосом от
другого вида). Такие формы называются
аллотетраплоид ами.
Полиплоидия приводит к изменению
признаков организма, а поэтому является
важным источником изменчивости в эво
люции и селекции, особенно у растений.
Это связано с тем, что у растительных
организмов весьма широко распростра
нены гермафродитизм (самоопыление),
апомиксис (партеногенез) и вегетативное
размножение. Поэтому около трети ви
дов растений, произрастающих на нашей
планете,— полиплоиды, а в резко конти
нентальных условиях высокогорного Па
мира произрастает до 85 % полиплоидов.
Почти все культурные растения тоже по
липлоиды, у которых, в отличие от их ди
ких сородичей, более крупные цветки,
плоды и семена, а в запасающих органах
(стебель, клубни) накапливается больше
питательных веществ. Полиплоиды легче
Норма
ДИПЛОИД'
2
3
4
Анеуплоидия
Моносомик
4
2
Т рисомик<
Т е тр асо м и к^
1
4
Моноплоидия
Моноплоид
( I
Полиплоидия
2
3
4
3
4
Т риплоид<
2
Тетраплоид<
2
Р ис. 158
269
приспосабливаются к неблагоприятным
условиям жизни, легче переносят низкие
температуры и засуху. Именно поэтому
они очень распространены в северных и
высокогорных районах.
В основе резкого увеличения продук
тивности полиплоидных форм культурных
растений лежит явление полимерии
(см. § 67).
У анеуплоидов, или гетероплоидов, нормальное число хромосом увели
чивается или уменьшается менее чем на
целый набор. Анеуплоиды возникают тог
да, когда не расходятся хроматиды от
дельных хромосом в митозе или отдель
ные гомологичные хромосомы в мейозе.
В результате нерасхождения хромосом
при гаметогенезе могут возникать поло
вые клетки с лишними хромосомами, и
тогда при последующем слиянии с нор
мальными гаплоидными гаметами они
образуют зиготу 2п + 1 ( т р и с о м и к ) по
определенной хромосоме. Если в гамете
оказалось меньше на одну хромосому, то
последующее оплодотворение приводит, к
образованию зиготы 2п — 1 ( мо н о с о м и к ) по какой-либо из хромосом. Кроме
того, встречаются формы 2п — 2, или
н у л л и с о м и к и , так как отсутствует па
ра гомологичных хромосом, и 2п+х, или
полисомики.
Анеуплоиды встречаются как у расте
ний и животных, так и у человека. Анеуплоидные растения обладают низкой жиз
неспособностью и плодовитостью, а у че
ловека это явление нередко приводит к
бесплодию и в этих случаях не наследует
ся. У детей от матерей старше 38 лет
частота анеуплоидии повышена (до
2,5 %). Кроме того, случаи полиплои
дии у человека вызывают хромосом
ные болезни, о которых будет сказано
позже.
Наличие анеуплоидных рядов у расте
ний и животных указывает на большое
значение анеуплоидии для видообразова
ния. Анеуплоидные формы часто исполь
зуются в селекции растений. Скрещивая
270
растения с нуллисомиками и моносомиками, в геном можно вводить определен
ную хромосому с желательными генами.
Таким путем получены новые формы
пшеницы, устойчив.ле к ряду заболе
ваний.
Полиплоидия у животных. Установ
лено, что у раздельнополых животных как
в естественных, так и в искусственных ус
ловиях полиплоидия встречается крайне
редко. Это обусловлено тем, что поли
плоидия, вызывая изменение соотноше
ния половых хромосом и аутосом, приво
дит к нарушению конъюгации гомологич
ных хромосом и тем самым затрудняет
определение пола. В результате такие
формы оказываются бесплодными и ма
ложизнеспособными. Это подтверждает
ся тем, что у гермафродитных животных и
животных, размножающихся партеногенетическим путем, полиплоидия может
осуществляться почти также, как у расте
ний. Например, у дождевых червей
встречаются ряды In, 2п, 3п Ап и
более.
Однако и у раздельнополых животных
встречаются случаи полиплоидии. Извес
тны полиплоиды у аскарид, тутового шел
копряда, бабочек, рыб, амфибий и неко
торых млекопитающих. Так, у серебрис
того карася встречается двуполая дипло
идная и однополая триплоидная формы.
У некоторых представителей лососевых
рыб и ящериц также встречаются полип
лоидные формы.
У млекопитающих обнаружены триплоидные зиготы. Однако триплоидные
эмбрионы у мышей доживают только до
середины эмбриогенеза. Случаи триплоидии и тетраплоидии обнаружены при
анализе материала спонтанных выки
дышей или мертворожденных у че
ловека.
В мире животных полиплоидия неред
ко встречается в соматических клетках
ряда тканей, т. е. в тех случаях, когда де
ление клетки осуществляется только пу
тем митоза.
■ 1. Почему полиплоидные формы культурных
растений отличаются высокой продуктивностью, а
их дикие сородичи легче приспосабливаются к не
благоприятным условиям жизни? 2. Почему поли
плоидия у животных встречается редко?
78. Хромосомные и генные
мутации
Хромосомные мутации. В процессе
эволюции организма изменяться может
не только число и величина хромосом, но
и их организация: отдельные участки хро
мосом могут менять свое расположение
внутри хромосомы и даже переходить от
одних хромосом к другим. Изменения в
структуре хромосом, которые можно
выявить и изучить под световым мик
роскопом, называют хромосомными
мутациями, перестройками или аберра
циями. Известны перестройки разных
типов (рис. 159, с. 272):
1) нехватки, или дефишенси, — это
потеря концевых участков хромосо
мы;
2 ) делеции — выпадение участка
хромосомы в средней ее части;
3) дупликации — двух- или много
кратное повторение набора генов, ло
кализованных в определенном учас
тке хромосомы;
4) инверсии — поворот участка хро
мосомы на 180°, в результате чего в
этом участке гены расположены в
последовательности, обратной по
сравнению с нормальной;
5) транслокации — изменение по
ложения какого-либо участка хромо
сомы в хромосомном наборе. К наи
более распространенному типу тран
слокаций относятся реципрокные,
при которых происходит обмен учас
тками между двумя негомологичными
хромосомами. Участок хромосомы
может также изменять свое положе
ние и без реципрокного обмена, оста
ваясь в той же хромосоме или вклю
чаясь в какую-то другую. Транслока
ции такого типа иногда называют
транспозициями.
При дефишенси, делециях и дупли
кациях изменяется количество генети
ческого материала. Степень фенотипи
ческого изменения зависит от того, на
сколько велики соответствующие участ
ки хромосом и содержат ли они важные
гены. Примеры дефишенси известны у
многих организмов, включая человека.
Тяжелое наследственное заболевание —
синдром «кошачьего крика» (назван так
по характеру звуков, издаваемых больны
ми младенцами) обусловлен гетерозиготностью по дефишенси в 5-й хромосоме.
Этот синдром сопровождается сильным
нарушением роста и умственной отста
лостью. Обычно дети с таким синдромом
рано умирают, но некоторые доживают до
взрослого возраста.
Дупликации играют существенную
роль в эволюции генома, поскольку могут
служить материалом для возникновения
новых генов, так как в каждом из двух ра
нее одинаковых участков могут происхо
дить различные мутационные процессы.
При инверсиях и транслокациях
общее количество генетического матери
ала остается прежним, изменяется толь
ко его расположение. Такие мутации то
же играют значительную роль в эволю
ции, так как скрещивание мутантов с ис
ходными формами затруднено, а их гиб
риды F{ чаще всего стерильны. Поэтому
здесь возможно только скрещивание ис
ходных форм между собой. Если у таких
мутантов окажется благоприятный фено
тип, они могут стать исходными формами
для возникновения новых видов.
Генные мутации. Генные, или точковые, мутации — результат изменения
нуклеотидной последовательности моле
кулы ДНК в определенном участке хро
мосомы. Такое изменение последователь
ности оснований в гене воспроизводится
при транскрипции в структуре иРНК и
271
a b c d e f g h
an
N
n
Делеция
a b c e
f g h
гтттхс т
11
A
M B
D C
^ D E F rGu Hn
m ii
1 t
ix m
A B O D E
c
т
у
t
FGH
A B C B C D E F G H
A B C
т
t
A B O D E
n
Дупликация
Инверсия
c
G H
3
F
F GH
E
cm i i>r
t
t
A B C D E
СП
t
l
1Ж
FGH
D С В
FGH
M N O C D E
FGH
mm i ixm
Г О
Транслокация
M N О P Q
R
А В P Q
A B C D E
FGH
CEEDXED
t
R
C E E 3X D
t
t
ED
Транслокация
t
ADE
F B C G H
сгххххгю
Рис. 159. Хромосомные перестройки, изменяющие расположение генов в хромосомах.
приводит к изменению последователь
ности аминокислот в полипептидной це
пи, образующейся в результате трансля
ции на рибосомах. В результате синтези
руется белок с измененными свойствами,
что ведет к изменению каких-либо приз
272
наков организма. Существуют разные ти
пы генных мутаций, связанных с добавле
нием, выпадением или перестановкой
нуклеотидов в гене. Это дупликации,
вставки лишней пары нуклеотидов, делеции (выпадение пары нуклеотидов), ин
версии или замены пар нуклеотидов (АТ
±5 ГЦ; АТ а* ЦТ или АТ ** ТА).
Эффекты генных мутаций чрезвычай
но разнообразны. Большая часть генных
мутаций фенотипически не проявляется,
поскольку они рецессивны. Это очень
важно для существования вида, так как в
большинстве своем вновь возникающие
мутации оказываются вредными. Однако
их рецессивный характер позволяет им
длительное время сохраняться у особей
вида в гетерозиготном состоянии без вре
да для них и проявиться в будущем при
переходе в гомозиготное состояние.
Вместе с тем известен ряд случаев, когда
изменение лишь одного основания в оп
ределенном гене оказывает заметное
влияние на фенотип. Одним из примеров
служит серповидноклет очная а н е
мия — заболевание, вызываемое у чело
века заменой основания в одном из генов,
ответственных за синтез гемоглобина.
Это приводит к тому, что в крови эритро
циты с таким гемоглобином деформиру
ются (из округлых становятся серповид
ными) и быстро разрушаются. При этом
развивается острая анемия и снижение
количества кислорода, переносимого
кровью. Анемия вызывает физическую
слабость и даже может привести к нару
шениям деятельности сердца и почек и к
ранней смерти людей, гомозиготных по
мутантному аллелю.
Закон гомологических рядов в нас
ледственной изменчивости. Крупней
шим обобщением работ по изучению из
менчивости в начале XX в. стал закон
гомологических рядов в наследствен
ной изменчивости. Закон был сформу
лирован выдающимся русским ученым
Н. И. В а в и л о в ы м в 1920 г. в докла
де на III Всероссийском селекционном
съезде в Саратове. Сущность закона сос
тоит в том, что виды и роды, генетически
близкие, связанные друг с другом един
ством происхождения, характеризуются
сходными рядами наследственной измен
чивости. Зная,какие формы изменчивос
ти встречаются у одного вида, можно
предвидеть нахождение аналогичных
форм у родственного ему вида.
В основе закона гомологических рядов
фенотипической изменчивости у род
ственных видов лежит представление о
единстве их происхождения путем дивер
генции (расхождения) от одного предка в
процессе естественного отбора. Посколь
ку общие предки имели специфический
набор генов, то и их потомки должны об
ладать примерно таким же набором.
Более того, у родственных видов, име
ющих общее происхождение, возникают
и сходные мутации. Это означает, что у
представителей разных семейств и клас
сов растений и животных со сходным на
бором генов можно встретить паралле
лизм — гомологические ряды мутаций по
морфологическим, физиологическим и
биохимическим признакам и свойствам.
Так, например, у разных классов позво
ночных встречаются сходные мутации:
альбинизм и отсутствие перьев у птиц,
альбинизм и бесшерстность у млекопита
ющих, гемофилия у многих млекопитаю
щих и человека. У растений наследствен
ная изменчивость отмечена по таким
признакам, как пленчатое или голое зер
но, остистый или безостый колос и др.
Закон гомологических рядов, отражая
общую закономерность мутационного
процесса и формообразования организ
мов, представляет широкие возможности
для его практического использования в
сельскохозяйственном производстве, се
лекции, медицине. Знание характера из
менчивости нескольких родственных ви
дов дает возможность поиска признака,
который отсутствует у одного из них, но
характерен для других. Таким путем были
собраны и изучены голозерные формы
злаков, односеменные сорта сахарной
свеклы, не нуждающиеся в прорывке, что
особенно важно при механизированной
обработке почв. Медицинская наука в ка
честве моделей для изучения болезней
человека получила возможность исполь
273
зовать животных с гомологическими за
болеваниями: сахарный диабет крыс;
врожденная глухота мышей, собак, мор
ских свинок; катаракта глаз мышей,
крыс, собак и ряд других дефектов.
Закон гомологических рядов позволя
ет также предвидеть возможность появ
ления мутаций, еще неизвестных науке,
которые могут использоваться в селек
ции для создания новых ценных для хо
зяйства форм.
?
в 1. При облучении рентгеновскими лучами иног
да возникают хромосомные перестройки, а иногда
— хроматидные. Как это можно объяснить? 2. Ка
кое влияние на конъюгацию хромосом и жизнеспо
собность продуктов мейоза оказывает гетерозигот ность по транслокации? 3. Какое значение имеет
рецессивный характер большинства генных мута
ций? 4. В чем суть и практическое значение закона
гомологических рядов в наследственной изменчи
вости?
79. Спонтанный и индуцированный
мутагенез
Спонтанными называют мутации,
возникающие под влиянием неизвестных
природных факторов, чаще всего как ре
зультат ошибок при воспроизведении ге
нетического материала (ДНК или РНК),
поскольку репликация не происходит с
абсолютной точностью, а процессы репа
рации (исправление повреждений в мо
лекуле ДНК) не обладают абсолютной
эффективностью. Частота спонтанного
мутирования у каждого вида генетически
обусловлена и поддерживается на опти
мальном уровне — 106— 107.
До 1925— 1927 гг. генетики имели
дело только со спонтанными мутациями.
Различные попытки повысить частоту
мутаций не приносили успеха. Создава
лось впечатление, что мутационный про
цесс не зависит от условий окружающей
среды.
Радиационный мутагенез. Впервые
повышение частоты наследственной из
менчивости под влиянием внешних аген
274
тов (ионизирующая радиация) обнаружи
ли в 1925 г. советские микробиологи
Г. А. Н а д с о н и Г. С. Ф и л и п п о в ,
проводившие исследования с дрожжами.
В 1927 г. американский генетик Г.
М е л л е р сообщил о действии рентге
новских лучей на мутационный процесс у
дрозофилы, а позже мутагенное воздей
ствие этих лучей подтвердилось на мно
гих объектах. В дальнейшем было уста
новлено, что наследственные изменения
обусловливаются также всеми другими
видами проникающей радиации, особен
но нейтронами и у-лучами. При этом воз
никают как разрывы хромосом, так и точковые мутации. Причиной разрывов хро
мосом являются некоторые особенности
процессов, происходящих в тканях при
действии излучений. Излучения вызыва
ют в тканях ионизацию, в результате ко
торой одни атомы теряют электроны, а
другие присоединяют их; при этом обра
зуются положительно или отрицательно
заряженные ионы. Подобный процесс
внутримолекулярной перестройки в сос
таве хромосом может привести к их фраг
ментации.
Одним из самых опасных последствий
облучения является образование свобод
ных радикалов — ОН или — 0 2Н из на
ходящейся в тканях воды. Эти радикалы
обладают высокой реакционнной способ
ностью и могут расщеплять многие
органические вещества, в том числе ДНК
и РНК.
Химический мутагенез. Приоритет
открытия химических мутагенов принад
лежит советским ученым. В 1933 г. В . В .
С а х а р о в , а затем М . Е. Л о б а ш е в и
Ф. А. С м и р н о в (1934) показали, что
некоторые соединения (йод, уксусная
кислота, аммиак) способны индуцировать
рецессивные летали в Х-хромосоме. В
1946 г. советский генетик И . А. Р а п о
п о р т обнаружил сильное мутагенное
действие формалина и этиленамина, а ан
глийские исследователи Ш . Ау э р б а х
и Д ж . Р о б с о н — азотистого иприта.
Позже были открыты многие другие хи
мические мутагены: колхицин, кофеин,
некоторые компоненты табака, лекар
ственных препаратов, пищевых консер
вантов и пестицидов. Некоторые из них
усиливают мутационный эффект в сотни
раз по сравнению со спонтанным; они по
лучили название супермутагенов.
Другие мутационные факторы. Есть
все основания для предположения, что
подавляющее большинство факторов
внешней среды может оказывать то или
иное влияние на наследственные свой
ства организма. Установлено, что мута
генным действием обладают такие физи
ческие факторы, как ультрафиолетовый
свет, фотоны видимого света и темпера
тура. Повышение температуры увеличи
вает число мутаций, хотя и незначи
тельно.
К биологическим мутагенам относят
ся вирусы и токсины ряда плесневых гри
бов. Во многих отечественных и зарубеж
ных лабораториях было показано появле
ние большого числа хромосомных пере
строек в культурах микроорганизмов и
клеток животных и человека, поражен
ных вирусами. При этом мутагенным дей
ствием обладают не только те вирусы, к
которым восприимчивы растения и жи
вотные, но и непатогенные для них виру
сы. Таким образом, роль вирусов в био
сфере заключается в том, что они явля
ются не только возбудителями ряда забо
леваний растений, животных и человека,
но и виновниками многих спонтанных му
таций.
Причины мутаций. Возможность из
менять скорость мутационного процесса
послужила решающим стимулом к выяс
нению причин спонтанных мутаций.
Современная точка зрения на причи
ны спонтанных мутаций сформировалась
в 60-е гг. благодаря выяснению механиз
мов воспроизведения, репарации и ре
комбинации генов и открытию фермен
тных систем, ответственных за эти про
цессы. Возникла гипотеза, согласно ко
торой генные мутации — это ошибки в
работе ферментов матричного синтеза
ДНК- Притягательность этой общеприз
нанной гипотезы заключается в том, что
она позволяет дать единое объяснение
причин спонтанных и индуцированных
мутаций, происходящих под генетическим
контролем. Были открыты гены, мутации
которых могут повышать или понижать
частоту как спонтанных, так и индуциро
ванных мутаций.
Мутация рассматривается как ре
зультат перестройки атомов в сложной
молекуле гена. Причиной такой пере
стройки является попадание в ген кванта
света или ионизирующей частицы или же
случайные колебания атомов. Причем
мутации возникают в результате процес
са, длящегося во времени, а не непосред
ственно в момент прохождения кванта
энергии или ионизирующей частицы че
рез ген.
Согласно физиологической гипотезе
М. Е. Лобашева (1946), появлению му
таций должно предшествовать предмутационное состояние или потенциальное
изменение, которое может быть устране
но в процессе репарации либо реализует
ся в виде мутаций. Связь мутаций с про
цессами репарации в настоящее время
доказана практически для всех исследо
ванных объектов.
Репарация ДНК и мутационный про
цесс. Под действием различных физичес
ких и химических агентов могут возник
нуть повреждения ДНК в виде димеров
тимина, сшивок между нитями или с бел
ком, разрывов ДНК, появления нукле
отидов аномальной структуры и т. д. По
вышенная частота этих повреждений
блокирует репликацию ДНК, а тем са
мым и деление ядра и клетки. Оказалось,
что клетка обладает различными меха
низмами исправления повреждений в ни
тях ДНК. Способность клеток всех орга
низмов к самовосстановлению повреж
денной первичной структуры ДНК полу
чила название репарации.
275
В настоящее время выявлены три ос
новных механизма репарации ДНК: фо
тореактивация, эксцизионная репа
рация и пострепликативная репара
ция. Последние два типа называют также
темповой репарацией, поскольку и та и
другая не зависят от света (рис. 160).
Лучше всего изучена репарация по
вреждений, вызванных ультрафиолето
выми лучами. При облучении ультрафио
летом между соседними пиримидиновыми
основаниями одной цепи возникают ди
меры, чаще всего димер Т — Т, т. е. вмес
то водородных связей между Т и А двух
комплементарных нуклеотидных цепей
образуются связи Т — Т внутри одной
цепи.
Фотореактивация происходит при
воздействии видимого света. При этом
репарирующий фермент связывается с
поврежденной ДНК, разделяет димер на
мономеры и водородные связи Т — А
между комплементарными цепями опять
восстанавливаются.
Если при фотореактивации исправля
ются повреждения, возникшие только
под воздействием ультрафиолетовых лу
чей, то при темновой репарации —
повреждения, появившиеся под влияни
ем других физических, химических и био
логических факторов.
При эксцизионной репарации (от лаг.
excisio — вырезание) вырезается по
врежденный участок ДНК (см. рис. 160).
Сначала фермент эндонуклеаза разреза
ет одну цепь. Следующий фермент — экзонуклеаза — удаляет измененную часть,
Рис. 160. Механизм темновой репарации (эта
пы вырезания и репарации поврежденного
участка молекулы ДНК): 1 — пиримидиновый
димер искажает структуру ДНК, 2 — репараци
онная эндонуклеаза разрезает поврежденную
цепь, 3 — репарационная эксцизия при дей
ствии экзонуклеазы, 4 — репарационный син
тез, 5 — ДНК-лигаза заменяет оставшийся од
ноцепочечный разрыв.
276
а ДНК-полимераза вновь синтезирует
недостающий участок, комплементарный
неповрежденной цепи ДНК. И наконец,
свободные концы старой части цепи со
единяются с концами вновь синтезиро
ванного участка при помощи лигазы. Фо
тореактивация и эксцизионная репара
ция коротких участков ДНК протекают
без ошибок и, следовательно, не ведут к
мутациям.
Если димеры не будут устранены, то
соответствующие основания не смогут
выполнять роль матрицы и в этих местах
во вновь синтезированной цепи ДНК
окажутся пропуски (пробелы, или бре
ши). Путем рекомбинации между двумя
двойными цепями ДНК — продуктами
репликации — возможно образование
одной нормальной двойной цепи (по
стрепликативная репарация).
Если повреждения лежат так тесно
друг возле друга, что пропуски перекры
ваются, то для их заполнения использует
ся другая «ремонтная» система (SOS-ре
парация), способная синтезировать но
вую цепь ДНК и на дефектной матрице.
При этом новые основания включаются
таким образом, что могут возникнуть му
тации. Таким образом, репарации в зна
чительной мере ограничивают количес
тво мутаций, что имеет огромное биоло
гическое значение.
Однако не все виды повреждений
ДНК репарируются, часть их проявляет
ся в виде мутаций. В частности, известно
несколько мутаций, проявляющихся как
тяжелые наследственные заболевания за
счет нарушения процесса репарации.
Примером может служить пигментная
ксеродерма — рецессивная аутосомная
мутация — сверхчувствительность к
ультрафиолетовым лучам под влиянием
которых на открытых частях тела
появляются пигментные пятна вначале в
виде веснушек, затем все более крупных
папиллом (родинок на ножках) различной
величины и формы и, наконец опухоли.
Для 70— 75 % больных эта болезнь
заканчивается летально к моменту
наступления половой зрелости.
Способность клеток осуществлять
эффективную репарацию генетического
материала играет существенную роль в
замедлении процессов старения.
Искусственное получение мутаций. В
настоящее время во многих странах мира
ведутся работы по созданию новых и со
вершенствованию уже существующих ме
тодов направленного мутагенеза для соз
дания организмов с качественно новыми
признаками. Например, химические мута
гены используются для получения мутан
тных форм плесневых грибов (таких, как
пеницилл), актиномицетов, бактерий, вы
рабатывающих в больших количествах
пенициллин, стрептомицин и другие анти
биотики. Получены десятки перспектив
ных мутантов культурных растений.
?
■ 1. Какие факторы среды могут оказывать наи
большее мутагенное воздействие? 2. Какие мутаге
ны называют супермутагенами? Приведите приме
ры. 3. Какова причина разрывов хромосом при их
облучении и каковы последствия облучения? 4. Как
осуществляется фотореактивация и темповая репа
рация ДНК? 5. Каковы возможности использова
ния мутагенов в селекции микроорганизмов, расте
ний и животных?
80. Генетика человека.
Анализ родословных
Генетика человека изучает явления
наследственности и изменчивости в раз
личных популяциях людей, особенности
наследования нормальных и патологи
ческих признаков,зависимость заболева
ний от генетической предетерминации и
условий окружающей среды, в том числе
от социальных условий жизни. Задачей
медицинской генетики является выявле
ние и профилактика наследственных бо
лезней.
Особенности генетики человека.
Часто общебиологические и генетичес
кие закономерности, открытые на живот
277
ных, противопоставляются таковым у че
ловека. Однако известно, что человека от
животных отличает вторая сигнальная
система и социальные условия жизни, но
биологические и генетические законо
мерности у них общие. Это означает, что
основные закономерности наследствен
ности и изменчивости имеют универсаль
ное значение и в полной мере приложимы
к человеку.
Изучение наследственности и измен
чивости человека связано с большими
трудностями, основными причинами ко
торых являются:
1) невозможность
направленных
скрещиваний для последующего ге
нетического анализа;
2 ) невозможность эксперименталь
ного получения мутаций;
3) позднее половое созревание;
4) малое количество потомков в каж
дой семье;
5) медленная смена поколений;
6) невозможность обеспечения оди
наковых и строго контролируемых ус
ловий для развития потомков от раз
ных браков;
7) недостаточная точность регистра
ции наследственных признаков и не
большая родословная;
8 ) сложный кариотип (2п = 46) с
большим числом групп сцепления.
Несмотря на все эти затруднения, ус
пехи в познании генетики человека в пос
ледние годы очень велики.
Невозможность направленных скре
щиваний компенсируется тем, что у ис
следователя есть возможность из огром
ной человеческой популяции выбрать из
многих тысяч брачных пар именно те, ко
торые необходимы для генетического ана
лиза. Метод гибридизации соматических
клеток позволяет экспериментально, из
учать локализацию генов в хромосомах,
проводить анализ групп сцепления и т. д.
Методы генетики человека. При из
учении наследственности и изменчивости
278
у человека используются следующие ме
тоды: генеалогический, близнецовый, по
пуляционно-статистический, дерматоглифический, биохимический, цитогене
тический, методы гибридизации сомати
ческих клеток и моделирования.Традици
онные (генеалогический и близнецовый)
и новейшие методы, используемые в ге
нетике человека, отражают его особен
ности как генетического объекта.
Генеалогический метод, предложен
ный в конце XIX в. Ф. Г а л ь т о н о м,
позволяет выяснить родственные связи и
проследить наследование нормальных
или патологических признаков среди
близких и дальних родственников в дан
ной семье на основе составления родос
ловной -— генеалогии. Если есть родос
ловные, то, используя суммарные данные
по нескольким семьям, можно опреде
лить тип наследования признака — до
минантный или рецессивный, сцеплен
ный с полом или аутосомный, а также его
моногенность или полигенность.
Генеалогическое изучение какой-либо
семьи начинается с установления носите
ля наследственного признака — пробан
да. В случае патологического проявления
признака пробанд является исходным па
циентом. Братья и сестры пробанда на
зываются сибсами. При составлении ро
дословной принято пользоваться соот
ветствующими обозначениями (рис.
161).
Анализ родословной, представленной
на рисунке 162, показывает, что полидак
тилия (лишние пальцы) передается из по
коления в поколение лицам любого пола,
т. е. наследуется как доминантный ауто
сомный признак. К числу признаков, нас
ледующихся по такому типу, относятся
«габсбургская губа» (толстая выпячен
ная нижняя губа, прослеженная, начиная
с XV в., в династии Габсбургов), брахидактилия, или короткопалость, ахондроп
лазия — карликовость, связанная с рез
ким укорочением конечностей, веснушки,
катаракта глаз, хрупкость костей и др.
□о<Ш«Мн©шйи
М у ж ч ина
Л
В ы ки д ы ш
Ж ен щ и н а
_|_
М ед иц инский аборт
Пол н е и з в е с т е н
□!
Лично обсл ед ован
О бладатели
и зу ч а е м о го
пр изнака
Ребенок с уро д ствам и
О-П
0
- 0
О О
Брак
Н еоф ициальны й брак
Р одственны й брак
Р од ите ли
[Г е т е р о з и го т н ы е
и зу ч а е м о го
] пр изнака
I (р е ц е с с и в н о г о г е н а )
J носители
Д ети
М и кр о пр и зн а к
Р а зн о я й ц е в ы е близнецы
У м е р д о го д а
Умер в
п
лет
М ер тво рож д е н ны е
О д н оя йц е вы е близнецы
Б е зд е тны й брак
Рис. 163. Родословная по фенилкетонурии (ре
цессивное наследование).
Основными признаками аутосомнорецессивного типа наследования являют
ся относительно небольшое число боль
ных в родословной, наличие больных по
«горизонтали» (болеют сибсы — род
ные, двоюродные). Родители больного
ребенка часто фенотипически здоровы,
но являются гетерозиготными носителя
ми рецессивного гена. Пример рецессив
ного наследования приведен на рисунке
163. К числу признаков, наследующихся
по такому типу, относятся рыжие волосы,
альбинизм, подверженность полиомие
литу, фенилкетонурия и др.
В случае близкородственных браков
вероятность рождения ребенка, страда
ющего рецессивно наследуемой болез
нью, сильно возрастает вследствие уве
личения вероятности встречи Тамет, не
сущих мутантные гены. Яркий пример
.проявления патологического рецессив
ного. Признака при родственном1браке
показан на рисунке 164. Из этой родо
словной видно, что в брак вступили сиб
сы четвертой степени родства. От двух
родственных браков появилось в одной
семье 4 ребенка из 8, а в другой — 2 из
280
5, страдающих тяжелым наследствен
ным заболеванием — идиотией ТеяСакса (причина этой болезни — накоп
ление липидов в нервных клетках мозга
и их разрушение). По-видимому, все
четвероюродные сибсы были гетерози
готными носителями этого рецессивно
го гена, полученного ими от общего
предка.
Существует тип наследования, сцеп
ленного с полом. Признаки, обусловлен
ные геном, локализованным в ^-хромосо
ме, могут быть как доминантными, так и
рецессивными. При доминантном насле
довании признак одинаково проявляется
как у мужчин, так и у женщин. Примером
такого признака является особая форма
рахита, устойчивого к лечению кальци
феролами (витамин D). У человека най
дено более 100 сцепленных с полом ре
цессивных признаков, причем около по
ловины из них связано с заболеваниями
глаз.
При рецессивном наследовании забо
леваниями, сцепленными с А-хромосомой, как правило, страдают мужчины. В
редких случаях эти признаки могут проя
виться и у женщины, если ее отцом был
больной мужчина, а мать была гете
розиготной поданному признаку. Приме
рами таких заболеваний являются даль
тонизм, гемофилия.
У человека обнаружены также груп
пы признаков, которые наследуются
сцепленно: фенилкетонурия и группы
крови системы АВО, цвет волос и кариес
зубов и др.
?
1. Каковы причины затруднений изучения на
следственности и изменчивости человека и как они
решаются? 2. Приведите пример родословной
семьи, в которой обнаружена аномалия, наследую
щаяся по типу рецессивного и сцепленного с полом
признака. 3. В чем основные отличия аутосомно-доминантного и аутосомно-рецессивного типов насле
дования у человека? 4. В чем заключается опас
ность близкородственных браков?
Рис. 164. Родословная по идиотии в двух родственных семьях.
81. Близнецы
Основоположником близнецового
метода в генетике человека был Ф. Гальтон, который в 1876 г. предложил срав
нивать похожих и различающихся между
собой близнецов для выяснения степени
влияния наследственности и среды на
проявление признаков.
Известно, что у человека близнецы
бывают однояйцевыми (монозиготными)
и разнояйцевыми (дизиготными). Одно
яйцевые, или идентичные, близнецы
развиваются из одной яйцеклетки, опло
дотворенной одним сперматозоидом. Они
всегда одного пола и поразительно похо
жи друг на друга, так как имеют один и
тот же генотип. Кроме того, у них одина
ковая группа крови, одинаковые отпечат
ки пальцев и почерки, их путают даже ро
дители и не различают по запаху собаки.
Только у идентичных близнецов на 100 %
удаются пересадки органов, поскольку
набор белков у них одинаков и переса
женные ткани не отторгаются. Доля од
нояйцевых близнецов у человека состав
ляет около 35—38 % от общего их числа.
Разнояйцевые, или дизиготные,
близнецы развиваются из двух разных
яйцеклеток, одновременно оплодотво
ренных различными сперматозоидами.
Дизиготные близнецы могут быть как од
ного, так и разного пола, а с генетической
точки зрения они сходны не больше, чем
обычные братья и сестры.
У человека чаще встречаются двойни,
реже тройни и совсем редко — четверни
и пятерни. Статистика показывает, что в
среднем частота рождения близнецов
составляет 1 %, а вот пять детей появля
ются не более чем один раз на 54 млн ро
дов. Это означает, что рождение близне281
цов у человека все-таки исключение (хо
тя, например, уроженка Австрии Шейнберг родила 69 детей за 27 родов —
4 четверни, 7 тройняшек и 16 двойня
шек).
Близнецовый метод основан на срав
нении пар однояйцевых близнецов с од
нополыми двуяйцевыми близнецами, а
также сравнении пар однояйцевых близ
нецов, воспитанных вместе и раздельно.
Если изучаемый признак (например, цвет
волос, глаз, форма носа, глаз, губ, ушных
раковин, группа крови и др.) проявляется
у обоих близнецов пары, их называют
конкордантными (от лат. concordari —
быть согласным, сходным), если только у
одного из них,— то дискордантными.
Степень конкордантности определяется
как отношение числа конкордантных пар
к общему числу всех изученных пар близ
нецов. Следовательно, конкордантность — это процент сходства по изучае
мому признаку, а дискордантность —
частота различий.
Для получения точной оценки степени
конкордантности необходимо изучить
очень большую группу пар близнецов по
ряду наследственных детерминированных
признаков. Это позволяет сделать опре
деленные выводы о степени влияния на
следственности и факторов среды на раз
витие какого-либо нормального или па
тологического признака. Поскольку у
идентичных близнецов одинаковые гено
типы, то все различия между ними опре
деляются только средой.
Большой интерес для решения ряда
вопросов имеют случаи, когда близнецы
по каким-либо причинам росли и воспи
тывались в разных условиях. Сохраняю
щаяся конкордантность по ряду призна
ков в этом случае объясняется влиянием
только генотипа.
Некоторые примеры сравнения одно
яйцевых и разнояйцевых близнецов (ОБ
и РБ соответственно), представленные в
таблице 9, показывают, что у ОБ конкор
дантность значительно выше, чем у РБ,
282
Т а б л и ц а 9. Конкордантность некоторых приз
наков у однояйцевых и разнояйцевых близнецов
человека.
Признаки
Нормальные
Группа крови
Форма бровей
Цвет глаз
Цвет волос
Папиллярные линии кистей рук
Патологические
Косолапость
«Заячья губа»
Синдром Дауна
Рахит
Паралитический полиомиелит
Бронхиальная астма
Корь
Туберкулез
Скарлатина
Дифтерит
Рак
Эпилепсия
Шизофрения
Конкордантность, %
ОБ
РБ
100
100
99,5
97
92
46
51
28
23
40
23
33
89
88
36
19
95
37
84
50
16
67
80
2
5
7
22
6
4.8
87
15
47
38
14
3
13
однако степень сходства для разных приз
наков варьирует. Это означает, что на оп
ределение группы крови, формы бровей,
цвета глаз и волос у ОБ среда почти не
оказывает влияния, а решающее воздей
ствие имеет генотип. Значительна роль
наследственных факторов в развитии у
детей рахита, шизофрении, туберкулеза и
др. Напротив, доля наследственности в
возникновении косолапости и бронхиаль
ной астмы очень мала. Следовательно,
признаки, характеризующиеся высокой
дискордантностью, в основном определя
ются влиянием условий среды.
Следует указать, что и монозиготные
близнецы могут отличаться по качествен
ным признакам. Причиной отличий могут
быть мутации соматических клеток и ва
риации в экспрессии генов на всех этапах
их индивидуального развития. Это лиш
ний раз подтверждает вывод, что все
признаки организма человека определя
ются в результате взаимодействия гено
типа и условий среды.
■. 1. Каким образом можно определить относи
тельную роль генотипа и условий среды в формиро
вании признаков у человека? 2. Приведите примеры
признаков или заболеваний, определенных близне
цовым методом. 3. Что такое конкордантность?
Когда и как используется этот показатель в ге
нетике человека?
82. Цитогенетический,
популяционный и другие методы
генетики человека
Цитогенетический метод. Этот метод
основан на микроскопическом исследо
вании структуры хромосом у здоровых и
больных людей.
В 1956 г. шведские ученые Д ж .
Тийо и А. Л е в а н установили, что
нормальный кариотип человека включает
46 хромосом, из них 22 пары аутосом и
одна пара половых хромосом (рис. 165).
Идентификация всех хромосом человека
стала возможной благодаря применению
till ни
ил
XX
и
XX
методики культуры тканей и метода диф
ференциальной окраски хромосом. Воз
можность культивирования клеток чело
века (чаще всего лейкоцитов перифери
ческой крови) позволяет получать доста
точно большой материал для описания
цитологических особенностей исследуе
мого человека.
Цитологический контроль применяют
при диагностике ряда наследственных за
болеваний, которые связаны с явлениями
анеуплоидии и различными хромосомны
ми перестройками. Он позволяет также
изучать старение тканей на основе иссле
дований возрастной динамики структуры
клеток, устанавливать мутагенное дей
ствие факторов внешней среды на чело
века и т. д.
В последние годы цитогенетический
метод приобрел большое значение в связи
с возможностями генетического анализа
человека, которые открыла гибридизация
соматических клеток в культуре. Получе-
ку»«
к
л
»
и
7
и mi их ¥
му ж ч ина
XX
м
Рис. 165. Набор хромосом человека: слева — 22 пары аутосом, справа— половые хромосомы.
283
ние межвидовых гибридов клеток (напри
мер, человека и мыши) позволяет в значи
тельной степени преодолеть проблемы,
связанные с невозможностью направлен
ных скрещиваний, локализовать ген в оп
ределенной хромосоме, установить группу
сцепления для ряда признаков и т. д. Объе
динение генеалогического метода с цитоге
нетическим, а также с новейшими метода
ми генной инженерии значительно ускори
ло процесс картирования генов у человека.
Популяционный метод. Этот метод
позволяет изучать распространение от
дельных генов или хромосомных анома
лий в человеческих популяциях. Он осно
ван главным образом на данных демогра
фической статистики, которая занимает
ся исследованием наследственной струк
туры населения. При этом обычно произ
водят непосредственное выборочное ис
следование части популяции либо прово
дят опрос путем анкетирования. Выбор
способа зависит от цели исследования.
Последний этап состоит в статистичес
ком анализе полученных результатов. В
результате становится возможным опре
делить частоту генов в различных группах
населения, частоту гетерозиготных носи
телей ряда наследственных болезней и
аномалий. Это имеет важное значение не
только для анализа распространения нас
ледственных болезней человека, но и для
оценки последствий родственных браков,
которые особенно часты в изолятах, а
также для выяснения генетической исто
рии человеческой популяции в целом.
Изучение распространенности генов
на определенных территориях показыва
ет, что в этом отношении их можно разде
лить на три категории: 1) имеющие уни
версальное распространение (к их числу
относится большинство известных ге
нов); 2 ) характеризующиеся широким
распространением (так, примерно каж
дый сотый житель Европы гетерозиготен
по типу амавротической идиотии, тогда
как заболевают этой болезнью из 1 млн
только 25 человек, у которых этот ген
284
проявляется в гомозиготном состоянии);
3) встречающиеся локально, преимущес
твенно в определенных районах (ген сер
повидноклеточной анемии распространен
в странах Тропической Африки и Среди
земноморья, а ген, определяющий врож
денный вывих бедра,— у коренных жите
лей северо-востока России).
Биохимические методы. Эти методы
изучения наследственности человека по
могают обнаружить ряд заболеваний об
мена веществ (углеводного, аминокис
лотного, липидного и др.) при помощи,
например, исследования биологических
жидкостей (крови, мочи, амниотической
жидкости) путем качественного или ко
личественного анализа. Причиной этих
болезней является изменение активности
определенных ферментов.
С помощью биохимических методов
открыто около 500 молекулярных болез
ней, являющихся следствием фенотипи
ческого проявления мутантных генов.
При различных типах заболеваний удает
ся либо определить сам аномальный бе
лок-фермент, либо промежуточные про
дукты обмена. По результатам биохими
ческих анализов устанавливается диагноз
болезни, на основании которого опреде
ляются методы лечения. Ранняя диагнос
тика и применение различных диет на
первых этапах постэмбрионального раз
вития позволяют излечить заболевание
или хотя бы облегчить состояние боль
ных с неполноценными наследственными
ферментными системами.
Дерматоглифический метод. Дерма
тоглифика (от греч. derma — кожа,
gliphe — рисовать) — это изучение
рельефа кожи на пальцах, ладонях и по
дошвах стоп. В отличие от других частей
тела здесь имеются эпидермальные выс
тупы — гребни, образующие сложные
узоры, причем строго индивидуальные.
На Земле нет двух людей с одинаковыми
рисунками на пальцах (кроме однояйце
вых близнецов). В 1892 г. Ф. Гальтон раз
работал классификацию этих узоров.
Дерматоглифика включает три разде
ла: дактилоскопия (изучение узоров на
подушечках пальцев), пальмоскопия
(рисунки на ладонях) и плантоскопия
(изучение дерматоглифики подошвенной
поверхности стопы).
Закладка узоров на подушечках паль
цев происходит между 10-й и 19-й неде
лями внутриутробного развития; у 20-недельного плода узор уже хорошо разли
чим. Полное формирование деталей
строения дактильных узоров отмечается к
шести месяцам, после чего они остаются
неизменными до конца жизни. При по
вреждении кожи их рисунок восстанавли
вается в своем первоначальном виде.
Дерматоглифические исследования
имеют важное значение в определении
зиготности близнецов, в диагностике не
которых наследственных заболеваний, в
судебной медицине, в криминалистике
для идентификации личности. В послед
ние годы метод дерматоглифики стали ис
пользовать в клинической генетике для
подтверждения диагноза хромосомных
синдромов у людей с изменениями карио
типа.
Как и любая другая дисциплина, сов
ременная генетика человека использует
методы смежных наук: физиологии, моле
кулярной биологии, генной инженерии,
биологического и математического моде
лирования и т. д. Значительное место в ре
шении проблем генетики человека и меди
цинской генетики занимает онтогенети
ческий метод, который позволяет рассмат
ривать развитие нормальных и патологи
ческих признаков в ходе онтогенеза.
. 1. Каковы возможности и перспективы исполь
зования цитогенетического метода генетического
анализа человека? 2. Какие категории распростра
ненности генов на определенных территориях уста
навливаются популяционным методом? 3. Какова
разрешающая способность биохимических методов
изучения наследственности человека? 4. Каково
практическое значение дерматоглифического мето
да генетики человека?
83. Наследственные б о л е з н и
человека, и х л е ч е н и е
и профилактика
К настоящему времени зарегистриро
вано более 2000 наследственных болез
ней человека, причем большинство из
них связано с психическими расстрой
ствами. Поданным Всемирной организа
ции здравоохранения, благодаря приме
нению новых методов диагностики, еже
годно регистрируется в среднем три но
вых наследственных заболевания, кото
рые встречаются в практике врача любой
специальности: терапевта, хирурга, нев
ропатолога, акушера-гинеколога, педи
атра, эндокринолога и т. д. Болезней, не
имеющих абсолютно никакого отноше
ния к наследственности, практически не
существует. Течение разных заболеваний
(вирусных, бактериальных, микозов и да
же травм) и выздоровление после них в
той или иной мере зависят от наслед
ственных иммунологических, физиоло
гических, поведенческих и психических
особенностей индивидуума.
Исследования,
проведенные А.
С т и в е н с о н о м в Северной Ирландии,
показали, что около 4 % новорожденных
отягощены наследственными болезнями.
Эта цифра не включает выкидыши и мертворождения, частота которых составля
ет около 14 % от зарегистрированных бе
ременностей, а также не учитывает такие
распространенные заболевания, как диа
бет и шизофрения.
Изучение и возможное предотвраще
ние последствий генетических дефектов
человека является предметом медицин
ской генетики. Условно наследственные
болезни можно подразделить на три
большие группы: болезни обмена ве
ществ, молекулярные болезни, которые
обычно вызываются генными мутациями,
и хромосомные болезни.
Генные мутации и нарушения обме
на веществ. Генные мутации у человека
являются причиной многих наследствен
285
ных заболеваний. Некоторые из них были
известны в клинике давно, но механизм
их возникновения стал понятен только
после создания молекулярной генетики и,
в частности, после экспериментального
обоснования ее основного положения:
один ген — один фермент (один белок,
или полипептидная цепь).
Мутация может выражаться в повы
шении или понижении активности фер
мента, вплоть до полного его отсутствия.
Фенотипически такие мутации проявля
ются как наследственные болезни обмена
веществ, которые определяются по от
сутствию или избытку продукта опреде
ленной биохимической реакции.
Генные мутации классифицируют по
их фенотипическому проявлению: болез
ни, связанные с нарушением аминокис
лотного, углеводного, липидного, мине
рального обмена, обмена нуклеиновых
кислот. Примером нарушения обмена
аминокислот является фенилкетонурия,
которая наследуется по аутосомно-рецессивному типу. В результате генной мута
ции возникает недостаточность фермента
фенилаланингидроксилазы, катализирую
щего превращение фенилаланина в тиро
зин. При этом фенилаланин не усваивает
ся организмом. Нарушаются и последую
щие звенья биохимической цепи реакций,
в ходе которых образуются не только ти
розин, но и адреналин, норадреналин,
пигмент меланин. Фенилаланин превра
щается в фенилпировиноградную и фенилмолочную кислоты, которые накапли
ваются в крови и выделяются с мочой. В
конечном итоге эти крайне токсические
вещества приводят к нарушению высшей
нервной деятельности, слабоумию, рас
стройству регуляции двигательных фун
кций. Своевременный перевод новорож
денного уже с первых недель жизни на
специальную диету, лишенную фенилала
нина, предотвращает развитие заболеванния, и дети развиваются нормально.
Другим примером нарушения амино
кислотного метаболизма является аль
286
бинизм — относительно безобидная бо
лезнь, встречающаяся в странах Запад
ной Европы с частотой 1:25 000. Причи
ной этого заболевания является дефект
фермента тирозиназы, в результате чего
блокируется превращение тирозина в ме
ланин. У альбиносов молочный цвет ко
жи, очень светлые волосы и отсутствует
пигмент в радужной оболочке глаз. Они
имеют повышенную чувствительность к
солнечному свету, который вызывает у
них воспалительные заболевания кожи.
Одним из наиболее распространен
ных заболеваний углеводного обмена яв
ляется сахарный диабет. Эта болезнь
связана с дефицитом гормона инсулина,
что приводит к нарушению процесса об
разования гликогена и повышению уров
ня глюкозы в крови; при этом часть глю
козы выводится с мочой. Лечение произ
водится путем введения в организм не
достающего гормона, получаемого обыч
но из поджелудочной железы крупного
рогатого скота, а в последнее время и из
бактерий, в которые введен инсулиновый
ген человека.
Ряд патологических признаков (ги
пертония, атеросклероз, подагра и др.)
определяются не одним, а несколькими
генами — явление полимерии. Это бо
лезни с наследственным предрасположе
нием, которые в большей степени зави
сят от условий среды: в благоприятных
условиях такие заболевания могут и не
проявиться.
Молекулярные болезни. Необходимо
отметить, что граница между болезнями
обмена веществ и молекулярными болез
нями условна. Она скорее всего отража
ет степень осведомленности о причинах
наследственной аномалии: дефект на мо
лекулярном уровне, нарушение в струк
туре молекулы ДНК, нарушения репли
кации и репарации ДНК.
Подробно изучено более 100 так на
зываемых гемоглобинопатий, т. е. бо
лезней, возникающих из-за наследствен
ного нарушения структуры гемоглобина
(уже приводился пример серповиднокле
точной анемии). В результате рецессив
ного аутосомного дефекта эксцизионной
репарации и самой репаративной ДНКполимеразы возникает уже упоминавша
яся пигментная ксеродерма. Механизмы
репарации нарушены и в случае прежде
временного старения.
Хромосомные болезни. Этот тип
наследственных заболеваний связан с из
менениями числа или структуры хромо
сом. Частота хромосомных аномалий у
новорожденных составляет от 0,6 до 1 %,
а на стадии 8— 12 недель их имеют около
3 % эмбрионов. В основном это анеуплоидии. Среди самопроизвольных выкиды
шей частота хромосомных аномалий рав
на примерно 30 %, а на ранних сроках (до
двух месяцев) — 50 % и выше. К леталь
ному исходу в этом периоде приводят по
липлоидии.
Характерным отличием большинства
хромосомных болезней от болезней, при
чинами которых оказываются генные му
тации, является их повторное возникно
вение, а не наследование от предшеству
ющих поколений. Хромосомные и геном
ные мутации возникают как в гаметогенезе родителей, так и непосредственно в зи
готе или на ранних стадиях ее дробления.
Любые хромосомные перестройки при
водят к патологии.
У человека описаны все типы хромо
сомных и геномных мутаций, включая по
ли- и анеуплоидию. Анеуплоидия может
быть двух типов — моносомия, когда
хромосомный комплект уменьшается (2я
— 1), и полисомия, когда число отдель
ных хромосом увеличено, например трисомия (2га + 1). Особенно тяжелы моносомии.
Моносомия всего организма описана
для Х-хромосомы. Это синдром Шерешевского — Тернера (44 + X), проявля
ющийся у женщин, для которых харак
терны патологические изменения тело
сложения (малый рост, короткая шея),
нарушения половой системы (отсутствие
большинства женских вторичных поло
вых признаков), умственная ограничен
ность (рис. 166). Частота встречаемости
этой аномалии 1:4000 —■5000.
Женщины-гагригалшкы (44 + XXX),
как правило, отличаются нарушениями
полового, физического и умственного
развития, хотя у части больных эти приз
наки могут не проявляться. Известны
случаи плодовитости таких женщин. Час
тота синдрома 1:1000.
Мужчины с синдромом Клайнфельтера (44 + XXY) характеризуются нару-
Рис. 166. Больная с синдромом Шерешевского — Тернера (девочка 7 л е т).
287
шением развития и активности половых
желез, евнухоидным типом телосложе
ния: более узкие, чем таз, плечи, оволо
сение и отложение жира на теле по жен
скому типу, удлиненные по сравнению с
туловищем руки и ноги. Отсюда и более
высокий рост. Эти признаки в сочетании
с некоторой психической отсталостью
проявляются у относительно нормально
го мальчика начиная с момента полового
созревания.
Синдром Клайнфельтера наблюдает
ся не только при полисомии по Х-хромосоме (XXY, XXXY, XXXXY), но и по F-xpoмосоме (XYY, XXYY, XXYYY). Частота син
дрома 1:1000.
Из числа аутосомных болезней наи
более изучена трисомия по 21 -й хромосо
ме, или синдром Дауна. Поданным раз
ных авторов, частота рождения детей с
синдромом Дауна составляет 1:500— 700
новорожденных, а за последние десяти
летия частота трисомии-21 увеличилась.
Типичные признаки больных с син
дромом Дауна — маленький нос с широ
кой плоской переносицей, раскосые гла
за с эпикантусом — нависающей склад
кой над верхним веком, деформирован
ные небольшие ушные раковины, полу
открытый рот, низкий рост, умственная
отсталость. Около половины больных
имеют порок сердца и крупных сосудов
(рис. 167).
Существует прямая связь между рис
ком рождения детей с синдромом Дауна и
возрастом матери. Установлено, что 22—
40 % детей с этой болезнью рождают ма
тери старше 40 лет, составляющие лишь
2—3 % женщин детородного возраста.
Здесь рассмотрены лишь некоторые
примеры генных и хромосомных болез
ней человека, которые, однако, дают оп
ределенное представление о сложности и
хрупкости его генетической организации.
Профилактика и лечение наслед
ственных болезней. Основным путем
предотвращения наследственных заболе
ваний у человека является их профилак288
Рис. 167 Больной с синдромом Дауна.
тика. Для этого во многих странах мира, в
том числе и в Беларуси, существует сеть
учреждений, обеспечивающих медико
генетическое консультирование населе
ния. В первую очередь его услугами дол
жны пользоваться лица, вступающие в
брак, у которых имеются генетически
неблагополучные родственники.
Генетическая консультация обяза
тельна при вступлении в брак родствен
ников, лиц старше 30—40 лет, а также
работающих на производстве с вредными
условиями труда. Врачи и генетики смо
гут определить степень риска рождения
генетически неполноценного потомства и
обеспечить контроль за ребенком в пери
од его внутриутробного развития. Следу
ет отметить, что курение, употребле
ние алкоголя и наркотиков матерью
или отцом будущего ребенка резко по
вышают вероятность рождения младен
ца с тяжелыми наследственными неду
гами.
В случае рождения больного ребенка
иногда возможно его медикаментозное,
диетическое и гормональное лечение.
Наглядным примером, подтверждающим
возможности медицины в борьбе с нас
ледственными болезнями, может слу
жить полиомиелит. Эта болезнь характе
ризуется наследственной предрасполо
женностью, однако непосредственной
причиной заболевания является вирусная
инфекция. Проведение массовой имму
низации против возбудителя этой болез
ни позволило избавить всех наследствен
но предрасположенных к ней детей от тя
желых последствий заболевания. Диети
ческое и гормональное лечение, как ска
зано нами выше, успешно применяется
при лечении фенилкетонурии и сахарного
диабета.
?■ . 1. Каковы
к
причины различных типов наслед
ственных заболеваний человека? 2. Каковы послед
ствия генных мутаций, моносомии и трисомии у че
ловека? Приведите примеры. 3. Мужчина, страда
ющий наследственной болезнью, женился на здоро
вой женщине. У них было 8 детей: 4 девочки и 4
мальчика. Все девочки (но ни один из мальчиков)
унаследовали болезнь отца. Каков тип наследова
ния этой болезни: а) аутосомно-рецессивный; б) аутосомно-доминантный; в) сцепленный с У-хромосо
мой; г) доминантный, сцепленный с X-хромосомой;
д) рецессивный, сцепленный с ^-хромосомой?
Обоснуйте ответ. 4. Каковы возможности лечения
наследственных аномалий у человека? 5. Каковы
задачи медико-генетических консультаций?
84. Проблемы генетической
безопасности
Интенсивное развитие научно-техни
ческой революции поставило человечес
тво перед проблемой долговременных и
необратимых изменений окружающей
среды, которые приняли угрожающие
масштабы. В последние десятилетия наб
людается резкое сокращение площади
лесов и естественных луговых угодий,
загрязнение водоемов, атмосферы и поч
вы. В частности, катастрофа на Черно
быльской атомной станции (апрель
1986 г.) привела к загрязнению радио
нуклидами пятой части территории Бе
ларуси и создала огромную опасность для
10. Общая биология, 10 кл.
биосферы в целом, частью которой явля
ется человек.
Особенно сильное воздействие на
среду оказывают следующие факторы ан
тропогенного происхождения: пестици
ды, тяжелые металлы, оксиды углерода,
серы, азота, разливы нефти, сточные во
ды промышленных предприятий, хими
ческие и радиоактивные отходы. Наряду с
экологическими последствиями и отрица
тельным воздействием на здоровье ны
нешнего поколения людей загрязнение
биосферы имеет еще и далеко идущие ге
нетические последствия. Например, даже
небольшое повышение естественного
фона радиации (на l /З ) в результате ис
пытаний ядерного оружия может привес
ти к появлению в каждом поколении до
полнительно 20 млн человек с тяжелыми
наследственными нарушениями. Нетруд
но представить себе, какую опасность не
только для населения Беларуси, Украины
и России, но и для всего человечества
представляют такие события, как авария
на Чернобыльской АЭС.
Результатом осознания этого обстоя
тельства было появление в 60— 70-х гг.
новой науки — генетической токсико
логии.
Генетическая токсикология. Рас
пространение таких мутагенов, как про
мышленные и радиоактивные отходы,
пестициды (инсектициды, гербициды,
фунгициды), многие лекарственные и
косметические средства, чревато повы
шением частоты мутаций, а следователь
но, увеличением генетического груза че
ловечества — рецессивных мутаций,
приводящих к развитию наследственных
болезней. Эти мутации остаются скрыты
ми до тех пор, пока не перейдут в гомози
готное состояние. Кроме того, рецессив
ные мутации могут длительно переда
ваться из поколения в поколение, причем
концентрация их постепенно увеличива
ется вследствие накопления повторных
мутаций (рис. 168, с. 290). Это ведет к
росту заболеваемости и ставит сложные
289
моральные и экономические проблемы
перед обществом.
Генетическая токсикология изучает
мутагенную активность факторов антро
погенной природы, прежде всего хими
ческих соединений, разрабатывает мето
ды и способы оценки их генетической ак
тивности. Эта наука ставит своей целью
свести к минимуму степень риска мута
генных воздействий, уменьшить генети
290
ческую опасность во всех областях чело
веческой деятельности прежде всего за
счет снижения контактов человека с му
тагенами. Новые химические соединения
и другие генетически активные агенты
изымаются из употребления или их при
менение строго ограничивается.
Антимутагенез. Этот процесс вклю
чает такие воздействия на клетку и орга
низм, которые блокируют или уменына-
ют вероятность возникновения спонтан
ных и индуцированных мутаций. Подоб
ные воздействия антимутагенов могут
проявляться по-разному: в одних случаях
происходит стимуляция системы инакти
вации или подавление системы актива
ции промутагенов, в других — стимуля
ция процессов безошибочной репарации
или непосредственное модифицирование
мутагена, «отвлечение» его от генетичес
кого материала.
Важнейший природный антимутаген,
образующийся в самом организме, —
фермент к а т а л а з а . Этот фермент при
останавливает мутагенное действие об
разующегося в клетках пероксида водо
рода (Н20 2) и таким образом поддержи
вает частоту спонтанных мутаций на низ
ком уровне.
Антимутагенное действие оказывает
низкая температура, видимый свет, а так
же некоторые витамины и серосодержа
щие аминокислоты — цистеин, цистин и
метионин. Названные аминокислоты
способны уменьшать летальный эффект
ионизирующей радиации. Ненасыщен
ные жирные кислоты, таниновая кислота
и катехин, содержащийся в чае и кофе, и
ряд других соединений также обладают
антимутагенной активностью по отноше
нию к отдельным мутациям.
Генетический мониторинг окружаю
щей среды. Мониторинг — это ком
плексная система наблюдений, оценки и
прогноза состояния природной среды, из
менения биосферы или ее отдельных эле
ментов под влиянием человеческой дея
тельности. Мониторинг может быть ло
кальным (в отдельном районе), глобаль
ным (для всей планеты), абиотическим
(геофизическим) и биологическим.
В последние годы интенсивно разви
вается генетический мониторинг —
служба регистрации числа мутаций и
сравнение темпов мутирования в после
дующих поколениях по сравнению с пре
дыдущими. Мониторинг включает приме
нение тестов для выявления возникаю
щих мутаций и анализа мутагенных фак
торов. В качестве тест-объектов исполь
зуются культуры микроорганизмов, лей
коциты и др.
Предстоит дальнейшая разработка
унифицированных методов оценки раз
личных соединений, вводимых в среду, и
параметров слежения за генофондом по
пуляции. Перспективная цель монито
ринга — анализ мутагенной обстановки
среды, наблюдение за мутагенными изме
нениями в отдельных регионах и в биос
фере в целом. Генетическая служба будет
иметь общегосударственное и меж
дународное значение.
? 1., Какова
,
опасность испытания ядерного ору
жия с позиций генетики? 2. Что такое генетическая
токсикология? 3. Какие факторы генетической
опасности вам известны? 4. Что такое антимутаге
нез? Приведите примеры антимутагенов? б. На ка
ких этапах возможно вмешательство в мутацион
ный процесс, изменяющее его результаты. 6. Что
такое генетический мониторинг и каковы его цель и
задачи?
ГЛАВА VII
Регуляция и координация
жизнедеятельности организмов
85. Виды регуляторных систем
у животных и растений
Упорядоченное и эффективное функ
ционирование сложного многоклеточно
го животного организма обеспечивается
согласованной работой двух тесно взаи
модействующих мещлу собой систем —
нервной и эндокринной. Они контроли
руют активность отдельных частей орга
низма, обеспечивают передачу информа
ции от одних частей к другим и между час
тями и центрами, которые сопоставляют,
интегрируют и отдают надлежащие ко
манды. Благодаря согласованной дея
тельности нервной и эндокринной систем
обеспечивается важнейшее свойство ор
291
ганизма — саморегуляция физиологи
ческих функций.
Несмотря на то что сферы влияния
двух регулирующих систем во многом пе
рекрываются, между ними есть два су
щественных различия: первое касается
быстроты действия, второе — величины
объекта (мишени). Быстрые реакции и
четкие локальные воздействия (напри
мер, на часть волокон определенной
мышцы, не затрагивая остальных) осу
ществляются нервной системой. Эндок
ринная система контролирует относи
тельно медленные и длительно протекаю
щие процессы, например, рост организ
ма, размножение, обмен веществ, под
держание постоянного содержания глю
козы в крови и т. п. Вырабатываемые же
лезами эндокринной системы биологи
чески активные вещества ;— гормоны —
доставляются органам-мишеням кровью,
одновременно оказывая воздействие на
многие органы и системы органов, т. е. их
действие генерализованно.
Однако резкой грани между нервной и
гуморальной регуляцией провести нель
зя. Нервная система не только участвует
в регуляции функций эндокринных же
лез, но и играет важную роль в выработ
ке гормонов. Это происходит тогда, когда
аксон нейрона не передает нервный им
пульс эффектору, например мышце или
железе, а заканчивается свободно, и ак
тивное вещество, выделяемое им при
возбуждении, поступает во внутреннюю
среду организма, в кровь или гемолимфу,
оказывая воздействие на эффекторы,
расположенные вдали от выделяющего
его окончания аксона. В этом случае ак
тивные вещества, выделяемые нервной
клеткой, называют нейрогормонами, а
нервную клетку, их выделяющую, —
нейроинкреторной клет кой. Тесная
анатомическая и функциональная связь
между нервной системой и железами
внутренней секреции наблюдается у чле
нистоногих и позвоночных животных. Эта
взаимозависимость двух систем дает ос
292
нование говорить о нейро-гуморальной
регуляции функций организма.
Животные, в отличие от растений,
имеют обе регуляторные системы —
нервную и эндокринную. У растений
нервной системы нет, так как при прик
репленном образе жизни отпадает необ
ходимость в быстрых механизмах регуля
ции. Координация деятельности различ
ных клеток и тканей растительного орга
низма осуществляется с помощью фитогормонов, перемещающихся по сосудам
из одной части растения в другую.
?я , I. В
R чем сходство
и различие нервной и гумо
ральной регуляций функций организма? 2. Как про
исходит регуляция жизнедеятельности в раститель
ном организме?
86. Нервная система животных
В процессе эволюции животного мира
нервная ткань образовалась на ранних ее
этапах. Как система она впервые присут
ствует у примитивного типа кишечнопо
лостных. В последующих эволюционно
более сложно устроенных типах живот
ных развитие нервной системы происхо
дит в направлении увеличения числа ней
ронов, дифференциации их формы и фун
кциональной специализации, усложне
ния межнейронных связей и в образова
нии группировок из нейронов, обладаю
щих общими или связанными между со
бой функциями, и, наконец, в ее центра
лизации.
По мере совершенствования нервной
системы усиливается ее интегрирующая
функция. Чем сложнее и совершеннее
нервная система, тем сложнее и разнооб
разнее реакции организма на внешние
раздражения. Совершенствование нерв
ной системы напрямую связано с актив
ной мышечной деятельностью животных.
Развитие нервно-мышечной деятельнос
ти, дальностных рецепторов (зрения,
обоняния) особенно стимулируется хищ
ным образом жизни, активной охотой.
Поэтому возврат к другим способам пи
тания ведет к регрессу в строении орга
нов чувств, движения, упрощению нер
вной системы, а у паразитических живот
ных к регрессу органов вегетативной
жизни.
Нервная система беспозвоночных. У
кишечнополостных она наиболее прими
тивна и представляет собой сетевидное
соединение сравнительно равномерно
размещенных по телу нервных клеток,
называемое диффузной нервной систе
мой (рис. 169). Однако уже у некоторых
групп этого типа наблюдается концен
трирование нейронов вокруг рта и на по
дошве, в частности у гидроидных полипов
и актиний, а у сцифоидных медуз — по
краю зонтика.
Примитивность диффузной нервной
системы состоит в отсутствии длинных
проводящих путей и неразделении ее на
центральную и периферическую части.
Сеть относительно медленно проводит
раздражение по всем направлениям от
Рис. 169. Нервная система гидры: 1 — рот, 2 —
щупальца, 3 — нервные клетки.
нейрона к нейрону. Так как нейроны свя
заны с эпителиально-мускульными клет
ками, то волна возбуждения от какой-ли
бо точки тела распространяется далее и
сопровождается волной мышечных со
кращений. Реакции организма имеют не
точный, расплывчатый характер.
Следствием такого устройства нер
вной системы является способность лю
бого вырезанного кусочка тела кишечно
полостных к самостоятельным рефлек
сам. Множество связей между элемента
ми диффузной системы обеспечивают
также их широкую взаимозаменяемость и
тем самым большую надежность функци
онирования.
У плоских червей усиливается кон
центрация нервных клеток. Это проявля
ется в образовании продольных и попе
речных сгущений, которые формируют
продольные и кольцевые нервные ство
лы — ортогон (рис, 170, с. 294). В про
межутках меяеду стволами остаются
только элементы периферического отде
ла нервной системы. Дальнейшее совер
шенствование ортогона связано с умень
шением числа стволов и смещением все
большего числа нейронов к переднему
концу тела, что постепенно приводит к
образованию мозгового ганглия. Сюда
же смещаются и органы чувств: осяза
ния, зрения, вкуса.
Близкий к ортогону тип строения нер
вной системы имеют круглые черви.
Еще большая централизация и функ
циональная специализация нервных кле
ток характерна для третьего типа нервной
системы — узловой, или ганглионар
ной, свойственной кольчатым червям,
моллюскам и членистоногим. Его
отличает концентрация тел нервных кле
ток в компактные, четко выраженные
ганглии (узлы) (рис. 171, с. 294), внутри
которых образуется сплетение отростков
и осуществляется контакт между отдель
ными нейронами. Промежуточные учас
тки нервных стволов, соединяющих два
ганглия, полностью лишены тел нервных
293
2
3
4
клеток. Стволы состоят из параллельно
идущих аксонов, быстро проводящих
нервные импульсы. По образному выра
жению известного зоолога В. Н. Бекле
мишева, ганглии — это телефонные
станции, в которых возможно пере
ключение, а промежуточные участки
нервных стволов — простые пучки про
водов.
Нейроны, сосредоточенные в гангли
ях, образуют центральную нервную
систему. Резко возрастает роль нервных
узлов головного отдела. Происходит дифференцировка нейронов в соответствии с
выполняемыми функциями. Так, нейро
ны, по которым информация поступает в
нервные центры, называются центро
стремительными, или чувствитель
ными. Нейроны, по которым информа
ция от нервных центров передается к ис
полнительным органам (мышцам, желе
зам), называются центробежными, или
двигательными. Нервные клетки, вос
принимающие импульсы от других нейро-
Рис. 171. Ганглионарный тип нервной системы: а —- дождевого червя: 1 — «мозг», 2 — окологлоточ
ное кольцо, 3 — подглоточный ганглий, 4 — нерв, 5 — нервный тяж, 6 — ганглий; б — брюхоногого
моллюска: 1 — головной ганглий, 2 — ножной, 3 — мантийный, 4, 5 — ганглии, иннервирующие
внутренности; b — ж ука: 1 — головной мозг, 2 — подглоточный ганглий, 3 — грудные ганглии, 4 —
брюшные ганглии нервной цепочки.
294
нов и передающие их нервным клеткам,
называются вставочными, или инт ер
нейронами. Благодаря специализации
нейронов нервный импульс проводится
по жестко определенным путям, что
обеспечивает быстроту и точность ответ
ных реакций организма.
Строение ганглионарного типа нер
вной системы у кольчатых червей, мол
люсков и членистоногих отличается лишь
частными деталями, касающимися прос
транственного расположения ганглиев.
Так, у дождевого червя (см. рис. 171, а)
парные ганглии сосредоточены над глот
кой, под глоткой и на брюшной стороне
каждого туловищного сегмента, что фор
мирует брюшную нервную цепочку. Са
мый передний надглоточный ганглий
крупнее остальных, и поэтому его иногда
называют «мозгом», хотя по сложности
организации его никак нельзя сравнивать
с мозгом хордовых животных. Благодаря
наличию в сегментах собственных пар
ных ганглиев достигается относительная
автономность каждого сегмента и воз
можность быстрой регенерации разре
занного на части червя.
У брюхоногих моллюсков пять пар
ганглиев расположены в жизненно важ
ных органах: голове, ноге, органах дыха
ния, мантии и внутренностях (см. рис.
171, б). В связи с редукцией головного
отдела у двустворчатых моллюсков отсут
ствуют головные ганглии.
В пределах типа Моллюски нервная
система достигает наибольшей сложнос
ти у класса головоногих. У них несколько
пар сильно развитых ганглиев сконцен
трированы в головном отделе и образуют
головной мозг, от которого отходят нервы
к различным органам. Мозг защищен
хрящевой капсулой (единственный при
мер хрящевого скелета у беспозвоноч
ных!). Высокое развитие и централиза
ция нервной системы связаны с более
высокой активностью и значительно бо
лее сложным поведением головоногих по
сравнению с другими моллюсками. Их
заслуженно называют «приматами»
моря.
У членистоногих строение нервной
системы чрезвычайно сходно с таковой у
кольчатых червей, что свидетельствует о
родстве этих групп (см. рис. 171, в). От
личием является более или менее выра
женное слияние ганглиев соседних сег
ментов друг с другом в связи с укорочени
ем тела или малыми размерами предста
вителей некоторых групп членистоногих,
а также в связи со слиянием головного и
грудного отделов тела, наблюдающимся у
ракообразных и паукообразных.
Достаточно высокого развития дости
гает нервная система насекомых (рис.
172). У общественных насекомых наибо
лее развит парный надглоточный ган
глий, преобразуемый в мозг с тремя отде
лами — передне-, средне- и заднемозгом. Он осуществляет иннервацию раз
нообразных органов чувств — осязания,
обоняния, равновесия, зрения.
Рис. 172. Головной мозг насекомого (кузнечи
ка): 1 — зрительные доли, 2 — нервы парных
простых глазков, 3 — переднемозг, 4 — нерв
непарного простого глазка, 5 — усиковый
нерв, 6 — среднемозг, 7 — дополнительный
усиковый нерв, 8 — лобногубный нерв, 9 — ос
нование кишечного нерва, 10 — перемычка, 11
— окологлоточное кольцо.
295
Несмотря на явные прогрессивные
преобразования нервной системы, наб
людаемые у беспозвоночных, они так и не
достигают столь сложной структуры, как
центральная нервная система позвоноч
ных животных. Поэтому беспозвоночным
свойственны в основном инстинкты и
врожденные рефлексы. Их способность к
обучению фактически равна нулю.
Нервная система позвоночных. Поз
воночные животные имеют трубчатую
нервную систему. В эмбриогенезе на пе
реднем конце нервной трубки образуются
три вздутия — передний, средний и зад
ний мозговые пузыри. Позднее перед
ний мозговой пузырь подразделяется на
два отдела, из которых образуется перед
ний и промежуточный мозг. Из сред
него мозгового пузыря формируется
средний мозг. Разделившись на два отде
ла, задний мозговой пузырь образует
мозжечок и продолговатый мозг, пе
реходящий в спинной. Таким образом, у
всех классов позвоночных животных го
ловной мозг состоит из вышеназванных
пяти отделов, степень развития которых
неодинакова.
a
б
в
У рыб сильно развиты обонятельные
доли переднего мозга, а также мозжечок,
координирующий движения (рис. 173, а).
Головной мозг земноводных отлича
ется от такового у рыб большим развити
ем переднего мозга, полным разделением
его на два полушария и слабо развитым
мозжечком. Крыша переднего мозга име
ет первичный мозговой свод — архипаллиум. Слабое развитие мозжечка
обусловлено малой подвижностью этих
животных и однообразием производимых
ими движений (рис. 173, б).
У пресмыкающихся полушария пе
реднего мозга развиты значительно силь
нее за счет увеличения мозгового свода.
Обособлены теменные доли, более диф
ференцированы обонятельные (рис. 173,
в). Лучше, чем у земноводных, развит
мозжечок в связи с активным образом
жизни. Впервые в процессе эволюции на
поверхности полушарий появляются за
чатки коры, что повышает приспособи
тельные возможности и обеспечивает бо
лее сложные формы поведения животных
этой группы.
Головной мозг птиц отличается от
мозга пресмыкающихся большими общи-
г
д
Рис. 173. Головной мозг различных позвоночных: а — костн ой рыбы, б — земноводного, в —
пресмыкающегося, г — птицы , д — млекопитаю щ его: 1 — обонятельные доли, 2 ■
— передний мозг
(большие полушария), 3 — промежуточный мозг, 4 — средний мозг, 5 — мозжечок, 6 — продолго
ватый мозг.
296
ми размерами, сравнительно большими
полушариями и зрительными долями, ог
ромным мозжечком и маленькими обоня
тельными долями (рис. 173, г).
У млекопитающих чрезвычайно раз
виты полушария мозга, покрывающие
средний мозг и мозжечок (рис. 173, д).
Разрастается нервное вещество крыши
мозга, образующее складчатую кору
переднего мозга, или неопаллиум.
В коре в соответствии с функциями нерв
ных клеток выделяются области трех ти
пов. Сенсорные (чувствительные) зоны
получают импульсы от большинства ре
цепторов тела (входные сигналы), обес
печивая определенные виды ощущений:
зрительные, слуховые, обонятельные,
осязательные и др. Ассоциативные зоны
осуществляют оценку, хранение, сопо
ставление вновь поступающей информа
ции с полученной ранее и хранящейся в
блоках памяти, сравнение информации,
полученной от одних рецепторов, с
информацией, полученной от других, и на
этой основе вырабатывают наиболее
подходящий ответ. Таким образом, в этой
зоне происходят процессы запоминания,
научения, мышления. Из двигательных
зон выходят нервные импульсы к эффек
торам, т. е. к тем или иным произвольным
мышцам, контролируя их активность.
Таким образом, кора больших полу
шарий, собирая информацию от всех сен
сорных и двигательных систем, осущест
вляет анализ и становится аппаратом
тонкой условно-рефлекторной деятель
ности, а у человека — органом его психи
ческой деятельности, сознательного
мышления (рис. 174). Зависимость функ
ционирования организма от работы коры
головного мозга усиливается по мере ее
усложнения. Так, после удаления боль
ших полушарий, лягушка продолжает
вести себя почти так же, как и до опера
ции. Голубь после удаления коры спосо
бен к полету и сохранению равновесия,
однако часами может оставаться непод
вижным. Под действием раздражения он
начинает бесцельно двигаться, не спосо
бен есть предлагаемую пищу. Собака
Д в и га те л ь н а я
с о м а т и ч е с к а я зона
П ре м отор ная зона
С енсорная
сом атическая
зон а
А ссоци а ти вн ая
зон а речи
Р5
М оторны й
ц е н т р р е чи
Зрительная
се н со р н а я зона
^о
^ ч 3 р ите л ьна я
-'г> а с с о ц и а т и в н а я
зона
С луховая
а с с о ц и а т и в н а я зон а
С луховая
се н со р н а я зона
Рис. 174. Схема расположения четырех главных долей коры головного мозга, а также
зон и центров, связанных со специфическими функциями и участками тела.
297
после удаления коры больших полушарий
сохраняет способность ходить и прогла
тывать вложенную ей в рот пищу, но не
проявляет страха или возбуяедения. Мла
денец, рожденный без коры больших по
лушарий, ничему не научается и не совер
шает произвольных движений. Такие дети
обычно умирают вскоре после рождения.
У рыб и земноводных от головного
мозга отходят 10 пар черепномозговых
нервов, а начиная с пресмыкающихся —
12 пар. Они осуществляют иннервацию
органов чувств, расположенных в голов
ном отделе, а также поперечно-полоса
той мускулатуры, управляющей движе
нием глазных яблок, языка, жевательных
мышц.
Второй отдел ЦНС позвоночных жи
вотных и человека — спинной мозг (рис.
175). От него отходят спинномозговые
нервы (у человека их 31 пара), иннерви
рующие мускулатуру различных частей
тела (рис. 176, а, б). Основными функци
ями спинного мозга являются: осущест
вление двигательных рефлексов, а также
проведение нервных импульсов от орга
нов в головной мозг (восходящие нерв
ные пути), а от него в обратном направ
лении (нисходящие нервные пути).
Отходящие от ЦНС нервы, их сплете
ния, узлы, рецепторные аппараты сос
тавляют периферическую нервную сис
тему. Узлами, или ганглиями, называют
скопления тел нервных клеток вне
спинного и головного мозга. По анато
мическим и функциональным особеннос
тям нервная система условно подразде
ляется на две взаимосвязанные друг с
Рис. 175. Поперечный срез спинного мозга: 1 — белое вещество, 2 — спинномозговой канал, 3 — зад
няя продольная борозда, 4 — задний корешок спинномозгового нерва, 5 — спинномозговой узел, 6
— спинномозговой нерв, 7 — серое вещество, 8 — передний корешок спинномозгового нерва, 9 —
передняя продольная борозда.
298
•1
Рис. 176. Иннервация поверхности тела 31-й парой спинномозговых нервов (а) и их расположение (б)
(С — шейные, Т — грудные, L — поясничные, S — крестцовы е): 1 — кора головного мозга, 2 — моз
жечок, 3 — продолговатый мозг, 4 — шейные позвонки, 5 — спинной мозг, 6 — грудные позвонки,
7 — поясничные позвонки, 8 — крестец, 9 — концевая нить, 10 — копчик, 11 — 1-й копчиковый нерв.
другом части — соматическую (СНС) и
вегет ат ивную (ВНС) (рис. 177,
с. 300). СНС иннервирует поперечно-по
лосатую скелетную мускулатуру и органы
чувств, обеспечивая произвольные дви
гательные и чувствительные функции.
Эта часть нервной системы осуществляет
связь организма с окружающей средой и
быстрые реакции на ее изменения. ВНС
иннервирует гладкую мускулатуру внут
ренних органов, кровеносных сосудов,
мышцу сердца и железы. Кроме того, ее
волокна оканчиваются в скелетных мыш
цах и стимулируют их работоспособ
ность. Вегетативная нервная система
управляет деятельностью внутренних
органов, участвующих в осуществлении
функций питания, дыхания, выделения,
299
циркуляции жидкостей, и приспосабли
вает их работу к потребностям организма
и условиям внешней среды. Вегетативная
нервная система, так же как и соматичес
кая, состоит из центральных и перифери
ческих образований. Центры (ядра) рас
положены в области головного и спинно
го мозга. Периферическая часть включа
ет отходящие от этих центров нервы,
узлы и сплетения. Таким образом, цен
тральная и периферическая нервная сис
темы, соматическая и вегетативная раз
деляются лишь условно, представляя со
бой функционально единое целое, ибо
процессы возбуждения и торможения в
самых различных ее частях всегда взаи
мосвязаны и взаимодействуют друг с
другом.
Итак, наиболее сложно устроенная
нервная система позвоночных животных
и человека обеспечивает наибольшую
точность, быстроту, локальность ответ
ных реакций, возможность быстрого нау
чения, образования приспособительных
реакций в меняющихся условиях среды,
что дает им значительные преимущества
в борьбе за существование.
Рис. 177 Общая схема вегетативной нервной
системы (пунктиром обозначены постганглио
нарные волокна симпатической системы, иду
щие к органам; непрерывной линией — волок
на парасимпатической и преганглионарные во
локна симпатической системы. Область цен
тров симпатической системы в спинном мозге
заш трихована)'. 1 — центры парасимпатичес
кой системы в головном мозге, 2 — центры
симпатической нервной системы, 3 — центры
парасимпатической системы в нижнем конце
спинного мозга (крестцовый отдел), 4 — поло
вые органы, 5 — мочевой пузырь, 6 — толстая
кишка, 7 — тонкая кишка, 8 — почка, 9 — над
почечник, 10 — поджелудочная железа, 11 —
печень, 12 — желудок, 13 — сердце, 1 4 —-л е г
кие, 15 — сосуды головы, 16, 17 — слюнные
железы, 18 — глаз.
300
?
1. Каковы последовательные этапы эволюци
онных преобразований нервной системы животных
разных таксономических групп? 2. Что такое ган
глий, нервный узел, нервный центр? 3. Что дает ос
нование считать организацию нервной системы поз
воночных животных наиболее совершенной?
87. Органы чувств у животных
Как уже указывалось, нервная систе
ма осуществляет регуляторную функцию
в ответ на изменения во внешней или
внутренней среде организма. Для того
чтобы обнаружить эти изменения, в ходе
эволюции развились специализирован
ные к определенным раздражителям сен
сорные системы (органы чувств). Они не
только различают и регистрируют появ
ление и исчезновение раздражителей, но
и определяют их величину (силу), ско
рость и направление действия.
Органы чувств — совокупность об
разований, которые обеспечивают вос
приятие того или иного вида раздражите
ля и производят его анализ в нервной
системе. Свойство органов чувств осу
ществлять качественный и количествен
ный анализ раздражителей послужило
основанием для того, чтобы академик
И. П. Павлов назвал их анализат ора
ми. Независимо от вида анализатор сос
тоит из трех элементов: рецептора (пери
ферическая часть), проводящих нервных
путей (проводник) и участка ЦНС (цен
тральная часть) (рис. 178).
Рецептор — это клетка или специ
альный чувствительный орган, способ
ный воспринять, т. е. возбудиться от оп
ределенного вида возбудителя. Возбуж
дение связано с электрическими процес
сами в мембране чувствительной клетки,
выражающимися в смене зарядов на
мембране и, таким образом, приводящи
ми к возникновению нервного импульса.
Основное назначение рецептора состоит
в кодировании полученной информации
из внешней среды или от внутренних
органов организма в форму нервного
импульса.
Во многих органах чувств прежде, чем
раздражитель достигнет чувствительных
клеток рецептора, он видоизменяется
(усиливается, ослабляется) с помощью
вспомогательных структур, связанных с
рецептором. Примером таких структур,
например, в органе зрения служат хрус
талик, радужная оболочка, зрачок и др.
Рис. 178. Схема строения зрительного анализатора: 1 — периферическая часть анализатора, 2 —
проводниковая часть, 3 — центральный отдел анализатора.
301
Функцией проводящих нервных пу
тей органа чувств является передача по
лученной от рецептора информации, за
кодированной в форме нервного импуль
са, в соответствующий отдел ЦНС.
В участке ЦНС происходит декодиро
вание (расшифровка) информации с язы
ка нервного импульса в форму соответ
ствующего ощущения (зрительного, слу
хового, вкусового и т. п.), которое ис
пользуется организмом для адекватного
действия.
В ходе прогрессивной эволюции жи
вотных органы чувств претерпели сущес
твенные изменения. Так, у протает вос
приятие раздражителя осуществляется
участками поверхностных рецепторов
мембран, вызывая кратковременные из
менения мембранного потенциала. В ре
зультате этого, например, осуществляет
ся активизация аппарата управления дви
жениями ложноножек или ресничек, бла
годаря чему амеба или инфузория при
ближаются или удаляются от раздра
жителя.
Начиная уже с примитивных много
клеточных животных, при дифференцировке тканей выделяются клетки, специ
ализирующиеся на рецепторных функци
ях. Так, у кишечнополостных это чувстви
тельные эпителиальные клетки, соеди
ненные с нервными элементами диффуз
ной нервной системы.
С возникновением центральной нерв
ной системы становится возможным осу
ществлять анализ раздражителей, т. к.
устанавливаются постоянные связи ре
цепторов с эффекторами.
У высших беспозвоночных и позво
ночных животных развиваются дистан
тные рецепторы, обнаруживающие ис
точник раздражения на расстоянии (это
зрение, слух, обоняние), в отличие от
контактных, вступающих в действие
лишь при соприкосновении с раздра
жителем (вкус, механорецепторы).
В зависимости от формы энергии раз
дражителя, воспринимаемого рецепто
302
ром, выделяют органы зрения, а также
органы механического, химического, теп
лового, электрического чувств.
Фоторецепция. Несмотря на разно
образие органов зрения, в основе про
цессов фоторецепции лежат сходные
первичные реакции: поглощение квантов
света светочувствительными пигмента
ми, например родопсином, и последую
щее расщепление его на составляющие
части — белок опсин и каротиноид ретиналь (альдегидная форма витамина А).
В темноте происходит обратный про
цесс — ресинтез пигмента.
У некоторых групп животных отдель
ные фоторецепторные клетки диффузно
расположены в эпидермисе (дождевой
червь, некоторые виды насекомых) или в
нервной ткани (ланцетник). Такие рецеп
торы обеспечивают восприятие лишь ин
тенсивности освещения.
У многих животных рецепторные
клетки образуют светочувствительный
эпителий (сетчатку) с системой линз,
осуществляющих
светопреломление.
Благодаря эволюции от плоского и ямкообразного к камерному глазу (у медуз,
кольчатых червей, брюхоногих и голово
ногих моллюсков) стало возможным не
только определение направления и
интенсивности падающего света, но и
получение четкого изображения (рис.
179). Четкость изображения достигалась
развитием механизмов аккомодации
(например, у головоногих моллюсков).
У членистоногих имеются глаза двух
типов — простые и сложные, или фа
сеточные. Простой глаз состоит из един
ственной линзы, под которой расположе
но небольшое количество светочувстви
тельных клеток. Он различает лишь свет
и темноту. У ракообразных и насекомых
наряду с простыми глазами развился
сложный фасеточный глаз. Он состоит из
множества отдельных, изолированных
пигментными клетками, конических глаз
ков — омматидиев. Каждый глазок
воспринимает часть предмета, а вместе
а
б
в
г
Рис. 179. Эволюция органа зрения: а — восприятие изображения в плоском, б — ямкооб
разном, в — камерном глазах и г — в глазу с линзой: 1 — зрительные рецепторы (сетчат
ка), 2 — пигментный слой. Черная стрелка — предмет, зеленая — изображение.
они обеспечивают его мозаичное изобра
жение. Чувствительность фасеточного
глаза значительно большая, чем просто
го, что позволяет улавливать движение
объектов.
Способность различать цвета отмече
на у насекомых. Для насекомых и голово
ногих моллюсков выявлено поляризаци
онное зрение, т. е. способность опреде
лять плоскость поляризации. Это дает
животным возможность дополнительно
ориентироваться по положению Солнца
на небосводе.
Наиболее совершенен орган зрения
позвоночных животных, в особенности
птиц, млекопитающих и человека (рис.
180). Рецепторные клетки (палочки и
колбочки) воспринимают свет различной
длины волны, отражаемый от объектов,
находящихся в поле зрения на разных
расстояниях. В зрительной зоне коры го
ловного мозга осуществляется точное
восприятие изображения. Наличие трех
типов колбочек, возбуждаемых волнами
разных длин, соответствующих красной,
зеленой и синей частям спектра, обеспе
чивает цветовое восприятие объектов.
Механорецепция.
Механические
раздражители воздействуют давлением,
растяжением, вибрацией, звуком на
специализированные сенсорные орга
ны. У различных типов животных во
всех группах механорецепторов раз-
Рис. 180. Схема строения глаза человека: 1 —
роговица, 2 — веко, 3 — цилиарный отросток,
4 — склера, 5 — сосудистая оболочка, 6 — сет
чатка, 7 — зрительный нерв, 8 — стекловидное
тело, 9 — водянистая влага, 10 — зрачок, 11 —
хрусталик, 12 — радужная оболочка.
303
дражение заключается в сдвиговой де
формации чувствительных волосковых
клеток.
В противоположность сходным пер
вичным механизмам раздражения вспо
могательные структуры, воспринимаю
щие раздражения, сильно различаются.
Выделяют несколько типов механорецепторных органов.
Так, звуковые колебания многие на
секомые (личинки комаров, мухи, жуки)
воспринимают с помощью связки, напо
минающей струну и натянутой под покро
вами животного. Чувствительные клетки
соединяются одним концом с покровами,
а другим — со связкой. У других насеко
мых (кузнечики, сверчки, цикады, терми
ты) восприятие звука осуществляется
тонкой кутикулярной мембраной, играю
щей роль мембранной перепонки, под ко
торой лежит резонирующий трахейный
пузырь. Группа чувствительных клеток
располагается между перепонкой и пузы
рем.
Наиболее сложно устроенным и чув
ствительным к восприятию звуковых
волн является орган слуха млекопитаю
щих и человека (рис. 181). Ухо имеет три
отдела — наружное, среднее и внут
реннее. Функцией наружного уха, пред1
Рис. 181. Строение уха человека: 1 — слуховой
канал, 2 — слуховые косточки, 3 — полукруж
ные каналы, 4 — улитка, 5 — слуховой нерв,
6 — евстахиева труба, 7 — барабанная пере
понка.
304
ставленного ушной раковиной и на
ружным слуховым проходом, является
улавливание и направление звуковых ко
лебаний на барабанную перепонку, от
деляющую наружное ухо от среднего.
В среднем ухе (объем 1 см3) находятся
три слуховые косточки — молоточек,
наковальня и стремечко, которые в 22
раза усиливают колебания барабанной
перепонки и передают их на мембранную
перепонку овального окна, отделяюще
го среднее ухо от улитки внутреннего
уха. Утитка внутреннего уха представлена
спиральным (2,5 оборота и длиной 35 мм)
костным каналом, разделенным двумя
продольными перепонками на три части.
На мембране среднего канала располага
ются чувствительные волосковые клетки,
в которых происходит преобразование
звуковых волн в нервные импульсы.
Ухо млекопитающих обладает чрез
вычайно высокой чувствительностью и
способностью различать разные звуко
вые оттенки.
Положение тела животного в прос
транстве определяется статоцистами.
Принципиальная схема структуры статоциста представлена пузырьком под по
кровом, в котором на отростках чувстви
тельных клеток лежит статолит (из
вестковый камешек) (рис. 182). Смеща
ясь при изменении положения тела, он
вызывает одностороннее раздражение
волосковых клеток чувствительного эпи
телия. Статоцисты у медуз располагают
ся на краю зонтика, у ракообразных — на
основном членике вторых усиков (ан
тенн), у моллюсков — в ноге.
Механические раздражения повер
хности тела (осязание) воспринимаются
у многих животных изолированными друг
от друга чувствительными клетками по
кровов. У членистоногих образовались
специальные купгикулярные сенсил
лы — шарнирно закрепленные щетинки,
связанные с дендритом чувствительной
клетки. Отгибания щетинки в разных
направлениях ведут к разному по силе
Рис. 182. Орган равновесия — с т а т о ц и с т —
кишечнополостного животного: 1 — отросток
чувствительных клеток, 2 — статолит, 3 — на
ружная оболочка статоциста, 4 — чувстви
тельные клетки.
возбуждению чувствительной клетки.
Совокупность таких кутикулярных сен
силл на суставах наружного скелета на
секомых дает информацию о взаимном
положении частей тела.
Хеморецепция широко распростра
нена у всех групп животных. Она прояв
ляется в восприятии химических раздра
жителей. Контактную хеморецепцию оп
ределяют как вкусовое восприятие, а
дистантную — как обонятельное.
Обонятельные раздражители у жи
вотных играют решающую роль при по
иске пищи, организма-хозяина, привле
чении полового партнера и др. Органы
обоняния хорошо развиты у насекомых.
Они расположены на усиках и обладают
поразительной остротой восприятия. За
пах для них — средство информации и
общения.
Разнообразны приспособления для
восприятия вкуса. Так, у насекомых
функцию органа вкуса выполняют сен
сорные клетки с волосками трубчатой
формы, расположенные на лапках (у ба
бочек) или на ротовых конечностях (у
мух). Они содержат группу сенсорных
клеток, дендриты которых тянутся до от
11. Общая биология, 10 кл.
верстия на вершине волоска. Эти рецеп
торы у мух избирательно раздражаются
сахарозой, поваренной солью или водой.
Предполагают, что решающим пара
метром раздражителя является прос
транственная конфигурация его молеку
лы, обеспечивающая субстратную специ
фичность, воспринимаемую акцепторным
участком мембраны хеморецептора.
Рецепция у растений. У растений от
сутствуют дифференцированные органы
чувств. Однако в состав клеточных мем
бран некоторых тканей входят хемо-, фо
то- и механорецепторы белковой приро
ды, чувствительные к действию химичес
ких и физических факторов.
Растения способны воспринимать
красный, инфракрасный и синий цвета.
Активность фоторецепторов использует
ся для осуществления фототаксисов, фотонастий и восприятия фотопериодических сигналов. Фотовосприятие осущес
твляют специальные рецепторные бел
ки — фитохромы. Наибольшее коли
чество фитохромов обнаружено в мерис
темах и в камбии.
Хеморецепция позволяет раститель
ным организмам, их тканям и клеткам ре
агировать на фитогормоны, осущест
влять хемотаксисы и хемотропизмы.
Механорецепция лежит в основе таких
явлений, как геотропизм, тигмотропизм.
Таким образом, эти рецепторы вос
принимают сигналы, поступающие из
внешней и внутренней среды, обеспечи
вая адаптивные ответы клеток на измене
ния условий существования.
и 1. Из каких элементов состоят сложные органы
чувств и каковы их функции? 2. Дайте классифика
цию рецепторов в зависимости от вида раздражите
ля. 3. Какие процессы лежат в основе фоторецеп
ции? 4. В каком направлении происходила эволю
ция фоторецепции у животных разных таксономи
ческих групп? 5. Какими структурами воспринима
ют звуковые колебания насекомые? 6. Как устроен
орган слуха человека? 7. Как осуществляется ре
цепция у растений?
305
88. Эндокринная система
и ее работа
В координации деятельности различ
ных органов животных и человека важ
ную роль играют гормоны. Они выделя
ются эндокринными железами непос
редственно В' тканевую жидкость или в
кровь, которой и разносятся ко всем ор
ганам, Их еще называют железами внут
ренней секреции.
Как уже указывалось, гормоны могут
образовываться и специализированными
нервными клетками, транспортироваться
по аксонам до их окончаний, образующих
нейрогемальные органы, и выделяться
из них в кровь. Таким образом, нейросек
реторные процессы, протекающие, нап
ример, в гипоталамусе позвоночных и пе
редней доле мозга насекомых, являются
связующим звеном между нервной и гу
моральной регуляцией. Все гормоны бес
позвоночных, за исключением некоторых
гормонов членистоногих — экдизона,
ювенильного гормона, — нейросекреты.
Гормоны — чрезвычайно активные
вещества, ничтожное количество кото
рых оказывает сильное воздействие на
строение и функции различных органов,
расположенных вдали от места их обра
зования. По своей химической природе
гормоны весьма разнообразны. Некото
рые из них являются аминокислотами или
производными аминокислот (тироксин,
адреналин), производными жирных кис
лот (простогландины, ювенильный гор
мон), белками (окситоцин, антидиуретический и адренокортикотропный гормо
ны, глюкагон, инсулин, гонадотропины,
гормон роста), стероидами — из группы
неомыляемых липидов (половые гормо
ны, гормоны коры надпочечников, экдизон).
Гормоны сравнительно быстро разру
шаются, поэтому для обеспечения
функций организма они должны выде
ляться в кровь постоянно. Только в этом
306
случае возможно поддержание в крови их
необходимой концентрации.
Наиболее полно изучены гормоны на
секомых и позвоночных животных. У на
секомых выделены, четыре вида желез
внутренней секреции (рис. \Ъ2>). Нейро
секреторные клет ки передней- доли
«головного мозга» выделяют м о з г о в о й
1
Рис. 183. Расположение желез внутренней
секреции и переднего отдела нервной системы
насекомых: 1 — кишечник, 2 — надглоточный
ганглий (мозг), 3 — нейросекреторные клетки
мозга, 4 — переднемозг, 5 — среднемозг, 6 —
заднемозг, 7 — глазные доли, 8 — подглоточ
ный ганглий, 9 — кардиальные тела, 10 — при
лежащие тела, 11 — переднегрудная железа,
12 — ганглий брюшной нервной цепочки.
гормон, который контролирует и активи
зирует деятельность переднегрудных
желез. При прекращении выделения
мозгового гормона у личинок и куколок
затормаживается рост и развитие, возни
кает диапауза (приостановка развития).
Кардиальные тела сходны по форме и
положению с прилежащими телами и
связаны нервами с «головным мозгом».
Их роль наименее изучена. Они регули
руют секреторную деятельность перед
негрудных желез. Прилежащие, или до
бавочные, тела в виде пары округлых
образований расположены над передней
кишкой позади «головного мозга». Они
выделяют л и ч и н о ч н ы й ( ю в е н и л ь
ный) гормон, или н е о т е н и и . Гормон
способствует развитию личиночных орга
нов и препятствует метаморфозу. В связи
с отставанием роста прилежащих тел от
роста тела личинки действие неотенина
прекращается после последней линьки
насекомого, так как концентрация неоте
нина в гемолимфе резко уменьшается.
При этом у насекомых с неполным мета
морфозом развивается взрослая форма, а
у насекомых с полным превращением
формируется куколка. Переднегрудные
железы, расположенные на брюшной
стороне переднегруди, выделяют в гемо
лимфу гормон л инь к и, или э кд из он.
Он прекращает диапаузу, вызывая у ли
чинок линьку и регулируя, следователь
но, развитие личинки.
У ракообразных нейросекреторные
клетки, расположенные в ганглиях глаз
ных стебельков брюшной нервной цепоч
ки и многих других участках нервной сис
темы, выделяют разнообразные гормоны,
которые управляют движением глазных
пигментов, пигментов наружных покро
вов, тормозят линьку, изменяют ритм ра
боты сердца, регулируют размножение и
другие процессы. Нейросекреторные
клетки околоротовой области гидры вы
деляют гормон роста. Гормон нейросек
реторных клеток головного ганглия
планарии стимулирует регенерацию,
а также размножение путем деления
особи на части. Для кольчатого червя
нереиды описана гормональная регу
ляция регенерации, роста и размно
жения.
Среди хордовых животных наиболее
полно изучена гормональная регуляция у
человека (см. таблицу 10).
Т а б л и ц а 10. Эндокринные железы человека, гормоны и их физиологическое действие
Железа
1
Щитовидная
Гормон
2
Тироксин
Трийодотиронин
Тиреокальцитонин
Паращитовидная
Паратгормон
П одж елудочная
(островки Лангерганса)
Место действия
3
Весь организм
Физиологический эффект
4
Ускоряет обмен веществ
и потребление кислорода в тканях
Регулирует обмен кальция и фос
фора
Скелет, почки, же
Регулирует обмен кальция и фос
лудочно-киш ечный фора
тракт
Инсулин
Весь организм
Глюкагон
Печень
Регулирует обмен углеводов, сти
мулирует синтез белков
Стимулирует синтез и распад гли
когена
Регулирует обмен углеводов
Кортикостероиды
Весь организм
Регулирует обмен электролитов и
Канальцы почек
(кортизон)
Альдостерон
воды
Адреналин
Мышцы
сердца,
б) мозговое вещееПовышает частоту и силу сокраще
гладкие мышцы артетво
риол
ний сердца, тонус артериол, артериНадпочечники:
а)корковый слой
307
1
2
3
4
альное давление, стимулирует сок
ращение многих гладких мышц
Норадреналин
Гипоталамус
Гипофиз
а) передняя доля
Либерины, статины
Соматотропин
(гормон роста)
Тиротропин
Кортикотропин
Печень, скелетные
мышцы
Жировая ткань
Артериолы
Стимулирует распад гликогена
Стимулирует распад липидов
Повышает тонус артериол и арте
риальное давление
Регулирует секрецию гормонов ги
пофиза
Гипофиз
Весь организм
Ускоряет рост мышц и костей. Стимулирует синтез белка. Оказывает
влияние на обмен углеводов и жиров
Щитовидная желе
Стимулирует синтез и секрецию
гормонов щитовидной железы
за
Кора надпочечни
Стимулирует синтез и секрецию
гормонов коры надпочечников
ков
Стимулирует развитие желтого теФолликул ости муЯичники, семеннила после овуляции и выработку про
лирующий гормон
ки
гестерона у женщин
У мужчин стимулирует развитие
ткани семенников и секрецию андро
генов
Молочная железа
Вызывает разрастание ткани, про
Пролактин
дукцию молока
б) задняя доля
Собирательные
Антидиуретический
гормон (вазопрессин) трубочки почек
Артериолы
Семенники(яички)
Яичники
Увеличивает тонус. Поввннает ар
териальное давление
Окситоцин
Гладкие мышцы,
Усиливает сокращения матки и из
особенно матки
гнание плода
Тестостерон
Мужские половые
Стимулирует нормальный рост,
органы
развитие и функционирование
Весь организм
Стимулирует развитие вторичных
половых признаков
Эстрон, эстрадиол
Женские половые
Стимулирует нормальный рост,
органы
развитие и циклическую функцию
Молочная железа
Стимулирует развитие протоков
Весь организм
Стимулирует развитие вторичных
половых признаков
Женские половые
Поддерживает течение беремен
органы
ности
Прогестерон
Такие железы, как поджелудочная и
половые, являясь одновременно железа
ми и внешней и внутренней секреции, от
носятся к смешанным.
Гормоны продуцируются также и не
которыми органами и тканями, выпол
няющими в организме, помимо эндок
308
Облегчает обратное всасывание
(реабсорбцию) воды
ринной, другие специализированные
функции. Это гормоны местного дей
ствия, например гастрин желудочнокишечного тракта повышает секрецию
желудочных и поджелудочной желез,
усиливает моторику желудка, тонкой
кишки и желчного пузыря; ренин вызы
вает сосудосуживающий эффект в поч
ках и т. п.
Выделяемые в кровяное русло гормо
ны доставляются во все части тела. Неко
торые гормоны, например тироксин или
гормон роста (соматотропин), влияют на
обмен веществ всех клеток тела. Дей
ствие же большинства гормонов избира
тельно. Например, циркулирующий в
крови секретин воздействует только на
клетки поджелудочной железы. Орган,
реагирующий на определенный гормон,
называют «органом-мишенью» этого
гормона. В настоящее время доказано,
что восприятие гормона осуществляется
гликокаликсом — надмембранным ком
плексом клеток.
Механизмы действия гормонов пол
ностью не выяснены. Считают, что они
разные. В одних случаях показано их
участие в регуляции активности генов.
Так, тироксин стимулирует при метамор
фозе земноводных генную активность,
ответственную за ряд процессов, проис
ходящих при переходе к наземному обра
зу жизни (рассасывание хвоста, синтез
ферментов, участвующих в метаболизме
мочевины, и др.).
В других случаях было обнаружено,
что гормоны влияют на свойства мем
бран. Так, например, инсулин повышает
их проницаемость для глюкозы.
?
1. Какова химическая природа гормонов?
2. Какие гормоны регулируют рост и метаморфоз
насекомых? 3. Каково физиологическое действие
гормонов щитовидной и поджелудочной желез?
4. Какое действие оказывает адреналин на работу
органов? 5. Как регулируются процессы роста орга
низма у позвоночных животных? 6. Как осущес
твляется регуляция полового развития человека?
7. Какие гормоны местного действия вам известны?
89. Нарушения деятельности
эндокринных желез у человека
Нарушения деятельности желез внут
ренней секреции вызывают заболевания,
называемые эндокринными. Эти нару
шения бывают двоякого рода: а) вызван
ные усилением деятельности железы —
ее гиперфункцией, когда гормоны обра
зуются и выделяются в кровь в избытке;
б) вызванные ослаблением деятельности
железы — ее гипофункцией, когда выде
ление гормонов ниже нормы.
Так, при гиперфункции щитовидной
железы возникает базедова болезнь
(рис. 184, а, с. 310). Ее признаки — уве
личение железы, пучеглазие, учащение
сердцебиения, повышенная возбуди
мость, эмоциональность, быстрая утом
ляемость и потливость. При гипофункции
щитовидной железы взрослые люди стра
дают микседемой, или слизистым оте
ком (рис. 184, б). Кожа у таких больных
сухая, грубая, отечная, выпадают волосы,
значительно понижается обмен веществ,
может нарушаться психическая деятель
ность. При гипофункции железы в дет
ском возрасте развивается кретинизм:
задержка роста, умственная отсталость.
Гипофункция паращитовидных желез ве
дет к тяжелому заболеванию — тетании.
Оно сопровождается судорогами, разру
шением зубов, похудением, резким сни
жением содержания кальция в плазме
крови. При гипофункции коры надпочеч
ников развивается аддисонова болезнь:
кожа приобретает бронзовую окраску,
больной худеет, теряет аппетит, содержа
ние сахара в крови понижается, кровяное
давление падает.
Одно из массовых тяжелых эндокрин
ных заболеваний — сахарный диабет —
связано с гипофункцией островковых зон
поджелудочной железы. Сопровождается
оно значительным снижением проницае
мости мембран мышечных и жировых
клеток для глюкозы. Содержание глюко
зы в плазме крови, а также в моче резко
повышается.
При выделении в кровь избыточных
количеств гормона роста передней доли
гипофиза наблюдается ускорение рос
та — гигантизм (рис. 184, в), при его не309
Рис. 184. Заболевания, вызванные нарушением деятельности эндокринных желез: а — базедова бо
лезнь, б — кретинизм и микседема (девочка 12 л е т : справа — до лечения, слева — через 6 месяцев
после лечения препаратами щитовидной железы), в — гигантизм.
достатке развивается карликовость.
Повышенная выработка гормона роста у
взрослого человека приводит к акроме
галии. При этом заболевании чрезмерно
увеличиваются кости лица, рук и ног. Од
новременно растут и мягкие ткани, губы и
щеки становятся толстыми, увеличенный
язык не помещается во рту, удлиняется
нос, увеличиваются внутренние органы
(печень, сердце и др.). Заболевание это
чаще наблюдается у мужчин.
9, 1. кКакие виды нарушении деятельности желез
внутренней секреции существуют? 2. Расскажите,
какие заболевания вызваны гиперфункцией эндок
ринных желез человека. 3. Какие болезни вызваны
гипофункцией желез внутренней секреции чело
века?
90. Поведение животных.
Врожденное поведение
Приспособление организмов к посто
янно меняющимся условиям существова
ния в значительной степени осуществля
ется их поведенческими реакциями. Под
поведением понимают проявления жиз
недеятельности, выражающиеся в сово
310
купных разнообразных действиях орга
низма, способствующих его выживанию.
Поведенческие реакции часто становятся
более очевидными, если внешние усло
вия подвергаются изменениям. Так, если
животное испугается при встрече с дру
гим животным, в его ответной реакции,
например бегстве, участвуют нервная,
эндокринная, дыхательная, мышечная,
кровеносная и другие системы. Это и есть
та суммарная ответная реакция организ
ма, которая называется поведенческой.
Однако поведенческие акты вызыва
ются не только внешними раздражителя
ми. Известно, что организм может нор
мально существовать только в том слу
чае, если его основные внутренние кон
станты находятся на относительно посто
янном уровне. Таковыми являются: опре
деленный уровень глюкозы в крови, ос
мотическое давление крови, кровяное
давление, урцвень кислорода в крови,
кислотность и щелочность крови и т. п.
При отклонении от константных ве
личин организм начинает испытывать
потребность в специальных веществах и
вынужден их активно искать и потреб
лять. Таким образом, потребность ор
ганизма — это любое значительное от
клонение важных жизненных кон
стант от уровня, обеспечивающего его
нормальные физиологические отправ
ления.
Реакции и средства, направленные на
удовлетворение возникающих потреб
Организм, испытывающий потребность
ностей, у разных живых существ различ
ны в зависимости от эволюционного по
ложения организмов. Однако для всех
общим является то, что возникшая пот
ребность вызывает действие, направлен
ное на ее удовлетворение. Это может
быть выражено следующей схемой:
Целенаправленное
поведение
Удовлетворение, потребности, а сле
довательно, восстановление нормального
уровня основных внутренних констант,
называется подкреплением. Например,
подкрепление — это прием пищи, воды,
уход в тень от солнца и т. п. Принцип
циклической саморегуляции (см. схему)
лежит в основе различных форм поведе
ния организмов.
Итак, в ответ на стимулы, восприни
маемые организмом из внешней среды
или от самого организма, последний, пу
тем адекватного поведения, обеспечивает
себе возможность выживания и размно
жения (оставления потомства). В случае,
если поведение организма не соответ
ствует измененным условиям, он погиба
ет. Населяющие нашу планету современ
ные виды живых организмов выжили при
всех изменениях среды потому, что они
сумели приспособиться к ним разнооб
разными способами, в том числе и адап
тивным поведением. Роль поведенческих
приспособлений в выживании животных
трудно переоценить, поскольку самые
быстрые реакции организма — поведен
ческие. Например, человек может ук
рыться в тени несравненно быстрее, чем
его кожа потемнеет под действием сол
нечных лучей.
У животных различают наслед
ственные формы поведения и приобре
тенные организмом в течение жизни.
Наследственные формы поведе
ния — это врожденные реакции, т. е.
Удовлетворение
потребности
безусловные рефлексы, с которыми ор
ганизм рождается, наследуя их от роди
телей, как и любой другой признак. Они
составляют основу нервной деятельности
низкоорганизованных животных, чрезвы
чайно разнообразны и характерны для от
дельных видов, а также родов, семейств и
даже отрядов.
Безусловные рефлексы могут прояв
ляться в виде простых стереотипных ре
акций и в виде более или менее сложных
стереотипных форм поведения.
Кчислу простых стереотипных реак
ций относят, например, движение и ори
ентацию тела по отношению к источнику
раздражения — таксисы. В зависимости
от действующего фактора различают’'тер
мо-, фото-, хемо- и другие таксисы. При
мером положительного термотаксиса
является скопление насекомых на теплых
местах: мух — на освещенных первым
весенним солнцем стенах домов, комнат
ной мухи — осенью после включения
отопления в доме. Пример фототакси
са — скопление насекомых в темное вре
мя суток у источников света и, наоборот,
укрытие ночных видов в темных местах
(щелях, под корой) в дневные часы. Хе
мотаксис играет большую роль в поиске
пищи, а также в отыскании особей проти
воположного пола.
Сложными врожденными стереотип
ными формами поведения являются ин
стинкты. Они не зависят от выучки и
проявляются лишь под воздействием
311
внешних и внутренних раздражителей и,
как правило, только в определенные пе
риоды жизни (например, инстинкт гнез
дования или кормления потомства).
Внутренними раздражителями, запуска
ющими инстинктивные поведенческие
реакции, могут выступать, например,
гормоны. Так, гипофиз и половые железы
вырабатывают гормоны, определяющие
поведение животного в период размно
жения, но выделение этих гормонов зави
сит, по крайней мере частично, от внеш
них событий, например от времени года.
У многих позвоночных, обитающих в уме
ренной зоне Северного полушария, при
искусственном содержании в условиях
характерного для зимы короткого свето
вого дня не обнаруживается нормальной
Рис. 185. Брачное поведение озерной чайки (последовательность событий указана цифрами).
312
весенней активности половых желез, да
же если весна давно наступила. Но если
тех же животных содержать при посте
пенно увеличивающейся длине дня, то их
гипофиз и половые железы начнут выде
лять гормоны и животные смогут присту
пить к размножению даже среди зимы.
Инстинкты возникли у животных в
процессе эволюции как приспособления
к типичным для данного вида условиям
существования. Благодаря их генетичес
кой обусловленности инстинкты могут
проявляться в соответствующий период
индивидуального развития, не требуя
специального обучения. Например, ново
рожденные млекопитающие сразу начи
нают сосать молоко матери, только что
вылупившиеся утята умеют плавать
и т. п.
Рефлексы, из которых складывается
тот или иной инстинкт, тесно взаимосвя
заны и проявляются в серии строго пос
ледовательных действий — в виде цеп
ного рефлекса. В такой цепи каждый
предшествующий рефлекс является неп
ременным раздражителем для последую
щих рефлексов. В результате формиру
ются достаточно консервативные врож
денные программы. Один из примеров
консервативного инстинктивного поведе
ния приведен в книге известного фран
цузского энтомолога Ж- А. Фа б р а . Оса
бембекс обычно роет свою норку в песке
и откладывает в нее яйцо. Особенностью
экологии этого вида роющих ос является
то, что они доставляют пищу своей ли
чинке через вход в песке, который всегда
безошибочно находят. Ж- А. Фарб рас
крывал крышу норки, и личинка оказыва
лась под открытым небом. Несмотря на
то что, казалось бы, новый путь к личин
ке прост и доступен, оса по-прежнему
подныривала к личинке через песок.
Пример демонстрирует, что инстинкты
осуществляются автоматически, без
осознания их цели,вслепую.
Сходство и устойчивость инстинктив
ных действий у особей конкретного вида
послужили основанием для использова
ния их в систематике так же, как и приз
наков строения организмов. Например,
все хищники семейства кошачьих имеют
сходное поведение при охоте и убивают
жертву укусом в шею. Все самцы озерной
чайки, ухаживая за самками, совершают
одну и туже последовательность брачных
движений (рис. 185). Неповторимость
поведения, свойственного тому или ино
му виду, — это, главным образом, след
ствие генетической уникальности вида.
Инстинкты составляют основу пове
дения практически всех видов животных,
но для низкоорганизованных их роль осо
бенно велика. Благодаря им животные
наделяются набором приспособительных
реакций, которые могут быть использо
ваны при первой же необходимости, по
этому они особенно важны для животных
с коротким сроком жизни или лишенных
заботы о потомстве. Эти приспособи
тельные реакции передаются из поколе
ния в поколение, выдерживая жесткий
естественный отбор, и, безусловно, име
ют важное значение для выживания.
?•
1. Что такое поведение? 2. Чем вызываются
поведенческие акты? 3. Какие формы поведения
свойственны животным? 4. Что такое инстинкт и
каково его значение в выживании животного?
91. Поведение,
связанное с научением
Врожденные программы поведения
обеспечивают, как правило, трафаретные
средства для удовлетворения основных
потребностей организма. Они эффектив
ны только при неизменных внешнихусловиях. Чтобы выжить при изменившихся
условиях, животное должно обладать
способностью перестраивать свои пове
денческие реакции. Эта способность на
прямую определяется уровнем эволюци
онного развития животных. Поэтому
млекопитающие и птицы с их высоко раз
витой нервной системой наделены этой
способностью в наибольшей степени. Ее
проявление выражается в форме приоб
ретенных условных рефлексов. В этом
случае животное научается что-то делать,
т. е. реагировать на стимул, который был
для него ранее безразличен. Так, при вы
работке условных рефлексов у собаки
академик И. П. П а в л о в одновременно
сочетал кормление (безусловный р а з
дражитель) со звонком (безразличный
условный раздражитель). После мно
гократного повторения эксперимента у
собаки наступало слюноотделение при
подаче только звукового сигнала.
Адаптивное изменение индивиду
ального поведения в результате пред
шествующего опыта называется науче
нием. Научение основано на способности
животных хранить и извлекать информа
цию о прошлом опыте — памяти. Физи
ологические основы памяти до сих пор не
выяснены, о ней существуют лишь гипо
тезы. Согласно одной группе гипотез, в
ее основе лежат изменения межнейрон
ных связей в центральной нервной систе
ме, согласно другой — образование спе
цифических макромолекул (нуклеиновых
кислот и белков). Эти гипотезы не явля
ются взаимоисключающими: возможно,
что для изменений в области синапса не
обходим синтез специфических макромо
лекул, и, наоборот, этот синтез может
вызвать перестройку синаптических
структур.
В процессах научения наибольшую
роль играет долговременная память, ко
торая может храниться всю жизнь.
В кратковременной памяти информа
ция хранится в пределах нескольких ми
нут. Ее образование резко снижается с
возрастом организма.
Одним из способов индивидуального
научения является метод проб и ошибок.
Так, например, молодая неопытная щука
будет пытаться заглотить колюшку до тех
пор, пока не убедится на собственном
опыте, что маленькая колючая рыбка
несъедобна для нее. Именно способность
314
к научению на основе жизненного опыта
делает многих животных — потенциаль
ных жертв — неприкосновенными. Так,
птицы научаются не трогать ядовитых на
секомых, обладающих предостерегаю
щей окраской, после первого же непри
ятного знакомства с ними. Точно так же
индивидуальный опыт «убеждает» ля
гушку отказаться от поедания яркоокрашенных жалящих насекомых.
У многих видов теплокровных живот
ных, особенно у птиц, имеется врожден
ная реакция следования молодняка за
матерью, что имеет большое значение
для выживания молоди. Оставшиеся без
матери птенцы будут следовать за любым
животным, даже неодушевленным дви
жущимся предметом, оказавшимся в по
ле их зрения. Следовательно, эта инстинк
тивная реакция не полностью запрограм
мирована генетически. Совершенно яс
но, что это случай научения, так как у
птицы может запечатлеться подобным
образом почти любой движущийся
объект. Этот вид научения называется
импринтингом (от англ, imprinting — за
печатление). Он формируется при осо
бом состоянии нервной системы на ран
ней стадии развития, так как если у птиц
эта реакция научения не возникла сразу
после вылупления, то она вообще может
никогда не появиться.
Одной из форм научения считают так
же привыкание, формирующееся при
продолжительном повторении стимулов,
не подкрепленных поощрением или нака
занием. Это приводит к постепенной ут
рате реакции на них. Например, птицы на
огороде перестают бояться пугала, т. е.
научаются не обращать на него внима
ния. Привыкание — важный элемент
формирования поведения молодого жи
вотного, научающегося распознавать
нейтральные элементы окружающей сре
ды и не реагировать на них.
Для животных с высокоразвитой нерв
ной системой, таких, как млекопитающие
и птицы, важна такая форма научения,
как подражание. Так, певчие птицы
«правильно» (характерно для данного
вида) поют лишь в том случае, если они в
период формирования этого умения име
ли возможность слышать песню других
особей своего вида. Некоторые воспи
танные в одиночестве певчие птицы, на
пример зяблик, способны научиться раз
ным песням, но легче всего — песне
своего вида, т. е. той песне, которая
предопределена врожденной програм
мой.
Приведенный пример свидетельству
ет, что многие сложные формы поведения
нельзя назвать ни всецело врожденными,
ни полностью приобретенными. Они
представляют собой комбинацию того и
другого. Соотношение же врожденной и
индивидуально приобретенной форм по
ведения животных различно: первая пре
обладает у низкоорганизованных живот
ных, вторая —- у более высокоорганизо
ванных.
Таким образом, в жизни животных
врожденные рефлексы, включая и такие
сложные их формы, как материнские ин
стинкты и забота о потомстве, не обяза
тельно должны быть строго неизменными
автоматическими реакциями, приводя
щими особей в изменившейся среде к ту
пику и гибели. На помощь этим врожден
ным, характерным и обязательным для
каждого вида рефлексам, приходят при
обретенные в течение индивидуальной
жизни условные рефлексы, через кото
рые и происходит приспособление к но
вым условиям.
В конечном счете сочетание раздра
жений внешней среды, физиологичес
кого состояния организма и явлений
возбуждения и торможения приводят
к тому, что жизнедеятельность живот
ного организма приобретает закон
ченный и биологически целесообраз
ный характер. Организм действует
как единое целое, и этим он обязан
регулирующей и объединяющей роли
нервной системы.
Научение в филогенезе. Способнос
ти к научению тесно связаны с уровнем
развития нервной системы.
Свидетельства возможности научения были по
лучены даже для плоских и кольчатых червей. Одна
ко только после 150— 200 повторений воздействи
ем слабого электрического тока планарии и дожде
вые черви научились избегать его, выбирая иной
путь передвижения по лабиринту, ведущему в тем
ную, влажную и безопасную камеру.
Имеются также убедительные факты, свиде
тельствующие о возможности научения осьминога
(класс головоногие моллюски), обладающего до
вольно развитым мозгом и органом зрения. Однако
способность к научению моллюсков других классов
достоверно не доказана.
Несравненно быстрее происходит научение у
членистоногих. Так, пчелы могут быстро научиться
подлетать к тарелкам, помещенным на голубой бу
маге, оставляя без внимания другие тарелки, нахо
дящиеся на серой бумаге, в том случае, если только
голубая бумага ассоциируется у них с сахарной во
дой. Они безошибочно летят в сторону голубой бу
маги, независимо от ее расположения, даже тогда,
когда все тарелки пустые. Хорошими «учениками»
являются муравьи, научающиеся определенным
действиям за 35 повторений.
Таким образом, несмотря на позитив
ное свидетельство возможности научения
беспозвоночных животных, они все же
имеют относительно слабо развитые спо
собности к нему. Их стимулы весьма ог
раничены, а в поведении доминируют сте
реотипные акты, диктуемые врожденны
ми таксисами, рефлексами и инстинк
тами.
Среди хордовых наилучшими способ
ностями к научению обладают птицы и
млекопитающие, имеющие наиболее раз
витый мозг, хотя и низшие хордовые (ры
бы, земноводные и пресмыкающиеся)
способны к выработке способом проб и
ошибок относительно простых ответов
типа «стимул — реакция».
Птицы и млекопитающие отличаются
от низших хордовых не только быстротой
315
и устойчивостью приобретенных новых
реакций, но также и сложностью проб
лем, которые они могут решать. У них
возникают новые особенности поведе
ния, существенно освобождающие жи
вотных от обучения по типу «стимул —
реакция» и по методу «проб и ошибок».
Высшей формой поведения, связан
ной с научением и присущей человеку,
является мышление — процесс обоб
щенного и опосредованного отражения
действительности. Мышление позволяет
решать сложные проблемы взаимосвязи
с внешним миром, не прибегая к методу
проб и ошибок. Способность к мышле
нию изучают с помощью «проблемы об
ходного пути». На рисунке 186 суть ме
тода очевидна: чтобы подойти к пище,
енот должен отойти от нее и обойти ко
лышек, чтобы удлинить шнурок. Среди
многих животных, испытанных в этих эк
спериментах, только обезьяны оказались
способными решить задачу без многочис
ленных проб и ошибок. Человек эту зада
чу решает с первого предъявления, так
как ему в несравненно большей степени,
чем другим приматам, свойственна спо
собность к мышлению. Более того, он
может научиться в довольно раннем воз
расте воспринимать информацию в виде
символов, например букв и слов. Они да
ют ему представление о внешнем мире,
обеспечивают возможность общения
Рис. 186. Проблема обходных путей в опытах с
енотами.
316
между людьми и накопление знаний в
различных областях. Слово и письмо ста
ли второй сигнальной системой для че
ловека, расширившей его возможности к
адаптивному поведению. Благодаря спо
собности к мышлению и разнообразным
формам адаптивного поведения человек
играет ведущую роль в живой природе.
0
1 1. Что такое научение? 2. Какие способы нау
чения выделяют у животных? 3. Какие формы пове
дения свойственны человеку?
92. Координация и регуляция
у растений
Целостность многоклеточного расти
тельного организма поддерживается
межклеточными (межтканевыми, межор
ганными) системами регуляции, среди
которых гормональная регуляция являет
ся главной. Осуществляется она фито
гормонами — активными низкомолеку
лярными органическими соединениями,
образующимися в различных тканях и ор
ганах и действующими в очень низких
концентрациях.
Впервые предположение о существо
вании у растений веществ, действующих
подобно гормонам животных, сделал
Ч. Дарвин при изучении явления гелио
тропизма — изгибания растения к ис
точнику света. Результаты своих много
численных и оригинально проведенных
опытов и гипотезы, объясняющие явле
ние гелиотропизма у растений, он описал
в книге «Сила движения у растений»
(1880). Экспериментируя с проростками
растений, Дарвин установил, что наибо
лее чувствительными к свету являются
верхушки проростков (сенсорные зоны),
а медленная двигательная ростовая реак
ция осуществляется в другом участке (в
двигательной зоне) (рис. 187). Он пред
положил, что передача раздражения из
сенсорной зоны в нижележащую двига
тельную осуществляется с помощью ка
ких-то веществ. Открыты эти вещества
Рис. 187 Опыт Ч. Дарвина с этиолированными
проростками пшеницы.
были в течение первой половины XX ве
ка. Они были названы фитогормонами,
функции которых заключались не только
в регуляции процессов роста, но и созре
вания, старения, явлениях стресса, тран
спорта веществ и многих других.
В отличие от животных, имеющих
обособленные эндокринные железы, гор
мональная система растений менее спе
циализирована. Регуляция различных
морфологических и физиологических
процессов обеспечивается одними и теми
же фитогормонами, но в разных соотно
шениях.
Ростовое вещество, выделяемое вер
хушками проростков растений, вызываю
щее их изгибание в сторону света, впер
вые было определено в 1935 г. и названо
индолил-3-уксусной кислотой (ИУК).
Соединения этой группы были названы
ауксинами (от греч. «аихапо» — расту),
и л и ростовыми гормонами.
ИУК была обнаружена во всех расте
ниях, причем наиболее высоко ее содер
жание в почках, листьях, камбии, форми
рующихся семенах, пыльце. Предшес
твенником ИУК чаще всего является
аминокислота триптофан.
Ауксин стимулирует деление и растя
жение клеток, формирование проводя
щих пучков и корней, разрастание око
лоплодника. Первостепенна роль ИУК в
ростовых движениях — тропизмах и настиях.
Вторая группа фитогормонов — цитокинины, ускоряющие деление клеток.
Природный цитокинин из незрелых зер
новок кукурузы — зеатин -— был выде
лен в 1963 г.
Цитокинины найдены у микроорга
низмов, водорослей, папоротников, мхов
и у большинства растений. Наиболее бо
гаты ими апикальные меристемы корней,
из которых по ксилеме они транспортиру
ются в надземные органы. Велико их со
держание также в семенах и плодах.
Действие цитокининов, однако, про
является лишь в присутствии ауксина.
Сильное вытягивание стебля у расте
ний происходит под влиянием фитогор
мона гиббереллина (рис. 188, с. 318),
впервые выделенного из фитопатогенно
го гриба Gibberella fujikuroi.
Гиббереллин обнаружен у грибов, во
дорослей и высших растений. Больше
всего его содержат незрелые семена.
Синтезируется этот фитогормон в листь
ях и корнях, а перенос осуществляется
ксилемным и флоэмным током. Места
действия гиббереллина — верхушечные
и вставочные меристемы, в которых про
исходит активация деления клеток.
Противоположное тормозящее дей
ствие на рост растений оказывает фито
гормон абсцизин, или абсцизовая кисло
та (от англ, abscission — отделение, опа
дение), впервые выделенный из сухих
зрелых коробочек хлопчатника.
Абсцизин вырабатывается у покрыто
семенных и голосеменных растений, па317
Рис. 188. Влияние гиббереллинов на рост растений:
а —- до обработки гиббереллином, б — после обра
ботки.
поротникообразных, хвощей, мхов. Им
богаты старые листья, зрелые плоды, се
мена, покоящиеся почки.
Абсцизин задерживает прорастание
семян и рост почек. Его накопление в
тканях приводит к формированию отде
лительного слоя в черешках и плодонож
ках, опадению листьев и плодов.
При неблагоприятных факторах
внешней среды, особенно при засухе, аб
сцизин быстро накапливается в тканях,
способствует закрыванию устьиц, что
снижает испарение воды растениями. Та
ким образом он участвует в механизмах
стресса.
Бактерии, грибы и многие раститель
ные организмы способны вырабатывать
318
газ этилен. Его синтез наиболее интен
сивно происходит в стареющих листьях и
в созревающих плодах.
Этилен приостанавливает удлинение
проростков, рост листьев у двудольных,
вызывает задержку митоза. Он также
усиливает процессы старения, созрева
ния и опадания плодов и листьев.
Таким образом пять классов фитогор
монов (ауксины, цитокинины, гиббереллины — стимуляторы; абсцизовая кисло
та и этилен — ингибиторы), взаимодей
ствуя друг с другом путем изменения их
количественного соотношения, осущест
вляют координацию и регуляцию морфо
генетических п физиологических процес
сов в растительных организмах.
Изучение природных гормонов, их хи
мических формул создало основу для воз
можности синтеза искусственных регуля
торов роста; Теперь их активно использу
ют фермеры и садоводы для контроля
роста и развития растений. Так, ауксины
могут продлить цветение растений, спо
собствуют укоренению черенков, приво
дят к более раннему плодоношению, вы
зывают образование плодов без опыле
ния. Обработка абсцизином предотвра
щает прорастание клубней, корнеплодов
и луковиц во время хранения; его также
применяют для борьбы с сорной расти
тельностью.
?
■ 1. Какие фитогормоны свойственны растени
ям? 2. Каковы особенности действия фитогормо
нов? 3. Какое действие оказывает этилен на расте
ния?
ГЛАВА VIII
Адаптация организмов
к условиям обитания
гт :
' .М д '
ЭЗ. Среда обитания и условия
существования организмов.
Экологические факторы
Среда обитания — это совокупность
всех элементов и явлений природы, ее
в ещ еств о и п р о ст р а н ст в о , к о то р ы е н е
п о с р ед с т в е н н о о к р у ж а ю т ж и в ы е о р г а
низм ы и ок а зы в а ю т на них п р я м о е или
к о св е н н о е вл и я н и е.
Составные части среды обитания
чрезвычайно разнообразны и изменчи
вы. Разные элементы среды оказывают
различное воздействие на организмы,
так как одни из них чрезвычайно важны,
другие менее либо вовсе не нужны. Так,
например, шляпочные грибы не нужда
ются в свете, но чрезвычайно чувстви
тельны к недостатку органических ве
ществ. Растительные организмы, нап
ротив, все нуждаются в свете. Поэтому
понятие «среда обитания» не следует
путать с понятием «условия существо
вания» — совокупностью жизненно не
обходимых факторов среды, без кото
рых живые организмы не могут сущес
твовать. К таким факторам принадле
жат свет, вода, тепло, воздух, почва
и др. В отличие от них другие факторы
среды хотя и оказывают заметное влия
ние, однако не являются для живых ор
ганизмов жизненно необходимыми
(например, ветер, атмосферное элек
тричество и др.).
Э л ем ен ты о к р у ж а ю щ е й ср ед ы , к о
т о р ы е вы зы ваю т у ж и в ы х о р г а н и зм о в
п р и с п о с о б и т е л ь н ы е р еа к ц и и (а д а п т а
ц и и ), н азы в аю тся эк ол оги ч еск и м и ф а к
торам и.
Экологические факторы разнообраз
ны как по своему происхождению, так и
характеру действия. Их подразделяют на
три группы.
-А б и о т и ч е с к и е факторы включают
элементы неорганической, или неживой,
природы, воздействующие на организм.
В свою очередь абиотические факто
ры подразделяют на физические, или
климатические (свет, температура,
влажность, давление, ветер, течения),
почвенно-грунтовые, или эдафические (механический состав почвы, ее про
ницаемость, влагоемкость), химические
(соленость воды, газовый состав воды и
воздуха), топографические (особеннос
ти рельефа местности).
Б и о т и ч ес к и е факторы — это сово
купность разнообразных воздействий од
них организмов на жизнедеятельность
других, а также на среду обитания. Одни
организмы могут служить пищей для дру
гих (например, хозяин для паразита),
способствовать размножению и расселе
нию (например, птицы и насекомые-опы
лители для цветковых растений), оказы
вать механические, химические и другие
воздействия.
А н троп оген н ы е
(антропические)
факторы — это все формы деятельности
человеческого общества, изменяющие
среду обитания живых организмов или
непосредственно влияющие на их жизнь.
Выделение антропогенного фактора в
отдельную группу было вызвано тем, что
закономерности развития человеческого
общества часто шли вразрез с законами
природы. Благодаря широкомасштабной
производственной деятельности людей
преобразовывался лик планеты: интен
сивно использовались ресурсы природы,
они перерабатывались с возвратом в
природную среду чуждых ей отходов, за
грязняли ее. Академик В. И. В е р н а д
с кий -— создатель учения о биосфере —
рассматривал деятельность человека как
мощную геологическую силу, оказываю
щую преобразующее влияние на окружа
ющую среду.
Действие на организм различных эко
логических факторов имеет свои особен
ности. Так, абиотические факторы пря
мо или косвенно воздействуют на орга
низм через обмен веществ. Например,
изменение температуры среды приводит
к изменению интенсивности метаболизма
как у растений, так и у большинства жи
вотных. Действие абиотических факторов
одностороннее, т. е. организм может к
ним приспособиться, но, как правило, не
в состоянии оказывать на них обратное
влияние.
319
Действие биотических факторов
принципиально другое, так как организ
мы обоюдно влияют друг на друга. Нап
ример, хищник приобретает приспособ
ления к наиболее эффективной поимке
жертвы. Однако и жертва развивает
приспособления, которые помогают ей
избежать поимки. В данном случае про
исходит совместная эволюция организ
мов разных видов.
Изменчивость разных элементов сре
ды не одинакова. Так, например, темпе
ратура, влажность, сила ветра, осадки,
наличие пищи, хищников, паразитов,
конкурентов чрезвычайно непостоянно
во времени и в пространстве. Диапазон
изменчивости во многом зависит от осо
бенностей среды обитания. Например, в
нижних горизонтах почвы температура
почти постоянна, тогда как на поверхнос
ти она сильно варьирует. Другие элемен
ты среды относительно постоянны на
протяжении больших отрезков времени.
Таковыми являются состав атмосферы,
сила земного притяжения, солнечная
постоянная, солевой состав океана и др.
Поэтому действие одних факторов среды
бывает постоянным, а других — регуляр
но-периодическим (например, суточный
и сезонный ритмы, смена приливов и от
ливов в океане); третьи же действуют не
регулярно, без четкой периодичности
(например, ливни, бури, смерчи).
Таким образом, конкретные организ
мы определенного вида подвергаются
разнообразным как по природе и интен
сивности, так и по периодичности воздей
ствиям факторов среды и живут только в
тех средах, к которым они сумели приспо
собиться.
• 1. В чем отличия понятий: среда обитания, ус
ловия существования, экологические факторы?
2. Чем объясняется выделение антропогенного
фактора в отдельную группу? 3. В чем заключается
отличительная особенность действия биотических
факторов?
320
94. Закономерности действия
внешних факторов
В природе невозможно найти расте
ние или животное, которое могло бы пе
реносить все условия жизни, существую
щие на Земле. Каждый организм живет в
относительно узком диапазоне темпера
тур, влажности, содержания кислорода и
других факторов среды. Более того, жи
вые организмы разных видов, а нередко и
одного вида, но на разных стадиях разви
тия, сильно отличаются друг от друга по
отношению и чувствительности к факто
рам среды. Тем не менее, в характере
воздействия факторов среды и в ответ
ных реакциях живых организмов выявлен
ряд общих закономерностей, которые
укладываются в некоторую общую схему
действия экологического фактора разной
дозы на жизнедеятельность организма
(рис. 189).
Так, на рисунке 189 внизу изображена
зависимость скорости роста растений от
температуры при поддержании на посто
янном уровне всех остальных жизненно
важных параметров среды. Была получе
на куполообразная кривая: по мере повы
шения температуры до некоторой вели
чины скорость роста растений возраста
ла, достигая максимального значения, а
затем снижалась.
Весь диапазон температур, при кото
ром возможен рост растений, ограничен
двумя пороговыми точками, соответ
ствующими минимальной и максималь
ной величинам температур, за пределами
которых растения гибнут. Эти точки на
зываются нижним и верхним пределами
выносливости (толерантности).
Точка на оси абсцисс, соот
ветствующая температуре, при которой
наблюдается наибольшая скорость роста
растений, называется точкой оптимума.
Для большинства организмов опреде
лить оптимальное значение фактора с
достаточной точностью бывает трудно,
поэтому принято говорить о зоне опти-
▲
Пределы выносливости
Верхний предел
Степень благоприятности фактора
Ч
Интенсивность фактора
Оптимум
8
18
28
38 Температура,°С
Рис. 189. Зависимость результата действия экологического фактора от его интенсивности.
321
м ум а. Слева или справа от нее располо
жена область температур, при которых
растения растут еще достаточно хорошо
жить при разных температурах как в
пресных, так и в солоноватых водоемах.
Некоторые растения не переносят даже
небольших заморозков (огурцы, томаты),
в то время как другим они не вредят (са
довая земляника, петрушка, овес и др.).
Одни виды луговых трав предпочитают
почвы с довольно узким диапазоном кис
лотности. Например, щавелек, белоус
торчащий растут на кислых почвах с pH
3,5—4,5. Другие растения (например,
сосна обыкновенная) хорошо растут на
почвах с широким диапазоном кислотнос
ти. В связи с этим организмы, для сущес
твования которых необходимы строго оп
ределенные, относительно постоянные
условия внешней среды, называются
стенобионтными (от греч. stenos —
узкий, Ыоп —■живущий), а те, которые
могут жить в широком диапазоне измен
чивости условий среды, — эврибионтными (от греч. eurys — широкий) (рис.
190).
В зависимости от конкретного фак
тора среды различают организмы стено и эвритермные по отношению к темпе
ратуре, стено- и эвригалинные по отно
шению к концентрации солей, стено- и
эврифотные по отношению к свету. При
этом организмы одного и того же вида
могут иметь узкий предел толерантно
сти по отношению к одному фактору и
широкий — к другому (например, быть
приспособленными к жизни в узком ди
апазоне температур и в широком диапа
зоне солености). Кроме того, одна и та
— зо н а норм альной ж и зн ед ея т ел ь н ос
ти. Крайние участки кривой, соответ
ствующие резкому недостатку (слева)
или избытку (справа) тепла, называются
зо н о й п есси м у м а , или ст р е с с а . В этой
зоне растения еще не погибают, но и не
растут.
В природных условиях нередко наб
людается нарушение симметрии куполо
образной кривой. Так, например,устрицы
растут лучше всего при умеренной соле
ности воды, но при низкой солености их
рост подавляется сильнее, чем при высо
кой. Оптимальное для речной форели со
держание кислорода, сдвинуто также в
правую сторону градиента фактора.
Повторяемость описанных тенденций
позволяет рассматривать их как фунда
ментальный биологический принцип: для
к а ж д о го вида р аст ен и й и ж и в о т н ы х с у
щ е ст в у ет о п т и м у м , з о н а н о р м а л ь н о й
ж и зн е д ея т ел ь н о ст и , ст р е с с о в ы е зо н ы й
п р е д ел ы в ы н о с л и в о с т и в о т н о ш е н и и
к а ж д о го ф а к т о р а среды .
Разные виды живых организмов за
метно отличаются друг от друга как по-положению оптимума, так и по пределам
выносливости. Так, например, рифообра- Л'
зующие коралловые полипы обитают в :
зоне Мирового океана с. соленостью но
чи. равной 3,5 %, и высокими (не ниже
20,5 °С) и малоизменчивыми температу- К
рами. Напротив, рачки дафнии могут.
J3
Б
О
X
ш
S
Стенобионты
опт.
I
/
Эврибионты
'
[/мин.
Стенобионты
опт.
;
МАКС.
- МИН. |
МАКСЛ
Температура и другие факторы
Рис. 190. Экологическая пластичность видов.
322
-------
же доза фактора может быть оптималь
ной для одного вида, пессимальной для
другого и выходить за пределы выносли
вости для третьего. Способность орга
низмов адаптироваться к тому или ино
му диапазону изменчивости факторов
среды называют экологической плас
тичностью. Следовательно, эврибионтные организмы являются экологи
чески пластичными, выносливыми, что
способствует их широкому распростра
нению. Напротив, к маловыносливым,
экологически непластичным принадле
жат стенобионтные организмы, имею
щие обычно ограниченные ареалы рас
пространения.
В за и м о д е й ст в и е ф а к т о р о в . О г р а н и
чиваю щ ий ф а к т о р . Экологические фак
торы воздействуют на живой организм
одновременно и совместно. При этом
действие одного фактора зависит от того,
с какой силой и в каком сочетании дей
ствуют одновременно другие факторы.
Эта закономерность получила название
взаимодейст вия факторов. Напри
мер, жара или мороз легче переносятся
при сухой, а не влажной погоде. Ско
рость испарения воды листьями растений
(транспирация) значительно выше при
высокой температуре воздуха с ветром,
чем в безветренную прохладную погоду.
В некоторых случаях недостаток одного
фактора частично компенсируется уси
лением другого. Явление частичной вза
имозаменяемости действия экологичес
ких факторов называется эффектом
компенсации. Например, увядание рас
тений можно приостановить путем как
увеличения количества влаги в почве,
так и снижением температуры воздуха,
уменьшающего транспирацию. В пусты
нях недостаток осадков в определенной
мере восполняется повышенной относи
тельной влажностью воздуха в ночное
время. В Арктике продолжительный све
товой день летом компенсирует недоста
ток тепла.
Вместе с тем ни один из необходимых
организму экологических факторов не
может быть полностью заменен другим.
Отсутствие света делает жизнь растений
невозможной, несмотря на самые бла
гоприятные сочетания других условий.
Фактор, величина которого оказывается
близкой или выходит за пределы толе
рантности (ниже минимума или выше
максимума), называется о г р а н и ч и в а ю
щ им. Вот почему, если значение хотя бы
одного из необходимых экологических
факторов приближается или выходит за
пределы пороговых величин, то, несмот
ря на оптимальное сочетание остальных
условий, особям грозит гибель. Изменить
результат можно только воздействием на
ограничивающий фактор.
Природа ограничивающих факторов
может быть различной. Например, угне
тение роста травянистых растений под
пологом буковых лесов, где при опти
мальном тепловом режиме, повышенном
содержании диоксида углерода, богатых
почвах возможности развития трав огра
ничиваются недостатком света. Ограни
чивающие факторы среды определяют
также географический ареал вида. Так,
продвижение вида на север может лими
тироваться недостатком тепла, а в рай
оны пустынь и сухих степей — недостат
ком влаги или слишком высокими темпе
ратурами. Сезонные изменения климата
обычно выходят за пределы диапазона ус
ловий, оптимальных для большинства ви
дов. Соответственно условия среды бла
гоприятствуют росту организмов лишь в
течение определенных периодов, что оп
ределяет сезонные циклы их численности
и активности. В большинстве областей
умеренного и арктического поясов глав
ным фактором, определяющим сезоны
роста и активности, служит температура,
и эти сезоны обычно приурочены к теп
лым летним месяцам.
Ограничивающим распространение
организмов фактором могут служить и
биотические отношения, например заня323
1
2
Рис. 191. Индикаторные организмы чистых вод: 1 — личинка поденки, 2 — личинка веснянки, 3 — ли
чинка ручейника.
тость территории более сильным конку
рентом или недостаток опылителей для
цветковых растений.
Выявление ограничивающих факто
ров и устранение ихдействия, иными сло
вами, оптимизация среды обитания жи
вых организмов составляет важную
практическую цель в повышении уро
жайности сельскохозяйственных куль
тур и продуктивности домашних жи
вотных.
Экологические индикаторы. По
скольку специфические факторы и их до
за строго определяют, какие организмы
могут жить в данной местности, можно
использовать обратную закономерность
и судить по организмам о типе и состоя
нии (качестве) физической среды. На
пример, присутствие в водоеме губок бадяг, личинок ручейников (рис. 191), поде
нок и веснянок свидетельствует о чистоте
324
водоема; наоборот, массовое развитие
малощетинковых червей (рис. 192) гово
рит о его сильном загрязнении. Лишай
ники чутко реагируют на присутствие в
воздухе даже ничтожных количеств за
грязняющих веществ, особенно диоксида
серы, и не растут в таких местообитаниях
(например, в крупных городах). Обнару
жено также, что видовой состав сорных
растений служит хорошим показателем
плодородия и кислотности почвы. Напри
мер, на кислых почвах произрастает ве
реск обыкновенный, щавелек, лютик ед
кий, погремок большой, на щелочных —
мать-и-мачеха, очиток едкий.
Таким образом, организмы, характер
ные для определенных территорий, назы
ваются экологическими индикаторами
среды обитания. Метод экологической
индикации широко используется в систе
ме мониторинга — длительного слеже-
95. Абиотические факторы
(климатические факторы).
Тепло
Среди абиотических факторов важ
нейшее значение в создании условий
жизненной среды имеют климатические
факторы —• тепло, влажность, свет.
Тепло. Важность теплового фактора
для большинства организмов определя
ется его значительным влиянием на ско
рость процессов обмена веществ, пове
дение, развитие, популяционную динами
ку, географическое распределение. Тем
пература — величина, характеризующая
тепловое состояние тела. Диапазон из
менчивости температур в водной и назем
но-воздушной средах весьма велик (таб
лица 11)
Таблица
1 1 . Температурный диапазон актив
ной жизни на Земле
Среда жизни
Суша
Океан
Пресные
водоемы
Рис. 192. Малощетинковый червь трубочник —
индикатор загрязненных вод.
ния за состоянием почвенной, воздушной
и водной сред обитания.
?
1. Каков общий характер действия на организм
экологического фактора разной дозы? 2. Что такое
экологическая пластичность организма? 3. Как вы
понимаете взаимодействие экологических факто
ров? 4. Что такое ограничивающий фактор и каково
его действие на организм? 5. Какую информацию
можно получить о среде по экологическим индика
торам?
Максимум Минимум Амплитуда
55 °С
35,6 “С
93 °С
- 7 0 °С
-3 ,3 °С
0°С
125 "С
38,9 °С
93 "С
Как видим, наиболее стабильной яв
ляется морская, а наиболее изменчи
вой — наземно-воздушная среда.
По обеспеченности наземных орга
низмов теплом различают четыре основ
ных термических пояса.
1. Тропический пояс. Температура
всегда выше О °С (за исключением высо
когорных областей). Средняя температу
ра самого холодного месяца 15— 20 °С.
Амплитуда колебания температур не
превышает 5 °С. Вегетация растений
круглогодичная (кроме засушливых об
ластей).
2. Субтропические пояса. Темпера
тура самого холодного месяца выше
+4 °С, самого теплого — выше 20 °С.
Возможны кратковременные морозы. Ус
тойчивый снежный покров зимой отсут
ствует. Для сельскохозяйственных куль
325
тур два вегетационных сезона — летний
и зимний.
3. Умеренные пояса. Хорошо выра
жен летний вегетационный сезон и про
должительный зимний период покоя рас
тений. Средняя длительность безмороз
ного периода 70—80 суток. Зимой устой
чивый снежный покров и устойчивые мо
розы. Заморозки весной и осенью.
4. Холодные пояса. Заморозки воз
можны в течение всего вегетационного
периода, продолжительность которого
всего 1,5— 2 месяца. Зима длинная,
очень морозная.
Каждый термический пояс заселен
только такими организмами, пределы вы
носливости которых не выходят за рамки
крайних значений температур, характер
ных для этих зон. Так, умеренные пояса
заселены в основном эвритермными ви
дами, тропический пояс — стенотермными теплолюбивыми видами, а холодные
пояса — стенотермными холодолюбивы
ми видами.
Организмы способны к жизни в ин
тервале таких температур, при которых
не происходит денатурации белковых мо
лекул, необратимого изменения коллоид
ных свойств цитоплазмы, нарушения ак
тивности ферментов, изменения гидроли
тических процессов, дыхания. Для боль
шинства организмов этот диапазон тем
ператур составляет 0— 50 °С. Однако у
некоторых организмов выработались
адаптации для жизни в большем диапазо
не изменчивости температур. К ним при
надлежат, в первую очередь, представи
тели царства Бактерий. Так, например,
некоторые виды цианобактерий способ
ны обитать в горячих источниках при тем
пературе + 80— 88 °С.
Высокой устойчивостью обладают и
некоторые виды наземных лишайников,
семена и вегетативные органы пустын
ных растений, находящиеся в верхнем
раскаленном слое почвы.
Немало видов растений способны
переносить отрицательные температу
326
ры, предварительно подготовившись к
ним. Такие организмы впадают в состоя
ние глубокого покоя, или анабиоза,
при котором процессы жизнедеятель
ности сведены к минимуму. У растений
подготовка к этому состоянию сопро
вождается опадением листьев, надзем
ных органов у травянистых многолетни
ков, образованием чешуек на почках, их
засмолением, развитием толстого слоя
кутикулы, коры на стеблях. В тканях и
клетках накапливаются гормоны, тор
мозящие ростовые и формообразова
тельные процессы, благодаря чему рас
тение неспособно к прорастанию во
время случайных осенних и раннезимних
потеплений.
Чтобы воспрепятствовать замерза
нию воды в межклетниках и внутри кле
ток, что привело бы к обезвоживанию и
механическому повреждению клеток, у
морозостойких растений выработался
ряд биохимических адаптаций. Они
заключаются ц синтезе веществ, в час
тности таких, как гидрофильные и водо
растворимые белки, моно- и олигосаха
риды и др., способных связывать значи
тельное количество воды. Связанная вода
в виде гидратных оболочек не замерзает и
не транспортируется, оставаясь при этом
в клетке. Повышение концентрации рас
творимых углеводов в клеточном соке
способствует понижению точки замерза
ния воды.
Примерами морфологических адап
таций, касающихся формы растений хо
лодных местообитаний, являются их не
большие размеры — к а р л и к о в о с т ь
(карликовые березы, карликовые ивы),
стелющиеся формы — с т л а н и к и (кед
ровый стланик, можжевельник, рябина);
п о д у ш к о в и д н ы е формы растений,
образуемые в результате усиленного вет
вления и крайне замедленного роста по
бегов (рис. 193). Благодаря таким призе
мистым и компактным формам, растения
мало подвергаются воздействию ветра,
используют припочвенное тепло в летнее
Рис. 193. Формы растений полярных и высоко
горных областей: а — кедровый стланик, б —
арктическое растение-подушка, в — схема вет
вления растения-подушки.
327
время, а зимой защищены снежным по
кровом.
Для некоторых видов растений на оп
ределенном этапе онтогенеза необходимо
воздействие низких температур, без ко
торого они не в состоянии завершить
свой жизненный цикл. Примером являет
ся яровизация — процесс, протекаю
щий у озимых форм однолетних и двулет
них растений под действием низких поло
жительных температур определенной
длительности, способствующих последу
ющему ускорению развития этих расте
ний. Растения, нуждающиеся в яровиза
ции, называются озимыми, а развиваю
щиеся без нее — яровыми. Озимые рас
тения без яровизации не зацветают.
Температурные адаптации живот
ных. Животные обладают большим раз
нообразием способов адаптации к воз
действию различных температур. Прежде
всего у них выработались два способа по
лучения тепла. Большая часть животных
получает тепло из внешней среды. Эта
группа животных называется эктотермными (пойкилотермными, хо ло д
нокровными). Температура их тела
близка к температуре внешней среды.
Эти животные лишены способности под
держивать температуру тела в узких гра
ницах, так как не обладают совершенны
ми собственными механизмами образо
вания и сохранения достаточного коли
чества тепла. Активность эктотермных
животных зависит от температуры окру
жающей среды. Однако эктотермные жи
вотные не лишены способности к термо
регуляции. Средством такой регуляции
могут служить поведенческие реакции, в
частности соответствующая ориентация
тела животного по отношению к внешне
му источрику тепла. Многие виды прес
мыкающихся способны удерживать тем
пературу тела в некоторых пределах пу
тем изменения активности, прячась в те
ни, закапываясь в песок или выползая на
солнце. Другие животные приобрели
способность изменять интенсивность
328
пигментации эпидермиса в зависимости
от температуры. Некоторые развили спо
собность повышать температуру тела за
счет активизации мышечных движений.
Так, за счет дрожи летательных мышц об
разуется тепло у многих видов насеко
мых, что позволяет им поддерживать от
носительно постоянный уровень актив
ности в определенных температурных
границах. Активно плавающие тунцы
способны на несколько градусов повы
шать температуру тела по сравнению с
температурой воды.
Низкие температуры для многих экто
термных организмов являются ограничи
вающим фактором. Способом выжива
ния для большинства из них при наступ
лении периодов с низкими температура
ми является резкое снижение метаболи
ческой активности (оцепенение, спяч
ка). Местообитания с постоянно низки
ми температурами освоены лишь теми
эктотермными организмами, которые в
ходе длительной эволюции приобрели
ряд адаптаций, препятствующих замерза
нию внутри- и межклеточной воды, дена
турации белков и изменению физическо
го состояния липидов. Устойчивость к за
мораживанию у них достигается механиз
мами, понижающими точку замерзания
жидкостей, например, повышением кон
центрации солей в крови арктических
рыб, замещением части воды органичес
кими растворителями (антифризами),
например глицерином и др. В наземной
среде низкие температуры успешнее пе
реносят животные с мелкими размерами
тела, благодаря которым они, обитая в
ограниченных пространствах, способны
использовать малейшие преимущества
микроклимата (насекомые, клещи, пау
ки, круглые черви и т. п.).
При низких температурах преимущес
твом для эктотермных животных являет
ся снижение энергозатрат, а следова
тельно, резкое снижение потребления
пищи. При высоких температурах экто
термные животные избегают излишних
потерь воды путем изменения уровня ак
тивности и мест локализации. Особенно
четко это проявляется у обитателей пус
тынь и полупустынь.
Вторая группа животных —- эндотермные (гомойотермные, или т еп
локровные) приобрела способность под
держивать температуру своего тела на
постоянном уровне вне зависимости от
температуры внешней среды за счет теп
ла, образуемого самим организмом в ходе
метаболических процессов. К ним при
надлежат птицы и млекопитающие. Фер
ментные системы эндотермных животных
адаптированы к функционированию в уз
ком диапазоне температур. Эндотермные
организмы выработали разнообразные
приспособления для терморегуляции.
Так, снижение теплоотдачи достигается у
них развитым перьевым или волосяным
покровами, жировым слоем в подкожной
клетчатке, сужением капилляров кожи.
Увеличение теплоотдачи происходит пу
тем потоотделения, уменьшения густоты
шерстного или перьевого покровов, рас
ширения капилляров кожи, тепловой
одышки. При необходимости увеличение
теплопродукции достигается повышени
ем потребления пищи, окислением буро
го жира, который служит исключительно
источником тепла. Благодаря высокой
интенсивности окислительных процес
сов, наличию мощного комплекса термо
регуляционных средств эндотермные жи
вотные шире осваивают разнообразные
географические зоны нашей планеты.
Ограничивающим фактором в холодных
районах для них является не температура,
а пища.
Важное значение для поддержания
температурного баланса у эндотермных
организмов имеет отношение повер
хности тела к его объему, так как коли
чество выработанного тепла зависит от
массы тела, а теплообмен осуществля
ется через покровы. По мере возраста
ния массы тела удельная поверхность
уменьшается. В связи с этим замечено,
3
Рис. 194. Соотношение размеров головы и
ушей у песца (1) — обитателя тундры, лисицы
обыкновенной (2) — жительницы умеренных
широт и лисицы фенека (3) — обитателя пус
тынь.
что виды теплокровных животных, оби
тающие в холодных климатических зо
нах, имеют большие размеры тела, чем
их сородичи в более теплых местообитани я х ( п р а в и л о Б е р г м а н а ), а высту
пающие части (ушные раковины, клю
вы, хвосты, конечности) увеличиваются
при продвижении к югу ( п р а в и л о А л
л е н а ) (рис. 194).
Таким образом, у животных двух при
веденных групп разная стратегия освое
ния жизненного пространства: у эктотермных — пассивный путь, т. е. подчинение
жизненных функций ходу внешних темпе
ратур, у эндотермных — активный путь,
связанный с большой затратой энергии,
что позволяет им заселять разнообраз
ные местообитания.
9
1. Каков диапазон активной жизни организмов
в разных средах нашей планеты? 2. Каковы адапта
ции у организмов холодных местообитаний? 3. Ка329
коды адаптации организмов к жизни в условиях
жаркого сухого климата? 4. Какие механизмы тер
морегуляции выработали экто- и эндотермные жи
вотные?
96. Свет
Свет — важнейший экологический
фактор. Он является основным источни
ком энергии как для создания первичного
органического вещества фототрофными
организмами биосферы, так и для под
держания теплового баланса планеты.
Биологическое действие солнечного све
та зависит от его спектрального состава
(табл. 12), интенсивности, суточной и се
зонной периодичности.
Табл и ца 12, Спектральный состав приходящего наземную поверхность солнечного излучения и его би
ологическое действие
Длина волн, нм
Спектральный состав
Ультрафиолетовые 290— 400
лучи
Доля, %
9 — 10
Биологическое действие
Бактерицидное, мутагенное. Способ
ствуют синтезу у растений некоторых вита
минов, пигментов, а у животных — вита
мина D. Многими насекомыми исполь
зуются в пространственной ориентации.
Видимый свет — 400— 710
физиологически ак
тивная
радиация
(ФАР)
41— 45
Источник энергии для фотохимических
реакций при синтезе первичного органи
ческого вещества. Сигнальное действие в
регуляции
многих
физиолого
биохимических процессов и поведения.
Оказывает формообразующее действие на
растения.
Инфракрасное из 710— 3000
лучение
45— 50
Основной источник тепла для подав
ляющего большинства организмов всех
царств.
Количество солнечной радиации, посту
пающее на земную поверхность, в пер
вую очередь определяется географичес
кой широтой местности. Однако особен
ности местного характера, такие, как сос
тояние атмосферы, рельеф местности,
вносят свои изменения в световой режим
растений и животных. Благодаря этому
разнообразие световых условий чрезвы
чайно велико: от сильно освещенных
местообитаний, таких, как высокогорья,
степи, пустыни, до сумеречного освеще
ния в пещерах и водных глубинах. Соот
ветственно разнообразны и приспособ
ления организмов к жизни при том или
ином световом режиме.
В зависимости от потребности в свете
растения подразделяют на следующие
экологические группы:
1. Светолюбивые (световые), или
гелиофиты, — растения открытых,
330
постоянно освещаемых местообита
ний;
2. Тенелюбивые (теневые), или сциофиты, — растения затененных мес
тообитаний;
3. Теневыносливые — растут в усло
виях разной световой обеспеченнос
ти, однако лучше — при высокой ос
вещенности.
Растения первых двух групп отлича
ются положением экологического опти
мума: у светолюбивых он находится в об
ласти полного солнечного освещения, а у
тенелюбивых сдвинут в область слабой
освещенности.
Под влиянием разной световой обес
печенности у растений выработались со
ответствующие приспособления. Прежде
всего они касаются величины листовых
пластинок: у световых растений они
обычно мельче, чем у тенелюбивых. Ори-
ентация листьев у растений-гелиофитов
вертикальная или под большим углом к
горизонтальной плоскости, т. е. они как
бы «отворачиваются» от избыточного
света. И напротив, листья теневыносли
вых растений ориентированы к свету всей
своей плоскостью и расположены так,
чтобы одновременно не затенять сосед
ние листья (листовая мозаика) (рис.
195).
Поверхность листа у многих гелиофи
тов способствует отражению световых
лучей. Так, у лавра, магнолии она блестя
щая, словно лакированная; у молоча
ев — покрыта светлым восковым на
летом.
Листья световых растений обычно
толстые за счет мощного развития асси
миляционных тканей и прежде всего па
лисадной паренхимы. Она состоит из
2— 3 рядов узких и длинных клеток, при
сутствующих не только на верхней, но и
на нижней стороне листа. Устьица мелкие
и многочисленные. Напротив, мезофилл
теневых растений слабо дифференциро
ван на палисадную и губчатую паренхиму.
Пластинка листа тонкая, широкая, с
большой удельной поверхностью, что
способствует лучшему проникновению
света в глубь листа.
У светолюбивых растений количество
хлоропластов в клетках листа на единицу
площади в несколько раз больше, чем у
тенелюбивых. Сами хлоропласта у гели
офитов более мелкие и светлые, способ
ные перемещаться в клетке: при сильном
освещении они становятся «ребром» к
направлению лучей, что защищает хло
рофилл от разрушения.
От условий освещенности зависит
состав пигментов листа и их соотноше
ние. У растений открытых и хорошо осве
щенных местообитаний хлорофилл а пре
обладает над формой b в 3— 5 раз, а в за
тенении отношение хлорофиллов а и b
составляет 3:2.
Особенно четко адаптивное измене
ние состава пигментов проявляется у
водных растений при изменении спек
трального состава света на разных глу
бинах. С нарастанием глубины теряется
оранжево-красная часть спектра и до
больших глубин доходят в основном зе
леные, голубые и синие лучи. В соответ
ствии с этим у обитающих на мелко
водье зеленых водорослей преобладают
зеленые пигменты, у более глубо
ководных бурых водорослей чаще
встречаются бурые пигменты (фукоксантин и др.), а у самых глубоководных
красных водорослей — красные пигмен
ты (фикоэритрин идр.).
Несмотря на отмеченные адаптации
растений к световой обеспеченности, их
морфологическая структура и особеннос
ти фотосинтезирующего аппарата могут
перестраиваться при изменении светово
го режима. Так, у лиственных теневынос
ливых древесных пород и кустарников
листья, расположенные на периферии
кроны, приобретают структуру, сходную
со структурой листьев световых рас
тений.
У животных световой фактор прежде
всего играет огромную роль в зрительной
ориентации в пространстве. Практически
331
у всех групп животных, начиная с под
вижных форм кишечнополостных — ме
дуз, развиваются светочувствительные
органы — глаза, способные различать
спектральный состав света (цветовое
зрение). Особенно велика их роль для
животных, обитающих в наземно-воз
душной среде. Именно у них орган зрения
достигает наибольшего совершенства.
Зрение у животных совершенствовалось
одновременно с развитием разнообраз
ных форм и окрасок, помогающих скры
ваться от врага или узнавать особей сво
его пола, обнаруживать яркие венчики
цветков с нектаром.
Ориентация животных на свет осу
ществляется в результате ф ото т а к си со в :
положительного (движение в сторону
большей освещенности) или отрица
тельного (движение в сторону наимень
шей освещенности). Общеизвестно дви
жение от света у почвенных животных —
дождевого червя, горшечного червя (бе
лого энхитрея), крота и др.
Некоторые насекомые, например
пчелы, способны ориентироваться в
пространстве путем восприятия поля
ризованного света. Для этого им доста
точно видеть хотя бы часть голубого не
ба. Такая же способность ориентации
выявлена у осьминогов и некоторых
рыб.
Свет играет большую роль в форми
ровании годичного и сезонного циклов
развития животных, а также в регуляции
активности в течение светлого и темного
времени суток. Например, у насекомых
одни виды активны днем (дневные бабоч
ки, стрекозы), другие — в сумеречное
время (бражники, хрущ майский),
третьи — ночью (сверчки, луговой мо
тылек).
Поскольку световой режим место
обитаний в разных широтах различен, то
он оказывает большое влияние на геог
рафическое распространение животных.
Например, в тундре практически отсут
ствуют ночные хищники, так как за ко
332
роткую летнюю ночь они не могут про
кормить ни себя, ни потомство.
?
1. Какие экологические группы растений по от
ношению к потребности в свете выделены? 2. Како
во воздействие светового фактора на организм жи
вотных?
97. Влажность
Все организмы нуждаются в воде,
так как она является основной средой
клетки, где осуществляются процессы
метаболизма. Особая роль воды для на
земных организмов заключается в необ
ходимости постоянного пополнения ее
из-за потерь при испарении. Вся эволю
ция наземных организмов сопровожда
лась выработкой приспособлений к
добыванию и сохранению влаги. Нако
нец, для большой группы растений,
животных, микроорганизмов вода яв
ляется непосредственной средой их
обитания.
Основной источник поступления воды
в наземные местообитания — атмосфер
ные осадки. Водообеспечение наземных
организмов зависит также от наличия во
доемов, уровня грунтовых вод, запасов
почвенной влаги.
Обеспеченность влагой разных геог
рафических районов земного шара неоди
накова. Так, в аридных (сухих, засушли
вых) областях растения и животные ис
пытывают недостаток влаги в течение
большей части вегетационного сезона.
Напротив, в гумидных (влажных) рай
онах во влаге нет недостатка.
Большое влияние на водный режим
местности оказывает температура и дви
жение воздушных масс — ветер. При
высоких температурах и ветре воздух и
почва становятся суше, что вызывает по
вышенные потери воды у наземных орга
низмов.
Соотношение температуры и влаж
ности характеризует микроклимат данной
местности и имеет существенное значе
ние при выборе организмами мест оби
тания.
Исключительную важность приобре
тает сезонный режим выпадения осадков
в течение года, а также суточные коле
бания.
Влажность воздуха и осадки обуслов
ливают периодичность активной жизни
организмов, распределение их по местам
обитания, влияют на смертность, плодо
витость и др. Так, при наступлении сухого
сезона в пустынях многие животные
(например, пустынные жуки чернотелки)
впадают в спячку и пробуждаются только
после осенних дождей.
При ливневых затяжных дождях часто
гибнет большое количество мелких жи
вотных, в особенности насекомых, вымо
кают и погибают однолетние растения.
Влажность воздуха и осадки оказыва
ют большое влияние на развитие гриб
ковых и бактериальных заболеваний рас
тений и животных и, следовательно, кос
венным образом на их урожайность и
численность. Так, в дождливое лето гриб
фитофтора наносит большой ущерб по
садкам картофеля, резко снижая его про
дуктивность.
Из-за постоянной прикрепленности к
субстрату и неспособности отыскивать
подходящую среду для жизни растения
обнаруживают более четкую зависимость
от водного режима, чем животные. По
предпочтениям к местообитаниям с раз
ными условиями увлажнения и выработ
ке соответствующих приспособлений на
земные растения подразделяют на три ос
новные экологические группы: гигрофи
ты, ксерофиты и мезофиты.
Гигрофиты — растения избыточно
увлажненных местообитаний с высокой
влажностью воздуха и почвы. Для них ха
рактерно отсутствие приспособлений, ог
раничивающих расход воды, и неспособ
ность переносить даже небольшой дефи
цит
влаги.
Типичные гигрофиты
(рис. 196) — растения влажных тропи
ческих лесов, полупогруженные при-
2
Рис. 196. Растения-гигрофиты: 1 — тростник, 2
— пушица.
333
3
б
в
Рис. 197 Рыхлые ткани с системой межклетников (аэренхима) на поперечных срезах прибрежного
растения: а — лист, б — стебель, в — корень.
брежные растения (рогоз, камыш, трос
тник), пушица, росянка, рис, папирус,
травянистые растения темнохвойных ле
сов (кислица, майник) и др. Характерны
ми анатомо-морфологическими чертами
гигрофитов являются тонкие листовые
пластинки с небольшим числом широко
открытых устьиц и крупными межклетни
ками, а также развитие у некоторых ви
дов специальной воздухоносной ткани —
аэренхимы (рис. 197). Водопроводящая
система растения развита слабо, корни
тонкие, часто без корневых волосков,
слаборазветвленные. К физиологическим
адаптациям гигрофитов следует также
отнести низкое осмотическое давление
клеточного сока, высокую интенсивность
транспирации, а Следовательно, незначи
тельную водоудерживающую способ
ность (быстрое увядание срезанных рас
тений).
Ксерофиты — растения сухих мес
тообитаний, способные переносить зна
чительный недостаток влаги. Это расте
ния пустынь, сухих степей, саванн, сухих
субтропиков. В зависимости от особен
ностей анатомо-морфологических и фи
зиологических адаптаций их разделяют на
две группы: суккуленты (рис. 198) и склерофиты.
334
С уккуленты запасают в сочных мя
систых листьях (агавы, алоэ) или стеблях
(кактусы, молочаи) большое количество
воды. Фотосинтез у стеблевых суккулен
тов осуществляется периферическим
слоем паренхимы стебля, содержащим
хлорофилл. Во время редких дождей их
поверхностные, слабые корневые систе-
Рис. 198. Кактусы.
мы интенсивно всасывают воду, которая
хранится в водоносных тканях. Эконом
ный расход воды обеспечивается защитой
эпидермиса растений восковым налетом,
погруженными в ткань листа или стебля
немногочисленными устьицами, которые
в дневное время обычно закрыты. Интен
сивность фотосинтеза низкая, поэтому
рост суккулентов и накопление вегета
тивной массы осуществляются медленно.
Особенно много суккулентов в пус
тынях Центральной Америки и Южной
Африки. В Беларуси произрастают еди
ничные виды, такие, как молодило и
очитки, встречаются на сухих песчаных
склонах.
С к л ероф и ты — это растения с мел
кими, жесткими листьями, покрытыми
толстой, препятствующей испарению,
кутикулой. Наиболее характерными рас
тениями этой группы являются степные и
пустынные полыни, ковыли, верблюжья
колючка, астрагалы, саксаулы, чертопо
лох. Они, в отличие от суккулентов, не
накапливают в тканях воду, а интенсивно
испаряют поглощаемую влагу. Сильно
развитая корневая система либо повер
хностная, способная улавливать редкие
атмосферные осадки или утреннюю росу,
либо проникающая на большую глубину
до грунтовой воды. Благодаря повышен
ному осмотическому давлению клеточно
го сока, склерофиты развивают большую
сосущую силу, поэтому их еще называют
растениями-насосами.
При достаточном водообеспечении
для склерофитов характерна интенсивная
транспирация. Однако в периоды дли
тельных летних засух интенсивность
транспирации резко снижается. Этому
способствуют анатомо-морфологические
приспособления: мелкие, сухие, часто в
виде игл или колючек листья, покрытые
восковым налетом, жесткими волосками,
погруженного типа устьица. К физиоло
гическим адаптациям относится большая
водоудерживающая способность клеток и
тканей, достигаемая благодаря высокой
вязкости и эластичности цитоплазмы,
большой доле связанной воды. Это поз
воляет склерофитам переносить глубо
кое обезвоживание тканей (до 3/ 4 всего
водного запаса) без потери Жизнеспособ
ности.
М е зо ф и т ы — растения местообита
ний с умеренной увлажненностью почвы
и воздуха. Они более требовательны к
влаге, чем ксерофиты, и менее, чем гиг
рофиты. Сюда относятся растения лугов,
травяного покрова лесов, лиственные
древесные и кустарниковые породы из
областей умеренного климата, большин
ство культурных растений. У растениймезофитов умеренно развита корневая
система, имеются корневые волоски.
Способность переносить недостаток вла
ги в воздухе и почве ограничена.
Мезофиты — очень разнообразная
по требованиям к факторам среды и
пластичная группа растений. Некоторые
из них по своим требованиям приближа
ются к гигрофитам, другие — к ксерофи
там.
Особое место среди мезофитов зани
мают эфемеры и эфемероиды — весен
ние растения степей и пустынь. Для них
характерна краткая вегетация (4— 6 не
дель) в увлажненный весенний период и
длительный период покоя в виде семян у
эфемеров-однолетников (маки, крупки,
вероники) или луковиц, клубней, корне
вищ у многолетних эфемероидов (тюль
паны, гусиные луки).
Подобно растениям, сухопутные жи
вотные также населяют местообитания с
разным водным режимом. Животные
влажных местообитаний — гигрофилы
— не обладают механизмами регуляции
водного обмена. К ним относятся мокри
цы, комары, наземные планарии влаж
ных тропических лесов, наземные брюхо
ногие моллюски и земноводные.
Животные сухих местообитаний —
ксерофилы — имеют хорошо развитые
механизмы регуляции водного обмена и
приспособления к удержанию воды в ор
335
ганизме. Так, многие пустынные насеко
мые никогда не пьют воду и довольству
ются влагой, поступающей с пищей. Ряд
животных приспособился жить за счет
метаболической воды, образуемой в про
цессе окисления накапливаемых запасов
жира (верблюд, жирнохвостые тушкан
чики, гусеницы платяной моли, рисовый
и амбарный долгоносики). Большинство
ксерофильных животных обладают рядом
приспособлений для удержания воды, та
кими, как водонепроницаемые покровы,
отсутствие кожных желез, выделение ко
нечных продуктов азотистого обмена в
виде мочевой кислоты, повышенное об
ратное всасывание воды в канальцах
нефронов почек, пониженное потоотде
ление. Большую роль играют разнооб
разные адаптивные поведенческие реак
ции: закапывание в грунт, рытье нор, уход
в тень, соответствующая ориентация те
ла по отношению к солнечным лучам,
летняя спячка. Для активно передвигаю
щихся животных важны передвижения к
водоемам, как источникам воды.
9
I. Какие экологические группировки растений и
животных различают по отношению к обеспечен
ности влагой?
98. Биотические факторы
Каждый организм живет в окружении
других живых существ, вступая с ними в
разнообразные отношения. Такая связь
является необходимым условием их вы
живания, так как она обеспечивает воз
можность питания, размножения, защи
ты, расселения. Не все отношения между
организмами в равной степени полезны
каждой стороне; некоторые формы взаи
модействия приводят к гибели одного из
организмов либо к его угнетению.
Живое окружение организма состав
ляет его б и оти ч еск ую ср ед у , а разнооб
разные отношения между живыми орга
низмами называются б и оти ч еск и м и .
336
Типы б и о ти ч еск и х о т н о ш ен и й .
куренция —
Кон
взаимодействие (соревно
вание) между особями за один и тот же
ресурс, который имеется в ограниченном
количестве. В качестве ресурса могут
выступать различные пищевые объекты
для животных, элементы минерального
питания или свет для растений, прос
транство для закрепления на субстрате,
места, удобные для устройства гнезд, нор
и т. д.
Если конкурирующие особи принад
лежат к одному виду, то взаимоотноше
ния между ними называют в н у тр и в и д о
вой к он к ур ен ц и ей ; если же они относят
ся к разным видам •— м еж в и д о в о й к о н
к у р ен ц и ей .
О наличии конкуренции судят по сни
жению скорости роста, уменьшению раз
меров особей, снижению плодовитости и
выживаемости совместно обитающих ор
ганизмов.
Во многих случаях конкурирующие
особи непосредственно не взаимодей
ствуют друг с другом, но реагируют на
снижение уровня ресурса из-за присут
ствия и жизнедеятельности других осо
бей. Так, на карпов, конкурирующих за
пищу в рыбоводных прудах, другие карпы
влияют не прямо, а через уменьшение ко
личества и качества пищи.
Среди подвижных животных конку
ренция нередко принимает такую форму,
когда одна особь непосредственно пре
пятствует захвату другой особью части
местообитания и использованию находя
щихся на ней ресурсов. Эта форма внут
ривидовой конкуренции проявляется в
охране особью своей индивидуальной
территории. У некоторых животных за
щита своего участка включает жестокие
конфликты между конкурентами, тогда
как у других существует более тонкое
взаимное распознавание конкурирующи
ми особями запрещающих сигналов.
Птицы в гнездовой период в качестве
маркировки территории используют ха
рактерную песню; хищные млекопитаю-
Рис. 199. Мечение территориальных участков: а — зубром и б — самцом антилопы.
щие семейства псовых метят границы
участка мочой, представители семейства
кошачьих — секретом пахучих желез и
царапинами на деревьях. Зубр маркирует
деревья, сдирая с них кору, а самец анти
лопы оставляет на дереве метку с по
мощью секрета пахучей железы, распо
ложенной перед глазом (рис. 199, а, б).
В борьбе за территорию из числа конку
рентов побеждают сильнейшие особи,
которые и оставляют потомство.
Формой конкуренции среди растений
является выделение в почву токсических
веществ, которые подавляют рост расте
ний других видов. Такие токсические во
здействия называют аллелопатией (от
греч. allelon — друг друга, взаимно и
pathos — страдание, болезнь). Так, на
пример, разлагающиеся листья каштанов
выделяют в почву токсические соедине
ния, подавляющие рост сеянцев многих
других видов деревьев. Лещина и дуб сво
ими выделениями часто угнетают рост
травянистых растений под кроной. Био
логически активные вещества — фи
тонциды, выделяемые растениями, и
антибиотики, вырабатываемые микро
организмами, используются ими для по
давления жизнедеятельности микроорга
низмов других видов.
12. Общая биология, 10 кл.
Изучение межвидовой конкуренции в
природных условиях — трудная задача.
Поэтому последствия конкурентных вза
имоотношений исследуются в лаборатор
ном эксперименте. Классическим приме
ром таких работ являются исследования
русского ученого Г. Ф. Гаузе (1934). Он
культивировал два близкородственных
вида инфузорий-туфелек на бактериаль
ном корме. При выращивании по отдель
ности оба вида процветали, но при сов
местном культивировании выживал толь
ко один из них (рис. 200, с. 338). Повто
рение подобных экспериментов с плодо
выми мушками, мышами, однолетними
растениями всегда приводило к сходному
результату. Это дало основание сформу
лировать закон конкурентного исключе
ния: два вида не могут долго существо
вать совместно, если они зависят от од
ного и того же лимитирующего их ре
сурса; один из видов будет вытеснен.
В результате конкурентного исключе
ния в биоценозе уживаются вместе толь
ко те виды, которые в ходе эволюции при
обрели различия в потребляемых ресур
сах в соответствии с их размерами и фор
мой, химическим составом, местообита
нием и сезонностью. Например, четыре
вида медоносной пчелы, обитающие сов337
Рис. 200. Рост популяций двух
видов инфузорий-туфелек при
раздельном (Л) и совместном
(Б) выращивании.
Время , дни
местно, избегают конкуренции благодаря
различиям в длине хоботка, позволяюще
го им собирать нектар и пыльцу с цветков
разного размера. Или, как видно из ри
сунка 201, каждый из пяти видов амери
канских славок, живущих в еловых лесах,
кормится на разных частях деревьев и ха
рактеризуется различиями в способах
отыскивания насекомых среди веток и
листвы.
Таким образом, у особей одного вида в
присутствии конкурирующих с ними осо
бей другого вида наблюдаются внешние
признаки или особенности поведения, от
личающиеся от таковых в отсутствие кон
курента. Это явление, известное под наз
ванием см ещ ен и я (р а с х о ж д е н и я ) п р и з
нак ов, служит доказательством того, что
конкуренция может вызвать эволюцион
ную д и в е р г е н ц и ю , как это видно в
случае изменения формы клювов у вьюр
338
ков в зависимости от пищевой специали
зации (рис. 202, с. 340).
Конкурентные отношения являются
одним из важнейших механизмов форми
рования видового состава биоценозов и
пространственного распределения осо
бей. Конкурентное исключение и эволю
ционная дивергенция, уменьшая межви
довую конкуренцию, обеспечивают эф
фективное использование ресурсов в би
оценозах.
На огромную роль конкурентных от
ношений в эволюционном развитии видов
указывал Ч. Дарвин.
Взаимоотношения хищник — жер
тва включают все взаимодействия, при
которых одни организмы — хищники —
используют в пищу других — жертв.
Хищ ничест во представляет собой
главную силу, обеспечивающую передви
жение вещества и энергии в биоценозах.
Рис. 201. Расположения мест кормежки пяти видов американских славок в еловых лесах штата Мэн
(Канада).
Существует два основных способа
классификации хищников. Наиболее ес
тественной и привычной является «так
сономическая» классификация: хищники
в собственном значении этого слова пое
дают животных, растительноядные
организмы — растения, а всеядные — и
тех, и других.
Иногда под хищничеством понимают
такие взаимоотношения двух групп
любых организмов, при которых одна
группа использует другую в пищу. Сог
ласно этой функциональной классифика
ции, которая позволяет понять сущность
явления хищничества, выделяют четыре
основных типа хищников: истинные хищ
ники, хищники с пастбищным типом пи
тания, паразиты и паразитоиды.
Истинные хищники (рис. 203, с. 341)
убивают за свою жизнь множество жертв
почти сразу же при нападении, поедая их
целиком, либо только часть. К ним при
надлежат тигры, волки, орлы, лисы,
божьи коровки, насекомоядные расте
ния, питающиеся семенами грызуны и
муравьи, фильтрующие планктон киты,
дафнии, циклопы.
Хищники с пастбищным способом
питания в течение всей жизни использу
ют большое число жертв, съедая, как
правило, только часть каждой из них.
Последствия для жертвы различны: чаще
всего вредны, однако редко приводят к
гибели. Характерными примерами слу
жат травоядные позвоночные, мухи, ку
сающие позвоночных животных, пиявки
и самки комаров, сосущие кровь.
Паразиты, как и предыдущая группа
хищников, поедают только часть своей
жертвы, называемой хозяином, с такими
же последствиями для жертвы. Однако, в
отличие от хищников с пастбищным ти
пом питания, паразиты в течение своей
жизни нападают всего на одну или на
очень немногих особей-хозяев, с которы
ми они тесно связаны почти всю жизнь.
Примерами таких паразитов являются
печеночный сосальщик, ленточные черви
(рис. 204, с. 341), аскариды, вирус кори и
туберкулезная палочка, паразиты расте
ний — вирус табачной мозаики, фитопа339
Рис. 202. Формы клювов у вьюрков в зависимости от пищевой специализации: виды первого рода —
светло-коричневы й фон, второго рода — ж е л ты й , третьего — коричневый', дробящие клювы — го
лубой ц в е т, хватательные — бордовый, долбящие — красный', растительная пища — зеленый цвет,
преимущественно животный корм — оранжевый, животная пища — тем но-зелены й цвет.
тогенные грибы (ржавчинные, головне
вые, спорыньевые), омела, тля идр.
Паразитоиды — организмы, ведущие
паразитический образ жизни только на
стадии личинки. Сюда относятся некото
рые насекомые отряда перепончатокры
лых и многие виды двукрылых. Взрослые
насекомые паразитоиды ведут свободный
образ жизни; яйца чаще всего откладыва
ют в тело другого животного, реже на его
поверхность. Личинки всегда развивают
ся в теле хозяина. Примерами широко из
вестных насекомых-паразитоидов могут
служить наездник трихограмма (рис.
205), личинка которого паразитирует в
340
яблонной плодожорке и вызывает ее ги
бель на ранних стадиях развития, а также
бычий овод, откладывающий яйца под ко
жу крупного рогатого скота, что приводит
к истощению животных.
Степень воздействия хищных живот
ных на своих жертв различна и во многом
определяется наличием или отсутствием
предпочтения к определенному виду жер
твы — избирательностью питания.
Животные, у которых полностью или час
тично отсутствует избирательность в пот
реблении пищевых объектов, называют
ся неспециализированными хищника
ми. К ним относится большинство хищ-
Рис. 203. Хищные животные: а — лиса; б —
хищные жуки: 1 — жужелица, 2 — божья ко
ровка.
Рис. 204. Свиной цепень.
Рис. 205. Наездник, откладывающий яйца в гу
сеницу.
ников с пастбищным типом питания. И
действительно, кролики, крупный рога
тый скот имеют практически нулевую из
бирательность питания. При достаточно
высокой численности особей они способ
ны поддерживать травостой обширных
пространств в состоянии низкорослого
дерна. Следствием такого безвыборочного питания будет поддержание высокого
видового разнообразия растительности
благодаря подавлению межвидовой кон
куренции. Так, исследования показали,
что защита травостоя от поедания живот
ными путем огораживания приводит к ги
бели 45 % видов растений, вытесненных
оставшимися более конкурентоспособ
ными видами.
Кроме поддержания высокого фло
ристического разнообразия, умеренное
потребление растительности приводит к
усилению интенсивности фотосинтеза за
счет уменьшения самозатенения расте
ний в плотном травостое, что повышает
продуктивность растений (жертв).
Однако значительная плотность хищ
ников с пастбищным типом питания, пот341
ребляющих не только надземные части
растения, но и корни, как это типично для
коз, приводит к полному истреблению
растительности.
Специализированные хищники пи
таются одним или немногими видами
жертв.
У истинных хищников выбор конкрет
ной жертвы, в первую очередь, определя
ется ее возможной доступностью. Вот по
чему хищники сначала отлавливают
больных, старых, травмированных, более
уязвимых молодых, либо не нашедших
себе территории особей низшего ранга.
Такого рода избирательный отбор повы
шает жизнеспособность популяции жер
твы. Эту роль хищников оценивают как
санитарную.
Поскольку наличие пищевой специа
лизации ограничивает возможность пе
реключения на другие виды пищи, то
следствием являются циклические коле
бания численности хищников и жертв. На
рисунке 206 представлены многолетние
данные по циклическим колебаниям чис
ленности канадской рыси и ее жертвы —
зайца. При низкой численности как хищ
ника, так и жертвы численность жертвы
быстро возрастает. По мере возрастания
численности особей жертвы начинает
увеличиваться и численность хищников,
но с некоторым отставанием. Наконец
отлов жертвы хищниками становится
чрезмерным и численность жертвы со
кращается. До последней особи жертва
не истребляется, так как это энер
гетически невыгодно хищнику. Кроме
того, уровень снижения численности
жертвы напрямую зависит от наличия
убежищ.
Так устанавливается динамическое
равновесие между хищником и жертвой,
что способствует поддержанию стабиль
ности в биоценозе.
Взаимные приспособления хищника
и жертвы. Стратегии хищника и жертвы
противоположны: первый стремится най
ти и поймать, а жертва — избежать по
имки. Поэтому происходит усовершен
ствование приспособлений как у хищни
ка, так и у жертвы, благодаря чему между
ними устанавливается некоторое эволю
ционное равновесие. Многие виды на
земных растений в качестве средств за
щиты, кроме шипов, колючек, толстой
коры, вырабатывают биологически ак
тивные вещества, делающие их несъе
добными или даже ядовитыми. Напри
мер, у астрагала вырабатывается мизеротоксин, вызывающий паралич дыхания и
Рис. 206. Колебания численности зайца (сплошная линия) и рыси (пунктир) по числу заго
товленных шкур.
342
даже гибель овец и крупного рогатого
скота. Ядовиты зверобой продырявлен
ный, вех ядовитый и др.
Некоторые растения приобрели спо
собность синтезировать вещества, обла
дающие высочайшей специфичностью
действия только по отношению к опреде
ленному виду потребителя. Примером мо
жет служить бальзамическая пихта, вы
рабатывающая ювабион, являющийся хи
мическим аналогом ювенильного гормона
насекомых. Поедая листья с содержа
щимся в них ювабионом, личинки насеко
мых-вредителей не заканчивают своего
развития, так как дополнительно линя
ют еще один-два раза, после чего поги
бают.
Защита животных от поедания истин
ными хищниками разнообразна. Это и
покровительственная окраска, и особая
форма тела, делающие их трудно разли
чимыми в среде обитания (рис. 207, с.
344), быстрая скорость передвижения,
помогающая жертве уйти от преследова
ния хищника. У многих животных, не
имеющих укрытий, своеобразным спосо
бом компенсации потерь от хищников яв
ляется интенсификация размножения.
Этот способ развился, например, у тлей,
планктонных ракообразных и коловра
ток, приобретших партеногенетический
способ размножения (рис. 208, с. 345).
Как средство защиты от поедания,
многие животные вырабатывают яды и
токсины. Так, некоторые виды многоно
жек в качестве оборонительного средства
используют синильную кислоту, вызы
вающую у нападающих на них животных
судороги, паралич, удушье. Жуки нарыв
ники и шпанские мушки продуцируют
кант аридин — вещество кожно-на
рывного действия. Батрахотоксин ядо
витых лягушек Колумбии нарушает у
употребивших их в пишу нормальную ра
боту натриевых каналов нервных клеток
и мышечных волокон, что приводит к де
поляризации мембран. Буфоталин —
токсин европейских видов жаб — оста
навливает у хищника работу сердца в
систоле.
У паразитов развились приспособле
ния, позволяющие потреблять пищу их
хозяина, не приводя его к гибели. К тако
го рода приспособлениям относятся сме
на хозяина в цикле развития паразита,
сезонность заражения. Адаптации хозяи
на (жертвы) направлены на повышение
иммунитета и устойчивости к воздей
ствию паразита. Благодаря подобным
приспособлениям взаимоотношения па
разита и хозяина приходят к некоторому
равновесию.
О
f. 1. Что такое конкуренция и какие ее формы вы
знаете? 2. Какова роль конкурентных отношений в
эволюционном развитии видов? 3. Назовите основ
ные способы классификации хищников. 4. В чем от
личие истинных хищников от хищников с пастбищ
ным способом питания? 5. В чем отличие паразитоидов от паразитов?
99. Другие виды биотических
взаимоотношений
Комменсализм (нахлебничество, сотрапезничество) — взаимоотношения на
основе пищевых связей, когда одно жи
вотное поедает остатки или избытки пи
щи другого. Выгоду от совместного су
ществования извлекает только коммен
сал, присутствие которого для партнера
остается фактически безразличным. Та
ковы взаимоотношения львов и гиен, бе
лых медведей и песцов. Комменсалами
крупных рыб, например акул, являются
рыбы-прилипалы, использующие остат
ки их пищи. С помощью специальных
присосок на голове они прикрепляются к
брюху акулы и таким образом без особых
усилий повсюду следуют за ней, при слу
чае деля с ней добычу. Акуле они не при
носят ни вреда, ни пользы.
Синойкия (квартирантство) — прос
транственное сожительство, когда один
организм использует другой как субстрат
для обитания или перемещения с его по
мощью в пространстве. Также, как и в
343
2
344
Ч| Рис. 207 Покровительственная и предостерегающая окраска различных насекомых: 1 — бабочка углокрыльница, 2 — та ж е бабочка со сложенными крыльями среди сухих листьев; 3 — бабочка голу
бая ленточница, 4 — она же на коре дерева; 5 — гусеница репницы на листе капусты (покровительс
твенная окраска съедобной гусеницы); 6 — гусеница капустницы (предостерегающая окраска несъе
добной гусеницы); 7 — гусеница волнянки (предостерегающая окраска); 8 — предостерегающая ок
раска луговой пестрянки.
Рис. 208. Планктонный рачок с партеногенетическими яйцами.
Рис. 209. Рак-отшельник с актиниями, сидящими на занятой им раковине.
случае комменсализма, выгоду получает
только один организм, а другому это без
различно. Так, в гнездах птиц, норах гры
зунов поселяется много видов членисто
ногих, находящих там подходящий для
жизни микроклимат. Некоторые виды
рыб на коралловых рифах живут среди
щупалец актиний и медуз. Различные ви
ды кишечнополостных (например, акти
нии) поселяются на раковинах моллюс
ков, используя их как средство передви
жения (рис. 209).
К синойкии относят также эпифитность, когда определенные виды про
тестов, лишайников, мхов и папоротни
ков поселяются на стволах деревьев.
К различным формам синойкии отно
сится обитание животных в дуплах, в рас
щелинах коры деревьев, перенос семян и
плодов растений животными.
Мутуализм (от лат. mutuus — взаим
ный) — форма обязательного сожи
тельства, при которой оба организма из
влекают выгоду друг от друга и ни одна
из сторон не может существовать без
другой.
Примером мутуалистических взаимо
отношений является сожительство клу
беньковых бактерий рода ризобиум
(Rhizobium) с корнями растений семей
ства бобовых, мимозовых и др. (рис. 210,
с. 346). Заражение бактериями происхо
дит через корневые волоски. Ткани корня
под влиянием ростового вещества, выде
ляемого бактериями, усиленно разраста
ются и образуют клубеньки. Бактерии в
клубеньках быстро размножаются, за
полняя растительные клетки. Чтобы бак
териям было достаточно кислорода для
роста и получения энергии, они совмес345
Рис. 210. Мутуалистическая связь клубеньковых бактерий с корнями бобовых
растений: а — корень гороха с клубеньками, б — клубеньки в разрезе, в — рас
тительная клетка, заполненная бактериями, г — внедрение бактерий через кон
чики корневых волосков.
тно с растительными клетками синтези
руют родственный гемоглобину пигмент
— леггемоглобин. В результате такого
тесного взаимодействия растение обес
печивает бактерий питательными вещес
346
твами (главным образом, сахарами) и
создает для их жизни оптимальные усло
вия. Бактерии, в свою очередь, на первых
порах живут за счет растения, а затем на
чинают фиксировать свободный азот из
воздуха с последующим образованием
аммиака, а из него — нитритов и нитра
тов, которых достаточно и для бактерий,
и для растений. Кроме того, часть нитри
тов и нитратов выделяется в почву, повы
шая ее плодородие.
Взаимовыгодное сожительство су
ществует также между некоторыми де
ревьями (ель, лиственница, сосна, дуб,
береза) и грибами и называется микори
зой. При этом мицелий гриба оплетает
корни растений и проникает только под
эпидермис или в клетки паренхимы кор
ня, где может образовывать клубки. Ми
коризный гриб увеличивает всасываю
щую поверхность корня в 10— 14 раз,
лучше поглощает фосфор, выделяет ви
тамины и ростовые вещества, стимулиру
ющие развитие корня. От высшего расте
ния гриб получает углеводы и другие безазотистые соединения, кислород и кор
невые выделения, способствующие про
растанию спор.
Лишайники также представляют со
бой мутуалистическую ассоциацию гри
ба с автотрофными организмами — циа
нобактериями, протистами, водорос
лями.
Обитающие в пищеварительном
тракте жвачных животных, термитов и
тараканов бактерии, инфузории и жгути
ковые помогают животному-хозяину пе
реваривать растительную пищу, выраба
тывая целлюлозолитические ферменты.
Животное-хозяин предоставляет организмам-симбионтам место обитания и
пищу.
Таким образом, многообразие форм
взаимоотношений между организмами
создает ту необходимую биотическую
среду, в которой возможно нормальное
существование живых существ.
» 1. Что общего и в чем отличия между коммен
сализмом и синойкией? 2. Какие взаимоотношения
называют мутуализмом? Приведите примеры.
100. Ритмические изменения
факторов среды
и их влияние на организмы
Климатические факторы (свет, тем
пература, влажность, приливы и отливы
и др.) подвержены регулярно повторяю
щимся изменениям, возникающим в ре
зультате вращения Земли вокруг своей
оси и Солнца, а также вращения Луны
относительно Земли. Амплитуда измен
чивости этих факторов связана с широт
ной принадлежностью того или иного ре
гиона, его расстоянием от океана, в ре
зультате чего и сформировались клима
тические зоны на нашей планете.
Чтобы выжить, живые существа в
процессе длительной эволюции развили
системы восприятия периодических из
менений в окружающей среде и вырабо
тали способность соответственно изме
нять свое физиологическое состояние.
Такие повторяющиеся с определенной
последовательностью состояния организ
ма называются биологическими ритма
ми. Повторяемость состояний организма
может осуществляться через разный про
межуток времени, называемый перио
дом. Например, через 24 часа — суточ
ная, один раз в сезон — сезонная, еже
годно повторяющаяся — ежегодная пе
риодичность. Роль биоритмов — по
мочь организму согласовать свою дея
тельность с меняющейся средой и опти
мально приспособиться к ней.
Наиболее четко выраженным перио
дическим изменениям подвержен важ
нейший абиотический фактор — свет, с
которым связана также изменчивость
температуры и влажности.
Реакция организмов на суточный
ритм освещения, т. е. на соотношение
длины светлого и темного времени суток,
выражающаяся в изменении их физиоло
гического состояния, называется ф о т о
периодизмом.
Одно из основных проявлений этой
реакции у растительных организмов —
347
фотопериодическая индукция (наве
дение) зацветания. Она служит несом
ненным приспособлением растений к ус
ловиям существования, так как обуслов
ливает их зацветание в самые благопри
ятные сроки. В ходе фотопериодической
индукции к зацветанию в листьях образу
ется гормон цветения, который по флоэ
ме переносится в цветочные почки, сти
мулируя формирование цветков. Если же
фотопериод неблагоприятен для расте
ния, то в листьях формируются гормоныингибиторы (подавители) цветения.
В зависимости от реакции на длину
дня, ускоряющей зацветание, растения
делятся на длиннодневные (свекла, кле
вер, кукуруза, картофель, хлебные злаки,
многие крестоцветные, укроп), корот
кодневные (астры, георгины, рис, соя,
конопля, перец) и нейтральные (оду
ванчик, томаты, гречиха, горох).
Короткодневные растения цветут
обычно ранней весной или поздним летом
и осенью. Им достаточно 12 ч светлого
времени суток и даже меньше.
Длиннодневные растения зацветают
только при фотопериоде продолжитель
ностью более 12 ч в сутки. Такие расте
ния обычно цветут поздней весной и в на
чале лета. Длиннодневные растения рас
пространены в основном в умеренных и
приполярных широтах, а короткоднев
ные — в субтропиках.
Периодичность физиологических про
цессов у организмов с периодом около су
ток называется циркадными ритмами.
С такой периодичностью, например, у
растений изменяется митотическая ак
тивность в образовательных тканях (пе
риод около 20 ч) и скорость роста (пери
од около 24 ч). Такие же ритмы обнару
живают процессы фотосинтеза и дыха
ния, транспирации, движения листьев,
открывания и закрывания цветков, усть
иц. Так, ритм раскрывания и закрывания
цветков приспособлен к суточной актив
ности определенных групп опылителей.
Суточные движения листьев в определен
348
ных условиях могут служить регулятором
теплового режима растений: вертикаль
ное положение листьев ночью снижает
потери тепла, а горизонтальное положе
ние днем увеличивает получаемое тепло.
Околосуточные ритмы тесно связаны с
суточными колебаниями освещенности,
температуры, влажности и других факто
ров среды. Поэтому благодаря периодич
ности изменения физиологических про
цессов организмы хорошо приспособле
ны к тем условиям, в которых они обита
ют, и мало зависят от случайных погод
ных условий.
Для животных также свойственна су
точная цикличность многих процессов.
Так, резко меняющаяся в течение суток
освещенность, температура и влажность
определяют разную приуроченность ко
времени суток активности у сухопутных и
полуводных видов животных. Жабы и бу
рые лягушки активны»ночью, когда влаж
ность выше, чем в дневное время суток.
Напротив, озерная и прудовая лягушки,
находящиеся в условиях высокой влаж
ности, активны днем. Будучи теплолюби
выми, большинство пресмыкающихся
умеренных широт — дневные, немногие
виды — сумеречные и только гекконы —
ночные виды. В тропических пустынях,
напротив, многие виды животных актив
ны в ночное время, так как днем очень
жарко.
Подавляющее большинство птиц —
дневные животные, засыпающие с захо
дом и просыпающиеся с восходом солнца.
Такая четкая суточная цикличность
активности связана с тем, что большин
ство птиц питается растительными кор
мами и дневными животными и при добы
вании корма использует исключительно
зрение. Сравнительно немногие виды
(совы, ряд куликов, козодой и др.), кор
мящиеся ночными животными, активны
ночью или в сумерки. В отыскании добы
чи, кроме зрения, они используют острый
слух или осязание. Суточный ритм птиц
сильно меняется в период размножения,
когда самцы многих дневных видов поют
чуть ли не круглые сутки, и во время пе
релета, который совершается ночью и
многими строго дневными птицами.
Суточная цикличность активности
млекопитающих определяется, как и у
птиц, главным образом, возможностью
добывать корм в то или иное время суток,
а не температурой и влажностью, пос
кольку они гомойотермные животные.
Растительноядные млекопитающие ак
тивны днем. Большинство хищных зве
рей, охотящихся за животными с разной
суточной активностью, могут добывать их
как днем, так и ночью. Летучие мыши ак
тивны в ночное время, поскольку они
специализируются на ловле ночных насе
комых.
Кроме суточной периодичности, ак
тивность организмов подвержена изме
нениям в течение сравнительно длитель
ных периодов, например сезонной пери
одичности. У многолетних растений и
пойкилотермных животных она связана с
различиями в скорости роста в разные се
зоны года, что выражается в образовании
годичных колец в древесине растений, на
чешуе рыб. и раковинах моллюсков (рис.
211 ).
При наступлении неблагоприятного
для жизнедеятельности зимнего периода
растения и животные приполярных ши
рот и умеренного пояса, а в тропических
регионах в засушливый период впадают в
период покоя. У листопадных деревьев к
этому периоду сбрасывается листва. Вер
хушечные, меристемы побегов прекраща
ют активный рост и покрываются почеч
ной чешуей, превращаясь в зимние поко
ящиеся почки. В таком состоянии покоя
растения гораздо морозоустойчивее, чем
при активном росте. У многих растений
этой же цели служат специальные покоя
щиеся органы — луковицы, клубни,
клубнелуковицы и корневища. Эти орга
ны могут.быть также приспособлены и
для перенесения перегрева, летней засу
хи, как это наблюдается у многих пустын
ных растений.
У животных разных видов сезонная
периодичность выражается в кормовых,
зимовочных, нерестовых перемещениях.
Четко выражены кормовые кочевки и
миграции у птиц (рис. 212, с. 350). Так,
например, стрижи и ласточки ежегодно
улетают зимовать за тысячи километров
на юг Африки, возвращаясь в Европу, как
правило, на одни и те же места. Такие
ежегодные миграции совершают и белые
аисты.
Многие млекопитающие при недос
татке пищи или невозможности ее добы-
349
Рис. 212. Осенняя и весенняя миграции цен
трально-европейской мухоловки-пеструшки.
чи из-за глубокого снежного покрова
также совершают миграции. Им подвер
жены, главным образом, копытные и
двигающиеся за ними хищники. Напри
мер, северные олени проводят зиму на
окраине таежной полосы, питаясь ягелем
и другими съедобными лишайниками,
растущими на стволах деревьев. Но уже
при первых признаках весны они боль
шими табунами двигаются в тундру, при
держиваясь из года в год одного и того же
маршрута.
Зимовочные миграции в дельты круп
ных рек свойственны многим видам рыб
(сазан, лещ, судак, сом и др.), где они
поздней осенью собираются в больших
количествах в глубоких местах («зимо
вальных ямах») и проводят зиму в мало
подвижном или неподвижном состоянии.
У осетровых рыб (осетр, белуга) от
мечаются ежегодные нерестовые мигра
ции из моря в реки.
Сезонная цикличность у животных
выражается в явлениях зимней и летней
спячки. Она свойственна тем животным,
которые в определенные периоды
350
времени не могут добывать достаточное
количество полноценного корма. Впада
ющие в зимнюю спячку млекопитающие
накапливают с осени большое количес
тво жировых запасов. В настоящую зим
нюю спячку, которая характеризуется
сильным снижением температуры тела,
приостановкой процессов пищеварения,
низким уровнем дыхания и кровообраще
ния, впадают летучие мыши (рис. 213),
ежи, некоторые грызуны (сурки, суслики,
хомяки, тушканчики).
Когда весенняя сочная трава выгора
ет, в летнюю спячку впадает среднеазиат
ский желтый суслик.
Таким образом, фотопериодизм —•
важнейший сигнальный фактор, который
регулирует и управляет ритмикой суточ
ной и сезонной жизнедеятельности орга
низмов, наилучшим образом приспосаб
ливая их к меняющимся условиям среды,
что и обеспечивает их выживание.
В искусственных условиях (парниках,
теплицах, оранжереях), регулируя фото-
Рис. 213. Летучая мышь в зимней спячке.
период, можно и в зимнее время успешно
выращивать овощные культуры, рассаду,
декоративные растения. Увеличением
светлого времени суток за счет искус
ственного освещения достигают повыше
ния яйценоскости кур.
?
« 1. Что такое биологический ритм? 2. Дайте оп
ределение понятия фотопериодизма. 3. Каково зна
чение явления фотопериодизма? 4. Приведите при
меры сезонной периодичности в жизни растений и
животных.
101. Среды жизни
Среда жизни — это часть природы, в
которой живут организмы. Живые орга
низмы освоили четыре основные среды
обитания: водную, наземно-воздушную,
почвенную и непосредственно сам живой
организм. Каждая из сред имеет свои
специфические особенности, к которым
организмы приспособились в результате
длительной эволюции.
Водная среда, или гидросфера. Гид
росфера была первой из сред жизни, ос
военной организмами. Это самая обшир
ная среда обитания, занимающая 71 %
площади нашей планеты. Воды морей и
океанов составляют 98 %. Большая часть
пресных вод заключена в ледниках, и
лишь менее 0,5 % сосредоточено в реках,
озерах, болотах.
Особенности гидросферы как среды
обитания водных организмов, называе
мых гидробионтами, можно понять,
лишь ознакомившись с физическими
свойствами воды по сравнению с возду
хом. К числу главнейших свойств воды
относится плотность, которая примерно в
800 раз превышает плотность воздуха.
Из-за высокой плотности воды на разных
глубинах водоемов существует значи
тельная разница давлений.
Плотность воды зависит от температу
ры. Ее величина максимальна при +4 °С и
равна 1 г/см3. При повышении или пони
жении температуры плотность воды
уменьшается. При замерзании вода рас
ширяется, увеличивая свой объем при
мерно на 11 %. Благодаря этому свойству
лед располагается на поверхности водое
ма, в то время как более плотная вода
(t > 0° С) находится подо льдом и не про
мерзает. Самая плотная вода с температу
рой +4 °С располагается в придонной об
ласти (рис. 214). Следствием высокой
плотности воды является ее большая
опорность) используемая живыми орга
низмами, а также выталкивающая сила и
вязкость.
У воды одна из самых высоких вели
чин удельной теплоемкости. Чтобы изме-
Лед
20°С
0°С
со
х
15°С
VO
>.
10°С
X
1°С
2°С
5°С
3°С
4°С
4°С
Лето
Зима
Рис. 214. Вертикальное распределение температуры в водоеме в разные
сезоны года.
351
нить температуру 1 г воды на 1 °С, нужно
затратить 4,19 Дж тепла (в 500 раз
больше, чем для воздуха). Благодаря это
му свойству вода, медленно нагреваясь и
медленно остывая, уменьшает амплитуду
суточных и сезонных колебаний темпера
тур, стабилизируя ее.
У воды высокая теплопроводность (в
30 раз выше, чем у воздуха), благода
ря чему обеспечивается равномерное
распределение температуры в водной
среде.
Замечательным и важным свойством
воды является ее способность растворять
разнообразные минеральные вещества.
В водах гидросферы обнаружены в том
или ином количестве все химические эле
менты таблицы Менделеева. В зависи
мости от количества растворенных солей
выделяют пресные (до 0,5 г/л), солоно
ватые (0,5— 16 г/л), морские (16—
47 г/л) и пересоленные (47—350 г/л)
воды.
Природным водам свойствен опреде
ленный химический состав. Так, в прес
ных водах преобладают карбонаты, в
морских — хлориды.
С повышением солености возрастает
плотность воды и понижается температу
ра ее замерзания.
В воде растворяются и газы. Однако
кислорода содержится в 30 раз меньше,
чем при той же температуре в равном
объеме воздуха. Более того, коэффици
ент диффузии кислорода в стоячей воде
примерно в 320 тыс. раз ниже, чем в воз
духе. Это означает, что при отсутствии
движения воды малоподвижный водный
организм может погибнуть от недостатка
кислорода, т. к. скорость поступления
кислорода из толщи воды будет меньше,
чем потребность в нем. Поэтому кисло
род в водной среде — ограничивающий
фактор.
Содержание кислорода в природных
водах подвержено значительным измене
ниям и суточным колебаниям: в светлое
352
=:
Рис. 215. Суточный ход содержания растворен
ного в воде кислорода (1) и углекислого га
за (2).
время оно повышается, в ночное — сни
жается (рис. 215).
Насыщение воды кислородом осу
ществляется в основном за счет фотосинтетической деятельности водорослей и
погруженных высших растений. Диффу
зия кислорода из воздуха при наличии
ветрового перемешивания также способ
ствует насыщению воды кислородом.
Напротив, углекислый газ растворя
ется в воде примерно в 35 раз лучше кис
лорода. В воде его почти в 700 раз боль
ше, чем в равном объеме атмосферного
воздуха. В дневное время С 0 2 активно
потребляется в ходе фотосинтеза, а
ночью — содержание его возрастает изза прекращения фотосинтеза и выделе
ния углекислого газа в процессе дыхания
гидробионтов (см. рис. 215).
Свет проникает в водную толшу водо
емов на разную глубину в зависимости от
содержания взвешенных частиц и окра
шивающих ее растворенных веществ, а
также угла наклона солнечных лучей, па
дающих на поверхность воды. Поэтому
прозрачность природных вод различна: от
0,1 до 66,5 м. Величину прозрачности оп
ределяют путем погружения белого диска
в воду до предельной глубины его види
мости.
Наибольшее количество света полу
чает поверхностный слой воды, в котором
интенсивно осуществляется фотосинтез.
Важным фактором жизни гидробионтов является также концентрация ионов
водорода (pH). Ее величины подвержены
значительным суточным колебаниям в
пресных водоемах с сильным развитием
водорослей и цианобактерий («цветение
воды»). В цветущих водоемах в светлое
время суток величины pH могут достигать
10— 11, а в ночное время снижаться до 6.
В морской воде величины pH постоянны
и равны примерно 8,2.
В водоемах гидросферы на основании
комплекса сходных условий выделяют две
экологические зоны —- толщу воды —
пелагиаль и донную область — бенталь
(рис. 216).
Наземно-воздушная среда. Живые
организмы в наземно-воздушной среде
окружены воздухом. В отличие от воды он
обладает низкой плотностью и как след
ствие — малой подъемной силой, незна
чительной опорностью и низкой сопро
тивляемостью при движении организмов.
Наземные организмы живут в условиях
сравнительно низкого и постоянного ат
мосферного давления, также обуслов
ленного малой плотностью воздуха.
Воздух обладает низкой теплоемкос
тью, поэтому он быстро нагревается и
столь же быстро охлаждается.
Легкие воздушные массы имеют
большую подвижность как в горизон
тальном, так и в вертикальном направле
ниях. Это способствует поддержанию на
постоянном уровне газового состава воз
духа. Содержание кислорода в воздухе
значительно выше, чем в воде, поэтому
кислород на суше не является ограничи
вающим фактором.
Рис. 216. Экологические зоны водоема (пруда): 1 — толща воды (пелагиаль) и ее обитатели, 2 — дно
водоема (бенталь) и его население, 3 — абиотические вещества.
353
Низкое содержание углекислого газа,
равное 0,03 %, — характерная особен
ность воздушной среды.
Свет в условиях наземного обитания
не является лимитирующим фактором в
отличие от водной среды.
Наземно-воздушная среда имеет раз
ные режимы влажности: от полного и
постоянного насыщения воздуха водяны
ми парами в некоторых районах тропиков
до практически полного их отсутствия в
сухом воздухе пустынь. Велика также из
менчивость влажности воздуха в течение
суток и сезонов года. Влага на суше выс
тупает в качестве ограничивающего фак
тора.
Для температурного режима суши ха
рактерна также, как и для влажности,
большая амплитуда колебаний. Особен
но велика суточная и межсезонная из
менчивость температуры в континенталь
ных районах и в пустынях, где воздух со
держит мало влаги. Так, в пустынях Сред
ней Азии суточный диапазон колебаний
температуры воздуха достигает 25 —
38 °С.
Различные сочетания температуры,
влажности, облачности,осадков, силы и
направления ветров у земной повер
хности создают разные погодные усло
вия, действию которых подвергаются
живые организмы. В различных геогра
фических районах складываются сход
ные погодные условия, формирующие
климат местности. На нашей планете
существует большое разнообразие ти
пов климата.
Большое значение для наземных ор
ганизмов, особенно мелких^ имеют ло
кальные микроклиматические условия
в конкретном местообитании. Напри
мер, температура воздуха на обращен
ной к югу стороне дерева намного выше,
чем на северной. Резко отличаются тем
пература, влажность, сила ветра, осве
щенность на открытых пространствах и
в лесу, а зимой на открытых участках
почвы и под снегом, под слоем опавших
354
листьев, в норах, дуплах, пещерах и т. п.
Все это создает большое разнообразие
условий жизни на суше в наземно-воз
душной среде.
Почва. Почва представляет собой
сложную систему, состоящую из твердых
частиц, окруженных воздухом и водой. В
зависимости от типа почвы — глинистая,
песчаная, глинисто-песчаная идр. — она
в большей или меньшей степени прони
зана полостями, заполненными смесью
газов и водными растворами. В почве, по
сравнению с приземным слоем воздуха,
сглажены температурные колебания, а
на глубине 1 м не ощутимы и сезонные
изменения температуры.
Самый верхний горизонт почвы со
держит некоторое количество перегноя,
от которого зависит продуктивность рас
тительного покрова. Расположенный под
ним средний слой содержит вымытые из
верхнего слоя и преобразованные вещес
тва. Нижний слой представлен материн
ской породой.
Вода в почве находится в крупных
пустотах, куда проникает под воздействи
ем силы тяжести (гравитационная во
да), либо в виде пленочной воды, удер
живаемой поверхностью почвенных час
тиц. Вода содержится также и в мельчай
ших пространствах — порах — и может
передвигаться по ним в разных направле
ниях, подобно движению ее в капилляре.
В почвенном воздухе присутствует и па
рообразная вода.
Состав почвенного воздуха резко ме
няется с глубиной: содержание кислорода
уменьшается, а углекислого газа возрас
тает. При затоплении почвы водой или
интенсивном гниении органических ос
татков возникают бескислородные зоны.
Таким образом, условия существова
ния в почве различны на разных ее гори
зонтах.
Организм как среда обитания. Раз
нообразные организмы, как раститель
ные, так и животные, и даже одни из са
мых мелких — бактерии служат средой
обитания для других организмов —■пара
зитов. Паразиты на протяжении всей
своей жизни или части ее пользуются ор
ганическими веществами другого орга
низма, называемого хозяином. Таким об
разом, хозяин и является средой жизни
паразита. Паразит может поселиться на
поверхности тела хозяина или внутри
него. В первом случае это явление назы
вается эктопаразитизмом, во втором
— эндопаразитизмом.
В отличие от свободноживущих орга
низмов, непосредственно сталкивающих
ся и взаимодействующих с факторами
внешней среды, паразиты «перекладыва
ют бремя регуляции отношений» с нею на
организм хозяина. Через хозяина регули
руется и получение паразитом пищи, яв
ляющейся важнейшим фактором внеш
ней среды.
Известный паразитолог Е. Н. П а в
л о в с к и й различал двоякую среду оби
тания паразитов: среду первого поряд
ка — непосредственное место обита
ния, т. е. организм хозяина, и среду
второго порядка — условия, окружа
ющие хозяина. Не следует считать, что у
паразита окружающая его среда посто
янна. Она может резко меняться на
протяжении короткого промежутка вре
мени (недели, дня), например, при забо
левании хозяина, выработке им имму
нитета и т. д. Для освоения этой особой
среды — организма хозяина — у пара
зитов в ходе эволюции выработался ряд
адаптаций.
?. 1. Какие главнейшие особенности воды как
среды обитания гидробионтов? 2. Назовите ограни
чивающие факторы водной среды. Ответ поясните.
3. Какие отличительные особенности наземно-воз
душной среды от водной? 4. Что такое климат и
микроклимат? 5. В чем заключаются особенности
почвы как среды жизни? 6. Что такое среда жизни
для паразита и его хозяина?
102. Адаптации организмов
к жизни в разных средах
как результат
их исторического развития
В процессе исторического развития
живых организмов ими был выработан
ряд адаптаций, позволивших заселить те
или иные местообитания. Адаптация (от
лат. adaptatio — прилаживание, приспо
собление) — процесс, а также резуль
тат приспособления строения и фун
кций организмов и их органов к услови
ям среды обитания. Благодаря адапта
ции обеспечивается выживание видов в
конкретных условиях среды.
Рассмотрим основные направления в
формировании адаптаций к жизни в че
тырех средах обитания.
Адаптации к жизни в водной среде.
Толщу воды заселили гидробионты, боль
шая часть которых относится к двум важ
нейшим экологическим группировкам:
нектону и планктону. Нектон (от греч.
nektos — плавающий) (рис. 217, с. 356)
— совокупность активно плавающих жи
вотных, способных преодолевать силу те
чения. Это пелагические рыбы, голово
ногие моллюски, ластоногие, китообраз
ные. Для них характерна обтекаемая
форма тела, развитая мускулатура, бла
годаря которым они могут передвигаться
с большой скоростью. Например, рыбамеч развивает скорость до 130 км/ч,
кальмары — до 45— 50 км/ч.
П ланкт он (от греч. planktos — па
рящий, блуждающий) (рис. 218, с. 357)
— совокупность пассивно парящих в
воде и переносимых течением бактерий
(бакт ериопланкт он), растений (фи
т опланкт он) и животных (зооплан
ктон), совершенно или почти не приспо
собленных к самостоятельному передви
жению на значительные расстояния. Что
бы удержаться в толще воды, они имеют
в основном небольшие размеры, нередко
с разнообразными и многочисленными
выростами наружных частей тела. Об355
Рис. 217 Организмы нектона: 1 — акула, 2 — осетр, 3 — щука, 4 — каракатица, 5 — кальмар, 6 —
осьминог.
356
6
Рис. 218. Организмы планктона: а — фитопланктон, б — зоопланктон.
легчения веса планктонные организмы
достигают за счет обильных жировых
включений, газовых вакуолей, полной
или частичной утраты утяжеляющих час
тей тела — раковин. К планктону отно
сятся разнообразные группы цианобак
терий, протист, водорослей, мелкие рако
образные, коловратки, медузы.
Донную область водоемов — б е н т а л ь — заселили бентосные организмы
(рис. 219, с. 358). Бент ос (от греч.
benthos — глубина) — совокупность ор
ганизмов, обитающих на дне водоемов,
адаптированных к соответствующей
структуре грунта. Например, бентосные
животные текучих водоемов (рек, кана
лов) имеют уплощенное тело и различ
ные приспособления для прикрепления к
грунту (присоски у пиявок, крючья у ли
чинок ручейников, массивные раковины у
беззубок, перловиц, ризоиды у водорос
лей).
357
Рис. 219. Население дна стоячих водоемов.
Адаптацией к добыванию пищи, взве
шенной в толще воды, является фильтра
ционный способ питания, свойственный
только водным животным. К фильтрате рам принадлежат животные разных
групп — двустворчатые моллюски (без
зубка, мидия, устрица и др.), ракообраз
358
ные (дафния, диаптомус), рыбы (сельдь,
белый и пестрый толстолобики, уклея и
др.), китообразные (беззубые киты).
Высокая плотность и большая вытал
кивающая сила воды способствовали
слабому развитию скелета водных живот
ных (хрящевой скелет акул, скатов, осет-
ровых рыб). По такому же пути шла эво
люция и у высших погруженных водных
растений. Их стебли и листья длинные и
тонкие, со слабо развитой механической
тканью (рис. 220). В их органах хорошо
развита воздухоносная паренхима, увели
чивающая плавучесть.
Низкая прозрачность природных вод
не способствовала эволюции органов
зрения водных животных на дальностное
видение. Так, глаза рыб, имеющие плос
кую роговицу и круглый хрусталик, прис
пособлены к видению предметов на рас
стоянии не более 10— 15 м.
Специализированным органом дыха
ния у первичноводных животных разных
систематических групп являются жабры
(многощетинковые черви, ракообразные,
двустворчатые моллюски, рыбы).
Адаптации к жизни на суше. Низкие
плотность и опорность, большая про
зрачность воздуха, низкое содержание
водяных паров и высокая концентрация
кислорода — главнейшие факторы, к ко-
Рис. 220. Погруженные растения: 1 — попереч
ный срез стебля урути (на срезе видна возду
хоносная паренхима); 2 — водокрас с зимую
щими почками, опускающимися на дно.
1
2
359
торым шло приспособление живых орга
низмов на суше.
Прежде всего у обитателей суши хо
рошо развиты опорные системы, поддер
живающие их тела. У растений —■ это
разнообразные механические ткани, осо
бенно мощно развитые у деревьев. Ж и
вотные выработали как наружный (чле
нистоногие), так и внутренний скелеты
(хордовые). Некоторые группы животных
приобрели гидроскелет (круглые и коль
чатые черви). Проблемы у наземных ор
ганизмов с поддержанием их тела в прос
транстве и с преодолением сил гравита
ции ограничили их предельную массу и
размеры. Самые крупные животные суши
(слон — до 5 т) уступают по размерам и
массе гигантам водной среды (синий
кит — до 150 т).
Низкая сопротивляемость воздуха
способствовала прогрессивной эволюции
систем передвижения наземных живот
ных. Так, наиболее высокие скорости
движения по суше приобрели млекопита
ющие, а птицы освоили воздушную среду,
развив способность к полету.
Большая подвижность воздуха в вер
тикальном и горизонтальном направле
ниях использовалась некоторыми назем
ными организмами либо отдельными ста
диями их развития для расселения с по
мощью воздушных потоков (молодь пау
ков, насекомые, споры, семена и плоды
растений, цисты протист). По аналогии с
водными планктонными организмами в
качестве приспособлений к пассивному
парению в воздушной среде насекомые
выработали сходные адаптации — мел
кие размеры тела, разнообразные вырос
ты, увеличивающие относительную по
верхность тела или некоторых его частей
(рис. 221). Семена и плоды, распростра
няемые ветром, имеют различные крыловидные и парашютовидные придатки,
увеличивающие их способность к плани
рованию (рис. 222).
Приспособления наземных организ
мов к сохранению влаги также разнооб
разны. Так, например, у насекомых — са
мого многочисленного класса наземных
животных — тело надежно защищено от
высыхания хитинизированной много
слойной кутикулой, в наружном слое ко
торой содержатся жиры и воскоподобные
вещества. Конечные продукты белкового
обмена выделяются в виде сухих экскре
тов — кристаллов мочевой кислоты.
Сходные водосберегающие приспо
собления развили и пресмыкающиеся,
сумевшие совместно с насекомыми ши-
2
Рис. 221. Адаптации животных к расселению ветром: 1 — комарик, 2 — личинка непарного шелкоп
ряда.
360
роко заселить безводные местообитания.
Выработанная у наземных животных
способность к внутреннему оплодотворе
нию сделала их независимыми от наличия
водной среды.
Приспособления растений к дефициту
влаги на суше наилучшим образом выра
жены у экологической группировки рас
тений, освоивших сухие местообита
ния — ксерофитов. Одни из них — сук
куленты — приобрели способность на
капливать воду в мясистых и сочных
361
стеблях либо в листьях. Другая группа —
склерофиты — пошла по пути приобре
тения приспособлений, направленных на
интенсивное добывание воды из почвы и
экономное ее расходование. Они облада
ют мощно развитой корневой системой,
позволяющей проникать на большую
глубину до грунтовой воды (верблюжья
колючка), либо имеют сильно развет
вленные, расположенные на небольшой
глубине корни, максимально использую
щие атмосферные осадки. Листья у скле-
рофитов игольчатые либо в виде чешуй,
покрытые восковым налетом либо опу
шенные. Устьица располагаются на ниж
ней, свернутой внутрь стороне листовой
пластинки.
Такое преимущество наземной среды,
как высокое содержание кислорода в ат
мосферном воздухе, было использовано
животными суши для интенсификации
обмена веществ. Именно у наземных жи
вотных (птиц и млекопитающих) была
выработана гомойотермия, которая обес-
Рис. 223. Обитатели почвы: 1 — личинка хруща майского, 2 — круглый червь, 3 — лжескорпион, 4 —
гамазовый клещ, 5 — инфузория, 6 — жгутиконосец, 7 — коловратка, 8 — дождевой червь, 9 — мок
рица, 10 — губоногая многоножка, 11 — личинка щелкуна, 12 — медведка.
362
печила им возможность повышения ин
тенсивности жизнедеятельности. Неза
висимость обменных процессов от коле
баний температуры внешней среды
расширила их возможности к рассе
лению.
А даптации к ж и зн и в п о ч в е. Мелкие
почвенные членистоногие (клещи, насе
комые, многоножки) используют для пе
редвижения по стенкам почвенных по
лостей коготки на лапках. Многие из них
имеют несмачивающиеся покровы, снаб
женные волосками, чешуйками, которые
позволяют переживать периоды затопле
ния почвы в пузырьках воздуха (рис.
223).
Более крупные почвенные животные,
такие, как личинки майского жука, дож
девые черви, при передвижении исполь
зуют сильное сопротивление со стороны
почвы. Поэтому у них выработались
приспособления для раздвигания почвен
ных частиц путем изгибания тела (круг
лые черви) либо использования гидрос
келета (дождевые черви).
Некоторые средние по размерам
(медведка) и крупные почвенные живот
ные (кроты, слепыши) выработали эф
фективный способ передвижения в поч
ве — рытье. Внешнее строение землероев отражает их приспособленность к ро
ющему подземному образу жизни
(рис. 224).
А даптации к ж и зн и в д р у го м о р г а
н и з м е — п а р а з и т и з м у . Главнейшие
приспособления следующие:
1. Наличие о р г а н о в п р и к р еп л ен и я
(присосок, кутикулярных крючков и
др.), обеспечивающих связь паразита
с организмом хозяина.
2. Развитие специализированных п о
кровны х обр азов ан и й
(кутикула
и др.), защищающих эндопаразитов от
воздействия пищеварительных фер
ментов хозяина.
3. Способность к а н а э р о б н о м у д ы х а
н и ю , обеспеченному ферментатив-
Рис. 224. Крот.
ным расщеплением запасных пита
тельных веществ в отсутствие кисло
рода. Невысокая эффективность
этого типа энергетического обмена
компенсируется практически неис
сякаемыми запасами углеводов, по
ставщиком которых является пища
хозяина, и отсутствием трат энергии
на движение.
4. Р е г р е с с и в н о е р а з в и т и е (упро
щение) н ер в н о й си стем ы и о р га н о в
ч ув ств. Укорачивается кишечник
(круглые черви), либо пищеваритель
ная система полностью утрачива
ется (ленточные черви), и раство
ренные питательные вещества вса
сываются всей поверхностью тела.
5. Интенсивное р а зв и т и е п о л о в о й
системы, появление у животного спо
собности размножаться уже на стадии
личинки(например, у печеночного со
сальщика) приводит к очень высокой
половой продуктивности.
6. Изменение форм размножения,
т. е. в о зн и к н о в е н и е г е р м а ф р о д и т и з
м а (плоские черви), обеспечивающее
гарантию размножения при наличии
даже единственной особи.
7. Эффективная за щ и т а о п л о д о т в о
р ен н ы х яи ц многослойными оболоч
ками и обеспечение питанием разви
вающегося в яйце зародыша.
363
8. Р а зв и т и е п р и с п о со б л ен и й для вы
хода личинок из яйца и тела хозяина
во внешнюю среду и их проникнове
ния в организм нового хозяина (желе
зы, выделяющие специальные фер
менты, колющие приспособления
и т. п.).
9. С м ен х о з я е в в жизненном цикле
паразита: в основном хозяине живут
взрослые особи, размножающиеся
половым способом; в промежуточном
хозяине развивается личинка и может
происходить бесполое размножение.
Это приспособление препятствует
чрезмерному накоплению паразитов в
организме хозяина и его гибели, а
также способствует расселению па
разитов.
На рисунке 225 представлены па
разитические круглые черви, вызы
вающие серьезные заболевания чело
364
века. Одно из таких заболеваний — сло
новая болезнь (рис. 226) — вызывает
ся червем-паразитом филярней Банк
рофта.
Таким образом, в результате длитель
ного действия естественного отбора про
исходило совершенствование приспособ
лений, пригодных для освоения живыми
организмами специфических условий
разных жизненных сред.
■ 1. Какие адаптации выработали планктонные
организмы? 2. Каковы приспособления, характер
ные для обитателей бентали водоемов? 3. Каковы
основные направления формирования приспособ
лений у наземных организмов? 4. Какие адаптации к
жизни в почве характерны для организмов разных
размеров? 5. Приведите примеры адаптаций у пара
зитов, которые обеспечивают им широкое распрос
транение.
ОГЛАВЛЕНИЕ
Введение .......................................................
3
Раздел первый
Клетка — структурная
и функциональная единица жизни
Клетка в системе живой природы. Клеточ
ная теория ...................................................
Гл а в а I. Молекулярные основы жизни . . .
1. Элементы и атомы ..............................
2. Химические соединения и молекулы
3. Вода и другие неорганические ве
щества. Их роль в клетке .................
4. Органические соединения .................
5. Липиды ................................................
6. Белки (протеины) ................................
7. Ферменты ................
8. Нуклеиновые кислоты .......................
9. АТФ и ее роль в клетке .....................
Г лав а II. Строение и системы жизнеобес
печения клетки ...............' ..............................
10. Строение кл етки ..................................
11. Биологические мембраны эукариоти
ческой клетки ......................................
12. Молекулярный транспорт через био
логическую мембрану .......................
13. Информационная система клетки.
Ядро, его строение и функции ...........
14. Хранение наследственной информа
ции. Генетический код .......................
15. Реализация наследственной ин
формации. Синтез белка на рибосо
мах .......................................................
16. Энергетическая система клетки . . . .
17. Фотосинтез ..........................................
18. Бактериальный фотосинтез ...............
19. Хемосинтез ..........................................
20. Клеточное дыхание ..............................
21. Дальнейшее превращение пировиноградной кислоты ..................................
22. Анаэробное дыхание. Брожение . . . .
23. Транспортная система клетки ...........
24. Внутриклеточное переваривание . . .
25. Вакуоли . , ............................................
26. Опорно-двигательная система клет
ки .........................................................
27. Клеточная стенка ................................
28. Система связи и взаимодействия меж366
6
8
9
14
15
17
20
22
27
30
35
36
37
41
43
46
49
50
53
55
64
66
67
69
72
75
78
79
82
85
ду клетками в многоклеточном ор
ганизме ................................................
Г лав а III. Воспроизведение клетки . . . .
29. Деление клетки. Клеточный цикл . . .
30. Митоз ...................................................
31. Амитоз. Клеточное деление прока
риот .......................................................
32. Мейоз и его биологическое значение
33. Клетка — структурная и функцио
нальная единица жизни .....................
91
93
—
95
99
—
103
Раздел второй
Организм — биологическая система
Основные свойства живых организмов ..
Г лава IV. Обмен веществ и энергии в жи
вых организмах ..................................................
34. Структурная организация живых орга
низмов ................................................
35. Растительные ткани ...............................
36. Животные ткани .....................................
37. Питание как способ получения ве
ществ и энергии из окружающей
среды ...................................................
38. Поступление и транспорт питатель
ных веществ в живых организмах . . .
39. Поступление питательных веществ в
животный организм (способы пи
тания) ...................................................
40. Пищеварение ......................................
41. Пищеварение у человека ....................
42. Всасывание питательных веществ ..
43. Витамины .............................................
44. Особенности пищеварения у расти
тельноядных животных .....................
45. Транспорт веществ в организмах жи
вотных и человека ..............................
46. Типы кровеносных систем .................
' 47. Кровь, тканевая жидкость и лимфа ..
48. Защитные функции крови .................
49. Иммунная система ..............................
50. Газообмен ............................................
51. Транспорт газов в организме чело
века. Перенос кислорода и углекисло
го газа кровью ....................................
52. Выделительная система у животных
и растений ..........................................
106
107
—
110
116
121
123
129
133
136
140
143
145
147
148
153
158
161
165
169
171
53. Гомеостаз — поддержание постоянст
ва внутренней среды .........................
54. Передвижение организмов. Опорные
системы и движение растений ...........
55. Опорные системы и движение жи
вотных .............................
Г лав а V. Размножение и индивидуальное
развитие организмов ....................................
56. Типы размножения организмов. Бес
полое размножение ............................
57. Половое размножение .......................
58. Оплодотворение ..................................
59. Индивидуальное развитие организма.
Онтогенез животных .........................
60. Онтогенез человека ...........................
61. Индивидуальное развитие растений
62. Старение и смерть организмов .........
Г л а в а VI. Наследственность и измен
чивость организмов ......................................
63. Гибридологический метод изучения
наследственности ................................
64. Моногибридное скрещивание. Пер
вый и второй законы Менделя .........
65. Аллельные гены и их взаимодействие.
Анализирующее скрещивание...........
66. Дигибридное и полигибридное скре
щивания. Третий закон Менделя . . . .
67. Неаллельные взаимодействия генов
68. Хромосомная теория наследствен
ности. Сцепленное наследование . . .
69. Генетические карты ...........................
70. Генетика пола ......................................
71. Наследование признаков, сцеплен
ных с полом ........................................
72. Цитоплазматическая наследствен
ность .....................................................
73. Современные представления о гене
74. Изменчивость организмов. Норма ре
акций ...................................................
75. Типы изменчивости. Модификационная изменчивость ................................
76. Комбинативная изменчивость. Му
тации .....................................................
77. Мутационная изменчивость. Геном
ные мутации ........................................
78. Хромосомные и генные мутации . . . .
179
185
187
192
—
195
200
204
209
211
216
218
221
226
230
234
238
241
245
247
251
255
259
262
265
267
271
79. Спонтанный и индуцированный му
тагенез ..............................................
80. Генетика человека. Анализ родослов
ных ........................................................
81. Близнецы .............................................
82. Цитогенетический, популяционный
и другие методы генетики человека
83. Наследственные болезни человека,
их лечение и профилактика ................
84. Проблемы генетической безопас
ности ......................................................
Гла в а VII. Регуляция и координация
жизнедеятельности организмов ..............
85. Виды регуляторных систем у живот
ных и растений ..................................
86. Нервная система животных................
87. Органы чувств у животных ................
88. Эндокринная система и ее работа . .
89. Нарушение деятельности эндокрин
ных желез у человека . . ....................
90. Поведение животных. Врожденное
поведение .............................................
91. Поведение, связанное с научением
92. Координация и регуляция у расте
ний ........................................................
Гл а в а VIII. Адаптация организмов к
условиям обитания .....................................
93. Среда обитания и условия существова
ния организмов. Экологические
факторы ............................................
94. Закономерности действия внешних
факторов ..........................................
95. Абиотические (климатические) фак
торы. Тепло ......................................
96. Свет ...................................................
97. Влажность ........................................
98. Биотические факторы .....................
99. Другие виды биотических взаимоот
ношений ............................................
100. Ритмические изменения факторов
среды и их влияние на организмы ..
101. Среды жизни ....................................
102. Адаптации организмов к жизни в
разных средах как результат их ис
торического развития .....................
274
277
281
283
285
289
291
—
292
300
306
309
310
313
316
318
320
325
330
332
336
343
347
351
355
(Н а з в а н и е и н о м ер ш к о л ы )
Учебный
год
19
/
20
/
20
/
20
/
Имя и фамилия ученика
Состояние
учебного пособия
при
получении
Оценка
ученику за
пользование
учебным пособием
У ч еб н о е п о с о б и е и зд а н о з а сч ет ср ед с т в го с у д а р с т в ен н о г о б ю д ж е т а
дл я ф о н д о в б и б л и о т е к п о з а к а з у М и н и с т е р с т в а о б р а з о в а н и я ,
св о б о д н о й п р о д а ж е н е п о д л еж и т
Учебное издание
ЛЕМЕЗА Николай Алексеевич
ЛИСОВ Николай Дмитриевич
КАМЛЮК Лилия Васильевна
ОБЩАЯ БИОЛОГИЯ
Учебное пособие для 10 класса
с углубленным изучением биологии
общеобразовательных школ
с русским языком обучения
Редактор Е. В. Литвинович
Художник Б. Г. Клюйко
Художественный редактор О. И. Романцов
Технический редактор 3. В. Романкевич
Корректоры С. А. Янович, Т. Н. Ведерникова
Сдано в набор 03.06.98. Подписано в печать
15.09.98. Формат 7 0x90'/,6. Бумага офс. № 1.
Гарнитура литературная. Офсетная печать. Уел.
печ. л. 26,91+0,29 форз. Уел. кр.-отт. 109,4. Уч,изд. л. 28,32 + 0,49 форз. Тираж 19 000 экз.
Заказ 347.
Издательское
государственное
предприятие
«Народная асвета» Государственного комитета
Республики Беларусь по печати. Лицензия ЛВ № 4
от 02.12.97. 220600, Минск, проспект Машерова, 11.
Набор сделан в типографии издательства
«Белорусский Дом печати». 220041, Минск,
проспект Ф. Скорины, 79.
Минская фабрика цветной печати. 220024, Минск,
Корженевского, 20.
ISBN 985-03-0814-1
9 789850 308146