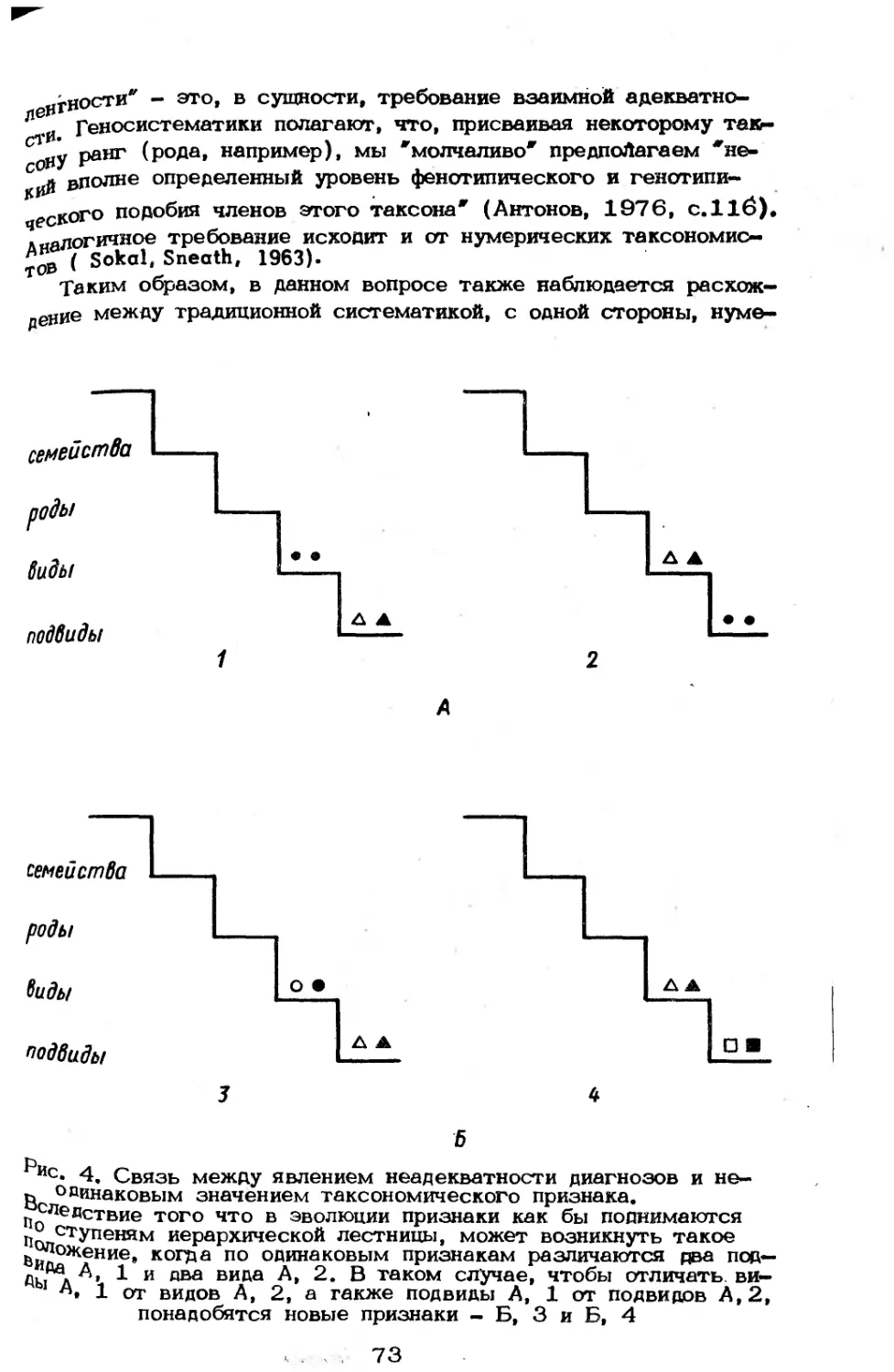
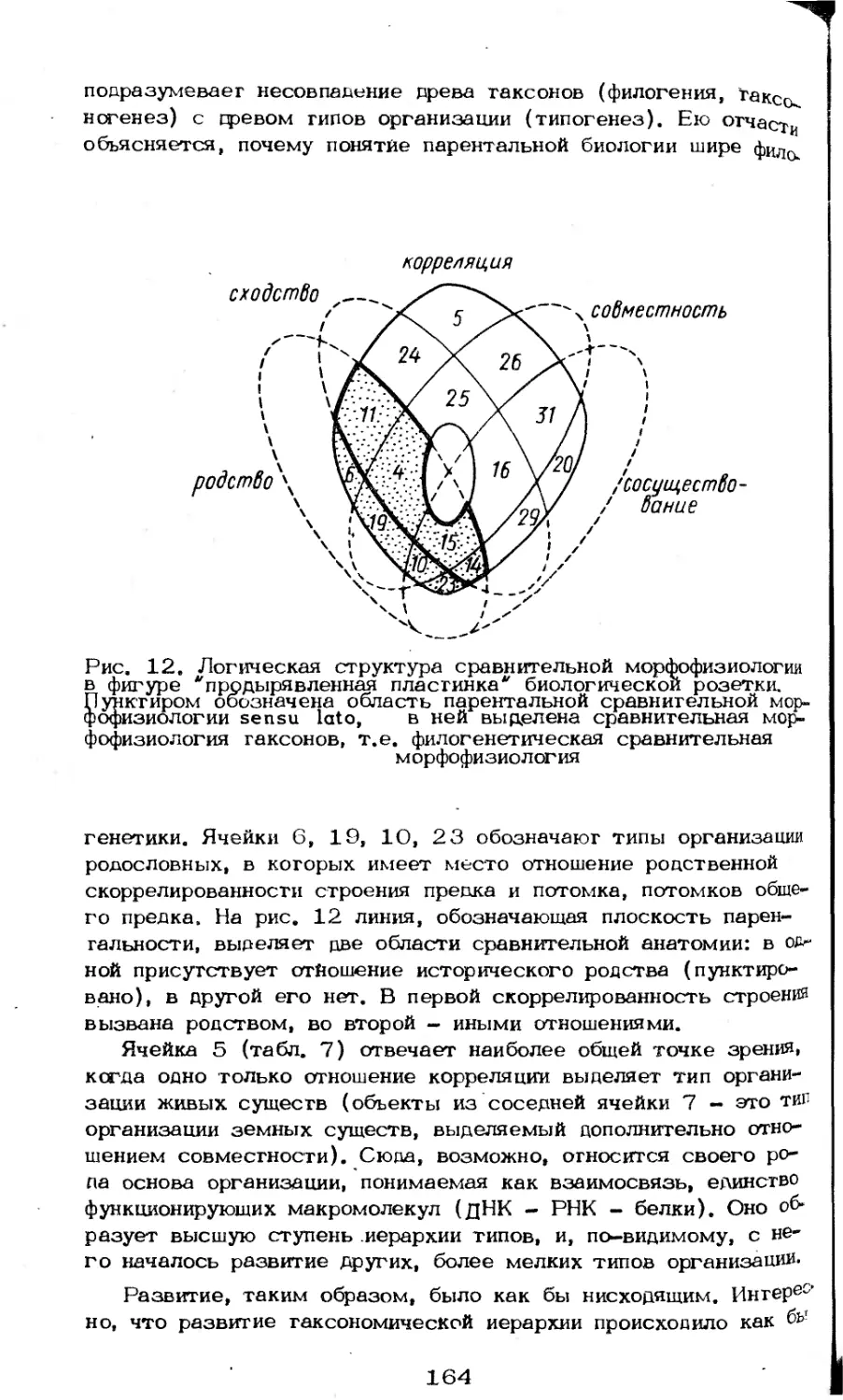
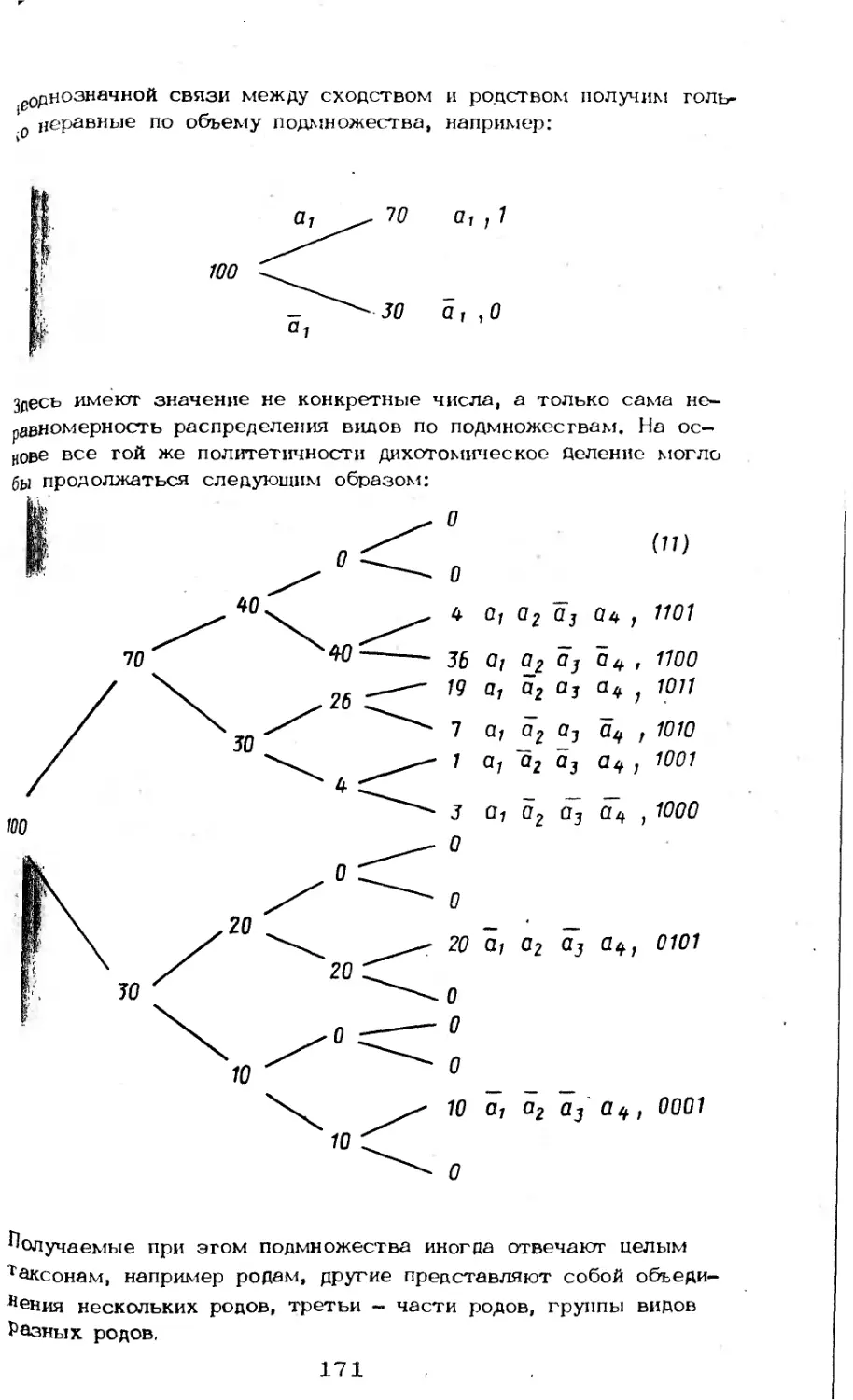
/






















































































































































































































Автор: Заренков Н.А.
Теги: общая экология биоценология гидробиология биогеография биология теоретическая биология
ISBN: 5-211-00138-9
Год: 1988




Текст
Н.АЗАРЕНКОВ
Издательство
Московского университета
Н.А.Заренков
Теоретическая биология
(Введение)
Издательство Московского университета 1988
УДК 574.8
Заренков Н.А. Теоретическая биология. — М.: Изя—во Моск, ун-та, 1988. - 216 с. - ISBN 5-211-00138-9.
В монографии рассматриваются вопросы структуры и единства биологического знания. Теоретические концепции и понятийный аппарат современной биологии анализируются с позиции, согласно которой носителем биологической специфичности является популяция. Широко привлекается философская и методологическая литература, в частности, в связи с проблемой единства и структуры естествознания. Освещается новый фактический материал, а также дается новая интерпретация традиционного материала.
Для широкого круга биологов, а также философов.
Табл. 7. Ил. 13. Библиогр.: 700 назв.
Рецензенты:
член-корреспондент АН. СССР А.В.Яблоков,
доктор биологических наук Я.И.Старобогатов, доктор философских наук З.В.Каганова, доктор биологических наук Г.Н.Симкин
Печатается по постановлению Редакционно-издательского совета Московского университета
2001000000-146 © Издательство Москов<
077(02) - 88 “ университета, 1988
ПРЕДИСЛОВИЕ
В настоящее время в отечественной философской и биологической литературе все острее ощущается недостаток работ, с единых методологических позиций освещающих наиболее острые проблемы биологии, т.е. работ, которые в 30-50-е гг. объединялись под эгидой "теоретической биологии". Такое положение, на наш взгляд, определяется стремительным нарастанием общего объема "биологической информации", невероятными трудностями сбора и обобщения гигантских материалов, а также как снежный ком нарастающей "разноголосицей" позиций и мнений. Есть и другая, более серьезная сторона. Стремительная разработка эмпирического знания все чаще приводит профессиональных биологов к пессимистическим выводам о самой возможности существования понятия "теоретическая биология" и его необходимости и, что самое главное, — о принципиальной целесообразности дедуктивных по своей природе научных схем, на поверку временем слишком часто оказывающихся поверхностными и быстротечными.
И тем не менее настоятельная потребность теоретических обобщений в наши дни очевидна в связи не только с чудовищным разрастанием абстрактных теоретических конструкций практически по всем основным направлениям современной биологии, но и с необходимостью профессионалов-исследователей сохранять широкую эрудицию, в том числе в сопредельных по профессии областях знания. Этот скачок теоретических построений по сравнению с 5О-ми и Даже 7О-ми годами по многим направде-1иям современной науки был совершен благодаря широкому про— <икновению в биологию математических приемов анализа, появ— 1ению достаточно сложных программ обработки биологических данных на ЭВМ.
Особенно актуальна "теоретическая биология" для студентов, спирантов и молодых исследователей, перед которыми методо-огические проблемы современного знания встают с предельной 'кСстротой. Все сказанное дает возможность высоко оценить тот зуд, который затратил автор на сбор, обработку и осмысление тактически необозримого и крайне разнородного материала. У эня нет сомнений в том, что даже при значительном количест-j несогласий с автором, книга должна быть опубликована хо-бы потому, что позволит читателям более внимательно вду— 1ться в состояние, тенденции развития и противоречия многих
вопросов современной биологии. Уровень аргументации автора достаточно высок. Его позиции по всем основным проблемам теоретической биологии, как в нашей стране, так и за рубежом, разделяют и отстаивают многие известные биологи и философы. Нельзя забывать, о том, что сама природа теоретической биологии, ее история и современное состояние столь противоречиво и столь различно трактуются отдельными школами и исследователями, что даже на обоснованную критику практически по всем направлениям может быть сформулирована не менее аргументированная антикритика. Такова на сегодняшний день сама природа теоретической биологии. Таково ее состояние в наши дни.
Г. Н. Симкин
ОТ АВТОРА
После того как книга написана и живет самостоятельной от автора жизнью, как будто бы становится ненужным знание тех обстоятельств, которые послужили побудительной причиной ее написания. Однако поскольку такое отчуждение нередко мешает правильному восприятию авторского замысла, представляется целесообразным рассказать об этом читателю.
Мне приходится читать лекции на биологическом факультете Московского университета по зоологии беспозвоночных, а также частные спецкурсы по зоогеографии, сравнительной анатомии и систематике. В процессе преподавания у меня сложилось впечатление, что знание спецкурса как системы фактов и умение передавать это знание слушателям не исчерпывают обязанностей лектора перед аудиторией. Представляется необходимым, чтобы при чтении лекций по отдельным биологическим дисциплинам между ними выявлялось очевидное соответствие, делающее возможным объединение их в системном знании общей биологии. Особенно важно проводить идею единства биологического знания преподавателям общей биологии. Именно единство биологии, ее внутренняя структура и положение среди других наук должны определять содержание учебной программы и методов обучения сту дентов-биологов.
В силу ряда причин вопросы преподавания общей биологии далеки от оптимального решения. Во-первых, система фактов, добытых в биологии и смежных науках, возросла настолько, что в лекциях уже невозможно охватить даже наиболее существенные из них достаточно детально. Факты, например строение какого-либо животного, имеют теперь значение не столько в связи со своей уникальностью, сколько возможностью показать на немногих из них то, что приложимо к другим фактам подобного рода, остающимся вне рамок учебной программы. Отсюда возникает необходимость внесения в спецкурсы теоретического подтекста, выявления внутренних связей между частными разделами биологии и преемственности между совершенно различными, на первый взгляд, понятиями, такими, как например,'тип и гаксон или классификация и районирование.
Во-вторых, в современной биологии сохраняется определенное разногласие относительно предмета биологии между натуралистами и представителями физико-химической биологии. В последние годы в высших учебных заведениях преобладает одностороннее понимание единства биологии на основе физико-химического знания. Его результатом является вытеснение из учеб-
5
ных программ ботаники и зоологии и расширение курсов физики, химии, математики. Возникает вопрос, насколько это оправдано и чему отдавать предпочтение - физической химии или собственно биологии?
В-третьих, требует осознания сущность современной трагедии живой природы,невольной причиной которой мы являемся. Необходимо осознать, для чего в действительности нужна человеку природа живая; для того ли только, чтобы оставались чистыми вода и воздух, чтобы сохранялись и возобновлялись пищевые и другие ресурсы? Положительный ответ на этот вопрос ле-, жиг на поверхности, но скорее всего он не может считаться исчерпывающим, потому что в духовном мире человека живая природа занимает несколько особое место, нежели неживая природа. Она будит в нем чувство милосердия. Именно поэтому биологическая культура, биологическое образование не могут оставаться нейтральными по отношению ко всему живому: милосердие, любовь к нему должны быть неотъемлемой частью преподавания биологии.
Предлагаемая читателю книга представляет собой попытку разобраться в этих вопросах с позиций традиционной биологии. Она, разумеется, не дает исчерпывающего ответа на них, но зато отличается от других изданий по аналогичной тематике следующей особенностью. Автор исходит из того, что натуралисты вполне объективно изучают живую природу, и это позволило традиционной биологии уже воссоздать объективную картину земной жизни. Поэтому задача науки в настоящее время состоит в том, чтобы 'извне привнести в биологию теорию биологии и жизни, на основе, например, термодинамики (Бауэр) или кристаллохимии (Шрёдингер), а в том, чтобы выделить эту теорию из са мой сущности биологии, ее "внутреннего мира". Для решения этой задачи не требуется какого-то иного (не биологического знания, однако необходимо новое понимание известных фактов, которое и определяет исходную познавательную позицию. Для биолога такой позицией будет признание самой возможности собственно биологической концепции жизни. Эта концепция в не явном вице содержится в теории естественного отбора Чарльза Дарвина, и свою задачу автор видит в том, чтобы изложить ее языком современной науки. Между тем, как показано в первых главах книги, уже длительное время в науке преобладает одностороннее обсуждение концепции жизни, главным образом с позиций физики.
В книге рассмотрены логическая структура биологии и пон« тийный аппарат ее частных разделов на основе биологической концепции жизни. Изложению этой части книги помогает диаграмма Венна, представляющая развертку на плоскости многомерной фигуры. Эта диаграмма понадобилась потому, что биологи фактически пользуются многомерной логикой. Последним
обстоятельством объясняются специфические трудности изложения. Дело в том, что в данной книге, как и в любом письменном источнике, изложение ведете в линейной последовательности и в этом смысле может считаться одномерным. Несоответствие линейной последовательности- изложения многомерной структуре объекта имеет своим следствием то, что многие понятия вводятся раньше, чем раскрывается их содержание. Эго несоответствие устраняется только после того, как завершается рассмотрение всех фигур диаграммы Венна.
Теоретический характер книги потребовал введения множества названий теорий, школ, направлений мысли, концепций: витализм, механицизм, редукционизм и пр. Автору очень не хотелось бы, чтобы читатель усмотрел в этом злоупотребление терминами, пресловутое "наклеивание ярлыков". Различать теории и школы — не означает порицать их, плюрализм - естественное состояние любой теории. Единственное, против чего решительно возражает автор - и в этом пафос книги — подмена сдних понятий другими, подмена биологической точки зрения на жизнь точкой зрения физики. Предлагаемая в книге биологическая концепция в свою очередь не отменяет физика листе кую хотя бы потому, что они относятся к разным системам знания. Они могут мирно сосуществовать, однако лишь при условии, что остаются самими собой и не подменяют друг друга.
Глава 1
О РАЗНЫХ ПОДХОДАХ К ОПРЕДЕЛЕНИЮ ПРЕДМЕТА ТЕОРЕТИЧЕСКОЙ БИОЛОГИИ
Выражение 'теоретическая биология* появилось в первой четверти нашего столетия (Reinke, 1901; Ehrenberg, 1928; Uexkull, 1928). Показательно, что тогда же выделилось понятие 'эмпирическая биология* (Schaxel, 1922). Поначалу пор. теоретической биологией понимался широкий круг вопросов: клеточная теория, обмен веществ в организме, понятие силы и организации, механика развития и дарвинизм, логико-методологические основы биологических теорий и концепций. По Л.Бер-таланфи ( Bertalanffy, 1932), теория биологии в первую очередь представляет собой теорию познания и методологию науки о жизни. Внимание к вопросам логики у Берталанфи — результат его контактов с Венским кружком, из которого вышли некоторые видные позитивисты ( Verhoog , 1975).
И все же внимание первых теоретиков привлекали прежде всего отношения между витализмом и механицизмом в вопросе о жизнедеятельности организма. Механицизм определяли как доктрину, '...согласно которой естественные процессы и особенно жизненные процессы детерминированы механистически и могут получить объяснение в законах физики и химии* (The Oxford English Dictionary, 1933, p. 1525). Противоположный механицизму витализм начала XX в. был охарактеризован Л.Плате следующим образом: 'Под общим названием виталистов в настоящее время объединились все противники физико-химического обоснования жизненных процессов* (1928, с. 16). Эта выдержка характеризует умонастроения в период зарождения теоретической биологии, но вследствие своего негативного характера не раскрывает содержание витализма, *...согласно которому, проявления жизни хотя и сводятся частично к механическим причинам, тем не менее полностью от них независимы и требуют признания непознаваемой жизненной силы (*vis vitalis " )* (Zoologisches Worterbuch, 1927, s. 764). М.Л.Левин (1936), возможно, был не так уж далек от истины, указывая, что исторически теория биологии связана с витализмом. Отмечено, в частности, что М.Экскюль, один из ее основателей, был виталистом (Geertde Klerk, 197 9). Все же более точным представляется заключение, что, возникнув как реакция на механицизм, теоретическая биология поначалу представляла собой арену борьбы двух взаимоисключающих подходов к пониманию жизнедеятельности организма.
В современной литературе выражение 'механицизм*, вызывающее ассоциации только с механикой (Savory, 1971), уступило место другому, имеющему более широкое значение, - *фи-
зико-химический редукционизм*. Что же касается классического витализма (Дриш, 1915), то он вытеснен из науки и под теоретической биологией понимают сейчас прежде всего теорию жизнедеятельности организма, излагаемую в понятиях физики и химии. При этом в центре внимания неизменно остаются три формулы, определяющие сущность жизни:
жизнь есть особая форма движения материи (1),
жизнь есть обмен веществ и энергии в организме (2),
жизнь есть жизнедеятельность организма (3).
Формула (1) принадлежит философии, ее автором является Ф.Энгельс (Кедров, 1971). Формула (2) принадлежит физике, потому что в ней использованы физические понятия. Формулу (3) можно было бы считать биологической, если бы ее содержание не исчерпывалось формулой (2). Здесь проходит граница между редукционизмом, настаивающим на сведении (3) к (2), и витализмом, отрицающим такую сводимость. Тем самым витализм “...утверждает неидентичность и автономность биологических феноменов как отличных от небиологических феноменов* . (Blandino, 1969,- с. 3), а следовательно, и самостоятельность биологии. Несводимость биологических процессов к небиологическим классический витализм основывал на заимствованной у Аристотеля энтелехии, наперед недоступной для научного изучения: ’'Энтелехия может быть только “'мыслима*'; она находится вне пространства и времени* (Дриш, 1915, с. 263).
Однако несводимость можно было бы понимать и как некий биологический принцип, пока не наполненный научным содержанием и поэтому еще недоступный научному (но не обязательно физико-химическому) исследованию. Как считал В.А.Фауссек (1903), витализм - это не решение, а постановка вопроса. В.С.Бек называет его “вербальным символом еще не решенных проблем* (Beck, 1957), и в этом смысле витализм можно считать приемлемой гипотезой. Поскольку вещество, энергия и поле, изучением которых ограничены физика и химия, не исчерпывают понятия материи, физикализм, как справедливо подчеркнула Л.М.Косарева (1977), не равновелик материализму, хотя действительно *...физическая картина мира довольно долгое время считалась синонимом материализма в естествознании* (Мещерякова, Пахомов, 1981, с. 163). Вопреки позитивизму, объявившему физику образцовым научным знанием, надо считать-ся с правомочностью разных систем знаний о природе. Подмена философского понятия материи естественнонаучными понятиями вещества, энергии, поля — грубая методологическая ошибка» потому что понятия философии не эквивалентны понятиям частных наук.
В последнее время витализм пережил своего рода обновление и вернулся в биологию в виде кибервигализма (Новик,
1975). В отличие от классического витализма, опиравшегося на непознаваемую энтелехию, он пользуется научными понятиями системы и информации. Вследствие этого чисто негативное определение современного витализма, согласно которому сущность жизненных явлений совершенно не связана с физическими законами... (Рашевски, 1966, с. 165), устарело, и формула (3) приобрела новый вид:
жизнь есть функционирование живой системы. (3*) Однако в силу общенаучного статуса системологии и кибернетики их принципы применимы и в,физической химии. Это обстоятельство отразила так называемая *организмическая* точка зрения Л.Берталанфи (1969, с. 28), согласно которой *... организмы есть организованные явления и их следует рассматривать в аспекте теории открытых систем и состояний подвижного равновесия, которая, по существу, является расширением обычной физхимии, кинетики и термодинамики*. В результате, вместо (2) мы имеем теперь новую формулу:
жизнь есть обмен веществ, энергии и информации, осуществляемый открытой системой (организмом). (2‘).
Итак, все вернулось на *круги своя*. Теперь неясно, можно ли считать организм открытой системой всего лишь в физика-листском понимании. Оппозиция * механицизм - витализм* теперь переросла в оппозицию *кибернетизованная биология — ор-ганицизм, если под органипизмом иметь в виду некий принцип целостности, не выявляемый аналитическими методами физхимии ( Hein , 1969). Иными словами, * жизнь есть способ организации материи* и таким * способом* является организм ( Muzller, 1955, с. 1). Расширение формул (2) до (2*), а (3) до (3*) произошло в принципиально неустранимом *зазоре' между философским понятием материи и естественнонаучными понятиями вещества и энергии. Противостояние витализма и редукционизма подеялось на новый, более высокий вещественно-энергетическо-информационный уровень. Именно на этом уровне с особой остротой встает вопрос о природе жизни и предмете теоретической биологии.
Могло бы показаться, что главное в этом вопросе — это равенство или неравенство между правыми частями естественнонаучных, а потому сопоставимых формул (2’) и (3*). На са мом же деле есть основания оспаривать правильность обеих этих формул: обе они имеют в виду организм, но можно ли считать организм первым и суверенным носителем жизни? Эгс вопрос все яснее вырисовывается по мере ознакомления с его предысторией, с миром тех понятий, которые хотя и сложилис в основном в античное время, но определились только в сере дине XIX в. в столкновении дарвинизма с типологией. В сов-ременной биологии типология выступает как *организмоцент-
10
ризм* (Завадский, 1968; Мамзин, 1974), дарвинизм (теория естественного отбора) - как популяционизм.
Типология как направление биологической мысли выросла из представлений Платона о предсущесгвующих идеях, только несовершенным образом воплотившихся в вещах реального мира (Хорев, 1979; Nordenskiold, 1928). Задача типолога состоит в преодолении несовершенства вещей и приобщении через них к вечным сущностям, типам. При таком подходе первой сущностью оставалось индивидуальное бытие вещей, организмов (Александров, 1939): "...душе есть причина и начало живого тела" (Аристотель, 1976; с. 402). По-видимому, отсюда следует витализм - вывод о том, что в каждом организме есть сущность жизни и что душа, энтелехия, есть биологический эйдос. По Аристотелю, "жизнью мы называем всякое питание, рост и упадок тела, имеющее основание в нем самом. Таким образом, всякое естественное тело, причастное жизни, есть сущность, притом сущность составная* (Там же, с.394). В этом утверждении — "зародыш* всей проблематики физико-химической биологии. Поскольку первая сущность - индивидуальный предмет (Аристотель, 1976, с. 396), а абсолютное подлежащее - субстрат предмета (Гайденко, 1980), постольку аристотелевская философия ориентирует биологов на изучение организменного субстрата. Получается, что труды Аристотеля могли бы служить отправной точкой для обоих направлений ор-ганизмоцентризма — для механицизма и витализма и при этом носителем жизни каждый раз остается единичный организм. Заметим, что дарвинизм на первое место выдвинул множество организмов — вид, подвид (популяцию).
Происхождение слова "типология* представляется следующим образом. Аристотель и Платон пользовались словом для обозначения образцовой формы, обобщения, очертания. Однако близкое значение имело в древнегреческом языке и другое слово - (ровный, равный, состоящий из одинако-
вых частей, соразмерно построенный). Дополнительно это слово могло бы означать и множество одинаковых организмов (одного вида). Аристотелевское слово принято пе-
реводить латинским "species * и русским "вид". Такое типологическое представление о виде исключает внутривидовую изменчивость биологического вида как ненужное, несущественное, вредное. Важно также, что и по звучанию и по происхождению «ес&о^» близко к «/№« » , больше того, иногда они считаются синонимами £ Boisacq, 1938; Prellwitz, 1892) и происходят от корня » глагола (видеть, созерцать)
(Луканин, 1984). Слово «iа » имело такие значения: внешний вид, красивая наружность, общее начало во многих вещах и, кроме того, архетип, идеальная форма ( Liddell, Scott, 1927; Дворецкий, 1958). Отсюда следует, что типология есть
эйдология, учение о существенном и неизменном в каждом живом организме, хотя сами организмы и могут чем-то несущественным отличаться друг от друга. Вероятно, что это существенное - жизнь. Аристотель и Платон пользовались этими словами в определенном контексте. У Платона идеи имели самостоятельное существование. Они ниоткуда не происходят, существуют всегда и, оставаясь чистыми и неизменными, противостоят изменчивой материи. В этом контексте «i&ca »— эго умопостигаемый образ сущего, первоначало мира, данное нам в чувственно воспринимаемой внешности, форме вещей. Аристотель же, как отметил В.Н.Карпов (1937), обладал развитым чувством природной действительности. Будучи не только философом, но и натуралистом, он превратил платоновский дуализм двух начал — идеального и материального — в дуализм в самих вещах: оформляющего » и оформляемой материи (точнее,
субстрата).В противоположность платоновскому «t8e<x» , аристотелевское «Ct&ot; » не существует отдельно от субстрата. Если у Платона воспринимаемая наблюдателем форма организма — только отблеск умопостигаемого «idea» , то у Аристотеля эта форма и есть сама сущность, надо только отбросить в сторону несущественные отклонения от типичной формы. Заметим, что именно эти отклонения, внутривидовая изменчивость, составляют сущность вида в теории естественного отбора.
Для европейской науки, чьи истоки лежат в христианской религии, существование типов изначально было гарантировано Творцом. Способом отыскания типов, сущностей биология, как и все классическое естествознание, обязана прежде всего экспериментальному методу. 'Самое лучшее из всех доказательств < сть опыт, если только он коренится в эксперименте' (Бэкон, 1978, с. 34). Какой современный исследователь не подпишется под этими словами? Поскольку экспериментирование с энтелехией невозможно, витализм с самого начала был отлучен от экспериментальной физико-математической науки, начавшейся с трудов Г.Галилея. С одной стороны, кажется очевидным, что 'наука о жизни может иметь только те же методы и основания, которые существуют и в науке о неодушевленных телах, и нет надобности делать какое бы то ни было различие между принципами наук физиологических и наук физико-химических' (Бернар, 1866, с. 31). С другой стороны, можно ли в организме отыскать что-либо, нё относящееся к физхимии, пока мы пользуемся методами физической химии? Разумеется, нельзя. Этим объясняется живучесть витализма, но в этом и его слабость, потому что он не предложил ничего позитивного, не объяснил, как надо изучать организм. Современный кибервитализм указывает, что изучать организм надо с позиций системологии и кибернетики, но и это не есть биологически специфический метод.
Почти современной, однако поначалу только механической
формой типологическая биология обязана Р.Декарту: 'Декарт, с его определением животных как простых машин, смотрит на цело глазами мануфактурного периоца в отличие от средних веков, когда животное представлялось помощником человека' . Точка зрения Декарта примечательна механистическим видением проблемы животного организма как 'живой машины'. В ней воздается должное механике, тогда наиболее развитой области знания. Именно ей было обязано своими успехами растущее мануфактурное производство. 'Для естествознания ХУЛ века природа - это машина в руках бога-механика...' (Гайденко, 1982, ’ с. 73). Природа - это 'machina mundi ' (машина мира), ее образ - 'хоры стройные светил'. 'Машинный подход' к проблеме жизни с неизбежностью приводил к мысли о том, что 'живая машина' и есть носитель жизни.
Выражениям Декарта'^ивая машина', 'автомат' (Декарт, 1950, с. 300) предстояла долгая (вплоть до современности) жизнь, однако уже не в узко механическом, а в кибернетизован-ном физико-химическом редукционизме. Разумеется, при атом осталось главное - модернизованная частная целевая установка минувшего периода естествознания. Что же касается Ч.Дарвина, то он заинтересовался приспособленностью, т.е. не тем, что находится внутри организмов, а тем, что существует между ними - борьбой за существование. В атом смысле он преодолел декартовский механицизм.. Надо отметить еще одно обстоятельство. Первоначально механицизм представлялся здоровой реакцией на экстерналистское видение жизни, понимание жизни как души, извне привнесенной- в неодушевленное само по себе тело. Механицизм предлагал внутренние естественные причины, т.е. существование механизма в самих организмах.
Развитию концепции 'живой машины' способствовало французское естествознание на рубеже ХУШ-ХТХ вв., в частности . взгляды П.А.Гольбаха и О.ЖЛаметтри (1976), но прежде всего А.Лавуазье. Дж.Лёб (1912, с. 7) считал, что работы Лавуазье положили 'начало научной биологии'. Если под наукой иметь в виду только физико-химию, то с этим невозможно не согласиться. 'С той поры, как дыхание, - эта, казалось бы, сущность жизни, было сведено гением Лавуазье на физико-химический процесс, витализму был нанесен роковой удар...' (Тимирязев., 1938, с. 171). В этой ретроспективной оценке открытия великого химика не учтено то обстоятельство, что самого Лавуазье интересовал процесс окисления (горения), а не сущность жизни. Им принималось, что этот процесс идет одинаково как в организмах, так и в неживой природе, и новое подтвержпе ние концепции 'живой машины' отнюдь не вызвало краха гипо—
п Ркс К.//Маркс К., Энгельс Ф. Соч. 2-е изд. Т.23.
тезы о таинственной сущности жизни в организме: даже сам термин 'витализм* появился, по-видимому, не раньше 1829 г. (The Oxford English Dictionary, 1933, p.2852).
Действительно, перечитывая Ж.Кювье, одного из наиболее ярких представигелей типологической школы, невозможно не заметить, что сущность жизни он по-прежнему связывает с загадкой жизнедеятельности, с таинственным вихрем (*1е tourbil -Ion" - Cuvier, 1817) вещества и энергии в организме. В этой связи представляется показательным и высказывание К.Бэра, относящееся к 1828 г.: 'Органические тела не только изменяемы, но являются единственными, которые изменяются сами' (Бэр, 1924, с. 94). Притягательность организма как вместилища загадки жизни еще больше возросла после доказательства Л.Пастером невозможности самозарождения. Благодаря этому, вопреки все большему усилению физико-химической концепции жизни, сделалось очевидным, что живые организмы не могут возникнуть только на физико-химической основе. По-видимому, этим можно объяснить двойственность позиции К.Бернара (1866). Он различал две 'причины' возникновения жизни: 'первоначальную', создававшую организм—машину, и 'ближайшую', 'отправляющую жизненные явления' - физико-химические свойства организма. Кажется, что К.Бернар указал тем самым на возможность примирения витализма и редукционизма.
Таким образом, типологический подход к сущности жизни представляется противоречивым единством дополняющих друг друга механицизма и витализма. Механицизм постепенно раскрывал понятие жизнедеятельности организма в терминах физхимии, а оттесняемый витализм хранил идею биологической специфичности. Дж.Вуцжер (Woodger, 1929, р. 205) обрисовал это единство в следующих словах: '...так называемая доказательность витализма совсем не есть доказательность витализма, но бездоказательность некоторых форм механицизма'. В этом поединке могло бы показаться, что дарвинизм и традиционная биология остаются сторонними наблюдателями.
Если для уяснения предмета современной теоретической биологии обратиться к тем публикациям, в которых встречается это название, то окажется, что она прежде всего продолжает искать различие между живым и неживым, ищет определение жизни и, следовательно, скорее является теорией жизни, нежели теорией биологии, т.е. теорией знания о жизни. Это означает, что сюда дополнительно относится значительная по объему литература по теории жизни, создаваемая совместным трудом биологизирующих философов и философствующих естественников, и среди последних - прежде всего химиков, физиков, специалистов в области физико-химической биологии. Общая черта этих публикаций состоит в том, что принимается справедливость формул (2‘) и (3*) и последняя сводится к первой. Проблема неизменно
связывается с организмом и решается в рамках организмоценг-ризма. Отвергнув ненаучную энтелехию виталистов-типологов, современные редукционисты-типологи ищут биологическую специфичность в организменном вещественно-энергетическом субстрате (субстратный подход), в особом способе его функционирования (функциональный подход). В принципе такой подход представляет собой развитие декартовской точки зрения на организм как на машину (теперь он называется открытой системой). Прогресс в теории биологии ставится в зависимость от успехов эмпирического изучения организмов ( Progress m Theoretical Biology, 1967) или от теории развития организма (Bertalanf-fy, 1933). Эти точки зрения находят отражение в "Журнале теоретической биологии" ( J - of Theoretical Biology).
Существуют едва ли не десятки естественнонаучных определений жизни. Их классификация удачно была осуществлена Л.Н.Серавиным (1973). Кроме того, имеется сводка Дж. Бландино "Теория природы жизни" ( Blandino, 1969), отличающаяся от книги С ера вина несколько большей философичностью. В книге В.Л.Кремянского (1969) проблема жизни рассмотрена с позиций концепции уровней организации и др. Несмотря на довольно широкий выбор определений жизни, согласно которым носителем жизни является организм, вопрос далек от разрешения. Об этом прежде всего свидетельствует неослабевающий интерес к проблеме. Нетрудно убедиться, что и в наши дни попытки ра— ’ зобраться в ней остаются в рамках додарвиновской типологии. Приведем несколько высказываний в этом плане. Жизнедеятельность организма — это "механизм жизни" ( Hill et all., 1971), и, наоборот, "выражение жизнь обозначает способ особой активности живых существ" ( Lataste, 1889, р. 1); "живые механизмы образуют один класс понятий с машинами" ( Polanyi, 1968); организм — эго "естественное тело", "живая машина* (Сент—Дьёрцьи, 1960); "разные организмы — различные химические лаборатории" (Ritter, 1919); "сущностью жизни занимается наука физиология" (Рейнке, 1903, с. 3). Особое внимание уделено клетке, хотя известно, что понятия клетка" и "организм" совпадают только частично. "В мельчайшей клетке лежат уже перед нами все загадки жизни" (Рейнке, 1903, с. 3); "Клетка — основной элемент жизни" (Гартман, 1936, с. 25); "элементарная биологическая структура" (Вер— мель, 1970). Более поздняя версия: "...комплекс атрибутивных признаков, которыми характеризуется жизнь, в полном объеме выражаемая организменным (клеточным) целым, формируется 'определенной кооперированностью свойств молекул, составляющих материальный субстрат живого, их физико-химическими взаимодействиями и превращениями" (Чепиков, 1976, с. 32). Выходит, например, что планетарная организованность жизни не есть необходимый атрибут жизни.
Разделяя точку зрения Н.К.Кольцова (1936), А.П.Руденко (1971) и А.А.Малиновский (1972) считают, что биологические явления отличаются от явлений косной природы способностью к ауторепродукции, к воспроизведению сложной каталитической системы. Между гем уже К.Бэр (1924) обратил внимание, что исходно размножение есть форма регулирования обмена веществ в процессе роста организма: "...размножение и образование особи представляет собой один и тот же процесс, и рост является лишь более общим его выражением" (с. 99). Представление об этом принято биологами в виде так называемого правила о ядерно-плазменном отношении (Де Робертис и др., 1967). Оно заключается в следующем. В клетке имеет место взаимодействие между ядром и цитоплазмой. По мере роста клетки соотношение между массой ядра и цитоплазмой может измениться в пользу последней, и тогда возникнут трудности для жизнедеятель ноет и клетки. В результате деления клетка возвращается к оптимальному соотношению между ядром и цитоплазмой. Таким образом, посредством размножения действительно осуществляется регуляция обмена веществ. Некоторые определения жизни довольно конкретны. Если оставить в стороне такие гипотезы, как, например, гипотезу Н.Чермака (1895) о "живых молекулах-вихрях", воскрешающие представления Ж.Кювье, то теперь мы имеем своего рода определения - программы. В таких случаях фактически принимается, что для понимания природы жизни традиционная биология ничего сделать не может. Естественно, что такие определения постоянно совершенствуются. Вот некоторые из них.
"Изучение структуры протеинов может помочь решению собственно биологических проблем" ( Whyte , 1949). "Живой белок — это однозначный целостный "носитель" биологической формы движения" (Букановский, I960, с. 52). В своей основе жизнь - это "некоторое состояние вещества (Ляпунов, 1964), прежде всего конформационные взаимодействия веществ (Курса-нов, 1979), и поскольку "живая система... скорее всего представляет собой агрегат молекул" (Кальвин, 1964), "примат" химических процессов остается бесспорным (Франк, Энгельгардт, 1959). Биологическое исследование надо начинать с составляющих организм химических элементов ( Doff in, 1953), Жизнедея тельность организма — это исключительно или преимущественно физико-химический процесс (Lillie, 1934; Жданов, 1964; Холум, 1965; Мелюхин, 1966). "На вопрос" Что такое жизнь?' призвана дать ответ молекулярная биология, и центральное место в этой науке занимает молекула ДНК" (Франк-Каменецкий, 1983).
Где проходит граница применимости молекулярно-биологического подхода к определению жизни? В.М.Волькенштейн (1981, с. 9) отвечает на этот вопрос так: "Биология имеет цело с
макроскопическими молекулами и надмолекулярными системами и, следовательно, не затрагивает ярерные процессы . Пожалуй, не все принимают это ограничение. Например, А.С^нг—Дьерди (1964, с. 24) допускает, что ю... более тонкие явления жизни заключаются в изменении формы и перераспределении... электронных облаков вокруг атомного ядра, а Дж.Бернал (1969, с. 213) полагал, что "жизнь есть частичная, непрерывная, многообразная и взаимодействующая со средой самореализация электронных состояний атомов*. По-видимому, здесь речь идет о каком-то особом, "живом* состоянии атомов, и поэтому у читателя не может не возникнуть ассоциации с живым веществом виталистов. Теперь посмотрим как трактуется происхождение жизни на внутримолекулярном уровне.
"...Жизнь никогда не происходила из неживой материи, но возникла самопроизвольно. Она скорее есть имманентная способность даже мельчайших индивидуальностей атомов и молекул, возведенных в особое качество посредством их ассоциации* (Bornemicza , 1955, р. 116). Дж.Бернал (1962, с. 17) допускает существование "...вдоль берегов, особенно в прибрежных глинах, так называемой жизни без организма, набора установившихся реакций в растворах и абсорбирующих слоях*. Глины могут образовывать так называемые кристаллиты, похожие в некоторых отношениях на мембраны. Причем катионы в таких структурах являются носителями информации, воспроизводимой в послойном росте кристаллитов (Cairus-Smith, 1981). Аналогичное предположение о "жизни в мертвой природе* было сделано и раньше (Гольдгаммер, 1922). И хотя при этом подразумевается всего лишь происхождение "преджизни* или "околожизни", тем не менее это знаменательные допущения. Во-первых, они как бы размывают границу между живым и неживым, подрывают надежность физикалистского представления о происхождении жизни. Во-вторых, это симптом самоотрицания организмоцентризма: если жизнь — эго физическая химия в организме, то можно обойтись без организма. Впрочем, многие с этой крайностью не согласны ( Beck, 1957; Buffaloe, 1964; Кузин, 1972). Наконец, у А.И.Опарина происхождение жизни есть прежде -всего проблема абиогенного синтеза организменного субстрата. Тем не менее В.И.Кремянский (1969) не без оснований заметил, что по А.И.Опарину вмесге с первичными формами "самой жизни одновременно возникли "первичные формы естественного отбора". С.Э.Щноль (197 9, с. 7), по-видимому, тоже склонен основывать биологическую специфику на естественном отборе. Здесь важно отметить, что "отбор" - это какое-то действие, состояние, г.е. совсем иная категория, нежели "вещество* или существо*. К сожалению, это интересное замечание А.И.Опарина и С.Э.Шнрля^осталось нераскрытым.
Изгнав иЬ организма..не^и^т^-еск^ю "живую силу* и нехими—
ческое "живое вещество", физико-химическая биология решила методологически важную задачу субстратного единства живой и неживой природы; нет какого-то особого "живого вещества", нег "таинственной "жизненной силы. Между тем задача биологии состояла в ином: понять, чем отличается живое от неживого. Эта задача осталась нерешенной и, по-видимому, этим обстоятельством вызвано появление системного кибернетического подхода (кибервитализма), о котором уже упоминалось.
Еще Т.Рейнке (1903) вслед за Декартом сравнивал организмы с автоматом, функционирующим под контролем "доминант" Биологическую специфичность иногда связывают со "стройностью" организмов (Умов, 1916), с упорядоченностью многочисленных, самих по себе химических , реакций (Кавецкий, Колпаков, 1962; Энгельгардт, 1977), с уникальностью самого сочетания этих реакций (Вирхов, 1866; Опарин, 1977).
Принципиальная сверхсложность живых организмов, не найденная в неживой природе, выражается в существовании внутри-организменных систематизирующих информационных связей и структур (Колмогоров, 1964; Юргеленас, 1967; Тюхгин, 1972; Украинцев, 1976; Анохин, 1977; Франц, 1977). Кибернетический подход с неизбежностью обязывает признать информацию не менее реальной, чем вещество, энергия, поле.
Существование в организме информационных связей оесспор-но. Вместе с тем Г.Гёсслер (1968) справедливо указывает, что понятие обмена веществ имплицитно включает в себя регуляцию обмена веществ. Это замечание др некоторой степени возвращает обсуждение к формуле (2), но теперь ударение смешается на "обмен". Впрочем, сторонники кибернетического подхода к сущности жизни понимают проблему регуляции обмена веществ достаточно широко, наделяя ее свойствами устойчивости, стационарности, метасгабильности и пр. (Штеренберг, 1967). Организмы - это системы со "встроенной" в них системой коммуникации (Лоусон, 1969), "...самоеохраняющиеся и саморазви-вающиеся посредством информационного и вещественно-энергетического обмена со средой..." (Сагаговский, 1967). Отсюда возникновение жизни не сводится только к абиогенному синтезу организменного субстрата: "...с появлением жизни поступающие извне воздействия приобретают для живых существ жизненно-приспособительную сигнальную информацию — они используются в процессах саморегулирования и самоуправления в целях самосохранения организма" (Тюхгин, 1972, с. 82). В данной цитате информация характеризует не только внутриорганизмен-ные процессы, но также подразумевает некоторую оценку организма со стороны внешней среды. Вообще же насчитывается пять видов информации, в том числе логическая (смысловая) и метрическая, исчисляемая в битах (Паск, 1966; Урсул, 1973). Важно подчеркнуть, что в цитированной литературе понятие ин
формации обсуждается в основном применительно к организму. Указывается, например, что при взаимодействии с внешней средой биологические самовоспроизводящиеся системы создают информацию в виде смысловых последовательностей нуклеотидов ДНК (Блюменфельц, 1971). Такие молекулы называются сема-тофорическими ( Zuckerkandall, Pouling, 1965). Более широкое понимание значения концепции информации для биологии принадлежит М.М.Камшилову (1966, с. 228): "Консервация информации в форме молекулярных, морфофизиологических, биоце-нологических структур в характерных чертах биотического круговорота являетсй наиболее важной особенностью эволюции".
Наконец, существует еще одно особое направление кибервитализма - термодинамическое. Своеобразие термодинамики живого организма настолько велико, что возможность особой биологической термодинамики не исключена (Михайловский, 1980). Хотя законы термодинамики справедливы как в живой, так и в неживой природе (Волькенштейн, 1980, с. 100), "... в энтропийной шкале живые системы не занимают исключительного положения" (Геодакян, 1970, с. 56) и для "...живых организмов несомненно справедливы второе начало термодинамики и понятие энтропии..." (Зотин, 1976, с. 22), тем не менее необходима "новая термодинамика", способная описывать необратимые нелинейные процессы в условиях автоматического регулирования в организме (Термодинамика необратимых процессов, 1976, с. 248), может быть, в виде "энергетико-информационного поля" ( G.Fischer, 1981, а,б). Таким образом, контуры "новой термодинамики* вырисовываются на пересечении классической термодинамики и кибернетики. Одним из любопытных следствий из кибервигализма является возможность воспроизведения жизни вне организма в кибернетических устройствах (Колмогоров, 1964), т.е. фактический выхоц из организмоцентризма, однако на иной, нежели у Дж. Бернала (1962), основе.
Итак, организм — носитель жизни. Движение научной мысли, отмеченное трудами Аристотеля, ЖКювье, Э.С.Бауэра, А.А.Гур-вича, Г.Дриша, Э.Шр ингера, продолжается в этом направлении уже более двух тысячелетий. Каковы же итоги? Выражая почти общепринятую и отнюдь не новую точку зрения (Шэфер, 1914) и как бы даже оправдывая давние ожидания виталистов (Бородин, 1894; Страхов, 1865), Г.М.Франк (1976, с. 12) заклю-чаег: ...в осуществлении Сорганизмом. — И.5.] жизненных функций ничего нет кроме химических и физических процессов". По словам С.Э.Шноля, приведенным к работе И.К.Лисеева и В.И.Тищенко (1975), "такие "сугубо биологические процессы", как ферментативный катализ, регуляция внутриклеточных биохимических процессов, биологическая подвижность, нервное возбуждение, наследственность, изменчивость, размножение и т.п., имеют в своей основе физико-химические процессы" (с. 168).
В организмоцен тризме "функциональное определение жизни не выполняет своей роли - отличить живое от неживого. В конечном счете его сторонники приходят к выводу, что между живым и неживым нет принципиального различия" (Ермолова, 1970, с. 157). М.С.Волькенштейн (1979) видит заслугу известного биохимика М.Эйгена в том, что он доказал возможность самоорганизации материи в процессе происхождения жизни на основе существующих принципов физики. Примерно в таком же положении оказались субстратные определения жизни.
Видимо, неудовлетворенностью субстратными и функциональными определениями жизни можно объяснить тот факт, что биологическую специфичность иногда усматривают в уникальной сверхсложности организма, упорядоченности и высокой специфичности протекающих в организме химических процессов, сложности клетки и ее органелл и пр. Обычно эта сверхсложность подразумевает высокоразвитую жизнь, современниками которой являемся мы ( Elsasser, 1969). В этом - слабость такого подхода. Действительно, если сверхсложность есть непременное свойство жизни, то она имелась и у только что возникших про-.тобионтов. Но тогда откуда ей взяться - ведь ее нет в неживой природе. Если же сверхсложность возникла постепенно, то она не может выполнить свою функцию критерия жизни. Шансы на благоприятное для возникновения жизни стечение случайных обстоятельств столь малы (Блюменфельд, 1967), что сверхсложность следовало бы признать чудом. По-видимому, здесь, выход только один: надо принять, что протобионгы были устроены много проще самых примитивных из известных нам существ. Но тогда мы попадаем в ситуацию, о которой писал К.Уоддингтон (1947, с. 214), имея в виду организмоцентризм: "...в биологии нет строгого определения жизни. Существование промежуточных ступеней между живым и неживым, таких, например, как вирусы, делает сомнительным плодотворность попыток сформулировать это понятие".
Широко распространено заимствованное из философии представление о том, что биологическая специфичность возникла скачком при переходе количества в качество (Ильин, 1966; Опарин, 1979). Однако предмет обсуждения в биологии составляет не философская формула (1), а естественнонаучные формулы, в которых особенности биологической формы движения материи должны быть раскрыты в научных терминах. А пока выражение "принципиально новое качество* служит только новым обозначением неизвестной науке биологической специфичности.
Сам по себе обмен веществ и энергии не может считаться чисто биологическим явлением. В сущности, это было доказано уже исследованиями А.Лавуазье и Пристли. Вполне определенно об этом писал Ф.Энгельс: "Обмен веществ как таковой имеет
место и помимо жизни*2. Это же было осознано в период становления теоретической биологии (Гольдгаммер, 1922) и остается очевидным для наших современников (Шкловский, 1962; Руденко, 1971). Часто приводимые ссылки на Ф.Энгельса, определившего жизнь как обмен веществ, осуществляемый белковыми телами, пока ничего не добавили для решения проблемы. Прежде всего, нельзя забывать о той научной атмосфере, в которой творил Ф.Энгельс. Эго был период, когда формула (2) выполняла функцию альтернативы витализму и, в частности, построениям Г.Дриша, впоследствии отвергнутым, например, Н.А.Холодковским (1923). Далее, в * Анти-Дюринге* сам Ф. Энгельс писал, что его собственная * дефиниция жизни* *...весьма недостаточна, поскольку она далека от того, чтобы охватить все явления жизни*, и обмен веществ *...в свою очередь нуждается в объяснении при посредстве жизни,..'3.
Обращая внимание на го, что в * Анти-Дюринге* Ф. Энгельс фактически отказался от того определения жизни, которое было дано им в 'Диалектике природы*, нельзя забывать об истории написания и публикования этих сочинений. Оба они были написаны в 70-80-е гг. прошлого века. Сднако *Анти-Дюринг* вышел из печати впервые в 1878 г. при жизни автора, имевшего, таким образом, возможность редактировать собственную рукопись. 'Диалектика приройы* осталась незаконченной и оставалась в рукописи до 1925 г., когда впервые была опубликована на русском языке. Естественно, что Энгельс не имел возможности подготовить ее к печати и устранить очевидные противоречия.
Естественный ход развития жизни на Земле вынуждает нас в эпоху назревающего экологического кризиса различать два представления о жизни: с одной стороны, как жизнедеятельности организма, а с другой - как планетарного явления. Идущее от В.И.Вернадского представление о жизни как свойстве планеты поддержано М.М.Камшиловым (1966, с. 283): 'Понятие жизни относится не к отдельным организмам, а ко всей совокупности живых существ...*. Между тем, по мнению К.М.Завадского (1966), для организмоцентризма все надорганизменные системы от популяции до биосферы вторичны и производны от организма. Согласно организмоцентризму, возникновение на Земле даже единичного организма означало бы зарождение жизни. Эта робинзонада заключала бы в себе не меньше сущности жизни, чем все население Земли. И хотя организмы от бактерии до человека очень разнятся, тем не менее для организмоцент—
^Энгельс Ф.//Маркс К., Энгельс Ф. Соч. 2—е изд. Т.20.
Там же. С. 82, 84.
ризма они остаются взаимозаменяемыми вместилищами одной и той же жизни. Формула А.Бергсона *...жизнь является как бы потоком, идущим от зародыша к зародышу при посредстве развитого организма* (1909, с. 24) только переносит организмо-центризм в историческое время и не предполагает органического разнообразия, потому что жизнь в каждый момент могла бы находиться в единственном организме.
Начавшись с Аристотеля, организмоцентризм долгое время развивался по расходящимся направлениям как витализм и механицизм. В настоящее время через понятие информации наблюдается схождение этих линий развития, что естественно: подлинной альтернативой типологии остается дарвинизм. Витализм упрекал механицизм в том, что тот сводил жизнь (жизнедеятельность) только к вещественно-энергетическим процессам. Сегодня этот упрек следует признать справедливым, потому что в определение жизни введено понятие информаций 3’). Механицизм отвергал витализм за то, что тот вывел загадку жизни (жизнедеятельности) за пределы науки, однако подразумевал под наукой редуцированный мир понятий классической физики. Сегодня для определения жизни редукционистам также недостаточно понятий вещества и энергии, и потому они привлекли, понятие информации. Сопоставление двух положений: 1) биология сводима к современной физике, 2) понимание жизни *не доступно человеческому разуму в науке биологии* (Kemeny, 1959) — могло бы означать не принципиальную непознаваемость, а непознаваемость для физикализма. Таким образом, противоречие между витализмом и редукционизмом спровоцировано подменой несопоставимых понятий: материи - веществом и энергией, знания вообще - ее частной формой, физической наукой, материализма — фи зикализмом. Редуцированное понятие материи в действительности не устраивало обе борющиеся стороны: редукционизм - потому, что знание жизнедеятельности не привело к пониманию специфичности жизни, витализм - потому, что признание существования в организме чего-то сверх вещества и энергии исключал! его из науки, считающейся синонимом собственно знания.
В этом вопросе важное значение имеет онтологический статус информации (Клаус, 1963). Она понимается по-разному. Одни считают ее свойством материи, другие признают существование информации только в системе управления в организме. Причем "...отсутствие информации в неорганической природе отнюдь не делает невозможным ее появление на уровне живого* (Коршунов, Мангатов, 1974, с. 31). "Информацию естественнее всего считать объективным свойством материи, связанным с предложенным В.И.Лениным всеобщим свойством материи -свойством отражения* (Баженов и др., 1963, с. 518). По В.И.Кремянскому (1977), информация обнаруживает сходство 1 энергией: последняя выражает не свойства носителя самого по
себе, а его способность совершать работу, т.е. отношения, внешние для носителей. Реальность информации оспаривается, например, М.И.Сетровым (1978) на том основании, что она не материальна. Болгарский исследователь М.Янков (197 9, с. 308) пишет по этому поводу: "Итак, информация не есть ни вещество ни энергия, она есть свойство материи. Формулы "Информация не есть ни вещество, ни энергия" и "Информация не есть НИ материя, ни энергия" были бы эквивалентны, если бы вещество исчерпывало объем понятия материи... Однако известно, что вещество - это лишь один вид или форма материи". В этой связи представляется двусмысленным замечание Н.Винера (1968, с. 201): "Информация есть информация, а не материя и не энергия. Тот материализм, который не признает этого, не может быть жизнеспособным в настоящее время". В этом замечании под материей, по нашему мнению, скорее всего подразумевается вещество, т.е. философская категория подменена естественнонаучным понятием. Информация, разумеется, не вещественна и не существует отдельно от вещественно-энергетического субстрата как своего носителя, как изображение - от зеркала и объекта; однако через субстрат она воздействует на органы чувств и через ощущение вызывает мысли и чувства и потому может считаться существующей. Если мы допустим, что информация не существует, то будем обязаны принять, что несуществующее воздействует на материальный мир.
Таким образом, в организмоцентризме "...противоречие между витализмом и механицизмом мертво... Аристотель приписал направленность и упорядоченность живых существ активности их душ, и лестница существ, которую он обнаружил в природе, была неотъемлемой иерархией таких душ. Сегодня мы объясняем эти особенности живых существ в терминах организации и описываем своего рода красную нить, которая протягивается через всю историю живого мира как непрерывное повышение уровня организации" ( Sommerhoff, 1954, с.1).
Приведенный в данной главе материал показывает, что обсуждение концепций жизни как основного вопроса теоретической биологии продолжается длительное время и имеет долгую предысторию. Несмотря на определенное разнообразие концепций, эго обсуждение характеризуется по меньшей мере двумя общими чертами. Во-первых, оно ведется с позиций физики, в последнее время укрепившихся благодаря кибернетическому подходу. Во-вторых, как само собой разумеющееся принимается, что жизнь — свойство единичного организма. Эти две особенности обсуждения имеют обратную сторону. В частности, полностью, на наш взгляд, игнорируется традиционная биология. Получается, что как наука о жизни традиционная биология за свою историю ничего не сделала для определения понятия жизни. Кроме того, несмотря на противоположность между классическим витализмом и физико
химическим редукционизмом (первый отказывает науке в правомочности изучения жизни, а второй отвергает витализм за йена учность), оба направления мысли фактически объединяются в организмопентризм. Следовательно, они вместе противопоставляются традиционной биологии, в которой изучаются такие надорганизменные объекты, как сообщества (экология), фауна и флора (биогеография), родословные (филогенетика) и пр. Известно, однако, что надорганизменные объекты изучает не только традиционная биология, но и теория видообразования - теория естественного отбора. Поэтому представляется полезным рассмотреть предмет теоретической биологии с точки зрения тех разделов науки, которым она призвана служить, с точки зрения традиционной биологии и теории эволюции.
Глава 2
ТРИ 'ОБРАЗА* БИОЛОГИИ
Объединение биологического знания следует считать своевременной задачей. Оно могло бы состояться на основе некоторого определения жизни, однако, как справедливо отмечено, этого пока не произошло (Фесенкова, 1976). Не в том ли причина, что господствующее определение жизни (2) и (2*) все-таки не является собственно биологическим? Может быть, та наука, которую мы называем биологией, представляет собой лишь искусственное объединение разных систем знания? Действительно, в связи со словом 'биология* в сознании возникают по меньшей мере три 'образа* этой области знания.
Во-первых, это физико-химическая биология. Эго быстро развивающаяся, очень важная в практическом отношении область знания. Она ставит своей программой замену, искусственное воспроизведение и улучшение получаемых ныне от живой природы ресурсов. Весьма актуальной задачей остается ддя нее управление протекающими в организме процессами жизнедеятельности организма. При решении этих проблем исключительное значение приобретают точные приборы, сверхчистые препараты, чистые линии лабораторных животных и растений. В живых организмах имеются только обычное вещество и уже известные виды энергии, для изучения которых чрезвычайно плодотворными оказались методы физики и химии. Вместе с ними физико-химическая биология усвоила математический язык, прежде всего дифференциальное исчисление. Однако обращает внимание тот факт, что множество тех объектов, которое подвергается изучению (кишечная палочка, дрозофила, лягушка, белая мышь и пр.), не образует целостного природного объекта. Эго не есть фауна (как, например, фауна Байкала), не есть сообщество (как, например, сообщество ельника— зеленомощнмка), не есть родословная (как, на
пример, родословная человека) и тл. Названные организмы выбраны в качестве объектов исследования всего лишь потому, что их удобно содержать в лаборатории. И тем не менее полученные при этом результаты используются для раскрытия сущности жизни как природного явления, поскольку считается, что в каждом организме есть жизнь.
Второй *образ* - традиционная биология, которую олицетворяют такие натуралисты, как Бюффон, Брэм, Фабр. Великих натуралистов прошлого восхищает преж е всего разнообразие живой природы. Им свойственно благоговение перед богодарованной природой: ''Природа не что иное есть, как непременный Создателя закон* (К.Линней), природа - эго *Храм Природы* (Эразм Дарвин). Разумеется, о какой-^либо переделке природы и мысли быть не может. Природа - эго самоценность, раскрытие тайн природы - это приобщение к премудрости Творца. Отношение к природе как к самоценности унаследовала современная традиционная биология. Ее интересует жизнь растений и животных, их особенности, расселение на планете, история и родословная и т.п. Довольно правильное представление о современной традиционной биологии создают такие издания, как * Жизнь животных* (1968-1973) и *Жизнь растений* (1974-1982). Типичнейшее занятие натуралиста — объективное наблюдение в природе и классифицирование, которое только в последнее время подвергается математизации. Натуралисты воспринимают природу, как гак называемые *деги* природы: *гора вижу — гора пою, река вижу — река пою*. То, что происходит сегодня с живой природой, - причина личной трагедии для современных натуралистов.
Третий "образ* - это теория естественного отбора. Ее автор Ч.Дарвин начинал как обычный натуралист: он коллекционировал, наблюдал, описывал, путешествовал, раскрывая тайны живой природы. С^нако основным результатом его работы стала теория, объясняющая органическое разнообразие. Для него не имели значения скромные в то время знания о химическом составе организмов, о процессе обмена веществ. Последние достижения биохимии, биофизики, физиологии только расширили предметную область теории естественного отбора, но они ничего не изменили в логике теории. Уже современники Дарвина отметили парадоксальность его теории. Было показано, что теория, созданная для объяснения разнообразия вцдов, сама в нем не нуждается: видовое разнообразие - только повод для разработки теории, и это разнообразие только вытекает из теории. Механизм видообразования, наследственная изменчивость и отбор могли быть раскрыты на примере даже одного вида. С одной стороны, это обстоятельство делает теорию отбора универсальным обобщением, объясняющим картину первичной жизни, когда не было видового разнообразия. С другой стороны, это обстоятельство отда—
ляет теорию отбора от натурализма, для которого именно видовое разнообразие есть объект изучения.
Перечисленные три "образа* биологии вместе составляют сво, его рода треугольник, предметную область теоретической биологии. Теперь сопоставим их друг с другом и посмотрим, как работают в этой предметной области существующие представления о предмете и задачах теоретической биологии. При этом будем иметь в виду, что основные теоретические расхождения в биоло-гии так или иначе выводятся из противоречий между типологией (организмопенгризмом) и дарвинизмом. *Как неоднократно подчеркивалось в биологической и философской литературе послодних лет, переход от типологического к популяционному образу мышления составил одну из характерных черт революционного преоб. разования биологического‘знания* (Карпинская, Хон, 1976, с. 374). Действительно, оппозиция типология - популяционизм, организм - популяция имеет множество аспектов и исключительно содержательна для теории: преформизм и эпигенез, инструктивный и селективный способ развития ( Danchin, 1971; Березин, 1981), лапласовский и вероятностный детерминизм, ламаркизм и дарвинизм (Завадский, Мамзин, 1970), возможность и невозможность наследования приобретенных свойств, нереальность и реальность вида и пр. Однако, если мы обратимся к сочинениям по общей биологии (Haas, 1964; Baker, Allen, 1979), то обнаружим, что "революционные преобразования* нисколько не затронули естественнонаучного мировоззрения современных типологов. В логической структуре современной типологии место энтелехии заняла вполне научная молекула ДНК, хотя остается фактом, что в литературе нет выражения "живая мо лекула*. Справедливо отвергая витализм за его принцип непознаваемости жизни для науки, современная типология ищет в субстрате живого организма что-то несводимое к физхимии, потому что ее задача состоит в установлении отличий живого от неживого. Тем не менее это искомое отличие понимается как нормальная проблема физхимии. В виде следствия из физико-химических предпосылок выводятся некоторые разделы традиционного натурализма, большей частью "эволюция организмов"и эко» логия. Такие взгляды объективно отражают преобладающее умонастроение научной общественности. Надо отметить, что такое ввдение биологии хронологически совпадает с распространением позитивизма, который провозгласил физико-математическое знание образцом собственно научного знания.
Что же собой представляют программы построения теоретической биологии, которые фактически не учитывают неоднород-
В действительности же "эволюция* - это вадообразование, т.е. надорганизменное понятие.
ность биологического знания и для которых понятие жизни укладывается в формулу (2*)?
Редакция журнала "Acta Вiotheoretica " (1935) в обращении к читателям определила его тематику следующим образом: теории биологического характера, формализованное логическое или математическое представление основ биологических наук (формулы графики, символы и пр.), исследования о содержании вдей, методов, разделение биологии на частные разделы и отношения между ними, эмпиризм, методы и тенденции мысли в этих разделах биологии и г.ц. Эго - своего рода программа-поиск, которая за 50 лет существования журнала еще не воплотилась в стройную теорию. Эго же надо заметить и о программе Г.Вер-хуга ( Verhoog , 1975).’ В ней дополнительно отмечена необходимость аксиоматизации и моделирования. Впрочем, об аксиоматизации пишут овольно часто ( Wood ger, 1929; Lagerspetz , 1959; Медников, 1982), априори допуская ее осуществимость.
Определяя биологию как "изучение организма", М.Маклин (Macklin 1969) оспаривает возможность оформления теоретической биологии по следующим причинам. Во-первых, теоретическая биология изучает физико-химические особенности организма и не объясняет поведения всего организма. Во-вторых, она не содержательна: если экспериментальные данные имеются, то они формализуются в аналитической модели, отражающей уже известное; если же таких данных нет, го теория пуста. В-третьих, нет и не может быть собственно биологических законов. Последний довод, как признает сам Маклин, вдет от Дж.Смарта (Smart, 1963). По Дж.Смарту, только физические науки располагают законами в собственном смысле этого слова. Законы физики справедливы везде и повсюду. В биологии же таких законов не может быть вследствие уникальности явления жизни. Как было замечено впоследствии ( Steen, von der, 1973), "Жизнь* по Смарту - имя собственное. Физика относится к биологии так же, как к электронике; биология же занимается естественной историей структур и объясняет, почему живые существа ведут себя так и не иначе. Обсуждая "естественную историю*, которая действительно отвечает понятию "имя собственное", Смарт, однако, остается на позициях организмоцентризма, который обязывает к признанию множественности жизни: сколько организмов - столько жизней. В этом - непоследовательность его позиции. Теперь обратимся к общим планам построения теоретической биологии, прежде всего в аспекте взаимосвязи традиционной и физико-химической биологии.
Г.Уотермен (1968), обсуждая перспективы теоретической иологии, высказал пожелание, чтобы "традиционалисты" научились больше использовать математику и теоретическое мышление. Этот призыв можно было бы принять в качестве программы, к сожаленщр, недостаточно конкретной. Он сохраняет свое
значение даже при учете того обстоятельства, что ''...развитие науки определяют содержательные теории, а* не математические методы и ЭВМ сами по себе* (Андреев, 1974, с. 4). С другой стороны, хотелось бы убедиться в том, что, например, изучение репликации, которая рассматривается О.М.Оленовым (1977) в качестве важной части теории, чем-то обогащает логику теории естественного отбора или проясняет вопрос о соотношении между жизненной формой и таксоном.
Любопытен подход Г.Моровица (1968), который из * дидактических соображений* подразделяет теоретическую биологию на формальную теорию, физическую теорию и теорию систем. С таким пониманием содержания теоретической биологии трудно согласиться. Физическая теория относится к физике и уже поэтому не может войти в искомую биологическую теорию, пока биология не сведена к физике. Теория систем есть общенаучная область, она охватывает самые разные области знания. В гаком случае ее плодотворность в биологии должна определяться спецификой биологических систем; однако Моровиц не указывает, в чем состоит их специфика, отличие от неживых систем. Что же касается формальной теории, то под нею подразумеваются математическое моделирование и экспериментирование, осуществляемые в генетике и экологии на основе некоторых постулатов. К сожалению, Моровиц не затрагивает вопроса о выборе постулатов и о перенесении имеющегося опыта, например, в биогеографию. Показательно, что одним из основателей теоретической биологии он называет Д’Арси Томпсона, автора замечательной книги *О росте и форме*. Между тем, как правильно указывает сам Моровиц, Д’Арси Томпсон считал проблему роста в основе своей физической, а форму - математической проблемой ( d'Arcy Thompson, 1945, р.15).
Программа, выдвинутая У.Эльзассером (Elsasset, 1981), опирается на 3 постулата: 1) конечности, 2) творческого отбора, 3) информационной стабильности. Не разъясняя сущности первого постулата, но отмечая его * особую природу*, Эльзассер пишет, что два оставшихся находятся внутри квантовой теории и биологическая теория соотносится с ней как здание с его чертежом. Особенность биологии состоит в том, что она изучает организмы - *негомогенные классы* (нет двух вполне одинаковых клеток*), тогда как физика имеет цело с 'гомогенными классами*. *Негомогенность* существенно ограничивает предсказательные возможности биологической теории, и '...общие законы биологии, не дедуцируемой из физики, будут иметь логическую структуру, совершенно отличную от привычной структуры физических наук* ( Elsasser, 1966, р. 4). Биология вообще *не картезианская наука*, поскольку структура живой материи не может быть выражена в числах, и целое не может быть разложено на части ( Elsasser, 1975, р. 64). Эти интересные
мысли перекликаются с идеями Г.Дриша, но, к сожалению, не сопровождаются анализом проблем традиционной биологии и явно тяготеют к органицизму.
* Энциклопедия биологических наук* ( The Encyclopedia of Biological Sciences, 1970, p. 112) в разделе "Биофизика* определяет содержание теоретической биологии так: 1) теория систем, 2) приложения для ЭВМ, 3) классическая математическая биология (по-видимому, биометрия), 4) приложения физики к биологии. Таким образом, биология оказывается как бы частью физики.
Близкая точка зрения высказана М.В.Волькенштейном (1981, с. 7): "Современная биофизика есть физика живой природы, и ее конечные задачи состоят в обосновании теоретической биологии". Об этом же пишет Ю.Н.Полянский (1964, 1981). Тем не менее считается, что биофизика - раздел биологии.
Создается любопытное положение. С одной стороны, "...принципиальных отличий живой природы от мертвой не существует и поэтому физических критериев живого предложить нельзя" (Блю-менфельд, 1971, с. 64). В таком случае, казалось бы, организменный подход в принципе не может вывести на биологически специфичную теорию. С другой стороны, названные авторы имеют в виду теоретическую биологию. Не было бы более логичным в данном случае иметь в виду прикладной раздел физики, физическую теорию жизнедеятельности организма? У.Эльзассер (1969) полагает, будто современные усилия в области теоретической биологии начались с работы Н.Бора по приложениям кван-. говой физики к биологическим исследованиям.
Развернутая программа теоретической биологии принадлежит А.А.Ляпунову (1971, с. 13): "Вся теоретическая биология должна состоять... из двух частей: физико-химической, назначение которой состоит в том, чтобы расшифровать физико-химическую природу актов жизнедеятельности на уровне макромолекул или клеточных органелл, и кибернетической, назначение которой состоит в том, чтобы понять функционирование живых систем, отправляясь от их структуры и сведений о свойствах их элементов*.
При таком подходе к проблеме теорией для биологии автоматически становятся законы физической химии и квантовой теории (Давыдов, 1979; Баженов, 1980; Elsasser, 1981) и, стало быть, не требуется знания филогенетики, экологии, биогеографии и других разделов традиционной биологии. Отсюда непонятно, как такая теория будет решать нерешенные вопросы традиционной биологии, такие как биогеографическое районирование и классифицирование. При обсуждении подобных, вопросов на— гУралистами может сложиться впечатление, что матричный син-все белков» величайшее открытие физико-химической биологии, еще не стал достоянием науки — настолько своеобразны
содержание и дух этих обсуждений. Известно, что большие надежды возлагаются на еще не открытые физико-химические явления (Семенов, 1959; Чепиков, 1976), на особые взаимодействия микро- и макрофизических явлений ( Jordan, 1959), которые, по существующему мнению, создадут предпосылки для окончательного сведения биологической теории к физической. В таком случае биология остается совокупностью вопросов, "пока*’ не разрешенных физхимией ( Commoner, 1961); если же какой-то особой физхимии в организме нет, то не существует и "автономной" науки биологии ( Roll-Hansen , 1962). Оставаясь фактически на позициях организмоцентризма, Б.П.Токин (1963) тоже допускает возможность плодотворного участия физиков (точнее; физического знания) в становлении теоретической биологии, но ничего не пишет об участии в этой разработке традиционной биологии, натурализма. Называя организмы "физическими системами", Г.Соммерхоф ( Sommerhoff, 1954) ставит вопрос о возможности математического описания активности организмов как отношений между физическими переменными. "Синтез жизни" (Игнатов, 1962) понимается как синтез организмов, о котором писал Н.Д.Зелинский (1952). Но если при таком синтезе всего лишь осуществлены законы физической химии, может ли сложившийся организм заключать в себе что-либо сверх нее, биологичек ли он и вообще живой ли? Аналогичным образом, если происхождение жизни есть абиогенный синтез соединений углерода, то откуда в организменной жизнедеятельности взяться собственно жизни? В этом контексте остается весьма привлекательной своей честностью формула, подразумевающая организм: "Жизнь имеет статус в физическом универсуме" (Wald, 1962). Таким образом, содержание теоретической биологии фактически задано организменной физико-химической концепцией жизни.
Большинство программ теоретической биологии примечательно засилием физико-химических методов и игнорированием традиционной биологии. Против математики как метода пока возражать не приходится, хотя бы потому, что ее можно считать по меньшей мере утонченной и развитой логикой ( Kemeny, 1959). Хотя, соблюдая традиционную строгость математической науки, ее применение в биологии было бы желательно предварить доказательством применимости математики в биологии. Тем не менее математика как наиболее абстрактный и развитый язык, по-ви-цимому, могла бы способствовать объединению всего естественнонаучного знания. Другая возможность применения математики в биологии, как полагает В.В.Налимов (устное высказывание),-это привитие биологам образного математического мышления. Эта идея очень привлекательна вследствие фантастического разнообразия математических образов, облегчающих самые неожиданные ассоциации. Известно, например (Мур, 1967), что идея геометрической и арифметической прогрессии оказалась очень
плодотворной В дарвинизме. В силу своей природы математика не претендует на содержательную сторону природных явлений. Поэтому не представляется целесообразным считать синонимами "теоретический* и "математический*, к чему склойен Н.Рашевский (Rashevsky» 1962); математизирование - это еще не теоретизирование.
В целом существующие программы теоретической биологии в качестве объекта называют организм, к изучению которого предлагается привлечь физико-химические методы. Само по себе это также еще не вызывает возражений. Известно, например, что материальные памятники истории и культуры изучают с помощью физических и химических методов и получают массу важных сведений не только о технике художника, но также о времени и месте создания художественно-исторических ценностей. Такие сведения позволяют разработать и методы реставрации памятников. Вместе с тем вполне очевидно, что физические и химические методы не могут выявить культурологического значения памятника, т.е. того, что составляет его сущность и отличает от природных тел. Точно так же и в биологии. Разумеется, организмы могут изучаться разными методами, и получаемые при этом сведения могут относиться к разным системам знания. Бесспорна исключительная плодотворность физико-химического подхода в медицине, сельском хозяйстве и других областях деятельности. Вмебте. с тем, очевидно, что если уж искать в организмах биологическую специфичность, то нужны какие-то специфические методы.
Существуют и другие программы построения теории биологии. По А.А.Малиновскому (1969), задача теоретической биологии — это наиболее полное использование информации, которую получают самыми различными методами во всех областях биологии. Б.Л.Астауров (1972, с. 62) определял теоретическую биологию кик ...упорядочение и осмысление фактических данных о жизни во всех ее проявлениях, позволяющие видеть и выразить наблюдаемые явления в виде возможно более общих и взаимосвязанных закономерностей или даже законов*. Считая ее частью общей биологии, Б.Л.Астауров, хотя и не указывает, какими принципами должен руководствоваться теоретик, зато перечисляет те области знания, которые составляют теорию биологии: теория наследственности, теория эволюции, молекулярная биология, цитология, теория онтогенеза, генетика. В этом перечне отсутствуют важные, на наш взгляд, разделы традиционной биологии: иогеография, экология, сравнительная анатомия и пр. М.С.Гиля-Р°в считает актуальной задачу анализа понятий и принципов огических исследований, таких как гомология, принципы сис-ики и ее отношение к филогенетике, понятие субстрата 197*2 \И Организменных систем (Цит. по Замилову, Лисееву,
)• Если Б.Л.Астауров предлагает программу объединения
отдельных областей знания, то М.С.Гиляров, кроме того, пред, лагает проанализировать понятийный аппарат частных разделов биологии. Думается, что такие программы, задающие цель изучения уже существующего знания, отвечают задачам теории био логии больше, чем теории жизнедеятельности. Игнорирование нв турализма, трацинйонной биологии стало, на наш взглад, дурной традицией и свойственно даже таким, казалось бы, наиболее теоретичным работам, как исследования /1. Берталанфи и Н.Рашевского.
Фактически в довольно обширной литературе, посвященной принципиальным вопросам биологической теории, обсуждаются разные области знания: теория жизни, теория жизнедеятельности организма, теория эволюции, ^Доктрина жизни* и "Наука о жизни* (Kladuw , 1966), логическая структура биологии в целом и т.д. Для того чтобы разобраться в этой сложной картине, было бы полезным обратить внимание на замечание А.В.Я&. локова о том, что теория объективно протекающего процесса жизни и теория знания о жизни, воссозданного биологами за ис. торическое время, не одно и то же (Цит. по Замилову, Лисееву, 1972). Определенное значение для обсуждаемого вопроса имеют сочинения, в которых предпринята попытка охватить разнообразие фактов и понятий в биологии, например, в виде Центральных концепций* ( Baer et al., 1971), а также публикации в которых сопоставляются теории жизнедеятельности и видообразования (Moment, 1962; Ayala, 1968; Смирнов, 1973а; Югай, 1976; Опарин, 1977). В общем они создают лишь видимость гармонии, якобы слитности организмоцентризма и популяционизма, теории жизнедеятельности и теории отбора.
Прежде всего, здесь имеются две предметные области: организм и популяция вида. Органицизму мы обязаны знанием жизне-деятельности организма, дарвинизм же представляет собой тео-^ рию видообразования. Для чего нужно теоретической биологии ( понятие вида, если загадка жизни - в организме? Теория эво- . люции, как подчеркнул А.А.Олицкий (1981), не обсуждает воэ-( можности существования биологических объектов и, по всей в и-, димости, организмов. Она лишь описывает механизм отбора и f объясняет видообразование. Поводом для постановки вопроса об, эволюции послужило разнообразие видов. Наоборот, для теории жизнедеятельности организма разнообразие видов - чуждое, как. бы лишнее явление. Знания о жизнедеятельности замечательно выросли после Дарвина, но они ничего не изменили в логической структуре теории отбора и теории видообразования в целом. Укрепив их новыми фактами на молекулярном уровне, они только значительно расширили предметную область дарвинизма. Формулы (2’) и (3*) не заключают в себе никаких намеков на тео* рию видообразования. Неудивительно, что их содержание успеШ' но раскрывает физико-химическое естествознание, вооруженное
математическим методом; но при этом раскрывается понятие физико-химического обмена веществ организма, а не жизни как особой формы движения материи (1).
Однако видовое разнообразие служит для дарвинизма только, природным феноменом, подлежащим объяснению на основе теории отбора, что же касается самой теории отбора, то она не предполагает видового разнообразия как элемента понятийного аппарата. Она приложима и к единственной популяции. Ввдовое разнообразие только следует с неизбежностью, если корректна теория отбора. Не случайно, А.И.Опарин (1979) связал зарождение жизни, когда явно не было видового разнообразия, с возникновением естественного отбора. Наоборот, для традиционной биологии видовое разнообразие есть сам объект изучения, а не повод для выработки какой-либо концепции вроде теории естественного отбора. Таким образом, при обсуждении отношений между организмоцентризмом и популяционизмом незаметно участвует "третье заинтересованное лицо" - традиционная биология, развитие которой связано прежде всего с изучением видового* разнообразия. Ее открытия, сделанные после 1859 г., новые виды и даже типы животных и растений тоже не оказали ни малейшего воздействия на логическую структуру теории видообразования и на ее понятийный аппарат.
Если организмоцентризм видит жизнь как живую машину в бесчисленных несущественных для него модификациях от бакте
рии до человека, теория эволюции - как множество неодинаково приспособленных видов (популяций) - и отсюда выводит разнообразие живых машин, т.е. организмов разных видов, то что вцдит традиционная биология?
В восприятии натуралистов - ботаников и зоологов - жизнь -это множество разных объектов: фаун и флор, типов организации, жизненных форм сообщества, родословных (филем), таксонов и пр. Множество таких надорганизменных объектов образу— эт многообразие, и разнообразие видов - это только сырой материал для воссоздания многообразия. Все ныне живущие и когда-либо существовавшие виды животных и растений вместе взятые перераспределены по таким объектам, как если бы одни и те же актеры были заняты в разных пьесах. Традиционная биология нуждается в теории, имеющей своим объектом именно мно-ообразие. Эго надо особо подчеркнуть потому, что, как нам сажется, в биологии имеет место подмена теоретической биоло— ни теорией эволюции (Борзенков, Северцов, 1980). И.Т.Фро-11973), например, указывает, что теоретиэацию биологии юнимаюг прежде всего как продвижение в исследовании эволюционных проблем.
Признавая заслуженно исключительное положение дарвинизма * биологии, его благотворное воздействие на все естествознание отдавая должное памяти Ч.Дарвина, великого труженика и вы
дающегося биолога, надо признать, что традиционная биология, имеющая дело с жизнью такой, какая она есть, богаче теории эволюции, освещающей пусть важнейший, но все же один из аспектов биологии. Знание механизма эволюции, как справедливо отметил А.И.Круликовский (1979), не есть решение таких проблем традиционной биологии, как классификация и районирование, экологическая ниша и жизненная форма, тип и таксон, таксон и града и пр. Изначально надо считаться с гой возможностью, что дарвинизм есть основа для решения этих и многих других задач. Возможно, что в неявном виде эти решения уже содержатся в дарвинизме; однако уже сама возможность оправдывает необходимость теоретической биологии. По-видимому, в центре внимания при обсуждении отношений между дарвинизмом и традиционной биологией должны находиться многообразие и разнообразие. Именно с этих позиций следует пересмотреть уже существующие системы биологии ( Haeckel , 1866; Tschulok, 1910; Меуег-Abich, 1963). При этом полезно иметь в виду, что сам принцип подхода к проблеме, план действий, внутреннее строение искомой теории должны считаться с присутствием всех "трех заинтересованных лиц", названных выше и составляющих треугольник.
В этой связи уместно обратиться к оценке некоторыми методологами существующих тенденций в теоретической биологии.
Как полагает А.И.Алешин (1980), процесс теоретизации биологии конструктивен постольку, поскольку он нацелен на создание гипотетико-дедуктивной теории. Он осуществляется в соответствии с логическими требованиями и заключается в физика-лизации и кибернетизации биологии, и уже поэтому, такая теория не может выявить "содержательно-физические" и "содержательно-кибернетические" отличия живого от неживого. Отсюда построение биологической по языку теории остается невозможным.
Более полно сложившееся положение описано в очень интересной статье Т.В.Лойта (1974, с. 89): "Дело не в том, что физико-химическая или логико-математическая модель рассматривается как модель научной теории вообще или как эталон развитой теоретической конструкции, а в том, что физика и математика превращаются здесь фактически в метатеорию биологического знания и тем самым из биологического исследования исключается вСе, что находится , за пределами физико-химического естествознания. Поэтому, видимо, более целесообразно ств вить задачу построения теоретической биологии вообще с. мета-теоретическим акцентом, т.е. рассматривать общую теорию биологии как метатеорию биологической науки, являющуюся познавательно-теоретической и общеметодологической областью в те? ретической системе биологического знания*. Фактически теория должна стать результатом обобщения теорий частных разделов биологии. В связи с вышеприведенной цитатой -становится понят
цым, почему "создание единого фронта биологических наук в результате выхода на молекулярный и субмолекулярный уровни исследования живого..." (Мирзоян, 1975, с. 17) состояться не может.
Таким образом, содержание понятия "теоретическая биология* оказалось, по нашему мнению, основательно скомпрометировано. Оно прежде всего обозначает область знания о строении и функционировании организма и ориентировано на физику и химию (Reed, 1978). Такая теория практически игнорирует традиционную (натуралистическую) биологию. Было бы наивностью ожидать, что такая "теоретическая биология" сделает что-то положительное для традиционной биологии.
Последнее время большое внимание уделяется концепции структурных уровней организации материи. Неувязки, возникающие при рассмотрении предметной области биологии (треугольник образов), становятся еще более отчетливыми, если обратиться к этой концепции.
Глава 3
УРОВНИ ОРГАНИЗАЦИЙ МАТЕРИИ
И ТЕОРИЯ БИОЛОГИИ
История учения о структурных уровнях организации материи описана М.В.Веденовым и соавт. (1972). Основные положения этого учения (Кремянский, 1969; Кедров, 1971а,б) состоят в следующем. В процессе саморазвития материя поднимается на все более высокие уровни организации, которые выделяются особенностями носителя: уровень элементарных частиц, ядерно-физический, атомарный, молекулярный, высокомолекулярный. Каж* дый последующий уровень получается объединением в систему предыдущих носителей: элементарные частицы объединяются в атомы, атомы в молекулы, молекулы в биополимеры. "То, что было целым на одном уровне, становится частью на более высоком уровне" ( Novikoff, 1969, р. 161). В.Ф.Сержантов (1972, с. 120) раскрывает понятия элементарности и сложности через отношения между ними: "Всякий материальный объект является атомарным (неделимым) только по отношению к данной системе и только в определенном отношении, будучи сам системой в каких-то иных отношениях и обладая своей структурой". Понятия элемента и структуры, следовательно, имеют реляционную природу (Мулуд, 1973). На каждом уровне организации материи имеются собственная структура носителя и особые законы его Функционирования. Эти законы описывают специфические для каждого уровня системные отношения, взаимосвязи элементов 6 системе. Общее значение реляционности можно оценить в све
те проблемы целого и части: *...гносеологический принцип структурного анализа выражает идеи когеренции истины, согласно которой, как известно, истинность единичного может быть установлена только в связи с общим, со всей системой* (Курсанов, 1973, с. 13).
Поскольку на каждом уровне имеются элементарные частицы, в субстратном отношении между этими уровнями сохраняется единство. Поскольку на каждом уровне осуществляются физические процессы, постольку на каждом уровне материальный носитель может изучаться методами физики (в биологии - биофизики).
Известны и разные формы движения материи. Во времена Ф. Энгельса казалось, что каждой форме движения отвечают стро го определенный структурный уровень организации материи и изучающая его наука: химии — атомы, физике — молекулы, биологии - белки. При этом, однако, Б.М.Кедров (1971а, с. 290) указывает, что * Энгельс исходил из неоднозначного соотношения между видом материи и формой движения*. В настоящее время эта неоднозначность сделалась очевидной. Например, к субстрату организмов, живой плоти, состоящей из макромолекул, возможны разные подходы: физический (биофизика), химический (биохимия), биологический (биология). В биологии, однако, господствует физико-химическая концепция жизни, формулы (2) и (2*), (3) и (3*); если эго так, то собственно биологический (не физико-химический) подход к изучению жизни в действительности отсутствует.
Что же собой представляет биологический уровень развития материи - предметная область биологии? М.И.Сетров (1971) понимает системную организованность на этом уровне следующим образом. В организме существуют уровни пространственной иерархии - органы, ткани, клетки, молекулы. Кроме того, имеются уровни надорганизменной генетической иерархии - царства, типы, классы, отряды и пр. Иерархической системе этих уровне! отвечает сложная структура биологического знания, системности жизни отвечает системность биологии. Последняя представляет собой как бы лестницу наук - от молекулярной биологии до популяционной биологии ( Monfent, 1962; Наумов, 1964; Бергаланфи, 1969; Иогансен, 1972; Шварц, 1972). Эта теоретическая конструкция отражает чрезвычайную сложность (многоплановость) реально существующей биологии, зафиксированной, например, в рубрикации * Реферативного журнала*. Вместе с тем представляется очевидным, что в картине первичн* й жизни не было такой сложности, которая сейчас дает основ, для выделения типов, классов, отрядов и пр. Сложная таксономическая иерархия предполагает материал - большое количество видов, а виды возникли постепенно в ходе исторического развития. Фактически обе у? даемые уровни организации относятся к современным нам высо
кОсЛожным существам, с которых не могла начаться жизнь, зародившаяся в Цировом бульоне*. Но в таком случае эти уровни организации не имеют общебиологического значения. Они свой ственны только высокосложным организмам, возникшим - в масштабах геологической истории - совсем недавно. Собственно биологическими, специфическими для жизни в целом следует считать такие уровни организации, которые свойственны жизни в тобой момент ее существования.
Что же касается поначалу отсутствовавшей клетки, которую часто считают * фундаментальной единицей* живых систем (Мс Elroy, 1962) (это равносильно признанию клеточной природы протобионтов), то она в действительности представляет особую трудность для теории структурных уровней. Сравним, например, одноклеточную8 бактерию и нервную клетку. Оставаясь клетками в структурном отноч/ении (цитоплазма, внутриклеточные органеллы, оболочка), они явно различаются функционально. Клетка бактерии функционирует как самостоятельный организм (осуществляет обмен веществ), и в этом она соответствует многоклеточному организму, несмотря на очевидные структурные различия между ними.
Таким образом, при обсуждении уровней организации материи в биологии полезно различать структурный и функциональный аспекты. Структура носителя жизни усложнялась чрезвычайно за время существования жизни (протобионт - клетка - многоклеточный организм). Этим обусловлена весьма ограниченная пригодность структурного критерия для определения жизни как особого уровня организации материи. Больше того, само понятие организма, в котором выделяются структурные уровни пространственной иерархии, лишено кажущегося очевидным статуса элементарного носителя жизни.
Обратимся, например, к общественным насекомым. Только вместе взятые рабочая пчела, трутень и матка более или менее отвечают привычному представлению об организме как самостоятельно осуществляющей жизненные функции единице жизни. Аналогичные трудности возникают при попытке применить понятие организма к таким колониальным существам, как сифонофоры, мшанки и др.
Совсем иное дело функционирование организма. Оно сохрани— ется неизменным в том смысле, что сводится к обмену веществом и энергией с внешней средой. Эта инварианта и называется в организмопентризме жизнью, и при этом не имеют значения ст©пень структурной сложности организма и наличие в нем уровней иерархии. В этой связи надо вернуться к формуле (1) и обратить внимание на то, что в ней сообщается о * движении*, а не о структуре (плоти) носителя биологической формы движе-Ния материи. Но если под * движением* понимать обмен веществом и энергией, то для биологической специфичности не останет
ся места: жизнедеятельность любого организма осуществляется в строгом соответствии с законами физхимии. Поэтому и функциональный подход не приводит физхимию к пониманию особенностей жизни как особой формы движения материи.
К.М.Завадский (1966) проницательно заметил, что в биологии "учение об уровнях организации" фактически имеет в виду Hfe уровни организации, а уровни изучения. В самом деле, высокосложные организмы можно изучать до уровня органов, тканей, клеток, органелл, молекул, атомов, элементарных частиц и т.д. Этим уровням изучения будут отвечать анатомия, гистология, цитология, молекулярная биология, атомная физика, физика элементарных частиц. Взятый в целом перечень наук отнюдь не составляет биологию. Количество таких "уровней организации" и число наук о них зависят от степени сложности организма. Очевидно, что в отличие от современной биологии в биологии первичной жизни не могло бы быть, например, гистологии, потому что у первичных протобионтов не было тканей.
Таким образом, теория уровней биологической организации в ее современном состоянии не может служить основой для теории биологии, потому что она не позволяет выявить собственно общебиологические Дисциплины, необходимые для изучения жизни в. целом, независимо от того, на какой стадии развития находится жизнь. Как для общего определения жизни не требуется сверхсложной многоуровневости современных организмов, так и Для разработки теории биологии нет необходимости анализировать понятийный аппарат тех дисциплин, которые заняты изучением сверхсложности.
Какие же общебиологические, инвариантные "уровни организации" (а следовательно, и дисциплины, изучающие эти уровни) могли быть у протобионтов, отделенных зыбкой границей от неживых фазово-обособленных систем физхимии? Смогли бы такие уровни послужить основой для биологического определения жизни? Не обернется ли этот вопрос окончательным крахом структурного и функционального критериев жизни, как они понимаются в организмоцентризме? Кажется, в организмоцентризме было бы правильнее считать организм безуровневым носителем жизни в том смысле, что независимо от структурной сложности любой организм, включая протобионтов, есть носитель жизни вслед' ствие способности самостоятельно осуществлять обмен веществ и энергии. Увы, обмен веществ и энергии имеет место и в неживой природе, и в нем нет ничего сверх обычной физхимии.
Совсем иная картина возникает на уровне выше организма. Традиционная биология, особенно не вникая в проблему жизнедеятельности, располагает классификациями множеств организмов, объединениями организмов в популяции, веды, подвиды. На" пример, существует классификация вадов в систематике, класса фикация жизненных форм в экологии, фаунистических элементов
в фаунистике и т.д. В таких классификациях гоже имеет место иерархическое соподчинение. Например, виды объединены в род, роды объединены в семейства и т.п. Над организменным уровнем занимается и теория отбора, основа дарвинизма. Для организмоцентризма такие уровни организации, как популяции, виды, фауны, сообщества, биологичны вследствие того, что они состоят нз организмов как элементарных носителей жизни. Вместе с тем, как справедливо отметил К.М.Завадский (1966), для организмоцентризма все над организменные единицы (включая биосферу) остаются вторичными, производными объектами. Но тогда, «согласно той же логике, вторичными и производными становятся теория отбора и традиционная биология.
До сих пор функционирование понималось как обмен веществ и энергии и было синонимом жизнедеятельности одного организма. Сднако под функционированием можно понимать и взаимодействие между организмами. Содержание понятия межорганизменных взаимодействий удобно начать рассматривать в связи с теоремами Н.А.Умова (1916, с. 188). Теорема I гласит: действие живой системы на неживую можно заменить действием неживой системы. Теорема II утверждает: действие неживой системы на живую можно заменить действием неживой системы на неживую. Смысл этих теорем состоит, по-видимому, в том, что, например, при соударении двух тел, живого и неживого, живой организм ведет себя как объект механики, имеющий массу, ускорение, центр тяжести и т.д., и в этом смысле живое и неживое неразличимы. Вместе они образуют механическую систему. . Можно было бы развивать теоремы Умова применительно к химии и вслед за Пристли рассмотреть взаимодействие живого организма с воздухом. Из классических опытов Пристли вытекает, что действие животного на воздух может быть заменено действием горящей свечи и растение "не заметит" этой подмены. Точно так же для А.Лавуазье живые животные оставались машинами, в которых осуществлялось горение (окисление)^-. Значит, организм проявляет свою двойственную природу в зависимости от той системы, в которой находится. В физической или химической системе (соударение, опыты Пристли) организм остается открытой системой, объектом физхимии. В какой-то другой системе тот же объект дополнительно мог бы функционировать как биологический организм. Иными словами, в согласии с формулой (1) надо установить особенность того способа существования, при котором открытые системы становятся организмами.
Я.Г.Дорфман (1962) сообщает, что в конце ХУ1П века под Впечатлением исследований Лавуазье выходил первый биофизический журнал "La medicine, eclaree par les sciences physiques" . Это наглядно свидетельствует о том, что биофизика, ныне претендующая на роль теоретической биологии, представляет собой °существление точки зрения физики на процессы жизнедеятельности. '
Этот особый способ существования Ч.Дарвин назвал борьбой за существование; особое свойство, отличающее организм от то* го, что мы называем сейчас открытой системой, получило у него название приспособленности, а совокупность организмов, в которой имеет место борьба за существование, является видом, разновидностью (популяцией). Приступая к раскрытию содержания этих положений, надо подчеркнуть, что они взаимосвязаны в едином понятийном аппарате теории естественного отбора и не могут быть поняты порознь.
Прежде всего приспособленность относительна, конкретна. Не существует абсолютной приспособленности, которая могла бы обеспечить выживание в любых условиях обитания. Каждый раз выражение "вид приспособлен* подразумевает какие-то конкретные условия обитания. Эго означает, что живая плоть, один и тот же вещественно-энергетический субстрат организма, является носителем приспособленности в одних условиях и он может лишиться данной приспособленности в иных условиях. Справедливо и другое: в одних и тех же условиях обитания различные по свойствам вещественно-энергетического субстрата организмы могут оказаться примерно одинаково приспособленными. Отсюда следует заключение, что приспособленность в принципе невозможно описать на языке физики и химии, созданном для описания, изучения вещества и энергии. Различающиеся свойствами вещественно-энергетического субстрата, совершенно непохожие друг на друга различные организмы именно потому на равных борются за существование, что в этой борьбе они сопоставляются по приспособленности, а не по свойствам субстрата.
Аналогию, поясняющую понятие приспособленности, можно найти в общенаучной проблеме знака и значения. Любые два или больше иностранных языка свидетельствуют о том, что носителем одного и того же значения могут быть разные знаки. И наоборот, один и тот же знак может иметь много значений. Хранение и передача одной и гой же смысловой информации возможны с использованием разных вещественных субстратов (бумага, чернила, магнитная лента, фотопленка и пр.), с неодинаковой затратой разных видов энергии. Разумеется, смысловая информация неотделима от субстрата-носителя, как значение неотделимо от знака, тем не менее информация и субстрат не одно и то же.
Известно, что, создавая понятийный аппарат теории отбора, Ч.Дарвин обратился к современной ему политэкономии. Следуя за ним, мы найдем еще одну поясняющую аналогию для понятия приспособленности. К,Марксуказывал, что даже тщательный физико-химический анализ не позволил бы отыскать в стоимости
Маркс К.//Маркс К., Энгельс Ф. Соч. 2-е изд. Т. 23.
С. 56.
товара /ни одного атома вещества природы*. Действительно, вспомним, например, историю открытия и производства алюминия. С точки зрения физики и химии он всегда был и остается сегодня одним и тем же веществом. Но его стоимость зависит оГ успехов производства: она представляет собой отношение алюминия-товара к другим товарам. Несмотря на то что стоимость невозможно изучать методами физики и химии (в товаре нет ни "атома* стоимости), стоимость - это реальность в том смысле, что мы с ней имеем дело в повседневной жизни и от закона • стоимости во многом зависит жизнь общества, взаимодействие общества с природой.
Как понятие стоимости относится к сфере идеального и в нем нет ничего мистического, в понятии приспособленности также нет ничего сверхъестественного: то и другое находится вне вещей (товар, организм), между ними, и не существует самостоятельно. Стоимость относится к идеальному, к сфере человеческой деятельности. Приспособленность относится к живой природе, за отсутствием более подходящего выражения ее приходится называть природным прообразом идеального.
Вернувшись к примеру с алюминием, надо заключить, что алюминий, оставаясь веществом в неживой природе, становится товаром в системе общественных отношений. Понятие приспособленности вообще близко к понятию значения, напоминает понятие семантической информации. Поскольку понятие приспособленности нельзя описать с помощью тех понятий, которыми пользуются физика и химия при изучении вещества и энергии, на приспособленности можно основывать биологическую специфичность, отличающую живое от неживого.
Другая особенность понятия приспособленности состоит в том, что оно относится в первую очередь к множествам организмов: к видам и разновидностям - в дарвинизме, к видам, подвидам и популяциям — в современной эволюционной теории. Действительно, критериями приспособленности служат численность организмов и обширность ареала вида. Естественно, что оба эти критерия не приложимы к единичному организму. Однако в работах Ч.Дарвина и других исследователей понятие приспособленности применяется и к единичным организмам. Приспособленными считаются такие организмы, которые способны успешно выживать, т.е. нормально осуществлять жизнедеятельность (включая размножение) в конкретных условиях обитания. В этом случае приспособленность напоминала бы "работоспособность* Машин и механизмов и не представлялась бы биологически специфичным свойством, если бы не одно дополнительное обстоя— Тельство. Оно заключается в следующем. По Дарвину, межорга— ^изменная (внутривидовая) борьба за существование, в которой °рганизмы проявляют неодинаковую приспособленность, возника— вследствие того, что чего-то недостает виду в целом. Она
возникает всего лишь как следствие чрезмерно высокой для дац ных условий численности вида в целом. Внутривидовая борьба только опосредует межвидовую борьбу за существование. Такая приспособленность отличается от машинной "работоспособности*', И если приспособленность-биологическая специфичность, составляющая сущность жизни, го носителем первой сущности остается вид (популяция), носителем второй сущности будет единичный организм, которому она как бы навеяна извне, от вида.
По терминологии теории уровней организации, вид есть га система, в которой организм остается неделимым (атомарным) элементом в биологическом отношении. В иной системе (опыты Пристли, теоремы Умова) он сам оказывается системой и тогда имеет уровни строения, по недоразумению называемые биологами "уровнями организации". Принцип "вне вида нет организ ма" мог бы выражать го обстоятельство, что в пределах вида происходит как бы оживление открытых систем, превращение их в живые организмы. Подчиненное по отношению к виду положение организма выражается еще и в том, что отбор, движущая сила эволюции, имеет дело только с множеством, на единичном организме ему отбирать нечего.
Теперь обратимся к межорганизменным отношениям. Вообще следует различать взаимодействие и отражение. В неживой природе они не отделены друг от друга (Штофф, 1969).
При физическом взаимодействии живых и (или неживых) тел изменяются их физическая структура, химический состав, содер жание энергии, форма, положение в пространстве, ускорение и многие другие свойства, которые находят свое место в понятий ном аппарате физики и химии. В живой природе взаимодействие отделено от отражения (Штофф, 1969). Например, взаимодействие хищника и жертвы в борьбе за существование неизбежно со провождается изменением физико-химических свойств плоти животных. На языке биологов они называются "травма", "пресыщение", "истощение", "заболевание", "упражнение", "привыкание и т.д. Такие ненаследуемые изменения плоти имеют значение для ламаркизма, согласно которому изменения организмов совершаются в генеалогиях, и это уже есть эволюция, механизм видообразования. При этом вид в вышеуказанном смысле, как система, остается излишним. Эти же изменения плоти только мешают увидеть дарвиновский механизм видообразования (не ор ганизмообразования) на основе приспособленности. В дарвиниЭ' ме такие ненаследуемые изменения плоти не имеют самостоятельного значения. Они только влияют на приспособленность (выживание и размножение), зависящую от конкретных условий обитания. Вследствие конкретного характера приспособленности качественная определенность изменений плоти организмов сама по себе не имеет значения для отбора. В живой природе такие
42
„ 3
взаимодействия отделены от отражения уже потому, что отражение совершается в статистической структуре популяции, а не в организме. Отражение выражается в изменении статистических показателей изменчивости признаков организмов популяции: численность, средняя, лимиты, дисперсия, обширности ареала и пр. Они относятся ко всей популяции и характеризуют совокупный субстрат популяции. Обособленность взаимодействий организмов П отражения для популяции хищника в популяции жертвы (и обратно) состоит еще в том, что изменение структуры популяции не предполагает с необходимостью изменений плоти организмов, достаточно лишь одного изменения, чтобы исходно различные организмы (неопределенная наследственная изменчивость) размножались с неодинаковой скоростью. Взаимодействие и отражение - это аспекты двойственного существования биологического организма. "Живая особь существует, так сказать, на двух уровнях: непосредственном индивидуальном и опосредованном, видовом, - образующих противоречивое единство. С выделением видового уровня отражение как бы получает новый, относительно самостоятельный "экран", способный запечатлевать то, что выходит за рамки возможностей отдельной особи и принципиально не может быть достигнуто в этих рамках - "время" вида как совокупный эволюционный опыт. Возникает и новый субъект отражения — не только единичная особь с ее весьма ограниченным пространственно-временным полем деятельности, но и вид с его более широкими пространственно-временными возможностями и более емкими резервуарами памяти* (Давыдова, 1979, с. 37).
Таким образом, содержание межорганизменных отношений оказывается довольно сложным. С одной стороны, организмы взаимодействуют как открытые системы, их физическое взаимодействие отличается наглядностью и доступно дпя непосредственного наблюдения. С другой стороны, это взаимодействие - только внешняя сторона отражения в статистической структуре популяции ее борьбы за существование. Отражение как бы надстроено над физическим взаимодействием. В борьбе за существо- . вание, когда изменяются численность и обширность ареала видов, их популяции участвуют как носители идеального свойства приспособленности. Поэтому она не сводится к физическому взаимодействию. Вследствие этого борьбу популяций и отдельных Пидов за существование следует считать особым, не физическим взаимодействием. Его можно было бы назвать семангико—информационным взаимодействием.
Соотношение разных форм движения материи поясняет рис. 1. ^начала имело место развитие неживой материи от элементарных
v Отражение понимается здесь как понятие естественно-научное, а не философское.
43
частиц к веществу. Моменту зарождения жизни предшествовало достижение неживой материей структурного уровня макромолекул и открытых фазовообособленных систем. Существование эт\
социальная форма движения материи
Рис. 1. Соотношение между разными формами движения магерШ Усложнение вещественно-энергетического субстрата, начавшееся в природе с уровня элементарных частиц (I), продолжается в организмах от момента зарождения жизни под. действием естественного отбора (1J). С появлением общества этот процесс идет как бы внутри социальной формы цвижения материи (III) под де4 ствием общественных отношений. При сохранении субстратного единства разные формы движения материи различаются движушл* ми силами развития (стрелки): естественный отбор и обществен' ные отношения
го уровня могло продолжаться длительное время. Когда в "мировом бульоне* снизилась концентрация необходимых для обмена
44
0ешеств, открытые системы вступили в борьбу за существование, и с этого момента представляли собой живые организмы, ромеит зарождения жизни отмечен появлением у открытых систем идеального свойства приспособленности. В этот момент физико-химический коллоид становится еще и живой плотью. Поскольку такие системы различаются идеальным свойством, различие между ними не может быть обнаружено методами физхи-в момент зарождения жизни между коллоидом, и плотью йет различий по химическому составу и физической структуре, дальнейшее усложнение плоти (вещественно-энергетического субстрата) уже протекало как бы внутри организмов. Здесь особенно впечатляет развивающаяся сверхсложносгь макроструктур (органов, клеток, их органелл) и достигшая кульминации у высших животных. Такие структуры действительно не встречаются в неживой природе, но их нельзя считать отличительным существенным признаком жизни уже потому, что сущность жизни относится к категории идеального. С возникновением человеческого общества развитие живой и неживой материи продолжалось как бы внутри социальной формы движения материи и привело к появлению сложных орудий груда и продуктов производства, в том числе синтезированных веществ, к искусственно выведенным сортам культурных растений и породам домашних животных. Все перечисленные объекты не встречаются в неживой и в живой природе, но было бы явной ошибкой считать эту невстречаемость особенностью, отличающей общество от природы. В этих веществах, вещах, породах животных и сортах растений интериоризо-ван, овещнен (сделался вещью) труд, система общественных отношений. Сходным образом семантико-чшформационные взаимодействия постепенно интериоризовались в сверхсложной живой плоти.
Между тремя разными формами движения материи сохраняется субстратное единство через структурные уровни, различаются же они особыми способами движения материи. 1. В неживой природе — это физические и химические взаимодействия природных тел как вещественно-энергетических образований. 2. В жи— н°й природе становится ведущим семантико-информационное взаимодействие, потому что организмы становятся носителями значения - приспособленности. 3. С появлением общества над живой и неживой природой надстраивается и становится определяющей система общественных отношений. Не нарушая основных законов природы, общество управляет природой.
Таким образом, формы движения материи выделяются не особенностями вещественно-энергетического субстрата и его функционированием, а природой движущих сил развития, способом существования субстрата. Борьба за существование между видами й внутри видов и есть особый биологический способ существова-Ния открытых систем, делающий их живыми организмами.
45
Представляется возможным примерно так понимать представ ления В.М.Кедрова (1959) и В.Ф.Сержантрва (1972) о том, что, начавшись в добиологическое время, развитие неживой ма-терии продолжалось как бы внутри биологической формы движения. В этой связи представляет интерес и замечание Е.Ф.Солопова (1966, с. 92): "Живое тело как живое тело всегда функционирует, всегда участвует в своей главной, высшей форме движения, все остальные изменения происходят как бы включенными в эту форму движения как ее составные части*.
В итоге весьма широкое и лишенное четких границ понятие "теоретическая биология* может быть расчленено следующим образом.
1. Теория жизнедеятельности организмов - область исключительно плодотворного применения физикалистской методологии в изучении функционирования природных открытых систем, достигших чрезвычайной структурной сложности, когда они стали существовать особым, биологическим способом. В био л от ии такие системы получили название организма. До недавнего времени все такие системы без исключения оставались результатом либо естественного, либо искусственного отбора. Молекулярная биология начинает создавать открытые системы без применения отбора, вне вида (не относящиеся к какому-либо виду), Такие открытые системы, не отличаясь структурно и функционально (как открытые системы) от организмов, смогут вести биологическое существование (сделаться организмами), как толь ко вступят в борьбу за существование. По своему происхождению (способу изготовления) искусственные организмы не отличаются от других творений человеческих рук, в них интериори-зованы общественные отношения.
2. Общая теория жизни должна представлять собой развернутый перевод философской формулы (1) на общенаучный язык. Теория жизни должна вскрыть отличия жизни от других форм движения материи. В этом заключается ее внешняя по отношению к биологии функция. Оформление этой теории послужило бы стимулом для разработки теории неорганической формы движения материи, не совпадающей с теоретической физикой (Кре-мянский, 1969). В биологии функции теории жизни фактически выполняет теория отбора (Заренков, 1984). Г.А.Югай (1976, 1984) предлагает теорию жизни именовать биотикой. По этимологическим соображениям это предложение вряд ли приемлемо. Дело в том, что в биогеографии биотой называют совокупность фауны и флоры (Бобринский, 1946, с. 8). Ради, единства слово' образования (фауна - фаунистика, флора - флористика) производ' ным от биоты будет биотика (биотистика) как обозначение иной нежели у Г.А.Югая, области знания.
3. Теория биологии - теория науки о жизни, специфичео' кая биологическая теория, знание о биологическом знании. Она
46
0Меет дело прежде всего с готовым знанием о жизни, которое дожилось в историческое время. Содержание теории в какой-то степени определяется содержанием ее предметно# области, совокупностью фактов, имеющихся обобщений. Инвариантность теории определяется тем, что она исходит из некоторой теории жизни, основы для изучения биологического знания. Она может льдеть своей целью выработку единого понятийного, аппарата и языка на основе понятий и языка частных разделов биологии. Эта цель является весьма актуальной для современно# чрезвычайно разросшейся биологии. Практика биологов разных специальностей исключительно богата, понятийный аппарат и язык частных разделов биологии находятся в движении, быстро развиваются. Поэтому теория биологии тоже должна представлять собой как бы процесс. По своему построению она могла бы считаться аксиоматической (на основе некоторой теории жизни), а по функции, как охватывающая разные частные теории (например, теории систематики), - метагеоретической.
Надо считаться с той возможностью, что по своему статусу, внутренней структуре и функциям теория биологии окажется теорией в несколько особом, непривычном смысле. Эту возможность не следует сбрасывать со счетов, потому что "разные вицы наук организованы различными способами" (Коллингвуд, 1980, с. 24). Возможно, что теория биологии не сможет служить руководством к практическому изучению живой природы, так как не добавит знания как руководства к деятельности. Скорее, теория биологии должна представлять собой определенное понимание знания, места биологии среди других естественных наук» Имея в виду теорию речи - языка, теорию мышления и, наконец, биологическую теорию популяций, Г.П.Щедровицкий (1976, с. 175) пишет: "...что это объекты совершенно иного категориального типа, нежели объекты физических теорий, что их нужно описывать принципиально иначе, нежели описывались объекты физических теорий, что сами эти объекты будут представлены в существенно иных онтологических картинках, нежели объекты классической физики, что описывающие их теории будут иметь логическую структуру иную, нежели физические теории...". Эта цитата наводит на мысль о том, что биология, принадлежа Через свой объект к семье естественных наук, могла бы примы— Кать к гуманитарной культуре своей внутренней логической стру» тУрой.
и В этих трех теориях, вместе образующих так называемую теоретическую биологию, нетрудно узнать три 'образа* биоло-гИи, которым была посвящена гл. 2.
Глава 4
БИОЛОГИЯ И ФИЗИКАЛИЗМ
Принципы физикализма, сформулированные Б.С.Флейшманом (1980), вкратце сводятся к следующему: 1) для изучения родо достаточно и необходимо экспериментирование и измерь ние, 2) первопричины всех явлений находятся в микромире, 3) все явления в природе протекают на основе закона сохране ния вещества и энергии. К этому можно было бы добавить, т-для физикализьАа в природе нет ничего, кроме вещества, энер^ гии и поля, что же касается идеального, существование которс го в обществе очевидно и принимается при разрешении основу го вопроса философии (Ильенков, 1979 а,б; Лифшиц, 1984), то оно оставлено физикализмом без внимания. Таким образом, физикализм имеет перед собой редуцированную картину мира в целом. В социокультурном знании идеальным являются значени смысл, человеческий дух, стоимость и пр. В биологии, изучаю щей жизнь, которая занимает как бы переходное место между социумом и неживой материей, им отвечает природный первооб раз идеального, который Дарвин назвал приспособленностью в контексте непривлекательной своей безжалостностью теории ес тественного отбора. Своеобразие приспособленности как первообраза идеального обнаруживается в связи с популяцией, а не организмом.
Общее определение популяции — не простая задача (Тимофе ев-Ресовский и соавт., 1973). В ней выделяется основной во прос, содержание которого состоит в следующем. С одной стороны, "популяция представляет собой группу живых существ, ограниченную во времени и пространстве" (Парк, 1939, с.-ЗО1 Сходных взглядов придерживается, например, А.В.Яблоков (1980), и вообще такой подход считается весьма обычным ( Jonckers, 1973). С другой стороны, на понятии популяции основывается популяционизм, на него опирается синтетическая теория эволюции - преемница дарвинизма (Huxley, 1942). К.М.Завадский (1968) пишет о едином "популяционно-видовой уровне организации. Значит, на популяцию должно быть перене сено то существенное в понятии вида, что делает неизбежной борьбу за существование и видообразование, стремление попу* ции к неограниченной численности. Но тогда популяцию не еле довело бы наделять пространственно-временными границами. Так же, как вид, популяцию следовало бы считать неопределе!1 но протяженным во времени и пространстве множеством орга-* низмов. Имеется еще один довод в пользу такого понимания пуляции: существуют виды, состоящие из единственной популй' ции. Принимая вслед за К.М.Завадским единый "популяционно* видовой* объект, противопоставим его единичному организму.
48
Организм, 'живая машина", выполняет работу по обме— веществом и энергией с внешней средой и потому с неизбежностью саморазрушается. Причины смерти организма внутренние-в его "машинной* природе. В организме жизнь не отделена они
смерти. Вполне отчетливо неотделенность жизни от смерти выявляется у одноклеточных организмов. У них окончание индивидуального существования наступает вследствие размножения делением. Такое деление, приводящее к увеличению (по сравнению с родительской клеткой) величины относительной поверхности, есть всего лишь особый способ регуляции обмена веществ. * Машинная* природа организма - в его пространственно-временной определенности, конечности. Эго делает возможным экспериментирование и измерение. Именно эти две познавательные операции необходимы и достаточны для изучения "живой машины*.
Популяция (вид) - это самовозобновляемое множество организмов. Оно не имеет пределов, предустановленных самой популяцией: виды вымирают в борьбе за существование вследствие неблагоприятного стечения случайных, впошник относительно самого вида обстоятельств. Такое существование есть прин
ципиально незавершаемое становление и полная противоположность смерти, т.е. жизнь (рис. 2). Применимость экспериментирования и измерения к изучению природных видов более чем со
мнительна: как экспериментировать с множеством организмов, не имеющим собственных границ; как измерять открытое, длящееся множество организмов?
Могло бы показаться, что само стремление вида к неограниченной численности есть скрытая причина его собственного вымирания. На самом же деле при этом подразумевается ограниченность земного пространства - внешнее по отношению к виду обстоятельство. То, что называется *само-Регупяцией* в популяциях, в Действительности подразумевает влияние внешней среды, 'Точнее ограниченность внеш-Ней среды. Кроме того, надо
Рис. 2. Два типа множества.
Обычное изображение множества (слева) создает впечатление законченного неподвижного множества организмов. Для потенциально бесконечной популяции как самовозобновляемого множества организмов больше подходит образ разомкнутого кольца (справа)
отличать неограниченность во времени от неограниченности в пространстве, и первое свойство бо-гаче второго. Действительно, для заполнения организмами бесконечного просгранеочм требуется бесконечное же время, тогда 4 - 1560 49
как бесконечное во времени существование возможно и в конеч. ном, например земном, пространстве. Поэтому даже в частной, земной, форме жизни сохраняется противоречие между конечны^ во времени организмом и потенциально бесконечной во времени популяцией.
В литературе понятия вида и организма действительно противопоставлены по их неодинаковой временной природе. Например, К.Бэр писал в 1834 г.: "Известные формы организации, эти связанные при помощи размножения ряды, являются постоянными идеями творения, а отдельные особи представляют из себя лишь преходящие изображения данных идей. Только особи и подлежат смерти, и только новообразование является постоял, ным" (Бэр, 1924, с. 96). По Дж.Симпсону, "эволюционный вид есть родословная (последовательность популяций предсов и потомков)" (Simpson, 1961, р. 96). В противоположность заведомо конечному организму существование вида понимается многими авторами (Шмальгаузен, 1935; В.Беклемишев, 1964; Завадский, 1968; Шварц, 1968) как "неопределенно долгое", что отвечает дарвиновскому представлению о веде. Наоборот, такие утверждения, будто "...вид имеет финитное распространение в пространстве и во времени" (Eldredge, Cracraft, 1980), выводят понятие вида в область финалистических концепций (Давиташвили, 1969). Интуитивно понимаемая бесконечность заняла видное место в понятийном аппарате эволюционной биологии, Вот как, например, Ч.Давенпорт (Davenport, 1930, р. 331) описывает механизм эволюции: "1) бесконечная способность к размножению, 2) бесконечная способность мутировать, 3) бесконечное разнообразие местообитаний, 4) обширные возможности для расселения мутантов.., 5) вымирание бесконечного числа других индивидов". Скорость размножения организмов вида Дарвин определил как "геометрическую прогрессию". Такое особое существование вида во времени представляется возможным понимать в связи с понятием потенциальной бесконечности в смысле Г. Кантора (1914).
Распространение идей Г.Кантора вызвало в свое время переворот в теории множеств ив содержательной логике и привело к парадоксам. Известно, что парадоксы возникли вследствие неправомочной подмены бесконечного очень большим, но конечны’ и применением обычной логики в принципиально иной области -в бесконечности (Харин, 1963; Библер, 1975; Бурова, 1976) Важно, однако, отметить, что у Кантора понятие бесконечности несколько иное, нежели у Аристотеля, оказавшего глубокое во3 действие на классическое естествознание, а значит, и на типологию; оно подразумевало актуальную бесконечность. В аристо* телевской, преимущественно онтологической логике, напротив, актуальная бесконечность оставалась чисто логическим понятМ" ем, не имевшим под собой природного- обоснования. Признавал^
50
только потенциальная бесконечность, в которой как бы совершали 'со-дление* исследователь и природа, и вне этого "со-дления ничего не было. 'Бесконечность чувственного мира не умещалась н сфере античного миросозерцания' (Лосев, 1923, с. 16). Наоборот, у Кантора потенциальная бесконечность существует постольку, поскольку принимается реальностью актуальная бесконечность, первая как бы имеет место для своего осуществления (Мелюхин, 1966; Попов, Стяжкин, 1974). Сам Кантор относил актуальную бесконечность к области иррационального - к богу; Современная наука в боге не нуждается, но задача соотнесения двух вадов бесконечности остается.
Своеобразное понятие бесконечности не может изучаться естественными науками, оно относится к философии и математике. В биологии же оно принуждает исследователей к особой форме представления результатов исследований, придающих ей своеобразие, неприемлемое для физикалистски настроенного исследователя. Такой спецификой биологической формулы является презаде всего слабая и поверхностная математизация, не затрагивающая натурализм и -традиционную биологию. Далее - неопределенность, текучесть суждений, явственно выступающие в наиболее развитом разделе биологии: в систематике. Наконец, сам язык биологии с его нечеткими понятиями, явное сходство с принципиально неформализуемым естественным языком. Эта сделавшаяся 'притчей во языцех* неопределенность могла бы считаться репрезентантом потенциальной бесконечности популяционных объектов, выражением их незавершаемости, открытости. Здесь уместно процитировать Н.Л.Лузина (1957, с. 472): 'Мы не владеем идеей бесконечного иначе, как беря конечное и заставляя его расти. Всегда, следовательно, имеется неопределенность, которая является истинной идеей бесконечности*. Замечательно, однако, что традиционная неопределенность в биологии выявлена не на природных длящихся видах, а всего лишь на конечных выборках.
Действительно, то, что обычно называют 'видом', фактически есть всего лишь выборка организмов из длящегося множества организмов вида. Казалось бы, противоположность между выборкой и видом можно преодолеть, расширяя выборку, вовлекая в исследование все большее количество организмов. На самом деле это была бы 'гонка за тенью*: любая, даже очень большая выборка организмов есть неподвижность, тогда как вид есть поток организмов, Этим важным обстоятельством объясняется особая, отличная от физикалистской соотнесенность во времени на-бДюдателя и природы.
В Исследованиях Г.Галилея, открывших эпоху современного ручного экспериментального изучения природы, свободное падение тела оставалось природным явлением, которое можно было ИаУЧать по частям. Членение целого природного явления исследо
51
вателем не приводило к искажению приростного закона, поскольку в каждом отрезке траектории падающего тела неизменно сохраняется закон свободного падения. В такого рода исследованиях нет расхочдаений между временной точкой (точнее, временным интервалом наблюдения и бесконечной природой. Отсюда сама возможность экспериментирования, правомочность изучениям лаборатории законов природы. Теперь попробуем представить себе наблюдение за природным видом. Такое наблюдение каждый раз осуществляется только во временной точке (интервале), оно относится к выборке, выделенной из природы наблюдателем. Удалим наблюдателя из природы. Тогда вместе с ним пропадет и выборка, но останется беспредельно длящийся вид. Следовательно, выборка отличается от живого вида наличием границ, созданных наблюдателем.
Возможность измерения и правомочность применения количественной математики также созданы самим исследователем. В этом смысле любая выборка есть артефакт. Имея в своем распоряжении выборку, исследователь должен считаться как с реальностью с существованием той области, которая никогда не будет исчерпана исследованием. Суждения о выборке могут относиться и к ввду при том условии, что суждения останутся открытыми, незавершенными, становящимися. Такое суждение могло бы претендовать на объективность по отношению к виду. Внешне непостоянство и неоднозначность суждений представляются неопределенностью и субъективизмом, что вызывает критику со стороны так называемого "точного естествознания". Между гем именно этими свойствами суждения о живых объектах отличаются от суждений о неодушевленных объектах. Так обстоит дело при мысленном движении от временной точки "вправо"-в будущее". Движение "влево", в "прошлое", не может утвердить неопределенность как выражение объективности биологичео ких суждений. В самом деле, пополняя выборку уже вымершими организмами, исследователь вынужден считаться с ограниченностью их числа. Расширение выборки возможно только до какого-то объективно существующего в природе предела. Такая расширенная выборка останется объективно конечной, а следовательно, суждения о такой выборке обязаны сделаться однозначными. Если они не становятся такими, то в этом "виноват" исследователь, а не природа. Если ограничиваться изучением только уже осуществившихся организмов и игнорировать потенциально осуществимые организмы, то ^определенность суждений в градини* онной биологии д олжна считаться субъективной.
В бесконечных множествах, как подчеркивал Г.Кантор (191^ нельзя применять привычное понятие количества; такие множесТ ва различаются неодинаковым порядком слагающих их элементе® Упорядочение - это одна из задач классифицирования. В свою очередь классифицирование не предполагает с необходимостью
52
измерения. Отметив пристрастие натуралистов к классификациям, мы должны понять и причины сравнительно малой распространенности измерения. Что же такое измерение?
"Измерением мы называем познавательный процесс, заключающийся в сравнении путем физического эксперимента данной величины с некоторым ее значением, принятым за единицу сравнения. Число, выражающее отношение измеряемой величины к единице, называется числовым значением измеряемой величины; оно может быть целым или дробным и есть число отвлеченное" ( Маликов, 1949, с. 18). Что же касается единиц измерения, таких как сантиметр, грамм, секунда, то они представляют собой конвенциональные единицы и существуют на основе договоренности между исследователями. Казалось бы, биологи должны поступить сходным образом и договориться о биологически специфичных единицах измерения популяций, носителей первой биологической сущности. Но что может дать сопоставление популяций, лишенных пространственно-временных границ? Отдельные популяции и их объединения потенциально бесконечны и как бы одинаковы по величине, к ним неприменимо понятие количества. Остается лишь одно: при сравнении выборок выявлять признаки и принимать, что такие признаки в каком-то смысле являются и признаками вида. В итоге напрашиваются следующие соответствия (табл. 1).
Таблица!
Соответствие биологических и физических понятий по их физиологическому назначению в биологии и физике
Биология
Сопоставление видов (популяций) через выборки организмов; сравнение
К пасс ифинирование
Признаки
Система
Физика
Сопоставление величин объектов через эталоны; экспериментирование
Измерение
Единицы измерения
Закон
Понятие развития, будучи приложенным к виду и организму, °Каэывается имеющим неодинаковое содержание. Онтогенез, индивидуальное развитие организма, протекает по наследственной пРограмме, и ничего нового сверх этой программы у организма появитьбя не может. Наоборот, в процессе эволюции у видов в°ЭДикают такие свойства, которых не было у их предков. Такие ^воприобрегения не содержатся в наследственности предков, их объяснить только собственными свойствами вида. Необ
53
ходимо учесть влияние внешней среды в естественном отборе. Поэтому совсем не без оснований Е.А.Седов (1976) склоняет^ ся к тому, чтобы собственно развитием, как это принято в д&ч лектике, считать только развитие жизни, а не онтогенез организма. Если мы к тому же вспомним, что онтогенез в конце концов есть производное филогенеза, как это принято в современной формулировке биогенетического закона, и что у протори, онтов не могло быть онтогенеза, то проблема развития оргаг ма потеряет общебиологическое значение. Так же как в вопр о специфике жизни йбпуляция преобладает н^ц организмом, т« же и онтогенетическое развитие организма представляет собой частную проблему по сравнению с историческим развитием вида. Но это лишь одна из сторон общего вопроса о соотношении меж, ду единичным элементом и множеством элементов, решаемом в традиционной биологии совсем не так, как в организмоцеНтриэ» ме.
У Аристотеля помимо индивидов (организмов) упоминают^ вид род и класс. Они не означают таксономических рангов < гематики и подразумевают только относительный объем логи ких понятий. Организм - это абсолютное подлежащее, первая сущность и индивидуальное бытие, — онтологически доминирует над вторыми сущностями, видом и родом. Онтологическая роль рода состоит в том, что он фиксирует пределы смены противоположностей. В отличие от платонизма в философии Аристоте -противоположности были только тем, гЧТо сказывается о суС страте (организме)" (Гайденко, 1980). В процессе классиф! рования, когда организмы’ объединяются в виды, а виды в рсь,,^ исследователь как бы удаляется от сущности. Сущности было больше в организме, меньше в ваде, еще меньше в роде. Аристотелевские понятия вида, рода и класса через натуралистов ХУП1-Х1Х вв. перешли в научную биологию, и содержание их при этом резко изменилось: из логических они сделались природными. Больше того, в дарвинизме понятие вида сделалось центральным. При этом оказалось, что вад и вообще любые по» пуля иконные объекты обладают неопределенной изменчивостью. У потенциально бесконечных видов она вместе с тем и беспредельна. Поэтому ни род, ни какая-либо другая ступень иерарх не могут фиксировать смены противоположностей. Признаком длящегося вида становится длящаяся же изменчивость. Знач? вопреки мнению Л.Агассица (Agassiz, 1962), полагавшему бдато каждый организм есть носитель признаков вида, к кот му относится организм, признаков рода, к которому относит вид и т.д., организм не может подменить собою вида.t Признак организма хотя и развиваются в онтогенезе, но их конечное развитие совсем не то же самое, что становление признаков в# да в погенциально бесконечном существовании. Далее в биолог^ чески специфическом способе существования, который Дарвиг
иазв^л борьбой за существование, организм выступает носите— яек1 только второй сущности. Образно выражаясь, теперь орга-виэм биологии как бы светит биологии отраженным светом (как Луна Земле). Поэтому коперниковский переворот', фактически осуществленный Дарвином, означает отход от канторовского по-
вйыания множества, которое только мыслится как единое целое.
Если длящийся незавершаемый вид реально существует в приро-
де, ми
то это — реальность особого рода с совершенно непривычны— свойствами. Возможно, что наша невосприимчивость к тако-
го род® природным объектам есть выражение подсознательно
усвоенного физикализма, в котором нет различий между временной точкой и природой. Давние споры о реальности вица вызва-
ны, по-ввдимому, тем обстоятельством, что физикалистские критерии реальности (возможность экспериментирования и измерения) непригодны для изучения вада. Если приспособленность есть "идеальная" биологическая сущность каждого' вада, то кри
терии реальности вида не могут относиться к вещественно-энергетическому субстрату.
В длящейся популяции нет предела, а следовательно, и середины разнообразия. В этом смысле понятие типа становится ложным, наше представление о типе как середине разнообразия будет изменяться в ходе становления вида.
Онтологическое доминирование множества над единичным элементом служит основанием для пересмотра отношений между длящимря видом и конечной выборкой. Любая выборка, как и единственный организм, обладает пространственно-временной определенностью. В этом смысле они эквивалентны и противопоставляются длящемуся виду, который невозможно характеризовать числом, потому что число есть неподвижность. Поскольку обычному биологическому, исследованию доступна только выборка, естественным образом возникает вопрос о том, насколько выборка показательна для вида. Эта задача вряд ли может быть решена, пока не известен способ взятия выборки. Обсуждая подобную проблему в логике, М.В.Попович (1975, с. 46) пишет: Из теории прямо следует, что при условии, что множество индивидов бесконечно, степень подтверждения гипотезы, говорящей обо всех индивидах и опирающейся на конечное число благоприятных испытаний, равна нулю". Столь пессимистический вывод противоречит практике науки, которой известно много эмпирически обоснованных закономерностей, и его обсуждение выходит за пределы возможностей самой логики. Во всяком случае, заключает М.В.Попович, "мы должны считаться с соображениями бесконечности самым серьезным образом...". Сложность, по-види-м°му, состоит в том, что в цитированном отрывке упоминается 0 всех индивидах", и смысл этого выражения для становящегося множества совсем неясен. Проблему, таким образом, представляет само выражение "весь вид". Отсюда следуют и другие трудности.
55
Возникают сомнения в применимости закона исключенного третьего классической логикиs который разработан Аристотеле^ на примере утверждения * завтра будет морское сражение° (Ар^ стотель, 1978, с. 102). Возможность в данном примере сое» тоит из Двух противоположных суждений о "сражении*. Пока "завтра* не наступило, противоположности соединены в возмо^. ности, и логическая истинность высказываний субъекта о "сражении" определяется соответствием знаний субъекта истинному положению вещей и его способностью логически мыслить. Неоп, реДе ценность суждений о будущем остается объединением про» г иъоп сложных суждений и проистекает из неосведомленности субъекта относительно всех обстоятельств, определяющих насту, пленке или ненаступление "сражения*. Такую неопределенность следует считать субъективной (Ахманов, 1960). Сама неопределенность суждения (допущение обеих противоположностей) ог. раничена во времени в силу самой природы "сражения", и выра.. женке "все обстоятельства" обозначает конечное число обстоятельств. Когда наступает "завтра", противоположности распада, ются и тогда произойдет одно из двух, третье исключено. Так соблкдается закон исключения третьего в его логическом аспек те. Совсем иное наблюдается в высказывании "возникнут млекопитающие, не вскармливающие детенышей молоком". Здесь вслед ствие особой природы объекта "млекопитающие", который состоит из потенциально бесконечных видов, противоположности соединены в бесконечном д л он ии, и такие противоположности могут быть разделены только логически. О "млекопитающих", пока они пребывают в длении, возможно только неопределенное суждение, своей неопределенностью репрезентующее неопределенно долгое существование видов. Если даже изучены все без исключения организмы в выборке, живые и вымершие, и установлено, что все они вскармливают детенышей молоком, то все равно, пока обсуждаются млекопитающие как живое становящееся множество, неопределенность суждения сохраняется как третье, как со^д^нение противоположностей. Сколь обширным не было бы фактическое обоснование некоторого свойства вида, оно остается принципиально незащищенным перед лицом новых открытий, потому что любая конечная выборка несоизмерима с потенциала но бесконечной совокупностью. Именно таким незащищенным (открытым) останется суждение о млекопитающих в современно, систематике. Балансирования между "да* и "нет", соединенные "да" и "нет", представляют собой третье суждение, с которым не приходится иметь дела классической логике.
Статистические методы и вероятностное видение природы в современной биологии весьма обычны, и потому может показать ся, что представление о неопределенности, по крайней мере в ее статистической форме, уже усвоено; например, теория ДарвИ' на "насквозь пропитана духом вероятностных идей* (Седов,
56
1076, с° 88)- Значение вероятностных идей для биологии ярко лазала А.И.Равикович (1977). Тем не менее статистические етоды большей частью применяются в предположении, будто в ироде существует неподвижный порадок, а неопределенность несена в результате изучения исследователем, и от нее надо збавиться. О таком отношении к неопределенности свидетельствует распространенное стремление к однозначности суждений, обычное среди систематиков. Достаточно сильно оно дает о себе знать в фаунистике, флористике, экологии, где всевозможные индексы уже своей числовой природой создают иллюзию об однозначном порядке в природе. В додарвиновской типологии гарантом порядка был творец и поэтому порядок в живой природе, выявляемый исследователем, не требовал оправдания своему существованию. Сегодня можно задаться вопросом: на чем основывается предполагаемый само собой разумеющимся неподвижный порядок в живой природе? Может быть, эта гипотеза создана в подражание классической физике? Нетрудно убедиться в том, что и статистический вероятностный подход может служить типологическим представлениям об однозначном неподвижном по
рядке в природе.
Пусть имеется статистическое распределение, характеризующее изменчивость некоторого вида (в действительности — выборки). Все статистические показатели изменчивости — это некоторые числа, любое число — неподвижность. Следовательно, все статистические показатели неподвижны, их коллективным носителем является изученная выборка. Нетрудно догадаться, что выборочный объект исследования своей выборочной природой не только делает возможным применение чисел, но и предопределяет соответствующую выборке неподвижность числового описания выборки. В этой связи полезно вернуться к истории концепции эволюции. Как известно, для решения вопроса о том, изменяются ли виды, во Франции в 1802 г. была создана комиссия, которой было поручено сравнить древние египетские мумии некоторых видов животных с ныне живущими представителями этих же видов. Комиссия заключила, что за несколько тысяч лет, Разделяющих эти две выборки организмов, ожидаемых различий Не возникло. Отсюда последовал вывод о неизменяемости видов Во времени, о ложности концепции эволюции (Амлинский,1955). Можно, однако, задаться вопросом о том, была бы доказана эволюция видов, если бы ожидаемые различия были бы обнаружены. Думается, что такого вывода не следовало бы делать. В самом деле, объединим данные о древних мумиях с данными о с°Временных животных. Такая операция правомочна, потому что Л1°бая временная точка наблюдения выделена исследователем, нет в природе. В результате объединения получим выборку, к°Торая характеризуется некоторыми неподвижными статистичео-11Ми показателями. Эти показатели описывают различия между
57
организмами одного вида в выборке, флуктуации во временно^ срезе'толщиною* в несколько тысяч лет. В принципе это явле». ние того же порядка, что и различия между единовременно со, существующими организмами, обитающими в разных условиях: ведь толщина временного среза устанавливается только иссле^ дователем. Какой бы ни была численность выборки, она уже своей выборочной природой принуждает исследователя к вычислениям и уже этим обстоятельством создает основание для не. объективного суждения о длящемся живом виде. Объединение вечного числа таких выборок не меняет положения, не отменяв] противоположности между конечным и бесконечным.
Таким образом, надо еще раз подчеркнуть вслед за исследи вателями истории эволюционизма, что представление об измену мости видов во времени сделалось убедительным благодаря пе. ренесению на природу опыта животноводства и растениеводства, т.е. опыта деятельности человека (Филиппченко, 1979). У дель, ный вес этого довода в пользу эволюционизма можно оценивать по-разному. Тем не менее остается несомненным, что учение Дарвина в какой-то степени действительно сохраняет сродство с общественными науками не только благодаря перенесению на природу понятий из области политэкономии, но также благодаря фактологии: факты из области человеческой деятельности и из жизни природы имели значение для обоснования теории эволюци!
В заключение надо обратить внимание на еще одно парадоксальное следствие из представления о популяции как потенциально бесконечного множества организмов. Разнообразие организмов в пределах только одной популяции не уступает разнооб разню организмов всего органического мира в том смысле, что оба потенциально бесконечны; однако при условии, что в общем случае в природе нет неразличимых организмов и имеет место неопределенная изменчивость. Значит, так же, как в теории бео конечных множеств, целое (органический мир) не больше своей части (популяция вида), и таких частей потенциально бесконечное множество. Этот вывод не согласуется с постулатом классической логики, согласно которому часть всегда меньше пело'
В биологии это могло бы иметь следующее значение. В прШ ципе все традиционное биологическое знание (систематика, био-география, экология и пр.) могло бы быть воссоздано даже на одном виде, даже на одной суперпопуляции протобионтов в пер-вичной жизни. Если бы была показана возможность такого вое* создания биологического знания, то тем самым была бы утвер* ждена неизменность структуры традиционной биологии, ее способность объективно отображать в структуре знания планетарную организованность земной жизни в любой момент ее суше^ вования, включая момент зарождения жизни.
Ю. А. Шрейдер и соавт. (1980) предлагают назвать дарвин*^ скую биологию классической, а современную 'критическую шке
58
> связанную с именем А.А.Любишева, "неклассической". С
Л им трУДно согласиться. Скорее, наоборот, дарвинизм, сменив-й классическую типологию, поставил биологию перед необхо-
Ш мостью отказа от классической логики в пользу логики раэ-^нваюпхихся множеств. Поэтому целесообразно именно дарвинов-сКУ*° биологию назвать неклассической биологией.
Глава 5
ЛОГИЧЕСКАЯ СТРУКТУРА биологического ЗНАНИЯ
Обычно живая природа представляется разнообразием организмов "от бактерии до человека". Более широкое видение свойственно традиционной биологии: живая природа есть многообразие таких объединений популяций видов, как фауна и флора, так— соны, родословные, жизненные формы сообществ и др. Такие объединения выделяются на разнообразии популяций, когда на нем задаются те или иные отношения. Например, если задано отношение родства, то получаются родословные. Если задано отношение совместности, то получаются фауны и флоры.
Установление отношений может считаться познавательной операцией: "Когда ум рассматривает какую-нибудь вещь так, что как бы приводит ее к другой веши, ставит ее с другой вещью и переносит свой взор с одной на другую, то это есть, как обозначают сами слова, отношение и сравнение* (Локк, 1960, с. 322). Вследствие своей логической природы такие отношения имеют неограниченные возможности. Ничто не мешает нам,- например, сравнивать медведицу с созвездием. Отношение как мысленное соотнесение позволяет нас связывать отдельные предметы или явления в целостную картину и таким образом воспроизвести в сознании картину мира в целом. Отношения - это формально-логические репрезентанты природных взаимодействий. С их помощью в знании можно воспроизвести те природные взаимодействия, благодаря которым природа остается целостной. В живой природе - это совокупность тех взаимодействий, которые Ч.Дарвин "широкой метафорически" назвал борьбой за существование. В этом смысле взаимодействия и отношения — это не Дмрка от бублика". Отношения - инструмент изучения первых.
Почему бы в таком случае не ограничиться непосредствен-НЬ1м изучением борьбы за существование, чем и занята эволюционная теория? Почему эволюционная теория подменяет собой гРеакционную биологию, которая вместо природного взаимодействия пользуется отношениями? Думается, все цело в том, что • «еявном виде дарвиновская теория выступает как теория жиз— И Ц гл
’ Рассматривая жизнь как особую форму движения материи, да как традиционная биология воссоздает картину земной
59
жизни - осуществленной на планете биологической формы двц^ ния материи. Очевидно, любое знание о жизни будет конкретно пока изучается жизнь на конкретной планете. Здесь такое же положение, как в исторической науке: с сдной стороны, суще^ в уют концепция социальной формы движения материи и учение о всемирном историческом процессе, с другой — локальные ци„ вилизации, которые обладают определенным своеобразием и ют собственную историю.
Примером использования отношений может служить матема^ тика. Ее логическая структура отличается наглядностью, сама математика в определенной мере является наукой об отношен^ ях (Бурбаки, 1963). Возьмем точку - элементарный идеальны; объект. Зададим такие отношения между точками, чтобы получу лась прямая линия - новый объект. Ее свойства раскрываются в отношениях с другими прямыми и с точками. Зададим, напри, мер, отношение пересечения. Получим единственную точку как общее место двух прямых линий, и новые объекты - углы, наделенные собственными свойствами. Зададим отношение Пересе, чения для трех линий так, чтобы получился треугольник. Этот новый объект обладает собственными свойствами, которые раскрываются в отношениях с другими треугольниками, прямыми и точками. Математика обладает целым набором отношений: равенство, конгруэнтность, больше, меньше и тщ. Теоремы обычно утверждают осуществимость тех или иных отношений объектов.
Какие отношения использует биология и что они дают? Можно выделить не меньше пяти отношений: сходство (в признаках) корреляция (строения), совместность (обитания), историческое родство (предка и потомка), сосуществование (единовремен-ность). Вполне возможно, что в действительности таких отношений больше пяти, но пока ограничимся только что названными, как имеющими наиболее широкое хождение. Далее примем, что такие отношения связывают целые популяции, а не отдельные организмы. При выполнении этого условия отношения могут сделаться столь же биологически специфичными, что и популяции -элементарные носители жизни. Теперь возьмем генеральную совокупность всех ныне существующих и когда-либо существовав-ших популяций растений и животных и зададим на такой совокупности отношение сходства. * Задать отношение сходства* -это значит упорядочить по сходству. Вообще объединения на основе сходства называются фенонами. Ими занимается фенетичео кая, или феноническая, систематика (табл. 2). Объединения фенонов по сходству в более крупные образуют в конце концов стему фенонов, обладающую иерархической структурой. Далее, на той же совокупности популяций зададим отношение исторического родства. Тогда популяции окажутся упорядоченными в элементарные родословные, образованные пре жом и потомком.
60
желтый зеленый голубой синий фиолетовый
красный
родство
сходство
корреляция совместность сосуществование
А
5
фенетическая(фенони-ческая) систематика; фенон; филогенетическая систематика;
таксон
биотика (фаунистика и флористика) , биотический (фаунистический и флористический ) элемент
историческая биология; ро -дословная
сравнительная морфофизиология; тип организации—'
5
3
24
26
25
31
18
22
экология; жизненная форма сооб -
щества
12
15
р - 17
*ГИс» 3. Разложение семантико-информационного взаимодействия.
& “ разложение солнечного света на составляющие цвета. Разложение природного семантико-информационного взаимодействия ^Фухконечная стрелка) на формально-логические отношения (F)
Упорядочение с помощью отношений и их сочетаний генераль-совокупности животных и растений (В). Г — разделы биоло» Яч йИ их °®ьекть1« Выделена филогенетическая систематика. с четным числом отношений имеют четные номера, число «ошений в ячейке передано штриховкой неодинаковой плотности
Частные разделы биологии и относящиеся к ним понятия
Таблица 2
Отношение Раздел биологии Объекты (уровни организации): элементарный, сложный, генеральный Геометрический образ
Сходство фонетическая (фено-ническая) систематика; нумерическая таксономия фенон, фенон высшего уровня, иерархическая система фенонов; операциональные таксономические единицы - ОТЕ (OTU) фенограмма, дендрограмма; неориентированный граф в многомерном пространстве
Историческое родство филогенетика таксонов, родство других объектов родословная предка и потомка, родословные предков и потомков, филогенетическое древо таксонов органического мира; родословные других объектов ориентированный граф-де— рево
Родственное сходство филогенетическая систематика подтаксон, таксон, надтаксон; иерархическая -система органического мира круги Эйлера
Корреляция сравнительная мор— фофизиология (сравнительная анатомия) тип организации, последовательность град Гексли; типы организации других биологических объектов
Совместность фаунистика и флористика (биотика) фаунистический, флористический элементы (биотические элементы), биота; мировая биота (фауна и флора) ?2
Сосуществование синэкология жизненная форма сообщества, сообщество живой покров Земли
Нумерическая таксономия выделяется из фонетической систематики только количественным методом.
Ф®унистика и флористика (биотика) становятся эоо— и фитогеографией ( биогеографией), когда осуще— - грогда^ичес^ой оболочки. Поэтому к£1р—
Объединение элементарных родословных есть древо органического мира. Задавая все на той же совокупности оставшиеся три отношения, получим жизненные формы сообщества (сосуществование). биотические элементы биоты (фауны и флоры) (совместно), типы организации сравнительной анатомии (корреляции). Значение этих операций поясняет следующая аналогия.
Изучая такое природное явление, как солнечный свет, иссле-
доваЗ?ель пропускает его через призму и устанавливает, что он состоит из семи составляющих цветов (рис. 3). По аналогии пять рассматриваемых отношений могли бы считаться составляющими природного семантико-информационного взаимодействия — борьбы за существование. В этой связи уместно вспомнить ф.Бэкона (1978, с. 109): ^Следует совершать разложение и разделение природы, конечно не огнем, но разумом, который есть как бы божественный огонь*. Логическая операция упорядочения разнообразия животных и растений отношениями приводит к классификациям; упорядочение могло бы считаться изучением, которое не вредит природе. Этот способ изучения природы традиционная биология унаследовала от старого натурализма.
После того как осуществлено "разложение и разделение* по ф.Бэкону, возникает задача, отчасти обратная уже решенной. Она состоит в том, чтобы из отдельных отношений воссоздать
некоторую целостность, систему отношений. Такая целостность отвечала бы природной целостности взаимодействия, служила бы репрезентантом системного единства биологического знания. Эту задачу можно решить с помощью диаграммы Венна — графического приема логики (Кузичев, 1968) (см. рис. 3). Пять фигур диаграммы - четыре овала и продырявленная посредине пластинка — отвечают каждая одной и той же генеральной совокупности популяций животных и растений, и каждый раз на одной и гой же совокупности задано одно из пяти отношений. При пересечении пяти фигур получается (2^ - 1) ячеек (единица -э"го окружающая плоскость). В каждой из получившихся таким образом ячеек по-прежнему подразумевается все та же генеральная совокупность, упорядоченная каким-либо сочетанием отношений из общего числа (2^ - 1). Сочетаемость отношений — это тоже своего рода отношение между отношениями. В центре по-лУченной таким образом биологической розетки имеется ячейка с пятью отношениями. В прочих ячейках неодинаковой плотно-сТью штриховки обозначено то или иное число отношений. Все-возможные постепенные переходы по числу отношений между яПейками напоминают постепенные переходы между соседними Юговыми полосами солнечного спектра. В отличие от послед-биологическая розетка не поляризована, порядок отношений
Ней не имеет ни малейшего значения. Она представляет со-систему всех возможных способов упорядочений генераль— совокупности животных и растений пятью отношениями и их
всевозможными сочетаниями и поэтому не стесняет нас в дад^ нейших рассуждениях: ненужные ячейки в дальнейшем можно дет вычеркнуть, если это понадобится.. Разнообразие ячеек мо» жно было бы перечислить в таблице. Тогда окружающая плос-, кость обозначалась бы как ООООО (нет отношений^ централь^ ячейка - как 11111 (все отношения), а оставшиеся ячейки -прочими сочетаниями нулей и единиц. Такая табличная форма записи была бы незаменимой при числе отношений больше пяти, Ее можно было бы использовать и для пяти отношений, но дна» грамма Венна достаточно наглядна и красива. По этим же при., чинам нет пока необходимости изображать систему отношений в виде 31—гранника, разверткой которого на плоскости являет» ся Диаграмма Венна.
На рис. 3 видно, что частные разделы биологии и их обьек» ты перекрываются, так же как соответствующие отношения. Эго обстоятельство не следует считать формальностью, нечаянным результатом. Оно могло бы считаться наглядным выражением взаимосвязанности отдельных разделов биологии, наглядным вы. ражением единства и системности биологического знания.
В самом деле, достаточно хорошо известен тот факт, что объект одного раздела биологии может служить объектом и в других ее разделах. Например, класс амфибии есть таксон в си-стематике, но он же есть жизненная форма хищника водных и околоводных материковых биотопов в экологии. Такой "двугранный*' объект называется "таксон как жизненная форма". Класс амфибий можно рассматривать и как фаунистический элемент тропической и умеренной фауны материков, полностью отсутствующий в высокоширотной приполярной зоне. В этом смысле было бы оправданным называть класс амфибий так: "таксон как фаунистический элемент". Дополнительно амфибий можно было бы рассматривать как тип организаций, как единство скоррелированных органов. Эго был бы объект, именуемый так: "таксон как тип организации". Условно оценим единство амфибий как таксона баллом 1, как жизненной формы - 0,9 балла (потому что помимо материковых водоемов некоторые лягушки обитают в морском биотопе мангров), как фаунистического элемента -0,6 балла (объединение тропической и умеренной зон едва ли более естественно, чем объединение умеренной зоны с высоко^ ротной), как типа организации - 0,5 балла (существуют хвоста* тые и бесхвостые амфибии, безногие червячки, и, таким образомг единство амфибий как типа организации не слишком велико). Ноте* перь можно задаться вопросом о "трехгракных"объектах (например-"таксон как тип организации жизненной формы"), о "четырехгранных" объектах ("таксон как тип организации жизненной фор* мы фаунистического элемента"). Наконец, надо обратить вним3' ние на сложность понятия таксона (см. рис. 3, габл. 2), полу* чаемого при пересечении фенона родословной ("таксон есть р0*
64
словная как фенон"). Тогда получится наиболее сложный, *пя-т11Гранный" объект: "родословная как фенон типа организации жизненной формы фаунистического элемента*. Этому объекту бу-дет отвечать центральная ячейка. Отнюдь не считая совершенным пример с амфибиями и принимая условными количественные оценки целостности сложных объектов, надо подчеркнуть два существенных в данном рассуждении обстоятельства. Во-первых, в языке биологов действительно имеются подобные понятия. Во-вторых, они подразумевают эмпирическую выборку, включающую известных науке вымерших и ныне живущих животных. Объекты, выделенные в эмпирической выборке, сама их целостность остаются незащищенными от новых открытий. Здесь, как и вообще в биологии, сохраняется место для третьего суждения, которое было рассмотрено выше на примере, допускающем "возможность существования млекопитающих, не вскармливающих детенышей молоком*. Неопределенность, возникающая в связи с третьим суждением, становится объективной только в том случае, если незащищенность эмпирических суждений вызвана потенциальной бесконечностью объектов суждения. В противном случае, когда
неопределенность ограничивается только уже осуществленными ныне живущими и вымершими организмами, она остается субъективной. В самом деле, кто, как не исследователь, "виноват* в том, что все еще остается неизученной уже осуществленная и доступная вследствие своей конечности природная совокупность организмов? В такой неподвижной природе есть основание для определенного однозначного суждения, и именно исследователь "виноват* в том, что такое суждение еще не сделалось достоянием науки.
Геометрический образ биологического знания, каким является биологическая розетка, мог бы создать неправильное представление, будто 31 объект биологии (по числу ячеек) разграничен между собой столь же четко, что и ячейки, разграниченные четкими линиями. На самом деле таких границ не может быть вследствие популяционной природы самих объектов.
Представим себе три вида одного рода. Они состоят из популяций и в предельном случае - из одной и пребывают в потенциально бесконечном длении. Пусть в какой-то момент они равнопохожи друг на друга, и это обстоятельство поясняет треугольник, у которого вершинами служат виды, а длина сторон про-п°рциональна сходству между видами-точками. Пока виды равно-похожи, треугольник остается равносторонним. Это' мгновенное, очечное наблюдение в действительности относится к выборке, к некоторому произвольно выбранному моменту наблюдения. В Рваные моменты наблюдения мы будем иметь различные треу— гольники, потому что сходство между видами не будет оставаться постоянным, треугольники будут пульсировать, для того что-ы от мгновенных точечных суждений о выборках перейти к су-
' 1560
65
жпению о длящихся видах, надо объединить отдельные суждения в одно неопределенное суждение, геометрическим образом кото> рого станет пятно с размытыми краями, а не треугольник. Пред, ставляется, что не только род, объект систематики, ведет себя так, как только что было описано, но и другие "многогранные* объекты из всех ячеек биологической розетки. Но это означало бы и отсутствие четких границ между отдельными ячейками.
Возвращаясь к примеру с родом из трех видов, надо заметить, что в отдельные моменты наблюдения некоторый вид может сделаться столь непохожим на два прочих (это означало бы сильную неравное героин ость треугольника), что его можно было бы вынести в самостоятельный род, противопоставить дау^ другим видам. В свою очередь это означало бы не только неопределенность границ рода, но и возможность перехода вида из одного рода в другой. Больше того, это означало бы перемещение вида из одной ячейки в другую. Тогда за достаточно длительное время все ячейки розетки были бы заполнены одинаково плотно. Более подходящим образом существования вида в потенциально бесконечном длении было бы случайное блуждание точки, и тогда 31 ячейка могла бы считаться множеством тех ипостасей одного вида, которые он принимает при сопоставлении с другими видами. Ясно, что такое бытие вида невозможно характеризовать числами, как это было проделано выше в примерах с амфибиями. Задача характеристики видов и их объединений вообще не могла бы решаться метрически с помощью чисел. Для правильного восприятия биологической розетки полезно вспомнить рисунки голландского художника Эшера, изобразившего постепенное превращение одних животных в другие. На биологической розетке потребовалось бы превратить изображение из центральной ячейки в 5 изображений в краевых ячейках таким образом, чтобы между любыми соседними изображениями сохранялись постепенные переходы. Каждый такой образ отвечал бы одной из 31 классификаций. Своей пластичностью такие "размытые* образы вызывают в сознании картины поздних импрессионистов. Таким образом, неопределенность в биологической розег ке подразумевает: 1) отсутствие определенных границ между объектами одного рода в каждой ячейке и, в частности, отсутствие однозначных границ между разными ступенями иерархической системы: 2) отсутствие определенных границ между объектами в разных ячейках. Биологическая розетка - это множество всевозможных точек зрения на живую природу. Когда, напри* мер, исследователя интересует отношение сосуществования, это не означает, что в выделенном этим отношением множестве популяций нет иных отношений. Эго означает лишь, что данного исследователя интересует пока только сосуществование. Здесь уместна аналогия с садом камней в Японии. Для того чтобы увидеть все камни, наблюдатель должен обойти площадку кругов
66
f/гоМУ чТО 113 одной точки все камни сразу не видны. В свою г‘ \₽дь между отношениями также имеют место отношения. На-очер^
имер, почему животные сходны? Потому что они родственны
(или) обитают совместно. Здесь причиной сходства служит от— ^ошение родства и (или) совместности. Теперь зададимся воп—
осо.м: почему животные обитают совместно? Последует ответ: ^стомУ что они сходны. В этом случае сходство выступает уже как следствие совместности, и таким образом причинно-следственны6 отношения между отношениями могут иногда быть обратимыми. Кажется, из всех пяти отношений одно только родство
относительно всех прочих выступает в качестве причины, а не следствия. Надо заметить, однако, что когда филогенетики делают заключения о родстве (причина), основываясь на сходстве (следствие), они поступают прямо противоположным образом.
Совсем нерешенным остается вопрос о числе отношений. В
наших построениях использованы только те отношения, которые удалось выделить из письменных или устных сообщений биологов. Если бы удалось выявить или открыть новые отношения, то это означало бы выявление или открытие новых разделов биологии и новых объектов, потому что, как заметил А.И.Уемов (1961), возникновение новых наук связано с нахождением общих свойств и отношений путем абстракции. В новой, расширенной системе уже имеющиеся отношения и их сочетания сохранились бы, хотя в целом вся система усложнилась бы. Отыскивая новые отношения, надо опираться на содержательные биологические соображения. Например, отношение скрещиваемости, по-видимому, нецелесообразно вводить в биологическую розетку уже потому, что оно имеется только у разнополых форм и потому не имеет общебиологического значения. Скрещиваемость - частный случай отношения совместного сосуществования.
Сложная картина современной биологии не может рассматриваться всего лишь как отражение объективно существующей живой природы. Эта картина сложилась под влиянием таких факторов, как традиции, конъюнктура и преходящие интересы практики, особенности методов и др. Поэтому основанная на отношениях» воспроизводящих только природное взаимодействие популяций, биологическая розетка не может с точностью воспроизвести структуру биологии.
На биологической розетке нет, например, иммунологии, кари-°Лог'ии, радиобиологии и других дисциплин, выделенных по методам исследования. По этой же причине отсутствуют морфология, биохимия, биофизика, биоэнергетика. Тем не менее кариологичес-Кие» иммунологические, морфологические, биохимические признаки используются для установления сходства, родства, корреля-ЦИЙ и поэтому неявно представлены в соответствующих фигурах ^^з&гки. далее, в неявном виде на розетке не представлены фи-аИолОГия> генегика> эмбриология. В действительности же физио
67
логия лежит в основе корреляций; сходство в функциях — эго обстоятельство, учитываемое в систематике, филогенетике. По^ тому неявно физиология все же представлена в некоторых час., тях розетки. В генетике пользуются отношением сходства (на> пример, при выделении фенотипов) и отношением родства. Вместе с гем генетика - это пример раздела биологии, выделивши ся в ответ на запросы практики. Сравнительная эмбриология при сравнительном изучении зародышей, личинок пользуется сгг< ношением 1сходства и корреляции, полученные ею данные имек^ важное значение для сравнительной анатомии, филогенетики, сйч стематики. Поэтому этот раздел эмбриологии фактически тоже представлен на розетке. Вообще, если некоторый раздел биоло. гии пользуется рассматриваемыми отношениями, для него найдется место на биологической розетке. От аналогичных построе. ний других авторов (Одум, 1975; Рюиз, 1977) она отличается тем, что основана на отношениях, и разделы биологии выделяют, ся постериорно.
Наконец, на биологической розетке нет таких разделов биологии, как альгология, ботаника, зоология, ихтиология, микробиология.
Каждый из названных разделов биологии пользуется теми же отношениями, что и вся биология в целом. Вследствие этого структуры знания отдельных частей и всей биологии в целом, изоморфны. Действительно, как и в ботанике, в зоологии есть экология (животных), систематика (животных). Фитогеографии и флористике отвечают зоогеография и фаунистика. Примерно гак же обстоит дело в альгологии, энтомологии и тл.
СВ этой связи следует напомнить о взглядах Л.А.Зенкевича (1966) на структуру университетской биологии. По Л.А.Зенкевичу, существовавшая в бО-ые годы структура биологического факультета Московского университета основана на "горизонта.^: ном* членении биологического знания с подразделением на мор* фологию, которой вынуждены ограничиваться кафедры зоологий и ботаники, физиологию , биохимию, биофизику. Таким образом выделяются и соответствующие кафедры. 'Вертикальное' членение позволило бы лучше выделить такие кафедры и дисциплина как зоология, ботаника, микробиология. Одновременно это были и системы знаний - от морфологии до биофизики, повторяющие0 в каждом подразделении 'вертикального* членения. Вследствие 'вертикальности* членения на каждой из таких кафедр сохрани лась бы собственно биологическая тематика и осталась бы не* изменной и изоморфной биологическому знанию в целом темат^ ка каждой кафедры. Фетишизация методов исследования, выде^ ние кафедр по методам изучения организменного субстрата, вели к надуманному противопоставлению по 'горизонтали' та*1 называемых 'экспериментальных' наук наукам 'описательны^1 к искусственному разобщению единого познавательного пропев
68
0 результате к настоящему времени на биологическом факуль-геТе усилилась разобщенность кафедр, сохранивших изоморфную сей биологии структуру знания (зоология, ботаника) - кафед-выделенным особенностями физико-химического метода
^учения субстрата (биоорганическая химия, биофизика). К на— сТОящему времени организационная структура биологического Факультета не столько отражает специфичность жизни как особой формы движения материи, сколько подаинена решению хотя л практически важных, но гем не менее преходящих задач биотехнологии, успешно решаемых отраслевыми и академическими институтами.]
В заключение надо пояснить, каким образом биологическая розетка могла бы выполнять свою роль образа неизменной струк* ’ турой биологического знания уже в первый момент возникновения жизни, датируемый началом борьбы за существование. Мож
но представить, что в этот момент на планете существовала суперпопуляция протобионтов и тогда совпадали бы два понятия: существование суперпопуляции и существование жизни; единственным носителем жизни была бы суперпопуляция протобионтов. Отличаясь друг от друга в той степени, в которой внутривидовая изменчивость остается материалом для естественного отбора, будучи подразделенной на тропические, субтропические, не
ритические, пелагические и др. популяции, такие популяции протобионтов могли бы изначала состоять в межпопуляционном родстве. Вследствие неопределенной изменчивости они могли бы отвечать разным фенонам и типам организации, выполнять в первичных сообществах несколько различные функции. На таком множестве можно было бы воссоздать биологическое знание, изоморфное знанию о современной высокоразвитой жизни. За геологическое время естественный отбор сделал первичную картину более зрелой и четкой. По сравнению с первичной жизнью современная высокоразвитая жизнь - это как бы "сверхжизнь*, потому что каждый раз возникновение нового вада означало бы возникновение новой жизни. Это усложнение жизни сопровождалось Усложнением организменного субстрата — "эволюцией организмов"; появляются все новые структуры (клетка, хромосомы, Ультраструктуры, органы, ткани, системы органов), новые более сложные химические соединения и процессы, в которых они участвуют. Биологов уже давно занимают причины того удивительного явления, что эволюция сопровождается именно усложнением, прогрессом организменного субстрата. Не исключено, что. Эт° смогла бы объяснить особая, нефизическая природа семантико-информационного взаимодействия.
Глава t>
ТРАДИЦИОННАЯ ТАКСОНОМИЧЕСКАЯ НЕОПРЕДЕЛЕННОСТЬ
Филогенетическая систематика, старейший и высокоразвитый раздел биологии, характеризуется неоднозначностью суждений, непостоянством выводов — тем, что в дальнейшем называется неопределенностью. Издавна неопределенность остается мишещ,; для критики в адрес традиционной систематики, поводом для в мнений в произволе и субъективизме. Чтобы повысить научное, систематики, разрабатываются объективные математические топы классификации (нумерическая таксономия), предлагаются особо надежные признаки (геносистематика). При этом у пуске, ется из виду, что проблема состоит не в неопределенности се. мой по себе, а в том, естественная ли она. Неопределенность могла бы пониматься как следствие способности видов осущес, влять эволюцию, сввдетельство "открытости*, "незавершенности видов. Какой должна была бы стать система эволюционирующих живых видов? Как бы живой, развивающейся. Действительно, си стема всегда в движении (Козо-Полянский, 1950), и хотя в ней что-то остается неизменным, она остается незащищенной перед возможным открытием новых фактов. Рассмотрим, однако неопределенность по существу. Она складывается не менее чем из пяти взаимосвязанных составляющих элементов.
(I). Непостоянное таксономическое значение, относительность одного и того же признака состою в том, что один и тот же признак позволяет выделить таксоны разных рангов (Комаров, 1940). Это касается биохимических признаков растений (Федоров, Пименов, 1967), наблюдается в кариологии ("Морфология хромосом, как и всякий другой систематический признак, является признаком относительным* - Розанова, 1916), в микро- и макроморфологии. И.И.Соколов (1965, с. 40) заключил, что ни один взятый отдельно признак не может служить показателем ранга обладающей им группы. Обширными и достоверными материалами по этому вопросу рао полагают животноводство и растениеводство. Те признаки, кото рыми различаются породы домашних животных и сорта культу? ных растений (внутривидовые единицы), пригодны и для разлив ния видов. При этом выясняется любопытная деталь. Различия между внутривидовыми единицами (породы, сорта) зачастую пр восходят различия между природными видами. В этом убеждав сравнение, например, таксы с догом - с одной стороны, и ша" кала с лисицей - с другой. Заметим, что в данном примере пс* признаком, имеющим неодинаковое значение, - внутривидовым (межпородным) и межвидовым - подразумевается размах изМ^ чивости экстерьера.
Нетрудно убедиться и в том, что на любом уровне иерархии
аэличия между соподаиненными видами не остаются неизменной величиной. Каждому систематику известно, что внутри не-кОторог° надвидового таксона имеются четко различимые ("хорошие*') и трудно различимые ("плохие") соподчиненные таксоны. Многочисленные факты подобного рода были хорошо известно Ч.Дарвину (1939). Еще раньше обсуждаемое явление было известно К.Линнею (1804). В диалоге с Гизике (Уэвелл,
1914) он высказал мысль, поднятую именно в данном контексте: не признаки создают род, а, наоборот, род создает признаки. А.Л.Тахгатян (1947) объяснял обсуждаемое явление не-
равномерной эволюцией признаков, как поднимающихся по иерархической лестнице. Более общее объяснение принадлежит Ч.Дарвину. Оно состоит в том, что признаки видов и более высоких
категорий закладываются внутри вида — предка надвидового таксона - сначала как различительные особенности организмов. Их можно было бы называть прообразами таксономических признаков (Заренков, 1976). При этом "...размер различий в разных
ветвях или группах, находящихся на одаой ступени кровного родства с общим предком,- может колебаться весьма значительно,
гак как он зависит от разных степеней изменений, пройденных этими группами" (Дарвин, 1939, с. 612). В этой цитате "размер различий" мог бы пониматься как признак в том значении, которое было придано этому выражению в примере с таксой и Догом, лисицей и шакалом; Несмотря на наличие объяснения, пе
рекрывание межвидовых серологических различий внутривидовы-
ми различиями считается "парадоксом" (Hunziker, 1969). Пра
вомочность традиционного для систематики непостоянного значения одного и того же признака оспаривают геносистематики (Антонов, 1976, с. 116). Ими становится задача "количественных критериев эквивалентности" для таксонов одного ранга. Эта задача якобы решена в нумерической таксономии: так называемые операциональные таксономические единицы (ОТЕ, OTU ) выделяются определенным значением сходства, выраженным в процентах, отдельно для каждого уровня системы этих единиц. Наконец, обсуждаемое явление вызывало недоумение у В. У эвелла (1914, с. 418), потому что оно нарушает принцип существенного признака в формальной логике: для выделения видов нужны °ДДи признаки, для родов - другие, для семейств - третьи и Т-Д. Итак, с одной стороны - формальная логика, нумерическая таксономия и геносистематика, с другой,- биологическая содержательность. Между ними существует принципиальное расхождение.
(II). В заимная неадекватность (несопоставимость, неэкв иваленгнос гь) диагнозов одного ран-г а состоит в том, что даже близкие одноранговые таксоны основаны на разных признаках и характеризуются неодинаковым
размахом изменчивости одного и того же признака (в послед-нем случае размах сам является признаком). Неоднократно отмечалась неодинаковая устойчивость одного и того же признак^ в разных таксонах одного ранга ( Crowson, 1970), несопосгавц. мость Диагнозов, основанных на разных признаках ( Banarescu , 1973). Например, класс насекомых характеризуется наличием трахей, а в классе паукообразных трахеи либо имеются, либо отсутствуют. Если у насекомых имеется признак "всегда есть трахеи", то у паукообразных — "иногда есть трахеи". В четырех царствах органического мира (Тахтаджян, 1974), пожалуй, не найдется ни одного признака, о котором сообщалось бы с определенностью, что он всегда имеется или всегда отсутствует. Например, гетеротрофность всегда есть у животных, но у бактерий она иногда имеется и иногда отсутствует. Чтобы четче выделить такой таксон, виды которого только иногда имеют дан. ный признак, систематики вынуждены вводить в диагноз, например для паукообразных или бактерий, новые относительно устойчивые признаки^. Отсюда и вытекает взаимная неадекватность диагнозов. Можно видеть, что это явление имеет своей основой непостоянное значение одного и того же признака (1). Действ», гельно, если опин и тог же признак не остается неизменным и позволяет выделять разные ранги (например, виды и роды), то для описания разных видов (родов) понадобятся различные признаки (рис. 4). Этим объяснением мы, в сущности, обязаны Ч.Дарвину. Оно содержится в его представлениях об единой форме эволюции, о происхождении признаков высших таксонов из внутривидовых различительных признаков организмов. К.Линней (1804) безуспешно стремился к тому, чтобы диагнозы таксонов одного ранга были взаимоадекватными, основывались бы на одних и тех же признаках. В сущности, это была попытка создать иерархию признаков, параллельную иерархии таксономических рангов. И все же среди 24 линневских классов растений нашлись такие, которые основывались не "на числе, соразмерности и соединении тычинок цветка", а на иных признаках. Вследствие этого взаимная неадекватность диагнозов классов осталась не достигнутой (Станков, 1958). То же наблюдалось в построениях других систематиков, современников К.Линнея (Кир* пичников, 1968). Обсуждаемое явление представляет собой нарушение аристотелевского принципа деления по противоположностям. Например, у птиц "перья бывают расщепленные и нерас-щепленные" (Аристотель, 1937, с. 43). Требование "эквива-
Надо обратить внимание на ставшее традицией неодинаковое понимание признака. С одной стороны, самостоятельными признаками будут: 1) "всегда есть трахеи", 2) "иногда есть гра' хеи". С другой стороны, под признаками можно понимать "гра" хеи?# и тогда мы получим два суждения о признаке: 1)"всегда есть , 2) "иногда есть*.
Юности* - это, в сущности, требование взаимной адекватно-.й Геносистематики полагают, что, присваивая некоторому так-ранг (рода, например), мы "молчаливо* предполагаем *не-вП°лне определенный уровень фенотипического и генотипи-еского подобия членов этого таксона* (Антонов, 1976, с.116). 1 ^логичное требование исходит и от нумерических таксономис-Sokol, Sneath, 1963).
Таким образом, в данном вопросе также наблюдается расхож» рение между традиционной систематикой, с одной стороны, нуме-
семейства роды
виды
подвиды
семейства
роды
виды
подвиды
5
Рис. 4. Связь между явлением неадекватности диагнозов и не-° Знаковым значением таксономического признака.
^©Дствие того что в эволюции признаки как бы поднимаются п ступеням иерархической лестницы, может возникнуть такое Врд°Жение» когда по одинаковым признакам различаются два под— дк, 1 и два вида А, 2. В таком случае, чтобы отличать, ви—
1 от видов А, 2, а также подвиды А, 1 от подвидов А, 2, понадобятся новые признаки - Б, 3 и Б, 4
73
рической таксономией, геносистематикой и классической логи-кой - с другой стороны.
(III). Неопределенность числа рангов (ступеней) таксономической иерархии состоит в том, что систем^ таксонов даже одного ранга, например классов насекомых и кообразных, содержит неодинаковое число ступеней иерархии. системе обширного класса насекомых к таким обычным рангам как отрад, семейство и род добавляются серии, порядки, секции и пр. Прямая зависимость между числом видов и степень^ сложности иерархии очевидна: чем больше видов, тем больше требуется степеней иерархии, чтобы отобразить широкий диапам зон отдаленности родотва между видами. В этой связи представ ляется справедливым утверждение Е.С.Смирнова (1967) о том, что система тем совершеннее, чем больше выделено рангов. Вместе с тем очевидна и зависимость меж /у степенью изучен-носги таксона и сложностью системы: по мере изучения система усложняется. Тем не менее совсем не редкость, когда авто, ры—современники, основываясь на одном и том же материале, выделяют в одном и том же таксоне неодинаковое число ступеней иерархии. Каких-либо правил установления числа рангов не существует (Джеффри, 1980); связанный с этой проблемой "...вопрос о ранге групп — один из основных и трудных вопросов* (Скарлато, Старобогатов, 1974). Фиксирование числа раь гов, к чему призывал Л.Агассиц, было раскритиковано Э.Гекке-лем (1900). По Э.Геккелю, фиксирование числа рангов проистекает из идей типологической школы и объясняется стремлением отобразить в системе последовательность творческих актов, во площение идей в строении животных. Было якобы шесть таких актов.
Наиболее общая идея воплощена в типе (Pembranchement -Кювье), более частная - в классе, еще более частная - в поря; ке и Т.Д., вплоть до вида. В настоящее время, основываясь на представлениях об исключительности признаков ДНК, геносисте-матик Б.М.Медников (1974) признает реальное существование у позвоночных животных только четырех рангов. Однако если для старых типологов гарантом столь строгого порядка был тв<>' рец, то в современную эпоху следовало бы указать иную причл-ну порядка. Может быть, .ею являются физико-химические свой' ства ДНК? В противном случае, обращаясь к собственно биологическим доводам, положение о фиксировании числа рангов защитить невозможно. Действительно, пока мы признаем неогран^ ченность исторического развития, тем самым мы допускаем во3 можность сколь угодно отдаленного исторического родства. Сйс' тему в целом и систему отдельных таксонов в частности сл^д0, вело бы наделить свойством неопределенности числа рангов для того, чтобы отобразить необычность популяционных объектов классифицирования. Такое свойство предотавляет собой ие1?
ность в связи со следующими обстоятельствами. Известно, что цикт° не наблюдал непосредственно происхождение одних гаксо-нов других. Исключением служит происхождение культурных
Рис. 5. Классифицирование по сходству на заведомо известных п родословных.
Поскольку между уровнем родства и степенью сходства нет однозначной связи, на одной и той же родословной видов можно выделять разные роды. Обозначения: 1 - границы семейства; < — границы рода (в том числе монотипического, состоящего из одного вида); 3 — следование во времени, 4 — отношение сестринского родства
Форм животных и растений, породообразование и сортообразование — своего рода внутривидовая филогения на самом низшем Уровне. Тем большую ценность представляют наблюдения животноводов и растениеводов и обобщение этих наблюдений, принад
75
лежащее Ч.Дарвину (1939, с. 612). Оно сводится к следую^ му: между уровнем "кровного родства" и величиной межвидов^ различий нет однозначной связи. Отсюда следует, что количе^ во рангов, выделяемых, например, в пределах класса, не можеу быть однозначным, пока таксоны остаются таксонами (см. тац 1), не родословными.
Эго заключение поясняет рис. 5. Пусть в природе осущест-вляется некоторая непосредственно не наблюдаемая родословная видов. В ней по гомологическому сходству (наблюдаемому) вы. делено три вида (более точно, это всего лишь выборки), обра^ зующие один род (рис. 5,1). Хотя уровень родства между общим предком и каждым из его потомков одинаков, сходство ме, жду ними может оказаться неодинаковым. Тогда в пределах р&. да можно выделить подрод с двумя видами и подрод с одним видом (рис. 5,2). В гаком случае получим иерархию вид - под. род — род. На той же родословной сходство межде видами может оказаться одинаковым, и тогда иерархия будет более простой; вид - род (см. рис. 5,2). На практике систематики неодинаково оценивают различия даже между одними и теми же ви. дами, и поэтому в литературе сосуществуют разные системы одних и тех же таксонов. Обычно считается, что эго плохо. Од. нако посмотрим теперь, что произойдет с системой, если от выборок мысленно перейти к неограниченно длящимся видам. Тогда вследствие неопределенной изменчивости сходство между видами в разных временных точках наблюдения не оставалось бы неизменным. Разноречивые суждения систематиков по поводу системы таксонов как бы воспроизводят в логических формах природную неопределенность. Поэтому обсуждаемое явление в действительности представляет собой достижение традиционной систематики и не может считаться ее недостатком.
(ТУ). Непостоянство структуры таксона выражается в том, что, например, в пределах одного рода разные авторы выделяют неодинаковое число подродов и по-разному распределяют виды между подродами. Вследствие этого виды не имеют определенного положения* в системе и у разных авторов отнесены к разным подродам. Эго так называемые виды неопределенного систематического положения. На рис. 5,1 показан только один вариант разбиения рода на подроды, но их могло бы быть не меньше грех; причем каждый вариант - эго один подрод с двумя видами и один подрод с единственным видом. Этот пример поясняет, что такое непостоянство структуры таксона как явление, наблюдаемое на любых уровнях таксономической иерар* хии. В основе этого явления лежит установленный Ч.Дарвином факт неоднозначной связи между степенью сходства и уровнем "кровного родства". Именно неоднозначность связи между эги>1,! отношениями делает возможным выделение на непрерывной род0* словной дискретных таксонов. Особенно четко это видно на рйс‘
3 4, где степень родства уменьшается плавно в направле-от предка. На такой родословной только потому и можно вы-елять дискретные таксоны, что в одинаковом с родством направлении, хотя и неравномерно, изменяется сходство.
Р дополнительно это явление можно рассматривать и с несколько иной точки зрения. Дело в том, что не только признаки поц-Н|Олаюгся по ступеням иерархии (см. рис. 5), вместе с ними со ступеньки на ступеньку поднимается и само явление изменчивости. И если в пределах вида существует неопределенная изменчивость в межорганизменных признаках—прообразах, то на более высоких ступенях иерархии она перейдет в неопределенную изменчивость таксономических признаков. Поэтому межвидовые (вообще межтаксонные) различия в принципе не могут оставаться постоянными. Таким образом, виды монофилетического таксона обладают межвидовой изменчивостью, унаследованной от вида-предка (Заренков, 1972). Сходные построения развивает Г.Х.Шапошников (1974). Неопределенность, выявляемая систематиками на выборах, воспроизводит ту неопределенность, которая наблюдалась бы на множестве длящихся видов. Следовательно, обсуждаемое явление непостоянства структуры таксонов представляется одним из наиболее важных открытий традиционной систематики и не может служить поводом тля критики в ее
адрес.
(У). Политетичность таксономических диагнозов (Beckner , 1959; Мауг, 1978) состоит в том, что в них соо&-шение о наличии (отсутствии) признака часто сопровождается уточнениями вроде "обычно", "иногда*. Вследствие этого однозначное установление принадлежности вида к роду, например, часто оказывается невозможным. В этой связи В.Г.Гептнер (1956) пишет о "субъективной трактовке* таксонов. Как уже фактически было отмечено, политетичность надо рассматривать в связи с неодинаковым значением одного и того же признака (I) и взаимной неадекватностью диагнозов одного ранга (И). Действительно, выражения "обычно*, "иногда" могут считаться качественными оценками размаха изменчивости признака, когда сам размах изменчивости используется как признак. Когда признак "всегда имеется", величина размаха изменчивости равна нулю. Вместе с тем политетичность — это своего рода аналог внутривидовой изменчивости, однако уже на таксономическом Уровне. Поэтому политетичность можно рассматривать в связи с 1У. Политетичность противоречит понятию класса в формальной логике: отдельный предмет, относимый к данному классу, Должен обладать признаком (признаками) класса (Минто, 1909). аРушение этого требования, как обычно полагают, компрометирует традиционную систематику. Тем не менее очевидна ее биопсическая содержательность, возможность истолкования в терминах дарвиновской теории видообразования.
Особенности использования признаков в традиционной систь, матике, взаимосвязь между отдельными проявлениями неопред^ ленности, выступавшая вполне отчетливо уже в процессе их смотрения, можно дополнительно пояснить на следующем приме, ре:
класс Aves : "всегда имеется яйцерождение", класс Mammalia : "обычно живорождение, иногда яйцерожде-ние*.
В этом примере содержится явление взаимной неадекватности таксонов одного ранга (II), потому что названия признаков, за, ключенные в кавычки, несопоставимы. Диагноз класса MammaljQ политетичен (1У). В подобных случаях некоторые систематики пытаются преодолеть политетичность и признак биологии размнь жения выводят из диагноза класса. При этом получают, например, подклассы. У млекопитающих это будут подклассы Proto-theria (яйцерождение") и Eutheria ("живорождение"). Но тогда возрастет неадекватность диагнозов классов: диагноз класса птиц сохраняет признак биологии размножения, а диагноз класса млекопитающих останется без этого признака. При этом получится к тому же, что признак "яйцерождение" позволяет выделять класс (Aves) и подкласс (Prototheria ), т.е. проявляется неодинаковое значение одного и того же признака (I). Иерар. хическая структура класса млекопитающих утратила бы неопределенность, поскольку не все авторы выделяют подклассы. Разумеется, дело не в самих млекопитающих и птицах (конкретность примеров не имеет значения).
Ранее было отмечено, что нормы биологического классифицирования противоречат правилам формальной логики и обычным правилам классифицирования (Пиаже, Инелъдер, 1976; Свиридов, 1976; Мейен, 1977; Навроцкий, 1981). Указывается, что логика биологической классификации есть особый случай логики классов ( Buck, НиП, 1966). К тому же примем во внимание давление на традиционную систематику со стороны естествоиспытателей, которые считают классическую логику единственной основой классифицирования вообще. Кто же прав? Думается, что права традиционная систематика. Именно натуралистическое классифицирование как способ видения живой природы, без привлечения стереотипов со стороны, обеспечило систематике объективность. Непредвзятый объективный подход к изучению живой природы, свойственный систематике как своеобразному общественному явлению, сТал возможным при условии субъективности отдельных натуралистов. Биологическая содержательность выводов традиционной систематики состоит, в частности, в том, что она не только филогенетическая, но и эволюционная в том смысле, что оказывается связанной с теорией эволюции. Что же касается логики, полезно обратить внимание на высказывания самих логиков о возможностях своей науки.
хТо, что классическая логика не является единственно воз— ной теорией дедукции, на сегодняшний день хорошо извест-м<^(Бочеров, 1984, с. 127). "В зависимости от изменения Н°^р2кания рассматриваемого предмета изменяется и способ С°о рассмотрения - меняются логические средства" (Марков, еГдб4, с. 340). "Применение законов формальной логики возможно лишь в предположении, что предметы, о которых мы рас-\жДЛ®м» в определенных отношениях не изменяются..." (Гор-
1961. с. 27).
Условие, которое оговаривается- в последней цитате, виды явно не выполняют: оставаясь самими собой в потенциально неограниченном длении, они изменяются, и отношение сходства меЖУ ними ПРИ этом изменяется гоже. Таким образом, формула (1) чрезвычайно ответственна. Особенности жизни как осо
бой формы движения материи таковы, что ее изучение предполагает особую логику. Поэтому следует отвергнуть представление о том, будто биологическое классифицирование аналогично классифицированию неживых объектов, как эго полагают некоторые авторы (Starr, Heise, 1969; Андреев, 1980), и что образцом для биологии может служить таблица химических элементов Д.И.Менделеева. Не допуская мысли о природном характере таксономической неопределенности, критики традиционной система-
тики принимают в качестве образца, достойного подражания, обобщения классической физики и подразумевают при этом де
картовский принцип ясности, о котором пишет, например, Б.Г.Кузнецов (1972). В сущности, в этом выражается неверие в биологию как самостоятельную область науки. Неудачные по
пытки преодолеть таксономическую неопределенность тем не менее весьма поучительны.
Еще Ч.Дарвин (1939, с. 606) подметил факт: признаки, очень важные для жизнедеятельности, обычно не представляют существенного значения для систематики. Впоследствии эго заключение было неоднократно подтверждено (Зенкевич, 1939; Ушаков, 1959; Bremekamp, 1962). Вслед за А.Кэйном (Cain, 1959) Э.Майр (1971) указал, что такое (в зависимости от значения для жизнедеятельности организма) априорное взвешивание признаков есть наследие типологической школы. В связи с этим надо вспомнить о методе К.Линнея и Ж.Кювье. Первый апРиорно принял главенствующее значение цветка для выделения классов растений, второй - важнейшей для систематики животных считал нервную систему, потому что без нее невозможно Функционирование животного организма. Этот метод был принят и -Б. Ламарком (1955, с. 220). Названные авторы разработали иерархию признаков, параллельную иерархии таксонов. В Наще Время этог принцип организмоцентризма пытался отстаи-Ва7Ъ, например, В.И.Кудрявцев (1954).
Обсуждая метод К.Линнея и Ж.Кювье, В.Уэвелл (1914) при
шел к выводу о том, что в зоологической системе априорный подход (Уэфелл назвал его "физиологическим'1’) проявил себя с большей ясностью, чем в ботанической системе. Как методе^ бэконианского естествознания, В. У эвелл одобрял априорный энологический подход за его связь с физиологией, куда боле^ близкой к физике, нежели морфология.
В систематике проведение априорного принципа невозможно Трудность не только в том, что функциональное значение пр^ знаков часто неизвестно. И даже не в том, что в разных усла виях обитания у одного и того же организма разные признаки имеют неодинаковое значение для жизнедеятельности. Главная трудность в том, что в отличие от типологов современные сис. тематики привлекают признаки, свидетельствующие о родстве, и чем надежнее признаки об этом свидетельствуют, тем боль, шее значение они имеют для систематики. Совсем не очевидно, что такие признаки к тому же имеют важное значение для жиа. недеягельности. В этой связи уместно вспомнить о рудимента^, ных структурах, иногда сохраняющих важное значение для сио тематики.
В традиционной систематике признаки хотя и существуют независимо от наблюдателя в природе, тем не менее вовлекают, ся в систематику только в операции сравнения: какие виды, та кие и признаки. Если бы из всех существ были известны толь ко млекопитающие, то их признаки, например способность вскар мливать детенышей молоком, были бы признаками органического мира. Поэтому признаки в систематике - не только природное, но и логическое понятие. Вопрос о том, как соотносятся между собой в понятии признака логическое и онтологические (Магомедмирзаев, 1976), очень интересен. Этот вопрос выводит обсуждение на проблему, которая называется так: "систему открывают или строят?". По—видимому, однозначного ответа на этот вопрос давать не следует: "строят" — потому что при построении системы натуралисты пользуются логикой, которой нет в природе, "открывают" - потому что система отражает природный порядок видов. Во всяком случае, основа для априоризма сужается еще больше, если мы учтем, что в своей работе систематик зависит от того, какие виды ему известны.
Априоризм в систематике иногда принимает неожиданные формы. В так называемой "биологической концепции* вица, р^ витой Э.Майром, априорно принято, что внутривидовая скрещиваемость и репродуктивная изоляция - наиболее надежные признаки таксона видового ранга. Справедливости ради надо отм6' тить, что сами сторонники этой концепции не считают этот знак вполне надежным и скорее принимают его как "необходимый" (Креславский, 1973), тем не менее априоризм здесь и5' лицо. Учитывая большое количество агамных форм, эту конНеГ1/ цию не следовало бы считать обще биологической. Понятие
Поскольку признаки ДНК первичны, ими следу-первую очередь. Основание для наиболее обьек—
Орряжено с понятием видообразования, а видообразование не лолагает с необходимостью именно половое размножение.
Необходимо лишь, чтобы имелось какое-то размножение вообще, пусгь даже бесполое. Принятие "биологической* концепции вида, сущности, приводит к отрицанию общебиологической природы понятия вида. Кроме того, нельзя упускать из виду гиногению 0 гибридаое видообразование у животных и растений ( Dubois , Gunther, 1982; Васильев и др., 1983), что составляет для об-срКдаем°й концепции Дополнительные трудности. Естественно, чго некоторые зоологи вполне определенно не приемлют "биоло-гцческую" концепцию вида (Мошковский, 1969; Полянский, 1982).
другая форма априоризма — это молекулярная таксономия и, в частности, систематика генотипов (геносистематика), получившая развитие в труцах А.Н.Белозерского (1970). Она исходит из того, что признаки ДНК определяют все прочие фенотипические признаки. ~ ег заниматься в гивных суждений дает молекула ДНК, в которой, как указывают многие авторы (Frelin, Vullenmeir, 197 9), содержатся сведения О происхождении вида.
Происхождение молекулярной таксономии от типологии вполне очевидно ( Rasmussen, 1969). Прежде всего эго следует из того, что она исходит из представлений об исключительном значении химических веществ и прежде всего ДНК в жизнецеятель-ности организма. "Наиболее .объективное суждение", возможность которого при этом допускается, если следовать нормальному словоупотреблению, есть в то же время и единственное суждение. Следовательно, допускается возможность однозначности -в пределах точности метода, т.е. предела, "...дальше которого решение филогенетической проблемы не может быть продвинуто на основании данных о рецентных видах* (Гершензон, 1974, с« 87). Иными словами, принимается, что систематику можно избавить от неопределенности и таким образом лишить ее биологической содержательности, прервать ее связи с теорией эволюции. Однако пока нет оснований обсуждать далеко идущие по-следствия из этих предположений. В действительности суждения РеНосистематиков относятся к выборкам, а не к видам; эти су— ^Дения принципиально незащигимы от новых открытий, пока ос— таюгся в силе положения о неопределенной наследственной иэ-Менчивости и о стремлении каждого вида к неограниченной чис-^лности. Поэтому рассуждение по индукции, которое фактичео-имеется в виду геносистематиками, в данном случае непри— Нимо: пусть у п организмов сохраняются неизменными &°йства ДНК и это есть мономорфный признак вида, однако о (п +• 1) - м организме на основании имеющегося опыта
Gr° заключить нельзя (Малиновский, Мина, 1976). Обосно
вывая исключительность признаков ДНК, геносистематики обр^ щаются к миру больших чисел, указывая на очень большое со, четание нуклеотидов в этой замечательной молекуле, основе для "наиболее объективных суждений". Между тем известно, что отень большое число и бесконечность - в принципе разные понятия: "...эти две области по своей природе не имеют между собой ничего общего" (Галилей, 1964, с. 145). При изучении очень больших чисел возникает только субъективная неопределенность; любое даже очень большое число есть неподвижность тогда как систематике нужны "живые", развивающиеся сужде-ния.
Вообще представители молекулярной биологии, по-видимому, представляют себе биологические виды как что-то завершенное, неподвижное, неживое. Отсюда и своеобразная оценка тех неу-дач, которыми сопровождаются поиски однозначных определенных суждений и мономорфных неизменных признаков. Литература о биохимических признаках весьма обширна, насчитывает сотни названий ( Vegotsky, Fox, 1962; Leone et al. , 1965; Благовещенский, 1966; Alston, 1967; Janine, 1972; и др,),
Известна широкая изменчивость и на хромосомном уровне (Орлов, 1970; White, 1973; Sharma, 1974). Поскольку хромосомы содержат ДНК, это уже есть свидетельство ее внутривидовой изменчивости. О хромосомном полиморфизме сообщают едва ли не в каждом своем номере такие журналы, как "Генетика" и "Chromosoma". По-вед имому, случаи внутривидовой изменчивости представляются биохимикам временными досадными неу-дачами в поисках неизменных мономорфных признаков видов. Чтобы уменьшить изменчивость, предлагается изучать только сравнимые по физиологическому состоянию организмы, особи одного пола (Stephen, 1956; Благовещенский; Александров, 1974). Разумеется, это возврат к типологии. Вместо безвыбо речного изучения организмов веда, выявления изменчивости к® видового признака предлагается ограничить исследование толь® немногими избранниками, "типичными" особями. Парадоксально, что многие исследователи обескуражены фактом изменчивости биохимических и кариологических признаков и не замечают, чт5 именно благодаря изменчивости такие признаки представляют особую ценность для собственно биологической систематики. В этой связи полезно еще раз вернуться к типологическому пер^ ду систематики.
А.И.Теряев (1809), излагая метод типологической сисге^' тики К.Линнея, называет "вариететы" ("varitates ") "выродка" ми", и эта эмоциональная оценка внутривидовой изменчивости красноречива сама по себе. Другой представитель типологичв^ кой школы Ж Кювье (Cuvier, 1817, р. 19) считал, что "вЯ" р нации есть случайные подразделения вида", изменчивость 3s",. ключена в довольно тесные лимиты и вариации "с начала вен1
ле переступали эти лимиты. *Вид заключает в себе особей, про-0СХ°^шИХ °^ни с*7 других или от общих родителей, а также от
тех. которые похожи на них настолько же, насколько они похо-ят ДРУГ на ДРУга* (Кювье, 1937, с. 144). Иными словами, в представлениях ЖКювье, так же как и типологически мысля-
щих биохимиков, неопределенная изменчивость заключена в не— кОторые предопределенные границы. Но тогда четко ограниченный размах изменчивости сам становится мономорфным призна-
ком.
При допущении корректности дарвиновской теории видообразования поиски таких надежных признаков, продолжаемые современными типологами, не имеют оснований. Причины неопределенности в систематике вызваны прежде всего отсутствием однозначной связи между приспособленностью и другими свойствами субстрата. На протяжении исторического развития организмы остаются носителями приспособленности, неизменно заинтересованы в выживании, в таких признаках, которые обеспечивают выживание в конкретных условиях. Систематиков же интересует сходство как свидетельство родства, и такое сходство сохраняется безотносительно к меняющимся условиям обитания. Совсем не обязательно, чтобы интересующая геносистематиков последовательность ДНК сохраняла неизменно свое приспособительное значение в течение длительного исторического развития. В нормальной жизнедеятельности животных имеются механизмы для утраты сегментов хромосом и частей молекулы ДНК, которые не имеют значения в данных конкретных условиях. Зго - диминуции, делении, дефишенси. Нет оснований считать, что организмы при этом 'помнят* об интересах систематиков и теряют только такие последовательности нуклеотидов, которые не представляют ценности для установления родства.
Из безуспешных попыток преодолеть таксономическую неопределенность следует сделать вывод о том, что биологическая специфичность неустранима в операции классифицирования, пока исследователь остается объективным наблюдателем свойств видов. Биологическая специфичность вынуждает исследователей классифицировать животных и растений особым образом, иначе, чем неживые объекты. В гаком случае надо изменить отношение к неопределенности. С ней не следует бороться, ее надо Вь1Являть и изучать. Систематика объективна неопределенностью своих суждений. Опыт систематики, имеющий дело только с одним родом популяционных объектов, таксонов, надо распространить на другие объекты других частных разделов биологии.
Глава 7
ОТНОШЕНИЕ СХОДСТВА
И ФЕНЕТИЧЕСКАЯ (ФЕНОНИЧЕСКАЯ) СИСТЕМАТИКА
Особенности отношения сходства, широко используемого в биологии, раскрываются в дискуссиях вокруг фонетической систематики. При этом главным остается вопрос о носителе признаков сходства — популяция или организм.
Благодаря в первую очередь В.Иогансену ( Johansen , 1909) в биологии сложилась следующая система терминов:
ген - генотипический - генотип - генофонд - генетика, фен — фенотипический — фенотип - фенофонд — фенетика. Параллелизм понятий здесь вполне ясен. Понятия генофонда и фенофонда относятся к популяции, под фенотипом же подразумевается и единичный организм, и некоторое множество фенотипически сходных организмов. А.В.Яблоков (1980) энергично развивает фенетику популяций, имея в виду под феном альтернативный признак относительно несложной генетической обусловленности.
Затем был введен термин "фенон* как обозначение фенотипически однородной выборки сходных организмов ( Camp, Gilly, 1943). По своему содержанию он напоминает фенотип и фенофонд. Еще позже было предложено понятие фонетической классификации ( Cain, Harrison, 1960) как такого способа упорядочения разнообразия, при котором надо привлекать любые признаки без взвешивания, в том числе "генетические*, априори считая их равноправными. Признание за разными признаками равного значения (веса) при установлении сходства имеет то основание, что сходство по таким генетическим признакам, как хромосомы или химические свойства молекулы ДНК, - это тоже сходство. Таким образом, с самого начала фенетическая систематика оказалась противопоставленной филогенетической систематике Э.Геккеля, использующей не любые, а только гомологические признаки ( O.Solbrig, D.Solbrig, 1979). В фонетичес-
кой систематике получили широкое развитие математические методы классификации, чго послужило поводом для названия *нУ' мерическая таксономия*. В ней "...филогенетическая спекуляция отделена от таксономической процедуры", "...филогенетические дедукции делаются впоследствии на основе фенетической классй-фикации, но не включены в процесс классифицирования* (SokaL Sneath, 1963, р. 52, 55). "Таксономический анализ* Е.С.Смирнова (1969) сходен с нумерической таксономией в том, что тоже отделяет классифицирование по сходству от ф»710* генетического подхода, но отличается от первого взвешивание^ признаков в зависимости от частоты встречаемости. Надо nojr черкнуть, что в действительности никто из систематиков не
ввОдиг в свои построения филогенетические данные потому, что е1Де никто не наблюдал непосредственно сам процесс филогении. gaK Уже было отмечено, разграничительная линия между нуме— о11Ческой таксономией и филогенетической систематикой прохо— в другом месте: филогенетическая систематика опирается
не да любые, а только на гомологичные признаки сходства и поэтому остается частным разделом фенетической систематики.
Поскольку фенегическая систематика пользуется любыми, в
тОм числе и гомологическими, признаками, получаемые ею результаты часто согласуются с традиционной системой и в том случае, когда привлекаются математические методы нумеричес— кой систематики. Опыты по межвидовой гибридизации, в которых нумерическими методами установлено, что по сходству гибридные формы редко занимают полагающееся им промежуточное место между родителями (Heiser et al., 1965;Sneath, 1968, 1969), не компрометируют фенетическую систематику и ее нумерический метсд, потому что установление родства не входит в ее задачи. Между прочим, это еще одно свидетельство неоднозначной связи сходства и родства.
Те феноны, которые выделены с помощью нумерических ме
тодов, получили название операциональных таксономических единиц (см. табл. 2). Вместе с фенонами такие единицы заменили
таксоны.
Слово 'фонетический₽ (в основном неправильно) понимается как синоним слова 'фенотипический*,, например, у А.К.Скворцова (1972). Против такой подмены, неправильной в контексте систематики, резко протестовал А.Кэйн ( Cain, 1968). Тем не менее, как установил А.В.Яблоков (1976), В.Иогансен действительно понимал фенетику и фенотип как сопряженные понятия, парные к генетике и генотипу (4). В этом случае 'фонетический' и 'фенотипический' могли бы оставаться синонимами. Но систематики, как уже было указано, придали слову 'фонетический' несколько иное значение. К сожалению, авторы последнего термина не обратили внимания на систему терминов В.Иоган-сена.
Выход из этой путаницы, по-ви дам ому, состоит в том, чтобы фенетическую систематику переименовать в 'феноническую' включив в нее как частность филогенетическую систематику. В п'еРвой установление сходства есть самоцель, во второй сходство есть свидетельство родства (рис. 6). Что же касается их °бьектов — фенона и таксона, то они перекрываются, соотносят— 2* как показано на рис. 3. Тогда 'систематика генотипов'
•Майра (1968) и А»Н.Белозерского (1970) будет отделена Фенотипической систематики того периода истории науки, Огйа еще не были известны признаки генов и хромосом.
Более глубокое основание для упорядочения систематик — их место в оппозиции 'организм - популяция'.
Главная задача нумерической таксономии — это выделение "стабильных фонетических групп", достижение воспроизводимое, ти результатов как свойства объективности (Sokal, Sneath, 1963; Смирнов, 1969; Leuschner, 1974). Такой подход, не выделяющий специфический объект исследования, обернулся не-специфичностью методов вычисления сходства. Они применимы не только в нумерической систематике, но и в экологии при оценке, например, сходства пищи рыб ( Laughlin, 1982), сход, ства отдельных организмов (Leuschner , 1977), сообществ и биот (Песенко, 1982), биогеографических регионов ( Pielou, 1979) и т.д. Довольно скоро выяснилось, что результаты вычислений зависят от выборки видов (Вайнштейн, 1968, 1972) и что разные математические методы приводят к неодинаковым результатам (Воусе, 1964; Sokal, Michener, 1967; Тамарин, 1971). Теперь возникла проблема субъективного выбора объективных методов. Выбор достаточно широк: среди 43 коэффициентов сходства насчитывается не менее 20 оригинальных
популяционная (дарвинизм)
типологическая':'-'
Цррганйзмрцёнтризм^
Рис. 6.
А - деление типологической (организменной) систематики по с отелю признаков сходства; Б — деление систематики в целом по целям классификации и истолкованию результатов: сходство есть самоцель (фенетическая систематика) или сходство есть свидетельство ропства (филогенетическая систематика); В -ление систематики в целом на основе разных концепций жизнб
Г — деление систематики в целом по разным видам признак
( Hubalek, 1982). Этот неожиданный результат довольно чигелен. Вновь возникшая неопределенность есть слерюгвие
1959).
что« игнорируя биологически содержательный вопрос о носителе признаков сходства, исследователи не ограничены в выборе ме— тодов изучения. В данном случае разработке методов классифицирования должны были бы предшествовать осознание и приня— TJIe той или иной концепции элементарного носителя жизни, выбор дарвинизмом и типологией. "Важно осознать, что
манипуляция данными или числами в любой процедуре не увеличивает объективности процесса, которыми эти данные обнаружены" (Inge.-,
Видение системы как неподвижной упоряд оченности - это градация типологии. И вполне справедливо И.И.Канаев (1966) относит к ней феноническую систематику не только потому, что об этом прямо заявил один из ее основателей (Sokal, 1962), но еше и потому, что из фенонической систематики исключено филогенетическое истолкование полученной системы даже в том случае, ческие. просе о ты тем
когда среди прочих признаков использованы гомологи-Но к этому нодо добавить, что, не разобравшись в во-носителе признаков сходства, нумерические таксономис-самым пренебрегли и вопросом о биологической специ—
фичности, которая заключена прежде всего в популяции, а не в организме, тем самым из системы фенонов вместе с внутривидовой изменчивостью (Daly> 1961; Minkoff, 1983) оказал
ся исключенным механизм происхождения органического разнообразия, который должен был бы присутствовать в системе в неявном виде. Отсюда и более общее возражение против нумери— ческого метода: вследствие популяционной природы носителя признаков сходства сама правомочность применения метрического метода представляется очень сомнительной.
Любое число — неподвижность. Между тем суждение о популяционных живых объектах само должно быть как бы живым, развивающимся. Идею развития, незавершенности и передает традиционная неопределенность, против которой направлены усилия нумерических таксономистов. Приступая к изучению этой неопределенности, следует подчеркнуть два обстоятельства. Во-первых, чистое сходство" — это частный случай сходства вообще (ячейка 3, табл. 3), в общем же случае сходство сопряжено с другими отношениями (см. рис. 3). "Чистое сходство" в сравнении с родственным сходством" изучено весьма слабо. Споры вокруг фенетической (фенонической) систематики ведутся в контексте ее отношений с филогенетической систематикой ("родственное сходство"), и это естественно сужает область споров. Остается ® стороне малоизученное сходство вследствие "совместности" Ячейка 28) и другие формы сходства. Ниже рассматривается |°Лько частный случай "сходство вследствие родства" в ячейке
(рис. 3). Во-вторых, изучение неопределенности может проводить только на конечной выборке организмов и конечной же &ь,борке их признаков. Между гем полученные результаты надо
Таблица 3
Объекты фенонической (фонетической) систематики sensu lato (номера ячеек см. на рис. 6)
Ячейка Филогенетическая систематика Ячейка феноническая систек^' тика sensu stricto
2 таксоны 3 феноны
11 таксоны как типы организации 24 феноны как типы opj низании
4 таксоны как типы организации и биотические элементы 25 феноны как типы орге>( низании и биотические элементы
13 таксоны как биотические элементы 28 феноны как биотические элементы
12 таксоны как биотические элементы и жизненные формы 27 феноны как биотические элементы и жизненные формы
15 таксоны как типы организации, жизненные формы и биотические элементы 16 феноны как биотические элементы, жизней ные формы и типы организации
14 таксоны как типы организации и жизненные формы 29 феноны как типы организации и жизненные формы
17 таксоны как жизненные формы 18 феноны как жизненш формы
отнести к природным видам, наделенным способностью потенциально бесконечного существования. Значит, надо принять, что не сама по себе количественная оценка неопределенности представляет собой цель исследования, а в первую очередь незавершенность количественных параметров этой неопределенности, потому что именно незавершенность, открытость суждения отр£‘ жает природу вида.
Признаком О./ будем называть такое состояние, когда нек0' торое свойство либо имеется — a.t , либо отсутствует — Qi •
Qifai, Q[J t где Qz , - противоположные состояний
признака, положительная и отрицательная формы; символ без Qi и не имеет значения.
Как формируется эмпирическая выборка видов и других так»’ сонов, носителей гомологических (т.е. обусловленных родство’4
Ар^наков? Из теории естественного отбора, неявно йыполняю-еЙ функции теории жизни, следует, что новые виды возникают на основе внутривидового разнообразия вида—предка. Такое разнообразие состоит из фенотипов как носителей сочетаний аллель-' лЫХ признаков; более точно, аллели — это положительная и отри— цаТельная формы признака. В классических опытах Г.Мендэля по скрещиванию гороха было получено следующее разнообразие фенотипов, различающихся цветом и строением эпидермиса горошин:
а2
желтый гладкий зеленый гладкий
желтый морщинистый зеленый морщинистый
Обозначая признак цвета как ‘ q1 н строения эпидермиса как , мы можем написать решетку Пеннета:
®2 @2
Qfa2
11 01
10 00
После нее получено заменой положительных форм признаков символом "l’’ и отрицательных символом "О". Известно, что Г.Менделю повезло потому, что ему встретились фены - Признаки относительно простой наследственной обусловленности. Известно также, что в конкретных природных популяциях под действием многочисленных, разнонаправленных и сложно взаимодействующих факторов разнообразие фенотипов представляет непростую картину. Вместе с тем установлено, что сложные случаи наследования объясняются на основе законов Г.Менделя, представляя собой их частности. Сложности в структуре разнообразия конкретных популяций, наблюдаемые в ограниченное время, также не имеют значения. Существенно другое: в длительное историческое время в пределах ареала неопределенной протяжен ности несогласованно действующие факторы, вызывающие отбор в разных и противоположных направлениях, обусловят появление всевозможных фенотипов. Численность организмов каждого фенотипа колеблется во времени, свидетельствуя о неодинаковой Приспособленности (значение) одного и того же фенотипа (суб-страт) в меняющихся условиях обитания. Выражение "одного и Того же фенотипа" относится к субстрату и означает здесь * одно и то же сходство с другими фенотипами". Чтобы пояснить ЭТо> упорядочим по сходству разнообразие фенотипов (7):
Классифицирование, т.е. выделение дискретных внутривидовых 6Диниц, в таком множестве невозможно в том смысле, что, пр^д-^ввляя собой континуум фенотипов численностью 2 \ оно со-^еРЖит постепенные переходы между совершенно непохожими фе— й°Типами из крайних столбцов. Фенотипы из среднего столбца -^^внопохожие с последними. Сходство плавно убывает по оси
абсцисс, и это обстоятельство сохраняется даже при полное сходстве между фенотипами в среднем столбце.
& ____________I_______L---------L
сходство, т.е число единиц, (нулей)
Структура упорядоченного множества (7) биномиальна, по. тому что множество представляет собой члены разложения би. нома Ньютона:
(*1* + *О*)2 = *1*2 + (*1О*-*1О*) + *О*2, или
(а + а)2 = а2 + 2аа + а2, (8)
гпе л «= 2; а, а - разные формы "обезличенных* признаков.
Переходя рт 2 к л признакам, при достаточно длительном наблюдении и неопределенным образом изменяющихся условиях обитания мы получим множество фенотипов вида | (а + а) | = «2 л. Оно представляет собой множество сочетаний прообразов таксономических признаков. Строчки символов, вообще говоря, это будущие таксономические диагнозы, обладатели прообразов — это зародыши* новых видов. Если бы все прообразы перевоплотились в таксономические признаки, то получился бы таксономический континуум | Л/к I в 2Л.Вряд ли он осуществим в полной мере. Поэтому с<1. Отбор, воздействуя только на приспособленность и не пользуясь отношением сходства, будет превращать выборки фенотипов в виды, а прообразы - в таксономические признаки. При этом исключено такое положение: в изменяющихся условиях обитания за исторически длительный период (который требуется для видообразования) отбор целеу^ ремленно вычленяет и превращает в виды фенотипы только и3 правой (левой) части распределения (7). Эго означало бы, 4Tt отбор пользуется категорией сходства (несходства) субстрат^ и благоприятствует сходным (несходным). По этим же причм* нам исключено и другое положение, по которому отбор благо* приятсгвовал бы равнопохожим из среднего столбца распре ния (7).
Выборки фенотипов имеют шансы на выживание независим0
QCI
от ' за ' ей ' нйй ра, эцака» )азНовидность
сходства *“ различия в признаках субстрата; исход борьбы существование, преимущественное размножение определяют— только приспособленностью. Эго лишь иносказание определе-[ приспособленности как конкретного, относительного свойст-переформулировка положения о независимости значения от
лежащего в основе определения жизни. Далее "если бы [здесь - эго выборка фенотипов - A/.3.J достиг— такой степени процветания, что превысила бы численность одоначального вида, то она сделалась бы видом, а вид превратился бы в разновидность* (Дарвин, 1939, с. 308). В таком случае совокупность вновь возникших видов сохранит биномиальную структуру, причем такая совокупность отвечает представлению о монофилетическом таксоне. Обозначим ее через . Она не приурочена к какому-либо отрезку времени, а пребывает в становлении. Точно гак же она не ограничена в какой—ли— области, а разворачивается в пределах пульсирующего ареа-монофилетического таксона.
Процесс пространственного обособления популяций, в котором созревают предпосылки для видообразования, остается в центре внимания многих эволюционистов. Для решения проблемы немало сделано Э.Майром. Важный аспект этого явления состоит в следующем: *...вновь возникшие популяции будут обладать
бо ла
специфическим генным составом, отличающим их от остальных популяций вида, ибо здесь при заселении. новой территории случайной [ подчеркнуто мной. - Н. 3. J группой особей происходит перестройка генного состава* (Дубинин, Ромашов, 1932). Позже у Э.Майра этот принцип случайности был раскрыт в * принципе основателя*, согласно которому *...основатели новой колонии (или популяции) несут в себе лишь небольшую часть общей генетической изменчивости родительской популяции (или вида)* (Майр, 1974, с. 424).
t Длящееся множество Nр, лишенное пространственно-временных границ в длении, непрерывно (но не обязательно равномер-к°) поддерживается, пополняется за счет внутривидового разно-°брааия випа-предка или какой-то его популяции. В свою очередь каждая популяция /у& есть источник вновь возникающего видового разнообразия. Эмпирическая выборка видов образованная выборками из длящихся популяций этих видов, состав-ЛЯет лишь часть А/в. Природная совокупность скорее всего составляет только часть таксономического континуума:
/V3c:>V6c/VK.
(9)
сходст-
Структура Л/р биномиальна, потому что основана на
На* КотоРое не замечает отбор. Структура также биномиаль-а (по построению). Что можно сказать о структуре N3 ? Ка-
^ось бы, для ответа на этот вопрос надо обратиться к конк-HbiM таксонам, что и было сделано ранее (Заренков, 1972,
1976). Однако такое опытное решение общего вопроса о ст( тической структуре таксона не представляется целесообразна Эмпирический материал может включать ошибки, например установлении гомологий, которые здесь имеют решающее зна^ ние: ведь подразумевается сходство, обусловленное родство^ Предпочтение должно быть отдано дедуктивному методу, crpyKs тура таксона должна быть выведена из биологически содержу тельных посылок и в конечном счете опираться на теорию жи^ ни. На процесс выделения из природы эмпирической выборки г ли бы влиять следующие обстоятельства.
1. Неодинаковое число организмов у разных видов, т.е. нси динаковое число носителей сочетаний признаков в природной вокупности N& . Представляется естественным, что чем больще организмов представляет данный вид, тем больше шансов у си. стематиков открыть такой вид. Теперь достаточно допустить, что виды с большим количеством организмов, "банальные*, по. хожи друг на друга больше, чем организмы редких видов, и тогда биномиальность окажется невозможной. Такой же вывод последовал бы из допущения о том, что редкие виды обязательно резко отличны от * банальных" и не похожи друг на друга. Думается, что эти предположения неприемлемы. Численность ор. ганизмов вида — это свидетельство его приспособленности. Пр», с пос облени ость в свою очередь не связана со сходством: пер» вое есть аналог значения, второе — знак. Несходные виды могут быть одинаково приспособленными в одних условиях, и, наоборот, сходные виды могут обладать неодинаковой приспособленностью - в зависимости от условий. Надо учесть к тому же, что численность одного и того же вида не постоянна и колеблется в зависимости от изменяющихся условий обитания, и нет оснований считать, что вымирающие виды сходны (несходны) между собой больше, чем процветающие виды.
2. Неодинаковая изученность отдельных подразделений географической оболочки. .Известно, например, что биота вод Евро® изучена полнее биоты вод Антарктики, а глубины океана изучены слабее фауны и флоры равнин Евразии. Если мы допустим, что слабо изученные биоты состоят из трудноразличимых (резко различимых) видов, тогда биномиальность эмпирической выборки тоже окажется невозможной. В свою очередь это означа* л о бы: что особые условия, например условия Антарктики, способствуют выживанию сходных (несходных) видов, т.е. в водах Антарктики отбор пользуется понятием сходства, а не приспособленности. Оснований для такого предположения в действите^ ноет и нет. Антарктическая фауна и флора включают в себя как трудно различимые, так и хорошо различимые виды.
3. Неодинаковая изученность органического мира во времени, слабая изученность вымерших фаун и флор в сравнении с современной биотой. Если, например, в кембрии виды различа-
слабее (резче), чем современные виды, биномиальность '^^нческой выборки, объединяющей и вымершие и ныне живу-
БцДы, оказалась бы невозможной. Эго предположение пред-^^ляется крайне сомнительным. Если в кембрии эволюция осу-сТсТвЛЯлась на основе приспособленности, а не сходства, то ^ддлчия между кембрийскими таксонами в среднем были такие Р что и в любую другую геологическую эпоху, процент труд-^азличимых (резко непохожих) видов оставался неизменным, вполне естественно, что в начале исторического развития орга-ндзмы были много проще современных и общее число видов по-началу оставалось незначительным. Представляется почти очевидным, что морфологические различия между древнейшими вирами были невелики, пока протекал первый этап биохимической эвОлюции. Однако вполне допустимо считать, что и в суперпо-нулипии протобионтов имелась неопределенная изменчивость по биохимическим признакам (исходный материал для естественного отбора), и, таким образом, изначала существовали весьма сходные и совсем несходные формы в пределах этого разнообразия.
В заключение представим себе такую ситуацию: на Земле исчез свободный кислород и создались анаэробные условия. Очевидно, что после такой катастрофы остались бы в живых только анаэробы. Разве это не означает, что отбор все-таки учитывает сходство, в данном случае в типе обмена веществ? Некорректность этого примера состоит ц том, что он подразумевает не таксон, а тип организации - "анаэробы*, который объединяет самые различные неродственные виды. Эго объединение основано на негомологичных признаках. Между тем здесь рассматривается частный случай сходства по гомологическим признакам ("родственное сходство*). Таким образом, если не упускать из виду различие между сходством и приспособленностью и не пугать "родственное сходство* с другими формами сходства (например, "сходство вследствие сосуществования", как в примере с анаэробами, или сходство вследствие совместности и пр.), то Нет оснований отвергать вывод о том, что трудноразличимые и сильно несходные виды попадают в поле зрения систематика с гой же частотой, с которой они встречаются в природе. В та-ком случае эмпирическая выборка /V
. и первая получена с... ___ _ г___________r_..
к°в» 1976). Кроме того, это означает, что принцип случайнос-Ги Дает нам возможность узнать кое-что о N&, основываясь На свойствах /Уэ . Иными словами, есть возможность вероят-н°стного предсказания сочетаний свойсгв, не известных науке.
С этой целью рассмотрим модель: таксономический контину-для четырех признаков ( п = 4) и модель эмпирической *Ь1б°РКи /уэ . причем согласно уже осуществленным рассужде— последняя могла бы считаться монофилетическими таксона— • и оба множества биномиальны (8).
э биномиальна так же, как случайным образом из второй (Зарен-
В каждом из 4 столбцов континуума Л/к в 2^ (каждый бец отвечает признаку) содержится поровну единиц и нулей. * Сделаем так, чтобы в столбцах эмпирического множества | ц , 8 имелась бы биномиальная структура и содержалось неравц^' число единиц и нулей, что свидетельствовало бы о политетич^ сти. Диагноз таксона, включающего разнообразие /Уэ выгляде„ бы так: "первый и четвертый признаки обычно имеются, втор^ и третий признаки обычно отсутствуют". Поскольку /Уэ есть случайная выборка из более обширной природной совокупности то у последней в каждом столбце должно наблюдаться такое соотношение между символами, что и у Мэ . Этому условию не отвечает континуум /Ук , и потому он неосуществим. Знач^ /Уэ получена случайным образом из какой-то другой совокупно, сти. Но численность ддэбой другой совокупности меньше ( например /Vg. Она имеет биномиальную структуру, и в каждое е.е столбце символы "1" и "О" могут находиться примерно в том же соотношении, что и в N3 . Такая /7Э может считаться случайной выборкой из N . Кроме того, отсюда следует прин. ципиальная возможность предсказания, перечисления всех стро. чек символов /УБ . Что же касается континуума /Ук, то он неосуществим, и это свидетельствует о корреляции между приз-наками.
Между прочим, зная численность строчек символов в множестве /УБ, можно получить простейший показатель достоверно" сти знания той природной системы, которая могла бы быть построена на Wg ио которой мы знаем только по эмпирической выборке N3 . Таким показателем могло бы служить отношение | /уэ| • Д*® °ДНО® группы креветок этот показатель оказался равным 31/403 (Заренков, 1976), т.е. систематикам в данном случае известна примерно одна тридцатая часть таксономического разнообразия группы, точнее носителей данных изучаемых признаков. Имеется в виду, что в природе среди Kps-веток есть еще и такие, которые различаются какими-то иными, еще не учтенными признаками. Такие животные могут быть неразличимыми по учтенным признакам. Разнообразие сеяно во времени и не обязательно должно считаться уже осуществившимся, оно может пребывать в становлении.
Полученные дедуктивным путем результаты позволяют следующим образом понимать проблему континуальности - дискрет* и ости. В принципе таксоны в эмпирической системе разграничены естественным образом, поскольку континуум неосуществим и между признаками имеет место корреляция. Однако относит^ но границ между конкретными таксонами и корреляцией между отдельными признаками можно высказать только вероятностное суждение, т.е. имеется некоторая вероятность того, что эта граница сохранится при пополнении эмпирической выборки до /УБ . Определенное однозначное суждение по этому поводу во-
94
невозможно. Разумеется, ° ос^кая ме₽а достоверности неодинакова для разных буде надо считаться с тем, что веро-
границ между таксонами в
таксонов.
а4 0000 N3 0000
0001 0001 0001
0010 4Q За 4 0100
1000 1000 1000
1100 1100 1100
ООН
0101 es2a2 1001 0101 1001 0101 (10) 1001
1010 1010 1010
оно ОНО
1110
1101 4 а а3 1011 1101 1011 1101 1011
0111
а4 ни 1111
Таким образом, в целом проблема решается в пользу вероятностной дискретности. В сущности, этот вывод в значитель-ной мере предопределен тем обстоятельством, что эмпирическая выборка и генеральная совокупность обладают конечным числом Цризнаков и конечным же числом сочетаний признаков. В потенциально бесконечном существовании таксонов осуществляется бесконечное же число признаков и их сочетаний, поэтому нет оснований противопоставлять дискретность континуальности. В п°тенциально бесконечном "со-диении" сходство — различие меж-ИУ видами не может оставаться постоянным, и здесь требуется гРегье суждение. В этой связи уже построенные системы, в которых одни таксоны разделены вполне четко, а другие соедине— Hbl переходными формами, показательны в следующем отношении: твкие системы дают представление об изменении степени сход-Сгва в длении. При этом разные виды, хорошо разделенные и
95
соединенные переходами, отвечают отельным временным точкам дления одних и гех же видов.
Необычность биологического классифицирования вызвана оС(х бой биологической природой объектов классификации. Она же нуждает биологов несколько особым образом пользоваться понятием времени.
Глава 8
ПОНЯТИЕ ВРЕМЕНИ В БИОЛОГИИ
Иногда наша повседневная речь с удивительной отчетливостью выявляет отдельные стороны даже весьма сложных понятий. К ним относится понятие времени. Показательно, что мы говорим "жизнь течет", но счастье — "длится*. Течение обыден, ной жизни воспринимается нами как поток событий, тогда как счастье — это скорее состояние, дление.
Четкое различение земной быстротекущей жизни и вечного блаженства или мук на небе присуще христианскому вероучению и другим религиям. Земная жизнь — это "суета сует", кратковрь менное греховное пребывание' Души в телесной оболочке. Жизнь небесная — непреходящее бытие души, освободившейся от бренного тепа. Это - мирское и сакральное время. Первое дано человеку частями последовательно одна за другой, во второй душа погружена в не имеющее начала и конца бытие после того, как память о земном существовании "канула,в Лету".
Эти две стороны времени осознаны в поэзии: Бесстрашно отгоните
Надежд самообман, С достоинством примите
Тот жребий, что нам дан: Отжив, смежим мы веки, Чтоб не восстать вовеки, Все, как не вьются реки, -Вольются в океан.
Созвездий мириады Сюда не шлют лучи, Молчат здесь водопады, Не пенятся ключи: Ни радости беспечной, Ни скорби быстротечной, Один лишь сон - сон вечны.' Ждет в вечной той ночи.
(А.Ч.Суинберн)
В донаучном мифологическом знании происхождение течения времени от дления поясняется следующим образом. Блаженное существование Адама и Евы на небесах (дление) было прервано грехопадением. Последовало наказание, изгнание из рая на грешную Землю. Эго наказание выразилось и в том, что в земном существовании жизнь не отделена от смерти, зарождение в организме жизни есть и начало смерти; это иное, нежели в раю, земное, время.
Эти отступления в область повседневной жизни, поэзии и р©" литии помогают ощутить нашу предрасположенность и внутренне
96
одготовленность к усвоению сложного научного понятия време-Они наводят на мысль, что представление о времени как
Р^цинении дления и течения, бытующее за пределами науки, ^огло бы служить образом биологического времени.
Обширная естественнонаучная литература о природе времени
основном создана физиками или ради физики (Вяльцев, 1965; доепюхин, 1966; Молчанов, 1977). Больше того, как считает р рейхенбах (1962, с. 32), *для решения проблемы времени ве сушествУет других способов, кроме методов физики*. Впол—. ве справедливо М.Э.-Омельяновский (1962) расценивает эту установку как позитивистскую. Ценность книги А. Грюнбаума 'Философские проблемы пространства и времени* (1969), как считает Э.М. Чудинов (1969, с. 568), состоит в том, что она '...вводит читателя в курс сложнейших философских проблем физико-математического знания...* [^подчеркнуто мной - Н.5.1. Часто отдельные проблемы времени и пространства объединяют^ ся в единой хроногеометрии, но и тогда, как отметил В.С.Лямин (1978), абсолютизируются физическое пространство и вре
мя. Даже сама идея хроногеометрии о времени как о четвертом измерении пространства - времени (Уитроу, 1964) исходит от физики: 'Идея четырехмерного пространства родилась как отражение связи между пространством и временем, которая была открыта теорией относительности* (Аронов, 1974, с. 275). Вместе с тем физическая теория относительности могла бы служить прецедентом, оправдывающим поиски биологического вре
мени, потому что признание относительности пространства и времени *...позволяет нам утверждать, что сами формы пространства и времени, будучи абсолютными условиями бытия движущейся материи, по своей качественной природе изменчивы вследствие изменчивости своего содержания - конкретных качеств материи в том или ином ее состоянии* (Свидерский, 1964, с. 44).
Субстанциальная концепция времени принадлежит истории. Если время не субстанция, а форма бытия материи (Аскин, 1971), то особым формам материи могли бы отвечать разные способы ее существования, т.е. неодинаковые по содержанию и структуре времена. В таком случае свойства времени, как и пространства (Мелюхин, 1966), определились бы спецификой взаимодействия материальных объектов. Особые взаимодействия 6 Разных формах движения материи могли бы оказаться связанными со специфическими формами причинения, если 'причинно— СлеЛственная структура связей объектов обусловливает фундаментальные свойства пространства и времени* (Кузнецов, 1967, с,40). Время, таким образом, определяется через причину (Пуанкаре, 1906). Действительно, понятие времени в физике свя— аано с определенным пониманием причины и следствия.
После того как абсолютное время Н.Ньютона было отвергну—
то и его место заняло реляционное время, в физике утвердИГ1& получившее признание в классической логике препставление 0 " временной последовательности одноместных событий как одн^ сторонней причинно-следственной связи (Молчанов, 1977). гласно концепции реляционного времени все те одноместные бытия, на которые Данное событие воздействует или может Действовать, считаются по отношению к нему "будущим*. Все те одноместные события, которые воздействуют или могут во&. действовать на данное событие, считаются по отношению к не» му "прошлым". Одновременными считаются одноместные собы» тия, которые принципиально-не могут взаимодействовать между собой и, таким образом, не состоят в причинно-следственной связи.
В биологии осознание сложности и специфичности понятия времени затруднено господством типологии в прошлом и преобладанием организмоцентризме в настоящем. Типология и ее преемник-современный организмоцентризм поместили биологическую специфичность в организм, и поэтому для них биологическое время остается организменным понятием. Именно к нему обращается прочная литературная традиция (Du Nouy, 1936; Биологические часы, 1964; Уитроу, 1964; Аскин, 197J.). Однако поскольку жизнедеятельность организма сводима к физхимии, постольку в организме нет особых причинно-следственных срязей и организменное время следует считать физическим временем. Для популяционизма искомая специфичность могла бы находиться в семантико-информационных взаимодействиях, в которых отражение отделено ог физического взаимодействия; следовательно, ему могла бы отвечать особая форма причинности и особая структура времени. Отражению отвечает "сигнально-информационная причинность", которая не отменяет физической причинности, но как бы "надстраивается* над последней (Тюхтин,1972). Таким образом, понятие причинности выводит теорию на проблему биологического времени.
Применим эти представления к пребывающим в борьбе за существование популяциям хищника и жертвы.
Взаимодействие между популяциями есть прежде всего изначальное непрерывное взаимоотражение их статистических струн* тур. При этом популяция хищника (жертвы) есть не только при* чина структуры популяции жертвы (хищника), но и следствие. В таком взаимоотражении каждая популяция выступает и как причина и как следствие, как будто одно зеркало отражается в другом. Существенно, что в таком взаимоотражении статистичее ких структур каждая популяция "ощущает* только приспособленность партнера, и совокупный вещественно-энергетический суб* страт организмов популяции остается всего лишь носителем приспособленности. Такой биологически специфичный способ су-шествования живой материи, живой плоти, имеет место в дден^
98
дление живых взаимодействующих популяций удобно назы-С°”ъ сосуществованием. Формально сосуществование есть огно— ВеНйе, поэтому оно нашло свое место на биологической розет-
Взаимодействие сосуществующих популяций происходит при рблюдении физических причинно-следственных связей не только внугри организма, но и в поединках между организмами, например между организмом хищника и жертвы. Такие связи как бы включены в межпопуляционные взаимодействия и сохраняют • при этом определенную независимость. Действительно, при сохранении физических взаимодействий между организмами популяции, к которым организмы принадлежат, могут сохранять нейтралитет. Эту принципиальную возможность предполагает следующая несбыточная ситуация.
Пусть вследствие избытка "мест под солнцем" наступила идиллия, когда "и волки сыты и овцы целы* и все организмы имеют равные возможности оставить потомство. Тогда прекратилась бы борьба за существование, но физическое взаимодействие между ними сохранилось бы как между объектами физики, например в виде притяжения. Такая идиллия создавала бы у наблюдателя впечатление жизни, потому что в организмах осуществлялся бы нормальный обмен веществ. Однако прекращение борьбы за существование и вызываемого ею отбора приспособленных означало бы прекращение жизни как особого способа существования материи. Прекратилось бы взаимоотражение статистических структур и сохранилось бы взаимодействие организмов-машин. В конечной географической оболочке неограниченное размножение организмов, как одно из проявлений жизнедеятельности, постоянно приближало бы окончание идиллии. Ее прекращение сделалось бы неотвратимым, если бы некий внешний фактор не заботился постоянно о расширении ресурсов планеты, необходимых для безбедного существования организмов. В далеком прошлом противоречие между потребностями неограниченно возрастающего числа протобионтов и ограниченными возможностями географической оболочки вызвало вспышку жизни, превращение пробионгов в протобионтов. Оно же отметило начало биологического времени, в котором взаимоотражение статистических структур борющихся за существование популяций как бы надстроилось над физическим взаимодействием организмов. По прошествии Достаточно длительного идиллического существования сторонний наблюдатель признал бы в таких видах "живых ископаемых", не изменившихся потому, что, пока сохраняется идиллия, нет и вызываемого естественным отбором исторического развития. Реальное существование "живых ископаемых", почему-то не эволюционировавших, хотя окружающие виды претерпели эволюцию, имеет в этой связи следующее значение.
Во-первых, представляя собой весьма отдаленное сходство с
99
идиллией, существование таких 'живых ископаемых* помогает уяснить понятие дления. Основное содержание рления - это пск вседневная борьба за существование, в которой популяция оста, ется носителем приспособленности. В таком длении независимо от различий в субстрате (внутривидовая изменчивость) отдель. ные организмы изначала равны перед отбором, потому что по определению разные организмы могут оказаться (впоследствии) равноприспособленными. Презумпция равноуспешного выживания постоянно отменяется отбором, и тем не менее среди различай... щихся субстратом организмов нет выдающихся, заведомо высо-коприспособленных, потому что приспособленность конкретна, постериорна. В этом смысле популяция однородна, несмотря на внутривидовую изменчивость. Больше того, поскольку отбор име» ет дело с приспособленностью популяции, тогда как приспособленность организмов вторична, производна от популяционной, постольку организмы не индивидуальны в биологическом смысле, т.е. как носители приспособленности. К тому же отбор изменяет статистические показатели изменчивости популяции (отражение), а не субстрат отдельных организмов, остающихся индивидуальными (Пасеков, Коростелев, 1971) только по свойствам субстрата. В этом, биологическом, смысле индивидуальность и целостность — достояние популяции, а не организма.
Поэтому в равной мере следовало бы отвергнуть претензии ныне живущих, уже вымерших или еще не народившихся организмов представлять популяцию. В таком непрерывном длении не;: 'краевых* организмов. В сосуществовании как со-длении нет разницы между прошлым и будущим. Только произвольно привнесенная наблюдателем временная точка ('настоящее) рассекает однородное и неподвижное дление популяции на 'прошлое' и 'будущее*. Такие привилегированные точки создаются наблюдателем, их нет в природе. В самом деле, мысленно удалим из природы наблюдателя. Вместе с ним пропадет и временная точка. Это обстоятельство незаметно в классической физике. При изучении свободного падения тела различия между сколь угодно малым отрезком траектории и сколь угодно протяженной траекторией отсутствуют в том смысле, что в любой отрезок времени закон свободного падения тела остается неизменным.
Из таких измеряемых отрезков состоит любая природная трй,“ ектория даже неограниченной протяженности. В этом залог успеха математического метода изучения неживой природы: вмешательство исследователя -в природу не приводит к искажению природных законов, в выделенном для опыта 'куске' природы законы осуществляются столь же строго, что и в природе в и®’ лом. В этом смысле неживая природа для классической физики* аддитивное целое, состоящее как бы из взаимозаменяемых тей, и это обстоятельство объясняет полную отчужденность ио*, следователя от природы, духовный 'нейтралитет', 'аморальное^ законов физики.
0ное дело в биологии. Оставаясь самими собой, длящиеся 0ды изменяются от одной временной точки к другой, что и о(>« н0руживает наблюдатель. Поэтому, например, сходство между оПуляииями также пульсирует во времени столь же неопределенным образом, сколь неопределенна внутривидовая изменчивость. Выбор временной точки в таких условиях будет влиять на результаты упорядочения органического разнообразия по сходству в признаках организменного субстрата. Причина такого странного положения - собственное время сравниваемых популяций, г.е. Доение.
Во-вторых, на противопоставлении 'живых ископаемых* эво
люционирующим вицам можно раскрыть содержание течения времени. Видообразование происходит в течении времени. Видообразование, как историческое развитие состоит из генеалогий. В генеалогиях возникают организмы с принципиально новыми свойствами, одни организмы порождаются другими. Явления на
следования и размножения - одна из сторон жизнедеятельности, которая может быть сведена к физико-химическим процессам. Однако при этом нельзя упускать из виду, что эволюция вызвана отбором приспособленных, т.е. нефизическим механизмом. Значит, понятие эволюции неприложимо к организму, эволюционируют виды (популяции). Образуя течение времени, виды—предки и их потомки пребывают в длении, пока остаются живыми. В течении времени сохраняются направленность и неоднородность
вследствие того, что предок и потомок — не взаимозаменяемые объекты, их нельзя менять местами. Поэтому в течении времени в отличие от дления прошлое отделено от будущего. Не выходя из со
стояния дления, виды осуществляют историческое развитие, ко— герое сопоставимо с эволюцией в общенаучном понимании (историческое развитие Земли, Вселенной и пр.), но отличается от развития неживой природы движущими силами, вызывающими развитие. Поэтому историческое развитие органического мира есть особая форма движения материи.
На биологической розетке течению времени отвечает отношение исторического (не генеалогического) родства. Важно поп-Черкнуть еще раз, что при этом подразумевается познавательная операция установления отношений, разнообразие точек зрения исследователя. Только этим и оправдана частичная отделенность течения от Дления. В природе же, пребывая в биологичео-Ком времени, виды состоят в длении и течении с начала возник^ новения жизни. Что же касается исследователя, то Для решения своих задач он останавливает внимание либо на одном длении -и Тогда выделяются живые сосуществующие популяции, связан— Ные экологическими отношениями; либо на . течении — и тогда выявляется историческое родство независимо от того, живы или вымерли предки и потомки (есть или нет сосуществования); ^ибо на длении и течении — и тогда обнаруживаются связанные
101
экологическими отношениями и родгтвом сосуществующие предки и потомки. Последняя задача - в пересечении обоих овалов.
Если в длении имеет место семантико-информационное взаимодействие при взаимоотраженйи статистических структур попу*, ляций, то в вызванном цлением течении имеет место физико-химическое воздействие совокупного вещественно-энергетического субстрата популяций вша—предка на субстрат вира-потомка. Эго воздействие называется "генетической обусловленностью", "наследственностью". Ими создается то, что называют генетическим единством органического мира во времени.
Особенности понятия времени в биологии обусловливают своеобразие понятия объективности, объясняют причины особой соотнесенности исследователя и объекта наблюдения. Чтобы разобраться в этом вопросе, надо обратиться к истории науки (Коллингвуд, 1980). Следует, однако, подчеркнуть, что в рефлексии, когда обсуждается знание о знании, наука и религия не только антагонисты, но еще и стереопара.
В условиях господства религии в средневековой Европе любая деятельность была либо богоугодным делом, либо богопротивной ересью. Больше того, религия оставалась не только пробным камнем для оценки деятельности вообще. Сама природа была богоцарованной природой. Не составляло исключения и время. Хронология событий велась от рождества Христова. И хотя дата светопреставления неоднократно переносилась (Ламонт, 1984), будущее, как и прошлое, представлялось достаточно протяженным и вместе с гем оставалось обозримым, заведомо конечным. Наше ощущение времени как независимой от бога беспредельной протяженности было бы чуждым человеку средневековья. Сходным образом отсчеты в пространстве подразумевали некий центр. Считалось, что в центре Вселенной находится благословенная Земля, а центром Земли служит гроб господень.
1 "Естественные науки, как только одолели религиозное мировоззрение, одолели и хронологический взгляд на мир. Основным условием научного мировоззрения стали объективность и независимость явления от наблюдотеля, одоление двойного центризма: времени и пространства. [Действительно, пока исследователь был занят богоугодным делом, он фактически представлял богоцентризм времени и пространства. - Н.3.\ Естественные науки достигли этого требования, когда выключили местонахождение наблюдателя из системы явления и сделали внутреннюю связь явления самодовлеющей^. Наблюдатель здесь - активная сила, своим вмешательством образующая новое явление" (Иоффе, 1927, с. 28).
Отказ от бога означал отказ от экстерналистского видения мира из божественного центра времени и пространства, отказ от конечных внешних причин, от души, привнесенной богом.
102
Конец цитаты можно было бы понимать в том смысле, что, оДОлев богоцентризм, наука как бы пришла к антропоцентризму, росшей ценностью сделалась земная личность. Здесь-то и обнаруживаются неоднороиность науки как системы знания, проги— ^ложность между биологической и классической физикой, от-лИч/е биологии от наук о неживой природе.
В классической физике антропоцентризм не дает о себе знать- потому, что законы физики '''...универсальны в том смысле, что они приложимы повсюду в пространстве и времени" ( Smart , 1963, р. 53). В связи с этим, следуя своему принципу членения природы, разлагая сложное на составные части, физики могут не опасаться тех искажений, которые могли бы возникнуть вследствие подмены беспредельно протяженного пространства - времени пространственно-временной точкой, в которой и осуществляется экспериментирование, заменившее пассивное созерцание натурфилософского периода истории естествознания. На прогулке в яблоневом саду законы механики Ньютона соблюдаются столь же строго, что и во всей вечной Вселенной. Поэтому выбор места и времени не имел значения для открытия и содержания закона свободного падения тел Г.Галилея. Наоборот, как уже было отмечено, в биологии выбор временной точки наблюдения для опред-еления свойств вида имеет значение вследствие того, что объекты наблюдения совершают собственное дление. Возникает вопрос о том, объективно ли выбрана временная точка для наблюдения, можно ли по результатам, полученным в одной временной точке, судйть о вице "в целом". По-видимому, этот вопрос должен решаться по-разному в различных разделах биологии. В систематике, которая опирается на отношение сходства, эмпирическая выборка во временной точке могла бы считаться случайной выборкой из генеральной совокупности, пока рассматривается выборка одного и того же конечного числа признаков (Заренков, 1984). В филогенетике, основанной на отношении исторического родства, эмпирическая выборка видов могла бы оказаться в каких-то иных отношениях с генеральной совокупностью таксонов. Во всяком случае, в систематике сама постановка вопроса о выборке и природной совокупности и есть вопрос относительной объективности. Он приводит к принципиальной неопределенности, которая воспроиз-
... _
в неодушевленные тела. Им противопоставлены естественные пнутренние причины, механизм в самих вещах и явлениях. Отсюда господство интернализма вообще и механицизма в биологии, Углубленный интерес к внутренней структуре вещей и явлений.
эти запросы ответила молекулярная биология, открывшая внутреннюю пружину понимаемой как жизнь жизнедеятельности Рганизмов. Ею оказалась двойная спираль ДНК. Дарвинизм же Подразумевает некоторый возврат к экстернализму, однако в со-®ершенно особом смысле: приспособленность как сущность жи-ых существ раскрывается в борьбе за существование между организмами.
103
водит в логике классифицирования природную неопределенное^] потенциально бесконечного дления популяционных объектов кд. сификации.
В биологии надежды на достижение объективности обычно возлагаются на аналитическую химию и математический метод Думается, однако, что проблема объективности состоит не в том, какими именно методами. изучается эмпирическая выбору организмов - традиционными морфологическими или методами аналитической химии и математики, а в том, представляет ли эмпирическая выборка природный длящийся вид.
Глава 9
ОТНОШЕНИЕ ИСТОРИЧЕСКОГО РОДСТВА,
ФИЛОГЕНЕЗ, ЭВОЛЮЦИЯ
Словом 'эволюция* биологи обозначают широкий круг явлений (Завадский, 1973). К ним относится филогенез (филогения Э. Геккель ( Haeckel , 1894) определил филогению как * племен ное развитие*, в результате которого развились 'бесчисленны формы организмов*, как 'генеалогическую историю', 'историк вида' (Геккель, 1906). Изобразив филогению в виде древа, ветви которого впоследствии ( Meyer-Abich » 1926) получили названия таксонов (например, класс Amphibia)» Геккель тем о мым пояснил, что содержанием филогенеза является историчес кое развитие таксонов (таксоногенез). Первое филогенетическое древо, напоминающее схему логического деления, как это отметил в 1876 г. И.И.Мечников (1943), было построено еиг раньше Ж.-Б. Ламарком (1955, с. 436), Так же, как Геккель. Ламарк еще не пользовался словом 'таксон*. Вообще филогения частная форма исторического развития, которое осуществляют не только таксоны, но также фауны и флоры, сообщества и типы организации ('фауногенез*, 'филоценогенез*).
Выделяются по меньшей мере три разных подхода в вопрос о соотношении филогенетической и эволюционной проблематик.
1. Эволюция и филогенез - синонимы. Примером такого подхода могут служить взгляды А.Н.Северцова ('Морфологические закономерности в эволюции*, 1939). Пожалуй, эта точка зрения преобладает. Действительно, если филогенез есть гаксоно-генез, а вид есть один из таксонов, то филогенез есть и видообразование, т.е. эволюция. Значит, в пересечении проблем филогенетики и эволюции находится понятие виДа. Если к тому же генезис таксонов высокого ранга (родов, семейств) в конце концов сводится к видообразованию, то эволюция есть вообще таксоногенез. Тем не менее эволюция фауны - тоже эволюция.
104
2, Микроэволюция, 'элементарное эволюционное явление*
длительное и векторизованное изменение генотипического ^рс-гава популяции (Тимофеев-Ресовский и др., 196 9) - тоже Сролюпия» однако не филогенез.
3, Теория эволюции описывает механизм, вызывающий фило-^
редез. В этом случае она выступает прежде всего как теория ртбора. В ней центральное место занимает понятие приспособленности и игнорируется организменный субстрат. Наоборот, то, чгО могло бы именоваться теорией филогенеза, имеет своим предметом субстрат, закономерности его развития в филогенезе. Сюда относится известное правило Э.Копа. По-видимому, наиболее значительным обобщением в теории филогенеза надо считать концепцию диффузионного барьера С.Э.Шноля (1979).
Неоднократно подвергавшийся критике номогенез Л.С.Берга (Завадский, Георгиевский, 1977) фактически имеет в виду закономерное развитие морфологических признаков организма в
филогенезе, противопоставляя закономерность вероятностному принципу дарвинизма. При этом Л.С.Берг, по-видимому, не учитывал то обстоятельство, что эмпирический материал, на кото
ром основывалось заключение о закономерном характере развития, представляет собой только выборку из природной совокупности таксонов и поэтому направленность развития могла бы считаться только следствием неполноты знания природы. Не учитывалось также, что фактически закономерное историческое развитие организменного субстрата изучает сравнительная анатомия, где корреляция может пониматься как статистическая связь, а потому направленйость исторического развития могла бы пониматься тоже в статистическом смысле слова. Л. С. Берг (1922) критиковал дарвинизм, основываясь на изучении субстрата (знак), тогда как дарвинизм опирается на понятие при-способленности (значение). Допустим, что некоторое филогенетическое построение отвергнуто (что бывает довольно часто). Вполне очевидно, что это обстоятельство нисколько не отражается на дарвинизме. Дарвинизм совершенно равнодушен к судьбе филогенетического древа (Рьюз, 1977). Любопытный факт в этой связи описан А.Эллегардом ( Ellegard , 1958). Он проанализировал отклики 'массового читателя' на дарвиновское Учение в 1858-1872 гг. и показал, что основное внимание чин гателя привлек филогенетический вывод из дарвинизма о том, Что человек произошел от обезьяны. За этот подрывающий устои Религии вывод частного характера дарвинизм называли 'обезьяньей теорией*. Для 'массового читателя* теория эволюции и Ге логико-методологические выводы, которые из нее вытекали, казались заслоненными частным следствием относительно возможной родословной человека. Наши представления о родословной Человека сильно изменились за последнее время, но это обстоя— Тельство никак не сказалось на дарвинизме.
105
Далее, дарвинизм неявно выполняет функции теории жизцр тогда как филогенетика остается частным разделом биолог^' Наконец, интересующие эволюционистов события происходят i жду сосуществующими популяциями, в длении, среди живых гЛ' ганизмов популяции. Наоборот, в родословной, объекте фидог^1 нетики, могут оказаться вымершими оба составляющих, пр6до^ и потомок, или один из них; филогенез - это течение времецй
Если филогенез такоонов сводится к превращениям одних г> I ганизмов в другие, то родословная таксонов есть всего лишь пучок сплетенных генеалогий. В генеалогиях порождение одно., го организма другим есть свершившийся факт, сам по себе свободный от неопределенности. Поэтому такое видообразован^ как сумма перевоплощений организмов предкового вида в орга„ низмы вида—потомка само по себе тоже свободно от неопреде, ценности. В таком случае фактическая неопределенность в от, веге на вопрос "кто от кого произошел?* могла бы считаться субъективной, в ней как бы "повинен* исследователь. Очевидно, это умозаключение устроило бы организмоцентризм. С точки зрения эволюционной (популяционистской) филогенетики дело об стоит иным образом. Эволюционируют популяционные объекты. Суждения о их родстве основаны на сходстве в признаках суб. страта. Субстрат - это именно совокупный субстрат организмов популяции. Он характеризуется неопределенной изменчивостью уже в выборке. Поэтому суждение о родстве по сходству будет неопределенным. Такой результат ценен изводит природную неопределенность длящихся
В эволюционной филогенетике однозначного "кто от кого произошел?* в принципе быть не
согласии с теорией отбора элементарной эволюционирующей е» ницей остается популяция. Из популяций состоят виды, роды и другие таксоны. Поэтому через популяцию в филогенетику про никает объективная неопределенность. Казалось бы, это недор» зумение: если один вид уже произошел от другого, то ретроспективное суждение об их родстве может быть только субъективно неопределенным, и в такой неопределенности "повинен исследователь. Однако вдумаемся в выражение "один вид про8, зошел от другого". Ведь изначала длящиеся виды как состоящие из популяций лишены однозначных границ. Следовательно, и суждение о родстве таких видов не может быть однозначна любое единичное однозначное суждение по этому вопросу су^ ективно уже вследствие своей единственности. Объективное ждение - это некоторое множество точечных суждений, свое® множественностью выражающее идею развития. Отношение рического родства — это длящееся, развивающееся стношени6.
Поскольку таксоны состоят из популяций, постольку, ост^ ясь в длении, они осуществляют потенциально неограниченней филогенез. В этом отношении филогенез противоположен онт01
тем, что воспро-популяций.
ответа на вопра может, пока в
106
организма-машины, осуществляемому по конечной наследст-программе. Генеалогическое родство организмов, примере^ которого может служить родословная породистых лошадей, Р^сдедует смешивать с филогенетическим родством таксонов, Не сдельном случае популяций видов (подвидов). Мерой генеа-₽ Некого родства могло бы служить число родственных орга-л°Ги0В, связывающих потомка с предком. При непосредствен-родстве это число равно нулю. По аналогии мерой филоге-^^ического родства, казалось бы, должно служить число пере-'оряЫ* Ф°РМ между родственными таксонами. Могло бы пока-^тьСя, что длина стрелок, соединяющих таксоны в обычных фи-логенетических схемах, пропорциональна числу переходных
м Скорее всего, это ошибка. Опираясь на принцип *вне ви— фор1*1* и
да нет организма , мы должны были бы отвергнуть понятие несходной формы как совокупности организмов, не относящихся к таксонам. Они не могут не иметь таксономического статуса.
Известно, что существуют две формы родства: предок—потомок и сестринское родство (см. рис. 5). Первое несимметрично, второе выглядит симметричным (га* родственно *б*, как *б* родственно *а*; стрелка с двумя остриями), но в действительности они состоят из двух несимметричных отношений пре
док - потомок. Элементарной родословной следовало бы считать непосредственное родство предка и потомка. Граф для сестринского родства есть объединение (не суммирование) двух элементарных родословных с общим предком. Стрелки на рис. 5 обозначают не переходные формы, а лишь следование во времени. Рис. 5, 6 более точно передает это обстоятельство, не оставляя места для так называемых переходных форм, которые не могут быть отнесены к каким-либо видам. Вместе с тем он
создает ошибочное впечатление о скачкообразном видообразова— «ИИ. Вообще проблема переходных форм и различение дискретных единиц в непрерывном историческом развитии остается камнем преткновения для филогенетиков и систематиков. Думается, что главная причина этому - игнорирование популяционной при— P°Qbi видов и других таксонов, что следует считать прочной традицией типологического мышления.
Отношение исторического родства объективно, независимо °т наблюдателя осуществляется в природе. Его можно было бы непосредственно видеть, если бы биология и жизнь были сверсг-ннцами и развивались параллельно. Тогда родство не представ-тцЛ0СЬ эагаАКО®» разрешения которой привлекаются дру— отношения, и прежде всего сходство как свидетельство род-
До 'таксонов. Но без сходства мы не увидели бы таксонов.
Уская, что сходство тем ближе, чем теснее родство, не еле— забывать о полностью подтвердившемся выводе Дарвина с ^зутствии однозначной связи между степенью сходства и
ПеНью родства.
107
Но если мы признаем сходство паллиативом, а усгано^ сходства с целью выяснения родства только вынужденной к (за невозможностью непосредственного наблюдения филогС1 ческого родства), тогда надо признать условным и само ц( тие таксона, по крайней мере в филогенезе. В таком cnyt{e сама филогенетика покажется условностью. Так поступать зя, потому что таксон по определению выделяется родстве! сходством, И тогда уместно поставить вопрос: что увидел г наблюдатель, для которого историческое развитие было бы ь' посредственно наблюдаемым явлением, а сходство и другие > ношения - в принципе недоступными; если не таксоны, то «г/ же? Очевидно, это были бы непрерывные родословные попу, и не было бы оснований даже для постановки задачи класс пирования таксонов. Такой поток популяции распался бы не соны только в тот момент, когда наблюдатель воспользова, бы отношением сходства. Поэтому сходство - не паллиатив филогенетика - не условность. Именно благодаря сходству, торое связано с родством лишь неоднозначной связью, на непр, рывных родословных популяций выделяются дискретные таксон: и тогда возникает задача классифицирования в привычном пои?, мании. Неоднозначность связи между отношениями, точнее, не» возможность мерой сходства точно выразить меру родства, я&. ляется благом для филргенетики, оправдывает само существом ние этого раздела биологии.
На рис. 5, 1—5 вследствие неоднозначной связи между схс;. сгвом и родством классифицирование привело бы к неодинаковым результатам. Причем из рис. 5.1,2,3 следует, что при этом были бы получены различные родословные и системы гар-
сонов.
На этой основе среди систематиков возникли разногласия (Любищев, 1968). Одни во главе с В.Хеннигом ( Hennig, 1965) придают при классифицировании первенствующее значев; характеру ветвления родословного древа, т.е. (ненаблюдаемом-родству. Допускается, что, изучая сходство, можно обнаружу узлы ветвления древа. Критические замечания в адрес этого пушения со стороны В.Эйхлера (1981) справедливы: "филсге^ тическая систематика" В. Хеннига филогенетична не больше диционной систематики. Методические новшества, предложение В.Хеннигом, не превратили сходство в родство. Эта трудности не преодолена и приверженцами математических методов Р6СЧ? та филогенетического ррева по биохимическим признакам (C^i li-Sforza, 1965). Большинство же исследователей не ра-; ют этой иллюзии и считают, что филогенетика располагает 'т°"
~ т Г- л.,рйЗ?
ко сходством. Сходство и родство - разные, не взаимозамен мые отношения, в этом — принципиальная трудность. Сходств' симметричное отношение (Шрейдер, 1971): если "а" похоже ’-' , "б" то*б"похоже на "а". Наоборот, родство - несимметрично6'
108
^е: если *a* происходит от 4 б ц то их нельзя поменять мео-йе меняя отношения. Например, сходство по составу ДНК гаМ0» СХодством, потому что оно не отделяет предка от по-
• дискретность - континуальность системы, это прежде всего \_с о соотношении между сходством и родством. С одной Э° оны, вслед за Ч. Дарвином систематики признают постепен-с'_, эволюции и континуальность множества таксонов, а слецо-^ательно» и конвенциональность границ таксонов. Впрочем, ос— Вователем идеи континуальности можно было бы считать t R Ламарка (Simpson, 1961). С другой стороны, на блюде— животноводов и растениеводов, хорошо известные Чо Дарвин свидетельствуют о том, что по крайней мере внутри вида континуальность наблюдается не всегда: культурные внутривидо» вне формы (один уровень родства) зачастую различаются резче природных видов (более отдаленное родство). Эго следует считать доводом в пользу дискретности внутри вида. Последняя, если следовать логике дарвина, могла бы перевоплотиться в дискретность видов и других таксонов, потому что внутривидовые единицы (разновидности) суть зарождающиеся виды, и между ними нет принципиальных различий.
Впрочем, автор теории естественного отбора признавал дискретность таксонов и в явной форме, указывая, что вымирание промежуточных форм только очертило группы, но никак не создало их* (дарвин, 1939, с. 621). Таким образом, взгляды Ч.Дарвина на дискретность и континуальность неоднозначны, и, по-видимому, в этом их объективность. Тем не менее вместе с другими авторами ( Cracraft, 1979) А.А.Любищев (1966), , по-видимому, выразил относительно более распространенное понимание взглядов Ч.Дарвина по этому вопросу, заключая: *Дио-кретность таксонов - следствие вымирания, эволюция непрерыв— На . Современное состояние проблемы континуальности - дискретности далеко от разрешения. Континуальность предполагает Переходные формы и общих предков дискретных таксонов—потом— к°в, однако палеонтология дала мало предков (Махотин, 1959).
•Н.Криштофович (1950) считает, что они не дошли до нас ^едствие малочисленности организмов и * быстротечности*.
*°Дные взгляды развивают Дж.Симпсон (1948) и Э.Майр у'1). Дисконтинуум у разнополых симпатрических видов жи— них объясняется репродуктивной изоляцией (Gisin , 1969), ГоТаКЖе экологической изоляцией и дискретностью эволюционно-н Родесса ( Darlington, 1940). Что же касается микроорга— ( Мов» то у них, скорее, наблюдается п -мерный континуум с arr”Heise, 1969). Впрочем, в- геномах микроорганизмов от— с^вуюг некоторые так называемые *запрещенные* сочетания
УКЛеогИдОВ1 что, по-видимому, нельзя объяснить недостатком Носких данных (Ашмарин, Харченко, 1971). Для обычной,
109
фенотипической систематики микроорганизмов 'запрет' на тания некоторых* признаков предлагается принять как пост,-, без обсуждения (Заварзин, Старк, 1965). '...Многообрази ших организмов ограничено прежде всего несовместимостью признаков. Отсюда следует, что для низших организмов стр^ тура системы существенно отличается от традиционного эв0(ъ ционного древа, явно вырисовывающегося у наиболее диффер^ цированных форм' (Заварзин, 1974, с. 3). В системе живо?/ ных, как полагает С.И.Огнев (1944), также нет континуа,-п ности.
Невозможность 'фонетического континуума' признана и г ими авторами ( Silvestri —Hill, 1964). А.И.Коробков (197*1 заключает, что дисконтинуум — 'первичное явление, неполноте геологической летописи здесь не причем'. Разумеется, это со. вместим© с тем фактом, что не все вымершие виды, как при* знает, например, В.А.Красилов (1977), сохраняются в некого мом состоянии. Напротив, Ш.Мичнер (Michener , 1970) подар. ет, что дискретность вызвана вымиранием и, кроме того, *эес, люционными потенциями' предков современных таксонов. По С.Я.Парамонову (1940, с. 254), 'гиатус является отражение! эволюционного процесса, а не его основой'. Н.И.Вавилов (1967 рассматривал виды в движении и поэтому считал естественной разграниченность одних видов и нерезкую обособленность других. Три последних автора как будто бы считают дисконтинуум свойством самих таксонов, а не следствием вымирания уже во» никших переходных форм. Б.М.Козо-Полянский (1924, с. 128), признавший симбиогенез в качестве источника разнообразия в>> дов, считал, что во многих случаях переходных форм не было потому, что 'между суммой, полученной в результате интеграции разнородных слагаемых, и самими слагаемыми переходов не существовало'. И, наоборот, отсутствие переходных форм между про- и эукариотами рассматривается как свидетельстве симбиогенного происхождения последних (Хахина, 1979). По поводу этой проблемы, выходящей за рамки филогенетики, Т.И.Тихоненко (1979) заметил, что трансформации и транса^ пии, которые состоят в усвоении геномом прокариот частии ДНК, представляют собой явление совершенно иной природы,-собственно симбиогенез, слияние воедино разных организмов. Что же касается вирусов, которые также могут быть усвоена клеткой, то это явление особого порядка, потому что, не собственного обмена веществ, вирусы могли бы и не считать самостоятельными организмами. Наконец, П.В.Терентьев (1969) понимает таксоны как 'сгущения в поле континуума и, таким образом, предлагает некоторый компромисс между крайностями.
Очень интересна точка зрения К.Бэра (1959, с. 400). ‘ ‘ суждая в 1825 г. систему Р.Окена, он заключает, что нек:
110
модификации совершенно невозможны, "...если бы наша пла-л должна была реализовать все возможные формы живот— строения, то это ни в коем случае не произошло бы вне-это произошло бы только путем последовательного из-'. Здесь имеется в виду не столько неосуществимость, только неосуществляемость континуума, т.е. дискретность рас-г1Л#тривается как результат незавершаемости жизни.
бесследное вымирание переходных форм, признаваемых ког-
йета наго Запн°— менения
да^го существовавшими, необнаруженность их в ископаемом состоянии, есть благо для классифицирования (Бобринский, 1927; Кэйн, 1958; Скарлато, Старобогатов, 1974). Оно позволит лзбежать безвыходных для классификации состояний, таких, как, например, изображенное на рис. 5. "...Лишь неполнота палеонтологических данных создает спасительный морфологический hiatus" (Соколов, 1965, с. 39). Обнаружение переходных форм -неприятность ( Delevryas , 1964). "Плохие таксоны" соединены между собой переходами (Мазуренко, Хохряков, 1981).
Эмоциональная оценка в этих высказываниях любопытна сама
по себе. Она свидетельствует о том, что некоторые исследователи находят в природе то, что хотят найти. Получается, что систематики ищут "зияния" между видами (Н.Кузнецов, 1917), и только незнание (фактор субъективной природы) когда-то существовавших или ныне существующих переходных форм позволяет проводить границы между таксонами. Замечательно, что
это допущение пользуется столь же широким признанием, что и утверждение реальности границ таксонов. В связи с этим не мешает вспомнить о замечании М.Неймайра (1919, с. 131): 'Различать виды в палеонтологии единственно только по присутствию или отсутствию переходов не более чем забава, и выводы, которые делаются при этом, имеют столь же цены, как если бы из большого ящика, заключающего все переходы между Двумя очень отдаленными друг от друга ископаемыми, выхватили бы горсть экземпляров и хотели бы по этому признаку Различать виды". Своим замечанием М.Неймайр справедливо Указывает, что в палеонтологии дискретность нельзя считать объективным явлением, если она вызвана незнанием. Однако он имеет в виду разграниченность во времени. Разобщение "вертикальной" (во времени) и "горизонтальной" (отдельно для сосу— “^твующего на временном срезе разнообразия) классификаций, к Чему склоняется, например, Е.С.Смирнов (1969), разумеется» облегчает работу систематиков: "...система, которая явля-еТся в основном континуумом, становится дисконтинуумом в с^ое современности, и это позволяет неонтологическим систематизм различать их таксоны" ( Clifford, Stephenson, Г975, p.l 1).
* Вместе с тем согласиться с правомочностью разделения ^Ртикальной" и "горизонтальной" классификаций невозможно.
^первых, у нас нет объективных критериев для определения
111
'толщины* временного среза. Во-вторых, нельзя подходить этому вопросу, преследуя достижение удобств для практичен работы. В-третьих, само противопоставление двух классифика пий идет не от природы. В филогении нет разграниченных ьрк менных срезов, есть непрерывное течение фелогенеза. Такое противопоставление целесообразно в экологии, когда только вне сосуществующие животные и растения могут состоять в экологических отношениях. Задача упорядочения органическою разнообразия по сходству, вызванному родством, не влечет , собой целесообразность разобщения горизонтальной и вертик ной классификаций. Зато в неявном виде эта задача содержи-проблему соотношения между эмпирической выборкой и более обширной биологически возможной генеральной совокупностью органического разнообразия. Систематик, как и филогенетик ( Bretsky , 1979), находится в положении следователя, раскрь-. веющего преступление: каковы факты, таковы и выводы. Наука располагает только эмпирической выборкой фактов, выводы же должны быть распространены на все филогенетическое древо (рис. 7). Совсем не очевидно, что все или хотя бы важнейшие части филогенетического древа представлены в эмпирической выборке фактов. Но допустим все же, что в эмпирической выборке представлены все части древа, и, больше того, пусть из-вестны все виды полностью. Теперь их надо упорядочить по сходству так, чтобы порядок отразил ненаблюдаемое непосредс?. венное родство. Здесь возникает все та же принципиальная тр$-ность: одно только сходство не выделяет следование во времени предков и потомков. Независимо от того, работает филогене» тик в палеонтологии или в неонтологии, сходство остается едда-ственным универсальным средством установления родства. Вместе с гем хорошо известно, что сходство вызывается не только родством, но также и совместностью* обитанием в одном биотопе (акула и фин). Каким образом в своей работе
Рис. 7. Родословное древо видов как его изображают филогенетики. Уже из-вестные науке живые ( • ) и вымершее ( В ) видь! в действительности пред-, ставляют собой выборки из гениально*1 совокупности организмов вида. Такая вокупность потенциально бесконечна fW® живых видов и конечна для вымерших видов (двойная линия). Суждения филоГ котика, представленные филогенетичео-ким древом, зависят не только от выборки организмов из длящегося множества организмов вида, но еще и от наружения еще не известных живых В и вымерших ( □ ) видов. Пунктирная линия обозначает следование видов в 1 чение времени
112
«сги
распознают именно то сходство, которое обусловлено родст-для этого используется метод гомологий.
Основы гомологического метода фактически заложены ЖСент-
илером еще в 1825 г. (Амлинский, 1955). Впоследствии д Я.Бляхер (1976) и А.Ремане (Remane, 1955) всесторонне ^смотрели это понятие.
Р ромологическими у разных организмов называются органы и
структуры, такие, которые расположены одинаков,о относительно других органов и структур - критерий положения (Owen, 1855); ^логические органы и структуры развиваются в онтогенезе кз одних и тех же зачатков, и, кроме того, они имеют примерно одинаковое строение. Если же они устроены по-разному, то мо-гуг быть соединены постепенном переходом из структур переход-ноГО строений, наблюдаемых у других организмов. Гомологический метод проник в нумерическую систематику ( N. Jardine, С.Jardine, 1967). Он был распространен на цитологические структуры и биохимические признаки, правда, без особого успеха (Grim-stone, 1959; Wooihouse , 1970), потому что одни и те же тонкие ультраструктуры клетки и вещества возникают независимо у разных организмов различными способами. Через гомоло
гии в операцию установления сходства вовлекается едва ли не вся организация всего организма. Вследствие этого систематики избавлены от грубых ошибок, когда сходство обусловлено совме» стным обитанием в сходных условиях. Тем не менее метод гомологий не всегда позволяет отделить глубокое сходство, обу
словленное родством, от сходства, вызванного другими причинами (совместность, сосуществование, корреляция), и возможности его применения во многих случаях ограничены (Е.Смирнов, 1959; Гиляров, 1964). Главное же состоит в том, что гомологическое сходство - эго по-прежнему лишь свидетельство родства. В этом смысле использование гомологического сходства в филогенетике не создает каких-либо особо благоприятных условий для установления родства. Гомологическое сходство по-преж» Нему симметрично и не позволяет отличать предка от потомка. Филогенез таксонов - только одна из форм исторического па» Разные формы исторического родства занимают большую часть овала Сродство* биологической розетки, а 'чистое*, непосредственно наблюдаемое родство, для установления которого не Понадобилось бы других отношений, занимает ячейку 1 (рис. 8).
В биологической розетке (рис. 8) овал 'родство* обозначает ГеЧение времени. В той его части, которая перекрывается овалом 'сосуществование', дополнительно имеется и дление време-Пунктирная часть овала — эго филогенетическая систематика Филогенетика таксонов. Ее можно рассматривать ео стороны °тНощеНИЯ родства, вызывающего сходство (эго будет филогенез гика таксонов), и со стороны отношения сходства, обусловленное Г° Родством (это будет систематика родословных). В первом слу»
^1560
113
чае ведущим отношением будет родство (табл. 4), во втор, сходство.
Объединение популяций в ячейке 1 есть родословная пог ний. Она разветвлена уже вследствие сестринской формы р(
Рис. 8. Логическая структура исторической биологии в овал 'родство биологической розетки.
Пунктиром обозначена область филогенетической систематик! sensu rato , Собственно филогенетической систематике sensu stricto отвечает ячейка 2
ва, и Для такого членения родословной на ветви не требует никаких других отношений. Ограничиваясь одним только он нием родства, наблюдатель не смог бы выделить, на родосл ной никаких других объектов, кроме узлов ветвлений и окот ний ветвей. Если бы наблюдатель дополнительно воспользоь< отношением сосуществования (ячейка 22), тогда он смог бы выделить на разветвленной родословной сосуществующие попу* ляции даже в линейной последовательности предок — потомок. Такое наблюдение могло бы быть кратковременным или прод0/? жаться длительное время в воображаемом сосуществовании сверстниц — биологии и жизни. В последнем случае выделились бы всевозможные случаи сосуществования, наблюдатель увиА611 бы разобщение в течение времени популяции живых, сосушесГ| вующих организмов. Например, в современных нам фауне и Ф’0' ре есть родословные, когда предки и потомки состоят из организмов. Каргина жизни сделается еще более содержателе *ной, если привлечь дополнительно и другие отношения. С п014^, щью отношения совместности наблюдатель распределит мноЖе<^ во родственных популяций по местообитаниям (ячейка 8).
114
исторической биологии ячеек см. на рис. 7)
Таблица 4
Объекты (цОмеРа
Ячейке Историческая биология sensu stricto родство типов организации, жизненных Форм, биотических элементов Ячейка Филогенетика (филогенетическая систематика) — родство таксонов
1 6 8 19 родословные; жизнь в течении времени родословные типов организации! родословные биотических элементов; биото-генез2 родословные типов организации как биоти- ческих элементов; ти-погенез как биотогенез 2 11 4 родословная таксонов (родственных фенонов); филогенез (таксоноге-нез)4 родословная таксонов как типов организации; филогенез, гипогенез родословная таксонов как типов организации и биотических элементов; филогенез, гипогенез, биотогенез
10 21 родословные типов организации как жизненных форм и биотических элементов; типоге— нез, биом орфоген ез, биотогенез родословные биотических элементов как жизненных форм; био-гогенез, биоморфогенез 13 12' родословная таксонов как биотических эле- ментов и жизненных форм; филогенез, биотогенез, биоморфогенез родословная таксонов как биотических эле- ментов и жизненных форм; филогенез, био-гогенез, биоморфогенез
23 22 родословные типов организации как жизненных форм; типогенез, биоморфогенез родословные жизненных форм; биоморфогенез 15 14 родословная таксонов как типов организации, биотических элементов и жизненных форм; фи- логенез, гипогенез, биотогенез, биоморфо- генез^ родословная таксонов как типов организации
1 t ел. , Близко по значению к градогенезу или анагенезу Дж.Гек-ааз 1 Huxley, 1958).2 Объединение фауногенеза и флорогене-
Близко по значению к филоценогенезу5 Поскольку таксон
Фейл. о&ьегмнение родственных фенонов, филогенез есть еще и
и«огенезр По-видимому, отвечает видообразованию. 115
Продолжение таблицы .
Ячейка Историческая биология sensu stricto родство типов организации, жизненных форм, биотических элементов Ячейка Филогенетика (филог. нетическая система! ка) - родство таксо
17 и жизненных форм; с. логенез, типогенез, морфогенез родословная таксоне как жизненных форм филогенез, биоморфс нез
подразделения родословной окажутся приуроченными к океану или к суше, другие - к тропикам или высоким широтам, тре * к высокогорьям или абиссали. С точки зрения географии эгс элементы географической оболочки - сектора, зоны, пояса. ( точки зрения биотики такие местообитания предков и погомк: есть филохор, а сами обитатели — элементы биоты. Таким о зом, история жизни, обособление всех новых предков и пого ков, есть в то же время воссоздание биологического простр ства в географической оболочке.
После того как будет привлечено отношение корреляции (’ ка 6), родословная популяции распадается на родословные г пов организации. Тогда течение жизни будет представляться процессом становления и обособления типов организации, rat например, как микрофауна и микрофлора. Водный и наземны? типы организации, тропический и высокоширотный, высокого} ный и абиссальный типы подразумевают дополнительно совм< ность (ячейка 19). Можно видеть, что эта система типов о низании фаунистических элементов не совпадает с той, kotoj отвечает ячейке 8. Дифференциация первоначального типа ор низании живых существ 'вообще* вызвана, в частности, осы нием географической оболочки (ячейка 19). Дифференциация пов вызывается неоднородностью не только пространства, нс времени (ячейка 23). Причины этой неоднородности — заме? ние скорости вращения Земли, колебания климата, глобально изменения орографии. Существенное значение в этом проае‘ имело прохождение 'точки Пастера', когда вследствие наког ния в земной атмосфере свободного кислорода анаэробное Дг"• ние сменилось аэробным.
Ячейки 10 и 21 аналогичны: первая - ячейке 19, втор®1 ячейке 8. Отличие первых от последних состоит в том, что них дополнительно использовано отношение сосуществований"
116
г -честно сосуществующие родословные в ячейке 21 состоят ь Ж»вых сосуществующих предков и потомков, обитающих сов-^стно. Возьмем, например, фауну Байкала. Наряду с другими 1 улисгическими элементами она включает эндемиков, обитаю-• х только в Байкале. Очень вероятно, что среди эндемиков эндемичные же предки и потомки, образующие родослов—
е Такие родословные состоят из длящихся во времени пред— л потомков. Они служат примером для ячейки 21. Тогда пимером для ячейки 10 могли бы служить совместно сосуще— ^руюшие типы организации жизненных форм опылителей и энто-мофильных растений. Такое объединение непохожих существ геМ не менее естественно, потому что не задано отношение сходства. Три отношения — родство, корреляция и сосущесгво— ваййе — выделяют в ячейке 23 объект, похожий на тот, который отвечает ячейке 10, однако здесь не требуется совместности. Примером может служить объединение живых опылителей и энтомофильных растений, однако разобщенных в пространстве,
обитающих, например, на разных континентах.
В таксономической части овала ячейка 2 содержит собственно таксоны, не объединенные никакими другими отношениями, кроме родственного сходства. Например, это прокариоты или эукариоты. Ячейка 11 содержит такие таксоны, которые к тому же и типы организации. Не всякий таксон есть еще и тип организации, во всяком случае класс насекомых представляет собой большее единство как тип (однообразное расчленение тепа), нежели класс ракообразных. В ячейке 4 подразумевается, что такие таксоны - типы обитают совместно. Например, вид белый медведь, обитающий в Арктике, вместе с тем может считаться и типом организации арктического хищника, сохраняющего активность в течение круглого года. Другой пример - это тип организации пещерного животного — протея, обитающего в темноте в чрезвычайно стабильных условиях. В ячейке 15 подразумеваются только живые, совместно обитающие популяции гаксона — типа. Оба примера — белый медведь и протей - отве-чак>т этому условию, если ограничиться только параллельно длящимися популяциями сосуществующих современников и исклю-'кИГь ранее вымершие популяции, не сосуществующие с данными. Для ячейки 14 подошел бы пример таксона - типа, образованно-10 изолированными, но сосуществующими популяциями. Сюда от--й°сятся пространственно обособленные сосуществующие популя— ^ии протея в пещерах Югославии. У белого медведя, по крайней в настоящее время, популяции, может быть, не изолирова»
К ячейке 17 относятся сосуществующие таксоны, вэаимо— '-Тйующие как жизненные формы; в свою очередь их объели— Ие также есть таксон — жизненная форма. Например, сосуще— млекопитаюшие ВКЛ£°чаюг жизненные формы травояц-и хищных. Вместе взятые они образуют класс млекопитаю-
117
ших. (если бы не существовало других млекопитающих, напр^ грызунов) и в то же время жизненную форму активно подвид6 НЫХ ЖИВОТНЫХ, противоположных неподвижным ЖИВОТНЫМ, мёр губкам. Ячейки 12 и 13 содержат объединения популяцщ в таксоны. Такие популяции либо сосуществуют (ячейки 12) либо лишены единства во времени (ячейка 13). Например, не живущие и вымершие триасовые популяции щигня, живого ископаемого, обитают (обитали) совместно на Европейском Кс„ тиненте. Этот пример относится к ячейке 13. В ячейке 12 Он’ представляют самостоятельные объекты. Популяции современна, и вымерших щитней по отдельности выполняли (выполняют) ции. эврифага в сообществах временных водоемов.
Формы исторического родства многообразны. В течение Bps. меня связаны родством не только таксоны, но и типы органа запии, жизненные. формы, биотические элементы и всевозмож-ные сложные объекты, наделенные свойствами основных обьек, тов. Многообразие исторического времени вызывается способ, ностыо популяций включаться в разные объединения популяций на основе различных отношений и их всевозможных сочетаний.
Глава 10
ОТНОШЕНИЕ СОСУЩЕСТВОВАНИЯ
И ЭКОЛОГИЯ
Если биологическая специфичность заключена в межпопуляш-онных взаимодействиях, то понятие биологического сосуществования следует применять к популяциям, а не к отдельным организмам. Сосуществование есть необходимое условие для того, чтобы между популяциями могла осуществляться борьба за су-шествование; недостаток жизненных ресурсов есть достаточное условие. Поэтому разногласия по поводу того, что представляет собой сообщество - объединение жестко взаимозависимых или, наоборот, максимально независимых в своей жизнедеягелк ности организмов (Шорыгин, 1955; Whittaker, 1972), не представляются непримиримыми. По-видимому, следовало бы и* лагать, что сила борьбы за существование в сообществах не постоянна и зависит от внешних условий.
В сообществах животные и (или) растения объединены био* ценологическими или симфизиологическими связями (БеклемИ** шев, 1951). Последним выражением удачно подчеркнуто то 0 стоятельство, что популяции в сообществе связаны через жи3* ненные функции обменом веществ и энергии, что, однако, рует семантико-информационные взаимодействия. СимфизиоД^ ческие связи осуществляются в хищничестве, паразитизме, биозе, комменсализме и пр. Вслед за А.А. Быковым (1968)
118
0М выражения "экологические связи*. Разнообразие таких аей довольно велико: топические, трофические, фабрические, се^0Ческие. Примером первых могут служить ели и березы в вторых - хищник и жертва, третьих - раки-отшельники и
юХ°ноГие моллюски« четвертых - некоторые лианы и пальмы, т кН®* связи могли бы считаться экологическим выражением 1 бы за существование. Экологические связи бывают прямые
ревенные. Благодаря последним, едва ли не каждая популя-
0 а связана с любой другой через посредников. Об этом свиде-цИ**
льствуюг так называемые пищевые цепи в море, примеры ко-«срых приведены в книге Л.А.Зенкевича (1947), или аналогич-Hbie схемы из многочисленных сводок по экологии (Макфедьен, 1965; Одум, 1975). Вследствие этого имеется достаточное основание для понятия единого живого покрова Земли.
для литературы последних десятилетий особенно характерно пОдклкнение к сообществам небиологического компонента, тако-г0| как почва, грунт, вода, атмосфера. Эго было уже у Р.Лин-цемана (1943). Такие объекты получили название биогеоценозов и экосистем. Внутренняя непротиворечивость и широкая распространенность этих понятий объясняются тем, что, участвуя в экосистемах своим совокупным вещественно-энергетическим субстратом, животные и растения действительно утрачивают биологическую специфичность, которая заключена в семантико-информационных взаимодействиях. В экосистемах через обмен ве
ществ и энергии животные и растения участвуют в вещественно-энергетических процессах наравне с небиологическим компонентом. Миграция вещества и энергии внутри биологического компонента экосистемы и между ним и небиологическим компонентом (его иногда называют биомом) подчиняется одним и тем же закономерностям физхимии. Поэтому сообщество, биологический компонент экосистемы, рассматривается прежде всего как объединение продуцентов, консументов и редуцентов, как, например, у Р. Уиттекера (1980). В истории экологии, как эго следует из обзора Г.А.Новикова(1980), этот подход приобрела все большее значение.
Думается, однако, что такой подход выводит экологию за пределы собственно биологии в область биосферологии (Гегамян, ®80), планетарную зону взаимодействия между биологической ^органической формами движения материи, осложнившегося Результате воздействия на нее социальной формы движения ^нтерии общества. Биологический вклад в эго небиологическое ^еление глобальной миграции вещества и энергии, начавшийся ^Рождения жизни, выражается в существенном убыстрении Усложнении таких миграций. Масштабы и характер биологиза-эГих явлений, явственно проступающие при сравнении Земли зисизненными планетами, ярко описаны В. И. Вернадским
119
Современная биосферология развивает представление о геохимической функции живого покрова Земли как одного из уровней организованности биосферы (Шипунов, 1980). В бд^ сферологии выражение "живое вещество" имеет вполне опред^ ленное биогеохимическое, но не собственно биологическое чение. Оно свободно от той двусмысленности, которая северу ся в этом же выражении в биологическом контексте (вещест^" не живое, но организмы состоят из веществ).
Понятие сообщества, биологического компонента экосистем^ сопряжено с понятием жизненной формы (биоморфы). Начиная с трудов А.Гумбольдта, вышедших в 1805 г. (Гумбольдт, 193g) оно в основном разрабатывается ботаниками (Варминг, 1902; Raunkiaer, 1934). Его история достаточно подробно рассмот, рена И.Г.Серебряковым (1962) и Ю.Г.Алеевым (1980).
Д.А.Криволуцкий (1967) отмечает, что понятие биоморфы име. ет своего рода предысторию в таких народных выражениях, как "норники" и "землерои", в которых отражены особенности о фаза жизни животных, в данном случае в роде грызунов. Ознакомление с историей этого понятия позволяет установить еле. дующее. Прежде всего, биоморфа — это структурный элемент сообществ (биоценозов), живых или уже вымерших (Попов; 1962; Мер клин, 1968). В последнем случае они изучаются м. леоэкологией (Геккер, 1934). Будучи соединенными экологическими связями, биоморфы есть к тому же и экобиоморфы сообщества. Поэтому сообщество имеет "экобиоморфный состав" (Алексеенков, 1971), представляя собой "спектр биоморф" (Акимов, 1954), или "биоэкологический спектр" (Бычков, 1967). "Не существует местообитания такого, которое характеризовалось бы единственной жизненной формой..." (Cain , 1950, р. 5). В сообществах биоморфы связаны не родственны ми, сходственными и (или) другими отношениями, а именно симфизиологическими связями, "... с ис тематике-морфолог ичесю отношения не играют гут никакой роли..." (Варминг, 1901). Они "ощущают" один другого как биоморфы, независимо от родства, сходства и таксономического положения.
"Причисляя организм к данной жизненной форме, мы характ®* ризуем его экологически со стороны отношения к биотическим и абиотическим факторам среды, а также в отношении места и роли его в биоценозе. В таком понимании биоморфы являются структурными элементами биоценоза, пригнанными отбором друг к другу" (Акимов, 1954, с. 27). По-видимому, эго опре" деление соответствует наиболее широко распространенным прей" ставлениям о биоморфе. Показательно, что в нем биоморфа в равной степени определяется через взаимодействие с биотическим и абиотическим факторами. Такой подход представляется оправданным потому, что симфизиологические связи в своей основе — это обмен веществ и энергии, который осуществляйся
120
каково между живыми и (или) неживыми телами. Показагель-°'4 таюке, что в нем ничего не говорится об организменном суб-fi° ате. ° жив°й плоти. Следуя определению М.П.Акимова (1954), с F считать биоморфой "хищника". Эго слово уже выделя-h<bI бломорфУ через указание ее роли в сообществе, но не созда— еГ наглядного чувственного образа. И только заинтересовавшись Дратом, мы можем обнаружить, что "хищник* - эго объеди-СеВце хишных бактерий, хищных грибов, хищных растений и жи-рОгНЬ1Х» Более точно, "хищное животное* или "хищное растение* есТЬ "жизненная форма 'таксона", потому что оно имеет в виду таКсоны Animalia и Plantae . Значит, в таких выражениях, как "хишное животное", помимо отношения сосуществования, родразумевается отношение родственного сходства.
Разные биоморфы обычно выделяются на основе двух, пона-
чалу кажущихся равнозначными, принципов: функционального и субстратного (структурного). В таких названиях жизненных форм, как, например, хищник, фитофаг, ксерофит, подразумеваются особенности образа жизни и функционирования в сообществе. Наоборот, "травянистые растения* и "микрофауна* - это примеры использования субстратного принципа, в данном случае в его узком морфологическом значении (без явного указания на особенности вещественного состава).
Могло бы показаться, что использование обоих принципов
приводит к выделению одних и тех же названных форм, потому что представляются равносильными два суждения: 1) если организмы выполняют данную функцию в сообществе, то они устроены гак, а не иначе (от функции к структуре), 2.) если организмы устроены так, а не иначе, то они выполняют данную функцию в сообществе (от структуры к функции). Известно, однако, что органы и вообще структуры мультифункциональны, т.е. один и тот же орган может выполнять разные функции. Вместе с гем одна и та же функция может выполняться разными структурами, т.е. имеет место мультиструктурность. Например, газообмен у высших животных может осуществляться не только через Легкие, но и через кожу. В свою очередь легкие служат не только для газообмена, но и для водообмена. Что же касается кожи, то она дополнительно выполняет защитную функпжч Пред-ставляется очевидным, что оба суждения не равносильны и не ^огут привести к одинаковым результатам. Исторически раЬсуж-Аение от структуры к функции предшествовало рассуждению от Функции к структуре. А.Гумбольдт выделил биоморфы растений наиболее приметным особенностям внешнего строения (палы
Мы» лианы и пр.). Назвав их "физионимическими типами", он Фактически отдал предпочтение субстрату (структуре), думается» что гумбольдтовская жизненная форма "лианы" или "кактусу Должна скорее считаться феноном. Если же под "лианой" п Уразумевать именно паразитическое растение, а под "какту
121
сом* именно ксерофита, то это будут примеры объекта *фецс как жизненная форма*. Теперь примем, что *лианы* и *какт сы* — это два таксона. Тогда получим два объекта, называе мых так: *жизненная форма таксона*, или в развернутой фор, * жизненная форма родственных фенонов*.
Как следует из обзора Т.И.Серебряковой (1972), на пра ке субстратный и функциональный подходы обычно не различ. ются. Между тем скорее всего только функциональный подхо М.П.Акимова (1951) выделяет жизненные формы в 'чистом де*. Когда привлекается дополнительно субстратный подход, лучаются разные частные приятия жизненной формы.
Еще одна трудность субстратного подхода состоит в том, что если ограничиться только особенностями внешнего строе как вслед за А.Гумбольдтом предлагают М.Т.Мазуренко и А.П.Хохряков (1981), то из понятия биоморфы придется вьп ти, например, жизненную форму 'эндопаразита'. Такое оскуг ние понятия биоморфы было бы нецелесообразным. Нельзя ут. кать из.виду того соображения, что симфизиологические свя. могут осуществляться не только с помощью наружных, но те
Рис. 9. Логическая структура экологии. .Жирная линия обозначает плоскость синтопности—диатопности
же и внутренних органов, в данном случае хозяина. Эндопар ты — это частная жизненная форма паразитов вообще. Выра» ние * внутриклеточный паразит* - это дальнейшее подразделе жизненной формы паразита. Таким образом, по-видимому, эд имеется иерархическая структура, как в систематике. Оооты шение функционального и субстратного подходов при выделен жизненных форм видно из рис. 9. Субстратный подход, т.е. н
122
еление жизненных форм по сходству строения (морфологические л биохимические признаки) и (или) по скоррелированности ганов (морфологические и биохимические структуры), занима-
ет только пунктированную часть овала * сосуществование". Но р этой части овала структурный подход не исключает функцио-
^льного, т.е, отношения сосуществования. Оставшаяся непунк-тцрованная часть овала отвечает экологии "в чистом виде", не нуждающейся в субстратном подходе. Действительно, одни и те же функции в сообществе могут выполняться резко различающиеся в субстратном отношении организмами, например, уже на
званными хищниками-животными и хищными растениями и грибами; слово "хищник" обозначает функцию в сообществе и ничего не говорит о субстрате. Иногда, когда сообщество, например, надо представить как отвлеченную систему связей продуцентов с консументами, субстратный подход не нужен, он приводит к
выделению частных жизненных форм.
Своеобразным "местообитанием" сообщества является временная точка, выделенная наблюдателем. В ней нет со-дления
в том смысле, что жизненные формы, объединенные в сообщест-
во, взаимодействуют при осуществлении жизненных функций организмов и связаны через обмен веществ. Здесь работают обычные физикалистские представления о времени. Сосуществование оказывается еще и со-длением, когда мы обращаем внимание на то обстоятельство, что жизненные формы могут "ощущать" еще и приспособленность. Тогда сосуществование становится биологически специфичным со-длением, в котором нет привилегированных временных точек. Тогда временные границы сообществ пропадают и становится несодержательным вопрос о том, где во времени кончается одно сообщество и где начинается Другое. Становится некорректным и вопрос о четко однозначной классификации жизненных форм. Иными словами, снимается вопрос, аналогичный тому, который обсуждается в систематике. В принципе эти вопросы решаются так же, как в систематике, надо только заменить слово "таксон" словом "жизненная форма".
Рассматривая овал "сосуществование" (см. рис. 9), мы находим в нем плоскость симметрии. Ее положение показано жирной линией. Справа от этой Линии находятся сообщества жизненных форм, объектов диатонической экологии, обитающие в био-т°пах. Слева размещены сообщества—биоты. Они состоят из биоморф как биотических элементов и населяют биотопы как биохоры (см. табл. 5). Эго объясняется гем обстоятельством, область слева от плоскости симметрии есть пересечение Овалов "совместность" и "сосуществование". Она в равной ме-принадлежит биотике и экологии и обозначает синтопическую эКологию. В этой связи уместно вспомнить о замечании У.Тью-Релла (Turrill, 1959, р. 172): "Нет ясной границы между
123
• Таблица
Объекты экологии. Номера ячеек см. рис. 9
Ячейка Синтопическая экология Ячейка Диатоническая экод^
30 биоморфы сообщества как биотический элемент; биотоп как биохор 9 20 биоморфы сообщества биотоп биоморфы как типы о] ганизации; биотоп ка»
31 биоморфы сообщества как типы организации; типотоп
биотоп как типотоп (типохор) 29 биоморфы как типы ор ганизации и феноны; биотоп как типотоп и
16 биоморфы как типы организации и биотичес— ' 18 фенотоп
кие элементы; биотоп биоморфы как феноны,
как типотоп (типохор) биотоп как фенотоп
К и биохор 14 биоморфы как таксоны
27 биоморфы как биотические элементы и феноны; биотоп как био- и типы организации; биотоп как ареал и т«. потоп
хор и фенотоп (фенохор) 17 биоморфы как таксоны: биотоп как ареал
12 биоморфы как биотические элементы, феноны и родословные (таксоны); биотоп как био- 22 биоморфы как родослс ные; биотоп как фило хор
хор; фенотоп и филотоп (филохор) - ареал 23 биоморфы как родослс ? ные и тины организации; биотоп как фило-
15 10 биоморфы как биотические элементы, феноны и родословные (таксоны), типы организации; биотоп как биохор; фенотоп и филотоп (ареал), типотоп (типохор) биоморфы как биотические элементы, родословные и типы организации; биотоп, филотоп (филохор), типотоп (ти— похор) топ и типотоп
124
Продолжение таблицы 5
Ячейка
21
Синтопическая экология Ячейка Диатопическая экология
биоморфы как биотические элементы и родословные; биотоп как биохор и филотоп (филохор) •
экологией растений и географией растений*. Эго отметил и зоолог Д.Н.Кашкаров (1938). С.И.Левушкин (1974, 1976) пишет об "экологическом фаунистическом комплексе*. По Р.Мак—Ар гуру (MacArthur, 1972), "географическая экология* - это примерно то же, что "экологическая география*. Объективно такие сочинения, как, например, книга А.П.Кузнецова "Экология донных сообществ Мирового океана" (1980) или "Растительный мир Земли* (1982), представляют собой вклад не только в биогеографию, но также и экологию. Благодаря своей двойственной природе, объекты синтопической экологии могут служить объектом биогеографического районирования (Воронов, 1976), т.е. выделяются как индивидуальные регионы (в биогеографии) и биохоры (в биотике).
Диатопическая экология имеет дело 'с типами сообществ безотносительно к их географической индивидуальности, т.е. к локализации на планете. Возьмем, например, сообщество леса. Конкретные формы сообщества этого типа можно встретить в разных уголках планеты. Само понятие "сообщество леса* предполагает отвлечение от географической индивидуальности разных лесов. Другие примеры типов сообществ - литораль, тундра, степь и пр. Упорядочение разнообразия таких сообществ есть содержание задачи типологизации. Процедура типологизации под названном "классификация сообществ* (Griffiths, 1974) приводит к выделению "параллельных сообществ* (Тhors on, 1956 )» *парал-Дельных биот* и викарирующих ассоциаций животных и растений (Coinrad iop о lus, 1981) на основе одного лишь сосуществования без совместности. Примерами таких Диатонических объектов могут служить, например, хвойные леса (Северной Америки, Евразии), сообщество литорали островов и материков обоих полушарий. Сюда же относятся высотные и поясные викариаты (Cain, 1944), а также такие понятия, как экологические эквиваленты (Одум, 197 5), морфофизиологические параллелизмы (Николаев, 1977) и другие аспекты широкого явления викарирования (Saste-Ге« 1981). Яркие примеры викариатов представляют фауна и Флора Австралии в сравнении с другими регионами: волк и сум-ч^тый волк, белка и сумчатая белка, крот и сумчатый крот. Мно-
125
гочи с ленные примеры подобного рода приведены у Э.Пианки (1981). Синтопическая экология имеет дело с конкретной ной совместностью* и рассматривает сообщества на конкретном физико-географическом фоне. Он представляет собой фактически наблюдаемое распределение материков и океанов, климатических зон, форм мегарельефа и пр. При таком рассмотрении живой щч кров Земли распадается на индивидуальные сообщества-биоты, в равной мере интересующие биогеографов и экологов. Выделение таких сообществ—биот и есть задача районирования.
Время в овале "сосуществование* представлено либо одним длением, либо в пересечении с овалом "родство*, а также течь, нием. В ячейке 9 (рис. 9) имеется только дление, сосуществование популяций. Они взаимодействуют экологически как жизненные формы сообщества и состоят в борьбе за существование. Такой представилась бы, картина жизни наблюдателю, лишенному возможности устанавливать дополнительно и другие межпопуляцц. онные взаимодействия. Разумеется, эта точка зрения малосодер. жательна. В конце концов она позволяет увидеть генеральное объединение популяций как нерасчлененный живой покров плане*, ты, либо разнесенный по разным временным точкам наблюдения (экологии как биосферологическая проблема), либо в длении популяций с прошлым, не отделенным от будущего. В ячейке 22 дополнительно задано течение времени, родство. Оно выделяет в живом покрове планеты родословные популяционных жизненных форм. Объединение таких родословных есть "жизнь биоценоза во времени" (Кашкаров, 1939).
В истории цветковых растений четвертичного времени установлено изменение спектра жизненных форм, таких, как лианы, эпифиты, гидрофиты (Hansen , 1956). Эго явление тоже относит ся к ячейке 22. Биотоп, местообитание таких развивающихся жизненных форм, есть одновременно и филотоп Ячейка 30 обозначает дление живого покрова Земли. Верхнему предельному зна чению совместности отвечает биосфера как предельно широкий биохор. Такая совместность отвечает общему представлению о жизни как планетарному явлению в конкретном земном воплощении, т.е. о земной жизни. Более частные значения совместности ведут к выделению жизненных форм сообществ материков и океанов, разных физико-географических пояс-'ч и других подразделений географической оболочки. В ячейг 21 биота и биохор приобретают дополнительно свойство филотопа, потому что в ней задано помимо сосуществования и совместности еще и отношение род' ства. В соединении с родством совместность выступает как род" ственная совместность (популяции обитают совместно потому, что они родственны). Эго означает, что живой покров Земли по1? разделен на такие отдельные сообщества, которые состоят из совместно обитающих родственных популяций жизненных форм. Они обитают в биотопах-биохорах, являющихся и филотопами. &
126
^рдяя в стороне вопрос о мере совместности и родства, надо Гц1иь отметить, что родство и совместность могут быть гесны-
И отдаленными. Такое видение жизни, когда заведомо извесг— сосуществующие родословные (при этом не требуется отно-нйя сходства) как бы вплетены в живой покров Земли и взаи— □действуют экологически, не встречается на практике. Тем не ^едее это не абстракция. Дело в том, что картина жизни, вос-с0здайная современной биологией, соотнесена с весьма непродолжительным существованием науки. Поскольку биология не была свидетельницей истории жизни, историческое родство не было не— росредственно наблюдаемым феноменом. При параллельном разви— т1щ жизни и биологии историческое родство как самостоятельная категория не нуждалось бы в обсуждении на основании косвенных свидетельств, например сходства. Тогда экологические взаимодействия сосуществующих родственных популяций не казались бы абстракцией. Такой подход и проводится в ячейках 21 и 22.
На практике же, устанавливая экологические отношения, био-лдри обычно обращаются к совокупному вещественно-энергетическому субстрату и таким образом привлекают отношения сходства (в признаках) и корреляции (структур); это - пунктированная часть овала. Ячейка 20 отвечает гой точке зрения, когда сообщества представляются состоящими из биоморф как типов организации. Такими типами организации могли бы быть, например, продуценты, редуценты, консументы. Все эго биоморфы как типы морфофизиологической организации, единства скоррелированных морфологических и биохимических структур. Носителем таких структур является совокупный вещественно-энергетический субстрат популяции. Их местообитание можно называть типотопом. Типы организации биоморф функционируют как единое сообщество, потому что тип организации продуцентов скоррелирован с таковым ксдсументов и оба они скоррелированы с редуцентами. В свою очередь каждый тип подразделяется на своего рода подти— пы. Например, коДсументы подразделяются на хищников, паразитов, некрофагов, фитофагов и т.д. Далее, хищники бывают макрофагами и микрофагами, пассивными и активными, и т.д.
Таким образом, биоморфы как типы организации характеризуйся иерархической структурой, впрочем, как и в Других ячейках. ® экологии типы организации рассматриваются преимущественно в трофологическом аспекте. Примерами могут служить "экологические группы" с разными типами питания, выделенные Т.Г.Мир-^инк и И.П.Бабьевой (1981). Трофические взаимоотношения меж-подобными группами у растений и их консортов Т.А.Работнов 4 78) называет микосимбиотрофией, бактериосимбиотрофией, ^икосимбиотрофией. Однако в сообществах пищевые связи — эго °Пько одни из видов симфизиологических связей. Не следует ^Ускать из виду и другие связи, например топические. В них скрываются новые грани понятия биоморфы как типа организа—
127
ции. Например, корреляция между древостоем, подростом и вянисгым покровом, между сестонофагами и детритофагами в обществах морского дна. Подчеркивая то обстоятельство,
экологические взаимосвязи отражаются в строении животных (или) растений, биологи пишут о коадаптациях. Ячейка 31 чается от ячейки 20 тем, что сообщества, образованные био^ фами как типами организации, дополнительно упорядочены и шением совместности. Выделение этих объединений в ячейке 2 есть типологизации, упорядочение в ячейке 31 - районирован^
Сопоставляя ячейки 20 и 31, мы сравниваем частные обье ты диатонической и синтопической экологии (см. табл. 5). э», сопоставление можно продолжить и на примере другой пары нч ек - 10 и 23. Они содержат родственные биоморфы как типы организации, объединенные отношением совместности (ячейка ; или без совместности (23). В биологии фактически имеются эти понятия. Согласно широко распространенным сейчас пред^ влениям, жизнь зародилась в океане. Следовательно, океанские
сообщества предшествовали наземным и пресноводным, и, знач/ тип организации морских биоморф предшествовал прочим. В эп смысле в свете современных представлений о родственных св», зях населения океана, суши и пресных вод (Бирштёйн, 1949; Хлебович, 1974) мы можем рассматривать понятия экологии! историческом аспекте. В литературе эта проблема обозначаете как "эволюционная экология", и в этой связи выражение "рели> товый биоценоз" (Воробьев, 1949) представляется вполне опрз
данным.
Считается, что в тропиках смешение пресноводной и океанической фаун совершается с большей легкостью, чем в высоких широтах. В тропиках же обитает немало амфибийных животных и имеется немало наземных животных морского происхождения. Существенное значение для смешения фаун имеет литораль, и в частности мангры. Все это дает основание для вопроса о месте происхождения типа организации наземных животных в экологобиогеографическом аспекте. Этой проблемы касается ячейка 10. Взятая сама по себе, без вопроса о локализации события, та проблема утрачивает биогеографический аспект и рассматривается ячейкой 23. В обеих ячейках имеется в виду ценозогенез (биоморфогенез), а в ячейке 10 еще и биотогенез (фауногенез и флорогенез). Корреляция выступает здесь как выводимость, с(? гласованность строения разных биоморф, в частности выводимо^’ дыхательной, выделительной и других систем органов наземных биоморф из соответствующих систем органов их водных предков
Остается та часть овала "сосуществование", которая пере# вается овалом "сходство". Область перекрывания подразделена надвое стороной овала "родство", и слева от нее находятся я#* ки 12, 15, 14, 17. Объекты этих ячеек относятся к эколог^ и к тому же принадлежат филогенетической систематике. В
128
етЫРеХ ячейках каждое объединение популяций остается биомор-^аксоном и, исключая ячейку 17, дополнительно наделено и
ругими свойствами.
М Таксономический аспект понятия биоморфы в чистом лиде от-
осится к ячейке 17. Иногда полагают, что каждый таксон до некоторой степени есть и определенная жизненная форма. Хотя г совпадения систематических групп и жизненных форм далеко не обязательны* (Серебряков, 1962), все же отсюда вытекает понимание сообщества как таксоцена (Николаев, 1977). В свою очередь' таксон представляется состоящим из жизненных форм, т е, имеющим экологическую структуру. Так, существуют (начи-ная с А.Гумбольдта) многочисленные системы жизненных форм царства Plantae и его подразделений (Серебряков, 1962) и сио-
*зообиоморф* для царства Animalia (Акимов, 1954). Существует система биоморф для класса насекомых (функционально-
биоценотипические *группы* Д.В.Панфилова, 1966), семейства
земляных червей (Перель, 1981), класса млекопитающих (Фор-
мозов, 1956) и т.д. В этой связи представляется полезным выражение * экосистематика* (Griffiths, 1974). * Эволюция жизненных форм злаков* (Серебряков, 1971) - это своего рода биоморфогенез, историческое развитие жизненных форм таксона Gra mine a.
Границы жизненных форм и таксонов иногда совпадают. Вообразим, что живой покров Земли состоит только из гетеротрофных животных и фотоавтотрофных растений. Наблюдающему эту картину только с эколого-таксономической точки зрения он будет предоставляться состоящим из двух таксонов как жизненных форм: Animality и Plantae. В действительности же существуют и низшие формы жизни (бактерии, синезеленые) с весьма разнообразными типами обмена веществ. Кроме того, среди высших растений, помимо фотоавтотрофов, есть .паразитические и хищные формы. Поэтому в природе однозначное совпадение границ таксонов и жизненных форм (типов организации) было бы невозможно. Надо отметить, что еще Е. Варминг (1901) фактически признал статистическое перекрывание этих объектов у цветковых расте-«ий. При этом он заметил, что одни семейства растений состоят Иа единственной биоморфы, а другие включают несколько биоморф,
К возникшей проблеме можно относиться по-разному. Можно ^Ыло бы, преодолевая неопределенность искать более надежные, гетеротрофность, признаки. Однако можно удовлетвориться достигнутым результатом, считая естественным явлени-
Несовпадение границ таксона и жизненной формы. В любом сПучае отношение к возникшей проблеме должно опираться на био-^ически содержательные посылки, выводящие его на теорию
Изни, Автору представляется приемлемым положительное отно-^еНие к проблеме, а попытки преодолеть неопределенность в- от °Щениях *таксон — жизненная форма* бесперспективными.
Если элементарным носителем биологической специфичное! действительно является популяция, то элементарным объекте*, "таксон как биоморфа" также должна быть популяция (подви. Теперь представим себе популяцию в длении, когда, занимая место в. филогенетической системе в качестве подвида или ы да, она в то же время выполняет определенную роль в сообщ стве. В силу неопределенной изменчивости такая длящаяся ь потенциально бесконечном времени популяция не может имен постоянных границ как фенон, а следовательно, не может зан мать строго постоянного места в системе и как таксон (othi сительно других видов и подвидов). Неопределенным образом флуктуирующее во времени сходство данной популяции с друг; ми означает непостоянство тех морфологических и биохимич* ких структур, посредством которых выполняются ее функции j сообществе, а стало быть, и изменение их роли жизненной фс мы в сообществе. Разбивая доение популяции на отдельные о резки, мы будем регистрировать некоторую динамику в огней ниях таксона с жизненной формой: иногда их границы совпади ют, иногда нет. Именно такую картину нам хрмонстрирует кге жество таксонов и жизненных форм, и эго наблюдение в фикс рованный отрезок времени есть образ той картины, которая имеет место в длении отдельной популяции. Как же отобрази' эту динамику в фактически существующей системе таксонов -биоморф? Думается, эта задача уже решена, и решение есть ма неопределенность системы. Собственно, с этой проблемой мы встречаемся в каждой ячейке биологической розетки, за i ключением первых пяти. Последние соответствуют такому ва ду на генеральную совокупность популяций, который не выде; ет объектов. ВШЗление объектов, а следовательно, и несовп дение разных классификаций, начинается с использования дву и больше отношений. Еще раз продемонстрируем явление несс падения классификаций на примере ячейки 14. По сравнению ячейкой 17 в ней проведена более сложная точка зрения на i неральную совокупность популяций, и вследствие этого карт® жизни еще больше детализируется.
Например, класс ныне существующих головоногих моллюск включает таксоны десятиногих, восьминогих и наутилусов. В первые входат кальмары и каракатицы, вторые включают осы ногов и раковинных аргонавтов, третьи - раковинных животш Все головоногие - хищники и, представляя одну жизненную Ф му, тем не менее не обнаруживают в этом отношении полног единства. Быстро плавающие кальмары - активные хищники, ! следующие своих жертв. Для прочих головоногих это менее к рактерно. Гре имущественно ползающие осьминоги скорее отне с я гея к хищникам-засадникам, и каракатицы по своему спосо охоты ближе к осьминогам, чем к своим ближайшим родичам кальмарам. Выходит, что, выделяя разные жизненные формы
^ллков, мы должны разрушать границы таксона десятиногих ринить каракатиц и осьминогов.
Теперь посмотрим на систему класса головоногих с точки ения сравнительной анатомии. Тогда обнаружится следующая дгина» Раковинные аргонавты и наутилусы образуют тип ор-^цзании раковинных головоногих. Задний конец тела у этих Гйв(угных запрятан в раковине и не может выполнять рулевой ^пии, как у кальмаров, каракатиц и некоторых осьминогов. Значит, теперь нарушены границы таксона восьминогих и создало объединение аргонавта и наутилуса, искусственное с точки зрения систематики. Еще сложнее с осьминогами, среди которых преобладают ползающие формы, однако имеется и некоторое количество пелагических видов, образующих особый тип организации. Обладая широкой перепонкой, натянутой вроде парашюта между щупальцами, они плавают, как медузы. Их следует отделить в особый тип организации. Итак, снова перемешаны все границы, другой пример - птицы. Таксономически разные птины, например пингвины и бакланы, образуют одну биоморфу хищника-ры бое да, тогда как один тип организации (птицы, имеющие длинные клюв, шею и ноги) охватывает таксономически разобщенных цапель, фламинго, аистов и журавлей. Они относятся к разным жизненным формам. Журавли всеядны, прочие преиму
щественно хищники.
Ячейка 12 из сингопической части овала похожа на ячейку 17 из Диатонической части. Жизненная форма таксона здесь выступает в качестве биотического элемента биоты в определенном регионе системы биогеографического районирования. Вернувшись к классу головоногих моллюсков и применяя к ним новое сочетание отношений, мы должны подразделить их на донных (большинство осьминогов, каракатицы) и пелагических хищников (прочие головоногие), т.е. на жизненные формы сообществ морского дна и толщи воды. Таким образом, через отношение совместности границы таксонов снова оказались нарушенными, ЭДнако иным способом, чем прежде. Каргина усложнится еще больше, если привлечь к ней вымерших головоногих, однако этого надо покинуть пределы овала "сосуществование*.
Ячейка 15 - самая сложная в биологической розетке. В ней йспользованы все пять отношений и выделено такое объединение п°пуляций, которое обладает свойствами всех 5 объектов и в Рапной степени принадлежит всем пяти частным разделам биотин. Возможно, этим требованиям отвечают следующие два Римера,, В термальных водах источника пещеры в Сгеловой го-близ Кейптауна обитает рачок спелеогрифус. Он предсгавля-собой оригинальный тип организации жизненной формы микро-и эндемичный только для этого района аустральный эле-фауны Южной Африки. Важно, однако, подчеркнуть, что Пелеогрифус есть единственный вид отряда спелеогрифовых
131
(Gordon , 1957). Другой пример - это один из видов рак^ тельника, обитающий на Бермудских островах. В отличие qj других видов этого же рода, которые относятся к гак назьц мой жизненной форме собирающих клешнями пищу подвижны} животных, этот вид ведет совершенно особый образ жизни, обитает в пустых раковинах брюхоногих, неподвижно закреп, ных в полипняке кораллового рифа, и добывает пищу взмахе головных антенн. Таким образом, один из всего таксона эг вид относится к биоморфе фильтратора.
Остаются четыре ячейки: 18, 29, 16, 27. В них не отношение родства. Наименее сложная из них ячейка 18, в торую входят объекты, выделяемые в живом покрове Земли ношением сходства, т.е. биоморфы как феноны (см. табл. 5 Таким объектом могло бы быть объединение микроскопичен продуцентов, характеризующихся округлой формой тела: нею рые бактерии, синезеленые водоросли и простейшие. В ячей 2 9 добавляется отношение корреляции, и тогда выявляется кой объект, как ‘’микроскопические продуценты и консумент фильтраторы*. Эго единство основано на отношении корреля консументы обладают приспособлениями для фильтрации пи микроскопических продуцентов и соответствующими химизму последних ферментативными системами, а продуценты облад способностью быстро размножаться, чтобы восстановить уб1 численности.
В ячейке 16 мы находим, например, фитоценоз мангроы лесов тропической зоны. Мангровые деревья обладают сход ми приспособлениями к обитанию на границе суши и моря, частности, у них имеются ходульные корни и жесткие, покр воском листья. Своеобразная физиология этих растений поз ляет им существовать при высоком содержании солей в гру и воде. Внешнее •'физиономическое* сходство между такими стениями весьма значительно, поэтому они образуют не т<я тип организации, но также и фенон. Однако скоррелированы строения совместно сосуществующих растений и (или) живс ных может и не сопровождаться сходством. Например, к ос нию в тех же манграх приспособлены рыбки периофгальмуы Они используют ходильные корни деревьев как субстрат дл5 пребывания на воздухе во время охоты за насекомыми. Пр* сутствии сходства между растениями и животными есть ос! ние для корреляции их строения. Этот пример следует огне к ячейке 31. Другой пример такого рода представляют оби щие в манграх креветки. Они способны использовать в пиш татки листьев мангровых деревьев, покрытые воском, толь! потому, что имеют необходимые для этого пищеварительны* ферменты.
Наконец, ячейка 27 подразумевает относительно несло# точку зрения на живой покров Земли, когда он представляв состоящим из биоморф-фенонов
0 экологии принято выделять жизненные формы по взрослым здозмам. Между тем понятие организма в разных отделах °Рг50Гйи неодинаково. Сравним, например, бактериальную клет»
И устрицу, имеющую личиночную стадию. У устрицы планк-най личинка отличается от взрослой стадеи, обитающей на
Т и, стало быть, они могли бы принадлежать к разным жиэ-формам. Этого не наблюдается у бактерий. Личинка ус—
* йЦЬ] относится к планктонному сообществу, взрослый мол-
т\сК - к донному сообществу. Поэтому, например, И.X.Шарова ц981) построила для жужелиц две параллельные системы жиз» ^еляых форм: одну для личинок, другую - для имаго. Фактичео-
кй это означает отход от организмоцентризма, потому что има-го и личинки - это суборганизменные единицы; вместе они об-
разуют один организм в разные периоды его существования. Многочисленные примеры, подобные устрице и жужелицам, тем ие менее не имеют всеобщего биологического значения, о чем свидетельствует пример с бактериями. Больше того, в только
что зародившейся жизни, которая на равных правах с высокоразвитой жизнью образует объект биологии, не было основания для постановки обсуждаемой проблемы: протобионты скорее походили на бактерий, нежели на устрицу. И все же проблема су-
ществует, и ее надо решать на основе общебиологических прин
ципов.
Понятие биологического типа, предложенное И.Г.Серебряко-вым (1962), можно считать важным шагом в этом направлении. Биологический тип - это совокупность разных стадий развития одного организма в разных биотопах, само развитие функций организмов в разных сообществах. Эго понятие напоминает онтогенетическое развитие организма в привычном смысле. По-видамому, в систематике биоморф следует учесть опыт систематики таксонов, в которой в равной мере принимается во внимание строение личинки и взрослого животного. Онтогенез организма в гаком случае будет представляться и онтогенезом био— м°рфы. Организм сохранит своюх целостность как структурной единицы популяции, хотя и здесь, как в примере с устрицами, бУДет утрачена граница меэоду биотопами личинки (толща воды) и взрослого животного (дно). Если, грубо говоря, "онтогенез й°вторяет филогенез*, то онтогенетическая изменчивость орга-Низма как биоморфы (плавающая личинка - взрослая донная ста-ЙИя) имеет своим источником историческое развитие вода как и°Морфы _ биоморфогенез. С этой точки зрения онтогенетическая изменчивость есть отражение исторического развития био— ^°Рф в видообразовании. Индивидуальный и исторический био-°Рфогенез надо считать параллельными, подобно тому как пепельны эмбриогенез и филогенез. В таком случае справедливо и ,
ij ” другое заключение: исторический биоморфогенез есть при-индивидуального биоморфогенеза. Но поскольку эволюцио-
133
ньфуЮт виды-биоморфы, а не организмы-биоморфы, популяц^ низм способен ввести свой вклад в проблему неопределенное границ биотопов и состава сообществ. I
Биоморфы состоят из длящихся популяций. Следовательно ! обсуждаемое явление неопределенности надо соотнести с п<л циально бесконечным длением популяций. Вследствие неопре ленной изменчивости структур (субстрата) в популяции функ биоморфы-вида в сообществе не могут быть четко очерчены же во временной точке. В длении они должны флуктуировать столь же неопределенным образом, что и структуры. Тогда границы между сообществами в Длении и состав сообществ
\ Должны оставаться неизменными. В итоге следовало бы счи что га неопределенность границ сообществ и их состава, ко рую мы наблюдаем в срезе современности вследствие онтог тической изменчивость биоморф-организмов, вызвана неопре ленноегью биоморф-видов в длении. Этот эргодический прин; еще раз напоминает о подчиненном по отношению к популящ статусе организма.
Дискретность и континуальность флор, фитоассоциаций (В.Шмидт, 1981) и вообще сообщества, а также биохоров . регионов — эго разные состояния живого покрова Земли, на' даемого в произвольно выбранной исследователем временно! точке. В длении эти противоположности сходятся, образуя н ределенность, третье состояние. Стирание границ между пре воположностями происходит, в частности, через изменяемое жизненных циклов и типов развития.
Эмбриогенез эволюционирует, так же как и другие призи видов (Г.Шмидт, 1968). Эволюция эмбриогенеза могла бы шествляться на основе внутривидовой изменчивости предка, которые- обобщения в области сравнительной эмбриологии пс ляют уяснить природу изменчивости эмбриогенеза. Известие например, что в крупных яйцах эмбрион обеспечен питатель ми веществами на весь период развития. Вследствие этого ет место прямое развитие до взрослого состояния, минуя с дню личинки. Наоборот» когда размеры яиц невелики, длите, ное время продолжается личиночное развитие. Все это вреь личинка добывает пищу самостоятельно (Иванова-Казас, 1-Почему явление внутривидовой изменчивости эмбриогенеза i могло бы возникнуть вследствие зависимости изменчивости меров яиц? Есть и другие причины разнообразия типов раз* тия. Известно, что у морских животных микроскопические i чинки имеются чаще, чем у пресноводных, имеющих прямое вигне. Возможно, это объясняется трудностью осуществления 9 пресной воде осморегуляции, поскольку микроскопические ли4 ки имеют большую относительную поверхность. У холодновоД" ных животных развитие тоже часто упрощено, тогда как У ’ге|1 ловодных длительный личиночный период жизни наблюдается 4
134
_ чем У холодноводных и глубоководных животных. На этой 1Де» -
рснове У экологически пластичных вадов также могла бы сло-jKjrrbCfl внутривидовая изменчивость типов развития. ,
Следовательно, можно предполагать, что разнообразие типов дзвития, наблюдаемое в таксоне, до известной меры могло бы ^читаться следствием разнообразия типов развития внутри прод-нового вида этого таксона. Аналогия с систематикой остается рри этом вполне содержательной, и неопределенность в экологии представляется тоже биологически содержательной. Вместе с тем это явление наводит на мысль о том, что, размещая в пространстве популяционные биоморфы сообщества, мы должны помнить об их популяционной природе и пользоваться пространством несколько особым образом. Эго обстоятельство уже выявилось в фаунистике и флористике, в которых сообщества жизненных форм понимаются как фауны и флоры.
Глава 11
БИОГЕОГРАФИЯ, ФАУНИСТИКА 1
И.ФЛОРИСТИКА И ПОНЯТИЕ
БИОЛОГИЧЕСКОГО ПРОСТРАНСТВА
Если биологическая специфичность заключена в межпопуля— ционных взаимодействиях, то биологическое пространство, в котором они осуществляются, есть проблема популяционной биологии, и оно изучается биотикой - фаунистикой и флористикой. Бистике особенно не повезло: теоретики обычно не замечают ее существования. В сильно разросшейся проблеме биосферы биотике также не заметна, потому что взятые в аспекте обмена веществ и энергии животные и растения не выделяются среди таких элементов биосферы, как атмосфера, гидросфера и литосфера. В биосферологии выражение "живое вещество* имеет, биогеохимическое, но не собственно биологическое значение.
Объектом биотики служит биота - объединение фауны и флоры (Бобринский, 1951) отношением совместности. Местообитанием биоты можно считать биохор Э.Геккеля (Haeckel , 1866). Соответственно выделены зоо- и фитохоры (хорионы) (Turrill , 1959), Биотика становится биогеографией, когда биота как население биохора находит свое место в географическом пространстве, т„е. когда проведено картирование. Тогда из общего биологического понятия, характеризующего жизнь как особую форму движения материи безотносительно к планете, биотика становится частным понятием земной жизни. Биохор именуется теперь биогеографическим регионом, и на него переходит имя собственное территории или акватории (например, биота Байкала). Биохор как регион занимает определенное место в систе
v
135
ме биогеографического районирования планеты. Поэтому естес-венно, что биогеография в равной мере принадлежит биологии географии (Гептнер, 1936): первой - через биоту, второй -через регион. Задача теперь заключается в том, чтобы в биог ографии выделить биологическую и географическую составляющие. Сложность этой задачи в том, что биологическое простри ство существует только в уникальной земной версии.
Самостоятельность географии как науки в значительной мере основывают на понятии географического пространства (Хар. вей, 1974). Иногда этому обстоятельству придается исключительное значение: "География - наука о местоположениях... Пр странственные связи — предмет изучения теоретической геогра фии" (Бунге, 1967, с. 203, 214). А.А.Григорьев (1966) и В.С.Лямин (1978) считают объектом географии географически оболочку, в которой осуществляется особая географическая ма движения материи (Кузнецов, 1970), свойственная не толь ко Земле, но и деугим планетам. Географическая оболочка есть прежде всего ландшафтная сфера Земли. Несмотря на однознач. ность этого понятия (Марков и соавт., 1968), ландшафт оста-» ется специфическим объектом географии, ее "индивидуумом", "Географический ландшафт есть такая совокупность или группировка- предметов или явлений, в которой особенности рельефа, климата, вод, почвенного и растительного покрова и животного мира, а также, до известной степени, деятельности человека, сливаются в одно гармоническое целое..." (Берг, 1947, с. 5), Понятие ландшафта применимо к "подводным областям океана* (Полынов, 1956) и пелагиали (К.Беклемишев, 1969). Безжиэ ненный ландшафт, подобный лунному, - это тоже объект reorps фии. Такие ландшафты существовали и на Земле в добиологичес кую эпоху. В современных нам земных ландшафтах биота участвует наравне с рельефом, водоемами, климатом и пр. Биологам неизвестны биоты вне ландшафтов географической оболочки. По тому понятно, что ландшафты изучаются и биологами и геогра-фами.
В ландшафте легко распознаются свидетельства его принадлежности к зоне, поясу и сектору. С другой стороны, своей це лостностью и структурностью географическая оболочка обязана ландшафтам, их принадлежностью к зоне, поясу, сектору. В отдельном ландшафте отражается структурность географической оболочки; в свою очередь своей целостностью географическая оболочка обязана закономерному размещению ландшафтов.
Зональность в географической оболочке обусловлена тем обстоятельством, что угол падения солнечных лучей на поверхность Земли зависит от географической широты и сходным образом изменяется от экватора к полюсам. При этом наблкдак/г ся неодинаковые в разных зонах сезонные изменения поступле-* ния солнечной энергии в зависимости от угла наклона £ зеМ*
136
экватора к плоскости эклиптики, когда Земля вращается УР Солнца по гелиоцентрической орбите. Показано, что в 6дорическое время угол £ не оставался постоянным и поэ— Г£>МУ система зональности претерпела существенные изменения тЛоедтеченский, 1950; Монин, 1977). Подтвердив-ранее выс-^занное предложение Л.С. Берга (1947), климатологи призна-* зональность изначальным свойством нашей планеты (Сини—
1981). Зоны располагаются не вполне симметрично отно-^ельно экватора. Поэтому некоторые географы (Исаченко, |971), выделяют явление полярной асимметрии и придают ему авНый с зональностью статус. Полярная асимметрия возникает оТтого, что северное материковое полушарие представляет про-глвоположность южному океанскому полушарию. Вследствие нео-дцяаковой теплоемкости суши и воды плоскость симметрии зональности находится южнее экватора примерно на 5° (Витвип-кий, 1980). Асимметрия распределения материков и океанов как форм мегарельефа наблюдается и на других небесных телах Солнечной системы (Флоренский, Забелин, 1971), однако лишенных гидросферы. Если физико-географическая зональность и жизнь есть планетарные явления, то жизнь к тому же остается и зональным явлением. К числу зональных факторов иногда относят и рельеф. Он развивается в связи с горообразованием, вызванным деформацией земного эллипсоида в шар при замедлении вращения планеты вследствие приливного трения. Горообразование больших масштабов наблюдается в так называемых критических зонах обоих полушариев на 35° и 62° на материках и на дне океанов (Личков, 1960). В результате при продвижении от экватора к полюсам набдодается закономерное изменение площади материков, занятой горами и низменностями (Розов, Строганова, 1979). Замедление вращения Земли вызывает к тому же замедление протекания сезонных явлений в высоких широтах. Зональность выражают с помощью таких терми-н°в, как арктическая, бореальная (северная, умеренная), север ная субтропическая, тропическая и т.д. Более детальная схема зональности разработана на основе типов растительности. Она включает зону тундры, тайги, степи и пр. (Григорьев, 1963). “ океане зональные ландшафты определяются океанскими круто— в°Ротами, вызывающими вертикальное перемешивание вод (Бур— Ков» 1966). Положение круговоротов хорошо соответствует раз-^ению физико-географических зон на материке на схеме’
•^.Рябчикова (i960).
Поясность - чередование ландшафтов на разновысотном ^ьеф5 Земли. Оно вызвано существованием двух форм мега-^Ьефа т океанов и материков. Граница между ними проходит
Уровня моря на отметке 2000-3000 м, где материки со-с ложем океана (Богданов и соавт., 1978). В свою Р6Дь существование материков и океанов объясняется неод&-
137
наковой толщиной литосферных плит земной коры: До 75 |
под материками и только 6 км - под ложем океана. Пояснс выражается в неодинаковом протекании суточных и годовых пений на разной высоте от уровня моря, в различиях атмос него и гидростатического давлений, в особенностях почво* осадкообразования, характере циркуляции водных и воздущ^ масс, особенностях солнечной экспозиции и пр. Разумеется ясность не проявляется отдельно от зональности. Например распределение горных ландшафтов будет неодинаковым в выс^ к их и низких широтах. В частности, пояс альпийских лугов в тропиках находится выше, чем в умеренной зоне. В умеренно^ зоне Европы система горных ландшафтов начинается с пятого пояса, а на севере Европы - с шестого (Греонер, 1914). ясность сохранилась бы на Земле и при отсутствии большог количества воды, но тогда океанскими ландшафтами следове бы называть ландшафты океанского ложа, ныне залитого во Пояса имеют следующие названия: высокогорный, горный, р нинный, литоральный, шельфовый, абиссальный и пр. Океань материки как формы мегарельефа имеются также на Луне, ] се, Венере. Тем не менее пока неясно, можно ли такое расч^ некие рельефа считать свойством всех планет. Следовательно, вопрос о поясности как свойстве биоты на любом подходящем Для жизни небесном теле надо считать открытым.
Секторность, или меридианальная зональность, обусловлена тем обстоятельством, что материки и океаны (при наблк> дении с полюбов) составляют на нашей планете фигуру трехлучевой звезды. Она образована материковыми (одним американским и двумя несовершенными - евро-африканским и азиатско-австралийским) и океанскими (одним несовершенным иидфоке-анским и двумя совершенными - тихоокеанским и атлантическим) секторами. Вследствие этого по экватору наблюдается закономерное чередование материковых и океанских ландшафтов. В океане с удалением от материков закономерно изменяются геоморфологические, климатические и гидрологические характеристики, состав осадков, прозрачность и цветность воды (Гы-марев, 1978). На материках с удалением от береговой линии влажный приморский климат сменяется континентальным. В при* морских ландшафтах и вдалеке от моря условия обитания для пещерных животных существенно различаются ( Howarth , 1980)*
В истории Земли расположение океанов и материков не оставалось неизменным (Лисицын, 1980). Согласно принципу билизма, развитому в концепции глобальной тектоники, совре*'е1 ное размещение океанов и материков установилось в результ8* те сложных перемещений литосферных плит, плавающих на р86* плавленной основе. В мезозое материки составляли вместе е^' ный праконтинент Пангею, окруженную океаном Панталасса. ким образом, в мезозое Землю характеризовала двусторонняя
138
^^етрия» что,, разумеется, не препятствовало существованию ^^орности. Неизвестно, однако, сохранялась ли секторность С более древней Земле. Например, как будто бы нет основа-й0Й отвергать такую картину: материки сосредоточены у одного полюсаs а океаны вокруг другого полюса. В такой картине н^г секторности и Земля имеет радиальную осевую симметрию, у дожившаяся еще в XIX в. система биогеографического райо-^ования суши (Beaufort, 1951) фактически отражает исто-литосферных плит, степень и давность их обособления. Со-
временная система, разработанная для суши (Старобогатов, 1970; Тахтаджян, 1978) и океана (Ekmant 1935;Briggs, 1974) не только учитывает секторность, зональность и поясность, но и отражает возможность и степень расселения животных и растений в изменявшейся в геологическое время географической оболочке Земли. Для описания секторности помимо имен собственных материков и океанов используются такие названия, как прибрежный, неритический, приморский, континентальный и др.
На пересечении зон, поясов и секторов, которые имеют соб
ственную иерархическую структуру, возникают регионы, также населенные иерархической структурой. Животные и растения, обитающие в таких регионах, характеризуются как "зональный тип", "зональная форма". В них отражается географический аспект биоты и ее элементов. Примером географического видения биоты, когда соподчиненные биоты выделяются через физико-географическое районирование, может служить исследование П.Д.Ярошенко (1975). Дилемма "от региона к биоте" или "от биотопа к региону" обычно затушевана. Вместе с ней затушевано и различие между географическим и биологическим подходами к членению биоты, потому что в процессе районирования биота неминуемо становится элементом того или иного ландшафта. Тем не менее собственно биологический подход сохраняет свою самостоятельность вплоть до картирования, когда биолог выделяет различные биотические элементы на основе разных отношений. Возможны разные точки зрения на биоту. Для систематики биота есть объединение таксонов. Биохор такой биоты надо считать объединением ареалов\ Поскольку таксон есть Родословная фенонов, ареал как местообитание таксона обладает свойствами филохора и фенохора. Местообитание тех или , иных типов организаций животных (растений) есть биохор как Типохор. При условии сосуществования биохор является еще и биотопом, местообитанием жизненных форм .сообщества. В общей сложности в овале "совместность" насчитываются 16 ви—
п Хорион ( Turrill, 1959; Тахтаджян, 1978) - это область “ерекрывания, сгущения ареалов таксонов растений. Среди зоо-flQroB понятием хориона фактически пользуется например В. Н.Се-м®нов (1975).
139
дов всевозможных объектов биотики разной степени сложно^ (рис. 10). Им отвечают 16 видов биохоров (табл. 6). ”
показать, что зональный, поясный и секторный принципы
Рис. 10.
А - положение биоты в четырехмерном концептуальном биологическом пространстве; Б - логическая структура биотики (фаунистики и флористики) в овале 'совместность* биологической розетки
ния мировой биоты не мешают биоте сохранять свою собственную биологическую структуру, оставаться системой таксонов, фенонов, типов организации и пр.
Биологическое содержание зональности, или 'органической полярности* (Уимпеннл, 1945), состоит, в частности, в том, что таксономическое разнообразие возрастает в направлении от полюсов к экваторам. Причины этого явления все еще недостаточно понятны (Фишер, 1964;Buzac, 1972; Rhode , 1978).
Тип организации пойкилотермных животных, обитающих в холод-ном климате высоких широт, характеризуется крупными размерами (правило Бергмана) и плавными очертаниями тела (правило Аллена); таким образом, такой тип есть еще и фенон. В Ус" ловиях холодного климата при наличии слоя вечной мерзлоты растения имеют приповерхностную корневую систему и предств0* лены в основном стелющимися формами. Такого не наблюдаете# в умеренной зоне. По этому поводу К.Бэр (1959, с. 385) W с ал еще в 1822 г.: *В различных климатах различны формы
140
Таблица 6
объекты биотики (фаунистики и флористики): б0(ГГ1»ческие элементы (номера ячеек см. на рис. 10)
Ячейка Синхронная биотика Ячейка Диахронная биотика
30 биотические элементы как биоморфы; биохор как биотоп 7 биотические элементы; биохор
31 биотические элементы как биоморфы и типы организации; биохор как биотоп и типохор (типотоп) 26 биотические элементы как типы организации; биохор как типохор (ти— потоп)
16 биотические элементы как биоморфы, типы организации и феноны; биохор как биотоп, типохор (типотоп), фенохор (фенотон) 25 биотические элементы как типы организации и феноны; биохор, типохор (типотоп), фенохор (фенотоп)
27 биотические элементы как биоморфы и фено» ны; биохор как биотоп, фенохор (фенотоп) 28 биотические элементы как феноны; биохор, фенохор (фенотоп)
12 биотические элементы как биоморфы, родословные и феноны (таксоны); биохор как биотоп; филохор (филотоп) как фенохор (фенотоп) - ареал 13 биотические элементы, феноны как родословные (таксоны); биохор, фенохор .(фенотоп) как фи-лохор (филотоп) - ареал
15 биотические элементы как биоморфы, родословные и феноны (таксоны), типы организации; биохор как биотоп; филохор (филотоп) как фенохор (фенотоп)-ареал; типохор (типотоп) 4 биологические элементы, феноны как родословные (таксоны); биохор, фено хор (фенотоп) как филохор (филотоп) - ареал
10 биотические элементы как биоморфы, родословные и типы организации; биохор как био- 19 биотические элементы как родословные и типы организации; биохор, фо лохор (филотоп), типохор (типотоп)
141
1
i
Продолжение таблиц
Ячейка Синхронная биотика Ячейка Диахронная биотйка''
21 топ, филохор (филотоп), гипохор (типотоп) биотические элементы как биоморфы, родословные; биохор как биотоп, филохор (филотоп) 8 биотические элемент как родословные; бис хор как филохор (фи., лотоп)
жизни. Животные и растения южных стран высоки, стремятся I вверх, а в высоких широтах они низки, вытянуты, прижимаются к земле. В сходных же климатах имеются сходные формы животных и растений*. В северных странах преобладают гравяни тые растения, в тропиках, наоборот, древесные виды (Алехин, 1944). Здесь имеются в виду жизненные формы, типы органи зации и феноны таксона Plantae.
Сезонные явления в жизни растений и животных протекают в океане неодинаково в зависимости от географической ширсит (Богоров, 1974), и это, в частности, объясняется особенности ми годового ритма освещенности (Паули, 1959). В высоких широтах резко выраженная сезонность (полярные день и ночь) обусловливает такие явления, как листопад и четко выражение сезон линьки, перезимовывание и спячку, одноразовое размноя ние и быстрое, в условиях короткого полярного лета, укорочен ное развитие личинок у морских беспозвоночных, приводящее иногда к живорождению (последнее вызывает снижение плодов тостр). Биологические особенности могут отражаться в орган! ческом вещественно-энергетическом субстрате, например у пе ч©ночных мхов высокоширотной Арктики (Ладыженская, Жукоы 1971) или у растений аридной зоны (Кочетов, 1985). Имеют ся данные о возрастании объема ядра и увеличении содержав® ДНК на клетку у хвойных растений с увеличением географичее кой широты ( Mergen, Thielgs, 1967; Miksche , 1971; Miks che, Hott a, 1973). У рыб, обитающих в высоких широтах, число хромосом больше, чем у рыб низких широт (Никольский Васильева, 1973).
Вообще зональность у животных проявляется не менее четко, чем у растений (Ходашева, 1966; Чернов, 197 5). Количество микроорганизмов в почве (грибы, акгиномицеты, водоросли, простейшие, бактерии) возрастает в направлении к эква"* тору (Почвоведение, 1982). Сходные черты зональности наблют даются в бактериальном населении океана (Крисс, 1976). Но-
142
паТура зональных биохоров разработана главным образом таксонов. Зональные ареалы называются циркумполярными, ^антарктическими. Чаще, орнако, обнаруживаются разор-е ареалы: материками - для океанских животных и океа-- ~ для наземных животных. Эго является следствием того,
;fl обычно один таксон представлен одним типом организации ^ценной формы: либо наземной, либо морской. Строго говоря ^щочая птиц), непрерывные зональные ареалы есть только рысоких широтах Арктики и Антарктики.
' Одмн из самых ярких примеров поясности — изменение сос— ава флоры эвфотического слоя в океане: на мелководье преоб-^адакт зеленые и бурые водоросли, на глубине - красные. Эго вызвано тем, что красный цвет длинноволновой части спектра быстро рассеивается в воде и в глубины проникает коротковолновый синий цвет. Последний дополнителен к красному пигменту водорослей багрянок. Поскольку четкого чередования таксонов при этом не прослеживается, в этом примере налицо типы организации жизненных форм водорослей.
С возрастанием глубины моря закономерно изменяются со
общества морского дна. На мелководье, в условиях подвижного придонного слоя воды, преобладают животные, питающиеся взвешенными органическими частицами. На выровненных участках цна, в условиях малоподвижного слоя воды, преобладают животные, питающиеся органическими частицами грунта (Кузнецов, Нейман, 197 5). Поясность означает не только расстояние от нуля глубин, но также и закономерное чередование крутизны и пологостей на ступенчатой гипсографической кривой. Поэтому естественно, что два названных варианта сообществ данных жи-
вотных закономерно чередуются в океане, хотя один и тот же вариант сообщества включает таксономически разных животных (Виноградов, 1977). У глубоководных животных пелагиали обычно наблюдается обесцвечивание или красная окраска, орга— иы зрения увеличены или, наоборот, редуцированы. Кроме того, им свойствен так называемый глубоководный гигантизм (Бир-щтейн, 1963). В этом случае можно говорить скорее о типе организации жизненной формы, чем о феноне.
При рассмотрении в обратном направлении, т.е. от литорали в глубь материка, мы обнаружим пояс галофильных растений, ^пример мангры, а также амфибийных животных, выдерживающих осушение во время отлива. Для деревьев прибрежной зоны *арактерны так называемые флаговые кроны, для трав — упругие стебли, для кустарников — обтекаемая форма. Все эго позволяет растениям успешно противостоять сильным ветрам. На-СеЛение высокогорий в отличие от обитателей низменностей йМеет приспособления, защищающие его от усиленного ультрафиолетового облучения. Обитатели горных круч, например муф— резко Отличаются от обитателей влажных равнин, лосей
143
строением копыт, телосложением, особенностями аллюра, 0 кой. Для размещения горной растительности .важное значец^* имеет солнечная экспозиция в условиях неодинаковой кру^и поверхности (Павлов, 1918). Пожалуй, в литературе назвав 11 поясов больше тяготеют к жизненным формам и типам орга|7 зации, чем к таксонам, хотя существуют и поясные характе 4 тики таксонов морских беспозвоночных (Беляев, 1977),
Традиционное деление биотики (биогеографии) на морскую (океанскую) и материковую подразумевает секторные биохорЬ| и два типа организации жизненных форм. Другое деление би^, тики - на водную и наземную - менее естественно, потому биота материковых водоемов во многих отношениях ближе к земной, чем к морской. В материковых водоемах велика доля двух групп, весьма чуждых морской среде — цветковых расте. ний и насекомых. Эти группы выделены в общем по таксонок®. ческому принципу, и в качестве жизненных форм (цветение у растений, связь жизненного цикла с серией у насекомых) более близки к наземной биоте, чем к океанской. Планктон материке, вых водоемов резко отличается от океанского. Планктонные м вотные океана обычно имеют личинок. В прибрежных водах к ним добавляются личинки донных животных. Наоборот, обитать, ли материковых водоемов часто лишены личинок, и различия между планктоном прибрежья и открытой частью водоема не очень велики. Типичные растения океанской биоты - это одноклеточные водоросли. Они резко преобладают над макрофитами, В материковых водоемах площадь, занятая освещенным солнцем мелководьем, относительно обширна и велико значение макрофитов. Если в океане среди макрофитов преобладают водоросли> то в пресноводной биоте, как и на суше, преобладают цветковые растения. Океанская биота в основном состоит из первич-н сводных растений и животных, в биоте материковых водоемов МНОГО вторичноводных форм, проникших В водоемы с суши. Срб’ ди них особенно много насекомых, за редкими исключениями отсутствующих в океане. На суше и в материковых водоемах основные продуценты - цветковые растения, тогда как в океане — водоросли. Преобладающие консументы в океане — рыбы> на суше ~ другие группы позвоночных животных. Основным Фк' гофагом в океане являются планктонные рачки, тогда как на суше — насекомые.
Беглый обзор биологического содержания зональности, пояс* ности и секторности позволяет заключить, что биологические объекты (см. табл. 6 и рис. 10) сохраняются как бы внутри тех ландшафтов, которые выделяются по физико-географическо* му принципу. Зональность и секторность не проявляются пор03^1 но и в этом случае биологический компонент ландшафтов сох?8* няется как объект биологии и отличается как объект районир0* вания от рельефа, водоемов, климата и пр. Например, верти4®
144
система поясов в горах в высоких широтах не только Сменена вниз по отношению к тропикам (зональность), но еще неодинакова в условиях гумидного приморского и аридного континентального климата (секторность) ( Dansereau, 1957). Система поясов "литоральные водоросли - галофильные цветковые растения - растения приморской низменности* в полной мере 0сражена в тропиках и умеренных широтах, ее нет в высокой Арктике и Антарктике. В обоих примерах подразумеваются ка— кне-либо биологические объекты, выделенные согласно зонам, поясам и секторам.
Ландшафтное пространство в целом, как и отдельные ландшафты, имеет три измерения. Они отвечают трем плоскостям симметрии биологической структуры океана, по Л.А.Зенкевичу (рис. 11,А). Ось вертикальной поясности (на рис. 11, Б) -это как бы расправленная гипсографическая кривая, ось секторности - это как бы срединная линия материков (океанов), равноотстоящая от побережий, ось зональности - это как бы распрямленный меридиан. Рис. 11, Б создает геометрический образ концептуального пространства, он может считаться плодом ландшафтной концепции географической оболочки.
А Рис. 11.
- плоскости симметрии биологической структуры океана, по н.А.Зенкевичу (1948)\с изменениями: вместо четырехмериди— Ильных плоскостей изображено три по числу океанских или материковых секторов; Б - положение ландшафта I в концептуальном географическом пространстве
По аналогии биота своим внутренним строением, сложением Иа биотических элементов, задает биологическое пространство (см. рис. 10, А). Оно имеет четыре измерения, в нем размещаются 16 всевозможных видов биохоров из овала "совмесг-10 - 156G 145
ность" (16—й есть совместность в "чистом виде", ей отвечл^ ячейка 22).
В каком смысле можно говорить о каком-то особом биолог песком пространстве? Скорее, правильнее было бы обсуждать
особый способ использования жизнью обычного пространства, для биоты пространство - это не просто вместилище орга„ низмов (обычное пространство), но прежде всего место обита„ ния. Л.А.Зенкевич и В.А.Бронкая (1937) назвали такое прост.
ранство "экологическим ареалом*. Впоследствии его стали на-
зывать "экологической нишей" (Одум, 1975). Этим выражением именуют многомерное (по числу факторов обитания) концеп, туальное условное пространство. Оси - измерения такого пространства — температура, влажность, освещенность и пр. Факто-
ры обитания закономерно изменяются в географической оболочке (зоны, сектора, пояса), и, следовательно, между экологичес-
кой нишей и регионом имеет место некоторое взаимное соответствие. Отражением экологической ниши в географическом пространстве будет регион, в котором имеются условия для жизнедеятельности (обмен веществ и энергии) растений и животных как биологического компонента ландшафтов. Такие регионы и схема биогеографического районирования в целом неподвижны во временной точке наблюдения.
Сходным образом отражение биологического пространства в географическую оболочку означает осуществление биохора в регионе. Приобретя такие свойства, как величины площади и очертания, заняв определенное место на географической карте сообразно с зонами, поясами и секторами, биохор тоже становится регионом. Биологичность такого региона состоит в том, что даже во временной точке наблюдения он обнаруживает движение, вызванное популяционной природой биотических элементов.
В самом деле, при наблюдении региона (биохора) из временной точки его свойства представляются неизменными, а схема районирования неподвижной. Такая картина сохраняется, пока региону отвечает биохор такой биоты, которая выделена каким-либо одним отношением или одним сочетанием отношений. Не покидая временной точки, исследователь может привлечь дополнительно другие отношения и их сочетания. В общей сложности из каждой временной точки можно увидеть 16 граней одной и той же мировой биоты, 16 разных биохоров биоты, и им будУ1" отвечать 16 разных схем биогеографического районирования. Например, карта ареалов таксонов мировой биоты будет иной, нежели карта сообществ. Разные карты — это как бы отдельнь16 моменты движения во временной точке. Оставаясь в такой точке, наблюдатель мог бы раскрывать эго разнообразие (а значит, и осуществлять движение) в любой произвольной последо-
вательности, отражая в географическое пространство одну за другой все 16 составляющих биоты (см. рис. 10, табл. 6).
146
было бы познавательной операцией. В природе все разнооб-азце членений мировой биоты и схем районирования стянуто во временной точке и осуществлено "разом*, поскольку биологическое пространство "разом* отражено в географической. Отражение, перевоплощение абстрактно-биологических биохоров в конкретно-географические регионы можно понимать как осуществление биологического в географическом, как "оживление* когда— т0 безжизненных ландшафтов планеты. Формально это будет ото
Сражением четырехмерного пространства в трехмерное.
до сих пор биологическое пространство рассматривалось из 0ременной точки, выделенной наблюдателем. Между тем биотические элементы состоят из длящихся популяций, и в длении нег привилегированных точек. Вследствие неопределенной изменчивости у таких биотических элементов изменяются свойства, определяющие их принадлежность к таксону (как биотическому элементу), к типу организации (как биотическому элементу) и т.д.; изменяются составляющие биотического элемента. Их множество и перечислено в табл. 6. Остановив свое внима-
ние на отдельном популяционном биотическом элементе в его длении, наблюдатель тоже обнаружил бы движение, действительно, в длении нет противоположности между точкой и протяжен
ностью.
Следовательно, то разнообразие граней мировой биоты и ее соподчиненных единиц (иерарзия), которое разворачивается в познавательной операции во временной точке наблюдения, и то разнообразие, которое разворачивается в длении, эквивалентны. Этот, в сущности, эргодический принцип приложим к биоте вследствие популяционной природы ее элементов, дополнительная сложность состоит в том, что длящиеся биоты пребывают в географической оболочке, существующей по собственным законам. В историческом развитии Земли зоны, сектора и пояса не оставались неизменными, и эго несомненно имеет значение для свойств регионов как географически осуществленных биохоров и Для системы биогеографического районирования в целом.
В этом положении можно увидеть еще раз две составляющие биогеографии: биологическую и географическую. Разумеется, свойства региона определяются физико-географическими факторами, но это обстоятельство имеет для биотики вполне определенное и частное значение. Дело в том, что физико-географи-Ческие факторы определяют биогеографические регионы через °РГ'анизменный субстрат, для нормальной жизнедеятельности которого необходимы соответствующие условия в географической °болочке. Если организменный субстрат не принимать во внимание (в нем только опосредована биологическая специфичность), г° в вопросе "биологическое — географическое" решающим ос— ^анется семантико-информационное взаимодействие популяций.
Тогда собственно биологическим в биогеографии будут не
147
очертания и величина площади ареала, не месторасположение региона, а сложение биоты из 16 биотических элементов, име^ щих иерархическое строение. Биологичность биогеографии сост^ ит в том, что любая биота во временных точке и протяженное-ти имеет неизменную структуру в собственном четырехмерно^ пространстве, и эта структура раскрывается в географической трехмерной оболочке как 16 типов карт биогеографического районирования. Что же касается свойств регионов на любой карте, то это сфера географии.
Жирная линия на рис. 10 обозначает плоскость синхронии -диахронии. Синхрония - это со—дление биотических элементов. Поскольку в длении нет противоположности между точкой и про, тяженностью, для синхронии не требуется установления *толщи. ны" временного среза. Объединение таких биотических элементов есть биота как сообщество: вымершее (когда объединены вымершие элементы) или ныне живущие. Диахрония — это совместность без сосуществения, биота, но не сообщество. Такая биота включает не только ныне живущие, но и вымершие биотические элементы.
Вымершие биотические элементы, точнее их ископаемые остатки, остаются объектом биологии потому, что в них воплотилось, сделалось плотью прошлое, история их особого способа существования как носителя приспособленности. Что же касается настоящего, то оно не отличается от такового, вмещающего ископаемые остатки горных пород.
В связи с синхронией надо рассмотреть два варианта биот. 1. В биоту объединены вымершие биотические элементы. Здесь основанием для объединения служит только субстрат, его прошлое.
2. В биоту объединены живые биотические элементы. Основанием для объединения здесь служит не только прошлое субстрата, но и особый способ его существования в настоящем.
К диахронии относится гот случай, когда объединяются вымершая и ныне живущая биоты. Очевидно, что, как и в первом варианте, основанием, для объединения служит только прошлое субстрата организмов.
Вымерший ихтиозавр (мезозой) и современные крупные быстро плавающие животные океанской пелагиали образуют биотический элемент как тип организации и фенон крупного пелагического пловца (ячейка 25). Совершая зеркальный перевод этой ячейки в ячейку 16 через плоскость синхронии — диахр0* нии, надо выделить ныне существующие типы организации. этого объект из ячейки 25 надо подразделить на два: 1) современные акулы и дельфины, 2) мезозойские акулы и ихгиоз^ ры. Взятые порознь, они относятся к синхронной биотике как самостоятельные фаунистические элементы. Здесь уместно от^6* тить, что, например, В. В.Алехин (1944) выделяет географий
148
аСТений мела, карбона и т.д» Каждая из них по отдельности -сИН5Фония» пока имеет место со—дление популяций. При отсут— сТВии со—дления разные эпохи вместе дают диахронию.
Слева от плоскости симметрии находятся синхронная биоти-экологическая география, справа от той же плоскости -длахронная бистика, в которой рассматривается историческое развитие биот региона (совместность). Эго - область фа уно- и флорогенеза в широком смысле слова. Собственно биотой регио-здесь становятся изменяющиеся во времени списки видов дли других объединений популяций, как, например, в работе Д.Буко (1979). Частичная синхрония (диахрония) как будто бы указывает на возможность иерархической структуры по отноше-0йю сосуществования. Если, например, в фенонике имеются раэ-цие степени сходства, то в биотике, возможно, будут разные степени сосуществования (синхрония) или разные степени диа
хронии.
Пунктирная линия на рис. 10 обозначает плоскость парентальности (parens (лат.) - родитель). Ниже этой линии находится область биотики, выделенная отношением исторического родства. Она имеет своим предметом фауногенез и флорогенез в узком собственном смысле слова, т.е. как генетическую связь предков и потомков при наличии совместности. Выше пунктирной линии на рис. 10 имеется в виду другой частный случай фауно- и флорогенеза - парагенетическая связь, два вида связи и составляют понятие фа уно— и флорогенеза в широком смысле слова. Вследствие многозначности слова *биота* биото-генез остается широким разделом знания, если даже ограничиться областью ниже плоскости парентальности. "Чистый* филогенез таксонов в условиях совместности подразумевается только в ячейке 13. В других ячейках (4, 12, 15) таксоны выступают в качестве жизненных форм, типов организации и т.д. В других ячейках имеется в виду не филогенез, а иные генетические процессы в историческом времени: типогенез (19), биотогенез (8), биотогенез биоморфы (21) и пр. Соотношение между двумя областями биотики, разграниченными пунктирной линией, можно пояснить на следующем примере. Ныне живущие, совместно сосуществующие формы дарвиновских вьюрковых Птиц на Галапагосских островах связаны родством и, следова-Тельно, образуют некоторую родословную. Наравне с родословными других животных и растений они образуют биотический ^мент биоты Галапагосских островов (ячейка 21). Отразив я<1ейку 21 в ячейку 30 через плоскость парентальности, мы По,1Учим иной биотический элемент, для выделения которого не ^Рвбуется отношения родства. В таком биотическом элементе объединены не только предки и потомки, но и совершен— 1,0 * безродные* животные и растения, которых называют мигран» ^Ми. Такой биотический элемент выделен отношением совмест—
149
ного сосуществования, и его структура выявляется классифиц^ рованием биотопов Галапагосских островов: биота каменистых осыпей* пляжей, холмов, зарослей кустарника и пр.
Разграничительные линии четырех областей биотики, переев каясь, образуют центр понятийного аппарата этого раздела би^ логии. Вокруг центра имеются четыре квадранта по четыре ячейки в каждом:
синхронный парентальный (ячейки 10, 12, 15, 21), диахронный парентальный (ячейки 4, 8, 13, 19), диахронный апарентальный (ячейки 7, 25, 26, 28), синхронный апарентальный (ячейки 16, 27, 30, 31). Квадранты, как и отдельные ячейки, можно совмещать посредством вращения вокруг центра или отражения через плоскости и таким образом устанавливать соответствия между разными понятиями. Ниже приводятся примеры, поясняющие содержание ячеек квадрантов.
Ячейка 7 (см. рис. 10, Б) выражает наиболее простую точку зрения на мировую биоту безотносительно ко времени: совместное обитание популяций. Популяции могут быть распределены по местообитаниям, биохорам, принимающим на географической карте свойства регионов в системе секторов, зон и поясов, Когда к мировой биоте применяется дополнительно отношение корреляции (ячейка 26), она распадается на типы организации, Предельно широкий тип организации земного существа есть единство скоррелированных биохимических и морфологических особенностей* которые позволяют ему жить в земных условиях. На какой-то иной планете с иной силой тяжести, с иным составом атмосферы был бы другой тип организации. Расчленяя мировой биохор, мы тем самым выделяем и все более мелкие биоты. Поэтому отношение корреляции переносится на все более мелкие структуры частного значения, и тогда выделяются все более частные типы организации (материковый, океанский и пр>)
Даже в относительно однородном биотопе океанской пелагиа-ли выделяются разные биохоры: водные массы разных географических широт, прибрежья и открытой части океана, поверхностная и глубинная водные массы. В результате складывается иерархическая система типов организации как следствие классификации ландшафтов (от географического к биологическому). Ес-ли на множестве организмов пелагиали дополнительно задать отношение сходства (ячейка 25), то в типе организации пелагических существ выделяются соподчиненные подтипы на основ£ еще и корреляции и совместности (точнее, корреляция вследствие совместности): 1) микроскопические существа с более менее длинными выростами тела, 2) полупрозрачные существ3 со студенистым телом, 3) относительно крупные подвижные 0Г ганизмы. Первые сохраняют плавучесть благодаря большой относительной поверхности, вторые не тонут потому, что их уд6^
150
яый вес мало отличается от такового воды, третьи - активные рловны, и многие из них имеют воздухоносный пузырь или заполненные воздухом легкие. Эго - феноны типов организации биотических элементов. Биологами такие объединения воспринимались бы скорее всего как нечто ненаучное, думается, однако, что в действительности такое отрицательное отношение к указанным объектам скорее всего объясняется широким распространением среди профессиональных биологов таксономического восприятия разнооиразия, которое надо считать частным подходом изучению жизни. Таксономический подход (ячейка 4) позволил бы распознать в этих объектах: 1) мелких планктонных ракообразных и личинок разных животных, а также одноклеточных водорослей, 2) медуз, сальп и ктенофор, 3) рыб, кальмаров и дельфинов. Все это - названия таксонов. Отражение ячейки 26 в ячейку 31 приведет к подразделению трех указанных объединений на более дробные, выделяемые отношением сосуществования.
Оставшиеся ячейки 27 и 28 похожи на ячейки 31 и 26 соответственно и отличаются от последних отсутствием отношения корреляции. Примером могли бы служить некоторые донные водоросли и колониальные полипы, образующие фенон в условиях совместности в ячейке 28. Отношением сосуществования они подразделяются на более частные объединения, включающие современные типы организации (ячейка 27).
Глава 12
ОТНОШЕНИЕ КОРРЕЛЯЦИИ
И СРАВНИТЕЛЬНАЯ МОРФОФИЗИОЛОГИЯ (СРАВНИТЕЛЬНАЯ АНАТОМИЯ)
Как свидетельствует история биологии, поначалу организменный субстрат изучался в основном морфологией. Разнообразие организмов воспринималось прежде всего как разнообразие их формы, органов и более мелких структур. В настоящее время Центр тяжести исследований сместился в биохимию, что вполне объективно отметил К.Паавер (1976). Морфология как будто бы устарела, хотя, как полагают некоторые авторы (Любищев, 1983), именно морфологическое разнообразие может служить выражением биологической специфичности. Как объяснить иначе, что в течение многих миллионов лет эволюции происходило совершенствование морфоструктур? В этой связи своевременной вредставляется работа С.Э.Шноля (1979), который показал, Что, хотя любые обменные процессы можно воспроизвести in vitro, морфоструктуры остаются необходимым условием просекания этих Процессов в организме.
151
В биоморфологии ( Schoute , 1949) выделяются два разд^ ла: учение об относительной поверхности организма как факГ(х ра для осуществления жизнедеятельности, и биосимметрика (у маниев, 1974), или учение о биологической изомерии (Касин^ 1973). Значение первого раздела состоит в том, что он в де^' сгвительности как бы физиологизирует морфологию (что естесг венно, поскольку функция неотделима от структуры), и в наибо, лее общем виде его сущность выражена в концепции Диффузи01и ного барьера С.Э.Шноля (1979). Основные положения этого
учения состоят в следующем.
В "мировом бульоне" в результате абиогенного синтеза воз. никли фазово-обособленные системы - тельца округлой формы. Через собственную поверхность посредством молекулярной дИф. фузии они осуществляли обмен веществ. Рост таких пробионтов в условиях недостатка питательных и других веществ привел к конкуренции между ними. Поначалу конкуренты "боролись за существование", вырабатывая наиболее совершенные физико-химические механизмы и структуры для осуществления обмена. Выживали те из них, у которых наиболее успешно осуществлялась молекулярная диффузия через поверхность тела. Однако надо иметь в виду, что при возрастании массы тела относительная поверхность быстро сокращается, и поэтому одной только молекулярной диффузии становится недостаточно для успешного осуществления обмена веществ. Так возникает явление диффузного барьера.
На первом этапе исторического развития преодоление диффузионного барьера осуществляется посредством выработки наиболее совершенного физико-химического механизма диффузии. Этот этап завершается выработкой универсального для всех живых существ организменного субстрата. Отголоском широких поисков такого совершенства можно считать разнообразие вещественно-энергетического субстрата, наблюдаемое у ныне живущих прокариот.
Второй этап совершается на основе достижений первого, остающихся относительно неизменными, и называется морфологическим. Теперь диффузионный барьер преодолевается морфологическими средствами благодаря усложнению формы тела, что позволяет увеличить относительную поверхность. Особенно показательны в этом отношении высшие растения с их корневой системой и кроной, сложной системой устьиц и сосудов. Поверхность сосудов, заполненных воздухом и водой, как бы продолжает наружную поверхность. Если на первом этапе "движителем" вещества была молекулярная диффузия, го на втором этапе ее сменяют капиллярные силы. Возможности второго этапа велики, и об этом свидетельствует разнообразие высших расте-ний. И все же вид чахлых сосенок на болоте и скрюченных березок, укоренившихся в кирпичной кладке старинного здания,
152
язь1вает на пассивный характер морфологии как средства пре— 'рдения диффузионного барьера.
совершенно новые и богатые возможности открывает движе— движущийся организм как бы увеличивает свою поверхность, ^^ррикасаясь со средой гем больше, чем выше скорость перед-Сй#ения. Выработка механизмов перемещения в пространстве посредством совершенствования движителей (плавников, ног, кры-^уев» жгутов) составляет содержание третьего этапа исгоричео-ксГ0 Развкгкя организменного субстрата. Он отвечает животному типу организации. Уместно отметить, что, по Л „А. Зенкевич у (1977), движение - ключ к пониманию организации животных. д.А.Зенкевич выразил эго положение в такой форме: невозможно вообразить неподвижное животное в неподвижной среде. Неподвижные животные (губки, кишечнополостные) либо селятся в подвижной воде, либо сами вызывают вододвижение.
Гранины между названными этапами условны. На каждом но-вОм этапе средства преодоления диффузионного барьера предыдущего этапа превращаются в дополнительное средство достижения одной и той же цели новым способом. Например, совершен
ствование двигательного аппарата у животных предполагает совершенствование источников энергии (появление АТФ), усложнение морфологии, преодоление диффузионного барьера.
Как и другие широкие обобщения, построения С.Э.Шноля имеют свою предысторию. Использованное им понятие относительной поверхности еще ранее получило признание в сравнительной анатомии (Беклемишев, 1964); представление о биохимическом и морфологическом этапах развития организменного субстрата было сформулировано Н.Пири ( Pine, 1954). Естественно, это нисколько не умаляет общебиологической значимости построений С.Э.Шноля. Теперь биология обладает понятием организменного субстрата, в котором стерта граница между формой и вещественно-энергетическим субстратом, между морфологией, биохимией и биофизикой. Взятое в аспекте исторического развития как усложняющееся понятие, оно соотнесено с естественным для жизни существованием в геологическом времени. Разногласия между морфологами и биохимиками относительно значимости разных сторон организменного субстрата теперь поза— йи« Поэтому не ради возвеличения морфологии, а. ради компенсации явной и преходящей недооценки морфологии в современной биологии следует перечислить некоторые свидетельства ее ис-к,5очительной содержательности.
У растений, как считает Ф.Боуэр (Bower , 1930), увеличе-Нйе размеров с неизбежностью влечет за собой усложнение фор— Mbl как средства регуляции величины относительной поверхнос-
Эго отражается не только во внешнем облике растений, но
С их внутреннем строении. Например, у относительно крупных ^Мидиевых водорослей форма хлоропласта сложнее, чем у бо—
153
лее мелких. По поводу проводящей системы сосудистых р^^ ний Боуэр замечает, что, начиная с некоторого предела, t;e нейшее возрастание абсолютных размеров делает невозможн U использование растением поверхностного натяжения жидкост^’ для транспортировки веществ.
Отсюда возникает необходимость иных, нежели у растение механизмов движения внутренней среды, например таких, имеются у животных. По Ф.Боуэру, "размер-фактор* имеет ное значение для понимания организации живых существ. Поэ ' тому в определенном смысле растения и животные противощх ложны. "Тело животных характеризуется... сосредоточением ганов внутри и слабым развитием наружной поверхности сра^ нительно с массой тела. У растений, наоборот, поверхность ла обыкновенно сильно расчленена и развита" (Талиев, 1931 с. 9). Различия эти не абсолютны. С одной стороны, существуют кактусы с относительно несложной формой тела, а с дру_ гой - "зоофиты", колониальные беспозвоночные (кишечнополощ, ные, мшанки, морские лилии), напоминающие своей формой р0Сш тения. В.И.Талиев указывает, что "...при водном существовании внешний механизм добычи питательных веществ и у растений и у животных может сильно сближаться..." (1931, с. Ю), потому что нужно иметь большую относительную поверхность для усвоения того питательного материала, который несет мимо неподвижно прикрепленного животного вода. Экспериментально доказано, что у некоторых водных животных происходит усвоение растворенных питательных веществ, что сближает их с растениями.
Если учесть, что между внутренними органеллами клетки и окружающей цитоплазмой осуществляется обмен веществ, то по аналогии с внешним диффузионным барьером следует принять понятие внутреннего диффузионного барьера (Заренков, 1979). Это обстоятельство объясняет, почему с увеличением размеров тела простейших наблюдаются усложнение формы ядра и увеличение его массы, появляются многоядерность и гетероморфизм ядерного аппарата (Осипов, 1981). Многоядерность в свою очередь шаг по направлению к многоклеточности (Иванов, 1968), т.е. тенденция к увеличению размеров тела. Почему она выгодна? Дело в том, что скорость перемещения в пространстве зависит от размеров тела. Помимо абсолютной скорое-ти перемещения (см/с) надо различать еще и относительную скорость (длина тела/с). Если животное преодолевает за сек^' ду две длины тела, то, значит, в течение секунды оно имело удвоенную относительную поверхность. Если простейшие облеРа' ют высокой относительной скоростью (десятки длин тела в се> кунду)» то они значительно уступают в абсолютной скорости позвоночным, в частности некоторым рыбам (десятки м/с). Очевидно, высокие значения абсолютной скорости доступны
154
-лум животным. Мелкие животные, имеющие высокую отно-^ельную поверхность, энергетически не в состоянии решить С е заДачУ» Между тем в борьбе за существование организмы ^оставляются именно по абсолютной, а не относительной ско-соСТи. Поэтому достижение высокой скорости возможно именно Р крупных животных. Однако размеры животных не представля— хаотической картины.
Одним из удивительных фактов в области морфологии яви-^сЪ обнаруженная Л.И.Численко (1981) статистическая закономерность в распределении размеров организмов, но природа ^гих закономерностей еще не раскрыта. Остаются также невыясненными причины распределения размеров тела в таксонах позвоночных животных (Стрельников, 197 3). Идея о том, что мелкие и крупные организмы отличаются не только размерами,
но, прежде всего, они представляют разные типы организации с множеством совершенно неожиданных особенностей, оказалась чрезвычайно плодотворной. Она фактически содержится в сочинениях Дж.Гексли (Huxley, 1932) и Д’Арси Томпсона ( d'Arcy Thompson, 1942) и в настоящее время наполнена богатым биологическим содержанием (Стрельников, 1970; McMahon, 1973; дольник, 1982; Schmidt-Nielsen, 1984). Другой раздел биоморфологии - это биосимметрика, в которой, в частности, рассматривается симметрия растений, например расположение листьев или частей цветка. А.П.Бердышев (1972), исходя из представлений о неодинаковой способности зачатковых клеток цвет-
ка к дальнейшему делению, выдвинул гипотезу, объясняющую.
количественные соотношения, которые подчиняются закономерности Фибоначчи.
История понятия симметрии в биологии (Касинов, 1973; Урманцев, 1974) начинается с открытия Л.Пастером в середине XIX в. нерацемичности веществ в организме. Сам Л. Пастер и многие его последователи расценили эго открытие как установление биологической специфичности (Абакумов, 1972). В этом отношении примечательна уже своим названием статья 'Стереохимия и витализм* ( Japp, 1898). Дж.Бернал (1969) связывал возникновение жизни с дисимметрическим синтезом биополимеров. Таким образом, биосимметрика пытается утвердить биологическую специфичность как специфичность особого организменного пространства. Вместе с тем известно, что встречаемость 1- и d- форм молекул и макроструктур неспецифич-для живой природы (Урманцев, 1971). Известно также, что симметрия организмов имеет экологическое значение и отвеча-е<г условиям их обитания (Гиляров, 1944). Следовательно, фи-аиологизация биосимметрики тоже не исключена. Разумеется, Эт° не означало бы, что сам феномен формы сводим к химическому составу; форма и химический состав - разные стороны 11 снятия вещества, имеющие значение для жизнедеятельности.
155
Наконец, надо отметить высказанное И.А.Акчуриным (1974) предположение относительно особой организованности жизни в организме, объясняемой *гротендиковыми структурами реальн^ го физико-биологического пространства*. Здесь не имеется в виду биосимметрика и сохраняется организменный подход. можности этого принципа для объединения биологического зна.. ния пока представляются сомнительными.
В свое время выживанию витализма способствовала механц. ка развития, предметно связанная с морфогенезом, становление ем в онтогенезе морфологии высших животных. Она получила развитие в работах В.Ру, Г.Шпемана, Г.Дриша, Ч.Чайльда. но на явлениях онтогенетического формообразования было осо.. бенно ясно видно, что жизнь является не только физико-химической проблемой, но и Проблемой формы, неразрешимой в понятиях физики и химии* (Бляхер, 1976, с. 233). При внимательном рассмотрении это обстоятельство не может не показаться странным: общебиологическая концепция пытается опереться на данные о морфогенезе высших форм жизни, прежде всего животных. Между тем очевидно, что для низших форм жизни, отдаленное представление о которых дают бактерии, прр» блема морфогенеза не имеет особого значения. Следовательно, морфогенез, понимаемый как проблема биологического пространства, вряд ли может вывести мысль на биологическую специфичность.
В новое время в поисках биологической специфичности орга-ницизм гоже обращается к морфологии (Sinnot, 1963). Проблема органической формы, в той или иной степени отделенная от физхимии, претендует на биологическую специфичность ( Waddington, 1951). Дж.Вуджером ( Woodger , 1945, р. 99) * жизнь рассматривается как иерархия пространственных частей*', Дж. Нидхем ( Needham, 1936) считает * плодотворным* предложение Вуджера (1929) считать витализмом *теорию, которая постулирует некоторую целостность живого организма, т.е. С, Н, N , О, Р и т.д. плюс организующие отношения*, и вместе с тем подчеркивает, что Центральная проблема биологии есть проблема формы* (Needham, 1936, р. 23). Сходных взглядов придерживается А.А.Любищев (1982). Надо заметить, однако, что морфогенез, высокая сложность формы и пространственного размещения частей организма — эти свойства отличают толь* ко высокоразвитые формы жизни и их, очевидно, не было у про* тобионтов; следовательно, форма, пространственная организация животных и растений, не составляет столь широкой биологической проблемы, сопоставимой с теорией жизни.
Еще А’Арси Томпсон ( d'Arcy Thompson, 1942, р.282) отметил: 'Исключая мелкие организмы, форма которых (подобно капле воды) создается действием молекулярных сил, мы можем рассматривать форму организма как функцию или прямое след-
156
^pjje роста, скорость которого различна в разных направлен»» Поскольку рост в конце концов есть результат обмена веществ, понятия формы и морфогенеза оказываются соединенны— с жизнедеятельностью организма. Теперь дело за некоторым мктором, задающим неодинаковые скорости роста зародыша в оазных направлениях (Medewar, 1945; Huxley, 1950), и задача объяснения появления сложной формы приблизилась бы к разрешению. Таким фактором могла бы быть наследственность, сожалению, механизм осуществления наследственности остается во многом еще нераскрытым ( Waddington , 1957; Тимо-феев-Ресовский, 1965; Преснов, Исаева, 1985), и вследствие этого сохраняется объяснение биологической специфичности с позиций организменной биологии и механики развития.
Если биологическая специфичность заключена в межпопуляционных взаимодействиях, то проблема биологического пространства относится к биотике. Форма организма, целесообразное пространственное размещение его частей, взаимная обусловленность строения разных частей организма изучаются сравнительной анатомией с помощью отношения корреляции, и здесь тоже необходим популяционный подход. Между тем понятие корреляции в биологии весьма многозначно (В.Шмидт, 1979, 1981), и это обстоятельство в известной мере отражает борьбу разных направлений биологической мысли - типологии и популяционизма. В общенаучном значении корреляция - это особый вид синхронной соотносительной связи, не выделяющей причины и следствия ная связь сованно и организма
ганизменными. Они введены в биологию прежде всего Ж.Кювье. Идет корреляции заимствована Ж.Кювье у Аристотеля, и, возможно, у И.Канта (Лункевич, 1960; Канаев, 1976). В согласии с аристотелевскими представлениями о доминировании единичного над множеством и в соответствии с типологической концепцией организма-машины Ж. Кювье и его современники соотнесли корреляцию с единичным организмом. Скоррелирован— Кость частей организма, организменная целостность проявляются прежде всего в целесообразном размещении частей организма. Впоследствии у А.Н.Северцова (1939) они получили название топографических корреляций. По Ж.Кювье, организменная Целостность как целесообразное размещение органов и есть План строения, по Ж.Сент—И леру это - коннексия органов ( Cuvier, 1817, р.25).
Вследствие скоррелированности органов становится возможным функционирование организма как целого: * Всякое организованное существо образует целое, единую замкнутую систему, Насти которой соответствуют друг другу путем взаимного влия— ft
(Аскин, 1978). Корреляция в организме - это взаим-органов и вообще структур, функционирующих согла-тем самым делающих возможным само существование как живой машины. Такие корреляции называются ор—
157
ния, конечной цели* (Кювье, 1937, с. 130). Пор 'конечной целью* подразумевалась жизнедеятельность организма как С{1 ноним жизни (Борисяк, 1937). Особенность жизнедеятельно," ги животных вызвана их способностью передвигаться. Исходя" из этого в отличие от растений организацию животных надо изучать, начиная с нервной системы. Ею начинается иерархия
структур и органов: на первом месте нервная система, на ВТ(к ром — мышечная система и опорные структуры, на третьем -пищеварительная система и т.д. Так же как в систематике признаков, в сравнительной анатомии разрабатывалась иерарху органов по их значению в жизнедеятельности - подход, свойст-венный типологической школе. Бесспорно, что у Кювье корре^ ции, в сущности, оставались морфофизиологическими. Физиоло-гичность таких корреляций была подаеркнута В.Гёте (1957, с. 155). Например, по Кювье, у травоядных живогных длинный кишечник скоррелирован с особым строением зубов. Эта корре, ляция объясняется тем, что для переваривания малопитательной растительной пищи требуются значительные физиологические усилия. Из этого примера видно, что расширение сравнительной анатомии в 'сравнительную морфофизиологию* отвечало бы духу построений Кювье. У А.Н.Северцова (1939) наряду с другими также рассмотрены морфофизиологические корреляции. Тот факт, что закрепилось все же другое выражение - "сравнительная анатомия*, - возможно, следует объяснять распространенной среди морфологов иллюзией, будто Морфология (*с большой буквы') может выразить биологическую специфичность и сохранить значение альтернативы физико-химическому редукционизму. Между тем имеются основания расширить предметную область сравнительной анатомии в сравнительную морфофизиологию и биохимию.
У растений 'биохимические свойства... коррелятивно связаны со многими морфологическими особенностями* (Благовещенский, Александрова, 1974). Интересные примеры самых разных корреляций у животных собраны И.Д.Стрельниковым (1970), множеством материала располагают ботаники (Г.Кецров, 1968; Went , 1971). Далеко не все организменные корреляции получили причинное физиологическое объяснение. Например, со времен Аристотеля (1937, с. 34) остается непонятным, почему 'большинство парнокопытных имеет рога для нападения, так как некоторые, однокопытные*. Хотя функциональное значение этих образований известно, их совместная встречаемость может быть объяснена в самой общей форме: такова наследственность. Тогда корреляция в данном случае - проявление плейот-ропизма, ее нац о рассматривать в рамках проблемы "ген -знак*. Для понимания организменных корреляций существенно,
что корреляция вообще может пониматься как состояние и как процесс. На первый взгляд представляется, что Ж.Кювье
158
~uvier , 1800) под корреляцией понимает процесс, в котором 3^еЯениЯ одних органов могут вызывать изменения ,других. Су-1 до всему, имеются в виду "изменения*' в двух направлениях, ^первых, это согласованные изменения строения животных в ^огенезе - организменный процесс, которые для эволюциониста де будут собственно изменениями. Во-вторых, изменение сдой'структуры вследствие преобразования другой в гипотетичес-опыте. При этом представление о согласованности структур возникало при сравнении разных организмов, и этой логи— веской операцией сравнения была обязана своим названием *сра— рЛ^гельная анатомия*. Важно отметить, что сопоставление в современной сравнительной анатомии (В.Беклемишев, 1964) каК бы воспроизводит природаый процесс исторического развития, отрицавшийся Ж.Кювье. Таким образом, корреляции у Ж,Кювье - это скорее состояния, их действительно можно считать "статическими* (Новиков, 1937).
Меж организменные корреляции в строении самки и ее детенышей, разных каст термитов и пр. С.А.Северцов (1951) назвал конгруэнциями. Б.С.Матвеев (1945) вслед за В.В.Васне-цовым (1938) называл их экологическими корреляциями, потому что благодаря им виды, состоящие из неодинаковых организмов, могут жить в сообществах. Корреляции структур организмов разных видов, например энтомофильных растений и насекомых-опылителей, Б.С.Матвеев (1945) предложил называть коа-даптациями. Филегические корреляции, названные А.Н.Северцо-вым (1939) координациями, представляют собой систему взаимосвязанных изменений морфоструктур в историческом развитии. Например, в процессе возникновения скатов ог акул осуществились согласованные изменения общей формы тела, плавников, их опорных структур, зубного аппарата и пр. По А.Н.Северцову, носителями координаций являются организмы разных видов, критерий координации — константное сосуществование структур у организмов.
Таким образом, система организменных понятий как бы встроена в понятие видообразования, относящееся к популяцион— Ио-видовому уровню. И.И.Шмальгаузен (1942) соединил организменные корреляции, осуществляемые в онтогенезе, с филети-Ческими. В результате исторический процесс интеграции организменной целостности И.И.Шмальгаузен рассмотрел как развитие организменных корреляций.
Исторически старшими следует считать внутриорганизменные Корреляции. Представление о них возникло в типологической сравнительной анатомии (Шмальгаузен, 1935; Канаев, 1963).
В.Н.Беклемишева (1964) историческое развитие беспозво-н°чных в определенной мере изучено с помощью корреляций. ^Ни могли бы считаться координациями. Однако, как и в других названных формах межорганизменных корреляций (между
159
организмами разных видов), понятие вида при этом фактичее не использовано. Историческое развитие таксонов, в которое разворачиваются корреляции между строением вида-предка и вида—потомка, представляется чем-то вроде развития насекОк. в ходе которого из яйца выходит личинка, личинка превращ^^1 ся в куколку, а из куколки получается взрослый организм, и, тривидовая изменчивость и механизм видообразования в целок/' только подразумеваются, но сами по себе не использованы в логике исследования и не присутствуют в получаемых результа г ах.
Через корреляции определяется важнейшее в сравнительной анатомии понятие типа организации ( " I'embranchement"). "ЕшЬ ranchement является также техническим словом плотницкого ремесла и означает: соединять и сглаживать балки и стропила... анатомический тип, общий образ, в котором, по возможно, сти, содержались бы формы всех животных... общая схема, ко. торой были бы подчинены как человек, так и животные, с помощью которой можно было бы сравнивать между собой классы, роды и виды и судить о них* (Гёте, 1957, с. 146, 155, 2 54). Впоследствии К.Бэр остановился на понятии типа в ело. дующем отрывке: "Идея животной организации... осуществлена в некоторых главных формах, которые опять распадаются на варианты низшей ступени... Типом я называю характер расположения органических элементов и органов* (цит. по Микулинско-му, 1959, с. 320). Значение типов состоит в следующем: *Раз все обычные представления о животной лестнице оказываются ошибочными, то приходится рассматривать каждую форму как модификацию какой-нибудь общей формы, а последнюю как мода-фиканию основного типа...* (Бэр, 1924, с. 62). В этих цитатах тип определен как некоторая обобщенная конструкция. Ее модификации образуют иерархическую структуру. Те или иные скоррелированные органы и морфоструктуры выделяют ступени иерархии сообразно с их значением в жизнедеятельности.
Поскольку функционирование органов возможно при условии их целесообразного размещения ( Cuvier , 1817), четыре предельно широких типа животных у Кювье оказались и планами строения. Названия для ступеней иерархии были заимствованы из систематики. Если при последовательном типологическом подходе в систематике такие названия, как тип, класс, отряд, вид, имели только логическое (не природное) содержание (уст#* новленное Аристотелем), то иерархические ступени в типолог»^ ческой сравнительной анатомии тоже должны были бы иметь логическое, но не природное содержание. Когда благодаря дар" винизму вид сделался природным объектом, природное содержа ние должно было бы наполнить и понятие вида в сравнительной анатомии. И если в систематике сходство между видами и ИХ объединениями в таксоны стало возможным объяснять род ст-
160
ofAt то в сравнительной анатомии скоррелированность призна-видов также можно было бы истолковать в связи с истории ьеским родством, далее, так же как в систематике,-степень с,содства отражает уровень родства, а иерархическая структура рлстемы таксонов — отдаленность родственного сходства, так в сравнительной анатомии следовало бы сходным образом поддать степень скоррелированности и природу иерархичности слстемы типов. Наконец, так же как в систематике возникла принципиальная неопределенность системы и вокруг нее вспыхнули споры, в которых открылась противоположность между типологией и популяционизмом, так и в сравнительной анатомии должно было бы происходить то же, что в систематике. В действительности же в области сравнительной анатомии царит относительное спокойствие.
Переход от организменных корреляций к корреляциям у вида да первый взгляд не представляет сложности. Если скоррелированные структуры константно сосуществуют у организмов вида, то корреляция тех же структур у вида - это многократное повторение одной и той же корреляции у многих организмов. Если же некоторая корреляция наблюдается у вида, то она же будет многократно повторена в объединении видов, относимых к какому-либо типу организации. В действительности же такое умоза-. ключение было бы ошибочным. Вид нельзя понимать как сумму одинаковых организмов, а надвидовую единицу - как сумму одинаковых по данным структурам видов.
Прежде всего, между понятием признака *в систематике и понятием структуры или органа в сравнительной анатомии имеет место определенное соответствие. Признаком может служить наличие-отсутствие какой-либо структуры (органа). С другой стороны, систематики выделяют- признаки, не интересуясь их Функциональным значением, тогда как органы выделяются сравнительными анатомами с помощью понятия функции ('органы пищеварения'). Вместе с тем далеко не всегда остается понятным функциональное значение многих органов или структур, и хотя это не есть довод против функциональной значимости таких органов или структур, констатация корреляции между ними вполне допустима. В общем как будто бы можно принять, что 6 сравнительной анатомии и в систематике основу познавательной деятельности составляет сравнение, и во многом этот ме— работает в них сходным образом. В таком случае Для понимания корреляции структур внутривидовая изменчивость име-611 то же значение, что и в систематике для понимания отношения сходства. В сравнительной анатомии имеют смысл понятия прообразов структуры, поднимающихся по иерархической лестнице типов организации, сохраняет значение внутривидовая изменчивость как источник для разнообразия типов организации Н® надвидовом уровне и т.д. Есть и другие параллелизмы. В 11 - 1560 161
систематике вследствие политетичности таксономического ДИаъ ноза единичный организм не может заменить собой типа орге " низании.
Так же обстоит дело и в сравнительной анатомии. Для т>ог чтобы разобраться в этом вопросе, воспользуемся уже опроб^ ванными обозначениями:
— название структуры или органа (например, плацент у млекопитающих);
— структура (орган) имеется (например, плацента егся);
- структура (орган) отсутствует (например, плацента отсутствует).
Сходным образом с помощью символов a.j {Qj, QjJ но обозначить наличие - отсутствие сосцов млечной железы у млекопитающих. Этим структурам отвечают особенности функционирования млекопитающих: у живородящих плацентарных самки имеют сосцы, у яйцекладущих клоачных вместо сосцов имеются млечные поля. Корреляцию между этими структурами можно
туры встречаются или отсутствуют вместе и порознь никогда не встречаются. Такая корреляция есть свойство целого класса млекопитающих, ее носителем не может быть ни один единичный организм. Корреляцией можно было бы считать отсутствие хотя бы одного сочетания символов в корреляционной решетке. Наоборот, единственное сочетание символов ay a.j и отсутствие символов в трех прочих клетках тоже обозначало бы корреляцию, однако всего лишь ее частный случай, "как константное сосуществование", по А.Н.Северцову (1939). Если заполнены все четыре клетки, корреляция отсутствует.
Любое заключение о корреляции, полученное на эмпирическом материале, ничем не защищено от новых открытий. Среди новых, не известных науке видов могут встретиться обладатели еще не известных сочетаний структур, и тогда будут заполнены все четыре клетки решетки. Это означало бы отсутствие корр6* ляции между признаками - для сравнительной анатомии и конги' дуальность - для систематики. Наоборот, скоррелированность признаков, незаполненность хотя бы одной клетки корреляционной решетки, означала бы дискретность таксонов, невозможное1* Таксономического континуума. Таким образом, проблема понятий типа и корреляции переходит в проблему границ таксонов систематики. Как в систематике, любые суждения о систем©
162
остаются принципиально незащищенными от новых открытий, так 0 в сравнительной анатомии представление о типе организации Л корреляциях между структурами остаются гоже незащищенными, открытыми. Скоррелированноегь структур, как и сходство в признаках, есть становящееся, развивающееся отношение меж— ру длящимися популяционными объектами. Историческая общность сравнительной анатомии и систематики не случайна. Понятийный аппарат этих разделов биологии обладает глубоким внутренним сродством. Вплоть до настоящего времени сохраняется тради—
°j
а О —
— 1 o' 1 Q
ция изучения типов организации как типов организации таксонов. Вообще говоря, эго частная форма типов организаций вообще, они подразумеваются в ячейках 4, 11, 14, 15 (риа12). Между таксонами и типом организации в действительности нет однозначной связи. Возьмем, например, класс насекомых. Его можно рассматривать как единство скоррелированных структур: определенный набор ротовых придатков, определенное число грудных сегментов с гремя парами ног, трахеи, особые органы выделения. Сравним его с классом ракообразных, который характеризуется значительно меньшей устойчивостью признаков (д>-статочно сравнигь речного рака и циклопа): непостоянный состав грудного отдела, неодинаковое число грудных придатков очень несходного строения, разнообразие органов дыхания (листовидные ноги, особого рода трахеи у мокриц).
Значит, как тип организации класс насекомых представляет собой большую целосгносгь, чем класс ракообразных. В первом имеет место более полное совпадение понятий таксона и типа, чем во втором. Соответственно в первом совпадают полнее, чем во втором, понятия признака (сходства) и корреляции (структур). Несовпадение таксономического признака сравни-тельн©анатомической структуры означает относительную самостоятельность сравнительноанатомического подхода от таксономического при изучении родословных. Эта самостоятельность
163
подразумевает несовпадение древа таксонов (филогения, такс^ нсгенез) с древом типов организации (типогенез). Ею отчасти объясняется, почему понятие парентальной биологии шире фц;-]Ск
Рис. 12. Логическая структура сравнительной морфофизиологии в фигуре "продырявленная пластинка" биологической розетки. Пунктиром обозначена область парентальной сравнительной морфофизиологии sensu lato, в ней выделена сравнительная мор-фофизиология таксонов, т.е. филогенетическая сравнительная морфофизиология
генетики. Ячейки 6, 19, 10, 23 обозначают типы организации родословных, в которых имеет место отношение родственной скоррелированности строения предка и потомка, потомков общего предка. На рис. 12 линия, обозначающая плоскость парен-гальности, выделяет две области сравнительной анатомии: в одной присутствует отношение исторического родства (пунктировано), в другой его нет. В первой скоррелированность строения вызвана родством, во второй — иными отношениями.
Ячейка 5 (табл. 7) отвечает наиболее общей точке зрения, когда одно только отношение корреляции выделяет тип организации живых существ (объекты из соседней ячейки 7 - это тип организации земных существ, выделяемый дополнительно отношением совместности). Сюда, возможно, относится своего рода основа организации, понимаемая как взаимосвязь, единство функционирующих макромолекул (ДНК - РНК - белки). Оно о& разует высшую ступень иерархии типов, и, по-видимому, с него началось развитие других, более мелких типов организаций-
Развитие, таким образом, было как бы нисходящим. Ингер60’ но, что развитие таксономической иерархии происходило как бь'
164
Таблица 7
Объекты сравнительной морфофизиологии
Л биохимии (сравнительной анатомии): гипы организации (номера ячеек см. на рис. 12)
Ячейка Парентальная сравнительная анатомия Ячейка Непарентальная сравнительная анатомия
11 тип организации как 5 тип организации живых
таксон (родословная и существ
фенон) 45 тип организации и фенон
4 тип организации как таксон и биотический элемент 25 тип организации, фенон и биотический элемент
15 тип организации как таксон, биотический 26 тип организации как биотический элемент
элемент и жизненная 31 тип организации как би-
форма огический элемент и
14 тип организации как жизненная форма сооб-
таксон и жизненная
форма 16 тип организации как фе-
19 тип организации как родословная и биотический элемент нон, биотический элемент и жизненная форма сообщества
10 тип организации как родословная, жизненная форма и биотический 20 тип организации как жизненная форма сообщества
элемент 29 тип организации как фе-
23 тип организации как родословная и жизнен— нон и жизненная форма
й ная форма
ь обратном направлении, снизу вверх: суперпопуляция протоби— онтов - вид - род - семейство и т.п. На низшей ступени иерархии типов организации находится вид. Он представляет собой объединение популяций, различающихся мелкими структурами. Оставаясь в ячейке 5 и пользуясь одним только отношением Корреляции, мы не смогли бы выделить в нем типы организации объединения ихищник и жертвы4' (и отдельно “'хищника* и Жертвы) - для этого понадобилось бы отношение сосуществования ; рака—отшельника и актинии - для выделения подобных объектов потребовалось бы дополнительно отношение совместного сосуществования; "человек и его гоминидных предков" - для
165
этого необходимо привлечь еще отношение родства; отдельно "человека" и отдельно его "гоминидных предков4' - для этого дополнительно понадобилось бы отношение сходства. Системы иерархии в каждом случае были бы неодинаковыми, поскольку они выделены разными отношениями.
Выделение и расчленение типов организации для экологии, биотики и фенонической систематики происходит в той части фигуры "корреляция", которая относится к овалам "сосуществование", "сходство", "совместность*. Здесь ячейки обозначают типы организации в биотике (ячейки 25, 26), биотике и экологии (ячейки 16 и 31) и собственно в экологии (ячейки 20 и 29). Они были рассмотрены в соответствующих главах. Дополнительно там находятся такие объекты, как "горные организмы", "полуводные (околоводные) гомойотермные животные*, "Гомойотермные животные бореальной климатической зоны" (Экологическая физиология животных, 1979, 1981). Впрочем, два последних объединения выделены с помощью отношения. Частично названные ячейки вместе с ячейкой 24 находятся еще и в овале "сходство". Примером соединения сходства и корреляции может служить так называемая "параллельная эволюция*. Она состоит в том, что у ряда не связанных родственными отношениями растений Австралии имеется так называемый эвкалиптовый тип листа, единство скоррелированных особенностей внешнего и внутреннего строения этой части растений. Не исключено, однако, что такое сходство объясняется обменом сегментами хромосом между видами (Went, 1971), что напоминает трансдукцию. Если трансдукция относится к числу мута-факторов мутагенеза, то рассматриваемое явление не следует относить к филогенетике. Тогда пример эвкалиптового листа свидетельствует о том, что отношение корреляции действительно может сопровождаться сходством, независимым от родства. Если дополнительно в рассматриваемом примере имеются в виду рёценгные растения одной флоры, то пример относится к ячейке 16. Если же подразумевается флора биохора в историческом развитии, в диахронии (рецентная и вымершая), то место этого примера будет в ячейке 25. Вообще надо заметить, что анализ конкретного материала из биологических публикаций обычно затруднителен вследствие того, что авторы особо не оговаривают пространственно-временных рамок своих исследований.
Другой пример - это сопряженная эволюция паразита и хозяина. Для выделения такого объекта, как, например, "собака и клещ", не требуется отношения сходства. Скоррелированность строения этих животных (волосяной покров собаки, цепляющиеся ноги клеша и пр.) могла бы пониматься по-разному. Пока рассматривается живой клещ на живой собаке, пример этот тается в ячейке 31. Далее оба животных могли бы рассматри-166
^ъся в диахронии (вымерший клещ, живая собака), и тогда ^тог пример принадлежал бы ячейке 26 при дополнительном ус— принадлежности к одной фауне (совместность). Можно бы— д0 бы представить себе хозяина (собаку) в составе фруны од-роро биохора—сообщества и отдельно клеща (потенциального паразита собаки, однако паразитирующего на другом животном) в рругом биохоре-сообществе. Тогда этот пример будет относитъ-сй к ячейке 20, если эти животные остаются в синхронии.
В примере с растениями организмы неразличимы по двум
„коррелированным признакам (которые выделяют тип), и разные растения находятся в одной клетке, образуя фенон - тип. Во ртором примере собака и клещ образуют тип по скоррелирован— рости некоторых признаков, и по гем же признакам они не сходны; это тип, но не фенон.
Заканчивая рассмотрение биологической розетки, надо отме-
тить следующее. На конкретных примерах показано, что выде-
ляемые биологами объекты образуют системное многообразие. Оно состоит из 31 классификации (по числу ячеек) одного и того же разнообразия "от бактерий до человека". Отдельные
классификации являются достоянием разных отделов биологии. Системность многообразия состоит в том, что для описания одного какого-либо раздела биологии понадобились понятия всех других разделов (см. табл. 3—7). Таким образом, в биологической розетке воссоздано единство знания о жизни, продемонстрирована пластичность языка биологии. Вследствие этого обычая линейная последовательность изложения непригодна для описания жизни: в каждом частном разделе биологии все понятия
нужны разом, изложение должно быть пятимерным, а не линейным.
Многообразие — эго неизменная структура жизни, она изначала могла бы быть у мировой суперпопуляции протобионтов. Многообразие не знало развития, оно может быть названо эво— люцией в собственном смысле этого слова, как развертывание. Здесь подходит сравнение эволюции с распусканием бутона цветка. Наоборот, каждая классификация вызревает в течение времени и в своей иерархической структуре (например, вид - род-семейство - ...) хранит следы течения времени.
Работоспособность теории оценивается по тому, насколько она способна объяснить имеющиеся факты. Под фактом автор Понимает картину жизни, воссозданную традиционной биологией, Многообразие ее объектов, плывучесть границ объектов. Для объяснения этой картины и привлечена биологическая розетка, основанная на той концепции жизни, которая в неявном виде содержится в дарвинизме.
Глава 13
НЕКОТОРЫЕ ПРИКЛАДНЫЕ ВОПРОСЫ
ТЕОРИИ БИОЛОГИИ
Для выявления биологической специфичности первостепенна значение имеет приспособленность, когорая отличается от вес прочих физико-химических свойств организменного субстрата Приспособленность лежит в основе представлений о биномиал^. ной структуре таксона, о случайной природе эмпирической blu борки в систематике. Как было показано выше, она же послу» жила отправной точкой при анализе понятия времени, логичен, кой структуры биологического знания и установления взаимосвязи понятийных аппаратов частных разделов биологии и пр. Этим возможности такого подхода не исчерпаны. Он сохраняет свое значение биологического принципа при истолковании эмпирически установленных закономерностей и способствует осмыс. ливанию общебиологического содержания отдельных разрозненных явлений, таких, как кривая Виллиса, учение А.Н.Северцова об ароморфозах, теория В.А.Догеля олигомеризации гомологичных органов.
1. Правило Виллиса - одно из наиболее широких эмпирически установленных закономерностей - отражает статистическое распределение подтаксонов в надтаксоне. 'Вогнутая кривая систематиков' - это графическое представление закономерности, установленной Виллисом (Willis, 1940). Она состоит в том, что имеется много родов с немногими видами в каждом роде и немного обширных родов. В итоге получается кривая, напоминающая гиперболу (рис. 13). История изучения этой закономерности описана В.Вильямсом (Williams, 1964). Сам Вильямс, как и впоследствии Э.Майр (1971), считал это явление биологически содержательным. Вслед за С.Райгом (Wright, 1941) допускалось, что обсуждаемая закономерность объясняется определенным соответствием между вымиранием и видообразованием. Противопоставив вымершие виды ныне живущим, С. Райт фактически нарушил естественность границ диахронных таксонов, состоящих из живых и вымерших видов. А.Н.Голиков (1976) привлекает экологическое объяснение для 'вогнутой кривой'. Между гем известно, что далеко не всегда таксоны обладают экологическим единством и не всегда занимают единую экологическую нишу. Таксоны выделены по сходству в гомологичных признаках.
С.Андерсон ( Anderson, 1974), оставив в стороне биологическую содержательность, выделил стохастический и детерминистский подходы в изучении кривой Виллиса. Отражая общую неудовлетворенность состоянием вопроса, А.И.Кафанов и В.В.СУ" ханов (1981) пришли к выводу о его биологической бессодер-
168
^ательности. Фактически этим выводом они признали произвольными границы и объем таксонов, устанавливаемых систематиками. Иногда (Мейен, 1974; Численко, 1977) "вогнутая кри
вая" обсуждается как проявление общебиологической и раже общенаучной закономерности в рамках чрезвычайно широкого закона Ципфа. д.Э.Филиппов (1984, с.410) выводив кривую Виллиса из дарвинизма как "...следствие случайного распределения адаптивных требований к признакам вида, налагаемых условиями существования в разных экологических нишах", хаотически распределенных в природе. Эго объяснение представляется наиболее правдоподобным, потому что использованный в нем принцип случайности подразумевает неоднозначное соотношение между субстратом как носителем
1 роды,ёоодще надтаксоны
Рис. 13. Условная "вогнутая кривая систематиков". Пояс-
нения см. в тексте.
гомологических признаков, с одной стороны, и адаптивностью (приспособленностью) - с другой стороны. Тем не менее следует заключить, что основанная на обширном фактическом материале "вогнутая кривая* еще не получила общепринятого объяснения. В целом сложившееся положение показывает различие между знанием и пониманием: зная о широкой природной
закономерности, мы не понимаем ее причин.
Предлагаемая гипотеза заключается в следующем. Формально "вогнутая кривая" напоминает гиперболу. Ветви математической гиперболы ограничивают две равновеликие в пределе площади, бесконечно возрастающие по мере асимптотического приближения к осям системы координат. На конкретных "вогнутых Кривых" нет асимптот, и обе площади ограничены, как показано на рис. 13. Грубо говоря, величины этих площадей представляют собой произведение числа видов на число родов. Возможно, Что величины этих площадей примерно равны, и это предположение можно было бы проверить расчетами на конкретном материале, имеющемся в литературе. Однако более привлекательным Представляется дедуктивный метод. Он позволяет получить выводы достаточной общности и избежать разного рода сложностей, присущих исследованиям эмпирического материала. Что же представляют из себя величины этих площадей?
169
Площадь на рис. 13 охватывает немного крупных родОв с большим количеством видов в каждом, площадь Sg - множе-ство мелких родов с небольшим количеством вадов в каждом. Обширные рода потому содержат много вадов, что виды относи, тельно слабо отличаются друг от друга. Мелкие роды бедны дами потому, что резко отличные виды выделены в самостоятельные рады. В пределах обширного рода различия между видами неопределенны, они могут быть значительными и исчезающе малыми (свойство.(I), с. 70 ). Когда систематик-*дроби-тель* ревизует такие роды, то вследствие политетичности (свойство (У), с. 77 ) из такого рода выделяются только немногие "оригиналы* и остаются более многочисленные относительно слабо различающиеся "заурядные* виды; в результате возникают Два рсда и снова неравного объема. Ревизия в обрат, ном направлении, в сторону объединения мелких родов, тоже не в состоянии изменить соотношения между мелкими и обширными родами. Чтобы убедиться в этом, достаточно мысленно про-делать ревизию в стиле систематика-"обьединителя", после того как завершил свою работу систематик~"дробитель*. Тогда в силу неоднозначного сходства между родственными видами таксона (свойство (1), с. 7 О ) немногие * сверхоригиналы" сохраняют свою самостоятельность как мелкие роды, а прочие относительно "заурядные* вады разных родов могут быть сведены в обширные, но немногие роды. Такие противонаправленные ревизии воспроизвели бы в классификации природное явление неоднозначного колеблющегося сходства в длении вадов.
Равенство площадей и $2 означало бы, что в процессе видообразования отбор игнорирует сходство: относительно сходных вадов в обширных родах примерно столько же, сколько относительно хорошо различимых видов в мелких родах. По-видимому, такой вывод есть следствие из теории отбора приспособленных. Отбор как бы *не замечает* сходства. В сущности, здесь наблюдается то же, что и в биномиальном по сходству распределении таксономического разнообразия: статистическая структура, основанная на сходстве, сохраняется вследствие того, что отбор имеет дело только с приспособленностью. Таким образом, оба статистических распределения основаны на общем принципе. Неудивительно, что между ними существуют и более гесные связи.
Гипотетическое объяснение "вогнутой кривой* может получить более основательное подтверждение, если кривую вывести из тех особенностей классифицирования, которые разобраны в гл. 6.'
Возьмем таксон, включающий 100 видов. Пусть это будет семейство. Руководствуясь признаками, названными в диагнозе этого таксона, разделим его на подмножества, как в дихотомическом ключе, для определения. ВследстЕие ,титетичности и
170
однозначной связи между сходством и родством получим голь-0 неравные по объему подмножества, например:
Здесь имеют значение не конкретные числа, а только сама неравномерность распределения видов по подмножествам. На основе все гой же политетичности дихотомическое деление могло
бы продолжаться следующим образом:
(П)
о, а2 а3 а4 , 1101
аг а_2 а3 , 1100
Gj а2 а3 а4 } 1011
а, а2 Qj ач , 1010
а1 ~а2 а3 , Ю01
а1 а 2 аз ач > 10°0
аг а2 а3 0101
а? а2 а3 , 0001
Получаемые при этом подмножества иногда отвечают целым таксонам, например родам, другие представляют собой объединения нескольких родов, третьи - части родов, группы видов Разных родов.
171
Естественным образом некоторые единичные виды могут вечать монотипическим родам. "Нулевые* подмножества могу,г? означать сочетания признаков, не попавшие во множество 1qq видов. Такие сочетания признаков могут оказаться в принциПе неосуществимыми или всего лишь временно неизвестными исследователю. Подробно этот ветвящийся процесс рассмотрен автором на конкретном примере (I) ракообразных (Заренков 1976, 1984). Более существенно иное. В силу того что по сходству эмпирическое разнообразие есть случайная выборка из природной совокупности, все рассуждения на качественном уровне можно осуществлять на примере. выборки так, будто эгс. природная картина. Поэтому на некоторой стадии ветвящийся процесс можно прервать и рассмотреть полученное множество сочетаний признаков (II). Такие сочетания признаков суть события в том значении, которое было фактически предано этому слову в гл. 6. Если хотя бы на некоторых стадиях ветвящего, ся процесса исходное множество видов подразделяется на неравные педмножества (принцип политетичности), то с неизбежностью, начиная с некоторого момента, события будут распределены в согласии с правилом Виллиса: будет множество * единичных* событий и мало событий и мало событий с большим числом видов в каждом.
Теперь осталось только одно: * довести* события до таксонов. В сущности, события - это взаимоадекватно описанные таксоны: в строчке символов, отвечающей своеобразному диагнозу, однозначно сообщается о наличии (*1*) или отсутствии (*0*) данного признака. Чтобы от таких взаимоад екватных диагнозов перейти к диагнозам, на которых основаны обычные таксоны и "вогнутая кривая", надо, во-первых, разъединить роды, если они соединены в одном событии, и, во-вторых, объединить разъединенные части родов, попавших в разные события. Для решения первой задачи надо ввести дополнительный разъединяющий признак которым реально и разли-
чаются данные роды. Выделенные таким образом самостоятельные роды будут иметь диагнозы, неадекватные прочим диагнозам сопохниненных родов семейства. Вследствие политетичности эти роды, скорее всего, будут состоять из неодинакового числа видов, для решения второй задачи надо в одном роде объединить события, невзирая на различия по некоторому уже использованному признаку. Диагноз восстановленного таким образом таксона снова будет политетическим по этому признаку. Обе задачи, в сущности, представляют собой два направления в систематике: одно представляют "дробители", другое - "объединители*. Но пока остаются явления политетичности признаков и взаимной неадекватности таксонов, имеющие биологическое обоснование, "вогнутая кривая* неуничтожимо.
172
Таким образом, есть основания заключить, что "вогнутая р^щая* ~ действительно биологически содержательное явление, рис объясняется особенностями отношения сходства популяцион-объектов и биологической специфичностью, которая состоит особом бытии живых существ как носителей приспособленно-
сТ0- „ „
Существует ли соответствие между вогнутой кривой и би-?о1лиальным по сходству распределением событий? Этот вопрос (дожно поставить иначе, если несколько изменить содержание биномиального распределения. Новое содержание состоит в следующем. На оси ординат ((8), с. 90 ) теперь будем откледы-вать общее число видов в событиях, образующих данный член биномиального разложения. Например, если член 2(10. биномиального разложения (& + Q состоит из двух событий -и 01 0.2 и если первому отвечает пять ко один вид, то общее число видов для данного члена разложения равно шести. Такие подсчеты были выполнены на примере некоторых ракообразных (Заренков, 1976, 1984). Они показали, что одновершинное биномиальное распределение видов в событиях эмпирической выборки как будто бы сохранилось. При
-1U2 видов, а второму толь-
знавая преимущество дедуктивного метода изучения эмпирического материала перед индуктивным, мы должны были бы прежде всего понять общебиологическое значение полученного результата. Оно, по—видимому, заключается в следующем.
Если количество видов в данной группе животных или растений свидетельствует о приспособленности данной группы, так же как количество организмов говорит о приспособленности ви
да, то число видов в таких группах не может зависеть от сходства данной группы с другими группами видов. Иными словами, нет оснований принимать гипотезу, будто непохожие события (крайние члены биномиального разложения (8)) наиболее богаты видами. Нет оснований принимать и другую гипотезу, будто сходные события состоят из минимального количества видов. Пока остается в силе представление о независимости приспособленности от сходства, распределение ведов по событиям следует считать случайным. Более точное представление о таком распределении дает завершающая стадия ветвящегося процесса (11, с. 171): немного событий с большим числом видов и мно-ро событий с малым числом видов. Возможно, такое распределение можно считать пуассоновским. От структурированного таким образом множества ведов теперь надо перейти к вогнутой кривой, от распределения видов — к распределению видов в ро-Аах.
Как было отмечено выше, объемы родов и событий совпадают не всегда. События — это взаимоадекватно выделенные объ
единения видов, лишенные политетичности порознь и сохраняющие политетичность только в совокупности. Было отмечено так-
173
же, что на фоне противонаправленной деятельности 'дробц?^ и "объединителей* объединение событий в роды приведет к литетическим Диагнозам родов (результат деятельности джителей*), а также родов, основанных на взаимно неадеква" ных диагнозах (результат деятельности 'дробителей*, привп^" кающих новые признаки). Поэтому система отдельных групп вотных и растений, которые являются продуктом коллективног^"* творчества 'дробителей* и 'объединителей' в историческое мя, не могут стереть ту статистическую структуру, которая задана биномиальным распределением. Если 'дробигели' и
'объединители' классифицируют на основе биологической логи-ки, создавая взаимно неадекватные ('дробители') и политети-ческие ('объединители') диагнозы, то структура биномиально, го распределения (8) представляет собой пример множества, упорядоченного на основе правил классификации формальной по-гики. Перераспределяя даже одни и те же виды различным об-
разом по разным родам, систематики разных направлений не могут стереть биномиальную структуру потому, что не уступают друг другу. Каждый систематик мог бы подобрать такие
примеры, что в пределах крупного таксона сосуществуют взаимно адекватно и неадекватно выделенные роды. Нетрудно отыскать политетичные диагнозы и их противоположность и тогда, когда имеются однозначные указания о наличии - отсутствии
признака.
В контексте высказанных соображений следует прийти к выводу о том, что между обоими типами статисгических распределений могла бы осуществляться взаимосвязь. В свою очередь отсюда проистекало бы следующее.
'Вогнутая кривая', представляющая эмпирический факт, могла бы получить общебиологическое обоснование. И наоборот, биномиальное распределение, обоснованное биологически, но опробованное на незначительном фактическом материале, могло бы теперь претендовать на тот значительный по объему фактический материал, на который опирается 'вогнутая кривая".
За пределами систематики 'вогнутые кривые' известны в экологии, фаунистике и флористике. Они изображают количественное распределение таксонов жизненных форм, типов организации. Широкая распространенность этого явления, инвариантность статистической структуры столь различных объединений, по-видимому, могли бы получить свое объяснение на основе статистической природы популяции. Если отдельные объекты биологии статистически перекрываются в длении, как это показывает биологическая розетка, то структура одного объекта могла бы передаться и другим объектам. Современное состою ние вадов и других таксонов, которые иногда отвечают (или 116 отвечают) какой-либо одной жизненной форме, удовлетворителг* но совпадают (или не совпадают) с типом организации, соот
174
ветствуют (или не соответствуют) фаунистическому элементу, ^ог'ЛО бы пониматься гак: на срезе современности мы видим то, что могл° бы иметь место в длении.
2. В свете представлений о приспособленности как особом свойстве вещественно-энергетического субстрата между теорией отбора Ч.Дарвина и учением А.Н.Северцова (1939) об аро-мсрфозах существует глубокая связь. Преуспевающие виды, по Ч./Зарвину, отличаются высокой численностью организмов и об-। ширным ареалом. Биологический прогресс систематической группы характеризуется дополнительно "распадением, по мере того как животные попадают в новые условия существования, на подчиненные систематические единицы (разновидности, группы и т.Д.)" (Северцов, 1939, с. 271). В историческом развитии биологический прогресс может достигаться двумя противоположными способами: с помощью ароморфозов, изменений морфофи-зиологии широкого значения, поднимающих организацию животных на более высокую ступень, и идиоадаптаций, частных приспособлений, не повышающих "общей энергии жизнедеятельности". Из этого можно видеть, что понятие биологического прогресса близко к понятию приспособленности; что же касается идиоадаптации и ароморфозов, то они относятся к организменному субстрату. А.Н.Северцов (1939, 'С. 332) указывал, что оба тина исторического развития свободно переходят один в другой. Впоследствии А.С.Северцов (1973) подчеркнул конкрет-ный характер, относительность ароморфозов. Вслед за Б.С.Мат— веевым А.С.Северцов (1973) пришел к выводу, что признаки, ароморфные для одной группы, для другой группы лишь идиоадап— тивны.
В этом выводе фактически содержится указание на то, что между степенью биологической прогрессивности (приспособленности), с одной стороны, и морфофизиологическим субстратом организма, с другой стороны, нет однозначной связи. Эго позволяет считать учение А.Н.Северцова творческим развитием Дарвиновской концепции приспособленности. Из представлений А.Н.Северцова об ароморфозе и идиоадаптации вытекает, что свойства организменного субстрата сами по себе, без учета условий обитания, еще не могут служить основой для оценки п регрессивности (приспособленности).
' Общебиологическое представление о неоднозначной связи органа и вообще морфоструктур с их функциями также напоминает проблему субстрат — приспособленность. Известно, что мор-фоструктуры вообще мультифункциональны и в свою очередь в одной функции могут участвовать разные структуры. Морфофункциональная целостность организма рассматривается как конкретизация понятия жизнедеятельности, жизнеспособности в подходящих условиях обитания. В каких-то иных условиях обитания организм может оказаться нежизнеспособным. Здесь то
175
же проглядывает проблема знака и значения. Поэтому необхсх. димо подчеркнуть особенность понятий знака и значения для понимания организма, отличие этих понятий на организменном уровне от тех же понятий, перенесенных на популяционный ур^ вень. Отличие состоит в том, что приспособленность на попу., ляционном уровне имеет значение для отбора. Наоборот, жизне. способность (приспособленность), соотнесенная с единичным организмом, значения Для отбора не имеет. Таким образом, проблема знака и значения, несмотря на любопытные параллели, имеет разное содержание на организменном и популяционном уровнях. Приспособленность, которая имеет значение для отбора, единичный организм приобретает в борьбе за существование. Такая приспособленность вторична от противоречия меж. ду численностью всей популяции и ограниченными возможностями внешней среды.
3. Олигомеризацией органов и структур В.А.Догель (1954; назвал уменьшение их числа в ходе эволюции. Примером может служить сокращение числа головных щупалец у головоногих моллюсков от нескольких десятков (наутилус) до 8-10 у прочих видов этих животных. К олигомеризации В.А.Догель отнес также случаи концентрации отдельных структур, такие, например, как слияние в мозговую массу нервных ганглиев у насекомых. Олигомеризации подвержены гомологичные структуры (щупащ-. цы головоногих, ганглии насекомых), а также гомодинамные образования. Если, например, часть двигательных конечностей членистоногих переключится на сенсорную функцию (смена функции), то число двигательных конечностей сократится, хотя структуры остаются и выполняют новую для них функцию.
В своем критическом рассмотрении теории В.А.Догеля Н.А.Ливанов (1960) обратил внимание на то, что концентрация структур не ведет к сокращению их количества. Она сводится лишь к изменению их взаиморасположения и поэтому не подходит под определение олигомеризации как процесса уменьшения числа структур. Явление концентрации, слияния таких структур, как ганглии насекомых, получило название литации (Подлипаев, Наумов, Бор кин, 1974). Тем не менее олигомеризация в узком смысле (без лигаций и без случаев смены функций) по-прежнему основана на обширном материале. Обсуждение этого явления (Значение процессов полимеризации и олигомеризации в эволюции, 1977) выявило сложность и многоплановость построений В.А.Догеля. Н.В.Тимофеев—Ресовский, Н.Н.Воронцов и А.В.Яблоков (1969), не обратив внимания на существенные замечания Н.А.Ливанова (1969), рассматривают олигомеризацию в качестве модуса эволюции.
Несколько иное понимание количественных изменений гомологичных счетных (т.е. тех, которые можно сосчитать) структур предложил Н. А. Заренков (1972). В построениях В.А.Догеля,
176
хак и Других филогенетиков, не учитывается то обстоятельство, что признаки видов таксона закладываются в качестве прообразов (различительных особенностей внутривидовых подразделений) вида-предка этого таксона (с. 70, (1)). Примем, что признаком соподчиненных таксонов служит количество щупалец: g - у осьминогов и 10 - у кальмаров. Тогда общий предок этих животных должен обладать признаком: *8-10 щупалец*. В одних популяциях предкового вида преобладают 8-щупальце-рые организмы, от них могли произойти осьминоги. От других популяций, состоящих из 10-щупальцевых организмов, могли возникнуть кальмары.' Разумеется, для решения вопросов филогенетики нельзя ограничиваться одним каким-либо признаком, и дело не в головоногих моллюсках, а в подходе к проблеме. Этот подход состоит в том, что время должно рассматриваться в Двух аспектах: помимо течения времени (филогенез) надо пользоваться длением времени. Длящийся предковый вид раскрывает во времени собственную внутривидовую изменчивость, на основе которой могут возникать виды—потомки.
Если рассматривать возникшее таким образом множество родственных видов без их общего предка (такие виды обычно остаются неизвестными), то вслед за В. А. Доге л ем можно сделать заключение о закономерном уменьшении числа гомологичных структур. Такая родословная в основном будет представлена филогенетическим рядом. Если же учесть, что родственные виды имели общего предка и счетные структуры потому и гомологичны, что унаследованы от общего предка, го схема родственных отношений скорее будет напоминать букет или пучок.
В первом случае изучаемые виды связаны родством 'предок -погомок*, во втором — сестринским родством (см. рис. 5). Обычное филогенетическое древо скорее представляет собой с ре да ее между двумя крайностями, но это означает, что противонаправленные процессы олигомеризации и полимеризации (увеличение числа счетных структур) в общем уравновешивают друг друга. Эта уравновешенность представляется наиболее правдоподобной.
Обсуждаемая теория имеет еще один аспект. В.А.Догель отметил, что 'процесс олигомеризации характеризует особенно ярко явление прогрессивной эволюции*, поскольку подвергнувшиеся олигомеризации группы многоклеточных животных отличаются 'видовым богатством*, 'широкой приспособленностью к окружающим условиям среды* (1954, с. 10). Здесь можно узнать даа из грех критериев биологического прогресса, который равнозначен* 'понятию победы в борьбе за существование* (Северцов, 1939, с. 271). Следовательно, по В.А.Догелю, намечается возможность морфологического, критерия биологического прогресса в понимании А.Н.Северцова: олигомеризованные Животные обычно остаются победителями в борьбе за существо-12 - 1560
вание. Если же взгляды А.Н.Северцова действительно представ, ляют собой творческое развитие учения Ч.Дарвина о приспособленности, то в перспективе как будто бы вырисовывается возможность морфологического критерия приспособленности: приспособленные животные обычно олигомеризованы.
Известно, однако, что приспособленность, как и биологическая прогрессивность, относительна, не связана однозначно с морфофизиологией, и различия между ними принципиальны: первые есть цель, а последняя - средство. Показательно вместе с тем, что в качестве примера ароморфоза А.Н.Северцов (1939 с.27 4) называет концентрацию нервных ганглиев у насекомых, т.е. пример, использованный В.А.Догелем. Этот ароморфоз, как указывает А.Н.Северцов, дал возможность насекомым осуществить биологический прогресс. В данном примере связь ме>цду морфофизиологией и биологической прогрессивностью (приспособленностью) как будто бы однозначна. Вспомним, однако, что насекомые преуспевают только на суше, а в океане их насчитывается всего лишь около 30 видов. Мы можем представить себе планету Океан, лишенную суши и тем не менёе населенную разными животными. Останутся ли прогрессивными и приспособленными насекомые на такой планете?
IJ
ЗАКЛЮЧЕНИЕ
Остается фактом, что осмысление Ч.дарвином современного ему биологического знания стало возможным благодаря перенесению на живую природу понятий из политэкономии. Скорее все-ро, это обстоятельство еще недостаточно оценено методологами, ро всяком случае, аналогия между стоимостью и приспособленностью как будто бы ранее еще не была замечена. Надо последовать примеру Ч. Дарвина и поставить вопрос более широко: об осмыслении современной биологии с позиций социологии и гуманитарной культуры. Основание для постановки этого вопроса состоит в том, что сущность живых существ, так же как и человека, относится к сфере идеального.
Наверное, мы никогда не узнаем, по какому поводу впервые прозвучало слово "жизнь*, понятие о которой занимает центральное место в биологии и социокультурной области. Можно только предполагать, что при той невыделенности из природы, которая была свойственна первобытному обществу, противопоставление жизни человека жизни животных и растений не было столь глубоким, как в средневековой Европе. Эго противопоставление окончательно сложилось в важнейшем догмате христианства о бессмертии человеческой души, т.е. о собственно жизни как противоположности смерти. Этот догмат бесконечно отдалил человека от *меньших братьев*, выделил его из природы и вместе с тем сгладил различие между живой и неживой природой. Естественные науки, сплотившиеся вокруг физики, уже своим предметом ('физическая реальность* - это вещество, энергия, Поле) запрограммированы на изучение такой однородной природы и чужды природному идеальному. Опираясь на физику, они Успешно освоили эту, казалось бы, однородную область, в которой физикалистская методология наперед исключила какие-либо Неожиданности. Неожиданностью оказалась традиционная биология, не поддающаяся ‘физикализации. Объяснение этого факта в рамках физики невозможно, потому что из сферы ее интересов Выпадает идеальное. Остается социокультурная область, из которой можно заимствовать некоторое подобие природного идеального. Эго заимствование успешно осуществил Ч.Дарвин в своей теории естественного отбора, и так появилась основа для сближения человека и природы; однако уже совсем иного, не Такого, как в первобытном обществе.
В традиционной биологии идеальное дает о себе знать уже в особенностях операции классифицирования. Целесообразно еще Раз вернуться к этому вопросу.
179
В качестве лучшего образца для биологической систематики предлагается таблица химических элементов Д.И.Менделеева. Эго предложение представляет особую ценность, потому что помогает полнее понять особенности биологической классификации
В таблице д.И.Менделеева химические элементы упорядоче»" ны в систему по свойствам атомов, выраженным с помощью чисел. Каждый элемент занимает только одну клетку и остается в ней, пока сохраняются неизменными его свойства. Если в результате радиоактивного распада из атомов одного элемента получатся атомы другого элемента с новыми свойствами, то вновь возникший элемент найдет место в другой клетке таблицы: изменение свойств приводит к изменению названия элемента. Это превращение элементов не предполагает изменения свойств одного элемента (так чтобы он оставался самим собой)
Теперь представим себе некоторое подобие таблицы Д.И.Менделееве!, клетки которой по аналогии с химическими элементами заполнены названиями биологических видов. Виды состоят из организмов, состав видов меняется в длении от одной временной точки, выделенной наблюдателем, к другой временной точке. Поскольку не существует и двух неразличимых организмов (неопределенная наследственная изменчивость) и дление потенциально бесконечно, неограниченно длящимися оказываются и свойства видов: оставаясь самим собой, вид изменяет свои свойства. Эго должно было привести к перемещению вида из одной клетки таблицы в другую. Столь странное поведение элемента классифицирования совершенно неприемлемо для исследователя, который достоинства классификации видит в ее четкости и неподвижности. Между тем именно эти странности свойственны биологической классификации, и эти странности биологически содержательны.
В биологической классификации систематическое положение вида раскрывается при сопоставлении с видами. Хотя на практике систематик располагает только неподвижными выборками, принцип объективности по отношению к природе обязывает систематика за выборками видеть длящиеся природные виды. Система таких видов в свою очередь сама обязана быть длящейся (живой), как и классифицируемые виды. Именно этими свойствами, кдк уже было отмечено (с.7О- 77(1)-(У)). и обладает система видов традиционной биологии: пульсируют границы видов и других таксонов, изменяются их объем и положение в системе, изменяется место таксона на иерархической лестнице и пр. Система живых видов сама как бы живая. Нечто подобное можно найти в социокультурной сфере.
В толковых словарях, своего рода системах упорядоченных слов, смысл о них слов раскрывается с помощью других слов. Поскольку смысл объясняющих слов непостоянен, непостоянным оказывается и смысл объясняемого слова. В данном случае
180
средство соответствует поставленной задаче, потому что смысл объясняемого слова не однозначен. Смысл слова представляет собой нечто живое и незавершенное, способное продолжить свою ^изнь в соединении с другими словами. Именно поэтому словео цая речь, литература остаются наиболее пригодными средствами для выражения духовного мира человека, своего рода идеальной сущности собственно человеческой жизни.
При изучении смысла слов сопоставляются слова со словами, при классифицировании видов одни виды сравниваются с другими видами. Сходство между этими двумя познавательными операциями состоит в том, что для изучения живых Длящихся объектов привлекаются живые же объекты. Действительно, в лингвистике, например, семантика слов "в живом развитии языка* изменчива, подвижна, многозначна (Дмитриева, 1962). Слово как носитель смысла (сущности) в устной речи и в письменных текстах не остается неизменным и смысл его Длится в развивающемся языке. Эго напоминает биологическую эволюцию, в которой родственные виды (течение времени) имеют собственную историю существования (Дление).
Стихийно сложившийся язык традиционной биологии близок к естественному языку своим полиморфизмом, общей нерегулярностью: *Нечегкие и неотчетливые по своему смыслу слова с неровными краями областей их значений, неясность разграничительных линий между понятиями — все это создает возможность для нарушения строго дедуктивных форм мышления... Полиморфизм языка - это один из способов допущения 'нестрогой* логики при 'внешнем* сохранении видимости дедуктивной строгости...* (Налимов, 1968, с. 72). Таким языком фактически пользуется биология с ее логикой длящихся понятий. 'Классические логики имеют дело только с абсолютно истинными и абсолютно ложными значениями, осуществляемыми только в узкой сфере высказываний; эти логики требуют дополнения в виде логики относительной истины и лжи, т.е. другими словами, в виде диалектической логики. Элементы этой логики, без сомнения, содержатся в многозначной логике '(Клаус, 1963, с.92). В биологии при классифицировании длящихся популяционных объектов исследователь как раз и находится в таком положении, когда абсолютно истинное (и абсолютно ложное) неподвижное суждение невозможно. Невозможность всеобщей жесткой логики и языка была показана теоремой Геделя, которая '...завершает собой целую эпоху глубокой и безусловной веры в жесткий детерминизм в сфере логики - той веры, последним всплеском Которой было появление логического позитивизма... Логические Позитивисты хотели построить новый 'совершенный язык' со строго однозначным значением слов...' (Налимов, 1968, с.69, 71). Показательно, что именно язык слов успешно послужил Ч .Дарвину при разработке теории эвойюции, оказавшейся теори
181
ей жизни. Попробуем, например, однозначно определить смысл слова "отбор*.
Числа, язык количественной математики обладают иными ВОа^ можностями. Еще в античной философии, в платонизме и пифа^о, реизме была осознана неподвижность, неизменяемость чисел. Поэтому именно числа оказались пригодным^ для выражения неподвижных эйдосов, идей (Лосев, 1923). Математическая теория чисел учит, что каждое число индивидуально, имеет уникальное внутреннее строение и нет ограничений для количества таких чисел. Поэтому каждой индивидуальной вещи можно было бы поставить в соответствие свое собственное число. Эго могло бы явиться не только нумерацией, потому что неподвижное внутреннее строение числа могло бы выразить неподвижную су-.щность каждой вещи. Они пригодны для описания превращения химических элементов, «но не пригодны для описания развития биологических видов, каждый из которых совершает собственное развитие (точнее, дление). Язык чисел - это язык типоле*, гии, и вполне естественно, что им воспользовалась нумерическая таксономия, относящаяся к типологической школе.
О внутреннем сродстве традиционной биологии с обществоведением свидетельствует следующее.
Многообразие в общественных науках, по-видимому, устроено, как в биологии. Вернемся в этой связи к диаграмме Венна и на одной и той же генеральной совокупности людей зададим несколько отношений общности. Отношение культурной, прежде всего языковой, общности выделяет культуры: европейскую, античную, славянскую и пр. Отношение генеалогического родства выделяет роды, фамилии, кланы, династии. Отношение общности исторических судеб выделяет народы, народности, племена. Отношение собственности на средства и орудия производства выделяет классы: рабов, рабовладельцев, феодалов, крепостных. Отношение подданства или гражданства приводит к таким общностям, как империи, республики, царства и пр. По-видимому, в науке об этносе (Бромлей, 1983) существует гораздо большее число отношений и общностей. Что же касается уже перечисленных отношений, то ими можно пользоваться порознь и во всевозможных сочетаниях. Получаемре при этом многообразие напоминает упорядоченное множество биологических классификаций. Несколько особое место в гаком социологическом многообразии занимает отношение морфофизиологического сходства, выделяющее расы и другие расовые единицы. Возможно, что через это отношение проводится биологическая точка зрения на человека. Расы остаются фенонами, пока в них не вкладывается собственно социологическое содержание (*раса господ*); Ра” зумеегся, выведение социального (право на господство) из биологического (морфофизиология) есть неправомочное биологиза-торство.
182
Соотношение между физическим и биологическим в организме остается центральной проблемой в теории жизни. Аналогичная проблема — это соотношение биологического и социального в природе человека. В этой связи поучительна точка зрения социологов на состояние биологии. "...Сущность живых индивидов, как доказала современная биология, определяется не только отдельной особью и ее внутренним строением, а также "совокупностью отношений", биологических, популяционно-видовых и био-ценогических. Поэтому 'сущность человека^не есть абстракт, присущий индивиду" [Маркс, Энгельс. Соч. Т. 3. С. 3. - Н.З.], а система особых отношений, в которую он включен" (Тарасов, Черненко, 1979, с. 31). В этой цитате положение в биологии оценивается слишком оптимистично. Господство физико-химического редукционизма свидетельствует о том, что сущность живых существ ищут прежде всего в них самих, в отрыве от "совокупности отношений". Что же касается "совокупности отношений", то из цитаты следует, что по аналогии с социологией отношения надо считать столь же реальным объектом, что и живую плоть организмов. Конечно, для изучения отношений непригодны такие'методы, как центрифугирование и измерение. Но изучение отношений означало бы дальнейший отход от редукционизма.
С исторической наукой биологию сближает особая соотнесенность во времени исследователя и исторического процесса как объекта исследования. В исторической науке "каждое новое настоящее открывает новые проблемы, новые стороны в вопросах, казавшихся в прежнем настоящим (ставшим теперь уже прошедшим) решенным. Очевидно, что даже при неизменности числа и объема источников информации историческое знание столь же неисчерпаемо, как историческое время... Не подлежит сомнению, что выразить непрерывную связь процесса с сохранением устойчивости различной длительности на разных уровнях целостности, в терминах традиционной (формальной) логики невозможно" (Барг, 1984, с. 94, 119). В приведенной цитате принимается неотделимость прошлого от движущегося настоящего и от будущего ("новое настоящее"). Это явление вызывает такие же логические трудности, какие в биологии вызваны выражением "весь вид" (нет привилегированных временных точек и противоположности между прошлым и будущим: все организмы в равной мере необходимы, чтобы существовало понятие вида).
Нитью, связывающей биологию и социо-культурную область, возможно, служит и психология. Рассматривая происхождение психики, А.Н.Леонтьев (1981) отделяет раздражимость от чувствительности. Раздражимость - это способность организма отвечать на физико-химическое воздействие, чувствительность -это форма раздражимости, обеспечивающая выполнение сигнальной, информационной функции. Чувствительность и раздражимость
183
находят свое место в таблице нарастания семиотических свойств знаковых сисгем (Степанов, 1971, с. 82). Способностью отвечать на физико-химические воздействия обладают и роботы, яв-но неживые объекты. Они функционируют на основе количественной информации, исчисляемой в битах. В этом смысле раздражимость — это свойство той стороны организма, которая получила у Р.Декарта название "живая машина". Чувствительность -не только особая форма раздражимости, но и более высокая по сравнению с ней форма отражения, она основана на сигнальной информации. Борьбу за существование автор назвал семантикоинформационным взаимодействием, подчеркивая этим самым, что в живой природе объективно существует некоторое подобие семантической информации; во всяком случае, важно, что имеется в виду не количественная информация. Разнообразие видов информации в природе и разнообразие способов их существования (чувствительность, раздражимость, борьба за существование и др.), по-видимому, надо рассматривать в связи с представлением философов об отражении как всеобщем свойстве материи. Эго разнообразие, возможно, облегчает саму постановку вопроса о том, откуда взялась биологическая и человеческая (психика) сущность. В самом деле, если сущность не предсу-ществовала, то откуда ей взяться.
Уже указывалось, что, создавая понятийный аппарат теории естественного отбора, Ч.Дарвин прибегал к заимствованиям из политэкономии: учение Мальтуса, "отбор", "конкуренция" и пр. В современном ему английском обществе викторианской эпохи эти выражения пользовались широким распространением (Малкей, 1983). Показательно, что при этом остались в небрежении понятия физики и химии и что впоследствии физико-химическая биология ничего не изменила в теории отбора. Она лишь открыла для нее новую предметную область - молекулярный уровень. Таким образом, учение о природе своими корнями связано с науками о человеческом обществе. В этой связи надо отметить следующие обстоятельства.
д.Халл (Hull , 1973) показал, что уже современники упрекали Ч.Дарвина в том, что вопреки его же заявлениям о приверженности бэконианскому методу и неприятии дедуктивизма он изменил традициям эмпирического бэконианского естествознания. Выводя из посылки ("отбор") следствия ("разнообразие видов"), Дарвин воспользовался дедукцией в принципе так же, как верующие, признав всемогущество творца, выводят из этой посылки картину мира. Дедуктивизм и аксиоматичность, отмеченные и другими авторами (Завадский, Галл, 1980), разумеется, не компрометируют дарвинизм. Они лишь еще больше отдаляют его от эмпирических наук индуктивного естествознания, вооруженных математическим методом.
В гл. 4 были изложены причины, по которым в биологии
184
классифицирование заменяет собой измерение. Но поскольку от измерения величин зависит проникновение математики в естествознание (Перминов, 1983), могло бы возникнуть впечатление, будто вместе с измерением отрицательно решается вопрос о применении в биологии математики вообще. На сходную мысль могла бы навести и следующая цитата: "В ряде наук очень важную роль играет классификация, в других науках она практически не нужна; в одних науках возможны и важны точные измерения, в других - то, что лежит в их основании, не поддается измерению* (Томсон, 1970, с. 16). Эго впечатление было ошибочным. Математическое знание не представляет собой застывшей схемы. Начавшие^ с геометрии и арифметики, оно еще в первой четверти прошлого века оставалось знанием о величине и измерении, той '’’всеобщей математикой* Р. Де карта, которая объясняет все "относящееся к порядку и мере* (Асмус, 1956). Современную математику особенно интересуют сама структура математического знания, структура количественных отношений, категории, бесконечность и другие идеальные объекты. Им невозможно поставить в соответствие какие-либо предметы и явления чувственного мира, они не наглядны и не имеют аналогов даже в той природе и в том опыте, на которых выросло физико-математическое естествознание. Современная математика-явление более богатое, чем наука о количественных отношениях, числах и величинах (Яновская, 1962). Г.И.Рузавин (1983, с.230) пишет: *...если для математизации наук о неорганической природе достаточно ограничиться метрическими понятиями и методами, то исследование более сложных процессов живой природы и общества требует обращения к неметрическим методам, основанным на абстрактных структурах современной мате* матики... именно посредством дальнейшего развития и усовершенствования новейших абстрактных структур и методов можно создать более адекватные методы для математического анализа биологических и социальных явлений. Во всяком случае, аппарат и методы классической математики, ориентированные на решение проблем физической науки, меньше подходят для указанной цели, чем структуры и методы новейшей математики*. Выходит, что способом добывания знания биология ближе к социологии, нежели к физике. Впрочем, это обстоятельство было замечено еще М. Гартманом (1936), который исходил из принадлежащего Виндельбанду деления наук на идиографические и,немоте гические. Номотетической наукой следует считать физику, и в биологии это направление представлено физиологией организма (включая механику развития). Оно руководствуется экспериментальным методом и *приводит физические процессы в систему логических законов, которые до известной степени стоят вне времени и не индивидуальны*. В .остальном же биология идиографична. Она изучает индивидуальные тела резко выражен
185
ной исторической природы, которые при всех переменах состояний устойчивы во времени. Поэтому биология якобы вынуждена заниматься описанием и классифицированием, хотя в целом ее природа остается двойственной. По М.Гартману, идиографичео-кое знание преходяще и 'рационализация мира явлений' остается за номотетической наукой. По-видимому, расцвет физико-химического редукционизма означал бы Для Гартмана расцвет биологии. А пока с гуманитарным знанием биология остается связанной, например, через явно идиографическую биогеографию.
Принадлежа через объект (жизнь) к наукам о природе, биология примыкает к социологии и гуманитарному знанию способом изучения жизни (воссоздание многообразия через сопоставление для классифицирования на основе разных отношений). В этой раздвоенности - причина своего рода драмы традиционной биологии, раздвоенности учебных программ, запутанности рубрикаций 'Реферативного журнала' и других подобных изданий. Физико-химический органицизм дает обществу технологическое знание и не принимает традиционную биологию, дающую понимание живой природы. В понимании жизни и биологии как знания о жизни теория биологии вслед за Ч.дарвиным должна обратиться к понятийному аппарату общественных наук и к гуманитарной культуре как источнику идей и образов.
Литература
Абакумов В.А. О специфике пространственно-временной орга— ‘ низации биосистем//'Развитие концепции структурных уровней , в биологии. М., 1972.
Акимов М.П. Биоморфический метод изучения биоценозов// Бюл. МОИП, Отд. биол. 1954. Т. 59. №3.
Акчурин ИЛ. Проблема адекватности фундаментальных понятий реальным структурам//Проблемы истории и методологии научного познания, М., 1974.
Алеев Ю. Г. Жизненная форма как система адаптаций//Усп.
1 совр. бйол. 1980. Т. 90, № 3/6.
Александров Г.В. Учение Аристотеля о категориях бытия//
1 Аристотель. Категории. М., 1939.
Алексеенко Л. Н. Экобиоморфный состав луговой флоры в связи с условиями среды//Учен. зап. ЛГУ. 1971. Т.360, №22.
F лехин В.В. География растений. М., 1944.
лешин А.И. О типологии теоретических исследований в современной биологии (логико-методологический аспект)//Биоло-
, гия и современное научное познание. М., 1980.
^млинский И.Е. Жоффруа Сент-Илер и его борьба против Кювье. М., 1955.
^ндреев В.Л. Системы-классификации в биогеографии и сис-тематике (детерминистские методы)//Иерархические класси—
| фикационные построения в географической экологии и систе-/ матике. Владивосток, 1979.
Андреев В.Л. Классификационные построения в экологии и систематике. М., 1980.
Анохин П. К. Теория функциональной системы и ее место в построении теоретической биологии//Эволюция типов индивидуального развития животных. М., 1977.
Антонов А.С. Геносистематика, ее достижения, проблемы и перспективы//Взаимодействие методов естественных наук в
i познании жизни. М., 1976.
Аристотель. О частях животных. М., 1937.
Аристотель. Метафизика//Соч. М., 1976. Т. 1.
Аристотель. Об истолковании//Соч. М., 1978. Т. 2.
Аронов Р. А. Пространство и время и пространство - время// Проблемы истории и методологии научного познания. М., 1974.
А скин Я. Ф. Направление времени и временная структура про-цессов//Пространство. Время. Движение. М., 1971.
187
А скин Я.Ф. Корреляция как виц синхронной детерминации// Методологические вопросы науки. Саратов. 1978. Вып. § Асмус В. Ф. Декарт. М., 1956.
Астауров Б. Л. Теоретическая биология и некоторые ее очередные задачи//Вопр. философии. 1972. № 2.
Ахманов А.С. Логическое учение Аристотеля. М., I960.
Ашмарин И. П., Харченко Е. П. "Запрещенные*' сочетания нуклеотидного состава и количество ДНК в геноме микроор-ганизмов//Журн. эвол. биохимии и физиол. 1971. Т.7, Nog, Баженов Л. Б. Принцип редукционизма и проблема взаимоог^ ношения физики и биологии//Биология и современное научное познание. М., 1980.
Баженов Л. Б., Бирюков Б. В., Спиркин А. Г. О философских вопросах кибернетики//Клаус Г. Кибернетика и философия. М.» 1963.
Баранцев Р. Г. Системные триады и классификация//Теория и методология биологических классификаций. М., 1983.
Барг М. А. Категории и методы исторической науки. М„ 198 5.
Беклемишев В. Н. О классификации биоценологииеских (симфизиологических) связей//Бюл. МОИП. Отд. биол. 1951. Т. 56, № 5.
Беклемишев В. Н. Основы сравнительной анатомии беспозвоночных. Т. 1. Проморфология. М., 1964. Т. 2. Органоло гия. М., 1964.
Беклемишев В. Н. Об общих принципах организации жизни// Бюл. МОИП, Отд. биол. 1964. Т. 69. № 2.
Беклемишев К.В. Экология и биогеография пелагиали. М., 1969.
Белозерский- А. Н. Молекулярная биология — новая студень познания природы. М., 1970.
Беляев Г. М. Ультраабиссальная (хадальная) фауна//В кн.: Биология океана. Т. 1. Биологическая структура океана. М., 1977.
Берг Л. С. Номогенез или эволюция на основе за коном ернос-тей//Тр. Геогр. ин-та. 1922. Т. 1.
Берг Л.С. Климат и жизнь. М., 1947.
Бергсон А. Творческая эволюция. М., 1909.
Березин В.А. Селективный и инструктивный принципы в био логии//Журн. общ. биол. 1981. Т. 42, № 5.
Бернал Дж. О возможной роли космоса в происхождении жизни на Земле//Тр. У Междунаропн. биохим. конг. Эвол. биохим. М., 1962.
Бернал Дж. Возникновение жизни. М., 1969.
Бернар К. Прогресс в физиологических науках//Обший вывод положительного метода. Спб., 1866.
Берталанфи Л. Общая теория систем. Критический обзор// Исследования по общей теории систем. М., 1969.
188
Киблер В. С. Мышление как творчество. М., 1975. биологические часы//Поцред. С.Э.Шноля. М.» 1964. Бирштейн Я.А. Некоторые проблемы происхождения и эволюции пресноводной фауны//Усп. совр. биол. 1949. Т. 27, №1.
бирштейн Я. А. Глубоководные равноногие ракообразные севере—западной части Тихого океана. М., 1963.
благовещенский А.В. Биохимическая эволюция цветковых растений. М., 1966.
Благовещенский А.В., Александрова Е. Г. Биохимичео-кие основы филогении высших растений. М., 1974.
Блюменфельц Л.А. Предисловие//Кастлер Г. Возникновение биологической организации. М., 1967.
Блюменфельц Л.А. Критерий живого и физика//Критерий живого. М., 1971.
Б ля хер Л. Я. Проблемы морфологии животных. М., 1976.
Бобринский Н. А. Зоогеография и эволюция. М.; Л., 1927.
Бобринский Н. А. География животных (курс зоогеографии). М.» 1951.
Богданов Ю.А., Каплин П.А., Николаев С.Д. Происхождение и развитие океана. М., 1978.
Богоров В.Г. Планктон Мирового океана. М., 1974.
Борзенков В. Г., Северцов А. С. Теоретическая биология: размышление о предмете. М., 1980.
Борисяк А. А. Ж.Кювье и его научное значение//В кн.: Ж.Кювье. О переворотах на поверхности земного шара. М.; Л.. 1937.
Бородин И. П. Протоплазма и витализм. Мир божий. ‘Спб., 1894.
Бочаров В.А. Аристотель и традиционная логика. М., 1984.
Бромлей Ю. В. Очерки истории этноса. М., 1983.
Букановский В.М. Принципы и основные черты классификации современного естествознания. Пермь, 1960.
Букс А. Эволюция и темпы вымирания. М., 1979.
Бунге В. Теоретическая география. М., 1967.
Бурбаки Н. Очерки по истории математики. М., 1963.
Бурков В.А. Структура течений Тихого океана и их номенкла^-тура//Океанология. 1966. Т. 6, № 1.
Бурова И.Н. Парадоксы теории множеств и даалектика, М., 1976.
Бычков А.А. Геоботаническая терминология. Алма-Ата, 1967.
Бэкон Ф. Сочинения: В 2 т. Т. 1. М., 1977. Т. 2. 1978.
Бэр К. Э. Всеобщий закон природы, проявляющийся во всяком развитии//Избранные работы. Л., 1924.
Бэр К. Э. Об истории развития животных//Избранные работы. Л., 1924.
Бэ^ К. Э. Неизданные рукописи//Анналы биологии. М., 1959.
[ 18 9
Вавилов Н.И. Линнеевский виц как система. Л., 1967.
Вайнштейн Б.А. О числовой таксономии//Журн. общ. биол 1968. Т. 29, № 2.
Вайнштейн Б. А. Систематика, классификация и числовая сономия//Журн. общ. биол. 1972.
Варминг Е. Ойкологическая география растений. Введение в изучение растительных сообществ. М., 1901.
Варминг Е. Распределение растений в зависимости от внеш», них условий (экологическая география растений). Спб., 1902.
Васильев В.П. Эволюционная кариология рыб. М., 1985. 300 с.
Васильев В. П., Васильева Е.Д., Осинов А.Г. Первое свидетельство в пользу основной гипотезы сетчатого видообразования у позвоночных//Докл. АН СССР. 1983. Т. 271, № 4.
Васнецов В.В. Экологические корреляции//Зоол. журн. 1938. Т. 17. № 4.
Веценов М.Ф., Кремянский В.И., Шаталов А. Т. Концепция структурных уровней в биологии//Развитие концепции структурных уровней в биологии. М., 1972.
В ер мель Е. М. История учения о клетке. М., 1970.
Вернадский В. И. Биосфера. Т. I—II. Л., 1926.
Визгин В.П. Генезис и структура квалитативизма Аристотеля, М., 1982.
Винер Н. Кибернетика. М., 1968.
Виноградова Н. Г. Фауна шельфа, материкового склона и абиссали//Биология океана. Т. 1. Биологическая структура океана. М., 1977.
Вирхов Р. О механическом понимании жизни//Сборник статей по естествознанию и медицине. Харьков, 1866.
Витвицкий Г. Н. Зональность климата Земли. М., 1980.
Волькеншгейн М.В. Предисловие редактора//Эйген М., Винклер Р. Игра жизни.1979.
Волькенштейн М.В. Физика и биология. М., 1980.
Волькенштейн М.В. Биофизика и теоретическая биология// Методологические и теоретические пробл. биофизики. М., 1981.
Воробьев В. П. Бентос Азовского моря. Симферополь, 1949.
Воронов А.Г. Общие вопросы биогеографии и ее основные направления- на современном этапе//Итоги науки и техники. Биогеография. Т. 1. М., 1976.
Вяльцев А. Н. Дискретное пространство-время. М., 1965.
Гайденко П.П. Эволюция понятия науки: становление и развитие первых научных программ. М., 1980.
Гайденко П. П. Культурно-исторический аспект эволюции науки//Методология. пробл. историко-научных иссл. М., 1982.
190
I а л ил ей Г. Избранные труды. Т. 2. М., 1964.
»арден Ж-К. Теоретическая археология. М., 1983.
Ьртман.М. Общая биология. Введение в учение о ^кизни.
|.М.; Л., 1936.
Кгамян Г. В. О биосферологии В.И.Вернадского//Журн.общ.
| биол. 1980. Т. 41, № 4.
Кеслер К. О сущности жизни. М., 1968.
fere И.-В. Избранные сочинения по естествознанию. М.; Л., | 1957.
Эккель Э. Трансформизм и дарвинизм. Популярное изложение общего учения о развитии. Спб., 1900.
еккер Р. Ф. К постановке исследований по палеобиологии (палеоэкологии и палеозоологии)//Вопр. экол. и бионенол. М.;
Л., 1934.
ьодакян В. А. Организация систем - живых и неживых //
L Системные иссл. М., 1970.
Ьптнер В. Г. Общая зоогеография. М., 1936.
1птнер В. Г. Предисловие//В кн.: Майр Э., Линсли Ж, Юзин-
[ rep Р. Методы и принципы зоологической систематики. М., 1956.
ершензон С.М. Молекулярная биология и теория эволюции (методологический аспект проблемы)//Философия и теория эволюции. М., 1974.
риляров М. С. О функциональном значении симметрии орга-низмов//3оол. журн. 1944. Т. 2 3, № 5.
[иляров М.С. Современные представления о гомологии/Усп. совр. биол. 1964. Т. 57, Ng 2.
оликов А. Н. О количественных закономерностях процесса дивергенции//Гидробиологические исследования самоочищения
; водоемов. Л., 1976.
Гольдгаммер Д. А. Процессы жизни в "мертвой* природе.
| Берлин, 1922.
борский Д.П. Вопросы абстракции и образования понятий.
F М., 1961.
|Гребнер П. География растений. М., 1914.
Григорьев А. А. Теоретические основы современной физической географии//Взаимодейсгвие наук при изучении Земли.
М., 1963.
Григорьев А.А. Закономерности строения и развития географической срецы//Избранные теоретические работы. М., 1966.
Грюнбаум А. Философские проблемы пространства и времени. М., 1969.
Гумбольдт А. География растений. М.; Л., 1936.
Давиташвили Л. ID. Причины вымирания организмов. М., 1969.
Давыдов А.С. Биология и квантовая механика. Киев, 1979.
Давыдова Г.А. О роли принципа отражения в исследовании процесса развития//Вопр.филос. 1979, № 12.
191
Дарвин Ч. Происхождение видов. Сон. Т. 3# М»; Л., 1939, Дворецкий И.Х. Древнегреческо-русский словарь.Т. 1.М., 1958. Декарт Р. Избранные произведения. М., 1950.
Джеффри Ч, Биологическая номенклатура. М., 1980.
Дмитриева Н.И. Изображение и слова. М., 1962.
Дольник В.Р.Аллометрия морфологии,функции и энергетики гомо&
отермных животных и ее физический контр оль/ДКурн. общ. би ол. Ig^j Дорфман Я.Г. Антуан Лоран Лавуазье (1743-1794). М», 1962, Дриш Г. Витализм, его история и система. М., 1915.
Дубинин Н.П. Эволюция популяций и радиация. М., 1966.
Дубинин Н.П.,Ромашов Д.Д. Генетические основы строения
вида и его эволюции//Биол.журн., 1932. Т. 1, No 5-6.
Ермолова Е.А. Методологическая роль категорий "структура", "система", "функция* в понимании природы живого//Методологии. пробл. совр. науки. М., 1970.
Жданов В.Н, К определению понятия "жизнь*// Вести. АМН СССР. 1964. Т.1.
Завадский К.М. Основные формы организации живого и их под-разделения//Философские пробл. совр. биол. М.; Л., 1966, Завадский К.М. Вид и видообразование. Л., 1968.
Завадский К.М. Развитие эволюционной теории после Дарви-
на (1859-1920-е годы). Л., 1968.
Завадский К.М. Галл Я.М. Основные этапы и закономерности познания борьбы за существование как фактора эволюции// Биологии и совр. науч, познание. М., 1980.
Завадский К.М., Георгиевский А.Б. К оценке эволюционных взглядов Л.С.Берга//Берг Л.С. Труды по теории эволюции. Л., 1977.
Заварзин Г.А. Фенотипическая систематика бактерий. М., 1974.
Заварзин Г.А., Старк Ю.С. Анализ запрещенных вариантов в
систематике микроорганизмов//Изв. АН СССР. Сер. биол.
1965. Вып, 5.
За милов Д.А., Лисе ев И.К. Методологические аспекты и пу-
ти формирования теоретической биологии//Вопр.филос. 1972, N? 3.
Заренков Н.А. Вопросы эволюции счетных признаков//Журн.
общ. биол. 1972. No 5.
Заренков Н.А. Лекции по теории систематики. М., 1976.
Заренков Н.А. Происхождение многоклеточных как проблема
общей морфологии//Состояние и перспективы развития морфологии. М., 1979 а.
Заренков Н.А. К вопросу об определении предмета биологии//
Журн. общ. биол. 1979 б. Т. 40, № 5.
Заренков Н.А. Некоторые следствия из теории естественного отбора Ч.Дарвина//Журн. общ. биол. 1984. Т. 45, № 1.
Заренков Н.А. Общебиологические принципы зоологической
систематики: Автореф. дис... д-ра биол. наук. М., 1984.
192
Зелинский Н. Д. Проблема белка//Вестн. АН СССР. 1962.
Т. 3.
Зенкевич Л. А. Система и филогения//Зоол. журн. 1939.
Т. 18. № 4.
Зенкевич Л. А. Фауна и биологическая продуктивность моря.
Ч. 2. Моря СССР//Сов. наука. 1947.
Зенкевич Л. А. Биологическая структура океана//3оол. журн. 1948. Т. 27, № 2.
Зенкевич, Л. А. О положении ботаники и зоологии в системе биологических наук//Бюл. МОИП. Отд. биол. 1966. Т. 71, № 3.
Зенкевич Л. А. Очерки по эволюции двигательного аппарата животных//Избранные труды. Т. 1. М., 1977.
Зенкевич Л. А., Бро икая В. А. Материалы по экологии руководящих форм бентоса Баренцева моря//Учен. зап. МГУ. 1937, № 3.
Зотин А. И. Второе начало, негэнтропия, термодинамика линейных необратимых процессов//Термодинамика биологичео- • ких процессов. М., 1976.
Иванов А. В. Происхождение многоклеточных животных. Л., 1968.
Иванова-Казас О.М. Анализ раннего развития ракообразных. 1. Основные варианты раннего развития//Биол. моря. 1977, № 1.
Игнатов А. И. Проблема происхождения жизни. М., 1962.
Ильенко Э. В. Проблема идеального//Вопр. философии.
1979 а. № 6, С. 128-140.
Ильенков Э. В. Проблема идеальногб//Вопр. философии. .1979 б. № 9.
Ильин А. Я, Об основных формах витализма в современной биологии//Вести. Моск, ун-та. Серия.8. Философия. 1966. № 6.
Иогансен Б. Г. К изучению надорганизменных систем//Раз-вигие концепции структурных уровней в биол. М., 197,2.
Иоффе И. И. Культура и стиль. Л., 1927.
Исаченко А. Г. Развитие географических идей. М., 1971.
Кавецкий Р. Е., Колпаков Е. В. О взаимоотношениях биологической, физической и химической форм движения мате-рии//Философские вопр. совр. биол. Киев, 1962.
Каганова 3. В. Проблемы философских оснований биологии. М„ 1979.
Кальвин М. Химическая эволюция//Пробл. эвол. и гехнич. биохимии. М., 1964.
К амшилов М. М. Круговорот органического вещества и проблема сущности жизни//Журн. общ. биол. 1966. Т. 27, №3.
Канаев И. И. Очерки из сравнительной анатомии до Дарвина. М.; Л., 1963.
Канаев И. И. Сверки из истории проблемы морфологического типа от Дарвина до наших дней. М.; Л., 1966.
Канаев И. И. Жорж Кювье. Л., 1976.
Кантор Г. Учение о множествах Георга Кантора//Новые идеи в математике. Спб., 1914, № 6.
Карпинская Р.С. Философские проблемы молекулярной биологии. М., 1971.
Карпинская Р. С., Хон Г. Н. Принцип редукции в системе методов исследования живого//Взаимоцействие методов ес-теств. наук в познании жизни. М., 1976.
Карпов В. Н. Аристотель и его научный метод//Аристотель. О частях животных. М.; Л., 1937.
Касинов В. Б. Биологическая изомерия. Л., 197 3.
Кафанов А.И., Суханов В.В. О зависимости между числом и объемом таксонов//Журн. общ. биол. 1981. Т. 42, № 3.
Каш каров Д. Н. Основы экологии животных. М.; Л., 1938.
Кедров Б.М. О соотношении форм движения материи в при-роце//философские пробл. совр. естествознания. М., 1959.
Кедров Б. М. Взаимосвязь форм движения материи и их классификации//Пространство. Время. Движение. М., 1971 а.
Кедров Б.М. Критерии различения высших и низших форм движения матерйи//Просгранство. Время. Движение. М., 1971 б.
Кедров Г. Б. Взаимосвязь некоторых признаков древесины двудольных и ее эволюционное значение//Морфология высших растений. М., 1968.
Кирпичников М. Э. К концепции рода у цветковых растений// Бот. журн. 1968. Т. 53, № 2.
Клаус Г. Кибернетика и философия. М., 1963.
Козо-Полянский Б. М. Новый принцип биологии. Очерк теории симбиогенеза. Л.; М., 1924.
Козо-Полянский Б.М. Значение различных методов в систематике растений, преимущественно Anthophyta (к оценке "новой" систематики)//Пробл. бот. 1950, № 1.
Коллингвуд Р.Дж. Идея истории. Автобиография. М., 1980.
Колмогоров А. Н. Жизнь и мышление с точки зрения кибер-нетики//Докл. теор. конф, философ, семинаров, научн. учреждений АН СССР. М., 1962.
Колмогоров А. Н. Жизнь и мышление как особые формы существования материи//О сущности жизни. М., 1964.
Кольцов Н. К. Организация клетки. М.; Л., 1936.
Комаров В. Л. Учение о виде у растений. М.; Л., 1940.
Коробков И. А. Палеонтологические описания. М., 1971.
Коршунов А.М., Мантатов В.В. Теория отражения и эвристическая роль знаков. М., 1974.
Косарева Л.М. Предмет науки. М., 1977.
194
Кочетов Ю. В. Особенности строения поверхности листьев растений аридной зоны//Изв. АН СССР. Сер. биол. 1985.
Красилов В.А. Эволюция и биостратиграфия. М., 1977.
Кремянский В.И. Типы отражения как свойства материи// Вопр. философии, 1963, № 8.
Кремянский В. И. Структурные уровни живой материи. М., 1969.
Кремнянский В.И. Методологические проблемы системного подхода к информации. М., 1977.
Креславский А. Г. Репродуктивная изоляция как критерий видд//Журн. общ. биол. 1973. Т. 34, № 4.
Криволуцкий Д. А. Понятие "жизненная форма" в экологии животных//Журн. общ. биол. 1967. Т. 37, № 2.
Крисс А. Е. Микробиологическая океанография. М., 1976.
Криштофович А. Н. Эволюция растений по данным палеоботаники//Пробл. бот. 1950. Вып. 1.
Круликовский А.И. О функциях и структуре теоретической биологии// Методология частных наук. Иркутск, 1979.
Кудрявцев В. И. Систематика дрожжей. М., 1954.
Кузин А. М. Клетка как единая упорядоченная на молекулярном уровне система//Развитие концепции структурных уровней в биологии. М., 1972.
Кузичев А. С. Диаграммы Венна. М., 1968.
Кузнецов А. П. Экология донных сообществ Мирового океана (трофическая структура морской донной фауны). М., 1980. ,
Кузнецов Б. Г. Идеи и образы Возрождения. М., 1979.
Кузнецов И. В. Категория причинности и ее познавательное значение//Теория познания и совр. наука. М., 1967.
Кузнецов Н.’Я. О "таксономических понятиях" и попытках их обоснования морфологическими данными//Русск. энтомол. обозрение. 1917, № 17.
Курсанов А. Л. Биология XX века//Вестн. АН СССР. 1979. Т. 8.
Курсанов Г. Современный структурализм//Мулуд Н. Современный структурализм-'/Размышления о методе и философии точных наук. М., 1973.
Кэйн А. Вид и его эволюция. М., 1958.
Кювье Ж. Размышления о переворотах на поверхности земного шара. М.; Л., 1937.
Ламарк Ж.—Б. Философия зоологии. Избр. произв. в 2 томах. М.» 1955.
Ламе гтри Ж.О. Человек-машина. Сочинения. М., 1976.
Ламонт К. Иллюзия бессмертия. М., 1984.
Левин М. Л. Предисловие//Гартман М. Общая биология. Введение в учение о жизни. М.; Л., 1936.
Леонтьев А. Н. Проблемы развития психики. М., 1981.
195
Лёб дж. Жизнь. Одесса, 1912.
Левушкин С. И. Об экологических фаунистических комплексах (на примере подземных фаун)//Журн. общ. биол. 1974.
Т. 36, № 6.
Левушкин С. И. К постановке вопроса об экологическом фаунистическом комплексе//Журн. общ. биол. 1976. Т. 35, № 5.
Линдерман Р. Трофико-динамическое направление в экологическом исследовании//Усп. совр. биол. 1943. Т. 16, № 5.
Линней К. Философия ботаники, изъясняющая первой оной основания... Спб., 1800.
Линней К. Система природы Карла Линнея... Царство Животных. Часть 1. Спб., 1804.
Лис ее в И. К., Тищенко В. И. Биология и современное научное познание//Вопр. философии, 1975. Т. 10.
Лисицын А. П. Палеоокеанология, история океанской среды, древнейшая история и эволюция океанов/УГеология океана. Геологическая история океана. М., 1980.
Лич ков Б. Л. Природные воды Земли и литосфера//3ап. Геогр. об-ва СССР. Нов. сер. 1960. Т. 19.
Л ой г Т. В. Теоретическая биология как метатеория биологичес кой науки//Вопр. философии. 1974, № 6.
Локк Дж. Избранные сочинения. М., 1960. Т. 1.
Лосев А. Ф. Диалектика числа у Плотина. М., 1928.
Лоусон И. А. Язык, коммуникация и биологическая организа-ция//Иссл. по общ. теории систем. М., 1969.
Лузин Н.fa. Аналогия между множествами, измеримыми ft и аналитическими множествами//Собр.соч. Т. II. М., 1958.
Луканин Р. К. "Органон" Аристотеля. М., 1984.
Лункевич В. В. От Гераклита до дарвина. Очерки по истории биологии. М., 1960.
Л ы маре в В. И. Основные проблемы физической географии океана. М., 1978.
Любишев А. А. Систематика и эволюция//'Внутривицовая изменчивость наземных позвоночных животных и микроэволюция: Тр. Всесоюзн. сов. Свердловск, 1966.
Любишев А. А. Проблемы систематики//Проблемы эволюции. Т. 1. Новосибирск, 1968.
Любишев А.А. Проблемы формы систематики и эволюции организмов. Сборник статей. М., 1983.
Лямин В. С. География и общество. М., 1978. 309 с.
Ляпунов А. А. Об управляющих системах живой природы//О сущности жизни. М., 1964.
Ляпунов А. А. В чем состоит системный подход к изучению реальных объектов живой природы?//Системные иссл. М., 1971.
196
Магомедмирзаев М. М. О проблемах морфологического измерения и счета с позиций фенетики растений//Журн. общ. биол. 1976. Т. 37, № 3.
Мазуренко М. Т., Хохряков. А. П. Биоморфологическая изменчивость и ее взаимосвязь с таксонообразованием у растений//Жизненные формы: структура, спектры и эволюция. М., 1981.
Майр Э. Зоологический вид и эволюция. М., 1968.
Майр Э. Принципы зоологической систематики. М., 1971.
Майр Э. Популяции, виды и эволюция. М., 1974.
Макфедьен Э. Экология животных. Цели и мегоды. М., 1965 Маликов М. Ф. Основы метрологии. М., 1949.
Малиновский А..А. Пути теоретической биологии. М., 1969.
Малиновский А. А. Общие особенности биологических уровней и чередование типов организации//Развитие концепции структурных уровней в биологии. М., 1972.
Малиновский А.А., Мина М. В. Рецензия: Алтухов Ю.П. Популяционная генетика рыб//Журн. общ. биол. 1976. Т. 37, № 5.
Малкей М. Наука и социология знания. М., 1984.
М а м з и н А. С. Очерки по методологии эволюционной теории. Л., 1974.
Марков А. А. Математическая логика//Философская энциклопедия. М., 1964. Т. 3.
Марков К.К., Бардин В.И., Лебедев В.Л. и др. Геогра— ч фическое районирование Ангарктиды//География Антарктиды.
М., 1968.
Матвеев Б.С. О соотносительных-изменениях функции и сре ды в эволюции животных//Зоол, журн. 1945. Т. 24, № 1.
Махотин А. А. О принципах составления филогенетических схем//Тр. Ин-та морфологии животных им.. А.Н.Северцова. 1959. Т. 27.
Медников Б. М. О реальности высших систематических категорий позвоночных живогных//Журн. общ. биол. 1974.
Т. 35, № 5.
Медников Б. М. Аксиомы биологии. М., 1982.
М е й е н С. В. О соотношении тихогенетического и номогенети-ческого в эволюции//Журн. общ. биол. 1974. Т; 35, № 3.
Me йен С. В. Таксономия и мерономия//Вопросы методологии в геологических науках. М., 1977.
Мейен С. В. Основные аспекты типологии организмов//Журн. общ. биол. 1978.
Мейен С.В., Соколов Б. С., Шрейдер Ю. А. Классическая и неклассическая биология. Феномен Любишева//Вести. АН СССР. 1977. Т. 11.
Мелюхин С. Т. Материя в ее единстве, бесконечности и развитии. М., 1966.
197
М ер к л ин Р. Л. , Жизненные формы и их значение для палеоэк(>. логического анализа//Проблемы палентологии (23-я сессия Междунар. геол, конгр. Докл. сов. геологов). М., 1968.
Мечников И. И. О дарвинизме. М.; Л., 1943.
Мещерякова Н. А., Пахомов Б. Я. Идеи кибернетики в со-здании концептуального аппарата биологии//Биология и сов* ременное научное познание. М., 1981.
Микулинский С. Р. Взгляды К.М.Бэра на эволюцию в додар. виновский период//Анналы биологии. 1959. Т. 1.
Минто В. Дедуктивная и индуктивная логика, М., 1909.
Мирзоян Э. Н. О некоторых тенденциях развития биологии в XX веке//История биологии с начала XX века до наших дней. М., 1975.
Мирч ин к Т. Г., Баб'ьева Н.П. Грибы, образующие мицелий, и дрожжи в природных экосистемах//Журн. общ. биол. 1981, Т. 42, № 2.
Михайловский Г. Е. Элементы биологической термодинамики Иркутск, 1980.
Молчанов Ю. Б. Четыре концепции времени в философии. М., 1977.
Монин А. С. История Земли. Л., 1977.
Моровиц Г. Исторический очерк//Теоретическая и математическая биология. М., 1967.
Мошковский ID. Д. Проблема вида в прогозоологии//Успехи протозоологии: Тез. ок. и сообщ., представленные II! Меж-дунар. конгр. протозоологов. Л., 1969.
Мул уд Н. Современный структурализм. Размышления о методе и философии точных наук. М.-, 1973.
Мур Э. ф. Математика в биологических исследованиях//Математика в современном мире. М., 1967.
Навроцкий Б. А. Структура биологической классификации// Науч. докл. высш, школы. Философ, науки. 1979, № 6.
Налимов Н.П. Вероятностная модель языка. М., 1979.
Наумов Н.П. О методологических проблемах биологии//Науч« докл. высш, школы. Философ, науки. 1964. Т. 1.
Николаев И. И. Таксоцен как экологическая категория. Экология. 1977. №‘5.
Неймайр М. Корни животного царства. М., 1919. 300 с.
Новик И. Б. Вопросы стиля мышления в естествознании. М., 1975. 143 с.
Новиков Г. А. Очерк истории экологии животных. Л., 1980. 287 с.
Новиков П.А. Комментарии к книге Ж.Кювье "О переворота* на поверхности земного шара". М., 1982.
Огнев С. И. Проблемы систематики. 11. Вопрос о виде в св€ те новейших данных. Зоол. журн., 1944. Т. 23, № 1.
198
О л ум Ю. Основы экологии. М., 1975. 740 с.
О л ено в Ю. М. Проблемы молекулярной генегики. Л., 1977.
Олицкий А. А. О традиционной биологии и биофизиН'е//А\етодологические и теоретические проблемы биофизики. М., 1981.
Омельяновский М. Э. Послесловие//Рейхенбах Г. Направление времени. М., 1962.
ЙОпжрин А. И. Жизнь и ее соотношение с другими формами i движения материи//О сущности жизни. М., 1964.
Опарин А. И. Материя, жизнь, интеллект. М., 1977.
Опарин А. И. О сущности жизни//Вопр. философии. 1979, №4.
(ррлов В. Н. Кариосисгематика млекопитающих. М., 1974.
О сипов Д. В. Проблемы гетероморфизма ядер у одноклеточных организмов. Л., 1981.
Jlaa в ер К. Вопросы синтетического подхода в биоморфологии. Таллин, 1976.
Павлов Н. В. Ботаническая география СССР. Алма-Ата, 1948.
Панфилов д.В. Географическое распространение функционально- биоценотических групп насекомых на территории СССР// Зональные особенности населения наземных животных. М., 1966.
Парамонов С. Я. Возможно ли построение единой биологической системы низших таксономических единип//Сов. бот.
1940, № 6/7.
|Пари Т. Аналитическое изучение популяций животных и его ‘ отношение к общей экологии//Усп. совр. биол. 1939. Т.10, № 2.
Пасеков В. П., Коростылев А. П. Об оценке параметров । эволюционного процесса дивергенции, популяций в результа— те случайного генного дрейфа//Усп. совр. биол. 1971. Т. 32, № 3.
Паск Г. Модель эволюции//Принципы самоорганизации. М., 1966.
Паули В. Л. Ритмика биологических процессов в океане//Тр. Севастопольской биол. станции. 1959. Т. 11.
Пере ль Т.С. Жизненные формы Lumbricidae //Журн. общ. биол. 197 5. Т. 36, № 2.
Перминов В. Я. Процесс математизации знания: предпосылки и структура//На пути к единству науки. М., 1983.
Песенко Ю. А. Принципы и методы количественного анализа в фаунистических исследованиях. М., 1982.
Пиаже Ж., Инельдер Б. Генезис элементарных логических структур. Классификации и сериации. М., 1963.
Пианка Э. Эволюционная экология. М., 1981.
Плате Л. Эволюционная теория. М.; Л.,1928.
Полынов Б. Б. Учение о ландшафтах: Избр. тр. М., 1956.
Полянский Ю, Н. Об определении предмета и метода совре—
менной биофизики//Вопр. философии. 1964, № 2.
Полянский Ю. Н. К проблеме познания современной биофизики (биофизика общая и теоретическая биология)//Методологические проблемы биофизики. М., 1981.
Полянский Ю. Н. Проблема вида и внутривидовой изменчивости в протозоологии//Весгн. АН СССР. 1982. Т. 3.
Попов М. Г. Основы флорогенетики. М., 1963.
Попов П.С., Стяжкин Н. И. Развитие математических идей от античности до эпохи Возрождения. М., 1974.
Попович М. В. Философские вопросы семантики. Киев, 1975.
П очвоведение/Под ред. И.С.Кауричева. М., 1982.
Предтеченский П. П. Динамика климата в связи с изменениями солнечной деятельности//Тр. Главк, геофизич. обсерватории им. Воейкова. 1950. Т. 19, № 81.
Преснов Е. В., Исаева В.В. Перестройки типологии при морфогенезе.
Пуанкаре А. Ценность науки. М., 1906.
Пшибыльский А. Структура стимула и биологическая эволю-ция//Проблема взаимосвязи организации и эволюции в биологии. М., 1978.
Работнов Т. А. Фитоценология. М., 1978.
Равикович А. И. Вероятностный образ мышления в естествен-ноисторических науках XIX века//Методология и история геологических наук. М., 1977.
Растительный мир Земли/Под ред. Ф.Фукарека. М., 1982. Т. 1-2.
Рашевски Н. Некоторые медицинские аспекты математической биологии. М., 1966.
Рейнке Г. Сущность жизни//Сушность жизни. Спб., 1903.
Рейхенбах Г. Направление времени. М., 1962.
Робертис Э. де, Новинский В., Саэс Ф. Биология клет-» ки. М., 1967.
Розанова М. А. Экспериментальные основы систематики растений. М.; Л., 1946. Г f
Розов Н. Н., Строганова М. Н. Почвенный покров мира.
М., 197 9.
Руденко А.П. Химическая добиологическая эволюция каталитических систем и критерий живого//Критерий живого. М., 1971.
Руза в ин Г. И. Философские проблемы оснований математики. М., 1983.
Рябчиков А. М. Общие закономерности географической зональности суши земного шара//Весгн. Моск, ун-та. Сер. Географ. 1960, № 4.
Сагатовский В. Н. Замечания по вопросу об определении жизни//Вопр. философии. 1967, № 3.
Свидервский В. И. О философском понимании конечного и бесконечного//Вопр. философии. 1964, № 6.
Свиридов А. В. Материалы по истории методов диагностики биологических объектов//Науч. докл. высш, школы. Биол. науки. 1976, № 8.
Северцов А.Н. Морфологические закономерности эволюции. М.; Л., 1939.
Северцов А.С. Становление аром орфоза//Журн. общ. биол. 1973. Т. 34, № 1.
Северцов С. А. О крнгруэнциях и понятии целостности вида// Проблемы экологии животных. 1951. Т. 1.
Седов Е.А. Эволюция и информация. М., 1978.
Семенов В. Н. Некоторые принципы биологического райониро-вания//Тез. докл. 1-го сов, океанологов. Вып. 2. М., 1977.
Семенов Н. Н. О соотношении химии и биологии//Вопр. философии. 1959, № 10.
Сенг-Дьерцьи А. Биоэнергетика. М., 1960.
Сенг-Дьерцьи А. Введение в молекулярную биологию. М., 1964.
С ер а в ин Л. И. Теория информации с точки зрения биолога. Л., 1973.
Серебряков И. Г. Экологическая морфология растений. М., 1962.
Серебрякова Т.Н. Морфология побегов и эволюция жизненных форм злаков. М., 1971.
Серебрякова Т.Н. Учение о жизненных формах растений на современном этапе//Итоги науки и техники. Ботаника.
Часть 1. М., 1972.
Сержантов В. Ф. Введение в методологию современной биологии. Л., 1972.
Сетров М. И. Организация биосистем. М., 1971.
Сетров М. И. Организация и эволюция информационных механизмов биосистем. М., 1978.
Симпсон Дж.Г. Темпы и формы эволюции. М., 1948.
Синицын В.М. Введение в палеоклиматологию. М., 1981.
Скарлато О. А., Старобогатов Я.И. Филогенетика и принципы построения естественной системы//Теоретические вопросы систематики и филогении животных. Л., 1974.
Скворцов А. К. Рецензия: Э.Майр. Принципы зоологической систематики//Бот. журн. 1972. Т. 57, № 11.
С л он им А. Д. Предмет, задачи, приемы и история развития эколого-физиологических исследований//Экологическая физиология животных. Ч. I. М., 1979.
Смирнов Е. С. Гомология и таксономия//Тр. Ин-та морфологии животных. 1959. Т. 27.
Смирнов Е.С. Систематика живого//Структура и формы материи-. Диалектический материализм и современное естествознание. М., 1967.
4 - 1560 201
Смирнов Е. С. Таксономический анализ. М., 1969.
Смирнов И. Н. Теоретическая биология: трудности создания и перспективы развития//Журн. общ. биол. 1973. Т. 34, № 2.
Смирнов И. И. О некоторых методах и принципах сисгемати-ки//Тр. Зоол. Ин—га АН СССР. 1965. № 35.
Солопов Е. Ф. Соотношение форм движения и видов материи и проблема определения жизни//Философские проблемы современной биологии. М., 1966.
Станков Н.С. Жизненный путь и труды Карла Линнея//Карл Линней. М., 1958.
С та робота г ов Я. И. Фауна моллюсков и зоогеографическое районирование континентальных водоемов земного шара. Л., 1970.
Степанов Ю.С. Семиотика. М., 1971.
Страхов Н. Н. О методе естественных наук и значении их в общем образовании. Спб., 1865.
Стрельников И. Д. Анатомо-физиологические основы видообразования позвоночных. Л., 1670.
Стрельников И. Д. Закон взаимосвязи видообразования животных с величиной их тела, мозга и энергией физиологических процессов 2. О возможности по одному из известных свойств и функций анатомо-физиологических взаимосвязей организма млекопитающих и птиц определять и предсказывать другие неизвестные//3оол.журн. 1973. Т. 52, № 3.
Та лиев В. И. Основа ботаники в эволюционном изложении. М.; Л., 1931.
Тамарин П. В. О противоположном характере результатов, получаемых с помощью весового и адансоновского принципов в таксономии//Журн. биол. 1971. Т. 32, № 4.
Тарасов К. Е., Черненко Е. К. Социальная детерминированность биологии человека. М., 1979.
Тахтаджян А.Л. О принципах, методах и символах филогенетических построений в ботанике//Бюлл. МОИП, отд. биол. 1947. № 52, № 5.
Тахтаджян А.Л. Растения в системе организмов//Жизнь растений. Т. 1. М., 1974.
Тахтаджян А. Л. Флористические области Земли. Л., 1978.
Терентьев П.В. В защиту вида как конечного таксона//3о-ол. журн. 1969. Т. 47, № 6.
Термодинамика биологических процессов. М., 1976.
Теряев А.М. Начальные основания ботанической философии. Спб., 1809.
Тимирязев К. А. Сочинения. Т. 5. М., 1938.
Тимофеев-Ресовский Н. В. Структурные уровни биологических систем//Системные иссл. М., 1970.
202
Тимофеев-Ресовский Н.В., Иванов В. И. Некоторые вопросы феногенетики//Актуальные вопросы совр. генетики. М., 1965.
Тимофеев—Ресовский Н.В., Воронцов Н. Н., Яблоков А. В. Краткий очерк теории эволюции. М., 1969.
Тимофеев—Ресовский Н.В., Яблоков А. В., Глотов Н.В. Очерк учения о популяции. М., 1973.
Тихоненко Т. И. О возможности межвидового обмена генетической информацией в биосфере//Вестн. АН СССР. 1979. № 8.
ТЬкин Б. П. Теоретическая биология и творчество Э.С.Бауэра. Л., 1963.
Толмачев А.И. Введение в географию растений. Л., 1974.
Томсон Д.П. Дух науки. М., 1970.
Тюхгин В.С. Отражение, системы, кибернетика. Теория отражения в свете кибернетики и системного подхода. М., 1972.
Уемов А. И. Некоторые тенденции в развитии естественных наук и принципы их классификации//Вопр. философ. 1961. № 8.
Уильямсон М. Анализ биологических популяций. М., 1975.
Уимпенни Р.С. Органическая полярность с экологической и физиологической точки зрения//Усп.совр. биол. 1945. Т. 19, № 2.
Уитроу Дж. Естественная философия времени. М., 1964.
Уиттекер Р. Сообщества и экосистемы. М., 1980.
Умов Н. А. Физико-механическая модель живой материи. Соч. Т. 3. М.» 1916.
Уоддингтон К.Х. Организаторы и гены. М„ 1947.
Уотермен Т. Проблема//Теоретическая и математическая биология. М., 1968.
Украинцев Б.С. Информационная форма причинности//Философские основания естественных наук. М., 1976.
Урманцев Ю. А. Симметрия природы и природа симметрии.
М.» 1974.
Урсул А.д. Отражение и информация. М., 1973.
Ушаков Б.П. Физиология клетки и проблема вида в зоологии// Цитология. 1959. Т. 1, № 5.
У эвелл В. История индуктивных наук от древнейшего до настоящего времени. Спб., 1914. Т. 2.
Фа ус с ек В. А. Сущность жизни//Сущность жизни. Спб., 1903.
Федоров А. А., Пименов М. Г. Современная хемосистематика растений и возможности использования химических данных на уровне вида и внутривидовых таксонов//Совещание по объему вида и внутривидовой систематике. Л., 1967.
Фесенкова Л. В. Методологические аспекты исследования жизни в космосе. М., 1976.
203
Филиппов А. Э. Дискретное видообразование и закон Виллиса//Журн. общ. биол., 1948.
Фишер А. Г. Широтные изменения в органическом разнообра— зии//Океанология. 1964. Т. 4, № 1.
Флейшман Б.С. Сложность, эффективность и осуществимость// Автоматика. 1980. № 4.
Флоренский П.В., Забелин Е.И. Полярная асимметрия планет//Природа. 1971. № 6.
Флоркэн М. Биохимическая эволюция. М., 1947.
Формозов А. Н. Биологические формы животных в аридных, полуаридных областях Средней и Центральной Азии//Вопросы географии. М.; Л., 1956.
Франк Г.М. О путях познания материальной сущности явлений жизни//Взаимодействие методов естественных наук в познании жизни. М., 1976.
Франк Г.М., Энгельгардт В.А. О роли физики и химии в исследовании биологических проблем//Философские проблемы современного естествознания. М., 1959.
Франк-Каменецкий М. Д. Самая главная молекула//Квант. 1983. Т. 25.
Франц П. К обоснованию общей концепции информации//Кибер-нетика и диалектика. М., 1978.
Фроло*в И. Т. Методологические принципы теоретической биологии. М., 1973.
Харвей Д. Научное объяснение в географии. М., 1974.
Харин Н. Н. Математическая логика и теория множеств. Рос-* вузиздат, 1963.
Хахина Л. Н. Проблема симбиогенеза. Историко-критический очерк. Л., 197 9.
Хлебович В. В. Критическая соленость биологических процессов. Л., 1974.
Ходашова К.С. О географических особенностях структуры населения наземных позвоночных животных//Зональные особенности населения наземных животных. М., 1966.
Холодковский Н. А. Аналитическая теория развития (по по-» воду книги Ганса Дриша)//Биологические очерки. М.; Пг., 1923.
Хорев Н. В. Философия как фактор развития науки. М., 1979.
Чернов Ю. И. Природная зональность и животный мир. М., 1975.
Численко Л.И. О структуре таксонов и таксономическом разнообразии//Журн. общ. биол. 1977. Т. 38, № 3.
Численко Л. И. Структура фауны и флоры в связи с размерами организмов. М., 1981.
Чудинов Э. Послесловие//Г^)юнбаум А. Философские проблемы пространства и времени. М., 1969.
204
Шапошников Г.X. Популяция, вид, род как живые системы и их структура у тлей//Тр.Зоол. ин-та АН СССР. 1974, №53.
Шарова И. X. Жизненные формы жужелиц (Coleoptera Carabi-dae ) М., 1981.
Шварц С. С. Единство жизни. Свердловск, 1972.
Шилин К.И. Экологичность японской лирики//Человек и мир в японской культуре. М., 1985.
Шипунов Ф. Я. Организованность биосферы. М., 1980.
Шкловский И.С. Вселенная, жизнь, разум. М., 1973.
Шмальгаузен И.И. Основы сравнительной анатомии. М., 1935.
Шмальгаузен И. И. Организм как целое в индивидуальном и историческом развитии. М.; Л., 1942.
Шмидт В.М. О корреляциях. I. Сущность, онтогенетический и филогенетический аспекты явления биологических корреляций// Вести. ЛГУ. Биол. 1979. № 3.
Шмидт В.М. О корреляциях П. Вопросы классификации биологических корреляций//Вестн. ЛГУ. Биол. 1981 а. Т. 15, № 3.
Шмидт В. М. Современные проблемы сравнительной флористики//Вестн. ЛГУ. Биол. 1981 б. Т. 21, № 4.
Шмидт Г. А. Типы эмбриогенеза и их приспособительное значение. М., 1968.
Шноль С.Э. Физико-химические факторы биологической эволюции. М., 1979.
Шорников Б.С. О некоторых проблемах эволюции и математической биологии//Системность и эволюция. М., 1984.
Шоры г ин А. А. О биоценозах//Бюлл. МОИП. Ога. Биол. 1955. Т. 60, № 6.
Шрейдер Ю. А. Равенство, сходство, порядок. М., 1971.
Шрейдер Ю. А. Теория познания и феномен науки//Гносеоло— гия в системе философского мировоззрения. М., 1983.
Штеренберг М.И. К вопросу о функциональном определении жизни//Вопр. филос. 1967. № 3.
Штофф В. А. Моделирование и философия- М.; Л., 1966.
Шэфер Э. А. Жизнь, ее сущность, происхождение и сохранение. Спб., 1914.
Щедровицкий Г.П. Проблема построения системной теории сложного гпопулятивного* объекта//Системные иссл. М., 1976.
Чепиков М. Г. Современная революция в биологии. М., 1976.
Чермак Н.К. О построении живого вещества. Гипотеза о живых молекул—вихрей Н.Чермака. Спб., 1895.
Цукер кэндл Э., Полинг Л. Молекулы как документы процесса эволюции/ХПроблемы эволюционной и технической био^ химии. М., 1964.
205
Эйхлер В. Некоторые тенденции развития современной эволюционной систематики//Журн. общ. биол. 1981. Т. 42*, № 1.
Экологическая физиология животных. Л., 1979 Ч 1 1981 Ч 2 1982. Ч. 3.
Энгельгардт В. А. О некоторых атрибутах жизни: иерархия, интеграция, узнавание//Современное естествознание и материалистическая диалектика. М., 1977.
Югай Г. А. Философские проблемы теоретической биологии. М., 1976.
Югай Г. А. Общая теория жизни. М., 198 5.
Юргеленас Д. И. Теория отражения и понятие информации// Теория познания и современная наука. М., 1967.
Яблоков А.В. Популяционная морфология как новое направление эволюционно-морфологических и популяционных исследо-ваний//Журн. общ. биол. 1976. Т. 37, № 5.
Яблоков А.В. Фенетика. Эволюция, популяция, признак. М., 1980.
Ян ко в М. Материя и информация. М., 1979.
Яновская С. Количество (в математике)//Философская энциклопедия. Т. 2. М., 1962.
Ярошенко П. Д. Общая биогеография. М., 1975.
Acta Biotheoretica. 1935- T.l.'
A gds s iz L. Essay on classification//Cambr., Mass. 1962 •
Alston R.E. Biochemical system'atics//E volutionary biology. 1967.
N 1.
Anderson S. Patterns of Jdunal evolution//Quart. Rev. Biol. 19’г4-.
T.49,N 4.
Ayala F.J. Biology as an autonomous science//Amer. Sci. 1968.
Vol. 56.
Baer A.S., Hazen W.E., J ameson D.L., S loan W.C. Central concepts of biology. N.Y., 1971.
Baker J.J.W., Allen G.E. A course in biology. 3 ed. Mass., 1979.
Banarescu P. Principiile si metodele zoologiel sistematice. Bu-curesti, 1973 •
Beck W.S. Modern sience and nature of life. N.Y., 1957.
Beckner M. The biological way of thought. N.Y., 1959 -
Berta lanffy L. Theoreticahe Biologie. I. Berlin, 1932.
Bertalanffy L. Modern theories of development; An introduction to theoretical biology. 1933.
Blandino G. Theories on the nature of life. N.Y., 1969. t
Boisacq E .Dictionnaire etymologique de la langue greque. Etudies dans ses rapports avec les autres langes indo - europees. Heidelberg; Paris, 1938.
Bornemisza S.Th. The explanation of life. Zurich, 1948.
Boy#er F.O Size and form in plants with special reference to the primary conducting tracts. London.
206
Воусе A.J. The value of some methods of numerical taxonomy with reference to Hominoid classification//Phenetic and Phylogenetic Classification. Systematics Ass. publ., 1964. Vol. 6.
Bremekamp C.E.B. The various aspects dL biology. Verh. Konkicle bfeder. acad. wetens. Afd. nat.( 1962. Reeks 2, Deel 54, no 2, Amsterdam.
Bretsky S.S. Recognition of ancestor -descendsnt relationships in invertebrate paleontology//Phylogenetic analysis and paleontology. 1979-
Briggs J'.S. Marine zoogeography. N.Y.
Buck R .C., Hull D.L. The logical structure of the Lhmean hierarhy// Syst. Zool. 1966. Vol. 15.
Buff aloe N.D. Principles of biology. New Delhi, 1964.
В uzas M.A. Patterns of apecies diversity and their explanation// Taxon. 1972. Vol. 21.
Cain A.J. Deductive and inductive methods in post - Linnean taxo-nomy//Proc. Linn. Soc. London, 1959.
Cain S.A. Foundations of plant geography. N.Y., 1944.
Cain S.A. Life - forms and phytoclimate//Bot. Rev. 1950. Vol. 16, N 1.
Cain A.J. The assessment of new types of character in taxonomy// Chemotaxonomy and Serotaxonomy. The Systematics Ass. 1968. Spec. Vol. 2»
Cain A.J. , Harrison G.A. Phyletic weighting//Proc. Zool. Soc.
L. I960. Vol. 135.
Cairns - Smith A.C. Beginnings of organic evolution//Study Time, vol. 4, pap. 4 th.Conf.Int.Soc.Study Times, Alpbach, 1979- N.Y.,, 1981, p. 15-33.
C a m p W.H., G i 11 у C.L. The structure and origin of species//Brit-tonia. 1943. Vol. 4-
Cavalli - Sforza L.L. Bull. Intern. Statist. Inst., 1965, 41, 2, p. 803-807.
C lifford H.T., Stephenson W. In introduction to numerical clas-sificatio. N.Y.; San Franc.; London, 1975.
Commoner B. In defence of biology; the integrity of biology must be properly applied to the problems of life//Science. 1961, N 133-
C on trand iopolus J. Quelques reflexions a propos de la vicarian
ce//C.R. Sec. Biogeogr. 1981. T.57, N 5-
Cracraft J. Phylogenetic Analysis. Evolutionary models and Paleontology. N.Y., 1979.
Crowson R.A. Classi fication and biology. L., 1971-
Cuvier G. Lemons d'anatomie comparee de G.Cuvier, recueillies et publiees sous les yeux par C.Dumeril, etc. T. I, 1800, Paris.
C u v ie r G. Le regne animal distribue d'apres son organisation. T .1.
P., 1817.
Daly H.V. Phenetic classification and typology//Syst. Zool. 1961 -Vol. 10, N 4.
Danchin A. Thimes de la biologie: theories instructives et theories
selectives. I.Theories instructives// Rev. quest, sci. 1979. Vol. 150, . N 2.
Dansereau P. Biogeography: an ecological perspective. N.Y., 1957.
Darlington P.J., Jr. Moderr taxon -ny, reality and usefulness//Syst. Zool. 1971. Vol. 20, N 3.
Davenport Ch.B. The mechanism of organic evolution// J. Wash. Acad. Sci. 1930. Vol. 20, N 14.
Delevoryas Th. The role of paleobotany in vascular plant classification/ Phenetic and phylogenetic classification//Systematics Ass. 1964, N 6.
Doff in H. Le roman de la molecule. L'explication de la vie. Saint-Germain-en-Laye. 1953.
Dubois A., G iintherR. Klepton and Synklepton: two new evolutionary systematics category in zoology// Zool. Jb.Syst. 1982. Vol. 109-
Ehrenberg R. Theoretische Biologie vom Standpunkt der Irreversibi-litat des elementaren Lebensvorganges. Berlin, 1923.
Ekman S. Zoogeography of the sea. L., 1935.
Eldridge N., CracraftJ. Phylogenetic patterns and the evolutionary process. N.Y., 1980.
E llegard A. Darwin and the general reader. The reception of Darwin's theory of evolution in the British periodical press. 1859-1872, Geteborg.
E 1 s as s er W.M. The physical foundation of biology. An analytical study. L., 1S66.
E Isasser W.M. Atom and organism. A new approach to theoretical • biology. Princeton.
E Isasser W.M. What is theoretical biology?//Theoretical physics and biology. Amsterdam; London. 1969.
Elsasser W.M. The chief abstractions of biology. N.Y., 1 975.
E Isasser W.M. Principles of a new biological theory: a summary // J.The or. Biol. 1981. Vol. 89.
Fischer von G. Das Energie-Informations-Kontinuum und eine These zur Evolution//Zool.Jb.Anat. 1 9 8 1 - a. Bd. 1 05. H.4 .
F ischer von G. Zur axiornatischen Begriindung der Biologie// Zool. Jb. Anat. 1981b. Vol. 105. H.3.
Flew A.G JN. The structure of darwinism//New Biol. 1959, Vol.28«
F r e 1 i n Ch., V ullenmier F. Biochemical methods and reasoning in systematics// Ztschr. Zool. Syst. Evolutionsforsch. 1979<,Bd. 17, N 1.
Geert J.M. de Klerk. Mechanism and vitalism. A history of the contro-versy//Acta .biothecretica. 1979. Vol. 28. H. 1.
Gisin H. Discussion//Principles and conception os Systematic biology. Systematic Biol. Proc. Int. Conf. Nat. Acad. Sci., 1969. Publ. 1962.
Gordon I. Spelaeogriphus, a new cavernicolous crustacean from
South Africa//BulLBrit.Mus. (Nat.Hist.) Zool. 1957.Vol.5, N 2.
Griffits G.C.D. On the foundations of biological systematics//Acta Biotheoretica. 1974. Vol. 23.
Grimstone A.V. Cytology, homology and phylogeny * a note on "organic design"//Amer. Naturalist. 1959. Vol. 93.
Haas J. An der Basis des Lebens. Berlin, 1964-
Haeckel E. Generelle Morphologic der Organismen. Allgemeine grun-dziige der organischen Formenwissenschaft, mechanisch begriin-det dutch die von Ch.Darwin reformierte Descendentheorie. I.
1866 • Berlin.
Haeckel E. Systematische Phylogenie der Protisten und Pflanzen.I. Berlin, 1984.
Hansen H. Life - forms as age indicators. Riuckjbing, 1956.
Hein H. Molecular biology vs organicism//Synthese. 1969. Vol. 20. 254.
Hennig W. Phylogenetic systematics. Urbane, 1966-
HillL., Bellanyl., Chester J. Integrated biology. L.,1057-
Howatrh F.G.The zoogeography of specialized cave animals; a bioclimatic model//Evolution. 1980. Vol.34, N 2.
Hubalek Z . Coefficient of association and similarity, based on binary (presence-absence) data: an evAlution//Biol. Rev. 1982. Vol. 57.
Hunziker J.H. Molecular data in plant systematics//Systematic Biol. Proc. Int. Conf .Nat. Acad. Sci. 1969 (Publ. 1962).
Huxley J.S. Problems of relative growth. L., 1932-
Huxley J.S. Toward a new systematics//The New Systematics. Oxford, 1940.
Huxley J.S. Relative growth and form transformation. A discussion on the measurement of growth and form// Pr. Roy. Soc. B. 1950. Vol. 137, N 889.
Huxley J.S. Evolutionary processes and taxonomy with special reference to grades. Uppsala, 1958. Arsskr., 1958/
Inger R.F. Reply to Sokal//Evolution. 1959, N 13.
Japp F.K. Stereochemistry and vitalism//Nature. 1898- Bd. 58-Jardine N., Jardine C. Numerical homology//Nature. 1967. Bd.’216. J ohannsen W. Elemente der exacten Erblichkeitslehre. Jena, 1909.
Jordan P. La physique et le secret de la vie organique. P., 1959.
Jouckers L.H.M. The concept of population in biology//Acta Biotheoretica. 1973- Vol. 22, N 2-
Ke men у J. A philosopher looks at science. New York; London, Toronto, 1959.
Lagers petz K. Teological explanations and terms in biology// Ann. Zool. Soc. Vanamo. 1959. Vol. 19, N 6.
Lataste F. Qu'est - ce que Petre vivant, Definition nouvelle// Compt. Rend. Sci. Soc. Roy., 1889, 5» 1889.
Laughlin R.A. Feeding habits of the blue crab, Callinectes sapi-dus Rathbun, in the Apalachicola estuary Florida//Bull. Mar. Sci.
209
1982. Vol. 32, N 4.
Leuschner D. Einfiihrung in die numerische Taxonomie. J., 1974.
Lillie R.S. The living and non living/ZAmer. Naturalist. 1934- Vol.68
Liddell H.G., S с о 11 R. A Greek - English lexicon. Oxford. 1 927.
Pt.3. 1930. Pt.5.
L other R. Die Beherrschung der Mannigfaltigleit. Philosophische Grundlagen der Taxonomie. Jena, 1972-
MacArthur R.H. Geographical ecology; patterns in the distribution of species. 1972. N.Y.
Macklin M. Theoretical biology. A statement and defenceZZSynthese. 1969. Vol. 20.
Mayr S. Origin and history of some terms in systematic and evolutionary biology//Syst. Zool. 1978. Vol. 27, N 1.
McE Iroy W.B. Energy and life//Frentiers of modern biology. Boston, 1962.
McMahon Th. Size and shape in biologyZZScience. 1973. Vol. 179, N 4079.
Medewar P.B. Size, shape and ageZZEssays on growth and Form presented to d'Arcy W.Thompson. Oxford, 1945.
Meyer - Abich A. Logik der Morphologie. Berlin, 1926.
Meyer - Abich A. Geistesgeschichtliche Grundlangen der Biologie. Stuttgart, 1963.
Michener Ch.D. Diverse approaches to systematics/ZE volutionary biology. 1970. Vol. 4.
Miksche J.P. Infraspecific variation of DNA per cell between Picea sitchensis (Bong). Carr. ProvenancesZZ Chromosoma. 1971. Vol.32, N 4.
Miksche J. P., Hotta Y. DNA base composition and repetitions DNA in several conifersZZChromosoma. 1973 .Vol. 41, N 1.
MinkoffE.C. Evolutionary biology. Massachusets, 1983•
Moment G.B. Biological sciences todayZ/Frontiers of modern biology.
Boston, 1962.
Muzzier H.J. LifeZZScience. 1955. Vol. 121, N 3132.
Needham J. Order and life. Yale Univ. Press., 1936.
N ordenskiold E. The history of biology. N.Y., 1928.
Novikoff A.B. The concept of integrative levels and biologyZZ Interrelations: the biological and physical sciences. Chicago, 1969.
N oiiy du, L. Le temps et la vie. Dijon, 1936.
Owen R. Principes d'osteologie comparee ou recherches sur 1'arche-type. P., 1855.
P i e 1 о u E .C. Biogeography. N.Y., etc. 1979-
Pirie N.W. On making and recognizing lifeZZNew Biol. 1954.
Polanyi M. Life's irreducible structure. Life mechanisms end information in DNA are boundary conditions with a sequence of boundaries above them// Science. 1968. Vol. 160.
P r e 11 w i t z W. E tymologisches Worterbuch der Griechischen Sprache. Gottingen, 1892.
Progress in theoretical biology. N.Y., 1967. Vol. 2.
R as hew s к у N. Mathematical foundations of general biology//Ann.
N.Y. Acad. Sci. 1962- Vol. 96, N 4<
Rasmussen D.I. Molecular taxonomy and typology/ZBioscience’.l969.
Vol. 19, N 5.
Raunkiaer C. The life form of plants and statistical plant geography. Oxford, 1934.
Reed E.S. Darwin's evolutionary philosophy:the laws of change//Acta biotheoretica. 1978. Vol. 27, N 3/4.
Reinke J. Einleitung in die theoretische Biologiel. Berlin, 1901.
Remane A. Morphologic und Homologieuforschung//Verh. Dtsch. Zool.
Ges. (1954), 1955. Suppl. Bd., N 18.
R itter W.E. The unity of the organism or the organismal conception of life. Boston, 1919.
Roll-HansenN. On the reduction of biology to physical science// Synthese. 1969 .Vol. 20.
Sastere Cl. La vicariance. Compt. rend. Sec. Biogeogr. 1981. Vol. 57, N 4.
Savory Th.H. The principles of mechanistic biology. Merrow, 1971.
Schaxel J. Grundziige der Theorien bildung in der Biologie. Jena, 1922.
Schmidt-Nielsen K. Scaling; why is animal size so important? Cambr., 1984.
Schoute J.C. Biomorphology in general//Verh. Kon. Nederl. Akad.
Wetensh. Afd. Natuur. 1949. Ser.2, Deel 46, N 1.
Sharma A.K. Plant cytogenetics//The cell and nucleus. 1974. Vol.2.
Silvestri L.G., Hill L.R. Some problems of the taxonomic approach// Phenetic and Phylogenetic Classification. Systematic Ass. 1964. Publ. N 6.
Simpson G.G. Principles of animal taxonomy. N.Y., 1961.
Sinnott E.W. The problem of organic form . L., 1963.
Smart J.J.C. Phylosophy and scientific realism. L., 1963.
Sneath P.H.A. Numerical taxonomy study of the graft chimaere Laburnocytiscus adamii (Cytisus purpureus Laburnum anagyroides)// Proc. Linn. Soc. London. 1968. Vol. 179.
Sneath P.H.A. Recent trends in numerical taxonomy//Taxont 1969.
Vol. 18.
Sokal R. Typology and empiricism in taxonomy//!.Theor. Biol. 1962. Vol .3.
Sokal R., Michener C.D. The effect of different numerical techniques on the phenetic classification of bees of the Hoplitis complex (Megachilidae)// Proc. Linn. Soc. L. 1967. Vol. 178.
Sokal R., Sneath P.H.A. Principles of numerical taxonomy’. San Francisco, 1963-
Solbrig O.T., Solbrig D.J. Introduction to population biology and evolution. Massachusets, 1979.
Starr M.P., Heise H. Discussion//; The construction of a classifi
cation//Systematic Biology .Proc. Int. Conf. Nat. Acad. Sci. 1969.
. Publ. 1692).
Steen van der, W.J. Review of M.A .Simon: The matter of life //Acta biotheoretica .1973. Vol. 2^. N 3-
Stephen W.P. Hemolymph proteins and their use in taxonomic studies// Proc. X Intern .Conf. Entomol .Montreal. 1956, N 1.
The Encyclopedia of biological sciences. N.Y., et al. 1970.
The Oxford English Dictionary. Oxford. 1932- Vol. 3. " V-Z" .
Thorson G. Parallel level - bottom communities, their temperature adaptation, and their " balance" between predators.and foodx animals//Adv. marine biol. 1956.
T schulok S. Das System der Biologie in Forschung und LehreKJena.
Thompson d'Arcy W. Essays on growth and form. Cambr., 1945.
T urrill W.B. Plant geography//Rollins R.C., Taylor G.(ed.); Vistas in botany., L., 1959.
U exkull J. Theoretische Biologie. Berlin, 1928.
VegotskyA., Fox S.W. Protein molecules: intraspecific and interspe-cific variations//Comparative Biochemistry. 1962. Vol. 4. B.
V e r h о о g H. Social and ethical aspects of biology part of theoretical biology? //Acta biotheoretica. 1975* Ser. A. Vol. 24, N 1/2.
Waddington C.H. The character of biological form//Aspects of
Form. Ed. L.L.Whyte, 1951.
Waddington C.H. The strategy of genes. L., 1957.
W aid G. Theories of the origin of life//Frontiers of modern biology Boston, 1962.
Went E.W. Parallel evolution//Taxon. 1971. Vol. 20, N 313.
White M.J.D. Animal cytology and evolution. Cambridge, 1973.
Whittaker R.H. Evolution and measurement of species diversity// Taxon, 1972. Vol. 21.
Whyte L.L. The unitary principle in physics and biology. 1949.
W illiams C.B. Patterns in the balance of nature and related problems in quantitative ecology. L.; N.Y., 1964.
W ill is J.C. The course of evolution by differentation or divergent mutation rather than selection. Cambridge, 1940.
Wood ger J.H. Biological principles. A critical study. L„; N.Y., 1929.
Wood ger J.H. The axiomatic method in biology. Cambridge. 1937.
W oodger J.H. On biological transformations//Essays on Growth and Form presented to d'Arcy W.Thompson. Oxford, 1945-'
W oolhouse H.W. Environment and enzyme evolution in plants// Phytochemical phylogeny. L.; N.Y., 1970.
Wright S. The " age and area" concept extended//Rev. Ecol. 1941.
Vol. 22.
Zoologische Worterbuch der zoologische Fachausdriicke. Jena, 1927.
Z uckerkandal P., Poul ing L. Evolutionary divergence and con* vergense in proteins// Evolving genes and proteins. N.Y., 1965.
СОДЕРЖАНИЕ
Предисловие .......................................... 3
Глава 1. О предмете теоретической биологии ........... 8
Глава 2, Три "образа" биологии....................... 24
Глава 3. Уровни организации материи и теория биологии 35
Глава 4. Биология и физикализм....................... 48
Глава 5. Логическая структура биологического знания 59
Глава 6. Традиционная таксономическая неопределенность 70
Глава 7. Отношение сходства и фенетическая (феноничес-кая) систематика.......................................84
Глава 8. Понятие времени в биологии.................96
Глава 9. Отношение исторического родства, филогенез, эволюция..............................................104
Глава 10. Отношение сосуществования и экология . . . .118
Глава 11. Биогеография, фаунистика и флористика и понятие биологического пространства.....................135
Глава 12. Отношение корреляции и сравнительная морфо-физиология (сравнительная анатомия)..................151
Глава 13. Некоторые прикладные вопросы теории биологии ..................................................168
Заключение...........................................179
Литература...........................................187
Научное издание
Заренков Николай Алексеевич
ТЕОРЕТИЧЕСКАЯ БИОЛОГИЯ
Зав. редакцией Н.М. Глазкова
Редактор Г.Г.Есакова
Художественный редактор М.Ф.Евстафиева
Обложка художника Е. Н.В о л к о в а
-Технический редактор Н.И.С ми рнова
Корректоры Л.А.А йд а рб е к о ва, М.А.М еренкова
ИБ № 3045
Сдано в набор 02.03.88. Подписано в печать 25.08.88.
Л-35721. Формат 60x90/16. Бумага офсет. № 2.
Гарнитура литературная. Офсетная печать.
У с л. печ. л. 13,5. Уч.-изд. л. 14,47.
Тираж 3460 экз. Заказ № 1560. Изд. № 4323.
Пена 1 р. 40 коп.
Ордена *3нак Почета* издательство Московского университет.-103009, Москва, ул. Герцена, 5/7.
Типография ордена *3нак Почета* изд-ва МГУ.
119899, Москва, Ленинские горы
УВАЖАЕМЫЕ ЧИТАТЕЛИ!
Ознакомитьс университета и в магазинах - :я с планами Издательства Московского оформить предварительный заказ можно опорных пунктах по изучению спроса;
в Москве - магазин № 110 *Университетская книжная лавка* ( 117296, Москва, Ломоносовский просп., 18);
в Киеве - магазин №12 *Книги* (252001, Киев, Крешатик, 44);
в Новосибирске -» магазин №2 Академгородка (630090, Новосибирск, ул. Ильиче^ 6).
Жители других тельный заказ Магазин № ул. Кржижановс казы и высыла< университета м городов также могут оформить предвари-в местных книжных магазинах. 93 *Книга - почтой* ( 117168, Москва, :кого, 14) принимает предварительные за-эт литературу Издательства Московского аложенным платежом.