






















































































































































































































































































































































































































































































Автор: Атраментова Л.А. Филипцова О.В.
Теги: развитие и способности психики сравнительная психология психология отдельных видов деятельности отраслевая (прикладная) психология социально-психологические проблемы некоторых отраслей психологии медицина биология психология генетика психогенетика учебное пособие
ISBN: 5-89349-656-6
Год: 2004




Л.А. Атраментова, О.В. Филипцова
ВВЕДЕНИЕ
ПСИХОГЕНЕТИКУ
Учебное пособие
Рекомендовано Редакционно-издательским советом
Российской Академии образования
использованию в качестве учебно-методического пособия
Москва
Издательство «Флинта»
Московский психолого-социальный институт
2004
?УДК 159.922(075.8) Ж'
"ББК 88.4
А92
Библиотека психолога
Главный редактор Д.И. Фельдштейн
Заместитель главного редактора
С.К. Бондырева
Члены редакционной коллегии:
А.А. Бодалев, В.П. Борисенков, С.В. Дармодехин, А.А. Деркач,
Ю.И. Дик, А.И. Донцов, И.В. Дубровина, М.И. Кондаков,
В.Г. Костомаров, В.С. Леднев, В.И. Лубовский, Н.Н. Малафеев,
Н.Д. Никандров, А.И. Подольский, В.А. Поляков, В.В. Рубцов,
Э.В. Сайко, В.М. Тиктинский-Шкловский
Атраментова Л.А.
Введение в психогенетику: Учебное пособие / Л.А. Атраменто-
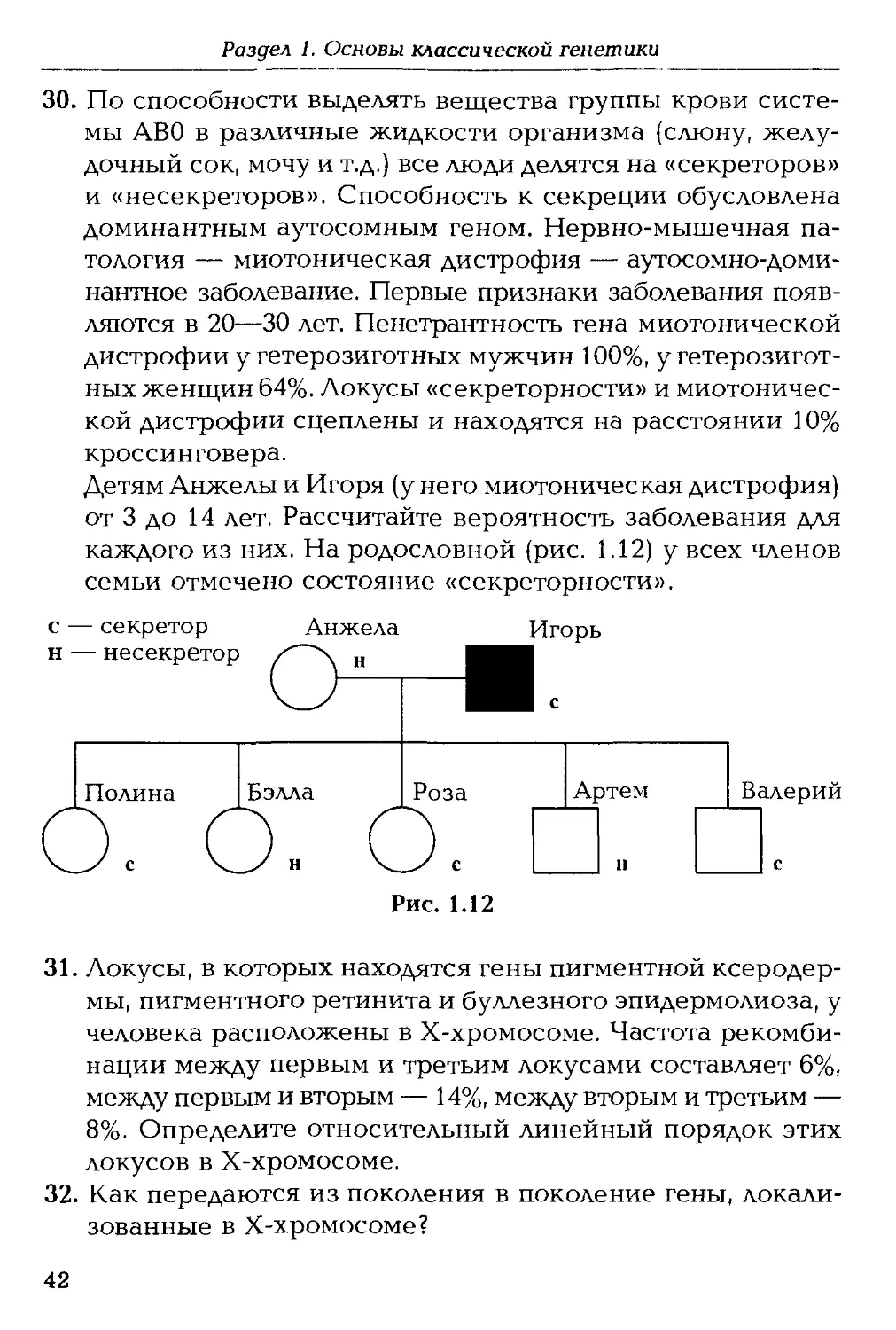
ва, О.В. Филипцова. — М.: Флинта: Московский психолого-соци-
альный институт, 2004. — 472 с.: ил.
ISBN 5-89349-656-6 (Флинта)
ISBN 5-89502-676-1 (МПСЦ)
В книге рассматриваются проблемы психогенетики — нового направ-
ления в науке, пограничной области знания между психологией и гене-
тикой. Представлены основные задачи современной психологической
генетики: изучать, как реагируют на новые ситуации люди с разными
генотипами, разрабатывать методы анализа поведения человека, его ин-
теллекта.
Для студентов-психологов.
ISBN 5-89349-656-6 (Флинта)
ISBN 5-89502-676-1 (МПСИ)
© Издательство «Флинта», 2004
Содержание
Введение.............................................9
Раздел 1. ОСНОВЫ КЛАССИЧЕСКОЙ ГЕНЕТИКИ............... 15
1.1. Основные понятия...................... 17
1.2. Моногенное наследование................21
Полное доминирование.....................21
Неполное доминирование ..................23
Кодоминирование..........................24
Решетка Пеннета..........................25
1.3. Независимое наследование...............26
1.4. Сцепление и кроссинговер...............28
1.5. Сцепленное с полом наследование........31
Х-сцепленное наследование................31
Y-сцепленное наследование................33
1.6. Наследование полигенных признаков......34
Контрольные вопросы и задания к разделу 1...37
Раздел 2. МАТЕРИАЛЬНЫЕ НОСИТЕЛИ
НАСЛЕДСТВЕННОСТИ....................................47
2.1. Химическая структура ДНК...............48
2.2. Генетический код.......................50
2.3. Транскрипция и трансляция..............52
2.4. Генные мутации.........................54
2.5. Динамические мутации...................55
2.6. Геном человека.........................56
2.7. Хромосомный набор человека.............58
2.8. Структурная организация хромосом ......60
2.9. Цитогенетическая символика.............62
2.10. Анеуплоидии............................63
2.11. Аутосомные синдромы....................65
Трисомия 21 (синдром Дауна)..............65
Трисомия 13 (синдром Патау)..............68
Трисомия 18 (синдром Эдвардса)...........68
2.12. Синдромы половых хромосом..............69
Моносомия по Х-хромосоме (45, ХО),
или синдром Тернера......................69
Трисомия по Х-хромосоме (47, XXX),
или синдром трипло-Х.....................70
Дисомия по Х-хромосоме у мужчин (47, XXY),
или синдром Клайнфельтера...............71
Дисомия по Y-хромосоме (47, XYY),
или синдром Жакоб.......................72
2.13. Полиплоидия ........................ 73
2.14. Псевдонормальный кариотип.............74
2.15. Однородительская дисомия..............75
2.16. Мозаицизм.............................77
2.17. Геномный импринтинг...................78
2.18. Хромосомные перестройки...............79
Делеции.................................82
Транслокации............................82
Инверсии................................83
Изохромосомы и кольцевые хромосомы......83
Аберрация по типу «ломкой» Х-хромосомы...84
2.19. Повторный риск хромосомных болезней ..86
Контрольные вопросы и задания к разделу 2...87
Раздел 3. НАСЛЕДСТВЕННОСТЬ И СРЕДА.................93
3.1. Модификационная изменчивость..........93
3.2. Норма реакции.........................97
, 3.3. Экспрессивность и пенетрантность.......98
3.4. Компенсация дозы гена
(инактивация Х-хромосомы)................ 102
3.5. Признаки, зависимые от пола
и ограниченные полом..................... 105
3.6. Мультифакториальные признаки........ 107
3.7. Закономерности клеточных делений.... 109
Митоз................................. 109
Мейоз................................. 110
Гаметогенез у человека................ 111
3.8. Материнский эффект................... ИЗ
3.9. Врожденные дефекты.................. 116
Тератогены............................ 118
ф10. От гена к поведению.................. 124
Контрольные вопросы и задания к разделу 3. 128
Раздел 4. АНАЛИЗ НАСЛЕДСТВЕННОСТИ ЧЕЛОВЕКА....... 135
4.1. Структура признака.................. 136
, 4.2. Генеалогический метод................ 140
Анализ большой родословной............... 146
Анализ малочисленных родословных....... 149
4.3. Компонентный анализ.................. 150
Фенотипические модели...................151
Коэффициенты корреляции................ 153
Модель главного гена................... 155
4.4. Метод генетических маркеров.......... 157
4.5. Моделирование на животных............ 159
Контрольные вопросы и задания к разделу 4. 161
Раздел 5. БЛИЗНЕЦЫ В ГЕНЕТИКЕ ЧЕЛОВЕКА............ 168
5.1. Типы близнецов....................... 170
5.2. Частота рождения близнецов........... 173
5.3. Близнецовый метод изучения наследственности .... 175
Формирование близнецовой выборки....... 177
Диагностика зиготности................. 177
Конкордантность........................ 180
Вычисление показателя наследуемости.....181
Семьи близнецов........................ 183
Контрольные вопросы и задания к разделу 5..184
Раздел 6. ПОЛОВЫЕ И ГЕНДЕРНЫЕ РАЗЛИЧИЯ.............187
6.1. Хромосомное определение пола......... 189
6.2. Половая дифференцировка и половые гормоны.. 190
6.3. Интерсексуальные состояния человека . 192
6.4. Гендерные различия в когнитивных
способностях и личностных свойствах....... 199
6.5. Роль среды в гендерных различиях.... 203
6.6. Воспитание и поло-ролевые установки ..204
6.7. Гомосексуализм........................206
Контрольные вопросы и задания к разделу 6..209
Раздел 7. СТРУКТУРА ПОПУЛЯЦИИ И ГЕНЕТИЧЕСКИЕ
ПРОЦЕССЫ...........................................211
7.1. Нахождение частот аллелей.............212
Сцепление с полом.......................213
Множественный аллелизм..................213
7.2. Уравнение Харди-Вайнберга.............214
7.3. Дрейф генов...........................216
7.4. Эффект основателя ....................218
7.5. Эффект «бутылочного горлышка».........220
' 7.6. Поток генов............................221
7.7. Естественный отбор....................223
7.8. Мутационный процесс...................225
7.9. Инбридинг.............................226
7.10. Аутбридинг............................229
7.11. Брачная ассортативность...............231
Контрольные вопросы и задания к разделу 7...232
Раздел 8. ЧЕЛОВЕЧЕСКОЕ РАЗНООБРАЗИЕ................239
8.1. Телосложение .........................242
8.2. Черты лица............................248
8.3. Пигментация...........................249
8.4. Дерматоглифика........................254
8.5. Скорость развития.....................258
8.6. Особенности темперамента..............259
8.7. Группы крови..........................259
8.8. Система HLA...........................261
8.9. Полиморфизм ДНК.......................265
8.10. Электрическая активность мозга........268
8.11. Функциональная асимметрия.............273
8.12. Реакция на фармакологические средства..276
Контрольные вопросы и задания к разделу 8...278
Раздел 9. ГЕНЕТИЧЕСКИЙ КОНТРОЛЬ ВОСПРИЯТИЯ..........282
9.1. Зрение................................282
9.2. Слух..................................288
9.3. Вкусовая чувствительность.............290
9.4. Восприятие запаха.....................298
Контрольные вопросы и задания к разделу 9. 299
Раздел 10. ГЕНЕТИЧЕСКИЕ ОСНОВЫ
ИНТЕЛЛЕКТУАЛЬНЫХ РАЗЛИЧИЙ..........................305
10.1. Проблемы тестирования.................307
10.2. Генетические исследования интеллекта..309
10.3. Моделирование на животных.............315
10.4. Эффект Флинна.........................316
10.5. Межпопуляционные различия в IQ........318
Контрольные вопросы и задания к разделу 10...324
Раздел И. ГЕНЕТИКА ЛИЧНОСТНЫХ ОСОБЕННОСТЕЙ .........326
11.1. Близнецовые исследования характеристик
личности...................................326
11.2. Дерматоглифика и личность.............330
11.3. Молекулярно-генетические исследования
личности...................................331
11.4. Наследственность и профессия..........332
11.5. Антисоциальное и криминальное поведение.334
11.6. Генетические основы агрессивности.....340
Контрольные вопросы и задания к разделу 11..343
Раздел 12. ГЕНЕТИЧЕСКИЕ ОСНОВЫ АНОМАЛИЙ
ПОВЕДЕНИЯ...........................................345
12.1. Генетическая природа психических болезней ... 346
12.2. Гипотеза о происхождении психических
болезней................................349
12.3. Дерматоглифические маркеры нервных
и психических болезней человека............352
12.4. Социальное значение пограничных
психических состояний .....................355
12.5. Расстройства настроения...............357
Униполярный, или депрессивный синдром...357
Биполярный синдром......................358
12.6. Шизофрения ...........................360
12.7. Психические расстройства в онтогенезе.363
Аутизм..................................364
Нарушения речевого развития и обучения..366
Заикание................................368
Синдром Туретта.........................370
Дислексия...............................371
Синдром дефицита внимания
и гиперактивности.......................372
12.8. Органические поражения мозга .........373
Болезнь Альцгеймера.....................373
Болезнь Паркинсона......................375
Хорея Гентингтона.......................379
Эпилепсия...............................381
Болезнь Феллинга (фенилкетонурия).......384
Болезнь Тэя—Сакса.......................385
Рассеянный склероз......................386
12.9. Химическая зависимость.................387
Генетика алкоголизма.....................387
Генетика наркомании......................392
Генетика курения.........................394
12.10. Расстройства самоконтроля.............395
Гэмблинг.................................396
Расстройства питания.....................398
Анорексия................................399
Булимия..................................401
Контрольные вопросы и задания к разделу 12...403
Ответы и решения.....................................412
Приложение...........................................440
Метод %2.............................................440
Литература...........................................442
Введение
Интерес к наследственным основам поведения возник в се-
редине XX в., а ранее считалось, что поведение детермини-
ровано исключительно средой. Это убеждение возникло из
наблюдений над изменчивостью и ситуативностью поведе-
ния. На этапе становления генетики исследователей интере-
совали легко выявляемые морфологические различия между
живыми организмами. Учет и анализ поведенческих призна-
ков представляет значительные трудности, поэтому генети-
ческий компонент в поведении обычно игнорировался.
Сама идея использовать генетический подход к анализу
поведения считалась неприемлемой, особенно если речь шла
о поведении человека. Генетики занимались анализом при-
знаков, которые были неизменными в течение жизни и пото-
му удобными для генетического анализа. Понимание того, что
наследуется не признак, а способность к его формированию
пришло позже. Поведение представляет собой реакцию орга-
низма на внешние воздействия. В связи с этим создавалось
впечатление, что оно бесконечно изменчиво и независимо от
генотипа. Между тем накапливались факты, которые можно
было объяснить, только допустив, что поведение имеет на-
следственную основу. Генетические исследования, выполнен-
ные на животных, дали убедительные доказательства того, что
поведение зависит от наследственности. Наличие генетичес-
кого компонента в поведении животных давно не вызывает
сомнения. Проблема заключается лишь в том, чтобы выяснить
роль наследственности в отдельных поведенческих актах у
разных видов.
Наследственные основы поведения человека представля-
ют не только академический интерес, но важны и в практи-
ческом отношении. С условиями, в которых живет современ-
ный горожанин, люди не сталкивались на протяжении всей
предыдущей истории. Появляются новые профессии, кото-
рые предъявляют все более жесткие требования к биологи-
ческим и психическим особенностям работников. Высокая
плотность населения порождает жесткую конкуренцию.
У одних людей на стрессовую ситуацию развивается болез-
Введение
ненная реакция, у других — социально не одобряемое пове-
дение. Адекватная реакция на окружающую обстановку во
многом определяет жизненный успех человека. Зная свои на-
следственные задатки, человек может избегать ситуаций, ко-
торые приведут к нежелательной реакции. Индивиды, наслед-
ственно предрасположенные к формированию химический
зависимости, должны активно избегать ситуаций, где им мо-
гут быть предложены алкоголь и наркотики.
Много внимания специалисты в области психологической
генетики уделяют наследственным основам интеллекта. От
уровня интеллекта зависит способность к обучению, овладе-
нию профессией и в конечном счете жизненный успех.
Среда обитания человека меняется очень быстро. Пове-
денческая реакция на новую культурную ситуацию у некото-
рых людей оказывается патологической. В последнее время
появилось много игровых заведений (казино, залы компью-
терных игр и т.п.). Вскоре обнаружилось, что у некоторых
людей развивается состояние, которое можно назвать «иг-
ральная наркомания». Широкое распространение компьюте-
ров породило ряд новых проблем. Появились «компьютерные
болезни». Отдельные компьютерные игры оказались опасны-
ми для детей, имеющих наследственную предрасположен-
ность к эпилепсии.
Новые реальности ставят перед психологическими гене-
тиками вполне определенные задачи: изучать, как реагиру-
ют на новые ситуации люди с разными генотипами, разраба-
тывать методы, с помощью которых можно было бы выявлять
индивидов с потенциально патологической реакцией и пре-
дупреждать развитие болезненного состояния.
Сегодня в школах работают профессиональные психоло-
ги. Им особенно необходимо знание психологической гене-
тики. Школьный психолог общается не только с ребенком, но
и с его семьей. Зная, какими чертами характера обладают ро-
дители, психолог может предсказать, в каком направлении бу-
дет развиваться психика ребенка, и при необходимости свое-
временно ее скорректировать. Говоря о необходимости ин-
10
дивидуального подхода в образовании, прежде всего надо'
иметь в виду наследственные задатки обучаемого. От педаго-
гов обычно требуют, чтобы они применяли к каждому ребен-
ку индивидуальный подход. Это возможно только при опоре
на знания законов генетики. До сих пор среди психологов и
педагогов ведутся дискуссии о том, следует ли тестировать
детей при распределении их по группам обучения. Есть мне-
ние, что ориентация на среднего ученика не дает возможнос-
ти особо одаренным детям развить свои задатки, а детям, ко-
торые успевают медленно, и такой уровень окажется непо-
сильным.
При изучении наследственности человека исследователь
сталкивается с рядом проблем, которые связаны рядом огра-
ничений и запретов. Особенно серьезные проблемы возника-
ют при генетическом анализе поведения человека. Человек при-
спосабливается к окружающей среде, не только обретая био-
логические особенности в ходе эволюции и индивидуального
развития, но и совершенствуя технологию, то есть через куль-
турное развитие. Биологические особенности наследуются
путем передачи наследственного материала в соответствии с
законами генетики. Наследование культуры происходит как
передача опыта. Иногда передача генов и трансляция куль-
турных особенностей имеют сходную картину и могут ими-
тировать друг друга. Однако у человека как объекта генети-
ческих исследований есть и преимущества, важнейшее из ко-
торых речь. С ее помощью можно получить показатели,
которые нельзя зарегистрировать прибором. Речь позволяет
сделать объектом генетического анализа признаки, которые
невозможно изучить ни у какого другого биологического вида.
Человек, в отличие от животного, может охарактеризовать
вкус вещества, сообщить, видит или нет отдельные цвета, раз-
личает ли высоту звука. Это существенно расширяет диапа-
зон признаков, которые могут быть включены в генетичес-
кий анализ.
Психологи, исследующие генетические основы поведе-
ния, используют самые разные, как классические, так и но-
11
Введение
вейшие методы. Эти методы созданы с таким учетом, чтобы
преодолеть ограничения, которые налагает на исследователя
работа с человеком. Наиболее доступным является генеало-
гический метод, или метод родословных, разработанный в
XIX в. Ф. Гальтоном. Еще один специфический метод генети-
ки человека — близнецовый. Его автор — также Ф. Гальтон.
С помощью близнецового метода решается проблема соотно-
шения наследственности и среды в формировании различий
между индивидами. Эта же проблема решается и при изуче-
нии приемных детей. Приемные дети — удобная модель для
выяснения того, насколько различия между людьми обуслов-
ливаются сходством их генотипов и насколько сходством сре-
ды, в которой они выросли. В обществах, где отсутствует тайна
усыновления, изучение приемных детей позволило многое
понять в наследовании психических болезней и некоторых
видов отклоняющегося поведения. Животные модели позво-
ляют связать между собой генетические, биохимические,
физиологические компоненты для разных признаков. Много
ценной информации о биологических основах поведения по-
лучено при воздействии фармакологическими средствами на
животных с разными генотипами. Психофармакологический
метод позволяет сравнивать поведение животных и человека
под действием одних и тех же веществ. После того как было
доказано, что носителями наследственной информации в
клетке являются хромосомы, их структуру стали изучать у
людей с морфологическими и поведенческими аномалиями.
Этот метод получил название цитогенетического. Оказалось,
что изменения в хромосомном аппарате приводят к грубым
нарушениям соматического и психического развития чело-
века. Прогресс в области молекулярной генетики позволяет
обнаружить различия между индивидами с вариантами по-
ведения на уровне молекулы ДНК. Популяционно-генетичес-
кий метод позволяет дать ответ на вопрос, в чем заключается
приспособительная ценность различных вариантов поведе-
ния. Любое исследование по генетике человека является кон-
кретно-популяционным. Генетические параметры, получен-
12
Введение
ные при изучении популяции, характеризуют именно эту по-
пуляцию и могут существенно отличаться от параметров, по-
лученных на другом населении.
Кроме перечисленных генетических методов, используют
и вспомогательные методики, с помощью которых удается
полнее охарактеризовать изучаемый признак. Так, электро-
физиологические методы дают возможность оценить био-
электрическую активность мозга. С помощью биохимических
методов определяют количество того или иного вещества в
крови и другом биологическом материале. Антропологичес-
кие методики дают возможность охарактеризовать телосло-
жение и т.д. Показатели, полученные перечисленными мето-
дами, сравнивают у индивидов, проявивших разные свойства
при психологическом тестировании, у больных и здоровых
и т.п. На материале, полученном с помощью биохимических,
цитогенетических и молекулярно-генетических методик ос-
нован метод генетических маркеров. Каждый из перечислен-
ных методов, используемых в психологической генетике, по-
зволяет решать специфические задачи, а все вместе они дают
цельное представление о роли наследственности в поведении
человека.
Поведенческие признаки сложны для генетического ана-
лиза, так как в сильной степени модифицируются условиями
среды. Даже используя модельные объекты, трудно контро-
лировать условия эксперимента. Различия в жизненном опы-
те, накопленные животными до эксперимента, влияют на по-
ведение во время исследования. При изучении человека
объективные измерения особенно затруднительны, кроме тех
случаев, когда исследуются простые сенсорные восприятия.
При анализе поведения человека исследователь сталкивает-
ся с многозначностью и комплексностью психологических
функций. В каждом исследовании возникает необходимость
ответить на вопрос, наследственная основа чего именно изу-
чается. Успех генетического анализа зависит от того, насколь-
ко исследователю удалось разложить сложные поведенчес-
кие акты на более простые элементы и проанализировать каж-
Введение
дый из них в отдельности. При измерении уровня интеллекта
или оценке личностных характеристик избежать субъектив-
ности особенно трудно, однако ее можно свести к минимуму,
совершенствуя приемы работы, привлекая объективные био-
логические методики. Все перечисленные проблемы являют-
ся предметом нового научного направления — психогенети-
ки — области знания, пограничной между психологией и ге-
нетикой.
Раздел 1
ОСНОВЫ КЛАССИЧЕСКОЙ ГЕНЕТИКИ
Зарождение генетики связано с именем Грегора Менделя
(1822—1884), монаха августинского монастыря, жившего в ав-
стрийском городе Брюнне (Брно, современная Чехия). Г. Мен-
дель (рис. 1.1), скрещивая различные сорта гороха, первым
установил основные закономерности наследования. Его глав-
ное научное достижение состоит в доказательстве того, что
наследственность является дискретной. Это означает, что
Рис. 1.1. Основоположник генетики Грегор Мендель
Раздел /. Основы классической генетики
наследственное вещество состоит из неделимых единиц —
атомов наследственности, которые Мендель назвал фактора-
ми. Впоследствии менделевские факторы наследственности
получили название генов.
До Менделя в биологии господствовала теория слитной
наследственности. Согласно этой теории наследственный
материал представлялся чем-то вроде жидкости. Обычно на-
следственность связывали с кровью, отсюда и термины «чис-
токровный», «полукровный». Считалось, что, переходя от ро-
дителей к потомкам, наследственное вещество проявляет
свойства жидкости: при скрещиваниях смешивается в любых
соотношениях и, смешавшись, в дальнейшем не может раз-
делиться на исходные «чистые» варианты. Такие представле-
ния о природе наследственного материала сформировались
благодаря многовековой сельскохозяйственной практике.
Размножая животных и растения, люди отмечали, что потом-
ки по внешнему виду напоминают родителей, родственные
организмы похожи больше, чем не родственные. Было также
замечено, что гибридное потомство имеет внешний вид, про-
межуточный по сравнению с родителями. Это и создавало
впечатление смешивания признаков, а значит, и наследствен-
ного материала. Хотя подкрепленная многочисленными на-
блюдениями теория слитной наследственности и была обще-
признанной, тем не менее она не могла объяснить некоторые
факты. Так, иногда у чистопородных животных рождались
потомки с необычными признаками, например, коротконо-
гий ягненок или бесхвостый котенок. Позднее таких живот-
ных стали называть мутантами. Мутантные признаки стой-
ко наследовались на протяжении многих поколений. С таки-
ми же фактами сталкивались и растениеводы. Примеры
стабильного наследования редких признаков известны и у че-
ловека.
Мендель, работая с чистыми линиями гороха, т.е. такими,
у которых в течение нескольких поколений проявлялись одни
и те же константные признаки, сформулировал и опублико-
вал в 1866 г. ряд положений, известных как законы Менделя:
16
1.1. Основные понятия
закон доминирования, закон расщепления и закон независи-
мого наследования признаков.
Работы Грегора Менделя и его последователей начала XX в.
заложили фундамент, на котором выстроилась генетика —
наука о наследственности и изменчивости. Официальной да-
той зарождения генетики как науки считается 1900 г., когда
законы Менделя были независимо друг от друга переоткры-
ты сразу тремя учеными — австрийцем Чермаком, немцем
Корренсом и голландцем де Фризом.
Сегодня объектом генетики являются самые разные орга-
низмы — от вирусов до человека. Предметом генетического
анализа стали разнообразные признаки — от морфологичес-
ких до поведенческих. Генетический анализ ведется на моле-
кулярном, организменном и популяционном уровне.
1.1. Основные понятия
Содержание важнейших понятий классической генетики —
наследственность, ген, признак, генотип, фенотип и других
раскрывается в следующих определениях.
Наследственность — это свойство организма сохранять
и передавать какой-либо признак от родителя потомку.
Изменчивость — это свойство организма приобретать ка-
кие-либо новые признаки, отличные от родительских.
Ген — участок ДНК, который определяет синтез клеткой
какого-то белка или контролирует активность другого гена.
Аллель — одна из двух или более форм рассматриваемого
гена, находящегося в определенном месте хромосомы, или ло-
кусе. Если ген существует в форме двух аллелей, то речь идет
о диаллельной системе. Если ген существует в форме более
двух аллелей, то тогда говорят о серии множественных алле-
лей или о множественном аллелизме. Как правило, аллели
одного гена обозначают латинской буквой или одним симво-
лом, доминантный обозначается большой буквой, а рецессив-
ный — маленькой (например Айа). Если мутантных состоя-
ний несколько, то используют символы, стоящие вверху обо-
Раздел 1. Основы классической генетики
значения гена (например, 1А и Р). За нормальный признак при-
нимают такой, который свойствен большинству особей в по-
пуляции. Мутантных состояний может быть одно или несколь-
ко. Например, при дальтонизме аллель, определяющий это
заболевание, считается мутантным, а аллель нормального зре-
ния — нормальным. Пример обозначений для диаллельной
системы — группа крови резус, где имеются два аллеля Rh и
rh. Группа крови АВО является примером множественного
аллелизма, где выделяют три аллеля IA, Р и i°.
Гомозигота — организм, у которого имеются одинаковые
копии рассматриваемого гена. Например, гомозиготный че-
ловек с резус-положительной группой крови обозначается
как RhRh.
Гетерозигота — организм, у которого имеются различные
копии рассматриваемого гена. Например, гетерозиготный
человек с резус-положительной группой крови обозначается
как Rhrh.
Генотип — набор всех генов организма.
Фенотип — внешнее проявление генотипа. Часто при раз-
ных генотипах фенотип проявляется одинаково. Например,
гомозиготный генотип RhRh и гетерозиготный генотип Rhrh
фенотипически проявляются резус-положительной группой
крови.
Моногибридное скрещивание — это скрещивание, в кото-
ром анализируется один признак. Например, у человека изу-
чается только наследование цвета глаз.
Полигибридное скрещивание — это скрещивание, в кото-
ром анализируется несколько признаков. Оно может быть ди-
гибридным (одновременно изучается два признака, напри-
мер, цвет глаз и цвет волос), тригибридным (одновременно
изучается три признака, например, цвет глаз, цвет волос и
способность ощущать запах мускуса) и т.д.
Доминантный аллель — аллель, который маскирует выра-
женность другого аллеля. Например, аллель ахондроплазии
(вид карликовости) является доминантным по отношению к
аллелю, определяющему нормальное развитие скелета. Так,
гетерозигота по ахондроплазии имеет аномальный скелет.
Понятно, что для проявления доминантного признака доста-
точно наличие хотя бы одного доминантного аллеля.
Рецессивный аллель — аллель, фенотипическое проявле-
ние которого маскируется другим аллелем. Так, для проявле-
ния рецессивного признака необходимо наличие у человека
обоих рецессивных аллелей, т.е. состояние гомозиготности
по данному аллелю. Например, альбинизм у человека — ре-
цессивный признак, проявляющийся лишь у рецессивных го-
мозигот.
Аутосома — любая из хромосом человека, кроме половой.
Половые хромосомы — это пара хромосом, различающая-
ся у представителей разных полов.
Аутосомный признак — признак, за который отвечает ген,
локализованный в аутосоме. Например, аутосомные призна-
ки — альбинизм, фенилкетонурия.
Сцепленный с полом, или Х-сцепленный, признак — при-
знак, находящийся под контролем гена Х-хромосомы. Напри-
мер, сцепленные с полом признаки — дальтонизм, гемофи-
лия, мышечная дистрофия Дюшенна.
Кариотип — это хромосомный набор. В норме в кариотип
человека входят 44 аутосомы и 2 половые хромосомы (XX —
у женщин, XY — у мужчин). В кариотипе все хромосомы пар-
ные или гомологичные (кроме Y-хромосомы, для которой нет
гомолога).
Гаплоидный набор хромосом — один экземпляр гомологич-
ных хромосом, обозначаемый как п. У человека гаплоидный
набор хромосом составляет 23, из которых 22 аутосомы и
1 половая хромосома. Гаплоидный набор хромосом характе-
рен для половых клеток — яйцеклеток и сперматозоидов.
Диплоидный набор хромосом — два экземпляра гомологич-
ных хромосом, обозначается 2п. У человека диплоидный на-
бор хромосом составляет 46, из которых 44 аутосомы и 2 по-
ловые хромосомы. Диплоидный набор хромосом характерен
для соматических (не половых) клеток.
Геном — совокупность генов в гаплоидном наборе.
19
Раздел 1. Основы классической генетики
Процесс передачи наследственного материала от родите-
лей потомкам называется наследованием. Закономерности на-
следования изучают с помощью генетического анализа —
совокупности методов, основанных на сравнении родствен-
ных организмов. Главный метод генетического анализа — гиб-
ридологический, или метод скрещиваний. Гибридологический
метод разработал Г. Мендель. Сущность менделевского подхо-
да к изучению наследственности состоит в следующем. Из
множества признаков, по которым различаются представите-
ли одного вида, для анализа отбирается их ограниченное коли-
чество (один-два, реже три). Для скрещивания берется родитель-
ская пара, которая различается по анализируемым признакам.
В ряду поколений ведут количественный учет потомков.
Для записи скрещиваний и их результатов используют спе-
циальные символы. Родительское поколение обозначают бук-
вой Р, буквой F обозначают потомство. Цифровой индекс при
букве F соответствует номеру гибридного поколения: F, F.
Записывая скрещивание, на первое место обычно ставят жен-
ский пол^1, зеркало богини Венеры, материнский организм),
на второе — мужской щит и копье бога войны Марса, от-
цовский организм). Скрещивание обозначают символом х.
Потомок от родителей, имеющих разную наследственность,
является гибридом.
Скрещивания, в которых материнский и отцовский орга-
низмы берутся с различными вариантами признака белая х
$ черный и черная х белый), называются реципрокными,
или взаимно обратными.
При анализе скрещиваний удобно пользоваться решеткой
Пеннета. Ее описание будет дано несколько позже.
Изучение наследования у человека начинается с состав-
ления родословной. Родословная — это графическое изобра-
жение родственных отношений. На родословной членов од-
ного поколения располагают на одном уровне. Старшие по-
коления размещают выше молодых. Поколения нумеруют
сверху вниз римскими цифрами. Членов одного поколения
20
1.2. Моногенное наследование
нумеруют слева направо арабскими цифрами. При таком обо-
значении каждый член родословной получает свой код — ряд/
место, или так называемые генеалогические координаты. Че-
ловек, для которого составляют родословную, называется
пробанд, на родословной его обозначают стрелкой. Индиви-
ды с альтернативными признаками обозначаются разным
цветом или штриховкой.
При составлении родословной используют символы, пред-
ставленные на рис. 1.2.
женщина
мужчина
женщина с изучаемым признаком
мужчина с изучаемым признаком
Рис. 1.2. Символика родословных
1.2. Моногенное наследование
Полное доминирование
На примере группы крови системы резус у человека рассмот-
рим, как происходит наследование одного признака. По это-
му признаку люди делятся на две группы: резус-положитель-
ные (Rh) и резус-отрицательные (rh). На эритроцитах резус-
21
Раздел 1. Основы классической генетики
положительных индивидов имеется белковый фактор, назы-
ваемый резус. У резус-отрицательных людей такого белка нет.
Различия по резус-группе зависят от гена, который находит-
ся в первой хромосоме. Аллель Rh обеспечивает присутствие
в крови группоспецифического вещества, аллель rh — его от-
сутствие. Аллель Rh является доминантным, rh — рецессив-
ным. Резус-положительные индивиды могут быть как гомо-
зиготными RhRh, так и гетерозиготными Rhrh, резус-отрица-
тельные всегда гомозиготны — rhrh.
Рассмотрим, какие группы крови возможны у потомков в
разных семьях. В записи скрещивания укажем генотипы ро-
дителей и генотипы образуемых ими гамет. Гаметы гаплоид-
ны, в половую клетку попадает только один ген. У гомозигот-
ной особи формируется один тип гамет, у гетерозиготной —
два типа гамет в соотношении 1:1.
Все потомки резус-положительных гомозиготных родите-
лей резус-положительны:
Р: RhRh Ч RhRh
Гаметы: Rh
F,: RhRh
Все потомство резус-отрицательных родителей имеет ре-
зус-отрицательную группу:
Р: rhrh Ч rhrh
Гаметы: rh
F(: rhrh
Потомки гомозиготных резус-положительных и резус-от-
рицательных родителей гетерозиготны по генотипу и резус-
положительны:
Р: RhRh Ч rhrh
Гаметы: Rh, rh
Fp Rhrh
Потомство гетерозиготных родителей расщепляется по ге-
нотипу в отношении 1:2:1, а по фенотипу в отношении 3:1:
Р: RhRh Ч RhRh
Гаметы: Rh, rh
Fjt 1/ARhRh, l/2Rhrh, 1/4 rhrh
22
1.2. Моногенное наследование
В семьях, где один из родителей резус-положительная ге-
терозигота, а второй резус-отрицательный, половина потом-
ков резус-положительна и половина резус-отрицательна:
Р: Rhrh Ч rhrh
Гаметы: Rh, rh
F,: l/2Rhrh, 1/2 rhrh
Для того чтобы определить генотип особи, имеющей до-
минантный признак, используют анализирующее скрещива-
ние. В этом случае анализируемую особь скрещивают с ана-
лизатором — рецессивной гомозиготой. Если в потомстве от
анализирующего скрещивания появляются особи с рецессив-
ным признаком, значит, анализируемый родитель был гете-
розиготным.
Допустим, в семье один из супругов резус-положительный,
а второй резус-отрицательный. Генотип резус-отрицательно-
го индивида rhrh, генотип резус-положительного неизвестен
и записывается в виде фенотипического радикала — Rh-. Фе-
нотипический радикал — это та часть генотипа, которая про-
является в фенотипе. В данном случае при полном доминиро-
вании генотипы RhRh и Rhrh в фенотипе проявляются одина-
ково, как признаки аллеля Rh. Фенотипическим радикалом
является Rh-. Если в такой семье рождается резус-отрицатель-
ный ребенок, это означает, что родитель с доминантным при-
знаком был гетерозиготным.
Неполное доминирование
В этом случае гетерозигота занимает промежуточное поло-
жение между доминантной гомозиготой и рецессивной гомо-
зиготой. У человека по типу неполного доминирования насле-
дуется гиперхолестеринемия (повышенный уровень холесте-
рина в крови). Гомозигота по нормальному аллелю дикого
типа АА имеет нормальное число рецепторов в клетках пече-
ни для холестерина, приносимого кровью. У гетерозиготы Аа
таких рецепторов вдвое меньше, а у рецессивной гомозиго-
ты аа их вообще нет. Рецессивные гомозиготы умирают в ран-
23
Раздел 1. Основы классической генетики
нем детстве от сердечного приступа, гетерозиготы умирают
по той же причине, но в юности. Сердце доминантных гомо-
зигот, имеющих два нормальных или диких аллеля, здорово.
По типу неполного доминирования человек наследует также
форму волос: гомозиготы имеют прямые (А;А;) либо курча-
вые (А.Д,) волосы, а у гетерозигот (A^J волосы волнистые.
В случае неполного доминирования по фенотипу особи мож-
но точно определить генотип и необходимость в анализиру-
ющем скрещивании отпадает.
Кодоминирование
При данном типе аллельного взаимодействия генов в фено-
типе гетерозиготы проявляются два признака, то есть присут-
ствуют белки двух молекулярных форм. Наиболее известный
пример кодоминирования у человека —- генетический конт-
роль групп крови системы АВО. Принадлежность человека к
этой группе обусловлена присутствием на его эритроцитах
веществ белковой природы А и/или В. За принадлежность
индивида к той или иной группе крови отвечает аутосомный
ген I, для которого известно три аллельных варианта: i°, 1А, 1В.
Аллели 1А и Is ответственны за выработку соответственно фак-
торов А или В. Аллель i°является «молчащим» и рецессивным
по отношению к аллелям 1Аи 1В. Генотипы РР и /А/А фенотипи-
чески не различимы и соответствуют второй (II) группе кро-
ви (она называется также группа А). У индивидов с генотипа-
ми IBi° и РР третья (III) группа крови (другое название — груп-
па В). Генотип i°i° формирует первую группу (I) крови, она же
нулевая — 0. Аллели 1А и Р по отношению друг к другу кодо-
минантны. Генотип 1А1В соответствует четвертой (IV) группе
крови, или АВ. В данном случае при сочетании двух различ-
ных аллелей возникает новый признак. По типу кодомини-
рования у человека наследуются также группы крови систе-
мы MN. Люди с группой крови М имеют генотип ММ, с груп-
пой крови N — генотип NN и с группой крови MN — генотип
MN.
24
1.2. Моногенное наследование
Решетка Пеннета
Для удобства учета возможных комбинаций признаков у по-
томства пользуются решеткой Пеннета. Решетка Пеннета
представляет собой графы, в которых отражено сочетание
генов отца и матери у предполагаемого потомства. С помо-
щью решетки Пеннета можно вычислить вероятность рожде-
ния потомка с тем или иным признаком. По вертикали записы-
ваются гаметы отца, а по горизонтали — матери. Сочетание
отцовских и материнских гамет указывается непосредствен-
но в графах таблицы. Так, у отца с группой крови АВ и у мате-
ри с группой крови 0 по системе АВО возможно рождение
детей с такими генотипами:
.0 1
1А ТА -0 1 1
1в ТВ-0 11
Видно, что при таком браке возможно рождение детей с
генотипами IAi° и Pi0 с равными вероятностями. В данном слу-
чае число возможных генотипов совпадает с числом возмож-
ных фенотипов. Если оба родителя имеют группу крови АВ,
то у них возможно рождение детей со следующими геноти-
пами:
1А1А 1А1В
1в 1А1В 1в1в
В таком браке могут родиться дети с тремя различными
генотипами 1А1А, РР и 1АР и фенотипами А, В и АВ. Вероятность
рождения ребенка с группой крови А составляет 1/4, с груп-
пой крови АВ — 1/2 и с группой крови В — 1/4,
25
Раздел 1. Основы классической генетики
1.3. Независимое наследование
Рассмотрим у человека два признака — цвет волос и структу-
ру волос. Признак «рыжие волосы» является рецессивным по
отношению к другим окраскам волос. Признак «курчавые
волосы» является доминантным по отношению к признаку
«прямые волосы», а признак «волнистые волосы» является
промежуточным и проявляется у гетерозигот. Таким образом,
окраска волос наследуется по типу полного доминирования
(а — аллель рыжих волос, А — аллель нерыжих волос), а струк-
тура волос — по типу неполного доминирования (В — аллель
курчавых волос, b — аллель прямых волос). Допустим, роди-
тели гетерозиготны по обоим генам. При мейозе у каждого
родителя формируется 4 типа гамет: АВ, АЬ, аВ и ab. Запишем
возможные сочетания родительских гамет в виде решетки
Пеннета:
АВ АЬ аВ ab
АВ ААВВ ААВЪ АаВВ АаВЬ
АЬ ААВЬ Aabb АаВЬ Aabb
аВ АаВВ АаВЬ ааВВ ааВЬ
ab АаВЪ Aabb ааВЬ aabb
Проанализируем возможные генотипы потомства отдель-
но по каждому признаку:
S цвет — 4 АА :8 Аа : 4 аа, или 1 АА :2 Аа :1 аа (3 не рыжих :
1 рыжий);
S структура — 4 ВВ : 8 Bb :4 bb, или 1 ВВ : 2 Bb: 1 bb (1 прямой :
2 волнистых : 1 курчавый).
Чтобы вычислить вероятность рождения ребенка с опре-
деленным сочетанием признаков, необходимо перемножить
многочлены фенотипов: (3 нерыжих : 1 рыжий) х (1 курчавый :
2 волнистых : 1 прямые) = 3 нерыжих курчавых : 6 нерыжих
волнистых : 3 нерыжих прямых : 1 рыжий курчавый : 2 ры-
жих волнистых : 1 рыжий прямой.
26
1.3. Независимое наследование
Среди потомков второго поколения обнаруживаются лица,
генотип которых напоминает родительский, а также лица с
новым сочетанием аллелей. Таким образом, в результате по-
лового размножения (скрещивания) возрастает генетическое
разнообразие. Новое сочетание признаков у потомков, полу-
чаемое в результате скрещивания, называется комбинатив-
ной изменчивостью.
В общем случае число генотипов при полигибридном скре-
щивании равно Зп, где п — число пар гетерозиготных генов.
Так, для моногибридного скрещивания Аа х Аа число возмож-
ных генотипов у потомства равно З1 = 3, т.е. 1 АА: 2Аа : 1 аа —
3 возможных генотипа. При дигибридном скрещивании, на-
пример, АаВЬ х АаВЬ, число генотипов потомков равно З2 = 9,
а при скрещивании ААВЬ х АаВЬ, число генотипов потомков
равно З1 = 3, т.к.:
АВ АЬ
АВ ААВВ ААВЬ
АЬ ААВЬ ААЬЬ
Число фенотипов при полном доминировании равняется
2п, где п — число пар гетерозиготных генов. Так, при скрещи-
вании Аа ? Аа, число фенотипов у потомства 21 = 2, т.к.:
А а
А АА Аа
а Аа аа
При скрещивании АаВЬ х АаВЬ, число фенотипов для по-
томства составит 22 = 4, т.к. (3 А : 1 а) х (3 В : 1 Ь) = 9 АВ : 3 АЬ:
3 аВ : 1 ab.
При неполном доминировании и кодоминировании число
фенотипических классов у потомства равняется числу гено-
типических классов, т.е. Зп, где п — число пар гетерозигот-
ных генов.
27
Раздел 1. Основы классической генетики
Зная закономерности наследования отдельных признаков,
можно рассчитать вероятность унаследования различных со-
четаний.
Допустим, у жены вторая группа крови, у мужа третья,
кроме того, они резус-положительны. Их резус-отрицатель-
ный сын имеет первую группу крови. Возникает вопрос, дети
с какой еще группой крови могут родиться у этих супругов и
с какой вероятностью?
Фенотипы членов семьи:
P:$IIRhx$IIIRh
F.r^Irh
Им соответствуют генотипы:
Р: $ IA— Rh— х /1В— Rh—
F^ i°i° rhrh
По генотипу потомка определяем генотипы родителей.
Муж и жена гетерозиготны по обоим генам:
Р: lAi° Rhrh х ^IBi° Rhrh
У них могут быть потомки с такими генотипами (феноти-
пами):
Р: $ IAi° х IBi° -> F(: 1/4 IAIB (IV), 1/4 IAi° (II), 1/4 p!°(III),
1/4 ;V(I)
P: $ Rhrh x $ Rhrh -> F,: 3/4 Rh— (Rh), 1/4 rhrh (rh)
Анализируемые признаки свободно комбинируются, так
как ген резус-фактора находится в первой хромосоме, а ген
группы АВО — в шестой.
Зная вероятность появления в потомстве отдельного при-
знака, можно рассчитать вероятность их различных сочетаний.
Так, вероятность рождения резус-положительного ребенка с
четвертой группой крови представляет собой произведение
вероятностей наследования каждого из этих признаков по
отдельности: 3/4 (Rh) х 1/4 1AIB (IV) — 3/16 RhTV.
1.4. Сцепление и кроссинговер
У живых организмов число генов превышает число хромосом.
Гены, расположенные в одной хромосоме, называются сцеп-
28
1.4. Сцепление и кроссинговёр
ленными и образуют группу сцепления. Число групп сцепле-
ния равно количеству хромосом в гаплоидном наборе. Сцеп-
ленные гены наследуются вместе, но могут разойтись в ре-
зультате кроссинговера.
Кроссинговер — это обмен участками между гомологич-
ными хромосомами. Он происходит в мейозе, когда гомоло-
гичные хромосомы, перед тем как разойтись в разные клет-
ки, обмениваются участками. Относительное расстояние
между генами находят по формуле:
количество кроссоверных особей . ппо/
-----—-------------------—s-----X 1UU/6.
общее количество особей
Это расстояние обозначается в процентах перекреста, и
1% перекреста в честь американского генетика Томаса Мор-
гана называется морганидой.
Кроссоверные особи имеют новое сочетание признаков.
Сцепленные гены могут быть в двух фазах (рис. 1.3):
1) цис-положение — два гена, сцепление которых определя-
ют, находятся в одном гомологе;
2) транс-положение — два гена находятся в разных гомоло-
гах.
АВ А b
-------•-----• « »..
------------------------------------------------•—-•———— -«-- »
а b---------------------------------------------а В
Гены А и В в цис-положении Гены А и В в транс-положении
Рис. 1.3. Фазы сцепления генов
Существуют несколько методов выявления сцепленности
генов. Один из них — метод сдвоенных сибсов Пенроза. При
этом анализируется сибство (потомки) из нескольких семей
или из одной большой семьи по двум каким-то признакам.
Всех сибсов (братьев и сестер) разбивают на четыре группы.
В первую группу входят сибсь(, сходные по обоим признакам,
во вторую группу — сходные по первому и не сходные по вто-
рому признакам, в третью группу — не сходные по первому и
сходные по второму признакам и в четвертую группу — не
к м
Раздел 1. Основы классической генетики
сходные по обоим признакам. Далее методом %2 устанавлива-
ют, существует ли связь между двумя признаками. Затем де-
лают вывод о сцепленности генов, лежащих в основе изучае-
мых признаков.
Так, у человека одна из аномалий скелета — хондродист-
рофия — наследуется как аутосомно-доминантный признак.
Чтобы проверить предположение, что ген, обусловливающий
эту аномалию, сцеплен с локусом, ответственным за группу
крови системы АВО, применим к данной ситуации метод Пен-
роза. В нашем распоряжении имеется родословная, представ-
ленная на рис. 1.4. 1 2
О В
Рис. 1.4. Родословная
Анализ потомства позволяет выяснить генотипы родите-
лей (1/1,2). Если обозначить ген хондродистрофии Н, его нор-
мальный аллель h, а аллели групп крови 0 и В как i° и Р соот-
ветственно, то, по-видимому, структура генотипа у 1/1 гено-
тип i°h/i°h, а у 1/2 генотип i°h/PH. Методом сдвоенных сибсов
Пенроза проверим наличие сцепления между аллелями Н и Р.
Среди сибсов фенотипы распределились так: 3 человека —
группа крови В и аномалия, 4 человека — группа крови 0 без
аномалии и 1 человек с группой крови 0 с аномалией. Попар-
ное сопоставление сибсов (всего 28 сопоставлений) дало та-
кой результат: 9 пар сходны по обоим признакам, 4 пары сход-
ны по первому и не сходны по второму признаку, 3 пары не
сходны по первому и сходны по второму признаку и 12 пар не
сходны по обоим признакам:
30
l.i>. Сцепленное с полом наследование
1 сибс 2 сибс сходные несходные
сходные 9 4 13
несходные 3 12 15
S 12 16 28
t = 5,03, df - 1, р < 0,05.
Таким образом, гипотеза о сцеплении верна, а фенотип
П/4 — результат кроссинговера.
Генетики используют связь между частотой кроссингове-
ра и расстоянием между генами для построения хромосом-
ных карт. Хромосомная карта — это схема, на которой пред-
ставлен порядок расположения генов на хромосоме. По доле
кроссоверных потомков устанавливают частоту кроссинго-
вера между двумя генами. Чем дальше расположены гены на
хромосоме, тем больше вероятность кроссинговера для этих
генов, и, наоборот, чем ближе, тем меньше вероятность раз-
рыва хромосомы между ними и обмена участками гомологич-
ных хромосом.
1.5. Сцепленное с полом наследование
Х-сцепленное наследование
Любой ген в Х-хромосоме мужчины проявляется в феноти-
пе, поскольку отсутствует второй аллель данного гена, кото-
рый маскировал бы его. Аллель в Х-хромосоме женщины мо-
жет проявляться, а может и не проявляться — это зависит от
jproro, доминантный он или рецессивный, и какова природа
второго аллеля. Женщина может быть гомозиготной (ХАХА,
ХаХа) или гетерозиготной (ХАХа). Мужчина является гемизи-
готой (XAY и XaY).
| Мужчина наследует Y-хромосому от отца и Х-хромосому
|от матери. Женщина наследует по одной Х-хромосоме от каж-
[дого родителя. Если мать является гетерозиготой по сцеплен-
|ному с полом гену, то ее сын имеет вероятность 1/2 унаследо-
вания любого из ее двух аллелей.
31
Раздел 1. Основы классической генетики
Наиболее известный пример рецессивного Х-сцепленно-
го наследования — генеалогическое дерево правящих динас-
тий Европы, восходящих к английской королеве Виктории.
В этой родословной встречается гемофилия. Гемофилия — бо-
лезнь, при которой замедлена свертываемость крови из-за от-
сутствия особого белка, который называется фактор сверты-
вания крови. Среди предков Виктории гемофилия не встре-
чалась, поэтому, по одной из версий, предполагается, что
аллель, ответственный за гемофилию, возник в результате му-
тации в гамете одного из родителей Виктории.
Закономерности наследования Х-сцепленных признаков
можно рассмотреть на примерах. Так, если мать — носитель
гена гемофилии (XHXh), а отец здоров (XHY), то у них могут
родиться дети:
У'
Л* У/У
У У'у Уу
Все дочери будут здоровы, а вероятность рождения боль-
ного или здорового сына одинакова, и равна 1/2.
Можно рассмотреть другой случай (рис. 1.5), когда у жен-
щины брат-гемофилик, здоровые родители, здоровый муж, и
она хочет знать вероятность рождения у нее больного ребен-
ка.
Рис. 1.5. Фрагмент родословной с Х-сцепленным
рецессивным заболеванием
32
1.5. Сцепленное с полом наследование
Изучив данный случай, можно установить, что мать жен-
щины гетерозигота, поскольку передала дефектный ген сыну.
Сама женщина могла получить с равной вероятностью 1 /2 ген
гемофилии от матери, и, следовательно, передать его своему
будущему сыну с вероятностью 1/2. Поэтому, риск рождения
больного сына составит 1/2х1/2 = 1/4.
Обычно женщины-носительницы не обнаруживают при-
знаков рецессивных болезней, сцепленных с Х-хромосомой.
Наследование Х-сцепленных доминантных генов проис-
ходит сходным образом с аутосомно-доминантным наследо-
ванием. Для проявления признака необходимо наличие хотя
бы одного аллеля у женщин и одного аллеля у мужчин. При-
мером доминантного Х-сцепленного признака у человека яв-
ляется коричневая окраска зубной эмали.
Y-сцепленное наследование
При Y-сцепленном наследовании признак передается от отца
к сыну, а ген такого признака находится в Y-хромосоме. На
рис. 1.6 приведена родословная с Y-сцепленным наследова-
нием.
Рис. 1.6. Родословная с Y-сцепленным наследованием
Классическим примером является признак «волосатые
уши» (гипертрихоз ушной раковины) у некоторых мужчин
Шри-Ланки, Индии, Израиля и аборигенов Австралии (рис. 1.7).
Рис. 1.7. «Волосатые уши» у человека
1.6. Наследование полигенных признаков
Наследование многих признаков не описывается схемой мо-
ногибридного скрещивания, так как они зависят от несколь-
ких генов (полигенный контроль). При этом наблюдаются нео-
бычные численные соотношения фенотипов, например, 15:1,
12:3:1, 9:3:4, 9:7, 9:6:1, 1:4:6:4:1. Эти соотношения являются
модифицированной формулой скрещивания, в котором рас-
щепляются две пары генов (9:3:3:1) и обусловлены тем, что
некоторые генотипы не различаются фенотипически.
Комплементарность — это такое взаимодействие генов,
при котором два или более гена вызывают развитие нового
признака. Например, синтез интерферона у человека зави-
сит от двух генов, один из которых находится во второй хро-
мосоме (ген А), а другой — в пятой хромосоме (ген В). Ин-
терферон будут синтезировать только те индивиды, у ко-
торых имеются оба доминантных аллеля. Так, у двух гетеро-
зиготных родителей (АаВЬ) возможно рождение такого потом-
ства:
Р: QAaBb х ААаВЬ
F,: (ЗА : 1а-) х (ЗВ- : 1b) = 9А-В- : ЗА-b- : За-В- : 1а-Ъ-
(по генотипу) и 9 :7 (по фенотипу)
Вероятность рождения ребенка, способного синтезиро-
вать интерферон, 9/16, а неспособность его синтезировать —
7/16.
34
1.6. Наследование полигенных признаков
Полимерией называется такой тип взаимодействия неал-
лельных генов, при котором гены действуют на признак оди-
наково. Известна некумулятивная полимерия (гены действу-
ют на признак одинаково, их действие не суммируется) и ку-
мулятивная (гены действуют на признак одинаково, их
действие суммируется).
По типу кумулятивной полимерии у человека наследует-
ся пигментация кожи. Генетические исследования показали,
что разнообразие людей на земле по цвету кожи зависит от
четырех-пяти пар генов. Цвет кожи определяется количе-
ством в ней пигмента меланина. Для простоты допустим, что
цвет кожи находится под контролем двух пар генов А;(а;) и
А2(а2). Доминантные аллели этих генов активны в отношении
синтеза меланина, их рецессивные аллели нет. Аллели Af и А2
обладают аддитивным (суммирующим) эффектом: количе-
ство пигмента зависит от числа этих аллелей в генотипе. При
накоплении доминантных аллелей увеличивается интенсив-
ность окраски. При отсутствии доминантных аллелей (а;а;а2а2)
формируется белая кожа. Наличие одного (любого) доминант-
ного гена (А;а;а2а2 или а ,0^0^ обеспечивает светлую окрас-
ку кожи, двух (А!А1а2а2плпА1а1А2а2) — смуглую, трех (АуАД^
или Apfl^) — темную, четырех (А/кДД.) — черную.
Считается, что способность к лактации у женщин опре-
деляется двумя полимерными генами Ц и L2. При генотипе
ЦЦ у женщин образуется оптимальное количество моло-
ка, которое необходимо ребенку, при генотипе 11111:J2 молока
не образуется вообще, при генотипе L1L1L2L2 молока образу-
ется слишком много.
Эпистаз — это подавление аллелью одного гена аллелей
другого гена. Другими словами, это своеобразное взаимодей-
ствие доминантного и рецессивного генов, но не на уровне
аллелей, а на уровне разных генов. Выделяют доминантный и
рецессивный эпистаз. При доминантном эпистазе В < А > bb,
фри рецессивном эпистазе В < аа > bb.
При эпистазе в потомстве двух гетерозигот также наблю-
дается модификация менделевского соотношения 9:3:3:1 —
может быть 13:3, 12:3:1 и 9:3:4.
Раздел 1. Основы классической генетики
Рис. 1.8. Альбинизм у человека и у мыши
У человека примером эпистатического наследования яв-
ляется так называемая «бомбейская» группа крови. «Бомбей-
ский» фенотип является результатом взаимодействия гена I
(система группы крови АВО) и гена Н. Аллель Н отвечает за
синтез фермента, который присоединяет молекулу сахара к
особому гликопротеину на поверхности эритроцита. Затем
антигены А и В присоединяются к этому сахару. Рецессив-
ный аллель h вырабатывает неактивную форму этого фермен-
та, который не может присоединить сахар. Если присутству-
ет хотя бы один аллель Н, то группа крови человека определя-
ется генотипом по системе АВО. Но у человека с генотипом hh
антигены А и В не могут присоединиться к поверхности эрит-
роцита. Поэтому у таких людей при определении группы кро-
ви по системе АВО всегда нулевая группа крови, хотя генотип
может быть любой (А, В, АВ и 0). «Бомбейская» группа крови
является примером рецессивного эпистаза — IA, IB < hh > И.
На примере цвета кожи также можно рассмотреть эпис-
таз. У человека известен признак, называемый альбинизмом
(рис. 1.8). Альбиносы встречаются среди представителей разных
рас с любым цветом кожи. Рецессивный ген с подавляет выра-
36
Контрольные вопросы и задания к разделу 1
ботку меланина, и если человек гомозиготен по этому гену (сс),
то независимо от того, какое количество доминантных генов,
ответственных за синтез меланина, он имеет (— — — — сс), ок-
раска кожи у него будет альбинотической.
Контрольные вопросы и задания к разделу 1
1. Почему Грегора Менделя считают основателем генетики?
2. Изложите основные положения теории слитной наслед-
ственности. Какие факты эта теория не могла объяснить?
3. Какая связь существует между генотипом и фенотипом?
4. Всегда ли между генотипом и фенотипом существует од-
нозначное соотношение?
5. Чем отличается наследование аутосомных признаков от
признаков, сцепленных с полом?
6. Какие задачи решаются при использовании реципрокных
скрещиваний?
7. Для каких целей используют анализирующее скрещива-
ние?
8. Приведите примеры доминирования и неполного домини-
рования у человека.
9. Перечислите рецессивные признаки у человека.
10. Какие признаки наследуются кодомининантно?
11. На основании ряда фенотипических признаков, наследу-
емых по аутосомной моногенной диаллельной схеме, ус-
тановите свой генотип. В скобках приведены обозначения
для доминантных и рецессивных аллелей.
Способность языка сворачиваться в трубочку (принимать
U-образную форму), доминантный признак (R и г):
Раздел 1. Основы классической генетики
Прикрепление мочки уха,
рецессивный признак (D и d):
Веснушки на лице, доминант-
ный признак (F и f):
Ямочки на щеках, доминант-
ный признак (Dud):
«Вдовья» линия волос, доми-
нантный признак (W и w):
12. У Веры и Андрея аутосомно-рецессивное заболевание.
Укажите вероятность носительства аутосомно-рецессив-
ного гена у членов родословной, изображенной на рис. 1.9.
13. У человека ген карликовости доминирует над геном нор-
мального роста. Мужчина нормального роста вступил в
брак с женщиной карликового роста, отец которой был
нормального роста. Какова вероятность рождения ребен-
ка карлика у этих супругов ?
14. Отсутствие эмали на зубах человека наследуется как ауто-
сомно-доминантный признак. У мужа нормальные зубы,
а у жены зубы без эмали. У ее отца были нормальные зубы.
Может ли их будущий ребенок унаследовать этот дефект?
15. Муковисцидоз — нарушение углеводного обмена, насле-
дуется по аутосомно-рецессивному типу. У здоровых суп-
ругов родился здоровый ребенок и ребенок, больной му-
ковисцидозом. Какова вероятность, что следующий ребе-
нок будет болен?
38
Контрольные вопросы и задания к разделу 1
Рис. 1.9
I
II
III
IV
V
16. Жена резус-отрицательна. Ее муж резус-положителен. Их
первый ребенок резус-отрицателен и здоров. Какова ве-
роятность, что следующие дети будут резус-положитель-
ными и поражены гемолитической болезнью из-за резус -
конфликта между резус-отрицательной матерью и резус-
положительным плодом?
17. Парагемофилия — аутосомно-рецессивное заболевание,
при котором имеется склонность к кожным и носовым
кровотечениям. В консультацию обратились супруги Ан-
тон и Жанна, у каждого из которых был брат с таким же
заболеванием. Каков риск рождения у них больного ре-
бенка (рис. 1.10)?
Рис. 1.10
39
Раздел 1. Основы классической генетики
18. У Виктории врожденная нечувствительность боли, кото-
рая наследуется по аутосомно-рецессивному типу (рис.
1.11). По этой причине у нее часто происходят несчастные
случаи, поэтому из-за травм ее тело, а особенно руки, изу-
родованы множеством шрамов. У Виктории есть троюрод-
ная сестра Анастасия, которая опасается, что у ее ребенка
не будет защитного болевого ощущения. Какова вероят-
ность рождения у Анастасии такого ребенка, если она за-
мужем за Юрием, который является единокровным бра-
том Виктории?
Рис. 1.11
20. Какие признаки наследуются независимо?
21. Какое расположение генов в хромосомах ограничивает
комбинативную изменчивость?
22. Сколько групп сцепления у человека?
23. Приведите примеры сцепленного наследования у челове-
ка.
24. Темные волосы — доминантный признак, светлые — ре-
цессивный. Кудрявые волосы не полностью доминируют
над прямыми — у гетерозигот волнистые волосы. Темно-
волосая женщина с кудрявыми волосами вступила в брак
с мужчиной, имеющим темные гладкие волосы. Супруги
гетерозиготны по гену цвета волос. Гены, контролирую-
40
Контрольные вопросы и задания к разделу 1
щие цвет волос и их структуру, находятся в разных хро-
мосомах. Какие волосы могут быть у их детей?
25. Карие глаза — доминантный признак, голубые — рецес-
сивный. Темноволосая кареглазая женщина вступила в
брак со светловолосым кареглазым мужчиной. Женщина
гетерозиготна по гену цвета волос, мужчина — по гену
цвета глаз. Какие глаза и волосы возможны у их детей, если
гены, отвечающие за цвет глаз и цвет волос, находятся в
разных хромосомах?
26. У человека карие глаза доминируют над голубыми, а тем-
ные волосы над светлыми, причем гены, ответственные за
цвет глаз и волос, находятся в разных хромосомах. Голу-
боглазый брюнет, отец которого был блондином, женился
на кареглазой блондинке из семьи, все члены которой в
нескольких поколениях имели карие глаза. Какие глаза и
цвет волос будут у детей этих супругов ?
27. Что такое хромосомные карты?
28. В каких единицах выражают расстояние между генами на
хромосомной карте?
29. В родословной на рис. 1.12 обозначены случаи аутосомно-
доминантной патологии эллиптоцитоза — аномалии эрит-
роцитов, обусловленной нарушением структуры гемогло-
бина. Решите вопрос о возможности сцепленного насле-
дования гена эллиптоцитоза с генами группы крови
системы резус.
Рис. 1.12
41
Раздел 1. Основы классической генетики
30. По способности выделять вещества группы крови систе-
мы АВО в различные жидкости организма (слюну, желу-
дочный сок, мочу и т.д.) все люди делятся на «секретеров»
и «несекреторов». Способность к секреции обусловлена
доминантным аутосомным геном. Нервно-мышечная па-
тология — миотоническая дистрофия — аутосомно-доми-
нантное заболевание. Первые признаки заболевания появ-
ляются в 20—30 лет. Пенетрантность гена миотонической
дистрофии у гетерозиготных мужчин 100%, у гетерозигот-
ных женщин 64%. Локусы «секреторности» и миотоничес-
кой дистрофии сцеплены и находятся на расстоянии 10%
кроссинговера.
Детям Анжелы и Игоря (у него миотоническая дистрофия)
от 3 до 14 лет. Рассчитайте вероятность заболевания для
каждого из них. На родословной (рис. 1.12) у всех членов
семьи отмечено состояние «секреторности».
31. Локусы, в которых находятся гены пигментной ксеродер-
мы, пигментного ретинита и буллезного эпидермолиоза, у
человека расположены в Х-хромосоме. Частота рекомби-
нации между первым и третьим локусами составляет 6%,
между первым и вторым — 14%, между вторым и третьим —
8%. Определите относительный линейный порядок этих
локусов в Х-хромосоме.
32. Как передаются из поколения в поколение гены, локали-
зованные в Х-хромосоме?
42
Контрольные вопросы и задания к разделу 1
33. Как передаются в поколениях признаки, сцепленные с
Y-хромосомой?
34. У здоровых родителей пятеро детей. Два их сына Анато-
лий и Михаил больны гемофилией, сын Максим и две до-
чери Зинаида и Клавдия здоровы (рис. 1.13). Определите
вероятность заболевания у внуков.
35. Николай и его дядя по матери Геннадий по причине даль-
тонизма не могут водить машину — они смешивают крас-
ные цвета с серыми, желтыми и темно-зелеными. Опре-
делите вероятность того, что у дочери Геннадия Марины
и у его племянницы Инны (рис. 1.14) будет ребенок-даль-
тоник. Дальтонизм определяется рецессивным геном,
сцепленным с Х-хромосомой.
Марина
Рис. 1.14
36. В семье отец и сын — дальтоники, а мать различает цвета
нормально. От кого из родителей сын унаследовал ген даль-
тонизма?
37. Для орофациодигитального синдрома характерны такие
I признаки: расщепление челюсти и языка, синдактилия,
43
Раздел 1. Основы классической генетики
иногда умственная отсталость. Синдром наследуется как
доминантный сцепленный с Х-хромосомой признак. В ге-
мизиготном состоянии этот ген летален. Несмотря на яв-
ные физические уродства больные женщины из родослов-
ной на рис. 1.15 всегда находили себе мужей, т.к. владели
огромным состоянием, переходящим из поколения в по-
коление, что и прельщало потенциальных женихов. Ни у
кого из этих женщин не было сыновей. Антонина — пос-
ледняя представительница этого рода — здорова и заму-
жем за Сергеем. Каков риск, что ее дети будут больны?
Рис. 1.15
38. Какие численные соотношения фенотипов могут наблю-
даться при взаимодействии неаллельных генов?
39. Приведите примеры плейотропного действия генов у че-
ловека.
40. Интерферон — белок, образующийся в клетках организ-
мов при вирусных инфекциях. В клетках человека интер-
ферон синтезируется, если присутствуют одновременно
два доминантных аллеля разных генов — А и В (компле-
ментарное взаимодействие). Родители гетерозиготны по
генам А и В. Какова вероятность рождения детей, не спо-
собных синтезировать интерферон?
44
Контрольные вопросы и задания к разделу 1
41. Белок гемоглобин состоит из двух полипептидов — а и Р,
синтез которых контролируется генами, расположенны-
ми в негомологичных хромосомах. Какова форма взаимо-
действия этих генов?
42. У пятерых сестер — Александры, Клавдии, Эльвиры, Ве-
роники и Владиславы — в один год родилось по ребенку.
Наибольшие трудности при грудном вскармливании воз-
никли у Александры и Эльвиры. У Александры было очень
много молока, что приводило к застойным, а как следствие,
и воспалительным процессам в молочных железах, из-за
чего ей пришлось прекратить кормление ребенка грудью.
Эльвира столкнулась с другой проблемой — молоко после
рождения ребенка у нее так и не появилось, несмотря на
многочисленные советы и «диеты», способствующие по-
вышению лактации. Веронике приходилось докармливать
своего ребенка детской смесью, потому что малыш часто
оставался голодным. Ребенок Владиславы быстро набирал
в весе из-за обилия молока у матери, хотя его было все же
меньше, чем у ее сестры Александры. Лишь у Клавдии ко-
личество молока соответствовало потребностям ребенка.
Какие генотипы могли быть у пятерых сестер, если извес-
Г тно, что секреция грудного молока находится под контро-
лем, по меньшей мере, двух доминантных полимерных ге-
нов {Lt и L2) с кумулятивным действием?
43. Кровь больных, страдающих гемофилией, в стеклянной
' пробирке сворачивается гораздо медленнее, чем кровь
i здоровых людей. При обследовании мальчика, оказалось,
что его кровь характеризуется плохой свертываемостью.
Это дало основание предполагать, что он болен гемофи-
лией, сцепленной с Х-хромосомой. При смешивании крови
этого мальчика с кровью другого гемофилика, достоверно
страдающего Х-сцепленной формой гемофилии, наблюда-
лось быстрое образование сгустка, характерное для свер-
тывания крови нормального человека. Объясните явление.
>44. Допустим, что различия в цвете кожи у человека обуслов-
। лено в основном двумя парами независимо расщепляю-
ll 45
Раздел 1. Основы классической генетики
щихся генов. Гены взаимодействуют по типу кумулятив-
ной полимерии с межаллельным эффектом полудомини-
рования: ААВВ — черная кожа, aabb — белая. Сочетание
любых трех аллелей черной окраски обусловливают тем-
ный цвет, двух — смуглый. Присутствие в генотипе толь-
ко одного аллеля (А или В) формирует светлую кожу. Ка-
ковы генотипы родителей, если а) оба смуглых родителя
имеют одного черного и одного белого ребенка?, б) оба
смуглых имеют только смуглых детей?
Раздел 2
МАТЕРИАЛЬНЫЕ НОСИТЕЛИ
НАСЛЕДСТВЕННОСТИ
Шведский врач и биохимик Фридрих Мишер первым провел
химический анализ содержимого ядра. В 1869 г. он выделил
ядра лейкоцитов, содержащихся в гное солдат, раненных во
время франко-прусской войны. В ядрах он обнаружил веще-
ство нуклеин. Впоследствии нуклеин получил название нук-
леиновая кислота. Открытие Мишера, как и Менделя, про-
шло незамеченным.
В 1929 г. ученые открыли дезоксирибозу и стали различать
две кислоты — рибонуклеиновую (РНК) и дезоксирибонуклеи-
новую (ДНК). Примерно в это же время, в 1928 г., английский
микробиолог Фредерик Гриффитс сделал первый шаг в изу-
чении ДНК в качестве носителя наследственной информации
клетки. Гриффитс заметил, что у животных с определенным
типом бактериальной пневмонии обнаруживался один из двух
типов бактерии Diplococcus pneumoniae. Поверхность коло-
ний бактерий штамма R (от анг. rough — грубый, шерохова-
тый) была шероховатой, а штамма S (от агл. smooth — глад-
кий) — гладкой, поскольку бактерии штамма S были окруже-
ны гладкой полисахаридной капсулой. Гриффитс обнаружил,
что инъекция мышам бактерий штамма R не вызывала у жи-
вотных пневмонию. Введение животным бактерий типа S при-
водило к пневмонии. Гриффитс предположил, что все дело
было в полисахаридной капсуле. Он подверг бактерии штам-
ма S термообработке, после чего они уже не вызывали пнев-
|Монию у мышей. Однако, после того как он смешал бактерии
штамма R с убитыми бактериями штамма S и ввел смесь мы-
шам, животные заболели пневмонией. После исследования в
трупах мышей были найдены живые бактерии штамма S в
полисахаридных капсулах.
47
Раздел 2. Материальные носители наследственности
Позже, в 1930-х гг., ученые Эйвери, МакЛеод и МакКарти
предложили возможное объяснение полученным Гриффит-
сом результатам. Они предположили, что какой-то фактор в
уничтоженных термообработкой бактериях штамма S транс-
формировал безвредные бактерии штамма R в болезнетвор-
ные. Эксперименты показали, что при введении мышам фер-
мента протеазы, разрушающего белки, штамм R все равно
оставался болезнетворным. Однако при введении фермента,
разрушающего ДНК, трансформация не происходила. Эйве-
ри, МакЛеод и МакКарти подтвердили, что именно ДНК вы-
зывала трансформацию бактерий при изоляции ДНК из раз-
рушенных после тепловой обработки бактерий штамма S и
введении ее вместе с бактериями штамма R мышам. Мыши
погибали, а их тела содержали активные бактерии штамма S.
Напрашивался следующий вывод: ДНК бактерий штамма S
вызывала нарушения бактерий штамма R, приводя к тому, что
последние начинали синтезировать полисахаридную капсу-
лу, необходимую для инфицирования животных.
В 1950 г. американские микробиологи Альфред Херши и
Марта Чейз подтвердили, что именно ДНК, а не белок, явля-
ется веществом наследственности. Для доказательства этого
они инфицировали бактерию E.coli вирусом. В вирусе были
ДНК и белковая капсула^ маркированные различными радио-
активными элементами: белковая капсула — радиоактивной
серой, ДНК — радиоактивным фосфором. Использовались два
штамма вирусов. У одного штамма маркированной была белковая
капсула, у другого — ДНК. Бактерии, зараженные штаммом виру-
са с маркированной белковой капсулой, не обнаружили радиоак-
тивности. В то же время у бактерий, зараженных вирусами с
маркированной ДНК, радиоактивность присутствовала. Это
означало, что именно ДНК, а не белок вируса, встраивалась в
бактерию и направляла размножение вируса.
2.1. Химическая структура ДНК
Материальным носителем наследственной информации яв-
ляется дезоксирибонуклеиновая кислота (ДНК). Гены пред-
48
2.1. Химическая структура ДНК
ставляют собой участки молекулы ДНК. Структура ДНК была
расшифрована английскими учеными Дж. Уотсоном и
ф. Криком в 1953 г.
Молекула ДНК состоит из двух полинуклеотидных цепей,
закрученных одна вокруг другой в виде спирали. Каждая по-
линуклеотидная цепь состоит из множества мономеров — нук-
леотидов. Нуклеотид — это химическое соединение, состоящее
из одной молекулы углевода (дезоксирибозы) —• остатка моле-
кулы фосфорной кислоты и молекулы азотсодержащего орга-
нического соединения — азотистого основания (рис. 2.1).
Фосфатная^
группа
О
Основание
Пятиуглеродный
сахар
Нуклеотид
Рис. 2.1. Схема строения нуклеотида
В состав ДНК входят нуклеотиды с четырьмя типами азо-
тистых оснований — двух пуриновых (аденин А и гуанин Г) и
двух пиримидиновых (тимин Т и цитозин С) (рис. 2.2). Сосед-
ние нуклеотиды одной цепочки соединены друг с другом ко-
валентными связями между остатками фосфорной кислоты
одного нуклеотида и молекулой углевода соседнего нуклео-
тида. Азотистые основания двух цепей обращены внутрь спи-
рали так, что аденин располагается против тимина, а гуанин
против цитозина. Такое соответствие азотистых оснований
называется комплементарностью. Азотистые основания двух
цепочек, взаимодействуют друг с другом водородными свя-
зями. Пара А-Т удерживается двумя водородными связями, а
пара Г-Ц — тремя.
к.
Раздел 2. Материальные носители наследственности
цитозин (Ц)
тимин (Т)
Рис. 2.2. Химическое строение азотистых оснований
При удвоении двойная спираль ДНК разделяется на две
одиночные цепочки, и против каждой из них в соответствии
с принципом комплементарности достраивается вторая цепь.
Это процесс называется репликацией.
2.2. Генетический код
В молекуле ДНК записана информация о последовательнос-
ти аминокислот в молекуле белка. Между последовательнос-
тью нуклеотидов ДНК и последовательностью аминокислот
в молекуле белка имеется соответствие — называемое гене-
тическим кодом (табл. 2.1). В последовательности нуклеоти-
дов ДНК закодирована генетическая информация, которая
наделяет организм присущими ему признаками. Генетичес-
кий код включает 64 триплета, которые еще называются ко-
донами. Триплет — это три последовательно расположенных
50
Генетический код (РНК)
1-й нуклео тид 2-й нуклеотид
У ц А
У Фенилаланин Фенилаланин Лейцин Лейцин Серин Серин Серин Серин Тирозин Тирозин Стоп-сигнал Стоп-сигнал
ц Лейцин Лейцин Лейцин Лейцин Пролин Пролин Пролин Пролин Г истидин Г истидин Глутамин Глутамин
А Изолейцин Изолейцин Изолейцин Метионин Треонин Треонин Треонин Треонин Аспаргин Аспаргин Лизин Лизин
Г Валин Валин Валин Валин Аланин Аланин Аланин Аланин Аспаргиновая кислота Аспаргиновая кислота Глутаминовая кислота Глутаминовая кислота
Раздел 2. Материальные носители наследственности
нуклеотида в цепочке ДНК. Три кодона не соответствуют ни-
какой аминокислоте и называются стоп-сигналами. Осталь-
ные 61 кодируют 20 аминокислот. Некоторые аминокислоты
кодируются одним триплетом, другие несколькими. Это свой-
ство генетического кода называется вырожденностью. Код не
перекрывается. Это означает, что в последовательности ДНК
каждый нуклеотид входит в состав только одного кодона. Код
не имеет запятых — за последним нуклеотидом одного кодо-
на стоит первый нуклеотид следующего кодона.
2.3. Транскрипция и трансляция
Реализация наследственной информации, или путь от гена к
признаку, осуществляется через ряд биохимических реакций.
Для многих признаков этот путь известен до мельчайших под-
робностей. Последовательность нуклеотидов ДНК находит
отражение в строении белка и таким образом определяет его
функцию. Начальные этапы реализации генетической инфор-
мации — транскрипция и трансляция. На стадии транскрип-
ции информация с молекулы ДНК переписывается (транскри-
бируется) на молекулу РНК (рибонуклеиновую кислоту).
Молекула РНК, так же как и ДНК, является полинуклеоти-
дом, но состоит из одной цепи. Нуклеотид РНК включает одно
из азотистых оснований (аденин, гуанин, цитозин, урацил),
углевод рибозу и остаток фосфорной кислоты. Переписыва-
ние информации с ДНК на РНК происходит в соответствии с
принципом комплементарности A-У, Т-А, Г-Ц, Ц-Г (первое
азотистое основание ДНК, второе — РНК).
Переписывание информации с ДНК (через РНК) на белок
происходит в соответствии с генетическим кодом, который
представляет собой соответствие между триплетами РНК и
отдельными аминокислотами.
В состав белка входит большое количество молекул ами-
нокислот, относящихся к 20 различным вариантам. От того,
какие аминокислоты содержатся в молекуле белка и в каком
порядке они расположены, зависят особенности белка. По-
52
2.3. Транскрипция и трансляция
рядок и вид аминокислот, из которых состоит молекула бел-
ка, определяется генетической информацией. Клетки чело-
века способны синтезировать около 100 тысяч различных бел-
ков. Функции белков разнообразны. Строительные белки
формируют различные структуры организма, определяя их
анатомо-морфологические и функциональные особенности.
Под контролем ферментов (белков-катализаторов) протека-
ют все биохимические процессы в организме. Тип реакции, а
также ее скорость зависят от особенностей этих белков.
Матричная молекула РНК содержит копию гена и вклю-
чает как информативные, так и не информативные нуклео-
тидные последовательности. Не информативные последова-
тельности вырезаются ферментами рестриктазами, а инфор-
мативные фрагменты соединяются с помощью фермента
лигазы. Молекула РНК, образованная в результате выреза-
ния—сшивания, содержит только те нуклеотидные последо-
вательности, в которых закодирована информация о после-
довательности аминокислот в белке. Эта РНК называется ин-
формационной (и-РНК). Синтез белка в клетке происходит в
цитоплазме на рибосомах по программе, записанной в моле-
куле и-РНК. Синтез белков кодируется наследственной ин-
формацией, заключенной в ДНК структурных генов. Другие
гены — регуляторные — контролируют скорость синтеза бел-
ковых продуктов и их количество.
Синтезированная в ядре и-РНК выходит в цитоплазму, где
на рибосомах происходит синтез белков из аминокислот, до-
ставляемых транспортными РНК (т-РНК), Последователь-
ность триплетов, или кодонов, и-РНК переводится в последо-
вательность аминокислот в соответствии с генетическим ко-
дом. Синтезируемые полипептидные цепи слагаются в
белковые молекулы. Белки имеют несколько уровней орга-
низации. Первичная структура -— это последовательность
аминокислотных остатков в полипептидной цепи, вторичная —
представляет собой спиральную конфигурацию белковой мо-
лекулы, третичная — трехмерную конфигурацию. Белки, со-
стоящие из нескольких полипептидных цепей, имеют и чет-
Раздел 2. Материальные носители наследственности
вертичную структуру. Примером белка с четвертичной струк-
турой является хорошо изученный нормальный человеческий
гемоглобин (НЬА), состоящий из двух а- и двух |3-полипептид-
ных цепей, контролируемых разными генами. Генетический
локус, кодирующий a-цепь, активен в течение всей жизни,
начиная с самых ранних стадий развития плода, |3-локус на-
чинает функционировать после рождения. Еще одна, у-цепь,
контролируемая другим гемоглобиновым геном, синтезиру-
ется в период внутриутробной жизни плода. При ассоциации
у-цепи с a-цепью образуется гемоглобин F, характерный для
плода. В крови новорожденных надолго HbF приходится око-
ло 80% всего гемоглобина. Гемоглобин, характерный для кро-
ви взрослого человека, обнаруживатся в эритроцитах плода
уже к 13-й неделе развития, а к концу первого года жизни
ребенка гемоглобин HbF полностью заменяется на гемогло-
бин НЬА. В крови взрослого человека присутствует неболь-
шое количество еще единого вида гемоглобина НЬА,, в кото-
ром цепь б объединяется с цепью 8, кодируемой локусом, ко-
торый становится активным после рождения.
2.4. Генные мутации
При копировании молекулы ДНК могут происходить ошиб-
ки, которые меняют последовательность нуклеотидов. Этот
процесс напоминает ошибки, которые возникают, например,
при переписывании текстов или при неточном их переводе с
одного языка на другой. Изменения наследственного материа-
ла — мутации — могут представлять собой выпадения, встав-
ки, замены, перестановки одного или нескольких нуклеоти-
дов. Изменения в ДНК могут возникать спонтанно (самопро-
извольно) в силу химической природы молекулы ДНК и
индуцировано — при действии специфических факторов,
называемых мутагенными. Мутагенами являются ультрафио-
летовое излучение, различные химические соединения (азо-
тистая кислота, некоторые красители, предметы бытовой
химии, инсектициды, гербициды, некоторые лекарственные
54
2.5. Динамические мутации
препараты и др.). События на уровне молекулы ДНК могут
изменить смысл генетической информации, что проявится
в структуре, а следовательно, и функции кодируемого бел-
ка.
2.5. Динамические мутации
Генетикам, изучающим наследственность человека, давно из-
вестно явление, получившее название антиципация, или уп-
реждение. Антиципация проявляется в том, что некоторые
наследственные заболевания у потомков начинаются в более
раннем возрасте и протекают тяжелее, чем у родителей. Дол-
гое время антиципация воспринималась как статистическая
погрешность, вызванная особенностями сбора генеалогичес-
кого материала. Для проверки этого были проведены специ-
альные исследования, исключающие методические ошибки.
Анализ этого материала доказал, что антиципация — реально
существующее явление. Механизм антиципации получил
объяснение после того, как с помощью молекулярно-генети-
ческих методов был открыт новый класс наследственных за-
болеваний, вызванный умножением числа нуклеотидных по-
второв ДНК. Было установлено, что в основе некоторых на-
следственных заболеваний лежит «динамическая мутация».
Динамическая мутация — это прогрессирующее изменение
наследственного материала, которое происходит при его пере-
даче из поколения в поколение. Этот вид мутаций обусловлен
изменением числатринуклеотидных повторов ДНК. В резуль-
тате такой мутации уменьшается или увеличивается число
тринуклеотидных повторов, причем умножение (экспансия)
происходит в 2,5 раза чаще, чем уменьшение.
Экспансия тринуклеотидных повторов представляет со-
бой последовательное (тандемное) умножение участков мо-
лекулы ДНК. При этом какой-то тип тринуклеотида много-
кратно повторяется. В молекуле ДНК имеются участки, склон-
ные к амплификации (умножению). При образовании половых
клеток этот участок многократно повторяется, и потомок по-
55
Раздел 2. Материальные носители наследственности
лучает хромосому с большим числом повторов, чем имел его
больной родитель. В связи с этим заболевание у потомка на-
чинается в более раннем возрасте и протекает тяжелее. Ди-
намические мутации повторов являются причиной некоторых
нервных и психических заболеваний с наследственным ком-
понентом (синдром ломкой Х-хромосомы, хорея Гентингто-
на, атаксия Фридрейха и др.).
2.6. Геном человека
ДНК, находящаяся в гаплоидном наборе хромосом данного
вида организмов, обозначается понятием геном. Размер гено-
ма выражают в парах оснований (п.о.). Ядерный геном чело-
века содержит примерно 3,3 миллиарда п.о.; длина всей ДНК —
более 1 м. Около 30% ДНК составляют повторяющиеся пос-
ледовательности. Количество копий этих последовательнос-
тей в геноме человека варьирует от нескольких единиц до
нескольких тысяч. 70% ДНК представляют собой уникальные
последовательности, присутствующие в виде одной или еди-
ничных копий. Геном человека включает около 30 тысяч ге-
нов. Гены человека сильно варьируют по размерам — от не-
скольких сотен до нескольких миллионов пар нуклеотидов.
Функционально ген разделяется на регуляторную и струк-
турную части. Первая представляет собой последователь-
ность ДНК, связанную с регуляцией генной активности, вто-
рая содержит информацию о последовательности аминокис-
лот в молекуле белка. Гены человека, как и других высших
организмов, устроены мозаично. Их структурные части состо-
ят из перемежающихся участков, называемых экзонами и ин-
тронами. В экзонах содержится информация о последователь-
ности аминокислот в кодируемом белке, интроны — неин-
формативные участки гена. Участки ДНК, разделяющие гены,
называются спейсерами. Некоторые гены собраны в группы,
называемые кластерами.
Последовательность ДНК у двух случайно взятых людей
различается примерно на 0,2%. Последовательности ДНК,
2.6. Геном человека
кодирующие белок, занимают около 10% генома, остальные
приходятся на долю некодирующих участков. В некодирую-
щих участках располагаются последовательности, регулирую-
щие активность генов, однако функция большей части неко-
дирующей ДНК не ясна.
Прогресс в области молекулярной генетики человека свя-
зан с исследованиями, проведенными в рамках международ-
ной программы «Геном человека». Благодаря этим работам
возникло новое научное направление — геномика. В рамках
этого направления выявлены многие особенности организа-
ции генома, обнаружены новые гены, расшифрованы мута-
ции, вызывающие болезни и аномалии поведения.
В цитоплазме клетки имеются органоиды, содержащие
собственную ДНК, — митохондрии. Признаки, которые на-
ходятся под контролем митохондриальных генов, получили
название цитоплазматических. Наследование цитоплазма-
тических признаков отличается от наследования признаков,
контролируемых ядерными генами. Цитоплазматические
признаки передаются потомкам только от матери. Это свя-
зано с тем, что материнская гамета (яйцеклетка) содержит
цитоплазму с большим количеством митохондрий, в то вре-
мя как отцовская гамета цитоплазмы практически не содер-
жит.
Митохондриальный геном человека (хромосома М, или
мтДНК) представляет собой кольцевую молекулу ДНК дли-
ной 16 569 пар нуклеотидов. В каждой митохондрии находит-
ся от 2 до 10 копий мтДНК, а всего в одной клетке присутству-
ет несколько тысяч таких копий. МтДНК включает 13 генов,
особенностью которых является отсутствие интронов. Ми-
тохондриальные гены передаются потомкам только от мате-
ри. Митохондриальная ДНК не подвержена рекомбинации и
наследуется как единый блок, или гаплотип. Это свойство
митохондриальной ДНК позволило исследователям реконст-
руировать генеалогию митохондриальных последовательно-
стей и выдвинуть предположение о том, что все митохондри-
альные ДНК человека могут иметь одного предка.
Раздел 2. Материальные носители наследственности
2.7. Хромосомный набор человека
В 1956 г. шведские ученые Тио и Леван опубликовали работу,
в которой было показано, что у человека 46 хромосом.
В начале 1960-х гг. в Денвере (США) состоялась конферен-
ция, где была предложена номенклатура хромосом человека.
С тех пор примерно через 3—5 лет на международной кон-
ференции в эту номенклатуру вносятся коррективы по мере
накопления новой информации. В настоящее время, соглас-
но денверской номенклатуре, хромосомы человека делятся на
7 групп. Группы обозначаются буквами от А до G, а в преде-
лах групп хромосомы нумеруются арабскими цифрами по
нисходящей величине: А — 1, 2, 3; В — 4, 5; С — 6—12, X; D —
13, 14, 15; Е — 16, 17, 18; F — 19, 20; G — 21, 22, Y. Половые
хромосомы чаще всего выносят в отдельную 23-ю пару или
же в ту группу, куда они подходят по размеру (рис. 2.3).
Ж Hl й* нг
19 20 21 22 __________
F G X ’ Y
Рис. 2.3. Кариотип человека
58
2.7. Хромосомный набор человека
Морфологию хромосом описывают, когда они максималь-
но спирализованы и хорошо видны в микроскоп — на стадии
клеточного деления, которая называется метафаза. Тело хро-
мосомы состоит из двух плеч, разделенных перетяжкой.
В первичной перетяжке расположена центромера — струк-
тура, состоящая из ДНК, к которой прикрепляются нити ве-
ретена деления. Теломеры, или концевые участки, ответствен-
ны за существование хромосом как отдельных образований.
Концы разорванных хромосом могут слипаться, теломеры не
слипаются. У некоторых хромосом есть спутник — участок,
соединенный с остальной частью хромосомы тонкой нитью —
вторичной перетяжкой.
Основным критерием, по которому хромосомы распреде-
ляются по группам, является положение центромеры, а точ-
нее, центромерный индекс — это отношение короткого пле-
ча к длине всей хромосомы. Центромерный индекс выража-
ется в процентах. У метацентрических, или равноплечих,
хромосом центромерный индекс равен 50%, у акроцентричес-
ких, или неравноплечих, хромосом — около 10—12%. В телоцен-
трических, или резко неравноплечих, хромосомах центромера
отделяет лишь совсем маленький участок. Длинное плечо хро-
мосомы обозначают буквой д, короткое — р (рис. 2.4).
Акроцентрическая Метацентрическая
Телоцентрическая
хромосома
21
хромосома
короткое
плечо
Р
длинное
плечо
Я
3
Рис. 2.4. Виды хромосом человека
в зависимости от положения центромеры
69
Раздел 2. Материальные носители наследственности
При специальном окрашивании хромосом видны темные
и светлые полосы, закономерно чередующиеся и создающие
специфический для каждой хромосомы рисунок. Более тем-
ные участки называются гетерохроматиновыми, светлые —
эухроматиновыми. Гетерохроматиновые участки более спи-
рализованы, и поэтому при окраске они захватывают больше
частиц красителя и выглядят более темными.
2.8. Структурная организация хромосом
Двуспиральная молекула ДНК непрерывно располагается
вдоль всей длины хромосомы. Комплекс ДНК и белка состав-
ляет хроматин ядра.
Основными белками, принимающими участие в укладке
ДНК, являются гистоны. Гистоны — это белковые молекулы,
содержащие много положительно заряженных аминокислот
(лизин и аргинин), которые соединяются с отрицательно за-
ряженной ДНК. Блок приблизительно из 200 нуклеотидов мо-
лекулы ДНК оборачивается вокруг сердцевины и образует
дисковидные нуклеосомы (рис. 2.5). Сердцевина, или кор, пред-
ставляет собой комплекс из восьми гистоновых молекул —
Н2А, Н2В, НЗ и Н4 (каждый гистон представлен двумя моле-
кулами) .
Сердцевина из
8 гистоновых молекул
Нуклеосома
Рис. 2.5. Строение нуклеосомы
во
Двойная
спираль
ДНК
Нуклеосомы
«—>1
2 нм
11 нм
Упаковка
в петли
<------->1
300 нм
Рис. 2.6. Различные степени укладки молеку
Раздел 2. Материальные носители наследственности
Нуклеосома относится к укладке первого порядка. Далее
нуклеосомы укладываются друг относительно друга и форми-
руют нити толщиной около 30 нм, которые называются соле-
ноиды (рис. 2.6).
Соленоиды, в свою очередь, упаковываются во множество
петель. В полностью конденсированной ДНК петли уклады-
ваются кольцами диаметром примерно в 700 нм, и именно в
таком виде они формируют хромосомы, которые выглядят
палочкообразными структурами и изучаются при установле-
нии кариотипа человека.
2.9. Цитогенетическая символика
Кариотип (набор хромосом) описывается числом хромосом,
дополнительно указываются половые хромосомы: 46,XX —
кариотип нормальной женщины, 46,ХУ — кариотип нормаль-
ного мужчины, 47,ХХУ — кариотип включает 47 хромосом,
половые хромосомы ХХУ. Аутосомы специально обозначают
только тогда, когда их количество изменено. Если число хро-
мосом увеличено (гиперплоидия), используют символ « + »,
если уменьшено (гипоплоидия) — символ « — »: 47.ХХ + 8 —
имеется добавочная хромосома № 8 (трисомия по 8-й хромо-
соме). Мозаичность обозначают косой линией: 46,XX/
47,ХХ + 21. Короткое плечо хромосомы обозначают буквой
«р», длинное «q». Уменьшение длины плеча обозначается сим-
волом « — », увеличение — « + ». Запись 46,XX 2р + означает,
что у женщины увеличено короткое плечо хромосомы 2.
Символ «г» с номером в скобках указывает на присутствие
соответствующей кольцевой хромосомы 46,XX,г (7). Изохро-
мосомы (хромосомы с идентичными плечами) обозначают
буквой «й>. Запись 46,X,i(Xq) означает, что у женщины с 46-ю
хромосомами одна из Х-хромосом нормальная, а вторая пред-
ставляет собой изохромосому, включающую два длинных пле-
ча этой хромосомы. Символ «t» обозначает транслокацию.
Например, запись 46,XX,t(4p — ; 13q+) свидетельствует об уко-
рочении короткого плеча хромосомы № 4 и увеличении ддин-
62
2.10. Аиеуплойдии
ного плеча 13-й хромосомы за счет переноса на нее участка
четвертой хромосомы.
При дифференциальном окрашивании хромосом выявляют-
ся серии полос (рис. 2.7). Чередование полос специфично для
каждой хромосомы и служит отправными точками при ее то-
пографировании. Линии разметки хромосом проводят через
центромеры и середины темных или светлых полос. Участки
хромосомы между двумя линиями разметки называются райо-
нами. Нумерация районов начинается от центромеры. Участки
внутри района делятся на полосы (темные и светлые), которые
нумеруются от центромеры к теломере. Так, запись lq25 озна-
чает полоску 5 во втором районе длинного плеча первой хромо-
сомы. Эту систему используют для обозначения места хромо-
сомной аномалии. Например, запись 46,ХХ, del (2)(q23) означа-
ет, что разрыв произошел во второй хромосоме в длинном плече
во втором районе на уровне полос-
ки 3. У этой женщины одна из хро-
мосом № 2 нормальна, а вторая име-
ет короткое плечо и часть длинного
плеча.
Робертсоновские транслока-
ции (центрические слияния) обо-
значают символом «гоЬ». В карио-
типе 46,XY,rob( 13q; 15q) имеется
центрическое слияние длинных
плеч 13-й и 15-й хромосом.
2.10. Анеуплоидии
В норме у человека 46 хромосом, из
которых 44 аутосомы и 2 половые
хромосомы (XX у женщин и XY у
мужчин). Такой хромосомный на-
|бор называется эуплоидным. При
’анеуплоидии в кариотипе отмеча-
ется недостающие или избыточ-
ные хромосомы.
Рис. 2.7. Топографиче-
ские особенности хромо-
сомы №1 человека
63
Раздел 2. Материальные носители наследственности
Анеуплоидия вызывает множественные врожденные по-
роки развития внутренних и наружных органов, аномалии в
строении лица, замедленное внутриутробное развитие, отста-
вание психического развития, нарушение функций нервной,
эндокринной и иммунной систем. Анеуплоидии часто приво-
дят к умственной отсталости. Анеуплоидии, связанные с по-
ловыми хромосомами, приводят к более легким нарушениям.
Избыток генетического материала, по-видимому, менее опа-
сен, чем его недостаток. У большинства детей, рожденных с
аномальным числом хромосом, отмечается избыточная хро-
мосома (трисомия). Трисомии и моносомии обозначаются по
вовлеченной хромосоме, а синдромы, вызванные ими, изна-
чально были названы по именам впервые описавших их уче-
ных. Например, трисомия по 21-й хромосоме (47,XY, + 21) на-
зывается также синдромом Дауна.
Анеуплоидия возникает в результате нерасхождением хро-
мосом. В норме пары гомологических хромосом расходятся,
и в каждом сперматозоиде или яйцеклетке содержится толь-
ко по одному гомологу каждой пары. Нерасхождение может
происходить как при первом, так и при втором мейотическом
делении. Результатом нерасхождения является гамета, содер-
жащая две копии хромосомы или вообще ни одной. У челове-
ка при слиянии этой аномальной гаметы с нормальной при
оплодотворении образуется зигота с 47 или с 45 хромосома-
ми.
Любая из 22 пар аутосом может иметь дополнительную или
недостающую хромосому, и, кроме того, половые хромосо-
мы могут оказаться в пяти аномальных, анеуплоидных ком-
бинациях (Y0, ХО, XXX, XXY и XYY). Однако, лишь немногие
встречаются у новорожденных, а в основном развитие пре-
кращается еще в эмбриональном периоде. В среднем избы-
точные или недостающие хромосомы вызывают спонтанные
аборты в 50% случаев.
64
г
2.11. Аутосомные синдромы
2.11. Аутосомные синдромы
Трисомия 21 (синдром Дауна)
В 1866 г. английский врач Джон Лэнгдон Даун опублико-
вал отчет, в котором были описаны несколько детей с умствен-
ной отсталостью, не похожих на других детей с низким ин-
теллектом. Даун провел свои исследования в психиатричес-
кой больнице в Сюррее (Англия), где он первым разграничил
детей с кретинизмом, вызванным гипотиреоидизмом, и детей,
заболевание которых он охарактеризовал как «монголизм».
Внешне у детей с «монголизмом» наблюдается маленькая
и круглая голова, наклоненный кзади лоб, слабо развитая пе-
реносица, низко прикрепленные с маленькой мочкой уши,
глаза с эпикантной складкой, приоткрытый рот с выступаю-
щим языком, единственная поперечная складка на ладонях,
большой промежуток между первым и вторым пальцами ног,
головной мозг меньшего веса (рис. 2.7).
Термин «монголизм» Даун применил на основании того,
что описанные им дети внешне напоминали представителей
монголоидной расы. В 1960-х гг. со стороны азиатских гене-
тиков поступил этический протест по поводу термина «мон-
голизм», и термин был изъят из научного обихода. Описан-
ную Дауном болезнь стали называть синдром Дауна.
| В первой половине XX в. в отношении причин синдрома
Дауна высказывалось много предположений. Первыми уче-
ными, которые высказали мнение относительно хромосом-
ных аномалий, лежащих в основе синдрома Дауна, были Ва-
арденбург и Блейер в 1930-х гг. Однако лишь в 1959 г. Джером
Лежен и Патрисия Жакоб независимо друг от друга устано-
вили, что причиной синдрома Дауна была трисомия по 21-й
хромосоме.
Классическая полная трисомия по 21-й хромосоме возни-
кает при слиянии двух гамет, в одной из которых произошло
нерасхождение указанной хромосомы. Аномальной гаметой
примерно в 90% случаев является яйцеклетка, и лишь в 10%
Iслучаев — сперматозоид. Одной из известных и доказанных
Г Введение в психогенетику 65
Раздел 2. Материальные носители наследственности
причин нерасхождения 21-й
хромосомы является возраст
матери, выходящий за рамки
репродуктивного оптимума.
Дети с синдромом Дауна
рождаются с частотой 1:700. У
молодых матерей вероятность
рождения ребенка с данным
синдромом составляет 1:2000, у
женщин после 40 лет — 1:22.
Дополнительный набор ге-
нов 21 хромосомы приводит к
их сверхэкспрессии, следстви-
ем чего является увеличенный
синтез определенных белков.
Ранее предполагали, что для
Рис. 2.8. Некоторые из внешних проявлений
синдрома Дауна
2.11. Аутосомные Синдромы
Рис. 2.9. Девочка с синд-
ромом Дауна
проявления неблагоприятных
последствий необходима была
тройная копия лишь небольшого
участка 21-й хромосомы, кото-
рый так и назывался — критичес-
кий участок для синдрома Дауна.
Коэффициент умственного
развития больных синдромом Дау-
на варьирует от 20 до 70 единиц
(в норме около 100). Дети с синд-
ромом Дауна в большинстве слу-
чаев вполне воспитуемы, могут
научиться говорить, иногда обу-
чаются чтению и письму. Как
правило, они не способны к абст-
рактному мышлению, поэтому не
могут научиться считать. Дети с
синдромом Дауна спокойны и
дружелюбны, послушны и терпе-
ливы. Терапией, направленной
на улучшение функции центральной нервной системы, и спе-
циальными методами обучения удается адаптировать их к
жизни. Многие люди с трисомией 21, получившие специаль-
ное воспитание, способны к самостоятельной жизни, овладе-
вают несложной профессией и создают семьи (рис. 2.9).
Часто отчаявшиеся родители детей с синдромом Дауна на-
ходятся перед так называемой дилеммой «Baby Doe». Само
понятие «Baby Doe» вошло в обиход после широкого освеще-
ния в прессе случая с ребенком, фигурировавшим под таким
псевдонимом. Baby Doe родился с заращенным пищеводом и
синдромом Дауна в Блумингтоне, штат Индиана (США) в 1982 г.
Родители Baby Doe решили, что ребенка следует лишить пи-
тания и медицинской помощи и оставить его умирать. Они
считали, что если бы врачи сделали хирургическую операцию
на пищеводе, все равно их ребенок остался бы неполноцен-
ным по причине синдрома Дауна. Ребенок умер в возрасте
67
Раздел 2. Материальные носители наследственности
нескольких месяцев. В последние годы последовали другие
случаи с так называемыми «Baby Doe». Президентская комис-
сия в США для разработки этических положений относитель-
но отказа в медикаментозной помощи сделала заключение,
что симптомы, сопутствующие синдрому Дауна, не являются
настолько тяжелыми, чтобы оправдывать отказ во врачебной
помощи новорожденным с этой болезнью.
Трисомия 13 (синдром Патау)
Рис. 2.10. Расщелина
неба и губы у ребенка
с синдромом Патау
Частота синдрома Патау составляет
около 1 на 5000—15000 новорожден-
ных. У таких детей имеется расще-
лина неба, которой часто сопутству-
ет расщелина губы (рис. 2.10). У них
маленькая голова, скошенный назад
лоб, крупный треугольный нос, от-
сутствуют или сращены глаза. Могут
быть дополнительные пальцы на ру-
ках и ногах, дополнительная селезен-
ка, аномалии печени и поджелудоч-
ной железы, нарушение изгибов ки-
шечника. Развитие больных детей
обычно не превышает шестимесяч-
ного возраста. Продолжительность
их жизни обычно не превышает од-
ного года, и лишь 20% живут несколько дольше. Смерть чаще
всего наступает в результате пороков развития сердца и по-
чек.
Трисомия 18 (синдром Эдвардса)
Синдром Эдвардса проявляется в целом комплексе аномалий
развития, которые приводят к летальному исходу. Внешние
признаки: ямкообразный подбородок, неправильной формы
уши, маленькие глаза и сжатые пальцы рук за счет повышен-
68
2.12. Синдромы половых хромосом
Рис. 2.11. Лицо и рука ребенка с синдромом Эдвардса
ного мышечного тонуса (рис. 2.11), Дети с синдромом Эдвар-
дса рождаются с частотой примерно в 1 на 4000—8000.
2.12. Синдромы половых хромосом
Моносомия по Х-хромосоме (45, Х0),
или синдром Тернера
В 1938 г. американский эндокринолог Генри Тернер опубли-
ковал отчет о нескольких бесплодных женщинах маленького
роста. В то время причину задержки их полового развития
обнаружить не удалось. В настоящее время известно, что опи-
санный Тернером синдром связан с присутствием в кариоти-
пе лишь одной, а не двух Х-хромосом. Классический синдром
Тернера встречается в 60% случаев заболевания, а у 40% жен-
щин наблюдается мозаицизм или делеция короткого плеча
К-хромосомы. У женщин с синдромом Тернера яичники за-
чаточны и молочные железы недоразвиты (рис. 2.12).
i Синдром Тернера часто диагностируется при рождении.
Ж пораженных новорожденных наблюдаются характерные
ВИейные складки кожи сзади («шея сфинкса»), отеки рук и ног
К очень низкая масса тела. Частота синдрома Тернера среди
69
Раздел 2. Материальные носители наследственности
Рис. 2.12. Женщина с синдромом Тернера
новорожденных девочек 1:2000. При синдроме Тернера ин-
теллект практически всегда находится в пределах нормы.
Трисомия по Х-хромосоме (47, XXX),
или синдром трипло-Х
Триплоидия по Х-хромосоме, или синдром трипло-Х, встре-
чается с частотой примерно 1 на 1000—2000 женщин. Внеш-
не такие женщины почти не отличаются от женщин с нор-
70
2.12. Синдромы половых хромосом
мальным хромосомным набором за исключением более вы-
сокого роста и нарушения месячного цикла. IQ женщин с
триплоидией по Х-хромосоме находится в пределах нормы,
однако может быть ниже, чем у остальных членов семьи. Боль-
шинство женщин с синдромом трипло-Х фертильны (могут
иметь детей).
Дисомия по Х-хромосоме у мужчин (47, XXY),
или синдром Клайнфельтера
Эта половая аномалия впервые была описана в 1942 г. амери-
канским врачом Гарри Клайнфельтером. В основе синдрома
Клайнфельтера лежит кариотип 47.XXY. У пораженных муж-
чин очень маленькие семенники (около 1/3 их нормального
размера) и увеличенные молочные железы (рис. 2.13).
Рис. 2.13. Увеличенные молочные железы у мужчины
с синдромом Клайнфельтера
Большинство больных стерильны (не могут иметь потом-
ство) и продуцируют мало тестостерона. Микроскопическое
исследование семенников обнаруживает дегенеративные
изменения в стенке семявыносящих канальцев. У людей с
синдромом Клайнфельтера наблюдаются длинные конечно-
ft
71
Раздел 2. Материальные носители наследственности
сти и слабая волосяная растительность на лице. У многих муж-
чин с данным заболеванием пассивный характер и низкая са-
мооценка. В большинстве случаев их IQ находятся в преде-
лах нормы. Небольшая часть мужчин с лишней Х-хромосо-
мой не имеет никаких клинических проявлений синдрома
Клайнфельтера за исключением бесплодия. В среднем частота
синдрома Клайнфельтера в популяции довольно высокая —
1 мальчик на 1000 новорожденных имеет данное заболевание.
У мужчин с синдромом Клайнфельтера в 12 раз повышен
риск развития рака молочной железы, что соизмеримо с час-
тотой этого заболевания у женщин. Зарегистрированы боль-
ные с синдромом Клайнфельтера, имеющие наборы XXXY,
XXXXY и XXXXXY. При большем числе Х-хромосом у муж-
чин сильно страдают интеллектуальные способности и их IQ
значительно ниже нормы.
Дисомия по Y-хромосоме (47, XYY),
или синдром Жакоб
Лишняя Y-хромосома у мужчин встречается с частотой при-
мерно 1:1000. В 1965 г. Патрисия Жакоб опубликовала резуль-
таты исследования, проведенные среди пациентов закрыто-
го учреждения для психических больных в Carstairs (Шотлан-
дия). Из 197 обследованных ею мужчину 12 были обнаружены
аномалии кариотипа, из них 7 имели дополнительную Y-хромо-
сому. Жакоб предположила, что наличие дополнительной Y-xpo-
мосомы могло быть причиной жестокости и агрессивности у
этих мужчин и совершения ими противоправных действий. Ког-
да и в других подобных учреждениях были проведены анало-
гичные исследования, обнаружилась сходная статистика. Это
привело к возникновению в обществе образа так называемых
«врожденных преступников». В 1968 г. адвокаты во Франции
и Австралии выдвигали в качестве довода для оправдания сво-
их клиентов описанный недавно «синдром Жакоб».
В начале 1970-х гг. в лечебных учреждениях Англии, Ка-
нады, Дании, а также Бостона (США) начался скрининг на ка-
72
2.13. Полиплоидия
риотип XYY. Социальные служащие и психологи подробно
инструктировали родителей как надо обращаться с детьми с
таким хромосомным набором.
2.13. Полиплоидия
Изменение числа хромосом в клетке происходит из-за нару-
шения их поведения во время клеточного деления. При этом
могут возникнуть полиплоидные клетки. При полиплоидии
число хромосом в клетке увеличивается в целое число раз —
кратно гаплоидному набору. Организмы с кратно умножен-
ным числом хромосом — полиплоиды. У триплоидов тройной
набор хромосом (Зп), у тетраплоидов — учетверенный (4п) и
т.д. Полиплоидия обычна в растительном мире, большинство
культурных растений — полиплоиды. Их дикие родственни-
ки имеют диплоидный хромосомный набор. У животных по-
липлоидия встречается гораздо реже и в основном среди бес-
позвоночных.
Генетики описали несколько случаев рождения триплоид-
ных детей, у которых было 69 хромосом. Триплоидность у че-
ловека не совместима с жизнью. Все новорожденные с трип-
лоидным набором хромосом имели множественные аномалии
развития и умерли через несколько дней.
Триплоидные зиготы возникают, если половая клетка, имею-
щая диплоидный набор хромосом, соединяется с нормальной
гаплоидной половой клеткой. Диплоидная половая клетка об-
разуется в том случае, если не расходятся дочерние клетки
только в одном из мейотических делений. Если нарушается
расхождение хромосом в обоих мейотических делениях, фор-
мируется тетраплоидная половая клетка.
Триплоидный хромосомный набор у человека может иметь
разное происхождение. Ребенок может получить два отцовс-
ких и один материнский хромосомный набор или два мате-
ринских и один отцовский. Плод, имеющий два материнских
и один отцоцский хромосомный набор, называется дигеничес-
кий триплоид, один материнский и два отцовских — диан-
Раздел 2. Материальные носители наследственности
дрический триплоид. Диандрические и дигенические трип-
лоиды, несмотря на то что у них численно одинаковый хро-
мосомный набор, различаются по характеру аномалий. У ди-
андрических триплоидных плодов сильно развита плацента
и задержано развитие собственно эмбриональных структур,
в то время как при дигенической триплоидии рост плацентар-
ных тканей сильно угнетен. Этот пример свидетельствует о том,
что хромосомы отцовского и материнского происхождения
оказывают неодинаковое влияние на развитие организма.
Если аномальное количество хромосом имеет небольшая
группа соматических клеток, то последствия для здоровья
могут быть незначительными. Например, клетки печени че-
ловека часто содержат участки полиплоидных (триплоидных)
клеток, однако они не оказывают существенного влияния на
функцию печени.
2.14. Псевдонормальный кариотип
В результате событий, нарушающих расхождение хромосом
при делении клетки, может образоваться зародыш с необыч-
ным диплоидным хромосомным набором. В таком диплоидном
наборе все хромосомы происходят от одного из родителей, а
хромосомы второго родителя отсутствуют. Такие наборы, хотя
и содержат 23 пары хромосом, являются псевдонормальны-
ми. Диплоидные эмбрионы, у которых все хромосомы имеют
отцовское происхождение, называются диандрическими, если
все хромосомы имеют материнское происхождение — диге-
ническими. У зародышей с псевдонормальным кариотипом
отцовского происхождения происходит злокачественное пе-
рерождение ткани одной из плодных оболочек, чего не на-
блюдается при материнском происхождении хромосом. Пе-
речисленные примеры свидетельствуют о том, что гаплоид-
ный набор отцовских и материнских хромосом у зародыша
функционирует неодинаково. Геном отца вносит существен-
ный вклад в развитие структур, через которые осуществля-
ется связь зародыша с организмом матери и его питание. Ге-
74
2.15. Однородительская дисомия
ном матери оказывает преимущественное влияние на разви-
тие самого зародыша.
2.15. Однородительская дисомия
Для нормального развития человеческого организма в его хро-
мосомном наборе должны присутствовать хромосомы мате-
ринского и отцовского происхождения. Нормальное разви-
тие обеспечивается сочетанием гаплоидных наборов матери
и отца. Если это условие нарушается, развитие организма от-
клоняется от нормы. Такие нарушения возникают при одно-
родительской дисомии. Явление однородительской дисомии
выражается в том, что при численно нормальном диплоидном
хромосомном наборе одна пара имеет только отцовское или
только материнское происхождение. При этом все остальные
хромосомы представлены парами, в которых один гомолог
получен от отца, а другой от матери. Существует два типа од-
нородительской дисомии — изодисомия и гетеродисомия.
Изодисомия возникает при нерасхождении хромосом во вто-
ром делении мейоза. В этом случае обе хромосомы являются
копиями и имеют идентичную последовательность ДНК с
полностью гомозиготным хромосомным материалом. Гетеро-
дисомия возникает при нерасхождении хромосом в первом
мейотическом делении. В этом случае парные хромосомы не
являются копиями.
Однородительская дисомия может возникнуть разными
путями. Один их них — комплементация (дополнение) гамет —
возможен при соединении нуллисомной и дисомной гамет. В
гаплоидном наборе нуллисомной гаметы отсутствует одна из
хромосом, а в гаплоидном наборе дисомной гаметы эта хро-
мосома представлена двумя гомологами. При соединении та-
ких гамет нехватка хромосомы первой гаметы компенсиру-
ется избыточной хромосомой второй гаметы. В диплоидной
зиготе все хромосомы, кроме описанной пары, происходят от
разных родителей, а одна пара происходит либо от отца, либо
от матери.
75
Раздел 2. Материальные носители наследственности
Другой способ происхождения однородительской дисо-
мии — коррекция трисомии. Происходит это следующим об-
разом. При слиянии нормальной гаплоидной гаметы с гаме-
той, у которой на фоне гаплоидного хромосомного набора
одна из хромосом представлена двумя гомологами (дисомия),
образуется трисомная зигота. В ходе одного из делений три-
сомная зигота может утратить лишнюю (третью) хромосому.
Однородительская дисомия возникает в том случае, если ут-
рачивается хромосома, полученная с нормальной гаплоидной
гаметой, и сохраняется пара хромосом, полученная от друго-
го родителя. С утратой одной из двух хромосом, полученных
от одного родителя, нормальный хромосомный набор восста-
навливается. При этом в части клеток сохраняется первона-
чальная хромосомная конституция зиготы и организм оказы-
вается хромосомным мозаиком.
Однородительская дисомия может возникнуть в резуль-
тате коррекции моносомии. В этом случае происходит дупли-
кация единственной хромосомы, полученной от отца, либо от
матери.
Однородительская дисомия может оказывать влияние на
развитие организма в том случае, если хромосома содержит
участки, подверженные импринтингу. В этом случае зародыш
получает две хромосомы, каждая из которых имеет одинаковым
образом модифицированные аллели, в то время как для нормаль-
ного развития необходимо, чтобы эти участки были модифици-
рованы различным образом. Если хромосома не содержит участ-
ков, подверженных импринтингу, то у индивидов с однороди-
тельской дисомией по данной хромосоме не возникает
аномалий фенотипа. К настоящему времени у человека эф-
фект импринтинга установлен для хромосом №15, 11, 7 и 14.
Об импринтинге на хромосомном уровне свидетельству-
ют два хорошо изученных наследственных заболевания: синд-
ром Прадера-Вилли и синдром Энгельмана. У ребенка с синдро-
мом Энгельмана отмечается малый размер мозга, отсутствие
речи, эпилептические припадки. Для синдрома Прадера-Вил-
ли характерны ожирение, аномалии лицевого черепа, ум-
2.16. Мозаицизм
ственная отсталость, маленькие кисти и стопы. Каждый из
этих синдромов обусловлен нехваткой в одном и том же уча-
стке хромосомы 15 (qll-ql2 ). При делеции в хромосоме мате-
ринского происхождения у ребенка обнаруживаются призна-
ки синдрома Энгельмана, при нехватке в хромосоме отцовс-
кого происхождения — признаки синдрома Прадера-Вилли.
2.16. Мозаицизм
Если нарушение в ходе митоза возникает в раннем эмбриоге-
незе, то от клетки с измененным хромосомным набором об-
разуется множество дочерних клеток, и тогда возникает орга-
низм-мозаик. Комплементарный мозаицизм — явление, при
котором мозаичный организм состоит из клеток, хромосомные
наборы которых как бы дополняют друг друга. При некомпле-
ментарном мозаицизме наборы разных клеточных клонов не
являются дополнительными. Комплементарные мозаики —
удвоение
нерасхождение
клон 45, X
клон 47, ХХУ
Рис. 2.14. Схема нерасхождения хромосом у мозаика
с клонами клеток с недостающей и лишней Х-хромосомами
Раздел 2. Материальные носители наследственности
результат нерасхождения хромосом, некомплементарные —
их утраты. Например, некоторые люди со сглаженной фор-
мой синдрома Дауна являются хромосомными мозаиками, т.е.
некоторые их клетки имеют нормальный хромосомный на-
бор, а другие — дополнительную 21 хромосому.
Для того чтобы понять, как получаются организмы-мозаи-
ки, можно привести следующий пример. Допустим, что цито-
генетический анализ выявил у женщины два типа клеток — 45, X
и 47,XXX — и нам необходимо объяснить такой феномен.
Зигота была нормальной (46,XX). При первом дроблении зи-
готы хроматиды одной из удвоенных Х-хромосом не разош-
лись и образовали два клона клеток — с одной Х-хромосомой
и с тремя Х-хромосомами (рис. 2.14).
2.17. Геномный импринтинг
За последние десятилетия накоплено много фактов наследо-
вания, в том числе и у человека, которые не укладываются в
схемы классической генетики. Так, долгое время считалось,
что проявление гена у потомства не зависит от того, кто из
родителей его передал. Однако постепенно накапливались
данные, которые ставили под сомнение это положение. Ста-
ло понятно, что генетический материал, полученный от мате-
ри и отца, в какой-то мере функционально различается.
У диплоидных организмов обычно функционируют (экспресси-
руются) оба аллельных гена — отцовский и материнский [биал-
лелъная экспрессия). Есть, однако, гены, проявление которых в
признаке обусловлено тем, от кого из родителей они получены.
Дело в том, что в хромосомах есть участки, которые при образо-
вании половых клеток подвергаются молекулярной модифика-
ции. Одни участки модифицируются при формировании мужс-
ких половых клеток, модификация других участков происходит
при образовании женских половых клеток. Молекулярная мо-
дификация приводит к тому, что материнская и отцовская
хромосомы становятся функционально различными. Это яв-
ление получило название геномный импринтинг.
2.18. Хромосомные перестройки
Гены, подверженные импринтингу (импринтированные
гены), проявляются у потомства по-разному, их активность
зависит от пола родителя, от которого они получены. В участ-
ках генома, подверженных импринтингу, экспрессируется
только отцовский или только материнский аллель. Это явле-
ние называется моноаллельной экспрессий.
При геномном импринтинге происходят изменения не
всей хромосомы, а отдельных ее участков. Аллель, находящий-
ся в импринтированном участке и полученный от одного ро-
дителя, функционально неравен аллелю, полученному от дру-
гого родителя. Гены, локализованные в участках, не затраги-
ваемых импринтингом, проявляются у потомства не зависимо
от того, кто из родителей их передал.
Точные механизмы, лежащие в основе геномного имприн-
тинга, до конца не известны, однако основную роль в этом
процессе отводят метилированию цитозиновых оснований
ДНК. Метилирование (присоединение метильных групп) ДНК
изменяет активность некоторых генов.
Геномный импринтинг иллюстрируется таким примером.
У женщин с синдромом Тернера присутствует только одна
Х-хромосома (набор половых хромосом ХО). Эта хромосома
может иметь материнское (Xм) или отцовское (Х°) происхож-
дение. Женщины с Х-хромосомой материнского и отцовско-
го происхождения различаются в психическом отношении.
У женщин Хм0 в большей степени, чем у пациенток Х°0 вы-
ражены трудности в социальной и познавательной деятель-
ности. Цитогенетический анализ показал, что эти особеннос-
ти поведения связаны с наличием в Х-хромосоме небольшо-
го метилированного участка, соответствующего одному гену
или небольшому числу генов.
2.18. Хромосомные перестройки
Причиной недостатка или избытка хромосомного материала
в клетке могут быть хромосомные перестройки (аберрации),
которые возникают при разрывах хромосом и их соединении
70
Раздел 2. Материальные носители наследственности
в ином порядке. Восстановление разрывов может привести к
перемещению участков хромосом, их удвоению, утрате или
повороту внутри хромосомы. Могут также возникать обме-
ны участками между негомологичными хромосомами (рис.
2.15). Хромосомные перестройки у человека обычно приво-
дят к множественным порокам развития. В результате этого
возникают так называемые хромосомные болезни. Некоторые
из них передаются потомству. В отличие от анеуплоидий, при
хромосомных перестройках количество хромосом не изме-
няется.
Структурные изменения хромосом имеют специальные
названия: нехватка (делеция), удвоение (дупликация), поворот
хромосомного фрагмента (инверсия), перестановка (инсер-
ция), перенос части генетического материала на негомологич-
ную хромосому (транслокация).
Структурные перестройки хромосом могут привести к час-
тичной трисомии или частичной моносомии. При частичной
трисомии одна из гомологичных хромосом нормальная, а в
другой один из участков дуплицирован (удвоен). При частич-
ной моносомии одна из хромосом нормальная, а в другой име-
ется нехватка. Влияние хромосомных перестроек на фенотип
зависит от величины хромосомного фрагмента, затронутого
аберрацией. Специфическая картина для синдрома Дауна
развивается при трисомии по сегменту 21 q22.1, синдрома ко-
шачьего крика — при моносоми 5р15, синдрома Эдвардса —
при трисомии 18gll.
Индивиды с хромосомными перестройками обычно про-
являют ту или иную степень умственной отсталости и имеют
множественные физические аномалии.
В настоящее время хромосомные аномалии встречаются
примерно у 0,6% новорожденных. Большинство эмбрионов с
хромосомными аномалиями нежизнеспособны и самопроиз-
вольно абортируются. Все хромосомные аномалии связаны с
нарушением интеллекта и поведения. Хромосомные болезни
не излечиваются, поэтому единственный способ их профи-
лактики — предупреждать рождение больных детей.
80
2.18. Хромосомные перестройки
a b ( d cf
X h i! k I m n
и b <: i! It i j k ! m >i
a be tie fd cf g h i i k I »t n
и beg fed hijkImn
Рис. 2.15. Хромосомные перестройки
нормальная
последовательность генов
делеция
дупликация
инверсия
Большинство хромосомных болезней не являются наслед-
ственными в том смысле, что они не передаются от родите-
лей к детям, так как больные, как правило, умирают в детстве,
а если доживают до репродуктивного возраста, то обычно
оказываются бесплодными. В связи с этим хромосомные бо-
лезни представляют собой новые мутации, возникающие в
половых клетках родителей.
Причины хромосомных нарушений разнообразны, но
чаще всего они индуцируются химическими мутагенами и ра-
диоактивным излучением. Поэтому профилактика хромосом-
ных болезней — это, прежде всего, сохранение здоровой эко-
логической обстановки. Серьезным фактором риска хромо-
сомных заболеваний у потомства является возраст матери.
Как у юных, так и у пожилых матерей вероятность рождения
ребенка с хромосомной аномалий повышена. Оптимальным
репродуктивным возрастом для женщины считается 20—35 лет.
В профилактике хромосомных болезней важную роль играет
дородовая диагностика. Существуют методы, при помощи
которых можно исследовать хромосомы эмбриона. Если об-
наруживается хромосомная аномалия, беременность реко-
мендуют прервать.
81
Раздел 2. Материальные носители наследственности
Делеции
Примером делеции у человека является синдром, «кошачьего
крика», при котором наблюдается утрата участка короткого
плеча в пятой хромосоме. Это наиболее распространенная
делеция у человека. У больных детей круглое, лунообразное
лицо, и они издают слабые, жалобные крики, сходные с мяу-
каньем кошки (рис. 2.16). Со временем такие мяукающие зву-
ки исчезают. IQ больных обычно менее 20, т.е. они в значи-
тельной степени являются умственно отсталыми.
Рис. 2.16. Лицо ребенка с
синдромом «кошачьего
крика»
При транслокации короткого
плеча двадцать второй хромосо-
мы на девятую (обозначается
22р —, 9р+) в 97% случаев наблю-
дается злокачественное заболева-
ние крови — миелоидная лейкемия.
В 7-й хромосоме обнаружена де-
леция, приводящая к развитию
группы аномалий развития, обо-
значаемых как синдром Вильямса.
Дети с этим синдромом друже-
любны и сохраняют способность
понимать состояние другого чело-
века по выражению его лица.
В 17-й хромосоме есть делеция,
приводящая к развитию синдро-
ма Смита—Маджениса. Поведе-
ние детей с этим синдромом отли-
чается агрессивностью, импуль-
сивностью, взрывчатостью.
Транслокации
При транслокации различные (негомологичные) хромосомы
обмениваются участками или объединяются. Некоторые ви-
русы (например, вирус, вызывающий свинку, лекарства (в
частности, противораковые) и радиация (рентген-облучение
или УФ-облучение) могут вызывать транслокации,
2.18. Хромосомные перестройки
При нереципрокной транслокации участок одной хромо-
сомы отрывается и присоединяется к другой хромосоме. На-
пример, у 97% больных хронической миелоидной лейкемией
конечный участок хромосомы 22 отрывается и присоединя-
ется к хромосоме 9.
При реципрокной транслокации две различные хромосо-
мы обмениваются участками. Если при хромосомном обмене
не происходит разрыва никаких генов, человек с двумя транс-
локационными хромосомами здоровый, хотя является транс-
локационным носителем. У такого человека нормальное ко-
личество генетического материала, но он перегруппирован.
Транслокационный носитель продуцирует некоторые «не-
сбалансированные» гаметы — сперматозоиды или яйцеклетки
с делециями или дупликациями генов в транслокационных хро-
мосомах. Одна из распространенных хромосомных транслока-
ций происходит между хромосомами 14 и 21. Когда в семье встре-
чаются характерные врожденные дефекты и эмбриональная
гибель, генетик обычно предполагает транслокационное носи-
тельство. Транслокация 21-й хромосомы вызывает, например,
около 6% случаев транслокационного синдрома Дауна.
Инверсии
При инверсии участок внутри хромосомы повернут на 180?.
Взрослый человек, являющийся гетерозиготой по инверсии
в одной хромосоме может оставаться здоровым, но иметь реп-
родуктивные проблемы. Это может произойти из-за того, что
инвертированная хромосома и ее неинвертированный гомо-
лог обернутся друг вокруг друга во время мейоза и при крос-
синговере произойдет образование хромосом с отсутствую-
щими или лишними генами. Недостаток или избыток генов
может вызвать гибель эмбрионов или врожденные дефекты.
Изохромосомы и кольцевые хромосомы
Еще одной мейотической ошибкой, которая нарушает баланс
генетического материала, является формирование изохромо-
h 83
Раздел 2 Материальные носители наследственности
Рис. 2.17. Образование
изохромосомы
Рис. 2.18. Кольцевая хромосома
Ломкий
участок
Рис. 2.19. Вид Х-хро-
мосомы при синдро-
ме фрагильной
Х-хромосомы
сом — хромосом с идентичными плеча-
ми. Такое явление происходит, когда
центромера расщепляется в другой плос-
кости (рис. 2.17).
Кольцевые хромосомы образуются
приблизительно с частотой 1:25 000.
Это случается при потере теломерных
участков, тогда «липкие» концы слива-
ются друг с другом (рис. 2.18). Кольце-
вые хромосомы могут возникать при
радиационных облучениях.
Аберрация по типу «ломкой»
Х-хромосомы
К особому типу хромосомных аберра-
ций относится так называемый синд-
ром ломкой, или фрагильной, Х-хромо-
84
2.18. Хромосомные перестройки
Рис. 2.20. Больные с синдромом фрагильной Х-хромосомы
(в детском и взрослом возрасте)
сомы. Синдром фрагильной Х-хромосомы распространен с ча-
стотой около 1:1000. При специальном культивировании Х-хро-
мосом больных людей в среде с низким содержанием фолие-
вой кислоты кончик Х-хромосомы свисает (рис. 2.19), хромо-
85
Раздел 2. Материальные носители наследственности
сома становится более подверженной к разрыву в сайте
Хр27.3.
Клиническое проявление болезни и ломкость Х-хромосо-
мы в локусе Xq27.3 обусловлены многократным увеличени-
ем в первом экзоне гена FMR-1 тринуклеотидного повтора
CGG. У нормальных индивидов число этих повторов колеб-
лется от 5 до 50. При возрастании числа CGG-повторов от 50
до 200 отмечается состояние премутации, а увеличение этого
числа свыше 200 приводит к клиническим проявлениям за-
болевания и выявлению фрагильной, или ломкой, Х-хромо-
сомы. Переход от состояния премутации к полной мутации
возникает только при передаче гена от матери. Отмечается
корреляция между числом амплифицированных CGG-повто-
ров и тяжестью клинических проявлений заболевания.
Человек с синдромом фрагильной Х-хромосомы имеет ха-
рактерные торчащие уши и удлиненную нижнюю челюсть
(рис. 2.20), а кроме того, генитальные аномалии.
У больных наблюдается умственная отсталость, их IQ ле-
жит в пределах 35—50. Большинство пораженных индивидов
социально адаптированы, контактны и доброжелательны,
могут выполнять несложную физическую работу. Тяжелая
умственная отсталость обнаруживается примерно у 15% боль-
ных. Мальчики болеют в 2—3 раза чаще, чем девочки, и забо-
левание у них протекает в более тяжелой форме. Заболева-
ние в последующих поколениях протекает в более тяжелой
форме, т.е. проявляется эффект антиципации.
2.19. Повторный риск хромосомных болезней
Если у родителей нормальные кариотипы, риск хромосомной
болезни для потомства оценивается по эмпирическим данным
для каждого типа аномалий. При этом учитывается возраст
матери, профессиональная вредность, экологическая обста-
новка и т.д. Если у супругов родился ребенок с хромосомной
аномалией, вызванной нерасхождением хромосом при гаме-
тогенезе родителей (такой синдром называется спорадичес-
86
Контрольные вопросы и задания к разделу 2
ким), то вероятность ее повторения у следующих детей нахо-
дится на уровне среднего популяционного. Эмпирический
риск хромосомных аномалий у потомства в зависимости от
возраста матери приведен в табл. 2.2.
Если причиной хромосомного синдрома у ребенка явля-
ется сбалансированная хромосомная перестройка у родите-
лей, то повторный риск рассчитывается на основе закономер-
ностей поведения хромосом в мейозе и эмпирических дан-
ных о выживаемости эмбрионов.
Доля клеток с аномальным хромосомным набором выра-
жается в процентах, который учитывается при вычислении
риска. В случае мозаицизма родителей вероятность рожде-
ния ребенка с хромосомной аномалией рассчитывают по фор-
муле р — Кх/(2—х), где х — доля мозаичного клона, К — ко-
эффициент элиминации гамет, зигот, эмбрионов.
Таблица 2.2
Суммарный популяционный риск трисомий 13, 18 и 21
хромосом в зависимости от возраста матери
Возраст матери Вероятность хромосомной аномалии у потомства, %
До 19 лет 20—24 года 25—29 лет 30—34 года 35—39 лет 40—44 года 45 лет и старше 0,08 0,06 0,10 0,20 0,54 1,60 4,20
Контрольные вопросы и задания к разделу 2
1. Как было доказано, что наследственным материалом яв-
ляется ДНК?
2, Перечислите свойства генетического кода.
3. В чем суть процессов транскрипции и трансляции?
В. >7
Раздел 2. Материальные носители наследственности
4. Перечислите молекулярные причины мутаций.
5. Чем отличаются митохондриальные гены от ядерных?
6. Чем отличается наследование митохондриальных призна-
ков от ядерных?
7. Какое количество аминокислот закодировано в цепочке
ДНК, состоящей из 1500 пар нуклеотидов, если в ней не
содержится неинформативных последовательностей?
8. Сколько аминокислот закодировано в цепочке РНК, со-
стоящей из 189 нуклеотидов, если два триплета являются
бессмысленными ?
9. Цепочка молекулы ДНК из 1444 нуклеотидов, кодирую-
щих белок, содержит 5 интронных участков длиной 100,
120, 135 и два по 150 нуклеотидов. Сколько аминокислот в
кодируемом белке?
10. Какие изменения произойдут в первичной структуре бел-
ка, если на интронном участке произойдет замена несколь-
ких нуклеотидов?
11. Какая последовательность образуется с матрицы АААААА
при участии фермента РНК-полимеразы?
12. Перечислите особенности, положенные в основу класси-
фикации хромосом человека.
13. Что представляют собой гетерохроматиновые районы хро-
мосом?
14. Сколько хромосом содержится в нормальных спермато-
зоидах человека?
15. Сколько хромосом в нормальных соматических клетках
человека?
16. Какой набор половых хромосом в лейкоцитах женщины?
17. Какой набор половых хромосом в эпителиальных клетках
мужчины ?
18. Какую информацию несет запись, сделанная с помощью
цитогенетической символики?
19. Перечислите аутосомные синдромы и синдромы половых
хромосом.
20. Какие клеточные процессы приводят к образованию ане-
уплоидных клеток?
88
Контрольные вопросы и задания к разделу 2
21. Какие клеточные процессы приводят к полиплоидизации
клеток ?
22. В результате каких процессов образуются псевдонормаль-
ные кариотипы?
23. Как возникает однородительская дисомия?
24. Перечислите процессы, приводящие к образованию орга-
низмов — хромосомных мозаиков.
25. В консультацию обратилась супружеская пара, первый
ребенок которых имел трисомию по 18-й хромосоме. Жене
36 лет, мужу 42. Кариотип обоих супругов нормальный. Ка-
кова повторная вероятность рождения у них ребенка с ане-
уплоидией?
26. У супругов первый ребенок здоров, у второго трисомия
по 13-й хромосоме. В настоящее время супругам по 28 лет.
Каков риск повторения трисомии у их следующего ребен-
ка, если при кариотипировании у них не выявлено анома-
лий хромосомного аппарата?
27. У женщины легкая форма синдрома Дауна — она мозаик
по клеткам трисомикам. Доля аномального клеточного кло-
на 0,25. У мужа нормальный хромосомный набор. Оцени-
те вероятность рождения у нее ребенка с синдромом Дау-
на, если коэффициент элиминации анеуплоидных зигот и
эмбрионов 0,5.
28. У женщины 15% клеток с трисомией по 18-й хромосоме.
Оцените вероятность рождения у нее ребенка с синдро-
мом Эдвардса, если примерно половина зигот-трисомиков
элиминируется.
29. У женщины обнаружено центрическое слияние двух 21-х
хромосом. Какова вероятность рождения у нее здорового
ребенка?
10. Дети синдромом Патау рождаются с частотой примерно
> 1:50 000. Какова вероятность повторения этого заболева-
ния у следующего ребенка, если у одного из супругов об-
наружена сбалансированная транслокация 13-й хромосо-
мы на 20-ю, а коэффициент элиминации анеуплоидных
; зигот 0,5?
к
89
Раздел 2. Материальные носители наследственности
31. У женщины, имеющей ребенка с синдромом Дауна, выяв-
лена траслокация 21-й хромосомы на 13-ю. Какова веро-
ятность рождения у нее здорового ребенка, если коэффи-
циент элиминации анеуплоидных эмбрионов 0,5?
32. Супружеская пара (женщине 30 лет, мужчине 55) обрати-
лась в консультацию по поводу рождения у них ребенка с
трисомией по 21-й хромосоме. Они считают, что причи-
ной аномалии у ребенка является возраст отца и хотят
уточнить вероятность рождения у них здорового ребенка.
Какова эта вероятность?
33. Используя цитогенетическую символику, напишите фор-
мулы хромосомных наборов клеток, с такой характерис-
тикой кариотипа: 1) трисомия по 14-й хромосоме; 2) моно-
сомия по 10-й хромосоме; 3) изохромосома, образованная
длинными плечами 15-й хромосомы; 4) центрическое сли-
яние 21-й и 13-й хромосом; 5) транслокация длинного пле-
ча 2-й хромосомы на короткое плечо 8-й.
34. Опишите особенности кариотипов, обозначенных следу-
ющими формулами:
a) 46,XX,inv(5p + q-);
б) 46,XX,t(13q;22q);
в) 46,XY,del(2)(q23);
г) 46,XY,del(l)(p34).
35. Во время митоза в культуре нормальных клеток человека
восемнадцатая хромосома, не разделившись на хромати-
ды, направилась к одному из полюсов. Какое количество
хромосом в дочерних клетках?
36. В культуре клеток человека с нормальным хромосомным
набором после облучения ее рентгеновскими лучами на-
рушилось расхождение хромосом. В одной из клеток че-
тырнадцатая и двадцатая хромосомы не разделились на
хроматиды и направились к противоположным полюсам
деления. Сколько хромосом и какие содержит каждая из
дочерних клеток?
37. Оцените потенциальную жизнеспособность эмбрионов че-
ловека со следующими хромосомными наборами;
Контрольные вопросы и задания к разделу 2
а) трисомия по 2 хромосоме;
б) моносомия по 5 хромосоме;
в) трисомия по 20 хромосоме;
г) тетрасомия по Х-хромосоме;
д) дисомия по Y-хромосоме.
38. На каждые 1000 новорожденных мальчиков приходится
один с синдромом Клайнфельтера (XXY), а на каждые 700 —
один с синдромом Жакоб (XYY). Объясните возможный
цитогенетический механизм происхождения данных син-
дромов.
39. В 1961 г. во Франции было зарегистрировано необычное
явление — рождение разнополых монозиготных близне-
цов. Один из близнецов был здоровым мальчиком, второй —
девочка с аномалиями развития. Какова причина этого яв-
ления? Какая болезнь у девочки?
40. У кого из родителей и на какой стадии мейоза произошло
нарушение, приведшее к появлению потомков с такими
хромосомными наборами: 1) XXX, 2) XXXXY, 3) XXY,
4) XYY?
41. При кариологическом исследовании у женщины обнару-
жены три типа клеток с хромосомными наборами: 45,X;
46,XX и 47,XXX. Объясните причину такого мозаицизма
женщины.
42. А) Объясните происхождение у людей следующих типов
мозаицизма, приводящих к интерсексуальному состоя-
нию: X0/XY, X0/XYY, X0/XXY, XX/XY, XX/XXYY, Х0/ХХ/
XY, XX/XXY/XXYYY.
Б) Каково происхождение у мужчин таких типов мозаициз-
ма: XY/XXY, XY/XXXY, XXXY/XXXXY, XXXX/XXXXY?
В) Как произошли женщины с таким типом мозаицизма: Х0/
XX, ХО/ХХХ, ХХ/ХХХ, ХХХ/ХХХХ, X0/XX/XXX, XX/
ххх/хххх?
43. Отцы с синдромом Дауна неизвестны. У женщин с синд-
ромом Дауна иногда рождаются дети — здоровые, с три-
сомией по 21-й хромосоме и мертвые. Почему у этих жен-
щин возможно рождение здоровых детей?
91
Раздел 2. Материальные носители наследственности
44. Половой хроматин — инактивированная Х-хромосома —
является у млекопитающих способом компенсации дозы
генов, сцепленных с Х-хромосомой. Сколько телец поло-
вого хроматина наблюдается в клетках мужчины с тетра-
сомией по Х-хромосоме?
45. Какой наиболее вероятный набор половых хромосом у
женщины, если в ее клетках не обнаружено полового хро-
матина?
46. Сколько телец полового хроматина содержится в клетках:
1) больного с синдромом Клайнфельтера; 2) женщины с
синдромом Тернера; 3) женщины с синдромом Дауна; 4) жен-
щины с нормальным хромосомным набором; 5) мужчины
с нормальным хромосомным набором; 6) мужчины с хро-
мосомным набором XYY; 7) женщины с тетрасомией по
Х-хромосоме?
47. У мужчины с умственной отсталостью в клетках эпителия,
взятого с внутренней стороны щеки, обнаружено тельце
Барра. Какова возможная причина его умственной отста-
лости?
48. У женщины с небольшими отклонениями в психике в клет-
ках эпителия обнаружено два тельца полового хроматина.
Какова возможная причина ее психических отклонений?
49. Каков молекулярный механизм геномного импринтинга?
50. У Анатолия сбалансированная дупликация в 5-й хромосо-
ме. Какова вероятность рождения у него ребенка с нор-
мальным хромосомным набором?
51. У Веры сбалансированная дупликация в 11-й хромосоме.
Какова вероятность рождения у нее ребенка с аномалией
по 11-й хромосоме, если у мужа нормальный хромосом-
ный набор?
52. Супружеская пара обратилась в медико-генетическую
консультацию по поводу неоднократного самопроизволь-
ного прерывания беременности. Был проведен цитогене-
тический анализ. У жены обнаружена транслокация 22-й
хромосомы на 15-ю. Возможно ли рождение здоровых де-
тей у этой супружеской пары?
Раздел 3
НАСЛЕДСТВЕННОСТЬ И СРЕДА
3.1. Модификационная изменчивость
Анализируя изменчивость любого организма, исследователь
убеждается в том, что многие различия между особями зави-
сят от окружающей среды. Даже при одинаковом генотипе
две особи, если они в процессе развития находились в раз-
ных условиях, могут быть фенотипически непохожими. Фе-
нотипические различия, вызываемые внешними факторами
у наследственно одинаковых организмов, называкУгся моди-
фикациями.
Понимание природы модификаций имеет большое прак-
тическое значение. Зная, как реализуется тот или иной гено-
тип в различных условиях среды, можно целенаправленно
влиять на формирование у организма полезных признаков и
создавать условия для того, чтобы нежелательные признаки
не проявились. Все это важно для специалистов, имеющих
дело с развитием человека, — медиков, педагогов, психоло-
гов. Для воспитания здоровых людей необходимо знать, как с
помощью гигиенических мероприятий и образовательных
программ вызывать у развивающегося организма полезные
модификации и как устранять возможность возникновения
вредных.
Модификации — это реакции организма на условия сре-
ды. Одно и то же воздействие вызывает вполне определен-
ную модификацию у всех наследственно схожих особей. Глав-
ная особенность модификационной изменчивости — ее впол-
не определенная направленность. Этим модификации
отличаются от мутаций, которые, как правило, лишены на-
правленности.
Раздел 3. Наследственность и среда
Большинство наиболее заметных модификаций, которые
встречаются у животных и человека, известны из повседнев-
ных наблюдений. К числу обычных модификаций относятся
фенотипические изменения, связанные с питанием. Измене-
ние количества пищи модифицирует вес, изменение ее каче-
ства различным образом сказывается на физическом и пси-
хическом здоровье. Некоторые болезненные состояния чело-
века вызваны недостатком в пище тех или иных витаминов:
это, к примеру, цинга при недостатке витамина С, куриная
слепота при недостатке витамина А и др. Наиболее частыми
модификациями являются фенотипические изменения, вы-
зываемые физическими нагрузками. В результате трениров-
ки увеличивается объем мышц, усиливается кровообращение.
При малоподвижном образе жизни возникают обратные из-
менения. В горных условиях в крови человека увеличивается
количество эритроцитов и содержание гемоглобина.
Модификации представляют собой полезную приспосо-
бительную реакцию организма на тот или иной внешний фак-
тор. Кожа человека отвечает загаром на солнечное облуче-
ние, а загорелая кожа лучше противостоит вредному дей-
ствию ультрафиолетовых лучей. Повышение содержания
гемоглобина у людей, попавших в горы, представляет собой
приспособление для лучшего использования кислорода в ус-
ловиях разреженности воздуха. Большинство модификаций
имеют адаптивное значение для организма и полезны для
приспособления в постоянно меняющихся условиях среды.
В отличие от мутаций, модификации не передаются по на-
следству, хотя это было очевидным не всегда. В течение дли-
тельного времени среди биологов шел спор о том, передают-
ся ли потомству признаки, приобретенные родителями в те-
чение жизни. Могут ли потомки унаследовать признаки,
которые сформировались у их родителей под действием спе-
цифических условий среды? Иными словами, разовьются ли
у потомков эти же признаки, даже если факторы, породив-
шие их у родителей, будут отсутствовать? Этот вопрос имеет
не только теоретическое, но и практическое значение. Ведь
94
3.1. Модификационная изменчивость
если модификации наследуются, то достаточно целенаправ-
ленно изменить свойства родителей, чтобы их многочислен-
ное потомство уже без специального воздействия имело же-
лаемые признаки. Понятно, какое значение имеет решение
этого вопроса для сельского хозяйства, медицины, а также в
психолого-педагогической практике. В последнем случае на-
следование приобретенных признаков сильно бы облегчило
воспитательную работу: «правильно» воспитав одно поколе-
ние, можно было бы не беспокоиться о последующих.
Проблема наследования результатов научения долго вол-
новала ученых. Сторонники идеи наследования приобретен-
ных признаков, считали, что условные рефлексы, выработан-
ные родителями, могут передаться потомкам. Первым, кто
сделал попытку экспериментально проверить эту идею, был
русский физиолог И.П. Павлов. Смысл этих опытов состоял в
следующем. У мышей путем многократных повторений вы-
рабатывали условный рефлекс по звонку бежать к кормуш-
ке. От обученных мышей получали потомство и выясняли,
через сколько повторений у него вырабатывается рефлекс.
Идея наследования результатов обучения не получила экспе-
риментального подтверждения. Это имеет важное практичес-
кое значение, так как опровергает утверждение, что при про-
чих равных условиях у более образованных родителей долж-
ны рождаться более одаренные дети, чем у родителей, не
получивших образования.
Это справедливо и в отношении морфологических призна-
ков. Немецкий ученый Август Вейсман (1834—1914) в тече-
ние 22 поколений отрезал мышам хвосты, а затем получал от
них потомство. Длина хвостов не менялась. Опыты Вейсма-
на, безусловно, дали важную научную информацию, однако
еще больше примеров, и, может быть, более убедительных,
могут привести собаководы. Щенкам некоторых пород обре-
зают хвосты и уши. Так делают в течение многих поколений,
а хвосты и уши все равно от рождения не соответствуют стан-
дарту. Интересные факты можно найти и у человека. К при-
зеру, обряд обрезания, принятый у иудеев, если судить по
Раздел 3. Наследственность и среда
религиозным книгам, существует, по крайней мере, 4000 лет.
Это значит, что в течение не менее чем 160 поколений, про-
водился «естественный эксперимент», а еврейские мальчи-
ки и до сих пор рождаются необрезанными. Если бы наследо-
вались приобретенные признаки, после войн от искалечен-
ных мужчин рождалось бы столько калек, что можно было бы
говорить об эпидемии увечий.
В настоящее время вопрос о наследовании модификаций
решен окончательно. Изменения организма, не затрагивающие
генотип, биологически не наследуются. Большинство факторов,
которые влияют на организм человека в течение его развития и
жизни (питание, тренировки, температурное воздействие, обу-
чение и др.), не изменяют генотип. Изменения генотипа (му-
тации) происходят под действием других причин.
Способность давать мутации (мутировать) представляет
собой универсальное свойство всех живых организмов, от
вирусов до человека. Мутация — это внезапно возникающее
изменение наследственного материала. Мутационные изме-
нения чрезвычайно разнообразны. Они могут затрагивать
любые морфологические, физиологические, биохимические
характеристики организма. Мутации могут быть доминант-
ными и рецессивными; вызывать резкие или едва заметные
отклонения от нормы; повышать и снижать жизнеспособ-
ность организма. Мутации, возникающие в половых клетках
(генеративные), передаются по наследству. Мутации, возни-
кающие клетках тела, которые не участвуют в размножении
(соматические мутации), потомкам не передаются.
Классифицируют мутации по характеру изменения гене-
тического аппарата, обозначая их как генные и хромосомные.
Генные мутации представляют собой переход генов из одно-
го аллельного состояния в другое. Этот процесс связан с из-
менениями на уровне молекулы ДНК. Все разнообразие ге-
нов существует благодаря мутационному процессу. Мутации
присущи самой молекулярной природе генов. При копиро-
вании молекул ДНК изредка происходят ошибки. Передача
любой информации происходит с ошибками: когда не было
96
3.2. Норма реакции
печатных станков, книги переписывались от руки и при пе-
реписке иногда случались ошибки, которые изменяли смысл
текста. Среди хромосомных мутаций выделяют численные и
структурные изменения хромосом. Первые называются ге-
номными мутациями, вторые — хромосомными перестрой-
ками, или аберрациями.
3.2. Норма реакции
Фенотип организма определяется генетической информаци-
ей, полученной от родителей, и теми конкретными условия-
ми среды, в которых эта информация реализуется. Среда мо-
дифицирует признаки. Степень модификации обычно про-
порциональна силе и продолжительности действия на
организм модифицирующего фактора. Иными словами, чем
интенсивнее и дольше действие фактора, тем сильнее изме-
няется фенотип. Однако изменчивость признаков не беспре-
дельна. Относительная роль генотипа и факторов среды в фор-
мировании различных признаков неодинакова. У человека
можно проследить всю гамму переходов от признаков, пол-
ностью определяемых генотипом (группа крови, цвет глаз),
через такие, на которые факторы среды налагают заметный
отпечаток (рост, вес), к признакам, полностью зависящим от
условий среды (навыки и умения).
У всех организмов характер фенотипических изменений,
вызываемых влиянием среды, обусловлен генотипом. Способ-
ность организма отвечать на действие факторов среды опре-
деленным образом, теми, а не иными модификациями, назы-
вается нормой реакции. Представление о норме реакции име-
ет важное значение в сельском хозяйстве. Зная генетически
обусловленную норму реакции растений того или иного сор-
та, животных той или иной породы, можно создавать специ-
альные условия для желаемого развития у них полезных че-
ловеку признаков.
Понимание нормы реакции важно и в генетике человека.
Например, отдельные люди отличаются наследственно обус-
Раздел 3. Наследственность и среда
ловленной непереносимостью некоторых лекарств. Многие
вещества, в том числе продукты питания, вызывают аллерги-
ческую реакцию, причем не у всех людей, а у индивидов с
определенными генотипами. В педагогической практике нор-
ма реакции проявляет себя в том, что разные люди, обучаясь
с помощью одних и тех же методик, достигают неодинаковых
результатов. Особенно наглядные примеры нормы реакции
дает спорт.
В зависимости от того, насколько подвержены признаки
действию внешней среды, их обозначают как признаки с уз-
кой или широкой нормой реакции. К первым относятся белки
крови, ко вторым вес, рост, артериальное давление, уровень
интеллекта. Норму реакции у человека хорошо иллюстриру-
ют разлученные в раннем детстве генетически идентичные
индивиды - монозиготные близнецы. Признаки с узкой нор-
мой реакции у партнеров идентичной близнецовой пары, вы-
росших в разных условиях, оказываются одинаковыми, а при-
знаки с широкой нормой реакции могут существенно разли-
чаться. Наиболее широкую норму реакции демонстрируют
поведенческие признаки.
3.3. Экспрессивность и пенетрантность
Один и тот же ген может проявляться по-разному у разных
особей. Особенности проявления гена в признаке русский
ученый Н.В. Тимофеев-Рессовский обозначил терминами эк-
спрессивность и пенетрантность.
Под экспрессивностью понимается степень выраженнос-
ти признака. Большинство генов проявляется в признаке у
всех носителей соответствующего генотипа. Это полностью
пенетрантные гены. Некоторые гены, они получили назва-
ние не полностью пенетрантных, проявляются не у всех их
обладателей. Пенетрантность как возможность гена про-
явиться в признаке имеет количественную выраженность и
представляет собой процент индивидов с признаком среди
всех обладателей соответствующего генотипа. Другими сло-
98
3.3. Экспрессивность и пенетрантность
вами, под пенетрантностью понимают долю индивидов, у ко-
торых признак выражен.
На пенетрантность и экспрессивность гена влияют наслед-
ственные и средовые причины — другие гены, пол, возраст,
специфические воздействия. У человека не полностью пенет-
рантными обычно являются доминантные гены, контролирую-
щие патологическйе признаки. Неполная пенетрантность на
родословной проявляется в виде «проскакивающих» поколе-
ний. В родословной на рис. 3.1 встречаются люди с полидак-
тилией (многопалостью). Цифры внизу обозначают количе-
ство пальцев на левой и правой руке (первая пара цифр) и
левой и правой ноге (вторая пара цифр). У человека полидак-
тилия наследуется по аутосомно-доминантному типу с непол-
ной пенетрантностью. «Проскакивающее» поколение на ро-
дословной видно на индивиде с генеалогическими координа-
тами III/1, поскольку у него больные дети.
У людей с геном полидактилии может быть по пять, шесть
или семь пальцев на разных руках и ногах, т.е. наблюдается
различная его экспрессивность (рис. 3.2). Этот ген может быть
выражен в признаке настолько слабо, что не проявляется при
обычном наблюдении: у некоторых людей лишний палец
представлен маленькой косточкой, полностью закрытой ко-
жей. При этом кисть выглядит обычной пятипалой и только
рентгенологический анализ позволяет обнаружить зачаток
шестого пальца.
Плейотропия — это действие одного гена на многие при-
знаки. При синдроме Марфана, аутосомно-доминантном за-
болевании с неполной пенетрантностью и варьирующей эк-
спрессивностью, у человека может наблюдаться высокий
рост, гиперподвижность суставов, нарушение работы сердца
и дефекты хрусталика глаза (рис. 3.3). Могут наблюдаться не
все из этих симптомов или же человек может быть феноти-
пически здоров, т.е. ген синдрома Марфана может не про-
явиться.
Существуют признаки, которые проявляются у человека
с возрастом. Например, витилиго (депигментация отдельных
к
99
100
Рис. 3.1. Родословная с полидактилией
ж
К Ж
3.3. Экспрессивность и пенетрантность
Рис. 3.2. Полидактилия рук
Рис. 3.3. Руки человека с синдромом Марфана
Рис. 3.4. Витилиго на лбу
Раздел 3. Наследственность и среда
участков кожи) — аутосомно-доминантная болезнь с пенет-
рантностью около 50% (рис. 3.4), проявляющаяся в возрасте
до 20 лет, хорея Гентингтона — аутосомно-доминантная бо-
лезнь, пенетрантность которой зависит от возраста, у обла-
дателей данного гена проявляется в основном после 30 лет.
3.4. Компенсация дозы гена
(инактивация Х-хромосомы)
Женщины имеют два аллеля каждого гена Х-хромосомы, в то
время как мужчины лишь один. Механизм, названный Х-инак-
тивацией, позволяет сгладить разницу в количестве сцеплен-
ных с полом генов у мужчин и женщин. В раннем эмбриональ-
ном развитии девочки одна Х-хромосома в каждой клетке
инактивируется. Это происходит примерно на 3-й неделе раз-
вития человека. «Выключение» одной из двух Х-хромосом
(полученных от матери или от отца) является случайным про-
цессом. В результате у женщины проявляются гены отцовс-
кой Х-хромосомы в одних клетках и материнской Х-хромо-
сомы — в других.
При изучении женщин, у которых присутствует не вся
Х-хромосома, а лишь ее фрагмент, обнаружен участок, вы-
ключающий Х-хромосому, и называемый X-инактивацион-
ным центром. Предполагается, что Х-инактивация происхо-
дит в одном гене в определенное время.
Женщина является генетическим мозаиком, поскольку в
одних клетках проявляется один аллель, а в других — другой.
Клетка с определенной инактивированной Х-хромосомой об-
разует дочерние клетки с такой же самой инактивированной
Х-хромосомой. Поскольку инактивация происходит в раннем
эмбриогенезе, взрослая женщина имеет фенотипически раз-
личные популяции клеток по выраженности сцепленных с
полом генов.
Гетерозиготность защищает женщину от Х-сцепленных
рецессивных болезней. Если женщина наследует один аллель,
определяющий выработку жизненно важного фермента, ко-
102
3.4. Компенсация дозы гена (инактивация Х-хромосомы)
торий отвечает за синтез этого же фермента в неактивном
состоянии, она, вероятно, останется здоровой, поскольку не-
которые из ее клеток будут вырабатывать данный фермент.
Мужчина с таким дефектным аллелем погибает. В редких слу-
чаях гетерозиготная носительница сцепленного с полом гена
может иметь признаки болезни. Например, у женщины-но-
сительницы гена гемофилии в случае инактивации Х-хрбмо-
сомы с нормальным аллелем фактора свертывания крови име-
ются сглаженные симптомы гемофилии, а кровь свертывает-
ся дольше, чем в норме.
Явление Х-инактивации используется для выявления но-
сительниц некоторых генов сцепленных с полом болезней.
Так, генетики могут определить носительниц мышечной дис-
трофии Дюшенна, летальной рецессивной Х-сцепленной бо-
лезни, проявляющейся в прогрессирующей деградации мышц
и мышечной слабости (рис. 3.5). У женщины с подозрением
на носительство проводят мышечную биопсию. Образец тка-
ни окрашивается для обнаружения белка дистрофина, который
отсутствует у больных. Если женщина является носительницей,
то одни ее клетки синтезируют дистрофин, а другие — нет. Та-
ким образом, у носительницы мышечной дистрофии Дюшен-
на по сути гибридный состав мышц. Такие женщины могут
испытывать легкие симптомы мышечной слабости.
Рис. 3.5. Симптомы мышечной дистрофии
Дюшенна у мальчика
103
Раздел 3. Наследственность и среда
а
Рис. 3.6. Тельце Барра в ядре женщины (а) и
его отсутствие в ядре мужчины (б)
Инактивацию Х-хромосомы можно наблюдать в клетках
женщины при цитологических исследованиях. Инактивиро-
ванная Х-хромосома окрашивается в намного большей сте-
пени, чем активная Х-хромосома. В интерфазе в ядре клетки
женщины можно обнаружить темно-окрашенную инактивиро-
ванную Х-хромосому, которая называется тельце Барра в честь
канадского ученого Мюррея Барра, впервые обнаружившего в
1949 г. такие тельца в нервных клетках кошек. В норме клетка
мужчины не содержит тельца Барра, поскольку его единствен-
ная Х-хромосома остается активной (рис. 3.6).
В 1961 г. английский генетик Мери Лайон предположила,
что тельце Барра представляет собой инактивированную
Х-хромосому и такая инактивация происходит в раннем эмб-
риогенезе и является необратимой. Она акцентировала свое
внимание на том, что для гомозигот по Х-сцепленным генам
Х-инактивация не имеет каких-либо эффектов. В этом слу-
чае не имеет значения, какая Х-хромосома выключена. Од-
нако при гетерозиготности Х-инактивация приводит к функ-
ционированию того или иного, но разных аллелей.
104
3.5. Признаки, зависимые от пола и ограниченные полом
3.5. Признаки, зависимые от пола
и ограниченные полом
В случае признаков, зависимых от пола, аллель является до-
минантным у одного пола, но рецессивным у другого пола. На
проявление признака могут оказывать влияние гормональные
различия.
Примером признака, зависимого от пола, является алопе-
ция, или облысение, по мужскому типу (рис. 3.7). Аллель об-
лысения (Н) является доминантным у мужчин, но рецессив-
ным у женщин, поэтому больше лысых мужчин, чем женщин.
Гетерозиготный мужчина лысый — (Hh), а гетерозиготная
женщина — (Hh) нет. Для того чтобы у женщины проявилась
лысина, ей необходимо иметь два аллеля, определяющие ало-
пецию (НН).
Рис. 3.7. Наследование алопеции у человека
105
Раздел 3. Наследственность и среда
Джон Адамс
(1735—1826)
Джон Кинси Адамс
(1767—1848)
Чарльз Фрэнсис Адамс
(1807—1886)
Генри Адамс
(1838—1918)
Рис. 3.8. Фрагмент генеалогического дерева семьи Адамсов,
на котором прослеживается наследование алопеции
106
3.6. Мулыпифакториальные признаки
Хорошо известны семьи, где прослеживается алопеция. На
рис. 3.8 приведены четверо мужчин из семьи Адамсов с ало-
пецией: Джон Адамс (второй президент США), Джон Кинси
Адамс (шестой президент США), Чарльз Фрэнсис Адамс (дип-
ломат) и Генри Адамс (историк).
Признаки, ограниченные полом, проявляются лишь у пред-
ставителей какого-то одного пола. Ген, который контролиру-
ет такие признаки, может быть сцепленным с полом или ауто-
сомным. Оба родителя могут передавать потомству гены при-
знаков, ограниченных полом. У человека, например, рост
бороды и размер молочных желез относятся к признакам, ог-
раниченным полом. У женщин не может расти борода, т.к. они
не продуцируют необходимых для этого гормонов. Однако
женщины могут передавать гены, обусловливающие рост бо-
роды, своим сыновьям. Другим примером признака, ограни-
ченного полом, является одна из форм рака груди. Мужчина
может передавать дочери такой ген, но сам при этом не боле-
ет, и такой тип наследования особенно важно учитывать при
диагностике рака груди.
3.6. Мультифакториальные признаки
Большинство признаков и болезней человека зависят не от
одного или двух генов, а от целого комплекса генов и в то же
время от условий внешней среды. Наследование таких муль-
тифакториальных болезней не поддается точному описанию,
как это имеет место в случае менделирующих признаков.
К мультифакториальным болезням относятся, например: псо-
риаз, сахарный диабет, язва, шизофрения, эпилепсия, различ-
ные формы рака, мигрень, ревматизм, ишемическая болезнь
сердца и другие. Для оценки вероятности болезни использу-
ют значения эмпирического риска. Величины эмпирического
риска устанавливают при эпидемиологических обследовани-
ях населения. Результатом таких исследований являются таб-
лицы, которые используют при оценке риска для разных ти-
пов семей. Одной из них является так называемая таблица
107
Раздел 3. Наследственность и среда
Смита (табл. 3.1). Для определения повторного риска при
мультифакториальном заболевании необходима информация
о популяционной частоте, характеризующей распространен-
ность гена в популяции, и о величине наследуемости, кото-
рая указывает на долю генетического влияния на признак.
Таблица 3.1
Повторный риск при мультифакториальном наследовании
(таблица Смита)
Популяционная частота, % Наследуемость, % Больные родители
0 1 2
Больные сибсы
0 1 2 0 1 2 0 1 2
1 80 1,0 6,5 14,2 8,3 18,5 27,8 40,9 46,6 51,6
50 1,0 3,9 8,4 4,3 9,3 15,1 14,6 20,6 26,3
20 1,0 2,0 3,3 2,0 з,з 4,8 3,7 5,3 7,1
0,1 80 0,1 2,5 8,2 2,9 9,8 17,9 31,7 37,4 42,4
50 0,1 1,0 3,2 1,0 3,4 6,9 10,9 15,3
20 0,1 0,3 0,7 0,3 0,7 1,3 0,8 1,4 2,3
Так, в формировании рака груди играют роль различные
факторы. У женщин, имеющих детей, риск снижен по срав-
нению с не рожавшими. Высококалорийная пища с содержа-
нием насыщенных жиров повышает вероятность рака. Если
у матери и тети рак сформировался до менопаузы, то риск
для пробанда составляет 50%. Если заболевание у родствен-
ниц появилось после менопаузы, то риск для пробанда очень
низкий. Для профилактики рака груди у женщин с высоким
риском развития этого заболевания в некоторых странах, в
том числе США, врачи рекомендуют замену пока еще здоро-
вой ткани груди на силиконовый имплантант, хотя материа-
лы последнего тоже могуг быть канцерогенно опасными.
108
3.7. Закономерности клеточных делений
3.7. Закономерности клеточных делений
Основой размножения и развития организма является деле-
ние клеток. Наиболее известны два типа деления — митоз и
мейоз.
Митоз
Биологическая роль митоза состоит в сохранении постоян-
ного числа хромосом в ряду клеточных поколений. При ми-
тозе после удвоения наследственного материала происходит
одно клеточное деление. Дочерние клетки, образующиеся в
результате митоза, содержат практически одинаковый на-
следственный материал, сходный с наследственным материа-
лом материнской клетки.
Митоз является одной из двух составляющих клеточного
цикла: совокупности стадий деления и интерфазы — фазы,
когда клетка не делится.
Профаза является обычно самой длительной фазой деле-
ния. Хроматиды укорачиваются до 4% первоначальной дли-
ны и утолщаются. Ядерная мембрана распадается и образу-
ется веретено деления.
В метафазе нити веретена прикрепляются к центромерам
и хромосомы располагаются вдоль экватора.
Анафаза является очень короткой стадией. Каждая цент-
ромера расщепляется на две, и нити веретена оттягивают до-
черние центромеры к противоположным полюсам, и хрома-
тиды отделяются и называются уже хромосомами.
В телофазе митотическое веретено разрушается, форми-
руется мембрана вокруг’ ядер дочерних клеток и заканчива-
ется деление цитоплазмы.
Максимальное число делений клетки называется пределом
Хейфлика, по фамилии ученого, который открыл этот фено-
мен в 1960-е гг. Так, клетка соединительной ткани фиброб-
ласт из ткани новорожденного делится 35—63 раза, фиброб-
ласт взрослого делится около 14—29 раз. Чем моложе чело-
век, тем больше у него число последующих клеточных
109
Раздел 3. Наследственность и среда
делений. В организме человека различные клетки делятся по-
разному, например, клетки эпителия тонкого кишечника всю
жизнь, нейроны мозга — никогда.
Мейоз
Мейоз — это клеточное деление, связанное с половым раз-
множением организмов. Биологическая роль полового раз-
множения заключается в создании генетически разнообраз-
ного потомства. Наиболее эффективно наследственное раз-
нообразие создается за счет комбинаторики уже имеющегося
генетического разнообразия. Этой цели служит оплодотво-
рение. Биологически этот процесс представляет собой слия-
ние женской и мужской половых клеток, а генетически —
объединение разнокачественного наследственного материа-
ла. Для того чтобы при оплодотворении сохранялся хромосом-
ный набор, при образовании половых клеток происходит
уменьшение числа хромосом вдвое. Редукция числа хромо-
сом обеспечивается двумя клеточными делениями, которые
следуют за одним удвоением наследственного материала.
Первое деление — редукционное, или мейоз I, при котором
вдвое уменьшается число хромосом. Второе деление — эква-
ционное, или мейоз И, при котором получаются четыре клет-
ки из двух от первого деления. В результате этих процессов в
четырех дочерних клетках, образовавшихся из одной мате-
ринской, обнаруживается половинное (гаплоидное) число
хромосом. В половой клетке человека каждая хромосома пред-
ставлена одним гомологом. При слиянии половых клеток дип-
лоидный набор хромосом восстанавливается и из оплодотво-
ренной яйцеклетки — зиготы развивается новый многоклеточ-
ный организм. Эти два связанных между собой процесса —
мейоз и оплодотворение — поддерживают постоянное число
хромосом в поколениях организмов, которые размножаются
половым путем.
ПО
3.7. Закономерности клеточных делений
Гаметогенез у человека
Сперматогенез
Во время сперматогенеза диплоидная клетка делится, и из нее
получаются дочерние клетки — сперматогонии. Несколько
сперматогониев соединяются друг с другом цитоплазматичес-
кими мостиками, и они вместе проходят мейоз. Сначала спер-
матогонии накапливают цитоплазму и реплицируют ДНК,
становясь первичными сперматоцитами, которые являются
все еще диплоидными клетками. Затем во время редукцион-
ного деления (мейоз I) каждый первичный сперматоцит де-
лится, формируя две гаплоидные клетки — вторичные спер-
матоциты. В мейозе II из вторичных сперматоцитов образу-
ются сперматиды. Затем сперматиды специализируются и у
них образуются жгутики. После формирования сперматид
цитоплазматические мостики разрушаются и образуются зре-
лые сперматозоиды.
Зрелый сперматозоид содержит три функциональные ча-
сти: гаплоидное ядро, систему передвижения и ферменты для
проникновения в яйцеклетку. Каждый сперматозоид состо-
ит из жгутика, тела и головки. На переднем конце содержит-
ся акросома — небольшой бугорок, где содержатся фермен-
ты для проникновения в яйцеклетку (для растворения внеш-
ней мембраны). В течение жизни у мужчин образуются
миллиарды сперматозоидов.
Сперматогенез обеспечивает защиту от повреждений ге-
нетического материала. Сперматогонии, подверженные воз-
действию токсинов, настолько разрушаются, что никогда не
превращаются в зрелые сперматозоиды. Более зрелые спер-
матозоиды, подверженные воздействию токсинов, теряют
подвижность. Но некоторые токсические лекарства перено-
сятся в семенной жидкости без вреда для сперматозоидов.
Они могут наносить вред матке или попадать в кровеносную
систему женщины и двигаться к плаценте — месту соедине-
ния матери и плода. Например, кокаин влияет на плод по-ино-
му. Он прикрепляется к рецепторам на поверхности сперма-
111
Раздел 3. Наследственность и среда
тозоида без вреда для сперматозоида и переносится в яйцек-
летку.
Оогенез
Развитие яйцеклетки начинается с диплоидной клетки — оого-
ния, которая находится в яичниках. Каждый оогоний растет,
реплицирует ДНК, накапливает цитоплазму и превращается
в первичный ооцит. При оогенезе, в отличие от сперматоге-
неза, клетки получаются разными по размеру. В мейозе I пер-
вичный ооцит делится на небольшое полярное тельце и вто-
ричный ооцит, каждая клетка гаплоидна. В мейозе II поляр-
ное тельце может делиться на две равные клетки, а может и
не делиться. Вторичный ооцит делится на две неравные час-
ти — маленькое полярное тельце и зрелую яйцеклетку, или
ооцит. Таким образом, каждый оогоний потенциально дает
4 клетки, но лишь одна становится яйцеклеткой. Затем поляр-
ное тельце разрушается и не играет роли в дальнейшем раз-
витии. Однако 1 из 100 спонтанных абортов вызван оплодот-
ворением именно полярного тельца.
В яйцеклетке находится запас питательных веществ для
нового организма. Временные рамки для развития яйцеклет-
ки сильно отличаются от временных рамок для развития спер-
матозоида. За б месяцев до рождения, т.е. в трехмесячном
внутриутробном возрасте плод женского пола содержит око-
ло 2 млн. первичных ооцитов. С этого времени их число умень-
шается. При рождении содержится около 1 млн., а к периоду
полового созревания остается около 400 000 ооцитов. При
рождении ооциты находятся в стадии профазы I. После поло-
вого созревания один или несколько ооцитов завершает мей-
оз I ежемесячно. Затем эти ооциты снова приостанавливают
мейоз, но уже на стадии метафазы II. Гормоны ежемесячно
стимулируют яичник к овуляции, или освобождению вторич-
ного ооцита. Если происходит оплодотворение, то мейоз за-
вершается. Таким образом, оогенез не завершается без опло-
дотворения, а неоплодотворенный вторичный ооцит покида-
112
3.8. Материнский эффект
ет организм женщины во время менструации. В течение жиз-
ни у женщины овуляция происходит около 400 раз (от перио-
да полового созревания до менопаузы).
Генетики пытаются установить некоторые наследствен-
ные болезни по изучению полярного тельца, удаляя яйцеклет-
ку и прикрепившееся к нему полярное тельце (поскольку обе
клетки имеют один генетический материал). После такого
исследования яйцеклетка снова вносится в организм после
оплодотворения in vitro, если анализ полярного тельца не об-
наружил дефектов.
3.8. Материнский эффект
Биологи накопили множество фактов, свидетельствующих о
том, что потомки по некоторым признакам больше напоми-
нают своих матерей, чем отцов. Это явление, получившее на-
звание материнский, эффект, обнаружено у растений, живот-
ных и у человека. Вот некоторые примеры. Длина тела и вес
новорожденного ребенка сильнее коррелирует с соответствую-
щими признаками матери, чем отца. Старение супругов уве-
личивает вероятность рождения у них ребенка с наследствен-
ными патологиями. При этом у пожилых матерей вероятность
рождения детей с хромосомным заболеванием повышается в
десятки раз, а у пожилых отцов увеличивается незначительно.
Материнский эффект обусловлен тем, что при развитии
потомства организм матери и среда, которую она создает,
оказывают большее влияние, чем организм отца и создавае-
мая им среда. Среда, влияющая на формирование признаков
человека, понимается широко. Это и внутриутробная среда,
и семейное окружение, и социальное положение родителей,
а также многое другое.
Уже при зачатии мать вкладывает в ребенка больше на-
следственной информации, чем отец, так как, кроме ядерных
хромосом, женская половая клетка содержит еще и тысячи
копий митохондриальных генов. В сперматозоиде митохонд-
рии располагаются лишь в середине и на конце жгутика, ко-
113
Раздел 3. Наследственность и среда
торый при оплодотворении не проникает в яйцеклетку. В го-
ловке же сперматозоида митохондрий не содержится.
Генетические особенности матери начинают сказываться на
ребенке еще в период его внутриутробного развития. Беремен-
ная женщина и плод физически связаны, поэтому особенности
организма матери влияют на развивающийся плод как комплекс
средовых факторов. Иногда это действие оказывается неблагоп-
риятным. Наиболее известный пример такого рода — гемоли-
тическая болезнь новорожденных. Это заболевание возника-
ет из-за иммунологической несовместимости матери и плода
по группе крови системы резус (рис. 3.10).
Болезнь возникаету резус-положительных детей, рожден-
ных резус-отрицательными матерями, и проявляется в виде
желтухи, обусловленной разрушением у ребенка красных
кровяных клеток — эритроцитов. Причина болезни состоит
в том, что в организме резус-отрицательной матери выраба-
тываются антитела против белка, называемого резус-факто-
ром. Этот белок имеется на мембране эритроцитов резус-по-
ложительного плода. Материнские резус-антитела разруша-
ют резус-положительные эритроциты ребенка еще до его
рождения. При этом страдают различные системы органов
ребенка, а особо тяжелые случаи болезни приводят к его ги-
бели. Путем срочного переливания крови можно избавить
ребенка от последствий иммунологического конфликта.
Женщинам, которые страдают некоторыми хронически-
ми заболеваниями —• эндокринными, сердечно-сосудистыми —
ы антитело; @ антиген.
Рис. 3.10. Возникновение резус-конфликта
114
' 3.8. Материнский эффект "
врачи не рекомендуют рожать детей. При этих заболеваниях
нарушается обмен веществ, ослабевают функции различных
систем организма. Эти нарушения нередко усиливаются при
беременности и неблагоприятно отражаются на развитии
плода. У женщин, больных сахарным диабетом, иногда рож-
даются дети с пороками развития скелета. Эти аномалии у
ребенка вызваны не его генотипом, а обусловлены наруше-
нием обмена веществ у матери в связи с ее заболеванием.
Дети с дефектами скелета нередко рождаются и у женщин,
страдающих заболеваниями щитовидной железы. В организ-
ме таких женщин нарушен обмен кальция, а при его недостат-
ке у плода происходит неправильное формирование костной
ткани.
Особенности материнского организма могут оказать влия-
ние и на умственное развитие ребенка. Наиболее изученный
пример — излеченная фенилкетонурия (болезнь Феллинга).
При этом тяжелом наследственном заболевании, связанным
с нарушением обмена аминокислоты фенилаланина, поража-
ется нервная система. В крови фенотипически здоровых
взрослых людей, излеченных в детстве от этой болезни, по-
вышено содержание токсических продуктов обмена фенила-
ланина. Плод беременных женщин, излеченных от фенилке-
тонурии, в течение всего периода внутриутробного развития
подвергается действию этих опасных для нервной системы ве-
ществ. Дети, рожденные такими женщинами, хотя сами не
имеют наследственной предрасположенности к заболеванию,
обычно обнаруживают пониженные умственные способнос-
ти. Для того чтобы развитие плода протекало нормально, жен-
щине, излеченной от фенилкетонурии, на весь период бере-
менности назначают диету с пониженным содержанием фе-
нилаланина.
На развивающийся плод оказывает влияние не только ге-
нотип матери, но и факторы среды, действию которых она
подвергается. Наиболее опасное действие на развивающий-
ся плод оказывают химические соединения, которые женщи-
на по тем или иным причинам принимает во время беремен-
115
Раздел 3. Наследственность и среда
ности. Некоторым женщинам для сохранения беременности
при угрозе ее самопроизвольного прерывания назначают син-
тетические гормоны — гестагены. Эти вещества оказывают
маскулинизирующее действие на плод женского пола. У ново-
рожденных девочек обнаруживаются признаки, характерные
для мужского пола. Это ненаследственное состояние напоми-
нает наследственную болезнь, известную как адреногениталь-
ный синдром.
Как видим, влияние, которое оказывает материнский орга-
низм на развивающийся плод, чрезвычайно разнообразно по
своим последствиям. Отсюда вытекают важные практичес-
кие выводы. Для того чтобы наследственные задатки потом-
ка проявились наилучшим образом, беременной женщине как
никому другому следует вести здоровый образ жизни. Ей ре-
комендуется полноценное питание, запрещается курение,
употребление алкоголя и наркотиков и т.д.
В первые годы жизни мать продолжает оказывать более
сильное влияние, чем отец. Генетические особенности мате-
ри могут отразиться на ребенке через ее молоко. Исследова-
ниями на животных доказано, что состав молока самки зави-
сит от ее генотипа. У некоторых женщин в молоке присут-
ствуют необычные вещества, которые вызывают у грудных
младенцев нарушение обмена веществ, порождающее раз-
личные заболевания.
3.9. Врожденные дефекты
Развитие эмбриона под воздействием патологического гена
или неблагоприятных факторов окружающей его среды при-
водит к врожденным аномалиям.
Специфическая природа врожденных дефектов обычно
зависит от того, какие структуры развиваются во время дей-
ствия каких-либо негативных факторов. Временной отрезок,
когда гены, токсические вещества, вирусы, излучение и ряд
других влияний могут нарушить структуру определенного
органа или системы, называется критическим периодом. Не-
116
3.9. Врожденные дефекты
которые части человеческого организма, такие, как пальцы
конечностей, чувствительны к подобным воздействиям в те-
чение короткого промежутка времени, а развитие централь-
ной нервной системы может быть нарушено в течение всего
пренатального периода, так же как и в первые два года пост-
натального периода. Следствием длительного критического
периода нервной системы в целом и головного мозга в част-
ности является множество врожденных дефектов, включаю-
щих разную степень умственной отсталости. Мозг ребенка
остается чувствительным к вредным воздействиям и после
рождения.
Около 2/3 всех врожденных дефектов происходит из-за
нарушений развития на эмбриональных стадиях (эмбрионом
считается развивающийся организм в первые три месяца
внутриутробной жизни). Более сглаженные дефекты, кото-
рые становятся заметными лишь по мере взросления ребен-
ка, такие, как затруднения в обучении, часто происходят из-
за нарушений в развитии уже не эмбриона, а плода (плодом
считается развивающийся организм после 3 месяцев внутри-
утробной жизни). Например, химические вещества, влияю-
щие на развитие мозга в первом триместре беременности,
могут вызвать тяжелую умственную отсталость ребенка. Ве-
щество, подействовавшее на седьмом месяце беременности,
может снизить способность к обучению.
Некоторые врожденные дефекты появляются из-за ано-
мального гена, включившегося в определенный период пре-
натального развития. При редком наследственном заболева-
нии фокомелии («ласты моржа») происходит остановка раз-
вития конечностей на третьей-пятой неделе эмбрионального
развития и ребенок рождается с редуцированными конечно-
стями.
Много врожденных дефектов появляется из-за воздей-
.ствия токсических веществ, употребляемых женщиной во
(время беременности. Аномалии развития этого рода не пере-
едаются по наследству, а могут возникнуть лишь при повтор-
ком воздействии повреждающего фактора. Такие факторы
117
Раздел 3. Наследственность и среда
носят название тератогены (от греч. «вызывающий урод-
ства»).
Тератогены
Талидомид
Идея о том, что плацента (орган, развивающийся во время бе-
ременности и связывающий мать и ребенка) защищает эмб-
рион и плод от воздействия вредных веществ, была опровер-
Рис. 3.11. «Талидомидный»
ребенок
гнута в 1957—1961 гг., ког-
да в Европе родилось око-
ло 10 ООО детей с симпто-
мами фокомелии. Было
обнаружено, что матери
всех дефектных детей
применяли в начале бере-
менности препарат тали-
домид. « Талид омидные»
дети рождались с недораз-
витыми конечностями
или вообще без конечно-
стей (рис. 3.11).
В США талидомид не
был разрешен, поскольку
ученые заметили некото-
рые побочные эффекты в
доклинических исследо-
ваниях на животных.
В Южной Америке в 1994
г. родилось несколько «та-
лидомидных» детей, по-
скольку там лекарство
употреблялось женщина-
ми в результате неофици-
ального ввоза его из стран
Европы.
118
3.9. Врожденные дефекты
Краснуха
Австралийские ученые впервые заметили тератогенные эф-
фекты вируса краснухи, вызвавшем в 1941 г. так называемую
«немецкую корь». В США внимание к вирусу краснухи было
привлечено в начале 1960-х гг., когда после эпидемии красну-
хи родилось примерно 20 000 детей с врожденными уродства-
ми и 30 000 мертвых детей. Если вирус краснухи поражает
беременных женщин в течение первого триместра, у них вы-
сокий риск рождения детей с катарактой, глухотой и сердеч-
ными аномалиями. Последствия вируса краснухи у плодов,
развивающихся во втором и третьем триместрах, менее ощу-
тимы. К ним могут быть отнесены нарушения в обучении,
расстройства речи и слуха и ювенильный сахарный диабет.
Алкоголь
Люди давно заметили неблагоприятное действие алкоголя на
потомство. Существует даже понятие «дети карнавала», ко-
торым обозначают неполноценных детей, зачатых во время
массовых праздников, когда спиртные напитки употребляют-
ся в особенно больших количествах. Результатом этих наблю-
дений у некоторых народов стала традиция не разрешать
жениху и невесте во время свадьбы пить спиртные напитки.
Эти наблюдения подтверждены научно. Накоплено множе-
ство фактов, которые показывают, что у людей, страдающих
алкоголизмом, повышен процент неполноценных детей.
Особенно тяжелы для ребенка последствия материнского
алкоголизма. Еще 23 столетия назад нарушения у детей, рож-
денных пьющими матерями, были замечены Аристотелем.
Беременные женщины, периодически употребляющие алко-
голь в умеренном количестве или однократно в большом ко-
личестве в период какого-либо особенно важного этапа в эб-
риогенезе, имеют риск рождения ребенка с фетальным ал-
когольным синдромом (ФАС). Поскольку ученые еще не
Установили последствия употребления малых количеств ал-
коголя беременными и поскольку генотипически разные жен-
119
Раздел 3. Наследственность и среда
щины по-разному метаболизируют это вещество, лучше все-
го полностью отказаться от алкоголя во время беременности
и в период кормления ребенка грудью.
Плод пьющих матерей развивается в среде с повышенным
содержанием алкоголя, поэтому высокая концентрация спир-
та становится для ребенка естественной. Метаболизм плода
адаптируется к высокому содержанию алкоголя. После того
как ребенок теряет связь с телом матери, его организм начи-
нает испытывать недостаток алкоголя. По наблюдениям вра-
чей, такие дети находятся в состоянии похмельного синдро-
ма. Они постоянно плачут и успокаиваются только после того,
как получат некоторое количество алкоголя. Такие дети обыч-
но рано становятся алкоголиками.
Ребенок с фетальным алкогольным синдромом имеет ха-
рактерную маленькую голову, анатомические нарушения
глаз, плоское лицо и плоский нос (рис. 3.12). Его рост замед-
лен как до, так и после рождения. У ребенка ухудшены ин-
теллектуальные способности: от легких нарушений в обуче-
нии до умственной отсталости. В одном исследовании было
показано, что если женщина во время беременности употреб-
ляла до трех смешанных алкогольных напитков ежедневно (в
виде коктейлей), IQ ее ребенка был в среднем на 5 баллов
ниже по сравнению с контрольным. В настоящее время в
США ФАС занимает третье место среди причин умственной
отсталости у детей, и на каждую 1000 новорожденных от 1-го
до 3-х приходится с этим синдромом.
Последствия ФАС проявляются и в более старшем возра-
сте. У подростков и взрослых с этим синдромом невысокий
рост и маленькие головы. Более 80% сохраняют черты лица,
характерные для детей с ФАС, включая нарушение формы
губ, неправильно сформированные и расположенные зубы и
широкий промежуток между верхней губой и основанием
носа. Такие черты лица делают людей с ФАС внешне схожи-
ми, хотя и без особых грубых нарушений.
Многие взрослые с ФАС имеют психическое развитие на
уровне детей младшего школьного возраста. У них часто воз-
120
3.9. Врожденные дефекты
короткий
нос
малая
окружность
головы
низкое
расположение
переносицы
складки
глаза
малое расстояние
от верхней губы
до носа
। тонкая
' ’ верхняя губа
Рис. 3.12. Лица детей с фетальным алкогольным синдромом
никают трудности в общении, им сложно предвидеть послед-
ствия своих поступков, они безынициативны и неправильно
понимают социальные роли в обществе.
Кокаин
Кокаин является чрезвычайно опасным веществом для еще
неродившегося ребенка. Он может вызвать сердечный при-
121
Раздел 3. Наследственность и среда
ступ у эмбриона. Выжившие, но подвергшиеся воздействию
кокаина дети имеют гораздо меньшую способность концент-
рировать внимание на окружающих объектах, чем нормальные
дети. По мере взросления проявляются другие проблемы с фи-
зическим здоровьем и психикой. Оценка последствий действия
кокаина на пренатальных стадиях часто затрудняется тем, что
помимо кокаина происходило воздействие и других веществ,
которые могли быть причиной появившихся симптомов.
Интересным обстоятельством является то, что употребле-
ние кокаина не только будущей матерью, но и будущим от-
цом, может повлиять на развитие ребенка. Дело в том, что
кокаин может быть внесен в организм женщины, как и ряд
других токсических веществ, в составе семенной жидкости,
не повреждая при этом сами сперматозоиды. В дальнейшем
кокаин может вызвать нарушения в матке или попасть в кро-
веносную систему женщины, затем в плаценту и оттуда к раз-
вивающемуся эмбриону или плоду.
Сигареты
Химические компоненты сигаретного дыма оказывают небла-
гоприятное действие на плод. Монооксид углерода проходит
через плаценту и присоединяется к молекулам гемоглобина,
тем самым не давая присоединиться кислороду. Другие токси-
ческие соединения сигаретного дыма затрудняют попадание
питательных веществ к плоду. Исследования, в которых срав-
нивались плаценты курящих и некурящих женщин, показали,
что в плацентах курящих женщин отмечается недостаток важ-
ных факторов роста, вызывая тем самым замедленный рост
ребенка до и после рождения. Курение во время беременно-
сти коррелирует с риском спонтанных абортов, мертворож-
дением, недоношенностью и малым весом при рождении.
Питание
Некоторые питательные вещества в больших количествах
действуют на человеческий организм подобно наркотикам.
3.9. Врожденные дефекты
Лекарство, используемое для лечения акне Accutane являет-
ся производным витамина А, которое вызывает спонтанные
аборты и нарушение развития сердца, нервной системы, при-
водит к лицевым аномалиям. Тяжелые последствия этого пре-
парата впервые были замечены спустя девять месяцев после
того, как они назначались дерматологами молодым женщи-
нам в начале 1980-х гг. В настоящее время лекарство сопро-
вождается соответствующими маркировками и никогда не
назначается беременным. Это лекарство на основе витамина А
также используется для лечения псориаза; и оно само, и из-
быток витамина А вызывают врожденные аномалии. Неко-
торые формы витамина А сохраняются в жировых отложе-
ниях тела в течение трех лет после его употребления.
Другим пищевым продуктом, способным повреждать плод,
является витамин С в избыточных количествах. Плод привы-
кает к большим дозам витамина, употребляемого беременной
женщиной. После рождения, когда ребенок употребляет го-
раздо меньше витамина, у него появляются симптомы недо-
статочности витамина С (цинга) и он становится сильно под-
верженным инфекциям.
Недостаток питания негативно сказывается на развиваю-
щемся плоде. Во время беременности и во время кормления
ребенка грудью женщина нуждается в избыточных калори-
ях. Акушерские записи по наблюдению беременных до, во
время и после Великой Отечественной войны прослеживают
связь между недостаточным питанием женщин на ранних
сроках беременности и повышением риска спонтанных абор-
тов. У абортированных плодов были отмечены слишком ма-
лые размеры головного мозга. Плохое питание на более по-
здних сроках беременности угнетает развитие плаценты, что
может приводить к небольшому весу при рождении, малень-
кому росту, в дальнейшем подверженности зубов кариесу,
задержке полового развития, нарушению в обучении и воз-
можной умственной отсталости.
123
Раздел 3. Наследственность и среда
Профессиональные вредности
Немало людей сталкивается с тератогенами на своем рабо-
чем месте. Ученые отмечают повышение частоты спонтанных
абортов и рождения детей с аномалиями у женщин, работаю-
щих с текстильными красками, свинцом, полупроводниковы-
ми материалами, ртутью и кадмием.
3.10. От гена к поведению
От гена к поведенческому признаку ведет сложная цепь раз-
личных биохимических реакций и физиологических процессов.
Генотип влияет на поведение через ряд уровней: биохимичес-
кий, морфологический, нейрофизиологический. У многоклеточ-
ных организмов отдельные группы клеток приспособлены для
выполнения разных функций. Развитие организма начинает-
ся с зиготы, из которой в ходе многочисленных митозов обра-
зуются специализированные клетки, в которых происходит эк-
спрессия определенного набора генов. Поведение связано с
нервной системы. Основой современных нейрогенетических
исследований является нейронная теория, разработанная
Рамон-и-Кахалем (рис. 3.13).
Автономной единицей нервной системы является нейрон —
нервная клетка с отростками. Нервная система человека сла-
гается из огромного количества нейронов, в коре головного
мозга их насчитывается около 14 миллиардов. Каждый ней-
рон возникает из одной зародышевой клетки — нейроблас-
та. Нейрон состоит из тела клетки, содержащего ядро, и от-
ростков, которые подразделяются на аксон и дендриты. По
дендритам к нейрону поступает нервный импульс, по аксону
нервный импульс распространяется от нейрона к другим клет-
кам. Аксон всегда один. Он может быть длинным, коротким,
разветвленным. Функциональный контакт окончания аксо-
на с другой клеткой получил название синапса. Их количе-
ство у разных нейронов разное. Один нейрон может образо-
вывать связи с тысячами других нервных клеток. Нейроны
124
3.10. От гена к поведению
Рис. 3.13. Рамон-и-Кахаль (а) и его схематическое
изображение нейронов (б)
б
складываются вместе наподобие отдельных звеньев, образую-
щих цепь. Каждый нейрон не сливается, а тесно соприкаса-
ется с соседним нейроном своими отростками. Нейрон с от-
ростками функционально поляризован — каждая цепь слу-
жит для проведения нервных импульсов только в одном
определенном направлении. Один конец цепи (рецепторный)
воспринимает импульс, другой (эффекторный) передает его
исполнительным элементам. Дендриты воспринимают нерв-
ные импульсы, аксон проводит импульсы от нейрона. Аксон
оканчивается на теле или дендритах других нейронов с помо-
щью синаптического контакта, который состоит из трех ком-
понентов: пресинаптической области, синаптической щели и
постсинаптической области. Важной частью синаптического
контакта является участок пресинаптической мембраны, где
происходит выброс медиатора в синаптическую щель. Содер-
жимое синаптической щели обеспечивает направленную
диффузию медиатора и его взаимодействие с рецепторами
(специализированными белковыми комплексами) постсинап-
125
Раздел 3. Наследственность и среда
тической мембраны. Каждый нейрон вырабатывает специфи-
ческий медиатор. Одним из медиаторов является вещество
ацетилхолин. Известны и многие другие химические передат-
чики нервного импульса с возбуждающим или тормозящим
эффектом.
Установлено, что нейронные системы дифференцируют-
ся еще до того, как они начали функционировать. Это так на-
зываемый дофункциональный период развития нервной си-
стемы. На ранних стадиях формирования мозга образуется
огромный избыток синаптических связей, число которых за-
тем постепенно уменьшается. Предполагается, что процесс
отмирания и стабилизации синапсов служит одним из основ-
ных механизмов, с помощью которых жизненный опыт из-
меняет структуру мозга при его формировании. Установле-
но, что способность к выполнению определенных поведен-
ческих актов и к обучению заложены в структуре нервных
сетей. Эти структуры формируются под генетическим конт-
ролем в ходе индивидуального развития. Гены оказывают вли-
яние на поведение, реализуя свое влияние в процессе инди-
видуального развития. Они создают специфическую органи-
зацию мозга, выстраивают нейронные ансамбли, формируют
межнейронные отношения и синаптические контакты. Сло-
жившаяся в онтогенезе морфофункциональная система как
бы подготовлена к осуществлению поведенческих актов та-
ким, а не иным способом. Гены, следовательно, детермини-
руют не поведение как таковое, а специфическую организа-
цию мозга, которая и определяет специфику поведения. При
осуществлении поведенческого акта не происходит актива-
ции специфических генов. Генетическая активность необхо-
дима лишь для восполнения запасов белков, которые утрачи-
ваются при функционировании клеток. Мутации генов, от ко-
торых зависит развитие нервной системы, приводят к
нарушению в соответствующих разделах головного мозга или
определенных типах нервных клеток. Так возникает анома-
лия форм поведения, зависящих от этих отделов мозга или
соответствующих типов клеток.
3.10. От гена к поведению
Генетические исследования функциональной деятельно-
сти мозга связаны с рядом трудностей, которые обусловлены
изменением поведения человека в онтогенезе и множеством
прямых и обратных связей организма со средой. Развитие и
функционирование мозга чувствительно к изменениям био-
химической среды. Нарушения метаболизма приводят к сни-
жению или увеличению различных веществ, появлению ано-
мальных соединений, которые влияют на работу мозга и, сле-
довательно, отражаются в особенностях поведения. У
человека описано множество генетически контролируемых
дефектов ферментных систем, которые приводят к измене-
нию поведения. При синдроме Леша-Найхана из-за недоста-
точности одного из ферментов в крови накапливается боль-
шое количество мочевой кислоты. Больные отличаются по-
вышенной возбудимостью, постоянно совершают какие-то
движения, кусают и калечат себя. Не только аномальные про-
дукты обмена, но также избыток или недостаток нормальных
метаболитов могут нарушать работу нервных клеток. У гете-
розиготных носителей рецессивных генов, ответственных за
болезни обмена веществ, активность соответствующих фер-
ментов примерно в два раза ниже, чем у нормальных гомози-
гот. У гетерозигот по гену фенилкетонурии отмечаются бо-
лее низкие показатели IQ, чем у индивидов с нормальным ге-
нотипом. У них повышена возбудимость коры головного
мозга, увеличен риск психических расстройств. Предполага-
ют, что эти аномалии вызваны увеличением фенилаланина в
нервных клетках.
Небольшие отклонения от нормы обнаруживаются и у ге-
терозиготных носителей по другим рецессивным патологи-
ческим генам. У них отмечаются небольшие отклонения в те-
стах IQ, связанных с выполнением действий, и в тестах на
пространственное восприятие, увеличены показатели по лич-
ностным опросникам, тестирующим психосоматические рас-
стройства, депрессию и эмоциональную лабильность. Причи-
ну этих отклонений усматривают в накоплении аномальных
метаболитов. Считается, что среди лиц, страдающих умствен-
127
Раздел 3. Наследственность и среда
ной отсталостью невыясненной этиологии, значительно чис-
ло гетерозиготных носителей вредных рецессивных генов.
Контрольные вопросы и задания к разделу 3
1. Зависит ли модификационная изменчивость от наслед-
ственности?
2. Полидактилия (многопалость) контролируется аутосомно-
доминантным геном с неполной пенетрантностью. При-
няв, что пенетрантность этого гена составляет около 80%,
рассчитайте вероятность рождения многопалых детей в
семье, где у жены нормальная кисть, а муж шестипалый.
3. Короткопалость (брахидактилия) контролируется аутосом-
но-доминантным геном с полной пенетрантностью.
У жены нормальное строение кисти, муж короткопалый.
Короткопалой была и мать мужа. Какова вероятность того,
что у супругов родятся короткопалые дети?
4. При синдроме Марфана (аутосомно-доминантное заболе-
вание) нарушен синтез коллагеновых волокон, формиру-
ющих соединительную ткань. Составляющие синдрома:
смещение хрусталика глаза, длинные, тонкие конечности,
длинные, «паучьи» пальцы (арахнодактилия), вогнутая
грудная клетка, ослабленная мускулатура тела. Наиболее
серьезное нарушение - ослабление стенки аорты, которое
иногда приводит к ее разрыву. У разных больных наблю-
дается неодинаковое сочетание и выраженность перечис-
ленных признаков. Для объяснения наследственной при-
роды этого заболевания были предложены две гипотезы.
Одна из них утверждает, что три составляющих синдрома
(поражение аорты, поражение хрусталика и скелетной
системы) находятся под контролем отдельных сцепленных
генов, которые могут рекомбинировать при кроссингове-
ре. Согласно второй гипотезе, заболевание контролирует-
ся одним плейотропным геном.
Проверьте обе гипотезы, используя родословную на рис. 3.14.
У Олега болезнь выражена в наибольшей степени по срав-
128
Контрольные вопросы и задания к разделу 3
нению с его сыновьями: у Игоря очень высокий рост, у
Андрея — отклонения в положении зрачков, а у Петра —
характерные для врожденного порока сердца шумы. Все
сыновья Олега женаты на здоровых женщинах. У Игоря —
две девочки, одна здорова, а у другой — Марии — арахно-
дактилия и врожденный порок сердца. У Андрея один здо-
ровый сын, а два других очень высокие с аномалией хрус-
талика. У Петра — здоровый сын и дочь Анна с арахнодак-
тилией и смещением хрусталика.
Олег
Рис. 3.14
5. Каково диагностическое значение тельца Барра?
6. 8% носительниц сцепленной с полом рецессивной мышеч-
ной дистрофии Дюшенна испытывают мышечную сла-
бость. Объясните возможную причину такого состояния
этих женщин.
7. Одна из форм рака груди наследуется как доминантный
признак, ограниченный полом. Зинаида не беспокоилась
по поводу рака, так как ее мать, сестра и родственницы по
материнской линии были здоровы. Однако тетя и двою-
родная сестра Зинаиды со стороны отца больны раком гру-
ди. Маммограмма выявила у Зинаиды начальные призна-
ки заболевания. Определите вероятность заболевания у
сестры Зинаиды Лидии, у ее двоюродной сестры Марии и
у племянниц — дочерей Веры и Петра (рис. 3.15).
Введение в психогенетику
129
Раздел 3. Наследственность и среда
Рис. 3.15
8. Перечислите мультифакториальные признаки человека
9. Какие меры следует принимать людям, у которых имеет-
ся большой риск рождения больного ребенка?
10. Риск развития ревматизма для сибсов и потомков боль-
ных составляет 10% и увеличивается вдвое, если больны
родитель и сибс. Оцените вероятность заболеть ревматиз-
мом у А, Б, В и Г в родословных на рис. 3.16.
Рис. 3.16
И. Гипертоническая болезнь относится к мультифакториаль-
ным заболеваниям. Фактором риска при этой болезни яв-
ляется наследственная отягощенность, избыточный вес
тела, повышенное артериальное давление. Укажите на ро-
дословной (рис. 3.17) лиц, относящихся к группе повышен-
ного риска.
130
Контрольные вопросы и задания к разделу 3
ож. — ожирение, | — повышенное артериальное давление
Рис. 3.17
12. Язвенная болезнь желудка является мультифакториаль-
ным заболеванием, чаще поражающим мужчин, чем жен-
щин. Сравните риск заболевания язвой желудка для чле-
нов III поколения родословной на рис. 3.18.
13. Язвенная болезнь относится к болезням с наследственной
предрасположенностью. Она встречается у 0,9% населе-
ния данной популяции, а наследуемость болезни состав-
ляет 60%. У консультирующегося дядя страдает язвенной
5*
131
Раздел 3. Наследственность и среда
болезнью, а родители и двое братьев здоровы. Какой риск
заболевания язвой у консультирующегося?
14. В чем биологическое значение митоза?
15. Назовите наиболее известные канцерогены.
16. С каким размножением связан мейоз?
17. В ооците I порядка появились два новых мутантных гена.
Укажите максимальное число зигот, которые смогут по-
лучить эти гены.
18. Почему у 45-летней женщины, если она забеременела,
меньше шансов родить здорового ребенка, чем у молодой?
19. Перечислите проявления материнского эффекта в потом-
стве?
20. В медико-генетическую консультацию обратились две суп-
ружеские пары, у которых родились дети с незаращени-
ем твердого неба («волчья пасть»). Изучив родословную
каждой семьи и выяснив их бытовые и производственные
условия, врачи пришли к заключению, что у супругов А
все последующие дети, а также внуки не должны иметь
этого дефекта, но у супругов Б существует большая веро-
ятность появления его у детей и внуков. Укажите возмож-
ные механизмы появления данного дефекта у ребенка в
семье Айв семье Б.
21. Врожденная глухота может возникнуть в результате на-
личия мутантного гена в генотипе плода, но она может раз-
виваться и у организма с нормальным генотипом. Чем мож-
но объяснить наличие глухоты при нормальном генотипе?
Нужно ли выяснять природу этой аномалии у пациента,
если лечение в обоих случаях одинаково?
22. Человек вошел в противоречие со своей биологией. Он хо-
чет иметь детей, не когда молод, а когда уверенно стоит на
ногах. Данная тенденция свойственна наиболее цивили-
зованным человеческим популяциям. Решением подобной
проблемы является эмбриональная криоконсервация, или
замораживание эмбрионов. В ряде исследований показа-
но, что замороженные эмбрионы не обнаруживают уве-
личения доли врожденных дефектов или аномалий разви-
132
Контрольные вопросы и задания к разделу 3
тия по сравнению со «свежими» эмбрионами. Также по-
казано, что возраст женщины, которой пересаживается
эмбрион, не имеет существенного значения. Как Вы счи-
таете, разумно ли будет, если большинство женщин будет
пользоваться достижениями репродуктивной медицины,
извлекая ооциты в молодости, оплодотворяя их и пересажи-
вая для развития в возрасте, выходящем за рамки репро-
дуктивного? Какие неблагоприятные социальные и биоло-
гические последствия могут возникать в результате этого?
23. Эмбриональная криоконсервация находится в фокусе при-
стального внимания широких кругов общественности в
связи с ее значительной биоэтической компонентой. Речь
идет о том, что замораживание эмбрионов всегда предпо-
лагает предшествующее оплодотворение in vitro. Не все
религиозные направления одобряют эту процедуру. Като-
лическая церковь, к примеру, выступает против многих
форм альтернативных репродуктивных технологий, а в
особенности против тех, где зачатие походит вне организ-
ма. Кроме того, перед извлечением яйцеклетки из тела
женщины, проводится гормональная стимуляция, приво-
дящая к множественной овуляциии. В результате множе-
ственной овуляции созревает более одной яйцеклетки. Это
необходимо для увеличения шансов наступления беремен-
ности, поскольку не все яйцеклетки могут быть успешно
оплодотворены и не все эмбрионы могут быть успешно
имплантированы или заморожены.
В странах, где развиты технологии по эмбриональной кри-
оконсервации, как правило, супружеская пара подписы-
вает соглашение с клиникой, где проводится in vitro опло-
дотворение и криоконсервация, о дальнейшей судьбе
«лишних» замороженных эмбрионов. Такие эмбрионы
могут быть использованы в дальнейшем этой же супружес-
кой парой, переданы людям с бесплодием для усыновле-
ния или же уничтожены. Однако далеко не все можно
предвидеть при заключении таких соглашений, и в миро-
вой практике описано много интересных случаев. Так, в
133
Раздел 3. Наследственность и среда
1985 г. в результате авиакатастрофы погибли родители
двух замороженных эмбрионов в Австралии. Взрослые
дети погибших оказались в весьма щекотливом положе-
нии по отношению к своим 8-клеточным родственникам
и возможности раздела имущества. В США известным де-
лом 1996 г. было «Дэвис против Дэвиса», когда после раз-
вода супругов осталось семь замороженных эмбрионов.
Опекунство над ними с целью дальнейших попыток забе-
ременеть хотела получить экс-жена, экс-муж был настро-
ен против этого. Решением суда было уничтожение эмб-
рионов.
Как Вы считаете, следует ли отдавать «лишние» эмбрио-
ны для научных целей?
24. Согласны ли вы с утверждением, что поведенческие акты
находятся под генетическим контролем?
Раздел 4
АНАЛИЗ НАСЛЕДСТВЕННОСТИ
ЧЕЛОВЕКА
Совокупность методов изучения наследственности называ-
ется генетическим анализом. Психолога, изучающего генети-
ческие основы поведения, интересуют самые разные призна-
ки, так или иначе связанные с поведением. Это и легко опре-
деляемые реакции сенсорного восприятия, и сложные
личностные характеристики. Для генетического анализа про-
стых признаков используют методы, разработанные основа-
телем генетики Г. Менделем. При анализе сложных количе-
ственных признаков используют методологию, разработан-
ную Ф. Гальтоном. В последнее время при анализе различных
поведенческих вариантов используют хромосомный и моле-
кулярно-генетический анализ. В основе генетического ана-
лиза лежит следующая зависимость: если признак находится
под контролем генотипа, то родственные организмы по это-
му признаку более похожи, чем организмы, не связанные
родством. Законы наследования универсальны, поэтому прин-
ципы генетического анализа едины для всех живых организ-
мов, но для работы с конкретными биологическими видами
существуют специфические приемы. Генетический анализ
адаптирован к различным организмам — лабораторным и
сельскохозяйственным животным и растениям с разным ти-
пом размножения, микроорганизмам, а также человеку. От-
дельные биологические объекты стали модельным: это муха
дрозофила, домовая мышь, кишечная палочка, бактериофа-
ги. Модельные объекты обычно не имеют практического зна-
чения, но удобны для исследований, так как обеспечивают
высокую эффективность генетического анализа. Разрешаю-
щая способность генетического анализа зависит от скорос-
ти, с которой появляются новые поколения, и от численности
135
Раздел 4. Анализ наследственности человека
потомства, получаемого от одного скрещивания. Всем этим
требованиям удовлетворяют виды с коротким жизненным
циклом и высокой плодовитостью.
Исходная задача генетического анализа — установить, яв-
ляется ли исследуемый признак наследственным или он пол-
ностью детерминирован средой. После того как наследствен-
ная природа признака считается доказанной, выясняют, ка-
кие именно наследственные структуры его контролируют:
ядерные или цитоплазматические гены, сцепленные с полом
или аутосомные, каково их количество и т.д. В зависимости
от того, сколько генов контролирует признак, его обознача-
ют как моногенный или полигенный.
Главный методический прием при анализе качественных
признаков — сравнение его частоты у родственников разных
степеней родства. Если наследственные факторы не участву-
ют в формировании признака, то его частота у родственни-
ков и у лиц, не состоящих в родстве, примерно одинакова.
Если признак имеет генетическую природу, наблюдается се-
мейное накопление — более высокая частота признака сре-
ди родственников, чем в целом у населения.
Основой для генетического анализа количественных при-
знаков являются коэффициенты корреляции между род-
ственниками. Чем больше влияние наследственности на при-
знак, тем выше коэффициент корреляции. Однако, как и в
случае с качественными признаками, сходство по количе-
ственным признакам родственников может быть обусловле-
но как общностью наследственности, так и условий жизни.
Специальные приемы генетического анализа позволяют раз-
делить генетическое и средовое влияние.
4.1. Структура признака
Признаки, с которыми имеет дело генетик, различны по сво-
ей природе и выраженности. Например, человек от рожде-
ния может быть слепым или глухим, но может ослепнуть или
потерять слух без видимых причин, либо в результате несча-
136
4.1. Структура признака
стного случая или после инфекционного заболевания. При-
знаки, которые присутствуют у индивида с момента рожде-
ния, называются врожденными. Признаки, которые незави-
симо от причин появляются у человека после рождения, на-
зываются приобретенными.
Умственная отсталость ребенка может быть вызвана тем,
что он унаследовал от родителей специфические гены, а мо-
жет быть следствием того, что мать во время беременности
злоупотребляла алкоголем или перенесла инфекционное за-
болевание. Если признак определяется генетическими при-
чинами, он называется наследственным, если возникает
вследствие действия специфических условий среды — не на-
следственным. Признак определяют как семейный, если он
встречается еще у кого-то из родственников. Если признак в
родословной встретился один раз, его обозначают как спора-
дический.
Понятие «врожденный» не равнозначно понятию «наслед-
ственный». Термин «приобретенный» не означает, что на раз-
витие признака не влияет генотип. Врожденный признак мо-
жет быть как наследственным, так и не наследственным. Ре-
бенок может родиться с недоразвитыми конечностями
вследствие полученных от родителей генов. Потомок с нор-
мальным генотипом также может от рождения иметь уродли-
вые конечности, если его мать во время беременности упот-
ребляла лекарственные препараты, обладающие тератоген-
ным (порождающим уродства) эффектом. В обоих случаях
уродство конечностей врожденное, но в первом случае при-
чина аномалии генетическая, во втором — средовая.
Многие хронические заболевания (эпилепсия, шизофре-
ния, сахарный диабет, гипертония, близорукость, сколиоз) не
являются врожденными, а возникают в течение жизни. В свя-
зи с этим нередко создается впечатление, что причина этих
болезней не генетическая, а средовая — неправильный об-
раз жизни, действие каких-то вредных факторов. В действи-
тельности же на развитие перечисленных заболеваний ока-
зывает влияние наследственная предрасположенность. Ре-
137
Раздел 4. Анализ наследственности человека
шить, является признак наследственным или нет, можно толь-
ко с помощью генетического анализа.
Понятия «семейный» и «спорадический» указывают лишь
на частоту признака в родословной, но ничего не говорят о
его происхождении. Спорадический признак может иметь
наследственную природу, а его генетической причиной мо-
жет быть новая мутация или редкая комбинация генов. В то
же время наличие повторяющихся семейных случаев не яв-
ляется доказательством генетической природы признака, так
как родственники могут быть похожими не только из-за об-
щей наследственности, но и в силу одинаковых семейных ус-
ловий, нездоровых привычек или сходной профессии с ее спе-
цифической вредностью. Семейные случаи дают основание
лишь предполагать наследственную природу признака. Дока-
зать, что признак имеет наследственное происхождение, мож-
но с помощью специальных исследований.
Генетики в своей работе могут встретиться с фенокопия-
ми. Это не наследственные признаки, которые имитируют
мутации. Так, коревая краснуха беременной женщины может
вызвать ненаследственную глухоту у ребенка. Врожденная
глухота генетически нормального ребенка является феноко-
пией наследственной глухоты. Лекарственный препарат та-
лидомид, принимаемый беременной женщиной, вызывает у
потомства такие аномалии конечностей, которые наблюдают-
ся при некоторых хромосомных болезнях. Отличать феноко-
пии от наследственных случаев важно для генетического про-
гнозирования. Наличие у родителя фенокопии не повышает
риск для потомства. В некоторых случаях при мутантном ге-
нотипе может сформироваться нормальный фенотип — нор-
мокопия. Это возможно, если создать условия (например, с
помощью лекарственных препаратов, специальной диеты)
невозможные для проявления мутантного гена.
Признаки, выраженность которых зависит от многих ге-
нов и на которые оказывают влияние многие факторы сре-
ды, получили название мулыпифакториальных. Некоторые из
них можно легко классифицировать по принципу «есть—нет»
138
4.1. Структура признака
(например, болен—здоров). Такие признаки еще называют
качественными, или альтернативными. Частоту качественно-
го признака в группе выражают долями единицы или процен-
тами. Альтернативные признаки, которые не меняются в те-
чение всей жизни и не модифицируются средой, наиболее
удобны для генетического анализа. Количественные призна-
ки численно выражают в соответствующих единицах изме-
рения (баллы, сантиметры, секунды) с различной точностью
(килограммы, граммы, миллиграммы). Числовые показатели
у большого количества людей формируют непрерывный ряд
значений, и по этим признакам обследуемых невозможно
распределить по четко разграниченным классам.
Разделение признаков на качественные и количественные
условно. Непрерывный ряд распределения можно превратить
в классы, обозначенные, как, например, в случае с кровяным
давлением — «норма» и «гипертония». В свою очередь каче-
ственный признак можно рассматривать как количествен-
ный, например, альтернативные состояния, обозначаемые как
«нормальный вес» и «ожирение», заменить весом в килограм-
мах.
Анализируя количественные признаки, используют поня-
тие порога, или порогового значения признака. Так, величина
кровяного давления, выше которой человек считается больным
гипертонией, является пороговой. В основу модели пороговых
признаков положено допущение о том, что их генетическая
основа представляет собой полигонную систему. Например,
при анализе умственных способностей в соответствии с при-
нятыми для данного теста пороговыми показателями всех
людей можно разделить на группы нормальных, умственно
отсталых и пограничных между ними.
Мы коснемся рассмотрения нескольких из наиболее важ-
ных методов генетического анализа, а именно: генеалогичес-
кого метода, компонентного метода, метода генетических
маркеров, метода моделирования на животных. Близнецовый
метод будет рассмотрен в отдельной главе.
139
Раздел 4. Анализ наследственности человека
4.2. Генеалогический метод
Генеалогический метод представляет собой изучение наслед-
ственности человека с помощью родословных. Метод был раз-
работан Ф. Гальтоном (рис. 4.1), о котором Карл Пирсон гово-
рил так: «Он знал математику и физику больше, чем девять
биологов из десяти, больше из биологии, чем девятнадцать
математиков из двадцати, и больше из патологии и физио-
логии, чем сорок девять из пятидесяти биологов и матема-
тиков его времен». Генеалогия — это наука о родословных
человека. Чем больше семья, тем более информативной яв-
ляется родословная. В 1826 г. была описана семья персидско-
го шаха, имевшего 66 сыновей, 53 замужних дочери, 271 вну-
ка и внучку. К 80-летию этого шаха в его семье было уже 860
детей, внуков, правнуков и праправнуков. Особенно важно
наличие большого числа детей в поколении для установления
типа наследования признака. Люди изображали внутрисемей-
ные отношения в графическом виде, начиная с XV в.
Рис. 4.1. Фрэнсис Гальтон
Слово родословная (англий-
ский эквивалент «pedigree»)
происходит от французского
выражения «ргё de дгие», кото-
рое буквально означает «лапа
журавля», поскольку линии, со-
единяющие родителей и детей
на родословных, напоминают
птичью лапу. До XX в. родослов-
ные использовались преимуще-
ственно в генеалогии для ото-
бражения родственных отноше-
ний. В XX в. родословные стали
использоваться в генетике для
прослеживания признаков в
ряду поколений.
В нашей стране до револю-
ции родословным придавалось
140
4.2. Генеалогический метод
большое значение. Издавались специальные родословные
книги именитых людей. Сейчас, к сожалению, эта традиция
утрачена. Во многих обществах, где от происхождения чело-
века зависит его социальное положение, генеалогии и сейчас
уделяется большое внимание. Генеалогический метод в гене-
тике человека выполняет ту же функцию, что и метод скре-
щиваний в генетике животных и растений. По сути, это по-
иск скрещиваний, поставленных природой. Генеалогический
метод универсален, составление родословной сопутствует
всем другим методам, с помощью которых изучают наслед-
ственность человека. Он используется для определения ха-
рактера наследования признака, определения генотипа чле-
на родословной, вычисления вероятности рождения больно-
го ребенка.
Сбор материала для генетического анализа проводят од-
ним из способов — по пробанду п сплошным методом. Выбор
метода определяется характером изучаемого признака и его
распространенностью в популяции. Если ставится цель изу-
чить редко встречающийся признак, например, психическое
заболевание, то регистрация семей осуществляют по пробан-
ду. Потенциальные пробанды — обладатели признака, могут
состоять на учете в специализированных лечебных или со-
циальных учреждениях. Если планируется изучить наслед-
ственную природу признака, который имеется у каждого че-
ловека, например, уровень интеллекта, сбор материала про-
водят сплошным методом. Намечают группу людей, которая
может быть изучена (генеральная совокупность) и из нее фор-
мируют выборку. При любом сборе данных объекты иссле-
дования должны включаться в выборку случайно, недопусти-
мо отбирать «наиболее интересные» или «типичные» случаи.
При составлении родословных могут возникать опреде-
ленные трудности. Обычно люди могут проследить свою ро-
дословную не более чем на 2—3 поколения назад и не помнят
всех родственников. Иногда они умышленно скрывают фак-
ты, которые для них неприятны или о которых они не хотят
упоминать (алкоголики, умственно неполноценные родствен-
141
Раздел 4. Анализ наследственности человека
ники и т.д.). В ряде случаев путаница в интерпретации родос-
ловных может наблюдаться из-за внебрачных детей, повтор-
ных браков, современных репродуктивных технологий (ис-
кусственное оплодотворение, суррогатное материнство).
Родители и дети представляют собой ядерную семью. Ро-
дословная обычно включает несколько ядерных семей. Роди-
тели и дети, родные братья и сестры (сибсы) являются род-
ственниками первой степени родства. У них половина общих
генов. Родственники второй степени родства — бабки/деды
и внуки/внучки, дядья/тетки и племянники/племянницы
имеют четверть общих генов. Двоюродные сибсы — это род-
ственники третьей степени родства, у них восьмая часть об-
щих генов. Дети, у которых только один общий родитель на-
зываются полусибсами, они родственники второй степени
родства. Дети, имеющие общую мать (единоутробные), гене-
тически более сходны, чем имеющие общего отца (единокров-
ные), так как в первом случае полусибсы имеют одну и ту же
цитоплазматическую наследственность, во втором случае ми-
тохондриальные гены получены от разных женщин (табл. 4.1).
Рис. 4.2. Свойственные отношения
между членами семей мужа и жены
142
4.2. Генеалогический метод
Генетическое родство
Таблица 4.1
Родственная связь (заштриховано) Степень родства Доля общих генов
Монозиготные близнецы — 1
л л t О ..ж Дизиготные близнецы 1 1/2
1 Сибсы 1 1/2
Родитель-ребе- нок 1 1/2
X в Дядя (тетка) — пле-мянница (племянник) 2 1/4
ж 1 — Полусибсы 2 1/4
Дважды двою- родные сибсы 2 1/4
— -I
Двоюродные сибсы 3 1/8
143
Раздел 4. Анализ наследственности человека
Продолжение табл. 4.1
Родственная связь (заштриховано) Степень родства Доля общих генов
i а Полутетя-полу- племянник (и анало- гичные случаи) 3 1/8
% Двоюродные дядя и племянница (и анало- гичные случаи) 4 1/16
1 Троюродные сибсы (и аналогичные случаи) 5 1/32
1 1 Троюродные дядя и племянница (и анало- гичные случаи) 6 1/64
144
4.2. Генеалогический метод
Окончание табл. 4.1
Интерес представляют и сводные сибсы. Эти дети теоре-
тически не имеют общих генов, но каждый из них имеет по-
ловину общих генов с одним из родителей. Общей у таких
«сибсов» является только семейная среда, которую создали
сводные братья и сестры
Рис. 4.3. Свойственные отношения
между сводными сибсами и их родителями
145
Раздел 4. Анализ наследственности человека
родители каждого из них, заключив брак. Генетик может
встретиться и с семьей, где есть приемные дети — не имею-
щие общих генов ни с одним из «родителей».
Свойство представляет собой отношения между людьми,
возникающими вследствие брака. Такие люди друг' по отно-
шению к другу называются свойственниками. На рис. 4.2 и
рис. 4.3 приведены различные отношения свойства.
Анализ большой родословной
Большие родословные, включающие несколько поколений и
большое число членов, удобны для анализа качественных при-
знаков. Просматривая родословную, обращают внимания на
то, как распределяется в родословной изучаемый признак: в
каждом поколении он встречается или имеются пропуски,
признак встречается у представителей одного пола или у обо-
их, кому из детей (сыновьям или дочерям) отец и мать пере-
дают признак. Общий вид родословной дает основание выдви-
нуть предварительную генетическую гипотезу. Ее проверяют,
анализируя каждую ядерную семью. Анализируя родословную,
руководствуются знанием закономерностей наследования, а
именно: аутосомный признак встречается с одинаковой час-
тотой у мужчин и женщин, сцепленный с полом — только у од-
ного или по преимуществу у представителей одного пола;
аутосомный ген мать и отец передают дочерям и сыновьям с
равной вероятностью, ген, сцепленный с Х-хромосомой, мать
с равной вероятностью передает как сыновьям, так и доче-
рям, а отец только дочерям; ген, локализованный в Y-хромо-
соме, отец передает только сыновьям; признак, который кон-
тролируется цитоплазматическими генами, дети наследуют
только от матери. Из сказанного вытекает ряд следствий. Если
признак в родословной встречается с одинаковой частотой у
мужчин и женщин, допускаем, что он контролируется ауто-
сомными генами. Если признак передается без пропусков по-
колений и ребенок с признаком рождается в семье, где у од-
ного из родителей был такой же признак, то он считается до-
146
4.2. Генеалогический метод
Рис. 4.4. Родословная с признаком
«белая прядь волос надо лбом»
минантным. Если признак передается с пропусками в поко-
лениях, считается, что он контролируется рецессивным геном.
Если признак встречается у представителей только одного пола
(обычно мужчин), считаем, что гены, его контролирующие,
локализованы в половой хромосоме. Доказательством того,
что признак находится под контролем аутосомного гена, слу-
жит его наследование сыновьями и дочерями как от матери,
так и отца. Отец не может передать сыну ген, локализован-
ный в Х-хромосоме, следовательно, передача признака от отца
к сыну отвергает гипотезу о его сцеплении с Х-хромосомой.
Отец не может передать дочери признак, который находится
под контролем гена, локализованного в Y-хромосоме. Пере-
дача признака от мужчины к дочери отвергает гипотезу о его
сцеплении с Y-хромосомой, Отец также не может передать
детям признак, который находится под контролем митохонд-
риальных генов, поэтому передача признака от мужчины де-
h 147
Раздел 4. Анализ наследственности человека
тям свидетельствует о его сцеплении с ядерными генами. Выд-
винутая на основе обзора родословной генетическая гипоте-
за требует математического доказательства.
В качестве примера рассмотрим родословную, представ-
ленную на рис. 4.4, где у некоторых членов имеется белая
прядь волос надо лбом, и определим тип наследования данно-
го признака.
Из родословной видно, что данный признак обнаружива-
ется во всех поколениях, что указывает на доминантный тип
наследования. Наличие признака как у мужчин, так и у жен-
щин опровергает сцепление с Y-хромосомой, а передача от
отцов к сыновьям делает невозможным сцепление с Х-хро-
мосомой. Выдвигаем гипотезу аутосомно-доминантного на-
следования и проверяем ее математически. Поскольку при-
знак редкий, его носители являются гетерозиготами Аа, а не
носители рецессивные гомозиготы аа. Выделим шесть семей,
в которых один из родителей имеет этот признак, а другой
нет (Аа х аа). (Это браки носителей признака, обозначенные
1/1, II/1, III/3, IV/1, IV/2, IV/3, их супруги, не имеющие при-
знака, в родословной не указаны.) Если генотипы супружс-
ких пар правильно отражают генетическую гипотезу, то по-
ловина потомков в таких семьях должны иметь признак, со-
отношение полов среди потомков с признаком и без признака
должно быть 1:1. Всего в этих семьях 17 потомков, из кото-
рых 11 имеют признак (7 мужчин и 4 женщины) и 6 не имеют
(4 мужчины и 2 женщины). С помощью критерия %2 (см. табл. 1
Приложения) проверяем имеются ли межполовые различия
в наследовании данного признака:
Фенотип Пол Всего
Мужчины Женщины
Здоровые 7 4 И
Больные 3 3 6
Всего 10 7 17
%2 = 0,0009, df = 1, р > 0,05.
148
4.2. Генеалогический метод
Расчет значения ?2 показывает, что в распределении при-
знака достоверной разницы между полами нет. Поскольку
предполагаемые генотипы родителей во всех анализируемых
семьях Аа ? аа, то теоретически ожидаемое соотношение
больных и здоровых потомков в них должно соответствовать
1:1, или для данной задачи 8,5:8,5 (дробность цифр не имеет
значения, поскольку для осуществления расчетов свойства
объектов, вкладываемых в имеющиеся данные, не важны).
Далее решаем вопрос о достоверности различий между тео-
ретически ожидаемым и фактическим количеством больных
и здоровых потомков:
Фенотип Фактические Ожидаемые
Больные 11 8,5
Здоровые 6 8,5
Всего 17 17
X2 = 1,47, df = 1, р > 0,05.
Различия между фактической и ожидаемой сегрегацией
больных и здоровых также недостоверны. Гипотеза об ауто-
сомно-доминантном наследовании белой пряди надо лбом
верна.
Анализ малочисленных родословных
В настоящее время исследователи имеют в своем распоряже-
нии в основном небольшие родословные. Для анализа такого
материала все данные суммируют и выполняют сегрегацион-
ный анализ. Сегрегационный анализ, с помощью которого
проверяется гипотеза о моногенной доминантной природе
признака, приведен в предыдущем примере. Проверка гипо-
тезы о моногенно-рецессивной природе признака сложнее,
потому что в этом случае возникает необходимость избежать
погрешностей, вызванных особенностями сбора генеалоги-
ческого материала. Источник этих погрешностей лежит в сле-
дующем. Например, в семье, где оба родителя гетерозиготны
(Да х Аа), теоретически 25% детей должны иметь рецессив-
149
Раздел 4. Анализ наследственности человека
ный признак. Однако, если семье немного детей, среди них
может не оказаться ребенка с рецессивным признаком и эта
семья не попадет в поле зрения генетика. При таком отборе
формируется смещенная выборка и генетические показате-
ли, полученные на ней, оказываются искаженными. Для того
чтобы учесть погрешности, вызванные особенностями сбора
материала, сегрегационные частоты (SF) рассчитывают по
формуле, Вайнберга:
= ^F(1^SF)
T-N' SF \ T-N '
где N — количество семей данного типа; Т — общее количе-
ство детей у семей данного типа с учетом пробанда; А — ко-
личество пораженных детей у семей данного типа; SF — сег-
регационная частота признака; SSF — статистическая ошиб-
ка.
Вычисленную сегрегационную частоту сравнивают с тео-
ретически ожидаемой используя статистический критерий
Стьюдента t:
t = (SF—SF )/S_
где SF— эмпирическая сегрегационная частота, SF . — тео-
ретически ожилаемая сегрегационная частота, SSF — статис-
тическая ошибка.
4.3. Компонентный анализ
Генетическая основа многих признаков настолько сложна, что
не может быть описана терминами менделевской генетики.
Для генетической характеристики таких признаков исполь-
зуют показатель наследуемости. Показатель наследуемости —
это математическая величина, выраженная в процентах, ко-
торая показывает, какая часть наблюдаемого внутригруппо-
вого фенотипического разнообразия определяется генетичес-
кими различиями между индивидами. Показатель наследуе-
мости находят с помощью компонентного анализа, который
150
4.3. Компонентный анализ
представляет собой процедуру, с помощью которой феноти-
пическое разнообразие в группе (Р) можно разложить на ге-
нетический (G) и средовый (Е) компоненты:
Р = G + Е.
Разнообразие признака в группе выражают с помощью ма-
тематической величины — показателя фенотипической дис-
персии. Причины фенотипического разнообразия могут быть
генетическими и средовыми: особи могут различаться пото-
му, что у них разные генотипы, и потому, что они развива-
лись в разных условиях. Для компонентного анализа исполь-
зуют коэффициенты корреляции г между родственниками.
Обычно вычисляют коэффициент корреляции между роди-
телями и потомками (гр), а также между сибсами (г.). Компо-
нентное разложение проводят по формулам:
G = 2г ;
о р'
G — 4(г — г );
d < s p/l
G=G +G-.
a d
Е= 1 — G
где G — общий генетический компонент, Ga — его адлитив-
ная и Gd — доминантная составляющие, Е — средовой компо-
нент.
Показатель наследуемости — групповая характеристика,
поэтому его величина зависит от структуры популяции, на
которой проводится исследование. При выборе формулы для
вычисления коэффициента корреляции учитывают природу
признака, руководствуясь теорией фенотипических моделей.
Фенотипические модели
В генетическом анализе принято разделять признаки на аль-
тернативные, непрерывные, квазинепрерывные и квазиаль-
тернативные.
К альтернативным признакам относятся пол (мужской и
женский), принадлежность к группе крови резус (положи -
151
Раздел 4. Анализ наследственности человека
тельный и отрицательный) и другие особенности дискретно-
го характера. Генетическая основа этих признаков также име-
ет альтернативную природу, например, X- или Y-хромосома,
аллель резус-положительности или резус-отрицательности
ит.д.
К непрерывным признакам относят рост, вес, артериаль-
ное давление, уровень интеллекта и другие особенности, ко-
торые можно выразить количественно. Принято считать, что
наследственная основа этих признаков представляет собой
систему полигенов — множество генов со слабым эффектом,
распределение которых в популяции подчиняется нормаль-
ному закону.
Примером квазинепрерывных признаков могут служить
хронические заболевания, в том числе и психические — ши-
зофрения, эпилепсия и др. Фенотипически эти признаки про-
являются как альтернативные (болен или здоров). Предпола-
гается, что генетической основой этих признаков является
система полигенов. Считается, что гены, ответственные за
признак, имеются у некоторой части, а может быть и у всех
представителей популяции. Но проявляется признак (заболе-
вание) только у отдельных лиц, у которых в генотипе присут-
ствует определенное — пороговое — количество этих генов.
Такие признаки еще называют мулыпифакториальными и
пороговыми.
Квазиалыпернативные — это признаки с непрерывной ко-
личественной выраженностью, но альтернативной генетичес-
кой основой. Примером такого признака может быть уровень
интеллекта, выраженный в баллах IQ, у индивидов с нормаль-
ным хромосомным набором и у больных с синдромом Дауна.
Наследственность нормальных и больных людей различает-
ся одной хромосомой, то есть альтернативно, но баллы IQ в
обеих группах распределяются как непрерывный признак: у
нормальных людей интеллект варьирует от 80 до 125 баллов,
а у больных синдромом Дауна с 40 до 70.
4.3. Компонентный анализ
Коэффициенты корреляции
При выборе формулы для вычисления коэффициента корре-
ляции между родственниками учитывают характер анализи-
руемого признака. При анализе непрерывных признаков вы-
числяют межклассовые и внутриклассовые коэффициенты
корреляции. Для квазинепрерывных признаков вычисляют
тетрахорический коэффициент корреляции. Анализ альтер-
нативных признаков проводят с использованием коэффици-
ента сопряженности. Для оценки связи по непрерывному и
квазинепрерывному признакам используют бисериальный ко-
эффициент корреляции. Связь между непрерывным и альтер-
нативным признаками оценивается с помощью точечно-би-
сериального коэфцииента корреляции. Формулы для вычис-
ления различных коэффициентов корреляции можно найти
в руководствах по статистике для исследователей.
Коэффициент корреляции для непрерывных признаков
вычисляют по формуле:
г _
v
ъху~
п
где г — коэффициент корреляции, х — значения первого при-
знака, у — значения второго признака, п — объем выборки
(число пар признаков), st — статистическая ошибка выбороч-
ного коэффициента корреляции.
153
Раздел 4. Анализ наследственности человека
Если признак рассматривается как квазинепрерывный,
используют формулы:
Хр ~ XR ~ (Х р ~ XR )0 ~ Х р / а р)
rR~
ap+XR<ap-Xp)
s 1 '
Л rR Д j
\ A apaR
аР =<
у/ 271
1 2
• ехр(—xR / 2) / Qr
у 271
SGa = ^Sr0P ' SGd ~ ^SrOP + SrSB '
где rR и srR — коэффициент корреляции между родственника-
ми и его стандартная ошибка; Qp — распространенность при-
знака в общей популяции; QR — частота признака среди
родственников пробандов; А — число родственников с при-
знаком среди всех родственников пробанда, sGa, sCd — стати-
стические ошибки. Значение хр и xR находят из величин <Эри
QRno таблице Фолконера (см. литературу к главе).
При анализе альтернативного признака коэффициент кор-
реляции и его статистическую ошибку вычисляют по формулам:
где rR и srR — коэффициент корреляции между родственника-
ми и его стандартная ошибка; Qp — частота признака в попу-
154
4.3. Компонентный анализ
ляции; Qr — частота признака среди родственников пробан-
дов; А — число родственников с признаком среди всех род-
ственников пробанда, sr — статистическая ошибка.
Величина коэффициента корреляции зависит от структу-
ры популяции. При положительной брачной ассортативнос-
ти корреляция между родителями и детьми выше, чем при
панмиксии, а при отрицательной ниже. Если по признаку, взя-
тому для генетического анализа, в популяции существует не-
случайный подбор супружеских пар, в формулу для вычисле-
ния коэффициента корреляции между родственниками не-
обходимо ввести поправку на величину коэффициента
корреляции между супругами
г' =г/(1+гс),
где г' — коэффициент корреляции с учетом поправки на брач-
ную ассортативность, г— коэффициент корреляции для пан-
миктичной популяции, гс— коэффициент корреляции меж-
ду супругами.
Модель главного гена
Если при компонентном анализе получены высокие значения
показателя наследуемости, делают предположение, что в си-
стеме генетического контроля признака присутствует глав-
ный ген. Этот ген проявляется не у всех его носителей (ген не
полностью пенетрантный). Считается, что пенетрантность
этого гена выше у гомозигот (АА), чем у гетерозигот (Аа). Для
нахождения частоты главного гена и его пенетрантности у
гомо- и гетерозиготных индивидов используют дисперсион-
ные уравнения:
Qp = р2П2 + 2/7(1 - р)П, + (1 - р)2 По
rp=QP(i-QP)
Va=Ga-Vp= 2р(1 -р)Ь(772 - 77,) + (1 -р)(77, -770)]2
155
Раздел 4. Анализ наследственности человека
^=с„-ур=р2(1-р)г [п2 -2П,+П0 Г,
где р — частота тестируемого аллеля, П2 — пенетрантность
гомозигот по данному аллелю, П1 — пенетрантность гетеро-
зигот, По — пенетрантность оппозитивных гомозигот.
Уравнения решаются алгебраическим путем. Все парамет-
ры, используемые для решения, вычисляются на основе ко-
эффициентов корреляции, рассчитанных по формулам аль-
тернативной модели наследования. Всего имеется четыре воз-
можных решения:
1. k. = 1, т = 1, п — 1,
р = (—с + п4с2 - 4ab)/2Ь
n2=(Qp+myiVd)/p
п, =(йр-р2772)/[2р(1-р)]
2. k = 1, m = 1, л = — 1,
и решение по предыдущим формулам.
3. к — — 1, m = 1, л = 1,
и решение по предыдущим формулам.
4. к = — 1, т = 1, п = — 1,
и решение по предыдущим формулам,
где к, т, п — коэффициенты, определяющие знак корня при
решении квадратных уравнений; а, Ь, с, — вспомогательные
величины, которые вычисляются по следующим формулам:
a = Q), b = a+ Vd + klQp-Jyd+2Va,
+ -2К„.
Среди полученных корней уравнения выбирают те, что
имеют биологический смысл (частота и показатель пенетран-
тности лежат в пределах от 0 до 1).
156
4.4. Метод генётических маркеров
4.4. Метод генетических маркеров
О генетической основе признака у человека, в том числе и
поведенческого, можно судить по статистической ассоциации
его с другим признаком, который выполняет роль генетическо-
го маркера. В качестве маркеров выбирают структуры изучен-
ной генетической природы: белки, хромосомные варианты,
полиморфизм ДНК, моногенные морфологические признаки,
которые находятся под контролем полностью пенетрантных
генов. Ассоциация генетического маркера с анализируемым
признаком может иметь разные причины. Одна из них — гене-
тическое сцепление между маркерным геном и геном, который
контролирует анализируемый признак. Еще одна причина ас-
социации — множественное действие гена. Анализируемый
признак может находиться под контролем того же гена, кото-
рый оказывает влияние на маркерный признак. В результате
этого может оказаться, что анализируемый признак и признак,
с которым он ассоциирован, контролируются одним и тем же
геном. Следует учитывать, что ассоциация может возникнуть
из-за методических ошибок в сборе материала исследования,
например, если популяция этнически неоднородна.
Маркеры не являются общезначимыми, а ассоциация аб-
солютной. Например, в одной семье признак ассоциирован с
группой крови М, в другой — с группой крови N. В ряду поко-
лений эта связь будет прослеживаться до тех пор, пока не бу-
дет разделена кроссинговером.
Использование генетических маркеров дает возможность
решать разные проблемы: подтвердить наследственную при-
роду признака, установить его генетическую гетерогенность.
Если одна и та же поведенческая особенность в одних случа-
ях ассоциируется с каким-либо одним генетическим марке-
ром, а в других случаях с другим, то это может свидетельство-
вать о том, что перед нами две генетически различные фор-
мы поведения —- фенотипически сходные, но имеющие
разные генетические причины. Маркерами признака могут
быть хромосомы, например, маркером синдрома Дауна яв-
ляется лишняя 21-я хромосома.
к
157
Раздел 4. Анализ наследственности человека
Ассоциация изучаемого признака с белком или последо-
вательностью ДНК служит одним из доказательств его наслед-
ственной природы. В течение долгого времени в качестве мар-
керов исследователи использовали полиморфные белки. Этим
маркеры стали называть классическими, чтобы отличить их
от появившихся в последнее время маркеров ДНК. Исполь-
зование в качестве маркеров молекул ДНК значительно рас-
ширяет возможности генетического анализа, так как инфор-
мационное содержание ДНК значительно выше белкового.
Генетическая структура или белок (назовем его кандидатом
в маркеры, или просто кандидатом) может считаться маркером,
если его ассоциация с анализируемым признаком статистичес-
ки доказана. Для этого подбирается группа лиц с интересующим
признаком (основная группа — О) и группа лиц без этого при-
знака (контрольная — К). В каждой группе учитывается число
индивидов, у которых предполагаемый маркер-кандидат при-
сутствует (+) и число индивидов, у которых кандидат отсут-
ствует (—). Как контрольная, так и основная группа включа-
ют индивидов с генетическим вариантом (О+ и К+) и без него
(О-и К”). Если кандидат одинаково часто встречается в обе-
их группах (О+ :О = К+:К_), он не может выполнять функцию
маркера. Если кандидат чаще (или реже) встречается в одной из
групп, его можно использовать как маркер. Чем сильнее ассо-
циация кандидата с признаком, тем больше выражено нера-
венство О+:О~#К+:К~, тем надежнее маркер. На степень ас-
социации указывает показатель х:
ОК
о к~
Величина х по-
казывает, во сколько раз маркер чаще (реже) встречается в одной
из групп по сравнению с другой. Достоверность отклонения х
от 1 оценивают с помощью критерия х2:
X2 — y2w у = 1пх
158
4.5. Моделирование на животных
4.5. Моделирование на животных
Исследования на животных проводят для моделирования си-
туаций, которые невозможно осуществить, работая с челове-
ком. Главная проблема, связанная с генетическими исследо-
ваниями на животных, состоит в корректности переноса на
человека данных, полученных при исследовании модельных
объектов. Правомочность использования животных в каче-
стве генетических моделей основана на их генетическом сход-
стве с человеком. Согласно молекулярно-генетическим дан-
ным, человек по составу генов имеет 60%-ное сходство с му-
хой дрозофилой и 90%-ное сходство с мышью, 223 гена
человека сходны с генами бактерии кишечной палочки, ДНК
шимпанзе на 99% совпадает с ДНК человека. Мутации, влияю-
щие на поведение человека, могут быть найдены у животных.
Поэтому в качестве моделей могут быть использованы раз-
личные виды животных, однако чем ближе они к человеку в
систематическом отношении, тем более адекватной является
экстраполяция результатов исследования.
Генетические эксперименты на животных позволяют дать
ответ на вопрос: имеет ли вообще отношение наследствен-
ность к тому или иному типу поведения. Для решения этих
задач скрещивают животных, различающихся определенны-
ми поведенческими особенностями, и анализируют потом-
ство. Один из приемов моделирования состоит в исследова-
нии генетического контроля метаболических путей, сходных
у животных и человека. Другой подход заключается в иссле-
довании участков генома человека, гомологичных геномам
модельных животных.
Первые экспериментальные доказательства того, что на
поведение оказывают влияние наследственные факторы, по-
лучены именно на животных. При наблюдениях за поведе-
нием животных разных генетических линий и пород были по-
лучены факты, которые свидетельствовали о влиянии гено-
типа на поведенческие особенности. Если генетически
.различные группы животных отличаются поведением, то это
Раздел 4. Анализ наследственности человека
дает основание считать, что эта особенность находится под
генетическим контролем. В распоряжении генетиков имеют-
ся экспериментальные модели многих признаков человека.
Среди лабораторных животных есть линии, где все или боль-
шинство особей на определенном этапе жизни заболевают ка-
кой-либо болезнью: сахарным диабетом, раком, и др. Суще-
ствуют линии животных, различающиеся по типу нервной дея-
тельности. Имеются линии, моделирующие психические
болезни человека, например, эпилепсию. Для многих линий
животных характерны такие поведенческие особенности, как
предпочтение алкоголя, морфина, кокаина. Эти линии служат
моделью для познания генетических механизмов, которые
формируют предрасположенность к наркомании и алкоголиз-
му у человека.
Еще одним доказательством того, что поведенческий при-
знак находится под генетическим контролем, является успеш-
ность отбора, направленного на его усиление или ослабление.
Если в результате отбора животных признак не меняется, это
указывает на то, что его разнообразие в группе животных
обусловлено не генетическими причинами. Успешный отбор
свидетельствует о том, что признак находится под генетичес-
ким контролем.
На лабораторных животных можно смоделировать прием-
ную семью. Для этого у одной самки забирают детенышей и
передают их на воспитание самке другой породы. В то же вре-
мя детенышей второй самки передают на воспитание первой,
то есть производят обмен детенышами. Когда животные вы-
растают, сравнивают их поведение с поведением животных
первой и второй породы. Для того чтобы исключить влияние
материнского организма на развивающийся зародыш, обме-
нивают не детенышей, а оплодотворенные яйцеклетки, и за-
тем сравнивают потомство, развивавшееся в организмах соб-
ственной и приемной матери.
160
Контрольные вопросы и задания к разделу 4
Контрольные вопросы и задания к разделу 4
1. О чем говорит показатель степени родства?
2. Могут ли свойственники иметь общие гены?
3. Пользуясь графической символикой родословных, со-
ставьте собственную родословную, родословную одного
из Ваших соседей и Вашего лучшего друга.
4. Какова доля общих генов у поименованных членов родос-
ловных на рис. 4.5?
Рис. 4.5
5. Обозначьте родственные связи между поименованными
членами родословных на рис. 4.6.
6 Введение в психогенетику
161
Раздел 4. Анализ наследственности человека
Рис. 4.6
6. Сколько родственниц женского пола у Софьи и сколько
родственников мужского пола у Ирины в родословных на
рис. 4.7?
Рис. 4.7
162
Контрольные вопросы и задания к разделу 4
7. Из родословных, изображенных на рис. 4.8, определите
сколько свойственников у Людмилы, сколько свойствен-
ников мужского пола у Ольги и сколько свойственниц жен-
163
Раздел 4. Анализ наследственности человека
8. У Анны Петровны и Александра Ивановича есть два сына
и пять внуков. На серебряную свадьбу к юбилярам при-
шли все сыновья, зятья и внуки. В доме имеется только де-
вять стульев. Всем ли хватит стульев?
9. Дарья и Дмитрий — жена и муж. Они собираются идти в
поход. Дарья хочет пригласить с собой своего брата и де-
веря, а Дмитрий — своего брата и шурина. Сколько чело-
век пойдут в поход?
10. Ирина и Игорь — молодожены. К себе в гости Ирина хо-
чет пригласить своих родителей и свекра со свекровью,
Игорь — своих родителей и еще тещу с тестем. Сколько
гостей будет у Ирины и Игоря?
11. В лес за грибами пошли две сватьи и два свата, зять и неве-
стка, сын и дочь, теща и тесть, свекровь и свекор, три мужа
и три жены. Сколько человек пошли в лес?
12. По дороге шли два отца, два сына, дед и внук. Сколько че-
ловек шли по дороге?
13. Какие задачи решаются с помощью генетического ана-
лиза?
14. На чем основан генетический анализ количественных при-
знаков?
15. Какая проблема решается при использовании генетичес-
ких маркеров?
16. Почему для изучения наследственности человека можно
использовать животных?
17. У представителей белой расы изредка встречаются лица с
шерстистыми волосами. Этот признак изучен в семьях, где
один из супругов (муж или жена) имел шерстистые, а дру-
гой — обычные волосы. Среди потомков этих браков было
145 детей (примерно поровну сыновей и дочерей) с шер-
стистыми и 130 детей (поровну сыновей и дочерей) с обыч-
ными волосами. В браках между людьми, которые имеют
обычные волосы, и людьми с обычными волосами, но имев-
шими родственников с шерстистыми волосами, всегда
рождаются дети с обычными волосами. Как наследуется
признак шерстистые волосы?
164
Контрольные вопросы и задания к разделу 4
18. Фаина и ее дочь Марина больны кератозом. У Марины три
здоровых дочери и два больных сына Иван и Григорий. У
Ивана трое здоровых сыновей и один больной, у Григория
два больных сына и здоровая дочь. Супруги всех упомяну-
тых лиц здоровы. Установите, как наследуется кератоз.
19. Определите генетическую основу признака в родослов-
ной на рис. 4.9.
20. Одна из форм рахита является наследственным призна-
ком. От браков 14 мужчин, больных рахитом, со здоровы-
ми женщинами родились 21 дочь и 16 сыновей. У всех до-
черей со временем стали обнаруживаться признаки рахи-
та, все сыновья были здоровы. Какова генетическая
природа этого заболевания?
21. У Тимофея большие зубы, такие же зубы у трех его сыно-
вей, а у двух дочерей зубы нормального размера. Все се-
меро внуков от сыновей имеют большие зубы, а трое вну-
чек — обычные. Внуки и внучки от дочерей имеют нор-
мальные зубы. Как наследуется признак большие зубы?
22. В основе брахидактилии (короткопалости) лежит укоро-
чение и сращение костей фаланг. Определите тип насле-
дования брахидактилии у членов родословной, представ-
ленной на рис. 4.10.
165
1
Рис. 4.10
Контрольные вопросы и задания к разделу 4
23. На рис. 4.11 приведена родословная, в которой встреча-
ются люди, у которых зубная эмаль коричневого цвета. Оп-
ределите, как наследуется этот признак.
11
167
Раздел 5
БЛИЗНЕЦЫ В ГЕНЕТИКЕ ЧЕЛОВЕКА
Появление на свет близнецов всегда завораживало умы и да-
вало обильный материал для создания мифов и легенд. Близ-
нецам приписывались сверхъестественные силы, они счи-
тались то виновниками несчастья, то предвестниками со-
бытий в природе и жизни, которых ожидали или боялись.
Очень часто подчеркивалась противоположность близнецов.
Один олицетворял добро, положительное начало, другой —
зло, отрицательное начало. Необъяснимым казалось появле-
ние близнецов на свет: ведь обычно один мужчина мог зачать
только одного ребенка, а если их двое, следовательно, в зача-
тии замешаны боги, демоны, духи, животные.
Во многих религиях боги являлись близнецами. В антич-
ной мифологии часто встречаются подкинутые близнецы,
вскормленные животными и спасенные людьми. Самыми из-
вестными из них являются Ромул и Рем. Они были вскормле-
ны волчицей и воспитаны пастухами. Согласно мифу, в 753 г.
до н.э. ими был основан город на том месте, где они были най-
дены. Ромул, стремясь к единовластию, убил своего брата
Рема и стал первым царем Рима. Близнецы увековечены в го-
родском гербе Рима.
Эмоции, связанные с близнецами, чрезвычайно многооб-
разны и противоречивы: от эйфорического почитания до де-
монического страха. Даже между соседствующими племена-
ми может наблюдаться резко противоположное отношение к
близнецам: радостные праздники у одного племени, пресле-
дования и уничтожение близнецов — у другого. У инков, жив-
ших на территории современного Перу, близнецы мужского
пола почитались как сыновья бога Молнии. У племени дакота
в Северной Америке близнецы тоже считались приносящими
168
Раздел 5. Близнецы в генетике человека ,
счастье сверхъестественными существами. Индейцы племе-
ни ноатка видели в рождении близнецов доброе предзнаме-
нование и связывали их появление с богатым уловом лососе-
вых рыб. Родители близнецов сооружали в стороне от дерев-
ни на берегу реки маленькую хижину и ставили вокруг нее
деревянные фигуры и маски. Здесь, вдали от своих соплемен-
ников, они должны были провести два года. Отец целый год
очищался купанием и красил лицо в красный цвет. Во время
купания он пел ритуальные песни, которые содержали хвалу
лососям, а также призыв явиться, чтобы увидеть близнецов.
Если рыба приходила, верили, что ее вызвали колдовством.
Если же она не появлялась, то близнецам и их родителям, ко-
нечно, могло не поздоровиться. В верховьях Конго мать близ-
нецов должна была точно соблюдать следующий ритуал: ре-
бенка, родившегося первым, она всегда носила на правой
руке, а рожденного вторым — на левой. Если к ней обраща-
лись с приветствием, она должна была дважды благодарить,
то есть за каждого ребенка отдельно. Если она сама привет-
ствовала кого-нибудь, то также должна была это делать дваж-
ды, чтобы оба ребенка были послушными. По той же самой
причине она должна была есть обеими руками, иначе один
ребенок отстанет в упитанности.
Но есть ритуалы поистине жестокие. У аборигенов Юж-
ной Америки рождение близнецов очень часто воспри-
нималось как дурной знак, как доказательство неверности
жены или ее связи со злым духом. Поэтому матерей близнецов
подвергали наказаниям, иногда даже убивали. У айнов, корен-
ных жителей японских островов, женщины, родившие близ-
нецов, изгонялись своими соплеменницами из боязни, что они
могут заразиться и тоже рожать близнецов. Самих близне-
цов избегали в течение всей жизни. Младшего большей час-
тью убивали или родственники прятали его, а потом бросали
в лесу. Рождение близнецов часто рассматривалось как по-
следствие нарушения супружеской верности и считалось, что
они отмечены роком. Так было у древних индусов, в Ассирии,
Вавилоне, Египте, Абиссинии. В древнем Перу родителей пос-
169
Раздел 5. Близнецы в генетике человека
ле рождения близнецов подвергали строгому посту, во время
которого они должны были по очереди лежать с одним согну-
тым коленом до тех пор, пока подложенный под колено боб
благодаря теплу и поту не начинал прорастать. После этого
супружескую чету с веревкой на шее еще водили напоказ.
Обоих близнецов или одного из них часто после рождения
убивали. В Европе вплоть до средневековья многих матерей
близнецов обвиняли в сожительстве с дьяволом и вместе с
детьми сжигали на костре. В Испании XV в. даже существо-
вал королевский указ, предписывающий сжигать всех жен-
щин, родивших близнецов.
Достоверными знаниями о близнецах стали располагать
сравнительно недавно. Всего немногим более 200 лет прошло
с тех пор, как в одном толковом словаре t> близнецах писали
следующее: «Близнецы — это двое детей, которые появляют-
ся на свет из материнского чрева путем родов, то есть двое
детей от одного бремени».
5.1. Типы близнецов
Близнецы — это дети, родившиеся от многоплодной беремен-
ности. Они могут быть однополыми — все мальчики или все
девочки, и разнополыми — и мальчики, и девочки. В зависи-
мости от количества одновременно родившихся близнецов их
называют двойня, тройня, четверня и т.д. По биологическо-
му происхождению близнецы делятся на монозиготных (М3)
и дизиготных (ДЗ).
Монозиготные близнецы происходят из одной оплодотво-
ренной яйцеклетки — зиготы. Число монозиготных партне-
ров определяется количеством делений зиготы, которые пред-
шествуют развитию самостоятельных организмов. Если после
первого дробления зиготы каждая из двух дочерних клеток —
бластомеров дает начало отдельному зародышу, то формиру-
ется монозиготная двойня. Если после двух делений один из
бластомеров начинает развиваться самостоятельно, а второй
дробится еще раз и уже из них развиваются отдельные эмб-
170
5.1. Типы близнецов
рионы, возникает монозиготная тройня. Монозиготные близ-
нецы — пример бесполого размножения у человека, есте-
ственного клонирования. Монозиготные близнецы являют-
ся генетическими копиями. Это единственный тип родствен-
ников, у которых сто процентов общих генов. Нормальные
монозиготные близнецы всегда одного пола. Они настолько
похожи, что часто их практически невозможно различить
(рис. 5.1).
Рис. 5.1. Монозиготные близнецы
Существуют два типа М3 близнецов — монохорионные и
дихорионные. В случае монохорионных М3 близнецов зигота
расщепляется после того, как она имплантируется в плацен-
ту, и у близнецов имеется общий хорион (зародышевая обо-
лочка). В случае дихорионных М3 близнецов зигота расщеп-
ляется в первые пять дней после оплодотворения, т.е. до того,
как она имплантируется в плаценту, и у каждого партнера
свой хорион. Примерно 2/3 М3 близнецов являются монохо-
рионными и 1/3 — дихорионными. «Сиамские» близнецы
получаются в случае, когда зигота расщепляется лишь спустя
около двух недель после оплодотворения, что приводит к час-
тичному объединению их тел (рис. 5.2.).
171
Раздел 5. Близнецы в генетике человека
Рис. 5.2. «Сиамские» близнецы
Дизиготные близнецы
возникают иначе. Их коли-
чество определяется тем,
сколько яйцеклеток одно-
временно созревает у жен-
щины. Из оплодотворенных
яйцеклеток развиваются от-
дельные эмбрионы. Генети-
чески они сходны, как обыч-
ные братья и сестры, и мо-
гут быть как однополыми,
так и разнополыми. Обычно
дизиготные двойни разли-
чаются настолько, что со-
мнения в их происхожде-
нии не возникает, но иног-
да они бывают настолько
похожи, что напоминают
монозиготных.
В некоторых исследова-
ниях обнаружено, что моно-
хорионные близнецы более
похожи друга на друга, чем дихорионные близнецы, по лич-
ностным качествам и по когнитивным способностям, в то вре-
мя как в других исследованиях таких различий найдено не
было. Дизиготные близнецы всегда являются дихорионными,
таким образом, большее сходство у М3 близнецов, чем у ДЗ,
может быть объяснено более сходной пренатальной средой.
Близнецы, как правило, рождаются раньше и весят мень-
ше, чем одиночные новорожденные. Также в постанатальный
период родители часто не могут уделять достаточного внима-
ния сразу двоим детям одновременно, поэтому у близнецов
речь развивается в среднем позже и они немного хуже вы-
полняют вербальные тесты. Большинство таких различий,
включая вербальный дефицит, обнаруживаются уже в ран-
ние школьные годы.
172
5.2. Частота рождения близнецов
С возрастом М3 близнецы становятся все менее сходны-
ми по физическим чертам и массе тела и более сходными по
способностям, отражающимся в вербальных и математичес-
ких тестах. В противоположность, ДЗ близнецы с возрастом
становятся все менее сходными по данным видам когнитив-
ных способностей. М3 близнецы доживают примерно до оди-
накового возраста, а ДЗ близнецы чаще имеют разную дли-
тельность жизни.
Давно замечено, что в некоторых семьях близнецы рож-
даются чаще, чем в среднем в популяции. Это касается как
М3, так и ДЗ близнецов.
Если природа предрасположенности к рождению М3 близ-
нецов не совсем ясна, то причины склонности к рождению
ДЗ близнецов более понятны. Так, если у женщины есть род-
ная сестра, родившая ДЗ близнецов, то для нее самой вероят-
ность рождения ДЗ близнецов в 1,7 раза выше, чем в среднем
в популяции. Если у женщины мать является ДЗ близнецом,
то вероятность рождения у нее ДЗ близнецов в 2,5 раза выше
среднепопуляционных значений. Предполагается, что у чле-
нов семей с неоднократным рождением ДЗ близнецов чаще
встречаются аллели гена, которые отвечают за повышенную
выработку фолликулстимулирующего гормона (ФСГ). В свою
очередь, повышенный уровень ФСГ приводит к тому, что у
женщины возможно одновременное созревание нескольких
яйцеклеток. Ген, способствующий повышению репродуктив-
ных шансов, был найден у овцы. Предполагают, что у челове-
ка подобный ген находится в хромосоме 4.
5.2. Частота рождения близнецов
Рождение близнецов — явление редкое: на каждые сто оди-
ночно рожденных детей приходится одна двойня, на сотню
двоен — одна тройня, на сотню троен одна четверня и т.д.
Однако для разных популяций имеются различные данные.
Так, в среднем в США среди каждых 60 родов одни приводят
к появлению двойни. В США количество двоен увеличилось
173
Раздел 5. Близнецы в генетике человека
примерно на 28 000 за последние пять лет. Данный факт мож-
но объяснить применением фармпрепаратов для повышения
вероятности забеременеть у женщин с бесплодием. Кроме
того, альтернативные репродуктивные технологии также при-
водят к повышению риска многоплодной беременности (на-
пример, пересадка сразу нескольких эмбрионов после ш vitro
оплодотворения).
Частоту рождения монозиготных и дизиготных близнецов
выражают коэффициентом близнецовости (число близнецо-
вых пар на 1000 родов). Для дизиготных близнецов этот ко-
эффициент (d) (коэффициент дизиготности) вычисляется по
формуле d = 2и/ 1000N, где и — число разнополых близнецо-
вых пар, N — общее число изученных родов. Коэффициент
монозиготности (т) находится по формуле т — (L — 2и)/
1000N, где L — общее число близнецовых пар в изученной вы-
борке, и и L обозначают то же, что и в предыдущей формуле.
Тестом на репрезентативность выборки служит соотно-
шение моно- и дизиготных близнецов, которое должно соот-
ветствовать их соотношению в популяции. В любой популя-
ции еще до этапа диагностики зиготности легко определить
ожидаемое соотношение моно- и дизиготных пар, исходя из
принципа, сформулированного В. Вайнбергом: для дизиготных
близнецов равновероятно оказаться одно- и разнополыми. Сле-
довательно, разнополые близнецы составляют половину всех
дизиготных пар. Отсюда примерное число дизиготных пар в
выборке равно удвоенному числу разнополых двоен в ней, а
число монозиготных пар — общему количеству близнецов ми-
нус удвоенное число разнополых пар. Для европейских по-
пуляций доля монозиготных близнецов составляет около 35%,
поэтому репрезентативной близнецовой выборкой является
такая, в которой соотношение моно- и дизиготных пар при-
мерно 1:2. А поскольку для генетического исследования из ди-
зиготных пар отбираются только однополые, то соотношение
тех и других в репрезентативной выборке должно быть 1:1.
Рассмотрим следующий пример. В сельских популяциях
Японии выявляли близнецов. Всего за год было зарегистри-
174
5.3. Близнецовый метод изучения наследственности
ровано 1420 пар, из них в 598 парах были оба мальчика, в 547 —
обе девочки и в 275 — мальчик и девочка. На основании этих
данных следует определить примерную долю моно- и дизи-
готных близнецов в исследованной популяции. Итак, рожде-
ние дизиготных близнецов можно представить как рождение
двух сибсов. Из двух детей могут родиться два мальчика, две
девочки и мальчик с девочкой. Если предположить равную ве-
роятность рождения мальчика и девочки (по 1/2), то при раз-
ложении бинома во второй степени (общее число детей рав-
но 2), где составляющими событиями являются рождение
мальчика (м) и рождение девочки (д), получим: (1/2(м) +
1/2(д))2 = 1/4(м)2 + 1/2(мд) + 1/4(д)2. Видно, что доля разно-
полых дизиготных близнецовых пар (1/2(мд)) составляет
столько же, сколько и суммарная доля однополых дизиготных
близнецовых пар (1/4(м)2 + 1/4(д)2). Зная общее число близ-
нецовых пар разного пола, можно найти число монозиготных
близнецов, которое будет равняться разности между числом
всех близнецовых пар и удвоенным числом разнополых пар.
Для исследованной популяции 1420 — 2 х 275 = 870. Доля мо-
нозиготных близнецовых пар 870/1420 — 0,61, а дизиготных
пар 1 — 0,61 = 0,39.
5.3. Близнецовый метод
изучения наследственности
Использовать близнецов для изучения наследственности че-
ловека предложил Ф. Гальтон. В настоящее время близнецо-
вый метод используют для анализа признаков, наследование
которых не подчиняется менделевским закономерностям.
Генетическая основа таких признаков сложна и, как правило,
мало изучена. Близнецовый метод позволяет решить, имеет ли
вообще наследственность отношение к анализируемому призна-
ку, а если имеет, то насколько велика ее роль. Этот метод осо-
бенно часто используется в психологической генетике.
Близнецы интересны для генетиков по ряду причин: не-
которые из них являются генетическими копиями, или кло-
175
Раздел 5. Близнецы в генетике человека
нами, Кроме того, используя близнецов, можно соблюсти усло-
вия, практически не выполнимые при работе с человеком, —
создать одинаковые условия для всех испытуемых. Пара близ-
нецов на протяжении длительного времени находится в оди-
наковых или очень сходных условиях. Вначале — это общая
внутриутробная среда. После рождения — одно семейное ок-
ружение, одна социальная среда (детский сад, школа).
Для генетических исследований представляют интерес не
любые близнецы, а только определенные их категории. Это
монозиготные близнецы, выросшие вместе, а также разлучен-
ные монозиготные близнецы. Среди дизиготных близнецов
для генетических исследований отбирают только однополые
пары, выросшие в одинаковых условиях. Разнополые и раз-
лученные дизиготные близнецы в генетических исследовани-
ях не используются.
В основе близнецового метода лежит сопоставление мо-
нозиготных близнецов с дизиготными, партнеров монозигот-
ной и дизиготной пары между собой, близнецовой выборки с
общей популяцией. Близнецы представляют собой генетичес-
кую модель, в которой можно учесть как генетические, так и
средовые факторы, которые оказывают влияние на форми-
рование признака. Монозиготные близнецы — это индиви-
ды с одинаковыми генотипами, которые обычно и развива-
ются в сходной среде. Имея в своем распоряжении монози-
готных разлученных близнецов, исследователь моделирует
ситуацию — одинаковые генотипы в разных условиях. Срав-
нение фенотипов монозиготных близнецов, выросших в раз-
ных условиях, дает ответ на вопрос: влияет ли генотип на раз-
витие признака или он целиком зависит от среды. Дизигот-
ные близнецы, выросшие в одинаковых условиях, позволяют
решить вопрос о том, как влияет одинаковая среда на разви-
тие признака у индивидов с разными генотипами.
Исследование на близнецах складывается из ряда этапов:
формирование близнецовой выборки, установление зиготно-
сти близнецовых пар, сопоставление пар и групп близнецов
по изучаемому признаку.
5.3. Близнецовый метод изучения наследственности
Формирование близнецовой выборки
Формирование группы близнецов проводят каким-либо из
двух способов — сплошным или пробандным. Выбор метода
определяется характером признака, который собираются ис-
следовать. Сплошным методом формируют выборку для изу-
чения признаков, которые имеются у каждого человека (рост,
вес, артериальное давление, уровень интеллекта). В этом слу-
чае в выборку включают все встретившиеся пары при сплош-
ном обследовании популяции. Если предстоит изучить ред-
кий признак, выборку близнецов формируют по пробанду.
Пробандом является индивид, зарегистрированный в специ-
ализированном медицинском или социальном учреждении по
поводу того признака, который интересует исследователя.
Среди зарегистрированных индивидов для исследования от-
бирают тех, у кого есть близнец того же пола. Весьма ценная,
но очень редкая генетическая модель — разлученные моно-
зиготные близнецы. Эти близнецы представляют особый ин-
терес для специалистов по психогенетике, так как партнеры,
имея одинаковые генотипы, развиваются в разных средах.
Чем в более раннем возрасте близнецы были разлучены, тем
более надежные результаты получаются при их исследовании.
Чем сильнее на признак влияет генетический фактор, тем
больше сходны по этому признаку разлученные партнеры
монозиготной пары. Однако к результатам, полученным на
таких близнецах, следует относиться с осторожностью. Дело
в том, что, будучи разлученными, близнецы нередко попада-
ют в семьи сходного социально-экономического положения,
что может приводить к неправильным выводам о соотноше-
нии наследственности и среды в формировании признаков,
сильно зависимых от социального окружения.
Диагностика зиготности
Монозиготные близнецы совпадают по всем генетически кон-
тролируемым полностью пенетрантным признакам, которые
не модифицируются средой. Наиболее удобны биохимичес-
177
Раздел 5. Близнецы в генетике человека
кие маркеры генов, например, группы крови. Дизиготные I
близнецы могут быть похожими, а могут и различаться. Раз- {
личие хотя бы по одному такому признаку дает основание
считать пару дизиготной, сходство даже по нескольким при-
знакам не дает абсолютной уверенности, что пара монозигот-
на. По этой причине диагностика дизиготности абсолютна, а
диагностика монозиготности всегда вероятна. При неизвест-
ном фенотипе родителей зиготность близнецов можно уста-
новить на основе частот анализируемых признаков в популя-
ции. Если зиготность устанавливается с использованием ро-
дословной, необходимо исключить неинформативные
признаки. Неинформативными являются признаки, по кото-
рым потомки данной супружеской пары не могут различать-
ся. Признаки, по которым потомки данной супружеской пары
могут различаться, являются информативными. Вероятность
того, что дизиготные близнецы будут иметь одинаковые по-
казатели по n-ному количеству признаков, равняется произ-
ведению вероятностей по каждому тесту: РДЗ — Р1*Р2>' —Рп' а
вероятность того, что близнецы монозиготны равняется
Рмз =7 — Рдд. Учет все новых генетических маркеров прово-
дят до тех пор, пока не достигается принятый уровень веро-
ятности, например, 95%. Наиболее надежный метод опреде-
ления зиготности близнецовой пары является анализ ДНК,
однако в обычных исследованиях для установления типа близ-
нецов ограничиваются их портретным сходством.
Например, нам нужно определить вероятность монозигот-
ности мужской близнецовой пары, если частота рождения мо-
нозиготных близнецов в данной популяции составляет 32%, а
фенотипы близнецов и их родителей следующие:
Система группы крови Отец Мать Близнецы
1. Система АВО 0 АВ А
2. Система MN MN MN М
3. Система Rh'rh Rh+ Rh4 rh’
178
Будем рассуждатьследующим образом. Близнецовая пара
может быть как монозиготной с вероятностью 0,32, так и ди-
зиготной с вероятностью 1 — 0,32 — 0,68. В дизиготной близ-
нецовой паре могут родиться или двое детей одного пола, или
же двое детей разного пола. Вероятность рождения двух де-
тей одного пола составляет (1/4)2 + (1/4)2 = 1/2.
Генотипы родителей по системе АВО и MN определяются
однозначно, а по системе Rh+rh- исходя из фенотипов их де-
тей. У родителей с группой крови 0 и АВ дети могут иметь ге-
нотип АО или ВО с вероятностью 1/2. Вероятность того, что
генотипы пары дизиготных близнецов по системе группы
крови АВО будут одинаковы, составляет (1/2)2 + (1/2)2 = 1/2.
У двух гетерозиготных родителей по системе группы крови
MN могут родиться дети с тремя возможными генотипами и
такими их вероятностями — 1/4ММ, 1/2MN и 1/4NN. Веро-
ятность рождения ребенка ММ и NN составляет 1/4, а ребен-
ка MN — 1/2, Следовательно, вероятность того, что у двух
дизиготных близнецов будет совпадение по какому-либо фе-
нотипу из этих трех возможных, составляет (1/4)2 + (1/2)2 +
(1/4)2 = 3/8. И, наконец, у двух гетерозиготных родителей по
системе группы крови Rh+rh“ возможно рождение детей с
двумя фенотипами — Rh+ и rh-, причем дети Rh+ могут ро-
диться с вероятностью 3/4 (сюда входит гомозигота Rh+Rh+
и гетерозигота Rh+rh-), а дети rh- — с вероятностью 1/4. От-
сюда, вероятность, что два дизиготных близнеца будут иметь
любую, но одинаковую группу крови по этой системе, равна
(3/4)2 + (1/4)2 = 5/8,
Ход этих рассуждений представлен в таблице (см. с. 178).
Исследованная однополая пара может быть дизиготной с
вероятностью 0,03984 или монозиготной с вероятностью 0,32.
Нормированная вероятность для этой пары быть дизиготной
составляет 0,03984/(0,03984 + 0,32) = 0,11, а монозиготной
соответственно 1 —0,11 = 0,89,
Раздел 5. Близнецы в генетике человека
Тип близнецов Дизиготные Монозиготные
1. Близнецы по рождению 1 - 0,32 = 0,68 0,32
2. Близнецы одного и того же пола (1/2)2 + (1/2)2 = 1/2 1
3. Близнецы из данной се- мьи, имеющие одинаковые антигены системы АВО (1/2)2 + (1/2)2 = 1/2 1
4. Близнецы из данной се- мьи, имеющие одинаковые антигены системы MN (1/4)2 + (1/2)2 + (1/4)2 = 3/8 1
5. Близнецы из данной се- мьи, имеющие одинаковые антигены системы RhTh (3/4)2 + (1/4)2 = 5/8 1
Совокупная вероятность 0,68 х 1/2 х 1/2 х 3/8 х 5/8 = 0,03984 0,32 х 1 х 1 х 1 х 1 = 0,32
Конкордантность
Для качественных признаков используют показатель конкор-
лднтности, для количественных — показатель внутрипарно-
го сходства. Пара близнецов считается конкордантной, если
изучаемый признак имеется у обоих партнеров. Если признак
имеется только у одного из них — пара дискордантна. Сте-
пень конкордантности выражается процентом пар, у которых
анализируемый признак совпадает. В зависимости от спосо-
ба сбора материала рассчитывают либо парную, либо пробанд-
ную конкордантность. Если конкордатные пары сформирова-
лись путем независимо выявленных партнеров, то рассчиты-
вают пробандную конкордантность. Если в конкордантной
паре выявлен только один, а второй зарегистрирован как его
партнер, то правильным будет расчет парной конкордантно-
сти. Парная конкордантность рассматривается в том случае,
если только один член из пары является пробандом. Если оба
близнеца рассматриваются как пробанды (выявлены незави-
симо), то должна рассчитываться пробандная конкордант-
ность. В данном случае следует рассчитывать пробандную
конкордантность (Кпроб.): Кцра..=100%2П/(2Пк+Пд) где Пк —
число конкордантных пар, П — число дискордантных пар.
Если второй партнер и в конкордантных и дискордантных
парах зарегистрирован по первому, то в этом случае правиль-
но рассчитывать парную конкордантность — это процент
конкордантных пар от общего числа. К =100%П/(Пк+Пд),
где Пк — число конкордантных пар, П — число дискордант-
ных пар. Если исследователь получает в свое распоряжение
данные и не знает, как формировалась близнецовая выбор-
ка, то правильно будет, если он вычислит и парную и пробанд-
ную конкордантность. Значения конкордантности использу-
ют для того, чтобы вычислить показатель наследуемости. Со-
поставление парной конкордантности у моно- и дизиготных
близнецов дает приблизительное представление о роли на-
следственных и средовых различий в формировании анали-
зируемого признака. Исходным является положение, что сте-
пень парной конкордантности должна быть выше у монозигот-
ных близнецов по сравнению с дизиготными, когда ведущую
роль в формировании фенотипических различий между инди-
видами играет генотип. Если в формировании фенотипичес-
ких различий ведущую роль играет среда, конкордантность
монозиготных близнецов будет приближаться к конкордант-
ности дизиготных близнецов.
Вычисление показателя наследуемости
Для вычисления наследуемости качественного признака ис-
пользуют формулу Хольцингера: Н = (Кмз — Кдз)/(100 — Кдз)
где: Н — показатель наследуемости, Кю — конкордантность
монозиготных близнецов в %, Кдз — конкордантность дизи-
готных близнецов в %.
При Н = 1 фенотипическая изменчивость в популяции обус-
ловлена только генотипическими различиями, при Н = 0 — толь-
ко средовыми. Доля средового влияния (Е) на фенотипическое
разнообразие популяции вычисляется по формуле: Е = 100 — Н.
Если признак контролируется моногенно с полной пенет-
рантностью, то Кмз = 1.
Раздел 5. Близнецы в генетике человека
Величина конкордантности у монозиготных близнецов по
моногенному признаку может служить мерой пенетрантнос-
ти гена, ответственного за этот признак.
Одним из вариантов близнецового метода является иссле-
дование монозиготных близнецов, воспитывавшихся раздель-
но. В этом случае возможно непосредственное вычисление
доли средовых влияний (Е) в развитии качественных призна-
ков: Е = (Кв — Кр)/(100 — Кр), где К иК — показатели кон-
кордантности вместе и раздельно воспитывавшихся близне-
цов.
Для оценки наследуемости количественных признаков ис-
пользуют разность в значениях признаков у партнеров одной
пары. Соотношение наследственности и среды в формирова-
нии различий по количественным признакам оценивают сле-
дующим образом. Поскольку монозиготные близнецы гене-
тически идентичны, считают, что фенотипические различия
между ними обусловлены средовыми факторами. Фенотипи-
ческие различия между дизиготными близнецами складыва-
ются из генетических и средовых причин. Если известна об-
щая фенотипическая дисперсия в популяции (V опуляц), то для
вычисления наследуемости используют формулу: Н = 2(V —
Умз)/У„опулЯи V рассчитываются как среднее квадратов разно-
стей фенотипических дисперсий для двух членов нескольких
пар близнецов V= 1/п (ф2 + d22 +... dn2), где d — внутрипарная
разница, п — количество пар близнецов. Если известна лишь
дисперсия между близнецами, наследуемость вычисляется по
формуле: Н = (V.., — V^f/V^
В качестве примера вычисления наследуемости можно
привести следующий. В 1976 г. американский психолог Джон
Лехлин изучал наследуемость ряда личностных качеств чело-
века. Исследование было проведено на близнецовых парах,
среди которых 514 были монозиготными и 336 дизиготными.
В целом 50% монозиготных и 28% дизиготных близнецовых
пар оказались конкордантными. Чтобы определить коэффи-
циент наследуемости этих признаков у человека, необходи-
мо воспользоваться формулой Хольцингера:.
182
5.3. Близнецовый метод изучения наследственности
Семьи близнецов
Семьи монозиготных близнецов представляют особый инте-
рес для генетиков. Рассмотрим два типа таких семей. Первый
тип — это семьи близнецов, супруги которых не состоят в
родстве. Дети этих супружеских пар формально являются
двоюродными сибсами (они дети родных братьев или сестер).
Фактически (генетически) они полусибсы, так как имеют 1/4
общих генов. Потомки монозиготных близнецов-сестер гене-
тически более сходны, чем потомки близнецов-братьев. Дети
сестер имеют одинаковую цитоплазматическую наследствен-
ность и генетическое сходство по ядерным генам, равное 1/4.
Дети братьев имеют разную цитоплазматическую наслед-
ственность, генетическое сходство по ядерным генам у них
также 1/4. Дети сестер имеют также сходную внутриутроб-
ную среду, а дети братьев разную. Материнское влияние пос-
ле рождения может усилить фенотипическое сходство детей
сестер.
Описано несколько случаев, когда в брак вступали моно-
зиготные сестры и монозиготные братья. Дети от таких бра-
ков, формально являются двоюродными сибсами, но генети-
чески они сходны, как родные братья и сестры: у них 50% об-
щих генов. Кроме того, как и родные сибсы, они имеют
одинаковую цитоплазматическую наследственность. Есть ос-
нования считать, что и среда, в которой происходит разви-
тие и формирование этих сибсов как личностей, также бо-
лее сходна, чем среда обычных двоюродных братьев и сес-
тер: она определяется сходством поведения матерей и отцов.
Семьи близнецов интересны еще в одном аспекте. Для спе-
циалистов по психогенетике это материал для изучения того,
насколько генотип индивида влияет на выбор брачного парт-
нера. Если супруги монозиготных близнецов более сходны,
чем супруги дизиготных близнецов, то это свидетельствует о
том, что на выбор брачного партнера оказывает влияние ге-
нотип.
183
Раздел 5. Близнецы в генетике человека
Контрольные вопросы и задания к разделу 5
1. Перечислите известные Вам типы близнецов.
2. Каково происхождение «сиамских» близнецов?
3. Какие проблемы в генетике человека решаются с помо-
щью близнецового метода?
4. Какие категории близнецов используются для генетичес-
ких исследований?
5. Какую информацию получают, изучая семьи близнецов?
6. По данным, приведенным в таблице, вычислите коэффи-
циент близнецовости в различных этнических группах:
Группа Количество родов Всего близнецо- вых пар Число разнополых близнецовых пар
Русские 50 000 375 НО
Финны 40 000 612 234
Французы 20 000 215 71
Австрийцы 4000 44 15
Японцы 10 000 75 И
Китайцы 60 000 492 63
7. Среди цветного населения США выявлено 3363 пар близ-
нецов, среди которых 1195 пар оказались разнополыми.
В данной популяции частота рождения мальчиков состав-
ляет 51,32%. Определите долю моно- и дизиготных близ-
нецовых пар в исследованной выборке.
8. Свидетельствует ли сходство однополых пар близнецов по
группе крови системы АВО в пользу их монозиготности?
№ пары Фенотип пары Фенотип матери Фенотип отца
1 0 0 0
2 0 А В
3 АВ А 0
4 А АВ АВ
5 В 0 АВ
6 АВ АВ АВ
9. В таблице приведены сведения о фенотипах однополых
близнецов и их родителей. Свидетельствует ли в пользу
184
Контрольные вопросы и задания к разделу 5
монозиготности пары близнецов их сходство по анализи-
руемым признакам?
№ близнецовой пары Фенотип близнецов Фенотипы родителей
1 rh Rh+, Rh*
2 rh rh, rh
3 Rh+ Rh, rh“
4 rh Rh+, rh
5 M MN, M
6 MN M, MN
7 N MN, MN
10. Определите вероятность монозиготности братьев, если
они происходят из популяции, где среди двоен 36% моно-
зиготных близнецов и 32% однополых дизиготных. Мужчи-
ны конкордантны по признакам, частота которых в данной
популяции следующая: способность ощущать ФТК — 0,12,
группа крови MN — 0,54, Хд(а +) среди мужчин — 0,69.
11. Обследование 256 пар близнецов по заболеванию тубер-
кулезом дало следующие результаты: монозиготные кон-
кордантные — 30 пар, монозиготные дискордантные — 34,
дизиготные конкордантные — 46, дизиготные дискордан-
тные — 146 пар. Оцените роль наследственности в фор-
мировании туберкулеза.
12. Проанализируйте результаты, приведенные в таблице, и
укажите, влияют или нет генетические факторы на забо-
леваемость инфекционными болезнями.
Заболева- ние Количество пар близнецов
Монозиготных Дизиготных
Всего Конкордантных Всего Конкордантных
Корь 1629 1587 1996 1910
Коклюш 1047 1017 1287 1184
Скарлатина 327 179 381 179
Дифтерия 282 141 340 128
Ангина 309 158 204 81
Пневмония 328 106 472 86
185
1
Раздел 5. Близнецы в генетике человека
13. Конкордантность монозиготных пар по смертности в воз-
расте от 20 до 60 лет составляет 30,1%, у дизиготных —
17,4%, по смертности от травм — у монозиготных 6,9%, ди-
зиготных — 3,9%. Используя эти данные, вычислите долю
генетического влияния на указанные признаки.
14. Приведены внутрипарные различия для монозготных (М3)
и дизиготных (ДЗ) близнецов мужского пола по росту
(в см) и по весу (в кг). Рассчитайте наследуемость роста и
массы тела по результатам анализа выборки моно- и дизи-
готных близнецов.
Масса (М3); 1, 2, 1, 2, 1, 3, 2, 0, 1, 2, 3, 4.
Масса (ДЗ): 4, 2, 1, 5, 3, 5, 6, 5, 0, 3, 2, 2.
Рост (М3); 3, 2, 2, 0, 4, 5, 1, 2, 3, 0, 1, 2. '
Рост (ДЗ); 5, 7, 8, 2, 1, 3, 4, 5, 6, 6, 7, 9.
Дисперсия веса у мужчин в данной популяции 21 кг2, рос-
та — 55 см2.
15. Оцените роль генетических факторов в развитии рака у
человека на основании одного из близнецовых исследо-
ваний, если имеются следующие данные:
Тип близнецов Всего пар Один болен раком,число пар Оба больны раком, число пар
затронут один и тот же орган затронуты разные органы
Монозиготные 1528 143 8 13
Дизиготные 2609 292 9 39
16. В августе 1993 г. в штате Индиана (США) родились «сиам-
ские» близнецы — Эми и Анджела. Через семь недель
близнецы были разъединены хирургическим путем, при
этом погибла Эми. В возрасте 10 месяцев умерла и Анджела.
Медицинские расходы на операцию и послеоперационный
уход составили около 1 млн. долларов, что вызвало негатив-
ную реакцию многих налогоплательщиков штата, которые
считали, чго подобные операции слишком дорогостоящи, а
результаты не оправдывают себя, поэтому «сиамских» близ-
нецов не следует разъединять вообще. Каково Ваше мнение ?
186
Раздел 6
ПОЛОВЫЕ И ГЕНДЕРНЫЕ РАЗЛИЧИЯ
Говоря о различиях между мужчинами и женщинами, исполь-
зуют два понятия: пол и гендер. Термин пол имеет отношение
к телесным различиям (анатомическим и физиологическим)
между мужчиной и женщиной, а гендер затрагивает их пси-
хологические, социальные и культурные особенности. Разгра-
ничение пола и гендера является фундаментальным, так как
одни различия между мужчиной и женщиной обусловлива-
ются биологическими причинами, а другие по своей природе
биологическими не являются. Это мужские и женские про-
фессии, мода, обряды. В какой степени различия в поведении
мужчин и женщин определяются биологическими различия-
ми? Иными словами, в каком соотношении находятся пол и
гендер? Как всегда, у ученых нет единого мнения.
Одни считают, что в поведении мужчин и женщин суще-
ствуют врожденные (биологические) различия, которые об-
наруживаются в том или ином виде во всех культурах. Для
доказательства приводится обширный антропологический
материал, показывающий, что практически у всех народов
мужчины чаще, чем женщины, принимают участие в охоте и
в войнах, следовательно, более агрессивны. Женщины зна-
чительную часть времени заняты воспитанием детей и не при-
нимают участия в охотах и войнах.
Биологи накопили большой фактический материал о про-
явлении полового диморфизма человека по различным морфо-
логическим и физиологическим признакам. Хорошо известен
половой диморфизм и в отношении индивидуально-психоло-
гических особенностей. Все личностные черты присутству-
ют у представителей как мужского, так и женского пола, раз-
личия имеют количественный характер.
к 187
Психологи Кэттел и Торндайк были первыми, кто пред-
положили, что гендерные различия между мужчинами и жен-
щинами объясняются редкостью чрезвычайно умных женщин.
Франсис Гальтон в своем исследовании о наследственной при-
роде интеллекта практически не обнаружил выдающихся
женщин. В то время женщина вряд ли могла проявить себя
интеллектуально, поскольку общество традиционно отводи-
ло ей роль в домашнем хозяйстве и в уходе за детьми. Равных
прав в образовательной и профессиональной сфере у жен-
щин по сравнению с мужчинами не было.
За последние десятилетия появились данные, свидетель-
ствующие об уменьшении различий в когнитивных способ-
ностях между мужчинами и женщинами. Даже различия в
спортивных достижениях, которые могут быть частично обус-
ловлены действием стероидных половых гормонов, снижают-
ся, что хорошо прослеживается во время Олимпийских игр.
Объяснения этому можно найти в высокой мотивации и хо-
рошей тренировке женщин.
Обнаруживается сходство когнитивных способностей
мужчин и женщин. Многие ученые отмечают, что в когни-
тивных способностях между мужчинами и женщинами боль-
ше сходства, чем различий. Оба пола практически одинаково
выполняют тесты IQ. В то же время в пределах каждого пола
существуют большие вариации, особенно большой размах
отмечается у мужчин. Так, среди мальчиков на одном полюсе
много лиц с умственной отсталостью, дислексией, задержкой
речевого развития, гиперактивным синдромом, на другом —
много одаренных, особенно в математике и естественных
науках. Среди девочек полюса подвинуты больше к средним
значениям, т.е. у них наблюдается меньшая дисперсия. Дей-
ствительно, среди женщин наблюдается меньше умственно
отсталых, однако и меньше одаренных.
В современную эпоху отмечается сглаживание историчес-
кого неравенства и отсутствия возможностей у женщин в
когнитивных сферах, хотя и не во всех популяциях. Напри-
мер, в Афганистане до настоящего времени женщины не име-
ют права на получение образования. Все же в современном
6.1. Хромосомное определение пола
мире имеется тенденция к тому, что девочки демонстрируют
более высокие показатели при обучении на всех этапах обра-
зовательного процесса, чем мальчики.
6.1. Хромосомное определение пола
У человека представители обоих полов имеют одинаковое чис-
ло аутосом, но у мужчин присутствует одна X- и одна Y-xpo-
мосомы, а у женщин — две Х-хромосомы. Пол с различными
половыми хромосомами называется гетерогаметным, а пол с
одинаковыми половыми хромосомами — гомогаметным. Та-
ким образом, у человека мужской полгетерогаметный, а жен-
ский — гомогаметный. Х-хромосома человека значительно
больше и содержит больше генов, чем Y-хромосома. Около
1/4 генов Y-хромосомы соответствует генам Х-хромосомы.
Доминирующее влияние на развитие мужского пола оказы-
вает Y-хромосома: наличие этой хромосомы при любом чис-
ле Х-хромосом направляет развитие зародыша по мужскому
типу. Индивиды только с одной Х-хромосомой всегда женщи-
ны (ХО). Ген, обеспечивающий маскулинизацию, находится
в коротком плече Y-хромосомы. Лица с изохромосомой по
короткому плечу Y-хромосомы являются фенотипическими
мужчинами, а с изохромосомой по длинному плечу феноти-
пическими женщинами.
Обнаружен участок Y-хромосомы, который определяет
мужской пол. Были описаны мужчины с двумя Х-хромосома-
ми и женщины с одной X- и одной Y-хромосомами. Более тща-
тельное изучение половых хромосом у таких людей выявило,
что мужчины с набором XX в действительности имели неболь-
шой участок Y-хромосомы. Этот же участок Y-хромосомы от-
сутствовал у женщин с набором XY. Этот участок занимает
всего около 0,5% всей Y-хромосомы, состоит из 300 000 пар
оснований ДНК. Внутри этого участка ген, определяющий
мужской пол SRY (sex-determining region of the Y).
Ген SRY отвечает за синтез белка, который включает дру-
гие гены, направляющие развитие у эмбриона мужских струк-
189
Раздел 6. Половые и гендерные различия
тур. До 6-й недели пренатального развития у эмбриона чело-
века имеются зачаточные протоки, которые могут развивать-
ся как в мужские, так и в женские репродуктивные органы.
Когда примерно на 7-й неделе включается ген SRY, зачаточ-
ные яички начинают синтезировать тестостерон — половой
гормон, который стимулирует к развитию мужские структу-
ры. Запускается каскадный механизм действия других генов,
определяющих развитие в сторону мужского пола, и пример-
но на 16-й неделе эмбрионального развития начинают фор-
мироваться незрелые сперматозоиды.
6.2. Половая дифференцировка
и половые гормоны
У человека анатомические различия между мужским и жен-
ским полом видны при рождении. В онтогенезе половые раз-
личия в строении тела увеличиваются. Взрослея, мальчики и
девочки начинают различаться и поведением, и эти различия
объясняют действием половых гормонов. Различают первич-
ные и вторичные половые признаки, внутренние и наружные
половые органы. Первичные половые признаки (половые орга-
ны) непосредственно связаны с производством потомства.
Вторичные половые признаки непосредственно не вовлече-
ны в размножение, а связаны с выращиванием потомства.
Внутренние половые органы — это половые железы: яичники у
женщин и семенники у мужчин. Наружные половые органы —
мужские и женские гениталии.
Формирование эмбриона начинается с недифференциро-
ванной пары гонад (половых желез). Присутствие Y-хромо-
сомы превращает гонады в семенники, при отсутствии Y хро-
мосомы гонады превращаются в яичники. Семенники выра-
батывают мужские половые гормоны — андрогены, яичники
продуцируют женские половые гормоны — эстрогены и про-
гестерон. Андрогены обусловливают развитие генитальных
тканей во внешние половые органы. Без андрогенного сти-
мула эти же самые ткани развиваются в женские гениталии.
6.2. Половая дифференцировка и половые гормоны
Иногда эмбрион с хромосомным набором XY оказывается
нечувствительным к влиянию тестостерона, который выра-
батывается его собственными семенниками. Тогда вместо
мужских внешних гениталий формируются женские. Нор-
мальное развитие пола зависит не только от хромосом и на-
личия гормонов, но и от способности развивающихся тканей
реагировать на сигналы гормонов. Способность тканей отве-
чать на действие гормонов является следствием предшествую-
щего развития. Таким образом, первоначальные различия в
наборах хромосом дают толчок организму к развитию по муж-
скому либо женскому типу.
Гонады (половые железы) мужчин и женщин выделяют
«женские» эстрогенные гормоны, но секреция их у женщин
сильнее, чем у мужчин. Эстрогены, не будучи белками, не яв-
ляются непосредственными продуктами генов. Однако они
синтезируются при участии белков-ферментов и поэтому
находятся под контролем генов опосредовано. И мужчины, и
женщины имеют гены, которые кодируют эти ферменты, и эти
гены активны у представителей обоего пола. Различие между
полами, по-видимому, лежит либо в количестве производимых
ферментов, либо в условиях, создаваемых в клетках для их
активности. Простого наличия гормонов, независимо от их
концентрации, недостаточно для того, чтобы вызвать половую
дифференциацию, ткани должны быть восприимчивы к тому
или иному гормону. Мужской половой гормон — тестосте-
рон также синтезируется у представителей обоего пола, но у
женщин в меньшем количестве. Под действием тестостеро-
на генитальные ткани плода развиваются в мужские половые
органы.
Есть предположение, что половые гормоны действуют на
развитие мозга, в частности, тестостерон и его побочные про-
дукты — дегидротестостерон и эстроген, поскольку они вза-
имодействуют с гипоталамусом, лимбической системой и ко-
рой больших полушарий. Эти гормональные воздействия вна-
чале делают мужской мозг чувствительным к воздействию
тестостерона, а позднее активируют мозг для осуществления
191
Раздел 6. Половые и гендерные различия
типичного для мужского пола стиля поведения. Половые раз-
личия в головном мозге особенно заметны в зонах гипотала-
муса, которые участвуют в регуляции типичного для мужчи-
ны сексуального поведения. Тестостерон обычно ассоцииру-
ется с агрессивностью и склонностью к насилию. О том, что
половые железы оказывают влияние на поведение, было из-
вестно давно. Еще древние животноводы кастрировали сам-
цов домашних животных, для того чтобы сделать их не агрес-
сивными и управляемыми. У приматов — ближайших род-
ственников человека, самцы также более агрессивны, чем
самки, хотя в зависимости от вида наблюдаются большие от-
клонения. Многие самки приматов в некоторых ситуациях
более агрессивны, чем самцы, например, когда их детенышам
угрожает опасность. Кастрированные самцы-обезьяны менее
агрессивны, чем обычные, а обезьяны самки, получившие
тестостерон, становятся более агрессивными. Вместе с тем
было обнаружено, что когда обезьянам создавали условия,
благодаря которым повышался их статус в группе, у них в
крови повышался и уровень тестостерона.
О действии гормонов на дальнейшее поведение человека
свидетельствует и такое наблюдение. Для сохранения беремен-
ности некоторым женщинам вводят синтетические вещества —
прогестины. По своей химической структуре они напоминают
мужские половые гормоны. Оказалось, что прогестины мас-
кулинизируют женский плод. Маскулинизирующее действие
гормона на женский плод после рождения прекращается.
Наблюдения за этими девочками показывают, что по своему
поведению они напоминают мальчиков сорванцов.
6.3. Интерсексуальные состояния человека
Под интерсексуальным состоянием подразумевается одно-
временная экспрессия мужских и женских половых призна-
ков у одного и того же человека. Медицинским термином ин-
терсексуального состояния является гермафродитизм. Само
слово происходит от имени одного из младших богов в древ-
192
6.3. Интерсексуальные состояния человека
негреческом Пантеоне Гермафродита, который был самым
настоящим двуполым существом. Согласно греческой мифо-
логии, Гермафродит был сыном бога торговли Гермеса и бо-
гини любви Афродиты, во время купания он объединился с
нимфой. На одной из античных статуй, найденной в Центу-
рине, Гермафродит представлен с мужским половым органом,
женскими молочными железами и бедрами и мягкими жен-
ственными чертами лица. Кроме этой статуи, со времен Древ-
ней Греции и Рима сохранилось много других статуй и фре-
сок, свидетельствующих об интересе людей к этому явлению.
Социальный и психологический удел людей с интерсек-
суальными состояниями далеко не радостен. Среди них часто
бывают депрессии, низкая самооценка и склонность к самоубий-
ству. В Древней Греции гермафродитов обычно убивали при
рождении. В средние века гермафродитов преследовали, хотя
само явление признавалось как гражданскими, так и религи-
озными законами. У народа хинди в определенные истори-
ческие периоды отмечалось поклонение гермафродитам из-
за веры в то, что Высшее Существо также обладало одновре-
менно мужскими и женскими признаками. В некоторых
культурах к гермафродитам относятся просто как к людям с
альтернативной половой принадлежностью.
Гермафродитизм распространен среди растений и беспоз-
воночных животных. Представители одного вида моллюска
всегда рождаются самцами и остаются ими до определенно-
го периода своей жизни, после которого примерно половина
из них превращается в самок. Рыбки гуппи могут менять свой
пол.
До XX в. определение гермафродитизма было довольно
простым: гермафродит — это человек, имеющий мужские и
женские половые органы различных полов (рис.6.1). Внастоя-
щее время в связи с развитием генетики и эндокринологии
простые анатомические определения более не являются дос-
таточными. По новой терминологии истинным гермафроди-
том считается человек, у которого одновременно имеются
ткани как яичников, так и семенников. Сочетания могут быть
7 Введение в психогенетику
193
Раздел 6. Половые и гендерные различия
самыми разными — один яичник и один семенник (около 34%
случаев истинного гермафродитизма), половые железы, со-
стоящие из обоих видов тканей (ovotestis, или яйцесеменник)
(около 20% случаев) и яичник или семенник на одной стороне
и овотестис на другой (около 46% случаев). Люди с опреде-
ленными половыми железами (два семенника или два яични-
ка), однако, с сомнительными внешними половыми призна-
ками, не соответствующими хромосомному полу или недораз-
витыми, в-настоящее время классифицируются как ложные
гермафродиты, среди которых можно выделить несколько
разновидностей.
На любой стадии эмбриогенеза может произойти наруше-
ние развития. Основными веществами, которые отклоняют
развитие плода от заложенной генетической формы, являют-
ся андрогены семенников. Без семенников и веществ, кото-
рые они секретируют, развитие плода будет идти по женско-
му типу.
Основными причинами женского ложного гермафродитиз-
ма является врожденная гиперплазия надпочечников, а муж-
ского ложного гермафродитизма — синдром андрогенной не-
чувствительности .
Рис. 6.1. Выраженность внешних половых признаков
обоих полов у гермафродита
194
6.3. Интерсексуальные состояния человека
Врожденная гиперплазия надпочечников, или адреногени-
тальный синдром, является аутосомно-рецессивным заболева-
нием, которое приводит к усиленной продукции андрогенов
корой надпочечников. При этом заболевании андрогенная сти-
муляция является слишком запоздалой для преобразования
зачаточных половых протоков и вирилизация затрагивает
лишь внешние половые органы, дефекты которых можно
скорректировать хирургическим путем.
Девочки, тем или иным образом маскулинизированные,
отличаются своим поведением от обычных девочек. Они пред-
почитают игры и игрушки мальчиков. Не любят играть в кук-
лы и возиться с детьми. Они уверенны и настойчивы, успеш-
но конкурируют с мальчиками, однако не так агрессивны.
Желание иметь поклонника возникает у них позже, чем у
сверстниц. Половая ориентация у таких девушек не наруше-
на, но на первом месте у них стоит профессия и карьера, се-
мейная жизнь на втором.
Особенности темперамента также сильно зависят от дей-
ствия гормонов. Исследования, проведенные на макаках ре-
зус, показали, что самки, которым вводили мужские половые
гормоны, чаще дрались и гонялись друг за другом, чаще при-
нимали угрожающие позы, чем обычные самки. Девочки с
врожденной гиперплазией надпочечников чаще, чем их здо-
ровые сестры, обнаруживают энергию и драчливость. Они
предпочитают играть с мальчиками и не проявляют интереса
к куклам, однако чувствуют себя девочками и не имеют гомо-
сексуальных наклонностей.
Синдром андрогенной нечувствительности, известный
также как синдром тестикулярной феминизации, или синд-
ром Морриса, является Х-сцепленным рецессивным заболе-
ванием, в результате которого лица с генотипом XY развива-
ются в фенотипических женщин. Это происходит из-за того,
что в клетках отсутствуют рецепторы для андрогенов, в част-
ности для тестостерона, хотя сами эти вещества вырабатыва-
ются в достаточном количестве. Поскольку внешние половые
органы вне сомнения являются женскими, то данная болезнь
7*
195
Раздел 6. Половые и гендерные различия
редко диагностируется вплоть до полового созревания, когда
у таких фенотипических женщин не наступают месячные.
Пациентки с тестикулярной феминизацией по внешнему
виду и поведению почти ничем не отличаются от нормальных
женщин, за исключением более высокого роста и небольшо-
го оволосения на теле, зависящего от андрогенов, а также
короткого влагалища (рис. 6.2). Женщины с этим синдромом
нередко имеют высокий интеллект, сильный («мужской») ха-
рактер, который помогает им достичь высоких результатов в
профессиональной деятельности. Частота фенотипа состав-
ляет около 1 на 40 000.
Интересная вариация синдрома андрогенной нечувстви-
тельности была открыта среди 23 родственных семей в Доми-
никанской Республике. Было изучено 38 человек, которые
после рождения без сомнения были отнесены к лицам женс-
кого пола и воспитывались соответствующим образом. На са-
мом деле, эти дети были мужчинами с генотипом XY, у кото-
рых отсутствовал фермент 5-а-редуктаза, необходимый в син-
тезе мужского полового гормона. Хотя у этих детей был
нормальный для мужчин уровень тестостерона, у них из-за
отсутствующего фермента совсем не синтезировался дигид-
ротестостерон. Во время полового созревания примерно в 12
лет эти дети подвергались физическому превращению в муж-
чин — у них формировались мужские гениталии и развива-
лись семенники. Таким мужчинам было довольно легко из-
менить социальную и физическую компоненту, несмотря на
12-летний женский социальный тип.
Точная этиология истинного гермафродитизма менее
ясна. Истинными гермафродитами считаются лица, у кото-
рых имеются железы, функционирующие как по типу ооге-
неза, так и сперматогенеза. К случаям истинного гермафроди-
тизма относятся химеры и мозаики. В организме химеры име-
ются клетки генетически различных зигот, например, XX/XY.
В организме мозаика присутствуют клетки одной зиготы, од-
нако в результате нерасхождения хромосом при делении об-
наруживается наличие клонов клеток с различными перерас-
196
6.3. Интерсексуальные состояния человека
пределениями генетического материала, например, XX/XXY,
XX/XXYY.
И все же большинство истинных гермафродитов (около
70%) имеют нормальный кариотип XX или XY. Возможно, они
представляют собой случаи невыявленного химеризма, и по-
следние исследования мужчин с генотипом XX и женщин с
генотипом XY подтвердили это. У мужчин с хромосомным на-
бором XX был найден специфический участок Y-хромосомы,
где локализован ген SRY, запускающий механизмы развития
по мужскому типу. С другой стороны, потеря этого участка
гена у людей с генотипом XY приводит к половой дифферен-
циации по женскому типу. В то же время были зарегистриро-
ваны случаи истинного гермафродитизма, которые нельзя
объяснить ни транслокацией, ни делецией этого гена.
В гражданско-процессуальных делах иногда возникают
проблемы, связанные с расторжением брака на основании не-
Рис. 6.2. Проявления синдрома
андрогенной нечувствительности
197
Раздел 6. Половые и гендерные различия
возможности нормального репродуктивного функциониро-
вания интерсексуала. В мировой литературе зафиксированы
единичные случаи беременности у истинных гермафродитов
с генотипом XX и у химеры XX/XY, однако случаи мужской
гермафродитной плодовитости науке пока неизвестны.
Огромный интерес к интерсексуальным состояниям чело-
века существует в мире спорта. Малоизвестным является тот
факт, что по требованию Международного Олимпийского
Комитета все женщины-спортсменки должны были прохо-
дить так называемую «верификацию пола», целью которой
было выявление подставных мужчин, имеющих преимуще-
ства в состязаниях. В реальности такие мужчины никогда об-
наружены не были. Однако процедуры «верификации пола»
привели к значительному подрыву репутации женщин-спорт-
сменок, рожденных со сравнительно редкими генетически-
ми аномалиями, влияющими на развитие половых гонад или
на выраженность вторичных половых признаков.
В 1992 г. на зимних играх в Альбервилле был введен ут-
вержденный Международным Олимпийским Комитетом, ос-
нованный на ДНК-анализе тест по определению специфичес-
кого участка Y-хромосомы — района SRY. В 1996 г. на летних
играх в Атланте у 8 из 3387 спортсменок тест на SRY-участок
оказался положительным. У 7 оказалась андрогенная нечув-
ствительность (у четырех неполная и у трех полная), а у од-
ной ранее были удалены гонады. Все они были допущены к
соревнованиям.
После игр в Атланте многие организации продолжали на-
стаивать на отмене процедуры «верификации пола», которая
была сложной, дорогостоящей, противоречивой и унизитель-
ной. К тому же мужчин, желающих предстать женщиной,
можно обнаружить при дополнительном исследовании обя-
зательного анализа мочи на допинг. Под значительным обще-
ственным давлением Международный Олимпийский Коми-
тет в 2000 г. запретил «верификацию пола».
198
6.4. Гендерные различия в когнитивных способностях
6.4. Гендерные различия в когнитивных
способностях и личностных свойствах
Наблюдения за поведением, исследование мозга живых лю-
дей и трупов, а также моделирование на животных разного
пола дают основание предполагать наличие различий в ког-
нитивных способностях мужчин и женщин.
Современная психология констатирует три основных по-
ловых различия в когнитивных способностях. Это касается
вербальных способностей, которые более развиты у женщин,
математических способностей, более выраженных у мужчин,
и пространственно-образного воображения, более характер-
ного для мужчин. На рис. 6.3 приведены примеры вербально-
го теста, в котором по первой предложенной букве каждого
слова надо составить предложения, и невербального теста
на проверку пространственно-образных способностей, в ко-
тором надо выбрать куб, соответствующий развертке.
1. М________В С________________А.
2. С Ф И К.
3. Ж О Е Б.
Рис. 6.3. Вербальный (а) и невербальный (б) тесты
на проверку когнитивных способностей
199
Раздел 6. Половые и гендерные различия
Различия между интеллектуальными особенностями полов
проявляются не в общем проявлении интеллекта, а в некото-
рых специфических когнитивных способностях. Особенно-
сти внимания и восприятия, которые являются наиболее пер-
вичными этапами переработки информации, имеют межпо-
ловые различия. Обнаружено, что у девочек младенческого
возраста взгляд дольше фиксируется на зрительных стиму-
лах, в то время как у мальчиков уже в это время диагностиру-
ются состояния, связанные с дефицитом внимания. Некото-
рые ученые считают, что различия в восприятии касается не
только зрительного восприятия, но и всех остальных сенсор-
ных систем. Мужчины легче адаптируются к динамическому
визуальному процессу, и они легче обнаруживают небольшие
движения в поле своего зрения. Мужчинам также более при-
суще «чувство времени», т.е. они лучше определяют течение
времени. Женщины более чувствительны к прикосновению,
запахам, вкусу и звукам.
Из-за того что у мужчин лучше развиты пространствен-
но-образные способности, они могут более точно прицеливать-
ся в неподвижные и движущиеся мишени, атакже бросать пред-
меты по определенной траектории. Мужчины лучше женщин
ориентируются на местности и больше полагаются на оцен-
ку направления, расстояния и геометрических форм при по-
иске пути, в то время как женщины лучше ориентируются по
сухопутным отметкам.
Мужчины превосходят женщин в количественных расче-
тах и мысленном вращении предметов, а также в заданиях,
требующих вовлечения оперативной памяти для сохранения
и манипуляции с визуальным изображением.
Была выдвинута теория, что в эволюции многие из этих
способностей были важны для выживания в обществах охот-
ников и собирателей, когда мужчины осваивали незнакомые
им территории во время охоты, а женщины больше времени
проводили в поиске растений, ягод, плодов в близлежащей
местности. Была даже предложена эволюционная гипотеза о
развитии синдрома дефицита внимания. В соответствии с
200
6.4. Гендерные различия в когнитивных способностях
этой теорией, способность быстро просматривать окружаю-
щую местность и в то же время постоянно находиться начеку
при новом стимуле, например, при приближении опасного
животного, давали неоспоримые преимущества древним
охотникам.
С помощью методов магнитно-резонансного анализа моз-
га показано, что между мужчинами и женщинами существу-
ют различия в функционировании зон головного мозга, от-
ветственных за речь. У женщин лучше, чем у мужчин, рабо-
тают механизмы, вовлеченные в формирование памяти.
В частности, у женщин лучше развита ассоциативная память —
они легче предлагают синонимы к определенному слову, со-
ответствующий цвет, перечень пунктов, начинающихся с за-
данной буквы. Женщины быстрее извлекают информацию
из памяти, они более ловкие, быстрее, чем мужчины, могут
подбирать подходящие объекты из набора предлагаемых.
Существует мнение, что в основе различий в когнитивных
способностях и личностных характеристиках мужчин и жен-
щин лежат нейроанатомические различия. Считается, что
гены играют ключевую роль не только в половой, но и в ген-
дерной дифференциации. Если не происходит включение
гена SRY на шестой-седьмой неделе эмбриогенеза, то разви-
тие плода идет по женскому пути. Именно в этот период на-
блюдается сильное гормональное влияние на плод, которое
затем прослеживается и в постнатальной жизни человека.
Гормональное влияние возрастает еще раз в период полово-
го созревания. Кроме того, гормональные перепады отмеча-
ются ежедневно, сезонно, а также связаны с менструальным
циклом и наступлением менопаузы. Изучение гормонально-
го влияния на поведение и обучение показало, что у девочек
с врожденной гиперплазией надпочечников, которые подвер-
жены значительному воздействию тестостерона внутриут-
робно, лучше, чем у других девочек, развито пространствен-
ное воображение и наравне с другими мальчиками выраже-
но агрессивное поведение. Предполагается, что естественное
повышение уровня эстрогенов в период полового созревания
201
Раздел 6. Половые и гендерные различия
у девочек с дислексией способствует улучшению способнос-
тей к чтению.
Мужской мозг по абсолютному весу примерно на 10% тя-
желее женского, однако по отношению к массе тела мозг жен-
щин больше. Существует много споров относительно адап-
тивной ценности более крупного головного мозга, поскольку
во многих исследованиях отмечается положительная корре-
ляция между размером головного мозга и интеллектом, вы-
раженным в баллах IQ.
Ядерно-магнитная томография головного мозга показала,
что мозг мужчин содержит больше белого вещества и спин-
номозговой жидкости, чем мозг женщин. Мозг женщин, на-
против, содержит относительно больше серого вещества. Счи-
тается, что увеличение белого вещества мозга у мужчин по-
зволяет более эффективно передавать информацию в
различные зоны мозга и положительно влияет таким образом
на пространственное воображение. Увеличение серого веще-
ства у женщин позволяет более эффективно обрабатывать по-
лученную информацию.
Исследования у высших животных обнаруживают поло-
вой диморфизм головного мозга, включая гиппокамп, гипо-
таламус и мозолистое тело, хотя для человека подобные ис-
следования пока находятся в стадии незавершенности. Так,
показано, что правое полушарие у мужчин больше и более
плотное. Внутриутробно правое полушарие развивается бы-
стрее, чем левое. Именно поэтому левое полушарие больше
подвержено вредному влиянию в эмбриогенезе. Предполага-
ется, что по этой причине среди мужчин высокая частота лю-
дей с проблемами в обучении речи, поскольку зоны, ответ-
ственные за речь, располагаются в левом полушарии. Также
выдвинута гипотеза, что за счет различного строения мозо-
листого тела у мужчин и женщин взаимосвязь между обоими
полушариями сильнее у женщин.
Если рассматриватьлпчносшные свойства человека, то по-
ловые различия более всего проявляются в таких характери-
стиках: агрессивность, сотрудничество, вербальное общение.
202
6.5. Роль среды в гендерных различиях
Известно, что мужчины более агрессивны, чем женщины.
Неодинаково проявление агрессивности: у девочек и женщин
она чаще вербальная, у мальчиков и мужчин — физическая.
В ситуациях, когда незнакомому человеку нужна помощь, ко-
торая может быть сопряжена с потенциальной опасностью,
мужчины в большей степени, чем женщины, оказывают эту
помощь, особенно если в ней нуждается женщина или же если
события разворачиваются на виду у других людей. Мужчины
в меньшей степени поддаются влиянию, чем женщины. Вер-
бальные коммуникативные особенности мужчин состоят в
том, что они больше склонны к иерархической организации
и общаются посредством направляющих и доминирующих
взаимоотношений. В противоположность им, женщины яв-
ляются сторонницами равенства и сотрудничества, а во вза-
имоотношениях стремятся выразить свои чувства, понять, обод-
рить и поддержать другого человека. В невербальном поведе-
нии женщины обладают большими способностями, чем
мужчины, в определении чувств другого человека по выра-
жению его лица, в умении скрывать свои собственные эмо-
ции на лице, в узнавании лица, которое они видели ранее, и в
умении маскировать свое лицо, голос и тело. Считается, что
девочки и женщины обладают более изысканными эстетичес-
кими вкусами.
6.5. Роль среды в гендерных различиях
Причины гендерных различий могут быть обусловлены соци-
умом, культурой и специальными упражнениями. Например,
женщины, набирающие высокие баллы при тестировании по
шкале маскулинности, лучше выполняют тесты на проверку
математических способностей и пространственно-образно-
го мышления. У эскимосов по выраженности пространствен-
но-образного воображения мало различий между мужчина-
ми и женщинами. Логичным считается то объяснение, что,
во-первых, арктическая среда для выживания требует хоро-
шей пространственной ориентировки независимо от пола, и,
203
Раздел 6. Половые и гендерные различия
во-вторых, родители предоставляют самостоятельность как
мальчикам, так и девочкам. У европейцев специальные про-
странственные тренировки в начальной школе сглаживают
и даже вовсе стирают различия между мальчиками и девоч-
ками по развитию пространственного воображения.
Выбирая профессию, юноши и девушки отдают неодина-
ковое предпочтение различным видам деятельности. Женщи-
нам легче даются вербальные науки, а мужчинам — матема-
тические и требующие пространственного воображения, по-
этому, видимо, они подбирают соответстветствующие
профессии. Существует точка зрения, что различия в выборе
профессии в какой-то мере биологически обусловлены.
Нельзя, однако, не учитывать, что на распределение профес-
сиональных ролей между мужчинами и женщинами сильное
влияние оказывают социокультурные установки и ценност-
ные ориентации общества. В 70-х гг. прошлого века в США
среди лиц с высшим образованием доля женщин составила
6%, в Великобритании — 16%, в СССР — 75%. Значение соци-
окультурных установок в проявлении полового диморфизма
в поведении иллюстрируется на примере одного из племен в
Новой Гвинее. У этого народа женщины занимают домини-
рующее положение в обществе, они активны, имеют сильный
и решительный характер, а мужчины слабохарактерны, не-
уравновешенны, подвержены эмоциям.
6.6. Воспитание и поло-ролевые установки
Каким образом развиваются различия по признаку гендера?
Изучение взаимоотношений матери и ребенка указывает на
то, что с мальчиками и девочками родители обращаются по-
разному, даже если они уверены, что не делают при этом ни-
каких различий. Когда взрослых просили охарактеризовать
незнакомого ребенка, их ответы отличались в зависимости от
того, к какому полу, по их мнению, принадлежал ребенок.
Наблюдения показывают, что если ребенка окружающие
воспринимают как мальчика, если к нему относятся как к
204
6.6. Воспитание и поло-ролевые установки
мальчику, то он и считает себя мальчиком. По этой причине у
него развивается соответствующая поло-ролевая установка.
То же самое справедливо и для женского пола. Осознание себя
как представителя того или иного пола обычно определяется
наружными гениталиями и зависит от того, какую роль опре-
деляют ребенку в результате регистрации. Имя, которое да-
ется ребенку, одежда, которую ему приобретают, формиру-
ют у него осознание своей половой принадлежности. Воспи-
танием у ребенка подчеркивают мужские либо женские
черты.
Известен такой пример. В результате ошибки, случившей-
ся при обрезании в 1967 г., один из канадских монозиготных
мальчиков-близнецов Джон лишился наружного полового
органа. Психолог Дж. Мани, считая, что ребенку до трех лет
можно привить любую гендерную роль, посоветовал провес-
ти хирургическую коррекцию пола травмированного маль-
чика, назначить соответствующее гормональное лечение эс-
трогенами и воспитать его как девочку. «Девочка» любила иг-
рать в куклы и помогала матери по дому. Его брат любил играть
с машинками и заявлял о своем желании избрать одну из муж-
ских профессий. В пятилетием возрасте эти идентичные близ-
нецы проявляли четкие различия в поведении. В целом вос-
питание имело успех, но у ребенка, воспитанного как девоч-
ка, со временем стал проявляться активный мужской
темперамент. Этот ребенок рвал предлагаемые платья, отвер-
гал куклу Барби, предпочитал мочиться стоя, хотел бриться,
подобно отцу, и накопил карманные деньги, чтобы купить иг-
рушечное ружье, которым с ним не хотел делиться брат. Хотя
операция была проведена, когда малышу было 17 месяцев, и
он не знал о происшествии, ему было все труднее приспосаб-
ливаться к жизни по женскому типу. Во время полового со-
зревания родители сообщили Джону о том, что произошло с
ним в раннем детстве. Джон выбрал жизнь мужчины и снова
подвергся операции по хирургическому восстановлению свое-
го исходного пола. Как оказалось, одним только воспитанием
сделать установку на ту или иную половую роль невозможно.
205
Раздел 6. Половые и гендерные различия
Описаны двое детей с хромосомным набором XX и с анд-
рогенитальным синдромом. Оба ребенка имели внешние ге-
ниталии промежуточного типа. У одного из них в двухмесяч-
ном возрасте был проведен цитогенетический анализ, обна-
ружен женский хромосомный набор, поэтому ребенка стали
воспитывать как девочку. Другой ребенок воспитывался как
мальчик. Когда дети выросли, каждый из них соответствовал
той роли, в которой его воспитывали.
Наблюдения девочек с синдромом врожденной гиперпла-
зии надпочечников показывают, что вопреки желанию роди-
телей и воспитанию, они остаются неисправимыми сорван-
цами. Все эти факты говорят о том, что пол, как и многие дру-
гие признаки у человека, в своем формировании испытывают
влияние как наследственных факторов, так и факторов среды.
6.7. Гомосексуализм
Гомосексуализм — это сексуальная активность, направленная
на представителей своего пола. Гомосексуализм встречается
у всех народов, но отношение к этому явлению в разных куль-
турах неодинаково. У батаков — народа, проживающего на
острове Суматра, гомосексуальные отношения между муж-
чинами до брака считаются нормальным явлением. После
женитьбы большинство из них прекращает гомосексуальную
активность и переходят к гетеросексуальным связям со свои-
ми женами. У африканского племени эторо почти двести дней
в году запретны для общения с женщинами, поэтому большая
часть сексуальных контактов у мужчин этого народа проис-
ходит друг с другом. У древних греков половые отношения
между мужчинами и юношами приветствовались, а у древ-
них иудеев осуждались. В европейской культуре, основанной
на христианстве, гомосексуалисты на протяжении большей
части истории подвергались гонениям. За однополые контак-
ты полагалось тюремное заключение и даже смертная казнь.
Однако ни убеждения, ни наказания не искоренили это явле-
ние.
206
у 6.7. Гомосексуализм
Опросы современного населения показывают, что гомо-
сексуализм распространен весьма широко. Оказывается,
только половина американских мужчин являются истинно ге-
теросексуальными. 8% мужчин имели исключительно гомо-
сексуальные связи, 10% практиковали как гомо-, так и гете-
росексуальные контакты. 37% мужчин имели хотя бы одну го-
мосексуальную связь, 13% имели желание гомосексуального
контакта, но не реализовали его. Полностью гомосексуальны
около 2% женщин, 13% имели гомосексуальный опыт, 15%
имели желание вступить в гомосексуальную связь, но не осу-
ществили ее. Относительно терпимое отношение к однопо-
лым связям в европейской культуре сформировалось только
в последние десятилетия. В некоторых европейских странах
браки между лицами одного пола уже узаконены.
Наличие однополых связей в разных человеческих куль-
турах, стабильный процент лиц, практикующих гомосексуа-
лизм в условиях свободного выбора, невозможность истре-
бить его репрессивными мерами навели на мысль, что у этого
явления имеется биологическая основа. Действительно, ис-
следования, проведенные на близнецах, показали, что склон-
ность к гомосексуализму обусловлена генетической предрас-
положенностью. По разным данным наследуемость мужско-
го гомосексуализма колеблется в пределах 0,20—0,70.
Биологические основания гомосексуализма пытались обна-
ружить в особенностях полового развития, однако анатоми-
ческие и физиологические исследования индивидов с гомо-
сексуальными наклонностями не выявили у них никаких от-
клонений в развитии половой системы. Принадлежность
гомосексуалиста к тому или иному биологическому полу ни-
когда не вызывала сомнения, и до сих пор не существует ни-
каких биологических тестов, по которым можно было бы вы-
явить человека с гомосексуальными наклонностями.
Сексуальное поведение обычно связывали с половыми
гормонами, поэтому одно время предполагалось, что мужчи-
ны гомосексуалисты отличаются низким содержанием анд-
рогенов. Эта гипотеза не выдержала экспериментальной про-
207
Раздел 6. Половые и гендерные различия
верки, прямой зависимости между выработкой андрогенов и
гомосексуальностью не обнаружили. Было выдвинуто пред-
положение, что причиной отклоняющегося поведения явля-
ется измененная чувствительность рецепторов к андрогенам.
При исследовании пациентов, скончавшихся от СПИДа,
выявлено, что у гомосексуалистов передний отдел гипотала-
муса, ответственного за сексуальную активность, отличается
по размерам от соответствующей структуры гетеросексуалов
и напоминает таковую женщин.
О биологической основе гомосексуализма свидетельству-
ет то, что это не чисто человеческое явление. Гомосексуаль-
ное поведение обнаружено и у многих животных. Некоторые
самцы дрозофилы проявляют половое поведение по отноше-
нию к другим самцам охотнее, чем к самкам. Оказалось, что
эта особенность поведения контролируется генетически и
является моногенным признаком. Соответствующий ген был
выделен и клонирован. У дрозофилы также обнаружена ано-
малия полового поведения, названная «лесбиянский фено-
тип». Он проявляется в том, что самки ухаживают за самка-
ми и за самцами. Это поведение также генетически детерми-
нировано.
Едва ли можно представить, что такое сложное явление,
как гомосексуализм у человека, контролируется лишь одним
геном. В 1993 г. Хамер исследовал роль генов в определенных
формах рака у мужчин-гомосексуалистов со СПИДом. В ре-
зультате построения родословных, ученый заметил, что в не-
которых семьях само явление гомосексуализма прослежива-
лось в трех поколениях. Поскольку некоторые из этих гомо-
сексуалистов были родственниками не по прямой, а по
боковой линиям (дяди и двоюродные сибсы), то Хамер пред-
положил, что роль среды, а именно воспитания, была невели-
ка, а роль генов значительна.
Хотя исследования Хамера широко освещались в научной
литературе, он был не первым, кто изучал генетическую пред-
расположенность к гомосексуализму. Пилар исследовал ряд
семей и обнаружил группы мужчин-гомосексуалистов. Он
208
1 Контрольные вопросы и задания к разделу 6
также обнаружил родственные связи между женщинами-лес-
биянками из одних и тех же семей, хотя они были слабее, чем
для мужчин-гомосексуалистов. Бейли изучал гомосексуаль-
ные пары близнецов. Для М3 близнецов он обнаружил, что
вероятность одному партнеру стать гомосексуалистом, если
у другого наблюдается подобное отклонение, равна 52%, для
ДЗ близнецов эта же величина была меньше — 22%. Он так-
же обнаружил значительный наследственный компонент лес-
биянства.
Проанализировав всю имеющуюся информацию, Хамер
в своем исследовании выделил генетические профили 40 муж-
чин-гомосексуалистов и обнаружил общий для всех генети-
ческий маркер в длинном плече Х-хромосомы (q28). Он пред-
положил, что другие гены также вовлечены в проявление го-
мосексуализма, а обнаруженный им маркер может лишь
частично свидетельствовать о склонности к гомосексуализ-
му.
Контрольные вопросы и задания к разделу 6
1. От чего зависит пол у человека?
2. Какова роль гена SRY в Y-хромосоме человеке?
3. Приведите примеры половых и гендерных различий.
4. Обсудите вопрос о том, насколько различия между муж-
чинами и женщинами имеют биологическую и насколько
культурную основу.
5. Чем определяется половая принадлежность человека?
6. В доказательство несостоятельности гипотезы о том, что
Y-хромосома дает определенные спортивные преимуще-
ства можно привести один из интересных исторических
фактов, касающихся спортивных успехов Стеллы Уолш,
двукратной Олимпийской чемпионки в беге на 100 метров
и участнице 11 мировых состязаний. В 1936 г. в Берлине
она завоевала серебряную медаль, уступив первенство
американке Хелен Стефенс. Группа поддержки Уолш под-
вергла сомнению половую принадлежность Стефенс, мо-
209
Раздел 6. Половые и гендерные различия
тивируя свою идею чересчур успешным для женщины вы-
ступлением последней. После осмотра Стефенс немецкие
врачи заявили об однозначной принадлежности ее к «сла-
бому» полу. Сама Стелла Уолш погибла от шальной пули
во время ограбления торгового центра в Кливленде в 1980 г.,
после чего была проведена аутопсия, выявившая ее не-
однозначную половую принадлежность. Стелла Уолш ока-
залась гермафродитом с кариотипом XY. Хелен Стефенс
умерла женщиной в 1994 г. Как Вы относитесь к мероприя-
тиям, касающимся «верификации пола» в спорте? Нуж-
ны ли такие тесты в гражданских ситуациях, связанных,
например, с репродуктивной неполноценностью супру-
гов?
7. Какие генетические факторы контролируют особеннос-
ти полового поведения?
8. Приведите факты, полученные при исследовании живот-
ных, которые свидетельствуют о биологической основе
гомосексуализма.
9. В основе суррогатного материнства лежит использование
женщины в качестве «инкубатора» для вынашивания ею
ребенка и последующего рождения и передачи другим лю-
дям. Суррогатное материнство бывает коммерческим (за
денежное вознаграждение) и альтруистическим (бесплат-
но). Двое мужчин-гомосексуалистов решили нанять сур-
рогатную мать, которая родила бы им ребенка. После сда-
чи спермы одного из мужчин в специализированный реп-
родуктивный центр и оплаты услуг суррогатной матери в
размере 10 000 долларов, через положенное время муж-
чины получили ребенка и стали его воспитывать. Как Вы
считаете, какие психические последствия в дальнейшем
для всех участников этих событий могут возникнуть — для
обоих мужчин, для суррогатной матери и для ребенка?
Стоит ли разрешать гомосексуальным парам заводить де-
тей путем суррогатного материнства или усыновления?
210
Раздел 7
СТРУКТУРА ПОПУЛЯЦИИ И
ГЕНЕТИЧЕСКИЕ ПРОЦЕССЫ
Любой вид живых существ распространен по занимаемому
ареалу не равномерно, а в той или иной мере обособленными
группами, которые называются популяциями. Такое распре-
деление живых организмов связано с тем, что условия жизни
в разных местах ареала неодинаковы и особи концентриру-
ются в наиболее благоприятных участках. Не всякая изоли-
рованная группа представляет собой популяцию. Наиболее
четкое определение популяции можно дать для видов, размно-
жающихся половым путем. У таких видов популяция — это
совокупность свободно скрещивающихся особей, в течение
большого числа поколений населяющая определенное про-
странство и в той или иной мере репродуктивно изолирован-
ная от других совокупностей. Сказанное справедливо и для
человека как биологического вида. У человека популяциями
могут считаться отдельные населенные пункты, этнические
группы, религиозные касты и другие виды сообществ. Все
человечество также можно рассматривать как огромную по-
пуляцию.
Любая популяция представляет собой сообщество генети-
чески различных особей и характеризуется своим собствен-
ным генофондом — совокупностью всех генов популяции.
В популяционной генетике важнейшими понятиями являют-
ся частота гена (аллеля), частота генотипа, частота фено-
типа, в то время как в менделевской генетике таковыми яв-
ляются понятия «ген», «генотип», «признак».
Среди задач, решаемых популяционной генетикой, важ-
нейшая — изучение генетической структуры различных по-
пуляций, а также генетических процессов, которые в них про-
текают.
211
Раздел 7. Структура популяции и генетические процессы
7.1. Нахождение частот аллелей
Вероятность сочетания разных аллелей в генотипах обуслов-
лена системой скрещивания, сложившейся в популяции. Ста-
тистически случайное (не зависящее от генотипа) образова-
ние родительских пар обозначают термином панмиксия. Если
какие-то сочетания генотипов в супружеских парах встреча-
ются чаще или реже, чем при случайном процессе, говорят
об ассортативности.
Описать структуру популяции, значит найти частоты ал-
лелей и вероятность различных типов скрещивания (у чело-
века — разных типов браков). Как это делается, рассмотрим
на примере моногенного признака — группы крови системы
MN у человека. Принадлежность к той или иной группе в этой
системе контролируется аутосомным локусом L, который су-
ществует в двух аллельных состояниях Iм и LN. Аллель Iм
обеспечивает выработку эритроцитарного фактора М, аллель
LN — фактора N. В популяции присутствуют индивиды с ге-
нотипами LMLM, LNLN и LMLN, которым соответствуют феноти-
пы (группы крови) М, N и MN. Допустим, среди 1000 человек,
взятых в случайном порядке из определенной популяции, 361
имел группу М, 159 группу N, а у 480 была группа MN. По-
скольку аллели кодоминантны, по фенотипу можно опреде-
лить генотип каждого человека. 361 человек имели генотип
LMLM, 159 — LNLNn 480 — LMLN. Всего в этой группе 2000 генов.
Определим, какую долю составляют аллели LM и какова доля LN.
Частоты альтернативных аллелей обозначают буквами р
и q (р + q = 1). Гомозиготный индивид вносит в генофонд
два одинаковых аллеля, а гетерозиготный два разных. Коли-
чество индивидов с генотипом LMLM обозначим символом D, а
количество индивидов с генотипом LNLN символом R, число
LM LN символом Н.
Частоты аллелей найдем с помощью простых арифмети-
ческих действий.
р = (2D + H)/2(D + R + H) =
= (2x361 + 480)72(360 + 159 + 480) = 0,6
212
7.1. Нахождение частот аллелей
Таким же образом находим частоту второго аллеля:
q = (2R + H)/2(D + R + H) =
= (2 х 159 + 480)/2(361 + 159 + 480) = 0,4
Правильность расчетов проверяется сложением:
р + q = 0,6 + 0,4 = 1.
Сцепление с полом
Частоты генотипов по генам, сцепленным с Х-хромосомой, у
гомогаметного пола (у человека это женский пол) подчиняет-
ся формуле Харди—Вайнберга (в случае двуаллельной систе-
мы) и формуле Бернштейна (для множественного аллелизма).
У гетерогаметного (мужского) пола частоты генотипов равны
частотам аллелей.
У человека 2/3 генов, сцепленных с полом, принадлежат
женской части популяции и 1/3 — мужской. Средняя часто-
та аллелей, сцепленных с Х-хромосомой, в популяции равня-
ется р = 2/3pz>+ 1/Зр^.
Например, если в популяции у 8% мужчин встречается ре-
цессивный сцепленный с Х-хромосомой дальтонизм, то доля
мужчин-дальтоников совпадает с частотой гена у них. Доля
женщин-дальтоников составляет q2 = 0,082 = 0,0064, или
0,64%, что в 12,5 раз меньше, чем мужчин.
Множественный аллелизм
Структура популяций для случая множественного аллелизма
описывается формулой Бернштейна. Если частоты аллелей А',
А", А'" и т.д. соответственно р,, р2, р3... рп (pt + р2 + р3 + ... +
pn = 1), то доля любой гомозиготы равняется квадрату часто-
ты соответствующего аллеля, а доля гетерозиготы — удвоен-
ному произведению соответствующих частот.
Так, группы крови системы AB0 определяются тремя ал-
лелями — ген 1А определяет наличие на поверхности эритро-
цитов антигена А, ген 1в — соответственно наличие антигена
В и ген i° не определяет выработку ни одного из антигенов.
Гены 1А и 1в кодоминантны по отношению друг к другу и до-
213
Раздел 7. Структура популяции и генетические процессы
минантны по отношению к гену i°. Частоты указанных алле-
лей обозначим соответственно р, q и г, и они легко определя-
ются графически из следующей таблицы возможных комби-
наций аллелей у индивидов в популяции:
\ $ \ Ра Чв Го
Ра Р2а РаЧв РаГо
Чв РаЧв Ч2в ЧвГо
Го РаГо ЧвГо Г2о
Можно привести следующий пример. При исследовании
выборки итальянцев с острова Сицилия по частоте у них групп
крови системы АВО оказалось, что 45,9% имели I группу кро-
ви, 33,4% — II группу, 17,3% — III группу и 3,4% — IV группу.
Для определения частоты генов группы крови этой системы
в изученной популяции поступим следующим образом. Обо-
значим частоту гена А — р,В — qni° — г. Частоту гена i° опре-
делим исходя из доли лиц с группой крови 0:
г = д/0,459 =0,678.
Суммарная доля лиц, имеющая группу крови 0 и В, равна
q2 + 2qr + г2, или же (q + г)2:
(q + г)2 = 0,632, где 0,173 = q2 + 2qr (доля фенотипов В и0);
q +г = 70,632 =0,795..
Зная г, получим q:
q = 0,795 — г = 0,795 — 0,678 = 0,117.
Из равенства р + q + г = 1, находим р:
р = 1 — q — г = 1—0,117 — 0,678 = 0,205.
7.2. Уравнение Харди-Вайнберга
Система скрещивания оказывает влияние на распределение
генотипов в популяции. При случайном скрещивании — пан-
214
7.2. Уравнение Харди-Вайнберга
миксии соотношение генотипов (в популяции) описывается
уравнением Харди—Вайнберга:
р2АА + 2pqAa + q2aa = 1.
Эта запись означает, что различные генотипы в панмик-
тической популяции представлены в таком соотношении:
доли гомозигот равняются квадратам частот соответствующих
аллелей, а доля гетерозигот — удвоенному произведению этих
частот. Такое состояние популяции называется равновесным.
Равновесие Харди—Вайнберга выполняется только в идеальной
популяции — бесконечно большой, генетически изолирован-
ной, в которой отсутствуют отбор и мутационный процесс.
В такой популяции соотношение генотипов (p2AA:2pqAa:q2aa)
сохраняется на протяжении бесконечного числа поколений.
Уравнение Харди—Вайнберга используют для нахожде-
ния частот аллелей непрямым путем. К нему прибегают, ког-
да признак контролируется доминантным аллелем и не все
генотипы можно идентифицировать по фенотипу. Используя
уравнение Харди—Вайнберга для нахождения частот аллелей,
необходимо быть уверенным, что по своим характеристикам
популяция приближается к идеальной, то есть она достаточ-
но большая и панмиктичная. Допустим, необходимо найти
частоту рецессивного гена альбинизма в городской популя-
ции, где один альбинос приходится на каждые 15 000 населе-
ния. Задачу решаем так. Обозначим гены, генотипы и фено-
типы: аллель альбинизма — а, аллель нормальной пигмента-
ции — А, частота нормального аллеля — р, частота аллеля
альбинизма — q, АА и Аа — нормально пигментированные
индивиды, аа — альбиносы.
Соотношение генотипов р2 : 2pq, : q2
Доля альбиносов q2 = 1/15000.
Отсюда q = 0,008, р = 1 — q = 0,992.
Зная частоты аллелей, можно рассчитать количество в
популяции гетерозиготных носителей гена альбинизма:
2pq = 2 х 0,992 х 0,008 = 1,6% .
215
Раздел 7. Структура популяции и генетические процессы
7.3. Дрейф генов
Теория дрейфа генов была разработана американским уче-
ным С. Райтом в 1930-х гг. Под дрейфом генов, или эффектом
Райта, подразумеваются случайные изменения частот генов
из поколения в поколение в популяциях малого размера. Из-
за ограниченного эффективно-репродуктивного объема по-
пуляции генофонд нового поколения может существенно от-
личаться от генофонда поколения родителей.
В малочисленных популяциях случайным образом одни ал-
лели, в том числе и вредные, могут размножиться, а другие
вообще утратиться. Дрейф генов приводит к снижению гене-
тического разнообразия, обедняет генофонд популяции. Осо-
бенно сильно дрейф генов проявляется в периоды резкого
снижения численности популяций — во время войн, эпиде-
мий, катастроф.
Оценку дрейфа генов проводят с помощью формулы
= yjpfl— p)/2N, где о — квадратическое отклонение,
р — частота аллеля, N— репродуктивный объем популяции.
Примером дрейфа генов может быть и высокая распрост-
раненность в США хореи Гентингтона. Считается, что груп-
па нескольких мигрантов из Англии, среди которых, по край-
ней мере, один был носителем этого патологического гена,
образовала поселение. Их многочисленные потомки унасле-
довали патологический ген.
Популяция данкеров — одна из религиозных сект в Пен-
сильвании (США) — основана 27 семьями, эмигрировавши-
ми из Германии в середине XVIII в. Они жили замкнутым со-
обществом, заключая браки внутри своей группы. Современ-
ные популяционно-генетические исследования населения
Германии и американцев немецкого происхождения показа-
ли, что эти популяции существенно различаются по частотам
генов, определяющих группы крови и ряд морфологических
признаков (рис. 7.1). Так, в популяции данкеров часто встре-
чаются левши с оволосением средних фаланг пальцев, при-
216
7.3. Дрейф генов
сверхподв ижность
большого пальца
прикрепленная
и свободная мочка уха
оволосение средних
фаланг пальцев леворукость
Рис. 7.1. Характерные наследственные
признаки популяции данкеров
крепленной мочкой уха и сверхподвижностью указательно-
го пальца руки (палец «голосующего»).
Среди индейцев Северной Америки встречаются высокие
частоты альбинизма. Так, в европейских популяциях частота
альбинизма составляет 1:20 000, а в некоторых индейских —
1:200. У индейцев гопи альбинизм получил широкое распрос-
транение в силу дрейфа генов, несмотря на отсутствие ада-
пативной ценности данного гена. Дело в том, что в традици-
онной культуре индейцев гопи альбиносы почитаются. Муж-
чины-альбиносы пользуются успехом у женщин, что ведет к
217
Раздел 7. Структура популяции и генетические процессы
увеличению числа потомков, получающих ген альбинизма.
Однако постепенно в индейских популяциях идолопоклон-
ничество по отношению к альбиносам уходит в прошлое, что в
конечном счете приведет к снижению в них гена альбинизма.
7.4. Эффект основателя
Эффект основателя является следствием дрейфа генов, ког-
да небольшая группа людей выходит из родительской попу-
ляции и обосновывается в новом месте, унося с собой нереп-
резентативную выборку для родительской популяции. В этой
группе одни аллели вообще могут быть утеряны, а другие —
представлены у большинства индивидов. Возникновение уни-
кальной частоты генов в новой популяции «основателей» соб-
ственно и является эффектом основателя.
Эффект основателя проявляется в необычайном распре-
делении ряда наследственных болезней на острове Тристан
да Кунья и в популяции амишей.
Тристан да Кунья был открыт в 1506 г. В 1817 г. выходец из
Шотландии обосновался на острове вместе с семьей, состоя-
щей из жены, двух детей, двух компаньонов и цветной слу-
жанки. Эта семья считается основателем современного по-
селения.
В течение последующих 130 лет на остров прибыло не-
сколько женщин. На острове высаживались жертвы кораб-
лекрушений. Таким образом, в 1946 г. 222 человека острова
были потомками 8 мужчин и 9 женщин и связаны друг с дру-
гом родством.
На Тристане распространенным наследственным заболева-
нием является пигментный ретинит, одна из форм рака глаз.
Считается, что одна из женщин, прибывших на остров, была но-
сительницей гена пигментного ретинита. В настоящее время
1 % населения острова являются носителями гена пигментного
ретинита по сравнению с менее чем 1/4000 в среднем в мире.
Половина жителей Тристан да Кунья страдают астмой,
которая является наследственной болезнью.
218
7.4. Эффект основателя
Популяция амишей представляет собой религиозный изо-
лят. Изначальная иммиграционная группа амишей в Амери-
ку состояла примерно из 200 поселенцев и в последующих по-
колениях браки заключались между их потомками. Эффект
основателя в популяции амишей сопровождался инбридин-
гом, поскольку при ограниченном числе брачных партнеров
кровнородственные браки неизбежны. В настоящее время
считается, что средняя степень кровнородственности браков
в округе Ланкастер в Пенсильвании находится на уровне бра-
ков между троюродными сибсами.
Среди наследственных болезней с повышенной частотой
у амишей встречаются кистозный фиброз, мышечная дис-
трофия верхнего пояса, гемолитическая анемия с недостаточ-
ностью пируват-киназы и ряд психических наследственных
расстройств. У амишей распространены наследственные бо-
лезни, которые редко встречаются или вообще отсутствуют
в других популяциях. Ген одного из типов карликовости, из-
вестного как синдром Эллиса-ван Кревельда, распространен
среди амишей Ланкастера с частотой 7%, в то время, как в
других популяциях его частота составляет менее 0,1%. Кроме
Рис. 7.2. Ребенок с синдромом Эллиса-ван Кревельда,
распространенного среди амишей
219
Раздел 7. Структура популяции и генетические процессы
карликовости, необычным симптомом указанного заболева-
ния является наличие зубов при рождении, полидактилия и
ряд других признаков (рис. 7.2).
Эффект основателя также заметен в популяции африка-
неров в Южной Африке. Африканеры являются потомками
небольшой группы датских, французских и немецких иммиг-
рантов. Эффект основателя привел к увеличению доли одно-
го из вариантов порфирии — аутосомно-доминантной болез-
ни обмена, эффекты которой заметны после приема барби-
туратов.
7.5. Эффект «бутылочного горлышка»
Если размер популяции быстро уменьшается, редкие аллели
могут быть утеряны при гибели их носителей. Так происхо-
дит снижение аллельного разнообразия и уменьшение час-
тоты гетерозигот. Оставшаяся популяция имеет более одно-
образный генофонд по сравнению с исходной. Происходит
так называемый эффект «бутылочного горлышка». Под этим
образным выражением подразумевается передача лишь не-
большого числа аллелей следующему поколению (рис. 7.3).
Эффект «бутылочного горлышка» можно продемонстри-
ровать на примере острова Пинджлеп островной цепи вос-
Рис. 7.3. Схематическое представление эффекта
«бутылочного горлышка»
7.6. Поток генов
точной Каролины. Среди населения острова у 4—10% наблю-
дается ахроматопсия, сочетающая в себе цветовую слепоту,
близорукость и катаракты. Предполагается, что огромная ча-
стота заболевания на острове является следствием событий
1780 г., когда сильный тайфун привел к гибели большого числа
островитян. В результате эффекта «бутылочного горлышка»
частота гена ахроматопсии значительно увеличилась в следую-
щих поколенях, когда численность населения восстановилась.
7.6. Поток генов
Богатое археологическое наследие дает информацию о пере-
движениях людей в древних популяциях. Некоторые мигра-
ции были спорадическими и затрагивали небольшие группы.
Другие были более или менее постоянными и включали ог-
ромное количество людей. Крупномасштабные миграции, за
которыми следует смешение людей в результате заключения
браков, вводят новый генетический материал в чужие попу-
ляции. Проникновение генов в популяции путем миграции но-
сит название поток генов, или просто миграция.
Своеобразное распределение аллеля Р системы АВО в Ев-
ропе является следствием движения монгол в западном на-
правлении от материнской популяции в период между VI и
XVI вв. В Центральной Азии отмечена высокая частота алле-
ля Р, составляющая 0,25—0,30. В Европе частота аллеля Р пос-
ледовательно снижается, начиная от границ с Азией с наи-
большими частотами и заканчивая Голландией, Францией,
Испанией и Португалией с наименьшими частотами (0,05 и
менее). У басков, населяющих область Пиренеев на границе
между Испанией и Францией, частота гена Р минимальна —
менее 0,03. С популяционно-генетической точки зрения по-
пуляция басков является эндогамной. Чрезвычайно низкую
частоту аллеля Р среди басков можно объяснить малой сте-
пенью брачных контактов с соседними популяциями. Воз-
можно, что несколько столетий назад аллель Р полностью от-
сутствовал в генетически закрытой популяции басков.
221
Раздел 7. Структура популяции и генетические процессы
Обмен генами между популяциями может иметь ощути-
мые медицинские последствия. До недавнего времени гемо-
литическая болезнь новорожденных, или резус-конфликт,
практически не встречалась в Китае. Все китаянки резус-по-
ложительны (RhRh). Однако межрасовые браки американских
иммигрантов с коренным населением Китая ввели в китайские
популяции резус-отрицательный аллель (rh). В первом поколе-
нии у потомства американских мужчин и китайских женщин
резус-конфликт не наблюдался. В противоположность, все
браки резус-орицательных американских женщин (rhrh) с
резус-положительными китайскими мужчинами (RhRh) пред-
ставляли собой группу риска для будущего потомства. Все
дети китайских отцов (RhRh) были резус-положительными
(Rhrh) и были потенциальными жертвами гемолитической бо-
лезни.
В то время как американские иммигранты вносят резус-
отрицательный аллель (rh) в китайские популяции, где его
ранее не было, китайские иммигранты (все RhRh) вносят все
больше резус-положительных аллелей (Rh) в американские
популяции, уменьшая таким образом частоту резус-отрица-
тельности. Изначально браки резус-положительных китайцев
(RhRh) с резус-отрицательными американками (rhrh) приве-
ли к возрастанию доли новорожденных с резус-конфликтом.
Однако в последующих поколениях частота резус-отрица-
тельных женщин в США снизилась ввиду того, что женщины
смешанного (американо-китайского) происхождения имели
генотип как RhRh, так и Rhrh (преимущественно последний
из указанных генотипов). Таким образом, для США долговре-
менным эффектом американо-китайских браков является
снижение частоты гемолитической болезни новорожденных
в популяции.
При смешении популяций А и В вследствие миграции оце-
нить долю вклада каждой популяции в образовавшейся попу-
ляции С можно следующим образом:
222
7.7. Естественный отбор
ь _дс-ди
Вклад популяции А: , _ , вклад популяции В:
о + а qb qa
а _ЯС- <к
Ь + a qa-qb
где а — число лиц в популяции А, включенных в популяцию С;
b — число лиц в популяции В, включенных в популяцию С;
qa — частота гена в популяции А;
qb — частота гена в популяции В;
qc — частота гена в популяции С.
Миграция изменяет частоту аллелей среди старожилов,
если у пришельцев частота аллелей была иной. Если т — доля
иммигрантов, то доля старожилов 1 — т. Если в популяции,
из которой происходит миграция, частота аллеля Р, а в анали-
зируемой локальной популяции Ро, то в поколении t частота
аллеля в локальной популяции станет р — (1 — т)‘х (Рд — Р) + Р.
Например, нам нужно определить, сколько могло быть бе-
лых среди предков среднего американского негра, если изве-
стно, что частота гена серповидно-клеточной анемии в попу-
ляциях американских негров 0,056, африканских негров 0,085,
а у белого населения данный ген не присутствует. Будем рас-
суждать следующим образом. Генетический вклад африкан-
ских негров в генофонд американских негров рассчитываем
исходя из частоты исследуемого гена в популяции американ-
ских (qa), африканских негров (qb) и белых (qc):
qc - qb _ о,°5б
Ча - Чь 0,085
- 0,66 = 66%
Следовательно, 34% предков американских негров не при-
надлежали к негритянской расе.
7.7. Естественный отбор
Естественный отбор представляет собой важнейший фак-
тор эволюции, который вызывает адаптивные изменения в ге-
223
Раздел 7. Структура популяции и генетические процессы
нетической структуре популяций. Отбор проявляется в том,
что особи с разными генотипами оставляют неодинаковое
количество жизнеспособных потомков, т.е. вносят неодина-
ковый генетический вклад в следующее поколение. Некото-
рые ученые полагают, что в связи с развитием цивилизации
естественный отбор у человека потерял свое значение. Бла-
годаря медицине люди, страдающие сахарным диабетом,
врожденным пороком сердца и другими тяжелыми заболева-
ниями, могут выживать и иметь детей. Снижением интенсив-
ности отбора, особенно в промышленно развитых странах,
объясняют высокую частоту наследственных болезней в со-
временных человеческих популяциях. Есть и другая точка
зрения: отбор в популяциях человека не исчез, но изменил
свою направленность.
У народов, ведущих примитивное натуральное хозяйство,
отбор проявляет себя на стадии после рождения. В этих по-
пуляциях высока плодовитость и детская смертность. Те, кому
удалось выжить и вырасти, обладают отменным здоровьем.
В индустриально развитых странах с хорошо развитой меди-
циной, рождаемость и детская смертность очень низкие, од-
нако практически все взрослое население страдает теми или
иными болезнями с наследственным компонентом. В этих
популяциях отбор проявляет себя в основном до рождения в
виде гибели эмбрионов на разных этапах развития.
Интенсивность отбора s связана с показателем генетичес-
кой приспособленности w таким уравнением: s = 1 — w. По-
казатель приспособленности представляет собой вероятность
передачи генов следующему поколению. Для вычисления
коэффициента отбора количество потомков, оставляемых наи-
более плодовитым генотипом, принимают за единицу (w = 1).
Приспособленность (w) других генотипов рассчитывается как
доля от приспособленности наиболее адаптивного генотипа.
Адаптивность большинства носителей хромосомных анома-
лий нулевая (ы = 0), так как они не передают свои гены по-
томству. Против таких генотипов действует отрицательный
отбор с интенсивностью, равной единице (s = 1). У людей с
224
7.8. Мутационный процесс
наследственной карликовостью детей в среднем в пять раз
меньше, чем у их нормальных братьев и сестер. Адаптивность
гена карликовости 0,2 (1/5 = 0,2), против него действует от-
бор с интенсивностью s = 0,8.
7.8. Мутационный процесс
Мутации — это ненаправленные, случайные изменения ге-
нетического материала, происходящие спонтанно или под
влиянием особых физических, химических и биологических
факторов. Спонтанный мутационный процесс — это главный
источник появления новых аллелей, приводящий к увеличе-
нию генетического разнообразия популяции. Большинство
мутаций не закрепляется в популяции, утрачиваясь в ряду
поколений. Мутации возникают очень редко — одна мутация
на сотни тысяч гамет за одно поколение. Вновь возникшие
мутации, как правило, вредны. Однако в редких случаях му-
тантный ген сообщает своим носителям некоторые преиму-
щества, и тогда он подхватывается естественным отбором.
В современных человеческих популяциях уровень мути-
рования повышен. Это связано с тем, что в среде обитания
человека появилось большое количество новых химических
веществ. Многие из них являются мутагенами, это ядохимика-
ты, красители и другие соединения. Мутагенным действием об-
ладает ионизирующее, рентгеновское, ультрафиолетовое излу-
чение. Мутагенным эффектом обладают и вирусы, которые,
встраиваясь в ДНК хозяина, вызывают в ней изменения.
Эффекты отбора и мутаций уравновешиваются, если чис-
ло вредных генов, элиминируемых отбором, совпадает с чис-
лом вредных генов, возникающих в результате мутаций.
Гены переходят из одного аллельного состояния в другое
(мутируют) с частотой примерно 10'5—10’6. Оценить скорость
мутирования при доминантных новообразованиях не пред-
ставляет труда — для этого необходимо знать количество по-
томков с доминантным признаком, родившихся от родителей
с рецессивными признаками.
8 Введение в психогенетику
225
Раздел 7. Структура популяции и генетические процессы
Равновесные частоты для доминантного (р) и рецессивно-
го (д) аллеля вычисляются по формулам р = u/s и q- -Ju/s , где
и — частота мутирования, s — коэффициент отбора. Для леталь-
ных генов (s = 1) формулы имеют вид р = и и q ~-Ju Допус-
тим, частота возникновения аутосомного рецессивного леталь-
ного аллеля, обусловливающего болезнь Тэя—Сакса, равна 10'5.
Равновесная частота этого аллеля равна q = 4s = 41СР = 0,0032.
Генетические процессы в популяции взаимосвязаны — му-
тационный и миграционный процессы создают генетическое
разнообразие, система скрещиваний формирует из них мно-
гообразие генотипов, а естественный отбор оценивает их,
решая судьбу отдельных генов и генотипов в дальнейших по-
колениях.
7.9. Инбридинг
Инбридинг — это браки между биологическими родственни-
ками.
При кровнородственном браке у потомков повышается ве-
роятность гомозиготного состояния. Такой потомок называ-
ется инбредным, а инбридинг измеряется коэффициентом
инбридинга F. Величина Рпоказывает вероятность гомозигот-
ности потомства по какому-либо случайно выбранному ауто-
сомному локусу. Значение F варьирует от 0 до 1. Коэффици-
ент инбридинга F может быть рассчитан для индивида на осно-
ве менделевских закономерностей с помощью родословной.
Указанный подход к расчету коэффициента инбридинга впер-
вые был разработан монахами в средние века для определе-
ния степени «греха» для людей, как правило, знатных и бога-
тых, которые хотели заключить брак с родственником. Для
того чтобы брак был разрешен, в церковную казну необходи-
мо было внести денежную компенсацию, пропорциональную
коэффициенту инбридинга в современном его понимании.
Для описания уровня инбридинга используют понятия как
коэффициента инбридинга, так и коэффициента родства. Ко-
226
7.9. Инбридинг
эффициент родства двух индивидов — это вероятность того,
что случайно выбранный ген, принадлежащий одному инди-
виду, идентичен гену того же локуса другого индивида. Ко-
эффициент инбридинга индивида равен коэффициенту род-
ства его родителей. Коэффициент родства определяется для
двух индивидов, которые могут иметь общих предков. Коэф-
фициент инбридинга определяется для одного индивида и
измеряется степенью генетического сходства между его ро-
дителями. Вычисление коэффициента инбридинга (F) можно
проводить с помощью путевого анализа, разработанного
С. Райтом. Метод основан на подсчете этапов передачи генов.
Вероятность передачи гена в одном поколении равняется 1/2,
поэтому F = г/2, где г — степень родства. Доля общих генов у
детей от кровнородственных браков равняется 2г.
В браках между генетическими родственниками повыше-
на вероятность появления потомства с рецессивными призна-
ками, что важно учитывать при генетическом прогнозирова-
нии наследственных болезней. Генетические последствия
кровнородственных браков можно проиллюстрировать на
примере ряда наследственных болезней в популяциях Евро-
пы и США. Например, среди белого населения США на кров-
нородственные браки между двоюродными сибсами прихо-
дится лишь 0,05%, однако 18—24% альбиносов и 27—53% слу-
чаев болезни Тэя—Сакса происходят именно от этих браков.
Не во всех популяциях последствия инбридинга неблагоп-
риятны. У сельского населения Индии, Китая и Японии кров-
нородственные браки довольно часты, но ожидаемые небла-
гоприятные последствия обнаружены в них не были, разли-
чий по частоте врожденных уродств, мертворождений,
смерти в течение первого года жизни в кровнородственных
браках и не родственных не выявлено. Этому факту дают та-
кое объяснение. Если в европейских странах и в Северной
Америке кровнородственные браки запрещены, то в странах
Востока, наоборот, они поощряются. Инбридинг в течение
многих поколений приводил к выщеплению рецессивных го-
мозигот, которые обладали пониженной жизнеспособностью
8’
227
Раздел 7. Структура популяции и генетические процессы
и не передавали свои гены потомству. Таким образом, инб-
редные популяции за много веков освободились от вредных
рецессивных генов, поэтому в настоящее время в этих попу-
ляциях кровное родство супругов не приводит к выщеплению
гомозигот по вредным рецессивным генам.
Примерами популяций, в которых уже многие поколения
практикуется инбридинг, являются арабские популяции тау-
регов и индийские популяции тамилов, где приветствуются
браки между двоюродными сибсами.
В некоторых популяциях, например, в Японии, кровнород-
ственные браки поощряются по ряду причин. Традиция тре-
бует, чтобы семьи жениха и невесты были знакомы. Также
желательно, чтобы эти семьи происходили из одной местнос-
ти и были одного социоэкономического статуса.
Другим примером кров-
Рис. 7.4. Королева Елизавета-
мать — глава британской
королевской семьи
неродственных браков яв-
ляются браки в высших
классах Европы, где брачные
партнеры должны иметь рав-
ные социальные уровни. По-
скольку представителей выс-
ших классов значительно
меньше, чем представите-
лей средних и низших, ста-
новится неизбежных заклю-
чение браков между членами
родственных семей. Край-
ним примером являются ев-
ропейские королевские ди-
настии (рис. 7.4).
Последствиями инбри-
динга на популяционном
уровне может быть значи-
тельное повышение доли
людей с редким аутосомно-
рецессивным заболевани-
228
7.10. Аутбридинг
ем. Так, если частота аллеля q какой-либо аутосомно-рецес-
сивной болезни равна 0,001, то частота пораженных людей в
панмиктической популяции (f = 0) будет 0,0012 = 0,000001
(1 на миллион). Если допустить, что f = 0,01, а частота рта же,
то доля больных составляет q2 + pqf = 0,000001 + 0,999 х 0,001 х
0,01 = 0,000011. ?идно, что инбридинг в популяции на уровне
1% вызывает 1100% увеличение частоты больных (11-кратное
увеличение).
7.10. Аутбридинг
Термин «аутбридинг» (как и термин «инбридинг») пришел
из сельского хозяйства, где им обозначают генетически отда-
ленные скрещивания животных или растений. Обычно это
скрещивание разных сортов растений, пород животных, а
также разных видов, а иногда и родов. Термином аутбридинг
у человека обозначают не родственные браки, когда хотят
противопоставить их кровнородственным. К ним можно от-
нести браки между выходцами из разных населенных пунк-
тов (обычно удаленных), между представителями разных на-
циональностей и рас.
По-видимому, в процессе эволюции выявлялись опреде-
ленные преимущества аутбридинга. Избегание у человека
близкородственных браков стало проявляться в виде инцест-
табу, особенно среди родственников первой степени родства.
Еще лет пятьдесят назад в бывшем СССР анализ биологи-
ческих последствий отдаленных браков, в особенности меж-
национальных и межрасовых, не проводился (в основном по
идеологическим причинам), а их влияние на потомство апри-
орно считалось благоприятным. Вывод о полезности отдален-
ных браков не основывался на результатах научных исследо-
ваний, а представлял собой как бы логическое продолжение
вполне справедливой идеи о вредности кровного родства.
Научным аргументом, приводимым для доказательства этого
положения, были данные генетики о том, что у потомства от
не родственных браков вредные рецессивные гены перехо-
229
Раздел 7. Структура популяции и генетические процессы
дят в гетерозиготное состояние и не оказывают влияния на
фенотип. Между тем рецессивными болезнями не исчерпы-
вается все разнообразие наследственной патологии челове-
ка. Последствия аутбридинга, обнаруженные при тщательно
спланированных исследованиях, оказались неоднозначными.
Аутбридинг повышает степень гетерозиготности потомства.
При этом повышается вероятность перехода вредных рецес-
сивных генов в гетерозиготное состояние. Действительно, у
потомства от аутбредных браков реже встречаются рецессив-
ные заболевания. Еще один эффект, который ожидается от
аутбидинга — гетерозис — повышенная жизнеспособность
потомства, полученного от неродственного скрещивания.
Отмечено, что высокая гетерозиготность у человека ускоря-
ет рост и половое созревание, но при этом сокращается про-
должительность жизни. Так, в течение последних 100 лет в
некоторых популяциях время полового созревания у девушек
снизилось по сравнению с данными предыдущих исследова-
ний примерно на 0,3 года за каждые 10 лет. К отрицательным
эффектам аутбридинга относят также разрушение историчес-
ки сложившихся адаптивных комплексов генов. Если потом-
ство первого поколения отдаленных браков действительно в
ряде случаев демонстрирует гетерозис, то во втором и после-
дующих поколениях выщепляются генотипы с пониженной
приспособленностью.
Популяционные процессы часто сопряжены с другими,
причем биологические аспекты переплетаются с социальны-
ми, что можно рассмотреть на примере усилившейся мигра-
ции и межрасовых браков.
В 1967 г. в США был снят запрет на заключение межрасо-
вых браков, хотя по данным социологического опроса 1965 г.
от 42 до 72% белого населения США высказались против.
С того времени доля межрасовых браков возрастала. В пери-
од с 1960 по 1990 гг. доля браков между белым населением и
азиатами возросла почти в 10 раз, между белым населением
и африканцами — почти в 4 раза.
230
Г 7.11. Брачная ассортативность
7.11. Брачная ассортативность
Выбор брачного партнера является одним из наиболее важ-
ных событий в жизни человека. Большинство исследований
сходится к тому, что браки формируются не случайно. Логич-
но предположить, что человек подбирает себе партнера по
интересам, личностным качествам, внешности и не обраща-
ет внимания на группы крови и другие биохимические
признаки. Так, умеренная положительная корреляция супру-
гов отмечается по антропометрическим характеристикам
(г —0,10—0,30) и IQ (г = 0,37), несколько большая по физичес-
кой привлекательности (г = 0,38—0,52) и по уровню образо-
Рис. 7.5. Отметки, используемые при измерении
лицевой асимметрии
231
Раздел 7. Структура популяции и генетические процессы
вания (г = 0,46). По личностным характеристикам сходство
супругов ниже (г = —0,23—0,47, в среднем г = 0,15), в то время
как по личностным ценностям несколько выше (г = 0,20—0,58).
Интересно, что наибольшая корреляция у супругов наблюда-
ется по таким чертам, как религиозность, консерватизм и ав-
торитарность (г = 0,40—0,70). Корреляция наблюдается так-
же и по возрасту вступления в брак. Показано, что люди с пси-
хическими нарушениями довольно рано вступают в первый
брак по сравнению с людьми, психически здоровыми.
При выборе брачного партнере значительное внимание
уделяется чертам лица. Черты лица развиваются на протя-
жении жизни человека и некоторыми учеными рассматри-
ваются даже как вторичные половые признаки. Асимметрия
черт лица, например, изучается по степени асимметрии меж-
ду соответствующими точками левой и правой стороны, обыч-
но их берут не менее 14 (рис. 7.5).
Многие телесные признаки (пальцы, запястья, уши) не яв-
ляются предметом полового выбора и прямо не влияют на при-
влекательность лица.
Контрольные вопросы и задания к разделу 7
1. Можно ли назвать коллектив большого предприятия по-
пуляцией?
2. Какая популяция описывается уравнением Харди—Вайн-
берга?
3. Перечислите свойства идеальной популяции.
4. Группа крови системы Ik контролируется аутосомным ло-
кусом с двумя аллелями 1кП и 1кь. Из 4200 человек, обследо-
ванных в одной из популяций, 1218 имели генотип 1кП1ка,
882 — генотип IkbIkb и 2100 — Ika 1кь. Определите часто-
ты аллелей Ika ulkb в популяции.
5. Поликистоз почек контролируется аутосомным доминант-
ным геном. Среди 1500 детей, которые родились в одном
из городов, у пятерых обнаружен поликистоз почек. Оп-
ределите частоту гена поликистоза.
232
Контрольные вопросы и задания к разделу 7
6. Гиподонтия (отсутствие одного или нескольких постоян-
ных зубов) у человека наследуется как аутосомно-доми-
нантный признак с варьирующей экспрессивностью. Этот
дефект в современных цивилизованных странах встреча-
ется примерно у 10% населения. Рассчитайте, как соотно-
сятся доли гетеро- и гомозигот по гену гиподонтии.
7. Аутосомный доминантный ген, вызывающий нефроблас-
тому (рак почек), локализован в 11-й хромосоме. Пенет-
рантность данного гена около 63%. Среди населения ис-
следованной популяции численностью 237 500 человек
обнаружено 7 больных. Определите частоту гена нефроб-
ластомы.
8. Акаталазия — заболевание, вызванное рецессивным ге-
ном. У гетерозигот по этому гену наблюдается понижен-
ное содержание каталазы в крови. Частота гетерозигот в
одной из популяций 1,4%. Каковы частоты аллелей гена,
ответственного за синтез фермента каталазы ?
9. Аниридия — отсутствие радужной оболочки — наследу-
ется как аутосомный доминантный признак и встречает-
ся в популяции с частотой 1:10 000. Определите частоту ал-
леля, контролирующего развитие данной аномалии.
10. У человека локус, ответственный за синтез макроглобу-
лина, сцеплен с Х-хромосомой. Наличие глобулина доми-
нирует над его отсутствием. В выборке одной популяции
было 114 макроглобулинположительных женщин и 46
мужчин, 88 макроглобулинотрицательных женщин и 154
мужчины. Каковы частоты аллелей этого гена у мужчин,
женщин и в целом в популяции?
И. При фронтометафизарной дисплазии (Х-сцепленное ре-
цессивное наследование) наблюдается ряд дефектов труб-
чатых костей, аномалии черепа, зубов, боли в суставах, а
также снижение слуха и зрения. Это заболевание выяв-
лено у 3% населения. Какова доля пораженных среди муж-
чин и женщин при соотношении полов 1:1? Изменится ли
ответ, если соотношение полов будет 0,9 мужчин :1 жен-
щина?
233
Раздел 7. Структура популяции и генетические процессы
12. Частота аллелей гена, определяющего группу крови АВО,
среди наслеления одного города составляет: А — 0,25, В —
0,189,0 — 0,561. Каков процент людей в данном городе име-
ет группы А, В, АВ и 0?
13. Перечислите факторы популяционной динамики.
14. В каких популяциях происходит дрейф генов?
15. Что такое антропологический изолят?
16. Для каких человеческих популяций характерен инбри-
динг?
17. Какое генетическое влияние оказывают на популяцию
миграции?
18. Частота гена R0 в Западной Африке составляет 0,63, у ев-
ропейцев 0,02, а у американских негров 0,44. В каком со-
отношении произошло смешение генов у американских
негров?
19. Потомство от смешанных браков негров с белыми в США
относят к негритянскому населению, поэтому такие бра-
ки можно рассматривать как поток генов от белых к нег-
рам. Предки негров США вывезены из Африки около 300
лет назад, что составляет примерно 12 поколений (1 поко-
ление ? 25 лет). Рассчитайте интенсивность потока генов,
если у негров США частота аллеля Fya составляет 0,094, у
негров Африки — 0,0, а у белых США — 0,422.
20. Частота гена резус-отрицательности в одной популяции
составляет 15%, в другой — 1%. В группе населения, сфор-
мировавшейся в результате браков между представителя-
ми этих популяций, частота резус-отрицательности 9%.
В каком соотношении в гибридной популяции представ-
лены генофонды каждой из предковых форм?
21. У 47 карликов в браке с людьми нормального роста всего
было 54 ребенка (25 карликов и 29 нормальных). У 56 их
нормальных сибсов, состоящих в браке с нормальными
людьми, было 322 ребенка нормального роста. Определи-
те приспособленность гена карликовости.
22. Определите приспособленность каждого из трех геноти-
пов по локусу серповидноклеточности, если среди 4130 об-
234
Контрольные вопросы и задания к разделу 7
следованных было 3122 нормальных гомозигот, 10 были го-
мозиготны по серповидно-клеточной анемии, 998 были ге-
терозиготны.
23. При исследовании ахондропластических карликов оказа-
лось, что за 15-летний период в одной популяции дети с
такой наследственной патологией появились в 15 случаях
из 105 555 изученных. При этом 5 больных родились от
браков, где один или оба родителя были поражены этой
болезнью, остальные 10 — от браков здоровых индивидов.
Определите частоту мутаций по гену ахондроплазии в дан-
ной популяции.
24. Частота новорожденных с фенилкетонурией, вызываемой
аутосомно-рецессивным летальным аллелем, составляет
4:100 000. Определите темп мутирования нормального гена
в аллель фенилкетонурии.
25. Некоторые хромосомные перестройки обладают доминан-
тным фенотипическим эффектом. Больные с синдромами
Дауна, Патау, Эдвардса (трисомики), как правило, не ос-
тавляют потомства. Дети с этими фенотипами рождаются
соответственно с частотой 1:700, 1:6000, 1:7000. Определи-
те частоты мутирования этих хромосомных болезней.
26. По каким признакам браки являются панмиктичными?
27. Какое влияние на структуру популяции оказывают инб-
ридинг и аутбридинг?
28. Перечислите признаки, по которым имеется положитель-
ная и отрицательная брачная ассортативность.
29. Рассчитайте коэффициент инбридинга в кровнородствен-
ных браках, указанных в родословных на рис. 7.6 (с. 236).
30. Супруги Стелла и Всеволод — двоюродные брат и сестра.
Их отцы — Тихон и Георгий — родные братья. У их деда
Михаила было аутосомно-рецессивное заболевание —
лейциноз (болезнь кленового сиропа), которое приводит
к нарушению процессов миелинизации в ЦНС. Опреде-
лите вероятность рождения больного ребенка у этих суп-
ругов (рис. 7.7) (с. 236).
31. Ипполит здоров, но его брат Игорь и сестра Елена больны
аутосомно-рецессивным заболеванием гистидинемией,
235
Раздел 7. Структура популяции и генетические процессы
вызываемой дефектом фермента гистидазы. Ипполит же-
нат на своей двоюродной сестре Зое (рис. 7.8). Рассчитай-
те риск рождения больного ребенка в этой семье.
236
Контрольные вопросы и задания к разделу 7
Рис. 7.9
32. Мукополисахаридоз — аутосомно-рецессивное заболева-
ние. Давид женат на своей двоюродной племяннице Ека-
терине. У его родной племянницы Симоны мукополиса-
харидоз (рис. 7.9). Каков риск рождения больного ребен-
ка в этом кровнородственном браке?
33. В выборке из 10 000 человек группы крови М, MN и N име-
ли 3944, 4512 и 1444 человек. Рассчитайте коэффициент
инбридинга в популяции.
34. Измерено кровяное давление и вычислены коэффициен-
ты корреляции между различными типами родственников.
Для систолического давления в парах родитель — потомок
г = 0,24, в сибсовых парах г = 0,33, для диастолического
давления в парах родитель — потомок г = 0,18, в сибсо-
вых — г = 0,27. Рассчитайте наследуемость кровяного дав-
ления в данной популяции.
35. В некоторых обществах существует положительная брач-
ная ассортативность по вероисповеданию. Вследствие это-
го возникает относительная репродуктивная изоляция
между разными группами верующих. Ниже представле-
ны две гипотетические популяции, включающие одинако-
вое число мужчин и женщин (по 500 человек), принадле-
жащих к различным конфессиональным группам и не со-
стоящих в браке. Определите, какая доля населения в
изученных выборках может остаться в безбрачии, если
237
Раздел 7. Структура популяции и генетические процессы
предположить, что браки между разными группами верую-
щих не заключаются.
Группа вероисповедания Мужчины, чел. Женщины, чел.
1-я попу- ляция 2-я попу- ляция 1-я попу- ляция 2-я попу- ляция
Иудаисты 15 1 11 4
Мусульмане 29 20 44 44
Христиане 213 205 202 452
Атеисты 205 256 216 0
Буддисты 38 18 27 0
Всего 500 500 500 500
Раздел 8
ЧЕЛОВЕЧЕСКОЕ РАЗНООБРАЗИЕ
Если мы вспомним, как ведут себя знакомые нам люди в раз-
ных ситуациях, то отметим, что их поведение отличается боль-
шим разнообразием. В сходных обстоятельствах разные люди
совершают разные поступки. От чего зависит индивидуаль-
ная реакция человека на конкретные обстоятельства? Пыта-
ясь найти ответы на эти вопросы, наука еще делает только
первые шаги на этом пути. Ясно, что поведение человека за-
висит от того, как он воспринимает окружающую среду, как
перебатывает информацию. Понять, почему поведение лю-
дей так разнообразно, невозможно без анализа биологичес-
кого разнообразия вида. Внутривидовое разнообразие чело-
века огромно и проявляется на всех уровнях биологической
организации — молекулярно-генетическом, физиолого-био-
химическом, морфологическом. Еще больше впечатляет куль-
турное разнообразие людей.
В человеческих популяциях наблюдается значительная ге-
нетическая изменчивость по множеству признаков: цвету
кожи, глаз, волос, росту, весу, группам крови и др. Люди раз-
личаются и по чертам характера, темпераменту, тому, как они
реагируют на раздражители. Это разнообразие представля-
ет собой внутрипопуляционную изменчивость. Существует
также и межпопуляционное разнообразие. Наиболее очевид-
ны различия между отдельными популяциями по пигмента-
ции, чертам лица, телосложению.
Все человечество условно разделяют на группы, отличаю-
щиеся по морфологическим признакам, — расы. Существует
несколько классификаций, по которым людей относят к той
или иной расе. Линней выделял четыре подвида в виде Ното
sapiens — H.europaeus, H.afer, H.asiaticus и H.americanus. He-
239
Раздел 8. Человеческое разнообразие
которые из систем включают несколько десятков расовых
групп. Расы можно рассматривать как большие популяции,
отличающиеся по морфологическим признакам. Обычно вы-
деляют три большие расы: негроиды, монголоиды и европео-
иды. Различия между расами определяются в пределах от 4%
до 34% по целому ряду признаков, причем, как правило, по
различным признакам европеоиды располагаются между нег-
роидами и монголоидами.
Термин «раса» несет биологическую нагрузку. Если в клас-
сификацию включаются культурные и исторические аспек-
ты, используют понятие этническая группа. Группы, связан-
ные генетически, обычно имеют и общую культуру, поэтому
понятия раса и этническая группа перекрываются. Расы раз-
личаются генофондами. Аллельное разнообразие большин-
ства генов одинаково представлено в генофондах разных рас,
различаются лишь частоты аллелей. Так, аллель серповиднок-
леточного гемоглобина часто встречается у негров и крайне
редко у народов Северной Европы. Известна роль расовых
различий в реакции на психоактивные вещества. Так, у лиц
монголоидной расы снижена переносимость этанола, негро-
иды и некоторые этнические группы Юго-Восточной Азии от-
вечают возбуждением на те опийные алкалоиды, которые
обладают успокоительным эффектом у европейцев.
Формирование рас было возможно благодаря изоляции
групп людей. Оно происходило под влиянием специфических
условий среды: климата, особенностей пищи, инфекций. Глав-
ным фактором эволюции рас считается естественный отбор,
который приспосабливал популяции к конкретным условиям
среды. Родина первых людей — Африка. После их выхода из
Африки, льды, образовавшиеся на евразийском континенте
вследствие всеобщего похолодания, разделили территорию,
на которой обитали первобытные люди, на три области. Это
создало условия для независимой эволюции рас: европеоид-
ной на западе, монголоидной на востоке и негроидной на юге.
В настоящее время считается, что в генофонде человече-
ства отражена как его прежняя история, так и следы адапта-
240
Раздел 8. Человеческое разнообразие
ции к современным условиям. Информация о генофонде мо-
жет уточнить историю народа, а исторические летописи по-
могают реконструировать направленность отбора и мигра-
[ цию популяций. Наблюдая человеческое разнообразие, уче-
ные пытаются объяснить его происхождение, рассматривая
вариации как приспособленность к конкретным условиям.
Между тем не всегда можно объяснить, почему в популяции
распространен этот, а не другой признак. Так, многие мута-
ции являются адаптивно нейтральными и различия по ним
могут быть не связаны с отбором. Генофонды популяций мо-
гут различаться благодаря дрейфу генов, в результате кото-
рого признак распространяется независимо от его адаптив-
ного значения. Кроме того, изменения окружающей среды в
связи с производственной деятельностью человека происхо-
дят чрезвычайно быстро, поэтому адаптивные в недавнем
прошлом признаки в новых условиях могут оказаться нейт-
ральными или даже вредными.
Межпопуляционные различия могут быть связаны и с миг-
рацией населения. Мигрируя, люди переносят как адаптив-
ные, так и неадаптивные признаки. Гены, контролирующие
эти признаки, переходят через поколения, даже если в новых
условиях они уже не имеют приспособительного значения.
В новых условиях естественный отбор начинает смещать ген-
ные частоты, однако процесс адаптации будет происходить
многие поколения.
Используя технологию, люди оказывают противодействие
естественному отбору. У человека в основном культура, а не
гены обеспечивают успех приспособления на новом месте.
Когда британские переселенцы впервые ступили на землю
Австралии, испанцы в Южную Америку, арабы в Восточную
Африку, маловероятно, чтобы биологически они были боль-
ше приспособлены к новым условиям, чем коренные обита-
тели этих мест. Генетически адаптированными были абори-
гены, которые на протяжении многих поколений путем есте-
ственного отбора приспосабливались к местным условиям.
Переселенцы, несмотря на меньшую биологическую при-
241
Раздел 8. Человеческое разнообразие
способленность, имели более выгодное преимущество на Запа-
де — развитую технологию, что оказалось важнее биологи-
ческой адаптации.
8.1. Телосложение
Тело человека по своему внешнему виду характеризуется
множеством разнообразных вариаций. Учеными делались
неоднократные попытки создать классификацию типов телос-
ложения. Наиболее известные из них — система Э. Кречме-
ра и система У. Шелдона. Кречмер выделил три типа телосло-
жения: пикнический (широкий, округлый, с большим количе-
ством жира), лептосомный (длинный, тонкий, вытянутый) и
атлетический (мускулистый, широкоплечий, узкобедрый).
лептосом
пикник
Рис. 8.1. Типы телосложения (по Кречмеру)
242
В действительности имеются переходные варианты и чаще
всего человека невозможно отнести к какому-то из классов
Кречмера (рис. 8.1).
Более совершенной является система Шелдона, в которой
учитывается непрерывность метрических показателей разме-
ров тела. Система основана на выделении трех основных
физических типов: эндоморфного, мезоморфного и эктоморф-
ного. Название типа телосложения зависит от того, какой из
зародышевых слоев эмбриона в наибольшей степени оказывал-
ся развитым у индивида. У эндоморфов больше развиты орга-
ны, которые возникают из эндодермального зародышевого слоя.
У людей с эндоморфным телосложением очертания тела мяг-
кие и округлые. У них слабо развит костно-мышечный аппа-
рат, низкое отношение поверхности тела к весу, хорошо раз-
виты органы пищеварения. Мезоморфы — это индивиды с
большим весом тела и угловатым телосложением, с преиму-
щественным развитием костно-мышечного аппарата. У них
сильнее развиваются органы, которые в эмбриогенезе закла-
дываются из мезодермального зародышевого слоя. Эктомор-
фы худощавы, тонкокостны, плоскогруды, имеют хрупкое
телосложение. Наиболее развитые ткани эктоморфов по пре-
имуществу закладываются в эктодермальном зародышевом
слое. Степень развития каждого из перечисленных компонен-
тов выражается баллами. Тип телосложения (соматотип)
представляет собой набор из трех цифр, характеризующий
степень развития каждого из перечисленных компонентов.
В разных условиях селективным преимуществом может об-
ладать тот или иной соматотип. Высокий худой тип негров-
нилотов сформировался благодаря преимуществам, которы-
ми обладает данный тип телосложения при поддержании
физиологических функций в условиях жаркого климата. Арк-
тический тип — коренастый, с большим жировым отложени-
ем лучше приспособлен к обитанию в холодном климате.
Давно замечено, что люди определенного телосложения
более склонны к некоторым заболеваниям. Смертность от
туберкулеза выше среди худых и высоких людей, чем среди
243
Раздел 8. Человеческое разнообразие
низкорослых и коренастых. Коронарная болезнь сердца, ин-
сулинонезависимый сахарный диабет чаще поражает эндо-
морфов. Отмечена связь телосложения и с психическими
заболеваниями. Так, шизофрения чаще встречается у лиц с
высоким эктоморфным компонентом, а маниакально-депрес-
сивный психоз у людей с низкой степенью эктоморфии и вы-
соким мезомофным и эндоморфным компонентами. Истерия
и депрессия более характерны для мезоморфного и эндоморф-
ного типов, симпотомы тревоги и страха — для эктоморфно-
го. К паранойе более склонны мезоморфные типы. Считает-
ся, что характер симптомов при неврозах также связан с те-
лосложением.
Умственное развитие, выражаемое в баллах IQ, слабо кор-
релирует с телосложением, однако хорошо выражена связь
между телосложением и темпераментом. Шелдон при описа-
нии темперамента использовал тот же компонентный подход,
что и для описания телосложения. Компоненты темперамен-
та получили название «висцеротония» (расслабленность в
движениях, легкость в общении и выражении чувств), «сома-
тотония» (уверенность в движениях, эмоциональная чер-
ствость), «церебротония» (заторможеность в движенях, не-
общительность, эмоциональная сдержанность). Шелдон об-
наружил сильную положительную корреляцию между
висцеротонией и эндоморфией, соматотонией и мезоморфи-
ей, церебротонией и эктоморфией. Приведенные примеры
могут свидетельствовать об общности генетического контро-
ля телосложения и психического типа. Предполагается, что
гены, ответственные за телосложение, контролируют также
развитие мозга и эндокринной системы, определяющей тем-
перамент. Возможно, что это отдельные тесно сцепленные
гены. Существует еще одно объяснение, которое главную
роль отводит среде: ребенок с выраженной степенью мезо-
морфии, растущий среди других детей, случайно осознает, что
он сильнее остальных и может их победить. Впоследствии он
привыкает выделяться из среды сверстников, и это становит-
ся ведущей чертой его поведения.
244
8.1. Телосложение
Замечено, что люди с разным телосложением нередко вы-
бирают различные жизненные пути. Специальные исследо-
вания показывают, что индивиды, занятые исследовательской
работой, в среднем более высокие и худые, чем работающие
на производстве. Водители транспорта на дальние расстояния
и летчики отличаются более высоким мезоморфным компо-
нентом. Американские студенты, изучающие естественные
и социальные науки, отличаются более атлетическим телос-
ложением по сравнению со студентами искусствоведами,
филологами, философами. У будущих инженеров более вы-
ражен мезоморфный компонент, а у студентов физиков и
химиков эктоморфный. Юноши-правонарушители отличают-
ся более выраженным мезоморфным компонентом.
Северо-Восточная Аф-
рика является родиной са-
мых высоких людей на Зем-
ле. В племенах нилотов, вы-
пасающих скот, высокий
рост является адаптивным
признаком, которому благо-
приятствовал естественный
отбор. Дело в том, что у вы-
соких худощавых людей
высокие отношения поверх-
ности тела к объему, что не-
обходимо для более инте-
сивного и быстрого излуче-
ния тепла с поверхности
тела. Представителем пле-
мени нилотов является Ма-
нут Бол, в прошлом пастух
из Судана, а в настоящем
известный баскетболист
NBA, чей рост составляет
231 см (рис. 8.2). Несмотря Рис. 8.2. Баскетболист NBA —
на то что родиной баскетбо- представитель племени нилотов
245
Раздел 8. Человеческое разнообразие
Рис. 8.3. Массивное тело — требование для борцов сумо
ла является США, много спортсменов, входящих в состав
сборной NBA, поставляет африканский континент.
Так же как белым американцам приходится мириться с
тем, что на их родине в баскетбольную лигу входят африкан-
цы, японцы признают, что для японского национального
спорта — сумо — в их популяции вряд ли преобладают люди
с необходимой для этого вида спорта массивностью тела. Так,
у борца сумо американско-полинезийского происхождения
Конишики масса тела состаляет около 272 кг (рис. 8.3).
Короткие ноги и руки являются преимуществом в мало-
весовой категории при поднятии тяжестей. Крепкое тулови-
ще и короткие конечности характерны для представителей
степных популяций Евразии, простирающейся от Болгарии
до Монголии, которые поставляют в спортивный мир много
знаменитых штангистов и борцов. Одним из них является ту-
рок Наим Сулейманоглу (рис. 8.4).
В современном модельном бизнесе красивые женщины с
изящным удлиненным телом из Северо-Восточной Африки
246
8.1, Телосложение
Рис. 8.4. Короткие конечности — атрибуты
спортсмен ов-штангистов
Рис. 8.5. Фотомодели из Северо-Восточной Африки — идеал
современного модельного бизнеса
247
Раздел 8. Человеческое разнообразие
(Сомали, Эфиопии, Судана) переходят из категории редких
экзотичных в категорию распространенных. Внешность та-
ких моделей представляет собой сочетание северо-восточных
африканских и западно-азиатских и европейских генов. Од-
ной из первых супермоделей из Сомали является Иман, став-
шая женой известного музыканта Дэвида Боуи (рис. 8.5).
8.2. Черты лица
Черты лица определяются формой лба, носа, глаз, челюстей
и других признаков. Существует предположение, что общее
строение лица эволюционно сформировалось под действием
таких климатических факторов, как температура и влаж-
ность. Для свободного дыхания наиболее приспособлены ши-
рокие носовые ходы с круглыми широко открытыми ноздря-
ми (признаки черных жителей Африки). Однако в холодном,
сухом или пыльном климате (такой климат был в Европе в
эпоху оледенения) более узкие носовые ходы и сплющенные
ноздри (признаки, свойственные представителям северо-за-
падных популяций Европы) могли иметь приспособительное
значение. Такая форма носовых ходов создает большую по-
верхность для воздушного потока и обеспечивает его увлаж-
нение, прогревание и фильтрацию. Характерные черты мон-
голоидного лица — высокие выдающиеся скулы, сочетающие-
ся с уплощенным носом, делают лицо плоским. Складка кожи,
покрывающая внутренний угол глаза (эпикант) и толстое веко
создают раскосую форму глаза (рис. 8.6). Монголоидность
больше выражена в холодных районах — Северном Китае,
Монголии, Сибири и меньше у населения Юго-Восточной и
Центральной Азии, на Урале, в Гималаях, Северной и Юж-
ной Америке. Считается, что монголоидные особенности лица
делают человека более устойчивым к экстремально холодно-
му климату. Монголоидный тип лица у коренных народов
Америки появился вследствие миграции: обитатели северной
Азии прошли через Берингов пролив в Америку и, постепен-
но расселяясь, донесли свои анатомические особенности до
248
8.3. Пигментация
а б в
Рис. 8.6. Нормальные глаза европеоида (в), монголоида (б)
и эпикант у человека (а)
Южной Америки. Монголоидные черты лица жителей тро-
пической Индонезии также обязаны миграции.
Современные популяции различаются по совершенству
зубо-челюстного аппарата. Коренные жители тропической
Африки, американские индейцы, австралийские аборигены
имеют правильно развитые здоровые зубы. Они хорошо при-
способлены к грубой пище, которая обычна у народов, веду-
щих образ жизни охотников и собирателей. Мягкая пища на-
родов, занимающихся растениеводством, не требует мощно-
го жевательного аппарата, поэтому отбор на массивность и
прогнатность (прогнатнъш — выступающий вперед) нижней
части лица человека ослаблялся. Наиболее часто зубо-челюст-
ные аномалии встречаются среди народов, которые давно
перешли на культивирование растений. В индустриально раз-
витых странах почти нет людей, которые бы не обращались к
врачам по поводу заболеваний зубов. Высокотехнологичная
стоматология практически полностью прекратила отбор, на-
правленный против мутаций, вызывающих аномалии в стро-
ении и функционировании зубо-челюстного аппарата.
8.3. Пигментация
Наиболее заметный признак, отличающий расы — пигмен-
тация кожи. Окраска кожи человека в основном зависит от
концентрации черного пигмента меланина и глубины его за-
легания в коже.
249
Раздел 8. Человеческое разнообразие
Меланин синтезируется одними из разновидностей кле-
ток кожи — меланоцитами. Пигмент в больших количествах
находится в любой коже — черного, желтого и белого цвета.
Кровь, циркулирующая в дерме, добавляет красные тона к
цвету кожи. Там, где эпидермис тонкий и много капилляров,
например, в губах, кожа более розовая.
Меланоциты находятся в нижнем слое эпидермиса. Ме-
ланосомы — органеллы, в которых формируется пигмент. Об-
разование меланина катализируется ферментом тирозина-
зой. Меланоциты секретируют меланосомы с пигментом пря-
мо в окружающие клетки.
Существует два типа меланина, который синтезируется в
меланоцитах: эумеланин — коричневый или черный и фео-
меланин — желтый. У блондинов и рыжих синтезируется
феомеланин. У взрослого человека любой расы примерно
1000—2000 меланоцитов на 1 мм2. Более темный цвет кожи
связан с повышенным синтезом меланина, а не с плотностью
меланоцитов. Таким образом, в черной коже содержится боль-
ше меланосом, чем в белой, но не самих меланоцитов.
При сравнении цвета кожи у представителей разных на-
родов, населяющих земной шар, выявляется его огромное раз-
нообразие. Распределение различно пигментированных рас
по земному шару совпадает с интенсивностью ультрафиоле-
тового излучения. Коренные обитатели северо-западной Ев-
ропы очень светлые. Наиболее темные народы обитают в не-
которых частях тропической Африки, Азии, Австралии и на
островах Тихого океана.
Считается, что различия по степени пигментации возник-
ли в ходе эволюции как реакция на разную интенсивность
солнечного облучения. Каким образом ультрафиолетовая ра-
диация производит селекцию по цвету кожи? Одна из гипо-
тез состоит в следующем. Меланин, находящийся в верхних
слоях кожи, рассеивает и поглощает ультрафиолетовое излу-
чение, предотвращая его проникновение в более глубокие
слои кожи. Избыточное ультрафиолетовое излучение вызы-
вает ожог, повреждающий кожу, вызывая опасность инфи-
8.3. Пигментация
цирования. Кроме того, ультрафиолетовое излучение облада-
ет мутагенным и канцерогенным эффектом — вызывает рак
кожи. Темная кожа жителей Африки защищает кожу от вред-
ного действия большой дозы ультрафиолета, что особенно
важно для детей, которым солнечный ожог причинят особый
вред. В доказательство этой гипотезы приводят факты о том,
что в Южной Африке рак кожи намного чаще встречается у
светлокожих выходцев из Европы, чем у чернокожих афри-
канцев. Кожное заболевание псориаз, при котором наблюда-
ется воспаление кожи, у негров отсутствует. Частота псориа-
за наиболее высокая на Фарерских островах (2,8—7,0%), в
Норвегии — 4,82%, Швеции — 2,3—6,25%, Дании — 3,29—
4,18%. От Северного полюса к экватору идет снижение час-
тоты псориаза. Различные частоты можно объяснить адапта-
ционными возможностями людей различных климатических
поясов к уровню солнечной радиации. Однако у лапландцев,
которые проживают на севере Норвегии, частота невысокая —
около 0,6%, т.к. они являются малой расой, происходящей от
смешения европеоидов и монголоидов. Псориаз в основном
встречается у людей с голубыми глазами, светлыми волосами
и светлой кожей. У европеоидов светлые глаза встречаются с
частотой до 75%, а светлые волосы — до 50%. Частота псориа-
за снижена от европеоидной расы к монголоидной и к негро-
идной.
Другой возможный фактор отбора связан с синтезом ви-
тамина D, который продуцируется в коже под действием уль-
трафиолета. Витамин D участвует в обмене кальция в орга-
низме, что важно для нормального развития скелета. При не-
достатке витамина у ребенка развивается рахит. Между тем
избыток витамина D токсичен для организма. Возможно, что
толстый меланиновый слой темнокожих людей в жарком сол-
нечном климате защищает кожу от избытка ультрафиолета
и, следовательно, от избыточной продукции витамина D. Эту
гипотезу подтверждают факты, что рахит чаще обнаруживал-
ся среди черных детей, живущих в северных городах, чем у
их белокожих сверстников. Чтобы исключить влияние раз-
Раздел 8. Человеческое разнообразие
личий в питании, сравнивались дети из семей с одинаковым
социальным и экономическим статусом. Эти различия исчез-
ли после того, как промышленность стала производить моло-
ко, обогащенное витамином D.
Дефицит витамина D в организме приводит к аномалии
скелета. При деформации тазовых костей становятся невоз-
можными роды. Смерть матери и ребенка вследствие небла-
гополучных родов, связанных с аномалией тазовых костей,
обусловленной недостатком витамина D, — сильный фактор
естественного отбора. Низкая температура воздуха в север-
ных районах земного шара на протяжении значительной ча-
сти года вынуждает жителей закрывать большую часть тела
одеждой, что ограничивает доступ к ней ультрафиолета. Кро-
ме того, в этих широтах ультафиолетовая радиация ослабле-
на. В условиях недосточности ультрафиолета в более выгод-
ном положении оказываются слабо пигментированные инди-
виды, способные более эффективно использовать его слабое
излучение.
Считается, что в первобытных африканских популяциях
отбор благоприятствовал генотипам, обеспечивающим более
темную кожу, защищенную от ультрафиолета, а при рассе-
лении людей из Африки отбор изменил направленность.
В районах с низкой освещенностью, особенно зимой, когда
большая часть поверхности тела покрыта одеждой, черная
кожа с большим трудом поглощает достаточное количество
ультрафиолетовой радиации, чтобы вырабатывать необходи-
мое количество витамина D. Светлая кожа имеет селектив-
ное преимущество в климате с низкой интенсивностью ульт-
рафиолетового излучения, так как позволяет более эффектив-
но поглощать слабое ультрафиолетовое излучение.
Выйдя из Африки, предки современных людей заселяли
более высокие широты, где не только понижена интенсив-
ность солнечного излучения, но происходят также резкие
сезонные изменения температурного и светового режима.
В меняющихся условиях более выгодна не просто слабо пиг-
ментированная кожа, а кожа, способная к изменению степе-
252
8.3. Пигментация
ни пигментации. Благодаря этой способности меняется защит-
ная функция кожи в соответствии со световым режимом.
Летом светлокожие индивиды загорают, предохраняясь от
солнечных ожогов, зимой светлеют, избегая дефицита вита-
мина D. У европеоидов, живущих в районах, которые оста-
ются облачными даже летом (например, Ирландия), отбор на
способность к загару настолько ослабился, что в этих райо-
нах часто встречаются люди с так называемой кельтской ко-
жей, которая вообще не способна к загару и реагирует на уль-
трафиолетовое излучение ожогом.
Полного соответствия между цветом кожи и интенсивно-
стью солнечной радиации нет, поэтому не все случаи распре-
деления пигментации по земному шару можно объяснить.
Индейцы тропической части Южной Америки имеют свет-
лую кожу, аборигены Тасмании, живущие в прохладном
влажном климате, темную. Эти факты можно объяснить не
адаптацией к конкретным условиям, а миграцией. Предста-
вители монголоидной расы — племена охотников и рыболо-
вов, живущие на Севере, имеют темную кожу, не способную
эффективно использовать слабое ультрафиолетовое излуче-
ние для синтеза витамина D. Считается, что у этих людей по-
требность в витамине D удовлетворяется за счет животной
пищи.
В популяциях человека существует также разнообразие
по цвету глаз и волос, однако оно менее выражено, чем вари-
ация по цвету кожи. Большинство людей на земном шаре тем-
новолосы и темноглазы, только на северо-западе Европы жи-
вут голубоглазые блондины. Встречается и иная комбинация
признаков — брюнеты могут иметь голубые глаза и светлую
кожу, а среди темнокожих австралийских аборигенов встре-
чаются блондины. В отличие от окраски кожи адаптивное зна-
чение полиморфизма по цвету волос и глаз менее понятно.
Предполагается, что окраска глаз является приспособлением
к различной освещенности. Цвет глаза зависит от количества
меланина в радужке — диафрагме, которая, расширяясь и
сужаясь, регулирует количество света, попадающего на сет-
253
Раздел 8. Человеческое разнообразие
чатку. Предполагается, что темные глаза более адаптирова-
ны к яркому солнечному свету.
8.4. Дерматоглифика
Большая часть поверхности тела человека покрыта волосяной
растительностью, за исключением поверхностей ладоней,
пальцев и стоп, где имеется специфический папиллярный
рисунок — дерматоглифика. Интерес к рисункам дермаль-
ной кожи можно проследить от древних времен до первых
научных публикаций Грю, Бидлу и Мальпиги XVII столетия.
Большой вклад внес в развитие дерматоглифики Фрэнсис
Гальтон, чьи отпечатки пальцев представлены на рис. 8.7.
При указании места расположения какого-либо признака
или направления линии часто употребляют общепринятые в
анатомии термины: дистальный (distalis), проксимальный
(proximalis), ульнарный (ulnaris) и радиальный (radialis). Ди-
стально ладонь ограничена пястно-фаланговыми сгибательны-
ми складками (Plicae flexoriae metacarpo-phalangeales —
P.f.m.-ph.), а проксимально — запястной, или браслетной, сги-
Рис. 8.7. Дерматоглифика Фрэнсиса Гальтона
254
8.4. Дерматоглифика
бательной складкой (Plica flexoria carparalis — P.f.c.). Вокруг
центрального углубления ладони располагаются шесть ладонных
подушечек: у основания большого пальца—тенар (Thenar — Th),
на проксимально-ульнарном крае ладони — гипотенар
(Hypothenar — Н), а проксимально от 1, 2, 3 и 4 межпальцевых
промежутков лежат I, II, III и IV межпальцевые подушечки.
Практически тенар и I межпальцевая подушечка — это еди-
ное возвышение, ограниченное постоянно присутствующей
на ладони сгибательной складкой большого пальца (Plica
flexoria policis). Из постоянных сгибательных складок ладони
можно назвать еще две — дистальную поперечную сгибатель-
ную складку (Plica flexoria transversa distal is), которая образу-
ется от сгибания III—V пальцев и ограничивает проксималь-
но III и IV межпальцевые подушечки, и проксимальную сги-
бательную складку (Plica flexoria transversa proximalis),
идущую наклонно от I межпальцевого промежутка по направ-
лению к проксимально-ульнарному краю ладони (рис. 8.8).
У человека существу-
ют три основных типа
пальцевых узоров — дуга,
петля и завиток, осталь-
ные являются их модифи-
кациями. Выделяют два
типа дуг — обычную и вы-
гнутую, или Т-дугу, два
типа петель — ульнарные
и радиальные, два типа за-
витков — концентричес-
кие и спиральные. К моди-
фикациям, относится, на-
пример, составной узор
(рис. 8.9). Дуга не имеет
трирадиуса, или дельты,
— места, где сходятся три
направления линий. Пет-
ля имеет одну дельту, за-
Рис. 8.8. Схема ладонной топогра-
фии (обозначения в тексте)
255
Раздел 8. Человеческое разнообразие
Дуга
Петля
Выгнутая дуга
Концентрический
завиток
Спиральный
завиток
Составной узор
Рис. 8.9. Основные пальцевые узоры
виток — две. В генетических исследованиях используют раз-
личные дерматоглифические характеристики, как качествен-
ные, так и количественные, однако наиболее распространен-
ной является гребневой счет. Его учитывают следующим об-
разом. От дельты до центра узора проводят прямую линию и
подсчитывают количество гребешков, отрезков гребешков и
точек, которые касаются или пересекают эту линию. Гребневой
счет не проводят в дуговых узорах, где нет дельты. В среднем
гребневой счет на одном пальце составляет 15—20 гребешков.
Особенно интересны для генетики редкие сочетания дер-
матоглифических признаков, к которым относят, например,
такие, как 8—10 дуг, 10 завитков, 10 ульнарных петель с низ-
ким гребневым счетом (75 и ниже), 10 ульнарных петель с вы-
256
8.4. Дерматоглифика
соким гребневым счетом (180 и более), 3 и более радиальных
петли на пальцах, единственную сгибательную складку ладо-
ни, любой узор одновременно тенара и 1-й межпальцевой
подушечки (Th/I), завиток в любой области ладони и другие.
Всего у человека насчитывается более 30 редких признаков
дерматоглифики, которые используются как информативные
варианты, указывающие на возможность хромосомной болез-
ни, менделирующих мутаций или тератогенного эффекта.
Ярким примером диагностических возможностей дермато-
глифики является работа Камминса, описавшего характерные
признаки дерматоглифики людей с синдромом Дауна.
Иногда у человека встречаются настолько редкие дерма-
тоглифические признаки, что за все время дерматоглифичес-
ких исследований они выявлялись лишь единично, например,
дополнительные межпальцевые трирадиусы (рис. 8.10) или
продольное направление главных ладонных линий (рис. 8.11).
Дуги редко встречаются у мужчин, чаще у женщин. В не-
которых семьях дуги довольно часты. Наиболее часто дуга
отмечается на указательном пальце и ее частота уменьшает-
ся в ульнарном направлении с самым низким процентом на
мизинце. Дуговые узоры часто встречаются при различных
формах брахидактилии. Преобладание дуг диагностичный при-
знак триплоидии, трисомии 18, полисомии X, синдрома XXY и
XYY, синдрома «ногти—надколенник», ломкой Х-хромосомы.
Радиальные петли чаще всего локализованы на указатель-
ном пальце. Локализация радиальных петель на 3—5 пальцах
может свидетельствовать в пользу следующих диагнозов: син-
дром Дауна, триплоидия, синдром фрагильной Х-хромосомы.
При брахидактилии и трехфаланговом первом пальце кисти
отчетливо повышена частота радиальных петель.
Ульнарные петли — частый узор. При синдроме Дауна (фе-
нотип 10 петель) и синдроме Клайнфельтера частота ульнар-
ных петель повышена.
Завитки — обычный узор, чаще локализуется на 1-м и 4-м
пальцах. Очень редко у человека встречаются и завитки, и
дуги (конкурирующие узоры).
9 Введение в психогенетику
257
Раздел 8. Человеческое разнообразие
Рис. 8.10. Дополнительные
межпальцевые
трирадиусы (а‘, b', с‘, d')
Рис. 8.11. Продольное на-
правление главных ладон-
ных линий
8.5. Скорость развития
Гестационный период (период вынашивания ребенка) в сред-
нем меньше у негроидов, чем у европеоидов. Так, в некото-
рых исследованиях показано, что к 39 неделям беременности
рождается 51% детей негроидов по сравнению с 33% детей
монголоидов. Несмотря на то что младенцы негроидов рож-
даются раньше, они физиологически более зрелые, чем мла-
денцы европеоидов, что можно установить по функциониро-
ванию легких и составу амниотической жидкости.
В дальнейшем онтогенезе также отмечаются межрасовые
отличия. Дети негроидов раньше начинают держать голову,
развивать различные моторные навыки и самостоятельно
одеваться. Дети монголоидов, наоборот, отстают от детей ев-
ропеоидов. В среднем монголоид начинает ходить примерно
в 13 месяцев, европеоид — в 12 месяцев и негроид в 11 меся-
цев. Развитие постоянных зубов у негроидов начинается в
258
8.7. Группы крови
5,8 лет и заканчивается в 7,6 лет, у европеоидов — в 6,1 года и
в 7,7 лет, у монголоидов — в 6,1 года и 7,8 лет соответственно.
8.6. Особенности темперамента
Межрасовые отличия в темпераменте отмечаются в следую-
щем. Европеоиды более несдержанны, чем негроиды. Самы-
ми спокойными являются монголоиды. Среди детей меньшая
агрессивность и большая социальная приспособляемость на-
блюдается у монголоидов по сравнению с негроидами, а ев-
ропеоиды занимают промежуточное положение. При изуче-
нии показателей экстраверсии и нейротизма по классичес-
кому тесту Айзенка на выборке из 25 стран было показано,
что у монголоидов наблюдается меньшая экстравертирован-
ность и больший нейротизм, чем у европеоидов. Наиболее эк-
стравертированными и наименее нейротичными являются
негроиды.
8.7. Группы крови
Разнообразие морфологических признаков человека несравни-
мо с разнообразием белков его тела. Как отдельный вид молеку-
лярного полиморфизма рассматривают группы крови. У чело-
века известно 14 различных систем групп крови, которые конт-
ролируются независимыми генными локусами: ABO, Lewis,
Lutheran, MNS, Rhesus, Kell, Kidd, Diego, Duffy, Xg, P и другие.
Принадлежность человека к группам крови большинства
систем определяется аутосомными генами, существующими
обычно в двух аллельных состояниях. Каждый из кодоминан-
тных аллелей ответствен за выработку соответствующего
эритроцитарного фактора.
Группа крови системы Хд(а) контролируется геном, сцеп-
ленным с Х-хромосомой. В Y-хромосоме соответствующего
аллеля нет. Ген имеет два аллельных состояния: Хд(а+) — эрит-
роцитарный фактор «а» присутствует и Хд(сг) — этот фак-
тор отсутствует.
9*
259
Раздел 8. Человеческое разнообразие
Представители одной популяции сильно различаются на-
борами генов, контролирующих принадлежность к группам
крови. Выявлены различия в частотах групп крови между по-
пуляциями земного шара. Процент резус-отрицательных ин-
дивидов особенно высок в европейских популяциях, но в по-
пуляциях монголоидов и негроидов эта группа крови встре-
чается крайне редко. Группа крови А чаще встречается у
европеоидов, группа В — у монголоидов. В некоторых попу-
ляциях американских индейцев вообще отсутствует группа В.
Есть предположение, что на различия по частоте групп крови
системы АВО оказали влияние инфекционные эпидемии, ко-
торые переживало человечество в прежние эпохи. Были сде-
ланы попытки исследовать сходство группоспецифических
факторов крови и веществ на поверхности болезнетворных
микроорганизмов — бактерий и вирусов. Нормальный орга-
низм не вырабатывает антител по отношению к белкам соб-
ственного тела. Если организм инфицирован микроорганиз-
мом, поверхностные белки которого напоминают белки кро-
вяных клеток или других тканей человека, то он не может быть
распознан иммунной системой как чужеродное тело и про-
тив него не вырабатываются защитные белки — антитела.
Показано, что поверхностные белки бактерий бубонной чумы
сходны с белком фактора первой группы крови. В связи с этим
индивиды с этой группой крови должны быть более чувстви-
тельны к чуме. При сопоставлении исторических сведений о
распространении бубонной чумы и распределении первой
группой крови возникло предположение о взаимосвязи этих
явлений. Бубонная чума появилась в Юго-восточной Азии и
распространилась на запад. Инфекция переносится блохами,
которые паразитируют на черных крысах. Крысы предпочи-
тают селиться вблизи зернохранилищ, поэтому чума поража-
ет популяции земледельцев. Чума, известная под названием
«черная смерть», поразила Европу в середине XIV столетия,
уничтожив четверть населения. В Центральной Азии бубон-
ная чума существовала в течение длительного времени. Это
считается причиной того, что на данной территории самый
260
8.8. Система HLA
низкий процент людей с нулевой группой крови, так как эта
болезнь была фактором отбора против гена, определяющего
нулевую группу крови. Сейчас эпидемий чумы нет, поэтому
не представляется возможным проверить, действительно ли
люди с нулевой группой крови более восприимчивы к этой
болезни. Статистический анализ показал, что индиивиды с
разными группами крови неодинаково подвержены различ-
ным болезням. Люди с группой 0 чаще болеют язвой желуд-
ка, а при группе А чаще бывает рак желудка.
8.8. Система HLA
Еще один вид белкового полиморфизма у человека представ-
лен антигенами тканевой совместимости (гистосовместимо-
сти) HLA. В 70-х гг. XX в. началось интенсивное изучение ге-
нов, ответственных за узнавание и отторжение транспланти-
рованных органов и тканей от одного индивида другому.
Главный комплекс гистосовместимости человека МНС
(major histocompatibility complex) известен еще как система
HLA (human leukocyte antigens), а ген этого комплекса лока-
лизован в 6-й хромосоме.
Продукты генов этого комплекса — антигены, препятствую-
щие обмену тканями между представителями одного и того
же вида. Продукты локусов А, В и С — гликопротеины, на-
званные молекулами класса I, экспрессирующие на поверх-
ности клеток организма, имеющих ядра, в то время как про-
дукты локуса D — гликопротеины класса II, кодирующие ан-
тигены иммунокомпетентных клеток (В- и Т-лимфоцитов и
макрофагов).
Очень близкое расположение в хромосоме генов системы
HLA благоприятствует их сцепленному наследованию. Дру-
гими словами, каждый человек наследует по одному набору
А, В, С и D локусов от каждого родителя совместно (без ре-
комбинации) . Эту тесно сцепленную группу называют гапло-
типом. Таким образом, в каждой семье сегрегируют лишь
4 гаплотипа (рис. 8.12).
261
Раздел 8. Человеческое разнообразие
Трансплантация является успешной, если у донора такие
же антигены главного комплекса гистосовместимости, как у
реципиента, или же, по крайней мере, если у донора нет ан-
тигенов, которых нет у реципиента (рис. 8.13).
Гены системы HLA полиморфны — известно, по меньшей
мере, 24 аллеля локуса А, 50 аллелей локуса В, 11 аллелей ло-
куса С и аллели DQ, DP и D/DR локуса D (Al, А2, АЗ, В5, В7,
В14, В15 и др.).
Статистически доказано, что определенные аллели систе-
мы HLA ассоциированы с повышенным риском развития оп-
ределенных болезней. В настоящее время с маркерами HLA
связывают около 50 заболеваний человека. Эти болезни за-
трагивают различные системы органов, однако все они, по-
видимому, происходят из-за нарушения работы иммунной
системы. К таким болезням, к примеру, относится анкилози-
рующий спондилит, или болезнь Бехтерева — воспаление и
деформация позвоночника, синдром Рейтера — воспаление
суставов, глаз и мочеполовых путей, ревматоидный артрит —
воспаление суставов (рис. 8.14), псориаз — чрезмерное ше-
лушение кожи, ювенильный сахарный диабет — нарушение
метаболизма сахара с результате развития аномалий [3-клет-
ками печени, зоб щитовидной железы — гормональная дис-
Рис. 8.12. Наследование HLA-гаплотипов
262
8.8. Система HLA
ЛОКУС: А С В 1 СИБСЫ 2 3 4
<7> CO N. ’Г- -г" Щ СП СО L- Т- 1О CM Oi 2 V- ’Г" Ш см см 2 СО < 03
ГЕНОТИП: А-1,8-5, С-1 А-9, В-7, С-3 А-1.В-5, С-1 А-9, В-7, С-3 А-1,8-5, С-1 А-2, В-13, С-2 А-3, В-8, С-4 А-2, В-13, С-2
Кожа пересаживается
от сибсоа 2, 3, 4
сибсу 1
Отторжение'
Успешная
трансплантация гпрансп'пантатов
Рис. 8.13. Трансплантация участка кожи между сибсами
функция щитовидной железы, миастения — прогрессирую-
щая атрофия мышц, хронический активный гепатит — вос-
паление печени и т.д. HLA-ассоциированные болезни просле-
живаются в семьях, однако рассчитать генетический риск для
них не представляется возможным, поскольку они не насле-
дуются в соответствии с менделевскими закономерностями.
Хорошо изученным примером HLA-ассоциированной бо-
лезни является анкилозирующий спондилит, когда воспаля-
ется и деформируется позвоночник. Если у человека имеется
антиген В27, тогда вероятность развития у него заболевания
в 100 раз выше, чем у человека без этого антигена. Такая вы-
сокая вероятность рассчитана на основе того, что более чем у
90% людей, страдающих анкилозирующим спондилитом, при-
сутствует антиген В27, хотя в популяции антиген В27 встре-
чается лишь у 5% населения. У черного населения Америки
антиген В27 встречается значительно реже, чем у белых. Ан-
килозирующий спондилит у американских негров также
встречается гораздо реже. Антиген В27, таким образом, явля-
263
Раздел 8. Человеческое разнообразие
рентгенологический снимок
Рис. 8.14. Ревматоидный артрит — клинические перспективы
диагностики по системе HLA
ется маркером анкилозирующего спондилита. Естественно,
что к такой статистике следует подходить осторожно — ведь
10% больных не имеют антигена В27, а некоторые люди с этим
антигеном не заболевают вовсе.
Можно указать еще на некоторые ассоциации. Риск забо-
леть сахарным диабетом при наличии антигена DR3 повышен в
3,3 раза, чем при его отсутствии. В четыре раза повышен риск
множественного склероза у индивидов с антигеном DR2. Веро-
264
8.9. Полиморфизм ДНК
ятность заболеть псориазом увеличивается в 13 раз при нали-
чии антигена Cw6. Если присутствует специфичность D/DR2,
в 2,4 раза повышается риск воспаления зрительного нерва.
Природа связи аллелей HLA с развитием определенных за-
болеваний до конца не установлена. Определенный гаплотип
может означать генетическую подверженность к болезням
или чувствительность к специфическим вирусам. Таким об-
разом, хотя наших знаний пока недостаточно, имеющиеся
статистические закономерности могут быть клинически по-
лезными для выявления групп повышенного риска.
8.9. Полиморфизм ДНК
Полиморфные белки (группоспецифические антигены, HLA-
антигены, ферменты и др.) получили название классических
маркеров. В последнее время появился новый инструмент
изучения наследственности человека — маркеры ДНК. Инфор-
мационное содержание ДНК значительно выше белкового, а
методы работы с молекулами ДНК проще, чем с белками. Наи-
более важное преимущество маркеров ДНК — наличие по-
лиморфизма (разнообразия нуклеотидных последовательно-
стей) разного типа. ДНК-полиморфизм обусловлен замена-
ми, вставками, выпадениями нуклеотидов.
В 1980-х гг. в Великобритании Алек Джеффрис с сотруд-
никами обнаружили высоковариабельные участки генома.
Эта группа ученых занималась исследованием гена, кодирую-
щего миоглобин. Они установили, что один из интронов в этом
гене содержит многократно повторяющуюся последователь-
ность пар оснований. Количество таких повторов у разных
людей варьировало. Джеффрис хотел использовать этот ва-
риабельный участок как маркер для локализации гена мио-
глобина. Участок ДНК, содержащий этот район был изолиро-
ван и использован как зонд в ДНК-анализе. ДНК-зонд — это
участок одноцепочечной молекулы ДНК, который может ком-
плементарно достраиваться к ДНК анализируемого гена.
ДНК-зонд является меченым веществом (химически или ра-
265
Раздел 8. Человеческое разнообразие
диоактивно) и, следовательно, способен обнаружиться при
анализе его, например, с помощью чувствительной фотогра-
фической пленки.
С помощью ДНК-зонда было обнаружено, что каждый ана-
лизируемый образец давал множество «полос» на фотоплен-
ке. Расположение полос оказалось строго индивидуальным,
и таким образом был открыт профиль ДНК, или генетический
профиль. В дальнейшем было открыто много районов с вари-
абельным числом тандемных повторов. Вариабельные учас-
тки тандемных повторов наследуются как простые менделиру-
ющие признаки. В генетическом смысле участок с вариабель-
ным числом тандемных повторов — это локус с определенным
количеством аллелей, каждый из которых характеризуется
определенной длиной. О таком локусе, содержащем множе-
ство аллелей, каждый из которых имеет довольно высокую
частоту, говорят, что он генетически полиморфный. В высо-
кополиморфном локусе с вариабельным числом тандемных
повторов более 95% популяции будет иметь аллели различной
длины и таким образом быть гетерозиготами по этому локу-
су. Вероятность того, что два не связанных родством челове-
ка будут иметь ту же самую комбинацию аллелей в опреде-
ленном локусе с вариабельный числом тандемных повторов,
часто гораздо меньше 1%.
Позже было открыто большое количество участков с ва-
риабельным числом тандемных повторов, состоящих всего из
2—4 пар оснований, повторяющихся 5—15 раз. Эти участки
называются короткие тандемные повторы.
Маркеры ДНК относят к трем группам: маркеры митохон-
дриальной ДНК, аутосомные маркеры и маркеры Y-хромосо-
мы. Особенностью полиморфизма митохондриальной ДНК
является отсутствие рекомбинаций и материнский тип насле-
дования. Y-хромосомный полиморфизм имеет отцовское на-
следование, обнаруживает практически полное отсутствие ре-
комбинации. Оба типа полиморфизма дополняют друг друга.
Полиморфизм ДНК используют для решения разных про-
блем генетики человека. Рассмотрим некоторые примеры.
266
8.9. Полиморфизм ДНК
Один из них — мутация в гене рецептора CCR5. Этот ген ло-
кализован в хромосоме 3 (3р.21.3). Рецептор CCR5 использу-
ется вирусом иммунодефицита для проникновения в клетки.
В гене обнаружена делеция размером 32 нуклеотидные пары.
Эта делеция препятствует взаимодействию рецептора с ви-
русом и тем самым определяет устойчивость человека к ВИЧ-
инфекции. Люди, гомозиготные по этой делеции, не могут
заболеть СПИДом. Мутантный аллель, сообщающий устой-
чивость к СПИДу, в европейских популяциях встречается у
2—15% населения, а среди коренного населения Африки и
Японии эта мутация не выявлена.
Маркеры ДНК используются и при изучении наследствен-
ных основ человеческого поведения. Одним из ключевых меди-
аторов нервной системы, участвующих в передаче возбужда-
ющих импульсов, является дофамин. Обнаружено функцио-
нальное, биохимическое и фармакологическое разнообразие
дофаминовых рецепторов (DI, D2, D3, D4, D5). Ген дофами-
нового рецептора находится в 11 хромосоме в районе q22-23.
Рецептор D2 отличается высоким сродством к психоактивным
веществам, особенно алкоголю, поэтому ген D2 рассматри-
вается в качестве одного из основных генов-кандидатов фор-
мирования алкогольной зависимости. Полиморфизм этого
гена может обусловливать неодинаковую чувствительность
к алкоголю разных людей. Еще один важный нейромедиатор
центральной и периферической нервной системы — серото-
нин. Он участвует в регуляции настроения, аппетита, сна, бо-
левого восприятия, а также ряда других сложных поведенчес-
ких реакций. Прекращение действия серотонина в мозге пос-
ле освобождения из нейронов осуществляется путем его
активного обратного всасывания с помощью переносчика се-
ротонина SERT. У человека ген переносчика серотонина
(hSERT) расположен на хромосоме 17 в области qll.l-ql2. При
анализе полиморфных вариантов гена обнаружены варьиру-
ющие тандемные повторы минисателлитного типа, локализо-
ванные во втором интроне. Считается, что тандемные повто-
ры, расположенные в регуляторных районах гена, могут вли-
ять на его экспрессию.
267
Раздел 8. Человеческое разнообразие
Разнообразие человеческих популяций по маркерам ДНК
используется в тех случаях, когда стоит задача реконструи-
ровать происхождение отдельных популяций, народов, рас.
В 1987 г. американский ученый А. Вилсон опубликовал ре-
зультаты сравнительного анализа митохондриальной ДНК,
взятой у представителей разных этносов всех человеческих
рас, заселяющих пять континентов. Оказалось, что все мито-
хондриальные ДНК возникли из одной предковой ДНК пу-
тем дивергенции. Считается, что предки современных людей
жили в Северо-восточной Африке примерно 200 тыс. лет на-
зад. Это следует из того, что африканские популяции отлича-
ются наибольшей гетерогенностью митохондриальной ДНК
по сравнению с популяциями других континентов, что объяс-
няют большей древностью африканских популяций. Данные
по маркерам Y-хромосомы также показывают, что наиболее
древние варианты обнаруживаются в Африке. Все эти резуль-
таты позволили подтвердить африканское происхождение
человека. Через 10 лет удалось расшифровать фрагмент ДНК
митохондрий из останков неандертальца, и оценить время
существования последнего общего предка человека и неан-
дертальца в 500 тыс. лет назад.
Маркеры ДНК применялись и для решения проблемы о
том, как происходило заселение различных территорий, к
примеру, американского континента. У американских индей-
цев в Y-хромосоме выявили гаплотип, который не встречает-
ся в других регионах мира. Анализ данных по митохондри-
альной ДНК позволил сделать предположение о том, что все
популяции американских индейцев происходят из одной
предковой популяции, проживавшей ранее в районе Монго-
лии и Северного Китая.
8.10. Электрическая активность мозга
Биоэлектрические процессы в нервных клетках связаны с
биохимическими механизмами, которые отражают функци-
онирование генома. Кора головного мозга находится в состо-
268
8.10. Электрическая активность мозга
янии постоянной электрической активности. Причиной этой
активности является генерация синаптических потенциалов
и импульсных разрядов в отдельных нервных клетках. Порож-
даемые мозгом электрические колебания можно записать на
бумаге в виде кривых. Для этого к определенным точкам го-
ловы прикладывают электроды, и электрические колебания
после прохождения через усилители записываются в виде
электроэнцефалограммы (ЭЭГ). ЭЭГ представляет собой
сложный признак из многих составляющих: распределение
частот и амплитуд в одном отведении, форма волн. Комбина-
ции различных элементов кривой ЭЭГ многообразны, и их
можно сравнивать. Различные параметры ЭЭГ хорошо опи-
саны и используются для оценки состояния различных обла-
стей коры головного мозга. У взрослого человека обнаружи-
вается несколько типов волн. В спокойном состоянии в коре
больших полушарий регистрируется регулярный ритм с час-
тотой около 8—13 колебаний в секунду (а-ритм).
ЭЭГ примерно 4% индивидов в популяции характеризует-
ся тем, что у них в затылочной области либо полностью отсут-
ствуют a-волны, либо выявляются только короткое время с
очень низкой амплитудой. В состоянии активной деятельно-
сти a-ритм сменяется частыми (более 13 в секунду) колеба-
ниями небольшой амплитуды — [3-ритмом. Во время сна он
сменяется медленными (0,5—3,5 в секунду) колебаниями —
8-ритмом. Электрофизиологам известны также у- , 0-, ц-ритм
и другие (рис. 8.15).
Особенности ЭЭГ связаны с различными психофизиоло-
гическими и психологическими характеристиками человека:
свойствами нервной системы, особенностями темперамента,
когнитивными способностями. Каждый человек имеет соб-
ственную, характерную только для него ЭЭГ. Становление
особенностей ЭЭГ происходит в детстве и юности. По мере
взросления картина ЭЭГ меняется, однако у одного человека
?-волны имеют постоянный характер.
Рисунок ЭЭГ складывается примерно к 15—18 годам и со-
храняется на протяжении жизни, изменяясь только в экстре-
269
Раздел 8. Человеческое разнообразие
а-ритм
$-ритм
б-ритм
Рис. 8.15. Различные волны в электроэнцефалограмме
человека
мальных ситуациях — при тяжелых психических заболева-
ниях, опухолях мозга, при сильном переутомлении, интокси-
кации. Из-за больших индивидуальных различий в скорости
развития детей у них велики и различия в ЭЭГ. Благодаря ЭЭГ
можно выявить области коры, которые на разных этапах он-
тогенеза подвергаются в той или иной мере генетическим или
средовым влияниям.
Индивидуальные различия нейрофизиологических пара-
метров, характеризующих состояние спонтанной активнос-
ти мозга, находят отражение в индивидуально-психологичес-
270
8.10. Электрическая активность мозга
ких различиях. Это было показано при параллельных иссле-
дованиях ЭЭГ и тестировании, позволяющем оценить уровень
интеллекта, скорость работы, концентрированность внима-
ния, установки личности, сенсорные и моторные способнос-
ти. Между личностными характеристиками и особенностя-
ми ЭЭГ обнаружены различные корреляции. Индивиды с
мономорфными a-волнами активны, стабильны и надежны.
Они точны в работе, устойчивы к стрессам, у них хорошая
кратковременная память, но процесс обработки информации
у них несколько замедлен. Индивиды с низкоамплитудной
ЭЭГ обладают низкой спонтанной активностью, склонны к
экстраверсии, у них хорошо развита пространственная ори-
ентация. Индивиды, обладающие низковольтной ЭЭГ, отли-
чаются расслабленностью и беззаботностью, экстравертиро-
ваны, малоинициативны, демонстрируют низкие показатели
в тестах на концентрацию. Лица с диффузными [3-волнами
отличаются низкой скоростью работы, у них удлинено время
реакции и при этом они делают много ошибок, неустойчивы
к стрессам, плохо выполняют тесты, которые оценивают про-
странственную ориентацию, допускают большое количество
ошибок в тестах на внимание и точность. У лиц с редким ва-
риантом затылочных быстрых a-волн обычно высокий интел-
лект, они способны к абстрактному мышлению, обладают
прекрасными моторными способностями, быстро обрабаты-
вают информацию.
Изучение электроэнцефалограммы позволяет понять ме-
ханизмы, лежащие в основе генетической регуляции поведе-
ния. Генетический анализ ЭЭГ затрагивает как ее целостный
рисунок, так и отдельные количественные характеристики.
Генеалогическим методом выявлены варианты ЭЭГ с простым
типом наследования. Использование таких вариантов может
связать биохимические механизмы, лежащие в основе их воз-
никновения, с индивидуальными особенностями психики.
Генетический контроль различных параметров ЭЭГ соответ-
ствует разным типам, как монотонным, так и полигонным
(табл. 8.1.).
271
Раздел 8. Человеческое разнообразие
Таблица 8.1
Наследование характеристик ЭЭГ
Вариант ЭЭГ Тип наследования
Низкоамплитудная Затылочные медленные в-волны Монотонные б-волны Диффузные в-волны Аутосомно-доминантный Аутосомно-доминантный Аутосомно-доминантный Полигенный
Данные, полученные на близнецах, свидетельствуют, что
в норме индивидуальные различия в ЭЭГ обусловлены в ос-
новном генетически. У монозиготных близнецов кривые ЭЭГ
совпадают в больше степени, чем у дизиготных. Так, монози-
готные близнецы генерируют примерно одно и то же количе-
ство a-волн в единицу времени, чего не наблюдается у дизи-
готных партнеров. Изучение разлученных близнецов пока-
зало, что их ЭЭГ совпадает так же, как и у близнецов,
воспитывавшихся вместе. Сходство по ЭЭГ пары монозигот-
ных близнецов сохраняется в течение всей жизни, а дискор-
дантной по ЭЭГ близнецовая пара становится лишь в случае
тяжелого психического заболевания или травмы одного из
партнеров. Скорость возрастных изменений ЭЭГ также нахо-
дится под генетическим контролем. Наиболее заметные изме-
нения в ЭЭГ происходят в детстве и в преклонном возрасте.
Известно, что элементы кожи, как и структуры централь-
ной нервной системы (ЦНС), являются производными обще-
го эмбрионального листка — эктодермы, и время формирования
кожных узоров совпадает с периодом эмбриональной диффе-
ренцировки ЦНС. Имеются также свидетельства, что более
сложному папиллярному рисунку на пальцах соответствует
и более сложная структура нервных окончаний.
Поэтому логично предположить, что отклонения в дерма-
тоглифическом рисунке при хромосомных заболеваниях, ког-
да имеют место грубые пороки развития мозга, в такой сте-
пени отчетливы и характерны, что могут быть использованы
272
8.11. Функциональная асимметрия
в диагностике этой патологии. Анализ папиллярных узоров
представляет значительный интерес и в исследованиях не-
рвно-психических расстройств не хромосомной этиологии.
В то же время дерматоглифика может быть использована и
при изучении индивидуальных особенностей строения и фун-
кции ЦНС у здоровых людей.
Известно, что картина ЭЭГ человека чрезвычайно разно-
образна у отдельных индивидуумов, и в ней имеются генети-
чески детерминированные варианты. Признаки дерматогли-
фической картины кисти человека коррелируют с определен-
ными паттернами ЭЭГ. Наиболее ярко это проявляется при
высокой сложности узора на пальцах рук — завитках или на-
личии узора в области тенара, чему соответствует увеличе-
ние мощности в высокочастотном диапазоне a-активности и/
или в диапазоне низкочастотной 0-активности.
Генетическая детерминация сложной эмбриональной
структуры эктодермы обусловливает и сложную структуру ее
производных, в частности — морфологию ЦНС и гребневой
кожи. На уровне папиллярных узоров это, по-видимому, ве-
дет к усложнению дерматоглифических узоров, а на уровне
ЦНС может проявляться в усложнении ее макро- или микро-
структуры (в повышении складчатости коры, в увеличении
числа нейронов или глиальных элементов, в усложнении вет-
вления дендритов, в росте количества синаптических контак-
тов) . На уровне ЭЭГ такая структурная сложность может оп-
ределять способность к более дифференцированному режи-
му деятельности, что, возможно, и проявляется в увеличении
доли высокочастотных компонентов ЭЭГ.
8.11. Функциональная асимметрия
У человека правое и левое полушария головного мозга функ-
ционально неравнозначны. Это явление получило название
межполушарная асимметрия. Она проявляется в различиях
восприятия и переработки информации и выявляется посред-
ством снятия электроэнцефалограммы. Левое полушарие от-
273
вечает преимущественно за абстрактную переработку ин-
формации. Правое полушарие «специализировано» на конк-
ретной, пространственно-образной переработке информа-
ции. Элекрофизиологические исследования показали, что
биоэлектрическая активность правого полушария мозга на-
ходится в большей зависимости от генотипа, чем активность
левого полушария.
С функциональной асимметрией мозга у человека связа-
но преимущественное использование правой или левой руки.
Некоторые исследователи приводят данные о неодинаковых
познавательных способностях правшей и левшей, других раз-
личий в поведении право- и леворуких индивидов не обнару-
живают.
Поиск генетических причин право- и леворукости прово-
дится давно, однако четкого представления о наследственных
механизмах двигательного доминирования до сих пор нет. О
наличии генетического компонента в формировании право-
и леворукости указывают генеалогические исследования.
Есть сведения, что праворукие индивиды, происходящие из
семей левшей, отличаются от праворуких, которые происхо-
дят из семей правшей, по способности к пространственной
ориентации и скорости восприятия. Вероятность появления
леворукого ребенка у праворуких родителей составляет 2%.
В семьях, где один из родителей леворукий, вероятность рож-
дения леворукого ребенка составляет 17%. Если оба родителя
леворукие, ребенок будет леворуким с вероятностью 47%. В
отношении леворукости обнаружен материнский эффект: ле-
ворукость матери оказывает большее влияние на формиро-
вание леворукости у ребенка, чем леворукость отца.
Для объяснения асимметрии владения рукой предложено
несколько генетических гипотез. Одна из моделей предпола-
гает существование одного локуса с двумя аллелями. Соглас-
но другой модели этот признак находится под контролем двух
независимых локусов: один локус с двумя аллелями опреде-
ляет доминирование речевого полушария, другой — соответ-
ствие доминирования руки доминированию речевого полу-
274
8.11. Функциональная асимметрия
шария. Еще одна модель постулирует существование гена
«правого сдвига». Наличие этого гена обусловливает предрас-
положенность индивида стать правшой. При отсутствии это-
го гена человек становится левшой или правшой в зависимо-
сти от обстоятельств. Хотя ни одна из моделей не объясняет
все случаи распределения право- и леворуких людей в семь-
ях, влияние генотипа на функциональную асимметрию не
вызывает сомнения.
У большинства людей доминирующей является правая
рука, левши составляют примерно 5% населения. В связи с
«дискриминацией» левшей, их переучиванием указанная ча-
стота по некоторым оценкам является заниженной в 2—3
раза. Практически во всех культурах существует предубеж-
дение против леворукости. Конструкция асимметричных ору-
дий производства и бытовых приборов рассчитана на прав-
шей. При массовом производстве музыкальных инструмен-
тов потребности левшей игнорировались. Ранее врожденную
леворукость считали чем-то вроде аномалии развития, и ле-
воруких детей учили работать правой рукой, за пользование
левой рукой, случалось, наказывали. У некоторых детей от
такой практики развивались неврозы и отвращение к учебе.
В последнее время предубеждение против леворукости ос-
лабло, практика переучивания леворуких детей ушла в про-
шлое. Для левшей стали изготавливать предметы особой кон-
струкции, которые продаются в специальных магазинах.
Антропологи установили, что левши чаще встречаются в
некоторых традиционных культурах по сравнению с индуст-
риальными. Они обратили внимание на то, что предметы быта,
которые они коллекционировали в антропологических экс-
педициях, устроены «под левую руку». Для объяснения меж-
популяционных различий в частоте право- и леворукости об-
ратились к явлению функциональной асимметрии мозга, ко-
торая связана с этими признаками. Было показано, что в
различных этнических группах функциональные способнос-
ти левого и правого полушария для восприятия и переработ-
ки информации используются неодинаково. В популяциях с
275
Раздел 8. Человеческое разнообразие
высоким развитием цивилизации преобладает левополушар-
ный фенотип, и, наоборот, в популяциях, относящихся к тра-
диционным, преобладает правополушарный фенотип. Дела-
ется допущение, что разные культуры основаны на разных
способах обработки информации и естественный отбор бла-
гоприятствует фенотипам, адаптивным в специфических ус-
ловиях среды.
8.12. Реакция на фармакологические средства
В 1902 г. английский врач Арчибальд Гэррод предположил, что
генетические факторы отвечают за химические превращения
в организме человека и лежат в основе межиндивидуальных
различий. В 1957 г. Арно Мотульски показал связь между по-
бочным действием лекарственных препаратов и особеннос-
тями генотипа.
Гены ответственны за синтез множества ферментов в пе-
чени человека. Некоторые из этих ферментов отвечают за то,
насколько быстро лекарственный препарат будет удален из
организма. Поэтому различия в активности ферментов будут
определять неодинаковую реакцию людей на лекарства.
Мужчины и женщины часто по-разному реагируют на вве-
дение одной и той же дозы лекарства. Так, женщины более
эффективно реагируют на антидепрессанты, которые дей-
ствуют на специфические ингибиторы обратного захвата се-
ротонина в пресинаптическую клетку. Женщины иначе, чем
мужчины, реагируют на некоторые обезболивающие сред-
ства, стероидные гормоны и средства, понижающие тревож-
ность.
Одним из первых открытий на генетическом уровне были
межпопуляционные различия в полиморфизме фермента N-
ацетил-трансферазы, который учавствует в метаболизме не-
которых лекарств. Фермент глутатион-Б-трансфераза, уча-
ствует в детоксикации вредных веществ. Люди, у которых
данный фермент отсутствует, в большей степени подверже-
ны различным формам рака. Среди черного населения США
276
8.12. Реакция на фармакологические средства
частота гена глутатион-Б-трансферазы примерно равна 0,31.
В разных европейских популяциях частота этого гена колеб-
лется в пределах от 0,39 до 0,54. Данный феномен может слу-
жить объяснением различной межпопуляционной заболева-
емости раком.
В зависимости от набора ферментов, одни люди являются
медленными инактиваторами, другие — быстрыми. Медлен-
ные инактиваторы могут быть подвержены более длительно-
му воздействию лекарства и значительно меньшему воздей-
ствию активных метаболитов. И наоборот, быстрые инакти-
ваторы дольше испытывают действие метаболитов и гораздо
меньше действие самого лекарственного вещества. 5—10%
людей в популяциях африканцев и европейцев относятся к
этим двум, а не к нормальным типам инактиваторов, в то вре-
мя как в азиатских популяциях доля «нетипичных» (быстрых
или медленных) инактиваторов меньше.
Некоторые гены у человека отвечают за то, будет ли чело-
век после приема кодеина чувствовать облегчение боли, или
же будет ли человек, больной определенной формой рака, ре-
агировать на определенное лекарственное вещество. Так, у
онкобольных с генетически обусловленной недостаточностью
дигидропиримидин дегидрогеназы наблюдаются нейротокси-
ческие эффекты при лечении широко используемым препа-
ратом 5-фторурацилом, и поэтому они нуждаются в подборе
других лекарств.
Другим механизмом, определяющим активность лекар-
ственных веществ, является присутствие или отсутствие спе-
цифических рецепторов данного лекарства. Поскольку лекар-
ственные препараты присоединяются к рецепторам, пред-
ставляющим собой белковые молекулы, то эффект лекарства
будет определяться числом этих белков, детерминируемых
генетически.
Действие широко распространенного антипсихотическо-
го лекарственного препарата галоперидола зависит от его
способности связываться с рецепторами дофамина D2. Коли-
чество данных рецепторов определяется генетически. Так, в
277
Раздел 8. Человеческое разнообразие
одном исследовании было обнаружено, что у 63% пациентов
с большим количеством указанных рецепторов отмечался
положительный эффект от лечения галоперидолом и только
у 29% пациентов с малым количеством рецепторов.
Другие исследования указывают на то, что люди по-раз-
ному реагируют не только на лекарства, но и на пищевые ве-
щества в силу генетических различий.
Контрольные вопросы и задания к разделу 8
1. Перечислите условия, которые были решающими при эво-
люции человеческих рас.
2. Объясните адаптивное значение цвета кожи у человека.
3. Чем обусловлено разнообразие людей по группам крови?
4. У супружеской пары потерялся ребенок. В течение не-
скольких лет они вели поиски и обнаружили мальчика, воз-
раст которого соответствовал возрасту их сына. Анализ по-
казал, что женщина имеет группу крови М, мальчик N. Мо-
жет ли ребенок быть ее сыном?
5. У Анастасии родился сын с первой группой крови. Муж
Анастасии заинтересовался, как это могло получиться,
если у них с женой вторая группа крови. Объясните.
6. В родильном доме перепутали двух мальчиков. Родители
одного из них имеют I (0) и IV (АВ) группы крови, родите-
ли второго I и III (В) группы. Анализ показал, что у первого
мальчика I группа крови, а у второго II (А) группа крови.
Определите, кто чей сын.
7. Две супружеские пары разыскивают сыновей, потеряв-
шихся в раннем детстве. Был найден ребенок с первой
группой ребенок, который достоверно принадлежал одной
из супружеских пар. У одной супружеской пары I и IV
группы крови, у другой — II и III. Чей это сын?
8. У мальчика группа крови MN. У его матери группа крови
М. Какая группа крови невозможна у отца?
9. Можно ли исключить отцовство, если у матери группа кро-
ви II, у ребенка III, а у предполагаемых отцов I и IV группы?
278
Контрольные вопросы и задания к разделу 8
10. Для установления родительства были исследованы груп-
пы крови системы ABO, MN и Rh. Получены такие резуль-
таты: первая супружеская пара АВ М гй"и АВ N Rh+; вто-
рая супружеская пара 0 М Rh+ и АВ MN rh“; мальчик АВ
MN Rh+ .Чей он сын?
11. Охарактеризуйте систему генетического контроля HLA.
12. Степень генетического родства учитывается при транс-
плантации тканей и органов. Трансплантация может быть
успешной, если донор не имеет антигенов, которых нет у ре-
ципиента. Важное значение при трансплантации имеет сис-
тема антигенов гистосовместимости. Могут ли родители с
антигенами гистосовместимости А11В15С1 и АЗЗВ10С7
быть донорами ткани для своих детей?
13. Могут ли быть донорами друг для друга сибсы, если их ро-
дители имеют такие антигены гистосовместимости:
A1B5C7D8 и A3B6C8D3?
14. У членов одной семьи обнаружены следующие антигены
гистосовместимости: женаА30В4В46С1, мужА30А36В4С2,
дети: 1) А30В4С1С2, 2) А30В4В46С1С2, 3) А30А36В4С1С2,
4) А30В4В46С1. У третьего ребенка обнаружено заболева-
ние почек и ему требуется трансплантация этого органа.
Почку могут дать родители и сибсы. Кто из них может быть
донором?
15. Члены одной семьи имеют наборы антигенов гистосовме-
стимости: жена АЗВ7В15С8, муж АЗА9В15С4С8, дети:
1) АЗВ7В15С8, 2) АЗА9В15С4С8, 3) АЗВ7В15С8. Третий ре-
бенок получил тяжелые ожоги и нуждается в пересадке
кожи. Родственники готовы дать ткань. Кто из них может
быть донором?
16. Какие виды полиморфизма ДНК известны в настоящее
время?
17. Какую информацию получает исследователь с помощью
электроэнцефалограммы?
18. Существует ли связь между ЭЭГ и личностными характе-
ристиками?
19. Приведите примеры наследования различных вариантов
ЭЭГ.
279
Раздел 8. Человеческое разнообразие
20. Человеку свойственна функциональная асимметрия по-
лушарий головного мозга. За абстрактную переработку ин-
формации отвечает преимущественно левое полушарие,
за конкретную, пространственно-образную — правое. На
данных, полученных при изучении популяций малочислен-
ных народов Севера, решите, играют ли какую-либо роль
генетические факторы в формировании функциональной
асимметрии мозга.
Среди 80 пар, в которых у одного из супругов был право-
полушарный фенотип, было обнаружено 88 детей с пра-
вополушарным фенотипом, 12 детей с левополушарным
фенотипом и 36 детей со смешанным фенотипом. В 38 бра-
ках, где один из родителей имел левополушарный фено-
тип, было обнаружено соответственно 24, 28 и 21 ребенок
с разными фенотипами, и, наконец, в 38 браках, где один
из родителей имел смешанный фенотип, были обнаруже-
ны 12, 9 и 30 детей с тремя разными фенотипами.
21. Петя хорошо владеет правой рукой. Его братья Юра и Ки-
рилл и сестра Таня — левши. Петина мать Нина — прав-
ша, отец Иван — левша. У Нины два брата: Артур — прав-
ша и Анатолий — левша. Мать Нины — правша, отец Па-
вел — левша. Артур женат на Татьяне — правше. У Артура
и Татьяны две дочери — Катя и Маша — обе левши. Со-
ставьте родословную этой семьи и определите характер
наследования право- и леворукости.
22. Активность ферментов определяется с помощью «дибу-
каинового числа». Это мера степени ингибирования фер-
мента с помощью дибукаина. У гомозигот по гену недоста-
точности холинэстеразы дибукаиновое число равняется
25, у гетерозигот — 50—70, у нормальных гомозигот боль-
ше 75. 100 человек, обследованных на активность холинэ-
стеразы, распределились следующим образом: 4 имели
дибукаиновое число 65, 58, 67 и 50, а у остальных этот по-
казатель был больше 80. Определите частоты аллелей это-
го гена.
23. По способности инактивировать путем ацетилирования
некоторые лекарственные средства все люди делятся на
280
Контрольные вопросы и задания к разделу 8
«быстрых» и «медленных». Фенотип «медленные инак-
тиваторы» контролируется гомозиготностью по соответ-
ствующему гену. Среди европейцев примерно 50% «мед-
ленных инактиваторов», а среди азиатов около 15%. Оп-
ределите частоту аллелей данного локуса.
24. Пищевая индустрия использует много искусственных до-
бавок в различных продуктах. Искусственные красители
и ароматизаторы дешевле натуральных. В некоторых слу-
чаях изначально используются натуральные экстракты из
растений и животных, однако их патентование усложня-
ется малой прибыльностью в будущем. Новые биотехно-
логические компании, например, американская компания
Senomyx, по-другому подходят к данному вопросу. При оп-
ределении точной структуры белка-рецептора, ответ-
ственного за вкус или обоняние, ученые в таких компани-
ях пытаются методом моделирования подобрать вещества,
которые имеют наибольшее сродство к данному рецепто-
ру. Например, химическое вещество 1-ацетил-4-метилбен-
зен имитирует вкус черешни, 4-метоксибензилацетат —
вкус ванили. Подобные вкусовые добавки добавляются и
в фармацевтические препараты для устранения неприят-
ного, чаще горького вкуса последних. Многие добавки
, могут иметь негативные последствия для здоровья чело-
века, в силу их новизны, возможно, еще неизвестные. Как
, Вы считаете, стоит ли в настоящее время, когда экологи-
ческая обстановка и так довольно сложная, подвергать
> человека дополнительному намеренному риску, сопря-
► женному с употребление искусственных веществ ? Как Вы
можете связать рост фармакогенетических осложнений,
i в том числе аллергических реакций у человека в последние
I десятилетия, с использованием искусственных добавок?
Раздел 9
ГЕНЕТИЧЕСКИЙ КОНТРОЛЬ
ВОСПРИЯТИЯ
Поведение человека в значительной мере определяется его
способностью воспринимать действительность. Органы
чувств дают первичную информацию об окружающем мире.
От того, в каком виде эта информация поступает в мозг, зави-
сит поведение индивида в той ли иной ситуации. Строение
зрительного, слухового и других анализаторов находится под
генетическим контролем. Функционирование органов чувств
определяется их структурными особенностями. Таким образом,
если мы говорим о влиянии наследственности на поведение, то
имеем в виду не прямое действие генотипа на поведенческий
акт, а последовательность событий, среди которых — развитие
и функционирование органов чувств. Не все в этой цепи со-
бытий известно, однако некоторые звенья изучены достаточ-
но хорошо.
9.1. Зрение
Орган зрения формируется под контролем множества генов,
мутация каждого из которых может вызвать те или иные на-
рушения в строении, а следовательно, и функционировании
зрительного анализатора. Эти нарушения проявляются как
аномалии зрения.
При нормальном зрении лучи света, попадающие в глаз,
после преломления в роговице и хрусталике собираются в
точку (фокусируются) на сетчатке, и человек четко воспри-
нимает изображения предметов. Если лучи фокусируются не
на сетчатке, изображение оказывается нечетким. Эти нару-
шения зрения обозначаются как аномалии рефракции. К ним
относят близорукость, дальнозоркость и астигматизм. Близору-
282
9.1. Зрение
кие люди плохо видят удаленные предметы, дальнозоркие —
предметы, находящиеся вблизи. Наиболее часто встречается
близорукость. Она связана с тем, что лучи света фокусиру-
ются не на сетчатке, а перед ней, в стекловидном теле. Такая
аномалия возникает при врожденном удлинении переднезад-
него диаметра глаз. У дальнозорких людей фокус находится
I за сетчаткой. Дальнозоркость бывает старческая и врожден-
ная. Врожденная дальнозоркость обусловлена укорочением
переднезаднего диаметра глаза, старческая — уплощением
хрусталика и его неспособностью изменять свою кривизну
из-за возрастной потери эластичности. Близнецовые иссле-
дования доказали наследственную предрасположенность к
развитию близорукости. При исследовании семей были по-
лучены данные, указывающие на аутосомно-рецессивный тип
наследования, однако наиболее правдоподобной считается
полигенная модель.
У некоторых людей кривизна хрусталика по вертикально-
му и горизонтальному диаметрам не одинакова. При таких
условиях лучи света после преломления не собираются в од-
ной точке, а рассеиваются по сетчатке. Это является причи-
ной астигматизма — невозможности ясно видеть. Сходство
родственников по наличию и по степени астигматизма ука-
зывает на его наследственную природу. Неодинаковая ост-
рота зрения может быть связана и с косоглазием, когда нару-
шается бинокулярное зрение. Семейные случаи также сви-
детельствуют о наследственной предрасположенности к этой
аномалии.
Описано несколько аномалий зрительного нерва. Одна из
них наследуется по аутосомно-доминантному типу и начина-
ет проявляться у взрослых, другая — начинающаяся в детстве,
передается как аутосомно-рецессивный признак. Еще одна
форма наследуется по цитоплазматическому типу {атрофия
зрительного нерва Лебера). Ген, ответственный за одну из
форм атрофии зрительного нерва, наследуемой по аутосом-
но-доминантному типу, был обнаружен в длинном плече хро-
мосомы 3.
283
Раздел 9. Генетический контроль восприятия
Некоторые люди плохо видят в сумерках. Эта особенность
может иметь наследственную и ненаследственную природу.
Анализ родословных показал, что слабое зрение при малой
освещенности (куриная слепота] наследуется как аутосомно-
доминантный признак. Однако это состояние может возник-
нуть и у генетически нормальных людей. Эта аномалия раз-
вивается при нехватке в рационе некоторых витаминов. Нор-
мализация питания у больных с ненаследственной формой
куриной слепоты улучшает способность видеть при слабом
свете. Если у человека куриная слепота вызвана генетичес-
ким дефектом, излечить болезнь с помощью витаминотера-
пии не удается. Предполагается, что необходимые для нор-
мального зрения витамины не усваиваются организмом из-
за того, что нарушен обмен веществ, вызванный генетическим
дефектом. Лечение наследственной формы куриной слепо-
ты возможно, если будет установлено, какой этап метаболиз-
ма нарушен.
Известно несколько генетически различных форм врож-
денной слепоты, вызванных мутациями разных генов. Боль-
шинство этих мутаций рецессивны. Допустим, что гены А и В
ответственны за развитие нормального зрения, а их мутации
а и Ь приводят к развитию слепоты и люди с генотипами ААЬЬ
и ааВВ слепые. В их браке рождаются дети с нормальным зре-
нием:
Р: АаЬЬ х ааВВ
Fp АаВЬ
В данном случае слепота наследуется по типу комплемен-
тарное™. Если у супругов слепота имеет одну генетическую
природу (ААЬЬ х ААЬЬ или ааВВ х ааВВ), то все их потомки
также будут слепыми. Одинаковые мутации более вероятны
у родственников, чем у людей, не состоящих в родстве. Этот
пример дает представление о генетической опасности для
потомства кровнородственных браков.
При катаракте происходит «затуманивание» в норме про-
зрачного хрусталика глаза. Человек видит предметы будто бы
через замерзшее стекло или запотевшие очки. Катаракта яв-
284
9.1. Зрение
ляется одной из наиболее частых причин нарушения зрения.
Причиной катаракты может быть травма глаза, сахарный диа-
бет, чрезмерная подверженность глаз УФ-лучам и, конечно,
гены. Молекулярно-генетические исследования выявили
гены, ответственные за врожденную форму катаракты, в хро-
мосомах Ip, 1 q, 13, 12q и 17р. Примерно у 1 из 250 новорож-
денных наблюдается врожденная катаракта, у других катарак-
та развивается позже, часто с лежащей в основе генетичес-
кой причиной.
Некоторые болезни глаз не имеют четкого генетического
механизма, и в этих случаях определяют эмпирический риск.
Например, при одной из форм глаукомы (повышенном глаз-
ном давлении) — первичной открытой угловой глаукоме —
симптомы наступают после 40 лет. Риск для родственников
первой степени родства пораженного составляет 5—16%. Ген,
ответственный за другую форму глаукомы, был обнаружен в
коротком плече хромосомы 6. При страбизме, или косогла-
зии, риск развития заболевания у сибса пораженного ребен-
ка составляет 15%. В семьях, где родитель и ребенок имеют
страбизм, риск болезни у следующего ребенка составляет
около 40%.
Дегенерация рецепторов сетчатки включает множество
отдельных генетически обусловленных заболеваний, отлич-
ных по механизму наследования и типу потери зрения. В Ве-
ликобритании, например, это заболевание встречается с час-
тотой 1:2000. Эти болезни являются наиболее важной причи-
ной, приводящей к потере зрения в юношеском возрасте, и
составляют большую долю слепоты у взрослых. Некоторые
люди с рецепторной дисфункцией обладают дефектами зре-
ния в средней области зрительного поля, имеют слабую адап-
тацию к темноте и входят в группу лиц с пигментным рети-
нитом (ПР). ПР в среднем встречается с частотой 1:5000. На-
пример, мутация гена пигмента родопсина обнаружена
примерно у 30% лиц с доминантной формой ПР. При ПР люди
с трудом ориентируются в темноте. ПР наступает, как прави-
ло, в первые 20 лет жизни человека. В течении длительного
285
Раздел 9. Генетический контроль восприятия
времени люди с ПР чувствуют ухудшение бокового зрения,
которое затем в большинстве случаев полностью пропадает.
Около 50% лиц с ПР имеет наследственную основу болезни.
Разновидности ПР отличаются типом наследования, тяжес-
тью течения болезни и возрастом начала. Имеется, по мень-
шей мере, три основных типа наследования ПР — Х-сцеплен-
ное рецессивное, аутосомно-доминантное и аутосомно-рецес-
сивное. Самой тяжелой формой является Х-сцепленный
рецессивный тип ПР. У больных с данной формой полная сле-
пота наступает к 40 годам. К наименее тяжелой относится
аутосомно-доминантный тип ПР. Аутосомно-рецессивный
тип ПР называется синдромом Ашера. При синдроме Ашера
наблюдается как ПР, так и врожденное не прогрессирующее
ухудшение слуха.
Нормальные люди (трихроматы) способны различать цве-
та, полученные путем смешения тонов трех основных участ-
ков спектра: красного, зеленого и синего. Если в сетчатке гла-
за отсутствуют чувствительные элементы для восприятия ка-
кого-либо из этих участков спектра, то возникает нарушение,
называемое дихромазией. Существуют две разновидности
дихромазии — протанопия (слепота на красный цвет) и дей-
теранопия (слепота на зеленый цвет).
Аномалии цветовосприятия были описаны английским
врачом Дальтоном и потому носят название дальтонизм. Ге-
нетическая основа дальтонизма состоит в следующем. Спо-
собность воспринимать цвет контролируется двумя тесно
сцепленными генами (R и G), локализованными в Х-хромосо-
ме. Рецессивный аллель одного из этих генов (г) формирует
невозможность различать красный цвет, другого (д) — зеле-
ный. Эти гены локализованы в Х-хромосоме, поэтому нару-
шение цветового зрения чаще встречается у мужчин, чем у
женщин. В европейских популяциях цветовая слепота встре-
чается примерно у 8% мужчин и у 0,64% женщин. Существу-
ет еще один редкий дихроматический эффект — тритано-
пия (или тританомалия), при котором нарушено различение
цветов в сине-зеленой области спектра. Предполагается, что
эта аномалия контролируется аутосомным геном.
286
Очень редко встречается аномалия цветного зрения, на-
зываемая монохромазией, при которой отсутствует восприятие
всех цветов. В этом случае человек различает только степень
яркости. Этот признак сочетается со светобоязнью, атрофи-
ей зрительного нерва и другими аномалиями. Наследуется
монохромазия по аутосомно-рецессивному типу.
Аномалии цветового зрения неодинаково часто встреча-
ются в различных популяциях. Это может свидетельствовать
о существовании генетического полиморфизма, который под-
держивается естественным отбором. Одним из факторов от-
бора по генам дальтонизма может быть цивилизация. Цвето-
вая слепота очень редко встречается в племенах, основное
занятие которых охота и собирательство, и довольно часто в
индустриальных странах. Эти различия свидетельствуют о
том, что с развитием цивилизации отбор на нормальное цве-
товое зрение ослабился. Для народов, живущих охотой, жиз-
ненно важно иметь не просто хорошее зрение, но и чувство-
вать цвет, чтобы различать на фоне зеленой растительности
коричневатую шерсть животных. Логично допустить, что
охотники, не способные успешно охотиться вследствие де-
фекта цветового зрения, а также их потомство имеют мень-
ше шансов выжить. В условиях цивилизации мутации по ге-
нам цветового зрения не приводят к таким серьезным послед-
ствиям, поэтому отбор против них ослаблен либо вообще
отсутствует. Можно даже считать, что в индустриальных стра-
нах отбор благоприятствует генам дальтонизма в связи с тем,
что в некоторых странах мужчин с дефектами зрения не при-
зывают на военную службу, что повышает вероятность их
выживания по сравнению с пригодными к военной службе
нормально видящими мужчинами. Изучая зрение у народов
традиционных культур, антропологи столкнулись с необыч-
ным явлением. Оказалось, что в некоторых австралийских
племенах, а также у индейцев Южной Америки частота даль-
тонизма необычно высока. Было сделано предположение, что
охотники и воины дальтоники обладают обостренным чув-
ством формы и способностью хорошо различать очертания
Раздел 9. Генетический контроль восприятия
предметов. Если среди охотников есть дальтоники, охота мо-
жет быть более успешной, поскольку такие индивиды способ-
ны обнаружить животное, имеющее маскирующую окраску.
Дальтоник-воин в отряде может быть полезен тем, что спосо-
бен различать камуфляж. История свидетельствует, что на
заре развития авиации в летчики-разведчики отбирали даль-
тоников, так как они легче обнаруживали замаскированные
военные объекты, чем разведчики с нормальным зрением.
Дефекты зрения серьезно ограничивают выбор профес-
сии. Особенно строго идет отбор на качество зрения работ-
ников транспорта. Однако оценка работы водителей-дальто-
ников показала, что частота совершаемых ими нарушений
дорожного движения достоверно не отличается от частоты
нарушений, совершаемых водителями с нормальным цвето-
вым зрением. По-видимому, у людей, имеющих те или иные
дефекты восприятия, развивается компенсаторное поведе-
ние, позволяющее преодолеть физический недостаток.
Генетически обусловленные особенности восприятия ок-
ружающего мира могут не только оказать решающее действие
в выборе профессии, но и определить успех в том или ином
виде деятельности. К примеру, индивиды с аномалией цвето-
вого зрения, занимающиеся живописью, могут использовать
необычное сочетание красок и создавать оригинальные, с
точки зрения обычных людей, произведения искусства. Та-
кой особенностью отличался художник Ван Гог, у которого
была наследственная болезнь зрительного нерва, поэтому он
видел краски не так, как большинство людей.
9.2. Слух
Врожденное отсутствие слуха приводит к глухонемоте, зат-
рудняющей общение. Средовые причины врожденных де-
фектов слуха хорошо известны. Главная из них — воздей-
ствие на эмбрион тератогенных факторов, когда происходит
закладка слухового анализатора — до 14-й недели беремен-
ности. Наиболее опасны для развития слуха будущего ребен-
288
9.2. Слух
ка инфекционные заболевания беременной женщины. Врож-
денная глухота может развиться у ребенка после принятия
беременной некоторых лекарственных препаратов, может
быть вызвана и родовой травмой.
В формировании органа слуха принимает участие большое
количество генов, и мутация любого из них может стать при-
чиной тугоухости. Ослабленный слух является составной ча-
стью многих наследственных синдромов, например, синдро-
ма Ашера. О генетической гетерогенности глухоты свидетель-
ствует разнообразие ее клинических проявлений. В одних
случаях глухота проявляется с рождения, другие ее формы
развиваются в течение жизни. Генеалогический анализ по-
зволил обнаружить несколько десятков рецессивных мута-
ций, приводящих к глухоте. Некоторые формы глухоты обус-
ловлены доминантными мутациями.
В связи с тем, что наследственная глухонемота — генети-
чески гетерогенное состояние (определяется мутациями раз-
ных генов), в семьях, где оба родителя глухонемые, могут ро-
диться дети с нормальным слухом. Это можно проиллюстриро-
вать таким примером. Допустим, С и D — гены, принимающие
участие в формировании нормального слуха. Мутация любо-
го из генов (с или d) нарушает нормальный процесс форми-
рования слухового анализатора и приводит к глухоте. В бра-
ке глухонемых индивидов, имеющих глухоту рецессивной
природы, но обусловленную мутациями разных генов (CCdd
х ccDD), потомство будет гетерозиготным по обоим генам
{CcDd) и за счет доминирования нормальных аллелей над му-
тантными иметь нормальный слух. В то же время, если у суп-
ругов нормальный слух, но они гетерозиготны по одному и
тому же гену, то у них могут родиться глухонемые дети: CcDD х
CcDD (генотип пораженного потомка ccDD) или CCDd х CcDd
(генотип пораженного потомка CCdd). Если родители гетеро-
зиготны по разным локусам {CcDD х CCDd), то среди потом-
ства не окажется гомозигот ни по одному из рецессивных ге-
нов. Вероятность того, что супруги являются носителями од-
ного и того же мутантного гена, значительно повышается, если
они состоят в родстве. Люди, имеющие те или иные анома-
10 Введение в психогенетику
289
Раздел 9. Генетический контроль восприятия
лии, нередко объединяются в общества инвалидов. Они вме-
сте работают и отдыхают, в своем узком кругу обычно нахо-
дят брачных партнеров. Такие люди особенно нуждаются в
генетическом консультировании. Генетик поможет опреде-
лить риск наследственных аномалий у будущих потомков и
даст рекомендации, которые позволят его снизить. Для этого
необходимо выяснить, не состоят ли потенциальные родите-
ли в кровном родстве, с помощью точной диагностики решить,
какова причина глухонемоты у супругов. Прогноз для потом-
ства будет благоприятным, если у супругов генетически раз-
ные рецессивные формы заболевания или если хотя бы у од-
ного из них заболевание имеет не наследственную природу.
Прогноз для потомства неблагоприятен, если у кого-либо из
супругов доминантная форма глухонемоты или оба они стра-
дают одной и той же рецессивной формой. Каков бы ни был
прогноз, решение о том, иметь или не иметь детей, принима-
ет сама супружеская пара.
9.3. Вкусовая чувствительность
Вкус является главным фактором выбора пищи. В понятие
вкуса пищи входит как собственно ее вкус и запах, так и внеш-
ний вид. В ряде случае аппетитная пища связана с ее энерге-
тической ценностью, которая, в свою очередь, зависит от со-
держания в ней жиров и углеводов.
Влияние вкуса пищи на вкусовые предпочтения наиболее
существенно в младенчестве и детском возрасте. Дети любят
сладкое и отвергают горькое. Такие врожденные вкусовые
качества заметны сразу после рождения. Традиционным
объяснением считалось то, что сладость указывает на энерге-
тическую ценность пищи, в то время как горечь — предуп-
реждение о пищевой опасности.
Отторжение ряда горьких веществ является генетически
обусловленным. К изученным примерам генетического поли-
морфизма вкуса у человека относится лишь реакция на горь-
кий вкус.
290
। 9.3. Вкусовая чувствительность
В 1931 г. химик Артур Фокс, смешивая порошкообразное
вещество, случайно просыпал небольшое его количество. Из
окружающего воздуха частички этого вещества попали в рот
Фоксу и его коллеге. Коллега Фокса почувствовал вещество
чрезвычайно горьким, сам Фокс — безвкусным. Затем Фокс
предложил попробовать порошок некоторым своим друзьям,
среди которых одни нашли его безвкусным, другие — горь-
ким, а третьи — очень горьким.
Порошок Фокса, названный фенилтиокарбамидом (ФТК),
широко использовался с тех пор для выявления различий во
вкусовой чувствительности. Исследования, которые были
проведены вскоре после случая в лаборатории Фокса, пока-
зали, что в основе чувствительности к ФТК лежат гены. Уче-
ные обнаружили, что люди с большей вероятностью находи-
ли ФТК горьким, если другие члены их семей также чувство-
вали горечь этого вещества. Генетическая природа ощущения
горького вкуса ФТК была настолько очевидной, что перед раз-
работкой современного ДНК-анализа тест на чувствитель-
ность к ФТК использовался в качестве теста на отцовство.
Спустя более чем 70 лет, в хромосоме 7 был локализован
ген, отвечающий за чувствительность к фенилтиокарбамиду.
В настоящее время установлено, что этот ген может находить-
ся в трех аллельных состояниях. Небольшие отличия в бел-
ках, которые они кодируют, в конечном счете влияют на функ-
ционирование вкусовых рецепторов языка. Изменения в
строении рецепторов приводят к тому, что человек оказыва-
ется неспособным воспринимать ФТК. Поскольку каждый
человек имеет по два аллеля данного гена, различные их ком-
бинации (1/1, 1/2, 2/2 и т.д.) определяют, будет ли человек
чувствовать ФТК очень горьким (такие люди называются су-
пертестерами), в какой-то степени горьким (этих людей на-
зывают тестерами) или безвкусным вообще (эти люди явля-
ются не тестерами).
Супертестеры не только в наибольшей степени ощущают
вкус определенных веществ, но также и в максимальном вы-
ражении воспринимают раздражающее действие веществ, и
291
Раздел 9. Генетический контроль восприятия
особенно велика у них тактильная чувствительность к жирам,
поступающим в составе пищи. Такой феномен можно объяс-
нить тем, что в грибовидных ворсинках находятся не только
нервные окончания, участвующие в восприятии вкуса, но и
окончания тройничного нерва, которые воспринимают ощу-
щение прикосновения и боли.
Генетические различия во вкусовом восприятии влияют
на пищевые предпочтения и, таким образом, на потребление
продуктов, а в конечном счете на состояние здоровья челове-
ка. Избегание овощей и фруктов с горьковатым вкусом сре-
ди супертестеров ставит этих людей в группу риска по болез-
ням, при которых потребление указанных продуктов снижа-
ет вероятность развития некоторых болезней, в частности,
отдельных форм рака. Снижение потребления пищи, содер-
жащей много жиров или же слишком сладкой, женщинами
не тестерами уменьшает риск развития других заболеваний,
в том числе сердечно-сосудистых.
Возможно, что межпопуляционные различия в частоте ге-
нов, ответственных за формирование вкусовой чувствитель-
ности, — одна из причин, обусловивших специфику нацио-
нальных кухонь. Специальные исследования показали, что
между способностью ощущать различные концентрации ФТК
и предпочтением определенных пищевых продуктов имеет-
ся хорошо выраженная корреляция: индивиды с пониженной
чувствительностью к ФТК предпочитают острые блюда, а
люди, чувствующие это вещество в минимальных концент-
рациях, предпочитают более пресную пищу. Не исключено,
что различия в остроте блюд у разных народов связаны с рас-
пределением в популяции признака высокой и низкой вкусо-
вой чувствительности к различным химическим соединени-
ям. Генетически детерминированная низкая вкусовая чув-
ствительность понуждает людей готовить более острую пищу,
в то время как люди с высокой вкусовой чувствительностью
способны получить удовольствие и от пищи, не приправлен-
ной острыми специями.
Другое вещество, 6-п-пропилтиоурацил (ПРОП), одни
люди также ощущают как горькое, а другие как безвкусное.
292
9.3. Вкусовая чувствительность
Способность ощущать вкус ПРОП не является альтернатив-
ным качеством к неспособности ощущать это вещество, а
представляет собой пример непрерывно распределяемого
признака. Женщины более чувствительны к ПРОП, чем муж-
чины. Редко, но все же встречаются люди, способные уловить
горечь этого вещества в растворах, разбавленных в концент-
рации 1 часть к 5 миллионам. Большая чувствительность к
ПРОП связана с неприятием ряда других веществ с горько-
ватым вкусом. Замечено, что тестеры ПРОП не любят черный
кофе, сок грейпфрута и овощи рода Brassica.
Способность идентифицировать и отвергать горькие ве-
щества является эволюционным преимуществом. У человека
насчитывается от 40 до 80 различных вкусовых рецепторов
на горькое, и имеются данные о множественных механизмах
преобразования горького вкуса. Химически различающиеся
горькие вещества, многие из которых ядовитые, в целом дают
ощущение горечи. Тот факт, что чувствительность к ПРОП
максимальна в молодом возрасте и во время беременности,
но снижается к старости, дает основание полагать, что вкусо-
вой ответ на горькое первоначально имел защитную основу.
Пожилые люди менее чувствительны к горькому, а некото-
рым из них горький вкус даже нравится.
По своему химическому строению ФТК относится к ве-
ществам, которые подавляют функцию щитовидной железы.
Она при этом разрастается и формируется зоб. Замечено, что
зоб, связанный с ослаблением функции щитовидной железы,
чаще встречается у людей не тестеров, чем у тестеров. Эти
наблюдения привели ученых к мысли, что способность ощу-
щать ФТК дает обладателям этого признака некоторые селек-
тивные преимущества. По-видимому, тестеры отвергают
пищу, в которой содержатся сходные с ФТК вредные соеди-
нения, а не тестеры этого делать не могут.
В настоящее время ученые обнаружили около 30 различ-
ных генов, которые кодируют рецепторы, воспринимающие
горькие вещества, у млекопитающих (семейство генов T2R).
Отмечено, что данные рецепторы реагируют на широкий
fa
293
Раздел 9. Генетический контроль восприятия
спектр горьких веществ. Кроме того, у человека обнаружены
три вариации гена T1R, связанные с рецепцией сладкого вку-
са — T1R1, T1R2, T1R3. Показано, что комбинация T1R2/T1R3
осуществляет узнавание различных натуральных и синтети-
ческих подсластителей.
В то время как горькие вещества отторгаются, углеводы и
жиры являются предпочтительной пищей. Новорожденные
предпочитают подслащенные растворы, нежели воду, а с воз-
растом все больше потребляют сладкого. Восприятие и вос-
произведение сладкого вкуса осуществляется более просты-
ми способами по сравнению с горьким вкусом. Человек, по-
видимому, имеет не более 2—3 различающихся рецепторов
на сладкое. К тому же сладкий вкус ощущается при гораздо
больших концентрациях, чем потенциально токсические
горькие вещества. В целом более сладкие сахара, например,
сахароза, являются более предпочтительными, чем менее
сладкие. Жировые предпочтения также вырабатываются на
очень ранних этапах жизни человека, сразу как только дети
начинают распознавать вкус, сопряженный с высокой энер-
гетической ценностью и увеличенным содержанием жира.
Жиры и сахара являются основными компонентами шоколад-
ных конфет, пирожных, сладких десертов, йогуртов и моро-
женого. Сочетание сахаров и жиров представляет собой наи-
более полный источник энергии. Кроме того, в некоторых ис-
следованиях показано, что потребление энергетически
ценной пищи, такой, как шоколад, сопряжено с выделением
эндорфинов.
Предпочтение углеводов и жиров может осуществляться
через ряд нейрофизиологических механизмов. В одних иссле-
дованиях показана роль серотонина, в то время как в других —
эндогенных опиатов в опосредованном положительном вос-
приятии сладкого вкуса и вкуса жирной пищи. В данном слу-
чае эффекты эндорфинов отличны от простого чувства голо-
да и сытости. Предполагается, что эндорфины влияют на пред-
почтение пищи путем формирования более приятных ответов
на эту пищу.
294
9.3. Вкусовая чувствительность
Вкусовые предпочтения сахара и жира, по-видимому, яв-
ляются врожденным, генетически обусловленным, свойством
человека, и они отмечаются среди всех рас, этносов и во всех
возрастных группах. В целом предпочтение отдается более
энергетически ценной пище. Так, шоколад содержит 6 ккал/г, в
то время как шпинат и фруктовые соки — 0,2—0,5 ккал/г. Пик
чувствительности на энергетически ценную пищу приходит-
ся примерно на 12 лет, а затем постепенно снижается. Это
можно объяснить усиленным ростом человека, половым со-
зреванием и эволюционно сформированной потребностью в
высококалорийных пищевых веществах особенно в данный
период.
Хорошо демонстрирует связь между генами и культурны-
ми особенностями отношение к свежему молоку у разных на-
родов. В свежем молоке содержится молочный сахар лакто-
за. Переваривание этого углевода происходит при участии
фермента — лактазы, который вырабатывается в клетках тон-
кого кишечника. Способность усваивать или не усваивать
молочный сахар контролируется парой аллелей одного гена.
Доминантный ген L активен в течение всей жизни человека,
его аллель 1 активен только в детстве и неактивен у взрослых.
У людей с генотипами LL и L1 лактаза вырабатывается в тече-
ние всей жизни, у людей с генотипом 11 — только в раннем
детстве, примерно до четырехлетнего возраста. Затем ген ут-
рачивает свою активность и продукция лактазы прекращает-
ся. В отсутствии фермента молочный сахар проходит по ки-
шечнику, не расщепляясь и утрачивая питательную ценность,
а сам процесс сопровождается кишечными расстройствами,
приводящими к заболеваниям, опасным для жизни. Ген лак-
тазы прекращает работу, когда в этом, «с точки зрения при-
роды», исчезает необходимость. Диким млекопитающим во
взрослом состоянии молоко «не положено», поэтому необхо-
димости в постоянной активности соответствующего гена нет.
Система генетического контроля устроена так, что лактазная
активность на определенном этапе онтогенеза подавляется.
За «выключение» лактазного гена ответственны другие гены,
295
Раздел 9. Генетический контроль восприятия
гены-регуляторы. Мутации, нарушавшие процесс выключе-
ния лактазного гена, возникали всегда, однако они не имели
приспособительного значения и, по-видимому, были нейт-
ральны. Предполагается, что у человека эта мутация подхва-
тилась отбором около 10 000 лет назад в скотоводческих по-
пуляциях. Эта мутация давала какие-то преимущества ее об-
ладателям и потому распространилась во многих популяциях
Северной Европы. Можно предположить, что дела обстояли
так. В северных районах с долгой и холодной зимой, когда
отсутствовала растительность, а охота не всегда была успеш-
ной, люди нередко переживали голод. Приручение животных,
как известно, происходило в эпоху неолита. Одомашненные
животные были стабильным источником мяса, и это страхо-
вало людей в случаях неудачной охоты. Содержавшиеся при
человеке животные размножались и выкармливали своих по-
томков молоком. Дети, играя с телятами и козлятами, вместе
с ними могли подкормиться молоком козы или коровы. Пи-
таться молоком домашних животных ребенок мог до тех пор,
пока ген лактазы сохранялся у него в активном состоянии.
Ясно, что мутанты с продленной лактазной активностью име-
ли преимущества, так как были лучше обеспечены высокока-
чественной пищей. В периоды голода, которые были неред-
ки, дети с мутантным геном лактазы оказывались в более вы-
годном положении. Таким образом, мутация продленной
лактазной активности подхватилась отбором и распростра-
нилась в северных районах. В южных районах планеты, где
растительная пища доступна почти круглый год, эта мутация,
по-видимому, не давала особых преимуществ и потому не рас-
пространилась. Позднее люди научились доить животных, а
способ употребления молока — свежее или в виде кисломо-
лочных продуктов — определялся генотипом животноводов.
В странах Северной Европы от 80% до 100% взрослого насе-
ления обладают геном L и усваивают свежее молоко. Корен-
ное взрослое население Америки, Австралии, Африки, Азии
и Южной Европы в основном имеют генотип 11 и способны
употреблять в пищу только кисломолочные продукты, в кото-
296
9.3. Вкусовая чувств1!гпельност1,
рых молочный сахар расщеплен бактериями. В популяциях,
где население по преимуществу не способно усваивать све-
жее молоко, развито мясное животноводство, а там, где ос-
новная часть населения усваивает его, — также и молочное.
В Западной Африке есть популяции пастухов, которые пьют
свежее молоко в большом количестве. В этих популяциях лак-
тазная недостаточность встречается очень редко. Рядом жи-
вут племена скотоводов, которые не пьют свежего молока и
имеют высокую частоту гена лактазного дефицита.
У некоторых народов, проживающих в районах Средизем-
номорья, строго запрещено употреблять в пищу бобы Vicia
faba. В этом регионе встречается тяжелое заболевание фа-
визм, которое возникает после съедания бобов (название болез-
ни происходит от латинского наименования этого растения) и
проявляется в виде лихорадки, рвоты, головокружения. За-
болевание известно с древности, но только современная на-
ука раскрыла его генетическую природу. Под действием ве-
ществ, имеющихся в бобах (таким же действием обладают
определенные лекарственные препараты), у некоторых лю-
дей разрушаются эритроциты. Повышенная чувствитель-
ность к химическим соединениям связана с недостаточнос-
тью фермента глюкозо-б-фосфат-дегидрогеназы (Г6ФД). Ген
Г6ФД локализован в Х-хромосоме. Недостаточность фермен-
та является рецессивным признаком, поэтому фавизмом чаще
болеют мужчины. Этот фермент необходим для стабилизации
мембраны эритроцита, и при его отсутствии она становится
неустойчивой к действию некоторых химических соедине-
ний. Люди, склонные к фавизму, невосприимчивы к малярии.
Климат Средиземноморья благоприятен для малярии, поэтому
генетическая устойчивость к этому тяжелому заболеванию под-
хватилась отбором и повысила частоту гена недостаточности
Г6ФД в этом регионе. Платой за устойчивость к малярии явля-
ется фавизм. В некоторых странах число носителей мутантного
гена достигает 20%. Патологическая реакция на конские бобы
значительной части населения заставила выработать прави-
ла, запрещающие употреблять в пищу это растение.
Раздел 9. Генетический контроль восприятия
9.4. Восприятие запаха
Способность различать запахи имеет приспособительное зна-
чение. У многих животных обоняние играет главную роль при
поиске пищи, внутри- и межгрупповых контактах. Люди раз-
личаются по способности воспринимать запах различных ве-
ществ, их оттенки и отдельные составляющие. Индивиды с
особенно тонким обонянием работают экспертами в парфю-
мерной промышленности. Обоняние, как признак, имеющий
адаптивное значение, находится под генетическим контро-
лем. У мыши имеется около 1000 генов, ответственных за обо-
няние. Очевидно, что у человека подобных генов не меньше.
У человека гены, ответственные за восприятие запаха, рас-
полагаются во многих хромосомах. Вот несколько примеров.
Ощущение запаха мускуса зависит от доминантного гена М,
а неспособность ощущать это вещество обусловлена его ре-
цессивным аллелем т. Локус, ответственный за это свойство,
является аутосомным, поэтому признак с одинаковой часто-
той встречается у мужчин и женщин и одинаково наследует-
ся от матери и отца детьми обоего пола.
Одни люди чувствуют запах синильной кислоты, другие
нет. Среди нечувствительных больше мужчин, чем женщин.
Это объясняется тем, что ген, ответственный за этот признак,
локализован в X хромосоме. Y-хромосома этого локуса не со-
держит. Неспособность чувствовать запах синильной кисло-
ты контролируется рецессивным геном, а способность — его
доминантным аЛлелем. Обозначим ген способности обонять
запах S, неспособности — s. Генотипы женщин: XsXsh X'sXs
(тестеры), XSXS (не тестеры). Мужчины имеют один ген — XSY
(тестеры) или XSY (не тестеры).
Среди европейцев лишь небольшая доля не чувствует за-
паха фрезий, а для большинства популяции данный запах яв-
ляется одним из наиболее сильных из известных им. Неспо-
собность чувствовать запах фрезий определяется аутосомно-
рецессивным геном. При исследовании более 1600 людей
было выявлено, что частота рецессивного фенотипа велика
298
Контрольные вопросы и задания к разделу 9
(около 10%) у людей восточно-европейского и британско-
кельтского происхождения и мала (около 4%) у людей преиму-
щественно скандинавского, английского, датского и герман-
ского происхождения.
На грызунах показано, что гены рецепторов, ответствен-
ных за обнаружение феромонов (веществ, привлекающих
особей противоположного пола), отличны от генов, ответ-
ственных за определение других запахов. У женщин обнару-
жено, что гены, ответственные за восприятие запаха муж-
ского тела, формирующегося с участием антигенов HLA, пе-
редаются лишь по отцовской линии, а передача генов по
материнской линии в восприятии данного запаха роли не иг-
рает.
Контрольные вопросы и задания к разделу 9
1. Какие факты свидетельствуют о том, что аномалии реф-
ракции имеют наследственную природу?
2. Как наследуются дефекты цветового восприятия?
3. Супруги нормально различают цвета, а брат жены — даль-
тоник. Какова вероятность рождения в этой семье ребен-
ка-дальтоника ?
4. У жены дальтонизм, у мужа нормальное зрение. Какова
вероятность дальтонизма у их детей?
5. Катаракты имеют несколько наследственных форм. Ка-
кова вероятность того, что у потомка разовьется катарак-
та, если оба его родителя страдают доминантной формой
болезни и гетерозиготны по соответствующему гену и еще
являются гетерозиготами по другому рецессивному гену
катаракты? Оба локуса являются аутосомными и не сцеп-
ленными.
6. На рис. 9.1 приведена родословная со случаями гемерало-
пии — неспособности видеть в сумерках. Как наследует-
ся этот признак?
7. Причинами врожденной слепоты могут быть аномалии
хрусталика и роговицы глаза. Аномалии хрусталика и ро-
299
Раздел 9. Генетический контроль восприятия
1
Рис. 9.1
300
Контрольные вопросы и задания к разделу 9
говицы передаются, как аутосомные рецессивные призна-
ки.
1) Какова вероятность рождения слепого ребенка в семье,
где отец слепой вследствие аномалии хрусталика, а
мать — вследствие аномалии роговицы, а по другому
гену они нормальны и гомозиготны?
2) Какова вероятность рождения слепого ребенка в семье,
где отец слепой вследствие аномалии роговицы, а мать —
вследствие аномалии хрусталика и роговицы?
3) Какова вероятность рождения слепого ребенка в семье,
где отец и мать здоровы, но гетерозиготны по генам ано-
малии хрусталика и роговицы?
8. Изолированное косоглазие (не связанное с рядом нервно-
мышечных болезней, при которых оно присутствует) мо-
жет возникнуть у потомства с вероятностью 15%, если оба
родителя здоровы, но имеют одного больного ребенка, и с
вероятностью 40%, если косоглазием страдает и один из
родителей. Оцените вероятность рождения больного ре-
бенка для текущей беременности у пар А, Б и В в родос-
ловных на рис. 9.2.
Рис. 9.2.
9. Приведите примеры генетической гетерогенности глухо-
ты.
10. В родословной, изображенной на рис. 9.3, в трех семьях
прослеживается глухонемота. Объясните, почему у глухо-
немых родителей из поколения III все дети здоровы (поко-
ление IV)?
301
Раздел 9. Генетический контроль восприятия
И. У человека одна из форм наследственной глухоты опреде-
ляется двумя рецессивными аллелями разных генов. Для
нормального слуха необходимо наличие двух доминант-
ных аллелей, один из которых определяет развитие улит-
ки, а другой — слухового нерва. В семье родители глухие,
а двое их детей слышат нормально. Определите генотипы
членов семьи.
12. Глухота — рецессивный признак. Молодые супруги глу-
хие. Анализ родословной показал, что у каждого из них глу-
хота наследуется по аутосомно-рецессивному типу, поэто-
му считалось, что все их дети также будут глухими. У них
родились дочь, сын и разнополая двойня. У всех детей был
нормальный слух. Объясните.
13. Охарактеризуйте генетическую систему чувствительнос-
ти к ФТК.
14. В чем состоит биологическое значение чувствительности
к горьким веществам?
15. Приведите примеры популяционных различий в перено-
симости лактозы и дайте им объяснение.
16. Почему в малярийном поясе высока частота гена, ответ-
ственного за недостаточность фермента Г6ФД?
302
Контрольные вопросы и задания к разделу 9
17. Способность ощущать запах синильной кислоты контро-
лируется доминантным геном, сцепленным с Х-хромосо-
мой. Неспособность ощущать это вещество — рецессив-
ный признак. Женщина не ощущает запах синильной кис-
лоты, ее муж ощущает запах синильной кислоты. Будут ли
чувствовать этот запах их дети?
18. 75% взрослых европейцев усваивают лактозу, 25% нет.
Способность к усвоению этого сахара контролируется
аутосомным доминантым геном. Найдите частоты аллелей
гена лактазы в популяции европейцев.
19. 7% европейцев не ощущают запах мускуса. Способность
ощущать это вещество — аутосомный доминантный при-
знак. Найдите частоты аллелей гена, ответственного за
ощущение мускуса.
20. Усвоение молочного сахара лактозы, имеющегося в све-
жем молоке, происходит при участии фермента лактазы.
Синтез фермента лактазы находится под контролем ауто-
сомного гена. Известны два аллеля этого гена. Доминант-
ный аллель активен в течение всей жизни, благодаря чему
его взрослые носители могут усваивать свежее молоко. Ре-
цессивный аллель активен только в раннем детстве, поэто-
му взрослые рецессивные гомозиготы не могут усваивать
свежее молоко. Среди шведов 97% взрослого населения
усваивают свежее молоко, среди китайцев таких людей 8%,
у народа йоруба 2%. Найдите частоты аллелей гена лакта-
зы у этих народов.
21. Для работников химической промышленности, связанных
с производством синильной кислоты, важным является
способность ощущать запах этого ядовитого вещества.
Способность ощущать синильную кислоту контролирует-
ся доминантным геном, сцепленным с Х-хромосомой. 18%
мужчин не чувствуют этого запаха. Какой процент жен-
щин не ощущает запах этого ядовитого вещества?
22. Рецессивный ген недостаточности глюкозо-6-фосфатде-
гидрогеназы (Г-6-ФДГ) локализован в Х-хромосоме. Эта
особенность генотипа проявляется при определенных ус-
303
Раздел 9. Генетический контроль восприятия
ловиях. Такими условиями является принятие некоторых
медицинских препаратов — сульфаниламида, примахина
стрептоцида, фенацитина, фенилгидразина и др. Эти пре-
параты, не вызывающие побочных эффектов у носителей
нормального гена Г-6-ФДГ, у индивидов, дефицитных по
Г-6-ФДГ, вызывают разрушение эритроцитов, что сопро-
вождается головной болью, пожелтением кожи, анемией
различной степени.
Недостаточность глюкозо-6-фосфатдегидрогеназы эрит-
роцитов наблюдается у 10% негров. Признак контролиру-
ется рецессивным, сцепленным с Х-хромосомой, геном.
Какова частота этого гена в популяции негров ?
23. В одном из районов Греции частота гена недостаточности
Г-6-ФДГ составляет 0,144. Где доля детей с недостаточнос-
тью Г-6-ФДГ будет больше — в браках между здоровыми
или между больными супругами? Как относится суммар-
ная доля больных детей от этих браков к доле всех боль-
ных в популяции?
24. Синтез фермента лактазы у человека контролируется по-
лиморфным аутосомным локусом, имеющим три аллель-
ных состояния: L, ^и 12. Аллель L обусловливает наличие
лактазы в детском возрасте и у взрослых; аллель Ц обе-
спечивает присутствие этого фермента только у детей; ал-
лель 12 не активен в отношении синтеза лактазы. Аллель L
доминирует над всеми остальными аллелями, 12 рецесси-
вен по отношению к другим аллелям этого гена. На каж-
дые 100 000 населения приходится 44 человека, которые
вообще не усваивают лактозу во взрослом состоянии, а
люди, не усваивающие молоко и в детстве, встречаются с
частотой 1:1 000 000. Определите частоты аллелей данного
гена в исследованной популяции.
Раздел 10
ГЕНЕТИЧЕСКИЕ ОСНОВЫ
ИНТЕЛЛЕКТУАЛЬНЫХ РАЗЛИЧИЙ
Под интеллектом обычно понимают мыслительную способ-
ность. Научный подход к измерению величины интеллекта че-
ловека начался в Англии во второй половине XIX в. и был свя-
зан с именем Фрэнсиса Гальтона, двоюродного брата Чарль-
за Дарвина. Он первым выполнил систематические семейные
и близнецовые исследования, изучил приемных детей, про-
слеживая поведенческие характеристики. Гальтон проанали-
зировал множество семей выдающихся людей и обнаружил,
что среди их родственников талантливых личностей больше,
чем в популяции. В родословной семьи Баха имеется большое
число крупных музыкантов, в семье Бернулли можно найти
знаменитых математиков, род Дарвина включает выдающих-
ся ученых. В то же время Бетховен, Гаусс и Франклин стоят
одиноко на высоте своей славы среди других родственников.
Гальтон разработал тест на изучение интеллекта, руковод-
ствуясь следующей упрощенной теорией. Поскольку вся инфор-
мация попадает к человеку через органы чувств, то у самых
умных людей органы чувств должны быть наиболее развиты-
ми. В его тесте были задания на восприятие, двигательную
активность и время реакции. Однако в начале XX в. обнару-
жилась несостоятельность и упрощенность такого подхода.
Отвечая на потребность времени, французский психолог
Альфред Бине в 1904 г. разработал первый удачный тест на
измерение интеллекта. Это было сделано по заказу француз-
ского правительства, которое хотело выявить способности
обучающихся для создания альтернативного образования.
Вместе со своим коллегой Теодором Симоном Бине подобрал
тест, который помог исключить из общей массы детей таких,
которые мало что получали от занятий по общей программе.
305
Раздел 10. Генетические основы интеллектуальных различий
Тест состоял из многих маленьких заданий на проверку па-
мяти и мышления. Акцент был сделан на вербальном, или язы-
ковом, развитии человека. Льюис Термен из Стенфордского
университета перевел и адаптировал тест Бине—Симона в
США. Появилось понятие IQ (intelligence quotient], или коэф-
фициента интеллекта.
По мнению разработчиков, с помощью тестов оценивают-
ся врожденные умственные способности, которые, как счи-
тается, не зависят от прежнего обучения и жизненного опы-
та. Задания для тестов подбираются с учетом возраста тести-
руемых. Чтобы исключить влияние пола в тесты не вводят
вопросы, на которые мальчики и девочки отвечают по-разно-
му, а чтобы на результаты тестирования не влияли языковые
различия, создаются невербальные тесты. При тестировании
ребенок набирает баллы, по которым устанавливают «ум-
ственный возраст». Этот «возраст» сравнивают с хронологи-
ческим возрастом ребенка и определяют степень его умствен-
ного развития — задержан он или продвинут. Эталоном для
данного возраста считается среднее количество баллов, на-
бираемых детьми соответствующей возрастной группы. Тес-
ты считаются надежными, если баллы IQ, полученные у од-
ного человека, через небольшой промежуток времени высо-
ко коррелируют друг с другом (г = 0,95). Такие тесты дают
высокую корреляцию (г — 0,80) и с ответами, полученными
через несколько лет после первого тестирования. Считается,
что общий интеллект по-разному отражается в различных его
составляющих, но сам он является устойчивой чертой инди-
вида. Корреляция между тестами и их частями рассматрива-
ется как доказательство того, что все они измеряют лежащий
в их основе общий интеллект. Различные компоненты IQ (сло-
варный запас, нахождение аналогий, распознавание образов,
вербальные и невербальные задания) также высоко коррели-
руют друг с другом.
Тест Стенфорда—Бине был опубликован в 1916 г. и оста-
вался наиболее признанным среди других тестов на интел-
лект почти полвека. Толчком к внедрению тестов IQ в США
306
10.1. Проблемы тестирования
было вступление Америки в Первую мировую войну в 1917 г.
Необходимость тестирования большого количества рекрутов
быстро привела к развитию групповых тестов для оценки вер-
бальных и невербальных способностей. Из этих разработок
был практический выход, который заключался в осознании
того, что тесты IQ полезны и для взрослых, а не только для
детей, а также достоверны, поскольку были проанализирова-
ны данные почти о 2 млн солдат. Позже Дэвид Векслер объ-
единил задания из ряда предшествующих тестов. Он был пер-
вым врачом, который увидел целесообразность тестов IQ для
исследования психических свойств человека. По его мнению,
эти тесты могли дать более ценную информацию, чем любые
психометрические приборы. Так, недостаток баллов по вер-
бальному IQ можно было объяснить нарушениями в левом
полушарии, а недостаток баллов по образному IQ — наруше-
ниями правого полушария.
Одним из наиболее известных ученых, занимавшимся ис-
следованием IQ, был Ганс Айзенк. Его исследования показа-
ли, что интеллект человека по меньшей мере на 70% обуслов-
лен наследственностью и только на 30% определяется окру-
жающей средой. Все, чего человек добивается в обучении,
профессии и всей социальной жизни, в значительной степе-
ни зависит от его наследственности.
В бывшем СССР запрет на развитие генетики и тестиро-
вание умственных способностей пришелся на 1936 г. Пример-
но в 1980 г. табу с тестов IQ в бывшем СССР попытались снять
московские ученые, когда был проведен ряд исследований на
близнецах и установлена наследуемость IQ, равная 0,78.
10.1. Проблемы тестирования
Использование тестов сопряжено с целым рядом проблем.
Корректное применение теста требует, чтобы тестирующий
был знаком с системой ценностей той группы, к которой при-
надлежит тестируемый. К примеру, тестирование австралий-
ских аборигенов с помощью бумаги и карандаша бессмыслен-
307
Раздел 10. Генетические основы интеллектуальных различий
но, если они никогда не видели этих предметов. Тестируемый
может не знать некоторых слов и выражений людей, кото-
рые составляли тесты. Результаты тестирования зависят и от
того, принадлежит ли тестирующий к той же социальной и
этнической группе, что и тестируемый. Интеллект бывает
трудно отделить от культуры и воспитания, поэтому при тес-
тировании могут возникать проблемы из-за того, что в тесте
заложены ценности его составителя. К примеру, на вопрос
«Что ты будешь делать, если сломаешь чужую вещь? » соглас-
но тесту правильным считается ответ «предложу уплатить ее
стоимость», а ответы «почувствую себя виноватым» или «при-
знаюсь, что это сделал я» — неправильными. Учитывая все
эти проблемы, психологи много внимания уделяют поискам
«первичного» или «базового» интеллекта, не зависящего от
особенностей культуры. Чтобы нейтрализовать культурные
факторы, разработчики направляют усилия на создание не-
зависимых от культуры тестов. Такие тесты не должны содер-
жать слов или понятий, известных представителям одной
культуры и неизвестных представителям другой. Следует,
однако, признать, что как нет независимого от культуры ис-
следователя, так, по-видимому, не может быть и независимых
от культуры тестов. Другой подход, с помощью которого ста-
раются сделать процедуру тестирования более объективной,
заключается в том, что сравнивают группы культурно сопос-
тавимых людей. Такими группами, к примеру, могут быть дети
разных национальностей, родители которых принадлежат
одному социальному слою и имеют сходные профессии. Од-
нако даже если семьи имеют одинаковый доход и бытовые
условия, а дети учатся в одной школе, на них все-таки не мо-
жет не оказать влияние то, что в сознании окружающих, а
также и их самих они принадлежат к разным этническим
группам. Различия в жизненном опыте, связанном с нацио-
нальной или расовой принадлежностью, также могут отра-
зиться на результатах тестирования. Надо признать, что до
сих пор еще не найдены способы, с помощью которых мож-
но было бы нейтрализовать этнические и социальные особен-
308
10.2. Генетические исследования интеллекта
ности и получить «интеллект в чистом виде», который лежит
в основе характеристик, оцениваемых с помощью тестов.
Значение интеллекта, получаемое по тестам IQ, представ-
ляет собой относительную характеристику, которая отража-
ет место индивида по этому показателю в группе людей одно-
го возраста, имеющих сходные условия жизни. Изменение
условий жизни приводит к повышению или снижению инди-
видуальных показателей IQ. При этом соотношение показа-
телей IQ у индивидов, испытавших одинаковые изменения в
условиях жизни, не меняется.
Индивиды с повышенным уровнем интеллекта обычно
проявляют его в профессиональной деятельности либо при
тестировании. В обычных условиях они могут ничем не вы-
деляться среди других людей. Нередко у известных ученых
не обнаруживались особенно высокие значения в тестах IQ.
В то же время у некоторых журналистов, актеров обнаруже-
ны на редкость высокие баллы IQ. Случается, что люди, имею-
щие высокие баллы IQ, оказываются малоприспособленны-
ми к обычной жизни, и в то же время индивиды с невысоким
значением IQ добиваются больших жизненных успехов.
В связи с этим все еще остается открытым вопрос, о том, что
именно измеряется баллами IQ. Принято считать, что интел-
лект — это то, что измеряется при тестировании.
10.2. Генетические исследования интеллекта
Изучение близнецов показало, что уровень интеллекта зави-
сит от генотипа, хотя результаты, полученные разными ис-
следователями, неоднозначны. В одних работах показано, что
конкордантность монозиготных близнецов по показателям IQ
с возрастом увеличивается, то есть роль генетического фак-
тора растет, авторы других работ утверждают, что с возрас-
том роль генотипа в формировании различий по IQ уменьша-
ется.
Существенное снижение уровня интеллекта обычно свя-
зано с генной или хромосомной мутацией. Если в отношении
309
Раздел 10. Генетические основы интеллектуальных различий
грубых нарушений умственного развития роль наследствен-
ности понятна, то до сих пор не ясно, чем вызваны отличия в
уровне интеллекта между людьми с нормальными генотипа-
ми. Не обнаружено также ни одного гена, который бы повы-
шал умственные способности.
Существует несколько теорий, объясняющих генетичес-
кие механизмы наследования интеллекта. Большинство ученых
склонны к полигенному типу наследования, согласно которому
на уровень интеллекта влияет не один, а много разных генов,
а варьирование признаков подчиняется нормальному распре-
делению. В пользу этой теории свидетельствует анализ потом-
ства от браков с разным уровнем IQ. В одном из таких иссле-
дований родители были разделены по усредненной величине
IQ на 6 групп: одаренные — с IQ 122, норма — с IQ 100 и в
разной степени отсталые: с IQ 78, с IQ 56, с IQ 34 и с IQ 12.
Главным методом, который используется для установле-
ния роли генетических факторов в определении IQ, является
исследование людей, отличающихся по степени родства. Роль
генов в определении IQ подтверждается теми фактами, что
коэффициент корреляция по IQ у монозиготных близнецов
выше (0,86), чем у дизиготных (0,60), у сибсов выше (0,47), чем
у полусибсов (0,31), а у полусибсов, в свою очередь, выше, чем
у двоюродных сибсов (0,15). Между биологическим родите-
лем и ребенком, живущими вместе, корреляция по IQ также
выше (0,42), чем между приемным родителем и ребенком
(0,19). В пользу большой генетической компоненты IQ также
свидетельствует его слабое изменение в течение жизни.
О значении внешней среды в формировании IQ у челове-
ка свидетельствует то, что IQ дизиготных близнецов корре-
лирует больше (0,60), чем IQ сибсов различного возраста
(0,47), несмотря на ту же самую степень генетического сход-
ства, что свидетельствует о влиянии общей внутриутробной
среды. Не связанные родством дети, воспитывавшиеся вмес-
те, имеют корреляцию по IQ больше (0,32), чем биологичес-
кие сибсы, воспитывавшиеся раздельно (0,24). Корреляция по
IQ между приемным родителем и ребенком, живущими вме-
310
10.2. Генетические исследования интеллекта
сте (0,19), близка к таковой между биологическим родителем
и ребенком, живущими раздельно (0,22). Кроме того, у сиб-
сов, воспитывавшихся вместе, корреляция по IQ выше (0,47),
чем у сибсов, воспитывавшихся раздельно (0,24). Аналогич-
ное заключение характерно и для случая биологического ро-
дителя и ребенка, живущих вместе (0,42) или раздельно (0,22).
В то же время существует одно обстоятельство, касающее-
ся корректности всех генетических исследований наследуе-
мости IQ на близнецах. Оценка наследуемости в большой
мере базируется на корреляции монозиготных и дизиготных
близнецов. Однако в большинстве генетических исследова-
ний такая важная составляющая, как хорионический эффект,
который ранее регистрировался при рождении лишь в Кана-
де и Франции, не учитывается.
Исследования, проведенные во Франции среди 8—12-лет-
них монозиготных близнецов, показали, что дихорионные
близнецы, составляющие 1/3 от всех монозиготных близне-
цов, различаются по весу при рождении, уровню холестери-
на в крови пуповины, личностным характеристикам во взрос-
лом возрасте и познавательным способностям. Монозиготные
близнецы обнаружили почти идентичные словарные способ-
ности вне зависимости от хорионного происхождения, одна-
ко сходные результаты в тестах по графике были зарегистри-
рованы лишь среди монохорионных монозиготных близне-
цов. Дихорионные близнецы при тестировании не показали
отличий по сравнению с дизиготными близнецами.
Если бы интеллект полностью зависел только от генотипа,
то, согласно генетической теории, коэффициент корреляции
между родственниками был бы равен показателю родства.
Некоторые типы родственников действительно демонстриру-
ют эту закономерность. Корреляция между другими типами
родственников более высокая, чем в случае, если бы интел-
лект определялся исключительно наследственными фактора-
ми. Можно назвать, по крайней мере, три причины, вслед-
ствие которых коэффициенты корреляции между родствен-
никами отклоняются от значений, которые предписывает
к
311
Раздел 10. Генетические основы интеллектуальных различий
генетическая теория. Одна из них — положительная брачная
ассортативность по уровню умственного развития. Люди
обычно вступают в брак с партнерами, примерно с таким же
уровнем интеллекта. Коэффициент корреляции между суп-
ругами по IQ довольно высок. Наследственное сходство ро-
дителей увеличивает коэффициент корреляции у потомков
по сравнению с тем его значением, которое ожидается, если
бы браки в отношении изучаемого признака были случайны-
ми. Другая причина повышенного коэффициента корреляции
между родственниками — общая семейная среда. Родствен-
ники, как правило, имеют сходное социально-экономическое
положение, одинаковый доступ к образованию, похожее вос-
питание. Все это развивает интеллект в одном направлении и
увеличивает сходство. О значительном средовом влиянии на
уровень интеллекта свидетельствуют высокие значения ко-
эффициента корреляции между детьми и их приемными ро-
дителями.
Величина коэффициента корреляции по IQ между род-
ственниками может зависеть и от особенностей методики
исследования. Интеллектуальное развитие разных людей про-
текает с неодинаковой скоростью и завершается в разном
возрасте. Скорость умственного развития может определять-
ся генетическими причинами. Обычно однократные измере-
ния IQ проводятся у родственников, когда они находятся в
разном возрасте, и это приводит к занижению коэффициен-
тов корреляции. Все эти примеры указывают на сложность
получения несмещенных коэффициентов корреляции, на
необходимость учитывать множество условий при организа-
ции корректного генетического исследования интеллекта.
В 60-х гг. XX в. в США были проведены широкие генети-
ческие исследования интеллекта. Были изучены разные эт-
нические и социальные группы людей. Результат был одно-
значный — доля генетического влияния (наследуемость) в
уровне интеллекта, оцениваемого в баллах IQ, довольно вы-
сока. Оказалось, что внутригрупповое разнообразие людей
по интеллекту на 50—80% зависит от генетических факторов
312
10.2. Генетические исследования интеллекта
и на 20—50% обусловлено негенетическими причинами —
особенностями воспитания, социального положения и др. Та-
ким образом, различия в интеллекте между индивидами оп-
ределяется как генетическими различиями между индивида-
ми, так и разнообразием условий жизни в самом широком
смысле.
Если интеллект находится под генетическим контролем,
то на его проявления должны оказывать влияние генетичес-
кие процессы, такие, как инбридинг, гибридизация, отбор.
Исследования, проведенные в различных популяциях, пока
не дали четкого ответа на вопрос о том, насколько инбридинг
и аутбридинг влияют на величину IQ. Так, в некоторых ис-
следованиях прослежено снижение 1Q у детей по сравнению
с родителями, если последние состоят в кровнородственных,
или инбредных, браках. И, наоборот, у детей от аутбредных
или отдаленных по происхождению родителей браков, напри-
мер, между европеоидами и монголоидами, IQ выше, чем у
детей от внутрирасовых браков. В других исследованиях по-
добной связи не обнаружено.
Люди, у которых по каким-то причинам нарушена целост-
ность миелиновых оболочек аксонов мозга, а следовательно,
и скорость передачи сигналов, отличаются невысокими зна-
чениями IQ. Установлена положительная связь между объе-
мом серого вещества мозга и величиной интеллекта. Длина
дендритов также положительно коррелирует с величиной IQ.
К средовым факторам, влияющим на величину IQ, отно-
сится наличие в пищевом рационе витаминов и минералов.
После приема этих веществ зафиксированы случаи возрас-
тания IQ в среднем на 4 балла, однако это касается в основ-
ном людей, которые получают их недостаточно. Люди, пита-
ющиеся полноценно, от приема дополнительных добавок в
баллах IQ не выигрывают.
Если в распоряжении нет тестов IQ, то примерную оцен-
ку умственных способностей человека дают нейрофизиоло-
гические показатели, такие, как электроэнцефалограмма и
позитрон-эмиссионная томография. Отмечено, что после по-
313
Раздел 10. Генетические основы интеллектуальных различий
314
10.3. Моделирование на животных
дачи раздражения конфигурация волн на энцефалограмме го-
раздо сложнее у людей с высоким IQ (рис. 10.1). Позитрон-
эмиссионная томография показывает, что потребление глю-
козы корой больших полушарий отрицательно коррелирует
с величиной интеллекта.
10.3. Моделирование на животных
Сложные поведенческие признаки способны изменяться при
целенаправленном воздействии. Воспитанием, обучением,
тренировкой можно изменить поведение в желаемом направ-
лении. Большинство количественных признаков, в том числе
и интеллект, сильно модифицируются средой. При этом сте-
пень модификации (норма реакции) обусловлена генетичес-
кой конституцией индивида. Особенность интеллекта как
количественного признака состоит в том, что он имеет очень
широкую норму реакции. Интеллект можно повышать тре-
нировкой, однако следует признать, что не все цели дости-
жимы. Генетические исследования на животных четко пока-
зали, как наследственность и условия жизни влияют на ум-
ственное развитие. По способности находить выход из
лабиринта лабораторные крысы делятся на умных и глупых.
Селекция на скорость выхода из лабиринта в течение несколь-
ких поколений позволила создать неперекрывающиеся линии
глупых и умных крыс: самая умная крыса из «глупой» линии
была глупее самой глупой из «умной» линии. Сам по себе факт
успешного отбора на умственные способности свидетельству-
ет о том, что они находятся под генетическим контролем. Из
крысят «умной» и «глупой» линий были сформированы три
группы. Молодняк одной группы выращивали в обычных ус-
ловиях — таких же, при которых проводили селекцию. Вто-
рая группа воспитывалась в условиях, неблагоприятных для
исследовательской и познавательной активности. Животные
третьей группы росли в информационно обогащенной среде.
В их клетках были туннели, мячи, зеркала. Эксперимент по-
казал, что если крысята обеих линий росли в бедных услови-
k 315
Раздел 10. Генетические основы интеллектуальных различий
ях, то, став взрослыми, они почти не различались по скорос-
ти решения задачи. Животные обеих линий, выращенные в
обогащенной среде, справлялись с задачей значительно луч-
ше, чем выращенные в обычных условиях, причем особых
успехов по сравнению с контролем (обычные условия) дос-
тигали животные «глупой» линии. Различия между «глупы-
ми» и «умными» животными сохранялись, но были менее вы-
ражены, чем в контроле. Этот эксперимент показал, что обед-
ненные условия не дают возможности проявиться хорошим
генетическим задаткам, а обогащенная среда улучшает пока-
затели животного с любым генотипом.
Для того чтобы нейтрализовать материнский эффект, но-
ворожденных крысят обменивали. Воспитание чужой самкой
не повлияло на скорость обучения взрослых крыс. Она по-
прежнему зависела от генетического происхождения живот-
ных и не зависела от того, какая мать — родная или приемная
их воспитывала. Чтобы исключить влияние материнского
организма на развивающийся зародыш, проводили экспери-
менты по пересадке оплодотворенной яйцеклетки в самку
другой линии животных. Оказалось, что способность к обу-
чению зависела от генотипа зиготы и не изменялась под влия-
нием организма «приемной» матери, в которую был имплан-
тирован зародыш.
10.4. Эффект Флинна
Во всем мире отмечается тенденция, которая проявляется в
том, что поновые поколения выполняют старые тесты IQ луч-
ше, чем их предшественники. Первым обнаружил данное яв-
ление Джеймс Флинн. Его исследования показали, что IQ в
популяции увеличивался за поколение примерно на 5—30 бал-
лов, т.е. примерно на 0,3 балла в год. В 1952 г. в исследовании
новобранцев-мужчин в возрасте 18 лет было показано, что
среди них 0,04% имели 1Q от 150 и выше. В 1982 г. среди такой
же категории лиц IQ выше 150 баллов имели уже 2,27%, что
составило по сравнению с прошлым периодом 57:1.
316
10.4. Эффект Флинна
Тем не менее нет подтверждений тому, что прибавки в
баллах IQ существовали на протяжении всей истории чело-
seKa. Одна из гипотез, которая объясняет прибавки IQ в пос-
леднее время, является предположение Ричарда Линна о том,
1то улучшение в питании, произошедшее в последнее время,
фивело как к увеличению роста, так и к повышению интел-
лекта человека, измеряемого в баллах IQ. Так, за последние
>000 лет средний рост британских мужчин практически ос-
тавался постоянным и составлял 172 см. Так обстояли дела
Таблица 10.1
IQ выдающихся людей (по Кокс и с поправкой Флинна)
Историческое лицо IQ (по Кокс) Q1 (с поправкой Флинна)
Гёте 210 188
Паскаль 195 173
Ньютон 190 168
Байрон 180 158
да Винчи 180 158
Гюго 180 158
Микеланджело 180 158
Спиноза 175 153
Штраусс 170 148
Вагнер 170 148
Бах 165 143
Бетховен 165 143
Дарвин 165 143
Гайдн 160 138
Мольери 160 138
Бальзак 155 133
Рембрандт 155 133
Линкольн 150 128
Наполеон 145 123
Вашингтон 140 118
Кромвель 135 113
311
Раздел 10. Генетические основы интеллектуальных различий
Таблица 10.2
Значения IQ в различных профессиональных группах (по Кокс)
Профессиональная группа IQ (по Кокс) QI (с поправкой Флинна)
Философы 180 158
Ученые 175 153
Писатели (кроме фантастов) 170 148
Религиозные лидеры 170 148
Писатели-фантасты 165 143
Революционные лидеры 165 143
Г осу дарственные деятели 165 143
Художники 160 138
Музыканты 160 138
Военные 140 118
примерно до когорты, рожденной в 1930 г. С этого времени
рост стал увеличиваться. Логично предположить, что увели-
чение роста привело к увеличению других антропометричес-
ких характеристик, в том числе и размеров головного мозга,
что, в свою очередь, привело к повышению интеллекта.
Исследовательница Кэтрин Кокс провела ретроспектив-
ное изучение 300 выдающихся людей, родившихся в период
с 1450 по 1850 гг. Используя архивные материалы учебных за-
ведений, где учились знаменитости, их биографические све-
дения, дневники, опубликованные труды и официальные до-
стижения, Кокс попыталась определить их примерные бал-
лы IQ (табл. 10.1).
Кокс распределила изученных ею людей по родам деятель-
ности и также обнаружила определенные закономерности
(табл. 10.2).
10.5. Межпопуляционные различия в IQ
' Тестирование представителей разных этнических групп про-
водились в основном в США. Эти исследования выявили ста-
318
10.5. Межпопуляционные различия в IQ
бильные различия только в двух случаях: были отмечены бо-
лее высокие средние значения IQ у евреев ашкенази по срав-
нению с нееврейским населением, среди которого они жи-
вут, и более низкий средний IQ у негритянского населения
Америки по сравнению с белым населением. Последний факт
породил многочисленные дискуссии, которые вышли за рам-
ки чисто научного обсуждения и захватили широкие слои об-
щественности.
Оказалось, что средние значения показателя IQ сильно
варьируют в зависимости от того, на каких группах белого и
черного населения проводились исследования. При этом во
всех случаях у черного населения средние показатели IQ ока-
зывались примерно на 15 единиц ниже, чем у белых того же
социального положения и образовательного уровня. Особен-
ность распределения баллов Ю состоит в том, что различия
между отдельными индивидами внутри любых этнических
групп выше, чем средние различия между группами. Эти ре-
зультаты неоднократно проверялись и сомнения не вызыва-
ют, однако при их обсуждении участники дискуссии, в зави-
симости оттого, в чем усматривались причины различий в IQ,
разделились на два лагеря. Одни считали, что различия в ве-
личине IQ между чернокожим и белым населением обуслов-
лены генетически, другие видели причину в условиях жизни.
Сторонники генетической природы различий делали упор на
то, что эволюция белой и черной рас в течение тысяч лет про-
исходила в состоянии репродуктивной изоляции, что могло
привести к случайным флуктуациям частот генов, которые
ответственны за развитие функциональных особенностей
мозга, участвующих в формировании интеллекта. Кроме того,
различные условия среды, в которых жили группы людей в
прошлом, могли направить отбор на сохранение комбинаций
генов, адаптивных в специфических условиях и определяю-
щих поведенческие признаки. Теоретическая посылка безуп-
речна, но ее невозможно проверить, так как до сих пор неиз-
вестно, какие и сколько генов формируют нормальный ин-
теллект. Другой аргумент, к которому прибегают сторонники
319
Раздел 10. Генетические основы интеллектуальных различий
генетической природы различий, вообще говоря, основан на
ошибочном понимании того, что такое наследуемость. Мно-
гочисленные исследования, проведенные на различных этни-
ческих группах, показали, что у всех народов наследуемость
показателя IQ находится в пределах 80%. На этом основании был
сделан неправильный вывод о том, что разница в IQ между
группами обусловлена генетическими причинами. Такое объяс-
нение методологически неверно, так как наследуемость — это
внутригрупповая характеристика, и какова бы ни была вели-
чина этого показателя, она ничего не говорит о причине меж-
групповых различий.
Сторонники средовой природы различий, со своей сторо-
ны, считают, что IQ американских негров ниже в силу того,
что по причине дискриминации они не имели такого же до-
ступа к образованию, как белые. Аргументом против этого до-
вода являются такие факты: в Америке дискриминации в не
меньшей степени, чем негры, подвергались представители и
других этнических групп, однако столь существенных разли-
чий в величине IQ между ними и потомками белых европей-
цев не обнаружено. Другой серьезный аргумент, который
приводят сторонники негенетических причин, касается ме-
тодики исследования. Они считают, что различия в IQ между
белыми и неграми возникают из-за того, что негры не так хо-
рошо, как белые, владеют английским языком и не совсем
точно понимают, что от них требуют тесты, составленные бе-
лыми, поэтому результат тестирования как бы предопреде-
лен заранее. Против этого довода свидетельствуют те факты,
что различия между белыми и неграми больше выражены в
тестах на выполнение действий, чем вербальных. Кроме того,
различия в IQ не устраняются, если в тестах используют спе-
цифический язык американских негров.
Худшие результаты негров пытались объяснить также и
психологическими причинами. Считалось, что испытания,
проводимые белыми инструкторами, тревожат чернокожих
детей, и это отрицательно сказывается на результатах тести-
рования. Для проверки этого предположения подбирали ин-
320
10.5. Межпопуляционные различия в IQ
структоров такой же этнической группы. При этом различия
в IQ между белыми и неграми хотя и уменьшались, но незна-
чительно.
Бесспорно, уровень умственного развития человека зави-
сит от условий жизни, а жизненный уровень белых амери-
канцев в среднем значительно выше, чем чернокожих. А в
целом средняя величина IQ как негритянских, так и белых
детей, проживающих в лучших условиях, выше, чем у детей
из бедных семей. Для того чтобы различия в условиях жизни
белого и черного населения не оказывали влияния на резуль-
таты исследования, прибегли к методу приемных детей. Были
выявлены чернокожие и белые дети, в раннем возрасте усы-
новленные белыми супругами. Контролем служили дети, вос-
питанные биологическими родителями. Исследование пока-
зало, что различия в IQ между белыми и черными детьми со-
храняются и располагаются в таком порядке. Наиболее
высокие значения Ю проявили белые дети, выросшие со свои-
ми биологическими родителями. Затем следовали белые дети
из приемных семей. Место после них заняли результаты тес-
тирования приемных черных детей с большой долей генов бе-
лых. За ними шли приемные черные дети с небольшой долей
генов белых.
Сторонники средовой гипотезы интеллектуальных разли-
чий приводят и такой факт: дети белых и черных американс-
ких солдат не выявили различий в IQ. Объяснить его можно
следующим образом. Набор в американскую армию прово-
дится по результатам тестирования. Из-за низких показате-
лей IQ отсеивается гораздо больше негров, чем белых. Дети
чернокожих американских военных просто были детьми бо-
лее интеллектуальной части негритянского населения.
Различиям между неграми и белыми в баллах IQ в свое вре-
мя придали слишком большое значение, считая, что эти бал-
лы однозначно и адекватно отражают какую-то очень важ-
ную сторону человеческой сущности. Наблюдения показали,
что, несмотря на плохое выполнение тестов и неважные ус-
пехи в школе, негритянские дети обнаруживают прекрасный
11 Введение в психогенетику
321
Раздел 10. Генетические основы интеллектуальных различий
ум в повседневных житейских ситуациях. Психологи объяс-
няют эти различия существованием двух уровней умственной
деятельности. На первом уровне информация обрабатывается
прямым способом, достаточным для большинства повседнев-
ных целей. Второй уровень — это обработка информации
способом, необходимым для абстрактного мышления. Сниже-
ние показателей IQ у негров относится ко второму, а не к пер-
вому уровню. Поэтому, интерпретируя результаты тестиро-
вания, правильнее говорить не об интеллекте вообще, а бал-
лах в определенных тестах.
Интеллект рассматривают как наиболее эффективное ус-
ловие биологической адаптации, выработанной в процессе
эволюции. Если допустить, что различия в среднем интеллек-
те американских негров и белых обусловлены генетически,
то имеет смысл охарактеризовать условия, к которым при-
шлось приспосабливаться американским неграм. Негры были
завезены в Америку как рабы 300 лет назад и более десяти
поколений они жили в неестественных для них природных и
социальных условиях. Можно допустить, что за это время
происходил отбор, отметавший наиболее развитых в умствен-
ном отношении индивидов. Высокие показатели наследуемо-
сти интеллекта указывают на то, что такой отбор, если он дей-
ствительно имел место, был эффективным. Очевидно, что сво-
бодолюбивым и интеллектуально развитым индивидам
труднее было смириться с условиями рабства. Возможно, вос-
ставая против рабовладельцев или убегая от них, они чаще
погибали, поэтому имели меньше шансов дожить до зрелого
возраста и оставить потомков.
Высокие средние значения IQ у евреев также поставили
вопрос об их причинах. Их попытались найти в истории на-
рода. В течение многих столетий евреи в европейских стра-
нах проживали в условиях тяжелой дискриминации. Им не
позволялось владеть землей, запрещались многие профессии,
разрешалось селиться только в строго определенных районах.
На рубеже XIX—XX вв. дискриминация ослабилась, и права
евреев увеличились. У них появилась возможность получать
322
10.5. Межпопуляционные различия в IQ
высшее образование, осваивать профессии, требующие вы-
соких интеллектуальных способностей. Значительно вырос
процент евреев среди врачей, адвокатов, университетских
профессоров, в несколько превысив их средний процент в на-
селении. Каким образом у этого народа накопился интеллекту-
альный потенциал, который проявил себя, как только появились
условия? Считается, что причина лежит в особенностях ев-
рейской культуры. Вся жизнь евреев была регламентирова-
на священным писанием. По всем житейским вопросам люди
обращались к раввину, и все проблемы он должен был решать
в соответствии с законами религии. Естественно, что в еврей-
ской общине ценились обладатели большого ума, способные
читать и толковать священные книги. Таким людям община
всячески содействовала: оказывала финансовую помощь, ус-
траивала выгодные браки. Прибегая к биологической терми-
нологии, можно сказать, что в еврейской среде происходил
отбор в пользу генов, обеспечивающих высокие умственные
способности. О том, что такой отбор может быть эффектив-
ным, свидетельствуют высокие значения наследуемости IQ.
Согласно статистике, в богатых семьях до репродуктивного
возраста доживало в 3—4 раза больше детей, чем в бедных.
Считалось, что тяжелые условия жизни евреев способство-
вали выживанию наиболее интеллектуально развитых инди-
видов. Возможно, что более сообразительные молодые люди
чаще избегали насильственной смерти и потому имели боль-
ше шансов передать свои гены потомкам, чем индивиды, ме-
нее развитые в умственном отношении. По-видимому, все эти
условия обеспечили в еврейской популяции накопление ге-
нов, формирующих высокие умственные способности. Эти
гены не могли проявиться в условиях дискриминации, но ког-
да дискриминация ослабилась, генетический потенциал это-
го народа не замедлил проявиться.
Объяснение приведенных фактов не претендует на окон-
чательную истинность. Правильно было бы считать, что еще
нет окончательного ответа на вопрос о причинах межгруппо-
вых различий. Хотя многочисленные работы по генетике ин-
и
323
Раздел 10. Генетические основы интеллектуальных различий
теллекта и не дали ответа на главный вопрос — в чем причи-
на межгрупповых различий, эти исследования позволили ре-
шить много других проблем. Было установлено, что при бла-
гоприятных условиях интеллект можно развить. Специальны-
ми приемами можно повысить успеваемость учащихся. Эти
выводы оказались полезными для разработки образователь-
ных программ. Ученые пришли к пониманию того, что мето-
ды обучения должны быть адекватны индивидуальным осо-
бенностям ребенка, независимо от того, детерминированы
они генетически или нет. Методика обучения должна быть
такой, чтобы максимально выявить благоприятные наслед-
ственные задатки. Если в образовании не учитывать особен-
ностей группы или индивида, оно не будет эффективным.
В настоящее время дискуссии о причинах межгрупповых
различий в IQ потеряли свою остроту. Ученые пришли к вы-
воду, что знание того, чем обусловлены межгрупповые раз-
личия в уровне интеллекта, имеет чисто академический ин-
терес и не должно влиять на цели образования. Какими бы
ни были причины различий по уровню развития интеллек-
та между представителями разных групп — средовыми или
генетическими, это никак не должно сказываться на отно-
шениях, определяемых правами человека. Даже если допу-
стить, что обнаруженная разница определяется генетичес-
кими факторами, для практических целей это не имеет ни-
какого значения, точно так же, как если бы было доказано,
что причиной всех различий в уровне интеллекта является
среда.
Контрольные вопросы и задания к разделу 10
1. Что отражают тесты IQ?
2. Приведите примеры, свидетельствующие в пользу гене-
тической природы интеллектуальных различий.
3. Приведите примеры, свидетельствующие в пользу средо-
вой обусловленности интеллектуальных различий. ч
4. Какую информацию дают исследования на животных в
324
Г"
Контрольные вопросы и задания к разделу 10
понимании наследственных основ интеллектуальных раз-
личий?
5. При помощи специальных тестов исследовали показатель
интеллектуального развития IQ. Установлено, что браки
по этому показателю у людей положительно ассортатив-
ны, коэффициент корреляции по IQ между супругами со-
ставляет г = 0,39. Коэффициент корреляции по IQ в па-
рах родитель—потомок г = 049, в парах сибсов г — 0,51.
Вычислите наследуемость показателя IQ в данной попу-
ляции.
6. По результатам измерения коэффициента интеллекта (ус-
ловные единицы) у моно- и дизиготных близнецов рассчи-
тайте наследуемость этого признака.
Монозиготные близнецы:
IQ (усл. ед.): 108—104, 98—105, 107—102, 97—94, 80—86,
115—124, 110—115, 118—120, 88—90, 101—105.
Дизиготные близнецы:
IQ (усл. ед.): 100—112, 99—104, 112—130, 89—103, 95—109,
85—100, 110—112, 94—99, 105—108, 114—120.
7. Используя данные, приведенные в таблице, обсудите зна-
чение наследственности и среды в уровне интеллекта.
Группа Коэффициент корреляции
Монозиготные выросли вместе 0,87
Дизиготные выросли раздельно 0,75
Дизиготные одного пола 0,56
Дизиготные разного пола 0,49
Братья и сестры 0,55
Не родственники 0,24
Раздел И
ГЕНЕТИКА ЛИЧНОСТНЫХ
ОСОБЕННОСТЕЙ
Характеризуя какого-либо знакомого человека, мы стараем-
ся применять по отношению к нему прилагательные, наибо-
лее полно описывающие его как личность, например, эгоис-
тичный, болтливый, добродушный, навязчивый, авторитар-
ный, озабоченный мелочами и т.д. Психологи определяют все
эти черты как личностные свойства и оперируют ими для
диагностики своих клиентов. Личностные свойства характе-
ризуют восприятие человека и его отношение к определен-
ным жизненным явлениям.
Личностные расстройства проявляются как выход опре-
деленного личностного свойства за рамки адаптивного, при
этом другие характеристики могут оставаться в пределах нор-
мы. К личностным расстройствам относятся антисоциальное
поведение, пограничные психические состояния, параноидаль-
ные, шизоидные и шизотипические состояния, избегание дру-
гих людей и зависимость от них, навязчивые состояния, не-
гативизм и другие.
11.1. Близнецовые исследования
характеристик личности
Многие ученые детально занимались генетическим анализом
личностных и когнитивных особенностей. Основным мето-
дом были близнецовый анализ. Тестирование проводили с по-
мощью опросников, среди которых Миннесотский многофак-
торный опросник (MMPI), калифорнийский личностный оп-
росник (CPI), 16-факторный опросник Кэттела и др. При
создании Миннесотского многофазного опросника его авто-
ры руководствовались клиническими представлениями о воз-
326
11.1. Близнецовые исследования характеристик личности
можных патологических изменениях личности. В связи с этим
каждая из клинических шкал MMPI дает оценку близости ис-
пытуемого к соответствующему варианту дисгарморническо-
го развития личности. Оцениваются такие показатели, как
ипохондрия, депрессия, истерия, психопатия, маскулин-
ность/фемининность, паранойяльность, психастения, шизо-
идность, гипомания, социальная интроверсия.
В своей выборке Томас Бушар, занимавшийся близнецо-
выми исследованиями, среди М3 близнецов, разлученных в
детстве, обнаружил чрезвычайно много интересных пар, силь-
но сходных между собой. Среди таких пар были «близнецы
Джимы» — Джим Льюис и Джим Спрингер, разлученные в
раннем детстве и встретившиеся 39 лет спустя. У обоих были
собаки по кличке Toy, оба служили помощниками шерифа,
водили «Шевроле», курили' одну и ту же марку сигарет и по-
строили круглую скамью вокруг дерева в своем саду.
В качестве М3 близнецов с большим внешним и поведен-
ческим сходством, но различными судьбами, можно привес-
ти братьев МакКьюсиков. Виктор МакКьюсик, отец медицин-
ской генетики, создал каталог всех известных наследствен-
ных болезней человека и в 1997 г. был награжден высшей
американской наградой за свою научную деятельность — пре-
мией Альберта Ласкера. Винсент МакКьюсик пошел по пути
юриспруденции, где также сделал блестящую карьеру.
Данные разных авторов, в модифицированном виде при-
веденные в табл. 11.1, свидетельствуют, что по большинству
характеристик монозиготные близнецы больше похожи, чем
дизиготные.
Результаты исследований с помощью шкал MMPI обычно
свидетельствуют о низкой наследуемости большинства пока-
зателей. Различные компоненты темперамента характеризу-
ются средней степенью наследуемости — 30—50%. Наиболее
высокая наследуемость отмечена для шкал доминантности,
толерантности и социабельности у мужчин, гибкости — у
женщин. Шкалы теста MMPI, характеризующие депрессию,
мнительность, асоциальность, интроверсию, имеют наследу-
Раздел 11. Генетика личностных особенностей
емость, равную 27—37%. Среди личностных черт наиболее
четкую зависимость от генотипа обнаруживает параметр эк-
страверсия—интроверсия. По показателям экстраверсии—
интроверсии, психотизма, нейротизма между женщинами и
их потомками были обнаружены корреляции, указывающие
на наличие материнского эффекта.
Таблица 11.1
Коэффициенты корреляции у близнецов
по психологическим характеристикам
Характеристика Близнецы
Монозиготные Дизиготные
Эмоциональность 0,64 0,03
Активность 0,51 0,50
Социабельность 0,61 0,04
Импульсивность 0,77 0,32
Реакция на еду 0,43 0,49
Способность успокаиваться 0,76 0,06
Е ровень активности 0,75 0,57
Страх 0,66 0,46
Настойчивость 0,68 0,59
Покладистость 0,60 0,25
Воображение 0,67 0,18
Рациональность 0,39 0,15
Экстраверсия 0,34 0,29
Нейротизм 0,37 0,23
В изменчивости содержательных характеристик личнос-
ти роль генотипа снижается и возрастает роль среды. Состав-
ляющие темперамента (эмоциональность, активность, социа-
бельность, импульсивность) формируют стиль поведения.
Более сложные по своей природе психические свойства лич-
ности контролируются генотипом менее жестко, чем свойства
темперамента. По большинству показателей коэффициент
внутрипарной корреляции по характеристикам темперамен-
та у монозиготных близнецов выше, чем у дизиготных Разлу-
328
11.1. Близнецовые исследования характеристик личности
ченные в детстве монозиготные близнецы обнаруживают
высокое сходство в мимической экспрессии, интонации, по-
ходке, однако оказываются мало похожими в интересах, со-
циальных воззрениях, способах межличностного взаимодей-
ствия, умении владеть собой в стрессовой ситуации.
На близнецах изучено много интересных феноменов, в ча-
стности, склонность к разводам. Так, исследования, проведен-
ные на основе Миннесотского регистра близнецов, показа-
ли, что на момент обследования частота разводов в Миннесо-
те составляла около 19%, а среди близнецов в целом — около
20%. Вероятность развода возрастала до 29%, если у человека
разводились родители, и до 31%, если родители супруга (суп-
руги) состояли в разводе. Вероятность расторжения брака для
ДЗ и М3 партнера близнецовой пары составляла 30% и 45%
соответственно, если их партнер был в разводе. Таким обра-
зом, риск развода для М3 близнецов был на 25% выше, чем
для ДЗ близнецов. По-видимому, в основе стабильности бра-
ка лежат какие-то наследуемые личностные черты.
Принято считать, что отношение человека к различным яв-
лениям вырабатывается в течение жизни, однако специаль-
ные исследования показали, что такие склонности и предпоч-
тения имеют генетическую компоненту. Сравнивая резуль-
таты анкетирования близнецов, ученые определили, что для
26 из 30 изученных склонностей был выявлен генетический
компонент, среди них чтение книг, занятия спортом и др.
Прямая связь между генами и личностными склонностя-
ми маловероятна, поэтому предполагается, что в их основе ле-
жат определенные высоконаследуемые личностные характе-
ристики, которые, в частности общительность, физическая
привлекательность и атлетические способности, значитель-
но генетически связаны с некоторыми из перечисленных
склонностей. Скорее всего, именно личностные свойства пре-
допределяют отношение человека к явлениям жизни. Напри-
мер, человек с врожденной хорошей координацией и физи-
чески сильный может больше преуспеть в спорте, чем менее
физически развитый. Поэтому у первого более вероятно
благоприятное отношение к спорту, чем у второго.
329
Раздел 11. Генетика личностных особенностей
11.2. Дерматоглифика и личность
В 1933 г. Ноэль Жакэн начал изучать связь между отпечатка-
ми пальцев и особенностями характера человека. Часто тако-
го для анализа берутся сохранившиеся отпечатки пальцев и
ладоней выдающихся личностей (рис. 11.1 и рис. 11.2).
Рис. 11.1. Отпечатки пальцев Владимира Маяковского
Рис. 11.2. Отпечатки ладоней Альберта Эйнштейна
330
F n ’
11.3. Молекулярно-генетические исследования личности
После Жакэна поиски связи между дерматоглификой и
личностными и когнитивными характеристиками человека
проводили многие ученые, в частности Берил Хатчинсон, ко-
торая первой провела глубокие исследования дерматоглифи-
ки в связи с психологическими проблемами. Берил Хатчин-
сон в 1967 г. высказала мнение, что личностные черты, свя-
занные с дерматоглификой, наследуемы.
11.3. Молекулярно-генетические
исследования личности
Изучение молекулярно-генетических основ темперамента и
личности направлено на поиск генов, участвующих в форми-
ровании психических характеристик. Определенные успехи
в этой области уже есть: открыты гены, которые кодируют
белки, участвующие в биохимических процессах мозга. Из-
вестны также гены, контролирующие синтез нейромедиато-
ров серотонина, дофамина, норадреналина, с помощью кото-
рых передается сигнал от одной нервной клетки к другой. Эти
гены в настоящее время рассматриваются как кандидатные
гены темперамента.
Считается, что темперамент, наследуемость которого со-
ставляет 30—60%, зависит от действия многих генов, обладаю-
щих слабым эффектом. Что это за гены, можно установить,
если известны биохимические процессы, которые влияют на
функционирование нервной системы. Психофизиологические
исследования показали, что склонность формировать теплые
межличностные отношения, отзывчивость, сентименталь-
ность зависят от норадреналиновой системы. Такие черты
темперамента, как тревожность, застенчивость, утомляе-
мость, страх перед неизвестностью связывают с серотонино-
вой системой мозга. Серотонин тормозит агрессивное пове-
дение. Этот нейромедиатор выделяется в синаптическую
щель, где воздействует на белки-рецепторы на поверхности
другого нейрона, порождая нервный импульс. Затем белком-
переносчиком серотонин переносится назад в клетку, из ко-
331
Раздел И. Генетика личностных особенностей
торой он был выделен. В настоящее время известен ген, ко-
дирующий структуру белка-переносчика серотонина. Этот
ген расположен в 17-й хромосоме. Обнаружены две аллель-
ные формы этого гена, различающиеся длиной нуклеотидных
последовательностей. Фермент моноаминоксидаза (МАО)
расщепляет серотонин. Ген, кодирующий фермент МАО (он
расположен в Х-хромосоме), рассматривается как кандидат-
ный «ген агрессивности». Нервная клетка синтезирует серо-
тонин из получаемой с пищей аминокислоты триптофана.
В этом процессе важную роль играет фермент триптофангид-
роксилаза. Поэтому другим кандидатным геном агрессивно-
го поведения стал ген триптофангидроксилазы, расположе-
ний в хромосоме 11. Известны два аллельных варианта этого
гена, более редкий вариант характеризуется точечной мута-
цией в седьмом интроне.
Импульсивность, раздражительность, склонность к нару-
шению правил связывают с дофаминовой системой. Имеют-
ся данные о том, что люди, у которых эти характеристики
сильно выражены, имеют более длинный аллель гена DRD4.
Длина гена зависит от числа нуклеотидных повторов в экзо-
не. Обычно в экзоне этого гена содержится от 2 до 10 повто-
ров, наиболее часто встречаются индивиды, у которых 4 по-
втора. Отмечены и межпопуляционные различия в частоте
этого гена. В частности, среди представителей монголоидной
расы чаще встречаются индивиды с двумя повторами.
11.4. Наследственность и профессия
Вопрос о том, чем обусловлена склонность к тому или иному
роду занятий, имеет не только академический, но и практи-
ческий интерес. Сейчас особенно актуальна возможность
определить, имеется ли генетическая предрасположенность
к тому или иному виду деятельности, а если имеется, каким
образом ее учитывать, так как современные профессии
предъявляют жесткие требования к индивидуальности работ-
ника. Известно несколько впечатляющих примеров, которые
332
11.4. Наследственность и профессия
вроде бы свидетельствуют о наследственных наклонностях к
определенной профессии. Антропологи проследили родослов-
ные сотен семейств на протяжении многих поколений и уста-
новили, что одни из них вообще не дали талантливых потомков,
в то время как в других число талантов было очень велико. Так, в
роду Баха в пяти поколениях было 16 композиторов и 29 про-
фессиональных музыкантов. Среди родственников Тициана
в восходящих линиях насчитывалось 8 художников.
О том, что успешная профессиональная деятельность за-
висит от биологических задатков, педагогам известно из по-
вседневного опыта. Что бы ни говорили теоретики по поводу
социальной обусловленности человеческой деятельности,
отбор детей в специальные школы — спортивные, художе-
ственные, музыкальные, балетные и др. ведут по их биологи-
ческим задаткам. Отбор на пригодность к профессионально-
му занятию музыкой, спортом проводился всегда и вопросов
не вызывает. Другое дело, когда речь идет о массовых про-
фессиях. Здесь дела обстоят сложнее, поскольку психичес-
кие особенности, требуемые, допустим, для работы бухгалте-
ра, учителя или медсестры, не так очевидны и не ясна их на-
следственная основа. Тем не менее и в выборе массовых
профессий биологические задатки нельзя сбрасывать со сче-
тов. Теоретически возможны разные точки зрения на то, что
является определяющим в выборе профессии. Одна из них
предполагает равную вероятность в выборе любой профес-
сии, другая — жесткую наследственную детерминацию. Бо-
лее реалистичной представляется третья точка зрения. Она
свидетельствует, что на выбор профессии человеком оказы-
вают влияние как наследственные, так и средовые факторы.
К первым относятся телосложение, особенности темперамен-
та, наличие специальных способностей, ко вторым — мате-
риальная возможность получить образование, наличие учеб-
ных заведений и др. Для каждого человека выбор одних про-
фессий более вероятен, чем других.
О наличии генетического компонента в формировании
специальных способностей свидетельствуют многочислен-
333
Раздел И. Генетика личностных особенностей
ные наблюдения. Музыкальные династии — самый яркий
пример. Однако нельзя забывать и о роли среды. Человек с
прекрасными музыкальными задатками, который появился на
свет и живет в обществе с неразвитой музыкальной культу-
рой, не сможет их проявить.
Если говорить о составляющих музыкальных способнос-
тей, то следует отметить, что важнейшей из них является фи-
зиологическая возможность различать частоту звуковых ко-
лебаний, которая связана с устройством слухового анализа-
тора. Наличие вокальных данных также дается природой, т.е.
обусловлено генетически. Как было показано в одном из ис-
следований, различия в певческих голосах — бас, баритон и
тенор у мужчин, сопрано, меццо-сопрано и альт у женщин —
находится под контролем одной пары генов. Бас у мужчин и
сопрано у женщин контролирует один и тот же генотип AtAv
а тенору у мужчин и альту у женщин — соответствует гено-
тип А^А2. Гетерозигота Д;Д2 проявляется в виде баритона у
мужчин и меццо-сопрано у женщин. Более поздние исследо-
вания показали, что различия в голосе контролируются не мо-
ногенно, а зависят от большого числа наследственных факто-
ров, однако вполне возможно, что описанный ген является
«главным геном», от которого зависит «положение» голоса в
вокальной классификации, а большое их разнообразие обус-
ловлено множеством генов с небольшим эффектом — поли-
генов, или генов модификаторов.
11.5. Антисоциальное и криминальное
поведение
Одним из наиболее изученных с генетической точки зрения
расстройств является антисоциальное и криминальное пове-
дение, в основе которого лежит такая личностная черта, как
агрессивность.
В течение многих лет ученые исследуют вопрос о том, яв-
ляется ли преступное поведение генетически обусловленным
и, следовательно, неизбежным или же на него влияют социум,
334
11.5. Антисоциальное и криминальное поведение
окружающая среда и другие внешние факторы. К настоящему
времени выявлено много факторов, предраспологающих к
криминальному поведению. Антисоциальное и криминальное
поведение является результатом действия нескольких фак-
торов.
Антрополог Чезаре Ломброзо, признанный многими отцом
криминологии, воодушевленный трудом Дарвина «Проис-
хождение человека», развил теорию, в соответствии с кото-
рой некоторые люди генетически ближе к своим более при-
митивным предкам и поэтому рождены с внутренней пред-
расположенностью к криминальному и антисоциальному
поведению.
Ломброзо считал, что в результате произошедшей мута-
ции преступник является отдельным видом Homo Delinquens,
не эволюционировавшим в «нормального» человека Ното
sapiens. Ломброзо собрал обширный фактический материал.
В его выборку входили итальянские заключенные и военнос-
лужащие. Он пришел к выводу, что физические черты пре-
ступников отличали их от обычных людей. Так, среди преступ-
ников преимущественно встречались люди с плоским лицом,
большими ушами, полными губами, крупной нижней челюс-
тью, выступающими скулами. Эти люди увлекались татуиров-
ками, жестокими играми и общались на особом, понятном им
сленге. Все указанные признаки как бы отодвигали преступ-
ников на более низкую ступень развития. Однако под описа-
ние Ломброзо подходили лишь около 1/3 преступников, дру-
гие 2/3 случаев он объяснить не мог.
Ломброзо был не единственным ученым, кто пытался свя-
зать определенные внешние черты человека со склонностью
к криминальному поведению. Гиппократ в свое время связы-
вал строение тела с темпераментом, а темперамент, в свою
очередь, с жидкостями тела. Уильям Шелдон искал корреля-
цию между соматотипом и склонностью к правонарушени-
ям. Шелдон считал, что у мезоморфов выше порог болевой
чувствительности, они более агрессивны и жестоки по срав-
нению с эктоморфами и эндоморфами. После восьмилетних
335
Раздел 11. Генетика личностных особенностей
исследований Шелдон пришел к выводу о существовании тес-
ной связи между мезоморфным строением тела и предраспо-
ложенностью к правонарушениям. Другие ученые также об-
наруживали, что среди людей с антисоциальным поведением
также больше мезоморфов, чем в остальном населении.
Более поздние исследования не обнаружили связи между
ростом, весом, физической силой и криминальным поведе-
нием. В некоторых работах, наоборот, констатировали факт,
что в среднем у преступников росто-весовые показатели ниже
этих же показателей у законопослушных граждан, и преступ-
ники позже достигают половой зрелости.
Исследователи часто прибегали к изучению родословных
семей, где было много преступников или, наоборот, таковых
не было, для доказательства генетических основ криминаль-
ного поведения. Позже, когда такие родословные стали ана-
лизировать количественно, было показано, что в криминаль-
ных семьях около 40% сыновей становились преступниками,
а в не криминальных — около 13%. Так, для доказательства
генетических основ преступности часто приводили генеало-
гическое дерево семьи Дюков из США, давшей 140 преступ-
ников на 1200 членов семьи, и семьи Эдвардсов. В роду Дж.
Эдварса — широко известного проповедника — не было пре-
ступников. В этом роду был президент Соединенных Штатов,
а также высшие судейские чиновники, писатели и религиоз-
ные деятели. Эти данные должны были продемонстрировать
генетическую предрасположенность к преступной деятель-
ности. Однако позже выяснилось, что среди предков Эдвар-
са были люди, осужденные за преступления. Постепенно ста-
ло понятно, что простое сопоставление генеалогий мало дает
для понимания того, как зависит криминальное поведение от
наследственности, поскольку трудно развести наследствен-
ное влияние и влияние среды.
Близнецовые исследования, проведенные в первой поло-
вине XX в., формально вроде бы выявили значительную ге-
нетическую предрасположенность к криминальному поведе-
нию. Так, в среднем у М3 близнецов конкордантность по пре-
336
11.5. Антисоциальное и криминальное поведение
ступности была 0,75, в то время как у ДЗ близнецов — лишь
0,24. Однако в ранних исследованиях численность выборок
была не очень велика. Кроме того, в то время зиготность близ-
нецов устанавливалась лишь на основе внешнего сходства.
В более поздних исследованиях, когда зиготность устанавли-
вали с большой вероятностью, также была получена высокая
генетическая предрасположенность к преступному поведе-
нию, хотя и ниже, чем в предыдущих исследованиях. У М3
близнецов конкордантность составляла около 0,48, а у ДЗ
близнецов — 0,20.
Было ясно, что не только генетические, но и средовые фак-
торы играют роль в склонности к криминальному поведению.
Кроме того, близнецовые исследования также имели некото-
рые недостатки, например, часто оба близнеца росли в оди-
наковой среде. И, наоборот, бывало, что близнецы настолько
не хотели походить друг на друга, что намеренно вели себя
отличным от своего партнера образом.
Исследования приемных детей и их приемных и биологи-
ческих родителей дали результаты, приведенные в таблице
11.1, которые свидетельствуют о том, что на формирование
преступного поведения оказывают влияние как среда, так и
наследственность.
Таблица 11.1
Процент преступников среди мужчин, выросших
в приемных семьях
Биологические родители Усыновители % преступников среди усыновленных мужчин
Преступники Законопослушные 20
Законопослушные Законопослушные 14
Преступники Преступники 25
Законопослушные Преступники 15
Одним из наиболее детальных исследователей генетичес-
ких основ криминального поведения был Ганс Айзенк. Он
предложил психологическую теорию, объяснявшую крими-
337
Раздел 11. Генетика личностных особенностей
нальное поведение. Айзенк считал, что криминальное пове-
дение является продуктом как биологических, так и соци-
альных факторов.
Ученый выделял три основных элемента личности — эк-
страверсию, психотизм и нейротизм, каждый из которых из-
мерялся и выражался в баллах от низкого до выского значе-
ния. Айзенк предполагал, что дети вырабатывают в себе уме-
ние управлять антисоциальным поведением сознательно,
посредством формирования условных рефлексов. Он считал,
что скорость, с которой условный рефлекс вырабатывается,
зависит от личностных свойств человека, в частности, от эк-
страверсии и нейротизма. Согласно его теории, у людей с
высокими значениями экстраверсии и нейротизма условные
рефлексы вырабатываются плохо. Таким людям недостает
умения управлять собой, следовательно., среди преступников
людей с такими психологическими характеристиками будет
больше всего. И, наоборот, люди с низкими значениями экст-
раверсии и нейротизма эффективно контролируют себя и с
малой вероятностью могут попасть в группу преступников.
Нейротизм отражает врожденную биологическую предрас-
положенность физиологически реагировать на стрессовые
ситуации. У нейротиков симпатическая система активирует-
ся слишком быстро, парасимпатическая — слишком медлен-
но для того, чтобы оказать противоположный тормозящий
эффект.
По расчетам Айзенка, в популяции 16% людей являются
типичными экстравертами, 16% — типичными интровертами
и 68% занимают промежуточное положение. Считается, что
у людей с высокими значениями экстраверсии кора больших
полушарий находится в состоянии недостаточного возбужде-
ния из-за врожденных особенностей функционирования ре-
тикулярной системы. Поэтому такие люди постоянно нахо-
дятся в состоянии поиска стимуляции коры. Таким образом,
экстраверты являются импульсивными людьми, которым тре-
буется дополнительное возбуждение для поддержки баланса
в коре головного мозга. У интровертов, наоборот, кора нахо-
338
11.5. Антисоциальное и криминальное поведение
дится в состоянии сверхвозбуждения под влиянием ретику-
лярной системы, поэтому они избегают стимуляции и возбуж-
дения для поддержания баланса и являются спокойными,
сдержанными людьми.
У людей с высокими значениями нейротизма вегетатив-
ная нервная система работает нестабильно. Таким людям
свойственны перепады настроения и тревожное состояние.
В противоположность им, у людей с низкими значениями ней-
ротизма вегетативная нервная система функционирует ста-
бильно, они спокойны и уравновешены. Считается, что вы-
сокие значения нейротизма нарушают образование условных
рефлексов из-за высокого проявления тревожности. Впослед-
ствии теория Айзенка была подвергнута критике со стороны
ряда ученых.
Генетики отдают себе отчет в том, что преступность не
может быть предметом генетического анализа. Отнесение
человека к категории преступника определяется законодатель-
ством и зависит от решения суда. Изменение закона может ис-
ключить человека из когорты преступников. Ясно, что в такой
ситуации генетический анализ невозможен, преступность —
предмет не генетического, а социологического анализа.
Идея о врожденной преступной программе человека ушла
в прошлое. В настоящее время уже не говорят о «генах пре-
ступности» и «преступной наследственной программе». В ге-
нах записана информация о структуре органических моле-
кул, взаимодействие которых обеспечивает рост и развитие
организма, формирует его нервную систему, особенности
психики, а следовательно, способ реагирования на окружаю-
щую среду. На всех этапах развития признаки испытывают
модифицирующее влияние среды. Чем дальше в своем раз-
витии признак отстоит от гена, тем сильнее он подвержен
влиянию среды.
При анализе преступного поведения может быть прило-
жима концепция подверженности и порога реагирования.
В контексте криминологической генетики можно говорить об
устойчивости и чувствительности к отдельным криминоген-
к
339
Раздел И. Генетика личностных особенностей
ным факторам, сопротивлению индивида влиянию кримино-
генной среды. Генетический анализ может быть применен
только к каким-то первичным, природным свойствам инди-
вида, ассоциация которых с антисоциальным поведением ста-
тистически доказана, например агрессивности.
11.6. Генетические основы агрессивности
Психологи выделяют разные типы агрессивности. С помощью
специальных тестов уровень агрессивности человека можно
выразить количественно в баллах. По выраженности агрес-
сивности люди различаются весьма сильно. Генетический
анализ, основанный на вычислении коэффициентов корре-
ляции между родственниками, а также близнецовый анализ
позволили сделать оценку наследуемости агрессивности. Ус-
тановлено, что наследуемость агрессивного поведения в сред-
нем составляет 40—54%. Эти данные свидетельствуют о том,
что различия между людьми по агрессивности примерно на-
половину обусловлены их наследственными особенностями
и наполовину средовыми причинами.
Соотношение наследственности и среды для разных ви-
дов агрессивности неодинаково. Так, наследуемость прямой
физической агрессивности (по отношению к людям) равна
0,47, непрямой физической агрессивности (по отношению к
предметам) — 0,40 и вербальной агрессивности (оскорбления,
угрозы) — 0,28.
Между видами агрессивного поведения имеются значи-
тельные корреляции (0,31—0,51). Это указывает на то, что
различные проявления агрессивности связаны с действием
общих генов. Анализ показал, что имеется общий генетичес-
кий контроль различных видов агрессивного поведения и не-
которых черт темперамента, таких, как эмоциональность и
импульсивность. Был выведен генеральный фактор агрессив-
ности, наследуемость которого составила 48%.
На генном уровне обнаружена связь между увеличением
агрессивности и некодирующими участками гена триптофан
340
11.6. Генетические основы агрессивности
гидроксилазы (ТРИ). Данный фермент участвует в синтезе
серотонина (5-НТ), который отвечает за проявление и регу-
ляцию импульсивной агрессивности. Ученые также связыва-
ют с агрессивностью нейромедиаторы норадреналин и дофа-
мин. Однако данные по всем этим веществам пока противо-
речивы и недостаточны. Отмечена роль тестростерона в
развитии агрессивного поведения, хотя результаты более оп-
ределенны у животных, а не у человека.
Соотношение наследственности и среды в проявлении аг-
рессивного поведения не остается неизменным в течение
жизни. У детей с возрастом влияние генетических факторов
на уровень агрессивности повышается, а роль факторов об-
щей среды снижается. Агрессивному поведению детей обыч-
но предшествуют высокие оценки эмоциональности и актив-
ности в более раннем возрасте, причем между агрессивнос-
тью и эмоциональностью имеются генетические корреляции
Изучению физической агрессивности отводится доволь-
но много места, поскольку именно данный вид агрессивнос-
ти особенно опасен для окружающих. Наиболее часто физи-
ческая агрессивность проявляется в 2—3-летнем возрасте.
Это можно объяснить тем, что примерно с конца первого до
конца второго годов жизни дети овладевают основными мо-
торными навыками — они могут ходить, бегать, захватывать,
толкать и бросать предметы. Дети этого возраста все в боль-
шей степени познают окружающее их пространство и дру-
гих людей. Большинство их открытий эмоционально положи-
тельные. Однако все чаще они сталкиваются с определенны-
ми конфликтами, большинство из которых касается желания
владеть либо предметом, либо территорией. Важным обстоя-
тельством является то, что в этом возрасте лингвистические
способности развиты хуже, чем моторные, поэтому ребенку
легче забрать понравившуюся игрушку, нежели попросить ее.
Подобные ситуации «учат» ребенка агрессивности и приме-
нению физической силы. В то же время происходит ответная
реакция — акт агрессивности со стороны другого ребенка —
владельца предмета. У большинства детей через некоторое
341
Раздел И. Генетика личностных особенностей
время формируется условный рефлекс, состоящий в понима-
нии того, что за их агрессивным поступком последует агрес-
сивный ответный поступок. К тому же взрослые также не
поощряют агрессивного поведения детей. Со временем у ре-
бенка вырабатывается умение ждать, пока игрушка не осво-
бодится, или же просить ее. В последующие годы дети все
больше прибегают к формам поведения, альтернативным
формам агрессивного поведения. Однако примерно у 4% де-
тей высокий уровень физической агрессивности прослежи-
вается вплоть до позднего юношеского возраста, а у некото-
рых и дальше. У таких детей наблюдается состояние, извест-
ное как хроническая физическая агрессивность. Одни входят
в группу риска по причинению физического вреда другим
людям и себе. Среди них повышена вероятность сопутствую-
щих умственных расстройств, ухода из школы, наркомании,
депрессии, безработицы, оскорбления членов семьи и буду-
щих супругов и детей, а также самоубийства. Кроме того,
вследствие нездорового образа жизни у них повышена веро-
ятность развития сердечно-сосудистых расстройств и рас-
стройств нервной системы.
Ученые обнаружили ряд факторов, предрасполагающих к
хронической физической агрессивности, среди которых име-
ются как генетические, так и средовые. Существуют, по мень-
шей мере, три группы таких факторов: индивидуальные осо-
бенности, физическое окружение и социальное окружение.
К индивидуальным особенностям относятся как родовые
осложнения, так и генетически наследуемые признаки — уро-
вень тестостерона и серотонина, масса тела, пол, раса, нацио-
нальность, темперамент, гиперактивность, тревожность,
склонность к противоречивости, импульсивность и наруше-
ние когнитивного развития. Известны исследования по гене-
тике физической агрессивности у детей, проведенные на
близнецах и приемных детях, в которых отмечается наличие
наследственной компоненты физической агрессивности. Ис-
следования в области молекулярной генетики позволили про-
следить связь определенных генов с физическим агрессив-
342
Контрольные вопросы и задания к разделу 11
ным поведением. К ним относятся гены, ответственные за
синтез ферментов метаболизма нейромедиаторов (МАО А) и
рецепторов серотонина.
К физическим факторам риска хронической физической
агрессивности относится большинство вредных веществ, на-
рушающих развитие нервной системы. Например, курение
и употребление алкоголя матерью во время беременности по-
вышает риск когнитивных и поведенческих проблем. Среди
вредных веществ постнатального действия наиболее опасным
из изученных является свинец, который вызывает ряд нару-
шений, в том числе может повышать риск развития хрони-
ческой физической агрессивности.
И, наконец, к социальным фактором риска относятся на-
рушения в стиле поведения и отношении к ребенку со сторо-
ны родителей, сибсов, других членов семьи, друзей, учителей
и соседей.
Контрольные вопросы и задания к разделу 11
1. Почему наследственные основы поведения человека удоб-
нее всего изучать на близнецах?
2. Приведите примеры того, что на выбор профессии могут
оказать влияние наследственные особенности человека.
3. Считается, что для успешного занятия спортом требуется
специальная одаренность. Среди спортсменов обследова-
ны 92 монозиготные пары, среди которых в 61 паре оба
партнера были спортсменами. Среди 227 дизиготных пар
оба были спортсменами в 59 парах. Обсудите эти данные.
4. В таблице приведены данные (число пар), отражающие
мнение родителей о характере близнецов. Проанализи-
руйте и обсудите их.
Тип близнецов Очень похожи Похожи как обычные сибсы Непохожи
Монозиготные Дизиготные 50 4 34 49 16 47
343
Раздел И. Генетика личностных особенностей
5. Как вы относитесь к показателю «наследуемости» преступ-
ного поведения, полученного близнецовым методом?
6. Может ли криминальное поведение быть предметом гене-
тического анализа?
7. Какие факты свидетельствует о том, что различия в агрес-
сивности имеют наследственную основу?
8. В исследованиях Джоанн Ланге (1929 г.) было установле-
но, что среди М3 близнецов конкордантность по преступ-
ности составляла 77%, асреди ДЗ близнецов лишь 12% были
конкордантны. Какова наследуемость криминального по-
ведения по результатам данного исследования?
9. Обсудите такие данные. Среди заключенных обнаруже-
но 27 мужчин, имевших монозиготного близнеца, и 17,
имевших дизиготного близнеца. 18 монозиготных и 3 ди-
зиготных брата также были преступниками.
Раздел 12
ГЕНЕТИЧЕСКИЕ ОСНОВЫ
АНОМАЛИЙ ПОВЕДЕНИЯ
Поведение характеризуется множеством особенностей, по-
этому разделить людей на четко различающиеся поведенчес-
кие классы затруднительно. Несмотря на то что варьирова-
ние поведения обычно имеет непрерывный характер, как
правило, можно указать индивидов, чьи поступки резко вы-
деляют их среди большинства людей. Поведение, резко от-
личающееся от ожидаемого, называется девиантным (откло-
няющимся). Понятие девиации имеет культурные корни. Бы-
вает так, что один и тот же поступок в одном обществе
считается нормой, в другом — преступлением.
Главная проблема состоит в том, чтобы решить, что явля-
ется причиной того или иного вида девиантного поведения —
патологическая наследственность или условия жизни. Реше-
ние этого вопроса имеет не только академический интерес,
но и практическое значение. Нередко человека, привлечен-
ного к уголовной ответственности, подвергают психиатричес-
кой экспертизе, для того чтобы решить вопрос, куда его на-
правлять — в клинику или в суд. Так, выявление лишней хро-
мосомы у человека, совершившего тяжкое преступление,
может коренным образом изменить его дальнейшую судьбу.
Понимание причин девиантного поведения важно для его про-
филактики. Показателен такой пример. Когда алкоголизм не
считали болезнью, борьбу с этим явлением вели исключитель-
но социальными мерами, например, введением «сухого зако-
на». Алкоголиков воспитывали, штрафовали, но не лечили. То
же и в отношении лиц с нетрадиционной сексуальной ориен-
тацией; на гомосексуалистов смотрели как на извращенцев,
социально опасных личностей, место которым в тюрьмах и
концентрационных лагерях. В древние времена у психичес-
Ъ 345
Раздел 12. Генетические основы аномалий поведения
Рис. 12.1. Трепанированные черепа с целью
изгнания злых духов
ких больных проводили трепанацию черепа для изгнания
злых духов из головы больного (рис. 12.1). В Европе в средние
века психически больных считали одержимыми дьяволом и
держали на цепи. В настоящее время отношение к перечис-
ленным видам отклоняющегося поведения становится более
гуманными, и во многом эти сдвиги обязаны генетике, кото-
рая выявила наследственную основу различных вариантов че-
ловеческого поведения.
12.1. Генетическая природа психических
болезней
Разновидностью отклоняющегося поведения, являются пси-
хические заболевания. Представление о том, что люди с пси-
хическими аномалиями являются больными, сформировалось
примерно двести лет назад, а до этого их рассматривали как
невменяемых. Признание сумасшедших больными было боль-
шим прогрессом, однако причины повреждения ума видели
во внешних обстоятельствах. Только в последние десятиле-
тия сложилось понимание того, что у психических отклоне-
ний есть генетическая основа.
12.1. Генетическая природа психических болезней
Исследования, выполненные разными методами, показа-
ли, что наследственная предрасположенность к психическим
болезням может быть различной: контролироваться одним
локусом (быть моногенной), небольшим числом локусов (оли-
гогенный контроль) или множеством локусом [полигенный
контроль). В последнем случае наследственная основа пред-
ставляется множеством локусов с аддитивным (суммирую-
щим) эффектом аллелей. Каждый из этих генов в отдельнос-
ти может не проявлять самостоятельного патологического
действия, но при их накоплении в определенной комбинации
в генотипах они приводят к развитию болезни. Существует
также модель «главного гена», согласно которой развитие бо-
лезни определяется действием гена с сильным эффектом, а
ее выраженность — множеством генов со слабым эффектом
(генов-модификаторов, или полигенов). С развитием молеку-
лярной генетики стала обнаруживаться сложная молекуляр-
но-генетическая природа психических заболеваний. В настоя-
щее время психические болезни относят к мультифактори-
альным, или болезням с наследственным предрасположением.
Если при моногенных и хромосомных болезнях людей
можно легко разделить на больных и здоровых, то подобная
классификация мультифакториальных болезней затрудни-
тельна, так как в пределах одной и той же формы заболевания
имеются непрерывные переходы. Для мультифакториальных
болезней характерен широкий клинический полиморфизм —
разнообразие в проявлении заболевания у разных лиц. Одно
и то же заболевание может быть вызвано различными гене-
тическими причинами. Это явление носит название генети-
ческая гетерогенность.
Распространенность мультфакториальных заболеваний в
популяции составляет несколько процентов, что значитель-
но выше, чем распространенность болезней моногенной или
хромосомной природы. Для мультифакториальных болезней
характерно семейное накопление — частота болезни у род-
ственников выше, чем в среднем в популяции. В родословных
с психическими болезнями повышена также частота индиви-
347
Раздел 12. Генетические основы аномалий поведения
дов с пограничными психическими состояниями и другими
отклонениями в поведении. Болезни с наследственным пред-
расположением неодинаково проявляются у мужчин и жен-
щин. Фактором повышенного риска является принадлеж-
ность человека к часто поражаемому полу и наличие больных
родственников редко поражаемого пола. Риск заболевания
тем выше, чем ближе генетическое родство с пораженным
родственником и чем выше процент больных. Проявление за-
болевания зависит от возраста, в котором появились его пер-
вые признаки. Обычно оно протекает тяжелее при раннем
начале и имеет стертые формы, если начинается в старшем
возрасте.
Для того чтобы выяснить, какие гены участвуют в форми-
ровании мультифакториальных (в том числе и психических)
заболеваний, используют биохимические, молекулярно-гене-
тические методы и данные генеалогического анализа. Они
позволяют определить, какие гены формируют подвержен-
ность заболеванию, какие биохимические реакции протека-
ют под их контролем, как биохимические продукты оказыва-
ют влияние на функции организма, как это проявляется в
поведении. Эта методология построена на концепции «кан-
дидатных генов». Кандидатными называют гены, которые
контролируют ферменты, участвующие в формировании
структур организма, нарушение которых приводит к заболе-
ванию. Используется также подход, получивший название
«геномный поиск». В этом случае сравнивают наличие вари-
антов ДНК в группе больных и группе здоровых людей.
Одними из средовых факторов риска развития психичес-
ких и нервных болезней являются вирусы. При исследовании
спинномозговой жидкости психиатрических и неврологичес-
ких пациентов на содержание вирусных антител и интерфе-
рона были обнаружены комплемент-зависимые нейтрализую-
щие антитела к вирусу герпеса (HSV1) у 52% пациентов со
старческим слабоумием (52%) ,51% больных энцефалитом, 44%
больных алкоголизмом, у 37% пациентов с повреждениями
периферических нервов, при множественном склерозе (42%),
348
| 12.2. Гипотеза о происхождении психических болезней
менингите (35%), шизофрении (32%), умственной отсталости
(11%). Интерферон в спинномозговой жидкости был найден
у 40% психиатрических и 35% неврологических больных.
В последующих главах будут описаны генетические при-
чины некоторых аномалий поведения. Эти аномалии поведе-
ния входят в группы психических болезней, классифицируе-
мых на основе определителей DSM (Diagnostic and Statistical
Manual of Mental Disorders), которые постоянно обновляются.
12.2. Гипотеза о происхождении
психических болезней
Генетиков всегда интересовал вопрос, почему, несмотря на
пониженную генетическую приспособленность психических
больных, психозы так широко распространены во всех чело-
веческих популяциях. Для объяснения высокой частоты пси-
хических болезней в популяциях была выдвинута эволюци-
онно-генетическая гипотеза. Согласно этой гипотезе психи-
ческие болезни представляют собой животное наследие
человека, а их высокая распространенность объяснялась тем,
что гены, их формирующие, в невысоких дозах, по-видимо-
му, полезны и благодаря этому сохраняются в популяции. Из-
вестно, что у низших животных поведение состоит из гене-
тически запрограммированных реакций, а поведение высших
позвоночных, особенно у млекопитающих, в мёныпей степе-
ни определяется генетической программой, а является резуль-
татом научения. Считается, что эволюция поведения идет не
путем полного исчезновения старых реакций и замены их
новыми, а путем перевода первых в скрытое состояние. В ре-
зультате гибридизации, селекции, а возможно и стрессовых
факторов, могут создаться условия для проявления филоге-
нетически древних реакций. Если по каким-то причинам рез-
ко снижается порог реагирования, то реакции могут возни-
кать не только в ответ на специфические раздражители, но и
на нейтральные. В таких случаях они становятся неадекват-
ными ситуации и приводят к аномалиям поведения.
Ь 349
Раздел 12. Генетические основы аномалий поведения
У животных описаны три типа нервных реакций, эволю-
ционно сформировавшихся как защитные: эпилептиформ-
ный, кататонический и аффективный, а у человека известны
три группы психозов: эпилепсия, шизофрения и аффектив-
ные психозы. Сходство этих состояний у человека и живот-
ных может свидетельствовать о том, что психические болез-
ни человека эволюционно произошли от защитных реакций
животных: эпилепсия от эпилептиформной реакции, аффек-
тивные психозы от аффективной реакции, шизофрения от ка-
татонической реакции. Из-за того что у человека резко сни-
зился порог реагирования, эти реакции утратили свою адап-
тивную роль и стали патогенными. Таким образом, в основе
шизофрении, эпилепсии и маниакально-депрессивного пси-
хоза лежат биологически древние защитные реакции.
Наиболее архаичной защитой у животных является судо-
рожная реакция. Она встречается у животных, стоящих на
различных ступенях эволюционной лестницы: рыб, амфибий,
птиц, млекопитающих, что и указывает на ее адаптивный ха-
рактер. Считается, что эпилептический припадок — это чрез-
мерное выражение свойства нервной системы готовности к
бурным защитным двигательным реакциям борьбы или бег-
ства. Его значение заключается в разрядке опасных для
нервной ткани очагов высокого напряжения и в освобожде-
нии мозга от токсинов. Генетические исследования на живот-
ных показали, что предрасположенность к судорожным при-
падкам имеет полигенный характер. Сохранение в популяции
животных особей с очень низким порогом судорожной реак-
ции позволяет предположить, что «эпилептические» гены свя-
заны с определенными биологическими преимуществами. Об
этом говорит более высокая подвижность нервных процес-
сов, в частности, более высокая скорость выработки услов-
ных рефлексов у крыс линии Крушинского—Молодкиной,
специально отселектированной на предрасположенность к
эпилепсии. Крысы этой линии лучше решают предлагаемые
им задачи, но имеют более низкий порог электросудорог, ко-
торые возникают у них при действии звука.
350
12.2. Гипотеза о происхождении психических болезней
Шизофрения рассматривается как проявление чрезмер-
но низкого порога кататонической реакции. Кататонические
реакции (явление, известное как животный гипноз) относят
к нормальным адаптивным реакциям. Хищники, как прави-
ло, обращают внимание на движущиеся предметы, а засты-
вание (кататония) делает жертву незаметной. В этом состоит
адаптивное значение кататонических реакций. Чрезмерная
выраженность таких реакций делает их патологическими.
Кататонические позы нередко встречаются у крыс низших
социальных рангов, биологически плохо приспособленных.
Вывести линию крыс с предрасположенностью к кататони-
ческой реакции не удалось из-за их низкой способности к раз-
множению. О параллели между шизофренией и кататоничес-
кой реакцией свидетельствует сходство в двигательной актив-
ности. Типичным проявлением «шизофренического облика»
является манерность и вычурность поведения. Причина это-
го — резко выраженные кататонические нарушения мотори-
ки, сочетание «застывания» и повышенной двигательной ак-
тивности (гиперкинезов). Из всех психозов кататония наибо-
лее характерна именно для шизофрении. У большинства
больных шизофренией при всех ее формах наблюдаются мик-
рокататонические симптомы — это пониженная чувствитель-
ность вестибулярного аппарата, мышечная заторможенность,
угловатость движений, стереотипная деятельность.
Аффективные психозы возникают у особенно эмоцио-
нальных лиц, поэтому их рассматривают как проявление па-
тологически низкого порога эмоциональных реакций. Ярко
выраженная суточная периодичность маниакально-депрес-
сивного психоза указывают на то, что аффективные психозы
являются реакцией мозга на биологические ритмы организ-
ма у лиц с патологически сниженным порогом эмоциональ-
ных реакций
Психоз рассматривают как «плату» за сохранение в попу-
ляции определенных генов, которые в других комбинациях
дают их носителям какие-то биологические преимущества.
Какие же преимущества могут давать людям умеренные дозы
351
Раздел 12. Генетические основы аномалий поведения
генов, ответственных за психические болезни? При попыт-
ках в эксперименте найти ответ на этот вопрос были обнару-
жены интересные факты. Получена информация о том, что у
больных шизофренией женщин высок процент музыкально
и художественно одаренных детей. У пилотов высокого клас-
са, обладающих очень быстрой реакцией, часто наблюдают-
ся, изменения в электроэнцефалограмме, соответствующие
эпилептиформному типу. В экспериментах на животных по-
казано, что индивидуальные характеристики наиболее ярко
проявляются при повышенной возбудимости. Известно не-
мало примеров того, что индивиды, одаренные специальны-
ми творческими способностями, не только сами имеют пси-
хические отклонения, но также и повышенный процент род-
ственников с психическими аномалиями.
12.3. Дерматоглифические маркеры нервных
и психических болезней человека
В качестве примера связи особенностей дерматоглифики с
нервной болезнью можно привести синдром Туретта — не-
врологическое заболевание, характеризующегося множе-
ственными моторными и вокальными тиками. Дерматоглифи-
ческий комплекс при синдроме Туретта является редким ва-
риантом нормальной дерматоглифики. Возможно, что такая
комбинация очень сложного (завиток на большом пальце) и
очень простых (дуги и петли с низким гребневым счетом на
указательном и среднем пальцах) дерматоглифических
признаков характеризует тип ЦНС, который, в силу ограни-
ченности компенсаторных возможностей, оказывается не-
способным нормально функционировать в условиях воздей-
ствия различных повреждающих факторов, что в данном слу-
чае оборачивается развитием гиперкинетических состояний.
Тонкая взаимосвязь топографии и характера кожных узо-
ров со структурами мозга неизвестна. Однако можно пред-
полагать, что узорность на тенаре, в отличие от пальцевых
узоров, характеризует какие-то эволюционно более древние
352
12.3. Дерматоглифические маркеры нервных и психических болезней
структуры мозга. Доказательства этому имеются в антропо-
логии. Известно, что узор на тенарной подушечке является
специфическим расовым признаком. Узорность в этой обла-
сти достаточно сходна у европеоидов и монголоидов, обладаю-
щих минимальными значениями признака, в то время как она
максимально выражена у коренных жителей Америки и Авт-
ралии. Значительная частота этого признака характеризует
негроидов, которые являются наиболее древней и специали-
зированной расовой ветвью человечества. С другой стороны,
повышенная узорность тенара — признак, характерный для
приматов. Отсюда есть все основания считать справедливым
предположение об эволюционной древности узора в области
тенара. Опираясь на эти представления, можно говорить о
схожести клинических проявлений синдрома Туретта с ост-
рым транзиторным психозом, возникающим после испуга,
характеризующимся двигательным возбуждением и вырази-
тельными движениями, имитирующими поведение окружа-
ющих, наблюдающийся у жителей островов Малайского ар-
хипелага.
В связи с этим у человека предполагается наличие двух раз-
личных, самостоятельных механизмов речи, причем, наряду
с эволюционно более молодым, имеется дополнительный, эво-
люционно более древний, ответственный за примитивные,
эмоционально окрашенные звуки. Именно этот механизм, в
норме не функционирующий, считают вовлеченным в работу при
синдроме Туретта. С узорностью в области тенара коррелирует
увеличение высокочастотной а- и ^-активности мозга. В срав-
нении с этим, преобладание на пальцах рук завитковых узо-
ров (наиболее сложный тип пальцевой дерматоглифики) кор-
релирует с возрастанием активности только в границах вы-
сокочастотного ос-диапазона. Таким образом, узорность в
области тенара отражает такую организацию ЦНС, которая
характеризуется очень сильным и едва ли не постоянным
возбуждением высших отделов мозга. По-видимому, это хро-
ническое возбуждение вносит весьма значительный вклад в
легкость невротических срывов у детей с болезнью Туретта.
12 Введение в психогенетику
353
Раздел 12. Генетические основы аномалий поведения
В распределении узорности тенара отмечены существенные
половые различия. У девочек с болезнью Туретта узор в этой
области чаще всего имеется на обеих ладонях, тогда как у
мальчиков — чаще только на левой. В соотношении полов при
болезни Туретта также имеются очень выраженные разли-
чия с преобладанием пораженных лиц мужского пола.
В качестве еще одного примера можно привести и заболе-
вание хромосомной природы, а именно — синдром 5р- (синд-
ром «кошачьего крика»), дерматоглифический тип при кото-
ром достаточно близок к представленным выше, хотя имеют-
ся и серьезные отличия, соответствующие большей тяжести
заболевания. Специфической особенностью этого заболевания,
кроме задержки роста и микроцефалии, является необычный
крик у детей раннего возраста, напоминающий мяуканье котен-
ка. Интересно, что в большинстве исследований никаких мор-
фологических изменений гортани не обнаружено и причиной
необычного крика считают поражение центральной нервной
Рис. 12.2. Отпечатки левой руки
шизофреника-правши
системы. Последнее обстоя-
тельство заслуживает осо-
бенного внимания, посколь-
ку вокализмы при синдроме
Туретта и специфический
крик при синдроме 5р- мо-
гут иметь общий патогенез,
связанный с дисфункцией
определенной области моз-
га.
В одном исследовании
показано, что у больных
шизофренией ладонные
линии нередко отличаются
от ладонных линий здоро-
вых людей (рис. 12.2). При
изучении М3 близнецов,
дискордантных по шизоф-
рении, также были отмече-
354
12.4. Социальное значение пограничных ДсиХйЧескиХ состояний
ны различия в дерматоглифике. Таким образом, данное забо-
левание могло быть следствием неблагоприятных внутриут-
робных факторов заболевшего близнеца.
12.4. Социальное значение пограничных
психических состояний
Психические особенности людей представляют неодинако-
вую ценность в разных культурах. В индустриально развитых
обществах люди с психическими аномалиями считаются боль-
ными, а в обществе с традиционным укладом люди с психи-
ческими отклонениями могут выполнять важную социальную
роль. Например, шаманы. У современных народов, занимаю-
щихся охотой и собирательством, шаман — центральная фи-
гура в религиозном культе. Шаманство выполняет структу-
рирующую и стабилизирующую роль в обществе. Шаман ру-
ководит религиозными обрядами, которые сплачивают людей
и усиливают дух коллективизма. Согласно верованиям охот-
ников и собирателей, обыкновенный человек не может не-
посредственно общаться с духами. Это может делать только
шаман, который является посредником между миром людей
и миром духов. По наблюдениям антропологов, изучавших
традиционные общества, человек не может стать шаманом по
желанию. Он становится шаманом помимо своей воли, а не-
редко и вопреки собственному желанию. Внезапно, обычно
при наступлении половой зрелости, у будущего шамана на-
чинается острое нервное заболевание, которое сопровожда-
ется истерическими припадками, обмороками, галлюцинация-
ми. Во время одного из припадков человеку «является дух-
покровитель», который «приказывает» ему стать шаманом,
предлагая свою помощь и руководств. Обычно больной отка-
зывается, колеблется, но затем покоряется и вступает в союз
со своим духом-покровителем. После этого припадки прекра-
щаются, больной выздоравливает и начинает выполнять фун-
кции шамана. Воздействие шаманов на психику соплеменни-
ков столь велико, что они во время шаманства видят те же
Раздел 12. Генетические основы аномалий поведения
галлюцинации, что и сам шаман. Изучая это явление, врачи и
антропологи пришли к выводу, что шаманами в основном ста-
новятся люди, отличающиеся особой болезненной организа-
цией. Это крайне возбудимые индивиды, с наклонностями к
экстатическим припадкам и разным видам галлюцинаций,
страдающие истеричностью. В родословных у многих шама-
нов кто-нибудь из предков по мужской или женской линии
также был шаманом. Как всегда, при анализе семейных дан-
ных возникает проблема о том, чем обусловлено сходство
между родственниками — общностью генов или общностью
семейной среды. В данном случае вопрос стоит о шаманстве
как семейной профессии, династии. Специальные исследо-
вания показали, что «выучиться на шамана» невозможно, что
отвергает идею о социальной преемственности, культурном
наследовании этого признака. Считается, что формирование
личности шамана представляет собой не овладение профес-
сией в результате научения, а является реализацией генети-
ческой склонности к определенному типу поведения. Обяза-
тельным условием «профессиональной пригодности» к это-
му занятию является склонность к нервно-психическим
расстройствам, у которых, как известно, имеется наследствен-
ная основа.
Приведенный пример свидетельствует об относительной
ценности различных вариантов человеческой психики. Если
в традиционных обществах индивиды, склонные к нервным
расстройствам, занимают почетное место в обществе тради-
ционного типа, то такие же люди в обществе индустриально
развитых стран нередко становятся пациентами психо-невро-
логических клиник. Приведенные факты можно рассматри-
вать как доказательство (возможно, несколько утрированное)
того, что к тому или иному виду профессиональной деятель-
ности, к той или иной роли, которую человек займет в обще-
стве, существует наследственная предрасположенность. Ка-
ким образом она реализуется, зависит от условий среды. А
что касается того факта, что индивиды с аномалиями поведе-
ния выполняют социально полезные функции, позволяет по-
356
12.5. Расстройства настроения
нять, почему в человеческих популяциях так распростране-
ны психические аномалии.
12.5. Расстройства настроения
Расстройства настроения включают нарушения эмоцио-
нального статуса, или настроения. По современной психиат-
рической нозологии на основе положения Карла Леонгарда у
человека могут проявляться два основных типа расстройств
настроения — униполярный, или депрессивный, и биполярный
синдромы. Основной характеристикой депрессии является
превалирование в эмоциональном состоянии человека дис-
фории, или грусти. При биполярных нарушениях происходит
превалирование эйфории, или радости. Термин «биполярный»
используется в связи с тем, что у многих людей с биполяр-
ным синдромом на самом деле наблюдаются колебания на-
строения между двумя полюсами — дисфорией и эйфорией.
Униполярный, или депрессивный синдром
Хотя у многих людей время от времени наступают депрессив-
ные состояния, лишь у немногих такие состояния проявля-
ются настолько сильно, что они диагностируются как депрес-
сивный, или униполярный, синдром. Униполярный синдром
встречается у 3,5% популяции и включает острые, хотя и ог-
раниченные во времени приступы депрессии. Среди женщин
униполярный синдром распространен чаще, чем среди муж-
чин.
При близнецовых исследованиях показано, что оптимис-
тический или пессимистический настрой человека обуслов-
лен в значительной степени генетически.
Риск развития депрессивного синдрома у родственников
первой степени родства с больным колеблется от 5% до 25%.
Конкордантность по заболеванию составляет Для М3 близне-
цов 40% и для ДЗ близнецов 17%, что свидетельствует в пользу
генетической предрасположенности к депрессивному синд-
Раздел 12. Генетические основы аномалий поведения
рому. Наследственную природу заболевания подтверждают
исследования приемных детей и их родителей. Наследуемость
униполярного синдрома в среднем составляет 0,75.
В развитии униполярного синдрома прослежен эффект ан-
тиципации, однако увеличения тринуклеотидных повторов
выявлено не было. В одних исследованиях обнаружена связь
депрессивного синдрома с геном переносчика серотонина на
хромосоме 17 q и с геном рецептора дофамина D4, однако в
других исследования подтверждения этому не найдено. В ряде
исследований предполагается мультифакториальная приро-
да депрессивного синдрома или же выдвигается модель глав-
ного гена.
Биполярный синдром
Ранее использовавшийся термин маниакально-депрессивный
синдром заменен в настоящее время термином биполярный
синдром. Дело в том, что при биполярном синдроме наблюда-
ются маниакальные приступы или происходит чередование ма-
ниакальных и депрессивных состояний. Под маниакальным
состоянием подразумевается приподнятое настроение, повы-
шенная активность и самооценка, раздражительность, умень-
шенная потребность во сне. Больные постоянно продуциру-
ют идеи, часто отвлекаются. Депрессивные симптомы включа-
ют снижение настроения, уменьшение интереса к различным
видам деятельности, бессонницу или сонливость, психомотор-
ное возбуждение или торможение, усталость, чувство ненуж-
ности и вины, неспособность концентрироваться и действовать
решительно, мысли о смерти или самоубийстве. Биполярный
синдром распространен реже, чем униполярный — примерно
у 0,3—1,5% популяции с равной вероятностью среди мужчин
и женщин.
У родственников лиц с биполярным синдромом риск раз-
вития заболевания выше, чем в среднем в популяции, что мо-
жет предполагать наследственную природу данного заболе-
вания. Действительно, конкордантность по биполярному син-
358
12.5. Расстройства надтроения
Рис. 12.3. Роберт Шуман (со
своей женой Кларой),
страдавший биполярным
синдромом
дрому среди М3 близнецов
колеблется от 33% до 80%, сре-
ди ДЗ близнецов — от 0% до
8%. Исследования приемных
детей также подтверждают
генетическую основу заболе-
вания. Механизм наследова-
ния биполярного синдрома
сложен и, скорее всего, вклю-
чает много локусов, ответ-
ственных за предрасполо-
женность к данному заболе-
ванию. При исследовании
маркеров биполярного синд-
рома предполагалось много
маркеров на разных хромосо-
мах (18р, 18q, 21q), однако в
более поздних исследованиях
результаты не всегда были
воспроизведены.
Многие известные лично-
сти, особенно музыканты,
страдали значительными перепадами настроения. Например,
у Генделя, Берлиоза, Шумана и Малера описаны особеннос-
ти поведения, которые в настоящее время характеризуются
как симптомы биполярного синдрома. У Берлиоза и Шумана
в музыкальных произведениях чередуются меланхолические
и оживленные фрагменты. Следует заметить, что Шуман за-
кончил свою жизнь в психиатрической лечебнице, где в воз-
расте 46 лет умер от сознательной голодовки (рис. 12.3). Ма-
лер, венский композитор, был пациентом Зигмунда Фрейда,
и также проявлял психическую нестабильность и перепады
настроения. Возможно, заболевание как-то связано с творчес-
кой активностью этих людей.
Сходное с биполярным синдромом состояние циклотомии
характеризуется менее выраженными перепадами между со-
Раздел 12. Генетические основы аномалий поведения
стояниями эйфории и дисфории. В данном случае легкую эй-
форию называют термином гипомания. Циклотомия обычно
начинается в возрасте около 20 лет. Как правило, люди вна-
чале не замечают признаков болезни и лишь спустя некото-
рое время отмечают значительные и неизбежные перепады
настроения.
12.6. Шизофрения
К шизофрении относят группу тяжелых хронических нару-
шений психики, проявляющихся в расстройстве мышления,
эмоций и поведения, которые часто начинаются без видимой
причины. Шизофрения характеризуется клиническим поли-
морфизмом. Некоторые больные беспрестанно хихикают,
другие большую часть времени неподвижно сидят, у третьих
наблюдаются периоды агрессивного поведения. Состояние
одних больных быстро улучшается, у других годами сохраня-
ется помраченное сознание. Одни больные теряют способ-
ность к концентрации внимания и логическому сопоставле-
нию фактов, у других в некоторых жизненных ситуациях от-
мечается хорошо выраженное логическое мышление, но при
действии самых незначительных причин оно утрачивается.
Некоторые больные в течение длительного времени могут
нормально жить и успешно работать и только временами впа-
дать в тяжелое состояние дезориентации.
В популяциях шизофрения встречается с частотой около
0,85%, а с учетом шизоидности — 4%. Шизофрения обнару-
живается как в промышленно развитых, так и в развивающих-
ся странах: в Северной Америке распространенность шизоф-
рении составляет 2,3—40 случаев на 1000 населения, на севе-
ре Европы — от 0,048 до 15, в Азии — 1,8—3,8, в Африке —
6—11,5. В больших городах шизофрения чаще встречается в
низших слоях населения. Это, возможно, связано с тем, что
предрасположенные к шизофрении люди из-за умственной
и волевой недостаточности опускаются на более низкие сту-
пени социальной лестницы. При этом отцы шизофреников и
12.6. Шизофрения
здоровых индивидов по социальному положению не разли-
чаются.
Высокая популяционная частота шизофрении при пони-
женной плодовитости больных (на 25—30% ниже, чем сред-
непопуляционная) может поддерживаться лишь в том случае,
если генотипы с мутантным аллелем придают их носителям
какие-либо селективные преимущества. Было выдвинуто
предположение, что пониженная плодовитость больных ши-
зофренией уравновешивается их повышенной устойчивос-
тью к некоторым заболеваниям. Результаты исследований
указывали на повышенную устойчивость больных шизофре-
нией к аллергиям, вирусным инфекциям и злокачественным
опухолям. Однако не все из перечисленных наблюдений уда-
лось подтвердить, и факт селективного преимущества гете-
розигот сейчас нельзя считать установленным.
Высказывалось предположение о существовании инфек-
ционного агента — бактерии или вируса, — вызывающего ши-
зофрению. Такая инфекция может быть получена от кого-то
из родственников и может сохраняться в ткани мозга в скры-
том состоянии долгое время. В настоящее время уже извест-
ны вирусы — возбудители мозговых инфекций с длительным
инкубационным периодом. Роль наследственного фактора
может состоять в том, что он делает некоторых людей осо-
бенно восприимчивыми к этой инфекции. В спинномозговой
жидкости некоторых больных шизофренией удалось обнару-
жить признаки инфекции цитомегаловирусом, а на препара-
тах мозга этих больных обнаруживаются структурные изме-
нения, которые можно считать результатом вирусной инфек-
ции
Генеалогические исследования показали, что у родствен-
ников больного шизофрения встречается чаще, чем в общей
популяции. Среди них часто наблюдается тип личности, от-
клоняющийся от нормы и классифицируемый как шизоид,
чаще, чем в среднем в популяции отмечаются депрессии. На-
следственностью определяется не только развитие шизофе-
нии как таковой, но и ее формы. Наличие у родственников
361
Раздел 12. Генетические основы аномалий поведения
разных форм шизофрении говорит о генетическом родстве
разных форм этого заболевания. Если оба родителя пробанда
здоровы, риск для его сибсов составляет около 10%. Риск по-
вышается до 15%, когда болен один из родителей, и достигает
40—60%, когда больны оба родителя. Риск заболеть шизоф-
рений выше у родственников ближних степеней родства,
даже если они живут в разных условиях. Чем тяжелее проте-
кает шизофрения у родителей, тем выше риск заболевания
для детей. Хотя пол не оказывает сильного влияния на часто-
ту шизофрении, у мужчин заболевание проявляется в более
раннем возрасте, чем у женщин. У близких родственников
больного риск развития болезни в 10—20 раз выше, чем в об-
щей популяции. Если пробанд имеет кататоническую или ге-
бефреническую симптоматику, то у его родственников риск
заболеть шизофренией выше, чем у родственников пробан-
дов с параноидной и простой шизофренией. Внутри семьи су-
ществует корреляция клинических форм. Родственники боль-
ных кататонической формой имеют больший риск заболеть
кататонической шизофренией, чем родственники больных
параноидной шизофренией.
О роли наследственности в формировании шизофрении
свидетельствуют также близнецовые исследования. То, что
более половины монозиготных пар дискордантны по шизоф-
рении, указывает на то, что формирование болезни при на-
личии наследственной предрасположенности провоцирует-
ся какими-то средовыми факторами, которым может подвер-
гаться один из близнецов. Следует, однако, отметить, что до
сих пор ни одного специфического средового фактора, вы-
зывающего шизофрению, не обнаружено. Конкордантность
по шизофрении монозиготных близнецов, воспитанных
врозь, такая же, как и конкордантность воспитанных вместе.
Конкордантность по шизофрении дизиготных пар в три раза
ниже, чем монозиготных. Если пробанд болен шизофренией,
то риск для его монозиготного партнера в 50 раз выше по срав-
нению со среднепопуляционым, а риск заболеть той же фор-
мой болезни, увеличен примерно в 30 раз. У дизиготных парт-
362
12.7. Психические расстройства в онтогенезе
неров больных шансы заболеть шизофренией примерно в три
раза меньше, чем у монозиготных, но при этом в 14 раз выше,
чем для человека, не имеющего больных родственников.
Одно время считалось, что родитель, больной шизофре-
нией, своим ненормальным поведением как бы индуцирует
расстройство поведения у детей, поэтому близкие родствен-
ники болеют шизофренией из-за сходства среды. Исследова-
ния семей с приемными детьми показали, что дети больных
шизофренией, очень рано отданные на воспитание в другие се-
мьи, болеют шизофренией с частотой значительно большей, чем
в целом в популяции. Частота шизофрении у этих детей при-
мерно такова, как и частота этого заболевания у детей, воспи-
танных своими больными родителями. При этом дети нормаль-
ных родителей, даже если они воспитывались в семье, где были
больные шизофренией, заболевают реже. Эти факты также ука-
зывают на ведущую роль наследственности в заболевании
шизофренией. Молекулярно-генетические исследования
шизофрении показали наличие у больных нескольких гене-
тических специфичностей в сайтах 5qrl 1.2-qrl3.3; 6р23; 22qrll.
12.7. Психические расстройства в онтогенезе
Некоторые психические расстройства, начинаются в детстве,
другие развиваются в первые годы жизни. К расстройствам
онтогенеза относят умственную отсталость, когда IQ ребен-
ка не превышает 70 баллов. Причинами умственной отстало-
сти могут быть фенилкетонурия, болезнь Тэя—Сакса, синд-
ром фрагильной Х-хромосомы, синдром Дауна, фетальный
алкогольный синдром и некоторые другие. Генетические при-
чины этих болезней уже обсуждалась в соответствующих гла-
вах. Онтогенетические расстройства включают также аутизм
и сходные состояния (синдром Ретта и Аспергера). В данную
группу болезней входят различные нарушения в обучении,
общении и двигательных навыках (дислексия, заикание и дру-
гие), синдром дефицита внимания и гиперактивности и не-
которые другие болезни. Известно много выдающихся людей,
363
Раздел 12. Генетические основы аномалий поведения
которые в детстве страдали расстройствами онтогенеза, в
частности, в обучении, например, Альберт Эйншейн, Томас
Эдисон, Вудро Вильсон, Нельсон Рокфеллер, Винстон Чер-
чиль, Чарльз Дарвин, Джон Кеннеди.
В данной главе мы рассмотрим наиболее интересные и изу-
ченные с точки зрения генетики расстройства онтогенетичес-
кого развития.
Аутизм
Для аутизма характерно отсутствие осознанности о суще-
ствовании и чувствах окружающих людей. Люди с аутизмом
полностью безразличны к комфорту. Они не могут подражать,
не понимают свою социальную роль или имеют искаженное
представление о ней, неспособны обзаводиться друзьями и
знакомыми и отвечать социальной и эмоциональной взаим-
ностью. У людей с данным заболеванием нарушено вербаль-
ное и невербальное общение, отсутствует воображение. Раз-
говорный язык вообще не развивается или развивается с за-
держкой. Речь людей с аутизмом лишена смысла, часто идет
повторение слов и предложений. Они малоразговорчивы, не
могут начинать и поддерживать разговор, при разговоре мо-
гут наблюдаться эхолалии (например, на вопрос «Как тебя
зовут?» они повторяют последнее слово, как эхо: «зовут, зо-
вут...»). Для аутизма характерен ограниченный набор видов
деятельности, к которым относятся, например, стереотипные
движения тела, концентрация на каком-нибудь одном пред-
мете или виде деятельности, постоянные занятия с частями
каких-либо предметов, волнение при минимальных измене-
ниях в окружающей обстановке, необъяснимая настойчи-
вость в точной последовательности действий, значительно
суженный круг интересов. Больные дети не отзываются на
свое имя, не любят, когда их ласкают и обнимают, не выра-
жают никаких эмоций на лице и не контактируют глазами,
плохо спят, у них развивается чувство страха. Около 66—75%
людей с аутизмом имеют IQ до 70 баллов.
364
12.7. Психические расстройства в онтогенезе
Аутизм распространен в популяции с частотой 2—5 на
10 ООО человек, причем среди мужчин он встречается в 3—4 раза
чаще, чем среди женщин. Различия касаются и возрастных
групп: так, среди детей 7—9 лет аутизм встречается с часто-
той 12,6 на 10 000, среди взрослых 18—20 лет — с частотой 0,4
на 10 000, что, по-видимому, объясняется различными диаг-
ностическими критериями и улучшением состояния у взрос-
лых. Действительно, с возрастом наблюдается сглаживание
симптомов аутизма. Так, в одном исследовании среди боль-
ных аутизмом с высоким IQ показано, что у 80% мужчин и
100% женщин улучшаются социальные и коммуникативные
умения, у 50% лиц обоего пола уменьшается повторяемость
действий. В целом в настоящее время в мире идет тенденция
к росту заболеваемости аутизмом.
В биологическом отношении предполагается, что при
аутизме недоразвита лимбическая система головного мозга,
область, которая отвечает за память и эмоции. У многих де-
тей с аутизмом отмечаются судорожные приступы. В одном
исследовании показано увеличение размеров головного моз-
га у людей с аутизмом, которое, однако, происходит не внут-
риутробно, а уже в постнатальном периоде на втором-третьем
годах жизни. У больных аутизмом наблюдается высокий уро-
вень серотонина в крови (в тромбоцитах). В то же время об-
наружена отрицательная корреляция между уровнем серо-
тонина в крови и вербальными способностями.
Ранее предполагали, что к средовым факторам риска раз-
вития аутизма относится вакцинация детей против кори, крас-
нухи и свинки, которая происходит в возрсте около 15 меся-
цев и сопровождается в ряде случаев симптомами аутизма.
Дело в том, что в вакцинах до недавнего времени содержался
консервант тимеросал на основе ртути. Известно, что боль-
шие дозы некоторых форм ртути негативно влияют на разви-
тие мозга, однако тимеросал, как показано в исследованиях,
таким эффектом не обладает. Кроме того, у детей, которые
не были привиты, аутизм развивается с такой же самой час-
тотой, как и у привитых, поэтому в настоящее время данная
Раздел 12. Генетические основы аномалий поведения
гипотеза отвергнута. Многие дети с аутизмом рождаются недо-
ношенными. В прошлом к средовым факторам риска развития
аутизма также относилось недостаточно внимательное отноше-
ние родителей к ребенку в раннем детском возрасте, особенно
матери, но сейчас эта гипотеза также не подтверждена.
Семейные исследования указывают, что риск для сибса
больного аутизмом составляет в среднем 4,2%. При близне-
цовых исследованиях показана большая конкордантность по
аутизму среди М3 близнецов, которая в среднем составляет
73%. Для ДЗ близнецов в ряде исследований конкордантность
была равна 0%, что указывает на значительный генетический
компонент аутизма. Высказывались различные мнения по
поводу механизма наследования аутизма (полигенный, ауто-
сомно-рецессивный и Х-сцепленный), однако в настоящее
время ученые более склонны к тому, что аутизм вызывается
множеством генов с относительно небольшим эпистатичес-
ким взаимодействием. В более ранних исследованиях была
прослежена связь между аутизмом и синдромом фрагильной
Х-хромосомы, но впоследствии наличие маркеров аутизма на
Х-хромосоме не подтвердилось. В одном исследовании пред-
положена связь аутизма с дупликацией в районе 15ql l-qrl3 и
предложен механизм наследования по типу генетического
импринтинга со стороны матери. В других исследованиях
указана связь аутизма с геном переносчика серотонина на
хромосоме 17 q и с гаплотипом системы HLA B44SC30DR4 на
хромосоме 6р. При моделировании на генетически модифи-
цированных животных, несущих человеческие гены, показа-
но, что участок 21 </22.2 ответствен за обучение и память, и на-
рушения в нем характерны для аутизма. Однако в целом роль
данного локуса в общем вкладе в предрасположенность к
аутизму у человека пока неизвестна.
Нарушения речевого развития и обучения
Способность членораздельно говорить, писать и читать —
важнейшие условия коммуникации. Речь является уникаль-
366
12.7. Психические расстройства в онтогенезе
ным свойством человека. Способность развивать артикуля-
ционную речь основана на таких качествах, как тонкий конт-
роль мышц гортани и рта, который отсутствует у шимпанзе и
других крупных человекообразных обезьян. FOXP2 был пер-
вым обнаруженным геном, имеющим отношение к способ-
ности человека развивать речь. Точковая мутация в данном
гене вызывала ухудшение речи в одной изученной семье, в
которой у половины членов были значительные сложности в
артикуляции, сопровождаемые лингвистическими и грамма-
тическими нарушенями. Ученые выделили комплементарный
участок ДНК с геном FOXP2 у шимпанзе, гориллы, орангутан-
га, макаки резус и мыши. Оказалось, что человеческий ген
FOXP2 отличен от таковых у животных по ряду кодируемых
им аминокислот, что указывает на тот факт, что ген FOXP2
был предметом отбора в течение предыдущей эволюции че-
ловека.
Изучая близнецов в возрасте двух лет, ученые пришли к
выводу, что генетические, а не средовые факторы играют ос-
новную роль в задержке начала разговорной речи у детей,
которые проявляют наибольшие трудности в овладении ре-
чью.
Для исследования речевых навыков двухлетних детей их
родителям был предложен список из 100 наиболее распрост-
раненных слов, в котором необходимо было отметить исполь-
зуемые близнецами слова. Варьирование было чрезвычайно
большим: одни дети использовали все 100 слов, другие — ни
одного. Среднее количество воспроизводимых из списка слов
равнялось 48 для близнецовой выборки в целом и лишь 4,2 сло-
ва для группы отстающих детей.
Было обнаружено, что как ДЗ, так и М3 близнецы имели
очень сходное речевое развитие. Однако результаты отстаю-
щей в речевом отношении группы значительно отличались.
Так, если в М3 близнецовой паре один ребенок был отстаю-
щим, то вероятность того, что и его партнер попадал в эту же
группу составляла 81 %. В то же время для ДЗ близнецовых пар
эта величина составляла 42%.
Раздел 12. Генетические основы аномалий поведения
Все указанное свидетельствовало в пользу значительного
генетического компонента в обучении речи отстающих детей.
Для неотстающих детей среда являлась определяющим фак-
тором. При быстром, среднем и даже слегка замедленном ре-
чевом развитии именно среда играла большую роль, чем гены.
Считается, что ранняя задержка в речевом развитии не оз-
начает, что ребенок обязательно столкнется с трудностями в
освоении речи или чтения. Половина этих детей догоняет сво-
их ровесников.
В настоящее время делается акцент на половые различия
в речевой задержке — мальчики чаще, чем девочки, сталки-
ваются с этой проблемой, и, таким образом, принадлежность
к мужскому полу является фактором повышенного риска в
нарушении речевого развития.
Заикание
Заикание характеризуется остановками речевого потока, ко-
торые выражаются в повторении звуков, слогов или слов, уд-
линенности звуков, в результате чего слова становятся «рас-
тянутыми». Заикание начинается в детстве. Примерно у од-
ной пятой части заикающихся это состояние сохраняется на
всю жизнь. Причины заикания до конца не ясны. Иногда на-
чало этого дефекта объясняют тем, что маленькие дети под-
ражают заикающимся старшим.
Как правило, степень заикания постоянно варьирует — в
спокойном состоянии человек говорит лучше, чем в тревож-
ном. В некоторых случаях при сильном волнении, когда от заи-
кающегося человека ждут быстрого ответа на вопрос, заика-
ние может значительно усиливаться. Заикание встречается у
1% взрослых. У детей в возрасте до 5 лет частота заикания 5%,
у школьников — с частотой 1,2%.
Заикание можно лечить у специальных врачей — логопе-
дов. Успех лечения выше у детей до 5 лет, их речь может стать
нормальной. У детей старшего возраста и у взрослых лече-
ние в основном направлено уже не на речевое улучшение, а
368
12.7. Психические расстройства в онтогенезе
Мэрилин Монро Брюс Уиллис
Рис. 12.4. Знаменитости-заики
на способы решения проблем, связанных с заиканием, в том
числе и психологических. Лечение заикания включает специ-
альные мероприятия, связанные с постановкой правильного
межреберно-диафрагмального дыхания, тренировкой говоре-
ния при широко открытом рте с целью большего расслабления
артикуляционных мышц (большинство заик разговаривают, еле
открывая рот) и техникой медленного разговора, когда дыха-
ние более свободно и в легкие поступает больше воздуха.
Многие известные люди с заиканием после соответствую-
щего лечения избавлялись от данного заболевания. К ним от-
носятся актеры Брюс Уиллис, Сэм Нейл, Мерилин Монро
(рис. 12.4).
Поиск генетических причин заикания проводили с помо-
щью генеалогического и близнецового методов. Анализ ро-
дословных показал, что в некоторых семьях заикание наблю-
дается в нескольких поколениях, у мальчиков в четыре раза
Раздел 12. Генетические основы аномалий поведения
чаще, чем у девочек. У заикающихся женщин процент род-
ственников с таким же дефектом речи выше, чем у заикаю-
щихся мужчин. Тот факт, что монозиготные близнецы не все-
гда конкордантны по заиканию, указывает на то, что в фор-
мировании этого дефекта речи определенную роль играют
средовые факторы. В некоторых исследованиях отмечается
аутосомно-доминантная модель наследования заикания. Од-
нако большинство наблюдаемых фактов укладываются в тео-
рию мультифакториальной природы этого признака. Счита-
ется, что генетическая подверженность заиканию представ-
ляет собой полигенную систему с пороговым эффектом,
неодинаковым у представителей разного пола. Более высо-
кий процент заикающихся среди мужчин и более высокая на-
следственная отягощенность заикающихся женщин указыва-
ет на то, что порог подверженности этому дефекту речи у
женщин выше, чем у мужчин. Практический вывод, который
делается из этого заключения, состоит в том, что мальчики
формируют группу более высокого риска по заиканию, чем
девочки. Другой важный вывод состоит в том, что потомство
заикающихся женщин формирует группу более высокого
риска, чем потомство заикающихся мужчин.
Синдром Туретта
В 1885 г. французский невропатолог Жиль де ля Туретт впер-
вые диагностировал женщину с заболеванием, впоследствии
получившим название синдром Туретта. Синдром Туретта
проявляется обычно до 18-летнего возраста и характеризует-
ся непроизвольными, быстрыми, повторяющимися, неритми-
ческими и стереотипными множественными двигательными
и вокальными тиками. Двигательные тики обычно затрагива-
ют область головы и других частей тела (туловище, верхние и
нижние конечности). К вокальным тикам относятся различ-
ные шумовые сопровождения, например, прищелкивание
языка, похрюкивание, рычание, лаяние, покашливание, про-
изнесение обычных слов. Популяционная частота синдрома
370
Г2.7. Психические расстройства в онтогенезе
Туретта составляет 5 больных на 10 000 человек, однако для
разных популяций данные варьируют. Так, в минессотском
исследовании частота синдрома Туретта на 10 000 человек
составляла 0,046, среди взрослого населения Северной Дакоты —
0,5, среди подростков Северной Дакоты — 5,2, среди подрос-
тков Новой Зеландии — 0,7, среди взрослых Израиля — 4,3.
В одном исследовании школьников эта цифра составила 23,4
на 10 000 человек. Среди мужчин синдром Туретта распрост-
ранен чаще, чем среди женщин (в 1,6—9,3 раза).
Риск заболеть синдромом Туретта для родственников пер-
вой степени родства больного по разным данным составляет
примерно от 2% до 51%, причем более подверженным, как уже
было отмечено, является мужской пол, для представителей
которого риск в 5 раз выше, чем для женщин. В одном иссле-
довании была получена конкордантность для М3 близнецов
в 53% и для ДЗ близнецов в 8%. Многие исследования предпо-
лагают моногенную аутосомно-доминантную модель с непол-
ной пенетрантностью и варьирующей в зависимости от пола
и возраста экспрессивностью. В некоторых исследованиях
указывается на мультифакториальный тип наследования син-
дрома Туретта. В ряде исследований при синдроме Туретта
обнаружены хромосомные перестройки — делеции в хромо-
сомах 9р и 18g и транслокацию 1(7; 18).
Дислексия
Дислексия определяется как специфическое и значительное
ухудшение способности к чтению, которую нельзя объяснить
снижением интеллекта, возможностей овладеть чтением,
мотивацией или сенсорными повреждениями. Дислексия яв-
ляется одной из наиболее частых аномалий, диагностируемых
в детстве, и представляет огромную образовательную и со-
циальную проблему. Несмотря на то что дислексия считается
аномалией, дислекеики часто имеют повышенные показате-
ли умственного развития. Среди детей, страдающих дислек-
сией, мальчики составляют около 80%.
Раздел 12. Генетические основы аномалии поведения
У 60% людей, страдающих дислексией, среди родственни-
ков обнаруживается задержка развития речи. В одном из ис-
следований показано, что среди детей с дислексией 88% име-
ли одного-двух родственников, которые также были не спо-
собны обучиться чтению. В исследованиях на близнецах
отмечается более высокая конкордантность монозиготных
пар по сравнению с дизиготными. В одном из исследований
на 36 парах монозиготных близнецов отмечена стопроцент-
ная конкордантность по дислексии, по другим данным — 68%,
в то время как конкордантность у ДЗ близнецов составила
32%. Эти данные говорят о наличии генетической компонен-
ты в формировании дислексии, а неполная конкордантность
монозиготных близнецов указывает на определенную роль
среды.
Установлено, что дислексия является высоконаследуемым
признаком. На основании исследований большого числа се-
мей из Великобритании и США была прослежена связь дис-
лексии с хромосомой 18р11.2 и указан маркер D18S53.
Синдром дефицита внимания и гиперактивности
Гиперкинетический синдром проявляется в повышенной
склонности отвлекаться. Гиперактивные дети не могут уси-
деть на одном месте и сконцентрироваться на каком-либо за-
нятии. Они легко переключаются с одного дела на другое,
ничего не доводя до конца. Они болтливы и рассеянны, им-
пульсивны и невнимательны. Гиперкинетический синдром
встречается у 3—9% детей, причем среди мальчиков в несколь-
ко раз чаще, чем среди девочек. Гиперкинетические дети часто
происходят из семей с какими-либо психическими наруше-
ниями. Примерно у 40% из них один или оба родителя страда-
ют психическим расстройством. Часто родители гиперактив-
ныхдетей являются алкоголиками. Клинико-генеалогические
исследования показали, что родственники пробандов с гипе-
рактивным синдромом чаще имеют депрессии, тревожность,
чаще употребляют наркотические вещества.
372
12.8. Органические поражения Moira
Риск развития гиперкинетического синдрома у ребенка
больной матери составляет от 4% до 38%, больного отца — от
15% до 44%. В другом исследовании показано, что риск разви-
тия гиперкинетического синдрома у сибса больного ребенка
составляет 25%. Близнецовые исследования показали, что
конкордантность по гиперактивности ДЗ близнецов состав-
ляет 29%, а конкордантность М3 близнецов в 2,5 раза выше.
Наследуемость данного признака, по данным разных авторов,
колеблется в пределах 0,64—0,88. Для изучения генетики дан-
ного заболевания также используются модельные животные —
крысы линии Вистар—Киото со специальным геном, локали-
зованным в хромосоме 8 и отвечающим за гиперактивность.
У мышей подобный ген локализован в хромосоме 9. Некото-
рые исследователи предполагают не полностью пенетрант-
ную аутосомно-доминантную модель наследования гиперки-
нетического синдрома у человека.
12.8. Органические поражения мозга
Болезнь Альцгеймера
Болезнь Альцгеймера, ранее известная также под названием
деменция, впервые была описана немецким врачом Альцгей-
мером в 1907 г. В США по статистике ею страдают 5% людей
после 65 лет и 25% после 85 лет. В настоящее время выявлена
наследственная природа болезни Альцгеймера в некоторых
семьях, однако наследственная основа выявлена лишь у 5%.
Болезнь Альцгеймера развивается начиная примерно с 60 лет.
Чаще болеют женщины. Болезнь Альцгеймера начинается
постепенно. После появления первых симптомов умственная
функция ухудшается в течение 3—10 лет. Отмечается двига-
тельное возбуждение, маниакального типа, суетливость, рас-
пад речи. Больные часто теряются и могут бесцельно бродить
в различных местах. Постепенно исчезают двигательные на-
выки, нарастает расстройство памяти. Больные не могут
вспомнить имен своих родственников, не знают свой возраст,
не помнят профессии, не понимают смысл речи. У них осла-
373
Раздел 12. Генетические основы аномалий поведения
бевает внимание, нарастает вялость и безразличие ко всему
окружающему. Наблюдаются бредовые идеи ущерба, ограб-
ления, преследования. На последних стадиях больные не мо-
гут говорить и принимать пищу и нуждаются в уходе. Смерть
часто наступает в результате инфекций, что обычно наблю-
дается у людей, прикованных к постели.
При патологоанатомическом исследовании отмечается ат-
рофия коры головного мозга. При гистологическом анализе
наблюдается жировое перерождение и дегенерация нервных
клеток (рис. 12.5). Типичным признаком болезни Альцгейме-
ра является отложение белка /3-амилоида вокруг кровеносных
сосудов мозга. Считается, что отложение амилоидного белка -—
главный момент в развитии этой болезни.
Описаны три мутации в кодоне 717 гена АРР, которые вы-
зывают болезнь Альцгеймера. Все они связаны с замещени-
ем аминокислоты валина на изолейцин, фенилаланин или гли-
цин. Эти мутации связаны с отложением белка |3-амилоида.
При вскрытии бляшки [3-амилоида содержатся в мозгу в обла-
сти центров обучения и памяти. Открыты еще три гена, ответ-
ственных за болезнь Альцгеймера. Один из них локализован в
14-й хромосоме (14q24.3) и кодирует мембранный белок S182
(белок may, или тау-протеин), который обусловливает насле-
дование болезни как аутосомного доминантного признака.
Белок тау прикрепляется к микротрубочкам в нервных клет-
ках, разрушая эти клетки. Локус AD3 детерминирует возник-
новение злокачественной формы болезни Альцгеймера. Было
найдено 5 различных мутаций этого гена, связанных с ран-
ним началом семейной болезни. Ген, кодирующий белок S182,
вызывает около 80% рано начинающихся случаев болезни Альц-
геймера. Его продукт имеет сходство с мембранным белком, об-
наруженным у одного из модельных объектов генетики пове-
дения — червя нематоды. В работе на нематоде показано, что
этот белок участвует в транспортировке белков внутри клетки.
Считается, что при мутациях гена S182 соответствующий де-
фектный белок способен прерывать транспорт АРР в клетку и
тем самым способствует накоплению амилоидных бляшек.
374
12.8. Органические поражения мозга
Рис. 12.5. Дегенеративные изменения мозга при болезни
Альцгеймера (увеличение желудочков мозга
и борозд за счет гибели нейронов)
Методы генной инженерии позволяют внести человечес-
кий ген болезни Альцгеймера в геном мыши. У генетически
трансформированью мышей также начинают откладываться
бляшки в тех же областях мозга, что и у человека, — в зоне
памяти и зоне обучения.
Болезнь Паркинсона
Болезнь Паркинсона, или дрожательный паралич, впервые
была описана английским врачом Джеймсом Паркинсоном в
1817 г. Болезнь Паркинсона является медленно прогрессирую-
щим дегенеративным заболеванием центральной нервной
системы, которое примерно одинаково встречается у мужчин
и женщин. Европеоиды чаще страдают болезнью Паркинсо-
на, чем представители других рас. Болезнь Паркинсона встре-
чается у 1 % популяции в возрасте старше 65 лет и у 0,4% попу-
ляции в возрасте старше 40 лет. В абсолютном выражении
число случаев болезни Паркинсона растет, что связано с рос-
Раздел 12. Генетические основы аномалий поведения
Рис. 12.6. Нарушение осанки при болезни Паркинсона
Рис. 12.7. Изменение почерка при болезни Паркинсона
том продолжительности жизни и неблагоприятными эколо-
гическими факторами.
Данное заболевание включает несколько симптомов, сре-
ди которых наиболее выражены тремор (дрожание), мышечная
376
12.8. Органические поражения мозга
ригидность (напряженность), гипокинезия (замедленность и
обеднение рисунка движений), деменция (утрата интеллек-
туальных способностей), депрессия, снижение обоняния и
нарушение сердечного ритма и кровяного давления. Заболе-
вание обычно начинается дрожанием рук в виде «счета мо-
нет» и нарушением походки в виде замедленных шажков,
наклона туловища вперед и перехода в бег (рис. 12.6). Умень-
шается громкость голоса, речь становится замедленной и мо-
нотонной, лицо часто представляет собой застывшую маску.
Люди с болезнью Паркинсона имеют проблемы со многи-
ми видами двигательной активности. Одна из наиболее нео-
бычных проблем впервые была обнаружена Арнольдом Пи-
ком в 1903 г. Он сравнивал образцы почерков людей до и пос-
ле появления у них болезни Паркинсона. В некоторых, хотя и
не во всех случаях он обнаружил, что размер букв значитель-
но уменьшился. Он назвал данный феномен «микрография»,
что в переводе с греческого означает «мелкий почерк». Не все,
у кого мелкий почерк, страдают болезнь Паркинсона. Речь
идет о произошедшем уменьшении почерка. На рис. 12.7 по-
казан почерк француженки до и после болезни Паркинсона.
При болезни Паркинсона существенно повреждаются
клетки черной субстанции головного мозга. Характерные
симптомы болезни появляются тогда, когда погибает 80—85%
клеток данной области. Болезнь Паркинсона происходит из-
за недостатка дофамина, который синтезируется в мозгу.
Существует несколько форм болезни Паркинсона. Две
наиболее важные — ранняя и поздняя формы. Ранняя форма
начинается до 50 лет и составляет 5—10% всех случаев, на
позднюю — после 50 лет приходится 90—95%.
Болезнь Паркинсона имеет семейный характер. На раз-
витие болезни оказывают влияние как гены, так и среда.
В одном исследовании на близнецах было показано, что бо-
лезнь Паркинсона, развивающаяся до 50 лет, имеет значитель-
ную генетическую компоненту. Болезнь Паркинсона, разви-
вающаяся после 50 лет, имеет средовую обусловленность.
К настоящему времени ученые открыли несколько генов
подверженности ранней, или ювенильной, форме болезни Пар-
Раздел 12. Генетические основы аномалий поведения
кинсона. К ним относятся гены SNCA (PARK1), parkin (PARK2)
и ген убиквитина CH-L1. Ген SNCA, локализованный в чет-
вертой хромосоме, был первым обнаруженным геном, кото-
рый, по мнению ученых, способствовал развитию болезни
Паркинсона. Данный ген был обнаружен у членов несколь-
ких семей из Италии и Греции, где болезнь прослеживалась
по аутосомно-доминантному типу наследования. Ген SNCA
отвечает за выработку альфа-синуклеина, который является
токсическим веществом и поражает клетки мозга. Интерес-
ным является тот факт, что тау-протеин и альфа-синуклеин
при взаимодействии могут образовывать амилоид, который
является причиной поражения мозга как при болезни Альц-
геймера, так и при болезни Паркинсона. Это отчасти объяс-
няет, почему у людей с одним заболеванием, появляются при-
знаки другого. Ген parkin, локализованный в шестой хромо-
соме, часто обнаруживается в семьях, где прослеживается
аутосомно-рецессивный тип наследования болезни. В одной боль-
шой изученной семье болезнь Паркинсона проявлялась у лиц от
7 до 58 лет под влиянием указанного гена. Ген убиквитина CH-L1,
расположенный в четвертой хромосоме, также отвечает за раз-
витие болезни Паркинсона, однако пока он обнаружен у од-
ной пары сибсов, страдающих этим заболеванием.
О влиянии факторов среды на развитие болезни Паркин-
сона говорит случай, произошедший в Калифорнии в 1931 г.
У группы наркоманов быстро развивались симптомы болез-
ни Паркинсона после внутривенных инъекций вещества
N-МФТП (2,3,6-тетрагидропиридин), который является побоч-
ным продуктом при синтезе «уличного» героина. Хотя дан-
ный случай продвинул исследования в области обнаружения
средовых факторов развития болезни Паркинсона, N-МФТП
не является значимой причиной развития болезни в популя-
ции в целом. Проживание в сельской местности также повы-
шает риск развития болезни из-за влияния сельхозудобрений,
в больших количествах присутствующих в питьевой воде. Это
объясняется тем, что некоторые из удобрений имеют хими-
ческую структуру, сходную с N-МФТП. Из 400 добровольцев,
12.8. Органические поражения мозга
которым вводили N-МФТП в 1980-х гг., лишь у немногих раз-
вились признаки болезни, а остальные остались невосприим-
чивыми.
Часто причиной развития болезни Паркинсона является
прием ряда нейролептиков, например галоперидола, а также
вещества резерпина, которые вызывают блокаду дофамино-
вых рецепторов. Реже причиной является отравление угар-
ным газом или марганцем, опухоли и инфаркты мозга. Болез-
ни Паркинсона чаще подвержены работники умственного
труда. Частые травмы головы также могут приводить к раз-
витию заболевания, что можно продемонстрировать на при-
мере боксера Мухамеда Али, у которого симптомы болезни
проявились в возрасте до 40 лет.
Курение и употребление кофе снижает риск развития бо-
лезни Паркинсона. Некоторые ученые считают, что в сигаре-
тах и кофе имеются некоторые защитные вещества. Другие
ученые полагают, что сами по себе данные вещества не име-
ют никакого значения, а все объясняют тем, что людям с при-
страстием к каким-либо веществам свойственны специфичес-
кие биохимические процессы в головном мозгу, что делает
их более устойчивыми к болезни Паркинсона.
Установлено, что у 75% лиц с болезнью Паркинсона в роду
никогда не было случаев данного заболевания. И, наоборот, в
семьях, где встречалось много лиц с данной болезнью, неко-
торые люди не болели вообще. Однако если у консультирую-
щегося лица в семье были больные, то у него самого риск за-
болеть выше среднепопуляционного.
В настоящее время основным лечением болезни Паркин-
сона является введение предшественника дофамина. В неко-
торых случаях больным проводят трансплантацию поражен-
ного участка мозга.
Хорея Гентингтона
Хорея Гентингтона — это заболевание, которое на началь-
ных этапах характеризуется нарастанием повышенной дви-
379
Раздел 12. Генетические основы аномалий поведения
гательной активности конечностей, усиливающейся при
эмоциональном напряжении. Впервые болезнь была описа-
на американским ученым Джорджем Гентингтоном в 1872 г.
Болезнь проявляется начиная с 30 лет и старше. Начало за-
болевания может быть связано с интеллектуальными рас-
стройствами, психозом и бредом. Отмечаются нарушения
речи, эмоциональная притупленность, эндокринные нару-
шения. В мозгу умерших больных обнаруживают тяжелые
изменения в лобных долях, сглаживание извилин, пораже-
ния подкорки.
Распределение случаев хореи Гентингтона в родословных
соответствует аутосомно-доминантному типу наследования.
Индивиды, имеющие признаки этого заболевания, обладают
генетической особенностью к тандемному умножению ДНК
в четвертой хромосоме. Передача из поколения в поколение
хромосомы с такой особенностью приводит к тому, что пото-
мок получает хромосому с более длинным участком ДНК, чем
имел его родитель. Чем большее число раз повторен один и
тот же участок ДНК, тем раньше начинается заболевание.
Болезнь Гентингтона, признаки которой у потомков начина-
ют проявляться раньше, чем у родителей, — результат амп-
лификации последовательности CAG в сайте 4р16.3. В норме
число повторов в этом участке составляет 11—34, при их чис-
ле больше 42 развиваются симптомы заболевания.
Хорея Гентингтона передается преимущественно от боль-
ного отца. При этом у детей наблюдается нарастание длины
повтора, что сопровождается более ранним появлением пер-
вых признаков болезни. Между числом тринуклеотидных
повторов в мутантном гене и возрастом появления первых
симптомов заболевания имеется обратная корреляция. Чис-
ло положительно коррелирует с темпом неврологической и
психической симптоматики.
Болезнь Гентингтона довольно распространена в популяции.
Ее частота в зрелом возрасте составляет примерно 1:10 000 и
в крупной психиатрической больнице обычно можно встре-
тить 2—3 человек, пораженных данной болезнью.
380
12.8. Органические поражения мозга
4Й.
Хореей Гентингтона в течение
15 лет страдал известный американ-
ский фольклорный певец и компо-
зитор Вуди Гатри (рис. 12.8).
Эпилепсия
Эпилепсия представляет собой
заболевание, которое проявляет-
ся сочетанием внезапных рас-
стройств сознания и судорог.
Различают генуинную, рефлек-
торную и симптоматическую эпи-
лепсию. При генуинной эпилепсии
припадки возникают без видимой
причины. При рефлекторной эпи-
лепсии припадок провоцируется
Рис. 12.8. Вуди Гатри,
пораженный болезнью
Гентингтона
звуком или мелькающим светом. Симптоматическая эпилеп-
сия обусловлена поражениями мозга в результате травмы,
инфекции, интоксикации или в связи с соматическими забо-
леваниями.
Заболеваемость эпилепсией (число вновь заболевших в
течение года) составляет в среднем 30—50 случаев на 100 ты-
сяч населения. При этом наблюдается широкая межпопуля-
ционная изменчивость — от 17 случаев на 100 тысяч населе-
ния в Японии до 1000 случаев на 100 тысяч населения в Авст-
ралии. Распространенность эпилепсии (фактическое число
больных) варьирует от 2 до 9 больных на 1000 населения в
индустриальных странах и от 7 до 21 в развивающихся. Веро-
ятность появления эпилепсии в возрасте до 20 лет составляет
примерно 1% и 2% до 40 лет. Риск заболеть эпилепсией до кон-
ца жизни составляет около 3,5%. Риск для родственника пер-
вой степени родства больного пробанда в 5—10 раз выше, чем
в среднем для популяции. Заболеваемость среди мужчин
выше, чем среди женщин. Эту разницу объясняли повышен-
ной подверженностью мужчин к травмам, однако в исследо-
Раздел 12. Генетические основы аномалий поведения
ваниях на лабораторных животных (крысах) обнаружена бо-
лее высокая судорожная готовность у самцов по сравнению с
самками, что свидетельствует о генетической природе отли-
чий.
О наследственной природе эпилепсии говорят результа-
ты генеалогического и близнецового анализов. Частота эпи-
лепсии среди близких родственников пробанда в несколько
раз выше, чем в популяции. Конкордантность монозиготных
пар по генуинной эпилепсии 89%, дизиготных 4%. Тяжелая
травма головного мозга может инициировать развитие судо-
рожных припадков, для порога возникновения таких припад-
ков характерна индивидуальная изменчивость. Лица, у кото-
рых судорожные припадки возникают как следствие травмы
головного мозга, действия высокой температуры и других
стрессовых факторов, могут в генетическом отношении от-
личаться от тех, у кого припадки после таких воздействий не
развиваются При симптоматической эпилепсии конкордант-
ность моно- и дизиготных близнецов 12% и 0%. Следователь-
но, симптоматическая эпилепсия развивается на благоприят-
ной наследственной почве. Это подтверждается также и тем,
что заболеваемость эпилепсией среди сибсов больных симп-
томатической формой этой болезни выше, чем в популяции.
Известно около сотни наследственных признаков, связанных
с судорожными припадками. К ним относятся нарушения
обмена аминокислот, заболевания, связанные с накоплени-
ем промежуточных продуктов обмена в клетках мозга, нару-
шения мозгового кровообращения, опухоли мозга.
Согласно одной из генетических гипотез эпилепсии, это
заболевание обусловлено аутосомным доминантным геном,
который в гетерозиготе вызывает церебральную дизритмию,
а гомозиготе эпилепсию. Эпилепсия считается генетически
гетерогенным заболеванием — в одних семьях она контроли-
руется доминантным геном с вариабельной экспрессивнос-
тью и высокой пенетрантностью, в других — рецессивным
геном, в третьих - находится под контролем полигенного ком-
плекса. Молекулярно-генетические исследования эпилепсии
382
12.8. Органические поражения мозга
показали наличие у больных нескольких генетических спе-
цифичностей в сайтах 6q23-q25; 8pter-q22; 8q; 8q24; 11 q.
Посредством элекроэнцефалографии были вскрыты цереб-
ральные механизмы эпилепсии. Главную роль в них играет су-
дорожная готовность мозга, которая обусловлена функциональ-
ными перестройками нервной ткани. Главный признак — на-
рушение электрического ритма мозга, или церебральная
дизритмия. Дизритмия проявляется вспышками острых и
медленных волн, наличием высокоамплитудных и островер-
шинных компонент. У больных отмечаются аномалии ритма
не только во время судорог, но и в межсудорожный период.
В обычных условиях дизритмия отмечается примерно у 10%
населения, а среди родственников больного количество таких
людей достигает 60%.
Генетический анализ дизритмии ЭЭГ показал, что она яв-
ляется именно тем наследуемым признаком, который адек-
ватно характеризует предрасположенность к эпилепсии. При
неоднократном отведении потенциалов дизритмия обнаружи-
вается примерно у 95% больных эпилепсией. У некоторых
людей судорожные припадки могут развиваться в ответ на
частые световые вспышки определенного цвета и частоты.
При этом возникают изменения в электроэнцефалограмме.
У сибсов таких людей подобная реакция возникает чаще, чем
в случайной популяции. Чем больше сходство в клинических
проявлениях эпилепсии у родственников, тем выше сходство
ЭЭГ.
Исследования на животных выявили, что генетически раз-
личные линии крыс отличаются по частоте судорог. У пасю-
ков частота аудиогенной (вызываемой звуком) эпилепсии со-
ставляет около 4%. Среди белых крыс лабораторной линии
Вистар к эпилептическим припадкам склонны 15% животных.
У крыс линии Крушинского—Молодкиной, специально отсе-
лектированной на предрасположенность к эпилепсии, при-
падки встречаются примерно у 99% особей. Скрещивание жи-
вотных из линий показало, что предрасположенность к судо-
рожным припадкам имеет полигенный характер.
383
У линий, имеющих более низкий порог электросудорог, об-
наруживается более высокая способность к решению задач.
Крысы линии Крушинского—Молодкиной имеют более вы-
сокую подвижность нервных процессов, чем крысы линии Ви-
стар, у них быстрее вырабатываются условные рефлексы.
Церебральная дизритмия у практически здоровых людей
рассматривается как «скрытая эпилепсия», которая может
проявиться в специфических условиях. Мелькающим светом
дизритмию можно вызвать более чем у 50% здоровых людей.
Подобным эффектом обладают и компьютерные игры. Поэто-
му люди (в особенности дети) с повышенной судорожной го-
товностью мозга, наличием дизритмий, страдающие эпилеп-
сией должны это учитывать, а производители игр обязаны
делать соответствующее предупреждение в инструкции.
Болезнь Феллинга (фенилкетонурия)
Рис. 12.9. Ребенок с фенил-
кетонурией
Болезнь Феллинга, или фенилке-
тонурия (ФКУ), была открыта в
1934 г. норвежским ученым Фел-
лингом. Это заболевание связан-
но с мутацией аутосомного гена,
расположенного в 12 хромосоме
и наследуется по рецессивному
типу. Сейчас открыто уже более
400 мутаций этого гена.
ФКУ — одно из наиболее рас-
пространенных заболеваний,
связанных с дефектом в метабо-
лизме аминокислоты фенилала-
нина. Заболевание проявляется в
первые месяцы жизни и выража-
ется задержкой моторного и пси-
хического развития. Причиной
заболевания является невозмож-
384
12.8. Органические поражения мозга
ность усваивать аминокислоту фенилаланин, которая посту-
пает в организм ребенка с молоком матери. При фенилкето-
нурии в крови накапливаются ядовитые продукты метаболиз-
ма. Они патологически влияют на нервные клетки, что при-
водит к постепенному нарастанию признаков умственной
отсталости (рис. 12.9). Если своевременно не приняты меры, дети
погибают. Особенно часто фенилкетонурия встречается в ев-
ропейских популяциях, поэтому в родильных домах проводит-
ся генетический скрининг (выявление) фенилкетонурии.
Дети, имеющие генетический дефект, с первых дней жизни пе-
реводятся на диету. Это пища, в которой содержится очень ма-
лое количество фенилаланина. Находясь на диете примерно до
восьмилетнего возраста, дети вырастают здоровыми.
Гетерозиготность женщины по гену фенилкетонурии мо-
жет оказать неблагоприятное влияние на умственное разви-
тие ребенка. Во время беременности у нее может повышать-
ся содержание в крови фенилаланина и продуктов его обме-
на, что может явиться причиной нарушения в развитии мозга
плода, хотя после рождения у ребенка обмен фенилаланина
будет нормальный.
Болезнь Тэя—Сакса
Болезнь Тэя—Сакса — наследственное нарушение метаболиз-
ма, вызываемое отсутствием фермента гексозаминидазы А.
При отсуствии этого фермента в нервных клетках накап-
ливается и разрушает их. Дети с таким заболеванием редко
живут дольше пяти лет. Болезнь Тэя—Сакса названа так по
имени английского доктора Уоррена Тэя и американского не-
вропатолога Бернарда Сакса. В 1881 г. Тэй описал изменения
глаз больного, а Сакс понял, что в клетках происходят опас-
ные изменения и причиной тому — наследственность.
Наследуется болезнь Тэя—Сакса по аутосомно-рецессив-
ному типу. У гетерозиготных носителей обычно не проявля-
ются симптомы заболевания. Болезнь наиболее распростра-
нена среди потомков евреев, ведущих свое происхождение
13 Введение в психогенетику
385
Раздел 12. Генетические основы аномалий поведения
из Центральной и Восточной Европы (ашкенази). Приблизи-
тельно 1 из 25 ашкеназских евреев является носителем. В дру-
гих популяциях это соотношение 1:250.
Дети с болезнью Тэя—Сакса в первые шесть месяцев жиз-
ни внешне развиваются вполне нормально. Затем они стано-
вятся чрезмерно чувствительными к шуму и у них постепен-
но ухудшается зрение. Приблизительно в год начинаются су-
дороги эпилептоморфного характера; мускулы становятся все
слабее, нарушается координация движений. Глотание проис-
ходит с трудом, легкие не справляются со своей функцией.
Приблизительно в возрасте пяти лет наступает летальный
исход, поскольку нервная система к тому времени оказвает-
ся полностью разрушенной.
Рассеянный склероз
Рассеянный склероз — это болезнь с аутоиммунным компо-
нентом, при которой происходит разрушение миелиновых
оболочек нервных волокон головного и спинного мозга. Мно-
жественный склероз трудно диагностировать, поскольку бо-
лезнь имеет много едва уловимых симптомов. Обычно они ус-
танавливаются при повторяющихся случаях мышечной сла-
бости, вызванной склерозом волокон в ЦНС, которые
утратили свои миелиновые оболочки. По-видимому, мишеня-
ми рассеянного склероза являются не сами нейроны, а олиго-
дендроциты, клетки нейроглии, выполняющие функции пита-
ния и обмена веществ нейронов. Макрофаги (разновидности
лейкоцитов) атакуют эти клетки и разрушают их миелиновые
оболочки, что замедляет и блокирует проведение импульса
по аксону. Результатом являются различные симптомы, такие,
как зрительное раздвоение, онемение и похолодание конеч-
ностей, утрата двигательной координации в руках и ногах,
приступообразная потеря речи и способности глотать.
20% случаев рассеянного склероза представляют собой се-
мейные случаи и ассоциированы с генами главного комплек-
са гистосовместимости HLA.
386
12.9. Химическая зависимость
12.9. Химическая зависимость
Генетика алкоголизма
Необходимым условием для развития алкоголизма является
доступность алкоголя. По-видимому, алкоголь был известен
людям еще с доисторических времен. С появлением земледе-
лия между X и V вв. до н.э. систематическое производство
алкоголя стало возможным во многих популяциях благодаря
сбраживанию ячменя, меда, молока и винограда.
Стимулом употребления алкоголя, по-видимому, были его
психотропные эффекты, позволяющие преодолеть чувство
страха, присущего первобытным людям. Возможно, именно
благодаря этому свойству алкоголь довольно рано был введен
в религиозные обряды. Изобретение метода дистилляции ал-
коголя произошло примерно в X в. н.э., и с этого времени ста-
ли производить концентрированные алкогольные напитки.
В течение XIII и XIV вв. эта технология распространилась в
Европе и положила начало алкоголизму.
Алкоголь распространен повсеместно, за исключением
стран, где он запрещен религией.
Алкоголизм является болезнью, которая проявляется а на-
рушении контроля над приемом спйртных напитков. Попу-
ляционно-генетические исследования показали, что алкого-
лизм развивается в среднем у 3—4% лиц, причем мужчины
болеют в десять раз чаще женщин.
По своей химической природе алкоголь относится к угле-
водам и является потенциальным источником энергии для лиц,
употребляющих его в больших количествах. У сильных алкого-
ликов данное вещество составляет половину их энергетических
нужд. Алкоголь всасывается в желудке через 5—10 минут после
его употребления на голодный желудок. Эффекты от употреб-
ления алкоголя длятся несколько часов и зависят от массы тела
человека. Именно поэтому одна и та же доза вызывает у жен-
щин, как правило, больший эффект, чем у мужчин.
Алкоголизм обнаруживает семейное накопление. Причи-
ны семейного накопления могут быть наследственность и
культурные особенности.
13’
387
Раздел 12. Генетические основы аномалий поведения
Многочисленные исследования четко указывают на то, что
алкоголиками чаще бывают люди, имеющими мать, отца или
более отдаленных родственников, страдающих алкоголизмом.
Так, если у человека страдает алкоголизмом родственник пер-
вой степени родства, то риск развития у него самого алкого-
лизма увеличен в 4 раза, если родственник второй степени
родства — в 2 раза по сравнению со среднепопуляционными
данными.
У М3 близнецов конкордантность по алкоголизму намно-
го выше, чем у ДЗ близнецов, причем как у мужчин, так и у жен-
щин. Также выявлено, что на каждой стадии алкоголизма М3
близнецы более сходны между собой, чем ДЗ близнецы.
Исследования приемных детей, проведенные в Дании и
Швеции, показали, что алкоголизм имеет генетическую ос-
нову.
Родственники женщин, лечившихся от алкоголизма, име-
ют более высокий риск развития данного заболевания, чем
родственники лечившихся мужчин. Некоторые исследования
предполагают наличие полоспецифических локусов предрас-
положенности к алкоголизму. Наиболее изученным фактором
предрасположенности к алкоголизму, является наличие у че-
ловека отца-алкоголика. Неодинаковая эмоциональная реак-
ция разных людей на алкоголь общеизвестна. У одних прием
алкоголя вызывает состояние эйфории, у других он вызыва-
ет «оглушение». Наркологи считают, что индивиды первого
типа входят в группу повышенного риска по алкоголизму.
Физиологи различают эндогенный и экзогенный алкоголь.
Эндогенный алкоголь — это продукт естественного обмена
веществ, экзогенный алкоголь — это напиток. Уровень эндо-
генного алкоголя у человека определяется особенностями
обмена веществ и находится под контролем генотипа. Отме-
чено, что индивиды с естественно низкой концентрацией эн-
догенного алкоголя реагируют на прием спиртных напитков
положительной эмоциональной реакцией. У людей, в крови
которых концентрация эндогенного алкоголя повышена, при-
ем спиртных напитков вызывает неприятные ощущения.
388
12.9. Химическая зависимость
Несмотря на то что генетическая компонента в развитии
алкоголизма очевидна, до сих не ясен механизм наследова-
ния данного заболевания.
Метаболизм алкоголя является одной из биологических
причин, которые влияют на склонность к алкоголизму. Об-
мен алкоголя в организме упрощенно можно отразить такой
схемой:
ADH ALDH
Алкоголь —> Альдегид —> Уксусная кислота —> Углекислый газ и вода
Первая реакция (превращение алкоголя в альдегид) конт-
ролируется ферментом алкогольдегидрогеназой (ADH), вторая
(превращение альдегида в уксусную кислоту) — ферментом
альдегиддегидрогеназой (ALDH}. По активности обоих фер-
ментов выявлен полиморфизм, связанный с этническими раз-
личиями. В печени человека фермент ADH2 существует в трех
вариантах — ADH2-1, ADH2-2 и ADH2-3, которые кодируют
соответственно субъединицы рр р2, и 03. В локусе ADH3 име-
ются два аллельных состояния — ADH3-1 и ADH3-2, кодиру-
ющие субъединицы gt и д2. В азиатских популяциях преобла-
дают аллели ADH2-2 и ADH3-1, а их продукты ферменты р2Р2
и gtgt в 40 и 2 раза соответственно более активны, чем формы
Р1Р1 и g2g2. В печени также присутствуют два фермента ALDH —
цитозольный ALDH 1 и митохондриальный ALDH2. Митохон-
дриальная форма гораздо более активна в метаболизме уксус-
ного альдегида, чем цитозольная. Примерно у 50% представи-
телей монголоидной расы форма ALDH2 является неактивной.
Хорошо изученным является тот факт, что частоты алле-
лей ADH2-2, ADH3-1 и ALDH2-2 значительно снижены у ал-
коголиков в сравнении с представителями азиатских попу-
ляций в целом. Альдегид — весьма токсичное вещество. Его
накопление в крови приводит к интоксикации организма, ко-
торое сопровождается неприятными ощущениями. Считает-
ся, что низкая склонность к потреблению алкоголя связана с
накоплением альдегида вследствие высокой активности фер-
ментов P2-ADH и gt-ADH и малой активности фермента
14 Введение в психогенетику
389
Раздел 12. Генетические основы аномалий поведения
ALDH2-2. Защитные эффекты ALDH2-2 значительно выше,
чем у ADH2-2 и ADH3-1.
Кроме алкогольдегидрогеназы и альдегиддегидрогеназы в
метаболизме алкоголя может участвовать один из подвидов
ферментов семейства цитохромов Р4502Е1 (CYP2E1).
Фермент моноаминоксидаза (МАО), которая катализиру-
ет окисление моноаминовых нейромедиаторов, таких, как
катехоламины, индоламины и другие биогенные амины, воз-
можно, играет роль в регуляции поведения и настроения. Две
формы фермента МАО А и МАО В — распределены преиму-
щественно в головном мозгу, печени и тромбоцитах крови.
В тромбоцитах возможно обнаружить лишь форму В данного
фермента. У алкоголиков обнаружена низкая активность фер-
мента МАО в тромбоцитах, что также может быть фактором
подверженности к алкоголизму.
Кроме того, у алкоголиков снижена активность фермента
аденилапщиклазы в лимфоцитах и тромбоцитах даже после
периодов лечения алкоголизма, что позволяет считать данное
свойство фактором риска развития алкоголизма.
Также у алкоголиков частота аллеля А; гена DRD2, кодирую-
щего рецептор дофамина D2, значительно повышена по срав-
нению с не-алкоголиками.
Недавно обнаружен ген предрасположенности к антисо-
циальному алкоголизму HTR1B , который снижает уровень
серотонина и увеличивает риск импульсивной агрессии. Му-
тантный ген был обнаружен в результате генетического ана-
лиза финских преступников-алкоголиков и одного юго-запад-
ного племени американских индейцев, среди которых высо-
кая частота алкоголизма.
Давно отмечено, что представители разных рас реагиру-
ют на алкоголь неодинаковым образом. Жители Европы, осо-
бенно славяне, в отличие от представителей монголоидной
расы, нередко употребляют алкоголь в очень больших коли-
чествах, но при этом не всегда становятся алкоголиками.
Чувствительность к интоксикации алкоголем, которая оце-
нивается по ряду поведенческих и физиологических показа-
тёлей ^покраснение кожи, частота пульса, кровяное давление,
сонливость, изменение походки) у представителей монголо-
идной расы встречается чаще, чем у европеоидов. Эти разли-
чия выявляются даже у новорожденных, что свидетельству-
ет о существовании генетического контроля в реакции на ал-
коголь.
Неблагоприятную физиологическую реакцию монголои-
дов на прием даже небольшого количества алкоголя объясня-
ют особенностями ферментных систем у этих народов. По-
пуляционные исследования показали, что примерно 40—50%
жителей Южного Китая и Японии отличаются резкой недо-
статочностью фермента ALDH, участвующего в метаболизме
алкоголя. Среди представителей других народов монголоид-
ной расы населения недостаточность этого фактора колеблет-
ся в пределах 5—57%. 40% представителей племен южноаме-
риканских индейцев характеризуется дефицитом ALDH, тог-
да как в племенах сиу и навахо, обитающих в Северной Америке,
частота ALDH-недостаточности составляет только 5%. Среди
популяций европейцев и африканцев дефицит ALDH прак-
тически не наблюдается.
Считается, что быстрое накопление и медленное расщеп-
ление весьма токсичного альдегида, который является про-
дуктом окисления алкоголя, сопровождается неприятными
субъективными ощущениями, и это препятствуют повторно-
му употреблению спиртных напитков, защищая человека от
алкоголизма. Реакция людей на алкоголь в значительной мере
зависит от предыдущей истории популяций. Малые северные
народы, проживавшие на территории бывшего СССР, оказа-
лись на грани вымирания, после того как столкнулись с пи-
тейными традициями русской культуры. Охотники и рыбо-
ловы никогда не занимались изготовлением спиртных напит-
ков, так как в их суровом климате не было соответствующих
условий. Поэтому предыдущая эволюция не выработала у
этих людей ферментные системы для детоксикации алкого-
ля. Когда представители малых народов получили практиче-
ски неограниченный доступ к спиртным напиткам, которые
Раздел 12. Генетические основы аномалий поведения
доставлялись им в обмен на продукты их труда, то вскоре об-
наружилось, что многие из них в течение нескольких меся-
цев погибали от злокачественного алкоголизма. Практику ис-
требления американских индейцев с помощью «огненной
воды» использовали и белые переселенцы. Устойчивость к
действию алкоголя выработалась у народов, в течение мно-
гих поколений изготовлявших и употреблявших спиртные на-
питки. Представители разных народов проявляют неодина-
ковую устойчивость и чувствительность и к другим наркоти-
ческим веществам растительного происхождения. Различия
в реакции на наркотическое вещество обычно связаны с тем,
присутствовало оно или нет в предыдущей истории.
Генетика наркомании
Из-за широкой доступности наркотических средств в настоя-
щее время наркомания стала серьезной социальной пробле-
мой.
Близнецовые исследования показали, что наследуемость
наркомании составляет 30%.
Пристрастие к разным наркотикам имеет неодинаковую
генетическую компоненту. Наследуемость подверженности
героиновой наркомании составляет 50%, к психоделическим
средствам 26%. Большое влияние (53%) на прием психодели-
ческих средств оказывает внесемейное окружение. Показа-
но, что одним из важных факторов предрасположенности к
наркомании является такая психологическая черта человека,
как поиск новизны.
Ученые обнаружили практически одну и ту же корреля-
цию между обоими партнерами М3 и ДЗ пар близнецов по
употреблению марихуаны и кокаина, в результате чего был
сделан вывод о том, что при пробовании наркотиков основ-
ную роль играет социальная среда. Однако М3 близнец парт-
нера-наркомана с большей вероятностью может стать нарко-
маном, чем ДЗ близнец партнера-наркомана, что свидетель-
ствует в пользу значительного генетического компонента в
392
12.9. Химическая зависимость
развитии наркомании. Таким образом, среда подталкивает
человека попробовать наркотики, а гены «подсаживают» его
на определенный наркотик.
Кофеин является наиболее доступным из легальных нар-
котических веществ. Он относится к легким стимуляторам.
Высокие дозы этого вещества препятствуют сонливости, а
очень высокие могут вызывать зрительные и слуховые гал-
люцинации. Кофеин повышает частоту сердечных сокраще-
ний и кровяное давление, учащает дыхание и оказывает сла-
бое диуретическое влияние. В большом количестве кофеин
может поступать в организм в виде кофе или в виде кока-колы
и аналогичных напитков. Стандартной дозой для стимулирую-
щего эффекта является 200 мг кофеина или две чашки креп-
кого кофе. Смертельные случаи от употребления кофеина
могут наблюдаться после принятия около 10 г кофеина, что
составляет около 100 чашек крепкого кофе. Высокие дозы
кофеина, действующие внутриутробно, влияют на размер
тела новорожденного.
Если женщина во время беременности употребляет мно-
го кофе или кока-колы, риск гибели плода вдвое выше, чем у
женщин, употребляющих мало кофеина. Дело в том, что ко-
феин стимулирует дыхательные центры, и при его отсутствии
дыхание плода может быть угнетено, особенно если в данное
время у плода наблюдается какая-либо инфекционная инток-
сикация. Малый вес новорожденных у матерей, злоупотреб-
ляющих кофе, объясняется тем, что, как и другие вредные
вещества, проходящие через плаценту (в частности, алкоголь
и никотин), кофеин затрудняет попадание к плоду кислорода
и питательных веществ, что приводит к недостаточному пи-
танию плода и замедленному его росту.'
Марихуана является наиболее употребляемым из нелегаль-
ных наркотиков. Активным ингредиентом марихуаны явля-
ется тетрагидроканнабинол (рис. 12.10).
Эффект от курения марихуаны проявляется практически
сразу и длится от одного до четырех часов. Под действием
марихуаны человек расслабляется и становится более разго-
393
Рис. 12.10. Молекула тетрагидроканнабинола
ворчивым. Марихуана является также депрессантом и поэто-
му людям с депрессивным синдромом от ее употребления ста-
новится хуже. У наркоманов после курения марихуаны цве-
та и звуки становятся более интенсивными, улучшается от-
ношение к другим людям и к жизни вообще. Они становятся
более терпеливыми и эмпатичными, у некоторых на какое-то
время появляется способность к творчеству. В то же время
марихуана в короткое время снижает способность человека
концентрироваться и уменьшает время реакции, поэтому явля-
ется фактором риска при дорожно-транспортных нарушениях.
Генетика курения
От последствий курения погибает больше людей, чем от
СПИДа, алкоголя, других наркотических веществ, автоката-
строф, убийств и самоубийств вместе взятых. Активным ве-
ществом сигарет является никотин. По степени пристрастия
никотин сходен с героином, а его негативные последствия для
здоровья популяции в целом гораздо серьезней.
Никотин является стимулятором и седативным средством,
снижая тревожность. Никотин проникает из легких в мозг
всего за несколько секунд, а его эффекты для организма длят-
ся около получаса. Повторный прием приводит к депрессии
и усталости, поэтому курильщикам требуется все больше си-
гарет.
12.10. Расстройства самоконтроля
Сигаретный дым содержит различные газы, а больше все-
го монооксида углерода. Кроме никотина в сигаретах содер-
жится смола. Смола вызывает рак, эмфизему и другие болез-
ни легких, монооксид углерода — сердечно-сосудистые бо-
лезни. У курящих матерей риск рождения недоношенных и
мертворожденных детей выше.
При изучении семей предполагают, что генетические фак-
торы влияют на склонность к курению, поскольку растет доля
курильщиков среди родственников людей, которые также
курят. Вероятность для человека стать курильщиком повыша-
ется в 2—3 раза, если курит его брат.
Результаты изучения приемных детей свидетельствуют о
наличии генетической предрасположенности к этой привыч-
ке.
На близнецовых выборках популяций Швеции, Дании,
Финляндии, Австралии и США было показано, что наследуе-
мость курения находится в пределах 37—84% для женщин и
28—84% для мужчин.
На моно- и дизиготных близнецах мужчинах показано, что
начало и длительность курения находятся под генетическим
контролем. Кроме того, у М3 близнецов конкордантность по
прекращению курения также была выше, чем у ДЗ близне-
цов, что свидетельствует о генетическом компоненте склон-
ности бросить курить.
12.10. Расстройства самоконтроля
Импульсивные расстройства самоконтроля проявляются в
нарушении способности управлять своими порывами. Люди
с расстройствами самоконтроля действуют импульсивно и
своим поведением наносят себе вред. Само по себе импуль-
сивное поведение неопасно, и многие люди иногда действу-
ют под влиянием внезапного порыва. Однако в ряде случаев
импульсивное поведение может привести к печальным по-
следствиям. Можно рассмотреть следующие примеры. Муж-
чина, не имеющий денег, но страстно желакиций приобрес-
Раздел 12. Генетические основы аномалий поведения
ти дорогой автомобиль, может пойти на все (кражу, убийство
другие действия, сопряженные с риском) ради достижения
своей цели. Женщина, которая не в состоянии регулировать
свои сексуальные порывы, может вступать в случайные кон-
такты, результаты которых также могут быть плачевными.
Среди импульсивных расстройств самоконтроля можно
назвать клептоманию (неспособность удержаться от воров-
ства), гэмблинг (пристрастие к азартным играм), пироманию
(тягу к поджигательству), сексуальную импульсивность (не-
способность контролировать сексуальное поведение), трихо-
тилломанию (вырывание собственных волос) и некоторые
другие.
В отдельную группу расстройств самоконтроля входят рас-
стройства питания — анорексия (отказ от еды) и булимия
(объедание).
Гэмблинг
Гэмблингом (от англ, gamble — азартная игра) называется при-
страстие к азартным играм.
Границы между гэмблингом как развлечением и патоло-
гией стерты. Во всем мире увеличивается частота случаев па-
тологического гэмблинга, что связано с легализацией заведе-
ний, которые привлекают любителей острых ощущений: лоте-
реи, казино, ипподром, игровые автоматы. На патологический
гэмблинг приходится примерно 25% всех нарушений самокон-
троля.
В гэмблинг вовлекаются самые разные слои населения.
Так, в конце 1980-х гг. американский спортивный мир был по-
трясен тем, что национальный кумир баскетболист Пит Роуз
оказался гэмблером, делавшим ставки в тысячах долларов на
баскетбольных играх. В настоящее время около 3% людей
страдают этой зависимостью, причем среди мужчин гэмбле-
ров больше, чем среди женщин.
Описано две стадии развития патологического гэмблин-
га. В течение первой стадии гэмблер чувствует эйфорию.
396
12.10. Расстройства самоконтроля
Даже если при этом он несет потери, то гордится своими ус-
пехами и игнорирует будущие проблемы. Втягиваясь в игру,
человек неуклонно деградирует, круг его интересов сужает-
ся, постепенно разрушается психика. Вторая стадия сопря-
жена с разочарованием. На этот период приходится большое
число самоубийств.
В США среди патологических гэмблеров около 25% явля-
ются безработными, что почтив пять раз больше, чем среди
обычных людей. Среди гэмблеров повышена преступность.
Для первичной профилактики гэмблинга необходимо оп-
ределить группу повышенного риска. Психологи отмечают,
что дети гэмблеры в раннем возрасте отличаются повышен-
ной общительностью, отсутствием стеснительности. Тяга к
игре чаще наблюдается у детей из неблагополучных семей, а
также из семей, где играют родители. Ребенок гэмблер посто-
янно находится в состоянии стресса. Это состояние — благо-
приятная почва для развития наркомании. Детей, которые
слишком увлекаются игровыми автоматами, следует не нака-
зывать, а лечить.
Среди личностных характеристик взрослых гэмблеров
мужчин преобладают такие, как недружелюбность, стремле-
ние повелевать другим человеком, агрессивность, неискрен-
ность, безответственность, среди гэмблеров женщин — обыч-
но зависимость, подчиненность и пассивная агрессивность,
стремление к успеху нетрадиционными методами. Патологи-
ческий гэмблинг коррелирует с расстройствами настроения.
У патологических гэмблеров отмечена генетически обуслов-
ленная повышенная активность нейромедиатора норадрена-
лина и аномалии ЭЭГ.
Патологический гэмблинг по своей генетической приро-
де относится к мультифакториальным и полигенным болез-
ням. Ученые обнаружили связь между этим состоянием и спе-
цифическими генами одного из рецепторов дофамина D2. Так,
у патологических гэмблеров аллель D2A1 встречается в два
раза чаще, чем у людей, не участвующих в азартных играх.
Нейромедиатор дофамин участвует в формировании чувства
боли, положительных и отрицательных эмоций, а также свя-
зан с так называемым «чувством вознаграждения». Предпо-
лагают, что гэмблерам присущ «синдром недостаточного воз-
награждения» , в результате нарушения функционирования спе-
цифических дофаминовых нервных путей. Именно потому эти
люди пытаются компенсировать недостаточное физиологическое
вознаграждение путем азартной игры. В таком состоянии у
гэмблеров изменяется сила и направленность биохимических
процессов в мозге, и они получают удовольствие.
Патологический гэмблинг является частой причиной се-
мейных разладов. Около 20% жен гэмблеров происходили из
семей, в которых патологическим гэмблером был один из ро-
дителей. С генетической точки зрения, такая ситуация явля-
ется отражением теории генотропизма венгерского ученого
Леопольда Зонди. Согласно его теории, пожительная брачная
ассортативность является следствием притяжения друг к дру-
гу людей со сходными генами или, как в данном случае, поис-
ке брачных партнеров, похожих на членов семьи (для жен-
щин — на отца, братьев, для мужчин — на мать, сестер).
Расстройства питания
Психологическое значение еды простирается далеко за пре-
делы ее пищевой ценности. Люди обычно уделяют много вре-
мени приготовлению пищи и сервировке стола. Человек за-
висит от пищи не только физически, но и эмоционально. Го-
лодный человек раздражен, и, наоборот, хороший обед ведет
к чувству удовлетворенности. Для некоторых людей прием
пищи имеет необычно большое значение, и они становятся
рабами странных и нездоровых ритуалов в отношении еды.
Люди с расстройствами питания находятся в постоянной
борьбе со своей зависимостью.
Наиболее хорошо изучены расстройства питания анорек-
сия и булимия. Расстройства питания часто прослеживаются
в семьях и особенно в сочетании с расстройствами настрое-
ния. Даже в случаях, когда родственники больных не были
398
официально диагностированы, как правило, в беседе о них
фигурируют выражения типа «страдала навязчивой идеей о
своем весе», «ела, как птичка» или «ненавидела свою внеш-
ность».
Анорексия
Распространенным мнением является то, что анорексия яв-
ляется болезнью, которую создает сама женщина в результа-
те следования некоему идеалу, навязываемому современным
обществом, где делается упор на худощавость. Анорексию
сложно лечить одними лишь психологическими средствами,
поскольку болезнь имеет в своей основе и биологические
причины. Анорексия определяется как болезнь, при которой
люди сознательно голодают, теряя, по меньшей мере, 15% сво-
ей нормальной массы тела. Анорексия обычно начинается в
период полового созревания и, как правило, распространена
среди европейских женщин среднего или высшего классов.
В некоторых популяциях частота анорексии невелика — 0,1—
0,7%, в некоторых высокая — 1—3%, причем до 95% всех боль-
ных могут составлять женщины. Так, в США анорексия об-
наружена у 8 млн человек (около 3% популяции), из них око-
ло 7 млн женщины.
Слово анорексия буквально означает «без аппетита». У
анорексиков развиваются все симптомы голодания — низкое
кровяное давление, замедленные или нерегулярные сердце-
биения, запоры, ощущение холода и др. От 15 до 21% случаев
анорексии заканчиваются летальным исходом.
Женщины, происходящие из семей с расстройствами пи-
тания, чаще, чем в среднем в популяции, сами страдают таки-
ми болезнями. Показано, что у М3 близнецов конкордант-
ность по анорексии выше, чем у ДЗ. В одном исследовании на
близнецах показано, что анорексия на 58% генетически обус-
ловлена. В другом исследовании показано, что если мать или
сестра женщины страдали анорексией, то риск для консуль-
тирующейся в 12 раз выше, чем в среднем в популяции.
Раздел 12. Генетические основы аномалий поведения
Средовыми факторами в развитии анорексии является
стремление к упомянутому выше стереотипу идеальной фи-
гуры. Кроме того, неосмотрительное поведение родителей,
состоящее в их сильной обеспокоенности в отношении внеш-
него вида и веса своих дочерей, также часто провоцирует рас-
стройства питания, имеющие генетические предпосылки.
Такой стиль поведения более характерен для семей среднего
и высшего класса.
На лабораторных крысах была показана связь между ано-
рексическим поведением животных и нарушениями в зоне
гипоталамуса, называемой центром насыщения. Частичное
разрушение указанной области приводит к уменьшенному
воздействию на нее нейромедиатора адреналина. Происхо-
дит постепенная адаптация к меньшему потреблению пищи,
поскольку насыщение начинает наступать при гораздо сни-
женном количестве еды. При увеличенном уровне серотони-
на у крыс в области центра насыщения происходит прекра-
щение потребления пищи и начинается голодание (в проти-
воположность ожирению, при котором уровень серотонина
снижен). Аналогичные эффекты наблюдались и при измене-
нии концентрации норадреналина.
Эффекты нейромедиаторов сходны у крыс и человека. Ус-
тановлено, что у анорексиков вдвое чаще наблюдаются измене-
ния в гене рецепторов серотонина, чем у обычных людей.
Нейромедиатор серотонин был открыт в 1950-х гг. Серо-
тонин оказывает влияние на многие проявления поведения
человека, включая голод, сон, половой ответ, контроль побуж-
дений, агрессию, гнев, депрессию, тревожность и восприя-
тие. Низкий уровень серотонина встречается у людей со
склонностью к суицидальному поведению, чрезмерно агрес-
сивных или чрезмерно депрессивных людей. Высокий уро-
вень серотонина обнаружен у людей, находящихся в состоя-
нии постоянной тревоги, страдающих бессонницей, со склон-
ностью к сверхточному выполнению заданий.
У людей с анорексией повышение серотонинового воздей-
ствия коррелирует с рядом психологических характеристик,
400
12.10. Расстройства самоконтроля
в частности, с состоянием нервозности, стремлением к совер-
шенству и порядку, низкой толерантностью к новым ситуа-
циям, страху взросления, низкой самооценке. Голодание, воз-
можно, является для таких людей попыткой избавиться от
этого дискомфорта. В одном исследовании, проведенном сре-
ди 322 женщин-анорексиков, установлена значительная связь
между степенью заболевания и степенью стремления к со-
вершенству. Для анорексиков характерны большие затраты
времени на такие пищевые ритуалы, как нарезка своей еды
тонкими кусочками.
У людей с анорексией повышен уровень гормона мозга
кортизола. Обычно кортизол выделяется в ответ на стресс.
Недавно открытые гормоны орексин А и орексин В связны
с пищевым поведением у крыс. Регулируя чувства голода и
сытости, ученые могут оказывать влияние на количество по-
требляемой крысами пищи. Инъекции указанных гормонов
в области центра насыщения в гипоталамусе приводят к тому,
что крысы сразу начинают съедать в 8—10 раз больше пищи,
чем в норме.
Помимо этого у анорексиков обнаружены повышенные
уровни опиоидов, энкефалинов и эндорфинов в спинномоз-
говой жидкости. Некоторые исследования показали, что уг-
нетение синтеза этих веществ приводило к тому, что люди с
анорексией начинали прибавлять вес.
Считается, что предрасположенность к анорексии имеет
полигенный характер. Уже обнаружен один из генов пред-
расположенности к анорексии. Он расположен в коротком
плече 1-й хромосомы. Кроме того, в этом участке был найден
соответствующий маркер анорексии — D1S3721, который
позволит генотипировать людей и выявлять группу риска.
Булимия
В соответствии со строгими критериями булимия встречает-
ся у 1—3% женщин и 0,2% мужчин. Некоторые симптомы забо-
левания характерны для значительно большего количества —
401
Раздел 72. Генетические основы аномалий поведения
от 5 до 15%. В основном это девушки-подростки и молодые
женщины. Симптомы булимии начинаются обычно в юношес-
ком возрасте и включают чрезмерное переедание, за которым
следует избавление от съеденного путем изматывающих за-
нятий спортом, рвоты или приема слабительных.
На близнецах установлено, что наследуемость булимии со-
ставляет 83%. Речь идет не об одном гене, а о множестве, т.е.
даное расстройство имеет полигенный контроль. Самыми рас-
пространенными факторами среды, которые провоцируют
развитие булимии, являются сложности в отношениях с парт-
нером, социальные проблемы и психологические травмы.
Ученые предполагают наличие связи между генами предрас-
положенности к депрессии и генами предрасположенности
к расстройствам питания. Так, в одном исследовании показа-
но, что 50% женщин с расстройствами питания сопутствуют
депрессивные состояния.
Показано, что в развитии булимии имеет место снижение
серотонинового воздействия. Предшественником серотони-
на является триптофан, который может образовываться при
многоэтапных метаболических превращениях углеводов, в
огромных количествах потребляемых булимиками и практи-
чески сразу избавляющихся от них, следствием чего являет-
ся низкий уровень серотонина.
Поскольку серотонин участвует в регуляции настроения,
то нарушение активности данного нейромедиатора головно-
го мозга вызывает тревожность и навязчивые идеи о совер-
шенстве, влияет на контроль аппетита и таким образом уве-
личивает подверженность к булимии. У людей с булимией
часто имеется чувство беспомощности, безнадежности и низ-
кая самооценка безотносительно к реальному положению
вещей. Женщины с булимией часто имеют искаженные пред-
ставления о своем теле. Им может сопутствовать алкогольная
или наркотическая зависимость. Установлено, что изменение
серотониновой активности не исчезает после лечения боль-
ных булимией. Поэтому полностью не исчезают симптомы и
булимии. В одном исследовании при изучении женщин, ле-
402
F Контрольные вопросы и задания к разделу Г2
I -------------------------------------------------------
чившихся по поводу булимии препаратом, специфически влия-
ющим на серотониновую систему и вызывающим гормональ-
ные и поведенческие эффекты, было установлено, что после
лечения у женщин все же остается повышенным уровень
метаболитов серотонина. Также у них наблюдается ухудше-
ние настроения и продолжаются навязчивые идеи о совер-
шенстве и утонченности. Уровни других нейромедиаторов
мозга, в частности, дофамина и норадреналина, находятся в
пределах нормы.
Контрольные вопросы и задания к разделу 12
1. Какие вам известны кандидатные гены психических за-
болеваний?
2. Какие особенности поведения животных сходны с психи-
ческими болезнями человека?
3. Приведите примеры отношения к индивидам с погранич-
ными психическими состояниями в различных культурах.
4. В 1973 г. в журнале «Science» была опубликована статья,
которая произвела огромное впечатление на читателей.
В ней говорилось о том, что восемь человек ловко одура-
чили медицинский персонал нескольких психиатрических
клиник США. Эти люди заявили, что у них наблюдались
галлюцинации, они слышали какие-то странные звуки
типа «пустота», «дыра» и т.д. В других отношениях все эти
люди были совершенно здоровы. Врачи не могли поста-
вить им диагноз, поскольку по отдельному симптому они
не подпадали ни под какую категорию психических боль-
ных. Тем не менее все они были помещены в больницы.
Став пациентами, они сразу стали вести себя «нормаль-
но», однако медицинские работники приняли изменение
их поведения за дальнейшее прогрессирование неизвест-
ной болезни. Лишь некоторые другие пациенты считали,
что новые «псевдопациенты», возможно, были либо жур-
налистами, либо научными работниками, проникшими в
больницы для детального исследования «внутреннего»
403
Раздел 12. Генетические основы аномалий поведёния
мира психиатрических учреждений. «Псевдопациенты»
провели в больницах от 7 до 52 дней и были выписаны с
диагнозом «шизофрения в стадии ремиссии». Когда было
обнародовано, что в данном случае имел место научный
эксперимент, последовали неоднозначные высказывания,
начиная с поддержки обманутых психиатров и заканчи-
вая недоверием в целом психиатрической диагностике.
Каково Ваше мнение по поводу сказанного выше? Возмож-
ны ли такие фальсификации, если для диагностики пси-
хических больных применять современные генетические
знания? Достаточно ли этого?
5. Известно, что настроение человека зависит от ситуации,
а наследственность остается неизменной в течение жиз-
ни. Как Вы относитесь к утверждению, что наследствен-
ность оказывает влияние на настроение?
6. Приведите факты, доказывающие роль наследственности
в формировании эмоциональных расстройств.
7. Изначально электрошок применялся для наказания людей.
Затем его стали использовать для лечения различных бо-
лезней, в частности, тяжелых депрессивных состояний че-
ловека. Больным назначают 6—8 сеансов электрошока. Не-
которые люди негативно относятся к этому лечению, по-
скольку его последствия часто непредсказуемы. Сами
ученые до сих пор не могут объяснить механизм происхо-
дящих во время электрошока изменений в головном моз-
ге. Каково Ваше отношения к подобным методам лечения
в отношении психики человека, в которых большая состав-
ляющая отводится «методу проб и ошибок» и «попадания
пальцем в небо»? Какие альтернативные методы Вы счи-
таете более целесообразными?
8. Как Вы считаете, можно ли людей, диагностированных би-
полярным или униполярным синдромом, допускать в ка-
честве педагогов в учебные заведения? Какие эффекты
для обучаемых (положительные и отрицательные) может
повлечь деятельность таких педагогов ?
9. Приведите факты, которые свидетельствуют о том, что ши-
зофрения — наследственная болезнь.
404
Контрольные вопросы и задания к разделу 12
10. Известны ли Вам факторы среды, вызывающие шизофре-
нию?
11. Риск заболевания шизофренией составляет примерно 10%,
если болен один родитель, 40%, если больны оба родителя,
и 16%, если болен брат или сестра. Определите пример-
ный риск заболевания шизофренией для индивидов А, Б и
В (рис. 12.11).
12. На рис. 12.12 представлена родословная, в которой встре-
чается шизофрения. Частота этого заболевания в иссле-
405
Раздел 12. Генетические основы аномалий поведения
дованной популяции составляет 1,1%, а наследуемость 85%.
Определите вероятность развития данного заболевания у
членов родословной в III поколении.
13. У 75 человек, больных шизофренией, был партнер — мо-
нозиготный близнец, среди которых 26 также были боль-
ны. 94 больных имели брата или сестру — дизиготного
близнеца, из которых шизофренией болели 7 человек.
Оцените роль генетических факторов в развитии шизоф-
рении.
14. Обсудите такое суждение: «поскольку способность на-
учиться читать имеет генетическую основу, учитель, выс-
тавляя ребенку оценку за чтение, оценивает не его уси-
лия, а родительские гены».
15. Известно, что заикание имеет наследственную основу. Оз-
начает ли это, что от заикания можно избавиться?
16. Как вы относитесь к идее не наказывать нарушителей дис-
циплины в классе, если известно, что гиперкинетический
синдром имеет наследственную природу?
17. У монозиготных близнецов конкордантность по гиперак-
тивности составляет 73%, у дизиготных 29%. На основании
этих данных решите, является ли гиперактивность резуль-
татом плохого воспитания или какую-то роль играет на-
следственность.
18. Среди 31 пары монозиготных близнецов по дислексии
была конкордантна 21 пара. У 97 пар дизиготных близне-
цов конкордантной была 31 пара. Вычислите показатель
наследуемости дислексии.
19. Для решения вопроса о наличии наследственной основы
аутизма у детей проведены близнецовые исследования.
Изучены 23 пары монозиготных близнецов и 17 пар дизи-
готных. Среди монозиготных близнецов конкордантными
оказались 22 пары, среди дизиготных — 4. По этим дан-
ным вычислите показатель наследуемости аутизма.
20. Из 276 детей с дислексией 88% имели 1—2 родственников,
также не способных обучиться чтению. Все изученные
пары монозиготных близнецов были конкордантны по дис-
406
Контрольные вопросы и задания к разделу 12
лексии. Из трех пар дизиготных близнецов конкордант-
ной была одна пара. Соотношение девочек и мальчиков
среди детей, страдающих дислексией, соответствовало 1:4.
Обсудите роль наследственности в формировании дислек-
сии.
21. Некоторые дети не могут осознанно регулировать свою
двигательную активность. Гиперкинетические дети невни-
мательны, недисциплинированны в школе, используют
свои интеллектуальные резервы не в полной мере. Хотя
никаких патологических нарушений у таких детей не об-
наружено, считается, что в основе гиперкинетического
синдрома лежат органические причины, поэтому его иног-
да называют «минимальная дисфункция головного мозга».
Среди американских младших школьников гиперкинез
встречается примерно у 3%. Амфетамины, которые ис-
пользуются для повышения возбуждения и улучшения на-
строения у взрослых, парадоксальным образом действу-
ют на гиперкинетических детей. В отличие от эффектов у
взрослых, амфетамины успокаивающе действуют на
сверхподвижных детей, повышая их желание работать и
снижая отвлекаемость. Сходные стимуляторы, такие, как
метилфенидат (выпускаемый под торговой маркой Ritalin)
и кофеин (назначаемый в виде двух чашек кофе ежеднев-
но), также оказывают благоприятные эффекты на гипер-
кинетических детей. Если бы у Вашего ребенка были про-
блемы с поведением и школьной успеваемостью, согласи-
лись бы Вы на прием ребенком амфетаминов и их аналогов
для того, чтобы разрешить сложившуюся ситуацию? Су-
ществует ли опасность некорректного использования ле-
карств или вызывания зависимости от них? Можете ли Вы
сделать предположения о связи между клиническим на-
значением амфетаминов у детей и последующей нарко-
тической привязанностью у взрослых?
22. У некоторых людей с аутизмом наблюдаются необычай-
ные способности. Так, в известном фильме «Человек дож-
дя» у героя, страдающего аутизмом, была развита фено-
407
Раздел 12. Генетические основы аномалий поведения
менальная память. Его брат, пользуясь этим, водил его в
казино, где они выигрывали огромные деньги. Как Вы счи-
таете, этично ли использование больных людей в подоб-
ных ситуациях? Или же их способности настолько уни-
кальны, что позволяют в определенных ситуациях изме-
нять положение дел настолько, что тогда их использование
оправдано?
23. Какие особенности в структуре ДНК выявлены при хорее
Гентингтона?
24. Определите вероятность развития хореи Гентингтона у
20-летней Анны, если ее 50-летний отец и 55-летний дядя
страдают данной болезнью. Мать этих братьев тоже была
больна хореей (рис. 12.18). Какова вероятность рождения
у Анны больного ребенка?
Рис. 12.13
25. Эпилепсия — мультифакториальное заболевание. Каков
риск для ребенка быть эпилептиком, если его мать и брат
страдают этим заболеванием? Частота эпилепсии в дан-
ной популяции 0,7%, наследуемость болезни 70%.
26. Консультирующийся — здоровый мужчина, имеет боль-
ных эпилепсией отца и мать. В популяции это заболева-
ние встречается с частотой 0,2%, а наследуемость его 45%.
Какова вероятность, что у данного мужчины разовьется
эпилепсия?
27. Аутосомно-рецессивная болезнь Тэя—Сакса широко рас-
пространена в популяции евреев ашкенази.
408
Контрольные вопросы и задания к разделу 12
Мириам, Яков и Иосиф (рис. 12.14) происходят из популя-
ции евреев ашкенази, где частота гетерозигот по болезни
Тэя—Сакса, определенная на основе скрининговой про-
граммы, составляет 4%. Мириам обратилась к семейному
врачу с просьбой помочь ей остановить свой выбор на од-
ном из двух кандидатов в мужья — Якове или Иосифе.
Какова вероятность рождения больного ребенка в каждом
из возможных браков Мириам с Яковом или с Иосифом,
если все трое пока не прошли тестирование на носитель-
ство гена болезни Тэя—Сакса?
Рис. 12.14
28. Скрининг на гетерозиготность по аутосомно-рецессивной
болезни Тэя—Сакса заключается в недорогом, но не даю-
щем 100%-ной уверенности анализе уровня гексозамини-
дазы А в сыворотке крови. У носителей уровень данного
фермента понижен, хотя фенотипически они не проявля-
ют никаких симптомов. Более точный и дорогой тест за-
ключается в определении уровня фермента в изолирован-
ных лейкоцитах. На популяционном скрининговом уров-
не (массовое просеивание) применяют в основном первый
тест из-за его дешевизны.
На рис. 12.15 изображена родословная, члены которой —
евреи ашкенази, частота носительства гена Тэя—Сакса в
популяции которых 4%. Из поименованных лиц только Аб-
рам прошел скрининг по второй, более дорогой методике,
15 Введение в психогенетику
409
Раздел 12. Генетические основы аномалий поведения
и оказался гетерозигным носителем. Михаил прошел скри-
нинг по первой методике и тоже оказался гетерозиготным.
В лаборатории, в которой проходили скрининг указанные
мужчины, первый тест гарантирует правильность резуль-
татов с вероятностью 99/100, во втором же тесте резуль-
таты однозначны. Определите риск рождения больного
ребенка у каждой из трех пар, третьего поколения родос-
ловной.
Рис. 12.15
29. Болезнь Тэя—-Сакса встречается среди евреев ашкенази
с частотой 11:10 000, а среди евреев сефардов с частотой
11:100 000. Какова вероятность рождения ребенка с этой
болезнью в браке между дядей и племянницей у евреев
ашкенази и сефардов?
30. В чем проявляются различия в реакции людей на прием
алкоголя?
31. Что лежит в основе различий в реакции людей на прием
наркотических средств?
32. Какие гены считаются кандидатными в предрасположен-
ности к алкоголизму?
33. Приведите примеры межпопуляционных различий в ре-
акции на алкоголь.
410
Контрольные вопросы и задания к разделу 12
Тип близнецов Оба курят Оба не курят Один курит
Дизиготные 30 20 50
Монозготные выросли вместе 48 23 29
Монозиготные выросли врозь 36 43 21
34. В таблице приведены данные о курении у близнецов (ука-
зано число пар). Обсудите гипотезу о наличии наслед-
ственной основы курения.
35. Среди курящих женщин были выявлены близнецовые
пары: 53 монозиготных и 69 дизиготных. Среди монози-
готных пар в 44 курили обе, в 9 одна. Среди дизиготных в
34 курили обе, в 35 — одна. Вычислите конкордантность
женщин по курению и обсудите результаты.
36. Как Вы относитесь к такому высказыванию: «поскольку
расстройство самоконтроля имее’!' наследственную осно-
ву, то с человека снимается ответственность за свои по-
ступки» ?
37. Приведите факты, которые доказывают генетическую
природу анорексии и булимии.
38. Среди гэмблеров можно выделить по меньшей мере четы-
ре типа лиц. «Гэмблер действия» стремится к соперниче-
ству, интересуется азартными играми, требующими сно-
ровки. Данная категория предпочитает спортивные пари,
казино, карточные игры и гонки. «Гэмблеры, уходящие от
действительности», наоборот,-играют в игры, в которых
не требуется специальных умений, например, игровые ав-
томаты, видео-покер. «Internet-гэмблеры» наслаждаются
собственностью своего персонального компьютера, за ко-
торым они могут проводить 24 часа в сутки, причем в тай-
не от других. И, наконец, «биржевые гэмблеры» специа-
лизируются в биржевых сделках. Какой тип гэмблера, по
Вашему мнению, наиболее опасен для общества? Какие
физические и психологические последствия ожидают гэм-
блеров каждого типа?
15*
411
ОТВЕТЫ И РЕШЕНИЯ
К разделу 1
12. К облигатным гетерозиготам относятся II/1,2; IV/1. Носи-
телями с вероятностью 1/2 являются 1/1,2; П/З; V/1, а но-
сителем с вероятностью 1/4 — 1/3.
13. 50%.
14. Да, с вероятностью 50%.
15. 25%.
16. 50%.
17. 1/9.
18. Родители Виктории — носители, что можно заключить из
фенотипа пораженной. Следовательно, отец Юрия мог
передать мутантный ген сыну с вероятностью 1 /2. Дедуш-
ка Виктории гетерозиготен с вероятностью 1/2, а его род-
ной брат — дедушка Анастасии — с вероятностью 1/2 х
1/2 = 1/4. Далее, мать Анастасии гетерозиготна с вероят-
ностью 1/4 х 1/2 = 1/8, а сама Анастасия с вероятностью
1/8 х 1/2 = 1/16. Отсюда, риск рождения больного ребенка
у консультирующихся составляет 1/2x1/16x1/4= 1/128.
29. Ген эллиптоцитоза обозначим Е, его нормальный аллель е,
аллель резус-положительности Rh, а резус-отрицательно-
сти rh. Тогда, у I/1 генотип rhe/rhe, а у 1/2 генотип rhe/RhE.
В потомстве 3 сибса имеют фенотип RhE, 5 сибсов фено-
тип rhe и один сибс фенотип rhE. Попарное сопоставле-
ние сибсов дало такие значения: сходных по обоим при-
знакам 13 пар сибсов, сходных по первому и не сходных
по второму признакам 5 пар, не сходных по первому и сход-
ных по второму признакам 3 пары сибсов и не сходных по
обоим признакам 15 пар (всего 36 сопоставлений):
'' ...... 1 сибс 2 сибс сходные несходные X
сходиыс 13 5 18
несходные 3 15 18
I 16 20 36
X2 = 9,11, df = 1, р < 0,001.
412
Сцепление имеется, фенотип П/6 — результат кроссинго-
вера.
30. Обозначим ген дистрофии D, его нормальный аллель d, ген
«секреторное™» S, аллель «несекреторности» s. Тогда,
исходя из анализа потомства, генотипы родителей тако-
вы: Анжела sd/sd, Игорь SD/sd. Таким образом, ген дист-
рофии (D) сцеплен с геном «секреторности» (с). Вероят-
ность кроссинговера между этими генами равна 0,1, сле-
довательно, вероятность того, что кроссинговер не
произойдет, равна 0,9. Учитывая состояние «секреторно-
сти» сибсов, вероятность кроссинговера и пенетрантность
гена дистрофии у разных полов, можно получить следую-
щие значения риска развития дистрофии у детей:
Полина — 0,64 х 0,9 = 0,576; Бэлла — 0,64 х 0,1 = 0,064;
Роза — 0,64 х0,9 = 0,576; Артем— 1,00x0,1 =0,1; Валерий —
1,00x0,9 = 0,9.
31. Очевидно, что ближе всего к гену пигментной ксеродер-
мы расположен ген буллезного эпидермолиза — между
ними кроссинговер происходит с наименьшей вероятно-
стью (6%). Дальше всего от гена пигментной ксеродермы
расположен ген пигментного ретинита — между ними
кроссинговер происходит наиболее часто (14%). Ген бул-
лезного эпидермолиза лежит между геном пигментной
ксеродермы и пигментного ретинина, что согласуется с
численными значениями кроссинговера между всеми тре-
мя генами— 6% + 8% — 14%.
34. По линии сыновей близка к 0, дочерей — 1/4 (для мальчи-
ков).
35. Геннадий передал дочери ген дальтонизма. Следователь-
но, у Марины, как облигатной носительницы, риск рож-
дения сына дальтоника равен 1/2. Наличие у Николая даль-
тонизма указывает на то, что его мать Нина — гетерози-
готная носительница этого гена. Отец Инны здоров,
следовательно не имеет гена дальтонизма. Нина с вероят-
ностью 1/2 могла передать Инне ген дальтонизма. Вероят-
ность, что сын Инны будет дальтоником 1/2 х 1/2 = 1/4,
риск для дочери равен нулю.
Ответы и решения
37. Поскольку Антонина здорова, то ген орофациодигиталь-
ного синдрома она не унаследовала. Следовательно, с рож-
дением Антонины ее род избавился от мутантного гена, и
естественно, риск рождения у Антонины больных детей —
как мальчиков, так и девочек — равен 0.
40. 7/16.
41. Комплементарное, т.к. только вместе они обеспечивают
образование качественно нового продукта.
42. Среди пятерых сестер мы можем наблюдать все возмож-
ные типы лактации — от максимальной ее выраженности
(у Александры) до полного отсутствия (у Эльвиры) с про-
межуточными состояниями у Владиславы, Клавдии и Ве-
роники. В предположении, что секреция молока зависит
от пары полимерных доминантных генов, действие кото-
рых суммируется, можно заключить, что генотип Александ-
ры — ЦЦЦЦ, Владиславы — L(L(L212 (или же ЬфЦЦ), Клав-
дии — Вероники — ЦЁЦг (или же и Эльви-
ры — 1,1,1^.
44. а) Оба родителя АаВЬ; б) могут быть различные варианты
ААЬЬ х ААЬЬ, ааВВ х ааВВ, ААЬЬ х ааВВ.
К разделу 2
7. 500.
8. 61.
9. 263.
10. Никаких.
11. YYYYYY.
14. 23.
15. 46.
16. XX.
17. XY.
25. 0,54%.
26. 0,1%.
27. 7,14%.
28. 4,1%.
414
К разделу 2
29. 0%.
30. Возраст отца не играет роли. Риск 0,2%.
32. а) 47, XX, + 14 или 47, XY, + 14; б) 45, XX, - 10 или 45, XY,
- 10; в) 46, XX, i( 15q) или 46, XY, i(l5q); г) 46, XX, rob(21q,
13q) или 46, XY, rob(21q, 13q); д) 46, XX, t(2q —, 8p + ) или
46, XY, t(2q —, 8p + ).
34. а) у женщины имеется инверсия в пятой хромосоме, при-
ведшая к увеличению короткого и уменьшению длинного
плеча; б) у женщины присутствует слияние длинных пле-
чей 13-й и 22-й хромосомы; в) у мужчины делеция длинно-
го плеча второй хромосомы, разрыв произошел на уровне
третьей полосы второго района; г) у мужчины делеция ко-
роткого плеча 1-й хромосомы с разрывом в третьем райо-
не на уровне 4-й полосы.
35. Одна клетка содержит 45 хромосом и является моносоми-
ком по 18 хромосоме, вторая содержит 47 хромосом и яв-
ляется трисомиком по 18 хромосоме.
36. Одна клетка является моносомиком по 14 и трисомиком
по 20 хромосоме, другая — трисомиком по 14 и моносоми-
ком по 20 хромосоме.
37. Наименьшие шансы на выживание у эмбрионов с трисо-
мией по аутосомам. Дисомики по Y-хромосоме имеют бо-
лее благоприятные шансы по сравнению с тетрасомика-
ми по Х-хромосоме.
38. Зигота с хромосомным набором синдрома Клайнфельте-
ра может получиться в двух случаях:
1) слияние материнской гаметы с неразошедшимися в пер-
вом или втором делении мейоза Х-хромосомами и нор-
мальной отцовской гаметы с Y-хромосомой:
2) слияние материнской гаметы с нормальным хромосом-
ным набором с отцовской гаметой, имеющей хромосом-
415
ный набор XY, который получился от нерасхождения хро-
Зигота с хромосомным набором синдрома Жакоб получа-
ется при слиянии нормальной материнской гаметы и от-
цовской гаметы с двумя Y-хромосомами. Такая гамета —
результат нерасхожденеия хромосом во втором мейоти-
ческом делении.
39. Хромосомный набор зиготы XY. В первом митотическом
делении зиготы одна из дочерних клеток утратила Y-xpo-
мосому, что и привело к формированию набора ХО, что со-
ответствует синдрому Тернера. В другой дочерней клетке
развитие пошло по обычному пути и привело к формиро-
ванию здорового мальчика XY.
40. 1) Нерасхождение хромосом в одном из мейотических де-
лений при оогенезе или в первом мейотическом делении
при сперматогенезе. 2) Нерасхождение Х-хромосом в пер-
вом и втором делениях мейоза при оогенезе. 3) Нерасхож-
дение Х-хромосом в одном из делений при оогенезе или
во втором делении мейоза при сперматогенезе. 4) Нерас-
хождение Y-хромосом во втором делении мейоза при спер-
матогенезе.
41. Можно предположить, что зигота имела набор хромосом
46,XX. В одном из делений произошло нерасхождение хро-
матид одной из удвоенных Х-хромосом. Одна из дочерних
клеток получила три Х-хромосомы, а другая — одну. Эти
416
удвоение
клон 46, XX
клон 45, X
клон 47, XXX
клетки далее делились нормальным митозом и дали нача-
ло соответствующим клонам:
Степень мозаицизма зависит от того, в каком по счету де-
лении произошло не расхождение. Чем позднее это случи-
лось, тем меньше доля мозаичного клона.
42. А. 1) Зигота XY. Утрата Y-хромосомы в одном из митоти-
ческих делений. 2) Зигота XY. Нерасхождение У-хромосо-
мы в митозе. 3) Зигота XXY или ХУ. Утрата или нерасхож-
дение половых хромосом. 4) Возможно, что в образовании
зиготы участвовали яйцеклетки и полярное тельце, кото-
рые были оплодотворены сперматозоидами X и Y. 5) Зиго-
та XXY. Нерасхождение У-хромосомы. 6) Возможно, зиго-
та XXY, либо ситуация 4 с последующей утратой хромо-
417
Ответы и решения
сом. 7) Возможен хромосомный набор зиготы XXY с пос-
ледующим и неоднократным нерасхождением и утратой
половых хромосом.
Б. 1) Зигота XXY. Утрата Y-хромосомы. 2) Зигота XXY. Нерас-
хождение Х-хромосомы. 3) Утрата Х-хромосомы в зиготе
XXXXY. 4) Зигота XXXXY. Утеря Y-хромосомы и нерас-
хождение Х-хромосомы.
В. 1) Зигота XX. Утрата Х-хромосомы. 2) Зигота XX. Нерас-
хождение Х-хромосомы. 3) Зигота XXX. Утрата Х-хромо-
сомы. 4) Зигота полисомна по Х-хромосомам. Утрата или
нерасхождение Х-хромосом. 5) Зигота XX. Первые мито-
зы нормальны. В последующих нерасхождение Х-хромо-
сомы.
43. При мейозе из диплоидной клетки с трисомией образуют-
ся гаметы, половина которых несет нормальный гаплоид-
ный набор хромосом, которые могут дать начало нормаль-
ной зиготе.
46. 1) Два. 2) Ни одного. 3) Одно. 4) Одно. 5) и 6) Ни одного.
7) Три.
50. 0%.
51. 100%.
52. Да.
К разделу 3
2. 40%.
3. 50%.
4. Предположим, что отдельные признаки, входящие в синд-
ром Марфана, контролируются сцепленными доминант-
ными генами: ген А отвечает за высокий рост и арахно-
дактилию, ген В — за смещение хрусталика, ген С — за
врожденный порок сердца (а, Ь, с — нормальные аллели
этих генов). Согласно этой гипотезе Олег, обладающий
всеми тремя доминантными генами (А-В-С-), передал каж-
дому сыну (исходя из симптомов болезни у них) по одно-
му гену из триады генов синдрома: Игорю — ген А, Анд-
418
К разделу 3
рею — ген В и Петру — ген С. Поскольку информация о
болезни матери братьев отсутствует, вероятнее всего, она
здорова, следовательно, ее генотип aabbcc. Информация
о сыновьях указывает на то, что Олег был гетерозиготным —
АаВЬСс. Генотип его сыновей —Игоря Aabbcc, Андрея
ааВЬсс, Петра ааЬЬСс — результат кроссинговера при га-
метогенезе у Олега. Однако информация о внуках Олега
опровергает эту гипотезу, поскольку у них имеются при-
знаки, которые отсутствовали у их отцов. Так, у Марии долж-
ны присутствовать гены А и С (у Игоря же нет гена С), у
Михаила и Руслана — гены А и В (у Андрея нет гена А), и,
наконец, у Анны — гены А и В (у Петра имеется лишь ген С).
Следовательно, более вероятно действие одного гена с
плейотропным эффектом. Его неодинаковое проявление
у родственников может быть связано с действием генов-
модификаторов или какими-то негенетическими причи-
нами.
6. Поскольку на ранних стадиях эмбриогенеза во всех клет-
ках женского зародыша происходит инактивация одной
из Х-хромосом, то вследствие этого женщина является
мозаиком по Х-хромосомам родителей. В одних клетках
активна материнская Х-хромосома (с мутантным геном),
а в других — отцовская (с нормальным геном). Выражен-
ность мышечной слабости будет зависеть от того, в какой
части клеток активна Х-хромосома с геном дистрофии —
чем эта доля выше, тем меньше у женщины выражена
мышечная слабость.
7. Ген предрасположенности к раку Зинаида получила от сво-
его отца Олега, который не проявился у него, так как огра-
ничен полом. Лидия могла получить ген от отца с вероят-
ностью 1/2. Вера могла передать мутантный ген дочери
также с вероятностью 1/2. Петр мог получить дефектный
ген от матери с вероятностью 1/2, но болезнь у него не про-
явилась, и он не информативное звено в данной родослов-
ной. Своей дочери Петр мог передать, таким образом, этот
аутосомный ген с вероятностью 1/2 х 1/2 = 1/4. Мария не
419
имеет родства с Олегом и семьей, из которой он происхо-
дит, поэтому риск заболеть этой формой рака у нее бли-
зок нулю. Лидия и дочь Веры имеют риск 1/2, дочь Петра —
1 /4, потому эти женщины должны находится под медицин-
ским наблюдением.
10. У А, Б и В вероятность заболеть ревматизмом определяет-
ся из данных условия задачи, а у Г эта вероятность повы-
шена (задействованы дополнительные факторы). Таким
образом, искомая вероятность для консультирующихся та-
кова:
1) для А— 10% (больная сестра);
2) для Б — 10% (оба родителя больны);
3) для В — 20% (мать и брат больны);
4) для Г — более 20% (оба родителя и сестра больны).
И. В целенаправленной профилактике гипертонической бо-
лезни в порядке снижения риска нуждаются: Ш/2 — три
фактора риска (больной родственник первой степени род-
ства, ожирение, повышенное артериальное давление), Ш/З
(два фактора риска), Ш/5 (один фактор риска).
12. Самый высокий риск у Ш/З и Ш/4. Риск для Ш/2 ниже.
Самый низкий риск среди братьев у Ш/8. Среди сестер
наиболее высок риск у Ш/5, наименьший у Ш/6 и Ш/7.
13. По таблице Смита консультирующийся имеет риск язвы
немного более 1% (никто из сибсов или родителей не бо-
лен).
17. Из ооцита I порядка может образоваться только одна зи-
гота.
18. Если беременность наступила у женщины 45 лет, ооцит
I порядка находился в яичнике 45 лет, за это время он мог
подвергаться многократным воздействиям мутагенов и
других неблагоприятных факторов. Поэтому у зиготы мо-
жет быть нарушен генотип.
20. В семье Б этот дефект имеет наследственную природу, по-
этому он может повториться у других детей и у внуков,
если они получат соответствующий ген. В семы1 А такого
гена нет, дефект возник под влиянием тератогена и не дол-
420
1 К разделу 4
жен повторяться, если исключить тератогенные воздей-
ствия в период беременности.
21. Наличие глухоты при нормальном генотипе можно объяс-
нить тем, что тератогенный фактор помешал генам, уча-
ствующим в развитии органа слуха, реализовать их нор-
мальную генетическую программу. Выяснять природу
этой аномалии у пациента нужно для генетического про-
гнозирования вероятности повторного рождения глухих
детей в данной семье, а потом и у пациента, когда он ста-
нет взрослым.
К разделу 4
4. Вероятность передачи определенного гена каждому сле-
дующему поколению равна 1/2. Чтобы определить долю
общих генов между любыми двумя родственниками, не-
обходимо установить степень их родства. Она определя-
ется на родословной как число связующих их звеньев.
Доля общих генов равна (1/2)п, где п — степень родства.
Для данного случая:
а) степень родства у Василия и Лидии равна 0, следователь-
но, у них нет и общих генов;
б) степень родства у Андрея и Дианы равна 5, поэтому доля
их общих генов (1/2)5 = 1/32.
5. а) Николай и Александр — троюродные братья; б) Ольга и
Татьяна — троюродные племянница и тетя.
6. а) У Софьи нет родственниц женского пола; б) у Ирины 3
родственника мужского пола (генеалогические координа-
ты: 1/1, II/1, П/2).
7. а) У Людмилы 6 свойственников (генеалогические коор-
динаты — 1/4, 1/5, 11/1, 11/5, 11/6, П/7); б) у Ольги 5 свой-
ственников мужского пола (генеалогические координаты —
I/4, II/1, II/4,11/5,111/2); в) у Сергея 4 свойственницы женско-
го пола (генеалогические координаты — 1/2, II/1,11/3, П/6).
8. Да.
9. 4.
421
10. 4.
11. 6.
12.3.
19. Из родословной видно, что больные дети рождаются от здо-
ровых родителей. Если же один родитель болен, а второй
здоров, то возможно рождение здоровых детей. Следова-
тельно, болезнь наследуется по рецессивному типу. Боле-
ют только мальчики, значит, признак сцеплен с половой
хромосомой. Передача признака от мужчины к мужчине
происходит через женщин, таким образом, признак сцеп-
лен с Х-хромосомой.
22. Из родословной обнаруживаем, что наследуемый признак
встречается во всех поколениях, как у женщин, так и у
мужчин. Пораженные потомки обоего пола рождаются от
браков обоих типов — с пораженной матерью или с пора-
женным отцом. Выдвигаем гипотезу об аутосомно-доми-
нантном наследовании брахидактилии. Для анализа отби-
раем семьи —1/1, П/8, III/1, Ш/7, Ш/9, III/10, III/11, III/12,
IV/1, IV/3, IV/5, IV/6, IV/9, IV/12, IV/15 (второй здоровый
партнер не указан). С помощью критерия проверяем на-
личие межполовых различий в разных группах больных и
здоровых потомков:
Фенотип Пол Всего
Мужчины Женщины
Здоровые 13 23 36
Больные 16 15 31
Всего 39 38 67
X2 = 0,799, df = 1, р > 0,05.
Критерий %2 показал отсутствие достоверных различий по
распределению случаев брахидактилии между мужчина-
ми и женщинами, следовательно, данный признак не сцеп-
лен с полом.
Аналогично определяем значение х2 для определения дос-
товерности различий между ожидаемым и фактическим
количеством больных и здоровых потомков. Ожидаемое
422
К разделу 4
соотношение в браках Аа х аа больных и здоровых детей
равно 1:1, или для наших данных 34:34. Тогда, расчет при-
нимает следующий вид:
Фенотип Фактические Ожидаемые
Больные 36 34
Здоровые 32 34
Всего 68 68
X2 = 0,235, df = 1, р > 0,05.
Недостоверность различий по критерию /2 позволяет при-
нять гипотезу об аутосомно-доминантном наследовании
брахидактилии.
23. Очевидно, что зубная эмаль коричневого цвета наследу-
ется по доминантному типу, поскольку у пораженных ро-
дителей бывают пораженные дети. Среди пораженных
встречаются лица обоего пола. Однако интересен тот факт,
что у больного отца (Ш/1) из 12 детей поражены лишь 8
дочерей, а 4 сына здоровы. Поэтому, мы может предполо-
жить, что признак сцеплен с Х-хромосомой, и как уже было
сказано, доминантен. Проверим с помощью критерия %2
гипотезу о доминантности анализируемого признака. Мы
имеем дело с 5 браками, где был болен один родитель и
здоров другой (1/1, II/1, Ш/1, IV/3, IV/4 — для обеспече-
ния меньшей «перегруженности» родословной здоровые
супруги в ней не отмечены). Всего в этих 5 семьях имеется
22 ребенка — 13 больных и 9 здоровых. При доминирова-
нии теоретически ожидается одинаковое количество боль-
ных и здоровых (по 11 человек), если предположить гено-
тип больных женщин Аа, а больных мужчин — А— (вто-
рого гомолога у мужчин нет, т.к. признак по нашему
предположению сцеплен с Х-хромосомой):
Фенотип Фактические Ожидаемые
Больные 1 3 9,5
Здоровые 6 9, 5
Всего 19 19
X2 = 2,58, df = 1, р > 0,05.
423
Видно, что рассчитанный критерий /2 свидетельствует о
недостоверности различий между фактическими и ожи-
даемыми классами больных и здоровых. Следовательно,
изучаемый признак доминантный, как мы и предполага-
ли.
Доказать сцепление с полом гена, ответственного за зуб-
ную эмаль коричневого цвета, опираясь на данную родос-
ловную, сложно, поскольку в ней имеется лишь один слу-
чай, когда у больного мужчины можно проанализировать
его потомство. Однако этот единичный случай весьма по-
казателен. Если рассуждать, что признак действительно
доминантен и сцеплен с полом, то вероятность рождения
любого количества дочерей у данного мужчины больны-
ми, равна 1.
Если же признак доминантный и аутосомный, то вероят-
ность его передачи как дочери, так и сыну равна и состав-
ляет 1/2. Вероятность рождения девочки равна 1/2, такая
же вероятность рождения мальчика, причем эти события
независимы. Вероятность, что девочка будет больна, со-
ставляет 1/2, такая же вероятность рождения здоровой де-
вочки. Для мальчика риск родиться больным составляет
также 1/2, как и вероятность оказаться здоровым — 1/2.
Независимыми событиями также являются рождение де-
вочки больной, а мальчика здоровым, что характерно для
рассматриваемого случая в нашей родословной. Таким
образом, мы может рассчитать вероятность совместного
события — рождения 8 девочек больными и рождение 4
мальчиков здоровыми в предположении аутосомно-доми-
нантного типа наследования коричневого цвета зубной
эмали как произведение вероятностей этих двух незави-
симых событий. В свою очередь вероятность каждого из
этих двух событий можно рассчитать на основе формулы
п! k к
Бернулли — Q(k)= _ iq/p) 1 гАе п — общее ко-
личество повторяющихся событий, к. — количество инте-
424
К разделу 5
ресующих нас событий, р — вероятность интересующего
нас события, q — вероятность противоположного ему со-
бытия.
Если обозначить количество всех рожденных девочек п(1
количество больных девочек к,, вероятность рождения
больной девочки р(, вероятность рождения здоровой де-
вочки q,, то вероятность рождения из 8 девочек 8 больных
составит
С8(8) = СЖ =
8! flYflT8
8!(8 -8)'.[ 2 Д 2)
Аналогично, обозначив количество всех рожденных маль-
чиков п2, количество больных мальчиков к2, вероятность
рождения здорового мальчика р2, вероятность рождения
больного мальчика q2, найдем, что вероятность рождения
из 4 мальчиков 4 здоровых составит
4' Л1 1V4 /1Y
С (4) = Cptq’ =---:-- - - = -
4 2 4!(4 - 4Д 2 J 2 ) ДД
Совместная вероятность рождения 8 больных девочек и 4
здоровых мальчиков у больного отца (Ш/1) в предположе-
нии аутосомно-доминантного типа наследования призна-
Y _ 1
д] ~ 4096'ВиА"
ка составит С8(8)хС4(4) =
2
но, что полученная вероятность 1/4096 слишком мала для
того, чтобы объяснить аутосомно-доминантным наследо-
ванием рождение всех 8 дочерей больными. Поэтому, более
вероятно, что признак «коричневый цвет зубной эмали» на-
следуется по доминантному, сцепленному с полом, типу.
К разделу 5
6. m — 3,1; 3,6; 3,7; 3,4; 5,3; 6,1; d — 4,4; 11,7; 7,1; 7,5; 2,2; 2,1.
7. Вероятность рождения мальчика р (0,5132), девочки
q (1 —0,5132 = 0,4868). Для определения вероятности рож-
425
Ответы и решения
дения разных дизиготных пар воспользуемся разложени-
ем бинома в степени 2 (2 детей), где двумя событиями, со-
ставляющими полную группу, будут рождение мальчика
(м) с вероятностью р и рождение девочки (д) с вероятнос-
тью q: (р(м) + ц(д))2 = р2(м)2 + 2рц(мд) + д2(д)2. Доля раз-
нополых среди дизиготных близнецов 2рц(мд)/(р2(м)2 +
2рц(мд) + ц2(д)2) = 2рц(мд)/1 = 2рц(мд). Доля дизиготных
пар (N ) равна отношению числа разнополых близнецовых
пар (NAp) к их ожидаемой доле — Мдр/2рц(мд). Остальные
пары монозиготные:
Nm = N — Ыд = N — (NAp/2pq(MA)).
Для изученного населения монозиготных и дизиготных
пар примерно:
NM = 3363 — (1195/(2x0,5132x0,4868)) = 971.
NM=N — N = 3363 — 971 = 2392.
Д м
Доля монозиготных пар 971/3363 = 0,29, дизиготных —
2392/3363 = 0,71.
8. 1 — нет, 2 — да, 3 — происхождение пары проблематично,
4 — да, 5 — да, 6 — да.
9. 1 — да, 2 — нет, 3 — да, 4 — да, 5 — да, 6 — да, 7 — да.
13. Для смертности в возрасте от 20 до 60 лет — 0,15, смертно-
сти от травм — 0,03.
15. Имеющиеся данные можно проанализировать в два эта-
па. Во-первых, при анализе пар, где имеется один больной
партнер, а также пар, где оба партнера больные, можно
установить связь заболевания с зиготностью (или, други-
ми словами, с генотипом). Во-вторых, в группе обоих боль-
ных близнецов обоих типов (моно- и дизиготных) можно
установить связь между зиготностью (или генотипом) и
локализацией рака (один и тот же орган или разные орга-
ны). Для удобства эти два расчета разместим в две табли-
цы, воспользовавшись критерием %2 для установления свя-
зи между признаками:
426
К разделу 5
1. Больные (конкордантные + дискордантные по болезни)
-к з Конкордантные Дискордантные Всего
Монозиготные 21 143 164
Дизиготные 40 292 332
Всего 61 435 496
3 — зиготность близнецов, К — конкордантность,
%2 = 0,009, df = 1, р > 0,05.
2. Оба больные (конкордантные + дискордантные
по локализации)
° 3 Конкордантные (одинаковая локализация) Дискордантные (разная локализация) Всего
Монозиготные 8 13 21
Дизиготные 9 39 48
Всего 17 52 69
3 — зиготность близнецов, О — орган, %2 = 1,995,
df = 1, р > 0,05.
На основании полученных значений %2 можно сделать сле-
дующие заключения. Из анализа первой таблицы следует
вывод, что болезнь раком одного партнера из обоих типов
близнецовых пар (моно- или дизиготных) не связана с бо-
лезнью или же ее отсутствием у другого партнера пары.
Иными словами, имеется слабое влияние генетических
факторов на развитие рака у человека. После анализа вто-
рой таблицы можно сделать вывод, что, если все же в близ-
нецовой паре оба партнера больны, то связь между нали-
чием болезни и ее локализацией в одном и том же органе
также отсутствует. Таким образом, и само заболевание
раком, и его локализация имеют слабые генетические
предпосылки, и, следовательно, имеют не генетическую,
а в большей степени средовую обусловленность.
427
К разделу 7
4. О, 54 и 0,46.
5. 0,00167.
6. Частоту нормального аллеля (q) находим, исходя из доли
’ лиц, у которых этого заболевания нет — q = д/0,9 = 0,95-
Тогда частота гена гиподонтии равна р = 1 — q — 1 — 0,95 =
0,05. Соотношение доли гетеро- и гомозигот по этому при-
2pq _ 2x0,05x0,95 _
знаку:
Таким образом, гетерозигот по гиподонтии в популяции в
38 раз больше, чем гомозигот.
7. Поскольку пенетрантность нефробластомы составляет
0,63, то людей, имеющих мутантный ген (один или два), в
популяции больше, чем обнаруженных больных 7 человек.
Всего в популяции И носителей этого гена (7:0,63 = 11).
Число людей, не имеющих гена нефробластомы, состав-
ляет 237 500 — 11 = 237 489. Обозначив частоту гена не-
фробластомы р, а частоту его рецессивного аллеля q, по
формуле Харди—Вайнберга найдем частоту рецессивно-
го гена как корень квадратный из доли здоровых людей в
популяции —
/237489
V 237500
- 0,999977. Соответственно,
р = 1 — q = 1 — 0,999977 = 0,000023.
8. 2pq = 0,014, р = 1— q, 2q(l — q) = 0,014, 2q2 —2q = 0,014.
Решаем квадратное уравнение 2q2 — 2q — 0,014 = 0. Корни
квадратного уравнения вида ах2 + Ьх + с = 0 находим по
-b±Vb? - 4ас 2±д/4-0,112 2±1,97
формуле х, 2 =-------------. q, 2 =---------= —-—,
2а 4 4
по условию q < 1, q = 0,0075.
Тот же результат можно получить и другим способом. Так
как 2pq очень мало (1,4% = 0,014), то мало и q, а р близко к
единице. Следовательно, 2pq = 2q = 0,014, q = 0,007.
9. Аниридия — АА и Аа, доля пораженных — р2 + 2pq, доля
нормальных (аа) q2 = 0,9999. q — 0,99995, р = 1 — q =
0,00005.
10. Частоту аллелей у мужчин находим, используя данные о
частоте фенотипов у этого пола: р^ = 46/(46 + 154) = 0,23;
q^= 154/(46 + 154) — 0,77. Частота аллелей у женщин рав-
на: q’2 = 88/(88 + 114) = 0,44, q? = 0,66; р^ = 1 — 0,66 =
0,34. Женщины имеют две Х-хромосомы, а мужчины одну,
поэтому, если соотношение полов в популяции 1:1, то 2/3
генов, сцепленных с Х-хромосомой, принадлежат женщи-
нам, а 1/3 — мужчинам. Частота аллелей в среднем в по-
пуляции р = 2/3 р£ + 1/3 р_7 = 2/3 х 0,34 + 1/3 х 0,23 =
0,30; q = 2/3 q?+ 1/3 q^= 2/3x0,66 + 1/3x0,77 = 0,70.
11. Доля больных мужчин q, больных женщин — q2. Дтя на-
хождения q при соотношении полов 1:1 составим и решим
следующее квадратное уравнение:
— х Nx q2 + — х N х q = 0,03х N
2 2 4
q2 + q = 0,06
q2 +q- 0,06 = 0
_ ~1±д/12 ~ 4x1 x(-0,06)
012 —* ,
2x1
q = 0,057
Доля больных мужчин 0,057, а доля больных женщин 0,0572 =
0,0032 (мужчин почти в 18 раз больше). Если соотношение
полов отклоняется от 1:1, вычисления будут аналогичны-
ми, лишь с учетом долей лиц обоего пола в популяции, ко-
торые найдем из соотношения: х + 0,9х = 1, откуда х =
0,53, где х — доля женщин в популяции. В этом случае квад-
ратное уравнение и его решение примет вид:
0,53xNxq2 + (l-0,53)xNxq = 0,03xN
0,53q2 + 0,47q = 0,03
0,53q2 + 0,47q - 0,03 = 0
- 0,47 ± Jo,472 - 4 x 0,53x(-0,03)
41,2 —
2x0,53
q = 0,060
При этом соотношении полов доля больных мужчин 0,060,
а доля больных женщин 0,0602 = 0,0036.
12. А = р2АА + 2pqA0 = 0,343 = 43,3%, В = q2BB + 2qrB0 =
0,248 = 24,8%, АВ = 2pq = 0,0945 = 9,5%, 0 = г2 = 0,315 =
31,5%.
18. Генетический вклад африканских негров в генофонд аме-
риканских негров рассчитываем исходя из частоты иссле-
дуемого гена в популяции американских (qa), африканс-
ких негров (qb) и белых (qc):
= °’44~0’02 = о 69 = 69%
qa-qb 0,63-0,02
Следовательно, 31% среди предков американских негров
не принадлежали к негритянской расе.
19. Интенсивность потока генов, или миграции, m можно оп-
ределить по формуле:
0^94-0^22
0,0-0,422
1-т = !Д78 = 0,9795;
m = 1 - 0,9795 = 0,0205 = 2,05%.
Таким образом, поток генов от белых к неграм шел со сред-
ней интенсивностью 2,05% за одно поколение. В результа-
те через 12 поколений доля генов африканских предков
составляет сейчас (1 —ш)12 = 0,78 общего числа генов нег-
430
К разделу 7
ров США. Около 22% генов (1 — 0,78 = 0,22) американс-
кие негры унаследовали от белых.
20. Определим частоту гена резус-отрицательности в двух ис-
ходных (qt и q2) и в гибридной (q3) популяциях:
q, = Т0Д5 = 0,38;q2 = Д01 = 0,1 ;q3 = 7W = 0,3-
Соотношение генофондов первой и второй популяций в
третьей составляет:
q3-q, _ 0,3-0,38 Q2
q2-qt 0,1-0,38 ’ '
Таким образом, в гибридной популяции генофонд первой
популяции (с частотой гена 0,38) составляет 28%, а второй
(с частотой гена 0,1) — 72% (1 — 0,28 = 0,72).
22. Частота нормального аллеля (А) и аллеля серповиднокле-
точности (S) составляют:
2x3122 +998 А
р, =-------------= 0,8768; q = 1 - р. = 0,1232.
Га 2x4130 4s Еа
Теоретически ожидаемые частоты генотипов для равно-
весного состояния составляют:
Р1а = 0,77; 2pqAS = 0,22; q*s=0,01.
Фактические частоты составляют соответственно: АА =
3122/4130 = 0,756; AS = 998/4130 = 0,242; SS = 10/4130 =
0,002.
Выживаемость рассчитываем как отношение наблюдае-
мых частот фенотипов к теоретически ожидаемым: АА =
0,756/0,77 = 0,98; AS = 0,242/0,22 = 1,12; SS = 0,002/0,01 =
0,2.
Относительную приспособленность рассчитываем при де-
лении этих величин на самую большую из них — 1,12, кото-
рую принимаем за единицу. Величины приспособленности
для трех генотипов составляют 0,88,1 и 0,18. Против нормаль-
ной гомозиготы действует отбор с интенсивностью 0,12, а
против гомозиготы по гену серповидно-клеточной анемии
интенсивность отбора составляет 0,82.
431
23. Прежде всего, исключим из рассмотрения 5 детей, рож-
денных от браков, где один или оба родителя страдали
ахондроплазией, поскольку вероятность передачи патоло-
гического гена потомку значительно выше, чем вероят-
ность возникновения мутации в гамете с нормальным ге-
ном. Частоту мутации гена ахондроплазии определяем
прямым подсчетом исходя из отношения гамет, в которых
возникла мутация, ко всем родительским гаметам:
(105555 - 5) х 2
24. Поскольку гомозиготы по гену фенилкетонурии не остав-
ляют потомства, против них действует отбор с интенсив-
ностью s = 1. Скорость мутирования составляет u = sq2 =
4х 10 s.
25. Поскольку хромосомный набор диплоидный, получим ско-
рость мутирования в отношении указанных синдромов
1/1400, 1/12000 и 1/14000 гамет.
29. а) (1/2)5 + (1/2)5 + (1/2)5 + (1/2)5 = 1/8; б) (1/2)5 + (1/2)5 +
(1/2)5 + (1/2)5 + (1/2)5 + (1/2)5 + (1/2)5 + (1/2)5 = 1/4.
30. 1/2 Ч 1/2 Ч 1/4 = 1/16.
31. 2/3 Ч 1/4 Ч 1/4 = 1/24.
32. Допустим, что заболевание Симоны — результат сегрега-
ции генов, а не новая мутация. Тогда считаем, что она по-
лучила по рецессивному гену от каждого гетерозиготного
родителя. Следовательно, Михаил гетерозиготный, а веро-
ятность того, что гетерозиготен Давид, как родной брат,
составляет 1/2. Такая же вероятность быть гетерозигот-
ным для Степана — 1/2, для Ибрагима — 1/2 х 1/2 = j и,
наконец, для Никона — 1/4 х 1/2 = 1/8. Таким образом,
сама Екатерина гетерозиготна с вероятностью 1/8 х 1/2 =
1/16. Поэтому, вероятность рождения больного ребенка у
Екатерины и Давида равна 1/16х 1/2 х 1/4 = 1/128.
35. Максимально возможное число браков равно сумме ми-
нимального числа лиц с одинаковым вероисповеданием.
Для 1-й популяции возможно заключение 474 браков, или
432
К разделу 8
94,8% (11 + 29 + 202 + 205 + 27), в то время как в безбра-
чии останется 5,2% населения. Во 2-й популяции возмож-
но заключение 226 браков, или 45,2% (1 + 20 + 205), а в
безбрачии останется 54, 8% населения.
К разделу 8
4. Нет.
5. Супруги гетерозиготны IAi°.
7. Второй пары.
8. М.
9. Исключается первый.
12. У родителей нет антигенов, которые не будут иметь их дети
(родители гомозиготны), поэтому они могут быть донора-
ми ткани для своих детей.
13. Родители гомозиготны, поэтому у сибсов будет одинако-
вый набор антигенов. Следовательно, они могут быть до-
норами друг для друга.
14. Донорами могут быть отец и старший ребенок.
15. Возможные доноры — мать и старший ребенок.
20. Нулевую гипотезу об отсутствии связи между фенотипа-
ми родителей детей проверяем с помощью критерия х2-
Фенотип родителя Фенотип детей, чел.
Право- полушарный Лево- полушарный Смешан- ный
1. Правополушарный (всего 80 браков) 88 12 36
2. Левополушарный (всего 38 браков) 24 28 21
3. Смешанный (всего 38 браков) 12 9 30
X2 = 52,16, df — 4, р < 0,001. Нулевая гипотеза отбрасыва-
ется, наследственные факторы оказывают влияние на
асимметрию мозга.
22. Среди обследованных 4 были гетерозиготы и 96 гомозиго-
ты. р = 0,98, q = 0,02.
433
23. A— быстрые, a — медленные инактиваторы. Европейцы:,
рА = 0,293. Азиаты:, рА = 0,613.
К разделу 9
3. Риск для сына 25%, для дочери 0%.
4. Вероятность для дочерей 0%, для сыновей 100%.
5. 13/16 = 81%.
6. Родословная дает следующую информацию: изучаемый
признак встречается во всех поколениях (потомству семьи
V/4 ген не передался) и у лиц обоего пола. Обозначим ген
гемералопии А, а его нормальный аллель а.
Предполагаем аутосомно-доминантный тип наследования
гемералопии. Анализируемые семьи — Аа хаа — 1/1, II/2,
11/3, 111/2, Ш/6, W/2, IV/12, V/3, V/4, VI/3, VII/3, VIII/2, IX/4
(здоровый родитель не отмечен). Вычисляем критерий %2
для проверки связи между признаком и половой принад-
лежностью:
Фенотип Пол Всего
Мужчины Женщины
Здоровые 13 7 20
Больные 14 15 29
Всего 27 22 49
X2 = 0,748, df = 1, р > 0,05.
Критерий %2 указывает на отсутствие ассоциации гемера-
лопии с полом, т.е. это аутосомный признак.
При выдвинутой гипотезе наследования в анализируемых
семьях Аа х аа ожидается соотношение больных и здоро-
вых 1:1, или для данного случая 24,5:24,5. Вычислим значе-
ние х2 для сопоставления количества ожидаемых и наблю-
даемых потомков разного типа:
Фенотип Фактические Ожидаемые
Больные 29 24,5
Здоровые 20 24,5
Всего 49 49
X2 = 1,65, df ~ 1, р > 0,05.
434
К разделу 9
Поскольку различия между фактическими и теоретичес-
ки ожидаемыми классами больных и здоровых недосто-
верны, то гипотеза о доминантном характере наследова-
ния признака принимается.
В целом математический анализ подтверждает аутосомно-
доминантный тип наследования гемералопии.
8. У пар А и Б вероятность рождения больного ребенка уста-
навливается из данных условия задачи. У пары В вероят-
ность повышена в связи с наложением дополнительного
риска. Вероятность рождения больного ребенка для пар:
1) для А — 15% (оба родителя здоровы и один сибс больной);
2) для Б — 40% (один родитель и один сибс больные);
3) для В — более 40% (оба родителя и один сибс больные).
10. Вероятно, механизм контроля глухонемоты во всех случа-
ях аутосомно-рецессивный моногенный. Однако если в
двух семьях (1/1, 2, II/3-7 и 1/3, 4, II/8-10) глухонемота кон-
тролируется одним и тем же локусом, о чем свидетельству-
ет рождение всех глухонемых детей в браке II/7 и П/8, то
в третьей семье (II/1, 2, III/1-6) этот признак контролиру-
ется локусом, отличным от двух предыдущих семей, в
пользу чего говорит рождение в браке Ш/б и Ш/7 всех здо-
ровых детей.
12. Генотипы супругов ААЬЬ и ааВВ, детей АаВЬ.
17. Все дочери будут чувствительны, все сыновья не чувстви-
тельны.
18. 0,5.
19. 0,26.
20. Шведы р — 0,83 и q = 0,17, китайцы р — 0,04 и q = 0,96,
йоруба р = 0,01 nq = 0,99.
21. Частота аллелей и фенотипов по генам, сцепленным с по-
лом, у мужчин совпадают, поэтому частота искомого
аллеля q — 0,18. Допустив, что популяция равновесна, счи-
таем, что такая же частота этого аллеля и у женщин.
Поэтому доля рецессивных гомозигот у них составляет
q2 = 0,182 = 0,0324 = 3,24%.
23. В браках здоровых возможны две комбинации: женщина,
гомозиготная по нормальному аллелю (А) и здоровый муж-
435
чина, а также женщина, гетерозиготная по нормальному
аллелю, и здоровый мужчина. От этих двух типов браков
больные дети могут родиться лишь от комбинации гетеро-
зиготной матери (Аа) и здорового отца (А—), причем боль-
ных будет 1/4 (все они мальчики). Доля браков, где жен-
щина гетерозиготна (Аа), а мужчина здоров (А—), в попу-
ляции составляет 2pq х р, т.к. частота здоровых и больных
мужчин совпадает с частотами у них нормального и му-
тантного гена соответственно. Доля больных детей от та-
ких браков составляет 2pq х р х 1/4 = p2q/2.
Другой тип браков, о котором идет речь в условии задачи
и в котором оба родителя больны (женщина аа и мужчина
а — ) распространен в популяции с частотой q2 х q = q3, и
все дети (обоих полов) от таких браков будут больными.
Отношение доли больных детей в браках между здоровы-
ми родителями к доле больных детей в браках между боль-
ными родителями:
? p2q/2 _ р2
“^Г-27-
Если q = 0,144, а р = 1 — q = 0,856, p2/2q2 = 0,8562/2 X
0,1442 = 17,67. Таким образом, в браках здоровых родите-
лей больных детей ожидается почти в 18 раз больше, чем в
браках больных родителей, в силу того, что доля браков
первого типа больше.
Суммарная доля больных детей в браках здоровых роди-
телей и в браках больных родителей составит p2q/2 + q3.
Всего же больных в популяции (q2 + q)/2 (для простоты
предполагаем равенство полов). Тогда, соотношение сум-
марной доли больных детей, рожденных в браках двух ука-
занных типов, к доле всех больных в популяции, равно:
p2q/2+q3 _ р2 + 2q2 _ 0,8562 + 2х 0,1442 _Q67
I (q2+q)/2~~ q + 1 0,144+1 " ’ ’
В данной популяции дети с недостаточностью Г-6-ФДГ,
рожденные в браках обоих здоровых и в браках обоих
К разделу 12
больных людей, составят около 2/3 от всех больных попу-
ляции.
24. Обозначим частоты аллелей L, lt и 12 как р, q и г соответ-
ственно. Частоту аллеля 12 определим исходя из доли лиц,
не усваивающих лактозу и в детском, и во взрослом пери-
оде:
r = J-----=0,001.
V1000000
Из суммарной доли лиц, вообще не усваивающих лактозу
(г2) и не усваивающих ее только во взрослом состоянии
(q2 + 2qr), найдем частоту гена q:
44 1
(q + г)2 = - -- +------= 0,0004401;
100000 1000000
q + r = V0,0004401=0,021; q = 0,021-0,001 = 0,02.
Частоту аллеля L найдем из соотношения: р + q + г = 1,
р = 1 — q — г = 1 — 0,02 — 0,001 = 0,979.
К разделу 11
8. Н = (77 — 12)/( 100 — 12) = 0,74.
К разделу 12
11. А — 10%, Б — 40%, В — 16%.
12. По таблице Смита можно определить, что риск для маль-
чика первой семьи (Ш/2) составляет немного более 6,5%
(болен сибс, оба родителя здоровы), для детей второй се-
мьи (Ш/4,5 — немного более 18,5% (больны сибс и один
родитель), для детей третьей семьи (Ш/7,8) — немного бо-
лее 46,6% (больны сибс и оба родителя).
13. Нулевую гипотезу об одинаковой конкордантности моно-
и дизиготных близнецов проверим с помощью критерия %2:
X2 = 17,975; df = 1; р < 0,001. Нулевая гипотеза отвергает-
Конкордантность Зиготность —____ Конкор- дантные Дискор- дантные Всего
Монозиготные 26 49 75
Дизиготные 7 87 94
Всего 33 136 169
ся. Конкордантность монозиготных близнецов (35%) дос-
товерно выше, чем дизиготных (7%), что указывает на роль
наследственных факторов в развитии этой болезни.
17. Наследуемость гиперактивности, определенная на осно-
вании формулы Хольцингера, составляет 62%. Следова-
тельно, в развитии гиперактивности генетические факто-
ры играют большую половину, чем средовые (в частности,
плохое воспитание).
24. Бабушка Анны передала мутантный ген своим сыновьям.
Отец Анны мог передать ген хореи дочери с вероятнос-
тью 1/2 (более точно определить нельзя, поскольку Анна
еще не дожила до критического возраста, в котором начи-
нается проявление болезни). Следовательно, риск рожде-
ния больного ребенка у Анны составляет 1/2 х 1/2 = 1/4.
25. И частота, и наследуемость болезни в данной популяции
высокая. По таблице Смита определяем, что риск ребенка
заболеть эпилепсией составит около 8,3% (один сибс и один
родитель больны).
26. Популяционная частота может быть определена как низ-
кая, наследуемость — средняя. При одном больном роди-
теле и одном больном сибсе вероятность заболевания для
консультирующегося составляет примерно 3,4% (по таб-
лице Смита).
27. Брак Мириам и Якова кровнородственный, а Мириам и
Иосифа не родственный. Риск рождения ребенка с ауто-
сомно-рецессивной патологией выше в кровнородствен-
ном браке. Рождение больного ребенка у двух фенотипи-
чески здоровых людей возможно в случае, если оба они
гетерозиготы, и тогда этот риск составит 1/4. Мириам и
Иосиф, как случайно взятые индивиды в популяции, име-
438
К разделу 12
ют шанс быть гетерозиготами с вероятностью 2pq, или по
условию 4% (0,04). Следовательно, риск рождения у них
больного ребенка равен 2pq х 2pq х 1/4 = 0,04 х 0,04 х 1/4 =
0,0004.
В кровнородственном браке, если один из будущих суп-
ругов (не имеет значения, кто именно) с вероятностью 2pq
имеет дефектный ген, то вероятность присутствия этого
гена у другого равна не 2pq, а коэффициенту родства.
Мириам — двоюродная племянница Якову, у них четвер-
тая степень родства, а коэффициент родства соответствен-
но равен (1/2)4. Следовательно, риск рождения больного
ребенка в браке Мириам и Якова составит: 2pq х (1/2)2 х
1/4 - 0,04 х 1/64 = 0,000625.
28. Носительство Абрама установлено абсолютно, носитель-
ство Сары составляет 2pq. Следовательно, рождение у них
больного ребенка составляет 2pq х 1/4 = 0,04 х 1/4 = 0,01
(1%).
Для Михаила с Ребеккой вероятность быть гетерозигот-
ной парой равна произведению вероятностей каждому из
них быть гетерозиготой — для Михаила эта величина рав-
на 99/100 (по результатам скрининга) и для Ребекки 2pq
(без скрининга). Тогда, вероятность рождения у них боль-
ного ребенка равна 99/100 х 2pq х 1/4 = 0,0099 (1%).
Поскольку Абрам — облигатная гетерозигота, то Семен,
будучи его двоюродным братом, имеет ген болезни Тэя—
Сакса с вероятностью (1/2)3 = 1/8 (у них третья степень
родства). Руфь гетерозиготна с вероятностью 2pq. Веро-
ятность того, что оба родителя гетерозиготны составляет
1/8 х 2pq = 1/8 х 0,04 — 0,005. Риск рождения больного
ребенка у этой пары равен 0,005 х 1 /4 = 0,00125 (0,125%).
35. 67%.
ПРИЛОЖЕНИЕ
Метод %2
Известно, что расщепления в скрещиваниях носят случайный
характер, поэтому практически всегда в опытах наблюдают-
ся отклонения от теоретически ожидаемого расщепления. Эти
отклонения носят случайный характер и в значительной мере
зависят от величины выборки. Кроме того, отклонения могут
быть следствием нарушения любого из условий менделевс-
кого наследования (гибель гамет или зигот определенного ге-
нотипа, сцепление генов и т.д.).
Поэтому при анализе расщепления прежде всего необхо-
димо оценить величину отклонений, их значимость, чтобы
понять причину отклонения. Чаще всего для этой цели при-
меняют метод %2. Суть метода сводится к сопоставлению тео-
ретически рассчитанного на основе нулевой гипотезы (Но)
расщепления с опытными данными по всем классам.
Данные опыта и теоретическое ожидание заносятся в таб-
лицу. Далее определяют величину отклонения, которую воз-
водят в квадрат, чтобы избежать отрицательных величин.
Квадрат отклонения делят на теоретически ожидаемую для
каждого класса величину. Сумма этих величин и представля-
ет собой величину %2. После этого по таблице с учетом степе-
ней свободы определяют вероятность случайности отклоне-
ния (р). Число степеней свободы df, или н определяется по
формуле н = (п—1 )(т— 1), где пит — количество строк и столб-
цов с ненулевыми значениями.
Если значение %2 превышает табличное при соответствую-
щих степенях свободы, то это означает, что отклонение в опы-
те неслучайно, оно не может быть объяснено причинами ста-
тистического характера. Следовательно, нулевая гипотеза
должна быть отвергнута. Допустимой границей вероятности
в биологии принято считать величину 0,05.
Иногда для таблиц 2x2 величину %2 рассчитывают с по-
правкой по Йейтсу, которая несколько отличается от описан-
ной выше методики. Поэтому в данном издании читатель мо-
440
Приложение
жет в ответах на ряд задач, где %2 рассчитывается для таблиц с
двумя строками и двумя столбцами, обнаружить результат
именно с поправкой по Йейтсу. В этом случае расчет %2 выг-
лядит следующим образом:
В следующей таблице приведены стандартные значения
%2 при уровне значимости 0,05, 0,01 и 0,001 для степеней сво-
боды от 1 до 10 (более полные таблицы можно найти в реко-
мендуемой литературе по статистике):
Стандартные значения критерия %2
df ч2 (р < 0,05) ч2 (р< 0,01) ч2 (р < 0,001)
1 3,8 6,6 10,8
2 6,0 9,2 13,8
3 7,8 11,3 16,3
4 9,5 13,3 18,5
5 П,1 15,1 20,5
6 12,6 16,8 22,5
7 14,1 18,5 24,3
8 15,5 20,1 26,1
9 16,9 21,7 27,9
10 18,3 23,2 29,6
Литература
К разделу 1
1. http://student.biology.arizona.edu/sciconn.
2. Lewis R. Life. — Wm. С. Brown Publishers, 1995. — 884 p.
3. McKusick V.A. Mendelian inheritance in man. Baltimore, 1990. —
2300 p.
4. Raven P.H., Johnson G.B. Biology. — Wm. C. Brown Publishers /
McGraw-Hill, 1996. — 1311 p.
5. Volpe P. Biology and Human Concerns. — Wm. C. Brown
Publishers, 1993. — 611 p.
6. Бердышев Г.Д., Криворучко И.Ф. Генетика человека. — Киев:
«Выща школа», 1979. — 448 с.
7. Бердышев Г.Д., Криворучко И.Ф. Медицинская человека. —
Киев: «Выща школа», 1990. — 336 с.
8. Бочков Н.П. Клиническая генетика. — М., Медицина, 1997. —
288 с.
9. Гмурман В.Е. Руководство к решению задач по теории вероят-
ности и математической статистике: Учебн. пособие для сту-
t дентов втузов. — М.: Высш, шк., 1979. — 400 с.
10. Гофман-Кадошников П.В. Задачник по общей и медицинской
генетике. — М.: Первый Московской медицинский институт
! им. И.М. Сеченова, 1980. — 155 с.
11. Дженкинс М. 101 ключевая идея: Генетика. — М.: Фаир-Пресс,
2002. — 240 с.
12. Дышловой В.Д., Пилявская С. М. Обучающие задачи по генети-
ке человека и генетическим механизмам индивидуального и
исторического развития. — Киев, 1988. — 100 с.
13. Медицинская генетика: Учебник / Н.П. Бочков, А.Ю. Асанов,
Н.А. Жученко и др.; Под ред. Н.П. Бочкова. — М.: Мастерство;
Высшая школа, 2001. — 192 с.
14. Инге-Вечтомов С.Г. Генетика с основами селекции. — М.: Выс-
шая школа, 1989. — 591 с.
15. НильДж., Шелл У. Наследственность человека. — М.: ИЛ, 1958.
442
К разделу 1
16. Харпер П. Практическое медико-генетическое консультиро-
вание. — М.: Медицина, 1984. — 302 с.
17. Штерн К. Основы генетики человека. — М.: ИЛ., 1965. — 687 с.
18. Эфроимсон В.П. Введение в медицинскую генетику. М.: Ме-
дицина, 1968. — 395 с.
К разделу 2
1. Delabar J.M., Theophile D., Rahmani Z. et al. Molecular mapping
of twenty-four features of Down syndrome on chromosome 21 //
Europ. J. Hum. Genet. — 1993. — Vol. 1. — P. 114—124.
2. FoxS.I. Human Physiology. —Wm. C. Brown Publishers / McGraw-
Hill, 1996. — 704 p.
3. Hernandez D., Fisher E.M.C. Down syndrome genetics: unravelling
a multifactorial disorder // Hum. Mol. Genet. — 1996. Vol. 5. —
P. 1411 — 1416.
4. Korenberg J.R. et al. Down syndrome phenotypes: The consequen-
ces of chromosomal imbalance // Proc. Natl. Acad. Sci. USA. —
1994. — Bull. 91. — P. 4997—5001.
5. Lewis R. Life. — Wm. C. Brown Publishers, 1995. — 884 p.
6. McCormick M., Schinzel A., Petersen M. et aL Molecular genetic
approach to the characterization of the Down syndrome region of
chromosome 21 // Genomics. — 1989. — Vol. 5. — P. 325—331.
7. Raven P. H., Johnson G.B. Biology. — Wm. C. Brown Publishers /
McGraw-Hill, 1996. — 1311 p.
8. Shapiro B.L. Whither Down syndrome critical regions? // Hum.
Genet. — 1997. — Vol. 99. — P. 421—423.
9. Volpe P. Biology and Human Concerns. — Wm. C. Brown
Publishers, 1993, — 611 p.
10. Айала Ф., Кайгер Дж. Современная генетика: В 3-х т. Т. 1. Пер.
с англ. — М.: Мир, 1987. — 295 с.
11. Айала Ф., Кайгер Дж. Современная генетика: В 3-х т. Т. 2. Пер.
с англ. — М.: Мир, 1988. — 368 с.
12. Айала Ф., Кайгер Дж. Современная генетика: В 3-х т. Т. 3. Пер.
с англ. — М.: Мир, 1988. — 335 с.
13. Дженкинс М. 101 ключевая идея: Генетика. — М.: фаир-Пресс,
2002. — 240 с.
14. Дышловой В.Д., Полянская С.М. Обучающие задачи по генети-
ке человека и генетическим механизмам индивидуального и
исторического развития. — Киев, 1988. — 100 С.
15. Инге-Вечтомов С.Г. Генетика с основами селекции. — М.: Выс-
шая школа, 1989. — 591 с.
6*
443
Литература
16. Ленц В. Медицинская генетика: Пер. с нем. — М.: Медицина,
1984. — 448 с.
17. Литвиненко А.И., Атраментова Л.А. Генетика: Сб. задач. —
К.: Выща школа, 1987. — 95 с.
18. Пузырев В.П., Степанов В.А. Патологическая анатомия гено-
ма. — Новосибирск: Наука. — 1997. — 224 с.
19. Тарантул В.З. Геном человека. Энциклопедия, написанная че-
тырьмя буквами. — М.; Языки славянской культуры, 2003. —
393 с.
20. Штерн К. Основы генетики человека. — М.: ИЛ., 1965. — 687 с.
21. Эфроимсон В.П. Введение в медицинскую генетику. М.: Ме-
дицина, 1968. — 395 с.
К разделу 3
1. Bird G. W. Another example of haemopoetic (twin) chimaerism in a
subject unaware of being a twin // Journal of Immunogenetics. —
1982. — Vol. 9. — P. 317—322.
2. Boklage C.E. Survival probability of human conceptions from
fertilazation to term // International Journal of Fertility. — 1990. —
Vol. 35. — P. 75—94.
3. FoxS.I. Human Physiology. —Wm. C. Brown Publishers / McGraw-
Hill, 1996. — 704 p.
4. Gazzaniga M.S., Ivry R.B., Mangun G.R. Cognitive neuroscience. —
W.W. Norton & Company, New York, London, 1998. — 550 p.
5. Halgin R., Whitbourne S. Abnormal psychology. — Brown &
Benchmark Publishers, 1994. — 520 p.
6. Lewis R. Life. — Wm. C. Brown Publishers, 1995. — 884 p.
7. Lindsay D.T. Functional human anatomy. — Mosby Year Book,
1996. — 822 p.
8. Nelson J.L. Microchimerism: incidental byproduct of pregnancy
or active participant in human health? // Trends in Molecular
Medicine. — 2002. — Vol. 8. — P. 109—113.
9. Raven P.H., Johnson G.B. Biology. — Wm. C. Brown Publishers /
McGraw-Hill, 1996. — 1311 p.
10. Strain L., Dean J.C.S., Hamilton M.P.R., Bonthron D.T.A. A true
hermaphrodite chimera resulting from embryo amalgamation after
in vitro fertilization // New England Journal of Medicine. — 1998. —
Vol. 338. — P. 166—169.
11. Volpe P. Biology and Human Concerns. — Wm. C. Brown
Publishers, 1993. — 611 p.
444
К разделу 4
12. Блум Ф., Лейзерсон А., Хофстедтер Л. Мозг, разум поведение:
Пер. с англ. — М.: «Мир», 1988. — 248 с.
13. Дышловой В.Д., Пилявская С.М. Обучающие задачи по генети-
ке человека и генетическим механизмам индивидуального и
исторического развития. — Киев, 1988. — 100 с.
14. Инге-Вечтомов С.Г. Генетика с основами селекции. — М.: Выс-
шая школа, 1989. — 591 с.
15. Корочкин Л.И., Михайлов А.Т. Введение в нейрогенетику. —
М.: Наука, 2000. — 274 с.
16. Крушинский, Полетаева ИИ. Проблемы генетической психофи-
зиологии человека // Вопросы психологии. —1979. — №4. —
С. 170—173.
17. Малиновский А.А. Физиология и одаренность // Природа. —
1976. — № 9. — С. 73—78.
18. Малых С.Б. Исследования генетической детерминации ЭЭГ
человека // Вопросы психологии. — 1997. — № 6. — С. 109—128.
19. Мерфи Э.А., Чейз Г.А. Основы медико-генетического консуль-
тирования: Пер. с англ. — М.: Медицина, 1979. — 389 с.
20. Роль среды и наследственности в формировании индивидуаль-
ности человека / Под ред. И.В. Равич-Щербо. — М.: Педагоги-
ка, 1989. — 335 с.
21. Фогель Ф., Мотульски А. Генетика человека. В 3—х т. Т.З: Пер.
с англ. — М.: Мир, 1990. — 366 с.
22. Штерн К. Основы генетики человека. — М.: ИЛ., 1965. — 687 с.
23. Эрман Л., Парсонс П. Генетика поведения и эволюция: Пер. с
англ. — М.: Мир, 1984. — 566 с.
К разделу 4
1. Wald J. Статистические методы, применяемые в генетике че-
ловека / Проблемы медицинской генетики. — М., «Медици-
на», 1970. — С. 130—154.
2. Айала Ф., Кайгер Дж. Современная генетика: В 3-х т. Т. 1. Пер.
с англ. — М.: Мир, 1987. — 295 с.
3. Айала Ф., Кайгер Дж. Современная генетика: В 3-х т. Т. 2. Пер.
с англ. — М.: Мир, 1988. — 368 с.
4. Айала Ф., Кайгер Дж. Современная генетика: В 3-х т. Т. 3. Пер.
с англ. — М.: Мир, 1988. — 335 с.
5. Гмурман В.Е. Руководство к решению задач по теории вероят-
ности и математической статистике: Учебн. пособие для сту-
дентов втузов. — М.: Высш, шк., 1979. — 400 с.
445
Литература
6. Закс Л. Статистическое оценивание: Пер. с нем. — М.: Стати-
стика, 1976. — 598 с.
7. Лильин Е.Т., Трубников В.И., Ванюков М.М. Введение в совре-
менную фармакогенетику. — М.: Медицина. — 1984. — 139 с.
8. Фогель Ф., Мотульски А. Генетика человека. В 3-х т. Т. 3: Пер.
с англ. — М.: Мир, 1990. — 366 с.
9. Фолконер Д. Введение в генетику количественных признаков. —
М.: Агропромиздат, 1985. — 486 с.
10. Харпер П. Практическое медико-генетическое консультиро-
вание. — М.: Медицина, 1984. — 302 с.
11. Штерн К. Основы генетики человека. — М.: ИЛ., 1965. — 687 с.
К разделу 5
1. Lewis R. Life. — Wm. С. Brown Publishers, 1995. — 884 p.
2. McKusick V.A. Mendelian inheritance in man. Baltimore, 1990. —
2300 p.
3. Volpe P. Biology and Human Concerns. — Wm. C. Brown
Publishers, 1993. — 611 p.
4. www.dermatoglyphics.com.
5. Айала Ф., Кайгер Дж. Современная генетика: В 3-х т. Т. 1. Пер.
с англ. — М.: Мир, 1987. — 295 с.
6. Айала Ф., Кайгер Дж. Современная генетика: В 3-х т. Т. 2. Пер.
с англ. — М.: Мир, 1988. — 368 с.
7. Айала Ф., Кайгер Дж. Современная генетика: В 3-х т. Т. 3. Пер.
с англ. — М.: Мир, 1988. — 335 с.
8. Ниль Дж., Шелл У. Наследственность человека. — М.: ИЛ, 1958.
— с.
9. Фолконер Д. Введение в генетику количественных признаков.
— М.: Агропромиздат, 1985. — 486 с.
10. ФридрихВ. Близнецы: Пер. с нем. — М.: Прогресс, 1985. — 216 с.
11. Штерн К. Основы генетики человека. — М.: ИЛ., 1965. — 687 с.
К разделу 6
1. Baity J.M., Pillard R.C. A genetic study of male sexual orientation
// Archives of General Psychiatry. — 1991. —Vol. 48. — P. 1089—
1096.
2. Baity J.M., Pillard R.C., Agyei Y. A genetic study of female sexual
orientation // Archives of General Psychiatry. — 1993. —Vol. 48. —
P. 1089—1096.
446
К разделу 6
3. Collaer M.L., Hines М. Human behavioural sex differences: a role
for gonadal hormones during early development? // Psychological
Bulletin. — 1995. — Vol. 118, No. 1. — P. 55—77.
4. FoxS.I. Human Physiology. —Wm. C. Brown Publishers / McGraw-
Hill, 1996. — 704 p.
5. Gazzaniga M.S., Ivry R.B., Mangun G.R. Cognitive neuroscience. —
W.W. Norton & Company, New York, London, 1998. — 550 p.
6. Gur R.C., Turetsky B.I., Matsui M. et al. Sex differences in brain
gray and white matter in healthy young adults: Correlations with
cognitive performance // The Journal of Neuroscience. — 1999. —
Vol. 19, No. 10. — P. 4065—4072.
7. Halpern D.F. Sex differences and cognitive abilities. — Mahwah,
NJ: Erlbaum, 2000.
8. Hamer D.H., Hu S., Magnuson V.L. et al. A Linkage Between DNA
Markers on the X chromosome and Male Sexual Orientation //
Science. — 1993. — Vol. 261. — P. 321—327.
9. Hartmann T. Hunter in a farmer's world: ADD/ADHD 2002 (http:/
/www.thomhartmann.com/home-add.shtml).
10. Herdt G., Davidson J. The Sambian «turnim man»: Sociocultural
and Clinical Aspects of Gender Formation in Male Pseudoher-
maphrodites With 5—alpha-reductase Deficiency in Papua // New
Guinea Archives of Sexual Behavior. — 1988. — Vol. 17. — P. 33—56.
11. Kimura D. Sex differences in the brain // Scientific American. —
1992. — Vol. 10, Summer Quarterly, — P. 26—31.
12. Kimura D. Sex, sexual orientation and sexual hormones influence
human cognitive function // Current Opinion in Neurobiology. —
1996. — Vol. 6. — P. 259—263.
13. Kottak С. P. Anthropology. — New York: McGraw-Hill Inc., 1991. —
496 p.
14. LeVayS.A Difference in Hypothalamic Structure Between Hetero-
sexual and Homosexual Men // Science. — 1991. — Vol. 253. —
P. 1034—1037.
15. MacIntyre F., Estep K.W. Sperm competition and the persistence
of genes for male homosexuality // Biosystems. — 1993. — Vol.
31, No. 2—3. — P. 223—233.
16. Orbach S., Schwartz J. Why are little boys slugs and snails while
little girls are all things nice? // The Guardian, 1997, 14th., June.
17. Pillard R.C., Weinrich J.D. Evidence of Familial Nature of male ho-
mosexuality // Archives of General Psychiatry. — 1986. — Vol. 43. —
P. 808—812.
18. Radford T. Genes say boys will be boys and girls will be sensitive //
The Guardian, 1997, 12th., June.
447
Литература
19. Raine М. Absence of Y Specific DNA Sequence in Two Siblings
With 46XX Hermaphrodism // Archives of Disease in Childhood. —
1989. — Vol. 64. — P. 1185—1187.
20. Rosman A., Rubel P. G. The tapestry of culture. — New York:
Random House, 1989. — 338 p.
21. Shaywitz B.A., Shaywitz S.E., Pugh K.R. et al. Sex differences in the
functional organization of the brain for language // Nature. — 1995.
— Vol. 373. — P. 607—609.
22. Shaywitz S., Shaywitz B., Pugh K. et al. Estrogen changes functional
organization of brain in postmenopausal women // Journal of the
American Medical Association. — 1999. — Vol. 281. — P. 1197—
1202.
К разделу 7
1. Barrett S.C., Charlesworth D. Effects of a change in the level of
inbreeding on the genetic load // Nature. — 1991. — Vol. 352,
No. 6335. — P. 522—524.
2. Contreras L„ Escobar T., Coupal N. et al. Comparative study of
height and age at menarche according to the socioeconomic level
in Venezuela//Arch. Latinoam. Nutr. — 1981. —Vol. 31, No. 4. — P.
740—757.
3. Couvet D., Ronfort J. Mutation load depending on variance in
reproductive success and mating system // EXS. — 1994. —
Vol. 68. — P. 55—68.
4. Eorthofer M.S., Kessler R.C., Story A.L., Gotlib I.H. The effects of psy-
chiatric disorders on the probability and timing of first marriage // J.
Health Soc. Behav. — 1996. — Vol. 37, No. 2. — P. 121 — 132.
5. Frances V., Frances A. The incest taboo and family structure //
Fam. Process. — 1976. — Vol. 15, No. 2. — P. 235—244.
6. Gangestad S.W., Thornhill R. Menstrual cycle variation in women's
preferences for the scent of symmetrical men // Proc. Roy. Soc.
bond. — 1998. — Bull. 265. — P. 927—933.
7. Guz K., Dedeoglu N., Luleci G. The Frequency and Medical Effects
of Consanguineous marriages in Antalya, Turkey // Hereditas. —
1989.—Vol. Ill, No. 1, —P. 79—83.
8. Hafez M., El-TahanH., Awadalla M. etal. Consanguineous Matings
in the Egyptian Population // J. Med. Genet. — 1983, — Vol. 20,
No. 1. — P. 58—60.
9. HedrickP. W. Evolutionary genetics of the major histocompatibility
complex // Am. Nat. — 1994. — Vol. 143. — P. 945—964.
448
К разделу 7
10. Hill A. Immunogenetics: Defence by diversity // Nature. — 1999. —
Vol. 398. — P. 668—669.
11. Hume D., Montgomerie R. Facial Attractiveness signals different
aspects of «quality» in women and men // Evolution and Human
Behavior. — 2001. Vol. 22. — P .93—112.
12. Ihara et al. HLA and human mate choices: Tests on Japanese
couples // Anthropological Science. — 2000. — Vol. 108. —
P. 199—214.
13. Imaizumi I. A Recent Survey of Consanguineous Marriages in Japan
// Clin. Genet. — 1986. — Vol. 30, No. 3. — P. 230—233.
14. Jorde L.B., Pitkanen K.J. Inbreeding in Finland // Am. J. Phys.
Anthropol. — 1991. — Vol. 84, No. 2. — P. 127—139.
15. Lewis R. Life. — Wm. C. Brown Publishers, 1995. — 884 p.
16. Little B.B., Malina R.M. Gene flow and variation in stature and
craniofacial dimensions among indigenous populations of southern
Mexico, Guatemala, and Honduras // Am. J. Phys. Anthropol. —
1986. — Vol. 70, No. 4. — P. 505—512.
17. Lykken D., TellegenA. Is Human Mating Adventitious or the Result of
Lawful Choice? // Journal of Personality and Social Psychology. —
1993. — Vol. 65, No. 1. — P. 56—68.
18. McGraw K. Environmental Predictors of Geographic Variation in
Human Mating Preferences // Ethology. — 2002. — Vol. 108. —
P. 303—317.
19. Milinski M., Wedeking C. Evidence for МНС-correlated perfume
preferences in humans // Behavioral Ecology. — 2001. — Vol. 12. —
P. 140—149.
20. Nikitiuk B.A. New data on the ambiguous nature of the effects of
exogamy on human growth processes // Arkh. Anat. Gistol.
Embriol. — 1979. — Vol. 77, No. 11. — P. 5—11.
21. Penn D., Potls W. MHC-disassortative mating preferences reversed
by cross-fostering// Proc. Roy. Soc. Lond. — Bull. 265. — P. 1299—
1306.
22. Rao P.S., Inbaraj S.G. Trends in Human Reproductive Wastage in
Relation to Long-term Practice of Inbreeding // Ann. Hum. Genet. —
1979. — Vol. 42, No. 3. — P. 401—413.
23. Raven P.H., Johnson G.B. Biology. — Wm. C. Brown Publishers /
McGraw-Hill, 1996. — 1311 p.
24. Roche A.F. Secular trends in human growth, maturation, and
development // Monogr. Soc. Res. Child. Dev. — 1979. — Vol. 44,
No. 3—4. — P. 1 — 120.
25. Sailer S. Is Love Colorblind? // National Review. — 1997.
26. Sensus. — 1990.
449
Л итература
27. Thompson Е.А., Roberts D.F. Kinship Structure and Heterozygosity
on Tristan da Cunha // American Journal of Human Genetics. —
1980. — Vol. 32. — P. 445—452.
28. Volpe P. Biology and Human Concerns. — Wm. C. Brown
Publishers, 1993. — 611 p.
29. Wahab A., Ahmad M. Biosocial Perspective of Consanguineous
Marriages in Rural and Urban Swat, Pakistan // J. Biosoc. Sci. —
1996. — Vol. 28, No. 3. — P. 305—313.
30. Wayn for th D., Dunbar R. Conditional male choice strategies in
humans — evidence from lonely hearts advertisements //
Behaviour. — Vol. 132, — P. 755—779.
31. Wedekind C., Seebeck F., Bettens F., Paepke A. MHC-dependent
mate preferences in humans // Proc. R. Soc. bond. — 1995. — Bull.
260. — P. 245—249.
32. Whaley M. Book review «Survival of the Prettiest: The Science of
Beauty» by N. Etcoff // Human Nature Review. — 2002. — Vol. 2. —
P. 398—401.
33. Wolanski N. Assortative mating in somatic traits and its conse-
quences // Stud. Hum. Ecol. — 1994. — Vol. 11. — P. 73—111.
34. Woolf C.M., Dukepoo F.C. Hopi Indians, Inbreeding and Albinism
// Science. — 1969. — Vol. 164. — P. 30—37.
35. Айала Ф. Введение в популяционную и эволюционную гене-
тику: Пер. с англ. — М., «Мир», 1984. — 232 с.
36. Айала Ф., Кайгер Дж. Современная генетика: В 3-х т. Т. 1. Пер.
с англ. — М.: Мир, 1987. — 295 с.
37. Айала Ф., Кайгер Дж. Современная генетика: В 3-х т. Т. 2. Пер.
с англ. — М.: Мир, 1988. — 368 с.
38. Айала Ф., Кайгер Дж. Современная генетика: В 3-х т. Т. 3. Пер.
с англ. — М.: Мир, 1988. — 335 с.
39. Алтухов Ю.П. Генетические процессы в популяциях. — М.:
Наука, 2003. — 431 с.
40. Дышловой В.Д., Пилявская С.М. Обучающие задачи по генети-
ке человека и генетическим механизмам индивидуального и
исторического развития. — Киев, 1988. — 100 с.
41. Инге-Вечтомов С.Г. Генетика с основами селекции — М.: Выс-
шая школа, 1989. — 591 с.
К разделу 8
1. Abramson P.R., Imari-Marque J. The Japanese-American: A cross-
cultural, cross-sectional study of sex guilt // Journal of Research
in Personality. — 1982. — Vol. 16. — P. 227—237.
450
К разделу 8
2. Aithal G.P., Day C.P., Kesteven P.J., DalyA.K. Association of poly-
morphisms in the cytochrome P450 CYP2C9 with warfarin dose re-
quirement and risk of bleeding complications // Lancet — 1999. — Vol.
353. — P. 717—719.
3. Alda M. Pharmacogenetics of lithium response in bipolar disorder
// J. Psychiatry Neurosci. — 1999. — Vol. 24, No. 2. — P. 154—158.
4. Allen G. Frequency of triplets and triplet zygosity types among U.S.
births, 1964 // Acta Geneticae Medicae et Gemellologiae. — 1988.
Vol. 37. — P. 299—306.
5. Andreasen N.C., Flaum M., Swayze V. et al. Intelligence and brain
structure in normal individuals // American Journal of Psychiatry. —
1993. — Vol. 150. — P. 130—134.
6. Armstrong E. Brains, bodies and metabolism // Brain, Behavior and
Evolution. — 1990. — Vol. 36. — P. 166—176.
7. Arranz M.J., Munro J., Birkett J. et al. Pharmacogenetic prediction
of clozapine response // Lancet. — 2000). — Vol. 355, No. 9215. —
P. 1615—1616.
8. Asayama S. Adolescent sex development and adult sex behavior in
Japan // Journal of Sex Research. — 1975. — Vol. 11. — P. 91—122.
9. Bayley N. Comparisons of mental and motor test scores for ages
1 —15 months by sex, birth order, race, geographic location, and
education of parents // Child Development. — 1965. — Vol. 36. —
P. 379—411.
10. Beals K.L., Smith C.L., Dodd S.M. Brain size, cranial morphology,
climate and time machines // Current Anthropology. — 1984. —
Vol. 25. — P. 301—330.
11. Bertilsson L., Dahl M.L., Sjoqvist F. et al. Molecular basis for rational
mega-prescribing in ultrarapid hydroxylators of debrisoquin //
Lancet. — 1993. — Vol. 341. — P. 363.
12. bogdanov@finger-print.ru.
13. Egan V., Chiswick A., Santosh C. et al. Size isn't everything: A study
of brain volume, intelligence and auditory evoked potentials //
Personality and Individual Differences. — 1994. — Vol. 17. —
P. 357—367.
14. Ellis L., Nyborg H. Racial/ethnic variations in male testosterone
levels // Steroids. — 1992. — Vol. 57. — P. 72—75.
15. G№siorowski A., Hajn V. A report on the occurence of accessory
triradii in the subdigital area, and carpal triradii on the palm in Po-
lish and Czech populations // Anthropol. Rev. — 2000. — Vol. 63. —
P. 127—132.
16. Hajn V., G№siorowski A. Extremely rare longitudinal direction of
all main lines on the human palm // Biologia. — 1998. —Vol. 36. —
P. 101 — 106.
451
Литература
17. Harvey Р. Н., Krebs J.R. Comparing brains // Science. — 1990. —
Vol. 249. — P. 140—145.
18. Hixson J.R. Benign prostatic hypertrophy drug to be tested in
prostate CA prevention. // The Medical Post. — 1992, October 20. —
P. 17.
19. Но K.C., Roessmann U., Straumfjord J.V., Monroe G. Analysis of
brain weight: I. Adult brain weight in relation to sex, race, and age //
Archives of Pathology and Laboratory Medicine. — 1980. —Vol. 104.
— P. 635—639.
20. Но K.C., Roessmann U., Straumfjord J.V., Monroe G. Analysis of
brain weight: II. Adult brain weight in relation to body height,
weight, and surface area // Archives of Pathology and Laboratory
Medicine. — 1980. — Vol. 104. — P. 640—645.
21. Imaizumi Y. Twinning rates in Japan, 1951 —1990//Acta Geneticae
Medicae et Gemellologiae. — 1992. — Vol. 41. — P. 165—175.
22. Iwawaki S., Wilson G.D. Sex fantasies in Japan // Personality and
Individual Differences. — 1983. —Vol. 4. — P. 543—545.
23. Jensen A.R., Johnson F.W. Race and sex differences in head size
and IQ // Intelligence. — 1994. — Vol. 18. — P. 309—333.
24. Johansson I., Lundquist E., Bertilsson L. et al. Inherited amplification
of an active gene in the cytochrome P450 CYP2D locus as a cause
of ultrarapid metabolism of debrisoquine // Proc. Natl. Acad. Sci.
(USA). — 1993. — Vol. 90. — P. 11825—11829.
25. Krynetski E.Y., Tai H.L.r Yates C.R. et al. Genetic polymorphism of
thiopurine S-methyltransferase: clinical importance and molecular
mechanisms // Pharmacogenetics. — 1996. — Vol. 6. — P. 279—290.
26. Lazarou J., Pomeranz B.H., Corey P. N. Incidence of adverse drug
reactions in hospitalised patients: a meta-analysis of prospective
studies // JAMA. — 1998. — Vol. 279. — P. 1200—1205.
27. Lennard L., LilleymanJ.S., Van Loon J., Weinshilboum R.M. Genetic
variation in response to 6—mercaptopurine for childhood acute
lymphoblastic leukaemia // Lancet. — 1990. — Vol. 336. — P. 225—
229.
28. Lerer B., Macciardi F. Pharmacogenetics of antidepressant and
mood stabilizing drugs: a review of candidate-gene studies and
future research directions // Int. J. Neuropsychopharmacol. —
2002. — Vol. 5, No. 3. — P. 255—275.
29. Lerer B., Segman R.H., Fangerau H. et al. Pharmacogenetics of
tardive dyskinesia: combined analysis of 780 patients supports
association with dopamine D3 receptor gene Ser9Gly polymor-
phism // Neuropsychopharmacology. — 2002. — Vol. 27, No. 1. —
P. 105—119.
Л52
К разделу 8
30. Lewis R. Life. — Wm. C. Brown Publishers, 1995. — 884 p.
31. Lynn R. Race differences in intelligence: A global perspective //
Mankind Quarterly. — 1991. — Vol. 31. — P. 255—296.
32. Lynn R. The intelligence of Ethiopian immigrant and Israeli adole-
scents // International Journal of Psychology. — 1994. — Vol. 29. —
P. 55—56.
33. Lynn R., Shigehisa T. Reaction times and intelligence: A comparison
of Japanese and British children // Journal of Biosocial Science. —
1991. — Vol. 23. — P. 409—416.
34. Mahgoub A., Idle J.R., Dring L.G. et al. Polymorphic hydroxylation
of Debrisoquine in man // Lancet. — 1977. — Vol. 2, No. 8038. —
P. 584—586.
35. McHenry H.M. How big were the early hominids? // Evolutionary
Anthropology. — 1992. — Vol. 1. — P. 15—20.
36. MotulskyA.G. Drug reactions, enzymes and biochemical genetics
//JAMA. — 1957. — Vol. 165. — P. 835—837.
37. Papiernik E., Cohen H., Richard A. et al. Ethnic differences in
duration of pregnancy // Annals of Human Biology. — 1986. —
Vol. 13. — P. 259—265.
38. Pare C.M., Mack J.W. Differentiation of two genetically specific
types of depression by the response to antidepressant drugs // J.
Med. Genet. — 1971. — Vol. 8, No. 3. — P. 306—309.
39. Raz N., Torres I.J., Spencer W.D. et al. Neuroanatomical correlates
of age-sensitive and age-invariant cognitive abilities: An in vivo
MRI investigation // Intelligence. — 1993. — Vol. 17. — P. 407—422.
40. Roses A.D. Pharmacogenetics and the practice of medicine //
Nature. — 2000. — Vol. 405, No. 6788. — P. 857—865.
41. Ross R., Bernstein L., Judd H. et al. Serum testosterone levels in
healthy young Black and White men // Journal of the National
Cancer Institute. — 1986. —Vol. 76. — P. 45—48.
42. Rushton J.P. Corrections to a paper on race and sex differences in
brain size and intelligence // Personality and Individual Differen-
ces. — 1993. — Vol. 15. — P. 229—231.
43. Rushton J.P. Cranial capacity related to sex, rank and race in a
stratified random sample of 6,325 U.S. military personnel //
Intelligence. — 1992. — Vol. 16. — P. 401—413.
44. Rushton J.P. Differential К theory and race differences in E and N
// Personality and Individual Differences. — 1985. — Vol. 6. —
P. 769—770.
45. Rushton J.P. Japanese inbreeding depression scores: Predictors of
cognitive differences between Blacks and Whites // Intelligence. —
1989. — Vol. 13. — P. 43—51.
453
Литература
46. Rushton J.P. Life history comparisons between Orientals and
Whites at a Canadian university // Personality and Individual
Differences. — 1992. — Vol. 13. — P. 439—442.
47. Rushton J.P. Mongoloid-Caucasoid differences in brain size from
military samples // Intelligence. — 1991. — Vol. 15. — P. 351—359.
48. Rushton J.P. Race and crime // Canadian Journal of Criminology. —
1990. — Vol. 32. — P. 315—334.
49. Rushton J.P. Race differences in behaviour: A review and
evolutionary analysis // Personality and Individual Differences. —
1988. — Vol. 9. — P. 1035—1040.
50. Rushton J.P. Race, evolution and behavior: A life history perspective
— New Brunswick, NJ: Transaction Publishers, 1995.
51. Rush ton J.P. Sex and race differences in cranial capacity from Inter-
national Labour Office data // Intelligence. — 1994. — Vol. 19. —
P. 281—294.
52. Rushton J.P. , Ankney C.D. The evolutionary selection of human
races: A response to Miller // Personality and Individual
Differences. — 1993. — Vol. 15. — P. 677—680.
53. Rushton J.P. , Bogaert A.F. Race differences in sexual behavior:
Testing an evolutionary hypothesis // Journal of Research in
Personality. — 1987. —Vol. 21. — P. 529—551.
54. Rushton J.P. , Bogaert A.F. Race versus social class differences in
sexual behavior: A follow-up of the r/K dimension // Journal of
Research in Personality. — 1988. — Vol. 22. — P. 259—272.
55. Rushton J.P. , Brainerd C.J., Pressley M. Behavioral development
and construct validity: The principle of aggregation // Psycholo-
gical Bulletin. — 1983. — Vol. 94. — P. 18—38.
56. Smeraldi E., Zanardi R., Benedetti F. et al. Polymorphism within
the promoter of the serotonin transporter gene and antidepressant
efficacy of fluvoxamine // Mol. Psychiatry. — 1998. — Vol. 3,
No. 6. — P. 508—511.
57. Sokal R.R., Harding R.M., Oden N.L. Spatial Patterns of Human
Gene Frequencies in Europe // Am. J. Phys. Anthropol. — 1985. —
Vol. 80, No. 3. — P. 267—294.
58. Steen V.M., LovlieR., MacEwan T., McCreadieR.G. Dopamine D3—
receptor gene variant and susceptibility to tardive dyskinesia in
schizophrenic patients // Mol. Psychiatry. — 1997. — Vol. 2, No. 2.
— P. 139—145.
59. Tremblay R.E., Baillargeon L. Les difficultes de comportement
d'enfants immigrants dans les classes d'accueil, au prescolaire //
Canadian Journal of Education. — 1984. — Vol. 9. — P. 154—170.
60. Vogel F. Moderne probleme der Humangenetik // Ergeb. Inn. Med.
Kinderheild. — 1959. — Vol. 12. — P. 52—125.
454
К разделу 9
61. Volpe Р. Biology and Human Concerns. — Wm. C. Brown
Publishers, 1993. — 611 p.
62. Willerman L., Schultz R., Rutledge J.N., Bigler E.D. In vivo brain
size and intelligence // Intelligence. — 1991. —Vol. 15. — P. 223—
228.
63. WolfC.R., Smith G. Pharmacogenetics // Br. Med. Bull. — 1999. —
Vol. 55. — P. 366—386.
64. Wolf C.R., Smith G., Smith R.L. Science, medicine and future:
Pharmacogenetics // BMJ. — 2000. — Vol. 320, No. 8. — P. 987—
990.
65. Атраментова Л.А., Федота AM. Изучение распространенно-
сти псориаза в различных климато-географических условиях
и этнических группах населения // Вестник проблем биоло-
гии и медицины. — 1998. — Вып. 17. — С. 3—5.
66. Гладкова Т.Д. Кожные узоры кисти и стопы обезьян и челове-
ка. — М.: 1966. — 151с.
67. Дымшиц Г.М. Сюрпризы митохондриального генома // При-
рода. — 2002. - № 6. — С. 54—61.
68. Кречмер Э. Строение тела и характер. — М.: НО Научный Фонд
«Первая Исследовательская Лаборатория имени академика
В.А. Мельникова», 2000. — 208 с.
69. Левонтин Р. Человеческая индивидуальность: наследствен-
ность и среда: Пер. с англ. — М.: Издательская группа «Про-
гресс» «Универе», 1993. — 208 с.
70. Лимборская С. А., Хуснутдинова Э.К., Балановская Е.В. Этно-
геномика и геногеография народов Восточной Европы. — М.:
Наука, 2002. — 261 с.
71. Рогинский Я.Я., Рычков Ю.П. Генетика расообразования у че-
ловека //Проблемы медицинской генетики. — М.: «Медици-
на», 1970. — С. 215—232.
72. Фогель Ф., Мотульски А. Генетика человека. В 3-х т. Т. 3: Пер.
с англ. — М.: Мир, 1990. — 366 с.
73. Харрисон Дж., Уайнер Дж., Тэннер Дж. и др. Биология челове-
ка: Пер. с англ. — М.: Мир, 1979. — 616 с.
74. Хрисанфова Е.Н., Перевозчиков И.В. Антропология. — Изд-во
Московского университета, 1991. — 319 с.
75. Штерн К. Основы генетики человека. М., ИЛ., 1965.
К разделу 9
1. Bartoshuk L.M. Comparing sensory experiences across individuals:
Recent psychophysical advances illuminate genetic variation in
455
Литература
taste perception // Chem. Senses. — 2000. — Vol. 25. — P. 447—
460.
2. Berry V., Francis P., Kaushal S. et al. Missense mutations in MIP
underlie autosomal dominant «polymorphic» and lamellar
cataracts linked to 12q // Nature Genetics. — 2000. — Vol. 25. —
P. 15—17.
3. Bhattacharya S.S., Wright A.F., Clayton J.F. et al. Close genetic
linkage between X-linked retinitis pigmentosa and a restriction
fragment length polymorphism identified by recombinant DNA
probe LI.28. // Nature. — 1984. — Vol. 309. — P. 253—255.
4. Birch LL. Children's preference for high-fat foods // Nutr. Rev. —
1992. — Vol. 50. — P. 249—255.
5. Drewnowski A. Taste preferences and food intake // Ann. Rev. Nutr.
— 1997. — Vol. 17. — P. 237—253.
6. Drewnowski A., Gomez-Garneros C. Bitter taste, phytonutrients and
the consumer: a review // Am. J. Clin. Nutr. — 2000. — Vol. 72. —
P. 1424—1435.
7. Drewnowski A., Krahn D.D., Demitrak M.A. et al. Naloxone, an
opiate blocker, reduces the consumption of sweet high-fat foods
in obese and lean female binge eaters // Am. J. Clin. Nutr. — 1995. —
Vol. 61. — P. 1206—1212.
8. FoxS-1. Human Physiology. —Wm. C. Brown Publishers / McGraw-
Hill, 1996. — 704 p.
9. Francis P., Berry V., Moore A., Bhattacharay S.S. Lens biology, deve-
lopment and human cataractogenesis // Trends in Genetics. —
1999. — Vol. 15. — P. 191 — 196.
10. Gazzaniga M.S., Ivry R.B., Mangun G.R. Cognitive neuroscience. —
W.W. Norton & Company, New York, London, 1998. — 550 p.
11. Inglehearn C.F, Carter S.A., Keen T.J. et al. A new locus for
autosomal dominant Retinitis Pigmentosa (adRP) on chromosome
7p // Nature Genetics. — 1993. — Vol. 4. — P. 51—53.
12. Inglehearn C.F., Keen T.J., Bashir R. et al. A completed screen for
mutations of the rhodopsin gene in a panel of patients with
autosomal dominant retinitis pigmentosa // Hum. Mol. Genet. —
1992. — Vol. 1. — P. 41—45.
13. Jacob S., McClintock M.K., Zelano B., Ober C. Paternally inherited
HLA alleles are associated with women's choice of male odor //
Nature Genetics. — 2002. — Vol. 30, No. 2. — P. 175—179.
14. Kim U., Jorgenson E., Coon H. et al. Positional cloning of the human
quantitative trait locus underlying taste sensitivity to phenylthio-
carbamide // Science. — 2003. —Vol. 299. — P. 1221 —1225.
15. Kottak C.P. Anthropology. — New York: McGraw-Hill Inc., 1991. —
496 p.
456
К разделу 9
16. Lane R.P. Cutforth T., Young J. et al. Genomic analysis of
orthologous mouse and human olfactory receptor loci // Proc. Natl.
Acad. Sci. USA. — 2001. — Vol. 98, No. 13. — P. 7390—7395.
17. Lewis R. Life. — Wm. C. Brown Publishers, 1995. — 884 p.
18. Li X., Staszewski L., Xu H. et al. Human receptors for sweet and
umami taste // Proc. Natl. Acad. Sci. USA. — 2002. — Vol. 99,
No. 7. — P. 4692—4696.
19. Mackay D., lonides A., Kibar Z. et al. Connexin-46 mutations in
autosomal dominant congenital cataract //Am. J. Hum. Genet. —
1999. — Vol. 64. — P. 1357—1364.
20. Mombaerts P. How smell develops // Nat. Neurosci. — 2001. —
Suppl.4. — P. 1192—1198.
21. Mombaerts P. Seven transmembrane proteins as odorant and
chemosensory receptors // Science. — 1999. — Vol. 286, No. 5440. —
P. 707—711.
22. Raven P.H., Johnson G.B. Biology. — Wm. C. Brown Publishers /
McGraw-Hill, 1996. — 1311 p.
23. Rosman A., Rubel P.G. The tapestry of culture. — New York:
Random House, 1989. — 338 p.
24. Shiels A., Mackay D., lonides A. et al. A missense mutation in the
human connexin50 gene (GJA8) underlies autosomal dominant
«zonular pulverulent» cataract, on chromosome Iq. //Am. J. Hum.
Genet. — 1998. — Vol. 62. — P. 526—532.
25. Sohocki M.M., Bowne S.J., Sullivan L.S. et al. Mutations in a novel
photoreceptor-pineal gene on 17p cause Leber congenital
amaurosis (LCA4) // Nature Genetics. — 2000. — Vol. 24. —
P. 79—83.
26. Stubbs J., Ferres S., Horgan G. Enegry density of foods: effects of
energy intake // Grit. Rev. Food Sci. Nutr. — 2000. — Vol. 40. —
P. 481—515.
27. Volpe P. Biology and Human Concerns. — Wm. C. Brown
Publishers, 1993, —611 p.
28. Zou Z., HorowitzL.F., MontmayeurJ.P. et al. Genetic tracing reveals
a stereotyped sensory map in the olfactory cortex // Nature. —
2001. — Vol. 414, No. 6860. — P. 173—179.
29. Инге-Вечтомов С.Г. Генетика с основами селекции. — М.: Выс-
шая школа, 1989. — 591 с.
30. Ниль Дж., Шелл У. Наследственность человека. — М.: ИЛ, 1958.
— с.
31. Харпер П. Практическое медико-генетическое консультиро-
вание. — М.: Медицина, 1984. — 302 с.
32. Штерн К. Основы генетики человека. — М.: ИЛ., 1965. — 687 с.
457
Литература
33. Эфроимсон В.П. Введение в медицинскую генетику. М.: Ме-
дицина, 1968. — 395 с.
К разделу 10
1. Сох С. The Early Mental Traits of Three Hundred Geniuses. —
Stanford University Press, 1926.
2. Flynn J. Massive IQ Gains in 14 Nations: What IQ Tests Really
Measure // Psychological Bulletin. — 1987. — Vol. 101, No. 2. —
P. 171—191.
3. http://www.nobel.se/index.html.
4. Kaufman A.S. Genetics of Childhood Disorders: II. Genetics and
Intelligence // J. Am. Acad. Child. Adolesc. Psychiatry. — 1999. —
Vol. 38, No. 4. — P. 487—488.
5. Kottak C.P. Anthropology. — New York: McGraw-Hill Inc., 1991. —
496 p.
6. Lewis R. Life. — Wm. C. Brown Publishers, 1995. — 884 p.
7. Lynn R. The role of Nutrition in Secular Increases in Intelligence //
Personality and Individual Differences. — 1990. — Vol. 11, No. 3.
— P. 273—285.
8. Raven P.H., Johnson G.B. Biology. — Wm. C. Brown Publishers /
McGraw-Hill, 1996. — 1311 p.
9. Rosman A., Rubel P.G. The tapestry of culture. — New York:
Random House, 1989. — 338 p.
10. Volpe P. Biology and Human Concerns. — Wm. C. Brown
Publishers, 1993. — 611 p.
11. Weiss V. It could be Neo-Lysenkoism, if there was over a break in
continuity! // Mankind Quarterly. — 1991. — Vol. 31. — P. 231—
253.
12. World Almanac and Book of Facts 2000 (for 1999). — Mahwan,
New Jersey, 1999.
13. Айзенк Г. Супертесты IQ. — M.: Изд-во ЭКСМО-Пресс, 2002. —
208 с.
14. Штерн К. Основы генетики человека. — М.: ИЛ., 1965. — 687 с.
К разделу 11
1. Archer J. The influence of testosterone on human aggression //
Br. J. Psychol. — 1991. — Vol. 82, Part 1. — P. 1—28.
2. Bohman M., Cloninger C.R., Sigvardsson S., von Knorring A.L.
Predisposition to petty criminality in Swedish adoptees. I. Genetic
<458
К разделу 11
and environmental heterogeneity // Arch. Gen. Psychiatry. —
1982. Vol. 39. — P. 1233—1241.
3. Bouchard T., Prusse L., Leblanc C. Using MZ twins in experimental
research to test for the presence of a genotype-environment
interaction effect // Acta Geneticae Medicae Gemellologicae
Roma. — 1990. — Vol. 39. — P. 85—89.
4. BowlbyJ. Some pathological processes set in train by early mother-
child separation // Journal of Mental Science. — 1953. — Vol. 2. —
P. 265—272.
5. BowlbyJ. The making and breaking of affectional bonds // British
Journal of Psychiatry. — 1977. — Vol. 130. — P. 201—210.
6. Brown G.L., Goodwin F.K., Ballenger J.C. et al. Aggression in
humans correlates with cerebrospinal fluid amine metabolites //
Psychiatry Res. — 1979. — Vol. 1. — P. 131 —139.
7. Caspi A., McClayJ., Moffitt T. et al. Role of genotype in the cycle
of violence in maltreated children // Science. — 2002. —Vol. 297. —
P. 851—854.
8. CoccaroE.F., Bergeman C.S., KavoussiR.J., Seroczynski A.D. Herita-
bility of aggression and irritability, a twin study of the Buss-Durkee
aggression scales in adult male subjects // Biol. Psychiatry. — 1997.
Vol. 41. — P. 273—284.
9. Dalgard K.E. A Norwegian twin study of criminality // Br. J.
Criminology. — 1976. — Vol. 16. — P. 213—232.
10. Farrington D. The Development of Offending and Antisocial
Behaviour from Childhood: Key Findings from the Cambridge
Study in Delinquent Development // Journal of Child Psychology
and Psychiatry and Allied Disciplines. — 1995. — Vol. 360. —
P. 929—964.
11. Farrington D.P. , Barnes G.C., Lambert S. The concentration of
offending in families // Leg. Criminol. Psychol. — 1996. — Vol. 1. —
P. 47—63.
12. Finkle D., McGue M. Sex differences and nonadditivity in the
heritability on the Multidimensional Personality Questionnaire
scales // J. Pers. Soc. Psychol. — 1997. — Vol. 72. — P. 929—938.
13. Gazzaniga M.S., Ivry R.B., Mangun G.R. Cognitive neuroscience. —
W.W. Norton & Company, New York, London, 1998. — 550 p.
14. Gjone FL, Stevenson J. The association between internalizing and
externalizing behaviour in childhood and early alolescence:
Genetic or environmental common influences? //J. Abnorm. Child
Psychol. — 1997. — Vol. 25. — P. 277—286.
15. Laub J.S., Sampson R.J. Unravelling families and delinquency: a
reanalysis of the Glueck data // Criminology. — 1998. — Vol. 26. —
P. 355—380.
459
Литература
16. Jones S. In The Blood: God, Genes and Destiny. — London: Harper
Collins, 1996. — P. 231.
17. Kreuger R.F., Moff it T.E., Caspi A. et al. Assortative mating for
antisocial behaviour: development and methodological implica-
tions // Behav. Genet. — 1998. — Vol. 28. — P. 173—186.
18. Lykken D. T., Bouchard T.D., McGue M., Tellegen A. The Minnesota
twin family registry: some initial findings // Acta Geneticae
Medicae Gemellologicae Roma. — 1990. — Vol. 39. — P. 35—70.
19. McKusick V.A. Mendelian inheritance in man. Baltimore, 1990. —
2300 p.
20. Mednick S.A., Gabrielli W.F., Hutchings B. Genetic factors in
criminal behaviour: evidence from adoption cohort // Science. —
1984. — Vol. 224. — P. 891—893.
21. Nielsen D.A., Goldman D., Virkkunen M. et al. Suicidality and 5—
hydroxyindoleacetic acid concentration associated with a trypto-
phan hydroxylase polymorphism // Arch. Gen. Psychiatry. — 1994.
— Vol. 51, —P. 34—38.
22. Olson J., Vernon A., Aitken J. The Heretability of Attitudes: A study
of Twins // Journal of Personality and Social Psychology. — 2001. —
Vol. 80, No. 6. — P. 845—860.
23. Orlebecke J.F., Knol D.L., Verhulst F.C. Child behavior problems
increased by maternal smoking during pregnancy // Arch. Environ.
Health. — 1999. — Vol. 54. — P. 15—19.
24. Rhee S.H., Waldman I.D. Genetic and environmental influences on
antisocial behavior: a meta-analysis of twin and adoption studies //
Psychol. Bull. — 2002. — Vol. 128. — P. 490—529.
25. Simon N.G., Cologer-Clifford A., Lu S.F. et al. Testosterone and its
metabolites modulate 5—НТ[Д and 5—HT]B agonist effects on
intermale aggression // Neurosci. Biobehav. Rev. — 1998. — Vol. 23.
— P. 325—336.
26. Tremblay R.E. Prevention of injury by early socialization of
aggressive behavior // Injury Prevention. — 2002. — Vol. 8. —
P. 17—21.
27. Virkkunen M., De Jong J., Bartko J., Linnoila M. Psychobiological
concomitants of history of suicide attempts among violent offenders
and impulsive fire setters // Arch. Gen. Psychiatry. — 1989. —
Vol. 46. — P. 604—606.
28. Wasserman G.A., Staghezza-Jaramillo B., Shrout P. et al. The effect
of lead exposure on behavior problems in preschool children //
Am. J. Public Health. — 1998. — Vol. 88. — P. 481—486.
29. www.dermatoglyphics.com.
30. Блум Ф., Лейзерсон А., Хофстедтер Л. Мозг, разум поведение:
Пер. с англ. — М.: «Мир», 1988. — 248 с.
460
К разделу 12
31. Крушинский, Полетаева И.И. Проблемы генетической психофи-
зиологии человека // Вопросы психологии. —1979. — №4. —
С. 170—173.
32. Малиновский А.А. Физиология и одаренность // Природа. —
1976. — №9. — С. 73—78.
33. Ниль Дж., Шелл У. Наследственность человека. — М.: ИЛ, 1958.
34. Роль среды и наследственности в формировании индивидуаль-
ности человека / Под ред. И.В. Равич-Щербо. — М.: Педагоги-
ка, 1989. — 335 с.
35. Трут Л.Н. Очерки по генетике поведения. — Новосибирск:
Наука, СО. — 1978. — 256 с.
36. Харпер П. Практическое медико-генетическое консультиро-
вание. — М.: Медицина, 1984. — 302 с.
37. Штерн К. Основы генетики человека. — М.: ИЛ., 1965. — 687 с.
К разделу 12
1. Agarwal D. The Genetics of Alctjhol Metabolism and Alcoholism //
Int. J. Hum. Genet. — 2001. —/Vol. 1, No. 1. — P. 1—9.
2. Allen M.G., Cohen S., Pollin W. et al. Affective illness in veteran
twins: A diagnostic review // American Journal of Psychiatry. —
1974. — Vol. 131. — P. 1234—1239.
3. Amrose N., Cox N., Yairi E. The genetic basis of persistence and re-
covery in sluttering // J. Speech Hear Res. — 1997. — Vol. 40. —
P. 567—580.
4. Apter A., Pauls D.L., Bleich A. et al. An epidemiological study of
Giles de la Tourette's syndrome in Israel // Archives of General
Psychiatry. — 1993. — Vol. 50. — P. 734—738.
5. Bandmann O„ Vaughan J., Holmans P. et al. Association of slow
acetylator genotype for N-acetyltransferase 2 with familial
Parkinson's disease // Lancet. — 1997. —Vol. 350, No. 9085. —
P. 1136—1139.
6. Baron M., Shapiro E., Shapiro A. Genetic analysis of Tourette
syndrome suggesting a major gene effect // American Journal of
Human Genetics. — 1981. —Vol. 33. — P. 767—775.
7. Belkovets A., Kurilovich S., Avkenstyuk A., Agarwal D. Alcohol
Drinking Habits and Genetic Polymorphism of Alcohol Metabolism
Genes in West Siberia // Int. J. Hum. Genet. — 2001. — Vol. 1,
No. 2. — P. 151 — 156.
8. Bertelsen A., Harvald B., Hauge M.A. A Danish twin study of manic-
depressive disorders // British Journal of Psychiatry. — 1977. —
Vol. 130. — P. 330—351.
461
Литература
9. Biederman J., Faraone S.V., Keenan K. et al. Family-genetic and
psychosocial risk factors in DSM-III attention deficit disorder //
Journal of the American Academy of Child and Adolescent
Psychiatry. — 1990. — Vol. 29. — P. 526—533.
10. Bonifati V., FabrizioE., VanacoreN. et al. Familial Parkinson«s disease:
a clinical genetic analysis // Canad. J. Neurol. Sci. — 1995. —
Vol. 22. — P. 272—279.
11. Bruun R.D. Gilles de la Tourette syndrome: an overview of clinical
experience // Journal of the American Academy of Child and
Adolescent Psychiatry. — 1984. — Vol. 23. — P. 126—133.
12. Bryson S., Clark B., Smith T. First report of a Canadian epidemio-
logical study of autistic syndromes // Journal of Child Psychology
and Psychiatry. — 1988. — Vol. 29. — P. 433—445.
13. Burd L., Kerbeshian J., Wikenheiser M., Fisher W. Prevalence of
Gilles de la Tourette's syndrome in North Dakota adults //
American Journal of Psychiatry. — 1986. — Vol. 143. — P. 787—788.
14. Burd L., Kerbeshian J., Wikenheiser M., Fisher W. Prevalence of
Gilles de la Tourette syndrome in North Dakota school-age children
// Journal of the American Academy of Child and Adolescent
Psychiatry. — 1986. — Vol. 25. — P. 552—553.
15. Cantwell D.P. Psychiatric illness in the families of hyperactive
children // Archives of General Psychiatry. — 1972. — Vol. 27. —
P. 414—417.
16. Carmelli D., Swan G.E., Robinette D., Fabsitz R. Genetic influence
on smoking — a study of male twins // The New England Journal
of Medicine. — 1992. — Vol. 327. — P. 829—833.
17. Chakravartti R., Roy A.K., Rao K.U., Chakravartti M.R. Hereditary
factors in stammering // J. Genet. Hum. — 1979. — Vol. 27, No. 4. —
P. 319—328.
18. Cohen I.L., Sudhalter V., Pfadt A. et al. Why are autism and the
fragile-X syndrome associated? // American Journal of Human
Genetics. — 1991. — Vol. 48. — P. 195—202.
19. Collier D. The genetics of heroin abuse. What are the implications
of new scientific findings for effective drugs treatment? // The
Observer. — 2002, May 12.
20. Comings D. The molecular genetics of pathological gambling //
CNS Spectrums. — 1998. — Vol. 3, No. 6. — P. 20—37.
21. Comings D.E., Comings B.G., Devor E.J. Detection of a major gene
for Gilles de la Tourette syndrome // American Journal of Human
Genetics. — 1984. —Vol. 36. — P. 586—600.
22. Comings D.E., Himes J.A., Comings B.G. An epidemiologic study
of Tourette's syndrome in a single school district // Journal of
Clinical Psychiatry. — 1990. — Vol. 51. — P. 463—469.
462
К разделу 12
23. Cook Е.Н., Courchesne R., Lord C. et al. Evidence of linkage
between the serotonin transporter and autistic disorder //
Molecular Psychiatry. — 1997. — Vol. 2. — P. 247—250.
24. Cook E.H., Lindgren V., Leventhal B.L. et al. Autism or atypical
autism in maternally but not paternally derived proximal 15q
duplication // American Journal of Human Genetics. — 1997. —
Vol. 60. — P. 928—934.
25. Coon H., Hoff M., Holik J. et aL Analysis of chromosome 18 DNA
markers in multiplex pedigrees with manic depression // Biological
Psychiatry. — 1996. — Vol. 39. — P. 689—696.
26. Cox N.J., Reich T., Rice J.P. et al. Segregation and linkage analyses
of bipolar and major depressive illness in multigenerational
pedigrees // Journal of Psychiatric Research. — 1989. — Vol. 23. —
P. 109—123.
27. Grice D.E., Halmi K.A., Fichter M.M. et al. Evidence for a
susceptibility gene for anorexia nervosa on chromosome 1 //
American Journal of Human Genetics. — 2002. — Vol. 70. —
P. 787—92.
28. Crowe R.R., Namboodiri K.K., Ashby H.B., Elston R.C. Segregation
analysis and linkage analysis of a large kindred of unipolar depres-
sion // Neuropsychobiology. — 1981. — Vol. 7. — P. 20—25.
29. Curtis D., Robertson M.M., Gurling H.M.D. Autosomal dominant
gene transmission in a large kindred with Gilles de la Tourette
syndrome // British Journal of Psychiatry. — 1992. —-Vol. 160. —
P. 845—849.
30. Daniels W.W., Warren R.P. , Odell J.D. et al. Increased frequency
of the extended or ancestral haplotype B44BSC30BDR4 in autism
// Biological Psychiatry. — 1995. — Vol. 32. — P. 120—123.
31. De Bruyn A., Souery D., Mendelbaum K. et al. Linkage analysis of
families with bipolar illness and chromosome 18 markers //
Biological Psychiatry. — 1996. — Vol. 39. — P. 679—688.
32. De Lisi L.E. Speech disorder in schizophrenia: review of the
literature and exploration of its relation to the uniquely human
capacity for language // Schizophr. Bull. — 2001. — Vol. 27,
No. 3. — P. 481—496.
33. Donnai D. Gene location in Tourette syndrome // Lancet.- 1987. —
Vol. 14. — P. 627.
34. Eapen V., Pauls D.L., Robertson M.M. Evidence for autosomal
dominant transmission in Tourette's syndrome: United Kingdom
Cohort Study//British Journal of Psychiatry.— 1993. — Vol. 162.—
P. 593—596.
35. Enard W., Przeworski M., Fisher S.E. et al. Molecular evolution of
463
Литература
FOXP2, a gene involved in speech and language // Nature. — 2002. —
Vol. 418, No. 6900. — P. 869—872.
36. Engstrom C., Thornlund A., Johansson E. et al. Anticipation in
unipolar affective disorder // Journal of Affective Disorders. —
1995. — Vol. 35. — P. 31—40.
37. Faraone S.V., Biederman J., Chen W.J. et al. Segregation analysis
of attention deficit hyperactivity disorder: evidence for single gene
transmission // Psychiatric Genetics. — 1992. — Vol. 2. — P. 257—
275.
38. Fisher S.E., Francks C., Marlow A. J. et al. Independent genome-
wide scans identify a chromosome 18 quantitative-trait locus
influencing dyslexia // Nat. Genet. — 2002. — Vol. 30, No. 1. —
P. 86—91.
39. Foroud T., Uniacke S.K., Liu L. et al. Heterozygosity for a mutation
in the parkin gene leads to later onset Parkinson disease //
Neurology. — 2003. — Vol. 60. — P. 796—801.
40. Foundas A.L., Bollich A.M., Corey D.M. et al. Anomalous anatomy
of speech-language areas in adults with persistent developmental
sluttering // Neurology. — 2001. — Vol. 57. — P. 207—215.
41. Frick P. J., Lahey B.B., Christ M.G. et al. History of childhood
behavior problems in biological relatives of boys with attention
deficit hyperactivity disorder and conduct disorder // Journal of
Clinical Child Psychology. — 1991. — Vol. 20. — P. 445—451.
42. Garr L.G., Yi I.S., Li T.K., Yin S.J. Cytochrome P4502E1 genotypes,
alcoholism, and alcoholic cirrhosis in Han Chinese and Atayal
Natives of Taiwan // Alcohol Clin. ExP. Res. — 1996. — Vol. 20,
No. 1. — P. 43—46.
43. Gasser T, Muller-Myhsok B., Wszolek Z.K. et al. A susceptibility
locus for Parkinson«s disease maps to chromosome 2pl3 // Nat.
Genet. — 1998. — Vol. 18, No. 3. — P. 262—265.
44. Gazzaniga M.S., Ivry R.B., Mangun G.R. Cognitive neuroscience. —
W.W. Norton & Company, New York, London, 1998. — 550 p.
45. Gershon E.S., Hamovit J.H., Guroff J. J. et al. A family study of
schizoaffective, bipolar I, bipolar II, unipolar, and normal control
probands // Archives of General Psychiatry. — 1982. — Vol. 39. —
P. 1157—1167.
46. Gillberg C. Infantile autism and other childhood psychoses in a
Swedish urban region: epidemiological aspects // Journal of Child
Psychology and Psychiatry. — 1984. — Vol. 25. — P. 35—43.
47. Gjone EL, Stevenson J., Sundet J.M. Genetic influence on parent-
reported attention-related problems in a Norwegian general
population twin sample // Journal of the American Academy of
Child and Adolescent Psychiatry. — 1996. — Vol. 35. — P. 588—596.
464
К разделу 12
48. Goldin L.R., Gershon E.S., Targum S.D. et al. Segregation and
linkage analyses in families of patients with bipolar, unipolar, and
schizoaffective mood disorders // Journal of Human Genetics. —
1983. — Vol. 35. — P. 274—287.
49. Goldstein L., Manowitz P., Nora R. et al. Differential EEG activation
and pathological gambling // Biological Psychiatry. — 1985. —
Vol. 20. — P. 1232—1234.
50. Goodman R., Stevenson J. A twin study of hyperactivity I: an
examination of hyper-activity scores and categories derived from
Rutter teacher and parent questionnaires // Journal of Child
Psychology and Psychiatry. — 1989. —Vol. 30. — P. 671—689.
51. Goodman R., Stevenson J. A twin study of hyperactivity II: the aetio-
logical role of genes, family relationships and perinatal adversity //
Journal of Child Psychology and Psychiatry. — 1989. — Vol. 30. —
P. 691—709.
52. Gurling H. Candidate genes and favored loci; strategies for
molecular genetic research into schizophrenia, manic depression,
autism, alcoholism, and Alzheimer disease // Psychiatric
Developments. — 1986. —Vol. 4. — P. 289—309.
53. Halgin R., Whitbourne S. Abnormal psychology. — Brown &
Benchmark Publishers, 1994. — 520 p.
54. Halgin R., Whitbourne S. Abnormal psychology. — Brown &
Benchmark Publishers, 1994. — 520 p.
55. Hall W„ Madden P., Lynskey M. The Genetics of Tobacco Use:
Methods, Findings and Policy Implications // Tobacco Control. —
2002. — Vol. 11. — P. 119—124.
56. Hess E.J., Rogan P. K., Domoto M. et a]. Absence of linkage of
apparently single gene mediated ADHD with the human syntenic
region of the mouse mutant coloboma // American Journal of
Medical Genetics. — 1995. — Vol. 60. — P. 573—579.
57. Heutink P., van de Watering Pakstis A.J. et al. Linkage stu-
dies on Gilles de la Tourette syndrome: what is the strategy of choice?
// American Journal of Human Genetics. — 1995. — Vol. 57. —
P. 465—473.
58. Hoshino Y., Kumashiro H., Yashima Y. et al. The epidemiological
study of autism in Fukushimaken // Folia Psychiatrica Neurologica
of Japan. — 1982. — Vol. 36. — P. 115—124.
59. jennings@cyberport.com.
60. Jorde L.B., Hasstedt S.J., RitvoE.R. etal. Complex segregation analysis
of autism // American Journal of Human Genetics. — 1991. —
Vol. 49. — P. 932—938.
61. Kessler R.C., McGonagle K.A. ZhaoS. et a]. Lifetime and 12-month
465
Литература
prevalence of DSM-II1-R psychiatric disorders in the United States
// Archives of General Psychiatry. — 1994. — Vol. 51. — P. 8—19.
62. Kottak C.P. Anthropology. — New York: McGraw-Hill Inc., 1991. —
496 p.
63. Kotulak R. Study Links Genetics and Addiction // The Toronto Star.
— 1999, March 26.
64. Kremer B., Goldberg P., Andrew S.E. et al. A worldwide study of the
Huntington's disease mutation. The sensitivity and specificity of
measuring GAG repeats // N. Engl. J. Med. — 1994. — Vol. 330,
No. 20. — P. 1401 — 1406.
65. Kreuz F.R. Attitudes of German persons at risk for Huntington's
disease toward predictive and prenatal testing // Genet. Couns. —
1996. — Vol. 7, No. 4. — P. 303—311.
66. Kunugi H., Tatsumi M., Sakai T. et al. Serotonin transporter gene
polymorphism and affective disorder // Lancet. — 1996. —
Vol. 347. — P. 1340.
67. LaBuda M.C., Maldonado M., Marshall D. et al. A follow-up report
of a genome search for affective disorder predisposition loci in the
Old Order Amish // American Journal of Human Genetics. — 1996.
— Vol. 59. — P. 1343—1362.
68. Lang A.E., Lozano A.M. Parkinson«s disease // New Engl. J. Med. —
1998. — Vol. 339 — P. 1044—1053.
69. Leroy E. et al. The ubiquitin pathway in Parkinson«s disease //
Nature. — 1998. — Vol. 395, No. 6701. — P. 451—452.
70. Lewis R. Life. — Wm. C. Brown Publishers, 1995. — 884 p.
71. Lindsay D.T. Functional human anatomy. — Mosby Year Book,
1996. — 822 p.
72. Locke, 1983, P. 172.
73. Lorenz V.C., Shuttlesworth D.E. The impact of pathological
gambling on the spouse of the gambler // Journal of Community
Psychology. — 1983. —Vol. 11. — P. 67—76.
74. Lotter V. Epidemiology of autistic conditions in young children: I.
prevalence // Social Psychiatry. — 1966. —Vol. 1. — P. 124—137.
75. Lbcking C.B. et al. Association between early-onset Parkinson«s
disease and mutations in the PARKIN gene // New Engl. J. Med. —
2000. — Vol. 342. — P. 1560—1567.
76. Manki EL, Kanba S., Muramatsu T. et al. Dopamine D2, D3 and D4
receptor and transporter gene polymorphisms and mood disorders
// Journal of Affective Disorders. — 1996. — Vol. 40. — P. 7—13.
77. McCarthy P. , Fitzgerald M., Smith M.A. Prevalence of childhood
autism in Ireland // Irish Medical Journal. -— 1984. — Vol. 77. —
P. 129—130.
466
К разделу 12
78. McGuffin Р., Katz R., Watkins S., Rutherford J. A hospital-based
twin register of the heritability of DSM-IV unipolar depression //
Archives of General Psychiatry. — 1996. — Vol. 53. — P. 129—136.
79. McMahon W.M., van de Wetering B.J.M., Filloux F. et al. Bilineal
transmission and phenotypic variation of Tourette's disorder in a
large pedigree // Journal of the American Academy of Child and
Adolescent Psychiatry. — 1996. — Vol. 35. — P. 672—680.
80. Mendlewicz J., Rainer J.D. Adoption study supporting genetic
transmission in manic-depressive illness //Nature. — 1977. —Vol.
268. — P. 326—329.
81. Moisan M.-P. , Courvoisier H., Bihoreau M.-T. et al. A major
quantitative trait locus influences hyperactivity in the WKHA rat
// Nature Genetics. — 1996. —Vol. 14. — P. 471—473.
82. Moldin S.O., Reich T., Rice J.P. Current perspectives on the genetics
of unipolar depression // Behavior Genetics. — 1991. — Vol. 21. —
P. 211—242.
83. Montgomery E.B., Baker K.B., Lyons K., Koller W.C. Abnormal
performance on the PD test battery by asymptomatic first-degree
relatives // Neurology. — 1999. — Vol. 52. — P. 757—762.
84. Morrison J.R., Stewart M.A. A family study of the hyperactive child
syndrome // Biological Psychiatry. — 1971. — Vol. 3. — P. 189—195.
85. Ogilvie A.D., BattersbyS., Bubb V.J. et al. Polymorphism in serotonin
transporter gene associated with susceptibility to major depression
// Lancet. — 1996. — Vol. 347. — P. 731—733.
86. Oruc L., Lindblad K., Verheyen G.R. et al. CAG repeat expansions
in bipolar and unipolar disorders // American Journal of Human
Genetics. — 1997. — Vol. 60. — P. 730—732.
87. Patel P.l. Invited editorial: quest for the elusive genetic basic of
Tourette syndrome // American Journal of Human Genetics. —
1996. — Vol. 59. — P. 980—982.
88. Pauls D.L., Cohen D.J., Heimbuch R.C. et al. The familial pattern
and transmission of Tourette syndrome and multiple tics // Annals
of Human Genetics. — 1981. — Vol. 38. — P. 1091 —1093.
89. Pauls D.L., Pakstis A.J., Kurlan R. et al. Segregation and linkage
analyses of Tourette's syndrome and related disorders // Journal
of the American Academy of Child and Adolescent Psychiatry. —
1990. — Vol. 29. — P. 195—203.
90. Pauls D.L., Raymond C.L., Stevenson J.M., Leckman J.F. A family
study of Gilles de la Tourette syndrome // American Journal of
Human Genetics. — 1991. — Vol. 48. — P. 154—163.
91. Pickles A., Bolton P. , MacDonald H. et al. Latent-class analysis of
recurrence risks for complex phenotypes with selection and
467
Литература
measurement error: a twin and family study of autism // American
Journal of Human Genetics. — 1995. — Vol. 57. — P. 717—726.
92. Piven J., ArndtS., Bailey J. et al. An MRI study of brain size in autism
//American Journal of Psychiatry.— 1995.—Vol. 12. — P. 1145—
1149.
93. Piven J., Harper J., Palmer P., Arndt S. Course of behavioral change
in autism: a retrospective study of high-IQ adolescents and adults
// Journal of the American Academy of Child Adolescent
Psychiatry. — 1996. — Vol. 35, No. 4. — P. 523—529.
94. Pomrock J.H., Ginath J. Differences between Schizophrenics and
Normal Controls Using Chirological (Hand) Testing // Israel
Journal of Psychiatry and Related Sciences. — 1982. — Vol. 19,
No. 1. — P. 5—22.
95. Price R.A., Kidd K.K., Cohen D.J. et al. A twin study of Tourette
syndrome // Archives of General Psychiatry. — 1985. — Vol. 42. —
P. 815—820.
96. Price R.A., Pauls D.L., Kruger S.D., Caine E.D. Family data support
a dominant major gene for Tourette syndrome // Psychiatry
Research. — 1987. — Vol. 24. — P. 251—261.
97. Reider C.R., Halter C.A., Casteluccio P.F. et al. Reliability of
Reported Age at Onset for Parkinson's Disease // Movement
Disorders. — 2003. — Vol. 18, No. 3. — P. 275—279.
98. Ritvo E.R., Freeman B.J., Mason-Brothers A. et al. Concordance of
the syndrome of autism in 40 pairs of afflicted twins // American
Journal of Psychiatry. — 1985. — Vol. 142. — P. 74—77.
99. Robertson M.M., Gourdie A. Familial Tourette's syndrome in a large
British pedigree. Associated psychopathology, severity, and
potential for linkage analysis // British Journal of Psychiatry. —
1990. — Vol. 156. — P. 515—521.
100. Robertson M.M., Verrill M., Mercer M. et al. Tourette's syndrome
in New Zealand: a postal survey // British Journal of Psychiatry. —
1994. — Vol. 164. — P. 263—266.
101. Rosenhan D.L. On being sane in insane places // Science.— 1973.—
Vol. 179. — P. 250—258.
102. Rosman A., Rubel P.G. The tapestry of culture. — New York:
Random House, 1989. — 338 p.
103. Ross G.W. et al. Association of coffee and caffeine intake with the
risk of Parkinson disease // JAMA. — 2000. — Vol. 283. —
P. 2674—2679.
104. Roy A., AdinoffB., Roehrich L. et al. Pathological gambling: A psycho-
biological study // Archives of General Psychiatry. — 1988. —
Vol. 45. — P. 369—373.
468
К разделу 12
Schachar R., Wachsmuth R. Hyperactivity and parental psycho-
pathology // Journal of Child Psychology and Psychiatry. — 1990. —
Vol. 31. — P. 381—392.
Silberg J., Michael R., Joanne M, et al. Genetic and environmental
influences on the covariation between hyperactivity and conduct
disturbance in juvenile twins // Journal of Child Psychology and
Psychiatry. — 1996. — Vol. 37. — P. 803—816.
Smalley S.L., Asarnow R.F., Spence M.A. Autism and genetics: a
decade of research // Archives of General Psychiatry. — 1988. —
Vol. 45. — P. 953—961.
Smith D.J., Stevens M.E., Sudanagunta S.P. et al. Functional
screening of 2 Mb of human chromosome 21q22.2 in transgenic
mice implicates minibrain in learning defects associated with Down
syndrome // Nature Genetics. — 1997. — Vol. 16. — P. 28—36.
Sommer M., Koch M.A., Paulus И7 et al. Disconnection of speech-
relevent brain areas in persistent developmental sluttering //
Lancet. — 2002. — Vol. 360. — P. 380—383.
Steffenburg S., Gillberg'C. Autism and autistic-like conditions in
Swedish rural and urban areas: a population study // British Journal
of Psychiatry. — 1986. — Vol. 149. — P. 81—87.
Steinhausen H.C., Gobel D., Breinlinger M., Wohlleben B. A
community survey of infantile autism // Journal of the American
Academy of Child and Adolescent Psychiatry. — 1986. — Vol. 25. —
P. 186—189.
StoberG., HeilsA., Lesch K.P. Serotonin transporter gene polymorp-
hism and affective disorder // Lancet. — 1996. — Vol. 347. —
P. 1340—1341.
Straub R.E., Lehner T., Luo У. et al. A possible vulnerability locus
for bipolar affective disorder on chromosome 21q22.3 // Nature
Genetics. — 1994. — Vol. 8. — P. 291—296.
Szatmari P., Jones M.B. IO and the genetics of autism // Journal of
Child Psychology and Psychiatry. — 1991. — Vol. 32. — P. 897—
908.
Tanaka F, Shiratori Y., Yokosuka O. et al. Polymorphism of alcohol-
metabolizing genes affects drinking behavior and alcoholic liver
disease in Japanese men // Alcohol Clin. ExP. Res. — 1997. —
Vol. 21, No. 4. — P. 596—601.
Tanner C.M., Ottman R., Goldman S.M. et al. Parkinson Disease in
Twins // JAMA. — 1999. — Vol. 281, No. 4. — P. 341—346.
Taylor L.D., Krizman D.B., Jankovic J. 9p monosomy in a patient
with Gilles de la Tourette syndrome // Neurology. — 1991. —
Vol. 41. — P. 1513—1515.
469
Литература
118. Thapar A., Hervas A., McGuffin Р. Childhood hyperactivity scores
are highly heritable and show sibling competition effects: twin
study evidence // Behavior Genetics. — 1995. — Vol. 25. —
P. 537—544.
119. Thapar A., McGuffin P. A twin study of depressive symptoms in
childhood // British Journal of Psychiatry. — 1994. —Vol. 165. —
P. 259—265.
120. Todd R. Genetics of Childhood Diorders: XXI. ADHD, Part 5: A
Behavioral Genetic Perspective //J. Am. Acad. Child. Adolesc.
Psychiatry. — 2000. — Vol. 39, No. 12. — P. 1271 — 1274.
121. Torgersen S. Genetic factors in moderately severe and mild affective
disorders // Archives of General Psychiatry. — 1986. — Vol. 43. —
P. 222—226.
122. Torrey E.F., Taylor E.H., Bracha H.S. et al. Prenatal origin of
schizophrenia in a subgroup of discordant monozygotic twins //
Schizophr. Bull. — 1994. — Vol. 20, No. 3. — P. 423—432.
123. Treffert D.A. Epidemiology of infantile autism // Archives of
General Psychiatry. — 1970. — Vol. 22. — P. 431—438.
124. Tsai L., Stewart M.A., August G. Implications of sex differences in
the familial transmission of infantile autism // Journal of Autism
and Developmental Disorders. — 1981. —Vol. 11. — P. 165—173.
125. Tsuang M. T., Bucher K.D., Fleming J.A., Faraone S. V. Transmission
of affective disorders: An application of segregation analysis to blind
family study data // Journal of Psychiatric Research. — 1985. —
Vol. 19. —P. 23—29.
126. Volpe P. Biology and Human Concerns. — Wm. C. Brown
Publishers, 1993. — 611 p.
127. Warren R.P., Singh V.K., Averett R.E. et al. Immunogenetic studies
in autism and related disorders // Molecular and Chemical
Neuropathology. — 1996. — Vol. 28. — P. 77—81.
128. Weissman M.M., Bland R.C., Canino G.J. et al. Cross-national
epidemiology of major depression and bipolar disorder // Journal
of the American Medical Association. — 1996. — Vol. 276. —
P. 293—299.
129. Weissman M.M., Gershon E.S., Kidd K.K. et al. Psychiatric disorders
in the relatives of probands with affective disorders // Archives of
General Psychiatry. — 1984. — Vol. 41. — P. 13—21.
130. Wender P. EL, Kety S.S., Rosenthal D. et al. Psychiatric disorders in
the biological and adoptive families of adopted individuals with
affective disorders // Archives of General Psychiatry. — 1986. —
Vol. 43. — P. 923—929.
131. Widiger T.A., Clark L.A. Toward DSM-V and the classification of
470
К разделу 12
psychopathology // Psychological Bulletin. — 2000. — Vol. 126,
No. 6. — P. 946—963.
132. Willerman L. Activity level of hyperactivity in twins // Child
Development. — 1973. — Vol. 44. — P. 288—293.
133. WingL., Yeates S.R., Brierley L.M., Gould J. The prevalence of early
childhood autism: comparison of administrative and epidemiolo-
gical studies // Psychological Medicine. — 1976. — Vol. 6. —
P. 89—100.
134. Winokur G., Tsuang M.T., Crowe R.R. The Iowa 500: Affective
disorder in relatives of manic and depressive patients // American
Journal of Psychiatry. — 1982. —Vol. 139. — P. 209—212.
135. Дженкинс M. 101 ключевая идея: Генетика. — M.: Фаир-Пресс,
2002. — 240 с.
136. Смоляк А.В. Шаман: личность, функции, мировоззрение: (на-
роды Нижнего Амура). — М.: Наука, 1991.
137. Солониченко В.Г., Богданов Н.Н., Остренко Т.Я., Малышев Ю.И.
Дерматоглифическии рисунок при синдроме Туретта // Фи-
зиология человека. — 1997. —• Т. 23, № 1. —С. 113—117.
138. Трут Л.Н. Очерки по генетике поведения.’— Новосибирск:
Наука, СО. — 1978. — 256 с.
139. Фогель Ф., Мотулъски А. Генетика человека. 3—й том. — М.:
Мир, 1990.
140. Штерн К. Основы генетики человека. — М.: ИЛ., 1965. — 687 с.
К приложению
1. Закс Л. Статистическое оценивание: Пер. с нем. — М.: Стати-
стика, 1976. — 598 с.
Учебное издание
Любовь Алексеевна Атраментова,
Ольга Владимировна Филипцова
ВВЕДЕНИЕ В ПСИХОГЕНЕТИКУ
Учебное пособие
Подписано в печать 14.04.2004. Формат 60x88/16. Печать офсетная
Усл. печ. л. 28,9. Уч.-изд. л. 28,1
Тираж 5 000 экз. Заказ № 2442. Изд. № 881
ИД № 04826 от 24.05.2001 г.
ООО «Флинта», 117342, г. Москва, ул. Бутлерова, д. 17-Б, комн. 332
Тел/факс 334-82-65; тел. 336-03-11. E-mail: flinta@mail.ru, flinta@flinta.ru
WebSite: www.flinta.ru
МПСИ, 113191, г. Москва, 4-й Рощинский пр., д. 9-а
Тел/факс: 234-43-15; 958-17-74 (доб. 111). E-mail: publish@col.ru
Отпечатано с готовых диапозитивов во ФГУП ИПК
«Ульяновский Дом печати». 432980, г. Ульяновск, ул. Гончарова, 14